
Printed by H. Howard, RA. Engraved by W. T. Fry.
William Kirby. M.A. F.R. L.S. &c
The publication of the concluding volumes of the "Introduction to Entomology" has been unavoidably delayed by the continued ill health of one of the Authors, which has devolved upon the other a considerable increase of labour, and demanded a greater expenditure of time than would otherwise have been required: for though Mr. Spence put every facility in Mr. Kirby's power, and had drawn up a rough copy of every Letter belonging to his department; yet, as most of them had been written several years ago, many curious facts, and a great variety of interesting information subsequently derived from various sources, were necessarily to be inserted, and the whole to be prepared for the press.
When the thousands of objects that were to be examined, and many of them repeatedly, in composing the Letters on the External Anatomy of Insects, are considered, it will not appear surprising[Pg iv] if some errors should have crept in; especially as Mr. Kirby was deprived of the effectual help formerly derived from the acumen, learning, and judgement of his esteemed coadjutor, by his lamented and protracted indisposition: but it is hoped that these errors will be found of minor importance, and not to affect any general principles advanced. The same remarks are also in part applicable to the Anatomical and Orismological Tables (Vol. III. p. 354-393, and Vol. IV. p. 257-354), which were drawn up by the Authors jointly many years ago, before any other portion of the work was composed, but which have, especially the former, required considerable alterations and additions in consequence of subsequent observations and information.
It will not be amiss here to state, in order to obviate any charge of inconsistency in the possible event of Mr. Kirby's adverting in any other work to this subject, that though on every material point the authors have agreed in opinion, their views of the theory of instinct do not precisely accord. That given in the second and fourth volumes is from the pen of Mr. Spence.
It was originally intended, as mentioned in the Preface, to have given a complete list of Entomological[Pg v] works, of which a large portion was drawn up; but the great length to which more important matters have extended, has rendered necessary the intire omission of this list,—an omission in some degree compensated by the catalogue of Authors quoted, which comprises most of the standard Entomological works.
| Page. | Line. | |
| 29 | 27, | for Pseudo-cordia read Pseudo-cardia. |
| 33 | 7, | for 1/12 read 1½. |
| 35 | 7, | and elsewhere, for Gigas read grandis. |
| 46 | 16, | for number and situation read in some respects. |
| 98 | 6, | for Furtina read Jurtina. |
| 121 | note252, | for c read c´. |
| 135 | note297, | for XXIV. read XXIII. |
| 137 | note301, | for 17 read 18. |
| 251 | 4, | for ten read nine. |
| 378 | 10, | } for frœnum read frænum. |
| 359 | 21, | |
| 422 | note1199, | for a´ read a´´. |
| 425 | note1212, | for b´ read b´´. |
| 471 | 1, | dele Pelecotoma. |
| 10, | for orbicular read subtriangular. | |
| 512 | antepenult. | After genera insert except in some Acridæ, as A. viridissima. |
| 562 | note1681, | for πνεω read πνυμι. |
| 606 | 5, | for Heteropterous read Homopterous. |
The Synoptical Table of the Nomenclature of the Parts of the External Crust of Insects should be placed opposite to page 354.
Plates VI-XX. should be placed in this Volume, and the remainder in the Fourth.
It is however suggested to Purchasers, that in binding complete Sets of the Work, a separate Volume may be formed of the Synoptical Table, the Plates and their Explanations, and the Indexes.
| Letter. | Page. | |
| XXVIII. | Definition of the Term Insect | 1-51 |
| XXIX. | States of Insects. Egg state | 52-104 |
| XXX. | The same Subject continued. Larva state | 105-237 |
| XXXI. | The same Subject continued. Pupa state | 238-290 |
| XXXII. | The same Subject continued. Imago state | 291-347 |
| XXXIII. | External Anatomy of Insects. Terms and their Definition | 348-393 |
| XXXIV. | The same Subject continued. The Head and its Parts | 394-528 |
| XXXV. | The same Subject continued. The Trunk and its Parts and Organs | 529-697 |
| XXXVI. | The same Subject continued. The Abdomen and its Parts | 698-720 |
It being judged expedient, since the publication of the last Edition of the first and second Volumes of this Work, to adopt a new plan with respect to the reference letters of the Plates, the Reader is requested to make the following corrections in those Volumes.
| Vol. I. | ||
| Page. | Note. | |
| 125 | 197, | for f, d read a´´. |
| 273 | 501, | for a read a. |
| 395 | 724, | for 29, 30 read 13. |
| Vol. II. | ||
| 244 | 343, | for a read a. |
| 319 | 492, | for 10 read 14. |
| 348 | 550, | for a read e´´´. |
| 353 | 564, | for 7—. read 16—. |
| 366 | 585, | for a read s´´, v´´´. |
| 586, | for b read t´´. | |
| 405 | 674, | for 1.8. aa read 18. c†. |
| 675, | for bb read q´´´. | |
| 406 | 681, | for bb read C´´. |
| 407 | 684, | for cc read C´´ C´´, which represent the bundles of muscles in connexion with the drums. In the above figure the mirror is the part directly under those bundles. |
What is an insect? This may seem a strange question after such copious details as have been given in my former Letters of their history and economy, in which it appears to have been taken for granted that you can answer this question. Yet in the scientific road which you are now about to enter, to be able to define these creatures technically is an important first step which calls for attention. You know already that a butterfly is an insect—that a fly, a beetle, a grasshopper, a bug, a bee, a louse, and flea, are insects—that a spider also and centipede go under that name; and this knowledge, which every child likewise possesses, was sufficient for comprehending the subjects upon which I have hitherto written. But now that we are about to take a nearer view of them—to investigate their anatomical and physiological characters[Pg 2] more closely—these vague and popular ideas are insufficient. In common language, not only the tribes above mentioned, but most small animals—as worms, slugs, leeches, and many similar creatures, are known by the name of insects. Such latitude, however, cannot be admitted in a scientific view of the subject, in which the class of insects is distinguished from these animals just as strictly as beasts from birds, and birds from reptiles and amphibia, and these again from fishes. Not, indeed, that the just limits of the class have always been clearly understood and marked out. Even when our correspondence first commenced, animals were regarded as belonging to it, which since their internal organization has been more fully explained, are properly separated from it. But it is now agreed on all hands, that an earthworm, a leech, or a slug, is not an insect; and a Naturalist seems almost as much inclined to smile at those who confound them, as Captain Cook at the islanders who confessed their entire ignorance of the nature of cows and horses, but gave him to understand that they knew his sheep and goats to be birds.
You will better comprehend the subsequent definition of the term Insect, after attending to a slight sketch of the chief classifications of the animal kingdom, more especially of the creatures in question, that have been proposed. That of Aristotle stands first. He divides animals into two grand sections, corresponding with the Vertebrata and Invertebrata of modern Zoologists: those, namely, that have blood, and those that have it not[1]:—by this it appears that he only regarded red blood as real blood; and probably did not suspect that there was[Pg 3] a true circulation in his Mollusca and other white-blooded animals. His Enaima, or animals that have blood, he divides into Quadrupeds, Birds, Fishes, Cetacea, and Apods or reptiles; though he includes the latter, where they have four legs, amongst the quadrupeds[2]; and his Anaima, or animals without blood, into Malachia, Malacostraca, Ostracoderma, and Entoma. The first of these, the Malachia, he defines as animals that are externally fleshy and internally solid, like the Enaima; and he gives the Sepia as the type of this class, which answers to the Cephalopoda of the moderns. The next, the Malacostraca, synonymous with the Crustacea of Cuvier and Lamarck, are those, he says, which have their solid part without and the fleshy within, and whose shell will not break, but splits, upon collision[3]. The Ostracoderma, corresponding with the Testacea of Linné, he also defines as having their fleshy substance within, and the solid without; but whose shell, as to its fracture, reverses the character of the Malacostraca. He defines his last class Entoma, in Latin Insecta, with which we are principally concerned, as animals whose body is distinguished by incisures, either on its upper or under side, or on both, and has no solid or fleshy substance separate, but something intermediate, their body being equally hard both within and without[4]. This definition would include the Annelida and most other Vermes of Linné, except the Testacea, which accordingly were considered as insects by those Zoologists that intervened between Aristotle and the latter author. The Stagyrite, however, in another place,[Pg 4] has expressly excluded all apods[5]. From other passages in his works, it appears that he regarded the Vermes, &c. either as larvæ, or as produced spontaneously and not ex ovo[6].
This definition of an insect, though partly founded on misconception, as well as his primary division of animals in general, is by no means contemptible. If you look at a bee or a fly, you will observe at first sight that its body is insected, being divided as it were into three principal pieces—head, trunk, and abdomen[7]; and if you examine it more narrowly, you will find that the two last of these parts, especially the abdomen, are further subdivided. And this character of insection, or division into segments, more or less present in almost every insect[8], is not to be found (with the exception of the Crustacea, which Aristotle[Pg 5] distinguishes by the nature of their integument and its contents) in any of the other classes into which he divided animals without blood. It was on account of this most obvious of their characters, that these little creatures were in Greek named Entoma, and in Latin Insecta; and from the former word, as you know, our favourite science takes the name of Entomology.
Pliny adhering to the definition of Aristotle, as far as it relates to the insection of the animals we are speaking of, expressly includes Apods, as well as Aptera, amongst them[9]; and in this was followed, without any attempt at improvement, by all the entomological writers that intervened between him and the great Aristotle of the moderns, Linné.
This illustrious naturalist, aware of the incorrectness of the primary divisions of the animal kingdom founded upon the presence or absence of blood, establishes his system upon the structure of the heart, and upon the temperature and colour of the circulating fluid. He divided animals into two great sections or sub-kingdoms, each comprising two classes. His first section included those having a heart with two ventricles, two auricles, and warm and red blood, viz. the Mammalia or beasts, and the Aves or birds. His second, those having a heart with one ventricle, one auricle, and cold and red blood, namely, the classes Amphibia, which included reptiles, serpents, &c. and Pisces or fish. His third, those having a heart with one ventricle and no auricle, and cold white sanies in the place of blood, namely, his classes Insecta et Vermes, including the Invertebrate animals of Lamarck.[Pg 6] Thus the first of Aristotle's great divisions he increased by the addition of a new and very distinct class, the Amphibia, by which some ground was gained in the science; but as much was lost by his compressing the four classes of which the last consisted into two, by which the natural classes of Cephalopoda and Crustacea merged under Insecta and Vermes. Linné was not aware of the extraordinary fact, that the Cephalopoda have three hearts; and that though the Crustacea and Arachnida have a circulation, Insects have none, or he would never have taken this retrograde step.
Indeed Linné's definition of an Insect is, in many most material points, inapplicable, not only to the Crustacea, but to many other animals included under that denomination. This will appear evident from a very slight examination. Thus it runs: "Polypod animalcula, breathing by lateral spiracles, armed every where with an osseous skin, whose head is furnished with movable sensitive antennæ[10]." Now of this definition only the first member can be applied to the whole class which it is meant to designate; for the entire genus Cancer L., which, with some others, forms the class Crustacea of the moderns, does not respire by spiracles at all, but by gills; and the same in some degree may be said of spiders, scorpions, &c. With the last member of the definition Linné himself must have been aware that a large number of what he conceived to be insects were at variance, as mites, spiders, and many other of his apterous tribes: though from some very recent observations of M.[Pg 7] Latreille[11], there seems some ground for thinking, that in these the antennæ are represented by the mandibles, palpi, &c.[12], and to the soft flexible, coriaceous or membranous skin of a vast number of insects, the term cutis ossea is by no means applicable.
Evident as these incongruities are, when the Herculean task which Linné imposed upon himself, and the vastness and variety of his labours, are considered, they become very venial. Indeed, unless he had divided his class Insecta into two or more, it was impossible to define it intelligibly to ordinary readers, otherwise than nearly in the terms which he actually employed; and these characters, restricted and amended by qualifying clauses, are still those to which recurrence must be had in a popular definition of the class, when separated as it ought to be from the Crustacea and Arachnida.
Pennant, Brisson, and other zoologists, who, attending to nature rather than system, saw the impropriety of uniting a crab or a lobster in the same class with a bee or a beetle, long since assigned the Crustacea their ancient distinct rank. "But these changes," as Latreille observes[13], "being only founded upon external characters, might be deemed arbitrary; and to fix our opinion, it was necessary to have recourse to a decisive authority—the internal and comparative organization of these animals.[Pg 8]" It results from the observations of the most profound comparative anatomist of our age, M. Cuvier, that the Crustacea and Arachnida differ from insects properly so called, and particularly from those that are furnished with wings, in having a complete system of circulation, a different mode of respiration, and that they have a more perfect organization. Influenced by these motives, both Cuvier and Lamarck have considered them as forming two classes separate from insects. Treviranus, led by considerations founded on the organs of circulation, of respiration, and of generation, is of opinion that spiders and scorpions ought to form one class with the Crustacea: he observes, however, that the nervous system of all three is very dissimilar; and that in an arrangement founded on this circumstance, the organs of motion, and the external shape, even spiders and scorpions must be placed in different classes[14].
It is to be observed with regard to the Arachnida of the French school, that the class as laid down by them includes several animals that have no circulation, and breathe by tracheæ, of which description are the mites (Acarus L.), and the harvest-men (Phalangium L.) &c.; and therefore it has been divided into two orders, Pulmonaria and Tracheana; but if the definition from the internal organization be adhered to, the latter should either remain with the class Insecta, or form a new one by themselves. Yet the animals that compose the Trachean order of Arachnida, their external form considered, are certainly much more nearly related to the spiders and[Pg 9] scorpions than to any members of the class Insecta at present known. This circumstance, perhaps, may seem to throw some doubt upon the modern system of classification.
I must further observe, that the assertion of Treviranus, which appears to intimate that the respiration of the pulmonary Arachnida is the same with that of the Crustacea, is not quite correct, since in the latter the branchiæ or gills are external, and in the former internal, the air entering by spiracles before it acts upon them[15].
It may not be amiss in this place to lay before you the principal points in which the Crustacea and Arachnida agree with Insecta, and also those in which they differ.
The Crustacea agree with Insecta in having a body divided into segments, furnished with jointed legs, compound eyes, and antennæ. Their nervous system also is not materially different, and they are both oviparous. They differ from them in having the greater insections of the body less strongly marked; in the greater number of legs on the trunk, the anterior ones performing the office of maxillæ: in their eyes usually on a moveable footstalk: their palpigerous mandibles; and their four antennæ at least in the great majority. But the principal difference consists in the internal organization and the fountains of vitality; for the Crustacea have a double circulation, the fountain of which is a heart in the middle of their thorax[16]. They have too a kind of gizzard and liver, at least the Decapods[17], and their respiration is by gills. Genuine insects terminate their[Pg 10] existence after they have laid their eggs[18]; but the Crustacea live longer, and lay more than once.
The Arachnida will be found to differ from insects more widely than even the Crustacea. They agree in their jointed legs and palpi; immoveable eyes; and in being covered with a coriaceous or corneous integument: but they differ in having a system of circulation; gills instead of tracheæ; their organs of generation double; and the females lay more than once in their lives. Their head also is not distinct from the trunk as in insects; they have no compound eyes; and their antennæ, if we admit the opinion on this head of MM. Latreille and Treviranus, that they have representatives of these organs, differ totally in structure, situation, and use, from those of the great body of insects. In the Araneidæ or Spiders, their body seems to have no segments or incisure but that which separates the abdomen from the trunk; and in the Scorpionidæ they are observable only in the abdomen. Other particulars might be enumerated in which these two classes differ from insects; but these will be sufficient to convince you that Aristotle and MM. Cuvier and Lamarck were justified in separating them.
The two last-mentioned authors made further improvements in Zoology. The latter, from the consideration of the general structure of animals, perceiving that Aristotle's Enaima were distinguished from his Anaima, by being built as it were upon a vertebral column, very judiciously changed the denomination, which was indeed improper, of "The Philosopher's" two sub-kingdoms, into[Pg 11] that of Vertebrata or animals that have a vertebral column, and Invertebrata or those that have no vertebral column. These he distributes into three primary divisions according to their supposed degrees of intelligence—Thus:
| * Apathetic Animals. | 1. Infusoria. |
| 2. Polypi. | |
| 3. Radiata. | |
| 4. Vermes. | |
| ** Sensitive Animals. | (Epizoaria.) |
| 5. Insecta. | |
| 6. Arachnida. | |
| 7. Crustacea. | |
| 8. Annelida. | |
| 9. Cirrhipeda. | |
| 10. Mollusca. | |
| *** Intelligent Animals. | 11. Pisces. |
| 12. Reptilia. | |
| 13. Aves. | |
| 14. Mammalia.[19] |
Profiting by the light afforded by the Aristotelian system, this eminent zoologist improved, we see, upon that of Linné, by resolving his Insecta into three classes, and his Vermes into seven, interposing the Linnean Insecta between the four first and three last, in which he was not so happy, since as to sense insects should certainly occupy the place he has here assigned to the Mollusca.
In the work from which I have taken this statement of Lamarck's system, that acute writer has given a sketch of another method of arrangement, in which he has made the first deviation from the beaten track of an unbroken[Pg 12] and unbranching series. In the Supplement to the first volume, he has distributed the Invertebrata in a double subramose series—one consisting of articulate, and the other of inarticulate animals[20].
Upon Lamarck's system, most of the modern ones, with some variation, are founded. There is one, however, by a learned countryman of ours, that is more unique, sui generis, and I may add profound, than any that has yet appeared. I am speaking of that, you will perceive, of which our friend Mr. Wm. MacLeay has given a detailed statement in his Horæ Entomologicæ. In this he goes even far beyond what Lamarck has attempted in the above sketch, and substantiates his claim to be considered as one of those original thinkers, rari nantes in gurgite vasto, that do not appear every day. The following are the principal bases of his system.
1. That all natural groups, whether kingdoms or any subdivision of them, return into themselves; a distribution which he expresses by circles.
2. That each of these circles is formed precisely of five groups, each of which is resolvable into five other smaller groups, and so on till you reach the extreme term of such division.
3. That proximate circles or larger groups are connected by the intervention of lesser groups, which he denominates osculant.
4. That there are relations of analogy between the corresponding points of contiguous circles.
This system he has represented by tables of circles inscribed with the five primary divisions of each group. His first table exhibits a general view of organized matter[Pg 13] as distributed in the animal and vegetable kingdoms—Thus:
Our learned author here divides the animal kingdom into what may be denominated five sub-kingdoms or provinces, in three of which (with the exception of the Crustacea and Arachnida belonging to his Annulosa) no circulation of blood is visible, but which obtains in the rest. These he names—
1. Acrita, consisting of the Infusory Animals, the Polypi, the Corallines, the Tæniæ, and the least organized of the Intestinal Worms.
2. Radiata, including the Jelly-fish, Star-fish, Echini, and some others.
3. Annulosa, consisting of Insecta, Arachnida, and Crustacea.
4. Vertebrata, consisting of Beasts, Birds, Reptiles, Amphibia, and Fishes.
5. Mollusca, including the numerous tribes of shell-fish, land-shells, slugs, &c., which, from their mucous or gelatinous substance, from their nervous system and the imperfection of their senses, return again to the Acrita,[Pg 14] though connected with the Vertebrata by having a heart and circulation.
His next set of circles shows the sub-division of these five sub-kingdoms into classes—Thus:
In this scheme the osculant classes are those placed between the circles. In the Mollusca circle two classes are still wanting to complete the quinary arrangement of that sub-kingdom. I am not sufficiently conversant[Pg 15] with the details of the animal kingdom at large to hazard any decided opinion upon Mr. MacLeay's whole system, or to ascertain whether all these classes are sufficiently distinct[21]. My sentiments with regard to those of the Annulosa I shall state to you hereafter.
Upon a future occasion I shall consider more at large the station to which insects seem entitled in a system of invertebrate animals, which will not accord exactly with that assigned by MM. Cuvier and Lamarck. But I am now in a field in which I have no intention to expatiate further, than as it is connected with the subject of the present letter. I shall therefore confine myself in what I have more to say to the definitions of Insecta that have been given by modern authors, beginning with that of the zoologist last mentioned. Insects form a part of his second group, which he terms sensitive animals (animaux sensibles), which group he thus defines: "They are sentient, but obtain from their sensations only perceptions of objects—a kind of simple ideas which they cannot combine to obtain complex ones. Charact. No vertebral column; a brain, and most commonly an elongated medullary mass; some distinct senses; the organs of movement attached under the skin: form symmetrical, by parts, in pairs[22]." This division of animals, from the[Pg 16] kind and degree of sense and intelligence that they possess, seems rather fanciful than founded in nature, since many insects show a greater portion of them than many vertebrate animals. Compare in this respect a bee with a tortoise[23]. Lamarck divides his group of animaux sensibles into two sections, namely, Articulated animals, exhibiting segments or articulations in all or some of their parts; and Inarticulatcd animals, exhibiting neither segments nor articulations in any of their parts. Insecta, Arachnida, and Crustacea, belong to the first of these sections, which he defines as "those whose body is divided into segments, and which are furnished with jointed legs bent at the articulations[24]." Insecta he defines—"Articulate animals, undergoing various metamorphoses, or acquiring new kinds of parts—having, in their perfect state, six feet, two antennæ, two compound eyes, and a corneous skin. The majority acquiring wings. Respiration by spiracles (stigmates), and two vascular opposite chords, divided by plexus, and constituting aeriferous tracheæ, which extend every where. A small brain at the anterior extremity of a longitudinal knotty marrow, with nerves. No system of circulation, no conglomerate glands. Generation oviparous: two distinct sexes. A single sexual union in the whole course of life[25]." Arachnida he defines—"Oviparous animals, having at all times jointed legs, undergoing no metamorphosis, and never acquiring new kinds of parts. Respiration tracheal or branchial: the openings for the entrance of the air spiraculiform (stigmatiformes). A heart and circulation beginning in[Pg 17] many. The majority couple often in the course of life[26]." I shall next add his definition of Crustacea: "Oviparous, articulated, apterous animals, with a crustaceous integument more or less solid, having jointed legs; eyes either pedunculate or sessile, and most commonly four antennæ, with a maxilliferous mouth seldom rostriform; maxillæ in many pairs placed one over the other; scarcely any under-lip; no spiraculiform openings for respiration; five or seven pair of legs; a longitudinal knotty marrow terminated anteriorly by a small brain. A heart and vessels for circulation. Respiration branchial with external branchiæ, sometimes hid under the sides of the shell of the thorax, or shut in prominent parts; sometimes uncovered, and in general adhering to particular legs or to the tail. Each sex usually double[27]."
I have given Lamarck's definitions of these three classes, all considered as Insecta by Linné, that by comparing them together you may be better enabled to appreciate the system of this author. On looking over the characters of the Arachnida as here given, you will see at once that it consists of heterogeneous animals—for in fact he includes in this class not only the Trachean Arachnida of Latreille, but the Ametabolia of Dr. Leach, or the Hexapod Aptera, and the Myriapoda.
I shall next copy for you Latreille's latest definition of Insecta and Arachnida.
"Insecta: A single dorsal vessel representing the heart: two trunks of tracheæ running the whole length of the body, and opening externally by numerous spiracles; two antennæ; very often upper appendages for flight, indicating the metamorphosis to which the animal[Pg 18] is subject when young; legs most commonly reduced to six. Arachnida: Distinguished from Crustacea by having their respiratory organs always internal, opening on the sides of the abdomen or thorax to receive the respirable fluid. Sometimes these organs perform the office of lungs, and then the circulation takes place by means of a dorsal vessel, which sends forth arterial, and receives venose branches. Sometimes they are tracheæ or air-vessels, which, as in the class Insecta, replace those of circulation. These have only the vestige of a heart, or a dorsal vessel alternately contracting and sending forth no branch. The absence of antennæ, the reunion of the head with the thorax, a simple trachea but ramified and almost radiating, serve to distinguish these last Arachnida, or the most imperfect of insects, which respire only by tracheæ[28]." Under this head he observes—"Of all these characters, the most easy to seize and the most certain would doubtless be, if there were no mistake in it, that of the absence of antennæ; but later and comparative researches, confirmed by analogy, have convinced me, that these organs, under particular modifications it is true, and which have misled the attention of naturalists, do exist[29]:" and he supposes, from the situation and direction of the mandibles of the Arachnida, corresponding with that of the intermediate pair of antennæ in Crustacea, that they really represent the latter organs. If this supposition be admitted, their use is wholly changed; the palpi, in fact, executing the functions of antennæ, which probably induced Treviranus to call them Fühlhörner (Feelinghorns).[Pg 19] Perhaps these last may be regarded as in some sort representing the external antennæ of the Crustacea? With regard to Insecta, their antennæ seem to disappear in the Pupiparæ Latr., or the genus Hippobosca L.
The above definitions of the Arachnida by these two celebrated authors, appear to me the reverse of satisfactory. When we are told of animals included in it, that some breathe by gills and others by tracheæ, that some have a heart and circulation and others not, we are immediately struck by the incongruity, and are led to suspect that animals differing so widely in the fountains of life ought not to be associated in the same class. A learned zoologist of our own country, Dr. Leach, seems to have made a nearer approach to a classification in accordance with the internal organization, by excluding from Arachnida the Acari and Myriapoda.
Sub-kingdom Annulata Cuv.
| * Gills for respiration. | Classes. | |
| Legs sixteen: | Antennæ two or four | 1 Crustacea. |
| ** Sacs for respiration. | ||
| Legs twelve: | Antennæ none | 3 Arachnöidea. |
| *** Tracheæ for respiration. | ||
| a. No Antennæ. | ||
| 4 Acari. | ||
| b. Two Antennæ. | ||
| Six thoracic legs: | Abdomen also bearing legs | 2 Myriapoda. |
| Six thoracic legs: | No abdominal legs | 5 Insecta[30]. |
Mr. MacLeay, on whose system I shall now say a few words, divides his sub-kingdom Annulosa into five classes, namely, Crustacea, Ametabola, Mandibulata, Haustellata, Arachnida. From the Crustacea he goes by the genus[Pg 20] Porcellio Latr. to Iulus[31], which begins his Ametabola: these he connects with the Mandibulata, by Nirmus, which he thinks approaches some of the corticarious Coleoptera[32]. This class he appears to leave by the Trichoptera Kirby, and so enters his Haustellata by the Lepidoptera[33], and leaves it again by the Diptera by means of the Pupiparæ Latr., especially Nycteribia, connecting this class with the Arachnida, which he enters by the Hexapod Acari L.[34], and these last he appears to leave by the Araneidæ, and to enter the Crustacea by the Decapods[35]: thus making good his circle of classes, or a series of Annulose animals returning into itself. Mr. MacLeay's whole system upon paper appears very harmonious and consistent, and bears a most seducing aspect of verisimilitude; but it has not yet been so thoroughly weighed, discussed, and sifted, as to justify our adopting it in toto at present: should it, however, upon an impartial and thorough investigation, come forth from the furnace as gold, and be found to correspond with the actual state of things in nature, my objections, which rest only upon some parts of his arrangement of Annulosa, would soon vanish. Some of those objections I will state here, and some will come in better when I treat of the Systems of Entomology. My first objection is, that his Ametabola, Mandibulata, and Haustellata, approach much nearer to each other than they do to the other two classes of his circle, or than even these last to each other; so that under this view it should primarily consist of three greater groups, resolvable, it may be, into five smaller ones. My next objection is, that he has also considered[Pg 21] the Trachean and Pulmonary Arachnida as forming one class. Whether an animal breathes by gills or tracheæ, or has a circulation or not, is surely as strong a reason for considering those so distinguished as belonging to different classes, as the taking of their food by suction or by manducation is, for separating others to the full as much or more nearly related as to their external structure. But of this more hereafter. I cannot help, as a last objection, lamenting that our learned author has rejected from his system a term consecrated from the most remote antiquity, and which, even admitting his arrangement, might have been substituted for Annulosa, a name borrowed by Scaliger from Albertus Magnus, neither of whom, in Entomology, is an authority to weigh against Aristotle, from whom we derive the term Insecta, in Greek Εντομα.
As Fabricius did not alter Linné's class Insecta, but merely broke up his orders into new ones, which he named classes, I shall give you a detail of the alterations he introduced into the science in a future letter.
Having stated what my predecessors have done in classification, I shall next proceed to lay before you my own sentiments as to—What is an insect. Since our correspondence commenced, the Arachnida, principally on account of their internal organization, have been excluded from bearing that name, carrying with them, as we have seen, several tribes, which as yet have not been discovered to differ materially in that respect from the present Insecta: for the sake, therefore, of convenience and consistency, that I may, as far as the case will admit, adhere to the Horatian maxim
I shall regard as Insects all those Annulosa that respire by tracheæ[36] and have no circulation, considering the Trachean Arachnida and the Myriapoda for the present as sub-classes, the one bordering upon the Arachnida, and the other upon the Crustacea. Some of these I am ready to own seem separated by an interval sufficiently wide from the Hexapods, which may be regarded as more peculiarly entitled to the denomination of Insects. The most striking differences will be found in the coalition of the head with the trunk in some (Phalangidæ), and the disappearance of the annulose form of the body in others (Acarus L.), so that the legs only are jointed[37]. Yet an[Pg 23] approach to such structure may be traced in some Hexapods; for instance, the coalition of the head and trunk in Melophagus, Latr., and that of the trunk and abdomen in Sminthurus, Latr.[38] The Myriapoda exhibit other remarkable differences; though their head and trunk are distinct, the former antenniferous, and their body annulose, the abdomen as well as the trunk is furnished with legs, sometimes amounting to hundreds; but even to this a tendency has been observed in some Hexapods[39]. If you examine a specimen of Machilis polypoda, an insect related to the common sugar-louse (Lepisma saccharina), you will find that the abdomen is furnished with a double series of elastic appendages, which, being instruments of motion, may be regarded as representing legs. It is worthy of notice, that the Myriapoda when first disclosed from the egg have never more than six legs[40], and keep acquiring additional pairs of them and additional segments to their abdomen as they change their skins: and it is equally remarkable, that many Hexapods are subject to a law in some degree the very reverse of this, having many abdominal legs in their first state, and losing them all in their last. The union of the head with the trunk in the Trachean Arachnida has been regarded as almost an unanswerable argument, in spite of their different internal organization, for including them in the same class with the Pulmonary Arachnida; but the case of Galeodes, which, though furnished with gills, (as an eminent Russian Entomologist Dr. G. Fischer is reported to have discovered,) implying also a circulation, and evidently belonging to the last-mentioned class, has nevertheless a distinct thorax consisting of more than one piece, to which[Pg 24] are affixed only six legs[41], proves that even this circumstance possesses no weight when set against the organization. If it was a difference in this respect, that proved the Crustacea classically distinct from Insecta—that likewise was the principal reason for the separation also of the Arachnida—it seems to follow that it ought also to furnish an argument equally cogent for considering the Trachean Arachnida, as well as the Myriapoda, distinct from the Pulmonary.
Another difference between the tribes in question is that of their metamorphosis; and this appears to have had great weight with Lamarck, inducing him to include in his Arachnida, not only the Tracheans and Myriapods, but even the apterous Hexapods, except Pulex, or the Anoplura and Thysanura of modern authors. But the metamorphosis alone, unless supported by the internal organization, will I think scarcely be deemed a sufficient reason for separating from each other tribes agreeing in that respect, and placing them with others with which they disagree. The metamorphosis in some of the Hexapods (Lepidoptera) consists in the loss of legs, the acquisition of wings, a great change in the oral organs and in the general form; in others (some Coleoptera), in the acquisition only of wings and a change of shape, the oral organs remaining much the same; in others again (Curculio L.), in the acquisition of six legs and wings and a change of form; in the flea, in the acquisition of six legs and a change of form only; in the Orthoptera, Hemiptera, &c. in the mere acquisition of wings; in the Libellulidæ, in the loss of the mask that covers the mouth and the acquisition of wings; in the Diptera, in the[Pg 25] acquisition of six legs, wings, a change of the oral organs and of the form; in some of the Octopods (Acarus L.), in the acquisition of a pair of legs; and in others (Phalangium and Aranea L.), solely in a modification of them as to their proportions; in the Myriapods, the alteration that takes place in this respect is considerable; a large number of pairs of legs is acquired and many additional abdominal segments, and the proportion which the abdomen bears to the whole insect is quite altered. In all these cases there is a change more or less, either partial or general, of the original shape or organs of the animal; and with regard to their metamorphosis, there is a greater difference between a young and adult Iulus than between a young and adult grasshopper or bug: so that if the metamorphosis, per se, be assumed as a principal regulator of the class, the grasshopper or bug have as little claim to belong to it as the Iulus.
M. Lamarck lays considerable stress upon another character—That Insecta engender only once in the course of their lives, and Arachnida more than once. But this, if examined, will be found to be confined chiefly to the Pulmonary Arachnida, the Tracheans following the law of Insecta in this respect[42].
You may perhaps object that the bringing of the Trachean Arachnida and the Myriapoda into the class Insecta will render the approximation of them to a natural arrangement more difficult, since it will be impossible at the same time to connect the Myriapods with the Crustacea, and the Trachean with the genuine Arachnida.[Pg 26] I admit the validity of your objection, but by no arrangement of insects in a simple series can we attain this object: the difficulty, however, may perhaps be obviated in this way. The distribution of organized matter, to adopt Mr. Wm. MacLeay's metaphor[43], begins in a dichotomy, constituting the animal and vegetable branches of the great tree of nature, and from these two great branches, by means of infinite ramifications, the whole system is formed, and, what is remarkable, these branches unite again so as to represent a series returning into itself, a discovery due to the patient investigation and acumen of our learned friend just mentioned. Now, in considering the Aptera order, we find at first setting out from the Hexapods, a dichotomy, where the Anoplura Leach branch off on the one side, and the Thysanura Latr. on the other—the former, by means of the Pediculidæ, taking their food by suction, particularly Phthirus Leach, or the Morpion (in which the segments of the trunk and abdomen become indistinct[44]) approach the Octopods by the hexapod Acari L.—the latter by Machilis polypoda tending towards the Myriapods. In the Octopod branch a further dichotomy takes place, from which you proceed on one side to the Araneidæ in the Arachnida, by Phalangium, &c.; and in the other by Chelifer, &c. to Scorpio. Again, the Myriapod branch also divides, going by the Iulidæ to one branch of the Isopod Crustacea, and by the Scolopendridæ to another.
But there is another view of this subject before alluded to, which may be repeated here, and which seems to[Pg 27] prove that the types of form in one natural group or class are reproduced in another; this appears to result from the following parallel series:
| Neuropterous Larvæ. | Aptera. | Arachnida. | Crustacea. |
| Psocus | Hexapoda | Galeodes | Larunda. |
| Myrmeleon | Phalangium | Aranea | Decapoda brachyura. |
| Octopoda | |||
| Panorpa? | Chelifer | Scorpio | Decapoda macroura. Thalassina Scorpio especially. |
| Ephemera | Myriapoda | ***** | Isopoda. |
No type representing the Myriapoda has yet been discovered in the Arachnida class; but I have little doubt of its existence. You will observe that the analogies between the larvæ of the winged orders and the Aptera were first noticed by Mr. W. MacLeay[45]. It is probable that these parallel series of representatives of each other might be increased, as well as the numbers in the respective columns.
What I have said will, I trust, sufficiently justify me for making at present no more material alterations in the classification I long since proposed to you[46]; I shall, therefore, now proceed to define the objects I consider as Insecta; but I shall first observe—that as Latreille considers the branchiopod Crustacea or Entomostraca of Müller as entitled to the denomination of Crustaceo-Arachnida[47]; so his Trachean Arachnida might be called Arachnida-Insecta, and his Myriapoda, Crustaceo-Insecta.
Sub-kingdom—Annulosa[48].
Class—Insecta.
First Definition—From their external Organization.
Second Definition—From their internal Organization.
In defining the Arachnida I shall only mention those particulars in which they differ from Insectæ in their external anatomy.
Class—Arachnida.
The external characters in this class are the same almost in every respect as those which distinguish the Phalangidæ, the whole difference consisting almost in the systems of circulation, respiration, and digestion. Perhaps some future anatomist may discover in the tribe just mentioned, that there is a nearer agreement between them and the Arachnida in these systems than is at present suspected, which would prove them true Arachnida. I am inclined to think that Phrynus and Gonyleptes, &c. breathe by branchial spiracles; but having no opportunity[Pg 32] of examining living specimens, I dare not speak with any confidence on the subject.
Having thus given you a view of the most important diagnostics by which what we have all along called Insects may scientifically be distinguished from other invertebrate animals, it may not be without use, if, under this head, I take a more popular and familiar view of the subject, and say something upon those distinctions which may attract the attention of the more common observer.
The notion of diminutive size, particularly as compared with vertebrate animals, seems more frequently attached to the idea of an insect than any other; and this notion is generally correct, for one insect that is bigger than the least of the above animals, thousands and thousands are vastly smaller: but there exist some that are considerably larger, whether we take length or bulk into consideration, and this in almost every order. To prove this most effectually, and that you may have a synoptical view of the comparative size of the larger insects of the different orders and tribes, I now lay before you a table of the dimensions of such of the largest as I have had an opportunity of measuring, including particularly those giants that are natives of the British isles.[Pg 33]
From this table you see that several insects included in it exceed some of the smallest Vertebrata in bulk. In the Mammalia, the Sorex Araneus, called by the common people here the Ranny, is not more than two inches long excluding the tail; and the Mus messorius, or harvest-mouse, peculiar to the southern counties of England, is still more diminutive: so that to these little animals, the larger Dynastidæ, Goliathi, and Prioni, &c., appear giants, and may compete with the mole in size. Even some of the beetles of our own country, as the great Hydrophilus, the stag-beetle, &c., are more bulky than the two first-named quadrupeds. Amongst the birds, many Picæ, Passeres, &c., yield to several insects in dimensions, and their wings when expanded do not extend so far as those of not a few Lepidoptera. The great owl-moth of Brazil (Erebus Strix) in this respect is a larger fowl than the quail. Those beautiful little creatures, the humming-birds (Trochilus L.), the peculiar ornament and life of tropical gardens, which emulate the most splendid butterflies in the brilliancy of their plumage, are smaller than a considerable number of insects in almost every order, and even than some of those that are natives of Britain. Various reptiles also are much inferior in size to many of the insects of the above table. The smallest lizard of this country would be outweighed by the great British beetles lately mentioned, and the mole-cricket (Gryllotalpa vulgaris); and some of the serpent tribe are smaller than the larger Scolopendræ and Iuli. Amongst the fishes also, though some are so enormous in bulk, others in this respect yield the palm to several insects. The minnow and the stickleback that frequent our own pools and streams are considerably inferior in size to some of our water beetles.
In looking over the table, and comparing the different species that compose it with each other, you will perceive that the largest insects of the two sections of Hemiptera, of the Lepidoptera as to their body merely, of the Hymenoptera and Diptera, in general size fall considerably short of those of the other orders; and that certain individuals of the Orthoptera and Aptera bear away the palm in this respect from all the rest. In the Coleoptera the giants, with the exception of the Goliathi, are chiefly to be found amongst the timber devourers in the Lamellicorn and Capricorn tribes. Of orthopterous insects the Phasmidæ present the most striking examples of magnitude; and in the Neuroptera, the Agrionidæ of great length.
It is worthy of remark here, that although the tropical species of a genus usually exceed those of colder climates in size, the Gryllotalpa of Brazil is very considerably smaller than that of Europe: whether this is the case with the rest of the cricket tribe I have not had an opportunity of ascertaining. The Lepidoptera, though often remarkable for the vast expansion of their "sail-broad vans," if you consider only their bodies, never attain to gigantic bulk. Even the hawk-moths (Sphinx L.), though usually very robust, make no approach to the size of the great beetles, or the length of some of the spectres (Phasma) and dragon-flies (Agrionidæ). With regard to the superficial contents of their wings, a considerable difference obtains in different species where they expand to the same length—for the secondary wings are sometimes smaller than the primary, and sometimes they equal them in size. In some instances, also, the latter although long are narrow, and in others they are nearly as wide as long: regard, therefore, should be had to their expansion[Pg 40] both ways. In the Hymenoptera and Diptera, the principal giants are to be found in the predaceous or blood-sucking tribes, as Scolia, the Sphecidæ, Pompilidæ, Vespidæ, &c., belonging to the former order; and the Asilidæ and Tabanidæ to the latter. The true and false humble bees (Bombus and Xylocopa) and the fly tribe (Muscidæ), though they sometimes attain to considerable size, scarcely afford an exception to this observation. Amongst the Aptera none of the Hexapods strike us by their magnitude, and few of the Octopods, though the legs of some of the Phalangidæ inclose a vast area. That in the table would with them describe a circle of six inches diameter, though its body is little more than a quarter of an inch in length. The Myriapods exceed most insects in the vast elongation of their body, which with their motion gives them no slight resemblance to the serpents. In the class Arachnida, the bird-spiders (Mygale) are amongst the principal giants, nor do the Scorpions fall far short of them—both of them when alive often alarming the beholder as much by their size as by their aspect.
But as I have before observed, generally speaking, one of the most remarkable characters of the insect world, is the little space they occupy; for though they touch the vertebrate animals and even quadrupeds by their giants, yet more commonly in this feature they go the contrary way, and by their smallest species reach the confines of those microscopic tribes that are at the bottom of the scale of animal life. I possess an undescribed beetle, allied to Silpha minutissima E. B.[50] which, though furnished[Pg 41] with elytra, wings, antennæ, legs, and every other organ usually found in the order it belongs to, is absolutely not bigger than the full stop that closes this period. In several other coleopterous genera there are also very minute species, as in Cryptophagus, Anisotoma, Agathidium, &c. I know no orthopterous insect that can be called extremely minute, except that remarkable one found on the Continent in the nests of ants, the Blatta Acervorum of Panzer[51], but now called, I believe, Myrmecophilus: nor indeed any in the Hemiptera, Neuroptera, and Diptera, that approach the extreme limits of visibility: but in the Lepidoptera, the pygmy Tinea occultella is almost invisible except in flight, being scarcely thicker than a horse's hair, and proportionably short; indeed, many others of those lovely Lilliputians, the subcutaneous Tineæ, decorated with bands of gold and silver, and studded with gems and pearls, that in larger species would dazzle the beholder's eye, are in size not much more conspicuous. In the Hymenoptera order, Ichneumon Punctum of Dr. Shaw, which forms so striking a contrast to his giant Phasma dilatatum, being placed together in the same plate; and another that I possess, under the trivial name of Atomos, would elude the searching eye of the entomologist unless when moving upon glass. Linné named the tribe of parasites to which these belong, Minuti, on account of their generally diminutive size. But these little minims, under the superintendence of Providence, are amongst the greatest benefactors of the[Pg 42] human race, since they keep within due bounds the various destroyers of our produce.
The number of minute species of insects seems greatly to exceed that of large ones, at least in Europe, of which it may be asserted probably with truth, that two-thirds are under a quarter of an inch in length, and one-third not exceeding much a duodecimal of it. It might hold good perhaps in Coleoptera, Hymenoptera, Diptera, and Aptera: but in Orthoptera, Hemiptera, Neuroptera, and especially Lepidoptera, a large proportion would be found to exceed three lines in length. Neither can it be affirmed of extra-European species, of those at least preserved in cabinets, amongst which it is rare to find an insect less than the fourth of an inch long. This, however, must probably be attributed to the inattention of collectors, who neglect the more minute species.
Though size forms a pretty accurate distinction between insects and the great bulk of vertebrate animals, it affords less assistance in separating them from the invertebrate classes, which are of every size, from the monstrous bulk of some Cephalopoda (cuttle-fish) and Mollusca (shell-fish, &c.) to the invisible infusory animalcule: but external characters, abundantly sufficient for this purpose, may be drawn from the general covering, substance, form, parts, and organs of the body. As I shall enter into pretty full details upon this subject when I come to treat of the external anatomy of insects, I shall here, therefore, only give such a slight and general sketch of the distinctions just mentioned, as will answer the end I have in view. I must here repeat what I have before observed, and what it is necessary that you should always bear in mind, namely,[Pg 43] that at the limits of classes and of every other natural group, the characters begin to change, those peculiar to the one group beginning gradually to disappear, and those of the other to show themselves; so that it is impossible almost to draw up a set of characters so precise as exactly in every respect to suit all the members of any natural group.
Whichever way we turn our eyes on the objects of creation, above—below—athwart, analogies meet us in every direction, and it appears clear, that the Book of Nature is a Book of Symbols, in which one thing represents another in endless alternation. And not only does one animal, &c. symbolize another, but even between the parts and organs of one set of animals there is often an analogy as to their situation and use, when there is little or no affinity as to their structure—or again, the analogy is in their situation, without affinity in either structure or use. Thus certain parts in one tribe represent other certain parts of another tribe, though as to their structure there is often a striking disagreement. This is particularly observable between the vertebrate and invertebrate animals. I shall therefore, in my remarks on the general and particular structure of insects, contrast it in its most important points with that of the first-mentioned tribe.
The first thing that strikes us when we look at an insect is its outside covering, or the case that incloses its muscles and internal organs. If we examine it attentively, we find that it is not like the skin of quadrupeds and other Vertebrata, covering the whole external surface of the body; but that in the large majority it consists of several pieces or joints, in this respect resembling the[Pg 44] skeleton of the animals just named; and that even in those in which the body appears to have no such segments, as in many of the Mites (Acarus L.), they are to be found in the limbs. This last circumstance, to have externally jointed legs, is the peculiar and most general distinction by which the Insecta of Linné, including the Crustacea, may always be known from the other invertebrate animals[52].
If we proceed further to examine the substance of this crust or covering, though varying in hardness, we shall find it in most cases, if we exclude from our consideration the shells of the Mollusca, &c., better calculated to resist pressure than that of the majority of animals that have no spine. In all the invertebrate tribes, indeed, the muscles, there being no internal skeleton, are attached to this skin or its processes, which of course is firmer than the internal substance; but in insects it is very often rigid and horny, and partially difficult to perforate, seldom exhibiting that softness and flexibility which is found in the cuticle of birds and most quadrupeds. From this conformation it has been sometimes said, that insects carry their bones on the outside of their body, or have an external skeleton. This idea, though not correct in all respects, is strictly so in this—that it affords a general point of support to the muscles, and the whole structure is erected upon it, or rather I should say within it. The difference here between Insects and the Vertebrata seems very wide; but some of the latter make an approach towards it. I allude to the Chelonian Reptiles (Testudo L.),[Pg 45] in which the vertebral column becomes external or merges in the upper shell. The cyclostomous fishes also are not very wide of insects as to their integument. But on this subject I shall be more full hereafter.
The forms of insects are so infinitely diversified that they almost distance our powers of conception: in this respect they seem to exceed the fishes and other inhabitants of the ocean, so that endless diversity may be regarded as one of their distinctions. But on all their variations of form the Creator has set his seal of symmetry; so that, if we meet with an animal in the lower orders in which the parts are not symmetrical, we may conclude in general that it is no insect.
But it is by their parts and organs that insects may be most readily distinguished. In the vertebrate animals, the body is usually considered as divided into head, trunk, and limbs, the abdomen forming no part of the skeleton; but in the insect tribes, besides the organs of sense and motion, the body consists of three principal parts—Head, Trunk, and Abdomen—the first, as was before observed, bearing the principal organs of sense and manducation; the second most commonly those of motion; and the third those of generation—the organs of respiration being usually common to both trunk and abdomen. These three primary parts,—though in some insects the head is not separated from the trunk by any suture, as for instance in the Arachnida; and in others, head, trunk, and abdomen form only one piece, as in some mites,—still exist in all, and in the great majority they are separated by incisures more or less deeply marked: this is particularly visible in the Hymenoptera and Diptera, which, in this respect, are formed upon a common model;[Pg 46] and in the rest, with the above exceptions, it may be distinctly traced.
The head of insects is clearly analogous to that of vertebrate animals, except in one respect, that they do not breathe by it. It is the seat probably of the same senses as seeing, hearing, smelling, tasting—and more peculiarly perhaps of that of touch. The eyes of insects, though allowed on all hands to be organs of sight, are differently circumstanced in many particulars from those of the animals last mentioned; they are fixed, have neither iris nor pupil, are often compound, and are without eyelids to cover them during sleep or repose; there are usually two compound ones composed of hexagonal facets, but in some instances there are four; and from one to three simple in particular orders. The antennæ of insects in some respects correspond with the ears of the animals we are comparing with them; but whether they convey the vibrations of sound has not been ascertained: that they receive pulses of some kind from the atmosphere I shall prove to you hereafter—so that if insects do not hear with them in one sense, they may, by communicating information, and by aëroscopy, to use Lehman's term, not directly in his sense[53], supply the place of ears, which would render them properly analogous to those organs. That in numbers these remarkable organs are tactors is generally agreed, but this is not their universal use. That insects smell has been often proved; but the organ of this sense has not been ascertained. What has improperly been called the clypeus, or the part terminating the face above the upper lip (labrum), is in the situation of the nose of the Vertebrata,[Pg 47] and therefore so far analogous to it, and in some cases even in form: I therefore call it the nose. Whether this part represents the nose by being furnished with what answer the purpose of nostrils, residing somewhere at or above the suture that joins it to the upper lip, I cannot positively affirm; but from the observations of M. P. Huber, with regard to the hive-bee, it appears that at least these insects have the organ of the sense in question somewhere in the vicinity of the mouth, and above the tongue[54]: analogy, therefore, would lead us to look for its site somewhere between the apex of the nose and the upper lip; and in some other cases, which I shall hereafter advert to, there is further reason for thinking that it actually resides at the apex of the nose. The organ of taste in insects, though some have advanced their palpi to that honour, is doubtless in some part within the mouth analogous in a degree to the tongue and palate of the higher animals. The organs of manducation, in what may be deemed the most perfect description of mouth, consist of an upper lip closing the mouth above, a pair of mandibles moving horizontally that close its upper sides, and a lower lip with a pair of maxillæ attached to it, which close the mouth below and on the under sides, both labium and maxillæ being furnished with jointed moveable organs peculiar to annulose pedate animals, called palpi. In some tribes these organs assume a different form, that they may serve for suction; but though in many cases some receive an increment at[Pg 48] the expense of others, and a variation in form takes place, none, as M. Savigny has elaborately proved, are totally obliterated or without some representative[55]. The organs now described, except the upper lip, are formed after a quite different type from those of Vertebrata, with which they agree only in their oral situation and use.
The second portion of the body is the Trunk, which is interposed between the head and abdomen, and in most insects consists of three principal segments, subdivided into several pieces, which I shall afterwards explain to you. I shall only observe, that some slight analogy may perhaps be traced between these pieces and the vertebræ and ribs of vertebrate animals, particularly the Chelonian reptiles. This is most observable in Gryllus L. and Libellula L., in which the lateral pieces of the trunk are parallel to each other[56]. In the Diptera and many of the Aptera most of these pieces are not separated by sutures. Each of the segments into which the trunk is resolvable bears a pair of jointed legs, the first pair pointing to the head, and the two last to the anus. These legs in their composition bear a considerable analogy to those of quadrupeds, &c., consisting of hip, thigh, leg, and foot; but the last of these, the foot or Tarsus, is almost universally monodactyle, unless we regard the Calcaria that arm the end of the tibia, as representing fingers or toes, an idea which their use seems to justify. Acheta monstrosa and Tridactylus paradoxus, however[57], exhibit some appearance of a phalanx of these organs. They differ from them first in number, the thoracic legs being[Pg 49] invariably six in all insects, with the exception of the Octopods or most of the Trachean Arachnida, which have usually eight. In the Myriapods, though there are hundreds of abdominal legs, only six are affixed to the trunk. Next they differ with regard to the situation of their legs; for though the anterior pair or arms are analogous in that respect, the posterior pair are not, since in quadrupeds these legs are placed behind the abdomen, but in insects before it—in fact, in the former the legs may be considered as placed at each end of the body, excluding only the head and tail, but in the latter in the middle. Though they correspond with those of quadrupeds in being in pairs or opposite to each other, yet their direction with respect to the body is different, the legs of quadrupeds, &c. being nearly straight, whereas in insects they are bent or form an angle, often very obtuse at the principal articulations, which occasions them to extend far beyond the body, and when long to inclose a proportionally greater space. The wings are the organs of motion with which the upper side of the trunk is furnished; and these, though they are the instruments of flight, are in no other respect analogous to those of birds, which replace the anterior legs of quadrupeds, but approach nearer, both in substance and situation, to the fins of some fishes, and perhaps in some respects even to the leaves of plants. M. Latreille is of opinion, That the four wings or their representatives replace the four thoracic legs of the decapod Crustacea[58]. Upon this opinion, which shows great depth of research and practical acumen, I shall have occasion to express my sentiments when I come to treat more at large on the anatomy[Pg 50] of the trunk and its members; at any rate they do not replace the two anterior pair of legs of the hexapod Aptera. When merely used as wings, they commonly consist of a fine transparent double membrane, strengthened by various longitudinal and transverse nervures, or bones as some regard them, accompanied by air-vessels, of which more hereafter, as well as of their kind and characters. I shall only observe, that insects are known from all other winged animals, by having four wings, or what represent them, and this even generally in those that are supposed to have only a pair. Another peculiarity distinguishes the trunk of insects that you will in vain look for in the vertebrate animals—these are one or two pair of lateral spiracles or breathing pores. Though the respiratory sacs, &c. of birds are almost as widely dispersed as the tracheæ and bronchiæ of insects[59], yet their respiration is perfectly pulmonary, and nothing like these pores is to be discovered in them.
The principal peculiarity of the third part of the body, the abdomen, is its situation behind the posterior pair of thoracic legs, and its rank as forming a distinct portion of what represents the skeleton. In most insects it is so closely affixed to the posterior part of the trunk as to appear like a continuation of it, but in the majority of the Hymenoptera and Diptera, and in the Araneidan Arachnida, or spiders, it is separated by a deep incisure; and in the first-mentioned tribe is mostly suspended to the trunk by a footstalk, sometimes of wonderful length and tenuity. In the Mammalia the male genital organs are partly external; but in insects as well as in many of the vertebrate animals, except when employed, they are[Pg 51] retracted within the body. This part is the principal seat of the respiratory pores or spiracles, many having eight in each side, while others have only one.
Such are the principal external characters which distinguish Insecta and Arachnida, or what we have heretofore regarded as insects, to which here may be added another connected with their internal organization. The union of the sexes takes place in the same manner as amongst larger animals; and the females with very few exceptions, more apparent than real, are oviparous. They are, however, distinguished by this remarkable peculiarity already alluded to, that, except in the case of the Arachnida, one impregnation fertilizes all the eggs they are destined to produce. In most cases, after these are laid, the females die immediately, and the males after they have performed their office, though they will sometimes unite themselves to more than one female. One other circumstance may be named here—that no genuine insect or Arachnidan has yet been found to inhabit the ocean.
Before I conclude this letter, it is necessary to apprize you, that every thing which it contains relative to the characters of insects, has reference to them only in their last or perfect state, not in those preparatory ones through which you are aware that the majority of them must pass. The peculiar characteristics of them in these states—in the egg, the larva, and the pupa, will be the subjects of my next letters, which will be devoted to a more detailed view of the metamorphosis of insects than I gave you before when adverting to this subject[60].
On a former occasion I gave you a general idea of what has been called, perhaps not improperly, the metamorphosis of insects[61]; but since that time much novel and interesting speculation on the subject has employed the pens of many eminent Physiologists; and besides this, the doctrine then advanced of successive developments has been altogether denied by a very able Anatomist, Dr. Herold, who, with a hand, eye, and pencil, second only to those of Lyonnet, has traced the changes that gradually take place in the structure of the cabbage-butterfly (Pieris Brassicæ) on passing through its several states of larva, pupa, and imago. It is necessary, therefore, that previously to considering separately and in[Pg 53] detail the states of insects, I should again call your attention to this subject, and endeavour to ascertain whether Dr. Herold's hypothesis rests upon a solid foundation; or whether that adopted from Swammerdam by all the most eminent Entomologists and Physiologists since his time can be maintained against it.
I shall first give you a short abstract of the new hypothesis.
According to Dr. Herold—The successive skins of the caterpillar, the pupa-case, the future butterfly, and its parts and organs, except those of sex which he discovered in the newly excluded larva, do not preexist as germes, but are formed successively from the rete mucosum, which itself is formed anew upon every change of skin from what he denominates the blood, or the chyle after it has passed through the pores of the intestinal canal into the general cavity of the body, where, being oxygenated by the air-vessels, it performs the nutritive functions of blood. He attributes these formations to a vis formatrix (Bildende Kraft).
The caul or epiploon (Fett-masse), the corps graisseux of Reaumur, &c., which he supposes to be formed from the superfluous blood, he allows, with most physiologists, to be stored up in the larva, that in the pupa state it may serve for the development of the imago. But he differs from them in asserting that in this state it is destined to two distinct purposes—first, for the production of the muscles of the butterfly, which he affirms are generated from it in the shape of slender bundles of fibres;—and secondly, for the development and nutrition of the organs formed in the larva, to effect which, he says, it is dissolved again into the mass of blood, and being oxygenated by the air-vessels,[Pg 54] becomes fit for nutrition, whence the epiploon appears to be a kind of concrete chyle[62].
Need I repeat to you the hypothesis to which this stands opposed—That every caterpillar at its first exclusion contains within itself the germe of the future butterfly and of all its envelopes, which successively presenting themselves are thrown off, till it appear in perfection and beauty, with all its parts and organs, when no further development takes place.
I believe you will agree with me, when you have read and considered the above abstract of Dr. Herold's hypothesis, that in it he substitutes a name for knowledge, talks of a vis formatrix because his assisted eye cannot penetrate to the primordial essence or state of the germes of being, and denies the existence of what he cannot discover[63]. From ancient ages philosophers have done the same, to conceal their own ignorance of causes under a sounding name, when they have endeavoured to penetrate within the veil of the sanctum sanctorum, which it is not permitted to vain man to enter. This has occasioned the invention, not only of the term in question, but of many others, as little meriting the appellation of Signs of ideas; such as Plastic Nature, Epigenesis, Panspermia, Idea seminalis, Nisus formativus, &c. But upon[Pg 55] this subject you cannot do better than consult what the learned Dr. Barclay has said in his admirable work On Life and Organization[64], in which he has placed the inanity, the vox et præterea nihil, of such high-sounding terms in their true light. The processes of nature in the formation and development of the fœtus in utero, of the chick in the egg, of the butterfly in the caterpillar, we in vain attempt fully to investigate; yet we can easily comprehend that pre-existent germes, by the constant accretion of new matter in a proper state, may be gradually developed, but we find it impossible to conceive how, by the action of second causes, without the intervention of the first cause, the butterfly should be formed in the caterpillar, unless it preexists there as a germe or fœtus. "Is it not clear," asks Dr. Virey in his lively manner, "as Blumenbach and other Physiologists maintain, that there is a formative power, a nisus formativus, which organizes the embryo? Admirable discovery!" says he, "which teaches us that the fœtus forms itself because it forms itself! As if you should affirm that the stone falls because it falls[65]!" Had Dr. Herold considered what Bonnet says with as much good sense as modesty, he would never have imagined that his discovering the organs of the butterfly one after the other at certain periods in the caterpillar, was any sound argument against their preexistence and coexistence as germes. "Organs," says that amiable and excellent Physiologist, "that have no existence as to us, exist as they respect the embryo, and perform their essential functions; the term of their becoming visible is that which has been[Pg 56] erroneously mistaken for the period of their existence[66]." This has been Dr. Herold's grand error; he mistook the commencement of the appearance of the organs of the butterfly for that of their existence, and yet the early appearance of the sexual organs ought to have led him to a conclusion the reverse of that which he has adopted.
Dr. Virey has observed with great truth—that "Every being has a peculiar and unique nature, which would be impossible if the body was composed of parts made at several intervals, and without a uniform power that acts by concert[67]:" and every Physiologist acquainted with the history of insects that undergo a complete metamorphosis will allow, that their developments and acquisition of new parts and organs take place according to a law which regulates the number, kind, and times of them, differing in different species, and which has had an invariable operation, since the first creation, upon every sound individual that has been produced into the world.
In consequence of this law, one species changes its skin only four times, and another five or six;—in some cases the first skins shall be covered or bristled with hairs or spines, and the last be naked and without arms;—that which forms the case of the pupæ shall differ in form and substance from the preceding skins, varying in both respects in different species; and finally the butterfly shall invariably follow, when no other change but the[Pg 57] last mortal one shall take place. Can this law, so constantly observed, be the result of a blind power? Or are we to suppose that the Deity himself is always at work to create the necessary organs in their time and place? Is it not much more consonant to reason and the general analogy of nature, to suppose that these parts and organs exist in embryo in the newly-hatched caterpillar, and grow and are successively developed by the action of the nutritive fluid? In the pupa of many Diptera the inclosed animal, even under the microscope, appears without parts or organs, like a mere pulp; but Bonnet tells us, that if boiled, all the parts of the pupa appear[68], which proves the preexistence of these parts even when not to be discerned, and that nothing but the evaporation of the fluids in which they swim is wanted to render them visible.
Mr. William MacLeay has with great truth observed: "The true criterion of animal as well as vegetable perfection is the ability to continue the species[69];" and in their progress to this state certain changes take place in the parts and organs of all animals and vegetables: there is, therefore, an analogy in this respect between them; and this analogy also furnishes another argument against Dr. Herold's hypothesis, as we shall presently see. These changes are of three kinds: In the vegetable kingdom, at least in the phænogamous classes, there is a succession of developments terminating in the appearance of the generative organs, inclosed in the flower; in this kind the integuments, or most of them, are usually persistent. In insects and other annulose and some vertebrate animals, there is a succession of spoliations, or[Pg 58] simultaneous changes of the whole integument, till the animal appears in its perfect form with powers of reproduction; in this kind the integuments are caducous.—In man and most of the vertebrate animals there is a gradual action of the vital forces in different organs till they are fitted for reproduction; accompanied, as progress is made to the adult state, by the acquisition of certain organs, &c. as of teeth, horns, pubes, feathers, &c.[70] Let us now consider a little in detail the analogies that appear to exist between the second and the first and third kinds. I shall first consider the latter as the least obvious. That able, judicious, and learned physiologist, Dr. Virey, has pointed out no inconsiderable resemblance between the metamorphosis of the insect, and the changes, which he denominates a metamorphosis by metastasis, to which most vertebrate animals are subject. In them, he observes, a state analogous to the larva state begins at the exclusion of the fœtus from the womb; it is deprived of teeth, and its viscera are only accommodated to milk: in the cornute species the horns are in embryo: the digestive system now preponderates, and the great enjoyment is eating. A second state, in a degree analogous to that of pupa, commences at the period of dentition—the teeth now produce another modification in the intestinal canal, which becomes capable of receiving and digesting solid food: during this period the vital forces are all tending to produce the perfect state of the animal; and in this state, in man especially, the individual is educated and fitted to discharge the duties of active life. Again, analogous to the imago state is the age of puberty, in which[Pg 59] the complete development of the sexual powers takes place in both sexes, and the animal has arrived at its acme, and can continue its kind[71]: now the digestive powers diminish in their activity, and love reigns paramount. When this state is fully attained, no further or higher change is to be expected, and the progress is soon towards decay and the termination of the animal's mortal career. So we see that in fact man and other mammalia, though they do not simultaneously cast their skins like the insect; or pass into a state of intermediate repose, before they attain the perfection of their nature, like the caterpillar; have their three states, in each of which they acquire new parts, powers, and appetites.
But a more striking analogy has been traced between the insects that undergo a complete metamorphosis and the vegetable kingdom; for though the primary analogy seems to be between the Polypus and the Plant, yet the secondary one with the Insect is not by any means remote. There are circumstances to which I shall have occasion hereafter to call your attention, which afford some ground for supposing, that the substance of the insect and the vegetable partakes of the same nature, at least approximates more nearly, than that of the insect and the vertebrate animal; and every one who has observed these little creatures with any attention, will have observed amongst them forms and organs borrowed as it were from the kingdom of Flora; and vice versâ the Botanist, if he makes the comparison, will find amongst his favourite tribes many striking resemblances of certain insects. But the analogy does not stop here; for the butterfly and the plant appear to have been created with a particular[Pg 60] reference to each other, both in the epoch of their appearance and the changes that take place in them. Thus, as Dr. Virey has observed, the caterpillar is simultaneous with the leaf of the tree or plant on which it feeds, and the butterfly with the flowers of which it imbibes the nectar[72]. Swammerdam, I believe, was the first who noticed the analogy between the changes of the insect and the vegetable, and has given a table in which he has contrasted their developments, including other animals that undergo a metamorphosis[73]: an idea which has been generalized by Bonnet[74], and adopted and enlarged by Dr. Virey[74]. A state analogous to that of the larva in the insect begins in the plant when it is disclosed from the seed, or springs from its hybernaculum in the bulb, &c., or is evolved from the gemma; integument after integument, often in various forms, as cotyledon, radical, cauline, or floral leaves, expands as the stem rises, all which envelopes incase the true representative of the plant, the fructification, as the various skins do the future butterfly. When these integuments are all expanded, the fructification appears inclosed by the calyx or corolla as the case may be, in which the generative organs are matured for their office—this is the bud, which is clearly analogous to the pupa state of the insect. Next the calyx and corolla expand, the impregnation of the germen takes place, and the seed being ripened, and dispersed by the opening of the seed-vessel or ovary of the plant, the individual dies: thus the imago state of the insect has its representative in the plant. "If we place," says Dr. Virey, "here the egg of the insect, next its caterpillar, a little[Pg 61] further the chrysalis, and lastly the butterfly—what is this but an animal stem—an elongation perfectly similar to that of the plant issuing from the seed to attain its blossoming and propagation?"[75]
There being, therefore, this general analogy in their progress to that state in which they can continue their species between every part of animated nature, it holds good, I think, that the same analogy should take place in their developments. If the adult man or quadruped, &c. is evidently an evolution of the fœtus, as from microscopical observations it appears that they are[76], if the teeth, horns, and other parts, &c. to be acquired in his progress to that state are already in him in their embryos, we may also conclude that the butterfly and its organs, &c. are all in the newly-hatched caterpillar. Again, if the blossom and its envelopes are contained in the gemma, the bulb, &c. where they have been discovered[77], it follows analogically that the butterfly and its integuments all preexist in its forerunner.
Perhaps after this view of the objections to Dr. Herold's hypothesis, it will not be necessary to say much with regard to the argument he draws from the change of organs—the loss of some and the acquisition of others—since this may readily be conceived to be the natural consequence of the vital forces tending more and more to the formation of the butterfly, and the withdrawing of their action more and more from the caterpillar; I shall not, therefore, enter further into the question, especially[Pg 62] since the change of organs will come more regularly under our notice upon a future occasion.
Winged insects, many branchiopod Crustacea, and the Batracian reptiles, have been observed by Dr. Virey to bear some analogy to the mammalia, aves, &c. in another respect. In leaving their egg, they only quit their first integument, answering to the chorion or external envelope of the human fœtus; they therefore still continue a kind of fœtus, so to speak, more or less enveloped under other tunics, and principally in their amnios, or the covering in which the fœtus floats in the liquor amnii[78]. This the butterfly does in the pupa case; and its birth from this, under this view, will be the true birth of the animal. In the human subject, the ova upon impregnation are said to pass from the ovary through the Fallopian tube into the uterus. In the insect world, upon impregnation, the eggs pass first from the ovaries into the oviduct, answering to the Fallopian tube, which in them terminates in the ovipositor, or the instrument by which the parent animal conveys the eggs to their proper station: there is, therefore, nothing properly analogous to the uterus in the insect, and the substance upon which the larva feeds upon exclusion answers the purpose of a placenta.
After this general view of the most modern theories with regard to the metamorphosis of insects, I shall in the present and some following letters, treat separately of the different states through which these little beings successively pass.
The first of these is the Egg state, the whole class of insects being strictly oviparous. Some few tribes indeed[Pg 63] bring into the world living young ones, and have on that account been considered as viviparous, but incorrectly, for the embryos of none of these are nourished, as in the true viviparous animals, within a uterus by means of a placenta, but receive their development within true eggs which are hatched in the body of the mother. This is proved by the observations of Leeuwenhoek, who found eggs in the abdomen of a female scorpion[79]; and of Reaumur, with regard to the flesh-fly (Musca carnaria) and other viviparous flies as they have been called[80]. A similar mode of production takes place in vipers and some other reptiles, which have hence been denominated ovo-viviparous, to distinguish them from the true viviparous animals—the class Mammalia. By far the larger portion of insects is oviparous in the ordinary acceptation of the term. The ovo-viviparous tribes at present known are scorpions; the flesh-fly and several other flies; a minute gnat belonging to Latreille's family of Tipulariæ[81]; some species of Coccus; some bugs (Cimicidæ)[82]; and most Aphides, which last also exhibit the singular fact of individuals of the same species being some oviparous and others ovo-viviparous, the former being longer in proportion than the latter.—Bonnet, however, is of opinion that the eggs of the first are not perfect eggs, but a kind of cocoon, which defends the larva, already formed in some degree, from the cold of winter[83].
When excluded from the body of the mother, or from the egg, as has been before observed, some insects appear nearly in the form of their parents, which, with a very slight alteration, they always retain; others, and the greater number, assume an appearance totally different from that of their parents, which they acquire only after passing through various changes. It is to these last, which have chiefly engaged the attention of Entomologists, that the title of metamorphoses has been often restricted. As, however, those insects which undergo the slightest change of form, as spiders do, undergo some change, and almost all insects cast their skins several times[84] before they attain maturity, Linné and most Entomologists, till very recently, have regarded the whole class as undergoing metamorphoses, and as passing through four different states, viz. the Egg—the Larva—the Pupa—and the Imago.
It is obvious, however, that in ovo-viviparous species three states of their existence only come under our cognizance, as these, being hatched in the body of the mother, come forth first under the form of larvæ. There is even one tribe of insects which presents the strange anomaly of being born in the pupa state. This is the Linnean genus Hippobosca (Pupipara fam. Latr.), to which our forest-fly belongs, the females of which lay bodies so much resembling eggs, that they were long considered as such until their true nature was ascertained by Reaumur (most of whose observations were confirmed by De Geer), who, from their size, which nearly equals[Pg 65] that of the parent fly—from their slight motion when first extruded—from spiraculiform points which run down each side of them—and lastly, from their producing not a larva, as all other insects' eggs do, but perfect flies in the winged state—inferred, and doubtless with reason, that they are not real eggs, but pupæ, or larvæ just ready to assume the pupa state, which, however strange it may seem, have passed the egg and larva states in the body of the mother[85].
Insects, therefore, as to their mode of birth, may be divided into—
I. Ovo-viviparous, subdivided into—
1. Larviparous, coming forth from the matrix of the mother in the state of larvæ, as the Scorpion (Scorpio), the Flesh-fly (Musca), the Plant-louse (Aphis), &c.
2. Pupiparous, continuing in the matrix of the mother during the larva state, and coming forth in that of pupa, as the Forest-fly (Hippobosca equina), the Sheep-louse (Melophagus ovinus), the Bat-louse (Nycteribia Vespertilionis), &c.
II. Oviparous. All other insects.
Our business for the remainder of this letter will be with the latter description of these little animals.
The unerring foresight with which the female deposits her eggs in the precise place where the larvæ, when excluded, are sure to find suitable food; and the singular instruments with which, for this purpose, the extremity of their abdomen is furnished, have been noticed in a former letter[86], and those last mentioned will be adverted to in a future one. I shall now, therefore, confine myself[Pg 66] to other circumstances connected with the subject, arranged for the sake of order under several distinct heads, as—their exclusion—situation—substance—number—size—figure—colour—and period of hatching.
i. Exclusion. The exclusion or extrusion of the impregnated eggs takes place, when, passing from the ovary into the oviduct, they are conducted by means of the ovipositor, in which it terminates, to their proper situation. By far the greater number of insects extrude them singly, a longer interval elapsing between the passage of each egg in some than in others. In those tribes which place their eggs in groups, such as most butterflies and moths, and many beetles, they pass from the ovaries usually with great rapidity; while in the Ichneumonidæ, Sphegidæ, Œstri, and other parasitic genera, which usually deposit their eggs singly, an interval of some minutes, hours, or perhaps even days, intervenes between the extrusion of each egg. One remarkable instance of the former mode I noticed in my letter on the Perfect Societies of Insects[87]; another may be cited, to which you may yourself be a witness—I allude to that common moth, vulgarly called the Ghost (Hepialus Humuli), which lays a large number of minute black eggs, resembling grains of gunpowder, and ejects them so fast that, according to De Geer, they may be said to run from the oviduct, and are sometimes expelled with the force of a popgun[88]. A Tetrapterous insect, the genus of which is uncertain, is said, when it is taken, to discharge its eggs like shot from a gun[89]. And a friend of mine, who had observed with attention the proceedings of a common crane-fly (Tipula),[Pg 67] assured me that several females which he caught projected their eggs to the distance of more than ten inches.
A few Diptera extrude them in a sort of chain or necklace, each egg being connected by a glutinous matter with that which precedes and follows it. In a small species of a genus allied to Psychoda (a kind of midge), which one season was abundant in a window of my house, this necklace is composed of eggs joined by their sides, not unlike those strung by children of the seeds of the mallow[90]. Other Tipulidæ on the contrary extrude their eggs joined end to end, so as to resemble a necklace of oval beads. Beris clavipes and Sciura Thomæ, two other flies, produce a chain about an inch long, consisting of oval eggs connected, in an oblique position, side by side; an arrangement very similar prevails in the ribband of eggs which drop from some of the Ephemeræ[91].
These eggs, like those of the insects first mentioned, though connected, are expelled in succession; but other tribes, as the Libellulidæ, with the exception of Agrion, many Ephemeræ, Trichopterous insects, &c. expel the whole at once, as it were in a mass. In those first mentioned they are gummed together in an oblong cluster[92]. In one Ephemera mentioned by Reaumur[93], they formed two oblong masses, each containing from three to four hundred eggs, and three and a half or four lines long. These animals as soon as their wings are developed eject these masses by two orifices, and are aided in the process by two vesicles full of air, wherever they happen to alight or to fall; in most instances it is the water, their proper element, that receives them, but the animal does not appear to know the difference between a solid and a liquid,[Pg 68] and seems only anxious how to free herself from a burthen that oppresses her; all has been contrived that an insect so short-lived may finish her different operations with the utmost celerity: the term of her existence would not have admitted the leisurely extrusion of such a number of eggs in succession[94]. Some Trichoptera, or May-flies, as Phryganea grandis L., exclude their eggs in a double packet, enveloped in a mass of jelly, (a circumstance often attending the eggs that produce aquatic larvæ,) upon the leaves of willows[95]. A similar double packet in the year 1810 I observed appended to the anus of a black species with long antennæ, probably Phryganea atrata F.[96] Upon taking several of the females I was surprised to find in the above situation a seemingly fleshy substance of a dirty yellow. At first, from its annular appearance, I conceived it to be some parasitic larva, but was not a little surprised upon pulling it away that it was full of globular transparent dusky eggs: it was about two lines and a quarter in length and nearly one in breadth. Being bent double it was attached to the animal by the intermediate angle, and when unfolded was constricted in the middle[97]. Each half, which was roundish, had about ten sharp transverse ridges, the interstices of which appeared as if crenated, an appearance produced by the eggs which it contained. Upon more than gentle pressure it burst and let out the eggs. Though resembling the packet of P. grandis in shape and other circumstances, it was nothing like[Pg 69] jelly, but had rather a waxy appearance, and seems to have been covered by a membrane: so that the excluded larvæ must probably have eaten their way out of it. I have still by me, in 1822, specimens of these egg-packets, which, after the lapse of so many years, retain their original form and colour. It is not improbable that other species extrude their eggs in a similar case. Scopoli says of P. bicaudata L., that the female carries about under her belly her eggs united into a globe, like Lycosa saccata[98]. The eggs of Geometra Potamogata F. are also enveloped in a gelatinous substance, and the mass is covered with leaves[99].
Insects of the Diptera order also, like frogs and toads, commit their eggs to the water imbedded in masses of jelly. Dr. Derham describes two different kinds of them, in one of which the eggs were laid in parallel rows end to end, and in another in a single row, in which the sides were parallel[100]. But the most remarkable and beautiful specimen of this kind that I ever saw was one that, many years ago, I took out of a pond at Wittersham in Kent, from which I requested a young lady to make the drawing I send you[101]. The mass of jelly, about an inch and a quarter long, and rather widest in the middle, was attached by one end to some aquatic grass, and from one end to the other ran a spiral thread of very minute eggs, the turns of the screw being alternately on each side.
The mode of exclusion of the eggs of the Blattæ, which are engaged for a whole week in the business of oviposition, is very singular: the female deposits one or two large suboviform capsules, as large as half their abdomen, rounded on one side, and on the other straight and serrated,[Pg 70] which at first is white and soft, but soon becomes brown and hard. This egg-case, as it may be called, contains sixteen or eighteen eggs arranged in a double series, and the cock-roaches when hatched make their escape through a cleft in its straight side, which shuts so accurately when they have quitted it, that at first it appears as entire as before[102]. The insects of the genus Mantis also, or what are called the praying insects, when they deposit their eggs, eject with them a soft substance, which hardens in the air and forms a long kind of envelope resembling parchment, in which the eggs are arranged also in a double series. And the Locusts (Gryllus Locusta L.) are said by Morier[103] to deposit in the ground an oblong substance, of the shape of their abdomen, which contains a considerable number of eggs arranged neatly in rows. The peristaltic motion observed in the females of some insects during oviposition has been before described[104].
ii. Situation. Under this head I include the situation in which the female insect places her eggs when extruded, whether she continues her care of them and carries them about till they hatch, or whether she entirely deserts them, placing them either without a covering within reach of their food, or enveloping them in hair or otherwise protecting them from accident or the attack of enemies. I shall consider them under two views: first, as depositing their eggs in groups, whether covered or naked; and secondly, as depositing them singly.
1. Those that deposit their eggs in groups are first to be considered. I shall begin with those that protect them with some kind of covering.
I have already mentioned in a former letter[105] the silken bag with which Lycosa saccata Latr., a kind of spider, surrounds her eggs, and in which she constantly carries them about with her, defending them to the last extremity. Many other spiders, indeed nearly the whole tribe, fabricate similar pouches, but of various sizes, forms, texture, and colours. Some are scarcely so big as a pea, others of the size of a large gooseberry; some globular, some bell-shaped; others, the genus Thomisus Walck. in particular, depressed like a lupine; some of a close texture like silk; others of a looser fabric resembling wool: some consisting of a single pellicle, but most of a double, of which the interior is finer and softer[106]; some white; others inclining to blue; others again yellow or reddish; most of them are of a whole colour, but that of Epeira fasciata is gray varied with black[107]. And while the parent spider of some kinds (the Lupi) always carries her egg-bag attached to her anus, others hold them by their palpi and maxillæ; and others suspend them by a long thread, or simply fasten them in different situations, either constantly remaining near them (the Telariæ), or wholly deserting them (the Retiariæ). The eggs of one of these last Lister describes as often fixed in a very singular situation—the cavity at the end of a ripe cherry; and thus, as he expresses it—"Stomachi maxime delicatuli quoties hanc innocuam buccam non minus ignoranter quam avide devorarunt[108]."
Herman informs us, that the species of the genus Chelifer carry their eggs in a mass under their belly[109].
Madam Merian gives an account of two species of Blatta, which she affirms carry an egg-pouch about with them—one species (B. gigantea?) she describes as carrying its eggs in a globular pouch of web like certain spiders, and the other in a brown bag, which, when alarmed, it drops and makes off[110]. But this admirable paintress of natural objects was not always correct in her statements[111]: it seems very improbable, from the habits of those species of which we know the history, that any of them should spin a pouch of web for their eggs.
The only insects certainly known to spin an egg-pouch like the spiders, are the Hydrophili, a kind of water-beetles. Some of these, as H. lividus, carry them about with them, like Lycosa saccata, attached to the under side of their body, as M. Miger observed[112]; and others when they are finished desert them. That of the great water-beetle (Hydrophilus piceus) was long ago described and figured by Lyonnet[113]; and a more detailed account of it has since been given by M. Miger[114]. In form it somewhat resembles a turnip when reversed, since it consists of a pouch of the shape of an oblate spheroid, the great diameter of which is three quarters of an inch; and the small, half an inch, from which rises a curved horn, about an inch long and terminating in a point[115]. The animal is furnished with a pair of anal spinners, which move from right to left, and up and down, with[Pg 73] much quickness and agility: from these spinners a white and glutinous fluid appears to issue, that forms the pouch, which it takes the animal about three hours to construct. The exterior tissue is produced by a kind of liquid and glutinous paste, which by desiccation becomes a flexible covering impermeable to water; the second, which envelops the eggs, is a kind of light down of great whiteness, that keeps them from injuring each other. The tissue of the horn is of a silky nature, porous and shining, and greatly resembling the cocoons of Lepidoptera. This part, contrary to what Lyonnet supposes, appears calculated to admit the air, the water soon penetrating it when submerged. At its base is the opening prepared for the egress of the larvæ, when hatched, which is closed by some threads, that, by means of the air confined in the cocoon or pouch, hinder the water from getting in[116]. This nidus does not float at liberty in the water till after the eggs are hatched, the parent animal always attaching it to some plant. By means of this anomalous process for a beetle, which this insect is instructed by Providence thus to perfect, the precious contents of its little ark are secured from the action of the element which is to be the theatre of their first state of existence, from the voracity of fishes, or the more rapacious larvæ of its own tribe, until the included eggs are hatched, and emerge from their curious cradle.
I shall next amuse you with a few instances, in which the Allwise Creator instructs the parent insect, instead of defending her eggs with a covering furnished by her internal organs, to provide it from without, either from[Pg 74] her own body or from some other substance. Most commonly, indeed, the female leaves her cluster of eggs without any other covering than the varnish with which in this case they are usually besmeared. Either they are deposited in summer and will soon be hatched, or they are of a substance calculated to encounter and resist the severities of the season. But many species, whose eggs are more tender or have to resist the cold and wet of winter, defend them in the most ingenious manner with a clothing of different kinds of substance.
Cassida viridis, a tortoise beetle, Rösel tells us, covers her group of eggs with a partially transparent membrane. Arctia Salicis F., a moth, common on willows, wholly conceals hers with a white frothy substance, which when dry is partly friable and partly cottony, and being insoluble in water effectually protects them from the weather[117]. The female of Lophyrus Pini (a saw-fly), having by means of her double saw made a suitable longitudinal incision in the leaf of a fir, and placed in it her eggs in a single row end to end, stops it up with a green frothy fluid mixed with the small pieces of leaf detached by her saws, which when dry becomes friable: a necessary precaution, since these eggs are extremely brittle[118]. Arctia chrysorhœa, Hypogymna dispar, and several other moths, surround theirs with an equally impervious and more singular clothing—hair stripped from their own bodies. With this material, which they pluck by means of their pincer-like ovipositor, they first form a soft couch on the surface of some leaf: they then place upon it successively layers of eggs, and surround them with a similar downy coating, and when the whole number is deposited cover[Pg 75] the surface with a roof of hairs, which cannot be too much admired; for those used for the interior of the nest are placed without order, but those employed externally are arranged with as much art and skill as the tiles of a roof, and as effectually keep out the water, one layer resting partly on the other, and all having the same direction, so that the whole resembles a well-brushed piece of shaggy cloth or fur. When the mother has finished this labour, which often occupies her for twenty-four hours, and sometimes even twice that period, her body, which before was extremely hairy, is almost wholly naked—she has stripped herself to supply clothing to her offspring, and having performed this last duty she expires. The female moths which thus protect their eggs are often furnished with an extraordinary quantity of hair about the anus for this express purpose; and Reaumur conjectures, that the singular anal patch of scales resembling those of the wings, but considerably larger, which is found in the female of Lasiocampa Pityocampa, is destined for the same purpose[119].
Reaumur had once brought to him a nidus of eggs clothed still more curiously: they surrounded a twig in a spiral direction, like those of Lasiocampa Neustria, but were much more numerous, and were thickly covered with fine down, not pressed close, but standing off horizontally, which assumed much the same appearance as a fox's tail would if twisted spirally round a branch[120].
A procedure nearly similar was observed by De Geer in some species of Aphides (A. Alni and A. Pruni), which covered their eggs with a white cottony down detached[Pg 76] from their belly by means of their hind legs[121]. In this case, however, the eggs were separately coated with the down, but there was no general covering to the group.
Several insects make the leaves and other parts of plants serve as coverings for their eggs. Tenthredo Rosæ L., a saw-fly, and other species of the same genus, with their saws make an incision in the green twigs of shrubs and trees, and fill it with a line of eggs placed end to end, taking care that, as the eggs grow after they are laid, they are placed at such distances as to leave room for their expansion[122]. Rhynchites Bacchus, a brilliant weevil, well known to the vine-dressers for the injury it does[123], rolls with much art the leaves of the vine, so as to form a cavity, in which it places its eggs; other species practise similar manœuvres; and some probably place their young progeny in the interior of twigs, making an opening for that purpose with their rostrum—at least, I once saw Rhynchites Alliariæ L. with its rostrum plunged up to the antennæ in the twig of a crab-tree. Others of this tribe, as we know, place their eggs in the interior of fruits and grain, as the nut, acorn, and common weevils.
It is probable that most of the above coverings serve another purpose besides the protection of the eggs from wet and cold—that of sheltering them from the action of too great light, which, as Dr. Michellotti by numerous experiments has ascertained, is fatal to the included germe[124]. On this account it is perhaps that so many insects fasten their eggs to the under side of leaves. Those[Pg 77] exposed in full day have usually an opaque and horny texture.
Some insects are spared all trouble in providing a covering for their eggs, their own bodies furnishing one in every respect adapted to this purpose. Not to mention the Onisci, or wood-lice, since they rather belong to the Crustacea, which have a four-valved cell under the breast, in which they carry their eggs, as the kangaroo does its young in its abdominal pouch, the whole body of the female of those strange animals the Cocci becomes a covering for her eggs, which it incloses on every side. To make this intelligible to you, further explanation is necessary. You must have noticed those singular immovable tortoise-shaped insects, which are such pests to myrtles and other greenhouse plants. These are the young of a species of Coccus (C. Hesperidum L.), and their history is that of the whole race. Part of them never become much bigger than the size of which you ordinarily see them, and when full-grown disclose minute two-winged flies, which are the males. The size of the females, which glue themselves to a twig or leaf as if lifeless, now augments prodigiously, and the whole body, distended with the thousands of eggs which it includes to the bigness of a large pea, without any vestige of head or limb, resembles a vegetable excrescence or gall-apple rather than an insect. If you remove one of them, you will perceive that the under part of its abdomen is flat and closely applied to the surface of the branch on which it rests, only a thin layer of a sort of cotton being interposed between them. In laying her eggs the female Coccus does not, like most insects, protrude them beyond her body into day-light; but as soon as the first egg has passed the orifice of her[Pg 78] oviduct, she pushes it between her belly and the cottony stratum just mentioned, and the succeeding eggs are deposited in the same manner until the whole are excluded. You will ask how there can be found space between the insect's belly and the cotton, to which at first it was closely applied, for so large a mass of eggs? To comprehend this, you must consider that nearly the whole contents of its abdomen were eggs; that in proportion as these are extruded a void space is left, which allows the skin of the under side of the body to be pushed upwards, or towards that of the back, affording room between it and the cottony web for their convenient stowage. If you examine the insect after its eggs are all laid, you will find that they have merely changed their situation; instead of being on the upper side of the skin forming the belly, and within the body, they now are placed between it (now become concave and nearly touching the back) and the layer of cotton. As soon as the female Coccus has finished her singular operation she dies; but her body, retaining its shape, remains glued upon the eggs, to which it forms an arched covering, effectually protecting them, until they are hatched, from every external injury. Some species lay so many eggs, that the abdomen is not sufficiently large to cover the whole mass, but merely one side of it, the remainder being enveloped in cottony web[125].
I am next to consider the situation of those eggs that are excluded by the mother in groups without any other covering than the varnish with which they are usually besmeared in their passage from the oviduct. The females[Pg 79] only place them upon or near the food appropriated to the young larvæ, to which they adhere by means of the varnish just mentioned. These groups consist of a greater or less number; and when the eggs are hatched by the heat of the sun, the larvæ begin to disperse and attack with voracity the food that surrounds them. It is thus that most butterflies and moths attach their eggs to the stems, twigs, and leaves of plants; that the lady birds (Coccinellæ), the aphidivorous flies (Syrphi &c.), and the lace-winged flies (Hemerobii), deposit them in the midst of plant-lice (Aphides); that the eggs of some flesh-flies are gummed upon flesh; those of crickets and grasshoppers buried in the earth; those of gnats and other Tipulidans set afloat upon, or submerged in, the water.
Frequently the whole number of eggs laid by one female is placed in one large group, more commonly, however, in several smaller ones, either at a distance from each other on the same plant, or on distinct plants. The object in the latter case seems to be, in some instances, to avoid crowding too many guests at one table, in others to protect the unhatched eggs from the voracity of the larvæ first excluded, which would often devour them if in their immediate neighbourhood.
In the disposition of the eggs which compose these groups much diversity prevails. Sometimes they are placed without order in a confused mass: more frequently, however, they are arranged in different, and often in very beautiful modes. The common cabbage-butterfly (Pieris Brassicæ) and many other insects place theirs upon one end, side by side, so as, comparing small things with great, to resemble a close column of soldiers, in consequence of which those larvæ which, on[Pg 80] hatching, proceed from the upper end, cannot disturb the adjoining eggs. Many indeed have a conformation purposely adapted to this position, as the hemisphærical eggs of the puss-moth (Cerura Vinula), which have the base by which they are gummed membranous and transparent, while the rest is corneous and opaque. The same ready exit to the larva is provided for in the oblong eggs of the emperor moth (Saturnia Pavonia), which are piled on their sides in two or more lines like bottles of wine in a bin[126].
Where the larva does not emerge exactly from the end of the egg other arrangements take place. The whirlwig-beetle (Gyrinus natator) and the saw-fly of the gooseberry &c. (Tenthredo flava L.) dispose theirs end to end in several rows; the former upon the leaf of some aquatic grass, the rows being parallel[127], the latter gummed to the main nerves of gooseberry or currant leaves, the direction of which they follow[128].
But the lackey-moths (Lasiocampa Neustria, castrensis, &c.) adopt a different procedure. As their eggs, which are laid in the autumn, are not to be hatched until the spring, the female does not, like most other moths, place them upon a leaf, with which they might be blown by the winter's storms far from their destined food, but upon the twig of some tree, round which she ranges them in numerous circles. If you examine your fruit-trees, you can scarcely fail to find upon the young twigs collections of these eggs, which are disposed with such admirable art, that you would take them rather for pearls, set by the skilful hand of a jeweller, than for the eggs of[Pg 81] an insect. Each of these bracelets, as the French gardeners aptly call them, is composed of from 200 to 300 pyramidal eggs with flattened tops[129], having their axes perpendicular to the circumference of the twig to which they are fastened, surrounding it in a series of from fifteen to seventeen close spiral circles, and having their interstices filled up with a tenacious brown gum, which, while it secures them alike from the wintry blast and the attack of voracious insects, serves as a foil to the white enamel of the eggs that it encompasses. It is not easy to conceive how these moths contrive to accomplish so accurately with their tail and hind feet an arrangement which would require nicety from the hands of an artist; nor could Reaumur, with all his efforts and by any contrivance, satisfy himself upon this head. He bred numbers of the fly from the egg, and supplied the females after impregnation with appropriate twigs; but these, as though resolved that imprisonment should not force from them the secret of their art, laid their eggs at random, and made no attempt to place them symmetrically[130].
This illustrious Entomologist was more successful in discovering the mode in which another insect, the common gnat, whose group of eggs is, in some respects, as extraordinary as that last described, performs its operations. The eggs of this insect, of a long phial-like form, are glued together, side by side, to the number of from 250 to 300, into an oblong mass, pointed and more elevated at each end, so as considerably to resemble a little boat in shape. You must not here suppose that I use the term boat by way of illustration merely; for it has all the essential properties of a boat. In shape it[Pg 82] pretty accurately resembles a London wherry, being sharp and higher, to use a nautical phrase, fore and aft; convex below and concave above; floating, moreover, constantly on the keel or convex part. But this is not all. It is besides a life-boat, more buoyant than even Mr. Greathead's: the most violent agitation of the water cannot sink it; and what is more extraordinary, and a property still a desideratum in our life-boats, though hollow it never becomes filled with water, even though exposed to the torrents that often accompany a thunder-storm. To put this to the test, I yesterday (July 25, 1811) placed half a dozen of these boats upon the surface of a tumbler half full of water; I then poured upon them a stream of that element from the mouth of a quart bottle held a foot above them. Yet after this treatment, which was so rough as actually to project one out of the glass, I found them floating as before upon their bottoms, and not a drop of water within their cavity.
This boat, which floats upon the surface of the water until the larvæ are disclosed, is placed there by the female gnat. But how? Her eggs, as in other insects, are extruded one by one. They are so small at the base in proportion to their length that it would be difficult to make them stand singly upright on a solid surface, much more on the water. How then does the gnat contrive to support the first egg perpendicularly until she has glued another to it—these two until she has fixed a third, and so on until a sufficient number is fastened together to form a base capable of sustaining them in their perpendicular position? This is her process. She fixes her four anterior legs upon a piece of leaf, or a blade of grass, and projects her tail over the water. She[Pg 83] then crosses her two hind legs, and in the inner angle which they form, retains and supports the first laid egg, as it proceeds from the anus. In like manner she also supports the second, third, &c., all of which adhere to each other by means of their glutinous coating, until she feels that a sufficient number are united to give a stable base to her little bark; she then uncrosses her legs, and merely employs them to retain the mass until it is of the required size and shape, when she flies away, and leaves it to its fate floating upon the water[131].
