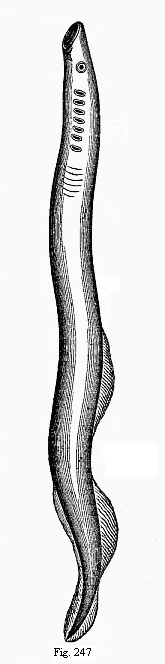
 Fig. 247—The large marine lamprey (Petromyzon marinus), much reduced. Behind the eye there is a row of seven gill-clefts visible on the left, in front the round suctorial mouth.
Fig. 247—The large marine lamprey (Petromyzon marinus), much reduced. Behind the eye there is a row of seven gill-clefts visible on the left, in front the round suctorial mouth.Our task of detecting the extinct ancestors of our race among the vast numbers of animals known to us encounters very different difficulties in the various sections of man’s stem-history. These were very great in the series of our invertebrate ancestors; they are much slighter in the subsequent series of our vertebrate ancestors. Within the vertebrate stem there is, as we have already seen, so complete an agreement in structure and embryology that it is impossible to doubt their phylogenetic unity. In this case the evidence is much clearer and more abundant.
The characteristics that distinguish the Vertebrates as a whole from the Invertebrates have already been discussed in our description of the hypothetical Primitive Vertebrate (Chapter XI, Figs. 98–102). The chief of these are: (1) The evolution of the primitive brain into a dorsal medullary tube; (2) the formation of the chorda between the medullary tube and the gut; (3) the division of the gut into branchial (gill) and hepatic (liver) gut; and (4) the internal articulation or metamerism. The first three features are shared by the Vertebrates with the ascidia-larvæ and the Prochordonia; the fourth is peculiar to them. Thus the chief advantage in organisation by which the earliest Vertebrates took precedence of the unsegmented Chordonia consisted in the development of internal segmentation.
The whole vertebrate stem divides first into the two chief sections of Acrania and Craniota. The Amphioxus is the only surviving representative of the older and lower section, the Acrania (“skull-less”). All the other vertebrates belong to the second division, the Craniota (“skull-animals”). The Craniota descend directly from the Acrania, and these from the primitive Chordonia. The exhaustive study that we made of the comparative anatomy and ontogeny of the Ascidia and the Amphioxus has proved these relations for us. (See Chapters XVI and XVII.) The Amphioxus, the lowest Vertebrate, and the Ascidia, the nearest related Invertebrate, descend from a common extinct stem-form, the Chordæa; and this must have had, substantially, the organisation of the chordula.
However, the Amphioxus is important not merely because it fills the deep gulf between the Invertebrates and Vertebrates, but also because it shows us to-day the typical vertebrate in all its simplicity. We owe to it the most important data that we proceed on in reconstructing the gradual historical development of the whole stem. All the Craniota descend from a common stem-form, and this was substantially identical in structure with the Amphioxus. This stem-form, the Primitive Vertebrate (Prospondylus, Figs. 98–102), had the characteristics of the vertebrate as such, but not the important features that distinguish the Craniota from the Acrania. Though the Amphioxus has many peculiarities of structure and has much degenerated, and though it cannot be regarded as an unchanged descendant of the Primitive Vertebrate, it must have inherited from it the specific characters we enumerated above. We may not say that “Amphioxus is the ancestor of the Vertebrates”; but we can say: “Amphioxus is the nearest relation to the ancestor of all the animals we know.” Both belong to the same small family, or lowest class of the Vertebrates, that we call the Acrania. In our genealogical tree this group forms the twelfth stage, or the first stage among the vertebrate ancestors (p. 228). From this group of Acrania both the Amphioxus and the Craniota were evolved.
The vast division of the Craniota embraces all the Vertebrates known to us, with the exception of the Amphioxus. All of them have a head clearly differentiated from the trunk, and a skull enclosing a brain. The head has also three pairs of higher sense-organs (nose, eyes, and ears). The brain is very rudimentary at first, a mere bulbous enlargement of the
fore end of the medullary tube. But it is soon divided by a number of transverse constrictions into, first three, then five successive cerebral vesicles. In this formation of the head, skull, and brain, with further development of the higher sense-organs, we have the advance that the Craniota made beyond their skull-less ancestors. Other organs also attained a higher development; they acquired a compact centralised heart with valves and a more advanced liver and kidneys, and made progress in other important respects.
|
Fig. 247—The large marine lamprey (Petromyzon marinus), much reduced. Behind the eye there is a row of seven gill-clefts visible on the left, in front the round suctorial mouth. |
We may divide the Craniota generally into Cyclostoma (“round-mouthed”) and Gnathostoma (“jaw-mouthed”). There are only a few groups of the former in existence now, but they are very interesting, because in their whole structure they stand midway between the Acrania and the Gnathostoma. They are much more advanced than the Acrania, much less so than the fishes, and thus form a very welcome connecting-link between the two groups. We may therefore consider them a special intermediate group, the fourteenth and fifteenth stages in the series of our ancestors.
