THE EVOLUTION OF MAN
Volume II
CHAPTER XXVIII
EVOLUTION OF THE VASCULAR SYSTEM
The use that we have hitherto made of our biogenetic law will give the reader an idea how far we may trust its guidance in phylogenetic investigation. This differs considerably in the various systems of organs; the reason is that heredity and variability have a very different range in these systems. While some of them faithfully preserve the original palingenetic development inherited from earlier animal ancestors, others show little trace of this rigid heredity; they are rather disposed to follow new and divergent cenogenetic lines of development in consequence of adaptation. The organs of the first kind represent the conservative element in the multicellular state of the human frame, while the latter represent the progressive element. The course of historic development is a result of the correlation of the two tendencies, and they must be carefully distinguished.
There is perhaps no other system of organs in the human body in which this is more necessary than in that of which we are now going to consider the obscure development—the vascular system, or apparatus of circulation. If we were to draw our conclusions as to the original features in our earlier animal ancestors solely from the phenomena of the development of this system in the embryo of man and the other higher Vertebrates, we should be wholly misled. By a number of important embryonic adaptations, the chief of which is the formation of an extensive food-yelk, the original course of the development of the vascular system has been so much falsified and curtailed in the higher Vertebrates that little or nothing now remains in their embryology of some of the principal phylogenetic features. We should be quite unable to explain these if comparative anatomy and ontogeny did not come to our assistance.
The vascular system in man and all the Craniotes is an elaborate apparatus of cavities filled with juices or cell-containing fluids. These “vessels” (vascula) play an important part in the nutrition of the body. They partly conduct the nutritive red blood to the various parts of the body (blood-vessels); partly absorb from the gut the white chyle formed in digestion (chyle-vessels); and partly collect the used-up juices and convey them away from the tissues (lymphatic vessels). With the latter are connected the large cavities of the body, especially the body-cavity, or cœloma. The lymphatic vessels conduct both the colourless lymph and the white chyle into the venous part of the circulation. The lymphatic glands act as producers of new blood-cells, and with them is associated the spleen. The centre of movement for the circulation of the fluids is the heart, a strong muscular sac, which contracts regularly and is equipped with valves like a pump. This constant and steady circulation of the blood makes possible the complex metabolism of the higher animals.
But, however important the vascular system may be to the more advanced and larger and highly-differentiated animals, it is not at all so indispensable an element of animal life as is commonly supposed. The older science of medicine regarded the blood as the real source of life. Even in the still prevalent confused notions of heredity the blood plays the chief part. People speak generally of full blood, half blood, etc., and imagine that the hereditary transmission of certain characters “lies in the blood.” The incorrectness of these ideas is clearly seen from the fact that in the act of generation the blood of the parents is not directly transmitted to the offspring, nor does the embryo possess blood in its early stages. We have already seen that not only the differentiation of the four secondary germinal layers, but also the first structures of the principal organs in the embryo of all the Vertebrates, take place long before there is any
[ 319 ]
trace of the vascular system—the heart and the blood. In accordance with this ontogenetic fact, we must regard the vascular system as one of the latest organs from the phylogenetic point of view; just as we have found the alimentary canal to be one of the earliest. In any case, the vascular system is much later than the alimentary.
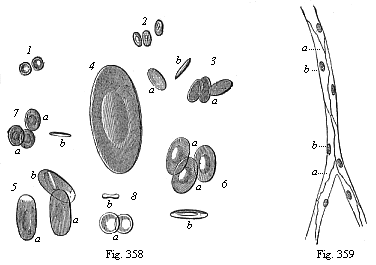 Fig. 358—Red blood-cells of various Vertebrates (equally magnified). 1. of man, 2. camel, 3. dove, 4. proteus, 5. water-salamander (Triton), 6. frog, 7. merlin (Cobitis), 8. lamprey (Petromyzon). a surface-view, b edge-view. (From Wagner.)
Fig. 358—Red blood-cells of various Vertebrates (equally magnified). 1. of man, 2. camel, 3. dove, 4. proteus, 5. water-salamander (Triton), 6. frog, 7. merlin (Cobitis), 8. lamprey (Petromyzon). a surface-view, b edge-view. (From Wagner.)
Fig. 359—Vascular tissues or endothelium (vasalium). A capillary from the mesentery. a vascular cells, b their nuclei. |
The important nutritive fluid that circulates as blood and lymph in the elaborate canals of our vascular system is not a clear, simple fluid, but a very complex chemical juice with millions of cells floating in it. These blood-cells are just as important in the complicated life of the higher animal body as the circulation of money is to the commerce of a civilised community. Just as the citizens meet their needs most conveniently by means of a financial circulation, so the various tissue-cells, the microscopic citizens of the multicellular human body, have their food conveyed to them best by the circulating cells in the blood. These blood cells (hæmocytes) are of two kinds in man and all the other Craniotes—red cells (rhodocytes or erythrocytes) and colourless or lymph cells (leucocytes). The red colour of the blood is caused by the great accumulation of the former, the others circulate among them in much smaller quantity. When the colourless cells increase at the expense of the red we get anæmia (or chlorosis).