It may not be out of place to mention here a remarkable circumstance which not seldom attends a kind of water-scorpion (Naucoris F.) occasionally to be met with in collections of Chinese insects. Its back is often covered with a group of rather large eggs, closely arranged; but whether these are its own eggs or those of some large species of water-mite (Hydrachna Maïll.) has not been clearly ascertained. On the former supposition, the ovipositor must be remarkably long and flexile to enable the animal to place the eggs on its back. In confirmation of the latter it may be observed, that the species of the genus Hydrachna usually attach their eggs to the body and legs of aquatic insects, as for instance H. abstergens to the water-scorpion (Nepa cinerea), &c.[132]
2. After having thus laid before you some of the procedures of those insects that usually deposit their eggs in groups, either naked or defended by coverings of various kinds, I next proceed to a rapid survey of those of the species that commonly deposit them singly. Some[Pg 84] of these, as for instance the Admiral Butterfly (Vanessa Atalanta), glue each egg carefully to its destined leaf by alighting on it for a moment. Another butterfly (Hipparchia Hyperanthus) whose caterpillar is polyphagous, drops hers at random on different plants. In general it may be observed, that all those larvæ which live in solitude, as in the interior of wood, leaves, fruits, grain, animals, &c., proceed from eggs laid singly by the female, which is usually provided with an appropriate instrument for depositing them in their proper situation. Thus the nut-weevil (Balaninus Nucum Germ.) and also that of the acorn (B. Glandium) pierce a nut or an acorn with their long beak, and then deposit in the hole an egg, from which proceeds the maggot that destroys those fruits. Leeuwenhoek asserts that the common weevil (Calandra granaria) adopts the same process, boring a hole in every single grain of corn before it commits an egg to it, and at the same time, by this manœuvre, prepares a small quantity of flour to serve for the food of the tender grub when it is first hatched[133]. It is probable that the Rhyncophorous or weevil tribe in general chiefly use their beaks for the purpose of depositing their eggs in different vegetable substances, and perhaps principally in fruit or grain. The tribe of gall-flies (Cynips) on the contrary, whose economy, detailed in a former letter[134], interested you so much, bore an opening for the egg with their spiral oviduct, which also conveys it.
Another large tribe of insects depositing their eggs singly, are those which feed upon the bodies of other animals, into the flesh of which they are either inserted, or placed so as speedily to find their way into it. Some[Pg 85] of these introduce them into living animals, and then leave them to their fate, as the Ichneumons and gad-flies: others deposit them along with the dead body of an insect interred in a hole, often prepared with great labour, as the different species of sand-wasps (Sphecidæ), spider-wasps (Pompilidæ), &c.: the manners of the latter of these tribes have been already adverted to[135], and those of the Ichneumonidæ will come more fully under consideration when I treat of the diseases of insects.
A similar labour in providing suitable habitations for their eggs is undergone by various other insects whose larvæ live chiefly on vegetable food, some inserting their egg within the substance the larva devours, as those that prey on timber, twigs, roots, or the like, and others on its surface. One would suppose at first, that the exceedingly small egg which produces the subcutaneous larvæ would, by the parent moth, be imbedded in the substance of the leaf which is to exhibit hereafter their serpentine galleries: but this is not the case, for she merely glues it on the outside; at least such was the situation of the only egg of these very minute moths Reaumur had ever an opportunity to observe[136].
Other insects, belonging to the tribe which lay their eggs singly, bury them in the ground. Of this description are many of the lamellicorn insects, the dung-chafers (Scarabæidæ MacLeay) particularly, which, inclosing their eggs in a pellet of dung, deposit them in deep cylindrical cavities. Concerning the proceedings of some of these, as well as of the whole race of bees, wasps, &c., which all lay single eggs, I have before detailed to you[Pg 86] many interesting particulars[137]. I must not conclude this subject without observing, that the female Pycnogonidæ, an osculant tribe between Insects and Crustacea, carry their eggs upon two pair of false legs[138].
iii. Substance. From this long dissertation on the situation of the eggs of insects and matters connected with it, I pass on to their substance or their external and internal composition, giving at the same time some account of the embryo included in them. The eggs of insects, like those of birds, consist in the first place of an external coat or shell, varying greatly, as to substance, in different genera. Most commonly, particularly in those which deposit their eggs in moist situations, as in dung, earth, and the like, it is a mere membrane, often thin and transparent, and showing, as in spiders, all the changes that take place in the inclosed embryo, as the formation of the head, trunk, and limbs[139]. This membrane is sometimes so delicate as to yield to the slightest pressure, and insufficient to protect the included fluids from too rapid an evaporation, if the eggs be exposed to the full action of the atmosphere. In most Lepidoptera, and several other tribes, this integument is considerably stronger, in those moths whose eggs are exposed throughout the winter, as Lasiocampa Neustria, &c., so hard as not to yield easily to the knife. Even in these, however, its substance is more analogous to horn or a stiff membrane than to the shell of the eggs of birds. Nothing calcareous enters into its composition, and it is not perceptibly acted upon by diluted sulphuric[Pg 87] acid. The eggs of birds are lined by a fine membrane; but I have examined several of those of insects, and have been able to discover nothing of the kind in them. I will not, however, affirm that it does not exist, though the shell of the insect egg appears more analogous to the membrane that lines that of the bird than to the outside shell itself.
Within this integument is included a fluid, on the precise nature of which, except that it is an aqueous whitish fluid, few or no observations have been made, or indeed are practicable; but it is reasonable to suppose, that like the white and yolk of the bird's egg, it serves for the development of the organs of the germe of the future insect.
But few observations are recorded that relate to the embryo included in the egg. It is stated, that it is invested with an extremely fine and delicate pellicle—supposed by some analogous to the Chorion and Amnios of the human fœtus, though others think the shell of the egg to correspond with the Chorion, and the successive integuments of the larva with the Amnios[140]. When the egg is first laid, nothing indeed is to be seen in it but the fluid just mentioned; the first change in this fluid is the appearance of the head of the embryo, more particularly in Coleoptera, of two points, the rudiments of the mandibles, and of those apertures into the tracheæ which I have called spiracles[141]; the little animal we may suppose[Pg 88] then assumes its form and limbs. The embryo is usually so folded in the egg that the head and tail meet[142], and the head, annuli, and other parts of the larva are often visible through the shell[143]. Swammerdam even saw the pulsation of the great dorsal vessel through the shell of the egg of Oryctes nasicornis.
Under this head I must notice another singular circumstance peculiar I believe to the eggs of insects, that sometimes, though rarely, they are covered with down or hair. Those of a singular little hemipterous insect, of a genus I believe at present undescribed, the ravages of which upon the larch have been before noticed[144], are covered by a downy web, as is the case with the animal itself. De Geer has described the eggs of a bug, not uncommon in this country (Pentatoma juniperina Latr.), which are reticulated with black veins, in which very short bristles are planted[145]. I possess also a nest of brown eggs, probably of a species of the same genus, found upon furze, which appear to be covered with very short downy hairs. The top of these is flat, and surrounded by a coronet of short bristles, each surmounted by a small white ball, so as to wear the appearance of a beautiful little Mucor. But hairy eggs are not confined to the Hemiptera Order, for, according to Sepp, those of the figure-of-eight moth (Bombyx cæruleocephala) are of this description[146].
iv. Number. The fertility of insects far exceeds that of[Pg 89] birds, and is surpassed only by that of fishes[147]. But the number of eggs laid by different species, sometimes even of the same natural family, is extremely various. Thus the pupiparous insects may be regarded as producing only a single egg; Musca Meridiana L., a common fly, lays two[148], other flies six or eight; the flea twelve; the burying beetle (Necrophorus Vespillo[149]) thirty; May-flies (Trichoptera K.) under a hundred; the silk-worm moth about 500; the great goat-moth(Cossus ligniperda) 1,000; Acarus americanus more than 1,000[150]; the tiger-moth (Callimorpha Caja) 1,600; some Cocci 2,000, others 4,000; the female wasp at least 30,000[151]; the queen bee varies considerably in the number of eggs that she produces in one season, in some cases it may amount to 40,000 or 50,000 or more[152]; a small hemipterous insect, resembling a little moth (Aleyrodes proletella Latr.) 200,000. But all these are left far behind by one of the white ants (Termes fatale F. bellicosus Smeath.)—the female of this insect, as was before observed[153], extruding from her enormous matrix not less than 60 eggs in a minute, which gives 3,600 in an hour, 86,400 in a day, 2,419,200 in a lunar month, and the enormous number of 211,449,600 in a year: probably she does not always continue laying at this rate; but if the sum be set as low as possible, it will exceed that produced by any other known animal in the creation.
v. Size. The size of the eggs is in proportion to that of[Pg 90] the insect producing them, though in some instances small ones produce larger eggs than those laid by bigger species. Thus the eggs of many Aptera, as those of that singular mite Uropoda vegetans, and of the bird-louse found in the golden pheasant, are nearly as large, it is probable, as the parent insect; while those of the ghost-moth (Hepialus Humuli) and many other Lepidoptera, &c. are vastly smaller. This circumstance perhaps depends principally on the number they produce: the majority of them, however, are small. The largest egg known, if it be not rather an egg-case, is that of a spectre insect (Phasma dilatatum), figured in the Linnean Transactions[154], being five lines in length and three in width, which probably approaches near the size of that of some humming-birds. The largest egg of any British insect I ever saw was that of the common black rove-beetle (Staphylinus olens) sent me by Mr. Sheppard—this is a line and half long by a line in width. But we do not often meet with insect-eggs exceeding a line in length. A vast number are much smaller: those of Ephemeræ are more minute than the smallest grains of sand[155], and some almost imperceptible, as those of the subcutaneous moths, to the naked eye. Commonly the eggs laid by one female are all of the same size; but in several tribes, those containing the germe of the female are larger than those that are to give birth to a male. This appears to be the case with those of the Rhinoceros beetle (Oryctes nasicornis[156]), and according to Gould with those of ants[157]. As the female in a vast number of instances is much bigger than the male, it is not improbable that this law may hold[Pg 91] very extensively. It is stated, however, by Reaumur[158], that the reverse of this takes place in the eggs of the hive-bee, those that are to produce males being larger than the rest.
Another peculiarity connected with the present head is the augmentation in bulk which takes place, after exclusion, in the eggs of the great tribe of saw-flies (Tenthredo L.), the gall-flies (Cynips L.), the ants (Formica L.) and the water-mites (Hydrachna Maïll. Atax F.). Those of the two former, which are usually deposited in the parenchymous substance of the leaves, or of the young twigs, of various plants, imbibe nutriment in some unknown manner, through their membranous skins, from the vegetable juices which surround them[159], and when they have attained their full size are nearly twice as large as when first laid. Except in the eggs of fishes, whose volume in like manner is said to augment previously to the extrusion of the young, there is nothing analogous to this singular fact in any other of the oviparous tribes of animals, the eggs of which have always attained their full size when they are laid.
It is to M. P. Huber that we are indebted for the knowledge of the fact that the eggs of ants grow after being laid, a circumstance favoured probably by the moist situation in which the workers are always careful to keep them. By an accurate admeasurement he found that those nearly ready to be hatched were almost twice as big as those just laid[160]. A similar observation was made on the red eggs of a water-mite (Hydrachna abstergens) by Rösel, who conjectured that they draw their means of increase from the body of the water-scorpions[Pg 92] (Nepæ), of which they form so singular an appendage[161], which opinion is confirmed by De Geer, who observes that when the water-scorpions are covered by an unusual number of the eggs of the water-mites, they grow weak and languid, and endeavour to rid themselves of their parasitic appendages[162]. It is most probable that the mite lately named (Uropoda vegetans), which is often found planted as it were upon the bodies of various beetles, by means of a long pedicle, through which, as the fœtus by an umbilical chord and placenta, it derives its nutriment from the above animals, is at first so fixed in the egg state, though before it is disengaged from the pedicle it is hatched, since it is often found with its legs displayed and quite active—this is the more probable, as the eggs of the water-mite are fixed by a pedicle to the animals to which they are attached[163]. I have met with a remarkable instance, in which pedunculated eggs seem to draw nutriment from the mother, which brings the pedicle still near to the nature of the umbilical chord. Those of the small hemipterous insect which infests the larch before alluded to, are attached to the anal end of the mother by a short foot-stalk not longer than the egg.
Dr. Derham seems to have observed, that the eggs of some Diptera, of the tribe of Tipulidæ, also increase in size before the larva is excluded[164]. It seems to me likely enough, that in this and many of the above cases in which the egg is supposed to grow, it is rather an extension of the flexile membrane that forms their exterior proportioned to the growth of the included embryo from food[Pg 93] it finds within the egg, than from any absorption from without.
vi. Shape. We are accustomed to see the eggs of different species of oviparous animals so nearly resembling each other in form, that the very term egg-shaped has been appropriated to a particular figure. Amongst those of birds, with which we are most familiar, the sole variations are shades of difference between a globular and oval or ovate figure. The eggs of insects, however, are confined by no such limited model. They differ often as much, both as to their shape, sculpture, and appendages, as one seed does from another; and it is not improbable that, if duly studied, they would furnish as good indications of generic distinctions as Gærtner has discovered in those of plants. Their most usual form indeed is globular, oval, or oblong, with various intermediate modifications. We meet with them ovate, or of the shape of the common hen's egg, flat and orbicular, elliptical, conical, cylindrical, hemispherical, lenticular, pyramidal, square, turban-shaped, pear-shaped, melon-shaped, boat-shaped, of the shape of an ale-stand, of a drum, &c.[165], and sometimes of shapes so strange and peculiar, that we can scarcely credit their claim to the name of eggs. Thus the eggs of the gnat are oblong and narrow, or nearly cylindrical, having at the top a cylindrical knob[166], so as to give them the precise form of the round-bottomed phial sometimes used by chemists: those of the common[Pg 94] water-scorpion (Nepa cinerea) are oblong, and at the upper end are surrounded by a sort of coronet, consisting of seven slender rays or bristles of the length of the egg[167], so as to resemble somewhat the seeds of Carduus benedictus (Cnicus acarna[168]) of the old botanists. One would think this spinous circlet a very awkward appendage to bodies which are to be gradually extruded through the fine membranous ovaries and oviduct which inclose them: but they are so admirably packed, the unarmed end of each egg fitting closely into the space inclosed by the spines of the one next below it, or, rather, the spines which are moveable, embracing it closely, that not only is no room lost, but the ovaries are perfectly secure from injury. The eggs of another species of this tribe (Ranatra linearis) have only two of these spines or bristles—they are inserted in the stem of a water-rush (Scirpus) or other aquatic plant, so as to be quite concealed, and are only to be detected by the two bristles which stand out from it[169]. The eggs of the beautiful lace-winged flies (Hemerobius), those golden-eyed insects so serviceable in destroying the plant-lice (Aphides[170]), are still more singular. Those of H. Perla are oval, and each of them attached to a filiform pedicle not thicker than a hair, and seven or eight times as long as the egg. By this pedicle (which is supposed to be formed by a glutinous matter attached to one end, which the female draws out by abstracting her ovipositor with the egg partly in it[Pg 95] from the leaf, to which she has previously applied it, to a proper length, when the gluten becoming sufficiently solid she wholly quits the egg,) the eggs are planted in groups of ten or twelve on the surface of leaves and twigs, from which they project like so many small fungi, to some of which they have a remarkable resemblance. When the included larva has made its way out of them by forcing open the top, they look like little vases, and were actually once figured by a Naturalist, as we learn from Reaumur, as singular parasitic flowers growing upon the leaves of the elder, for the origin of which he was extremely puzzled to account[171]. Eggs similarly furnished with a pedicle are also laid by other insects; but as most of these have been before alluded to, it is not necessary to describe them here[172]. The cause of these differences of form is for the most part concealed from us: in many instances it may perhaps be referred to that will to vary forms, and so to glorify his wisdom[173] and power, independently of other considerations, which, as Dr. Paley has well remarked[174], seems often to have guided the Great Author of Nature. But in some cases the end to be answered is sufficiently evident. The long footstalks of the eggs of the Hemerobius just mentioned, there can be little doubt, are meant to place them out of the reach of the hosts of predaceous insects which roam around them, from whose jaws, thus elevated on their slender shaft, they are as safe as the[Pg 96] eggs of the tailor bird in its twig-suspended nest from the attack of snakes. Reaumur has described the eggs of a kind of fly, common upon the excrements of the horse and other animals (Scatophaga vulgaris Latr.), or one related to it, that requires to be immersed in the dung to which it is committed, on which the future grubs are to feed. He found that if not thus surrounded with moisture, they infallibly shrivelled up and came to nothing; but it is equally necessary that they should not be wholly covered: if they were, the young larva would be suffocated at its first exit from the egg. In what way is this nice point secured? In this manner. Each egg is provided at its upper end, at which the animal when hatched comes out, with two diverging horns[175]; these prevent it from being stuck into the excrement, in which the female deposits the eggs one by one, more than three-fourths of its length: and when examined they resemble not badly, as Reaumur remarks (except that their colour is white), a parcel of cloves stuck into a pudding, as they are neatly inserted at due distances in the disgusting mass[176]. The French Naturalists found these eggs in swine's dung; I have observed them in cow-dung. Latreille thinks that the bristles above described attached to the eggs of Nepa and Ranatra have a similar use, as the female plunges them all but these bristles into the stems of aquatic plants[177]: but may not this have something to do with their oxygenation? Reaumur has figured another egg of a dipterous insect which has a longitudinal wing or lateral margin attached to it, giving[Pg 97] it the form of an oblong square, the object of which, he conceives, is to give a greater surface by which it may be more firmly fixed to the substance against which the fly attaches it[178].
Besides these more striking variations in figure, their surface, though often smooth, is frequently curiously and most elegantly sculptured, a circumstance that distinguishes the eggs of no other oviparous animals. Some, as the margined egg just mentioned, are only sculptured on one side, the other being plain; or, as those of the Tusseh silk-worm[179] (Attacus Paphia) and other Bombyces, which have orbicular depressed eggs with a central cavity above and below, have their circumference crossed with wrinkles corresponding with the rings of the inclosed embryo[180]. Others again are sculptured all over. Of these, in some, the sculpture of the two sides is not symmetrical, as in those of a fly figured by Reaumur[181]: but in general there is a correspondence in this respect between the different parts of the egg. In those elegant ones before alluded to of some bird-louse attached to the golden pheasant, the shell resembles the purest wax, and is scored with longitudinal striæ, each distinguished by a series of impressed points, which give it a beautiful appearance of net-work. In the others, as in a common butterfly (Hipparchia Ægeria) and moth (Geometra cratægata), the whole surface is covered with hexagonal reticulations[182]. Others, as those of another butterfly[Pg 98] (Hipparchia Hyperanthus), are beset with minute granules or tubercles[183]. Others again, like those of the cabbage and hawthorn butterflies (Pieris Brassicæ and Cratægi), are remarkable for beautiful longitudinal ribs, often connected by elevated lines crossing them at right angles[184]; and in some, as in another butterfly (Hipparchia Jurtina), crowned by imbricated scales[185]. Many other minor differences in this respect might be noticed, but these will suffice to give some idea of the infinite variety exhibited in this respect by these little atoms. If the Creator has wrought them with so much art and skill, can it be beneath his reasonable creatures to examine and admire them, that they may glorify those attributes which they serve to illustrate?
Some eggs after exclusion occasionally become slightly corrugated: Malpighi supposed that this occurs only when the eggs are barren, having observed that those of the moth of the silk-worm which preserved their plumpness always produced caterpillars, while those which lost their original rotundity and became wrinkled were constantly unprolific. Bonnet, however, found exactly the reverse take place in another moth[186], so that these appearances are scarcely to be depended upon. Kuhn asserts, that a virgin female of the puss-moth (Cerura Vinula) having begun to lay eggs, which were yellow above, green below, and depressed, he introduced to her an hour afterwards a male, and some minutes subsequently to the union, she again deposited eggs, which were wholly of a dark brown and convex[187].
vii. Colour. The colour of the eggs of insects is as various as their shape and sculpture. They are very often white, those of some spiders like minute pearls[188]; some are yellow, as those of the silk-worm; others orange, such are the eggs of the bloody-nosed beetle (Timarcha tenebricosa); others again of a golden hue; sometimes they are of a sanguine red. I remember once being much surprised at seeing the water at one end of a canal in my garden as red as blood: upon examining it further I found it discoloured by an infinite number of minute red eggs, belonging probably to some dipterous insect of the Tipulidan tribe. There are also eggs of every intermediate shade between red and black; some again are blue and others green. They are not always of whole colours, for some are speckled like those of many birds, of which I can show you specimens, that are also shaped like birds' eggs; these I think were laid by a common moth (Odenesis potatoria); others are banded with different colours—thus the blue eggs of the lappet-moth (Gastropacha quercifolia) are encircled by three brown zones[189]; others are brown with a white zone[190].
Many eggs assume a very different colour after being laid a few days. In general upon their first exclusion they are white. Those of the chameleon-fly (Stratyomis Chamæleon) which I once found in great numbers, arranged like tiles on a roof one laid partly over another, on the under side of the leaves of the water-plantain, from white become green, and then change to olive green. Those of the hemipterous enemy of the larch, more than[Pg 100] once mentioned in this letter, are first mouse-coloured, then they assume a reddish hue, and lastly a blackish one. Those of the gnat from white in a short time assume a shade of green, in a few hours they are entirely green, and at length become gray[191]. Those of the silk-worm, which at first are of a yellow or sulphur colour, acquire a violet shade. The eggs of that rare moth Endromis versicolor, are at first sulphur-coloured, then green, next rose-coloured, and lastly blackish. The colour of almost all eggs changes when they are near hatching; but this change depends more frequently upon the colour of the included larva, which appears through the transparent shell of the egg, than upon any actual alteration in the egg itself.
viii. Period of hatching. The general rule for the hatching of the eggs of insects is the absorption by the embryo of all the superabundant moisture included in them; but the time varies according to the state of the atmosphere, to the action of which they are subjected. Like those of other animals, they require a certain degree of heat for the due evolution of the included larva. This heat in much the greater number of instances is derived from the temperature of the air, but often also from other sources. The eggs of the gad-fly tribe are hatched principally by the heat of the body of the animal to which they are committed; and doubtless the vital heat of various larvæ, small as it may be, must contribute something to the hatching of the eggs deposited in them by various Ichneumons. In the fermenting bark in which the instinct of the rhinoceros beetles (Oryctes nasicornis &c.)[Pg 101] impels them to place theirs, the dung which the Scarabæidæ select for that purpose, and the decaying vegetables chosen by many other insects, a degree of artificial heat must exist: and the eggs, or rather egg-like pupæ, of the spider-fly of the swallow (Ornithomyia Hirundinis) are hatched by the heat of those birds which sit upon them along with their own eggs.
Fabricius says, "Insects never sit upon their eggs[192];" but certainly, as I formerly related to you[193], the female earwig does this, and one would be induced to suppose, from the circumstance of the young ones following their mother, as chickens do the hen, that Pentatoma grisea (Cimex Linn.), formerly mentioned, may do the same[194].
With these exceptions, the eggs of all insects are hatched by atmospheric heat alone, the variations in which determine the more speedy or more tardy disclosure of the included insect. The eggs of such species as have several broods in the year, as the nettle butterfly (Vanessa Urticæ) when laid in summer are hatched in a few days; but if not laid till the close of autumn, they remain dormant through the winter, and are only hatched at the return of spring. That this difference is to be attributed to the influence of heat has been often proved by experiment: the autumnal eggs if brought into a warm room may be hatched as soon as those laid in the height of summer. Silk-worms' eggs naturally are not hatched till they have been laid six weeks, but in countries where they are reared, the women effect their exclusion in a much shorter period by carrying them in their bosoms: yet to retard their hatching with particular[Pg 102] views is in many circumstances impossible. When the heat of the atmosphere has reached a certain point, the hatching cannot be retarded by cellars; and M. Faujas has remarked, that in June the silk-worm's eggs would hatch in an ice-house[195].
The period of exclusion does not, however, depend solely upon temperature: the hardness or softness of the shell, and possibly differences in the consistence of the included fluid, intended to serve this very purpose, cause some eggs to be hatched much sooner than others exposed to the same degree of heat. Thus the eggs of many flesh-flies are hatched in twenty-four hours[196]; those of bees and some other insects in three days; those of a common lady-bird (Coccinella bipunctata) in five or six days; those of spiders in about three weeks; those of the mole-cricket in a month; while those of many Lepidoptera and Coleoptera require a longer period for exclusion. The hard eggs of Lasiocampa Neustria and castrensis, noticed above, remain full nine months before being hatched[197], as do those of another moth (Hypogymna dispar), which, though laid in the beginning of the warm month of August, do not send forth the included caterpillar till the April following[198]. We know no more of the cause of[Pg 103] this difference than of that which takes place in the period of exclusion of the eggs of the different species of birds.
Some eggs change considerably both their form and consistence previously to being hatched. M. P. Huber found that those of different species of ants when newly laid are cylindrical, opaque, and of a milky white; but just before hatching their extremities are arched, and they become transparent with only a single opaque whitish point, cloud, or zone, in their interior[199]. An analogous change takes place in the eggs of many spiders, which just before hatching exhibit a change of form corresponding with that which the included spider receives when its parts begin to be developed, the thin and flexible skin of the egg moulding itself to the body it incloses[200].
In proportion as the germe included in the egg is expanded, it becomes visible through the shell when transparent: this is particularly the case with spiders, in which, as was before observed, every part is very distinctly seen. At length, when all the parts are consolidated so as to be capable of motion, which in spiders takes place in four or five days after they begin to be visible in the egg, the animal breaks the pellicle by the swelling of its body and the movement of its legs, and then quits it, and disengages all its parts one after the other[201]. In general, at least where the shell is harder than that of spiders, insects make their way out by gnawing an opening with their mandibles in the part nearest their head, which, when the shell is very strong (as in Lasiocampa Neustria, &c.), it is often several[Pg 104] hours in accomplishing[202]. In many instances, however, the larva is spared this trouble, one end of the egg being furnished with a little lid or trap-door, which it has but to force up, and it can then emerge at pleasure: such lids are to be found in the eggs of several butterflies and moths, as Satyrus Mæra, Saturnia pavonia major, &c. and the common louse[203]. In those exquisitely elegant eggs, before described, of some kind of bird-louse (Nirmus) found adhering to the base of the neck feathers of the golden pheasant[204], there is a lid or cap of this kind of a hemispherical form terminating in a tortuous style. Those of a species of bug (Pentatoma Latr.), found by our friend the Rev. R. Sheppard, besides a convex lid are furnished with a very curious machine, as it should seem, for throwing it off. This machine is dark-brown, of a corneous substance, and of the shape of a cross-bow[205], the bow part being attached to the lid or pushing against it, and the handle, by means of a membrane, to the upper end of the side of the egg.
When the included animal has made its way out of the egg, it enters upon a new state of existence, that of Larva, to which I shall direct your attention in the following letter.
The Larva state is that in which insects exist immediately after their exclusion from the egg (or from the mother in ovo-viviparous species), in which they usually eat voraciously, change their skin several times, and have the power of locomotion, but do not propagate.
Almost all larvæ, at their birth, are for a time in a very feeble and languid state, the duration of which differs in different species. In most it continues for a very short time, a few minutes or perhaps hours, after which they revive and betake themselves to their appropriate food. In others, as in the generality of spiders, this debility lasts for seven or eight days, and in some species even a month, during which the young ones remain inactive in the egg-pouch[206], and it is not till they have cast their first skin that their active state of existence commences.
All larvæ may be divided into two great divisions:—
I. Those which in general form more or less resemble the perfect insect.
II. Those which are wholly unlike the perfect insect.
I shall begin by calling your attention to the characters of the first of these divisions: the second, which is by far the most numerous, will be afterwards considered.
I. The first division includes the larvæ of Scorpions, Spiders, Cockroaches, Grasshoppers, Lanthorn-flies, Bugs, &c.; or generally, with the exception of the Flea and Crustacea, the whole of the Linnean Orders Aptera and Hemiptera. All these larvæ, however remotely allied in other respects, agree in the general similarity which they bear to the perfect insects which proceed from them. The most acute entomologist, untaught by experience, could not even guess what would be the form of the perfect insects to be produced from larvæ of the second division, while they can recognise the form of the spider, the cricket, the cockroach, the bug, and the frog-hopper, in that of the larvæ. There are, however, differences in the degrees of this resemblance, according to which we may, perhaps, divide this tribe in their second state as follows:—
i. Those that resemble the perfect insect, except in the relative proportions and number of some of their parts.
ii. Those which resemble the perfect insect, except that they are apterous, or not yet furnished with organs of flight.
i. Spiders, Phalangia, scorpions, lice, Poduræ, sugar-lice (Lepisma), mites, centipedes, millepedes, &c. come under the first subdivision. The larvæ of the first six tribes here mentioned differ at their birth from the perfect insect, only in size and the proportions of their parts.[Pg 107] Thus the larvæ of spiders have their legs of a different relative length from that which they subsequently acquire; and the palpi in the males, which previously to the discoveries of Treviranus were regarded as their sexual organs, are not yet fully developed[207]: and a similar difference takes place in the legs of Phalangia. The general form too of the body undergoes slight alterations, and the colour very considerable ones, with each change of the skin—a change to which all these tribes are subject.
The larvæ of the three last-mentioned tribes (the mites, centipedes, and millepedes) differ from the perfect insect not only in the proportion but also in the number of their parts. Leeuwenhoeck states (and De Geer confirms his assertion, extending it to other species of mites[208]), that the common cheese-mite, which in its perfect state has eight legs, when first excluded from the egg has but six, the third pair being wanting[209]. Some however are born with eight legs, for instance A. eruditus of Schrank, which he saw come from the egg itself with that number[210]. Others again have never more than six legs: this is the case with Latreille's genera—Caris, Leptus, Atoma, and Ocypetes of Dr. Leach[211]. In the centipedes (Scolopendridæ) and millepedes (Iulidæ) differences still more remarkable, as I have stated in a former letter, have been observed by De Geer; these animals, in their progress to the perfect state, not only gain several additional pairs of legs, but also several additional segments of the body. This illustrious Entomologist found that Pollyxenus lagurus (Scolopendra L.) was born a hexapod,[Pg 108] with but three segments and as many pairs of feet, but successively acquired five additional segments with other appendages, and nine more pairs of feet[212]. A species of millepede (Iulus terrestris L.), which he also traced from its birth, and which begins the world at first with only eight segments and six feet, by a successive development at length acquires, in its perfect state, 50 segments and not less than 200 feet[213]. The nature of these very singular accretions, which Latreille and Mr. Wm. MacLeay have also observed in the centipedes[214], seems not well understood. If, as is most probable, though De Geer could not find any exuviæ[215], the larvæ cast a skin before each change, they do not essentially differ from the metamorphosis of other insects. The legs that these insects thus acquire are affixed to the abdomen, the six that they set out with being attached to the part representing the trunk, so that the former may be regarded as analogous to the prolegs of caterpillars. These animals therefore, as I have before intimated, invert the order of Nature, and from perfect degenerate into imperfect insects.
ii. If you examine the cockroach, cricket, or grasshopper, in different stages of their growth, you will find that the larva does not vary essentially from the perfect insect, except in wanting wings and elytra. The case is the same in almost all the Linnean genera of the modern order—Hemiptera; and with Raphidia, Termes, and Psocus, in the Neuroptera. Some of these, however, exhibit[Pg 109] slighter discrepancies in the proportion of some of their parts, but without affecting the general resemblance. Thus the larvæ of the common ear-wig have at first only eight, and subsequently nine joints to their antennæ, whereas the perfect insect has fourteen[216]; and the forceps is quite different, resembling rather two straight styles than what its name implies. In those also of many bugs (Coreus marginatus F. &c.), the joints of the antennæ are of a shape dissimilar to that which obtains in the perfect insect. In that of the common water-scorpion, the anal air-tube, which is so long in the imago, is scarcely visible[217]. In the Cicada tribe, so celebrated for their song[218], neither the larva nor the imago have the enormous thigh armed below with strong teeth, the tibiæ terminating in a fixed incurved claw, probably for the purpose of digging the holes into which they retire till they disclose the fly, which distinguish the pupæ of some species, and is particularly conspicuous in one commonly brought from China[219]. These often exhibit also other minor differences.
II. In treating of the second great division of larvæ, those that are wholly unlike the parent insect,—which includes, with few exceptions[220], the whole of the Linnean[Pg 110] orders, Coleoptera, Lepidoptera, Hymenoptera, Diptera, the majority of the Neuroptera, Coccus and Aleyrodes in Hemiptera, and the genus Pulex in Aptera,—I shall advert to their characters, under several distinct heads; and to avoid unnecessary circumlocution, I shall in what follows wholly leave out of consideration the first division already explained, and use the term larvæ with reference only to those of the second. The heads under which I propose to treat of them are: The substance of their body, its parts, shape, or figure, clothing, colour. Also the Economy or mode of life of these creatures: their food, moultings, growth, age, sex, and their preparations for assuming the Pupæ.
i. Substance, with the exception of the head and six fore-feet, which are usually corneous, the exterior integument or skin of larvæ is commonly of a membranous texture, and the body is of a much softer consistence than in the perfect insect. In those, however, of some Staphylinidæ and other Coleoptera, the dorsal part of the three first pieces, which represent the trunk of the perfect insect, is hard and horny. Some also have their whole skin coriaceous, as the tortoise-shell butterfly (Vanessa polychloros); and some few, as the wire-worm (Elater segetum), and other Elateres, very hard. I possess a very remarkable larva from Brazil, from the extreme flatness of its body, and from its having cavities to receive its legs when unemployed, probably living under bark, the skin of which is still harder than that of the grub of the Elaters. Perhaps it has to resist great pressure; and on that account is gifted with this quality, so seldom to be met with in other kinds of larvæ. The[Pg 111] interior of the body of these animals is generally of a softer consistence than in the perfect insect. Their intestines, and other internal organs, are usually wrapped in a voluminous substance of a fatty nature, which is regarded as analogous to the epiploon, omentum, or caul, which envelops the viscera of quadrupeds, &c., and is called by Reaumur the corps graisseux. The use of this general flexibility of larvæ is obvious; for, their bodies being mostly long and narrow, a hard rigid covering would have been very inconvenient, and a considerable impediment to their motions. When a caterpillar is feeding, it has occasion to apply its body to any part of the margin of a leaf so as to support itself by its prolegs, and when moving it wants to give it all the curves that are necessary to enable it to avoid obstacles, and thread its way through the sinuous labyrinths which it must often traverse. On the other hand, the hardness of the substance of its head affords a strong fulcrum to the muscles which keep its powerful jaws in constant play. The larvæ, indeed, of some Diptera have a membranous head; but their mandibles, which serve also as legs, are not grinders, but merely claws, the muscles of which require less powerful support[221]. Under this head it may be proper to observe, that generally larvæ are opaque; but some, as those of ants, and a few Lepidoptera[222], are diaphanous. That of Corethra crystallina (Tipula De Geer) is so beautifully transparent as to resemble[Pg 112] a piece of crystal, and scarcely to be distinguished from the water in which it lives[223].
ii. Parts. The body of each larva consists of the head, including its different organs, and of the succeeding segments, of which the three first may usually be denominated the trunk, and have the six anterior feet, when present, attached to their under side: the remainder is the abdomen. The latter includes in some species a variable number of membranous feet, as well as various appendages affixed usually to its tail and sides. No larva is ever furnished with wings[224]. Each of these greater divisions, and the organs which they include, require separate consideration.
1. Head. This, as was lately observed, is exteriorly of a horny substance, or at least harder than the rest of the body, in most larvæ; and on this account, though rarely separated from it by any visible distinct neck[225], is, if the[Pg 113] larva be of a tolerable size, distinguished at the first view. In those of many Dipterous insects, however, the head is covered with the same flexible membranous skin with the rest of the body, from which it is often scarcely to be distinguished. In these, except that it contains the organs of manducation, it wears no more the appearance of a head than any other segment of the body, and scarcely so much as the last or anal one. The head of these larvæ is also remarkable for another peculiarity,—that it is capable of being extended or contracted, and assuming different forms at the will of the insect: a property which the head of no superior animal can boast. It is probable that there is a considerable variety in the shape and circumstances of the heads of larvæ; but since, with the exception of those of Lepidoptera, they have had less attention paid to them than they deserve (indeed in a vast number of cases, from the difficulty of meeting with them, these variations, except in a few instances, have not been described), I will here mention a few of the most remarkable. The head of the young larva at its first exclusion from the egg is usually the most dilated part of the body, but it does not often continue so. In that of Cicindela campestris, however,—the beautiful green beetle sometimes found in sandy banks,—and also in several caterpillars of Lepidoptera, it is much larger than any of the following segments[226], which, in conjunction with the animal's formidable jaws, gives it a most ferocious appearance. In some lepidopterous larvæ the head is of the same diameter with the rest of the body, but in insects in general it may, I think, be stated as less; and[Pg 114] occasionally it bears no proportion whatever to it. This is the case with the subcortical one from Brazil lately mentioned. It is more commonly longer than broad; but in some, as in the larvæ of carrion beetles (Silphæ), the reverse of this takes place. Its shape varies from triangular to orbicular, the mouth of the animal forming the vertex of the triangle. In some larvæ of Hemerobii, however, the head is narrowest behind. That of the grub of a gnat noticed above (Corethra crystallina) forms a kind of sharp horn or claw, terminating the body anteriorly[227]. The contour of the head of larvæ is usually intire and unbroken; but in the caterpillars of some Lepidoptera, as the butterfly called the grand admiral (Vanessa Atalanta), the Glanville fritillary (Melitæa Cinxia), &c. it is divided into two lobes[228]. In the Brazil flat larvæ it is trilobed, each lateral lobe being divided into three smaller ones: in which circumstance it somewhat resembles the head of some subcortical Cimicidæ. Although the part we are treating of is generally without horns, yet in some tropical butterflies of the tribe of Nymphales, it is singularly armed with them. Thus Papilio Anchises is distinguished, according to Madame Merian[229], by two in the occiput, which it has the power of retracting. In the purple highflier (Apatura Iris), a British species, the[Pg 115] two lobes of the head, I am informed, terminate behind in two horns; as they do likewise in the brilliant Morpho Menelaus[230], the lobes assuming the form of a pear, and the horn representing the stalk. In a caterpillar I found amongst Mr. Francillon's larvæ, the head is bilobed, with a very long recurving subcapitate subramose spine. In Satyrus Cassiæ, the head is armed with three occipital stout spines[231]. The larva of Nymphalis Amphinome Latr. (Limenitis F.) is crowned with a coronet of eight occipital stout acute spines, the intermediate ones being the longest[232]; and that of Morpho Teucer has a similar coronet, consisting of only seven blunt rays, seemingly, rather than spines[233]. With regard to the articulation of the head with the trunk, it is generally by its whole diameter; but in some instances, only by a part of it. This is the case with one of a sphinx figured by Mad. Merian[234]; and I have another, probably belonging to the nocturnal Lepidoptera (Phalæna L.)[235]. In both these, the head is vertical and triangular; and in the latter (which is a remarkable creature, the tail itself being more like a head, and furnished with what resemble two prominent black eyes) the vertex of the triangle is considerably higher than the back of the animal. Whatever may be the clothing of the body, the head is usually naked. Sometimes, however, it is itself beset with very small simple spines, as in the butterfly of the mallow (Hesperia Malvæ); or with longer compound ones, such as are found on the rest of the body.[Pg 116] This is the case with one of a butterfly named by Rösel Papilio morsa. The most common colour of the head of larvæ, where it differs from the rest of the body, is a darker or lighter reddish brown, or piceous. This is particularly observable in those of Coleopterous insects, but it is very commonly in other tribes of the same hue. Sometimes, amongst the Lepidoptera, the head is of a different colour from the rest of the body; especially where a contrast renders it striking. I can show the caterpillar of some insect, probably of the hawk-moth tribe (Sphingidæ), from Georgia, remarkable for the length of its anal spine, in which the body is black, and the head red: another has a white head and a brown body. In the larvæ of some Lepidoptera, Coleoptera, and Diptera, the head can be wholly or nearly withdrawn within the first segment of the body. This may be readily seen in that of the common glow-worm; and that of a small gnat (Tipula replicata De Geer) withdraws it so completely that the anterior margin of that segment closes the orifice, so that the animal appears to have no head[236].—The parts of the head which require distinct consideration are, the eyes, antennæ, and the mouth: consisting of various organs, which will be specified. Some of these parts and organs are peculiar to larvæ of one order, others to those of another, and some are furnished with them all.
Eyes. The larvæ of many insects have no eyes. Those with antennæ which terminate in a lamellated clava (Scarabæus L.), and capricorn beetles also (Cerambyx L.),[Pg 117] amongst the Coleoptera, are without them, and probably several others; and amongst the Diptera, all those with a membranous or variable head. Those of the remaining orders, with the exception, perhaps, of some Hymenoptera and Lepidoptera, are furnished with these organs; and in the Coleoptera all the predaceous tribes, as well as most of those that are herbivorous or granivorous, and the Gnats and other Tipulidans (Tipulariæ Latr.) in the Diptera, are also distinguished by them. In the larvæ of the dragon-flies (Libellula L.), and other Neuroptera, they are composed of many facets as in those of the perfect insect, from which they differ chiefly in being smaller. But in the other insects of this description they are simple, and resemble those of the Arachnida, and many aptera. These simple eyes vary in their number, in different genera and tribes, from one to six on each side of the head. Thus the larva of Telephorus, and the saw-flies, has only one[237]; that of Cicindela three, the two posterior ones being large with a red pupil surrounded by a paler iris, which adds to the fierce aspect of this animal; and the anterior one very minute. Those of the tortoise-beetles also (Cassida) have three[238]; of Staphylinus, four; of Timarcha (the bloody-nosed beetle) five; of Carabus, and the Lepidoptera in general, six. In the last they are of different sizes, and generally arranged in a circle: in that of Hemerobius there are five in a circle, with one central one[239]. The appearance of these[Pg 118] globules, which are often not visible but under a powerful lens, is so different from that of the eyes of a butterfly or moth, or other perfect insect, that it has been doubted whether they actually perform the office of eyes, but without reason. They occupy the usual station of those organs, being situated in many instances upon a protuberance which appears to incase them; and seem of a construction closely analogous to that of the eyes of spiders, and the stemmata or ocelli of Hymenoptera, which have been satisfactorily proved to be organs of vision. In the larva of a moth not yet ascertained to exist in this country, Attacus Tau, and probably other species, the eyes, after the skin has been changed a few times, are no longer to be seen[240].
Antennæ. Most larvæ are provided with organs near the base of the mandibles, which from their situation and figure may be regarded as antennæ. Fabricius has asserted that the larvæ of the saw-flies (Tenthredo L.) have no antennæ; but in this he was mistaken, for though very short, they are discoverable in them, as he might have learned by consulting De Geer[241]. In the majority of Neuropterous larvæ, they almost precisely resemble those of the perfect insect. In all the rest they are very different. The antennæ of Coleopterous larvæ are usually either filiform or setaceous, consisting of four or five joints, nearly equal in length. Those of Lepidopterous larvæ are commonly conical, as are those likewise of Chrysomela and Coccinella &c. amongst the Coleoptera, and very short, composed of two or three joints, of which the last is much thinner than the first, and ends in one or[Pg 119] two hairs or bristles. These antennæ the larva has the power of protruding or retracting at pleasure. Lyonnet informs us, that the caterpillar of the great goat-moth (Cossus ligniperda) can draw the joints of its antennæ one within the other, so as nearly to conceal the whole[242]. The larva of the common gnat has two long incurved setaceous antennæ, fringed with hairs at some distance from their apex, which consist only of a single joint[243]. The greater number of Dipterous larvæ, however, all indeed except the Tipulidans (Tipulariæ Latr.), and many belonging to the Coleoptera and Hymenoptera orders (as those of Curculio, Apion, Apis, &c.), are wholly deprived of antennæ. It is a general rule, that the antennæ of larvæ are shorter than the same organs in the perfect insect, the tribe Ephemerina perhaps affording the only example in which the reverse of this takes place[244].
Mouth. All larvæ have a mouth situated in the head, by which they receive their food, and furnished with one or more instruments for the purpose of mastication and deglutition. These instruments, in all the orders except Lepidoptera, some Neuroptera and Diptera, bear a general resemblance to the same parts in the perfect insect. In larvæ of the Coleopterous, Lepidopterous, and Hymenopterous orders, we can distinguish for the most part an upper and under lip; two pairs of jaws answering to the mandibulæ and maxillæ; and two, four, or six palpi[245]: and some of these instruments may be found in most Diptera. Each of these parts require separate notice.
Upper-lip (Labrum). The mouth of almost all larvæ,[Pg 120] except some of the order Diptera, are provided with a distinct upper-lip, for retaining their food during mastication. As the construction of this part does not widely differ from that of the perfect insect, which will hereafter be more fully described, it is only necessary to observe, that it is usually a transverse moveable plate, attached posteriorly to the nasus (clypeus F.), and situated just above the mandibles[246].
Upper-jaws (Mandibulæ). The most usual figure of these, which are of a hard horny consistence[247], is that of two slightly concave, oblong, or triangular plates, often at their lower extremity of considerable thickness, and of very irregular form, the base of which is filled with powerful muscles, and planted in the side of the mouth so as to move transversely. The other extremity can be made to meet or diverge like the claws of pincers, and are divided into one or more tooth-like indentations, by means of which the food of the larva is cut[248]. This is their construction in the larvæ of all Lepidoptera, and in many of those of the other orders. They frequently, however, assume a different form, though their situation is always the same. Thus in the larvæ of the capricorn beetles (Cerambyx L.) and of other wood-boring species, they are shaped like the half of a cone, the inner sides of which, applying close to each other, form a pair of powerful grindstones, capable of comminuting the hardest[Pg 121] timber[249]. M. Cuvier has observed, with regard to the mandibulæ of those of stag-beetles (Lucanus), that besides their teeth at the extremity, they have towards their base a flat striated molary surface; so that they both cut and grind their ligneous food[250]. It seems to have escaped him, that a similar structure takes place in many perfect insects of the lamellicorn tribe, as I shall hereafter show you. In the larvæ of the water-beetles (Dytiscus L.), ant-lions (Myrmeleon L.), and lace-winged flies (Hemerobius L.), they resemble somewhat the forceps at the tail of an ear-wig, being long and incurved; and, what is more remarkable, hollow and perforated at the end, so as to serve as a channel for conveying into the larva's mouth the juices of the prey which by their aid it has seized. Reaumur even asserts, that the larva of Myrmeleon has no other entrance into its throat than through these tubular mandibles[251]. That of the rove-beetles (Staphylinus L.), and of many other Coleopterous genera, have these organs of this forcipate construction, without being perforated[252]. In the larva of the carnivorous flies, and many other Diptera, are two black incurved subulate parts, connected at the base, and capable of being protruded out of, and retracted into, the head, through the skin of which they are usually visible. As I informed you in a former letter[253], these mandibles are used for walking as well as feeding: they are parallel to each other, and are neither formed for cutting nor grinding[Pg 122] like the mandibles of other insects, but merely detach particles of food by digging into it and tearing the fibres asunder. In this operation they are probably assisted by an acutely triangular dart-like instrument of a horny substance, which in some species (Musca vomitoria) is placed between the two. In others this part is wanting. Some Dipterous larvæ have two similar mandibles, but instead of being parallel, they are placed one above the other; others (Musca domestica and meridiana) have but one such mandible, and some have no perceptible mandible of any kind. The mandibles of the larva of the crane-flies (Tipula), which are transverse and unguiform, do not act against each other, but against two other fixed, internally concave and externally convex, and dentated pieces[254].
Under-jaws (Maxillæ). These are a pair of organs, usually of a softer consistence, placed immediately under the upper-jaws; but as they are usually so formed and situated as not to have any action upon each other, it is probable that in general they rather assist in submitting the food to the action of the mandibulæ, than in the comminution of it. In Lepidopterous larvæ they appear to be conical or cylindrical (at least in that of the cossus so admirably figured by Lyonnet[255]), and to consist of two joints; which may, I imagine, be analogous to the upper and lower portions of which the maxillæ of perfect insects usually consist. The last of these joints is surmounted by two smaller jointed palpiform organs. If any part of the maxillæ can act upon each other, it is these organs or palpi; but it is evident they are not calculated for mastication, although they may assist in the retention of the[Pg 123] substance to be masticated. In a figure given by Reaumur of the under side of the head of another lepidopterous larva (Erminea Pomonella), the maxillæ consist of a single joint, and appear to be crowned by chelate palpi[256]: a circumstance which is also observable in that of a common species of stag-beetle (Lucanus parallelipipedus), the weevil of the water-hemlock (Lixus paraplecticus[257]), and other insects. In general the maxillæ of larvæ are without the lobe or lobes discoverable in those of most perfect insects, this part being usually represented by a kind of nipple, or palpiform jointed process, strictly analogous to the interior maxillary palpi of the predaceous coleoptera; but in most of the lamellicorn beetles the lobe exists in its proper form[258], as it does likewise in that of the capricorn-beetle before noticed (Callidium violaceum[259]). In the former instance, it is armed with spines or claws; but in the latter it is unarmed, and rounded at the end. In the larva of Cicindela campestris, the base of the maxilla runs in a transverse direction from the mentum, to which, as is usually the case, it is attached. From this at right angles proceeds the lobe, from the outer side of which the feeler emerges; and the inner part terminates in an unguiform joint, ending in two or three bristles. The structure in the larvæ of water-beetles (Dytiscus L.) is different, for they appear to be without maxillæ[260]; but the case really seems to be, that these organs are represented by the first joint of what M. Cuvier calls their palpi[261]; from which proceed the real palpi, the interior[Pg 124] one being very short, and consisting only of a single joint. These maxillæ of larvæ were regarded by Reaumur and other writers as parts of the under-lip, on each side of which they are situated; and indeed, as well as those in the perfect insect, they form a part of the same machine, being connected by their base with the mentum, which is part of the labium, but they are clearly analogous to the maxillæ of the imago. They are not to be found in the larvæ of many Dipterous insects, and perhaps in some species belonging to other orders. In some Neuropterous larvæ, as those of the Libellulina MacLeay, the maxillæ are of a substance quite as solid and horny as the mandibles, which in every respect they resemble[262].
Under-lip (Labium). Between the two maxillæ in the larvæ of most of the insects under consideration is a part termed by Reaumur the middle division of the under-lip, but which is in fact analogous to the whole of that organ in the imago. This organ varies in shape, being sometimes quadrangular, often conical, &c. Interiorly it is frequently connected with a more fleshy protuberance, called the tongue by Reaumur[263], and supplying the place of the ligula in the perfect insect. On each side of the apex of the under-lip is a minute feeler, and in the middle between these in the Lepidoptera and many others, is a filiform organ, which I shall call the spinneret (Fusulus), through which the larva draws the silken thread employed in fabricating its cocoon, preparatory to assuming the pupa state, and for other purposes[264]. This organ is[Pg 125] found only in those larvæ which have the power of spinning silk; that is, in all Lepidoptera, most Hymenoptera, Trichoptera, some Neuroptera, and even a Dipterous insect[265]. This tube, Lyonnet had reason to believe, is composed of longitudinal slips, alternately corneous and membranous, so as to give the insect the power of contracting its diameter, and thus making the thread thicker or smaller. There is only a single orifice at the end, which is cut obliquely, somewhat like a pen, only with less obliquity, and without a point, the opening being below, so as to be conveniently applicable to the bodies on which the larva is placed. Reaumur conceived that this spinneret had two orifices; but Lyonnet ascertained this to be a mistake, the two silk tubes uniting into one before they reach the orifice. From the contractile nature of the sides and the form of the orifice, combined with the power the insect has of moving it in every direction, results the great difference which we see in the breadth and form of the threads, some being seven or eight times as thick as others, some cylindrical, others flat, others channelled, and others of different thickness in different parts[266]. In the larvæ of many Diptera the under-lip is merely a small tubercle, which can be protruded from the insect's mouth by pressure[267].
One of the most remarkable prepensile instruments, in which the art and skill of a Divine Mechanician are singularly conspicuous, and which appears to be without a parallel in the insect world, may be seen in the under-lip of the various species of dragon-fly (Libellula L.). In[Pg 126] other larvæ this part is usually small and inconspicuous, and serves merely for retaining the food and assisting in its deglutition; but in these it is by far the largest organ of the mouth, which when closed it entirely conceals; and it not only retains but actually seizes the animal's prey, by means of a very singular pair of jaws with which it is furnished. Conceive your under-lip (to have recourse, as Reaumur on another occasion[268], to such comparison,) to be horny instead of fleshy, and to be elongated perpendicularly downwards[269], so as to wrap over your chin and extend to its bottom,—that this elongation is there expanded into a triangular convex plate[270], attached to it by a joint[271], so as to bend upwards again and fold over the face as high as the nose, concealing not only the chin and the first-mentioned elongation, but the mouth and part of the cheeks[272]: conceive, moreover, that to the end of this last-mentioned plate are fixed two other convex ones, so broad as to cover the whole nose and temples[273],—that these can open at pleasure, transversely like a pair of jaws, so as to expose the nose and mouth, and that their inner edges where they meet are cut into numerous sharp teeth or spines, or armed with one or more long and sharp claws[274]:—you will then have as accurate an idea as my powers of description can give, of the strange conformation of the under-lip in the larvæ of the tribes of Libellulina; which conceals the mouth and face precisely as I have supposed a similar construction of your lip would do yours. You will probably admit[Pg 127] that your own visage would present an appearance not very engaging while concealed by such a mask; but it would strike still more awe into the spectators, were they to see you first open the two upper jaw-like plates, which would project from each temple like the blinders of a horse; and next, having by means of the joint at your chin let down the whole apparatus and uncovered your face, employ them in seizing any food that presented itself, and conveying it to your mouth. Yet this procedure is that adopted by the larvæ provided with this strange organ. While it is at rest, it applies close to and covers the face. When the insects would make use of it, they unfold it like an arm, catch the prey at which they aim by means of the mandibuliform plates, and then partly refold it so as to hold the prey to the mouth in a convenient position for the operation of the two pairs of jaws with which they are provided. Reaumur once found one of them thus holding and devouring a large tadpole;—a sufficient proof that Swammerdam was greatly deceived in imagining earth to be the food of animals so tremendously armed and fitted for carnivorous purposes. Such an under-lip as I have described is found in the tribe of dragon-flies (Libellulina); varied, however, considerably in its figure in the different genera. In the larva of Libellula Fab., such as Libellula depressa, &c. it is of the shape above described; so exactly resembling a mask, that if Entomologists ever went to masquerades, they could not more effectually relieve the insipidity of such amusements and attract the attention of the demoiselles, than by appearing at the supper table with a mask of this construction, and serving themselves by its assistance. It would be difficult, to be sure, by mechanism to supply the place of the muscles[Pg 128] with which in the insect it is amply provided: but Merlin, or his successor, has surmounted greater obstacles. In the larva of the Fabrician Æshnæ (Libellula grandis, &c. L.), this apparatus is not convex but flat: so that, though it equally conceals the face, it does not so accurately resemble a mask; and the jaws at its apex are not convex plates, but rather two single conical teeth[275]. It is, as to its general shape, similarly constructed in Agrion Fab. (L. Virgo, &c. L.); but the first joint is more remarkably elongated, the jaws more precisely resemble jaws than in any of the rest, and are armed with three long, very sharp teeth: between them also there is a lozenge-shaped opening, through which, when the apparatus is closed, is protruded a circular sort of nipple, apparently analogous to the ligula[276]. Libellula ænea, L., which is the type of another tribe (Cordulia Leach), has a mask somewhat different from all the above, the jaws being armed with a moveable claw and an internal tooth[277]. You will admire the wisdom of this admirable contrivance, when you reflect that these larvæ are not fitted to pursue their prey with rapidity, like most predaceous animals; but that they steal upon them, as De Geer observes[278], as a cat does upon a bird, very slowly, and as if they counted their steps; and then, by a sudden evolution of this machine, take them as it were by surprise, when they think themselves safe. De Geer says, it is very difficult for other insects to elude their attacks, and that he has even seen them devour very small fishes[279].[Pg 129] As these animals are found in almost every ditch, you will doubtless lose no time in examining for yourself an instance of so singular a construction.
Feelers (Palpi). In the orders Diptera and Hymenoptera are many larvæ in which these organs have not been certainly discovered; yet Reaumur in that of a common fly (M. meridiana L.) found four retractile nipples[280] which seem analogous to them; and Latreille has observed, that below the mandibles of those of ants are four minute points, two on each side[281]: but in all other larvæ their existence is more clearly ascertained. The maxillary palpi vary in number, many having two on each maxilla and others only one. In the perfect insect the former is one of the distinguishing characters of the predaceous beetles (Entomophagi Latr.), but in the larvæ it is more widely extended; since even in the caterpillars of Lepidoptera the inner lobe of the maxilla which represents this feeler is jointed, which is precisely the case with the beetles just named. Cuvier has observed this circumstance in the larva of the stag-beetle[282]; and it belongs to many other Coleoptera that have only a pair of maxillary palpi in the perfect state. The labial palpi are always two, emerging usually one on each side from the apex of the under-lip. With regard to the form of the palpi, those of the Lepidoptera are mostly conical; in other orders they are sometimes setaceous and sometimes filiform. Their termination is generally simple, but sometimes the last joint is divided. They are for the most part very short, and the labial shorter than the[Pg 130] maxillary. The latter never exceed four joints[283], which seems the most natural number; and the former are limited to three. Both vary between these numbers, and one joint. The joints, though commonly simple, are sometimes branched. This is the case with one I met with in considerable numbers upon the Turnip, in October 1808, the second joint of the palpi of which sends forth near the apex an internal branch. In the larva of the Cossus, as Lyonnet informs us[284], the joints of the palpi are retractile, so that the whole of the organ may be nearly withdrawn.
After thus describing the head of larvæ, and its principal organs, we must next say something upon the remainder of the body, or what constitutes the
2. Trunk and Abdomen: which I shall consider under one article. These are composed of several segments or rings, to which the feet and other appendages of the body are fixed. The form of these segments, or that of their vertical section, varies considerably: in many Lepidoptera, the wire-worm, &c., it would be nearly circular; in others a greater or less segment of a circle would represent it; and in some, perhaps, it would consist of two such segments applied together. Their lower surface is generally nearly plane. Their most natural number, without the head and including the anal segment, is twelve: this they seldom exceed, and perhaps never fourteen. The three first segments are those which represent[Pg 131] the trunk of the perfect insect, and to which the six anterior legs when present are affixed. In general, they differ from the remaining segments only in being shorter, and in many cases less distinctly characterized; but in Neuropterous larvæ, those of Dytisci, and some other Coleoptera, they are longer than the succeeding ones, and pretty nearly resemble the trunk of the animal in its last state. The surface of the trunk and abdomen will be considered under a subsequent head; I shall not, therefore, describe it here. The conformation of the different segments varies but little, except of the terminal one, or tail, which in different larvæ takes various figures. In most, this part is obtuse and rounded; in others acute or acuminate; in others truncate; and in others emarginate, or with a wider sinus, and with intermediate modifications of shape which it would be endless to particularize. In some, also, it is simple and unarmed; in others beset with horns, spines, radii, and tubercles of different forms, some of which will come under future consideration. The parts connected with the trunk and abdomen which will require separate consideration, are the legs, the spiracles, and various appendages.
Legs. It may be stated generally that the larvæ of the orders Coleoptera, Lepidoptera, and Neuroptera, have legs; and that those of the orders Hymenoptera and Diptera have none. This must be understood, however, with some exceptions. Thus the larvæ of some Coleoptera, as the weevil tribes (Curculio L.) have no legs, unless we may call by that name certain fleshy tubercles besmeared with gluten, which assist them in their motions[285];[Pg 132] while those of Tenthredo and Sirex in the order Hymenoptera are furnished with these organs. At present I know no Dipterous larva that may be said to have real legs, unless we are to regard as such certain tentacula formed upon a different model from the legs of other larvæ[286]. Rösel has, I think, figured a Lepidopterous apode. No Neuropterous one has yet been discovered.
The legs of larvæ are of two kinds; either horny and composed of joints, or fleshy and without joints[287]. The first of these, as I observed in a former letter[288], are the principal instruments of locomotion, and the last are to be regarded chiefly as props and stays by which the animal keeps its long body from trailing, or by which it takes hold of surfaces; while the other legs, or where there are none, the annuli of its body, regulate its motions. The former have been commonly called true legs (pedes veri), because they are persistent, being found in the perfect insect as well as in the larva; and the latter spurious legs (pedes spurii), because they are caducous, being found in the larva only. Instead of these not very appropriate names, I shall employ for the former the simple term legs, and for the latter prolegs (propedes)[289].
The legs, when present, are always in number six, and attached by pairs to the underside of the three first segments of the trunk. They are of a horny substance, and consist usually of the same parts as those of the perfect[Pg 133] insect; namely, coxa, trochanter, femur, tibia, and tarsus, suspended to each other by membranous ligaments: these parts are less distinctly marked in some than in others. Thus in the legs of a caterpillar, or the grub of a capricorn-beetle, at first you would think there were only three or four joints besides the claw; but upon a nearer inspection, you would discover at the base of the leg the rudiments of two others[290], in the latter represented indeed by the fleshy protuberance from which the legs emerge. In the larvæ of the predaceous Coleoptera, the hip and trochanter are as conspicuous nearly as in the perfect insect; and the tarsus, which still consists of only a single joint, is armed with two claws[291]. In those of the Neuroptera order, in which all the joints are very conspicuous, the tarsi are jointed, as well as two-clawed[292]. The legs of larvæ are usually shorter than those of the perfect insect, and scarcely differ from each other in shape, for they all gradually decrease in diameter from the base to the apex. This is the most usual conformation of them in Lepidopterous, Hymenopterous, and some Coleopterous larvæ, (those of the capricorn-beetles are very short and minute, so as to be scarcely visible,) in which they are so small as to be concealed by the body of the insect[293]. In Neuropterous larvæ, however, and[Pg 134] several Coleoptera, as those of Dytiscus, Staphylinus, Coccinella, &c., they more resemble the legs of the perfect insect, the joints being more elongated, and the femoral one projecting beyond the body[294].
You will find no other than true legs in most Coleopterous, Neuropterous, and Hymenopterous larvæ. But those of the saw-flies (Tenthredo L.), and all caterpillars, have besides a number of prolegs: a few Dipterous larvæ also, are provided with some organs nearly analogous to them. These prolegs are fleshy, commonly conical or cylindrical, and sometimes retractile protuberances, usually attached by pairs to the underside of that part of the body that represents the abdomen of the future fly[295]. They vary in conformation and in number; some having but one, others as many as eighteen.
With regard to their conformation, they may be divided into two principal sections: first, those furnished with terminal claws; and secondly, those deprived of them. Each of which may be divided into smaller sections, founded on the general figure of the prolegs, and arrangement of the claws or hooks.
i. The prolegs of almost all Lepidopterous larvæ are furnished with a set of minute slender horny hooks, crotchets, or claws, of different lengths, somewhat resembling fish-hooks; which either partially or wholly surround the apex like a pallisade. By means of these claws, of which there are from forty to sixty in each proleg, a short and a long one arranged alternately, the insect is enabled to cling to smooth surfaces, to grasp the smallest twigs to which the legs could not possibly adhere: a circumstance which the flexible nature of the prolegs greatly facilitates[296]. Claws nearly similar are found on the prolegs of some Dipterous larvæ[297], but not in any of those of the other orders. These last, however, are seldom either so numerous, or arranged in the same manner, as in caterpillars. When the sole of the foot is open, the claws with which it is more or less surrounded are turned outwards, and are in a situation to lay hold of any surface; but when the animal wishes to let go its hold, it begins to draw in the skin of the sole,[Pg 136] and in proportion as this is retracted, the claws turn their points inwards, so as not to impede its motion[298].
The prolegs with claws may be further divided into four different kinds.
1. In the larvæ of the great majority of butterflies and moths they assume the form of a truncated cone, the lower and smaller end of which is expanded into a semicircular or subtriangular plate, having the inner half of its circumference beset with the claws above mentioned; and, from its great power of dilating and contracting, admirably adapted for performing the offices of a foot. Jungius calls these legs pedes elephantini[299]; and the term is not altogether inapplicable, since they exhibit considerable resemblance to the clumsy but accommodating leg and foot of the gigantic animal he alludes to.
2. The larvæ of many minute moths, particularly of the Fabrician genera Tortrix and Tinea—those which live in convoluted leaves, the interior of fruits, &c., as well as the Cossus, and some other large moths,—have their prolegs of a form not very unlike those of the preceding class, but shorter, and without any terminal expansion; the apex, moreover, is wholly, instead of half, surrounded with claws[300]; the additional provision of which, together with a centrical kind of nipple capable of being protruded or retracted, in some measure, though imperfectly, supplies the place of the more flexible plate-like expansion present in the first class.
3. The third class is composed of a very few Lepidopterous larvæ which have their prolegs very thick and conical at the base, but afterwards remarkably slender,[Pg 137] long, and cylindrical, so as exactly to assume the shape of a wooden leg[301]. These, as in the first class, are expanded at the end into a flat plate: but this is wholly circular, is surrounded with claws, and has also in the middle a retractile nipple, as in the preceding class. In Cossus, at least in an American species (Cossus Robiniæ), described by Professor Peck[302], the anal prolegs have the claws only on their exterior half.
4. The remaining description of unguiferous prolegs, if they may not rather be deemed a kind of tentacula, are those of certain Diptera, provided with no true legs; which differ from the three preceding classes, either in their shape, or the arrangement of their claws. In one kind of those remarkable larvæ, which from their long respiratory anal tubes Reaumur denominates "rat-tailed," that of Elophilus pendulus, there are fourteen of these prolegs, affixed by pairs to the ventral segments, the twelve posterior ones of which are subconical, and truncate at the apex, which is surrounded with two circles of very minute claws, those of the inner being much more numerous and shorter than those of the exterior circle; while the anterior pair terminate in a flat expansion, and in shape almost exactly resemble those of a mole[303]. The prolegs of the larvæ of a kind of gnat called by De Geer Tipula amphibia, and of Syrphus mystaceus F., (Musca plumata De Geer,) are nearly of a similar construction, but in the last are armed with three claws only[304]. Long moveable claws also distinguish the singular[Pg 138] prolegs before described[305] of another gnat (Tanypus maculatus Meig., Tipula De Geer). The case-worms (Trichoptera K.) and some others, have two prolegs at the anus, each furnished with a single claw[306].
ii. The prolegs deprived of claws are found in the larva of the Hymenopterous tribe of saw-flies (Tenthredo L.), in those of some Lepidoptera (Hepialus F. &c.), and in some few Coleopterous and Dipterous genera. Those of the former are of the shape of a truncated cone, and resemble the second class of unguiculate prolegs, except in the defect of claws. In the latter they are a mere retractile nipple-like protuberance, in some species so small as scarcely to be perceptible. In all they aid in progressive motion; but it is by laying hold of surfaces, and so enabling the body more readily to push itself forward by annular contraction and dilatation, and not by taking steps, of which all prolegs are incapable: to assist in this purpose the protuberance sometimes secretes a gluten[307], which supplies the place of claws. Some larvæ have the power of voluntarily dilating certain portions of the underside of their body, so as to assume nearly the shape and to perform the functions of prolegs. In a Coleopterous (?) subcortical one from Brazil, before alluded to, there are four round and nearly flat areas in each ventral segment of the abdomen, but the last very little raised above the surface, and rough, somewhat like a file; and besides these, the base of the anal segment has ten of these little rough spaces, but of a different shape, being nearly linear, placed in a double series, five on each side. Doubtless these may be regarded as a kind of prolegs,[Pg 139] which enable the animal to push itself along between the bark and the wood[308].
In considering, in the next place, the number and situation of the prolegs, it will contribute to distinctness to advert to these circumstances as they occur in the different orders furnished with these organs.
To begin with the Lepidoptera.—Lepidopterous larvæ have either ten, eight, six, or two prolegs, seldom more[309], and never fewer. Of these, with a very few exceptions, two are attached to the last or anal, and the rest, when present, to one or more of the sixth, seventh, eighth, and ninth segments of the body: none are ever found on the fourth, fifth, tenth, or eleventh segments.
1. Where ten prolegs are present, as is the case in by far the greatest proportion of Lepidopterous larvæ, there is constantly an anal pair, and a pair on each of the four intermediate segments just mentioned.
2. In caterpillars, which like those of a few species of the genera Sphinx, Pyralis, and of the Bombycidæ, &c. have eight legs, they are placed in three different ways. In those which have an anal pair, the remaining six are in some fixed to the sixth, seventh, and eighth; in others, to the seventh, eighth, and ninth segments. In those which, like Cerura Vinula, and several other species of the same family, have no anal prolegs; the whole eight emerge from the sixth, seventh, eighth, and ninth segments.
3. The Hemigeometers, as Noctua Gamma, &c. have[Pg 140] only six legs: namely, an anal pair, and two ventral ones, situated on the eighth and ninth segments.
4. The larvæ of the Geometers (Geometræ F.) have but four prolegs; of which two are anal, and two spring from the ninth segment. It should be observed, however, that the larvæ of Hemigeometers, and even of some of those that have ten prolegs, where the four anterior ones are much shorter than the rest, move in the same way as the Geometers. This even prevails in a few where these organs are all of equal length.
5. Many of the larvæ of Tinea L. which live in the interior of fruits, seeds, &c., have but one pair of prolegs, which are attached to the anal segment.
6. The larvæ of Haworth's genus Apoda (Hepialus Testudo and Asellus F.), remarkable for their slug-like shape and appearance, move by the aid of two lateral longitudinal pustule-like protuberances, which leave a trace of a gummy slime in their course.
Hymenoptera.—The larvæ of the different tribes of Tenthredo L., almost the only Hymenopterous insects in which prolegs are present, have a variable number of these organs; some sixteen, as the saw-fly of the willow (T. lutea L.), and this is the most numerous tribe of them, including the modern genera, Cimbex F., Pterophorus, &c. Others have fourteen, as that of the cherry (T. cerasi L.); and many others with only nine joints to their antennæ. A third class have only twelve, as that of the rose (T. Rosæ L.), but this contains but few species. The last class contains those that have no prolegs at all, but only the six horny ones appended to the trunk. Of this tribe, the caterpillars of which have a very different aspect from the preceding, are those of the genus Lyda F. (T. crythrocephala[Pg 141] L.)[310]. Two of the prolegs are anal, and the rest intermediate, and none are furnished with claws. This circumstance, in conjunction with the greater number of prolegs, except in the case of Lyda, will always serve as a mark to distinguish these fausses chenilles, as the French call the larvæ of saw-flies, from true caterpillars. The dorsal prolegs of a species of Cynips described by Reaumur have been before noticed.
Coleoptera.—The larvæ of insects of this order are so little known or attended to, that no very accurate generalization of them in this respect is practicable. Many of them, in addition to their six horny legs, have a proleg at the anus; which in many cases appears to be the last segment of the abdomen, forming an obtuse angle with the remainder of it, so as to support that part of the body, and prevent it from trailing; and in some instances, as in Chrysomela Populi, a common beetle, secreting a slimy matter to fix itself[311]. In the larvæ of Staphylinidæ this proleg is very long and cylindrical; in that of Cicindela it is shorter, and in shape a truncated cone rather compressed; it is very short, also, in those of the Silphæ that I have seen. In the wire-worm (Elater Segetum) it is a minute retractile tubercle, placed in a nearly semicircular space, shut in by the last dorsal segment, which becomes also ventral at the anus. This space is in fact[Pg 142] the last ventral segment. This seems characteristic of the genus[312]. From the underside of the body of the common meal-worm (Tenebrio Molitor), at the junction of the two last segments, when the animal walks, there issues a fleshy part, furnished below with two rather hard, long, and moveable pediform pieces, which the animal uses in walking[313]. In the larva of another beetle, whose ravages have been before noticed, under the name of the cadelle[314] (Trogosita mauritanica), a pair of prolegs are said to be found under the anal segment; and in that of the bloody-nose beetle (Timarcha tenebricosa), that segment is bifid. That of the weevil of the common water-hemlock (Lixus paraplecticus F.) exhibits a singular anomaly: prolegs occupy the usual station of the true legs, being attached to the three segments representing the trunk[315]. This insect, however, does not appear to use them in moving. A pair in each of the twelve segments of the body are found in the grub of another weevil (Hypera Rumicis Germ.), the nine last pair being the shortest, which all assist the insect in walking[316]. But the greatest number of prolegs is to be found in the Brazil subcortical larva lately mentioned. Besides the six horny legs of the trunk, this remarkable animal has four prolegs on each of the seven intermediate abdominal segments, and five on each side of the base of the last, making the whole number of prolegs, if so they may be called, amount to forty-four: a far greater number than is to be found in any larva at present known. When I wrote to you upon the motions of insects, I informed[Pg 143] you that some larvæ moved by means of legs upon their back[317], but I was not then aware that any were furnished with them both on the back and the belly at the same time. By the kindness of Mr. Joseph Sparshall of Norwich, a very ardent and indefatigable entomologist, I am in possession of the larva of Rhagium fasciatum, a timber-feeding beetle. This animal on the ten intermediate segments of the underside of the body, which in the centre form a fleshy protuberance, has on it a double series of rasps, as it were, consisting each of two rows of oblique oblong prominences; and on the seven intermediate dorsal segments there are also in the centre seven rasps of three or four rows each, of similar prominences: so that this animal at the same time can push itself along both by dorsal and ventral prolegs. It is worthy of observation, that a pair of these rasps is between the second and third pair of true legs.
Diptera.—The larva of a little gnat, Tipula stercoraria De Geer[318] (Chironomus Meig.?), drags itself along by the assistance of a single tubercle, placed on the underside of the first segment of the body, which the animal has the power of lengthening or contracting[319]. That of another beautiful Chironomus (C. plumosus), remarkable for the feathered antennæ of the male[320], has two short prolegs, or pediform but not retractile tentacula in the same situation[321]. Others, as that of Tanypus maculatus,[Pg 144] &c. have two pairs, one attached to the anal and the other to the first segment[322]. Tipula amphibia De Geer in this state has ten prolegs, placed by pairs on the fourth, fifth, eighth, ninth, and tenth dorsal segments[323]; and Scæva Pyrastri F., one of the aphidivorous flies, has not fewer than forty-two, arranged in a sextuple series, seven in each row[324].
It may not be useless to close this long description of the legs of larvæ with a tabular view of them, founded chiefly upon these organs; which afford very obvious marks of distinction.
I should next say something upon the spiracles, or breathing-pores, or any other external apparatus for the purpose of respiration, in larvæ; but I think it will be best to reserve the consideration of these for a subsequent Letter. We will therefore conclude this detailed description of their parts in their first state, with some account of their other.
iii. Appendages. The generality of larvæ have no other external organs than those already described; but in several of them we observe various kinds of retractile ones and others—protuberances—horn-like processes—rays, &c.; which, though not properly coming either under any of the above parts, or under the clothing of these animals, yet require to be noticed. Upon these I shall now enlarge a little.
You must have observed upon the back of the last segment but one of the caterpillar of the silk-worm a horn-like process, rising at first nearly perpendicularly, and then bending forward. A similar horn, though confined in the genus Bombyx to the silk-worm and a few others, if we may believe Madame Merian, who, however, often makes great mistakes, is found in the beautiful caterpillar of one of the largest and finest moths that we know (Erebus Strix[325]), the glory of the Noctuidæ, and in most of those of the hawk-moths (Sphinx F.) [S. Porcellus, Vitis, and a few others excepted; in some of which, as S. Labruscæ, &c., this anal horn is replaced by a gibbosity, and in others, as S. Œnotheræ, by a callous eye-like plate[326]] in the same situation, but much longer[327], and[Pg 146] commonly curving backwards over the tail[328]. Sometimes, however, as in S. ocellata and S. Stellatarum, it is perfectly straight. These organs towards the apex are horny, and often end in a sharp point; nearer the base they are fleshy. They are without any true joint[329], yet the insect can elevate or depress them at pleasure. Under a lens, they usually appear covered with spinous eminences, arranged like scales. The use of these horns is quite unknown: Goedart fancies that they secrete a potent poison, and are intended as instruments of defence; but both suppositions are altogether unfounded. It has been remarked, that the body of those caterpillars which have these horns, is firmer, and yields less to the touch than that of those which have no such appendages[330]. The larva of a small timber-devouring beetle (Lymexylon dermestoides F.) has, like the above caterpillars, a long horn, and in the same situation: it has also a singular protuberance on the first segment[331]. Upon some other caterpillars, as in Bombyx Stigma F., a singular pair of horn-like appendages arises from the back of the second segment of the body, excluding the head. In a tawny-coloured one from Georgia, with a transverse row of short black spines on each segment, these horns are half an inch long, black, covered with spinous eminences, rather thickest at the base, and terminate in a little knob. They appear to articulate with the body at the lower extremity. I have another species, black, with narrow longitudinal yellow[Pg 147] stripes, in which these horns are of equal thickness at base and apex, but with the same terminal knob. Danais Archippus has a pair of tentacula at the head, and another pair, but shorter, at the tail; and D. Gylippus has, besides these, two in the middle of the body[332].
We are equally ignorant of the use of the upright horn found upon the back of the fourth segment in the larva of some moths (Noctua Psi, and tridens F.) which is of a construction quite different from that of those last described. It is cylindrical, slightly thinner at the apex, which is obtuse, fleshy, incapable of motion, of a black colour, and about two lines long. On the same segment, also, in the case-worms (Trichoptera K.) are three fleshy conical eminences, which the animal can inflate or depress, so that they sometimes totally disappear, and then in an instant swell out again. When retracted, they form a tunnel-shaped cavity, varying in depth[333]. Reaumur conjectured that these eminences were connected with respiration, and one circumstance seems in favour of this conjecture, that this segment has not the respiratory threads observable in the subsequent ones. Latreille mentions certain fleshy naked eminences placed upon the ninth and tenth segments of some hairy caterpillars, which, like those just mentioned, the animal can elevate more or less. They are often little cones; but when it would shorten them, the summit is drawn in, and a tunnel appears where before there was a pyramid[334].
In a former Letter I gave you a short account of the[Pg 148] remarkable Y-shaped, as it should seem, scent-organs (Osmateria) of the beautiful caterpillar of the swallow-tailed butterfly (Papilio Machaon L.), and others of the Equites[335]; I will now speak of them more fully. That found in the former is situated at the anterior margin at the back of the first segment, close to the head, from which at first view it seems to proceed. At the bottom it is simple, but divides towards the middle, like the letter Y, into two forks, of a fleshy substance[336], which it can lengthen, as a snail does its horns, to five times their ordinary extent, or retract them within the stalk, so as wholly to conceal them. Sometimes it protrudes one fork, keeping the other retracted; and often withdraws the whole apparatus for hours together under the skin, and its place is only marked by two tawny-coloured dots, so that an ordinary spectator would not suspect the existence of such an instrument[337]. Unfortunately this larva is rare in this country, so that I can scarcely flatter you with the hope of seeing this curious organ in a living specimen[338], unless you choose to import a parcel of its eggs from the south of Europe, where it is common. This you will think rather a wild proposition; but why should not Entomologists import the eggs of rare insects, as well as botanists the seeds of rare plants? But if you will be satisfied with the dissection of a dead specimen, I have several,[Pg 149] done by the ingenious Mr. Abbott of Georgia, in which this part is well exhibited[339].
Another small caterpillar, as it should seem, of a geometer, prepared by the same gentleman, exhibits a pair of similar horns on the fifth and sixth segments: in these the common base from which the fork proceeds is very short and wide, and each branch grows gradually more slender from the base to the apex, where it is involute. Whether these are retractile, or whether they correspond with those of P. Machaon in their nature and use, cannot be ascertained from a dead specimen: as they belong to a larva of a quite different tribe of Lepidoptera, the probability is, that they essentially differ. Two globose retractile vesicles issue from the ninth and tenth segments of those of Arctia chrysorrhea, &c.[340]
A great number of Lepidopterous larvæ, particularly those which are smooth and of a moderate size, have between the under-lip and fore-legs a slender transverse opening, containing a teat-like protuberance of the same construction as the furcate horn of the caterpillar of the beautiful mountain-butterfly, Parnassius Apollo; and, like that, can either be wholly retracted and concealed, or by pressure be extended to the length of one of the legs. In some larvæ this part is of a subhemispherical figure, generally single, but sometimes double. It is commonly, however, more slender and conical; and when of this shape, it is sometimes quadruple[341]. The use of this part is not very clearly known: some have supposed it to be a second spinneret, and to be of use in fabricating the cocoon; but[Pg 150] it is more probable that it secretes some other kind of fluid, and is connected with defence.
The singular organ in a similar situation, evidently for that purpose, with which the puss-moth endeavours to annoy its assailants, has been described in a former Letter, to which I refer you[342]. Bonnet, who was the first that discovered this organ, ascertained that it might be cut off without injury to either larva or imago. He also remarked in a caterpillar found in the wild succory (Cichorium Intybus) another short, black, needle-shaped organ between the conical part just described and the under-lip[343]. De Geer mentions a remarkable fleshy horn-like style, which issues from the lower side of the first segment, between the head and the legs of the case-worms (Trichoptera): he does not describe it as retractile, or it might be regarded as analogous to those of Lepidoptera similarly situated, that I have just noticed[344]. In that of the emperor-moth (Saturnia Pavonia), there are perforated tubercles, which when the animal is molested spirt forth a transparent fluid[345].
The horn-like appendage of the puss-moth (Cerura Vinula) is situated at the tail of the insect, and is composed of two distinct cylindrical diverging branches, each about four lines long, not united at the base. Each of these is hollow, and includes a smaller cylindrical piece, which can be protruded at pleasure, and withdrawn again, as a pencil within its case; or, rather, as the horns of a snail. The two outer horns are tolerably firm, moveable at their base, and beset with black spines; the interior tentacula are fleshy, moveable in every direction, and in full-grown[Pg 151] larvæ of a rose colour. The animal seldom protrudes them, unless in some way disturbed; and frequently it approximates the two outer cases so closely that they resemble a single horn. It appears to use these inner horns, when protruded, as a kind of whip to drive away the flies, especially the Ichneumons, that alight upon its body. When touched in any place, it will unsheath one of them, and sometimes both, and with them strike the place where it is incommoded[346]. A similar organ is found in some other Bombycidæ, as B. Tau and Furcula F. Reaumur mentions a caterpillar that to this kind of tail added the resemblance of two ears, or two cylindrical bodies, terminating in a point, which emerged from the first segment behind the head[347]. In another observed by the same author, the legs were replaced by a single horn, but which did not appear to send forth an internal one: from the back of its fourth segment also emerged a single conical or pyramidal fleshy eminence or cleft, terminating in two points[348]. Some of the tropical butterflies also, as may be seen in the figures of Madame Merian, have two diverging anal horns instead of anal prolegs; but it does not appear that they incase tentacula[349]. Wherever these caudal horns are found, the above prolegs are wanting[350].[Pg 152] Two conical anal horns also distinguish the caterpillar of one of the moths called Prominents, Notodonta camelina; but these are not terminal, but on the back of the last segment but one[351]. In that of another British moth, N. ziczac F., there are three dorsal prominences, one near the anus, and two more in the middle[352]. Some Geometers (G. fuliginosa, &c.) have two erect horns on the eleventh segment, and others (G. syringaria, &c.) two recurved ones on the eighth[353]. I must not here omit to mention the curious hooks emerging from two tubercles on the back of the eighth segment of the ferocious larva of that beautiful tiger-beetle, the Cicindela campestris L., not uncommon on warm sunny banks. This animal with incessant labour, as we are informed by M. Desmarets, digs a cylindrical burrow, to the enormous depth, the size of the animal considered, of eighteen inches. To effect this, it carries out small masses of earth upon its large concave head; and having often occasion to rest in ascending this height, by means of these hooks[354] it fixes itself to the sides of its burrow, and, having finally arrived at its mouth, casts off its burthen. When these insects lie in wait for their prey, their head, probably in conjunction with the first segment of the body, accurately stops the mouth of the burrow, so as to form an exact level with the surrounding soil; and thus careless insects, walking over it without perceiving the snare, are seized in a moment and devoured[355].
Another kind of appendage, which is found in some[Pg 153] larvæ, is the organ employed by them to carry the excrement; with which, instead of letting it fall to the ground, they form a kind of umbrella to shelter and probably conceal them. All the tortoise-beetles (Cassida L.) have instruments for this purpose, as well as an Indian genus (Imatidium Latr.) very nearly related to them. This instrument is a kind of fork, half as long as the body, consisting of two branches, growing gradually smaller from the base to the summit, where they terminate in a very fine point, of a substance rather horny, and attached to the body near the anal orifice. They are armed on the outside with short spines, from the base for about a third of their length. When this fork, as it usually is, is laid parallel to the back, with its points towards the head, the anal aperture points the same way. When the animal walks, the fork points the other way, and is in the same line with the body, and the anus assumes a prone position[356].
The larvæ of a genus of flies (Volucella Geoffr.) remarkable for inhabiting the nests of humble bees, are distinguished on their upper side by six long, diverging, pointed, membranous radii; placed in a semicircle round the anus[357]: what the particular use of these organs may be, has not been conjectured. Another in my collection has only four upper radii, but below the anus are two fleshy filiform tentacula. One of a Tipulidan described by Reaumur, has also four upper teeth; but instead of two subanal tentacula, has six[358]. The singular larva of another of this tribe (Chironomus plumosus) has on the[Pg 154] two last segments four long, fleshy, filiform, flexible tentacula, often interlaced with each other; which, according to the same illustrious author, are used by the animal to fix its caudal extremity, like the geometers, that the other end may be at liberty. Besides these organs round the anus, it has also four other oval ones, of uncertain use: not to mention the two prolegs, which M. Latreille thinks are air-tubes[359]. Jointed anal organs are observable in other larvæ: those in that of a saw-fly described by De Geer (Lyda F.) consist of three joints[360]; in that of Hister cadaverinus, a carnivorous beetle, of two[361]. The larva as well as the pupa and imago of Ephemera is furnished with three long diverging multiarticulate tails, which are probably useful as a kind of rudder to assist and direct their motions. That of the smaller dragon flies (Agrion F.) is furnished with three long vertical laminæ, by moving which, as fish do their tails, from side to side, the animal makes its way in the water[362]. That singular one, also, with a hooked head, figured by Reaumur, has a single swimming lamina, or fin, shaped like a fan, and placed in a vertical position under the tail[363].
The whole circumference of the body in some coleopterous larvæ,—for instance, in that of the tortoise-beetle lately mentioned,—is surrounded with appendages like rays. These are sometimes simple, rough with very short spinous points[364]; but I have a dipterous larva, in[Pg 155] which these radii themselves are beautifully pinnated by a fringe of longish spines on each side. Reaumur has described the grub of a beetle, the genus of which is uncertain, and which feeds upon the larva of Aleyrodes Proletella, whose body is margined on each side by eight triangular fleshy mammular processes, terminating each in a bristle, which give it a remarkable aspect[365]. The curious scent-organs with which the larva of Chrysomela Populi is fringed have been before fully described; and therefore I shall only mention them here[366].
In the larvæ of the lace-winged flies (Hemerobius), and ant-lions (Myrmeleon), the anus is furnished with a small fleshy retractile cylinder, from which proceeds the silken thread that forms the cocoon inclosing the pupa[367]. Providence has many different ways of performing the same operation. From the structure of the oral organs of these animals, the silk could not conveniently be furnished by the mouth; the Allwise Creator has therefore instructed and fitted them to render it by a spinneret at the other extremity of the body.
The respiratory anal appendages of many Dipterous larvæ will be fully described in a subsequent Letter: I shall therefore now only further observe upon this subject, that although there is seldom any alteration in the form of these appendages &c. in the same species, the caterpillars of two moths (Cerura Vinula and Attacus Tau), however, are exceptions. The former, when young, has two hairy projecting ear-like protuberances, which it entirely loses, as I have myself observed, before it assumes the pupa; and the latter, in like manner, after its third[Pg 156] change of skin, is deprived of its bent thorn-like points which attend it when young[368]. It is remarkable that these last larvæ, when just excluded from the egg, are also entirely destitute of these appendages; they soon, however, appear, from slight elevations which mark their situation, and rapidly acquire their usual form[369]. Changes of a similar kind, hitherto unobserved, may probably take place in other species.
iii. Figure. I am next to consider the general figure or shape of larvæ. All of them, with but few exceptions[370], agree in having a body more or less constricted at intervals into a series of rings or segments; usually in number, twelve; often nearly equal in length, but sometimes in this respect very dissimilar[371]. The general outline or shape of the body is extremely various: most frequently it approaches to cylindrical, as in most of the caterpillars of Lepidoptera, and of the Hymenopterous tribe of saw-flies (Tenthredo L.). The next most common figure is that more or less oblong or oval one, sometimes approaching to conical, found in many of the larvæ usually called grubs; such as those of the weevil (Curculio L.) and[Pg 157] of the capricorn (Cerambyx L.), and other coleopterous tribes; of bees, and all Hymenopterous insects but the saw-flies; and also of a large number of flies (Diptera). In some the figure approaches to fusiform, as in most of the moths of the Fabrician genus Lithosia. In others, as in those of the water-beetles (Dytiscus, &c.), it approaches to an obovate shape, being widest towards the head, and terminating in a point at the anus. In others, again, it is linear; an example of which is that of Staphylinus. Some are convex, and others gibbous, above, and flat underneath; as those of Silpha, Chrysomela and many other beetles. Others are flat, both above and below, and depressed like a leaf; a remarkable instance of which has been before noticed[372]. Some are very long, as those of most Lepidoptera; others very short, as that of the ant-lion (Myrmeleon). Many other peculiarities of form in individuals might be instanced; but a dry enumeration of these would be of no great use to you. They can only be advantageously learned by the study of good figures, and by watching the actual metamorphosis of the singularly-formed larvæ that you meet with.
Instead, therefore, of any further specification of individual forms, I shall now endeavour to give you, as far as my own knowledge of them and the information I can collect from other sources will enable me, a larger and more general view of the kinds of larvæ; for analytical inquiries lose half their value and importance unless we proceed to apply them synthetically, by forming, if possible, into groups the objects with which we are individually acquainted.
Partial attempts at a synthetical arrangement with regard[Pg 158] to the larvæ of Lepidoptera and the saw-flies (Tenthredo L.) have been made both by Reaumur and De Geer. M. Latreille also has recently given a Tableau méthodique et général of articulated animals furnished with jointed legs, considered in their first state[373]. The former of these is chiefly founded upon the number of the prolegs, and the latter upon the metamorphosis, prolegs, habits, head, and parts of the mouth, without any other notice of the configuration. Mr. Wm. MacLeay, who, though young in years, is old in science and critical acumen, has started a perfectly new hypothesis upon this subject. In the progress of his inquiries into the natural arrangement of animals, particularly of insects in the Linnean sense, he has been the first to observe, that the relation which organized objects bear to each other is of two kinds; one of real affinity, and the other only of analogy, or resemblance. This important distinction, upon which I shall enlarge in a future Letter, when I come to treat of Systems of Entomology, he has applied, in a way quite original, to larvæ in general, but more particularly to those of the Coleoptera order. For the basis of his system he assumes a relation of analogy between the larvæ of Insects that in the progress of their metamorphosis assume wings, and those that do not, which form his class Ametabola, so that the prototypes of the former shall be found amongst the latter[374]. But though Mr. MacLeay appears to consider the analogy between these two as primary, he extends it in a secondary sense to the Crustacea, at least in several instances[375]. Upon this occasion[Pg 159] he very judiciously remarks, that "in terming larvæ Chilognathiformes or Chilopodiformes, it is not meant that they are Scolopendræ or Iuli, or even near to them in affinity; but only that they are so constructed that certain analogical circumstances attending them strongly remind us of these Ametabola[376]." This remark you will bear in mind while I am treating of this subject. It should seem from another part of the same paragraph, that the comparison which our learned Physiologist recommends, is between the young of the Ametabola and the larvæ of the corresponding groups of Coleoptera. This must be understood to refer chiefly to the young of the Chilopoda and Chilognatha, since they approach nearer to them in that state, having then only six legs; but the rest of the Ametabola should certainly be brought to this comparison in their adult state: and even the two former orders in that state more strongly resemble numerous coleopterous larvæ, than when they are young and much shorter. I before called your attention to the remarkable circumstance that contrasts very many larvæ of Hexapod insects that become winged in their perfect state with adult Myriapoda: namely, that in one the progress to this state is by losing their prolegs and shortening their body; while in the other, the reverse of this takes place, numerous prolegs and additional segments being gained before they arrive at maturity[377]. As the multiplication of organs is a sign of imperfection, it may be affirmed of the former of these tribes, that their progress is towards greater perfection; while that of the other may be called a degradation. As larvæ may be regarded as a stepping-stone by which approach is made[Pg 160] from the apterous to the winged tribes of Insects, it seems most consistent with general analogy that each should connect with the other in that state in which the resemblance is greatest. Now the Myriapoda resemble larvæ, as we have just seen, most when in their adult state; therefore the comparison should be between larvæ and adult Myriapoda.
Mr. MacLeay divides coleopterous larvæ into five tribes thus characterized:—
1. A carnivorous hexapod larva, with an elongate linear flattened body, having a large head armed with two sharp falciform mandibles, and furnished with six granular eyes on each side. This kind he denominates Chilopodiform, as having for its type in the Ametabola, Scolopendra L. The examples he gives are Carabus and Dytiscus.
2. A herbivorous hexapod larva, with a long and almost cylindrical body, so fashioned that the posterior extremity being curved under the breast, the animal when at rest necessarily lies like an Iulus on its side. This tribe he denominates Chilognathiform, from Iulus L. His examples are, the larvæ of Petalocerous insects, as Scarabæus L., Lucanus L. &c.
3. An apod larva, having scarcely the rudiments of antennæ, but which is furnished instead of feet with fat fleshy tubercles; which, when continued along the back and belly, give the animal a facility of moving in whatever way it may be placed. These he denominates Vermiform, from certain of the Vermes intestina and Mollusca of Linné which he has associated with his Annulosa[378]. His examples are, Curculio L. and Cerambyx L.
4. A hexapod and distinctly antenniferous larva, with a[Pg 161] subovate rather conical body, of which the second segment is longer and of a different form from the others, so as to give the appearance of a thorax. His denomination for these is Anopluriform, from Pediculus L., forming Dr. Leach's Anoplura. His examples are, Coccinella and Chrysomela L.
5. A hexapod antenniferous larva of an oblong form, having like the former vestiges of a thorax, besides two or more articulated or inarticulated setaceous or corneous appendages to the last segment of the abdomen. This tribe he calls Thysanuriform, from Lepisma and Podura L., forming M. Latreille's order Thysanura. His example is Meloe with a note of interrogation[379].
