
Painted by J. J. Masquerier. Engraved by W. T. Fry.
William Spence, Esqr., F.L.S.
Published by Longman & Co. London, July 1825.
| Letter. | Page. | |
| XXXVII. | Internal Anatomy and Physiology of Insects. Sensation | 1-33 |
| XXXVIII. | Internal Anatomy and Physiology of Insects continued. Respiration | 34-80 |
| XXXIX. | Internal Anatomy and Physiology of Insects continued. Circulation | 81-101 |
| XL. | Internal Anatomy and Physiology of Insects continued. Digestion | 102-126 |
| XLI. | Internal Anatomy and Physiology of Insects continued. Secretion | 127-151 |
| XLII. | Internal Anatomy and Physiology of Insects continued. Reproduction | 152-173 |
| XLIII. | Internal Anatomy and Physiology of Insects concluded. Motion | 174-203 |
| XLIV. | Diseases of Insects | 204-240 |
| XLV. | Senses of Insects | 241-264 |
| XLVI. | Orismology, or Explanation of Terms | 265-363 |
| XLVII. | System of Insects | 364-428 |
| XLVIII. | History of Entomology | 429-485 |
| XLIX.[Pg iv] | Geographical Distribution of Insects; their Stations and Haunts; Seasons; Times of Action and Repose | 486-527 |
| L. | On Entomological Instruments; and the best Methods of collecting, breeding, and preserving Insects | 528-559 |
| LI. | Investigation of Insects | 560-573 |
| Appendix | 575-584 | |
| Authors quoted | 585-602 | |
| Explanation of the Plates | 603-614 | |
| Indexes | 615-683 |
Having given you this full account of the external parts of insects, and their most remarkable variations; I must next direct your attention to such discoveries as have been made with regard to their Internal Anatomy and Physiology: a subject still more fertile, if possible, than the former in wonderful manifestations of the power, wisdom and goodness of the Creator.
The vital system of these little creatures, in all its great features, is perfectly analogous to that of the vertebrate animals. Sensation and perception are by the means of nerves and a common sensorium; the respiration of air is evident, being received and expelled by a particular[Pg 2] apparatus; nutrition is effected through a stomach and intestines; the analogue of the blood prepared by these organs pervades every part of the body, and from it are secreted various peculiar substances; generation takes place, and an intercourse between the sexes, by means of appropriate organs; and lastly, motion is the result of the action of muscles. Some of these functions are, however, exercised in a mode apparently so dissimilar from what obtains in the higher animals, that upon a first view we are inclined to pronounce them the effect of processes altogether peculiar. Thus, though insects respire air, they do not receive it by the mouth, but through little orifices in the sides of the body; and instead of lungs, they are furnished with a system of air-vessels, ramified ad infinitum, and penetrating to every part and organ of their frame; and though they are nourished by a fluid prepared from the food received into the stomach, this fluid, unlike the blood of vertebrate animals, is white, and the mode in which it is distributed to the different parts of the system, except in the case of the true Arachnida, in which a circulation in the ordinary way has been detected, is altogether obscure.
In order that you may more clearly understand the variations that occur in insects, and in what respects they differ amongst themselves, and from the higher animals, in the vital functions and their organs, I shall consider them as to their organs of sensation, respiration, circulation, nutrition, generation, secretion, and muscular motion.
Organs of Sensation.—The nervous system of animals is one of the most wonderful and mysterious works of[Pg 3] the Creator. Its pulpy substance is the visible medium by which the governing principle[1] transmits its commands to the various organs of the body, and they move instantaneously—yet this appears to be but the conductor of some higher principle, which can be more immediately acted upon by the mind and by the will. This principle, however, whatever it be, whether we call it the nervous fluid, or the nervous power[2], has not been detected, and is known only by its effects. The system of which we are speaking may therefore be deemed the foundation and root of the animal, the centre from which emanate all its powers and functions.
Comparative anatomists have considered the nervous system of animals as formed upon four primary types, which may be called the molecular, the filamentous, the ganglionic, and cerebro-spinal[3]. The first is where invisible nervous molecules are dispersed in a gelatinous body, the existence of which has only been ascertained by the nervous irritability of such bodies, their fine sense of touch, their perceiving the movements of the waters in which they reside, and from their perfect sense of the degrees of light and heat[4]. Of this description are the infusory animals, and the Polypi. The nervous molecules in these are conjectured to constitute so many ganglions, or centres of sensation and vitality[5]. The second, the filamentous, is where the nervous system consists of nervous threads radiating from the mouth, as in the[Pg 4] Radiata, or star-fish and sea-urchins[6]. The third, the ganglionic, is where the nervous system consists of a series of ganglions connected by nervous threads or a medullary chord, placed, except the first ganglion, below the intestines, from which proceed nerves to the various parts of the body. This system may be considered as divisible into two—the proper ganglionic, in which it is ganglionic with the ganglions arranged in a series with a double spinal chord. This prevails in the classes Insecta, Crustacea, Arachnida, &c., and the improper ganglionic, in which it is ganglionic with the ganglions dispersed irregularly, but connected by nervous threads, as in the Mollusca[7]. In the fourth, the cerebro-spinal, the nervous tree may be said to be double, or to consist of two systems—the first taking its origin in a brain formed of two hemispheres contained in the cavity of the head, from which posteriorly proceeds a spinal marrow, included in a dorsal vertebral column. These send forth numerous nerves to the organs of the senses and the muscles of the limbs. The second consists of two principal ventral chords, which by their ganglions, but without any direct communication, anastomose with the spinal nerves and some of those of the brain, and run one on each side from the base of the skull to the extremity of the sacrum. This system consists of an assemblage of nervous filaments bearing numerous ganglions, from which nervous threads are distributed to the organs of nutrition and reproduction[8]. Its chords are called the[Pg 5] great sympathetic, the intercostal, or trisplanchnic nerves[9]. While the first of these two systems is the messenger of the will, by means of the organs of the senses connects us with the external world, and is subject to have its agency interrupted by sleep or disease[10]; the latter is altogether independent of the will and of the intellect, is confined to the internal organic life, its agency continues uninterrupted during sleep, and is subject to no paralysis. While the former is the seat of the intellectual powers, the latter has no relation to them, but is the focus from whence instincts exclusively emanate: from it proceed spontaneous impulses and sympathies, and those passions and affections that excite the agent to acts in which the will and the judgement have no concern[11].
It is probable, though the above appear to exhibit the primary types of nervous systems, that others exist of an intermediate nature, with which future investigators may render us better acquainted[12]: but as our business is solely with that upon which insects in this respect have been modelled, without expatiating further in this interesting field, I shall therefore now confine myself to them.
We have before seen[13] that the nervous system of insects belongs to the ganglionic type: but it requires a more full description, and this is the place for it. It originates[Pg 6] in a small brain placed in the head, and consisting almost universally of two lobes, sometimes extremely distinct. It is placed over or upon the œsophagus or gullet, and from its posterior part proceeds a double nervous chord, which embracing that organ as a collar dips below the intestines, and proceeds towards the anus, forming knots or ganglions at intervals, in many cases corresponding in number with the segments of the body, and sending forth nerves in pairs, the ramifications of which are distributed to every part of the frame. In the perfect insect the bilobed ganglion of the head or the brain is usually of greater volume than in the larva, and the ganglions of the spinal chord are fewer, which gives a more decided character of centricity to the whole nervous system[14]. This may be considered more particularly with respect to its substance and colour; its tunics, and parts.
I. Substance and Colour.—The nervous apparatus of insects is stated by those who have examined it most narrowly, though consisting of a cortical and medullary part, the latter more delicate and transparent than the former, to be less tender and less easy to separate than the human brain[15]. It has a degree of tenacity, and does not break without considerable tension; in general, it is clammy and flabby, and under a microscope a number of minute grains are discoverable in it, and when left to dry upon glass, it appears to contain a good deal of oil, which does not dry with the rest[16]. That of the ganglions differs from the substance of the rest of the spinal[Pg 7] chord, in being filled with very fine aërial vessels, which are not discoverable in the latter[17]. With regard to colour, Lyonet states that the chords of the spinal marrow in the larva of the great goat-moth are of a blueish gray, and have some transparence[18]; Malpighi and Swammerdam observed that the cortical part of the ganglions of that of the silk-worm and the hive-bee had a reddish hue, while the medullary part was white[19]; Cuvier relates that the brain and the third ganglion in Hypogymna dispar, with us a scarce moth, differed in colour from all the rest, being quite white, while the others were more or less tinted, and examined under a lens appeared variegated by reddish sinuous markings, resembling blood vessels as they are seen in injected glands[20].
II. Tunics.—The coats that inclose the various branches of the nervous system in insects seem analogous to those of vertebrate animals. The first thing that strikes the eye, when these parts in a recent subject are submitted to a microscope, is a tissue of very delicate vessels, which ramify beyond the reach of the assisted sight; these are merely air-vessels or bronchiæ derived originally from the tracheæ of the animal: but besides these is an exterior and an interior tunic; the first corresponding with the dura mater of anatomists; and the other, which is[Pg 8] the most delicate and incloses the cortical and medullary parts, with the pia mater[21].
III. Parts.—The nervous system of insects consists of the brain; the spinal marrow and its ganglions; and the nerves.
i. Brain.[22] Linné denied the existence of a brain in insects, and most modern physiologists seem to be of the same opinion. A part however, analogous to this important organ—at least in its situation, and in its emission of nerves to the principal organs of the senses, in which respect it certainly differs very materially from the upper cervical ganglion, which Dr. Virey regards as its analogue[23]—is certainly to be found in them; and as Messrs. Cuvier and Lamarck distinguish this part by the name of brain, we may continue to call it by that name without impropriety. The brain of insects, then, is distinguished from the succeeding ganglions of the spinal chord by its situation in the head, the middle of the internal cavity of which it occupies, and by being the only ganglion above the œsophagus. It is usually small, though in some cases larger than they are[24]. It consists of two lobes, more or less distinct and generally of a spherical form. In Oryctes nasicornis and Pontia Brassicæ the lobes are separated both before and behind[25]; while in the larva of Dytiscus marginalis, but not in the imago, in which there are two large hemispheres separated by a furrow, the brain is undivided[26]. Cuvier mentions the[Pg 9] larva of a saw-fly in which this part is formed of four nearly equal spherical bulbs[27]: in the Scorpion (to judge by the figure of Treviranus[28]) the two lobes represent an equilateral triangle, the exterior angle of which terminates in several lesser spherical bulbs; in Acrida viridissima, Nepa cinerea, Clubiona atrox, and the common Louse, the lobes are pear-shaped[29].
ii. The spinal marrow and its ganglions[30]. From the posterior part of the brain of insects, but in the ground and water beetles (Eutrechina and Eunechina) from its sides below[31], issue two chords which diverging embrace the œsophagus, and dipping below it and the intestines,—a situation they maintain to the end of their course,—and in their further progress uniting at intervals and dilating into several knots or ganglions, compose their spinal marrow. This part is so named, from a supposed analogy to the spinal marrow of vertebrate animals, which however admits of some degree of doubt; yet, since it mixes the functions of that organ with those of the great sympathetic nerves, the denomination is not wholly improper, and may be retained. Though this chord is usually double when it first proceeds from the brain, and surrounds the œsophagus like a collar, yet in some insects it may be called a single chord. This is the case with that of the common louse, in which Swammerdam could perceive no opening for the transmission of the part just named[32]; if he was not mistaken in this, the brain, as well as the rest of the spinal marrow in[Pg 10] that animal, would be below the intestines; from the figures of Treviranus it should seem that the spiders, at least Clubiona atrox, are similarly circumstanced[33]; in the cheese-maggot, which turns to a two-winged fly (Tyrophaga Casei), the chord is also single, but it has a small orifice through which the gullet passes[34]. At the union of the chords in other cases below that organ, a knot or ganglion is usually formed, and an alternate succession of internodes and ganglions commonly follows to the end. The internodes also may generally be stated to consist of a double chord, though in many cases the two chords unite and become one, or are distinguished only by a longitudinal furrow, and even where they are really distinct and separable, in the body of the insect they lie close together[35]. In the rhinoceros beetle (Oryctes nasicornis) and Acrida viridissima &c. all the internodes consist of a double chord[36]; but in many other insects numerous variations in this respect occur.—Thus in the stag-beetle the last internode is single[37]; in the caterpillar of the cabbage butterfly (Pontia Brassicæ) the five first are double, and the six last single[38]; in that of the great goat-moth (Cossus ligniperda) the three first only are double, but the others terminate in a fork[39]; in the cockroaches (Blatta) the four first, in Hydrophilus piceus the three first, and in Eristalis tenax the two first only are double, the rest being all single[40]. A singular variation takes place in Hypogymna dispar; all the internodes are[Pg 11] single, except the second, the chords of which at first are separate, and afterwards united[41]; and, to name no more, in Clubiona atrox there is only one internode, which is single, with a longitudinal furrow[42]. In some, as in the louse, the grub of Oryctes nasicornis, and the cheese-maggot, there are no internodes, the spinal marrow being formed of knots separated only by slight or deep constrictions[43].
I must next say something of the ganglions[44]. Lyonet has observed that, in the caterpillar of the great goat-moth, these in one respect differ remarkably from the chords that connect them; in the latter the air-vessels or bronchiæ only cover the outside of the tunic, while in the former they enter the substance of the ganglion, which is quite filled with their delicate and numberless branches[45]. Every ganglion may be regarded in some degree as a centre of vitality or little brain[46], and in many cases, as well as the brain, they are formed of two lobes[47]. I shall now consider them more particularly as to their station, number, and shape.
1. With regard to the first head, their station, they are most commonly divided between the trunk and abdomen; but in some cases, as in Hydrophilus piceus and Acrida viridissima, the first ganglion is in the head[48]; in others, as in the louse, the water-scorpion, and the grub of the rhinoceros-beetle, they are confined to the trunk, their functions in the abdomen being supplied[Pg 12] by numerous radiating nerves[49]; in others again, as in the scorpion, they are all abdominal. The ganglions vary also in their situation with respect to each other. Thus in some, as in the larva of the Chamæleon-fly (Stratyomis Chamæleon), they are so near as to appear like a string of beads[50]; in that of the ant-lion (Myrmeleon) the two ganglions of the trunk are separated by an interval from those of the abdomen, which are so contiguous as to resemble the rattle of the rattle-snake[51]. In others the internodes are longer, and the ganglions occur at nearly equal intervals, as in the larva of the Ephemeræ[52]; but in the majority they are unequal in length: thus in the scorpion the three first ganglions are the most distant[53]; in the hive-bee the third and fourth[54]; and in the spider the last[55].
2. The ganglions also in different species, and often in the same insect in its different states, vary in their number. Thus in the grub of the rhinoceros-beetle the whole spinal marrow appears like a single ganglion divided only by transverse furrows[56]; in the water-scorpion there are two[57]; in the louse there are three[58]; in the rhinoceros-beetle there are four[59]; five in the stag-beetle[60];[Pg 13] seven in the hive-bee and some Lepidoptera[61]; eight in the grub of the stag-beetle[62]; nine in the great Hydrophilus[63]; ten in Dytiscus[64]; eleven in the grub of the great Hydrophilus[65]; twelve in the grub of Dytiscus and the caterpillars of Lepidoptera[66]; thirteen in the larva of Æshna[67]; and twenty-four in Scolopendra morsitans[68]. You must observe that, generally speaking, the number of ganglions is less in the imago than in the larva. With regard to the distribution of these knots to the different primary parts of the body, the following table will exhibit it, as far as I am acquainted with it, at one view. I omit those in which the ganglions are only in one of these parts.
| Head. | Trunk. | Abdomen. | |
| Acrida viridissima | 1 | 3 | 6[69] |
| Hydrophilus piceus | 1 | 6 | 2 |
| Clubiona atrox | 0 | 2 | 1 |
| Gryllotalpa vulgaris | 0 | 2 | 7[70] |
| Myrmeleon, Larva | 0 | 2 | 8[71] |
| Eristalis tenax | 0 | 3 | 2[72] |
| Apis mellifica | 0 | 3 | 4 |
| Ephemera, Larva | 0 | 3 | 7 |
| Æshna, Larva | 0 | 6 | 7 |
3. I am next to say a few words upon the shape of the ganglions. Most commonly it approaches to a spherical figure, but in many instances, as I said before, they, as[Pg 14] well as the brain, consist of two lobes: they are, however, seldom all precisely of the same shape. In the Dytisci, and Carabi, the last is marked with a transverse furrow, which seems to indicate the reunion of two[73]; in the stag-beetle, the first ganglion is oval or elliptical, the second hexagonal; the third and fourth shaped like a crescent, and the last like an olive[74]; in the caterpillar of the great goat-moth the first is oblong and constricted in the middle, and the seven last are rhomboidal[75]; in the great Hydrophilus the second, and in the silk-worm all the ganglions are quadrangular[76]; in Hypogymna dispar the third is heart-shaped[77]; the great ganglion which forms the spinal marrow of the cheese-maggot is pear-shaped[78]; that of the grub of the rhinoceros-beetle is fusiform[79]; and in the scorpion all the ganglions are lenticular[80]. But the most remarkable in this respect are those of a spider (Clubiona atrox): in this insect the brain sits upon a bilobed ganglion of the ordinary form, which is immediately followed without any internode by another bilobed one, terminating on each side in four pear-shaped processes or fingers, which give it a very singular appearance[81].
iii. The nerves[82] of insects, as of other animals, are white filaments running from the brain and spinal marrow to every part of the body which they are destined to animate; and their numerous ramifications, when delineated,[Pg 15] form no unpleasing picture[83]. In the caterpillar of the goat-moth the accurate Lyonet counted forty-five pairs of them, and two single ones, making in all ninety-two nerves; whereas in the human body anatomists count only seventy-eight[84]. From the brain issue several pairs, which go to the eyes, antennæ, palpi, and other parts of the mouth: sometimes those that render to the mandibles issue from the first ganglion, as in the larva of Dytiscus marginalis, the stag-beetle, &c.[85]; those both of mandibles and palpi in the great Hydrophilus[86]; and in Blatta some which act also upon the antennæ[87].
The optic are usually the most conspicuous and remarkable of the nerves. In some insects with large eyes, as many Neuroptera, Hymenoptera, and Diptera, their size is considerable; in the hive-bee they present the appearance of a pair of kidney-shaped lobes, larger than the brain[88]; in the dragon-flies, whose brain consists of two very minute lobes, these nerves dilate into two large plates of a similar shape, which line all the inner surface of the eyes[89]; in the stag-beetle they are pear-shaped, and terminate in a bulb, from which issue an infinity of minute nerves[90]; it is probable that this takes place in all cases, and that a separate nerve renders to every separate lens in a compound eye[91]; the optic nerve in Dytiscus and Carabus is pyramidal, with the base of the pyramid at the eye and the summit at the brain[92]; in Eristalis tenax it[Pg 16] is very large, cylindrical, and of a diameter equal to the length of the last-mentioned part, upon the side of which it is supported; it terminates in a very large bulb corresponding to the eye[93]: in Scolopendra morsitans the optic nerves divide into four branches long before they arrive at the eyes, and in this insect the nerves which render to the antennæ are so thick as to appear portions of the brain, which they equal in diameter[94]. Swammerdam discovered in the grub of the rhinoceros-beetle and in the caterpillar of the silk-worm, a pair of nerves which he regarded as analogous to the recurrent nerves in the human subject, and therefore he distinguishes them by the same name[95]: they issue from the lower surface of the brain, or that which rests on the œsophagus, and at first go towards the mouth, but afterwards turn back, and uniting form a small ganglion; this produces a single nerve, which passing below the brain follows the œsophagus to the stomach, where it swells into another ganglion, from which issue some small nerves that render to the stomach, and one more considerable which accompanies the intestinal canal, producing at intervals lateral filaments which lose themselves in the tunics of that tube[96]. Lyonet afterwards discovered these nerves in the caterpillar of the goat-moth[97], and Cuvier in other insects[98].
The other nerves which issue from the brain exhibit no remarkable features. Those which originate in the spinal marrow are mostly derived from the ganglions, and[Pg 17] are sometimes interwoven with the muscles, as the woof with the warp in a piece of cloth[99]; those from the three or four first commonly rendering to the muscles of the legs, wings, and other parts of the trunk, and those from the remainder to the abdomen. After their origin they often divide and subdivide, and terminate in numerous ramifications that connect every part of the body with the sensorium commune. A pair of nerves is the most usual number that proceeds from each side of a ganglion[100]; but this is by no means constant, since in the louse, the hive-bee, and several other insects, only a single nerve thus proceeds[101]; and in the larva of Ephemeræ, while two pairs issue from the six first ganglions, only a single one is emitted by the five last[102]. In the spinal marrow of the rhinoceros-beetle, both larva and imago, the nerves consist of simple filaments which diverge like rays in all directions[103]: the same circumstance distinguishes the cheese-maggot, only some of the nerves appear to branch at the end[104]: in the louse, the last ganglion sends forth posteriorly three pairs of nerves which render to the abdomen[105]. Sometimes, though rarely, nerves originate in the internodes of the spinal marrow. Cuvier indeed has asserted that in invertebrate animals all the nerves spring from the ganglions, and never immediately from the spinal marrow; but Swammerdam, in describing those of the silk-worm, mentions and figures four pairs as proceeding from the four anterior internodes, excluding[Pg 18] the first[106]; and at the same time he gives it as his opinion, that all the nerves in insects really originate from the marrow itself, and not from the ganglions, which he asserts are of a different substance, and are inclosed in the marrow for the sake of giving it greater firmness[107]. In this opinion, however, he seems singular[108]. Those remarkable nerves described by Lyonet under the name of spinal bridle (bride épinière) also take their origin, not from the ganglions, but from a bifurcation of the spinal marrow. Of these, in the caterpillar of the goat-moth there are ten, the first issuing from the bifurcation of the internode between the fourth and fifth ganglions, and the remainder from the succeeding ones. After approaching the succeeding ganglion, these nerves form a pair of branches that diverge nearly at right angles from the bridle, and producing several lesser branches, lose themselves in the sides of the animal[109]. Besides the nerves above mentioned, two generally issue from the posterior part of the last ganglion, diverging in opposite and oblique directions: some of these render to the parts of generation; and in the silk-worm, and probably other species, the innermost pair is perforated for the passage of the vasa deferentia[110].
After duly considering this general outline of the nervous system of insects, the question will continually occur to you,—is then what you have called the brain the sensorium commune of these animals, in the same manner as it is in those with warm blood? To this query a negative must be returned. In the latter, the brain is the common centre to which, by means of the nerves and spinal marrow, all the sensations of the animal are conveyed, and in which all its perceptions terminate. The nerves and spinal marrow are merely the roads by which the sensations travel; and if their communication with the brain, by any means be cut off at the neck, the whole trunk of the animal becomes paralytic, evidently proving that the organ by which it feels is the brain. This, however, is so far from being the case in insects, that in them, if the head be cut off, the remainder of the body will continue to give proofs of life and sensation longer than the head: both portions will live after the separation, sometimes for a considerable period; but the largest will survive the longest, and will move, walk, and occasionally even fly, at first almost as actively without the head, as when united to it. Lyonet informs us, that he has seen motion in the body of a wasp three days after it had been separated from the head; and that a caterpillar even walked some days after that operation; and when touched, the headless animal made the same movements as when intire[111]. Dr. Shaw has observed—an observation confirmed in Unzer's Kleine Schriften,—that if Geophilus electricus be cut in two, the halves will live and appear vigorous even for a fortnight afterwards; and what is more remarkable, that the tail part always survives the[Pg 20] head two or three days[112]. The sensorium commune of insects, therefore, does not, as in the warm-blooded animals, reside in the brain alone, but in the spinal marrow also. It was on this account probably that Linné denied the existence of a brain in insects, regarding it merely as the first ganglion of the spine.
Cuvier and other modern physiologists, from the ganglionic structure of this organ, are of opinion that it is not the analogue of the cerebro-spinal system of vertebrate animals, but rather of their great sympathetic nerves. Indeed, considering solely the external structure of the nervous system of insects, a great resemblance strikes us between it and these nerves; for besides its general ganglionic structure, there is also in them an upper ganglion in the neck, seemingly corresponding with what we have named the brain of insects, from which the nervous chord dips to the lower part of the neck, where it forms a second ganglion, which appears to correspond with what we have considered as their second ganglion[113]. We may observe, however, that at least in one respect there is even an external resemblance between the brain of insects and that of vertebrate animals:—it most commonly consists, as has been stated, like them, of two lobes, often very distinct; a circumstance which not unfrequently distinguishes the other ganglions[114], and is not borrowed from the ganglions of the great sympathetics. With respect[Pg 21] to the internal structure of the ganglions and spinal marrow of insects, we know little to build any theory upon, except that the internal substance of the former is filled with air-vessels; at least so Lyonet, as has been already observed, found in the goat-moth, while only the tunics of the latter are covered by them. Taking the above resemblance to the brain of vertebrates into consideration, there appears ground for thinking that the nervous system of insects, like some of their articulations[115], is of a mixed kind, combining in it both the cerebro-spinal and the ganglionic systems; and this will appear further if we consider its functions.
That learned and acute physiologist Dr. Virey, assuming as an hypothesis, that the structure of the system in question is simply ganglionic, and merely analogous to the sympathetic system of vertebrate animals, has built a theory upon the assumption, which appears evidently contradicted by facts. Because, as he conceives after Cuvier, insects are not gifted with a real brain and spinal marrow, he would make it a necessary consequence that they have no degree of intellect, no memory, judgement or free will; but are guided in every respect by instinct and spontaneous impulses,—that they are incapable of instruction, and can superadd no acquired habits to those which are instinctive and inbred[116]. This consequence would certainly necessarily follow, was their nervous system perfectly analogous to the sympathetic of warm-blooded animals. But when we come to take into consideration the functions that in insects this system confessedly discharges, we are led to doubt very strongly[Pg 22] the correctness of the assumption. Now in these animals the system in question not only renders to the nutritive and reproductive organs, which is the principal function of the great sympathetic nerves in the vertebrates; but by the common organs maintains a connexion with the external world, and acquires ideas of things without, which in them is a function of the cerebral system: from the same centre also issue those powers which at the bidding of the will put the limbs in action, which also belongs to the cerebral system. That insects have memory, and consequently a real brain, has been before largely proved, as also that they have that degree of intellect and judgement which enables them to profit by the notices furnished by their senses[117]. What can be the use of eyes,—of the senses of hearing, smelling, feeling, &c. if they are not instructed by them what to choose and what to avoid? And if they are thus instructed—they must have sufficient intellect to apprehend it, and a portion of free will to enable them to act according to it. With regard to the assertion that they are incapable of instruction, or of acquiring new habits; few or no experiments have been tried with the express purpose of ascertaining this point: but some well-authenticated facts are related, from which it seems to result that insects may be taught some things, and acquire habits not instinctive. They could scarcely be brought from their wild state, and domesticated, as bees have been so universally, and both ants and wasps occasionally[118], without some departure from the habits of their wild state; and the fact of the corsair-bees, that acquire predatory habits[Pg 23] before described[119], shows this more evidently: but one of the most remarkable stories to our purpose upon record, is that of M. Pelisson, who, when he was confined in the Bastile, tamed a spider, and taught it to come for food at the sound of an instrument. A manufacturer also in Paris, fed 800 spiders in an apartment, which became so tame that whenever he entered it, which he usually did bringing a dish filled with flies but not always, they immediately came down to him to receive their food[120].
All these circumstances having their due consideration and weight, it seems, I think, most probable, that as insects have their communication with the external world by means of certain organs in connexion with their nervous system, and appear to have some degree of intellect, memory, and free will, all of which in the higher animals are functions of a cerebral system, and at the same time in other respects manifest those which are peculiar to the sympathetic system,—it is most probable, I say, as was above hinted, that in their system both are united.
I must bespeak your attention to a circumstance connected with the subject of this letter, which merits particular consideration: I mean the gradual change that takes place in the nervous system when insects undergo their metamorphoses; so that, except in the Orthoptera, Hemiptera, and Neuroptera Orders, in which no change is undergone, the number of ganglions of the spinal chord is less in the imago than in the larva. There seems an exception indeed to this rule in the case of the rhinoceros-beetle,[Pg 24] in the larva of which there is only one ganglion, while in the imago there are four[121]. But as this one ganglion occupies the whole spinal marrow, it is really of greater extent than the four of the imago; so that even in this case there is a concentration of the cerebral pulp. In some cases, as in Dytiscus marginalis, and Hydrophilus piceus[122], the imago has only one ganglion less than the larva, but more generally it loses four or five. Dr. Herold has traced the gradual changes that take place in the spinal marrow of the common cabbage-butterfly (Pontia Brassicæ), from the time that it has attained its full size to its assumption of the imago. Of these I shall now give you some account.
In the full-grown caterpillar, besides the brain there are eleven ganglions, the chords of the four first internodes being double, and the rest single: from each ganglion proceed two pairs of nerves, one from each side. In this the lobes of the brain form an angle with each other[123]. In two days the double chords mutually recede, so as to diminish the interval between the ganglions, and the single ones have become curved: thus the length of the spinal marrow is shortened about a fourth, and the fourth and fifth ganglions have made an approach to each other[124]. On the eighth day, when the insect has assumed the pupa but remains still in the skin of the caterpillar, the flexure of the internodes is much increased; the first ganglion is now united to the brain, and the fourth and fifth have joined each other, though they are still distinct; the spinal marrow has now lost considerably[Pg 25] more than a third of its length[125]. On the fourteenth day, the internodes, except the double ones, have become nearly straight again; the fourth and fifth ganglions have coalesced so as to form one, and the sixth and seventh have each lost their pairs of nerves[126]. Shortly after this, these last ganglions have nearly disappeared, and the chords of the three first internodes have again approached each other[127]. The next change exhibited is the absorption of the first ganglion by the brain, the union of the chords of the first internode, which is now straight, the approximation of the second and third ganglions, and the enlargement of the one formed by the union of the fourth and fifth, at the expense perhaps of the sixth and seventh, which have now intirely disappeared, and in their place is a very long internode. These united ganglions retain the pairs of nerves they had when separate[128]. Just before the assumption of the imago, the direction of the lobes of the brain becomes horizontal, the second and third ganglions unite, and the internode between the third and fourth is shortened[129]. Lastly, when the animal is become a butterfly, the second and third ganglions have coalesced, and are joined to that formed by the union of the fourth and fifth; a short isthmus or rather constriction, with an orifice, being their only separation: each of these united ganglions send forth laterally four pairs of nerves[130]. In his figure, Dr. Herold has not represented the orifice for the passage of the gullet, but doubtless one exists, which for an animal that imbibes only fluid food is probably[Pg 26] very minute. In Hypogymna dispar, we learn from Cuvier, this orifice is of that description, and of a triangular shape[131].
It can admit of no reasonable doubt that one of the principal intentions of these changes is to accommodate the nervous system to the altered functions of the animal in its new stage of existence, in which the antennæ, eyes, and other organs of the senses, as well as the limbs and muscles moving them, and the sexual organs, being very different from those of the larva, and if not wholly new, yet expanded from minute germs to their full size, may well demand corresponding changes in the structure of the nervous system by which they are acted upon.
But are these changes also concerned, as Dr. Virey conjectures, in producing that remarkable alteration which usually takes place between the instincts of the larva and imago? In order to answer this question, it will be requisite first to quote the ingenious illustration with which this able physiologist elucidates his ideas on this point. "The more readily," he observes, "to comprehend the action of instinct, let us compare the insect to one of those hand-organs in which a revolving cylinder presents different tunes noted at its surface, and pressing the keys of the pipes of the organ, gives birth to all the tones of a song: if the tune is to be changed, the cylinder must be pulled out or pushed in one or more notches, to present other notes to the keys. In the same manner let us suppose that nature has impressed or engraved certain determinations or notes of action, fixed in[Pg 27] a determinate series in the nervous system and the ganglions of the caterpillar, by which alone she lives, she will act according to a certain sequence of operations; and, so to speak, she will sing the air engraven within her. When she undergoes her metamorphosis into a butterfly, her nervous system being, if I may so express myself, pulled out a notch, like the cylinder, will present the notes of another tune, another series of instinctive operations; and the animal will even find itself as perfectly instructed and as capable of employing its new organs, as it was to use the old ones. The relations will be the same; it will always be the play of the instrument[132]."
This illustration is doubtless at the first glance very striking and plausible: but a closer examination will, I think, show, that, as in so many other instances in metaphysical reasoning, when fanciful analogies are substituted for a rigid adherence to stubborn facts, it is satisfactory only on a superficial view, and will not stand the test of investigation; and as this is a question intimately connected with what I have advanced on the subject of instinct in a former letter, I must be permitted to go somewhat into detail in considering it.
To prove his position, Dr. Virey ought at least to be able to show that, whenever a change takes place in the instincts of insects in their different states of larva and imago, a corresponding change takes place in the external structure of the nervous chord. But what are the facts? In three whole orders, viz. Orthoptera Hemiptera, and Neuroptera, as mentioned above[133], the structure[Pg 28] of the nervous chord is not changed; and yet we know that many tribes of these orders acquire instincts in their imago state altogether different from those which directed them in their state of larvæ. A perfect Locust, for instance, acquires the new instincts of using its wings; of undertaking those distant migrations of which so many remarkable instances were laid before you in a former letter[134]; and, if a female, of depositing its eggs in an appropriate situation. But if such striking changes in the instinct of these tribes can be effected without any perceptible alteration in the structure of the nervous chord, it is contrary to the received rules of philosophical induction to refer to this alteration the changes in the instincts of other tribes where it is found. Is it not far more probable that this alteration has in fact no connexion with the changes of instinct, but is solely concerned with those remarkable changes in the organs of sense and motion, which occur in the larva and imago states of the orders in which it is observed? In a common caterpillar, the form of the body, the legs, the eyes, and other organs of the senses, all strikingly differ from those of the imago; whereas, with the exception of the acquisition of new wings, a perfect locust differs little from its larva: so that we may reasonably expect a corresponding change, such as we find it, in the structure of the nervous chord of the lepidopterous insect, not called for in that of the neuropterous species, in which accordingly it does not take place.
This reasoning, in opposition to Dr. Virey's theory, that the changes of instinct depend on the altered structure of the nervous system, becomes greatly strengthened[Pg 29] when we advert to the higher classes of animals, which surely in any investigation of the nature of instinct ought to be closely kept in view; for the faculty, though often less perfect in them than in insects, is still of the same kind, and may consequently be expected to follow the same general laws. In a young swallow, for example, all its instincts are not developed at once any more than in an insect. The instinct which leads it to migrate does not appear for some months after its birth, and that of building a nest still later. But we have not the slightest ground for believing that these new instincts are preceded by any change in the structure of the great sympathetic nerve, or of any other portion of the nervous system: and the same may be said as to the sexual instincts developed in quadrupeds some years subsequent to their birth. If, then, these remarkable changes in the instinct of the higher classes of animals can take place independently of any visible change in the nerves, what substantial reason can be assigned why they may not also in the class of insects?
On the whole, I think you will agree with me, that there is nothing in Dr. Virey's hypothesis which should lead me to alter the opinion I have already so strongly expressed in a former letter[135], as to the insufficiency of the mechanical theories of instinct hitherto promulgated, adequately to explain all the phenomena; and unless they do this they are evidently of small value. Such theories as I have there adverted to may often seem to be supported by a few insulated facts, but with others, far more numerous, they are utterly at variance; and, to omit many other instances, I am strongly inclined to[Pg 30] doubt the possibility of satisfactorily explaining the variety of instincts exercised by a bee[136], or the extraordinary development of new ones in particular circumstances only[137], on any merely mechanical grounds.
And after all, even suppose it could be demonstratively shown that every instinct is as clearly dependent on secondary causes, as I have formerly admitted that some doubtless seem to be, yet what would this teach us as to the essential nature of instinct? We have advanced indeed a step; but still, as I have before observed in referring to the theories of Brown and Tucker, we have only placed the world upon the tortoise, and instinct, as to its essence, which is what we want to detect, is as mysterious as ever: just as, though we can clearly prove that the mind is acted upon by the senses, yet this throws no light upon the essential nature of the mind, which we are forced to admit is inscrutable, as if to teach us humility, and prevent our vainly fancying, that though allowed to discover some of the arcana of nature, we shall ever be able to penetrate into her inmost sanctuaries.
That Dr. Virey should regard instinct in insects as purely mechanical was the natural consequence of his denying them any portion of intellect; but his opinion cannot I think be consistently assented to, if it be the fact, as I have just shown[138], that they are not wholly devoid of the intellectual principle. Whatever is merely mechanical, must, under similar circumstances, always act precisely in the same way. An automaton once constructed, whilst its machinery remains in order, will invariably perform the same actions; and Des Cartes, when[Pg 31] he had constructed his celebrated female automaton, imagined that he had irrefragably proved his principle, that brutes are mere machines. But if, instead of losing himself in the wilds of metaphysical speculation, he had soberly attended to facts, he would have seen that the instinct of animals can be modified and counteracted by their intellect, and consequently cannot be regarded as simply mechanical. Though the instinctive impulse of an empty stomach powerfully impel a dog to gratify his appetite, yet, if he be well tutored, the fear of correction will make him abstain from the most tempting dainties: and in like manner a bee will quit the nectary of a flower, however amply replenished with sweets, if alarmed by any interruption. The ants on which Buonaparte amused himself with experiments at St. Helena, though they stormed his sugar-basin when defended by a fosse of water, controlled their instinct and desisted when it was surrounded with vinegar[139]: and in the remarkable instance communicated to Dr. Leach by Sir Joseph Banks, the instinct of a crippled spider so completely changed, that from a sedentary web-weaver it became a hunter[140]. There is evidently, therefore, no analogy between actions strictly mechanical and instincts, which, though they may often seem to be excited by mechanical causes, are liable to be restrained or modified by the connexion of the instinctive and intellectual faculties[141]; and while we are ignorant how this connexion takes place, it is obviously impossible to reason logically on the subject.
In thus denying that any existing mechanical theory of instinct is satisfactory, I by no means intend to assert[Pg 32] that instinct is purely intellectual. I have already given you my opinion[142], that it is not the effect of any immediate agency of the Deity; nor am I prepared to assent to the doctrine of a writer, who has in some respects written ably on the subject in question, who says, that "the Divine Energy does in reality act not immediately, but mediately, or through the medium of moral and intellectual influences upon the nature or consciousness of the creature, in the production of the various, and in many instances truly wonderful, actions which they perform[143]." The same objection applies to this as to so many other metaphysical theories, that it is not adequately supported by facts; and all theories not so supported are injurious to science in proportion as their plausibility is greater, by leading the student to relax in that observation of nature and attentive study of the instincts of animals, on which alone sound hypothesis on this subject can be ultimately founded.
I shall conclude these remarks on the nature of instinct with a few observations as to the circumstances in which insects may be supposed to be guided by this faculty, and those in which intellect seems to direct them. The bee, when it takes its flight to a field where flowers abound, is governed by intellect in the use of its senses; for these are given to it as guides: and when it arrives there, they direct it to the flowers, and enable it to ascertain which contains the treasures it is in search of; but having made this discovery, its instinct teaches it to imbibe the nectar and load its hind legs with pollen.—Again: its senses, aided by memory, enable it to retrace[Pg 33] its way to the hive, where instinct once more impels it in its various operations. So that when we ascribe a certain degree of intellect to these animals, we do not place them upon a par with man; since all the most wonderful parts of their economy, and those manipulations that exceed all our powers, we admit not to be the contrivance of the animals themselves, but the necessary results of faculties implanted in their constitution at the first creation by their Maker. I may further repeat, that the mere fact of being endowed with the external organs of sense, proves a certain degree of intellect in insects. For if in all their actions they were directed merely by their instinct, they might do as well without sight, hearing, smell, touch, &c. but having these senses and their organs, it seems to me a necessary consequence, that they must have a sufficient degree of intellect, memory, and judgement, to enable them advantageously to employ them.
There is this difference between intellect in man, and the rest of the animal creation. Their intellect teaches them to follow the lead of their senses, and make such use of the external world as their appetites or instincts incline them to,—and this is their wisdom; while the intellect of man, being associated with an immortal principle, and being in connexion with a world above that which his senses reveal to him, can, by aid derived from heaven, control those senses, and bring under his instinctive appetites, so as to render them obedient to the το ἡγεμονικον, or governing power of his nature: and this is his wisdom.
I am, &c.
"Life and flame have this in common," says Cuvier, "that neither the one nor the other can subsist without air; all living beings, from man to the most minute vegetable, perish when they are utterly deprived of that fluid[144]." The ancients, however, not perceiving insects to be furnished with any thing resembling lungs, took it for granted that they did not breathe; though Pliny seems to hesitate on the subject[145]. But the microscopic and anatomical observations of Malpighi, Swammerdam and Lyonet, and the experiments of more modern physiologists, have incontestably proved that insects are provided with respiratory organs, and that the respiration of air is as necessary to them as to other animals. They can exist indeed for a time in irrespirable air; and immersion in hydrogen or carbonic acid gases is not, as I have often ascertained, so instantly fatal to them as it would be to vertebrate animals; but like them, they[Pg 35] speedily perish in air altogether deprived of its oxygen, or placed in situations to which all access to this essential element is excluded. Their respiration too of atmospheric air produces the same change in it with that of the vertebrate animals, the oxygen disappearing, and carbonic acid gas being produced in its place. Boyle had long since ascertained, that when bees, flies, and other insects were placed under an exhausted receiver, they often perished[146]: and the same effect was even observed by the ancients to ensue, when their bodies were by any means covered with oil or grease, which necessarily closed the orifices of their respiratory organs[147].
But for the first series of experiments ascertaining the necessity of a supply of air to insects, and their conversion of it into carbonic acid, we are indebted to the illustrious Scheele[148]; and his experiments have been repeated and confirmed by Spallanzani, Vauquelin, and other chemists. The former found, that when caterpillars and maggots were confined in vessels containing only about eleven cubic inches of atmospheric air, though furnished with sufficient food, they soon died, and sooner when the space was more confined[149]. He ascertained too, that a larva weighing only a few grains consumed, in a given time, as much oxygen as an amphibious animal a thousand times as voluminous[150]. A male grasshopper (Acrida viridissima) in six cubic inches of oxygen lived but eighteen hours, and the female placed in eight cubic inches of atmospheric air, only thirty-six hours. The[Pg 36] usual tests in both instances detected the conversion of the oxygen present into carbonic acid[151]. Precisely the same result was obtained by Sorg and Ellis, who, having placed a number of flies in nine cubic inches of atmospheric air, found them all dead by the third day, the oxygen intirely vanished, and a quantity of carbonic acid nearly equal in bulk produced[152].
It is ascertained too, that insects like other animals require in the process of respiration not merely oxygen, but such a mixture of it with nitrogen or azote as composes atmospheric air: for Vauquelin found that a grasshopper placed in six cubic inches of oxygen lived only half as long (eighteen hours) as another placed in eight inches of atmospheric air; its breathing was much more laborious, and it died when not more than one-twentieth of the oxygen had been converted into carbonic acid[153]. That a large quantity of oxygen penetrates all parts of insects, is evident also from the acid prevalent in the fluids of most of them, as likewise from the wonderful power of their muscles. That azote is also received, seems probable from the ammonia which has been extracted from the fluids of many, and from the rapid putrescence of these animals[154].
The mode, however, in which the respiration of insects is carried on, differs greatly from that which obtains in the higher animals. They have no lungs, no organs confined to a particular part of the body, by means of which the whole of the blood is regularly exposed to the[Pg 37] action of the inspired air. They do not breathe through the mouth, but through numerous orifices called spiracles, and the respiratory vessels connected with these are conducted to every part of the body. In some indeed, that we have included under the denomination of insects, as the Arachnida, an approach is made to the branchial respiration of fishes.
The respiratory apparatus of insects may be considered under two principal heads:—viz. the orifices or spiracles, and other external organs by which the air is alternately received and expelled; and the internal ones, by which it is distributed. Each of these is well worthy of your attention.
I. The external respiratory organs of insects may be divided into three kinds. Spiracles; Respiratory plates; and branchiform and other pneumatic appendages.
i. Spiracles[155] (Spiracula), or breathing pores, are small orifices in the trunk or abdomen of insects, opening into the tracheæ, by which the air enters the body, or is expelled from it[156]. They may be considered principally as to their composition and substance; shape; colour; magnitude; situation; and number.
1. Composition and substance. Perhaps you may not be aware that the structure of these minute apertures is not so simple as at the first view it may seem; but when you recollect that by them the insect breathes, you will suspect that provision may be made for their opening and shutting. A spiracle therefore, speaking analogically,[Pg 38] may be regarded in numerous cases as a mouth closed by lips. In caterpillars and many other insects, the substance of the crust where it surrounds the spiracle, is elevated so as to form a ring round it. The lips, properly speaking, are formed of a single cartilaginous piece or platform, with a central longitudinal cleft or opening, when closed often extending the whole length of the piece[157]; but in some appearing always open and circular: of the former description are those covered by the elytra in the common cockchafer; and of the latter, those that are not so covered: in some, as in the antepectoral pair of the mole-cricket, there appear to be no lips, the orifice being merely closed with hairs[158]. Though the aperture is usually in the middle of the platform, in the female of Dytiscus marginalis, it is nearer the posterior side, the anterior or upper lip being the longest. In the majority, the mouth or cleft is nearly as long as the spiracle; yet in the puss-moth (Cerura Vinula) it is shorter[159]. Some spiracles, however, are unilabiate, or have only one lip. This is the case with Gonyleptes and perhaps others[160]. The lips are usually horizontal, but sometimes they dip so as to make the spiracle appear open.
With regard to the substance of these organs, it is more or less cartilaginous, and probably elastic; the surface frequently appears to be corrugate or plaited; this is very distinctly seen in the stag-beetle and the cockchafer: in the last insect, under a powerful magnifier, we are told that the lips appear to consist of parallel cartilaginous processes, separated by a cellular web[161]. In some species[Pg 39] of Copris the corrugations form a perplexed labyrinth; in the caterpillar of the puss-moth the plaits are so narrow as to look like rays[162]; and in some Dynastidæ the lips approach to a lamellated structure. Again, in Hydrophilus caraboides the upper lip, and in Dytiscus circumflexus, both lips seem formed of elegant plumes[163]: a similar ornament distinguishes the inner edge of the lips in the caterpillar of the great goat-moth (Cossus ligniperda) and others[164]. In the grub of the rhinoceros-beetle (Oryctes nasicornis) the margin of the lower or inner lip is decorated by pinnated rays, which enter the cellular membrane that covers the upper lip[165]: in this larva, and that likewise of the cockchafer, the two lips are formed of different substances; in the last the upper or outer one consists of a perforated cellular membrane, through which the air can pass, while the lower or inner one is a cartilaginous valve that closes the orifice[166]: in the former this valve is surmounted by a boss[167]. In the pupa of Smerinthus Populi, a hawk-moth not uncommon, and of some dragon-flies (Libellula depressa), the margin of the two lips is crenated, probably with notches which alternate, that the mouth of the spiracle may shut more accurately[168]. The substance is unusually thick in the spinose caterpillars of butterflies; and in the pupa of one, Uria Proteus, it is villose.
Under the present head I may observe, that in some cases, as in the puss-moth, and the larva of the common[Pg 40] water-beetle (Dytiscus marginalis), the spiracles are closed by a semifluid substance, which however, according to Sprengel, is permeable to the air[169]. The animal, where these organs are furnished with lips, has doubtless, by means of a muscular apparatus, the power of opening and shutting them: this is done, we are told, by elevating and depressing, or rather by contracting and relaxing them. Sorg counted in one case (Oryctes nasicornis) twenty, and in another (Acrida viridissima) fifty, of these motions to take place in little more than two minutes[170]: but the quickness and force of this motion is not always uniform; for the same physiologist observed, that in Carabus auratus, when feeding or moving its body rapidly, the contraction of the spiracles took place at very short intervals; but when it was fasting, and its motions were slow, the intervals were longer[171]: it is probable also, that the temperature may accelerate or retard the motion. In the summer I examined a specimen of Phyllopertha horticola, that had indeed been somewhat injured, with this view: the pulses of the abdomen, which alternately rose and fell, were at about the rate of the pulse of a man in health, sixty in a minute, and the spiracles appeared to me to keep pace with this motion: later in the year, when the temperature was lower, as I was walking, I took a specimen of some grasshopper (Locusta).[Pg 41] Upon viewing it under a lens, I observed one of the convex pectoral spiracles open and shut, and the interval between two breathings appeared nearly half a minute.
2. With regard to their shape, spiracles vary considerably. In general we may observe that the abdominal ones are usually flat, while those of the trunk are often convex[172]. Sometimes they are very narrow and nearly linear, as in many pupæ of Lepidoptera, and those in the metathorax of the sand-wasps (Ammophila) and affinities; at others they are wider and nearly elliptical, as in Lucanus and many Lamellicorn beetles: again, in Copris they are circular; in Cordylia Palmarum ovate; in Dytiscus oblong[173]; in Goerius olens lunulate; in Gonyleptes nearly of the shape of a horse-shoe[174]; and probably many other forms might be traced, if a thorough investigation with this view were undertaken.
3. The colour of spiracles will not detain us long. In the caterpillars of Lepidoptera this is often so contrasted with that of the rest of the body, as to produce a striking and pleasing effect. Thus when the body is of a dark colour, they are usually of a pale one[175]; or if the body is pale, they are dark[176], or surrounded with a dark ring[177]. This contrast is often rendered more striking by their position with regard to the partial colours that often ornament caterpillars: in those whose sides are decorated by a longitudinal stripe, the spiracles are often planted in it[178]; or just above it[179]; or between two[180]: in[Pg 42] some hawkmoths the intermediate ones are set in white or pale spots, which gives great life to the appearance of the animal. In general, in perfect insects the most prevalent colour is buff, or reddish-yellow. In the larva of the great water-beetle these organs resemble the iris of the eye, being circular with concentric rings alternately pale and dark[181].
4. The size of spiracles varies considerably. Those in the larva last mentioned are so minute as to be scarcely visible except under a lens, while those behind the fore-legs in the mole-cricket are a full line in length, and those in the pleura of Acrocinus accentifer, a Brazilian Capricorn beetle, are more than twice as long. In the same species they are often found of different sizes;—thus the anal pairs in the water-beetle lately alluded to, I mean in the perfect insect, are much larger than the rest[182], probably that the animal may imbibe a larger quantity of air when it rises to the surface of the water, where it suspends itself by the tail. In those Lamellicorn beetles in which the terminal part of the abdomen is not protected by the elytra, the covered spiracles are the largest.
5. Under the next head, the situation of spiracles, I shall not only consider the part of the body in which they are situated, but likewise their position in the crust; to which last, as it will not detain us long, I shall first call your attention. Their position in this respect is most commonly oblique: but in the abdomen of the above water-beetle they are transverse, and in a larva I possess, probably of an Elater, they are longitudinal. In spinose[Pg 43] caterpillars these organs are generally planted between two spines, one being above and the other below. The lateral line of the body most commonly marks their situation; but in many cases they become ventral, and in others dorsal. The most important circumstance, however, connected with the present head is their appropriation to particular segments or parts of the body, for, like the ganglions of the spinal marrow, they are distributed to almost every segment. Let us take a summary view of their arrangement in this respect.
No insect has any spiracle in the head; but in caterpillars and many other larvæ there is a pair in the first segment of the trunk. This is also to be found in the other states, but is not easily detected in the pupæ of Lepidoptera: in the Coleoptera order, in the grub of the Lamellicorn beetles, it is extremely conspicuous, and planted in the side of the first segment[183]; in other Coleopterous grubs it is not so readily found, but probably its station is somewhere behind the base of the arms, where it is very visible in that of the Staphylinidæ. In the imago of insects of this order, this antepectoral spiracle has been overlooked, and indeed is not soon discovered: to see it clearly, the manitrunk should be separated from the alitrunk; and then if you examine the lower side of the cavity, you will see a pair of, usually, large spiracles planted just above the arms, in the ligament that unites these two parts of the trunk to each other: in the common rove-beetle, however, (Goerius olens)you may easily see it without dissection[184]. In the Orthoptera it is situated behind the arms, as in Gryllotalpa: or between them[Pg 44] and the prothorax, as in Blatta: in the Hemiptera and Neuroptera probably the situation is not very different. In the Lepidoptera this pair of spiracles is planted just before the base of the upper or primary wings[185]: a similar situation, I suspect, is appropriated to it in the Trichoptera, but covered by a tubercle or scale. Something similar has been noticed by M. Chabrier, in the same situation and circumstances, in the collar of Hymenoptera[186]. In numerous Diptera this breathing pore is planted on each side between the collar and the dorsolum above the arms[187], and in Hippobosca in the collar itself[188].
In Lepidopterous, Coleopterous, and some other larvæ, the two segments of the body corresponding with the alitrunk in the perfect insect, are without spiracles, neither have they in this state, though pneumatic organs have been discovered[189], any real ones in that part: but not so the remaining orders, all of which have these organs in that section of the trunk. To begin with the Orthoptera:—in Blatta there seems to be a long narrow one behind the intermediate leg; in the Gryllotalpa there is one in the posterior part of the pleura; and in Locusta, above both the intermediate and hind legs[190]. It is probable, that in general those that have no spiracles in the manitrunk have four in the alitrunk, which seems the natural number belonging to the trunk. In many of the Heteropterous Hemiptera in the parapleura[Pg 45] there is an open spiracle without lips[191], to which, as in that beautiful bug Scutellera Stockeri, a channel sometimes leads. The space in which this spiracle is planted in other genera of bugs (Pentatoma &c.) is covered with a kind of membranous skin, often much corrugated[192]. In the aquatic insects of this section, and many terrestrial ones, as Reduvius, &c. this spiracle is obsolete. There is another circumstance, possibly connected with their respiration, relating to many of the bugs, which may be mentioned here. If you examine Pentatoma rufipes, a very common one, you will find between the scapula and parapleura a long orifice or chink; this upon a closer inspection, under a good magnifier, you will see completely filled with minute stiff hairs or bristles, which fringe the posterior margin of the scapula[193]. In a Brazilian species of Lygæus (sexmaculatus K. M. S.) with incrassated posterior thighs, these hairs are replaced by lamellæ which have the aspect of gills. A red, vertical, convex spiracle, with its orifice towards the head, and terminating posteriorly in a kind of conical sac, is situated towards the hinder part of the pleura in the giant water-scorpion (Belostoma grandis[194]); this seems analogous to one lately mentioned in the mole cricket. In the other section of this Order it is not easy to decipher the parts of the under side of the alitrunk. In Fulgora, Cicada, and many others of its genera, there appears to be more than one opening into the chest; but whether they are of a pneumatic nature or not, can only be ascertained by an inspection of the living animal. There is a very visible[Pg 46] spiracle over each of the four last legs of the Libellulina[195], but in the remainder of the Neuroptera Order they have eluded my search. In the Hymenoptera and Diptera they are nearly in the same situation, being placed behind the wings on each side of the metathorax; in the latter Order with the poiser near them on the inner side[196]: in this also, the spiracles of the trunk are without lips, except in the larvæ, but are often merely an orifice, sometimes fringed with hairs; this is particularly conspicuous in Syrphus, in which these orifices are very large, and in some species closed by an elegant double fringe of white hairs. This is doubtless to prevent the entrance of any particles of dust or the like.
We are next to consider the situation of the spiracles of the abdomen: these which are supposed to be appropriated exclusively to inspiration, are usually more numerous than those of the trunk, by which it is probable that expiration is performed, and have principally attracted the notice of Entomologists: they are either dorsal, lateral, or ventral. In Dytiscus, Copris, &c. amongst the beetles, all the spiracles are dorsal; in the larvæ of Coleoptera and Lepidoptera they are lateral; and in the Heteropterous Hemiptera they are usually ventral: in Dynastes they are commonly found of all three descriptions;—the three first being dorsal, the two next lateral, and the last pair ventral[197]. In some instances, as in Perga Kirbii, and probably other Hymenoptera, these organs are planted in that portion of the dorsal segment which turns under, as was observed in a former letter[198],[Pg 47] and becomes ventral. Generally there is a pair of spiracles to each segment, and in those insects that have a hypochondriack joint[199] there is often a spiracle in it. The last segment of the abdomen is always without these orifices, as is the basal one in Velia, Ranatra, and some other bugs. A singular anomaly distinguishes the Libellulina: they appear to have no abdominal spiracles[200], yet I have seen the abdomen of Libellula depressa when reposing, contract and dilate alternately, from whence it follows that this part is concerned in respiration. Sprengel says that the larvæ in this tribe have seven or nine on each side[201], and Reaumur speaks of them as discoverable in the pupa[202]. I have carefully examined the pupa-skin of most of the genera of Libellulina, under a powerful magnifier, but have not succeeded in discovering any thing like these organs in the abdomen. The Ephemera and probably the other Neuroptera have abdominal spiracles[203]. M. Latreille observed one on each side of the base of the scale on the footstalk of the abdomen in ants[204]. Generally the abdominal spiracles may be described as planted in the crust of the insect; but in many cases their station is in the membranous folds, which I have therefore named the pulmonarium, that sometimes separate the dorsal from the ventral segments: these folds allow of a considerable distention of the abdomen, which is probably necessary when all the air-vessels are full. In a gravid Ichneumon I once saw it enlarged to more than twice its natural size by means of this membrane, through which the eggs were distinctly[Pg 48] visible.—Before I bid adieu to this subject, I must say a few words upon the situation of the organs in question in the myriapods. In Iulus, in each segment is a pair of orifices which have usually been regarded as spiracles, but M. Savi found that these orifices opened into vesicles containing a fetid fluid, and upon a very close examination he discovered the real spiracles above the base of the legs, in connexion with tracheæ[205]. In some of the larger species of Scolopendræ large open spiracles in the same situation are extremely visible[206]. Cermatia presents a singular anomaly:—a single series of spiracles of the usual form, each planted in a cleft of the posterior margin of the dorsal scuta, runs along the back of the animal[207]: unless we may suppose that, like the seeming spiracles of Iulus just mentioned, these are merely orifices by which it covers itself with some secretion.
6. A few words upon the number of spiracles.—If you examine the common dog-tick (Ixodes Ricinus), you will find only one of these organs on each side of the abdomen[208]; the Libellulina, as we have seen, have only four, all in the trunk; in the Dynastidæ, Melolontha, and the larva of Dytiscus, there are fourteen; sixteen in the Copridæ; eighteen in Dytiscus, and probably the majority of Coleoptera, both larva and imago, and Lepidoptera; and a pair to each segment except the last, in the Myriapods.
ii. Respiratory plates (Respiratoria). The nearest approach[Pg 49] to spiracles is made by those remarkable plates that are found in such larvæ of Diptera, as in that state inhabit substances that might impede or altogether stop the entrance or exit of the air by the ordinary spiracles, such as dead or living flesh, dung, or the like. The Creator therefore, as he has seen it good for wise reasons[209] to commission certain insects to feed on unclean food, has fitted them for the offices that devolve upon them, and has placed their orifices for breathing in plates at each extremity of the body. There are usually two of these plates at the head, and two at the tail. In the grub of the common flesh-fly (Sarcophaga carnaria), at the junction of the first segment of the body with the second, two of these plates are planted, which are concave and circular, with a denticulated margin; in the cavity near the lower side is a round spiracle. These plates the animal can withdraw within the body, so as to prevent this spiracle from being stopped up by any greasy substance[210]. The posterior extremity of this grub is truncated, and has a large and deep cavity surrounded by several fleshy prominences: at the bottom of this are two oval brown plates, in each of which are three oval spiracles, placed obliquely: by the contraction of the fleshy prominences, this cavity also can be closed at the will of the animal[211]. In some cases, several stiff rays or[Pg 50] spines replace the prominences[212]. In Echinomyia grossa and others the anal plates appear not to be perforated, being surmounted only by a central boss[213]; but this, most probably, as in the case of Œstrus Ovis[214], is a valve that closes the respiratory orifices. In the gad-fly of the ox (Œ. Bovis) there are no plates at the anterior extremity of the body; but those planted in the other end are very remarkable, and demand particular attention. Each is separated by a curved line into two unequal portions; the smallest of which is contiguous to the convex belly, and the largest to the concave back of the animal. This last is distinguished by two hard, brown, kidney-shaped pieces, a little elevated with the concave sides turned towards each other: in this sinus is a single, small, white spot, which appears to be a spiracle: in the smallest portion are eight minute circular orifices, arranged in a line[215]. As the only communication which this grub has with the atmosphere is at its anal extremity, it has no occasion for respiratory organs at the other. The gad-fly of the horse (Gasterophilus Equi, &c.) which has no communication at all with the external air, breathing that which is received into the stomach, has these plates at both ends of the body.
iii. Respiratory Appendages[216]. These may be divided into two kinds; those by which the animal has immediate communication with the atmosphere, and those by which it extracts air from water.
1. To begin with the first. These are often found in insects which, during their two first states, live in the water. No better example, nor one more easy to be examined, of this structure, can be selected, than the gnat (Culex). You must have occasionally observed in tubs of rain-water, numerous little wriggling worm-like animals, which frequently ascend to the surface; there remain a while, and then bending their head under the body rapidly sink to the bottom again. These are the larvæ of some species of the genus just named; and if you take one out of the water and examine it, you will perceive that it is furnished near the end of its body with a singular organ, which varies in length according to the species, and forms an angle with the last segment but one[217]. The mouth of this organ is tunnel-shaped, and terminates in five points like a star; and by this it is usually suspended at the surface of the water, and preserves its communication with the atmosphere: in its interior is a tube which is connected with the tracheæ, and terminates in several openings, visible under a microscope, at the mouth of the organ. The points or rays of the mouth when the animal is disposed to sink in the water, are used to close it, and cut off its communication with the atmosphere. When the animal is immersed, a globule of air remains attached to the end of the tube, so that it is in fact of less specific gravity than that element, and it is not without some effort that it descends to the bottom; but when it wishes to rise again, it has only to unclose the tube, and it rises without an effort to the surface, and remains suspended for any length of time. Its anal extremity is clothed with bunches of[Pg 52] hairs, which are furnished with some repellent material which prevents their becoming wet[218]: it is this repellent quality that probably causes a dimple or depression of the surface, which if you look narrowly you will discover round the mouth of the tube[219].
When the gnat undergoes its first change and assumes the pupa, instead of a single respiratory appendage it is furnished with a pair, each in shape resembling a cornucopia, and, what is remarkable, placed near the opposite extremity of the body, for they proceed from the upper side of the trunk[220]. By these tubular horns, which Reaumur compares to asses' ears[221], they respire, and are suspended at the surface.
Other respiratory tubes or horns are more complex. The rat-tailed grub of a fly (Helophilus pendulus), like the gnat, breathes by a tube: but as if the Creator willed to show those whose delight it is to investigate his works, by how many varying processes he can accomplish the same end, this respiratory organ is of a construction totally different from that we have been considering. It is not fixed to the side of the tail, but is a continuation of the tail itself, and is composed of two tubes, the inner one, like the tube of a telescope, being retractile within the other[222]. The extremity, which is very slender, and through which the air finds admission by a pair of spiracles, terminates in five diverging hairs or rays, which probably maintain it in equilibrio at its station at the surface[223]. As these larvæ seek their food[Pg 53] amongst the mud at the bottom of shallow pools, in which they are constantly employed, they require an apparatus capable of being lengthened or shortened, to suit the depth of the water, that they may maintain their necessary communication with the atmosphere; and for this purpose a single tube would not have been sufficient: therefore Providence has furnished them with two, and both are extremely elastic, consisting of annular fibres, so as to admit their being stretched to an extraordinary length. Reaumur found that these animals could extend their tails to near twelve times their own length. The mechanism by which the terminal piece is pushed forth or retracted, is very curious, though extremely simple. Two large parallel tracheæ, the direction of which is from the head[224] of the grub to its tail, occupy a considerable portion of its interior: near the origin of the tail, where they are very ample, they suddenly grow very small, so as to form a pair of very slender tubes, but so long that, in order to find room in a very contracted space, they form numerous zigzag folds attached to the terminal tube; when this issues from the outer tube they consequently begin to unfold, and when it is intirely disengaged, they are become quite straight and parallel to each other. Reaumur has figured them as being united at the base of the inner tube[225]; most probably, however, they do not here stop short, but, as in[Pg 54] other instances, proceed to the end, and terminate in the two spiracles mentioned above: he conjectures that when the animal has occasion to push forth its respiratory apparatus, it injects into these vessels part of the air contained in the body of the tracheæ, which of course would cause them to unfold and push forth the tube[226]. When this insect assumes the pupa, instead of its anal respiratory organ it has four respiratory horns in the trunk near the head[227].
The larva of the chamæleon-fly (Stratyomis Chamæleon) is furnished with a respiratory organ of a still different and more elegant structure, exhibiting some resemblance to the tentacula of what are called sea anemones. In this larva the last joint of the body is extremely long, and terminates in an orifice to receive the air, which is surrounded by a circle of about thirty diverging rays, consisting of beautifully feathered hairs or plumes[228]. This apparatus serves the same purpose with that above described of the larva of the gnat. The feathery hairs are so prepared as to repel the water, and thus to suspend the animal by its tail at the surface, and preserve a constant access of air. When it has occasion to sink, it turns these hairs in and shuts the orifice, carrying down with it an air-bubble that shines like quicksilver, and which Swammerdam conjectures enables it again to become buoyant when it wants to breathe[229].
In the red aquatic larva of a small gnat (Chironomus plumosus) there are two anal respiratory subcylindrical horns, with the orifice fringed with hairs[230]; and in another[Pg 55] gnat Reaumur discovered four[231]. The larva of Tanypus maculatus, whose remarkable legs I formerly noticed[232], exhibits in the interior of its trunk two long, oval, opaque bodies, which De Geer conjectures may be air-reservoirs; these, when the animal assumes the pupa, according to every appearance become external, and are placed on the back, precisely where the respiratory horns of aquatic pupæ are usually situated,—they appear to terminate in a transparent point[233]. The pupa of a Tipula observed by Reaumur, instead of two has only one of these respiratory organs, in the form of a very fine hair proceeding from the anterior end of the trunk, and considerably longer than the animal itself[234].
It is observable that aquatic insects that come to the surface of the water for air, receive it at the anus, often carrying it down with them as a brilliant bubble of quicksilver. This is generally done by means of spiracles in perfect insects, but in the water-scorpion tribe in that state respiration is by means of a long hollow tube, consisting of two concavo-convex pieces which apply exactly to each other. This is found in both sexes, and therefore cannot be an ovipositor, as some have thought[235].
These respiratory organs, however, are not invariably confined to aquatic larvæ and pupæ, for those of some aphidivorous flies have anal ones, and the pupa of Dolichopus nobilitatus, or a fly nearly related to it, which is terrestrial, has likewise a pair of long sigmoidal ones on the back of the trunk[236]. The pupa also of the rat-tailed[Pg 56] larva just noticed as having four horns, resides under the earth, the insect being only aquatic in its grub state.
2. I am next to consider those respiratory appendages by which aquatic insects, since they do not come to the surface for that purpose, appear to extract air for respiration from the water; so that they may be looked upon in some degree as analogous to the gills of fishes: there is, however, this difference between them—in fishes, the blood is conveyed in minute ramifications of the arteries to the surface of the branchial laminæ, through the membranes of which they abstract the air combined with the water; but as insects have no circulation, the process in them must be different, and their branchiform appendages may be regarded as presenting some analogy rather than any affinity to those of fishes. The first approach to this structure is exhibited by the pupa of a gnat lately mentioned (Chironomus plumosus); for on each side of the trunk this animal has a pencil consisting of five hairs elegantly feathered, which, when they diverge, form a beautiful star; its anus also is furnished with a fan-shaped pencil of diverging hairs[237].
On most of the abdominal segments of the larvæ and pupæ of the Trichoptera are a number of white membranous floating threads, arranged in bundles, four on each segment, two above and two below, and traversed longitudinally by several air-vessels or bronchiæ, which run in a serpentine direction, growing more slender as they approach the extremity, and in some places sending forth very fine ramifications,—these are their respiratory organs[238]. The caterpillar also of a little aquatic[Pg 57] moth (Hydrocampa stratiotata) at first sight appears to be covered on each side with hairs, but which examined under a microscope are found to be branching flattish filaments, each furnished with tubes from the tracheæ. These caterpillars have also the semblance of spiracles, but apparently found in the usual situation[239]. The larva of a little beetle often mentioned in my letters (Gyrinus Natator), is furnished on each side of every abdominal segment with a long, hairy, slender, acute, conical process, of the substance of the segment, through each of which an air-tube meanders; the last segment but one has four of these processes, longer than the rest[240].
Laminose or foliaceous respiratory appendages distinguish the sides of the abdomen of the larvæ and pupæ of the Ephemeræ, whose history you found so interesting[241]. In them these organs wear much the appearance of gills. In the different species they vary both in their number and structure. With regard to their number, some have only six pair of them, while others have seven. In their structure the variations are more numerous, and sometimes present to the admiring physiologist very beautiful forms[242]. They usually consist of two branches, but occasionally are single, with one part folding over the other, as in one figured by Reaumur, which precisely resembles the leaf of some plant, the air-vessels or bronchiæ in connexion with the tracheæ branching and traversing it in all directions, like the veins of leaves[243]. The double ones[Pg 58] differ in form. In the larva and pupa of Ephemera vulgata there are six of these double false gills on each side of the abdomen, the three last segments being without them; each branch consists of a long fusiform piece, rather tumid and terminating in a point, which is fringed on each side with a number of flattish filaments, blunt at the end. An air-vessel from the trachea enters the gill at its base; is first divided into two larger branches, each of which enters a branch of the false gill. These branches send forth on each side numerous lesser ramifications, one of which enters each of the filaments[244]. In another species (E. vespertina) each false gill presents the appearance of a pair of ovate leaves with a long acumen, and the air-vessels represent the midrib of the leaf, with veins branching from it on each side[245]; and, to name no more, in E. fusco-grisea, one branch represents the leaf of a Begonia, the sides not being symmetrical, with its veins, while the other consists only of numerous branching filaments[246]. In other aquatic larvæ, as in that of the common May-fly (Sialis lutaria), these appendages consist of several joints[247].
By the above apparatus these aquatic animals are enabled to separate the air from the water, as the fish by their gills; but how this separation is made has not been precisely explained. The false gills in many species are kept in continual and intense agitation. When they move briskly to one side, Reaumur conjectures they may receive the air, and when they return back they may[Pg 59] emit it[248]. This brisk motion probably disengages it from the water. In many species, when in repose, they are laid upon the back of the animal[249], but in others they are not[250].
The larvæ of the Agrionidæ appear to respire like those of the Ephemeræ, &c. by means of long foliaceous laminæ or false gills filled with air-vessels; but instead of being ventral, they proceed from the anus. They are three in number, one dorsal and two lateral, perpendicular to the horizon, of a lanceolate shape, beautifully veined, with a longitudinal middle nervure, from which others diverge towards the margin, which are probably bronchiæ. They are used by the animal, which swims like a fish, as fins, but it does not appear to imbibe the water like the other Libellulinæ, nor to propel itself by ejecting it,—a circumstance which furnishes an additional argument for the more received opinion, that this action in them is for the purpose of respiration as much as for motion[251].
The larvæ and pupæ of the Libellulinæ, receive the water and air that they respire by a large anal aperture, which is closed at the will of the animal by five hard, moveable, triangular, concavo-convex pieces, all very acute and fringed with hairs. These pieces are placed so that there is one above, which is the largest of all; one on each side, which are the smallest, and two below; when these are closed they form together a conical point[252]. Sometimes only three of these pieces are conspicuous[253]:[Pg 60] three other cartilaginous pieces, resembling the valves of a bivalve shell, close the passage within the pointed pieces[254]. At this orifice the water is received; and when, by an internal process to be described afterwards, it has parted with its oxygen, is again expelled.
Under this head I shall mention a fact which may be connected with respiration of the insects concerned. In dissecting a moth related to Catocala Pronuba, but I do not recollect the particular species,—at the base of the abdomen of the male I discovered two bunches of long fawn-coloured parallel hairs, planted each in an oval plate, plane above, but below convex and fleshy; while the plates remained attached to the insect, they appeared to have a distinct pulsation. The hairs, which are about half an inch long, diverge a little, and form a tuft not very unlike a shaving-brush[255]. I have not since met with this species, but I have preserved the brush and scale. Somewhere in Bonnet's works, but I do not recollect where, I have since found mention of a similar fact in another moth.
II. Having considered the external respiratory organs of insects, by which the air is received, we are next to consider the internal ones, by which it is distributed. These are gills; tracheæ and bronchiæ; and sacs or pouches[256].
i. Gills (Branchiæ[257]). Having lately described what[Pg 61] may be denominated false gills, or branchiform appendages, I shall now call your attention to what may be denominated true ones, which are peculiar to the Arachnida Class: but what is remarkable, the animals that breathe by them are very rarely inhabitants of the water, so that their functions cannot be perfectly analogous to those of fishes.
In the Scorpion, on each side of the four first ventral segments a spiracle may be discovered, which has no lip as in other insects, but is merely a circular orifice. These orifices do not lead to tracheæ or vesicles, but to true gills, which are situated below a muscular web which clothes the internal surface of the crust. Each gill consists of many semicircular very thin plates, of a dead milky white, which are connected together at the dorsal end like the leaves of a book. There appear to be more than twenty of these leaves, which when strongly magnified look transparent and destitute of any vessels. Each gill is fastened at the back to the spiracle[258]. In the spiders also, gills are discoverable, but differently circumstanced. On the under side of the abdomen, near the base, is a transverse depression, on each side of which is a longitudinal opening leading to a cavity, which is covered from above by a cartilaginous plate. In this cavity is situated a true gill, which is white, triangular, and covered with a fine skin; the leaves of this gill are far more numerous and much finer and softer than those of the gills of the scorpion. On account of their softness they have often the appearance of a slimy skin; but their[Pg 62] laminated structure shows itself very clearly in old specimens, and in such as have been immersed in boiling water[259].
ii. Tracheæ and Bronchiæ[260]. Parallel with each side of the body of most insects and extending its whole length, run two cylindrical tubes[261], which communicate with the spiracles[262], and from which issue, at points opposite to those organs, other tubes which ramify ad infinitum, and are distributed to every part of the body[263]. The first of these tubes are called the tracheæ and the latter the bronchiæ. This structure appears, however, not to be universal: it is to be found in caterpillars and many Dipterous larvæ; but in that of the rhinoceros-beetle and other Lamellicorns, the bronchiæ branch directly from the spiracle, the bottom or interior mouth of which is lined by a membrane from which they proceed[263]: something similar has been observed to take place in many insects in other states, as the common cockchafer[264]; in the pupa of Smerinthus Populi[265]; in the Cicadæ[266]; in the Locust tribe[267]; and many others. In the Cossus, or larva of the great goat-moth, the trachea commences with the first spiracle, and finishes a little beyond the last, after which it diminishes considerably in diameter, and terminates in several branches or bronchiæ, which proceed to the anal extremity of the body[268]. The bronchiæ which originate from the tracheæ in the[Pg 63] vicinity of each spiracle, may be considered as consisting in general of three packets;—dorsal ones, which are distributed to the back and sides of the animal; visceral ones, which enter the cavity of the body, and are lost amongst the viscera and the caul; and ventral ones, which dipping from the tracheæ overrun the lower part of the sides and belly[269].
The tracheæ and bronchiæ consist of three tunics[270]: the first or external one is a thickish membrane, strengthened by a vast number of fibres or vessels, which form round it a number of irregular circles; the second is a membrane more thin and transparent, without a vascular covering[271]; the third is formed of a cartilaginous thread running in a spiral direction, which may be easily unwound[272]. This structure gives a great elasticity to these organs, so that they are capable of considerable tension, after which they return to their usual length[273]. The Bronchiæ are cylindrical or slightly conical, insensibly diminishing in size as they leave the trunk, in which they originate. In larvæ, after losing their spiral fibre, they appear to terminate in membrane, but in perfect insects they pass into vesicles[274]. In the Cossus the trachea is flattened, and in every segment, except the first and two last, is bound by a fleshy cord four or five times as thick as its threads. Where this occurs, there is a slight constriction,—probably here is a sphincter, by the contraction[Pg 64] of which Lyonet supposes the trachea may be shut when it is necessary to stop the passage of the air, and direct it to any particular point[275]. The structure here described is admirably adapted for the purpose it is intended to serve; for had these vessels been composed of membrane, they could not possibly have been prevented from collapsing; but by the intervention of a spiral cartilaginous thread this accident is effectually guarded against, and the necessary tension of the tubes provided for. However violent the contortions of the insect, however small the diameter of these vessels, they are sure to remain constantly open, and pervious to the air. And by this circumstance they may be always distinguished from the other organs of the animal, and likewise by their pearly or silvery hue, for from being constantly filled with air, these tubes, when viewed under a powerful microscope in a recently dissected insect, present a most beautiful and brilliant appearance, resembling a branching tree of highly polished silver or pearl:—though sometimes they are blue, or of a lead colour, and sometimes assume a tint of gold. In the dead insect the larger tubes soon turn brown, but the finer ones preserve their lustre several weeks[276]. The ramifications of the tracheal tree may be seen without dissection through the transparent skin of the common louse[277] and most of the thin skinned larvæ.
You will not expect to view in this way the minuter ramifications of the bronchiæ, when I have mentioned their number and incredible smallness. Nothing but[Pg 65] the scalpel of a Lyonet and the most powerful lenses are adequate to trace the extremities of these vessels; and even with every help, they at last become so inconceivably slender as to elude the most piercing sight. That illustrious anatomist found that the two tracheæ of the larva of the Cossus gave birth to 236 bronchial tubes, and that these ramify into no less than 1336 smaller tubes, to which, if 232, the number of the detached bronchiæ, be added, the whole will amount to 1804 branches[278]. Surprising as this number may appear, it is not greater than we may readily conceive to be necessary for communicating with so many different parts. For, like the arterial and venous trees, which convey and return the blood to and from every part of the body in vertebrate animals, the bronchiæ are not only carried along the intestines and spinal marrow, each ganglion of which they penetrate and fill, but they are distributed also to the skin and every organ of the body, entering and traversing the legs and wings, the eyes, antennæ, and palpi, and accompanying the most minute nerves through their whole course[279]. How essential to the existence of the animal must the element be that is thus anxiously conveyed by a thousand channels, so exquisitely formed, to every minute part and portion of it! Upon considering this wonderful apparatus we may well exclaim, This hath God wrought, and this is the work of his hands.
Though in general there is only a pair of tracheæ, yet in some larvæ a larger number have been discovered.[Pg 66] In those of the Libellulinæ there are six. According to M. Cuvier, Reaumur, who mentions only four, overlooked the two lateral ones that are connected with the spiracles[280]. The reason of this and other parts of their internal structure I shall explain under the next head. In the grub of the gad-flies of the horse (Gasterophili,) Mr. B. Clark discovered eight longitudinal tracheæ,—six arranged in a circle and two minute ones, which appeared to him to terminate in a pair of external nipples (spiracles) in the neck of the animal[281]. This is a singular anomaly, as the other Œstridæ have only a pair of tracheæ[282].
iii. Respiratory Sacs or Pouches. Besides their tracheæ and bronchiæ, many insects are furnished with reservoirs for the air, under the form of sacs, pouches, or vesicles. These are commonly formed by the bronchial tubes being dilated at intervals, especially in the abdomen, into oblong inflated vesicles; from which other bronchial tubes diverge, and again at intervals expand into smaller vesicles, so as to exhibit no unapt resemblance—as Swammerdam has observed with respect to those of the rhinoceros-beetle—to a specimen of Fucus vesiculosus. Cuvier compares them in the Lamellicorn beetles in general to a tree very thickly laden with leaves[283]; and Chabrier observes that they particularly occur in the intestinal canal[284]. This structure of the pulmonary organs may be seen also in the common hive-bee, and other Hymenoptera;[Pg 67] but the vesicles are less numerous, and those at the base of the abdomen much larger than the rest[285]. These vesicles, by a very rough dissection, may be distinctly seen in the abdomen of the cockchafer, which appears to be almost filled with them. Not being composed of cartilaginous rings like the air-tubes, but of mere membrane, if a pin pierces one, the air that inflates it escapes, and it collapses. In the larva of a little gnat (Corethra culiciformis) the tracheæ appear to proceed from a pair of oblong vesicles of considerable size[286] in the trunk, and towards the anus they form two other smaller ones[287],—upon piercing the former, De Geer observed a considerable quantity of air to make its escape[288]. Another species, probably of the same genus, described by Reaumur, exhibits something similar[289].
But one of the most remarkable structures, in this respect, is to be seen in the larva and pupa of the dragon-flies (Libellulina). I have before noticed the number of their tracheæ, but I shall here describe their whole internal respiratory apparatus. I must observe that Reaumur, Cuvier, and most modern writers on the physiological department of Entomology, have affirmed that they respire the water, and that they receive it for that purpose at their anal extremity: but M. Sprengel, from having observed in the larvæ abdominal spiracles, is unwilling to admit this as a fact[290]; and De Geer also seems to hesitate upon it, especially as he discovered that the animal seemed to absorb the water to aid it in its motions[291].[Pg 68] But when we consider that it is by the action of a pneumatic apparatus that the absorption and expulsion of the water takes place, and that the animal when it has been taken out of that element, upon being restored to it, immediately has eager recourse to this action[292], we shall feel inclined rather to adopt the opinion of those great physiologists Reaumur, Lyonet, and Cuvier, and admit that it absorbs water for the purpose of respiration. I shall now explain how this takes place. The pieces both internal and external that close the anal orifice have been before described; the others employed in the admission and expulsion of the water are evidently respiratory organs. When this orifice is opened, the parts that are above it are drawn back in an opposite direction, so that the five last segments of the abdomen become entirely empty, and form a chamber to receive the water that enters by it. When the water is to be expelled, the whole mass of air-vessels which had receded towards the trunk, is pushed forwards, and forms a piston that again expels the water in a jet. It consists of an infinite number of bronchiæ, entangled with each other, which proceed from the middle and posterior end of the tracheæ. M. Cuvier in the interior of the rectum of the larva discovered twelve longitudinal rows of little black spots, in pairs, which exhibited the resemblance of six pinnated leaves. These are minute conical tubes, of the spiral structure of tracheæ, which decompose the water, and absorb the air contained in it. He also discovered that each of these tubes gave birth to another outside the rectum, which connected itself with one of the six great longitudinal tracheæ; two of which are of enormous size,[Pg 69] and appear to serve as reservoirs, since they furnish air by transverse branches to two other tubes; they have each a recurrent branch, which follows the course of the intestinal canal, and furnishes it with an infinity of bronchiæ[293]. These tracheæ are found in the perfect insect. The principal ones in some send forth many branches, terminating in vesicles, which in shape resemble the seed-vessels of some species of Thlaspi, while others appear to form a file of oblong ones[294]. Near each of their spiracles also is a vesicle which appears to be a reservoir[295].
But this kind of structure is not confined to insects strictly aquatic. Even such species of terrestrial ones as live upon aquatic plants, and are, consequently, necessarily or accidentally often a considerable time under water, are furnished with some apparatus by means of which they can exist in this element for a considerable period. For example, most of the Weevils (Rhyncophora) die in a short time if immersed in water; yet the species of the genera Tanysphyrus, Bagous, and Ceutorhynchus which feed on aquatic plants, can exist for days under water, as I have ascertained by experiment. C. leucogaster and another of the same tribe, swims like a Hydrophilus, and will live a long time in a bottle filled with water and corked tight. Other insects also, that are not at all aquatic, have pneumatic pouches. A striated or channeled vesicle I have found under the lateral angles of the collar in the humble-bee, where Chabrier supposes the vocal spiracles are situate; and also at the mouth of the spiracles of the metathorax in Vespa, &c.[296] In Sphinx[Pg 70] Ligustri the bronchiæ terminate in oblong vesiculoso-cellular bodies, almost like lungs[297]; in Smerinthus Tiliæ these are preceded by a simple vesicle bound with spiral fibres[298]. M. Chabrier thinks that these air-bladders of insects, amongst other functions, give more fixity and force to the muscles for flight[299].
Many physiologists have seen an analogy between the spiral vessels of plants and the tracheæ of insects; and some of great name, as Comparetti, Decandolle, and Kieser, have thought that in some instances they terminated in the oscula or cortical pores: but Sprengel contends that they are not accurate in this opinion[300]. In fact, the principal analogy seems to be in the spiral structure of both these vessels.
Having considered the different organs of respiration both external and internal, I shall make a few further observations upon this function. We know little more respecting the mode in which insects respire, except that they breathe out the air by the same kind of organs by which they receive it,—namely, the spiracles, or their representatives. This has been satisfactorily proved by Bonnet, who showed that the experiments by which Reaumur thought it established that insects inspire by their spiracles, but expire through the mouth, anus, or pores of the skin, are founded on an erroneous assumption.[Pg 71] This physiologist, having observed on the surface of submerged insects numerous bubbles of air, concluded that they had passed through the above orifices[301]: but Bonnet found by various experiments carefully conducted, that this appearance was caused by air which adhered to the skin and its hairs, and that when the access of this was precluded by carefully moistening the skin with water previously to immersion, this accumulation of air-bubbles on its surface did not take place[302]. And in a variety of instances he observed large ones issue from all the spiracles, especially the anterior ones. These bubbles sometimes were alternately emitted and absorbed without quitting the spiracle[303], and at others were darted with force to the surface of the water, where they appeared to burst with noise[304]. This author is of opinion that the first and last pair of these organs are of most importance to respiration[305]. Reaumur subsequently owned that Bonnet's arguments had shaken his opinion[306]; and some observations of his own, with respect to the respiration of the bot of the ox, go to prove that expiration and inspiration are not by the same spiracles; for he found that the air in this animal was expired by the eight little lower orifices before mentioned[307], from which he clearly saw the air-bubbles issue—the upper one he conjectures receives the air[308]. As the only communication that this grub has with the atmosphere is by its posterior extremity, it follows, reasoning from analogy, that the anterior respiratory plates of Dipterous larvæ, which may be regarded as representing the spiracles of the trunk in[Pg 72] insects in general, are destined for the escape of the air, after it has parted with its oxygen, received by the anal ones[309]. So that there seems very good ground for M. Chabrier's opinion that inspiration is ordinarily by the abdominal spiracles, and expiration by those of the trunk of insects[310]. He seems to have been led to the adoption of this opinion, not so much by experiments similar to that of Reaumur just stated, but by observing that in many instances these two sets of spiracles differ from each other, the latter having a convex and the former a concave mouth or bed[311]. In some cases, however,—for instance during flight,—he supposes the spiracles of the trunk may receive as well as emit the air[312]: he likewise is of opinion, and it seems not improbable, that by means of these openings in the trunk, from the rush of the superfluous air through them, insects produce those sounds for which they are remarkable,—as the humming of bees and flies. In the former he thinks the sound is produced by the pneumatic apparatus covered by the ends of the collar; while in the latter he attributes it to the spiracles in the metathorax behind the wings attended by a poiser[313]. I incline, however, to M. Dufour's opinion[314],—that the vocal spiracles in the Hymenoptera, as well as in the Diptera, are those behind the wings. Perhaps both theories may be right; for if you take any common humble-bee, you will find that, in the hand, it produces one kind of sound when its wings are motionless,[Pg 73] and another more complex and intense when they vibrate. In numerous instances, however, there is no very striking external difference between the spiracles of the trunk and those of the abdomen: this observation applies more particularly to the caterpillars of Lepidoptera; but whether these receive the air by those of the abdomen, and return it by those of the trunk, has not yet been ascertained; and indeed, too little is at present known upon the subject, and too few facts have been collected, to admit of dogmatizing.
The external signs of respiration in insects are not universally to be discovered. The alternate contraction and expansion of the abdomen is, however, very visible in some beetles, bees, the larger dragon-flies, and grasshoppers. In one of the latter, Acrida viridissima, Vauquelin observed that the inspirations were from fifty to fifty-five times in a minute in atmospheric air, and from sixty to sixty-five when in oxygen gas[315]. But M. Chabrier has given the most satisfactory account of these signs: The abdomen, says he, is the principal organ of inspiration; it can dilate and contract, lengthen and shorten, elevate and depress itself. In flight, in elevating its extremity at the same time with the wings, it contracts itself, pushes the air into the trunk, and diminishes the weight of the body by the centrifugal ascending force[316]. In the majority of insects perhaps the dilatation of the abdomen takes place by the recession of the segments from each other by means of the elastic ligaments that connect them; in others, as the Dynastidæ, Galeodes, &c. by the longitudinal folded membrane that unites the dorsal[Pg 74] and ventral segments—in the Libellulina by similar ventral folds; and in Cimbex by membranous pieces in the first dorsal segment, which De Geer observed was elevated and depressed at the will of the animal[317].
Air is as essential to insects in their pupa as in their larva or perfect states. Lyonet, however, Musschenbroek, Martinet, and some other physiologists, have doubted whether quiescent pupæ breathed[318]; but Reaumur and De Geer seem to have proved that they do[319]: and if thrown into water, the same proof of respiration, by the emission and retraction of a bubble of air takes place, as in the larvæ; and De Geer found that if one be transferred under water from one spiracle to another, it will be absorbed by it[320]. Indeed, unless these pupæ had breathed, where would have been the necessity for the spiracles with which all are furnished? It is remarkable, however, that all these spiracles do not seem of equal importance in this respect. Reaumur found that if the posterior spiracles only were closed with oil, the insect suffered no injury; but that if the anterior ones were similarly treated, it infallibly died[321]. The respiration however of pupæ seems more perfect in those that have recently assumed that state, than in those that are more advanced towards the imago; in which at first, from Reaumur's experiments[322], it appears that the posterior spiracles were stopped; and in others still older, from Musschenbroek's[323], even the anterior ones. Those quiescent pupæ that during that state remain submerged, respire[Pg 75] air. De Geer has given an interesting record of this, in the case of Hydrocampa stratiotata. This insect spins a double cocoon, the outer one thin, and the inner one of a close texture. In the pupa there are three pair of conspicuous spiracles on the second, third, and fourth segments of the abdomen, which are placed on cylindrical tubes, and they appear to have no other air-vessels. The respiratory gills of the larva having vanished, like some others of the same genus, they know how to surround themselves with an atmosphere of air in the midst of the water, so that the interior of their inner cocoon is impervious to the latter element—how they renew the air has not been ascertained. Though they respire air, water is equally necessary, for the animal died when kept out of water[324].
The great majority of insects respire in much the same manner in all their states, particularly as to their external organs; for when the larva breathes by the lateral spiracles, the pupa and imago usually do the same. The converse of this, however, by no means holds; for it not unfrequently happens that the two latter breathe by means of lateral spiracles, though they received the air in their larva state by an apparatus altogether different. Thus the larvæ of many Diptera breathe by an anal tube, while the pupa and imago follow the general system. Sometimes a tribe of insects breathe by an apparatus quite different in all their states, as we have seen to be the case with the common gnat[325], which has an anal respiratory tube in its first state, thoracic respiratory horns in its second, and the ordinary lateral spiracles in its third.
Changes also take place in their internal organs. In the larvæ the respiratory apparatus, especially the tracheal tubes, is often much larger and more ramified than in the imago; and as the former is the principal feeding state, there seems good ground for Mr. B. Clark's opinion—that the respiration is intimately connected with the conversion of the food[326]. In the imago, there appears to be more provision for storing up the air in vesicular reservoirs, than in the larva. Wonderful is the mode in which some of the changes in the internal structure, which these variations indicate, must necessarily take place. They are, however, probably not more singular than those which less obviously occur in the air-vessels of all insects in their great change out of the larva into the pupa state. But having before enlarged on this subject, I need not repeat my observations[327].
The access of air is as necessary to insects even in their egg state[328], and in many cases its presence seems provided for with equal care, by means as beautiful as those Sir H. Davy and Sir E. Home have shown to occur in the oxygenation of the eggs and fœtuses of vertebrate animals[329]. It is only necessary to view the admirable net-work of air-vessels which Swammerdam discovered spread over the surface of the eggs of the hive-bee while in the ovaries[330],—a provision which, from analogy, we may conclude obtains generally; from the importance[Pg 77] which nature has attached to the oxygenation of the germ while in the matrix. And judging from analogy, we may infer that the access of this element is as carefully secured after the egg is laid, as before. The eggs of most insects being of a porous texture, often attached to the leaves of plants, and some of them embedded in the very substance of a leaf or twig[331], are in a situation for the abundant absorption of oxygen: and the pouch of silk in which the eggs of spiders and Hydrophili are deposited, may probably, from Count Rumford's experiments, be of utility in the same point of view. In the case of the Trichoptera and other insects[332] whose eggs are dropped into the water enveloped in a mass of jelly, this substance perhaps serves for aërating the included embryo, in the same way with the jelly surrounding the eggs of the frog, dog-fish, &c. It would be desirable to ascertain whether the former jelly be of the same nature as the experiments of Mr. Brande have shown the latter to be[333]. It is not improbable that the singular rays that terminate the eggs of Nepa[334] may in some way be connected with the aëration of the egg.
To what I have before remarked with regard to the vital heat of insects[335], I may under this head very properly add a few further observations. I there stated, that the temperature of these animals is usually that of the medium they inhabit, but that bees, and perhaps other gregarious ones, furnish an exception to this rule[336]. A confirmation of this remark is afforded by Inch, a German[Pg 78] writer, who, upon putting a thermometer into a bee-hive in winter, found it stand 27° higher than in the open air; in an anthill, he found it 6° or 7° higher; in a vessel containing many blister-beetles (Cantharis vesicatoria,) 4° or 5° higher. A thermometer, standing in the air at 14° R., put into a glass vessel with Acrida viridissima, in nine minutes rose to 17°, and a similar result was observed with respect to other insects[337]. Dr. Martine says that caterpillars have but two degrees of heat above that of the air they live in[338]. Coleopterous insects are said to move slowly and with difficulty when the thermometer sinks to 36°, to become torpid at 34°, and to lose muscular irritability at a lower degree[339]. I have before observed that some insects will bear to be frozen into an icicle, and yet survive[340]: they share this power with reptiles, fishes, and amphibia. But, however small the excess of it in some insects above that of the medium they inhabit, it proves that they possess the power of generating heat. Whether, like the warm-blooded animals, they generally possess that of resisting heat by perspiration, &c. is not so clear. Yet the heat to which some can bear to be exposed, basking at noon, as Dr. Clarke informs us[341], on rocky and sandy places, exposed to the full action of the sun, appears sufficient, if not resisted by some principle of counteraction, to roast them to a cinder. That bees perspire is well known, but probably not singly.
When the respiration of insects is suspended by immersion[Pg 79] in any fluid, it is often resumed, even when it has been long and they are apparently dead, if they be brought into contact with the atmosphere. Reaumur found this to be the case with bees[342]; and Swammerdam tells us that the maggot of the cheese-fly (Tyrophaga Casei) lived six or seven days in rain-water[343]: he found it so difficult to kill the larva of Stratyomis Chamæleon, which he first immersed twenty-four hours in spirits of wine, and then put them several days in water, without killing them,—that he lost his patience, and dissected them alive. He tried to drown them also in vinegar, in which they held out more than two days[344].
That the suspended animation and subsequent death of most terrestrial insects when thrown into water is caused by the want of air, is evident from this,—that the same effect ensues if the spiracles be covered with any oily or fatty matter. In this case too, their vital powers soon become suspended: they revive, if the suffocating matter be soon removed; and if this be not done, infallibly perish. This fact was known to the ancients, for Pliny observes that bees die if dipped in oil or honey[345]. One exception to this law has been before mentioned[346]: a similar contrivance secures the cheese-maggot from having its respiration interrupted by its moist and greasy food; the grub also of Sarcophaga carnaria, and of other Muscidæ probably, has its posterior spiracles placed in a plate at the bottom of a kind of fleshy pouch, which has the shape of a hollow, truncated, and[Pg 80] reversed cone. This pouch the grub can close whenever it pleases, so as to cover its spiracles[347]. And numerous other larvæ, both of Diptera and Coleoptera that devour unclean and oily food, have doubtless some protection of this kind for their spiracles and respiratory plates.
I am, &c.
We learn from the highest authority, that the blood is the life of the animal[348]: every object of creation, therefore, that is gifted with animal life, we may conclude, in some sense, has blood, which in this large sense may be defined—The fluid that visits and nourishes every part of a living body[349]. But the Great Author of nature has varied the machinery by which this nutritive fluid is formed and distributed, gradually proceeding from the most simple to the most complex structure; in which he seems to have seen it fit to invert the process observable in the systems of sensation and respiration, where the ascent is from the most complex, to the most simple structure. In the lowest members of the animal creation, the blood seems the portion they imbibe of the fluid medium in which they reside, which when chylified, distributes new molecules to all parts of their frame[350]. In others, as in insects, it is formed by the chyle that transpires[Pg 82] through the intestinal canal into the general cavity of the body, where it receives oxygen from the air-vessels, and is fitted for nutrition[351]. In these animals it is accompanied by a long dorsal vessel, the first step towards a heart, which alternately contracts and dilates with an irregular systole and diastole, but appears to have no vascular system connected with it, though in their preparatory states it has an extra-vascular circulation which ceases in the perfect insect. Again: in others, as the Tubicoles, Annelida, &c., a real circulation has been discovered; that is to say, a system of veins and arteries, but unaccompanied by a muscular heart[352]. In the Arachnida and Branchiopod Crustacea the long dorsal vessel is also found; but in these it is connected with an arterial and venous system, which receives, distributes, and returns the blood[353]. It has therefore now become a true heart, and there is a regular circulation; and in the Decapod Crustacea the dorsal vessel is contracted into an oval form, and placed nearly in the centre of the trunk[354]. In the great majority of invertebrate animals the blood is white, but in the Annelida, to which Class the common dew-worm belongs, a curious anomaly takes place—for it is red[355]. Thus a gradual ascent is made to the circulating system of the vertebrate and red-blooded animals. In all, however, the blood is the principal instrument of nutrition and accretion; and is on that account properly so denominated, though not connected with a circulating system.
Having given you this general outline of the means[Pg 83] by which the blood is distributed in the different Classes of animals, I shall now confine myself to the case of insects and Arachnida, beginning with the former.
I. If you examine attentively the back of any smooth caterpillar with a transparent skin, you will perceive in that part an evident pulsation, as though a fluid were pushed at regular intervals towards the head, along a narrow tube which seems to run the whole length of the body. Accurate dissections have proved that this appearance is real, that there is actually present in the back of most insects, placed immediately under the skin and furnished with numerous air-vessels, a longitudinal vessel[356] originating in the head near the mouth[357], running parallel with the alimentary canal nearly to the anus, containing a fluid which is propelled in regular pulsations of from 20 to 100 per minute, more or less as the weather is colder or warmer[358], causing a sensible alternate systole and diastole from the anal extremity towards the head. In the Cossus these pulses were observed by Lyonet to begin in the eleventh segment, from which they passed from segment to segment, till they arrived at the fourth, where they terminated[359]. This vessel is what Malpighi, who first discovered it, termed a heart, or rather series of hearts[360]; but which Reaumur, who injected it, regarded as a simple artery without striking contractions[361]: but to steer clear of any hypothesis, I shall merely call it the dorsal vessel (Pseudo-cardia). When carefully taken out of the body it is found to be a membranous tube, appearing to be closed[Pg 84] at each end[362], in many larvæ of equal diameter every where, but in perfect insects usually widest at the anal extremity[363], and attenuated into a very slender filament towards the head. In some insects, however, as in the larva of the chamæleon-fly (Stratyomis Chamæleon), it is attenuated at both ends, and in the Ephemera is alternately constricted and dilated as Malpighi describes that of the silkworm[364], a dilated portion belonging to each segment[365]. In the Cossus, and probably others, after the third segment, it is furnished with nine pair, the three posterior pair being the largest, of triangular transverse bundles of muscular fibres, which Lyonet denominates its wings[366], the action of which produces its systole and diastole, and their propagation from the tail towards the head[367]. Under the last pair of these wings it is strengthened by a large number of circular muscular fibres[368]. I have stated it as appearing to be closed at each extremity, because Cuvier and most writers have so regarded it, and probably it is so closed in the perfect insect; but from Lyonet's words it should seem that, in the larva of the Cossus, he considered it as open and expanded at its anterior end[369]. He seems also to suspect, that, by means of what he calls the frontal ganglions, a fluid is derived from the dorsal vessel to the spinal marrow. He likewise describes a large nerve as passing through it and becoming recurrent[370]. Carus, as we shall soon see, has also proved that this tube is not closed in larvæ.
The fluid which this vessel contains is very abundant; in the animal it appears colourless and transparent like water, but when collected in drops it becomes more or less yellow, and even orange[371]. Examined under the microscope it appears filled with a prodigious number of transparent globules, of incredible minuteness[372]. When mixed with water, which it does readily, its globules lose all their transparency, and coagulate into small clammy masses. After evaporation it becomes hard, and cracks like gum, as blood does also. This gummy substance is so abundant, that the fluid contained in the dorsal vessel of the caterpillar of the Cossus yields a mass of it of the size of a grey pea[373].
From the situation of this dorsal vessel, which is precisely the same with that of the heart in Arachnida and the Branchiopod Crustacea, and from the systole and diastole which keep its fluid contents in constant motion, who can wonder that the physiologists who first discovered it, reasoning analogically, maintained that it was a true heart? But modern comparative anatomists, and those of the highest name, from the absence of a vascular system for a circulation, have contended that it is not a true heart, but an organ appropriated to other purposes: a third hypothesis, and intermediate between these two, has very recently been promulgated, that the organ in question, namely, is a real heart, and in the preparatory states of insects, the centre of a real circulation, which, in the imago state, ceases with the full development of the wings; but that this circulation is[Pg 86] extravascular, or without peculiar vessels analogous to veins and arteries.
I shall now enlarge a little upon each of these hypotheses, beginning with the first or original one.
No one will deny that the argument from analogy is strongly in favour of this: I need not therefore dwell upon it, but proceed to others. Swammerdam, to whose exactness in observing, and scrupulous accuracy, every reader of his immortal work will bear testimony, expressly asserts that he has seen vessels issuing from the dorsal vessel in the silkworm, and even succeeded in injecting them with a coloured fluid[374]. Now it seems extremely improbable that so practised and expert an anatomist should have been deceived, especially upon a point which would naturally excite his most earnest and undivided attention. Without this recorded experiment, perhaps, it might be thought, though this was very unlikely, that he had mistaken bronchiæ for veins and arteries: but how could they have been injected from the supposed heart? Another great physiologist, Reaumur, in the caterpillar of the saw-fly of the rose (Hylotoma Rosæ) observed, besides the dorsal vessel, a ventral one of similar form, in which also was a pulsation, but slower than that of the other. This he supposes may be the principal trunk of the veins[375]. Bonnet thought he discovered a similar vessel in a large caterpillar, but with[Pg 87] all his attention could perceive no motion in it[376]. Reaumur also fancied he perceived in the grub of Musca vomitoria, in which he in vain looked for the dorsal vessel, a fleshy part which exhibited alternate pulsations; and when with a pair of scissors he made a lateral incision in the insect, amongst other parts that came out, there was one that had movements of contraction and dilatation for several minutes,—this experiment was repeated with the same result upon several grubs[377]. De Geer, whose love of truth and accuracy no one will call in question, saw the appearance of blood-vessels in the leg of the larva of a Phryganea L. (as Lyonet did in those of a flea[378]); and in the transparent thigh of Ornithomia avicularia he discovered a pulse like that of an artery[379]. Baker, whose only object was to record what he saw, speaks of the current of the blood being remarkably visible in the legs of some small bugs[380]: what he meant by that term is uncertain, but they could not be spiders, which he had just distinguished. This author has likewise seen a green fluid passing through the vessels of the wings of grasshoppers[381]; and M. Chabrier is of opinion that insects possess the power of propelling a fluid into the nervures of their wings and withdrawing it at pleasure, as they are elevated or depressed[382]; but this last fact may be independent of a circulation.
But though these arguments, which I have stated in their full force, appear strong, and at first sight conclusive,[Pg 88] those which may be urged for the more modern opinion—that no circulation exists in insects, properly so called,—appear to have still greater weight. Lyonet, whose piercing eye and skilful hand traced the course of so many hundred nerves and bronchiæ long after they became invisible to the unassisted eye, and which were a thousand times smaller than the principal blood-vessels, opening into so large an organ as the supposed heart of insects, might be expected to be, could never discover any thing like them. His most painful researches, and repeated attempts to inject them with coloured liquors, were unable to detect the most minute opening in the dorsal vessel, or the slightest trace of any artery or vein proceeding from or communicating with it[383]. And Cuvier, whose unrivalled skill in Comparative Anatomy peculiarly qualified him for the investigation, repeated these inquiries, and tried all the known modes of injection, with equal want of success; and is thus led to the conclusion, that insects have no circulation, that their dorsal vessel is no heart, and therefore ought not to be called by that name: that it is rather a secretory vessel, like many others of that kind in those animals. As to the nature of the fluid that it secretes, and its use, he thinks it impossible, from our present information on the subject, to form any satisfactory conclusion[384]. Marcel de Serres informs us—which further seems to prove that it can be no real heart—that this vessel may be totally removed without causing the immediate death of the insect[385]. This opinion receives additional confirmation from the mode in which respiration is performed in insects. In[Pg 89] those animals that have a circulation, this takes place by means of lungs or gills;—thus we find, even in the Crustacea and Arachnida so nearly related to insects, that the organs of this function are true gills; whereas in insects, though in some of their states their respiratory tubes are branchiform, yet they are not gills, and the respiration is by tubes and spiracles. And these tubes, as you have seen, are so numerous and so infinitely ramified and dispersed, as to occupy the place of arteries and veins, and to imitate their distribution,—and thus to oxygenate what may be deemed the real analogue of the blood, which bathes every internal part of the body of an insect. Those animals likewise that have a circulation are furnished with a liver, as is the case with the Arachnida and even many aggregate animals that have a heart; but in insects there are only hepatic ducts. M. Cuvier has also proved that the conglomerate glands, which exist in all animals that have a heart and blood-vessels, do not exist in insects, in which they are replaced by long slender secretory tubes, which without being united float in the interior of the body: from this circumstance, he is led to conclude that their nutrition is by imbibition or immediate absorption, as in the Polypi and other zoophytes, the chyle transpiring through the alimentary canal, and running uniformly to all parts of the body[386].
These arguments appear so satisfactory, that Physiologists in general seem to have been convinced by them that no circulation, at any time, takes place in insects, and that their supposed heart is merely a secretory vessel, though of what kind they were at a loss to[Pg 90] conjecture[387]. But, convincing as they seem, they appear to have been founded in error, and on the idea that a circulation, as well as a heart, necessarily implies a vascular[Pg 91] system consisting of veins and arteries; for by the recent discoveries of M. Carus, it has been satisfactorily proved that insects in their preparatory states, have an extravascular circulation, the arterial and venose currents not being confined by parietes. The observations upon which M. Carus' hypothesis is founded, were made in the Autumn of 1826; and an abstract of their results presented to the Union of German Naturalists and Physicians, which then held its meeting at Dresden, many of the members of which, as MM. Oken, Husche, Heyne, Purkinje, Otto, Weber, and Müller, had ocular proofs of the reality of the phenomena.
His first observations were made on the larva of Agrion Puella, which swims by means of three vertical laminæ attached to the tail; which, when the wings first appear as rudiments, begin to be exsiccated and are finally detached. Each of these laminæ, in its natural vertical position, presents an inferior abdominal and a superior dorsal edge, has two tracheæ running along its centre with ramifying bronchiæ, and consists of granular substance contained between two strata of the external integuments. A current of blood-globules enters each[Pg 92] lamina somewhat nearer to its abdominal than to its dorsal edge, and running through the greater part of its length suddenly turns and bends its course back towards the body, somewhat nearer to the dorsal than to the abdominal margin of the lamina. The channel thus formed in the midst of the granular substance is perfectly transparent, except where it is occupied by the blood-globules, or crossed by the bronchiæ. The parietes of the channel are not strictly defined, nor formed by any thing like the coats of a vessel, the blood circulating through the granular Parenchyma; a circumstance however which is not peculiar to this case, but also occurs generally in the first states of the circulation, as it presents itself for instance in the embryo of Fishes, and in the figura venosa of the incubated egg[394]. The blood-globules are elongated like a grain of wheat, considerably larger than those of the human blood, and float in a fluid which is invisible because of its transparency, but the existence of which is proved by the variations in the position of the globules in the current, sometimes following its direction, at others crossing it transversely, or more or less obliquely.
When the animal is vigorous, the current is uninterrupted, although its velocity is accelerated at regular intervals; and that not only in the excurrent (arterial), but also in the recurrent (venous) part of its course through the lamina. When the animal becomes exhausted, or the laminæ exsiccated, the circulation is interrupted, and in the same manner, as under the same circumstances, in the larvæ of frogs and lizards; the disturbance displaying itself not merely by a cessation of the process, but also[Pg 93] by retrograde movements of the currents, or by oscillatory motions of the blood-globules.
In proportion as the wings are developed, the circulation in the laminæ diminishes, and ultimately ceases, preparatory to the detachment of the laminæ themselves. At the same time, however, it presents itself under a new form in the wings. In these the excurrent or arterial stream takes its course along the inner margin of the wing, and the recurrent or venous returning along the outer; whilst, occasionally, other transverse currents take their course through the net-work of the wing from its inner to its outer margin. As the wings are further developed, the circulation in them, like that in the caudal laminæ, gradually becomes weaker and ultimately ceases[395].
The next observations were made on the transparent larva of a neuropterous insect (probably a Semblis or Sialis), in which the pulsations of the dorsal vessel were distinctly seen at its posterior extremity, from which they were propagated towards the anterior; these two divisions of that vessel appearing to bear to each other the relation of a heart and aorta. There were no traces of other vessels, though regular and rapid currents of blood-globules, exterior to the tracheæ, proceeded from the head towards the posterior extremity of the body, where each of these currents entered the heart, which again propelled its contents with accelerated velocity through the anterior part of the dorsal vessel towards the head. The lateral currents also were accelerated upon each contraction of the heart, proving that they must communicate with the dorsal vessel at the anterior part of the body, though the[Pg 94] opacity of the head rendered it impossible to ascertain the mode of anastomosis. An excurrent and returning current were also traced to each of the legs[396]. But the phenomena of the circulation was most distinctly visible in the larva of Ephemera vulgata, even more distinctly than it is possible to trace it in the larvæ of frogs and newts. In this animal the circulation, with the help of the microscope, is at once visible in the three last segments of the body; and with a little attention is discoverable not only in the three terminal caudulæ, and in the upper joints of the legs, but also in the head, and particularly the roots of the antennæ. In the posterior part of the body there are on each side two currents of blood, not bounded by parietes, situate on each side of the intestinal canal, the inner one being the most considerable. The external one communicates with the internal by several intermediate branches; from this probably the streams are detached, which in the form of loops are seen at the upper joints of the legs, though it is not possible precisely to ascertain this, nor even whether these lateral currents continue distinct in the thorax, which probably they do. At the ninth abdominal segment these currents which flow posteriorly from the head, change their direction, and are inflected so as to enter the pulsating heart, from which the current again flows towards the head. Before they enter the heart they give off three streams, one for each of the three caudulæ. The currents in these caudulæ present the phenomena of the circulation with peculiar distinctness, and are particularly remarkable from the circumstance, that the excurrent and recurrent streams, though closely approximated without any visible separation,[Pg 95] flow without disturbing each other. The excurrent stream is accelerated in correspondence with the pulsations of the heart; the recurrent on the contrary being always somewhat more sluggish, and the first to stagnate and cease when the strength of the animal is impaired. In the anterior part of the head currents can be discovered, forming loops like those of the legs, at the roots of the antennæ; each current proceeding from the cranial surface, and in returning taking its course towards the region of the larynx[397].
M. Carus has likewise observed currents of blood in the larvæ of water-beetles (Hydrophilus and Dytiscus)[398]; but at present he appears to have detected it in no terrestrial larva. Whether this is occasioned by their opacity, or it exists only in the ovum, as he seems to suspect[399], must be left for determination to future observers; it is scarcely probable, however, that the larvæ of Dytisci and Hydrophili should differ from other Coleoptera in their circulation.
The endeavours of M. Carus to discover any proofs of a circulation in insects in their last state, except in the wings at their first development, were without success[400]. He observes that the fact of the currents of fluids in larvæ not being defined by vascular parietes, enables us to comprehend the rapidity and facility with which the traces of the circulation are lost in the perfect insect. On the other hand, that the existence of a circulation at one period, and its cessation at another, elucidate many circumstances connected with the physiology of these animals: for instance, the contrast between the[Pg 96] rapid growth and transformations of the larvæ, and the stationary existence of the imago, &c. Lastly he remarks, that the phenomena of this circulation do not throw any light on the obscure subject of the mode of nutrition in perfect insects; which therefore must still be supposed to be effected according to the idea of Cuvier, without the intervention of vessels[401].
Whatever be the functions of the dorsal vessel, this seems the most proper place to state to you what further is known respecting it. Its construction is nearly alike in insects in all their states, except that in the imago it is shorter and narrower. Reaumur has affirmed, and before him Malpighi made a similar observation, that in chrysalises newly disclosed from the larva, and yet transparent, the motion of the included fluid is the reverse of what it has been in that state, it being propelled from the head to the tail, which he found to be the case also in the imago[402]. If this be true, and there is no reason to doubt his accuracy, when they are more advanced, it resumes its old course, as Lyonet observed, from the tail to the head[403]. But probably it is not always uniformly in the same direction, since Malpighi states that a very slight cause will change its course, and that the pulsations differ in quickness in different portions of the heart[404]. If its course were really always the same, and in one direction, without any reflux, it would seem to follow that the fluid must be absorbed at one end, and, if there was no outlet, transpire at the other, which would be a kind of circulation. In Syrphus Pyrastri and other aphidivorous[Pg 97] flies, this dorsal vessel, instead of the usual form which it had in the larva, assumes a very peculiar appearance. If, taking one of these flies by the head and wings and holding it up to the light, you survey under a lens the base of the lower part of its abdomen, you will see through its transparent skin, which exactly forms such a window as physicians have sometimes wished for in order to view the interior of their patients, a flask-shaped vessel having its long end directed towards the trunk, in which there is a manifest pulsation and transmission of some fluid. This vessel extends in length from the junction of the trunk with the abdomen to about the termination of the second segment. The included fluid does not run in the dorsal vessel in a regular course, but is propelled at intervals by drops, as if from a syringe, first from the wide end towards the trunk, and then in the contrary direction, forming a very interesting and agreeable spectacle. One circumstance led Reaumur to conjecture that the neck of this vessel, which he at first regarded as simple, is in fact composed of two or more approximated tubes, and that the blood is conveyed forward by the outward ones, and backward by the intermediate one[405]: he even thinks that he saw a kind of secondary heart, at the extremity next the trunk, for the purpose of causing the reflux. This illustrious author observed the above remarkable structure not only in the Syrphi, but in many of their affinities, and thinks that it is also widely diffused amongst the Muscidæ[406].
I must now say something upon what I conceive to be the real blood of insects; for I think no one will object to that name being given to their nutritive fluid, especially[Pg 98] in the larva, though it does not circulate by means of a vascular system. The chyle that is produced in the intestines of animals from the food, is that fluid substance from which their blood is formed: in insects it is not absorbed by the lacteals, but transpires through the pores of the intestinal canal into the general cavity of the body, where, being exposed to the influence of the oxygen in the air-vessels, it becomes, though retaining its colour, a different fluid from what it was before, and analogous to blood in its use and office[407]; only that in these animals, as Cuvier has observed, at least in their perfect state, the blood, for want of a circulating system, not being able to seek the air, the air goes to seek the blood[408]. The dispersion of this fluid appears to be universal, so that all the parts and organs contain it in a greater or less degree[409]. In many insects, if you break only an antenna or a leg, a drop of fluid flows out at the wound. In larvæ, the fluid which bathes[410], or visits, all the internal parts and organs is not only sufficient for their nutriment, but a large quantity of seemingly superfluous blood remains that is not wanted for this purpose. This is expended in the production of the caul or epiploon (Corps graisseux[Pg 99] Reaum.), which laps over and defends all the viscera of the animal, and goes principally to the formation of the imago[411]. I have said that Cuvier conceives nutrition in insects to take place by imbibition or immediate absorption; that is, I suppose, the different parts and organs thus constantly bathed in the blood, imbibe from it the particles necessary for their constant accretion. M. Chabrier seems to think that it is the compression and dilatation of the trunk that duly distributes the nutritive fluid[412]; Lyonet compares the nutrition of insects by their fibres from this fluid, when formed into the corps graisseux, to that of plants that draw their support by their roots from the earth[413]. Much obscurity, however, at present rests upon this subject—much for future investigation to explore; but in all the works of the Most High there is always something inscrutable, something beyond the reach of our senses and faculties, which teaches us humbly to adore his infinite perfections.
