There is a substantial agreement throughout the animal world in the first changes which follow the impregnation of the ovum and the formation of the stem-cell; they begin in all cases with the segmentation of the ovum and the formation of the germinal layers. The only exception is found in the protozoa, the very lowest and simplest forms of animal life; these remain unicellular throughout life. To this group belong the amœbae, gregarinæ, rhizopods, infusoria, etc. As their whole organism consists of a single cell, they can never form germinal layers, or definite strata of cells. But all the other animals—all the tissue-forming animals, or metazoa, as we call them, in contradistinction to the protozoa—construct real germinal layers by the repeated cleavage of the impregnated ovum. This we find in the lower cnidaria and worms, as well as in the more highly-developed molluscs, echinoderms, articulates, and vertebrates.
In all these metazoa, or multicellular animals, the chief embryonic processes are substantially alike, although they often seem to a superficial observer to differ considerably. The stem-cell that proceeds from the impregnated ovum always passes by repeated cleavage into a number of simple cells. These cells are all direct descendants of the stem-cell, and are, for reasons we shall see presently, called segmentation-cells. The repeated cleavage of the stem-cell, which gives rise to these segmentation-spheres, has long been known as “segmentation.” Sooner or later the segmentation-cells join together to form a round (at first, globular) embryonic sphere (blastula); they then form into two very different groups, and arrange themselves
in two separate strata—the two primary germinal layers. These enclose a digestive cavity, the primitive gut, with an opening, the primitive mouth. We give the name of the gastrula to the important embryonic form that has these primitive organs, and the name of gastrulation to the formation of it. This ontogenetic process has a very great significance, and is the real starting-point of the construction of the multicellular animal body.
The fundamental embryonic processes of the cleavage of the ovum and the formation of the germinal layers have been very thoroughly studied in the last thirty years, and their real significance has been appreciated. They present a striking variety in the different groups, and it was no light task to prove their essential identity in the whole animal world. But since I formulated the gastræa theory in 1872, and afterwards (1875) reduced all the various forms of segmentation and gastrulation to one fundamental type, their identity may be said to have been established. We have thus mastered the law of unity which governs the first embryonic processes in all the animals.
Man is like all the other higher animals, especially the apes, in regard to these earliest and most important processes. As the human embryo does not essentially differ, even at a much later stage of development—when we already perceive the cerebral vesicles, the eyes, ears, gill-arches, etc.—from the similar forms of the other higher mammals, we may confidently assume that they agree in the earliest embryonic processes, segmentation and the formation of germinal layers. This has not yet, it is true, been established by observation. We have never yet had occasion to dissect a woman immediately after impregnation and examine the stem-cell or the segmentation-cells in her oviduct. However, as the earliest human embryos we have examined, and the later and more developed forms, agree with those of the rabbit, dog, and other higher mammals, no reasonable man will doubt but that the segmentation and formation of layers are the same in both cases.
But the special form of segmentation and layer formation which we find in the mammal is by no means the original, simple, palingenetic form. It has been much modified and cenogenetically altered by a very complex adaptation to embryonic conditions. We cannot, therefore, understand it altogether in itself. In order to do this, we have to make a comparative study of segmentation and layer-formation in the animal world; and we have especially to seek the original, palingenetic form from which the modified cenogenetic (see p. 4) form has gradually been developed.
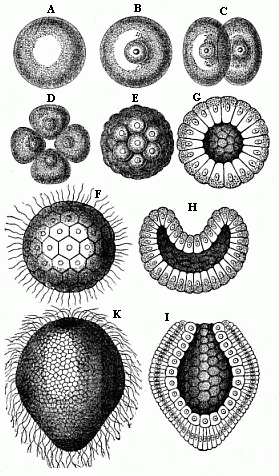
This original unaltered form of segmentation and layer-formation is found to-day in only one case in the vertebrate-stem to which man belongs—the lowest and oldest member of the stem, the wonderful lancelet or amphioxus (cf. Chapters XVI and XVII). But we find a precisely similar palingenetic form of embryonic development in the case of many of the invertebrate animals, as, for instance, the remarkable ascidia, the pond-snail (Limnæus), and arrow-worm (Sagitta), and many of the echinoderms and cnidaria, such as the common star-fish and sea-urchin, many of the medusæ and corals, and the simpler sponges (Olynthus). We may take as an illustration the palingenetic segmentation and germinal layer-formation in an eight-fold insular coral, which I discovered in the Red Sea, and described as Monoxenia Darwinii.