The system here stated, of naming and characterizing larvæ from the resemblance and analogy, in many cases very striking, that they bear to the apterous tribes, is a very happy and original one, and does its author great credit; yet I think in some instances, as I shall soon have occasion to point out to you, the application of it is not so happy as the first idea. But this is always the case when a new law of nature is discovered; the proper application of it is gradually developed, and it does not at all detract from the merit of the first discoverer, that all the bearings of such law do not strike him as it were intuitively.
Having, however, got the vantage-ground afforded by this discovery of my friend, let us see if by standing upon it we cannot get a tolerable generalization of the larvæ of all orders of insects that undergo a metamorphosis. But first I must observe, that as in the perfect animals, so in their larvæ, the different groups are connected by certain transition species, exhibiting characters common to two or more of them; and likewise that in many cases, which you will see as we proceed, the analogy is as strong or stronger between them and the Crustacea (and in a few instances Arachnida, and even Mollusca) than the Ametabola. My denominations, therefore, will be taken from those tribes where the analogy appears to me the most striking, and not from the Ametabola alone.
I shall begin by drawing up for you a list of the Primary forms that I seem to have observed, and their characters; and then going through the orders, shall give you the examples of each, with such observations upon them as the case may require.
Primary Forms of Larvæ.
| APTERA. | ARACHNIDA. | CRUSTACEA. | MOLLUSCA. |
| Anopluriform. | Araneidiform. | Isopodiform. |
Limaciform. |
| Thysanuriform. | Onisciform. |
||
| Chilopodiform. | Idoteiform. |
||
| Chilognathiform. | Amphipodiform. |
ANNELIDA. | |
| Stomapodiform. |
Vermiform. | ||
| Decapodiform. |
|||
| Branchiopodiform. |
Characters.
1. Anopluriform. Carnivorous; hexapod; antenniferous: with a shortish oblong depressed body, and distinct[Pg 163] thoracic shield. Example: Psocus, Coccinella, most Hemiptera[380].
2. Thysanuriform. Polyphagous; hexapod; antenniferous: body with segments of trunk distinctly marked; anus often furnished with setæ or mucro. Ex. Meloe[381]? Thrips, Aphis.
3. Chilopodiform. Carnivorous; subhexapod; antenniferous: body depressed, elongate, linear, with falcate acute mandibles, a distinct thoracic shield, and an anal proleg. Ex. Cicindela[382]? Carabus L.
4. Chilognathiform. Herbivorous; body subcylindrical, elongate, linear; no thoracic shield; often many prolegs, sometimes a retractile one, and sometimes none.—Ex. Elater, Petalocera, most Lepidoptera, Tenthredo L.[383]
5. Vermiform. Polyphagous; apod or hexapod: with very short legs; antennæ nearly obsolete; body fleshy, plicate, with sides often plicato-papillose;[Pg 164] no distinct thoracic shield. Ex. Curculio L., Cerambyx L., Musca L., and many other Diptera[384].
6. Araneidiform. Carnivorous; hexapod: body very short; mandibles long, suctorious; animal lying in wait for its prey in a pitfall it has prepared; motion retrogressive. Ex. Myrmeleon L.[385] Cicindela?
7. Isopodiform. Saprophagous; hexapod; antenniferous, with longer antennæ: body oblong; thoracic shield distinct; anus styliferous or laminiferous. Ex. Blatta L. Silpha L.?
8. Onisciform. Herbivorous; polypod; antenniferous: body short, oblong, depressed, margined. Ex. Erycina, Lycæna, in the Lepidoptera, and some species of Tenthredo L.[386]
9. Idoteiform. Subcortical; hexapod; antennæ obsolete: body much depressed, with the last segment elongate, terminating in three or more mucros. Ex. Larva from Brazil. Perfect insect at present unknown.
I have placed this larva, which was described above[387], amongst crustaceous forms, because of the remarkable resemblance which the last segment of the body bears to that of the Idoteidæ; but I do it with considerable hesitation, since in other respects its type seems to be in the Ametabola. In its want of antennæ, very short legs, and ventral asperities, it resembles some of the Vermiform larvæ; in its small head, distinct thoracic shield, and oblong shape, it approaches the Anopluriform; and in its[Pg 165] very depressed body, but not at all in other respects, the Chilopodiform. At any rate, it appears of a primary Type.
10. Læmodipodiform. Herbivorous; hexapod; antenniferous, with long antennæ: body elongate, subcylindrical; second segment of the trunk the longest; anterior pair of legs distant from the other two. Ex. Phasma.
11. Amphipodiform. Herbivorous; hexapod; antenniferous, with long antennæ: body shortish, compressed, saltatorious. Ex. Gryllus L.[388]
12. Stomapodiform. Carnivorous or saprophagous; hexapod; antenniferous, with long antennæ: body elongate, subdepressed, with raptorious hands, and abdomen wider than the trunk; in aquatic species furnished with lateral gills. Ex. Mantis, Ephemera? Sialis?
13. Decapodiform. Carnivorous; hexapod; antenniferous: body elongate, narrow, convex, compressed, tapering towards the tail; tail with natatorious laminæ. Ex. Dytiscus L., Agrion F.[389]
14. Branchiopodiform. Carnivorous?; aquatic; apod; antenniferous: head distinct: body transparent, flexile, furnished with a respiratory tube just above the tail. Ex. Culex[390].
15. Limaciform. Herbivorous; apod, or with very short legs: body ovate or obovate, slimy. Ex. Apoda Haw., Tenthredo Cerasi L.
The above are the principal forms that appear to me Primary (though some doubt may rest upon the ninth and tenth); and probably others will hereafter be discovered, since at present our knowledge of the larvæ of most of the Orders is very limited. And now having given you this generalization of them, as far as they are known to me, I shall next, in a slight survey of those of the different orders, lay before you what I have further to observe upon this subject.
Coleoptera. The Anopluriform coleopterous larvæ, according to Mr. W. MacLeay's view of them, include both those of Coccinella L., Chrysomela L., and Cassida L.; but this appears to me to admit of further consideration. With regard to the two former—those of Coccinella are carnivorous, those of Chrysomela herbivorous; the first is also usually more flat and depressed. As to the latter, Cassida[391], it seems to me to belong to a peculiar type, distinguished not only by its radiated margin, but by the remarkable deflected anal fork on which it carries its excrement. At present I know no analogous form amongst the apterous tribes; I must therefore leave this without a denomination. Perhaps the larva of Hispa or Alurnus, when known, will throw light upon this subject. The larva of Endomychus agrees with that of Coccinella.
There are very few known larvæ that approach to a true Thysanuriform type in this order: that most celebrated is the one supposed to belong to Meloe; but the claim of this to be so considered, is, as we have seen, rather dubious. Should this point at last be satisfactorily ascertained, it[Pg 167] will probably carry with it the whole tribe of vesicatory beetles. But even this animal in its general structure is anopluriform: the only circumstance that gives it any analogy to the Thysanura being its anal setæ. Mr. William MacLeay is inclined to regard some of the larvæ of the Malacodermi Latr., but which of them he does not state, as probably belonging to the tribe in question[392]. Those of Lampyris and of Telephorus, as described and figured by De Geer[393], appear to me intermediate between the Anopluriform and Chilopodiform Types: they have no anal setiform or styliform appendages, their mandibulæ are falcate, and their habits seem carnivorous.
Examples of Chilopodiform coleopterous larvæ are more numerous. Of this description are those of Gyrinus, Cicindela, Carabus, and Staphylinus. That of the first, indeed, appears to be the most perfectly Scolopendriform of any yet known; yet the gills or respiratory laminæ, a pair of which issues from each abdominal segment, and two pair from the last[394], prove that there is no slight analogy between it, and indeed many other aquatic larvæ, and the Stomapoda amongst the Crustacea. A remarkable instance of analogy with the Decapoda of the same Class is presented by the larva of Dytiscus, &c. which Mr. MacLeay considers as Chilopodiform, but which exhibits no other resemblance to Scolopendræ than[Pg 168] in its predaceous habits and threatening aspect. Its convex, compressed, tapering body, terminating in setæ or laminæ, is certainly much more like that of a shrimp or a prawn; to which the older Entomologists thought it was akin[395], and after which they named it. As Mr. MacLeay's object was, to take all his forms from the Ametabola, perhaps these larvæ will best fall in with his Chilopodiform type; though in the general form of their body they most represent a section (Lepisma L.) of the Thysanura.
Chilognathiform forms are equally numerous in the Coleoptera with the preceding. The wire-worm, or larva of Elater Segetis, as to shape best represents the full-grown Iulus[396], and those of the Petalocera (Scarabæus L., Lucanus L.) the young one.
The most abundant of all forms in this order, is, I think, the Vermiform, upon which I have nothing further to remark.
With regard to Crustaceous forms in Coleoptera, besides the Decapodiform just noticed, I possess two specimens of larvæ of Silphidæ which seem to exhibit a considerable analogy with the Isopodous Crustacea, one rather[Pg 169] convex and the other flatter, so as to give the idea of an Armadillo and of an Oniscus.
Strepsiptera. Larva Vermiform.
Dermaptera. Larva Thysanuriform. Type Podura or Sminthurus.
Orthoptera. Mr. MacLeay considers the larvæ of this Order as primarily Thysanuriform[397], though he allows the resemblance between them and Amphipoda to be particularly striking[398]. For my own part, their prototypes appear to me to be in the Crustacea, and their analogical relations to the Thysanura much more distant. I trust this will appear to you the reverse of dubious in a progress through the Crustaceous Orders. I begin with the Isopoda. Take the larva of a Blatta, and place it between a Lepisma, or Machilis, and an Oniscus, or Porcellio; you will find that in shape and width, and the form of its anal styles, it resembles the latter much more than it does the former, with which it possesses scarcely any character in common, except its multiarticulate antennæ. It is remarkable, that amongst the Blattidæ we meet with species that represent both the Oniscidæ and Armadillo or Glomeris[399], the latter being more convex than the former. In their habits the Blattæ certainly agree with Lepisma; and Dumeril, who thought the latter and Podura subject to a metamorphosis, imagined they were related[400].
The Spectres of Stoll (Phasma F.) are so strikingly analogous to another crustaceous tribe, the Læmodipoda, particularly the genus Caprella, that Montagu gave one species the Trivial name of Phasma[401]. The jumping[Pg 170] Amphipodiform Crustacea are represented extremely well by Gryllus L., and the Stomapodiform, particularly Squilla Mantis, by Mantis. The resemblance in this last instance is so very striking, that it cannot escape the eye of the least intelligent observer. Orthopterous insects may perhaps one day be discovered analogous to the two other crustaceous orders, the Decapods and Branchiopods; but at present I know of none of that description.
Hemiptera. The larvæ of this order, which in general resemble the perfect insect, except that they have no wings, seem most commonly to belong to the Anopluriform type[402]; but the Aphides, Chermes, and Thrips may, I think, be regarded as more analogous to the genera Podura and Sminthurus in the Thysanura[403]. I have some suspicion that the Nepidæ, Naucoris, and the remipedes, Notonecta, Sigara, &c. may find their prototypes amongst the Crustacea; but my confined knowledge of the latter does not enable me to point to any individual genera or tribes that they may be presumed to represent.
Neuroptera. As the kinds of larvæ of the different tribes composing this order, as it now stands, are very various, it is to be expected that the analogical forms they represent are equally so. The Libellulina MacLeay (whose metamorphosis that gentleman has denominated subsemicomplete, a term warranted by their losing in their perfect state the mask before described[404]) in their oral organs, particularly by their galeate maxilla and distinct ligula[405], have some relation to the Orthoptera, the prototypes of whose larvæ we have found amongst the[Pg 171] Crustacea: probably, therefore, those of the tribe in question lurk in that class; a suspicion that receives strong confirmation from the larva of Agrion[406], which in its tapering body and anal natatorious laminæ represents a shrimp. The larvæ of that very peculiar and distinct tribe, the Ephemerina, appear to be intermediate between the Stomapodiform and Thysanuriform types. Their natatory respiratory abdominal laminæ seem copied from the former, and their anal diverging setæ from the latter[407]. The Myrmeleonina, as well in their general form as in their motions and habits, present a most singular analogy with the tribe of spiders, as does also in some respect that of Cicindela. With regard to Panorpa, which Mr. MacLeay remarks is related to Myrmeleon[408], and is a most ferocious insect[409], as its larva has not yet been discovered, nothing certain respecting its analogical form can be asserted; but should it, like the male fly, represent the scorpion, both orders of Arachnida will have their representatives in the class we are considering. The Corydalina, as far as the larva of Hemerobius instructs us, is Chilopodiform, but with a tendency to the Araneidiform Type. The Ametabola also furnish the prototype of the next tribe, the Termitina, which, as is evident both from Psocus and Termes, are perfectly Anopluriform. The Sialina, or Plicipennes of Latreille, excluding Trichoptera Kirby, appear to me to be intermediate between the Chilopodiform and Stomapodiform Types, and not without some relation to the Thysanuriform. Their pediform, jointed, respiratory abdominal appendages, their head and falcate mandibles, seem copied from the first tribe. The same appendages[Pg 172] considered as organs of respiration, and their taper forks, are moulded upon the plan of the Stomapodiform Crustacea, and the long seta which terminates the abdomen is upon the Thysanuriform plan[410].
Trichoptera. The larvæ of this order appear also to be constructed upon a double plan. The respiratory threads observable in both the upper and under sides of the abdomen connect it with the Stomapoda, and its cylindrical elongate body with Chilognathiform types in the Lepidoptera[411].
Lepidoptera. The great majority of larvæ in this order are Chilognathiform, but there are exceptions to this remark. Those of the Geometræ recede from this type, both in their motions and the distance and number of their legs. In both these respects they represent the Læmodipoda in the Crustacea[412]. Other caterpillars are Onisciform; and a third sort seem to leave the Annulose type, and imitate that of the Mollusca, and one is figured by Madame Merian[413] which appears to tend even to the Chilopodiform type.
Hymenoptera. In this order the larvæ of the saw-flies, Tenthredo L., are in general Chilognathiform, though some are Onisciform, others Limaciform, and those of Lyda F. (Cephaleia Jur.) and Sirex[414] have a Vermiform tendency,[Pg 173] and are a stepping-stone to those of the rest of the order, which are all Vermiform and apods.
Diptera. The majority of this order may be set down as Vermiform, though it is not improbable that some of them bear an analogy to animals that appear far removed from the Annulosa. Thus, the larva of Stratyomis Chameleon seems to exhibit no small resemblance to some of the Polypi vaginati in the Acrita subkingdom of Mr. W. MacLeay[415]. That of Culex and some others is constructed on a quite different type from the rest, and seems to possess some analogy to the Branchiopod Crustacea.
Though some of these analogies are more striking than others, yet in almost all that I have stated there is that kind of resemblance that could not be the result of what is called mere chance; and Mr. MacLeay, by first pointing out this plan of the All-wise Creator, and by laying down the doctrine of analogies in general, as distinguished from affinities in the animal kingdom, has furnished the believer with a new argument against those attacks of the infidel, that would render null those proofs of the wisdom and goodness of the Author of nature with which the animal and vegetable creation furnish us; by affirming most absurdly, and under the most stultifying blindness of mind, that the creatures were in a manner their own creators, their wants under local circumstances stimulating them to efforts that in a long course of years produced all the different forms and organizations that are now to be found in our globe. The affinities and close connexion of beings with each other, so that the ascent from low to high is usually by the most gentle gradations, is the[Pg 174] circumstance on which they build this strange and impious theory. But the fact, that certain animals of one tribe were created with a view to certain animals of another, so as to present a striking aspect of correspondence, parallel almost with that of type and antitype, without any real affinity or approximation;—this triumphantly proves a Power above and without them, who has associated them not only in a complex chain of affinities, but has caused them to represent and figure each other, even when evidently far removed, so as to give a mutual correspondence and harmony to the whole, which could be produced only by a Being infinite in power and wisdom, who made all things after a general preconceived plan and system.
iv. We are now to consider the clothing with which larvæ are furnished. Many are quite naked, and smooth or rough only with granular elevations or tubercles orderly arranged; but a very considerable number, especially of the Lepidoptera order, are clothed with hair or bristles of different kinds, in greater or less abundance, and arranged in different modes; and a proportion still smaller have their skin beset with spines or a mixture of spines and hairs. Lyonnet found that the hairs of the caterpillar of the great goat-moth (Cossus ligniperda) were hollow, though not to the apex: probably this is the case with those of other larvæ, as well as with their spines. In this instance they were set, he observes, in a corneous ring, or very short cylinder, elevated a little above the skin. The hair passes through this ring, and appears to be rooted in a soft integument, which clothes the skin within, and upon which the nerves form a reticular tissue,[Pg 175] some of which he thinks he has even seen enter the root of the hairs, which perhaps are organs of touch[416].
Of the pilose larvæ, some, like most of those of the smaller moths (Geometra, Tortrix, Pyralis, &c.), have merely a few scattered short hairs, scarcely perceptible except through a lens: others (Odenesis potatoria, Lasiocampa Rubi) are covered with down more or less thick: in others (Eriogaster lanestris, Lasiocampa Neustria) the hair is slenderer, and more like wool; the body of two species which I purchased from the collection of Mr. Francillon is covered with woolly hairs, so long as to give them the appearance of a shock-dog; and Madam Merian has figured a similar one, which she could not bring to the perfect state[417]. The hairs of many Bombycidæ, known commonly by the name of hairy caterpillars, as Arctia erminea, &c. are stiffer, resembling bristles; sometimes, as in Arctia chrysorhea, mixed with shorter ones. The hairs either spring immediately from the skin (Noctua Aceris, leporina), or, as is more general, proceed only from certain tubercular elevations, usually subhemispherical, but sometimes conical; of which a number varying from four to twelve are found on each segment of different species. They seem to issue from these tubercles, as little diverging streams from the rose of a watering-pot. In both cases, they form a coating usually so dense as to conceal the body, but sometimes more thinly set, and admitting the skin to be seen more or less between them. In a caterpillar of the beautiful Arctia ocularia, the hairs are set upon tubercles alternately nearer the anterior and posterior margin of each segment, so as to form a dense[Pg 176] band, the rest being naked; and in the lovely green and black one of Saturnia Pavonia, each tubercle bears but six hairs, diverging like a star, the central one being the longest and capitate, so that the chief part of the body appears naked. This diverging position of the hairs is most common in the thick-clothed larvæ also, but many have them placed differently: thus, in those of Callimorpha Caja and Arctia villica[418] they are all directed towards the tail, like the quills of a porcupine: in some others the anterior ones point towards the head: in that of Eriogaster Quercus half of the tuft of hairs of each tubercle is directed downwards, the other half upwards: in that of Arctia Salicis all the hairs point downwards, so that the belly is thickly covered, while the back is bare. Another variation is, that the hairs of half the tubercle are sometimes very long, while those of the other half are very short, and even of a different colour[419]. In the larva of Tussuck moths (Laria pudibunda, fascelina, &c.) the hairs are collected into tufts of a singular appearance, those on the intermediate segments of the back being quite level at the top, so as to resemble so many brushes; while those on the first and last segments are longer, and composed of feathered hairs converging to a point at their extremity, like a common camel-hair-pencil[420]. This last mode of arrangement prevails also in the larva of Noctua Aceris; but in this the pencils are shorter, exactly wedge-shaped, and distinguished by another particularity, that of springing directly from the[Pg 177] skin, and not from a tubercle. This is also the case with the large caterpillars of Odenesis potatoria, which has a double row of short bundles of black hairs on the back, intermixed with larger ones: at each end of the body is a pencil of converging hairs, and the sides are spotted with bundles of white ones, which with longer tawny ones are bent downwards, so as to cover the sides of the creature[421]. Some have the anterior aigrettes disposed like the arms of a cross, of which the body of the caterpillar is the stem[422]. But not only is there considerable variety in the general arrangement of the hairs that clothe our little larvæ, the hairs themselves differ much in their kind and structure, of which I will now, before I proceed to consider spines, give you some account. Several of them are feathered like the plumes of a bird: this is the case with those of Morpho Idomeneus, on each segment of the body of which are three blue tubercles, like so many little turquois beads, from each of which proceeds a long black plume[423]. Other hairs terminate in a club; those of the larva of Noctua Alni, a specimen of which I possess taken in England, are flat and incrassated at the apex, something like the antennæ of some Sphingidæ. Mad. Merian has figured the caterpillar of another moth which feeds upon the Papaw-tree (Carica Papaya) with similar hairs[424]. But the most remarkable larva for the shape of its hairs is that of Anthrenus Musæorum, the little pest of our cabinets,[Pg 178] which I noticed in a former letter[425]. All the hairs of its body are rough with minute points; but those of six diverging long tufts or aigrettes, laid obliquely on the anal extremity of the body, which the animal when alarmed erects as a porcupine does its quills, are of a most singular structure: every hair is composed of a series of little conical pieces, placed end to end, the point of which is directed towards the origin of each hair, which is terminated at the other extremity by a long and large conical mass, resembling somewhat the head of a pike[426].
Besides the one lately mentioned, other caterpillars are rendered striking by the brilliant colour of the tubercles from which their hairs emerge. A remarkable instance of this is the thick large caterpillar of a Bombyx, which feeds upon the Psidium pyriferum, or white Guava, figured by Madame Merian. This caterpillar, which is white, with transverse black stripes, and which has two singular long converging curved bunches of hairs near the tail, is splendidly adorned on each side with fifty red tubercles, shining like coral, from which proceed six or seven long diverging hairs. Leeuwenhoeck took these tubercles for eyes[427]. Another figured by the same lady, who mistakes it, with her usual inaccuracy, for the larva of a Lygæus F., and which seems by her description to be between the onisciform and limaciform types, has the apparently fleshy mamillæ that project from its sides and back crowned with little hairy red globes, which give the animal a most singular and unique appearance[428]. Having thus described some of the principal modes in which[Pg 179] the All-wise Creator has decked and defended these creatures with hairs, I shall next give you a short account of the spines with which he has armed others. The spinous larvæ are principally lepidopterous, and more particularly conspicuous in some tribes of the genus Papilio L., though some saw-flies and Diptera are also distinguished by them. Vanessa Io[429], Atalanta and Urtica, Argynnis Paphia, Urania Leilus, and many other Butterflies, &c. are clothed with long sharp points, which claim the denomination of spines, rather than that of hairs or bristles; being horny and hard, and so stiff at the point as readily to pierce the skin. Those of the last-mentioned species, Madame Merian says, are as stiff as iron-wire[430]. They are sometimes entirely simple, and look like spikes rather than spines, as in the caterpillar of Nymphalis Amphinome and Morpho Menelaus[431]; but ordinarily they are beset with hairs, or more commonly with shorter spines, which often give them the appearance of plumes, as in Urania Leilus just mentioned: sometimes these lateral spines are so long as to have the appearance of a branch of a tree; this is strikingly the case with a small caterpillar which Captain Hancock brought from Brazil; its body is so thickly planted with spines of this description, that it absolutely wears the appearance of a forest or thicket in miniature. A singular circumstance attends the spines of this species: in many cases a smaller and very slender hair-like spine issues from them, resembling a sting; and this accounts for an observation of Abbott's, that many American caterpillars sting like a nettle, raising little white blisters on the skin[Pg 180] when accidentally or slightly touched[432]. Lewin has described the caterpillar of a moth found in New Holland, which he names Bombyx vulnerans, that, like these Americans, has also the power of wounding, but in a different way. It darts out, he says, when alarmed by the approach of any thing, from as many knobs or protuberances in its back eight bunches of little stings, with which it inflicts a very painful and venomous wound[433]. The caterpillar of Papilio Protesilaus F., if Madame Merian's account and figure of it are correct, has its body armed with hairy spines, the extreme point of which is surmounted by a star-shaped appendage[434]. Those of a few saw-flies (Tenthredo Pruni L.), and another figured by Reaumur[435], are covered with a little forest of spines without lateral branches, but divided into a fork at the apex. Some spines are merely rough, with very short points, as those round the head, which give so terrific an appearance to the caterpillar of the Bombyx regalis, of some proceedings of which I gave you an account in one of my former letters[436].
I must now say something upon the arrangement of these spines. Though in a few instances so thickly set as entirely to conceal the body of the animal, as in the case of the Brazil one lately mentioned, yet generally speaking, even when they are most numerous, they permit the skin to be distinctly seen. Their arrangement is various, though always orderly: in the majority they[Pg 181] are planted singly, but in some caterpillars in bundles. In that of Saturnia Io, on each segment there are six bundles of longish, quill-shaped, sharp, slender, diverging spines, which also appear to sheath aculei. Madame Merian has figured this larva, or one very near it, as the grub of a Euglossa[437], with which, though she affirms she traced it to the fly, it can have no connection. With regard to number, some larvæ have only four spines on each segment; others five, others again six, and others seven, or even eight: they are planted on the sides and back only, never on the belly. They are often more numerous on the intermediate than on the anterior and posterior segments; but sometimes the reverse of this takes place; in that of Attacus Erythrinæ only the head and tail are armed with spines, the rest of the body being without any[438]; and in that of Morpho Teucer there is only a single spine on the four intermediate segments[439]. They are usually all nearly of equal length; but in some cases those of the head and tail are much longer than the rest, and remarkably so in the caterpillar of Urania Leilus, also beautifully plumose, and gracefully waved[440]. Those in the second and third segments are much longer than any of the rest in that of Bombyx regalis; which circumstance gives it the terrific appearance lately alluded to. In the family to which Argynnis Paphia belongs, the larva is adorned with two on the back of the first segment twice as long as the rest, and resembling at first sight two antennæ.
The spines, as well as the hairs of the new skin, are concealed under the old one, and not incased in its spines;[Pg 182] but Bonnet ascertained, that if cut off very closely, the larva sometimes died in consequence, whilst no such result followed a similar operation on hairy larvæ. We learn from Reaumur[441], that some spinous larvæ of saw-flies (Tenthredo L.) lose their spines at the last change of their skin; and from Madame Merian, that that of Attacus Erythrinæ before mentioned loses also at the same period the six tremendous black spikes that arm its black and yellow larvæ. The grubs of ants that are destined to pass the winter in the larva state are hairy, but are not so in summer[442]. The spines found in the grubs of some gad-flies (Œstrus L.) are of a different kind from those above described, being very minute triangular flat plates, arranged in different and contrary directions[443], and serving the insect merely to change its place and fix itself[444].
Two other kinds of clothing, if so they may be called, neither coming under the description of hairs nor spines, are found in some other larvæ, not only amongst the Lepidoptera, but also in some of the other orders. Nymphalis Populi and others of the same family have larvæ furnished on the back of each segment with cylindricoconical processes of a fleshy substance, obtuse at the apex and surrounded with capitate hairs. In that of N. Sybilla, which has on each segment two fleshy protuberances, they are bifurcate or trifurcate, and also encircled at the base with a hairy tuft[445]. Others, as those of Melitæa Artemis, Cynthia, &c. have each segment beset on the back with from seven to nine fleshy, pubescent, wedge-shaped protuberances; two larger ones projecting[Pg 183] over the head. Under this head, too, may be noticed, the glutinous secretion which clothes the grub of Cionus Scrophulariæ, a little weevil; and of Tenthredo Cerasi L. a saw-fly, and that waxy or powdery substance which transpires through the skin of the larvæ of several Aphides, Chermes, Cocci, Hylotoma ovata F., &c. The Aphis, whose extensive ravages of our apple-trees (A. lanata) were before described to you[446], is covered and quite concealed by this kind of substance, so that the crevices in the bark which they inhabit look as if they were filled, not with animals, but with cotton. The insect, also, that forms those curious galls produced upon the spruce fir, and which imitate its cones (Chermes Abietis L., Aphis De Geer) secretes a similar substance. In these and other cases of the same kind, this matter seems to be, if I may so speak, wire-drawn through numerous pores in certain oval plates in the skin, more depressed than the rest of the back, arranged regularly upon the segments, and exhibiting minute tuberosities. When young, these animals have more of this secretion than when more advanced: it then hangs from their anal extremity in locks[447].
But the insects most remarkable for a covering of this nature are those Coccidæ of which Bosc has made a genus under the name of Dorthesia. De Geer is the first author that notices them, and has given a description and figure of one species under the name of[Pg 184] Coccus floccosus[448]. It was discovered by Modeer upon some sere fir-leaves in a thick bed of moss. Panzer has figured a second found upon Geranium sanguineum, which from the figure appears distinct from De Geer's, under the name of Coccus dubius[449]. Fabricius regards this as synonymous with the Dorthesia characias of Bosc, inhabiting Euphorbia characias in South Europe[450]. Olivier found a species upon the bramble[451]. I once took one, which appears to differ in some respects from the preceding species, upon Melampyrum cristatum, and our indefatigable friend Mr. Sheppard has sent me another, on what plant found I do not remember, which does not agree with any that I have mentioned. The body of the animals of this genus is covered by a number of cottony or waxy laminæ which partly cover each other, and are arranged usually in a triple series: in De Geer's figure the series appears quadruple, the lateral ones being placed obliquely. The anterior one in my specimen covered the head, and they are all canaliculate. Above the anus are four diverging ones: the whole are of the most dazzling whiteness. When these laminæ are removed, the body appears divided into segments.
With respect to those larvæ which imitate slugs by the viscid covering that besmears them and issues from their pores, we learn from Professor Peck that this exudation takes place as soon as they are hatched; that the animal retains its humidity although exposed to the fiercest heat of the sun, and that at the last moult the skin becomes[Pg 185] quite clean, and free from all viscidity[452]. It is probable that the other limaciform larvæ are similarly circumstanced. Madame Merian has figured an onisciform one, the legs of which, she says, are covered with a viscid skin: this produced a Noctua. Those of Papilio Anchises also are slimy, and adhere to each other[453].
v. Amongst other qualities which attach to larvæ, we must not omit to say something concerning their Colour. For though those which live in darkness, in the earth, in wood, in fruits, &c. are, with few exceptions[454], of an uniform whitish colour, yet such as are exposed to the influence of the light are usually adorned with a vast variety of tints, sometimes the most vivid that can be imagined. That the white colour of the former may be attributed to the absence of light is proved by an experiment of M. Dorthes, who having forced some to live under glasses, exposed to the light, found that they gradually became brown[455]. To attempt any classification of coloured larvæ would be in vain, since they are tinged with almost every possible shade that can be conceived, of many of which it would be difficult to find examples elsewhere; and infinitely diversified as to the arrangement and figure of their multiform markings and spots. A few general remarks, therefore, are all that you will expect on this head. Many are of one uniform colour; while a variety of tints, very different, and very vivid and distinct, ornament others. Sometimes they are distributed in[Pg 186] longitudinal rays or bands, at others in transverse ones. Sometimes they are waved or spotted, regularly or irregularly; at others they are sprinkled in dots, or minute streaks, in every possible way. Various larvæ are of the colour of the plant on which they feed, whence they are with difficulty discovered by their enemies. Thus, a large proportion of Lepidoptera are green of different shades, sometimes beautifully contrasted with black bands; a circumstance which renders the caterpillars of two of our finest insects of this order as lovely as the fly: I mean that of Papilio Machaon and Saturnia Pavonia. Very frequently the larvæ of quite different species resemble each other so exactly, in colour as well as shape, as scarcely to be distinguishable: this sometimes takes place even where they belong to different genera, as in those of Bombyx versicolor a moth, and Smerinthus Populi a hawk-moth. And it sometimes happens, very fortunately for distinguishing allied species, that where the perfect insects very nearly resemble each other, the larvæ are altogether dissimilar. Thus, the female of Pieris Rapæ is so much like the same sex of Pieris Brassicæ, that it might be taken for a variety of it, did not the green caterpillar of the one, and the spotted one of the other, evince the complete distinction of these butterflies. Noctua Lactuca, N. umbratica, and several other species of the same tribe, which includes N. Absinthii, Verbasci, Chamomillæ, Abrotani, are so extremely alike, that the most practised eye can scarcely discover a shade of difference between them, though their larvæ in colour and markings are constantly distinct[456]. The markings of[Pg 187] species belonging to the same family are usually different; but in some cases the latter may be prejudged from the former. The larvæ of many of the genus Sphinx L., for example, have their sides marked by oblique streaks running from the back in a direction towards the head; and by this last circumstance they are distinguished from those of Bombyx versicolor, Attacus Tau, and others of the same tribe, which have also lateral oblique striæ, but running from the back towards the tail[457]. The colours of individual larvæ of the same species are usually alike, but in Sphinx Elpenor and some others they vary exceedingly. Many, like those of Lasiocampa Rubi, Saturnia minor, &c., are of one colour when first disclosed, and assume others quite different in riper age. Just previously to changing their skin, the tints of most larvæ become as dull and obscure, as they are fresh and vivid when the change has fully taken place; and in some instances the new skin is quite differently marked from the old one. This is remarkably the case with the last skin of some of the larvæ of the genus Tenthredo L., which is entirely different from all the preceding ones. As people when they advance far in years usually become more simple in their dress than when they were young, so the larvæ in question change an agreeably variegated skin for one of a uniform and less brilliant colour[458]. Madame Merian has observed with respect to Attacus Erythrinæ, that its caterpillar is at first yellowish, with nine black striæ on each side: when arrived at one third of its size, they become orange; the striæ are obliterated, and in their place a round black spot appears on each of the[Pg 188] eight intermediate segments[459]. Mr. Sheppard has remarked to me, that the skin of that of Sphinx Ligustri, after being under ground four days, was changed from a vivid green to a dull red. Very rarely, however, it becomes of a more brilliant hue just before entering the pupa state: thus, that of another hawk-moth (Smerinthus Tiliæ) changes to a bright violet; and the yellow hairs of that of Laria pudibunda then become of a lovely rose colour. And here I may observe, that the hairs and spines also, of larvæ, vary greatly in colour. They are to be met with brown, black, red, yellow, violet, white, &c. De Geer found, that in the larva of Cimbex nitens the two sides of the body were of a different colour, the left being of a deep green, whilst the right side and the rest of the body were paler[460]; but as he saw only a single individual, this was probably an accidental circumstance. Though the caterpillars, as I lately said, of one of the most beautiful butterflies and moths that inhabit Britain contend with the perfect insect in loveliness, yet in general no judgement can be formed of the beauty of the future fly from the colour of the larva; and the young Aurelian must not flatter himself always with the hope, because the caterpillar excites admiration by its colours and their arrangement, that the butterfly or moth it is to produce will do the same; nor ought he to despise and overlook a sombre or plain-coloured individual of the former, under the idea that it will produce one equally plain of the latter, for it often happens that the splendid caterpillar gives a plain butterfly or moth, and vice versâ. De Geer, however, gives us two instances of conformity[Pg 189] between the colours of the caterpillar and those of the future moth; the one is that of the common currant-moth (Phalæna G. grossulariata L.), the caterpillar of which is white, ornamented with several black spots varying in size. At the two extremities it is yellowish, with a longitudinal ray of the same colour on each side, the head and legs being black. These colours are all to be found in the fly, the ground of its wings being white ornamented with many black spots of different sizes. Its upper wings are traversed by a yellowish band; and towards their base is a spot of the same colour. Its body is yellowish, with black spots; but the head and legs are black[461]. The other is that of a green caterpillar, which gives a green moth, figured by Reaumur (Pyralis prasinaria Fab.)[462]. Sometimes, also, the sex of the future perfect insect may be predicted from the colour it exhibits in its first state: thus, the brown caterpillars of Noctua Pronuba produce males, and the green ones females[463]. The sexes, also, of N. exoleta and Persicariæ differ in that state.
vi. To the full account of the Food of insects given in a former letter[464], which had reference chiefly to their larva state, it is only necessary in this place to add a few particulars not there noticed. Many larvæ when first excluded, as those of Pieris Cratægi, &c. devour the shells of the eggs from which they have proceeded[465]; and[Pg 190] others (Cerura Vinula, Sphinx Euphorbiæ, Noctua Verbasci), though their usual food is of a vegetable nature, eat with great apparent satisfaction the skins which they cast from time to time, not leaving even the horny legs. This strange repast seems even a stimulating dainty, which speedily restores them to vigour, after the painful operation by which they are supplied with it. Under this head it will not be out of place to mention, that some larvæ of insects, which feed only on the juices of animals, or the nectar and ambrosia of flowers, have no anal passage, and of course no feces. This is said to be the case with the grubs of bees, wasps, the larvæ of Myrmeleon, &c.[466]
vii. You will require no stimulus to induce you to attend to the subject I am next going to enter upon,—the Moulting, namely, of Larvæ; or their changes of skin. This, indeed, is a subject so replete with interest, and which so fully displays the power, wisdom, and goodness of the Creator, affording at the same time such large occasion for nice investigation, that a pious and inquisitive mind like yours cannot but be taken with it. In the higher orders of animals, though the hairs of quadrupeds and the feathers of birds are in many cases annually renewed, the change, or scaling and increment of the skin, is gradual and imperceptible; no simultaneous rejection[Pg 191] of it, in which it is stripped off by the animal itself like a worn shirt, being observable, till you descend in the scale to the Serpent tribe[467], which at certain periods disengage themselves from their old integument, and start forth with that new and deadly beauty so finely described by the Mantuan bard:—
In these the new skin, I imagine, is formed under the old from the rete mucosum; but in insects, as I formerly stated[469], since the time of Swammerdam it has generally been believed by entomologists, that the larva includes a series of cases or envelopes, one within the other, containing in the centre the germe of the future perfect insect, whose development and final exclusion take place only when these cases have been successively cast off. This hypothesis, as was explained to you on a former occasion[470], has been controverted by a late writer, Dr. Herold; who affirms that the skins of caterpillars are also successively produced out of the rete mucosum. I have however, I hope, satisfied you that the old system is most consonant[Pg 192] to nature and probability: but as we are now to enter at large upon the Moults of insects, it will not be without use if I add a few additional reasons which seem to me still further to prove the correctness of Swammerdam's system, as far as it relates to that subject. With regard to the mere formation of the skin from the rete mucosum, were this the whole question few would hesitate to adopt the sentiments of M. Herold; but when we come to consider further—that the number of moults of individuals of the same species is always the same, and that it varies in different species, and takes place at certain periods,—we begin to suspect that something more than the mere formation of a new skin upon an old one being cast is to be accounted for; and that the law which prescribes its own definite number of skins to each species, must begin to act in the primordial formation of the larva. Again, the hairs observable in the higher animals do not take their origin from the epidermis solely, but are planted below it in the rete mucosum, or deeper[471]; so that the change of skin does not affect them; but in the larvæ of insects they are a continuation of that integument, since, when the moult takes place, they always remain on the rejected skin[472]: this is the case, also, even with spines. If you shave a caterpillar ready to change its skin, either partially or generally, you will find that the parts in the new skin that correspond with those that are denuded, are equally hairy with those that were not[473]; and if you pay attention to the new-clad animal, you will find further, that the hairs never grow after a moult. From[Pg 193] hence it follows, that the hairs have their place and take their whole growth between the new skin and the old[474]. Whether the spines, simple or compound, lately described to you, that arm some larvæ are similarly circumstanced, seems not as yet to have been ascertained; but as the spinous ones of certain Tenthredines L. and Lepidoptera at their last moult have no spines, the presumption is, that, whether incased or not, they are mere appendages of the skin on which they appear. A new set of hairs, therefore, and probably of spines in spinous larvæ, accompanying each skin, and these varying very much in size, composition, &c. though a new membrane may be admitted to be formed from an action in the rete mucosum without a pre-existent germe of it, it seems not easy to conceive how these hairs and spines can spring up and grow there, each according to a certain law, without existing previously as a kind of corculum or punctum saliens; and that the germes of the tubercles, in which the hairs are so generally planted, according to a certain arrangement and in a given number, should also pre-exist, seems most consonant to reason. These and the several skins may all co-exist in their primordial germes, and remain beyond the discovery of our highest powers of assisted vision, till a certain period when they may first enter the range of the microscope-aided eye. It does not therefore follow, because these primordia semina rerum are not discoverable, that therefore they may not exist. Our faculties and organs are too limited and of too little power to enable us to see the essences of being.
Upon the supposition that the hypothesis of Swammerdam[Pg 194] is the true one, we may imagine that the envelope that lies within all the rest is that which covers the insect in its pupa state. Above this are placed several others, which successively become external integuments. These changes or casting of the skin in larvæ, analogous, as before observed, to that of serpents, are familiar to every breeder of silk-worms, in which four such changes occur: the first at the end of about twelve days from its birth, and the three next each at the end of half that time from the moulting which preceded it. With some exceptions[475], similar changes of the skin take place in all larvæ, not however in the same number and at the same periods. Most indeed undergo this operation only three or four times; but there are some that moult oftener, from five up to eight (Arctia villica), nine (Callimorpha Dominula), or even ten times; for so often, M. Cuvier informs us, the caterpillar of the tiger-moth (Callimorpha Caja) casts its exuviæ. It has been observed that the caterpillars of the day-flying Lepidoptera (Papilio L.) usually change only three times, while those of the night-flying ones (Phalæna L.) change four[476]. The periods that intervene between each change depend upon the length of the insect's existence in the larva state. In those which live only a few weeks or months, they are from eight to twenty days; while in those that live more than a year, as the cockchafer, &c. they are probably proportionably longer: though we know very little with[Pg 195] regard to the moult of any insects besides the Lepidoptera.
A day or two previously to each change of its skin, the larva ceases eating altogether; it becomes languid and feeble, its beautiful colours fade, and it seeks for a retreat in which it can undergo this important and sometimes dangerous and even fatal operation in security. Here, either fixing itself by its legs to the surface on which it rests, or, as is the case with many caterpillars, by its prolegs, to a slight web spun for this purpose, it turns and twists its body in various directions, and alternately swells and contracts its different segments. The object of these motions and contortions seems to be, to separate the exterior skin, now become dry and rigid, from the new one just below it. After continuing these operations for some hours, resting at intervals without motion, as if exhausted by their violence, the critical moment arrives: the skin splits in the back, in consequence of the still more violent swelling of the second or third segment: the opening thus made is speedily increased by a succession of swellings and contractions of the remaining segments: even the head itself often divides into three triangular pieces, and the inclosed larva by degrees withdraws itself wholly from its old skin. All larvæ, however, do not force their way through this skin in precisely the same place. Thus, those of the hawthorn butterfly (Pieris Cratægi), according to Bonnet[477] make their way out by forcing off what may be called their skull, or the horny part of their head, without splitting the skin, which remains entire; others have been observed to make their way out at the side and the belly. Reaumur[Pg 196] noticed the larva of Zygæna Filipendulæ, previously to its last moult, actually biting off and detaching several portions of its old skin; and before this, drops of a fluid resembling water were seen to exude from it[478].
The skin when cast is often so entire, that it might be mistaken for the larva itself; comprising not only the covering of the main trunk with the hairs which clothed it, but of the very skull, eyes, antennæ, palpi, jaws, and legs; which, if examined from within, are now found to be hollow, and to have incased, like so many sheaths, similar parts in the new skin. That the feet of the newly-coated larva were actually sheathed, as fingers in a glove, in the same parts of the exuviæ, may be proved by a very simple experiment: if a leg of one just ready to cast its skin be cut off, the same limb will be found mutilated when that change has ensued. The anal horns, also, of the larvæ of the hawk-moth (Sphinx L.) and other similar protuberances, are incased in each other in like manner; but hairs are laid flat between the two skins, and contribute considerably towards their more easy separation. Thus, if you saved the skins cast by the larva of Callimorpha Caja, for instance, you would appear to have ten different specimens of caterpillars, furnished with every external necessary part, and differing only in size, and the colour perhaps of the hairs, and all representing the same individual.
But further changes than this take place. Swammerdam says, speaking of the moult of the grub of Oryctes nasicornis, a beetle common in Holland, but not satisfactorily ascertained to inhabit Britain, "Nothing in all nature is in my opinion a more wonderful sight than the[Pg 197] change of skin in these and other the like worms. This matter, therefore, deserves the greatest consideration, and is worthy to be called a specimen of nature's miracles; for it is not the external skin only that these worms cast, like serpents, but the throat and a part of the stomach, and even the inward surface of the great gut, change their skin at the same time. But this is not the whole of these wonders; for at the same time some hundreds of pulmonary pipes within the body of the worm cast also each its delicate and tender skin. These several skins are afterwards collected into eighteen thicker, and, as it were, compounded ropes, nine on each side of the body, which, when the skin is cast, slip gently and by degrees from within the body through the eighteen apertures or orifices of the pulmonary tubes before described, having their tops or ends directed upwards towards the head. Two other branches of the pulmonary pipes that are smaller, and have no points of respiration, cast a skin likewise." ... "If any one separates the cast little ropes or congeries of the pulmonary pipes with a fine needle, he will very distinctly see the branches and ramifications of these several pipes, and also their annular composition[479]."—Bonnet makes a similar observation with regard to caterpillars; but he appears to have observed it more particularly, at least the change of the intestines, previously to the metamorphosis of the insect, when he says with the excrements it casts the inner skin of the stomach and viscera[480]. Both these great men appear to have recorded the result of their own actual observations with regard to the proceedings of two very different[Pg 198] kinds of insects; the one the grub of a beetle, and the other the caterpillars of Lepidoptera. The account of the former is given quite in detail, as that of a person who is describing what he has actually seen: yet by a later and very able physiologist, Dr. Herold, it is affirmed that the inner skin of the intestinal canal is never cast, that canal constantly retaining its two skins. He further affirms, that they are only the large trunks of the Tracheæ that cast their skins, none being detached from their smaller ramifications[481]. When men so eminent for their anatomical skill and nicety, and for their depth and acumen, disagree, the question must be regarded as undecided till further observations throw sufficient weight into one scale or the other.
The larva which has undergone this painful process is at first extremely weak: all its parts are soft and tender; even the corneous ones, as the head and the legs, are then scarcely more than membranous, and are all bathed with a fluid, which, before the moult, intervenes between the two skins, and facilitates their separation[482]: and it is only after some hours, or in some cases even days, during which it lies without motion, that this humidity evaporates, all its parts become consolidated, and it recovers its strength sufficiently to betake itself to its wonted food. Its colour, too, is usually at first much paler than before, and its markings indistinct, until their tints have[Pg 199] been enlivened by exposure to the air, when they become more fresh, vivid, and beautiful to appearance than ever. When a few meals have invigorated its languid powers, the renovated animal makes up for its long abstinence by eating with double voracity. . A similar preparatory fast, and succeeding state of debility, accompany every change of the larva's skin. Each time except the last, the old skin is succeeded by a new one, with few exceptions, similar to the one it has discarded. Previously to the final change, which discloses the pupa, it quits the plant or tree on which it had lived, and appears to be quite unsettled, wandering about and crossing the paths and roads, as if in quest of some new dwelling. It now abstains from food for a longer time than before a common moult, empties itself copiously, and as I have just said, if Swammerdam and Bonnet are to be depended upon, casts the skin that lines the stomach and intestines, as well as that of the Tracheæ.
I have observed above, that all larvæ, with few exceptions, change their skins in the manner that I have described. These exceptions are principally found in the order Diptera, of which those of the Linnean genera Musca, Œstrus, and probably all that, like the maggot of the common flesh-fly, have membranous contractile heads, never change their skin at all, not even preparatory to their becoming pupæ. The skin of the pupa, though often differing greatly in shape from that of the larva, is the same which has covered this last from its birth, only modified in figure by the internal changes that have taken place, and to which its membranous texture readily accommodates itself. The larvæ of the Dipterous genera Tipula, Culex, and those which have corneous[Pg 200] heads, like other larvæ change their skins several times previously to becoming pupæ[483]. The grubs, also, of bees, wasps, ants? and probably many other Hymenoptera, do not change their skin till they assume the pupa, nor the larva of the female Coccus[484].
If you feel disposed to investigate the reasons of that law of the Creator which has ordained that the skins of the higher animals shall be daily, and imperceptibly, and as it were piece by piece renewed, while those of insects are cast periodically and simultaneously,—the proximate cause must be sought for in the nature of the two kinds of skin, the one being more pliable and admitting a greater degree of tension than the other, and being so constructed as to scale off more readily. If, ascending higher, you wish to know why the skins of insects are so differently circumstanced from our own, the most apparent reason is, to accommodate the skin to the very rapid growth of these animals, which a gradual and slower change would have impeded too much, or the skin have suffered constant dilapidation and injury; therefore their Beneficent Creator has furnished them with one which will stretch to a certain point, and during a certain period, and then yield to the efforts of the inclosed animal, and be thrown aside as a garment that no longer fits the wearer.
viii. And this leads me to a subject to which I am desirous[Pg 201] now to bespeak your attention,—the Growth, I mean, and size of Insects in this state. As to size, larvæ differ as much as insects in their perfect state: these last, however, never grow after their exclusion from the pupa, while larvæ increase in bulk in a proportion, and often with a rapidity, almost without a parallel in the other tribes of animals. Thus Lyonnet found, that the caterpillar of the great goat-moth (Cossus ligniperda F.) after having attained its full growth is at least 72,000 times heavier than when it was first excluded from the egg[485]; and of course had increased in size in the same proportion. Connected with the size of larvæ, is the mode in which their accretion takes place. This, with respect to the more solid parts, as the head, legs, &c., is not, as in other animals, by gradual and imperceptible degrees, but suddenly and at stated intervals. Between the assumption of a new skin and the deposition of an old one, no increase of size takes place in these parts, while the rest of the body grows and extends itself, till, becoming too big for these solid parts, nature restores the equilibrium between them by a fresh moult[486], in which the augmentation of bulk, especially in these parts, is so great, that we can scarcely credit the possibility of its being cased in so small an envelope. Malpighi declares, that the head of a silk-worm that has recently cast its skin is four times larger than before the change[487]. It is very probable, also, that when the outer skin becomes rigid, it confines the body of the larva within a smaller compass than it would expand to if unconfined, so that, when this compression is removed, the soft and elastic new integument[Pg 202] immediately swells out, and the animal appears all at once much larger than it was before the moult. In fact, the proximate cause of the rupture and rejection of the old skin is the expansion of the included body, which at length becomes so distended as to split its envelope, aided, indeed, as before described, by the contortions of the creature itself.
The larvæ most notorious for the rapidity of their growth are those of Musca carnaria and other flesh-flies: some of which Redi found to become from 140 to more than 200 times heavier in twenty-four hours[488]: an increase of weight and size in so short a time truly prodigious, but essential for the end of their creation—the rapid removal of dead and putrescent animal matter. As the skins of these larvæ are never changed, we may conclude, if the cause of the change of skin in other larvæ above surmised be accurate, that their skins are more contractile and capable of a greater degree of tension than those of larvæ that are subject to moulting. And two peculiarities observable in them confirm this idea: in the first place, their head is not hard and corneous, as that of the others, but capable of being shortened or lengthened; and in the next, their breathing-pores are not in the sides, but at the extremities of the body, while in the moulting larvæ there are two in almost every segment, which must form so many callous points that impede the stretching of the skin to the utmost. The hairs, spines, and tubercles, that are so often found on caterpillars, must also form so many points of resistance that prevent that full extension of the integument which it might otherwise admit.
There is not always that proportion between the size of larvæ and of the insects that proceed from them that might have been supposed, some small larvæ often producing perfect insects larger than some of those proceeding from such as are of greater size.
ix. As insects often live longest in the state we are treating of, I shall say something next upon the age of larvæ, or the period intervening between their exclusion from the egg and their becoming pupæ. This is exceedingly various, but in every case nicely adapted to their several functions and modes of life. The grubs of the flesh-fly have attained their full growth, and are ready to become pupæ, in six or seven days; the caterpillar of Argynnis Paphia, a butterfly, in fourteen days; the larvæ of bees in twenty days; while those of the great goat-moth (Cossus ligniperda) and of the cockchafer (Melolontha vulgaris) live three years, or at least survive three winters, before the same change. That of another lamellicorn beetle (Oryctes nasicornis F.) is said to be extended to four or five; that of the wire-worm (Elater segetum) to five. That of the stag-beetle (Lucanus Cervus) is affirmed by Rösel to be extended to six years; but the most remarkable instance of insect longevity is recorded by Mr. Marsham in the Linnean Transactions[489]. A specimen of Buprestis splendida, a beautiful beetle never before found in this country, made its way out of a deal desk in an office in London in the beginning of the year 1810, which had been fixed there in the year 1788 or 1789; so that according to every appearance it had existed in this desk[Pg 204] more than twenty years. Ample allowance being made for its life as a pupa, we may conclude that it had existed as a larva at least half the above period. The grubs of the species of the genus Cynips L. attain their full size in a short time; but they afterwards remain five or six months in the gall before they become pupæ[490].
With few exceptions it may be laid down, that those larvæ which live on dead animals, in fungi, in dung, and in similar substances, are of the shortest duration in this state; and that those which live under the earth, on the roots of grass, &c. and in wood, the longest: the former becoming pupæ in a few days or weeks, the latter requiring several months, or even years, to bring them to maturity. The larvæ which live on the leaves of plants seem to attain a middle term between the one and the other,—seldom shorter than a few weeks, and rarely longer than seven or eight months. Aquatic larvæ appear to be subject to no general rule: some, as the larvæ of Gnats, becoming pupæ in two or three weeks; and others, as those of the Ephemeræ, which are thus compensated for their short life as flies, in as many years[491]. The cause of all these differences is obviously dependent on the nature of the food, and the purposes in the economy of creation to which the larvæ are destined.
x. The last part of the history of larvæ relates to their Preparations for assuming the pupa state. When they have acquired their full size, after having ceased to take[Pg 205] food, by a copious evacuation they empty the intestinal canal, even rejecting the membrane that lines it and the stomach[492]; their colours either change totally, or fade; and they make themselves ready for entering upon a new stage of their existence. Some merely rest in a state of inactivity in the midst of the substances in which they feed, as if conscious of their inability to select any safer abode. Of this description are most Coleopterous, Hymenopterous, and Dipterous larvæ, that feed under ground, or in the interior of trees, fruits, and seeds.
But a still larger tribe, those which feed on leaves, animals, &c. act as if more sensible of the insecurity of this to them important epoch. They are about to exchange their state of vigour and activity for a long period of deathlike sleep. The vigilant caution which was wont to guard them from the attack of their enemies will be henceforward of no avail. Destitute of all the means of active defence, their only chance of safety during their often protracted night of torpor must arise from the privacy of their place of repose. About this, therefore, they exhibit the greatest anxiety. Many, after wandering about as if[Pg 206] bewildered, retire to any small hole on the surface of the earth, covering themselves with dead leaves, moss, or the like, or to the chinks of trees, or niches in walls and other buildings, or similar hiding-places. Many penetrate to the depth of several inches under ground, and there form an appropriate cavern by pushing away the surrounding earth; to which they often give consistence by wetting it with a viscid fluid poured from the mouth. The larvæ of other insects undertake long and arduous journeys in search of appropriate places of shelter. Those of flesh-flies, now satiated with the mass of putridity in which they have wallowed, leave it, and conceal themselves in any adjoining heap of dust. The grubs of the gad-fly (Œstrus) creep some of them out of the backs of cattle, in tumours of which they have resided, and suffer themselves to fall to the earth; while others, which have fed in the stomach of horses, quit their hold, and by a still more extraordinary and perilous route are carried through the intestines the whole length of their numerous circumvolutions, and are discharged at the anus. And without enumerating other instances, various aquatic larvæ, as that of a common fly (Elophilus pendulus), &c. leave the water, now no longer their proper element, and betake themselves to the shore, there to undergo their metamorphosis.
Most of these, having reached their selected retreat, require no other precaution; but another large tribe of larvæ have recourse to further manœuvres for their defence before they assume the pupa. Those of the aphidivorous flies (Syrphus F. &c.), of the various lady-birds (Coccinella L.), and tortoise-beetles (Cassida L.), &c. fix themselves by the anus with a gummy substance to the[Pg 207] leaves or twigs under which they propose to conceal themselves during their existence in that state. Others previously suspend themselves by a silken thread fixed to the tail, or passing round the body; by which also, when become pupæ, they are afterwards pendent in a similar position; and lastly, a very great number of larvæ wholly inclose themselves in cases or cocoons, composed of silk and various other materials, by which during their state of repose they are protected both from their enemies and the action of the atmosphere. As these two last-mentioned processes are extremely curious and interesting, I shall not fear tiring you by entering into some further detail respecting them: explaining first the mode by which larvæ suspend themselves, both before and after they are become pupæ, by silken threads; and next, the various cases or cocoons in which others inclose themselves, and their manner of operating in the formation of them.
1. The larvæ which suspend themselves and their pupæ, with the exception of the tribe of Alucitæ, and some Geometræ of the family of G. pendularia, punctaria, &c. are almost all butterflies[493]. No others follow this mode. They may be divided into two great classes—those which suspend themselves perpendicularly by the tail, and those which suspend themselves horizontally by means of a[Pg 208] thread girthed round their middle. In both cases it should be observed, that the suspension of the pupa is the object in view; but as the process is the work of the larva, this seems the proper place for explaining it. To begin with the first case.
You are aware that all lepidopterous larvæ have the faculty of spinning silk threads from their mouths, and it will readily occur to you that it is by means of these threads that they suspend themselves. But how? How is a caterpillar to hang itself by the tail to threads spun from the mouth? Even suppose this difficulty overcome, others still greater remain. Suppose the caterpillar to be suspended by its tail,—this is but a preparatory operation,—what is required is, that the pupa shall hang in the same position: now when you take into consideration that it is incased within the skin of the larva, and without feet or other external organs; that it has to extricate itself from this skin; to hang itself in its place, and to detach the skin from the threads which hold it—this will appear no trifling task. Indeed at first view it seems impossible. Country-fellows for a prize sometimes amuse the assembled inhabitants of a village by running races in sacks: take one of the most active and adroit of these, bind him hand and foot, suspend him by the bottom of his sack with his head downwards, to the branch of a lofty tree; make an opening in one side of the sack, and set him to extricate himself from it, to detach it from its hold, and suspend himself by his feet in its place. Though endowed with the suppleness of an Indian juggler, and promised his sack full of gold for a reward, you would set him an absolute impossibility: yet this is what[Pg 209] our caterpillars, instructed by a beneficent Creator, easily perform. Their manœuvres I shall now endeavour to explain.
When the caterpillar has selected the under-side of the leaf or other object to which it purposes suspending itself, its first process is to spin upon it a little hillock of silk consisting of numerous loosely interwoven threads; it then bends its body so as to insinuate the anal pair of prolegs amongst these threads, in which, by a slight exertion, the little crochets which surround them[494] become so strongly entangled as to support its weight with ease. It now suffers the anterior part of the body to fall down, and it hangs perpendicularly from its silken support with its head downwards. In this position it remains often for twenty-four hours, at intervals alternately contracting and dilating itself. At length the skin is seen to split on the back near the head, and a portion of the pupa appears, which by repeated swellings acts like a wedge, and rapidly extends the slit towards the tail. By the continuance of these alternate contractions and dilatations of the conical pupa, the skin of the caterpillar is at last collected in folds near the tail, like a stocking which we roll upon the ancle before withdrawing it from the foot. But now comes the important operation. The pupa, being much shorter than the caterpillar, is as yet at some distance from the silken hillock on which it is to be fastened; it is supported merely by the unsplit terminal portion of the latter's skin. How shall it disengage itself from this remnant of its case, and be suspended in the air while it climbs up to take its place? Without arms or legs to[Pg 210] support itself, the anxious spectator expects to see it fall to the earth. His fears, however, are vain; the supple segments of the pupa's abdomen serve in the place of arms. Between two of these, as with a pair of pincers, it seizes on a portion of the skin; and bending its body once more, entirely extricates its tail from it. It is now wholly out of the skin, against one side of which it is supported, but yet at some distance from the leaf. The next step it must take is to climb up to the required height. For this purpose it repeats the same ingenious manœuvre, making its cast-off skin serve as a sort of ladder, it successively with different segments seizes a higher and a higher portion, until in the end it reaches the summit, where with its tail it feels for the silken threads that are to support it. But how can the tail be fastened to them? you ask. This difficulty has been provided against by Creative Wisdom. The tail of the pupa is furnished with numerous little hooks pointing in different directions[495], as well adapted to the end in view as the crochets of the larva's prolegs, and some of these hooks are sure to fasten themselves upon the silk the moment the tail is thrust amongst it. Our pupa has now nearly completed its labours; it has withdrawn its tail from the slough, climbed up it, and suspended itself to the silken hillock—manœuvres so delicate and perilous, that we cannot but admire that an insect which executes them but once in its life, should execute them so well: nor could it, as Reaumur has well and piously observed, had it not been instructed by a Great Master. One more exertion remains: it seems to have as great an antipathy to its cast-off skin,[Pg 211] as one of us should, when newly clothed after a long imprisonment, to the filthy prison garments we had put off. It will not suffer this memento of its former state to remain near it, and is no sooner suspended in security than it endeavours to make it fall. For this end—it seizes, as it were with its tail, the threads to which the skin is fastened, and then very rapidly whirls itself round, often not fewer than twenty times. By this manœuvre it generally succeeds in breaking them, and the skin falls down. Sometimes, however, the first attempt fails: in that case, after a moment's rest, it makes a second, twirling itself in an opposite direction; and this is rarely unsuccessful. Yet now and then it is forced to repeat its whirling, not less than four or five times: and Reaumur has seen instances where the feet of the skin were so firmly hooked, that after many fruitless efforts the pupa, as if in despair, gave up the task and suffered it to remain[496]. After these exertions, it hangs the remainder of its existence in this state until the butterfly is disclosed.
We are now to consider the second mode of suspension, in which larvæ by means of a silken girth round their middle, fix themselves horizontally under leaves, &c. These follow the same process with that of those last described, in spinning a small hillock of silk to which they fasten their hind legs; and if the operation concerned the larva state alone, this would be sufficient, as by means of this support, and of their prolegs, they could easily retain themselves in a horizontal position. But these larvæ[Pg 212] act as if they foresaw the assumption of a state in which they will be deprived of legs. It is the suspension of the forthcoming pupa that is the object in view; and though this can be hung by the tail in the same way with those of the first class, yet it is plain that it cannot be retained in a horizontal position, which for some unknown reason is essential to it, without some support to its anterior extremity. It is necessary for the larva, therefore, not only to fix its posterior legs amongst a collection of silken fibres, but to spin a girth of the same material round its body. This girth, though apparently of a single thread, will be found on examination to be composed of several, often as many as fifty or sixty; and is fastened on each side of the body of the larva about the middle, to the surface under which it is placed. Three different modes of fixing these girths are adopted by the caterpillars of different butterflies. Some, as those of the common cabbage-butterfly (Pieris Brassicæ), which have remarkably pliable bodies, bend them almost double on one side, then fix the thread and carry it over to the other in the same position, repeating this operation as often as is necessary. Others, as that of Lycæna Argus and many more of the Papiliones Rurales and Urbicolæ L., which have a short and more rigid body, after having bent the head on one side so as to fix one end of the thread, bring themselves into a straight position, and, by a manœuvre not easily described, contrive to introduce the head under the thread, which they then bend themselves to fasten on the other side, pushing it to its proper situation by the successive tension and contraction of their segments. But the most curious mode, though indeed that which seems most natural, is adopted[Pg 213] by the caterpillar of the beautiful swallow-tail butterfly (Papilio Machaon) and others of the same family. This first forms the loop which is to serve for its girth, and then creeps under it. But the difficulty it has to surmount is, to keep itself from being entangled in the fifty or sixty fine distinct threads of which the girth is composed, and to preserve them all extended so as to be able to introduce its body beneath them. For this purpose it makes use of the two first pair of its fore-legs, employing them as a woman does her hands in winding a skein of cotton, to collect and keep all the threads of its card unentangled and properly stretched; and it is often with great difficulty, towards the end of the process, that it prevents them from slipping off. When a sufficient number of threads is completed, the animal bends its head between its legs, and insinuates it under the collected loop, which by its annular contraction it easily pushes to the middle of the body.
In about thirty hours after the larvæ which girth themselves have finished their operations, the skin splits, and the pupa disengages itself from it by those contractions and dilatations of its segments which have been before described, pushing the exuviæ in folds to the tail, by different motions of which it generally succeeds in detaching them. One would have thought there would be considerable difficulty in slipping the skin past the girth; but this, according to Reaumur, seems to be easily effected[497].
If you are desirous of witnessing for yourself the manœuvres by which these curious modes of suspension are[Pg 214] effected, you may be readily gratified. It is only necessary to collect and feed until their metamorphosis the black spinous caterpillars of the common peacock-butterfly (Vanessa Io), which in most places may be found upon nettles, or those of the Pieris Brassicæ, which swarm in cabbages or brocoli in every garden. The former will exhibit to you a specimen of vertical, the latter of horizontal suspension. It should be observed, however, that to hit the precise moment when these processes are going on, it is necessary to feed a considerable number of the larvæ of each kind; some one of which, if you watch them narrowly when they have attained their full growth, you will scarcely fail to surprise in the act.
I must observe here, that although the vertical and horizontal are the two principal positions in which caterpillars suspend themselves, yet that others are inclined at various angles; and some are attached with less art, appearing only to be fastened by some part of their abdomen to the body upon which they are fixed[498].
2. The larvæ whose procedures I am in the next place to describe, are those which, previously to assuming the pupa state, inclose themselves in cases or cocoons of different materials. For the sake of method, I shall divide these into two great classes: First, those which form their cocoons entirely or principally of silk; and secondly, those which form them chiefly of other substances.
To begin with the first. The larvæ which inclose themselves in silken cocoons are chiefly of the Lepidopterous tribes of Bombycidæ and Noctuidæ; but a few[Pg 215] Geometræ (G. papilioniaria, lactearia, &c.); most of the Hymenoptera; some Coleoptera, as certain of the weevil tribe (Hypera Arator, Rumicis Germ.), and those brilliant beetles frequenting aquatic plants constituting the genus Donacia F.; the Neuropterous genera Hemerobius and Myrmeleon; Mycetophila and a few others in the Diptera; and Pulex in the Aphaniptera fabricate coverings of the same material. In all, with the exception of Myrmeleon and Hemerobius (and perhaps Hypera Rumicis, &c.?) which have their spinning apparatus at the extremity of the abdomen, the silken thread employed in forming these coverings proceeds from the middle part of the under-lip, as before explained; and is in fact composed of two threads gummed together as they issue from the two adjoining orifices of the spinner.
Of the larvæ which inclose themselves in silk, the most familiarly known is the silk-worm: the cocoon of this consists exteriorly of a thin, transparent, gauze-like coating, through the interstices of which can be seen an inner, smaller, oval ball of a more close and compact texture. The whole is in fact composed of one single thread, but arranged in two distinct modes. To form the exterior envelope, which is merely the scaffolding by means of which the inner and more solid covering is constructed, the caterpillar, after fixing upon a space between two leaves or twigs or angles suitable for its purpose, begins by glueing one end of its thread to one of the adjoining surfaces. This thread it next conducts to another part and then fastens, repeating this process and interlacing it in various directions, until it has surrounded itself with a slight and loosely spun netting. In the centre of this, when contracted into a space sufficiently small,[Pg 216] it lays the foundation of the interior cocoon. Fixing itself by its prolegs to some of the surrounding threads, it bends its body, and by successive motions of its head from side to side spins a layer of silk on the side opposite to it: when this is of the requisite thickness, the larva shifts its position, and repeats the same process in another quarter, covering each layer in turn with a new one until the interior cavity is reduced to the size desired. Thus, the silken thread which forms this new cocoon is not, as might have been supposed, wound circularly as we wind the thread of a ball of cotton; but backwards and forwards in a series of zigzags, so as to compose a number of distinct layers. Malpighi could distinguish six of these layers[499], and Reaumur suspects there is often a greater number[500]. The former found the length of the thread of silk composing them when wound off, without including the exterior case, to be not less than 930 feet[501]; but others have computed it at more than a thousand[502]: consequently the threads of five cocoons united would be a mile in length. Estimating by the weight,—the thread of a pound of cocoons, each of which weighs about two grains and a half, would extend more than 600 miles[503], and such is its tenuity, that the threads of five or six cocoons require to be joined to form one of the thickness requisite in the silk manufacture. It is the continuous thread of the inner cocoon which is most valuable; the outer loose coating from its irregularity cannot be[Pg 217] wound off, and is known in commerce by the name of floss silk.
Manœuvres in their general principle similar to those of the silk-worm are followed by most of those larvæ which inclose themselves in silken cocoons. Many species, however, adopt variations in the mode of procedure all of which it would be tedious to particularize, but some of them are worth mentioning. The larvæ of Tortrix prasinana, and other species of moths which form cocoons resembling a reversed boat, arrange their threads in layers, so as to construct two parallel walls gradually inclining towards the top and ends, where they finally force them to approach each other by means of an apparatus of silken cables[504]. And the larva of Saturnia Pavonia, though it forms the base of its flask-shaped cocoon by spinning like the silk-worm a number of interwoven zigzags, places the threads which compose the interior funnel-like opening of the apex nearly straight, parallel to each other, and converging towards the same point in the centre[505].
These last, as well as almost all larvæ, constantly remain in the inside of the cocoon during its construction. But De Geer has given us the history of a minute caterpillar of a species of moth (Tinea L.) which feeds on the under side of the leaves of the Rhamnus Frangula, or Black Alder, that actually weaves half of its cocoon on the outside. This cocoon, which is very small, is beautifully fluted, consisting of several longitudinal cords, with the intervals filled by fine net-work, and shaped like a reversed boat[506]. The animal begins by laying the foundations[Pg 218] of one of the ends of her cocoon, she adds new threads to this small beginning, and so proceeds. As the work advances she retreats backwards, and her body is situated nearly in the same line with the cocoon she has begun, and quite out of it; she only touches with her head and legs its anterior margin. When half the cocoon, or rather of its exterior layer, is finished, she suspends her operations for some moments. She then for the first time introduces her head into this demi-cocoon, and turns herself in it by doubling her supple body, and passing one part over the other, so that at last she manages to bring her tail into the pointed end of the cocoon, the head and the anterior half of her body remaining without. Thus situated, she commences her operations afresh. At a distance from the margin of the demi-cocoon, equal to its length, she begins to spin the pointed end of the other moiety, the length of her body serving her as a measure that enables her to begin at the proper distance from it. This new portion she spins in the same manner as the other; but as she is prevented by the demi-cocoon in which the posterior part of her body is lodged from retreating backwards, she contracts her body more, which answers the same purpose. When the new work is so advanced that she can no longer contract her body, she bends the anterior part of it considerably, and reverses her head. When the distance between the margin of the two halves of the cocoon is very small, so as no longer to admit the head between them, in order to unite them she is obliged to have recourse to another manœuvre. Withdrawing her head, she extends silken longitudinal threads between the two margins, and thus unites them. This part is more clumsy, and not so regular as[Pg 219] the rest of the cocoon, so that the point of union is always discoverable. These caterpillars do not always divide the cocoon into two equal portions, for often they will finish three quarters of the cocoon before they enter it, and begin at the other end[507].
The general rule is,—that each larva spins for itself a separate cocoon; but amongst those of Arctia chrysorhea and others which live in society, two or three sometimes begin their operations so close together that they are under the necessity of forming one common cocoon, which serves for a covering to the whole number. The same thing happens to silk-worms, the double or treble cocoons of which are called Dupions by the breeders. The larvæ of some Ichneumons, besides forming each its separate cocoon, spin a joint cottony covering for the whole[508], which is effected thus:—After they leave the caterpillar they have devoured, they fix themselves side by side at a little distance from it, and begin to spin each a cocoon; and in order to defend its end and side that is not covered by others, they spin further an envelope of loose silk, and thus this exterior covering is formed.
The size, figure, colour, substance, and texture of silken cocoons are extremely various. Their size indeed is usually proportioned to that of the included larva or pupa; yet it is by no means always so. Some large caterpillars spin cocoons so small, that the observer can hardly conceive how they can be contained in so narrow a compass: Eriogaster Catax is a moth of this description[509]. And others smaller in size lodge themselves in[Pg 220] apartments apparently much more spacious than necessary. The transparent hammock-like cocoons of Hepialus Humuli and Arctia villica, two other moths, would contain several of their pupæ. I possess one in which the pupa is suspended in the centre, that is ten times its size, and not very short in dimensions of that of Attacus Paphia, a giant silk-moth. The largest cocoon I ever read or heard of, is that thus described by Mr. Hobhouse in his Travels: "Depending," says he, "from the boughs of the pines, near the Attic mountain Parnes, and stretching across from tree to tree so as to obstruct our passage, were the pods, thrice as big as a turkey's egg! and the thick webs of a chrysalis, whose moth must be far larger than any of those in our country."[510] If this statement is correct, and I am not aware that there is any reason for doubting it, the cocoon must be vastly larger than the pupa, or the moth it produced would far exceed in size any yet known. Perhaps, however, as this gentleman is probably no entomologist, what he took for a cocoon might be a nidus, in which many larvæ were associated, of the nature of those formerly described[511].
With regard to figure, the majority are like those of the silk-worm, of a shape more or less oval or elliptic: some, however, vary from this. That of Lasiocampa Rubi is oblong. I have one from New Holland somewhat resembling an acorn, fixed to the twigs of some tree or shrub, of a very close silk, and covered by a circular operculum, which the animal pushes off when it assumes the imago; this is ovate or conico-ovate; others again are globose[512]; others are conical[513], as that of Gastropacha[Pg 221] quercifolia; others almost fusiform[514] (Odenesis potatoria). Reaumur received one from Arabia which was nearly cylindrical[515]. Those of T. prasinana before noticed, and many other Tortrices, are shaped like a reversed boat[516]; that of Saturnia Pavonia and others of the same tribe, like a Florence flask with a wide and short neck. The cocoon of Lygæna Filipendulæ resembles a grain of barley. Another cocoon in my cabinet, of very slight network, is shaped something like an air-balloon. But the most remarkable one for its form and characters, is one that I received from the rich cabinet above quoted. This, which is of an unusually close texture, is suspended by a thread more than two inches long from the point of a leaf; it then swells into a perfect cone, at the base about four-fifths of an inch in diameter and half an inch in length, and covered with scattered setiform appendages: from the centre of the base projects a large hemispherical protuberance, which terminates in a long stalk, much thicker than the thread that suspends the cocoon. There is commonly no difference between the shape of cocoons spun by larvæ which are to give birth to different sexes of the same species. The silk-worm cocoons, however, which will produce male moths, have more silk at the ends, and consequently are more round than those which are to produce females: but the difference is not striking.
The most usual colour of silken cocoons is white, yellow, or brown, or the intermediate shades. The whites are very pure in the general envelope of some species of Ichneumonidæ, and yellows often very brilliant. But[Pg 222] besides these more general colours, some cocoons are black[517], some few blue or green, and others red[518]. Sometimes the same cocoon is of two different colours. Those of certain parasites of the tribe of Ichneumones minuti L. the motions of one of which I noticed on a former occasion[519], are alternately banded with black or brown and white, or have only a pale or white belt in the middle, which gives them a singular appearance. In both cases the difference in colour depends upon the different tints with which the silky gum is imbued in the reservoirs: the first portion of it is white, and with this the larva first sketches the outline of its cocoon, and then thickens the layers of silk considerably in those parts where the white bands appear: when these are finished, its stock of white silk is exhausted, and the remainder of the interior of the cocoon is composed of brown silk[520]. The circular operculum above mentioned as covering an acorn-shaped cocoon, is paler than the latter, and also ornamented by a zone within the margin of deep brown. The pale cocoon also of Attacus Paphia is veined with silk of a deep red.
I have very little to say on the substance of the silk of cocoons. Though that of the silk-worm is composed of such a slender thread, that of many others is still finer, scarcely yielding in tenuity to the spider's web. On the other hand, the silk of the cocoons of Saturnia Pavonia and of several foreign species is as thick as a hair.
With regard to the texture of their cocoons—in some, as in that of the silk-worm, the threads are so slightly[Pg 223] glued to each other, as to separate with facility; but in that of the emperor-moth just mentioned they are intimately connected by a gummy matter, furnished, as Reaumur conjectures, from the anus[521], with which the whole interior of the cocoon is often plastered. Some, as that of the silk-worm, are composed of an exterior loose envelope, and an inner compact ball; others have no exterior covering, the whole cocoon being of an uniform and thick texture. The larva of Cossus Robiniæ Peck, in spinning its cocoon, makes the end next the opening to the air, by which the imago is to emerge, of a slighter texture than the rest of it[522]. The exterior case is sometimes, as in Laria pudibunda, very closely woven, so as to resemble a real cocoon[523]: its form is usually adapted to that of the inner one; but in some which fix them under flat surfaces (Laria fascelina, Callimorpha Caja,) it resembles a hammock[524]. Cocoons of a close texture have generally no orifice in any part; but that of Eriogaster lanestris is spun with openings, as if bored from without, the use of which, however, does not seem to have been ascertained[525].
Many silken cocoons are of so close a fabric, as, when finished, entirely to conceal the included insect; but a very considerable number are of a more open texture, composed of a much smaller quantity of silk, and that woven so loosely, that the larva or pupa may always be discovered through it. Of this description are the cocoons of Hypogymna dispar, Arctia Salicis, &c., which consist only of a few slight meshes. Those of some others[Pg 224] resemble gauze or lace[526]. Of the first description is one in my cabinet before alluded to, shaped somewhat like an air-balloon; the meshes are large and perfectly square. The pupa hangs in the centre, fixed by some few slight threads which diverge from it to all parts of the cocoon—so that it looks as if it was suspended in the air, like Mahomet's coffin, without support. Of the second description is a black one with very fine and nearly circular meshes: the threads that form these are thick, and seem to be agglutinated. In our own country, the cocoons of some beetles, as of Hypera Arator, Galeruca Tanaceti, and of some little Tineæ, also resemble gauze. Many of the larvæ, however, which spin these cocoons, whose thinness is probably attributable to the smallness of their stock of silk, seem anxious for a more complete concealment; and therefore commonly either hide them between leaves tied together, in some with a certain regularity, in others without art[527]; or thicken their texture, and render it opaque, by the addition of grains of earth[528], or of other materials with which their bodies supply them. These are principally of two kinds. The larvæ of Lasiocampa Neustria, Arctia Salicis, &c. after spinning their cocoons, cast from their anus three or four masses of a soft and paste-like matter, which they apply with their head all round the inside of the cavity; and which, drying in a short time, becomes a powder that effectually renders it opake. This is not, as might be conjectured, an excrement, but a true secretion, evidently[Pg 225] intended for this very purpose: and, according to Reaumur, a similar powder, but white, derived from the varicose intestines, is used by the caterpillars of Gastropacha quercifolia, &c.[529] The other material, which is still more frequently employed, and which is occasionally mixed with the former, is the hair which everyone has observed to cover so thickly the bodies of some caterpillars. This, after spinning a sufficient envelope, they tear, or in some instances cut off with their mandibles, and distribute all round them, pushing it with their head amongst the interstices of the silk, so as to make the whole of a regular thick texture. After this process, which leaves the body completely denuded, and often seems to give them great pain, they conclude by spinning another tissue of slight silk, in order to protect the forthcoming pupa from the surrounding prickly points. It should be observed, however, that though many hairy larvæ, as those of Noctua Aceris, Arctia Caja, and others, employ their hairs in the composition of their cocoons, the rule is not general, several never making any such use of them. Nor do all that do so employ them distribute them in the same manner as those above described, which rarely attempt to arrange them in any regular position. Reaumur has noticed a small hairy caterpillar that feeds on lichens, which is more methodical: this actually places its hairs upright, side by side, as regularly as the pales in a palisade, in an oval ring around its body, connecting them by a slight tissue of silk, which forces them to bend into a sort of roof at the top; and under this curiously-formed cocoon assumes its state of pupa[530]. Some larvæ make so much[Pg 226] hair and so little silk enter into the composition of their cocoons, that on the first inspection they would be pronounced wholly composed of it[531]; others, thickening the interior of their cocoon with hair, line the whole with a viscid matter like varnish[532].
The larvæ of some saw-flies (Tenthredo L.) are remarkable for inclosing themselves in a double cocoon, in which the inner is not, as in the silk-worm &c., connected with the outer, but perfectly distinct from it. Some species, as T. Rosæ (Cryptus Jur.), which have but a small stock of silk, compose the outer cocoon of thick silken cords crossing at right angles, and forming an oval net; which at the same time that it protects them effectually from the ants, which are always ready to attack them, demands much less silk than a covering of a closer texture. But the tender nymph itself requires to be inclosed in a case of a softer and more delicate substance; and accordingly the inner cocoon is composed of fine silk, woven so closely that the threads are scarcely perceptible under a microscope[533]. Reaumur mentions a hymenopterous larva belonging to Latreille's Fossores (Sphex L.) which thickened its cocoon with the legs, wings, and other relics of the flies which it had devoured[534]: trophies—like the drinking-cups of some savages, made of the skulls of their enemies, or the skull pyramid near Ispahan—of its powers of devastation.
It is a general rule, that those larvæ which spin cocoons, never in ordinary circumstances become pupæ without having thus inclosed themselves. An exception, however, is met with in the larva of a species of ant noticed[Pg 227] by De Geer (Formica fusca L.), some of the individuals of which inclose themselves in cocoons; while others neglect this precaution, and undergo their metamorphosis uncovered[535]. Rösel also made nearly the same observation on the larva of the flea[536].
I must say something with regard to the situation, often very remote from their place of feeding, in which larvæ fabricate their cocoons. A very considerable number, probably the majority, form them either partially (Arctia lubricipeda) or wholly under ground; others beneath dead leaves, moss, or in the chinks of the trees; others within the wood in substances on which they have fed; the larva of Cossus leaves in these a communication with the open air by which the imago emerges; and a large number attach them to the leaves and branches of trees and plants; the cocoon of Donacia fasciata(?) is fastened by one side to the roots or surculi of Typha latifolia. There is usually nothing very remarkable in the mode of fixing them, the exterior threads being merely gummed irregularly to different portions of the objects which support them. But some effect this with greater art. I have one from New Holland, very long, which is suspended from a twig by a long riband, as it were, which entirely girths the twig. The larva of the magnificent silk-moth, Attacus Paphia, actually forms a solid silken stalk to its cocoon, an inch and half in length[Pg 228] and a line in diameter, fastened by the other extremity to a twig, which it closely surrounds as if with a ring, at first sight resembling a fruit of a very singular appearance[537]. I have specimens of this cocoon with both stalk and ring. A bell-shaped cocoon fastened by a foot-stalk, but of softer consistence, to a blade of grass, found by Mr. Sheppard, I can also show you; and my friend Mr. Wilkin had a similar one out of the late Mr. Hudson's collection. Most larvæ spin their cocoons in solitude: some of those, however, which live in society do it close together under their common tent.
There are other cocoons that should be noticed here, such as those formed by the larva of Zygæna Filipendulæ, and some Bombyces, saw-flies (Tenthredo L.), and beetles (Curculio, Donacia F.), &c. These are formed of a substance which seems more analogous to gum than silk, yet furnished from the silk reservoirs, and usually present the appearance externally of parchment or membrane. That of the insect first mentioned is coated, however, with a slight interior silken lining; as indeed are almost all cocoons, of whatever substance.
The second class, into which I have divided larvæ that inclose themselves in cocoons, includes those which form their coverings not solely or principally of silk, but in which other materials are mixed more or less. The cocoons of some of these larvæ are merely composed of a few leaves slightly tied together, either irregularly, or arranged, particularly when they are of a linear figure, with considerable symmetry. The grubs of many beetles, as of[Pg 229] the rose-beetle, Cetonia aurata, &c., prepare themselves a cocoon, composed of earth, pieces of rotten wood, and any substances within their reach: which they fasten together with a glutinous secretion. The same material is employed by others in forming a cocoon wholly of earth; which is sometimes, as that of the stag-beetle, Lucanus Cervus, exceedingly hard; at others, as that of some moths, Noctua ambigua, &c., so slight as to fall to pieces as soon as touched[538]. Other cocoons are formed of grains of earth. Reaumur has given a very interesting account of the procedures of a larva in repairing one of these cocoons, from which he had broken off the top when just completed. Without quitting the interior of the walls that remained, it put out its head from the breach, and for more than an hour employed itself in selecting one by one grains of earth, which it conveyed with its mandibles and deposited within its case: it next spun all round the opening threads of silk, to which it attached grains of earth taken from the previously-stored heap, uniting them compactly by means of other silken threads. After employing three hours in this laborious process, the industrious little mason had reduced the diameter of the breach to a few lines. Reaumur was very curious to know how it would fill up this orifice, which would no longer admit the protrusion of its head outside the walls, as in its previous operations. He concluded, that while the rest of the cocoon was exteriorly formed of earth, this opening would be merely closed with silk. He was mistaken, however: the artist knew how to vary its[Pg 230] manœuvres, and make its vault of one uniform texture. It spun across the opening a little net of silk, between the meshes of which it thrust grains of earth so dexterously that they projected as far as the outer surface, retained there probably by silken lines previously attached and fastened within. It then finished its habitation by fortifying the inside of the orifice with another layer of earth[539]. The ant-lion (Myrmeleon) spins a globular cocoon with its anus, which it covers with grains of sand[540]. One that I took in the forest of Fontainebleau, in the quarry that produces the crystallized sandstone called the Fontainebleau fossil, was covered with large and shining grains. Instead of the grains of earth or sand employed by these larvæ, those of another tribe substitute grains of stone detached from the softer walls, upon whose lichens they previously feed, which they unite into solid oval cocoons[541]. Those of a fourth form their cocoons of patches of short moss arranged with the roots downwards, and forming a vault, as it were, of verdant turf, admirably adapted for concealment[542]. The larvæ of some moths form their cocoons of irregular pieces of bark tied together with silk, and resembling when completed a knotty protuberance of the twig on which they are fixed. That of Pyralis tuberculana constructs a pannier-shaped one of the parenchyma of the leaves of plants[543].
All these cocoons, however, must yield in point of singularity of construction, materials, and ingenuity, to one formed by a small caterpillar, described by the illustrious naturalist lately quoted, which feeds upon the oak. This cocoon is wholly composed of small rectangular[Pg 231] strap-shaped pieces of the fine upper skin, or epidermis of the twig upon which it rests, regularly fastened to each other in a longitudinal direction with very slender silken cords. But the mode of its construction is even more remarkable than the substance of which it is fabricated. The caterpillar's first process is to form its slips of bark into two flat triangular wing-like pieces, projecting opposite to each other from each side of the twig, somewhat like the feathers of an arrow. It does not, perhaps, require any great degree of intelligence in a larva to give its cocoon the usual oval form, when it begins to arrange its materials in that shape from the very first, and round so good a mould as its own bent body; but we surely must admit that it is a task to which no stupid artist would be competent, to form first a multitude of strap-shaped laminæ into two triangular plates, and then to bend these plates into a case resembling the longitudinal section of a cone, with an elliptical and protuberant base,—the figure which the cocoon of this insect assumes. All the minutiæ of the manœuvres which it employs in this nice operation could not be comprehended without a more diffuse explanation than I have here room to give: suffice it to say, that the caterpillar fastens silken lines to each exterior opposite and longer side of the laminæ, and by applying all the weight of its body forces them to bend and approach each other, in which position it secures them by other shorter lines. It next repeats the same process with the upper and shorter sides of the plates; which when joined form the base of the cocoon. Both these tasks are accomplished in less than an hour, and the seams are so nicely joined as to be imperceptible. A fine inner tapestry of silk, covering all the asperities of the exterior[Pg 232] walls, concludes its labours[544]. It is to be lamented that Reaumur was unacquainted with the moth that proceeds from the pupæ inclosed in these ingenious cocoons; which being small, and precisely of the same colour as the bark of the twig that supports them, are not to be discovered but by a very narrow inspection. It would seem, however, to be Noctua Strigula of Berkhausen, Pyralis strigulalis of Hubner[545]. The larva, he informs us, is found in May: its body is flatter than common, of a yellowish flesh-colour, clothed with tufts of red hair on each segment, and furnished with fourteen feet. Should this description enable you to detect it upon your oaks, a view of its ingenious procedures would amply repay you for the trouble of seeking for it. The larvæ of Cerura vinula, Stauropus Fagi, and several other moths, form their cocoons of grains of wood gnawed from the trees on which they feed. These grains they masticate, mixed with a glutinous fluid secreted from the mouth, into a paste, which forms a covering of an uniform smooth texture, and so hard as not readily to yield to a knife. Of a substance apparently nearly similar is composed the cocoon of a weevil related to Liparus Pini; which with its inhabitant was given me by the ingenious Mr. Bullock. A little moth, whose ravages have been before noticed[546], lines the interior of the grain of barley, of which it has devoured the contents, with silk; divides it into two apartments, into one of which it pushes the excrement it had voided, and in the other assumes the pupa[547].
These, and the other larvæ mentioned above, commonly[Pg 233] form their cocoons of the substances I have indicated; but when by any cause they are prevented from access to them, they often substitute such other materials as are at hand. Reaumur fed a larva that formed its cocoon of minute fragments of paper, which with its mandibles it had cut from the piece that covered the glass vessel that contained it[548]: and the same circumstance happened to Bonnet.
Upon a former occasion I described to you the cases of various kinds formed and inhabited by the insects of the Trichoptera Order (Phryganea L.) commonly called case-worms[549]. As these serve for the pupa as well as the larva, they may be regarded as a kind of cocoon. I shall not repeat here what I then said; but having purchased from the collection of the late Mr. Francillon some that seem to belong to this or some cognate tribe, that are of a curious construction, I shall give you some account of two or three of them in this place. The first is not quite three inches long, of a sublanceolate shape, but rather widest towards one end. It consists of an internal tough and thick bag or cocoon, of a silk resembling fine wool of a dirty white colour, which is closely covered transversely by pieces of the stalk of a plant, about three-fourths of an inch in length, and crossing each other at an obtuse angle. The next is thicker and shorter: the internal bag is just covered with small fragments of wood like sawdust; over these are fastened irregularly, short stout pieces of a pithy stick or stalk, and the whole is clothed with a very close-woven ash-coloured web. It seems difficult to conceive how the inclosed animal could contrive to cover her habitation with[Pg 234] this web without going wholly out of it. The third is the most curious and remarkable of all. It is nearly six inches long, and about four-fifths of an inch in diameter. It consists of a bag of thick cinereous silk web, to which are fastened, in a sextuple series, pieces of stick about an inch long, the end of one mostly resting upon the base of another: between each series a space of about three-tenths of an inch intervenes, but at the apex they all converge. This probably imitates the branch or stem of some tree or plant, in which the leaves are linear, and diverge but little from the stem. A label upon it states its country to be New Holland. I suspect the inhabitants of the two last cocoons to be terrestrial animals: the first is probably a true aquatic case-worm.
The same purpose for which the cocoons above described serve, is answered in the case of numerous Dipterous insects, by a humble and less artificial contrivance—the skin, namely, of the larva; which, as was before observed[550], is never cast, but, when the insect is about to enter into the pupa state, assumes a different form and colour; becomes of a thicker and more rigid texture; and defends the included pupa, which is separate from it, till its exclusion. In this case the mouth of the larva is constantly different from that of the perfect insect, or at least has not with it those relations as to number and kind of organs, which have been observed in the mouth of other larvæ compared with the insects that they produce. The animal, immediately after it is clothed with this skin, if it is opened, exhibits only a soft gelatinous[Pg 235] pulp, in the surface of which the exterior organs of the adult insect cannot yet be detected. Nature requires more time for their elaboration, or at least for the appearance of their outline, and to consolidate them. This pulp first takes an oblong form (Boule allongée Reaum.), and afterwards that of the insect it is destined to give birth to[551]. The skin of the larva also serves for a cocoon to the pupæ of male Cocci[552]. The grub of the genus Anthrenus, so destructive to our cabinets of natural objects[553], when it assumes the pupa does not quit its skin, but only splits it open longitudinally on the back, and when it becomes an imago makes its exit through the orifice[554]. Some Lepidopterous larvæ even (Alucita pentadactyla, Callimorpha rosea, &c.) assume the pupa state within their last skin[555].
When a larva has finished its cocoon,—which with[Pg 236] some species, that proceed so earnestly as though they had not a moment to lose, is the work of a few hours, of others about two or three days,—after a certain interval it casts its last skin, which is usually suffered to remain in the cocoon (but which one moth, Geometra lacertinaria, ejects through an opening purposely left in its bottom), and the pupa makes its appearance[556]. This interval is exceedingly various. Most larvæ assume the pupa state within a few days after they have formed their cocoons; but some not for several weeks, or even months. The caterpillar of Bombyx cæruleocephala, according to Rösel, lies three weeks in the cocoon before this change is effected; those of many Pupivora and Diplolepariæ Latr., according to Reaumur, six months[557]; that of Phalæna urticata nine months[558]; and that of Cimbex lutea, according to De Geer, sometimes eighteen months[559]. Brahm observes, that such larvæ of the double-brooded moth, Hepialus Testudo, as form their cocoons in autumn, do not become pupæ until the following spring; while those which form them in summer undergo this change in a few days[560]. From this fact it might be conjectured, that the degree of heat prevailing at the time the insect incloses itself determines the period of the pupa's appearance; but this supposition seems contradicted by what Reaumur observed of a brood of the larvæ of Phalæna urticata, just mentioned, which, though they formed themselves cocoons in September, did not become pupæ till the June following[561]. I am unable, therefore, to assign any plausible cause for these extraordinary variations.[Pg 237] The difficulty of comprehending how animals before so voracious can live so long without food may be partly surmounted, by adverting to the circumstance of its having attained its full growth, and laid up a store of nutriment for the development of the perfect insect. It is consequently no more wonderful that it should not have need of any further supply without casting off its upper integument, than that it should not eat after having done so and become a pupa.
We have now traced our little animals through their egg and larva states, and have arrived at the third stage of their existence, the Pupa State. This, to include all, can only be defined,—that state intervening between the larva and imago, in which the parts and organs of the perfect insect, particularly those of sex, though in few cases fully developed, are prepared and fitted for their final and complete development in the last-mentioned state; and in which the majority of these animals are incapable of locomotion, or of taking food.