II. The circulation of the Arachnida is next to be considered; and the term applied to these becomes strictly proper. Two great tribes, in our view of the subject, constitute this Class,—the spiders (Araneidea) and scorpions (Scorpionidea): I shall give you some account of the circulating vessels of each.—In spiders, the heart in general is a long dorsal vessel as in insects, but supposed to be confined to the abdomen, growing slenderer towards each extremity, particularly the anal. In some also, as in Aranea domestica, like that of insects, it has lateral muscular appendages; but in others, as in[Pg 100] Clubiona atrox, it is without them[414]. It exhibits a pair of vessels that appear to connect with the gills, by which the oxygenation of the blood takes place, and a number of others that ramify minutely and are lost in the analogue of the epiploon, supposed to be their liver[415]. Whether these last are to be regarded merely as veins, has not been ascertained; they seem rather to convey the blood outwards, than to return it back to the heart: but this question must be left for future investigation. I may observe, however, that though the heart of the spider has been traced only in the abdomen, it may probably extend into the trunk.
The heart of the scorpion has been examined both by Treviranus and Marcel de Serres; but I shall principally confine myself to the description of the latter, as the most clear and intelligible. The heart, then, of these animals is elongated, almost cylindrical, but attenuated at each end; it is extended from the head to the extremity of the tail, and appears to have four pairs of lateral muscles. On each side are four pairs of principal vessels, which go to the pulmonary pouches, and there ramify. These may be assimilated to veins. Besides these, there are four other vessels that cross them, forming with them an acute angle, and which, with four branches of smaller size, receive the blood from the pulmonary pouches, and distribute it to the different parts of the body,—these are the arteries. Before it enters the tail, the heart throws out two vascular branches which do not go to the gills, but distributing the blood[Pg 101] to different parts, ought to be considered as arteries[416]. Treviranus mentions bunches of reticulated vessels, concerning the use and origin of which he seems uncertain[417]; but as they approach the gills they are probably the branching extremities of what M. de Serres considers as the veins.
I am, &c.
"The immense Class of insects," says the immortal Cuvier, "in the structure of its alimentary canal exhibits as many variations as those of all the vertebrate animals together: there are not only the differences that strike us in going from family to family and from species to species; but one and the same individual has often a canal quite different, according as we examine it in its larva or imago state; and all these variations have relations very exact, often easily estimable, with the temporary or constant mode of life of the animals in which it is observable. Thus the voracious larvæ of the Scarabæi and butterflies have intestines ten times as large as the winged and sober insects—if I may use such an expression—to which they give birth[418]."
In the natural families of these creatures, the same analogy takes place with respect to this part that is observable in the rest of the Animal Kingdom; the length and complication of the intestines are here, as in the[Pg 103] other Classes, often an index of a less substantial kind of nutriment; while their shortness and slenderness indicate that the insect lives by prey[419].
In considering therefore the parts connected with the digestive functions of the insect world, it will not be amiss to have reference to their food, and their mode of taking it; but first it will be proper to state and define the parts of this important organ.
In general the alimentary canal[420] is composed of the same essential tunicks as that of the vertebrate animals, consisting of an interior epidermis, a papillary and cellular tunick, and an exterior muscular one[421]. The first is usually tender, smooth, and transparent; but not always discoverable, perhaps on account of its tender substance[422]. Ramdohr does not notice the papillary and cellular tunicks; they are probably synonymous with what he denominates—the flocky layer (Die flockige lage), and which he describes, when highly magnified, as appearing to consist of very minute globules or dark points, and as being of a cellular structure[423]. The exterior tunick is thicker and stronger than the interior, and composed of muscular fibres, running either longitudinally, or transversely so as to form rings round the canal. This tunick mostly begins at the mouth, and goes to the anus, changing its conformation in different parts of the above intestine. Sometimes however it originates only at the beginning of the stomach[424]. With respect to its general disposition, that canal—in its relative length, in[Pg 104] the size of its different parts, in the number and form of its dilatations, and particularly of its stomachs and its cœcums, and in the folds of its interior—exhibits variations altogether analogous to those of vertebrate animals, and which produce similar effects[425]. As to its parts, it may be considered as consisting of two larger portions, between which the biliary or hepatic vessels form the point of separation. In the first, the most universal parts are the gullet and the stomach; and in the second, the small intestine and the large intestine[426].
1. The gullet (Œsophagus[427]) is that portion of the intestinal canal which, receiving the food from the pharynx, or immediately from the mouth, conveys it to the stomach. Though it often ends just behind the head[428], it is usually continued through the trunk, and sometimes even extends into the middle of the abdomen[429]; it therefore seldom much exceeds in length half the body. It is constantly long when the head is connected with the trunk by a narrow canal—as in the Hymenoptera, Neuroptera, Lepidoptera, &c.; but is frequently short when these parts are more intimately united[430]. It often ends in a kind of sac analogous to the crop of birds. Under this head I must mention a part discovered by Ramdohr, which he calls the food-bag (Speisesack), as he thinks, peculiar to Diptera[431]. From the mouth in these proceeds a narrow tube into the abdomen, where[Pg 105] it expands into a blind sac having no connexion with the stomach; so that the fluid food, as blood, &c. stored in it, must be regurgitated into the mouth before it can pass into that organ[432]. Thus these animals, besides their stomach, have a reservoir in which to store up their food; the product therefore of a single meal will require several days to digest it.
2. The stomach (Ventriculus[433]) is that part of the intestinal canal immediately above the bile-vessels, which receives the food from the gullet for digestion, and transmits it when digested to the lower intestines[434]. By its admixture with the gastric juice, the food acquires in the stomach a quite different colour from what it had in the gullet. In herbivorous insects it contains no acid, but, like the gastric juice of herbivorous quadrupeds, is of an alkaline nature[435]. The chyle is forced through this organ, probably in part by the pressure of the muscular fibres during the peristaltic motion; and being pressed through the inner skin, is first collected in the intermediate cellular part, and ultimately forced through the outer skin[436]. At its posterior end it terminates in the pylorus, a fleshy ring or sphincter formed of annular muscular fibres[437]. The stomach often consists of two or more successive divisions, which are separated from each other, and are often of an entirely different conformation and shape[438]. In the Orthoptera, Predaceous Coleoptera, and several other insects, an organ of this[Pg 106] kind precedes the ordinary stomach, which from its structure Cuvier denominates a second stomach or gizzard[439]; Posselt improperly calls it Cardia[440]; and by Ramdohr it is named the plaited-stomach (Falten-magen[441]). It is a short fleshy part consisting of two skins, placed above the opening of the stomach, and perhaps rather belongs to the gullet. The inner skin is formed into longitudinal folds, and sometimes armed with horns, teeth, or bristles. Its cavity is very small and compressed, so as to admit only small masses of food, and yet present them to a wide surface for the action of the teeth or bristles;—in this stomach therefore, as in the gizzard of birds, to which it seems clearly analogous[442], the food is more effectually comminuted and rendered fit for digestion. The muscles, by which its action upon the food is supported, in some species amount to many thousands[443]. Rudiments of a gizzard are sometimes found concealed in the gullet of many insects[444]. The idea of Swammerdam, Cuvier, &c. that grasshoppers and other insects that have this kind of stomach, chew the cud[445], Ramdohr affirms is entirely erroneous[446]. Besides its divisions, the stomach has other appendages that require notice. In most Orthoptera, a pair or more of blind intestines or cœca may be found at the point of union of the gizzard with the stomach[447], which have been regarded as forming a third stomach: they also begin the stomach in the louse[448]; they form a coronet round[Pg 107] the apex of that organ, in the grub of the cockchafer[449]; and in that of the rose-beetle, there is one at the apex, one in the middle, and a third at the base[450]. Besides these appendages, which are formed of the skin of the stomach, there are others that are not so. In the Predaceous and some other beetles, the whole external surface of this organ is covered with small blind appendages opening into the space between its two skins, which cause it to resemble a shaggy cloth; these Ramdohr calls shags (zotte[451]), and Cuvier, hairs[452] (villi). These appendages the latter author seems to regard as organs that secrete the gastric juice and render it to the stomach[453]; but the former thinks their use uncertain[454].
3. The small intestines (Intestina parva) are the portion of intestines next the stomach, and consist often of three distinct canals;—the first is supposed to be analogous to the duodenum; it is found only in the Coleopterous genera Silpha L. and Lampyris L., and is distinguished from the succeeding intestine by being perfectly smooth[455]. Next follows the thin intestine (Dünndarm), which in the above insects is wrinkled; it most commonly immediately follows the stomach. Sometimes it is wholly wanting, as in Agrion, the Hemiptera[456], &c. Ramdohr conjectures that it is not solely destined for conveying the excrement, but that probably some juices are separated in it from the food especially for the nutrition of the gall-vessels, as their principal convolutions are mostly near this intestine[457]; which perhaps[Pg 108] may in some cases be regarded as analogous to the jejunum in vertebrate animals. The third pair of the small intestines, which perhaps represents the ileum, Ramdohr distinguishes by the name of club-shaped (Keulförmigen Darm[458]). It may generally be regarded as only a continuation of the former thickened at the end so as to resemble a club reversed. It is however sometimes separated from the thin intestine, as in Cerambyx moschatus[459].
4. The large intestines (Intestina magna) consist sometimes of two portions. The thick intestine (Dicken-darm), which may be regarded as a kind of cœcum, is found only in the larvæ of the Lamellicorn beetles, but never in the perfect insect. In shape it is oval and folded; whence it is thicker than the rest of the intestinal canal, and is constantly filled with excrement[460]. The second portion of these intestines is the rectum (Mastdarm), which terminates in the anal passage. This part is scarcely ever wanting, except when the insect evacuates no excrement, which is the case with the grubs of bees, wasps, and the antlion (Myrmeleon). In the imago of Telephorus, at least in T. fuscus, it is also obsolete[461]: in most cases, however, it is very distinct from the preceding intestine. Sometimes it consists of only one tunick composed of muscular fibres[462]. When the gullet is wide, the rectum is usually so likewise; but when it follows a club-shaped or thick intestine, it is narrow[463]. It generally may be termed short[464]. When[Pg 109] wide, it often contains a great quantity of excrement, as the gullet does of undigested food; but when narrow, the excrement seldom remains long in it. This intestine also in a few cases has a lateral enlargement or cœcum (Blind-darm), being a continuation of the same skin; but perhaps this enlargement is really analogous to what Ramdohr calls the thick intestine, though in these cases he regards it as an appendage of the rectum[465].
I must now call your attention to the bile-vessels of insects. These, by Malpighi[466] and the earlier physiologists, who regarded them as a kind of lacteals, were denominated varicose vessels: but Cuvier—and his opinion after some hesitation has been adopted by Ramdohr—considers them as vessels for the secretion of bile, and as analogous to the liver of animals that have a circulation[467]. As the want of blood-vessels prevents insects from having any gland, the bile is produced with them, as all their other secretions, by slender vessels that float in their nutritive fluid, and from thence secrete the elements proper to form that important product, which usually tinges them with its own yellow hue; though in the Lamellicorns and Capricorns they are of an opaque white, and in the Dytisci of a deep brown colour[468]. Their bitter taste further proves that they contain the bile[469]. They are long, slender, filiform, tortuous or convoluted, and mostly simple vessels; sometimes gradually smaller toward the base[470], at others towards the apex[471]. In[Pg 110] some, screw-shaped[472]: in one larva, with hemispherical elevations[473]: in the cockchafer, part of them are fringed on each side with an infinity of short, blind, minute, setiform tubes, while the rest are naked[474]; they are composed of a single, thin, transparent membrane, according to Ramdohr[475]; but Cuvier thinks their texture is spongy[476]. They appear to contain a number of small, irregular, dark granules, which float in a peculiar fluid, with which, however, they are not always filled throughout, nor are they constantly permeable from one end to the other. Thus in the meal-worm beetle (Tenebrio Molitor), the common trunk by which they are attached to the intestinal canal is composed of gelatinous granules[477]. The place of their insertion is generally a little below the pylorus, but in the common cockroach they are inserted into the stomach just above that part[478]. Usually each vessel opens singly into the intestinal canal, which the whole number surround at an equal distance from each other[479]. Sometimes, however, they are connected with it by a common tube in which they all unite, as in the asparagus-beetle (Lema Asparagi[480]), and the mole-cricket (Gryllotalpa vulgaris[481]); in the house-fly (Musca domestica), and other Muscidæ, each pair unites so as to form a single branch on each side of the canal previously to their insertion[482]; in the field-cricket (Gryllus campestris) they are all inserted in one spot[483];[Pg 111] and when numerous, they are generally attached singly though irregularly[484]. These vessels at their base do not open into the cavity of the intestinal canal, but merely into the space between its outer and inner tunicks, the last being constantly imperforated[485].
With regard to their apex, the bile-vessels are sometimes fixed singly or connectedly to the intestine merely by a few muscular fibres; for they do not enter it, their ends having no orifice. This structure is mostly to be met with in the Coleoptera[486]. In caterpillars, the tops of these vessels perforate the outer skin of the rectum, and proceeding in dense convolutions to the anus, become at last so fine that their terminations cannot be discovered[487]. In other cases, the extremities of a pair of these vessels unite so as to form a double one: this may be seen in those of Philonthus politus[488], and probably other rove-beetles: and lastly, in others the bile-vessels are free, hanging down by the intestinal canal, without being attached to it or to each other. This structure is constantly found in the Orthoptera and Hymenoptera Orders, &c.[489].
With regard to their number, the bile-vessels vary from two to upwards of one hundred and fifty, yet so that their whole amount is constantly the product of the number two,—at least as far as they have been counted: and even when those on one side are not alike, a similar variation takes place in the other, as may be seen in Galleruca Vitellinæ, where on each side are two long ones and one shorter[490]; the most usual numbers are, four—six—or many, that is, more than twenty—
| Two bile-vessels are found in | the larva of Cetonia aurata[491]. |
| Four | most Coleoptera, Diptera, and Hemiptera[492]. |
| Six | Lepidoptera, some Coleoptera[493], &c. |
| Eight | Myrmeleon, Hemerobius[494]. |
| Fourteen | Formica rufa[495]. |
| Twenty | larva of Clavellaria Amerinæ[496]. |
| Many | Libellulina, Orthoptera, and Hymenoptera[497]. |
The bile-vessels vary considerably in length: in many cases where they are free they are short[498]; they are often very long, and perhaps those that are fixed may be generally stated as the longest. In the Lamellicorn beetles they are remarkable for their great length[499].
Having given you this general account of the intestinal canal and its parts and appendages, I shall now state some of the peculiarities that in this respect distinguish particular tribes and families.
The Coleoptera alone, exhibit as many variations in the structure of the alimentary tube as all the other Orders of insects together:—to particularize these would occupy too large a portion of this letter, I shall therefore only notice a few of the most remarkable. In general they may be stated as having universally a stomach, a[Pg 113] small intestine and rectum, and not more than three pairs of fixed or united bile-vessels. In the Predaceous beetles, the gullet mostly widens at the base into a considerable crop, followed by a gizzard, a shaggy stomach, and two pairs of united bile-vessels. The whole alimentary canal in these, is never less than double, and sometimes treble the length of the body[500]. In the carnivorous beetles, at least the Staphylinidæ and Silphidæ, there is little or no crop, and the gizzard is hidden: in the former, the whole length of the intestinal canal is not twice, while in the latter it is more than four times that of the body[501]. In these also the intermediate portion of the large intestine is singularly annulated[502]. In the Petalocera the stomach is usually longer than all the rest of the intestines together, and often convoluted: in the cockchafer the whole intestinal canal is nearly five times the length of the body, four parts of which is occupied by the stomach[503]. In the grub the canal scarcely exceeds the length of the animal[504]. In Lampyris the stomach exhibits a remarkable appearance, having on each side a series of spherical folds or vesicles[505]. Have these any thing to do with the secretion of its phosphoric matter? Tenebrio has a gizzard armed internally with calluses, and a shaggy stomach, and Blaps does not differ materially; their entire canal is more than twice the length of the body[506]. In the vesicatory beetles (Cantharis, Meloe, &c.) there is no gizzard, and the canal is less than twice the length of the body[507]. Little is known with regard to the alimentary canal of the beetles distinguished by a rostrum (Rhyncophora).[Pg 114] In the only two that appear to have been examined, Rhynchites Betuleti and Cryptorhynchus Lapathi, that canal is moderately long, the stomach partially shaggy, and the small intestine inversely claviform; but in other respects they differ materially[508]. In the former there is no crop or gizzard, the stomach is fringed on each side, except at its upper extremity, with a series of small cœca or shags, and there are three pairs of bile-vessels[509]; while in the latter the gullet is dilated into a crop which includes a gizzard in which the skill of a Divine artist is singularly conspicuous:—though so minute as scarcely to exceed a large pin's head in size, it is stated to be armed internally with more than 400 pairs of teeth, moved by an infinitely greater number of muscles[510]. A transverse section of this gizzard represents two concentric stars, with nine rays each[511]: the object of this structure is, the comminution of the timber which this beetle has to perforate and probably devour[512]. The stomach is very slender, but dilates in the middle into a spherical vesicle[513], and there are only two pairs of bile-vessels[514]. In the Capricorn beetles, the part we are considering varies much: in general we may observe that it is more than double the length of the body, that the stomach is long and slender, and usually naked, that the gullet terminates in a crop without a distinct gizzard, and that there are three pairs of bile-vessels[515]. In the Herbivorous beetles (Chrysomela, Cassida, &c.) the canal[Pg 115] is more than double the length of the body, and in some much longer[516], the stomach is long, and commonly naked; but in Chrysomela violacea it is covered with hemispherical prominences[517], and in Chrysomela Populi it is shaggy[518]; in the insect last named and Galleruca Vitellinæ the rectum consists of two pieces[519]. In this tribe the intestines of the larva resemble those of the perfect insect[520].
In the Orthoptera the alimentary canal, which continues the same in every state, is short, or only moderately long; the gullet has one or two lateral pouches or crops[521], and terminates in a gizzard of curious construction, with singular folds and teeth[522]; then follows a short stomach, usually with a pair or more of cœca at its upper extremity[523]; the lower intestines are not distinct, and the bile-vessels numerous, short and free[524].
In the Neuroptera, many of the genera are distinguished by the remarkable length of the gullet, and by the lower intestines forming one short piece[525]. In the Libellulina the bile-vessels are numerous, short, and free, as in the Orthoptera[526]. In Hemerobius and Myrmeleon there is a gizzard[527], and just above it a cœcum, in the former very remarkable, is connected with the gullet[528].
The Hymenoptera appear all to be distinguished by a long slender gullet, terminating in a dilated crop forming the honey-bag; their stomach is variable, their small intestine slender, and the rectum dilated;—their bile-vessels,[Pg 116] like those of the two preceding Orders, are numerous, short, and free[529]. In the ants and ichneumons there is an approach to a gizzard[530]. In the wasp and humble-bee the stomach is very long, with muscular rings surrounding it[531]. In this Order the larvæ at first have no lower intestines and void no excrement[532], but as they approach to the pupa state one begins to appear[533].
The next insects whose alimentary canal we are to consider, are those which, taking their food by suction, have no occasion for masticating organs: this may in part be predicated of the preceding Order, in which most of the tribes in their perfect state imbibe fluid food, and use the ordinary organs of mastication principally in operations connected with their economy; and their crop, in which the honey in many is stored up for regurgitation, may be regarded in some degree as analogous to the food-bag of the Diptera and other suctorious insects.
The two sections of the Hemiptera Order differ widely in the canal we are considering, and I shall therefore give a separate account of each. In the Heteropterous section, appended to the gullet by a long convoluted capillary tube, besides the usual saliva-reservoirs there is often a double vessel, which Ramdohr regards as discharging the same function, but which in many respects seems rather analogous to the food-reservoir of the Diptera[534]. As I have had no opportunity of examining this vessel, I shall content myself with stating this idea, and describe the vessel more fully hereafter. The gullet, in[Pg 117] these, usually terminates in an ample crop consisting of many folds[535], followed by a long, slender, cylindrical tube, dilated at its base into a spherical tumour; these two may be said to form the first stomach: to this succeeds a second[536], which Ramdohr denominates the bug-stomach (Wanzen-magen), which varies in its figure, and in Pentatoma consists of four demi-tubes, so as to form a quadrangular canal[537]. In the Homopterous section of this Order Ramdohr seems to have examined but few; Chermes however and Aphis exhibit one remarkable feature; they have no bile-vessels, at least he could discover no trace of these organs[538]. Their intestinal canal is very simple, their stomach very long, widest above, and somewhat convoluted, with a very slender gullet[539]. In Cercopis spumaria the structure is more complex, and extremely singular. It has two or rather three stomachs; the two first of a horny substance, and the last a slender somewhat convoluted membranous tube, which becoming reversed, is attached by what should be deemed its lower extremity to the first stomach, from the other side of which emerge the lower intestines, terminating in a thick pear-shaped rectum. At the same point of the first stomach the four bile-vessels are attached, they grow gradually thicker for about a third of their length, when they become twisted like a cord, and taper towards the rectum, to which also they are attached[540]. From this structure it should seem that the food has to pass twice through the first stomach, before the process of digestion is complete, and it is rejected at the anus.
The next suctorious Order is the Lepidoptera: in these the gullet is long and slender, surrounded at the beginning with a loose transparent skin, and at the base furnished with a pair of lateral sacs, forming the honey-stomach, and probably analogous to the food-reservoirs of the Diptera, which when blown up are of an oval form; the stomach, as in the bugs, consists of two portions, the first being the longest[541]. There are three free bile-vessels on each side, proceeding from a single branch[542]. It will not be uninteresting here to abstract from Herold the progressive changes which take place in the intestinal canal in this Order, during the transition of the animal from the larva to the imago state. In the larva, the gullet, the small intestine, and the rectum, are short and thick[543], there are a pair of silk reservoirs (sericteria), as well as vessels for the secretion of saliva (sialisteria): if you examine it two days after its first change, you will find the gullet and the small intestine much lengthened and become very slender; the stomach contracted both in length and size; the rectum also changed, and the silk vessels contracted[544]. These in a pupa eight days old have wholly disappeared; the gullet is become still longer, its base is dilated into a crop or food-reservoir; the stomach is still more contracted, and instead of a cylinder represents a spindle; the small intestine also is lengthened[545]: at a still more advanced period, when it is near appearing under its last form, the gullet and small intestine are still more drawn out; and the honey-bag, though very minute, has become a lateral[Pg 119] appendage of the gullet[546]; and lastly, in the butterfly it appears as a large vesicle[547]; the small intestine is grown very long[548]; and the rectum has changed its form and acquired a cœcum[549]. When we consider the adaptation of all these changes of form, the loss of old organs and the acquisition of new ones, to the new functions and mode of life of the animal, we see evidently the all-powerful hand of that Almighty Being who created the universe, upholding by his providence, and the law that he has given to every creature, the system that he at first brought into existence.
We now come to the Diptera. These have a very slender gullet, to which is attached on one side a long filiform tube, terminating in the food-reservoir, which in some instances is simple[550], but most generally consists of two or more vessels[551], collapsing when empty, but varying in shape and size when inflated with food: the mouth of the stomach in many cases is dilated into a kind of ring[552]; sometimes there is on each side a blind appendage or cœcum opening into it, in Bombylius covered with shags, which though not connected with the mouth by a tube, Ramdohr regards as saliva-reservoirs[553]; in Musca vomitoria the beginning of this organ below the mouth is covered with hemispherical prominences, and in Tipula it is dilated and marked with transverse folds. There are usually two pairs of bile-vessels; in the Muscidæ pedunculate and free[554]; in Tipula, Bombylius, and Leptis,[Pg 120] sessile and united[555]; and in Tabanus sessile and fixed[556]. It is remarkable that in some of this Order—the reverse of what usually happens—the alimentary canal appears to be much longer in the larva than it is in the imago; in Musca vomitoria, its length in the former is two inches and a quarter, while in the latter it is only one inch and one third[557]. A singular organ distinguishes the imago of this species, the use of which appears not to be discovered. It succeeds the rectum, and has on each side two short club-shaped appendages, open at the end, which receive tracheæ, and terminate in a short piece that opens into the anus[558].
In Hippobosca and its affinities the canal in question differs from that of other Diptera, in having no food-reservoir; in other respects it resembles it[559].
From the above statement it appears that the principal character which distinguishes those that take their food by suction, from those that masticate it, is the faculty with which they are furnished by means of an ample crop, honey-stomach, or food-reservoir, of regurgitating the food they may have stored up. Another distinction still more striking, which will appear more evidently hereafter, is to be seen in the saliva-secretors with which the suctorious tribes are furnished, to be found in very few masticators, by which they are enabled to render the juices more fluid and fit for suction.
The only insect amongst the Aptera whose alimentary[Pg 121] canal I shall notice, is the common harvest-man (Phalangium Opilio): in this, though the stomach and lower intestine are remarkably simple, yet their cœcal appendages are numerous and singular: the former, which has no distinct gullet, is pear-shaped[560]; and the latter, tapering downwards, and truncated at the end[561]; connected with it above are no less than twenty-three cœca or blind appendages, of various forms and dimensions; the last pair but one of which is very remarkable, being bent like a bow, and furnished externally with four short clavate processes[562]. It is probable that some of these organs are analogous to the bile-vessels of other insects.
When the Creator in his wisdom fixed the limits of the various tribes of animals, he united them all into one harmonious system by means of certain intermediate forms, exhibiting characters taken some from those that were to precede, and others from those that were to follow them, and this not only in their external structure, but likewise in their internal organization; so that we are not to wonder if in the same individual we meet with organs that belong to two distinct tribes, or if, remaining nearly the same in their prima facie appearance, they begin to exercise new functions. An instance of this we have seen in the dorsal vessel of insects, which in the Arachnida, though not materially different in situation or general form, by the addition of a small apparatus of arteries and veins becomes the centre and fountain of a regular system of circulation[563]. From the circumstances here alluded to, physiologists have been led to entertain[Pg 122] very different sentiments with regard to the structure of the alimentary organs of the Class we are now to enter upon, the Arachnida: what some regard as a real liver, others look upon as an epiploon or caul; and what the last denominate bile-vessels are by some of the former considered as appropriated to the secretion of chyle[564]. Yet both these opinions have some foundation in nature. When, in the Arachnida, we discover a lobular substance consisting of granules filling the whole cavity of the body and wrapped round the intestines, every one will see in it no small analogy to the epiploon which in insects performs the same function: but when, upon a further examination, we detect certain vessels communicating with this substance and the intestinal canal[565], the idea that these may be hepatic ducts, and this substance analogous to the liver, immediately strikes us as not improbable. Again: when we discover pairs of other capillary and tortuous vessels connecting with the intestinal canal either at the pylorus[566] or below it[567], which in appearance strikingly resemble the bile-vessels which we so constantly find in insects, we seem warranted in concluding that they are of the same nature and use: but when a nearer inspection enables us to detect the hepatic ducts just mentioned in the scorpion, and we find that these capillary vessels in the spider are in a very different situation from those in insects which we suppose them to represent, it occurs to us as not unlikely, that their function may be different.
Let us now consider how the intestinal canal is circumstanced in the two sections into which the Class Arachnida is divided; the Scorpionidea, and Araneidea. In the Scorpions, this organ proceeds from the mouth to the anus without any flexure or convolution, so that its length is scarcely equal to that of the body[568]; it is slender, and its diameter, with the exception of an irregular dilatation here and there, is nearly the same in its whole extent; the gullet is short; the stomach long, and nearly cylindrical; the duodenum shorter and thicker than the stomach, from which, as well as from the rectum, it is separated by a valve; the latter is cylindrical, and opens at the anus above the insertion of the vesicle that secretes the poison[569]. With regard to the biliary system and its organs: The liver is of a pulpy granular consistence and of a brownish colour, fills the whole cavity of the trunk and abdomen, and serves as a bed for the other intestines. It is divided longitudinally into two portions, by the channel in which the heart reposes—its anterior part is formed into many irregular lobes, by the sinuosities of the trunk; at the other extremity, it terminates in two acute ends, which enter the first joint of the tail; its surface presents a reticular appearance, the result of the approximation of polygonous lobuli; its interior is a tissue of infinitely minute glands: in Scorpio occitanus there are about forty pyramidal lobuli detached from each other, the summits of which, by their union, form bunches that have their excretory canals, varying in number in different species, which convey the bile to the alimentary[Pg 124] tube; in the above insect there are six pairs, three in the trunk and three in the abdomen, and in S. Europæus a smaller number[570]; these vessels run transversely from the liver, or aggregation of conglomerate glands, to the intestinal canal[571]; the bunches consist of an infinite number of spherical glands, generally filled with a brown thick fluid[572]: besides the transverse vessels, from the base of the stomach there issue two pairs of very slender tortuous ones, seemingly analogous to the common bile-vessels; one pair of which runs upwards, one on each side that organ towards the mouth, forming here and there some ramifications which enter the liver; and the other runs nearly transversely to it[573]. As the fluid contained in these vessels is different from that contained in the glands of the liver, M. Marcel de Serres supposes they may be chyliferous[574].
In the Araneidea also the alimentary canal is nearly straight, and scarcely exceeds the length of the body: the gullet is rather thick and cylindrical[575]; the stomach is distinguished anteriorly by two pairs of sacs, the upper pair being much the largest and nearly triangular, the lower linear[576]; from these sacs a narrow tube runs towards the rectum, but which is so entangled with the liver, muscles, &c., as not to be easily made out[577]; the rectum is rather tumid, and has a lateral cœcum[578]. The disposition of the liver or conglomerate glands is stated to be similar to that of the scorpion[579]; it is usually white,[Pg 125] but in some species it is yellowish, or reddish, and its lower surface has sometimes regular excavations[580]; no transverse hepatic ducts connecting it with the alimentary canal, as in the scorpion, appear to have been at present discovered: two pairs of capillary free vessels are attached to the base of the rectum on one side, which, except in their situation, seem analogous to the bile-vessels of insects[581].
From the above detailed account of the alimentary canal of the animals whose internal anatomy we are considering, it appears that M. Cuvier's observation—that the length and complication of the intestines indicate a less substantial kind of nutriment—does not hold universally: thus, in Necrophorus and Silpha, carnivorous insects, the intestinal canal in its length and convolutions exceeds those of most herbivorous ones, and in Cassida viridis and some others of the latter tribe are not longer than those of the predaceous beetles. In herbivorous larvæ also, in general, the length of the alimentary canal does not exceed that of the body, but in those of some flesh-flies (Musca vomitoria) it very greatly exceeds it[582]. So true is the observation—that there is no general rule without exceptions.
In this letter it may not be out of place to say a few words upon the excrements of insects; which, strange as the observation may seem, but it is no less true than strange, are sometimes pleasing to the eye, from their symmetry, and to the taste, from their sweetness. In those that masticate their food they are solid, and in those that take it by suction, fluid or semi-fluid. In the[Pg 126] caterpillars of Lepidoptera they are of the former description, and every grain wears some resemblance to an insect's egg: as the passage in many of these consists of six fleshy parts separated by channels, so the excrement represents six little prisms separated by six channels[583]. The Aphides all secrete a fluid excrement as sweet as honey, of which the ants are so fond[584], which is ejected not only at the anal passage, but, in many, by two little siphonets also above it[585]. A semi-fluid excrement is produced by some species of Chermes, as that which inhabits the Box, which often comes from the animal in long convoluted strings resembling vermicelli. Reaumur says its taste is agreeable, much more so than that of manna[586]. Under this head should be included the abundant spume with which the larva of Cercopis spumaria envelopes itself[587].
I am, &c.
Having given you so full an account of the system of digestion in insects, I am now to say something concerning their secretions, and the organs by which they are elaborated. Though no individual amongst them perhaps secretes so many different substances as the warm-blooded animals; yet in general the Class abounds in secretions perhaps as numerous and extraordinary as in the last-mentioned tribes, to some of which a few of them are analogous, while others are altogether peculiar. We know little or nothing of the mode in which the process of secretion in insects is accomplished; in most cases we cannot even discover, except in general, whence the secreted substance originates; and in others, though we are able to trace the vessels that contain it, we are often in the dark as to their structure.—Cuvier, as has been before hinted, from not being able to detect any thing in them like glands, and from their being constantly bathed in the blood or nutritive fluid, conceives that they separate the peculiar substances they contain, by imbibition[Pg 128] or infiltration, through the pores of the skin[588]; a circumstance which seems to indicate a certain conformation of the pores both as to size and figure, so as to enable them to admit only one peculiar product.
In treating on this subject, I shall first consider the organs of secretion, and next their products.
I. Organs of Secretion. In general, these are membranous vessels that float in the blood or nutritive fluid, and secrete from it a peculiar substance. They may be denominated according to their products—Silk-secretors, Saliva-secretors, Varnish-secretor, Jelly or Gluten-secretor, Poison-secretor, and Scent-secretors.
i. Silk-Secretors (Sericteria). These organs are most remarkable in the caterpillars of the nocturnal Lepidoptera or moths, especially in that tribe called Bombyces, to which the silkworm belongs: but this faculty is not confined to these insects, but is shared by many other larvæ in different Orders; and in one instance at least, by the imago. In general, the outlet of the silk-secretors is at the mouth; sometimes, however, as in the larva of Myrmeleon and the imago of Hydrophilus, its exit is at the anus. The first is the organ which in the silk-worm provides for us that beautiful substance from which the animal takes its name. There are always two of these vessels, which are long floating tubes, growing slender towards the head of the insect, where they unite to form the spinneret (fusulus) before described[589], which renders the silk. Their lower extremity also is commonly more slender than the middle, and is closed at the end. These organs are usually very much convoluted and twisted[590].[Pg 129] According to Ramdohr[591], they consist of two transparent membranes, between which is found a yellow or transparent jelly. The greater the quantity of silk employed by the caterpillar in the construction of its cocoon, &c., the longer are the silk-secretors. Those of the silkworm are a foot long[592], while those of the larva of the goat-moth are little more than three inches[593].
Other insects spin silk with the posterior extremity of their body. In the great water-beetle (Hydrophilus piceus) the anus is furnished with two spinnerets, with which it spins its egg-pouch[594]; these are in connexion, probably, with the five long and large vessels containing a green fluid, described by Cuvier[595], which surround the base of each branch of the ovaries. The larva of Myrmeleon, which also spins a cocoon with its anus, differs remarkably in this respect from other insects, since its reservoir for the matter of silk is the rectum; this is connected with a horny tube, which the animal can protrude, and thus agglutinate the silk and grains of sand that compose its cocoon[596].
The web of spiders is also a kind of silk remarkable for its lightness and extreme tenuity. It is spun from four anal spinnerets, which never vary in number; two longer organs peculiar to some species have been mistaken for additional ones, but Treviranus affirms that they are merely a kind of anal feeler. Their structure, as far as known, has been before described[597]. The web is secreted in vessels varying in form. In some (Clubiona[Pg 130] atrox) they consist of two larger and two smaller ones, at the base of which lie many still more minute[598]. The four larger vessels are wide in the middle, branching at top, and below terminating in a narrow canal leading to the spinnerets[599]. Treviranus thinks the fluid contained in the lower minute vessels different from that furnished by the larger ones—but for what purpose it is employed has not been ascertained.
ii. Saliva-secretors (Sialisteria). These are organs, rendering a fluid to the mouth or stomach, that are found in many insects, especially those that take their food by suction, as the Hemiptera, Lepidoptera, and Diptera, though they are not confined to the perfect insect, being also in some cases visible in the larva. Swammerdam was one of the first that discovered them, and he suspects that they may be salival vessels; though he, as well as Ramdohr, thinks they are the same with the silk vessels of the caterpillar[600]; an opinion which Herold has sufficiently disproved, by showing that at one period of the insect's life they co-exist[601], and Lyonet discovered a very conspicuous pair in the caterpillar of the Cossus, co-existent with the silk-secretors[602]. But the physiologist who has given the fullest account of these organs is Ramdohr:—I shall therefore extract chiefly from him what I have further to communicate with respect to them.
They are variously constructed blind vessels, that are present in almost all insects that take their food by suction, but are mostly wanting in those that masticate it. They have been found, however, in Cryptorhynchus Lapathi,[Pg 131] Chrysopa Perla, and Iulus terrestris. The most usual number of the saliva-secretors is two[603]; but sometimes, as in the first of the last-named insects, there is only one[604]; in others (Pentatoma Baccarum) there are three, the exterior one consisting of a pair of reservoirs connecting with the gullet by a single capillary tube[605]; in Pentatoma prasina there appear to be four[606]; in Nepa cinerea, even six—the exterior double pair in this insect, under a powerful lens, is found to consist of spherical vesicles, resembling somewhat a bunch of currants[607]; and in Syrphus arcuatus they are covered with four rows of similar ones[608]. In the flea they consist of two pair of spherical reservoirs, each of which is connected with a short tube, which uniting with that of the other forms a common capillary one connecting with the mouth or gullet[609]; these organs sometimes terminate below in slender vessels;—thus, in Nepa, the inner pair terminates in a single vessel of this description[610], and in Tabanus and Hemerobius apparently in many[611]. It admits of a doubt however, as was lately observed, whether in the Hemiptera, which have usually more than a pair of these organs, some are not rather food-reservoirs as in the Diptera.
The saliva-secretors open either into the instruments of suction themselves (Tabanus, Musca); or into the entrance of the gullet (Pentatoma, &c.); or, lastly, into that of the stomach (Syrphus, Bombylius). Those which lie at the entrance of the stomach consist only of a blind uniform[Pg 132] tube[612]; but there is commonly to be distinguished in those that open into the mouth, a reservoir, varying in shape in different species, and terminating in a capillary tube, or tubes, at one or both extremities[613]. In Bugs, two pair of these vessels are often present, one of which opens into the stomach (Reduvius), or gullet (Pentatoma), but the other into the instruments of suction[614]. In the Diptera they open into the stomach when the insect feeds only upon the nectar of flowers (Syrphus), and into the proboscis when it feeds upon both animal and vegetable juices (Tabanus, Musca). The function of the fluid secreted by these organs is to moisten or dilute the food before it is received by the instruments of suction and passed to the stomach[615]. When a common house-fly applies its proboscis to a piece of sugar, it is easy to see that it moistens and dissolves it by some fluid.
iii. Varnish-secretor (Colleterium). In butterflies, moths, and several other insects, one or more vessels called blind vessels open into the oviduct, concerning the use of which, physiologists are not agreed. In the cabbage butterfly there is a pair of ovate ones, or rather a bilobed one, each lobe of which externally terminates in long perplexed convolutions, not easily traced, filled with a yellow fluid, which Reaumur and Herold think is used for varnishing or gumming the eggs, so that they may adhere to the leaves on which they are deposited: it may probably serve likewise for other uses[616]. Another vessel is also to be found in the above butterfly, which enters[Pg 133] the oviduct above this, filled with a thick white fluid, the function of which is, probably, to lubricate the passage[617]. A similar organ is found in Phryganea grandis[618].
iv. Jelly-secretor (Corysterium). This is a remarkable organ, related to the preceding, which secretes the jelly of Trichoptera, some Diptera, &c.; this organ in the former, at least in Phryganea grandis, is of an irregular shape, with four horns or processes[619].
Poison-secretor (Ioterium). This organ, which is most conspicuous in the Hymenoptera Order, has not received much notice, except in the case of the Hive-bee and the Scolia: in the former, it is an elliptical membranous vesicle or reservoir, furnished at its lower extremity with a tube which renders to the sting, and at the other by a blind, long, filiform, secretory, vessel, which according to Swammerdam divides into two terminal blind branches[620], though Reaumur could detect but one[621]; in this vessel the poison is secreted and stored up. In Scolia there are two secretory vessels, which enter the reservoir in the middle on each side[622]. In the Scorpion, we learn from Marcel de Serres that the poison-secretor is clothed externally with a horny thickish membrane, containing two yellowish glands, composed of an infinity of spherical glandules, terminating in a canal, enlarged towards its base so as to form a reservoir, and leading to the extremity of the sting[623]. Connected by a slender tube with each mandible in spiders is a vessel with spiral folds, which seems properly to belong to this head—though[Pg 134] Treviranus calls it a saliva-vessel[624]—since in the Mygale avicularia and other spiders, the effect of the bite is said to be so venomous as to occasion considerable inflammation, and sometimes death[625].
v. Scent-secretors (Osmateria). Amongst other means with which insects are gifted for the annoyance of their foes and pursuers, are the powerful scents which many of them emit when alarmed and in danger. Concerning the internal organs by which these effluvia are secreted we possess but little information, but more notice has been taken of the external ones by which they are emitted. We may conclude in general, that the secretory organs are membranous sacs or vesicles, perhaps terminating in longer or shorter blind filiform vessels, sometimes secreting a fetid fluid, and at others a fetid gaseous effluvium. The Iulidæ, at least Iulus and Porcellio[626], cover themselves when alarmed, with a fluid of this kind, or emit one, for this faculty is not peculiar to the species noticed by Savi. I observed early in the year, when I handled Iulus terrestris, that it was covered with a slimy secretion, of a powerful scent, which stained my fingers of an orange colour. The spiraculiform pores that mark the sides of the animal are the outlets by which this fluid is emitted, and not spiracles as has been supposed: each of these orifices, as we learn from Savi, terminates internally in a black vesicle, which is the reservoir of the fluid[627]. The most remarkable insect for its powers of annoyance in this way, is one on that account called the bombardier (Brachinus crepitans), which can fire numerous[Pg 135] volleys of stinking vapour at its assailants before its ammunition is exhausted[628]. M. Dufour has given a very particular account of the organ that secretes this vapour;—it consists of a double apparatus, one on each side, in the cavity of the abdomen, both formed of two distinct vessels. The first, which is the innermost, presents itself under two different aspects, according as it is contracted or dilated: in the former case it is a whitish, irregularly rounded, soft body, apparently glandular, placed under the last abdominal segments; communicating at one end with the reservoir, and terminating constantly at the other in a very long and slender filament: in the second case, or when it is dilated, it resembles an oblong, membranous, diaphanous sac, filled with air, then occupying the whole length of the abdomen, and appearing free except where it communicates with the reservoir. The second vessel or reservoir is a small, spherical, brown or reddish body, constant in its form, internally hollow, placed under the last dorsal segment, precisely above the rectum, and opening by a small pore into the anus[629]: so that the tail of this little beetle may be regarded as a battery mounted with two pieces of cannon, which our alert bombardier fires alternately without intermission till all his ammunition is expended. The ground-beetles (Eutrechina) in general have a pair of these anal scent-secretors, which discharge an acrid and caustic fluid, and sometimes a volatile one[630]. The external organ of the scent-secretors in Gyrinus consists of two minute hairy cylindrical retractile tubes, of a red colour[631]. Numerous insects of other tribes and genera[Pg 136] emit scents from their anus, and from various other parts of the body, of which having before given you a very full account[632], I shall proceed to the consideration of the secretions themselves: but first I must observe, that in many cases, as in some of the cottony and powdery Aphides, Chermes, &c., the substance secreted appears to be a transpiration through the pores of the body, a kind of excretion from the superabundance of its fluid contents[633]. In many, however, this secretion transpires through appropriate orifices: thus in Chermes Abietis, which produces those curious galls resembling the cone of a fir[634], the flocoons of seeming cotton that cover it proceed from little oval concavities on its back, four of which are arranged in a transverse line on each dorsal segment of the abdomen: these concavities have minute tubercles probably terminating in a pore[635]. In Aphis Fagi the cottony flocoons are almost an inch long[636].
The secretions of insects may be considered under the following heads—Silk; Saliva; Varnish or Gum; Jelly; Oils; Milk; Honey; Wax; Poisons and Acids; Odorous fluids and Vapours; and Luminous matter.
i. Silk. This valuable product of insects, while in the silk-secretor, assumes in the Lepidoptera the appearance of a viscid gum, but the moment it is exposed to the air it hardens into a silken thread. It is remarkable for the following qualities:—it dries the instant it comes in contact with the air; it is then insoluble not only in water[Pg 137] but in the most active solvents, and even heat has no effect upon it to melt or soften it: indeed, without these qualities it would be of no use to us[637]. As soon as it leaves the spinneret it becomes the thread we call silk, which being drawn through two orifices is necessarily double through its whole length. This thread varies considerably in colour and texture, as has been before stated[638], and sometimes resembles cotton or wool rather than silk. In spiders it is of a much softer and more tender texture than that of other spinning insects; and Mr. Murray seems to have proved that it is imbued, in the case of the gossamer, with negative electricity: in the sericterium the fluid that produces it is sometimes white or grey, and at others yellow[639]. A remarkable gnat (Ceroplatus tipuloides), living on an agaric, carpets its station of repose and its paths with something between silk and varnish, which it spins, not in a thread, but in a broad riband[640].
ii. Saliva. Many insects have the power of discharging from their mouth a fluid which seems in some degree analogous to the saliva of larger animals. Thus many, as Lepidoptera, Hemiptera, Diptera, &c., can dilute their food, and render it fitter for deglutition. I have seen a common fly when not employed in eating, emit a globule of fluid as big as a grain of mustard-seed from its proboscis, and retract it again. On a former occasion[Pg 138] I observed to you that many predaceous, carnivorous, and some herbivorous beetles, when alarmed emit a drop of coloured acrid fluid from the mouth[641]. That this is not secreted in any of the ordinary salival vessels is evident from Ramdohr's dissections of those beetles[642], who, had there been such an organ, would doubtless have discovered it: but as the stomach of all of them is distinguished by those minute cœca or blind vessels, which he denominates shags (zotten)[643], perhaps these may be the secretors of this fluid, probably analogous to the gastric juice[644]; in which case its primary office would be the digestion of the food. We are not however warranted in considering every fluid effused from the mouth as saliva. The glutinous material with which wasps cement the woody fibres for their paper edifices[645]; that with which some sand-wasps moisten the sand which they scrape away, of which they form the singular tubes that lead to their nests[646]; and that with which the aphidivorous larvæ fix themselves previously to their becoming pupæ[647],—may be a secretion distinct from saliva; possibly intermediate between it and gum or the matter of silk, and secreted by peculiar organs. In the wasp, however, Ramdohr discovered nothing of the kind[648]; and in Syrphus, as before observed, the saliva-secretors are very peculiar in their structure, as if appropriated to the secretion of a peculiar fluid[649]. Something[Pg 139] similar has been observed by Reaumur with regard to the larva of Crioceris merdigera, which forms its cocoon with a kind of froth produced from the mouth[650].
iii. Varnish or Gum. The eggs of various insects, when they leave the oviduct, are covered with a kind of varnish or gum by which they adhere to the substances that the young larvæ are to feed upon, or are placed in a proper position for their hatching in an appropriate station. Several instances of this have been already mentioned[651]; I shall therefore not enlarge further upon the subject. With regard to the secretion itself, little has been recorded except its colour, which has been before noticed. Some Lepidoptera also as we learn from Reaumur and Bonnet[652], use a varnish in the construction of their cocoons.
iv. Jelly or Gluten. This secretion is particularly conspicuous in the Trichoptera and some Diptera, serving as a bed or nidus for those eggs that are committed to the water,—upon which I have nothing to add to what has been before said[653]. Under this head also may be noticed the fluid, secreted in peculiar vesicles, that lubricates the oviduct and the passages of the sexual organs[654].
v. Oils. Oily substances are sometimes produced by insects. The common oil-beetle (Meloe Proscarabæus) when touched sends forth a drop of this kind of fluid, of an orange colour, from each joint of its legs[655]: something similar I have observed in Coccinella bipunctata:[Pg 140] Ray mentions a locust taken in Spain which emits a yellow oleaginous fluid from between the claws of its fore legs[656]; but the precise nature of these substances has not been ascertained, nor whether they are secreted by peculiar organs.
vi. Milk. A milky fluid is produced by the larva of Chrysomela Populi. Willughby observed a similar effusion from pores in the upper surface of the body of Acilius sulcatus; and other insects emit it from other parts of their body[657].
vii. Honey. It is certain that honey is not an animal secretion; yet the saccharine matter collected from the nectaries of flowers, from which it is derived, seems to undergo some alteration in the stomach; for the consistence of honey is greater than that of any vegetable nectar, and its taste does not vary greatly, while that of the nectar in different plants is probably not the same. Reaumur also has observed, that each honey-cell in a bee-hive is always covered by a cream-like layer of a thicker consistence than the rest, which apparently serves to prevent the more liquid honey, which from time to time is introduced under it, from running out[658]. Now if honey were the unaltered nectar of plants, it is difficult to conceive how this cream could be collected in proper proportions. The last-mentioned naturalist likewise ascertained, that if bees, in a season in which the fields afford a scarcity of food, be supplied with sugar, they will from this substance fill their cells with honey which differs in no respect from the common sort, except that its flavour is a little heightened[659]:—a similar[Pg 141] argument may be deduced from the circumstance of the bees imbibing the juices of fruits of various kinds as they are well known to do[660]. It seems therefore evident that the honey collected by bees undergoes some modification in their honey-stomach before it is regurgitated into the cells, and therefore may be regarded in some degree as a peculiar secretion.
Huber says that he has ascertained by a great number of observations that electricity is singularly favourable to the secretion of the substance of which honey is formed by flowers; the bees never collect it in greater abundance, nor is the formation of wax ever more active, than when the wind is in the south, the air humid and warm, and a storm gathering[661].
viii. Wax generally transpires through the pores of the skin of those insects that produce it, either partially or generally, and it is secreted from honey or other saccharine substances taken into the stomach. In the hive-bee, as has been before stated, it is produced partially[662], but in many other insects it is a general transudation of the body. This is particularly the case with a large number of the Homopterous Hemiptera; and those flocoons that look like cotton, and cover the body of several Chermes and Aphides, if closely examined will be found of the nature of wax: this I have particularly noticed with respect to Chermes Fagi, in which the cotton-like flocoons are often so long as to cause the insect to look like a feather, and a leaf covered by them exhibits a very singular appearance, as if clothed with the[Pg 142] fine down of a swan[663]. Probably the white powder or threads that appear to transpire through the skin of many other insects is of a waxy nature. In the larva of a beetle described by Reaumur, the flocoons are so arranged as to give the animal some resemblance to a hedgehog, and when rubbed off they are reproduced in twelve hours[664]. Gyllenhal, speaking of Peltis limbata, observes, that when alive it is covered with a white powder resembling mould, which if rubbed off returns again as long as the animal lives[665].
It will not be improper to include under this head what further account I have to give of Lac, which though regarded as a resin, since Cocci sometimes certainly produce wax[666], probably has some analogy with the latter substance. When the females of this Coccus (C. Lacca) have fixed themselves to a part of the branch of the trees on which they feed (Ficus religiosa and indica, Butea frondosa, and Rhamnus Jujuba[667]), a pellucid and glutinous substance begins to exude from the margins of the body, and in the end covers the whole insect with a cell of this substance, which when hardened by exposure to the air becomes lac. So numerous are these insects, and so closely crowded together, that they often entirely cover a branch; and the groups take different shapes, as squares, hexagons, &c., according to the space left round the insect which first began to form its cell. Under these cells the females deposit their eggs, which after a certain period are hatched, and the young ones eat their way out. Though indisputably an animal[Pg 143] secretion, many of the properties of lac are not very different from those of the juices of the trees on which the animal feeds, and which therefore would seem to undergo but little alteration.
Wax seems also to form a constituent part of some insects which are not found to secrete it. The yellow substance deposited in vessels containing spiders in alcohol is said to be a true wax, and may be obtained from these animals by gently heating them[668].
ix. Poisons and Acids. The bite as well as the sting of many insects is followed by inflamed tumours, so that the sialisteria of some bugs, Diptera, Aptera and spiders, may be regarded as producing a poisonous fluid; but we know nothing of the real nature of it, nor of that of other venomous insects, except the ant—whose celebrated acid may be considered under the present head,—the bee, the wasp, and the scorpion.
Contrary to the once received doctrine that no acid was to be found in any animal, except as the effect of disease in the alimentary canal, many insects secrete peculiar and powerful ones. I have on a former occasion related an instance in which an acid of this description, secreted in its sialisteria, is employed by a moth to soften its cocoon[669]; and Lister mentions a species of Iulus which produced one resembling that of ants[670]; but this last is the most powerful of all. The fact that blue flowers when thrown into an ant-hill become tinged with red has been long known; but Mr. Fisher of Sheffield, about 1670, seems to have been the first who ascertained that this effect is caused by an acid with which ants abound, and which may be obtained from them by distillation or[Pg 144] infusion in water[671]. Margraff and other chemists confirmed this discovery[672]; and concluding that this acid was of a peculiar kind, they gave it the name of the Formic acid. This name, however, is now exploded; the subsequent experiments of Deyeux, Fourcroy and Vauquelin having ascertained that the acid of ants is not of a distinct kind, but a mixture of the Acetic and Malic[673]. These acids are in such considerable quantities, and so concentrated in these animals, that, when a number of Formica rufa are bruised in a mortar, the vapour is so sharp that it is scarcely possible to endure it at a short distance. It also transpires from them, for they leave traces of it on the bodies which they traverse: and hence, according to the experiments of Mr. Coleridge, the vulgar notion that ants cannot pass over a line of chalk is correct; the effervescence produced by the contact of the acid and alkaline being so considerable, as in some degree to burn their legs[674]. The circumstance of much of the food of ants being of a saccharine nature may account for this copious secretion of acid, the use of which is probably to defend themselves and their habitations from the attack and intrusion of their enemies: if a frog be put into a nest of Formica rufa that has been deranged, it will be suffocated in five minutes[675]. That which they ejaculate from their anus when attacked, as formerly stated[676], must be secreted in an ioterium; but their very blood seems of an acid nature. It is very probable, as Dr. Thomson has observed[677], that acids may be obtained from many other insects, and that they are various modifications of the acetic.
From the circumstance that water is absorbed by greasy moths, that crystals of a salt are occasionally found adhering to them, that they change blue litmus paper red,—it has been inferred that their supposed oiliness is in fact an acid or acid salt, having the property of attracting moisture from the air, the infected moths being in fact not greasy, but wet; hence the application of chalk and clay, usually recommended in this case, can have only a temporary and superficial effect. The only effectual remedy, is steeping the body in spirits of wine till all the acid is extracted[678]. This acid is probably the same as Chaussier obtained from silkworms, since called Bombic Acid[679].
The poison of bees and wasps, as to its chemical qualities, is a transparent fluid, at first sweet to the taste, but immediately afterwards hot and acrid like the milky juice of the spurge[680]; soluble in water, but not in alcohol; and separable from the former in the state of white powder, when the latter is added giving a slight red tinge to paper stained with vegetable blue, and when dry and chewed appearing tenacious, gummy and elastic. This last property, as well as solubility in water and not in alcohol, is common also to the poison of the viper, which however differs in being tasteless, and not affecting vegetable blues. From hence Fontana concludes that this fluid is united with an acid, but in a very small proportion, and not with an alkali[681]. The venom of bees is extremely active; a grain in weight, it is conjectured, would kill a pigeon in a few seconds[682]. It is[Pg 146] remarkable, however, that while in some constitutions the sting of a single bee or wasp is sufficient sometimes to induce alarming symptoms, in others numerous punctures will produce little or no pain or inflammation. That this fluid, and not the puncture of the sting, is the sole cause of the inflammation that usually follows the wound inflicted by one of these animals, is proved by the facts, that if it be introduced into one made by a needle, the same effect ensues, and that when the whole contents of the poison-bag have been exhausted by the insect's stinging three or four times in succession, its weapon then becomes harmless[683].
The venom of scorpions, though much more potent, probably resembles that of bees, &c., in many of its chemical qualities: it issues from two pores in the sting before described[684], where, when the animal is irritated, it accumulates under the form of two little drops of a whitish colour: spread upon paper this fluid produces a spot like what would be caused by oil or grease, and this part of the paper becomes by desiccation firmer and transparent[685].
x. Odorous fluids and Vapours[686]. The powerful scents which different insects emit are extremely numerous, much more so indeed than the generality of Entomologists have been aware, for there is scarcely a scent odious or agreeable that may not be met with in the insect world. This you will be convinced of, by following a practice which I would recommend to you—that of smelling[Pg 147] the insects you take. Some of these scents are peculiar to particular parts or organs, and some are exhaled generally by the whole body; some are emitted by a fluid secretion, and others are gaseous effluvia. On a former occasion I gave you a rather full account of these scents and their organs[687]; I shall relate here only what I there omitted. To begin with sweet odours. Many beetles emit an agreeable scent. The rose-scented Capricorn or musk-beetle (Cerambyx moschatus) has long been noted for the delicious scent of roses which it exhales; this is so powerful as to fill a whole apartment, and the insect retains it long after its death. Captain Hancock also informed me that another species of the same genus, C. sericeus, has in a high degree a scent resembling that of the cedar[688] on which they feed. Though most of the micropterous tribes (Brachyptera) have a fetid smell, yet there are some exceptions to this amongst them. One species (Philonthus suaveolens K. MS.) related to P. micans, which I once took, smelt precisely like a fine high-scented ripe pear; another, Oxytelus morsitans, like the water-lily; a third, O. rugosus, like water-cresses; and lastly, a fourth (P. fuscipes) like saffron[689]: Trichius Eremita, one of the Petalocerous beetles, is stated to have the scent of Russia leather; Geotrupes vernalis, in spite of its stercorarious food, of lavender-water[690]. Mr. Sheppard has observed that Dytiscus marginalis when recently taken smells not unlike liquorice: Bonnet mentions a caterpillar that had the scent of new hay. A little gall-fly (Cynips Quercus Ramuli) has[Pg 148] the remarkable odour of Fraxinella: the larva of another species of this genus (C. Rosæ) has an odour which seemed to Reaumur as attractive to cats as that of Nepeta cataria or Teucrium Marum[691]: some Phalangia smell like walnut leaves[692]; and the various species of the genus Prosopis (Melitta * b. K.) have a very agreeable scent of Dracocephalum moldavicum[692].
We next come to fetid odours. These in numerous cases are known to be secreted and emitted by appropriate vessels and organs; they are often exhaled from a fluid secretion, of which, in the letter lately referred to, I gave you almost all the known instances. Savi, in his history of Iulus fœtidissimus, informs us that it emits a yellow fetid fluid from its supposed spiracles, which if applied in sufficient quantity imparts a red colour to the skin, to be removed neither by friction nor washing, but only disappearing by time; when removed from the black vesicles in which it is stored, it shoots into very transparent octahedral crystals[693].
I have before mentioned the coloured fluid which some insects emit when they are disclosed from the pupa, and that it probably exhales some powerful odour which attracts the males[694].
The great Hydrophilus, in its larva state, when first taken into the hand remains without motion; in a minute afterwards it renders itself so flaccid as to appear like a cast skin. Taken by the tail it contracts itself considerably, it then agitates itself briskly, and ejaculates with a slight noise a fetid and blackish fluid[695].
In other cases these odours are produced by gaseous vapours. That of the Bombardiers (Brachinus) is the most celebrated and remarkable. It is whitish, of a powerful and stimulating odour, very like that exhaled by nitrous acid. It is caustic, producing upon the skin the sensation of burning, and forming instantly upon it red spots which soon turn brown, and which, in spite of frequent lotions, remain several days. It turns blue paper red[696]. That amiable, intelligent, and unfortunate traveller Mr. Ritchie,—whose premature death, when attempting to penetrate to the interior of Africa, all lovers of Natural History so deeply lamented, and whose ardour in the pursuit of that science I had an opportunity of witnessing, when, in company with him, Messrs. Savigny, Du Fresne, and W. S. MacLeay in 1817, I visited the forest of Fontainebleau,—in a letter to the last-mentioned gentleman[697], relates that his companion M. Dupont, near Tripoli took a nest consisting of more than a thousand of a species of this genus. "I am making a few experiments," says he, "on the substance which they emit when they crepitate, but do not know whether I can collect enough to arrive at any conclusion. It made Dupont's fingers entirely black when he took them. It is neither alkaline nor acid, and it is soluble in water and in alcohol." From this we may conjecture that it formed crystals.
xi. Phosphorus. On this remarkable secretion I have so fully enlarged on a former occasion[698], that here I shall merely add a few observations which Mr. Murray obligingly[Pg 150] communicated to me. He remarks that in a box in which glow-worms were kept—five luminous specks were found secreted by the animal, which seemed to glow and were of a different tinge of light. One put into olive oil at eleven p. m. continued to yield a steady and uninterrupted light until five o'clock the following morning, and then seemed, like the stars, to be only absorbed by superior effulgence. The luminous spherical matter of the glow-worm is evidently enveloped in a sac or capsule perfectly diaphanous, which when ruptured discloses it in a liquid form, of the consistency of cream. M. Macaire, he observes, in the Bibliothèque Universelle, draws the following conclusions from experiments made on the luminous matter of this animal;—that a certain degree of heat is necessary to their voluntary phosphorescence—that it is excited by a degree of heat superior to the first, and inevitably destroyed by a higher—that bodies which coagulate albumen take away the power—that phosphorescence cannot take place but in a gas containing no oxygen—that it is not excited by common electricity, but is so by the Voltaic pile—and lastly, that the matter is chiefly composed of albumen.
xii. Fat. There is one product found in the body of insects most copiously in their larva state, but more or less also in the imago, which may be called their fat. In the former it is a many-lobed mass, occupying the whole of the interior, except the space that is required for the muscles and the internal organs, which it wraps round and protects. It is contained in floating membranes, very numerous, which fill all the interstices, and assume the appearance sometimes of small globules, and sometimes of a thickish mucilage, which easily melts and inflames;[Pg 151] in colour it is most commonly white, but sometimes yellow or green. It is imagined to be a kind of epiploon or caul, and is accumulated in the larva as a store of nutriment for the growth and development of the organs of the perfect insect while in the pupa state[699]. The blood in which the different organs float that is not required for their nutriment, is supposed to be expended in the formation of this substance. Marcel de Serres is of opinion that it is secreted from the chyle by passing through the pores of the dorsal vessel, formerly called the heart of insects[700].
Under this head I may mention what little is known with regard to the perspiration of these animals[701]. That a considerable quantity of fluid passes off from them when in the pupa state, is sufficiently proved by the loss of weight which they undergo, and by the experiments of Reaumur, who collected the fluid in closed glass tubes; and that in their perfect state they are constantly passing off perspirable matter by the pores of their skin or crust, is not only rendered probable by the succulent nature of their food and the absence of any urinary discharge, but is proved by what takes place in a swarm of bees. These insects, when crowded together in hot weather in a large mass, become heated to such a degree, and perspire so copiously, that those near the bottom are quite drenched with the moisture it produces, which so relaxes their wings that they are unable to fly[702].
I am, &c.
The reproductive organs of insects in their general denominations and functions correspond with those of the higher classes of animals; but as to number, proportions, and other particular details of their structure, they differ from them very considerably. I shall not now, however, enter at large upon this subject, but confine myself principally to the consideration of those organs in the female which are appropriated to the formation, fecundation, maturation, exclusion and deposition of their eggs, and other circumstances relating to that subject. The organs connected with this function are the Sperm-reservoir; the Oviduct; the Ovaries; and the Ovipositor.
I. The Sperm-reservoir (Spermatheca) is an organ connecting the vagina with the oviduct, which, according to Herold, receives the male sperm as into a reservoir[703], and fecundates the eggs in their transit through that passage. This vessel, which consists of a double tunic, in the cabbage-butterfly terminates the vagina, and is connected with the oviduct by a lateral undulating tube: in shape it is a rather irregular oblong, and is surmounted by a[Pg 153] small orbicular vesicle, connected by a short tubular footstalk with the main reservoir[704]. A similar organ was discovered by Malpighi in the imago of the silkworm, who denominates it the uterus; to which indeed it seems analogous, and which he also regards as a reservoir for the sperm for the gradual fecundation of the eggs[705]. But in that fly the organ is of a rather different shape, and the interior vessel terminates in several spherical vesicles[706]. John Hunter by the most decisive experiments, such as covering the eggs of the unimpregnated moth, after exclusion, with the liquor taken from the spermatheca in those which had been impregnated, and rendering them fertile, he demonstrated that this organ was a reservoir for the spermatic fluid, to impregnate the eggs as they were ready for exclusion, and that coition and impregnation were not simultaneous[707]. It is not improbable that in all insects whose eggs are gradually laid, this provision for their gradual fecundation, if carefully sought for, might be detected[708]. Rifferschweils is of opinion, that[Pg 154] in these cases the eggs are fertilized in their transit through the oviduct by sperm adhering to the folds of the cloacæ[709]: but this opinion seems less analogous to what takes place in other cases, with regard to the due preparation of the eggs for a safe and effectual transit[710].
II. The Oviduct (Oviductus) is the canal, always separate from the vagina, which receives the eggs from the ovary, transmitting them, often by a peculiar and complex instrument in which it terminates, to their proper station. This canal sometimes opens into the anal passage or cloaca, and at others, as in the cabbage-butterfly[711], is distinct, and lies between the sexual organ and the anus. In the Arachnida there are two oviducts[712].
III. The Ovaries (Ovaria) in insects are the viscera in which the eggs are generated and grow till they arrive at maturity, when they pass through the oviduct, and are extruded or deposited in their appropriate station. They vary considerably in their structure. In all however, except the Iulidæ, in which there is only a single ovary[713], the oviduct at its upper or inner extremity terminates in two branches, usually further subdivided into a number of smaller conical ones, which several ramifications constitute the ovaries, or egg-tubes as they are sometimes called: these tubes generally consist of a[Pg 155] single membrane, and are joined to the oviduct by membranous rugose cloacæ[714]: in the Phalangia, however, there are two tunics; the outer one of a cellular substance, and the inner one consisting of spiral fibres like tracheæ—a kind of structure which renders them capable of great extension[715]. Rifferschweils considers the ovaries as formed upon two primary types.—First, flagelliform ovaries, consisting of conical tubes equal in length, and inserted at the same place at the end of the primary branches as in the Lepidoptera, the Bee, &c. Secondly, racemose ovaries, consisting of short conical tubes, so proceeding from the primary branches as to render the ovary racemose or pinnated, as in certain Neuroptera, Coleoptera, and Diptera[716]: but perhaps their structure will be better understood if they are divided into agglomerate ovaries and branching ovaries: in the first the egg-tubes form two bundles, in which the branches are not discernible, as in the Ephemera, the chamæleon-fly, and spiders[717]: and in the second the branches are distinct, as in the Lepidoptera and the majority of insects.
The number of branches varies in different genera and species. In Echinomyia grossa, a large fly, there are only the two primary branches[718]; in the common dung-beetle (Geotrupes stercorarius) these appear divided at their apex into fingers[719]: in Scolia, a Hymenopterous genus, and the butterfly of the nettle, there are three secondary branches on each side[720]: in many other[Pg 156] Lepidoptera and the humble-bee there are four[721]; in the common louse there are five[722]; in the rhinoceros-beetle and the cockchafer, six[723]; in the wasp, seven[724]; eight in the cockroach[725]; twelve in the Carabi and the mealworm-beetle[726]; thirty in the large green grasshopper (Acrida viridissima[727]); thirty-two in the cheese-maggot-fly[728]; and in the hive-bee more than a hundred and fifty[729].
The number of eggs also contained in the ovaries varies. In Echinomyia grossa there is only one egg in each, and only two at once in the matrix[730]: in another fly produced by the cheese-maggot there are four[731]; in the louse there are five; in the cockchafer six[732]; in the hive-bee sixteen or seventeen are visible at the same time[733]; and in the silkworm-moth sixty or seventy[734]. Besides the eggs, the tubes contain a pellucid mucus, and at their upper extremity the eggs are lost in a granular mucous mass, in which, however, they may still be discovered with a microscope[735]. With regard to the termination of the ovaries or egg-tubes internally,—in those that have agglomerated ones it is not to be traced, the whole appearing like an oblong obtuse or acute body[736]: but in the branching ones it is more easily traced; at first they converge in most cases to a point; this is seen to advantage in the caterpillar of some butterflies, when near assuming the pupa, in which they are readily discovered, and represent with great truth and elegance the[Pg 157] bud of some blossom[737]; but in time they diverge, and sometimes become convoluted[738]; they generally terminate in a slender simple filament, but in the louse in a fork[739]; they are sometimes extremely long, as in the wasp and Lepidoptera[740]; in the hive-bee they appear to be shorter[741].
IV. We are next to consider the Ovipositor, or instrument by which numerous insects are enabled to introduce their eggs into their appropriate situations, and where the new-born larva may immediately meet with its destined food. As this instrument is one of the most striking peculiarities with which the wisdom of the Creator has gifted these little animals, and in many cases is extremely curious and wonderful, both in its structure and modes of operation—though on a former occasion I gave you a brief account of several kinds of them[742], I shall now enter more at large into the subject, and describe these often complex machines, as they are exhibited in most of the different Orders of insects.
With regard to the Coleoptera Order, there are doubtless numerous variations in the structure of this organ; but very few have been noticed, and those chiefly belong to insects whose grubs feed on timber. In these it is usually retractile one part within another, like the pieces of a telescope: in Buprestis it consists of three long and sharp laminæ, the two lateral ones forming a sheath to the intermediate one, which probably conveys the eggs[743]: in Elater it is a cylindrical organ, terminating in a pair of conical joints, which seem to form a forceps, and including a tube probably conveying the egg to the forceps,[Pg 158] which perhaps introduces it[744]. The Ovipositor of Prionus coriarius differs from that of Callidium violaceum, and many Capricorns before described[745]: it consists merely of a long bivalve piece ending in a kind of forceps, and hollowed above into a channel for the passage of the eggs[746].
In the Orthoptera the instrument of oviposition is more simple; in Locusta consisting merely of four robust three-sided pieces, two above and two below, the former pair at the end curving upwards and the latter downwards[747], these pieces seem calculated when they have entered the earth to enlarge the burrow, and the animal appears able to separate them very widely from each other[748]. The ovipositor of Acrida viridissima, which like that of many Hymenopterous insects forms a kind of appendage or tail to the body, has been described both by De Geer and Latreille as consisting of two valves only[749]; but in reality it consists of six, two upper and four lower, as you may ascertain by means of a pin or the point of a penknife, which will readily separate them. This is confirmed by a figure of Stoll's of a species which seems to connect Conocephalus with Gryllus. In this the ovipositor is considerably longer than the body of the animal, and is composed of six distinct pieces; viz. two external ones stouter than the rest, and within these four others finer than a hair and convolute at the apex[750]. There is a considerable variety in the shape of the ovipositors of the Acridæ and the cognate genera:—thus in[Pg 159] A. viridissima this organ is straight, in A. verrucivora bent like a sabre, and in Pterophylla citrifolia and some others, the whole machine is short and boat-shaped; in Scaphura Vigorsii it is also rough with sharp little tubercles[751]. I had an opportunity of observing, with respect to the first of these insects, that in boring, as is the case with the Cicadæ and saw-flies, the motion of the valves was alternately backwards and forwards. It appeared also to me that the two outer pieces of each of the apparent valves were fixed in a groove in the margin of the intermediate one. I saw this clearly with respect to the upper pieces, and it is most probable that the lower are similarly circumstanced. In the cricket tribe (Gryllus) the ovipositor is as long as the abdomen, very slender, terminating in a knob[752]. It is apparently bivalve like that of Acrida, but I believe is resolvable into the same number of pieces.
In the Homopterous Hemiptera there seems to be more than one type on which the ovipositor is constructed. In an insect very common with us, the froth froghopper (Cercopis spumaria), some approach is made to the ovipositors last described, at least the number of pieces is the same—for it has a pair of external valves forming a sheath, which includes three sharp laminæ resembling the blades of a lancet, the middle one of which can be separated into two; this instrument De Geer had reason to think was scored transversely like a file[753]. In the insects of this Order so noted for their song[754] (Cicada), there are only five pieces; namely, two valves forming the[Pg 160] sheath, two augers or borers, and an intermediate piece upon which they slide, each being furnished with an internal groove for that purpose, and the middle piece with a ridge to fit; a contrivance of Divine Wisdom, to prevent their dislocation when employed in boring; the augers terminate in a knob which is externally toothed[755]. This structure approaches that of the Hymenoptera, especially the saw-flies. With regard to the Heteropterous section of this Order—as they usually do not introduce their eggs into any substance, they have no call for any remarkable ovipositor, and therefore are not so furnished. A remark which will also apply to the Lepidoptera Order.
In the Libellulina amongst the Neuroptera, an organ of this kind is sometimes discoverable. In Agrion, Reaumur noticed a part which he conjectured to be an ovipositor; it consists of four laminæ or lancets, the interior pair slender, the exterior wider, and all externally serrated[756].
The insects of the Hymenoptera Order have long been celebrated for the organs we are describing, whether used as saws, augers, or darts. I formerly gave you a very general account of the saws,—I shall now give you a very interesting one in detail copied from an admirable little essay of Professor Peck. "This instrument," says he, "is a very curious object; and in order to describe it it will be proper to compare it with the tenon-saw used by cabinet-makers, which being made of a very thin plate of steel, is fitted with a back to prevent its bending. The back is a piece of iron, in which a narrow and deep groove is cut to receive the plate,[Pg 161] which is fixed: the saw of the Tenthredo is also furnished with a back, but the groove is in the plate, and receives a prominent ridge of the back, which is not fixed, but permits the saw to slide forward and backward as it is thrown out or retracted. The saw of artificers is single, but that of the Tenthredo is double, and consists of two distinct saws with their backs: the insect in using them, first throws out one, and while it is returning pushes forward the other; and this alternate motion is continued till the incision is effected, when the two saws receding from each other, conduct the egg between them into its place. In the artificial saw the teeth are alternately bent toward the sides, or out of the right line, in order that the fissure or kerf may be made sufficiently wide for the blade to move easily. To answer this purpose in some measure, in that of the Tenthredo the teeth are a little twisted, so as to stand obliquely with respect to the right line, and their point of course projects a little beyond the plane of the blade, without being laterally bent; and all those in each blade thus project a little outwards: but the kerf is more effectually made, and a free range procured for the saws, by small teeth placed on the outer side of each; so that while their vertical effect is that of a saw, their lateral effect is that of a rasp. In the artificial saw the teeth all point outward (towards the end) and are simple; but in the saw of the Tenthredo they point inward, or toward the handle, and their outer edge is beset with smaller teeth which point outwards (towards the end)[757]." Valisnieri, Reaumur, and De Geer describe the groove as being in the back; but in Mr. Peck's insect, if there is no error in his account, it is, as[Pg 162] in the Cicadæ, in the saw itself[758]. In the genus Cimbex, belonging to the same tribe, the saw differs in shape, being somewhat sigmoidal or resembling the letter S, while in that of other saw-flies it is cultriform with a concave edge: other minor differences distinguish them, which need not be particularized.
A similar structure, with regard to the organ in question, obtains in the rest of the Hymenoptera, even those that use it as a weapon of offence; but the backs of the saws in them, composed of a single piece, become a sheath for the darts. The valves, however, vary. In most of those with an exerted sting, as Pimpla, they are linear, exerted, and as long as the aculeus itself[759]. In Proctotrupes they appear to be united so as to form a tube for the ovipositor, and are produced by a prolongation of the last abdominal segment. The darts usually run in two grooves of the sheath, and at their apex are retroserrulate[760]. In some cases the sheath itself is serrated[761]. The shanks of the darts are connected with the valves; so that when these open they are pushed out: sometimes on their outer side they have a triangular plate towards the base, which prevents their being pushed out too far[762].
In Sirex and many ichneumons, in which the ovipositor is too long to be withdrawn within the abdomen, it remains always exerted; but in general it is retracted within that part when unemployed. In the gall-fly (Cynips) this instrument is really as long as in Pimpla, &c.;[Pg 163] but as it is infinitely more slender, when in repose it is rolled up spirally and concealed within the abdomen. It is the puncture of this minute organ that produces the curious galls formerly described to you[763]. But the most anomalous ovipositor in this Order appears to be that of Chrysis (C. ignita, &c.), which is covered by several demi-tubes or scales enveloping and sliding over each other: when these scales are removed, the true ovipositor appears, which is of a structure similar to that of the rest of the Order, but the valves are long and slender with their summit generally visible without the anus[764].
Though the ovipositor of the majority of Dipterous insects is a tube with retractile joints[765], in the crane-flies this organ is different, and, like that of Acrida above described, consists of what at first sight appear two valves, but each of which is formed of two pieces, the upper ones sharp and longer, and the lower pair blunt. The upper pair forms the auger that bores a hole in the ground, and the lower conducts the eggs into it after it is bored[766].
In the Aptera and Arachnida in general there seems no remarkable instrument of this kind; but Treviranus has described one in spiders for extruding the eggs of a singular construction. It is an oval plate lying between the external genitals and spinning organs, and is composed of a number of small screw-shaped cartilages, connected together in the most wonderful manner. There are few organs, he observes, in the animal kingdom which[Pg 164] for their artificial mechanism can be compared with this. Each cartilage inosculates very closely in the adjoining one, and all are besides bound together by a strong skin[767].
The manner in which the eggs of insects are fecundated by the male sperm is one of those mysteries of Nature that are not yet fully elucidated and understood. We can readily conceive that all the eggs may be fertilized by a single intercourse in the case of insects which, like the Ephemeræ and Trichoptera, exclude the whole mass at once; or like many moths and butterflies, in a very short time afterwards; but the subject becomes much more difficult to explain when we advert to the female of the hive-bee, the whole number of whose eggs, deposited in two years, are, as Huber has demonstrated, in like manner fertilized by a single act[768]:—if you bear in mind, however, what I have lately observed with regard to Malpighi's discovery of a sperm-reservoir in insects, you will more readily comprehend how in this case a gradual fecundation may take place. The principal objection to this solution of the difficulty in the case before us, is derived from the very small size of the organ supposed to be destined for this purpose—it being scarcely bigger than the head of a pin[769]: it seems therefore incredible that it should retain any portion of an extraneous fluid at the end of twelve or eighteen months, and still more unlikely that the fluid should in the interval have sufficed for the slightest moistening of not fewer than 30,000 or 40,000 eggs. The only hypothesis that seems at all to square with this fact, is that of Dr. Haighton,—that impregnation is the result not of any[Pg 165] actual contact of the sperm with the eggs, but of some unknown sympathetic influence[770], or rather perhaps of some penetrating effluvia or aura seminalis, which, though small in quantity, it may retain the power of emitting for a long period.
Certain female moths, of the species of that family which, from the remarkable cases or sacs the larvæ inhabit, the Germans call sack—träger, before noticed[771], have been supposed to have the faculty of producing fertile eggs without any sexual intercourse; and various observers, after taking great pains, appeared to have satisfactorily proved the fact; so that some doubted whether these insects produced any males at all[772]. The enigma was at length explained by the accurate Von Scheven. At first his experiments were attended with the same result as those of his predecessors; but upon making them more carefully, and separating what he conceived to be the female from the male pupæ, he ascertained not only the existence of a female in the species he examined (Psyche vestita), but that when thus secluded she laid barren eggs; evidently proving that in the contrary instances above alluded to, an unperceived sexual intercourse must have taken place[773]. Though he thus ascertained that these insects do not in this respect deviate from the general rule, he remarked or confirmed several facts in their economy sufficiently anomalous and striking;—as that the female is not only without wings, but with scarcely any feature of a moth, much more closely resembling a caterpillar; and that in ordinary circumstances[Pg 166] she never attempts to leave the pupa-case in which she has been disclosed, but that being there impregnated by the male, she there also, apparently after the manner of the female Cocci, deposits her eggs, which hatching produce young larvæ that make their way out of the case, and thus seem to originate without maternal interference[774].
But the most remarkable fact bearing upon this head, though as relating to a viviparous insect it does not strictly belong to it, is the impregnation of the female Aphides, or plant-lice, before alluded to[775]. If you take a young female Aphis at the moment of its birth, and rigorously seclude it from all intercourse with its kind, only providing it with proper food, it will produce a brood of young ones: and not only this; but if one of these be treated in the same way, a similar result will ensue, and so on, at least to the fifth generation!! to which period Bonnet, who first made an accurate series of observations on this almost miraculous fact, successfully carried his experiments, till the approach of winter and the want of proper food forced him to desist[776]; and Lyonet extended it still further[777]. It is now generally admitted as an incontestible fact, that female Aphides[Pg 167] have the faculty of giving birth to young ones without having had any intercourse with the other sex. How are we to explain this most extraordinary fact? Are we to suppose with Bonnet that these insects are truly androgynous, as strictly uniting both sexes in one? This supposition, however, is completely overturned by the circumstance, that there are actually male as well as female Aphides, and that these, as was first observed by Lyonet, are united towards the close of the summer in the usual manner[778]. The most likely supposition therefore is, that one conjunction of the sexes suffices for the impregnation of all the females that in a succession of generations spring from that union. It is true that at the first view this supposition appears incredible, contradicting the general laws and course of nature in the production of animals. But the case of the hive-bee, stated above, in which a single intercourse with the male fertilizes all the eggs that are laid for the space of two years, and in the case of a common spider mentioned by Audebert[779], for many years, shows that the sperm preserves its vivifying powers unimpaired for a long period, indeed a longer period than is requisite for the impregnation of all the broods that a female Aphis can produce; and if immediate contact with the fluid be not necessary, who can say that this is impossible? It is, however, one of those mysteries of the Creator that human intellect cannot fully penetrate. But this anomaly in nature is not wholly confined to the Aphides; since Jurine has ascertained that the same thing takes place with Daphnia pennata Müll (Monoculus Pulex L.), one[Pg 168] of Branchiopod Crustacea[780]. It is worth observing whether the female Aphides in their natural state, I mean those of the summer or viviparous broods, have intercourse with the male. I think I have noticed males amongst them; but they seem to become most numerous in the autumn, preparatory to the impregnation of the oviparous females. The object of this law of the Creator is probably the more ready multiplication of the species[781].
As to the period of gestation, most insects begin to lay their eggs soon after fecundation has taken place: but in some Arachnida, as the Scorpion, which seems to be both oviparous and ovo-viviparous, nearly a year intervenes, and the eggs increase to four times the size which they had attained at that period, before they are extruded[782]. The time that is required to lay the whole they are to produce, varies also in insects. In this respect they may be divided into two great classes:—those namely which deposit the whole at once, as Ephemerina, Trichoptera, &c., and those which deposit them in succession, occupying in this operation a longer or shorter period. Many in the first class, as the Trichoptera or caseworm-flies, envelope their eggs in a gelatinous substance[783], which renders their extrusion in a mass more easy. Of the second class, which includes by far the[Pg 169] greater proportion of insects, some exclude the whole number in a very short period, others require two or three days or a week, as the cockroach[784]; and others, as the queen-bee, not less than two years. The eggs in the ovaries of the last vary infinitely in size; those that have entered the oviduct have arrived at maturity, while the rest grow gradually smaller as they approach the capillary extremity of the tubes, where they become at length invisible to the highest magnifier[785]. In many insects the eggs seem nearly to have reached their full growth previously to the exclusion of the female from the pupa; and this exclusion and the impregnation and laying of the eggs rapidly succeed each other. One moth (Hypogymna dispar), which is remarkable for the number of eggs she contains, sometimes deposits them, even before they are fecundated, in the pupa-case[786]. But in other cases the sexual union is not so immediate, and some time, longer or shorter, is requisite for the due expansion of the eggs; and the ovaries of the animal swell so much, as often to enlarge the abdomen to an extraordinary bulk: this is seen in a very common beetle (Chrysomela Polygoni) that feeds upon the knot-grass; but in no insect is it so striking as in the female of the white ants, whose wonderful increase of size after impregnation I have related to you on a former occasion[787].
I shall conclude this subject with a few observations upon ovo-viviparous insects; supposed neuters, and hybrids, which, though they do not fall in regularly under any of the foregoing heads, may very well have a place in this letter.
1. It has already been observed that there are a few ovo-viviparous insects[788], the young of which exist in the ovaries at first as eggs, but are hatched within the body of the mother, and come forth in the living form of a larva and sometimes even of a pupa. Of the first description are certain Diptera, the Aphides, and the Scorpion.
Reaumur has described two modes in which the larvæ of the first are arranged in the matrix of the mother. In some they are heaped together without much appearance of order, being placed merely parallel to each other[789]; but in others they are arranged in a kind of riband—the length of the little animals, which are also parallel, forming its thickness—rolled up like the mainspring of a watch[790]. These larvæ in general are not divided into two masses corresponding with the pair of ovaries in other insects, but form only a single one[791]. You must not suppose that these little fetuses lie naked in the womb of the mother; each has its own envelope formed of the finest membrane, which, however, is not entirely divided from that of those adjoining to it, but appears to be one tube, which becomes extremely slender between each individual, so as when drawn out to look like a chain[792]. Reaumur seems to have thought that in these flies the larvæ were never confined in any other case or egg[793]; but De Geer sometimes found eggs in the body of Sarcophaga carnaria, though most generally larvæ, from which he conjectures that it is really ovo-viviparous, the eggs being hatched in the body of[Pg 171] the mother[794]. As these flies are all carnivorous, and their office is to remove putrescent flesh, you may see at one glance the object of Providence in this law of nature—that no time may be lost, and the animal exercise its function as soon as it is disclosed from the matrix.
The Aphides, so fruitful in singular anomalies, are ovo-viviparous, as I have before hinted[795], at one period of the year, that is during the summer, but strictly oviparous at its close. From the experiments of De Geer, however, upon Aphis Rosæ, it would appear that this faculty is not conferred upon the same individuals, but only upon those of different generations of the same species; all the generations being ovo-viviparous except the last, which is oviparous[796]: nor does it appear, as has been sometimes imagined, that it is common to the whole genus. De Geer observed a species in the fir, which makes curious galls resembling a fir cone (Aphis Abietis), which appeared never to be ovo-viviparous[797].
With regard to scorpions, it does not seem clear that they are always ovo-viviparous: M. Dufour twice found in the midst of the eggs nearly mature, a young scorpion which appeared to him at large in the cavity of the abdomen; it was so large that it was difficult to comprehend how it could possibly be excluded from the animal, without an extraordinary operation[798]. The pupiparous insects (Hippobosca, &c.) have been sufficiently noticed before[799].
2. I have already in several of my former letters stated[Pg 172] to you what the modern doctrine of physiologists is with respect to certain individuals, usually forming the most numerous part of the community with insects living in society, that were formerly supposed to be neuters, or as to their sex neither male nor female—that they are in almost every instance a kind of abortive females, fed with a different and less stimulating food than that appropriated to those whose ovaries are to be developed, and in consequence in most instances incapable of conception[800]. Upon these sterile females, you also heard, devolve in general the principal labours of their respective colonies, showing the beneficent design of Providence in exempting them from sexual cares and desires, and meriting for them the more appropriate name, now generally used, of workers. The differences in the structure of the female bee and the workers were also then accounted for; and similar reasoning may be had recourse to with regard to those of ants, in which the worker and the female differ still more materially. My reason for introducing this subject here, is to observe to you that I have some grounds for thinking that this system extends further than is usually supposed, and that to each species in some Coleopterous and other genera there are certain individuals intermediate between the male and female; this I seem to have observed more especially in Copris and Onthophagus. For in almost every British species in my cabinet of these genera I possess such an individual, distinguished particularly by having a horn on the head longer than that of the female, but much shorter[Pg 173] than that of the male. I once observed a pair of Pentatoma oleracea, a very pretty bug, in coitu, both sexes being ornamented with white spots, and by them stood a third distinguished from them by red ones. I do not, however, build on this circumstance, though singular; but mention it merely that you may keep it in your eye. It would be curious should it turn up, that, to answer some particular end of Providence, in some tribes of insects there are two kinds of males, as in the gregarious ones two descriptions of females.
I am, &c.
We have seen upon a former occasion the great variety of movements that insects can perform, and of the external organs with which they perform them[801]: but we are now to consider the internal apparatus, by the immediate action of which they take place—their system of muscles. When we reflect upon the wonderful velocity, their size considered, with which many insects move, and the unparalleled degree of muscular force that many exert[802], we feel no small degree of curiosity to know something of that part of their internal structure that produces these almost incredible effects. I shall in the present letter endeavour in some degree to gratify that curiosity, and give you an account of the muscles of these little animals,—first considering them in general; and then, as far as my information goes, adverting to those in particular that move the different parts and organs of an insect's body.
I. The muscles of insects may be considered in general as to their Origin; Substance and Parts; Shape; Colour; Kinds; Attachment; and Motions.
i. Origin. The origin of the muscular fibre in the higher animals is from the blood, the globules of which, by their coagulation in a series, appear to form it[803]; and in insects it is derived from the same universal source of nutrition and accretion, but not till it has been concreted into the adipose tissue or epiploon before noticed[804]. In the pupa of the cabbage-butterfly, Herold observed that this substance first assumed a fine flocky appearance and a blue-green colour, and that from it so changed were produced tender bundles of muscular fibres, extending in various directions, the epiploon itself decreasing in proportion as they were formed[805].
ii. Substance and Parts. The muscular fibre in vertebrate animals appears to consist of globules arranged in a series, and of no larger diameter than those of the blood,—the mean diameter of which in the human subject, when measured under the microscope by a micrometer, is found to be about 1⁄5000th part of an inch[806]. When Cuvier published his immortal work in 1805, the powers of any magnifier then constructed were not sufficient to enable this great physiologist to arrive at the simple fibre[807]; but Mr. Bauer, by the use of improved glasses, amongst other discoveries that will immortalize his name, was the first to detect, under the directions of Sir E. Home, the ultimate thread of which the muscular bundles are composed[808]. Chemists distinguish the substance[Pg 176] of which we are speaking, by the name of fibrine. By the abundance of azote or nitrogen that enters into its composition, it possesses a character of animalization more marked than any other animal substance; and its elements are so approximated in the blood, that the slightest stagnation causes them to coagulate: and the muscles are without doubt, in the living subject, the only organs that can separate this matter from the mass of blood, and appropriate it to themselves[809]. The primary bundles of muscles are formed of the simple fibres, and the secondary are the result of an aggregation of the primary. The smaller bundles are not always exactly parallel to each other, but must in many cases diverge more or less, to produce those variations in shape observable in the muscles themselves: there are intervals therefore between the bundles, which in some animals are filled by a cellular substance[810]. Probably much of this statement will apply in most instances to the muscles of insects, but we may conclude that the globules that form them are infinitely smaller[811]. Lyonet has given some interesting observations with regard to those of the caterpillar of the Cossus: he describes them as of a soft transparent substance, capable of great extension, covered and filled by silver tubes of the bronchiæ, penetrated by the nerves, and containing oily particles. Each muscle was enveloped in membrane, and was composed of many parallel bands, consisting of bundles of fibres enveloped likewise in separate membranes. The fibres themselves, (but it is doubtful whether he arrived at the ultimate term of muscular fibre,) in a favourable light and under a good magnifier,[Pg 177] appeared to be twisted spirally[812]. In spiders the muscles seemed to him to consist of two substances, the one soft and the other hard, the last forming a kind of stiff twisted filament[813]. A muscle thus composed of different bundles of fibres may be stated as to its parts, in insects, to consist of base, middle, and apex: the base is that part by which they are fixed to any given point of the internal surface of the crust, or of one of its processes, which serves as their fulcrum; the apex is that part by which they are fixed, either mediately or immediately, to the organ to be moved; and the middle is the remainder of the muscle. We usually discover in them no inflation of the middle corresponding with the belly of the muscles in vertebrate animals; they occasionally, however, terminate in a tendon, as those of the thighs and legs; but these tendons are of a different nature from the fibrous ones of warm-blooded animals; for they are hard, elastic, and without apparent fibres: the fleshy ones of the muscle envelope them, and are inserted in their surface[814].
iii. Shape. The muscles of insects are usually linear, with parallel sides; some are cylindrical, as those of the wings of the Libellulina[815]; and others, as those that move the legs in the caterpillar of the Cossus, are triangular[816]. In the suctorious mandibles of the grub of a common water-beetle[817] they are penniform, or shaped like a feather; and some in the Cossus are forked[818]. Under[Pg 178] this head I may also observe, that the muscles are sometimes extremely slender threads, crossing each other, and often curiously interwoven in various directions, so as to resemble lace or fine gauze, as may be seen in the alimentary canal of some caterpillars[819]; sometimes also they surround part of this organ, like a series of minute rings[820].
iv. Colour. The most usual colour of the muscles of insects is white: those for flight however, according to Chabrier, differ from the rest, by being of a deeper and reddish colour[821]; and I have observed likewise that those in the head of the stag-beetle, when dried at least, are red, and look something like the flesh of warm-blooded animals.
v. Kinds and Denomination. In general, muscles may be regarded as divided into primary and secondary—the primary being the muscles by which the principal movements of any organs are effected, and the secondary their auxiliaries which are the cause of subordinate movements[822]. Every muscle almost has its antagonist, the action of which is in an opposite direction; so that when it is equal, the organ to which they are attached remains without motion; but when that of one preponderates, a movement in proportion takes place[823]. The principal antagonist muscles that may be found in insects are the following. 1. Levator muscles that raise an organ, and Depressors that depress it. 2. Flexors that bend an organ, and Extensors that unbend or extend it.[Pg 179] 3. Abductors that draw an organ back, and Adductors that draw it forwards. 4. Constrictors that contract an opening, and Laxators that relax it. 5. Supinators that turn the underside of an organ upwards, and Pronators that return it to its natural situation. Some of these muscles in insects, like some of their articulations and their spinal chord[824], seem to exercise a double function,—thus the levators and depressors of the wings are constrictors and laxators of the trunk[825]. At first it may seem that insects, not having the power of turning up the hand, cannot have the Supinator and Pronator muscles; but some muscle of this kind must be in the Gryllotalpa, and in those that have a versatile head[826].
v. Attachment and Insertion. The attachment and insertion of the muscles in insects in general is to the interior of the crust, or to some of its internal processes as a fulcrum, and to the organ to be moved. In some cases, however, the muscles act upon the organ by the intervention of other bodies. Thus, those that move the wings are often attached to little bones, as Chabrier calls them[827], which are connected with the base of the wings by ligaments. In the Dynastidæ and other Lamellicorns, and the Libellulina, &c., a remarkable provision is made for giving a vast increment of force to the muscles of the wings, by means of caps or cupules surmounted by a tendon, which receive their extremity; the tendon terminating in a fine point attached to the wing, and thus more muscles are brought to bear upon it[828].[Pg 180] Chabrier seems to think that, in some cases, the back that intervenes between each pair of wings is the medium by which the muscles act upon it[829].
vi. Motions. Irritability is the universal distinction of the muscular fibre,—when put in action by the will or involuntarily, it causes it to contract or become shorter; and the intermediate agents of the will and other causes are the nerves, which, as galvanic experiments seem in some degree to prove, are the conductors of an invisible fluid or power which immediately causes that action. If a nerve is divided, the muscles to which it renders obey it no longer, evidently proving that the nerves cause muscular irritability[830]. How this contraction is immediately effected,—whether the fibre, as some suppose, undergoes any crispation, or becomes zigzag[831], or whether there is any sudden change in their chemical composition that rapidly and strongly augments their cohesion, as Cuvier hints[832], cannot be clearly ascertained, unless a Bauer could submit the living fibre to his glasses. All that we know certainly on the subject is, that muscles alternately contract and relax at the bidding of the will or involuntarily, and so occasion all the movements of animal bodies.
II. Having considered the muscles of insects in general, I must next make a few observations, as far as my means of information will enable me, upon those that move their different parts and organs—at least the principal ones; since to descend to minutiæ would be an[Pg 181] endless and unprofitable labour. As larvæ, except those whose metamorphosis is semicomplete[833], differ widely in their system of muscles from perfect insects, I shall begin my observations with them.
We owe by far the most accurate and detailed account of the muscles of larvæ to the illustrious Lyonet, who, with incredible labour and patience without example, dissected the caterpillar of the Cossus, and has described every air-vessel, every nerve, and every muscle that could be detected by the microscope. Cuvier also has given a description of the muscles not only of caterpillars, but of the larvæ of the Lamellicorn beetles, the Hydrophili, and the Capricorn beetles[834]. From these sources are derived what I have now to lay before you. If you look at one of Lyonet's plates[835], the layers of longitudinal muscles look like so many parallel ribands, others run in an oblique, and others again in a transverse direction[836]. He divides them into dorsal, ventral, and lateral muscles[837], terms which sufficiently explain themselves. Of the longitudinal muscles there are four principal rows[838], the others are more numerous. The principal object of these muscles, which are flexors and extensors, is to shorten or lengthen the body, or to act on any particular segment as the circumstances of the animal may require. I shall not here notice the muscles of the head and legs, as they are not remarkably different from those of perfect insects. The prolegs are moved by two muscles—the anterior one covering in part the posterior—of a remarkable structure: one of their points[Pg 182] of attachment is by many branches or tails to the sole of the foot, and by several heads to the skin of the animal; so that they can draw the proleg within the body or push it out, and perform other necessary movements[839].
I shall now call your attention to the muscles of the perfect insect, as they move the head and its organs; the Trunk; the Abdomen; and the Viscera.
i. The Head. This part in insects moves upwards, downwards, inwards, to right and left, is pushed forth or drawn in, is often capable in part of a rotatory movement, and is sometimes versatile, turning as it were upon a pivot. All these movements are of course produced by an appropriate apparatus of muscles, which have their attachment in the anterior part of the trunk, mostly in the manitrunk, while their insertion is in the posterior part of the head, in the margin of the occipital cavity. To enumerate and describe them all would be tedious and uninteresting—I shall only mention some of the principal ones. The levators of the head are usually a pair of muscles situated in the manitrunk, to the upper side of which they are attached, and perhaps in Coleoptera and some others to the phragma, which probably Cuvier means by the anterior part of the scutellum[840]; they are inserted in the posterior margin of the upper part of the head, in Coleoptera in a pair of notches (Myoglyphides[841]), or a single one[842]. In Cordylia Palmarum these muscles as they approach the head, to judge from the dead animal, divide into two branches or a fork:[Pg 183] thus, as the muscle-notches are wide in this insect, the muscle acts upon each extremity of the sinus—these branches appear to be tendinous[843]. The depressors of the head are the antagonist muscles to the above, and have their attachment to the antepectus and its antefurca[844]. A circumstance distinguishes these muscles in many Coleoptera, that seems hitherto to have been overlooked. If you take the common dung-beetle (Geotrupes stercorarius), and carefully extract the head with its muscles from the trunk, you will see on each side of the depressors a subovate corneous scale, of a pitch colour[845], which is attached only to the muscle, and designed to strengthen it: if you then examine the anterior cavity of the manitrunk, you will perceive on each side, just within the lower margin, a minute triangular scale, of a similar substance; these ligaments, like the pax-wax, or ligamenta nuchæ, in mammalia, though in a lower situation, are doubtless intended to sustain the action of the muscles.
With regard to the moveable organs of the head—the antennæ, maxillæ, palpi, tongue, mandibulæ, &c., have each their appropriate apparatus of muscles: but I shall only notice those of the last, the mandibulæ. These are principally abductors and adductors to open and shut them: from the work that the jaws of some insects have to do, you may conjecture that they must be furnished with powerful muscles. In caterpillars and other larvæ, in which state the action of the mandibles is most in requisition, the muscles are what Cuvier calls penniform[846], and are attached on each side to a tendinous lamina or[Pg 184] cartilage. In the grub of Dytiscus the power and magnitude of the adductor muscle is wonderful[847]. In the Orthoptera this structure of the mandibular muscles takes place also in the imago[848]; but in the Coleoptera, at least in the stag-beetle and some others that I have examined, these muscles in this state have no cartilage or tendon. Their attachment is always to the parietes of the head, of the cavity of which the adductors, in some cases, occupy a considerable portion[849]. As to their insertion—these last, in some Orthoptera, enter more or less the interior of the mandible[850]; but commonly they are inserted at or near the interior angle of the mandibular basal cavity, and the abductors at the exterior.
ii. The Trunk. We have little information with regard to the muscles of the parts of the trunk itself, by which, in some insects, the manitrunk is enabled to move independently of the alitrunk: it is more probable that the levators have in part at least their attachment to the anterior surface of the prophragm[851], than that the levators of the head should be there fixed, as Cuvier seems to think; since both the phragma and the ligament that appears in many cases to close the cavity of the manitrunk round the viscera[852], would prevent all communication between those muscles and any part connected with the scutellum: probably the depressors have their attachment partly on the anterior face of the medifurca[853]. These points, however, must be left to future investigators.
With regard to the organs of the trunk, we have more certain and satisfactory information;—the muscles of the legs having been described by Lyonet and Cuvier, and those of the wings most particularly by Chabrier. In caterpillars, the muscles are situated in the interior of the articulations that form the legs: they consist of several bundles appropriated to each, which have their attachment in the parietes of the preceding joint, near the margin, and are inserted in the margin of that they move[854]. Lyonet counted twenty-one muscles in the leg of the caterpillar of the Cossus; but eight of these were appropriated to the claw, or rather formed a pair of semipenniform muscles, having their insertion at the inner angle of its base[855]. In perfect insects, according to Cuvier, each joint of the legs is furnished with a pair of antagonist muscles—a flexor and extensor, the former being the lower, and the latter the upper muscle; and this pair has its insertion in the joint it moves, and its attachment usually in the preceding one: but those of the coxæ—which are rotators, causing it to turn backwards or forwards—and the extensor of the thigh, have their attachment in the parietes of the trunk, and to the endosternum; one of the rotators of the anterior coxa, and the extensor of the anterior thigh to the antefurca; of the intermediate pairs to the medifurca, and of the posterior to the postfurca[856]. Every joint of the tarsus has also its flexor and extensor. In the ground- and water-beetles (Eutrechina and Eunechina), &c., whose posterior coxæ are immoveable, the thigh includes two pair of[Pg 186] antagonist muscles[857]. In extracting the posterior leg of Necrophorus Vespillo I observed more than a single pair of muscles that had their attachment in the coxa; and probably many other variations in this respect exist.
Little was known with respect to the most interesting part of the muscular apparatus of insects, that by which such wonderfully rapid and varied motions are imparted to their organs of flight, till Chabrier undertook to elucidate it; which he has done in a manner that will confer a lasting honour upon his name, as one of the most able successors to Swammerdam and Lyonet in their peculiar department. He has given a most admirable account of the internal anatomy of the trunk of insects in general, as far as it relates to their flight; particularly of that of the cockchafer (Melolontha vulgaris), of one of the Libellulina (Æshna grandis), and of a bumble-bee (Bombus); and I believe he has thus illustrated insects of some of the other Orders, but his memoirs on these I have not had an opportunity of consulting. What I have to say on this subject, therefore, will be principally derived from what he has communicated with respect to the above insects.
A considerable difference in the volume of the muscles of the wings takes place in insects according to the force of their flight. Where it is rapid and powerful, the alitrunk is nearly filled by them, and the alimentary canal is much attenuated; but in those whose flight is feeble, they occupy less space, and the alimentary canal is proportionally enlarged[858]. In the Lepidoptera, Hymenoptera and Diptera, the principal muscles of both[Pg 187] wings have their attachment in the anterior portion of the alitrunk[859]; in the Coleoptera, in the posterior[860]; and in the Libellulina, those of the anterior wings are confined to the anterior portion, and those of the posterior pair to the posterior[861]. The muscles for flight in general differ from others by their mass, length, and colour; the bundles of fibres are very distinct, strong, and parallel; their direction is uniform, according to the motion they are to produce; their fibres are either attached to the solid parts to be moved, or to cupules, but they never terminate in a tendon; the muscles are perfectly independent of each other, and the wings can be moved by them separately[862]. As to their denomination and kind—the principal ones are the levators and depressors, which with respect to the trunk, as was before observed, are constrictors and laxators. The levator muscles form several distinct bundles in Coleoptera, Lepidoptera, &c.; in the Diptera there are three[863]; in the Libellulina they seem to be single, are all environed with a blackish pellicle, with numerous aërial vesicles, symmetrically arranged, filling the interstices[864]. The most common number is a levator to each wing; there are often, however, as in the cockchafer and the dragon-fly, two depressors[865]: but in the Hemiptera, Lepidoptera, and saw-flies (Serrifera) amongst the Hymenoptera, the secondary wings have distinct levators, but not depressors[866]; the other insects of that Order have only a pair of each[867].[Pg 188] The other wing-muscles are of a secondary description, and auxiliary to the above. Their office is to extend and close the wings: so that though the denomination of extensor will suit the former, that of flexor is not so proper for their antagonists; their office being not so much to bend, as to bring back the wing to its station of repose. The folding of certain wings, as those of Coleoptera, Dermaptera, the Vespidæ, &c., seems more the function of the abdomen than of the wing-muscles; this you may easily see, as I have often done, if you attend to any Staphylinus, when after alighting from flight it proceeds to fold up its wings under the elytra. Perhaps the term retractor might not be inapplicable to the muscles in question. Both these and the extensors are usually small slender muscles, but sometimes numerous[868]. They are larger in the Coleoptera, Lepidoptera, and saw-flies[869]. The muscles that open and shut the elytra of Coleoptera, and probably of Heteropterous Hemiptera, and which also aid their movements during flight, are very slender[870]. With regard to the attachment and insertion of the wing-muscles, it is according to two very distinct types, one of which appertains to insects in general, and the other is peculiar to the Libellulina. In insects in general, the principal muscles for flight have not their insertion in the wings, but act upon their bases by the intervention of small long pieces. The depressors occupy the middle and upper region of the alitrunk, and are inserted anteriorly and posteriorly upon the concave surfaces of two transverse horny semi-partitions, adapted by their elasticity to dilate the trunk—and thus acting[Pg 189] the part of both diaphragm and ribs[871]: but in the Libellulina, as in birds, these muscles are placed on each side of the point of support of the humerus[872]; the depressors being attached immediately to the wings without it, and the levators within it, with this sole difference, that they are connected to the internal extremity of the base of the wing by the intervention of a cupule terminating in a tendon; all are disposed perpendicularly to the arms of the levers on which they act, and all incline more or less outwards, the one to dilate, and the other to contract the trunk[873]. It may be observed in general, that in insects formed upon the first type, the great action of these muscles is the dilatation and contraction of the alitrunk, the main tendency of which is to depress and raise the wings[874]. I shall add here a few words upon the attachment of the wing-muscles in the different Orders: but first I must request you to read what I have said on the partitions and chambers of the alitrunk in a former letter[875]. In most insects of the first type, the depressors are longitudinal dorsal muscles that have their posterior point of attachment in the metaphragm (costale Chabr.); but the anterior varies:—in those that have elytra, tegmina, or hemelytra, the muscles for them seem to be contained in the chamber, varying in size, that lies between the prophragm and mesophragm; and the anterior point of attachment of their depressor muscles is the mesophragm: they are also attached in some to the metathorax or back of the posterior portion of the alitrunk[876].[Pg 190] The levator muscles in Coleoptera, at least in the cockchafer, by a long tendon have their posterior attachment in the lower part of the posterior coxæ[877], their anterior attachment to the solid parts to be moved. In the Cockchafer and the Dynastidæ, but not in Geotrupes, on each side of the cavity of the metathorax under the base of the wing is a large and small cupule, which from their lateral situation one would think must receive the levator muscles—apparently unnoticed by M. Chabrier; but as there is a pair of these cupules on each side, there must have been also a pair of muscles attached to them, which does not agree with his statement[878]. In the Hymenoptera and Diptera the anterior attachment of the depressors is to the back of the alitrunk and to the prophragm, and the levators to the breast, and the sides of the back of the trunk[879]. In the Libellulina the depressors and levators that terminate, by a tendon surmounting a cupule, in the base of the wings, have their posterior attachment in the breast. These cylindrical muscles with their cupule and tendon look like so many syringes[880].
Having thus described to you the powerful muscular apparatus by which, either mediately or immediately, the wings of insects are moved, it will not be out of place if I add a few words upon their flight itself. The great object in this is to generate a centrifugal force which may counteract the weight of the body. Its wings are the external organs by which the insect as it were takes hold of the air when they fall, and is impelled by it when[Pg 191] they rise; its head makes way for it; its abdomen, as a rudder, steers it; and by alternately increasing and diminishing in volume, and rising and falling, enables it to win an easy way through the fluctuations of the atmospheric sea. The trunk by its elasticity admits the internal action of antagonist muscles, which by turns compress and dilate it; an action promoting the elevation and depression of the wings, and keeping up the elasticity of the internal air, which is thus now rarified and now condensed: in the former state flowing like a tide, accompanied by the blood, into the nervures of the wings[881], and thus increasing their tension and centrifugal force;—in the latter ebbing and receding to the trunk, thus relaxing the one and diminishing the other. The spiracles by which the air enters or is expelled, open and shut at the animal's pleasure[882]; and besides, many insects are furnished, as we have seen[883], with numerous vesicles or reservoirs, which can give out a supply of internal air when wanted: and thus they can vary their aërial motions, diminish or increase the counteracting centrifugal force; rise and fall, and move onwards and in different directions, as their occasions demand.
iii. The Abdomen is perhaps capable of the greatest variety of motions of the three primary sections of the body. Even when the insect is reposing, a constant dilatation and contraction usually takes place in it[884]; and from its annular structure, its parts capable of separate motion are numerous:—it expands and contracts; it[Pg 192] rises and falls; it bends in various directions; and its segments can often be lengthened or retracted. Besides all this, its spiracles open and shut, and its reproductive and other anal organs have their appropriate motions. In numerous Coleoptera, however, and some Hemiptera, the upper-side of the abdomen is almost the only part that is moveable, especially near the trunk; the under-side, having its first segments soldered together, is only capable of motion near the tail[885]. The muscles that produce the various motions of this part must be entitled to all the denominations stated above[886]. I have on a former occasion explained to you how, in insects that have a petiolate abdomen, that part is elevated and depressed[887]. In those with a sessile one the base is attached to the metaphragm by strong ligaments[888], and the muscles that move the first piece act from one segment to another. The partial movements of the segments of this part, where they have place, are produced by muscular fibres which extend from the whole anterior margin of one to the whole posterior one of that which precedes it. If those, for example, of the back contract, the abdomen becoming shorter above, bends upwards; and if those of the sides or belly, it bends sideways or downwards[889]: this is a beautiful as well as simple contrivance.
The alternate rush of air from the abdomen into the alitrunk, and from the atmosphere into the abdomen, is attended by the constriction or expansion of that part[Pg 193] as it rises or falls in flight[890], which seems to require the action of constrictor and laxator muscles.
iv. The Viscera. Having before had occasion sufficiently to notice the muscles by which the systole and diastole of the dorsal vessel of insects is maintained[891], I shall now only mention those that are woven round their alimentary canal, by which the peristaltic motion of that organ, causing its contractions and the propulsion of its contents, takes place. One would at first think that a view of the intestines of any animal could under no circumstances afford any very pleasing spectacle to the eye of any but a scientific spectator; but any lady who is fond of going to Disons to be tempted with an exhibition of fine lace, would experience an unexpected gratification could she be brought to examine those of a caterpillar under a microscope: with wonder and delight she would survey the innumerable muscular threads that in various directions envelope the gullet, stomach, and lower intestines of one of these little animals; some running longitudinally, others transversely, others crossing each other obliquely, so as to form a pattern of rhomboids or squares; others again, surrounding the intestine like so many rings, and almost all exhibiting the appearance of being woven, and resembling fine lace,—one pattern ornamenting one organ; another, a second; and another, a third. This will suffice to give some idea of this part of the muscular structure of these little animals[892].
Lyonet counted the muscles contained in the body of the caterpillar of the Cossus. In the head he found 228; in the body, 1647; and enveloping the intestines,[Pg 194] no less than 2186; which, after deducting 20 that are common to the gullet and the head, gives a total of 4061[893]. In the human subject only 529 have been counted[894]: so that this minute animal has 3532 muscles more than the Lord of the creation!
The muscles of the Arachnida seem less numerous than those of insects. In the Scorpionidea they appear to be robust, formed of simple straight fibres, of a whitish gray colour: a muscular web, rather strong, clothes the parietes, but rarely adheres to them, of the abdomen, and envelopes the viscera, with the exception of the lungs, and probably of the heart. The dorsal part of this web gives birth to seven pairs of filiform muscles, which traverse the liver, and are attached to a muscular riband which, passing above the lungs, runs the whole length of the ventral parietes. These muscles when exposed to view resemble extended cords. The abdominal segment preceding the tail is filled with a powerful muscular mass which moves that organ[895]. Treviranus discovered two longitudinal muscles in Scorpio europæus, running from the breast to the tail, which above and below each gill were connected by another running transversely across the heart, thus forming a quadrangular area in which the gills are situate[896]. The heart appears to be moved by muscles not very dissimilar to those of the Cossus[897], as is likewise that of the Araneidea; in Clubiona atrox the wider part of this organ is muscular, and incloses a considerable cavity[898]. In this tribe the muscles of the[Pg 195] abdomen, the skin of which is soft and unfit to act as a lever to them, are attached to a cartilage, and thus their action is better sustained[899].
Having thus laid before you all of importance that I can collect with regard to the apparatus of muscles discoverable in insects, I shall next say something upon a few other points connected with that subject. When I enlarged upon their motions, I related a few instances of the extraordinary power of that apparatus[900] in leaping ones; but this power is not confined to that circumstance. The flea, not more remarkable for its compressed form, enabling it to glide between the hairs of animals, and its elastic coat of mail, by which it can resist the ordinary pressure of the fingers, than for its muscular strength, has attracted notice on this account from ancient times. Mouffet relates that an ingenious English mechanic, named Mark, made a golden chain of the length of a finger, with a lock and key, which was dragged by a flea;—he had heard of another that was harnessed to a golden chariot, which it drew with the greatest ease[901]. Another English workman made an ivory coach with six horses, a coachman on the seat with a dog between his legs, a postillion, four persons in the coach, and four lacqueys behind—which also was dragged by a single flea. At such a spectacle one would hardly know which most to admire, the strength and agility of the insect, or the patience of the workman. Latreille mentions a flea of a moderate size dragging a silver cannon on wheels, that was twenty-four times its own weight, which being charged with powder, was fired[Pg 196] without the flea appearing alarmed[902]. Many caterpillars are accustomed to extend their bodies from a twig, supported merely by the four hind feet, in one fixed attitude, either in an oblique, horizontal, or vertical direction, either upwards or downwards, and that for hours together. We may conceive what prodigious muscular force must be exerted upon this occasion, by reflecting that the most expert rope-dancer, though endued with the power of grasping with his feet like a bird with its claws, could not maintain himself in a horizontal position even for an instant. Bradley asserts that he has seen a stag-beetle carry a wand half a yard long and half an inch thick, and fly with it several yards[903]. Some insects have the faculty of resisting pressure in a wonderful degree. If you take a common dung-chafer (Geotrupes) in your hand and press it with all your strength, you will find with what wonderful force it resists you; and that you can scarcely overcome the counteraction, and retain the insect in your hand: was it not for this quality, the grub of the gad-fly must be crushed probably in passing through the anal sphincter of the horse[904]. But that of Eristalis tenax affords a more surprising instance of this power of counteraction:—an inhabitant of muddy pools, it has occasionally been taken up with the water used in paper-making, and strange to say, according to Linné, has resisted without injury the immense pressure given to the surrounding pulp[905]; like leather-coat Jack mentioned by Mr. Bell[906], who, from a similar force of[Pg 197] muscle, could suffer carriages to drive over him without receiving any injury. Almost as remarkable is the state of extreme relaxation into which the muscles of some larvæ fall, when their animation is suspended; and the revived tension to which a subsequent resumption of the vital powers restores them. Bonnet having suspended the animation of the caterpillar of Sphinx Ligustri by keeping it submerged, squeezed it between his fingers, until it had wholly lost its cylindrical form and was as flat and supple as the empty finger of a glove; yet in less than an hour the very same caterpillar became as firm, as compact, as cylindrical, and in short, as well, as though it had never been submitted to treatment so rough[907].