The few surviving species of the Cyclostoma are divided into two orders—the Myxinoides and the Petromyzontes. The former, the hag-fishes, have a long, cylindrical, worm-like body. They were classed by Linné with the worms, and by later zoologists, with the fishes, or the amphibia, or the molluscs. They live in the sea, usually as parasites of fishes, into the skin of which they bore with their round suctorial mouths and their tongues, armed with horny teeth. They are sometimes found alive in the body cavity of fishes (such as the torsk or sturgeon); in these cases they have passed through the skin into the interior. The second order consists of the Petromyzontes or lampreys; the small river lamprey (Petromyzon fluviatilis) and the large marine lamprey (Petromyzon marinus, Fig. 247). They also have a round suctorial mouth, with horny teeth inside it; by means of this they attach themselves by sucking to fishes, stones, and other objects (hence the name Petromyzon = stone-sucker). It seems that this habit was very widespread among the earlier Vertebrates; the larvæ of many of the Ganoids and frogs have suctorial disks near the mouth.
The class that is formed of the Myxinoides and Petromyzontes is called the Cyclostoma (round-mouthed), because their mouth has a circular or semi-circular aperture. The jaws (upper and lower) that we find in all the higher Vertebrates are completely wanting in the Cyclostoma, as in the Amphioxus. Hence the other Vertebrates are collectively opposed to them as Gnathostoma (jaw-mouthed). The Cyclostoma might also be called Monorhina (single-nosed), because they have only a single nasal passage, while all the Gnathostoma have two nostrils (Amphirhina = double-nosed). But apart from these peculiarities the Cyclostoma differ more widely from the fishes in other special features of their structure than the fishes do from man. Hence they are obviously the last survivors of a very ancient class of Vertebrates, that was far from attaining the advanced organisation of the true fish. To mention only the chief points, the Cyclostoma show no trace of pairs of limbs. Their mucous skin is quite naked and smooth and devoid of scales. There is no bony skeleton. A very rudimentary skull is developed at the foremost end of their chorda. At this point a soft membranous (partly turning into cartilage), small skull-capsule is formed, and encloses the brain.
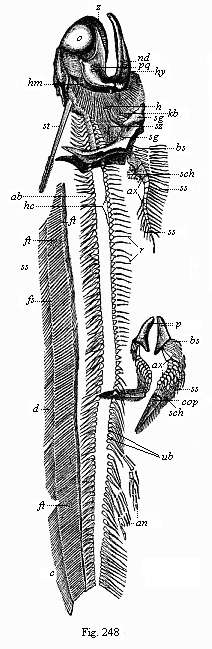 Fig. 248—Fossil Permian primitive fish (Pleuracanthus Dechenii), from the red sandstone of Saarbrücken. (From Döderlein.) I Skull and branchial skeleton: o eye-region, pq palatoquadratum, nd lower jaw, hm hyomandibular, hy tongue-bone, k gill-radii, kb gill-arches, z jaw-teeth, sz gullet-teeth, st neck-spine. II Vertebral column: ob upper arches, ub lower arches, hc intercentra, r ribs. III Single fins: d dorsal fin, c tail-fin (tail-end wanting), an anus-fin, ft supporter of fin-rays. IV Breast-fin: sg shoulder-zone, ax fin-axis, ss double lines of fin-rays, bs additional rays, sch plates. V Ventral fin: p pelvis, ax fin-axis, ss single row of fin-rays, bs additional rays, sch scales, cop penis.