The impregnated ovum of this coral (Fig. 29 A, B) first splits into two equal cells (C). First, the nucleus of the stem-cell and its central body divide into two halves. These recede from and repel each other, and act as centres of attraction on the surrounding protoplasm; in consequence of this, the protoplasm is constricted by a circular furrow, and, in turn, divides into two halves. Each of the two segmentation-cells thus produced splits in the same way into two equal cells. The four segmentation-cells (grand-daughters of the stem-cell) lie in one plane. Now, however, each of them subdivides into two equal halves, the cleavage of the nucleus again preceding that of the surrounding protoplasm. The eight cells which thus arise break into sixteen, these into thirty-two, and then (each being constantly halved) into sixty-four, 128, and so on.1 The final result of this
1. The number of segmentation-cells thus produced increases geometrically in the original gastrulation, or the purest palingenetic form of cleavage. However, in different animals the number reaches a different height, so that the morula, and also the blastula, may consist sometimes of thirty-two, sometimes of sixty-four, and sometimes of 128, or more, cells.
repeated cleavage is the formation of a globular cluster of similar segmentation-cells, which we call the mulberry-formation or morula. The cells are thickly pressed together like the parts of a mulberry or blackberry, and this gives a lumpy appearance to the surface of the sphere (Fig. E).1
When the cleavage is thus ended, the mulberry-like mass changes into a hollow globular sphere. Watery fluid or jelly gathers inside the globule; the segmentation-cells are loosened, and all rise to the surface. There they are flattened by mutual pressure, and assume the shape of truncated pyramids, and arrange themselves side by side in one regular layer (Figs. F, G). This layer of cells is called the germinal membrane (or blastoderm); the homogeneous cells which compose its simple structure are called blastodermic cells; and the whole hollow sphere, the walls of which are made of the preceding, is called the blastula or blastosphere.2
In the case of our coral, and of many other lower forms of animal life, the young embryo begins at once to move independently and swim about in the water. A fine, long, thread-like process, a sort of whip or lash, grows out of each blastodermic cell, and this independently executes vibratory movements, slow at first, but quicker after a time (Fig. F). In this way each blastodermic cell becomes a ciliated cell. The combined force of all these vibrating lashes causes the whole blastula to move about in a rotatory fashion. In many other animals, especially those in which the embryo develops within enclosed membranes, the ciliated cells are only formed at a later stage, or even not formed at all. The blastosphere may grow and expand by the blastodermic cells (at the surface of the sphere) dividing and increasing, and more fluid is secreted in the internal cavity. There are still to-day some organisms that remain throughout life at the structural stage of the blastula—hollow vesicles that swim about by a ciliary movement in the water, the wall of which is composed of a single layer of cells, such as the volvox, the magosphæra, synura, etc. We shall speak further of the great phylogenetic significance of this fact in Chapter XIX.
A very important and remarkable process now follows—namely, the curving or invagination of the blastula (Fig. H). The vesicle with a single layer of cells for wall is converted into a cup with a wall of two layers of cells (cf. Figs. G, H, I). A certain spot at the surface of the sphere is flattened, and then bent inward. This depression sinks deeper and deeper, growing at the cost of the internal cavity. The latter decreases as the hollow deepens. At last the internal cavity disappears altogether, the inner side of the blastoderm (that which lines the depression) coming to lie close on the outer side. At the same time, the cells of the two sections assume different sizes and shapes; the inner cells are more round and the outer more oval (Fig. I). In this way the embryo takes the form of a cup or jar-shaped body, with a wall made up of two layers of cells, the inner cavity of which opens to the outside at one end (the spot where the depression was originally formed). We call this very important and interesting embryonic form the “cup-embryo” or “cup-larva” (gastrula, Fig. 29, I longitudinal section, K external view). I have in my Natural History of Creation given the name of depula to the remarkable intermediate form which appears at the passage of the blastula into the gastrula. In this intermediate stage there are two cavities in the embryo—the original cavity (blastocœl) which is disappearing, and the primitive gut-cavity (progaster) which is forming.