It is fortunate that animals of a large size, as has been well remarked, especially noxious ones, have not been endowed with a muscular power proportionable to that of insects. A cockchafer, respect being had to their size, would be six times stronger than a horse; and if the elephant, as Linné has observed, was strong in proportion to the stag-beetle, it would be able to pull up rocks by the root, and to level mountains[908]. Were the lion and the tiger as strong and as swift for their magnitude as the Cicindela and the Carabus, nothing could have escaped them by precaution, or withstood them by strength. Could the viper and the rattlesnake move with a rapidity and force equivalent to that of the Iulus and Scolopendra, who could have avoided their venemous bite? But the Creator in these little creatures has manifested his Almighty power, in showing what he could have done had he so willed; and his goodness in not creating the[Pg 198] higher animals endued with powers and velocity upon the same scale with that of insects, which would probably have caused the early desolation of the world that he has made. From this instance we may conjecture, that after the resurrection, our bodies by a change in the structure and composition of their muscular fibre—for we know that their locomotive powers and organs, as far as the muscle is concerned, will then be of a very different nature[909]—may become fitted for motions and a potent agency of which we have now no conception.
This wonderful strength of insects is doubtless the result of something peculiar in the structure and arrangement of their muscles, and principally their extraordinary power of contraction, excited by the extent of their respiration: for animals that respire but little, as the fœtus in the womb and the pullet in the egg, have very little contractile muscular power[910]. To get some idea from facts of this extraordinary contractile power in insects,—extract the sting of a bee or a wasp, with its muscles, which appear to be attached to powerful cartilaginous plates[911], and you will find it continue for a long time to dart forth its spicula, almost as powerfully as when moved by the will of the animal. A still more extraordinary instance of irritability is exhibited by the antlia, or instrument of suction of the butterfly. If this organ, which the insect can roll up spirally like a watchspring or extend in a straight direction, be cut off as soon as the animal is disclosed from the chrysalis, it will continue to roll up and unroll itself as if still attached[Pg 199] to its head: and if after having apparently ceased to move for three or four hours it be merely touched, it will again begin to move and resume the same action. This surprising irritability and contractility of muscle doubtless depends upon the peculiar structure of the antlia, which is composed of an infinite number of horny rings, acted upon by muscles, more numerous probably than those which move the trunk of the elephant. The motion only ceases when the muscles become dry and rigid.
I have already, under another head[912], considered the annual sleep, or winter state of torpidity of insects, during which an intermission for the most part of muscular motion and action takes place. I shall now make a few observations with respect to their diurnal sleep, which may very properly have its place in the present letter. That insects, usually so incessantly busy and moving in every direction, require their intervals of repose, seems to call for no proof. We see some that appear only in the day, and others only in the night, others again only at certain hours; which leads to the conclusion, that when they withdraw from action and observation, it is to devote themselves to rest and sleep. The cockchafer flies only in the evening; but if you chance to meet with it roosting in a tree in the earlier part of the day, you will find it perfectly still and motionless, with its antennæ folded and applied to the breast:—we cannot indeed say that its eyes are shut; for as insects have no eyelids, that sign of sleep can never be found in them. Again, if a Lepidopterist goes into the wood to capture moths in[Pg 200] the day-time, he finds them often perched on the lichens that cover the north side of the trunk of a tree, with their wings and antennæ folded, and themselves without motion, and insensible of his approach and their own danger. Thus it was that I captured that rare insect the lobster-moth (Stauropus Fagi) in the New Forest. Some, however, have asserted that the caterpillar of the silkworm, except when they moult, never intermits feeding day or night, and consequently does not sleep: but the accuracy of this statement, both from analogy and observation, admits of great doubt. Malpighi informs us that these caterpillars for an hour and more, twice a day, remain immoveable with their heads bent down as though asleep, and even if disturbed, resume again the same inactive posture[913]; and other larvæ in great numbers certainly seem to have regular intermissions from eating of considerable duration: those called Geometers, for hours together remain motionless projected from a twig, to which they adhere by their posterior prolegs alone; and the processionary caterpillars make only nightly sorties from their nests, passing the day in inaction and repose[914]. Bees have been often seen by Huber, when apparently wearied with exertion, even in the middle of the day, to insert the half of their bodies into an empty cell, and remain there, as if taking a nap, without motion for half an hour or longer[915]; and at night they regularly muster in a state of sleep-like silence. Mr. Brightwell once observed an individual living specimen of Haltica concinna, which appeared to remain motionless on the same spot of a wall for three successive days.
Before concluding these remarks on the Internal Anatomy and Physiology of Insects, I shall explain to you, as you will probably feel inclined occasionally to pursue the subject, the best mode of dissecting them.—By far the most useful dissecting instruments for this purpose are very fine-pointed and sharp scissors, as these will enable you to divide the integument and separate other parts with much less risk of injuring their delicate structure than any knife. These scissors are what Swammerdam chiefly used; and he had some so extremely small and fine, that he was necessitated to employ a lens when he sharpened them. If to these be added a sharp and fine-pointed knife or two, some needles fixed in handles, also fine-pointed—(you will find them more convenient than any other instrument for detaching minute parts and fibres,) a pair of fine and accurately adjusted pliers, and an assortment of camel's-hair brushes,—you will be nearly set up as an Entomological dissector. You will still, however, require a small dissecting table, with a projecting and moveable arm for lenses of various descriptions, so as to admit both the hands to be employed upon the subject under examination; and for this purpose probably no contrivance can be better adapted than that of Lyonet, of which the figure in Adams On the Microscope will convey a better idea than any description[916].
Previously to dissecting any insect, it must be killed by plunging it into boiling water, which is recommended by Lyonet, or spirits of wine or of turpentine; and it is often useful to let larvæ remain a few days in the latter,[Pg 202] by which means the vessels become firmer and stronger. The parts of pupæ become much more distinct if they are boiled for a few minutes: and the same mode may be adopted in the examination of spiders.
The most convenient mode of proceeding, which was that also of Lyonet, is to dissect the insect in water, or, to avoid putridity, in diluted spirits,—if small, upon a concave glass, to which it should be fastened by means of a little melted wax; if larger, in the bottom of a common chip box, surrounded with a border of wax to retain the fluid. The integuments of the insect, being carefully divided longitudinally with scissors, should if flexible be turned back, and fixed by small pins stuck in by a fine pair of pliers, while the skin at the same time is stretched by another. After making such observations as present themselves without further dissection, the viscera must be cautiously extracted, washing away the fat which surrounds them with spirits of turpentine, in which it is soluble, applied by camel's-hair pencils. After separation they may conveniently be examined by putting them into water, and gently shaking them so as to cause the parts to unfold. If endowed with the patience of Swammerdam, you may even arrive at injecting these minute parts with wax or coloured fluids, conveyed by delicate glass tubes having one end as fine as a hair, which he also employed to fill the viscera with air; and afterwards drying them in the shade, and anointing them with oil of spike in which a little resin had been dissolved, he succeeded in preserving them. If it is not convenient to finish the dissection of an insect at once, it should be covered with spirits of wine. Swammerdam found a mixture of spirits and distilled vinegar[Pg 203] very useful for keeping caterpillars previously to dissecting them, as it consolidated the parts[917].
And now having brought to a close my long wanderings in this ample and intricate field, and having threaded, as well as my slender powers and limited knowledge enabled me, the infinite turnings and convolutions of this Dædalean labyrinth—the Anatomy and Physiology of insects,—will you not own that the volume of wonders I have laid before you proves irrefragably that, though these minims of nature apparently rank so low in the scale of being, yet in their structure, instead of being, as might be expected, more simple, they are infinitely more complex and highly wrought than those animals that are placed the nearest to ourselves? the Creator in the latter doing every thing by a beautiful simplicity; while in the former, the more to magnify his power and skill, because they afford no apparent space for it, by a wonderfully curious and intricate multiplicity: and whether we study the one or the other, we shall in both trace the footsteps of that adorable Love which has shown attention to the comfort and well-being of the lowest insect, as well as of the highest of his creatures.
I am, &c.
Having laid before you what observations I thought might sufficiently explain all the principal features of the Anatomy of insects both external and internal, you will next expect to be informed whether, like the higher animals, they are subject to have the admirable order observable in their frame interrupted by Disease; and you will perhaps imagine, from the multiplicity of their organs and vessels, that they must be peculiarly exposed to derangements of the vital and other functions. That they have their diseases is certain; but, except in the case of their appropriate parasitic assailants, which is a part of their economy, it does not appear that their maladies are more numerous and frequent than those of other animals. The same Almighty Power which endowed them with so complex a structure, generally upholds them in health during their destined career, until they have fulfilled the purpose of their creation, when they die and return again to their dust[918].
But perhaps I may seem to you as making too great a parade about these little insignificant creatures if I assign a separate letter to the consideration of their diseases: but when you recollect that Aristotle has a chapter on[Pg 205] this subject[919], and that the learned Willdenow has devoted a distinct portion of his excellent introductory work on Botany to the diseases of Plants[920],—you will perhaps be of a different mind: indeed, some facts I shall have to communicate are so remarkable and interesting, that I am sure, when you have read this letter, you will not think the subject one that deserves to be slighted.
Insect diseases may, I think, be divided into two great classes; those resulting, namely, from some accidental external injury or internal derangement, and those produced by parasitic assailants.
I. Under the first head we may begin with wounds, fractures, mutilations, and other extraneous causes of disease. To these—insects are peculiarly subject; and though they are not, like the Crustacea and Arachnida[921] and some other invertebrate animals, endowed with the power of reproducing a mutilated limb, yet their wounds appear to heal very rapidly, and at the time they are inflicted to produce little pain[922]. But if those important members, their antennæ, are mutilated, insects seem to suffer a kind of derangement; the great organ of their communication with each other, and in various respects with the external world, being removed, all their instincts at once fail them. I formerly related how the amputation of these affects the queen-bee[923]. A similar result, as Huber[Pg 206] tells us[924], follows, when the same experiment is repeated on the workers or drones: they immediately become unable to take any further part in the labours of the hive; they can no longer guide themselves except in the light; if they petition one of their fellow-citizens for honey, they are unable to direct their tongue to its mouth to receive it; they remain near the entrance of the hive, and when the light is intercepted they rush out of it to return no more.
Insects occasionally are subject to tumours or a preternatural enlargement of their parts and organs. The antennæ of bees sometimes swell at their extremity so as to resemble the bud of a flower ready to open, becoming at the same time very yellow, as does the fore part of the head[925]. I once saw a specimen of a Hydrobius—agreeing with H. fuscipes in every other respect even to the most minute punctum—which had a large tumour on each side of the prothorax, evidently accidental, occasioned probably by the stoppage of the pores by which the superfluous moisture and air escape when it undergoes its last change. The converse of this I have observed to take place sometimes in the same part of Geotrupes foveatus, the ordinary lateral foveæ becoming very considerably enlarged;—this was the case with the specimen from which Mr. Marsham made his description of that insect. The species is, however, very distinct in other respects, and may always be known by its small size. It happens now and then also, that these tumours represent blisters. I saw one once on one elytrum of a beetle and not on the other. Those of Serropalpus (as[Pg 207] Mr. MacLeay, on the authority of M. Clairville, informs me) are particularly subject to this disease. But, of all the organs, the wings are most exposed to derangements of this kind. De Geer, in a specimen of Pieris Cratægi just excluded from the chrysalis, observed that one of these was distended by a considerable quantity of extravasated green fluid—two or three large drops following an incision. This disease appeared to arise from the lower membrane not adhering to the upper; so that the nervures—which are rather longitudinal channels, being open below, than tubes—were not closed to confine the fluid to its proper course. The malady, which might be called a dropsy of the wing, carried off the insect the day after its exclusion[926]. Reaumur observed that the wings of some flies were affected by an air-dropsy, as he calls it, which appeared to arise from the air escaping from its natural channels, and thus separating, the two membranes that form the wing, and filling the cavity produced by their separation[927].
Sometimes also monstrosities are to be met with in these animals, or variations from a symmetrical structure in organs that are pairs. I have a beetle in which the terminal joint of one of the maxillary palpi is short, ovate, and acute; and that of the other, long, semiovate, and rather obtuse. A specimen of Blaps mortisaga in my cabinet, taken by Mr. Denny, besides the terminal mucro of the elytra, has a long diverging lateral one. Goeze had the larva of a Semblis brought to him in which one of the two fore-legs, though perfect in all its parts, was only half the length of the other[928]; which he regarded[Pg 208] as a reproduction, but it seems rather a malformation. Müller mentions a most extraordinary fact of one of the Noctuidæ, which when disclosed from the pupa retained the head of the larva[929]. One of the most remarkable instances of this kind that have fallen under my own observation, may be seen in a specimen of Chrysomela hæmoptera in the cabinet of our friend Curtis; in which one of the thighs produces a double tibia, but only one of these is furnished with a tarsus.
The diseases of insects which arise from some internal cause are not very numerous. The first that I shall mention is a kind of vertigo. "Ants have also their maladies," says M. P. Huber: "I have noticed one extremely singular; the individuals attacked by it lose their power of guiding themselves in a straight line, they can walk only by turning round in a circle of small diameter and always in the same direction. A virgin female shut up in one of my glasses was seized on a sudden with this distemper; she described a circle of an inch in diameter, and made about a thousand turns in an hour, or not quite seventeen in a minute. She continued constantly turning round for seven days, and when I visited her in the night I found her still in motion. I gave her honey—and I think that she ate some of it." He observed that some workers were attacked by a similar disease: one of these, however, had the power of walking from time to time in a straight line; when placed upon its head it continued its gyrations[930]. Similar motions of a little moth, mentioned on a former occasion[931], may perhaps have been produced by the same cause. Bees are also[Pg 209] subject to vertigo, which has been attributed to their eating poisonous honey[932]—but may not this disease in all these cases arise from some derangement of the nervous system? One of the ants which was so affected had lost one of its antennæ; but as this was not the case with the others, no great stress is to be laid upon the circumstance. Huber does not inform us whether those attacked by this disease recovered or not.
I have observed more than once, that the flesh-fly and some others of the same tribe are subject in particular seasons to a kind of convulsions. When thus attacked, they kick and struggle, and seem unable to fly. Sometimes they lie upon their backs without motion, but if a finger be placed near them their convulsive motions are renewed. When thrown into the air, instead of flying, they fall to the ground. Had this distemper occurred earlier or later in the year I should have attributed it to the benumbing effects of cold; but as my observations were made one year (1816) in May, and in another (1811) in the latter end of June, this could scarcely be the case. In the year last mentioned I observed that many flies died under its influence. In wet seasons this tribe is subject to another disease, which proves fatal to many of them, and indeed to other Diptera. A white crust appears to be formed upon the abdomen both above and below, of a granular appearance, much resembling fine moist sugar. On the back of that part this crust does not cover the margins of the segments, which gives it the appearance of white bands; so that deceived by it, I have often at first flattered myself that I had met with[Pg 210] some new species. The under-side of the abdomen is wholly covered by it, divided in the middle into two longitudinal masses, the anal segment being bare. De Geer has noticed this or a similar disease, which, when flies are attacked by it, causes the abdomen to swell so as even to burst, and the segments become dislocated. Upon opening the abdomen it is found filled with a white unctuous substance, which often accumulates (as above described) on its external surface[933]. Dr. Host says that in this disease when the animal is dead, the wings, which were before incumbent, become extended, and its almost invisible pubescence grows into long hairs[934]. De Geer seems to think that these flies are thus affected in consequence of having eaten some poisonous food[935]; but I rather suspect, as I have observed it become prevalent chiefly in wet seasons, that it arises from a superabundance of the nutritive fluid, or of the fat, so that it seems to be a kind of plethora. I once observed a fly fixed to a pane of glass, round which was a semicircle of what appeared to be merely vapour, whose radius was nearly three-fourths of an inch. Taking it for an aqueous fluid that had transpired from the dead animal, I paid no further attention to it at that time: but observing from day to day that the moisture did not evaporate, after two or three months had elapsed, I had the curiosity to examine it more closely, and, upon scraping some of it off with a penknife, I found it was a white substance of a fatty nature. In this case, then, the fat must have exploded on all sides with considerable violence from half the body or the abdomen. Probably this was[Pg 211] a more intense degree of plethora. When I examined this appearance the fly had fallen off, and I could not find it.
Mr. Sheppard once brought me a panicle of grass, the glumes of which were rough with hairs, or small bristles, to which several specimens of a fly related to Xylota pipiens adhered by their proboscis. At first I thought that having been entrapped by the bristles, and unable to extricate themselves, they had perished from want of food; but since when touched they readily dropped from the glumes, some other cause, perhaps disease, probably occasioned this singular suspension of themselves.
The maladies to which bees and silkworms are subject are more interesting to us than those of flies, on account of their utility as cultivated insects. One of the worst distempers which attacks the first of these animals is a kind of looseness or dysentery: this happens early in the year, when they are fed with too much honey without any portion of bee-bread[936], and sometimes destroys whole hives. Their excrements, instead of a yellowish red, then become black, and the odour they emit is insupportable; the bees no longer observe their usual neatness, inducing them to leave the hive when they void their excrements, but they defile it, their cells, and each other. Several remedies have been prescribed for this disease. To prevent it, a syrup made by an equal mixture of[Pg 212] good wine and honey is recommended; and as a cure, to place in the hive combs containing cells filled with bee-bread[937]. But one of the worst maladies to which these useful animals are subject, is that called by Schirach Faux Couvain. It originates with the larvæ; and is caused either by their being fed with unwholesome food, or when the queen, as sometimes happens, lays her eggs so that the head of the grub is not in a proper position for emerging from the cell when the period for its disclosure is arrived:—the consequence is, that in both cases it dies and becomes putrid, which sometimes produces a real pestilence in a hive. The remedy for this evil is to cut away the infected combs, and to make the bees undergo a fast of two days[938]. The hive should be cleaned and fumigated, by burning under it aromatic plants.
The cultivators of the silkworm in France have given names to several diseases to which that animal is subject. One is called La Rouge, and is supposed to be occasioned either by too great heat, or by too sudden a transition from cold to heat. It takes place when the caterpillar is first hatched; which lives perhaps, but in a very sickly state, till it should spin its cocoon and assume the pupa, when it expires. Another degree of the same disease is called Les Harpions or Passis. A second distemper of this animal is Des Vaches, Le Gras or La Saune: this is a mortal disease, supposed to be of a putrid nature, and produced by mephitic air; it shows itself after the second moult, but rarely after the subsequent ones. When a caterpillar is first attacked, changing the air may prove a remedy; but when the disease has made[Pg 213] progress, it is best to burn or bury them, since if the poultry pick them up they might be poisoned by them. A third disease of silkworms is called Les Morts Blancs, or Tripes, which is also occasioned by impure air, when the leaves the animal feeds upon are heaped so as to produce fermentation. The caterpillars attacked by it die suddenly, and preserve after their death the semblance of life and health. Too great heat, whether artificial or natural, occasions La Touffe, a fourth, which, when the heat continues long, destroys all those that are arrived at their last stage of existence in their larva state. Black points scattered over different parts of the body, or livid and blackish spots in the vicinity of the spiracles, followed by a yellowish or reddish tint, are symptoms of a fifth malady, called La Muscardine. After this the animal soon dies, and becomes mouldy, but does not stink. This disease is not contagious, and is thought to be caused by a moist heat, attended by pernicious exhalations. La Luzette, Luisette, or Clairène, is another malady, which shows itself most commonly after the fourth moult. It seems to arise from some original defect in the egg. The caterpillars attacked by it may be known by their clear red and afterwards dirty white colour; their body becomes transparent, and the matter of silk exudes in drops from their spinnerets; consequently, though as voracious as the rest, they are never able to construct a cocoon, and should be destroyed. Les Dragées is the name given to cocoons which include a larva that never becomes a pupa. The cause of this disorder has not been ascertained, and whole broods are sometimes subject to it, which, as in the last, seems to imply some defect in the eggs. But as the caterpillar spins its cocoon, and the silk is as good as usual,[Pg 214] it is a malady of no great importance. Lastly, sometimes the mulberry leaves have a gummy rather acrid secretion, which purges the silkworms; their excrement is no longer solid; they become weak and languid; and if the secretion is abundant, their transpiration is impeded, and at the time of moulting they are become so feeble as to be unable to cast their skin[939].
In the case of many caterpillars of Lepidoptera that died, Bonnet found by dissection that the disease was remotely occasioned by a diarrhea, which taking place immediately before they became pupæ, prevented the inner membrane of their intestines from being rejected, as it would have been if no extraordinary cause had prevented it, attached to the hard excrement. He found this membrane converted into a jelly occupying great part of the stomach, which he conjectured was the proximate cause of their death[940].
To conclude this head—spiders are reputed to be subject to the stone: I do not say Calculus in Vesica; but we are informed by Lesser that Dr. John Franck having shut up fourteen spiders in a glass with some valerian root, one of them voided an ash-coloured calculus with small black dots[941].
II. I now come to that class of diseases which appears to prevail almost universally amongst insects—I mean those resulting from the attack of parasitic enemies. Thus millions and millions annually perish before they have arrived at their perfect state. Diseases of this kind proceed either from vegetable or animal parasites. I shall begin with the first, which will not occupy us long.
i. As insects pass often no small portion of their life[Pg 215] in a state of torpidity, in which they remain chiefly without motion, it will not seem wonderful, should any partial moisture accidentally accumulate upon them, that it affords a seed plot for certain minute fungi to come up and grow in. Persoon observes with regard to his genus Isaria, that one species grows upon the larvæ of insects (I. truncata), and another upon pupæ (I. crassa[942]):—as he does not say upon dead larvæ and pupæ, as upon a former occasion[943], perhaps in these cases these plants may constitute an insect disease; but I lay no stress upon it, and only mention the circumstance here as connected with the history of these animals. Mr. Dickson has described a Sphæria under the name of entomorhiza that grows upon dead larvæ; it has a slender long stipes and spherical granulated head: on the pupa of a species of Cicada in my cabinet, another kind of Sphæria, with a twisted thickish stipes and oblong head, springs up in the space between the eyes. I observed something similar but longer, in the grub of some large beetle in M. Du Fresne's museum at Paris; and I have a memorandum of having noticed something of the kind on the rostrum of a Calandra. Bees and humble-bees have been sometimes thought to have some species of mucor or other Fungilli occasionally growing upon them; but Mr. Brown is of opinion that stamina which they have filched from flowers have been mistaken for these Fungilli, since he has detected those of Orchideæ in some of this tribe, and upon a beetle shown to him by Mr. MacLeay, one which he knew to be the stamen of an Aristolochia. I once observed a bunch of what I mistook for a singular mucor that adorned the vertex of a humble-bee, between the antennæ, which doubtless were[Pg 216] of the same description; and I even saw one upon its wing. Upon a former occasion I mentioned a parallel circumstance with respect to a species of Xylocopa[944].
ii. The animal parasites that infest insects are either themselves insects; or worms.
1. Their insect infesters, as far as we know at present, are confined to the Orders Strepsiptera, Hymenoptera, Diptera, and Aptera: they attack them sometimes in their egg state, most frequently when they are larvæ, occasionally when pupæ, and very rarely in their perfect state. Upon many of these I have formerly enlarged[945], and I shall now add such further circumstances as I then omitted. The Strepsiptera Order, as at present known, consists only of two genera, Stylops and Xenos; the first being appropriated to the imago of Andrena, a kind of bee, and the latter to that of the wasps. Their eggs appear to be deposited in the abdomen of these insects in which they feed, till having attained their full growth they perforate the membrane that connects its segments; and at the proper time their pupa-case bursts, they emerge, and take their flight. Sometimes four or five infest a single bee. Whether the latter dies upon their quitting it I have not been able to ascertain, but from their flying, when the little parasite is very near leaving them, with their usual activity, it should seem that this disease is not mortal; but it probably prevents their breeding: I do not recollect observing the exuviæ of one in a male bee[946].
The great body of insect parasites, however, belong to the Hymenoptera Order, and chiefly to the Linnean genus Ichneumon. The insects of this order have been[Pg 217] denominated Principes, because of the wonderful instincts of ants, wasps, bees, and other gregarious tribes that belong to it; and they merit a name of honour not less for the benefits that they confer upon mankind, by keeping within their proper limits the various insect-destroyers of the produce of the globe. It deserves notice that when these latter increase to a degree to occasion alarm, their parasites are observed to increase in a much greater, so as to prevent the great majority of them from breeding[947]. Though these benefactors of the human race constitute numerous genera, at present not well ascertained, I shall speak of most of them under the common name of Ichneumon.
The appearance of these little four-winged flies puzzled much the earlier naturalists:—that a caterpillar usually turning to a moth or butterfly should give birth to myriads of flies, was one of those deep mysteries of nature which they knew not how to fathom[948]: even the penetrating genius of our great Ray, though he ultimately ascertained the real fact[949], was at one time here quite at fault; for he seems at first to have thought, when from any defect or weakness nature could not bring a caterpillar to a butterfly, in order that her aim might not be entirely defeated, that she stopped short, and formed them into more imperfect animals[950].
Before I detail more particularly the proceedings of Ichneumons, I shall make a few general remarks upon them. The structure of the instrument by which they are enabled to deposit their eggs in their appropriate station has been before sufficiently described[951]; it is long[Pg 218] or short according to the situation and circumstances of the larva which receives them: if this lives in the open air, and the access to it is easy, it is usually short and retracted within the body; but if it lies concealed in deep holes or cavities, or shuns all approach, it is often very long. Thus in Pimpla Manifestator, which commits its eggs to the grub of a wild bee inhabiting the bottom of deep holes bored in posts and rails, the ovipositor is nearly an inch and half in length, and in some extra-European species three inches. How the egg is propelled so as to pass in safety from the oviduct, along this extended and very slender instrument to the grub for which it is destined, has not been certainly ascertained: but from an observation of Reaumur's[952] it should seem that it is aided in its passage by some fluid ejected at the same time with it, or is so lubricated as to slide easily without being displaced. The flies we are speaking of, by some authors are called Muscæ vibrantes, because when searching for the destined nidus of their eggs their antennæ vibrate incessantly, and it is by the use of these wonderful organs that they discover it wherever it lurks. Bergman observed that Fœnus Jaculator searches for the latent grub of certain bees and other Hymenoptera with its antennæ[953]: and from Mr. Marsham we learn that Pimpla Manifestator, before it inserts its ovipositor in the nest of the grub of Chelostoma maxillosa, explores it first with one antenna and then with the other, plunging them all the while intensely quivering up to the very root[954]. With respect to their size, Ichneumons vary greatly; some being so extremely minute as to be invisible to the naked eye, unless moving upon glass; while others, as to[Pg 219] their length, emulate the giants amongst insects. The former, unless appropriated to the eggs themselves, usually commit many eggs to a single larva, while the latter are directed by their instinct to introduce into them only one. Some of the former description are endowed with the faculty of leaping[955]. The food of Ichneumons, and indeed of other internal parasites, is chiefly the epiploon or fat of the larva, but they never touch any vital organ; so that it continues to feed, and probably more voraciously, grow, cast its skin, and often it changes to a chrysalis, although at the same time inhabited by an army of these little devourers.
Ichneumons, as far as has been at present ascertained, are parasitic upon other insects chiefly in their three first states, a solitary instance only having been observed of their inhabiting an imago; but from their first exclusion as eggs from the ovary till their assumption of that state they give them no rest. I shall therefore first treat of those that infest the eggs; next those appropriated to larvæ; and lastly those that devour pupæ.
Vallisnieri appears to have been the first naturalist who discovered that Ichneumons were appropriated to the eggs of other insects. He observed one proceed from those of the emperor-moth (Saturnia Spini): finding two holes in each egg, one larger than the other, he conjectured that one was made when it entered, and the other when it emerged. In this case the egg of the Ichneumon must be fixed on the outside of the egg it[Pg 220] was to feed upon; though some appear to pierce it with their ovipositor, and consequently introduce their egg within: for he says afterwards; "I have seen with my own eyes a certain kind of wild flies deposit their eggs upon other eggs, and bore and pierce others with an aculeus—by which they have introduced the egg[956]." Count Zinanni, a correspondent of Reaumur's, saw an Ichneumon pierce the eggs with her ovipositor repeatedly; which in about fifteen days were filled with the pupa, and in six more produced the imago[957]. I. Ovulorum L. is the only known species of egg-devourers; but most likely there are many, varying in size, according to the size of the egg they inhabit. Probably I. Atomus L., and I. Punctum Shaw, are of this description[958]. It is wonderful what a number these little flies destroy:—out of a mass of more than sixty eggs which was brought to De Geer, not one had escaped the Ichneumon[959]. But the most extraordinary thing is, that even these little creatures we are told are destroyed by another still more minute[960].
Though the animals we are speaking of usually destroy only a single egg, yet some appear not so to confine themselves. Geoffrey informs us that the larva of one of the Ichneumons whose females are without wings (Cryptus) devours the eggs of the nests of spiders, and from its size—it is nearly a quarter of an inch long—it must require several of them to bring it to maturity[961]. One of those also which destroys the gnat infesting the wheat (I. inserens) appears to devour them in[Pg 221] their egg state, and could not be brought to perfection by the food that a single one would furnish[962].
The Ichneumons that are parasitic upon larvæ are the most numerous of all. Some of them are deposited by the parent fly on the outside of their prey, and others introduced into its interior. Ophion luteum is one of the former tribe; it plants its eggs in the skin of the caterpillar of the puss-moth (Cerura Vinula). Each egg is furnished with a footstalk terminating in a bulb[963], which is so deeply and firmly fixed that it is impossible to extract it without detaching a portion of the animal with it, and even when the caterpillar changes its skin it is not displaced. After it is hatched, the grub, while feeding, keeps its posterior extremity in the egg-shell, to which it adheres so pertinaciously, that it is scarcely possible to disengage it without crushing it. It fixes itself by its mandibles to the skin of the caterpillar, and keeps constantly sucking the contents of its body till it dies: sometimes nine or ten of these larvæ inhabit a single caterpillar[964]. Reaumur has given an account of other external Ichneumons. Upon one caterpillar that he examined, they were so numerous as to render the poor animal quite a spectacle, and they underwent their metamorphosis attached to it[965]. One species of this description avenges the cause of insects upon their most pitiless foes, the all-devouring spider—for in the midst of her toils and lines of circumvallation it makes her its prey. De Geer, meeting one day with a young spider of a common kind, observed with surprise, engaged in sucking it, a small white grub, which was firmly attached to the abdomen near the trunk. Putting it by in a[Pg 222] glass, after some days he examined it again; when he observed that it had spun the outline of a vertical web, had stretched threads from the top to the bottom of the glass and from one side to the other, and had also spun the radii that meet in the centre, and this was all;—but what was remarkable, the larva that had fed upon it was suspended in the centre of this web, where it was engaged in spinning its own cocoon, while the spider, exhausted by this last effort, had fallen dead to the bottom of the glass. It cannot be asserted positively that this suspension of the larva of the Ichneumon in the centre of the web always takes place; but if it does, as seems most probable, it shows that this little parasite is endowed with an instinct which causes it so to act upon the spider as may induce it to spin a web so nicely timed as to be sufficiently complete at the period of its death and of the change of the Ichneumon, for the latter to cast it down and assume its station[966].
But the great bulk of the parasitic Hymenopterous devourers of larvæ have their assigned station within the body. As Entomologists in breeding insects have paid their principal attention to Lepidoptera, it necessarily follows that their Ichneumon infesters must be most generally known; but doubtless the larvæ of the other Orders are not wholly liberated from this scourge: they also require to be kept within due limits, and have their appropriate parasites. Some, however, in most of them have been detected: of which I shall now proceed to state to you the most interesting examples, beginning with the Coleoptera.
Alysia Manducator[967], remarkable for having mandibulæ[Pg 223] that do not close, and toothed at the end, usually attends masses of dung, both of man and cattle, probably for the purpose of depositing its eggs in some of the Coleopterous larvæ that inhabit it. Mr. Stephens, one of the most accurate observers as well as one of the best Entomologists of the present day, informs me that he once captured three specimens of Timarcha tenebricosa, from each of which forty or fifty minute Ichneumons emerged. An insect also of this Order, that is a great benefactor to mankind, as a destroyer of the plant-lice,—I mean the lady-bird (Coccinella), in its larva state is itself subject to the attack, as we learn from De Geer, of one of these small parasites[968]. He detected them also in that of two species of weevils: and in the pupa of some large grub of a beetle inhabiting the wood of the elm, perhaps that of the stag-beetle, he found the pupa of one of those Ichneumons that have an exserted ovipositor[969]. Doubtless, did we know their history, we should find that numberless species have their internal assailants belonging to this tribe.
Orthopterous larvæ seem not to have been yet announced as affording a pabulum to these animals: but the late Dr. Arnold, whose tact for observation with regard to the manners and economy of insects has rendered his loss irreparable, discovered that the remarkable parasitic genus Evania was appropriated to the all-devouring Blatta. Whether it attacked it in its egg or larva state I have not been informed. This little benefactor is here extremely rare, at least in the country; perhaps in towns, where the cock-roach abounds, it may be more common.
The observations of naturalists have chiefly been confined to the Hemipterous genus Aphis; but these early attracted their notice. Leeuwenhoek has given a particular and entertaining account of the proceedings of I. Aphidum. As soon as the little flies approached their prey, they bent their abdomen, which is rather long, between their legs, so that the anus projected beyond the head; then with their ovipositor they pierced the body of the Aphis, at the same time carefully avoiding all contact with it in every other part: whenever they succeeded in their attempt, a tremulous motion of the abdomen succeeded. Only a single egg is committed to one Aphis: when hatched, the latter becomes very smooth and appears swelled; it is, however, full of life, and moves when touched. Those that are thus pricked separate themselves from their sound companions, and take their station on the underside of a leaf. After some days the inclosed grub pierces the belly of the Aphis, and attaches the margin of the orifice to the leaf by silken threads; upon this it dies, becomes white, and resembles a brilliant bead or pearl[970]. De Geer observed also an Ichneumon on the Coccus of the elm, I. Coccorum[971].
Amongst the Neuropterous tribes likewise, probably the Ichneumonidæ commit their usual ravages; but their exploits, as far as I recollect, have met with no historian. I have a small species related to Chelonus, which a memorandum made when I took it tells me was obtained from Æshna viatica; yet I do not remember ever tracing that species to its final change, so that I must have taken this Ichneumon from the perfect insect. It suffices, however, to prove that this tribe is also exposed to the attack[Pg 225] of these parasites. Where larvæ and pupæ are aquatic, it seems probable, if any attack is made upon them, that it must take place after they have quitted the water.
In the Hymenoptera Order itself, almost every genus has been ascertained to have its Ichneumon parasites. Not even the fortified habitations of the gall-flies (Cynips) can escape them, almost every species becoming their prey; a circumstance which puzzled not a little some of the older naturalists, when they at one time saw a fly not remarkable for its colours or brilliancy emerge from the curious moss-like Bedeguar of the wild rose, and at another were struck by the appearance of one of those splendid minims of nature which almost dazzle the sight of the beholder[972]. Immunity, however, from this pest seems to have been granted to the gregarious Hymenoptera; at least none has yet been discovered to attack the ant, the wasp, the humble-bee, or the hive-bee; in which last, had there been one appropriated to it, it could never have escaped the notice of the Reaumurs and the Hubers. The solitary bees, however, as we have seen above[973], do not escape; and Epipona spinipes, a solitary wasp which feeds its own young with a number of green caterpillars[974], is itself, when a larva, though concealed in a deep burrow, the prey of the grub of an Ichneumon, which by means of a long ovipositor introduces its egg into its body[975]. Even these parasites, whose universal office it is in their first state to prey upon insects, are themselves subject to the same malady. Ichneumonidan devourers are kept in check by other[Pg 226] Ichneumonidan devourers. These in some cases are so numerous as to destroy the tithe of the kinds they attack[976]. Thus an ever-watchful Providence prevents these parasites from becoming so numerous as to annihilate in any place the species necessary for the maintenance of the general economy and proportion of animal and vegetable productions. Amongst the assailants of the Hymenoptera, none seem to have a more laborious task assigned them than those that pierce the various galls in which the larvæ of the Cynips tribe are inclosed. To look at an oak-apple, we should think it a work of difficulty, requiring much sagacity and address, for one of our little flies to discover the several chambers lurking in its womb, and to direct their ovipositor to each of them. Its Creator, however, has enabled it instinctively to discover this, and furnished it with an appropriate elongated instrument, which will open a way to the deep and hidden cells in which the grubs reside, penetrate their bodies, and to each commit an egg. When it prepares to perforate the gall, the Ichneumon begins by depressing this organ, that it may extricate it from its sheath; it next elevates its body as high as possible, and bending the instrument till it becomes perpendicular to the body and to the gall, so as to touch the latter with its point, it then gradually plunges it in, till it is quite buried[977]. A very remarkable Hymenopterous parasite (Leucospis), which when unemployed turns its ovipositor over the back of its abdomen, so that its end points to its head, is said to deposit its eggs in the nest of the mason-bee, most probably in the larva: but the curious observations that are stated to have been[Pg 227] made by M. Amédée Lepelletier upon its history have not yet been given to the public[978].
Dipterous insects, likewise, do not escape from these pests of their Class: but few observations, however, have been recorded as to the species assailed by them. We learn from De Geer, that a gnat (Cecidomyia Juniperi), which forms galls upon the juniper is devoured by an external Ichneumon[979]; that which injures the wheat in the ear, whose ravages I formerly mentioned to you[980], affords food to three of these parasites,—one I lately mentioned as probably devouring its eggs; another pierces the glumes of the floret, where its destined prey is concealed; and the third enters it. I once placed a number of the larvæ of the gnat upon a sheet of paper, at no great distance from each other, and then set down one of these last Ichneumons in the midst of them. She began immediately to pace about, vibrating her antennæ very briskly: a larva was soon discovered, upon which she fixed herself, the motion of her antennæ increasing intensely; then bending her abdomen obliquely under her breast, she inserted her ovipositor, and while the egg was depositing these organs became perfectly motionless. The larva when pricked gave a violent wriggle. This operation was repeated with all that had not already received an egg, for only one is committed to each larva. I have often seen it mount one that was already pricked, but it soon discovered its mistake, and quitted it untouched[981]. The Hessian Fly also (Cecidomyia Destructor) related to the preceding, whose alarming ravages I formerly described to you[982], has a peculiar parasite[Pg 228] attached to it, which keeps it in check. The only other Dipterous insects that I have seen mentioned as affording pabulum to an Ichneumon, are—one of the aphidivorous flies mentioned by De Geer, who does not note the species, to the larva of which the Ichneumon commits only a single egg, producing a grub that intirely devours its interior[983];—and two described by Scopoli, one, the larva of a fly frequenting hemp; and the other, which feeds on a Boletus, that of a gnat[984].
The Lepidoptera, however, is the Order over the larvæ of which the Ichneumons reign with undisputed sway; attacking all indiscriminately, from the minute one that forms its labyrinth within the thickness of a leaf, to the giant caterpillar of the hawk-moth. The most useful of all, however, the silkworm, appears at least with us, exempted from this scourge. De Geer, out of fifteen larvæ that were mining between the two cuticles of a rose-leaf, belonging to the first tribe here alluded to, found that fourteen were destroyed by one of these parasites, only one coming forth to display itself in all its brilliancy and miniature magnificence[985]. One of the most useful to us is that which destroys the clothes-moth, which the same writer also traced[986]. Another, equally serviceable, takes up its abode in the caterpillar that ravages our cabbages[Pg 229] and brocoli (Pontia Brassicæ) which perish by hundreds from its attacks. As this falls frequently under our notice, it will not be uninteresting to give a fuller account of it. Reaumur has traced and related its whole history. One of these little flies that he observed, was so intent upon the business in which she was engaged, that she suffered him to watch her motions under a lens, without being discomposed. She pursued nearly the same plan of proceeding with that of the Ichneumon of the wheat-gnat just described; except that she repeated her operations frequently on the same caterpillar in different parts, alternately plunging in and extracting her ovipositor. She seemed to prefer the spot where the segments of the body are united, particularly where the eighth meets the ninth, and the ninth the tenth. When the fly had completed its work and quitted the caterpillar, Reaumur gave it food, and it did not seem less lively and vigorous than others of its kind; in less than a fortnight it assumed the pupa; and in four days the whole of its interior being devoured, it died: but its parasites, perhaps not finding a sufficient supply of nutriment in it, never came to perfection[987]. Sometimes, however, these little grubs arrive at maturity before the caterpillar has become a chrysalis, when they pierce the skin and begin to emerge. First appears a little white tubercle, which gradually elevates itself in a direction perpendicular to the body; while this is doing, a second appears in another place; and so on, till fifteen or sixteen are seen on each side, giving the caterpillar a very grotesque appearance. By the alternate contraction and relaxation of their bodies the grubs effect their complete liberation, which takes[Pg 230] place with respect to the whole in less than half an hour. When entirely disengaged, they place themselves close to the sides of the caterpillar: even before this they begin spinning, and draw unequal threads in different directions, of which they form a cottony bed, which serves as the base of the separate cocoon of each individual, which they next construct of a beautiful silk thread of a lovely yellow, which, if it could be unwound and in sufficient quantity, would yield a silk unrivalled in lustre and fineness[988].
De Geer has recorded a very singular fact which deserves your notice. An Ichneumon, appropriated to one of the Tortrices, had deposited its eggs in two of their caterpillars; each produced a considerable number; but those that emerged from one were all females, and those from the other, males[989]. He observed a similar fact take place with Misocampus Puparum[990]. One might conjecture from this circumstance, that as in the queen-bee[991], so in these Ichneumons, the eggs producing the two sexes were arranged separately in the ovaries. Reaumur has related, that in one instance three or four males were produced to one female; and in another four or five females to one male[992].
But though the great majority of insects are subject to this Scolechiasis[993] in their larva state, yet sometimes they are not attacked by the Ichneumon till they have become pupæ. Of this kind is one just mentioned (M. Puparum), which commits its eggs to the chrysalis of the butterfly of the nettle, Vanessa Urticæ: the moment this caterpillar[Pg 231] quits its skin to assume that state, while it is yet soft they pierce it and confide to it their eggs[994]. De Geer and others have supposed that this same Ichneumon attacks the Cocci and Coccinellæ[995]; but this probably is an erroneous supposition. Cryptus Compunctor also attacks the pupæ of butterflies[996].
If we consider the great purpose of Providence in giving being to this tribe of destroyers—the keeping of insects within their proper limits,—we may readily conceive that this purpose is more effectually answered by destroying them in their preparatory than in their ultimate state, since at that time the laying of their eggs and a future progeny could not so effectually be prevented;—this will account for there being few or no Ichneumons appropriated to them in their latter state.
The next tribe of insect parasites are to be found in the Diptera Order. The species that has been particularly noticed as such is the Tachina Larvarum; its larva is polyphagous, laying its eggs upon the bodies of caterpillars of different kinds. Sometimes a pair is placed on the first segment, sometimes on the head itself, and sometimes near the anus. These eggs are very hard, convex, of an oval figure, polished and shining like a mirror. They are fixed so firmly that if you attempt to remove them with a penknife the skin comes off with them. When hatched, they enter the body and feed on the interior, and, undergoing their metamorphosis within it, do not emerge till they enter their perfect state. The caterpillar thus attacked lives long enough to spin its cocoon, when it dies[997]. Sometimes, however, these animals[Pg 232] quit their prey sooner. Reaumur saw a grub of one of the Muscidæ come out of a caterpillar, and then become a pupa, which was so large that he wondered how it could have been contained in the animal it had quitted[998].
We have now done with those parasites that produce in insects the disease I have called Scolechiasis[999]: the rest, which belong to the Aptera Order, will afford us examples both of Phthiriasis and Acariasis[1000].
I begin with the first. Mr. Sheppard once brought me a specimen of a bird-louse (Nirmus) which he took upon a butterfly (Vanessa Io): and should such a capture be more than once repeated, it would afford a certain instance of the first of these diseases amongst insects;—but most probably the specimen in question had dropped from some bird upon the butterfly. The only remaining animal belonging to the apterous hexapods that is parasitic on insects, is by many supposed to be the larva of a giant-beetle (Meloe Proscarabæus). I have before alluded to this animal[1001], and shall now resume the subject. Gœdart, Frisch, and De Geer, observed that it deposited in the earth one or two considerable masses, containing an infinite number of very minute orange-coloured eggs adhering to each other, which in about a month were hatched, and produced a number of small hexapods distinguished by two pairs of anal setæ and a proleg, by means of which they could move readily upon glass, as I have myself seen: these little animals precisely corresponded with one found by the latter author upon[Pg 233] Eristalis intricarius; and when that fly was placed amongst them, they immediately attached themselves to it, so as to leave no doubt of their identity[1002]. A congenerous species had been detected upon wild bees, and described by Linné under the name of Pediculus Apis. De Geer is so thoroughly to be depended upon for his veracity and accuracy of observation, that we cannot suppose there is any incorrectness in his statement. If the mass of eggs be, as he represents it, of the size of a hazel-nut, it must have been the product of a very large insect: in confirmation of this opinion it may be further observed, that the larva of the kindred genus Cantharis agrees with it in having anal setæ, though it appears to differ in having only two conspicuous segments in the trunk[1003]. Those which infest wild bees make their first appearance upon acrid plants, which the Meloe likewise feeds upon; from whence with wonderful agility they leap upon the Andrenæ, &c. that visit these flowers. Strong, however, as all these facts appear, still we cannot help exclaiming with the illustrious Swede last named, Who could ever have imagined that the larva of this great beetle would be found upon the body of flies,—and we may add, or bees? Who could ever imagine that it would feed like a bird-louse and resemble it so closely? that in the insertion of its palpi it should exhibit a character exclusively belonging to that tribe[1004]? Another circumstance seems to indicate that these hexapods at the time that they take their station in bees or flies are perfect insects—they do not vary in size, at least not materially. Where, we may also ask, if they are to become[Pg 234] large beetles, where do they take their principal growth? It cannot be as parasites on the little bees or flies that they are usually found upon; they must soon desert them, and like their kindred blister-beetles, as is most probable, have recourse to vegetable food. What an anomaly in rerum natura! It is much to be wished that some skilful insect-anatomist would carefully dissect the Meloe; or perhaps by digging round the roots of the ranunculuses and other acrid plants the larva of that beetle might be discovered in a later stage of growth, and so this mystery be cleared up. I should observe here, that Scopoli has described three parasites as Pediculi; viz. P. rostratus, coccineus, and Cerambycinus; the first of which Fabricius has adopted under the name of P. Gryllotalpæ, but which are all evidently hexapod Acarina[1005].
Acariasis seems a disease almost as universal amongst insects as Scolechiasis; with this difference however, that Acari most commonly take their station upon them in their perfect state. You have doubtless often observed the common dung-beetles (Geotrupes) covered on the underside of their body with small mites (Gamasus Coleoptratorum) which look as if they were engaged in suction—they are often so numerous that no part is uncovered; they also attack other beetles[1006], and are sometimes found on humble-bees. They are easily disturbed, run with great swiftness, and may often be seen in hotbeds and fermenting dung prowling in search of the stercorarious beetles. But the most remarkable insect of this kind is the Uropoda vegetans: it derives its nutriment from the insects it assails not by its mouth, but[Pg 235] by means of a long anal pedicle by which it is attached to them. De Geer found these in such numbers upon a species of Leptura, that its whole body was almost covered with them; they hung from the legs and antennæ in bunches, and gave the animal a most hideous and disgusting appearance. Under this load of vermin it could scarcely walk or move, and all its efforts to get rid of them were in vain: many were attached to its body and to each other by their anal pedicles, but others had cast them off and were walking about. When put into a glass with earth, they began to abandon their prey, so that in a few days it was quite freed from its plagues. He found that these parasites lived long in alcohol[1007].
If you inquire—How are these mites originally fixed by their pedicles? it seems most probable, that as the Hemerobii, when they lay their eggs, know how to place them upon a kind of footstalk, so the parent Uropoda has the same power; and this pedicle appears to act the part of an umbilical chord, conveying nutriment to the fœtus not from a placenta, but from the body of the insect to which it is attached; till having thus attained a certain maturity of growth and structure, it disengages itself and becomes locomotive. Many eggs of the aquatic Acarina (Hydrachna, &c.) are also furnished with a short pedicle by which they are fixed to Dytisci and other water insects. De Geer found some of this description on the underside of the water-scorpion, so thickly set as to leave no void space: they were oval, of a very bright red, and of different sizes on different individuals; whence it was evident that they grow when thus[Pg 236] fixed: when hatched or released—for perhaps they may be regarded as fœtuses in their amnios rather than eggs—they cease to be parasitical. Let us admire on this occasion, (piously observes this great Entomologist,) the different and infinitely varied means by which the Author of Nature has endowed animals, particularly insects, for their propagation and preservation: for it is a most extraordinary sight to see eggs grow, and pump as it were their nutriment from the body of another living animal[1008]. As these mites are fixed to the crust as well as its inosculations, they must have some means of forcing their nutriment through its pores.
Another insect, remarkable for its resemblance in some respects to the scorpion—called in this country the book-crab (Chelifer cancroides), from its being sometimes found in books—occasionally is parasitic upon flies, especially the common blue-bottle-fly (Musca vomitoria). They adhere to it very pertinaciously under the wings; and if you attempt to disturb them, they run backwards, forwards, or sideways, with equal facility.
Spiders also are infested by mites. Mr. Briggs once found a very small Theridion, to the thorax of which were attached four oblong bright scarlet mites, each of which was as large as the thorax itself. He afterwards met with another spider still smaller, attacked by two of these swoln parasites, one of which appeared to him nearly equal to the spider in size. This mite was probably either Leptus Phalangii, or Astoma parasiticum.
2. We now come to a perfectly distinct tribe of insect parasites, which belong to that section or order of[Pg 237] intestinal worms which Rudolph has denominated Entozoa nematoidea, and Lamarck Vers rigidules[1009]. To this tribe belong the Gordius of Linné and the Filaria of modern zoologists, which from the experiments and observations of De Geer, Dr. Matthey, &c. appear to have been too hastily separated, being really congenerous, and living indifferently in water and in the intestines of insects and other animals[1010]. To this genus belong the guinea-worm (Gordius medinensis[1011]), the Furia infernalis, and several others that are found in various vertebrate animals. These little worms have been discovered in insects of almost every Order; and their attack generally produces the death of the animal, though they appear not to devour those parts that are essential to life[1012]. I once took a specimen of Pœcilus azureus, and upon immersing it in boiling water I was surprised to see what at first I mistook for an intestine, thrust itself forth; but upon a nearer inspection, to my great surprise I found it was one of these worms, thicker than a horse-hair and of a brown colour. Mr. W. S. MacLeay also once found one in Abax Striola. It still remains in my specimen, making it appear as if it had a long tail. De Geer long ago found these worms in grasshoppers[1012]; but Dr. Matthey has given the fullest account of one which infested Acrida viridissima. A friend of his noticing one of these insects which had not strength enough to leap and could scarcely even walk,[Pg 238] being struck with the circumstance, caught the animal, upon which its hind legs were immediately detached from it. His surprise was greatly increased when he saw issue from its body a cylindrical worm about two feet and a half in length. Upon being called, Dr. M. soon recognised it for a Gordius or Filaria; and on his putting it into water, it moved in it with great velocity, twisting its long and slender body in all directions. Upon opening the body of the grasshopper, nothing appeared within it but the intestine shrunk up to a thread. A few days after, another was brought, which appeared in full vigour, but its abdomen was enormously distended, and from it another worm was extracted, which remained without motion rolled in a spiral direction: intending to preserve this in spirits of wine—as it had become flat he first immersed it in water, that it might recover if possible its cylindrical form. Upon immersion a movement took place in the animal, and it gradually recovered its plumpness; but it still remained without motion, as if dead, for nearly five days, when another living specimen being brought and placed with it, as soon as water was poured on them, the seemingly dead one began to show by a slight oscillation in its extremities that life was not extinct in it. Fresh water being poured upon it, at the end of the day it had recovered all its strength and agility. He afterwards often repeated the same experiment with a similar result[1013]. From this account it appears that the Gordius or Filaria has a property resembling that of the Vibrio Tritici, so well described and so beautifully figured by M. Bauer[1014], of apparently dying and being resuscitated by immersion in water.[Pg 239] How long it can retain this property remains to be ascertained.
De Geer states that he had seen them of the length of two feet[1015]; but they vary considerably in this respect. In ants, in which Gould detected them, he states their length to be not more than half an inch[1016]. In caterpillars, which they sometimes infest, they are longer; in that of Notodonta Ziczac, De Geer found one three inches and a half long[1017]; and Rösel three, of six inches, in that of Deilephila Euphorbiæ[1018]; and in Phalangium cornutum, according to Latreille, they extend to more than seven inches[1019]. In the larva of a Phryganea L. the author first named found one which was more than a foot long, corresponding exactly with the Gordius aquaticus of Linné; being forked at one extremity, brown above, gray below, and black at each end[1020]. These animals appear to die as soon as they leave the body[1021] they have preyed upon; except this happens in water, when their activity has no repose. In this element they give their bodies every possible inflexion, often tying themselves in knots in various places, interlacing and twisting themselves in a hundred different ways; so that when confined in the body of an insect, from their extreme suppleness and power of contortion they find sufficient space wherein to pack their often enormous length[1022]. Linné makes one of their habitats clay; and Mr. W. S. MacLeay finds them very common at Putney in clay at the bottom of pools.
Dr. Matthey asks—How does the Gordius get into Acrida viridissima[1023]? And De Geer—Why do they die after having quitted a caterpillar? and where do they perpetuate their species[1024]? These questions, without further observations, cannot easily be answered. However, it may be supposed that carnivorous insects, such as Harpali, &c. may swallow them when found apparently dead in clay, where the water has been evaporated, or when they have been ejected by other insects; and they may revive in their bodies, as Dr. Matthey found them to do in water. It is not difficult to conjecture that the larvæ of Phryganeæ may meet with them when young in the water, and sometimes unluckily swallow them with their food. Why they become as dead when they emerge from their prey we cannot at present conjecture; but no doubt to answer some wise purpose;—in rainy seasons they probably revive and get into little hollows full of rain-water. Upon De Geer's last question—How they perpetuate their species—at present I can offer no conjecture.
I am, &c.
At first one would think that the senses of insects might be described in very few words, and scarcely afford matter for a separate letter; but when we find that physiologists are scarcely yet agreed upon this subject, and that the use of some of their organs, which appear to be organs of sensation, has not yet been satisfactorily ascertained—we shall not wonder that it requires more discussion than at the first blush we were aware of. In treating on this head I shall first say something on the senses in general, and then confine myself to those of insects.
Touch, taste, smell, hearing, and sight, I need not tell you, is the usual enumeration of the senses: but as the term includes every means of communication with the external world, the list perhaps might be increased; and there is ground for thinking that the number seven, so signalized as a sacred number[1025], may also here have place. Dr. Virey, an eminent physiologist, whose sentiments on various subjects I have before noticed with approbation[1026], appears to be of opinion that there are[Pg 242] really seven senses; which he divides into those that are altogether physical, and those that are more connected with the intellect. The first of these divisions contains four senses,—touch, love, taste, and smell;—the second three,—hearing, sight, and the internal sense of thought, or the brain[1027]. That he is right in adding love to the list seems to me evident, because it is as distinct from touch, as smelling and taste are. With regard to the other, though it may be expected that there should be a transitive sense connecting the intellect (if I may so speak) with the external organ of sense, and as a medium by which the former can receive the notices of the external world furnished by the latter; yet it seems improper to make the entire brain itself a sense. We know that the agent between the common sensory and the sense is the consciousness or perception of the impression. "Seeing we may see and not perceive, and hearing we may hear and not understand." The picture may be painted upon the retina of the eye, the sound may strike upon the tympanum of the ear; but neither the one nor the other be received by the intellect, unless the internal power or faculty of perception be in action and mediate between them. This is what I mean by the internal sense, which, to use a term of Mr. W. S. MacLeay's[1028], is osculant between intellect and sense, or forms the transit from one group of powers to the other.
Of the ordinary senses, sight holds the first rank: it[Pg 243] can dart to the region of the stars, and convey by the perceiving sense, to the sensory, ideas of innumerable objects. Next in rank is hearing, which can receive sounds from a great distance; but the ideas it remits are confined only to one object, the variations of tones. In the other organs the sensitive power is much more confined. There is another difference between the intellectual and physical senses:—the former are the only ones that receive and convey sensations of the beautiful and sublime; of harmony and discord,—the latter, though they minister more to our sensual enjoyments, add little to our intellectual; and therefore too devoted an indulgence in them debases our nature, and levels us with the brutes, which use their eyes and ears only for information, not for pleasure[1029].
In man the ordinary five senses are usually in their greatest perfection, although in some animals particular senses have a greater range. The Vertebrates in general are also gifted with the same number, though there are some exceptions. But in the Invertebrates they are seldom to be met with all together in the same object. The Cephalopods have no smell. Several Gasteropods can neither hear nor see. The animals of bivalve shells have neither eyes, nor ears, nor smell; and the zoophytes and the races below them have, it is affirmed, only the single sense of touch, which in them is so extremely delicate as to be acted upon even by light[1030].
Not so our insects. These, there is good reason to[Pg 244] believe, possess all the ordinary senses. That they can see, touch, taste, and smell, no one denies. Linné and Bonnet, however, thought them deprived of hearing[1031]; but numerous observations prove the contrary. That they hear in their larva state, is evident from facts stated by the latter physiologist. He found that the sound of his voice evidently affected some caterpillars; which he attributes, but surely without reason, to the delicacy of their sense of touch: at another time, when some caterpillars of a different species were moving swiftly, he rang a small bell; upon which they instantly stopped and moved the anterior part of their body very briskly[1032]. That they possess this faculty in their imago state is confirmed still more strongly by facts. I once was observing the motions of an Apion under a pocket microscope: on seeing me it receded. Upon my making a slight but distinct noise, its antennæ started: I repeated the noise several times, and invariably with the same effect. A Harpalus, which I was holding in my hand, answered the sound in the same manner repeatedly. Flies, I have observed, at brisk and distinct sounds move all their legs; and spiders will quit their prey and retire to their hiding places. Insects that live in society give notice of intended movements, or assemble their citizens for emigration by a certain hum[1033]. But the most satisfactory proof of the hearing of these animals is to be had from those Orthoptera and Hemiptera whose males are vocal. Brunelli kept and fed several males of Acrida viridissima (a grasshopper with us not uncommon) in a closet, which[Pg 245] were very merry, and continued singing all the day; but a rap at the door would stop them instantly. By practice he learned to imitate their chirping: when he did this at the door, at first a few would answer him in a low note, and then the whole party would take up the tune and sing with all their might. He once shut up a male in his garden, and gave the female her liberty; but as soon as she heard the male chirp, she flew to him immediately[1034].
But although physiologists are for the most part agreed that insects have the ordinary five senses of vertebrate animals, yet a great variety of opinions has obtained as to their external organs; so that it has been matter of doubt, for instance, whether the antennæ are for smell, touch, or hearing; and the palpi for smell, taste, or touch. Nor has the question, as it appears to me, been satisfactorily decided: for though it is now the most general opinion that the primary use of antennæ is to explore as tactors, yet by the most strenuous advocates of this opinion they are owned not to be universally so employed; so that granting this to be one of their principal functions, yet it seems to follow that there may be another common to them all, which of course would be their primary function. We are warned, however, not to lay any stress upon the argument to be drawn from analogy; and told that we might as well dispute about the identity of the nose of a man, the proboscis of the elephant, the horn of the rhinoceros, the crest of the cock, or the beak of the toucan[1035]. But this is merely[Pg 246] casting dust in our eyes: for though three of these are nasal organs, bearing nostrils; the two others have no relation to the question, the horn of the rhinoceros and the crest of the cock being merely appendages, and have no more analogy to the nose and nostrils, which co-exist with them, than they have to the eyes or ears. I have on a former occasion observed, that a gradual change sometimes takes place in the functions of particular organs; but still, generally speaking, this observation regards secondary functions—the primary usually remaining untouched. We may say, for instance, with regard to the primary use of the legs of animals, that it is locomotion; while the secondary is either walking, running, jumping, flying, or swimming, according to the circumstances and nature of the animal. Thus the fore-legs of the Mammalia, in birds become wings, and both pair in fish are changed to fins. Observe, I do not say always and invariably, but in most cases,—that analogous parts have analogous uses, at least as far as primary uses are concerned. When, therefore, we cannot have demonstrative evidence concerning the function of an organ discoverable in any animal, we may often derive satisfactory probable arguments from the analogies observable in their structure compared with that of other animals, concerning the nature of whose organs we have no doubt. In fact, the chief evidence we have with regard to the office of the organs of sense in the animals immediately below ourselves, is that of analogy;—because we see with our eyes, hear with our ears, &c., we conclude, with reason, that they do the same.
In inquiring therefore into what may be the most general[Pg 247] use of the antennæ of insects, I shall endeavour to discover whether there is any part in the higher animals to which they may be deemed to exhibit any analogy. And here I must refer you to what I have said on a former occasion upon the present subject; where I made it evident, I hope, that the great bulk of the parts and organs of insects, in this particular differing from the majority of Invertebrates, are, some in one respect, some in another, and some in many, really analogous to those of the higher animals[1036]; and that a great many of them, though varying in their structure, have the same functions. Thus the analogues of the eyes of Vertebrates are for seeing; of the jaws for masticating; of the lips for closing the mouth; of the legs for walking, &c. We have seen also very recently, that a similar analogy, more or less strongly marked, holds also in their internal organs[1037]; so that it may be safely affirmed, that if all the invertebrate insects, though gifted with numerous peculiarities, present the most striking picture of those animals that have an internal skeleton, and more particularly of the Mammalia,—we may assume it as a probability, the above circumstances being allowed their due weight, that where facts do not prove the contrary, the function of analogous organs is more or less synonymous, though perhaps the structure and modus operandi may be different.
In the letter lately referred to, I observed that the antennæ of insects are analogous to ears in Vertebrates[1038]. Their number corresponds; they also stand out from the[Pg 248] head; and what has weighed most with me, unless they are allowed as such, no other organ can have any pretension to be considered as representing the ear. If we reflect, that in every other part and organ, the head of insects has an analogy to that of Mammalia, we must regard it as improbable that these prominent organs should not also have their representative. Admitting then that they are the analogues of ears, it will follow, not as demonstratively certain, but as probable, that their primary function may be something related to hearing. I do not say direct hearing, or that the vibrations of sound are communicated to the sensorium by a complex structure analogous to that of the internal ear in Mammalia—but something related to hearing. I conceive that antennæ, by a peculiar structure, may collect notices from the atmosphere, receive pulses or vibrations, and communicate them to the sensorium, which, though not precisely to be called hearing, may answer the same purpose. From the compound eyes that most of them have, the sense of seeing in insects must be very different from what it is in vertebrate animals; and yet we do not hesitate to call it sight: but since antennæ, as we shall see, apparently convey a mixed sensation, I shall have no objection, admitting it as their primary function, to call it after Lehmann Aëroscepsy[1039]. I lately related some instances of sound producing an effect on the antennæ of insects: I will now mention another that I observed, still more remarkable. A little moth was reposing upon my window; I made a quiet, not loud, but distinct noise: the antenna nearest to me immediately[Pg 249] moved towards me. I repeated the noise at least a dozen times, and it was followed every time by the same motion of that organ; till at length the insect being alarmed became more agitated and violent in its motions. In this instance it could not be touch; since the antenna was not applied to a surface, but directed towards the quarter from which the sound came, as if to listen. Bonsdorf made similar observations, to which Lehmann seems not disposed to allow their proper weight[1040]. It has been used as an argument to prove that antennæ are primarily tactors, or instruments of touch, that Fœnus Jaculator, before it inserts its ovipositor, plunges its antennæ into the hole forming the nidus of the bee, to the grub of which it commits its egg[1041]. But had those who used this argument measured the antennæ and the ovipositor of this ichneumon, they would have discovered that the latter is thrice the length of the former: and as these insects generally insert it so that even part of the abdomen enters the hole, it is clear that the antenna cannot touch the larva; its object therefore cannot be to explore by that sense. Others suppose that by these organs it scents out the destined nidus for its eggs; but Lehmann has satisfactorily proved that they are not olfactory organs. We can therefore only suppose, either that by means of its antennæ it hears a slight noise produced by the latent grub, perhaps by the action of its mandibles; or else that by its motions it generates a motion in the atmosphere of its habitation, which striking upon the antennæ of the Fœnus, are by them communicated[Pg 250] to its sensory. A similar disproportion is observable between the antennæ and ovipositor of Pimpla Manifestator, before signalized[1042]. Bees, when collecting honey and pollen, first insert the organs in question into the flowers which they visit; but, as I have more than once observed, they merely insert the tip of them. If anthers are bursting, or the nectar is exuding, these processes probably are attended by a slight noise, or motion of the air within the blossom, which, as in the last case, affects, without immediate contact, the exploring organs.
If the structure of antennæ be taken into consideration, it will furnish us with additional reasons in favour of the above hypothesis, with regard to their primary function. We shall find that these organs, in most of those insects which take their food by suction, are usually less gifted with powers of motion, than they are in the mandibulate tribes; so that in the majority of the Homopterous Hemiptera and Diptera, as is generally acknowledged, they cannot be used for touch. Under this view, they may be divided into active antennæ and passive antennæ: of the former, the most active and versatile are those of the Hymenoptera. By means of them, as was before observed[1043], their gregarious tribes hold converse, and make inquiry—frequently without contact—in the pursuit and discharge, if I may so speak, of the various duties devolved upon them by Providence. Amongst active antennæ, some are much more complex in their structure than others—a circumstance which is often characteristic[Pg 251] of the male insect[1044]: but if we examine such antennæ, we shall find that their most sensitive parts cannot come in contact with the earth or other bodies for exploring their way; but having thus a greater surface exposed to the action of the atmosphere, they have more points to receive vibrations, or any pulses or other notices communicated to it. It is thus, probably, that in their flights, when they approach within a certain distance, they discover the station of the other sex. Even the plumose antennæ of male gnats may in some respects thus be acted upon. In the Lamellicorn beetles, the knob of these organs in both sexes consists of laminæ, the external ones on their outside, of a corneous substance; while their internal surface, and the inner laminæ—which are included between them, as an oyster between the valves of its shell—are covered with nervous papillæ. If you examine the proceedings of one of these little animals, you will find before it moves from a state of repose that its antennæ emerge, and the laminæ diverge from each other; but that it does not apply them to surfaces to explore its way, but merely keeps them open to receive notices from the atmosphere. Even simple antennæ are often employed in this way, as well as for touch. I once noticed a species of Leptocerus, a trichopterous genus, in which these organs are very long, that was perched upon a blade of grass; its antennæ vibrated, and it kept moving them from side to side in the air, as if thus by aëroscepsy it was inquiring what was passing around it. Dr. Wollaston has an observation bearing so[Pg 252] precisely upon this question, and in general so extremely similar to what is here advanced, that I must copy it for your consideration. "Since there is nothing in the constitution of the atmosphere," says he, "to prevent vibrations much more frequent than any of which we are conscious, we may imagine that animals like the Grylli, whose powers appear to commence nearly where ours terminate, may have the faculty of hearing still sharper sounds, which at present we do not know to exist; and that there may be other insects, hearing nothing in common with us, but endued with a power of exciting, and a sense that perceives, vibrations indeed of the same nature as those which constitute our ordinary sounds, but so remote, that the animals who perceive them may be said to possess another sense, agreeing with our own solely in the medium by which it is excited, and possibly wholly unaffected by these slower vibrations of which we are sensible[1045]." That insects, however, hear nothing in common with us, is contrary to fact; at least with respect to numbers of them. They hear our sounds, and we theirs; but their hearing or analogous sense is much nicer than ours, collecting the slightest vibratiuncle imparted by other insects, &c. to the air. In inquiring how this is done, it may be asked—How know we that every joint of some antennæ is not an acoustic organ, in a certain sense distinct from the rest? We see that the eyes of insects are usually compound, and consist of numerous distinct lenses;—why may not their external ears or their analogues be also multiplied, so as to[Pg 253] enable them with more certainty to collect those fine vibrations that we know reach their sensory, though they produce no effect upon our grosser organs? I propose this merely as conjecture, that you may think it over, and reject or adopt it, in proportion as it appears to you reasonable or the contrary; and in the hope that some anatomist of insects, who, to the sagacity and depth of a Cuvier and a Savigny adds the hand and eye of a Lyonet, may give to the world the results of a more minute dissection and fuller investigation of the antennæ of these animals, than has yet been undertaken.
But besides receiving notices from the atmosphere, of sounds, and of the approach or proximity of other insects, &c., the antennæ are probably the organs by which insects can discover alterations in its state, and foretel by certain prognostics when a change of weather is approaching. Bees possess this faculty to an admirable degree. When engaged in their daily labours, if a shower is approaching, though we can discern no signs of it, they foresee it, and return suddenly to their hives. If they wander far from home, and do not return till late in the evening, it is a prognostic to be depended upon, that the following day will be fine: but if they remain near their habitations, and are seen frequently going and returning, although no other indication of wet should be discoverable, clouds will soon arise and rain come on. Ants also are observed to be excellently gifted in this respect: though they daily bring out their larvæ to sun them, they are never overtaken by sudden showers[1046]. Previously to rain, as you well know,[Pg 254] numberless insects seek the house; then the Stomoxys calcitrans, leaving more ignoble prey, attacks us in our apartments, and interrupts our studies and meditations[1047]. The insects of prey also foresee the approach of wet weather, and the access of flies, &c. to places of shelter. Then the spiders issue from their lurking-places, and the ground-beetles in the evening run about our houses. Passive antennæ, which are usually furnished with a terminal or lateral bristle, and plumose and pectinated ones, seem calculated for the action of the electric and other fluids dispersed in the atmosphere, which in certain states and proportions may certainly indicate the approach of a tempest, or of showers, or a rainy season, and may so affect these organs as to enable the insect to make a sure prognostic of any approaching change: and we know of no other organ that is so likely to have this power. I say electric fluid, because when the atmosphere is in a highly electrified state, and a tempest is approaching, is the time when insects are usually most abundant in the air, especially towards the evening; and many species may then be taken, which are not at other times to be met with: but before the storm comes on, all disappear, and you will scarcely see a single individual upon the wing. This seems to indicate that insects are particularly excited by electricity[1048].—But upon this head I wish to make no positive assertion, I only suggest the probability of the opinion[1049].
From all that has been said, I think you will be disposed[Pg 255] to admit that the primary and most universal function of the antennæ is to be the organs of a sense, if not the same, at least analogous to hearing, and answering the same end; something perhaps between it and touch. In some, however, as has been found in the Crustacea, an organ of hearing, in the ordinary sense, may exist at the base of the antennæ, which may act the part in some measure of the external ear, and collect and transmit the sound to such organ[1050].
That numerous antennæ, as a secondary function, explore by touch, is admitted on all hands, and therefore I need not enlarge further upon this point; but shall proceed to inquire whether insects do not possess some other peculiar organs that are particularly appropriated to this sense. First, however, I must make some general observations upon it. Of all our senses, touch is the only one that is not confined to particular organs, but dispersed over the whole body: insects, however, from the indurated crust with which they are often covered, feel sensibly, it is probable, only in those parts where the nerves are exposed, by being covered with a thinner epidermis, to external action. Not that they cannot feel at all in their covered parts; for as we feel sufficiently for walking, though our feet are covered by the thick sole[Pg 256] of a boot or shoe, so insects feel sufficiently through the crust of their legs for all purposes of motion. Besides, the points that are covered by a thinner cuticle are often numerous; so that touch, at least in a passive sense, may be pretty generally dispersed over their bodies; but active or exploring touch is confined to a few organs, as the antennæ, the palpi, and the arms. The two last I shall now discuss.
Various opinions have been started concerning the use of the palpi. Bonsdorf thought that they were organs of smell; Knoch, that this sense was confined to the maxillary ones, and that the labial ones were appropriated to taste[1051]: but the most early idea, and that from which they derive their present name of palpi (feelers), is, that they are organs of active touch; and this seems to me the most correct and likely opinion. Cuvier, himself a host, has embraced this side of the question[1052], and Lehmann also admits it[1053]. The following observations tend to confirm this opinion. The palpi of numerous insects when they walk, are frequently, or rather without intermission, applied to the surface on which they are moving—this you may easily see by placing one upon your hand; which seems to indicate that they are feelers. In the Araneidæ they are used as legs; and by the males at least, as exciting if they be not really genital organs[1054]. In the Scorpionidæ they answer the purpose of hands: besides being usually much shorter than antennæ, they[Pg 257] are better calculated to assist an insect in threading the dark and tortuous labyrinths through which it has often to grope its way, and where antennæ cannot be employed. I have noticed that Hydrophili—in which genus the palpi are longer than the antennæ—when they swim, have their antennæ folded; while the former are stretched out in front, as exploring before them. As these are attached to the under-jaws and under-lip, we may suppose they are particularly useful to insects in taking their food; and upon this occasion I have often observed that they are remarkably active. I have seen Byturus tomentosus, a beetle which feeds upon pollen, employ them in opening anthers; and the maxillary pair appear to me to assist the maxillæ in holding the food, while the mandibles are at work upon it.
The arms or fore-legs of some insects are also organs of active touch, being used, as we have seen, for cleaning the head, digging, repairing their dwellings, and the like[1055]. By the Ephemeræ, which have very short antennæ, the fore-legs, when they fly, are extended before the head, parallel with each other and quite united—probably to assist in cutting the air. The Trichoptera use their antennæ for the same purpose.
Another sense of which the organ seems uncertain is that of smelling, and various and conflicting opinions have been circulated concerning it. Christian thought that insects smell distant objects with their antennæ, and near ones with their palpi[1056]. Comparetti has a most singular[Pg 258] opinion. He supposes in different tribes of insects that different parts are organs of smell: in the Lamellicorns he conjectures the seat of this sense to reside in the knob of the antennæ; in the Lepidoptera in the antlia; and in some Diptera and Orthoptera in certain frontal cells[1057]. At first sight, one of the most reasonable opinions seems to be that of Baster, adopted by Lehmann, and which has received the sanction of Cuvier[1058],—that the spiracles are organs of smell as well as of respiration. Lehmann has adduced several arguments in support of this opinion. Because we both respire and smell with our nostrils, he concludes that neither the antennæ nor any other part of the head of insects can serve for smell, since they are not the seat also of respiration; and that there can be no smell where the air is not inspired[1059]. Again, because nerves from the ganglions of the spinal chord terminate in bronchiæ near the spiracles, they must be for receiving scents from those openings. Though it was necessary, in the higher animals, that the organ of scent should be near the mouth, because they are larger than their food; yet the reverse of this being the case with insects, which often even reside in what they eat, it is to them of no importance where their sense of smelling resides[1060]. By exposing antennæ, by means of an orifice in a glass vessel, to the action of stimulant odours, they appeared quite insensible to it: but he does not name the result of any experiment in which he exposed the mouth to this action; nor at all distinctly how the insect was affected when the spiracles were exposed to it[1061].
But though some of these arguments appear weighty, there are others, I think, that will more than counterbalance them, making it probable that the seat of this sense is in the head, either in its ordinary station at the extremity of what I call the nose, between it and the upper-lip, or under those parts. That the nose corresponds with the so-named part in Mammalia, both from its situation and often from its form, must be evident to every one who looks at an insect[1062]; and when we further consider the connexion that obtains between the senses of smell and taste, how necessary it is that the seat of the one should be near that of the other, and that it really is so in all animals in which we certainly know its organ[1063]; we shall feel convinced that the argument from analogy is wholly in favour of the nose, and may thence consider it as probable that the sense in question does reside there. Lehmann seems to be of opinion, because an insect is usually smaller than what it feeds upon, that it makes no difference whether it smells with its head or with its tail: but one would think that a flying insect would be more readily directed to its object by smelling with the anterior part of the body than with the posterior; and that a feeding one would also find it more convenient in selecting its food. As to the argument,—that smell must be the necessary concomitant of the respiratory openings, and that there can be no smell where the air is not inspired,—this seems asserting more than our knowledge of these animals will warrant: for the organs of the other senses, though the senses themselves seem analogous, are so different in their structure, and often in the mode in[Pg 260] which they receive the impressions from external objects, that analogy would lead us to expect a difference of this kind also in the sense of smell. Besides, smell does not invariably accompany respiratory organs even in the higher animals,—for we breathe with our mouths, but do not smell with them. Cuvier says that the internal membrane of the tracheæ being soft and moist, appears calculated to receive scents[1064]. But here his memory failed him; for it is the external membrane alone that answers this description; the internal consisting of a spiral elastic thread, and seeming not at all fitted to receive impressions, but merely to convey the air[1065]. That nerves penetrate to the bronchiæ, does not necessarily imply that they are connected with the sense in question, since this may be to act upon the muscles which are every where distributed.
I shall now state some facts that seem to prove that scents are received by some organ in the vicinity of the mouth, and probably connected with the nose. M. P. Huber, desirous of ascertaining the seat of smell in bees, tried the following experiments with that view. These animals, of all ill scents, abominate most that of the oil of turpentine. He presented successively to all the points of a bee's body, a hair-pencil saturated with it: but whether he presented it to the abdomen, the trunk, or the head, the animal equally disregarded it. Next, using a very fine hair-pencil, while the bee had extended its proboscis, he presented the pencil to it, to the eyes and antennæ, without producing any effect; but when he pointed it near the cavity of the mouth, above the insertion[Pg 261] of the proboscis, the creature started back in an instant, quitted its food, clapped its wings, and walked about in great agitation, and would have taken flight if the pencil had not been removed. On this, it began to eat again; but on the experiment being repeated, showed similar signs of discomposure: oil of marjoram produced the same effect, but more promptly and certainly. Bees not engaged in feeding appeared more sensible of the impression of this odour, and at a greater distance; but those engaged in absorbing honey might be touched in every other part without being disturbed. He seized several of them, forced them to unfold their proboscis, and then stopped their mouth with paste. When this was become sufficiently dry to prevent their getting rid of it, he restored to them their liberty: they appeared not incommoded by being thus gagged, but moved and respired as readily as their companions. He then tempted them with honey, and presented to them near the mouth, oil of turpentine, and other odours that they usually have an aversion to; but all produced no sensible effect upon them, and they even walked upon the pencils saturated with them[1066].
These experiments incontestibly prove that the organ of scent in bees—and there is no reason to think that other insects do not follow the same law—is in or near the mouth, and above the proboscis. It remains, therefore, that we endeavour to discover its precise situation: and as insects cannot tell us, nor can we perceive by their actions, in what precise part the sense in question[Pg 262] resides, the only modes to which we can have recourse to form any probable conjecture, are analogy and dissection. At first, the opinion noticed above, that the palpi are its organs, seems not altogether unreasonable; but as the argument from analogy, except as to their situation near the mouth, is not in favour of them, and there seems no call, were smell their function, for the numerous variations observable in their structure, I think we must consider them, as I have endeavoured to prove, rather as instruments of touch. Let us now inquire, whether there be not discoverable upon dissection, in the interior of the head of any insects, some organ that may be deemed, from its situation, under what we have called the nose and nostrils, the seat of the sense we are treating of. The common burying-beetle (Necrophorus Vespillo) is an insect remarkable for its acuteness of smell, which enables it to scent out and bury, as was formerly related to you[1067], the carcases of small animals. Take one of these insects, and kill it as formerly directed,—examine first its nose: in the middle of the anterior part you will see a subtrapezoidal space, as it were cut out and filled with a paler piece of a softer and more membranous texture. Next divide the head horizontally; and under the nose, and partly under this space, which I call the rhinarium or nostril-piece[1068], you will find a pair of circular pulpy cushions, covered by a membrane transversely striated with beautifully fine striæ. These are what I take to be the organs of smell, and they still remain distinctly visible in a specimen I have had by me more than fifteen years. A similar organ may be discovered[Pg 263] in the common water-beetle (Dytiscus marginalis), but with this peculiarity, that it is furnished with a pair of nipples. I have before described an analogous part covered with papillæ, in Æshna viatica, and you will find it in other insects[1069]. Perhaps at first this part may seem merely a continuation of the palate; but if you consider the peculiarities in its structure just noticed, it is evidently a sensiferous organ; and as the sense of smell appears to reside in the head, this is its most probable seat. But by what channel scents act upon it,—whether they are transmitted through the pores of the part representing the nostrils, or received by the mouth,—I will not venture to assert positively: but from the circumstance of their being membranous in some insects remarkable for acute scent, as in Necrophorus, Staphylinus, &c., there seems some ground for the former opinion, which receives further confirmation from an observation of an eminent Comparative Anatomist, M. Carus, with respect to Acrida verrucivora, in which under the nose and rhinarium, as appears from his description, he found some tracheæ, and two lobes of the cerebral ganglion, which caused him to regard this as the seat of the sense of smell[1070]. He also tells us that Rosenthal, in the blue-bottle-fly (Musca vomitoria) places the sense of smell partly in a delicately folded membrane observable in its head[1071]. As the sense of smell in these little beings is extremely acute, as well as their hearing, the perception of odours may reach their sensory through the above pores;[Pg 264] and even those in the hard rhinarium of an Anoplognathus may receive and transmit them; and besides the upper-lip and nose are often united by membrane, perhaps representing the rhinarium, as in Goerius, &c.[1072] which may facilitate such transmission.
That insects taste, no one hesitates to believe, though some have supposed the palpi to be the organ of that sense; but as they have a tongue, as we have shown, we may with Cuvier conclude, that one of its primary functions is to taste their food[1073]. I shall not therefore launch out further upon this head.
I have now placed before you a picture, or rather sketch, of the insect world. And whether we regard their general history and economy, their singular metamorphoses, the infinite varieties and multiplicity of their structure both external and internal, and their diversified organs both of sense and motion—I think you will be disposed to own, that in no part of his works is the hand of an Almighty and All-wise Creator more visibly displayed, than in these minutiæ of creation; that they are equally worthy of the attention and study of the Christian Philosopher with any of the higher departments of the animal kingdom; and that all praise is due to Him, for placing before our eyes, for our entertainment and instruction, such a beautiful moving picture of little symbols and agents, perpetually reflecting his glory and working his will.
I am, &c.
It was by the language of terms that he invented and employed, as well as by his system and methods of arrangement, that Linné smoothed the way to the study of Natural History;—having therefore led you through a large portion of the flowery fields of the Science of Entomology, I must now conduct you into that arid but not barren or unprofitable region. To enable you to understand descriptions of insects, or to describe them yourself, you must have a knowledge of the technical language by which their parts and characters are expressed. Much of this you already know from the definitions of external parts, furnished in a former letter[1074]: I shall now give you a more full and general explanation of terms, adding many new ones for unnoticed characters, that may be conveniently employed.
The science of terms, which I shall call Orismology[1075], may be divided into two branches—General Orismology, and Partial Orismology; the first containing general definitions, and the last those relating to particular parts and organs.
A. GENERAL ORISMOLOGY.
I. SUBSTANCE.
1. Membranous (Membranacea). A fine, thin, transparent substance. A Membrane.—Ex. Wings of Hymenoptera and Diptera.
2. Pergameneous (Pergamenea). A thin, tough, and less transparent substance, somewhat resembling parchment.—Ex. The Tegmina of the Orthoptera[1076].
3. Coriaceous (Coriacea). A thicker, flexible substance, resembling leather.—Ex. Elytra of Telephorus and the Malacodermi.
4. Corneous (Cornea). A hard inflexible substance resembling horn.—Ex. Elytra of Lucanus Cervus and many other Coleoptera.
5. Crustaceous (Crustacea). A rigid calcareous substance.—Ex. The Shell of a Lobster or Crab.
6. Callous (Callosa). A substance without pores, harder than the surrounding matter, and usually elevated above it.—Ex. Elevated parts of the Collar in Nomada. (Mon. Ap. Angl. Apis * b.)[1077] Spots on the elytra of Stenocorus (Tylostagmus K. MS.) bimaculatus and affinities.
7. Cartilagineous (Cartilaginea). A gristly substance between bone and ligament.—Ex. The Tongue of many Hymenoptera.
8. Subereous (Suberea). A soft elastic substance[Pg 267] somewhat resembling cork[1078]. The galls of some species of Cynips when mature approach to this substance.
9. Spongiose (Spongiosa). A soft elastic substance resembling sponge.—Ex. The Pulvilli of Thanasimus, Buprestis, &c.
10. Ligneous (Lignosa). A hard unelastic substance like wood.—Ex. Galls of some species of Cynips.
11. Carnose (Carnosa). A soft, fleshy substance.—Ex. Caterpillars and Grubs.
12. Tubulose (Tubulosa). When the interior is hollow or empty.
13. Solid (Solida). When the interior is full.
II. RESISTANCE.
1. Rigid (Rigida). Hard, which does not bend or yield to pressure.—Ex. The weevils (Rhyncophora).
2. Flexile (Flexilis). Which easily bends, or yields to pressure without breaking.—Ex. Elytra of Telephorus.
3. Soft (Mollis). Flexile and retaining the marks of pressure.—Ex. Elytra of Meloe.
III. DENSITY.
1. Foliaceous (Foliacea). Very thin and depressed, scarcely thicker than a leaf.—Ex. Aradus corticalis and Coreus paradoxus.
2. Depressed (Depressa). When the vertical section is shorter than the transverse.—Ex. Trogosita mauritanica.
3. Compressed (Compressa). When the transverse section is shorter than the vertical.—Ex. Centrotus cornutus: Abdomen in Cynips.
4. Plump (Pinguis). Naturally and proportionably plump.—Ex. Thylacites, &c. Most of the Cicadæ.
5. Obese (Obesa). Unnaturally enlarged and distended, as if from disease or too much food. Ex. Chrysomela Polygoni ♀, Galeruca Tanaceti ♀, Brachycerus.
6. Ventricose (Ventricosa). Bellying out as if filled with air.—Ex. Pneumora.
IV. PROPORTION.
1. Thick (Crassa). Disproportionably thick throughout.—Ex. Copris.
2. Incrassate (Incrassata). Disproportionably thick in part.—Ex. Base of the Abdomen of Æshna and many Libellulina. Plate IX. Fig. 9.
3. Slender (Tenuis). Disproportionably slender throughout.—Ex. Lixus paraplecticus.
4. Attenuate (Attenuata). Disproportionably slender in part.—Ex. Tail of Scorpio, Raphidia ♂, &c.
5. Broad (Lata). Disproportionably broad throughout.
6. Dilatate (Dilatata). Disproportionably broad in part.—Ex. Elytra of Lycus fasciatus, &c. Plate XIII. Fig. 20.
7. Narrow (Angusta). Disproportionably narrow throughout.—Ex. Abdomen of Agrion.
8. Angustate (Angustata). Disproportionably narrow in part.—Ex. Elytra of Sitaris humeralis. Plate XIII. Fig. 19.
9. Long (Longa). Disproportionably long throughout.—Ex. Scolopendra.
10. Elongate (Elongata). Disproportionably long in part.—Ex. Abdomen of Libellulina.
11. Short (Brevis). Disproportionably short throughout.—Ex. Copris.
12. Abbreviate (Abbreviata). Disproportionably short in part.—Ex. Elytra of Staphylinidæ, Atractocerus, &c.
V. FIGURE[1079].
1. Circular (Circularis). Having the diameter every way equal. Plate XXIX. Fig. 16, 17.
2. Rotundate (Rotundata). Rounded at the angles or sides. Plate XXIX. Fig. 19.
3. Oval (Ovalis). Having the longitudinal diameter twice the length of the transverse, and the ends circumscribed by equal segments of a circle. Plate XX. Fig. 6.
4. Elliptic (Elliptica). Oval, but having the longitudinal diameter more than twice the length of the transverse. Plate XX. Fig. 19.
5. Oblong (Oblonga). Having the longitudinal diameter more than twice the length of the transverse, and the ends varying, or rounded. Plate XX. Fig. 3, 9.
6. Ovate (Ovata). Oval, but having the ends circumscribed by unequal segments of circles. Plate XX. Fig. 12, 13.
7. Cordate (Cordata). Heart-shaped. Ovate or subovate and hollowed out at the base, without posterior angles. Plate IX. Fig. 22.
8. Sagittate (Sagittata). Arrow-shaped. Triangular, hollowed out at the base with posterior angles. Plate XXVII. Fig. 41. w´´´.
9. Hastate (Hastata). Halberd-shaped. Triangular, hollowed out at the base and sides with the posterior angles spreading.—Ex. Horn of the prothorax of Dynastes hastatus. Postfurca in many Coleoptera. Plate XXII. Fig. 5. b †.
10. Triangular; Quadrangular; Quinquangular; Sexangular (Triangula; Quadrangula; Quinquangula; Sexangula). Having three, four, five, or six angles.
11. Turbinate (Turbinata). Top-shaped, triangular with curved sides. Plate XXV. Fig. 18.
12. Ensate (Ensata). Gradually tapering till it ends in a point.—Ex. Ovipositor of Acrida viridissima. Plate XV. Fig. 19.
13. Lanceolate (Lanceolata). Oblong and gradually tapering towards each extremity.—Ex. The Cerci in Blatta. Plate XV. Fig. 23. Q´´.
14. Sigmoidal (Sigmoidea). S-shaped. Lanceolate and concave on one side at the base, and on the other at the apex.—Ex. Ovipositor of Cimbex, Plate XV. Fig. 21. H´´.
15. Cuneate (Cuneata). Wedge-shaped. Having the longitudinal diameter exceeding the transverse,[Pg 271] and narrowing gradually downwards. Plate X. Fig. 11.
16. Acinacicate (Acinacicata). Falchion-shaped. Curved with the apex truncate, and growing gradually wider towards the end.—Ex. Abdomen of Ophion, Fœnus, and other Ichneumonidæ[1080].
17. Lunulate (Lunulata). Crescent-shaped. Curved with both ends acute, like the moon in her first quarter.—Ex. Last joint of the labial palpi of Oxyporus. Plate XIII. Fig. 4. a.
18. Falcate (Falcata). Sickle-shaped. Curved with the apex acute.—Ex. Ovipositor of Acrida varia. Antennæ of Atractocerus. Plate XI. Fig. 8.
19. Linear (Linearis). Narrow and of the same width throughout.—Ex. Wings of Pterophorus monodactylus.
20. Arcuate (Arcuata). Linear and bent like a bow.—Ex. Rostrum of Balaninus Nucum. Plate XIII. Fig. 12.
21. Cultrate (Cultrata). Coulter-shaped. Straight on one side and curved on the other.—Ex. Ovipositor of some Saw-flies. Under-wing of many Ichneumonidæ.
22. Spatulate (Spatulata). Spatula-shaped. Broader and rounded at the apex, linear and narrow at the base.—Ex. Abdomen of Ichneumon amictorius Panz.
23. Clepsydrate (Clepsydrata). Hour-glass-shaped.[Pg 272] Broader at the base and apex—Ex. The Prosternum of many Capricorn beetles.
24. Clavate (Clavata). Club-shaped. Linear at the base, but towards the apex growing gradually broader. Plate XI. Fig. 4.
25. Quadrate (Quadrata). Square. Quadrilateral with the sides equal and the angles right angles.
26. Rhomboid (Rhomboidea). Quadrilateral with the sides equal, but with two opposite angles acute, and two obtuse. Plate XXVII. Fig. 62. t´´.
27. Trapezate (Trapezata). Quadrilateral with the four sides unequal, and none of them perfectly parallel. Plate XIV. Fig. 4.
28. Trapezoid (Trapezoidea). Quadrilateral, with two sides unequal and parallel[1081]. Plate XXVI. Fig. 34. b´.
29. Parallelogramical (Parallelogramica). Quadrilateral, with all the angles right angles, and all the sides parallel, but two longer than the others.
VI. FORM[1082].
1. Spherical (Sphærica). The shape of a globe. A body whose diameter every way is equal. Plate XX. Fig. 5.
2. Orbiculate (Orbiculata). A depressed globe, whose horizontal section is circular, and vertical oval. Plate XX. Fig. 10, 11.
3. Lenticular (Lenticularis). Lens-shaped. Whose[Pg 273] horizontal section is circular, and vertical lanceolate.—Ex. Abdomen of Cynips aptera.
4. Ovaliform (Ovaliformis). Whose longitudinal section is oval, and transverse circular. Plate XX. Fig. 6.
5. Ellipsoid (Ellipsoidea). Whose longitudinal section is elliptical, and transverse circular. Plate XX. Fig. 19.
6. Oviform (Oviformis). Whose longitudinal section is ovate, and transverse circular. Plate XX. Fig. 12, 13.
7. Cucumiform (Cucumiformis). Cucumber-shaped. Whose longitudinal section is oblong, and transverse circular. Plate XX. Fig. 18, excluding the neck.
8. Cordiform (Cordiformis). Oviform and hollowed out at the base without posterior angles. Plate IX. Fig. 22.
9. Conical (Conica). Whose vertical section is triangular, and horizontal circular.—Ex. Abdomen of Cœlioxys conica (Apis * * b. K.). Plate XX. Fig. 7.
10. Turbiniform (Turbiniformis). Whose vertical section is turbinate, and horizontal circular.—Ex. Joints of antennæ of Aleochara socialis, and many others of that genus.
11. Pyramidal (Pyramidalis). Whose vertical section is triangular, and horizontal quadrangular.
12. Cuneiform (Cuneiformis). Whose vertical section is cuneate, and horizontal parallelogramical.
13. Triquetrous (Triquetra). Whose horizontal sections are equilateral triangles. Plate XI. Fig. 6.
14. Ensiform (Ensiformis). Whose horizontal sections are acute-angled triangles gradually diminishing in diameter from the base to the apex, and propagated in a straight line. Plate XI. Fig. 7.
15. Acinaciform (Acinaciformis). Whose horizontal sections are acute-angled triangles gradually increasing in diameter from the base to the apex, and propagated in a curved line.
16. Cultriform (Cultriformis). Whose horizontal sections are equal acute-angled triangles, or a three-sided body with two equal sides large and the third small.
17. Deltoid (Deltoidea). Short with the horizontal section triangular and decreasing in diameter towards the base.—Ex. Apex of the posterior tibia in Copris lunaris.
18. Trigonal; Tetragonal; Pentagonal; Hexagonal; Polygonal (Trigona; Tetragona; Pentagona; Hexagona; Polygona). Whose horizontal section is triangular; quadrangular; quinquangular; sexangular; multiangular.
19. Triedral; Tetraedral; Pentaedral; Hexaedral; Polyedral (Triedra; Tetraedra; Pentaedra; Hexaedra; Polyedra). That hath three sides; four sides; five sides; six sides; many sides.
20. Prismoidal (Prismoidalis). Having more than four sides and whose horizontal section is a polygon[1083]. Plate VI. Fig. 13. a, b, d´.
21. Trapeziform (Trapeziformis). Whose horizontal section is a Trapezium.
22. Trapezoidiform (Trapezoidiformis). Whose horizontal section is trapezoid.
23. Rhombiform (Rhombiformis). Whose horizontal section is rhomboidal. Plate VIII. Fig. 11.
24. Two-edged (Anceps). Whose horizontal section is lanceolate.
25. Cylindrical (Cylindrica). Whose horizontal sections are all equal circles. Plate XXI. Fig. 4.
26. Fusiform (Fusiformis). Spindle-shaped. Whose vertical section is lanceolate or lineari-lanceolate, and horizontal circular. Plate XXIII. Fig. 12.
27. Columnar (Teres). Whose vertical section is cuneate, and horizontal circular. Plate XVI. Fig. 2, 3.
28. Claviform (Claviformis). Whose vertical section is clavate, and horizontal circular. Plate XI. XII. Fig. 4.
29. Cubical (Cubica). Six-sided, with sides quadrate.
30. Parallelopipedous (Parallelopipeda). Six-sided, with four parallelogramical and two quadrate sides.
31. Pyriform (Pyriformis). Pear-shaped. Whose vertical section is spatulate, and horizontal circular.—Ex. Apion, &c.
32. Infundibuliform (Infundibuliformis). Funnel-shaped. Whose horizontal sections are circular, at first equal and then progressively larger and larger. Plate XXII. Fig. 12. c.
33. Fornicate (Fornicata). Convex above and concave beneath. Plate XIII. Fig. 18. a.
34. Coarctate (Coarctata). When the diameter of[Pg 276] the middle is less than that of the ends.—Ex. Posterior thigh of Locusta. Plate XIV. Fig. 5.
35. Calceoliform (Calceoliformis). Oblong, and somewhat coarctate in the middle.—Ex. Abdomen of Chelonus.
36. Lageniform (Lageniformis). Bellying out and then ending in a narrow neck, something like a bottle.—Ex. Sperm-reservoir attached to the oviduct in Pontia. Plate XXX. Fig. 12. d.
37. Constrict (Constricta). Suddenly and disproportionably smaller at one end. Plate XXII. Fig. 15.
38. Luniform (Luniformis). Whose longitudinal section is lunate. Plate XIII. Fig. 4.
39. Nodose (Nodosa). Having one or more knobs or swellings. Plate XII. Fig. 5.
40. Geniculate (Geniculata). Bent so as to form a knee or angle. Plate XII. Fig. 7.
VII. SUPERFICIES.
i. PARTS.
1. Disk (Discus). The middle of a surface.
2. Limb (Limbus). The circumference.
3. Margin (Margo). The extreme sides.
4. Apex (Apex). The summit.
5. Base (Basis). The bottom.
6. Supine Surface (Pagina superior). The upper surface.
7. Prone Surface (Pagina inferior). The under surface.
ii. ELEVATION and DEPRESSION.
1. Navicular (Navicularis). When two sides meet and form an angle like the outer bottom of a boat.—Ex. Notonecta glauca.
2. Convex (Convexa). An elevation the arc of which is the segment of a circle.—Ex. Upper Surface of the body of most Coleoptera.
3. Gibbous (Gibba). An elevation the arc of which is not the segment of a circle[1084].—Ex. Shoulders of the elytra of Prionus coriarius, and of many other Coleoptera.
4. Plane (Plana). Flat. When the disk is not higher than the limb, nor the limb than the disk.
5. Concave (Concava). A depression the arc of which is the segment of a circle.
6. Excavate (Excavata). A depression the arc of which is not the segment of a circle.—Ex. Prothorax of Sinodendrum cylindricum.
iii. SCULPTURE.
1. Equate[1085] (Æquata). Without larger partial elevations or depressions.
2. Smooth (Lævis). Without smaller partial elevations or depressions.
3. Levigate (Lævigata). Without any partial elevations or depressions.
4. Pore (Porus). A minute impression that perforates the substance.
5. Porose (Porosa). Beset with many pores.—Ex. Elytra of most Apions.
6. A Point (Punctum). A minute impression upon the surface, but not perforating it.
7. Punctate (Punctata). Beset with many points.—Ex. Impression on the Head and Prothorax of Phyllopertha Horticola, &c.
8. Variole (Variola). A shallow impression like a mark of the small-pox.
9. Variolous (Variolosa). Beset with many varioles.—Ex. Scarabæus variolosus.
10. Umbilicate (Umbilicata). When a variole, tubercle, granule, &c. has a depression in its centre.—Ex. Thorax of Pachygaster scabrosus.
11. Foveolet (Foveola). A roundish and rather deep depression, larger than a variole.
12. Foveolate (Foveolata). Having one or more foveolets.—Ex. Prothorax of Geotrupes stercorarius.
13. Fossulet (Fossula). A somewhat long and narrow depression.
14. Fossulate (Fossulata). Having one or more fossulets.—Ex. Oxytelus rugosus, &c.
15. Unequal (Inæqualis). Having very slight and indeterminate excavations.—Ex. Prothorax of Silpha thoracica, Cerambyx moschatus, &c.
16. Lacunose (Lacunosa). Having a few scattered, irregular, broadish but shallow excavations.—Ex. Elytra of Donacia vittata, Sagittariæ, &c.
17. Rimose (Rimosa). Chinky, resembling the bark of a tree. Having numerous minute, narrow and[Pg 279] nearly parallel excavations, which run into each other.—Ex. Elytra of Colymbetes adspersus ♀, and Cybister Rœselii.
18. Undose (Undosa). Having undulating nearly parallel broader depressions which run into each other, and resemble the sand of the sea-shore when left by the tide.—Ex. Cymatodes[1086] undosus K. MS.
19. Vermiculate (Vermiculata). Having tortuous excavations as if eaten by worms.—Ex. Prothorax of Colymbetes Hybneri and transversalis.
20. Reticulose (Reticulosa). Having a number of minute impressed lines which intersect each other in various directions like the meshes of a net.—Ex. Prothorax of Cybister Rœselii.
21. Acuducted (Acuducta). Scratched across very finely as if with the point of a needle or pin.—Ex. Colymbetes acuductus.
22. Striate (Striata). Having rather slightly impressed longitudinal parallel lines.—Ex. Amara communis, &c.
23. Sulcate (Sulcata). Having deeper impressed longitudinal parallel lines.—Ex. Dytiscus marginalis ♀.
24. Clathrose (Clathrosa). When strias or furrows cross each other at right angles.—Ex. Abdomen of Micropeplus porcatus.
25. Rivose (Rivosa). When furrows do not run in a parallel direction and are rather sinuate.—Ex. Prothorax of Elophorus stagnalis, &c.
26. Interstice (Interstitium). The space between elevations and depressions running in lines.
27. Interval (Intervallum). The space between irregular and scattered elevations and depressions.
28. Complanate (Complanata). A convex or irregular surface having a plane slight depression.—Ex. Sides of the Prothorax of Prionus cervicornis.
29. Canaliculate (Canaliculata). Having a longitudinal impressed line or channel.—Ex. Prothorax of Geotrupes, Broscus cephalotes, &c.
30. Carinate (Carinata). Having a longitudinal elevated line.—Ex. Rostrum of Curculio nebulosus. Bicarinate, Tricarinate, &c., having two or three such lines.—Ex. Elytra of Silpha recta.
31. Cristate (Cristata). Having one or two very elevated lines usually crenate.—Ex. Prothorax of Pterophylla laurifolia.
32. Porcate (Porcata). Having several parallel elevated longitudinal ridges.—Ex. Onthophilus striatus.
33. Costate (Costata). Having several broad elevated lines.—Ex. Brachinus bimaculatus, &c.
34. Clathrate (Clathrata). Having several elevated lines which cross each other at right angles.—Ex. Abdomen of Micropeplus porcatus.
35. Reticulate (Reticulata). Having many small elevated lines which intersect each other in various directions like the meshes of a net.—Ex. Lycus reticulatus. Wings of the Libellulina.
36. Rugose (Rugosa). Wrinkled. Intricate with approximating elevations and depressions whose direction is indeterminate.—Ex. Elytra of Prionus coriarius.
37. Cicatricose (Cicatricosa). Having elevated spots of a different colour from the rest of the surface, resembling scars.—Ex. Elytra of Ptomaphila lachrymosa K. MS[1087].
38. Embossed (Cælata). Having several plane tracts of a different shape higher than the rest of the surface.—Ex. Prothorax of Prionus damicornis, maxillosus, &c.
39. Gibbose (Gibbosa). Having one or more large elevations.—Ex. Sides of the Prothorax of Brachycerus barbarus.
40. Tubercle (Tuberculum). A pimple-like knob.
41. Tuberculate (Tuberculata). Having several tubercles.—Ex. Apoderus gemmatus. Base of Prothorax of Cerambyx moschatus.
42. Verruca. A small flattish wart-like prominence.
43. Verrucose (Verrucosa). Having several verrucæ.—Ex. Pimelia muricata.
44. Muricate (Muricata). Armed with sharp thick, but not close, elevated points like a Murex.—Ex. Bronchus Tribulus, quadridens[1088], &c.
45. Echinate (Echinata). Armed with sharp spines like a hedgehog or Echinus.—Ex. Hispa atra.
46. Rugged (Salebrosa). When a surface is rough[Pg 282] with mucros, spines and tubercles intermixed.—Ex. Numerous species of Bronchus.
47. Granule (Granulum). A very minute elevation.
48. Granulate (Granulata). Beset with many granules like shagreen.—Ex. Otiorhynchus sulcatus. Prothorax of Copris Molossus.
49. Scabrous (Scabra). Rough to the touch from granules scarcely visible.—Ex. Elytra of Otiorhynchus Ligustici.
50. Papillule (Papillula). A tubercle or variole with an elevation in its centre.
51. Papillulate (Papillulata). Beset with many papillules.—Ex. Elytra of Dynastes Hercules ♀.
52. Catenulate (Catenulata). Having a series of elevated oblong tubercles resembling a chain.—Ex. Carabus catenulatus.
53. Spherulate (Sphærulata). Having one or more rows of minute tubercles.—Ex. Trox lutosus, Limnius tuberculatus.
54. Consute (Consuta). Having very minute elevations in a series at some distance from each other, of a different colour from the rest of the surface, and somewhat resembling stitching.—Ex. Elytra of Oryctes Sylvanus (Cœlosis K. MS.).
55. Intricate (Intricata). When depressions or elevations so run into each other as to be difficult to trace.—Ex. Elytra of Carabus intricatus.
56. Corrugate (Corrugata). When a surface rises and falls in parallel angles more or less acute.—Ex. Front of Nothiophilus aquaticus.
57. Obliterate (Obliterata). Applied to impressions and elevations when almost effaced.
iv. CLOTHING.
a. general.
1. Scutate (Scutata). Covered with large flat scales.—Ex. Machilis polypoda.
2. Squamose (Squamosa). Covered with minute scales.—Ex. Lepidoptera.
3. Pulverulent (Pulverulenta). Covered with very minute powder-like scales.—Ex. Ceutorhynchus Sisymbrii.
4. Pollinose (Pollinosa). Covered with a loose mealy and often yellow powder resembling the pollen of flowers.—Ex. Lixus paraplecticus.
5. Farinose (Farinosa). Covered with a fixed mealy powder resembling flour.—Ex. Spots on the Elytra of Cetonia aurata, variegata, &c.
6. Lutose (Lutosa). Covered with a powdery substance resembling mud or dirt, which easily rubs off.—Ex. Trox lutosus.
7. Rorulent (Rorulenta). Covered like a plum with a bloom which may be rubbed off.—Ex. Peltis limbata.
8. Stupeous (Stupea). Covered with long loose scales resembling tow.—Ex. The Palpi of Lepidoptera. Antennæ of some Diptera. Plate XII. Fig. 23.
9. Pilose (Pilosa). Covered with long distinct flexible hairs.—Ex. Thorax of Vespa Crabro.
10. Villose (Villosa). Covered with soft flexible hairs thickly set.—Ex. Prothorax of Amphimalla solstitialis.
11. Lanate (Lanata). Covered with fine, very long,[Pg 284] flexible and rather curling hairs like wool.—Ex. Melolontha lanigera F.
12. Lanuginose (Lanuginosa). Covered with longish very soft fine down.—Ex. Prothorax of Trichius fasciatus. Thorax and base of the Abdomen of Megachile circumcincta (Apis **. c. 2. α K.).
13. Hirsute (Hirsuta). Covered with long stiffish hairs very thickly set.—Ex. Bombus.
14. Plumulose (Plumulosa). When the hairs branch out laterally like feathers.—Ex. Hair on the base of the Maxilla of Eucera (Apis ** d. 1. K.).
15. Hairy (Hirta). Covered with short stiffish sub-distinct hairs.—Ex. Genus Lagria.
16. Tomentose (Tomentosa). Covered with short interwoven inconspicuous hairs.—Ex. Acanthocinus Ædilis.
17. Pubescent (Pubescens). Covered with very fine decumbent short hairs.—Ex. Harpalus ruficornis, &c.
18. Stupulose (Stupulosa). Covered with coarse decumbent hairs.—Ex. Elytra of Melolontha vulgaris.
19. Velutinous (Velutina). Covered with very thick-set upright short hairs or pile, resembling velvet.—Ex. Trombidium holosericeum. Scutellum of Staphylinus hybridus.
20. Holosericeous (Holosericea). Covered with thick-set shining short decumbent hairs, resembling satin[1089].—Ex. Under side of the body of Elophorus stagnalis, Argyroneta aquatica, &c.
21. Setose (Setosa). Bristly. Sprinkled with stiff scattered hairs like bristles.—Ex. Echinomyia grossa.
22. Setulose (Setulosa). Setose with the bristles truncated.—Ex. Thylacites setosus.
23. Hispid (Hispida). Rough from minute spines, or very stiff rigid bristles.—Ex. Hispa atra. Phoberus horridus, &c.
24. Rough (Aspera). Rough from pubescence in general.
25. Bald (Calva). A part of a surface with little or no hair, when the rest of it is very hairy.—Ex. Vertex of Melitta and Apis Kirby.
26. Glabrous (Glabra). Without any hair or pubescence.
27. Lubricous (Lubrica). Slippery as if lubricated.—Ex. Dynastes Centaurus.
b. partial.
1. Cirrus (Cirrus). A lock of curling hair.
2. Cirrose (Cirrosa). Having one or more cirri.—Ex. Antennæ of Acanthocinus araneiformis.
3. Fascicule (Fasciculus). A bundle of thick-set hairs often converging at the apex. Plate XIX. Fig. 6. c.
4. Fasciculate (Fasciculata). Having one or more fascicules.—Ex. Catenulated lines in the Elytra of Trox arenosus. Buprestis fascicularis.
5. Penicil (Penicillus). A small bundle of diverging hairs. Plate XIX. Fig. 6. a.
6. Penicillate (Penicillata). Having one or more penicils.—Ex. Larva of Orgyia antiqua.
7. Verricule (Verriculum). A thick-set tuft of parallel hairs. Plate XIX. Fig. 6. b.
8. Verriculate (Verriculata). Having one or more verricules.—Ex. Larva of Dasychira pudibunda. Under side of Abdomen of Megachile ♀. (Apis **. c. 2. α. K.).
9. Barbate (Barbata). When any part is clothed with longer hairs, resembling a beard.—Ex. Anus of Macroglossa stellatarum. Antennæ of Cerambyx Ammiralis L. Plate XII. Fig. 26.
10. Ciliate (Ciliata). When the margin is fringed with a row of parallel hairs.—Ex. The base and apex of the Prothorax of Lucanus Cervus.
11. Fimbriate (Fimbriata). When a part is terminated by hairs or bristles that are not parallel.—Ex. Anus of many Andrenæ[1090]. (Melitta **. c. K.).
12. Comate (Comata). When very long flexible hairs thickly cover a space in the upper surface.
13. Crinite (Crinita). When very long hairs thinly cover any space.
14. Jubate (Jubata). Having long pendent hairs in a continued series.—Ex. Intermediate Legs of Anthophora pilipes (Apis **. d. 2. α. K.).
15. Furred (Pellita). When shorter decumbent hairs thickly cover any space, as in the Bombyces dorso cristato L.
v. COLOUR.
1. Niveous (Niveus). The pure unblended white of snow.—Ex. Arctia chrysorhea.
2. White (Albus). White less intense than niveous. The colour of chalk.—Ex. Spilosoma mendica ♀.
3. Lacteous (Lacteus). White with a slight tint of blue. The colour of milk.—Ex. Minoa lactearia.
4. Cream-coloured (Lactifloreus). White with a proportion of yellow.—Ex. Pale part of the Primary wings of Euprepia Caja.
5. Flesh-coloured (Carneus). White tinted with red. The colour of young and healthy flesh.—Ex. Secondary wings of Sphinx Ligustri.
6. Hoary (Incanus). White with a small proportion of black. The colour of a gray head. N.B. This term is usually confined to pubescence.—Ex. Curculio sulcirostris.
7. Cinereous (Cinereus). White with a shade of brown.—Ex. Sitona diffinis, Dasychira pudibunda.
8. Griseous (Griseus). White mottled with black or brown.—Ex. Curculio nebulosus.
9. Yellow (Flavus). Pure yellow.—Ex. Bands on the Abdomen of Nomada (Apis *. b. K.), Crabro, &c.
10. Straw-coloured (Stramineus). Pale yellow with a very faint tint of blue.—Ex. Ennomos cratægata.
11. Sulphureous (Sulphureus). Yellow with a tint of green. The colour of brimstone.—Ex. Gonepteryx Rhamni ♂.
12. Luteous (Luteus). Deep yellow with a tint of red. The colour of the yolk of an egg.—Ex. Primary wings of Colias Edusa.
13. Orange (Aurantius). Equal parts of red and yellow.—Ex. Apex of Wings of Pontia Cardamines.
14. Saffron-coloured (Croceus). The colour of saffron.—Ex. Yellow in the Elytra of Trichius fasciatus.
15. Miniatous (Miniatus). The colour of red lead.—Ex. Secondary wings of Euprepia Caja.
16. Fulgid (Fulgidus). A bright fiery red.—Ex. Lycæna Virgaureæ and Hippothoe.
17. Rufous (Rufus). A pale red.—Ex. Apion frumentarium.
18. Testaceous (Testaceus). The colour of a tile, a dull red.—Ex. Chrysomela Populi.
19. Scarlet (Coccineus). A bright pale red.—Ex. Elytra of Pyrochroa coccinea.
20. Red (Ruber). Pure red.—Ex. Under Wings of Hypercampa Dominula.
21. Sanguineous (Sanguineus). Red with a tint of black. The colour of blood.—Ex. Spots in Chilocorus Cacti, and Prothorax of Locusta morbillosa.
22. Rose-coloured (Roseus). Colour of the rose.—Ex. Parts of the Wings and Body of Deilephila Elpenor.
23. Crimson (Puniceus). A bright red with a tint of blue.—Ex. Base of the Under Wings of Catocala Sponsa.
24. Purple (Purpureus). Equal parts of blue and red.—Ex. Sagra purpurea. Vitta on the Elytra of Donacia fasciata.
25. Violet (Violaceus). Blue with some red. The colour of Viola odorata.—Ex. Chrysomela Goettingensis, Abdomen of Geotrupes vernalis.
26. Lilac (Lilacinus). Colour of the flowers of the lilac.—Ex. Part of the Iris of the Ocellus, in the Wings of Vanessa Io.
27. Blue (Cyaneus). Pure blue. Colour of Centaurea Cyanus.—Ex. Disk of the Wings of Papilio Ulysses. Callidium violaceum.
28. Azure (Azureus). A paler and more brilliant blue.—Ex. Wings of Morpho Menelaus, Telemachus, &c.
29. Sky-Blue (Cæruleus). A paler blue. The colour of the sky.—Ex. Polyommatus Adonis.
30. Cæsious (Cæsius). Very pale blue with a little black. The colour of blue eyes.—Ex. Under side of the Wings of Polyommatus Argiolus.
31. Green (Viridis). Equal parts of blue and yellow.—Ex. Cicindela campestris.
32. Æruginous (Æruginosus). Green with a blue tint. The colour of the rust of copper, verdigris.—Ex. Polydrosus Cnides.
33. Prasinous (Prasinus). Green with a mixture of yellow. The colour of the leaves of leeks or onions.—Ex. Pentatoma prasina. Under side of Wings of Thecla Rubi.
34. Glaucous (Glaucus). Pale blueish green. Sea green.—Ex. Elytra of Dynastes Hercules, Alcides, Tityus, &c.
35. Mouse-coloured (Murinus). Black with a small proportion of yellow. The colour of the common mouse.—Ex. Base of the abdominal segments of Cossus ligniperda.