Fig. 248—Fossil Permian primitive fish (Pleuracanthus Dechenii), from the red sandstone of Saarbrücken. (From Döderlein.) I Skull and branchial skeleton: o eye-region, pq palatoquadratum, nd lower jaw, hm hyomandibular, hy tongue-bone, k gill-radii, kb gill-arches, z jaw-teeth, sz gullet-teeth, st neck-spine. II Vertebral column: ob upper arches, ub lower arches, hc intercentra, r ribs. III Single fins: d dorsal fin, c tail-fin (tail-end wanting), an anus-fin, ft supporter of fin-rays. IV Breast-fin: sg shoulder-zone, ax fin-axis, ss double lines of fin-rays, bs additional rays, sch plates. V Ventral fin: p pelvis, ax fin-axis, ss single row of fin-rays, bs additional rays, sch scales, cop penis. |
The brain of the Cyclostoma is merely a very small and comparatively insignificant swelling of the spinal marrow, a simple vesicle at first. It afterwards divides into five successive cerebral vesicles, like the brain of the Gnathostoma. These five primitive cerebral vesicles, that are found in the embryos of all the higher vertebrates from the fishes to man, and grow into very complex structures, remain at a very rudimentary stage in the Cyclostoma. The histological structure of the nerves is also less advanced than in the rest of the vertebrates. In these the auscultory organ always contains three circular canals, but in the lampreys there are only two, and in the hag-fishes only one. In most other respects the organisation of the Cyclostoma is much simpler—for instance, in the structure of the heart, circulation, and kidneys. We must especially note the absence of a very important organ that we find in the fishes, the floating-bladder, from which the lungs of the higher Vertebrates have been developed.
When we consider all these peculiarities in the structure of the Cyclostoma, we may formulate the following thesis: Two divergent lines proceeded from the earliest Craniota, or the primitive Craniota (Archicrania). One of these lines is preserved in a greatly modified condition: these are the Cyclostoma, a very backward and partly degenerate side-line. The other, the chief line of the Vertebrate stem, advanced straight to the fishes, and by fresh adaptations acquired a number of important improvements.
The Cyclostoma are almost always classified by zoologists among the fishes; but the incorrectness of this may be judged from the fact that in all the chief and distinctive features of organisation they are further removed from the fishes than the fishes are from the Mammals, and even man. With the fishes we enter upon the vast division of the jaw-mouthed
or double-nosed Vertebrates (Gnathostoma or Amphirhina). We have to consider the fishes carefully as the class which, on the evidence of palæontology, comparative anatomy, and ontogeny, may be regarded with absolute certainty as the stem-class of all the higher Vertebrates or Gnathostomes. Naturally, none of the actual fishes can be considered the direct ancestor of the higher Vertebrates. But it is certain that all the Vertebrates or Gnathostomes, from the fishes to man, descend from a common, extinct, fish-like ancestor. If we had this ancient stem-form before us, we would undoubtedly class it as a true fish. Fortunately the comparative anatomy and classification of the fishes are now so far advanced that we can get a very clear idea of these interesting and instructive features.
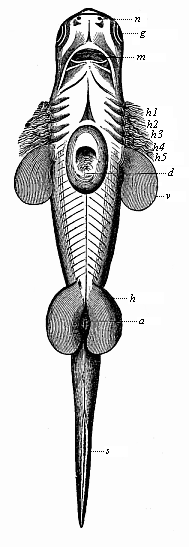 Fig. 249—Embryo of a shark (Scymnus lichia), seen from the ventral side. v breast-fins (in front five pairs of gill-clefts), h belly-fins, a anus, s tail-fin, k external gill-tuft, d yelk-sac (removed for most part), g eye, n nose, m mouth-cleft.
Fig. 249—Embryo of a shark (Scymnus lichia), seen from the ventral side. v breast-fins (in front five pairs of gill-clefts), h belly-fins, a anus, s tail-fin, k external gill-tuft, d yelk-sac (removed for most part), g eye, n nose, m mouth-cleft. |
In order to understand properly the genealogical tree of our race within the vertebrate stem, it is important to bear in mind the characteristics that separate the whole of the Gnathostomes from the Cyclostomes and Craniota. In these respects the fishes agree entirely with all the other Gnathostomes up to man, and it is on this that we base our claim of relationship to the fishes. The following characteristics of the Gnathostomes are anatomic features of this kind: (1) The internal gill-arch apparatus with the jaw arches; (2) the pair of nostrils; (3) the floating bladder or lungs; and (4) the two pairs of limbs.
