 Fig. 305—Head of a shark (Scyllium), from the ventral side. m mouth, o olfactory pits, r nasal groove, n nasal fold in natural position, n′ nasal fold drawn up. (The dots are openings of the mucous canals.) (From Gegenbaur.)
Fig. 305—Head of a shark (Scyllium), from the ventral side. m mouth, o olfactory pits, r nasal groove, n nasal fold in natural position, n′ nasal fold drawn up. (The dots are openings of the mucous canals.) (From Gegenbaur.)The sense-organs are indubitably among the most important and interesting parts of the human body; they are the organs by means of which we obtain our knowledge of objects in the surrounding world. Nihil est in intellectu quod non prius fuerit in sensu. They are the first sources of the life of the soul. There is no other part of the body in which we discover such elaborate anatomical structures, co-operating with a definite purpose; and there is no other organ in which the wonderful and purposive structure seems so clearly to compel us to admit a Creator and a preconceived plan. Hence we find special efforts made by dualists to draw our attention here to the “wisdom of the Creator” and the design visible in his works. As a matter of fact, you will discover, on mature reflection, that on this theory the Creator is at bottom only playing the part of a clever mechanic or watch-maker; all these familiar teleological ideas of Creator and creation are based, in the long run, on a similar childlike anthropomorphism.
However, we must grant that at the first glance the teleological theory seems to give the simplest and most satisfactory explanation of these purposive structures. If we merely examine the structure and functions of the most advanced sense-organs, it seems impossible to explain them without postulating a creative act. Yet evolution shows us quite clearly that this popular idea is totally wrong. With its assistance we discover that the purposive and remarkable sense-organs were developed, like all other organs, without any preconceived design—developed by the same mechanical process of natural selection, the same constant correlation of adaptation and heredity, by which the other purposive structures in the animal frame were slowly and gradually brought forth in the struggle for life.
Like most other Vertebrates, man has six sensory organs, which serve for eight
different classes of sensations. The skin serves for sensations of pressure and temperature. This is the oldest, lowest, and vaguest of the sense-organs; it is distributed over the surface of the body. The other sensory activities are localised. The sexual sense is bound up with the skin of the external sexual organs, the sense of taste with the mucous lining of the mouth (tongue and palate), and the sense of smell with the mucous lining of the nasal cavity. For the two most advanced and most highly differentiated sensory functions there are special and very elaborate mechanical structures—the eye for the sense of sight, and the ear for the sense of hearing and space (equilibrium).
Comparative anatomy and physiology teach us that there are no differentiated sense-organs in the lower animals; all their sensations are received by the surface of the skin. The undifferentiated skin-layer or ectoderm of the Gastræa is the simple stratum of cells from which the differentiated sense-organs of all the Metazoa (including the Vertebrates) have been evolved. Starting from the assumption that necessarily only the superficial parts of the body, which are in direct touch with the outer world, could be concerned in the origin of sensations, we can see at once that the sense-organs also must have arisen there. This is really the case. The chief part of all the sense-organs originates from the skin-sense layer, partly directly from the horny plate, partly from the brain, the foremost part, of the medullary tube, after it has separated from the horny plate. If we compare the embryonic development of the various sense-organs, we see that they all make their appearance in the simplest conceivable form; the wonderful contrivances that make the higher sense-organs among the most remarkable and elaborate structures in the body develop only gradually. In the phylogenetic explanation of them comparative anatomy and ontogeny achieve their greatest triumphs. But at first all the sense-organs are merely parts of the skin in which sensory nerves expand. These nerves themselves were originally of a homogeneous character. The different functions or specific energies of the differentiated sense-nerves were only gradually developed by division of labour. At the same time, their simple terminal expansions in the skin were converted into extremely complex organs.
The great instructiveness of these historical facts in connection with the life of the soul is not difficult to see. The whole philosophy of the future will be transformed as soon as psychology takes cognisance of these genetic phenomena and makes them the basis of its speculations. When we examine impartially the manuals of psychology that have been published by the most distinguished speculative philosophers and are still widely distributed, we are astonished at the naivete with which the authors raise their airy metaphysical speculations, regardless of the momentous embryological facts that completely refute them. Yet the science of evolution, in conjunction with the great advance of the comparative anatomy and physiology of the sense-organs, provides the one sound empirical basis of a natural psychology.
|
Fig. 305—Head of a shark (Scyllium), from the ventral side. m mouth, o olfactory pits, r nasal groove, n nasal fold in natural position, n′ nasal fold drawn up. (The dots are openings of the mucous canals.) (From Gegenbaur.) |
In respect of the terminal expansions of the sensory nerves, we can distribute the human sense-organs in three groups, which correspond to three stages of development. The first group comprises those organs the nerves of which spread out quite simply in the free surface of the skin itself (organs of the sense of pressure, warmth, and sex). In the second group the nerves spread out in the mucous coat of cavities which are at first depressions in or invaginations of the skin (organs of the sense of smell and taste). The third group is formed of the very elaborate organs, the nerves of which spread out in an internal vesicle, separated from the skin (organs of the sense of sight, hearing, and space).