36. Lurid (Luridus). Yellow with some mixture of brown. Dirty yellow.—Ex. Elytra of Aphodius luridus and nigrosulcatus.
37. Livid (Lividus). A pale purplish brown. The colour of a bruise.—Ex. Berosus luridus.
38. Tawny (Fulvus). A pale dirty orange.—Ex. The pale parts of the Wings of Hipparchia Pamphilus.
39. Fawn-coloured (Cervinus). A reddish brown.—Ex. Lasiocampa Rubi.
40. Olive (Olivaceus). A brownish green. The colour of olives.—Ex. Dytiscus marginalis.
41. Fuscous (Fuscus). A dull brown.—Ex. Hipparchia Semele. Prionus scabricornis.
42. Ferruginous (Ferrugineus). A yellowish brown with some red. The colour of the rust of iron.—Ex. Base of Under Wings of Smerinthus Populi. Gastropacha quercifolia.
43. Cinnamon-coloured (Cinnamomeus). A yellowish brown. The colour of cinnamon.—Ex. Prionus cinnamomeus.
44. Brown (Brunneus). Pure brown.—Ex. Dark parts in the Primary Wings of Euprepia Caja.
45. Bay (Badius). Bright red brown of the chestnut.—Ex. Elytra of Melolontha vulgaris when the hairs are rubbed off.
46. Chestnut (Castaneus). Colour of the dark part of the chestnut.—Ex. Elytra of Lucanus Cervus.
47. Piceous (Piceus). Shining reddish black. The colour of pitch.—Ex. Prionus coriarius.
48. Fuliginous (Fuliginosus). The opaque black of soot.—Ex. Wings of Lithosia rubricollis.
49. Black (Niger). A dull black with some brown.—Ex. Pachygaster niger.
50. Atrous (Ater). Pure black of the deepest tint.—Ex. Liparus anglicanus.
vi. SPLENDOUR.
a. gemmeous.
1. Margaritaceous (Margaritaceus). Glossy, white with changeable tints of purple, green, and blue.[Pg 291] The splendour of pearls.—Ex. The drums in Cicada capensis.
2. Opaline (Opalinus). A blueish white reflecting the prismatic colours. The splendour of the opal.—Ex. Wings of Notonecta glauca and some Nepæ.
3. Crystalline (Crystallinus). The white splendour of crystal or glass.—Ex. Stemmata of many Hymenoptera, &c.
4. Topazine (Topazinus). The yellow splendour of the topaz.—Ex. Many Stemmata of Hymenoptera, and Eyes of Spiders.
5. Rubineous (Rubineus). The red splendour of the ruby.
6. Smaragdine (Smaragdinus). The green splendour of the emerald.
7. Amethystine (Amethystinus). The purple splendour of the amethyst.
b. metallic.
1. Argent (Argenteus). The splendour of silver.—Ex. The spots on the under side of the Wings in Argynnis Lathonia, &c.
2. Golden (Aureus). The splendour of gold.—Ex. Entimus imperialis. Spot in the Wings of Plusia Festucæ.
3. Orichalceous (Orichalceus). A splendour intermediate between that of gold and brass.—Ex. Upper Wings of Plusia Chrysitis.
4. Æneous (Æneus). The splendour of brass.—Ex. Elytra of Carabus clathratus.
5. Cupreous (Cupreus). The reddening splendour of copper.—Ex. Carabus nitens.
6. Chalybeous (Chalybeus). The blue splendour of steel case-hardened, or of the mainspring of a watch.—Ex. Helops chalybeus. Legs of Lithosia Quadra.
7. Plumbeous (Plumbeus). The colour of lead.—Ex. Prothorax of Clytra dentata?
8. Inaurate (Inauratus). When striæ or other impressed parts have a metallic splendour.—Ex. Margin of Prothorax and Elytra of Carabus violaceus. Striæ of Elytra &c. of Phanæus Mimas.
9. Deaurate (Deauratus). A metallic hue which looks as if the gilding was worn off.—Ex. Donacia ænea, &c.
c. bombycine.
1. Sericeous (Sericeus). The splendour of silk.—Ex. Cryptocephalus sericeus.
2. Tramosericeous (Tramosericeus). The splendour of satin.—Ex. Chlamys Bacca, monstrosa, &c.
d. reflected.
1. Resplendent (Splendens). Reflecting the light intensely.—Ex. The Head and Thorax of Philonthus splendens, æneus, politus, &c.
2. Shining (Nitidus). Reflecting the light, but less intensely.—Ex. Dytiscus marginalis.
3. Pruinose (Pruinosus). When the splendour of the surface is somewhat obscured by the appearance of a bloom upon it like that of a plum, but which cannot be rubbed off[1091].—Ex. Elytra of Serica ruricola and brunnea.
4. Obscure (Obscurus). A surface which reflects the light but little.—Ex. Pælobius Hermanni.
5, Opaque (Opacus). A surface which does not reflect the light at all.—Ex. Trox sabulosus, arenarius. Silpha opaca.
vii. TRANSPARENCE.
1. Hyaline (Hyalina). The clear transparency of glass.—Ex. The Wings of many Neuroptera, Hymenoptera, and Diptera.
2. Diaphanous (Diaphana). Transparent, but less purely than hyaline. Semitransparent.—Ex. The Wings of many Coleoptera.
3. Adiaphanous (Adiaphana). Which does not transmit the light at all.—Ex. Elytra of Coleoptera.
viii. PAINTING.
1. Atom (Atomus). A very minute dot.
2. Irrorate (Irrorata). Sprinkled with atoms, as the earth with dew.—Ex. Onthophagus Vacca. Papilio Paris.
3. Gutta (Gutta[1092]). A roundish dot intermediate in size between an atom and a macula.
4. Guttate (Guttata). Sprinkled with guttæ.—Ex. Coccinella.
5. Macula (Macula). A larger indeterminately shaped spot.
6. Maculate (Maculata). Painted with such spots.—Ex. Abraxas grossulariata.
7. Litura (Litura). An indeterminate spot growing paler at one end, as if daubed or blotted.
8. Liturate (Liturata). A surface painted with one or more such spots.—Ex. Aphodius conflagratus.
9. Plaga (Plaga). A long and large spot.—Ex. Aphodius plagiatus.
10. Islet (Insula). A spot of a different colour, included in a plaga or macula.—Ex. The Ocelli in the Primary Wings of Hipparchia Semele. A spot in the middle fascia of the under side of the Primary Wing in Papilio Podalirius.
11. Crepera (Crepera). A gleam of paler colour upon a dark ground.—Ex. Elytra of Dytiscus marginalis.
12. Shadow (Umbra). A slight shade, not easily perceptible upon a paler ground.—Ex. Elytra of Acanthocinus Ædilis. Wings of Plutella asperella.
13. Signatures (Signaturæ). Markings upon a surface resembling in some degree letters and characters.
14. Signate (Signatus). Marked with signatures.—Ex. Elytra of Acrocinus longimanus.
15. Inscribed (Inscriptus). When the surface is marked with the resemblance of a letter of any language.—Ex. Plusia Gamma. Vanessa C. album.
16. Hieroglyphic (Hieroglyphicus). Painted with characters somewhat resembling hieroglyphics.—Ex. Acrocinus longimanus. Schizorhina Australasiæ.
17. Annulet (Annulus). A ring-shaped spot.—Ex. Cyclophora omicronaria, &c. Plate XIV. Fig. 1. o.
18. Lunulet (Lunula). A small crescent-shaped spot.—Ex. Marginal spots above and below the Secondary Wings in Melitæa Artemis, &c.
19. Reniculus (Reniculus). A small kidney-shaped spot.—Ex. Upper Wings of Mamestra Persicaria.
20. Ocellus (Ocellus). An eye-like spot in the Wings of many Lepidoptera, consisting of annuli of different colours, inclosing a central spot or pupil.
a. Pupil (Pupilla). The central spot of the ocellus. Plate XIV. Fig. 1. t. An ocellus is called bipupillate, tripupillate, &c., when there are two, three, &c. of these spots.—Ex. Primary Wing of Hipparchia Tithonus, &c. Plate XIV. Fig. 1. p.
a. Hastate Pupil (Pupilla hastata). When the pupil is a halberd-shaped spot.—Ex. Pupil of Ocellus of Aglia Tau. Plate XIV. Fig. 1. k.
b. Suffulted Pupil (Pupilla suffulta). When the pupil shades into another colour.—Ex. Primary Wing of Vanessa Io.
b. Iris (Iris). The circle which incloses the pupil. Plate XIV. Fig. 1. u.
c. Atmosphere (Atmosphæra). The exterior circle of the ocellus. Plate XIV. Fig. 1. v.
21. Blind Ocellus (Ocellus cæcus). An ocellus without the pupil.—Ex. Hipparchia Davus.
22. Spurious Ocellus (Ocellus spurius). A circular spot without any defined iris or pupil.—Ex. Spot in the Disk of the Primary Wings of Colias Helice.
23. Simple Ocellus (Ocellus simplex). When the ocellus consists only of iris and pupil.—Ex. Ocelli on the under side of Primary Wings of Hipparchia Semele. Plate XIV. Fig. 1. t, n, u.
24. Compound Ocellus (Ocellus complexus). When the ocellus consists of three or more circles.—Ex. Saturnia Spini. Plate XIV. Fig. 1. l.
25. Nictitant Ocellus (Ocellus nictitans). When the ocellus includes a lunular spot of a different colour.—Ex. Under side of Wings of Morpho Perseus. Plate XIV. Fig. 1. m.
26. Fenestrate Ocellus (Ocellus fenestratus). When an ocellus has a transparent spot.—Ex. Attacus Paphia and Cytherea.
27. Dioptrate Ocellus (Ocellus dioptratus). A fenestrate ocellus divided by a transverse line.—Ex. Attacus Polyphemus.
28. Double Ocellus (Ocellus geminatus). When two ocelli are included in the same circle or spot.—Ex. Under side of Secondary Wing of Morpho Perseus. Plate XIV. Fig. 1. r, v.
29. Twin Ocellus (Ocellus didymus). When such ocelli join each other.—Ex. Under side of Secondary Wing of Hipparchia Hyperanthus.
30. Sesquialterous Ocellus (Ocellus sesquialterus). An ocellus with a smaller near it, called also Sesquiocellus.—Ex. Under side of Secondary Wing of Colias Edusa. Plate XIV. Fig. 1. q.
31. Supercilium (Supercilium). An arched line resembling an eyebrow, which sometimes surmounts an eyelet.—Ex. Under side of Secondary Wing of Morpho Achilles. Plate XIV. Fig. 1. i.
32. Nebulose (Nebulosus). Painted with colour irregularly darker and lighter, so as to exhibit some resemblance of clouds.—Ex. Curculio sulcirostris, nebulosus; Catocala nupta.
33. Testudinarious (Testudinarius). Painted with red, black and yellow, like tortoise-shell.—Ex. Elytra of Aphodius testudinarius.
34. Consperse (Conspersus). Thickly sprinkled with minute irregular dots often confluent.—Ex. Biston Betularia.
35. Achatine (Achatinus). Painted with various concentric, curved, or parallel lines, resembling the veining of an agate.—Ex. Cossus labyrinthicus. Cerura vinula.
36. Ustulate (Ustulata). So marked with brown as to have the appearance of being scorched.—Ex. Wings of Ennomos dolabraria.
37. Marmorate (Marmorata). So painted with streaks, veins, and clouds, as to resemble marble.—Ex. Under side of the Wings of Vanessa Io. Marmarina marmorata.
38. Tessellate (Tessellata). Painted in checquerwork.—Ex. Abdomen of Sarcophaga carnaria and Musca maculata.
39. Fascia (Fascia). A broad transverse band.
a. Pyramidate Fascia (Fascia pyramidata). A band which juts out into an angle on one side.—Ex. Wing of Apatura Iris. Argynnis Paphia. Plate XIV. Fig. 1. h.
b. Macular Fascia (Fascia macularis). A band consisting of distinct spots.—Ex. Wings of Abraxas grossulariata. Plate XIV. Fig. 1. b.
c. Articulate Fascia (Fascia articulata). A band consisting of contiguous spots.—Ex. Under side of Wings of Melitæa Dictynna. Upper side of[Pg 298] Primary Wing of Morpho Menelaus. Plate XIV. Fig. 1. a.
d. Dimidiate Fascia (Fascia dimidiata). A band traversing only half the wing.—Ex. Primary Wing of Papilio Turnus. Plate XIV. Fig. 1. f.
e. Abbreviate Fascia (Fascia abbreviata). A band traversing less than half the wing.—Ex. Primary Wing of Papilio Podalirius, Ajax, &c. Plate XIV. Fig. 1. g.
f. Sesquialterous Fascia (Fascia sesquialtera). When both wings are traversed by a continued band, and either the primary or secondary by another.—Ex. Endromis versicolor. Plate XIV. Fig. 1. d, c.
g. Sesquitertious Fascia (Fascia sesquitertia). When both wings are traversed by a continued band, and more than half of either the primary or secondary by another; or, when a wing or elytrum contains a band and the third of a band.—Ex. Tortrix Avellana. Plate XIV. Fig. 1. d, e.
40. Striga (Striga). A narrow transverse streak.
41. Strigose (Strigosa). Painted with several such streaks.—Ex. Ennomos prunaria.
42. Line (Linea). A narrow longitudinal stripe.
43. Lineate (Lineata). Painted with several such stripes. N.B. If with two, we say bilineata, with three, trilineata, &c.—Ex. Elater lineatus.
44. Vitta (Vitta). A broad longitudinal stripe.
45. Vittate (Vittata). Painted with several such stripes.—Ex. Chrysomela fastuosa, cerealis, &c.
46. Undulate (Undulata). When fasciæ, strigæ, lines,[Pg 299] &c. curve into alternate sinuses resembling the rise and fall of waves.
47. Sinuato-Undulate (Sinuato-Undulata). When the sinuses are obtuse.—Ex. Boarmia repandaria.
48. Anguloso-Undulate (Anguloso-Undulata). When they go in a zigzag direction, or with alternate acute sinuses.—Ex. Acidalia undulata.
49. Radiate (Radiata). When a dot, spot, &c. appear to send forth rays.—Ex. The large blue area common to all the Wings of Papilio Ulysses.
50. Venose (Venosa). Painted with lines that branch like veins.—Ex. Under side of Wings of Pontia Napi.
51. Cancellate (Cancellata). Painted with transverse lines crossing longitudinal ones at right angles.—Ex. Macaria clathrata.
52. Areolate (Areolata). Painted with lines which intersect each other in various directions, so as to exhibit the appearance of net-work.—Ex. Wings of Tetanocera marginata and Cossus ligniperda.
53. Limbate (Limbata). When the disk is surrounded by a margin of a different colour.—Ex. Dytiscus marginalis.
54. Armillate (Armillata). When a leg, antenna, &c. is surrounded by a broad ring of a different colour.—Ex. Posterior Tibia of Prosopis annulata (Melitta * b. K.).
55. Annulate (Annulata). When a leg, antenna, &c. is surrounded by a narrow ring of a different colour.—Ex. Antennæ of many Ichneumons.
56. Cingulate (Cingulata). When the abdomen or the trunk is wholly surrounded by one or more[Pg 300] belts of a different colour.—Ex. Abdomen of many Nomadæ (Apis *. b. K.).
57. Decolorate (Decolor). When the colour appears to be discharged from any part.—Ex. Margin of the Abdominal segments in Stelis punctulatissima Latr. (Apis ** c. 1. β. K.).
58. Unicolorate (Unicolor). When a surface is of one colour.
59. Concolorate (Concolor). Of the same colour with another part. If speaking of Lepidoptera, when the upper and under sides of the wings are of the same colour.—Ex. Hesperia Linea, Paniscus.
60. Discolorate (Discolor). Of a different colour from another part. When the upper and under sides of the wings of Lepidoptera are of a different colour.—Ex. Polyommatus Corydon, Argiolus, &c.
61. Versicolorate (Versicolor). When a surface changes its colour as the light varies.—Ex. Apatura Iris ♂.
62. Iridescent (Iricolor). When a surface reflects the colours of the rainbow.—Ex. Mesothorax of Xylocopa iricolor. Wings of Hymenoptera, &c.
63. Infuscate (Infuscata). When a colour is darkened by the superinduction of a brownish shade or cloud.—Ex. Apex of the Upper Wings of Cossus ligniperda.
ix. DISTINCTION.
1. Distinct (Distincta). When spots, puncta, granules, &c. do not touch or run into each other, but are completely separate.—Ex. Under side of Wings of Lycæna Hippothoe.
2. Ordinate (Ordinata). When spots, puncta, &c. are placed in rows. Thus we say ordinato-punctate, ordinato-maculate, &c.—Ex. Spots on the Abdomen of Spilosoma lubricipeda, erminea, &c.
3. Contiguous (Contigua). When spots, &c. are so near that they almost or altogether touch each other.—Ex. Spots in the margin of the Wings of Argynnis Aglaia.
4. Confluent (Confluens). When spots, &c. run into each other.—Ex. Apex of the Primary Wings and Under side of the Secondary in Pontia Daplidice.
5. Obliterate (Obliterata). When the borders of spots fade into the general ground-colour; and when elevations and depressions, &c. are so little raised or sunk from the general surface, as to be almost erased.—Ex. Streak in the Wings of Hipparchus papilionarius, &c. Striæ in the Elytra of Sphodrus leucophthalmus.
6. Obsolete (Obsoleta). When a spot, tubercle, punctum, &c. is scarcely discoverable. Ex. Lycæna Hippothoe ♂ and ♀.—N.B. This term is often employed where one sex, kindred species, or genera, want, or nearly so, a character which is conspicuous in the other sex, or in the species or genus to which they are most closely allied.
7. Geminous (Gemina). When there is a pair of spots, tubercles, puncta, &c.—Ex. Head of one sex of Dorcus parallelopipedus. Upper Wings of Odenestis potatoria.
8. Didymous (Didyma). When this pair of spots, &c. touch or are confluent.—Ex. Spots in Elytra of Tylostagmus quadrimaculatus.
9. Connivent (Connivens). The meeting of two lines so as to form an angle.—Ex. Streaks on the Under side of Secondary Wings of Thecla Pruni.
10. Common (Communis). Common to two. When a spot for instance is partly on one elytrum and partly on the other.—Ex. Coccinella septempunctata.
VIII. MARGIN.
1. Crisp (Crispa). When the Limb is disproportionably larger than the Disk, so as to render the margin uneven with irregular rises and falls.
2. Undulate (Undulata). When the surface rises and falls obtusely, not in angles.—Ex. Margin of Wings of Hipparchia Semele.
3. Corrugate (Corrugata). When the surface rises and falls acutely in angles.—Ex. Acidalia luteata, &c.
4. Plicate (Plicata). Longitudinally or transversely folded; or so impressed with striæ as to have that appearance.—Ex. Abdomen of Staphylinus.
5. Dilatate (Dilatata). Dilated disproportionably with respect to the Disk.—Ex. Prothorax of Necrophorus.
6. Filate (Filata). When the edge is separated by a channel, often producing a very slender threadlike margin.—Ex. Elytra of Choleva.
7. Incrassate (Incrassata). When the margin is disproportionably thick.—Ex. Mr. Marsham's Family of Chrysomela "thorace utrinque incrassato."
8. Intire (Integra). When the margin has neither teeth, serratures, nor other incisions.
9. Channel (Canalis). An impressed line more or[Pg 303] less wide, which attends the edges, and is usually produced by its reflexion.
10. Edge (Acies). The extreme termination of the margin.
IX. TERMINATION.
1. Summit (Fastigium). The tip or extreme termination of the upper part.
2. Apex (Apex). The top or upper termination of any part.
3. Bottom (Fundus). The extreme termination of the lower part.
4. Base (Basis). The lower termination of any part.
5. Acute (Acuta). Terminating in an acute angle. Plate XV. Fig. 17.
6. Obtuse (Obtusa). Terminating bluntly, but within the segment of a circle. Plate XIV. Fig. 1. f.
7. Rotundate (Rotundata). Terminating in the segment of a circle.—Plate VI. Fig. 1. e.
8. Truncate (Truncata). Terminating in a transverse line. Plate XIII. Fig. 5. a´´´.
9. Premorse (Præmorsa). Terminating in an irregular truncate apex, as if bitten off.—Ex. Elytra of Lebia, Dromia, Lomechusa, &c.
10. Retuse (Retusa). Terminating in an obtuse sinus.—Ex. Back part of the Head in Cimbex.
11. Emarginate (Emarginata). When the end has an obtuse notch taken out.—Ex. Nose of Pedinus arenosus.
12. Exscinded (Excisa). When the end has an angular notch taken out.—Ex. Nose of Opatrum sabulosum.
13. Producted (Producta). Disproportionably long.
14. Mucronate (Mucronata). Terminating suddenly in a strong point.—Ex. Elytra of Lixus paraplecticus. Abdomen of Sirex Gigas ♀.
15. Acuminate (Acuminata). Terminating gradually in a sharp point.—Ex. Abdomen of Sirex Juvencus ♀.
16. Apiculate (Apiculata). Terminating suddenly in a small filiform truncate apex.—Ex. Abdomen of Thelyphonus.
17. Cuspidate (Cuspidata). Terminating in a long setiform point.—Ex. Tail of Scorpio.
X. INCISION.
1. Incised (Incisa). Cut into equal marginal segments.
2. Cleft (Fissa). Cut into equal and deep segments, but not reaching the base. Plate XIV. Fig. 3. a.
a. Bifid (Bifida). Cut into two segments.
b. Trifid (Trifida). Cut into three.
c. Quadrifid (Quadrifida). Cut into four.
d. Multifid (Multifida). Cut into more than four.
3. Laciniate (Laciniata). Cut into unequal, irregular, and deep segments.
4. Squarrose (Squarrosa). Cut into laciniæ that are elevated above the plane of the surface.
5. Partite (Partita). Divided to the base. Plate XIV. Fig. 3. b.
a. Bipartite (Bipartita). Divided thus into two parts.
b. Tripartite (Tripartita). Divided into three parts.
c. Quadripartite (Quadripartita). Divided into four parts.
d. Multipartite (Multipartita). Divided into more than four parts.
6. Lobate (Lobata). Divided to the middle into parts with convex margins, which recede from each other.—Ex. Acanthia paradoxa. Bilobate, with two lobes. Trilobate, with three lobes, &c.
7. Cruciate (Cruciata). Divided to the middle into four opposite arms, the angles being either four right ones, or two obtuse and two acute.—Ex. Prothorax of many Locustæ.
8. Sinuate (Sinuata). Having large curved breaks in the margin resembling bays. Plate XIV. Fig. 1.
9. Erose (Erosa). Sinuate, with the sinuses cut out into smaller irregular notches as if gnawed.—Ex. Wings of Vanessa C. album.
10. Crenate (Crenata). Cut into segments of small circles.
11. Serrate (Serrata). Cut into teeth like a saw, with teeth whose sides are unequal.—Ex. External margin near the Apex of the Elytra of many species of Buprestis.
12. Dentate (Dentata). Cut into teeth, with teeth whose sides are equal or nearly so.—Ex. The Wings of many Butterflies.
13. Repand (Repanda). Cut into very slight sinuations, so as to run in a serpentine direction. Plate XXII. Fig. 11. s.
XI. RAMIFICATION.
1. Dichotomous (Dichotoma). Dividing regularly in pairs.
2. Furcate (Furcata). Dividing into two. Plate XVIII. Fig. 11.
3. Ramose (Ramosa). Furnished with lateral branches. Plate XI. Fig. 18.
4. Decussate (Decussata). Sending forth lateral branches which alternately cross each other.
5. Divaricate (Divaricata). Standing out very wide.
XII. DIVISION.
1. Segment (Segmentum). The great inosculating joints of the body.
2. Joint (Articulus). The joints of a limb or member.
3. Incisure (Incisura). A deep incision between the segments, when they recede from each other.
4. Suture (Sutura). The line of separation of any two parts of a crust which are connected only by membrane or ligament, but do not inosculate.
a. Spurious Suture (Sutura spuria). An impressed line in any part of a body, which resembles a suture, but does not really divide the crust.
XIII. DIRECTION.
1. Longitudinal (Longitudinalis). Running lengthwise.
2. Transverse (Transversa). Running across: when the longitudinal line is cut through at right angles.
3. Oblique (Obliqua). Running sideways. When the longitudinal line is cut through at acute angles.
4. Horizontal (Horizontalis). Parallel with the horizon.
5. Erect (Erecta). Nearly perpendicular.
6. Vertical (Verticalis). Perpendicular.
7. Sloping (Declivis). A gentle descent.
8. Descending (Descendens). A steeper descent.
9. Acclivous (Acclivis). A gentle ascent.
10. Ascending (Ascendens). A steeper ascent.
11. Reclined (Reclinata). Leaning towards any thing as if to repose upon it.
12. Recumbent (Recumbens). Leaning or reposing upon any thing.
13. Reflexed (Reflexa). Bent back or upwards.
14. Inflexed (Inflexa). Bent inwards.
15. Recurved (Recurva). Curving outwards.
16. Incurved (Incurva). Curving inwards.
17. Revolute (Revoluta). Rolled outwards.
18. Involute (Involuta). Rolled inwards.
19. Forwards (Antrorsum).
20. Backwards (Retrorsum).
21. Upwards (Sursum).
22. Downwards (Deorsum).
23. Outwards (Extrorsum).
24. Inwards (Introrsum).
25. Straight (Recta).
26. Porrect (Porrecta). Reaching forth horizontally as if to meet something advancing.
27. Broken (Fracta). Bent with an elbow, as if broken.
28. Converging (Convergens). Tending to one point from different parts.
29. Diverging (Divergens). Tending to different parts from one point.
XIV. SITUATION.
1. Obverse (Obversa). When an object is viewed with its head towards you.
2. Reverse (Reversa). When an object is viewed with its anus towards you.
3. Resupine (Resupina). When an object lies upon its back.
4. Prone (Prona). When an object lies upon its belly.
XV. CONNEXION.
1. Colligate (Colligata). Adhering, or so fixed to any part as to have no separate motion of its own.
2. Free (Libera). Having a motion independent of that of the part to which it is affixed.
3. Connate (Connata). When parts that are usually separated, are, as it were, soldered together, though distinguished by a suture.—Ex. Elytra of Gibbium.
4. Coalite (Coalita). When parts usually separate are distinguished neither by incisure, segment, nor suture.—Ex. Trunk in Mutilla.
5. Distinct (Distincta). When parts are separated from each other by a suture.—Ex. Parts of the Trunk in Coleoptera, &c.
6. Distant (Distans). When they are separated by an incisure.—Ex. Head, Trunk, and Abdomen, in Hymenoptera.
7. Inosculating (Inosculans). When one part is inserted into the cavity of another.—Ex. Head in Buprestis.
8. Suspended (Suspensa). When one part is joined to another by a ligature, without being inserted in it.—Ex. Legs of Orthoptera.
XVI. ARMS.
1. Tooth (Dens). A short flattish process, somewhat resembling a tooth.
2. Horn (Cornu). A longer process, resembling a horn.
a. Laminate Horn (Cornu laminatum). A horn dilated at its base into a flat plate.—Ex. Onthophagus nutans.
b. Nodding Horn (Cornu nutans). When a horn bends forwards.—Ex. Onthophagus nutans.
3. Spine (Spina). A fine, long, rigid, pointed process.—Ex. Those on Elytra of many Hispæ, and the Posterior Tibiæ of Locusta.
4. Mucro (Mucro). A short, stout, sharp-pointed process.—Ex. Elytra of Lixus paraplecticus.
5. Spur (Calcar). A spine that is not a process of the crust, but is implanted in it.—Ex. Those on the lower side of the Tibiæ of Acrida.
XVII. APPENDAGES.
1. Auricle (Auricula). An appendage resembling an ear.—Ex. Thorax of Ledra aurita.
2. Caruncle (Caruncula). Having fleshy excrescences somewhat resembling the caruncles of birds.—Ex. Prothorax of Malachius.
XVIII. MOTION.
1. Vertical (Verticalis). When it is up and down.
2. Horizontal (Horizontalis). When it is from side to side.
3. Compound (Composita). When a part is capable of both vertical and horizontal motion.
4. Versatile (Versatilis). When it moves partly round as if upon a pivot.—Ex. Head of Hymenoptera and Diptera.
5. Vibratile (Vibratilis). When there is a constant oscillation of any part.—Ex. Antennæ of the Chalcidites. Legs of Tipula when reposing.
6. Rotatory (Rotatoria). When a body or a part of it turns wholly round, or describes a circle.—Ex. Ants and Moths in a certain disease[1093].
XIX. SCENT.
1. Acid (Acidus). A pungent acid scent.—Ex. Many Formicæ.
2. Moschate (Moschatus). A scent of musk.
3. Alliaceous (Alliaceus). A scent of garlic.—Ex. Some Andrenæ.
4. Cimicine (Cimicinus). A scent like that of the Bed-bug.—Ex. Cimex.
5. Rosaceous (Rosaceus). A scent of roses.—Ex. Cerambyx moschatus.
6. Aromatic (Aromaticus). A pungent scent of spices.—Ex. Oxytelus rugosus.
7. Balm-scented (Melissæus).—Ex. The species of Prosopis (Apis *. b. K.).
8. Sweet-scented (Odoratus). An undefined sweet scent.—Ex. Philonthus suaveolens.
9. Fetid (Fœtidus). A disagreeable scent.—Ex. Goerius olens. Chrysopa Perla.
GENERAL RULES.
In the above tables no notice is taken of diminutives, compounds, and similar terms, because it seemed best, with respect to these, to lay down only some general rules which may include the whole.
Rule I.
Terms in English ending in cle, ule, or let, and which in Latin add lus, la, or lum, to a word, diminish its sense. As, Denticle, a little tooth; Setule, a little bristle; Eyelet, a small eye: Denticulus, a little dens; Guttula, a little gutta; Punctulum, a little punctum. N.B. Where length or breadth are concerned, the diminutive implies a diminution in the length of the predicate. As, Lineola, Lineolet, a short line; Striola, Striolet, a short stria; Fasciola, Fasciolet, a short fascia.
Rule II.
The preposition sub prefixed to any word reduces the sense of it. As, Subpunctate, not fully punctate; Subhirsute, not fully hirsute, &c.
Rule III.
The termination culus in Latin words added to a comparative implies the state of the object comparatively. As, Convexiusculus, rather convex than not; Majusculus, rather large than not. This is usually denoted in English by the termination ish, or the adverb rather; as, largish, rather large, &c.
Rule IV.
The participle present used instead of the adjective[Pg 312] implies a tendency to the quality expressed by it. As, Cinerascens, cinerascent, tending to cinereous, &c.
Rule V.
The preposition ob prefixed to a term reverses it. As, Obconical, Obcordate, a conical or heart-shaped body, of which the narrowest part is the base.
Rule VI.
In compound terms the last member indicates the preponderating character. For instance, when it is said of a body that it is nigro-æneous, it means that the æneous tint prevails: but if, vice versâ, it is termed æneo-nigrous, the black tint is predominant.—N.B. In Sculpture the terms punctato-striate, or punctato-sulcate, signify that striæ or furrows are drawn with puncta in them.
Exception 1.
Some compound terms only indicate the union of two characters in one subject. As, when we say of wings that they are cruciato-incumbent, we mean both that they cross each other and are incumbent upon the body.
Exception 2.
Compound terms are sometimes employed very conveniently to restrict the application of a character to particular circumstances. As, when we say hirsuto-cinereous, we mean that the hirsuties only of a body is cinereous.
Rule VII.
When the term ordinary (ordinarius) is added either to terms expressing impressed puncta, lines, spots, &c., it signifies that such puncta, lines, or spots are common[Pg 313] to a particular section in any genus or tribe. As, the impressed lateral puncta on the thorax of Scarabæidæ; the lateral furrows and dorsal channel in the ground beetles (Eutrechina), and the spots in the primary wings of Xylina Polyodon and affinities[1094].
SYMBOLS.
Male ♂. Female ♀. Neuter ⚲. Egg θ. Larva ⊕. Pupa ☽. Imago ⊙. Head △. Trunk □. Abdomen ▽[1095].
B. PARTIAL ORISMOLOGY.
I. BODY (corpus).
1. Disjunct (Disjunctum). When head, trunk, and abdomen are separated by a deep incisure.—Ex. Hymenoptera, Diptera. Plate IV. Fig. 2, 3, 5.
2. Compact (Compactum). When head, trunk, and abdomen are not separated by a deep incisure, but inosculate in each other.—Ex. Buprestis, Elater, and many other Coleoptera, Orthoptera, and Hemiptera.
3. Bisect (Bisectum). When the head and trunk are not separated by a suture, so that the insect consists only of two pieces.—Ex. Araneidea. Plate V. Fig. 4.
4. Coalite (Coalitum). When neither head, trunk,[Pg 314] nor abdomen are separated by any incisure or suture.—Ex. Many Acari L., Phalangium, &c.
5. Multisect (Multisectum). When an insect appears to have no distinct trunk and abdomen, but is divided into numerous segments.—Ex. Scolopendra; Iulus, &c. Plate V. Fig. 6.
6. Cymbiform (Cymbiforme). When the margin of the thorax and elytra are recurved so as to give a body the resemblance of the inside of a boat.—Ex. Heleus, Cossyphus.
II. HEAD (caput).
i. DIRECTION.
1. Prominent (Prominens). When the head is in the horizontal line, and forms no angle with the trunk.—Ex. Carabus. Plate I. Fig. 1.
2. Porrected (Porrectum). When the head is prominent and elongate.—Ex. Cychrus.
3. Nutant (Nutans). When the head forms downwards an obtuse angle with the horizontal line, or trunk.—Ex. Harpalus.
4. Cernuous (Cernuum). When the head forms downwards a right angle with the trunk.—Ex. Most Gryllina and Locustina.
5. Inflexed (Inflexum). When the head forms inwards an acute angle with the trunk.—Ex. Blatta. Plate II. Fig. 3.
6. Turreted (Turritum). When the head is producted into a kind of columnar recurved turret or rostrum, in the sides of which, towards the end, the eyes are fixed.—Ex. Truxalis.
ii. INSERTION.
1. Retracted (Retractum). When the head is wholly withdrawn within the trunk.—Ex. Parnus.
2. Intruded (Intrusum). When the head is nearly withdrawn within the trunk.—Ex. Melasis.
3. Inserted (Insertum). When the head is partly withdrawn within the trunk.—Ex. Buprestis.
4. Exserted (Exsertum). When the head is quite disengaged from the trunk.—Ex. Tenebrio, Blaps.
5. Amplected (Amplexum). When the head is received into a sinus of the thorax.—Ex. Hister.
6. Recondite (Reconditum). When the head is wholly covered and sheltered by the shield of the thorax.—Ex. Cassida, Lampyris.
7. Semirecondite (Semireconditum). When the head is half covered by the shield of the thorax.—Ex. Silpha, Cyphon.
8. Retractile (Retractile). When an insect can at pleasure exsert its head, or withdraw it within the trunk.—Ex. Hister, Larva of Lampyris.
9. Versatile (Versatile). When the head can turn nearly round.—Ex. Hymenoptera, Diptera.
10. Pedunculate (Pedunculatum). When the head is constricted behind into a distinct neck.—Ex. Apoderus Coryli, &c.
11. Sessile (Sessile). When the head does not move in the socket of the trunk, but is attached to it by a kind of ligament.—Ex. Hymenoptera, Diptera.
iii. TERMINATION.
1. Clypeate (Clypeatum). When the Nasus, Genæ, &c. are dilated so as to shelter and overshadow the mouth.—Ex. Scarabæus, Copris, &c. Plate XIII. Fig. 14.
2. Capistrate (Capistratum). When the anterior part of the head is attenuated and subelongated into a kind of flat rostrum or muzzle.—Ex. Nitidula. Plate XIII. Fig. 13.
3. Rostrate (Rostratum). When the anterior part of the head is elongated and attenuated into a cylindrical or many-sided rostrum or beak.—Ex. The weevils (Rhyncophora). Plate XIII. Fig. 12.
4. Buccate (Buccatum). When the Nasus and anterior part of the head are inflated.—Ex. Conops and other Diptera. Plate XIII. Fig. 16.
iv. APPENDAGES.
1. Umbraculate (Umbraculatum). When there is upon the head an umbrella-shaped process.—Ex. Gryllus umbraculatus.
2. Lychnidiate (Lychnidiatum). When the Vertex, Frons, and Postnasus are porrected so as to form a kind of rostrum which gives light in the night.—Ex. Fulgora. Plate XIII. Fig. 15.
v. MOUTH (os).
1. Terminal (Terminale). When the mouth terminates the head.—Ex. Coleoptera, &c.
2. Prone (Pronum). When the mouth is wholly under the head.—Ex. Truxalis, Proscopia.
3. Perfect (Perfectum). When the mouth is furnished with all the Trophi. Viz. Labrum; Labium;[Pg 317] Mandibulæ; Maxillæ; Maxillary and Labial Palpi; and Tongue.—Ex. The Masticating Orders.
a. Feeders retracted (Trophi retracti). When, in a perfect mouth, the Trophi are not capable of being much pushed out or drawn in.—Ex. Most Coleoptera, Orthoptera, &c.
b. Feeders retractile (Trophi retractiles). When, in a perfect mouth, the Trophi can be considerably pushed forth or drawn in.—Ex. Stenus, Apis, &c.
4. Imperfect (Imperfectum). When the mouth wants any of the Trophi, or they exist in it only as rudiments.—Ex. The Suctorious Orders.
5. Elabrate (Elabratum). When an imperfect mouth has Mandibulæ, Maxillæ, Labium, and Maxillary Palpi, or what perform their office, but no Labrum.—Ex. Araneidea.
6. Emandibulate (Emandibulatum). When an imperfect mouth has all the Trophi but the Mandibulæ.—Ex. Trichoptera.
7. Bipalpate (Bipalpatum). When an imperfect mouth has only either Labial or Maxillary Palpi.—Ex. Tabanus, &c.
8. Expalpate (Expalpatum). When an imperfect mouth has no Palpi.—Ex. Hemiptera.
9. Stomapodous (Stomapodum). When the Legs and Sternum act the part of Maxillæ, Labium, and Palpi.—Ex. Araneidea, Scolopendra, &c.
a. upper lip (Labrum).
1. Whiskered (Mystacinum). When the upper lip is furnished with whiskers (Mystax), or bearded.—Ex. Creophilus hirtus. Plate XXVI. Fig. 30.
b. upper jaws (Mandibulæ).
1. Chelate (Chelatæ). When the upper jaws are furnished at the end with a chela or thumb.—Ex. Scorpio, Phalangium.
2. Unguiculate (Unguiculatæ). When they are armed with a moveable claw.—Ex. Araneidea. Plate VII. Fig. 10. c´.
3. Buried (Sepultæ). When they are covered and quite concealed by the upper lip.—Ex. Colliuris.
4. Open (Apertæ). When they are not quite concealed by the upper lip.—Ex. Most Coleoptera.
5. Toothless (Edentulæ). When they are not armed with teeth.—Ex. Apogonia gemellata. Plate XXVI. Fig. 22.
6. Toothed (Dentatæ). When they are armed with teeth.—Ex. Cicindela. Plate XXVI. Fig. 19.
7. Suctorious (Suctoriæ). When they have an orifice by which they imbibe their food.—Ex. Larva of Dytiscus, Myrmeleon, &c. Plate XIII. Fig. 6.
c. under jaws (Maxillæ).
1. Simple (Simplices). When the under jaws have but one lobe.—Ex. Hymenoptera. Plate VII. Fig. 2, 3. d´.
2. Compound (Compositæ). When they have more than one lobe.—Ex. Staphylinus and many other Coleoptera. Plate XXVI. Fig. 9, 10. d´´´. e´´´.
3. Adnate (Adnatæ). When they adhere to the lower lip through their whole length.—Ex. Trichoptera. Plate VII. Fig. 1. d´.
4. Adherent (Adhærentes). When they adhere to it[Pg 319] only at their base.—Ex. Coleoptera, Hymenoptera, &c. Plate VI. VII. Fig. 3. d´.
5. Spinose (Spinosæ). When they are armed at the apex with spines.—Ex. Libellulina. Plate VI. Fig. 12. f´´´.
6. Dentate (Dentatæ). When they are armed with teeth.—Ex. Melolonthidæ. Plate XXVI. Fig. 15. g´´.
7. Pectunculate (Pectunculatæ). When the stipes below the feeler has a row of minute spines set like the teeth of a comb.—Ex. Apis[1096].
8. Disengaged (Liberæ). When they do not adhere to the lower lip at all, or are only connected by membrane or ligaments.—Ex. Apis, &c. Plate VII. Fig. 3. d´.
9. Mandibuliform (Mandibuliformes). When they are hard and horny and shaped like the upper jaws.—Ex. Melolonthidæ, Anoplognathidæ, &c. Plate XXVI. Fig. 13, 15.
10. Unguiculate (Unguiculatæ). When they terminate in a moveable claw.—Ex. Cicindela.
d. feelers (Palpi).
1. Maniform (Maniformes). When they are chelate or furnished with a finger and thumb.—Ex. Scorpio, Chelifer. Plate XV. Fig. 7.
2. Pediform (Pediformes). When they resemble the legs either in structure or use.—Ex. Araneidea, Acarina. Plate VII. Fig. 10. h´´.
3. Antenniform (Antenniformes). When they are very long resembling antennæ.—Ex. Hydrophilus, Bryaxis, Culex ♂.
4. Unguiculate (Unguiculati). When they are armed with a claw at the end.—Ex. Gonyleptes. Plate XIII. Fig. 1.
5. Securiform (Securiformes). When the last joint of the feeler is triangular, and the preceding joint is connected with the vertex of the triangle.—Ex. Cleridæ. Plate XIII. Fig. 2. a.
6. Lunulate (Lunulati). When the last joint is shaped like a half-moon or crescent.—Ex. Oxyporus. Plate XIII. Fig. 4. a.
7. Fasciculate (Fasciculati). When the feeler terminates in a bunch of very slender laminæ.—Ex. Lymexylon flavipes. Plate XXVI. Fig. 3.
8. Lamellate (Lamellati). When the last joint is divided into transverse lamellæ.—Ex. Atractocerus. Plate XXVI. Fig. 1.
9. Inflated (Inflati). When the last joint of the feeler is very large and tumid.—Ex. Araneidea ♂. Plate XIII. Fig. 3.
10. Appendiculate (Appendiculati). When from one of the joints there issues an accessory joint or appendage.—Ex. Atractocerus, Trombidium. Plate XXVI. Fig. 1. a. Plate XXIII. Fig. 13. a.
11. Mammillate (Mammillati). When the last joint is very short, smaller than the preceding one, and retractile within it.—Ex. Pæderus[1097].
12. Subulate (Subulati). When the last joint is short, and vastly smaller than the preceding one.—Ex. Bembidium, Aleochara. Plate XXVI. Fig. 7.
13. Fusiform (Fusiformes). When the two last joints are[Pg 321] conical, and the base of the cones forms the point of union.—Ex. Trechus. Plate XXVI. Fig. 8.
14. Heteromorphous (Heteromorphi). When the two intermediate joints are vastly larger than the first and the last.—Ex. Cerocoma ♂. Plate XXVI. Fig. 2.
e. tongue (Lingua).
1. Linguiform (Linguiformis). When the tongue is quite distinct from the labium, usually retracted within the mouth, short and shaped something like a vertebrate tongue.—Ex. Gryllina, Libellulina. Plate VI. Fig. 6, 12. e´.
2. Liguliform (Liguliformis). When it emerges from the labium, is short, flat, and not concealed within the mouth.—Ex. Vespa and many Hymenoptera. Plate VII. Fig. 2. e'.
3. Tubulose (Tubulosa). When it emerges from the labium, is long and tubular, and capable of inflation.—Ex. Apis. Plate VII. Fig. 3. e´.
4. Setiform (Setiformis). A short minute sharp tongue discoverable between the scalpella of a promuscis.—Ex. Cimex L. Plate VII. Fig. 14. e´.
5. Palatiform (Palatiformis). When the tongue forms the inner surface of the Labium, but is not separate from it.—Ex. Most Coleoptera.
vi. NOSE (nasus).
1. Included (Inclusus). When the nose is included between the two sides of the Postnasus which run towards the upper lip.—Ex. Geocorisæ.
2. Vaulted (Fornicatus). When the nose is elevated, convex and hollow underneath.—Ex. Vespa.
vii. CANTHUS.
1. Entering (Intrans). When the Canthus takes a little angle or sinus out of the eye.—Ex. The Capricorn beetles, Mylabris. Plate VI. Fig. 1. h´.
2. Cleaving (Findens). When the Canthus cleaves the eye half through or more.—Ex. The Petalocerous beetles.
3. Dividing (Dividens). When the Canthus passes right through the eye and divides it into two.—Ex. Gyrinus, Tetraopes.[1098] Plate XXVI. Fig. 36.
4. Septiform (Septiformis). When the Canthus forms an elevated ridge or septum.—Ex. Many Petalocerous beetles.
viii. EYES (oculi).
1. Simple (Simplices). Eyes which do not consist of an aggregate of hexagonal lenses.—Ex. Araneidæ, Scorpio, Phalangium. Plate VII. Fig. 9. h.
a. Scattered (Sparsi). When simple eyes are separate from each other and not arranged in a certain order.—Ex. Eyes of Caterpillars, and some Scolopendræ.
b. Ordinate (Ordinati). When simple eyes are arranged in a certain order.—Ex. Araneidea. Plate XXVI. Fig. 37.
c. Conglomerate (Conglomerati). When a number of simple eyes are collected together so as to exhibit the appearance of a compound one.—Ex. Iulus. Plate XIII. Fig. 11.
d. Dorsal (Dorsales). When they are placed on the[Pg 323] back.—Ex. Phalangium. Plate XXVI. Fig. 43. h.
2. Compound (Compositi). Eyes which consist of an aggregate of hexagonal lenses.—Ex. All the Winged Orders. Plate XIII. Fig. 10. and XXVI. Fig. 38-42. h.
a. Sessile (Sessiles). Eyes that do not sit upon a footstalk.—Ex. Most insects. Plate XXVI. Fig. 40, 41.
α. Superior (Superiores). When they are placed in the upper part of the head.—Ex. Libellulina.
β. Lateral (Laterales). When they are placed in the side of the head.—Ex. Apis.
γ. Inferior (Inferiores). When they are placed in the lower side of the head.—Ex. The lower pair in Gyrinus.
δ. Posterior (Posteriores). When placed in the posterior part of the head.—Ex. Locusta.
ε. Anterior (Anteriores). When placed in the anterior part of the head.—Ex. Crabro, Goerius olens, &c.
ζ. Medial (Medii). When placed in the middle part of the head.—Ex. Harpalus, &c.
η. Belting (Cingentes). When the eyes nearly meet both above and below the head, so as to form a kind of belt round it.—Ex. Culex pipiens, Cordylia Palmarum.
θ. Immersed (Immersi). When they are quite imbedded in the head.—Ex. The Melasoma or Darkling beetles.
ι. Prominent (Prominuli). When they stand out from the head.—Ex. Cicindela.
b. Columnar (Columnares). When they sit upon a[Pg 324] short footstalk or pillar.—Ex. Strepsiptera, Ephemera ♂. Plate XXVI. Fig. 38, 39. h.
c. Pedunculate (Pedunculati). When they sit upon a long footstalk which also bears the antennæ.—Ex. Diopsis. Plate XIII. Fig. 9.
d. Operculate (Operculati). When the eyes are covered by an operculum.—Ex. Xylina conspicillaris[1099].
e. Ciliate (Ciliati). When the margin of the socket of the eye is fringed with hairs, so as to resemble an eyelash.—Ex. Apion vernale and Malvarum.
ix. STEMMATA.
1. Vertical (Verticalia). When they are placed in the Vertex.—Ex. Reduvius personatus. Plate XXVI. Fig. 40. i.
2. Frontal (Frontalia). When placed in the Frons.—Ex. Hymenoptera. Plate VII. Fig. 2. i.
3. Intraocular (Intraocularia). When placed in the space between the eyes.—Ex. Cercopis, Ledra, &c. Plate XXVI. Fig. 42. i.
4. Subocular (Subocularia). When placed in the space below the eyes.—Ex. Fulgora laternaria. Plate XXVI. Fig. 41. i.
5. Spurious (Spuria). A flat subdiaphanous space above the base of the antennæ, which seems to represent them.—Ex. Blatta, Gryllus.
x. ANTENNÆ.
a. number.
1. Dicerous (Dicera). Insects that have two antennæ.—Ex. Insects in general.
2. Acerous (Acera). Insects that have no antennæ.—Ex. The Acarina, &c.
b. situation.
1. Preocular (Præoculares). When antennæ are inserted before the eyes.—Ex. Chrysis.
2. Interocular (Interoculares). When inserted any where between the eyes.—Ex. Leptura, Haliplus.
3. Inocular (Inoculares). When inserted in the Canthus of the eyes.—Ex. The Capricorn beetles.
4. Subocular (Suboculares). When inserted under the eyes.—Ex. Fulgora, Nepa. Plate XXVI. Fig. 41. k.
5. Extraocular (Extraoculares). When inserted without the eyes.—Ex. Notonecta, Delphax.
6. Rostral (Rostrales). When seated on a rostrum.—Ex. The Rhyncophorous beetles.
7. Superior (Superiores). When inserted in the upper surface of the head.—Ex. Most insects.
8. Inferior (Inferiores). When inserted under the head.—Ex. Copris, &c.
c. approximation.
1. Distant (Distantes). When remote at their base.—Ex. Buprestis rustica.
2. Approximate (Approximatæ). When they approach each other at their base.—Ex. Donacia, Galeruca.
3. Contiguous (Contiguæ). When they nearly or altogether touch each other at their base.—Ex. Imatidium MacLeayanum.
4. Connate (Connatæ). When united at their base.—Ex. Ceria. Plate XII. Fig. 13.
d. proportion.
1. Very short (Brevissimæ). When shorter than the head.—Ex. The Muscidæ, &c.
2. Short (Breves). When as long as the head.—Ex. Hister.
3. Shorter (Breviores). When longer than the head and shorter than the body.—Ex. Dytiscus.
4. Mediocral (Mediocres). When of the length of the body.—Ex. Callidium violaceum.
5. Longer (Longiores). When longer than the body.—Ex. Monochamus Sutor.
6. Very long (Longissimæ). When much longer than the body.—Ex. Acanthocinus ædilis.
e. direction.
1. Intire (Integræ). When they have no elbow or angle.—Ex. Antennæ of most Coleoptera.
2. Broken (Fractæ). When the Clavola forms an angle with the Scapus.—Ex. Curculio, Apis, &c. Plate XXV. Fig. 15.
3. Geniculate (Geniculatæ). When they form an elbow in the middle but not with the Scapus.—Ex. Meloe. Plate XII. Fig. 7.
4. Straight (Rectæ). When they are without any angle, convolution, or curvature. Plate XI. Fig. 5.
5. Porrect (Porrectæ). When they are placed parallel with each other, and in the same line with the body.—Ex. Trichoptera in flight.
6. Excurved (Excurvæ). When they curve outwards.
7. Incurved (Incurvæ). When they curve inwards.
8. Decurved (Decurvæ). When they curve downwards.
9. Recurved (Recurvæ). When they curve upwards.
10. Reflexed (Reflexæ). When they are bent back over the body.
11. Deflexed (Deflexæ). When they are bent downwards.
12. Convolute (Convolutæ). When they roll inwards. Plate XII. Fig. 6.
13. Revolute (Revolutæ). When they roll outwards.
14. Spiral (Spirales). When they are convoluted spirally. Plate XXV. Fig. 31.
15. Rigid (Rigidæ). When they are very stiff and inflexible.—Ex. Libellulina, Fulgora. Plate XII. Fig. 12, 15.
f. reposition.
1. Hidden (Receptæ). Antennæ which when the animal reposes, are hidden under the head or trunk.—Ex. The Lamellicorns, Elater, Belostoma.
2. Exposed (Apertæ). Antennæ which when the animal reposes are not concealed.—Ex. Cerambyx.
g. figure and size.
1. Setaceous (Setaceæ). Long flexile antennæ which taper somewhat from the base to the apex. Plate XI. Fig. 1.
2. Setiform (Setiformes). Short rigid antennæ which taper from the base to the apex like a bristle. Plate XII. Fig. 14-16.
3. Capillary (Capillares). Antennæ nearly as slender as a hair. Plate XI Fig. 2.
4. Filiform (Filiformes). Antennæ every where of an equal thickness. Plate XI. Fig. 3.
5. Thick (Crassæ). Antennæ disproportionably thick. Plate XII. Fig. 29.
6. Incrassate (Incrassatæ). Antennæ disproportionably thick in any part: at the base, middle, or apex. Plate XXV. Fig. 34, 19, 7.
a. Gradually Incrassate (Sensim Incrassatæ). When they grow gradually thicker from the base to the apex. Plate XXV. Fig. 10.
b. Suddenly Incrassate (Subito Incrassatæ). When they grow suddenly thicker in any part. Plate XXV. Fig. 18, 19, 24.
7. Broad (Latæ). Antennæ disproportionably wide. Plate XXV. Fig. 24.
8. Dilated (Dilatatæ). When they are disproportionably wide in any part; base, middle, or apex. Plate XXV. Fig. 12. Plate XII. Fig. 1, 20.
9. Slender (Tenues). When they are disproportionably slender. Plate XI. Fig. 2.
10. Attenuate (Attenuatæ). Antennæ disproportionably slender in any part; base, middle, or apex. Plate XXV. Fig. 8, 21, 34.
a. Gradually Attenuate (Sensim Attenuatæ). When they grow gradually more slender from the base to the apex. Plate XI. Fig. 7.
b. Suddenly Attenuate (Subito Attenuatæ). When they grow suddenly slender in any part. Plate XII. Fig. 1. Plate XXV. Fig. 18, 34.
11. Fusiform (Fusiformes). Antennæ thickest in the middle and tapering more or less towards each[Pg 329] extremity. Plate XI. Fig. 5. Plate XXV. Fig. 8.
12. Prismatic (Prismaticales). Antennæ with three nearly equal sides. Plate XI. Fig. 6.
13. Ensiform (Ensiformes). Antennæ compressed and three-sided, with one side much narrower than either of the others. Plate XI. Fig. 7.
14. Falciform (Falciformes). When the Clavola of the Antennæ grows gradually narrower towards the apex, and is arcuate or incurved so as to resemble a sickle. Plate XI. Fig. 8.
15. Nodose (Nodosæ). When antennæ have one, two, or more joints larger than those which precede or follow them. Plate XII. Fig. 5.
16. Moniliform (Moniliformes). Antennæ consisting of oval or globular joints so as to resemble a necklace of beads. Plate XI. Fig. 9.
17. Dentate (Dentatæ). Toothed with teeth whose sides are equal. Plate XI. Fig. 10.
18. Serrate (Serratæ). Toothed with teeth whose sides are unequal like those of a saw. Plate XI. Fig. 11. Plate XXV. Fig. 8.
19. Biserrate (Biserratæ). So toothed on each side. Plate XXV. Fig. 18.
20. Imbricate (Imbricatæ). When the summit of each joint is incumbent upon the base of that which precedes it. Plate XI. Fig. 12.
21. Distichous (Distichæ). When the joints in general terminate in a fork. Plate XI. Fig. 13.
22. Cirrate (Cirratæ). When the joints terminate in a pair of curling hairy branches resembling tendrils. Plate XXV. Fig. 4.
23. Flabellate (Flabellatæ). When the antennæ on one side send forth from the joints, except those at the base, long flat flexile branches, which open and shut like the sticks of a fan. Plate XI. Fig. 17.
24. Biflabellate (Biflabellatæ). When they are flabellate on both sides. Plate XXV. Fig. 11.
25. Pectinate (Pectinatæ). Antennæ furnished on one side with a number of parallel stiff branches, resembling somewhat the teeth of a comb. Plate XXV. Fig. 25. Plate XI. Fig. 14.
26. Bipectinate (Bipectinatæ). Pectinate on both sides. Plate XXV. Fig. 22.
27. Duplicato-pectinate (Duplicato-pectinatæ). Bipectinate with the branches on each side alternately long and short. Plate XI. Fig. 15.
28. Ramose (Ramosæ). Antennæ furnished on one side with two or three irregular longish branches. Plate XI. Fig. 18.
29. Furcate (Furcatæ). Antennæ divided at the end into two prongs or branches. Plate XI. Fig. 19. Plate V. Fig. 3.
30. Bipartite (Bipartitæ). When they are divided to the base into two nearly equal branches. Plate XXV. Fig. 20.
31. Palmate (Palmatæ). Very short antennæ which send forth externally a few long finger-shaped branches, giving them some resemblance of a hand. Plate XI. Fig. 24.
32. Irregular (Irregulares). When the joints of the antennæ vary so much in size and shape that they cannot well be defined. Plate XI. Fig. 22.
h. termination.
α. versatile antennæ.
1. Subulate (Subulatæ). When they terminate in a minute joint, much slenderer than the preceding one. Plate XII. Fig. 16.
2. Setigerous (Setigeræ). When they terminate in a bristle. Plate XII. Fig. 14, 15. Plate XXV. Fig. 29.
3. Capillaceous (Capillaceæ). When they terminate in a fine capillary joint. Plate XII. Fig. 1.
4. Mucronate (Mucronatæ). When they terminate in a short point or mucro. Plate XII. Fig. 2.
5. Uncinate (Uncinatæ). When their apex is incurved so as to form a kind of hook. Plate XII. Fig. 3.
6. Unguiculate (Unguiculatæ). When they terminate in a hard horny incurved sharp claw resembling those of the tarsi of insects. Plate XXV. Fig. 16. a.
7. Clavate (Clavatæ). When their apex grows gradually thicker. Plate XII. Fig. 4. Plate XXV. Fig. 7, 14.
8. Capitate (Capitatæ). When they terminate suddenly in a larger knob of one or more joints. Plate XII. Fig. 8-10, and XXV. 1-3, 5, 6.
a. Fissile Knob (Capitulum fissile). When it is divided into several laminæ which the insect can open and shut. Plate XXV. Fig. 1-3, 5.
b. Tunicate Knob (Capitulum tunicatum). When the laminæ, at least on one side, appear to inosculate[Pg 332] or to be imbedded in each other. Plate XII. Fig. 8. Plate XXV. Fig. 5, 6.
c. Perfoliate Knob (Capitulum perfoliatum). When the joints of the knob are connected by a pedicle, which has the appearance of passing through them. Plate XII. Fig. 10.
d. Solid Knob (Capitulum solidum). When the knob consists of a single joint, or if of more, exhibits very faint traces of their separation. Plate XII. Fig. 9. Plate XXV. Fig. 33.
e. Inflated Knob (Capitulum inflatum). When the knob is disproportionably large, and looks as if blown out. Plate XII. Fig. 28. Plate XXV. Fig. 9.
β. inversatile antennæ.
1. Setigerous (Setigeræ). Antennæ furnished with a terminal bristle. Plate XII. Fig. 14-16, 21, 22. Plate XXV. Fig. 29.
a. Globiferous (Globiferæ). When the setigerous joint is larger than the preceding one, and globose. Plate XII. Fig. 12.
b. Angustate (Angustatæ). When the setigerous joint is not conspicuously larger than the preceding one. Plate XII. Fig. 14, 15.
2. Aristate (Aristatæ). Antennæ terminated by a variously shaped flat joint longer and usually larger than the preceding one, laterally setigerous. Plate XII. Fig. 21, 22.
a. Setarious (Setariæ). When the awn or bristle is naked. Plate XII. Fig. 21. a.
b. Plumate (Plumatæ). When the awn is feathered. Plate XII. Fig. 22. a.[Pg 333]
3. Filate (Filatæ). When inversatile antennæ have neither a terminal nor a lateral bristle. Plate XII. Fig. 17-20.
a. Simple (Simplices). When the last joint is exarticulate. Plate XII. Fig. 17, 18, 20.
b. Compound (Compositæ). When the last joint is itself obsoletely jointed. Plate XII. Fig. 19. a.
i. pubescence.
1. Verticillate (Verticillatæ). Antennæ beset with hair in whorls. Plate XII. Fig. 27.
2. Plumose (Plumosæ). Antennæ feathered on all sides with fine long hair. Plate XII. Fig. 24.
3. Ciliate (Ciliatæ). Antennæ fringed with parallel hairs on each side. Plate XI. Fig. 16.
4. Fimbriate (Fimbriatæ). Antennæ fringed with parallel hairs on one side.
5. Barbate (Barbatæ). Antennæ hairy on one side. Plate XII. Fig. 26.
6. Fasciculate (Fasciculatæ). Antennæ having several bundles of hair. Plate XXV. Fig. 32.
7. Scopiferous (Scopiferæ). When they are furnished with one or more dense brushes of hair. Plate XII. Fig. 25. a. Plate XXV. Fig. 17.
k. articulation.
1. Exarticulate (Exarticulatæ). Without visible articulations.
2. Biarticulate (Biarticulatæ). Consisting of two joints.
3. Triarticulate (Triarticulatæ). Consisting of three joints.[Pg 334]
4. Quadriarticulate (Quadriarticulatæ). Consisting of four joints.
5. Multiarticulate (Multiarticulatæ). Consisting of many joints.
l. joints.
1. Campanulate (Campanulatæ). Bell-shaped. When the joints are obconical, with the vertex of the cone rounded.
2. Pateriform (Pateriformes). When the joints are somewhat dilated and very short, shaped something like a shallow bowl.
3. Patellate (Patellatæ). When the whole joint is dilated and shaped something like a patella or platter.—Ex. Prosopis dilatata (Melitta *. b. K.) Plate XXV. Fig. 12. a.
4. Lobate (Lobatæ). When they are expanded at the tip into a lobe.—Ex. Belostoma, Cerocoma. Plate XI. Fig. 21, 22.
5. Torulose (Torulosæ). When they are a little tumid.
m. appendages.
1. Auriculate (Auriculatæ). When they have an ear-like process at their base.—Ex. Gyrinus, Parnus. Plate XII. Fig. 29. a. Plate XXV. Fig. 28. a.
2. Appendiculate (Appendiculatæ). When they have one or two antenniform processes at their base.—Ex. Otiocerus. Plate XXV. Fig. 29. b.[Pg 335]
III. TRUNK (truncus).
1. Monomerous (Monomerus). When the trunk has no suture or segment.—Ex. Araneidea.
2. Dimerous (Dimerus). When the trunk consists of two greater segments.—Ex. Coleoptera, &c.
3. Trimerous (Trimerus). When the trunk consists of three greater segments.—Ex. Neuroptera, &c.
4. Isthmiate (Isthmiatus). When an isthmus is formed between the Prothorax and Elytra, either in consequence of the former being constricted behind so as to form a neck, or the scutellum not being interposed between the elytra at their base, or the chief part of the mesothorax not being covered by the prothorax.—Ex. Clerus, Passalus, and Spondylis.
i. MANITRUNK (manitruncus).
a. prothorax.
1. Clypeiform (Clypeiformis). When the prothorax by its magnitude and distinct separation forms one of the most conspicuous pieces of the upper side of the trunk, so as nearly to represent the whole thorax; the mesothorax and metathorax being mostly hidden by the elytra and other organs for flight.—Ex. Coleoptera, Orthoptera, &c. Plate VIII. Fig. 1, 10.
2. Colliform (Colliformis). When the prothorax is short and narrow, and not so conspicuous as the other pieces of the trunk.—Ex. Libellulina. Plate IX. Fig. 6.[Pg 336]
3. Cerviculate (Cerviculatus). When the prothorax is elongate, attenuate, and distinguished from the Antepectus by no suture; so as to form a distinct and usually long neck. Plate III. Fig. 6.
4. Evanescent (Evanescens). When no distinct prothorax is discoverable or it is only represented by membrane.—Ex. Most Hymenoptera, Diptera, &c.
5. Marginate (Marginatus). When an impressed line or channel separates the edge of the prothorax from the rest of its surface, and so forms a margin.—Ex. Harpalus, &c.
6. Immarginate (Immarginatus). When it has no such margin.—Ex. The Rhyncophorous beetles.
7. Explanate (Explanatus). When its sides are so depressed and dilated as to form a broad margin.—Ex. Necrophorus, Silpha.
8. Emarginate (Emarginatus). When a segment of a circle is taken out of its anterior part for the reception of the head.
9. Ambient (Ambiens). When this sinus is so large as to receive the whole head.—Ex. Chilocorus.
10. Circumambient (Circumambiens). When its sides are elongated anteriorly and curve inwards, their ends lapping over each other and the head, so as to form a circle round the posterior part of the latter, and leave a space open for the eyes to see objects above them.—Ex. Heleus.
11. Clypeate (Clypeatus). When it quite covers and overshadows the head.—Ex. Lampyris, Cassida, Cossyphus.
12. Cucullate (Cucullatus). When it is elevated into a kind of ventricose cowl or hood which receives[Pg 337] the head.—Ex. Dictyonota crassicornis. Plate XIII. Fig. 18. a[1100].
13. Alate (Alatus). When its sides are expanded into a kind of wing.—Ex. Dictyonota crassicornis.
14. Auriculate (Auriculatus). When it expands on each side into two processes resembling ears.—Ex. Ledra aurita.
15. Angulate (Angulatus). When its sides or base jut out into one or more angles.—Ex. Copris.
16. Cruciate (Cruciatus). When it has two elevated longitudinal obtusangular lines, the angles of which approach each other in its middle, so as nearly to form a St. Andrew's cross.—Ex. Locusta. Plate XIII. Fig. 17.
17. Obvolving (Obvolvens). When there are neither ora nor suture to separate it from the antepectus.—Ex. Stenus, Curculio L.
18. Pulvinate (Pulvinatus). When in consequence of being depressed in one place, it seems to puff out in another.—Ex. Aleochara canaliculata, picea, &c.
19. Producted (Productus). When behind it terminates in a long scutelliform process which covers the Mesothorax, Metathorax, and great part of the Abdomen.—Ex. Acrydium, Centrotus.
b. antepectus.
1. Trachelate (Trachelatum). When of itself it forms a neck, the prothorax being represented only by membrane.—Ex. Xiphydria.
2. Unarmed (Inerme). When it has no prosternum.—Ex. The Rhyncophorous beetles. [Pg 338] 3. Armed (Armatum). When it has a prosternum. N.B. These two last terms may be extended also to the Medipectus and Postpectus. And also to the whole together. Thus, if there was no Sternum at all, it should be called Pectus inerme; and if one existed in all these divisions of the breast, it would be Pectus armatum.
c. arm (Brachium).
α. cubit (Cubitus).
1. Clypeate (Clypeatus). When a concavo-convex plate is affixed to the outside of the cubit.—Ex. Crabro clypeatus, scutatus, &c. ♂. Plate XV. Fig. 3. a.
2. Palmate (Palmatus). When towards the apex the cubit is armed laterally with several divaricate spiniform teeth.—Ex. Scarites, Clivina. Plate XV. Fig. 5.
3. Digitate (Digitatus). When the apex of the cubit is divided into several long teeth or fingers.—Ex. Gryllotalpa. Plate XV. Fig. 6.
4. Dolabrate (Dolabratus). When the apex of the cubit is dilated and shaped something like the head of a hatchet. Plate XV. Fig. 4.
β. hand (Manus).
1. Patellate (Patellata). When several joints of the hand are dilated so as to form an orbicular patella furnished underneath either with suckers, or a dense brush of hairs.—Ex. Dytiscus ♂, Staphylinus. Plate XV. Fig. 9.
2. Scutate (Scutata). When a single joint of the[Pg 339] hand is dilated into a broad scutiform plate.—Ex. Hydrophilus piceus ♂. Plate XV. Fig. 8.
3. Strigilate (Strigilata). When on the inner side of the first joint of the hand or palm the segment of a circle is taken out at the base opposite to the spur, the sinus being often pectinated with spines.—Ex. Apis[1101]. Plate XXVII. Fig. 36. a.
4. Auriculate (Auriculata). When any of the joints are externally dilated into an auriform process.—Ex. Gryllotalpa. Plate XV. Fig. 6. t´´.
ii. ALITRUNK (alitruncus).
1. Buried (Sepultus). When its upper surface is wholly or nearly covered and hidden by the thorax, elytra or other organs of flight[1102].—Ex. Coleoptera, Orthoptera.
2. Revealed (Revelatus). When it is not so covered, but is equally conspicuous with the Prothorax, or even more so.—Ex. Neuroptera, Hymenoptera, Diptera. Atractocerus in Coleoptera.
3. Coalite (Coalitus). When it is not separable into two segments, the Medipectus and the Postpectus forming one piece.—Ex. Cimex L.
4. Bisected (Bisectus). When it is separable into two segments.—Ex. Lamellicorn beetles.
a. mesothorax.
α. collar (Collare).
1. Uncovered (Apertum). When it is not concealed[Pg 340] by the shield of the prothorax.—Ex. Hymenoptera.
2. Covered (Tectum). When it is quite concealed by the prothorax.
3. Areate (Areatum). When it is larger than the prothorax, and terminates towards the wings in two oblique areas, inclosed by a ridge often crowned anteriorly with little teeth.—Ex. Libellulina. Plate IX. Fig. 7. g., a. N.B. The space between these areas is fitted with a membrane capable of tension and relaxation, which in flight causes them to approach to and recede from each other.
4. Amplectent (Amplectens). When posteriorly it is so curved as to form a large sinus which embraces the dorsolum.—Ex. Vespa L. Plate IX. Fig. 11. g..
5. Phonetic (Phoneticum). When its posterior angles, approaching the wings, cover the vocal spiracles[1103].—Ex. Hymenoptera.
β. dorsolum.
1. Thoraciform (Thoraciforme). When it forms the principal part of the upper surface of the trunk.—Ex. Bombus, Apis, Vespa &c. Plate IX. Fig. 11. i..
γ. scutellum.
1. Distinct (Distinctum). When it is separated from the dorsolum by a suture.—Ex. Hymenoptera, Diptera. Plate IX. Fig. 11, 19, &c. k´.
2. Coalite (Coalitum). When it is not separated[Pg 341] from the dorsolum by a suture.—Ex. Coleoptera, &c. Plate VIII. Fig. 3. k´.
3. Scutellate (Insectum scutellatum). An insect having a visible scutellum.—Ex. Melolontha.
a. Rejected (Rejectum). When, though visible, it does not intervene between the elytra at their base.—Ex. Passalus.
b. Received (Receptum). When it intervenes between the elytra at their base.—Ex. Most scutellate Coleoptera.
4. Exscutellate (Insectum exscutellatum). When an insect has no visible scutellum, it being wholly covered by the Prothorax.—Ex. Copris.
5. Ascending (Ascendens). When it curves upwards from the dorsolum.—Ex. Sagra.
6. Tabular (Tabulare). When it is elevated on a footstalk above the dorsolum, and forms a tabular or flat surface.—Ex. Elater.
7. Obumbrant (Obumbrans). When it overhangs the metathorax.—Ex. Musca. Plate IX. Fig. 19. k´.
δ. base-covers (Tegulæ).
1. Conchiform (Conchiformes). When they are a semicircular concavo-convex scale something resembling the valve of a bivalve shell.—Ex. Hymenoptera. Plate IX. Fig. 11, 12. g´´.
2. Laciniform (Laciniformes). When they are long, of an irregular shape, and appear like lappets on each side of the trunk.—Ex. Lithosia, &c. Plate IX. Fig. 5.[1104] [Pg 342]
ε. elytra.
1. Base (Basis). The part next the Prothorax.
2. Apex (Apex). The part next the Anus.
3. Humeral Angle (Angulus Humeralis). The exterior basal angle.
4. Scutellar Angle (Angulus Scutellaris). The interior basal angle.
5. Coleoptra (Coleoptra). The two elytra spoken of together.
6. Spinigerous (Spinigera). When the Coleoptra have a spine common to them both.—Ex. Cassida bidens.
7. Auriculate (Auriculata). When the Elytra have an elongate process at the shoulders.—Ex. Cassida bicornis, Taurus, &c.
8. Intire (Integra). When they have neither abbreviations nor incisions.
9. Fastigiate (Fastigiata). When of equal or greater length than the abdomen, and transverse at the end.
10. Truncate (Truncata). When they are shorter than the abdomen, and transverse at the end.
11. Abbreviate (Abbreviata). When they are shorter than the abdomen, but cover more than half its length. Plate I. Fig. 4.
12. Dimidiate (Dimidiata). When they are about half the length of the abdomen. Plate I. Fig. 5.
13. Very Short (Brevissima). When they are not half the length of the abdomen. Plate I. Fig. 2, 3, 7.
14. Mutilate (Mutilata). When they appear unnaturally short or curtailed as if mutilated.—Ex. Acrydium.[Pg 343]
15. Subulate (Subulata). When they are attenuated towards the end.—Ex. Sitaris humeralis.
16. Elongate (Elongata). When they extend beyond the anus.—Ex. Trox.
17. Obvolving (Obvolventia). When their Epipleuræ cover a considerable portion of the sides of the alitrunk. Plate XXVIII. Fig. 7.
18. Complicant (Complicantia). When they lie a little over each other.—Ex. Meloe. Plate I. Fig. 6.
19. Dehiscent (Dehiscentia). When they diverge a little at the apex.—Ex. Pyrochroa.
20. Ampliate (Ampliata). When they are disproportionably wide at the end.—Ex. Lycus fasciatus. Plate XIII. Fig. 20.
21. Plicate (Plicata). When they have two or three contiguous abbreviated furrows which exhibit the appearance of folds.—Ex. Pselaphidæ.
22. Perforate (Perforata). When a little hole appears drilled through them.—Ex. Cassida perforata.
N.B. Many of the above terms will apply to Tegmina, Hemelytra, Wings, &c.
A. Side-covers (Epipleuræ).
1. Marginal (Marginales). When they are only an inflexed continuation of the margin.—Ex. Buprestis.
2. Discoidal (Discoidales). When they are a process from the disk of the under surface of the elytra.—Ex. Lampyris, Cossyphus, Cassida, Notoclea[1105]. [Pg 344]
ζ. tegmina.
1. Fenestrella (Fenestrella). A transparent eye-like spot in the Anal Area of the Tegmina of Acrida ♂[1106].
2. Convolvent (Convolventia). When the Anal Area is horizontal, incumbent on the back of the insect, and forms a right angle with the rest of the tegmen, which is vertical and covers the sides.—Ex. Locusta. N.B. In this case the Anal Area of one Tegmen covers that of the other.
3. Aliform (Aliformia). When their substance approaches to membrane, and they nearly resemble Wings.—Ex. Most Homopterous Hemiptera.
η. hemelytra.
1. Obtected (Obtecta). When the Hemelytra are covered by a scutelliform mesothorax.—Ex. Scutellera.
2. Detected (Detecta). When they are not so covered.—Ex. Most Heteropterous Hemiptera.
θ. wings (Alæ).
A. Denomination.
1. Anterior (Anticæ). The fore or upper wings.
a. Superior (Superiores). The anterior wings are so denominated if when at rest they are placed upon the posterior wings.—Ex. Hymenoptera.
b. Primary (Primores). The anterior wings are so denominated if when at rest they are not placed[Pg 345] upon the posterior.—Ex. Lepidoptera diurna, Libellulina.
2. Posterior (Posticæ). The hind or lower wings.
a. Inferior (Inferiores). The posterior wings are so denominated if the anterior wings, when at rest, are placed upon them.
b. Secondary (Secundariæ). The posterior wings are so denominated if the superior wings, when at rest, are not placed upon them.—Ex. Lepidoptera diurna, Libellulina.
B. Magnitude.
3. Equal (Æquales). When the four wings are of equal length.—Ex. Libellulina.
4. Unequal (Inæquales). When they are not of equal length.—Ex. Hymenoptera.
C. Complication.
5. Plane (Planæ). Flat wings that are neither plicatile nor tumid.—Ex. Apis.
6. Tumid (Tumidæ). When the membrane between the nervures that form an areolet is bigger than the areolet, which gives it convexity.—Ex. The Serrifera or saw-flies. N.B. The object of this structure is to expose a larger surface to the action of the air.
7. Plicatile (Plicatiles). When the wings at rest are folded in one or more longitudinal plaits.—Ex. Vespa L.
8. Duplicatile (Duplicatæ). When they are folded transversely.—Ex. Coleoptera.
9. Convolute (Convolutæ). When the wings so envelope the body as to give it a cylindrical form.—Ex. Crambus.[Pg 346]
10. Incumbent (Incumbentes). Wings which when at rest cover the back of the insect.—Ex. The Noctuidæ, Geometra.
11. Cruciato-complicate (Cruciato-complicatæ). Wings crossed and folded.—Ex. Pentatoma, &c.
12. Cruciato-incumbent (Cruciato-incumbentes). Wings crossed but not folded, and covering the back.—Ex. Apis.
13. Extended (Extensæ). Wings that when at rest do not lie upon the body.—Ex. Libellula, Æshna, &c.
a. Expanded (Patentes). Wings that when at rest are horizontally extended and do not cover each other.—Ex. Libellula, &c.
b. Horizontal (Horizontales). Very narrow wings which when at rest are extended horizontally forming a right angle with the body, and covering the posterior wings.—Ex. Pterophorus[1107].
c. Erect (Erectæ). Wings which when at rest are extended vertically.—Ex. Vanessa, Agrion.
d. Erecto-patent (Erecto-patentes). When the primary wings at rest are erect and the secondary horizontal.—Ex. Hesperia.
e. Connivent (Conniventes). When erect wings are so closely applied to each other that the corresponding margins touch.—Ex. Vanessa.
f. Divaricate (Divaricatæ). When wings at rest are somewhat erect but diverge from each other.
14. Patulous (Patulæ). When wings at rest partly cover each other.
15. Applicant (Applicantes). When wings at rest are parallel with the abdomen.—Ex. Tipula.
16. Divergent (Divergentes). When wings at rest recede from the abdomen.
17. Deflexed (Deflexæ). When wings at rest covering each other are so bent downwards as to imitate a roof, of which their interior margin forms the ridge.—Ex. Homopterous Hemiptera.
18. Reversed (Reversæ). When wings at rest are deflexed, but so that the anterior margin of the inferior projects beyond the anterior margin of the superior.—Ex. Gatropacha quercifolia. Plate XIV. Fig. 2.
19. Broad (Latæ). When the interior margin is shorter than the posterior.—Ex. Papilio.
20. Narrow (Angustæ). When the posterior margin is shorter than the interior.—Ex. Heliconius.
D. Shape.
1. Falcate (Falcatæ). Wings having their posterior margin concave, and the posterior angle acute and curved.—Ex. Attacus Atlas. Plate XIV. Fig. 4.
2. Digitate (Digitatæ). Wings cleft to the base into several subdivisions.—Ex. Pterophorus. Plate XIV. Fig. 3.
a. Radius (Radius). A single subdivision of a digitate wing.
3. Caudate (Caudatæ). When wings terminate in a tail-like process.—Ex. Papilio Machaon. Plate XIV. Fig. 1. s.
a. Bicaudate (Bicaudatæ). Having two such tails. Tricaudatæ having three, &c.
E. Surface.
1. Squamate (Squamatæ). Wings covered with minute[Pg 348] scales.—Ex. Lepidoptera. Plate XXII. Fig. 16. a, b, c, d, &c.
2. Denudate (Denudatæ). When the wings of Lepidoptera appear more or less as if the scales had been rubbed off, either partially or generally.—Ex. Heliconius, Sesia, Zygæna, Nudaria.
3. Fenestrate (Fenestratæ). When one or two definite spaces in a Lepidopterous wing are denuded of scales.—Ex. Attacus Atlas, &c.
4. Bare (Nudæ). When wings have neither perceptible hairs nor scales.—Ex. Coleoptera.
F. Margin.
1. Anterior or Exterior (Anterior or Exterior). The outer margin of the wing, or that from the body. Plate XIV. Fig. 1. a.
2. Interior (Interior). The inner margin of the wing, or that next the body. Plate XIV. Fig. 1. b.
3. Posterior (Posterior). The terminal margin of the wing, or apex. Plate XIV. Fig. 1. c.
4. Plectrum (Plectrum). A marginal bristle stronger than the rest, observable about the middle of the costa and standing out from it.—Ex. Many Muscidæ.
G. Angles.
1. Humeral (Humeralis). Basal angle next the head. Plate XIV. Fig. 1. d.
2. Scutellar (Scutellaris). Basal angle next the scutellum or its region. Plate XIV. Fig. 1. e.
3. Posterior (Posterior). Outer apical angle. Plate XIV. Fig. 1. f.[Pg 349]
4. Anal (Analis). Inner apical angle. Plate XIV. Fig. 1. g.
H. Nervures.
1. Nervulet (Nervulus). A little nervure diverging obliquely from the costal into the disk of the wing towards the apex.
a. Simple (Simplex). When the nervulet does not terminate in a round punctum.—Ex. Eulophus.
b. Coronate (Coronatum). When it terminates in a round punctum.—Ex. Ichneumon penetrans[1108].
2. Neurose (Neurosæ). Wings that have nervures besides the marginal ones.
3. Aneurose (Aneurosæ). Wings that have no nervures besides the marginal ones.—Ex. Psilus.
4. Circumsepted (Circumseptæ). Wings whose margin is every where strengthened by a nervure.—Ex. Tabanus.
5. Varicose (Varicosæ). When the nervures are disproportionably swelled in any part.—Ex. Forficula auricularia. Plate X. Fig. 5.
6. Serpentine (Serpentinæ). Nervures that run in a serpentine direction.—Ex. Strategus Aloeus K. M.S[1109]. Plate X. Fig. 4.
7. Insulate (Insulatæ). Discoidal nervures that are entirely unconnected with any others, or with the base of the wing.—Ex. Strategus Aloeus. Plate X. Fig. 4. a, b.
[Pg 350]8. Uncinate (Unicinatæ). Nervures, that after running from the base towards the apex, turn back, and running a little towards the base, form a hook.—Ex. Strategus Aloeus. Plate X. Fig. 4. i..
9. Recurrent (Recurrentes). When a nervure, or a branch of it, after running towards the apex of the wing, turns back and runs towards the base.—Ex. Strategus Aloeus, &c. Plate X. Fig 4. l..
10. Connecting (Connectentes). Nervures that running transversely or obliquely connect the longitudinal ones, and so form the areolets.
I. Areolets.
1. Radiated (Radiatæ). When the areolets are chiefly formed by radiating longitudinal nervures.—Ex. Forficula, Psychoda. Plate X. Fig. 5, 13.
2. Areate (Areatæ). Radiated with a large basal area.—Ex. Papilio and many other Lepidoptera[1110]. Plate X. Fig. 6.
3. Areolate (Areolatæ). When the surface of the wing is divided into various areolets.—Ex. Diptera, Hymenoptera, and most Neuroptera. Plate X. Fig. 7-14.
4. Reticulate (Reticulatæ). When the areolets are extremely small and infinitely numerous.—Ex. Libellulina. Plate III. Fig. 5.
5. Open (Apertæ). Areolets that terminate in the margin of the wing, or that are not surrounded on all sides by nervures.
a. Marginal (Marginales). Open areolets that[Pg 351] terminate in the margin.—Ex. Tenthredo. Plate X. Fig. 8.
b. Incomplete (Incompletæ). Open areolets that terminate short of the margin.—Ex. Apis.
6. Radiant (Radiantes). When a small roundish areolet is a centre from which several long ones diverge.—Ex. Stratyomis. Plate X. Fig. 15.
7. Petiolate (Petiolatæ). When an areolet is connected with another by a stem like a footstalk[1111]. Plate X. Fig. 8.
8. Ramulose (Ramulosæ). When an areolet sends forth a little unconnected branch.—Ex. Pompilus, Sphex, &c.[1112]
9. Angular (Angulatæ). When an areolet juts out on one side into an angle from which no nervure proceeds, to form another areolet.—Ex. Eristalis, Cerceris.[1113] Plate X. Fig. 14.
10. Didymous (Didymæ). When areolets are nearly divided into two by a nervure.—Ex. Gyrostoma.
11. Sesquialterous (Sesquialteræ). When a minute areolet is appended to a large one.—Ex. Postcostal areolet of Hylotoma[1114].
* Areolets of the Costal Area.
1. Costal (Costales). Areolets, one or more, below the stigma. Plate X. Fig. 14. 15. a, b.
2. Postcostal (Postcostales). Areolets, one or more, above the stigma. Plate X. Fig. 8, 9. a, b.
* * Areolets of the Intermediate Area.
1. Protomesal (Protomesæ). First series of the middle areolets (Areolæ mediæ), often consisting of three, and then divided into upper, middle, and lower, areolets. Plate X. Fig. 8, 9. a.
2. Deuteromesal (Deuteromesæ). Second series of the same, often consisting of two, and then divided into upper and lower. Ibid. b.
3. Tritomesal (Tritomesæ). Third series of the same. Ibid. c.
K. Stigma.
1. Blind (Cœcum). When the stigma is wholly opaque, and neither begins nor terminates in a minute areolet.—Ex. Most Hymenoptera.
2. Fenestrate (Fenestratum). When the stigma begins or terminates in a minute areolet. Plate X. Fig. 11. m´´´.
L. Number.
1. Apterous (Aptera). Having no wings.
2. Dipterous (Diptera). Having two wings.
3. Tetrapterous (Tetraptera). Having four wings.
ι. legs (Pedes).
A. Number.
1. Tetrapod (Tetrapus). An insect having only four perfect legs.—Ex. Vanessa.
2. Hexapod (Hexapus). An insect having six legs.—Ex. Insects Proper in general.
3. Octopod (Octopus). Having eight legs.—Ex. Araneidea.[Pg 353]
4. Polypod (Polypus). Having more than eight legs but under fifty.—Ex. Glomeris, Cermatia.
5. Centipede (Centipes). Having more than fifty legs but under two hundred.—Ex. Scolopendra.
6. Myriapod (Myriapus). Having two hundred legs or more.—Ex. Iulus.
B. Situation.
1. Antepectoral (Antepectorales). The fore-legs or arms, affixed to the Antepectus.
2. Medipectoral (Medipectorales). The mid-legs, affixed to the Medipectus.
3. Postpectoral (Postpectorales). The hind-legs, affixed to the Postpectus.
4. Distant (Distantes). When the pairs of legs are remote from each other at their base.—Ex. Intermediate legs of Copris.
5. Approximate (Approximati). When they are near each other at the base.—Ex. Posterior legs of Copris.
6. Equidistant (Æquidistantes). When all the three pair are equally distant at the base.—Ex. Cassida.
C. Duration.
1. Persistent (Persistentes). Legs which the insect has in all its states.—Ex. The legs attached to the trunk. N.B. These are called Legs (Pedes). Plate XVIII. Fig. 11. a.
2. Deciduous (Decidui). Legs which the insect has not in all its states.—Ex. Membranous legs of Caterpillars. Plate XVIII. Fig. 11. b. N.B. These are called Prolegs (Propedes).[Pg 354]
3. Acquired (Acquisiti). Legs which the insect has not in its first state, but which it acquires subsequently.—Ex. Abdominal legs in Scolopendra, Iulus, &c.
D. Denomination.
1. Fore-Legs (Antici). The first pair. Taken by themselves called Arms (Brachia).
2. Anterior (Anteriores). The two first pair of legs.
3. Mid-legs (Intermedii). The middle pair of legs.
4. Hind-legs (Postici). The last pair of legs.
5. Posterior (Posteriores). The two last pair of legs.
6. Abbreviate (Abbreviati). Legs with an imperfect tarsus.—Ex. Vanessa.
7. Ambulatory (Ambulatorii). When the tarsi have a spongy sole.—Ex. Chrysomela.
8. Cursorious (Cursorii). When, the fore tarsi of some males excepted, they have not a spongy sole.—Ex. Carabus, Cicindela. Plate XIV. Fig. 7.
9. Saltatorious (Saltatorii). When the hind-legs have strong incrassated thighs formed for leaping.—Ex. Haltica, Orchestes, the Locustina. Plate XIV. Fig. 5.
10. Natatorious (Natatorii). When the legs are compressed or ciliated, and formed for swimming.—Ex. Dytiscus, Gyrinus, Notonecta. Plate XIV. Fig. 6.
11. Motatorious (Motatorii). Legs, which when the insect is at rest, are in a perpetual vibratory motion.—Ex. Tipula.
12. Fossorious (Fossorii). Leg with either palmate or digitate tibiæ.—Ex. Scarites, Clivinia, Gryllotalpa. Plate XV. Fig. 5, 6.
13. Raptorious (Raptorii). When the strong porrected[Pg 355] thighs, usually of the fore-leg, have a channel for the reception of the tibiæ, which are inflexed, and both armed with a double series of spurs.—Ex. Mantis, Nepa.
14. Prehensorious (Prehensorii). When the thighs of the hind-legs converge and the tibiæ diverge so as to form an angle which is armed with spines.—Ex. Gonyleptes. Plate XIV. Fig 8[1115].
E. Hip (Coxa).
1. Fixed (Fixæ). When they are not moveable.—Ex. Dytiscus, Gonyleptes.
2. Free (Liberæ). When they are moveable.—Ex. Hymenoptera, most Coleoptera.
3. Laminate (Laminatæ). When the posterior coxæ form a broad thin plate which covers the trochanter and the base of the thighs.—Ex. Haliplus. Plate XV. Fig. 1. p´´.
4. Flocculate (Flocculatæ). When the posterior coxæ are distinguished by a curling lock of hair (Flocculus).—Ex. Andrena[1116].
F. Trochanter (Trochanter).
1. Fulcrant (Fulcrans). When the trochanter merely props the thigh below at the base, but does not at all intervene between it and the coxa.—Ex. Carabus.
2. Intercepting (Intercipiens). When the trochanter intervenes between the thigh and the coxa, so as[Pg 356] intirely to separate them.—Ex. The Petalocerous beetles, Hymenoptera, &c.
3. Monomerous (Monomerus). When it consists of only one joint.—Ex. Coleoptera, &c.
4. Dimerous (Dimerus). When it consists of two joints.—Ex. Ichneumon.
G. Thigh (Femur).
1. Simple (Simplex). When it is no where particularly thick.
2. Incrassate (Incrassatum). When it is very thick, either partially or generally, and formed for leaping.—Ex. Haltica, &c.
3. Loricate (Loricatum). When the disk of the thigh appears covered with a double series of oblique scales like a coat of mail.—Ex. Locusta. Plate XIV. Fig. 5.
H. Shank (Tibia).
1. Alate (Alata). When the posterior tibia on each side is furnished with a dilated process which probably assists it in flight.—Ex. Petalopus phyllopus, &c. Plate XV. Fig. 2. a.
2. Foliaceous (Foliacea). When the tibia is laterally dilated into a thin plate for carrying pollen.—Ex. Euglossa cordata, &c.
3. Corbiculate (Corbiculata). When it is fringed with incurved hairs calculated for carrying kneaded pollen.—Ex. Apis, Bombus[1117].
4. Scopate (Scopata). When it is quite covered with a brush of hairs with which it brushes off the[Pg 357] gross pollen, and in which it carries it.—Ex. Andrena[1118].
5. Calcarate (Calcarata). When it is armed with one or more spurs (Calcaria).—Ex. The majority of insects.
6. Excalcarate (Excalcarata). When it has no such spurs.—Ex. Apion.
I. Foot (Tarsus).
1. Scopulate (Scopulatus). When the first joint on the under side is covered with a dense brush of rigid hairs.—Ex. Apis, Andrena, &c.[1119]
b. metathorax.
1. Simulant (Simulans). When the mesothorax is covered by the prothorax, and the Metathorax only is visible, under the form of an elongated or enlarged scutellum.—Ex. The Geocorisæ. Plate XXVIII. Fig. 12.
α. postdorsolum.
1. Latent (Latens). When it is covered by the mesothorax; it is then usually a mere membrane.—Ex. Most Coleoptera.
2. Exposed (Apertus). When it is not so covered.—Ex. Atractocerus, Hymenoptera, &c.
β. postscutellum.
1. Distinct (Distinctum). When the postscutellum is distinct from the postdorsolum.—Ex. Locusta. Plate VIII. Fig. 12 u´.
[Pg 358]2. Coalite (Coalitum). When it is not distinct.—Ex. Blatta.
3. Scutelliform (Scutelliforme). When it is a triangular elevated prominence resembling a scutellum.—Ex. Locusta.
4. Canaliform (Canaliforme). When it is a deepish elongate channel running from the postdorsolum to the abdomen.—Ex. Coleoptera. Plate VIII. Fig. 3. u´. XXVIII. Fig. 10. u´.
5. Obliterate (Obliteratum). When this channel is nearly or altogether obliterated.—Ex. Hymenoptera.
γ. postfrænum.
1. Tabulate (Tabulatum). When it forms a broad pannel or table on each side the postscutellum.—Ex. Most Coleoptera.
2. Funiculate (Funiculatum). When it forms a narrow ridge.—Ex. Pentatoma, Fulgora, Libellulina. Plate XXVIII. Fig. 11, 12. v´.
3. Cruciate (Cruciatum). When there are two funicular ridges diverging on each side, which inclosing a pannel form together a St. Andrew's cross, and are connected with the base of the wings.—Ex. Libellulina[1120]. Plate IX. Fig. 7. v´.
4. Adnate (Adnatum). When a funicular Postfrænum is closely adjacent to the sides of the metathorax till it nearly reaches the wings.—Ex. Pentatoma.
5. Transcurrent (Transcurrens). When a postfrænum[Pg 359] is at first adnate to the sides of the postscutellum, and then diverges across the pannel to the base of the wings.—Ex. Belostoma grandis.
IV. ABDOMEN.
1. Coalite (Coalitum). When the abdomen is not divided into segments.—Ex. Araneidea, Chelonus.
a. Plicate (Plicatum). When it consists of transverse folds.—Ex. Gonyleptes, Carkinodes[1121]. Plate XV. Fig. 11.
b. Tense (Tensum). When it is not folded.—Ex. Most Araneidea.
2. Insected (Sectum). When it is divided into segments.—Ex. Most insects.
3. Sessile (Sessile). When it has no footstalk, but is closely united to the trunk.—Ex. Coleoptera.
4. Petiolate (Petiolatum). When the first segment, or more, is longer and much narrower than the subsequent ones, so as to form a footstalk.—Ex. The Sphecidæ, Ichneumon.
5. Adjoined (Adjunctum). When it is connected with the trunk by a very short petiole.—Ex. Vespa, Apis.
6. Superimposited (Superimpositum). When the footstalk of the abdomen is inserted in the upper part of the postscutellum, so as to leave a considerable space between it and the postpectus.—Ex. Evania. Plate IV. Fig. 2.
7. Retracted (Retractum). When it is nearly withdrawn within the trunk.—Ex. Gonyleptes. Plate XV. Fig. 11. [Pg 360] 8. Obumbrate (Obumbratum). When it is overshadowed by the trunk and concealed under it.—Ex. Carkinodes. Plate XV. Fig. 10.
9. Saltatorious (Saltatorium). When the ventral segments or the anus are furnished with elastic processes which enable the animal to leap.—Ex. Machilis, Podura. Plate XV. Fig. 14.
10. Natatorious (Natatorium). When the abdomen is terminated by flat foliaceous appendages, or the tail is ciliated on each side with dense parallel hairs, which assist the insect in swimming.—Ex. Larva of Agrion, and Dytiscus.
i. CAUDA.
1. Uncinate (Uncinata). When the tail is inflected so as to form a kind of hook.—Ex. Dolichopus ♂.
2. Aduncous (Adunca). When it is crooked.—Ex. Chelostoma maxillosa ♂. (Apis ** c. 2. γ. K.)
3. Distinct (Distincta). When it is distinct from the abdomen.—Ex. Scorpio.
4. Cheliferous (Chelifera). When it is terminated by a very thick forceps somewhat resembling a lobster's claw.—Ex. Panorpa ♂. Plate XV. Fig. 12.
5. Papilliferous (Papillifera). When at the last segment but one the tail exerts two soft fleshy organs, which secrete a milky fluid and yield a powerful scent.—Ex. Staphylinus.
ii. OVIPOSITOR.
1. Ensate (Ensatus). When it is long, compressed, and shaped like a sword.—Ex. Acrida.[Pg 361]
2. Navicular (Navicularis). When it is shaped like a boat.—Ex. Cicada, Scaphura.
3. Telescopiform (Telescopiformis). When it consists of several tubes retractile within each other like the pieces of a telescope. Plate XVI. Fig. 2, 3.
4. Aculeiform (Aculeiformis). The ovipositors of Hymenopterous insects, which consist of the same parts, with the exception of the poison-bag (Ioterium), whether used as weapons or merely in oviposition.
a. Exerted (Exertus). When the vagina unemployed is partly out of the body.—Ex. Cleptes.
b. Extricated (Extricatus). When the valves and vagina unemployed are wholly out of the body.—Ex. Pimpla. Plate XVI. Fig. 1.
c. Reflexed (Reflexus). When the ovipositor is turned up and lies upon the back of the abdomen.—Ex. Leucospis.
APPENDIX.
Terms particularly applicable to Larvæ and Pupæ.
LARVÆ.
1. Spinneret (Fusulus). The organ which spins the silk. Plate XXI. Fig. 9.
2. Forcipate Lip (Labium Forcipatum). Mask of larvæ and pupæ of Libellulina[1122]. Plate XVI. Fig. 5. a. [Pg 362] 3. Unguiform Mandibles (Mandibulæ unguiformes). The parallel claw-shaped mandibles of many Diptera. Plate XX. Fig. 1, 2. c´.
4. Prop (Ereisma). A bipartite retractile glutinous organ exerted from between the legs of the genus Sminthurus, and employed by the animal to support itself when its legs fail it[1123].
5. Fecifork (Fæcifurca). The anal fork on which the larva of Cassidæ, &c. carry their feces. Plate XVIII. Fig. 2. a.
6. Mastigia (Mastigia). Two anal organs in the larvæ of Cerura Vinula, exerting from their apex a retractile flexible thread, with which they endeavour, by lashing their sides, to drive away the Ichneumons. Plate XIX. Fig. 2. a.
7. Syringes (Syringia). Organs situated in various parts of larvæ, from which they ejaculate a watery fluid to annoy or drive away their enemies[1124].
8. Rumules (Rumulæ). Teat-like fleshy protuberances observable on the bodies of various larvæ[1125].
9. Aeriducts (Aëriductus). Respiratory organs often foliaceous, with which the sides of the abdomen, the tail, and sometimes the trunk of aquatic larvæ and pupæ are often furnished. Plate XXIX. Fig. 3-7.
10. Prolegs (Propedes). Fleshy exarticulate pediform often retractile organs, which assist various larvæ in walking and other motions, but which disappear in the perfect insect. Plate XVIII. Fig. 11, 12. b.
[Pg 363]a. Coronate Prolegs (Propedes coronati). Prolegs that have an intire coronet of crotchets. Plate XXIII. Fig. 1.
b. Semicoronate Prolegs (Propedes semicoronati). Prolegs that have a semicoronet of crotchets.
c. Unarmed (Inermes). Prolegs that have no crotchets.
d. Stilt Prolegs (Propedes grabati). Prolegs that are unnaturally long, and elevate the animal. Plate XXIII. Fig. 7. a.
α. Coalite Stilt Prolegs (Propedes grabati coaliti). When stilt prolegs unite so as to form only one leg bifid at its apex. Plate XXIII. Fig. 7. b.
PUPÆ.
1. Adminicula (Adminicula). Semicoronets of minute teeth which arm the back of the abdomen of subterraneous pupæ, by which they are enabled to emerge from under the earth. Plate XVI. Fig. 13. e.
2. Cremastræ (Cremastræ). The anal hooks by which many pupæ suspend themselves. Plate XXIII. Fig. 8. a.
3. Cocoon (Folliculus). The silken case in which the pupæ of many insects are inclosed. Plate XVII. Fig. 5-8.
N.B. Other terms for Pupæ are explained Vol. III. p. 249.[Pg 364]
Having considered insects as to their History, Anatomy and Physiology, we must next enter a new and ample field, in which, like most of our predecessors, we shall often be perplexed and bewildered by the infinite variety of devious paths which traverse it, and by the mazy labyrinths in which the more we wander the less ground we seem to gain.—You will easily perceive I am speaking of the System of Insects. System is a subject which has engaged the attention of Naturalists from the time of Aristotle to the present day; and even now that it has been so much and so ably discussed, they are far from being agreed concerning it. In our own country a clue has, however, of late been furnished, which upon the whole seems better calculated to enable us to thread the intricate labyrinth of nature, than any thing previously excogitated.
There are two words relating to this subject concerning which Naturalists seem not to have very precise ideas—Method and System. They have often been confounded and used indifferently to signify the same thing. Thus we hear of a Natural Method and a Natural System. Linné seems to have regarded the former of these terms as representing the actual disposition of objects in[Pg 365] nature[1126], while by System he understands their classification and arrangement by Naturalists[1127]. But if we consider their real meaning,—a Method should signify an Artificial, and a System a Natural arrangement of objects[1128]. As many systematists, however, have aimed at giving a natural arrangement, though with various success,—some, as the French school, (to which we are principally indebted for the progress already made,) approximating nearer to the true idea than others,—and none having a perfect conception of it, of which probably in our present state, our minds, from its intricacy, are incapable,—it might perhaps be as well to call every arrangement whose object is confessedly artificial, a Method; and that which aims at the plan of nature, a System. Under this view system-makers would be divided into two classes,—the Methodists and Systematists.
The system of nature, which we are now to consider, may be viewed under a double aspect; for with regard to all created objects there is a System of Distribution, and a System of Correlation, which appear to be quite independent of each other. The former will best fall under our notice when we are treating of the Geography of insects: I shall therefore now confine myself to the latter.
When the Almighty Creator willed to bring into existence this mundane system, he formed it according[Pg 366] to a preconcerted plan, with all its parts beautifully linked together and mutually corresponding. All things were ordered in measure, and number, and weight[1129]. There was nothing deficient, nothing superfluous; but the whole in the strictest sense "was very good[1130]," and calculated in the highest degree to answer the purpose of its Great Author. I call it a system of Correlation, because there is discernible in it, in the first place, a concatenation of its parts, by which, as to their forms and uses, objects are linked together in groups by a chain of affinities; so that we pass from one to the other by gentle gradations, without having to overleap any wide interval. We see also a gradual ascent from low to high, from less to more excellent. And this leads us to another kind of relationship between natural objects, by which, though placed in distinct groups or in a different series, they in some sort represent and symbolize each other. Examples of this relationship by analogy are to be found in every kingdom of nature, and often form an ascending series from the lowest to the highest; for, as we shall see hereafter, these resemblances appear to maintain a certain correspondence with each other as to their relative situations; so that, for instance, in the animal kingdom they ascend step by step, without being linked by affinity or having any real juxtaposition, from the lowest groups, towards man, who stands alone at the head, or in the centre of all.—I shall say something on each of these kinds of relationship.
I. The relation of affinity may be considered as to its series and groups. A series, of course, consists of parts[Pg 367] either concatenated like a chain, or placed separately at small intervals from each other. It may run either in a right line, or deviate from it in various ways. It appears to be the opinion of most modern Physiologists, that the series of affinities in nature is a concatenated or continuous series; and that though an hiatus is here and there observable, this has been caused either by the annihilation of some original group or species in consequence of some great convulsion of nature, or that the objects required to fill it up are still in existence but have not yet been discovered[1131]: and this opinion is founded on a dictum of Linné, Natura ... saltus non facit[1132]. If this dictum be liberally interpreted, according to the evident meaning of the word saltus, few will be disposed to object to it; since both observation and analogy combine to prove that there must be a regular approximation of things to each other in the works of God; and that could we see the whole according to his original plan, we should find no violent interval to break up that approximation: but if it be contended, that in this plan there is no difference in the juxtaposition of the nearest groups or individuals, and never any interval between them, I think we are going further than either observation or analogy will warrant. Were this really and strictly the case, it seems to follow that every group or individual species must on one side borrow half its characters from the preceding group or species, and on the other impart half to the succeeding[1133]. But one of the[Pg 368] most evident laws of creation is variety; and if we survey all the works of the Most High, we shall no where discover that kind of order and symmetry that this strict interpretation implies. The general march of nature therefore seems to say, that there must be varying though not violent intervals in the series of beings: or in other words, some conterminous species or groups have more characters in common than others.
It was the opinion of Bonnet (in this field himself a host) and many other Naturalists, that the series of beings was not only continuous, but undeviating, ascending in a direct line from the lowest to the highest[1134]. Others, finding that this theory could not be made to accord with the actual state of things in nature, thought that a scale of the kingdoms of nature must represent a map or net[1135]; thus abandoning a continuous series: and Lamarck, as was before observed[1136], for the solution of the difficulty, arranged Invertebrate animals in a double subramose one. Mr. W. S. MacLeay and (without consultation nearly at the same time) Professor Agardh, Mr. Fries, &c. have given to the learned world an opinion which approximates more nearly to what we see in nature: viz. That the arrangement of objects is indeed in a continuous series, but which in its progress forms various convolutions, each of which may be represented by a circle, or a series that returns into itself[1137]. According to this opinion,—which seems the most consistent of any yet advanced, and which reconciles facts which upon no other plan can be reconciled,—the series of beings is[Pg 369] involved in the highest degree, rolling wheel within wheel ad infinitum, and revolving, if I may so speak, round its centre and summit—man[1138]: who, though not including in himself all that distinguishes them, is still the great Archetype in which they terminate, and from which they degrade on all sides.
It is by this convolving series that the various groups into which the kingdoms of nature seem resolvable are formed. We are instructed by the highest authority that every thing was created "after its kind;" and the common sense of mankind in all ages has imposed classic, generic, and other names implying sections, as well as specific ones, upon natural objects: and though many modern Physiologists have asserted that species form the only absolute division in nature; yet as all seem to allow that there are groups, and many that these are represented by a circle or group returning into itself[1139], the most absolute division in nature, we will not contend for a term[1140]. We now come to consider these groups themselves, and may notice them under various denominations.
It is customary to consider all the substances of which our globe consists as divided into three kingdoms,—the[Pg 370] Mineral, Vegetable, and Animal; but strictly speaking the primary division is into organized and inorganized matter; the former resolving itself into the two kingdoms last mentioned. These, like England and Scotland of old, have their "Land Debateable;" occupied by those Productions moyennes, (to use a term of Bonnet's[1141],) which are as it were partly animal and partly vegetable. From this territory common to both, the two kingdoms are extended in a nearly parallel direction till they reach their extreme limits, without any incursion from either side upon their mutual boundaries, but each showing its kindred with the other by certain resemblances observable between opposite points; so that valley corresponds with valley, mountain with mountain, river with river, sea with sea[1142]; not, however, so as to form an exact counterpart, but only in some general features. But to leave metaphor;—as the vegetable kingdom is distinguished from the mineral by its organization and life, by its circulation of sap, and by its powers of reproduction by seed or otherwise; so is the animal from the vegetable by its powers of volition and locomotion[1143], by its nervous systems and organs of sensation, and the senses to which they minister, by its muscular irritability, and by its instinctive endowments.
Having made these observations with regard to the primary division of natural objects in general,—what I have further to say will be confined to the animal kingdom, and ultimately to the branch of which we are treating.
[Pg 371]i. Lamarck divided the animal kingdom into two provinces, or subkingdoms as they are now called; the one consisting of all those animals whose skeleton is internal and built upon a vertebral column, which are denominated Vertebrates; and the second, of those whose skeleton or its representative is for the most part external, including the muscles,—these are called Invertebrates[1144]. Though this distinction is so marked as in general to form a most striking characteristic, yet when these two provinces approach each other, it begins to disappear. Thus the vertebral column, forming one piece with the shell[1145], becomes almost external in the Chelonian reptiles, or tortoises and turtles, and almost disappears in the cyclostomous fishes; and there is the beginning of an internal one in the Cephalopoda, or cuttle-fish belonging to the Invertebrates. Dr. Virey, assuming the nervous system as his basis, long since divided the animal kingdom, without assigning names to them, into three subkingdoms[1146]; M. Cuvier has four—Vertebrata; Mollusca; Articulata; Radiata[1147]: and Mr. MacLeay, finding five variations of that system, divides animals into five provinces or subkingdoms, of which I formerly gave you some account[1148];—viz. Vertebrata, in which the nervous system has only one principal centre; Annulosa, in which it is ganglionic, with the ganglions arranged in a series, with a double spinal chord; Mollusca, in which it is ganglionic, with the ganglions dispersed irregularly but connected by nervous threads; Radiata, in which it is filamentous, with the nervous threads radiating[Pg 372] from the mouth; and Acrita, in which this system is molecular[1149]. And to this division of the kingdom, as founded on a satisfactory basis, I should recommend you to adhere: still however we may speak of vertebrate and invertebrate animals, as forming the primary subdivision of them, taken from a striking character and obvious to every one who sees them.
If you inquire into the rank of each of these subkingdoms, of course you will assign the principal station to the Vertebrates, which are the most perfectly organized, to which man belongs, and over which he immediately presides. If we form the scale according to the nervous system of each province, that in which the organ of sensation and intellect is most concentrated will stand first; and in proportion as this organ is multiplied and dispersed will be the station of the rest, which will place them in the order in which I have mentioned them; and the Annulosa, to which insects belong, will precede the Mollusca, which Cuvier and Lamarck had placed before them on account of their system of circulation. But when we reflect that a heart and circulation occur in some of the conglomerate Polypi[1150], animals that approach the vegetable kingdom; that some of the acephalous Mollusca have no visible organs of sense, except that of taste, whose substance is little better than a homogeneous gelatinous pulp, and who seem from their inert nature to have very slight powers of voluntary motion[1151], we shall be convinced that a heart and circulation alone, unaccompanied by a more concentrated nervous system and more[Pg 373] perfect structure, cannot place an animal above those which in every other respect so obviously excel them. With regard to insects particularly, we may further ask—Who that considers how man employs his powers and organs even in his most degraded state, or that contemplates the wonderful works that he is enabled to accomplish when his faculties receive their due cultivation and direction, can avoid regarding him as superior to the rest of the animal creation? And what unsophisticated mind, not entangled in the trammels of system, when it surveys the industry, the various proceedings, and almost miraculous works that have been laid before you, the waxen palaces of the bee,—the paper cottages of the wasp and hornet,—the crowded metropolis of the white ants,—the arts, the manufactures, and stratagems of other insects,—the associations and labours for the common good of those that are gregarious;—will not at once conclude that they must be a superior race to the slug, the snail, and others, which live only to eat and propagate their kind?
Or who, that considers the wonderful structure of the animals whose cause I advocate,—the analogy that exists between their organs of manducation, of motion, and of sensation, and between various other parts of it[1152], with those of the higher animals,—the acuteness of their senses, their wonderful strength of muscle[1153], and powers of locomotion[1154],—but will think them superior to the headless and almost inanimate oyster or muscle, or the conglomerate Alcyonia, though they have a heart and circulation?
[Pg 374]Who again, that observes that in proportion as pedate animals approach to the human type, their motions are accomplished by fewer organs,—that man walks ore sublimi upon two legs; the majority of quadrupeds upon four; insects upon six: the Arachnida apparently upon eight; most Crustacea upon ten; and the Myriapods and others upon many,—but will thence conclude that insects must precede the Arachnida and Crustacea?
Who, once more, that reflects that if any of the superior animals are deprived of a limb it can never be reproduced, and that in insects the same circumstance occurs; while spiders and Crustacea if they lose a leg have the power of reproducing it, and the Mollusca if they are decapitated can gain a new head,—will consent to their being placed after any of these animals[1155]?
Lastly, who that recollects that the Mollusca are hermaphrodites, like most plants, bearing both male and female organs in the same body,—but will allow that insects, in which the sexes are separate as in the Vertebrates, must be more perfect, and of a higher grade[1156]?
ii. We now come to the Classes into which the Annulosa are divided. This term appears first to have been employed by Tournefort, and was adopted by Linné[1157]. As the nervous system of animals furnishes the most prominent distinction of a subkingdom, so the circulation of their fluids, and their respiration necessarily connected with it, seems best to point out the classes into which it may next be resolved. But having fully explained my[Pg 375] ideas on this subject in a former letter, I need not here repeat what I then said[1158].
iii. As we have subkingdoms, so we may also have subclasses, or such large divisions of a class—not founded upon internal organization or any of the primary vital functions, but upon different modes of taking their food, or such other secondary characters—as include more than one Order. To this description Clairville's Mandibulata and Haustellata appear to me to belong, which I think are by no means entitled to the rank of Classes; for whoever compares these two tribes together will at the first glance be convinced, by the numerous characters they possess in common, notwithstanding the different mode in which they take their food, that they form one connected primary group. This circumstance, therefore, only furnishes a clue for their further subdivision into two secondary groups, separated by distinctions certainly of a lower value than those which separate the Crustacea and Arachnida from Insecta. This is further confirmed by the variations that take place in their mode of feeding in their different states; some from masticators becoming suctorious (Lepidoptera), and others from being suctorious becoming masticators (Myrmeleon, Dytiscus, &c.),—which shows that this character does not enter the essential idea of the animal.
iv. Next to Classes and Subclasses we are to consider those groups of insects that are denominated Orders. The characters of these at first were taken principally from the instruments of flight or the absence of them; and the name appropriated to each Order by Linné, after[Pg 376] Aristotle, had reference to this circumstance. But this alone does not afford characters sufficiently discriminating: for though to an accurate observer a difference in these organs appears to be characteristic of most of the Orders, yet in some it is not easily detected or defined. In the Neuroptera there are as many different types of wings as there are of tribes or suborders. So that it seems not possible so to construct the definition of every Order, as to take its character from the organs of flight alone. Linné was sensible of this, and was compelled to have recourse to subsidiary characters in the majority of his: his observation therefore with regard to Genera,—that the character does not give the genus, but the genus the character[1159],—applies equally to Orders; and the characters included in the definition of an Order, should be the result of a careful examination of its component groups.
On a former occasion I named to you the Orders into which it appeared to me the Class Insecta might be divided[1160]; they were these. Coleoptera; Strepsiptera; Dermaptera; Orthoptera; Hemiptera; Trichoptera; Lepidoptera; Neuroptera; Hymenoptera; Diptera: Aphaniptera; Aptera. I then briefly explained them merely for the sake of illustration, and that you might know what description of insects were meant when these Orders were mentioned in my letters, without intending to affirm that I had arranged them in a natural series, or that all of them were perfectly natural. I shall[Pg 377] now consider them separately, and conclude with giving my sentiments as to which should be placed first.
* orders in which the ordinary Trophi all occur, or the Mouth is perfect[1161]. (Mandibulata.)
1. Coleoptera[1162] (Eleutherata F.). Aristotle may be called the founder of this Order, since he both named and defined it[1163]. Both his name and definition were adopted by Linné; and the former (with the exception of Fabricius and his school) by all succeeding Entomologists. To his definition Wings in a sheath[1164], other characters have been added; as the folding of the wings, and the straight suture by which the elytra are united[1165]. Aristotle's character, though to be found in the great majority of the Order, is not universal, since there are some beetles that have neither wings nor sheath, as the female glow-worm; and many that though they have the sheath have no wings, as Meloe, many Carabi, &c. To the transverse folding of the wings there are also exceptions; as in Buprestis, Molorchus, &c. The straight suture by which one elytrum exactly coincides with the other without lapping over, fails in Meloe: so that no one of these characters can be called universal in the Order; but as an exception or two does not invalidate a rule, and these are sufficiently universal for the purpose of pointing it out, they may be retained. Perhaps it will be an improvement to add the kind of the metamorphosis, which, as far as known, prevails universally.
Def. Metamorphosis incomplete[1166].
Legs inosculating, posterior coxæ usually transverse.
Elytra corneous, or coriaceous, without veins, united by a straight suture, so as mostly to cover the wings completely[1167].
Wings longitudinally and transversely folded[1168]: neuration simple[1169].
2. Strepsiptera[1170] (Rhiphiptera Latr.) The characters of this Order were first given in the Linnean Transactions, and it has been adopted by Latreille, who however, without sufficient reason, has changed the name originally imposed to Rhiphiptera[1171]. Rossi, who was the first that discovered an insect of this Order, concluded that because it was parasitic it must be Hymenopterous; and it is certainly more nearly related to that Order than to the Diptera, amongst which M. Lamarck has arranged it, and with which it has no character in common, except having two wings. This is one of those Orders, consisting of few genera and species, which, from their connecting two circles, Mr. MacLeay has called osculant, who places it between the Hymenoptera and Coleoptera[1172].