The peculiar formation of the frame work of the branchial (gill) arches and the connected maxillary (jaw) apparatus is of importance in the whole group of the Gnathostomes. It is inherited in rudimentary form by all of them, from the earliest fishes to man. It is true that the primitive transformation (which we find even in the Ascidia) of the fore gut into the branchial gut can be traced in all the Vertebrates to the same simple type; in this respect the gill-clefts, which pierce the walls of the branchial gut in all the Vertebrates and in the Ascidia, are very characteristic. But the external, superficial branchial skeleton that supports the gill-crate in the Cyclostoma is replaced in the Gnathostomes by an internal branchial skeleton. It consists of a number of successive cartilaginous arches, which lie in the wall of the gullet between the gill-clefts, and run round the gullet from both sides. The foremost pair of gill-arches become the maxillary arches, from which we get our upper and lower jaws.
The olfactory organs are at first found in the same form in all the Gnathostomes, as a pair of depressions in the fore part of the skin of the head, above the mouth; hence, they are also called the Amphirhina
(“double-nosed”). The Cyclostoma are “one-nosed” (Monorhina); their nose is a single passage in the middle of the frontal surface. But as the olfactory nerve is double in both cases, it is possible that the peculiar form of the nose in the actual Cyclostomes is a secondary acquisition (by adaptation to suctorial habits).
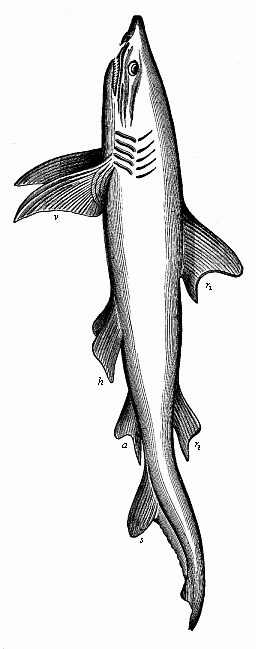 Fig. 250—Fully developed man-eating shark (Carcharias melanopterus), left view. r1 first, r2 second dorsal fin, s tail-fin, a anus-fin, v breast-fins, h belly-fins.)
Fig. 250—Fully developed man-eating shark (Carcharias melanopterus), left view. r1 first, r2 second dorsal fin, s tail-fin, a anus-fin, v breast-fins, h belly-fins.) |
A third essential character of the Gnathostomes, that distinguishes them very conspicuously from the lower vertebrates we have dealt with, is the formation of a blind sac by invagination from the fore part of the gut, which becomes in the fishes the air-filled floating-bladder. This organ acts as a hydrostatic apparatus, increasing or reducing the specific gravity of the fish by compressing or altering the quantity of air in it. The fish can rise or sink in the water by means of it. This is the organ from which the lungs of the higher vertebrates are developed.
Finally, the fourth character of the Gnathostomes in their simple embryonic form is the two pairs of extremities or limbs—a pair of fore legs (breast-fins in the fish, Fig. 250 v) and a pair of hind legs (ventral fins in the fish, Fig. 250 h). The comparative anatomy of these fins is very interesting, because they contain the rudiments of all the skeletal parts that form the framework of the fore and hind legs in all the higher vertebrates right up to man. There is no trace of these pairs of limbs in the Acrania and Cyclostomes.
Turning, now, to a closer inspection of the fish class, we may first divide it into three groups or sub-classes, the genealogy of which is well known to us. The first and oldest group is the sub-class of the Selachii or primitive fishes; the best-known representatives of which to-day are the orders of the sharks and rays (Figs. 248–252). Next to this is the more advanced sub-class of the plated fishes or Ganoids (Figs. 253–5). It has been long extinct for the most part, and has very few living representatives, such as the sturgeon and the bony pike; but we can form some idea of the earlier extent of this interesting group from the large numbers of fossils. From these plated fishes the sub-class of the bony fishes
or Teleostei was developed, to which the great majority of living fishes belong (especially nearly all our river fishes). Comparative anatomy and ontogeny show clearly that the Ganoids descended from the Selachii, and the Teleostei from the Ganoids. On the other hand, a collateral line, or rather the advancing chief line of the vertebrate stem, was developed from the earlier Ganoids, and this leads us through the group of the Dipneusta to the important division of the Amphibia.