The lymph-cells (leucocytes), commonly called the “white corpuscles” of the blood, are phylogenetically older and more widely distributed in the animal world than the red. The great majority of the Invertebrates that have acquired an independent vascular system have only colourless lymph-cells in the circulating fluid. There is an exception in the Nemertines (Fig. 358) and some groups of Annelids. When we examine the colourless blood of a cray-fish or a snail (Fig. 358) under a high power of the microscope, we find in each drop numbers of mobile leucocytes, which behave just like independent Amoebæ (Fig. 17). Like these unicellular Protozoa, the colourless blood-cells creep slowly about, their unshapely plasma-body constantly changing its form, and stretching out finger-like processes first in one direction, then another. Like the Amoebæ, they take particles into their cell-body. On account
[ 320 ]
of this feature these amoeboid plastids are called “eating cells” (phagocytes), and on account of their motions “travelling cells” (planocytes). It has been shown by the discoveries of the last few decades that these leucocytes are of the greatest physiological and pathological consequence to the organism. They can absorb either solid or dissolved particles from the wall of the gut, and convey them to the blood in the chyle; they can absorb and remove unusable matter from the tissues. When they pass in large quantities through the fine pores of the capillaries and accumulate at irritated spots, they cause inflammation. They can consume and destroy bacteria, the dreaded vehicles of infectious diseases; but they can also transport these injurious Monera to fresh regions, and so extend the sphere of infection. It is probable that the sensitive and travelling leucocytes of our invertebrate ancestors have powerfully co-operated for millions of years in the phylogenesis of the advancing animal organisation.
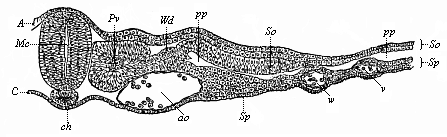 Fig. 360—Transverse section of the trunk of a chick-embryo, forty-five hours old. (From Balfour.) A ectoderm (horny-plate), Mc medullary tube, ch chorda, C entoderm (gut-gland layer), Pv primitive segment (episomite), Wd prorenal duct, pp cœloma (secondary body-cavity). So skin-fibre layer, Sp gut-fibre layer, v blood-vessels in latter, ao primitive aortas, containing red blood-cells.
Fig. 360—Transverse section of the trunk of a chick-embryo, forty-five hours old. (From Balfour.) A ectoderm (horny-plate), Mc medullary tube, ch chorda, C entoderm (gut-gland layer), Pv primitive segment (episomite), Wd prorenal duct, pp cœloma (secondary body-cavity). So skin-fibre layer, Sp gut-fibre layer, v blood-vessels in latter, ao primitive aortas, containing red blood-cells. |
The red blood-cells have a much more restricted sphere of distribution and activity. But they also are very important in connection with certain functions of the craniote-organism, especially the exchange of gases or respiration. The cells of the dark red, carbonised or venous, blood, which have absorbed carbonic acid from the animal tissues, give this off in the respiratory organs; they receive instead of it fresh oxygen, and thus bring about the bright red colour that distinguishes oxydised or arterial blood. The red colouring matter of the blood (hæmoglobin) is regularly distributed in the pores of their protoplasm. The red cells of most of the Vertebrates are elliptical flat disks, and enclose a nucleus of the same shape; they differ a good deal in size (Fig. 358). The mammals are distinguished from the other Vertebrates by the circular form of their biconcave red cells and by the absence of a nucleus (Fig. 1); only a few genera still have the elliptic form inherited from the reptiles (Fig. 2). In the embryos of the mammals the red cells have a nucleus and the power of increasing by cleavage (Fig. 10).
The origin of the blood-cells and vessels in the embryo, and their relation to the germinal layers and tissues, are among the most difficult problems of ontogeny—those obscure questions on which the most divergent opinions are still advanced by the most competent scientists. In general, it is certain that the greater part of the cells that compose the vessels and their contents come from the mesoderm—in fact, from the gut-fibre layer; it was on this account that Baer gave the name of “vascular layer” to this visceral layer of the coeloma. But other important observers say that a part of these cells come from other germinal layers, especially from the gut-gland layer. It seems to be true that blood-cells may be formed from the cells of the entoderm before the development of the mesoderm. If we examine sections of chickens, the earliest and most familiar subjects of embryology, we find at an early stage the “primitive-aortas” we have already described (Fig. 360 ao) in the ventral angle between the episoma (Pv) and hyposoma (Sp). The
[ 321 ]
thin wall of these first vessels of the amniote embryo consists of flat cells (endothelia or vascular epithelia); the fluid within already contains numbers of red blood-cells; both have been developed from the gut-fibre layer. It is the same with the vessels of the germinative area (Fig. 361 v), which lie on the entodermic membrane of the yelk-sac (c). These features are seen still more clearly in the transverse section of the duck-embryo in Fig. 152. In this we see clearly how a number of stellate cells proceed from the “vascular layer” and spread in all directions in the “primary body-cavity”—i.e. in the spaces between the germinal layers. A part of these travelling cells come together and line the wall of the larger spaces, and thus form the first vessels; others enter into the cavity, live in the fluid that fills it, and multiply by cleavage—the first blood-cells.
But, besides these mesodermic cells of the “vascular layer” proper, other travelling cells, of which the origin and purport are still obscure, take part in the formation of blood in the meroblastic Vertebrates (especially fishes). The chief of these are those that Ruckert has most aptly denominated “merocytes.” These “eating yelk-cells” are found in large numbers in the food-yelk of the Selachii, especially in the yelk-wall—the border zone of the germinal disk in which the embryonic vascular net is first developed. The nuclei of the merocytes become ten times as large as the ordinary cell-nucleus, and are distinguished by their strong capacity for taking colour, or their special richness in chromatin. Their protoplasmic body resembles the stellate cells of osseous tissue (astrocytes), and behaves just like a rhizopod (such as Gromia); it sends out numbers of stellate processes all round, which ramify and stretch into the surrounding food-yelk. These variable and very mobile processes, the pseudopodia of the merocytes, serve both for locomotion and for getting food; as in the real rhizopods, they surround the solid particles of food (granules and plates of yelk), and accumulate round their nucleus the food they have received and digested. Hence we may regard them both as eating-cells (phagocytes) and travelling-cells (planocytes). Their lively nucleus divides quickly and often repeatedly, so that a number of new nuclei are formed in a short time; as each fresh nucleus surrounds itself with a mantle of protoplasm, it provides a new cell for the construction of the embryo. Their origin is still much disputed.