There is little to be said of the development of the lower sense-organs. We
have already considered (p. 268) the organ of touch and temperature in the skin. I need only add that in the corium of man and all the higher Vertebrates countless microscopic sense-organs develop, but the precise relation of these to the sensations of pressure or resistance, of warmth and cold, has not yet been explained. Organs of this kind, in or on which sensory cutaneous nerves terminate, are the “tactile corpuscles” (or the Pacinian corpuscles) and end-bulbs. We find similar corpuscles in the organs of the sexual sense, the male penis and the female clitoris; they are processes of the skin, the development of which we will consider later (together with the rest of the sexual parts, Chapter XXIX). The evolution of the organ of taste, the tongue and palate, will also be treated later, together with that of the alimentary canal to which these parts belong (Chapter XXVII). I will only point out for the present that the mucous coat of the tongue and palate, in which the gustatory nerve ends, originates from a part of the outer skin. As we have seen, the whole of the mouth-cavity is formed, not as a part of the gut-tube proper, but as a pit-like fold in the outer skin (p. 139). Its mucous lining is therefore formed, not from the visceral, but from the cutaneous layer, and the taste-cells at the surface of the tongue and palate are not products of the gut-fibre layer, but of the skin-sense layer.
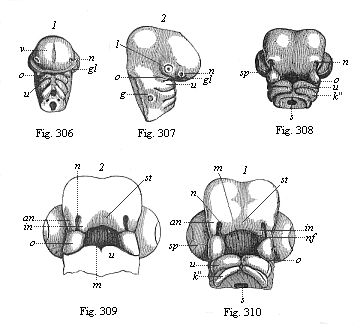 Fig. 306 and 307—Head of a chick embryo, three days old: 2.306 front view, 2.307 from the right. n rudimentary nose (olfactory pits), l rudimentary eyes (optic pits), g rudimentary ear (auscultory pit), v fore brain, gl eye-cleft, o process of upper jaw, u process of lower jaw of the first gill-arch.
Fig. 306 and 307—Head of a chick embryo, three days old: 2.306 front view, 2.307 from the right. n rudimentary nose (olfactory pits), l rudimentary eyes (optic pits), g rudimentary ear (auscultory pit), v fore brain, gl eye-cleft, o process of upper jaw, u process of lower jaw of the first gill-arch.Fig. 308—Head of a chick embryo, four days old, from below. n nasal pit, o upper-jaw process of the first gill-arch, u lower-jaw process of same, k″ second gill-arch, sp choroid fissure of eye, s gullet. Fig. 309 and 310—Heads of chick embryos: 309 from the end of the fourth, 310 from the beginning of the fifth week. Letters as in Fig. 308, except: in inner, an outer, nasal process, nf nasal furrow, st frontal process, m mouth. (From Kölliker.). |
This applies also to the mucous lining of the olfactory organ, the nose. However, the development of this organ is much more interesting. Although the nose seems superficially to be simple and single, it really consists, in man and all
other Gnathostomes, of two completely separated halves, the right and left cavities. They are divided by a vertical partition, so that the right nostril leads into the right cavity alone and the left nostril into the left cavity. They open internally (and separately) by the posterior nasal apertures into the pharynx, so that we can get direct into the gullet through the nasal passages without touching the mouth. This is the way the air usually passes in respiration; the mouth being closed, it goes through the nose into the gullet, and through the larynx and bronchial tubes into the lungs. The nasal cavities are separated from the mouth by the horizontal bony palate, to which is attached behind (as a dependent process) the soft palate with the uvula. In the upper and hinder parts of the nasal cavities the olfactory nerve, the first pair of cerebral nerves, expands in the mucous coat which clothes them. The terminal branches of it spread partly over the septum (partition), partly on the side walls of the internal cavities, to which are attached the turbinated bones. These bones are much more developed in many of the higher mammals than in man, but there are three of them in all mammals. The sensation of smell arises by the passage of a current of air containing odorous matter over the mucous lining of the cavities, and stimulating the olfactory cells of the nerve-endings.
Man has all the features which distinguish the olfactory organ of the mammals from that of the lower Vertebrates. In all essential points the human nose entirely resembles that of the Catarrhine apes, some of which have quite a human external nose (compare the face of the long-nosed apes). However, the first structure of the olfactory organ in the human embryo gives no indication of the future ample proportions of our catarrhine nose. It has the form in which we find it permanently in the fishes—a couple of simple depressions in the skin at the outer surface of the head. We find these blind olfactory pits in all the fishes; sometimes they lie near the eyes, sometimes more forward at the point of the muzzle, sometimes lower down, near the mouth (Fig. 249).
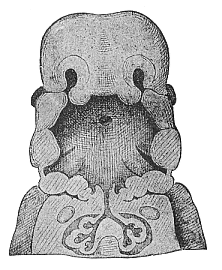 Fig. 311—Frontal section of the mouth and throat of a human embryo, neck half-inch long. “Invented” by Wilhelm His. The vertical section (in the frontal plane, from left to right) is so constructed that we see the nasal pits in the upper third of the figure and the eyes at the sides: in the middle third the primitive gullet with the gill-clefts (gill-arches in section); in the lower third the pectoral cavity with the bronchial tubes and the rudimentary lungs.
Fig. 311—Frontal section of the mouth and throat of a human embryo, neck half-inch long. “Invented” by Wilhelm His. The vertical section (in the frontal plane, from left to right) is so constructed that we see the nasal pits in the upper third of the figure and the eyes at the sides: in the middle third the primitive gullet with the gill-clefts (gill-arches in section); in the lower third the pectoral cavity with the bronchial tubes and the rudimentary lungs. |
This first rudimentary structure of the double nose is the same in all the Gnathostomes; it has no connection with the primitive mouth. But even in a section of the fishes a connection of this kind begins to make its appearance, a furrow in the surface of the skin running from each side of the nasal pit to the nearest corner of the mouth. This furrow, the nasal groove or furrow (Fig. 305 r), is very important. In many of the sharks, such as the Scyllium, a special process of the frontal skin, the nasal fold or internal nasal process, is formed internally over the groove (n, n″). In contrast to this the outer edge of the furrow rises in an “external nasal process.” As the two processes meet and coalesce over the nasal groove in the Dipneusts and Amphibia, it is converted into a canal, the nasal canal. Henceforth we can penetrate from the external pits through the nasal canals direct into the mouth, which has been formed quite independently. In the Dipneusts and the lower Amphibia the internal aperture of the nasal canals lies in front (behind the lips); in the higher Amphibia it is right behind. Finally, in the three higher classes of Vertebrates the primary mouth-cavity is divided by the formation of the horizontal
palate-roof into two distinct cavities—the upper (secondary) nasal cavity and the lower (secondary) mouth-cavity. The nasal cavity in turn is divided by the construction of the vertical septum into two halves—right and left.