Def. Metamorphosis subincomplete[1173]?
Pseudelytra twisted, attached to the anterior leg[1174].
Wings not covered by the elytra, longitudinally folded, forming nearly the quadrant of a circle[1175]: neuration simple.
Anus styliferous[1176].
3. Dermaptera[1177] (Ulonota F. Orthoptera Oliv.). This is another osculant Order, evidently connecting the Coleoptera with the Orthoptera. The elytra are of a coriaceous substance, have a straight suture, and are not veined, and the wings are folded longitudinally as well as transversely,—circumstances which connect it with the former Order,—while the shape of its wings, its oral organs, and its metamorphosis, show its affinity to the latter. It was established at the same time and in the same work with the preceding Order, in pursuance of a suggestion of Dr. Leach, and consists solely of the Linnean genus Forficula.
Def. Metamorphosis semicomplete.
Elytra coriaceous, without veins, united by a straight suture, so as partly to cover the wings.
Wings longitudinally and transversely folded, each forming nearly the quadrant of a circle: neuration radiating[1178].
Anus forcipate.
4. Orthoptera[1179] (Ulonota F.). This Order, which Linné at first regarded as belonging to the Coleoptera[1180], and afterwards improperly added to the suctorious Hemiptera, was very judiciously separated from both by De Geer, under the name of Dermaptera, a name not[Pg 380] improper, and which ought to have been retained. Its present name was, I believe, assigned to it by Olivier; and as this is generally in use, I shall not attempt to disturb it. Dr. Leach divided the Order into two, separating the Blattina from it, under the name of Dictyoptera[1181]. He was led to this by the tegmina decussating or lapping obliquely over each other, whereas in the rest the horizontal portion of one tegmen lies longitudinally over that of the other; he also probably took their depressed body into consideration;—these circumstances, however, rather indicate a tribe or suborder; and as such Mr. MacLeay regards it.
Def. Metamorphosis semicomplete.
Legs suspended.
Tegmina generally pergameneous[1182], reticulated with nervures, more or less incumbent, covering the wings.
Wings longitudinally folded, ample: neuration reticulated.
5. Neuroptera[1183] (Synistata, Odonota F.). Of all the Linnean Orders this appears to consist of the most discordant tribes; so that it seems next to impossible to construct a definition that will include them all, unless indeed we admit M. Latreille's idea, adopted by Mr. MacLeay[1184], that a varied metamorphosis is its essential character; or, to speak more largely, variety itself seems the characteristic of the insects composing it, in every state; and there is scarcely a common distinctive character in their perfect state, upon detecting which in any[Pg 381] individual you may exclaim—This is a Neuropterous insect. The only one that I have been enabled to seize is, that their scapulæ and parapleuræ are parallel and placed obliquely[1185]. Whether, with all this puzzling variation and dissonance between the different tribes of which it is now composed, this Order can be considered as a natural group, in the present state of our knowledge it would be rash to decide. I shall observe, however, that the Libellulina,—whether we regard their metamorphosis and the singular character before described that distinguishes their larva and pupa[1186], their oral instruments[1187], the remarkable position of their legs[1188], their general form, the wonderful and peculiar machinery by which their wings are moved[1189], and other circumstances of their internal anatomy,—if any are to be regarded as forming a separate Order, are the first entitled to that distinction. At present, with our friend Mr. MacLeay, I shall consider it as not further divisible, and as consisting of five principal forms. I must not omit to observe, that in the Ephemerina the parts of the mouth, except the labrum and palpi, appear to be mere rudiments[1190].
Def. Metamorphosis varying. Larva a hexapod.
Wings four in most, and reticulated with numerous areolets.
Prothorax distinct.
Scapulæ and Parapleuræ parallel and oblique.
Tail of the female without a terebrant, or pungent multivalve ovipositor[1191].
6. Hymenoptera[1192] (Piezata F.). Mr. MacLeay considers Sirex L. as being osculant between the Order we are now entering upon and the Trichoptera, and Tenthredo, L. as belonging to the latter. He appears to ground this opinion chiefly upon a consideration of their larvæ and a slight difference in their ovipositor. As the Order, as settled by Linné, has always been deemed one of the most natural ones, and all the great Entomologists of the present æra have agreed with him in thinking it so; it seems to me that to prove them mistaken in this opinion, the question should have been discussed at more length, and that it requires arguments of more weight than any Mr. MacLeay has at present produced to set it aside. He appears in general to lay great stress upon an agreement in larvæ and the kind of metamorphosis; and I am ready to acknowledge that it forms a strong presumption in favour of any hypothesis of affinity between certain tribes. But when it is had recourse to as fundamental and infallible, I think it is pushed far beyond what it will bear, or is warrantable. I may be wrong; but in my apprehension, a striking agreement in their general structure in the perfect state, which is the acme of their nature, affords a much more satisfactory reason for keeping two tribes together, than any difference observable in their larvæ or metamorphosis, for separating them. Let any one compare the structure of these two tribes with the Trichoptera on one side, and the Hymenoptera on the other, and it will require but a glance to convince him of their greater affinity to the latter; and the simple inspection only of Jurine's plates of the wings of Hymenoptera is calculated to produce[Pg 383] the same effect. With regard to their larvæ, the resemblance between the case-worms and the pseudo-caterpillars of the saw-flies seems to me very distant, and the numerous prolegs of the latter have scarcely a legitimate representative in the former. The larvæ of the genus Lyda lose the prolegs intirely, and in one species, which much resembles the vermiform larvæ of Hymenoptera, the real legs are so extremely short as to be scarcely discernible[1193]; so that it requires no great stretch of faith to believe that saw-flies or Sirices may exist in whose larvæ the legs disappear[1194]. But it is this very tribe, whose larvæ thus approach to those of the other Hymenoptera, in which Mr. MacLeay finds the greatest external resemblance to the Trichoptera[1195]. In fact the difference between the saw-flies and Siricidæ, and the remainder of the Hymenoptera, amounts to little more than what takes place in the Diptera Order between the Tipulidæ, Asilidæ, Muscidæ, &c., in which also the metamorphosis differs.
Another argument upon which Mr. MacLeay seems to lay some stress, is taken from the number of parts into which the ovipositor of the saw-flies is resolvable, which he finds to consist of four pieces; while in what he considers as the genuine Hymenoptera, it is formed only of three[1196]: but in fact, in these last there are two spiculæ, answering to the two saws of Tenthredo, so that the vagina[Pg 384] in which these move may be considered as a double sheath: only, as these were to be pushed out at the same time, and the others alternately, it was necessary that in the latter each sheath should be separate, to admit of this motion; but as to its composition, the weapon in both is essentially the same. At any rate this structure could furnish a reason only for the formation of a separate group in the same Order, but none for the transfer of such group to another, which had no such instrument at all; since, as we have seen, the Trichoptera extrude their eggs at once in a mass[1197]. I do not mean, however, that it should be inferred from what I have here said, that there is no tendency in the saw-flies towards a Trichopterous type, for in them nature seems pointing that way, but the distance is too great, and the number of types of form necessary to fill up the interval too many, to warrant in my opinion their removal from the one Order to the other.
Def. Metamorphosis incomplete[1198].
Trophi in most not used for mastication[1199].
Wings four: neuration generally areolate[1200].
Prothorax obsolete, giving place to an ample collar.
Tarsi pentamerous.
Ovipositor 5-6-valved, the vagina darting forth two retroserrulate spiculæ.
** orders in which all the ordinary Trophi do not occur, or the Mouth is imperfect[1201] (Haustellata).
7. Hemiptera[1202] (Ryngota F.). Linné at first confined this Order to those insects which have a promuscis, which he denominated a rostrum[1203]; but afterwards, convinced that the Orthoptera of the moderns could not be associated properly with the Coleoptera; instead of forming them into a distinct Order, as nature would have dictated—perhaps to avoid the multiplication of Orders and without altering his definition—with equal infelicity he added them to this. Subsequent Entomologists, who saw the impropriety of masticating insects thus herding with suctorious ones, restricted the Order to its old limits; but Latreille very judiciously altered its arrangement, and divided it into two Sections, separating those whose hemelytra terminate in membrane, from those in which they are mostly tegmina, or of a substance intermediate between that of the elytra of Coleoptera and that of the wings of the Tetrapterous Orders. He denominated the first of these sections, or rather suborders, Heteroptera, and the last Homoptera[1204]. Dr. Leach, observing that very considerable differences take place both in the economy and structure of Heteropterous and Homopterous insects, followed De Geer in considering them as separate Orders, which he has called Hemiptera and Omoptera, and in which he has been followed by Mr. MacLeay;[Pg 386] who, however, with his usual accuracy and judgment, has restored the aspirate to the latter name[1205]. Their agreement in having a promuscis, or instrument of suction, with a jointed sheath, at present induces me to hesitate as to the propriety of their separation, and to consider them as forming secondary rather than primary sections of the Class. That you may be enabled to judge for yourself upon this subject, I will state the principal features in which they differ. In the first place, the Heteropterous section usually sucks the juices of animals, and the Homopterous, those of plants; in the former, the Hemelytra, besides their different substance, as well as the wings, cross each other; while in the latter, the organs of flight are deflexed, and do not lap over each other at all. The antennæ also of the one are often long, and do not terminate in a bristle; while in the other, with few exceptions, they are very short and setigerous. In the Heteroptera the body is depressed and flat, in the Homoptera convex and thick. In the former, the scutellum is one of the principal features of the trunk; in the latter, not at all remarkable[1206]. Other differences in the structure, both of head, trunk, and abdomen, might be pointed out; but these you will chiefly find noticed in my letters on the External Anatomy of Insects, where I treated of those parts. I shall here, therefore, only further mention the ovipositor also as forming a most striking distinction[1207].
Def. Metamorphosis semicomplete in almost all.
Mouth promuscidate[1208].
Wings covered by Hemelytra or Tegmina[1209].
Tarsi mostly trimerous, rarely dimerous or monomerous[1210].
8. Trichoptera[1211] Kirby (Synistata F. Neuroptera Latr.). MM. Latreille and MacLeay are of opinion that Semblis F. and Phryganea L. ought to be associated in the same group; and the latter gentleman has backed his opinion by some apparently cogent arguments[1212]: there are others, however, that seem to me more cogent, for considering them as belonging to different Orders. Whoever examines the several tribes into which Mr. MacLeay has divided the Neuroptera, will observe in all of them a distinct prothorax, a circumstance which they possess in common with those Orders that use their mandibles for mastication; whereas in those that do not use them for mastication, as the Hymenoptera, or that take their food by suction, this part is replaced by a mostly narrow collar, forming a part of the alitrunk[1213]. The existence then of the prothorax in the Perlidæ, and of the collar in the Trichoptera, affords no slight presumptive evidence that they belong to different Orders. Another circumstance that weighs much with me is, that the type[Pg 388] of the neuration of the wings in Perla is taken from the Neuroptera, in the Trichoptera from the Lepidoptera; the same observation extends to the legs of both[1214], and likewise to the abdomen. Even in their oral organs, as far at least as relates to their mandibles, those of Perla, though membranaceous—a circumstance occurring even in Coleoptera—are of a Neuropterous type; while the angular termination of the cheeks in the Phryganeæ approaches to the Lepidopterous mandibular rudiments. The principal argument on which Mr. MacLeay's opinion seems to rest, is, that the larvæ of both are aquatic, and clothe themselves in cases formed of various materials: but though this circumstance shows that they approximate in the system, it does not prove that they belong to the same order, since the general habit and appearance of the two animals when arrived at perfection contravenes it. The larvæ of Myrmeleon and of Leptis Vermileo form pitfalls of sand for their prey, and when they become pupæ, cover themselves with it[1215]; but this in them does not even prove an affinity, but only an analogy. The larva of Perla is carnivorous[1216], that of Phryganea mostly herbivorous[1217]: so that they are not precisely similar in their habits. Whether they resemble each other altogether, in their form, does not clearly[Pg 389] appear. The above reasons will, I trust, justify me for considering them at present as belonging to different Orders; but if further discoveries should confirm the opinion Mr. MacLeay espouses, I shall have no hesitation in yielding to it.
Def. Metamorphosis incomplete[1218].
Mouth emandibulate.
Prothorax replaced by a collar.
Wings four, upper pair mostly hairy, lower ample, folded: neuration branching.
Anus without setæ. Eggs extruded in a gelatinous mass[1219].
9. Lepidoptera[1220] (Glossata F.). Concerning this Order, no difference of opinion exists amongst Entomologists. Besides the scales that cover their wings, they are distinguished by the peculiar instrument of suction formerly described: neither of these characters, however, is perfectly universal; some of the Order (Nudaria) having no scales upon their wings, and others being without any antlia (Aglossa). Other peculiar characters are to be found in them; for instance, the patagia, or tippets, that adorn their evanescent thorax[1221], and the tegulæ, or base-covers, of a shape quite dissimilar to those of Hymenoptera, which cover and defend the base of their wings[1222]. As in the last Order, their legs are located all together with scarcely any space intervening between them; and they often agree also in their spurs.
Def. Metamorphosis obtected[1223].
Mouth antliate[1224].
Prothorax very short, covered by a pair of tippets.
Wings four, covered partially or generally with minute scales: neuration branching, often with a central areolet.
10. Diptera[1225] (Antliata F.). This Order likewise appears indebted for its name to the philosopher of Stagyra, who distinguishes the members of it from their counter-parts—the Hymenoptera—by their having an oral, while these have an anal sting[1226]: and we may add, that while the last, on account of their wonderful economy and the benefits which by them Providence confers upon mankind, have been justly regarded as the princes of the winged insect world,—the former, when we consider the filthy and disgusting habits of their grubs, and the annoyance, both from their numbers and incessant assaults, of them, in their fly-state, may very properly be considered as its canaille. Almost all the tribes of Hymenoptera, from the saw-flies to the ants, have their representatives in this Order. Though the number of wings is its prominent feature, yet there are two-winged insects in other Orders, as some Ephemeræ: and the Eproboscidea of Latreille seem rather a kind of winged Aptera, if we consider their trophi, than real Diptera; or they may form an osculant group, partly winged and partly apterous, between the two. I have before remarked, that though, apparently, the insects of[Pg 391] this Order have only two wings, yet the under or secondary wings of the other Orders have in them their representative[1227]. Their poisers also, I formerly observed to you, are probably more connected with their respiration than with their flight[1228].
Def. Metamorphosis incomplete, or coarctate.
Mouth proboscidate[1229].
Prothorax replaced by a collar. Sutures of the trunk mostly spurious[1230].
Wings two, with winglets attached to them: neuration various[1231]. Poisers.
Tarsi pentamerous.
Ovipositor various[1232].
11. Aphaniptera[1233] (Aptera L. Lamarck. Rhyngota F. Suctoria Latr.) This is an osculant Order, and is distinguished from the other Aptera L. in undergoing a regular metamorphosis. The larva is vermiform, the pupa incomplete, and inclosed in a cocoon. Probably the common flea and the chigoe would form distinct genera. The number of species of fleas is greater than has been supposed. I have been informed that Dr. Leach is acquainted with fourteen British species alone. Besides their metamorphosis, they are distinguished from the Aptera by the number of segments into which their body is divided, and by their pentamerous tarsi. Something[Pg 392] like elytra and a scutellum appear to distinguish these insects.
Def. Metamorphosis incomplete.
Body apterous, compressed.
Mouth rostrulate[1234].
Tarsi pentamerous.
We are now come to those insects which, though they change their skin in their progress to their state of perfection, and some of them, as we have seen[1235], gain additional segments and pairs of legs, yet none of them acquire wings or wing-cases: these I have considered as forming one Order, under the denomination of
12. Aptera[1236] (Synistata, Antliata, Unogata, Mitosata F.). I do not give this as a natural Order. Our knowledge, however, of the internal organization of its groups, is not at present sufficiently matured to warrant the formation of them into new Classes[1237]: till that is more fully ascertained, it seems to me therefore best to consider these groups as forming three Suborders: the first consisting of the Hexapods; the second of the Octopods; and the third of the Polypods. It will be better, I think, instead of giving a general character of the Order,—which principally consists in the insects composing it being Apterous, or never acquiring organs of flight,—to define each of these groups.
Hexapods (Ametabolia Leach, Ametabola McL.). Six legs may be regarded as the natural number in all the[Pg 393] insect tribes[1238]: but our business now is with those Aptera whose body consists of three greater segments, and which in none of their states have ever more or less than six legs, and consist of the three Linnean genera Pediculus, Lepisma, and Podura (Thysanura and Anoplura). Some of the mites (Acarus L.) are hexapods, but their body has no distinction of head, trunk, and abdomen. The metamorphosis of most female Blattæ, and of some other Orthoptera that are apterous, cannot be regarded as materially different from that of the Hexapods. Amongst the Anoplura,—the Pediculi, or lice, are suctorious, and the Nirmi, or bird-lice, masticators,—a circumstance which in them does not appear to indicate even a different Order, and proves that undue stress ought not to be laid, independently of general characters, on the mode in which insects take their food.
Def. Metamorphosis complete.
Body consisting of three principal segments.
Mouth perfect, or rostellate[1239].
Antennæ distinct.
Legs six, in every state.
Octopods. This suborder consists of the Trachean Arachnida of Latreille, excluding the Pycnogonida; of the Acaridea, Sironidea, Phalangidea, and part of the Scorpionidea of Mr. MacLeay, and, with some exceptions, of the Linnean genera Acarus and Phalangium. This last tribe (for with Linné, I include Chelifer and Obsidium in the Phalangidea,) on one side approaches Scorpio by Thelyphonus, and on the other the Aranidea[Pg 394] by Gonyleptes; or, according to Mr. MacLeay, the transit is to both by Galeodes[1240]. But as there is reason for thinking that this last belongs to the Pulmonary Arachnida[1241], and forms a peculiar type in that Class, I consider the transit from the one to the other as above stated. The folded abdomen of Gonyleptes seems much to correspond with that of the cancriform spiders (Carkinodes cancriformis, &c.).
Def. Metamorphosis complete.
Body consisting of one or two principal segments.
Mouth various[1242].
Antennæ obsolete, or represented by mandibles.
Legs mostly eight, but in a few six only[1243].
Polypods. This suborder consists of Dr. Leach's Class Myriapoda, or the Chilognatha and Chilopoda of Latreille, corresponding with the Linnean genera Iulus and Scolopendra. Mr. MacLeay has arranged them in the same Class with the Hexapods, and connects them with the Anoplura by means of certain intestinal worms of an indistinct annulose structure[1244] (Entozoa Nematoidea Rud.), in which the sexes are diœcious, and some of which are furnished with lateral spinulæ,—thus, as he supposes, connected with the Polypods; and with the Anoplura by others (Epizoaria Lam.) in which appendages appear somewhat analogous to the legs of Hexapods, as in Cecrops Leach, and which like them are parasitic animals[1245]. But the right of these worms to be[Pg 395] considered as members of the same Class with the Hexapods and Polypods at present appears rather problematical, and requires further examination.
Def. Metamorphosis subcomplete[1246].
Body consisting of numerous segments.
Mouth perfect[1247].
Eyes compound or aggregate.
Antennæ distinct.
Legs six on the trunk, many on the abdomen.
I must next say something on the Orders of the Arachnida. Every one, at first sight, sees that spiders and scorpions are separated by characters so strongly marked, that they look rather like animals belonging to different Classes than to the same: these form the two primary Orders of the Arachnida, and they appear to be connected by two secondary or osculant ones,—on the one side by Galeodes, and on the other by Thelyphonus and Phrynus[1248]. This Class, although there is an appearance of eight legs, is, strictly speaking, of a Hexapod type; for the anterior pair, ordinarily regarded as legs and performing their function, are really the analogues of the maxillary palpi of perfect insects. This will be evident to you if you examine any species of Galeodes. These animals, if we look at them cursorily, we should regard[Pg 396] as Decapods; but when we trace the two anterior pairs of apparent legs to their insertion, we find that both proceed from the head, which in that genus is distinct from the trunk; while the three last pairs, which alone are furnished with claws, are planted, as legs usually are, in the latter part. The first pair represent the ordinary palpi of Arachnida, are analogous to the labial ones of Hexapods, and, as likewise in Phrynus and Thelyphonus, are more robust than what are usually taken for the first pair of legs; but they differ in being considerably longer, and instead of terminating in a chela are furnished with a retractile sucker[1249]. The second pair are more slender and shorter than the first; they correspond precisely with what are deemed the first pair of legs of Octopods and Arachnida, and are clearly analogous to the maxillary palpi of perfect insects. Whether the base of the first pair of these palpi is in any respect analogous to the labium of insects, (as that of the second seems to be to their maxillæ,) I am not prepared to assert: it will therefore be most advisable to name these palpi anterior and posterior: but as they evidently proceed from the head in Galeodes, and in that genus are clearly analogous to those of the Phrynidea, (which in their turn as clearly represent those of the Aranidea,) it follows that in all they are organs of the part representing the head, and therefore not in a primary sense legs; although in a secondary, as M. Savigny has proved, they may be so called[1250].
1. Araneidea McL. (Aranea L., Araneidæ Latr.) The Araneidea, or spiders, seem resolvable into two[Pg 397] suborders,—the Sedentaries and the Wanderers; thus forming, perhaps, what Mr. MacLeay would denominate the normal groups of a circle of Arachnida.
Def. Mandibles armed with a perforated claw.
Head and Trunk coalite.
Palpi pediform, anterior pair without claws.
Abdomen without segments or elongated tail.
Spiracles two[1251].
Anus furnished with an apparatus for spinning[1252].
2. Scorpionidea McL. (Scorpio L. Latr.)
Def. Mandibles chelate.
Head and Trunk coalite.
Anterior Palpi chelate[1253].
Posterior Palpi pediform.
Pectens two[1254].
Abdomen divided into segments and terminating in a jointed tail, armed at the end with a sting[1255].
Spiracles four pairs.
3. Galeodea.
Def. Head distinct[1256].
Eyes two.
Mandibles chelate with dentated chelæ.
Palpi pediform, the anterior pair thickest with a retractile sucker.
Trunk consisting of two principal segments, with a minute supplementary posterior one[1257].
Spiracles two placed in the trunk[1258].
Pseudo-pectens two[1259].
Abdomen divided into segments.
Anus unarmed and without a spinning apparatus[1260].
4. Phrynidea.
Def. Mandibles unguiculate.
Anterior Palpi chelate or unguiculate[1261], very robust.
Posterior Palpi pediform, very long and slender.
Abdomen divided into segments.
Spiracles two pairs.
Anus terminating in a mucro, and sometimes in a filiform jointed tail without a sting at the end.
v. Having considered the Orders into which Insecta and Arachnida may be divided, I am next to give you some account of the groups into which each is further resolvable. To draw out, however, a complete scheme of these would be deviating from my province, and extend[Pg 399] this letter to an enormous length. Indeed, to give the natural primary and subordinate sections of every Order, would require a knowledge of the subject to which no Entomologist has yet attained. I shall therefore only say something general upon them, and refer you to an example of each kind of group.
Previously to the groups themselves their nomenclature claims our attention. M. Latreille in his last arrangement of Annulose animals has divided his Orders into Sections; Families; Tribes; and Genera: his tribes he has often further subdivided into lesser sections, represented by capital and small letters, &c.[1262]. Mr. MacLeay, discarding the term section, has Tribes; Races (Stirps); Families; Genera, and Subgenera[1263]. But as in descending from the Order to the lowest term, or the species, a series of groups gradually diminishing in value, which require a greater number of denominations than have yet been employed by Entomologists, often occur, I think we may with benefit to the science add to the list. I would therefore propose the following primary and subordinate divisions of an Order: 1. Suborder; 2. Section; 3. Subsection; 4. Tribe; 5. Subtribe; 6. Stirps; 7. Family; 8. Genus; 9. Subgenus. I would further propose that each of these successive groups should have a name always terminating alike, so that the value of the group when spoken of might always be known by the termination:—thus if a subclass end in ata, a suborder might end in ita; a section in ana, a subsection in ena; a tribe in ina, a subtribe in ona; a stirps in una; and a family in idæ; the genera being left free.
[Pg 400]With regard to their characters, we are not to place our groups upon Procrustes' bed, and lop or torture them to accommodate them to every standard we may have fixed for them: assuming one set of characters for suborders, another for tribes, and so for every other group; for the value of characters varies,—those that in some cases are common to an Order, in others indicate only sections, or tribes, or genera and species, or sometimes even sexes. What is constant in one group is not so in another, and vice versâ; so that it is a vain labour to search for a universal character. If it is our wish really to trace the labyrinth of nature, we can only accomplish it by a careful perusal and examination of her various groups. It is singular how much and how far various Entomologists, and those of the very highest class, have been misled by a kind of favouritism to give too universal a currency to certain characters for which they have conceived a predilection. Some have been the champions of the antennæ; others of the trophi; others again of the wings; and others of the metamorphosis. These are all characters which within certain limits lead us right, and are an index to a natural group; but if we follow them further, we leave the system of nature, and are perplexed in the mazes of a method[1264].
Let us now see whether we can pitch upon any suborder which will afford an example of every group that we have lately named. Mr. MacLeay, from a consideration of the larvæ of that Order, has divided the Coleoptera into five primary groups that may be denominated Suborders. Whether these are all natural groups[Pg 401] has not yet been made sufficiently evident. It answers my present purpose, however, to assume it as proved. I select therefore his Chilopodimorpha for my suborder, altering the name as above proposed to Chilopodimorphita: for my Section I take the Predaceous beetles, or Adephaga of M. Clairville, distinguished by having the upper lobe of their maxillæ biarticulate and palpiform;—these I would denominate Adephagana, or devourers. They consist of two groups forming two subsections, the one terrestrial and the other aquatic; which I would name, following Mr. MacLeay, Geodephagena and Hydrodephagena. These two subsections are each resolvable into two Tribes constituted by Linné's four genera Cicindela and Carabus; Dytiscus and Gyrinus. The first tribe, remarkable for the swiftness of their flight, I would name Eupterina, or fliers; the second, equally noted for running, Eutrechina, or runners; the third Eunechina, or swimmers; and the fourth Gyronechina, or swimmers in a circle. The second of these groups, the Eutrechina, are resolvable into two other groups or Subtribes; one distinguished by having the cubit or anterior tibia notched, (which, from their being in general not very brilliant in colour, I would call Amaurona, or obscure); the other having the cubit without a notch, (which, from the brilliancy of many of them, I would name Lamprona, or splendid). These subtribes are both further resolvable into two or more races (Stirpes). I select that to which the crepitant Eutrechina belong, containing those which from their usually truncated elytra MM. Latreille and Dejean have named Truncatipennes[1265]: these, to shorten the name, I call Truncipennuna.[Pg 402] This brings us down to the lowest group formed out of genera and subgenera: or the family, which from its principal genus is named Brachinidæ, and which leads us to the genus Brachinus, and the subgenus Aptini. Thus we get the following scale, expressing every division of an Order, till we arrive at its lowest term, or the species that compose it.
Suborder
Chilopodimorphita McL.
Section
Adephagana Clairv.
Subsection
Geodephagena McL.
Tribe
Eutrechina
Subtribe
Amaurona
Stirps
Truncipennuna Latr.
Family
Brachinidæ
Genus
Brachinus
Subgenus
Aptini.
In the construction of this scale I have endeavoured to steer clear of being led by any system, but, with the exception of the Suborder, which I assume, to resolve it into natural groups gradually decreasing in value, or tending to the lowest term, which appear all of them to[Pg 403] have been considered as such by preceding Entomologists. The four Tribes into which the two subsections Geodephagena and Hydradephagena appear resolvable, are not only distinguished by the characters of the perfect insect, but likewise by those of their larvæ, which are constructed on four distinct types; those of the Gyronechina being the most perfectly Chilopodimorphous of the whole, and those of the Eunechina the least so[1266]. The former appear rather to form an osculant tribe, or one without the circle, than one within it; and to be going off towards another section, including Hydrophilus, Sphæridium, &c. I must observe, that between Dytiscus and Hydrophilus there is a striking agreement both in their form and habits in the larvæ[1266], and even in several characters in the perfect insect; so as in many respects to generate a doubt whether they ought not to enter the same circle and to follow each other. Yet the change of habits in the latter, which from a carnivorous larva becomes a herbivorous beetle; the consequent change of structure in their oral organs, their antennæ, and other striking differences; and the evident intervention of the Gyronechina and some other osculant tribes between the two, forbid their union in one and the same circle.
vi. I need not say more on those larger groups of an Order which conduct us to what are denominated its[Pg 404] genera; but upon these last it will not be a waste of your time to enlarge a little. In the last edition of the Systema Naturæ, and in its appendixes, Linné has described 2840 species of Insecta and Arachnida, which he divided into 83 genera, allowing upon an average nearly 35 species to each genus. From the paucity of the materials, therefore, of which his system was constructed, there was no loud call upon him for numerous genera. But now more than thirty times that number are said to have found a place in the cabinets of collectors[1267], and there is good reason for thinking that perhaps half that are in existence are as yet undiscovered;—this makes it a matter of absolute necessity to subdivide the Linnean genera, which in fact, with regard to the majority of them, were the primary groups of his Orders, rather than an approximation to the ultimate. But this principle may be carried too far: for it is the nature of man to pass from one extreme to the other: and this seems to me to be the case when it is proposed to make genera the extreme term of subdivision before you arrive at species. But it is argued by a very acute Zoologist, that simplicity, perspicuity, and room for necessary variations are best preserved by distinguishing these subdivisions each by an appropriate name[1268]:—Granted. But still it is only a choice of evils. It would require probably more than 10,000 names to designate them, were every extreme group distinguished by a name: but if Mr. MacLeay's admirable pattern exhibited in his genus Phanæus[1269] were followed, it would not call for more than 2000—could[Pg 405] the trifling difficulty occasioned sometimes by the discovery of a new group, be set against the advantage of having only 2000 names to commit to memory instead of 10,000[1270]? But if, after all, it is judged best to name subgenera, M. Savigny's excellent plan of distinguishing them by a plural termination would diminish the weight of the above objection, and might be used with advantage.
When the component parts of any minor group differ from another,—for the most part in important characters, indicating some tangible difference in their habits and economy, and confirmed by peculiarities in their larvæ; and these differences run through the whole, except that as usual they grow weaker as it is passing off to another; especially where they are striking in the centre or type of the group,—this is always a legitimate genus: but where the characters assumed are very slight, and nothing peculiar in its habits, economy or larva, warrant such distinction, it ought not to be conferred.
vii. I must next say a word concerning species and varieties. A species is a natural object whose differences from those most nearly related to it had their origin when it came from the hands of its Creator; while those that characterize a variety, have been produced since that event. As we do not know the value and weight of the momenta by which climate, food, and other supposed fortuitous circumstances operate upon animal forms, we cannot point out any certain diagnostic by which in all cases a species may be distinguished from[Pg 406] a variety;—for those characters that in some are constant, in others vary. In general, where there is no difference in form, appendages and organs, sculpture, proportions and larvæ,—colour alone, especially in insects inhabiting the same district, only indicates a casual variety. Thus Aphodius luridus has sometimes pale elytra with the striæ black (Scarabæus nigro-sulcatus Marsh.): at others it has black spots between the striæ, as in the type: in a third variety the elytra are black at the base and pale at the apex (Sc. varius Marsh.); and lastly, in a fourth they are intirely black (Sc. gagates Marsh.);—yet all these in every other respect precisely correspond. But the converse of this will scarcely hold good; for doubtless minor differences of structure are sometimes produced by a different food and climate: which may probably account for some variations observable in the individuals apparently of the same species obtained from different countries.
Having considered the kind and value of the groups into which Annulose animals, and more especially insects, may be divided, I shall next call your attention to their composition. There are five numbers and their multiples which seem more particularly to prevail in nature: namely, Two—Three—Four—Five and Seven. But though these numbers are prevalent, no one of them can be deemed universal. The binary number, which affords the most simple, and for that reason perhaps not the least valuable, mode of arrangement, we see exemplified when two branches, so to speak, diverge from a common stem,—as in the Vegetable and Animal kingdoms; the terrestrial and aquatic Predaceous beetles;[Pg 407] in the thalerophagous and saprophagous Lamellicorn ones; in the Anoplura and Thysanura; the Chilopoda and Chilognatha amongst Apterous insects; in the Scorpionidea and Aranidea amongst the Arachnida; and in the Macrura and Brachyura amongst the Decapod Crustacea. Again, in other cases three seems to be the most prominent number: this takes place sometimes with regard to the primary groups of an Order, or what I denominate the Suborders. Thus we have the Diurnal, Crepuscular, and Nocturnal Lepidoptera[1271]; the Linnean genera Blatta, Mantis, and Gryllus constitute the Orthoptera; and other instances of this number might be produced in some minor groups. But that which appears[Pg 408] to prevail most widely in nature is what may be called the quaterno-quinary; according to which, groups consist of four minor ones; one of which is excessively capacious in comparison of the other three, and is always divisible into two; which gives five of the same degree, but of which, two have a greater affinity to each other than they have to the other three[1272]. Mr. W. S. MacLeay, in the progress of his inquiries to ascertain the station of Scarabæus sacer, discovered that the thalerophagous and saprophagous Petalocerous beetles resolved themselves each into a circle containing five such groups. And having got this principle, and finding that this number and its multiples prevailed much in nature, he next applied it to the Animal Kingdom in general: and from the result of this investigation, it appeared to him that it was nearly, if not altogether, universal[1273]. Nearly at the same time a discovery almost parallel was made and recorded by three eminent Botanists, MM. Decandolle, Agardh, and Fries, with regard to some groups of the Vegetable Kingdom[1274]; and more recently Mr. Vigors thinks he has discovered the same quinary arrangement in various groups of birds[1275]. This is[Pg 409] a most remarkable coincidence, and seems a strong argument in favour of Mr. MacLeay's system. I should observe, however, that according to that system, as stated in his Horæ Entomologicæ, if the osculant or transition groups are included, the total number is seven[1276]:—these are groups small in number both of genera and species, that intervene between and connect the larger ones. Each of these osculant groups may be regarded as divided into two parts, the one belonging to the upper circle and the other to the lower; so that each circle or larger group is resolvable into five interior and two exterior ones, thus making up the number seven. Though Mr. MacLeay regards this quinary arrangement of natural objects as very general, it does not appear that he looks upon it as absolutely universal,—since he states organized matter to begin in a dichotomy[1277]: and he does not resolve its ultimate groups into five species; nor am I certain that he regards the penultimate groups as invariably consisting of five ultimate ones. In Copris I seem in my own cabinet to possess ten or twelve distinct types[1278]; and in Phanæus, the fifth type, which Mr. MacLeay regards as containing insects resembling all the other types[1279], appears to me rather divided into two; one formed by P. carnifex, Vindex, igneus, &c., and the other by P. splendidulus, floriger, Kirbii, &c.
The great point which demands our attention in[Pg 410] considering a numerical arrangement of the Kingdoms of Nature is the value of the component members of each group. It is by no means difficult to divide a Kingdom, a Class, or an Order into two, or three, or five, or seven or more groups, according to any system we may be inclined to favour; but it is not so easy to do this so that the groups shall be of equal rank. Yet it seems requisite that in grouping our objects, as we descend towards the lowest term we should resolve each only into its primary elements, and of them form the next group; and so on till we come to species. When I say of equal rank, I do not mean an exact parity between the members into which a group is primarily resolvable,—because there will always be a degradation in descensu from the perfection of the type; but merely that parity (to use a metaphor) that there is between children of the same mother, differing in their relative ages and approach to the perfection of their nature. Perhaps it may be observed with respect to the quinary system, that this condition is not complied with, since two of the groups taken per se appear really to form one group; or to be much nearer to each other than to the remaining groups. But when it is taken into consideration that this great group, always resolvable into two, is the typical group, and that the two are really equal, or rather superior in value to the three others, the objection seems to vanish.
With regard to all numerical systems we may observe, that since variation is certainly one of the most universal laws of nature, we may conclude that different numbers prevail in different departments, and that all the numbers above stated as prevalent are often resolvable or reducible into each other. So that where Physiologists[Pg 411] appear to differ, or think they differ, they frequently really agree.
II. The Almighty Creator, when he clothed the world that he had made with plants, and peopled it with animals, besides the manifestation of his own glory, appears to have had two most important purposes in view;—the one to provide a supply for the mutual wants of the various living objects he had created, for the continuance of the species, and for the maintenance of a due proportion, as to numbers, of each kind, so that all might subserve to the good of the whole; and the other, that by them he might instruct his creature man in such civil, physical, moral and spiritual truths, as were calculated to fit him for his station in the visible world, and gradually prepare him to become an inhabitant of that invisible one for which he was destined. The first of these purposes was best promoted by creating things "according to their kind," with sexes monœcious or diœcious; that groups of beings related to each other, and agreeing in their general structure, might discharge a common function. This we see to be the case generally in nature; for where there is an affinity in the structure, there is usually an affinity in the function. The last,—or the instruction of man in his primeval state of integrity and purity,—was best secured by placing before him for his scrutiny a book of emblems or symbols, in which one thing either by its form or qualities, or both, might represent another. If he was informed by his Creator that the works of creation constituted such a book, by the right interpretation of which he might arrive at spiritual verities as well as natural knowledge,[Pg 412] curiosity and the desire of information concerning these high and important subjects would stimulate him to the study of the mystic volume placed before him; in the progress of which he would doubtless be assisted by that Divine guidance, which even now is with those who honestly seek the truth. Both divines and philosophers have embraced this opinion, which is built upon the word of God itself[1280].
This last purpose of the Creator was the root of the analogies, connecting different objects with each other that have no real affinity, observable in the works of creation: so that from the bottom to the top of the scale of being, there is many a series of analogous forms, as well as of concatenated ones; and the intire system of nature is representative, as well as operative: it is a kind of Janus bifrons, which requires to be studied in two aspects looking different ways. To what degree of knowledge the primeval races of men attained after the fall, by the contemplation and study of this book of nature, we are no where informed; but we learn from the highest authority that the revelation that God thus made of himself was in time corrupted, by those that professing themselves to be wise became fools, to the grossest idolatry, which sunk men in the lowest depths of sensuality, vice, and wickedness[1281].
[Pg 413]In no country was this effect more lamentably striking than in Egypt, whose gods were all selected from the animal and vegetable kingdoms.
This species of idolatry doubtless originally resulted from their having been taught that things in nature were symbols of things above nature, and of the attributes and glory of the Godhead. In process of time, while the corruption remained, the knowledge which had been thus abused was lost or dimly seen. The Egyptian priesthood perhaps retained some remains of it; but by them it was made an esoteric doctrine, not to be communicated to the profane vulgar, who were suffered to regard the various objects of their superstitious veneration, not as symbols, but as possessed of an inherent divinity: and probably the mysteries of Isis in Egypt, and of Ceres at Eleusis, were instituted, that this esoteric doctrine, which was to be kept secret and sacred from the common people, might not be lost.
But this kind of analogy is of a higher order than that of which I am here principally to speak,—that, namely,[Pg 414] which the various objects of nature bear to each other. This, however, though of a lower rank, is essentially connected with the other, and leads to it; for it establishes the principle, that created things are representative or symbolical: and we find, when we view them in this light, that as we ascend from the lowest beings in the scale of creation, we are led from one to another till we reach the summit or centre of the whole, and are thus conducted to the boundaries of this visible and material system; from whence we may conclude that we ought not here to stop, but go on to something invisible and extra-mundane, as the ultimate object intended to be reflected from this great speculum of creation—the Creator himself, and all those spirits, virtues, and powers that have emanated from him.
The analogies which the various objects of the animal kingdom mutually exhibit, have for the most part been either overlooked by modern Physiologists, or have been mistaken for characters that indicate affinity; a circumstance that has often perplexed or disrupted their systems. Dr. Virey appears to have been one of the first who obtained a general idea of the parallelism of animals in this respect[1282]; and M. Savigny has contrasted the Mandibulata and Haustellata of the insect tribes as presenting analogies to each other[1283]. But a countryman of our own (often mentioned with honour in the course of our correspondence), peculiarly gifted by nature, and qualified by education and his line of study for such speculations, and possessing moreover the invaluable[Pg 415] opportunity of consulting at his ease one of the first Entomological cabinets in Europe, in a work that will for ever couple his name with the science that he cultivates[1284],—has first taught the Naturalist the respective value and real distinctions of the two kinds of relationship that I am now discussing. He has opened to the philosopher, the moralist and the divine, that hitherto closed door by which our first parents and their immediate descendants entered the temple of nature, and studied the symbols of knowledge that were there presented to them: and in addition to his labours (in numerous respects successful), in endeavouring to trace out the natural groups of beings connected by affinity, has pointed out how they illustrate each other by analogy; thus affording, as was before observed[1285], a most triumphant reply to the arguments of those modern sophists, who, from the graduated scale of affinities observable in creation, were endeavouring to prove that animals, in the lapse of ages, were in fact their own creators[1286].
For the more satisfactory elucidation of the subject before us, I shall consider, first, how we are to distinguish affinities from analogies; and then mention some of the various instances of the latter that occur between insects and other animals, and between different tribes of insects themselves.
To know what characters denote affinity and what are merely analogical, it must be kept in mind that the former being predicated of beings in a series (whether that series has its gyrations that return into themselves, or proceeds in a right line, or assumes any other[Pg 416] intermediate direction, it matters not), it cannot be satisfactorily ascertained but by considering attentively the gradual approximation or recession of the structure to or from a certain type in any point of such series. If, therefore, you wish to ascertain whether the characters, in which any given object resembles other objects in certain groups, indicate affinity or only analogy, you must first make yourself acquainted with the common features which distinguish the animals known to belong to that group,—either those relating to their structure, or to their habits and economy. If the object under your eye partakes in these characters more or less, in proportion as it approaches the type or recedes from it, the relation it exhibits is that of affinity; but if, though it resembles some members of it in several points of its structure, it differs from the whole group in the general features and characteristic marks that distinguish it, the relation it bears to those members is merely that of analogy. Thus, for instance, Ascalaphus italicus in its antennæ, the colouring of its wings, and its general aspect, exhibits a striking resemblance to a butterfly; yet a closer examination of its characters will satisfy any one that it is in quite a different series, and has no affinity whatever to that genus. A departure, however, in only one respect from what may be called the normal characters of its group, does not annul the claim of any tribe of insects to remain in it; since this very often only indicates a retrocession from the type, and not a disruption of its ties of affinity. Thus the saw-flies (Serrifera) differ from the other Hymenoptera, though not in their pupæ, yet more or less in their larvæ; but this alone cannot countervail their agreement with that Order in their organs of manducation[Pg 417] and motion, in their ovipositor, and in the other details of their structure[1287].
I have on a former occasion pointed out many of the analogies which take place between insects and other parts of the animal kingdom, and even between insects and the mineral and vegetable kingdoms[1288]: I shall now resume the subject more at large, but without recurring to those last mentioned. In considering the analogies which connect insects with other animals, or which they exhibit with respect to each other, we may have recourse to two methods. We may either consider them as placed somewhere between the two extremes of a convolving series, from which station we may trace these analogies upwards and downwards towards each limit; or we may conceive them and other animals in this respect arranged in a number of series that are parallel to each other, in which the opposite points are analogous. The first mode will perhaps best explain the analogies that exist between insects and other animals, and the last those between different groups of insects themselves. I shall give an example or two of each method, beginning with the first.
There are two tribes in the animal kingdom that seem placed in contrast to each other, both by their habits and by their structure. One of these is carnivorous, living by rapine and bloodshed, and can seldom be rendered subservient to our domestic purposes; while the other is herbivorous or granivorous, is quiet in its habits, and easily domesticated. Amongst insects we find the representatives of both: those of the first tribe are distinguished by their predaceous habits, by the open attacks, or by the various snares and artifices which they employ[Pg 418] to entrap and destroy other insects. They may usually be known by their powerful jaws or instruments of suction; by their prominent or ferocious eyes; by the swiftness of their motions, either on the earth, in the air, or in the water; by their fraud and artifice in lying in wait for their prey. Amongst the Coleoptera, the Predaceous beetles,—including the Linnean genera Cicindela, Carabus[1289], Dytiscus, and Gyrinus,—are of this description; and they symbolize those higher animals that by open violence attack and devour their prey:—for instance, the sharks, pikes, &c., amongst the fishes; the eagles, hawks, &c., amongst the birds; and the whole feline genus amongst the beasts. Similar characters give a similar relation of analogy to the Mantidæ and Libellulina amongst the Orthoptera and Neuroptera. The whole family of Arachne, the larvæ of the Myrmeleonina, &c., portray those animals that to ferocity add cunning and stratagem, or suck the blood of their victims. The Myriapods symbolize in a striking manner the Ophidian reptiles. Look at an Iulus, and both in its motions and form you will acknowledge that it represents a living serpent; next turn your eyes to a centipede or Scolopendra, and you will find it nearly an exact model of the skeleton of a dead one, the flat segments of its body resembling the vertebræ, its curving legs the ribs, and its[Pg 419] venomous maxillæ the poison-fangs. The great body of the Orthoptera, the Homopterous Hemiptera, the Lepidoptera, and Trichoptera, afford no example of Predaceous insects. All the analogies I have here particularized, ascending from the insect, terminate in races of a corresponding character and aspect amongst the Mammalia, and thus lead us towards man himself, or rather to men in whose minds those bad and malignant qualities prevail, which, when accompanied by power, harass and lay waste mankind; and thus ascending from symbol to symbol, we arrive at an animal who in his own person unites both matter and spirit, and is thus the member both of a visible and invisible world: and we are further instructed by these symbols,—perpetually recurring under different forms,—in the existence of evil and malignant spirits, whose object and delight is the corporeal and spiritual ruin of the noble creature who is placed at the head of the visible works of God.
The other tribe of animals that I mentioned of a milder character, may be looked upon as represented by many herbivorous, or not carnivorous, insects; amongst others, the Lamellicorn beetles imitate them by their remarkable horns, so that they wear the aspect of miniature bulls, or deer, or antelopes[1290], or rams, or goats, whether these horns are processes of the head or of the upper jaws. The gregarious Hymenoptera, some of which form part of our domestic treasures, may be regarded in some degree as belonging to this department. From insects the ascent upwards, with regard to form, is by some of the branchiostegous fishes, which symbolize the horns of[Pg 420] cattle; with regard to character, by the various species of Cyprinus and other similar genera.—Whether any of the reptiles may be looked upon as falling into this division, I am not sufficiently conversant with them to assert; but if any, the Chelonians, or tortoise and turtle tribes, are entitled to that distinction. Amongst the birds, the Gallinæ and Anseres,—from which Orders we derive our domestic poultry, whether terrestrial or aquatic,—and our game, form the step next below the ruminants, or cattle: and we are thus again led towards man, and are symbolically instructed in those domestic and social qualities which endear us to each other, best promote the general welfare, and render us most like good spirits and the Divinity himself; of whom the perpetual recurrence of animals exhibiting these amiable and useful qualities is calculated to impress upon us some notion. I might mention many more instances of ascending analogies; as from some of the Diptera by the parrots, to the Quadrumanes or monkey tribes—or from some of the Iulidæ that roll themselves into a ball, to the Armadillo; but these are sufficient to set your mind at work upon the subject, so that you may trace them for yourself. Nor shall I occupy your time by pointing out how analogies may be traced from insects downwards towards the lowest term in the scale of animal life, but proceed to consider the analogies observable between insects themselves; in which I shall follow the second method lately mentioned, and consider them as arranged in parallel series.
In studying the analogies that take place between insects themselves, we should always bear in mind that our inquiry is not concerning an affinity which demands a correspondence in various particulars that are not necessary to constitute an analogy; as, for instance, that there[Pg 421] should be a mutual imitation in all the states of any two insects. Wherever we discover a marked resemblance between two perfect insects, there is a true analogy, though their metamorphosis may differ; and where there is not that resemblance, though the metamorphosis may agree, there is no analogy. In fact, insects are sometimes analogous in their first state and not in their last; and at other times analogous in their last and not in their first; but the analogy is most perfect when it holds in all their states: it then, indeed, almost approaches to an affinity. They may also be analogous to each other in their habits and economy, when there is little or no resemblance in their form; and, vice versa, be analogous in their form and not in their habits. So that different sets of analogies may be assumed as foundations for different systems. Thus Mr. MacLeay assumes the metamorphosis as the basis of analogy between the corresponding Orders of Mandibulata and Haustellata[1291], while M. Savigny compares the perfect insects[1292]: the result therefore differs in some instances. I shall now lay before you in a tabular view their plans and my own.
Savigny.
| Mandibulata. | Haustellata. | |||
| Neuroptera | ... | Lepidoptera | ||
| Ascalaphus | Papilio | |||
| Hymenoptera | ... | Diptera | ||
| Eucera | Tabanus | |||
| Orthoptera | ... | Homoptera | ||
| Locusta L. | Cicada | |||
| Aptera | ... | Aphaniptera | ||
| Nirmus | Pulex. | |||
MacLeay.
| Mandibulata. | Haustellata. |
| Trichoptera | Lepidoptera |
| Hymenoptera | Diptera |
| Coleoptera | Aptera |
| Orthoptera | Hemiptera |
| Neuroptera | Homoptera. |
| K. and S. | |
| Coleoptera | Hemiptera Leach |
| Orthoptera | Homoptera Leach |
| Neuroptera | Lepidoptera |
| Hymenoptera | Diptera. |
In these two last columns, you see, I differ little from M. Savigny: I merely exclude the Aphaniptera as forming an osculant Order, and I have added the Coleoptera and Heteropterous Hemiptera for reasons I shall soon assign. From Mr. MacLeay I differ more widely, which has resulted from our different ideas as to the mode of tracing analogies; his theory leading him to the metamorphosis, and mine leading me[1293] to the perfect insect, for the foundation of our several systems. It remains that I show how each of the pairs in my columns represent each other: but I must observe, that the analogies exhibited by insects in the corresponding Orders of these columns are not equally striking in all their respective members; but only in certain individual species or genera, more or less numerous, by which the nearest approach is made to the contrasted forms.
To begin with the Coleoptera and Heteropterous Hemiptera.—Both are distinguished by having an ample[Pg 423] prothorax, a conspicuous scutellum, the neuration of their wings, the substance of the hard part of their hemelytra, which, as in Coleoptera, sometimes imitates horn and sometimes leather, and is occasionally, like elytra, lined with a hypoderma[1294]; the articulation of the head with the trunk is likewise the same in both[1295]: and some Heteropterous species so strikingly resemble beetles (Lygæus, brevipennis &c.), having little or no membrane at the end of their hemelytra, that they might easily be mistaken for them. These circumstances prove, I think, that this suborder is more analogous to the Coleoptera than to the Orthoptera, with which it agrees in scarcely any respect but its metamorphosis. The counterparts of this last Order indeed, instead of the Heteropterous, are to be sought for amongst the Homopterous Hemiptera, various species of which exhibit a most marked and multifarious analogy with numerous Orthoptera. Many of both Orders (Cicada, Locusta), as you have heard long since, are signalized by possessing the same powers of song, and produced by an analogous organ[1296]: a large proportion also of both are endued with wonderful saltatorious powers, and their posterior tibiæ are similarly armed; their legs in general likewise are longitudinally angular, and the head in both articulates with the trunk in the same manner[1297]. In both Orders too, the upper organs of flight are most commonly tegmina, but sometimes in both they are nearly membranous, like wings. In Centrotus and Acrydium, the one Homopterous and the other Orthopterous, the front is bilobed, the eyes are small; there are only two[Pg 424] stemmata between the eyes; the prothorax is conspicuous, and behind is producted into a long scutelliform process, under which all the parts also are analogous; the abdomen articulates with the trunk in the same way, is similar in shape in both, and consists of short inosculating segments. Some Fulgoridæ and Truxalides agree also in their producted front. Other analogous characters might be named between these tribes, but these are sufficient to confirm M. Savigny's opinion. That the Neuroptera present analogies to the Lepidoptera, though they differ so widely from them in their metamorphosis and habits, is evident from the instance lately adduced of Ascalaphus italicus, which was described as a butterfly by Scopoli[1298]; and many of the Libellulina, by their wings, partly transparent and partly opaque, and by the shape of those organs and of their bodies, imitate the Heliconian butterflies: and this resemblance is much more striking than any that occurs between the perfect insects in the Neuroptera and Homopterous Hemiptera. With regard to the Hymenoptera and Diptera the analogy is undisputed, and must strike every beholder; and one would almost say it was a real affinity, were it not that the resemblance is not only general between Order and Order, but that almost every Hymenopterous tribe has its counterpart amongst the Diptera; the saw-flies[1299] for instance, the ichneumons, the various false-wasps[1300], the false-bees[1301], the bees, the humble-bees, the ants, &c., severally find there a representative that wears[Pg 425] its livery and general aspect: a circumstance which evidently proves that it was part of the plan of the Creator to place them in contrast with each other. Were I to pursue this subject further, it might not be difficult to show that were the tribes of Mandibulata or of Haustellata also arranged in columns, analogies would be discoverable between their corresponding points: this seems to be Mr. MacLeay's opinion[1302]; and it is worth your pursuing the subject further, which cannot but prove very interesting.
But though the general analogy of these columns is that of Order to Order, yet individual species in each Order sometimes find their representatives in a different one from that with which they generally are contrasted;—thus some Diptera, as Culex, by the scales on the veins and other parts of their wings, are analogous to Lepidoptera rather than Hymenoptera[1303]; as is also the genus Psychoda by its form.
We come now to the consideration of a question not easy to be decided,—I mean, which Order of insects is to have the precedency, and which is the connecting link that unites them to Vertebrate animals.
Linné (and Mr. MacLeay seems in this to coincide with him) considered the Coleoptera as at the head of the Class of insects; De Geer thought the Lepidoptera entitled to that honour; Latreille and Cuvier begin with the Aptera: Marcel de Serres favours the Orthoptera[1304]; and others, on account of their admirable economy, have made the Hymenoptera the princes of the insect world[1305].[Pg 426] If the claim to priority was to be decided by the exquisiteness of instincts and the benefits conferred upon the human race, doubtless it would be in favour of the last-mentioned insects. If the power to do mischief carried it, and to lay waste the earth, the Orthoptera would be entitled as much as any to the bad pre-eminence. If beauty, and grace, and gaiety, and splendour of colours were the great requisite, and the law enjoined, Detur pulchriori,—the Lepidoptera would doubtless win the throne. But if perfection and solidity of structure, as they ought, are to regulate this point; we must, I think, with the illustrious Swede, assign the palm to the Coleoptera. If we consider these in all their parts, the organs for flight only excepted, they seem more perfectly formed and finished than the insects of any other order. But which of the Coleopterous tribes are entitled to the precedency? Linné placed the Lamellicorn beetles at the head of the order, beginning with the Dynastidæ, probably led by some characters which seem to connect these with the Branchiostegous fishes. In this he was followed by Fabricius. But Latreille and most modern Entomologists have begun with Cicindela and the other Predaceous beetles. I am not certain what are Mr. MacLeay's sentiments on this subject; but from what he says in the Annulosa Javanica[1306], it does not appear that he is a convert to the latter opinion. Bulk and strength seem the most striking characteristics of the former tribe, which represent the cattle or ruminants amongst Vertebrate animals.—Strength united with agility and a considerable portion of grace and symmetry evidently confers[Pg 427] a degree of pre-eminence upon the latter, symbolizing the feline race, which seems to throw no small weight into their scale.
There are two Classes of Vertebrate animals with which insects may appear to claim kindred. The fishes, and the reptiles. Fishes in their fins exhibit no small resemblance to insects; the pectoral and ventral ones representing their arms and legs, and the dorsal ones their wings: Pegasus Draco in this last respect is not unlike a butterfly[1307]. In some genera (Ostracion, Pegasus, &c.), like insects the animal is covered with a hard shell or crust, formed by the union of its scales. The oral cirrhi of many fishes seem analogous to the palpi of insects; and in some a pair longer than the rest represent their antennæ[1308]. Another circumstance in which insects and fishes correspond, is the wonderful variety of forms, often in the greatest degree eccentric, that occurs in both Classes. Some of the cyclostomous fishes, as Ammocœtus, Gastrobranchus, are supposed to connect the fishes with the Annulosa, by means of the Annelida as an osculant Class[1309], which Mr. MacLeay regards as the passage to the Chilopoda[1310]: his Mandibulata he considers as passing into the Anoplura by means of some osculant Order as yet unknown[1311]. But I must confess I can see no good ground for this last transition:—the Anoplura appear much more nearly related to the Psocidæ, especially by the apterous Atropos pulsatoria[1312] than to any[Pg 428] Coleopterous insect. But having stated these opinions, I shall leave you to draw your own conclusions, as the question is still perplexed with many difficulties. I am ready to admit that some Vertebrates approach near to the Annelida; but that it is through them alone that they are connected with insects, is not at present clear.
With regard to reptiles, they seem to be connected with insects by several characters. In the Chelonians, the skeleton merges in the external carapace or shell; the Ophidians change their skin like larvæ; the Batrachians undergo metamorphoses; some of the Saurians also have their changes: and the Draco volans has wings somewhat analogous to those of insects[1313]. Were I to be asked what Order of insects could connect with reptiles, I should point to the Orthoptera, especially Gryllus L., which by their noise and saltatorious powers not a little resemble frogs; and the larvæ of some strikingly imitate their form[1314]: and of others even that of a lizard[1315]. But these resemblances, after all, may only indicate analogies.
After the very general idea that I have attempted to embody for you of the System of Insects; of the groups in which nature has arranged them, and their mutual relations; it will not be out of place, if I next state to you what has been effected by Entomologists towards reducing them to order: or, in other words, if I give you some account of the various Methods and Systems[1316], beginning with the earliest, that have appeared and had their day, which will include a history of the progress of our science from its commencement to its present era.
In writing the history of any science, two modes present themselves. We may either give a chronological review of all the circumstances and publications connected with it; or content ourselves with a rapid survey, dwelling only on the principal epochs, and those lights of the science who by their immortal labours gave birth to them. The latter is that on every account best suited to our present purpose, which I shall therefore here adopt.
There seem to me to be seven principal epochs into which the History of Entomology may be divided: viz.[Pg 430] 1. The Era of the Ancients. 2. The Era of the revival of the science after the darkness of the middle ages. 3. The Era of Swammerdam and Ray, or of the Metamorphotic System. 4. The Era of Linné, or of the Alary System. 5. The Era of Fabricius, or of the Maxillary System. 6. The Era of Latreille, or of the Eclectic System. And 7. The Era of MacLeay, or of the Quinary System. All of these appear to form important points, or resting-places, in the progress of the science towards its acme; and of each of these I shall now proceed to give you a brief account.
1. The Era of the Ancients. To ascertain what attention was paid to insects in the earliest ages, we must have recourse to the most ancient of records, the Old Testament. In this sacred volume we are informed that after the Creation God brought the creatures to Adam that he might name them[1317]. Now the first man, in his unimpaired state of corporeal, mental, and spiritual soundness, under the divine guidance, doubtless imposed upon them names significant of their qualities or structure; which according to Plato was a work above human wisdom, and on account of which the ancient Hebrews deduced that Adam was a philosopher of the highest endowments[1318]. Whether on this great and interesting occasion he gave names to individual species, or only to natural groups, does not clearly appear. But probably as they were created, so were they brought before him "According to their kinds[1319]."
Subsequently Moses will be thought to have possessed no ordinary knowledge of insects, if we suppose, as the[Pg 431] ingenious remarks of Professor Lichtenstein[1320] render probable, that he distinguishes as clean insects the Fabrician genera Gryllus, Locusta, Truxalis, and Acheta, which a person unobservant of these animals would have confounded together. This discrimination presupposes this knowledge of their general characters, not only in the Jewish lawgiver, but also in the people themselves to whom the precept was addressed, to whom it would otherwise have been de ignotis.
Allusion is made in Holy Writ to insects of almost every one of the modern Orders[1321]. They are represented as employed divinitùs sometimes to annoy the enemies of the Israelites, and at others to punish that people themselves when they apostatized from their God. The prophets frequently introduce them as symbols of enemies that lay waste or oppress the church: as the fly of the Ethiopians or Egyptians; the bee of the Assyrians; and the locust of the followers of Mahomet and other similar destroyers[1322]. That Solomon, amongst other objects to the investigation of which his divinely inspired wisdom directed him, did not deem insects, those "Little things upon the earth[1323]," unworthy of his attention, we know from Scripture[1324]; but as his physical writings are lost, we are ignorant whether he treated of their natural arrangement, their economy and history, or of the instruction they afford analogically considered. Where[Pg 432] he has referred to them incidentally, it is generally with this latter view.
If we turn from the word and people of God to the Lovers of Wisdom (as they modestly styled themselves) of the heathen world, and their writings; we shall discern amongst them a great light shining, the beams of which illuminate even our own times. In the illustrious Stagyrite we recognize—"The father of philosophy, at least of our philosophy, who, rising superior to the darkness in which he lived, darted his penetrating glance through all nature, and established principles which a long course of ages of inquiry have but confirmed. With Aristotle begins the real History of science: and how much soever he may have erred upon particular points, the greatness of his conceptions and the justness of his ideas, on the whole entitle him to our high veneration. His labours in the investigation of the Animal Kingdom have laid the foundation of the knowledge we now possess[1325]." This language of the lamented and learned President of the Linnean Society is particularly applicable to what this great and original genius has effected in Entomology. We have seen upon a former occasion[1326], that Linné himself had not those precise ideas of the limits of the Class Insecta, which Aristotle so many centuries before him had adopted. In stating the obligations of Entomology to this true sçavant, I shall begin by laying before you a tabular view of what may be called his system, as far as I have been able to collect it from his works, especially his History of Animals.
| Coleoptera[1327]. | ||||||||
| Pedetica = Orthoptera saltatoria Latr.[1328] | ||||||||
| Astomata = Hemiptera Latr.[1329] | ||||||||
| Pterota vel Ptilota[1331] | Psychæ = Lepidoptera[1330]. | |||||||
| Tetraptera | majora = Neuroptera L. Orthoptera cursoria Latr.[1332]? | |||||||
| Opisthocentra = Hymenoptera[1333]. | ||||||||
| Diptera[1334] | minora = Musca, Tipula, &c. | |||||||
| Emprosthocentra = Culex, Stomoxys, Tabanus, &c. | ||||||||
| Insecta | Pterota simul et Aptera[1335] | Myrmex = Formica L. | ||||||
| Pygolampis = Lampyris L. | ||||||||
| Aptera[1336]. | ||||||||
It may be further stated, that Aristotle perceived also the distinction between the Mandibulata and Haustellata of modern authors: for he observes, that some insects having teeth are omnivorous; while others, that have only a tongue, are supported by liquid food[1337]. He appears to have regarded the Hymenoptera, or some of them, as forming a third subclass; since he clearly alludes to them, when he says that many have teeth, not for feeding, but to help them in fulfilling their instincts[1338].
From the above statement it will appear that this great philosopher had no contemptible notion,—though he[Pg 434] has only distinguished three of them as larger groups by appropriate names,—of the majority of the Orders of Insects at present admitted. His Coleoptera, Psychæ, and Diptera are evidently such. His idea of Hemiptera seems taken solely from the Cicada or Tettix: but the manner in which he expresses himself concerning it, as having no mouth, but furnished instead with a linguiform organ resembling the proboscis of Diptera[1339], proves that he regarded it as the type of a distinct group. Since he considers the saltatorious Orthoptera as forming such a group, it is probable that he included the cursorious ones with the Neuroptera in his majora section of Tetraptera; and the resemblance of many of the Mantidæ to the Neuroptera is so great, that this mistake would not be wonderful. His division of the Diptera is quite artificial.
How far Aristotle's ideas with regard to genera and species attained to any degree of precision, is not easily ascertained: in other respects his knowledge of insects was more evident. As to their anatomy, he observes that their body is usually divided into three primary segments,—head, trunk, and abdomen; that they have an intestinal canal,—in some straight and simple, in others contorted,—extending from the mouth to the anus; that the Orthoptera have a ventricle or gizzard[1340]. He had noticed the drums of Cicada, and that the males only are vocal. Other instances of the accurate observation of this great man might be adduced, but enough has been said to justify the above encomiums. His principal error was that of equivocal generation.
[Pg 435]Little is known with regard to the progress of other
Greek Naturalists in entomological science. It appears probable, from an epithet by which Hesiod distinguishes the spider—air-flying[1341], that the fact of these insects traversing the air was at that time no secret. Apollodorus, as we learn from Pliny[1342], was the first monographer of insects, since he wrote a treatise upon scorpions, and described nine species. But like many other Zoologists, by mistaking analogy for affinity, he has included a winged insect, probably a Panorpa, amongst his scorpions. From the time of Aristotle, however, to Pliny, no writer is recorded, with the exception of those before alluded to[1343], that appears to have attended much to insects. They are indeed incidentally noticed by Theophrastus, Dioscorides, Virgil, Ovid, &c., but without any material addition to the stock of entomological knowledge bequeathed to us by the Stagyrite. Even Pliny's vast compendium, as it professed to be, of the natural history of the globe, was in many respects little more than a compilation from that great philosopher. Still, however, though he does not appear to have paid much practical attention to insects,—which indeed, considering the extent of his views, was scarcely to be expected,—yet as a guide to the then state of entomological knowledge, and as an advocate for the study, which in the exordium of his eleventh book he has so eloquently and with so much animation defended from the misrepresentations of ignorance, Pliny has conferred a lasting obligation on the science. The last zoological writer of note was Ælian, who amongst other animals[Pg 436] often mentions insects. He has, however, few original observations. One was, that scorpions are viviparous[1344]. From him we learn incidentally that artificial flies were sometimes used by Grecian anglers[1345].
2. The Era of the Revival of the Science. From the time of Pliny and Ælian 1400 years rolled away, in which scarcely any thing was done or attempted for Entomology or Natural History in general. During that long night the glimmer of only one faint luminary appeared to make a short and feeble twilight. In the middle of the thirteenth century Albertus Magnus (so called from his family name of Groot, and justly, if incredible labour could entitle a man to the appellation), devoted one out of twenty-one folio volumes to Natural History. In this work he professes not so much to give his own opinions, as those of the Peripatetic philosophers[1346]. He occasionally, however, relates the result of observations made by himself, which prove him to have been no inattentive student of nature. He mentions a voyage that he made for the purpose of collecting marine animals, and that he found of them ten different tribes or genera, and several species of each. Amongst these he particularizes the Cephalopoda, the Crustacea, the testaceous Mollusca, and some of the Radiata and Acrita, &c.[1347] He gives a very correct account of the pitfalls of Myrmeleon. Insects he distinguishes, excluding the Crustacea, by the denomination of Anulosa (Annulosa), which he appears to employ as a known term[1348]. He also calls them worms, describing butterflies as flying[Pg 437] worms, flies as fly-worms, spiders as spider-worms; and what is still more extraordinary, the toad and the frog, which he includes amongst his Anulosa, he calls quadruped-worms[1349]!! Though it may appear so absurd to speak of these animals as insects, yet he had perhaps a deeper and more philosophical reason for this than we may at first be disposed to give him credit for. This would be the case if he separated these from the other reptiles and placed them amongst insects on account of their metamorphoses, mistaking perhaps an analogical character for one of affinity[1350]. Some of the Annelida, as Filaria and Lumbricus[1351], he also regarded as insects. I cannot gather from his desultory pages that he had any notion of a systematical arrangement of his Anulosa.
After the taking of Constantinople by the Turks in the middle of the fifteenth century, the light of learning, kindled by those of its professors who escaped from that ruin, appeared again in the West. The Greek language then began to be studied universally; and in consequence of the coeval invention of the art of printing, various editions of the great works of the ancients were published: amongst the rest those of the fathers of Natural History. From the perusal of these, the love of the sciences of which they treated revived in the West, and the attention of scientific men began to direct itself to the consideration and study of the works of their Creator. In the latter part of that century, a work entitled the Book of Nature appeared in the German language, in which animals and plants were treated of[Pg 438] and rudely figured; as they were likewise most miserably in Cuba's Ortus Sanitatis, published in 1485, in which insects and Crustacea were described under the three different denominations of Animals, Birds, and Fishes; so that but little profit was at first derived from the writings of Aristotle, Invertebrate animals not being then even honoured with
This unpromising and apparently hopeless state of the science proved, however, the dawn of its present meridian brightness.
The first attempt at a separate and systematical arrangement of insects subsequent to the times of Aristotle, was made in the ponderous volumes of Ulysses Aldrovandus, who, disregarding the Stagyrite, arranged insects according to the medium they inhabit, as you will see in the subjoined table:
| Membranacea | Favifica. | |||||||||||
| Anelytra | Non Favifica. | |||||||||||
| Alata | Farinosa. | |||||||||||
| Pedata | Elytrota. | |||||||||||
| Terrestria | Aptera | Paucipeda. | ||||||||||
| Multipeda. | ||||||||||||
| Insecta | Apoda. | |||||||||||
| Pedata | Paucipeda. | |||||||||||
| Aquatica | Multipeda. | |||||||||||
| Apoda. |
This artificial and meager system, which mixed insects with Annelida, was adopted by Charlton and other authors; and even in the eighteenth century had a patron of great eminence, who, endeavouring to improve upon[Pg 439] it, has rendered it still more at variance with nature and Aristotle: I mean the celebrated Vallisnieri, to whom in other respects, though in this he fell behind his age, the science was under great obligations. He divides insects into, 1. Those that inhabit vegetable substances living or dead. 2. Those that inhabit any kind of fluid and in any state. 3. Those that inhabit any earthy or mineral substances, dead bones, or shells. And 4. Those that inhabit living animals[1352].
The work that is usually called Mouffet's Theatrum Insectorum was produced in the present era, and was the fruit of the successive labours of several men of talent. Dr. Edward Wotton and the celebrated Conrade Gesner laid the foundation; whose manuscripts falling into the hands of Dr. Thomas Penny,—an eminent physician and botanist of the Elizabethan age[1353], much devoted to the study of insect,—he upon this foundation meditated raising a superstructure which should include a complete history of these animals; and with this view he devoted the leisure hours of fifteen years of his life to the study of every book then extant that treated of the science either expressly or incidentally, and to the description and figuring of such insects as he could procure; but before he had reduced his materials to order, in 1589 he was snatched away by an untimely death. His unfinished manuscripts were purchased at a considerable price by Mouffet, a contemporary physician of singular learning[1354], who reduced them to order, improved the style, added new matter, and not less than 150 additional figures; and[Pg 440] thus having prepared the work for the press, intended to dedicate it to queen Elizabeth[1355]. Fate, however, seemed still to frown upon the undertaking, for before he could commit his labours to the press he also died, and his book remained buried in dust and obscurity till it fell into the hands of Sir Theodore Mayerne, baron d'Aubone, one of the court physicians in the time of Charles I., who at length published it, prefixing a Dedication to Sir William Paddy, baronet, M.D., in 1634; and it was so well received that an English translation appeared twenty-four years afterwards. The work thus repeatedly rescued from destruction was indisputably the most complete entomological treatise that had then appeared. And though the arrangement (in which there is scarcely any attempt at system) is extremely defective, the figures very rude, often incorrect, and sometimes altogether false,—yet as an introduction to the study of insects its value at that day must have been very considerable; and as a copious storehouse of ancient entomological lore, it has not even at present lost its utility.
One of the most remarkable works of the era we are upon was published at Lignitz in the year 1603, by Caspar Schwenckfeeld, a physician of Hirschberg, under the title of Theriotrophium Silesiæ. This was probably the first attempt at a Fauna that ever was made. In it animals are divided into quadrupeds, reptiles, birds, fishes, and insects. The Crustacea, Mollusca, and Zoophytes, are included under fishes. He says of the Spongiæ that they are moved by animalcula which inhabit them[1356]. Did he borrow this observation from Aristotle,[Pg 441] or was it made by himself[1357]? It is singular that Linné should never allude to this work. Goedart, who belongs also to this era, is stated to have spent forty years of his life in attending to the proceedings of insects[1358]. But after this long study, his principal use to the science was the improvement he effected in the drawing and engraving of them,—for his figures, though sometimes incorrect and sometimes fabulous, were far superior to those of his predecessors.
3. The Era of Swammerdam and Ray, or of the Metamorphotic System. The great men whose names are here united, as they were cotemporary, so they agreed in founding their respective systems of insects on the same basis. To the former, however, is due the merit of being the first who assumed the metamorphoses of these animals as the basis of a natural arrangement of them; upon which the latter, in conjunction with his lamented friend Willughby, erected that superstructure which opened the door for the present improved state of the science. Swammerdam's system may be thus expressed in modern language:
| Class i. | Metamorphosis | complete[1359] = Aptera L.[1360] | ||||
| ii. | ——————— | semicomplete | Orthoptera, Hemiptera. Libellulina, Ephemerina[1361]. | |||
| Insects | iii. | ——————— | incomplete | Coleoptera, Hymenoptera, part of Neuroptera and Diptera[1362]. | ||
| obtected | Lepidoptera[1363]. | |||||
| iv. | ——————— | Ichneumones minuti L.[1364] | ||||
| Muscidæ, &c[1365]. | ||||||
[Pg 442]It was a great point gained in the science to introduce the consideration of the metamorphosis, and to employ it in the extrication of the natural system: for though when taken by itself it will, as in the table just given, lead to an artificial arrangement, it furnishes a very useful clue when the consideration of insects in their perfect state is added to it. The tables contained in the Prolegomena to Ray's Historia Insectorum divide insects into those which undergo no change of form, and those which change their form. The arrangement of the former Αμεταμορφωτα was made by Willughby, who subdivided them into Apoda and Pedata. As the only insects included in the former section were the grubs of Œstri, the remainder being Annelida, they need not be included in our table. I have endeavoured to compress these tables into as small a space as possible, by using the Linnean terms for metamorphosis, and reducing Ray's tribes of Orthoptera, Hemiptera, and Neuroptera to their modern denominations.
Ray details at considerable length the various tribes belonging to the four classes of metamorphosis established by Swammerdam[1366]. Most of his tribes indicate natural groups of greater or less value: but some of his larger groups are artificial, as you will see by the mere inspection of the table.
| Apoda | Terrestia[1367]. | |||||||||||||||
| Aquatica. | ||||||||||||||||
| Ametamorphota | Terrestia[1368] | Majora[1368]. | ||||||||||||||
| Hexapoda | Minora[1369]. | |||||||||||||||
| Aquatica[1370]. | ||||||||||||||||
| Pedata | Octopoda | Caudata[1371]. | ||||||||||||||
| Non caudata[1372]. | ||||||||||||||||
| 14-poda. | ||||||||||||||||
| 24-poda. | ||||||||||||||||
| 30-poda. | ||||||||||||||||
| Terrestia | Cylindrica[1373]. | |||||||||||||||
| Insecta | Compressa[1374]. | |||||||||||||||
| Polypoda | ||||||||||||||||
| Corpore tereti. | ||||||||||||||||
| Aquatica[1375] | ——— plano. | |||||||||||||||
| Bicaudata. | ||||||||||||||||
| Orthoptera. | ||||||||||||||||
| Heteroptera. | ||||||||||||||||
| Metamorphosis semicompleta[1376] | Homoptera. | |||||||||||||||
| Libellulina. | ||||||||||||||||
| Ephemerina. | ||||||||||||||||
| Metamorphumena | Coleoptera. | |||||||||||||||
| Metamorphosis incompleta vel obtecta | Alis farinaceis[1377]. | |||||||||||||||
| Anelytra | Diptera. | Gregaria et Favifica | Mellifica[1378]. | |||||||||||||
| Non Mellifica[1379]. | ||||||||||||||||
| Alis membranaceis | ||||||||||||||||
| Metamorphosis coarctata | Muscidæ et Ichneumones minuti L.[1385] | Apiformia[1380]. | ||||||||||||||
| Vespiformia | Breviora[1381]. | |||||||||||||||
| Tetrapter | Solitaria non Gregaria et Favifica | Augustiora[1382]. | ||||||||||||||
| Papilioniformia[1383]. | ||||||||||||||||
| Seticaudæ, seu Tripilia[1384]. |
[Pg 444]This era produced several great and original geniuses, who enriched the science with a vast increment of real knowledge. The illustrious Zoologists whose names it bears,—the one by his dissections and anatomical researches, and the other by his concise and well drawn descriptions of numerous insects, by various interesting observations on their manners and characters, and by the purity of his latinity,—contributed greatly to its progress towards perfection. Leeuwenhoek also, the compatriot of Swammerdam, and Hooke of Ray, amongst other objects submitted to their powerful microscopes, did not neglect insects.—To the former we are indebted for the remarkable discovery that the flea belongs to those that undergo a metamorphosis. Ray had besides two coadjutors whose names ought not to be forgotten,—Willughby and Dr. Martin Lister. The former is characterized by his lamenting friend as one of the profoundest of naturalists, as well as one of the most amiable and virtuous of men. What advantage Entomology would have reaped from his labours may be inferred from the eminent services that he rendered that science, amongst other branches of Zoology, during his short life. It appears from Ray's Letters[1386], that he drew up a history of insects and exsanguia, which probably formed the groundwork of the posthumous Historia Insectorum of that author; concerning which he says, "The work which I have now entered upon is indeed too great a task for me: I rely chiefly on Mr. Willughby's discoveries and the contributions of friends[1387]." And indeed Willughby's name and initials occur so frequently in that work, that it may be esteemed their joint production.[Pg 445] Lister by his various writings elucidated many points relating to insects; and he may be regarded as the first modern who observed that spiders can sail in the air. But the most important of his works, and that on which his fame as an Entomologist is principally founded, is his admirable treatise De Araneis; in which his systematic arrangement of these animals leaves far behind all former attempts, and rivals that of the best modern Arachnologists. His specific descriptions are drawn with a precision till then unknown; and each is headed by a short definition of the species, which he calls the Titulus, synonymous with the Nomen specificum of Linné, whose canon of twelve words it rarely exceeds.
One of the most important events of this era was the complete exposure and refutation of the absurd doctrine of equivocal generation, which had maintained its ground in the schools of philosophy from the time of Aristotle. Our own immortal Harvey was the first who dared to controvert this irrational theory: and his dictum—Omnia ex ovo—was copiously discussed and completely established by two of the ablest physiologists that Italy has produced, Redi and Malpighi.
Previously to the publication of the Historia Insectorum, no other works of eminence, with the exception of Madam Merian's beautiful illustration of the metamorphosis of the insects of Surinam, made their appearance: but in the interval of twenty-five years, which elapsed between the publication of that work and of Linné's first outline of his Systema Naturæ, Entomologists became more numerous and active. In England the pious and learned author of the Physico and Astro-Theology[Pg 446] was celebrated for the assiduity with which he studied insects; and in the former of these works has concentrated a vast number of interesting observations connected with their anatomy and history. No Englishman contributed more to the progress of Natural History, both as a writer and collector, than that disinterested physician and naturalist Sir Hans Sloane, whose extensive and valuable library and well-stored cabinets formed the original nucleus of the present vast collection of the British Museum. Amongst other departments, that of insects was not overlooked by him; and it is to be regretted that those which he had accumulated have either perished from neglect or are not accessible. Other Entomologists were eminent at this period in Britain. The principal of these were Petiver, Dale (to whom Ray bequeathed his collection of insects), Bobart, Bradley, and Dandridge; the last of whom, as Bradley tells us, delineated and described 140 species of spiders.
I must not omit here to observe that our Royal Society, the origin of which took place in this era, communicated a new and powerful impulse to the public mind in favour of Physical Science, and greatly accelerated the progress of Natural History. It acted not only as a centre of excitement which stimulated to exertion, but also as a focus to collect the scattered rays of light before they were dissipated. Insulated observations in every department of nature were thus preserved; and communications from the most eminent naturalists in various parts of Europe ornamented its Transactions. So that from the establishment of this illustrious Society, the triumphant march of Physical Science of every kind towards its acme may be dated.[Pg 447]
4. Era of Linné, or of the Alary System. We are now arrived at that period in the history of Natural Knowledge, especially of Entomology, in which it received that form, with respect to its general outline, which, amidst many lesser mutations, has been preserved ever since. Swammerdam had altogether deserted the system of Aristotle, and Ray mixed it with that of his predecessor. But a brilliant star soon appeared in the North[1388], which was destined to be the harbinger of a brighter day than had ever before illuminated the path of the student of the works of God. The illustrious philosopher whose name distinguishes this new era, imbibed a taste for Entomology almost as early as for Botany[1389]; and though the latter became his favourite, and absorbed his principal attention, he did not altogether neglect the former. In the first edition of his Systema Naturæ, published in 1735, and contained in only fourteen folio pages[1390], he began to arrange the three kingdoms of nature after his own conceptions. But this initiatory sketch, as might be expected, was very imperfect; and with respect to insects, instead of an improvement upon his predecessors, was extremely inferior to what Ray had effected; for he puts into one Order (to which he gives the name of Angioptera) the Lepidoptera, Neuroptera, Hymenoptera, and Diptera. In this work, however, Generic Characters were first given. In successive editions he continued to improve upon this outline: in the fourth he finally settled the number and denominations of his Orders; and[Pg 448] in the twelfth (uniting the Orthoptera, which he had at first considered as of a Coleopterous type, to the Hemiptera) also their limits. His system, being founded upon the absence or presence and characters of the organs for flight, is in some degree a republication of the Aristotelian, and may be called the Alary System.
| Superior | crustaceous with a straight suture | Coleoptera | 1. | ||||||||
| 4. | semicrustaceous, incumbent | Hemiptera | 2. | ||||||||
| All | imbricated with scales | Lepidoptera | 3. | ||||||||
| Wings | membranous—Anus | unarmed | Neuroptera | 4. | |||||||
| aculeate | Hymenoptera | 5. | |||||||||
| 2. | Poisers in the place of the posterior pair | Diptera | 6. | ||||||||
| 0. | Or without either wings or elytra | Aptera | 7. | ||||||||
In considering this table, it must strike every one acquainted with the subject, that although the assumption of a single set of organs whereon to build a system can scarcely be expected to lead to one perfectly natural, yet that the majority of the groups here given as Orders merit that character. The second indeed and the last require further subdivision, and concerning the fourth no satisfactory conclusion has yet been drawn. With regard to his series of the Orders, it is mostly artificial. Linné has the advantage of all his predecessors in giving clearer definitions of his Orders, and in their nomenclature; in which he has followed the path first trodden by Aristotle.
One of his most prominent excellencies, which led the way more than any thing else to a distinct knowledge of natural objects, was his giving definitions of his genera, or the groups that he distinguished by that name, since all preceding writers had merely made them known by the imposition of a name. His generic characters of insects were of two kinds: A shorter, containing the[Pg 449] supposed essential distinction of the genus, given at the head of the Class; and another, generally longer, and including non-essentials, given at the head of the Genus. The first he denominated the essential, and the latter the factitious or artificial character. He did not do for insects what he did for Botany,—draw up what he has called the natural character of a genus, which included both the others, and noticed every other generic distinction[1391].
The older Naturalists used to treasure in their memories a short description of each species, by which when they wished to speak or write of it they made it known. Thus, in speaking of the common lady-bird they would call it "the Coccinella with red coleoptra[1392] having seven black dots." This enunciation of any object was at first called its Title (Titulus), and afterwards its Specific Name (Nomen specificum), and by Linné was restricted to twelve words[1393]. But as the number of species increased to remember each definition was no easy task; that he might remedy this inconvenience, he invented what is called the Trivial Name (Nomen triviale), which expressed any species by a single term added to its generic appellation, as Coccinella septem-punctata; and thereby conferred a lasting benefit on Natural History. This convenient invention has rendered it less necessary to restrict the Nomen specificum to twelve words: it is desirable, however, that the definition of a species should be as short as possible, and contain only its distinctive characters. In his definitions and descriptions Linné was often very happy; but sometimes, in studying to avoid prolixity, he forgets Horace's hint,
and makes his definitions of species, without adding a description, so extremely short as to suit equally well perhaps a dozen different insects. The minor groups into which he has divided some of his Orders and Genera are sometimes natural, sometimes artificial. Those of the Coleoptera, from characters drawn from their antennæ (as is evident from his arrangement of the genera in that Order), are of the latter description; while those of his Aptera are more natural. The genera that he has most happily laboured in this respect are his Hemipterous ones of Gryllus, Cicada, and Cimex, and all his Lepidoptera. He had such a tact for discovering natural groups in general, that in him it seems almost to have been intuitive.
But in no respect were the labours of Linné more beneficial to the science and to Zoology in general, than when he undertook to describe the animals of his own country. His Fauna Suecica is an admirable exemplar, which ought to stimulate the Zoologists of every country to make it one of their first objects that its animal productions shall no longer remain unregistered and undescribed. Botanists have almost every where been diligent in effecting this with respect to plants, but other branches of Natural History have been more neglected. In his Systema Naturæ Linné attempted this for all the productions of our globe. The idea was a vast one; and the execution, though necessarily falling far short of it, did him infinite honour: and in it he has laid a foundation for his successors to build upon till time shall be no more.[Pg 451]
Such were the services rendered to Entomology by the labours of the immortal Swede; services so extensive as well as eminent, that had they been the fruit of a whole life devoted to this single object, they would have entitled him to a high rank amongst the heroes of the science. But how much more astonishing are they when considered but as gleanings from his hours of relaxation, snatched from labours infinitely greater, the produce, as he himself tells us, of moments consumed by others in "venationibus, confabulationibus, tesseris, chartis, lusibus, compotationibus[1394]." It is not so much in original discovery that the merits of Linné lie,—though considered in this view they are pre-eminent,—as in the unrivalled skill with which he sifted the observations of his predecessors, separating the ore from the dross, and concentrating scattered rays of light into one focus.
This era produced other systematists who adopted various methods, but none that merit particular notice except Geoffroy and De Geer. The former in this view is principally celebrated as the author of the method generally adopted by modern Entomologists, of dividing the Coleoptera into primary sections, according to the number of the joints of their tarsi. This method, though in many instances, as was formerly observed[1395], it leads to artificial results, in others affords a clue to natural groups; it can only therefore be applied subject to frequent exceptions. Geoffroy's work[1396], which was published in 1764, was further serviceable by indicating many genera not defined by Linné.
| GENERAL CLASSES. | ORDERS. | CLASSES. | ||||
| I. Wings covered with scales. Tongue spiral. Lepidoptera. | ||||||
| II. Wings membranous, naked. Mouth without teeth or tongue. Trichoptera. Ephemerina. | ||||||
| I. Four Wings without wing-cases | III. Wings membranous, equal, reticulated. Mouth with teeth. Rest of Neuroptera. | |||||
| IV. Wings membranous unequal, nervures mostly longitudinal. Mouth with teeth. A sting or borer in the female. Hymenoptera. | ||||||
| V. Wings membranous. Tongue bent under the breast. Homoptera. | ||||||
| I. Having wings | VI. Elytra half coriaceous and half membranous, crossed. A pair of membranous wings. Tongue bent under the breast. Hemiptera Leach. | |||||
| II. Two Wings covered by two wing-cases | VII. Elytra coriaceous or semicrustaceous, aliform. A pair of membranous wings. Mouth with teeth. Orthoptera. | |||||
| VIII. Elytra hard and crustaceous. A pair of membranous wings. Mouth with teeth. Coleoptera. | ||||||
| Insects | III. Two Wings uncovered | IX. A pair of membranous wings. A pair of poisers. Mouth with a tongue without teeth. Diptera. | ||||
| X. A pair of membranous wings. No poisers, tongue, or teeth in the male. No wings but a tongue in the breast of the female. Coccus L. | ||||||
| IV. Undergoing a metamorphosis | XI. No wings. Six legs. Mouth with a tongue. Aphaniptera. | |||||
| II. Without wings | ||||||
| XII. No wings. Six legs. Head and Trunk distinct. Hexapod Aptera, Termes, Psocus. | ||||||
| V. Undergoing no metamorphosis | XIII. No wings. 8 or 10 legs. Head united to the trunk. Octopod Aptera, Arachnida, Crustacea. | |||||
| XIV. No wings. 14 Legs or more. Head separated from the trunk. Polypod Aptera. Crustacea. |
[Pg 453] We next come to one of the greatest names in Entomology, the celebrated De Geer, who united in himself the highest merit of almost every department of that science. Both as a systematist, anatomist, and physiologist, and as the observant historian of the manners and economy of insects, his Mémoires pour servir à l'Histoire des Insectes are above all praise. His system[1397] is contained in a posthumous volume published in 1778[1398].
This system, though built upon the instruments of flight; in its ternary groups, equivalent to the Orders of Linné, adds likewise the instruments of manducation, and is thus intermediate between that of Linné and Fabricius, who perhaps from the consideration of it might derive the first idea of assuming the last-mentioned organs as the basis of a new method. But, though partaking of both, it is nearer to nature than either; and had its illustrious author laid less stress upon the number and substance of the organs of flight, it would probably have been as near perfection in this respect as most that have succeeded it. But following too strictly these characters, he has been led to place in different Classes, or rather Orders, insects that ought not to have been so separated,—as in the case of the two sections of the Hemiptera, and the Coccidæ. In other respects the whole of De Geer's Mémoires are a storehouse of valuable observations, in which he has furnished many a clue for threading the labyrinth of nature, and given most complete and interesting histories of the whole economy and habits[Pg 454] of many tribes and genera,—as of the Trichoptera, Aphides, Ephemerina, &c.
In this latter department of the science a light shone during part of the era we are now considering, which eclipsed every one that appeared before it, and has scarcely been equalled by any one that succeeded it. The date of its first appearance, indeed, was a year before that of Linné's first outline of his Systema Naturæ before alluded to; but it may properly be regarded as belonging to his era, since it did not disappear till some years after that had begun. A volume indeed would scarcely suffice to do justice to the preeminent merits of Reaumur, as exhibited in his admirable Mémoires pour l'Histoire des Insectes[1399]: I must therefore content myself with observing, that in judgement and ingenuity in planning his experiments; in patient assiduity in watching their progress; in the elegance of his language, and the felicity of his illustrations, he has rarely, if ever, been equalled. Every subject that he undertook was thoroughly investigated, and in the true spirit of philosophical inquiry. Every where you see him the same unprejudiced and profound observer, attached to no system, anxious only for truth and the advancement of science. If he has any fault, it is, perhaps, that of being sometimes too prolix; but we must recollect that from the nature of his subject much diffuseness was often necessary to render his meaning clear. A greater objection is his total inattention to all system, except with regard to Lepidoptera and their larvæ[1400], so that it is often difficult[Pg 455] to ascertain the insects whose history he gives. But with these exceptions, no observer of nature, who wishes his discoveries to be at once profound and interesting, can copy a better model or one nearer to perfection.
Next to that of Reaumur, the name of his admiring correspondent Bonnet may be mentioned. This great physiologist, though still more deficient in systematical knowledge[1401], was also an admirable observer of the economy and manners of insects. In this sense he became an Entomologist before he was seventeen years of age, in consequence of an impression made upon him by the account of the Antlion in that attractive work the Spectacle de la Nature. From verifying its wonderful history with his own eyes, he entered with enthusiasm upon the study of other insects, his observations on which he regularly communicated to Reaumur. Amongst other interesting inquiries, his experiments on that singular anomaly in nature the generation of Aphides[1402] do him the highest credit, and have set that question perfectly at rest[1403].
In another department of the science this period was distinguished by a work which may almost be deemed a prodigy. I am speaking of Lyonet's admirable treatise on the anatomy of the caterpillar of the Cossus,—a work which will uphold his reputation as long as Entomology shall be cultivated as a science, or the comparative Anatomist be delighted to trace the footsteps of Divine Wisdom in the gradually varying structure of animals. The plates to this publication, executed by the hand of its[Pg 456] excellent author, are as wonderful as the work itself; and together, to use Bonnet's words, form a demonstration of the existence of God. It is infinitely to be regretted that the author of this incomparable monument of scientific ardour and patient industry should have died before the full completion of his anatomical description of the pupa and imago of the same insect; of which he had prepared a considerable portion of the manuscript, and engraved upwards of twenty of the plates[1404].
Numerous other writers in various departments of the science appeared during this era; but it would be useless to enter into a particular detail of their works and merits. I cannot however omit noticing, on account of his inimitably accurate and chastely coloured representations of Lepidoptera, Sepp's beautiful Nederlandsche Insecten, in which the whole history of these animals, from the egg to the fly, is described and portrayed. In our own country this era was distinguished by no entomological work of any great eminence. Albin, Wilks, and Harris produced the principal. Gould, however, without having any thing of system, gave an admirable account of English ants, which I formerly noticed[1405].
One of our first poets, the celebrated Gray, was also much devoted to Entomology. From his interleaved copy of the Systema Naturæ, that venerable and able naturalist, Sir T. G. Cullum, Bart. copied the following characters of the genera of insects of Linné, drawn up in Latin Hexameters, which he kindly communicated to me.
Coleoptera.
Alas lorica tectas Coleoptera jactant.
*
**
***
Hemiptera.
Lepidoptera.
Squamam alæ, linguæ spiram Lepidoptera jactant.
Neuroptera.
Rete alæ nudum atque hamos Neuroptera caudæ.
Hymenoptera.
Diptera.
Diptera sub geminis alis se pondere librant.
Aptera.
Aptera se pedibus pennarum nescia jactant.
During this era, and by the influence of Linné, in the year 1739 the Royal Academy of Sciences at Stockholm was established, which did for Natural History in Sweden what our own Royal Society had done for it in England. Other societies, with a similar object, were formed in different parts of Europe, and were attended by similar good effects. At Paris, at Berlin, at St. Petersburg, at Moscow, at Turin, at Lisbon, &c., the lovers of Nature, at that time and subsequently, have associated for this purpose; and I may mention here, that I may not revert to the subject, the great Natural History association of our own country, The Linnean Society, named after the illustrious Swede, which was first instituted in 1788, and incorporated by royal charter in 1802. In the Transactions of this learned body, the Zoologist in general, and particularly the Entomologist, will find much useful information and many interesting observations connected with his science. This flourishing[Pg 460] society consists at this time of above 600 members, of whom more than 500 are Fellows;—a gratifying proof how widely Natural History is cultivated in the British Empire[1406].
5. Era of Fabricius, or of the Maxillary System.—We are now arrived, if its consequences be considered, at one of the most important epochs of the science. Fabricius, a pupil of Linné, who highly estimated his entomological acquirements[1407], thinking that the system of his master was not built upon a foundation sufficiently fixed and restricted[1408], conceived the idea of doing for Entomology what the latter had done for Botany. As the learned and illustrious Swede had assumed the Fructification for the basis of his system in that science, so the emulous and highly-gifted Dane, observing how happily those organs were employed as characters in extricating the genera of Vertebrate animals, assumed the instruments of manducation, far more numerous and various in insects, for the basis of a new system of[Pg 461] Entomology; which, from the maxillæ being principally employed to characterize the Classes or rather Orders, may be called the Maxillary System. De Geer, indeed, as we have seen above, had, in the majority of his Classes, to the organs of flight added the parts of the mouth: but Fabricius pursued the idea much further, and made the Trophi[1409], or Instrumenta Cibaria as he called them, the sole corner-stone of his whole superstructure. Though nothing seems to have been further from his intention than to follow Nature, since he complains that Linné by following her too closely had lost the Ariadnean thread of system[1410], yet it is singular that, by building upon this seemingly narrow foundation, he has furnished a clue, by the due use of which, instead of deserting her, his successors have been enabled with more certainty to extricate her groups: since the parts in question being intimately connected with the functions and economy of these animals, where they differ materially, indicate a corresponding difference in their character and station.
The first outline of his System, I believe, appeared in his Systema Entomologiæ published in 1775; and the last, in his Supplement to his Entomologia Systematica in 1798. In this the series and characters of his Classes (for so, after De Geer, he denominates his primary groups) were as follows:—
*
1. Eleutherata[1411]. (Coleoptera L.) Maxilla naked, free, palpigerous.
[Pg 462]2. Ulonata[1412]. (Orthoptera Oliv.) Maxilla covered by an obtuse galea or lobe.
3. Synistata[1413]. (Neuroptera L., excluding the Libellulina, and taking in Termes L. and Thysanura Latr.) Maxilla geniculate at the base and connate with the labium.
4. Piezata[1414]. (Hymenoptera L.) Maxilla corneous, compressed, often elongate.
5. Odonata[1415]. (Libellulina McL.) Maxilla corneous, toothed, two palpi.
6. Mitosata[1416]. (Myriapoda Leach.) Maxilla corneous, vaulted, not palpigerous.
**
7. Unogata[1417]. (Pulmonary Arachnida Latr.) Maxilla corneous, armed with a claw.
***
8. Polygonata[1418]. (Isopod and Branchiopod Crustacea Latr.) Palpi mostly six; Maxillæ many within the labium.
9. Kleistognatha[1419]. (Brachyurous Decapod Crustacea Latr.) Many Maxillæ without the labium, closing the mouth.
10. Exochnata[1420]. (Macrurous Decapod Crustacea Latr.) Maxillæ many without the labium, covered by palpi.
****
11. Glossata[1421]. (Lepidoptera L.) Mouth with a spiral tongue between reflexed palpi.
12. Ryngota[1422]. (Hemiptera Latr.) Mouth with a rostrum, having a jointed sheath.
13. Antliata[1423]. (Diptera L., Anoplura Leach., Trachean Arachnida Latr. &c.) Mouth with a haustellum without joints.
The Orders of Fabricius are equivalent usually to the primary groups of the Linnean Orders, and are regulated chiefly by the antennæ.
In estimating the value of the above system, we must bear in mind that, according to the statement of its author, it was intended to be partly artificial and partly natural: artificial as to its Classes and Orders; natural as to its genera, species, and varieties[1424]. He admitted, however, that natural Classes, &c. do exist; but he contended that artificial ones should be substituted for them, till further discoveries had cleared the way for their satisfactory development[1425]. As therefore his system, in its primary and secondary groups, was confessedly artificial, and the only use of an artificial system being to facilitate the study of any department of Natural History, its value must be estimated by the facilities it affords to the entomological student. But here, it must be allowed, that instead of enlarging the entrance to the temple of his science, it has made it narrower, and has placed most discouraging impediments in his way.
[Pg 464]If you examine the definitions of his Classes, you will find them in a variety of cases calculated rather to mislead than to instruct a learner. Thus that of the Eleutherata would equally well suit the Piezata and several others: that of the Piezata is scarcely to be found in it; since in this the maxilla, instead of being corneous, is usually coriaceous[1426], and its lobe sometimes nearly membranous. In the Unogata he even mistakes the mandibles for maxillæ. Let any young Entomologist endeavour to make out the Fabrician class of a Cicindela for instance; and finding its maxillæ corneous and armed with a claw, he would conclude that it belonged to the Unogata rather than to the Eleutherata. Besides all this, the necessity of examining minute parts not easily come at without dissection, is very discouraging to a beginner.
From hence it is evident, that the system of Fabricius, considered as an artificial one or a method, was no improvement upon the classification of his master Linné, but rather a retrograde movement in the science.
As to that part of his system in which he professes to take nature for his guide, his genera,—though even with respect to them he seems fearful of following her too closely[1427],—he certainly has rendered most essential services to Entomology, and laid the foundation of all that has since been done for its improvement. But it must be observed, that the series of his genera is often altogether artificial; as where he separates and places far asunder the Saprophagous and Thalerophagous Petalocerous beetles.
[Pg 465]Entomology, however, in other respects was deeply indebted to this great man. He first, as was lately observed, directed the attention of her votaries to parts which enabled them better to follow the chain of affinities, and to trace out natural groups. In his Philosophia Entomologica, drawn up on the plan of Linné's Philosophia Botanica, he bequeathed to the science a standard work that ought to be studied by every Entomologist. His incredible labours in defining new genera and describing new species, with which view he travelled into various parts of Europe, and seven times into Britain, have been of infinite service[1428], and placed the science upon a footing much nearer to that of Botany than it had ever before attained.
6. Era of Latreille, or of the Eclectic System. The system of Fabricius, though generally adopted in Germany and Switzerland, did not meet with a universal reception. It seems to have gained no permanent footing in the North of Europe, Britain, or France. In the latter country the Linnean phraseology and characters of the Orders were retained by the celebrated Olivier; while at the same time his definitions of genera were constructed, after the Fabrician model, upon the antennæ and the oral organs. But a new and brilliant genius had now appeared in France, whose indefatigable labours and singular talents have thrown more light over entomological science than those of all his predecessors. In 1796, about two years after Fabricius had completed his Entomologia Systematica emendata et aucta, M. Latreille published his Précis de Caractères Génériques des Insectes;[Pg 466] in which important work, walking in the steps of his great compatriot Bernard de Jussieu, he disregarded all artificial systems of Entomology, and attempted to construct one upon a natural basis: and to this end, uniting the consideration of the instruments of manducation with that of the organs for flight and motion, and of other external characters,—or the system of Linné with that of Fabricius,—he became the founder of the modern or Eclectic system[1429]; for he judiciously adopted that sensible dictum of Scopoli, "Classes et Genera naturalia, non sola instrumenta cibaria, non solæ alæ, nec solæ antennæ constituunt, sed structura totius, ac cujusque vel minimi discriminis diligentissima observatio[1430]." His object has been in the above and subsequent works, by dividing his Classes into natural Groups, from the Order to the Genus, to trace out in all its windings, to its inmost recesses, the perplexing labyrinth of the true system of the Creator:—of what he has effected, the subjoined tables will give you a sufficient idea[1431].
1817.
| Class. | Order. | Family. | Tribe. | Subtribe. | ||||||||
| I. Crustacea. | ||||||||||||
| Territelæ. | ||||||||||||
| Tubitelæ. | ||||||||||||
| Sedintariæ | Inequitelæ. | |||||||||||
| Orbitelæ. | ||||||||||||
| Araneides | Laterigradæ. | |||||||||||
| Pulmonariæ | Vagantes | Citigradæ. | ||||||||||
| Saltigradæ. | ||||||||||||
| Pedipalpæ | ||||||||||||
| II. Arachnida | Scorpioides. | |||||||||||
| Pseudoscorpiones. | ||||||||||||
| Entoma | Tracheariæ | Phalangita. | ||||||||||
| Holetra | Trombidites. | |||||||||||
| Acaridia | Riciniæ. | |||||||||||
| Hydrachnellæ. | ||||||||||||
| Microphthiræ. | ||||||||||||
| 1. Myriapoda | Chilognatha. | |||||||||||
| Chilopoda. | ||||||||||||
| 2. Thysanura | Lepismenæ. | |||||||||||
| Podurellæ. | ||||||||||||
| 3. Parasita | Mandibulata. | |||||||||||
| Edentula. | ||||||||||||
| III. Insecta | ||||||||||||
| 4. Suctoria. | ||||||||||||
| 5. Coleoptera. | ||||||||||||
| 6. Orthoptera. | ||||||||||||
| 7. Hemiptera. | ||||||||||||
| 8. Neuroptera. | ||||||||||||
| 9. Hymenoptera. | ||||||||||||
| 10. Lepidoptera. | ||||||||||||
| 11. Rhiphiptera. | ||||||||||||
| 12. Rhiphiptera. |
1825.
| Class. | Order. | Family. | Section. | Tribe. | ||||||
| I. Crustacea. | ||||||||||
| Pedipalpi | Scorpionides. | |||||||||
| Tarentulæ. | ||||||||||
| Pulmonariæ | ||||||||||
| Tetrapneumones. | ||||||||||
| Hyperhexapi. | Araneides | Tubitelæ. | ||||||||
| Inæquitelæ. | ||||||||||
| Dipneumones | Orbitelæ. | |||||||||
| Laterigradæ. | ||||||||||
| II. Arachnides. | Citigradæ. | |||||||||
| Saltigradæ. | ||||||||||
| Pycnogonides. | ||||||||||
| Pseudoscorpiones. | ||||||||||
| Phalangita. | ||||||||||
| Tracheariæ | Acarides. | |||||||||
| Hydrachnellæ. | ||||||||||
| Riciniæ. | ||||||||||
| Microphthira. | ||||||||||
| Condylopa. | Chilognatha | Anguiformia. | ||||||||
| III. Myriapoda. | Penicillata. | |||||||||
| Inæquipedes. | ||||||||||
| Æquipedes. | ||||||||||
| Thysanoura | Lepismenæ. | |||||||||
| Podurellæ. | ||||||||||
| Aptera. | ||||||||||
| Parasita | Mandibulata. | |||||||||
| Siphunculata. | ||||||||||
| Siphonaptera. | ||||||||||
| Hexapoda. | IV. Insecta. | |||||||||
| Coleoptera | ||||||||||
| Orthoptera | Elytroptera. | |||||||||
| Hemiptera | ||||||||||
| Alata | Neuroptera | |||||||||
| Hymenoptera | Anelytra quadripennia. | |||||||||
| Lepidoptera | ||||||||||
| Rhiphiptera | ——— bipennia. | |||||||||
| Diptera |
Having given you these tables of the Orders, from a comparison of which you will be able to trace the improvements in his system made by this learned Entomologist in the interval of eight years, I shall proceed to give those of his subordinate groups arranged under each. This I have already done, to save space, in the Arachnida and Insecta aptera.[Pg 469]
| Order. | Section. | Family. | Subfamily. | Tribe. | Subtribe. | |||||
| Cicindeletæ. | ||||||||||
| Terrestres | Truncatipennes. | |||||||||
| Bipartiti. | ||||||||||
| Adephagi | Carabici | Thoracici. | ||||||||
| Abdominales. | ||||||||||
| Subulipalpi. | ||||||||||
| Aquatica | Hydrocanthari. | |||||||||
| Gyrinites. | ||||||||||
| Fissilabres. | ||||||||||
| Brachyptera | Longipalpi. | |||||||||
| Depressi. | ||||||||||
| Microcephali. | ||||||||||
| Sternoxi | Buprestides. | |||||||||
| Elaterides. | ||||||||||
| Serricornes | ||||||||||
| Cebrionites. | ||||||||||
| Lampyrides. | ||||||||||
| Malacodermi | Melyrides. | |||||||||
| Clerii. | ||||||||||
| Pentamera | Xylotragi. | |||||||||
| Ptiniores. | ||||||||||
| Histeroida. | ||||||||||
| Peltoides. | ||||||||||
| Clavicornes | Palpatores. | |||||||||
| Dermestini. | ||||||||||
| Byrrhii. | ||||||||||
| Macrodactyli. | ||||||||||
| Palpicornes | Hydrophilii. | |||||||||
| Sphæridiota. | ||||||||||
| Coprophagi. | ||||||||||
| Arenicolæ. | ||||||||||
| Coleoptera | Scarabæides | Xylophili. | ||||||||
| Phyllophagi. | ||||||||||
| Lamellicornes | Anthobii. | |||||||||
| Melitophili. | ||||||||||
| Lucanides. | ||||||||||
| Pimeliariæ. | ||||||||||
| Melasoma | Blapsides. | |||||||||
| Tenebrionites. | ||||||||||
| Dioperiales. | ||||||||||
| Taxicornes | Cossyphenes. | |||||||||
| Crassicornes. | ||||||||||
| Heteromera | Helopii. | |||||||||
| Cistelides. | ||||||||||
| Stenelytra | Securipalpi. | |||||||||
| Œdemerites. | ||||||||||
| Rhyncostoma. | ||||||||||
| Lagriariæ. | ||||||||||
| Pyrochroides. | ||||||||||
| Trachelides | Mordellonæ. | |||||||||
| Anthicides. | ||||||||||
| Horiales. | ||||||||||
| Cantharidiæ. |
| Order. | Section. | Family. | Tribe. | |||
| Bruchelæ. | ||||||
| Anthribides. | ||||||
| Rhynchophora | Altelabides. | |||||
| Brentides. | ||||||
| Curculionites. | ||||||
| Scolitarii. | ||||||
| Xylophagi | Bostrichini. | |||||
| Paussili. | ||||||
| Trogossitarii. | ||||||
| Platysoma. | ||||||
| Prionii. | ||||||
| Tetramera | Cerambycini. | |||||
| Longicornes | Necydalides. | |||||
| Lamiariæ. | ||||||
| Lepturetæ. | ||||||
| Eupoda | Sagrides. | |||||
| Criocerides. | ||||||
| Coleoptera | ||||||
| Cassidariæ. | ||||||
| Cyclica | Chrysomelinæ. | |||||
| Galerucidæ. | ||||||
| Clavipalpi. | ||||||
| Trimera | Aphidiphagi. | |||||
| Fungicolæ. | ||||||
| Pselaphii. | ||||||
| Monomera. | ||||||
| Forficularia. | ||||||
| I. | Blattariæ. | |||||
| Mantides. | ||||||
| Spectra. | ||||||
| Orthoptera | ||||||
| II. | Gryllides. | |||||
| Locustariæ. | ||||||
| III. | Acridites. | |||||
| Longilabra. | ||||||
| Membranaceæ. | ||||||
| Geocorisæ | Nudicolles. | |||||
| Oculatæ. | ||||||
| Heteroptera | Ploteres. | |||||
| Hydrocorisæ | Nepides. | |||||
| Notonectides. | ||||||
| Hemiptera | ||||||
| Stridulantes. | ||||||
| Cicadariæ | Fulgorellæ. | |||||
| Membracides. | ||||||
| Cicadellæ. | ||||||
| Homoptera | ||||||
| Psyllides. | ||||||
| Hymenelytra | Physapi. | |||||
| Aphidii. | ||||||
| Gallinsecta. |
| Order. | Section. | Family. | Tribe. | Subtribe. | ||||
| Subulicornes | Libellulina. | |||||||
| Ephemerina. | ||||||||
| Panorpatæ. | ||||||||
| Neuroptera | Myrmeleonides. | |||||||
| Hemerobini. | ||||||||
| Planipennes | Psoquillæ. | |||||||
| Termitinæ. | ||||||||
| Filicornes | Raphidinæ. | |||||||
| Semblides. | ||||||||
| Perlides. | ||||||||
| Plicipennes. | ||||||||
| Securifera | Tenthredinetæ. | |||||||
| Urocerata. | ||||||||
| Terebrantia | ||||||||
| Evaniales. | ||||||||
| Ichneumonides. | ||||||||
| Pupivora | Gallicolæ. | |||||||
| Chalcidites. | ||||||||
| Chrysides. | ||||||||
| Oxyuri. | ||||||||
| Hymenoptera | Heterogyna | Formicariæ. | ||||||
| Mutillariæ. | ||||||||
| Scolietæ. | ||||||||
| Sapygites. | ||||||||
| Pompilii. | ||||||||
| Fossores | Sphegides. | |||||||
| Bembecides. | ||||||||
| Aculeata | Larratæ. | |||||||
| Nyssonii. | ||||||||
| Crabronites. | ||||||||
| Diploptera | Vespariæ. | |||||||
| Masarides. | ||||||||
| Andrenetæ. | ||||||||
| Mellifica | Solitariæ. | |||||||
| Andrenoides. | ||||||||
| Apiariæ | Dasygastræ. | |||||||
| Cuculinæ. | ||||||||
| Scobulipedes. | ||||||||
| Sociales. | ||||||||
| Hexapoda. | ||||||||
| Papilionides | Perlata. | |||||||
| Diurna | Argus. | |||||||
| Hesperides. | ||||||||
| Hesper-sphinges. | ||||||||
| Crepuscularia | Sphingides. | |||||||
| Lepidoptera | Zygænides. | |||||||
| Bombycites. | ||||||||
| Pseudo-Bombyces. | ||||||||
| Tineites. | ||||||||
| Nocturna | Noctuælites. | |||||||
| Tortrices. | ||||||||
| Phalænites. | ||||||||
| Crambites. | ||||||||
| Pterophorites. |
| Order. | Section. | Family. | Tribe. | Subtribe. | ||||
| Culicides. | ||||||||
| Nemocera | Culiciformes. | |||||||
| Gallicolæ. | ||||||||
| Tipulariæ | Terricolæ. | |||||||
| Fungivoræ. | ||||||||
| Florales. | ||||||||
| Tabanii. | ||||||||
| Sicarii. | ||||||||
| Mydasi. | ||||||||
| Leptides. | ||||||||
| Dolichopoda. | ||||||||
| Tanystoma | Asilici. | |||||||
| I. | Hybotina. | |||||||
| Empides. | ||||||||
| Anthracii. | ||||||||
| Bombyliarii. | ||||||||
| Vesiculosa. | ||||||||
| Notacantha | Xylophagei. | |||||||
| Stratyomides. | ||||||||
| Diptera | Syrphiæ. | |||||||
| Conopsariæ. | ||||||||
| Œstrides. | ||||||||
| Athericera | Cryptogastræ. | |||||||
| Creophilæ. | ||||||||
| Carpomyzæ. | ||||||||
| Muscides | Dolichoceræ. | |||||||
| Gonocephalæ. | ||||||||
| Scathophilæ. | ||||||||
| Apteræ. | ||||||||
| II. | Pupiparæ | Coriaceæ. | ||||||
| Phthiromyiæ[1432]. |
If you examine the Orders as here given, you will find that they mostly represent natural primary groups of his Classes, though with regard to their distribution you may perhaps feel disposed to differ from him. You will also think that his secondary and minor groups[1433], with the exception of some of his sections, merit the same character. Indeed, he has left far behind all his predecessors[Pg 473] in the progress that he has made towards extricating the true system. Setting out from a common centre he holds on his unwearied course, endeavouring to trace every set of objects that branches from it to its extreme term. But though he studied insects analytically with unrivalled success, he was not always equally happy in his synthetical arrangement of them. I do not here so much speak of the result which must necessarily follow from any arrangement in a series, and which cannot well be avoided; but I allude particularly to his intire adoption of the Geoffroyan system in the Coleoptera, which has prevented him in many instances from seeing the natural distribution of his groups.
In 1798, two years after the publication of Latreille's first enunciation of his system, M. Clairville, a very acute and learned Swiss Entomologist, drew up the following analytical table of insects.
| Sections. | ||||||
| 1. Elytroptera (Coleoptera). | ||||||
| Mandibulata | 2. Deratoptera (Orthoptera). | |||||
| 3. Dictyoptera (Neuroptera). | ||||||
| Pterophora | 4. Phleboptera (Hymenoptera). | |||||
| Insecta | 5. Halteriptera (Diptera). | |||||
| Haustellata | 6. Lepidioptera (Lepidoptera). | |||||
| 7. Hemimeroptera (Hemiptera). | ||||||
| Aptera | Haustellata | 8. Rophoteira. | ||||
| Mandibulata | 9. Pododunera. |
Every one will think that the change of the received names of the Orders, here denominated Sections, is perfectly needless. The principal merit of this system is the division of insects, tacitly pointed out by Fabricius, into[Pg 474] two groups or subclasses, from the mode in which they take their food.
Lamarck,—whose merits as a Zoologist, except in one point[1434], are of the highest order,—in his Système des Animaux sans Vertèbres, which was published in 1801, adopts the above division of insects; but, after Aristotle[1435], he makes the Hymenoptera an intermediate Order between the masticators and those that take their food by suction; he places the Lepidoptera at the head of the latter, and the Aphaniptera, which he denominates Aptera, at the end[1436]: the Hexapod, Octopod, and Polypod Aptera he considers as Arachnida[1437]. In his last great work (Histoire Naturelle des Animaux sans Vertèbres) he includes the Hymenoptera amongst the masticators, and reverses the disposition of his Orders, beginning with his Aptera and ending with the Coleoptera[1438].
M. Le Baron Cuvier, in his Anatomie Comparée (1805) divided Insecta into two subclasses, from the presence or absence of maxillæ: thus—
| With Maxillæ. | Without Maxillæ. |
| 1. Gnathaptera. | 1. Hemiptera. |
| 2. Neuroptera. | 2. Lepidoptera. |
| 3. Hymenoptera. | 3. Diptera. |
| 4. Coleoptera. | 4. Aptera. |
| 5. Orthoptera. |
His Gnathaptera include the Isopod Crustacea, the Arachnida, the Polypod, and some of the Octopod and Hexapod Aptera; and his Aptera—Pulex, Pediculus, and[Pg 475] the Acarina, with the exclusion of Hydrachna[1439]. It is remarkable enough that his Class as it stands, with a slight alteration, returns into itself, thus forming a circle; for his first Order (Gnathaptera) contains Hydrachna and the Thysanura, and his last (Aptera) ends with the Anoplura, and Acarina.