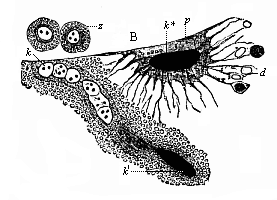 |
Fig. 361—Merocytes of a shark-embryo, rhizopod-like yelk-cells underneath the embryonic cavity (B). (From Ruckert.) z two embryonic cells, k nuclei of the merocytes, which wander about in the yelk and eat small yelk-plates (d), k smaller, more superficial, lighter nuclei, k′ a deeper nucleus, in the act of cleavage, k* asterisk chromatin-filled border-nucleus, freed from the surrounding yelk in order to show the numerous pseudopodia of the protoplasmic cell-body. |
Half of the twelve stems of the animal world have no blood-vessels. They make their first appearance in the Vermalia. Their earliest source is the primary body-cavity, the simple space between the two primary germinal layers, which is either a relic of the segmentation-cavity, or is a subsequent formation. Amoeboid planocytes, which migrate from the entoderm and reach this fluid-filled primary cavity, live and multiply there, and form the first colourless blood-cells. We find the vascular system in this very simple form to-day in the Bryozoa, Rotatoria, Nematoda, and other lower Vermalia.
The first step in the improvement of this primitive vascular system is the formation of larger canals or blood-conducting tubes. The spaces filled with blood, the relics of the primary body-cavity, receive a special wall. “Blood-vessels” of this kind (in the narrower sense) are found among the higher worms in various forms, sometimes very simple, at other times very complex. The form
[ 322 ]
that was probably the incipient structure of the elaborate vascular system of the Vertebrates (and of the Articulates) is found in two primordial principal vessels—a dorsal vessel in the middle line of the dorsal wall of the gut, and a ventral vessel that runs from front to rear in the middle line of its ventral wall. From the dorsal vessel is evolved the aorta (or principal artery), from the ventral vessel the principal or subintestinal vein. The two vessels are connected in front and behind by a loop that runs round the gut. The blood contained in the two tubes is propelled by their peristaltic contractions.
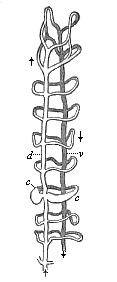 Fig. 362—Vascular system of an Annelid (Sænuris), foremost section. d dorsal vessel, v ventral vessel, c transverse connection of two (enlarged in shape of heart). The arrows indicate the direction of the flow of blood. (From Gegenbaur.
Fig. 362—Vascular system of an Annelid (Sænuris), foremost section. d dorsal vessel, v ventral vessel, c transverse connection of two (enlarged in shape of heart). The arrows indicate the direction of the flow of blood. (From Gegenbaur. |
The earliest Vermalia in which we first find this independent vascular system are the Nemertina (Fig. 244). As a rule, they have three parallel longitudinal vessels connected by loops, a single dorsal vessel above the gut and a pair of lateral vessels to the right and left. In some of the Nemertina the blood is already coloured, and the red colouring matter is real hæmoglobin, connected with elliptical discoid cells, as in the Vertebrates. The further evolution of this rudimentary vascular system can be gathered from the class of the Annelids in which we find it at various stages of development. First, a number of transverse connections are formed between the dorsal and ventral vessels, which pass round the gut ring-wise (Fig. 362). Other vessels grow into the body-wall and ramify in order to convey blood to it. In addition to the two large vessels of the middle plane there are often two lateral vessels, one to the right and one to the left; as, for instance, in the leech. There are four of these parallel longitudinal vessels in the Enteropneusts (Balanoglossus, Fig. 245). In these important Vermalia the foremost section of the gut has already been converted into a gill-crate, and the vascular arches that rise in the wall of this from the ventral to the dorsal vessel have become branchial vessels.
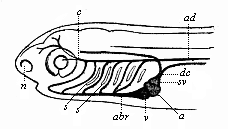 Fig. 363—Head of a fish-embryo, with rudimentary vascular system, from the left. dc Cuvier’s duct (juncture of the anterior and posterior principal veins), sv venous sinus (enlarged end of Cuvier’s duct), a auricle, v ventricle, abr trunk of branchial artery, s gill-clefts (arterial arches between), ad aorta, c carotid artery, n nasal pit. (From Gegenbaur.
Fig. 363—Head of a fish-embryo, with rudimentary vascular system, from the left. dc Cuvier’s duct (juncture of the anterior and posterior principal veins), sv venous sinus (enlarged end of Cuvier’s duct), a auricle, v ventricle, abr trunk of branchial artery, s gill-clefts (arterial arches between), ad aorta, c carotid artery, n nasal pit. (From Gegenbaur. |
We have a further important advance in the Tunicates, which we have recognised as the nearest blood-relatives of our early vertebrate ancestors. Here we find for the first time a real heart—i.e. a central organ of circulation, driving the blood into the vessels by the regular contractions of its muscular wall, it is of a very rudimentary character, a spindle-shaped tube, passing at both ends into a principal vessel (Fig. 221). By its original position behind the gill-crate, on ventral side of the Tunicates (sometimes more, sometimes less, forward), the head shows clearly that it has been formed by the local enlargement of a section of the ventral vessel. We have already noticed the remarkable alternation of the direction of the blood stream, the heart driving it first from one end, then from the other p. 190. This is very instructive, because in most of the worms (even the Enteropneust) the blood in the dorsal vessel travels from back to front, but in the Vertebrates in the opposite direction. As the Ascidia-heart alternates steadily from one direction to the other, it shows us permanently, in a sense, the phylogenetic transition from the earlier forward direction of the dorsal current (in the worms) to the new backward direction (in the Vertebrates).