Pupæ, like larvæ, may be separated into two great divisions:—
I. Those which, in general form, more or less resemble the larvæ from which they have proceeded.
II. Those which are wholly unlike the larvæ from which they have proceeded.
I. To the first division belong, with some exceptions[562],[Pg 239] the Dermaptera, Orthoptera, Hemiptera, and most Aptera, with the neuropterous tribes of Libellulina, Ephemerina, and the genus Termes, in the class Insecta; and the majority of the Arachnida. This, like the first division of larvæ, may be subdivided into two corresponding smaller sections; the first including those pupæ which resemble the larvæ, except in the relative proportion and number of some of their parts; and the second those that resemble them, except in having the rudiments of wings, or of wings and elytra.
i. The first subdivision will include the pupæ, if they may be so called[563], of insects of the Aptera order, and of the class Arachnida: as, lice, Poduræ, Lepismidæ, centipedes, millipedes, mites, harvest-men, spiders, scorpions, &c. These mostly differ from their larvæ only in that the relative length or number of their legs, the number of the segments of the body in some, or the development of their palpi, more nearly approach the characters of the perfect insect; and in that while in their larva state they have two or more skins to cast, previously to their assumption of the imago, in their pupa state they have but one. In fact, this last circumstance is the only one which, strictly speaking, characterizes the pupæ of this subdivision; as the changes which take place in the number and proportion of the organs are partly produced with each change of the larva's skin. And hence, as it is not easy to ascertain what number of skins a spider, for example, has yet to cast, and as both the larva and pupa differ so little from the perfect insect, it is very difficult to determine in what state insects of this division are. From[Pg 240] this difficulty has probably arisen the too great multiplication of species in some of these tribes, particularly the Arachnida, the larva and pupa having been mistaken for perfect insects. The pupæ of this subdivision were named by Linné complete, from the near resemblance which they bear to the imago.
ii. The second subdivision will include the pupæ of the Dermaptera, Orthoptera and Hemiptera orders, with few exceptions; as likewise the Libellulina, Ephemerina, and Termitina? amongst the Neuroptera: including the well-known tribes of earwigs, cockroaches, crickets, grasshoppers, locusts, lanthorn-flies, froghoppers (Cicada L.), bugs, plant-lice, dragon-flies, day-flies, white ants, &c. Of these, as in the former subdivision, the pupæ are equally capable of eating and moving with the larvæ, which they resemble, except in having the rudiments of wings, or of wings and elytra. The pupæ of the three orders first enumerated differ from those of the Neuroptera in resembling the perfect insect in most instances, both as to shape and the organs for taking their food; and in all other respects, except in not having their wings and elytra fully developed[564]. The resemblance of the pupæ of the Libellulina and Ephemerina to the perfect insects is more distant, and the above organs in the two states are very dissimilar; for the pupæ of the former are furnished with a prehensory mask similar to that of the larvæ before described[565], which the perfect insect has[Pg 241] not; and those of the latter with the usual oral organs of masticating insects, of which the imago has scarcely the rudiments.
I have applied the term rudiments to the wings and elytra in this state, not in a strict sense, but merely to denote their appearance; for in fact the wings, &c. are complete, but only folded up longitudinally and transversely, and inclosed in membranous cases, which when the last change takes place remain attached to the puparium or pupa-case. The tegmina or hemelytra in this state usually cover the wings, and the upper wings the under; but in the Libellulina both are usually visible. Though commonly very small compared with the instruments of flight in the perfect insect, some of these rudiments, contrasted with the majority, are of considerable magnitude. This is the case with those of some species of Chermes, as we learn from De Geer[566].
II. The second grand division comprises by far the largest number of pupæ: those of all coleopterous, strepsipterous, lepidopterous, hymenopterous, dipterous, and aphanipterous, and by far the majority of neuropterous insects, as well as the hemipterous genus Aleyrodes, and one sex of Coccus of the same order. These pupæ, however, though agreeing in the circumstance of being unlike the larvæ from which they proceed, differ from each other in several respects, and require to be divided into three great sections, as under:—
i. Those pupæ in which the parts of the future insect, being folded up under a membranous skin closely applying to each, are distinctly visible. To this head belong[Pg 242] generally, the pupæ of coleopterous[567] and hymenopterous insects; those of the neuropterous genera Myrmeleon and Hemerobius, &c.; the Trichoptera; amongst the Diptera, Culex, Tipula L., Tabanus, Bombylius, &c.; and that of the flea (Pulex). These were the incomplete pupæ of Linné.
ii. Those pupæ in which the parts of the future insect, being folded up under a harder skin, are less distinctly discoverable. To this subdivision belong the pupæ of all Lepidoptera, and of them alone. These are what Linné termed obtected pupæ.
iii. Those pupæ which are inclosed in the thick and opaque skin of the larva, through which no trace of the perfect insect can be discovered. These, which Linné termed coarctate pupæ, include a large proportion of the dipterous genera; as Œstrus L., Musca L., Empis L., Conops L., &c. &c.[568]
I shall next advert, chiefly to the pupæ of the grand division last described, under the distinct heads of substance, figure, and parts; colour, age, sex, motions, and extrication of the perfect insect.
i. As to their substance—at first interiorly all pupæ consist of a milky fluid, in which the unformed members of the future perfect insect may be said to float, and in[Pg 244] which they may be discerned, and separated with the point of a pin[569]. In proportion as these acquire consistency, and are more and more developed by the absorption of the surrounding fluid, they occupy its place, and fill up the cavity of the puparium. The rest of this fluid passes off by transpiration[570]. Reaumur is of opinion that it is from the epiploon, or corps graisseux, that this matter is prepared, which he regards as analogous to the white of an egg[571]. In coarctate pupæ the included animal, or the pulp that contains its germes (in which the limbs and body at first are not discernible), fills at this period the whole skin-cocoon; but in proportion as the above evaporation takes place, and the consolidation of the body and parts proceeds, it shrinks at each end, so that when near assuming the imago, a considerable cavity appears both at the head and tail of the cocoon[572]. At this period of its existence, from the quantity of fluid included in the puparium, the animal weighs usually considerably more than it does when become a perfect insect[573].
The exterior integument or skin of pupæ, which is usually lined with a very thin white pellicle, is of different consistence in different orders. In the Coleoptera and Hymenoptera it is, with a few exceptions, of a soft and membranous texture; in the Lepidoptera (especially those that are not defended by cocoons), and Diptera, it is more rigid and harder, being either coriaceous or corneous. Lepidopterous pupæ, however, are not excluded from[Pg 245] the last skin of the larvæ with this hard covering. At the moment of this change the envelope is nearly as soft and membranous as in the order first mentioned. But they are besides covered with a viscous fluid, which appears to ooze out, chiefly from under the wings, and which very soon drying, forms the exterior hard shell[574]. At first the antennæ, wings, and legs, like those of Coleoptera and Hymenoptera, can be each separated from the body; and it is only after these parts have been glued together by the fluid just mentioned, which takes place in less than twenty-four hours[575], that they are immoveably attached to the body of the pupa, as we usually see them. In fact, the essential difference between incomplete and obtected pupæ seems to be, that in the former the limbs and body are only covered each with a single membranous integument, whereas in the latter they are besides glued together by a substance which forms an additional and harder envelope. It is not easy to explain the alteration that takes place in the texture of the skin of such dipterous pupæ as retain the skin of the larva. In the latter this is generally a transparent and very fine membrane: yet the very same integument becomes to the pupa an opaque and rigid case.
The surface of the skin of the greater number of pupæ is smooth, but in those of many Papilionidæ it is rugose and warty: this you may see, particularly in that of Papilio Machaon. In many of the hawkmoths (Sphinx L.) it is covered with impressed puncta. In Attacus Io the upper side of the channels that separate the intermediate segments of the abdomen are curiously striated with transverse[Pg 246] striæ, formed of very minute granula, the lower side being transversely sulcated. In some few instances, as in Arctia Salicis, Laria pudibunda and fascelina, the skin of the pupa is clothed with hair[576]: as is also that of Hesperia Bixæ, according to Madame Merian[577]. De Geer has described a little beetle under the name of Tenebrio lardarius (Latridius Latr., Corticaria Marsh.), the pupa of which is beset with very fine hairs, terminating in a spherical or oval button[578].
ii. I shall include under the same head both the figure or shape, and parts of pupæ, as the latter in most kinds are either the same or nearly the same as those of the larva, or merely incasing those of the imago, so as not to require that detailed notice that I judged necessary when treating of the parts of larvæ.
With regard to incomplete pupæ, nothing further can be said of their extremely various figure, than that it has a general resemblance to that of the perfect insect. The head, trunk, abdomen, and their respective external organs, are alike visible in both; but in the pupæ, the latter, instead of occupying their natural situation, are all closely folded under the breast and abdomen: or, as in the case of the long ovipositors of some Ichneumons, laid along the back. In a specimen of some coleopterous insect now before me, the following is the order of the arrangement of the parts:—The head is inflexed; the mandibulæ are open; between them are seen the labium and labial palpi; these appear to cover and conceal the maxillæ, and the maxillary palpi extend on each side beyond them; the[Pg 247] antennæ pass above the thighs of the two anterior pair of legs, and then turning down over the breast between them and the posterior legs, repose upon the base of the wings; which also are turned down between the intermediate and posterior pair of legs, and rest upon the latter; the tibiæ are bent in and folded upon the thigh, and the tarsi turn outwards[579]. In another coleopterous species, the wings and elytra are placed under the hind-legs. In Hymenopterous pupæ the antennæ appear usually to lie between the legs[580]. In many Tipulæ the long legs are bent into three folds in the pupæ; but the tarsi are extended, and lie close to each other, the anterior pair being the shortest[581]. In a specimen belonging to this tribe in my cabinet, which I think contained Ctenocera pectinicornis, the six leg-cases are of the same length, exactly parallel and adjacent, and being annulated wear the appearance of tracheæ[582]. These parts have each their separate case, so that a pin may be introduced between them and the body: which cases, as well as the general envelope, are usually formed of a fine soft transparent membrane; but sometimes, as in the lady-bird (Coccinella), the tortoise-beetle (Cassida), the crane-fly (Tipula), &c. it is harder and more opaque, so that though it is usually easy for a practised Entomologist from an examination of the pupa, particularly in the Hymenoptera, to predict to what genus the insect to be disclosed[Pg 248] from them will belong, yet in these cases the organs being not so conspicuous, a less experienced examiner might be perplexed, and unable to come to a conclusion.
Although hymenopterous pupæ have usually no parts but what are afterwards seen in the perfect insect, this is not the case with several coleopterous and dipterous ones, which are furnished with various temporary appendages, indispensable to them to bring about their final change, or for other purposes. Thus, the pupa of the male of Lucanus Cervus has two short, jointed anal processes[583]. That of Hydrophilus caraboides has a pedunculated lunulate one; and moreover, the sides of the abdominal segments, and the top of the thorax, are beset with hairs, which are not seen in the perfect insect[584]. The abdomen of many, also, is armed with spines. That, the arrangement of whose organs I lately described, has a quadruple series in the back of this part; viz. on each of the first five segments, 3, 2, 2, 3. The five first ventral segments also have on each side three spines; the inner are incurved, the intermediate nearly upright, and the outer one recurved. These spines, except those of the innermost ventral series, terminate in a bristle. In another coleopterous species the back part of the head is armed with a pair of lateral spines, and that of the thorax with three processes, the external ones armed with a single spine, and the intermediate one with a pair. De Geer has figured the pupa of an Asilus, the head of which is armed with eight spines—two robust ones in front, and three smaller ones, connected at the base on each side. The abdominal segments, also, are fringed with spines[585]. The abdomen of the pupa of[Pg 249] Ctenocera pectinicornis is armed with several strong conical spines, pointing mostly towards the tail, which is likewise the case with that of Tipula lunata[586]. As the above pupæ are usually subterranean or subcortical, the spines assist in pushing them out of the ground, &c. The respiratory horns that proceed from the thorax of the pupæ of many of the aquatic gnats will be noticed in another place. Those of Corethra culiciformis and of some other aquatic gnat-like Diptera, have their anus furnished with a pair of oars, or natatory laminæ, by which they rise to the surface[587].
The figure of obtected pupæ, or chrysalises, is more uniform. They are commonly obtuse at the anterior extremity, and gradually contracted to a point at the posterior, or tail. The outline usually inclines to a long oval or an ellipse; but in some, as Attacus Io and Luna, the pupa is shorter and more spherical. In Geometra sambucaria it represents an elongated cone, and in Hepialus it is nearly cylindrical. In the butterfly tribe (Papilio L.) the outline is frequently rendered angular by various protuberances.
In all these pupæ may be distinguished the following parts:—first, the Head-case (Cephalo-theca), or anterior extremity; secondly, the Trunk-case (Cyto-theca), or intermediate part; and thirdly, the Abdomen-case (Gastro-theca).
1. The Head-case covers and protects the head of the inclosed imago. From its sides behind proceed the[Pg 250] antennæ-cases (Cera-theca); and before from the middle, the tongue-case (Glosso-theca). Just below the base of the antennæ-case you may discern the eye-cases (Ophthalmo-theca), surrounded on their inner side by a crescent-shaped lævigated piece, which may perhaps transmit some light to the inclosed prisoner.
2. The Trunk-case, divided into the thorax, or upper surface, extending from the head to the dorsal segments of the abdomen, and consisting of three pieces, answering to the prothorax, mesothorax, and metathorax of the perfect insect: the first answering to the prothorax small, the second covering the mesothorax very large, and the two next representing the metathorax, at first appearing to belong to the abdomen, but having no spiracle; and the breast (pectus) or under-surface reaching from the head to the ventral abdominal segments, from which proceed the wing-cases (Ptero-theca) and leg-cases (Podo-theca), which organs, with the antenna-cases and tongue-case, entirely cover, or rather form, the breast. The arrangement of the whole is as follows:—The wing-cases, which are more or less triangular, and exhibit the larger nervures of the wings, are a lateral continuation of the mesothorax, which turn downwards from the sides of the breast, and cover, or replace, the three first ventral segments of the abdomen. The antenna-cases, united to the anterior portion of the head just behind the eye-cases, repose immediately next to those of the wings running parallel with their inner margin. Then follow the legs, the tibiæ forming an angle with the thigh, and the case of the anterior pair being innermost, and representing the breast-bone in the pupa. The tongue lies over the forelegs,[Pg 251] except in the case of some sphinxes, which I shall notice afterwards: so that the glosso-theca covers both them and it.
3. The abdomen-case consists of nine segments when viewed on the back, and of only six when viewed below; so that it might be said to have ten dorsal and six ventral segments: but the fact is, that the place of the three anterior ventral segments, or rather ventral portions of the segments, (for they form complete rings without any lateral suture,) are replaced by the wings and other organs: in consequence of this, the fourth segment, which is less covered than the three first, at its posterior margin forms an annulus or ring. In counting the abdominal segments of a pupa, you must be careful not to include the piece that represents the metathorax, which looks as if it belonged to the abdomen[588]. In the pupæ of butterflies you will discover evident traces of ten dorsal segments; but in many moths, and some hawk-moths, you will perceive at first only eight, or even seven, but a closer examination will enable you to discover the line that marks out the others; and if you divide the puparium longitudinally, and inspect its internal surface, you will see very visible sutures between them. The intermediate segments are sometimes separated from each other and the preceding and subsequent ones by deep channels. In the pupa of Papilio Machaon there is one such channel between the third and fourth segments. In Bombyx[Pg 252] regalis the channel is between the sixth and seventh, and in B. imperatoria there are three, namely, a channel between the third and fourth, and fourth and fifth, and fifth and sixth segments. The way in which insects with an exserted sting fold it in the pupa seems not to have been noticed; but from an observation of De Geer upon one species of Ichneumon, it appears to be turned up over the back of the abdomen[589].
These little animals, thus swathed and banded, exhibit no unapt representation of an Egyptian mummy; though Lamarck applies the term Mumia to incomplete pupæ[590], to which it seems less happily applicable.
Chrysalises, as to the modifications of their general figure, may be conveniently divided into two great classes: first, those that have no angular projections, the anal mucro of some excepted, on different parts of their body; and secondly, those which have such projections. Each of these classes affords variations in its peculiar characters which require to be noticed.
1. The first of these are called angular pupæ[591], and are confined to the Butterfly or diurnal tribes. In some the head projects into one short conical protuberance: this you may see in the chrysalis of the common cabbage butterfly (Pieris Brassicæ), and others of the same genus[592]; in the brimstone-butterfly (Colias Rhamni[593]), and in the beautiful purple emperor or high-flier (Apatura Iris F.[594]): though in this last it is not conspicuous. But[Pg 253] the most remarkable instance of a single eminence from the head is exhibited by the pupa of a tropical butterfly (Morpho Idomeneus Latr.), figured by Madame Merian. In this the head projects into a long incurved obtuse horn[595]. In others the head is armed with two mucros, or conical eminences. This is the case with the common butterfly of the nettle (Vanessa Urticæ F.[596]), and with that of the beautiful Papilio Machaon[597]. In these the prominences are trigonal. These processes, which in some, as in the peacock-butterfly (Vanessa Io), stand upright[598], and in others diverge (Papilio Machaon), form the eye-cases of the included imago; and in their outer base is planted the crescent-shaped piece I lately mentioned, which seems intended to convey light into it. In many the prothorax, besides a lateral angular projection, has in the middle another triangular or trigonal one, somewhat resembling a Roman nose; on each side of which is a smaller elevated black point: so that it requires no great stretch of imagination to find out in it a sort of resemblance to the human face, which, though not quite so striking as honest Goedart figures it[599], is however very considerable. In the pupa of Morpho Menelaus, figured by Madame Merian[600], this nasiform prominence of the prothorax is extended into a long arched horn, reaching to the middle of the abdomen. The pupa of the silver-washed fritillary (Argynnis Paphia F.), and others of the same genus, exhibit beneath this nasiform prominence[Pg 254] a very deep depression, itself beset with one or more series of smaller angular elevations. The back of the abdomen is often furnished with two rows of protuberances, in some species larger, in others smaller[601]; sometimes sharp and conical, and sometimes flat, and in some instances resembling the fins of fishes[602]. These bosses usually decrease in size towards the tail.
2. The second kind of chrysalises are denominated conical[603]. These, which include the crepuscular and nocturnal Lepidoptera, and the butterflies with onisciform larvæ, have no protuberances, and are less variable in their form—their anterior extremity being almost constantly oval and rounded, and their posterior conical and acute. An exception to this form is met with in the pupa of a moth long celebrated (Lasiocampa Pithyocampa)[604], which has the head acute and the tail obtuse, and armed with two points[605]. Another occurs in that of the Cossus, which has two points on the head, by which it makes an opening in its cocoon: when it assumes the imago, one of these is placed below the other[606]. And some few have the anterior end nearly flat instead of rounded. The pupa of the orange-tip butterfly (Pieris Cardamines) seems intermediate between the angular and conical kinds: it is somewhat boat-shaped, and distinguished by a fusiform process from the head and tail[607]. Other modifications of the usual figure are met with, but are for the most part so slight as not to require notice. One[Pg 255] or two, however, should not be passed over. The pupæ of many hawk-moths (Sphinx L.) have the anterior piece of the head-case elongated into a sort of cylindrical proboscis, which is incurved beneath the breast: you will find this formation in S. Convolvuli and Ligustri[608]. In some, as in a species figured by Madame Merian, that feeds upon the Annona squamosa, it is rolled up like a serpent in many folds[609]. In Noctua Linariæ the tongue-case turns upwards, and is prominent laterally beyond the body[610]. This singular appendage is one of those beautiful instances of compensating contrivances, as Dr. Paley calls them, which perpetually occur in the insect tribes. The tongue of these hawk-moths is of very great length, often three inches, while the pupa itself is scarcely two; it could not possibly, therefore, have been extended at length, as it is in common cases, but is coiled up within the above protuberance. When the tongue is but a little longer than the breast, the ordinary plan is adhered to, but the apex of the breast projects a little over the abdomen into a sort of nose, in which the end of the tongue is contained. This conformation may be seen in the pupa of Noctua Gamma, Verbasci, and many other species. Sometimes, as in N. Linariæ F., this projection is recurved into a short horn.
I have before adverted to the adminicula or short spines looking towards the anus, with which the dorsal segments of the abdomen of some pupæ are armed; and by which, when the time for their exclusion is arrived, they are enabled to push themselves upwards or outwards from[Pg 256] their several places of confinement[611]: you will find these in the pupa of the great goat-moth (Cossus ligniperda); and in the cylindrical pupa of the moth called the ghost (Hepialus Humuli F.) there are two rows of sharp triangular spines on the back of each segment. These are not laid flat, but, as they do also in the Cossus, form an acute angle with the body; which gives them greater power of resistance. Those that constitute the row nearest the base of the segment are longer than the anterior row, the middle spines than the lateral ones. The first and last segment are without them, and the last segment but one has a sharp ventral transverse ridge, armed with many sharp teeth[612]. The abdominal spines lately mentioned, of semicomplete pupæ, are also adminicula.
The tail of this description of pupæ is in many instances armed with a mucro, or sharp point, emerging from its upper side. You will see this in most hawk-moths. In the pupa of Hesperia Proteus the mucro is truncate at the apex; in that of Bombyx imperatoria it is long, and terminates in two diverging points. In the majority of chrysalises of both descriptions the tail is acute, and usually furnished with hooks of different kinds. These are so various in shape and number, &c. that they would probably afford good characters for discriminating many allied species. In some there are but two or three, in others five or six, in others they are more numerous[613]. Sometimes they are quite straight[614], but most commonly recurved, so as to form a hook. The hawk-moths, and a[Pg 257] few others, as Bombyx Pini, Cerura Vinula, &c., have no anal hooks whatever. Under this head I shall observe, that in many conical pupæ below the anal angle or mucro, is the appearance of a vertical foramen or passage: this is particularly conspicuous in Hepialus, in which it is surmounted by a bifid ridge, and has under it a pair of minute black tubercles.
A pretty accurate judgement of the division to which the perfect insect when disclosed will belong, may usually be formed from the figure of its chrysalis. All the angular ones, with scarcely any exception, inclose butterflies. The converse, however, does not hold; for some that are not angular, as those of Parnassius Apollo and Mnemosyne, and most of the Linnean Plebeii urbicolæ, also inclose flies of that description. With these exceptions, all conical chrysalises give birth to moths or hawkmoths. An idea even of the family or genus under which the perfect insect will arrange, may be generally formed from the figure of the chrysalis; less distinctly, however, in the conical or rounded, than in the angular kinds, in which the prominences of the head and trunk, as before explained, usually vary in different families. Even the sex of some moths may be judged from the pupæ: those of females being thicker; and those also of the females that have no wings, or only the rudiments of them, will of course vary somewhat from the ordinary form: but there is a still more striking difference in that of Callimorpha? vestita F., and others of the singular tribe before noticed[615], called by the Germans Sacktrager (sack-bearers), from the sack-like cases in which the larva resides. The[Pg 258] females of these having not only no wings, but no antennæ, and legs not longer than those of the larva, their pupa more resembles that of a dipterous than of a lepidopterous insect, it being not easy to determine which is the head and which the tail[616].
In these too we can often learn from the outline of the wing-cases, whether the inhabitant of the chrysalis has these organs indented or intire. If the former, the margins of these cases are sinuate, as in that of Vanessa C. album; if the latter, they are intire, as in Pieris Brassicæ. Even in conical pupæ,—the size, the shape of the antennæ, which may be distinguished through the skin that covers them, and slight modifications of the ordinary form,—give indications of the genus of the included insect sufficiently conclusive to a practised eye.
The true figure of coarctate pupæ when they are mature, the parts of the future fly being very visible, and each being included in a separate case[617], is that of those that belong to the incomplete division; but as this is a character not cognizable without dissection, it is customary, in speaking of pupæ of this description, to refer solely to the shape of the exterior covering, which is in fact a cocoon formed of the dried skin of the larva moulded into a different form. In this sense the figure of coarctate pupæ is extremely various. The majority of them are more or less oval or elliptical, without any distinct parts, were it not that they usually retain traces of the segments which composed the larva's body[618]. Of this figure are the pupæ of the common cheese-maggot[619], and many other flies. Others[Pg 259] (Sepedon Latr.) have the pupa shaped like a boat. That of Scæva Pyrastri F. assumes the figure of a flask; or, according to Reaumur's more accurate comparison, of a tear[620]. The tail of many of these pupæ, particularly of aquatic species, is elongated into a sort of beak, either simple or forked, or is beset with spines variously arranged. The pupa of Stratyomis Chamæleon, and other allied species, differs from all the rest of this subdivision in retaining the exact form of the larva[621]; and hence constitutes an exception to the general character of our second great Division.
iii. There is much less variety in the colour of pupæ than in that of larvæ. The majority of coleopterous and hymenopterous pupæ are white, or whitish; of lepidopterous and dipterous, brown of various shades, often verging on black in the former and on red in the latter. The angular lepidopterous ones, however, are more gaily decorated. Some, Pieris Brassicæ, are of a greenish yellow, marked with spots of black; others are of a uniform green, Apatura Iris, Pieris Cardamines; others, reddish, Vanessa C. album; others again red with black spots, Urania Leilus[622]. A still greater number shine as though gilded with burnished gold—either applied in partial streaks, Vanessa Cardui; or covering the entire surface, Vanessa Urticæ. It was from this gilded appearance in some obtected pupæ that the terms Chrysalis and[Pg 260] Aurelia were applied to the whole. The alchemists mistook this for real gold; and referred to the case as an argument in favour of the transmutation of metals. But Reaumur has satisfactorily shown, that in this instance the old proverb is strictly applicable—"All is not gold that glitters." He found that this appearance is owing to the shining white membrane immediately below the outer skin, which being of a transparent yellow gives a golden tinge to the former; in the same way that tinfoil, when covered with a yellow varnish, assumes the metallic appearance which we see in gilt leather. He mentions, too, that for the production of this effect—it is essential that the inner membrane be moist: whence may be explained the disappearance of the gilding as soon as the butterfly is ready to escape from the pupa. The shade of colour in these gilded chrysalises is various: some are of a rich yellow, like pure gold; others much paler; and some nearly as white as silver. That of Hipparchia Cassiæ F. is red with silver spots[623].
Though by far the greater number of the chrysalises of moths are of an uniform chestnut, brown, or black,—a few are of other colours; as that of Geometra alniaria, which is of a glaucous blue; of Noctua sponsa, lilac; and of Noctua pacta, of a lovely blue, caused by a kind of bloom, like that of a plum, spread upon a brown ground. A similar bloom is found on that of Parnassius Apollo, and on the anterior part of that of Platypterix cultaria and sicula; in which last, Kliemann observed it to the[Pg 261] be renewed when rubbed off[624] Many pupæ have the sheaths of the wings of a different colour from that of the rest of the body; a few are variegated with paler streaks or bands, as Clostera Anastomosis, which has two red longitudinal stripes down its dark-brown back; and that of the common gooseberry and currant moth, which may be found in every garden, has alternate rings of black and yellow[625].
A few pupæ vary in their colour, as the painted lady-butterfly (Vanessa Cardui), some of which are light-brown with gray streaks and golden dots, others wholly of a golden yellow or brown, others of a light green[626].
Almost all at their first assumption of the pupa state have a different colour from that which they take a few days afterwards. This last they retain until the disclosure of the perfect insect; except some that have transparent skins, which a few days previously to this period exhibit the colours of the included animal.
iv. There is as great variety in the length of the age of Insects in their pupa as in their larva state. Some species continue in it only two or three days (Aleyrodes Chelidonii Latr., Tinea proletella L.); others, as many weeks, or months, or even years. Each, however, has in general a stated period, which in ordinary circumstances it neither much exceeds nor falls short of. The only general rule that can be laid down is—that small pupæ continue in that state a shorter time than those of larger bulk. Thus, amongst coleopterous genera, the more minute species of Curculio L.; amongst the Hymenoptera,[Pg 262] the Ichneumones minuti L.; amongst the Lepidoptera, the subcutaneous tribes; and the majority of the Diptera,—remain as pupæ only a few days or weeks: while the larger species in all these orders commonly exist in the same state several months—many even upwards of two years. There are, however, numerous exceptions to this rule; for some large pupæ are disclosed in a much shorter time than some others not a twentieth part of their bulk.
The reasons both of the rule and of the exceptions to it are sufficiently obvious. And first, as to the rule:—If you open a pupa soon after its assumption of that state, you will find its interior filled with a milky fluid, in the midst of which the rudiments of its future limbs and organs, themselves almost as fluid, swim. Now the end to be accomplished during the pupa's existence is, the gradual evaporation of the watery parts of this fluid, and the development of the organs of the inclosed animal by the absorption and assimilation of the residuum. Reaumur, by inclosing a pupa in a stopped glass tube, collected a quantity of clear and apparently of pure water, equal to eight or ten large drops, which had evaporated from it, and was condensed against the sides of the tube, and it was found to have lost an eighteenth part of its weight[627]. It is plain, therefore, that this necessary transpiration, other circumstances being alike, must take place sooner in a small than in a large pupa. Next, as to the exceptions:—Since the more speedy or more tardy evaporation of fluids depends upon their exposure to a greater or less degree of heat, we might à priori conclude, that pupæ[Pg 263] exposed to a high temperature would sooner attain maturity, even though larger in bulk, than others exposed to a low one:—and this is the fact. The pupa of a large moth, which has assumed that state in the early part of summer, will often disclose the perfect insect in twelve or fourteen days; while that of an Ichneumon, not one hundredth part of its size, that did not enter this state till late in autumn, will not appear as a fly for seven or eight months. But this is not the whole. The very same insect, according as it has become a pupa at an earlier or later period of the year, will at one time live but a few weeks, at another several months, in that state. Thus, if the caterpillar of Papilio Machaon, one of those which has annually a double brood, becomes a pupa in July, the butterfly will appear in thirteen days: if not until September, it will not make its appearance until the June following; that is, not in less than nine or ten months: and the case is the same with the pupæ of Noctua Psi, and of a vast number of other insects. To put beyond all doubt the dependence of these remarkable variations on temperature merely, it was only necessary that they should be effected, as Lister long ago advised[628], by artificial means. This Reaumur accomplished. In the month of January he placed the chrysalises of several moths and butterflies, which would not naturally have been disclosed until the following May, in a hothouse: the result was, that the perfect insects made their appearance in less than a fortnight, in the very depth of winter; and by other numerous and varied experiments he ascertained, that in this heated atmosphere five or six days[Pg 264] hastened their maturity more than as many weeks would have done in the open air. The disclosed insects were in every respect perfect, and the females, after pairing, laid their eggs, and then died, just as if they had not been thus prematurely forced into existence. The converse of this experiment equally succeeded:—by keeping pupæ the whole summer in an icehouse, Reaumur caused them to produce the fly one full year later than their ordinary period[629].
This extraordinary fact leads us to a very singular and unexpected conclusion—that we have the power of lengthening or shortening the life of many insects at pleasure; that we can cause one individual to live more than twice as long as another of the same species, and vice versâ. Had Paracelsus made this discovery, it would have led him to pursue his researches after the elixir of immortality with redoubled confidence, and would have supplied him with an argument for the possibility of prolonging the life of man beyond its usual term, which his sceptical opponents would have found some difficulty in rebutting. Even the logical Reaumur seems inclined to infer from it, that this object of the alchemists was not so chimerical as we are wont to conclude[630]. He confesses, however, if it were to be attained only by the same process as effects the extension of an insect's life—by prolonging its state of torpor and insensibility,—that few would choose to enjoy it on such conditions. The man of pleasure, blunted by excess of use to all modern stimuli, might perhaps not object to a sleep of a hundred years, in the hope of finding something new under the[Pg 265] sun when he waked; and an ardent astronomer would probably commit himself with scientific joy to a repose as long and as sound as that of the seven sleepers, for the chance of viewing his predicted return of a comet, on stepping out of his cave: but ordinary mortals would consign themselves to the perils of so long a night with reluctance, apprehending a fate no better than what befel the magician, who ordered himself to be cut in small pieces and put in pickle, with the expectation of becoming young again[631].
The duration, then, of an insect's existence in the pupa state, depends upon its bulk, upon the temperature to which it is exposed, and upon a combination of these two circumstances. This experiment appears very simple. We seem to ourselves to have accomplished what is so often undertaken in vain—to have found an entrance into the cabinet of Nature, and to have made ourselves masters of the contents of one of the pages of her sealed and secret book. We deceive, ourselves, however: this book, when it seems most legible, is often interlined with sympathetic inks, if I may so speak, which require tests unknown to us for their detection. If you lay up a considerable number of the pupæ of a moth now called Eriogaster lanestris, the larva of which is not uncommon in June on the black-thorn, selected precisely of the same size, and exposed to exactly the same temperature, the greater number of them will disclose the perfect insect in the February following; some not till the February of the year ensuing, and the remainder not before the same[Pg 266] month in the third year[632]. Mr. Jones of Chelsea, a most acute lepidopterist, in one of his excursions captured a female of Arctia mendica, another moth, which laid a number of eggs, thirty-six of which produced caterpillars: all these fed, spun their cocoons, and went into the pupa state in the usual manner, but at the proper season only twelve produced the fly. As this was no uncommon circumstance, he concluded that the rest were dead: to his great astonishment, however, in the next season twelve more made their appearance; and the following year the remainder burst into life, equally perfect with the foregoing[633]. In this extraordinary result, which also occasionally has been observed to take place in the emperor-moth (Saturnia pavonia), the privet-hawkmoth (Sphinx Ligustri), and that of the spurge (S. Euphorbiæ)[634], and other species,—it is clear that something besides mere size and temperature is concerned: for, these circumstances being precisely alike, one pupa arrives at maturity in six months, and another of the same brood requires between two and three years. We can guess, that the end which the All-wise Creator has in view, in causing this remarkable difference, is the prevention of all possibility of the destruction of the species. Eriogaster lanestris and Arctia mendica, &c., for instance, are doomed, for some reason[Pg 267] unknown to us[635], to be disclosed from the pupa in the cold and stormy months of February and March, almost every day of which in certain years is so ungenial that few insects could then survive exposure, much less deposit their eggs and ensure the succession of a progeny. Now, were all these to make their appearance in the perfect state in the same year, it might happen that the whole race in a particular district would be destroyed. But this possibility is effectually guarded against by the beautiful provision under consideration, it being very improbable that three successive seasons should be throughout unfavourable; and without such occurrence, it is clear that some of the race of this moth will be preserved. In the case of other moths, whose pupæ though disclosed in the summer are governed by the same rule, the prevention of the extinction of the species, by any extraordinary increase in a particular year of their natural enemies, seems the object in view[636]. But though the intention be thus obvious, the means by which it is effected are impenetrably concealed. What physiologist would not be puzzled with the eggs of a bird, of which one-third should require for their hatching to be sat upon only a fortnight, another third a month, and the remainder six weeks? Yet this would be an anomaly exactly analogous to that observed by Mr. Jones with respect to the pupæ of A. mendica. Reaumur found that when the skin of pupæ was varnished, so as to prevent absorption, the appearance of[Pg 268] the fly happened nearly two months later than in ordinary circumstances. Are we to conjecture that those of the moth just mentioned, or of E. lanestris, that are latest matured, from a greater degree of viscidity in the fluid that forms them[637], have thicker and more impervious skins than those disclosed at an earlier period? Or are we to refer the difference to some unknown peculiarity of organization? On any supposition, the fact remains equally wonderful; and I know of none the illustration of which is more worthy of the patient investigation of the physiologist.
As the period of maturity of the perfect insect is thus in some cases not fixed even to years, and as in many it seems dependent upon such variable causes; nothing appears more improbable than that it should ever be so strictly determined, that even the week in which the fly will leave its pupa-case can be pretty accurately predicted. Such, however, is the fact with regard to the Ephemera so interestingly described by Reaumur, the myriads of which that issue from the banks of the Seine all appear in two or three days, somewhere between the 10th and 18th of the month of August[638] in every year; at which time the fishermen regularly expect them. A like regularity attends the appearance of those described by Swammerdam, which every year, for three days about the feast of St. John, issue in clouds from the Rhine[639]—Not only is the week fixed, but in several instances even[Pg 269] the hour. The Ephemeræ observed by Reaumur appear at no other time than between eight and ten o'clock in the evening; and so unalterably is their exclusion fixed, that neither cold nor rain can retard it. Between these hours, in the evenings on which they appear, you may see them fill the air, but an hour before or after, you will in vain look for one[640]. So also the silkworm-moth and the hawkmoth of the evening primrose (Sphinx Œnotheræ) constantly break forth from the pupa at sunrise: and the hawkmoth of the lime (Smerinthus Tiliæ) as certainly at noon[641]. Schroeter states, that of sixteen specimens of the death's-head-hawkmoth (S. Atropos) which he bred, every one was disclosed between four and seven o'clock in the afternoon[642].
Before I conclude this head, I must observe, that after a caterpillar or gnat has spun its cocoon, it sometimes remains for a considerable period before it incloses itself in the pupa-case, and casts off the form of a larva. Thus the little parasite (Ichneumon glomeratus L.) that destroys the caterpillar of the common cabbage-butterfly, remains a larva in its cocoon for many months, but it becomes a perfect insect a few days after it has put on its puparium[643]; and the caterpillars of the great goat-moth (Cossus ligniperda), if they spin their cocoon in the autumn, remain in it through the winter in the larva state; whereas, if they inclose themselves in the month of June, they assume the pupa, so as to appear as flies in three or four weeks[644]. It is not therefore easy to state precisely[Pg 270] the age of those pupæ which are produced from larvæ that spin cocoons.
v. I have not much to say with regard to the sex of pupæ. The male is probably to be distinguished from the female by being smaller; but in the first great division of pupæ, those which resemble the larvæ, and are locomotive, the female in numerous cases may be known by the Ovipositor, or instrument for depositing her eggs in their proper station: and the male also has his anal instruments. Sometimes in this state the animal is so matured, as to be capable of continuing its kind. I have found the pupæ both of a Gryllus L. and of a Cimex L. in coitu.
vi. Though the pupæ of the second great division are usually not locomotive, yet I must not omit some notice of their motions. As the legs of insects in this state are folded within a common or partial integument, of course none of the pupæ now under consideration, with the exception of those of the Trichoptera order, can walk: coarctate ones are even incapable of the slightest motion, and exhibit no symptom whatever of animation. Some of those that are termed incomplete, however, and most chrysalises, have the power of communicating to their bodies a slight movement, extending more or less in different species, which is effected by the abdominal segments solely. The latter, during the first twelve hours of being pupæ, when their skin is soft, frequently turn[Pg 271] themselves, that the side on which they lie may not be flattened; afterwards by far the majority merely wriggle or twist their abdomen when touched, or in any way incommoded or disturbed. We learn from De Geer, that the pupa of the ghost-moth (Hepialus Humuli), the cocoon of which is more than twice the length of the chrysalis, moves in it from one end to the other[645]. Bonnet observed one of a moth (perhaps Lasiocampa Quercus), which alternately fixed itself at the top and bottom of its spacious and obliquely-fixed cocoon; descending slowly, but ascending as quickly, and almost in the same manner, as a chimney-sweeper in a chimney[646]. The pupa of the weevil of the water-hemlock (Lixus paraplecticus) will move from one end of the interior of a branch to another by means of its adminicula, aided by the motion of its abdominal segments[647]. But the most locomotive of pupæ of the second division are those of gnats, and many Tipulidans, which pass this state in the water. These will move from the bottom to the surface, and back again, with great facility and velocity. I have before mentioned several other motions of pupæ[648], which I shall not repeat here, by which they extricate themselves from their several places of intermediate repose, before they leave the puparium: if the imago were to be disclosed in the interior of a tree, or in the earth, its wings would be materially injured in forcing its way out. The object of several of the above motions may be to alarm insects that might attack these defenceless beings. The twirling motion in particular, formerly noticed[649], in some species, by[Pg 272] causing a rustling against the sides of the cocoon, makes a considerable noise—so singular in that of a red underwing-moth (Noctua pacta), that Rösel tells us, (who by the by was more timid than becomes a philosopher,) that the first time he heard it, he had nearly thrown away the box that contained it, in his fright[650].
vii. We are next to consider The extrication of the perfect insect from the puparium, or pupa-case, and from the cocoon. The period when the pupa has attained maturity, and the inclosed insect is ready to burst the walls of its prison, may be often ascertained. Just at this time the colour frequently undergoes an alteration, the golden or silver tint of the gilded chrysalises vanishes; and those which are transparent, usually permit the form and colours of the insect within and the motions of their limbs to be distinctly seen through them. In the Libellulina the eyes become more brilliant[651]. The mature pupæ of the moth lately mentioned (Eriogaster lanestris) have a particular swell of the abdominal segments, not apparent in those that are to continue till another season, or longer[652]. Those of the case-worms (Trichoptera) push off the grates from the cases which they have hitherto inhabited, and swim about[653]. Other signs and motions doubtless predict the approach of this great change in other species, which have not been recorded.
The mode in which insects make their way out of the puparium differs in different orders. In obtected pupæ, the struggles of the included butterfly or moth first effect a longitudinal slit down the middle of the thorax, where[Pg 273] there is usually a suture for the purpose. The slit rapidly extends along the head, and down the parts which compose the breast, and the insect gradually withdraws itself from its case. It is not, however, from the outer skin merely that it has to disengage itself, but also from a series of inner membranous cases, which separately inclose the antennæ, proboscis, feet, &c., as a glove does the fingers; and similar cases inclose the parts of the perfect insect in pupæ of all the other orders. This is sometimes a work of difficulty, but ordinarily it is effected with ease.
Incomplete and semicomplete pupæ undergo nearly the same process, save that in them the body is not swathed up in a common case; and therefore they have only to liberate themselves from the partial cases that envelop the several parts of their body.
In coarctate pupæ, as those of Muscidæ, Syrphidæ, Œstridæ, &c., the process is different. Their outer-case is ordinarily more rigid and destitute of the sutures, which in the former tribes so easily yield to a slight effort. Yet in these, at the anterior end under which the head of the fly lies, and from which it always issues, there is commonly a sort of lid, joined by a very indistinct suture to the rest, which can be pushed off, leaving a sufficient opening for the egress of the insect. In the pupæ of many of this tribe this lid is composed of two semicircular pieces, which can be separately removed. Many species seem to be able to force off the lid of their puparium, by merely pushing against it with their heads: but the common flesh-fly and many other Muscidæ, which are perhaps too feeble to effect this, or whose puparia are stronger than ordinary, are furnished with a very remarkable[Pg 274] apparatus for this express and apparently sole purpose. They are gifted with the power of introducing air under the middle part of the head, to which the antennæ are fixed, and of inflating that part into a sort of membranous vesicle as big as the head itself; by the action of which against the end of the pupa-case, the lid is soon forced off. So powerful is this singular lever, that it is even sufficient to rupture the fibrous galls in which the pupæ of the gay-winged Tephritis Cardui[654] are inclosed. That it is designed by Creative Wisdom to answer this sole purpose seems proved, from its disappearing soon after the disclosure of the fly, whose head shortly becomes all alike hard. Reaumur suspects that it may also be intended to promote the circulation of the insect's fluids; but to me his reasons appear not conclusive[655]. In one instance a mode still more unexpected obtains. The illustrious naturalist just named found that the fly which proceeded from one of the rat-tailed grubs (Elophilus Latr.) had actually the power of completely reversing its situation in its narrow case; and that it then employed its tail in pushing off the lid, which other species remove by means of their heads[656].
The extrication of insects whose pupæ are above ground, like those of butterflies, many beetles, flies, &c., is comparatively a simple operation. But what, you will ask, becomes of those species whose pupæ are concealed deep in the earth, or in the heart of the trees on which their larvæ have fed? Of this you shall be informed.—Coleopterous insects disclosed from pupæ thus circumstanced,[Pg 275] wait until their organs have acquired strength, and their elytra are sufficiently hardened to protect their filmy wings from damage in forcing their way through the earth or wood which covers them. Thus Oryctes nasicornis, a rhinoceros beetle common on the Continent, is a full month before it reaches the surface of the earth, after quitting its puparium. But it is evident that no delay would enable lepidopterous or dipterous insects, which are without elytra, to make their way out of such situations, without irreparable injury to their delicate wings. Many of these, therefore, while still within the hard case of the pupa, have the precaution, a few days previously to their exclusion, to force themselves up to the surface of the earth, or, when they reside in the interior of trees, to the entrance of their hole. This is effected by a successive wriggling of the abdominal segments, which in several species, of the Coleoptera, Lepidoptera, and Diptera orders, for this purpose, as has been more than once observed[657], are furnished with sharp points (adminicula), admitting a progressive, but not a retrograde motion. The puparia of the great goat-moth (Cossus ligniperda) may be often seen projecting from orifices in willow-trees; and those of the common crane-fly (Tipula oleracea) from the surface of the earth, to which they have thus made their way from a depth of several inches.
In all the preceding instances the exclusion of the perfect insect is complete, as soon as it has withdrawn itself from the puparium. But to a very large number, even after this is effected, the arduous task still remains of[Pg 276] piercing the cocoons of leaves, of thick silk, of tough gum, or even of wood, in which the pupæ are incased. We can readily conceive how the strong jaws of coleopterous and hymenopterous species may be employed to release them from their confinement. But what instruments can be used for this purpose by moths in a state of great debility, whose mouth has nothing like jaws—merely a soft membranous proboscis? How shall the silkworm-moth (B. Mori) force its way through the close texture of a silken ball, through which the finger could not be easily pushed? Or the puss-moth (Cerura Vinula) pierce the walls of its house of glue and wood, which scarcely yield to the knife? You will not doubt that these difficulties have been foreseen by Infinite Wisdom, and provided against by Infinite Power. The egress of moths from their cocoons is secured in two ways;—either by some peculiarity in the first construction of the cocoon by the caterpillar, or by some process which the pupa or perfect insect is instructed to perform. As examples of each, several curious instances may be cited.
The larva of the moth which about 1760 made such havoc in the province of Angoumois in France, becomes a pupa in the interior of the grain of wheat which it has excavated; but the opening by which it first entered is not bigger than a pin's point, and is quite insufficient for the egress of the moth. How, then, is the latter to force its way through the tough skin which surrounds it? The larva, previously to assuming the pupa state, gnaws out a little circular piece at that end of the grain where the head of the future moth would lie, taking care not to detach it entirely. At this little door, which is sufficient to protect it from intruders, the moth has but to push, when it[Pg 277] falls down, and leaves a free passage for its exit. A contrivance almost similar is adopted by a caterpillar which feeds in the interior of the heads of a species of teazel (Dipsacus L.), for a minute and interesting history of which we are indebted to Bonnet. This caterpillar previously to its metamorphosis actually cuts a circular opening in the head, sufficiently large for the egress of the future moth; but to secure this sally-port during its long sleep, it artfully closes it with fibres of the teazel, closely but not strongly glued together[658]. Another small caterpillar described by the same author, resides in the leaf of an ash curiously rolled up into a cone, and then assumes the pupa, which is inclosed in a silken cocoon, ingeniously suspended by two threads like a hammock in the middle of its habitation, and of so slight a texture that it presents no obstacle to the extrication of the moth. It is the closely-joined sides of its leafy dwelling that form a barrier, which, were it not for the precaution of the larva, would be impenetrable to so small and weak an animal. The little provident creature, before its change to a pupa, gnaws in the leaf a round opening, taking care not to cut through the exterior epidermis. This door is to serve the moth for its exit, like that formed by the wheat-caterpillar. But in proportion to its bulk its verdant apartment is of considerable size. How then shall the moth know the exact place where its outlet has been traced? How, without a clue, shall it discover in its dark abode the precise circle which requires only a push to throw it down? Even this is foreseen and provided against. Out of twenty positions in which its hammock[Pg 278] might have been slung, the caterpillar has been directed so to place it, that the silken cord that suspends the head is fastened close to the side of the door which it has previously constructed; and the moth, guided by this filum ariadneum, at once makes its way out of an apartment which, but for this contrivance, might have been to it a labyrinth as inextricable as that of Minos[659].
The mode in which other caterpillars provide for their extrication, when become moths, from their silken cocoons, is not less ingenious. Those of Eriogaster lanestris (of which I have lately said so much,) and others, form oblong cocoons, which, viewed externally, you would at the first glance assert were of one solid piece: but on examining them more narrowly, you perceive one end of them to be a distinct lid, of a size large enough to permit the moth to issue out; and that it is kept in its place by a few slight threads, easily broken by pressure from within[660]. A few pages back[661] I mentioned a cocoon formed by the larva of Tortrix prasinana, of the shape of a boat reversed, composed of two inclined walls fastened together at the top and ends. In constructing this cocoon, it firmly glues to each other the top and one end, so as to form an impermeable suture; but the other end, at which the moth is to issue, though externally it seems as strong as the rest, is merely drawn close by a slender thread or two fastened on the inside, and easily broken from within. And, what is particularly singular in the construction of this ingenious habitation, the sides forming the end last mentioned, though originally requiring force to draw[Pg 279] them into their required position, become so elastic as to close again when the moth has passed between them and made her escape; the cocoon preserving its usual shape, even when deprived of its inhabitant[662]. A similar cocoon is constructed by another leaf-rolling caterpillar, that of Tortrix chlorana[663]. Many similar proofs of contrivance in the construction of silken cocoons might be adduced, but I shall confine myself to one more only—I mean that furnished by the flask-shaped brown one of Saturnia Pavonia, and some other moths. If you examine one of these cocoons, which are common enough in some places on the pear-tree or the willow, you will perceive that it is generally of a solid tissue of layers of silk almost of the texture of parchment; but at the narrow end, or that which may be compared to the neck of the flask, that it is composed of a series of loosely-attached longitudinal threads, converging, like so many bristles, to a blunt point, in the middle of which is a circular opening[664]. It is through this opening that the moth escapes. The silk of its cocoon is of so strong a texture and so closely gummed, that had both ends been similarly closed, its egress would have been impracticable; it finds, however, no difficulty in forcing its way through the aperture of a sort of reversed funnel, formed of converging threads that readily yield to pressure from within. But an objection will here probably strike you. You will ask, Is not this facility of egress purchased at too dear a rate? Must not a chrysalis in an open cocoon be exposed to the attacks of those ichneumons of which you have said so much, and of numerous other enemies, which will find[Pg 280] admittance through this vaunted door? Our caterpillar would seem to have foreseen your dilemma; at least, under heavenly guidance, she has guarded against the danger as effectually as if she had. If you cut open the cocoon longitudinally, you will see that within the exterior funnel-shaped end, at some distance she has framed a second funnel, composed of a similar circular series of stiff threads, which, proceeding from the sides of the cocoon, converge also to a point, and form a sort of cone exactly like the closed peristome of a moss; or, to use a more humble though not less apt illustration, like the wires of certain mousetraps[665]. In this dome not the slightest opening is left, and from its arched structure it is impenetrable to the most violent efforts of any marauders from without; whilst it yields to the slightest pressure from within, and allows the egress of the moth with the utmost facility. When she has passed through it, the elastic threads resume their former position, and the empty cocoon presents just the same appearance as one still inhabited. Rösel relates with amusing naïvété how this circumstance puzzled him the first time he witnessed it: he could scarcely help thinking that there was something supernatural in the appearance of one of these fine moths in a box in which he had put a cocoon of this kind, but in which he could not discover the slightest appearance of any insect having escaped from it, until he slit it longitudinally[666]. But from an observation of Meinecken, it appears that these converging threads serve[Pg 281] a double purpose; being necessary to compress the abdomen of the moth as it emerges from the cocoon, which forces the fluids to enter the nervures of the wings, and give them their proper expansion. For he found, that when the pupa is taken out of the cocoon, the moth is disclosed at the proper time, but remains always crippled in its wings; which never expand properly, unless the abdomen be compressed with the finger and thumb, so as to imitate the natural operation[667].
I am next to give you some account of the second mode in which the release of the perfect insect from its cocoon is effected—that, namely, wherein its own exertions chiefly accomplish the work. I shall from a large number select only a few instances. The texture of the cocoon of the silkworm-moth is uniform in every part, and the layers of silk are equally thick at both ends. The moth makes its way out by cutting or breaking these threads at the end opposite to its head: an operation which, as it destroys the continuity of the silk, those who breed these insects are particularly careful to guard against, by exposing the cocoon to heat sufficient to destroy the included pupa. The question is—What instruments does the moth employ to effect this? And this we are not able to answer satisfactorily. Malpighi asserts that the animal first wets the silk with a liquid calculated to dissolve the gum that connects the threads, and then employs its lengthened head to push them aside and make an opening[668]. But, as Reaumur has observed, besides that so obtuse a part as the head of a moth is but ill fitted to act as a wedge, we find[Pg 282] the threads not merely pushed to each side, but actually cut asunder. He therefore infers that the eyes, which are the only hard organs of the head, are the instruments by which the threads are divided—their numerous minute facets serving the purpose of a fine file[669]. It should be observed, however, that Mr. Swayne confirms Malpighi's assertion, that the silkworm does not cut, but merely pushes aside, the threads of its cocoon; and he informs us that he has proved the fact, by unwinding a pierced cocoon, the thread of which was entire[670]. Yet Reaumur's correctness cannot be suspected: and he affirms, that from observation there can scarcely be a doubt that most of the threads are broken[671]; which is further confirmed in an account of the breeding of silk-worms published in the American Philosophical Transactions: in which it is expressly stated, that cocoons out of which the fly has escaped, cannot be wound[672]. Analogy, it must be confessed, is against Reaumur's opinion; since other kinds of silkworms make their escape by means of a fluid. Thus we are informed by Dr. Roxburgh, that Attacus Paphia, when prepared to assume the imago, discharges from its mouth a large quantity of liquid, with which the upper end of the case is so perfectly softened, as to enable the moth to work its way out in a very short space of time,—an operation which, he says, is always performed in the night[673]. Perhaps the two opinions may be reconciled, by supposing the silkworm first to moisten and then break the threads of its cocoon. In those that are of a slighter texture, a mere push against the[Pg 283] moistened end is probably sufficient: and hence we find in so many newly disclosed moths the hair in that part wet, and closely pressed down[674]. If it be apparently difficult for the silkworm-moth to effect an opening in its cocoon, how much harder must seem the task of the puss-moth (Cerura Vinula) to pierce the solid walls of its wood-thickened case? Here the eyes are clearly incompetent; nor could any ordinary fluid assist their operation, for the gum which unites the ligneous particles is indissoluble in aqueous menstrua. You begin to tremble for the fate of the moth incarcerated in such an impervious dungeon—but without cause: what an aqueous solvent cannot effect, an acid is competent to: and with a bag of such acid our moth is furnished. The contents of this she pours out as soon as she has forced her head through the skin of the chrysalis, and upon the opposite end of the cocoon. The acid instantly acts upon the gum, loosens the cohesion of the grains of wood, and a very gentle effort suffices to push down what was a minute ago so strong a barrier. How admirable and effectual a provision! But there is yet another marvel connected with it. Ask a chemist, of what materials a vessel ought to be to contain so potent an acid: he will reply,—of glass. Yet our moth has no glass recipient: her bottle is a membranous bag; but of so wonderful a fabric as not to be acted upon by a menstruum which a gum, apparently of a resinous nature, is unable to resist! This fact can only be explained by the analogous insensibility of the stomach to the gastric juice, which in some animals can dissolve bone,—and it is equally worthy of[Pg 284] admiration. In both cases, the vitality of the membranous or fleshy receptacle secures it from the action of the included fluid; but how—who shall explain?
Ordinarily it is the moth that breaks the cocoon; but in the goat-moth and many Tortrices it is the pupa itself that performs the work, either wholly or partially. The pupa of the former is for this purpose furnished with sharp points upon the head, capable of effecting this object[675]. The locust-moth, another species of Cossus (C. Robiniæ Peck), whose history has been admirably detailed by Professor Peck, has a different process. "In the silk-moth," says he, "and all others which I have had opportunity to observe, the chrysalis bursts in the cocoon, and the fluid which surrounded the new insect in it escaping at the same time, so weakens or dissolves the fibre and texture of the silk, that the moth is able to extricate itself, leaving the chrysalis behind it; but this is not the manner in the locust-moth. After remaining till all its parts are fully grown and it is ready to quit its prison, a certain quantity of exercise is necessary, to break the ligaments which attach the moth to the shell of the chrysalis, and to loosen the folds of the abdomen. In taking this exercise, it can only move the abdomen in various directions: as one side of the rings is moved forward, the hooks in the serrated lines above mentioned (the adminicula) take hold of the silk, and prevent their sliding back; the next flexure brings forward the opposite side of the rings, which are prevented by the points on that side from slipping back in the same manner, and the chrysalis is forced out of the slightly woven extremity[Pg 285] of the cocoon, and through the silk-lined cavity, till it is protruded for about one-third of its length out of the opening in the bark, and into the air[676]."
An exception to the general rule—that the rupturing of the cocoon is the business of the inclosed insect itself—is met with amongst ants; the workers of which not only feed the young, but actually make an aperture in their cocoons, cutting the threads with their mandibles with admirable dexterity and patience, one by one, at the time they are ready to emerge, the precise period for which these indefatigable nurses are well aware of, that they may meet with no obstacle. Without this aid, the young ant would be unable to force its way through the strong and dense coating of silk that infolds it[677]. And a proceeding somewhat akin to this was observed by the Hon. Captain Percy, R.N., who himself related it to me. Being fond of the study of insects, he was in the habit of attending to their motions; and in the beginning of September 1821 noticed those of a number of female Tipulæ, probably T. oleracea L., busily engaged in depositing their eggs amongst the roots of grass. While observing these proceedings, he at the same time saw one quitting its pupa-case, which had already by its own efforts got its head, thorax, and anterior legs out of it. It was then joined by two male flies; which, with their anal forceps and posterior legs taking hold of the pupa-case, appeared with their mouths and anterior legs to push the little prisoner upwards, moving her backwards and forwards; and as they kept raising her, shifting their hold of the[Pg 286] skin till she was entirely extricated, when they left her to recover her strength by herself. Probably the extreme length of the two pair of hind-legs of these animals may render such assistance necessary for their extrication.
There remains yet to be explained under this head the manner in which the perfect insect is excluded from certain aquatic pupæ; such as those of Phryganeæ, gnats, and one of those Tipulidæ that resemble gnats. These pupæ (perhaps that they may be safe from the attack of birds) are destined to remain during the greater part of their existence in this state at the bottom of the water. But it is obvious that if the perfect insects were there to be disclosed, their wings would be wetted, and they would be drowned. It is the provision by which this result is obviated that now calls for your attention.
You have already been told that the larvæ of Phryganeæ inclose themselves in cases of different materials, open at each end[678]. You have also learned, that in becoming pupæ, they secure each end of their cases with a grating of silk[679]. When that change has occurred, they remain motionless at the bottom of the water. Now how are these pupæ, encased in tubes of a greater specific gravity than the surrounding fluid, to make their way to the surface when the time has arrived for their becoming denizens of the air? This they accomplish in the following manner:—The pupa is furnished with two strong exterior moveable mandibuliform processes, and has the power of moving its four anterior legs and antennæ while in the pupa-case. With these temporary jaws it makes an opening in one of the silken doors of its case, forces its[Pg 287] way out at that end, and then by moving its legs, the cases of which in some species are ciliated for this very purpose, swims to the surface, where its skin splits, and discloses the included insect. That these jaws are given for the express and exclusive purpose of being thus applied, seems undeniable. The pupa eats nothing—they are therefore in every other point of view superfluous. They are given to it alone of all other similar pupæ, because unnecessary to all others; and they are cast off along with the rest of the puparium, the perfect insect having no vestige of jaws[680].
The gnat has to undergo its change on the surface of the water—How is it to accomplish this without being wetted? In the pupa state they usually remain suspended with the posterior end of the body turned downwards: but when the period for its change is arrived, it stretches it out upon the surface, above which its thorax is elevated. Scarcely has it been a moment in this position, than, swelling out the interior and anterior parts of the thorax, it causes it to split between the two respiratory horns. Through this opening the anterior part of the gnat then emerges. As soon as the head and trunk are disengaged, it proceeds with its labour, and gets out more and more; elevating itself so as to appear in the puparium like a mast in a boat. As it proceeds, the mast is more and more elevated and lengthened, till it becomes nearly perpendicular—just as the mast of a boat is gradually raised from a nearly horizontal to a vertical position: at this period a very small portion of the abdomen remains in the puparium. Neither its legs nor wings are of any use[Pg 288] in maintaining it in this position. The latter are too soft, and, as it were, folded; and the former are stretched out along the abdomen—the segments of this last part are the only agents. The observer who sees how the little boat gradually sinks, and how its margin approaches the water, forgets the mischievous insect it contains, which at another time he would crush without remorse, and becomes interested for its fate; especially should wind agitate the water. A very little is sufficient to drive about rapidly the little voyager, since it catches the wind in some degree as a sail. If it should be upset, it would be all over with it;—and numbers do thus perish. The gnat, after having fixed itself thus perpendicularly, draws first its two anterior legs out of their case, and moves them forward, and next the two intermediate ones; then inclining itself towards the water, it rests its legs upon it, for water is to them a soil sufficiently firm and solid to support them, although surcharged with the weight of the insect's body. As soon as it is thus upon the water, it is in safety; its wings unfold themselves and are dried, and it flies away. All this is the work of an instant[681].
The pupæ of Chironomus plumosus proceed from those red worm-like larvæ so common throughout the summer in tubs of rain-water, &c., described by Reaumur[682]. They are not inclosed in cases, but are of a greater specific gravity than the water at the bottom of which they reside, until within a few hours of the exclusion of the fly. They have the power of swimming, however; and by moving the tail alternately backwards and forwards, can slowly raise themselves to the top of the water. But[Pg 289] here occurs a difficulty. For the extrication of the imago it is necessary that they should remain quietly suspended at the surface; and moreover that the thorax, in which the opening for its exit is to be made, should be at least level with it: and this is precisely what takes place. If you watch one of these pupæ when it ascends from the bottom, you will see that as soon as it has reached the top it remains suspended there motionless; and that its thorax is the highest part of the body, and level with the surface. Now the question is, in what way this is accomplished? How can a pupa of greater specific gravity than water, remain suspended without motion at its surface? and how can its thorax, which is at its heaviest end, be kept uppermost?—By a most singular and beautiful contrivance, which I shall explain; the more particularly because it has escaped Reaumur, and, as far as I know, all other entomological observers. The middle of the back of the thorax has the property of repelling water—apparently from being covered with some oily secretion. Hence, as soon as the pupa has once forced this part of its body above the surface, the water is seen to retreat from it on all sides, leaving an oval space in the disk, which is quite dry. Now though the specific gravity of the pupa is greater than that of water, it is but so very slightly greater, that the mere attraction of the air to the dry part of the thorax, when once exposed to it, is sufficient to retain it at the surface; just as a small dry needle swims under similar circumstances. That this is a true solution of the phænomenon, I am convinced by the result of several experiments. If, when the pupa is suspended at the surface, a drop of water be let fall upon the dry portion of the thorax, it instantly sinks to the[Pg 290] bottom,—the thorax, which belongs to the heaviest half, being the lowest; and if the pupa be again brought to the surface, so that the fluid is repelled from its disk, it remains suspended there without effort, as before. Just previously to the exclusion of the fly, the dry part of the thorax is seen to split in the middle. The air enters, and forms a brilliant stratum resembling quicksilver, between the body of the insect and its puparium; and the former pushing forth its head and forelegs, like the gnat, rests the latter upon the water, and in a few seconds extricates itself wholly from its envelope.
Before I close this letter, I must state a fact connected with the subject of it that deserves to be recorded. It is a general rule, that one pupa-case incloses only one insect; but Kleesius, a German entomologist, asserts that he had once two specimens of Gastropacha quercifolia produced from one pupa; which was large, being full two inches long, and one thick.
When the insect has quitted the exuviæ of the pupa, it has attained the last stage of its existence. It is now termed an Imago, or perfect insect; and is capable of propagation.
Just after its exclusion, it is weak, soft, and languid: all its parts are covered with moisture; and, if a winged insect, its wings have so little the appearance, either in shape, size, or colour, which they are about to assume, that it might be taken for a mutilated abortion, rather than an animal in the most vigorous stage of life. If it be a beetle, its elytra, instead of covering the back of the abdomen, are folded over the breast: their substance is soft and leathery, and their white colour exhibits no traces of the several tints which are to adorn them. If the insect be a butterfly or a moth, the wings, instead of being of their subsequent amplitude, and variegated and painted with a variety of hues and markings, are in large species scarcely bigger than the little finger nail, falling over the sides of the trunk, and of a dull muddy colour, in which no distinct characters can be traced.[Pg 292] If the excluded insect be a bee or a fly, its whole skin is white and looks fleshy, and quite unlike the coloured hairy crust which it will turn to in an hour or two; and the wings, instead of being a thin, transparent, expanded film, are contracted into a thick, opaque, wrinkled mass.
These symptoms of debility and imperfection, however, in most cases speedily vanish. The insect, fixing itself on the spoils of the pupa, or some other convenient neighbouring support, first stretches out one organ, and then another: the moisture of its skin evaporates, the texture becomes firm, the colours come forth in all their beauty; the hairs and scales assume their natural position; and the wings expanding, extend often to five or six times their former size—exhibiting, as if by magic, either the thin transparent membranes of the bee or fly, or the painted and scaly films of the butterfly or moth, or the coloured shells of the beetle. The proceedings here described I witnessed very recently with regard to a very interesting and beautiful butterfly, the only one of its description that Britain has yet been ascertained to produce—I mean Papilio Machaon. The pupa of this being brought to me by a friend early in May this year (1822), on the sixteenth of that month I had the pleasure to see it leave its puparium. With great care I placed it upon my arm, where it kept pacing about for the space of more than an hour; when all its parts appearing consolidated and developed, and the animal perfect in beauty, I secured it, though not without great reluctance, for my cabinet—it being the only living specimen of this fine fly I had ever seen. To observe how gradual, and yet how rapid, was the development of the parts and organs, and[Pg 293] particularly of the wings, and the perfect coming forth of the colours and spots, as the sun gave vigour to it, was a most interesting spectacle. At first it was unable to elevate or even move its wings; but in proportion as the aërial or other fluid was forced by the motions of its trunk into their nervures, their numerous corrugations and folds gradually yielded to the action, till they had gained their greatest extent, and the film between all the nervures became tense. The ocelli, and spots and bars, which appeared at first as but germes or rudiments of what they were to be, grew with the growing wing, and shone forth upon its complete expansion in full magnitude and beauty.
To understand more clearly the cause of this rapid expansion and development of the wings, I have before explained to you that these organs, though often exceedingly thin, are always composed of two membranes, having most commonly a number of hollow vessels, miscalled nerves, running between them[683]. These tubes, which, after the French Entomologists, I would name nervures, contribute as well to the development of the wings, as to their subsequent tension. In the pupa, and commonly afterwards, the two membranes composing the organs in question do not touch each other's inner surface, as they afterwards do: there is consequently a space between them; and being moist, and corrugated into a vast number of folds like those of a fan, but transverse as well as longitudinal, and so minute as to be imperceptible to the naked eye, the wings appear much thicker than in the end. Now as soon as the insect is disclosed, a fluid enters[Pg 294] the tubes, and being impelled into their minutest ramifications, necessarily expands their folds; for the nervures themselves are folded, and as they gradually extend in length with them, the moist membranes attached to them are also unfolded and extended. In proportion as this takes place, the expanding membranes approach each other, and at last, being dried by the action of the atmosphere, become one. To promote this motion of the fluid, seems the object of the agitations which the animal from time to time gives to its unexpanded wings. That a kind of circulation, or rather an injection of an aqueous fluid into these organs, actually takes place, may be ascertained by a very simple experiment. If you clip the wings of a butterfly during the process of expansion, you will see that the nervures are not only hollow, but that, however dry and empty they may subsequently be found, they at that time actually contain such a fluid[684]. Swammerdam, who appears to have been the first physiologist that paid attention to this subject, was of opinion that an aëriform as well as an aquiform fluid contributes to produce the effect we are considering. He had observed that, if a small portion be cut off from the wing of a bee, a fluid of the latter kind exuded from its vessels in the form of pellucid globules, becoming insensibly drops—which he concluded proved the action of the latter; and he noticed, also, that the wings were furnished with tracheæ, which were at that time distended by the injected air; whence he justly surmised, that the action of the air was also of great importance to produce the expansion of the wing[685]. And Jurine found that every nervure contains[Pg 295] a trachea, which, proceeding from the interior of the trunk in a serpentine direction, follows all the ramification of the nervure, though it does not fill it[686]. Though Reaumur attributes the expansion of the wings chiefly to an aqueous fluid, yet he suspects that the air on some occasions contributed to it[687].
The wings of the other tribes of insects probably differ from the Lepidoptera in the manner in which they are folded. It should seem from Reaumur's description, that those of some flies, instead of the straight transverse folds of the former, have angular or zigzag folds[688]; which equally shorten the wing. Many Hymenoptera have wings without any nervures except the marginal. We may conjecture that these are more simply folded, so as to render their expansion more easy; but even in these wings there are often tracheæ, which appear as spurious nervures, and help to effect the purpose we are considering.
The operation of expanding their wings, in by far the larger number of insects, takes place gradually as described above; and, according to their size, is ended in five, ten, or fifteen minutes; in some butterflies half an[Pg 296] hour, in some even an hour. A few species, such as Sphinx Œnotheræ F., require several hours, or even a day, for this operation; and, from the distance to which they creep before it has taken place, a considerable degree of motion seems requisite for causing the necessary impulse of the expanding fluids[689]. In a few genera, however, as the gnat, the gnat-like Tipulidæ, and the Ephemeræ, this process is so rapid and instantaneous, that the wings are scarcely disengaged from the wing-cases before they are fully expanded and fit for flying. These genera quit the pupa at the surface of the water, from which, after resting upon it for a few moments, they take flight: but this would evidently be impracticable, and immersion in the fluid, and consequent death, would result, were not the general rule in their case deviated from.
Some species of the last of these genera, Ephemera, are distinguished by another peculiarity, unparalleled, as far as is known, in the rest of the insect world. After being released from the puparium, and making use of their expanded wings for flight, often to a considerable distance, they have yet to undergo another metamorphosis. They fix themselves by their claws in a vertical position upon some object, and withdraw every part of the body, even the legs and wings, from a thin pellicle which has inclosed them, as a glove does the fingers; and so exactly do the exuviæ, which remain attached to the spot where the Ephemera disrobed itself, retain their former figure, that I have more than once at first sight mistaken them for the perfect insect. You can conceive without difficulty how the body, and even legs, can be withdrawn[Pg 297] from their cases; but you must be puzzled to conjecture how the wings, which seem as thin, as much expanded, and as rigid as those of a fly, can admit of having any sheath stripped from them; much less how they can be withdrawn, as they are, through a small opening at the base of the sheath. The fact seems to be, that though the outer covering is rigid, the wing inclosed in it, notwithstanding it is sometimes more than twenty-four hours before the change ensues, is kept moist and pliable. In proportion, therefore, as the insect disengages itself from the anterior part of the skin, the interior or real wings become contracted by a number of plaits into a form nearly cylindrical, which readily admits of their being pulled through the opening lately mentioned; and as soon as the insect is released from its envelope, the plaits unfold, and the wing returns to its former shape and dimensions. Thus our little animal, having bid adieu to its shirt and drawers, becomes, but in a very harmless sense, a genuine descamisado and sansculotte. It does not seem improbable, that the pellicle we have been speaking of is analogous to that which, in addition to the outer skin, incloses the limbs of Lepidoptera, &c. in the pupa state, but which they cast at the same time with the puparium, and leave adhering to it[690].
The body of newly-disclosed insects commonly appears at first of its full size; but the aphidivorous flies (Syrphus F. &c.), and some others, in about a quarter of an hour after leaving the pupa become at least twice as large as they were at their first appearance: this apparent sudden growth, which is also noticed by Goedart,[Pg 298] Reaumur found to depend upon the expansion of the previously compressed segments of the animal by means of the included air[691]. Both in this instance and in that of insects whose wings only require expansion, the size of the imago often so greatly exceeds that of the pupa, that we can scarcely believe our eyes that it should have been included in so contracted a space. The pupa of one of the beautiful lace-winged flies (Hemerobius Perla) is not so big as a small pea, yet the body of the fly is nearly half an inch long, and covers, when its wings and antennæ are expanded, a surface of an inch square[692].
When the development of the perfect insect is complete, and all its parts and organs have attained the requisite firmness and solidity[693], it immediately begins to exercise them in their intended functions; it walks, runs, or flies in search of food; or of the other sex of its own species, if it be a male, that it may fulfill the great end of its existence in this state—the propagation of its kind. Previously to thus launching into the wide world, or at least immediately afterwards, almost all insects discharge from their intestines some drops of an excrementitious fluid, often transparent, and sometimes red. I have before related to you the alarm that this last circumstance has now and then produced on the minds of the ignorant and superstitious[694]. Whether this excrement[Pg 299] is produced indifferently both by males and females I cannot positively assert; but a circumstance related by Jurine affords some ground for a suspicion that it is peculiar to the latter. A specimen of a female of Lasiocampa Rubi, when killed emitted some of this fluid, which dropped upon the floor: this appeared to attract the males to the apartment in which it happened, and to the very spot—from whence it may be conjectured, that the scent of the fluid brought them there, and that the use of it is to bring the sexes together soon after exclusion from the pupa[695].
The colour, sculpture, and other peculiarities which distinguish insects in this state I shall consider at large in another letter, when I treat of their external parts and organs. Under the present head I shall confine myself to pointing out the characters by which the sexes of many species are distinguished from each other; as likewise the duration of their life in their perfect state; together with the circumstances on which this duration depends.
I. Sexual Distinctions. The first general rule that may be laid down under this section is,—That among insects, contrary to what mostly occurs in vertebrate animals, the size of the female is almost constantly larger than that of the male. Even in the larva and pupa states, a practised eye can judge, from their greater size, which individuals will become females. There are, however, some exceptions to this rule. Thus amongst the Coleoptera, the male Dynastidæ, remarkable for their horns,[Pg 300] as you may see in D. Alocus, Antæus, Actæon, &c., as likewise those of Lucanus, are larger than the unarmed females[696]. In the Neuroptera the female Libellulidæ are sometimes sensibly smaller, and never larger, than their males[697]. In the Hymenoptera the male of the hive-bee, but more particularly that of Anthidium manicatum and other bees of that genus, is much more robust than the other sex[698]. In the Diptera, the same difference is observable in Syrphus Ribesii, and some other aphidivorous flies, and also in Scatophaga stercoraria[699]. And amongst the apterous tribes, we are informed by De Geer that the male of Argyroneta aquatica, which builds an aërial palace in the bosom of the waters[700], usually exceeds the female in bulk[701]. The reason of this rule seems in some degree connected with the office of the female as a mother, that sufficient space may be allowed for the vast number of eggs she is destined to produce; and it is when impregnation has taken place, and the eggs are ready for extrusion, that the difference is most sensible. In the majority of cases this sexual disproportion is not very considerable, but in some few it is enormous. Reaumur mentions a beetle, of which he intended to give the history, the male of which is so small compared with the female, that a bull not bigger than a sheep, or even a hare, set by the side of the largest cow, would aptly contrast with them. This little beetle, he says, has wings[Pg 301] and elytra, while the giant female has no vestige of either, having the upper surface of its body naked and membranous[702]. The species to which this illustrious Naturalist here alludes, does not appear to have been ascertained. The female of many gall-insects (Cocci) is so large in comparison with the male, that the latter traverses her back as an ample area for a walk[703]. But this is nothing compared with the prodigious difference between the sexes of Termes fatale, and other species of white ants, whose males are often many thousand times less than the females, when the latter are distended with eggs[704]. Accidental differences in the size of the sexes sometimes arise: as when the female larva has, from any cause, been deprived of its proper supply of food, it will occasionally be less than the male. De Geer has stated a circumstance with respect to the Aphides that produce galls, that should be mentioned under this head—the first, or mother female, is larger than any of her progeny ever become[705].
The second observation that may be generally applied to the sexes of insects is, that, size excepted, there is a close resemblance between them in other respects. But to this rule the exceptions are very numerous, and so important that it is necessary to specify examples of each under distinct heads.
i. In some species the sexes are either partly or wholly of a different colour. Thus, in the order Coleoptera, the elytra of the male of Rhagium meridianum F. are testaceous, and those of the female black. Leptura rubra of[Pg 302] Linné, with red elytra, is the female of his L. testacea, in which they are testaceous. Cantharis dermestoides of the same author is the other sex of his Meloe Marci; one of which is chiefly testaceous, and the other black: which seems to have so misled Linné, that he placed them in different genera. One more instance in this order, the female of Cicindela campestris, as was first observed to me by our friend Sheppard, has a black dot on each elytrum, not far from its base near the suture, which the male has not.
Amongst the Orthoptera, the male Locustæ F., as Professor Lichtenstein has informed us[706], have a fenestrated ocellus, which is not to be found in the other sex. I was once attending to the proceedings of a Hemipterous species, Pentatoma oleracea Latr., which I found in union: the paired insects had white spots, but another individual was standing by them, in which the spots were of a sanguine hue. I mention this by the way only—the spots in the prolific sexes being of the same colour: but might not the red spotted one be a neuter?
The sexes of many Lepidoptera likewise differ in their colour. I must single out a few from a great number of instances. The males of Lycæna Argus F. have the upper surface of their anterior wings of a dark blue, while in the female it is wholly brown. The wings of the former sex of Hypogymna dispar are gray, clouded with brown; but those of the latter are white, with black spots. In the brimstone butterfly (Colias Rhamni), which is one of the first that appear in the spring, the wings of the male are yellow—of the female whitish. In the common[Pg 303] orange-tip (Pieris Cardamines F.), one sex has not the orange tip to the upper wings: and, to name no more, the male of Lycæna dispar, one of our rarest and most beautiful butterflies, has only a single black spot in the disk of its fulgid wings; while in the other sex, the primary pair have nine, and the secondary are black, with a transverse orange fascia near the posterior margin. But the most remarkable difference in this respect observable in the insects of the order in question, takes place in a tribe, of which only one species is certainly known to inhabit Britain—I mean the Papiliones Equites of Linné: what he has called his Trojani and Achivi in some instances have proved only different sexes of the same species. Mr. MacLeay's rich cabinet affords a singular instance confirming this assertion;—a specimen of a Papilio is divided longitudinally, the right hand side being male, and the left hand female. The former belongs to P. Polycaon, a Grecian, the latter to P. Laodocus, a Trojan. An instance of two Grecians thus united is recorded in the Encyclopédie Méthodique, as exhibited in a specimen preserved in the Museum of Natural History at Paris; which on the right hand side is P. Ulysses, on the left P. Diomedes[707].
In the Neuroptera, the Libellulidæ are remarkable for the differences of colour in the sexes. In the common Libellula depressa, which you may see hawking over every pool, the abdomen of the male is usually slate-colour, while that of his partner is yellow, but with darker side-spots. Reaumur, however, noticed some males that were of the same colour with the females[708]. Schelver[Pg 304] observed, when he put the skins of Libellula depressa into water, that the colours common to both sexes were in the substance of the skin, and remained fixed; while those that were peculiar to one could be taken off with a hair-pencil, and coloured the water: which therefore were superficial, and, as it were, laid on[709]. The yellow males, therefore, that Reaumur observed, were probably such as had the superficial blue colour which distinguishes them washed off. In Calepteryx Virgo Leach, the former are of a lovely silky blue, and the latter green. In Agrions F. nature sports infinitely in the colours of the sexes.
In the order Hymenoptera there are often differences equally great; the sexes of many of the Ichneumons and Saw-flies are of quite different colours. The former tribe Linné has divided into sections, from the white annulus observable in the antennæ of some, and from the colour of their scutellum: but these are often merely sexual characters[710]. The male of Anthophora retusa Latr., a kind of wild bee, is wholly black, the female wholly gray, and of so very different an aspect that they were long regarded as distinct species; a mistake which has likewise occurred with regard to the sexes of Osmia cærulescens, another bee, of which the male has a bronzed and the female a violet abdomen[711]. The nose of male Andrenæ Latr. is often yellow, or white, as in A. hæmorrhoidalis—when that of the female is black[712]. The labrum also is often of a different colour in the sexes, as in Ceratina Latr.
In the Diptera, Aptera, Arachnida, &c., I am not aware of any striking difference in the colours of the sexes.
ii. The sexes of insects vary (but more rarely than in colour) in their sculpture also, and pubescence. Thus the elytra of the females of many of the larger water-beetles (Dytiscus) are deeply furrowed, while those of the males are quite smooth and level[713]. The thorax of the female in several species of Colymbetes of the same tribe, as C. Hybneri and transversalis, on each side has several tortuous impressed lines or scratches, like net-work, which are not to be discovered in the male. Hyphydrus gibbus Latr., which differs solely from H. ovalis (Dytiscus ovalis Illig.) in being thickly covered with minute impressed puncta, is, from the observation of the Rev. R. Sheppard, the other sex of this last, with which he has taken it coupled; and it is by no means improbable that Hydroporus picipes (Dytiscus punctatus Marsh.) and H. lineatus,—between which, as Gyllenhal has justly observed, the same difference only exists,—are in like manner sexual varieties. With respect to pubescence, I have not much to say. Another aquatic beetle, Acilius sulcatus Leach, has not only its elytra sulcated, but the furrows of these, and a transverse one of the thorax, are thickly set with hair; while the male is smooth, and quite naked. Particular care seems to have been taken by the Creator, that when all the above inhabitants of the water are paired, the male should be able to fix himself so firmly, by means[Pg 306] of his remarkable anterior tarsi, (which I shall afterwards describe,) and these asperities, &c. in the upper surface of his mate, as not to be displaced by the fluctuations of that element, the reluctance of the coy female, or any other slighter cause.
In a moth called the ghost (Hepialus Humuli), the posterior tibia of the male is densely bearded, but not of the female[714].—Some Hymenoptera, as Ammophila Kirb. and Stigmus Jurine, have the upper lip of the male clothed with silver pile, while that of the female is not so ornamented. The legs of some bees are distinguished in the sexes by a difference in their clothing. That observable in those of the hive-bee has been before noticed[715]. In Andrena of Latreille[716] the posterior tibia of the female is covered externally with a dense brush of hairs, for collecting the pollen; and the posterior legs at their base have a curled lock of hair—which are not to be found in the male[717]. In Dasypoda, Melecta, Anthophora, Centris, Epicharis, &c. of the same author, the first joint of the tarsus of the female, and in Xylocopa almost the whole tarsus, is also similarly signalized from that of the other sex. In Bombus, as in the hive-bee, the posterior tibiæ of the females and neuters are furnished with a basket of hairs for carrying their pollen paste, which you will in vain look for in the male[718]. The latter, however, in some species of this tribe are distinguished from the former by the longer hairs of their legs, but not in the posterior ones. Thus, in Anthophora retusa the first joints of the[Pg 307] intermediate tarsus are bearded internally with a thin fringe of long hairs, and the first externally with a triangular one of short ones at the apex: but what is most remarkable, the last or unguicular joint, which in almost every other bee is naked, is on both sides fringed with long hairs[719]. In that remarkable genus Acanthopus Illig., of which the male only is known, the first and last joint of the intermediate tarsus have a dense external brush of stiff hairs, which probably is also a sexual character[720]. Another sexual kind of clothing is exhibited by the females of those bees that have their labrum or upper-lip inflexed (Megachile Latr.)[721]. Their abdomen is covered underneath with a brush of stiff hairs, involved in which they carry the pollen they collect. In the males of some of this tribe, as of M. Willughbiella, the first four joints of the anterior tarsus on their inner side have a long dense fringe of incurved hairs[722]: a circumstance also to be found in the same sex of Xylocopa latipes, in which the claw-joint also is bearded[723]. In Andrena Latr. the last dorsal segment of the abdomen of the same sex is fringed, while that of the male is naked[724]. In the humble-bees (Bombus), the mandibles of the male are bearded with curled hairs, while those of the females and neuters are without them. Some bees, as Andrena and Halictus Latr., have the anus of the female bearded, and that of the male naked: in some Bombyces the reverse takes place.
iii. With regard to the general shape of their body, the male and female usually resemble each other: but there are some exceptions to this rule. The male of the hive-bee is much thicker and more clumsy than either the female or the worker; but in Halictus Latr. the males are nearly cylindrical in shape, and very narrow; while the other sex are oblong or ovate, especially their abdomen: and in Andrena Latr. the former are much slenderer than the females, and of a more lanceolate shape. But a still more striking difference in this respect between the sexes is exhibited by some species of the genus Ptinus F., in which the male is long and slender, and the female short and thick. This, in more than one instance, has occasioned them to be mistaken for distinct insects: thus, P. Lichenum and P. similis, P. ovatus and P. testaceus, of Mr. Marsham, are mere sexual varieties. But the most entire abalienation of shape at present known, is that which distinguishes the male from the female Coccus; these are so completely dissimilar as scarcely to have any part in common. In Bombyx vestita F., and others of the same family, while the males are of the ordinary conformation of the order, the females are without even the slightest rudiments of wings; they have no antennæ, the legs are extremely short, not longer than those of the caterpillar; and the body is entirely destitute of scales, so that they altogether assume the exact appearance of hexapod larvæ[725]. A conformation nearly similar takes place in the female of Tinea Lichenella; but in this the feet are longer, and the anus is furnished with a long retractile ovipositor[726].
iv. In many cases, the structure of particular parts and organs of the body differs in the sexes. As the facts connected with this part of our present subject are extremely numerous and various, it will be convenient to subdivide it, and consider the sexual characters that distinguish—the Head, Trunk, and Abdomen of insects, and their several appendages.
1. The Head. This part in some females is considerably larger than it is in the male. This is the case with the ants, and several other Hymenoptera; while in some Andrenæ, as A. hæmorrhoidalis, and Staphylinidæ, as St. olens, that of the male is the largest. But in none is the difference more conspicuous than in the stag-beetle (Lucanus); in which genus the male not only exceeds the female in the length of his mandibles, but also greatly in the size and dimensions of his head. In the Apion genus, the rostrum of the female is generally longer and slenderer than that of her mate; and in Brentus, the rostrum of one sex (probably the male) is long and filiform, while in the other it is thick and short. This is particularly visible in B. dispar and maxillosus[727], &c.
One of the most striking distinctions of the males in this part of their body, are those threatening horns, usually hollow, with which the heads of many of the male lamellicorn insects and some others are armed, and which give them some resemblance to many of the larger quadrupeds. Many are unicorns, and have their head armed with only a single horn; which in some, as in Oryctes Illig., Dynastes Endymion[728], &c. is very short; in others,[Pg 310] very long, as in Dynastes Enema, Pan, Elephas[729]. In one, again, it is thick and robust; as in the clumsy Dynastes Actæon[730]: in another very slender, as in Onthophagus spinifer[731]. With respect to its direction in Elephastomus proboscideus MacLeay, it is horizontal[732] and straight; in Phaleria cornuta horizontal and broken, or the apex turning outwards and forming an angle with the base[733]; in Dynastes Hercules horizontal, and recurved at the apex[734]; in D. Actæon, Elephas, and Typhon, recurving from the base. In Geotrupes dispar it is recurved, so that its point exactly coincides with that of the porrected thoracic horn, with which it forms a kind of forceps[735]. In Copris lunaris F. and Diaperis horrida, the horn is nearly upright[736]. In Onthophagus Xiphias it is dilated at the base, and reclining upon the thorax; and at the apex attenuated, and bending forwards, or nodding. In Passalus cornutus it rises a little, and then bends wholly forwards. In Dynastes Milon, a most remarkable beetle, it slopes backwards in a waving line[737]; and in Onthophagus spinifer it is recurved and reclining.—In speaking of the direction of the horn, you must recollect that it will vary in proportion as the head varies from a horizontal position: so that an upright horn will become inclined or reclined, as the head bends forwards or backwards; but I speak of it as it appears when the head is horizontal.[Pg 311] Again, it varies in its teeth or branches. In Dynastes Hercules it is armed with several teeth. In D. Elephas and Actæon it has only one large one at its upper base[738]. In D. Milon it is serrated above. In D. Alcides, Tityus, Ægeon, Copris lunaris, &c. the horn is unarmed and simple at the apex. In D. Oromedon, Gedeon, Enema, Actæon and congeners, it is bifid. In some the horn is at first a broad lamina or ridge, which terminates in two branches, as in Onthophagus Vacca. In this the branches are straight; but in another undescribed species in my cabinet (O. Aries Kirby, MS.) they are first bent inwards, and then at the apex a little recurved: and in D. dichotomus it is divided into two short branches, each of which is bifid[739]. Other males emulate the bull, the he-goat, or the stag, in having a pair of horns on their head. In Onthophagus Taurus, these arms in their curvature exactly resemble those of the first of these animals[740]. In Goliathus pulverulentus, the straight, robust, diverging, sharp horns are not unlike those of some of the goat or gazel tribe. I have a beautiful little specimen in my cabinet, (I believe collected by Mr. Abbott of Georgia,) in which the horns have a lateral tooth, or short branch, like those of a stag; and which I have therefore named O. cervicornis. In O. Vacca, Camelus, &c. the horns are very short, and nearly perpendicular. In the male of[Pg 312] Copris Midas, the two longer perpendicular horns have a deep cavity between them, which, together with its black colour, give it a most demoniac aspect; so that you would think it more aptly representative of a Beelzebub or Beelzebul than a Midas[741], or than Phanæus Beelzebul MacL. A similar cavity is between the occipital horns of Diaperis hæmorrhoidalis Payk. Some species of Rynchænus, as R. Taurus, have a pair of long horns upon the rostrum of the male, the rudiments only of which are to be traced in the female[742]. Other species go beyond any known quadrupeds in the number of horns that arm their heads. Thus Ditomus calydonius Bonelli, belonging to Carabus L., has three equal horns[743]. The same number distinguishes Onthophagus Bonasus; but the intermediate one is very short. In Goliathus Polyphemus the middle horn, on the contrary, is much longer and thicker than the lateral ones, and forked at the apex; so that it looks as if it had four of these weapons[744]. A little Diaperis (D. viridipennis F.), a native of Carolina, has four horns upon the head of the male; namely, two long ones on the occiput, and two short dentiform ones on the nose. In a species nearly related to this, sent me by Professor Peck from New England, there is a cavity between the two occipital horns. The same number distinguishes Onthophagus quadricornis (Copris F.). The situation also of the horns varies: In some it is in the middle of the head, as Oryctes nasicornis, Copris lunaris, &c.: in others, as in Onthophagus nuchicornis, Xiphias, &c. it is a process[Pg 313] of the occiput or hind-head; and in O. Oryx F. the two horns proceed from the anterior part of the head. In the other sex, in insects the head of whose males is armed with horns, they are usually replaced by mere tubercles, or very short elevations, as you may see in the female of Copris lunaris; or by transverse ridges, as in the Onthophagi: or else the head is without arms, and quite smooth, as in Diaperis, Phaleria, &c. What may be the use of these extraordinary appendages, as well as those on the thorax, and in some cases on the abdomen, (which I shall mention afterwards), to the males, has not yet been ascertained. Whether the individuals of this sex are more exposed to the attack of birds and other enemies, in consequence of being more on the wing than the females, and are therefore thus provided with numerous projecting points for defence, is a question worth considering[745]. It is the only probable conjecture on the cui bono of these arms that I can at present make. Under this head I ought to notice the remarkable membranous process of an obovate shape, which like an umbrella covers the head of Acheta umbraculata F.[746] Whether the sharp curved horns which arm this part in another Acheta figured by Stoll[747], in an incumbent posture, with their point towards the mouth, are a sexual distinction, we are not informed,—probably they are.
The organs of the head also present many sexual[Pg 314] distinctions. The upper lip (labrum) in Halictus Latr., a tribe of wild bees, in the female is furnished with an inflexed appendage, which is not discoverable in that of the male[748]; and the shape of this lip in Sphecodes Latr. differs in the sexes[749]. Perhaps the horn or tubercle observable on this part of some female Nomadæ F.[750] may be wanting in the male.
The under-lip (labium)—taken in a restricted sense for that central part from which emerge the labial palpi, and which is often considered as the mentum,—does not offer any striking variations in the sexes. One, however, is of importance, as it helps to prove which are the true female Lucani. In the male the labium is emarginate, in the female it is intire. This may be seen both in L. Cervus and femoratus, and probably in other species. The sculpture also is different, the lip being smooth in the former and covered with excavated puncta in the latter. The tongue (lingua or ligula) of the sexes is usually the same; except in the hive-bee, in which that of the neuters is longer than that of the male and female.
The upper-jaws (mandibulæ), however, often afford striking sexual characters. The enormous protended ones of the common stag-beetle (Lucanus Cervus) attract the attention of the most incurious observer; and these are now generally allowed to be of this description. Geoffroy and Mr. Marsham, indeed, have asserted that they have taken in coitu those with long mandibles: but as these males are pugnacious, and attack each other with great fury, as Mr. Sheppard informs me, it is not improbable that these gentlemen may have mistaken a battle[Pg 315] for an amour: since not only have those with long mandibles been often taken united with those that have short ones[751], but the same difference obtains in the sexes of other species. This is particularly observable in Lucanus femoratus, of which I received from Brazil many specimens agreeing in every respect except in this, that one had short and the other very long mandibles. These organs vary in different specimens, as to the number of their teeth and branches. They are singularly robust in L. Alces[752]; but in none more threatening than in L. Elephas[753], in which they curve outwards and downwards. In Mr. W. MacLeay's genus Pholidotus, they are almost parallel to each other, and curve downwards; in Lucanus nebulosus Kirby, they assume a contrary direction[754]; as they do likewise in Lamprima Latr.[755] In Lucanus Capreolus the points close over each other[756]. In Lethrus F. in the female, but not the male, the mandible is armed below with a long incurved horn. In Lucanus serricornis they form a complete forceps[757]. In Siagonium quadricorne Kirby[758] the mandible is furnished at its base with an exterior horn, which is probably a sexual distinction. The male of Synagris cornuta, a kind of wasp, is still more conspicuous in this respect; for from the upper side of the base of its straight slender mandibles proceed a pair of crooked, decurved, tortuous, sharp horns, not only longer than[Pg 316] the mandible, but than the head itself[759]. Many sexual differences are observable in the mandibles of the various tribes of bees (Anthophila Latr.). Thus, in Colletes Latr. the male mandible is more distinctly bidentate at the apex than the female[760]: in Sphecodes Latr. and others, the reverse of this takes place[761]. Where these organs in both sexes are toothed at the apex, they often vary in the number of teeth. Thus, the female of Megachile centuncularis Latr. has four teeth at the apex of its mandible, while the male has only two[762]. In M. Willughbiella, though the mandibles of both sexes have four teeth, yet those of the male are sharp, and the two external ones the longest; while those of his mate are obtuse, and all nearly equal in length[763]. In Anthidium manicatum Latr., the former has only three teeth, while the latter has five[764]. The differences in this respect in the hive-bee have been before noticed[765]; those of the humble-bees (Bombus Latr.) are strikingly distinguished from each other; the female mandible being very stout and wide, constricted in the middle, and furrowed on its outer surface; and the male, on the contrary, very slender at the apex, dilated at the base, and without furrows[766].