I regard the gastrula as the most important and significant embryonic form in the animal world. In all real animals (that is, excluding the unicellular protists) the segmentation of the ovum produces either a pure, primitive, palingenetic gastrula (Fig. 29 I, K) or an equally instructive cenogenetic form, which has been developed in time from the first, and can be directly reduced to it. It is certainly a fact of the greatest interest and instructiveness that animals of the most different stems—vertebrates and tunicates, molluscs and articulates, echinoderms and annelids, cnidaria and sponges—proceed from one and the same embryonic form. In illustration I give a few
1. The segmentation-cells which make up the morula
after the close of the palingenetic cleavage seem usually to be
quite similar, and to present no differences as to size, form, and
composition. That, however, does not prevent them from
differentiating into animal and vegetative cells, even during the
cleavage.
2. The blastula of the lower animals must not be confused with the
very different blastula of the mammal, which is properly called the
gastrocystis or blastocystis. This cenogenetic
gastrocystis and the palingenetic blastula are sometimes
very wrongly comprised under the common name of blastula or
vesicula blastodermica.
pure gastrula forms from various groups of animals (Figs. 30–35, explanation given below each).
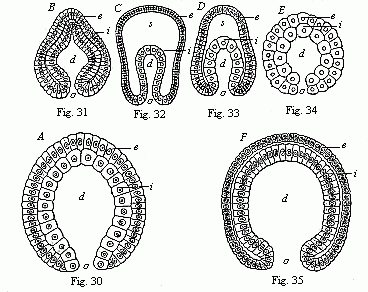 Fig. 30 (A)—Gastrula of a very simple primitive-gut animal or gastræad (gastrophysema). (Haeckel.) Fig. 31 (B)—Gastrula of a worm (Sagitta). (From Kowalevsky.) Fig. 32 (C)—Gastrula of an echinoderm (star-fish, Uraster), not completely folded in (depula). (From Alexander Agassiz.) Fig. 33 (D)—Gastrula of an arthropod (primitive crab, Nauplius) (as 32). Fig. 34 (E)—Gastrula of a mollusc (pond-snail, Linnæus). (From Karl Rabl.) Fig. 35 (F)—Gastrula of a vertebrate (lancelet, Amphioxus). (From Kowalevsky.) (Front view.) In each figure d is the primitive-gut cavity, o primitive mouth, s segmentation-cavity, i entoderm (gut-layer), e ectoderm (skin layer). |
In view of this extraordinary significance of the gastrula, we must make a very careful study of its original structure. As a rule, the typical gastrula is very small, being invisible to the naked eye, or at the most only visible as a fine point under very favourable conditions, and measuring generally 1/500 to 1/250 of an inch (less frequently 1/50 inch, or even more) in diameter. In shape it is usually like a roundish drinking-cup. Sometimes it is rather oval, at other times more ellipsoid or spindle-shaped; in some cases it is half round, or even almost round, and in others lengthened out, or almost cylindrical.
I give the name of primitive gut (progaster) and primitive mouth (prostoma) to the internal cavity of the gastrula-body and its opening; because this cavity is the first rudiment of the digestive cavity of the organism, and the opening originally served to take food into it. Naturally, the primitive gut and mouth change very considerably afterwards in the various classes of animals. In most of the cnidaria and many of the annelids (worm-like animals) they remain unchanged throughout life. But in most of the
higher animals, and so in the vertebrates, only the larger central part of the later alimentary canal develops from the primitive gut; the later mouth is a fresh development, the primitive mouth disappearing or changing into the anus. We must therefore distinguish carefully between the primitive gut and mouth of the gastrula and the later alimentary canal and mouth of the fully developed vertebrate.1
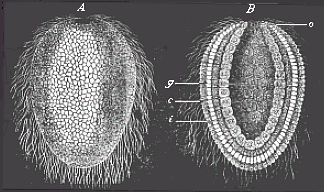
Fig. 36—Gastrula of a lower sponge (lynthus). A external view, B longitudinal section through the axis, g primitive-gut cavity, a primitive mouth-aperture, i inner cell-layer (entoderm, endoblast, gut-layer), e external cell-layer (outer germinal layer, ectoderm, ectoblast, or skin-layer). |
The two layers of cells which line the gut-cavity and compose its wall are of extreme importance. These two layers, which are the sole builders of the whole organism, are no other than the two primary germinal layers, or the primitive germ-layers. I have spoken in the introductory section (Chapter III) of their radical importance. The outer stratum is the skin-layer, or ectoderm (Figs. 30–35e); the inner stratum is the gut-layer, or entoderm (i). The former is often also called the ectoblast, or epiblast, and the latter the endoblast, or hypoblast. From these two primary germinal layers alone is developed the entire organism of all the metazoa or multicellular animals. The skin-layer forms the external skin, the gut-layer forms the internal skin or lining of the body. Between these two germinal layers are afterwards developed the middle germinal layer (mesoderma) and the body-cavity (cœloma) filled with blood or lymph.