As the new direction became permanent in the earlier Prochordonia, which gave rise to the Vertebrate stem, the two vessels that proceed from either end of the tubular heart acquired a fixed function.
[ 323 ]
The foremost section of the ventral vessel henceforth always conveys blood from the heart, and so acts as an artery; the hind section of the same vessel brings the blood from the body to the heart, and so becomes a vein. In view of their relation to the two sections of the gut, we may call the latter the intestinal vein and the former the branchial artery. The blood contained in both vessels, and also in the heart, is venous or carbonised blood—i.e. rich in carbonic acid; on the other hand, the blood that passes from the gills into the dorsal vessel is provided with fresh oxygen—arterial or oxydised blood. The finest branches of the arteries and veins pass into each other in the tissues by means of a network of very fine, ventral, hair-like vessels, or capillaries (Fig. 359).
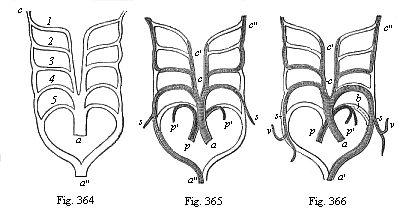 Fig. 364—The five arterial arches of the Craniotes (1–5) in their original disposition. a arterial cone or bulb, a″ aorta-trunk, c carotid artery (foremost continuation of the roots of the aorta). (From Rathke.)
Fig. 364—The five arterial arches of the Craniotes (1–5) in their original disposition. a arterial cone or bulb, a″ aorta-trunk, c carotid artery (foremost continuation of the roots of the aorta). (From Rathke.)
Fig. 365—The five arterial arches of the birds; the lighter parts of the structure disappear; only the shaded parts remain. Letters as in Fig. 364. s subclavian arteries, p pulmonary artery, p′ branches of same, c′ outer carotid, c″ inner carotid. (From Rathke.)
Fig. 366—The five arterial arches of mammals; letters as in Fig. 365. v vertebral artery, b Botall’s duct (open in the embryo, closed afterwards). (From Rathke.) |
When we turn from the Tunicates to the closely-related Amphioxus we are astonished at first to find an apparent retrogression in the formation of the vascular system. As we have seen, the Amphioxus has no real heart; its colourless blood is driven along in its vascular system by the principal vessel itself, which contracts regularly in its whole length (cf. Fig. 210). A dorsal vessel that lies above the gut (aorta) receives the arterial blood from the gills and drives it into the body. Returning from here, the venous blood gathers in a ventral vessel under the gut (intestinal vein), and goes back to the gills. A number of branchial vascular arches, which effect respiration and rise in the wall of the branchial gut from belly to back, absorb oxygen from the water and give off carbonic acid; they connect the ventral with the dorsal vessel. As the same section of the ventral vessel, which also forms the heart in the Craniotes, has developed in the Ascidia into a simple tubular heart, we may regard the absence of this in the Amphioxus as a result of degeneration, a return in this case to the earlier form of the vascular system, as we find it in many of the worms. We may assume that the Acrania that really belong to our ancestral series did not share this retrogression, but inherited the one-chambered heart of the Prochordonia, and transmitted it directly to the earliest Craniotes (cf. the ideal Primitive Vertebrate, Prospondylus, Figs. 98–102).
The further phylogenetic evolution of the vascular system is revealed to us by the comparative anatomy of the Craniotes. At the lowest stage of this group, in the Cyclostomes, we find for the first time the differentiation of the vasorium into two sections: a system of blood-vessels proper, which convey the red blood about the body, and a system of lymphatic vessels,
[ 324 ]
which absorb the colourless lymph from the tissues and convey it to the blood. The lymphatics that absorb from the gut and pour into the blood-stream the milky food-fluid formed by digestion are distinguished by the special name of “chyle-vessels.” While the chyle is white on account of its high proportion of fatty particles, the lymph proper is colourless. Both chyle and lymph contain the colourless amœboid cells (leucocytes, Fig. 12) that we also find distributed in the blood as colourless blood-cells (or “white corpuscles”); but the blood also contains a much larger quantity of red cells, and these give its characteristic colour to the blood of the Craniotes (rhodocytes, Fig. 358). The distinction between lymph, chyle, and blood-vessels which is found in all the Craniotes may be regarded as an outcome of division of labour between various sections of our originally simple vascular system. In the Gnathostomes the spleen makes its first appearance, an organ rich in blood, the chief function of which is the extensive formation of new colourless and red cells. It is not found in the Acrania and Cyclostomes, or any of the Invertebrates. It has been transmitted from the earliest fishes to all the Craniotes.
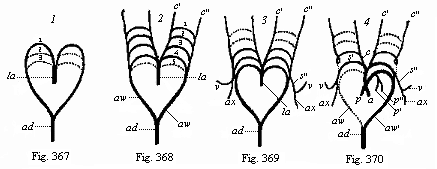 Figs. 367–70—Metamorphosis of the five arterial arches in the human embryo (diagram from Rathke). la arterial cone, 1, 2, 3, 4, 5 first to fifth pair of arteries, ad trunk of aorta, aw roots of aorta. In Fig. 367 only three, in Fig. 368 all five, of the aortic arches are given (the dotted ones only are developed). In Fig. 369 the first two pairs have disappeared again. In Fig. 370 the permanent trunks of the artery are shown; the dotted parts disappear, s subclavian artery, v vertebral, ax axillary, c carotid (c′ outer, c″ inner carotid), p pulmonary.