Of all the organs of the head, none seem so little subject to sexual variation as the under-jaws (maxillæ)[767]. I[Pg 317] can bring forward only one striking instance of it, and some degree of doubt rests even upon that. In the genus Nemognatha of Illiger, the maxillæ of the male are elongated, narrow, setiform, and often involute or spiral, like those of a bee or a butterfly. But that this is peculiar to the males is at present only surmised[768]. I possess several species of the genus, all of which are distinguished by long maxillæ; though in some they are as long as the body, and in others scarcely half that length. Gnathium Kirby is similarly characterized[769].
The maxillary palpi occasionally differ in the sexes. In Cerocoma those of the female are filiform, while the two intermediate joints of those of the other sex are much thicker than the first and the last[770]. In Hylæcetus and Lymexylon, those of the male are still more remarkable: they are pendent, the last joint very large, and laciniated so as to form a tuft[771]. The female ones grow gradually larger towards the end, but are not at all divided there[772]. The palpi of male spiders are of a very different structure from those of the other sex, terminating in a very complex incrassated piece, which has been supposed to contain the organ of generation; but this, according to Treviranus, is a mistaken idea—that organ being, as usual, to be found in the abdomen[773]. In the common gnat the palpi of the male are as long as the proboscis, consist of five joints, and at the end are tufted with hairs; while those of the female are scarcely one-fourth of its[Pg 318] length, have only three joints, and are not tufted. Whether the labial palpi in any genus differ in the sexes, I cannot affirm with certainty: I have not, however, observed any such variation in them.
I shall next mention some organs of the head, in which the difference between the sexes is often very striking and peculiar. You will readily conjecture that I am speaking of the antennæ. And here the advantage seems wholly on the side of the males: since in them these wonderful instruments of unknown sensations are not only more complex, but usually more elegant, than those of the other sex. You will pardon me, therefore, if I enlarge a little more than ordinary upon a subject so full of interest, and say something upon the differences observable between the sexes—in the shape, magnitude, and length, number of articulations, ramification and plumage, and individual joints of their antennæ.
With regard to their shape, variations are sometimes observable between the antennæ of the sexes; but this principally occurs in the Hymenoptera order. For instance, those of Chelostoma maxillosa, a small bee that deposits its eggs in little holes in posts and rails, are clavate in the female and filiform in the male[774]—a circumstance that distinguishes in some degree those of Sphecodes, Halictus, and Andrena of Latreille, three other genera of wild-bees[775]. In Dinetus Jur. the male antennæ are moniliform at the base, and filiform at the apex; the female, on the contrary, are entirely filiform[776].
The antennæ of the sexes also sometimes differ in magnitude and length. This is the case in the three genera of wild bees just mentioned; those of the female being thicker than those of the male, while these last are longer than the former. But in this tribe the males of the Fabrician genus Eucera are most remarkable for their long antennæ[777]. With regard to the different length of these organs in the sexes, no insects are more distinguished than some species of the capricorn-beetles (Cerambyx L.). In Lamia Sutor the male antennæ are twice the length of the female; and in another Brazilian species in my cabinet, related to L. annulata (Stenocorus F.), they are thrice their length. Some of the Anthribi F. approach the Cerambycidæ, not only in some other characters, but also in this circumstance:—thus the antennæ of A. albinus, a native of Britain, are vastly longer in the male than in the female; and in A. cinereus (Macrocephalus Oliv.)[778], which I suspect to be of the former sex, they are as long nearly as is usual in the tribe just named, called in France capricorn-beetles.
I may here observe, that sometimes in the sexes a difference is also to be found in the direction or flexure of their antennæ. Thus in Scolia F., Pepsis F. &c., in the males the antennæ are nearly straight, but in the females convolute or subspiral. The reverse of this takes place in Epipone spinipes, a kind of wasp, and its affinities; and Systropha Illig., a kind of bee: for in these the male antenna is convolute at the apex[779], and the female straight. In the various tribes of bees (Anthophila Latr.), these[Pg 320] organs in the latter are what is denominated broken, the main body of the antenna forming an angle with the first joints: but in the former this does not take place.
The antennas of the sexes do not always agree in the number of joints. In the bees, and many other Hymenoptera, the male has one more joint than the female; as is the case also in Œdemera notata (Cantharis acuta Marsh.). In Pteronus Laricis, a kind of saw-fly, the latter has only sixteen joints in its antennæ, while the former has twenty-four[780]. In Rhipicera marginata, a beetle, the beautiful antennæ of the male consist of thirty-two joints, while the female has no more than eleven! In Chelonus Jur. the male, on the contrary, has the smallest number of joints, namely sixteen; while the female has twenty-five[781].
In nothing do the sexes differ more materially than in the ramification of these organs, and their plumage. By attending to this, you may often detect the sexes in an instant; since the antennæ of the males in numerous instances are much more complex than those of the females. For what end the Creator has so distinguished them is not quite clear; but most probably this complex structure is for the purpose of receiving from the atmosphere information of the station of the female. A tendency to branching will be found in the antennæ of some males, in tribes where these organs are usually perfectly simple in both sexes. Thus, in the male of Chelostoma maxillosa,—mistaken for another species by Linné, which he names Apis florisomnis,—the intermediate joints on their inner side project into an angle[782]; and those of the same sex of the common[Pg 321] hornet, by means of a central sinus, have two obtuse teeth on each. With regard to more direct ramifications, some male antennæ terminate in a fork, or two branches. This is the case with Hylotoma furcata Latr., a saw-fly[783]; and the peacock-louse (Nirmus Pavonis Herm.)[784]. Others, again, have three lateral branches, as in Eulophus Geoffr. a little parasite, the male antennæ of which send forth a hairy external and rather long branch, from the base of the fourth, fifth, and sixth joints[785]. In Elater flabellicornis L., the eight last joints are flabellate, or elongated and flat, resembling the sticks of a fan in the male[786]; in the female they are shorter, and more properly may take their denomination from the teeth of a comb. In Lampyris Latreillii Kirby, the antennæ of the former are flabellate on both sides, while those of the latter are little more than serrate[787]. These organs are extremely beautiful in the males of the Rhipiceræ of Latreille. In R. marginata K. they consist of thirty-two joints, from thirty of which issues a branch, the first very short, but the rest gradually increasing in length as they approach the middle of the antenna; then gradually decreasing to the end, so as to represent an expanded fan[788]. But in none are they altogether so remarkable as in those moths that Linné denominates Bombyces Attaci, and some others. In these, in the males, these organs in their contour are lanceolate, and every joint is furnished with a couple of parallel equal branches on each side[789]. In the females these branches are shorter[Pg 322] on the whole, and alternately one long and one short; but in some, as Saturnia Pavonia, there is only one short branch or tooth on each joint in this sex[790]. In Bombyx regalis &c. only the first part of the antenna is so branched; and those of the female are setaceous and without branches. In B. versicolor, &c. there is only one branch from each side on every joint; those of the female being much shorter than those of the male. The latter sex of Pteronus Laricis Jur., a saw-fly, afford an example of a different structure, the antennæ on one side sending forth a branch from every joint but the two first; but on the other side, the nine or ten last joints also are without a branch. The female antenna is serrated[791]. In another of this tribe, Pterygopterus cinctus Klug, the male antenna resembles a single-toothed comb, being branched only on one side: that of the female, like the former instance, is serrated[792]. Whether the remarkable antennæ that distinguish the known individuals of the genus Phengodes (Lampyris plumosa F.) is a sexual character has not been ascertained; but it is not improbable that it may be, as in other Lampyridæ. A pair of delicate flexile and almost convolute plumose branches proceeds from the apex of each joint except the basal ones, which have something the air of cirri, and give a more than usual degree of lightness and elegance to these organs[793]. Other antennæ, especially in the Diptera order, assume an appearance of plumes—not from the branches that proceed from them, but from the fine long hairs that beset and adorn them. These are universally indications of the[Pg 323] male sex, those of the females being generally comparatively naked. If you take the common gnat, you will find that the antennæ of one individual are thickly fringed on each side, and tufted at the end with fine long hairs, while in the other only four or five placed at intervals in a whorl are to be perceived[794]. In Chironomus Meig., a kind of Tipula L., resembling a gnat, the male antennæ are beset on all sides with the finest hairs, and resemble a beautiful plume[795], while the females to the unarmed eye appear naked. Even in some Hymenoptera, the antennæ of the males are thus feathered, in a less degree: for instance, in Hylotoma Latr.[796] Whether the tufts and fringes which ornament, in a remarkable manner, the antennæ of many Cerambycidæ[797], are sexual characters, is not certainly known.
We are now to consider other sexual differences in these organs, resulting from the size or configuration of one or more individual joints. To begin with the first joint, or scapus. In many of the Hymenoptera, particularly the Anthophila Latr., this is elongated, and the remaining joints form an angle with it in the females: while in the other sex it is much shorter, and in the same line with the rest of the antennæ; and in Hylæus dilatatus (Melitta dilatata Kirby) the first joint in the male is dilated and shaped something like a patella[798]. In Malachius bipustulatus, &c. the sex just mentioned is peculiarly distinguished by a white excrescence on the first[Pg 324] four joints of the organs in question, most conspicuous in the second and fourth. The antennæ of male Cerocomæ are not very different[799]. Mr. Marsham has described a little Haltica under the name of Chrysomela nodicornis, from a peculiarity of the same sex not to be found in the other. The fourth joint is very large and obtriangular; in the female it is merely longer than the rest. In H. Brassicæ and quadripustulata the fifth joint is larger and longer than all but the first in the male, in their females it is only longer. In some moths (Herminia Latr., Crambus F.) there is also a knot in the middle of the male antennæ[800]. In Noterus, a water-beetle, the six intermediate joints are thicker than the rest, beginning from the fourth, and the last but one ends internally in a truncated tooth. The fifth and two following joints in the male antennæ of Meloe are larger than the rest, which distinguishes them, as well as a remarkable bend observable at that part[801].
Variations of the kind we are considering are also observable in the clava, or knob, in which antennæ often terminate. You have doubtless observed that the lamellated clava of the antennæ of the common cockchafer is much longer and more conspicuous in some individuals than in others—the long clava belongs to the male[802]. In another species, M. Fullo, that of this sex is nine or ten times the length of that of the other. In Colymbetes serricornis, a water-beetle, the male has a serrated clava of four joints. In Dorcatoma dresdensis[803], and also Enoplium damicorne, two beetles, it is nearly branched in the[Pg 325] male, but much less so in the female. In a little destructive beetle, common in our houses (Attagenus Pellio), in the latter it is very short, but in the former it is very long, and nearly formed by a single joint. In Eurhinus Kirby, a New Holland genus of the weevil-tribe, in the male the last joint, also, is much longer than it is in the female[804]. These examples will give you some idea of the principal variations that take place in the antennæ of the sexes, and of the wonderful diversity of forms in this respect to which mere sexuality gives rise amongst insects.
In the eyes, or stemmata, this diversity is less remarkable. Latreille has described two ants, Formica contracta and coeca, in the neuter of which he could discover no eyes[805]: in the former, the female, however, had large ones. The male he appears not to have known, but it probably was not destitute of these organs; of the latter he was acquainted only with the workers. The neuter of Myrmica rubra, another ant, has no ocelli or stemmata, although the male and female are provided with them[806]. They are discoverable only in the former sex of that singular insect related to the ants, Mutilla europæa. Other insects differ in the size of the eyes of their sexes. In the hive-bee, and some Ephemeræ, the eyes of the drone or male are much larger than those of the worker and female, and also meet at the vertex, having their stemmata below the conflux; whereas in these latter they are widely distant[807]. In Stratyomis, Tabanus, and many other[Pg 326] two-winged flies, the male eyes meet at some point below the stemmata, and above the antennæ. In the former they touch more at an angle; for the vertex forming a narrow isosceles triangle, and for the anterior part of the face one nearly equilateral: while those of the female are separated by a considerable interval. In Heptatoma and Hæmatopota in that sex, a similar interval obtains; while in the other, after forming a minute short triangle, they unite for a considerable space, and then diverging, form the face. This is also the case in Tabanus; but in the female, the space that intervenes between the posterior part of the eyes is much narrower than in these two cognate genera of the horse-flies. In some others of this order, as Musca Latr., the eyes of the male do not touch, but approach posteriorly much nearer to each other than those of the other sex. In a few instances the sexes vary even in the number of their eyes, as well as the size. This occurs in some species of Ephemera L. (E. diptera, &c.), in which the male, besides the common lateral ones, has two large and striking intermediate eyes, that sit upon vertical pillars or footstalks[808].
2. The Trunk. The thorax of many coleopterous males, especially of the Dynastidæ and Copridæ amongst the petalocerous tribes, exhibits very striking differences from that of the female. In many Lucani the lateral angle is more prominent. In Anthia it is bilobed posteriorly, while in the last-mentioned sex it is entire[809]. In Phanæus carnifex MacLeay (Copris F.) it is elevated into a plane triangular space, with the vertex of the triangle[Pg 327] pointing to the head; but in the female it is convex, with an anterior abbreviated transverse ridge[810].
In a large proportion terrific horns, often hollow, like those of the head lately noticed, arm the thorax of the male, of which you will usually only discover the rudiments in the other sex. In the first place, some are unicorns, or armed only with a single thoracic horn, which frequently, in conjunction with the thorax itself, not a little resembles a tunnel reversed: of this description are Dynastes Hercules, Tityus, Gedeon, Enema, &c.[811] In the three first this horn is porrected, or nearly in the same line with the body; but in the last, and D. Pan, it forms an angle with it; and in D. Ægeon it is nearly vertical[812]. In D. Hercules it is very long; in D. Alcides[813] and Tityus very short; in the two last, and in Oxytelus tricornis which is similarly armed, it is undivided at the apex; but in D. Gedeon, Pan, bilobus, &c.[814] it is bifid or bilobed. It is usually rather slender, but in D. Chorinæus[815] and bilobus, it is very stout and wide. In D. claviger it is hastate at the apex[816]. In D. hastatus it is short and truncated[817]. Others, again, have two thoracic horns. In Copris nemestrinus these are discoidal, diverging, and inclining forwards[818]. In Phanæus floriger[819] they are lateral, triangular, and incline towards each other, with, as it were, a deep basin between them. In P. splendidulus they sink into two longitudinal ridges, most elevated[Pg 328] posteriorly, with an intervening valley[820]. In P. bellicosus they are posterior, compressed, truncated, and emarginate at the apex, and include a basin[821]. In Copris Sabæus they are merely two acute prominences[822].—Three horns distinguish the thorax of many. In D. Aloeus[823] and its affinities, they are arranged in a triangle, whose vertex is towards the head. In D. Antæus[824] these horns are nearly equal in length, and undivided at the apex. In D. Titanus[825] the anterior horn is longer than the rest, and bifid at the apex; in D. Atlas and Endymion[826], both of which have a horn on the head, it is much shorter. In others, as in Megasoma Kirby, the vertex of the triangle is towards the anus. In M. Typhon[827] it is longer than the anterior ones, and bifid at the apex; in M. lanigerum they are equal in length[828]. In M. Elephas and Actæon[829] it is merely an elevation of the thorax; in the last almost obsolete. In Geotrupes Typhæus, common on our heaths, the anterior of this part is armed by three horizontal horns, the intermediate one being the shortest[830]. Copris lunaris also, another of our own beetles, has three short posterior thoracic horns, two lateral and triangular ones, and a transverse intermediate elevation, with a notch in the middle[831]. In Dynastes Neptunus the horns are porrected, the middle one being very long, and the lateral ones short[832]. In D. Geryon the point of the lateral[Pg 329] horns is towards the anus, and the base of the intermediate one covers the scutellum[833]. Others have four of these singular arms: this is the case with one of our rarest beetles, Bolbocerus mobilicornis K., which has four dentiform horns, the intermediate pair being the shortest, arranged in a transverse line on the anterior part of the thorax[834]. In B. quadridens these are merely teeth. In Phanæus Faunus[835] it has two lateral, elongated, compressed, truncate, horizontal horns, and two intermediate teeth. Dynastes Milon has a still greater number of horns on the thorax of the male, there being two lateral anterior ones and three posterior ones—the intermediate being the longest[836]; and Copris Antenor Fabricius and Olivier describe as having a many-toothed thorax; and from the figure of the latter[837], the male appears to have seven prominences.
But the males of insects are not only occasionally distinguished by these dorsal arms—in a few instances they are also furnished with pectoral ones. The illustrious traveller Humboldt found in South America a species of weevil (Cryptorhynchus Spiculator Humb.), the breast of which was armed with a pair of long projecting horns; and I possess both sexes of four species, three at least from Brazil, that exhibit in one individual the same character. One, concerning the country of which I am uncertain, recedes somewhat from the type of form of the rest, and comes very near that of Rynchænus Strix F.[838] In the individual which I take to be C. Spiculator, the pectoral horns are very long, curving upwards at the[Pg 330] apex, and nearly in a horizontal position; while in the three others they are much shorter, and inclined towards the horizon. The males of some species of Rynchites, as R. Bacchus and Populi[839], are also armed with a pair of lateral horns or spines, which may be termed pectoral rather than dorsal.
I shall now advert to the sexual characters that are to be found in the instruments of motion attached to the trunk—beginning with those for flight. In the female of the common glow-worm (Lampyris noctiluca) not the slightest vestige of elytra or wings is visible, and it resembles a larva rather than a perfect insect; yet its mate is a true beetle furnished with both. The same circumstance distinguishes the female cockroach (Blatta) and is more universally prevalent in that genus than in Lampyris, in which a large number of females have both elytra and wings. The males of Bombyx antiqua and Gonostigma, and of many other moths, have wings of the usual ample dimensions, while those of their females are merely rudiments. This is the case, also, with some of the Ichneumonidæ[840]. In the tribes of Ants, Termites, &c. the neuters or workers are without wings. Amongst the plant-lice (Aphides) there are individuals of both sexes, some of which have wings, and others not[841]. Amongst the Coleoptera, the female of Tenebrio Molitor, the common meal-worm, has elytra and no wings; while the male has both[842].—Sometimes these organs vary in size in the sexes: thus in Aradus Betulæ F., a kind of bug, the hemelytra and wings are narrower and shorter in the[Pg 331] female than in the male[843]. In the genus Blaps F., the mucro that arms the apex of each elytrum is longer in the former sex than in the latter. In Ateuchus gibbosus F., a dung-beetle, the elytra have a basal gibbosity near the suture in one sex that does not obtain in the other. In the Orthoptera order, the sexes are often to be known, almost at first sight, by a difference in the veining and areolets of the wings; but upon this I enlarged so fully when I treated of the sounds produced by insects, that it is not necessary to repeat what I have said; which observation also applies to the drums which distinguish the male Cicadæ[844]. The wings of some butterflies, and of most moths and hawkmoths (Sphinx L.), are furnished with a singular apparatus for keeping them steady, and the under-wing from passing over the upper in flight. This appears to have been first noticed by Moses Harris, and was afterwards more fully explained by M. Esprit Giorna[845]. From the base of the under-wing proceeds a strong bristle, received by an annulus or socket, which springing between the two principal nervures of the upper-wing terminates in the disk of the wing: in this annulus the bristle moves to and fro, and prevents the displacement of the under-wing. This apparatus is perfect only in the males, which alone have occasion for long flights; the females, though they have often several bristles, having no annulus[846].
The other instruments of motion, the legs, also differ in the sexes. In some instances they are disproportionably long. This is particularly the case with the anterior pair[Pg 332] of some beetles, as Macropus longimanus, Scarabæus longimanus L., in which they are so long as to make the males of these individuals rather inconvenient in a cabinet. Amongst British beetles Clytra longimana and Curculio longimanus Marsh. are also remarkable in this respect. In some other males the middle pair are the longest; as in Anthophora retusa Latr., a kind of wild-bee[847]. There are two known instances of remarkably long posterior legs in the Capricorn tribe, which I suspect belong to the present head. One is Saperda hirtipes Oliv.[848], in which the hind-legs are longer than the whole body, and adorned with a singular tuft of hairs; and the other a Clytus, I think, which Mr. MacLeay purchased from the late Mr. Marsham's collection, in which the hind-legs are not only very long, but have tarsi convolute, like some antennæ. From analogy I should affirm that these were the characters of male insects.
To come to the parts of legs. Sometimes the coxæ of the last mentioned sex are distinguished from those of the female by being armed by a mucro or spine. Thus the male of Megachile Willughbiella, and others of that tribe, have such a spine on the inner sides of the anterior coxa[849]. The Trochanter also of some differs sexually; and you will find that the posterior one of the male in Anthidium manicatum is of a different shape from what it is in the female[850]. In Sphodrus leucopththalmus, one of the beetles called black dors, in one sex the same trochanter[Pg 333] terminates in a long mucro or spine[851], and in the other it is rounded at the apex.
Peculiar characters in their thighs also often indicate different sexes. In Prionus damicornis there is a short spine at the apex of the anterior ones in the female that is not in the male; while in Macropus longimanus, at their base externally the male is armed with a mucro, which I cannot find in the female[852]. In Scarabæus longimanus L. this thigh is furnished with two teeth[853].—The intermediate thighs also sometimes differ. In an Onitis from China, a variety perhaps of O. Sphinx, those in the male are dolabriform, and in the other sex of the ordinary shape. In Odynerus spinipes they have on their lower side two sinuses, so as to give them the appearance of being toothed. The posterior thighs are sometimes incrassated in the male, and not in the female. This you will see in a weevil, not uncommon, Apoderus Betulæ, and also in many species of Cimbex F., a kind of saw-fly; and the same circumstance distinguishes the latter sex in many species of Lygæus F., a kind of bug: I discovered this from L. cruciger, of which I have both the sexes; and from Stoll's figure of L. Pharaonis[854]. In some of these the female thighs are enormously large. A remarkable variation in this respect is observable in the coleopterous genus Œdemera (Necydalis L.). In Œ. Podagrariæ these limbs are incrassated in one sex and not in the other[855]; in Œ. cærulea they are so in both sexes; and in Œ. ceramboides[Pg 334] in neither. In Pelecinus Polycerator F., one of the Ichneumon tribe, or an insect very near it from Brazil, these thighs in the female are armed with two spines underneath, which are not in the male.
The anterior tibiæ in Scarabæus longimanus L. differ remarkably in the sexes. In the female they are of the ordinary shape, and serrated externally; but in the male they are very long, incurved, and without teeth or serratures[856]. In the males of the genus Onitis F. they are bent like a bow, and acute at the end; but in the females they are formed on the common type[857]. In Hispa spinipes F. they are armed internally with a crooked spine[858]. But the most extraordinary sexual variation of this joint of the leg may be seen in the male of Crabro cribarius F. and several other species of the same family, in which these tibiæ are dilated externally into a concavo-convex plate, or rather have one fixed to them and part of the thigh, of an irregular and somewhat angular shape[859], with numerous transparent dots, so as not badly to resemble a sieve: whence the trivial name of the species. Rolander, who first described it, fancied that this plate was really perforated, and that by means of it the animal actually sifted the pollen; but it is most probably for sexual purposes. In another species, the plate is ornamented with transparent converging streaks. In the bee-tribes (Anthophila Latr.) the posterior tibia of the working sex is generally bigger than the corresponding part in their more idle partners: this is particularly[Pg 335] conspicuous in the genus Euglossa, in the females of which this part is triangular, very broad towards the apex, and fitted for carrying a large mass of pollen paste. The tibiæ of the males of some Lepidoptera are remarkable in this respect. That of Hepialus Humuli is much more hairy; but in H. Hectus it is a dilated mis-shapen mass, without a tarsus, and with long scales pendent from the disk[860]. Differences of this kind also occur in the calcaria or spurs that arm the apex of the tibiæ of a large number of insects. Thus in Acanthopus Klug, a singular bee, in the male the spur of the intermediate leg is dilated at the apex, and armed with six strong spines, the inner one larger than the rest[861].
But the part of the leg in which the sexes most vary is the tarsus; and this variation takes place both in the number of the joints, and their form and circumstances. The first case has been observed only with regard to certain species of Cryptophagus Herbst, as C. fumatus, &c. in which the female is pentamerous, or having five joints in all the tarsi; and the male heteromerous, or having five joints in the two anterior pairs, and only four in the posterior[862]. With respect to the form of the tarsal joints, the sexes more frequently differ; and by inspecting this part, especially in the predaceous and carnivorous Coleoptera, you may often, without further examination, ascertain whether any individual is male or female. Even in the slender-footed Cicindelidæ, the three first joints of the anterior tarsus of the male are more dilated than the two last, and covered underneath with a brush[Pg 336] of stiffish hair; in the female all are equally slender, and not so hairy. In Carabus, Feronia, &c. Latr. the four first joints of these tarsi in the males are dilated, and furnished with a brush or cushion: in the Silphidæ, also, the same circumstance takes place. In Harpalus Latr., and Silpha americana, the four anterior ones are similarly formed in this respect. But one of the most remarkable sexual characters, in this tribe of insects, that distinguish the males, are those orbicular patellæ, furnished below with suckers of various sizes, and formed by the three first joints of the tarsus, which are to be met with in the Dytiscidæ, &c.; but as I shall have occasion to treat of these more fully in another Letter, I shall only allude to them now. The second pair of tarsi have in these also the three first joints dilated and cushioned[863]. In Hydrophilus piceus, another water-beetle, the fifth joint of the tarsus is dilated externally, so as to form nearly an equilateral triangle[864]. Christian, a German writer on the Hymenoptera, has described some very singular appendages which he observed on the first joint of the four posterior tarsi of Xylocopa latipes F. These were battledore-shaped membranaceous laminæ, with a reticulated surface, of a pale colour; which were fixed in pairs by the intervention of a footstalk to the above joint, on which they sometimes amounted to more than a hundred: the use of which, he conjectures, is the collection of pollen[865]. I possess two specimens of this bee; one has none of these appendages, and on the other I can discover them only in one of the tarsi—from which circumstance I am[Pg 337] led to conjecture that, like the supposed Clavariæ that were imagined to grow on some humble-bees, but which are now ascertained to be the anthers of flowers—these also belong to the kingdom of Flora, and are spoils which the bee in question has filched from the blossom of some plant. The individuals that have been thus circumstanced are males; whether the female is guilty of similar spoliations is not known. In my specimen there are no traces of them. In many bees, the first joint of the posterior tarsi is much larger in the females and workers than in the males; but in the hive-bee this joint is largest in the latter[866]. In Beris clavipes and Empis nigra, two flies, the joint in question is large and thick in the male, but slender in the female. The penultimate tarsal joint in the posterior legs is dilated internally, and terminates in a mucro in one sex of Anoplognathus Dytiscoides of Mr. W. MacLeay[867]. In some insects the anterior tarsus of the males has been supposed to be altogether wanting: I allude to the petalocerous genus Onitis F.; but I have a specimen of Onitis Apelles of this sex, or a species nearly related to it, in which one of these tarsi is to be found[868]; which, though very slender, consists of five joints, and is armed with a double claw: from which circumstance it may, I think, be concluded, that although, as in Phanæus, these tarsi are very minute, they are not wanting. What renders this more probable is, a circumstance which every collector of insects, who has many specimens of Mr. W. MacLeay's Scarabæidæ in his cabinet, must have noticed: namely, that in all, except Copris[Pg 338] and Onthophagus, the anterior tarsi are usually broken off. Out of seventeen individuals of Scarabæus MacLeay in my own, not a single one has a relic of an anterior tarsus; and scarcely one in a much greater number of Phanæi. The tarsus in question in the nobler sex in Crabro, at least in C. cribrarius and its affinities, is also very short, especially the three intermediate joints; but at the same time very broad and flat. In the species just named, the external claw forms a kind of hook; and in the rest it is considerably longer than the other[869]. The claws, indeed, occasionally vary in the sexes in other Hymenoptera: thus in Melecta Latr., a kind of bee, in the female they are intire, but in the male they are furnished with an internal submembranaceous tooth or process[870]. In Cœlioxys conica and others, those of the latter sex are bifid at the apex, but those of the former acute[871]. In Megachile, the male claw is as in the instance just mentioned, while the female has a lateral tooth[872]; and a similar character distinguishes the sexes in the hive-bee[873].
3. The abdomen. This part affords many external sexual characters, whether we consider its general shape; the number of segments that compose it; its base, middle, or extremity.
In general shape it often differs in the sexes. Thus, the abdomen of female Tipulæ is lanceolate; that of the male cylindrical, and thickest at the extremity[874]. In[Pg 339] Molorchus F. it is convex above in the former, and flat in the latter,—the female of this beetle not unaptly representing some female Ichneumons in this respect, and the male their males[875]. In Andrena it is oblong in the one, and lanceolate in the other. In the hive-bee the drones have a thick, obtuse, and rather long abdomen; in the females it is long, and nearly represents an inverted cone; and in the workers a three-sided figure, or prism.
The number of segments, also, is generally different in the two sexes—the male having one more than the female; but in Dytiscus marginalis, &c. the reverse of this takes place: the female, if you reckon the bipartite half-concealed anal segment as one, having seven ventral segments, and the male only six. She has also eight dorsal, and the male seven.—In the ant tribes (Formica L.), the little vertical scale, at the base of the abdomen in one description of them, or the double knot in another, is less in the male than in the female. In a very singular male insect belonging to the Vespidæ, and related to Synagris, (which I purchased from the late Mr. Drury's cabinet,) the second ventral segment sends forth from its disk two remarkable parallel very acute and rather long spines. The same sex of Chelostoma maxillosa has likewise on the same segment a concave elevation, opposite to which on the fifth is a cavity which receives it, when the animal rolls itself up to take its repose[876]. In another species, C. Campanularum, the segment in question has only a tubercle[877].
On the second segment of the abdomen of some specimens,[Pg 340] probably males, of the remarkable African genus Pneumora before alluded to[878], there are thirteen little elevated ridges, placed rather obliquely in an oblique series; and gradually, though slightly, diminishing in size towards the belly: on their upper side they are flat, forming nearly a horizontal ledge, but on the lower they slope to the abdomen. The posterior thigh in its natural position covers the three first of them, and, if moved downwards, would strike them all[879]. I conjecture, therefore, that these are the animal's instruments of sound, imitating the harp or violin rather than the drum; and that the thigh acts the part of the hand or bow. The abdomen of these insects being blown out like a bladder, and almost empty[880], must emit a considerable sound when the thigh of the animal passes briskly over these ridges; and their different length would produce a modulation in the sound. When struck with a pin, they emit a grating noise.
In Staphylinus splendens, the penultimate ventral segment is very deeply cleft, and the antepenultimate emarginate in one sex, and intire in the other. In S. laminatus, an allied species, the penultimate segment is cleft, less deeply, however; but the antepenultimate is very short and intire; while the fourth is extremely long, and rounded at the margin, appearing as if it was only an elevated part of the last-mentioned segment; for which it was mistaken by Gravenhorst[881], while it is of the usual form in the other sex.
The extremity of the abdomen or its anal segments and organs furnish a variety of sexual characters. Sometimes the last dorsal segment is emarginate in the male, and not in the female; as in Megachile ligniseca, one of the leaf-cutter bees, Cimex hæmorrhoidalis, &c.[882] At other times little lateral teeth are added to this notch, as in another of the same tribe, M. Willughbiella[883]. Again, in other males, both the ventral and dorsal anal segment are armed each with a pair of teeth or mucros, as in Chelostoma maxillosa[884]. In Anthidium manicatum, another bee, the anus terminates in five spines[885]. In Cœlioxys conica of the same tribe, in which this part in the female is very acute, that of the male is armed with six points[886]. In that singular Neuropterous genus Panorpa, while the abdomen of the female is of the ordinary form, with a pair of biarticulate palpiform organs attached to the last retractile joint, or ovipositor, that of the male terminates in a jointed tail, not unlike a scorpion's, at the end of which is an incrassated joint armed with a forceps[887]. In the common earwig (Forficula auricularia) the two sexes differ considerably in their anal forceps: in one it is armed with internal teeth at the base, and suddenly dilated, above which dilatation it is bent like a bow: in the other it is smaller, without teeth, grows gradually narrower, is very minutely crenulate from the base to the end, and is straight, except at the very summit, where it curves inwards. Misled by these and similar differences, Mr. Marsham[Pg 342] has considered them (the sexes both of F. auricularia and F. minor) as distinct species.
The tail of some species of the genus Ephemera is furnished with three long, jointed, hairy bristles. We learn from Reaumur with respect to one, that though in the female these are all equal in length, yet in the male there is only a rudiment of the third. On the belly near the anus these males have four fleshy appendages, the posterior ones setaceous and long, and the anterior pair filiform and shorter. They are supposed to represent the anal forceps of other insects[888]. In Ephemera vulgata, described by De Geer, both sexes have three bristles, but those of the male are the longest; and he describes the forceps as consisting of only a pair of jointed pieces, forming a bow not unlike the forceps of an earwig[889].
v. All the differences I have hitherto noticed between the sexes of insects occur in their bodily structure; but there are others of a somewhat higher description observable in their character. You may smile at the idea of character in beings so minute; but if you recollect what I formerly related to you when treating upon the societies of insects, you will allow that something of this kind does take place amongst them. In general the males are more fitted for locomotion and more locomotive; and the females, on the contrary, are necessarily more stationary. And this for an obvious reason:—the law is, that the male shall seek the female, and therefore he is peculiarly gifted for this purpose, both in his organs of sensation and motion: while his partner in many cases has very simple antennæ, he has very complex ones; and[Pg 343] while she has either no wings or only rudiments of them, he is amply provided with them. Again: amongst the insects that suck the blood of man or beast, such as the gnat (Culex) or horse-flies (Tabanidæ), it is the female alone that is bloodthirsty, the males contenting themselves with the nectar of flowers[890]. But the difference of character in the sexes is most conspicuous, at least it has been more noticed, in those that live in societies, and is quite the reverse of what takes place in the human species. While the females and workers (which are now generally considered as sterile females, in which the ovaries are not developed) are laborious and active, diligent and skilful, wise and prudent, courageous and warlike;—the males, on the contrary, take no part in promoting the common weal, except merely a sexual one. Though till a certain period they are supported at the expense of the community, they take no part in its labours, either in collecting and forming the public stores, or in feeding and attending the young. They are idle, cowardly, and inactive; have neither art nor skill of any kind, and are unprovided with the usual offensive weapons of their species. These observations in their full force apply particularly to the hive-bee, and partially to the other social insects; amongst which, if you consult my former communications, there are some exceptions to this slothful character in the males[891].
II. Age. There is less diversity in the duration of the lives of insects in their perfect than in their larva or pupa[Pg 344] state. Some, like several species of Ephemeræ, live only a few hours; some never even see the sun[892]: others, as flies, moths, and butterflies, and indeed the majority of insects, a few days or weeks; and a comparatively small number, such as some of the larger Coleoptera, Orthoptera, &c., six, nine, twelve, or fifteen months—a period beyond which the life of perfect insects rarely extends. Some, however, certainly enjoy a longer existence in the perfect state. Mr. Baker kept one of the darkling beetles (Blaps Mortisaga) alive under a glass upwards of three years. The rose-beetle (Cetonia aurata), Rösel informs us he fed with fruit and moist white bread for as long a period[893]. Esper kept our most common water-beetle (Dytiscus marginalis) in water in a large glass vessel, feeding it with meat, for three years and a half[894]. With regard to the Arachnida, from the very slow growth of Scorpio europæus, Rösel suspects that it must live two or three years; and Audebert is stated to have kept a spider for several[895]. In this respect insects follow a law very different from that which obtains amongst vertebrate animals. In these the duration of their life is in proportion to the term of their growth: those which attain to maturity the latest, in almost every case living the longest. In insects, on the contrary, we often meet with the very reverse of this rule. Thus the larva of the great[Pg 345] goat-moth (Cossus ligniperda) is three years, that of the cabbage-butterfly (Pieris Brassicæ) not three months, in attaining maturity; yet the perfect insects live equally long. Melolontha vulgaris, which in its first state lives four years, as a beetle lives only eight or ten days[896]. And some Ephemeræ, whose larvæ have been two years in acquiring their full size, live only an hour; while the flesh-fly, whose larva has attained to maturity in three or four days, will exist several weeks.
There is yet another anomaly in the duration of the life of perfect insects. This is not, as in larger animals, a fixed period liable to be shortened only by accident or disease, and incapable of being prolonged; but an indeterminate one, whose duration is dependent on the earlier or later fulfilment of a particular animal function—that of propagation. The general law is, that a few days, or at most weeks, after the union of the sexes, both perish, the female having first deposited her eggs. If, therefore, this union takes place immediately after the disclosure of the insect from the pupa, their existence in the perfect state will not exceed a few days or weeks, or in some cases hours, as in that of the Ephemera, and likewise of the Phalænæ Attaci L. &c., which fall down dead immediately after oviposition[897]. But if by any means it be put off or prevented, their life may be protracted to three or four times that period. Gleditsch asserts, that by keeping apart the sexes of a grasshopper, their lives were prolonged to eight or nine weeks, instead of two or three, their ordinary length; and under similar circumstances Ephemeræ, which usually perish in a day, have[Pg 346] been kept alive seven or eight. It is in consequence of this very curious fact, which has not received from physiologists the attention that it merits, that many butterflies and other insects, which, when excluded from the pupa in summer, perish in less than a month, live through the winter, if excluded late in the autumn, and the union of the sexes does not ensue. It is probable that the great age to which Baker's Blaps, Rösel's Cetonia, and Esper's Dytiscus attained, was owing to their being virgins when taken, and subsequently kept from any sexual intercourse. A parallel case happens in the vegetable kingdom:—if annual plants are kept from seeding, they will become biennial; as, likewise, if they are sown too late in the year to produce seeds.
In the case, however, of the earlier or later exclusion of the imago, another agent has probably some influence. Buffon found that, other circumstances being alike, the silkworm-moths placed in a northern, lived longer than those exposed to a southern aspect: whence it appears that the stimulus of heat shortens the lives of insects, and consequently that cold tends to lengthen them.
It must be observed too, that as the death of the female insect does not take place until all the eggs are excluded, the term of her life, though usually short in the majority of species, which lay their whole number at once, is proportionably long in those which, like the queen-bee, have a longer period assigned them for this important office. Huber affirms, that he had certain proofs that she was engaged for two years in laying eggs, all impregnated by a single sexual union[898]; and in the[Pg 347] females of most insects that live in society, several months are required to mature the last eggs that are in the ovary. There is one tribe of insects, however, the females of which are affirmed to survive this operation: I mean Dorthesia Bosc; after which they even moult, though not so often as before[899].
I formerly related to you the singular fact, that the drones in a beehive at a certain period are without mercy slaughtered by the workers[900]. A fact the reverse of this is recorded by Morier with respect to the locusts: he affirms that the female, when she has done laying her eggs, is surrounded and killed by the males. He says that he never himself witnessed this extraordinary circumstance; but that he heard it from such authority that he gave full credit to it[901]. It is a fact, however, that seems to require further evidence to entitle it to such credit. These are instances in which, by a law of nature, the life of these insects is shortened by violence. It does not appear to have been ascertained how long those drones live that, under particular circumstances, as stated in a former letter[902], are exempted from the usual slaughter.
I am, &c.
Having shown you our little animals in every state, and traced their progress from the egg to the perfect insect, I must next give you some account of their structure and anatomy. And under this head I shall introduce you to a microcosm of wonders, in which the hand of an Almighty workman is singularly conspicuous. One would at first think that the giant bulk of the elephant, rhinoceros, or hippopotamus, must include a machine far more complicated, a skeleton more multifarious in its composition—covered by muscles infinitely more numerous—instinct with a nervous system infinitely more ramified—with a greater variety of organs and vascular systems in play, than an animal that would scarcely counterpoise a ten-millionth portion of it. Yet the reverse of this is the fact; for the Creator, the more to illustrate his wisdom, power, and skill, has decreed that the minute animals whose history we are recording, shall be much more complex in all the above respects than these mighty monarchs of the forest and the flood. Of this in the present and subsequent letters you will find repeated[Pg 349] and scarcely credible instances, which in every rightly constituted mind are calculated to excite, in an extraordinary degree, those sensations of reverence and love for the Invisible Author of these wonders, and that faith and trust in his Power and Providence, which an attentive survey of the works of Creation has a natural tendency to produce. And you will not only be struck by this circumstance, but equally by the infinite variations in the structure that will present themselves to your notice; and that not sudden and per saltus, but by approaches made in the most gradual manner from one form to another. And all along, where the uses of any particular organ or part have been ascertained, if you consider its structure with due attention, you will find in it the nicest adaptation of means to an end: a circumstance this, which proves most triumphantly, that the Power who immediately gave being to all the animal forms, was neither a blind unconscious power, resulting from a certain order of things, as some philosophists love to speak[903]; nor a formative appetency in the animals themselves, produced by their wants, habits, and local circumstances, and giving birth, in the lapse of ages, to all the animal forms that now people our globe[904]; but a Power altogether distinct from and above nature, and its Almighty Author[905].
I trust that what I have here advanced will excite your attention to the subject I am now to enter upon; and I flatter myself, that although at first sight it may promise nothing more than a dry and tedious detail of parts and organs, you will find it not without its peculiar interest and attraction.
This department of the science—the Anatomy of Insects—may still be regarded as in its infancy; and considering[Pg 351] the almost insuperable difficulties which, from the minuteness of the objects, oppose themselves to the skill and instruments of the entomological anatomist, we can scarcely hope that it will ever attain to that certainty and perfection to which, as far as the larger animals are concerned, anatomy has arrived. Yet infinitely more has been accomplished than might have been expected, and new accessions of light are daily thrown upon it. When[Pg 352] we consider what has been done by Malpighi, Leeuwenhoeck, and especially Swammerdam, we admire the patience, assiduity, and love of science, that enabled them, in spite of what seemed insurmountable obstacles, to ascertain, the first with respect to the silk-worm, and the latter in numerous instances, the internal organization of these minute creatures, as well as their external structure. Reaumur, and his disciple De Geer, extending their researches, have also contributed copiously to our knowledge in this branch of our science.
But in this field no one has laboured so indefatigably and with so much success as the celebrated Lyonnet; and though his attention was confined to one object—the caterpillar of the goat-moth (Cossus ligniperda F.),—every one who studies his immortal work must admire the patient and skilful hand, the lyncean eye, and keen intellect, that discovered, denuded, and traced every organ, muscle, and fibre of that animal. Much is it to be regretted that his proposed works on the pupa and imago of the same insect, which, he informs us, were far advanced[906], were never finished and given to the world. Our regret, however, is in some degree diminished by the elaborate work of M. Herold on the butterfly of the cabbage (Pieris Brassicæ), before eulogized[907]; in which he has done much to supply this desideratum.
In more modern times, besides Herold, MM. Latreille, Illiger, Marcelle de Serres, Savigny, Ramdohr, Treviranus[Pg 353] Sprengel, Audoin, Chabrier, and, above all, M. Cuvier in his celebrated Lectures on Comparative Anatomy, have considerably extended the boundaries of our knowledge in this department: and much of what I have to say to you in my letters on this subject, will be derived from these respectable sources. In the exterior anatomy of insects, I flatter myself that I shall be enabled to make some material additions to the discoveries of my predecessors; though few have occurred to me with respect to their internal organization.
In treating of the anatomy of the vertebrate animals, it is usual, I believe, to consider, first, the skeleton and its integuments, whether of skin or muscle, and their accessories; and afterwards the organs of the different vital functions and of the senses. But in considering the anatomy of Insects, the difference before stated[908], observable between them and the sub-kingdom just mentioned, as to their structure, renders it advisable to divide this subject into two parts—the first treating of their external anatomy, and the second of their internal.—I shall begin by drawing up for you a Table of the Nomenclature of the parts of their external crust; its appendages and processes[909], external or internal, accompanied by definitions of them; and followed by such observations respecting them as the subject may seem to require for its more full elucidation.
Anatomists have divided the human skeleton into three[Pg 354] greater sections—the Head, the Trunk, and the Limbs. That of insects, likewise, is resolvable into three primary sections, but without including the limbs (which, as being appendages, and therefore secondary, had best be considered under the section of which they form a part), for the abdomen in insects, as well as the rest of the body, being covered with a crust, and forming a distinct part, may be properly regarded as a primary section. And in fact these three parts may be received as primary under another view—the head, as containing the principal organs of sensation; the trunk, as containing those of motion; and the abdomen, as containing those of generation[910]. Under each of these primary sections, I shall consider its respective organs, members, and parts.
You are not to expect to find every part included in the following Table in every insect; since it has been my aim to introduce into it, the most remarkable of those that are peculiar to particular tribes, genera, &c. With respect to these, I shall generally refer you to the individuals in which they may be found.
DEFINITIONS.
Corpus (the Body). The whole crust of the insect; consisting of the Exoderma or external covering, and the Esoderma or internal cuticle that lines it[911]. It is divided into three primary parts, or sections—Caput, Truncus, Abdomen.
INSERT FOLDOUT HERE
INSERT FOLDOUT HERE
I. CAPUT (The Head).
The Head is the anterior section of the body; consisting of a kind of box without suture or segment, which receives the organs of sensation and manducation. It includes the Os, Facies, Subfacies, and Collum.
i. Os (the Mouth). That part of the head which receives and prepares the food for passing into the stomach. It includes the Trophi[912].
1. Trophi (the Trophi). The different instruments or organs contained in the mouth, or closing it, and employed in manducation or deglutition. They include the Labrum, Labium, Mandibulæ, Maxillæ, Lingua, and Pharynx.
A Labrum (the Upper-lip). A usually moveable organ; which, terminating the face anteriorly, covers the mouth from above, and is situate between the Mandibulæ[913]. It includes the Appendicula.
a Appendicula (the Appendicle). A small piece sometimes appended to the upper-lip[914]. Ex. Halictus ♀ Walck. (Melitta **. b. K.)
B Labium (the Under-lip). A moveable organ, often biarticulate, which terminating the surface anteriorly, covers the mouth from beneath, and is situate between the Maxillæ[915]. It includes the Mentum, and Palpi Labiales.
a Mentum (the Chin). The lower joint of the Labium,[Pg 356] where it is jointed; in other cases its base. It is usually seated between the base of the Maxillæ[916].
b Palpi Labiales (the Labial Feelers). Two jointed sensiferous organs, the use of which is not clearly ascertained, which emerge, one on each side, from the Labium, mostly near its summit[917].
C Mandibulæ (the Upper-jaws). Two transverse lateral organs, in most insects used for manducation; generally corneous, moving horizontally, and closing the mouth above, under the Labium[918]. They include the Prostheca, Dentes, and Mola.
a Prostheca (the Prostheca). A subcartilaginous process attached to the inner side, near the base, of the Mandibulæ of some Staphylinidæ[919]. Ex. Ocypus similis K., Creophilus maxillosus K., &c.
b Dentes (the Teeth). The terminating points of the Mandibulæ. They include the Incisores, Laniarii, and Molares[920].
A Incisores (the Cutting-teeth). Teeth somewhat wedge-shaped, externally convex and internally[Pg 357] concave[921]. Ex. Gryllotalpa Latr., Gryllus Latr. (Acheta F.), &c. &c.
B Laniarii (the Canine-teeth). Very sharp and usually long conical teeth[922]. Ex. Forficula L., Mantis L., Libellula L.
C Molares (the Grinding-teeth). Teeth that terminate in a broad uneven surface, fit for grinding the food[923]. Ex. the herbivorous Orthoptera.
c Mola (the Mola). A broad, flat, subrotund space, transversely grooved or furrowed, observable on the inner side of some mandibles that have no grinding-teeth at their apex[924]. Ex. Euchlora MacLeay, Anoplognathus Leach, Larva of Lucanus[925].
D Maxillæ (the Under-jaws). Two organs moving subhorizontally, fixed on each side at the base of the Labium, and often parallel with it—which in masticating insects seem primarily designed to hold the food[926]. They include the Cardo, Stipes, Lobi, and Palpi maxillares.
a Cardo (the Hinge). A small, transverse, usually triangular, corneous piece, upon which the Maxilla commonly sits[927].
b Stipes (the Stalk). The corneous base of the Maxilla, below the Palpus[928].
c Lobi (the Lobes). The parts of the Maxilla above the Palpus[929]. They include the Lobus superior, the Lobus inferior, and the Ungues.
A Lobus Superior (the Upper-lobe). The outer lobe of the Maxilla, incumbent on the inner one. In the Predaceous Beetles this lobe is biarticulate and palpiform[930]; and in Staphylinus olens, &c. it also consists of two joints[931]. It is called the Galea by Fabricius, in Orthoptera, &c.[932]
B Lobus Inferior (the Lower-lobe). The inner lobe of the Maxilla, covered by the outer one[933].
C Ungues (the Claws). One or more corneous sharp claws which arm the lobes of the Maxilla[934]. In the Predaceous Beetles there is only one terminating the lower lobe, with which, in Cicindela, it articulates; in the Orthoptera and Libellulina there are several.
d Palpi Maxillares (the Maxillary Feelers). Two jointed sensiferous organs, the use of which is not clearly ascertained, emerging from an exterior lateral sinus of the Maxilla[935].
E Lingua (the Tongue). The organ situated within the Labium or emerging from it, by which insects in many cases collect their food and pass it down to the Pharynx, situated at its roots above. It varies considerably in different orders and tribes. In the Orthoptera, Libellulina, &c. it is linguiform, and quite distinct from the Labium[936]; it appears also distinct in the lamellicorn beetles, &c.[937] In many[Pg 359] Hymenoptera it emerges from the Labium, and is fitted to collect liquids and pass them downwards[938]. In Formica it appears to be retractile[939]. In a considerable proportion of insects it seems connate with the Labium, and forming its inner surface? According to circumstances it might perhaps be denominated Lingua or Ligula. It includes the Paraglossæ.
a Paraglossæ (the Paraglossæ). Lateral and often membranous processes observable on each side of the tongue in some Hymenoptera, &c.[940]
F Pharynx (the Pharynx). The opening into the gullet[941]. It includes the Epipharynx and Hypopharynx.
a Epipharynx (the Epipharynx). A small valve under the Labrum, that in many Hymenoptera closes the Pharynx, and is an appendage of its upper margin[942].
b Hypopharynx (the Hypopharynx). An appendage[Pg 360] of the lower margin of the Pharynx, observable in Eucera F.[943]
The seven organs of the mouth above defined, viz. the Labrum, Labium, the two Mandibulæ, the two Maxillæ, and the Lingua, constitute what may be denominated a perfect mouth, peculiar to those insects that masticate their food[944]. In those that take it by suction, the Trophi, to adapt them for that purpose, assume a variety of forms, and should be distinguished by as many appellations. In almost every case, however, the rudiments or representatives of the above organs have been detected by the elaborate researches of that learned and able zoologist, M. Savigny[945]. I shall next subjoin definitions of the principal kinds of suctorious mouths.
2. Promuscis (the Promuscis). The oral instrument of Hemiptera, in which the ordinary Trophi are replaced[946] by a jointed sheath, covered above at the base by the Labrum, without Labella (Liplets) at the end, and containing four long capillary lancets, and a short tongue[947]. It includes the Vagina, and Scalpella.
A Vagina (the Vagina). The jointed sheath of the Promuscis, representing the Labium in a perfect mouth[948].
B Scalpella (the Lancets). Four pieces adapted for perforating the food of the insect, which when united form a tube for suction. The upper pair represent the Manidibulæ[949], and the lower the Maxillæ[950].
3. Proboscis (the Proboscis). The oral instrument of Diptera, in which the ordinary Trophi are replaced by an exarticulate sheath, terminated by Labella, and containing one or more lancets covered by a valve[951]. It includes the Theca, and Haustellum.
A Theca (the Theca). The sheath or case of the Proboscis, representing the Labium in a perfect mouth[952]. It includes the Basis, and Labella.
a Basis (the Base). The whole lower part of the Theca, from the mouth of the insect as far as the Labella, probably to be regarded as representing the Mentum?
b Labella (the Liplets). A pair of tumid lobes, often corrugated and capable of tension and relaxation, which terminate the Theca, and perhaps represent the termination of the Labium[954]?
B Haustellum (the Haustellum). The instrument of suction contained in the Theca[955]. It includes the Valvula, Cultelli, and Scalpella.
a Valvula (the Valvule). A corneous piece which covers the instruments of suction above, representing the Labrum in a perfect mouth[956].
b Cultelli (the Knives). The upper pair of the instruments of suction, which probably make the first incision in the food of the insect; they represent the Mandibulæ of the perfect mouth[957].
c Scalpella (the Lancets). A pair of instruments, usually more slender than the Cultelli, which probably enter the veins or sap-vessels, and together with them form a tube for suction[958].
4. Antlia (the Antlia). The oral instrument of Lepidoptera, in which the ordinary Trophi are replaced by a spiral, bipartite, tubular machine for suction, with its appendages[959]. It includes the Solenaria, and Fistula.
A Solenaria (the Solenaria). The two lateral subcylindrical air-tubes of the Antlia[960].
B Fistula (the Fistula). The intermediate subquadrangular pipe, formed by the union of the two branches of the Antlia, which conveys the nectar to the Pharynx[961]. These two branches represent the Maxillæ of the perfect mouth.—N. B. M. Savigny discovered the rudiments of the remaining Trophi in this kind of mouth[962].
5. Rostrulum (the Rostrulum). The oral instrument[Pg 363] of Aphaniptera (Pulex L.), in which the ordinary Trophi are replaced by a bivalve beak, between the valves of which there appear to be three lancets[963]. It includes the Laminæ, Scalpella, and Ligula.
A Laminæ (the Laminæ). Two corneous plates which are laterally affixed to the mouth of a flea, probably representing the Mandibulæ of the perfect mouth, which somewhat resemble the beak of a bird[964].
B Scalpella (the Lancets). The two upper or outer instruments, probably for making an incision in the skin; these are flat and acute, and seem to represent the Maxillæ of the perfect mouth[965].
C Ligula (the Ligula). A capillary instrument between the lancets; probably representing the tongue of the perfect mouth[966].
6. Rostellum (the Rostellum). The oral instruments of Pediculus and some other Aptera, in which the ordinary Trophi are replaced by an exarticulate retractile tube, which exerts a retractile siphuncle. It includes the Tubulus and Siphunculus.
A Tubulus (the Tubulet). The tube or retractile base of the Rostellum.
B Siphunculus (the Siphuncle). The real instrument of suction, which when unemployed is retracted within the tubulet.
Besides the above variations from the type of what I call a Perfect Mouth, there are others in which the parts of the Trunk appear to aid in the conversion of the food,[Pg 364] and become a kind of accessory Labium, Maxilla, &c. Thus in the Myriapods, the anterior pair of legs assume a Maxillary form and office[967]; the Prosternum those of a Labium[968]: in the Arachnida, also, the anterior Coxæ are accessory Maxillæ. In this Class, likewise, as has been more than once observed[969], the representatives of the interior pair of Antennæ of the Crustacea, are thought to assume the form and the functions of suctorious Mandibles[970].
ii. Facies (the Face). The upper surface of the head[971]. It includes all the parts that lie between its junction with the Prothorax and the Labrum: viz. Nasus, Postnasus, Frons, Vertex, Occiput, Genæ, Tempora, Oculi, Stemmata, and Antennæ.
1. Nasus (the Nose). That portion of the face, often elevated and remarkable, situated between the Labrum, Postnasus, and Genæ, and with which the Labrum articulates; called by Fabricius the Clypeus[972]. It includes the Rhinarium.
A Rhinarium (the Nostril-piece). The space between the anterior margin of the Nasus and the Labrum, in which, in vertebrate animals, the nostrils are often situated[973].—N. B. This is remarkable in some Lamellicorn beetles, as Anoplognathus Leach. In Necrophorus, and some others, it is membranous.
2. Postnasus (the Postnasus). That part of the Face immediately contiguous to the Antennæ, that lies[Pg 365] behind the Nasus, when distinctly marked out.—Ex. Sagra, Prosopis.
3. Frons (the Front). That part of the Face which lies behind the Postnasus, and usually between the posterior part of the eyes. This is sometimes the region of the Stemmata; or they are partly in this or partly in the Vertex[974].
4. Vertex (the Vertex). The horizontal part of the Facies, next the front, that lies behind the eyes and between the temples[975]. This also is often the region of the Stemmata.
5. Occiput (the Occiput). The back part of the head when it is vertical, or nearly so, to its point of junction with the trunk[976].—Ex. Meloe, Ripiphorus, Hymenoptera, Diptera.
6. Genæ (the Cheeks). Those parts which lie on the outside of the anterior half of the eyes, and intervene also between them and the Mandibulæ[977].
7. Tempora (the Temples). Those parts which lie on the outside of the posterior half of the eyes, between which the Frons and Vertex intervene[978].
8. Oculi (the Eyes). The principal organs of sight, most commonly two in number, placed in the sides of the head. In the majority they are compound, consisting of hexagonal lenses. In the Arachnida they are simple[979].
A Canthus (the Canthus). A process of the face, which enters the notch or sinus of the eye[980].—Ex. Scarabæus L., Cerambyx L.
9. Stemmata (the Eyelets). Two, or more commonly[Pg 366] three, convex, crystalline, simple eyes, observable in the Frons or Vertex, or common to both[981].—Ex. Orthoptera, Hemiptera, Hymenoptera.
10. Antennæ (the Antennæ). Two moveable and jointed sensiferous organs, situated in the space between or before the eyes, but in no instance behind them[982]. They include the Torulus, Scapus, Pedicellus, and Clavola.
A Torulus (the Bed). The cavity or socket in which the base of the Antenna is planted[983].
B Scapus (the Scape). The first and in many cases the most conspicuous joint of the Antennæ[984]. It includes the Bulbus.
a Bulbus (the Bulb). The base of the Scapus, by which it inosculates in the Torulus, often subglobose, and looking like a distinct joint[985]. It acts the part of a Rotula, being the pivot upon which the Antenna turns.
C Pedicellus (the Pedicel). The second joint of the Antenna[986]: in some insects acting also the part of a Rotula in the socket of the Scapus, to give separate motion to the Clavola.
D Clavola (the Clavolet). The remaining joints of the Antenna taken together[987]. It includes the Capitulum.
a Capitulum (the Knob). The last joints of the Clavola when suddenly larger than the rest[988].
iii. Subfacies (the Subface). The lower surface or underside of the head[989]. It includes the Lora and Jugulum.
1. Lora (the Lora). A corneous angular machine observable in the mouth of some insects, upon the intermediate angle of which the Mentum sits, and on the lateral ones the Cardines of the Maxillæ; and by means of which the Trophi are pushed forth or retracted[990].—Ex. Hymenoptera.
2. Jugulum (the Throat). That part of the subface that lies between the temples[991].
iv. Collum (the Neck). The constricted posterior part of a pedunculate head, by which it inosculates in the trunk[992]. It includes the Nucha, Gula, and Myoglyphides.
1. Nucha (the Nape). The upper part of the neck[993]. It includes the Myoglyphides.
A Myoglyphides (the Muscle-notches). Notches in the posterior margin of the neck, usually two in number, observable in Coleopterous insects, to which the levator muscles are attached[994].
2. Gula (the Gula). The lower part of the neck[995].
v. Cephalophragma (the Cephalophragm). A Y-shaped partition that divides the head internally in Locusta Leach, into two chambers, an anterior and posterior.
II. TRUNCUS (The Trunk).
The Trunk is the intermediate section of the body, which lies between the Head and the Abdomen[996]. It includes the Manitruncus, and the Alitruncus[997].
i. Manitruncus (the Manitrunk). The anterior segment of the trunk, in which the head inosculates, or on which it turns[998]. It includes the Prothorax and Antepectus.
1. Prothorax (the Prothorax). The upper part or the shield of the manitrunk, in Coleoptera, Orthoptera, &c. called by way of eminence the Thorax[999]. It includes the Ora, Patagia, Umbones, and Phragma.
A Ora (the Ora). The inflexed or inferior lateral margin of the Prothorax, separated in many genera from the Antepectus by a suture[1000].
B Patagia (the Patagia). Two corneous scales observable in Lepidoptera, fixed on each side of the trunk, just behind the head, and covered with a long tuft of hair[1001].
C Umbones (the Bosses). Two moveable bosses surmounted by a spine, with which the Prothorax of the Coleopterous genus Macropus is armed.
D Phragma (the Phragm). The Septum that closes the posterior orifice of the Prothorax in Gryllotalpa Latr.
2. Antepectus (the Forebreast). The underside or breastplate of the manitrunk, and the bed of the Arms[1002]. It includes the Spiracula Antepectoralia, Prosternum, Antefurca, and Brachia.
A Spiracula Antepectoralia (the Antepectoral Spiracles). A pair of breathing-pores fixed in the membrane[Pg 369] that connects the Antepectus with the Medipectus[1003].
B Prosternum (the Forebreast-bone). A longitudinal or other elevation of the Antepectus between the Brachia[1004].
C Antefurca (the Antefurca). An internal vertical process of the Antepectus, consisting usually of two branches, which afford a point of attachment to muscles of the Brachia[1005].
D Brachia (the Arms). The first pair of legs of Hexapods, the direction of which is usually towards the head; when spoken of with the other legs, called the Forelegs[1006]. They include the Clavicula, Scapula, Humerus, Cubitus, and Manus.
a Clavicula (the Clavicle). The first joint of the Brachium, answering to the Coxa in the legs.
b Scapula (the Scapula). The second joint of the Brachium, answering to the Trochanter in the legs.
c Humerus (the Humerus). The third and elongated joint of the Brachium, answering to the Femur in the legs.
d Cubitus (the Cubitus). The fourth and elongated joint, answering to the Tibia in the legs. It includes the Coronula and Calcaria. . A Coronula (the Coronula). A coronet or semicoronet of spines, observable at the apex of the Cubitus[Pg 370] or Tibia of some insects.—Ex. Dilophus Latr., Fulgora L.
B Calcaria (the Spurs). See the definition under Pedes Postici. They include the Velum.
a Velum (the Velum). A membrane attached to the inner side of the cubital spur in Apis L.[1007]
e Manus (the Hand). The terminal jointed portion of the Brachium, answering to the Tarsus in the legs[1008]. It includes the Pulvilli, Palma, and Digitus.
f Pulvilli (the Pulvilli). See definition under Pedes Postici.
g Palma (the Palm). The first joint of the Manus, when longer and broader than the subsequent ones, or otherwise remarkable; answering to the Planta in the legs[1009].
A Digitus (the Finger). See definition under Pedes Postici. It includes the Ungula.
a Ungula (the Claw-joint). See definition under Pedes Postici. It includes the Pollex, Unguiculi, and Palmula.
α Pollex (the Thumb). A small accessory joint, attached to the Ungula of the Manus in Mantis F.
β Unguiculi (the Claws). See definition under Pedes Postici.
γ Palmula (the Palmlet). A minute accessory joint between the claws, answering to the Plantula in the legs. It includes the Pseudonychia.
* Pseudonychia (the Spurious Claws). See definition under Pedes Postici.
ii. Alitruncus (the Alitrunk). The posterior segment of the trunk to which the abdomen is affixed, and which bears the legs and wings[1010]. It includes the Mesothorax and Medipectus, and the Metathorax and Postpectus.
1. Mesothorax (the Mesothorax). That segment of the alitrunk which bears the Elytra, or the anterior pair of wings, and the intermediate pair of legs[1011]. It includes the Collare, Prophragma, Dorsolum, Scutellum, Frænum, and Pnystega.
A Collare (the Collar). The first or anterior piece of the Mesothorax. In most insects that have a conspicuous Prothorax, as the Coleoptera, this piece appears scarcely to have a representative; but in the Libellulina it co-exists with it, and is more conspicuous[1012]. It is particularly remarkable in Hymenoptera and Diptera.
B Prophragma (the Prophragm). A partition of an elastic substance, rather horny, connected posteriorly with the Dorsolum, which passes down into the anterior cavity of the alitrunk, of which it forms the[Pg 372] upper separation from that of the manitrunk. It affords a point of attachment to several muscles of the wings, &c.[1013]
C Dorsolum (the Dorslet). The piece which lies between the Collare and Scutellum, to which the prophragm is anteriorly attached, and which bears the upper or anterior organs of flight[1014]. It includes the Pteropega, Elytra, Tegmina, Hemelytra, Alæ Superiores, and Tegulæ.
a Pteropega (the Wing-socket). The space in which the organs for flight are planted. That for the secondary or under-wings is in the Metathorax[1015].
b Elytra (the Elytra). The upper organs for flight, when they are without nervures, and uniformly of a thicker harder substance than membrane whether corneous, or coriaceous; lined by a fine membrane; and when closed, united by the longitudinal suture[1016]. They include the Axis, Sutura, Epipleura, Alula, and Hypoderma, and are peculiar to the Coleoptera and Dermaptera.
A Axis (the Axis). A small, prominent, irregular process of the base of the Elytrum, upon which it turns, and by the intervention of which it is affixed to the Dorsolum, in the anterior wing-socket[1017].
B Sutura (the Suture). The conflux of the sutural or inner margins of the two Elytra, where when closed they unite longitudinally[1018].
C Epipleura (the Epipleura). The inflexed accessory margin observable underneath in many Elytra, which covers the sides of the alitrunk and abdomen[1019].
D Alula (the Winglet). A small, membranous, wing-like appendage, attached to the Elytrum on one side and the Frænum on the other; which probably serves to prevent the dislocation of the former[1020].—Ex. Dytiscus. N. B. A similar organ for a similar purpose is to be found in Blatta and the Diptera.
E Hypoderma (the Hypoderma). The skin, in some species beautifully coloured, that lines the Elytra[1021]. N. B. This skin is also found in some Hemelytra, but not in Tegmina.
c Tegmina (the Tegmina). The upper organs of flight, when of a uniform coriaceous or pergameneous texture, veined with nervures, and lapping over each other[1022]. Ex. Orthoptera[1023].
d Hemelytra (the Hemelytra). The upper organs of flight, when they are corneous or coriaceous at the base and membranous at the apex[1024].—Ex. The heteropterous Hemiptera. They include the Corium and Membrana.
A Corium (the Corium). The corneous or coriaceous part of the Hemelytrum[1025].
B Membrana (the Membrane). The membranous part of the same[1026].
e Alæ Superiores vel Primariæ (the Upper or Primary Wings). The upper or anterior organs of flight when formed of membrane, or of the same substance with the under-wings[1027]. They include the Axes, Areæ, Areolæ, Neuræ, Stigma, Parastigma, and Lobuli.
A Axes (the Axes). Several osseous or horny pieces, by which the wing is connected with the Dorsolum[1028]. One usually to each area.
B Areæ (the Areas). The larger longitudinal spaces into which the wing may be divided[1029]. They include the Area Costalis, Intermedia, and Analis.
a Area Costalis (the Costal Area). That part of the wing lying between the anterior margin and the post-costal nervure[1030]. In Hymenoptera and Diptera it includes all the space bounded by the nervures that spring from the postcostal.
b Area Intermedia (the Intermediate Area). That part of the wing lying between the costal area and the interno-medial nervure, in Diptera; or the Anal in Orthoptera, Hemiptera, Hymenoptera, &c.[1031]
c Area Analis (the Anal Area). All that part of the wing which in Diptera lies between the internomedial[Pg 375] nervure; or in Orthoptera, &c. between the anal nervure and the posterior margin[1032].
C Areolæ (the Areolets). The smaller spaces into which the wing is divided by the nervures. They include the Areolæ Basilares, Mediæ, and Apicales.
a Areolæ Basilares (the Basal Areolets). The parallel areolets of the base of the wing[1033].
b Areolæ Mediæ (the Middle Areolets). The areolets of the wing that lie between the basal areolets and the apical[1034].
c Areolæ Apicales (the Apical Areolets). Those areolets of the wing that terminate in or very near the apex[1035].
D Neuræ (the Nervures). Corneous tubes, for expanding the wing and keeping it tense, and to afford protection to the air-vessels—commonly called the Nerves. They include the Neura Costalis, Postcostalis, Mediastina, Externo-media, Interno-media, Analis, Axillaris, and Spuria.
a Neura Costalis (the Costal Nervure). The first principal nervure of the wing, close to or forming the anterior margin in Lepidoptera, Hymenoptera, and Diptera; but sometimes remote from it in Tegmina[1036]. It includes the Phialum and Hamus.
α Phialum (the Phial). A little bag to receive fluid at the will of the insect, by which the weight of the wing is increased. It is found also in the under-wings in Coleoptera[1037].
β Hamus (the Hook). A Hook fixed to the Costal Nervure, near its base on the under-side, in the wings of some Lepidoptera, in which the tendon runs[1038].
b Neura Postcostalis (the Postcostal Nervure). The second principal, and often strongest, nervure of the wing[1039]. It includes the Neuræ Subcostales.
α Neuræ Subcostales (the Subcostal Nervures). Nervures springing from the under-side of the postcostal nervure, or from each other; called the first, second, third, &c. in the order of their occurrence[1040].
c Neura Mediastina (Mediastinal Nervure). A usually slender nervure, springing from near the base of the postcostal; between which and the costal it intervenes. In the Lepidoptera Diurna, however, it is often a strong nervure[1041].
d Neura Externo-media (the Externo-medial Nervure). The third principal nervure of the wing[1042]. It includes the Neura Subexterno-media.
α Neura Subexterno-media (the Subexterno-medial Nervure). A nervure that in some cases intervenes between the externo-medial and interno-medial[1043].
e Neura Interno-media (the Interno-medial Nervure). The fourth principal nervure[1044]. It includes the Neura Subinterno-media.
α Neura Subinterno-media (the Subinterno-medial Nervure). A nervure that sometimes intervenes between the externo-medial and the anal[1045].
f Neura Analis (the Anal Nervure). The principal nervure nearest the interior or posterior margin, with which it includes a space often subtriangular, traversed in most Diptera and many Hymenoptera by another nervure; and in many Tegmina and Hemelytra by several[1046]. In these kinds of upper-wing it is in many cases accompanied by a fold; and the part between it and the interior margin seems often capable of separate motion.
g Neura Axillaris (the Axillary Nervure). The short nervure, where there is only one, intervening between the anal nervure and the interior margin; replaced in some Muscidæ by a spurious nervure[1047].
h Neuræ Spuriæ (the Spurious Nervures). Very obsolete nervures, sometimes found in addition to those usually occurring; as in Syrphus[1048].
E Stigma (the Stigma). A corneous spot or plate, supposed to contain fluid, in the anterior margin of the upper wings; often produced by the conflux of the costal and postcostal nervures[1049].
F Parastigma (the Parastigma). A corneous spot between the costal and postcostal nervures, distinct from the Stigma observable in the Libellulina.
G Lobuli (Lobuli). One or more rounded portions of the base of the wing, separated from the rest by fissures peculiar to the Muscidæ, and the under-wings of some Hymenoptera[1050].
f Tegulæ (the Tegulæ). Small corneous concavo-convex[Pg 378] scales, which in many Orders, particularly Hymenoptera, cover and defend the base of the Upper-Wings[1051].
D Scutellum (the Scutellum). A piece, usually triangular, which follows the Dorsolum; and in Coleoptera is often only a continuation of it—placed between the base of the Elytra or upper-wings[1052].
E Frænum (the Frænum). A piece that lies under the lateral margin of the Scutellum and Dorsolum, or is adjacent to it; and which in many cases connects with the base of the upper organs of flight, so as to prevent their dislocation, by being pushed too far outwards[1053].
F Pnystega (the Pnystega). A corneous scale or plate, which covers certain pneumatic vessels, usually supported by the Scapularia, in Libellulina, &c. becoming dorsal[1054].
2. Medipectus (the Mid-breast). The underside of the first segment of the alitrunk[1055]. It includes the Peristethium, Scapularia, Mesosternum, Medifurca, and Pedes Intermedii.
A Peristethium (the Peristethium). The anterior piece of the Medipectus, which intervenes between the Brachia and mid-legs[1056].
B Scapularia (the Scapulars). Two pieces, one on each side the Medipectus, which succeed the Peristethium, and lie between the midlegs and the Pteropega[Pg 379] or wing-socket[1057]. It includes the Spiracula Scapularia.
a. Spiracula Scapularia (the Scapular Spiracles). Two spiracles observable, one in each scapular, in Acrida laurifolia, &c.
C Mesosternum (the Mid-breastbone). The elevated and central part of the Medipectus, between the midlegs, often terminating anteriorly in a mucro; sometimes, as in Elater, in a cavity, receiving the mucro of the Prosternum[1058].
D Medifurca (the Medifurca). A branching vertical process of the Endosternum, which serves as the point of attachment to the muscles that move the midlegs[1059].
E Pedes Intermedii (the Mid-legs). The intermediate pair of legs, consisting of the same parts as the posterior legs[1060].
3. Metathorax (the Metathorax). The posterior segment of the Alitruncus[1061]. It includes the Mesophragma, Postdorsolum, Postscutellum, Postfrænum, Pleuræ, and Metaphragma.
a Mesophragma (the Mesophragm). A partition of a firm consistence, connected by its posterior margin with the Postdorsolum, and passing down vertically into the mid-chest; serving as a point of attachment to several of the muscles that move the wings[1062]. This, with the prophragm, forms the anterior[Pg 380] cavity of the alitrunk, and with the metaphragm it forms the posterior cavity.
b Postdorsolum (the Postdorsolum). The middle-piece between the mesophragm and the Postscutellum. In Coleoptera it consists of a tense elastic membrane, which is quite covered by the Mesothorax[1063].
c Postscutellum (the Postscutellum). A narrow channel running from the Dorsolum to the Abdomen in Coleoptera, forming an isosceles triangle reversed. In other orders it is either a triangular elevation of the middle of the posterior part of the Postdorsolum, or a distinct triangular piece[1064].
d Postfrænum (the Postfrænum). In Coleoptera the part of the Metathorax in which the Postscutellum lies, at first nearly horizontal, but posteriorly it takes a vertical direction towards the abdomen. In general it may be defined, the part that intervenes between the Postscutellum and the Abdomen; and which in many cases is connected with the posterior basal margin of the under-wings, and prevents their being pushed too far forwards[1065].
e Pleuræ (the Pleuræ). The space behind the scapulars, on which the lower organs of flight are fixed[1066]. They include the Alæ Inferiores.
A Alæ Inferiores (the Under-wings). The lower or secondary pair of organs for flight[1067]. They include the Commissura, Tendo, Hamuli, Pterygium, Alula, and Halteres.
a Commissura (the Commissura). A joint in the costal nervure of the wings of Coleoptera, where they bend to take a transverse fold[1068].
b Tendo (the Tendon). A strong bristle, or bristles observable at the base underneath in the under-wings of many Lepidoptera, which plays in the Hamus of the upper-wings[1069].
c Hamuli (the Hooklets). Very minute hooks in the middle of the anterior margin, observable in some Hymenoptera, by which the under-wing is fixed to the upper, to cause both to act as one organ in flight[1070].
d Pterygium (the Pterygium). In under-wings this is a small wing-like appendage, fixed at the base of the wing in some Lepidoptera[1071].
e Alula (the Winglet). A small concavo-convex scarious appendage, fixed behind the wings at their base, in many Diptera[1072].
f Halteres (the Poisers). Small capitate processes or organs, observable under the wings of Diptera, attended by a spiracle[1073].
B Metapnystega (the Metapnystega). A corneous scale or lamina that covers the pneumatic organs in the Metathorax, situated sometimes in the Pleuræ, as in the Coleoptera; at others in the Postfrænum, as in Tenthredo L.; and sometimes, as in the Libellulina, between that part and the abdomen[1074].
C Metaphragma (the Metaphragm). A nearly vertical septum or partition, attached behind to the Postfrænum, and usually deeply cleft at its apex in Coleoptera, of a rather horny consistence, which forms the upper separation of the second cavity of the Alitrunk from that of the Abdomen[1075]. It affords a point of attachment for many muscles of both alitrunk and abdomen. It includes the Septula.
a Septula (the Septula). The lesser ridges and partitions raised on the surfaces of the metaphragm, and on those of other parts of the cavities of the trunk, serving as points of attachment to various muscles[1076].
4. Postpectus (the Postpectus). The underside of the second segment of the alitrunk[1077]. It includes the Mesostethium, Parapleuræ, Metasternum, Postfurca, Opercula, and Pedes Postici.
A Mesostethium (the Mesostethium). A central piece between the intermediate and posterior legs, and bounded laterally in Coleoptera by the Parapleuræ—along the middle of which, where it exists, the Metasternum runs[1078].
B Parapleuræ (the Parapleuræ). Two pieces, one on each side of the Postpectus, included between the Scapularia, Mesostethium, and Pleuræ[1079]. They include the Spiracula Parapleuritica.
a Spiracula Parapleuritica (the Parapleuritic Spiracles). Two spiracles, one in each of the Parapleuræ of Tetyra[1080].
C Metasternum (the Metasternum). The central and often elevated part of the Mesostethium. Its anterior mucro, in Coleoptera, often meets the posterior one of the Mesosternum, and sometimes appears to form one piece with it, as in Hydrophilus, and many Lamellicorn beetles. Sometimes, as in Cetonia vitticollis, it even passes between the arms, and covers the Prosternum, or supplies its place. Behind, it often terminates in a bifid mucro. It is not present in many Orders: as in the Hymenoptera, Diptera, &c.[1081] It includes the Pectines.
a Pectines (the Pectines). Two moveable processes, fixed one on each side by its base below the posterior legs to the Metasternum in Scorpio: on the lower side is fixed a series of parallel biarticulate processes, resembling the teeth of a comb[1082].
D Postfurca (the Postfurca). A process of the Endosternum, terminating in three subhorizontal acute branches, resembling the letter Y, and forming an acute angle with the Endosternum, to which the muscles that move the hind-legs, &c. are affixed[1083].
E Opercula (the Opercula). Plates that cover the vocal spiracles in humming insects; and likewise two large cartilaginous plates fixed to the posterior part of the Postpectus, which cover the Tympana in male Tettigonia F.[1084] Perhaps these may be regarded as a kind of Metapnystega in a new situation.
F Pedes Postici (the Hind-legs). The pair of legs[Pg 384] affixed to the postpectus[1085]. They include, the Acetabulum, Coxa, Trochanter, Femur, Tibia, and Tarsus.
a Acetabulum (the Socket). The socket in the Postpectus in which the leg is planted[1086]. It includes the Pessella.
A Pessella (the Pessella). Two little acute processes, fixed one in each, in the socket of the hind-legs in male Tettigoniæ, which appear designed to keep down the Opercula[1087].
b Coxa (the Hip). The first joint of the leg which plays in the socket[1088].
c Trochanter (the Trochanter). The second joint of the leg, by which the thigh inosculates in the Coxa. It appears to have no motion separate from that of the thigh. It is sometimes biarticulate[1089].
d Femur (the Thigh). The third joint of the leg, long and usually compressed[1090]. It includes the Gonytheca.
A Gonytheca (the Knee-pan). A concavity at the apex of the thigh, underneath, to receive the base of the Tibia[1091].
e Tibia (the Shank). The fourth joint of the leg, very long, and usually triquetrous[1092]. It includes the Epicnemis, Molula, Talus, Calcaria, and Coronula.
A Epicnemis (the Epicnemis). An accessory joint at the base of the Tibia in many Arachnida, which does not appear to have separate motion[1093].
B Molula (the Knee-ball). The convex and sometimes bent head of the Tibia, armed with a horny process on each side, by which it is attached to the thigh[1094].
C Talus (the Ankle). The apex of the Tibia, where it is united to the Tarsus[1095].
D Calcaria (the Spurs). One, two, or more moveable spines, inserted usually at the apex of the Tibia; and in many Carabi L., Lepidoptera L., and Trichoptera K., in the middle also. They may be regarded as a kind of fingers auxiliary to the Tarsus, and furnish often an important character in the discrimination of genera[1096].
E Coronula (the Coronula). A coronet or semicoronet of fixed spines observable at the apex of the posterior Tibia in Fulgora candelaria, &c.
f Tarsus (the Tarsus). The fifth principal portion of the legs; consisting in the majority of insects of 1-5 joints, but in the Phalangidæ of sometimes as many as 50[1097]. It includes the Planta, Digitus, and Solea.
A Planta (the Instep). The first joint of the Tarsus is so called when it is remarkably long and broad[1098]. It includes the Calx.
a Calx (the Heel). The curving part of the Planta, by which it inosculates with the Tibia.
B Digitus (the Toe). The remaining joints of the Tarsus taken together[1099]. It includes the Allux and Ungula.
a Allux (the Toe-ball). The last joint but one of the Tarsus, when remarkable, as in Rhyncophorous beetles (Curculio L.)[1100].
b Ungula (the Claw-joint). The last joint of the Tarsus, which bears the claws[1101]. It includes the Arthrium, Unguiculi, and Plantula.
α Arthrium (the Arthrium). A very minute joint at the base of the claw-joint, in most Tetramerous and Trimerous beetles[1102].
β Unguiculi (the Claws). One or two pair of moveable incurved claws, which usually arm the apex of the Ungula[1103].
γ Plantula (the Plantula). A minute accessory joint, sometimes attached within the claws to the apex of the Ungula[1104]. Ex. The Lucanidæ. It includes the Pseudonychia.
* Pseudonychia (the Spurious Claws). Two stiff clawlike bristles, that terminate the Plantula[1105].
C Solea (the Sole). The underside of the Tarsus[1106]. It includes the Pulvilli.
a Pulvilli (the Pulvilli). Cushions of short hairs very closely set; or of membrane, capable of being inflated,[Pg 387] or very soft; or concave plates, which cover the underside, or their apex, of the four first joints of the Manus or Tarsus, and sometimes even of the ends of the Calcaria, as in Cimbex; which act so as to produce a vacuum, and enable the animal to suspend itself, or walk against gravity[1107]. Ex. Timarcha, Buprestis, Priocera K., the Gryllina, Muscidæ, &c.
III. ABDOMEN (the Abdomen).
The Abdomen is the third or posterior section of the body which follows the Truncus[1108]. It includes the Tergum, Venter, Petiolus, Cauda, and Anus.
i. Tergum (the Tergum). The upper or supine surface of the abdomen[1109]. It includes the Segmenta Dorsalia, and Pulmonaria.
1. Segmenta Dorsalia (the Dorsal Segments). Transverse segments of the back, the sides of which often lap over and cover those of the ventral segments[1110].
2. Pulmonaria (the Pulmonary Space). Two longitudinal soft spaces, capable of tension and relaxation, one on each side of the back of the abdomen, in which, where they exist, the dorsal spiracles are planted[1111]. They include the Spiracula Dorsalia.
a Spiracula Dorsalia (the Dorsal Spiracles). Lateral breathing-pores observable in the dorsal segments, often covered by the preceding segment[1112].
ii. Venter (the Belly). The lower or prone part of the abdomen[1113]. It includes the Hypochondria, Epigastrium[1114], Segmenta Ventralia, and Elastes.
1. Hypochondria (the Hypochondria). Two portions of segments, one on each side; which in some genera[1115] (Carabus L., &c.) intervene between the first intire ventral segment and the posterior part of the Postpectus.
2. Epigastrium (the Epigastrium). The first intire ventral segment[1116]. It includes the Mucro and Tympana.
A Mucro (the Mucro). The central posterior point of the Epigastrium observable in many of the Orders, which reposes between the posterior legs; and, according to M. Chabrier, is useful to the insect during flight[1117].
B Tympana (the Drums). Two deep cavities, containing a complex machinery on each side of the Epigastrium in male Tettigoniæ, which are the instruments of sound[1118].
3. Segmenta Ventralia (the Ventral Segments). Transverse sections of the belly[1119]. In Elytrophorous insects they are usually of a firmer consistence than those of the back. They include the Spiracula Ventralia.
A Spiracula Ventralia (the Ventral Spiracles).—Breathing-pores[Pg 389] observable in some genera in the intermediate ventral segments, one on each side[1120]. Ex. Dynastes Aloeus, &c.
4. Elastes (the Elastes). The elastic organs on the ventral segments of Machilis polypoda which assist this insect in leaping.
iii. Petiolus (the Footstalk). A slender part by which the abdomen of many Hymenoptera is united to the trunk, in some genera very long, in others very short, and in others wanting[1121]. It includes the Funiculus, Foramen, Squama, and Nodus.
1. Funiculus (the Funiculus). A small cartilaginous cord, passing through a minute orifice of the Postfrænum, just above the point where the footstalk is fixed, to an opposite hole above it, which enables the animal the better to elevate or drop the abdomen[1122].
2. Foramen (the Foramen). The orifice in the abdomen, through which the above cord passes[1123].
3. Squama (the Scale). A vertical flat scale, observable on the footstalk of the genus Formica, &c.[1124]
4. Nodi (the Knots). One or more subrotund protuberances of the footstalk in the genus Myrmica[1125].
iv. Cauda (the Tail). Where the abdomen grows suddenly slenderer, and terminates in a long jointed tail, as in Scorpio and Panorpa[1126]. It includes the Centris.
1. Centris (the Centris). The last inflated joint of the tail, terminating in the Sting.
v. Anus (the Anus). The termination of the abdomen, consisting of the two last segments. It includes the Podex, Hypopygium, Culus, Ovipositor, and Appendices.
1. Podex (the Podex). The last dorsal segment of the abdomen[1127].
2. Hypopygium (the Hypopygium). The last ventral segment of the abdomen[1128].
3. Culus (the Culus). The orifice at the end of the anus.
4. Ovipositor (the Ovipositor). The instrument of oviposition, by which the insect conducts the eggs to their appropriate nidus, and often bores a way to it; the same instrument is by some genera used as a weapon of offence, when it is called the Aculeus[1129]. It includes the Unci, Tubulus, Valvæ, Vaginula, and Terebellæ.
A Unci (the Unci). Two pair of robust organs, the upper incurved and the lower recurved, with which the anus of Locusta Leach is furnished[1130].
B Tubulus (the Tubulus). A tubular ovipositor, consisting of several pieces often retractile within each other, like the tubes of a telescope[1131].
C Valvæ (the Valves). Two lateral laminæ, often coriaceous, by which the ovipositor when unemployed is covered[1132].
D Vaginula (the Sheath). A corneous case, with[Pg 391] two grooves, in which the Terebella or Spicula play[1133].
E Terebellæ (the Terebellæ). Instruments by which the insect saws or bores a passage for its eggs to the place in which her instinct directs her to deposit them[1134].
5. Aculeus (the Sting). The above instrument, when fitted for an offensive weapon[1135]. It includes, besides the Valvæ and Vaginula before defined, the Spicula.
A Spicula (the Darts). The proper stings which inflict the wound: retractile within the sheath, externally serrulate at the apex[1136]. They include the Retinaculum.
a Retinaculum (the Retinaculum). A minute horny moveable scale or plate with which the darts are furnished, which prevents their dislocation by being shot forth too far[1137].
vi. Appendices (the Appendages). Other instruments and organs, with which the anus of various insects is furnished. They include—the Forceps, Forfex, Furca, Styli, Foliola, Flosculus, Caudulæ, Fila, Mammulæ, Papillæ, and Siphonuli.
1. Forceps (the Forceps). A pair of anal organs that open and shut transversely, and meet at their inner margin, or at the apex. Ex. Forficula.
2. Forfex (the Forfex). A pair of anal organs, which open or shut transversely, and cross each other[1138]. Ex. Male of Raphidia Ophiopsis.
3. Furca (the Fork). An inflected elastic anal organ, ending in a fork, by which the animal is enabled to leap[1139]. Ex. Podura.
4. Styli (the Styles). Rigid, exarticulate, long and narrow anal organs[1140]. Ex. Staphylinus.
5. Foliola (the Leaflets). Rigid, exarticulate, dilated, leaf-like anal organs[1141]. Ex. Libellulina.
6. Flosculus (the Floret). A small, tubular, lunulate anal organ, with a central style[1142]. Fulgora candelaria, &c.
7. Cerci (the Cerci). Two short, flattish, sublanceolate, jointed, lateral anal organs[1143]. Ex. Blatta.—N.B. Analogous organs are observable in the Gryllina, but usually conical and without joints[1144]. In Gryllus Latr. they are setiform[1145].
8. Caudulæ (the Caudulæ). Two or more slender, filiform or setaceous, jointed, flexile anal organs[1146]. Ex. Lepisma, Machilis, Ephemera.
9. Fila (the Threads). Two exarticulate, slender, filiform anal organs[1147]. Ex. Machilis.
10. Mammulæ (the Mammulæ). Anal protuberances, containing instruments for spinning web[1148]. Ex. Araneidæ. They include the Fusi.
11. Fusi (the Spinners). Organs, consisting of two retractile pieces, issuing from the Mammulæ, and rendering the threads[1149].
12. Siphonuli (the Siphonets). Truncated, fistular, setiform anal organs, emitting a saccharine fluid[1150]. Ex. Aphis.
You will observe, that when the whole upper-side of the Truncus is spoken of, it is called the Thorax; and as in Coleoptera, and some other Orders, the whole of the Mesothorax except the Scutellum is covered by the Thorax, and the whole of the Metathorax by the Mesothorax and Elytra—the Thoracic shield, may without danger of mistake be denominated the Thorax, as it has always been. When the whole under-side of the Trunk is spoken of, it is called the Pectus. When the three Sternums are spoken of together, they may be called the Sternum; and the whole interior elevation of the Pectus may be called the Endosternum.
Before I confine my observations to the head of insects, which I propose to consider separately in the present letter, I must premise a few words upon their body in general, or rather its crust, or external integument. In this we may notice its substance, general form, sculpture, pubescence, and composition.
i. I have already noticed the substance of this integument in the preparatory states of insects[1151]; I shall not, therefore, here repeat what I then said, but restrict myself chiefly to the consideration of it as it is found in their last state, in which it is usually firmer than in their previous stages of existence. In this respect, however, it varies much in the different Orders, and even in the different genera of the same Order. In some Coleopterous insects, for instance, it is very hard, and difficult to perforate; while in others it is soft, flexible, and a pin easily passes through it[1152]. And in general, from a substance[Pg 395] in hardness resembling horn or shell, it passes through the intermediate degrees of that of leather and parchment, almost to a thin membrane. Yet in all cases there is enough of rigidity and hardness to answer the principal uses of a skeleton—to afford, namely, a sufficient point of attachment for the muscles, and to support and defend the interior organization; so that the play and action of the vital and secretory systems may not be interrupted or impeded.
With respect to the principles which enter into the composition of this integument, very little seems to be known at present; but few insects having been submitted to a chemical analysis. The blister-beetle (Cantharis vesicatoria), from its importance in medicine, has, however, been more than once analysed; and though the products have not been very precisely stated, yet we find amongst them phosphate of lime, albumen, and some other usual components of the substance of vertebrate animals[1153]. But which of these products belong to the integument, and which to its contents, cannot be ascertained, without a separate process for each; which would not, I conceive, be very feasible. The substance, however, of the integument of insects, though we know not its precise contents, which probably vary in different genera, &c., appears not to be exactly of the nature of any of those substances after which it has usually been denominated:[Pg 396] it is not properly analogous either to real horn, shell, skin, or leather, &c. This seems to result from the following circumstance:—Most of the excretions of vertebrate animals, as horn, skin (at least when tanned), feathers, wool, hair, &c. when exposed to the action of fire liquify, more or less, before they incinerate; emitting at the same time a peculiar and disagreeable scent: but upon applying this test to the parts of insects of the different Orders, I found, in every instance, that incineration took place without liquefaction, and was unaccompanied by that peculiar scent which distinguishes the others. Even the claws, which to the eye appear, as to their substance, exactly like those of Mammalia, birds, &c. burn without melting, and retain their form after red heat. That the insect integument is not calcareous like that of the Crustacea, and the shells of Molluscæ, you may easily satisfy yourself, by immersing them in an acid test. I made this experiment upon portions of insects of several of the Orders, in an equal mixture of muriatic acid and water, and the result was, not only that all hexapods, but octopods, Arachnida, and even Scolopendridæ, upon immersion only emitted a few air-bubbles; while, when the other myriapods, Polydesmus, Iulus, Glomeris, &c. and the Oniscidæ, were immersed, a violent effervescence took place; proving the different nature of their substance. It is remarkable that the two great branches of the Myriapods, the Scolopendridæ and Iulidæ (Chilopoda and Chilognatha Latr.), should in this respect be so differently circumstanced—the latter having a calcareous integument, and the former not.—A further difference distinguishes these two tribes: old specimens of the Iulidæ usually lose their colour and turn[Pg 397] white, like Oniscidæ; while those of the Scolopendridæ retain it.
ii. The form of insects is so variable, that it can be reduced to no other general rules—than that, for the most part, the length exceeds the breadth, and the breadth the depth, and that the upper surface is usually convex. But to these rules there are numerous exceptions. Thus many Tetyræ F. (Scutellera Latr.), a kind of bug, are as broad as they are long[1154]; in the genus Gonyleptes K.[1155] amongst the Aptera, and Epeira cancriformis, a crab-shaped spider, the breadth exceeds the length; in Cynips, and several other Hymenoptera, in Acrida K.[1156] (Locusta F.), and other Orthopterous insects, the depth exceeds the breadth; and in that singular beetle, Eurychora; the cockroach (Blatta), &c. the upper surface is flat.
iii. The sculpture of the integument of insects is often very remarkable; but as this will call for attention hereafter, I shall only here observe in general, that ornament and variety seem not to be the sole object of those elevations and depressions which form so prominent a feature of many of the animals in question; for by means of these, many important purposes, that at first sight do not strike the observer, may be served: such as giving firmness to the crust in those places where it is most wanted; diminishing[Pg 398] its powers of resistance in others, so that it may yield somewhat to the action of the muscles; increasing or deducting from the weight of the body, so as to produce a proper equipoise during its motions, whether on the earth, in the air, or in the water. The depressions of the outer surface of the crust, in many instances, produce an elevation of it in the interior, and so afford a useful point of attachment to certain muscles. This observation seems more especially applicable to those excavations that are common to particular tribes or genera: thus the dorsal longitudinal channel to be met with on the prothorax of most of the Carabi of Linné on the inside of the crust have a corresponding ridge. In Locusta Dux, also, (a Brazil locust,) the same part has four transverse channels, corresponding with which on the inside are as many septa, or ridges, to which muscles are attached; and those larger impressed puncta denominated puncta ordinaria, which distinguish the same part in Geotrupes and many of the Scarabæidæ, within are elevated, so as to form a kind of ginglymous articulation with the base of the anterior coxæ. The other impressed puncta so often to be seen on the different parts of various insects, which sometimes so intirely cover the surface that scarcely any interval is discoverable between them, though in many cases they appear to be mere impressions that attenuate but do not perforate the crust—yet in others, perhaps equally or more numerous, they are real pores, which pass through the integument. If, for instance, you take the thoracic shield of the cockchafer (Melolontha vulgaris), and after removing the muscle &c., hold it against the light, with the inner side towards the eye, you will see the light[Pg 399] through every puncture: or take the elytra of Geotrupes stercorarius, or any common beetle in which these organs have punctate striæ, and examine them under a lens on the inside, and you will see distinctly that the punctures pass through the elytrum, and the membrane that lines it[1157]. It is not improbable that in the case last mentioned these pores may be of use, as the spiracles are usually closely covered by the elytra, for the better transmission of the air to those respiratory organs. Whether the pores in the other parts of the body are for transpiration, is more than I shall venture to affirm; but as insects sometimes perspire, at least this has been ascertained with respect to the hive-bee[1158], this must be by the means of some pores.
iv. The integument of insects is often clothed, either partially or generally, with pubescence, or hairs of various kinds—a circumstance which seems to have more than one object. In Parnus, Heterocerus, Gerris, Argyroneta aquatica, and some other aquatic insects, the end in view seems to be to keep the water from wetting the crust; and in this case the covering of hairs is dense, silky, and decumbent. Another object is preventing friction from being injurious: thus humble-bees, that from their mode of nidification[1159], are usually more particularly exposed to it, are well clothed with hair; and in those articulations of insects where much friction takes place, we may often observe a dense fringe or coating of the same substance. This you may see in the common[Pg 400] stag-beetle (Lucanus Cervus), where the thorax receives the head; and very remarkably at the same point in the Hercules-beetle (Dynastes Hercules MacLeay): but besides these uses, there is probably one more universal, which will apply as well to those thinly scattered bristles and hairs, here and there one, to be noticed in many insects: but concerning this I can only throw out a conjecture, as I do not recollect ever to have seen any experiments with regard to this use of animal hairs. But may they not act as conductors, either to introduce some invisible fluid into the body in a positive state, or to convey it out, when received by other means, in a negative state? Every one knows that the fur of a cat has electric properties, and there may be an important general use of this kind attached to the fur and hairs of animals[1160]. But, as I said, I give this as a mere conjecture; and only wish it may excite your attention to the subject, and put in exercise your natural tact for experiment.