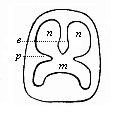 Fig. 312—Diagrammatic section of the mouth-nose cavity. While the palate-plates (p) divide the original mouth-cavity into the lower secondary mouth (m) and the upper nasal cavity, the latter in turn is divided by the vertical partition (e) into two halves (n, n). (From Gegenbaur.)
Fig. 312—Diagrammatic section of the mouth-nose cavity. While the palate-plates (p) divide the original mouth-cavity into the lower secondary mouth (m) and the upper nasal cavity, the latter in turn is divided by the vertical partition (e) into two halves (n, n). (From Gegenbaur.) |
Comparative anatomy shows us to-day, in the series of the double-nosed Vertebrates, from the fishes up to man, all the different stages in the development of the nose, which the advanced olfactory organ of the higher mammals has passed through at various periods in the course of its phylogeny. It first appears in the embryo of man and the higher Vertebrates, in which the double fish-nose persists throughout life. At an early stage, before there is any trace of the characteristic human face, a pair of small pits are formed in the head over the original mouth-cavity; these were first discovered by Baer, and rightly called the “olfactory pits” (Figs. 306 n, 307 n). These primitive nasal pits are quite separate from the rudimentary mouth, which also originates as a pit-like depression in the skin, in front of the blind fore end of the gut. Both the pair of nasal pits and the single mouth-pit (Fig. 310 m) are clothed with the horny plate. The original separation of the former from the latter is, however, presently abolished, a process forming above the mouth-pit—the “frontal process” (Fig. 309 st). Its outer edge rises to the right and left in the shape of two lateral processes; these are the inner nasal processes or folds (in). Opposite to these a parallel ridge is formed on either side between the eye and the nasal pit; these are the outer nasal processes (an). Thus between the inner and outer nasal processes a groove-like depression is formed on either side, which leads from the nasal pit towards the mouth-pit (m); this groove is, as the reader will guess, the same nasal furrow or groove that we have already seen in the shark (Fig. 305 r). As the parallel edges of the inner and outer nasal processes bend towards each other and join above the nasal groove, this is converted into a tube, the primitive nasal canal. Hence the nose of man and all the other Amniotes consists at this embryonic stage of a couple of narrow tubes, the nasal canals, which lead from the outer surface of the forehead into the rudimentary mouth. This transitory condition resembles that in which we find the nose permanently in the Dipneusts and Amphibia.
A cone-shaped structure, which grows from below towards the lower ends of the two nasal processes and joins with them, plays an important part in the conversion of the open nasal groove into the closed canal. This is the upper-jaw process (Figs. 306–310 o). Below the mouth-pit are the gill-arches, which are separated by the gill-clefts. The first of these gill-arches, and the most important for our purpose, which we may call the maxillary (jaw) arch, forms the skeleton of the jaws. Above at the basis a small process grows out of this first gill-arch; this is the upper-jaw process. The first gill-arch itself develops a cartilage at one of its inner sides, the “Meckel cartilage” (named after its discoverer), on the outer surface of which the lower jaw is formed (Figs. 306–310 u). The upper-jaw process forms the chief part of the skeleton of that jaw, the palate bone, and the pterygoid bone. On its outer side is afterwards formed the upper-jaw bone, in the narrower sense, while the middle part of the skeleton of the upper jaw, the intermaxillary, develops from the foremost part of the frontal process.
The two upper-jaw processes are of great importance in the further development of the face. From them is formed, growing into the primitive mouth-cavity, the important horizontal partition (the palate) that divides the former into two distinct cavities. The upper cavity, into which the nasal canals open, now develops into the nasal cavity, the air-passage and the organ of smell. The lower cavity forms the permanent secondary mouth (Fig. 312 m), the food-passage and the organ of taste. Both the upper and lower cavities open behind into the gullet (pharynx). The hard
palate that separates them is formed by the joining of two lateral halves, the horizontal plates of the two upper-jaw processes, or the palate-plates (p). When these do not, sometimes, completely join in the middle, a longitudinal cleft remains, through which we can penetrate from the mouth straight into the nasal cavity. This is the malformation known as “wolf’s throat.” “Hare-lip” is the lesser form of the same defect. At the same time as the horizontal partition of the hard palate a vertical partition is formed by which the single nasal cavity is divided into two sections—a right and left half (Fig. 312 n, n).
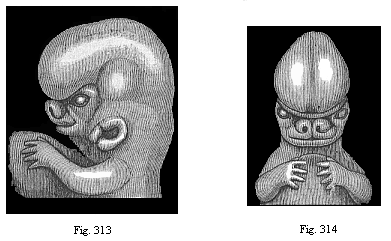 Figs. 313 and 314—Upper part of the body of a human embryo, two-thirds of an inch long, of the sixth week; Fig. 313 from the left, Fig. 314 from the front. The origin of the nose and the upper lip from two lateral and originally separate halves can be clearly seen. Nose and upper lip are large in proportion to the rest of the face, and especially to the lower lip. (From Kollmann.)