All the French Entomologists have followed Olivier and Latreille in adopting, with some variation, Geoffroy's system with regard to the Coleoptera, which has rendered them all more or less artificial. Dumeril has constructed a table of the Order, arranged differently from that above given[1440] of Latreille; but not more natural, for the very same reason.
Our learned countryman, Dr. Leach, by his zoological labours has thrown much light on the natural distribution of the Animal Kingdom, and no department of that kingdom is more indebted to him than the Annulosa; of which I have before stated to you his Classes[1441]. I shall now give a table of his Orders of Arachnida and Insecta Latr. and also his families, &c. of his Classes Myriapoda and Arachnides[1442].
| Class. | Order. | Family. | ||
| Glomerides. | ||||
| Chilognatha | Iulides. | |||
| Polydesmides. | ||||
| Myriapoda | ||||
| Cermatides. | ||||
| Syngnatha | Scolopendrides. | |||
| Geophilides. | ||||
| [Pg 476] | Podosomata | Pycnogonides. | ||
| Nymphonides. | ||||
| Sironides. | ||||
| Polymerosomata | Scorpionides. | |||
| Tarantulides. | ||||
| Arachnides | ||||
| Solpugides. | ||||
| Dimerosomata | Phalangides. | |||
| Araneïdes. | ||||
| Trombidides. | ||||
| Gammasides. | ||||
| Monomerosomata | Acarides. | |||
| Cheyletides. | ||||
| Eylaïdes. | ||||
| Hydrachnides. | ||||
| Ametabolia | Thysanura. | |||
| Anoplura. | ||||
| Coleoptera. | ||||
| Insecta | Dermaptera. | |||
| Orthoptera. | ||||
| Dictyoptera. | ||||
| Hemiptera. | ||||
| Omoptera. | ||||
| Metabolia | Aptera. | |||
| Lepidoptera. | ||||
| Trichoptera. | ||||
| Neuroptera. | ||||
| Hymenoptera. | ||||
| Rhiphiptera. | ||||
| Diptera. | ||||
| Omaloptera. |
I have before expressed my sentiments upon several of these Orders[1443]: I shall not here repeat them, but shall merely observe, with respect to those I have not adopted, that, though perhaps not entitled to rank as Orders, most of them form natural groups. His Orders, however, of Arachnida must be excepted from this remark, since they are evidently artificial. His analyses of his Orders, though in general they give natural groups, are usually not carried so far as those of M. Latreille, so as seldom to indicate what may properly be denominated families. He has made his nomenclature for his so-called families[Pg 477] more uniform and satisfactory than that of the French Entomologist: and we may say, with respect to the extent and effect of his zoological labours,—Nihil non tetigit, et omnia quæ tetigit ornavit.
7. Era of MacLeay, or of the Quinary System. I have more than once stated to you in my former letters the bases upon which the system which I am in the last place to explain to you is built. You know the Sub-kingdoms and Classes into which its learned and ingenious author, upon a novel and most remarkable plan, has divided the Animal Kingdom[1444]. I shall now copy for you his diagram of the Annulosa.
[Pg 478] I have before sufficiently noticed these Classes, or Orders as Mr. MacLeay terms them, of the Sub-kingdom Annulosa: I shall here therefore only throw out a few remarks on their composition. With regard to their circular distribution in the Crustacea, Mr. MacLeay thinks the series runs from the Branchiopods or Monoculus L. to the Decapods or Cancer L.; and so on, till by means perhaps of the genus Bopyrus, which Fabricius regards as a Monoculus, it returns to the Branchiopods again. This circle, through Porcellio, a kind of wood-louse, &c., which has only a pair of antennæ and at first but six legs, is connected with the Ametabola Class, which beginning with Glomeris goes by the other Chilognatha (Iulus L.), having also six legs at first, and certain Vermes to the Anoplura, and terminates in the Chilopoda (Scolopendra L.) their cognate tribe[1445]. From the Ametabola Mr. MacLeay proceeds to the Mandibulata, between which two groups he has discovered no osculant one, but he takes the Anoplura of the former as the transit to the Coleoptera in the latter; from whence passing to the Orthoptera, &c., he finally returns by the Hymenoptera. Between the Mandibulata likewise and Haustellata he finds no osculant class: but as the affinity between the Trichoptera and Lepidoptera is evident, proceeding by the Homoptera he returns to the Lepidoptera by certain Diptera, as Psychoda, &c. From the Aptera Lam. or Pulex L. he passes by the osculant class Nycteribida to the Arachnida; and beginning with the Acaridea, he goes to the Scorpionidea, and so to the Aranidea or spiders, which he connects with the Decapod Crustacea;—thus[Pg 479] forming his great circle of five smaller ones, each of which, as well as that which they form, returns into itself[1446].
We next take his Circles of Mandibulata: thus—
[Pg 480] In this arrangement of the tribes, as he calls them, of Mandibulata, Mr. MacLeay sets out from the Coleoptera, which he distributes, according to the supposed typical forms of their larvæ, into five minor groups, sufficiently noticed on a former occasion[1447]. From this tribe or Order he proposes to pass by Atractocerus to the osculant Order Strepsiptera, and from thence by Myrmecodes and the Ants to the Hymenoptera. From hence he next proceeds to his Trichoptera; in which, as we have seen[1447], he places not only Phryganea L., but also Tenthredo L. and Perla Geoffr., making his transit by Sirex L.; forming an osculant Order which he denominates Bomboptera. From this his way to the Neuroptera is by the Perlides, with Sialis as an osculant Order under the name of Megaloptera: he enters by Chauliodes, and leaves it by Panorpa or Raphidia by means of Boreus, forming also an osculant Order (Raphioptera) for the Orthoptera; which he enters by Phasma, Mantis, &c., and leaves by Gryllus, entering the Coleoptera again by the osculant Order Dermaptera formed of Forficula L.: and thus returning to the point from which he set out[1448]. He has not, however, made this return of the series into itself so clear in each order, excepting in the Orthoptera, as he has done in the whole Class or Sub-class. Thus in the Coleoptera there appears no particular affinity between the Predaceous and Vesicant beetles, his first and fifth forms[1449], or his Chilopodimorphous Coleoptera, and his Thysanurimorphous.
To enter fully into his doctrine of Analogies would lead us into a very wide field, and occupy a larger space[Pg 481] than I can afford; I must therefore refer you to his work for more particular and detailed information on that subject. With regard to the analogy between opposite points of contiguous circles, you may get a very good idea of it from his diagram of Saprophagous and Thalerophagous Petalocerous beetles, which I here subjoin.
It is a very singular circumstance that in these two circles we have two sets of insects,—one impure in its habits and feeding upon putrescent food, and the other clean and nourished by food that has suffered no decay,—set in contrast with each other, and that in each of the opposite groups, the one has its counterpart in some respect in the other. In none is this more striking than the Scarabæidæ and Cetoniadæ, both remarkable for having soft membranous mandibles unfit for mastication, and both living upon juices, the one in a putrescent and the other in an undecayed state[1450]. [Pg 482] Our learned author in subsequent works has stated every circle to be resolvable into two superior groups, which he denominates normal or typical, and three inferior ones, which he calls aberrant or annectent[1451].
Before I conclude this account of the various general systems that have distinguished the different entomological eras, i must say a few words on those partial ones which have been founded on the neuration of the wings of insects. Frisch, who died in 1743, attempted something in this way[1452]: Harris, in his Exposition of English Insects published in 1782, had arranged his Hymenoptera and Diptera according to characters derived from this same circumstance[1453]: Mr. Jones in the Linnean Transactions had made good use of it in dividing the Diurnal Lepidoptera into groups[1454]: and in the Monographia Apum Angliæ, the characters exhibited by the various groups into which Linné's genus Apis was resolvable, as to the neuration of their wings, were described[1455]. But M. Jurine was the first Entomologist who made that circumstance the keystone of a system; which indeed he restricted to Hymenopterous and Dipterous insects, but which might be extended much further. As this system has been before sufficiently enlarged upon[1456], I need here only mention it.
To particularize the various entomological works in every department of the science, that have appeared since the commencement of the era of Fabricius, would[Pg 483] require a volume. Such was its progress and spread, that in every corner of Europe the pens and pencils of able and eminent men, whose works have almost all been quoted in the course of our correspondence, have been employed to illustrate it[1457]. [Pg 484] I may observe, however, that the Internal Anatomy of Insects, a branch of Entomology which on account of its difficulty, from the extreme nicety required in dissecting them, had before been cultivated by scarcely more than a single student in an age, has now attracted numerous votaries. In Germany—Carus, Gaede, Herold, Posselt, Ramdohr, Rifferschweils, Sprengel, and others, have distinguished themselves in this arena: and in France, besides the illustrious Baron Cuvier (himself a host), Marcel de Serres, Leon Dufour, and very recently, by his elaborate essays On the Flight of Insects and its wonderful[Pg 485] apparatus, one of the most acute of anatomical physiologists, M. Chabrier,—have all contributed greatly to the elucidation of this interesting part of the science. In our own country very little has hitherto been effected in this line; but a learned Oxford Professor (Kidd) has presented to the Royal Society an account of the anatomy of the Mole-cricket, which entitles him to an eminent station amongst the above worthies.
I may likewise further observe, that the pictorial department of Entomology was, during the period I am speaking of, carried to its greatest perfection. Painters of insects formerly were satisfied with giving a representation generally correct, without attempting a faithful delineation of all the minor parts, particularly as to number;—for instance, the joints of the antennæ and tarsi, the areolets of the wings, &c.: but now no one gives satisfaction as an entomological artist unless he is accurate in these respects.
I am, &c.[Pg 486]
Though no subject is more worthy of the attention of the Entomologist than the Geographical Distribution of insects, yet perhaps there is none connected with the science, for the elucidation of which he is furnished with fewer materials. The geographer of these animals sitting by his fireside, even supposing his museum as amply stored as that of Mr. MacLeay, and the habitats of its contents as accurately indicated, still labours under difficulties that are almost insuperable; so that it is next to impossible, with our present knowledge of the subject, to give satisfactory information upon every point which it includes. Had he the talents and opportunities of a Humboldt, and could, like him, traverse a large portion of the globe, he would endeavour to note the elevation, the soil and aspect, the latitude and longitude, the mean temperature and meteorological phænomena, the season of the year, the kind of country, and other localities connected with the insects he captured, and so might build his superstructure upon a sure basis. But these are[Pg 487] things seldom registered by travellers that take the trouble to collect insects; who, if they specify generally the country in which any individual was found, think they have done enough. But to say that an insect was taken in India, China, New Holland, and North or South America,—when we consider the vast extent of those regions,—is saying little of what one wishes to know even with respect to its habitat. You must regard therefore, after all, what I have been able to collect,—and for which I am greatly indebted to the labours of my few but able precursors in this walk,—as merely approximations to an outline, rather than as a correct map of insect Geography.
Amongst the numerous obligations that he conferred upon Natural History, Linné was the first Naturalist who turned his attention to the Geographical Distribution of its objects, especially that of the Vegetable Kingdom[1458]: and the accomplished traveller Baron Humboldt, by the observations he made on this subject in the course of his peregrinations in tropical America, has furnished the Botanist with a clue which, duly followed, will enable him to perfect that part of his science; an end to which the learned observations of Messrs. R. Brown and Decandolle have greatly contributed[1459]. With regard to animals, Mr. White, so long ago as 1773, had observed that they, as well as plants, might with propriety be arranged geographically[1460]: and in 1778 Fabricius in his Philosophia Entomologica applied the principle to insects[1461]. Nearly forty years elapsed before[Pg 488] any improvement or enlargement of this last department was attempted; when in 1815 M. Latreille, stimulated by what had been effected in Botany, in a learned and admirable memoir[1462] endeavoured to place Entomology in this respect by the side of her more fortunate sister: and subsequently Mr. W. S. MacLeay, in the memorable work so often quoted in our correspondence, has viewed the subject in another light, and added some important information to what had been before collected[1463].
The point now under consideration naturally divides itself into two principal branches;—the numerical distribution of insects, and the topographical.
I. By the numerical distribution of insects I mean not only the number which Providence has employed to carry on its great plan on this terraqueous globe, or any given portion of it; or of the species of which each group or genus may be supposed to consist; or of the comparative number of individuals furnished by each species,—points of no easy solution: but more particularly their distribution according to their functions, whether they prey upon animal or vegetable matter, and in its living or decaying state.
We have no data enabling us to ascertain with any degree of accuracy the actual number of species of insects and Arachnida distributed over the surface of the globe; but it is doubtless regulated in a great degree by that of plants. We should first then endeavour to gain some just though general notion on that head. Now Decandolle conjectures that the number of the species[Pg 489] of plants, 60,000 being already known, may be somewhere between 110,000 and 120,000[1464]. If we consider with reference to this calculation, that though the great body of the mosses, lichens, and sea-weeds are exempt from the attack of insects, yet as a vast number of phanerogamous plants and fungi are inhabited by several species, we may form some idea how immense must be the number of existing insects; and how beggarly does Ray's conjecture of 20,000 species[1465], which in his time was reckoned a magnificent idea, appear in comparison! Perhaps we may obtain some approximation by comparing the number of the species of insects already discovered in Britain with that of its phanerogamous plants. The latter,—and it is not to be expected that any large number of species have escaped the researches of our numerous Botanists,—may be stated in round numbers at 1500, while the British insects, (and thousands it is probable remain still undiscovered,) amount to 10,000; which is more than six insects to one plant. Now though this proportion, it is probable, does not hold universally; yet if it be considered how much more prolific in species tropical regions are than our chilly climate, it may perhaps be regarded as not very wide of a fair medium. If then we reckon the phanerogamous vegetables of the globe in round numbers at 100,000 species, the number of insects would amount to 600,000. If we say 400,000, we shall perhaps not be very wide of the truth. When we reflect how much greater attention has been paid to the collection of plants than to that of insects, and that 100,000 species of the latter may be supposed already to have a place[Pg 490] in our cabinets[1466], we may very reasonably infer that at least three fourths of the existing species remain undiscovered.
Certain groups and genera are found to contain many more species than others: for instance, the Coleoptera and Lepidoptera Orders than the Orthoptera and Neuroptera; the Rhincophora than the Xylophagi: the Dytiscidæ than the Gyrinidæ; Aphodius than Geotrupes; Carabus than Calosoma. Again, some insects are much more prolific than others. Thus the Diptera Order, though not half so numerous with respect to species as the Coleoptera, exceeds it greatly in the number of individuals, filling the air in every place and almost at every season with its dancing myriads. We rarely meet with a single individual of the most common species of Calosoma or Buprestis; whilst the formicary, the termitary, the vespiary, and the bee-hive send forth their thousands and tens of thousands; and whole countries are covered and devastated by the Aphides and the Locusts. An all-wise Providence has proportioned the numbers of each group and species to the work assigned to them. And this is the view in which the numerical distribution of insects is most interesting and important: and we are indebted to Mr. W. S. MacLeay for calling the attention of Entomologists more particularly to this part of our present subject.
With regard to their functions, insects may be primarily divided into those that feed upon animal matter and those that feed upon vegetable. At first you would be inclined to suppose that the latter must greatly exceed[Pg 491] the former in number: but when you reflect that not only a very large proportion of Vertebrate animals, and even some Mollusca[1467], have more than one species that preys upon them, but that probably the majority of insects, particularly the almost innumerable species of Lepidoptera, are infested by parasites of their own class, sometimes having a different one appropriated to them in each of their preparatory states[1468], and moreover that a large number of beetles and other insects devour both living and dead animals,—you will begin to suspect that these two tribes may be more near a counterpoise than at first seemed probable. In fact, out of a list of more than 8000 British insects and Arachnida taken several years ago, and furnished chiefly by Mr. Stephens, I found that 3894 might be called carnivorous, and 3724 phytiphagous[1469]; so that, speaking roundly, they might be denominated equiponderant.
Carnivorous and phytiphagous insects may be further subdivided according to the state in which they take their food,—whether they attack it while living, or not till after it is dead. To adopt Mr. W. S. MacLeay's phraseology, the former may be denominated thalerophagous, and the latter saprophagous. The British saprophagous carnivorous insects, compared with those that are thalerophagous, are about as 1:6; while the phytiphagous ones are as 1:9. The thalerophaga in both tribes may be further subdivided as they take their[Pg 492] food by suction or mastication: in the carnivorous ones, the suckers to the masticators in Britain are nearly as 1:6; but with respect to the phytiphagous tribe you must take into consideration that some insects imbibing their food by suction in their perfect state (as the great body of the Lepidoptera), masticate it when they are larvæ: deducting therefore from both sides the insects thus circumstanced, the masticators will form about three fourths of the remaining British thalerophagous insects. Another circumstance belonging to this head must not be passed without notice:—there are certain insects feeding upon liquid food that do not suck, but lap it. This is the case with the Hymenoptera, who, though they are mandibulate, generally lap their food (the nectar of flowers) with their tongue, and may be called lambent insects: nor is this practice confined to that order, but all the mandibulate insects that feed on that substance merit the same appellation. The absorption of this nectar is so important a point in the economy of nature, that a very large proportion of the insect population of the globe in their perfect state, are devoted to it. Considerably more than half the species indigenous to Britain fulfill this function, and probably in tropical countries the proportion may be still larger.
To push this analysis still further—Amongst our carnivorous thalerophaga, aphidivorous insects are about as 1:14; and amongst the phytiphagous, the fungivorous ones form about a twentieth; and the granivorous about a twenty-fifth part of the whole. Again: in the saprophaga the lignivorous tribes form more than half, and the coprophagous ones more than a third.
If you wish to know further the relative proportions[Pg 493] of the different Orders to each other—The Coleoptera may be stated as forming at least 1:2 of our intire insect population; the Orthoptera and Dermaptera as about 1:160; the Hemiptera as 1:15; the Lepidoptera as more than 1:4; the Neuroptera with the Trichoptera as 1:29; the Hymenoptera as about 1:4; the Diptera as not 1:7; and the Aptera and Arachnida as perhaps amounting to 1:19[1470].
To extend this inquiry to exotic and more particularly to extra-European insects, in the present state of our knowledge, would lead to no very satisfactory results. The lists we have are so imperfect, that those which tell most in this country,—I mean the more minute insects and the Brachyptera—have hitherto formed a very small, if any part, of the collections made out of Europe. Mr. W. S. MacLeay however, who, besides his father's (particularly rich in Petalocera), has had an opportunity of examining the Parisian and other cabinets, finds that the species of coprophagous insects within the tropics, to those without, are nearly in the proportion of 4:3; and that the coprophagous Petalocera, to the remainder of the saprophagous ones, may be represented by 3:2[1471]. It may be inferred, from the superabundance of plants and animals in equinoctial countries, that the number of species of insects in general is greater within than without the tropics: the additional momentum produced by the vast size of many of the tropical species must also be taken into consideration.
II. There are three principal points that call for[Pg 494] attention under the second branch of our present subject—the topographical distribution of insects; namely, their Climates, their Range, and their Representation.
i. Entomologists, taking heat for the principal regulator of the station of insects, have divided the globe into entomological climates. Fabricius considers it as divisible into eight such climates, which he denominates the Indian, Egyptian, Southern, Mediterranean, Northern, Oriental, Occidental, and Alpine. The first containing the tropics; the second, the northern region immediately adjacent; the third, the southern; the fourth, the countries bordering on the Mediterranean sea, including also Armenia and Media; the fifth, the northern part of Europe interjacent between Lapland and Paris; the sixth, the northern parts of Asia where the cold in winter is intense; the seventh, North America, Japan, and China; and the eighth, all those mountains whose summits are covered with eternal snow[1472]. M. Latreille objects to this division, as too vague and arbitrary and not sufficiently correct as to temperature; and observes, with great truth, that as places where the temperature is the same, have different animals, it is impossible, in the actual state of our knowledge, to fix these distinctions of climates upon a solid basis. The different elevations of the soil above the level of the sea, its mineralogical composition, the varying quantity of its waters, the modifications which the mountains, by their extent, their height, and their direction, produce upon its temperature; the forests, larger or smaller, with which it may be covered; the effects of neighbouring climates upon it,—are all[Pg 495] elements that render calculations on this subject very complicated, and throw a great degree of uncertainty over them[1473]. This learned Entomologist would judiciously consider entomological climates under another view,—that which the genera of Arachnida and insects exclusively appropriated to determinate spots or regions would supply[1474]. Linné's dictum with regard to genera will here also apply; "Let the insects point out the climate, and not the climate the insects." If you expect invariably to find the same insects within the same parallels of latitude, you will be sadly disappointed; for, as our author further observes, "The totality or a very large number of Arachnida and insects, the temperature and soil of whose country are the same, but widely separated, is in general, even if the countries are in the same parallel, composed of different species[1475]." The natural limits of a country,—as mountainous ranges, rivers, vast deserts, &c.,—often also say to its insect population, "No further shall ye come;" interposing a barrier that it never passes[1476]. Humboldt observes, with respect to the Simulia and Culices of South America, that their geographical distribution does not appear to depend solely on the heat of the climate, the excess of humidity, or the thickness of forests; but on local circumstances that are difficult to characterize[1477]: and Mr. W. S. MacLeay makes a similar observation upon that of Gymnopleurus[1478]. So that the real insect climates, or those in which certain[Pg 496] groups or species appear, may be regarded as fixed by the will of the Creator, rather than as certainly regulated by any isothermal lines. Still, however, under certain limitations, it must be admitted that the temperature has much to do with the station of insects. The increase of caloric is always attended with a proportional increase in the number and kind of the groups and species of these beings. If we begin within the polar regions of ice and snow, the list is very meager. As we descend towards the line, their numbers keep gradually increasing, till they absolutely swarm within the tropics. Something like this takes place in miniature upon mountains. Tournefort long since observed at the summit of Mount Ararat the plants of Lapland; a little lower, those of Sweden; next, as he descended, those of Germany, France, and Italy; and at the foot of the mountain, such as were natural to the soil of Armenia. And the same has been observed of insects. Those that inhabit the plains of northern regions have been found on the mountains of more southern ones; as the beautiful and common Swedish butterfly Parnassius Apollo, on the mountains of France, and Prionus depsarius on those of Switzerland[1479].
M. Latreille, having given a rapid survey of the peculiar insect-productions of different countries, next attempts a division of the globe into climates, which he thinks may be made to agree with the present state of our knowledge, and be even applicable to future discoveries. He proposes dividing it primarily into Arctic and Antarctic climates, according as they are situated[Pg 497] above or below the equinoctial line; and taking twelve degrees of latitude for each climate, he subdivides the whole into twelve climates. Beginning at 84° N. L. he has seven Arctic ones, which he names polar, subpolar, superior, intermediate, supratropical, tropical, and equatorial: but his antarctic climates, as no land has been discovered below 60° S. L., amount only to five, beginning with the equatorial and terminating with the superior. He proposes further to divide his climates into subclimates, by means of certain meridian lines; separating thus the old world from the new, and subdividing the former into two great portions,—an eastern, beginning with India, and a western, terminating with Persia. He proposes further that each climate should be considered as having 24° of longitude, as well as 12° of latitude[1480]. In this chart of insect Geography he states that he has endeavoured to make his climates agree with the actual distribution of insects[1481]; and it should seem that in many cases such an agreement actually does take place: yet the division of the globe into climates by equivalent parallels and meridians, wears the appearance of an artificial and arbitrary system, rather than of one according with nature.
He has also pointed out another index to insect climates, borrowed from the Flora of a country. Southern forms in Entomology, he observes, commence where the vine begins to prosper by the sole influence of the mean temperature; that they are dominant where the olive is cultivated; that species still more southern are compatriots of the orange and palmetto; and that some[Pg 498] equatorial genera accompany the date, the sugar-cane, the indigo and banana[1482]. The idea is very ingenious, and, under certain limitations, supplies a useful and certain criterion. For though none of these plants are universal in isothermal parallels of latitude; yet, as plants are more conspicuous than insects, the Entomologist, furnished with an index of this kind, may by it be directed in his researches for them; and in all countries in which there is a material change of the climate, as in France, there will be a proportional change in the vegetable accompanied by one in the insect productions.
ii. In considering the range of insects I shall first advert to that of individual species. At the extreme limits of phanerogamous vegetation we find a species of humble-bee (Bombus arcticus), which, though it is not known to leave the Arctic circle, has a very extensive range to the westward of the meridian of Greenwich, having been traced from Greenland to Melville Island; while to the eastward of that meridian it has not been met with. In Lapland its place appears to be occupied by B. alpinus and lapponicus, with the former of which, though quite distinct, it was confounded by O. Fabricius; but whether these range further eastward of that meridian has not been ascertained. From its being found in the Lapland Alps[1483], it may be conjectured that B. alpinus ranges as high on this side as B. arcticus on the other, and may perhaps be found in Nova Zembla. Some species that have been taken in Arctic regions are not confined to them. Of this kind is Dytiscus marginalis,[Pg 499] which appears common in Greenland, abundant in Britain, and is dispersed over all Europe; while D. latissimus is more confined, neither ranging so far to the north or south; and though found in Germany, not yet discovered in Britain. Other species have a still more extensive range, and are common to the old world and the new. Thus Dermestes murinus, Brachinus crepitans, Tetyra scarabæoides[1484], Pentatoma juniperina, Cercopis spumaria, Vanessa Antiopa, Polyommatus Argiolus, Hesperia Comma, Vespa vulgaris, Ophion luteus, Helophilus pendulus, Oscinis Germinationis, and many besides, though sometimes varying slightly[1485], inhabit both Britain and Canada: and though vast continents and oceans intervene between us, New Holland, and Japan; yet all have some insect productions in common. With the former we possess the painted-lady butterfly (Cinthia Cardui), with scarcely a varying streak: and Thunberg, in his list of Japan insects, has mentioned more than forty species that are found also in this country. Whether any species has a universal range may be doubted, unless indeed the flea and the louse may be excepted. On the other hand, some are confined within very narrow limits. Apion Ulicis for instance, abundant upon Ulex europæus in Britain, has not, I believe, been found upon that plant on the continent.
The geographical distribution of groups, is, however,[Pg 500] far more interesting than that of individual species: for in considering this we see more evidently how certain functions are devolved upon certain forms, and can scan the great plan of Providence, in the creation of insects, more satisfactorily than by confining our attention to the latter. Groups, according to their range, may be denominated either predominant, dominant, sub-dominant, or quiescent.
1. M. Latreille has observed, that where the empire of Flora ceases, there also terminates that of Zoology[1486]. Phytiphagous animals can only exist where there are plants; and those that are carnivorous and feed upon the former, must of necessity stop where they stop. Even the gnat, which extends its northern reign so high[1487], must cease at this limit; while, where vegetation is the richest and most abundant, there the animal productions, especially the insect, must be equally abundant. I call that, therefore, a predominant group, members of which are found in all the countries between these points, or from the limits of animal-depasturing vegetation in the polar regions to the line.
Generally speaking, the carnivorous insects, whether thalerophagous or saprophagous, are of this description. Calosoma, which devours Lepidopterous larvæ, though poor in species and individuals, is widely scattered. Captain Frankland found C. calidum in his Arctic journey; C. laterale and curvipes inhabit tropical America[1488]: C. Chinense, as its name indicates, is Chinese[1489]; Mr. MacLeay[Pg 501] has an undescribed species from New Holland; and C. retusum was taken in Terra del Fuego. Another genus, equally universal and richer in numbers, is the lady-bird (Coccinella), which keeps within due limits the Aphides of every climate from pole to pole. The Libellulina pursue their prey both in Greenland and New Holland. The saprophagous carnivora are also similarly predominant;—the Silphidæ, the Dermestidæ, the Brachyptera, the Muscidæ, prey on carcases wherever the action of the solar beam causes them to become putrid. Many of the above insects have probably their capital station, or that where the species are most numerous, in or near the tropics; but the metropolis of the Brachyptera, at least as far as we can judge from our present catalogues, is within the temperate zone, particularly in Britain[1490]. The coprophagous Petalocera are most abundant in the hottest climates; but the Aphodiadæ form a predominant group: Professor Hooker took one species in Iceland[1491], and it probably ascends higher; others are found in India and China: but the metropolis of the group is within the temperate zone. Perhaps no genus is more completely universal than Bombus (Bremus Jur.), which, although its centre or metropolis is likewise in the northern temperate zone, extends from Melville Island to the line. It is remarkable that some of the tropical Bombi wear the external aspect of Xylocopæ, the kindred genus most prevalent in warm climates; and, vice versâ, some Xylocopæ resemble Bombi. I have a Brazilian undescribed species of the latter genus, whose[Pg 502] black body and violet-coloured wings would almost cause it to be mistaken for a variety of X. violacea; and B. antiguensis and caffrus F., (though their aspect belies it,) which misled Fabricius, are true Xylocopæ. I shall mention only one other predominant group, but that one of no common celebrity, formed of the gnats, or genus Culex. These piping pests, with their quiver—"venenatis gravida sagittis"—annoy man almost from the pole to the line. What remarkably distinguishes them, (as was formerly observed[1492],) and also the Simulium or true mosquito,—they appear to prevail most in the coldest and the hottest climates, and the Laplander and the tropical American are equally their prey; while the inhabitants of the temperate zone, with some exceptions, suffer but little from them: so that they may be stated to have both an arctic and a tropical metropolis.
2. There are other groups which, though their empire extends to the tropics, fall short of the polar circles:—these I call dominant groups. Of this description are some of the Scarabæidæ. Onthophagus is found both in the old world and in the new, and in the temperate and torrid zones. Its principal seat appears to be within the tropics, but it may almost be said to have also a northern metropolis. More than one species have been taken in New Holland. In general, tropical insects exceed those of colder climates in size; but in the genus we are speaking of, the European species are usually larger than the Indian. Copris seems more abhorrent of cold than its near relation Onthophagus. C. lunaris, which ranges northward as far as Sweden, is the only[Pg 503] recorded species found in Europe out of Spain. Latreille says, that all the large species of this genus are equinoctial: but C. Tmolus, described and figured by Fischer[1493], found in Asia near Orenburg, north of 50° N. L., is as big as C. Gigas or bucephalus. Another dominant group of Petalocera, remarkable for the bulk and arms of its tropical species, are the mighty Dynastidæ, the giants and princes of the insect race. Though their metropolis is strictly tropical, yet the scouts of their host have wandered even as far as the south of Sweden, where one of them, Oryctes nasicornis, is extremely common. O. Grypus[1494] and some other species are found in South Europe; but though in a torpid state they can endure unhurt the severity of a Scandinavian winter, they cannot when revived stand the cold that often pinches Britons in the midst of summer, and therefore are unknown in our islands[1495]. The Sphæridiadæ, whose metropolis is within the northern temperate zone, extend from thence beyond the line, since Dr. Horsfield found two species in Java[1496]. It is probable, indeed, that this group is predominant. Some dominant groups begin at a lower latitude. Of this description are the carpenter-bees (Xylocopa), whose larvæ are preyed upon by that of the Horiadæ[1497] under two forms, which extend from the tropics to about 50° N. L. Others are not common to both worlds. Thus, while Cantharis is the gift of Providence to America as well as the old world, Mylabris is confined[Pg 504] to the latter, where its range is very extensive;—in Europe, from South Russia to Italy and Spain; in Asia, from Siberia to India; and in Africa, from the shores of the Mediterranean to the Cape of Good Hope; which last continent, to judge from our present lists, especially the vicinity of the Cape, may be called the metropolis of the group[1498]. On the other hand, the Rutelidæ and Chlamys, which have a range from Canada to the tropics, (within which is their metropolis,) are purely American groups. Many more might be named under this head, but these will suffice for examples.
3. I call those subdominant groups, which either never enter the tropics, or those tropical ones whose range does not exceed 50° of N. L. in the old world, or 43° in the new. I make this difference because, as M. Latreille observes, the southern insects which in Europe begin between 48° and 49° N. L., in America do not reach 43°.[1499] But though the winters in Canada, within the same parallel as France, are longer and more severe than those even of Great Britain or of Germany, yet the summers are intensely hot; so that though tropical species do not range so high, those of a tropical structure, as Mr. W. S. MacLeay has intimated[1500], may be found at a higher latitude in the new world than in Europe.
The genus Melöe affords an instance of a subdominant group of the first description. It ranges from Sweden to Spain and the shores of the Mediterranean, and seems a tribe almost confined to Europe, where it is not[Pg 505] very unequally distributed. Of registered species Britain possesses the largest proportion; but Mr. W. S. MacLeay is of opinion that Spain is its true metropolis[1501]. I have a species of this genus, taken in North America by Professor Peck. The splendid genus Carabus ranges still further north than Melöe[1502]. A very fine species (C. cribellatus) inhabits the polar regions of Siberia[1503]; but the metropolis of the group appears to be the temperate zone: some, however, have been found in northern Africa; and Sir Joseph Banks captured one in Terra del Fuego. Of those whose range is between the tropics and 50° N. L. we may begin with Cicada. One species, indeed, has been found by Mr. Bydder and others, a little higher, near the New Forest, Hampshire. We may take Scolia for an example of a subdominant group beginning more southward. Its species first appear about 43° N. L., and abound in warm climates. In general most of those insects which M. Latreille denominates meridional,—such as Scarabæus, Onitis, Brentus, Scarites, Mantis, Fulgora, Termes, Scorpio, &c.—come under the present head, and in fact all tropical forms that wander to any distance within the above limits from their metropolis.
4. By quiescent groups I mean those that have none, or no high range as to latitude, from their centre or metropolis.[Pg 506] I say as to latitude, because these groups have often an extensive one as to longitude. Thus, Mr. W. S. MacLeay has remarked to me, that Goliathus appears to belt the globe, but not under one form. The types of the genus are the vast African Goliaths (G. giganteus, &c.), which, as well as G. Polyphemus, and another brought from Java by Dr. Horsfield, have, like Cetonia[1504], the scapulars interposed between the posterior angles of the prothorax and the shoulders of the elytra[1505]: while the South American species (G. micans, &c.) have not this projection of the scapulars; in this resembling Trichius. Mr. MacLeay further observes, that the female of the Javanese Goliathus is exactly a Cetonia, while that of the Brazilian is a Trichius. But quiescent groups have not generally this ample longitudinal range. Thus, Euglossa, in both its types,—one represented by Eu. cordata, and the other by Eu. surinamensis,—is confined to the tropical regions of America. Doryphora, likewise American, seems equally confined. Asida, though a southern genus, is not found to enter the tropics; and Manticora and Pneumora are in nearly the same predicament.
Under the present head we may consider what may perhaps be denominated without much impropriety endemial groups; by which I mean those groups that are regulated, as to their limits, not so much by the temperature, or the northing and southing of the latitude, as by the general aspect and circumstances of the country. Thus, the vast and nearly insular continent of Africa,[Pg 507] almost as wide as it is long, and situated in or near the tropics, instead of inland seas or sea-like rivers, is intersected by parched sandy deserts, extending far and wide; circumstances which, though in the vicinity of its streams it is humid, impart an unusual degree of aridity as well as heat to its general atmosphere; so that it well merits the poet's epithet, Leonum arida nutrix; and is also peculiarly fitted for all such animals, especially insects, as delight in a dry, sandy, hot country, particularly such as are predaceous in their habits. America, on the other hand, exhibits quite an opposite character. It is long, and comparatively narrow; surrounded, and almost divided into two continents, by immense circumfluent oceans; watered every where by rivers and lakes that emulate seas: in some parts covered by interminable forests; in others, intersected by ridges of the loftiest mountains. These circumstances, except in its Llanos, Pampas, or table-land, give a general character of humidity to its atmosphere, and fit it particularly for the production of a vast variety of peculiar plants, and for the residence of numerous and peculiar phytiphagous insects and other animals[1506]. Midway between these two continents lies a third (for so the vast island of New Holland may be denominated), which presents new features in its general aspect, and consequently new forms both in its Flora and Fauna, mixed with many old ones parallel to those both of the new world and the old. Perhaps Europe and Asia, with several that are peculiar, agree more in their animal productions than the continents just described.
[Pg 508] Let us next particularize a few of the peculiar types that distinguish particular continents and countries. The genera Manticora, Graphipterus, Glaphyrus, Eurychora, Pneumora, Masaris, and many others, are peculiar to Africa. In Asia alone we find Mimela[1507], Euchlora[1508], Colliuris, Catascopus[1509], Apogonia[1510], a peculiar type of Horia, &c. In America, Agra, Galerita, Nilio, another type of Horia, Tetraonyx, Rutela, Doryphora, Alurnus, Erotylus, Scotinus[1511], Cupes, Corydalis, Labidus, Heliconius, Castnia, &c. And in New Holland, Helluo, Elephastomus, Anoplognathus, Diphucephala[1512], Cerapterus, Heleus, Adelium, Notoclea, Achilus, Thynnus, Nycterobius, &c.
The countries bordering upon the Mediterranean, the Black, and the Caspian seas, agree in producing similar insects. These countries, and the Cape of Good Hope though so distant from them, appear to be the principal seat of Heteromerous Coleoptera, of the genera Lixus and Brachycerus, and of the conical Buprestes[1513]. But the insects of Guiana, on one side the Cordilleras, differ from those of New Granada and Peru on the other; and similar differences are observed in other neighbouring countries separated by natural boundaries.
iii. Another head connected with the topographical distribution of insects relates to their representation of each other. Here we may observe, that some insects represent each other only in their form; others also in their[Pg 509] function; and others in both. I shall give some instances of each. In Brazil there is a group of petalocerous beetles (Chasmodia), one of the Rutelidæ, which in New Holland has a representative, as to form, in one of the Cetoniadæ (Schizorhina[1514]), which, having soft mandibles, must have a different function:—it is to be observed, however, that these insects appear to approach each other in the series of affinities. Again, the Carabidæ may in the same country be said to have a representative in the remarkable heteromerous genus Adelium[1515], which is altogether an analogy. Others are representative only in their function. The general function of insects is to remove nuisances and to check redundances,—the saprophagous tribes do the one, and the thalerophagous the other. In going from the poles to the line,—in proportion as the heat increases, the quantum of work of both kinds increases; and new forms are either added to the old ones, so as to increase their momentum; or new ones, more powerfully talented, replace the old ones, and act in their stead: thus we see a gradual and interesting change take place in proportion as we approach the maximum of heat and of insect population. At the Cape, the universal Cicindelæ are aided by Manticora; in North America, the Silphidæ by a new group, the type of which is Silpha Americana (Necrophila, K.MS.); in South America, Copris by Phanæus. Again: Colliuris and Drypta of the old world, in the new give place to Ctenostoma and Agra. The honey and wax of Europe, Asia, and Africa, is prepared by bees congenerous with our[Pg 510] common hive-bee (Apis); while in America this genus is not found as a native, but is replaced by Melipona and Trigona[1516]; and in New Holland by a still different but undescribed type. The Melolonthidæ and Rutelidæ of the old and new world appear to have their work done in that country by the brilliant and numerous Anoplognathidæ. The Rhipicera of Brazil is of a different type from that of New Holland. The singular genus Cremastocheilus of North America has its representative in Africa in Genuchus[1517]. The Lucani of the rest of the world give place in New Holland to Lamprima and Ryssonotus.—I could produce a much greater number of examples, but these are sufficient to explain my meaning.
Having thus given you some, though an imperfect account, of the geographical distribution of insects, I am next to say something concerning their local distribution in any district, or their favourite haunts; a knowledge of which, with respect to those of our own country, is indispensable to the collector.
The surface of a country consists either of mountains, hills and valleys, or of plains. It is diversified by forest, wood, or copse; and watered by rivers, rivulets, lakes, and pools. Those parts that are not clothed with wood are either open or inclosed, forming grassy downs, heaths, pastures, meadows, morasses, and arable land. The soil also is equally various:—we find clay, loam, marl, chalk, vegetable mould, moor, sand, &c. The mountains and hills are either covered with a stratum of soil, or are[Pg 511] rocky and bare; the arable lands are divided by living or dead fences, the latter formed of various materials,—or else they are open, and the property only marked out by grassy balks, &c. All these places abound in shrubs and plants; some local, and some generally distributed. But besides the land and its fresh waters, we must look also to the sea, and its sandy, pebbly, or rocky shores, and the sea-wrack that is cast up upon them; the estuaries that receive its tides; the brackish waters and saline marshes in its vicinity. All the above places, when opportunity serves, the Entomologist should explore, for in almost all he will find peculiar kinds of insects.
As mountains and hills have usually their own Flora, the insects appropriated to alpine plants can only be met with where the pabulum is found. Here also those northern insects that are impatient of a warmer climate will take their station, if they migrate to the southward[1518]. The predaceous beetles likewise sometimes frequent a mountainous district. Carabus glabratus was first taken by Professor Hooker on Ingleborough; and probably, if the Welsh and Scotch mountains were duly investigated by an Entomologist, many novelties would reward his toils. The valleys and plains, especially those of a sunny exposition, abound in insects. When the heat of the atmosphere indisposes you for motion, you will find it no unprofitable or unpleasant employment, lying on the grass, to search for minute beetles, which you will there find coursing about amongst the tufts and roots of the herbage. Thus you may procure many of the Pselaphidæ, which you would not otherwise meet with. Even[Pg 512] when the grass is grown up, insects are fond of alighting upon its spikes, and thence drop or run to the ground. Should circumstances ever carry you abroad to the steppes or grassy plains of Tartary, or to Hungary, you would find there two or three species of the singular genus Lethrus, which burrows in the soil. Every hole is inhabited by a male and female;—from it they issue to attack the plants or vines; and having cut out the heart of a plant, go backwards like a crab with the prize to their burrow. At the time of pairing, sometimes violent battles, encouraged by the female, take place between the male and a stranger of that sex desirous of admission, which cease only with the death or flight of the stranger[1519]. The vicinity and borders of woods generally abound in insects of every Order; and if you proceed, as hereafter directed, will furnish you with numerous prizes, especially of Lepidoptera. Here alone you can meet with the purple emperor butterfly (Apatura Iris); and if properly equipped you may readily secure him.
The waters you will find nearly as prolific in insects as the land. In them, amongst the beetles, you may expect to meet with Dytiscus, Haliplus, Pælobius, Hyphydrus, Hydroporus, Noterus, Colymbetes, and other Dytiscidæ; the Gyrini, Hydrophili, Hydrænæ, Elophori, &c.: under stones, the Elmis; and in the mud, the Parni and Heteroceri. Some Sphæridiadæ are also aquatic: I have taken more than once Cercyon hæmorrhoidale from the under side of a piece of wood immersed in a canal[1520]. Even a few of the weevil tribes are to be met with in[Pg 513] water. Lixus paraplecticus, Tanysphyrus Lemnæ, Bagous atrirostris, are of this description. A species of Ceutorhynchus of Germar's third family (C. Natator K.) swims well. On aquatic plants you must look for Helodes and the splendid Donaciæ, which, living on submerged shoots and roots of these plants in their larva state, continue to attend them when perfect. Amongst the Eutrechina[1521],—Elaphrus, Notiophilus, and Bembidium frequent humid places, as the banks of rivers and ponds; and in such a station, under the roots of Potentilla anserina, Polygonum, &c. if you should be fortunate enough to find Omophron limbatum, which connects the Eutrechina with the Eunechina, you will make a valuable addition to the list of British insects. In the waters also you will meet with many Heteropterous Hemiptera; as Gerris, Hydrometra, and Velia, and all the Hydrocorisæ or water-bugs. On aquatic plants the larvæ of some Lepidoptera feed, as Hydrocampa stratiotata, potamogata, &c. Those also of the Trichoptera must be sought for in the water: and if you should feel inclined to see an interesting collection of their very curious cases, Mr. Sheppard of Wrabness can gratify your curiosity. Though few or no Hymenoptera frequent this element, vast numbers of Diptera are there alone to be met with in their preparatory state, particularly the gnats. We learn from Humboldt a curious fact with respect to those of South America, or the Zancudos; that, with some exceptions, these pests do not frequent those rivers called by the natives black waters, but only those which they name white waters[1522]. Of the Aptera, the genera[Pg 514] Hydrachna, Eylaïs and Limnochares are purely aquatic. Several spiders will walk over the water; and one species (Argyroneta aquatica) inhabits it[1523]. The stagnant waters in your vicinity will produce different species from running ones. Thus Haliplus elevatus, &c. inhabits only the latter, while the majority of the Dytiscidæ abound most in the former: the more minute ones may be sought for with success amongst the duckweed that covers a pool. I do not recollect finding any insect in waters absolutely salt[1524]; but brackish waters produce peculiar species: in these only, Hydræna marina occurs; and many of those large-eyed Cimicidæ (Acanthia), as A. saltatoria, littoralis, and Zosteræ occur in places where salt water has been. Latreille observes, that the genus Pimelia is to be met with only where the soil is impregnated with saline particles, or where the species of the genus Salsola abound[1525].
Heaths, though they do not afford numerous insects, have their rarities. Cicindela sylvatica, Carabus nitens and arvensis, frequent them, and are not elsewhere to be seen. Curculio nebulosus is also to be found on them, in places where the turf has been peeled; and some scarce Lepidoptera. In their vicinity, in sunny sandy banks, some of the rarer Ammophilæ and Pompili may be taken; and it is here only that I have ever met with Panurgus[1526]. Meadows and pastures are not to be neglected. Early in the year, when they are yellow with the blossoms of Ranunculus bulbosus, Leontodon Taraxacum, &c., many[Pg 515] minute beetles, and not a few Hymenoptera and Diptera, frequent them. Morasses also have their peculiar insects. In these you will meet with some of the scarcer Eutrechina; as Chlænia holosericea and nigricornis, Blethisa multipunctata, various Bembidia, &c. In this kind of district in the Isle of Ely Aphodius plagiatus has been taken, and that scarce and beautiful butterfly Lycæna Virgaureæ. Where land is cultivated the Entomologist as well as the farmer may expect a harvest. Insects in general are fond of perching on the summit of a blade of grass or corn; and many minute ones may be taken coursing about in the ears of the latter: some to devour the fungilli that infest the grain, as Phalacrus corruscus in Reticularia Segetum; others to attack the grain itself, as Cecidomyia Tritici; others to destroy these destroyers, as three little parasites belonging to the Chalcidites[1527]. But I have already mentioned most of those insects that are to be expected in such situations[1528]: I shall therefore only further observe, that upon barley particularly you will meet with the species of Latreille's genus Cephus.
With respect to soils, those that are light appear to be most prolific in insects. Warm sandy banks are frequented by Cicindela campestris, Opatrum sabulosum, Helops quisquilius, &c.: in them (when of a southern aspect) Ammophilæ, Pompili, and numerous Hymenoptera nidificate. Chalk also attracts various insects. Latreille observes, that the Licini, Papilio Cleopatra, several species of Dasytes, and some Lamiæ, delight in this kind of soil[1529]:—in my own neighbourhood I have observed Polyommatus Corydon principally in chalk-pits. One of these[Pg 516] pits, under a wood in an adjoining parish, has produced me several valuable insects. Here I took Apion ebeninum, Orobitis globosus, a new species of Evæsthetus, several of the rarer Pselaphidæ and Cholevæ, and Chætophorus cretifer before noticed[1530]. I do not mean, however, that all these are properly chalk insects; but they fall into these pits, where they are readily discerned, from the contrast of their colours with the whiteness of the chalk. By watching attentively the bottom of one, vast numbers in a warm day may be taken when they fall or are climbing upwards. Of all soils clay offers the fewest inducements to the Entomologist, who will lose both his time and labour in a clay-pit; while in one of sand, chalk, or marl, they will usually not be mispent. Vegetable earth also affords a harbour to various larvæ, and the pupæ of many nightfliers amongst the Lepidoptera, by digging in it, especially under trees, may be obtained. Even the bare rocks have their insect frequenters that take shelter in their fissures; and in the early part of your career especially you should always turn over large stones, as beneath them many of the Harpalidæ and other Eutrechina frequently lie hid: and in this situation, both in Suffolk and Sussex, Lomechusa emarginata, one of our scarcest Brachyptera, has been taken. Old trees also, and planks that have laid long without being moved, often afford a shelter to many of the minute Coleoptera; as Pselaphidæ, Aleocharidæ, Cryptophagidæ, Scymnidæ, &c. Live fences, especially when the hawthorn is in blossom, and where trees are also intermixed, are attended by innumerable insects of almost every description; and even the black-thorn will present you with one of[Pg 517] our most splendid weevils (Rhynchites Bacchus). Dead fences are almost as fertile in insects as living ones. In gates, posts, rails, and other timber when felled, the timber-devouring tribes take their station:—between the bark and the wood are the Bostrichidæ; in the wood itself, the Anobidæ and the Capricorn beetles. Here also you may meet with many Hymenoptera, which either devour timber or nidificate in it,—as the Siricidæ, Chelostoma, Trypoxylon, Sapyga, and several Diptera. In the decaying hedgestakes and sticks, where the Sphæria decorticans has turned off the bark, you may meet with Anthribus brevirostris; with A. latirostris, and other beetles, in S. fraxinea; and A. albinus, which I have more than once captured as it was emerging from the fissure of a gate-post, probably feeds on some internal fungus. The grassy balks that separate open fields usually abound in umbelliferous plants, which are attended by numerous Hymenoptera and Diptera, particularly by the various species of the splendid tribe of Chrysidæ: and the grassy banks of fences, where the aspect is sunny, are generally bored by a variety of insects of the former Order, to prepare a nest for their young. Andrenidæ and Nomadidæ particularly select this situation, the latter probably depositing their eggs in the burrows of the former[1531]. By watching these places in the spring, you may perhaps have the good fortune to meet with a Stylops. It is singular, that some insects choose, for their own residence or that of their young, the hardest and most trodden pathways. Thus, some ants will build their subterranean apartments under[Pg 518] gravel walks; and so do many species of the genus Halictus[1532], the habits and economy of which have been so ably detailed by M. Walckenaër[1533]: Cerceris also, and other Hymenoptera, will choose such places, however public, for the site of their nests or burrows. The ground is so consolidated by the constant foot, that they, probably find such situations spare them a world of labour, and therefore in their choice balance one inconvenience by another.
Though the sea itself, I believe, produces no true insects, yet there are many that constantly or occasionally haunt its shores. On the sand-hills of the Norfolk coast I found Ægialia globosa and Cicindela hybrida. Ceutorhynchus horridus inhabits thistles that grow near the sea. Under the Zostera and Fuci, (cast up both on its beach and the shores of estuaries,) many peculiar species of Cercyon, several Aphodii, and numerous Brachyptera, may often be found. In this situation the rare and singular Bledius armatus has been taken. At certain seasons of the year the beach and environs of the sea are covered by many species of Coccinella, which seem to bend their course thither from the inland country, as if they were about to emigrate[1534]. When the weather is fine and the tide begins to retire, at the line of its highest rise I have taken on the eastern coast a variety of insects, and amongst the rest Anomala Frischii. The inundations of rivers, except in the depth of winter, always bring a number of these little creatures, which float on the surface on bits of stick, weeds, &c.; and where they deposit these articles when the water begins[Pg 519] to subside, you may generally reap a plentiful harvest of various kinds.
You see, now, how varied is the scenery to which the diversion of the Entomologist introduces him; that he is never out of his way: whether on hill or in valley; on upland or plain; on the heath or in the forest; on the land or on the water; in the heart of a country or on its shores;—still his game is within his reach. But in order to enable him to pursue it with greater prospect of success, he must recollect that not only is every face of the country to be explored, but both the plants and the animals that it produces; and that he must not turn with disgust from even the carcase or the excrement of the latter. As numerous species of herbivorous insects feed only on one kind of plant, the Entomologist, when he discovers a scarce one, should examine it with the hope of finding upon it a scarce insect. Sometimes it happens that only a single opportunity occurs in a man's life of seeing certain plants growing wild: such opportunities should never be neglected. Some insects also inhabit a plant in one district or season, and not in another. Thus the most beautiful of the Apions, A. Limonii[1535], though the plant it feeds upon usually abounds near the sea, I have discovered only on the northern coast of Norfolk; and another scarcely less beautiful, but more minute (A. Astragali[1536]), though I have sought for it year after year, Astragalus glyciphyllus being abundant near me, I never found but once. The blossoms of plants as well as the leaves must be inspected. In those of the rose, the Cetonia aurata is often taken[1537]; and in the bells of the[Pg 520] different species of Campanula various bees may be captured enjoying a luxurious repose[1538]. No vegetable productions abound more in insect inhabitants than the Fungi. In Agarics several Diptera are to be taken, many Aleocharæ, Oxypori, &c.; in Boleti, the various species of Mycetophagus; in the arboreous ones, and under bark, more than one kind of Ips; and in Auricularia, as well as Boletus, the whole genus Cis. Upon living Vertebrate animals you must look for Pulices, Pediculi, Nirmi, Acari, and many Diptera, as Œstrus, Tabanus, Stomoxys, and the Pupipara of Latreille; and on the garden-snails for that curious genus Drilus, and some Acari[1539]. The caterpillars and pupæ of Lepidoptera, &c. will, as you have heard, furnish you with numerous ichneumons[1540]. On dead animals you will find the various species of Silphidæ, Nitidulidæ, Dermestidæ, Byrrhidæ, Chlolevidæ, Staphilinidæ, Muscidæ, &c.; and in excrement, various Scarabæidæ, Histeridæ, Aphodiadæ, Sphæridiadæ, the Brachyptera in general, and several Diptera[1541]. In putrescent roots and fruits, as the turnip, the cucumber, &c., you may also occasionally meet with rare Coleoptera.
I must next say something upon the seasons of insects, and their times of appearance. Those that collect honey and pollen are generally among the first that proclaim the approach of spring; and their appearance may be dated from the blossoming of certain trees and plants of common occurrence. Other plants, accompanied by peculiar[Pg 521] insects, blossom later; and so on till we arrive at the autumn. The earliest insect-season commences with the flowering of the sallow (Salix Caprea) usually accompanied in the garden by that of the crocus and the gooseberry. Then is your time to collect many species of wild bees and Diptera not afterwards to be met with: and various other insects now begin to emerge from their winter-quarters, or are produced from the pupa. Another and later season is marked by the general blossoming of the butter-cup (Ranunculus bulbosus), accompanied by the marsh-marygold (Caltha palustris) and ladies'-smock (Cardamine pratensis); when you may hunt the pastures, meadows, and marshes with success, and take some insects that do not show themselves later. The coprophagous insects are now abundant. Amongst others, Aphodius testudinarius, a perfectly vernal species, is now only to be taken, and usually flying. A third insect-season indicated by Flora, and a very prolific one, commences with the blossoming of the hawthorn, when you must desert the meads for the inclosures. At this time all nature begins to put on her gayest attire, and all her insect tribes are now on the alert, and fill the air. They are almost universally attracted by the sweet and lovely blossoms of the plant just named: so that by examining them you may entrap some of every Order, and many that during the year will appear no more. Even many of the saprophagous insects will sip nectar from these flowers. The umbelliferous plants proclaim the fourth season of insects, particularly the wild carrot and parsnip. You will scarcely ever fail to find, if the weather is genial, Hymenopterous and Dipterous insects of various genera,—especially such as have a short tongue,—engaged[Pg 522] in collecting the honey from those plants. Here you may take some of the rarer Chrysidæ, Crabronidæ, Cercerides, &c., and occasionally even Coleoptera. The last insect-season may be dated from the general flowering of the thistle tribe. When these are in blossom is the best time of all to collect the humble-bees (Bombus[1542]), the leaf-cutter bees (Megachile[1543]), and many other Apiariæ, which alone by their long tongues can imbibe the honey and collect the pollen of these flowers. The male humble-bees frequent them to the last, and often seem as if they were intoxicated with their sweets.
But perhaps you may prefer considering the whole summer appearance of insects as divided into three principal seasons. This may thus be done. Their vernal season may commence Florente Caprea, and end Florente Oxyacantha; their summer, Florente Oxyacantha and Florentibus Umbellatis; their autumn, Florentibus Umbellatis and Florente Carduo. In the first, the number of insects will be daily increasing; in the second (which is the harvest of the Entomologist, when his eyes and his hands ought to be every where), they will reach their utmost complement; and in the third, they will be gradually decreasing in number, till they generally die, or go into winter-quarters. At this time many minute Diptera and Ichneumons take shelter from the weather in the windows of our apartments. These seasons will not always exactly correspond with our usual reckoning, and take place at the same time; since, being regulated by our varying temperature, they will be sometimes sooner and sometimes later, sometimes longer and sometimes shorter. Though I have not named a brumal season, because insects are in[Pg 523] winter usually torpid,—yet some, as Diurnea Novembris, Cheimatobia brumata, and many Tipulariæ, even then make their appearance.
If you ask, Whether it be not possible to regulate our Entomological seasons by the appearance of insects themselves? I should answer, that probably this might be done; but that further observations seem wanted to enable us to do it satisfactorily. Perhaps the appearance of Formica rufa beginning the business of the year might form the commencement of one season; the flight of the orange-tip butterfly (Pontia Cardamines[1544]), of a second; a third might be indicated by the swarming of Melolontha vulgaris; a fourth, by that of Amphimalla solstitialis; and the last, by the appearance in numbers of Aphodius ciliaris, which in the autumn fills every horse-dropping.
Some insects are so ephemeral, that they are to be found in numbers only for a few days, and then disappear for that season. Of this description are the Ephemeræ, much of whose history has been detailed to you. Those of which De Geer has given an account (E. vulgata) appeared about the end of May or the beginning of June, and continued about a fortnight[1545]; while those which Swammerdam observed did not come forth till the middle of June, and lasted only three days[1546]. The same period distinguished those of which Reaumur has compiled so interesting a history, but they did not show themselves before the middle of August[1547]. My kind friend Mr. Marsham not long before his death copied[Pg 524] for me some memoranda he had made with respect to the sudden appearance of Cercopis bifasciata. On one occasion the white dress of a lady sitting upon a haycock was covered by these insects; but on the following day the same steps were taken at the same time to procure some, when after the most diligent search not a single one could be found. The same circumstance was observed a few years afterwards by another friend of his. He himself was of opinion that the insects in question were then migrating[1548].
I may here observe, that the London amateurs have particular seasons for collecting moths. For the imago they go into the woods in April, May, June, and October. For the larvæ they take the beginning of April, June, the beginning of July, and September. They dig for pupæ late in July, and in January and February.
I shall lastly make a few observations upon the times of action and repose of insects, the knowledge of which, as far as it can be obtained, is of essential use to the industrious collector. Entomologists have divided the Lepidoptera, with a view to this circumstance, into diurnal, crepuscular, and nocturnal; or butterflies (Papilio L.), hawkmoths (Sphinx L.), and moths (Phalæna L.). These terms may be applied to insects in general.
i. Diurnal insects are abundant. Butterflies in particular fly generally at no other time: they accompany the sun in his course, and before he sets disappear. Some other Lepidoptera, though not so named, are day insects:—such are the Zygænidæ, &c. amongst the hawkmoths;[Pg 525] and amongst the moths, Plusia Gamma, the Phytometræ solares of Haworth[1549], and some others. Numberless Coleoptera belong to this section. The Donaciæ fly only when the sun is out and the air is warm; they are then extremely agile and difficult to take. Some Hopliæ swarm in the day before noon, and then disappear[1550]: most of the tetramerous beetles also appear to be diurnal. The Libellulina and many other Neuroptera may also be so termed; and the Hymenoptera almost universally, with the sole exception of the Formicidæ[1551]. Amongst the Diptera, if we leave out the Tipulariæ, the rest will be found for the most part to belong to the present section.
ii. Crepuscular insects, strictly speaking, are those that appear only during the twilight, whether in the morning or evening; but the term may be understood, with some latitude, to signify all those insects that are seen only in the morning and evening, though after sunrise and before sunset. Of these, some come forth only in the morning, others only in the evening, and others both morning and evening. My memory only furnishes me with a single instance of an insect whose principal appearance and flight are in the morning. Catocala nupta I have often seen flying at this time, about six or seven o'clock, and never at any other: I am not however prepared to assert that it does not appear in the evening or night, but I have then never met with it. In the evening more particularly you hear the hum of the dung-beetle (Geotrupes), which Linné thought the prognostic of a following fine day; and of the swarms of Melolontha vulgaris[Pg 526] and Amphimalla solstitialis. Then also many other Coleoptera are in the air; especially before a thunderstorm, a state of the atmosphere that particularly excites insects[1552]: Ptinus imperialis and germanus I have never taken except under these circumstances. Then the Ephemeræ sport in the air, and lead their mystic dance. The majority of the hawkmoths are then too on the wing, with their long tongues imbibing the nectar of the flowers while they hover over them, both morning and evening.
iii. In the night the main body of the moths take their flight, as well as a vast number of Coleoptera and insects of other orders. At this time the Blattæ and crickets leave their hiding-places and run about: but the other Grylli L., though they sing in the night, fly only in the day. Then also the Carabi, like beasts of prey, leave their dark retreats,—in this differing from the Cicindelæ, which are diurnal,—and prowl about to entrap other unwary insects. Then, likewise, the female glowworm hangs out her lamp of love, and the male, led by it, wings his way to her: and then the water-beetles (Dytisci, Gyrini, &c.) forsake the waves and become tenants of the air.
Could we with certainty discover the stations in which insects after their excursions take their repose, we might capture many that we now search for in vain. Several of these stations were pointed out in a former part of this letter where I detailed their usual haunts. I may here add, that numbers of them, when reposing, conceal themselves from their enemies on the under side of the leaves[Pg 527] of trees and plants. Moths, especially the Noctuidæ, may often be met with in woods, as before observed[1553], on the north side of the trunks of trees. Mr. Marsham related to me, that once a little before sunset, observing over his head a number of insects on the wing moving on in one direction, he caught some of them, and they proved to be Labia minor. Struck with the circumstance, he watched them several evenings; and on one, as he was looking about a melon-pit for insects, he saw these little animals alight on the frame, hastily fold up their wings, and entering under the glasses, run down its sides and bury themselves in the loose earth. This he observed repeatedly. The onward flight of these insects was therefore evidently their return from their diurnal cruise to their nocturnal station.—This happened in September.
I am, &c.
Having in my last letter given you some account of the haunts of insects, I now proceed to describe the various instruments with which you ought to be provided, to enable you to collect them; and the best mode of employing each. The Entomologist when he makes an excursion should have three principal objects in view, for which he ought to be duly prepared. The first is to find insects, the next is to catch them, and the last when taken to bring them safe home. In exploring their haunts he must also recollect that some will be reposing; others feeding; others walking or running; others flying; others swimming; others lurking in various places of concealment, and in different states of existence; and that he must be prepared with means of coming at and capturing them under all these circumstances.
1. First furnish yourself with a strong knife or other instrument with which you can raise the bark or penetrate the wood of any tree, when circumstances indicate that insects are busy below the one or within the other. There[Pg 529] is no better tool for this and other purposes than Mr. Samouelle's digger, which consists of an iron five inches long, rather more than one-third of an inch in diameter, forming a curve towards the extremity, terminating in a lozenge-shaped point, and strongly fixed in a wooden handle[1554]. With this you may not only explore the interior of timber-trees, but grub up the turf under them, and examine the earth for the pupæ of Lepidoptera. When your object is merely this latter purpose, a potato-fork—which is better than a spade, as it will seldom injure the pupæ—will be your best implement.
2. Next have a stick, to resemble a common walking-stick, sufficiently stout to beat the branches of the trees and shrubs, fitted at one end with a male screw, and at the other with a female, with a brass cap to screw over each to keep the dirt from them. Besides this, you may carry with you a spare piece or two about a foot long, properly equipped to screw to it when you want to lengthen it.
3. Another implement must be a bag-net[1555]. This consists of a hoop of stout brass wire about nine inches or a foot in diameter, with a socket to receive the end of your stick, or, what is more secure, a screw to fix it to it, with a bag of gauze, muslin, or fine canvass, about twelve inches deep, sewed round it. The French collectors use a net of this kind, in which the hoop is formed of two semicircular pieces of iron or brass wire hooked together at one end, and at the other made to lap over the corresponding piece, and pierced to receive the screw at[Pg 530] one end of your stick. When not employed, they double the hoop and conceal it under the vest; they fix to it a muslin bag of two feet long. This net is made to serve various purposes. With it they catch Lepidoptera and other flying insects; and an adroit collector by giving it a certain twist completely closes the mouth, so as to prevent the escape of his captives. Fixed to a very long pole (Mr. Haworth says it should be twenty or thirty feet long[1556]), it is the best net for the purple emperor butterfly (Apatura Iris). It is also used with success to push before you through the grass of meadows, woods, &c., and thus often displaces numerous insects, which fall into it;—every now and then it is examined, and the valuable captures secured. The common bag-net will perform the same operations, but is not deep enough for flying insects. If you lengthen your stick before you screw it on, it enables you to brush with it the weeds at the sides and bottom of ditches. This employment of brushing the grass, &c. may be carried on if you are walking with any friend not interested in Entomology, without much interruption of conversation. For this last operation—sweeping the grass, &c.—if you wish at any time to devote a morning wholly to it, you will find a net invented by the late Mr. Paul, of Starston in Norfolk, and which he employs to clear his turnips of Haltica Nemorum[1557], a very useful implement. The accompanying figure will give you a better idea of it than any description[1558]; you may make it large or small according to your convenience: the wider it is, the greater space it will brush at[Pg 531] once. When your object is a more general investigation, the bag-net just described is preferable.
4. Scarcely any implement seems a greater favourite with British collectors than what may be called the fly-net[1559]. This is universally employed by them for capturing flying insects, especially Lepidoptera. It is similar to what is called a bat-fowling net, and should be made of green or white gauze or coarse muslin. The former colour, as being less visible, is most proper for mothing in the night; but the latter is best for the day, as this net is useful to hold under the branches of trees and shrubs to receive the insects that fall when they are beaten. The rods for the net we are considering,—which should be about five feet long, half an inch in diameter at the base, and gradually tapering to the end,—must be made of some tough wood; each should consist of about four joints for the convenience of carriage, and each joint should be fitted with a socket at the lower extremity, to receive the top of the joint below it: the terminal joint must either be bent into a curve, or fitted with an angular socket or ferrule, so as to form an obtuse angle with the rest of the rod[1560]. The gauze which is to form the net, being cut into the requisite shape, should be welted round, except at the bottom, where it should have a deep fold or a bag for preventing the escape of the included insects—in order to form a slide for the rods to slip in. At the apex where they meet, a few stitches should be set, or a piece of leather sewed in, to prevent their going too far. At the bottom, on each side, two strings must be sewed on the net, to receive[Pg 532] which there must be a hole in each rod about six inches from the bottom: these must be tied, which will keep the net from slipping upwards. When you go after moths and other insects that fly in the night, a plan, as I am told, of some of the London collectors may be adopted with advantage. Cause a lanthorn to be made with a concave back, and furnished with a reflector: this must be fastened, by means of a strap, upon the stomach. If you hold your expanded fly-net before this (as nocturnal insects fly to the light), you may thus entrap a considerable number. In sultry summer nights also, if you place a candle on a table in a summer-house, or even in a common apartment, and open the windows, you will often have excellent sport, and take insects you might otherwise never meet with.
When you use your fly-net, you must take the rods one in each hand, so as to keep it extended; and when you have brought it fairly beyond the insect you are pursuing,—to accomplish which you must be upon the alert,—you must bring the two sticks together, which, if you are commonly dexterous, will capture your prey. This net is likewise useful in taking winged insects when at rest upon the ground, by simply spreading it over them. When you use it to beat into, as above recommended, you must take both the sticks in one hand, and extend it by crossing them as much as you can. In the absence of this, a common umbrella, or even a sheet of stiff paper which you may carry folded in your pocket, are no bad substitutes. When your object is beating the bushes, bring your fly-net, &c. rapidly under the branches you mean to operate upon, or the insects will fall from them to the ground before you are prepared.[Pg 533]
Under this head I may mention a very ingenious net for taking Lepidoptera, particularly butterflies, invented by Dr. Maclean of Colchester, which I would call Maclean's elastic net. It is constructed of two pieces of stout, split cane, connected by a joint at each end and with a rod which lies between them, in which a pulley is fixed; through this a cord fastened to the canes passes; a long cane with a ferrule receives the lower end of the rod and forms a handle; and to the canes is fastened a net of green gauze. Taking the handle in your right hand, and the string in your left, when you pull the latter the canes bend till they form a hoop, and the net appended to them is open; when your prey is in it, relax the cord, and the canes become straight and close the mouth of the net, keeping them close with your left hand, you may soon disable your prey with your right. Dr. Maclean has scarcely ever found this net fail.
5. Another instrument which should be constantly in the hands of the Entomologist is the forceps[1561]. This is particularly useful for catching Diptera and Hymenoptera chiefly while at rest on the leaves and flowers of plants. Both these tribes are usually too agile to be taken by the hand alone, which besides without this contrivance would be exposed to the stings of many of the latter. The leaves of the forceps should be octagonal, five or six inches in diameter, and covered with green gauze, or rather very fine catgut, which will enable the head of a lace-pin to pass through it. You must direct your artisan to make the joint of the handle nearer the rings for the finger and thumb than to the leaves, or the instrument will not open well. An old pair of curling-irons[Pg 534] might be made into very good handles; but the hoop to which the catgut is fastened should be brass, or if iron it ought to be painted to secure it from rust. Some make the leaves of the forceps round; but when an insect is perched on a wall or any vertical surface, it has less chance of escape if you can apply a straight side to its station. The Germans use a much longer and larger instrument of this kind, having leaves of ten or twelve inches in diameter, which they use to catch Lepidoptera when settled on plants. When you aim at an insect with your forceps, you must expand the leaves as much as possible, and cautiously approach your prey; and when within reach, close them upon it suddenly, including the leaf or flower on which it rests. As these are sometimes bulky, and prevent the instrument from shutting closely,—that the included insect may not escape, it is often necessary to use the other hand to bring them together, when the pressure of the finger and thumb soon disables it.
6. As the waters, whether running or stagnant, as well as the earth and the air, teem with insects, you must likewise be provided with a net of a different description from any of the preceding, that you may fish them out. It may be made of fine canvass, just deep enough to prevent the insect from jumping out, and fastened to a brass hoop five or six inches in diameter, not perfectly circular, but having the segment of a circle cut off anteriorly, so that it will apply well to a flat vertical surface; and fitted posteriorly with a socket, to receive the end of your stick; or, what is better, with a screw, which will securely fasten it to it[1562]. In using this net, different[Pg 535] modes may be adopted. You may either watch the motions of an individual insect, and secure it by darting the net beyond it and drawing it towards you; or by placing it quietly under it, and then elevating it suddenly; or you may push your net at random along the margins of the pools and rivers amongst the weeds, &c.; amongst the duck-weed (Lemna) on their surface, or the mud at the bottom; and when you examine its contents, you will often find valuable captures. I have thus sometimes got rich booty in the most unlikely places;—such as Hydræna longipalpis, and an allied nondescript species, &c.; and by fishing amongst Zanichellia palustris, Hæmonia Zosteræ. If at any time you do not happen to have your water-net with you, with a common rake you may take the duck-weed from the surface of a pool, and upon examination you will often detect amongst it many minute water-beetles.
But besides all these implements you will find your finger and thumb a very handy forceps when insects are stationary or walking upon the ground; and even when flying, minute ones that you would not otherwise meet with may be taken by the palm of your hand, wetted with saliva, if, when you see them swarming in the air, you pass it to and fro amongst them. When such are stationary, or moving on the ground, on rails, the trunks of trees, &c., the fore-finger, so wetted, will often best secure them: but if they are perched on a summit or a vertical surface, before you approach near enough to alarm them bring forward quietly your bag-net, and hold it so that they may fall into it, if they attempt by falling to escape you. Other methods of entrapping insects may also be pursued with success. A table-cloth[Pg 536] spread on the grass in the open parts of a wood I have known allure several scarce insects: a lady's white dress is equally attractive. An old mattress, laid at night upon a grass-plat, if suddenly reversed in the morning, will supply the Entomologist occasionally with good Coleoptera. No better trap for the Silphidæ, Dermestidæ, &c., than a piece of carrion, a frog, or mole, &c. The numerous insects that inhabit excrement of every kind, especially that of the cow and the horse, may be best taken by immersing their pabulum in water: for this purpose, let a boy carry a spade and pail to the scene of action, and filling the pail nearly full of water begin the operation, and all the insects lurking in the submerged dung will come to the surface, and may be easily taken.
Another object of the collector of insects, when he has once entrapped them, is to bring them safe home. The Entomologists on the Continent, I believe, generally transfix their prey, of whatever Order, with a pin, as soon as they are caught: but as hard ones, such as Coleoptera, Hemiptera, &c., may be destroyed without injury by immersion either in spirits of wine or boiling water; and as large beetles, if transfixed (not to mention the unnecessary cruelty of so serving them), are apt to whirl round upon the pin in spite of any precaution, and injure themselves, and destroy other insects that are in their way, it seems best to kill them by other more effectual methods. With regard to those that would be injured by immersion in any fluid, as the Lepidoptera, Hymenoptera, Diptera, &c., they must be secured as soon as taken; and after having disabled them as much as you can without injuring them, by pressing the trunk below the wings with the finger and thumb, they should[Pg 537] be transfixed and put into a pocket-box lined with cork. Some use an oblong deep chip one, with paper pasted over it, and lined at top and bottom, the top being convenient for setting small moths. But this you will find not easy to open when you have an insect in one hand; and it is too deep for the pocket. I generally use a mahogany one, about 7½ inches by 4½ and 1¼ deep in the clear, corked only at the bottom, and opening by pressing a spring, which can be done with one hand. This will contain as many of the above insects as you will usually take in a day's excursion. When travelling, you should provide yourself with larger store-boxes, to receive at night the fruit of the day's hunt. These may be 18 inches square and 2½ deep, corked at top and bottom; which should be of equal depth, and fit very closely, to keep out Acari, &c. Entomologists have recourse to various ways of bringing home insects for immersion. For the larger ones, you must be provided with a number of small boxes, the lids of which are not liable to come off in the pocket. If it can be done, it is best to have only a single insect in a box. If you have several, those that are predaceous in their habits will probably devour the rest: and besides, if you open a box to put in other insects, generally one or two of those before imprisoned in it will make their escape. It is best to put the boxes containing an insect in one pocket, and the empty ones in another. If your boxes are numbered, in a small memorandum-book, which you should carry for the purpose, you may make any remarks as to the food, station, and habits of any insect you may take, inserting against them the number of the box or phial that contains it, and it will be ready for future use. For the smaller[Pg 538] beetles, &c. a number of phials, with their rims ground down and the mouths well fitted with corks, must be provided; but for those you do not wish to keep separate, a wide-mouthed phial filled with spirits of wine, which soon kills them, is the best receptacle. I have found, when at a loss, a piece of elder, with the pith taken out to a sufficient depth at each end and each mouth stopped with a wooden plug, a useful insect box. As numerous insects inhabit the various species of Boleti, if you go where these are to be found, unless you are a very agile person and expert at climbing, a boy with a short light ladder will be no useless accompaniment.
Something may be said in this place upon the dress with which the Entomologist should provide himself. I shall not recommend to you, in imitation of the insect-hunters in the vignette to Reaumur's second volume, to put on a bag-wig and a velvet court-dress; but the plain fustian jacket with side and other pockets used by English sportsmen will very well suit your purpose; only let the pockets be sufficiently ample: have also an inside one fixed on the left-hand side to receive your forceps. You may also have a bag like a shooter's, or an angler's basket, which may contain your nets till you want to use them. With all your implements about you, you will perhaps at first be stared and grinned at by the vulgar; but they will soon become reconciled to you, and regard you no more than your brethren of the angle and of the gun. Things that are unusual are too often esteemed ridiculous; and the philosopher whose object is to collect and study the wonderful works of his Creator, is often regarded by the ignorant plebeian as little short of a madman.[Pg 539]
Such is the apparatus to be provided by the entomological Nimrod: it is not often, however, that it will be necessary, except in distant excursions, to encumber and disfigure yourself with the whole. Even in this pursuit more may be effected by a judicious division of labour, than by grasping at every thing at once; and your acquisitions will in the end be more numerous, and your acquaintance with them more intimate, if at one time you devote yourself to the woods and hedges, another to the plains and meadows, a third to any heaths in your vicinity, and a fourth to the collection of aquatic insects whether from stagnant or running waters:—having thus chosen the scene of action, you may equip yourself accordingly. You will of course, though in pursuit of a particular description of game, not neglect to seize any other insects that fall in your way; but for this purpose it is unnecessary to be always provided with a certain instrument. Dr. Franklin used to say that a man would never make a Natural Philosopher, who, in performing his experiments, could not saw with a gimblet or bore with a saw; and so we may say, he will never make an expert collector of insects, who on occasion cannot fish with his hand or forceps, use his hat or an old letter to beat his game into, or, in the absence of boxes or bottles, contrive to secure his captures in small pieces of paper twisted up. Sparrman, when at the Cape, was wont,—to the no small amazement of the wondering natives, who took him for a conjurer,—to stick his impaled insects round the outside of his hat[1563]: and though I should not recommend such an exhibition in a civilized region, it[Pg 540] has often struck me that the cavity of a modern hat, if lined with cork, might be made a very useful receptacle for these animals in a long excursion. Indeed, an active Entomologist is never at a loss for an apparatus, but often makes his most valuable captures when unprovided with other instruments than his hands and eyes. A careful survey of the trunk and branches of trees and shrubs, particularly of the underside of their leaves, seldom fails to detect many a lurking moth or beetle, which may be transfixed or otherwise captured with little trouble by an expert hand. In this way an ingenious collector, who scarcely knew what a net of any kind was, told me he had made his whole collection, which was rather extensive. It is, in fact, only by thus detecting them when reposing, and adroitly shutting them up along with the leaf on which they sit, in a box, that minute moths (whose beauty and freshness the slightest handling destroys) can ordinarily be taken without being injured. The boxes containing them should afterwards be exposed to the action of heat, a low degree of which will destroy them.
Enough has been said upon the best modes of catching insects:—I shall next attempt to give you some further instructions as to the most effectual one of destroying them when caught, and to point out how you are to proceed with them after they are dead. As I sufficiently rebutted the charge of cruelty in a former letter[1564], it will not be necessary to enter here into that subject.
I have before recommended to you the use of spirits[Pg 541] of wine, and shall here repeat my recommendation; for after several years trial, I am of Böhm's opinion, who had tried it nine years[1565], that it is superior to any other method; particularly, because it not only effectually kills the insects, and they may be put together into it while you are collecting, if you have no reason for keeping them separate, of all sorts and sizes, in a wide-mouthed phial, without danger of their devouring each other: but when you come home wearied with a long day's hunt, you may let your insects remain in it without injury till the next morning. In collecting beetles abroad, when there is a want of store-boxes the readiest way is to put them into a wide-mouthed bottle or jar filled with any spirit, and send them home in it: some few may lose their colours, or become greasy; but in general they will receive little injury. This method saves room, and avoids the risk of breakage. The derangement which some hairy species sustain from this method may be readily repaired by brushing them with a dry camel's-hair pencil.
When you wish to take the insects you have immersed in spirits out of the phial, you must strain its contents through a piece of muslin, return the spirit into it for future use, and spread the insects separately upon blotting-paper, to absorb the moisture remaining about them. With regard to such as you have in boxes or phials without spirit, these must be immersed in a basin of boiling water. First empty into it the contents of your boxes,[Pg 542] and next, those of your phials; giving each, before you take out the cork, a smart rap, that the insects adhering to the latter may drop to the bottom: or you may immerse the phial itself, with the cork in, which soon destroys them, and is the safest plan. This done, with a camel's-hair pencil or feather take them out of the water, lay them upon blotting-paper to dry, and put them by for a few hours till you have leisure to impale and set them.
Those insects that are caught by the forceps would for the most part escape you, were you to attempt to get them out before you had transfixed them. You must therefore do this while the leaves of the instrument are closed; and then opening them, and taking the pin by the point, the head will readily pass through the catgut; and thus you may safely take, and more effectually kill your specimen by pressing it as before directed. With respect to Lepidoptera, it is necessary to disable them while yet in the fly-net, immediately after their capture. To effect this, while one hand holds both the rods of the closed net, with the other stretch the gauze so as to confine your insect within a narrow space; bring its wings into an erect position, and prevent its fluttering: which being done, with your finger and thumb give its breast a strong pinch below the wings; and then unfolding your net, and taking it up by one of its antennæ, place it between the finger and thumb of your left hand, stick a pin through it, and deposit it in your pocket-box.
But though nipping the breast will kill many small Lepidoptera, the larger ones will live long after it; as will likewise many Neuroptera, Hymenoptera, and Diptera: and besides this, in some Bombycidæ the thorax presents a very conspicuous and interesting character, which[Pg 543] renders it desirable, in order to avoid the damage or derangement occasioned by pressure, to transfix them without it. To dispatch these effectually, you will find the following apparatus very convenient. Fix in a small tin saucepan[1566] filled with boiling water, a tin tube consisting of two pieces[1567] that fit into each other; cover the mouth of the lower one[1568] with a piece of gauze or canvass, and place your insects upon it; then fix the upper one[1569] over it, and cover also the mouth of this with gauze, &c.; and the steam from the boiling water will effectually kill your insects without injuring their plumage. There is another more simple mode of doing this, the apparatus for which may be met with every where. Fix a piece or two of elder, willow, or any soft wood, with the bark on, across the bottom of a mug, and on this stick your impaled insects; invert the mug in a deep basin, into which pour boiling water till it is covered, holding it down with a knife, &c., that the expansion of the included air may not overturn it. In two minutes, or less, all the insects will be found quite dead, and not at all wetted. If the sticks do not exactly fit, they may be wedged in with a piece of cork. Professor Peck, who used to put minute insects into the hollow of a quill stopped with a piece of wood made to fit, killed them instantaneously by holding it over the flame of a candle.
Having killed your insects, your next object should be to prepare them for your cabinet. First, place by you a pincushion well stored with lace-pins of various magnitudes and lengths: for most insects those nearly an inch in length, for large ones, those that are thicker[Pg 544] and longer, but for Lepidoptera, a stouter kind, as short whites, are best. Next, take the Coleoptera and Hemiptera that, as before directed, you have laid by on blotting-paper after immersion, and begin your operations, selecting the largest first. The pin should be stuck through the middle of the right-hand elytrum[1570], and about one third of its whole length should emerge above the insect. Some foreign collectors, probably having in view its more convenient examination with a microscope under the glass of a drawer, bring it nearer the head of the pin: while the English ones, on the contrary, studying the most ornamental position of their specimens, leave only enough of the point free to fix them safely in their drawers[1571]. Both these methods are open to objection. When the insect is too near the head of the pin, it is difficult to fix it in your cabinet without bending the wire; and there is danger, without great care, of injuring the specimen when you put it in or take it out. Again: When the legs of your insect rest on the surface they collect the dust and dirt, are very liable to be broken, and the length of the pin above it is inconvenient when you have occasion to examine any one under a lens. Lepidoptera, however, which are never thus examined, may always be transfixed in this way, which sets them off to the greatest advantage.
Some insects, especially of the beetle tribe, are so extremely minute that it is next to an impossibility to get a pin through them without injuring, and often destroying them. By using fine needles, or very slender pins[Pg 545] manufactured on purpose, this difficulty might perhaps be surmounted; but the needles will be subject to rust, and the pins, I know by experience, cannot be fixed in cork without difficulty. For such minute insects, therefore, by far the best mode is to gum them on small pieces of card, which may be stuck upon a pin. Talc, which admits the underside of an insect to be seen through it, has been used for this purpose; and where you have only a single specimen, a thin small lamina of it would answer well; but ordinarily I should recommend the former mode. Your pieces of card, which must be small, may be either oblong and cut at the corners for neatness, with a couple of specimens gummed upon each, one on its belly and the other on its back; or you may cut little narrow card wedges, about four lines long and terminating in a point, upon which you may so gum your insects as to show the principal part of the under side, as well as the upper side of its body. Common gum-water made rather thin, with a very little glue mixed with it, will answer your purpose very well: it should be thinly spread on the card with a camel's-hair pencil, and then the insect placed upon it. With the same implement, if it has not been killed too long, before the gum is dry you may expand its antennæ, palpi, legs, and wings, &c. If you want to remove a specimen gummed on a card for any purpose, it is easily effected by plunging it into hot water.
Other insects may be transfixed through the thorax or upper side of the trunk; as also those Coleoptera, Orthoptera, and Hemiptera, whose wings you are desirous of expanding; only you should be careful that your pin passes through them behind the prothorax.[Pg 546]
Having impaled your insects, the next thing to be done is to set them. The best time for doing this is not till they have begun to stiffen, but before they are become quite stiff. If attempted soon after they are killed, the parts, unless you keep them in the intended position by means of pins or braces, will not retain it; and if after they are become too stiff, they are liable to be broken. Not only should the antennæ and palpi be extended so as to be readily seen; but the legs, and often the wings, ought to be placed in their natural position; all of which tends much to the beauty of your specimens, and adapts them for more ready examination. But as this operation requires time, and beauty and regularity may be purchased too dear if at the price of hours called for by science, you may be left to your own discretion in this business, only you should always with a pin expand the antennæ and palpi if possible. You might, however, both save your time and have your insects neatly set, if you would take the trouble to instruct some acute and handy youth in your neighbourhood in the modus operandi, and devolve this department upon him: and as none are quicker and more expert in capturing insects than boys, he might also assist you in your hunting expeditions.
I do not mean, however, to leave you at liberty with regard to the setting of Lepidopterous insects, which not only have a much worse appearance than those of other Orders if their wings be not regularly and uniformly expanded, but require it for the proper display of their characters. The necessary apparatus consists of a piece of cork about nine inches long, four broad, and half an inch thick, which should be made perfectly smooth, with[Pg 547] a piece of white paper pasted over it; and of several narrow slips of card or braces, tapering gradually to a point, of different lengths, from half an inch to two inches or more, with a pin fixed in each at the broadest end. Thus provided, you may proceed to action. But you must first decide whether, like the continental Entomologists, you will set your Lepidoptera horizontally; or, like the British, with their wings declining obliquely from the body. If you prefer the former method, the body must be let into a groove, and the wings expanded as flatly as possible, the anterior margin of the primary pair being brought forward so as to project beyond the head. But as this usually gives the insect an unnatural and formal appearance, I apprehend a man of your taste will prefer the mode adopted by your compatriots, the collectors of Britain, who in setting make the wings form an angle, varying according to the size and characters of the insect, with the body, and do not bring the anterior wings so forward. The wings of butterflies however, in order to appear at all natural, should be set more horizontally. Which fashion soever you prefer, the mode of operating is nearly the same; only that the English plan, except in the case of some large-bodied moths or hawk-moths, requires no groove in the setting-board. After you have stuck the insect upon the cork so as to bring its body close to its surface, stretch the anterior wing with a needle fixed in a handle, or a camel's-hair pencil, applied to the joint at the base, sufficiently forward, and then confine it by means of one of your card braces:—next, do the same by the opposite wing. Afterwards expand the posterior wings, which must not be separated from the anterior so as to leave any interval between them, and fix[Pg 548] them with braces. When you are become expert, you will find, if the fly is not large, that a single brace will be sufficient for each pair of wings[1572]: but sometimes, if the card be not sufficiently stiff, you may confine it by a pin near the point. You must be careful in expanding the wings that each is brought equally forward. Lastly, give the antennæ their proper position, and if necessary confine them with braces; and leave your specimen in an airy situation to dry and stiffen. In a few days the braces may be removed, and the specimen transferred to the cabinet. When you put them away to become stiff, you must be careful to place them and your other insects at night where earwigs cannot come at them; for in sultry weather these animals will often then attack and spoil them.
It is obvious that this process can only be performed while the joints and ligaments of the insect are still flexible; so that small species, in warm weather, will often be immoveably rigid before you can have an opportunity of setting them. On this account collectors usually set minute moths as soon as taken, which can be readily done on the lid of a cork-lined box. But fortunately both these, and specimens which have been dried for years, may be relaxed and rendered pliable by a very simple process. Fill a basin more than half full of sand, and saturate it with water; pour off the superfluous water, and cover the sand with blotting-paper: into this stick the insects you wish to relax, and covering the basin closely, leave them there for two or three days, according to their size; and the evaporation will render them sufficiently flexible for expansion or any[Pg 549] other purpose. Beetles may be relaxed by plunging them for a short time in warm water or spirits of wine[1573].
Many moths of the tribe of Tinea L. are so extremely minute, that it is almost impossible to set them without defacing their characters: indeed, the trunk of some is so small as not to admit being pierced by a pin. These, therefore, it is adviseable merely to gum upon card, expanding their wings (which the gum will easily retain in their proper situation) with a camel's-hair pencil. If you have two specimens, you may fix one in the natural position when at rest,—a method I should recommend with respect to other Lepidoptera, and indeed insects in general. Pezold advises that, by way of contrast, white card should be used for dark-coloured species of these little moths, and black for such as are pale. As the wings of different Coleopterous groups, as well as those of Hymenoptera, Diptera, &c., vary in their neuration[1574], you should, whenever you can, set open the elytra and expand the wings of one specimen at least in each group, which will be very important to you in making out the characters of your genera.
When sufficiently dried, your insects should be transferred from the setting-boards, either to their place in your cabinet or to the store-box before described, till you have leisure to investigate them.
However tedious some of the foregoing manipulations may seem, they are in fact much less so than those required in several other branches of Natural History, where, in addition to the labour of catching, the nice and[Pg 550] difficult task of clearing the skeleton of its muscular covering, and its internal cavity of its contents, and then of stuffing it and replacing its perished eyes by glass ones of the proper colour, is a necessary process with every individual. Happily the Entomologist, from the smallness of his game and the nature of their integument, is usually spared this labour. There are some few insects, however, in which a process in some degree analogous is requisite, if the beauty of the specimens be a consideration. Thus the abdomen of dragon-flies is very apt to lose its colour, and that of the Meloës to shrink up, if left in their natural state: these therefore should be eviscerated; which may be done by slitting the abdomen longitudinally on the under side, then carefully removing its contents, and stuffing it with cotton. In the former, a small straw or stalk of hay may be used, which will prevent the fractures to which that part, when dry, is so liable. Spiders, and a few apterous genera, as well as almost all larvæ, as they usually shrink up, in drying, into a shapeless mass, destitute of every character dependent on colour or form, require to be preserved in a different manner. They may all be very well kept in rectified spirits of wine mixed with water, in the proportion of three parts of the former to one of the latter. Each, suspended by a thread, should be put in a separate very small labelled phial. Larger spiders, such as Mygale aviculare, &c., when suffered to dry, though the abdomen shrinks, do not wholly lose their characters, and are often kept in cabinets: but if preserved in spirits, they may be put into larger wide-mouthed bottles, suspended at different heights, with a label on the outside opposite to each species. Mr. Abbott of Georgia had an excellent method[Pg 551] of preserving caterpillars, so that his specimens retain their colours and other attributes, and look as if they were alive. I am not acquainted with his process, but the following will answer very well.—The animal must first be killed by immersion in spirits of wine; next you must eviscerate it, which is best effected by gradual pressure of the finger and thumb. You must begin at the head, and so proceed till all the fluid contents of the body have passed out at the anus, which you may enlarge with a fine pair of scissors, being careful not to injure the anal prolegs. When you have cleared the skin as much as possible, introduce a fine glass tube, or a piece of hay or slender straw into the anus, round which, as near to the extremity as may be, pass loosely a fine thread: then blowing through the tube, when the skin is fully inflated withdraw it, at the same time pulling the thread tight and securing it by a knot. The caterpillar will now exhibit its proper shape and colours; to retain which, all that is necessary is to hold it near the flame of a lamp until perfectly dry, which will be in a few minutes, when it may be placed in the cabinet along with the imago to which it belongs[1575].
Although a very large proportion of the insect inhabitants of any country may be captured in their perfect state by the active Entomologist, yet there is no small number of them that probably he may never meet with in that state, and to secure which he must have recourse to other methods. He can procure pupæ by digging for[Pg 552] them in woods, under trees, &c., as above directed[1576], keeping them in some of their native earth till they are disclosed; or he must collect larvæ, and breed them; for which I shall now give you some instructions.—The insects we are particularly concerned with under this head are the caterpillars of Lepidoptera and of the saw-flies (Serrifera). If, however, in our entomological rambles we discover the larvæ of insects of other Orders upon their appropriate food, we may often attempt to breed them with success: but as you will seldom thus get species that you will not also meet with in their imago state, and the general directions for breeding will include almost all, I shall principally consider the best mode of breeding caterpillars, and pseudo-caterpillars. The first thing is to collect them. In beating the trees, bushes, and plants, while hunting for Coleoptera, &c., the Entomologist will often displace caterpillars, which, if unknown, he should put into a pill-box with a portion of their food: but Lepidopterists often sally into the woods, &c., for the express purpose of collecting these only. When engaged in this employment, the best plan is to take a sheet with you, and when you mean to beat the branches of any tree, place it as near them as you can, upon four or more sticks fastened in the ground, so as to leave the upper surface concave, and it will receive the falling caterpillars when you beat. If you aim at the pseudo-caterpillars of the Cimbicidæ, you must turn your attention principally to the different species of sallows and willows (Salix). Your spoils you will put into boxes with their food, as above directed, to bring them home.
[Pg 553] There are several kinds of boxes recommended to receive them and breed them in. If your only object is to get the perfect insect, a cubical box of moderate dimensions, glazed in front or on one side to enable you to watch their proceedings, with the other sides and top fitted with fine canvass for the admission of air, will very well answer this purpose; or your box may be canvassed all round, with a door in front[1577]. In this you may place a small garden-pot filled with earth, with a phial of water plunged in it to receive the insects' food. This may be moved, when you wish to change the water, without disturbing the earth, which should be kept somewhat moist. The earth is for those caterpillars whose pupæ are subterranean. But as you will probably wish to proceed scientifically, and ascertain precisely the moth that comes from each caterpillar, I should strongly recommend to you a box invented by Mr. Stephens, which he describes in a letter to me in nearly these words:—"The length of the box is 20 inches, height 12, and breadth 6; and it is divided into five compartments. Its lower half is constructed intirely of wood, and the upper of coarse gauze stretched upon wooden or wire frames: each compartment has a separate door, and is moreover furnished with a phial in the centre for the purpose of containing water, in which the food is kept fresh; and is half-filled with a mixture of fine earth and the dust from the inside of rotten trees; the latter article being added for the purpose of rendering the former less binding upon the pupæ, as well as being highly important for the use of such larvæ as construct their cocoons of rotten wood. The chief advantages of a breeding cage of the above[Pg 554] construction are, the occupation of less room than five separate cages, and a diminution of expense; both important considerations when any person is engaged extensively in rearing insects. Whatever be the construction of the box, it is highly necessary that the larvæ be constantly supplied with fresh food, and that the earth at the bottom should be kept damp. To accomplish the latter object, I keep a thick layer of moss upon the surface, which I take out occasionally (perhaps once a week during hot weather, and once a fortnight or three weeks in winter), and saturate completely with water, and return it to its place: this keeps up a sufficient supply of moisture, without allowing the earth to become too wet, which is equally injurious to the pupæ with too much aridity. By numbering the cells, and keeping a register corresponding with the numbers, the history of any particular larva or brood may be traced."
In attending to your insects in their cells, your expectations will sometimes be disappointed, when, instead of a butterfly or moth, you find only an Ichneumon. But this you must not regard as all misfortune; for by this means you will be better instructed in the history of each species, and learn to the attack of what enemies it is exposed: and thus you may get many species of these parasitic devourers of insects that you would not elsewhere meet with. If your caterpillars, however, appear to be of a rare kind, you must watch, and often examine them; and if you discover black specks upon any one, that appear unnatural or like nits, they may be extracted, Mr. Haworth assures us[1578], by a pair of small pliers; and if the operation is adroitly performed, the caterpillar will recover[Pg 555] and do well. You will often meet Lepidopterous larvæ travelling over roads and pathways: at such times they have usually done feeding, and are seeking a spot in which they may assume the pupa with safety. These you may place in one of your cells, and they will select a station for themselves. You must be careful frequently to examine the boxes in which you have pupæ, that you may take the imago as soon as it appears, and before it has had time to injure itself in attempting to escape. I mentioned to you on a former occasion Reaumur's experiments to accelerate the appearance of the butterfly[1579];—there is another still more remarkable, to which he had recourse for this purpose: it was by hatching his pupæ under a hen!! You will wonder, perhaps, how this could be effected, and be disposed to maintain that the pupæ must be crushed by the weight of the brooding animal. How did the ingenious and illustrious experimentalist prevent this? He prepared a hollow ball of glass, open at one end, about the shape and size of a turkey's egg. Having several chrysalises of the nettle-butterfly (Vanessa Urticæ) suspended to a piece of paper, he cut out some of these singly, with a square portion of the paper attached to them, and covered with paste the side opposite to that from which the chrysalis was suspended: these he introduced into the ball through the aperture, placing them as near to each other as possible, taking care so to apply the pasted surface to the inside of the ball, that when the side to which they were fixed was uppermost they all hung as from a vault. This being done, he stopped the aperture with a linen plug, but not[Pg 556] so completely as to cut off all communication with the atmosphere: he next placed the egg under a hen that had been sitting some days, who always kept it at the side of the nest, where it nevertheless derived benefit from her incubation. After the first day its interior was covered with vapour transpired by the chrysalises. Upon this Reaumur took the egg, and removing the linen plug it soon became dry again: he replaced it under the hen, and no vapour afterwards appeared. In about four days the first butterfly ever hatched under a hen made its appearance; it would probably have required fourteen under ordinary circumstances. He tried the same experiment with some Dipterous pupæ; but the heat was too great for them, and they all perished[1580].
Having properly prepared and set your specimens as above directed, the next step, when they have remained a sufficient time to be perfectly dry, is to place them in your cabinet. If you collect foreign insects as well as British, you may either preserve the latter in a separate cabinet, or keep both in the same, distinguishing the indigenous species by a particular mark. The letter B in red ink, if the pin which transfixes the insect be run through it, or, in the case of Lepidoptera, placed before the specimen, would be a very distinct and sufficient indication of them. The drawers of your cabinets should be about 18 inches square, and from the glass to the corked bottom about an inch and a half in depth: but the larger Dynastidæ, as Megasoma Actæon, &c., will require two inches. The frame of the glass should be[Pg 557] rabbeted underneath; and parallel with the sides of the drawer, but a little lower, there should be inner side-pieces fixed, so as to form a cavity all round of a proper width to closely receive the rabbet, and likewise to contain the camphor for preserving your insects from the attack of mites, &c.; to emit the scent of which, many holes should be bored in the side-pieces. Each cabinet may contain forty of these drawers in a double series, protected by folding doors; and you may place one cabinet upon another, if your space admits it. You will find a tool used by bell-hangers for cutting their wire very convenient to behead or otherwise curtail the pins, as those with which foreign insects are transfixed are often too long. If you cut them off below the insect, cut them obliquely, which will leave a point that will enter the cork.
When your drawers are smoothly corked[1581] and neatly papered, first divide each transversely by a full black line; parallel with this, on each side, draw a line with red ink: then, for arranging your insects, draw pencil lines, which are easily obliterated, at right angles with the others, according to the general size of the insects that are to occupy them. Insects look better thus arranged in double columns, than if the pencil lines traversed the whole width of the drawers. In arranging them, you may either place them in a straight line between the pencil lines,—which I think is best,—or upon them. You will begin your columns from the red lines in the middle, and not from the sides of the drawer; thus the heads of those on one side of it will be in an opposite[Pg 558] direction to those on the other. Where your pins are very fine and weak, you must make a hole first with a common lace-pin; otherwise, in forcing them into the cork, they will bend. In labelling your specimens, you should stick the appellation of the genus or subgenus with a pin before the species that belong to it. As to the species themselves, you may either number them 1, 2, 3, &c., sticking the pin they are upon through the number, and denoting them by a corresponding one in your catalogue; or you may at once write the trivial name, with the initial of the genus upon a label transfixed in the same manner. Lepidoptera cannot easily be arranged in columns. Perhaps if squares, corresponding with the size and number of the specimens of any given species you wish to preserve, were made with pencil, a label of the trivial name of the species, or a number being placed at its head, it would be as good a way as any other. But every one must be left to his own taste in these matters. Wherever you can, procure a specimen of each sex of an insect, and where important characters require it, let some of your Lepidopterous specimens exhibit the under side of the wings.
In arranging insects in your cabinet, if you wish to have it scientific, as much as the nature of the subject will admit, follow the series of affinities; but you may reserve a few drawers to place in contrast analogous forms. As your numbers of species increase you will have to alter your arrangement; but as pencil lines are easily rubbed out, this will occasion you less trouble than if they were drawn with ink. You should always be careful under each genus to leave space for new species.
As certain Acarina, Tineidæ, Ptinidæ, &c., prey upon[Pg 559] dead insects, you will of course wish to know how they may be kept out of your drawers, or banished when detected there. Camphor is the general remedy recommended. The cavity closed by the rabbet of the glass frame affords a good receptacle for this necessary article: put some roughly powdered into each side, and be careful to renew it when evaporated. This will generally preserve your insects, as will be seen from the result of the following experiment.—Some insects in a chip box having become much infested by mites and Psocus pulsatorius, I placed under a wine-glass several of each along with roughly-powdered camphor: at the end of twenty-four hours the mites were alive; but at the end of forty-eight they were all apparently dead, and did not revive upon the removal of the camphor. The specimens of Psocus all appeared dead in an hour, and never revived. If the camphor be put only into one side of a drawer, and in a lump, though perhaps it may keep out mites, &c., it will not expel them.
I am, &c.[Pg 560]
An entomologist who aspires to more than the character of a mere amateur, will not be content with filling his cabinet with nameless objects for the sole amusement of the eye; but will also be anxious to acquire some knowledge of what he has collected, and to ascertain by what names, whether indicating their genus or species, they have been distinguished by scientific writers who have described insects either in general or those of particular districts. Thus only can he himself derive profit from any discoveries he may make, or contribute to the further progress of the science[1582].
But in order to accomplish this object effectually, you must remember and practise the Onslow motto—Festina lente:—you must not be too eager to name your species, but begin first with grouping your collection. The only way to acquire, in any degree, a correct knowledge of the Natural System, or of the general plan of the Creator, which is the primum and ultimum of true science, is by studying groups. The knowledge of species is indeed indispensable for the registry of facts and other practical purposes, but the knowledge of groups leads[Pg 561] to a higher wisdom; and indeed it is through these that we best descend to the study of species.
I will suppose you have made yourself master of so much of the technical language, particularly the names and most important attributes of the principal organs of insects, as will suffice for understanding descriptions, or knowing these parts when you see them. I will also further suppose that what was formerly said on these subjects has been sufficiently studied, to enable you without much difficulty or hesitation to say whether any given object belongs to the Class Insecta or Arachnida, or to which of their respective Orders[1583]. You are therefore qualified to arrange your collection into its primary groups. But you have seen that many others intervene between the Order and the genus or species. As the genera of Linné are mostly primary groups of Orders, perhaps, setting aside such insects included in them by him as your eye and their apparent characters convince you have no claim to a place there, your next best step would be to make yourself thoroughly acquainted with them. When you have accurately marshalled and intimately studied these groups, you will probably have acquired an eye and a tact, experto crede, for grouping without book, and may proceed by analysis to resolve your whole collection, as nearly as possible, into as many as nature seems to indicate to you. In doing this you will doubtless at first fall into many errors; but these, practice and a closer examination will in time enable you to rectify. Having thus got your groups as near to nature as you can, you may now have recourse to those[Pg 562] authors, particularly Fabricius and Latreille, who have subdivided the genera of Linné; and you will see which of your groups agree with theirs, detect your own errors, and often theirs, and be enabled to label each of your genera and higher groups, if already known, with its modern appellation. You are now qualified also to enter scientifically into the study of the characters that distinguish groups, and may proceed, wherever opportunity is afforded, to examine the trophi, which may often be displayed sufficiently by the means recommended in my last letter[1584]. In this way you may learn also to know your groups as well by character as by habit, and be qualified to trace the gradual progress of nature from form to form; and may look upon yourself as duly prepared to put the last hand to your labours, and proceed to the examination of species.
It will have occurred to you, in making out your genera or lowest groups, that some consist of a vastly greater number of species than others. It seems advisable therefore, when you apply yourself seriously to ascertain what described ones your cabinet contains, to begin with those genera which appear to be poor in them; for here your labour will be comparatively light, from the small number you will have to examine; and you will become practised in the employment before you are called upon to attack those that overflow. Had Fabricius and other describers of species taken the trouble to subdivide the larger groups, as might easily have been done, into more genera and subgenera, the student would have been spared a most discouraging labour. To be obliged[Pg 563] to compare a single individual with the descriptions of from 100 to 300 species[1585], to ascertain its name, seems enough to make you start aside with horror from the employment, and be content that your species should remain unnamed, rather than expose yourself to such a waste of time and patience. But to lessen your alarm and encourage you to proceed, I must observe to you, though in a few instances it may be necessary to advert to the description of every single species in a section, yet that this is seldom requisite; and where it is, there are many helps to diminish the labour and abridge the process. A large number of insects are characterized by their colour; and it is the practice of all good describers to begin their definition of the species with that which predominates, and then to enumerate the variations from it. Thus, if an insect be all black except the thorax, antennæ, and legs, you will find it thus characterized, "Black: with thorax, antennæ, and legs ferruginous"; and so on. Hence, having noticed the predominant colour of your unknown species, in many genera you may compare it with the descriptions contained in a whole page at a single glance, and only read the further descriptions when the colour agrees. A practised Entomologist will thus investigate his insects with a rapidity which to an unlearned bystander would seem impossible. Though I have instanced colour as being the character most commonly employed in describing species of insects, you will readily conceive that in some tribes other characters afford more prominent distinctions. Thus in[Pg 564] the Dynastidæ and many other Petalocerous beetles, the principal specific character is derived from the horns or tubercles that arm the head and thorax: in Lucanus from the mandibulæ; and in Prionus from the marginal teeth of the thorax. If the insect, then, you want to name belongs to any of these genera, having observed its peculiar characters in this respect, you may ascertain in a very few minutes whether any already described exhibit the same. This facility of investigation can be better acquired by practice than precept, and cannot be attained all at once. The above hints, however, may be of some use; and cannot fail to be so, if you always endeavour to make yourself acquainted by a previous careful examination with the characters of every new insect you acquire,—whether those of form, colour, or sculpture,—before you attempt to discover its name in Fabricius or any other author.
When you have made such proficiency in the study as to be familiar with a few species of each section of an extensive genus, the labour of investigation will sometimes be greatly facilitated by attending to that conformity between the proportions, general aspect, and figure of a known and an unknown insect, which Naturalists express by the name of habit, and which, though easily perceived by a practised eye, is described with such difficulty. Scientific Entomologists in their descriptions have usually taken care to place near to each other, species agreeing in habit. When therefore you know the name of one species, and find another of the same general habit, you may commonly take it for granted that if described at all by your author, it will be placed near that already known to you. Thus, supposing[Pg 565] you are acquainted with that common weevil Cionus Scrophulariæ, and find its near relation C. Blattariæ; instead of comparing it one by one with the 161 species which compose the Longirostres femoribus dentatis of the Fabrician genus Rhynchænus in the Systema Eleutheratorum, you would at once turn to the former, very near which you would without further trouble discover it. Fortunate, would it be, could the Entomologist always depend on thus finding descriptions of allied species in the neighbourhood of each other; but unhappily the most distinguished authors have sometimes violated this important rule, so that we cannot always be certain that any given species is not elsewhere described than in its right place. Fabricius in many instances often removes widely asunder insects not merely related, but which are in reality scarcely more than varieties of the same species[1586]. In fact, the attention of this celebrated author was so distracted by the immensity of the materials he had to arrange, by the distance of the cabinets, in many cases, from each other, the new species of which he undertook to describe, and the rapidity with which they necessarily passed under his eye, that he seems never to have attained any nice perception of the affinities of insects.
You must not conclude, however, that the investigation of a new insect is even to an adept always a work of ease and dispatch. Often, when seemingly ascertained by the rapid process above indicated, a further inquiry[Pg 566] will be requisite; the more detailed description must be read, and figures consulted, before its name can be indisputably determined. In addition to the difficulty arising from the insufficient characters frequently given by Fabricius and the older authors, obstacles arising from their errors not seldom intervene. Thus they have sometimes selected for a specific character,—as in the case of Megachile centuncularis, Nomada ruficornis, and various other insects,—what really only indicates a family. At other times sexual characters common to many,—as in Eucera longicornis, Locusta perspicillata, &c.,—have been had recourse to. In these cases, in order satisfactorily to ascertain your species, you must further consult the synonyms and habitat given by the original describer, especially the figures he has referred to. When all these fail, as they sometimes will, the dernier resort is a reference to the cabinet containing the original specimen from which the description was drawn. British Entomologists possess an invaluable privilege, which their continental brethren may well envy them, in having the most liberal access, indulged to them by the learned President of the Linnean Society, to Linné's collection of insects, from which a large proportion of the species he described may be ascertained[1587]. Several of the cabinets, especially the Banksian,—now the property of the Linnean Society,—from which Fabricius described his[Pg 567] insects, may also still be consulted; and thus many mistakes rectified, which would otherwise greatly mislead[1588].
Though sometimes the limits that separate good species appear at first very slight, and require a practised eye to catch them, yet it occasionally happens that considerable apparent differences may safely be disregarded. The colour of insects,—to which unhappily for want of better characters we are so generally forced to have recourse,—though usually constant, is in some species very variable[1589]. This is the case sometimes with whole colours. Thus Carabus arvensis, Pœcilus cupreus, &c., are sometimes of a copper colour; at others, resemble brass; at others, they are green or blue, and even black. The colour of spots also often varies. In some individuals of Pentatoma oleracea they are pale, and in others red. The number and shape of spots are also often inconstant. Many of the species of Coccinella so abound in these variations, that nothing short of the most careful examination can enable you to distinguish the species from the variety. Insects vary also in size: but as this is never assumed as a specific character, it will not occasion you much trouble. Where the difference in this respect between two specimens is very great, the presumption is that they are specifically distinct. Differences in sculpture and proportion do not always indicate different species; this being sometimes, as we have seen[Pg 568] above, only a sexual character[1590]. Authors also in their descriptions, in this respect sometimes mislead the young student. When Linné calls the thorax of Aphodius erraticus smooth (lævis) he would not expect to find it covered with impressed puncta, and with a longitudinal posterior impressed line. Likewise in describing Chlænius vestitus and nigricornis, Fabricius passes without notice their punctate surface, so different from that of other Harpalidæ. Errors of this kind however, it is but fair to observe, are chiefly to be attributed to the circumstance that both Linné and Fabricius rarely employed a microscope in making descriptions; though no one now attempts this, except where insects are large, without such an aid.
If you ask, How am I to acquire this delicacy of tact which is to decide when the terms of a specific character are to be rigidly adhered to, and when taken with a certain latitude? I answer, In the same way in which a connoisseur attains the faculty of discerning the works of different masters in painting;—by such careful study of your author as will make you master of his style. Thus you will soon perceive in what cases expressions are to be taken literally and strictly, or with some allowance and abatement.
There yet remains more distinctly to be adverted to, the assistance that may be derived in the investigation of insects from figures. Generally speaking, these should never be referred to in the first instance, but be regarded as a resource when the ordinary methods leave the subject of inquiry doubtful. Those who begin their[Pg 569] entomological studies by turning over figures usually end them there, and never attain to that nameless tact in making out insects that can only be the result of patient study. Indeed figures, though often very useful, and sometimes indispensable, can scarcely ever exhibit those nice characters, particularly as to sculpture, that distinguish some insects. Our modern artists, indeed, are remedying this defect of the art, by giving in many cases the thorax or elytrum apart, with all its sculptural peculiarities: but this is not, and cannot be, done so as to represent every one. But though in general figures should be your last resort, I know not whether an exception to the rule may not be advisable with respect to the Lepidoptera, which are more difficult to be intelligibly described than any other order of insects; while a good figure exhibits to the eye all those markings and shades, that scarcely any description can place clearly before the mind.
When every attempt to investigate the name of your unknown species fails, and you have consequently reason to believe that it is undescribed, the best mode you can pursue for retaining that knowledge of its characters, which from your long investigation you must have acquired, is to note them down in your entomological journal, inserting it under its proper genus with a trivial name of your own. Such a journal you will find almost a sine qua non for containing a catalogue of your insects, and to register any observations concerning individuals you may have had an opportunity of making. With regard to this journal, I should recommend to you to get two blank books. One a duodecimo of 200 or 300 pages, to contain the mere catalogue of your insects,[Pg 570] their habitat and localities, or the source from which you derived them. In this you should number the genera in Roman capitals, and the species under each by a figure; leaving considerable space at the end of each genus for the insertion of new species. The other book should be of an octavo size, containing 400 or 500 pages. Under the number of each genus and species you might describe and figure it, if undescribed; if described, note in what it varies from the description, and what characters are overlooked: and in general, insert such observations, with regard to its economy and habits, as you may have had an opportunity of making.—As to foreign insects, wherever you can, upon good authority, be particular in indicating the country and station of each specimen.
I need not say much to you concerning the microscopes you should use for the examination of insects, a common pocket one of three glasses of different powers will answer every ordinary purpose[1591].
We have treated hitherto of insects as we find them now inhabiting our globe: but I must not conclude our correspondence without taking some notice of those that are found in a fossil state. Fossil insects may be divided into those that are found in amber, and those that are found in other substances.
It has been observed with respect to insectiferous amber, that the greater part of the insects found in it exist no longer in the countries that produce that amber, and that in every different locality the insects found in it are[Pg 571] different. Thus the amber of Sicily contains various species of Coleoptera not to be met with in other ambers, while that of the Baltic is rich in Diptera and Neuroptera[1592]. It is further observed, that the insects inclosed in the amber of Prussia, and those figured by Sendelius in his Historia Succinorum, all belong to genera at this time found in Europe[1593]. Insects of the following genera are recorded as having been found in this singular substance: Platypus, Elater, Atractocerus; Gryllus, Mantis; larvæ of Lepidoptera; Trichoptera; Ephemera, Perla, Termes; Formica; Tipula, Bibio, Empis; Scolopendra; and various Arachnida[1594]. In a piece of amber in my collection I find Evania, Formica, Chironomus, and some Arachnida.
Fossil insects have also been found in other substances. Parkinson figures larvæ of Libellulina found in limestone[1595]; some Melolonthæ in slate; a Polistes in schistus; Carabi and Necrobia in vegetable debris: but some of these rather belong to a comparatively modern formation[1596].
I observed in the outset of our correspondence, that we were entering an august temple, exhibiting in its inmost sanctuary the symbols of the Divine Presence[1597]. In proportion as we have penetrated, glory from that Shechinah has more and more shone forth: and whether we have considered the uses of insects, their ways and instincts, their forms and structure, and their arrangement[Pg 572] in a wondrous and complex system, the Wisdom, Power and Goodness of their and our Creator have every where been marvellously conspicuous, and calculated to awaken in us every devotional feeling. If, indeed, we admire and study these little creatures, or any other department of nature, without reference to their Creator, and collect and love them merely for themselves, we shall be in some sense idolaters, and, like the ancient world, put the works of God in his place. But if, while we admire them and store them up and study them, we see in them his glory reflected, and in the creature love the Creator, the study of them, in conjunction with that of the written Word, will be highly beneficial to us, and at the same time that it ministers to our temporal enjoyment will promote our eternal interests.
Taking this view, I cannot better close our correspondence on the subject that has so long occupied us, than in the pious words of one of our most admired poets:
Inter tot et tanta Optimi Creatoris miracula, quæ Regnum Animale tantopere illustrant, vix ulla sunt majori admiratione digna, et Physiologi eruditi introspectione, quam quæ ad generationem insectorum spectant. Quamvis enim inter sexûs organa vertebratorum animalium et insectorum analogia haud parva locum habet; numero tamen, figura et proportione partium, miro modo sæpius differunt; et organa insuper plura in insectis reperiuntur quorum in vertebratis exempla frustra quæsiveris.
Hoc argumentum tractando duo sunt imprimis consideranda, genitalia nempe ipsa utriusque sexûs, et coitus.