Figs. 367–70—Metamorphosis of the five arterial arches in the human embryo (diagram from Rathke). la arterial cone, 1, 2, 3, 4, 5 first to fifth pair of arteries, ad trunk of aorta, aw roots of aorta. In Fig. 367 only three, in Fig. 368 all five, of the aortic arches are given (the dotted ones only are developed). In Fig. 369 the first two pairs have disappeared again. In Fig. 370 the permanent trunks of the artery are shown; the dotted parts disappear, s subclavian artery, v vertebral, ax axillary, c carotid (c′ outer, c″ inner carotid), p pulmonary. |
The heart also, the central organ of circulation in all the Craniotes, shows an advance in structure in the Cyclostomes. The simple, spindle-shaped heart-tube, found in the same form in the embryo of all the Craniotes, is divided into two sections or chambers in the Cyclostomes, and these are separated by a pair of valves. The hind section, the auricle, receives the venous blood from the body and passes it on to the anterior section, the ventricle. From this it is driven through the trunk of the branchial artery (the foremost section of the ventral vessel or principal vein) into the gills.
In the Selachii an arterial cone is developed from the foremost end of the ventricle, as a special division, cut off by valves. It passes into the enlarged base of the trunk of the branchial artery (Fig. 363 abr). On each side 5–7 arteries proceed from it. These rise between the gill-clefts (s) on the gill-arches, surround the gullet, and unite above into a common trunk-aorta, the continuation of which over the gut corresponds to the dorsal vessel of the worms. As the curved arteries on the gill-arches spread into a network of respiratory capillaries, they contain venous blood in their lower part (as arches of the branchial artery) and arterial blood in the upper part (as arches of the aorta). The junctures of the various aortic arches on the right and left are called the roots of the aorta. Of an originally large number of aortic arches there remain at first six, then (owing to degeneration of the fifth arch) only five, pairs; and from these five pairs (Fig. 364) the chief parts of the arterial system develop in all the higher Vertebrates.
The appearance of the lungs and the atmospheric respiration connected therewith, which we first meet in the Dipneusts, is the next important step in vascular evolution. In the Dipneusts the auricle of the heart is divided by an incomplete partition into two halves. Only the right
[ 325 ]
auricle now receives the venous blood from the veins of the body. The left auricle receives the arterial blood from the pulmonary veins. The two auricles have a common opening into the simple ventricle, where the two kinds of blood mix, and are driven through the arterial cone or bulb into the arterial arches. From the last arterial arches the pulmonary arteries arise (Fig. 365 p). These force a part of the mixed blood into the lungs, the other part of it going through the aorta into the body.
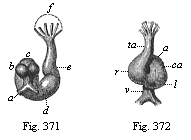 Fig. 371—Heart of a rabbit-embryo, from behind. a vitelline veins, b auricles of the heart, c atrium, d ventricle, e arterial bulb, f base of the three pairs of arterial arches. (From Bischoff.)
Fig. 371—Heart of a rabbit-embryo, from behind. a vitelline veins, b auricles of the heart, c atrium, d ventricle, e arterial bulb, f base of the three pairs of arterial arches. (From Bischoff.)
Fig. 372—Heart of the same embryo (Fig. 371), from the front. v vitelline veins, a auricle, ca auricular canal, l left ventricle, r right ventricle, ta arterial bulb. (From Bischoff.) |
From the Dipneusts upwards we now trace a progressive development of the vascular system, which ends finally with the loss of branchial respiration and a complete separation of the two halves of the circulation. In the Amphibia the partition between the two auricles is complete. In their earlier stages, as tadpoles (Fig. 262), they have still the branchial respiration and the circulation of the fishes, and their heart contains venous blood alone. Afterwards the lungs and pulmonary vessels are developed, and henceforth the ventricle of the heart contains mixed blood. In the reptiles the ventricle and its arterial cone begin to divide into two halves by a longitudinal partition, and this partition becomes complete in the higher reptiles and birds on the one hand, and the stem-forms of the mammals on the other. Henceforth, the right half of the heart contains only venous, and the left half only arterial, blood, as we find in all birds and mammals. The right auricle receives its carbonised or venous blood from the veins of the body, and the right ventricle drives it through the pulmonary arteries into the lungs. From here the blood returns, as oxydised or arterial blood, through the pulmonary veins to the left auricle, and is forced by the left ventricle into the arteries of the body. Between the pulmonary arteries and veins is the capillary system of the small or pulmonary circulation. Between the body-arteries and veins is the capillary system of the large or body-circulation. It is only in the two highest classes of Vertebrates—the birds and mammals—that we find a complete division of the circulations. Moreover, this complete separation has been developed quite independently in the two classes, as the dissimilar formation of the aortas shows of itself. In the birds the right half of the fourth arterial arch has become the permanent arch (Fig. 365). In the mammals this has been developed from the left half of the same fourth arch (Fig. 366).
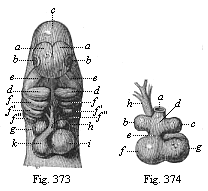 |
Fig. 373—Heart and head of a dog-embryo, from the front. a fore brain, b eyes, c middle brain, d primitive lower jaw, e primitive upper jaw, f gill-arches, g right auricle, h left auricle, i left ventricle, k right ventricle. (From Bischoff.)