Figs. 313 and 314—Upper part of the body of a human embryo, two-thirds of an inch long, of the sixth week; Fig. 313 from the left, Fig. 314 from the front. The origin of the nose and the upper lip from two lateral and originally separate halves can be clearly seen. Nose and upper lip are large in proportion to the rest of the face, and especially to the lower lip. (From Kollmann.) |
The double nose has now acquired the characteristic form that man shares with the other mammals. Its further development is easy to follow; it consists of the formation of the inner and outer processes of the walls of the two cavities. The external nose is not formed until long after all these essential parts of the internal organ of smell. The first traces of it in the human embryo are found about the middle of the second month (Figs. 313–316). As can be seen in any human embryo during the first month, there is at first no trace of the external nose. It only develops afterwards from the foremost nasal part of the primitive skull, growing forwards from behind. The characteristic human nose is formed very late. Much stress is at times laid on this organ as an exclusive privilege of man. But there are apes that have similar noses, such as the long-nosed ape.
The evolution of the eye is not less interesting and instructive than that of the nose. Although this noblest of the sensory organs is one of the most elaborate and purposive on account of its optic perfection and remarkable structure, it nevertheless develops, without preconceived design, from a simple process of the outer germinal layer. The fully-formed human eye is a round capsule, the eye-ball (Fig. 317). This lies in the bony cavity of the skull, surrounded by protective fat and motor muscles. The greater part of it is taken up with a semi-fluid, transparent gelatinous substance, the corpus vitreum. The crystalline lens is fitted into the anterior surface of the ball (Fig. 317 l). It is a lenticular, bi-convex, transparent body, the most important of the refractive media in the eye. Of this group we have, besides the corpus vitreum and the lens, the watery fluid (humor aqueus) that is found in front of the lens (at the letter m in Fig. 317). These three transparent refractive media, by which the rays of light that
enter the eye are broken up and re-focussed, are enclosed in a solid round capsule, composed of several different coats, something like the concentric layers of an onion. The outermost and thickest of these envelopes is the white sclerotic coat of the eye. It consists of tough white connective tissue. In front of the lens a circular, strongly-curved, transparent plate is fitted into the sclerotic, like the glass of a watch—the cornea (b). At its outer surface the cornea is covered with a very thin layer of the epidermis; this is known as the conjunctiva. It goes from the cornea over the inner surface of the eye-lids, the upper and lower folds which we draw over the eye in closing it. At the inner corner of the eye we have a rudimentary organ in the shape of the relic of a third (inner) eye-lid, which is greatly developed, as “nictitating (winking) membrane,” in the lower Vertebrates (p. 5). Underneath the upper eye-lid are the lachrymal glands, the product of which, the lachrymal fluid, keeps the outer surface of the eye smooth and clean.
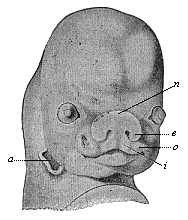 Fig. 315—Face of a human embryo, seven weeks old. (From Kollmann.) Joining of the nasal processes (e outer, i inner) with the upper-jaw process (o), n nasal wall, a ear-opening.
Fig. 315—Face of a human embryo, seven weeks old. (From Kollmann.) Joining of the nasal processes (e outer, i inner) with the upper-jaw process (o), n nasal wall, a ear-opening. |
Immediately under the sclerotic we find a very delicate, dark-red membrane, very rich in blood-vessels—the choroid coat—and inside this the retina (o), the expansion of the optic nerve (i). The latter is the second cerebral nerve. It proceeds from the optic thalami (the second cerebral vesicle) to the eye; penetrates its outer envelopes, and then spreads out like a net between the choroid and the corpus vitreum. Between the retina and the choroid there is a very delicate membrane, which is usually (but wrongly) associated with the latter. This is the black pigment-membrane (n). It consists of a single stratum of graceful, hexagonal, regularly-joined cells, full of granules of black colouring matter. This pigment membrane clothes, not only the inner surface of the choroid proper, but also the hind surface of its anterior muscular continuation, which covers the edge of the lens in front as a circular membrane, and arrests the rays of light at the sides. This is the well-known iris of the eye (h), coloured differently in different individuals (blue, grey, brown, etc.); it forms the anterior border of the choroid. The circular opening that is left in the middle is the pupil, through which the rays of light penetrate into the eye. At the point where the iris leaves the anterior border of the choroid proper the latter is very thick, and forms a delicate crown of folds (g), which surrounds the edge of the lens with about seventy large and many smaller rays (corona ciliaris.)
 Fig. 316—Face of a human embryo, eight weeks old. (From Ecker.)
Fig. 316—Face of a human embryo, eight weeks old. (From Ecker.) |
At a very early stage a couple of pear-shaped vesicles develop from the foremost part of the first cerebral vesicle in the embryo of man and the other Craniotes (Figs. 155 a, 297 au). These growths are the primary optic vesicles. They are at first directed outwards and forwards, but presently grow downward, so that, after the complete separation of the five cerebral vesicles, they lie at the base of the intermediate brain. The inner cavities of these pear-shaped vesicles, which soon attain a considerable size, are openly connected with the ventricle of the intermediate brain by their hollow stems. They are covered externally by the epidermis.