I. De genitalibus in genere prima observatio erit, "quo minor horum, habita corporis ratione, moles, eo magis nervorum systema, et cephalicum imprimis ganglium, predominans fit; eo major igitur intellectûs facultas (instincto naturali consociata) reperitur," ut in principibus, Apibus nempe, Formica, &c.[1599] In Hymenopteris, iterum, Dipteris, et Neuropteris, hæc organa maxime retracta sunt; dum in Lepidopteris, Coleopteris, et Orthopteris (quorum insuper mascula et feminea insigniter[Pg 576] inter se congruunt[1600]), magis exserta jacent[1601]. Genitalia plerumque in extremitate postica abdominis sub ano sita sunt[1602], sed in Arachnidis et Libellulinis masculis in basi ventris, in Phalangio sub ore, et in Chilognathis in anteriore corporis parte subtus latitant[1603]. Ubi organa duplicantur, ut testes, semper symmetrica sunt. Non obliviscendum est quòd in diversis generibus habitu externo persimili consociatis, imò in diversis unius generis speciebus genitalia diversa interdum reperiuntur[1604]: sic in Lamellicornibus stercorariis (Scarabæus, Copris, &c.), testes tantummodo sunt duo; in arboreis (Melolontha, &c.) duodecim, et in floralibus (Cetonia, &c.) viginti-quatuor.
Genitalia sunt vel mascula vel feminea.
i. Genitalia mascula, sunt penis; canalis excretorius; vesiculæ seminales; vasa deferentia; testes; prehensores; et semen.
1. Penis[1605] quoad substantiam plerumque membranaceus, at interdum corneus est, et intus cavernosus[1606]; in Coleopteris apice vagina bivalvi vulvam aperiente instructus est[1607]: figura variat admodum, sæpius tamen cylindricus vel subcylindricus est; in Blattis apicem versus sensim attenuatus[1608]; in Cherme Pyri capitatus[1609]; in Vespa vulgari cochleariformis[1610]; in Crabrone bilobus[1611]; in Poliste gallica? incurvus et apice bicornis[1612]; in Sarcophaga carnaria apice spinosus[1613]; in Megachile muraria[Pg 577] difformis[1614]; in Tyrophaga Casei et quibusdam aliis Muscidis, spiralis[1615]; in Cordulia ænea et Phalangio biarticulatus[1616]. Utplurimum nudus est, sed in Tephrite fimbriatus. In insectis proprie dictis simplex est hoc organon, in Scorpionibus autem duplex evadit; quod fit etiam in quibusdam reptilibus, Serpentibus nempe et Lacertis[1617].
2. Canalis excretorius e concursu vesicularum seminalium formatur, et a pene excipitur in quo terminat et cui semen reddit; interdum brevissimus est, ut in Blatta[1618], et interdum iterum prælongus, ut in Blapte Mortisaga, Tyrophaga Casei, et aliis[1619]. Plerumque cylindricus est, musculosus, compactus, et externe tracheis pertextus[1620].
3. Vesiculæ seminales conniventes formant, ut jam dictum est, canalem excretorium communem cujus prolongatio bifida esse videntur; vasa deferentia hinc excipiunt. Interdum vasa hæc ac vesiculæ seminales eodem loco in canali excretorio communi terminant, unde canalis hic tumidior fit[1621]. Vesiculæ supradictæ maxime variant: modo canalem exhibent ventricosum, tortum, implexum, longissimum; modo rectum, breviorem. In plerisque duæ sunt vesiculæ seminales, etiam in Lepidopteris monorchidis; in quibusdam (Tenebrione Molitore, Hydrophilo piceo) quatuor[1622]; in aliis (Dytisco marginali) sex[1623]; et, in Locustis et Blatta, plurimæ[1624]. Breves admodum sunt in Orthopteris et quibusdam Coleopteris[1625]; sed in aliis longissimæ; in Orycte nasicorni vicies, et in Cetonia aurata[Pg 578] ter decies corpus longitudine superant[1626]. In hisce organis semen e testibus per vasa deferentia acceptum ante emissionem elaboratur.
4. Vasa deferentia ita appellantur quia semen e testibus acceptum ad vesiculas seminales deferunt. Ex utroque teste unum vas deferens exit, et si utrinque plures sint testes, ut in Melolontha[1627], Cetonia, &c., omnia ad unicum utrinque canalem formandum confluunt, qui vesiculis supradictis semen reddit: interdum, ut in Lepidopteris[1628], ab his nullo modo separantur, unum canalem aut tubum formantia; sed in aliis penitus sunt distincta[1629]. Ex eodem filo quo contexuntur testes vasa deferentia sæpius deducuntur.
5. Testes organa sunt semen primum secernentia: variant compositione, numero, et figura. In quibusdam (Lepidopteris et Hymenopteris) sunt compacti vasculis visui se subducentibus; in aliis (Orthopteris, Neuropteris, Dipteris, et quibusdam Coleopteris) e vasculis brevibus cæcis variique voluminis conformati sunt, atque tunica densa tenaci vel rete tantum mucoso obducti[1630]; vel iterum ex unico variisque modis tecto canali varie contorto et implexo, qui deduci potest et haud raro massam ovalem trachearum ope contextam refert, conflantur, ut in Coleopteris Prædaceis tam aquaticis quam terrestribus[1631].
Numero etiam variant testes. Quædam Lepidoptera, ut Pontia Brassicæ, item Iulidæ[1632], unico gaudent; pleraque tamen insecta animalia vertebrata hic æmulantur, et testibus instruuntur duobus; in Nepa cinerea et reliquis Hemipteris quatuor vel quinque[1633], in Melolontha vulgari sex[1634], et in Cetonia aurata duodecim[1635], utrinque deteguntur. Interdum ex acinis[Pg 579] pluribus compacti videntur, et bacciformes appellari possunt. In Lamia duodecim glandulæ in utroque teste coalitæ inveniuntur[1636], et in Tenebrione Molitore plurimæ[1637].
Quoad figuram, interdum, ut in Pontia Papilionum genere, spherici evadunt[1638]; in Gryllo pyriformes[1639]; in Ape mellifica oblongi[1640]; lineares et longissimi in Procruste coriaceo, in quo decies longitudine corpus superant[1641]; in Nepa cinerea sub-ovati, et singuli filamento longo varie convoluto et contorto terminati[1642].
In larvis etiam hæc organa detegere est. Sic in eruca Pontiæ quatuor testes sunt utrinque, vel potius unicus ex quatuor serie ordinatis, conflatus[1643]. Hi sensim coacervantur donec in sphæricum testem antea descriptum coalescant.
6. Prehensores[1644] sunt organa figura varia quibuscum mas in coitu feminæ anum corripit et comprimit. Quoddam analogum in quibusdam Mammaliis, Avibus, Piscibus, et Reptilibus[1645] invenitur, sed in insectis maxime conspicui. Eorum situs, numerus; et forma, sunt notandi.
Quoad situm—circa foramen per quem prodit penis sub ano plerumque sunt inserti, sed in Conope cornu prehensorium in segmento ventrali antepenultimo deprehenditur[1646]; et in Libellulinis, præter prehensores anales, par est aliud anum spectans, in secundo ventris segmento pone penis ipsius situm[1647]. Prehensorum numerus minime constans: plerumque duo sunt, sed in Cicada, unicus furcatus tantummodo videre est[1648]; in Lepidopteris variis, Conope, Libellulidis, tres anum armant, difformes tamen[1649]; duo paria Culicem signant[1650], Megachilem murariam[1651],[Pg 580] et Agrionidas[1652]; in Locustis veris intra abdomen retracta sunt hæc organa; in pupa tamen L. morbillosæ, in nostro musæo asservata, quinque apparent; sex in Formicis De Geerius detexit, sed in cognato genere Myrmica, duo tantum[1653]; quatuor paribus postremo Tipula oleracea instructa est. Prehensorum forma multifarie variat, imò haud raro in specie eadem: interdum enim prehensioni soli hujusmodi instrumenta sunt adaptata, aliis diversæ figuræ compressionem efficientibus; interdum et utroque munere funguntur. In Pontia Brassicæ, in qua par unicum, concavo-convexi sunt, deltoidei, intus setis rigidis fimbriati, et apice dente incurvo armati[1654]; in Acrida varia tenues, simplices, recurvi; in Spilosomate lubricipeda, quæ tribus gaudet, laterales sunt concavo-convexi, ovati, dum intermedius brevior est, triangularis et unguiculo armatus[1655]; in Cordulia ænea, et affinibus, duo superiores sunt lineares et undulati, et inferior unicus profunde bifidus[1656]; in Vanessa Urticæ exteriores duo sunt conchiformes, par autem interius unguiforme[1657]; in Culice superiores longiores conici hirsuti, inferiores breviores et ut in præcedente unguem referunt[1658]; in Tipula oleracea, in qua octuplici prehensore anus armatus, valvulæ omnes figura diversæ—par exterius nempe concavum membranaceum reliquos includens, secundum unguiculatum, tertium subclavatum, et ultimum fere lunatum[1659]; in Megachile muraria, inter alios diversos, unum par literæ T formam habet[1660]; in Bombo forceps analis bivalvis est intus ramosus[1661]; et in Panorpa cheliformis[1662].
7. De semine ipso insectorum paucula sunt notanda. Fluidum[Pg 581] est spissum, lacteum, granulis repletum; sub lente punctula numerosa, nigra, oblonga, incurva, in illo deteguntur. Quoad analysin ejus, neque alkalinum neque acidum est, sed quoddam neutrum inter hos intermedium. Ex sanie vel sanguine deoxydato, et durante coitu copiosissime secernitur: in aqua tepida solvitur, et conquassatum fundum petit: spiritu vini rectificato superfuso flocculi quidam formantur[1663].
ii. Genitalia feminea vulva excepta antea tractavi[1664], hæc est tubus subcylindricus, foramine ovali vel lunato ab ano distincto, cum matrice connexus, et per quem semen in coitu transmittitur. In Scorpionibus duplicem esse vulvam affirmatur duobus ovariis connexum[1665].
II. Coitus.—Coitum insectorum tractaturo paucula de lenociniis amatoriis, et aliis ejusmodi, quæ antecedunt, sunt prædicenda. Olfactu mares Phalænarum interdum feminam latentem, uti canis leporem, odorantur[1666]; splendore phosphorico Lampyrides et quorundam aliorum insectorum feminæ maritum ad lectum gramineum prælucent; et huc referri forsan debet plurium cæcus ardor lumina circumvolandi, vel etiam in lumen irruendi; sonus excitat feminas Cicadarum et Gryllinarum[1667], &c. ad amores, et cantu stridulo querelisque amatoriis diem ducit mas cupidus, donec sponsa advolat, et tori foliosi fit haud invita particeps. Sonitu etiam uterque sexus formidati Anobii mutuo sese provocant ad venerem[1668].
In plurimis tamen insectis femina fit modestiæ et pudicitiæ exemplar, et non nisi difficillime et capite averso maris ardori se tradit. In insectorum moribus et œconomia virtutum plurimarum typum quendam et delineationem nobis proposuit Deus O. M., quem imitari nos voluit, interdum jussit[1669]. Sic[Pg 582] excitare nos ad laborem indefessum, ad prudentiam item et amorem erga prolem Formicæ dedit[1670]: Api ad devotam sui consecrationem, et omnium facultatum et virium ad reipublicæ emolumentum, ad obsequium quoque verum erga parentes et regem[1671]; atque ita, ut jam dictum est, in re amatoria insectorum feminæ sæpe speciem præ se ferunt pudoris et castitatis, et virginibus verecundiam, virtutum omnium custodem, et sexûs sui ornamentum maxime proprium, moribus suis prædicant. Hujus modestiæ exemplar insigne præbent Libellulinæ. Œstro amoris concitus, mas feminæ collum prehensore anali triphyllo arripit et avolat, illam quasi prædam secum gerens; sponsæ sic electæ, persuadere in animo est ut caudam suam inflecteret, et ad coitum se daret, quod, illa invita, fieri nequit; maris enim genitalia, ut antea dictum est, in basi ventris sita, feminæ vero in extremo ano; hinc, nolente illa, vix fit coitus, et sæpissime longo et vano labore, huc illuc volando virginem protervam frustra solicitat; sed tandem lacessitus aquas petit, quas sponsæ cauda longa, me teste, sæpius flagellat, donec defatigata, et quasi ex frigido calorem concipiens, demum et sensim caudam inflectit, et se reddit amori[1672]. Araneam ferocem, sævam etiam in amoribus, mas caute appropinquat, et, si blanditiis ejus minus propitiam sese ostendat, cito resilit, ne osculorum loco morte donetur: coitu etiam peracto, pede veloci ab uxore se subducit, quæ illum, imo post Veneres, aliàs forsan voraret[1673]. In genere mares feminas antennarum et abdominis motibus et frictione lenocinantur et ad coitum provocant.
Insecta sunt alia, ut Phalænæ, Muscidæ quædam, et Apis mellifica, in quibus inversa est hæc naturæ lex casta; harum enim feminæ marem petunt, vel blanditiis alliciunt ad amores.
Nunc de coitu ipso tractabimus, in quo hæc sunt præcipue notanda—modus, statio relativa, locus, et duratio.
[Pg 583]i. Plerisque insectis penis intrans est, sed in Muscidis quibusdam inversa est lex, et feminæ tubus retractilis analis, foramen sub ano maris penetrat et ita coëunt[1674]. Araneidis singulari et mirabili prorsus modo fit coitus; organi enim masculi functio partim palpis et partim membro ventrali delegatur: prioribus includitur glans quæ pudendum femineum penetrat, et sic in utroque sexu, palpis ambobus alternis vicibus huic officio inservientibus, orgasmus venereus producitur, cui insequitur fœcundatio, ab organo ventrali masculo; femina tubercula duo supra genitalia sita in rimas totidem inter branchias maris immittente, et in temporis momento omnia peracta sunt[1675]. Listerus, De Geerius, et alii in zootomia periti, in palpis latere organum masculum crediderunt, sed ex observationibus et dissectionibus Trevirani patet, testes et vesiculas seminales in abdomine locum habere[1676]; sed exitus horum solummodo in orificio[1677]; in palpis e contra est organum exsertile penem referens, quod in coitu erigitur et fere glandiforme est: hinc deduci potest, ut videtur, quod utrumque organum pro genitale habendum, et fœcundationem feminæ ab utroque pendere.
ii. Statio relativa. In plerisque insectis, durante coitu, maris statio superior est, et feminæ inferior, in hujus dorsum conscendente illo; interdum tamen hæc lex inversa est, et marem femina ascendit, quod ipse vidi in Vespa vulgari, et Scatophaga; in Pulice etiam femina superior, sed more humano os ori[1678]; quod fit etiam in aliis quibusdam masculo prædominanti, nempe in Cryptophago quodam minuto, nostris sub oculis, in Zygæna, Culice, et Phalangio[1679]. In insectis Orthopteris et pluribus Hemipteris sexus in coitu sibi invicem a latere paralleli stant[1680]; sed in aliis Hemipteris, saltem in Pentatomate,[Pg 584] more canum capitibus aversis, quod fit etiam in quibusdam Tipulidis, res venereas peragunt[1681].
iii. Locus. Interdum in terram et inter gramina; interdum inter arborum et fruticum ramos, et sub foliis; interdum iterum super aquas; et in ipso aëre demum haud raro amoris gaudiis ultimis fruuntur; hîc Ephemeræ caducæ in ipso venere choreas ducunt; sursum et deorsum, memetipso teste, alternatim volitantes[1682]: hîc etiam Apum regina et mater in sublime fertur maritum infelicem petens, qui voluptatem brevem vita emat[1683]: Phalænarum feminæ apteræ hue illuc per aërem inter arbores trahuntur a mare alato[1684]; et quarundam Tipularum mares a feminis tracti, per aërem item durante coitu rapiuntur. Modeste satis coëunt insecta, utplurimum plantarum sub umbra latitantes; et plura insuper, ut quædam Tipulæ, Tineidæ, et Bombycidæ, sub cortina alarum abdomen omnino tegente, veneri se tradunt[1685].
iv. Duratio. Coitus horum animalium duratio varia, interdum, ut in Araneidis, spatio perbrevi conficitur, in quibusdam tamen plus uno die opus est. Plures feminas interdum aggreditur idem mas, hoc in Bombyce, Chrysomela Polygoni, et Musca domestica obtinet. Aphidem masculum cum quinque feminis successive copulantem De Geerius videbat[1686].
N.B. Inter pupas Orthopterorum et Hemipterorum coitus interdum locum habet, quod maturiorem organizationem in his analogis, quam in aliis insectis probat.
[N. B. Those works in the following list to which an Asterisk is prefixed are useful to the Entomologist. The abbreviations of the titles of the works used in the text and notes of the Introduction to Entomology, in the list are put in Italics.]
Acerbi (Joseph) Travels through Sweden, Finland, and Lapland, to the North Cape, in 1798 and 1799. London 1802. 4to.
Adams (Joseph, M.D.) Observations on morbid poisons. London 1807. 4to.
Ælianus. De Natura Animalium.
Ahrens (Augustus) Fauna insectorum Europæ. Halæ 1812—. 12mo.
Aldrovandus (Ulysses) De animalibus insectis. Bononiæ 1602. fol.
Amoreux (P. J.?) Notice des insectes de la France reputés venimeux. A Paris 1789. 12mo.
Anderson (James, LL.D.) Recreations in Agriculture, natural history, the arts, and miscellaneous literature. 6 vols. London 1799—. 8vo.
Andrews (James Pettit) Anecdotes ancient and modern, with observations, and supplement. London 1789—. 8vo.
Angelinus (Fulvius), &c. De verme admirando per nares egresso. Ravennæ 1610.
Anonymous. A description of the island of St. Helena, containing observations on its singular structure and formation, and an account of its climate, natural history, and inhabitants. London 1808. 8vo.
Aristoteles. Tom. iv. Lutet. Paris. 1629. fol.
Azara (Felix de) Voyage dans l'Amerique Meridionale. Paris 1809. 8vo.
Bacon (Lord Verulam) Works of, by Mallet. 4 vols. London 1740. fol.
Baker (Henry) Of Microscopes and the discoveries made thereby. 2 vols. London 1785. 8vo.
Bancroft (Edward, M.D.) Experimental researches concerning the philosophy of permanent colours, &c. London 1794. 8vo.
Banks (The Right Hon. Sir Joseph, K.B. P.R.S., &c.) A short account of the cause of the disease in corn called by the farmers the blight, the mildew, and the rust. London 1805. 4to.[Pg 586]
Barclay (John, M.D.) An inquiry into the opinions, ancient and modern, concerning life and organization. Edinburgh 1822. 8vo.
Barrow (John) Account of travels into the interior of Southern Africa in the years 1797, 1798, &c. London 1801. 4to.
Bartram (William) Travels through N. and S. Carolina, Georgia, E. and W. Florida, &c. Philadelphia 1791. 8vo.
Beckmann (Johann.) Physikalisch-ökonomische bibliothek, &c. Göttingen 1778—.
Bell (Charles, M.D.) Essays on the anatomy of expression in painting. London 1806. 4to.
Belon (Pierre) Les observations de plusieurs singularités et choses memorables trouvées en Grèce, &c. Paris 1554, 12mo.
Berk (Van F. H.) Verhandeling ten bewijze, &c. Haarlem 1807. 8vo.
Berkhausen Naturgeschichte der Europaïschen schmetterlinge. Frankfurt 1784. 8vo.
Berneaud (Thiebaut de) Voyage to the isle of Elba. E. Tr. London 1814. 8vo.
Bewick (Thomas) The history of British Birds. London 1797. 8vo.
Bilberg (Gustavus Johannes) * Monographia Mylabridum. Holmiæ 1813. 8vo.
Billardiere. Relation du Voyage de la recherche de la Perouse pendant les années 1791-1794. 2 tom. A Paris, An. viii. 4to.
Bingley (William) Animal Biography, or anecdotes of the manners and economy of the animal creation, arranged according to the system of Linnæus. 3 vols. London 1803. 8vo.
Bochart (Samuel) Hierozoicon, sive bipartitum opus de animalibus S. Scripturæ. Francofurt: ad Mœn. 1675. fol.
Bonner (James) Plan for speedily increasing the number of beehives in Scotland. London 1795. 8vo.
Bonnet (Charles) * Œuvres d'histoire naturelle et de philosophie. 18 vols. à Neuchatel 1779—. 8vo.
Bonomo (Giovan. Cosim.) Osservazioni intorno a pellicelli del corpo umano. Firenze 1687. 8vo.
Bradley (Richard P. Bot. Cant.) A Philosophical account of the works of nature, &c. London 1721. 4to.
Brahm (Nikol. Jos.) Insekten kalender für sammler und œkonomen. Mainz 1790. 8vo.
Brez (Jacques) La Flore des insectophiles precédé d'un discours sur la utilité des insectes et de l'étude d'insectologie. Autrecht 1791.
Broughton (Thomas Duer) Letters written in a Mahratta camp in 1809, descriptive of the manners, &c. of the Mahrattas. London 1813. 4to.
Browne (Patrick) The civil and natural history of Jamaica. London 1756. fol.
Bruce (James) Travels to discover the source of the Nile in the years 1768-1773. 5 vols. Edinburgh 1790. 4to.[Pg 587]
Brunnich (Martin Thrane) Entomologia, sistens insectorum tabulas systematicas—Latine et Danice. Hafniæ 1764. 8vo.
Butler (Charles) The feminine monarchie or the history of bees. Oxford 1634. 4to.
Campbell (John) Travels in S. Africa, undertaken at the request of the Missionary Society. 2nd ed. London 1815. 8vo.
Carus (C.G.) Introduction to the Comparative Anatomy of Animals, compiled with constant reference to Physiology, and elucidated by twenty copper-plates. Translated from the German by R. T. Gore. 2 vols. London 1827. 8vo.
Catesby (Mark) The natural history of Carolina, Florida, and the Bahama islands. 2 vols. London 1731—. fol.
Charleton (Gualterus) Onomasticon Zooicon. London 1668. 4to.
Christ (J. L.) Naturgeschichte, klassification und nomenclatur der insekten, vom bienen, wespen, und ameisengeschelecht, &c. (Hymenopt.) Francfurt am Main 1791. 4to.
Clairville. * Entomologie Helvetique, ou catalogue des insectes de la Suisse, rangées d'apres une nouvelle methode. tom. 2. Zuric 1798—. 8vo.
Clark (Bracy) * An essay on the bots of horses and other animals. London 1815. 4to.
Clarke (Edward Daniel, LL.D.) Travels in various countries of Europe, Asia and Africa. 8 vols. London 1816—. 8vo.
Consett (Matthew) Tour through Sweden, Swedish Lapland, Finland, and Denmark, &c. London 1789. 4to.
Cook (James, Capt.) Account of the voyages undertaken by order of his present Majesty for making discoveries in the S. Hemisphere by John Hawkesworth, LL.D. &c. 3 vols. London 1773. 4to.
Coquebert (Anton. Johann.) * Illustratio iconographica insectorum quæ in musæis Parisinis observavit et in lucem edidit Joh. Christ. Fabricius, &c. Decas. 1-3. Parisiis 1779—. 4to.
Cramer (Peter) * Papillons exotiques des trois parties du monde, L'Asie, L'Afrique et L'Amerique. Utrecht 1779—. 4to.
Cuba? (M.D.) Ortus sanitatis. De herbis et plantis, de animalibus et reptilibus, de avibus et volatilibus, de piscibus et natatilibus, de lapidibus, &c. 1485. fol.
Curtis (John) * British Entomology, being illustrations and descriptions of the genera of insects found in Great Britain and Ireland, &c. London 1824. 8vo.
Curtis (William) A short history of the brown-tailed moth, &c. London 1782. 4to.
Cuvier (G. L. C. F. D. Baron) * Leçons d'Anatomie comparée. 5 vols. Paris 1805. 8vo.
——— Le Règne Animal distribué d'après son organisation, &c. tom. 4. Paris 1817. 8vo.
Darwin (Erasmus, M.D.)
——— Zoonomia, or the laws of organic life. London 1794. 4to.[Pg 588]
Darwin (Erasmus, M.D.) Phytologia, or the philosophy of agriculture and gardening. London 1800. 4to.
Davy (Sir Humphry, Bart., P.R.S.) Elements of agricultural Chemistry, in a course of lectures for the Board of Agriculture. London 1813. 4to.
De Geer (Baron Charles) * Mémoires pour servir à l'histoire des insectes. tom. 7. A Stockholm 1752—. 4to.
De Jean (M. le Baron) * Catalogue de la collection des Coléoptères de M. le B. de J. A Paris 1821. 8vo.
———– * Species Général des Coléoptères, de la Collection de M. Le Comte De Jean. tom. 2. A Paris 1825, 1826. 8vo.
Derham (William, D.D.) Physico-theology, or a demonstration of the being and attributes of God from his works of creation. 13th ed. London 1768. 8vo.
Detharding (George Christoph.) Disputatio de insectis coleopteris Danicis. Buetzovii 1763. 4to.
Donovan (Edward) * The natural history of British insects, explaining them in their several states, illustrated with coloured figures. London 1792—. 8vo.
———— Epitome of the natural history of the insects of China. London 1798. 4to.
———————————————————————— of India. 1800. 4to.
———————————————————————– of N. Holland. 1802. 4to.
Douce (Francis) Illustrations of Shakespeare and of ancient manners. 2 vols. London 1807. 8vo.
Drury (Dru) * Illustrations of natural history, wherein are exhibited figures of exotic insects. 3 vols. London 1770—. 4to.
Dufour (Leon) Description de six Arachnides nouvelles et d'une nouvelle espèce de Galeode. Extrait de la 4e tom. des Annal. Génér. des Scienc. Phys. A Bruxelles 1820. 8vo.
Duftschmidt (Gaspard) Fauna Austriaca. 2 tom. Lintz et Leipzig 1805—. 8vo.
Dumeril (A.M. Constant) Traité élémentaire d'histoire naturelle. tom. 2. 2nde ed. A Paris 1807. 8vo.
Ellis (Daniel) An inquiry into the changes induced in atmospheric air by the germination of seeds, the vegetation of plants, and the respiration of animals. Edinburgh 1807. 8vo.
Esper (Eugen J. Christoph.) * Die schmetterlingen in abbildungen nach der natur mit beschreibungen. Erlangen 1777—. 4to.
Escholtz. Beiträge sur naturkund.
Fabricius (Johann. Christian.) * Philosophia entomologica, sistens scientiæ fundamenta adjectis definitionibus, &c. Hamburgi et Kilonii 1778. 8vo.
———— * Systema Entomologiæ sistens, insectorum classes, ordines, genera, species, adjectis synonymis, locis, descriptionibus, observationibus. Flensburgi et Lipsiæ 1775. 8vo.[Pg 589]
Fabricius (Johann. Christian.) * Entomologia systematica emendata et aucta, secundum classes, &c. tom. 4. Hafniæ 1794—. 8vo.
———— * Supplementum Entomologiæ systematicæ. Hafniæ 1798. 8vo.
———— * Systema Eleutheratorum secundum ordines, &c. tom. 2. Kiliæ. 8vo.
———— * Systema Rhyngotorum secundum ordines, &c. Brunsvigæ 1803. 8vo.
———— * Systema Piezatorum secundum ordines, &c. Brunsvigæ 1804. 8vo.
———— * Systema Antliatorum secundum ordines, &c. Brunsvigæ 1805. 8vo.
———— Resultate natur historischer vorlesungen. Kiel. 1804. 8vo.
Fabricius (Otho) Fauna Groenlandica, systematice sistens animalia Groenlandiæ occidentalis hactenus indagata, &c. Hafniæ et Lipsiæ 1780. 8vo.
Fischer (Gotthelf) * Entomographia Imperii Russici, &c. vol. 1. Mosquæ 1820—. 4to.
Fischers. Beschreibung eins huhns mit menschenähnlichen profile. St. Petersburg 1816. 8vo.
Fleming (John, D.D.) * The Philosophy of Zoology, or a general view of the structure, functions, and classification of animals. 2 vols. Edinburgh 1822. 8vo.
Fontana (Felice) On Poisons. (E. Tr.) London 1787. 12mo.
Forbes (James) Oriental Memoirs, from a series of familiar Letters written during 17 years residence in India. 4 vols. London 1813—. 4to.
Forster (John Reinhold) Novæ species insectorum centuria I. London 1771. 8vo.
Forsyth (William) Observations on the diseases, defects, and injuries in all kinds of fruit and forest trees, with an account of a particular method of cure. London 1791. 8vo.
Franklin (William) Military memoirs of General Thomas, who rose from an obscure situation to the rank of a general in the service of the native princes in the N. W. of India, &c. Calcutta 1803. 4to.
Fuessli (Jean Gaspar) Archives de l'histoire des insectes traduites en François. Winterthour 1794. 4to.
Gaede (Heinrich Moritz) Beyträge zur anatomie der insekten, &c. Altona 1815. 4to.
Geoffroy * Histoire abregée des insectes dans laquelle ces animaux sont rangées suivant un ordre methodique, tom. 2. (Ins. Par.) A Paris 1764. 4to.
Germar (E.F.) * Insectorum species novæ aut minùs cognitæ descriptionibus illustratæ. Vol. I. Coleoptera. Halæ 1824. 8vo.
Gleditsch (Johan. Gottlieb.) Physicalisch-botanisch-œconomische abhandlungen. Halle 1765—. 8vo.
Goedartius (Johannes) De insectis in methodum cum notularum additione. Opera M. Lister, &c. cum Scarabæorum Anglicanorum quibusdam tabulis mutis. Londini 1685. 8vo.[Pg 590]
Goexe (Johann. Aug. Ephr.) Natur menshenleben verschung.
Goldsmith (Oliver, LL.D.) History of the earth and animated nature. 8 vols. London 1774. 8vo.
Good (John Mason, M.D.) Anniversary Oration, delivered March 8th, 1808, before the Medical Society of London.
Gould (William) An account of English ants. London 1747. 12mo.
Gravenhorst (J. L. C.) Coleoptera Microptera Brunsvicensia nec non exoticorum quotquot extant in collectionibus Brunsvicensium. Brunsvigæ 1802. 8vo.
Grew (Nehemiah) Musæum Regalis Societatis, or a catalogue and description of the natural and artificial rarities belonging to the R. Society, and preserved at Gresham College, &c. London 1681. fol.
Grosier (Abbé) A general description of China. (E. Tr.) London 1788. 8vo.
Gyllenhul (Leonard) * Insecta Suecica descripta. Classis I. Coleoptera sive Eleutherata. tom. 4. Scaris 1808-27. 8vo.
Hagenbach (Joh. Jacob.) Mormolyce. Novum Coleoptrorum Genus. Norimbergæ. 1825. 8vo.
Haworth (Adrian Hardy) * Lepidoptera Britannica, sistens digestionem novam insectorum Lepidopterorum quæ in Magna Britannia reperiuntur, &c. Londini 1803—. 8vo.
Hermann (J. F.) Memoire apterologique. Strasbourg 1804. fol.
Herold (Mauritius) * Entwickelungsgeschichte der schmetterlinge anatomische und physiologisch. Cassel und Marburg 1815. 4to.
Hobhouse (John Cam) Some account of a journey into Albania, Romelia, and other provinces of Turkey in 1809 and 1810. London 1812. 4to.
Hooke (Robert, M.D.) Micrographia, or some physiological descriptions of minute bodies made by magnifying-glasses, &c. London 1665. fol.
Hooker (Professor) Journal of a tour in Iceland in the summer of 1809. Yarmouth 1811. 8vo.
Horne (Thomas Hartwell) Introduction to the study of Bibliography; to which is prefixed a memoir on the publick libraries of the ancients, &c. 2 vols. London 1814. 8vo.
Huber (Francis) New observations of the natural history of bees. 2nd ed. (E. Tr.) Edinburgh 1808. 12mo.
——— * Nouvelles observations sur les abeilles, tom. 2. A Paris et à Genève 1814. 8vo.
Huber (P.) * Recherches sur les mœurs des fourmis indigènes. A Paris 1810. 8vo.
Hubner (Jacob) * Der gamlung Europaischen schmetterlinge. Augsburg 1796. 4to.
Hughes (Griffith) The natural history of Barbados. London 1750. fol.
Humboldt (Alexander, Baron de) and Bonpland (Aimé) Personal travels to the equinoctial regions of the new continent during the years 1799-1844. (E. Tr.) 5 vols. London 1818—. 8vo.[Pg 591]
Humboldt (Alexander, Baron de) and Bonpland (Aimé) Essais sur la géographie des plantes. Paris 1805.
————– Political essay on new Spain. (E. Tr.) London 1811. 8vo.
————– Recueil d'observations de zoologie et d'anatomie comparée. A Paris 1805. 4to.
Hunter (John, M.D.) Observations on certain parts of the animal economy. London 1786. 4to.
Jablonsky (Carl. Gustaf.) * Natursystem aller bekannten insekten. Ed. Herbst. (Herbst Jablonsk.) Berlin 1785—. 8vo.
Jackson (James Grey) An account of the empire of Morocco and the district of Suez; compiled from miscellaneous observations made during a long residence in those countries, &c. London 1809. 4to.
Jacquin (Nicolaus Joseph) Collectanea ad botanicam, chemiam, et historiam naturalem spectantia. Vindobonæ 1786.
Jermyn (Lætitia) * The Butterfly Collector's Vade Mecum, or A synoptical table of English Butterflies. 2nd ed. Ipswich 1827. 12mo.
Illiger (J. K. W.) * Versuch einer systematischen volständigen Terminologie das thierreich und pflanzenreich. Helmstadt 1800. 8vo.
Inch (C. W.) Ideen zu einer zoochemia. 1800.
Jungius (Joachimus) Historia vermium. Hamburgi 1691. 4to.
Jurine (L.) * Nouvelle methode de classer les Hyménoptères et les Diptères. tom. 1. Hyménoptères. A Genève 1807. 4to.
Kalm (Peter) Travels into N. America. (E. Tr.) 3 vols. Warrington 1772. 8vo.
Kæmpfer (Engelbert) The history of Japan. (E. Tr.) 2 vol. London 1728. fol.
Keys (John) A treatise on the breeding and management of bees. London 1814. 12mo.
Kirby (William) * Monographia Apum Angliæ, or an attempt to divide into their natural genera and families such species of the Linnean genus Apis as have been discovered in England, &c. 2 vols. Ipswich 1802. 8vo.
Kleemann (Christ. Fried. Karl.) Beiträge zur natur-oder insecten geschichte. Nurnberg 1761. 4to.
Knight (Thomas Andrew) A Treatise on the culture of the apple and pear, and on the manufacture of cider and perry. London 1797. 8vo.
Knoch (August. Wilhelm) Beiträge zur insecten geschichte. Leipzig 1781—. 8vo.
Knox (Robert) an historical relation of the island of Ceylon. London 1681. fol.
Lafontaine. Traité de la chirurgie. Leipzig. 1792.
La Lande de Lignac (Joseph Albert) Mémoire pour servir à commencer l'histoire des araignées aquatiques. Paris 1749. 8vo.
Lamarck (Jean Baptiste) Système des animaux sans vertèbres, ou tableau général des classes, des ordres et des genres de ces animaux, &c. A Paris 1801.[Pg 592]
Lamarck * (Jean Baptiste) Histoire naturelle des animaux sans vertèbres présentant les caractères généraux et particuliers de ces animaux, leur distribution, leur classes, &c. tom. 7. A Paris 1815—. 8vo.
———— Philosophie zoologique. A Paris 1809. 8vo.
Lambert (John) Travels through the United States of America, Canada, and Georgia in the years 1806-8. 3 vols. London 1810. 8vo.
Latham (John) A general synopsis of birds. 3 vols. London 1781—. 4to.
Latreille (Pierre André) * Histoire naturelle générale et particulière des crustacés et des insectes, tom. 14. Paris 1802—. 8vo.
———— * Histoire naturelle des fourmis et recueil des mémoires et d'observations sur les abeilles, les araignées, les faucheurs, et autres insectes. A Paris 1802. 8vo.
———— * Genera Crustaceorum et Insectorum secundum ordinem naturalem in familias disposita, iconibus exemplisque plurimis explicata. tom. 4. Parisiis et Argentorati 1806—. 8vo.
———— Familles naturelles du règne animal, exposés succinctement et dans un ordre analytique, avec l'indication de leurs genres. A Paris 1825. 8vo.
———— et De Jean (Baron) * Histoire naturelle et iconographie des insectes Coleoptères d'Europe. A Paris 1822—. 8vo.
Leach (William Elford, M.D.) * The Zoological Miscellany, being a description of new and interesting animals, illustrated with coloured figures, &c. London 1817—. 8vo.
Leeuwenhoek (Antonio) Arcana naturæ detecta. Delphis Batavor. 1695. 4to.
————— Arcana naturæ opes et beneficio exquisitissimorum microscopiorum detecta variisque experimentis demonstrata, &c. Lugd. Batav. 1696. 4to.
————— Epistolæ physiologicæ super compluribus naturæ arcanis, &c. Delphis 1719. 4to.
————— * Select works of, containing his microscopical discoveries in many of the works of nature, translated from the Dutch and Latin editions published by the author, by Samuel Hoole. 2 vols. London 1798. 4to.
Lehmann (Mart. Christ. Gottl.) De sensibus externis animalium exsanguium, insectorum scilicet et vermium, commentatio. Gottingæ 1798.
———— De antennis insectorum dissertatio prior fabricam antennarum describens; Dissertatio posterior usum antennarum recensens. Londini 1799—. 12mo.
Lempriere (William) Practical observations on the diseases of the army in Jamaica during the years 1792-7. 2 vols. London 1799. 8vo.
Lesser (Friedr. Christ.) Théologie des insectes, ou demonstration des perfections de Dieu dans tout ce que concerne les insectes, &c. Avec des ramarques de M. P. Lyonnet. (Lesser L.) tom. 2. A la Haye 1742. 8vo.
Lewin (William) * The insects of Great Britain systematically arranged, accurately engraved, and painted from nature, &c. London 1795.[Pg 593]
Lewin (William) Prodromus Entomology, or natural history of Lepidopterous insects of New South Wales. London 1805. 4to.
Lichtenstein (Aut. Aug. Hen.) Catalogus rerum naturalium rarissimarum auctionis lege distrahendarum. (Catal. Ham.) Hamburgi 1794.
Ligon (Richard) A true and exact history of the island of Barbados. London 1657. fol.
LINNE' (Carl. Von) * Systema Naturæ per regna tria naturæ secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Ed. 13. tom. 3. Vindobonæ 1767—. 8vo.
——— * Fauna Suecica. Sistens animalia Sueciæ regni, Mammalia, Aves, Amphibia, Pisces, Insecta, Vermes distributa per classes, &c. Ed. altera. Stockholmiæ 1761. 8vo.
——— Flora Lapponica, exhibens plantas per Lapponiam crescentes, secundum systema sexuale, collectas in itinere &c. anno 1732 instituto, &c. Ed. J. E. Smith. Eq. Londini 1792. 8vo.
——— Skanska resa, fürrättad ar 1749. (It. Scan.) Stockholm 1751. 8vo.
——— Lachesis Lapponica, or a tour in Lapland, now first published from the original MS. Journal of the celebrated Linnæus, by Sir J. E. Smith. 2 vols. (Lach. Lapp.) London 1811. 8vo.
——— Amœnitates Academicæ, seu dissertationes variæ physicæ, medicæ, botanicæ antehac seorsim editæ, nunc collectæ et auctæ, &c. tom. 7. Lugd. Bat. 1749—. 8vo.
——— * Philosophia Botanica in qua explicantur fundamenta botanica, cum definitionibus partium, exemplis terminorum, observationibus rariorum, &c. ed. 2da. Viennæ 1783. 8vo.
Lister (Martin, M.D.) * Historiæ animalium Angliæ tractatus de Araneis. Londini 1678. 4to.
Lyonet (Pierre) * Traité anatomique de la chenille qui ronge le bois de saule, &c. A la Haye 1762. 4to.
Mac Kinnen (Daniel) A Tour through the British W. Indies during the years 1802-3, giving a particular account of the Bahama islands. 2nd ed. London 1813. 8vo.
MacLeay (William Sharp) * Horæ Entomologicæ, or essays on the annulose animals.
Vol. 1. Part I. containing general observations on the geography, manners, and natural affinities of the insects which compose the genus Scarabæus of Linnæus, &c. London 1819. 8vo.
Vol. 1. Part II. containing an attempt to ascertain the rank and situation which the celebrated Egyptian insect, Scarabæus sacer, holds amongst organized beings. London 1821. 8vo.
————— * Annulosa Javanica. London 1824. 4to.
Magnus (Albertus) Opera. tom. 21. De natura animalium. fol.
Malpighius (Marcellus) Opera omnia. tom. 2. (De Bombyc.) Londini 1686. fol.
Marsham (Thomas) * Entomologia Britannica (E. B.) sistens insecta Britanniæ indigena secundum methodum Linnæanam disposita. tom. 1. Coleoptera. Londini 1802. 8vo.[Pg 594]
Martin (George, M.D.) Essays and observations on the construction and graduation of thermometers, and on the heating and cooling of bodies. Edinburgh 1792.
Martyr (Peter) The decades of the New World, or West Indies. (E. Tr.) London 1555. 4to.
Matthiolus (Petr. Andr.) Commentarii in libros Dioscoridis de Medica Materia. Venetiis 1554. fol.
Mead (Richard, M.D.) Medicina Sacra, seu de morbis insignibus qui in Bibliis memorantur; or a commentary on the diseases mentioned in Holy Scripture. London 1749. 8vo.
Meigen (Johann. Wilhelm.) * Nouvelle classification des mouches à deux ailes (Diptera L.) d'après un plan tout nouveau. A Paris 1800. 8vo.
———— * Systematische beschreibung der bekannten Europäischen zweiflügeligen Insekten. Vol. 5. Aachen. Hamm. 1818-1826. 8vo.
Merian (Maria Sibylla) * Metamorphosis insectorum Surinamensium. In qua erucæ et vermes ad vivum delineantur et describuntur, &c. Amstelod. 1705. fol.
———— Erucarum ortus, alimentum et paradoxa metamorphosis. Amstelæd. 1718. 4to.
Mills (John) An essay on the management of bees, &c. London 1766. 8vo.
Moldenhawer (Joh. Jac. Paul) Beyträge zur anatomie der pflanzen. Kiel 1812—. 4to.
Molina (I. Ignatius) The geographical, natural, and civil history of Chili. 2 vols. (E. Tr.) London 1809. 8vo.
Monro (Alexander, M.D.) The anatomy of the human bones and nerves, &c. Ed. Kirby, M.D. Edinburgh 1820. 12mo.
Morier (James) A second journey through Persia to Constantinople between the years 1810-16. With a journal of the voyage by the Brazils and Bombay to the Persian gulf. London 1818. 4to.
Mouffet (Thomas) * Insectorum sive minimorum animalium theatrum. Londini 1634. fol.
Müller (Otto. Frideric.) Zoologia Danica seu animalium Daniæ et Norvegiæ rariorum ac minùs notorum descriptiones et historia. vol. 2. Hafniæ et Lipsiæ 1779—. 8vo.
Olivier (Guill. Antoin.) * Entomologie, ou histoire naturelle des insectes, avec leur caractères génériques et specifiques, leur description, leur synonymie, et leur figure enluminée. Coleoptères, tom. 8. A Paris 1789—. 4to.
——— Travels in the Ottoman empire, Egypt and Persia. (E. Tr.) 2 vols. London 1813. 8vo.
Osbeck (Peter) A voyage to China and the E. Indies. Toreen (Olof) A voyage to Suratte, and an account of the Chinese husbandry by Capt. C. G. Ekeberg. (E. Tr.) 2 vols. London 1771. 8vo.
Paley (William, D.D.) Natural Theology, or evidences of the existence and attributes of the Deity collected from the appearances of nature. 11th ed. London 1807. 8vo.[Pg 595]
Pallas (Peter Simon) Spicilegia zoologica, quibus novæ imprimis atque obscuræ animalium species describuntur, et observationibus iconibusque illustrantur. Hagæ Com. 1766—. 4to.
——— Neue nordische beyträge zur physikalischen und geographischen erd und volkerbeschreibung, naturgeschichte und oekonomie. (N. Nord. Beytr.) St. Petersburg und Leipzig 1781—, 8vo.
——— Travels through the southern provinces of the Russian empire in the years 1793, 1794. (E. Tr.) 2 vols. Leipsig 1801. 4to.
Panzer (Georg. Wolfgang Franz. M.D.) * Faunæ insectorum Germanicæ initia. Deutschland's insekten. Heft l—109. (Fn. Germ.) Nurnberg.
Park (Mungo) Travels in the interior districts of Africa, performed under the direction and patronage of the African association, in the years 1795-7, &c. 2nd ed. London 1799. 4to.
—— The journal of a mission to the interior of Africa in the year 1805; together with other documents, official and private, &c. 2nd ed. London 1815. 4to.
Parkinson (James) Organic remains of a former world. 2 vols. London. 4to.
Peck (Professor) Natural history of the slug-worm. Boston, N.E. 1799. 8vo.
Percival (Robert, Capt.) An account of the island of Ceylon, containing its history, geography, natural history, with the manners and customs of its inhabitants. London 1803. 4to.
Persoon (Christian. Henric.) Synopsis methodica fungorum, &c. Gottingæ 1801—. 12mo.
Pezold (Ch. Ph.) Lepidopterologische anfangsgründe zum gebrauch augehender schmetterlingssommler. Coburg 1796. 12mo.
Piso (Gulielmus, M.D.) De Indiæ utriusque re naturali et medica, &c. Amstelædami 1558. fol.
Plinius (C. Secundus) Historia mundi naturalis. Francofurti ad Mœn. 1582. fol.
Posselt (Carl Friederich) Beytræge zur anatomie der insekten. Tubingen 1804. 4to.
Preysler (Johann. Daniel) Verzeichniss Böhmischer insekten. Prag. 1790. 4to.
Pulteney (Richard, M.D.) Historical and Biographical Sketches of the progress of botany in England, from its origin to the introduction of the Linnean System, vol. 2. London 1790. 8vo.
Purchas (Samuel) His Pilgrimes. London 1625. fol.
Ramdhor (Karl. August.) * Abbildungen zur anatomie der insecten. Halle 1809—. 4to.
Ray (John) Catalogus Plantarum circa Cantabrigiam nascentium, &c. Londini 1660. 12mo.
—— Historia Plantarum, species hactenus editas aliasque noviter inventas et descriptas complectens. Londini 1686. fol.
—— The wisdom of God manifested in the works of creation. London 1692. 8vo.[Pg 596]
Ray (John) * Historia Insectorum, cui subjungitur appendix de Scarabæis Britannicis autore M. Lister. Londini 1710. 4to.
—— Philosophical Letters between the late learned Mr. Ray and several of his ingenious correspondents; to which are added those of F. Willughby, Esq. published by W. Derham. London 1718. 8vo.
Reaumur (René Antoine Ferchault de) * Mémoires pour servir à l'histoire des insectes. tom. 6. A Paris 1734—. 4to.
Redi (Franciscus) Opusculorum pars prior sive experimenta circa generationem insectorum, &c. Amstelæd. 1686. 18mo.
—— Opusculorum tomus alter. Experimenta circa varias res naturales speciatim illas quæ ex Indiis afferuntur. Amstelæd. 1686. 18mo.
Reeve (Henry, M.D.) An essay on the torpidity of animals. London 1809. 8vo.
Rifferschweils (Johann. Jacob Hegetschweiler) * Dissertatio inauguralis zootomica de insectorum genitalibus. Turici 1820. 8vo.
Roemer (Johann. Jacob) * Genera insectorum Linnæi et Fabricii iconibus illustrata. Vitodur. Helvetor. 1789. 4to.
Rösel von Rosenhoff (August. Johann.) * Der monatlich-herausgegeben insecten belustigung. Nurnberg 1746—. 4to.
Rossi (Petrus) Fauna Etrusca sistens insecta quæ in provinciis Florentina et Pisana præsertim colligit. Liburni 1790. 4to.
Rudolphi (C. A.) Entozoa, seu vermium intestinalium historia naturalis. vol. 2. Amsterd. 1808. 8vo.
Sabine (Edward, Capt.) An account of the animals seen by the late Northern Expedition whilst within the Arctic Circle, &c. London 1821. 4to.
St. Pierre (James Hen. Bernardin) A voyage to the Mauritius or Isle of France, the Isle of Bourbon, the Cape of Good Hope, &c. (E. Tr.) London 1775. 8vo.
———— Studies from Nature. (E. Tr.) 4 vols. London 1796. 8vo.
Samouelle (George) * The Entomologist's useful compendium, or an introduction to the knowledge of British insects, &c. London 1819. 8vo.
Sandwith (Thomas) * An introduction to anatomy and physiology for the use of medical students and men of letters. London 1824. 12mo.
Sauvages (François Boisier de) Observations sur l'origin de miel. Nismes 1763.
Savi (Paolo) Osservazioni per servire alla storia di una specie de Iulus communissima nella pianura Pisana. Bologna 1817.
Savigny (Jules César) * Mémoires sur les animaux sans vertèbres. Parties I. II. A Paris 1816. 8vo.
———– Système des Annelides principalement de celles des côtes de l'Egypte et de la Syrie[1687]. fol.
Scheele (Charles William) Chemical observations and experiments on air and fire. (E. Tr.) London 1780. 8vo. [Pg 597] Schellenberg (J. R.) * Cimicum in Helvetiæ aquis et terris degentium genus in familias redactum observationibus et iconibus ad naturam delineatis illustratum. Turici 1800. 8vo.
———— * Genres de mouches Diptères representés en xlii planches. Zuric 1803. 8vo.
Schneider (David Heinrich) Systematische beschreibung der Europaischer schmetterlinge. Halle 1787. 8vo.
Schönherr (C. J.) * Synonyma insectorum, oder versuch einer synonymie aller bisher bekannten insecten nach Fabricii Systema Eleutheratorum geordnet, tom. 3. Stockholm 1806. 8vo.
———— * Curculionidum dispositio methodica cum generum characteribus, descriptionibus atque observationibus variis. Lipsiæ. 1826. 8vo.
Schrank (Franciscus de Paula) Enumeratio insectorum Austria, indigenorum. Augustæ Vindelicorum 1781. 8vo.
Schwenckfeeld (Caspar, M.D.) Theriotropheum Silesiæ, in quo animalium, h.e. quadrupedum, reptilium, avium, piscium, insectorum natura, vis, et usus sex libris perstringuntur. Lignicii 1603. 4to.
Scopoli (Johann. Anton.) Entomologia Carniolica exhibens insecta Carnioliæ indigena et distributa in ordines, genera, species, varietates methodo Linneana. (Ent. Carniol.) Vindobonæ 1763. 8vo.
———— Anni historico-naturales. Lipsiæ 1769—. 8vo.
Scriba (Ludwig Gottlieb.) Beiträge zu der insekten geschichte. Frankfurt 1790—. 4to.
Sendelius (Nathanel) Historia succinorum corpora aliena involventium, &c. Lipsiæ 1742. fol.
Sepp (Christian) * Beschowing der wonderen Gods inde minstgrachte schepgelen of Nederlandsche-insecten, &c. tom. 2. Amsterdam 1762. 4to.
Shaw (Thomas, D.D.) Travels and observations relating to several parts of Barbary and the Levant. 2nd ed. London 1757. 4to.
Shaw (George) The naturalist's miscellany, or coloured figures of natural objects, drawn and described immediately from nature. London 1789—. 8vo.
Shirach (Adam Gottl.) Histoire naturelle de la reine des abeilles, avec l'art de former les essaims, &c. (F. Tr.) A la Haye 1771. 8vo.
Smeathman (Henry) Some account of the Termites which are found in Africa and other hot climates. (From the Philosophical Transactions). London 1781. 4to.
Smellie (Wm.) The philosophy of natural history. Edinburgh 1790. 4to.
Smith (Sir J. E.) Sketch of a tour on the continent in the years 1786 and 1787. 3 vols. London 1793. 8vo.
——— * The natural history of the rarer Lepidopterous insects of Georgia, collected from the observations of John Abbott. 2 vols. London 1797. 4to.
——— Tracts relating to natural history. London 1798.[Pg 598]
Smith (Sir. J. E.) Introduction to physiological and systematic botany. London 1807. 8vo.
Sorg (F. L. A. W.) Disquisitiones physiologicæ circa respirationem insectorum et vermium. Rudolst. 1805. 12mo.
Southey (Robert, LL.D.) History of Brazil. London 1810—. 4to.
Sowerby (James) * British miscellany, or coloured figures of new, rare, or little-known animal subjects, many not before ascertained to be natives of the British Isles, &c. London 1806—. 8vo.
Spallanzani (Lazaro) Opuscoli di fisica animale e vegetabile. Modena 1776. 8vo.
————— Dissertations relative to the natural history of animals and vegetables. (E. Tr.) London 1784. 8vo.
————— Memoirs on respiration. (E. Tr.) London 1804. 8vo.
Sparrman (Andrew) A Voyage to the Cape of Good Hope, towards the Antarctic Circle and round the world, but chiefly into the country of the Hottentots and Caffres. 2 vols. London 1785. 4to.
Spence (William) Observations on the disease in turnips termed in Holdernesse fingers and toes. Hull 1812. 8vo.
Sprengel (Christian Conrad) Das entdeckte geheimniss der natur in bau und in der befrucktung der blumen. Berlin 1793. 4to.
Sprengel (Curtius) * Commentarius de partibus quibus insecta spiritus ducunt. Lipsiæ 1805. 4to.
Staunton (Sir George) An authentic account of the embassy from the king of Great Britain to the emperor of China. 3 vols. London 1797. 8vo.
Stedman (J. C. Capt.) Narrative of a five years expedition against the revolted negroes of Surinam in Guiana, on the wild coast of S. America, from 1772 to 1777. 2 vols. London 1796. 4to.
Stephens (James Francis) * Illustrations of British Entomology. London 1827, 1828. 8vo.
Stickney (William) Observations respecting the grub. Hull 1800. 8vo.
Stillingfleet (Benjamin) Miscellaneous tracts relating to natural history, husbandry, and physick, translated from the Latin. 2nd ed. London 1762. 8vo.
Stoever (D. H.) The life of Sir Charles Linnæus; to which is added a copious list of his works, and a biographical sketch of the life of his son. (E. Tr.) London 1794. 4to.
Stoll (Caspar) * Représentation exactement colorée d'après nature des cigales qui se trouvent dans les quatre parties du monde, &c. Amsterdam 1788. 4to.
——– * Représentation, &c. des punaises ditto. Amsterdam 1788. 4to.
——– * Représentation, &c. des spectres ou phasmes, et des mantes ou des feuilles ambulantes, des sauterelles, des grillons, des criquets et des blattes, &c. Amsterdam 1813. 4to.
Strack. Neue schriften der naturforschenden. Halle 1810.[Pg 599]
Sturm (Jacob) * Deutschlands Fauna in abbildungen nach der natur mit beschreibungen. tom. 5. Nurnberg 1805—. 12mo.
Swammerdam (John, M.D.) * The Book of Nature, or the history of insects reduced to distinct classes, confirmed by many particular instances displayed in the anatomical analysis of many species, and illustrated with copper-plates. (E. Tr.) [Swamm. Bibl. Nat. Hills. Swamm.] London 1758. fol.
Thenard (L. J.) Traité de Chimie élémentaire théorique et pratique. tom. 2. A Paris 1813. 8vo.
Thomson (Thomas, M.D.) System of chemistry. London 1802—.
Thorley (John) Μελισσηλογια, or the female monarchy, being an inquiry into the nature, order, and government of bees, &c. London 1744. 8vo.
Thunberg (Charles Peter) Travels in Europe, Africa, and Asia, performed between the years 1770 and 1779. (E. Tr.) 4 vols. London 1795. 8vo.
Treviranus (G.R. Von) * Ueber den innern bau der Arachniden. Nurnberg 1812. 4to.
Trost (P.) Kleinen Beyträge zur Entomologie, &c. Erlangen 1801. 8vo.
Tulpius (Nicolaus) Observationes medicæ. Amstelæd. 1652. 8vo.
Tusser (Thomas) Five hundredth points of good husbandry, united to as many of good houswifery, first devised, and now lately augmented with diverse approved lessons concerning hopps and gardening. London 1573. 4to.
Ulloa (George Juan de) A voyage to S. America, describing at large the Spanish cities, towns, provinces, &c. on that extensive continent, &c. 2 vols. London 1760. 8vo.
Vaillant (Le) Travels into the interior parts of Africa by the way of the Cape of Good Hope, in the years 1780-1785. (E. Tr.) 2 vols. London 1790. 8vo.
Valentia (George, Lord) Voyages and Travels to India, Ceylon, the Red Sea, Abyssinia, and Egypt, in the years 1802-1806. 3 vols. London 1809. 4to.
Vallisnieri (Antonio) * Esperienze ed osservazioni intorno all' origine, sviluppi, e costumi di vari insetti, con altre spettanti alla naturale e medica storia. Ed. 2. In Padova 1726. 4to.
Voet (Johannes Eusebius) Icones insectorum Coleopterorum. Ed. Panzer. Erlangen 1793. 4to.
Walckenaer (C. A.) * Tableau des Aranéïdes, ou caractères essentielles des tribus, gènres, familles, et races que renforme le gènre Aranea de Linné, avec la designation des espèces comprises dans chacun de ces divisions. Paris 1805. 8vo.
————— * Mémoires pour servir à l'histoire naturelle des abeilles solitaires qui composent le genre Halicte. Paris 1817. 8vo.
Walton (Izaac) The universal angler, made so by three books of fishing: the first written by Mr. Izaak Walton, the second by Charles Cotton, Esq., and the third by Col. Robert Venables. London 1676. 12mo.
Walton. Present state of the Spanish colonies, including a particular report[Pg 600] of Hispaniola, &c., with a general survey of the settlements on the S. continent of America, &c. 2 vols. London 1810. 8vo.
Waterton (Charles) Wanderings in South America, the North West of the United States, and the Antilles, in the years 1812, 1816, 1820, and 1824; with original instructions for the perfect preservation of birds, &c. for cabinets of natural history. 2nd ed. London 1828. 4to.
Weld (Isaac) Travels through the states of N. America and the provinces of Upper and Lower Canada in the years 1795 and 1797. London 1799. 4to.
White (Gilbert) * The natural history of Selborne. New ed. by Marckwick. 2 vols. London 1813. 12mo.
Wiedemann (C. R. W.) Archiv. für zoologie & zootomie. Berlin und Brunschweig. 1800—.
—————– Diptera exotica: Pars I. Kiliæ 1821. 12mo.
Wildman (Thomas) A treatise on the management of bees. 2nd ed. London 1769. 4to.
Willan (Robert, M.D.) Description and treatment of cutaneous diseases. London 1801. 4to.
Willdenow (Carl. Ludwig.) Grundriss der kräturkunde. Berlin 1792. 8vo.
———— The principles of botany and of vegetable physiology. (E. Tr.) Edinburgh 1811. 8vo.
Young (Arthur) Annals of agriculture and other useful arts. 40 vols. Bury St. Edmunds 1790—. 8vo.
——— Travels during the years 1787-1789, undertaken more particularly with a view of ascertaining the cultivation, wealth, resources, and natural prosperity of the kingdom of France. 2 vols. Bury St. Edmunds 1792. 4to.
Transactions (Philosophical) of the Royal Society of London, with the Abridgements of Lowthorp and Dr. Shaw. 4to.
———— of the Linnean Society of London. 4to.
———— of the Horticultural Society. 4to.
———— of the Society for the encouragement of arts, manufactures and commerce. 8vo.
———— (Asiatic Researches) of the Society instituted in Bengal for inquiring into the history and antiquities, the arts, sciences, and literature of Asia. 4to.
———— of the American Philosophical Society, held at Philadelphia, for promoting useful knowledge. 4to.
Memoirs of the Wernerian Society. 8vo.[Pg 601]
Mémoires de l'Academie royale des sciences. A Paris. 4to.
———— nouveaux de l'Academie de Dijon pour la partie des sciences et arts. 8vo.
———— de l'Academie royale des sciences à Turin. 4to.
Annales du Muséum national d'histoire naturelle. 4to.
Mémoires du Muséum national d'histoire naturelle. 4to.
Acta physico-medica Academiæ Cæsareæ naturæ curiosorum. (Nova Acta, Ephemerides, &c.) 4to.
Kongl. Vetenskaps Academiens nya handlingar. Academia regia scientiarum Suecana. (Act. Stock.) 8vo.
Communications to the Board of Agriculture. 4to.
Annals of botany (König and Sims). 4to.
——— of Philosophy (Thomson).
Annales de chimie, ou recueil de mémoires concernant la chimie et les arts qui en dependent. Paris. 8vo.
Annali di chimica.
Bulletin des sciences naturelles et de géologie (De Ferussac). 8vo.
Journal of natural philosophy, chemistry, and the arts (Nicholson's).
——— Edinburgh medical and surgical.
——— Zoological. 8vo.
——— Massachusetts Agricultural. 8vo.
——— de physique (Abbé Rozier, &c.). 4to.
——— für die liehaber der entomologie (Scriba). 8vo.
——— Loudon's Gardener's and Register of rural and domestic improvement.
Magazine, Tilloch's Philosophical.
———— Philosophical and Annals of Philosophy, by Taylor and Phillips. N. Series.
Magazin Berlinisches (Martini, Berlin natural history Societies).
———– Neuestes für die liehabers der entomologie (Schneider). 8vo.
———– Neu. entomolog. (Fuessli, Entomologische bemerkungen.) 8vo.
———– für das neueste aus der physik und naturgeschichte, &c. (Lichtenberg & Voigt). 8vo.
———– * für insektenkunde (Illiger). 8vo.
———– * der entomologie (Dr. Germar und Dr. Zicken Gennant Sommer). 8vo.
———– Encyclopédique, ou Journal des sciences, des lettres et des arts. 8vo.
Naturforscher der (The naturalist).
Review. London medical.
Systematisches verzeichniss der schmetterlinge der Wienergegend, &c. 4to.[Pg 602]
Dictionary of Chemistry (Messrs. Aikin).
——— medical (Dr. Hooper's). 8vo.
Dictionnaire Physique.
——— * Nouveau d'Histoire Naturelle. tom. 36. 8vo.
——— des sciences naturelles. 8vo.
Encyclopædia Britannica. 4to.[Pg 603]
PLATE XXI.[1688]
| FIG. | |
| 1. | Part of the interior of Cossus ligniperda. (Lyonet.) a, b. The spinal chord and its ganglions. d. The nerves. e. The bronchiæ connected with the trachea. |
| 2. | One of the labial palpi of ditto. (Ibid.) |
| 3. | Another view of the interior of ditto. (Ibid.) a. Trachea. b. Bronchiæ. c. Œsophagus. d. Ventricle or stomach. e. The lower intestines, f f. The bile vessels. g. Sericterium, or silk reservoir. h. Sialisterium, or saliva vessel. |
| 4. | Part of one of the tracheæ of ditto, to show its coats and spiral thread. (Ibid.) Vol. IV. p. 63. |
| 5. | A portion of the interior of ditto, to show the epiploon, or fat. (Ibid.) a a. Epiploon. Vol. IV. p. 150. |
| 6. | Leg of ditto laid open. (Ibid.) a a a. Semipenniform muscles of ditto. b. Their lower point of insertion in the claw. c. Muscles of the coxa. Vol. IV. p. 185. |
| 7. | Nervous system of the grub of Oryctes nasicornis. (Swamm.) a. The first ganglion or brain. c. The remaining ganglions, forming a thick spinal chord. d. The nerves issuing from them. |
| 8. | —————— of the louse. (Ibid.) a. The brain. c c c. The ganglions. d. Nerves. |
| 9. | The spinneret, or organ that renders the silk of the Cossus. (Lyonet.) Vol. III. p. 124. |
PLATE XXII.[1689]
| FIG. | |
| 1. | Male genital organ of a Bombus. a. The male organ. b. The prehensor. Vol. IV. p. 576, 580. |
| 2. | Female ditto of the louse. (Swamm.) a. The oviduct with an egg passing through it. b c. The ovaries. d. The colleterium, or varnish secretor. e. The lower extremity of the oviduct. |
| 3, 4. | Larvæ as arranged in the body of two species of viviparous flies. (Reaum.) Vol. I. p. 257; IV. p. 170. |
| 5. | Interior of postpectus. Dynastes. a. Parapleura. b b b. Points of the postfurca. c. Its stalk. |
| 6. | The medifurca of ditto, a. Base. b. Apex. |
| 7. | Antefurca of ditto, a. Base. |
| 8. | Mesothorax. Calandra. |
| 9. | Mesophragm and appendage. Dynastes. a a. Pieces adjacent. b. Septula. c. The notch for the transmission of the intestines. |
| 10. | Part of metaphragm, ditto, a. A kind of cupule affording a point of attachment to muscles. d. A deep notch for the intestines. |
| 11. | Interior of the upper side of alitrunk of ditto. a. The cavity of the chest between the prophragm and mesophragm. b. Ditto between the mesophragm and metaphragm. c. Cupules that afford a point of attachment to some of the wing-muscles. d. Notch of the metaphragm. |
| 12. | Portion of the alitrunk of Melolontha vulgaris. c. Cupule attached to the axis of the wings. |
| 13. | Part of the postpectus of Dytiscus marginalis, to show the operculum. Vol. III. p. 578. |
| 14. | Part of the metathorax of Melolontha vulgaris, to show the metapnystega. Ibid. 572. |
| 15. | The pseudocardia, or dorsal vessel of Stratyomis Chamæleon. (Swamm.) |
| 16.[Pg 605] | a.-w. Specimens of scales from the wings of various Lepidoptera. (Reaum., De Geer.) Vol. III. p. 644—. |
| FIG. | |
| 1. | One of the prolegs of a caterpillar. Cossus. (Lyonet.) a. Its coronet of spines. Vol. III. p. 134. |
| 2. | One of the spiracles of ditto. (Ibid.) Vol. IV. p. 37—. |
| 3. | Three of the hexagonal lenses of a bee's eye, with their prisms. (Swamm.) Vol. III. p. 496. |
| 4. | Trunk of a flea with the head removed, showing that all the legs are attached to the former. Ibid. p. 656. |
| 5. | Alitrunk of Dytiscus marginalis, exhibiting the wings as they are folded when unemployed. |
| 6. | Part of ditto, with the scutellum and apex of an elytrum, to show the alula. Vol. II. p. 343; III. 557. |
| 7. | Anterior and posterior prolegs of Tanypus maculatus. (De Geer.) a. Posterior proleg. b. Anterior ditto. Vol. II. p. 275; IV. p. 363. |
| 8. | Posterior extremity of a pupa, to show the cremastræ, or hooks by which it is suspended. a. Hooks. Vol. III. p. 209, 255; IV. p. 363. |
| 9. | Another specimen, in which the hooks are more numerous. a. Hooks. |
| 10. | Pulex penetrans, or the Chigoe. Vol. I. p. 49, 102. |
| 11. | Mandible of the larva of Myrmeleon Formicaleo. (Reaum.) Vol. III. p. 121. |
| 12. | Anal spinneret of ditto. (Ibid.) |
| 13. | Branching palpus, or feeler of Trombidium holosericeum. |
| 14. | Part of the tarsus of a spider, to show the simple and pectinated claws. (De Geer.) Vol. III. p. 690. |
| 15. | A pair of spinners of a spider. (Leeuwenh.) Vol. III. p. 392. |
| 16. | A mammula or teat of ditto. Vol. III. p. 391. |
| 17. | Anus of ditto. |
| 18.[Pg 606] | Stilt-legs of a dipterous larva. (De Geer.) |
| FIG. | |
| 1. | The bag-net. Vol. IV. p. 529. |
| 2. | The landing-net. Ibid. p. 534. |
| 3. | Mr. Paul's net. Ibid. p. 530. |
| 4. | The fly-net. Ibid. p. 531. |
| 5. | The forceps. Ibid. p. 533. |
| 6. | The breeding-cage. Ibid. p. 553. |
| 7. | Apparatus for effectually killing large moths, &c. a. The upper piece of the tube. b. The lower. c. The saucepan. Ibid. p. 543. |
| 8. | A beetle transfixed by a pin. Ibid. p. 544. |
| 9. | A butterfly, ditto, with the wings set out by card braces. Ibid. p. 546. |
| 10. | A scale of two inches, with one subdivided into lines, twelve to the inch. |
| FIG. | |
| 1. | Antenna with a lamellate knob. |
| 2. | Ditto Ditto |
| 3. | Antenna with a pectinate knob. |
| 4. | ———– cirrate. |
| 5. | ———– with a tunicate knob. Inside view. |
| 6. | ————————————– Outside view. |
| 7. | ———– clavate, with clava solid. |
| 8. | ———————————— serrate. |
| 9. | ———– with an inflated knob. |
| 10. | ———– gradually incrassate. |
| 11. | ———– biflabellate. |
| 12. | ———– with a patellate scape. a. Scape. |
| 13. | ———– with a solid knob. |
| 14. | ———– clavate, with last joint elongated. |
| 15. | ———– broken. |
| 16. | ———– unguiculate. a. Claw. |
| 17.[Pg 607] | ———– scopiferous. a. Stellated brush. |
| 18. | ———– suddenly incrassate, and biserrate. |
| 19. | ———– capillaceous, and suddenly incrassate. (Stöll.) |
| 20. | ———– bipartite. |
| 21. | ———– clavate, with clava subramose. |
| 22. | ———– bipectinate. |
| 23. | ———– broken. |
| 24. | ———– suddenly incrassate. |
| 25. | ———– pectinate. ♂. |
| 26. | ———– serrate. ♀. |
| 27. | ———– filiform, and submoniliform. |
| 28. | ———– auriculate. a. Auricle. |
| 29. | ———– appendiculate. a. Antenna. b. Appendicles. |
| 30. | ———– capitate, with a multiarticulate knob. |
| 31. | ———– spiral. |
| 32. | ———– fasciculate. |
| 33. | ———– capitate, with a transverse solid knob. |
| 34. | ———– capillaceous, or suddenly attenuated. (Latr.) |
| 35. | ———– embracing the eye. Vol. III. p. 523. |
| FIG. | |
| 1. | Feeler, maxillary, lamellate and appendiculate. a. Appendicle. b. Last joint lamellated. c. Second joint. Vol. III. p. 449, noted. Atractocerus. |
| 2. | ———————– heteromorphous. Cerocoma. |
| 3. | ———————– fasciculate. Lymexylon. |
| 4. | ———————– incrassate. |
| 5. | ———————– clavate. |
| 6. | ———————– conical. |
| 7. | ———————– subulate. |
| 8. | ———————– fusiform. |
| 9. | Maxilla compound, with the lobes spinous. |
| 10. | ——————————————– unarmed. |
| 11.[Pg 608] | ———————— with the upper lobe biarticulate. Vol. III. p. 442. |
| 12. | ———————— with the lobes penicillate. |
| 13. | ——— simple, mandibuliform, lobe unarmed. (MacLeay.) |
| 14. | —————— lobe penicillate. (Ibid.) |
| 15. | ————————— toothed. (Ibid.) |
| 16. | Mandibula. Cyphus Hancocki. |
| 17. | ————– Rhipicera marginata. |
| 18. | ————– Eurhynchus lævior. |
| 19. | ————– Manticora Gigas. |
| 20. | ————– Euchlora viridis. |
| 21. | ————– Macraspis tetradactyla. |
| 22. | ————– Apogonia gemellata. |
| 23. | Labium, &c. of Stenus. |
| 24. | —————— Stomis. (Clairv.) a. Lateral lobes of tongue. b. Intermediate lobe. |
| 25. | —————— Geotrupes. External view. |
| 26. | ———————————– Internal view. |
| 27. | —————— Hister maximus. (MacLeay.) |
| 28. | —————— Leistits. (Clairv.) |
| 29. | —————— Lepidiota Stigma K.MS. |
| 30. | Labrum whiskered. Halictus. a. Appendicle. |
| 31. | ———————— Megachile. |
| 32. | ———————— Pelecium. |
| 33. | ———————— Chasmodia viridis McL. |
| 34. | Labrum, &c. of Genuchus. |
| 35. | –—————— Cremastochilus. Vol. III. p. 422. |
| 36. | Lateral view of the head of Tetraopes, to show the eye wholly divided by the canthus. |
| 37. | Part of the trunk of a spider, to show the position of its simple eyes. |
| 38. | Eyes compound, columnar. Xenos. |
| 39. | ———————————— Ephemera. |
| 40. | Eyes compound and stemmata of Reduvius personatus. |
| 41. | ——————————————– Fulgora laternaria. Stemmata subocular. |
| 42.[Pg 609] | Eyes compound and stemmata of Cercopis. Stemmata intraocular. |
| 43. | Eyes simple, dorsal. Phalangium. |
| 44, 45. | Claw-joint of tarsus of Lamia, to show the arthrium. |
| 46. | ————————— Vespa Crabro. |
| 47. | Tarsus of Entimus imperialis. a. The rotula or ball received by the socket of the tibia. |
| 48. | The penultimate bilobed joint of ditto, with the arthrium separated from the claw-joint. |
| 49. | Part of the claw-joint so separated. a. Muscles which enter the arthrium. |
| FIG. | |
| 1. | Head of Cordylia Palmarum. a a. Muscles fixed in the myoglyphides or muscle-notches. |
| 2. | ———– Apoderus Coryli. |
| 3. | ———– Buprestis acuminata. |
| 4. | ———– Copris. a. Muscles. |
| 5. | ———– Elater. a a. Corneous scales analogous to pax-wax, attached to the depressor muscles, like those of Geotrupes. Vol. IV. p. 183. |
| 6. | ———– tibia. Cordylia Palmarum. |
| 7. | End of thigh, ditto, next the tibia. |
| 8. | ——————— Dynastes. Vol. III. p. 671. note2027. |
| 9. | Head of tibia of ditto. |
| 10. | ——————— Copris bucephalus. Vol. III. p. 670. |
| 11. | End of thigh, ditto, next the tibia. |
| 12. | Middle coxa. Melolontha vulgaris. a. The open part which receives the muscles. |
| 13. | Posterior ditto, ditto. a. The open part. |
| 14. | Head of posterior thigh of ditto. |
| 15. | End of ditto next the tibia. Gryllus. |
| 16. | Head of tibia of ditto. Vol. III. p. 669—. |
| 17. | Lateral view of ditto. |
| 18. | ————— of the head of the coxa of Lamia. a. The[Pg 610] point of attachment with the body, or the orifice through which the muscles pass. |
| 19. | Back view of ditto. |
| 20. | Posterior trochanter of Ichneumon, biarticulate. Vol. III. p. 665. |
| 21. | Tibia of Arachnida, to show the epicnemis. |
| 22. | Multiarticulate spiral antenniform tarsus of Cermatia. |
| 23. | Armed thigh. Scaurus. |
| 24. | ——— tibia. Hispa spinipes. |
| 25. | Auriculate posterior tarsus. Dasytes ater. |
| 26. | Armed anterior ditto. ditto. |
| 27. | ——— anterior coxa. Megachile Willughbiella. |
| 28. | ——— trochanter. Necrophorus. |
| 29. | Calcar or spur, thumb-shaped. Anterior tibia of Acherontia Atropos. |
| 30. | Calcar of posterior tibia. Œnas afer. |
| 31. | ———— anterior ditto. Zabrus gibbus. |
| 32. | ———— intermediate ditto. Acanthopus splendidus. |
| 33. | ———— posterior ditto. Ammophila vulgaris. |
| 34. | –—————————— Acanthopus splendidus. |
| 35. | ———— intermediate tibia. Cimbex Vitellinæ. |
| 36. | Calcar and velum of anterior ditto. Apis mellifica. a. The notch in the first tarsal joint. |
| 37. | Claws of Anomala Frischii. |
| 38. | –——— Macraspis quadrivittata. |
| 39. | –——— Serica brunnea. |
| 40. | –——— Melolontha vulgaris. |
| 41. | Posterior tarsus. Gryllus monstrosus. |
| 42. | Part of tarsus of Scolopendra. |
| 43. | ——————— Lebia, to show pectinated claws. |
| 44. | ——————— Phanæus. |
| 45. | Part of tibia of Onitis Apelles ♂, to show its very minute tarsus. Vol. III. p. 336. |
| 46. | Double claws of Oxypterum. |
| 47. | Claws. Anoplognathus. |
| 48. | ——— Hoplia. Anterior tarsus. |
| 49.[Pg 611] | ——— Macrodactylus subspinosus. |
| 50. | Pecten of Scorpio europæus. |
| 51. | Claw of Hoplia, posterior tarsus. |
| 52. | ———– Meloe variegatus. |
| 53. | Pulvilli and claws of the Asilidæ. |
| 54. | —————————– Tabanus. |
| 55. | —————————– Hive-bee. |
| 56. | Pulvilli and pseudonychia. Lucanus Cervus. |
| 57. | ——————————— Marmarina Lanius. |
| 58. | Segment of the body of Iulus. Showing that two pairs of legs are attached to each segment[1693]. |
| 59. | Tarsus of Priocera, with involute pulvilli. |
| 60. | ———— Nirmus Anseris. |
| 61. | ———— Xenos Peckii. |
| 62. | ———— Melittophagus K. Vol. I. p. 163. IV. p. 232. |
| 63. | ———— Ixodes Ricinus. (De Geer.) |
| FIG. | |
| 1. | Inside of elytrum of Dytiscus marginalis. |
| 2. | ———————— Strategus Aloeus. Part of hypoderma peeled off. |
| 3. | Part of ditto of Buprestis vittata, to show the axis. |
| 4. | —————— Passalus. |
| 5. | —————— Dynastes. |
| 6. | —————— Elater sulcatus, to show the epipleura. |
| 7. | —————— Blaps lethifera, ditto. |
| 8. | —————— Dynastes quadrispinosus, ditto. |
| 9. | Alitrunk and part of tegmina and wings of Locusta. a. Space marked by the transverse nervure in which all the nervures of the anal area terminate. Vol. III. p. 618. |
| 10. | Alitrunk and part of elytra and wings of Phyllopertha horticola. |
| 11.[Pg 612] | Alitrunk and part of tegmina and wings of Fulgora laternaria. Postfrænum funiculate, with an elastic part marked a. Vol. III. p. 558. |
| 12. | –———————— hemelytra and wings of Pentatoma. |
| 13. | –———————— wings of Panorpa. |
| 14. | ———————————— Trichoptera. |
| 15. | ———————————— Lepidoptera. Geometra. |
| 16. | ———————————— Lepidoptera. |
| 17. | ———————————— Diptera. |
| 18. | Wing of Chermes Fraxini. |
| 19. | Tegmen of Locusta. |
| 20. | ————– Issus. |
| 21. | Wing. Cercopis sanguinolenta. |
| 22. | ——– Locusta. |
| 23. | Hemelytrum of Reduvius? |
| FIG. | |
| 1. | Gills of a spider. (Treviran.) |
| 2. | ———– Scorpio europæus. (Ibid). |
| 3. | Aëriducts of larva of Ephemera fusco grisea. (De Geer.) Vol. IV. p. 58. |
| 4. | —————————————– vespertina. (Ibid.) Ibid. |
| 5. | —————————————– vulgata. (Ibid.) Ibid. |
| 6. | ————————– Sialis lutaria. (Ibid.) Ibid. |
| 7. | Thread-like ditto of the upper and under side of the larvæ of Trichoptera. (Ibid.) Ibid. p. 56. |
| 8. | Part of the body of the larva of a Libellula, laid open to show the tracheæ. (Reaum.) a. a. a. Tracheæ. Vol. IV. p. 66. |
| 9. | Part of the imago of ditto. (Ibid.) a. Vesicles that terminate the tracheæ. b. Oblong ditto. Vol. IV. p. 68. |
| 10. | Pupa of Corethra culiciformis. (De Geer.) a. b. Vesicles connected with the tracheæ. c. Tail. Vol. IV. p. 67. |
| 11. | Part of the head of Armadillo zonatus. a. Pseudo-spiracle. Vol. III. p. 493. |
| 12.[Pg 613] | Part of the trunk of Goerius olens, to show its antepectoral spiracle. Vol. IV. p. 43. |
| 13. | Part of the abdomen of Pneumora. a. The series of ridges, by striking the hind leg over which they probably produce their noise. Vol. II. p. 391; III. p. 339. |
| 14. | Underside of part of the alitrunk of Lygæus sexmaculatus K. MS. b. Branchiform apparatus between the scapula and parapleura. Vol. IV. p. 45. |
| 15. | —————————————————— Pentatoma rufipes. a. Corrugated membrane. b. Apparent fringe of hairs or bristles. Ibid. |
| 16. | Spiracle of larva of Oryctes nasicornis. (Sprengel.) |
| 17. | ———————— Dytiscus marginalis. (Ibid.) |
| 18. | Part of the trachea and bronchiæ of the pupa of Smerinthus Populi. (Ibid.) |
| 19. | Spiracle of the imago of Oryctes nasicornis. a. The boss. (Ibid.) |
| 20. | Dorsal spiracle of Cermatia. |
| 21. | Pencil of hairs attached to a supposed respiratory plate in certain Noctuidæ. Vol. IV. p. 60. |
| 22. | Part of the back of the abdomen of the pupa of a Pentatoma. a. Pseudo-spiracle. b. Connecting corrugations. Vol. III. p. 713. |
| 23. | Unilabiate spiracle of Gonyleptes. |
| 24. | Portion of ventral segments of abdomen of Aradus laminatus K. a. Tobaccopipe-shaped organ near the ventral spiracles. Vol. III. p. 713. |
| 25. | Part of the back of the alitrunk of Belostoma grandis, to show the metapnystega? or rather spiracle. Vol. III. p. 572; IV. p. 45. |
| 26. | Pseudo-spiracle of Carkinodes cancriformis. Vol. III. p. 714. |
| 27. | Part of the ventral segments of the abdomen of Lygæus compressipes. a. Pseudo-spiracles. |
| 28. | Portion of dorsal segments of Dytiscus marginalis, to show the large anal spiracles. Vol. IV. p. 42. |
| 29.[Pg 614] | Plumiform gills in the mouth of the spiracles of the larva of Cossus ligniperda. (Sprengel.) |
| FIG. | |
| 1. | Brain, spinal chord and ganglions of a full-grown caterpillar of Pontia Brassicæ. a. The brain. b. The double spinal chord. c d. Ganglions with a portion of their nerves. |
| 2. | Brain, spinal chord and ganglions, after two days, when the chord is shortened. |
| 3. | ——————————————— when the animal is become a pupa. |
| 4. | ——————————————— when it has been a pupa six days. |
| 5. | ——————————————— just before it assumes the imago. |
| 6. | ——————————————— when it has become a butterfly. Vol. IV. p. 24—. |
| 7. | Intestinal canal of the caterpillar. a. Saliva vessel. b. Silk reservoir. c. Gullet or œsophagus. d. Stomach. e e e. Bile vessels. f. Large intestine. g. Rectum. |
| 8. | —————————————— after it has assumed the pupa two days. |
| 9. | —————————————— after eight days. a. Crop or honey-stomach, first showing by the dilatation of the base of the œsophagus. |
| 10. | —————————————— a. Honey-stomach become a lateral appendage of the œsophagus b. |
| 11. | ——————- of the butterfly. a. Honey-stomach. b. Œsophagus. c. Small intestine become very long. d. Rectum. Vol. IV. p. 118. |
| 12. | Anal portion of the interior of the female butterfly. a. Ovaries. b. Oviduct. c. Colleterium or varnish secretor. d. Spermatheca or sperm receptacle. e. Part of the spinal chord. f. Rectum. g. A secretory organ filled with a thick white fluid, which is supposed to lubricate the passage. Vol. IV. p. 132, 152. |
N.B.—The reader is requested to observe that in the preceding Index, one object has been to point out, wherever it was possible, to what Linnean or Fabrician genus each of the genera included in it may be referred, though no species of it may have been known to those authors. Where a Synonym of any genus is printed in Roman letters, it indicates that such genus is so denominated in the former Editions of this work. The first namer of a genus, together with any author who has written a monograph upon it, or on any tribe, and the more eminent systematists, are usually only quoted. If the reader finds any difference, as is sometimes the case, between the text and the Index, he will be pleased to correct the former by the latter.—For instance, it was not discovered till too late, that the name Uria was pre-occupied in Ornithology;—in the Index this name is altered to Uriata. Doubtful species are printed in Italics.[Pg 669]
END OF THE FOURTH AND LAST VOLUME.
Published by the same Authors.
By Mr. Kirby,
MONOGRAPHIA APUM ANGLIÆ:
Or, An Attempt to divide into their natural Genera and Families such Species of the Linnean Genus Apis as have been discovered in England. In two volumes 8vo, with plates. Price 1l. 1s.
By Mr. Spence,
In one closely printed Volume, 8vo, price 7s. Boards,
TRACTS on POLITICAL ECONOMY, viz.
1. Britain independent of Commerce. Seventh Edition.
2. Agriculture the Source of the Wealth of Britain. Third Edition.
3. The Objections against the Corn Bill refuted. Fifth Edition.
4. Speech on the East India Trade. With Prefatory Remarks on the Cause and Cure of our Present Distresses, as originating from Neglect of Principles laid down in these Works.
THE MAGAZINE OF NATURAL HISTORY, AND JOURNAL OF ZOOLOGY, BOTANY, MINERALOGY, GEOLOGY, AND METEOROLOGY. Conducted by J. C. Loudon, F.L.S. H.S. &c. Vol. I. in 8vo. Price 18s. boards: (to be continued in Numbers every Two Months, alternately with the Gardener's Magazine,) price 3s. 6d. The different Departments edited by Gentlemen eminent in each. The Drawings of Botany and Conchology, by Sowerby;—of Animals, by Harvey;—of Trees, by Strutt: and the Engravings on Wood by Branston.
The objects of this work are—To record every new fact belonging to the subject; to render every part of the subject interesting to the amateur and general reader; to lead on the reader by degrees from the more elementary details to higher views and discussions; and to translate the technical terms, and Latin or Greek words used in Natural History, as they occur, and to give the derivation and accentuation of all systematic names.
By the same Author,
THE GARDENER'S MAGAZINE, AND REGISTER OF RURAL DOMESTIC IMPROVEMENT. In 8vo. No. XVIII. (to be continued every Two Months, alternately with the Magazine of Natural History) price 3s. 6d. Also may be had Vols. 1 to 4. price 21. 19s. 6d. boards.
AN ENCYCLOPÆDIA OF GARDENING; comprising the Theory and Practice of Horticulture, Floriculture, Arboriculture, and Landscape Gardening; including all the latest Improvements, a general History of Gardening in all Countries, and a Statistical View of its present State, with Suggestions for its future Progress in the British Isles. Illustrated with many hundred Engravings on Wood. A New Edition, complete in 1 large Vol. 8vo. price 2l. extra boards.
AN ENCYCLOPÆDIA OF AGRICULTURE; comprising the Theory and Practice of the Valuation, Transfer, Laying-out, Improvement, and Management of Landed Property; and the Cultivation and Economy of the Animal and Vegetable Productions of Agriculture, including the latest Improvements; a general History of Agriculture in all Countries; and a Statistical View of its present State, with Suggestions for its future Progress in the British Isles. In 1 large Vol. 8vo. closely printed, with upwards of 800 Engravings on Wood, 2l. 10s. boards.
AN ENCYCLOPÆDIA OF PLANTS; comprising the Scientific Name, its Authority, Derivation, and Accentuation, Synonym, French, German, Italian, and Spanish Appellation, &c. &c. &c. The whole in English, illustrated with Pictorial Types on a new principle, with Engravings of one or more species of every genus included in the work. The specific Characters by an eminent Botanist, the Drawings for the Figures by Sowerby, and the Engravings on Wood by Branston. In 1 large Vol. 8vo. closely printed, with upwards of Seven Thousand Engravings on Wood. Edited by J. C. Loudon, F.L.S. H.S. &c. (Nearly ready.)
A NEW SYSTEM OF GEOLOGY, in which the great revolutions of the Earth and Animated Nature are reconciled at once to Modern Science and to Sacred History. By Andrew Ure, M.D. F.R.S. Member of the Geological and Astronomical Societies of London, &c. &c. &c. (Nearly ready.)
This work will be illustrated by Copper-plate Engravings of Shells, characteristic of the Strata and Superposition of the Bone-Caverns, and of Casts of Fossil Plants; besides about 50 Wood Engravings, representing the most curious Animal Inhabitants of the primeval World described by Cuvier, and other Fossil Zoologists.
THE BOOK OF NATURE; being a Popular Illustration of the general Laws and Phænomena of Creation, in its Unorganized and Organized, its Corporeal and Mental Departments. By John Mason Good, M.D. F.R.S. F.R.S.E. 2nd Edit, in 3 Vols. 8vo. 1l. 16s. boards.
"The work is certainly the best philosophical digest of the kind which we have seen."—Monthly Review.
THE ENGLISH FLORA. By Sir James E. Smith, M.D. F.R.S. President of the Linnæan Society, &c. &c. A new Edition in 4 Vols. 8vo. 2l. 8s. boards.
By the same Author,
A GRAMMAR OF BOTANY, illustrative of Artificial, as well as Natural Classification, with an Explanation of Jussieu's System. 2nd Edition, 8vo. with 277 Figures of Plants, and their various Parts and Organs, price 12s.; or coloured, 1l. 11s. 6d.
AN INTRODUCTION TO THE STUDY OF PHYSIOLOGICAL AND SYSTEMATICAL BOTANY. In 8vo. 6th Edition, with 15 Plates, price 14s. plain, or coloured, 1l. 8s. boards.
COMPENDIUM FLORÆ BRITANNICÆ, 7s. 6d.
MUSCOLOGIA BRITANNICA; containing the Mosses of Great Britain and Ireland, systematically arranged and described; with Plates, illustrative of the characters of the Genera and Species. By William Jackson Hooker, LL.D. F.R.A. and L.S. &c.; and Thomas Taylor, M.D. M.R.I.A. and F.L.S. &c. The 2nd Edition, corrected and enlarged, in 8vo. 1l. 11s. 6d. plain, and 31. 3s. coloured plates.
AN ILLUSTRATED INTRODUCTION TO LAMARCK'S CONCHOLOGY, contained in his "Histoire Naturelle des Animaux sans Vertèbres;" being a Literal Translation of the Descriptions of the recent and Fossil Genera, accompanied by Twenty-two highly-finished Lithographic Plates, in which are given Instructive Views of the various Genera and their Divisions, drawn from Nature, from characteristic and generally well-known Species. By Edmund A. Crouch, F.L.S. Royal 4to. Price 1l. 11s. 6d. plain, or 3l. 3s. coloured.
Also, just published,
AN EPITOME OF LAMARCK'S ARRANGEMENT OF TESTACEA: with illustrative Observations and comparative Tables of the Systems of Linnæus and Lamarck. By C. Dubois, F.L.S. and F.H.S. 8vo. Price 14s. boards.
THE LINNÆAN SYSTEM OF CONCHOLOGY; describing the Orders, Genera, and Species of Shells, arranged into Divisions and Families, with a view to facilitate the Student's Attainment of the Science. By J. Mawe. In 8vo. with a Plate to each Genera (37); plain, 1l. 1s.; coloured after Nature, 2l. 12s. 6d.
By the same Author,
NEW DESCRIPTIVE CATALOGUE OF MINERALS, greatly extended, with Diagrams of their Simple Forms. 7th Edition. Price 6s.
CONVERSATIONS ON MINERALOGY; with Plates, engraved by Mr. and Miss Lowry, from Original Drawings, comprising upwards of 400 Figures of Minerals, including 12 beautifully coloured Specimens. 2nd Edition, in 2 Vols. 12mo. Price 14s. boards.
"The plan of these Conversations is happily conceived, and it is executed with ability and taste. The Author has studiously avoided all unnecessary parade of technical Diction, has rendered the Doctrine of Crystallography more familiar than heretofore to the Tyro in Mineralogy, and has included some of the most recently discovered Substances. We may, therefore, unhesitatingly characterise this Work as one of the most desirable Text Books that have issued from the British Press."—Monthly Review, Dec. 1822.
LECTURES ON BOTANY; containing the Descriptive ANATOMY of those Organs on which the Growth and Preservation of the Vegetable depend. By Anthony Todd Thomson, M.D. 8vo. plates, 1l. 8s. boards.
FIRST STEPS TO BOTANY, intended as popular Illustrations of the Science, leading to its Study as a Branch of general Education. By James L. Drummond, M.D. Professor of Anatomy and Physiology in the Belfast Academical Institution. In 12mo. with 100 Wood-Cuts, comprising 200 Figures. 2nd Edition. Price 9s. boards.
"This answers more completely to the proper notion of an Introduction to Botany than any work we have seen."—Eclectic Review, April.
CONVERSATIONS ON BOTANY, with Twenty-one Engravings. The Sixth Edition, enlarged, in 1 Vol. 12mo. Price 7s. 6d. plain, or 12s. coloured.
The object of this Work is to enable young persons to acquire a knowledge of the vegetable productions of their native country; for this purpose the arrangement of Linnæus is briefly explained, and a native plant of each class (with a few exceptions) is examined, and illustrated by an engraving; and a short account is added of some of the principal foreign species.
ELEMENTS OF AGRICULTURAL CHEMISTRY. By Sir Humphry Davy, Bart. In 8vo. (the 4th Edition) illustrated with 10 Engravings, price 15s. boards.
In the Press, and speedily to be Published, in Monthly Volumes.
THE CABINET CYCLOPÆDIA, conducted by the Rev. Dionysius Lardner, LL.D. F.R.S. L. and E. M.R.I.A. M.Z.S. Hon. F.C.P.S. M.A.S. &c. &c. Professor of Natural Philosophy and Astronomy in the University of London, assisted by numerous Eminent Literary and Scientific Characters.—1. Each volume will contain about 400 pages, foolscap 8vo. price 6s.—2. When plates are necessary, a corresponding reduction will be made in the letter-press. Each volume will be accompanied by its own plates, and no others. Principal Divisions of the Work:—I. The Cabinet of Natural Philosophy. Eight Volumes.—II. The Cabinet of Arts. Ten Volumes.—III. The Cabinet of Natural History. Fourteen Volumes.—IV. The Cabinet of Geography. Six Volumes.—V. The Cabinet of Philosophy. Twelve Volumes.—VI. The Cabinet of Physiology and Medicine. Six Volumes.—VII. The Cabinet of Literature. Ten Volumes.—VIII. The Cabinet of History and Antiquities. Twenty-two Volumes.—IX. The Cabinet of Biography. Twenty Volumes.—X. The Cabinet of Lexicography. Ten Volumes.
The order of publication will not follow the above scheme. The progress of the several divisions of the work will be contemporaneous, so as to sustain an interest by variety during the period of publication.
[1] Το Ἡγεμονικον.
[2] See Hooper's Medical Dictionary, under Nervous Fluid, and Mr. Sandwith's useful Introduction to Anatomy and Physiology, 83.
[3] N. Dict. d'Hist. Nat. xvi. 305—.
[4] Cuv. Anat. Comp. ii. 362. Compare MacLeay Hor. Entomolog. 215—.
[5] N. Dict. d'Hist. Nat. ubi. supr.
[6] Cuv. Anat. Comp. ii. 360. MacLeay Hor. Ent. 201.
[7] N. Dict. d'Hist. Nat. xvi. 306. MacLeay Hor. Ent. 200—.
[8] Ibid. 307. The great sympathetic nerves in fishes are said to have no ganglions. Cuv. ubi. supr. 297.
[9] They are called trisplanchnic because they render to the three cavities of the viscera:—viz. the thoracic, the abdominal and the pelvic. N. Dict. d'Hist. Nat. xxii. 524. 527.
[10] In Hemiplegia, &c.
[11] N. Dict. d'Hist. Nat. xvi. 307.
[12] Thus in the Molluscæ there must be a great difference in this respect, since in some of these the brain or cerebral ganglion has been cut off with the head, and another reproduced. Ibid. xvi. 306. Comp. v. 391.
[14] Comp. Plate XXX. Fig. 1. and 6. and Carus. Introd. to Comp. Anat. i. 64.
[15] Lyonet Anatom. 100.
[16] Ibid. 101.
[17] Lyonet Anatom. 100. In man and the vertebrate animals, the medullary pulp is every where homogeneous; under the microscope it appears to consist of a number of minute conglomerated globules. M. Vauquelin has analysed it, and found it to contain, of water 80 parts; of albumen in a state of demicoagulation 7·0; of phosphorus 1·50; of osmazone 1·12; of a white and transparent oily matter 4·53; of a similar red do. 0·75; of a little sulphur and some salts 5·15. N. Dict. d'Hist. Nat. xxii. 531—.
[18] Anat. 99.
[19] Malpigh. de Bombyc. 20. Swamm. Bibl. Nat. i. 224. a.
[20] Anat. Comp. ii. 348.
[21] Lyonet Anat. 100. t. iv. f. 6. Sandwith Introd. 59—.
[22] Plate XXI. Fig. 1. 7. 8. a.
[23] N. Dict. d'Hist. Nat. xxii. 527.
[24] Ibid. v. 591.
[25] Cuv. Anat. Comp. ii. 318. Swamm. Bibl. Nat. t. xxix. f. 7. Herold Schmetterl. t. ii. f. 1-10. a.
[26] Cuv. Ibid. 322. 337.
[27] Cuv. Anat. Comp. 324.
[28] Arachnid. t. i. f. 13. m.m.
[29] Cuv. ubi supr. 343. 346. Treviranus Arachnid. t. v. f. 45. a. Plate XXI. Fig. 8. a.
[31] Cuv. ubi supr. 337.
[32] Plate XXI. Fig. 8. Swamm. Bibl. Nat. i. 36. b.
[33] Arachnid. t. v. f. 45.
[34] Swamm. ubi supr. t. xliii. f. 7.
[35] Ibid. 112. a.
[36] Cuv. Anat. Comp. ii. 337. 343—.
[37] Ibid. 336.
[38] Herold Schmetterl. t. ii. f. 1.
[39] Lyonet Anat. 98.
[40] Cuv. ubi supr. 342. Gaede N. Act. Acad. Cæs. XL. ii. 323. Cuv. Ibid. 351.
[41] Cuv. ubi. supr. 348.
[42] Treviranus Arachnid. t. v. f. 45.
[43] Plate XXI. Fig. 7. 8. Swamm. Bibl. Nat. t. xliii. f. 7.
[45] Lyonet Anat. 100.
[46] N. Dict. d'Hist. Nat. xxii. 522—.
[47] Lyonet ubi supr. t. ix. f. 1-4.
[48] Cuv. Anat. Comp. ii. 339. 343.
[50] Swamm. ubi supr. t. xl. f. 5. Cuvier (ii. 332.) accuses Swammerdam of representing the spinal marrow in this grub as producing nerves only on one side; whereas he expressly states (ii. 50. b.) that a considerable number spring on each side from the eleven ganglions, but that to avoid confusion he had omitted some.
[51] Cuv. ubi supr. 325.
[52] Swamm. Bibl. Nat. t. xv. f. 6.
[53] Treviran. Arachnid. t. l. f. 13. 1-4.
[54] Swamm. ubi supr. t. xxii. f. 7.
[55] Treviran. ubi supr. t. v. f. 45.
[57] Cuv. Anat. Comp. ii. 346.
[59] Cuv. ubi supr. 337.
[60] Ibid. 335—.
[61] Cuv. ubi supr. 348.
[62] Ibid. 320—.
[63] Ibid. 340—.
[64] Ibid. 338—.
[65] Gaede ubi supr.
[66] Cuv. ubi supr. 323—. 327—. Mr. Bauer (Phil. Trans. 1824. t. ii. f. 1.) has figured only seven, excluding the brain, in that of the silk-worm, and Malpighi (De Bombyc. t. vi. f. 2.) ten,—Swammerdam (Bibl. Nat. t. xxviii. f. 3.) however has twelve.
[67] Ibid. 326.
[68] Ibid. 352.
[69] Ibid. 343—.
[70] Ibid. 345.
[71] Ibid. 325—.
[72] Ibid. 351.
[73] Cuv. ubi supr. 339.
[74] Ibid. 335—.
[75] Lyonet Anat. 190.
[76] Cuv. ubi supr. ii. 340. Malpigh. de Bombyc. t. vi. f. 2.
[77] Cuv. Ibid. 348.
[78] Swamm. Bibl. Nat. t. xlviii. f. 7.
[79] Cuv. Ibid. 319.
[80] N. Dict. d'Hist. Nat. xxx. 420.
[81] Treviran, Arachnid. t. v. f. 45. m.
[82] Plate XXI. Fig. 1. 7. 8. d.
[83] Lyonet ubi supr. t. x. f. 5. 6.
[84] Ibid. 192.
[85] Cuv. ubi supr. 323. 335.
[86] Ibid. ii. 339.
[87] Ibid. 342.
[88] Swamm. Bibl. Nat. t. xxii. f. 6. m.m.
[89] Cuv. ubi supr. 350.
[90] Ibid. 335.
[91] Vol. III. p. 495—. Lyonet. Anat. 581.
[92] Cuv. ubi supr. 337.
[93] Cuv. ubi supr. 351.
[94] Ibid. 352.
[95] Cuvier (Ibid. 319.) seems not to have been aware that Swammerdam was the first discoverer of these nerves, since he attributes their name to Lyonet.
[96] Bibl. Nat. i. 138. b. t. xxviii. f. 2. a, b, c. f. 3. g.
[97] Ubi supr. 578.
[98] Ubi supr. 320. 339, &c.
[99] Cuv. ubi supr. 349.
[100] Lyonet Anat. t. ix. x.
[101] Plate XXI. Fig. 8. Swamm. Bibl. Nat. t. xxii. f. 6.
[102] Ibid. t. xv. f. 6.
[104] Swamm. ubi supr. t. xliii. f. 7. h, h.
[106] In Mr. Bauer's figure (Philos. Trans. 1824. t. ii. f. 1.) no less than eighteen pairs of nerves are represented as issuing from the internodes; but it should seem as if in the specimen from which his figure was taken, several of the ganglions, perhaps from some injury received in the dissection, had become obliterated, while their nerves remained: yet still, even making allowance for these, many pairs will appear to take their origin from the spinal chord.
[107] Comp. Cuv. Anat. Comp. ii. 102-123.; with Swamm. Expl. of Plates XXXII. t. xxviii. f. 3. k.
[108] Malpighi seems, however, to agree with him. De Bombyc. t. vi. f. 1.
[109] Lyonet ubi supr. 201. t. ix. f. 1, 2. n. 1, 2. &c.
[110] Swamm. ubi supr. 1. 139. a. t. xxviii. f. 3. s, s.
[111] In Lesser Insecto-theol. ii. 84. note *.
[112] Linn. Trans. ii. 8. Aristotle had observed this vitality of insects, and that that of the myriapods is greatest. Hist. Animal. l. iv. c. 7. De Respiratione, c. 3. Reptiles have also this faculty. N. Dict. d'Hist. Nat. xxix. 161.
[113] Cuv. Anat. Comp. ii. 283—. These are named "the upper and lower cervical ganglions."
[114] Lyonet Anat. t. ix. x. Plate XXI. Fig. 1. a. b.
[116] N. Dict. d'Hist. Nat. ii. 47—. v. 592. xvi. 308—.
[118] Huber Fourmis, 260—. Reaum. vi. 172—.
[120] N. Dict. d'Hist. Nat. ii. 279—.
[121] Cuv. Anat. Comp. ii. 319. 337.
[122] Ibid. ii. 322, 323—; 338. 339—.
[126] Herold Schmett. t. ii. f. 6.
[127] Ibid. t. ii. f. 7.
[131] Anat. Comp. ii. 348.
[132] N. Dict. d'Hist. Nat. xvi. 313. Comp. i. 420.
[139] Antommarchi's Last Days of Napoleon.
[140] Linn. Trans. xi. 393.
[143] Zoological Journal, no. i. 5.
[144] Anat. Compar. iv. 296.
[145] Plin. Hist. Nat. l. xi. c. 3. Even Aristotle seems to have given into the common opinion. De Respirat. c. 3, 9. &c.
[146] Philos. Trans. v. 2011. Works, 4to. i. 79, 112.
[147] Aristot. Hist. Animal. l. viii. c. 27.
[148] On Air and Fire, 148, 155.
[149] Tracts, 208.
[150] Mem. on Respirat. 75.
[151] Ann. de Chimie, xii. 273.
[152] F. L. A. Sorg, Respirat. Insect. et Verm. Ellis, Inquiry into Chang. prod. on Atmosph. Air by Respirat. &c. 69.
[153] Ann. de Chimie, xii. 273.
[154] Sprengel, Commentar. &c. 27—.
[155] Plate XXIII. Fig. 2. and Plates VIII. IX. XVI. XXIX. c´,h´´, m´´, A´´, D´´.
[156] Moldenhawers (Anat. der Pflanz. 314—.) affirms that the spiracles of most insects are quite closed: but Sprengel (Commentar. § 8.) has satisfactorily refuted that opinion.
[158] Sprengel, Commentar. § 7.
[159] Ibid. t. iii. f. 30.
[162] Sprengel, 7. t. iii. f. 30.
[163] Ibid. t. ii. f. 22. t. iii. f. 29.
[165] Ibid. Fig. 16. Sprengel, Ibid. 9. t. 1. f. 4-6.
[166] Ibid. 9. t. i. f. 9.
[168] Sprengel, Ibid. t. iii. f. 27.
[169] Sprengel, Commentar. 7—.
[170] Sprengel, from whom I have borrowed this quotation, expresses the time by "scripulo horæ." This word is of uncertain meaning, being scarcely ever applied to time; but as it means the twenty-fourth part of an ounce, Faber conjectures it may mean the same portion of an hour.
[171] Sorg, Disquisit. circa respirat. insect. 27, 46, 66. Sprengel ubi supr. 11—.
[172] Chabrier sur le Vol des Ins. c. l. 454.
[173] Plate XXIX. Fig. 28. A´´.
[175] Sepp. I. iv. t. ii. f. 3.
[176] Ibid. t. xiv. f. 3.
[177] Ibid. t. v. f. 6, 7.
[178] Ibid. t. i. f. 7, 8.
[179] Ibid. t. x. f. 6, 7.
[180] Ibid. v. t. i. f. 3.
[181] Sphinx Labruscæ Merian Surinam. 34.
[182] Plate XXIX. Fig. 28. A´´.
[183] Swammerd. Bibl. Nat. t. xxvii. f. 5. Compare Sturm Deutsch. Fu. i. t. v. f. r.
[184] Plate XXIX. Fig. 12. c´.
[185] De Geer, i. 81. t. v. f. 10. f.
[186] Sur le Vol des Ins. c. i. 459.
[187] Reaum. iv. 246. t. xix. f. 8. s.
[188] In this tribe, which I forgot to remark before, (see Vol. III. p. 549—.) there seems both prothorax and collar.
[189] Vol. III. p. 550, 559. &c.
[190] Plate VIII. Fig. 14. h´´.
[191] Plate XXIX. Fig. 14, 15. m´´.
[195] Chabrier sur le Vol des Ins. c. iii. t. vi. f. 4. Sa, Sp.
[200] Sprengel, Comment. 3.
[201] Ibid.
[202] vi. 398.
[203] De Geer, ii. 635.
[204] Fourmis, 22.
[205] Osservaz. &c. sullo Iulus fœtid. 14—.
[206] They are particularly visible in an undescribed East Indian species, (S. alternata K. M. S.) with scuta alternately black and yellow.
[207] Plate XXIX. Fig. 20. A´´.
[208] De Geer, vii. t. vi. f. 3.
[210] De Geer vi. 67. t. iii. f. 10. ss. 14. Mr. W. S. MacLeay (Philos. Mag. N. Ser. No. 9. 178.) says that in this grub the longitudinal trunks of the Tracheæ send off at equal distances lateral branches just as if there were spiracula to correspond with them. This is evidently a preparatory step to the formation of those that ultimately appear in the perfect insect.
[211] De Geer 66. t. iii. f. 13.
[213] Reaum. iv. 375—. t. xxvi. f. 7, 8.
[214] Ibid. 555. t. xxxv. f. 10. ss.
[215] Ibid. 519—. t. xxxvii. f. 3, 4.
[216] Plates XVI. Fig. 9. a b. XIX. Fig. 9, 10, 12, 13. a. XXIX. Fig. 3-7.
[219] Compare Swamm. Bibl. Nat. i. 154. t. xxxi. f. 5. Reaum. iv. 601—. t. xliii. De Geer vi. 317—. t. xvii. f. 2-8.
[220] Swamm. Ibid. t. xxxi. f. 7, 8.
[221] Reaum. iv. 607.
[223] Reaum. iv. t. xxxii. f. 2. e.
[224] Mr. W. S. MacLeay (Philos. Mag. N. Ser. n. 9. 179.) asserts that what Reaumur (iv. 487. t. xxx. f. 6. ll) calls the first pair of legs of this grub, are the usual palmated stigmata which occur on the humerus of the larvæ of Muscidæ. It does not appear whether he has himself examined this grub, but Reaumur (443) states that it has seven pairs of legs all armed with claws. If this is correct, it is not properly a palmated organ.
[225] Reaum. iv. t. xxx. f. 10.
[226] Reaum. iv. t. xxx. f. 447—.
[227] Ibid. 456. t. xxxi. f. 1-7.
[229] Bibl. Nat. ii. 44.
[231] Reaum. v. t. iv. f. 6. s, u.
[233] De Geer vi. 395—. t. xxiv. f. 16. 18. d.
[234] v. t. vi. f. 1, 2.
[235] De Geer iii. 367. t. xviii. f. 1, 2, 9.
[236] Ibid. vi. 36. 194—. t. ii. f. 2, 3. s.
[237] Plate XVI. Fig. 9. a. b.
[238] De Geer ii. 539—. t. xi. f. 12, 16, &c.
[239] De Geer i. 526—. t. xxxvii. f. 2-6.
[240] Ibid. iv. 362—. t. xiii. f. 16-19.
[241] Vol. I. p. 282—. II. 365—.
[242] See Reaum. vi. t. xlii.—xlvi. and Plate XXIX. Fig. 3-5.
[243] Reaum. Ibid. t. xlv. f. 2.
[244] Plate XXIX. Fig. 5. De Geer ii. 624—.
[245] Ibid. Fig. 4. De Geer Ibid. 647—.
[246] Ibid. Fig. 3. De Geer Ibid. 653—.
[247] Ibid. Fig. 6. De Geer Ibid. 727—.
[248] Reaum. vi. 465.
[249] Ibid. t. xlii. f. 4, 5. De Geer ii. 623.
[250] Ibid. 648. t. xvii. f. 11, 12.
[251] Vol. III. p. 154. De Geer ii. 697—. t. xxi. f. 4, 5, 12.
[252] De Geer Ibid. 666—. t. xix. f. 6.
[253] Reaum. vi. 393. t. xxxvi. f. 8, 9. t. t.
[254] Reaum. vi. 395. t. xxxvi. f. 8-9. c. c.
[256] Marcel de Serres (Mem. du Mus. 1819. 137, &c.) calls the tubular tracheæ that receive the air, arterial tracheæ, and the vesicular ones which act as reservoirs, pulmonary tracheæ.
[258] Treviranus Arachnid. 7—. t. l. f. 1. r. f. 10. Comp. N. Dict. d'Hist. Nat. xxx. 419. Latreille calls these gills Pneumo-branches.
[259] Treviranus Ibid. 24. Plate XXIX. Fig. 1.
[263] Sprengel Commentar. t. i. f. 1.
[264] Ibid. f. 10.
[265] Ibid. t. ii. f. 15.
[266] Malpigh. De Bombyc. t. iii. f. 3.
[267] Ibid. t. iv. f. 1.
[268] Lyonet Anat. 101.
[269] Lyonet Anat. 101.
[270] Sprengel (ubi. supr. 16.) says that he never found more than two; but as Lyonet affirms that he has very often separated them (102), his accuracy cannot be questioned.
[271] Lyonet Anat. 103.
[272] Ibid. Cuv. Anat. Comp. iv. 438. This author says that the intermediate tunic is the spiral thread (437).
[273] Lyonet 102.
[274] Ibid. 104. Sprengel Commentar. 17.
[275] Lyonet 104. Sprengel Commentar. 17.
[276] Lyonet 102. Malpigh. De Bombyc. 12. Reaum. i. 130.
[277] Swamm. Bibl. Nat. t. ii. f. 7.
[278] Lyonet 411.
[279] Professor Kidd (Philos. Trans. 1825. 235.) conjectures that the tracheæ, as well as air-vessels, may possibly be blood-vessels; but this hypothesis is inconsistent with the fact recently discovered by Dr. Carus, of a circulation, by other means, in larvæ. See Carus Introd. to Comp. Anat. &c. ii. 400.
[280] N. Dict. d'Hist. Nat. xvii. 541. Reaum. vi. 397. Plate XXIX. Fig. 8. shows three of them at a.
[281] Essay on the Bots, &c. 23. t. i. f. 7, 32, &c.
[282] Ibid. 49. Valisnieri i. 101. t. vi. f. 4. &c.
[283] Bibl. Nat. i. 149. a. t. xxix. f. a. Cuv. Anat. Comp. iv. 439. Malpigh. De Bombyc. t. iii. f. 2.
[284] Sur le Vol des Ins. c. ii. 336. note 1.
[285] Swamm. Bibl. Nat. t. xvii. f. 9. Cuvier Ibid. 440.
[288] De Geer vi. 374.
[289] Reaum. v. 40. t. vi. f. 4, 7.
[290] Sprengel Comment. 4.
[291] De Geer ii. 667, 675.
[292] Reaum. vi. 394—.
[293] Reaum. vi. 394—. Cuv. Anat. Comp. iv. 440—. N. Dict. d'Hist. Nat. xvii. 540—.
[294] Plate XXIX. Fig. 9. a, b. Reaum. vi. 418—. 450.
[295] Cuv. Anat. Comp. iv. 441.
[297] Sprengel Comment. 17. t. iii. f. 24.
[298] Ibid. t. i. f. 11.
[299] Sur le Vol des Ins. c. ii. 336. note 1.
[300] Sprengel Comment. 13—. These oscula or pores in the straw of Triticum hybernum, as figured by Mr. Bauer's admirable pencil, (Sir J. Banks On the Blight, &c. t. ii. f. 3.) exactly resemble the spiracles of insects.
[301] Reaum. i. 136.
[302] Bonnet Œuvr. iii. 39—.
[303] Ibid. 43.
[304] Ibid. 50.
[305] Ibid. 69.
[306] De Geer ii. 117.
[308] Reaum. iv. 520.
[309] Mr. B. Clark thinks that he has discovered spiracles in this larva in the usual situation, (Essay on the Bots, &c. 48. t. ii. f. 3.) but they are probably analogous to the spiraculiform tubercles of Œ. Ovis. Reaum. iv. 566. t. xxxv. 17-19. t. Vallisnieri (Esperienz. &c. 136) notices them.
[310] Sur le Vol des Ins. c. i. 423.
[311] Ibid. 454. and c. iv. 66. note 1.
[312] Ibid. c. i. 453.
[313] Ibid. 459, 456.
[314] Ibid. 459.
[315] Annal. de Chim. xii.
[316] Sur le Vol des Ins. c. i. 423, 454. c. iii. 344. c. iv. 66.
[317] De Geer ii. 946—.
[318] Lesser, L. i. 124. note *. Lyonet Anatom. pref. xii. De Geer ii. 132.
[319] Reaum. i. 399—. De Geer i. 37—.
[320] Ibid. 40.
[321] Reaum. i. 400.
[322] Ibid.
[323] De Geer ii. 129.
[324] De Geer i. 531—. t. xxxvii. f. 13. s. Compare Reaum. ii. 396—.
[326] In Linn. Trans. iii. 302.
[328] Spallanzani found that the eggs of insects placed under the exhausted receiver of an air-pump, or in any small closed vessels, did not hatch, though every other condition for their development was present. Opusc. de. Phys. i. 141.