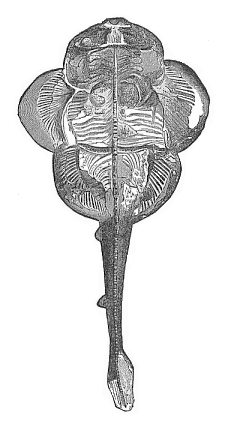 Fig. 251—Fossil angel-shark (Squatina alifera), from the upper Jurassic at Eichstätt. (From Zittel.) The cartilaginous skull is clearly seen in the broad head, and the gill-arches behind. The wide breast-fin and the narrower belly-fin have a number of radii; between these and the vertebral column are a number of ribs.
Fig. 251—Fossil angel-shark (Squatina alifera), from the upper Jurassic at Eichstätt. (From Zittel.) The cartilaginous skull is clearly seen in the broad head, and the gill-arches behind. The wide breast-fin and the narrower belly-fin have a number of radii; between these and the vertebral column are a number of ribs. |
The earliest fossil remains of Vertebrates that we know were found in the Upper Silurian (p. 201), and belong to two groups—the Selachii and the Ganoids. The most primitive of all known representatives of the earliest fishes are probably the remarkable Pleuracanthida, the genera Pleuracanthus, Xenacanthus, Orthocanthus, etc. (Fig. 248). These ancient cartilaginous fishes agree in most points of structure with the real sharks (Figs. 249, 250); but in other respects they seem to be so much simpler in organisation that many palæontologists separate them altogether, and regard them as Proselachii; they are probably closely related to the extinct ancestors of the Gnathostomes. We find well-preserved remains of them in the Permian period. Well-preserved impressions of other sharks are found in the Jurassic schist, such as of the angel-fish (Squatina, Fig. 251). Among the extinct earlier sharks of the Tertiary period there were some twice as large as the biggest living fishes; Carcharodon was more than 100 feet long. The sole surviving species of this genus (C. Rondeleti) is eleven yards long, and has teeth two inches long; but among the fossil species we find teeth six inches long (Fig. 252).
From the primitive fishes or Selachii, the earliest Gnathostomes, was developed the legion of the Ganoids. There are very few genera now of this interesting and varied group—the ancient sturgeons (Accipenser), the eggs of which are eaten as caviare, and the stratified pikes (Polypterus, Fig. 255) in African rivers, and bony pikes (Lepidosteus) in the rivers of North America. On the other hand, we have a great variety of specimens of this group in the fossil state, from the Upper Silurian onward. Some of these fossil Ganoids approach closely to the Selachii; others are nearer to the Dipneusts; others again represent a transition to the Teleostei. For our genealogical purposes the most interesting are the intermediate forms between the Selachii and the Dipneusts. Huxley, to whom we owe particularly important works on the fossil Ganoids, classed them in the order of the Crossopterygii. Many genera and species of this order are found in the Devonian and Carboniferous strata (Fig. 253); a single, greatly modified survivor of the group is still found in the large rivers of Africa (Polypterus, Fig. 255, and the closely related Calamichthys). In many impressions of the Crossopterygii the floating bladder seems to be ossified,
and therefore well preserved—for instance, in the Undina (Fig. 254, immediately behind the head).
Part of these Crossopterygii approach very closely in their chief anatomic features to the Dipneusts, and thus represent phylogenetically the transition from the Devonian Ganoids to the earliest air-breathing vertebrates. This important advance was made in the Devonian period. The numerous fossils that we have from the first two geological sections, the Laurentian and Cambrian periods, belong exclusively to aquatic plants and animals. From this paleontological fact, in conjunction with important geological and biological indications, we may infer with some confidence that there were no terrestrial animals at that time. During the whole of the vast archeozoic period—many millions of years—the living population of our planet consisted almost exclusively of aquatic organisms; this is a very remarkable fact, when we remember that this period embraces the larger half of the whole history of life. The lower animal-stems are wholly (or with very few exceptions) aquatic. But the higher stems also remained in the water during the primordial epoch. It was only towards its close that some of them came to live on land. We find isolated fossil remains of terrestrial animals first in the Upper Silurian, and in larger numbers in the Devonian strata, which were deposited at the beginning of the second chief section of geology (the paleozoic age). The number increases considerably in the Carboniferous and Permian deposits. We find many species both of the articulate and the vertebrate stem that lived on land and breathed the atmosphere; their aquatic ancestors of the Silurian period only breathed water. This important change in respiration is the chief modification that the animal organism underwent in passing from the water to the solid land. The first consequence was the formation of lungs for breathing air; up to that time the gills alone had served for respiration. But there was at the same time a great change in the circulation and its organs; these are always very closely correlated to the respiratory organs. Moreover, the limbs and other organs were also more or less modified, either in consequence of remote correlation to the preceding or owing to new adaptations.