The two primary germinal layers were first distinguished by Pander in 1817 in the incubated chick. Twenty years later (1849) Huxley pointed out that in many of the lower zoophytes, especially the medusæ, the whole body consists throughout life of these two primary germinal layers. Soon afterwards (1853) Allman introduced the names which have come into general use; he called the outer layer the ectoderm (“outer-skin”), and the inner the entoderm (“inner-skin”). But in 1867 it was shown, particularly by Kowalevsky, from comparative observation, that even in invertebrates, also, of the most different classes—annelids, molluscs, echinoderms, and articulates—the body is developed out of the same two primary layers. Finally, I discovered them (1872) in the lowest tissue-forming animals, the sponges, and proved in my gastræa theory that these two layers must be regarded as identical throughout the animal world, from the sponges and corals to the insects and vertebrates, including man. This fundamental “homology
1. My distinction (1872) between the primitive gut and mouth and the later permanent stomach (metagaster) and mouth (metastoma) has been much criticised; but it is as much justified as the distinction between the primitive kidneys and the permanent kidneys. Professor E. Ray-Lankester suggested three years afterwards (1875) the name archenteron for the primitive gut, and blastoporus for the primitive mouth.
[identity] of the primary germinal layers and the primitive gut” has been confirmed during the last thirty years by the careful research of many able observers, and is now pretty generally admitted for the whole of the metazoa.
As a rule, the cells which compose the two primary germinal layers show appreciable differences even in the gastrula stage. Generally (if not always) the cells of the skin-layer or ectoderm (Figs. 36 c and 37 e) are the smaller, more numerous, and clearer; while the cells of the gut-layer, or entoderm (i), are larger, less numerous, and darker. The protoplasm of the ectodermic (outer) cells is clearer and firmer than the thicker and softer cell-matter of the entodermic (inner) cells; the latter are, as a rule, much richer in yelk-granules (albumen and fatty particles) than the former. Also the cells of the gut-layer have, as a rule, a stronger affinity for colouring matter, and take on a tinge in a solution of carmine, aniline, etc., more quickly and appreciably than the cells of the skin-layer. The nuclei of the entoderm-cells are usually roundish, while those of the ectoderm-cells are oval.
When the doubling-process is complete, very striking histological differences between the cells of the two layers are found (Fig. 37). The tiny, light ectoderm-cells (e) are sharply distinguished from the larger and darker entoderm-cells (i). Frequently this differentiation of the cell-forms sets in at a very early stage, during the segmentation-process, and is already very appreciable in the blastula.
We have, up to the present, only considered that form of segmentation and gastrulation which, for many and weighty reasons, we may regard as the original, primordial, or palingenetic form. We might call it “equal” or homogeneous segmentation, because the divided cells retain a resemblance to each other at first (and often until the formation of the blastoderm). We give the name of the “bell-gastrula,” or archigastrula, to the gastrula that succeeds it. In just the same form as in the coral we considered (Monoxenia, Fig. 29), we find it in the lowest zoophyta (the gastrophysema, Fig. 30), and the simplest sponges (olynthus, Fig. 36); also in many of the medusæ and hydrapolyps, lower types of worms of various classes (brachiopod, arrow-worm, Fig. 31), tunicates (ascidia), many of the echinoderms (Fig. 32), lower articulates (Fig. 33), and molluscs (Fig. 34), and, finally, in a slightly modified form, in the lowest vertebrate (the amphioxus, Fig. 35).
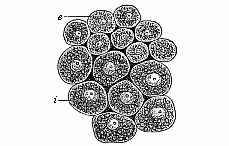 Fig. 37—Cells from the two primary germinal layers of the mammal (from both layers of the blastoderm). i larger and darker cells of the inner stratum, the vegetal layer or
entoderm. e smaller and clearer cells from the outer
stratum, the animal layer or ectoderm.