[329] Philos. Trans. 1820. 213.
[330] Bibl. Nat. i. 204. b. t. xix. f. 5.
[331] Vol. I. p. 446—. III. p. 76.
[333] Philos. Trans. 1820. 218.
[337] Inch, c. iv. Ideen zu Einer Zoochemie, 68—.
[338] On Thermom. 141.
[339] Carlisle in Philos. Trans. 1805. 25.
[341] Travels ii. 482.
[342] Reaum. v. 540.
[343] Swamm. Bibl. Nat. ii. 65. a.
[344] Ibid. 48. a.
[345] Hist. Nat. l. xi. c. 19.
[346] Swamm. Bibl. Nat. ii. 64. a.
[347] Reaum. iv. 428. t. xxix. f. 2. c, s.
[348] Genes. ix. 4.
[349] N. Dict. d'Hist. Nat. xxx. 130.
[350] Cuv. Anat. Comp. iv. 167.
[351] Herold Schmetterl. 25. note *. Vol. III. p. 53.
[352] N. Dict. d'Hist. Nat. vii. 313. Cuv. Anat. Comp. iv. 411.
[353] Ibid. 419, 407.
[354] Ibid.
[355] Ibid. 410.
[357] Lyonet Anat. 105.
[358] Ibid. 425.
[359] Ibid. 105—.
[360] De Bombyc. 15—.
[361] Reaum. i. 160—.
[362] Cuv. Anat. Comp. iv. 418.
[363] Marcel de Serres Mem. du Mus. 1819. 69.
[364] Swamm. Bibl. Nat. t. xl. f. 4. t. xv. f. 4.
[365] De Bombyc. t. iii. f. 4.
[366] Ubi supr. 414.
[367] Ibid. 425—.
[368] Ibid. 419.
[369] Ibid. 412.
[370] Lyonet Anat. 413.
[371] Lyonet Ibid. 426. Cuv. Anat. Comp. iv. 419.
[372] Lyonet says (426), "au-delà de trois millions de fois plus petits qu'un grain de sable"!!
[373] Ibid.
[374] His words are—"In silkworms I have clearly seen various small vessels spring from and approaching to the heart, which I have even filled with a coloured liquid. But whether they were veins or arteries I cannot yet affirm." i. 112. a. 176. a. According to Cuvier (Anat. Comp. iv. 418), but I cannot find the passage, Swammerdam also mentions having seen a red fluid issue from small vessels in grasshoppers.
[375] Reaum. v. 103.
[376] Bonnet ii. 309. Perhaps in both cases the alimentary canal was the organ seen.
[377] Reaum. iv. 171—.
[378] Lesser L. ii. 84. note.
[379] De Geer ii. 505—. vi. 287.
[380] On the Microscope. i. 130.
[381] Ibid.
[382] Sur le Vol des Ins. 325—.
[383] Lyonet Anat. 427—.
[384] Cuv. Anat. Comp. iv. 418—.
[385] Mem. du Mus. 1819. 71.
[386] N. Dict. d'Hist. Nat. xvi. 208.
[387] Marcel de Serres, in his Observations on the Dorsal Vessel of Insects[388], endeavours to prove that the principal use of that vessel is the more perfect animalization of the chyle that, transuding through the pores of the intestinal canal, is imbibed by it. In insects, he observes, that undergo metamorphoses, in which the growth or development of parts is often very rapid, it is requisite that a considerable portion of the chyle should be in reserve for this purpose. On this account it is that the Epiploon or adipose tissue is so abundant in larvæ to what it is in the perfect insect. That the importance also of this part to insects is proved by the circumstance, that all their interior parts communicate by fibrils with this tissue, and that probably their various organs derive the nutriment from it by their means. He then asks by which of the viscera is the fat elaborated, or by what means does the chyle which transudes from the intestinal canal pass to the state of fat? Facts seem to indicate, says he, that the function of the dorsal vessel is to pump up the chyle, and to cause it then to transude through the meshes of the adipose tissue, where it finishes by elaborating that mass of fat so abundant in larvæ and certain perfect insects, which are thus enabled to sustain the effects of a long fast. So that this vessel is only a secretory organ, analogous to so many others that exist in insects; but the secretion which it has to produce is the most important of all, since the support of the vital powers depends upon it: it is, in effect, that vessel which completes the function of animalization, and which itself prepares the nutritive fluid[389]. He observes, amongst other reasons he brings to support his theory, that the colour of the fluid which it contains is always analogous to that of the adipose tissue that surrounds it, and that the colour of that tissue never changes without that of the fluid undergoing a corresponding alteration,—that when, as in many perfect insects, the quantity of fat diminishes, the dorsal vessel also diminishes in size, and that the same reagents which coagulate the fat, coagulate equally the fluid in the dorsal vessel, which seems to indicate an identity between them[390].
But there are circumstances that militate against this hypothesis. The analysis which Lyonet has given of the fluid contained in the dorsal vessel of the Cossus[391], seems to prove that it is more analogous to gum or varnish. He saw indeed a few globules, which appeared ten times as big as the others, which swam upon the water, but which he did not regard as component parts of the fluid, but as little drops of grease extravasated by dissection. The fluid of the vessel itself easily mixed with water, and appeared to sink in it to the bottom[392]. This proves that it is not of a fatty or oleaginous nature. But the strongest objection is stated by M. Carus, who judiciously observes[393], That it is contradictory to suppose that a canal should absorb or exude fluids by its parietes in a different form. Further experiments however seem necessary to ascertain the nature of the fluid and its object.
[388] Mem. du Mus. 1819.
[389] Ibid. 68—.
[390] Ibid. 69—.
[392] Lyonet Anat. 426—.
[393] Introd. to Comp. Anat. ii. 277. Engl. Trans.
[394] This seems some confirmation of Dr. Virey's opinion, that insects in their first states are still a kind of fœtus. See above, Vol. III. p. 61—.
[395] Introd. to Comp. Anat. ii. 393—. Engl. Trans.
[396] Introd. to Comp. Anat. ii. 395—. Engl. Trans.
[397] Introd. to Comp. Anat. ii. 396—. Engl. Trans.
[398] Ibid. 398.
[399] Ibid. 399.
[400] Ibid. 398.
[401] Introd. to Comp. Anat. ii. 399—. Engl. Trans.
[402] Reaum. i. 409, 643—. Malpigh. De Bombyc. 38.
[403] Lesser L. ii. 87 note *.
[404] Ubi supra.
[405] Reaumur iv. 264.
[406] Ibid. 260—.
[407] Herold Schmetterl. 24.
[408] Anat. Comp. iv. 165.
[409] Marcel de Serres (p. 67.) speaks of this fluid as being, after it has transuded through the intestinal canal, a fluid in repose, which seems to indicate that it is perfectly stagnant; but when we consider that it is not only incessantly entering the body and making its way to every part, but is also, by means of the various secretory organs, constantly converted into new products, and so going out again in many cases, it will appear evident that it cannot be considered as a stagnant fluid, since there must be a constant though probably slow motion towards the points of absorption or imbibition.
[410] Dr. Kidd (Philos. Trans. 1825. 236.) did not find the abdominal viscera of the mole-cricket thus circumstanced, nor more lubricated than the intestines of the higher animals.
[411] Cuv. Anat. Comp. iv. 158. Herold Schmetterl. 28.
[412] Sur le Vol des Ins. c. iv. 88. note 1.
[413] Anat. 428.
[414] Treviranus Arachnid. 28. t. iii. f. 28, 29.
[415] Ibid. 29. t. iii. f. 30, 31.
[416] N. Dict. d'Hist. Nat. xxx. 420. Comp. Treviran. Arachnid. 10—.
[417] Ibid. 9—.
[418] Anat. Comp. iv. 129.
[419] Cuv. Anat. Comp. iv. 129.
[420] Plate XXI. Fig. 3. c, d, e, is the intestinal canal of the larva of the Cossus.
[421] Cuv. Ibid. 112.
[422] Ramdohr Anat. der Ins. 6.
[423] Ibid. 25.
[424] Ibid. 6.
[425] Cuv. ubi supr. 113.
[426] Comp. Ramdohr Anat. 7.
[428] Tenebrio Ramdohr, ubi supr. 9. t. iv. f. 1.
[429] Agrion. Ibid. t. xv. f. 4. a, b.
[430] Ibid.
[431] Many other insects that live by suction have something similar, as the honey-bag of butterflies, Plate XXX. Fig. 10, 11. a. Ramdohr t. xviii. f. 2. with t. xix. f. 1-3. and xxi. 1, 3, &c.
[432] Ramdohr Anat. 11—.
[434] Ramdohr Ibid. 28—.
[435] Herold (Schmetterl. 24) says that Ramdohr is mistaken here, and denies the existence of this juice in insects; but as Ramdohr's researches were so widely extended, he is most likely to be right.
[436] Ramdohr Ibid. 29.
[437] Ibid. 31.
[438] Ibid. 28.
[439] Anat. Comp. iv. 135. Comp. Dr. Kidd in Philos. Trans. 1825. 223. t. xv. f. 6, 7.
[440] Ramdohr Anat. 15.
[441] Ibid. 15.
[442] Ibid. 18.
[443] Ibid.
[444] Ibid.
[445] Swamm. Bibl. Nat. i. 94. b. Cuv. Anat. Comp. iv. 134.
[446] Ubi supr. 18.
[447] Ibid. t. i. f. 1. e. 5. c. 9. g, h.
[448] Ibid. t. xxv. f. 4. bb.
[449] Ramdohr Anat. t. viii. f. 3. cc.
[450] Ibid. t. vii. f. 2.
[451] Ibid. 20.
[452] Anat. Comp. iv. 132.
[453] Ibid. and 136.
[454] Ubi supr. 30.
[455] Ibid. 31. t. iv. f. 2. c. t. v. f. 1. d. f. 4. D.
[456] Ibid. 32.
[457] Ibid. 34.
[458] Ramdohr Anat. 35.
[459] Ibid. t. xxiv. f. 1. F.
[460] Ibid. 36. t. vii. f. 2. kk. t. viii. f. 3. g, hh.
[461] Ibid. t. xii. f. 1. t. xvii. f. 1. t. vii. f. 5.
[462] Ibid. 37.
[463] Ibid. 38.
[464] Ibid.
[465] Ramdohr Anat. 40.
[466] De Bombyc. 18—.
[467] Anat. Comp. iv. 153.
[468] Ibid.
[469] Ibid.
[470] Ramdohr 43. Cicindela campestris, t. iii. f. 1. K.
[471] Phryganea grandis, Ibid. t. xvi. f. 2.
[472] Notonecta glauca, Ibid. t. xxiii. f. 5.
[473] Of Musca vomitoria, Ibid. t. xix. f. 5.
[474] Ibid. t. viii. f. 1. H. and G. f. 2.
[475] Ibid. 50.
[476] Ibid.
[477] Ibid.
[478] Ibid. 44. t. i. f. 9.
[479] Ibid.
[480] Ibid. t. vi. f. 5. H.
[481] Kidd in Philos. Trans. 1825. t. xv. f. 6.
[482] Ibid. t. xix. f. 1. N, N, O, f. 2. P, P, O.
[483] Ibid. t. 1. f. 1. kkk.
[484] Ramdohr, t. xiii. f. 1-3.
[485] Ibid. 44.
[486] Ibid. 45.
[487] Ibid. 45. Plate XXI. Fig. 3. f. f.
[488] Rhamdohr, Ibid. t. iii. f. 6. E.
[489] Ibid. t. i. f. 1. 5. 9. t. xiv. f. 1-3.
[490] Ibid. 46. t. vi. f. 3.
[491] Ramdohr, t. vii. f. 2.
[492] Ibid. t. ii. iii. &c. t. xx. f. 1, 2. 6. t. xxii. f. 1-5. &c.
[493] Ibid. t. xviii. f. 1. 5. t. iv. f. 1. See also t. vi. f. 1. 3.
[494] Ibid. Anat. t. xvii. f. 1, 2. 6.
[495] Ibid. t. xiv. f. 3.
[496] Ibid. t. xiii. f. 4.
[497] Ibid. t. xv. f. 3, 4. t. 1. f. 1. 5. 9. t. xii. f. 4, 5, 6, &c.
[498] Ibid. t. xi. f. 4. t. xii. f. 4-6. t. xiii. f. 2-4, &c.
[499] Ibid. t. vii. f. 1. t. viii. f. 1, &c.
[500] Ramdohr Anat. t. ii. iii. xxv.
[501] Ibid. t. iii. f. 6. t. iv. f. 2. t. v. f. 1.
[502] Ibid. f. l. e. f. 3.
[503] Ibid. 122.
[504] Ibid. 123.
[505] Ibid. t. v. f. 4. B.
[506] Ibid. 94.
[507] Ibid. 96—.
[508] Ramdohr t. x. f. 1. 8.
[509] Ibid. f. 8. b. c.
[510] Ibid. 98. t. x. f. 2-4. From Ramdohr's figure, compared with the size of the insect, it appears that the gizzard could scarcely have been of greater diameter.
[511] Ibid. f. 2.
[512] See W. Curtis in Linn. Trans. i. 88.
[513] Ramdohr t. x. f. 1. d.
[514] Ibid. l l.
[515] Ibid. t. ix. f. 1, 2. t. xi. f. 3. t. xxiv. f. 1, 2.
[516] Ramdohr 103.
[517] Ibid. 104. t. vi. f. 4. D.
[518] Ibid. f. 2. B.
[519] Ibid. t. vi. f. 3. E.
[520] Ibid. 101.
[521] Ibid. t. i. f. 1. 5. 9.
[522] Ibid. f. 2, 3, 4. 7, 8. 12.
[523] Ibid. f. 1. e, f. 5. c. f. 9. g h.
[524] Ibid. f. 1. 9. k.
[525] Ibid. t. xv. f. 3, 4. t. xvii. f. 2. 6.
[526] Ibid. t. xv. f. 3, 4, f.
[527] Ibid. t. xvii. f. 2. c. f. 6. d.
[528] Ibid. f. 2. b. f. 6. c.
[529] Ramdohr t. xii. f. 6. H. t. xiii. f. 1. f.
[530] Ibid. t. xiv. f. 2, 3, C.
[531] Ibid. t. xii. f. 6. D. t. xiii. f. 1. b.
[532] Ibid. 133. t. xii. f. 1-3.
[533] Ibid. f. 4.
[534] Comp. Ramdohr t. xxii. f. 3. M. Fig. 4. 3. with t. xxi. f. 1. I.
[535] Ramdohr t. xxii. f. 1. c. f. 3, 4. B—.
[536] Ibid. f. 1. D E. f. 3. C D.
[537] Ibid. t. xxii. f. 1. D, E. f. 3. C, D. f. 4. C.
[538] Ibid. 198.
[539] Ibid. t. xxvi. f. 2. 4.
[540] Ibid. t. xxxiii. f. 3.
[541] Ramdohr t. xviii. f. 1. F, G.
[542] Ibid. L, K.
[550] Ramdohr, Ibid. t. xx. f. 1. E. f. 6. C.
[551] Ibid. t. xix. f. 2. C. f. 3. CCD. t. xx. f. 2. E.
[552] Ibid. t. xix. f. 2. D.
[553] Ibid. t. xx. f. 2. FF. f. 6. DD. 184. 180.—
[554] Ibid. t. xix. f. 1. ON. f. 2. OP. f. 3. F. t. xxviii. f. 1, 2. p. q.
[555] Ramdohr, Ibid. t. xx. f. 1. G. f. 2, 3. L.
[556] Ibid. t. xxi. f. 1. D.
[557] Ibid. 172.
[558] Ibid. t. xix. f. 2. K L. This organ seems analogous to that with four retractile fleshy horns, observed by Reaumur and De Geer in other species of Muscidæ. Reaum. iv. t. xxviii. f. 13. a, s. De Geer vi. t. iii. f. 18. c, d.
[559] Ramdohr t. xxi. f. 6.
[560] Ramdohr t. xxix. f. 1 *. A.
[561] Ibid. and f. 3. B, D.
[562] Ibid. f. 2, 3. 5. &c.
[564] Treviranus and Ramdohr are of the former opinion; and Meckel, Cuvier, Marcel de Serres, and Leon du Four, of the latter.
[565] Treviran Arachnid. t. 1. f. 6. v.
[566] Ibid. n.
[567] Ibid. t. ii. f. 24. β.
[568] Treviran Arachnid. f. 6. B B.
[569] N. Dict. d'Hist. Nat. xxx. 423—. Comp. Treviranus, Arachnid. t. i. f. 6.
[570] Treviranus, Ibid. v.
[571] N. Dict. d'Hist. Nat. xxx. 421—. Comp. Treviran. Ibid.
[572] N. Dict. d'Hist. Nat. Ibid.
[573] Treviran. Ibid. t. i. f. 6. i i, c c.
[574] N. Dict. d'Hist. Nat. Ibid.
[575] Treviran. Ibid. t. ii. f. 24. a.
[576] Ibid. v, b.
[577] Ibid. c, d, f.
[578] Ibid. g, n.
[579] N. Dict. d'Hist. Nat. Ibid.
[580] Treviran. Ibid. 28.
[581] Ibid. t. ii. f. 24. β.
[582] Ramdohr, t. xix. f. 1.
[583] Reaum. i. 143. t. v. f. 9.
[585] De Geer iii. 26.
[586] Reaum. iii. 357. t. xxix. f. 6-10.
[588] Cuv. Anat. Comp. iv. 163—.
[590] Malpigh. De Bombyc. t. v. f. 2. Swamm. t. xxxiv. f. 5. Lyonet, t. v. f. 1.
[591] Anat. der Ins. 59.
[592] Ibid. 60. Malpigh. 20.
[593] Lyonet Anat. 111.
[594] N. Dict. d'Hist. Nat. xv. 483.
[595] Anat. Comp. v. 198.
[596] Ramdohr, 60. t. xvii. f. 1. f, g, h, r.
[597] Vol. I. p. 403—. Treviran. Arachnid. 42.
[598] Treviran. Arachnid. 43. t. iv. f. 42. o. p. 9.
[599] Ibid. α, y.
[600] Swamm. ii. 21. a. t. xxxvi. f. 1. abcd. Ramdohr 58.
[601] Schmet. t. iii. f. 1.
[602] Lyonet—. 112. t. v. f. 1. P, Q, R, S.
[603] Ramdohr Anat. t. xviii. f. 1. M. f. 5. F.
[604] Ibid. t. x. f. 1. m.
[605] Ibid. t. xxii. f. 3. M L. Ramdohr regards the double one as a pair; but as they terminate in a single tube, they ought to be reckoned as one.
[606] Ibid. f. 4.
[607] Ibid. f. 2. K, L, M, N. t. xxiii. f. 6.
[608] Ibid. 177. t. xxi. f. 3. F. F.
[609] Ibid. f. 2. G, H.
[610] Ibid. t. xxii. f. 2. L.
[611] Ibid. t. xxi. f. 1. O. t. xvii. f. 6. n.
[612] Ramdohr Anat. t. xx. f. 6. D.
[613] Ibid. t. xxii. f. 1. K, L. f. 2. I, K, L.
[614] Ibid. f. 3, 4, 5.
[615] Ibid. 57—.
[616] Reaum. ii. 81. Herold Expl. of Plates, x. Malpigh. De Bombyc. 37. Plate XXX. Fig. 12. c.
[617] Herold Ibid. x. t. iv. f. 1. p, u, y. Marcel de Serres Mem. du Mus. 1819. 141.
[618] Gaede Anat. t. i. f. 3. d.
[619] Ibid. 17. t. i. f. 4.
[620] Bibl. Nat. t. xix. f. 3. β.
[621] Reaum. v. 377. t. xxix. f. 7. s.
[622] N. Dict. d'Hist. Nat. xxx. 388.
[623] Ibid. 427—.
[624] Arachnid. 31. t. ii. f. 21. p. 9.
[625] N. Dict. d'Hist. Nat. xxii. 114. 117. comp. Vol. I. p. 127.
[626] Ibid. xxviii. 6.
[627] Osservazioni, &c. 13—.
[628] Vol. II. p. 243—. N. Dict. d'Hist. Nat. iv. 308.
[629] Ibid. iv. 309.
[630] Ibid. v. 252.
[631] De Geer iv. 358. t. xiii. f. 9. m.
[632] Vol. II. p. 241—. III. p. 147—.
[633] De Geer iii. 41.
[634] Vol. I. p. 451, where by mistake it is represented as the work of Aphis Abietis.
[635] De Geer iii. 111.
[636] Reaum. iii. t. xxvi. f. 4-6.
[637] N. Dict. d'Hist. Nat. vi. 305.
[639] Treviran. Arachnid. 44. In Paraguay a spider is found which makes spherical cocoons of yellow silk, which are spun because of the permanence of the colour. This operation occasions a flow of water from the eyes and nose of the spinners. Azara Voyag. 212. See also Murray in Werner. Trans. 1823. 8—.
[640] Reaum. v. 24.
[642] Ramdohr Anat. t. ii.-vi.
[643] Ibid. 20. See above, p. 107. As some of the Sialisteria render to the stomach (see above, p. 131), there seems no small affinity between these shags and those organs.
[644] Cuv. Anat. Comp. iv. 132, 136.
[645] Reaum. vi. Pref. xxviii. 177—.
[646] Ibid. 253—.
[647] Ibid. iii. 375.
[648] Anat. t. xii. f. 6.
[649] Ibid. xxi. f. 3. I I.
[650] Reaum. iii. 230.
[652] Reaum. iii. 215. Bonnet ix. 182.
[654] Marcel de Serres Mem. du Mus. 1819. 133, 141.
[655] De Geer, v. 6.
[656] Rai. Hist. Ins. 62.
[657] Vol. II. p. 242—. 248. Rai. Hist. Ins. 94, 382.
[658] Reaum. v. 448.
[659] Ibid. v. 722.
[660] Vol. I. p. 196. II. p. 176.
[661] Encyclop. Britan. viii. 205. from Journ. de Phys.
[663] Reaum. iii. 318—. t. xxvi. f. 1-6.
[664] Ibid. 396—. t. xxxi. f. 20-29.
[665] Insect. Suec. i. 257.
[667] N. Dict. d'Hist. Nat. xvii. 189.
[668] Nicholson's Journ. i. 298—.
[670] Philos. Trans. 1670.
[671] Philos. Trans. Ibid. Ray's Lett. 74.
[672] Amoreux Ins. Venim. 236—.
[673] N. Dict. d'Hist. Nat. xii. 94.
[674] Southey's Brazil, i. 645.
[675] N. Dict. d'Hist. Nat. ubi supr.
[677] Syst. of Chemist. 533.
[678] Germar Mag. der Ent. iii. 445—.
[679] Mem. Dijon 1783. ii. 70.
[680] Reaum. v. 354.
[681] On Poisons, i. 265—.
[682] Ibid. 269.
[683] Reaum. ubi supr.
[684] Vol. I. p. 124. III. p. 716—.
[685] N. Dict. d'Hist. Nat. xxx. 427.
[686] I use the term odorous, not in the same sense as odoriferous, but to include both sweet and fetid scents.
[687] Vol. II. p. 238—. III. p. 147—.
[688] A Brazilian wood so called, but differing from the common cedar.
[689] Dotharding Insect. Coleopt. Danic.
[690] Sturm Deutsch. Fn. i. 27.
[691] Reaum. iii. 494.
[692] Mon. Ap. Angl. i. 136.
[693] Osservaz. sullo Iulus, &c. 14.
[695] N. Dict. d'Hist. Nat. xv. 487.
[696] N. Dict. d'Hist. Nat. iv. 308.
[697] Dated Tripoli in the West, January 21, 1819.
[699] Reaum. i. 145. Lyonet Anat. 106—. N. Dict. d'Hist. Nat. xvi. 224. Plate XXI. Fig. 5. a.
[700] See above, p. 90. note387.
[702] Huber i. 273.
[703] Herold Schmetterl. tab. expl. vii.
[704] Herold Schmetterl. t. iv. f. 1. x. &c. Plate XXX. Fig. 12. d.
[705] De Bombyc. 36.
[706] Ibid. t. xii. f. 1. I. and, f. 2. O. M.
[707] Philos. Trans. 1792. 186.
[708] Swammerdam, in dissecting the female of Oryctes nasicornis, discovered a blind vessel opening into the vagina, and at the other or inner extremity not terminated by any secretory tube, containing a yellowish matter, that seems analogous to the organ mentioned in the text; and in the hive-bee he found a similar organ covered with air-vessels, which he supposes to be connected with the Colleterium (see above, p. 132.), and which he states to contain a slimy matter. Bibl. Nat. i. 151. b. t. xxx. f. 10. g. 204. b. t. xxix. f. 3. t. Perhaps likewise the organ discovered by M. L. Dufour in Scolia,—which he imagines to belong to the poison-secretor, and which he describes as a sac consisting of a double tunic, the exterior one muscular and the interior membranous, and filled with a blueish-green gelatinous matter (N. Dict. d'Hist. Nat. xxx. 388.)—may be a spermatheca.
[709] De Insector. Genital. 17.
[710] I allude to those organs above described (p. 132.) for the secretion of matter for varnishing the eggs or lubricating the oviduct. It seems most probable, if the fecundation of the eggs takes place gradually, that upon their passing into the oviduct, a special reservoir should be appropriated to the reception of the male sperm, adapted to maintaining in due activity the vivifying principle, or aura seminalis.
[711] Herold Schmett. t. iv. f. 2. m n.
[712] Treviran. Arachnid. 36. t. iv. f. 32. aa. Marcel de Serres in Mém. du Mus. 1819. 89.
[713] Marcel de Serres, Mém. du Mus. 1819. 115.
[714] Rifferschw. De Genital. Ins. 11.
[715] Marcel de Serres in Mém. du Mus. 1819. 109. Plate XXX. Fig. 12. a.
[716] Rifferschw. ubi supr. 23—. Swamm. Bibl. Nat. t. xlii. f. 8. a, f, g, h.
[717] Ibid. i. 104. t. xv. f. 3. ii. 62. t. xii. f. 8. Treviran. Arachnid. t. iv. f. 32.
[718] Reaum. iv. 391.
[719] Posselt Anat. der Ins. t. i. f. 28, 29.
[720] N. Dict, d'Hist. Nat. xxx. 387—. Swamm. ubi supr. ii. 23. t. xxxv. f. 3.
[721] Ibid. i. 203.
[723] Swamm. ubi supr. i. 151. Gaede Anat. der Ins. t. ii. f. 3.
[724] Swamm. i. 203.
[725] Gaede Anat. der Ins. 20. t. i. f. 9.
[726] Ibid. 25, 28. t. ii. f. 10.
[727] Ibid. 32.
[728] Swamm. ii. 74.
[729] Ibid. 203. t. xix. f. 3.
[730] Reaum. iv. 391—.
[731] Swamm. t. xliii. f. 19.
[732] Gaede 22.
[733] Swamm. Bibl. Nat. i. 203.
[734] Ibid.
[735] Rifferschw. 11—.
[736] Swamm. t. xlii. f. 8. Gaede, t. i. f. 3. cc.
[737] Herold Schmett. t. v. f. 10. 12.
[740] Swamm. t. xix. f. 4. b.
[741] Ibid. f. 3.
[743] De Geer iv. 127. t. iv. f. 17.
[744] De Geer iv. 143. t. v. f. 15.
[746] De Geer. v. 62. t. iii. f. 12.
[748] Stoll Sauterel. t. xxii. b. f. 87, &c.
[749] De Geer iii. 418. t. xxi. f. 10, 11. Latr. Gen. Crust. et Ins. iii. 98.
[750] Stoll ubi supr. t. xiii. a. f. 51.
[751] This insect, which connects Conocephalus, Acrida, &c. with Locusta, is also distinguished by antennæ at first filiform and then setaceous.
[752] De Geer iii. t. xxiv. f. 1, 12.
[753] Ibid. 176. t. xi. f. 19.
[755] Reaum. v. 177—.
[756] Ibid. vi. 435. t. xl. f. 6, 7.
[757] Natural History of the Slug-worm, 12—. f. 12, 13.
[758] Valisn. Esperienz. &c. Musca dé Rosai. Reaum. v. 100—. De Geer ii. 916—. The last writer thought he saw in the back of the saw itself a longitudinal cavity (918), which applied to the groove would form an open canal.
[761] Reaum. v. 347. t. xlix. f. 10. d, f.
[762] See above, Vol. III. 390. a.
[763] See above, Vol. I. 448—.
[764] De Geer ii. 835. t. xxviii. f. 20, 21. Plate XV. Fig. 22. This figure was drawn by a friend—the organ seems more exerted than in De Geer's. I cannot make out the little appendage at the end.
[766] Reaum. v. 19—. t. iii. f. 3-6.
[767] Arachnid. 40.
[768] Huber Nouvel. Observ. i. 106.
[769] Swamm. Bibl. Nat. t. xix. f. 2.
[770] Philos. Trans. 1797. 80.
[772] Compare Reaum. iii. 153. Pallas Act. Nat. Cur. 1767. iii. 430. Wien. Verzeich. 292.
[773] Naturfor Stk. xx. 59—.
[774] It does not appear to be clearly decided whether the eggs are extruded from the female, or whether dying immediately after fecundation they are hatched within her body. As the young larvæ certainly are hatched in the pupa (not merely within the exterior case of bits of grass, &c., which includes it) which the body of the insect must fill, it does not seem easy to conceive how she can find room for oviposition; and yet Von Scheven expressly says that one female of Ps. vestita—which being kept from all access to the male actually left the pupa-case and wandered about the glass which contained them—laid unfruitful eggs.
[776] Bonnet i. 19—.
[777] Reaum. vi. 551.
[778] Reaum. vi. 552.
[779] N. Dict. d'Hist. Nat. ii. 284.
[780] N. Dict. d'Hist. Nat. ix. 125. Bonnet and Jurine both found that the female Aphides and Branchiopods that were fertile without the usual intercourse of the sexes were less fruitful than their mother, and those of the last generation less so than the first. Latr. Hist. Nat. des Crust. et Ins. xi. 292.
[781] See more on the subject of fecundation, Vol. II. p. 154—. 169—.
[782] N. Dict. d'Hist. Nat. xxx. 426.
[784] De Geer iii. 533.
[785] Swamm. i. 203. b. t. xix. f. 3.
[786] Reaum. ii. 66.
[791] Reaum. iv. 414.
[792] Ibid. t. xxviii. f. 14, 15.
[793] Ibid. 404.
[794] De Geer vi. 63—.
[796] De Geer iii. 70—.
[797] Ibid. 128.
[798] N. Dict. d'Hist. Nat. xxx. 426—.
[800] Vol. II. p. 50, 110—, 118—, 125—, 130—. The neuters of the Termites, however, (p. 33.) seem to be a distinct sex, if I may so speak—and to merit that name.
[801] Vol. II. Letter XXII. Vol. III. Letters XXXIV.-XXXVI.
[802] Vol. II. p. 280, 295—, 306, 310—. &c.
[803] Philos. Trans. 1818. 174. t. viii. f. 4-6.
[805] Schmetterl. 105.
[806] Philos. Trans. 1819. 172, 174, 187.
[807] Anat. Comp. i. 90.
[808] Philos. Trans. 1819. 175.
[809] Cuv. ubi supr. 90—.
[810] Cuv. Ibid. i. 89—.
[812] Lyonet Anat. t. iv. f. 3.
[813] Ibid. 93—.
[814] Cuv. Anat. Comp. i. 134.
[815] Chabrier Sur le Vol des Ins. c. i. 445.
[817] De Geer iv. t. xv. f. 11. m n, o p.
[818] Lyonet Anat. 93.
[819] Lyonet Anat. t. xiii. f. 1, 2.
[820] Ramdohr Anat. t. v. f. 1. e. f. 3.
[821] Chabr. ubi supr. 440—.
[822] Ibid. 442, &c.
[823] N. Dict. d'Hist. Nat. xxii. 80.
[824] Vol. III. p. 663, 670. See above p. 21.
[825] Chabrier Sur le Vol des Ins. c. i. 446.
[827] Ubi supr. 437, 439.
[828] Plate XXII. Fig. 11, 12. c. Chabrier ubi supr. c. iii. t. xi. viii. f. 9. S. D. i, k. c. i. 440—.
[829] Plate XXII. Fig. 11, 12. c. Chabrier Sur le Vol des Ins. c. iii. t. xi. viii. f. 9. S. D. i, k. c. i. 440—.
[830] Cuv. Anat. Comp. i. 94—.
[831] N. Dict. d'Hist. Nat. xxii. 80.
[832] Ubi supr. 101—.
[834] Anat. Comp. i. 432—.
[835] Anat. t. vii. f. 2. left hand.
[836] Ibid. right hand.
[837] Ibid. 115—.
[838] Cuv. ubi supr.
[840] Anat. Comp. i. 447.
[841] Vol. III. p. 366. Plate XXVII. Fig. 1, 4. n´.
[842] Ibid. Fig. 3. n´.
[844] Vol. III. p. 367—, 541, 584. Plate XXII. Fig. 7. Cuv. ubi supr. 448.
[846] Anat. Comp. i. 136.
[847] De Geer iv. t. xv. f. 11. o, p.
[848] Marcel de Serres Comparaison, &c. 3—.
[849] Ibid. 4.
[850] Ibid. 5.
[851] Plate XXII. Fig. 11. h´.
[853] Plate XXII. Fig. 6. Vol. III. p. 585—.
[854] Cuv. Anat. Comp. i. 436. Plate XXI. Fig. 6.
[855] Ibid. a, b. Lyonet Anat. 37.
[856] Cuv. ubi supr. 458—. Vol. III. p. 368, 378, 382.
[857] Cuv. ubi supr. 459.
[858] Chabr. Sur le Vol des Ins. c. i. 441.
[859] Chabr. Sur le Vol des Ins. c. i. 415.
[860] Ibid.
[861] Ibid. c. iii. 344. t. viii. f. 8, 9.
[862] Ibid. c. i. 440.
[863] Ibid. 444.
[864] Ibid. 445. c. iii. 359.
[865] Ibid. c. ii. 332. c. iii. 359.
[866] Ibid. c. i. 445.
[867] Ibid. c. iv. 78.
[868] Chabr. Sur le Vol des Ins. c. i. 415, 442. c. iv. 80.
[869] Ibid. c. i. 442.
[870] Ibid. 439—.
[871] Chabrier Analyse, 28. The latter part of this passage is copied from a MS. note of the author's in my copy.—W. K.
[872] Chabrier Analyse, Ibid. Sur le Vol des Ins. c. i. 445. Vol. III. p. 617.
[873] Analyse ubi supr.
[874] Sur le Vol des Ins. c. i. 448, c. ii. 336.
[876] Chabr. Ibid. c. i. 443. ii. 316, 332.
[877] Chabr. Sur le Vol des Ins. c. ii. 333.
[878] Ibid. 332. Plate XXII. Fig. 11, 12. c. A cupuliform process is also observable at the side of the metaphragm. Ibid. Fig. 10. a.
[879] Chabr. Ibid. c. iv. t. xi.-4. f. 14.
[880] Ibid. c. i. 445. xi.-8. f. 8, 9.
[881] Chabr. Sur le Vol des Ins. c. ii. 336. note 1. Vol. III. p. 292—.
[882] Chabr. Ibid. c. i. 447.
[885] Chabrier Sur le Vol des Ins. c. i. Addend. 298.
[888] Chabr. ubi supr. c. i. 422.
[889] Cuv. Anat. Comp. i. 451.
[890] Chabr. Analyse 25. Sur le Vol des Ins. c. i. 423, 452. Addend. 301.
[892] Lyonet Anat. t. xiii. f. 1, 2.
[893] Lyonet Anat. t. xiii. 188—, 584.
[894] Ibid. 189.
[895] N. Dict. d'Hist. Nat. xxx. 421.
[896] Arachnid. 9. t. i. f. 7. r.
[897] Ibid. o.
[898] Ibid. 10.
[899] Arachnid. 45. t. iii. f. 31. m, n, q, r, t.
[901] Mouffet Theatr. 275.
[902] N. Dict. d'Hist. Nat. xxviii. 249.
[903] Phil. Acc. of Works of Nat. 144.
[904] Clark in Linn. Trans. iii. 309.
[905] Fn. Suec. 1799.
[906] Anatomy of Expression in Painting, 170.
[907] Bonnet Œuvr. ii. 124.
[908] N. Dict. d'Hist. Nat. xxii. 81.
[909] 1 Cor. xv. 50—.
[910] N. Dict. d'Hist. Nat. ubi. supr.
[911] Swamm. Bibl. Nat. t. xviii. f. 2. l, m, n, o. Reaum. v. t. xxix. f. 7. m, n, o, p, q.
[913] De Bombyc. 5.
[914] Reaum. ii. 185—.
[916] t. vi. f. 3.
[917] These directions for dissecting are chiefly taken from Swammerdam, Life xiv.— and Lyonet Anat. 7—.
[918] Ps. civ. 29.
[919] Hist. Animal. l. viii. c. 27.
[920] The Principles of Botany and of Vegetable Physiology, § 310-353.
[921] Dr. Leach, from a communication of Sir Joseph Banks, has given a very interesting history of a spider which, having lost five of its legs, from a web-weaver had become a hunter; these legs it afterwards reproduced, though shorter than the others. Linn. Trans. xi. 393. Comp. N. Dict. d'Hist. Nat. ii. 282.
[924] Huber Abeilles ii. 409.
[925] N. Dict. d'Hist. Nat. i. 42.
[926] De Geer i. 72—.
[927] Reaum. iv. 342.
[928] Naturf. xii. 224. t. v. f. 8.
[929] Naturf. xvi. t. iv. f. 1-3.
[930] Huber Fourmis, 174. note 1.
[932] N. Dict. d'Hist. Nat. i. 42.
[933] De Geer vi. 75. Latr. Hist. Nat. xiv. 371.
[934] Jacquin Collectan. iii. t. xxiii. f. 7.
[935] De Geer ubi supr.
[936] Dr. Bevan asserts (The Honey-bee, 197) "that we have no evidence that pollen constitutes any part of the food of adult bees." Had he consulted Reaumur (v. 418) he would have found that this great man examined the proceedings of a bee with a magnifying glass, and distinctly saw her devour very deliberately the masses of pollen on her hind legs. He says also (Ibid. 419.) that if the stomach and intestines are opened, they will be found filled with that substance.
[937] Schirach Hist. &c. 54. Reaum. v. 713. N. Dict. d'Hist. Nat. i. 42.
[938] N. Dict. d'Hist. Nat. i. 42. Schirach Hist. 56.
[939] Latr. Hist. Nat. xiv. 163—. N. Dict. d'Hist. Nat. iv. 134—.
[940] Œuvr. ii. 48—.
[941] Lesser L. ii. 121.
[942] Synops. Meth. Fung. 687. g. 63. n. 1, 2.
[943] Ibid. 4. g. 1. n. 4.
[946] Mon. Ap. Angl. ii. 111. Linn. Trans. xi. 90—.
[947] Reaum. ii. 439.
[948] Ibid. 415. Mouffet 57.
[949] Hist. Ins. Præf. xv.
[950] Cat. Cant. 137.
[952] Reaum. vi. 306.
[953] Fn. Suec. 1626.
[954] Linn. Trans. iii. 26.
[955] De Geer i. 608. Linné has made a mistake with regard to the Ichneumon here alluded to, in calling De Geer's saltatorious Ichneumon I. Muscarum, and referring for it to t. xxxii. f. 19, 20 of that author; whereas the Ichneumon that preys upon the aphidivorous flies does not jump, and is figured by De Geer 605. t. xxxiv. f. 26-29. The jumping one feeds on the larva of a Coccinella.
[956] Vallisnieri Lettere, &c. 80.
[957] Reaum. vi. 296—.
[958] Linné evidently has described another species under I. Ovulorum, in Fn. Suec. 1644.
[959] De Geer i. 593—.
[960] N. Dict. d'Hist. Nat. vi. 10.
[961] Geoffr. Hist. Ins. Par. ii. 361.
[962] Linn. Trans. v. 102—.
[964] De Geer ii. 850—.
[965] Reaum. ii. 444—.
[966] De Geer ii. 863—.
[967] Panzer Fn. Germ. Init. lxxii. 4.
[968] De Geer, i. 583—. ii. 822—. 907.
[969] Reaum. vi. 312.
[970] Leeuwenh. Epist. Oct. 6, 1700. De Geer ii. 869.
[971] Ibid. i. 604.
[972] Rai. Hist. Ins. 259—.
[973] See above, p. 217; and Vol. I. p. 356.
[974] Ibid. 348.
[975] Reaum. vi. 303—.
[976] Reaum. ii. 454—.
[977] De Geer ii. 879—.
[978] N. Dict. d'Hist. Nat. xvii. 513.
[979] De Geer vi. 411—.
[981] Linn. Trans. iv. 236.
[982] Vol. I. p. 50—. 170—. Since that Volume of the present Edition was printed, Say's account of the Hessian Fly has been met with, where he distinguishes it by the above name. (Journal of the Acad. of Nat. Sciences of Philadelphia 1817.) The Ichneumon he calls Ceraphron Destructor.
[983] De Geer, i. 605. This, as before observed, is not the I. Muscarum of Linné; but it ought to have that name, and the other instead to be named, I. Coccinellæ.
[984] Ent. Carn. 760, 761.
[985] De Geer i. 587.
[986] Ibid. ii. 876.
[987] Reaum. ii. 417—.
[988] Reaum. ii. 419—.
[989] De Geer i. 583—.
[990] Ibid. ii. 884.
[992] Reaum. vi. 312.
[994] De Geer ubi supr.
[995] Ibid. 883.
[996] Linn. Fn. Suec. 1609.
[997] Reaum. ii. 443. De Geer i. 196—, 550—. vi. 24.
[998] Reaum. ii. 440—.
[1001] Vol. I. p. 163. note260. Vol. III. p. 162. note379.
[1002] De Geer v. 8—.
[1003] Naturf. xxiii. t. i. f. 8.
[1004] N. Dict. d'Hist. Nat. xx. 110—.
[1005] Ent. Carn. 1052-4.
[1006] Hister particularly.
[1007] De Geer vii. 126—.
[1008] De Geer vii. 144—.
[1009] Lamarck Anim. sans Vert. iii. 196.
[1010] De Geer ii. 554—. Pictet Bibliotheq. Univers. num. ult.
[1011] The existence of this animal has been satisfactorily ascertained by M. de Blainville, who had a specimen, extracted from a human body, sent him by M. Girard, a surgeon of Guadaloupe.
[1012] De Geer ii. 555.
[1013] Matthey ubi supr.
[1014] Philos. Trans. 1823. 8. t. i. ii.
[1015] De Geer ii. 556.
[1016] Gould Ants, 63.
[1017] De Geer i. 551.
[1018] Rösel I. iii. 20.
[1019] Latr. Fourmis, 373.
[1020] De Geer ii. ubi supr. t. xiv. f. 12-14.
[1021] Ibid. i. 553.
[1022] Ibid. ii. 556. t. xiv. f. 12, 13.
[1023] Ubi supra.
[1024] De Geer i. 553.
[1025] Vol. III. p. 15. note21.
[1026] Ibid. 58—. See above, p. 26.
[1027] N. Dict. d'Hist. Nat. xxx. 584.
[1028] Hor. Entomolog. 37.
[1029] N. Dict. d'Hist. Nat. xxx. 584—.
[1030] Cuv. Anat. Comp. ii. 362.
[1031] Syst. Nat. i. 535. Bonnet Œuvr. ii. 36.
[1032] Ibid.
[1034] Lehmann De Sens. Extern. Animal. Exsang. 22—.
[1035] Ibid. De Antenn. Insect. ii. 79.
[1039] De Antenn. Insect. ii. 65—.
[1040] De Antenn. Insect. ii. 42.
[1041] Ibid. 26.
[1045] Philos. Trans. 1820. 314.
[1046] Lehmann De Usu Antenn. ii. 66—.
[1048] Compare what is said above (p. 141) with respect to bees.
[1049] See, for further arguments, Lehmann ubi supr. c. ix.
[1050] Marcel de Serres thinks he has discovered an organ of hearing in most insects, but he does not state its situation. Mém. du Mus. 1819. 99. Treviranus, with regard to the Blattina, suspects it to be situated between the eye and the base of the antennæ, perhaps alluding to the spot noticed above. (Vol. III. p. 505.) Carus, who mentions the above, says, "Is it not reasonable to ask if the sense of hearing may not reside in the membrane which connects the antennæ with the head?" Introd. to Comp. Anat. i. 80—.
[1051] Lehmann De Sens. Extern. Anim. Exsang. De Olfactu.
[1052] Cuv. Anat. Comp. ii. 675.
[1053] Ubi supr.
[1054] Marcel de Serres says they are connected with testes seated in the trunk (Mem. du Mus. 1819. 95); but Treviranus denies this (Arachnid. 36—. t. iv. f. 33).
[1055] Vol. II. p. 361—. III. p. 544—.
[1056] Lehmann De Sens. Extern. &c. De Olfactu.
[1057] Lehmann ubi supr. &c. 27.
[1058] Ibid. and De Usu Antenn. ii. 24—. Cuv. Anat. Comp. ii. 675.
[1059] Lehmann De Usu Antenn. ii. 28.
[1060] Ibid. 31.
[1061] Ibid. 35—.
[1063] N. Dict. d'Hist. Nat. xxiii. 210.
[1064] Ubi supr.
[1065] See above, p. 63. Sprengel Commentar. 14—.
[1066] Huber Abeilles ii. 375—. Dr. Bevan in his interesting work on the Honey-Bee adopts the opinion here stated with respect to the organ of smell in that animal. 265, 303.
[1070] Introd. to Comp. Anat. i. 76. The part he alludes to, is figured Plate VI. Fig. 4. a. g´.
[1071] Ibid. This membrane likewise represents the Nose and Rhinarium in that fly.
[1073] Cuv. Anat. Comp. ii. 682—.
[1076] The elytra of this Order in general differ so materially both from membrane and corium, that it was requisite to invent a term to distinguish them.
[1077] Mon. Ap. Angl. i. t. v. f. 8. b, c.
[1078] We use this term because subcrosa is employed in a quite different sense.
[1079] We restrict the term Figure, to the shape of a superficies.
[1080] The term falcate has usually been applied to signify this figure, as well as that to which we have restricted it; but as the truncate and sharp extremity forms a striking difference, we thought it best to invent a new term.
[1081] We have departed from the more usual definition of trapezoid, "An irregular figure whose four sides are not parallel," because the above is best suited to forms in insects.
[1082] We use this term to denote the shape of solid bodies.
[1083] The word employed in Botany to denote a Polygon is prismatical; but since, properly defined, this term is synonymous with triquetrous, we thought it best to use an adjective derived from prismoid, which implies a body that approaches to prismatical.
[1084] This term in Anatomy denotes any unnatural protuberance or convexity of the body, as a person hunched, or hump-backed. In Astronomy it is used in reference to the enlightened parts of the moon, whilst she is moving from the first quarter to the full, and from the full to the last quarter; for all that time the dark part appears horned or falcated, and the light one hunched out, convex or gibbous.
[1085] We employ the term æquatus instead of æqualis commonly used in this sense, because æqualis is also applied to magnitude, to which we would restrict it.
[1086] I do not find in Schönherr (Curculionid. Method. Disp.) any genus or subgenus of Rhyncophorous beetles the characters of which correspond with those of the insect here alluded to, which I once thought might be a Cyphus Germ. but it is not. It appears common in Brazil, and I have at least two species of it.
[1087] Linn. Trans. vi. 194. t. xx. f. 5.
[1088] Germ. Insect. Spec. Nov. 332—. To this genus Curculio Tribulus and quadridens appear to belong.
[1089] This kind of pubescence has usually been denominated sericeous (sericea); but it certainly does not resemble silk; and is very different from the proper sericeous splendour, exhibited by Cryptocephalus sericeus.
[1090] Mon. Ap. Angl. 1. t. iv. ** c. f. 1. a.
[1091] See above, p. 283. no 7.
[1092] Linné in Coccinella has employed the term Gutta for a white or yellow spot in a darker ground, and Pustula for a red spot in a black ground. We thought one term sufficient to express spots bigger than atoms.
[1094] As this work is intended for general readers as well as for the learned, the above rules, &c. it is hoped will not be deemed without use.
[1095] These symbols are inserted here, because they may be very conveniently adopted in a correspondence on the subject of Entomology.
[1096] Mon. Ap. Angl. i. t. xii. * *. e. 1. neut. f. 6. c. and t. x. * *, d, 1, f. 1. c.
[1097] Oliv. Ins. No. 44. Pæderus. t. i. f. 1. e.
[1098] Germ. Insect. Spec. 486—.
[1099] Fn. Suec. 1183. Fabricius has not admitted this moth among his Noctuæ, I know not why.
[1100] Curtis, Brit. Ent. t. 154.
[1101] Monogr. Ap. Angl. i. 97. t. xii. Apis **. e. 1. Neut. f. 21. d.
[1102] This term may be applied to the Mesothorax in heteropterous Hemiptera, in which that part lies buried under the Prothorax. Plate VIII. Fig. 20. i, k.
[1103] Chabrier Sur le Vol des Insectes. Mém. du Mus. t. viii. 55.
[1104] In many moths, particularly Spilosoma ocularia, and affinities, the insect looks as if its neck was ornamented with a beautiful tippet formed by the Patagia, and its shoulders by these lappets.
[1105] Neither Chrysomela nor Imatidium have a discoidal Epipleura; which furnishes a further proof that Notoclea is distinct from Chrysomela, and Imatidium from Cassida.
[1106] For the reason of this change of the name of Locusta F., see Zool. Journ. No iv.
An Acrida with this spot is figured by Professor Lichtenstein. Linn. Trans. iv. t. v. A.
[1107] Reaum. i. t. xx. f. 12-15.
[1108] Linn. Trans. v. t. iv. f. 10, 11. From my specimens, which are not in a very good state, I cannot ascertain whether this belongs to any of the modern genera into which the Ichneumones minuti of Linné are now divided.
[1109] To this genus or subgenus Scarabæus Syphax, Antæus, Titanus, &c. belong.
[1110] Jones. Linn. Trans. ii. t. viii. f. 1, 3-6, 8.
[1111] Jurine Hymenopt. t. i. f. 3. b.
[1112] Ibid. t. iii. Gen. 4, 5.
[1113] Ibid. t. x. Gen. 23, 24.
[1114] Ibid. t. vi. Gen. 2.
[1115] See Kirby in Linn. Trans. xii. 450—. t. xxii. f. 16.
[1116] Mon. Ap. Angl. i. t. iv. Melitta **. c. f. 10. a.
[1117] Mon. Ap. Angl. i. t. xii. f. 19. a, b.
[1118] Mon. Ap. Angl. t. iv. **. c. f. 14 *. a. f. 12.
[1119] Ibid. t. xxi. f. 20.
[1120] Chabrier Sur le Vol des Insectes. Ann. du Mus. xiv. t. viii. f. 1. K. n.
[1123] De Geer vii. 38—. t. iii. f. 10. rr.
[1125] De Geer ii. 507. t. xi. f. 16. m. n.
[1126] Philos. Botan. 97. n. 153.
[1127] Ibid. 98. n. 155, &c.
[1128] Μεθοδος is rendered "An artificial and compendious mode of doing any thing; a mode of teaching or learning:" Μεθοδευω is "To overcome by artifice." Συσεμα applied to music is "A full and harmonious assemblage of tones." So that in fact, System should express the actual disposition of objects, or a Natural arrangement; and Method, an Artificial one.
[1129] Wisdom. xi. 20.
[1130] Genes. i. 31.
[1131] W. S. MacLeay in Linn. Trans. xiv. 54.
[1132] Linn. Syst. Nat. i. 11.
[1133] Qu. Whether every real species or group has not some one or more peculiar characters which it neither derives from its predecessor nor imparts to its successor in a series?
[1134] Œuvres vii. 51—.
[1135] N. Dict. d'Hist. Nat. xx. 485.
[1137] W. S. MacLeay. Hor. Entomolog. passim; and in Linn. Trans. ubi supr. 53—.
[1138] N. Dict. d'Hist. Nat. xx. 485.
[1139] The idea of a continuous series militates somewhat against that of a circle returning into itself. The progression of the series may be in a circle; but at the point of contact where the second circle meets the first, the lines must cut each other; and at this point of intersection of the two circles are of course the osculant groups constituting the first and the last of each circle, which in their intervention come in contact with each other, or rather forming transition groups. If each circle is regarded as absolute, the series is broken, though the osculant groups connect the circular ones.
[1140] Mr. MacLeay almost admits that there are natural genera. Hor. Ent. 492.
[1141] Œuvr. vii. 52.
[1142] N. Dict. d'Hist. Nat. ii. 34—.
[1143] Even those animals that like the Spongiæ and Alcyonia are aggregate, and fixed by a common base, have a partial degree of voluntary locomotion in their cells.
[1145] Cuv. Anat. Comp. i. 173.
[1146] N. Dict. d'Hist. Nat. ii. 25.
[1147] Ibid. 26—.
[1149] Hor. Entomolog. 200—. See above, p. 3—.
[1150] Savigny Mém. sur les Anim. sans Vertèbr. II. i. 3.
[1151] MacLeay Hor. Ent. 204.
[1152] Vol. III. p. 46—, See above, p. 247.
[1155] In this respect insects excel many reptiles, which can reproduce some of their parts.
[1156] See MacLeay Hor. Entomolog. 203, 206—. 298—.
[1157] Linn. Philos. Botan. n. 155, 160.
[1158] Vol. III. Letter XXVIII.
[1159] Scias Characterem non constituere Genus, sed Genus Characterem; Characterem fluere e Genere, non Genus e Charactere; Characterem non esse ut Genus fiat, sed ut Genus noscatur. Philos. Botan. m. 169.
[1162] Derived from κολεος, a sheath, and πτερον, a wing.
[1163] Hist. Animal. l. iv. c. 7. l. v. c. 20.
[1164] Ὁσα το πτερον εχει εν κολεῳ.
[1165] Latr. Gen. Crust. et Ins. i. 169. Oliv. Ins. i. Introd. v.
[1167] In some genera, as Molorchus, &c., they do not completely cover the wings. Plate X. Fig. 1. Plate I. Fig. 4, 5.
[1168] In Buprestis, Molorchus, &c., they are only longitudinally folded.
[1170] From σρεψις, a turning or twisting, and πτερον.
[1171] Vol. III. p. 589. note1769.
[1172] Hor. Entomolog. 371—.
[1173] Linn. Trans. xi. 96—.
[1174] Ibid. t. ix. f. 1. d.
[1176] Linn. Trans. Ibid. f. 15. b.
[1177] From δερμα, a skin.
[1179] From ορθος, straight.
[1180] Fn. Suec.
[1181] From δικτυον, a net.
[1183] From νευρον, a nerve.
[1184] Her. Entomolog. 433.
[1187] Ibid. p. 423, 441—, 451, 454—.
[1190] N. Dict, d'Hist. Nat. x. 344.
[1191] The ovipositor of Raphidia seems merely calculated to introduce its eggs under bark; it seems incapable of boring.
[1192] From ὑμην, a membrane.
[1193] De Geer ii. 1035.
[1194] Since this was written, Mr. Stephens has showed me a remarkable Hymenopterous insect taken by him in Hertfordshire, which appears to have the antennæ of one of the Ichneumonidæ and the wings and abdomen of a Tenthredo L., so as to form a link connecting the two tribes or suborders. This may probably have a vermiform larva.
[1195] Hor. Entomolog. 431.
[1196] Hor. Entomolog. 429.
[1197] Vol. III. p. 67. See above, p. 160.
[1198] Whoever consults De Geer ii. 941—. t. xxxiii. f. 14, 15. t. xxxvi. f. 27. and t. xxxix. f. 7, 8, will be convinced that the metamorphosis of Tenthredo L. is incomplete rather than obtected.
[1199] The Hymenoptera, though they have all the usual oral organs, cannot be denominated masticators generally; these organs, especially the mandibles, being chiefly used in their economy.
[1202] From ἡμισυ, the half.
[1203] Vol. III. p. 463—. Linn. Syst. Nat. Ord. II.
[1204] If considered as suborders, their denomination should not terminate precisely as that of Orders. Perhaps Hemipterita and Heteropterita might be an improvement.
[1205] Hor. Entomolog. 374—.
[1209] Vol. III. p. 611—. 604—.
[1211] From θριξ, τριχος, hair. Mr. MacLeay, thinking it indisputable that the Perlidæ should be included in this Order, suggests the propriety of changing its name, both as inapplicable, and as being preoccupied by a Dipterous genus. As I do not think the Perlidæ belong to the Order, and as the great body of the Trichoptera are distinguished by hairy upper wings, I cannot think the name improper: but to apply a name to a Genus which terminates like the denominations of Orders, I think leads to mistakes, and should not be tolerated.—K.
[1212] Hor. Entomolog. 430—.
[1214] The location of the legs together, their long coxæ, and their calcaria, are analogous also to those of the Lepidoptera.
[1215] Reaum. vi. Mem. x. t. xxxii. f. 13. t. xxxiv. f. 1-6. De Geer vi. 169—. t. x. f. 7, 8.
[1216] N. Dict. d'Hist. Nat. xxv. 286.
[1217] De Geer ii. 511—. He however observes, that they often attack other insects: but the form of their mandibulæ, like that of the caterpillars of Lepidoptera, which also on some occasions become carnivorous (Vol. I. p. 386), is fitted for a vegetable diet. De Geer, Ibid. 505.
[1218] This is evident from De Geer's account. Ibid. 516. t. xii. f. 14. t. xv. f. 4.
[1220] From λεπις, a scale.
[1221] Vol. III. p. 537. Plate IX. Fig. 4.
[1225] From δις twice, or double.
[1226] Hist. Animal. l. iv. c. 1, 27.
[1233] From αφανης; inconspicuous; so named because something like elytra appear.
[1236] From α, priv. and πτερον.
[1240] Hor. Entomolog. 381.
[1241] Vol. III. p. 22. note36.
[1245] Hor. Entomolog. 286.
[1246] The number of segments and legs acquired by these insects in their progress to their last state, distinguishes their metamorphosis from that of other Aptera, and requires a distinct name.
[1248] When I said (Vol. III. p. 31.) that Phrynus probably belonged to the true Arachnida, it escaped my recollection that Latreille had placed that genus there.
[1249] L. Dufour Six Nouvell. Arachnid. &c. Ann. Gen. des Scienc. Physiq. IV. iii. 17. t. lxix. f. 7, b.
[1250] Mém. sur les Anim. sans Vertèbr. I. i. 57—.
[1252] Plate XV. Fig. 10. T´´. Plate XXIII. Fig. 15. 17. T´´.
[1255] Called the Centris. Vol. III. p. 388, 716.
[1256] M. Latreille thinks that in Galeodes the prothorax is coalite with the head (N. Dict. d'Hist. Nat. xii. 370.); but that it is not so, is evident from the six real legs being affixed to the pieces behind it. See also Vol. III. p. 23. note40.
[1257] L. Dufour ubi supr. IV. iii. 18.
[1258] Ibid. 19.
[1259] Ibid. t. lxix. f. 7. d.
[1260] When the characters of the Class Arachnida were drawn up (Vol. III. p. 30.) I had not seen a Galeodes: they should be thus amended:
Palpi four: anterior pair pediform, cheliform, or unguiculate; posterior pediform.
Trunk Legs six, &c.
[1262] Familles Naturelles du Règne Animal.
[1263] Annulosa Javanica. 5.
[1265] Coléopt. d'Europe i. 75.
[1266] Vol. III. p. 167—. I formerly hinted (Ibid. p. 163.) that the larva of Cicindela may be regarded as Araneidiform: this is further confirmed by its having eight eyes, (and not six,) as I have since discovered, and by the aspect of its large head and prothorax. The other larvæ of the Adephagana have twelve eyes.—Mr. Stephens (Illustrations of British Entomology, no. xv. p. 175.) has confirmed the above statement, as to the number of eyes of the larva of Cicindela.
[1267] Mr. MacLeay says that more than 100,000 Annulosa exist in collections.—Hor. Ent. 469.
[1268] Vigors in Zoolog. Journ. I. ii. 188.
[1269] Hor. Entomolog. 125—.
[1270] See Bicheno in Linn. Trans. xv. 491.
[1271] Dr. Horsfield, in his very ingenious and generally admirable Descriptive Catalogue of the Javanese Lepidoptera in the Museum of the Honourable East India Company, has divided that Order into five primary groups, apparently to accommodate it to Mr. W. S. MacLeay's quinary system. I trust he will pardon me for observing, that in this arrangement he seems to me rather to force than to follow nature; and that though he adheres to the above system as to the number, he forsakes it in the construction of his groups.
The obvious primary sections of the Lepidoptera, which have been evident to almost every one who has at all studied the Order, are the three named in the text, corresponding with Linné's genera Papilio, Sphinx, and Phalæna. The groups of the last or nocturnal section, which Dr. Horsfield has elevated to the same rank with the two first, are evidently not of equal value, nor to be placed upon the same platform; for the Bombycidæ, Noctuidæ, and Phalænidæ, are clearly of a secondary rank. Indeed this section is resolvable into more groups of equal value than the learned Doctor has assigned to it; for the Tortricidæ, Tineidæ, &c. are not so united to the Geometers, or genuine Phalænidæ, as to form with them a primary group of the Nocturnal Lepidoptera, but are themselves entitled separately to that distinction. This will be evident to every one who will take the trouble to compare the larvæ and their habits, of the two tribes, as well as the perfect insects.
In the construction of his groups, he seems not to have discovered in the Lepidoptera a great typical group resolvable into two, or at least he has not built his system on this foundation, which appears an essential part of the quinary arrangement. (See Mr. W. S. MacLeay in Linn. Trans. xiv. 56—.) As to value, the Papilionidæ constitute the typical group or centre of the Order, though the Phalænidæ prevail as to numbers: but neither of these are resolvable into two primary groups.
[1272] Linn. Trans. xiv. 56—. It is to be observed, however, that what Mr. MacLeay calls the aberrant groups are usually also resolvable into two.
[1273] Hor. Entomolog. 318, et passim.
[1274] Linn. Trans. ubi supr. Mr. W. S. MacLeay informs me that M. Agardh has found that the distribution of Fuci is regulated by the same law.
[1275] Zool. Journ. iii. 312—.
[1276] Vol. III. p. 15. note21.
[1277] Hor. Entomolog. 199.
[1278] Viz. 1. Copris Hesperus; 2. C. reflexa; 3. C. Sabæus; 4. C. lunaris; 5. C. Carolina; 6. C. Œdipus; 7. C. Midas; 8. C. capucina; 9. C. Bucephalus; 10. C. Molossus; 11. C. Eridanus; 12. C. sexdentata K.
[1279] Hor. Entomolog. 518.
[1280] The most natural and consistent interpretation of 1 Cor. xiii. 12, Βλεπομεν γαρ αρτι δι' εσοπτρου εν αινιγματι, is, that "we see now as it were in a mirror the glory of God reflected enigmatically by the things that he has made." Comp. Rom. i. 20—. Our Saviour (Luke x. 19.) calls serpents and scorpions the power of the enemy; which can only mean that they are figures or symbols of the enemy.
[1281] Rom. i. 20, to the end of the chapter.
[1282] N. Dict. d'Hist. Nat. xx. 484. comp. ii. 30—.
[1283] Mém. sur les Anim. sans Vertèbr. I. i. 20—.
[1284] Horæ Entomologicæ.
[1289] A most singular insect belonging to this tribe, and which seems to form a link, having a notched cubit, between the Amaurona and the Lamprona, has been described and figured by Hagenbach under the name of Mormolyce phyllodes. It exhibits such a striking resemblance to a Mantis or Phasma, that it might be mistaken for one. It was found on the western side of the island of Java. Mr. Samouelle showed me a second species of this genus from China, belonging to a lady, who put it into his hands, it being broken, to put together.
[1290] A remarkable imitation of an antelope's horn, a process of the mandible of an insect, in the possession of R. D. Alexander, Esq. F.L.S., is figured in the fifth Number of the Zoological Journal.
[1291] Hor. Entomolog. 456. Comp. Linn. Trans. xiv. 67—.
[1292] Mém. sur les Anim. sans Vertèbr. I. i. 20—.
[1294] Vol. III. pp. 372, 598.
[1298] Ent. Carn. 168. n. 446.
[1299] Meigen has figured a Dipterous insect exactly resembling a Cimbex, which he calls Aspistes berolinensis (Dipt. i. 319. t. xi. f. 16, 17.)
[1300] Prædones Latr., &c.
[1301] Andrena F., &c.
[1302] Hor. Entomolog. 437.
[1304] Mém. du Mus. 1819. 136.
[1305] Rifferschw. de Ins. Genital. 9.
[1306] Annulos. Javan. i. 1.
[1307] N. Dict. d'Hist. Nat. xxv. 115—. xxvii. t. M. 8. f. 1.
[1308] Piso Hist. Nat. 63. Curui 1. Jundia v.
[1309] N. Dict. d'Hist. Nat. xxvii. 235. Hor. Entomolog. 203.
[1310] Ibid. 281—.
[1311] Ibid. 354, 390, 397.
[1312] This insect, except in its antennæ, so nearly resembles a Nirmus, that it might be mistaken for one. See Coquebert Illustr. Icon. i. t. ii. f. 14.
[1314] Fuessl. Archiv. t. lii. f. 5.
[1315] Stoll Saut. de Pass. t. xx. b. f. 79.
[1317] Genes., ii. 19—.
[1318] Pol. Synops. on Genes. ii.
[1319] Genes. i. 25.
[1320] Linn. Trans. iv. 51—. See Levit. xi. 20—.
[1321] The Neuroptera appears to be the only Order not so signalized. It is worthy of notice that insects are usually noticed generically and not specifically in Scripture. On the insects of Scripture see Bochart Hierozoic. ii. 1. iv.
[1322] Isai. vii. 18. Joel ii. Rev. ix. 3.
[1323] Prov. xxx. 24—.
[1324] 1 Kings iv. 33.
[1325] Linn. Trans. i. 5.
[1327] Ibid. l. i. c. 5.
[1328] Ibid. l. iv. c. 7.
[1329] Ibid.
[1330] Ibid. l. v. c. 19.
[1331] Aristotle calls winged insects Pterota when he would distinguish them from those that are apterous, and Ptilota when he contrasts them with birds. (Comp. Hist. Anim. l. iv. c. 1. with l. i. c. 5.) Sometimes he calls birds thus contrasted Schizoptera, and insects Holoptera. De Anim. Incess. c. 10.
[1332] Ibid. l. i. c. 5.
[1333] Ibid. and l. iv. c. 7.
[1334] Ibid.
[1335] Hist. Anim. l. iv. c. 1.
[1336] Ibid.
[1337] Ibid. l. viii. c. 11.
[1338] Gr. Ον τροφης χαριν εχει οδοντας αλλ' αλκης. Αλκη means Strength of mind, Fortitude, Strenuousness, also Help:—it here probably signifies their strenuous use of their oral organs in fulfilling their instincts. De Partib. Anim. l. iv. c. 5.
[1339] Hist. Anim. l. iv. c. 7.
[1340] Ibid.
[1341] Gr. Αερσιποτητος αραχνη. Dies. lin. 13.
[1342] Hist. Nat. l. xi. c. 25.
[1343] Vol. I. p. 481. Vol. II. p. 121—.
[1344] De Natur. Animal. l. vi. c. 20.
[1345] Ibid. l. xv. c. 1.
[1346] Opera vi. 683.
[1347] Ibid. 153—.
[1348] Ibid. 154, 233, 265, &c.
[1349] Opera vi. 676, 679, 680.
[1351] Opera vi. 682—.
[1352] Esperienz. ed Osserv. i. 42—.
[1353] Pultency's Sketches of Botany in England, i. 86.
[1354] Theatr. Insect. Epist. Ded. i.
[1355] Theatr. Insect. Epist. Ded. i.
[1356] Theriotroph. Siles. 455.
[1357] Aristotle (Hist. Anim. l. i. c. 1.) says, "The sponge seems to have some sensation: as a proof, it is not easily plucked up, unless, so they say, the attempt is concealed."
[1358] Lister's Goedart, Præf. ii.
[1359] See Vol. I. p. 65—, where these terms are explained.
[1360] Swamm. Bibl. Nat. i. 38—.
[1361] Ibid. 92—.
[1362] Ibid. 119—.
[1363] Ibid. ii. 1—.
[1364] Ibid. 31—.
[1365] Ibid. 30.
[1366] Hist. Ins. Prolegom. ix.—
[1367] These are all Annelida.
[1368] Larvæ.
[1369] Various Aptera and the Bed Bug.
[1370] Nymphon.
[1371] Scorpio.
[1372] Spiders, Phalangia, and Mites.
[1373] Iulus.
[1374] Scolopendra.
[1375] Annelida.
[1376] This section is divided by the author into thirteen tribes.
[1377] Lepidoptera.
[1378] Apis, Bombus, &c.
[1379] Vespidæ.
[1380] Andrena, Halictus, Nomada, &c.
[1381] Crabro, Philanthus, Cerceris, &c.
[1382] Serifera? Ichneumon, &c.
[1383] Trichoptera.
[1384] Pimpla Manifestator, and other Ichneumonidæ, with a long ovipositor.
[1385] Our author has followed Swammerdam in this unnatural separation of those Diptera whose metamorphosis is coarctate from the rest; and in associating with them the Chalcidites, whose metamorphosis is really different. Into this error both were led by system.
[1386] Philos. Lett. &c. 141.
[1387] Ibid. 343.
[1388] Ray died in 1705, and Linné was born in 1707.
[1389] When a boy he attempted to introduce wasps and bees into his father's garden, to the great annoyance of the old gentleman.—Stœver's Life of Linnæus, 4.
[1390] Ibid. 75.
[1391] Linn. Philos. Botan. n. 87, 188, 189.
[1392] See above, p. 342, n. 5.
[1393] Linn. n. 291.
[1394] Fn. Suec. Præf.
[1396] Histoire abrégée des Insectes.
[1398] The first volume of his Mémoires was published in 1752.
[1399] The first volume of this work was published in 1734, the sixth and last in 1742.
[1400] Reaum. i. Mém. vi. vii. and Mém. ii. 68—.
[1401] Smith's Tour, iii. 150.
[1402] Vol. I. p. 175. Also see above, p. 166—.
[1403] Bonnet i. 19—.
[1404] We have been informed that these valuable remains are at length likely to be rescued from oblivion, and given to the public.
[1405] Vol. II. p. 48, note51.
[1406] Since the former edition of these volumes was published, another and most important association has been formed, having for its object the Animal Kingdom solely; which not only has a museum to receive specimens of dead animals (by the liberal donation of its present learned secretary, of his own rich collection, and from other sources, already most interesting both as a spectacle and to the student), but also a Vivarium, in which a considerable and curious assemblage of living animals may be seen. This association, which is named The Zoological Society, is principally indebted for its formation to the efforts of a great, amiable, and lamented character, the late Sir Thomas Stamford Raffles, whose merits were equally conspicuous both as a Politician and a Naturalist, and who was its first President.
[1407] Linné is recorded to have said, "Si Dominus Fabricius venit cum aliquo Insecto, et Dominus Zoega cum aliquo Musco, tunc ego pileum detraho et dico: Estote doctores mei." Stœver's Life of Linnæus. 186.
[1408] Fab. Philos. Entomolog. Præf.
[1410] Philos. Entomolog. vi. §. 2. Syst. Ent. Prolegom.
[1411] From Ελευθερος, Free.
[1412] Derivation uncertain. Perhaps Αυλων, A long and narrow space or tract.
[1413] Συνιστημι, To stand together.
[1414] Πιεζω, To press.
[1415] Οδους, A tooth.
[1416] Μιτος, A thread.
[1417] Unogata is probably a mistake for Onychata; from Ονυξ, A claw.
[1418] Doubtless for Polygnatha; from Πολυς, Many, and Γναθος, A jaw.
[1419] Κλειστος, Closed, and Γναθος.
[1420] Εξω, Without, and Γναθος.
[1421] Γλώσσα, A tongue.
[1422] Ῥυγχος, A rostrum.
[1423] Αντλια, A pump.
[1424] Dispositio insectorum sistit divisiones s. conjunctiones eorum, et est artificialis quæ Classes et Ordines, et naturalis quæ genera, species, et varietates docet. Philos. Entomol. vi. §. 2.
[1425] Ibid. §. 7.
[1426] Latreille Gen. Crust. et Ins. iii. 214.
[1427] With respect to Natural Genera he says—"Cavendum tamen ne nimis imitando naturam systematis amittamus filum Ariadneum." Ibid. § 6.
[1428] Fab. Entomolog. Syst. em. et auct. i. Præf. iv.
[1429] Fabricius calls this a chaos, and threatens to prove it, but he never fulfilled his threat. See Fab. Supplem. Præf. i.
[1430] Introd. ad Hist. Nat. 401.
[1431] See N. Dict. d'Hist. Nat. x. article Entomologie; and Familles Naturelles du Règne Animal 262—.
[1432] These tables, except the first, are taken from the Familles Naturelles du Règne Animal. As a new edition of M. Le Baron Cuvier's Règne Animal is preparing, M. Latreille will doubtless give in it a still more improved arrangement of the Crustacea, Arachnida, and Insecta.
[1433] Several of the minor groups given in the table he has further resolved before he arrives at his genera.
[1434] Vol. III. p. 348, note903.
[1436] Syst. des Anim. sans Vertèbr. 185.
[1437] Ibid. 171.
[1438] Anim. sans Vertèbr. iii. 332—.
[1439] Anat. Comp. i. t. viii.
[1440] Expos. d'une Meth. Nat. 17.
[1442] Linn. Trans. xi. 376. N. B. I have transferred from the Arachnida his suborder Notostomata, as he subsequently placed it at the end of Insecta, under the Omaloptera.
[1443] See above, pp. 378, 380, 385, 390.
[1445] See Vol. III. p. 25—. and above, p. 394—.
[1446] Hor. Entomolog. c. vi.
[1448] Hor. Entomolog. 420—.
[1449] Ibid. 422.
[1450] Other systems or methods have been promulgated by various authors, as by Schæffer, Scopoli, Geoffroy, &c. Walckenaer and Blainville have proposed one founded on the number of the legs of insects; but those in the text are the principal and best known.—N. Dict. d'Hist. Nat. xvi. 277.
[1451] Linn. Trans. xiv. 59—. Annulos. Javan. 6. See above, p. 408.
[1452] Latreille Gen. Crust. et Ins. iii. 226. note 1.
[1453] Præf. ii.
[1454] Linn. Trans. ii. 63—.
[1455] Mon. Ap. Angl. i. 211—.
[1456] Vol. III. p. 620. n. 3.
[1457] It may not be unprofitable here to mention those works which the Entomologist may find it most useful to consult in various departments of the science. For descriptions of the Genera and Species of insects in general, he must have recourse to the Entomologia Systematica emendata et aucta of Fabricius, and its Supplement; to the volumes he subsequently published under the titles Systema Eleutheratorum, Rhyngotorum, Glossatorum, Piezatorum, and Antliatorum; to the Genera Crustaceorum et Insectorum of Latreille; to the same department of the Règne Animal of Cuvier; and to the Animaux sans Vertèbres of Lamarck. He will find the genera of Linné and Fabricius illustrated by figures, in Rœmer's Genera; and many of the species described by the latter in Coquebert's Illustratio Iconographica. In our countryman Drury's beautiful Illustrations of Natural History, a large number of new and rare insects are depicted; and in Mr. Donovan's Insects of China, India, and New Holland, some of the most brilliant and interesting that have been imported from those countries. Panzer's Faunæ Insectorum Germanicæ Initia has little short of 3000 figures of insects of every Order (a considerable number of which are found to inhabit Britain), by the celebrated Sturm; and the latter, in his Deutschlands Fauna, has illustrated many Coleopterous genera analytically (as has also M. Clairville the weevils and Predaceous beetles of Switzerland in his Entomologie Helvétique) by his admirable pencil. Beetles in general are well figured and described in Olivier's splendid Entomologie; as are those of Europe in a beautiful work now in course of publication, under the title of Coleoptères d'Europe, by MM. Latreille and Dejean. The latter author has also begun a work on this Order under the title of Species général des Coléoptères de la Collection de M. Le Comte Dejean; two volumes of which have appeared, containing part of the Carabici Latr. but I fear it has stopped for want of encouragement. Had the descriptions been less verbose it would have had a better chance of success. For the Orthoptera and Hemiptera, the student must have recourse to Stoll's Spectres, Mantes, Sauterelles, Grillons, Blattes, Cigales, and Punaises. To a knowledge of the species of Lepidoptera, the admirable figures of Cramer (Papillons Exotiques de trois Parties du Monde), Esper (Schmetterlinge, Tagschmetterlinge), Hübner (Schmetterlinge, &c.), and Ochsenheimer's valuable Schmetterlinge von Europa, with the continuation by Treitschke, will afford a useful avenue. Meigen also, author of a most valuable work on the Europæan Diptera, is publishing at this time a work on Lepidoptera under the title of Europäische Schmetterlinge. To the Hymenoptera Jurine and Christian are the best guides, and to the Diptera Meigen.
With regard to works in British Entomology in general—Donovan's Natural History of British Insects, and Samouelle's Entomologist's Useful Compendium, will be found very excellent helps to the student. For the British Genera, the most important work that has yet appeared is Mr. John Curtis's British Entomology, in which not only are the insects admirably represented, but their trophi correctly delineated, accompanied by able descriptions. For the Coleoptera of our country, Mr. Marsham's Entomologia Britannica should be consulted: for the Lepidoptera, the Butterflies of Lewin, Mr. Haworth's useful Lepidoptera Britannica, and Miss Jermyn's Butterfly-Collector's Vade Mecum; and for the English species of Linné's genus Apis, the Monographia Apum Angliæ. A British Fauna Insectorum, under the title of Illustrations of British Entomology, has at length been happily begun by a gentleman (J. F. Stephens, Esq.) who both by his accurate knowledge of the subject, and the extent of his collection of British Insects, is best qualified to undertake it. As far as it has proceeded, it is ably executed, and possesses this advantage, (an advantage seldom to be obtained in works published periodically,) that it finishes, as far as possible, as it goes.
[1458] Linn. Philos. Botan. § 334.
[1459] Linn. Trans. x. 20—. &c. Dict. des Scienc. Nat. xviii.
[1460] Selborne i. 173.
[1461] Philos. Entomolog. ix. § 20.
[1462] Mém. du Mus. 1815.
[1463] Hor. Entomolog. 42—. 518.
[1464] Essai Elément. de Géograph. Botan. 62.
[1465] Wisdom of God, &c. 2d edit. 9.
[1466] Hor. Entomolog. 469. This calculation includes the Crustacea.
[1467] It has lately been discovered that the larva of Drilus flavescens, a beetle, feeds upon the common snail. (Bulletin des Scienc. Nat. 1824. iii. 297; v. 110; vi. 221.) I have found an Acarus on the same animal.
[1469] We employ this term, because the more common one, herbivorous, does not properly include devourers of timber, fungi, &c.
[1470] If we consider the number of species of Acari, Nirmi, Poduræ, and Araneidæ, this proportion will appear moderate.
[1471] Hor. Entomolog. 48.
[1472] Philos. Entomolog. ix. § 20.
[1473] Géograph. Génér. des Ins. 5.
[1474] Ibid.
[1475] Ibid. 7—.
[1476] Ibid. 8, 11.
[1477] Personal Narrat. E. T. v. 88. He says also that each stream almost has its peculiar species (Ibid. 98), and that they sometimes emigrate to stations they had not infested before. Ibid. 106—.
[1478] Hor. Entomolog. 519.
[1479] Latr. ubi supr. 3.
[1480] Géographie, &c. 22—.
[1481] Ibid. 27.
[1482] Géographie, &c. 20—.
[1484] As this insect is the type of a distinct genus amongst the Scutelleridæ, I have distinguished it by the name Fabricius gave the whole tribe.
[1485] M. Latreille (Géographie, &c. 8.) seems to regard these varieties as distinct; in which case they would be the representatives of the species named in the text: but the variations are mostly so slight, as not to afford any satisfactory distinctive characters.
[1486] Géogr. Génér. des Ins. 2.
[1487] When I described the Melville Island insects for Captain Sabine, I received from him no Culices; but I afterwards saw in his possession a genuine one from thence.—K.
[1488] Linn. Trans. xii. 380—. n. 6, 7.
[1489] Ibid. n. 5.
[1490] Dejean in his catalogue gives only 434 species; while Mr. Stephens, four years ago, had 550, and has since increased the number to above 600.
[1491] Journal of a Tour in Iceland, 272.
[1493] Entomogr. Russ. Coleopt. t. xiii. f. 1.
[1494] Ahren's Fn. Europ. i. 1.
[1495] Hor. Ent. 47—.
[1496] Annulosa Javanica, 36.
[1497] See the Rev. L. Guilding's admirable History of Xylocopa Teredo and Horia (Cissites Latr.) maculata, Linn. Trans, xiv. 313—.
[1498] Out of 51 species described by Bilberg, 28 are African, and 19 of these are from the Cape.
[1499] Géogr. Génér. des Ins. 18.
[1500] Hor. Entomolog. 45.
[1501] Dr. Leach has described 8 British species (Linn. Trans. xi. 37.); Dejean has 7 Spanish ones.
[1502] I have a very splendid species of this genus taken by C. C. Elwes Esq. on the Pyrenees, which is undescribed, and falls under none of the count Dejean's Families, having its elytra perfectly smooth, without striæ, punctures, &c. It is of a brilliant golden green. It stands in my cabinet under the name of C. lævigatus. K.
[1503] Fischer Entomogr. Russ. 90—. t. viii. f. 13.
[1505] Major General Hardwicke gave me one of this description from Nepal.
[1506] Latr. Géograph. &c. 18—.
[1507] Linn. Trans. xiv. t. iii. f. 4.
[1508] Hor. Entom. 147.
[1509] Linn. Trans. ubi supr. f. 1.
[1510] Ibid. xii. t. xxi. f. 9.
[1511] Ibid. f. 14.
[1512] To this genus belong Melolontha aurulenta. Ibid. 400; and M. sericea. Ibid. 463.
[1513] Latr. Géograph. 7.
[1514] Cetonia atropunctata and Brownii of Linn. Trans. (xii. 464. t. xxiii. f. 6.) belong to this genus.
[1515] Linn. Trans. xii. t. xxii. f. 2; t. xxiii. f. 7.
[1516] Latreille, Géograph. &c. 10.
[1517] Linn. Trans. xiv. 569.
[1519] Fischer, Entomogr. Russ. i. 135.
[1520] From finding it in water, Fabricius considered this insect as a Hydrophilus, but it is a true Cercyon.
[1522] Personal Narrat. E. T. v. 91—.
[1524] A species of Gyrinus (G. Viola aquatica), described by Modeer (Linn. Syst. Nat. Ed. Gmel. i. 1612. n. 9.), is said to inhabit salt water.
[1525] Géograph. &c. 6.
[1526] Apis *., a. Mon. Ap. Angl. ii. 178—.
[1527] Linn. Trans. iv. 30—. v. 96—. t. iv.
[1529] Géograph. &c. 6.
[1531] These, as well as Melecta, are probably a kind of Cuckow-bee. Mon. Ap. Angl. i. 150.
[1532] Melitta * *. b. Mon. Ap. Angl. i. 138—.
[1533] Mémoires sur le gènre Halicte.
[1535] Linn. Trans. ix. 78—. t. i. f. 20.
[1536] Ibid. 55. t. i. f. 12.
[1537] This insect does not, I believe, eat the petals of the rose, but laps the nectar it produces. I have seen it employed upon wounded trees lapping the sap.
[1538] Mon. Ap. Angl. ii. 172. 257.
[1539] See above, p. 491, note1467.
[1540] Ibid. p. 219; and Vol. I. p. 267—.
[1541] Ibid. p. 256—.
[1542] Apis * *. e. 2. K.
[1543] Apis * *. c. 2. α. K.
[1544] Butterfly Collector's Vade Mecum, 66, noted.
[1545] De Geer ii. 638—. 641—.
[1546] Swamm. Bibl. Nat. i. Conf. 114 with 103.
[1547] Reaum. vi. 480—.
[1549] Lepidopt. Britann. 263—.
[1550] Linn. Trans. v. 256.
[1553] Vol. II. p. 217. See above, p. 200.
[1554] Entomologist's Useful Compendium. t. xi. f. 5.
[1556] Lepidopt. Britann. 20.
[1560] Samouelle's Compendium. t. xi. f. 1, 2.
[1562] Plate XXIV. Fig. 2. N.B. The net is represented too shallow in this figure.
[1563] Voyage to the Cape. i. 63. Eng. Trans.
[1565] Illig. Mag. iii. 222. Mr. Stephens however, whose experience is great in the best modes of collecting, is of opinion that insects that have been immersed in spirits of wine are apt to become mouldy. We have not ourselves observed this.
[1571] In the figure just quoted the artist has represented the insect as transfixed in this way.
[1573] Mr. Samouelle (Useful Compendium, 321) recommends a somewhat different method.
[1575] Some other methods are recommended by Mr. Samouelle, which the reader will find in his Useful Compendium, 318.
[1578] Lepidopt. Britann. 87.
[1580] Reaumur ii. 12—.
[1581] See Mr. Samouelle's Compendium, 311.
[1582] Compare what is said Vol. I. p. 47—.
[1583] Vol. III. p. 28—. See above, p. 377—.
[1585] In Elater, Fabricius describes 137 species; in Melolontha, 149; in one section of Rhynchænus, 161; of Curculio, 183; and in his Papiliones Heliconii, 300.
[1586] Thus he places Chlænius holosericæus and nigricornis, which might pass for varieties, far asunder; and Dromius agilis is even put in a different section from D. quadrimaculatus, truncatellus, &c.
[1587] The continuance of this important privilege, by the lamented death of the learned President, is now rendered uncertain; but I trust we may anticipate, that by the liberality of the members of the Linnean Society, and if necessary of the public, this invaluable treasure, by being fixed in the Metropolis, will be more than ever accessible to the British Naturalist.
[1588] It may not be amiss to mention a few:—Sphæridium dytiscoides is a Hydrophilus related to H. fuscipes. S. glabratum is heteromerous, probably one of the Helopii Latr. Carabus retusus and Maderæ both belong to Calosoma. Cistela angustata is a true Choleva. See Linn. Trans. xi. 138.—S.
[1591] For dissections the one recommended above, p. 201, may be used. Sometimes a watchmaker's eye-glass, which also sets the hands at liberty, will be found useful.
[1592] N. Dict. d'Hist. Nat. xxxii. 264.
[1593] Ibid. xvi. 281.
[1594] Ibid.
[1595] Organic Remains iii. t. xvii. f. 2.
[1596] Ibid. 281—.
[1598] Cowper's Retirement.
[1599] Rifferschweils De Insect. Genital. 9.
[1600] De Orthopteris hoc præcipue notavit D. Marcel. de Serres (Mém. du Mus. 1819. 113—.) in quibus vesiculæ seminales, colleterio; testes, ovariis; vasa deferentia, oviductui; canalis seminalis, ovipositori, &c., mutuò adamussim respondent.
[1601] Rifferschw. De Insect. Genital. 9.
[1602] Reaum. ii. 79. Herold. Schmetterl. t. iv. f. 2, 3.
[1603] Treviranus Arachnid. 11, 36—. Reaum. vi. 436. N. Dict. d'Hist. Nat. xi. 82. Marcel. de Serr. ubi supr. 104. Latreille Fam. Nat. 324.
[1604] Rifferschw. ubi supr.
[1606] Rifferschw. 10. N. Dict. d'Hist. Nat. xvi. 242.
[1607] Ibid. & xxxv. 412.
[1608] Gaede Anat. der Ins. t. i. f. 9. a.
[1609] De Geer iii. t. ix. f. 11. t.
[1610] Reaum. vi. t. xvi. f. 6, 7. g.
[1611] Ibid. t. xviii. f. 4, 5. g.
[1612] Ibid. t. xxvii. f. 16. c.
[1613] De Geer vi. t. iii. f. 17. d, e, f.
[1614] Reaum. vi. t. viii. f. 5. d, e, m.
[1615] Swamm. Bibl. Nat. t. xliii. f. 17. a, b, c.
[1616] De Geer ii. t. xix. f. 11. f. N. Dict. d'Hist. Nat. xi. 82.
[1617] Ibid. xxx. 41; xxix. 177.
[1618] Gaede Anat. t. i. f. 9.
[1619] Ibid. 18. Swamm. ubi supr. t. xliii. f. 17. e, d.
[1620] Rifferschw. 10.
[1621] Ibid. 22.
[1622] Gaede t. ii. f. 9. d, e. N. Dict. d'Hist. Nat. xvi. 241.
[1623] Swamm. ubi supr. i. 223. t. xxii. f. 5. h, i. Hoc insecto et Hydrophilo supradicto organa insunt quæ pro Prostatis habentur.
[1624] N. Dict. d'Hist. Nat. xvi. 242. Gaede t. i. f. 9. d d.
[1625] Ibid. etiam t. ii. f. 9. 14. d d.
[1626] Cuv. Anat. Comp. v. 192.
[1627] Gaede t. ii. f. 2. c.
[1628] Herold. Schmett. t. xxxii.
[1629] Gaede t. ii. f. 9.
[1630] Rifferschw. 19.
[1631] Ibid. 20.
[1632] Marcel. de Serres Mém. du Mus. 1819. 115.
[1633] Ibid. 128. Comp. Cuv. Anat. Comp. v. 195. cum Swamm. Bibl. Nat. i. 102.
[1634] Cuv. Ibid. 191.
[1635] Ibid.
[1636] Rifferschw. 22.
[1637] Gaede t. ii. f. 9. b b.
[1638] Herold. Schmett. t. iv. f. 8, 9.
[1639] Gaede t. ii. f. 14. b b.
[1640] Swamm. ubi supr. t. xxi. f. 1. a.
[1641] Rifferschw. 21.
[1642] Swamm. t. iii. f. 6. f.
[1643] Herold, ubi supr. t. v. f. 1, 9. &c.
[1645] Cuv. ubi supr. v. 115.
[1646] De Geer vi. t. xv. f. 8. d.
[1647] Ibid. ii. t. xix. f. 11. e.
[1648] Reaum. v. t. xix. f. 9.
[1649] Ibid. ii. t. xxvi. f. 10, 11. ll. De Geer ii. t. xix. f. 9.
[1650] Reaum. iv. t. xl. f. 8. c, e.
[1651] Ibid. vi. t. viii. f. 4. c, b.
[1652] De Geer ii. t. xxi. f. 20. b, c.
[1653] Ibid. t. xlii. f. 11. b, c, d; t. xliii. f. 13. p.
[1654] Herold. Schmett. t. iv. f. 3. x x.
[1655] Reaum. ii. t. iii. f. 2. c. l.
[1656] De Geer ii. t. xix. f. 9. b, c; f. 10. c.
[1657] Reaum. ii. t. iii. f. 3. c. l.
[1658] Ibid. iv. t. xl. f. 8. c. e.
[1659] Ibid. v. t. iii. f. 7, 8.
[1660] Ibid. vi. t. viii. f. 4. b, c.
[1662] Plate XV. Fig. 12. L´´.
[1663] Rifferschw. 12.
[1664] Vide supra, Letter XLII.
[1665] N. Dict. d'Hist. Nat. xxx. 16. 425. Marcel. de Serres Mém. du Mus. 1819. 89.
[1666] Rai. Hist. Ins. 177. Jurine Hymenopt. 9. not.
[1668] N. Dict. d'Hist. Nat. xxxvi. 255.
[1669] Prov. vi. 6; xxx. 25.
[1672] Reaum. vi. 432—.
[1673] De Geer vii. 179—.
[1674] Reaum. iv. 385.
[1675] De Geer vii. 249. Treviran. Arachnid. 41.
[1676] Marcel. de Serres penem in palpis cum teste pyriformi in thorace connexum esse affirmat, Mém. du Mus. 1819. 95.
[1677] Treviran. Ibid. 37. t. iv. f. 33.
[1678] De Geer vii. 10.
[1679] Reaum. ii. 72. t. ii. f. 2. De Geer vi. 314; vii. 165. Rai. Hist. Ins. 40.
[1680] De Geer ii. 24; iii. 132.
[1681] De Geer iii. 242. t. xiii. f. 15.
[1682] Ibid. iii. 642.
[1683] Huber Nouv. Observ. i. 37—.
[1684] De Geer ii. 276.
[1685] Reaum. ii. 65—.
[1686] De Geer iii. 62.
[1687] This memoir, which was sent me by its learned author, is stated as part of the first volume of his Histoire Naturelle.—K.
[1688] Vol. IV. Letters XXXVII and XL.
[1689] Vol. IV. Letter XLII.; III. p. 580—.
[1691] Vol. IV. p. 317-324. Vol. III. p. 415-455, 490, viii. 680. e.
[1693] N.B. The transverse lines in the figure are merely impressed, and do not represent a segment.
[1694] Vol. IV. Letter XXXVIII.
[1695] N.B. Where the volume is not indicated, the third is to be understood.
[1696] Where the volume referred to is not indicated, the fourth is to be understood. The reader will also be pleased to observe that the references to Partial Orismology are accompanied by the characteristic mark of each part, as given in the table, Vol. III.
[1697] It is to be observed, that the individuals of this Order belong some to Mandibulata, as Nirmus; others to Haustellata, as Pediculus; and others, as the Polypoda, in which the seven organs of a perfect mouth are multiplied, properly to neither of these subclasses.













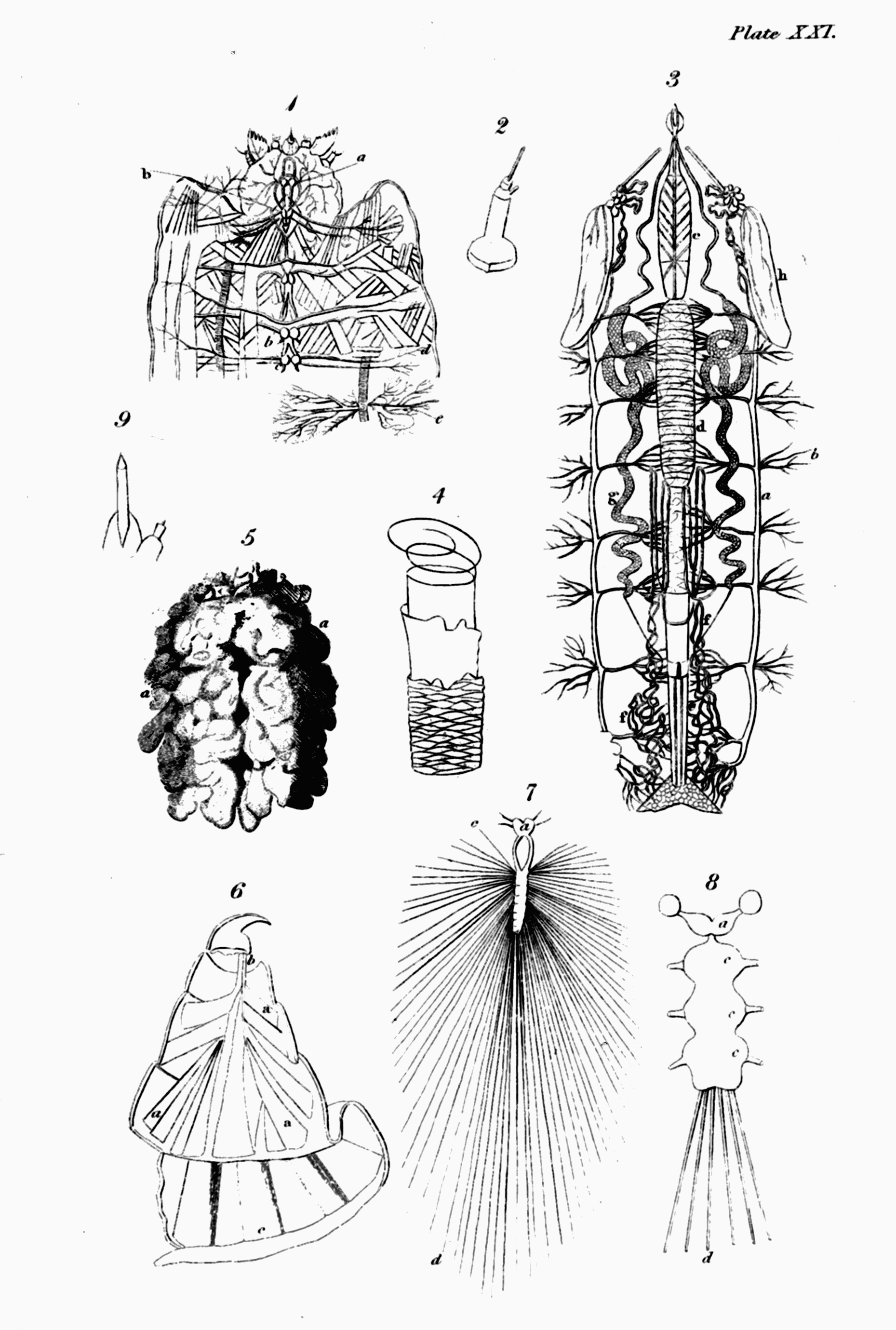{kind=link}
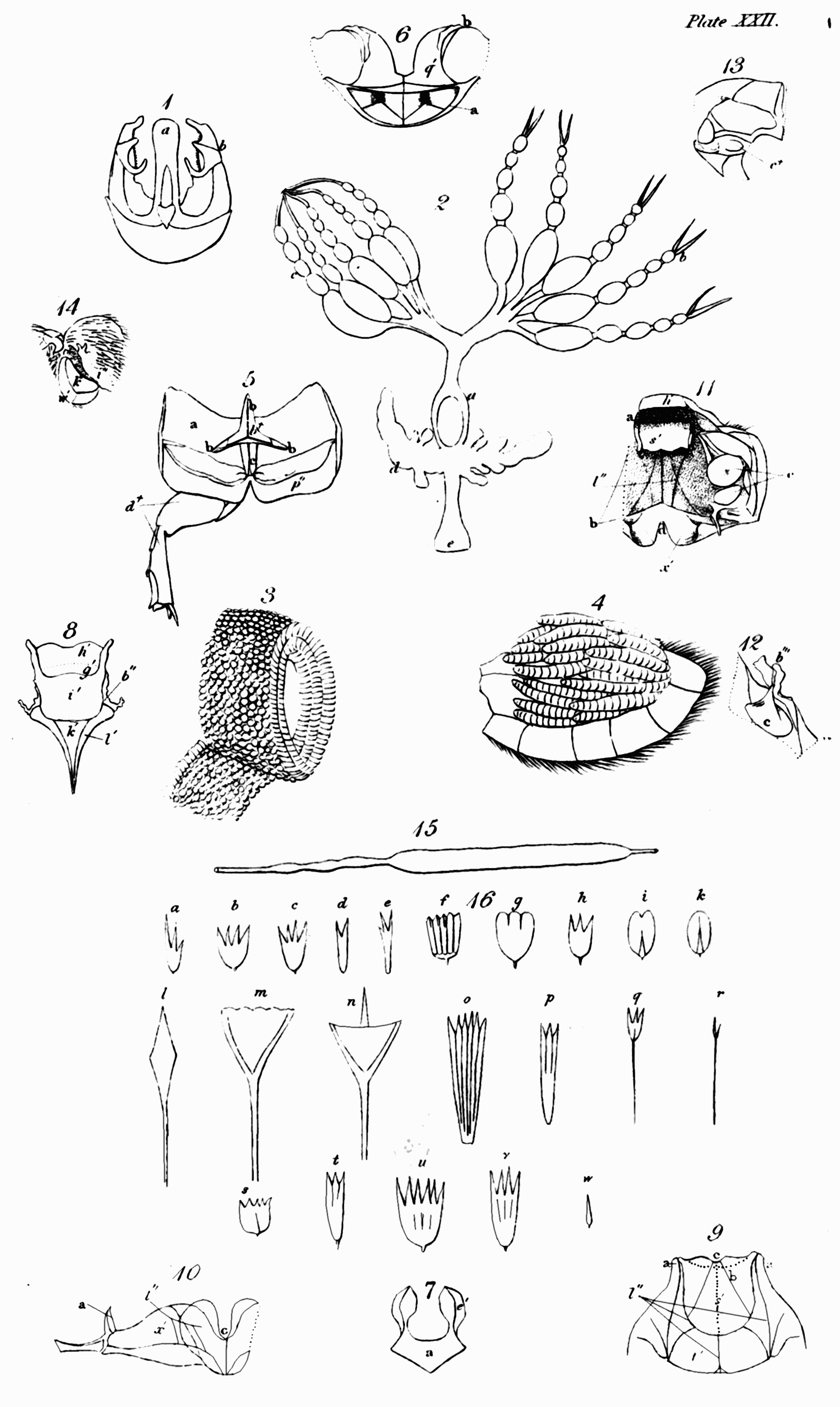{kind=link}
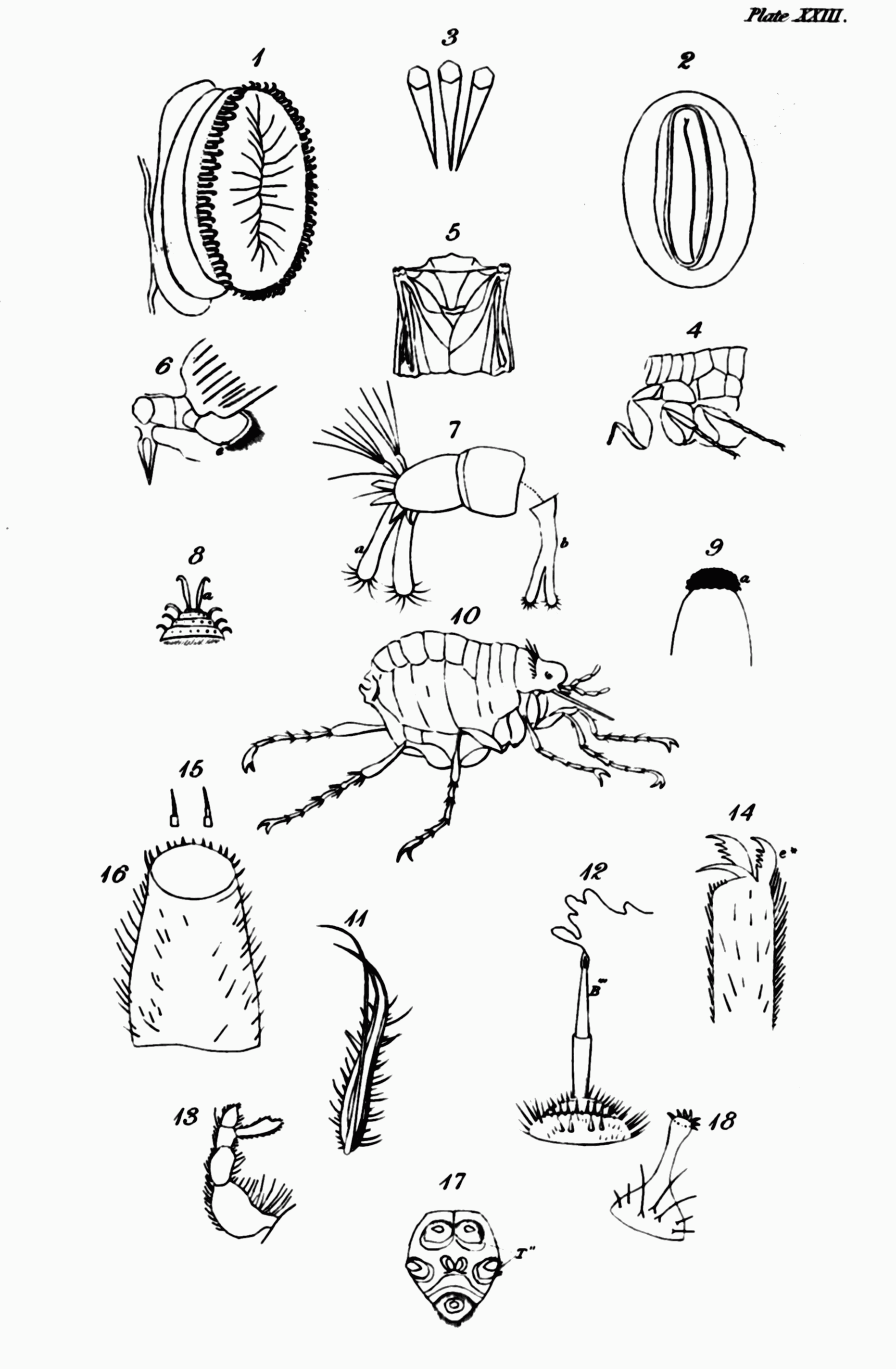{kind=link}
{kind=link}
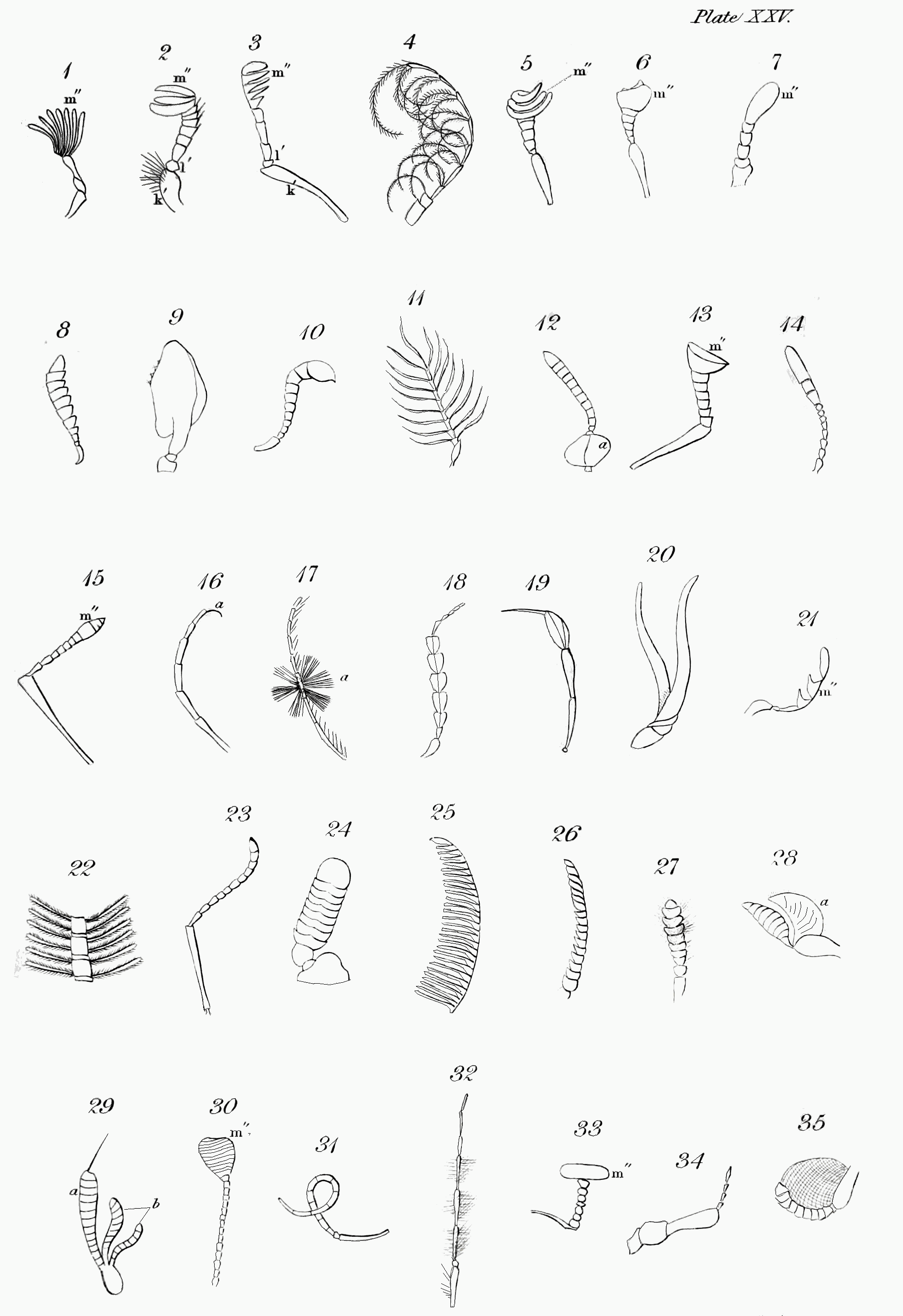{kind=link}
{kind=link}
{kind=link}
{kind=link}
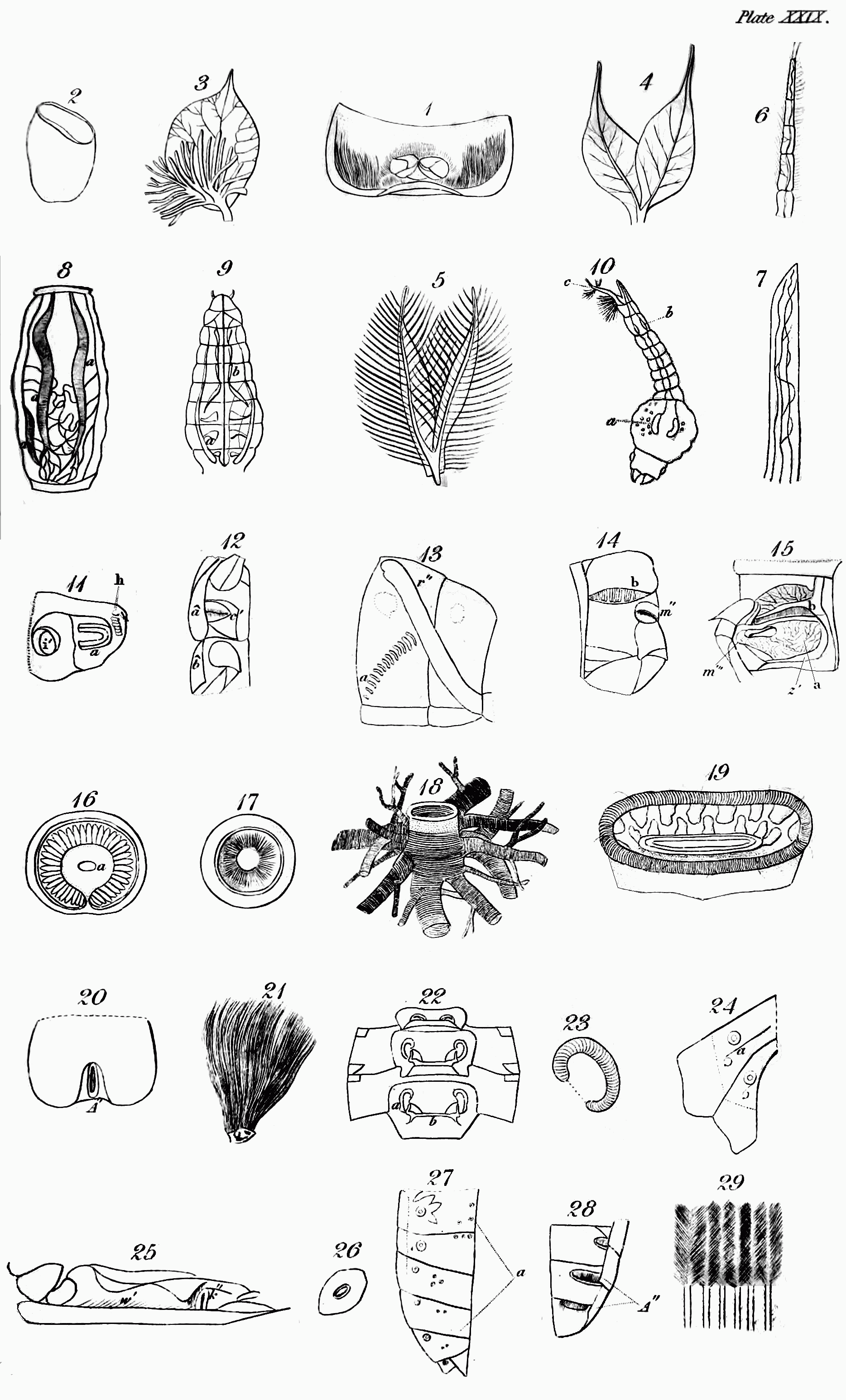{kind=link}
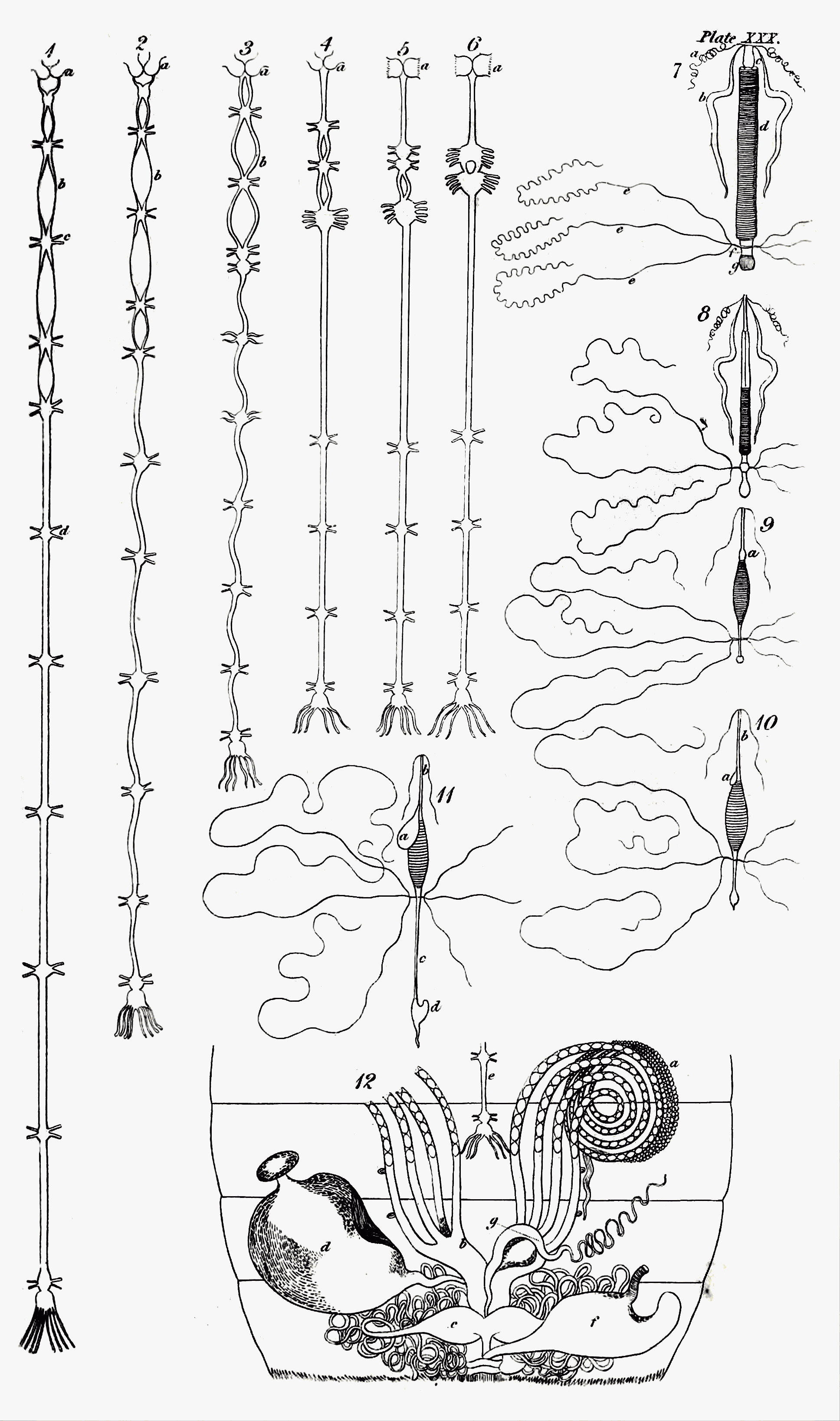{kind=link}