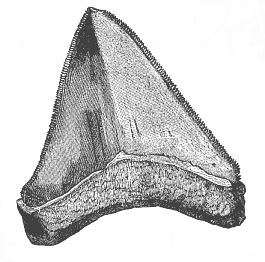 Fig. 252—Tooth of a gigantic shark (Carcharodon megalodon), from the Pliocene at Malta. (From Zittel.)
Fig. 252—Tooth of a gigantic shark (Carcharodon megalodon), from the Pliocene at Malta. (From Zittel.) |
In the vertebrate stem it was unquestionably a branch of the fishes—in fact, of the Ganoids—that made the first fortunate experiment during the Devonian period of adapting themselves to terrestrial life and breathing the atmosphere. This led to a modification of the heart and the nose. The true fishes have merely a pair of blind olfactory pits on the surface of the head; but a connection of these with the cavity of the mouth was now formed. A canal made its appearance on each side, and led directly from the nasal depression into the mouth-cavity, thus conveying atmospheric air to the lungs even when the mouth was closed. Further, in all true fishes the heart has only two sections—an atrium that receives the venous blood from the veins, and a ventricle that propels it through a conical artery to the gills; the atrium was now divided into two halves, or right and left auricles, by an incomplete partition. The right auricle alone now received the venous blood from the body, while the left auricle received the venous blood that flowed from the lungs and gills to the heart. Thus the double circulation of the higher vertebrates was evolved from the simple
circulation of the true fishes, and, in accordance with the laws of correlation, this advance led to others in the structure of other organs.
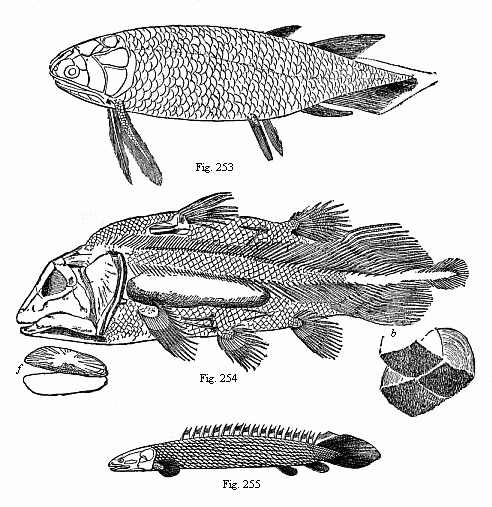 Fig. 253—A Devonian Crossopterygius (Holoptychius nobilissimus), from the Scotch old red sandstone. (From Huxley.)
Fig. 253—A Devonian Crossopterygius (Holoptychius nobilissimus), from the Scotch old red sandstone. (From Huxley.)Fig. 254.—A Jurassic Crossopterygius (Undina penicillata), from the upper Jurassic at Eichstätt. (From Zittel.) j jugular plates, b three ribbed scales. Fig. 255—A living Crossopterygius, from the Upper Nile ((Polypterus bichir). |
The vertebrate class, that thus adapted itself to breathing the atmosphere, and was developed from a branch of the Ganoids, takes the name of the Dipneusts or Dipnoa (“double-breathers”), because they retained the earlier gill-respiration along with the new pulmonary (lung) respiration, like the lowest amphibia. This class was represented during the paleozoic age (or the Devonian, Carboniferous, and Permian periods) by a number of different genera. There are only three genera of the class living to-day: Protopterus annectens in the rivers
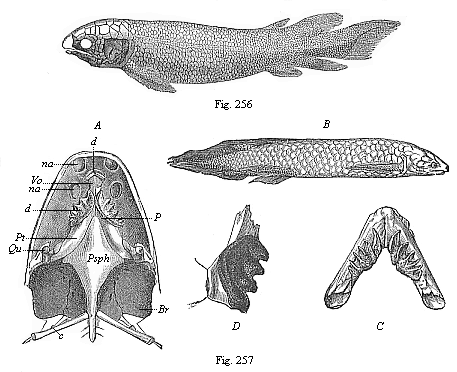 Fig. 256—Fossil Dipneust (Dipterus Valenciennesi), from the old red sandstone (Devon). (From Pander.)