Fig. 37—Cells from the two primary germinal layers of the mammal (from both layers of the blastoderm). i larger and darker cells of the inner stratum, the vegetal layer or
entoderm. e smaller and clearer cells from the outer
stratum, the animal layer or ectoderm. |
The gastrulation of the amphioxus is especially interesting because this lowest and oldest of all the vertebrates is of the highest significance in connection with the evolution of the vertebrate stem, and therefore with that of man (compare Chapters XVI and XVII). Just as the comparative anatomist traces the most elaborate features in the structures of the various classes of vertebrates to divergent development from this simple primitive vertebrate, so comparative embryology traces the various secondary forms of vertebrate gastrulation to the simple, primary formation of the germinal layers in the amphioxus. Although this formation, as distinguished from the cenogenetic modifications of the vertebrate, may on the whole be regarded as palingenetic, it is nevertheless different in some features from the quite primitive gastrulation such as we have, for instance, in the Monoxenia (Fig. 29) and the Sagitta. Hatschek rightly observes that the segmentation of the ovum in the amphioxus is not strictly equal, but almost equal, and approaches the unequal. The difference in size between the two groups of cells continues to be very noticeable in the further course of the segmentation; the smaller animal cells of the upper hemisphere divide more quickly than the larger vegetal cells of the lower (Fig. 38 A, B). Hence the blastoderm, which forms the single-layer wall of the globular blastula at the end of the cleavage-process, does not consist of
homogeneous cells of equal size, as in the Sagitta and the Monoxenia; the cells of the upper half of the blastoderm (the mother-cells of the ectoderm) are more numerous and smaller, and the cells of the lower half (the mother-cells of the entoderm) less numerous and larger. Moreover, the segmentation-cavity of the blastula (Fig. 38 C, h) is not quite globular, but forms a flattened spheroid with unequal poles of its vertical axis. While the blastula is being folded into a cup at the vegetal pole of its axis, the difference in the size of the blastodermic cells increases (Fig. 38 D, E); it is most conspicuous when the invagination is complete and the segmentation-cavity has disappeared (Fig. 38 F). The larger vegetal cells of the entoderm are richer in granules, and so darker than the smaller and lighter animal cells of the ectoderm.
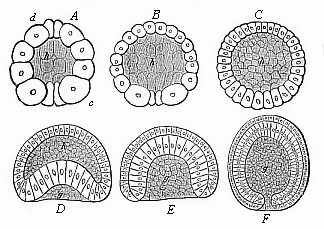 Fig. 38—Gastrulation of the amphioxus, from Hatschek (vertical section through the axis of the ovum). A, B, C three stages in the formation of the blastula; D, E curving of the blastula; F complete gastrula. h segmentation-cavity. g primitive gut-cavity. |
But the unequal gastrulation of the amphioxus diverges from the typical equal cleavage of the Sagitta, the Monoxenia (Fig. 29), and the Olynthus (Fig. 36), in another important particular. The pure archigastrula of the latter forms is uni-axial, and it is round in its whole length in transverse section. The vegetal pole of the vertical axis is just in the centre of the primitive mouth. This is not the case in the gastrula of the amphioxus. During the folding of the blastula the ideal axis is already bent on one side, the growth of the blastoderm (or the increase of its cells) being brisker on one side than on the other; the side that grows more quickly, and so is more curved (Fig. 39 v), will be the anterior or belly-side, the opposite, flatter side will form the back (d). The primitive mouth, which at first, in the typical archigastrula, lay at the vegetal pole of the main axis, is forced away to the dorsal side; and whereas its two lips lay at first in a plane at right angles to the chief axis, they are now so far thrust aside that their plane cuts the axis at a sharp angle. The dorsal lip is therefore the upper and more forward, the ventral lip the lower and hinder. In the latter, at the ventral passage of the entoderm into the ectoderm, there lie side by side a pair of very large cells, one to the right and one to the left (Fig. 39 p): these are the important polar cells of the primitive mouth, or “the primitive cells of the mesoderm.” In consequence of these considerable variations arising in the course of the gastrulation, the primitive uni-axial form of the archigastrula in the amphioxus has already become tri-axial, and thus the two-sidedness, or bilateral symmetry, of the vertebrate body has already been determined. This has been transmitted from the amphioxus to all the other modified gastrula-forms of the vertebrate stem.