At the point where this comes into direct contact with the most curved part of the primary optic vesicle there is a thickening (l) and also a depression (o) of the horny plate (Fig. 318, I). This pit, which we may call the lens-pit, is converted into a closed sac, the thick-
walled lens-vesicle (2, l), the thick edges of the pit joining together above it. In the same way in which the medullary tube separates from the outer germinal layer, we now see this lens-sac sever itself entirely from the horny plate (h), its source of origin. The hollow of the sac is afterwards filled with the cells of its thick walls, and thus we get the solid crystalline lens. This is, therefore, a purely epidermic structure. Together with the lens the small underlying piece of corium-plate also separates from the skin.
As the lens separates from the corneous plate and grows inwards, it necessarily hollows out the contiguous primary optic vesicle (Fig. 318, 1–3). This is done in just the same way as the invagination of the blastula, which gives rise to the gastrula in the amphioxus (Fig. 38 C–F). In both cases the hollowing of the closed vesicle on one side goes so far that at last the inner, folded part touches the outer, not folded part, and the cavity disappears. As in the gastrula the first part is converted into the entoderm and the latter into the ectoderm, so in the invagination of the primary optic vesicle the retina (r) is formed from the first (inner) part, and the black pigment membrane (u) from the latter (outer, non-invaginated) part. The hollow stem of the primary optic vesicle is converted into the optic nerve. The lens (l), which has so important a part in this process, lies at first directly on the invaginated part, or the retina (r). But they soon separate, a new structure, the corpus vitreum (gl), growing between them. While the lenticular sac is being detached and is causing the invagination of the primary optic vesicle, another invagination is taking place from below; this proceeds from the superficial part of the skin-fibre layer—the corium of the head. Behind and under the lens a last-shaped process rises from the cutis-plate (Fig. 319 g), hollows out the cup-shaped optic vesicle from below, and presses between the lens (l) and the retina (i). In this way the optic vesicle acquires the form of a hood.
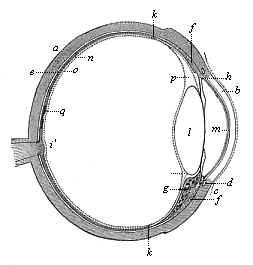 Fig. 317—The human eye in section. a sclerotic coat, b cornea, c conjunctiva, d circular veins of the iris, e choroid coat, f ciliary muscle, g corona ciliaris, h iris, i optic nerve, k anterior border of the retina, l crystalline lens, m inner covering of the cornea (aqueous membrane), n pigment membrane, o retina, p Petit’s canal, q yellow spot of the retina. (From Helmholtz.)
Fig. 317—The human eye in section. a sclerotic coat, b cornea, c conjunctiva, d circular veins of the iris, e choroid coat, f ciliary muscle, g corona ciliaris, h iris, i optic nerve, k anterior border of the retina, l crystalline lens, m inner covering of the cornea (aqueous membrane), n pigment membrane, o retina, p Petit’s canal, q yellow spot of the retina. (From Helmholtz.) |
Finally, a complete fibrous envelope, the fibrous capsule of the eye-ball, is formed about the secondary optic vesicle and its stem (the secondary optic nerve). It originates from the part of the head-plates which immediately encloses the eye. This fibrous envelope takes the form of a closed round vesicle, surrounding the whole of the ball and pushing between the lens and the horny plate at its outer side. The round wall of the capsule soon divides into two different membranes by surface-cleavage. The inner membrane becomes the choroid or vascular coat, and in front the ciliary corona and iris. The outer membrane is converted into the white protective or sclerotic coat—in front, the transparent cornea. The eye is now formed in all its essential parts. The further development—the complicated differentiation and composition of the various parts—is a matter of detail.
The chief point in this remarkable evolution of the eye is the circumstance that the optic nerve, the retina, and the pigment membrane originate really from a part of the brain—an outgrowth of the intermediate brain—while the lens, the chief refractive body, develops from the outer skin. From the skin—the horny
plate—also arises the delicate conjunctiva, which afterwards covers the outer surface of the eyeball. The lachrymal glands are ramified growths from the conjunctiva (Fig. 286). All these important parts of the eye are products of the outer germinal layer. The remaining parts—the corpus vitreum (with the vascular capsule of the lens), the choroid (with the iris), and the sclerotic (with the cornea)—are formed from the middle germinal layer.
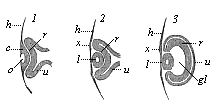 Fig. 318—Eye of the chick embryo in longitudinal section (1. from an embryo sixty-five hours old; 2. from a somewhat older embryo; 3. from an embryo four days old). h horny plate, o lens-pit, l lens (in 1. still part of the epidermis, in 2. and 3. separated from it), x thickening of the horny plate at the point where the lens has severed itself, gl corpus vitreum, r retina, u pigment membrane. (From Remak.)
Fig. 318—Eye of the chick embryo in longitudinal section (1. from an embryo sixty-five hours old; 2. from a somewhat older embryo; 3. from an embryo four days old). h horny plate, o lens-pit, l lens (in 1. still part of the epidermis, in 2. and 3. separated from it), x thickening of the horny plate at the point where the lens has severed itself, gl corpus vitreum, r retina, u pigment membrane. (From Remak.) |
The outer protection of the eye, the eye-lids, are merely folds of the skin, which are formed in the third month of human embryonic life. In the fourth month the upper eye-lid reaches the lower, and the eye remains covered with them until birth. As a rule, they open wide shortly before birth (sometimes only after birth). Our craniote ancestors had a third eye-lid, the nictitating membrane, which was drawn over the eye from its inner angle. It is still found in many of the Selachii and Amniotes. In the apes and man it has degenerated, and there is now only a small relic of it at the inner corner of the eye, the semi-lunar fold, a useless rudimentary organ (cf. p. 32). The apes and man have also lost the Harderian gland that opened under the nictitating membrane; we find this in the rest of the mammals, and the birds, reptiles, and amphibia.