Fig. 256—Fossil Dipneust (Dipterus Valenciennesi), from the old red sandstone (Devon). (From Pander.)Fig. 257—The Australian Dipneust (Ceratodus Forsteri). B view from the right, A lower side of the skull, C lower jaw. (From Gunther.) Qu quadrate bone, Psph parasphenoid, Pt P pterygopalatinum, Vo vomer, d teeth, na nostrils, Br branchial cavity, C first rib. D lower-jaw teeth of the fossil Ceratodus Kaupi (from the Triassic). |
of tropical Africa (the White Nile, the Niger, Quelliman, etc.), Lepidosiren paradoxa in tropical South America (in the tributaries of the Amazon), and Ceratodus Forsteri in the rivers of East Australia. This wide distribution of the three isolated survivors proves that they represent a group that was formerly very large. In their whole structure they form a transition from the fishes to the amphibia. The transitional formation between the two classes is so pronounced in the whole organisation of these remarkable animals that zoologists had a lively controversy over the question whether they were really fishes or amphibia. Several distinguished zoologists classed them with the amphibia, though most now associate them with the fishes. As a matter of fact, the characters of the two classes are so far united in the Dipneusts that the answer to the question depends entirely on the definition we give of “fish” and “amphibian.” In habits they are true amphibia. During the tropical winter, in the rainy season, they swim in the water like the fishes, and breathe water by gills. During the dry season they bury themselves in the dry mud, and breathe the atmosphere through lungs, like the amphibia and the higher vertebrates. In this double respiration they resemble the lower amphibia, and have the same characteristic formation of the heart; in this they are much superior to the fishes. But in most other features
they approach nearer to the fishes, and are inferior to the amphibia. Externally they are entirely fish-like.
In the Dipneusts the head is not marked off from the trunk. The skin is covered with large scales. The skeleton is soft, cartilaginous, and at a low stage of development, as in the lower Selachii and the earliest Ganoids. The chorda is completely retained, and surrounded by an unsegmented sheath. The two pairs of limbs are very simple fins of a primitive type, like those of the lowest Selachii. The formation of the brain, the gut, and the sexual organs is also the same as in the Selachii. Thus the Dipneusts have preserved by heredity many of the less advanced features of our primitive fish-like ancestors, and at the same time have made a great step forward in adaptation to air-breathing by means of lungs and the correlative improvement of the heart.
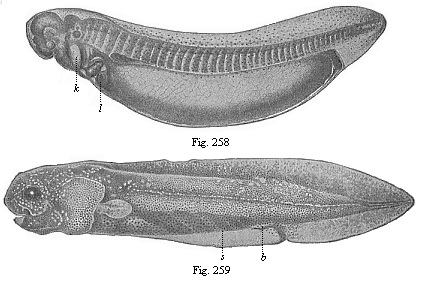 Fig. 258—Young ceratodus, shortly after issuing from the egg, magnified. k gill-cover,
Fig. 258—Young ceratodus, shortly after issuing from the egg, magnified. k gill-cover,l liver. (From Richard Semon.) Fig. 259—Young ceratodus six weeks after issuing from the egg. s spiral fold of gut, b rudimentary belly-fin. (From Richard Semon.) |
Ceratodus is particularly interesting on account of the primitive build of its skeleton; the cartilaginous skeleton of its two pairs of fins, for instance, has still the original form of a bi-serial or feathered leaf, and was on that account described by Gegenbaur as a “primitive fin-skeleton.” On the other hand, the skeleton of the pairs of fins is greatly reduced in the African dipneust (Protopterus) and the American (Lepidosiren). Further, the lungs are double in these modern dipneusts, as in all the other air-breathing vertebrates; they have on that account been called “double-lunged” (Dipneumones) in contrast to the Ceratodus; the latter has only a single lung (Monopneumones). At the same time the gills also are developed as water-breathing organs in all these lung-fishes. Protopterus has external as well as internal gills.
The paleozoic Dipneusts that are in the direct line of our ancestry, and form the connecting-bridge between the Ganoids and the Amphibia, differ in many respects
from their living descendants, but agree with them in the above essential features. This is confirmed by a number of interesting facts that have lately come to our knowledge in connection with the embryonic development of the Ceratodus and Lepidosiren; they give us important information as to the stem-history of the lower Vertebrates, and therefore of our early ancestors of the paleozoic age.
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XX
Chapter XXII
Figs. 1–209
Figs. 210–408