Apart from this bilateral structure, the gastrula of the amphioxus resembles the typical archigastrula of the lower animals (Figs. 30–36) in developing the two primary germinal layers from a single layer of cells. This is clearly the oldest and original form of the metazoic embryo. Although the animals I have mentioned belong to the most diverse classes, they nevertheless agree with each other, and many more animal forms, in having retained to the present day, by a conservative heredity, this palingenetic form of gastrulation which they have from their
earliest common ancestors. But this is not the case with the great majority of the animals. With these the original embryonic process has been gradually more or less altered in the course of millions of years by adaptation to new conditions of development. Both the segmentation of the ovum and the subsequent gastrulation have in this way been considerably changed. In fact, these variations have become so great in the course of time that the segmentation was not rightly understood in most animals, and the gastrula was unrecognised. It was not until I had made an extensive comparative study, lasting a considerable time (in the years 1866–75), in animals of the most diverse classes, that I succeeded in showing the same common typical process in these apparently very different forms of gastrulation, and tracing them all to one original form. I regard all those that diverge from the primary palingenetic gastrulation as secondary, modified, and cenogenetic. The more or less divergent form of gastrula that is produced may be called a secondary, modified gastrula, or a metagastrula. The reader will find a scheme of these different kinds of segmentation and gastrulation at the close of this chapter.
By far the most important process that determines the various cenogenetic forms of gastrulation is the change in the nutrition of the ovum and the accumulation in it of nutritive yelk. By this we understand various chemical substances (chiefly granules of albumin and fat-particles) which serve exclusively as reserve-matter or food for the embryo. As the metazoic embryo in its earlier stages of development is not yet able to obtain its food and so build up the frame, the necessary material has to be stored up in the ovum. Hence we distinguish in the ova two chief elements—the active formative yelk (protoplasm) and the passive food-yelk (deutoplasm, wrongly spoken of as “the yelk”). In the little palingenetic ova, the segmentation of which we have already considered, the yelk-granules are so small and so regularly distributed in the protoplasm of the ovum that the even and repeated cleavage is not affected by them. But in the great majority of the animal ova the food-yelk is more or less considerable, and is stored in a certain part of the ovum, so that even in the unfertilised ovum the “granary” can clearly be distinguished from the formative plasm. As a rule, the formative-yelk (with the germinal vesicle) then usually gathers at one pole and the food-yelk at the other. The first is the animal, and the second the vegetal, pole of the vertical axis of the ovum.
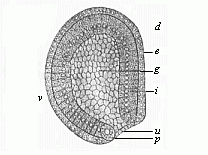 Fig.
39—Gastrula of the amphioxus, seen from left side
(diagrammatic median section). (From Hatschek.) g
primitive gut, u primitive mouth, p peristomal
pole-cells, i entoderm, e ectoderm, d dorsal
side, v ventral side.
Fig.
39—Gastrula of the amphioxus, seen from left side
(diagrammatic median section). (From Hatschek.) g
primitive gut, u primitive mouth, p peristomal
pole-cells, i entoderm, e ectoderm, d dorsal
side, v ventral side. |
In these “telolecithal” ova, or ova with the yelk at one end (for instance, in the cyclostoma and amphibia), the gastrulation then usually takes place in such a way that in the cleavage of the impregnated ovum the animal (usually the upper) half splits up more quickly than the vegetal (lower). The contractions of the active protoplasm, which effect this continual cleavage of the cells, meet a greater resistance in the lower vegetal half from the passive deutoplasm than in the upper animal half. Hence we find in the latter more but smaller, and in the former fewer but larger, cells. The animal cells produce the external, and the vegetal cells the internal, germinal layer.
Although this unequal segmentation of the cyclostoma, ganoids, and amphibia seems at first sight to differ from the original equal segmentation (for instance, in the monoxenia, Fig. 29), they both have this in common, that the cleavage process throughout affects the whole cell; hence Remak called it total segmentation, and the ova in question holoblastic, or “whole-cleaving.” It is otherwise with the second chief group of ova, which he distinguished from these as meroblastic, or “partially-cleaving ”: to this class belong the familiar large eggs of birds and reptiles, and of most fishes. The inert mass of the passive food-yelk is so
large in these cases that the protoplasmic contractions of the active yelk cannot effect any further cleavage. In consequence, there is only a partial segmentation. While the protoplasm in the animal section of the ovum continues briskly to divide, multiplying the nuclei, the deutoplasm in the vegetal section remains more or less undivided; it is merely consumed as food by the forming cells. The larger the accumulation of food, the more restricted is the process of segmentation. It may, however, continue for some time (even after the gastrulation is more or less complete) in the sense that the vegetal cell-nuclei distributed in the deutoplasm slowly increase by cleavage; as each of them is surrounded by a small quantity of protoplasm, it may afterwards appropriate a portion of the food-yelk, and thus form a real “yelk-cell” (merocyte). When this vegetal cell-formation continues for a long time, after the two primary germinal layers have been formed, it takes the name of the “after-segmentation.”