Fig. 374—Heart of the same dog-embryo, from behind. a inosculation of the vitelline veins, b left auricle, c right auricle, d auricle, e auricular canal, f left ventricle, g right ventricle, h arterial bulb. (From Bischoff.) |
If we compare the fully-developed arterial system of the various classes of Craniotes, it shows a good deal of variety, yet it always proceeds from the same fundamental type. Its development is just the same in man as in the other mammals; in particular, the modification of the six pairs of arterial arches is the same in both (Figs. 367–370). At first there is only a single pair of arches, which
[ 326 ]
lie on the inner surface of the first pair of gill-arches. Behind this there then develop a second and third pair of arches (lying on the inner side of the second and third gill-arches, Fig. 367). Finally, we get a fourth, fifth, and sixth pair. Of the six primitive arterial arches of the Amniotes three soon pass away (the first, second, and fifth); of the remaining three, the third gives the carotids, the fourth the aortas, and the sixth (number 5 in Figs. 364 and 368) the pulmonary arteries.
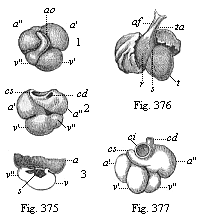 |
Fig. 375—Heart of a human embryo, four weeks old; 1. front view, 2. back view, 3. opened, and upper half of the atrium removed. a′ left auricle, a″ right auricle, v′ left ventricle, v″ right ventricle, ao arterial bulb, c superior vena cava (cd right, cs left), s rudiment of the interventricular wall. (From Kölliker.)
Fig. 376—Heart of a human embryo, six weeks old, front view. r right ventricle, t left ventricle, s furrow between ventricles, ta arterial bulb, af furrow on its surface; to right and left are the two large auricles. (From Ecker.)
Fig. 377—Heart of a human embryo, eight weeks old, back view. a′ left auricle, a″ right auricle, v′ left ventricle, v″ right ventricle, cd right superior vena cava, ci inferior vena cava. (From Kölliker.) |
The human heart also develops in just the same way as that of the other mammals (Fig. 378). We have already seen the first rudiments of its embryology, which in the main corresponds to its phylogeny (Figs. 201, 202). We saw that the palingenetic form of the heart is a spindle-shaped thickening of the gut-fibre layer in the ventral wall of the head-gut. The structure is then hollowed out, forms a simple tube, detaches from its place of origin, and henceforth lies freely in the cardiac cavity. Presently the tube bends into the shape of an S, and turns spirally on an imaginary axis in such a way that the hind part comes to lie on the dorsal surface of the fore part. The united vitelline veins open into the posterior end. From the anterior end spring the aortic arches.
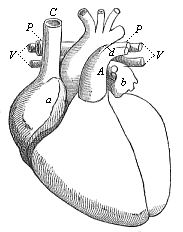 Fig. 378—Heart of the adult man, fully developed, front view, natural position. a right auricle (underneath it the right ventricle), b left auricle (under it the left ventricle), C superior vena cava, V pulmonary veins, P pulmonary artery, d Botalli’s duct, A aorta. (From Meyer.)
Fig. 378—Heart of the adult man, fully developed, front view, natural position. a right auricle (underneath it the right ventricle), b left auricle (under it the left ventricle), C superior vena cava, V pulmonary veins, P pulmonary artery, d Botalli’s duct, A aorta. (From Meyer.) |
This first structure of the human heart, enclosing a very simple cavity, corresponds to the tunicate-heart, and is a reproduction of that of the Prochordonia, but it now divides into two, and subsequently into three, compartments; this reminds us for a time of the heart of the Cyclostomes and fishes. The spiral turning and bending of the heart increases, and at the same time two transverse constrictions appear, dividing it externally into three sections (Figs. 371, 372). The foremost section, which is turned towards the ventral side, and from which the aortic arches rise, reproduces the arterial bulb of the Selachii. The middle section is a simple ventricle, and the hindmost, the section turned towards the dorsal side, into which the vitelline veins inosculate, is a simple auricle (or atrium). The latter forms, like the simple atrium of the fish-heart, a pair of lateral dilatations, the auricles (Fig. 371 b); and the constriction between the atrium and ventricle is called the auricular canal (Fig. 372 ca). The heart of the human embryo is now a complete fish-heart.
[ 327 ]
In perfect harmony with its phylogeny, the embryonic development of the human heart shows a gradual transition from the fish-heart, through the amphibian and reptile, to the mammal form, The most important point in the transition is the formation of a longitudinal partition—incomplete at first, but afterwards complete—which separates all three divisions of the heart into right (venous) and left (arterial) halves (cf. Figs. 373–378). The atrium is separated into a right and left half, each of which absorbs the corresponding auricle; into the right auricle open the body-veins (upper and lower vena cava, Figs. 375 c, 377 c); the left auricle receives the pulmonary veins. In the same way a superficial interventricular furrow is soon seen in the ventricle (Fig. 376 s). This is the external sign of the internal partition by which the ventricle is divided into two—a right venous and left arterial ventricle. Finally a longitudinal partition is formed in the third section of the primitive fish-like heart, the arterial bulb, externally indicated by a longitudinal furrow (Fig. 376 af). The cavity of the bulb is divided into two lateral halves, the pulmonary-artery bulb, that opens into the right ventricle, and the aorta-bulb, that opens into the left ventricle. When all the partitions are complete, the small (pulmonary) circulation is distinguished from the large (body) circulation; the motive centre of the former is the right half, and that of the latter the left half, of the heart.
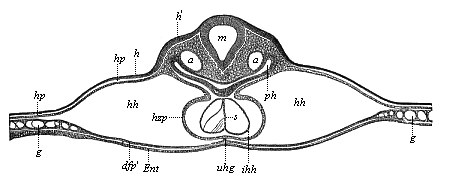 Fig. 379—Transverse section of the back of the head of a chick-embryo, forty hours old. (From Kölliker.) m medulla oblongata, ph pharyngeal cavity (head-gut), h horny plate, h′ thicker part of it, from which the auscultory pits afterwards develop, hp skin-fibre plate, hh cervical cavity (head-cœlom or cardiocœl), hzp cardiac plate (the outermost mesodermic wall of the heart), connected by the ventral mesocardium (uhg) with the gut-fibre layer or visceral cœlom-layer (dfp*prime;), Ent entoderm, ihh inner (entodermic?) wall of the heart; the two endothelial cardiac tubes are still separated by the cenogenetic septum (s) of the Amniotes, g vessels.