The peculiar embryonic development of the vertebrate eye does not enable us to draw any definite conclusions as to its obscure phylogeny; it is clearly cenogenetic to a great extent, or obscured by the reduction and curtailment of its original features. It is probable that many of the earlier stages of its phylogeny have disappeared without leaving a trace. It can only be said positively that the peculiar ontogeny of the complicated optic apparatus in man follows just the same laws as in all the other Vertebrates. Their eye is a part of the fore brain, which has grown forward towards the skin, not an original cutaneous sense-organ, as in the Invertebrates.
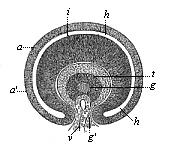 Fig. 319—Horizontal transverse section of the eye of a human embryo, four weeks old. (From Kölliker.) t lens (the dark wall of which is as thick as the diameter of the central cavity), g corpus vitreum (connected by a stem, g, with the corium), v vascular loop (pressing behind the lens inside the corpus vitreum by means of this stem g), i retina (inner thicker, invaginated layer of the primary optic vesicle), a pigment membrane (outer, thin, non-invaginated layer of same), h space between retina and pigment membrane (remainder of the cavity of the primary optic vesicle).
Fig. 319—Horizontal transverse section of the eye of a human embryo, four weeks old. (From Kölliker.) t lens (the dark wall of which is as thick as the diameter of the central cavity), g corpus vitreum (connected by a stem, g, with the corium), v vascular loop (pressing behind the lens inside the corpus vitreum by means of this stem g), i retina (inner thicker, invaginated layer of the primary optic vesicle), a pigment membrane (outer, thin, non-invaginated layer of same), h space between retina and pigment membrane (remainder of the cavity of the primary optic vesicle). |
The vertebrate ear resembles the eye and nose in many important respects, but is different in others, in its development. The auscultory organ in the fully-developed man is like that of the other mammals, and especially the apes, in the main features. As in them, it consists of two chief parts—an apparatus for conducting sound (external and middle ear) and an apparatus for the sensation of sound (internal ear). The external ear opens in the shell at the side of the head (Fig. 320 a). From this point the external passage (b), about an inch in length, leads into the head. The inner end of it is closed by the tympanum, a vertical, but not quite upright, thin membrane of an oval shape (c). This tympanum separates the external passage from the tympanic cavity (d). This is a small cavity, filled with air, in the temporal bone; it is connected with the mouth by a special tube. This tube is rather longer, but much narrower, than the outer passage, leads inwards obliquely from the anterior wall of the tympanic cavity, and opens in the throat below, behind the nasal
openings. It is called the Eustachian tube (e); it serves to equalise the pressure of the air within the tympanic cavity and the outer atmosphere that enters by the external passage. Both the Eustachian tube and the tympanic cavity are lined with a thin mucous coat, which is a direct continuation of the mucous lining of the throat. Inside the tympanic cavity there are three small bones which are known (from their shape) as the hammer, anvil, and stirrup (Fig. 320, f, g, h). The hammer (f) is the outermost, next to the tympanum. The anvil (g) fits between the other two, above and inside the hammer. The stirrup (h) lies inside the anvil, and touches with its base the outer wall of the internal ear, or auscultory vesicle. All these parts of the external and middle ear belong to the apparatus for conducting sound. Their chief task is to convey the waves of sound through the thick wall of the head to the inner-lying auscultory vesicle. They are not found at all in the fishes. In these the waves of sound are conveyed directly by the wall of the head to the auscultory vesicle.
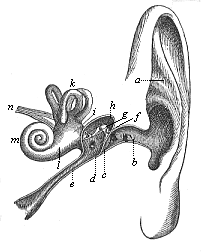 Fig. 320—The human ear (left ear, seen from the front), a shell of ear, b external passage, c tympanum, d tympanic cavity, e Eustachian tube, f, g, h the three bones of the ear (f hammer, g anvil, h stirrup), i utricle, k the three semi-circular canals, l the sacculus, m cochlea, n auscultory nerve.
Fig. 320—The human ear (left ear, seen from the front), a shell of ear, b external passage, c tympanum, d tympanic cavity, e Eustachian tube, f, g, h the three bones of the ear (f hammer, g anvil, h stirrup), i utricle, k the three semi-circular canals, l the sacculus, m cochlea, n auscultory nerve. | 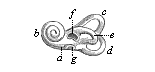 Fig. 321—The bony labyrinth of the human ear (left side). a vestibulum, b cochlea, c upper canal, d posterior canal, e outer canal, f oval fenestra, g round fenestra. (From Meyer.)