M. Cuvier regards the hairs of insects as merely a continuation of the epidermis, with which they fall when the animal changes its skin[1161]; but this will apply only to the hairs of larvæ: for the hairs of perfect insects in many cases are implanted in a pore, and pass through epidermis or crust to the membrane that lines it, in which they terminate.
v. We are now to consider the composition of the integument; under which term I would include the different layers of which it consists, and its articulation.
1. With respect to the first of these circumstances, the layers of which the integument consists, it seems to exhibit some, although not an exact, analogy with the skin, rather than the skeleton, of the vertebrate animals[1162]. In these last, the skin is stated to consist of four layers. Of these the exterior one is the epidermis, or scarf-skin: under this is the rete mucosum, or mucous tissue, which gives its colour to the skin; next follows the papillary tissue formed by the extremities of the nerves, and in which the sense of touch principally resides; the last and innermost layer is the skin proper, or leather, called Dermis, Derma, or Corium[1163]. Two of these layers M. Cuvier assigns to insects. They have, he observes, in every state, a true epidermis[1164]; and in their state of larva he finds that the infinite variety of colours that so adorn many of them is produced by a mucous substance observable between the epidermis and the muscles[1165]: this seems analogous to the rete mucosum. To this, dried and mixed with their horny substance, he attributes also the colours of the perfect insect: "for," says he, "when the Lepidoptera are in the chrysalis, the little coloured scales which are to ornament their wings, are then in a state of mucosity similar to that which is found under the skin of the caterpillar. The colours of the Arachnida," he goes on, "are also due to this mucosity: it is discoverable under the skin, and has the appearance of minute glandular points of which the shades vary considerably. But in the Coleoptera, and many other Orders, the[Pg 402] colours of the skin are mixed in its horny tissue, nearly as those of the Testacea are in their calcareous shells"[1166]. In the perfect insects, therefore, in most cases, we may consider the epidermis and rete mucosum as together forming the exterior and coloured integument of insects—that part which in the table, since it is not properly an epidermis, I have distinguished by the name of Exoderma.
The learned author just quoted has observed nothing under the skin of white-blooded animals that he regards as analogous to nervous papillæ[1167]. In some parts of insects, as in the lamellæ of the antennæ of the Petalocera, and the extremities of the joints, especially the last, of many palpi, there is however an appearance of them; and it seems reasonable to suppose that where the sense of touch resides, there must, even in insects, be something of a papillary tissue.
With regard to the innermost integument of the vertebrate animals,—the leather, or real skin,—this learned comparative anatomist finds nothing analogous to it in the integuments of insects[1168]; but as he does not notice it, he appears to have overlooked the substance that lines the outer crust, or exoderma, in the Coleoptera and most other orders. This is not always easily detected; but it may generally be discovered by breaking, or rather tearing (not cutting), after having cleared away the muscles, any part of the body of an insect. It is always very visible on the under side of elytra[1169], but is not discoverable[Pg 403] in tegmina. It appears to consist, in many cases, of several layers of a whitish membrane, and generally breaks into fibres. In some elytra of the larger Dynastidæ, towards the sides the exterior layer is separated from the rest by a kind of cellular substance. The fibrous structure of this inner skin (which I call the Esoderma) seems to give it some affinity to the skin of vertebrate animals[1170]. In many parts of the body, however, it appears to be merely a thin pellicle. A medical friend, to whom I showed specimens of it, thinks it a kind of cellular membrane.
2. A few words are next necessary with regard to the articulation of the integument, or the mode by which the several pieces of which it and its members consist, are united to each other. In some, as in several of the parts of the head, the occiput, vertex, temples, cheeks, &c.—the line of distinction is merely imaginary; in others an impressed line separates a part from its neighbours, as is the case with the nose in Vespa, &c. the head in the Arachnida. But in the majority of instances the parts are separated by a suture, or form a real joint. The kinds of articulation observed by anatomists in vertebrate animals do not all occur in insects, and they seem to have some peculiar to themselves. Thus, for instance, they have no proper suture; for though they exhibit the appearance both of the harmonic and squamose (ecailleuse Cuv.) sutures[1171], yet these parts being all limited by[Pg 404] the esoderma, or skin, above noticed as lining the integument, and all admitting a degree of motion more or less intense, rather afford examples, as the case may be, of other kinds of articulation[1172]. Again, they have no proper Enarthrosis, or ball and socket; though the anterior coxæ of the Capricorn-beetles (Cerambyx L.) approach very near to this kind of articulation, as will be shown more fully in another place. The inosculating segments or rings, which distinguish the abdomen, and sometimes other parts of insects, are an example of a kind of articulation not to be met with in the Vertebrata. The ginglymous articulation, in which the prominences of the ends of two joints are mutually received by their cavities, and which admits only of flexion and extension, often prevails in the limbs, &c. of insects; but in many cases the joints are merely suspended to each other by a ligament or membrane; and, in fact, the integument of insects, with regard to its articulation, even where the joints ginglymate, may be said in general to consist of pieces connected by the internal ligament, membrane, or skin that lines it; for even in the legs, where the ginglymous articulation is sometimes very remarkable and complex, as will be shown to you hereafter, the joints are also connected by this substance, as you may see if you examine the legs of any Coleopterous insect.
The number of articulations or pieces that form the integument and its members in these animals, varies greatly in different tribes, genera, &c. Thus, in the common louse (Pediculus humanus) they scarcely reach fifty, while in some cockroaches (Blatta) they amount to more than eight times that number.
Having premised these observations on the external anatomy of the body in general, in the remainder of the present letter I shall confine myself to the consideration of the head and its parts.
I. The Head of insects, as the principal seat of the organs of sensation, must be regarded in them, as well as in the vertebrate animals, as the governing part of the body. It may be considered with respect to its substance, figure, composition, superficies, proportion, direction, articulation with the trunk, motions—and more particularly as to its parts and appendages.
i. With regard to its substance—the head may be said in general to be the hardest part of the crust: and it is so for very good reasons. In the first place, as it has to make way for the rest of the body when the animal moves, it is thereby best fitted to overcome such resistance as may be opposed by the medium through which it has to pass; in the next, as it bears the organs of manducation, it was requisite that it should be sufficiently firm and solid to support their action, which is often upon very hard substances; and besides this, as no motion of its parts inter se, as in the case of the trunk, is requisite to facilitate the play of its organs, a thin integument was not wanted.
ii. The most general law relative to the figure or shape of the head seems to be, that it should approach to that of an equilateral triangle, with its angles rounded, and the vertex being the mouth; and that the vertical diameter should be less than the horizontal, whether longitudinal or transverse. But the infractions of this law are numerous and various. Thus, in some insects an isosceles triangle is represented by the head, the length being greater than the breadth; in others, instead of being flat it is compressed, so that the horizontal diameter is less than the vertical; in others, again, it is orbicular, or round and depressed; in others nearly spherical: occasionally it is rather cylindrical. In many instances it is very long; in others the width exceeds the length. Though often narrowest before, in some cases the reverse takes place. Its anterior end is often attenuated into a long or short snout or rostrum, and its posterior into a long or short neck. Its contour, though usually regular, is sometimes either cut into lobes, or scooped out into sinuosities. But to enumerate minutely all the variations of form which take place in the head of insects would be endless; I shall therefore proceed to the next particular.
iii. The composition of the head is very simple; for, exclusive of its organs, it consists only of a single piece or box, without suture or segment, with an aperture at the end below to receive the instruments of manducation, others for the eyes and stemmata when present, and also for the antennæ. In the Arachnida, &c., in which the head is not separated from the thorax, it is merely a plate, the under-side or cavity of which is occupied and filled by the above instruments.
iv. With regard to its superficies, the head of insects is generally more or less uneven, though in some cases it presents no inequalities. In many of the Lamellicorn tribes, and a few other individuals, in one sex at least, as has been before observed[1173], it is armed with long horns, or prominent tubercles; it is often covered with numerous puncta, or pores; and some of its parts, as the nose, after-nose, &c. in particular groups, marked out by an impressed line[1174]. In many Hymenoptera, Diptera, &c. its upper surface is convex, and the lower concave; in others both surfaces are convex.
v. It is the most general rule, as to its proportion, that it shall be smaller than either trunk or abdomen; but in some instances, as in the S. American ant, Atta megacephala, it is much larger than either.
vi. By the direction of the head, I mean its inclination with respect to the prothorax. The most natural direction, or that which obtains most generally, is for it to form an angle more or less obtuse with the part just mentioned. This seems to obtain particularly in Coleoptera; but in some, as Mylabris, it is inflexed, forming an acute angle with it. In the Heteropterous Hemiptera (Cimex L. &c.) it is generally in the same line with the body, or horizontal; and in many Diptera it is vertical.
vii. We now come to a circumstance which will detain[Pg 408] us longer, namely, its articulation with the trunk, or rather with its anterior segment, the prothorax.—M. Cuvier makes two principal kinds of articulation of the head upon the prothorax, in one of which the points of contact are solid, and the movement subordinate to the configuration of the parts; in the other, the articulation is ligamentous, the head and the thorax being united and kept together by membranes.
1. The first of these kinds of articulation—that by the contact of solid parts—takes place, he says, in four different ways. "In the most common conformation, in the part that corresponds to the neck, the head bears one or two smooth tubercles, which receive corresponding cavities of the anterior part of the prothorax observable in the Lamellicorn and Capricorn beetles. In this case the head can move backwards, and the mouth forwards and downwards. The second mode of solid articulation takes place when the posterior part of the head is rounded, and turns upon its axis in a corresponding cavity of the anterior part of the prothorax; as may be seen in Curculio, Reduvius, &c. The axis of motion is then at the centre of articulation, and the mouth of the insect moves equally backwards and forwards, upwards and downwards, to right and left.—The third sort of articulation, by solid surfaces, takes place when the head, truncated posteriorly, and presenting a flat surface, is articulated, sometimes upon a tubercle of the thorax, and sometimes upon another flat and corresponding surface, as in almost all the Hymenoptera and the majority of the Diptera. The disposition of the fourth kind of articulation allows the head only the movement of the angular hinge (le seul mouvement de charnière angulaire).[Pg 409] The only examples at present known are in some species of Attelabus F. The head of these insects terminates behind in a round tubercle, received in a corresponding cavity of the thorax: the lower margin of this cavity has a notch, and permits the movement of the head only in one direction[1175]."
2. The second kind of articulation, the ligamentous, he affirms takes place only in orthopterous and some neuropterous insects: "The head in this kind of articulation is only impeded in its movements towards the back, because it is stopped there by the advance of the prothorax; but below it is quite free. The membranes or ligaments extend from the circuit of the occipital cavity to that of the anterior part of the prothorax, which gives a great extent to the movement[1176]."
When I consider the well-deserved celebrity of the great author whose words I have here quoted, and the great and useful light that the genius and learning which conducted his patient labours and researches have thrown over every department of comparative anatomy,—a science he may be almost said to have founded,—I feel the most intire reluctance to differ in any thing from an authority so justly venerable to all lovers of that interesting study. But, however great my diffidence and hesitation to express an opinion at all opposed to his, the interests of truth and science require that I should state those particulars in which my own observations, made upon a careful examination of various insects of every Order, have led to results in some respects different from the[Pg 410] above. "Aliquando bonus dormitat Homerus;" and if the Genius of Comparative Anatomy ever nodded, it sometimes happened when he was examining the structure of insects. An instance of this with regard to the mouth of the bee has been noticed by Mr. Savigny[1177]; and indeed it is not wonderful that in so extensive an undertaking, in which the number of examples to be examined upon every branch of his subject must be immense, that he did not always scrutinize minutely those that seemed less important. Every writer on every department of Natural History, especially where the objects of research, as in the insect world, are so infinite in number, will be liable to such mistakes; but these will meet with due allowance from every candid mind—
and I shall express my trust that you will overlook any errors of mine; and doubtless I shall not be free from them—
The two kinds of articulation of the head which our learned author has stated as the principal ones, will, I think, be found upon examination not so widely distant as his expressions seem to indicate; for in fact in all insects, as well as the Orthoptera, this part is suspended by a membrane or ligament which unites the margins of the occipital cavity with those of the anterior one of the prothorax: thus forming all round some protection to the[Pg 411] organs that are transmitted from the head through the latter part to the rest of the body. Though the head in most Orthoptera is not partly received into the cavity of the prothorax, as is the case in the Order Coleoptera, but is rather suspended to it, yet in some instances, for example in the mole-cricket (Gryllotalpa vulgaris), it is partially inserted.
Again: when, in his first mode of articulation by contact of solid parts, he speaks of one or two smooth tubercles of the neck, with their corresponding cavities in the prothorax, as forming the most common conformation, you would expect to find examples of this in very many insects; yet upon a close examination, unless in Oryctes nasicornis[1178], and perhaps in others of the Dynastidæ MacLeay, you would scarcely meet with any thing that could be called a tubercle and its corresponding cavity in the neck or prothorax of any Lamellicorn or Capricorn beetle that you might chance to examine. You would find, indeed, that the occiput was usually smooth and very slippery, as if lubricated; that in its margin were one or two notches (Myoglyphides), with muscles attached to them; that in the former of these tribes, the Lamellicorns, it projected on each side so as to form a more or less prominent angle; and that the throat (jugulum) was very convex, and lodged in a cavity of the lower margin of the prothorax: but further appearances of tubercles &c. you would in vain look for even in this[Pg 412] tribe; and in the Capricorns you would find that the general conformation in this respect belonged to our learned author's second mode of solid articulation, resembling that of the majority of the weevils (Curculio L.), in which the head has no projecting angles or tubercles, or other elevation, but is received usually into the circular cavity of the prothorax.
His third mode of this articulation, that of the Hymenoptera and Diptera, is so peculiar, that it ought to be considered as a primary kind; since in this the head moves upon the prothorax as upon a pivot, and has a kind of versatile motion.
With regard to his fourth mode, which from his description appears to be that of Apoderus Oliv., he allows motion to the head only in one direction, observing that the lower margin of the prothoracic cavity has a notch. But M. Latreille calls the articulation of the head in this genus an Enarthrosis[1179], which admits motion in every direction; and if you examine the common species (A. Coryli), you will find that the prothorax has a sinus taken out of its upper margin, as well as out of its lower one—which at any rate will allow a motion upwards.
I merely mention these little inaccuracies, with due diffidence, as some apology for giving you a different and at least a more popular and general view of this part of my subject, which I shall now proceed to state to you. It seems to me most convenient for the Entomologist, and most consonant to nature, to divide insects, with respect to the articulation of the head with the trunk, into three primary sections, each admitting one or more subdivisions.
1. The first consists of those whose head inosculates more or less in the anterior cavity of the chest; and whose articulation, therefore, seems to partake in a greater or less degree of the ball and socket (Enarthrosis). The head, however, is often capable of being protruded from this cavity. If you take into your hand any common Harpalus that you may find under a stone, you will see, if pressed, that it can shoot forth its head, so as to be entirely disengaged from the prothorax: a neck of ligament intervening between them[1180]: of course this power of protruding the head enables the animal to disengage it at its will from the restriction imposed upon its motions by the surrounding margin of the prothoracic cavity. To this section belong all the Coleoptera, the Heteropterous Hemiptera (Cimex L., &c.), and some of the Neuroptera (Raphidia, Semblis, &c.).—It may be further divided into two subsections—those, namely, whose head inosculates in the prothorax by means of a neck: as for instance Latreille's Trachelides, Apoderus, and the Staphylinidæ, amongst the beetles; the Reduviadæ amongst the Heteropterous insects, and Raphidia in the Neuroptera; and those whose head inosculates in the prothorax without the intervention of a neck; as, the Petalocera, the aquatic beetles (Dytiscus, Hydrophilus, &c.), and most of the genus Curculio L. in the first of these orders, the great body of the Cimicidæ in the second, and Semblis, Corydalis, &c. in the third.
2. The second section consists of those insects whose head does not inosculate in the chest, but is merely suspended[Pg 414] to it by ligament or membrane. To this belong most of the tribes of the Orthoptera Order, with the exception of the Mantidæ, the Dermaptera, the Homopterous Hemiptera, and such of the Aptera as have the head distinct from the prothorax.—This section admits of a triple subdivision: those, namely, whose head is wholly covered by the shield of the prothorax, as in Blatta L.; those whose head is partly covered by it, as Gryllotalpa, and other Gryllina; and those whose head is quite free, not being at all impeded in its motion by the prothorax, as the Dermaptera, Nirmus, Pediculus, &c.
3. The third section consists of those whose head is truncated posteriorly, and flat or concave, with a very small occipital aperture, and is attached to a neck of the prothorax upon which it turns, or is merely suspended to that part. This includes the Lepidoptera, Hymenoptera, Diptera, the Libellulina, &c. in the Neuroptera, and the Mantidæ in the Orthoptera. Three subsections at least, if not more, present themselves in this section: the first is, of those whose head is united to the prothorax, without the latter forming any neck. To this belong the Lepidoptera, Trichoptera? The second is of those the upper side of whose thoracic neck is ligamentous; and here you may range most of the Hymenoptera. The third is of those in whom it is a continuation of the ordinary integument. In this subsection the Diptera, Libellulina and Mantidæ will find their place. In this last section the head appears to turn upon the thorax as upon a pivot.
Before I finish what I have to say on the articulation of the head, I must direct your attention to the analogies[Pg 415] that hold in this respect between the different tribes. Thus the Coleoptera are analogous to the Heteropterous Hemiptera; the Orthoptera, with the exception of the Mantidæ, to the Homopterous Hemiptera; the Mantidæ to the Libellulina; the Lepidoptera to the Trichoptera; the Hymenoptera to the Diptera, with a slight variation, and probably others might be traced.
viii. A word or two upon the motions of which the head of insects is capable. M. Cuvier, in the extracts lately laid before you, speaks of different powers of movement as the result of different configurations of the parts of the head. This probably is correct with regard to many cases; but a great deal will depend upon the power the insect has of protruding its head and disengaging its base from the restriction of the prothorax; for where, like the Harpali, Staphylini, &c. it is able to do this, it can probably move its head in every direction. It is only where the ligaments are less elastic, or allow of little tension, that its movements are confined; and few living insects have been sufficiently examined to ascertain how far this takes place. In those cases belonging to the third section of articulations, in which the head moves upon the thorax as upon a pivot, as is the case with Hymenoptera and Diptera, the movement is nearly versatile. I have seen a fly turn its head completely round, so that the mouth became supine and the vertex prone; and from the form and fixing of the head, it should seem that those of the Mantidæ were endued with the same faculty.
ix. The parts and appendages of the head are now in[Pg 416] the last place to be considered. I shall begin with the Mouth, or rather the orifice in which the trophi or organs of manducation are inserted. In some insects, as was before observed, they occupy all the under-side of the head, as in the Arachnida, Myriapoda, &c; but in the great majority they fill an orifice in its anterior part, which in some instances, as in Lampyris, the Lepidoptera, Cimex L., Truxalis, appears to be nearly under the head; but in general it terminates that part, though it extends further below than above. In Chermes, a Homopterous genus, the promuscis is stated to be in the Antepectus, and consequently the mouth; but, as I shall endeavour to prove to you hereafter, this is a fallacy. In the males of the species of Coccus there is no mouth at all. In that of the elm (C. Ulmi) in lieu there are ten little shining points, arranged two before and two behind in a line, and three on each side in a triangle[1181]. It is to be observed that the orifice of which I am speaking is usually much smaller in those insects which take their food by suction, the Hemiptera, Lepidoptera, Diptera, &c., than in the masticating tribes. With regard to the real mouth, or that through which the food enters, I have nothing at present to observe, except that it lies between the upper-lip and tongue, is sometimes covered by a valve, as in Apis, Vespa[1182], &c., and is different in the masticators and suckers.
I shall next offer a few observations seriatim, as they stand in the Table, upon the organs of manducation; which, to avoid circumlocution, instead of Instrumenta[Pg 417] cibaria, the name Fabricius gave them, I shall call trophi or feeders. It is upon these parts, you are aware, that the system of the celebrated Entomologist just mentioned is founded; and could they always, or even for the most part, be inspected with ease, they would no doubt afford characters as various and discriminative as those of the vertebrate animals. Differences in these parts indicate a difference in the mode in which the animal takes its food, and often in the kind of food, and sometimes in its general economy and habits,—circumstances which are powerful and weighty in supporting the claim of any set of animals to be considered as forming a natural genus or group. Trifling variations, however, of these parts, unless supported by other characters and qualities, ought not to have much stress laid upon them, since, if we insist upon these, in some tribes almost every species might be made a genus.
With respect to their trophi in general, insects of late have been divided into two great tribes[1183], masticators and suckers; the first including those that are furnished with instruments to separate and masticate their food; namely, an upper- and under-lip (labrum and labium), upper- and under-jaws (mandibulæ and maxillæ), labial and maxillary palpi, and a tongue (lingua): and the second those in which these parts are replaced by an articulate or exarticulate machine, consisting of several parts and pieces analogous to the above, which pierce the food of the animal, and form a tube by which it sucks its juices. If, however, the mode in which insects take their food be[Pg 418] strictly considered, it will be found that in this view they ought rather to be regarded as forming three tribes; for the great majority of the Hymenoptera order, and perhaps some others, though furnished with mandibles and maxillæ, never use them for mastication, but really lap their food with their tongue: these, therefore, might be denominated lappers.
When a mouth is furnished with the seven ordinary organs used in taking the food and preparing it for deglutition—I mean the upper-lip and the two upper-jaws; the under-lip and the two under-jaws, including the labial and maxillary palpi; and the tongue—I denominate it a perfect mouth; but when it is deficient in any of these organs, or they exist merely as rudiments, or when their place is supplied by others, (which, though they may be analogous parts, have little or no connection with them in their use or structure,) I denominate it an imperfect mouth. This last I would further distinguish, according to the nature of its trophi, by other and more distinctive terms, as I shall presently explain to you.
1. Labrum[1184].—I shall first consider the organs present in a perfect mouth, beginning with the upper-lip (labrum). This part, which Fabricius sometimes confounded with the nose, miscalling it clypeus, is usually a moveable[1185] piece, attached by its base to the anterior margin of the part last named, and covering the mouth, and sometimes the mandibles, from above. In insects in their last state it is usually of a horny or shelly substance; yet in some cases, as in Copris and Cetonia, beetles that[Pg 419] imbibe juices, it is membranous. In form and shape it varies greatly, being sometimes nearly square, at others almost round; in some insects representing a parallelogram, in others a triangle, and in many it is oblong. In some instances it is long and narrow, but more generally short and wide. It is often large, but occasionally very minute. In the majority it has an intire margin, but it is not seldom emarginate or bilobed, or even dentate. Its surface is commonly even, but it is sometimes uneven, sometimes flat, at others convex, and in a few species armed with a short horn or tubercle[1186]. As to its pubescence, it is often naked, but now and then fringed or clothed with down or hairs, or sprinkled with bristles. It consists in almost every instance of a single piece; but an exception to this occurs in Halictus, a little bee, in the females of which it is furnished with a slender appendage[1187].—The direction of the upper-lip is various. It is rarely horizontal, or in the same line with the nose, often vertical; it sometimes forms an obtuse angle with the anterior part of the head, and occasionally an acute one, when it is more or less inflexed. The use of this part is ordinarily to close the mouth from above, to assist in retaining the food while undergoing the process of mastication; but in many Hymenopterous insects its principal use seems to be, to keep the trophi properly folded; and in some cases where it is inflexed, as in the leaf-cutter bees (Megachile Latr.), to defend its base, while the mandibles are employed, from injury by their action[1188].
2. Labium[1189].—On the under-side of the head, and opposed to the upper-lip, the mouth is closed by another moveable organ, concerning the nomenclature and analogies of which Entomologists have differed considerably. At the first view of it, this organ seems a very complex machine, since the under-jaws or maxillæ are attached to it on each side, and the tongue is often seen to emerge from it above, so as to appear merely a part of it; but as the former answer to the upper-jaws, and the latter is the analogue of the part bearing the same name in the vertebrate animals, I shall consider these as distinct and primary organs, and treat of the under-lip (labium) of which I am now speaking, by itself. Linné takes no notice of this part, but his illustrious compatriot and cotemporary, De Geer, did not so overlook it: he appears to consider the whole apparatus, including the maxillæ, as the labium[1190]; but sometimes he distinguishes the middle piece by that name[1191]; and the tongue, in the case of the stag-beetle, he denominates a proboscis (trompe)[1192]. In the Hymenoptera he calls this part tongue, under-lip, and proboscis: but seems to prefer the last term[1193]. Fabricius originally regarded the whole middle piece as a labium[1194]; but afterwards (though his definition is not accurate, since he assigns the palpi to the ligula, which he affirms is covered by the labium—circumstances by no means universal in Coleoptera) he considers this as consisting of ligula and labium[1195]. Latreille at first regarded[Pg 421] the ligula of Fabricius as the labium, and called the labium of that author the mentum[1196]; but afterwards he gave the name of labium to the whole middle piece of the lower apparatus of the mouth—calling the upper piece, with Fabricius, the ligula, and retaining the denomination of mentum for the lower[1197].
If the circumstances of the case are duly considered, I think you will be convinced that the term under-lip, or labium, should be confined to that part which the learned Dane so named. For I would ask, Which is the part on the under side of the head that is the antagonist, if I may so speak, of the upper-lip or labrum? Is it not that organ which, when the mouth is closed, meets that part, and in conjunction with it shuts all in? Now in numerous insects, particularly the Lamellicorn beetles (Scarabæus and Lucanus L.), this is precisely the case. In the Predaceous beetles, indeed, (Cicindela, Carabus, Dytiscus L. &c.) the under-lip does not meet the upper, to close the mouth and shut in the tongue; neither can the tongue be said so to do, but only, from some circumstances connected with its manner of taking its food, it is requisite that the last-mentioned organ should not be retractile or covered; but its miscalled mentum is still the analogue of that part which helps to close the mouth in the former tribe. Should not this, therefore, which so often performs the office, be distinguished by the name, of a lip? Again, is it not rather incongruous to consider that organ which confessedly more or less performs the office of a tongue, as a part of the lip? Though it often wears that appearance, yet I believe, if the matter[Pg 422] is thoroughly and patiently investigated, it will be found that on their upper side its roots are attached to the interior of the upper side of the head, as well as on their lower side to the labium; so that it may be regarded as common to the two lips, and therefore not properly considered as an appendage of the under-lip alone.
Having assigned my reasons for preferring the name given to the part in question by Fabricius, rather than that of Latreille, I shall next make my observations on the part itself. In many cases the labium, or the middle piece of the lower oral apparatus, appears to consist of two joints: this you may see in the great water-beetle (Hydrophilus piceus), the burying-beetles (Necrophorus), the Orthopterous tribes[1198], the Hymenoptera[1199], and others. In this case the upper or terminal piece is to be regarded as the labium, and the lower or basal one (which Mr. MacLeay calls the stipes) as the mentum or chin—at other times, as in some Lamellicorn beetles, the only separation is a transverse elevated line, or an obtuse angle formed by the meeting of the two parts, and very frequently there is no separation at all, in which case the whole piece, the mentum merging in it, may be denominated the labium.
With respect to its substance, the labium in most Coleopterous insects is hard and horny, in Necrophorus it is membranous, and the mentum harder; in Prionus coriarius, our largest Capricorn-beetle, both are membranous; in the bee-tribes, Apis L., the labium rather resembles leather, while the mentum is hard. Its surface is often convex, sometimes plane, and sometimes even[Pg 423] concave; as for instance in Melolontha Fullo, a rare chafer occasionally found on the coast of Kent. In some it is covered with excavated points; in others it is quite smooth. In numbers, as in the Predaceous beetles, both labium and mentum are perfectly naked; in others, as in the common cockchafer, they are hairy; in Geniates barbatus Kirby, another chafer in the male insect, the labium is naked, while the mentum, which forms a piece distinct from that part, is covered with a dense rigid beard[1200]. In shape the whole labium varies considerably, much more than the labrum; for in addition to most of the forms I enumerated when I described that organ, which I shall not here repeat, you may meet with examples of many others. Thus, to instance in the Petalocerous tribes (Scarabæus L.), in some, as in the Rutelidæ, the labium is urceolate, or representing in some degree the shape of a pitcher[1201]; in others it is deeply concave, and not a little resembles a basin or a bowl[1202]; this form is peculiar to the labium of Cremastocheilus Knoch, a scarce North American beetle; in another related to this, but of an African type (Genuchus Kirby MS. Cetonia cruenta F.), it is a trapezoid plate, which is elevated from the head, and hangs over the throat like a chin[1203]. In the Hymenoptera it is extremely narrow and long, and embraces the sides of the tongue, as well as covering it from below; so that it wears the appearance of a kind of tube[1204]. Generally speaking, the length of the labium exceeds its breadth; but in the Predaceous beetles the reverse of[Pg 424] this takes place, it being very short and wide, and usually terminating towards the tongue in three lobes or teeth which form two sinuses varying in depth[1205].
The mentum taken by itself affords no very striking characters to which I need call your attention: I shall only observe, that in Hymenoptera it is generally of a triangular shape[1206]; but before I proceed to consider the labial palpi, it will be proper to notice the remarkable labium of Orthopterous insects, and of the Libellulina, between which there is no little analogy. At first you would imagine the terminal part of this organ in the former to be the analogue of the tongue, or ligula F.; as it is indeed generally regarded by modern Entomologists[1207]. It seems, like the tongue of the Carabi L., Dytisci, &c., to be a distinct piece, which has below it both labium and mentum; but when you look within the mouth, you will find a linguiform organ[1208], which evidently acts the part of a tongue, and therefore ought to have the name; and the piece just alluded to must either be regarded as the termination of the lip, or as an external accompaniment of the tongue, analogous, it may be, to the paraglossæ in bees[1209]. In a specimen of Acrida viridissima (Locusta F.) which I dissected, the tongue is as long as the appendage of the under-lip, and by its upper surface seems to apply closely to it. In the Libellulina a similar organ is discoverable[1210], which on its upper-side terminates in the pharynx, like that of one of the Harpalidæ before mentioned. In the Orthoptera, therefore, I regard the labium as consisting of three articulations, the upper one[Pg 425] divided into two, three, or more lobes[1211]; the intermediate one more directly answering to the labium of other insects, and the basal one or mentum. This organ in the Libellulina is of a different structure: it has only two articulations representing labium and mentum; but the former consists of three parallel pieces, the two exterior ones rising higher than the intermediate one, and at their inner angle having an acute sinus from which the palpi emerge; and the intermediate piece, which is longitudinally channelled, lapping over the inner side of the lateral pieces. From the angle of the covered part of these pieces, a subulate short horizontal horn points inwards towards the tongue, which it must keep from closing with the labium[1212].
3. Palpi Labiales[1213].—The last-mentioned organs, the labial palpi, next claim our attention; but before I advert particularly to them, it will be proper to premise a few words upon palpi, or feelers, in general. These are usually jointed moveable organs, of a corneous or coriaceous substance, attached by ligaments to the labium and maxillæ, and in the Crustacea even to the mandibulæ. Their joints, which are usually more or less obconical, articulate also in each other by ligaments, with perhaps some little mixture of the ball and socket. Their ends, the last joint especially, seem furnished with nervous papillæ which indicate some peculiar sense, of which they are the instrument. What that sense is has not been clearly ascertained, and concerning which I shall enter more into detail in another place. Their motion seems[Pg 426] restrained, at least in some, to two directions, towards and from the mouth. They were called palpi or feelers, because the insect has been supposed to use them in exploring substances. There seem to be no organs in the vertebrate animals directly analogous to the palpi of insects and Crustacea, unless, perhaps, the cirri that emerge from the lips of some fishes, as the cod, red mullet, &c. which Linné defines as used in exploring (prætentantes). Whether the vibrissæ, miscalled smellers, of some quadrupeds and birds have any reference to them, I will not venture to affirm; but the feline tribe evidently use their bristles as explorers, and they are planted chiefly in the vicinity of the mouth.
Having made these general remarks, I shall now confine myself to the labial palpi. I call them labial palpi, because that term is in general use, and because in many cases they really do emerge from what I consider as the labium, as in most of the chafers; but they might with equal propriety be denominated lingual palpi, since they sometimes appear to emerge from the tongue as in the stag-beetle (Lucanus Cervus). In some instances, as in the Predaceous beetles, they seem to be common to both labium and tongue, being attached at their base on the upper side to the former, and on the under side to the latter. As to their situation: they emerge from the base of the labium in the locusts (Locusta Leach)[1214]; from its middle in Hister maximus[1215]; from its summit in Amblyterus MacLeay[1216]; and from its lateral margin in Dynastes MacLeay, &c. They consist of from one to four[Pg 427] joints; which, I believe, they never exceed. They vary in length; though generally shorter than the maxillary palpi, yet in the ferocious tiger-beetles (Cicindela L.) they equal them in length; and in the hive-bee and humble-bees, and many other bees, they are considerably longer[1217]. The two first joints of these palpi, however, in these bees are different in their structure from the two last, being compressed and flat, or concave; and the two last joints, instead of articulating with the apex of the second, emerge from it below the apex: so that these two first joints seem rather elevators of the palpi than really parts of them[1218]. With respect to the relative proportions of their joints to each other: in some cases the first joint is the longest and thickest, the rest growing gradually shorter and smaller[1219]; in others, the second is the longest[1220]; in others, again, the third[1221], and sometimes the last[1222]; and often all are nearly of the same size and length[1223]. They are more commonly naked, but sometimes either generally or partially hairy. Thus in Cicindela, the last joint but one is usually planted with long snow-white bristles in a double series, while the rest of the joints have none; and in Copris Latr. all of them are extremely hairy. In shape they do not vary so much as the maxillary palpi, being most frequently filiform or subclavate, and sometimes setaceous; the last joint varies more in shape than the rest, and is often remarkably[Pg 428] large, triangular, and shaped like the head of a hatchet[1224]; and at others it resembles the moon in her first quarter[1225]. In the great dragon-fly, or demoiselle if you prefer the gentler French name (Æshna F.) the labial palpi, which are without any visible joints, are terminated by a minute mucro or point[1226]. With regard to their direction and flexure, they frequently, as in the instance just mentioned, turn towards each other, and lie horizontally upon the end of the labium. Sometimes, as in the Cicindelidæ, they appear to point towards the tail of the insect, the last joint rising, and forming an angle with the rest of the feeler. In other instances they diverge laterally from the labium, the last joint turning again towards it at a very obtuse angle.
4. Mandibulæ[1227].—Having considered the analogues of the lips in our little beings, I must next call your attention to the representatives of the jaws. The vertebrate animals, you know, are mostly furnished with a single pair of jaws, one above and the other below, in which the teeth are planted and which have a vertical motion. But insects are furnished with two pair of jaws, a pair of upper-jaws and a pair of under-jaws, which have no teeth planted in them, and the motion of which is horizontal.—I shall begin with an account of the upper-jaws. These by modern Entomologists, after Fabricius, are[Pg 429] denominated mandibles (mandibulæ): a term appropriated by Linné to the beaks of birds. The upper-jaws of insects this great naturalist named maxillæ—and not improperly, since the office of mastication is more peculiarly their office than that of the under-jaws, which Fabricius has distinguished by that name: as the term mandible, however, is generally adopted, I shall not attempt to disturb it.
The mandibles close the mouth on each side under the labrum or upper-lip. They are generally powerful organs, of a hard substance like horn; but in the Lamellicorn beetles of Mr. MacLeay's families of Scarabæidæ and Cetoniadæ, they are soft, membranous, and unapt for mastication. In Coleopterous insects they commonly articulate with the head by means of certain apophyses or processes, of which in many cases there are three discoverable at the exterior base of the mandibles; one, namely, at each angle, and one in the middle. That on the lower side is usually the most prominent, and wears the appearance of the condyle of a bone: it is received by a corresponding deep socket (or cotyloid cavity) of the cheek, in which, being perfectly smooth and lubricous, it moves readily, but without synovia, like a rotula in its acetabulum. The upper one projects from the jaw, forms the segment of a circle, and is concave also on its inner face. A corresponding more shallow, or, as anatomists speak, glenoid cavity of the cheek, where it meets the upper-lip, receives it, and the concave part admits a lubricous ball projecting from the cheek, upon which it turns[1228].[Pg 430] This structure you will find in the stag-beetle, and some other timber-devourers. Other Coleoptera have only a process of a similar structure at each of the dorsal angles of the base of the mandible, the intermediate one being wanting; and the articulation does not materially differ, as far as I have examined them, in the Hymenoptera and Neuroptera. In the Orthoptera, the structure approaches more nearly to that of the stag-beetle, since there are three prominences: it is thus well described by M. Marcel de Serres: "This articulation," says he, "takes place in two ways. At first, in the upper surface of the mandible, and at its base, may be observed two small prominences and a glenoid cavity; these prominences are received in two glenoid cavities excavated in the shell of the front, as the cavity of the mandible receives a small prominence of the same part. Below the mandible, and at its base, there is a kind of condyle, which has its play in a cotyloid cavity excavated in the shell of the temple, far below the eye, and at the extremity of the coriaceous integument of the head[1229]." Within the head in this order, at least in Locusta Leach, is a vertical septum, which divides the head into two chambers, as it were, an occipital and a frontal, consisting of a concave triangular stem, terminating in two narrower concave triangular branches, so as to resemble the letter Y, and forming three openings, an upper triangular one, and two lateral subquadrangular ones, which last are the cavities that receive the base of the mandibles. This partition, which I would name Cephalophragma, doubtless affords a point of attachment to many of the muscles of the head. It does[Pg 431] not appear to have been noticed, unless it be synonymous with the intermaxillary arcade of Marcel de Serres[1230]. Probably a corresponding support to the muscles, &c. may exist, as we have seen it does in Vespa L.[1231], in many other heads of the different Orders, which have not yet fallen under examination. Many mandibles, as those of the hornet &c., appear to be suspended to the cavity of the head on the inside by a marginal ligament sufficiently relaxed to admit of their play: those of the Orthoptera, M. Marcel de Serres informs us, are united to the head by means of two cartilages, the outermost being much the shortest, to which their moving muscles are attached. These he considers as prolongations of the substance of the mandible[1232]. The bottom of mandibles, when cleared of the muscles &c., inclines almost universally to a triangular form; but in some cases, as in the stag-beetle, it is nearly a trapezium. I cannot conclude this subject without noticing the motions of the mandibles. What the author lately quoted has said with regard to those of the Orthoptera, will, I believe, apply equally well to all the mandibulate orders. "The articulation of mandibles with the skull appears to take place by two points solely; and as these parts only execute movements limited to a certain direction, they may be referred to ginglymus[1233].—The movements of mandibles are limited to those from within outwards, and from without inwards[1234]." Whether they are restricted from any degree of vertical motion, has not yet been proved, as the jaws of vertebrate animals move horizontally as[Pg 432] well as vertically—so those of insects may have some motion vertically as well as horizontally; and it seems necessary for some of their operations that they should. I am not anatomist enough to speak with confidence on the subject, but the ball and socket articulation at the lower part of the mandible, and the curving one at the upper, though a kind of ginglymus, seems to imply a degree of rotatory movement, however slight.
I must next say something upon the general shape of these organs. Almost universally they incline to a triquetrous or three-sided figure, with their external surface convex, sometimes partially so, and their internal concave. Most frequently they are arched, curving inwards; but sometimes, as in Prionus octangularis[1235], a Capricorn beetle, and others of that genus, they are nearly straight; and in Rhina barbirostris[1236], a most remarkable Brazilian weevil, their curvature is outwards. In Pholidotus lepidotus MacLeay, and Lucanus Elephas, two insects of the stag-beetle tribe, they are bent downwards; and in Lucanus nebulosus K. (Ryssonotus MacLeay) they turn upwards[1237]. They are usually widest at the base, and grow gradually more slender to the apex, but in the hornet (Vespa Crabro) the reverse takes place, and they increase in width from the base to the apex; and in the hive-bee, and others of that tribe, they are dilated both at base and apex, being narrowest in the middle; others are nearly of the same width every where. In those insects that use their mandibles principally for purposes connected with their economy, they are often more broad[Pg 433] in proportion to their thickness, than they are in those which use them principally for mastication. In the locust tribes (Locusta Leach), they are extremely thick and powerful organs, and fitted for their work of devastation; but in the glow-worm (Lampyris), they are very slender and minute. In those brilliant beetles, the Buprestes, they are very short; but in the stag-beetles, and those giants in the Capricorn tribe, the Prioni, they are often very long[1238]. They either meet at the summit, lap over each other, cross each other, or are protended straight from the head; as you have doubtless observed in the stag-beetle, whose terrific horns are mandibles of this description. These organs are usually symmetrical, but in some instances they are not: thus in Hister lævus, a kind of dung-beetle, the left hand mandible is longer than the right; in Creophilus maxillosus K. (Staphylinus L.), a common rove-beetle, in the left hand mandible the tooth in the middle is bifid, and in the right hand one intire; and in Bolbocerus K. the mandible of one side, in some species the dexter, and in others the sinister, has two teeth, and the other none.
The next circumstance with respect to these organs which demands our attention, is the teeth with which they are armed. These are merely processes of the substance of the mandible, and not planted in it by gomphosis[1239], as anatomists speak, as they are in vertebrate animals. They have, however, in their interior, at the base[Pg 434] at least, in the Orthoptera, a coriaceous lamina that separates them in some sort from the body of the mandible[1240]. Many insects, however, have mandibles without teeth; some merely tapering to a sharp point, others obtuse at the end, and others truncated[1241]. Of those that have teeth, some have them on the inside at the base, as Manticora, an African tiger-beetle[1242]; others in the middle, as Staphylinus olens, a rove-beetle, Lethrus cephalotes, &c.[1243]; others at the end, as many weevils (Curculio L.)[1244]; others again on the back, as the Rutelidæ, a tribe of chafers[1245], and Lethrus, a beetle just named; others once more on the lower side of the base, in the form of a tooth or spine, as in Melitta spinigera, a species of wild-bee, and some of its affinities[1246]; and lastly, others on the upper side of the base in the form of a long tortuous horn, as in that singular wasp Synagris cornuta F. before noticed as a sexual character[1247]. In the stag-beetle tribes (Lucanus L.) these teeth are often elongated into short lateral branches, or a terminal fork[1248]. They are sometimes truncated, sometimes obtuse, and sometimes acute.
But with regard to their kind, it will be best to adopt the ideas of M. Marcel de Serres; for though his remarks are confined to the Orthoptera, they may be applied with advantage to the teeth that arm the mandibles[Pg 435] of insects in general. He perceives an analogy between those of this Order and the teeth of quadrupeds; and therefore divides them into incisive or cutting, laniary or canine, and molary or grinding teeth. He denominates those incisives that are broad, having in some degree the shape of a wedge, their external surface being convex, and their internal concave; whence they are evidently formed for cutting. The laniaries are those which have a conical shape, are often very acute, and in general the longest of any; and in some insects, as the carnivorous Orthoptera (and the Libellulina), they cross each other. The molaries are the largest of all, and their purpose is evidently to grind the food. There is never only a single one to each mandible, while the number of the incisives and laniaries is very variable. As the molaries act the principal part in mastication, they are nearer the inner base of the mandible or point of support: they serve to grind the food, which has been first divided by the incisives or torn by the laniaries. The carnivorous tribes are destitute of them; in the omnivorous ones they are very small, and in the herbivorous ones they are very large[1249]. So that in some measure you may conjecture the food of the animal from the teeth that arm its mandibles. Of incisive teeth you may find an example in those that arm the end of the mandibles of most grasshoppers (Locusta), and of the leaf-cutter-bees (Megachile Latr.)[1250]; of the laniary or canine teeth, you will find good examples in the mandibles of the dragon-flies (Libellulina); the two external teeth of the[Pg 436] apex of those of the leaf-cutter bees may be regarded as between the incisives and laniaries; and the pointed mandibles without teeth may be deemed as terminating in a laniary one[1251]. The lower part of the inner or concave surface of the mandibles of grasshoppers will supply you with instances of the molary teeth, and the apex, also, of those of some weevils, as Curculio Hancocki K.[1252] But the most remarkable example of a molary organ is exhibited by many of the Lamellicorn beetles, especially those that feed upon vegetables, whether flower or leaf.—Knoch, who indeed was the first who proposed calling mandibles according to their teeth, incisive, laniary, or molary, but who does not explain his system clearly, observed that the mandibles of some Melolonthæ have a projection with transverse, deep furrows, resembling a file, for the purpose of bruising the leaves they feed upon[1253]: and M. Cuvier, long after, observed that the larvæ of the stag-beetle have towards their base a flat, striated, molary surface; though he does not appear to have noticed it in any perfect insect[1254]. This structure, with the exception of the Scarabæidæ and Cetoniadæ, seems to extend very generally through the above tribe; since it may be traced even in Geotrupes, the common dung-chafer,[Pg 437] in which at the base of one mandible is a concave molary surface, and in the other a convex one, but without any furrows: a circumstance that often distinguishes those that have furrows.—In the Dynastidæ the affinity of structure with the Melolonthidæ &c. is more pronounced, the furrows to which ridges in the other mandible correspond being reduced to one or two wide and deep ones; whereas in some of the latter tribe they are very numerous. These mandibles, in many cases, at their apex are furnished with incisive teeth to cut off their food, and with miniature mill-stones to grind it[1255]. The part here alluded to I call the Mola.
Were I to ask you what your idea is with regard to the use of the organs we are considering, you would perhaps reply without hesitation, "Of what possible use can the jaws of insects be but to masticate their food?" But in this you would in many instances be much mistaken; as you will own directly if you only look at the mandibles of the stag-beetle—these protended and formidable weapons, as well as those of several other beetles, cannot be thus employed. "Of what other use, then, can they be?" you will say. In the particular instance here named, their use, independent of mastication, has not been satisfactorily ascertained; but in many other cases it has. Recollect, for instance, what I told you in a former letter, of those larvæ that use their unguiform mandibles as instruments of motion[1256]. Again: amongst the Hymenopterous tribes, whose industry and varied economy have so often amused and interested you,[Pg 438] many have no other tools to aid them in their various labours and mechanical arts: to some they supply the place of trowels, spades, and pick-axes; to others that of saws, scissors, and knives—with many other uses that might be named. In fact, with the insects of this intire Order mastication seems merely a secondary, if it is at any time their use. Still comprehending in one view all the mandibulate Orders, though some use their mandibles especially for purposes connected with their economy, yet their most general and primary use is the division, laceration, and mastication of their food; and this more exclusively than can be affirmed of the under-jaws (maxillæ). This will appear evident to you, when you consider that insects in their larva state, in which universally their primary business is feeding, with very few exceptions use the organs in question for the purpose of mastication, even in tribes, as the Lepidoptera, that have only rudiments of them in their perfect state—while the maxillæ ordinarily are altogether unapt for such use. The exceptions I have just alluded to are chiefly confined to the instance of suctorious mandibles; or those which, being furnished at the end with an orifice, the animal inserting them into its prey, imbibes their juices through it. This is the case with the larvæ of some Dytisci, Hemerobius, and Myrmeleon[1257]; and spiders have a similar opening in the claw of their mandibles, which is supposed to instil venom into their prey[1258].
Under this head I must not pass without notice an appendage of the mandibles, to be found in some of the[Pg 439] rove-beetles (Staphylinidæ), as in Ocypus, Staphylinus, and Creophilus Kirby. In the first of these it is a curved, narrow, white, subdiaphanous, submembranous, or rather cartilaginous piece, proceeding from the upper side of the base of the mandible[1259]; in the second it is broader, straighter, and fringed internally and at the end with hairs; and in this at first it wears the appearance of being attached laterally to the mandible under the tooth[1260], but if closely examined, you will find that it is separate: in Creophilus maxillosus it is broader. This is the part I have named prostheca. It is perhaps useful in preventing the food from working out upwards during mastication.
5. Maxillæ[1261]. The antagonist organs to the mandible in the lower side of the head, are the under-jaws, or maxillæ—so denominated by the illustrious Entomologist of Kiel. Linné appears to have overlooked them, except in the case of his genus Apis, in which he regards them, and properly, as the sheath of the tongue. De Geer looked upon them in general as part of the apparatus of the under-lip or labium; and such in fact they are, as will appear when we consider them more particularly. Fabricius has founded his system for the most part upon these organs, the principal diagnostic of ten out of his thirteen Classes (properly Orders) being taken from them; and in the modern, which may be termed the eclectic, system, although the Orders are not founded upon them, yet the characters of genera, and sometimes of large tribes, are derived from them: and as they appear less[Pg 440] liable to variation than almost any other organ, as Mr. W. S. MacLeay has judiciously observed, there seems good reason for employing them—it is therefore of importance that you should be well acquainted with them.
Their situation is usually below each mandible, on each side of the labium; towards which they are often somewhat inclined, so that their tips meet when closed. In some cases, as in the Predaceous beetles (Carabus L. &c.), they exactly correspond with the mandibles; but in others their direction with respect to the head is more longitudinal, as in the Hymenoptera, &c. In substance they may be generally stated to be less hard than those organs; yet in some instances, as in the Libellulina, Anoplognathidæ, &c. they vie with them, and in the Scarabæidæ and Cetoniadæ exceed them, in hardness. In the bees, and many other Hymenoptera, they are soft and leathery. Their articulation is usually by means of the hinge on which they sit: it appears entirely ligamentous, and they are probably attached to the labium at the base, or mentum—at least this is evidently the case with the Hymenoptera, in which the opening of the maxillæ pushes forth the labium and its apparatus. In that remarkable genus related to the glow-worms, now called Phengodes (Lampyris plumosa F.), and in the case-worm flies (Trichoptera K.), the maxillæ appear to be connate with the labium, or at least at their base.—As to their composition, these organs consist of several pieces or portions. At their base they articulate with a piece more or less triangular, which I call the hinge (Cardo)[1262]. This on its inner side is often elongated towards the interior[Pg 441] of the base of the labium, to which it is, as I have just observed, probably attached. This elongate process of the hinge in Apis, Bombus, &c. appears a separate articulation; and the two together form an angle upon which the mentum sits[1263], and by this the maxilla acts upon the labial apparatus.
The next piece is the stipes or stalk of the maxilla. This is the part that articulates with the hinge, and may be regarded in some cases, as in the Orthoptera &c., as the whole of the maxilla below the feeler; and in others, as in the Geotrupidæ, Staphylinidæ &c., as only the back of it, the inside forming the lower lobe. This piece is often harder and more corneous than the terminal part, is linear, often longitudinally angular, and in the bee-tribes (Apis L.) is remarkable on its inner side for a series of bristles parallel to each other like the teeth of a comb[1264]. In Pogonophorus Latr., a kind of dor or clock-beetle, it is armed on the back with four jointed spines, the intermediate one being forked[1265]. M. Latreille has thus described the stipes of the maxillæ of Coleoptera: "Next comes the stalk," says he, "which consists of three parts: one occupies the back and bears the feeler; the second forms the middle of the anterior face, and its figure is triangular; the third fills the posterior space comprised between the two preceding; and is that which is of most consequence in the use of the maxilla; the anterior feeler, where there are two, the galea, and the other appendages that are of service in deglutition, are part of that piece[1266]."
The third and terminal portion of the maxilla is formed by the lobe, or lobes (Lobi). This may be called the most important part of the organ, since it is that which often acts upon the food, when preparing for deglutition. When armed with teeth or spines at the end, its substance is as hard as that of the mandibles; but when not so circumstanced, it is usually softer, resembling leather, or even membrane[1267]; and sometimes the middle part is coriaceous, and the margin membranous. This part is either simple, consisting only of one lobe, as you will find to be the case with the Hymenoptera, Dynastidæ, Nemognatha, and several other beetles; or it is compound, consisting of two lobes. In the former case, the lobe is sometimes very long, as in the bee tribes, and the singular genus of beetles mentioned above[1268], Nemognatha; and at others very short, as in Hister, &c. The bilobed maxillæ present several different types of form. Nearest to those with one lobe are those whose lower lobe is attached longitudinally to the inner side of the stalk of the organ, above which it scarcely rises. Of this description is the maxilla in the common dung-beetle (Geotrupes stercorarius), and rove-beetle (Staphylinus olens).[1269] Another kind of formation is where the lower lobe is only a little shorter than the upper: this occurs in a kind of chafer (Macraspis tetradactyla MacLeay).[1270] A third is where the upper lobe covers the lower as a shield; as you will find in the[Pg 443] Orthoptera order, and the Libellulina, and almost in Meloe[1271]. A fourth form is where the upper lobe somewhat resembles the galeate maxilla just named; but consists of two joints. This exists in Staphylinidæ, &c.[1272] The last kind I shall notice is when the upper lobe not only consists of two joints, but is cylindrical, and assumes the aspect of a feeler or palpus[1273]. This is the common character of almost all the Predaceous beetles (Entomophagi Latr.). This lobe, which has been regarded as an additional feeler, is strictly analogous to the upper lobe in other insects, and therefore should rather be denominated a palpiform lobe than a palpus. Where there are two lobes, the upper one is most commonly the longest; but in many species of the tribe last mentioned the lower one equals or exceeds it in length[1274].
The lobes vary in form, clothing, and appendages. The upper palpiform lobe in those beetles just mentioned, in general varies scarcely at all in form; but the genus Cychrus (which is remarkable for a retrocession from the general type of form of the Carabi L. making an approach towards that of those Heteromera which, from their black body and revolting aspect, Latreille has named Melosomes,) affords an exception, the upper joint being rather flat, linear-lanceolate, incurved, and covering the lower lobe[1275], which it somewhat resembles. The lower[Pg 444] lobe also in this tribe varies as little as the upper, being shaped like the last joint of that lobe in Cychrus just described, except that in Cicindela it is narrowest in the middle[1276]. In other tribes the upper valve is sometimes linear and rounded at the apex, and the lower truncated, as in Staphylinus olens[1277]; sometimes the upper one is truncated or obtuse, and the lower acute, as in Trogosita and Parnus[1278]. In Ptinus, another tribe of beetles, before noticed as injurious to our museums[1279], the reverse of this takes place, the upper-lobe, which is the smallest and shortest, being acute, and the lower truncated[1280]. In Blaps both are acute[1281]. In Rhipiphorus and Scolytus the lobes are nearly obsolete. The lower lobe is bifid in Languria, a North American genus of beetles, so as to give the maxilla the appearance of three lobes[1282]; and in Erotylus, a South American one, the upper is triangular[1283]: it is often oblong, quadrangular, linear, &c. in others.—In those that have only one lobe the shape also varies. In Gyrinus, the beetle that whirls round and round on the surface of every pool, which, though it belongs to the Predaceous tribe, has only one lobe, the lobe represents a mandible in shape of the laniary kind, being[Pg 445] trigonal and acute[1284]; and in the Anoplognathidæ, a New Holland tribe of chafers, in which it is, as it were, broken, the lobe forming an angle with the stalk, it is concavo-convex and obtuse, and somewhat figures a molary tooth[1285]. In the first tribe into which the bees (Apis L.) have been divided (Melitta Kirby), the lobe is often linear or strap-shaped, and bifid at the apex; and in the second (Apis K.) lanceolate and intire[1286]. In Cerocoma it is long and narrow[1287]. More variations in form might be named, but these are sufficient to give you a general idea of them in this respect. With regard to their clothing, I have not much to observe—in examining the Predaceous beetles you will observe, that the interior margin of the lower incurved lobe is fringed with stiff bristles or slender spines, and in many other beetles either one or both lobes have a thick coating or brush of stiffish hairs[1288]; but in several cases only the apex of the lobe is hairy. In the Orthoptera order, and many of the Melolonthidæ or chafers, the whole maxilla is without hairs, or nearly so.
The appendages of the maxillæ are next to be noticed. These are principally their claws, or laniary teeth; for they are seldom armed with incisive or molary teeth. The whole tribe of Predaceous beetles, with few exceptions, have the inner lobe of their maxilla armed with a terminal claw, which in the Cicindelidæ articulates with the lobe, and is moveable, but in the rest of the tribe is[Pg 446] fixed[1289]. In Phoberus MacLeay the lower lobe has two spines[1290]. In Locusta this lobe has three or four spines or laniary teeth, and in Æshna there are six, which, like the claw of Cicindela, are moveable[1291]. In others both lobes terminate in a single spine or claw: this is the case with Paxillus MacLeay[1292]. In Passalus, nearly related to the last genus, the upper lobe is armed with a single spine, and the lower one with two[1293]. Those maxillæ that terminate in a single lobe are also often distinguished by the spines or teeth with which it is armed; thus in a nondescript chafer belonging to the Dynastidæ (Archon K. MS.) it terminates in two short teeth; in that remarkable Petalocerous genus Hexodon Oliv. in three truncated incisive ones[1294]; in Dynastes Hercules in three acute spines[1295]. Four similar ones arm the apex of the maxilla in that tribe of Rutelidæ which have striated elytra; and five that are stout and triquetrous those of Melolontha Stigma F. Many others have six spines, sometimes arranged in a triple series[1296]. Besides teeth or spines, in some cases the lobes of maxillæ terminate in several long and slender laciniæ or lappets fringed with hairs. At least those of a Leptura (L. quadrifasciata L.) described by De Geer, appear to be thus circumstanced. He conjectures that this beetle uses its maxillæ to collect the honey from the flowers[1297].
As the principal use of the mandibles is cutting and masticating, so that of the organs we are considering seems to be primarily that of holding the food and preventing it from falling while the former are employed upon it. I say this is their primary use; for I would by no means deny that they assist occasionally in comminuting or lacerating it. In fact, were there no organs appropriated to this use, and if both mandibles and maxillæ were employed at the same time in comminuting the food, it seems to me that it must fall from the mouth. In a large proportion of insects the lobes of the maxillæ are not at all calculated for laceration or comminution; and in those tribes—as the Melolonthidæ, Rutelidæ, Dynastidæ—in which they seem most fitted for that purpose, the mandibles have incisive teeth at their apex, and at their base a powerful mola or grinder: circumstances which prove, that even in this case the business of mastication principally devolves upon them.
6. Palpi Maxillares[1298]. There is one circumstance that particularly distinguishes the maxillæ from the mandibles—they are palpigerous, as well as the under-lip. The feelers, or palpi, emerge usually from a sinus observable on the back of the maxillæ where the upper lobe and stalk meet. Their articulation does not materially differ from that of the labial palpi. Each maxilla has properly only one feeler; but, as was lately observed[1299], in certain tribes the upper lobe is jointed and palpiform, which has occasioned it to be considered as a feeler, and these tribes have been regarded as having six feelers. The most general rule with regard to the length of the palpi[Pg 448] is, that the maxillary shall be longer than the labial; but the reverse often takes place. In many bees the maxillary consist only of a single joint, and are very short; while the labial consist of four, and are very long[1300]: and in some insects (as in Pogonophorus Latr.) the four palpi are of equal length[1301]. The antennæ are most commonly longer than the palpi; but in several aquatic beetles, as Elophorus, Hydrophilus, &c., whose antennæ in the water are not in use, the organs we are considering are the longest.—As to the number of their articulations, it varies from one to six; which number they are not known to exceed. In each of the Orders a kind of law seems to have been observed as to the number of joints both in the maxillary and labial palpi, but which admits of several exceptions. Thus in the Coleoptera, the natural number may be set at four joints for the maxillary, and three for the labial palpi: yet sometimes, as in Stenus, Notoxus, &c., the former have only three joints, and the latter, as in Stenus and Tillus, only two. In the Orthoptera the law enjoins five for the maxillary, and three for the labial; and to this I have hitherto observed no exception. In the Hymenoptera, the rule is six and four, but with considerable exceptions, especially as to the maxillary palpi, which vary from six joints to a single one: thus in the hive-bee and the humble-bee, the labials, including the two flat joints or elevators, have four joints, while the maxillaries are not jointed at all[1302]. In Chrysis, in which the latter consist of five, the former are reduced to three. The Libellulina may almost be regarded[Pg 449] as having no maxillary palpi, since they exhibit no organ that is distinctly palpiform. It seems to me that the upper lobe of their maxilla, which articulates with the stalk in the same manner as a feeler, may be regarded as an instance in which that lobe and the feeler coalesce into one; and the mucro that proceeds from the lobe has the aspect of an emerging feeler, and corresponds somewhat with the labial one above noticed[1303]. In the remainder of the Neuroptera and the Trichoptera, the prevailing number is five and three. In the latter there are exceptions, which will furnish good characters for genera. In the Lepidoptera we find two, and sometimes three, the maxillary being very minute[1304]. The Diptera Order presents two tribes in this respect quite distinct from each other. The most natural number of joints in the maxillary palpi of the Tipulidæ, Culicidæ, &c. is four or five: the last joint, however, in Tipula, Ctenocera, &c. like that of the antennæ in Tabanus L., appears to consist of a number of very minute joints[1305]; but in the Asilidæ and Muscidæ, &c., the number two seems to be most prevalent[1306]. The labial palpi in this order are obsolete.—As to shape, the maxillary palpi, as well as the labial, are usually filiform; but in the weevil tribes (Curculio L.) they are most commonly very short and conical[1307]; in the chafers (Scarabæus L.) they usually are thickest at the apex[1308]; in Megachile and Euglossa, wild bees, they are setaceous, growing gradually[Pg 450] more slender from the base to the summit[1309]: a tribe of small water-beetles (Haliplus), the saw-flies (Tenthredo L.), and several other Hymenoptera, have them thickest in the middle[1310]. Their most important part, however, and that which varies most in form, is the terminal joint:—of this I have already related some singular instances[1311], and shall now describe a few more. This joint is sometimes acute, at others blunt, at others truncated: in figure it is ovate, oblong, obtriangular, hatchet-shaped, lunate, transverse, conical, mammillate, subulate, branched, chelate, laciniate, lamellate, &c. &c.[1312]: terms which I shall more fully explain to you hereafter, and which I only mention here to show the numerous variations[Pg 451] as to figure, of which this joint exhibits examples. The palpi in general at their vertex are often rather concave; and this concavity is formed by a thin papillose membrane, which it is supposed the animal has the power of pushing out a little, so as to apply it to surfaces. The primary use of the palpi of insects will be considered when I treat of their senses; but they probably answer more purposes than one. For instance, when I was once examining, under a lens, the proceedings of a species of Mordella, which was busily employed in the blossom of some umbelliferous plant, it appeared to me to open the anthers with its maxillary palpi, and they often held the anther between them: when not so employed, they were kept in intense vibration, more than even its antennæ; and at the same time, as far as I could judge, an Elater made the same use of them.
7. Lingua[1313].—This name was applied by Linné to the part in insects representing the tongue in vertebrate animals; and as it performs most of the common offices of a tongue, and the pharynx is situated with respect to it, as we shall presently see, nearly as it is in those animals, there seems no more reason for giving it a new name, than there is for giving a new name to the head or legs of insects, because in some respects they differ from those of the higher animals. I shall not therefore call it Ligula, with Fabricius and Latreille, nor Labium, with Cuvier and others, but adhere to the original term, which every one understands.
The tongue lies between the two lips—the labrum and labium. On its upper side, at the base, it meets the palate[Pg 452] or roof of the mouth, below which it is attached, it may be presumed, by its roots to the crust of the head, on each side the pharynx or swallow; and on its lower side, in many cases, it is attached to the labium, and that very closely, so as to appear to be merely a part of it, and to form its extremity: but in the Orthoptera and Libellulina, it is more free, and in form somewhat resembling the tongue of the quadrupeds[1314].—In substance the tongue varies. In general it seems something between membrane and cartilage; but in the Predaceous beetles, in which it is not covered by the labium, it approaches nearer to the substance of the general integument, and in Anthia F. it is quite hard and horny:—that just mentioned of the Orthoptera and Libellulina is more fleshy[1315]. With regard to its station, in many cases, as in the instance just named, in the Lamellicorn tribe (Scarabæus L.) and others, it is, when unemployed, concealed within the mouth; the lips, mandibles, and maxillæ all closing over it. The tongue of some Hymenoptera also is retractile within the mouth. "When ants are disposed to drink," says M. P. Huber, "there comes out from between their lower jaws, which are much shorter than the upper, a minute, conical, fleshy, yellowish process, which performs the office of a tongue, being pushed out and drawn in alternately: it appears to proceed from the lower-lip.—This lip has the power of moving itself forwards in conjunction with the lower jaws: and when the insect wishes to lap, all this apparatus moves forward; so that the tongue, which is very short, does not require to lengthen itself much to reach[Pg 453] the liquid[1316]." M. Lamarck thinks that the labium of insects has a vertical motion (de haut en bas ou de bas en haut)[1317]. This it certainly has in some degree; but it has also, as in the above case, a more powerful horizontal one, which is produced, in Hymenoptera at least, by the opening of the maxillæ—as I have already observed[1318].
I have little to say with respect to the structure of the tongue: it generally seems to be without articulations; but in many bees it articulates with the labium where it enters it, so as when unemployed to form a fold with it. In the hive-bee it terminates in a kind of knob or button, which has been falsely supposed to be perforated for imbibing the honey by suction. The upper part of this tongue is cartilaginous, and remarkable for a number of transverse rings: below the middle, it consists of a membrane, longitudinally folded in inaction, but capable of being inflated to a considerable size: this membranous bag receives the honey which the tongue, as it were, laps from the flowers, and conveys it to the pharynx[1319]. In Stenus this organ is retractile, and consists of two joints[1320].
The shape of the tongue of insects probably varies as much as any other part; but as it is apt to shrink when dried[1321], and is not easy to come at, we know but little of its various configurations:—in the bees it is very long, in most other insects very short. Though frequently simple and undivided, in many cases it presents a different conformation. Thus in the saw-flies (Tenthredo L.)[Pg 454] it terminates in three equal lobes[1322]; in Stomis and Geotrupes in three unequal ones, the intermediate being very short[1323]; in Carabus, in three short teeth[1324]; in Pogonophorus it represents a trident[1325]; in the wasp it is bifid, each lobe being tipped with a callosity[1326]; in Melolontha Stigma it is bipartite[1327]; in Elaphrus, the analogue of the tiger-beetles, it terminates in a single tooth or point; in the aquatic beetles, Dytiscus L., it is quadrangular and without teeth[1328]; in some Ichneumonidæ it is concavo-convex, and forms a demitube; and in others it is nearly cylindrical[1329].
In many insects it has no hairs, but in the Predaceous beetles it generally terminates in a couple of bristles[1330]. In the hive- humble- and other bees, it is extremely hairy[1331]; a circumstance which probably enables it more effectually to despoil the flowers of their nectar. In Geotrupes stercorarius, the common dungchafer, and Melolontha Stigma lately mentioned, the lobes of the tongue are fringed with incurved hairs[1332]; and in Æshna it is hairy on the upper side, each hair or bristle crowning a minute tubercle. In many cases the tongue is attended, and sometimes sheathed at the base, by two usually membranous appendages:—these the learned Illiger has denominated[Pg 455] paraglossæ; and I shall adopt his term. You will find them frequently attached to the tongue of the Predaceous beetles[1333], and to that of many Hymenoptera. In the hive-bee and humble-bee they are short, and take their origin within the labial feelers[1334]: in Euglossa, another bee, they are long, involute at the tips, and, what is not usual with them, very hairy[1335]: in the wasp, like the lobes of the tongue, they are tipped with a callosity.
Under this head I may observe to you, that the insects whose oral organs we are considering besides a tongue appear likewise to be furnished with a palate (Palatum). This, though a part of the roof of the mouth, is not precisely in the situation of the palate of vertebrate animals, since it seems rather the internal lining of the labrum. If you take the common dragon-fly (Æshna viatica), you will find that the under side of this part and of the rhinarium is lined with a quadrangular fleshy cushion, beset, like the upper surface of the tongue, with minute black tubercles, crowned with a bristle. This cushion is divided transversely into two parts by a depression; the anterior or outer piece being attached to the labrum, and the other piece to the rhinarium. The former has a central longitudinal cavity, black at the bottom, on the sides of which the tubercles are flat and without a bristle. From its base on each side a spiniform process emerges, forming a right angle with it. These processes seem the antagonists of those mentioned above[1336], that emerge from the labium. The posterior or[Pg 456] inner piece has on each side a roundish space, attached to the under surface of the two sides of the rhinarium, beset also with bristle-bearing tubercles. You will find something similar lining the labrum and nasus of some Coleoptera,—say Geotrupes, Necrophorus, and Dytiscus. The first piece I regard as the analogue of the palate, and the second as connected with the sense of smelling. In Necrophorus the circular pieces are covered with a finely striated membrane, and in Dytiscus each has a little nipple.
8. Pharynx[1337].—On the upper side of the tongue, usually at its base or root, is the pharynx, or aperture by which the food passes from the mouth to the œsophagus. This orifice, which is situated with respect to the tongue of the Orthoptera and Libellulina nearly as in those insects (at least as far as I have been able to examine them), whose tongue is called a ligula or labium,—of course exists in all the mandibulate Orders whose mouth we are now considering. In the Hymenoptera it is covered by a valve, the Epipharynx of Savigny; and it appeared to me to be so likewise in one of the Harpalidæ that I examined. The formation seems different in Geotrupes, as far as I can get an idea of it; but it is so difficult to examine the interior of the mouth without laceration of some of the parts, that I can only tell you what the appearances were in one instance, upon removing the labrum from the mandibles; and in another, separating the whole apparatus of the labium, including the maxillæ, from the mandibles and labrum. In the former case, the mandibles coincided[Pg 457] at the base, the two molary plates (molæ), which in this genus are narrow, transverse and not furrowed, are so applied as evidently to have an action upon each other, as the mandible opens and shuts, proper for trituration. Within these is the base of the tongue, under the form of a ventricose sack. The upper part of this last organ, which forms the internal covering of the labium, appears to consist of three (in the recent insect fleshy) lobes, the middle one being bent downwards internally, so as to form a kind of sloping cover to an orifice in the part I call the base. After two or three days, the tongue shrinks and dries to a hard substance;—between the mandibles and the base of the tongue I could not discover the pharynx. The above apparent opening covered by the tongue was the only one I could perceive. In the latter case, the form and structure of the base of the tongue is more visible: it is an oblong ventricose tubular sack, projecting above anteriorly into an acute angle formed by a fine white membrane, most beautifully and delicately striated with oblique striæ, to be seen only under a powerful lens: on the anterior side of this sack are two parallel cartilaginous ridges close to each other, fringed with short hairs, which take their origin from the angle. I could not be certain whether the orifice covered by the intermediate lobe was only apparent, or real; but I did not succeed in my endeavour to find any other pharynx, though from the molary structure of the base of the mandibles one may conjecture that there must be one situated at the base of this sack to receive the food they render after trituration. The excrement of this animal is not fluid. In the Libellulina the pharynx seems closed by two valves meeting. This part in Hymenoptera,[Pg 458] and probably in other Orders, has the aspect of being cartilaginous and fitted to sustain the action of the substances that have to pass through it[1338].
The Epipharynx is a valve, called by M. Latreille sublabrum (sous labre[1339]), attached by its base to the upper margin of the pharynx, or that next the labrum. In the bees it is said by Reaumur to be of a fleshy substance, and capable of changing its figure. He seems to think it the real tongue of the bee[1340]; but as it does not appear to have any of the uses of a tongue, and merely closes the orifice of the mouth, it surely does not merit that name. M. Savigny calls it a membranous appendage which exactly closes the pharynx[1341]. De Geer has examined the epipharynx of the wasp, which he describes as of a scaly substance, and regards merely as the cover of the part just named[1342].
With regard to the Hypopharynx, which Latreille considers as a support and appendage of the epipharynx, I have little to add to the definition I have given of it above. In the Libellulina the base of the tongue terminates towards the pharynx in a fleshy cushion, armed at each angle next to that part with a short hard horn or tooth of a black colour. This cushion, I suppose, may be analogous to the hypopharynx of M. Savigny[1343]. On the opposite side the pharynx is closed by another fleshy cushion (epipharynx?), which appears to line the nose, behind those two mammillæ before described[1344], which form the internal covering of the rhinarium.
Before I call your attention to what I would denominate an imperfect mouth, in which some one or more of the seven organs above enumerated exist under another form, or only as rudiments,—I must say something upon the mouth of the Myriapods and Arachnida, in which there seem to be redundant organs of manducation.—M. Latreille, in the Essay lately quoted, in which, though some of his notions seem fanciful, he has shown a vast depth and range of thought and research, has asserted,—from the admirable and curious observations of M. Savigny, and those which since their publication he has made himself,—that the masticating organs of annulose animals (called by him condylopes) are a kind of legs[1345]. And M. Savigny, whose indefatigable labours and unparalleled acuteness have opened the door to a new and vast field in what may be denominated analogical anatomy,—has observed, that with certain Apiropods[1346] the organs that serve for manducation do not differ essentially from those which, with the other Apiropods and the Hexapods, serve for locomotion[1347]: and the unguiform mandibles of the larvæ of certain Diptera, you have before been told, are used not only in manducation, but also as legs[1348]. These remarks will satisfactorily prove to you, that organs which at first sight possess no visible affinity or analogy—as for instance, jaws and legs—may, if traced through a long series of beings, exhibit a very great one;—and will lessen your surprise[Pg 460] when you find, that in certain tribes such commutations of organs and their use take place.
The following is the structure, as to its organs, of the mouth of the myriapods, as exhibited by the centipedes (Scolopendridæ). The part which appears to perform the office of the upper lip (but which M. Savigny regards as the nose, calling it the chaperon,) is a transverse piece with a deep anterior sinus, in the centre of which is a minute tooth[1349]. This piece is separated from the forepart of the head by a suture; but it probably is not moveable: however, it covers the mouth, and may be regarded rather as analogous to the labrum. Below this are two mandibles, armed at their end with five sharp triangular teeth[1350], under which are the maxillæ, terminating in a moveable concavo-convex lobe, resembling the valve of a bivalve shell[1351]; and between them is the labium, of a rhomboidal shape, divisible into two lobes, attached laterally to the maxillæ: these lobes M. Savigny terms the second maxillæ, forming with the others, according to him, the labium[1352]. Affixed to the base of this labium, or covering it on the outside, are a pair of pediform palpi, which he considers as the first auxiliary labium, and representative of the first pair of legs of hexapods and Iuli[1353]. I imagine them to be also the analogues, in some degree, of the labial palpi of a perfect mouth. The last of the organs in question is a large rhomboidal plate affixed to the first apparent segment of the trunk, crowned at its vertex with two truncated denticulated teeth, and from[Pg 461] the upper sides of which emerge a pair of moveable organs terminating in a powerful incurved claw, and which entirely covers all the other parts of the mouth[1354]. This, M. Savigny deems as a second auxiliary labium, and the lateral organs of prehension,—which may be regarded each as a kind of maxillary hand, and as the only representatives in this tribe of the maxillary palpi, though widely different,—he looks upon as really analogous to the second pair of legs in Iulus and the hexapods[1355]. These two pairs of pedipalpes (to use an expressive French term) show their relation to legs by their general structure, and their analogy with palpi by their use as oral organs, though belonging to the trunk: so that here we see the legs and their appendages assume a material function in manducation, forming a singular contrast to what we had observed before with regard to mandibles becoming instruments of locomotion. The mouth of the Iulidæ, with little variation, is upon the same plan[1356] with those here described.
The next type of form with regard to the oral organs is that of the Arachnida. In these, as you know, the head is confounded with the trunk; so that they are a kind of Blemmyes in the insect world. Their organs of manducation, amongst which there is no labrum or upper lip, are, in the first place, a pair of mandibles planted close and parallel to each other in the anterior part of the head, which they terminate. In the spiders they consist of two tubular joints, of which the first is much the largest, more or less conical or cylindrical, and armed[Pg 462] underneath with a double row of stout teeth; and the terminal one is more solid and harder, in the form of a very sharp crooked claw, which in inaction is folded on the first joint between the teeth. Under its extremity on the outside is a minute orifice, destined to transmit a venomous fluid, which is conducted there by an internal canal from the base of the first joint, where is the poison-bag[1357]. In the scorpion and harvest-man (Phalangium) the mandible consists of two joints terminated by a chela or double claw, the exterior one being moveable[1358].—M. Latreille, as has been before observed, regards these not as representatives of the mandibles of hexapods, but as replacing the interior pair of antennæ, in the situation of which they are precisely placed, of the Crustacea[1359]: and M. Savigny is of opinion that the Arachnida may in some sort be defined as Crustacea without a head, and with twelve legs, of which the two first pair are converted into mandibles and maxillæ[1360]. From the situation of the organs in question, the first of these opinions seems preferable; but the conversion of the legs in other cases, at least the coxæ, into organs of manducation, gives some weight to the last. With regard to their use, it is said to be to retain the insect which the animal has seized, and to facilitate the compression which the maxillæ exercise upon it for the extraction of the nutritive matter[1361]. If this be correct, in this respect the mandibles may be said to represent the maxillæ of the mandibulate hexapods; and, vice versa, the sciatic maxillæ, as they have[Pg 463] been denominated[1362], of the Arachnida, their mandibles. The palpi are pediform, and the first joint of the coxa, or hip, acts the part of a maxilla:—this is composed of a single piece or plate, more or less oval or triangular, sometimes straight and sometimes inclined to the labium, with the interior extremity very hairy. The labium consists also of a single piece, and is only an appendage of the anterior extremity of the breast. The interior of the mouth, or palate, presents a fleshy, hairy, linguiform piece, which is usually applied to the internal face of the labium. An opening is supposed to exist in its sides, for the transmission of the alimentary juices[1363]. If you examine the under side of the body of a scorpion, you will find that not only the palpi, but the two anterior pair of legs, by means of their coxæ, are concerned in manducation: so that these insects have in fact three pairs of maxillæ—a circumstance that M. Savigny has observed to take place also in the harvest-men (Phalangium L.)[1364]. The palpi of the scorpion, which may be called its hands, like the anterior legs of the lobster and crab, terminate in a tremendous chela or forceps, consisting of a large triangular joint, armed at the end with a double claw internally toothed; the exterior one of which, contrary to what takes place in the animals just named, is moveable, and not the interior[1365].
Having given you this full account of the trophi of those animals that have all the organs of manducation developed, I must next advert to those in which one part[Pg 464] receives an increment at the expense of others, and the whole oral machine is fitted for suction; or where some parts appear to be deficient, so that this may be called an imperfect mouth. At first sight one would regard the trophi of a bee as of this description; but this is not the case, since it has all the ordinary organs, though the tongue is unusually long, and looks as if it was made for suction; which, however, as you have been informed, is not the case.
There are five kinds of imperfect mouth to be met with in insects that take their food by suction, each of which I shall distinguish by a separate denomination. The first is that of the Hemiptera Order:—this I term the Promuscis; the second is that of the Diptera, which with Linné I call Proboscis; the third, peculiar to the Lepidoptera, is with me an Antlia; the fourth, which I name Rostrulum, is confined to the Aphaniptera order, or genus Pulex L.; and the last is Rostellum, which I employ to denote the suctory organs of the louse tribe (Pediculidæ).
i. Promuscis[1366].—The organ we are first to consider has usually been denominated Rostrum: but since that term is likewise in general use for the snout of insects of the weevil tribes (Curculio L.), I think you will concur with me in adopting the one here proposed, for the very different oral instruments of the Hemiptera. Illiger has employed promuscis to denote those of bees[1367]: but since, as I have just observed, they consist of all the ordinary organs, they seem to require no separate denomination:[Pg 465] the term, therefore, may be applied to represent a different set of trophi, without any risk of producing confusion. This part consists of five pieces: viz. a minute, long, conical piece, commonly very slender, which covers the base of the promuscis, and represents the labrum[1368]; a jointed sheath (vagina), consisting of either three or four joints, the analogue of the labium, and four slender rigid lancets (scalpella), the two exterior ones, according to M. Savigny, representing the mandibles, and the intermediate pair the maxillæ[1369]. By the union of these four pieces a suctorious tube is formed, which the animal inserts into the substance, whether animal or vegetable, the juices of which form its nutriment. These pieces are dilated at their base, and serrated at their apex; and the two central ones, though at their origin they are asunder, form one tube, which has often been mistaken for a single piece. A pharynx and tongue have been discovered by M. Savigny in this apparatus; who thinks that in Nepa there are also rudiments, but very indistinct, of labial palpi: so that the maxillary palpi seem to be the only part absolutely wanting[1370].
The Promuscis when at rest is usually laid between the legs; but when employed, in most cases its direction is outward. In the genus Chermes L. (Psylla Latr.) the origin of the promuscis has been supposed to be in the breast; but if closely examined, this anomaly in nature will be found not to exist. If you take one of these insects, the first thing that strikes you upon inspecting the head, is a pair of remarkable conical processes into which[Pg 466] the front appears to be divided. Look below these, and you will there discover the upper-lip: and from this you may follow the promuscis till it gets beyond the forelegs, when it takes a direction perpendicular to the body[1371]; a circumstance which has given rise to the above false notion. Though in Coccus, Chermes, &c. this instrument is short, in some Aphides it is longer in proportion than in any other insect. In A. Quercus it is three times the length of the body; so that when folded, it stretches out beyond it, and looks like a long tail[1372]; and in A. Abietis it even exceeds that length[1373].
ii. Proboscis[1374].—Linné long since, and after him Fabricius, has employed this term to designate the oral instruments, or rather their sheath, in the Muscidæ and some others, calling the same organ, when without fleshy lips, rostrum and haustellum: but as the parts of the mouth in all true Diptera (for Hippobosca and its affinities can scarcely be deemed as co-ordinate with the rest), are analogous to each other; although in some they are stiff and rigid, in others flexile and soft, and in Œstrus (except the palpi) mere rudiments,—the same appellation ought to designate them all. I am happy to find that M. Latreille agrees with me in this opinion; and to his sensible observations on this head, if you wish for further information, I refer you[1375]. The mouth of Dipterous insects appears to vary in the number of pieces that it presents;[Pg 467] but in all, the theca or sheath is present, which represents the labium (including the mentum) of the mandibulate Orders[1376]. It consists of three joints, the last of which is formed by the liplets (Labella). Those in the Muscidæ are large, turgid, vesiculose, and capable of dilatation; in the Bombylidæ and other tribes they are small, slender, long and leathery, and sometimes recurved. The second joint or stalk, which may be said to represent the mentum, the liplets being properly in a restricted sense the analogue of the labium, its sides being turned up, forms a longitudinal cavity, which contains the haustellum. The upper piece of this, the valvula, is long, rigid, and very sharp, representing the labrum[1377]. Beneath this cover, in the above cavity, are the lancets; which, as far as they are at present known, vary in number and form: sometimes there are five of them, sometimes four, sometimes two, and sometimes, it should seem, only one[1378]. In the gnat (Culex) they are finer than a hair, very sharp, and barbed occasionally on one side[1379]; in the horse-fly (Tabanus L.) they are flat and sharp like the blade of a knife or lancet[1380]. In this tribe the upper pair, or the knives (Cultelli), represent the mandibles; the lower pair, or the lancets (Scalpella), usually palpigerous, the maxillæ; and the central one the tongue. In the horse-fly Reaumur has figured only four, exclusive of the labrum and labium; but in a specimen I have preserved[Pg 468] there appear to be five, one of which, as slender as a hair, I regard as the analogue of the tongue[1381].—When the lancets are reduced to two, they probably represent the maxillæ, the mandibles being absorbed in the labrum; and where there is only one, the maxillæ also are absorbed by the labium, which then bears the palpi, the lancet representing the tongue[1382]. The lancets are so constructed in many cases, as to be able by their union to form a tube proper for suction, or rather for forcing the fluid by the pressure of the lower parts to the pharynx[1383]. Labial palpi appear not usually present in the proboscis; but M. Savigny thinks he has discovered vestiges of them in Tabanus[1384]. In this genus the maxillary ones are large, and consist of two joints[1385]. The proboscis is often so folded, as to form two elbows; the base forming an angle with the stalk, and the latter with the lips, so as in shape to represent the letter Z, only that the upper angle points to the breast, and the lower one to the mouth: this is the case with the flesh-fly and many others. In other flies, as Conops and Stomoxys, whose punctures on our legs so torment us[1386], there is only a single fold, with its angle to the breast. The proboscis is received in a large oblong cavity of the underside of the anterior part of the head.
It may here be observed, that in the promuscis the elongation of the organs seems to be made chiefly at the expense of all the palpi, but in the proboscis at that of the labial only; and in some cases at that also of the mandibles or maxillæ;—the former merging in the labrum and the latter in the labium.
iii. Antlia[1387].—The third kind of imperfect mouth is that of the Lepidoptera, which I have called Antlia. Fabricius denominates it lingua: but as this organ has no analogy with the real tongue of insects, this is confessedly improper, and it appeared necessary therefore to exchange it for another denomination: I have endeavoured to apply a term to it that indicates its use—to pump up, namely, the nectar of the flowers into the mouth of the insect. On a former occasion I described to you the structure of this instrument[1388]; but further discoveries with regard to it having since been made by MM. Savigny and Latreille, I shall here give you the result of their observations. The former of these able physiologists has detected in the mouth of the Lepidoptera rudiments of almost all the parts of a perfect mouth. Of the correctness of this assertion you may satisfy yourself, if you consult his admirable elucidatory plates, and compare them with the insects. Just above the origin of the spiral tongue or pump, the head is a little prominent and rounded; and immediately below the middle of this prominence there is a very minute, membranous, triangular or semicircular piece; which from its position, as covering the base of the antlia, may be regarded as the rudiment[Pg 470] of the upper-lip (labrum)[1389]. On each side of the outer base of the antlia is another small immoveable piece, resembling a flattened tubercle, the end of which is internally hairy or scaly: these pieces appear to represent the mandibles[1390]. Near the base of each half of the antlia, just below a sinus, may be distinctly seen the minute, usually biarticulate rudiment of a maxillary palpus[1391]; demonstrating to a certainty that these spiral organs, at least their lateral tubes or Solenaria, are real maxillæ[1392]. The rudiment of the under-lip (Labium) is the almost horny triangular piece united by membrane to the two stalks of the maxillæ, and supporting at its base the recurved labial palpi; which are so well known that I need not enlarge upon them[1393]. Amongst these parts there seems at first sight no representative of the tongue; but M. Latreille has advanced some very ingenious, and I think satisfactory arguments[1394], which go to prove that this part, at least the tongue of Hymenoptera, has its analogue in the intermediate tube or Fistula formed by the union of the two maxillæ, and which conveys the fluid aliment of this Order to the pharynx. As in Diptera the maxillæ sometimes merge in the labium, so here the tongue (as it were divided longitudinally) merges in the maxillæ. He further observes, that in a transverse section of the maxilla of the death's-head hawk-moth (Sphinx Atropos), the lateral tube appeared[Pg 471] to be divided into two by a membranous partition, and to contain in the upper cavity a small cylindrical tube, which seemed to be a trachea[1395]. To animals that are without lungs, and breathe by tracheæ, suction must be performed in a very different way from what it is by those that breathe by the mouth: and as in the very extended organs in question the fluid has a long space to pass before it reaches the pharynx, in some way or other these lateral tubes may have the power of producing a vacuum in the middle tube, and so facilitate its passage thither. We see, in the antlia, that the maxillæ receive their vast elongation at the expense of all the other organs, except the labial palpi.
iv. Rostrulum[1396].—An animal very annoying to us affords the type of the next kind of imperfect mouth—I mean the flea. Its oral apparatus, which I would name rostrulum, appears to consist of seven pieces. First are a pair of triangular organs, the laminæ, which together somewhat resemble the beak of a bird, and are affixed, one on each side of the mouth, under the antennæ: these represent the mandibles of a perfect mouth[1397]. Next, a pair of long sharp lancets (Scalpella), which emerge from the head below the laminæ: these are analogous to maxillæ[1398]: a pair of palpi, consisting of four joints, are attached to these near their base[1399], which of course are maxillary palpi. And lastly, in the midst of all is a slender setiform organ (ligula), which is the counterpart of the tongue[1400]. Rösel, and after him Latreille, seem to[Pg 472] have overlooked this last piece, since they reckon only six pieces in the flea's mouth[1401]: but the hand and eye of our friend Curtis have detected a seventh, as you see in his figure. From this account it appears, that the elongation of the organs of the Aphaniptera Order is at the expense of the labium and its palpi.
v. Rostellum.—So little is known of the composition of the next kind of imperfect mouth, that I need not enlarge upon it. It is peculiar to the louse tribe (Pediculidæ), and it consists of the tubulet (Tubulus), and siphuncle (Siphunculus). The former is slenderer in the middle than at the base and apex, the latter being turgid, rather spherical, and armed with claws which probably lay hold of the skin while the animal is engaged in suction. When not used, the whole machine is withdrawn within the head; the siphuncle, which is the suctorious part, being first retracted within the tubulet, in the same way as a snail retracts its tentacula[1402]. This apparatus seems formed at the expense of all the other organs.
There are some other kinds of imperfect mouth, which, though they seem not to merit each a distinct denomination, should not be passed altogether without notice. The first I shall mention is that of the family of Pupipara Latr. (Hippobosca L.). It consists of a pair of hairy coriaceous valves, which include a very slender rigid tube or siphuncle, the instrument of suction, which Latreille describes as formed by the union of two setiform[Pg 473] pieces[1403]. In Melophagus, the sheep-louse, the union of the valves of the sheath is so short, that they appear like a tube; but if cut off they will separate, and show the siphuncle, as fine as a hair, between them. This organ is of a type so dissimilar, as was before observed, to that of the Diptera in general, and approaches so near to that of the dog-tick (Ixodes), that they may be deemed rather apterous insects with two wings, than to belong to that Order; and the circumstance that some of the family are apterous confirms this idea. In fact they are a transition family that connects the two Orders, but are nearest to the Aptera. In Nycteribia the oral organs differ from those of the other Pupipara in having palpi. This also is the case with those of the genus Ixodes, the palpi of which are placed upon the same base with the instrument of suction, than which they are longer: they appear to consist of two joints, the last very long and flat. The instrument of suction itself is formed by three hard rigid laminæ; two shorter parallel ones above, that cover the third, which is longer and broader, and armed on each side with several teeth like a saw, having their points towards the base[1404]. Many of the other Acari L. have mandibles, and several have not: but their oral organs have not yet been sufficiently examined; and from the extreme minuteness of most of them, this is no easy task; nor to ascertain in what points they differ or agree.
If you consider the general plan of the organs of manducation in the vertebrate animals, how few are the[Pg 474] variations that it admits! An upper and a lower jaw planted with teeth, or a beak consisting of an upper or a lower mandible with a central tongue, form its principal features. But in the little world of insects, how wonderful and infinite is the diversity which, as you see, in this respect they exhibit! Consider the number of the organs, the varying forms of each in the different tribes, adjusted for nice variations in their uses:—how gradual, too, the transition from one to another! how one set of instruments is adapted to prepare the food for deglutition by mastication; another merely to lacerate it, so that its juices can be expressed; a third to lap a fluid aliment; a fourth to imbibe it by suction—and you will see and acknowledge in all the hand of an almighty and all-bountiful Creator, and glorify his wisdom, power, and goodness, so conspicuously manifested in the structure of the meanest of his creatures. You will see also, that all things are created after a pre-conceived plan; in which there is a regular and measured transition from one form to another, not only with respect to beings themselves, but also to their organs—no new organ being produced without a gradual approach to it; so that scarcely any change takes place that is violent and unexpected, and for which the way is not prepared by intermediate gradations. And when you further consider, that every being, with its every organ, is exactly fitted for its functions; and that every being has an office assigned, upon the due execution of which the welfare, in certain respects, of this whole system depends, you will clearly perceive that this whole plan, intire in all its parts, must have been coeval with the Creation; and that all the species,—subject to those variations only that climate[Pg 475] and different food produce,—have remained essentially the same, or they would not have answered the end for which they were made, from that time to this.