Fig. 379—Transverse section of the back of the head of a chick-embryo, forty hours old. (From Kölliker.) m medulla oblongata, ph pharyngeal cavity (head-gut), h horny plate, h′ thicker part of it, from which the auscultory pits afterwards develop, hp skin-fibre plate, hh cervical cavity (head-cœlom or cardiocœl), hzp cardiac plate (the outermost mesodermic wall of the heart), connected by the ventral mesocardium (uhg) with the gut-fibre layer or visceral cœlom-layer (dfp*prime;), Ent entoderm, ihh inner (entodermic?) wall of the heart; the two endothelial cardiac tubes are still separated by the cenogenetic septum (s) of the Amniotes, g vessels. |
The heart of all the Vertebrates belongs originally to the hyposoma of the head, and we accordingly find it in the embryo of man and all the other Amniotes right in front on the under-side of the head; just as in the fishes it remains permanently in front of the gullet. It afterwards descends into the trunk, with the advance in the development of the neck and breast, and at last reaches the breast, between the two lungs. At first it lies symmetrically in the middle plane of the body, so that its long axis corresponds with that of the body. In most of the mammals it remains permanently in this position. But in the apes the axis begins to be oblique, and the apex of the heart to move towards the left side. The displacement is greatest in the anthropoid apes—chimpanzee, gorilla, and orang—which resemble man in this.
As the heart of all Vertebrates is originally, in the light of phylogeny, only a local enlargement of the middle principal vein, it is in perfect accord with the biogenetic law that its first structure in the embryo is a simple spindle-shaped tube in the ventral wall of the head-gut. A thin membrane, standing vertically in the middle plane, the mesocardium, connects the ventral wall of the head-gut with the lower head-wall. As the cardiac tube extends and detaches from the gut-wall, it divides the mesocardium into an upper (dorsal) and lower (ventral) plate (usually called the mesocardium anterius and posterius in man, Fig. 379 uhg). The
[ 328 ]
mesocardium divides two lateral cavities, Remak’s “neck-cavities” (Fig. 379 hh). These cavities afterwards join and form the simple pericardial cavity, and are therefore called by Kölliker the “primitive pericardial cavities.”
The double cervical cavity of the Amniotes is very interesting, both from the anatomical and the evolutionary point of view; it corresponds to a part of the hyposomites of the head of the lower Vertebrates—that part of the ventral cœlom-pouches which comes next to Van Wijhe’s “visceral cavities” below. Each of the cavities still communicates freely behind with the two cœlom-pouches of the trunk; and, just as these afterwards coalesce into a simple body-cavity (the ventral mesentery disappearing), we find the same thing happening in the head. This simple primary pericardial cavity has been well called by Gegenbaur the “head-cœloma,” and by Hertwig the “pericardial breast-cavity.” As it now encloses the heart, it may also be called cardiocœl.
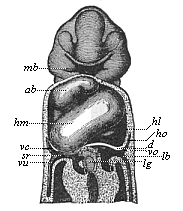 Fig. 380—Frontal section of a human embryo, one-twelfth of an inch long in the neck; “invented” by Wilhelm His. Seen from ventral side. mb mouth-fissure, surrounded by the branchial processes, ab bulbus of aorta, hm middle part of ventricle, hl left lateral part of same, ho auricle, d diaphragm, vc superior vena cava, vu umbilical vein, vo vitelline space, lb liver, lg hepatic duct.
Fig. 380—Frontal section of a human embryo, one-twelfth of an inch long in the neck; “invented” by Wilhelm His. Seen from ventral side. mb mouth-fissure, surrounded by the branchial processes, ab bulbus of aorta, hm middle part of ventricle, hl left lateral part of same, ho auricle, d diaphragm, vc superior vena cava, vu umbilical vein, vo vitelline space, lb liver, lg hepatic duct. |
The cardiocœl, or head-cœlom, is often disproportionately large in the Amniotes, the simple cardiac tube growing considerably and lying in several folds. This causes the ventral wall of the amniote embryo, between the head and the navel, to be pushed outwards as in rupture (cf. Fig. 180 h). A transverse fold of the ventral wall, which receives all the vein-trunks that open into the heart, grows up from below between the pericardium and the stomach, and forms a transverse partition, which is the first structure of the primary diaphragm (Fig. 380 d). This important muscular partition, which completely separates the thoracic and abdominal cavities in the mammals alone, is still very imperfect here; the two cavities still communicate for a time by two narrow canals. These canals, which belong to the dorsal part of the head-cœlom, and which we may call briefly pleural ducts, receive the two pulmonary sacs, which develop from the hind end of the ventral wall of the head-gut; they thus become the two pleural cavities.
The diaphragm makes its first appearance in the class of the Amphibia (in the salamanders) as an insignificant muscular transverse fold of the ventral wall, which rises from the fore end of the transverse abdominal muscle, and grows between the pericardium and the liver. In the reptiles (tortoises and crocodiles) a later dorsal part is joined to this earlier ventral part of the rudimentary diaphragm, a pair of subvertebral muscles rising from the vertebral column and being added as “columns” to the transverse partition. But it was probably in the Permian sauro-mammals that the two originally separate parts were united, and the diaphragm became a complete partition between the thoracic and abdominal cavities in the mammals; as it considerably enlarges the chest-cavity when it contracts, it becomes an important respiratory muscle. The ontogeny of the diaphragm in man and the other mammals reproduces this phylogenetic process to-day, in accordance with the biogenetic law; in all the mammals the diaphragm is formed by the secondary conjunction of the two originally separate structures, the earlier ventral part and the later dorsal part.