Fig. 321—The bony labyrinth of the human ear (left side). a vestibulum, b cochlea, c upper canal, d posterior canal, e outer canal, f oval fenestra, g round fenestra. (From Meyer.) |
The internal apparatus for the sensation of sound, which receives the waves of sound from the conducting apparatus, consists in man and all other mammals of a closed auscultory vesicle filled with fluid and an auditory nerve, the ends of which expand over the wall of this vesicle. The vibrations of the sound-waves are conveyed by these media to the nerve-endings. In the labyrinthic water that fills the auscultory vesicle there are small stones at the points of entry of the acoustic nerves, which are composed of groups of microscopic calcareous crystals (otoliths). The auscultory organ of most of the Invertebrates has substantially the same composition. It usually consists of a closed vesicle, filled with fluid, and containing otoliths, with the acoustic nerve expanding on its wall. But, while the auditory vesicle is usually of a simple round or oval shape in the Invertebrates, it has in the Vertebrates a special and curious structure, the labyrinth. This thin-membraned labyrinth is enclosed in a bony capsule of the same shape, the osseous labyrinth (Fig. 321), and this lies in the middle of the petrous bone of the skull. The labyrinth is divided into two vesicles in all the Gnathostomes. The larger one is called the utriculus, and has three arched appendages, called the “semi-circular canals” (c, d, e). The smaller vesicle is called the sacculus, and is connected with a peculiar appendage, with (in man and the higher mammals) a spiral form something like a snail’s shell, and therefore called the cochlea (= snail, b). On the thin wall of this delicate labyrinth the acoustic nerve, which comes from the after-brain, spreads out in most elaborate fashion. It divides into two main branches—a cochlear nerve (for the cochlea) and a vestibular nerve (for the rest of the labyrinth). The former seems to have more to do with the quality, the latter with the quantity, of the acoustic sensations. Through the cochlear nerves we learn the height and timbre, through the vestibular nerves the intensity, of tones.
The first structure of this highly elaborate organ is very simple in the embryo of man and all the other Craniotes; it is a
pit-like depression in the skin. At the back part of the head at both sides, near the after brain, a small thickening of the horny plate is formed at the upper end of the second gill-cleft (Fig. 322 A fl). This sinks into a sort of pit, and severs from the epidermis, just as the lens of the eye does. In this way is formed at each side, directly under the horny plate of the back part of the head, a small vesicle filled with fluid, the primitive auscultory vesicle, or the primary labyrinth. As it separates from its source, the horny plate, and presses inwards and backwards into the skull, it changes from round to pear-shaped (Figs. 322 B lv, 323 o). The outer part of it is lengthened into a thin stem, which at first still opens outwards by a narrow canal. This is the labyrinthic appendage (Fig. 322 lr). In the lower Vertebrates it develops into a special cavity filled with calcareous crystals, which remains open permanently in some of the primitive fishes, and opens outwards in the upper part of the skull. But in the mammals the labyrinthic appendage degenerates. In these it has only a phylogenetic interest as a rudimentary organ, with no actual physiological significance. The useless relic of it passes through the wall of the petrous bone in the shape of a narrow canal, and is called the vestibular aqueduct.
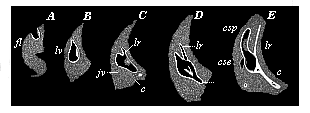 | Fig. 322—Development of the auscultory labyrinth of the chick, in five successive stages (A–E). (Vertical transverse sections of the skull.) fl auscultory pits, lv auscultory vesicles, lr labyrinthic appendage, c rudimentary cochlea, csp posterior canal, cse external canal, jv jugular vein. (From Reissner.) |
It is only the inner and lower bulbous part of the separated auscultory vesicle that develops into the highly complex and differentiated structure that is afterwards known as the secondary labyrinth. This vesicle divides at an early stage into an upper and larger and a lower and smaller section. From the one we get the utriculus with the semi-circular canals; from the other the sacculus and the cochlea (Fig. 320 c). The canals are formed in the shape of simple pouch-like involutions of the utricle (cse and csp). The edges join together in the middle part of each fold, and separate from the utricle, the two ends remaining in open connection with its cavity. All the Gnathostomes have these three canals like man, whereas among the Cyclostomes the lampreys have only two and the hag-fishes only one. The very complex structure of the cochlea, one of the most elaborate and wonderful outcomes of adaptation in the mammal body, develops originally in very simple fashion as a flask-like projection from the sacculus. As Hasse and Retzius have pointed out, we find the successive ontogenetic stages of its growth represented permanently in the series of the higher Vertebrates. The cochlea is wanting even in the Monotremes, and is restricted to the rest of the mammals and man.
The auditory nerve, or eighth cerebral nerve, expands with one branch in the cochlea, and with the other in the remaining parts of the labyrinth. This nerve is, as Gegenbaur has shown, the sensory dorsal branch of a cerebro-spinal nerve, the motor ventral branch of which acts for the muscles of the face (nervus facialis). It has therefore originated phylogenetically from an ordinary cutaneous nerve, and so is of quite different origin from the optic and olfactory nerves, which both represent direct outgrowths of the brain. In this respect the auscultory organ is essentially different from the organs of sight and smell. The acoustic nerve is formed from ectodermic cells of the hind brain, and develops from the nervous structure that appears at its dorsal limit. On the other hand, all the membranous, cartilaginous, and osseous coverings of the labyrinth are formed from the mesodermic head-plates.
The apparatus for conducting sound which we find in the external and middle ear of mammals develops quite separately from the apparatus for the sensation of sound. It is both phylogenetically and ontogenetically an independent secondary formation, a later accession
to the primary internal ear. Nevertheless, its development is not less interesting, and is explained with the same ease by comparative anatomy. In all the fishes and in the lowest Vertebrates there is no special apparatus for conducting sound, no external or middle ear; they have only a labyrinth, an internal ear, which lies within the skull. They are without the tympanum and tympanic cavity, and all its appendages. From many observations made in the last few decades it seems that many of the fishes (if not all) cannot distinguish tones; their labyrinth seems to be chiefly (if not exclusively) an organ for the sense of space (or equilibrium). If it is destroyed, the fishes lose their balance and fall. In the opinion of recent physiologists this applies also to many of the Invertebrates (including the nearer ancestors of the Vertebrates). The round vesicles which are considered to be their auscultory vesicles, and which contain an otolith, are supposed to be merely organs of the sense of space (“static vesicles or statocysts”).