The meroblastic ova are only found in the larger and more highly developed animals, and only in those whose embryo needs a longer time and richer nourishment within the fœtal membranes. According as the yelk-food accumulates at the centre or at the side of the ovum, we distinguish two groups of dividing ova, periblastic and discoblastic. In the periblastic the food-yelk is in the centre, enclosed inside the ovum (hence they are also called “centrolecithal” ova): the formative yelk surrounds the food-yelk, and so suffers itself a superficial cleavage. This is found among the articulates (crabs, spiders, insects, etc.). In the discoblastic ova the food-yelk gathers at one side, at the vegetal or lower pole of the vertical axis, while the nucleus of the ovum and the great bulk of the formative yelk lie at the upper or animal pole (hence these ova are also called “telolecithal”). In these cases the cleavage of the ovum begins at the upper pole, and leads to the formation of a dorsal discoid embryo. This is the case with all meroblastic vertebrates, most fishes, the reptiles and birds, and the oviparous mammals (the monotremes).
The gastrulation of the discoblastic ova, which chiefly concerns us, offers serious difficulties to microscopic investigation and philosophic consideration. These, however, have been mastered by the comparative embryological research which has been conducted by a number of distinguished observers during the last few decades—especially the brothers Hertwig, Rabl, Kupffer, Selenka, Rückert, Goette, Rauber, etc. These thorough and careful studies, aided by the most perfect modern improvements in technical method (in tinting and dissection), have given a very welcome support to the views which I put forward in my work, On the Gastrula and the Segmentation of the Animal Ovum [not translated], in 1875. As it is very important to understand these views and their phylogenetic foundation clearly, not only as regards evolution in general, but particularly in connection with the genesis of man, I will give here a brief statement of them as far as they concern the vertebrate-stem:—
1. All the vertebrates, including man, are phylogenetically (or genealogically) related—that is, are members of one single natural stem.
2. Consequently, the embryonic features in their individual development must also have a genetic connection.
3. As the gastrulation of the amphioxus shows the original palingenetic form in its simplest features, that of the other vertebrates must have been derived from it.
4. The cenogenetic modifications of the latter are more appreciable the more food-yelk is stored up in the ovum.
5. Although the mass of the food-yelk may be very large in the ova of the discoblastic vertebrates, nevertheless in every case a blastula is developed from the morula, as in the holoblastic ova.
6. Also, in every case, the gastrula develops from the blastula by curving or invagination.
7. The cavity which is produced in the fœtus by this curving is, in each case, the primitive gut (progaster), and its opening the primitive mouth (prostoma).
8. The food-yelk, whether large or small, is always stored in the ventral wall of the primitive gut; the cells (called “merocytes”) which may be formed in it subsequently (by “after-segmentation”) also belong to the inner germinal layer, like the cells which immediately enclose the primitive gut-cavity.
9. The primitive mouth, which at first lies below at the lower pole of the vertical axis, is forced, by the growth of the yelk, backwards and then upwards,
towards the dorsal side of the embryo; the vertical axis of the primitive gut is thus gradually converted into horizontal.
10. The primitive mouth is closed sooner or later in all the vertebrates, and does not evolve into the permanent mouth-aperture; it rather corresponds to the “properistoma,” or region of the anus. From this important point the formation of the middle germinal layer proceeds, between the two primary layers.
The wide comparative studies of the scientists I have named have further shown that in the case of the discoblastic higher vertebrates (the three classes of amniotes) the primitive mouth of the embryonic disc, which was long looked for in vain, is found always, and is nothing else than the familiar “primitive groove.” Of this we shall see more as we proceed. Meantime we realise that gastrulation may be reduced to one and the same process in all the vertebrates. Moreover, the various forms it takes in the invertebrates can always be reduced to one of the four types of segmentation described above. In relation to the distinction between total and partial segmentation, the grouping of the various forms is as follows:—
| I. Palingenetic (primitive) segmentation. |
1. Equal segmentation (bell-gastrula). |
A. Total
segmentation (without independent food-yelk). |
| II. Cenogenetic
segmentation (modified by adaptation). |
2. Unequal segmentation (hooded gastrula). |
|
| 3. Discoid segmentation (discoid gastrula). |
B. Partial
segmentation (with independent food-yelk). |
|
| 4. Superficial segmentation (spherical gastrula). |
The lowest metazoa we know—namely, the lower zoophyta (sponges, simple polyps, etc.)—remain throughout life at a stage of development which differs little from the gastrula; their whole body consists of two layers of cells. This is a fact of extreme importance. We see that man, and also other vertebrates, pass quickly through a stage of development in which they consist of two layers, just as these lower zoophyta do throughout life. If we apply our biogenetic law to the matter, we at once reach this important conclusion. “Man and all the other animals which pass through the two-layer stage, or gastrula-form, in the course of their embryonic development, must descend from a primitive simple stem-form, the whole body of which consisted throughout life (as is the case with the lower zoophyta to-day) merely of two cell-strata or germinal layers.” We will call this primitive stem-form, with which we shall deal more fully later on, the gastræa—that is to say, “primitive-gut animal.”