Sometimes the blending of the two diaphragmatic structures, and consequently the severance of the one pleural duct from the abdominal cavity, is not completed in man. This leads to a diaphragmatic rupture (hernia diaphragmatica). The two cavities then remain in communication by an open pleural duct, and loops of the intestine may penetrate by this “rupture opening” into the chest-cavity. This is one of those
[ 329 ]
fatal mis-growths that show the great part that blind chance has in organic development.
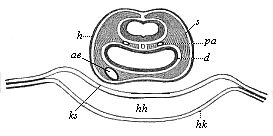 |
Fig. 381—Transverse section of the head of a chick-embryo, thirty-six hours old. Underneath the medullary tube the two primitive aortas (pa) can be seen in the head-plates (s) at each side of the chorda. Underneath the gullet (d) we see the aorta-end of the heart (ae), hh cervical cavity or head cœlom, hk top of heart, ks head-sheath, amniotic fold, h horny plate. (From Remak. |
Thus the thoracic cavity of the mammals, with its important contents, the heart and lungs, belongs originally to the head-part of the vertebrate body, and its inclusion in the trunk is secondary. This instructive and very interesting fact is entirely proved by the concordant evidence of comparative anatomy and ontogeny. The lungs are outgrowths of the head-gut; the heart develops from its inner wall. The pleural sacs that enclose the lungs are dorsal parts of the head-cœlom, originating from the pleuroducts; the pericardium in which the heart afterwards lies is also double originally, being formed from ventral halves of the head-cœlom, which only combine at a later stage. When the lung of the air-breathing Vertebrates issues from the head-cavity and enters the trunk-cavity, it follows the example of the floating bladder of the fishes, which also originates from the pharyngeal wall in the shape of a small pouch-like out-growth, but soon grows so large that, in order to find room, it has to pass far behind into the trunk-cavity. To put it more precisely, the lung of the quadrupeds retains this hereditary growth-process of the fishes; for the hydrostatic floating bladder of the latter is the air-filled organ from which the air-breathing organ of the former has been evolved.
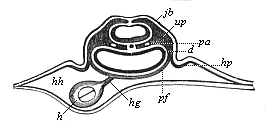 Fig. 382—Transverse section of the cardiac region of the same chick-embryo (behind the preceding). In the cervical cavity (hh) the heart (h) is still connected by a mesocard (hg) with the gut-fibre layer (pf). d gut-gland layer, up provertebral plates, jb rudimentary auditory vesicle in the horny plate, hp first rise of the amniotic fold. (From Remak.)
Fig. 382—Transverse section of the cardiac region of the same chick-embryo (behind the preceding). In the cervical cavity (hh) the heart (h) is still connected by a mesocard (hg) with the gut-fibre layer (pf). d gut-gland layer, up provertebral plates, jb rudimentary auditory vesicle in the horny plate, hp first rise of the amniotic fold. (From Remak.) |
There is an interesting cenogenetic phenomenon in the formation of the heart of the higher Vertebrates that deserves special notice. In its earliest form the heart is double, as recent observation has shown, in all the Amniotes, and the simple spindle-shaped cardiac tube, which we took as our starting-point, is only formed at a later stage, when the two lateral tubes move backwards, touch each other, and at last combine in the middle line. In man, as in the rabbit, the two embryonic hearts are still far apart at the stage when there are already eight primitive segments (Fig. 134 h). So also the two cœlom-pouches of the head in which they lie are still separated by a broad space. It is not until the permanent body of the embryo develops and detaches from the embryonic vesicle that the separate lateral structures join together, and finally combine in the middle line. As the median partition between the right and left cardiocœl disappears, the two cervical cavities freely communicate (Fig. 381), and form, on the ventral side of the amniote head, a horseshoe-shaped arch, the points of which advance backwards into the pleuro-ducts or pleural cavities, and from there into the two peritoneal sacs of the trunk. But even after the conjunction of the cervical cavities (Fig. 381) the two cardiac tubes remain separate at first; and even after they have united a delicate partition in the middle of the simple endothelial tube (Figs. 379 s, 382 h) indicates the original separation. This cenogenetic “primary cardiac
[ 330 ]
septum” presently disappears, and has no relation to the subsequent permanent partition between the halves of the heart, which, as a heritage from the reptiles, has a great palingenetic importance.
Thorough opponents of the biogenetic law have laid great stress on these and similar cenogenetic phenomena, and endeavoured to urge them as striking disproofs of the law. As in every other instance, careful, discriminating, comparative-morphological examination converts these supposed disproofs of evolution into strong arguments in its favour. In his excellent work, On the structure of the Heart in the Amphibia (1886), Carl Rabl has shown how easily these curious cenogenetic facts can be explained by the secondary adaptation of the embryonic structure to the great extension of the food-yelk.
The embryology of all the other parts of the vascular system also gives us abundant and valuable data for the purposes of phylogeny. But as one needs a thorough knowledge of the intricate structure of the whole vascular system in man and the other Vertebrates in order to follow this with profit, we cannot go into it further here. Moreover, many important features in the ontogeny of the vascular system are still very obscure and controverted. The characters of the embryonic circulation of the Amniotes, which we have previously considered (Chapter XV), are late acquisitions and entirely cenogenetic. (Cf. pp. 170–171; Figs. 198–202.)
Title and Contents
Vol. II Title and Contents
Glossary
Chapter XXVII
Chapter XXI
Figs. 1–209
Figs. 210–408