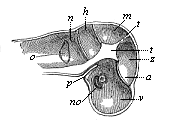 Fig. 323—Primitive skull of the human embryo, four weeks old, vertical section, left half seen internally. v, z, m, h, n the five pits of the cranial cavity, in which the five cerebral vesicles lie (fore, intermediate, middle, hind, and after brains), o pear-shaped primary auscultory vesicle (appearing through), a eye (appearing through), no optic nerve, p canal of the hypophysis, t central prominence of the skull. (From Kölliker.)
Fig. 323—Primitive skull of the human embryo, four weeks old, vertical section, left half seen internally. v, z, m, h, n the five pits of the cranial cavity, in which the five cerebral vesicles lie (fore, intermediate, middle, hind, and after brains), o pear-shaped primary auscultory vesicle (appearing through), a eye (appearing through), no optic nerve, p canal of the hypophysis, t central prominence of the skull. (From Kölliker.) |
The middle ear makes its first appearance in the amphibian class, where we find a tympanum, tympanic cavity, and Eustachian tube; these animals, and all terrestrial Vertebrates, certainly have the faculty of hearing. All these essential parts of the middle ear originate from the first gill-cleft and its surrounding part; in the Selachii this remains throughout life an open squirting-hole, and lies between the first and second gill-arch. In the embryo of the higher Vertebrates it closes up in the centre, and thus forms the tympanic membrane. The outlying remainder of the first gill-cleft is the rudiment of the external meatus. From its inner part we get the tympanic cavity, and, further inward still, the Eustachian tube. Connected with this is the development of the three bones of the mammal ear from the first two gill-arches; the hammer and anvil are formed from the first, the stirrup from the upper end of the second, gill-arch.
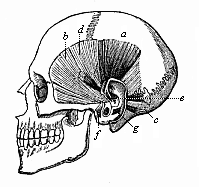 Fig. 324—The rudimentary muscles of the ear in the human skull. a raising muscle (M. attollens), b drawing muscle (M. attrahens), c withdrawing muscle (M. retrahens), d large muscle of the helix (M. helicis major), e small muscle of the helix (M. helicis minor), f muscle of the angle of the ear (M. tragicus), g anti-angular muscle (M. antitragicus). (From H. Meyer.)
Fig. 324—The rudimentary muscles of the ear in the human skull. a raising muscle (M. attollens), b drawing muscle (M. attrahens), c withdrawing muscle (M. retrahens), d large muscle of the helix (M. helicis major), e small muscle of the helix (M. helicis minor), f muscle of the angle of the ear (M. tragicus), g anti-angular muscle (M. antitragicus). (From H. Meyer.) |
Finally, the shell (pinna or concha) and external meatus (passage to the tympanum) of the outer ear are developed in a very simple fashion from the skin that borders the external aperture of the first gill-cleft. The shell rises in the shape of a circular fold of the skin, in which cartilage and muscles are afterwards formed (Figs. 313, 315). This organ is only found in the mammalian class. It is very rudimentary in the lowest section, the Monotremes. In the others it is found at very different stages of development, and sometimes of degeneration. It is degenerate in most of the aquatic mammals. The majority of them have lost it altogether—for instance, the walruses and whales and most of the seals. On the other hand, the pinna is well developed in the great majority of the Marsupials and Placentals; it receives and collects the waves of sound, and is equipped with a very elaborate muscular apparatus, by means of which the pinna
can be turned freely in any direction and its shape be altered. It is well known how readily domestic animals—horses, cows, dogs, hares, etc.—point their ears and move them in different directions. Most of the apes do the same, and our earlier ape ancestors were also able to do it. But our later simian ancestors, which we have in common with the anthropoid apes, abandoned the use of these muscles, and they gradually became rudimentary and useless. However, we possess them still (Fig. 324). In fact, some men can still move their ears a little backward and forward by means of the drawing and withdrawing muscles (b and c); with practice this faculty can be much improved. But no man can now lift up his ears by the raising muscle (a), or change the shape of them by the small inner muscles (d, e, f, g). These muscles were very useful to our ancestors, but are of no consequence to us. This applies to most of the anthropoid apes as well.
We also share with the higher anthropoid apes (gorilla, chimpanzee, and orang) the characteristic form of the human outer ear, especially the folded border, the helix and the lobe. The lower apes have pointed ears, without folded border or lobe, like the other mammals. But Darwin has shown that at the upper part of the folded border there is in many men a small pointed process, which most of us do not possess. In some individuals this process is well developed. It can only be explained as the relic of the original point of the ear, which has been turned inwards in consequence of the curving of the edge. If we compare the pinna of man and the various apes in this respect, we find that they present a connected series of degenerate structures. In the common catarrhine ancestors of the anthropoids and man the degeneration set in with the folding together of the pinna. This brought about the helix of the ear, in which we find the significant angle which represents the relic of the salient point of the ear in our earlier simian ancestors. Here again, therefore, comparative anatomy enables us to trace with certainty the human ear to the similar, but more developed, organ of the lower mammals. At the same time, comparative physiology shows that it was a more or less useful implement in the latter, but it is quite useless in the anthropoids and man. The conducting of the sound has scarcely been affected by the loss of the pinna. We have also in this the explanation of the extraordinary variety in the shape and size of the shell of the ear in different men; in this it resembles other rudimentary organs.
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XXIV
Chapter XXVI
Figs. 1–209
Figs. 210–408