According to this gastræa-theory there was originally in all the multicellular animals one organ with the same structure and function. This was the primitive gut; and the two primary germinal layers which form its wall must also be regarded as identical in all. This important homology or identity of the primary germinal layers is proved, on the one hand, from the fact that the gastrula was originally formed in the same way in all cases—namely, by the curving of the blastula; and, on the other hand, by the fact that in every case the same fundamental organs arise from the germinal layers. The outer or animal layer, or ectoderm, always forms the chief organs of animal life—the skin, nervous system, sense-organs, etc.; the inner or vegetal layer, or entoderm, gives rise to the chief organs of vegetative life—the organs of nourishment, digestion, blood-formation, etc.
In the lower zoophyta, whose body remains at the two-layer stage throughout life, the gastræads, the simplest sponges (Olynthus), and polyps (Hydra), these two groups of functions, animal and vegetative, are strictly divided between the two simple primary layers. Throughout life the outer or animal layer acts simply as a covering for the body, and accomplishes its movement and sensation. The inner or vegetative layer of cells acts throughout life as a gut-lining, or nutritive layer of enteric cells, and often also yields the reproductive cells.
The best known of these “gastræads,” or “gastrula-like animals,” is the common fresh-water polyp (Hydra). This simplest of all the cnidaria has, it is true, a crown of tentacles round its mouth. Also its outer germinal layer has certain special modifications. But these are secondary additions, and the inner germinal layer is a simple stratum of cells. On the whole, the hydra has preserved to our day by heredity the simple structure of our primitive ancestor, the gastræa (cf. Chapter XIX).
In all other animals, particularly the vertebrates, the gastrula is merely a brief transitional stage. Here the two-layer stage of the embryonic development is quickly succeeded by a three-layer, and then a four-layer, stage. With the appearance of the four superimposed germinal layers we reach again a firm and steady standing-ground, from which we may follow the further, and much more difficult and complicated, course of embryonic development.
The animal stems are indicated by the letters
a–g: a Zoophyta. b Annelida.
c Mollusca. d Echinoderma. e Articulata.
f Tunicata. g Vertebrata.
| I. Total Segmentation. Holoblastic ova. Gastrula without separate food-yelk. Hologastrula. |
I. Primitive Segmentation. Archiblastic ova. Bell-gastrula (archigastrula.) |
a. Many lower zoophyta (sponges, hydrapolyps, medusæ, simpler corals). b. Many lower annelids (sagitta, phoronis, many nematoda, etc., terebratula, argiope, pisidium). c. Some lower molluscs. d. Many echinoderms. e. A few lower articulata (some brachiopods, copepods: Tardigrades, pteromalina). f. Many tunicata. g. The acrania (amphioxus). |
| II. Unequal Segmentation. Amphiblastic ova. Hooded-gastrula (amphigastrula). |
a. Many zoophyta (sponges,
medusæ, corals, siphonophoræ, ctenophora). b. Most worms. c. Most molluscs. d. Many echinoderms (viviparous species and some others). e. Some of the lower articulata (both crustacea and tracheata). f. Many tunicata. g. Cyclostoma, the oldest fishes, amphibia, mammals (not including man). |
|
| II. Partial Segmentation. Meroblastic ova. Gastrula with separate food-yelk. Merogastrula. |
III. Discoid Segmentation. Discoblastic ova. Discoid gastrula. |
c. Cephalopods or cuttlefish. e. Many articulata, wood-lice, scorpions, etc. g. Primitive fishes, bony fishes, reptiles, birds, monotremes. |
| IV. Superficial Segmentation. Periblastic ova. Spherical-gastrula. |
e. The great majority of the articulata (crustaceans, myriapods, arachnids, insects). |
Title and Contents
Glossary
Chapter VII
Chapter IX
Figs. 1–209
Figs. 210–408