Having given you this particular account of the trophi or organs of the mouth of insects, I must now make some observations upon the other parts of the head. I have divided it, as you see in the Table, into face and subface; the former including its upper and the latter its lower surface. Strictly speaking, some parts of the face, as the temples and cheeks, are common to both surfaces; but I do not therefore reckon them as belonging to the subface, which, exclusive of the mouth and its organs, consists only of the throat, and where there is a neck, the gula.
i. Nasus[1405].—I shall consider the parts of the face in the order in which they stand in the Table, beginning with the nasus or nose. Fabricius has denominated this part the clypeus, in which he has been followed by most modern Entomologists. You may therefore think, perhaps, that I have here unnecessarily altered a term so generally adopted, and expect that I assign some sufficient reasons for such a change. I have before hinted that there is good ground for thinking that the sense of smell in insects resides somewhere in the vicinity of this part; and when I come to treat of their senses, I shall produce at large those arguments that have induced me to adopt this opinion: and if I can make out this satisfactorily, you will readily allow the propriety of the denomination. I shall here only state those secondary reasons for the[Pg 476] term, which, in my idea, prove that it is much more to the purpose than clypeus. This last word was originally applied by Linné in a metaphorical sense to the ample covering of the head of the Scarabæidæ, and the thoracic shield of Silpha, Cassida, Lampyris, and Blatta: in all which cases there was a propriety in the figurative use of it, because of the resemblance of the parts so illustrated to a shield. But when Fabricius (though he sometimes employs the term, as Linné did, merely for illustration,) admitted it into his orismological table, as a term to represent universally the anterior part of the face of insects to which the labrum is attached (though in some cases he designates the labrum itself by this name), it became extremely inappropriate; since in every case, except that of the Scarabæidæ, the part has no pretension to be called a shield;—so that the term is rather calculated to mislead than illustrate. This impropriety seems at length to have struck M. Latreille, since in a late essay[1406] he has changed the name of this part to Epistomis, a term signifying the part above the mouth. But there are reasons, exclusive of those hereafter to be produced concerning the sense of smell, which seem to me to prove that nasus is a preferable term; not to mention its claim of priority, as having been used to signify this part a century ago[1407]. When we come to consider the terms for the other parts of the head, as lips, jaws, tongue, eyes, temples, cheeks, forehead, &c. the concinnity, if I may so speak, and harmony of our technical language, seem to require that the part analogous in point of situation to the nose of vertebrate[Pg 477] animals should bear the same name. And any person who had never examined an insect before, if asked to point out the nose of the animal, would immediately cast his eye upon this part: so that one of the principal uses of imposing names upon parts—that they might be more readily known—would be attained. If it is objected, that calling a part a nose that has not the sense of smell, supposing it to be so, might lead to mistakes—I would answer, that this objection is not regarded as valid in other cases: for instance, the maxillæ are not generally used as jaws, and yet no one objects to the term; because, from their situation, they evidently have an analogy to the organs whose name they bear. But enough on this subject—we will now consider the part itself.
To enable you to distinguish the nose of insects when it is not separated from the rest of the face by an impressed line, you must observe that it is the terminal middle part that sometimes overhangs the upper-lip, and at others is nearly in the same line with it; that on each side of it are the cheeks, which run from the anterior half of the eyes to the base of the mandibles. Just below the antennæ is sometimes another part distinct from the nose, which I shall soon have to mention; so that the nose must not be regarded as reaching always nearly to the base or insertion of the antennæ, since it sometimes occupies only half the space between them and the upper-lip, which space is marked out by an impressed line. But you will not always be left at such uncertainty when you want to ascertain the limits of the nose; for it is in many cases a distinct piece, separated by an elevated or impressed line from the rest of the face. This separation is either partial or universal. Take any species of[Pg 478] the genera Copris, Onitis, or Ateuchus, and you will see the nose marked out in the centre of the anterior part of the face by two elevated lines, forming nearly a triangle and bounded by the horn[1408]. Or take a common wasp or hornet, and you will find a similar space, though approaching to a quadrangular figure, marked out by impressed lines[1409]. In Rhagio and Sciara, two Dipterous genera, this impression is so deep as to look like a suture. Between these lines, in those cases, is included what I call the nose. As to substance, in general it does not differ from the rest of the head; but in the Cleridæ it is almost membranous. You must observe, that in all these, what at first sight appears to be the termination of the front, is not the nose, but the narrow depressed piece that intervenes between it and the lip. With regard to its clothing, it is most commonly naked, but in some genera it is covered with hair; in Crabro F. often with golden or silver pile, which imparts a singular brilliance to the mouth of the insects of that genus: M. Latreille supposes that the brilliant colours of the golden-wasp (Chrysis L.) may dazzle their enemies, and so promote their escape[1410]; the brilliance of the mouth of the Crabro may on the contrary at first dazzle their prey for a moment, so as to prevent their escape. The form of the nose, where distinct from the rest of the face, admits of several variations: thus in the Staphylinidæ and Cleridæ it is transverse and linear; in Copris it is triangular, with the vertex of the triangle truncated; in Vespa Crabro it is subquadrate and sinuated. In many Heteromerous[Pg 479] beetles[1411] it is rounded posteriorly: in Pelecotoma, a new genus in this tribe, related to Asida, there is a deep anterior sinus; in Blaps the anterior margin is concave; in Cetonia[1412] Brownii, and atropunctata (forming a distinct subgenus), it is bifid: it varies in the Scarabæidæ, in some being bidentate, in others quadridentate, and in others again sexdentate, including the cheeks: in Mylabris, a kind of blister-beetle, it is transverse and nearly oval; in Lamia, a capricorn-beetle, it represents a parallelogram; and in most Orthoptera it is subtriangular: in Tettigonia F. it is prominent, transversely furrowed, and divided by a longitudinal channel: in Otiocerus K. it presents the longitudinal section of a cone[1413]: in the Diptera Order, with the exception of the Tipulidæ and some others, in which it unites with the cheeks, &c. to form a rostrum, the nose in general, as to form, answers to its name, resembling that of many of the Mammalia: in some of the Asilidæ it is very tumid at the end, and terminates in a sinus, to permit the passage of the proboscis to and fro: in many of the Syrphidæ, &c. it is first flat and depressed, and then is suddenly elevated, so as to give the animal's head the air of that of a monkey: in some tribes, as Rhingia, Nemotelus, Eristalis, &c., in conjunction with the cheeks it forms a conical rostrum: in Tabanus bovinus, and other horse-flies, it terminates in three angles or teeth. Many more forms might be mentioned, but these will suffice to give you a general idea of them. In size and proportions the nose also varies.[Pg 480] It is frequently, as in Tettigonia, the most conspicuous part of the face, both for size and characters; but in the Staphylinidæ it is very small, and often scarcely discernible, being overshadowed by its ample front: and it may be observed in general, that when the antennæ approximate the mouth, as in this genus and many others, the front becomes ample, and the nose is reduced to its minimum: but when they are distant from the mouth, the reverse takes place; and the nose is at its maximum and the front at its minimum. Mutilla, Myrmecodes, Scolia, &c. in the Hymenoptera, are an example of the former; and the Pompilidæ, Sphecidæ, Vespidæ, &c. of the latter. In Myopa buccata, &c. its length exceeds its width; but more commonly the reverse takes place. The circumscription of the nose also deserves attention. It is usually terminated behind by the front (frons), or, where it exists, by the postnasus, in the sides by the cheeks, and anteriorly by the labrum. But this is not invariably the case; for in the Cimicidæ, in which the cheeks form the bed of the Promuscis, the front embraces it on each side by means of two lateral processes, that sometimes meet or lap over each other anteriorly, which gives the nose the appearance of being insulated; but it really dips below these lobes to join the labrum. This structure you may see in Edessa F., and many other bugs. This part sometimes has its arms. Thus in Copris, and many Dynastidæ, the horns of the head seem, in part at least, to belong to this portion of it; in Tipula oleracea (the crane-fly), &c. it terminates before in a horizontal mucro. In Osmia cornuta, a kind of wild-bee, each side of the nose is armed with a vertical horn. The margin of the nose in most Lamellicorn insects, though mostly level, curves upwards.
I am next to mention a part of the nose which merits a distinct name and notice, which I conceive in some sort to be analogous to the nostrils of quadrupeds, and which I have therefore named the Rhinarium or nostril-piece. I had originally distinguished it by the plural term nares, nostrils; but as it is usually a single piece, I thought it best to denote it by one in the singular. When I treat of the senses of insects, I shall give you my reasons, as I have before said, for considering this part as the organ of scent, or connected with it, which you will then be able to appreciate. I shall only here observe, that the piece in question is in the usual situation of the nostrils—between the nose and the lip. In a large number of insects this part may be regarded as nearly obsolete; or at least it is merely represented by the very narrow membranous line that intervenes between the nose and the lip and connects them; which, as in the case of the head of Harpali before noticed, may be capable of tension and relaxation, and so present a greater surface to the action of the atmosphere. But I offer this as mere conjecture. In the lady-bird (Coccinella) this line is a little wider, and becomes a distinct Rhinarium; as it does also in Geotrupes. With respect to its insertion, the rhinarium is a piece that either entirely separates the nose from the lip, or only partially: the former is the most common structure. It is particularly remarkable in a New Holland genus of chafers (Anoplognathus Leach). In A. viridiæncus it is very ample, and forms the under side of the recurved nose, so that a large space intervenes between the margin of the latter and the base of the labrum. In Macropus Thunb., of the Capricorn tribe (Cerambyx L.), the nostril-piece, which forms a[Pg 482] distinct segment, is narrower than the nose, and the upper-lip than the nostril-piece, forming as it were a triple gradation from the front to the mouth. Again, in others the part in question is received into a sinus of the nose. This is the case with the dragon-flies (Libellulina), in which this sinus is very wide; in the burying-beetle (Necrophorus)[1414], in some species of which it is deep but narrow; and in a species of Tenebrio from New Holland, which perhaps would make a subgenus. If you examine with a common glass any of the larger rove-beetles (Staphylinidæ), you will find that the nose itself seems lost in the nostril-piece, both together forming a very narrow line across the head above the labrum, without any apparent distinction between them; but if you have recourse to a higher magnifier, you will find this divided into an upper and lower part, the former of the hard substance of the rest of the head, and the latter membranous. I once was of opinion that the prominent transversely furrowed part, so conspicuous in the face of Tettigonia F.[1415], was the front: but upon considering the situation of this, chiefly below the eyes and antennæ, and comparing it with the analogous piece in Fulgora laternaria and other insects of the Homopterous section of the Hemiptera, I incline to think that it represents the nose, and that the longitudinal ridge below it is the nostril-piece[1416]. In the Heteropterous section it is merely the vertical termination of their narrow nose. In other insects again, this part approaches in some measure to the common idea of nostrils; there being two, either one on each side the nose, or two approximated ones. If you catch the first humble-bee that you see busy upon a flower, you will[Pg 483] discover a minute membranous protuberance under each angle of the nose. Something similar may be observed in some species of Asilus L. In the Orthoptera, especially in Blatta, Phasma, and some Locustæ, two roundish or square pieces, close to each other on the lower part of the nose, represent the nostrils[1417].—With regard to substance, in the chafer-tribes, at least those that feed on leaves or living vegetable matter, as the Melolonthidæ, Anoplognathidæ, and in many other insects, the rhinarium is of the same substance with the rest of the head; but in Macropus Thunb., Staphylinus, Necrophorus, &c., it consists of membrane.
ii. Postnasus[1418].—This is a part that appears to have been confounded by Entomologists with the front of insects; in general, indeed, it may be regarded as included in the nose, and does not require separate notice: but there are many cases in which it is distinctly marked out and set by itself, and in which it forms a useful diagnostic of genera or subgenera. There is a very splendid and beautiful Chinese beetle, to be seen in most collections of foreign insects (Sagra purpurea), in which this part forms a striking feature, and helps to distinguish the genus from its near neighbour Donacia. If you examine its face, you will discover a triangular piece, below the antennæ and above the nasus, separated from the latter and from the front by a deeply-impressed line: this is the postnasus or after-nose. Again: if you examine any specimens of a Hymenopterous genus called by Fabricius Prosopis (Hylæus Latr.), remarkable for its scent of baum, you will find a similar triangle marked out in a[Pg 484] similar situation[1419]. In many Coleopterous insects, besides Sagra, you will discover traces of the part we are considering: as in Anthia, Dytiscus, and several others of the Predaceous beetles. In Cistela it is larger than the nose itself; but it is more conspicuous in the Orthoptera, particularly in Locusta (Gryllus F.), in which it is the space below the antennæ, distinguished by two or four rather diverging ridges[1420]. In the Libellulina, Myrmeleonina, &c. it is a distinct transverse piece. In Dasyga Latr., a kind of bee, it is armed with a transverse ridge or horn—But enough has been said to render you acquainted with it; I shall therefore proceed to the next piece.
iii. Frons[1421].—The Front of insects may be denominated the middle part of the face between the eyes, bounded anteriorly by the nose, or after-nose, where it exists, and the cheeks; laterally by the eyes; and posteriorly by the vertex. Speaking properly, it is the region of the antennæ; though when these organs are placed before the eyes, under the margin of the nose, as in many Lamellicorn and Heteromerous beetles, they seem to be rather nasal than frontal. This part is often elevated, as in the elastic beetles (Elater), whose faculty of jumping, by means of a pectoral spring, has been related to you[1422]. In Anthia, a Predaceous beetle, it has often three longitudinal ridges. In many of the Capricorn beetles (Cerambyx L.), it is nearly in the shape of a Calvary cross, with the arms forming an obtuse angle, and then terminating at the sinus of the eyes in an elevation for the site of the antennæ. In the ants also (Formicidæ),[Pg 485] the front is often elevated between those organs. In Ponera, one tribe of them, this elevation is bilobed, and receives between its lobes the vertex of the postnasus. In the hornet (Vespa Crabro) the elevation is a triangle, with its vertex towards the mouth. In Sagra it is marked out into three triangles, the postnasus making a fourth, with the vertexes meeting in the centre. In the Dynastidæ and Scarabæidæ the horns are often frontal appendages, as is that of Empusa Latr., a leaf-insect, and probably those of Sphinx Iatrophæ F., which affords a singular instance of a horned Lepidopterous one. Sometimes it is an ample space, reducing the nose to a very narrow line, as in the Staphylinidæ, or sending forth a lobe on each side, as before mentioned, which embraces the nose. In a species of bug from Brazil, related to Aradus F., these lobes are dilated, foliaceous, and meet before the nose, so as to form a remarkable extended frontlet to the head. In others this part is extremely minute: thus in many male flies and other insects, as the Libellulina, where the eyes touch each other, the front is cut off from the vertex and reduced to a small angle. In the female flies the communication with the vertex is kept open, and the front consequently longer. In the horse-flies (Tabanidæ), in Hæmatopota, and Heptatoma, the frontal space is wider than in the rest of that tribe. Many of these are distinguished by a levigated area behind the antennæ in the part we are treating of. In the Libellulina, and in the drone-bee, whose eyes are confluent, the stemmata are in the front. In many Orthoptera also, as Locusta Leach, one of them is below the antennæ; and in the lanthorn-fly tribe (Fulgoridæ), both these organs, which are situate between them and[Pg 486] the eyes, as they do also in Truxalis, appear to be in it[1423]. In this tribe the rostrum is an elongation of the part in question; and perhaps you would think at first that what I have considered as the nose in Tettigonia F. was also a tendency to this kind of rostrum; but if you examine the great lanthorn-fly (Fulgora laternaria), you will find besides, at the lower base of the lanthorn, a triangular piece analogous to the nose of Tettigonia, and below it another representing its nostril-piece:—the horizontal part of the nose in that genus may perhaps be regarded as part of the front. In Truxalis F. the face consists of a supine and prone surface, and the latter is composed of the front, after-nose, nose, and organs of the mouth. I may notice here a most remarkable and singular tribe of bugs, of which two species have been figured by Stoll[1424]: in these the head, or rather those parts of it that we have now been describing, the nose, namely, the after-nose, and front, are absolutely divided longitudinally in two, each half having an eye and antenna planted in it; or perhaps, as it is stated to be divided in one instance to the commencement of the promuscis, the nose is left intire, and dips down, as in cases before alluded to: so that in this the nose appears to leave the lobes of the front, which in others embrace its sides.
iv. Vertex[1425].—We now come to the vertex, or crown of the head; which is situated behind the front, and, except where the communication is intercepted by confluent eyes, adjoins it. It is laterally bounded by the hind part of the eyes and the temples; and posteriorly,[Pg 487] where that part exists, by the occiput. The vertex may be denominated the ordinary region of the stemmata: for though in several cases, as we have just seen, one or more of them are planted in the front; yet this in the great majority, especially in the Hymenoptera, is their natural station. In Blatta and some other Orthoptera the posterior angle of the head is the vertex. In many dung-chafers of Latreille's genus Onthophagus, which are said to have occipital horns, as O. nutans, nuchicornis, Xiphias, &c., the horn really arms the part I regard as the vertex. In Locusta Leach, this part is very ample, and in Truxalis very long; but more generally it is small, and not requiring particular notice.
v. Occiput[1426].—The occiput, or hind-head, is that part of the face that either forms an angle with the vertex posteriorly, or slopes downwards from it. It has for its lateral boundaries the temples, and behind it is either terminated by the orifice of the head, or in many cases by the neck. In those beetles that have no neck, as the Lamellicorn and Capricorn, the hind-head is merely a declivity from the vertex, usually concealed by the shield of the thorax, very lubricous, to facilitate its motion in the cavity of that part, and at its posterior margin distinguished by one or two notches, which I shall notice hereafter, for the attachment of the levator muscles: but in those beetles or other insects that have a neck, or a versatile head, the occiput forms an angle with the vertex, often rounded, and sometimes acute. This structure may be seen in Latreille's Trachelides, and several other beetles. In the Hymenoptera, Diptera, and others with a[Pg 488] versatile head, the part now under consideration curves inwards from the vertical line, so as with the temples and under parts of the head to form a concavity adapted to its movement upon the trunk.
vi. Genæ[1427].—The cheeks of insects (Genæ) usually surround the anterior part of the eyes, and lie between them and the mandibles or their representatives. Where they approach the latter, as in the Predaceous beetles (Cicindela, Carabus L. &c.), they are very short, and of course longer where the eyes are further removed from the mouth; as in the Rhyncophorous beetles (Curculio L.), where they form the sides of the rostrum, and often contain a channel which receives the first joint of the antennæ, when they are unemployed. In the Scarabæidæ and many other Lamellicorn beetles, their separation on each side from the nose is marked by a ridge[1428]; and in the wasps (Vespa) by an impressed line or channel. In an African tribe at present arranged with Cetonia F., to which C. bicornis Latr.[1429] and another, which he has named, I believe, C. vitticollis, belong, the cheeks are porrected on each side of the mouth into a horizontal horn. These horns have at first the aspect of a pair of open mandibles. In the magnificent Goliathi Lam., the horns of the male are rather a process of the cheek than of the nose. In Alurnus, Hispa, and other beetles, these parts, by their elevation and conjunction with the lower side of the head, form a kind of fence which surrounds and protects the oral organs; in many Cimicidæ, by a similar elevation of the cheeks, the bed of the promuscis is formed.[Pg 489] In the Homopterous Hemiptera they run parallel nearly with the rhinarium or nostril-piece. In the Hymenoptera they are almost always ample, but they are confined to the lower side of the eye. In Sirex grandis, and others of that genus, the cheek at the base of the mandible is dilated so as to form a rounded tooth below it. In the Capricorn-beetles it is considerable, and sometimes terminates, at the base of the mandible, in two or three notches. In Scaurus and Eurychora, darkling-beetles, the cheek below projects into a lobe that covers the base of the maxilla. But the animal distinguished by the most remarkable cheeks is a species of Phryganea L. (Phryganea personata Spence); for from this part projects a spoon-shaped process, which curves upwards, and uniting with that of the other cheek, forms an ample mask before the face, the anterior and upper margin of which, in the insect's natural state, are closely united; and the posterior part being applied to the anterior part of the eye, causes the face to appear much swoln. It looks as if it was a single piece; but upon pressing the thorax it opens, both above and in front, into two parts, each convex without and hollow within, and each having attached to its inside a yellow tuft of hair resembling a feather. The use of this machinery at present remains a mystery[1430].
vii. Tempora[1431].—The temples (Tempora) are merely a continuation of the cheeks to the posterior limit of the[Pg 490] head, forming its sides and posterior angles, and including the hinder part of the eyes, the vertex, and the occiput. They seldom exhibit any tangible character, except in certain ants (Atta Latr.), in which their angle terminates in one or two strong spines, giving the animal a most ferocious aspect; and in that remarkable genus Corydalis they are armed below with a tooth or point, which was not overlooked by De Geer[1432].
viii. Oculi[1433].—I must now call your attention to organs of more importance and interest, and which indeed include a world of wonders: I mean the eyes (Oculi) of insects. These differ widely from those of vertebrate animals, being incapable of motion. They may be regarded as of three descriptions—simple, conglomerate, and compound.
1. Simple Eyes[1434]. We will consider them as to their number, structure, shape, colour, magnitude, situation, and arrangement.
As to their number, they vary from two to sixteen. In the flea, the louse, the harvest-man (Phalangium), there are only a pair; in the bird-louse of the goose (Nirmus Anseris), and probably in others of the same genus, there are four[1435]; in some spiders (Scytodes, Dysdera, and Segestria Latr.[1436]), and some scorpions[1437], there are six. In the majority of spiders and Scolopendra morsitans,[Pg 491] Scorpio maurus, &c. there are eight; and in Podura and Sminthurus Latr. there are sixteen[1438].
As to their structure, nothing seems to have been ascertained; probably their organization does not materially differ from that of one of the lenses of a compound eye; which I shall soon explain to you.
Their colour in the many is black and shining, but in the bird-louse of the goose they are quite white and transparent. In spiders they are often of a sapphirine colour, and clear as crystal. In Scolopendra morsitans and many spiders, scorpions, and phalangia[1439], they appear to consist of iris and pupil, which gives them a fierce glare, the centre of the eye being dark and the circumference paler. In the celebrated Tarantula (Lycosa Tarantula), the pupil is transparent, and red as a ruby; and the iris more opaque, paler, and nearly the colour of amber.
Where there are more than two, they vary in magnitude. In the enormous bird-spider (Mygale avicularia) the four external eyes are larger than the four internal[1440]; but in the Tarantula and Sphasus, the two or four internal are the largest. In Clubiona and Drassus they are all nearly of the same size[1441]; and in the Micrommata family they are very small[1442].
They vary also in shape. In Scolopendra morsitans the three anterior ones are round, and the posterior one transverse, and somewhat triangular. In Mygale calpeiana, a spider, the two smallest are round and the rest[Pg 492] oval[1443]. In the trapdoor or mason spider (Mygale cæmentaria), the four small internal ones are round, and the large external ones oval[1444]; and those that are circumscribed posteriorly with an impressed semicircle, are shaped like the moon when gibbous[1445].
The situation and arrangement of simple eyes are also various. In many they are imbedded, as usual, in the head; but in the little scarlet mite, formerly noticed[1446], (Trombidium holosericeum), they stand upon a small foot-stalk[1447]: the hairiness of this animal might otherwise have impeded its sight. In spiders they are planted on the back of the part that represents the head, sometimes four on a central elevation or tubercle, and the remaining four below it—as in Lycosa; sometimes the whole eight are on a tubercle, as in Mygale; and sometimes, as in the common garden-spider (Epeira Diadema), upon three tubercles, four on the central one and two on each of the lateral ones. Other variations in this respect might be named in this tribe. In the scorpions a pair are placed one on each side, on a dorsal tubercle, and the other four or six on two lateral ones of the anterior part of the head[1448]. In the Phalangidæ the frontal eyes of the scorpion cease, and only a pair of dorsal ones are inserted vertically in the sides of a horn or tubercle, either bifid or simple, often itself standing upon an elevation which emerges from the back of the animal[1449]. If their eyes were not in a vertical and elevated position, the sight of[Pg 493] these insects would be very limited; but by means of the structure just stated, they get a considerable range of surrounding objects, as well as of those above them. With regard to the arrangement of the eyes we are considering, it varies much. Sometimes they are placed nearly in the segment of a circle, as in those spiders that have six eyes only, before noticed[1450]; sometimes in two straight lines[1451]; at others in two segments of a circle[1452]; at others, in three lines[1453], and at others in four[1454]. Again, in some instances they form a cross, or two triangles[1455]; in others, two squares[1456]; in others, a smaller square included in a large one[1457]; in others, a posterior square and two anterior triangles[1458]; sometimes a square and two lines. Though generally separate from each other, in several cases two of the eyes touch[1459]; and in one instance three coalesce into a triangle[1460]. But it would be endless to mention all the variations, as to arrangement, in the eyes of spiders.
2. Conglomerate Eyes[1461] differ in nothing from simple
eyes, except that instead of being dispersed they are collected
into a body, so as at first sight to exhibit the appearance
of a compound eye:—they are, however, not
hexagonal, and are generally convex. They occur in[Pg 494]
Lepisma, the Iulidæ, and several of the Scolopendridæ.
In Scolopendra forficata the eye consists of about twenty
contiguous, circular, pellucid lenses, arranged in five
lines, with another larger behind them, as a sentinel or
scout, placed at some little distance from the main body.
In the common millepede (Iulus terrestris) there are
twenty-eight of these eyes, placed in seven rows, and
forming a triangle, thus  —the posterior row containing
seven lenses, the next six, and so on, gradually
losing one, till the last terminates in unity. Each of
these lenses is umbilicated, or marked with a central depression.
In Craspedosoma Leach, you will find a similar
formation. In Glomeris zonata, a kind of wood-louse
that rolls itself into a ball, the lenses are arranged in a
line curved at the lower end, with a single one by itself
at the posterior end on the outside; they are oblong and
set transversely, and their white hue and transparency
give them the appearance of so many minute gems, especially
as contrasted with the black colour of the animal[1462].
Between these eyes and the antennæ is another transverse
linear white body, but opaque, seemingly set in a
socket, and surrounded by a white elevated line, like the
bezel of a ring. Whether it is an eye, or what organ, I
cannot conjecture[1463]. Its aspect is that of a spiracle.
—the posterior row containing
seven lenses, the next six, and so on, gradually
losing one, till the last terminates in unity. Each of
these lenses is umbilicated, or marked with a central depression.
In Craspedosoma Leach, you will find a similar
formation. In Glomeris zonata, a kind of wood-louse
that rolls itself into a ball, the lenses are arranged in a
line curved at the lower end, with a single one by itself
at the posterior end on the outside; they are oblong and
set transversely, and their white hue and transparency
give them the appearance of so many minute gems, especially
as contrasted with the black colour of the animal[1462].
Between these eyes and the antennæ is another transverse
linear white body, but opaque, seemingly set in a
socket, and surrounded by a white elevated line, like the
bezel of a ring. Whether it is an eye, or what organ, I
cannot conjecture[1463]. Its aspect is that of a spiracle.
3. Compound Eyes[1464].—These are the most common kind of eye in hexapod insects, when arrived at their perfect state; in their larva state, as we have seen, their eyes being usually simple[1465]; except, indeed, those whose metamorphosis is semicomplete, which have compound eyes[Pg 495] in every state.—In considering compound eyes, I shall advert to their structure, number, situation, figure, clothing, colour, and size.
As to their structure,—when seen under the microscope they appear to consist usually of an infinite number of convex hexagonal pieces. If you examine with a good glass the eye of any fly, you will find it traversed by numberless parallel lines, with others equally numerous cutting them at right angles, so as apparently to form myriads of little squares, with each a lens of the above figure set in it. The same structure, though often not so easily seen, obtains in the eyes of Coleoptera and other insects. When the eye is separated and made clean, these hexagons are as clear as crystal. Reaumur fitted one eye to a lens, and could see through it well, but objects were greatly multiplied[1466]. In Coleopterous insects they are of a hard and horny substance; but in Diptera, &c. more soft and membranous. The number of lenses in an eye varies in different insects. Hooke computed those in the eye of a horse-fly to amount to nearly 7,000[1467]; Leeuwenhoeck found more than 12,000 in that of a dragon-fly[1468]; and 17,325 have been counted in that of a butterfly[1469]. But of all insects they seem to be most numerous in the beetles of Mr. W. S. MacLeay's genus Dynastes. In the eyes of these the lenses are so small as not to be easily discoverable even under a pocket microscope, except the eye has turned white[1470]: it is not, therefore, wonderful, that Fabricius[Pg 496] should call these eyes simple[1471]. In some insects, however, as in the Strepsiptera Kirby, the lenses are not numerous: in Xenos they do not exceed fifty, and are distinctly visible to the naked eye[1472]. These lenses vary in magnitude, not only in different, but sometimes in the same eyes. This is the case in those of male horse-flies and flies, those of the upper part of the eye being much larger than those of the lower[1473]. The partitions that separate the lenses, or rather bezels, in which they are set, are very visible in the eyes just mentioned, and those of Xenos; but in many insects they are only discernible at the intersecting lines of separation between the lenses. In hairy eyes, such as those of the hive-bee, the hairs emerge from these septa. Every single lens of a compound eye may be considered as a cornea, or a crystalline humour, it being convex without and concave within, but thicker in the middle than at the margin: it is the only transparent part to be found in these most remarkable eyes. Immediately under the cornea is an opaque varnish, varying according to the species, which produces sometimes in one and the same eye spots or bands of different colours. These spots and bands form a distinguishing ornament of many of the Tabani and other flies. And to this varnish the lace-winged flies (Hemerobius, &c.) are indebted for the beautiful metallic hues that often adorn them. When insects are dead, this varnish frequently loses its colour, and the eye turns white: hence many species are described as having white eyes which when alive had black ones. The consistence[Pg 497] of this covering is the same with that of the varnish of the choroid in the eyes of vertebrate animals; but it entirely covers the underside of the lens, without leaving any passage for the light. Below this varnish there are numbers of short white hexagonal prisms[1474], every one of which enters the concavity of one of the lenses of the cornea, and is only separated from it by the varnish just described: this may be considered as the retina of the lens to which it is attached; but at present it has not been clearly explained how the light can act upon a retina of this description through an opaque varnish. Below this multitude of threads (for such the bodies appear), perpendicular to the cornea, is a membrane which serves them all for a base, and which consequently is nearly parallel with that part. It is very thin, of a black colour, not produced by a varnish; and in it may be seen very fine white tracheæ, which send forth branches still finer, that penetrate between the prisms of the cornea: this membrane may be called the choroid. Behind this is a thin expansion of the optic nerve, which is a true nervous membrane, precisely similar to the retina of red-blooded animals. It appears that the white pyramidal threads which form the retina of each lens are sent forth by this general retina, and pierce the choroid by a number of almost imperceptible holes[1475]. From this description it appears that the eyes of insects have nothing corresponding with the uvea or humours of those of vertebrate animals, but are of a type peculiar to themselves.
Having explained to you the wonderful and complex[Pg 498] structure with which it has pleased the Creator to distinguish the organs of vision of these minute beings, proving, what I have so often asserted, that when animals seem approaching to nonentity, where one would expect them to be most simple, we find them in many cases most complex, I shall now call your attention to the next thing I am to consider—the number of the eyes in question. Most insects have only two; but there are several exceptions to this rule. Those that have occasion to see both above and below the head, the eyes of all being immovable, must have them so placed as to enable them to do this. This end is accomplished in many beetles, for instance Scarabæus L., Helæus Latr., &c., by having these organs fixed in the side of the head, so that part looks upward and part downward; but in others four are given for this purpose. If you examine the common whirlwig (Gyrinus Natator) that I have so often mentioned[1476], which has occasion, at the same time, to observe objects in the air and in the water, you will find it is gifted with this number of eyes. Lamia Tornator (Cerambyx tetrophthalmus Forst.) and some others, of which I make a genus, under the appellation of Tetrops, are also so distinguished. In these insects, one eye is above and the other below the base of the antennæ; in fact, in these the canthus, instead of dividing the eye partially, as in the other Capricorn-beetles, runs quite through it at considerable width[1477]. In Ryssonotus MacLeay[Pg 499] (Lucanus nebulosus K.) the eye appears also to be divided in two by the canthus. In the Neuroptera Order there is more than one instance of the same kind. In Ascalaphus there are two considerable eyes on each side of the head, which, though clearly distinct, meet like those of many male flies and the drone. The male, likewise, of more than one species of Ephemera, besides the common lateral eyes and the stemmata on the back of the head, have a pair of compound eyes on the top of a short columnar process[1478]. In the Hemiptera Order, also, an instance occurs of four eyes in the genus Aleyrodes[1479]. Amongst the vertebrate animals, there is an example of eyes with two pupils in Anableps, a genus of fishes[1480], but no vertebrate animal has four of these organs. That many insects should have more than two eyes, will not seem to you so extraordinary as that any should be found that, like the Cyclops of old, have only one. There is, however, an insect, before celebrated for its agility[1481] (Machilis polypoda Latr.), which has a single eye in its forehead; or we may say, its eyes are confluent, without any line of distinction between them except a small notch behind. Now that I am treating of the number of eyes, I must not forget to observe to you, that in some insects no eyes at all have been discovered. In Polydesmus complanatus, on each side of the head there is an eye-shaped portion separated by a suture, in which under a powerful lens I cannot satisfy myself that I can discern any thing like the facets that usually distinguish compound eyes. In Geophilus electricus, another myriapod, they[Pg 500] certainly do not exist[1482]. Whence we may conclude, as was before observed[1483], that the faculty of emitting light is rather given it as a means of defence than to guide it in its path.
The situation of compound eyes differs in different tribes. In some, as in the Staphylinidæ, they are planted laterally in the anterior part of the head; in others, the Carabi &c., in the middle; in others again, Locusta Leach &c., in the posterior part. In some, their station is more in the upper surface, either before or behind; so that a very narrow space separates them, or perhaps none at all. Instances of this position of the eyes occur in a minute weevil (Ramphus Clairv.[1484]), and many Diptera, &c. Of those that form an union on the top of the head, some are placed obliquely, so as to leave a diverging space below them, as in many Libellulina[1485], the drone[1486], &c. Others, as Atractocerus, in which the eyes occupy nearly the whole head, and unite anteriorly, have this diverging space above their conflux. In Rhina barbirostris Latr., another kind of weevil, they are confluent below the head, at the base of the rostrum, and a very narrow interval separates them above. In a large number of the Heteromerous beetles, they are set transversely, in the Capricorn ones longitudinally. Their surface, when they are lateral, has usually two aspects, one prone to see below, the other supine to see above. In general the eyes are situated behind the antennæ, so that their position, whether it shall be anterior or posterior, depends upon that of those organs. Often, indeed, as in[Pg 501] the last-named beetles, part of the eye is behind and part before the antennæ; but except where there are four eyes, as in Tetrops, they are never placed before or below them.
Though the eyes of insects are generally sessile, yet to give them a wider range they are sometimes, but it rarely occurs, placed, like those of many Crustacea, on a footstalk, but not a moveable one. An instance of this in certain male Ephemeræ has already been mentioned. In the Hemiptera De Geer has figured two species of bugs (Cimicidæ) that are so circumstanced[1487]; as are also all the known Strepsiptera K., though in these the footstalk is very short[1488]: but the most remarkable example of columnar eyes is afforded by that curious Dipterous genus Diopsis, in which both eyes and antennæ stand upon a pair of branches, vastly longer than the head, which diverge at a very obtuse angle from its posterior part[1489].
In their figure eyes vary much. Sometimes they are so prominent as to be nearly spherical: this is the case with some aquatic bugs, as Ranatra, Hydrometra, and several male Ephemeræ[1490]. Very often they are hemispherical, as in the tiger-beetles (Cicindela L.), and the clocks or dors (Carabus L.); but in a large number of insects they are flat, and do not rise above the surface of the head.—With regard to their outline, they are often perfectly round, as in many weevils; oval, as in various bees;[Pg 502] ovate, as in other bees (Andrena F.); triangular, as in the water-boatman (Notonecta). They are also often oblong, and occasionally narrow and linear; as in that singular beetle Helæus. In many of the Muscidæ they form nearly a semicircle, or rather, perhaps, the quadrant of a sphere. The eyes of the Capricorn-beetles (Cerambyx L.) have a sinus on their inner side, as it were, taken out of them; so that they more than half surround the antennæ, before which is the longest portion of them. An approach to this shape is more or less observed in the darkling-beetles (Tenebrio L.); but in these the sinus is not so deep. I may under this head observe, that in those Mantidæ that represent dry leaves, and some others, these organs usually terminate in a spine[1491].
Though not distinguished by the beauty and animation that give such interest to the eye of vertebrate animals, and exhibiting no trace of iris or pupil, yet from the variety of their colours the compound eyes of insects, though most commonly black or brown, are often very striking. Look at those of one of the lace-winged flies that commit such havoc amongst the Aphides[1492], and it will dazzle you with the splendour of the purest gold, sometimes softened with a lovely green. The lenses of those of Xenos blaze like diamonds set in jet[1493]. You have often noticed the fiery eyes of many horse-flies (Tabanus L.) with vivid bands of purple and green[1494]. Others are spotted[1495]; and Schellenberg has figured one (Thereva hemiptera)[1496], that exhibits the figure of a flower[Pg 503] painted in red on a black ground. These colours and markings are all most vivid and brilliant in the living insect, and often impart that fire and animation to the eyes for which those of the higher animals are remarkable. Take one of the large dragon-flies that you see hawking about the hedges in search of prey, examine its eyes under a lens, and you will be astonished at the brilliance and crystalline transparency which its large eyes exhibit, and by the remarkable vision of larger hexagons which appear in motion under the cornea, being reflected by the retina—all which give it the appearance of a living eye. This moving reflexion of the hexagonal lenses in living insects was noticed long since in some bees (Nomada F., Cœlioxys Latr.)[1497]
Compound eyes differ greatly in their size. In some insects, as Atractocerus, the drone-bee, many male Muscidæ, &c., they occupy nearly the whole of the head; while in others, as numerous Staphylinidæ, Locusta Leach, &c., they are so small as to be scarcely larger than some simple eyes of spiders: and they exhibit every intermediate difference of magnitude in different tribes, genera, and species.
Under this head I must say something of the Canthus of the eye; by which I mean an elevated process of the cheek, which in almost all the genera of the Lamellicorn beetles enters the eye more or less, dividing the upper portion from the lower. Though usually only a process of the cheek, yet in the Scarabæidæ the whole of that part forms the canthus[1498]. It only enters the eye in the Rutelidæ, Cetonidæ, &c.; it extends through half of it in[Pg 504] Copris; it goes beyond the half in Ateuchus; and in Ryssonotus MacLeay (Lucanus nebulosus K.) it quite divides the eye into two[1499], as I before observed. In Lucanus, Passalus &c. it projects before the eye into an angle; in Lucanus femoralis nearly into a spine; but in Lamprima and Œsalus it does not exist. The part, also, that enters the eye in the Capricorn-beetles may be regarded as a kind of canthus, though it is merely a dilatation of the front.
4. Stemmata[1500].—Having given so full an account of the kinds and structure of the ordinary eyes of insects, you may perhaps expect that I should now dismiss the subject: you would, however, have great cause to blame me, did I not make you acquainted with a kind of auxiliary eyes with which a large portion of them are gifted; I mean those pellucid spots often to be found on the posterior part of the front of these animals, or upon the vertex, frequently arranged in a triangle. These, Linné, from his regarding them as a kind of coronet, called Stemmata. They have been of late denominated Ocelli; but as this latter term is also in general use for the eyelets on the wings of Lepidoptera, I have adhered to that of the illustrious Swede. Neither he nor Fabricius has expressed any opinion as to the use of these organs; but Swammerdam and Reaumur were aware that they were real eyes. The former found that there are nerves that diverge to them though not easily traced, and that they[Pg 505] have a cornea, and what he takes for the uvea[1501]; and the latter has supposed that the compound eyes and these simple ones have, the one the power of magnifying objects much, and the other but little, so that the former are for surveying those that are distant, and the latter those that are near[1502]. The same author relates some experiments that he tried with the common hive bee, by which he ascertained that the stemmata, as well as the compound eyes, were organs of vision. He first smeared the latter over with paint, and the animals, instead of making for their hive, rose in the air till he lost sight of them. He next did the same with the former, and placing the bees whose stemmata he had painted within a few paces of their hive, they flew about on all sides among the neighbouring plants, but never far: he did not observe that these ever rose in the air like the others[1503]. From this experiment it seems as if the compound eyes were for horizontal sight, and the stemmata for vertical.
The definition of them by Linné and Fabricius as smooth, shining, elevated or hemispheric puncta, conveys a very inadequate idea of them; for, except in a very few instances, they are perfectly clear and transparent, and their appearance is precisely the same as that of the simple eyes of Arachnida &c., under which head they might very well have been arranged; but as the last are primary eyes, and the stemmata secondary, it seemed to me best that they should stand by themselves. The structure of both is probably the same, and their internal organization that of one of the lenses of a compound eye, and both are set in a socket of the head.
Though a large number of insects have them, they are by no means universal, since some Orders, as the Strepsiptera, Dermaptera, and Aptera, are altogether without them. The Coleoptera, also, have been supposed to afford no instance of species furnished with them; but in the last number of Germar and Zincken Sommer's Magasin, it is affirmed that they are discoverable in Gravenhorst's genus Omalium, but not in the kindred genera Micropeplus and Anthophagus[1504]. Upon examining the former genus, I find, that although Omalium planum and affinities, O. striatulum, and some others, appear not to have them, yet with the aid of a good magnifier they may be discovered in most species of that genus; as likewise in Evœsthetus Grav. I find them also very conspicuous in A. Caraboides and other Anthophagi, but some species appear to want them. In these insects they are two in number, situated in the vertex a little behind the eyes but within them, and either at each end of a transverse furrow, or at the posterior termination of two longitudinal ones. Nor are they found in all the genera of the other Orders. In the Orthoptera, the Blattidæ, unless a white smooth spot on the inner and upper side of the eyes may be regarded as representing them, have them not; but in all the other genera of that Order they are to be found[1505]. In the Hemiptera all the Cicadiadæ are gifted with them; as are likewise Tetyra, Pentatoma, with many other[Pg 507] Cimicidæ, and the Reduviadæ very remarkably; but many others in both sections of this order, as Thrips, Coccus, Aphis, Capsus, Miris, Naucoris, Nepa, and Notonecta, &c. are deprived of them[1506]. Of the Neuroptera the Libellulina add stemmata to their large eyes, in the anterior angle of which they are stationed[1507]; but many other genera of that Order are without them; as Myrmeleon, Ascalaphus, Hemerobius, &c. The Trichoptera and Lepidoptera universally have them; though in the latter, except in Castnia and the Sphingidæ, they are not easily seen. In the Hymenoptera they are usually very conspicuous, but in Larra and Lyrops, two genera of this order, the posterior pair are scarcely discernible; and in the neuter ants they are quite obsolete. In the Diptera, though many genera are furnished with them, yet many also want them; amongst the rest Latreille's Tipulariæ, and all the horse-flies (Tabanus L.). The Pupiparæ (Hippobosca L.) usually have none; but in Ornithomyia avicularia, one of that tribe, though extremely minute they are visible, arranged in a triangle, in the polished space of their vertex.
As to the Number of the stemmata, three appears to be most universal. Reaumur mentions an instance in which he counted four in a fly with two threads at its tail; but great doubt rests upon this statement[1508]. Some Orthopterous genera, as Gryllotalpa, and many Hemipterous,[Pg 508] as Tetyra, Pentatoma, Reduvius[1509], Cercopis, Fulgora[1510], &c., have no more than two; and in Larra and its affinities, as just observed, the posterior ones are obsolete, so as to leave only one discernible.
Where there are three of these organs, they are usually arranged in an obverse triangle in the space behind the antennæ, at a greater or less distance from them. In those male flies (Muscidæ) whose eyes are confluent, the stemmata are in a little area behind their conflux; but, as before observed, in the drone-bee and the Libellulina they are before it. This triangle is in some cases nearly equilateral, as in Perla related to the may-flies, and many Hymenoptera; in others it is acutangular, as in Locusta &c., in which the stemma forming the vertex of the triangle is before the antenna[1511]: in others, again, it is obtusangular, as you will see in Pepsis and various Hymenoptera. In the humble-bees (Bombus), a line drawn through them would form a slight curve. Their situation also varies. In insects that have only two, they are sometimes placed a little behind the eyes, or in the back part of the space between them: this is the case with most of the bugs (Cimex L.) that have them.—They are often distant, as in Tetyra F., Edessa F.; and sometimes approximated, as in Reduvius F.[1512] In many of the Homopterous Hemiptera, as Cercopis, Ledra, &c. they are planted in the upper part of the head[1513], but in Iassus their situation is on the under part; and in a North American subgenus, as yet without a name, they are exactly[Pg 509] between the two, being placed in the frontal angle. In Fulgora their station is between the eyes and antennæ[1514]. They are most commonly sessile, and as it were set in the head; but in some, as Fulgora candelaria, they stand on a footstalk. The stemmata are set in the side of a frontal tubercle in that four-winged fly of threatening aspect, Corydalis, which in its perfect state has mandibles, but longer and more tremendous, like those that distinguish the larva only of the kindred genus Hemerobius[1515]. These organs differ little in shape, being usually perfectly round and somewhat convex; but occasionally they vary in this respect. In Fulgora serrata they are oblong, with a longitudinal depression; in F. Diadema they are also umbilicated, but the umbilicus is circular; in Corydalis they are oval; in other insects they are ovate; in some semicircular, and in a few triangular. They vary much in size: in some of these animals being so minute as to be scarcely visible, while in others, as Corydalis, Dorylus, Vespa pallida F., Reduvius, &c.[1515], they are as large as some compound eyes. They differ also in colour, though often black: in Fulgora laternaria they are of a beautiful yellow; in F. candelaria they are white; in many Hymenoptera they are crystalline, in others red: the fierce look of Reduvius personatus is rendered more hateful by its stemmata having a pale iris round a dark pupil[1516].
Let us here stop and adore the goodness of a beneficent Creator, who, though he has deprived these little beings of the moveable eyes with which he has gifted the[Pg 510] higher animals, has made it up to them by the variety and complex structure of their organs of vision, where we have only two points of sight, giving them more than as many myriads.
5. Antennæ.—But of all the organs of insects, none appear to be of more importance to them than their Antennæ, and none certainly are more wonderful and more various in their structure, and probably uses. Upon this last particular I shall enlarge hereafter. Their structure, as far as it differs in the sexes, I fully discussed in a former letter[1517]; and the most remarkable kinds of them will be included in a set of definitions which I shall draw up for you before our correspondence on this part of my subject closes: I shall therefore now confine myself to the following particulars—namely, their number, insertion, substance, situation, proportion, general form and structure, clothing, expansion, motions, and station of repose.
As to their Number, in the majority of crustaceous animals the antennæ amount to four, but no insect has more than two. A genus recently established (Otiocerus Kirby[1518]) seems to afford an exception to this rule, since the species composing it at first sight appear to have four, and in some instances even six antennæ; but as only two of them terminate in a bristle, the other, though proceeding from the same bed of membrane, may perhaps be regarded as merely appendages. Germar, who has described a species of this genus[1519] under the name of Cobax Wintheri, considers these appendages as analogous[Pg 511] to palpi: but as they do not proceed from the oral organs, but from the bed of the antennæ at the base of the nose[1520], they ought certainly to be regarded rather as accessories to the latter, than as representing the former. In the Aptera order the mites (Acacus L.) appear to be without these organs. In the pupiparous tribe Hippobosca they seem about to disappear; and in the Arachnida &c., as has been more than once observed[1521], the mandibulæ have been thought to represent, not indeed the antennæ of insects, but the inner pair of those of the Crustacea.
In considering the insertion of antennæ, by which I mean their articulation with the head, we must advert first to the orifice (Torulus) that receives them[1522]. This is a perforation of the crust of the head; commonly, though not invariably, circular: in Coleopterous insects often with concave lubricous sides, forming an acetabulum, with processes usual in ginglymous articulations, larger than the bulb or root of the antennæ; and which is commonly covered, except the central space occupied by the bulb, with a tense membrane. Though not in general remarkable, in some cases it merits attention. In the genus Rhipicera Latr., the elegant antennæ of whose males I have described in a former letter[1523], particularly the Brazilian species, it is a long process on each side of the nose, and might be mistaken for the first joint: in another Coleopterous genus, Priocera K.[1524], it has somewhat of the shape of a trumpet: in Cupes a tubercle rises[Pg 512] just above the base of the antenna: a circular process forms the torulus in Fulgora and others. It is also often placed in a cavity of the front, as in several wild-bees, Melitta K., and in Locusta Leach on the sides of an elevation of that part[1525]. In a large majority of insects the bulb (Bulbus) or ball which is received by the bed, wears the appearance, especially in the Hymenoptera, of a distinct joint; but if you carefully examine it, you will clearly see that it is merely the base of the scape swelled out into a spherical or other kindred form[1526]; and often marked, as in the Cicindelidæ, with impressed points: as it is the piece by which the antenna moves in its socket, this form of a rotula was doubtless given for its more ready motion in all directions. This structure is principally conspicuous in the Coleoptera and Hymenoptera Orders: in the others the base is not so distinguished from the rest of the scape. If you carefully extract the antennæ of a beetle, say a Copris or Lamia, and examine its base or bottom, you will find that it is open for the transmission of muscles and nerves; that in its upper margin it has a deep notch or sinus, on each side of which is a smaller notch; and that all round the margin, which is very lubricous, a membranous ligament is attached, by which it was affixed in the torulus. Its articulation, therefore, seems of a mixed kind, like that of most other organs and parts of insects, partaking of the ligamentous, ginglymous, and ball and socket. In the[Pg 513] Orthoptera, Hemiptera, &c. the articulation seems more purely ligamentous.
With regard to their substance—these organs are regulated, in some degree, by the nature of the integument of the animal of which they are appendages; in the softer insects being of a softer substance than they are in hard ones. The vertex of the joints, where they receive the succeeding one, appears in many cases to be softer than the rest of it, and especially towards the apex, often papillose. The antennæ are generally opaque; but in Nebria complanata, a beetle common on the sea-coast in Wales and Lincolnshire, they are semitransparent.
The situation of antennæ must next be considered. In this respect it seems necessary that they should be so situated as to be under the direction of the eyes: for if you examine ten thousand insects (except, as was before observed[1527], where there are four eyes), you will not find one in which these organs are situated either above or immediately behind them; their station being always either somewhere in the space between the eyes or that below them. In Ptinus F. they are placed near the vertex; but in Gibbium, which is so nearly related to that destructive genus[1528], they are beneath them. In many Melittæ K. they are in the middle of the space between the eyes; and in many other Hymenoptera and Coleoptera (Staphylinus &c.), in the anterior part of it. In many Lamellicorn genera (except in some Acridæ, as A. viridissima) as Melolontha, Cetonia, Lucanus, &c. they may be regarded as planted in the lower surface of the cheek before the eyes; but in Copris &c., in which they are inserted further under the[Pg 514] shield of the head, they are properly in the prone surface of the front. In the Capricorn-beetles (Cerambyx L.) and Cnodalon F. they may be termed inocular, or placed in a sinus of the eye; in the former tribe in its interior, and in the latter its anterior side. In the Rhynchophorous or rostrum-bearing beetles (Curculio L.) they vary in their situation. Thus in Macrocephalus Oliv. they are inserted at its apex; in Anthribus in its middle, and in Calandra at its base[1529]. In the water-scorpions (Nepa, Belostoma, &c.) they may be called extraocular, being placed under the head in its prone part, outside the eyes[1530]. In Nirmus Fringillæ, a kind of bird-louse, they appear to be oral, being situated, according to De Geer, under the head near the mouth, at a great distance from the eyes[1531].
In their proportions, both as to length and thickness, antennæ vary extremely. Thus sometimes they are very short—much shorter than the head; as in the aquatic beetles Gyrinus, Parnus, and the water-scorpion; and some land-beetles, as Anthrenus, &c. At other times they far exceed the length of the insect: the males of many Capricorn-beetles are so distinguished. In that of Lamia ædilis they are more than four times as long as the body; and every intermediate length between these two may be found amongst them. They vary also greatly in thickness: in Paussus, whose antennæ emit light in the night[1532], and Cerapterus, they are nearly as thick,—at least their knob, which forms the chief part of them,—as[Pg 515] the body of the insect[1533]; while in Mantis, Acrida K. and Psocus, they are as slender as a hair. The antennæ in many of the Prioni, especially in P. imbricornis, are thick from base to tip; while in other Capricorn-beetles they are quite the reverse.
It will not be necessary to enlarge here upon the general form of these organs: I shall therefore only notice the two principal divisions of them in this respect.—Antennæ, regard being had to one of their uses, may be divided into two sections, distinguished by forms extremely different: those, namely, that are employed by insects as tactors to explore their way, and those that cannot be so employed. The great majority are of the former kind; but those that may be denominated setigerous,—as the antennæ of the Libellulina, Ephemerina, of the Homopterous Hemiptera, and of many Diptera, the last joint of which terminates in a bristle, or is furnished with a lateral one, and of some gnats that have short feathered antennæ,—appear not fitted to be used as tactors to explore by touch, and form the latter description. This difference in these organs, as I shall have occasion to prove more at large hereafter, furnishes a strong presumption that their primary function is not touch. Were this the case, it would be common to them all.
As to their structure, antennæ consist in general of a number of tubular joints; each of which having separate motion, the animal is thereby enabled to give them every flexure necessary for its purposes. The scape, or first joint, by means of the bulb inosculates in the torulus, or is suspended to it; and the others, sometimes by a similar,[Pg 516] though less pronounced knob at their base, inosculate in the preceding one; but in some cases the inosculation seems not so perfect, the joints being simply suspended by ligament. In pectinated or lamellated antennæ, the branch is usually a lateral process of the joint from which it issues; but in Phengodes (Lampyris plumosa L.) its involute plumose branches appear to articulate with the apex of each joint[1534]. I have a specimen of one of the Cleridæ, of a genus undescribed, in which each branch is forked. In some tribes of the Capricorn-beetles (Stenocorus, &c.) the antennæ are often armed at their apex with spines, sometimes on the upper side and sometimes below. In some aquatic beetles (Gyrinus, Parnus) they are furnished with an auricle at their base, which, like the lid of a box, shuts them in when unemployed, and protects them from the water[1535].
The portions into which antennæ may in general be considered as divided, have been sufficiently explained to you above; but it may not be amiss to add here a few words on the principal variations in their structure that I have had an opportunity of observing. The scapus[1536] or first joint, which includes the bulbus, is usually the most conspicuous joint in the antenna (exclusive, I mean, of the capitulum, in those in which that organ terminates in a knob), it being thicker and often longer than the succeeding ones. In the Capricorn and Darkling beetles, indeed (Cerambyx and Tenebrio L.), the third joint is the longest, but the scape is still the thickest; and in the stag-beetles (Lucanus L.), many of the weevil tribes[Pg 517] (Curculio L.), and those of the bees (Apis L.), except in the males, it is as long nearly as the remainder of the antennæ, which forms an angle with it. In shape it is generally somewhat curved and subclavate, or increasing in size from the base to the summit; but it is sometimes straight and filiform, at others oblong or square, at others again triangular, in several instances three-sided: in one (Cetonia cruenta F. Genuchus K.) it is, as it were, broken, the upper part forming nearly a right angle with the lower; in Cerocoma Schæfferi it is foliaceous; and it is occasionally suborbicular: and probably many other forms might be enumerated.
The Pedicellus[1537] is the second, and may be deemed the least conspicuous joint of the antennæ. Though more slender than the scape, it is generally thicker than that which immediately follows it. In broken antennæ it is the hinge or pivot on which the clavola or upper member turns: it is usually very short, campanulate or bell-shaped, or obconical; but in a species of bug (Tetyra, from New Holland—T. pedicellata Kirb. MS.) it is nearly as long as all the rest of the joints taken together. In those species of Lycus, a genus of beetles related to the glow-worm, that have flattened antennæ (as L. reticulatus, fasciatus, &c.), this joint is almost received into the socket of the scape, so that their antennæ appear at first to have only ten joints, but in those which have those organs filiform (as L. minutus, Aurora, &c.) it is more conspicuous.
The Clavola[1538], or remaining joints of the antennæ taken together, constitutes the principal part of the organ, which, especially at its extremity, exercises its functions[Pg 518] of touch, or any other sense. The principal variations, as to form and structure, that occur in this part will be mentioned in another place. I shall only here observe, that in many instances the first joint of this part is longer than the rest; but in Tetyra pedicellata just mentioned, it is by far the shortest, and shaped like the pedicel of most insects. In the Libellulina, the Homopterous Hemiptera, and those flies whose antennæ terminate in a bristle, the clavolet is represented by the bristle. But in the flies which have a lateral bristle, on the last joint, and those with triarticulate antennæ that have no bristle, the terminal joint represents it. The clavolet often terminates in a knob, or in several joints thicker than that which precedes them. This varies greatly, not only in its form, but also in the number of joints of which it is composed. Thus in Paussus, Platypus, and many Calandræ, it consists of only a single joint[1539]; in Anthrenus, Ditoma, &c. of two; in Nitidula, Geotrupes, &c. of three[1540]; in Tetratoma, the Silphidæ, of four[1541]; of five in Scaphidium[1542]; of six in one species of Languria, of seven in the common cockchafer (Melolontha vulgaris[1543]); of eight in Diaperis Boleti, in which the whole clavolet forms the club[1544]; of nine in Oenas; and ten in Cerapterus[1545]. All the above, you will observe, are beetles. In the other orders there are eleven joints in the knob of some butterflies; twelve in that of Ascalaphus[1546] and Myrmeleon; and lastly, fourteen in Trachelus[1547].
Under structure also, the number of joints of which antennæ in general consist, should be considered. If you examine the insects belonging to the different orders, you will find remarkable variations in this respect. Let us run through them:—In the Coleoptera the natural number of joints is eleven; but this rule is not without many exceptions. Thus, many have fewer than the prescribed number: Paussus has only two[1548], Claviger and Platypus five, Dorcatoma and Calandra eight[1549], Geniates K. and Phanæus MacLeay nine[1550], and lastly Melolontha ten[1551]. Others, again, have more than eleven joints: Cebrio grandis, Chrysomela stolida, some Saperdæ, and several others, have twelve. In Prionus imbricornis the female has nineteen, and the male twenty[1552]. Rhipicera marginata has thirty-two; and in a New Holland species of this genus I counted thirty-eight. In the Orthoptera I can trace no general law in this respect. In Locusta Leach in some species you may count fourteen joints, in others sixteen, and in others twenty-five. In one, which appears to be a pupa, I found only thirteen. In Mantis they exceed thirty; but in Blatta, from between thirty and forty, they reach nearly one hundred and fifty; often varying in number in different individuals of the same species. The order Hemiptera exhibits two peculiar types of antennæ, which, with some exceptions, distinguish the two natural sections into which M. Latreille has judiciously divided it. In the Heteropterous section they are without a bristle at their end; and in the Homopterous one, with the exception of Aphis, Thrips, &c.[Pg 520] they have one. In the genera of both these tribes, the number of joints varies in these organs. Thus, exclusive of the seta, in Flata and Cixius there are only two joints; in Galgulus, Fulgora, and Cercopis, there are three; in Lygæus, Coreus, &c. there are four; in Tetyra, Pentatoma, Tettigonia, there are five[1554]; in Aleyrodes there are six; in Aphis seven; in Thrips eight; in Psylla ten, the last of which is terminated by two bristles[1555]; and in Coccus eleven. The Neuroptera order, as it stands at present, is regulated by no general rule with regard to the number of joints in the antennæ of the insects that compose it. Several types of form in these organs distinguish its discordant tribes. The first is that of the Ephemeræ, in which the antennæ consist of two short joints, crowned by a short, tapering, unjointed bristle. The second is that of the Libellulina, similar to the above, but with a jointed bristle. The third is that of Psocus, in which the antenna has two short thick joints at the base, terminated by a long filiform bristle, consisting of seven or eight joints, and finer than a hair. Perhaps these three may be regarded as belonging to a common type. The fourth type is presented by the short filiform antennæ of Termes; the fifth by the setaceous ones of Corydalis, Hemerobius, &c.; and the sixth and last by the clavate and capitate ones of Myrmeleon and Ascalaphus. In the Lepidoptera and Trichoptera orders the antennæ, though varying in their general form in the three tribes of which Linné formed his genera Papilio, Sphinx, and Phalæna, with the exception of Hepialus, in which the joints are[Pg 521] few, are always multiarticulate:—we will therefore, without further delay, proceed to the Hymenoptera. In Latreille's tribe Aculeata the general rule is, that the females shall have twelve joints and the males thirteen. In his Ichneumonides the law seems to be, that the antennæ shall be multiarticulate and setaceous; but in most of the other tribes of the order, even those that in other respects are most nearly related,—as in his Tenthredinetæ,—the number of joints of these organs varies without end. Thus in Hylotoma there are only three joints[1556]; in Cimbex læta[1557] five; in C. axillaris and Perga Leach[1558], six: and so on to twenty-five or more[1559]. The same fluctuation in this respect runs throughout the rest of the order. In the Diptera there are two general types of antennæ:—those of the Tipulariæ Latr., consisting usually of from fourteen to sixteen joints, in the males often resembling beautiful plumes; and those of the remainder of the order, in which they do not exceed three joints[1560]: though the last, or patella, is often further divided into obsolete or indistinct ones[1561]. These antennæ may be further subdivided into filatæ and aristatæ, or those without and those with a bristle, either lateral or terminal.
The clothing of antennæ also merits attention, since it is often not a little remarkable. By clothing I understand the down or hairs of every kind with which they are either generally or partially covered. A great number of filiform and setaceous antennæ of Predaceous beetles (Cicindela L., Carabus L.) have the first two, three, or[Pg 522] four joints naked, and the rest covered with a fine down. In insects that have a knob at the end of these organs, whether lamellated or perfoliate, this down is often confined to it, or to its intermediate joints, and seems intermixed with nervous papillæ. These are particularly visible in the flabellate antennæ of Rhipicera, Lampyris Latreillii[1562], Elater flabellicornis[1563], &c. covering both surfaces of the processes of the joints. In some male bees these papillæ are inclosed in hexagonal spaces into which the antennæ are marked out[1564]. It is to be observed, that in many antennæ the joints of the clavolet have one or two bristles or more at their apex, one above perhaps, and one below; the lower angle in those of the serrated antennæ of Elater is usually so furnished, and sometimes the upper. In many Capricorn-beetles and various insects the antennæ are clothed, instead of down, with stiffish hairs or short bristles. Other insects have these organs, at least the clavolet, beset with longer hairs standing out from them on all sides: of this kind are those of a singular beetle (Sarrotrium muticum) sometimes found in this country[1565]. Again, there are some that have only their underside bearded with longer hairs; as Lamia curculionoides, speculifera K., and other Capricorns[1566]. In another of this tribe, Saperda hirsuticornis, the three intermediate joints are ornamented with branches of long black hairs, which give them an elegant and feathery appearance[1567]. In Callichroma alpina the[Pg 523] apex of the slate-coloured joints of its antennæ is bearded with black hairs. In Lamia reticulata, and Saperda fasciculata and plumigera, all also Capricorns, a single bunch of hairs, resembling the brush of a bottle-cleaner, signalizes the middle of the antenna[1568]: in Saperda scopulicornis K. this is star-shaped[1569]. Sometimes the scape is externally bearded, as in Trox, a beetle found in horns and bones; and in many other Lamellicorns[1570]. In this last tribe the two exterior leaves of the knob of the antennæ are often set with short bristles[1571]; and in a minute beetle called by De Geer Dermestes atomarius, the hairs of this part are said to form a brush[1572].
When insects, I mean more particularly Coleoptera, are about to move from any station where they have been at rest, the first thing they usually do, before they set a step, is to bring forward and expand their antennæ, which have either been carefully laid up in a cavity fitted to receive them, or back upon the body: if they terminate in a lamellated knob, they separate the lamellæ as far as possible from each other; or if it is perfoliate, the joints of it mutually recede. The object of this is evidently to collect notices from the atmosphere, since the papillose part of these joints cannot be applied to surfaces. When the animal begins to move, in many cases the antennæ do the same, and continue their motion till it stops and returns to a state of repose. In the parasitic tribes of the Hymenoptera (Ichneumon L.) they are kept in an almost constant vibration. Many other insects move them in all directions without any order or regularity;[Pg 524] and others, when they elevate one depress the other, and so proceed as if balancing themselves by means of these organs like a rope-dancer. I have before stated to you how by motions of their antennæ, ants and bees communicate their wants or discoveries to each other, or make inquiry concerning any thing they wish to know[1573]. But as I shall have occasion to make some further remarks upon this subject, when the senses of insects are under discussion, I shall for the present take my leave of it.
I shall conclude what I have to communicate to you relative to the organs of which we are treating, with a few observations with respect to their station when the insect reposes. In the Capricorn-beetles, Eucera and other insects with long antennæ, they are merely turned back or on one side with no particular cavity for their reception when unemployed, but probably the apex passes under the body. In the Predaceous and Darkling beetles (Carabus L., Tenebrio L.) their station is usually under the sides of the prothorax, and in the Tortoise beetles (Cassida), under its anterior margin. In the Elastic beetles (Elater) they are received into a groove between the under margin of that part and the fore-breast (antepectus). In Anthrenus, when the animal reposes or counterfeits death, the antennæ are concealed in a cavity of the underside of the prothorax, at right angles with the throat[1574]. In the kindred genus Byrrhus, another simulator of death, a large cavity is excavated under the same part, to receive both the forelegs and antennæ, a narrow space being left between the angle of the prothorax[Pg 525] and fore-breast exactly admitting the base of the latter, which are quite concealed under the former. In Cryptocephalus and Chlamys, kindred beetles, when at rest they are withdrawn, except their scape and pedicel, with the head within the cavity of the prothorax. In others they are turned under the head, without any particular cavity for their reception; as in many moths, Apion, &c. In most of the Lamellicorn beetles their station is in the cavity formed by the eye and the throat, the knob forming an angle with the rest of the antenna. In Heterocerus they follow the contour of the eye[1575]. In Brentus, a genus of weevils remarkably long and slender, they are turned back and received by a slight longitudinal cavity of the rostrum; but in those of this tribe (Curculio L.) in which the clavolet forms an angle with the long scape, this latter part, bending back, is laid up in an oblique channel of that part; and the former, pointing in the contrary direction, is folded upon it. In many flies (Muscidæ) a vertical frontal cavity receives the antennæ, which point downwards during repose[1576]. Cryptocerus, a very remarkable ant, has on its head a singular square plate, the sides of which form a deep longitudinal cavity: in this cavity the antennæ, quite concealed, repose in safety. A cavity equally remarkable is exhibited by the water-scorpions, particularly Belostoma, in which is a very deep kidney-shaped box, between the eye and throat, to receive and defend its singular antennæ[1577]; which, when they are reposing, is closed by the exterior harder joints, and from which it seems as if they turned out, like a sentinel out of his box.[Pg 526] In some aquatic genera of beetles, as Gyrinus, Parnus, &c. they are withdrawn within a lateral cavity of the same part, and are defended from the water externally by the auricle at their base[1578]. The flabellated and lamellated antennæ, previous to their being folded for repose, close all their plates; which in action are as widely expanded as possible, so as to form a knob; and in some the middle piece is entirely concealed, as if in a box. In broken antennæ, or those in which the clavolet forms an angle with the scape, the former is folded upon the latter, with its point downwards.
II. Subfacies.—Having dispatched the Facies, or upper side of the head, I am next to consider the Subfacies, or under side: but as the principal parts that occupy this side have been already considered, I shall have no occasion to detain you long.
i. Jugulum[1579].—This part, which may be regarded as analogous to the throat in vertebrate animals, lies between the cheeks; from which it may usually be distinguished by being more lubricous and tumid, and often separated by an impressed line. It is particularly conspicuous and elevated in the Lamellicorn beetles, and calculated by its lubricity for easy motion in the lower side of the cavity of the chest. Its apex is the base in which the mentum sits. It is not necessary to enlarge further upon it, as it seldom exhibits striking characters.
III. Collum[1580].—In a large proportion of insects the head inosculates in the trunk without the intervention[Pg 527] of a neck, or a constriction of the head behind. In the Orders Orthoptera, Trichoptera, Lepidoptera, Hymenoptera, and Diptera, no instance of it that I recollect occurs: in the Coleoptera there are many. In the Predaceous beetles, though several have no distinct neck, yet others, as Anthia, &c. have a short and thick one; and some few, as Colliuris, Agra, &c. one more pronounced. Latreille has named a tribe in this Order Trachelides, from the circumstance of their having a neck: in this tribe you will find the blister-beetles (Cantharis and Mylabris) both of the moderns and the ancients. In the Hemiptera order the water-scorpions Nepa, &c. have a thick short neck; and Zelus, (a kind of bug,) one longer and more slender; and, like Raphidia, the snake's-head fly, which is similarly circumstanced, has the air of a serpent. Other Neuroptera, likewise, have a neck; as Hemerobius, Corydalis, &c. This part presents no other features that merit notice.
IV. Myoglyphides[1581].—The Myoglyphides, or muscle-notches, are sinuses, some shallow and some deeper, in the posterior margin of the upper side of the head, to which the levator muscles are affixed. They seem principally confined to the Coleoptera; though, in some cases at least, they may be traced in the Heteropterous Hemiptera. These notches vary in number and depth in different insects. Thus in Buprestis there is only one deep one[1582]: in Copris there are two shallow ones, in a deep sinus separated by a small prominence[1583]: in Elater and Lamia there are also two not in a sinus; and in[Pg 528] Calandra Palmarum there are four, two on each side, with a prominent lobe between them[1584]. To each of these notches, at its under margin, below the ligament that unites the occiput to the trunk, a muscle to raise the head is usually attached.
As the head of insects is the principal seat of the organs of sensation, so is the trunk of those of motion; and in it are contained the muscles by which they are moved: it may therefore be regarded as the great centre of motion, and as the main support and prop of the two other primary sections of the body—the head and abdomen, between which it is situated—it may be deemed the most important part of the insect, the key-stone of the whole structure. In treating upon it, for the greater clearness, I shall consider its substance, general form, proportions, composition, internal processes, and members. It will first, however, be necessary to assign my reasons for the nomenclature of its parts that I have adopted.
Had the entomological world been universally agreed upon this subject, and there was an established system of Orismology[1585], I should have proposed no alteration without great reluctance, and the fullest conviction of the absolute necessity of some change; but as the standard[Pg 530] of language in our science is still unsettled, and different terms are used by different writers, there seems full liberty left to me to select those that appear upon the whole most appropriate; and where proper and significant terms seem wanting, to invent new ones. M. Latreille, in a late Essay[1586], has proposed many changes of this kind, and seems to hesitate concerning the adoption of some of those recently coined in France for the parts of the trunk[1587]; it may therefore, I think, be permitted me to labour a little in this hitherto imperfectly cultured field, and to suggest such improvements as the subject may seem to require or admit.
Linné called the part we are now considering the trunk, its upper-side he usually denominated the thorax, and its under-side the breast: he notices also the scutellum and sternum[1588]. As the prothorax and scutellum are the only apparent parts of the back of the trunk in his first Orders (Coleoptera, Hemiptera L.), the rest being covered, in noticing these he puts the part for the whole, calling the prothorax the thorax, but which strictly was not synonymous with what he called by the same name in the other Orders. Linné's phraseology with regard to the trunk of insects was adopted by Fabricius and other Entomologists, till Illiger employed the term thorax to designate the whole of the trunk[1589], calling the upper part thorax superior and the lower thorax inferior. M. De Blainville, M. Latreille, and other French writers, improved[Pg 531] upon this, naming the upper part the back (dorsum), and the lower the breast (pectus); and dividing the trunk, or according to them thorax, into three sections, each bearing a pair of legs. But I see no sufficient reason for this alteration—the terms trunk, thorax, and breast, in the common acceptation are well understood, and lead to no confusion or glaring impropriety; I shall therefore adhere to the old phraseology, especially as French Entomologists in popular language still do the same.
As to the division of the trunk into segments by M. Latreille and others, it has been regarded as consisting of three primary ones, which have been called in the order of their occurrence, reckoning from the head—prothorax, mesothorax, metathorax. The first of these segments, however—and the learned Entomologist just named seems to hint as much[1590]—is usually more distinct from the other two, than they are from each other. If this idea be correct, the trunk is properly resolvable into two primary segments, the first bearing the arms or fore-legs, and the other the proper legs and the organs of flight. M. Chabrier calls the latter tronc alifère, or wing-trunk;—a happy term, which I have adopted and latinized, calling it the alitrunk (alitruncus): the first segment, because it bears the fore-legs, I have named manitrunk (manitruncus). I adopt likewise the terms above mentioned, prothorax, mesothorax, metathorax, to signify the three segments into which the thorax of Linné, or the upper side of the trunk, is resolvable; and those of the breast I denominate antepectus, medipectus, and postpectus. If terms be thought necessary to designate the two intire[Pg 532] segments into which the alitrunk is resolvable, the first may be the meditrunk (meditruncus), and the other the potrunk (potruncus).
I. Substance.—With regard to its substance, the trunk in general is softer than the head, and harder than the abdomen, especially as to its upper surface; but in some cases, where it is not protected by the elytra, as in the rove-beetles (Staphylinus L.), the abdomen appears as hard as the trunk. Though usually not very different from the elytra in this respect, in Meloe, Lytta, and other vesicatory beetles, it is of a firmer consistence.
II. General Form.—In the Coleoptera Order the only part of the trunk that is visible on its upper-side is the prothorax: the mesothorax, with the exception of the scutellum, and the metathorax, being entirely concealed by it and the elytra; so that, with regard to shape, it may nearly be considered as merging in the prothorax. Below it is more visible, and may be stated as more or less quadrangular; in oblong beetles inclining to a parallelogram, and in shorter or hemispherical ones to a square. In the majority it is more convex below than above, except in the case of the hemispherical or gibbous beetles (Coccinella, Erotylus, &c.), in which the under-side is flat and the upper-side very convex. In the Diurnal Lepidoptera the trunk approaches to a cubical shape, in the Nocturnal it is more spherical. A similar difference obtains in the Hymenoptera and Diptera: in the bees, wasps and flies, the trunk approaching to the figure of a sphere; in the ants, Scoliæ, crane-flies, &c. to that of a cube. The upper part of it in many Ichneumonidæ,[Pg 533] female ants, &c. is very elevated, forming an arch, and sloping towards the abdomen. In general it may be observed with respect to the remaining Orders, that the form of the trunk merges in that of the whole body, the tendency of which is often to a three-sided figure.
III. Proportions.—The trunk is usually longer and larger than the head and longer than the abdomen, but not wider: but there are exceptions to both these rules. In Colliuris, Mantis, &c., it is more slender; and in Atta megacephala and some neuter ants, it is shorter than the head; in Atractocerus, many Staphylinidæ, Phasma, the Libellulina, the Lepidoptera, and various Hymenoptera, it is shorter, and in the Mantidæ more slender than the abdomen. The greatest disproportion between it and the last part is exhibited by the genus Evania, parasitic upon the Blattæ, in which the abdomen appears merely as a minute and insignificant appendage of the trunk. The vertical diameter of this part, almost without exception, is greater than that of either head or abdomen. When we consider that it contains the muscles that move both the organs of flight and the legs, we see clearly the reason why the Creator gave it greater volume.
IV. Composition.—I lately intimated to you that the trunk, though resolvable into three segments, in most cases properly consists of only two primary ones. Whoever examines the perfect insects of every Order, except the Aptera[1591], will find this distinction strongly pointed[Pg 534] out, not only by the different direction of the first pair of legs from that of the two last, but also in a large proportion by a deep incisure; and in all it is further manifested by the anterior segment having a motion distinct from that of the rest of the trunk, and separating readily from it; and this not only where it is large, as in insects that have a thoracic shield, but also in those in which the prothorax is less apparent: whereas the other two pedigerous segments have little or no distinct motion, will not readily separate from each other, and in some cases exhibit no pectoral suture between them. Sometimes, however, these two last segments are more prominently distinguished: in Lytta, Mylabris, and other vesicatory beetles, they are separated below by an incisure, or rather the first or mid-leg segment, is not nearly so elevated as that of the hind-legs. In some ants (Atta Latr.), in the neuters, there is no distinction of segments in the trunk; but in others (Formica Latr.) it follows the general law, and consists of three. In the Arachnida, with the exception of Galeodes, in which the head is distinct, and the three segments of the trunk may be traced, these parts together form only a single segment. Induced by these reasons, I consider the trunk as consisting in general of two primary segments, the manitrunk and alitrunk: the latter resolvable into two secondary ones.
* Manitruncus[1592].—The manitrunk, then, is the anterior section of the trunk, which bears the arms and contains the muscles that move them. This part has free motion, or a motion independent of that of the rest of the[Pg 535] trunk. This indeed seems a necessary result of the direction and uses of the arms. It consists of an upper and lower part—the prothorax and antepectus.
i. Prothorax[1593].—The upper part of the manitrunk in the Coleoptera, Orthoptera, and Hemiptera, is by far the most conspicuous part of the trunk, but in the other Orders it is less so. With respect to it, insects may be divided into two classes—those that have and those that have not a prothorax. In the Coleoptera Order it is remarkable both for size and variations in its shape and sculpture. In the Orthoptera, though less various, it is almost equally conspicuous, especially in Blatta. In the Homopterous section of the Hemiptera, in many genera it has become extremely short; while in the Heteropterous section its dimensions are not much reduced. In the majority of the Neuroptera, likewise, it is comparatively large; in the Libellulina much shorter, and in the Trichoptera and Lepidoptera nearly evanescent[1594].—In the Hymenoptera and Diptera, with very few exceptions, the thoracic shield altogether disappears, at least if I am correct in an idea, which I shall hereafter explain, that the collar usually regarded as the analogue of the prothorax, is really a part of the alitrunk. In these last Orders, though there is no true prothorax, the manitrunk still remains under the form of an antepectus, bearing the fore-legs, and containing the muscles that move them.
The prothorax of insects may in general be considered[Pg 536] with respect to its parts, margin, appendages, shape, sculpture, clothing, and proportions.
1. The prothorax, regarded as a whole, distinct from the antepectus or fore-breast, consists commonly of two pieces—the shield, or upper part[1595], and the ora, or under part[1596]. In the shield you are to observe its apex[1597], base[1598], sides[1599], limb[1600], and disk[1601]. The apex is the part next the head; the base that next the abdomen; the limb the circumference, and the disk the central part. In many Orthoptera and Heteropterous Hemiptera, the shield appears further to consist of two pieces, an anterior and posterior one. The ora is a continuation of the shield below the lateral margin, turned downwards and inwards towards the fore-breast and the legs, but separated from the former in most cases by a suture, as in Carabus L.; and in others merely by an impressed line, as in Blaps F.; but in Curculio and Cerambyx L., &c. there is no ora, the shield being without a lateral margin, and forming one piece with the antepectus. The part we are now considering varies in different genera. Sometimes it is very narrow, as in Scarites; at others very broad, as in Buprestis, Nepa, &c. In Lampyris, except L. italica, and affinities, it projects posteriorly into a lobe or tooth, which forms a right angle with the rest of the ora, and becomes the lower part of the cavity that receives the head; and in Dermestes this part is excavated into an anterior and posterior one which admits the antennæ and arms when folded for repose.
2. The margin of the prothorax is a ridge, either defining[Pg 537] its sides or whole circumference. In many cases this margin is broad and dilated, but in others it is merely a thread or bead that separates the shield from the ora. Though generally terminating the upper surface, it sometimes, as in Staphylinus, dips below it. In many insects, however, as I just observed, the thoracic shield has no lateral margin whatever.
3. Various and singular are the appendages with which the prothorax of numerous insects is furnished. Many of these are sexual distinctions, and have been before described to you[1602]; but there are others common to both sexes, the most remarkable of which I shall notice.—Some are distinguished by a long horn which overhangs the head, as Membracis cultrata, ensata, &c.[1603]; in others it stands upright, as in Centrotus spinosus[1604]; C. Taurus has a pair of thoracic horns like those of a bull, only dorsal[1605]; in Ledra aurita they are flat, and represent ears[1606]; in some species of Tingis (T. Echii, Pyri, &c.) a kind of reticulated hood, resembling lace, is elevated from the anterior part of the prothorax, which receives and shelters the head[1607]. In Centrotus globularis and clavatus F., especially the former, the part in question is armed by a most singular and wonderful apparatus of balls and spines,—in one case standing erect[1608], and in the other being horizontal[1609],—which gives these animals a most extraordinary appearance. In many of the species here quoted[Pg 538] the prothorax is producted posteriorly into a long scutelliform horizontal horn, which more or less covers the wings and abdomen; a circumstance which also distinguishes the genus Acrydium F. (Tetrix Latr.). This horn seems to have been sometimes regarded by Linné and Fabricius as a real scutellum, and sometimes only as a process of the prothorax: but that it is merely the latter will be evident to you, if you examine carefully any insect furnished with this appendage; for if you remove that part, you will discover the true scutellum and other parts of the trunk concealed beneath it. A very remarkable prothoracic appendage is exhibited by some species of Mantis. In general the part we are treating of in this tribe is very slender; but in M. strumaria, gongyloides, &c., it appears dilated to a vast width, and assumes, either partially or generally, a subrhomboidal form; but if it is more closely examined, it will be found that the form of the prothorax is really similar to that of the rest of the tribe, but that this part is furnished on each side, either on its whole length or anteriorly, with a large membranous flat subtriangular appendage resembling parchment[1610]. Perhaps this kind of sail may be useful to the animal in flight. In Prionus coriarius &c. its sides are armed with teeth, and in many Lamiæ, Cerambyces, and other Capricorn beetles, and often in various bugs (Pentatoma Latr.) with sharp fixed spines. But the prothorax has moveable as well as fixed appendages; of this kind are those spines (umbones), whose base is a spherical boss moving in an acetabulum of the thoracic shield of the Capricorn subgenus Macropus Thunb. If I might[Pg 539] hazard a conjecture, I should say that these organs were given to this animal by an all-provident Creator, to enable it to push itself forward, when in the heart of some tree it emerges from the pupa, that it may escape from its confinement. Another kind of moveable appendages are attached to the thorax of Lepidoptera, usually in the form of a pair of concavo-convex scales covered externally with a tuft of hairs[1611]. M. Chabrier, who examined these scales in recent insects, describes them as vesicles, which appeared to him full of a liquid and of air, and capable of being alternately inflated and rendered flaccid; he regards them as accessories to a pair of spiracles, which he looks upon as vocal[1612], opening into the manitrunk just above the insertion of the arms. These organs are quite distinct from the tegulæ that cover the base of the primary wings of insects of this Order[1613], and are what, borrowing a term from Mouffet[1614], I have called in the table patagia, or tippets. Under this head I may include the caruncles at the anterior angles of the prothorax of a genus of beetles with soft elytra, named by Fabricius Malachius. When pressed, says De Geer of these insects, a red inflated soft vesicle, of an irregular shape, and consisting of three lobes, emerges from the thorax and from each side of the anterior part of the abdomen, which re-enters the body when the pressure is removed[1615]. M. Latreille seems to think that these vesicles have some analogy with the poisers of Diptera and[Pg 540] the pectens of scorpions; and that they are connected with the respiration[1616].
4. We are next to say something upon the shape of the prothorax. The forms of the thoracic shield, especially in the Coleoptera, are so various, that it would be endless to aim at particularizing all; but it may be useful to notice a few of the most remarkable. The prothorax of Moluris, a darkling-beetle, approaches the nearest of that of any insect to a spherical form, from its remarkable convexity; in the wheel-bug (Reduvius serratus) it is compressed, and longitudinally elevated into a semicircular serrated crest: it is crested, also, in many Locustæ and Acridæ, in some having two parallel ridges; but, generally speaking, its surface is more depressed. In Necrodes it is nearly circular, in Blatta petiveriana semicircular, in Nilion and some Coccinellidæ crescent-shaped, in Carabus obcordate, in Cantharis and Sagra approaching to a square, in Languria to a parallelogram; in many Cimicidæ, Belostoma, &c., it is triangular, with the vertex truncated; it is trapezoidal in Elater, in Ateuchus rather pentagonal, and exhibiting an approach to six angles in some other beetles[1617]: but the prothorax most singular in form is that of some species of M. Latreille's genus Helæus[1618], as H. perforatus, Brownii, &c.: in these its anterior angles are producted, and curving inwards, lap at the end one over the other, so as to form a circular orifice for the head, which otherwise would be[Pg 541] quite covered by the shield. Thus the upper portion of the eyes can see objects above, as well as their lower portion those below. I might enumerate many other forms, but these are sufficient to give you some notion of the variations of this part.
5. The prothorax is equally various in its sculpture; but since in the Orismological table almost every instance of it has its place, I shall here only notice it as far as it is common to the whole tribes, genera, or subgenera. The Scarabæidæ of Mr. W. S. MacLeay are distinguished by a small excavation on each side of this part, which, as has been before remarked[1619], furnishes an elevated base for an internal process with which the anterior coxæ ginglymate. In Onitis and Phanæus, to these excavations are superadded a pair impressed in the base of the prothorax, just above the scutellum; in Carabus L. a longitudinal channel divides the thoracic shield into two equal portions; and many genera of that great tribe have in addition, at the base on each side, one or two excavations or short furrows. Elophorus F. has on this part several longitudinal channels, alternately straight and undulated. Generally speaking, in Carabus L. the prothorax has no impressed points; but in one or two subgenera of Harpalidæ (Chlænia &c.) it is thickly covered with them. In numbers of Locusta Leach, the part we are considering is what Linné terms cruciate, being divided into four longitudinal portions by three elevated lines, the intermediate one being straight, and the lateral ones diverging from it both at their base and apex, so as to form a sinus or angle[1620]. In certain Acridæ K.[Pg 542] (Locusta F.) there are only two of these lines or ridges, but notched or toothed; and in some of the genus first named only one[1621]; in Locusta Dux and affinities the prothorax has several transverse channels or rather folds[1622], with corresponding ridges on its internal surface.
6. With respect to the clothing of the prothorax, I have not much to say: in Coleopterous insects this part is commonly naked; but in some genera, as Byrrhus, Anthrenus, Dermestes, and many weevils (Curculio L.) it is partially or totally covered with hairs or scales. In the other thoracic Orders it is usually naked, but in some Neuroptera, the Myrmeleonina, &c., it is hairy; and in the Libellulina it is fringed posteriorly with hairs.
7. As to its relative proportions, the prothorax is sometimes rather wider than the rest of the trunk and the head, as in Onitis, Pasimachus, &c.; it is considerably narrower in Collyuris and Odacantha; and of the same width in those Scaritidæ with striated elytra[1623]. Again, it is sometimes of the same width with the elytra, but wider than the head, as in Hydrophilus, Dytiscus, &c.; in some instances it is of the same width with the head, and narrower than the elytra, for instance in Anthia and Brachinus. In most Coleoptera it is longer than the head and shorter than the elytra; but in Manticora, the vesicatory beetles, &c., it is shorter than either. In Gnoma longicollis[1624], it is nearly as long as the elytra; in many Staphylinidæ, Atractocerus, &c., longer; in Phanæus carnifex, bellicosus, &c., it is longer than the elytra and the rest of the body. With regard to itself, it is sometimes[Pg 543] very wide in proportion to its length—Dytiscus, Helæus; at others very long in proportion to its width—Colliuris, Brentus, Mantis, &c. In Flata, and many other Homopterous Hemiptera, it is extremely short; extremely long in Gnoma; in Sagra and Donacia its width about equals its length; in Elater, Dytiscus, and many Heteropterous Hemiptera, it is narrowest before; in Languria it is every where of equal width; in Anthia, Carabus, &c., it is widest before; and, lastly, in the Scarabæidæ MacLeay it is usually widest in the middle.
ii. Antepectus[1625].—The antepectus, as was before observed[1626], in some tribes forms one piece, without any kind of separation, with the prothorax; but very often this is not the case. In Carabus L. it occupies almost the whole under-side of the manitrunk; but in Elater, in which the ora is very wide, the antepectus is merely the middle portion of that part. In Carabus F. &c. between the ora and the base of the arms is a convex triangular piece, distinguished from the rest of the antepectus by a spurious suture; and in Pentatoma and other Heteropterous Hemiptera a similar piece is observable, which terminates in a convex bilobed subtriangular sheath, receiving the base of the clavicle[1627]. This piece seems a prop to that part, and analogous to the scapula of the medipectus and parapleura of the postpectus. I shall say no more upon the antepectus, as it is seldom remarkable. In the mole-cricket, however, one peculiarity distinguishes it: it is in this of an elastic leathery[Pg 544] substance, while the prosternum is hard, resembling a bone. In other instances these parts are both of the same substance.
1. The sternum or breast-bone of insects consists mostly of three distinct pieces; in this resembling the human sternum, which is described by anatomists as composed originally of three bones[1628]. Each of these pieces is appropriated to a pair of legs, and each of them at times has been called the sternum: thus in Elater the prosternum, in the Cetoniadæ the mesosternum, and in Hydrophilus the metasternum, have been distinguished by this name. Our business is now with the first of these pieces, the sternum of the antepectus or prosternum[1629]: this is the middle longitudinal ridge of the fore-breast, which passes between the arms, when elevated, extended, or otherwise remarkable. It is most important in the Coleoptera Order, to which my remarks upon it will be chiefly confined. In these it is sometimes an elevation, and sometimes a horizontal process of the fore-breast. If you examine the great Hydrophilus (H. piceus), at first you will think that there is only a single sternum common to all the legs; but if you look more closely, you will perceive between the head and the arms a triangular vertical process, with a longitudinal cavity on its posterior face, which receives the point of the mesosternum that passes between the arms[1630]: this vertical piece is the real prosternum, and not the other, which really belongs to the alitrunk. In this case the elevation of the prosternum is before the arms; in others it is between them, as you[Pg 545] may see in a Chinese chafer (Mimela K.), which imitates the external appearance of a quite different tribe[1631]; in others again it is behind them, as in most of the Lamellicorn beetles. In the common dung-chafer (Geotrupes stercorarius), it is a hairy process, which, when the head is bent downwards, is received by a deep cavity of the mesosternum. The Dynastidæ MacLeay may always be known by a columnar prosternum rising vertically between the arms and the medipectus. Lastly, in other tribes there is a prosternal elevation both before and behind the arms, as in Cerambyx thoracicus, dimidiatus, and affinities. Of the second description, those that have a less elevated horizontal prosternum, the point in most is to the anus, but in some to the head: thus in Carabus L. it is generally a subspathulate flat piece, the point of which slides over the mesosternum, or covers it; but in Harpalus megacephalus Latr.[1632], one of this tribe, though similarly shaped, its point is to the head. These horizontal prosterna vary in their termination. In that of Carabus L. the apex is obtuse; in that of Elater, above described[1633], and Dytiscus it is acute; in Prionus lineatus, Spencii K., &c., it is bilobed; and in Buprestis variabilis, attenuata, &c., obsoletely trilobed. With regard to the other Orders no striking features of this part are observable, except in some Orthoptera. In Acrida viridissima K. (Locusta F.) it is represented by two long filiform vertical processes; and in Locusta Leach by a single conical horn[1634], mistaken by Lichtenstein for a process of the[Pg 546] throat[1635]. In one instance, Gryllotalpa, this part is a long piece between the arms, shaped like the human thighbone or tibia, being more slender in the middle and widest at the ends, and which is of a much harder substance than the rest of the antepectus, and forms the lower termination of a singular machine which will before long be noticed. In many bugs (Cimicidæ), instead of being elevated, the three portions of the sternum are hollowed out into a longitudinal groove, in which the promuscis when unemployed reposes.
The most conspicuous and remarkable appendages of the manitrunk, are the brachia or arms. I shall not, however, enter into the full consideration of these, as they consist numerically of the same parts, till I treat of the legs in general. Here it will only be necessary to assign my reasons for calling them by a distinct denomination. In this I think I am authorized, not only by the example of Linné, who occasionally found it necessary to do this[1636], and more particularly by the ancient notion that this pair of organs in insects were not to be reckoned as legs[1637], but likewise from their different position and functions. They are so inserted in the antepectus as to point towards the head, whereas the other two pair point to the anus. With regard to their functions, besides being ambulatory, and supporting the manitrunk in walking, they are applied to many other purposes independent of that office,—thus they are eminently the scansory or climbing legs in almost all insects; in most Carabi L., by means[Pg 547] of the notch and calcar[1638], they are prehensory legs; in Scarites belonging to that tribe, the Lamellicorn beetles, and the mole-cricket, they are fossorious legs, or proper for digging[1639]; in Mantis, Nepa, and some Diptera, they are raptorious, or fitted to seize and dispatch their prey[1640]: they are used also by many insects to clean their head, eyes, and antennæ, &c. For many of these purposes they cannot be fit without a structure different from that of the other legs, which renders it a matter of as great convenience in descriptions to speak of them and their parts under different names from those of the legs, as it is of the arms of man; on this account it is that I propose to give to the fore-leg and its part the names by which the analogous parts, or what are so esteemed, in the human species are distinguished;—when spoken of in common with the other legs, they may still be called the forelegs.
** Alitruncus. The alitrunk is the posterior segment of the trunk, which below bears the four true legs, and above the organs for flight or their representatives. In treating of this part we may consider its insertion or articulation, its shape, composition, substance, motions, and organs.
i. With regard to its insertion, or articulation with the manitrunk and abdomen, it may be observed that it is attached to both by its whole circumference by means of ligament; in the Coleoptera, Orthoptera, and Heteropterous Hemiptera being received by the posterior cavity of the prothorax, the shield of which in these Orders, especially[Pg 548] the last, almost covers and conceals it; but in the remaining ones it is merely suspended to it. In the former also, especially in the Coleoptera, it seems more separate and distinct from the manitrunk than from the abdomen, and more independent of its motions than of those of the latter part: but in the Hymenoptera and Diptera its greatest separation is from the abdomen in both respects. In many insects, as in the Lamellicorn beetles, the mole-cricket, &c., the manitrunk terminates posteriorly, drawing a line from the base of the prothorax to the antepectus, in an oblique section; in other tribes, as in the Cerambyx L., the Predaceous beetles, &c., the section here is often vertical, but in the alitrunk the anterior one is always vertical, while the posterior, by which it articulates with the abdomen, in the Orders with an ample thoracic shield, is oblique, so that the pectoral portion is more ample than the dorsal.
ii. As to its composition, the alitrunk is usually much more complex than the manitrunk; for, besides the instruments of motion, it consists of numerous pieces. It may be regarded as formed of two greater segments, the first bearing the elytra, or the primary wings, and the intermediate legs; and the second, the secondary wings and the hind legs.
1. Collare[1641]. The first segment of the alitrunk is the middle piece of the whole trunk, and therefore, when spoken of per se, may be called the meditruncus. It consists primarily of an upper and lower part, which in the table are denominated the mesothorax and the[Pg 549] medipectus. The first piece in the former that requires notice is the collar. I formerly regarded this piece, which is peculiar to the Hymenoptera, Diptera, and one tribe of the Neuroptera, as the representative of the prothorax in the other Orders, and this opinion seems at this time very generally adopted, but subsequent observations have caused me to entertain considerable doubts of its correctness. Many other Entomologists have thought it improper to distinguish these parts by the same name[1642]. Much, however, may be said on both sides of this question, and I shall now lay before you the principal arguments that may be adduced in defence of each opinion, beginning with those that seem to prove that the collar is the analogue of the prothorax. First, then, the collar, like the prothorax, is placed precisely over the antepectus, and being placed in the same situation, on that account seems entitled to the same denomination; especially as in some genera, for instance Chlorion F., it assumes the very semblance and magnitude of a thoracic shield, and is separated from the mesothorax by a considerable incisure. Again, in some cases that have fallen under my own observation, the collar is endued with some degree of motion distinct from that of the alitrunk, since in Pompilus and Chrysis the animal can make the former slide over the latter in a small degree. A third and last argument is, that no prophragm is formed from the collar: insects that have a thoracic shield are generally distinguished by having the anterior margin of the dorsolum deflexed so as to form a septum, called in[Pg 550] the table the prophragm, which enters the chest and separates the cavity of the mesothorax from that of the prothorax; now in Hymenoptera this septum is a process of the piece behind the collar, and excludes it from having any share in that cavity. These arguments at first sight seem to prove satisfactorily the identity of the collar and prothorax. But audi alteram partem, and I think you will allow that the scale containing the claims of the collar to be considered as a piece sui generis, dips much the lowest. And, first, I must observe, that though in Hymenoptera the collar seems to replace the prothorax by its situation, yet it is in fact a part of the alitrunk; for, if the manitrunk be separated from the latter, the collar remains, in most cases, attached to it[1643], while the antepectus and arm, with the ligament that covers its cavity above, the real representative of the prothorax, are easily removed, and this in recent individuals: as a further proof of this, I must request you will examine a neuter Mutilla; you will see that in this the collar is not separated from the alitrunk in any respect, but forms one piece with it, while the antepectus is distinct and capable of separate motion: further, the action of the collar is upon the alitrunk, it being of essential importance in flight, whereas the prothorax is of no other importance than as a counterpoise to that part[1644]. A further argument to prove the distinction of these parts may be drawn from the case of Xylocopa, a kind of bee. In this genus the collar forms a complete annulus or segment of[Pg 551] the body: now, if it really represented the prothorax, the under side of the segment, as in those Coleoptera in which no suture separates the upper from the lower part of the manitrunk[1645], should represent the antepectus, and have the arms inserted in it; but in the case before us there is a distinct antepectus bearing the arms received by the socket formed by this annulus. But the most powerful argument is the fact that some insects have both the prothorax and collar, a circumstance that completely does away every idea of their identity. If you examine the common hornet (Vespa Crabro), or any saw-fly (Tenthredo L.), you will find, as was before intimated, that the real covering of the cavity of the manitrunk is a ligamentous membrane, which properly represents the prothorax. In another genus of the same order (Xiphydria Latr.), the sides of the antepectus turn upwards and nearly form a horny covering distinct from the collar[1646], the ligamentous part being reduced to a very narrow line, and in Fœnus the dorsal fissure is quite filled up, so that in this the manitrunk is perfectly distinct, and exhibits both prothorax and antepectus of the usual substance. In Nomada likewise, N. Goodeniana K. was the species I examined, there is a short minute prothorax besides the collar. Next let us turn our attention to the Diptera; if you examine the common crane-fly (Tipula oleracea), you will find, first, a regular short prothorax, to which the antepectus, with the arms, is attached; and behind this also is a short collar embracing the alitrunk anteriorly. The next insects that I shall mention, as exhibiting both prothorax and collar, are the[Pg 552] Libellulina. These are generally admitted to have the former of these parts[1647], but besides this they have also the latter, which is the most ample and conspicuous piece in the whole trunk[1648]; intervening, as the collar should do, between the prothorax and those parts of the trunk to which the wings are attached. There is one circumstance connected with the subject which should not be overlooked. In the Hymenoptera, usually under a lateral process of the posterior part of the collar, is a spiracle or respiratory apparatus; in the Diptera there is also one, though not covered by the part in question, in the same situation; now this you will find precisely so situated with respect to the second piece in the thorax of Tipula oleracea, proving that this piece is the real representative of the collar. Enough, I think, has been said to satisfy you that I have not changed my sentiments on this subject upon slight grounds. Probably traces of the part in question might be detected in the thoracic Orders in general, in connexion with some vocal or respiratory organ[1649]; but having had no opportunity,[Pg 553] by an extended examination of living subjects, to verify or disprove this suspicion, I shall merely mention it, and conclude this head by observing, that the collar varies most in the Hymenoptera order, and that its most remarkable form is in Vespa, Cimbex, Dorylus, &c., in which it bends into an ample sinus that receives the dorsolum[1650].
2. Dorsolum[1651]. Where there is no apparent collar, the dorsolum (dorslet) is the first piece of the mesothorax, and where there is one, the second; it bears the elytra or other primary organs of flight. It varies in the different Orders, particularly with respect to its exposure. In Coleopterous insects it is most commonly, but not invariably[1652], covered entirely by the shield of the prothorax, the scutellum alone being visible; as it is also in the Orthoptera (with the exception of Mantis and Phasma, in the first of which it is partially, and in the latter intirely exposed), and the Heteropterous, and most of the Homopterous section of the Hemiptera. The scutellum is likewise covered in Gerris, Hydrometra, and Velia, and the whole of the back of the alitrunk by a process of the prothorax in Acrydium F., Centrotus, &c. But in the remaining Orders, and the tribe of Tettigonia in the Homopterous Hemiptera, the dorsolum is not hidden by the thoracic shield. It is usually less elevated than the scutellum; in Necrophorus, and some other beetles, however, the latter is most depressed. With regard to its substance, it is generally not so hard[Pg 554] and rigid as the scutellum, but in most Coleoptera harder than in the other Orders in which it is covered; in the Hemiptera, except in Tettigonia, it approaches to membrane. As to shape and other circumstances, it varies in the different Orders. In the beetle tribes it has generally a sinus taken out of its anterior margin, and it approaches more or less to a trapezium; in Blatta it is transverse and somewhat arched; in Gryllotalpa it is nearly square, and distinguished besides on each side by a minute aperture, fitted with a tense membrane, which perhaps covers a respiratory apparatus. In the locusts it is more or less triangular, and in Mantis and Phasma long and slender. In the Hemiptera the dorsolum appears to consist of several pieces, variously circumstanced, separated by sutures, corresponding with which are as many ridges on the inside of the crust[1653]. In the Libellulina it is rhomboidal[1654]; in Panorpa nearly hexagonal; in the Ephemerina it is ample and oblong; in Sialis and the Trichoptera this part is represented by three subtriangular pieces, the scutellum constituting a fourth, with the vertices of the triangles meeting in the centre[1655]; in the Lepidoptera the part in question is large, and receives the scutellum into its posterior sinus[1656]. The Hymenoptera usually exhibit a very ample dorsolum, mostly subtriangular with the vertex rounded or truncated, and pointing in some (Vespa L.) to the head[1657], and in others (Apis) to the anus; in the Diptera, except in Tipula, the parts of the mesothorax are not separated by any suture, but only indicated by impressed lines or channels;[Pg 555] in the genus last mentioned, however, the dorsolum is distinct, subrhomboidal, and received by an angular sinus of the scutellum, which last, I think, is not the part that has usually been regarded as entitled to that denomination; for this opinion I shall soon assign my reasons.
3. Scutellum[1658]. Some writers on the anatomy of insects, looking, it should seem, only at the Coleoptera and Orthoptera, have regarded the dorsolum and scutellum as forming only one piece[1659], and others have affirmed that the Lepidoptera and subsequent Orders have no scutellum[1660]. But as we proceed in considering the scutellum in all the Orders, we shall see that both these opinions are founded on partial views of the subject, and that all winged insects have a scutellum, more or less distinctly marked out or separated from the dorsolum. In the Coleoptera the scutellum is usually the visible, mostly triangular, piece that intervenes between the elytra at their base[1661], and which terminates the dorsolum. Some Lamellicorn beetles, &c. (Scarabæidæ MacLeay) are stated not to have the part in question (exscutellati): but this is not strictly correct, for in these cases the scutellum exists as the point of the dorsolum covered by the prothorax, though it does not intervene between the elytra: in others of this tribe, as Cetonia chinensis, bajula, &c., it separates these organs at their base, though it is covered by the posterior lobe of the prothorax: in Meloe F., the elytra[Pg 556] of which are immoveable, there seems really to be no scutellum. Generally speaking, as was lately observed, but not always, it is distinguished from the dorsolum by being more elevated: this is particularly conspicuous in the genus Elater, in which it is a flat plate elevated from the dorsolum by a pedicle; in Sagra the latter part is horizontal, while the scutellum is vertical: and even in cases where the distinction is not so striking, these parts are separated either by a line, or some difference in their sculpture and substance. In this Order this part varies greatly, and often in the same tribe or genus, both in size and shape; being sometimes very large[1662], and sometimes very minute; sometimes very long, and sometimes very short; sometimes nearly round, at others square; now oval or ovate, heart-shaped, triangular, acuminate, intire, bifid, &c. In the Orthoptera, though less conspicuous, it still is present as a triangular elevation of the middle of the posterior part of the dorsolum, with the vertex either pointing towards the head, as in Blatta, or towards the tail, as in Locusta Leach[1663]. In the Heteropterous section of the Hemiptera (which, in columns of Mandibulata and Haustellata, appear to bear the same reference to the Coleoptera, that the Hymenoptera do to the Diptera, and the Homopterous Hemiptera to the Orthoptera[1664]) the part we are considering is mostly very large and conspicuous, quite distinct from the[Pg 557] dorsolum, and in some (Tetyra F.) covering the whole abdomen, as well as the Hemelytra and the wings; it is most commonly, as in the Coleoptera, obtriangular[1665], but in the last-mentioned genus it often approaches to a pentagonal shape. Though usually so striking a feature in this tribe, in the aquatic bugs (Gerris &c.) it is covered by the prothorax. In some species of Reduvius F. (R. biguttatus, mutillarius, lugens, &c.) it is armed with one or more dorsal or terminal spines. In the Homopterous section, where the dorsolum, as in Tettigonia F., is not covered by the prothorax, the scutellum, which is merely a continuation of that part, bears some resemblance to a St. Andrew's cross, and terminates in a fork[1666]; in Fulgora, in which it is partly covered, it is merely the triangular point of the dorsolum: in the Cercopidæ, &c., whose dorsolum is wholly covered, the triangular scutellum is distinct from it; in Centrotus, Darnis, and Membracis, in which the prothorax is producted, and covers the abdomen more or less, the scutellum is a short transverse distinct piece. In the Lepidoptera, from the difficulty of abrading sufficiently the scales and hairs without injury, it is difficult to obtain a correct idea of the part in question; in the cabbage butterfly (Pieris Brassicæ) it appears to be triangular: in the humming-bird hawk-moth (Macro-glossum Stellatarum) it approaches to a rhomboidal shape[1667]; and in the eggar-moth (Lasiocampa Quercus) it is completely rhomboidal. In the Libellulina, in the Neuroptera Order, it seems to be represented by the posterior point of the dorsolum, which terminates[Pg 558] in something like a St. Andrew's cross[1668]. In most of the other tribes of this Order the scutellum is a triangular piece, with the vertex to the head, received between two pieces of the dorsolum; in Psocus it is nearly like that of Tettigonia before described. In the Hymenoptera the scutellum is separated from the dorsolum, which it often embraces posteriorly, as the collar does in front, by a suture; it varies occasionally in shape in the different tribes, most commonly it is crescent-shaped, but in many Ichneumonidæ and others it is triangular[1669]; in the hive bee, &c., it overhangs the succeeding piece of the alitrunk; in Melecta, Crocisa, &c., it is armed with a pair of sharp teeth[1670]; in others (Oxybelus uniglumis, &c.) with one or more spines, and in some with a pair of long horns[1671]. Before I describe this part in the Diptera, it will be proper to assign my reasons for considering a different piece as its representative, from what has usually been regarded as such, and which at first sight seems the analogue of what I admit to be the scutellum in the Hymenoptera. The dorsolum, and its concomitant the scutellum, belong to the first pair of the organs of flight, which are planted usually under the sides of the former, and in the case of wings, by their Anal Area, connected either mediately or immediately with the latter. Now, if you trace the sides of the piece that I have considered as the part in question in Hymenoptera, you will find that they lead you not to the base of the lower but to that of the upper wings[1672], and in the saw-flies (Tenthredo L.)[Pg 559] you will see clearly that the Anal Area of these wings is attached to a process of it, a proof that it belongs to the mesothorax, or region of that pair. But in the Diptera, the part that has been usually called the scutellum is not at all connected, either by situation or as a point of attachment, with the wing itself, but with the lower valve of the alula, which is with reason thought to be the representative of the secondary wing of the tetrapterous Orders. You may see this even in the common crane-fly (Tipula), in which there is a real alula, connected by means of a lateral process, terminating in ligament, with this supposed scutellum. If you examine further the same insect, you will easily find what I regard as the true one in the bilobed piece which receives the dorsolum, situated between the wings, and to the sides of which they are attached. In Asilus, Tabanus, &c., this part is transverse, and only distinguished on each side by an oblique impressed line; in the Muscidæ it is square, and marked by a straight transverse one.
4. Frænum[1673]. This appendage to the scutellum and dorsolum varies considerably in the different Orders, and in many cases, as you will see, is a very important part, being the process by which the former is mostly connected with the elytra or upper wings. In the Coleoptera, the elytra of which are nearly stationary in flight, and therefore less require any counteraction to prevent their dislocation, this part is commonly merely a process or incrassation of the under margin of the scutellum, which towards the base of the dorsolum is dilated to form the socket for the elytra. Its use as a countercheck[Pg 560] in this Order is best exemplified in the common water-beetle (Dytiscus marginalis). This at the inner base of the elytra has a membranous fringed alula resembling those of Diptera; to the lower fold of this the extremity of the frænum is attached, which forms a right angle with the scutellum, and the upper fold is attached to the base of the elytrum[1674]. The object of this appendage is probably to prevent the dislocation of these organs, which seems to indicate that they are used more in flight than those of other beetles. The Blattæ also, in the next Order, have a winglet attached to the anal area of the tegmina. The frænum, as in the preceding Order, lies under the margin of the scutellum and dorsolum, but which here forms one uninterrupted transverse line; it is nearly vertical, and is attached to the alula. The structure is not very different in the other Orthoptera[1675], but the frænum is surmounted or strengthened by one or two ridges; in Mantis it runs from the scutellum in an angular or zigzag direction—but in all it is attached immediately to the tegmen. In the Heteropterous Hemiptera it is represented by the narrow bead adjacent to the scutellum on each side[1676], which dilates into a flat plate as it approaches the Hemelytrum, with the Anal Area of which it is connected. But the Homopterous section of the Order in question furnishes examples of the most remarkable structure of this countercheck, which proves that it is really, what its name imports, a bridle. If you examine the great lanthorn-fly (Fulgora laternaria), or any species of Tettigonia, &c., you will find adjacent to the scutellum[Pg 561] or parallel with it, on each side a flat plate; and from the angle of that part in the first case, and from one of its processes in the last, you will further perceive a ridge or nervure which runs along this plate, in one forming an angle, and in the other being nearly straight, to the base of the tegmen, where it becomes a marginal nervure to a membrane that is attached to the posterior part of the base of the Anal and Costal Areas; and that this marginal nervure, like a trachea, consists of a spiral thread, or rather of a number of cartilaginous rings connected by elastic membrane[1677], and consequently is capable of considerable tension and relaxation, as the tegmen rises and falls in flight. In the Lepidoptera it appears to be a short piece overhung by the scutellum, which as it approaches the base of the wing is dilated. In the Libellulina, to go to the Neuroptera, it has the same kind of elastic nervure connected with the Anal Area of the wing which I have just described in the Homopterous Hemiptera; another nervure, in Æshna at least, appears to diverge upwards from the scutellar angle to the Intermediate Area[1678]: a structure little different distinguishes the rest of the Neuroptera, and even the Trichoptera. In the Hymenoptera this part varies somewhat; in the majority perhaps of the Order, as well as in the Diptera, it appears to be merely the lateral termination of the scutellum where it joins the wing; but in some tribes, as in Tenthredo L. (especially Perga Leach), Sirex L., and the Ichneumonidæ, a ridge, and sometimes two, runs from the scutellum to the wing; the upper one, where[Pg 562] there are two, as in Perga, being the stoutest, and connecting with the Costal Area, and the lower one with the Anal.
5. Pnystega[1679]. We learn from M. Chabrier, that in the common dragon-fly, a space, consisting of three triangles, which immediately succeeds the frænum, affords attachment to no muscles, but merely covers aërial vesicles[1680]. This is the part I have called the pnystega[1681]. An analogous piece may be discovered in Phasma and Mantis in a similar situation; but I cannot trace it in Locusta Leach, or in the other Orders.
Having considered the parts that constitute the mesothorax, we will next say something upon those, as far as they require notice, that compose the medipectus or mid-breast. But first I must observe, in general of the medipectus and postpectus taken together, or the whole underside of the alitrunk, that though usually they are in the same level with the antepectus or under side of the manitrunk, yet in several instances, as the Scarabæidæ MacLeay, the Staphylinidæ, &c. they are much more elevated than that part; they are also usually longer, very remarkably so in Atractocerus, but in Elater sulcatus and many others they are shorter. These parts are also commonly rather more elevated than the abdomen,—much so in some, as Molorchus; but scarcely at all in others, as Buprestis, the Heteropterous Hemiptera, &c. In some of the latter (Tetyra F.) the abdomen seems the most prominent. Another observation relating to[Pg 563] this part must not be omitted, namely, that though in many cases the medipectus and postpectus are perfectly distinct and may be separated, yet in others, as for instance the Lamellicorn beetles, the Hymenoptera and Diptera, &c., no suture separates them; so that though the upper parts, the mesothorax and metathorax, are separable, the lower ones just named are not so.
6. Peristethium[1682]. The first piece of the medipectus is what I have called, after Knoch, the peristethium[1683]. This immediately follows the antepectus; on each side it is limited by the scapulars, and behind by the mid-legs and mesosternum. Its antagonist above is usually the dorsolum. In the Coleoptera Order it varies occasionally, both in form and magnitude, but not so as to merit particular notice, except that both are regulated by the scapulars—if these are small, the peristethium is ample; and, vice versa, if they are large it is small. In all the following Orders, except the Hymenoptera, it is equally inconspicuous, but in them it is often more remarkable. I have a Brazilian species of Cimbex (C. mammifera K. MS.) which appears undescribed, in which this part swells into two breast-like protuberances, terminating posteriorly in membrane, as if it had separate motion: in the golden-wasps (Chrysis L.) it is anteriorly concave to receive the coxæ of the mid-legs; and in Stilbum, of the[Pg 564] same tribe, it is armed with one or more conical obtuse teeth.
7. Scapularia[1684]. The scapulars are situated between the coxæ of the mid-legs and the base or axis of the upper organs of flight, and they seem to act as a fulcrum to each. In the Coleoptera Order they are most commonly quadrangular or subquadrangular, often divided diagonally, and sometimes transversely, by an impressed line; the posterior part, which is usually the most elevated and often has an uneven angular surface, is that which intervenes between the coxæ and elytra: where the former are short, as in the Capricorn beetles, the scapulars are long; and where they are long, as in the Petalocerous ones, the latter are short. The anterior part is that which forms the lateral limit of the peristethium, upon which it often encroaches: this part, in conjunction with the dorsolum above, and the last-named part below, forms the kind of rotula that plays in the posterior acetabulum of the manitrunk, as the head does in the anterior one. In the flower-chafers (Cetonia F.) the scapulars are very thick and elevated, and interpose between the posterior angles of the prothorax and the shoulders of the elytra, which is one of the distinguishing characters of that tribe: in this case the lower angle of the scapular connects with the coxa of the mid-leg, and the upper angle with the axis of the elytra; and the most elevated and thickest part of the scapular is about midway between the two. This robust structure seems to indicate that the scapular has to counteract a powerful action both of the leg and elytrum. In the Orthoptera the scapulars[Pg 565] are usually divided into two parallel pieces, corresponding probably, though more distinct, with the two parts lately noticed of those of the Coleoptera: the upper side of the socket of the mid-leg is common to the base of both these pieces, but the articulation of the tegmen is chiefly with the anterior one. In the grasshoppers, locusts, &c. (Gryllus L.) in which tribe this leg is nearly opposite to that part, the scapular inclines but little from a vertical position[1685]; but in the praying-insects (Mantis), spectres (Phasma), and cockroaches (Blatta), in which the insertion of the mid-legs is behind that of the tegmina, it is nearly horizontal. In the Heteropterous Hemiptera the anterior part of the scapular is covered by the antepectus, and separated by a ridge, more or less pronounced, from the open part; the whole is of an irregular shape, and nearly parallel with the parapleura. In the Homopterous section it likewise consists of two pieces, and sometimes of more. Thus in Tettigonia F. it is bilobed, and between it and the coxa two small pieces are inserted[1686]. In some others, Iassus Lanio F., &c., it is not very unlike the scapular in Coleoptera, being subquadrangular and divided diagonally. In the Neuroptera this part and the parapleura are parallel, and placed obliquely[1687]. In the common dragon-fly (Æshna viatica) the former forms nearly a parallelogram[1688], which is not divided by any ridge or channel, but its lower half is separated into two unequal parts by a black longitudinal line, opposed to which on the inside is a ridge. The mid-leg in these is connected with the scapular by the intervention of a[Pg 566] triangular transverse anterior piece, which in fact seems only marked by a black channel, to which also interiorly a ridge is opposed[1689]. In the rest of the Order it is divided longitudinally into two parallel pieces. In Panorpa the posterior piece is longer than the anterior and props the coxa behind; in Myrmeleon and Perla, &c., it appears to consist of three pieces. I have not been able to obtain a clear idea of them in the Lepidoptera, except that they have more than one piece. Hymenopterous and Dipterous insects for the most part have no scapular distinct from the peristethium; but in Cimbex, Perga, and other saw-flies, it seems represented by its posterior depressed and sometimes membranous part: in Vespa, &c. a small subtriangular piece, just below the base of the upper wing, is probably its analogue[1690].
8. Mesosternum[1691]. The central part of the medipectus, or that which passes between the mid-legs when elevated, protended, or otherwise remarkable, is called the mesosternum or mid-breast-bone. In the Coleoptera Order it exhibits the most numerous variations, and is usually the most strongly marked of any of the three portions of the sternum, affording often important characters for the discrimination of genera and subgenera. It may be said to be formed upon three principal types—the first is, where it is a process of the posterior part of the peristethium, and points towards the anus or the head;—the second, where it is a process of the anterior part of the mesostethium, and points only towards the head: in this case there is no suture to separate the[Pg 567] medipectus from the postpectus;—the last type is where it is a ridge formed by a process both of the peristethium and mesostethium meeting between the legs; an example of this you will see in the common dung-chafer (Geotrupes Latr.). Upon the two first of these cases I shall offer a few remarks;—the last affording no variation need only be mentioned.
If you examine the terrestrial Predaceous beetles (Cicindela and Carabus L.) you will find that the peristethium is usually flat, terminating towards the postpectus in a kind of fork, the sinus of which receives the anterior point of the mesostethium—this is the mesosternum; but in the aquatic insects of this tribe, at least in Dytiscus marginalis, &c. the structure at first sight seems different, for apparently the prosternum is received by the anterior fork of the mesostethium; but if you proceed to separate the manitrunk from the alitrunk, you will find that the true mesosternum of the usual form is quite covered by this point, which curves towards the breast, is longitudinally concave to receive the point of the prosternum, and permit its motion in the groove. In some Heteromerous beetles, as the Helopidæ, &c. this part is anteriorly bilobed, so as to form a cavity which receives the point of the prosternum when the head is bent down: in Helops nitens (Tenebrio Oliv.) this sinus represents a crescent; in Cistela Ceramboides it is shaped like the Greek letter γ; in the Lady-bird (Coccinella L.) it assumes nearly the shape of a Saint Andrew's cross; in Spheniscus K.[1692] the mesosternum is wide, concave and wrinkled, with an anterior and posterior sinus; while in[Pg 568] the analogous genus Erotylus[1693] it is convex anteriorly, and posteriorly more or less rounded; in Doryphora it is a long, robust, subconical horn, often standing at an angle of about 45°, overhanging the prosternum.
In the genus last named, though its mesosternum in its direction and appearance resembles that of many Petalocerous beetles, yet it is separated by an evident suture from the mesostethium; but in the last-mentioned tribe its representative is a process of the latter part: yet as the peristethium and mesostethium are separated by no suture, though in some cases a transverse channel, and in others merely a coloured line, marks the point where they may be considered as soldered together, in these cases the mesosternum may perhaps be said to be common to both. In this great family, which includes within its limits some of the most singular and wonderful in their structure and armour, as well as some of the most brilliant and beautiful of the beetle tribes,—the part in question, in a vast number of cases, will enable the Entomologist satisfactorily to trace its numerous groups, not only where it rises or stretches out into a horn or ridge, but even often where it is merely a flat space between the mid-legs. I shall notice some of its most striking variations in this tribe. In Phanæus festivus, and in Macraspis and Chasmodia MacLeay, it is elongated horizontally, with the apex curving upwards; in[Pg 569] Anoplognathus it is horizontal, straight, and figures an isosceles triangle; in Cetonia suturalis, vitticollis, &c. it is very long, passing between the arms and nearly reaching the head; in C. marmorea, Lanius, &c. it is a lofty, robust, conical prominence; and in many Rutelidæ, especially those with striated elytra, it is pyramidal or four-sided; it varies also in its termination, particularly in the Cetoniadæ; and even where there is little or no elevation of it, as in the Scarabæidæ MacLeay, it is often terminated anteriorly by lines that vary in their angle or curvature. The genus Copris, as restricted by Mr. W. S. MacLeay, may from an inspection of this circumstance be divided into several families. Thus in C. Molossus and affinities its termination represents the letter ⅄ reversed, or a triangle surmounted by a mucro; in C. orientalis, &c., it ends in an acute-angled triangle; in C. lunaris, &c. in an obtuse-angled one; and in C. Iacchus, &c., in the segment of a circle.
The part we are considering is not so important in the other Orders. In the Orthoptera, however, it is occasionally remarkable. In Acrida viridissima (Locusta F.) attached to the anterior margin of the peristethium are two long triangular pieces which appear to represent this part; in the kindred subgenus, Conocephalus[1694], it is a single piece bifid at the apex; in Gryllotalpa it is a very elevated hairy ridge; and in Locusta Leach, it is a flat anterior process of the mesostethium. In the Heteropterous Hemiptera this part is often merely a portion of the channel in which the promuscis reposes; but sometimes, as in Edessa F., it is an elevated piece varying[Pg 570] in its termination. In the remaining Orders, as far as I have had an opportunity to examine them, it can scarcely be said to exist separately from the medipectus, except that in Tipula Latr. a bipartite subtriangular membranous piece seems to be its analogue.
We are now to consider the last segment of the alitrunk, which, as a whole, may be denominated the potruncus; it bears the second pair of the organs of flight, and the last pair of legs. The upper side of this is the metathorax, and its lower side the postpectus.
9. Postdorsolum[1695]. The first external piece of the metathorax is the postdorsolum, which presents itself under very different forms and circumstances in the different Orders. In the Coleoptera it is intirely covered by the dorsolum and scutellum; it is generally more or less of a membranous substance, or partly membranous and partly corneous, which enables it to yield more to the action of the wings in flight; it is usually an ample transverse piece with tumid sides[1696]; but in the Scarabæidæ MacLeay, it is short though very wide; and in Cychrus, and probably other apterous beetles, it is extremely minute and almost obsolete. In the Orthoptera Order, I observe once for all, the part in question, as well as the postscutellum and postfrænum are mere counterparts of the dorsolum, scutellum, and frænum, except that in some cases they are larger[1697]. In the Heteropterous Hemiptera at first sight it would appear that all the parts of the metathorax were altogether wanting or absorbed[Pg 571] in the ample scutellum; but if you remove this with care, you will find under it their representatives, its lower surface being hollowed out to receive them. The postdorsolum appears in these as a transverse obtusangular band; in the Nepidæ, Notonectidæ, &c. the three parts of the metathorax seem united into a single plate, emerging laterally from under the scutellum below the frænum; in which, however, some traces of a distinction between them may be discovered. In the Homopterous section the Fulgoridæ exhibit these pieces very distinctly, covered only at the base by the mesothorax: but in Tettigonia they are not so easily detected; they exist however as a narrow strip or band, almost concealed by that part. As to the Lepidoptera Order, in Pieris Brassicæ at least, the postdorsolum is represented by a pair of nearly equilateral triangles whose vertexes meet in the centre of the metathorax, and between which and the scutellum is a deep cavity; but in Macroglossum Stellatarum and Lasiocampa Quercus, there appears to be also a central transverse piece between them. In the Neuroptera there is no material or striking difference between the parts of the mesothorax and metathorax[1698]. In the Hymenoptera more variety occurs in this part. In the saw-flies, &c. (Tenthredo L.) the postdorsolum is a transverse piece covered by the scutellum; in the Ichneumonidæ it is smaller, but not covered; in the Vespidæ it is apparent, transverse, and with the postscutellum obtusangular[1699]; in Apis it is overhung by the scutellum. The Diptera exhibit some variations in this part. In Tipula it consists of three pieces placed transversely, the central one[Pg 572] quadrangular, and the lateral ones roundish; in the Asilidæ and most others of this Order, with the postscutellum, it forms a segment of a circle[1700], sometimes armed with a pair of spines, as in Stratyomis F., and is what has been usually regarded as the real scutellum, though, as I have endeavoured to show, not correctly[1701].
10. Postscutellum[1702]. The postscutellum bears the same relation to the postdorsolum that the scutellum does to the dorsolum, but it is seldom, if ever, a distinct piece. In the Coleoptera it is represented by the longitudinal narrow channel that terminates the postdorsolum towards the anus[1703]: this usually figures an isosceles triangle with the vertex truncated or open; but in Copris the triangle is equilateral. In the other Orders it is little more than the central posterior point of the postdorsolum[1704].
11. Postfrænum[1705]. The part now mentioned is much more important than the preceding one, and must not be passed over so cursorily. In the Coleoptera it usually presents itself under the form of two large and usually rather square panels, the disk of which is convex, but the rest of their surface unequal, which are situated one on each side of the postscutellum[1706]; under the anterior outer angle of these is the socket or principal attachment of the secondary wings, and their basal margin is attached to their outer side; posteriorly behind the vertex of the postscutellum the postfrænum is crowned with a ridge or bead, below which it descends vertically or obliquely to the abdomen; this ridge often turns upwards,[Pg 573] and proceeds towards the middle of the basal margin of the wing. In the Petalocerous beetles the part in question is usually more or less hairy; but in many others, as the rose-scented Capricorn (Callichroma moschatum), &c. it is naked. At its side you will commonly observe several plates and tendons (osselets Chabr.) connected inter se and with the base of the wing by elastic ligaments, which are calculated to facilitate the play of those organs. In the Orthoptera, Neuroptera, and Homopterous Hemiptera, the postfrænum does not differ materially from the frænum[1707]. In the Heteropterous section of the last Order it is usually a transverse ridge terminating the postdorsolum, with a bifurcation where it unites with the wing; but in Tetyra F. (at least so it is in Tetyra signata,) it is a nearly vertical piece, marked in the centre with an infinity of very minute folds, which probably by their alternate tension and relaxation let out and pull in the wings. Amongst the Lepidoptera it is not remarkable. In the Hymenoptera Order it is mostly represented, I think, by a double ridge or fork, sometimes however obsolete, but very conspicuous in the saw-flies, which laterally terminates the postdorsolum; the upper branch, usually the thickest, going to the anterior part of the base of the underwing, and the lower one to the posterior. You may observe something similar in the crane-flies (Tipula Latr.) and Asilidæ. A tendon proceeding from the point of the postscutellum forms a fork near its end, the upper branch of which connects with the anterior and the lower with the posterior valve of the winglet; the structure is a little, but not essentially, different in other Diptera.
12. Pleura[1708]. By this name I would distinguish the part which laterally connects the metathorax and postpectus. It includes in it the socket of the secondary wings. In the Coleoptera this is a two-sided piece lying between the postfrænum and the parapleura, with the upper side horizontal and the lower vertical[1709]—a tendon usually proceeds from its anterior extremity to the base of the wing. In the Orthoptera, Neuroptera, and other Orders, it is merely the longitudinal line of attachment of that part; but in the genus Belostoma Latreille, related to the water-scorpion, it presents a peculiar structure, being a deep channel or demitube, filled at its posterior extremity by a spiracle and its appendages[1710].
13. Metapnystega[1711]. This part, although in the table I have placed it as an appendage of the pleuræ, is not always confined to them, as you will soon see. It either covers aërial vesicles, or is the seat of a spiracle. In the Order Coleoptera it is of the former description. If you examine the metathorax of the common dung-chafer (Geotrupes stercorarius), in the horizontal part of the pleura you will see a sublanceolate or subelliptical rather membranous silky tense plate, with its point towards the head,—this is the part we are considering; something similar you will find in most beetles; but in some, as Callichroma moschatum, it is less conspicuous. This part, as far as I have observed, is not so situated in any other Order, except in some Heteropterous Hemiptera: in Belostoma the channel lately mentioned is filled up at its posterior end by a red organ with an anterior vertical fissure, terminating[Pg 575] behind in a conical bag: in Notonecta the pleura has something of a plate like that of Coleoptera, but of a horny substance. In the Orthoptera and Neuroptera this part changes its situation, if it be indeed synonymous; and as the pnystega follows the frænum, so the metapnystega succeeds the postfrænum. In the Libellulina M. Chabrier found that this as well as the other covered aërial vesicles[1712], and it probably does the same in the other cases in which it occurs. In Mantis and Phasma in the Orthoptera it is very minute; but in Locusta Leach, it is more conspicuous under the form of a tense membrane, the surface of which is depressed below that of the abdomen: in Acrida viridissima K. it fills the sinus of the postfrænum, and is vertical, as it is in Æshna. It is worthy of remark that this piece bears some analogy to that below the ridge of the part just named in Coleoptera, which descends either vertically or obliquely to the abdomen[1713]. A similar space, though often nearly obsolete, may be seen in the Hemiptera and Lepidoptera. But the Orders in which this part is most conspicuous are the Hymenoptera and Diptera, and in these its aërial vessels are connected with a spiracle. In Tenthredo L. and Sirex L., what Linné named grana, from their situation, should be regarded as belonging to the pnystega, and whether there is any part representing the metapnystega I am not quite satisfied; perhaps the membrane at the base of the abdomen in Tenthredo, and the bipartite piece, apparently its first segment, in Sirex[1714], may be its analogues: but in the great majority of the Order, the convex or flat piece that[Pg 576] intervenes between the postdorsolum and its adjuncts and the abdomen, and which bears a spiracle on each side, is the metapnystega[1715]. This part is often remarkable, not only for its size, but for the elevated ridges that traverse it, as in Ichneumon, Chlorion, &c. In the last genus it is of a pyramidal shape, with the anterior part horizontal and the posterior vertical; it is altogether vertical in Vespa, Apis, &c. Amongst the Diptera, in Tipula it is nearly horizontal, and shaped like a cushion; but in general in this Order it is vertical, and concealed under the postdorsolum[1716].
We are now to consider the parts that constitute the postpectus or under-side of the metathorax, and which bears the posterior pair of legs.
14. Mesostethium[1717]. This part in Coleoptera is terminated anteriorly by the peristethium, scapulars, and mesosternum, laterally by the parapleuræ[1718], and behind by the coxæ of the posterior legs[1719], which generally are inserted transversely between it and the abdomen. It is commonly very wide; but in Dytiscus L., Carabus L., &c., in which the coxæ and parapleuræ are dilated, it is proportionally reduced: its length is regulated by the distance of the intermediate and posterior legs; where these are far asunder, as in the rose-scented Capricorn (Callichroma moschatum), &c. it is long: but where they are near each other, as in the Scarabæidæ MacLeay, it is short; its width, however, generally exceeds its length. In shape it is generally subquadrangular[1720], though sometimes[Pg 577] rhomboidal, and other forms of it occur. Between the hind-legs it generally terminates in a notch or bifurcation distinct from the metasternum, as in Hydrophilus, &c.; in Hister there is no notch, and in many Scarabæidæ it projects between the hind-legs in a truncated or rounded mucro; in the Vesicatory beetles, Meloe L., it is more elevated than the medipectus, towards which it descends almost vertically; in Dytiscus L., Carabus L., &c., this part is usually divided into two by a transverse sinuous channel, and in Elater by a longitudinal straight one. In many Orthopterous genera, Gryllotalpa, Acrida K., Locusta Leach, &c., the mesostethium consists of two pieces[1721]. It is remarkable that in many of these genera, in this part, as likewise in the medipectus and antepectus, are one or more perforations which appear to enter the chest, the use of which I shall explain hereafter. In the Libellulina, as I shall soon have occasion to shew, there is a peculiar arrangement of the legs and wings, in consequence of which this part is placed behind the posterior ones. In the remaining Orders, the mesostethium, though it exists, exhibits no peculiarities worthy of particular notice, except in some Aptera and Arachnida: thus, in Nirmus Anseris it is terminated posteriorly by a pair of transverse membranous appendages which cover the base of the posterior coxæ; in Scorpio it consists of two pieces, the pectines[1722] being attached to the sides of the posterior one.
15. Parapleura[1723]. The parapleura, speaking generally, is that piece of the postpectus which, intervening between the pleura, mesostethium, and scapulars, is attached[Pg 578] by its posterior extremity to the coxæ of the hind-legs; by means of the pleura, from which it does not appear to be separated by any suture, it connects the secondary or under-wings with the hind-legs, as the scapular does the primary ones with the mid-legs; so that the direction of the parapleura depends upon the relative situation of the legs and wings. In Coleopterous insects its direction is horizontal, it being generally a narrow subquadrangular piece that runs straight from the posterior coxæ to the scapular[1724], and usually divided into two unequal portions by an elevated or impressed line. In the palm-weevil (Calandra Palmarum) this part is wider than usual; in Dytiscus marginalis,—in which genus, as likewise in Carabus L., the coxæ are incapable of separate motion,—it is nearly a right-angled triangle, and is divided longitudinally into two unequal portions. In the Orthoptera Order this part usually consists of two equal portions, and its direction is sometimes nearly horizontal, as in Mantis and Phasma; sometimes forming an angle with the horizon, as in Blatta; and sometimes nearly vertical, as in Locusta Leach. In the two first cases the wings are before the legs, and in the last their position is over them. In the Heteropterous Hemiptera it is parallel with the scapular, is divided into two unequal portions, and its direction is more or less inclined to the horizon[1725]. As to the Homopterous section—in Fulgora it is of a very irregular shape with an angular surface, and its direction from the leg to the wing is first nearly vertical and then horizontal: in Tettigonia it is almost vertical, and consists of two nearly equal portions. To[Pg 579] come to the Neuroptera—in the Libellulina it consists of two pieces, like those of the scapulars, but smaller[1726], and its inclination is towards the head: in Panorpa also it resembles the scapulars both in form and other circumstances[1727]. In the remaining Orders it exhibits no very remarkable features.
16. Metasternum[1727]. The central part of the mesostethium when elevated or porrected, or otherwise remarkable, is called the metasternum. In the Coleoptera, in those cases, as we have seen above[1728], in which the medipectus and postpectus form one piece, its anterior point becomes the mesosternum; but in others, as the Predaceous and Capricorn-beetles, &c., it is received in a sinus or fork of that part, or meets it. It is usually neither so remarkable nor important as the mesosternum. In Bolbocerus K. it is a rhomboidal elevation: in Gyrinus a ridge; as also in many Hydrophili, in which it passes between the hind-legs to the abdomen, and terminates in a sharp point[1729]; and in Dytiscus its two diverging lobes cover the base of the posterior trochanters[1730]. In the Orthoptera Order this part is not remarkable; but in Acrida viridissima K. it consists of three triangular pieces, the lateral ones being erect, and the intermediate one horizontal: in Locusta Leach it resembles the mesosternum[1731]. In the Heteropterous Hemiptera the whole mesostethium is elevated, and terminates at both ends in a fork, the anterior one receiving the point of the promuscis, and the posterior one that of the epigastrium: in the Homopterous section, the Tettigoniæ F. have usually a distinct[Pg 580] metasternal point between their hind-legs. In the remaining Orders there is no metasternum, or no remarkable one, except in one singular Hymenopterous genus, Evania, the parasite of the Blattæ[1732], in which there is a forked posterior process of the mesostethium with recurved points.
17. Opercula[1733]. By this term I distinguish those plates, before largely described[1734], which cover the drums of male Tettigoniæ F.; and likewise those called also by the same name by M. Chabrier[1735], which cover, in many cases, the vocal apparatus of the trunk of insects: those of Melolontha vulgaris he describes as situated below the wings, and between the two segments of the alitrunk[1736]; and if you take this insect and remove the elytra, the mesothorax and scapulars, under the latter and below the wing you will find an oval convex plate, which is probably the part he is speaking of;—but it is better exemplified, I think, in the common Dytiscus marginalis, in which it is very distinct as a convex subtriangular plate connected with the metathorax by membranous ligament, covering a kind of pouch, and appearing to open and shut at the vertex[1737].
I must here observe, with regard to the Aptera and Arachnida, that the trunk in them is much more simple than in those insects that are furnished with wings. In the hexapods, in the former Orders, though there are[Pg 581] usually three pedigerous segments, there is no distinction of dorsolum, scutellum, &c. In the Scolopendridæ and Scutigera amongst the Myriapods, according to the acute observations of M. Savigny[1738],—on which, however, some doubt at present rests,—there is a remarkable formation, the whole thorax being represented by the single plate that follows the head, to the under-side of which are attached the first and second pair of palpi or pedipalpi, and the first pair of legs, representing the three pairs of legs of hexapods. In the Iulidæ the three segments that follow the head, each bear a single pair of legs, while all the rest bear a double one: from whence it should seem to follow, that these segments and their legs represent the trunk and legs of Hexapods. In the Octopod Aptera and the Arachnida the trunk consists of a single piece, not separated from the head, and sometimes not distinct from the abdomen.
V. Internal processes[1739]. Perhaps you will think that this head would be better considered when I treat of the Internal Anatomy of Insects; but as the parts included under it are really processes of the external integument of the trunk, it seemed to me best to treat of them under that head. They are of two descriptions; processes of the thorax or upper part of the trunk, and processes of the breast or its under part.
i. Processes of the thorax[1740]. These are the phragma, prophragma, mesophragma, and metaphragma. The first belongs to the prothorax, the second to the mesothorax,[Pg 582] and the two last to the metathorax; each forming a kind of chamber of the under-side of each segment of the thorax.
1. Phragma. The phragm, or septum of the prothorax, is most conspicuous in the mole-cricket (Gryllotalpa), in which it is a hairy ligament attached to the inside of the upper and lateral margins of the base of that part: inclining inwards, it forms the cavity which receives the mesothorax. It is not, however, without a representative in many Coleoptera, though in these it is less striking, from its being smaller and taking a horizontal direction. In Elater, by means of some prominent points received by corresponding cavities of the vertical part of the base of the elytrum, it forms a kind of ginglymous articulation, which probably keeps them from dislocation in repose, and, by the sudden disengagement of these points from the cavities, assists the animal in jumping[1741].
2. Prophragma[1742]. This is a piece usually almost vertical, but in Elater horizontal; of a substance between membrane and cartilage, descending anteriorly from the dorsolum, and forming the first partition of the chest of the mesothorax; it is generally much shorter than the mesophragm. Though very visible in Coleoptera and the Heteropterous Hemiptera, in the other Orders it is less easily detected, and is sometimes obsolete. It may be observed here, that in the Hymenoptera, at least in the wasp, the hive-bee, the humble-bee, and the Diptera mostly, the interior of the upper-side of the alitrunk, instead of two, seems at first to be divided into four chambers, formed by septula: but as these ridges merely mark out the internal limits of the dorsolum, scutellum, postdorsolum,[Pg 583] and metapnystega, the last but one of these being usually less distinct, they seem not analogous to the three partitions of the alitrunk in other Orders; so that in these the mesophragm at least seems to have no representative, and the prophragm and metaphragm include between them only one ample chamber. In the Diptera, wherever there is an external depression or suture there is a corresponding internal ridge or seam, so that the parts seem more distinctly marked out on the inside than on the outside of the crust.
3. Mesophragma[1743]. This piece also, which forms the middle partition of the upper part of the cavity of the alitrunk, dividing it into two chambers, is most conspicuous in Coleoptera. It is usually in them a vertical piece, resembling the prophragm in substance, but twice its height, of a quadrangular shape with a notch in the middle; it fills the sinus of the postdorsolum, the sides of which sometimes descend below it[1744]. In this Order the chamber that it forms with the prophragm is very small[1745], the motions of the elytra requiring no powerful apparatus of muscles; but that which it forms with the metaphragm, which is appropriated to the muscles moving the wings, is very large[1746]. In the Orthoptera the anterior chamber is larger than in the preceding Order, which proves that tegmina are more moved in flight than elytra. In the Heteropterous Hemiptera a remarkable variation takes place—the anterior being larger than the posterior chamber; which last, in fact, consists of two, one for each wing: in these the mesophragm towards the abdomen forms an angle, which in Pentatoma, &c., is acute; in[Pg 584] Belostoma a right angle, and in Notonecta an obtuse one. In the two first the angle of the mesophragm sends two short diverging ridges to the metaphragm; and in the last only a single one: in this also the posterior chambers together are nearly as large as the anterior. From this structure it should seem that in flight the Hemelytra are more important than the wings. In the Homopterous section the anterior chamber is the smallest, at least in Fulgora candelaria; and the mesophragm is lofty and bipartite. In the Lepidoptera the anterior chamber is the largest, and the part in question conspicuous[1747]. In the Libellulina and Hymenoptera it is merely represented by a low ridge, and in the Diptera it seems evanescent.
4. Metaphragma[1748]. This, in many cases, is the largest and most remarkable of the three partitions of the upper portion of the cavity of the alitrunk, which separates it from that of the abdomen; it is attached to the posterior margin of the metathorax, and is nearly vertical: in substance it may be stated as rather firmer than the two preceding partitions. In the Coleoptera it is commonly of the width of the posterior orifice of the alitrunk; and its centre is cleft so as to form a deep sinus[1749] for the transmission of the intestines,—a circumstance which also, though less conspicuously, distinguishes the mesophragm[1750]: from this sinus it slopes gradually towards the sides, and is sometimes armed with an intermediate process on each side[1751]. This structure you will find exemplified[Pg 585] in the common cock-chafer and many others of the Order. I have not, however, discovered traces of it either in the Silphidæ, Staphylinidæ, or the vesicatory beetles (Meloe L.); or even in such species of Carabus L. and Cicindela L. that I have examined; while in Dytiscus it is very visible. In the Orthoptera it is nearly obsolete; but in Locusta Leach, under the metapnystega, one on each side, is a pair of seemingly pneumatic pouches which may be mistaken for it. It is almost equally inconspicuous in both sections of the Hemiptera. As to the Lepidoptera,—in Pieris Brassicæ, it resembles in some degree, though in miniature, the metaphragm of the Coleoptera; but in Sphinx Stellatarum and Lasiocampa Quercus it has a sinus on each side, but no middle one. In Panorpa it nearly closes the posterior orifice of the trunk, but in the Libellulina it is a mere ridge. In some Hymenoptera, as Cimbex sericea, the drone-bee at least, &c., it is a large convex bifid piece. In the wasps, under the spiracle of the metapnystega on each side, as in the Locusta, is what I also take to be a pneumatic pouch, which might easily be mistaken for a metaphragm. In the Diptera Order this part is very conspicuous. If you remove the abdomen of any common Tipula, you will find that the posterior orifice of the trunk is closed above by a pair of oblong, vertical, convex, diverging plates;—do the same by any fly (Musca L.), and you will detect in the same situation a very large convex or gibbous one notched below, which occupies almost the whole orifice: this is the metaphragm.
5. Septula[1752]. These are the smaller ridges of the interior of the alitrunk, which afford a point of attachment to[Pg 586] many muscles, and run in various directions both on the interior of the crust and of the metaphragm. These little seams are not to be found so generally in the other Orders; but very frequently, as has been before observed, where there is an exterior impression of the crust, or a suture, one of these forms its internal base.
ii. Processes of the pectus[1753]. We are next to consider the internal processes of the breast of insects: these consist for the most part of the endosternum, or internal sternum, and its branches. As the principal feature of this are the processes which rising from it serve as points of attachment to the muscles that move the legs, &c., I shall confine myself to them—they are, the antefurca, the medifurca, and the postfurca.
1. Antefurca[1754]. The first portion of the endosternum, or the internal prosternum, branches into the antefurca. In the Coleoptera a plate varying in shape and direction[1755] sends forth a pair of mostly vertical processes of a cartilaginous substance[1756], differing in height in different genera. In Carabus L. there is neither this plate nor its processes; but in Dytiscus the latter are very visible. A very singular and complex machine represents the part we are considering in that extraordinary insect the mole-cricket (Gryllotalpa Latr.). When we look at its prodigious arms and consider their office[1757], we may imagine that the requisite apparatus for moving them must be very powerful and peculiar. Their Creator has accordingly provided them with a machine for this purpose more than usually complex, extending from the prothorax to the[Pg 587] prosternum; the former being its base, and the latter its vertex. The cavity of the manitrunk is divided longitudinally by a double cartilaginous partition surmounted by a bony frame, with an anterior condyle or tuberosity, with which the inner part of the base of the clavicle of the arm appears to ginglymate; and the manitrunk is preserved from the injury the powerful action of the arm might occasion, by the counteraction of this machine, to describe which fully, would demand more space than I can afford[1758]. I mentioned under the mesostethium, the apertures visible in the breast of Locusta Leach and Acrida K. Each of these apertures opens into an internal, tubular, horny, process, which arching off is attached at the other extremity to the sides of the trunk—a pair being appropriated to each segment; the first analogous to the antefurca, the second to the medifurca, and the last to the postfurca. In the medipectus and postpectus of Acrida viridissima there is only a single aperture, terminating in a single tube, which after rising vertically a little way sends off a branch on either hand to the sides of the trunk. Where there are three of these holes, as in the antepectus and medipectus of Locusta Dux, there are three of these processes, the intermediate one being vertical. In the subsequent Orders the processes of the endosternum are not sufficiently remarkable to require particular notice: my further observations upon them will therefore be confined to the Coleoptera Order.
2. Medifurca[1759]. This part, which belongs to the mid-legs, is in many cases more conspicuous than the antefurca.[Pg 588] In Copris Molossus the endosternum of the medipectus is represented by a transverse zigzag ridge[1760] between the sockets of the mid-coxæ, from which proceeds a pair of branches wide at the base and growing gradually more slender to the extremity[1761], which is attached to the sides of the trunk; in Dytiscus marginalis a pair of slender, vertical, straight processes, fitted with a broad cartilaginous plate at their apex, rises from the endosternum, and sends forth a lateral one to the side of the medipectus: and lastly, in Carabus the medifurca is represented by a pair of subtriangular laminæ attached to the sides of the trunk.
3. Postfurca[1762]. This, which belongs to the hind-legs, is the most remarkable of the pectoral processes, and has been noticed by more than one writer[1763]. It is a kind of trident, the branches[1764] of which are acute, and on their upper surface longitudinally concave, elevated on a footstalk[1765] inclined towards the medifurca, consisting of two plates, a posterior one supporting the lateral branches, and an anterior or interior one forming a right angle with the other, supporting the intermediate one. This footstalk rises from between the posterior coxæ, which appear in the Lamellicorns to ginglymate with it at its base. The middle branch of the trident dips to the sinus of the medifurca. In Dytiscus marginalis the form is different; for the intermediate branch consists of two parallel pieces, and the lateral ones are dilated into broad vertical plates: the stalk of this is triquetrous, and a triple cartilaginous partition appears to go from its base anteriorly, the lateral ones diverging to the sides of the[Pg 589] trunk, and the intermediate one running straight to the base of the medifurca.
It may not be without interest to state here some of the several objects and uses of this structure of the trunk. When our Saviour says to his disciples, "But even the very hairs of your head are all numbered"[1766]—he taught them that the attention and care of the Deity were not confined to the mighty and the vast, but directed to every atom of his creation—that he not only decreed the number and magnitude of the planets and planetary systems, and of their various inhabitants, but that the most minute and apparently insignificant part of each individual, both as to its number and form, was according to the law by him laid down; and whoever studies them with attention will find that insects furnish a very interesting homily upon this text; since in various instances I think I have made it clear, that parts seemingly of the least importance—as a hair, a pore, or a slight impression—have their appropriate use[1767]. At first, it would seem that the various pieces of which we have seen the second primary segment of the trunk of these animals to be composed, would be of little importance; but when we reflect that this multiplicity of parts is usually not to be found in those that have no wings, whether they be apterous sexes or tribes[1768], a suspicion arises in the mind that they must be of more consequence than their prima facie appearance seems to warrant:—and this is really the case. The manitrunk, which is destined principally to incase the muscles that move the arms, did not require to be so complex as the part that had to support the action of wings as well as legs. In those that[Pg 590] have a large prothorax, as the Coleoptera, it may, indeed, be useful in flight as a counterpoise to the abdomen; and since when the wings descend it rises, and vice versa, it may be of some service by its vibrations[1769]; but for this it required no complexity of structure. But not so the alitrunk: it consists of parts much more numerous, and this number of parts is of great importance to the animal in its flight. All of them are so put together, being lined by a common elastic ligament[1770], as to be capable of a certain degree of tension and relaxation, which enables the animal to compress or dilate the trunk as its necessities require. To cause the elevation of the wings, it must be compressed or have its longitudinal diameter increased, and its vertical and transverse diminished: this compression is produced by the condensation of the internal air, which parts with some of its caloric, and by the action of the levator muscles. To cause the depression of the wings, it must be dilated, or have its longitudinal diameter diminished, and its vertical and transverse increased, which is effected by the rarefaction of the internal air, and the action of the depressor muscles[1771]. In some Orders, the Coleoptera, &c., this effect is promoted by the segments of the trunk, which are attached by loose ligamentous membranes, and received, one or more of them, into each other, which facilitates the above action[1772]. Thus much for the general use of these parts. I shall further here mention a partial one of two of them which seems indicated by a particular[Pg 591] circumstance, and upon which a theory may be built. In some insects the primary and secondary wings or their analogues are placed before the legs, in others over the legs, and in others behind the legs: but whatever their position, the pieces which I have named the scapularia and parapleuræ invariably connect the one with the other; the former, the primary wings with the mid-legs, and the latter, the secondary wings with the hind-legs. This circumstance seems to prove that the wings by the intervention of these pieces have an action upon the legs, and the legs upon the wings; and this is further proved in one case by an observation of M. Chabrier with regard to Melolontha vulgaris,—that the levator muscles of the wings, by means of a long tendon, are attached to the lower part of the posterior coxæ[1773]. Now, more than one medical friend has suggested to me, that what are called the coxæ in insects are really analogous to the thighs of vertebrate animals[1774]: consequently these parts must represent the coxæ; whence it would seem that the wings are really appendages of the legs. It must, however, be observed, that were this opinion admitted, in the Aptera, Hymenoptera, and Diptera, or even in the prothorax of other insects, there would scarcely be any analogue of the coxæ at all distinct from the trunk itself, of which even in the other Orders these pieces are component parts. An instance occurs in the Strepsiptera K., and in which the arms are furnished with an alary appendage, and the metathorax has none[1775].
VI. Organs of Motion. We are next to consider those organs attached to the trunk of insects which are instruments of motion. These are principally those by which they are transported through the air, and those by which they move on the earth or in the water—their wings and their legs. I shall begin with the first, the wings[1776]. These are not formed precisely after any type at present discovered in vertebrate animals: in some respects they have an analogy to those of birds[1777]; in others, to the dorsal fins of fishes: but, perhaps, altogether they approach the nearest to those of the dragon or flying-lizard (Draco volans L.), which do not, as in birds, replace the fore-legs, are kept expanded by diverging bony rays, and are connected with the hind-legs[1778]. As the Divine Creator appears in his works to proceed gradually from one type of structure to another, it has been supposed by a learned physiologist of our own country, that in winged insects, four of the legs of the Decapod Crustacea[Pg 593] are represented by the four wings[1779]: this opinion, however, is not yet fully proved; a remark which may also be applied to a more recent one of a celebrated French writer, who seems to think their origin and structure aërostatic, that they are auxiliary to the legs, and borrowed in part from the respiratory organs[1780]. Were I disposed to enter into these subtile speculations, I might here recall your attention to the analogy that, in their metamorphoses, exists between the Saurian Reptiles or lizard tribe and insects, and conjecture that the wings of the Draco are really representatives of the mid-legs of Hexapods, thus preparing to disappear altogether; but I shall content myself with throwing out this hint, which you are welcome to pursue. The organs of flight in general may be considered as to their number, kinds, and composition.
i. Number. The most natural number is four, for this obtains in the majority. In almost every Order, indeed, there occur instances of insects that have solely a single pair or none[1781].
These, however, are only exceptions to the rule; but in the Diptera, unless we consider the alulæ, the representatives of the secondary wings[1782], as a distinct pair, there are never more than two wings, and one instance is known in which an insect of this Order has none[1783]. Certain genera or individuals of the Tetrapterous Orders are also furnished with alulæ: besides Dytiscus, Blatta, Phalæna hexaptera, which have been before noticed[1784], they may be detected in miniature in Ammophila K. and affinities; these all may be regarded in some slight degree as insects with six wings.
ii. Kinds. Under this head we may consider the organs of flight as to their situation and as to their substance. As to their situation, usually the first pair are attached to the mesothorax, and the second to the metathorax; but in one instance, as has been before observed[1785], in the Strepsiptera K., the anterior pair belong to the manitrunk, and the posterior to the mesothorax. As to their substance, they take the several denominations of elytra, tegmina, hemelytra, and wings, for the most part according to its variations, as will be seen more at large hereafter. Under this head I shall only further observe, that in many instances the organs of flight appear to be mere abortions or rudiments, which serve to exemplify what has been more than once stated, that the CREATOR has seen it good to approach to new organs gradually as well as to new forms. Thus elytra are mere rudiments that do not serve to protect the wings in Atractocerus; tegmina in some species of Phasma, Acrydium,[Pg 595] &c.; hemelytra in the bed-bug[1786]; wings in many female moths, in Cryptus hemipterus a Hymenopterous insect, &c.
iii. Composition. The structure of wings has been before explained to you[1787], and I shall again have occasion to allude to it; but here I wish to call your attention to a circumstance that has not hitherto, that I recollect, been adverted to; I mean that all kinds of organs of flight, and it may be traced as we shall soon see even in elytra, are divided longitudinally into three areas or folds; the first or external one I call the Costal Area[1788] from its beginning with the costal nervure; the second is the Intermediate Area[1789]; and the third is the Anal Area[1790].
Having made these observations with respect to the organs of flight in general, I shall now proceed to consider more at large the elytra, tegmina, hemelytra, and wings.
i. Elytra. These are the wing-covers of the Coleoptera Order, distinguished from tegmina by the absence of nervures, from hemelytra by the want of the membrane at the apex, and from both by their uniting in almost every instance at the suture. I shall consider them as to their substance; articulation with the trunk; expansion; parts; shape; appendages; sculpture; clothing; colours, and uses.
1. Substance. The firmness of the substance of elytra is usually regulated by that of the crust of the insect to which they belong; in hard insects they are hard, and[Pg 596] in soft ones they also are soft. The most impenetrable ones that occur to my recollection are those of Illiger's genus Doryphora, and the softest and most flexile those of Telephorus, Meloe and affinities. With regard to individuals, they are mostly as hard as the prothorax, and harder than the back of the abdomen. Elytra also, as far as my observation goes, are never diaphanous.
2. Articulation with the trunk. This is by means of a process of the base of the elytrum which I call the axis[1791] or pivot, attached by elastic ligaments, and certain little bony pieces (osselets Chabr.) in the socket under the side of the anterior angle of the dorsolum[1792]. You may easily remove the elytra attached to the mesothorax from Geotrupes stercorarius, which will enable you to see the mode of articulation with little trouble[1793].
3. Expansion. It is by means of the bony pieces just mentioned that the organs in question are opened and shut[1794] under the action of the antagonist muscles. In opening for flight the two elytra recede from each other, and are elevated so as not to retain their horizontal position, which would interfere probably with the play of the wings, but form an angle with the body. When they return to a state of rest, the sutures usually meet and coincide longitudinally; but in some cases when closed, as in Necydalis, &c., they diverge from each other at the apex; and in Meloe, like the Orthoptera, to which that genus approaches, one laps over the other.
4. Parts. The parts to be considered in an elytrum are the areas, the axis, the suture, the margin, the[Pg 597] epipleura, the base and apex, the angles, and the hypoderma. At first it should seem as if an elytrum was not like other wings divided into areas; but I think upon examination it will be found that, though often nearly obsolete, these are represented in it; for the epipleura[1795] with the recurved part of the external margin seems to me analogous to the Costal Area; the inflexed part adjoining the scutellum and often going beyond it to the Anal, and the rest of the organ to the Intermediate. All this you may see in the dung-chafer, Geotrupes stercorarius. The axis[1796] or pivot by which the elytrum articulates with the trunk is generally placed about the middle of its base, but nearer the scutellar than the humeral angle, and varies in length and shape in the different tribes, but not so as to merit particular notice; it may be regarded as composed of three parallel pieces, one belonging to each area, that of the costal being the longest. In many these pieces are marked by no line of distinction, but in Macropus, &c., they may be readily traced[1797]. The suture[1798] is the internal margin of the elytrum from the point of the scutellum to the end. In many beetles the right hand suture, looking from the anus to the head, has a lower ledge or margin, and the other, one more elevated, which when they are closed lies upon the former; in some Dynastidæ there seems a kind of ginglymous structure in this part, each suture being fitted with a kind of ridge which is received by a channel of the other; in these the suture is generally marked out by an adjacent channel: but the most remarkable structure of this part distinguishes the genuine species of the genus Chlamys, in which both the sutures,[Pg 598] except at their base, are armed with little teeth, alternating with each other like the cogs of a mill-wheel. In apterous beetles the elytra are often connate, or have both sutures as it were soldered together. The margin[1799] or external edge of the elytra is generally formed by a bead or ridge, which, except in the case of the truncated ones, in which it is straight, curves more or less from the base to the apex; this ridge is often recurved so as to form a kind of channel between it and the disk of the elytrum, as may be seen in the Dynastidæ; in some there are two parallel ridges, as in Copris; in Silpha the margin is dilated; in Helæus and Cossyphus it is remarkably so and recurved, so that, in conjunction with those of the prothorax which are similarly circumstanced, they give the animal some resemblance to a small model of a barge. Though the margin of elytra is most commonly intire, yet in some beetles, as Gymnopleurus Illig., a sinus is taken out of it; in Cetonia it often projects at the base, and in Cryptocephalus in the middle, into a lobe; in Phoberus MacLeay it is denticulated, and in many Buprestes more or less serrulated; sometimes it terminates before it reaches the apex of the elytrum in a tooth, as in many Carabi Latr. The epipleura[1800] or side-cover is that part of the organ in question, below the margin, with which it usually forms an angle, being more or less inflexed, that covers the sides of the body. It varies in different tribes, being sometimes obsolete, as in the weevils (Curculio L.); in the Capricorn beetles it is very narrow; in Carabus, &c., dilated at the base; in many Heteromerous beetles, as Blaps, Pimelia, &c., it is very wide and conspicuous; in[Pg 599] Cossyphus it stands out a little from the abdomen, so as to form a kind of fence round it. Its shape generally approaches that of a scythe, being incurved and growing more slender towards the apex[1801]; but it is sometimes straighter and shorter. In Geotrupes and many other Lamellicorns, the base of the elytrum is nearly vertical, forming a right angle with the rest of it; it is usually transverse and straight; but in Calandra Palmarum and many Cassidæ it slants to the scutellum; in Chlamys it is sinuate, and in Elater it has a deep cavity above the axis which receives the points of the phragma mentioned before[1802]. The apex of elytra is usually acute, the angle being formed by the confluence of a curving and straight line: but there are many exceptions; for instance, in Mylabris it is rounded; in Hister obliquely, and in Necrophorus transversely, truncated; in many Capricorns it is emarginate; in others, as Macropus longimanus, it is bidentate; in some Prioni, P. cinnamomeus, &c., it terminates in a mucro at the internal angle; and in Cerambyx Batus, horridus, &c., at the external; and, to name no more, in some species of Necydalis it ends in a long acumen. The scutellar angle in insects that have a large scutellum, as Macruspis MacLeay, is obliquely truncated to admit it, but where it is small it is generally rectangular, with the angle rounded; in Buprestis vittata it is obtusangular; and in Dytiscus marginalis, &c., it is emarginate. In Cassida spinifex, perforata, &c., the humeral angle is producted into an acute lobe that stretches beyond the head, and in C. bicornis and Taurus it forms a horn at right angles with the elytrum. In general it[Pg 600] is either rectangular or rounded, with a prominence of the elytrum within it. The sutural and anal angles exist only where the elytra are truncated at the apex. In this case the sutural is generally rectangular, and the anal rather obtusangular or rounded. The Hypoderma is the fine soft membrane before noticed[1803] that lines the underside of the elytra, the use of which is probably to prevent injury to the wings from friction with their usually hard substance; this membrane is commonly of either a pallid or brownish colour; but in some insects, as Staphylinus hybridus, murinus, &c., Buprestis grandis, it is of a beautiful green or blue; and it exhibits the puncta, striæ, and other modes of sculpture of the elytra very distinctly, the pores of which usually perforate this membrane[1804]. Just under the shoulders of these organs you may observe an oblong and sometimes roundish spot, occasioned by the hypoderma in that part being particularly tense, and covering a cavity or pocket which appears to be connected with the axis by the hollow part, which I regard as representing the Costal Area; this pocket is evidently the analogue of a part in the wings noticed by M. Chabrier[1805], and named by me the phialum: from its connexion with the axis by a channel, this part in elytra should also seem destined to receive a fluid to add to the weight of the margin and its means of resistance.
5. Shape. The shape of elytra is various; taken together, in which case, in describing insects, they are denominated coleoptra, their most common form is more or less oblong, or forming more or less a considerable[Pg 601] portion of an ellipse; taken separately, it inclines to that of an isosceles triangle, with the exterior side curvilinear: truncated elytra are generally quadrangular, sometimes presenting a trapezium, at others nearly a parallelogram, and at others a square. With regard to their proportions they vary considerably, but the most general law seems to be that the length shall exceed twice the width; in some, as Buprestis grandis, it is more than thrice; in many Staphylinidæ they are as wide as they are long and sometimes wider; they are generally narrower at the apex than at the base, but in some species of Lycus, as L. fasciatus, &c., the reverse takes place; in Telephorus they are nearly of the same width every where: with regard to their surface they are sometimes very convex, as in Moluris; at others very flat, as in Eurychora, Akis, &c.
6. Appendages. These, though not so remarkable as those of the head and prothorax of beetles, ought not to be overlooked. In many Capricorns, as Lamia Tribulus, speculifera, &c., the disk and sides are armed with short sharp spines; in others (Stenocorus, &c.) the sutural and anal angles or one of them terminate in a spine or tooth; sometimes the whole surface, as in Hispa atra, &c., is covered, like a porcupine, with a host of slender spines, or its sides defended by spinose lobes, as in H. erinacea: the humeral prominence is armed with a spine pointing to the head in Macropus longimanus, and forming a right angle with the elytrum in some Curculionidæ, as Rhynchites spinifex; but the most remarkable appendage of this kind is exhibited by Cassida bidens and its affinities,—from the centre of the sutures of the elytrum rise perpendicularly a pair of long, slender, sharp processes[Pg 602] internally concave, which both apply exactly to each other, so as together to form a single horn which rises, like a mast from a ship, from the body of the animal[1806]. Besides the appendages here mentioned, the elytra exhibit a variety of tubercles and other elevations of various form and size, which it would be endless to particularize.
7. Sculpture. The sculpture of the organs in question is very various and often very ornamental: but as almost every kind of it will be noticed in the orismological tables, it will not be necessary to enlarge upon it here, especially since I have endeavoured upon a former occasion to explain how it may be useful and important as well as ornamental to the animal[1807]. I shall therefore only notice a few instances, amongst many, in which a particular kind of sculpture distinguishes particular tribes. Amongst those that are Predaceous the Cicindelidæ have elytra without striæ or furrows, while the majority of the subsequent terrestrial tribes of this section are distinguished by them: the Dynastidæ in the Lamellicorn section are remarkable for a single crenated furrow next the suture; in the weevil tribes the numerous species of the genus Apion are ornamented by furrowed elytra with pores in the furrows, which give them the appearance of neat stitching; in many of those beetles that have soft elytra, as the glow-worms (Lampyris), the blister-beetles (Cantharis, Mylabris), and still more in Œdemera, two or three slight ridges generally run longitudinally from the base to the apex, and are visible also on the under-side; as the furrows probably[Pg 603] lighten a hard elytrum, these ridges may serve to strengthen a soft one, and it is by these that the first approach is made to the reticular structure of tegmina or the wing-covers of Orthoptera: Lycus palliatus, &c., in its elytra exhibits a direct resemblance of the reticulations of nervures.
8. Clothing. To what I have before said on this subject in general[1808] I shall here add a few remarks, which, though they more properly belong to elytra, may in many cases be extended to the whole body of a beetle. In various instances it happens that the beautiful markings of these organs, as in Macropus longimanus, whose elytra when denuded are black, are produced by short decumbent hairs; in some these variegations are the effect of scales resembling those of Lepidoptera, often of a metallic lustre; from these scales is derived all the brilliancy of the diamond-beetle (Entimus imperialis, Germ.); in some the scales are so minute as to resemble the pollen of flowers, as the white marks observable on the green elytra of the rose-chafer (Cetonia aurata).
9. Colour. The organs of flight in the majority of the Orders with respect to colour are usually the most gaily decorated part of insects; I therefore deferred the notice of that subject till I came to treat of them. In general the colour of insects is either inherent in the substance of their crust, or produced by the hairs or scales that either partially or totally cover it. To confine myself to the Coleoptera, of whose elytra we are treating, it may be observed, I think, in general, that the majority of those that feed upon putrescent substances, the saprophagous tribes of Mr. W. S. MacLeay, are commonly of a[Pg 604] more dark and dismal aspect and colour than those which feed upon such as are living and fresh, denominated thalerophagous by the same learned author; this you may see exemplified in his Scarabæidæ and Cetoniadæ. Again, in the Predaceous beetles a similar contrast of colours is often observable. How brilliant and gay are the fierce Cicindelæ! those tigers of insects, as Linné calls them; how black as to colour, how horrible in aspect is their near relation the Manticora: what difference exists in the economy of these animals is not known, except, as I learn from Mr. Burchell, that the latter is subterraneous, whereas the former seek the sunbeam and fly rapidly. I shall now point out a few instances in which the colours of their elytra distinguish tribes or families. Amongst the Predaceous beetles a large family of the Cicindelidæ are distinguished by a middle angular white band, and several white dots on their green or brown elytra, as in C. sylvatica; a family of Brachinus, and the majority of Mylabris, Lamia capensis and fasciatus, &c., by black elytra, with yellow or red bands; Carabus violacea and affinities by the violet margin of these organs; Calliochroma Latreille by their sericeous, and Eumolpus by their metallic, lustre. These instances will be sufficient to turn your attention to this subject, which though not of primary importance in discriminating genera &c., is not without its use in a secondary view.
10. Uses. I must not quit this subject without saying something upon the ends which elytra seem designed to serve. Their first and most obvious use is the protection of the wings when unemployed, that they may not be lacerated or soiled, and rendered unfit for flight in the various retreats to which these animals betake themselves[Pg 605] either for food, repose, or to lay their eggs; to promote this purpose more effectually, the wings are usually curiously folded and laid up under them; and where the elytra are very short, as in the Staphylinidæ, these folds are very numerous and complex. In some instances, however, as in Molorchus F., Atractocerus, &c., the wings are only partially protected by the elytra and not folded under them; probably they are less in danger of laceration from their peculiar habits than the generality. Another use is to protect the upper-side of the alitrunk, which for reasons before assigned is usually softer than the under-side, and also of the abdomen, often above nearly membranous, from the injury to which they would otherwise be exposed; in the latter part also the spiracles in Coleoptera are not covered by the inosculations of the segments, as is the case in most other Orders, and therefore probably require some covering when the insect is not flying. In the Apterous beetles this appears to be their principal use; where these organs are connate, or as it were soldered together, the back of the abdomen is a thin membrane; the appearance of two elytra in these cases is given, doubtless, for the sake of symmetry and beauty, a subordinate attention to which may be traced in all the works of creation. If we consider the bulk and weight of many flying beetles, we may imagine that they want some assistance, more than the extent and dimension of their wings seem to promise, to support them in the air, and to enable them to move more readily in it; and although it seems clear from the state of their muscular apparatus that elytra do not move much in flight, yet by giving a broad and concave surface to the air, for then they are usually nearly vertical, they may assist in[Pg 606] some measure as sails, and help them in flying traversely and before the wind[1809].
ii. Tegmina[1810]. By this name the learned Illiger has distinguished the upper organs of flight of the Orthoptera and Homopterous Hemiptera[1811]. They may be considered under the same heads nearly as elytra.
1. Substance. Tegmina differ very materially from elytra in their substance, being generally more or less diaphanous, though in Blatta Petiveriana the dark parts are as opaque as elytra, and those of the Mantes that resemble dry leaves are only semidiaphanous. These organs are also of a less dense substance than elytra, something between coriaceous and membranous, which I shall express by the term pergameneous, as somewhat resembling parchment or vellum. Another circumstance relative to this head also distinguishes them,—they are not lined with membrane. In some instances, as in B. Petiveriana just named, they approach nearly to the substance of elytra, and in B. viridis, some Mantes, and Tettigonia, &c., they are little different from wings in their substance; but this does not diminish their right to be considered as tegmina, since their structure is altogether the same.
2. Articulation with the trunk. I observed above that the axis of elytra may be regarded as formed of three parts, one appertaining to each of the areas or their representatives[1812]; in tegmina, and indeed in wings in general,[Pg 607] these parts are separate and may be more distinctly traced, the axis of the Costal Area being generally the longest, and that of the Intermediate often the shortest; these axes are suspended in the wing-socket by elastic ligaments, intermixed with hard bony plates, the principal one of which, called by M. Chabrier the humerus[1813], is connected both with the tegmen and the trunk, and in some a little resembles the head and neck of a swan. This structure permits the animal to move the lateral areas in some degree separately, so that each, especially the anal, shall form an angle with the intermediate; as the motion of the latter is not wanted, its axis often falls short of the base, or is obsolete, as in Blatta.
3. Composition. The three areas, traces of which we had discovered in elytra, are particularly visible in tegmina. If you take any cockroach (Blatta), you will at first sight see that in it they are divided into three larger portions by stronger nervures or folds; and if you also take a Mantis, or Locusta Leach, a Fulgora or Tettigonia, the same circumstance will strike you, only you will see that in these the intermediate portion terminates also in an axis; these are what I call the three areas. The external one or Costal is usually the longest and narrowest[1814]; the Intermediate one is commonly triangular, with its inner side curvilinear[1815]; and the interior one, or Anal area, in the Orthoptera is rather oblong; in Fulgora angular, and in Tettigonia it presents an isosceles triangle; with its vertex to the apex of the wing[1816]. The first of these may be defined as that portion[Pg 608] of the wing that lies between the costal and postcostal nervures; and perhaps, in some cases, as in Mantis, for there is the fold of the tegmen, the mediastinal may be regarded as its limit; the Intermediate Area is that which lies between the postcostal or mediastinal nervure and the anal fold of the wing; and the Anal Area is the remainder. These areas may perhaps best be made out by tracing each to its axis. To study them carefully in tegmina and hemelytra is of considerable importance; for in them we find the first outline of the general plan upon which the wings of insects are constructed, and which, as we shall see hereafter, more or less enters into the composition of them all.
4. Position, and folding in repose. With regard to their position when not expanded, tegmina vary somewhat in the different tribes. In the Coleoptera we have seen that, except in a few instances, the elytra unite at their suture. Something like this takes place in Fulgora, Cercopis and affinities, in the Homopterous Hemiptera; in these, though the union is not near so exact, yet the tegmina do not lap over each other; they are usually more or less deflexed, with scarcely any portion in a horizontal position: in Tettigonia F., Chermes, Aphis, &c., the middle part only of these organs meets, from which point they diverge both towards their base and apex[1817]. In the Orthoptera the position is quite different, for one tegmen more or less lies over the other. In Blatta, in which the tegmina are nearly horizontal, the left hand one covers almost half the other[1818]: in the other tribes of the Order, with little variation, the Anal Area of the tegmen[Pg 609] is horizontal, and covers the back of the animal, and the Intermediate and Costal are vertical and cover its sides; the former, however, in some cases, only forms the angle between them. Sometimes in these the right-hand one is laid upon the left, as in Acheta; and sometimes the reverse of this takes place, as in Acrida K. With regard to the folding of the tegmina, the most remarkable instance that occurs is that of Acheta monstrosa, in which the ends of both these organs and the wings, in repose, are folded like a fan, and then rolled up like a serpent[1819].
5. Shape. The shape of tegmina is various. In the Blattæ and some Mantes they are more or less oblong; in Mantis precaria, strumaria[1820], and others, they incline to elliptical; in Phasma grandis and Acheta monstrosa they are rather panduriform[1821]; in M. gongyloides they are semi-cordate[1822]; in Pterophylla trapeziformis they are rhomboidal[1823]; in Conocephalus erosus they are sinuated; in Locusta Leach they are usually linear or linear-oblong[1824]; in Pterophylla K. they generally terminate in a short mucro[1825]; and in some of those Mantidæ whose tegmina simulate arid leaves, in a recurved one[1826]. In the Homopterous Hemiptera the shape of these organs is less various. In the Fulgorellæ Latr. they incline to a trapezium, sometimes to a pentagon[1827]; in the Tettigoniæ F. they approach to an obtuse-angled triangle;[Pg 610] and in others of the tribe they are nearly wedge-shaped[1828].
6. Neuration. The circumstance that most strikingly distinguishes tegmina from elytra is their neuration or veining; which adds much to their strength, without increasing their weight so much as to render them unapt for flight. To look at these organs in Blatta Petiveriana, you would imagine them at first to be deprived of this distinction; but if you observe them attentively, particularly their white spots, you will soon detect their nervures; and if you further examine their lower surface, you will find them very visible. The gibbous Blattæ also, Blatta picta and affinities, the analogues of Erotylus amongst the Coleoptera, have tegmina which, except at their apex, exhibit but faint traces of the nervures of their tribe, and approach to elytra besides by the innumerable minute impressed points that cover them. In the Orthoptera and some Homopterous Hemiptera the nervures may be divided into longitudinal ones more or less ramified, and traversing ones. In the Blattæ the traversing nervures cut the longitudinal ones nearly at right angles, but not at regular intervals, so as to cover the tegmen with quadrangular areolets; in Mantis precaria and affinities the longitudinal nervures of the Anal Area diverge from the base, and are traversed nearly as in Blatta, while those of the Costal diverge from the mediastinal nervure, but the traversing ones form innumerable irregular reticulations; in Mantis sinuata K.[1829] the whole tegmen has such reticulations but less numerous; in Locusta Leach it is regularly reticulated at the base, but the areolets of[Pg 611] the apex are quadrangular; in the Mantes, with oblong wings, all are quadrangular; in Pterophylla K. the longitudinal diverging nervures are not numerous, and the traversing ones cut them into quadrangular and triangular areolets, besides which they are covered by innumerable impressed points, so as altogether to exhibit a most exact resemblance of the leaf of some evergreen: in Gryllotalpa the longitudinal nervures of the Anal Area rather converge towards the apex, are traversed by few transverse nervures, and those of the Costal Area which diverge from the mediastinal nervure by still fewer; the neuration of Acheta F. has been before described[1830]; I shall only observe here, that the constructors of stringed instruments of music might, perhaps, from the tegmina of the male, the nervures of which probably modulate the sounds which it produces, take a hint for giving the strings in them a serpentine or convolute direction, and so might produce something new in that department, corresponding with the serpents and French-horns in wind instruments. Of the Homopterous Hemiptera in the Fulgorellæ Latr., which are most analogous to the Orthoptera of all that tribe, the longitudinal nervures are more numerous and branching, more especially toward the apex of the tegmen, and are traversed as much by transverse ones, sometimes reticulating the wing with roundish areolets, as in F. laternaria, and at others with quadrangular ones, as in F. candelaria; in some of these however, as Otiocerus K., Flata F., &c.[1831], there are no traversing nervures; and these lead to the Cercopidæ and others in which the longitudinal nervures become[Pg 612] few, and some are without any[1832], and these terminate those of this section of the Order in which the nervures in question are continued to the margin of the wing. We next come to those, Darnis, Centrotus, Membracis, &c., in which they are circumscribed a little within the apex by a traversing nervure, so that the tegmen ends in a margin of pure membrane, and thus some approach seems to be made to the Hemelytra, from Tettigonia, the most conspicuous genus of this tribe, in which the areolets, few in number, like those of Lepidoptera, are not formed, except the terminal ones, by traversing nervures, but by the ramifications of the longitudinal ones; in Chermes the Intermediate Area, which is connected with the base of the wing by a single nervure, is the only part that has any areolets[1833].
7. Colour. Orthopterous insects are seldom remarkable for tegmina of brilliant colours; there is in them none of that gilding or metallic lustre which so often distinguishes elytra: they are also frequently less ornamented in this respect than the wings, with which they usually form an agreeable contrast. Their reticulations and nervures, which are sometimes of a different colour from the rest of the tegmen, decorate them considerably: a remarkable circumstance belonging to this head attends the black tegmina of Blatta Petiveriana; one has four white spots, and the other only three; but as one laps over the other, the symmetry of the arrangement is preserved: the Homopterous Hemiptera are more distinguished in this respect, and some of the Fulgoridæ imitate the Lepidoptera both by their ocelli and spots: Fulgora laternaria,[Pg 613] Candelaria, serrata, and Diadema, sufficiently exemplify this remark, as do several Flatæ likewise[1834].
We may observe here—that tegmina are more calculated for flight than elytra, both from their thinner substance, and from the angle that their Anal Area, and often the Costal, forms with the rest of the tegmen; a circumstance which, in wings, M. Chabrier thinks presents some facilities in that kind of motion.
iii. Hemelytra[1835]. The next species of wing-covers, which though varying in the substance of their base, terminate in a part distinct from the three areas, consisting in almost every case of mere membrane, peculiar to the Heteropterous Hemiptera, are called hemelytra, or half-elytra:—this term was also formerly employed, but certainly incorrectly, to denote tegmina. I shall consider them with respect to such of the particulars noticed under the former heads as apply to them, but without repeating them formally.
1. As to their substance, they must be separately considered with regard to their base and apex. In various instances the base, or part consisting of the three areas, is almost corneous, as in Cydnus Morio and bicolor, bugs not uncommon with us, and many others[1836]; in these cases it is lined with a hypoderma like elytra; and in many the points, which are impressed upon it, also perforate the hemelytrum, and seem to act as pores: but in Lygæus, Reduvius, Capsus, Miris, and the majority of the Heteropterous Hemiptera, the organs in question being soft and flexible,[Pg 614] may be stated as rather resembling leather than horn;—on this account this part of a hemelytrum is denominated the corium. In Scutellera the portion covered by the scutellum is membranous; and in Acanthia paradoxa, and the cucullated species of Tingis, the wing-covers are entirely so. The apex of these organs is almost universally either membranous or coriaceo-membranous, on which account it is called the membrana. I say almost, because in Aradus and the Hydrocorisæ Latr., this part, though rather thinner than the rest of the Hemelytrum, is also coriaceous; in the latter tribe usually with a very narrow membranous edge; and in many Reduvii and Zeli there is scarcely any difference in the substance of the base and apex.
2. As to the articulation of Hemelytra with the trunk, it seems not strikingly different from that of tegmina: the point or base of the Intermediate Area, which falls short of that of the lateral areas, seems connected by a slender ligamentous piece, with its axis, which is thick; and I do not discern Chabrier's humerus shaped like a swan's head and neck[1837].
3. The composition of these organs differs from that of tegmina in more respects than one: in the first place, they consist, as was lately observed, of four instead of three areas; in the next, they appear to have, at least several of them, a part, which I suspect to be analogous to that above described in Coleoptera, supposed to represent the phialum of wings[1838]. I shall first speak of the areas. In some apterous species related to the bed-bug, Lygæus brevicollis Latr.[1839], &c., there is no trace of the usual areas,[Pg 615] and the membrana is a very narrow strip; in L. apterus the former are very faintly traced out, but they are present in all those that are furnished with wings; whence we may conjecture that they are of the same importance in flight with the folds observable in those organs[1840]. The three basal areas may be said most commonly to present three isosceles triangles, the Costal one being narrow and curvilinear[1841], the Intermediate the most ample[1842], and the Anal one the narrowest and shortest[1843], with its vertex towards the apex of the Hemelytrum, while in the two former it is at its base. In Lygæus compressipes (Rhinuchus K. MS.) the Anal Area is cultriform; and in most of the Hydrocorisæ it has an angle in the middle of its posterior margin. The proportion that the membrana or apical area bears to the rest of the wing varies in the different tribes. In some, as before stated, it is obsolete, in others nearly so; in the majority, perhaps, it occupies about a third of the hemelytrum; in Lygæus compressipes, cruciatus, &c., full half; in Alydus calcaratus, two-thirds; in Reduvius, nearly three-quarters[1844]; and in Aradus depressus the corium,—divided, however, though indistinctly, into the three areas,—is driven to the base of the wing: two ends are answered by this structure—as this insect lives under bark, its thin hemelytra take less room; and as it flies, though it has only rudiments of wings, they are more fit to supply their place: the part we are speaking of usually[Pg 616] runs obliquely from the vertex of the Anal Area to the base of the Costal.
4. As to their position and folding in repose, Hemelytra are usually nearly or altogether horizontal; but in Notonecta and Plea they are deflexed and cover the sides of the body; and the apical area of one wing precisely covers that of the other; where the scutellum does not intervene, as in Scutellera, Pentatoma, &c., the vertical angles of the Anal Area meet in the middle of the back, so as to exhibit the appearance of a cross. In Notonecta, in which the hemelytra are deflexed, at the apex of the membrana is a fissure which permits the two sides to form an angle with each other, and to apply exactly to the body. In Plea, in which there is no apical area, the posterior margins of the tegmina, as they ought rather to be termed, unite, but do not lap over each other. With regard to the appearance of something like a phialum, if you examine the hemelytra of most species of bugs on the underside, you will see that the costal nervure at the base is inflexed and covers a kind of channel; if you next take one of Belostoma grandis, where the structure is most conspicuous, or even the common Nepa cinerea, you will find in the same situation, adjacent to the inflexed costal nervure, a hollow tube running from the base of the wing, and terminating, after proceeding about one-fourth of its length, in a hollow cavity, which, as it is covered by a membrane, appears to me to be a collapsed pouch. This circumstance is worthy of further and more general investigation.
5. In their shape, with few exceptions, hemelytra more or less represent a wedge, being wider at their apex, where[Pg 617] they are usually obliquely truncated, than at the base; but in Plea Leach they are obtusangular, with the angle in the sutural margin; in Notonecta, on the contrary, an obtusangular sinus distinguishes that part; in Naucoris they are curvilinear and every where of equal width; in Ranatra they are linear and straight; in Aradus they are oblong, usually with an external lobe or dilatation at their base: a remarkable instance of the intention of this is observable in a nondescript Brazilian species, in which the head, prothorax, and abdomen, are edged with a number of broad foliaceous appendages; if the base of the hemelytrum had not been furnished with a similar appendage, the symmetry of the whole body would have been destroyed by the hiatus between the prothorax and abdomen, as may be seen by removing the hemelytra; but by this compensating contrivance of Providence, the gap is filled, the above lobe fitting exactly into it.
6. The neuration of these organs will not occupy us long, since the corium or harder part, though in some species there are traces of nervures, is often without them. Those of the cucullated species of Tingis resemble many tegmina in being ornamented by them with a kind of network, which looks like the finest lace; in several Lygæi, Edessa, and some Reduvii, there are a few diverging longitudinal nervures which occasionally by a ramification here and there form an areolet[1845], but there are seldom any traversing nervures. The Apical Area is usually most distinguished by nervures, in some forming several areolets, as in Aradus, in others running parallel to each other, nearly to the end of this area, as in Belostoma grandis,[Pg 618] where they are met by a traversing nervure; the object of this is doubtless to strengthen the membrane.
7. Both tegmina and hemelytra are most commonly naked, yet very short hairs are found on those of some species of Cercopis, and in many more instances in those of the latter description, as in Notonecta, several Lygæi and Reduvii, &c.
8. Colours in hemelytra are very various, and in many instances are peculiar to families; in certain Lygæi (L. Hyoscyami, &c.) black and red; in Lygæus compressipes and affinities a dingy black; in some Reduvii black with a large white spot;—but it is needless to enlarge further on this subject.
9. That hemelytra are used in flight is evident not only from the large space allowed for their muscles[1846], but likewise from a circumstance noticed by M. Chabrier, that in flight, in the Pentatomæ Latr., the corium of the hemelytrum is fixed to the wing[1846]; in which case both must describe the same arc.
iv. Wings. We are next to consider organs which are exclusively appropriated to flight, and therefore are properly denominated wings. These in the Orders that have elytra, tegmina, or hemelytra, are the pair that correspond with the secondary wings of the other Orders. It may be said, indeed, that in several instances both tegmina and hemelytra do not differ at all in substance or use from the wings that they cover. This is true; but as their structure in other respects is the same with that of those that are more solid and less apt for flight, it was convenient to consider them under the same name.
1. To begin with the articulation of these organs with the trunk; in general it may be stated that this, as in tegmina and hemelytra, is usually by the intervention of three axes, formed by the conflux of the nervures of the three areas at the base of the wing, which either immediately or by other pieces are implanted in the trunk, so as to receive from it the aërial and other fluids, necessary for its expansion and motions[1847]. Having given this general statement, I shall next apply it to the wings in some of the different Orders. If you carefully extract one from the stag-beetle (Lucanus Cervus) or any large species of the Dynastidæ, in the Coleoptera; the first thing that will strike you, upon examining the base, will be the plate before mentioned called by Chabrier the humerus, which is a stout transverse corneous piece, with a deep sinus towards the wing, filled with ligament: if you again follow the costal, mediastinal, and postcostal nervures, you will find them unite to form an axis, consisting of three parallel pieces, which connects by its intermediate internal piece with one end of this plate. The nervures of the Intermediate Area terminate also in a corneous axis at a greater distance from the base than the other two, which connects with Chabrier's humerus by means of the ligament of the sinus just named. Those of the Anal Area are received by a ligament attached to a transverse plate, widest at its anterior end, which connects with the posterior part of the said humerus; and at its posterior end is united to the postfrænum[1848], with which it forms a right angle. In the Orthoptera Order the structure is not very different, but the axes and other plates of the base of the[Pg 620] wing are less distinct and rather cartilaginous; the nervures of the Anal Area often terminate in a transverse one that there forms the segment of a circle[1849]; the inner base of this circle is ligament connected with the postfrænum[1850]. In the Homopterous Hemiptera the three axes may be readily traced, but the humeral plate, with which they all are connected, is more irregular in shape, and in Fulgora longitudinal, with an angular surface; in this Order the nervure, in some cases consisting of cartilaginous rings[1851], in which the frænum and postfrænum terminate in the tegmina and wings, is attached posteriorly to the ligament of the Anal Area. In the Heteropterous section the three axes are evident, but the humeral plate is not easily made out. In the Libellulina the axes of the Costal and Intermediate Areas are the coloured broad plates at their base, formed by the dilatation of their nervures; that, however, of the Anal is not dilated, but forms one nervure, in the primary wing, with the frænum, and in the secondary with the postfrænum. Having given you this clue to trace the axes in those tribes in which they are most conspicuous, it will assist you in searching for them in the remaining Orders, in all of which they may be traced, except perhaps in those minute Hymenoptera whose wings have solely the costal nervures; probably in these there is only one axis. In the Lepidoptera and Hymenoptera a circumstance connected with the present head is observable, which is not to be discovered in the other Orders: these are the tegulæ or base-covers, which appear intended to defend the base of the anterior wings. They are concavo-convex scales, which in the Lepidoptera are large[Pg 621] and of an irregular shape[1852], but in the Hymenoptera are smaller and semicircular[1853].
2. Wings, with regard to their substance, may generally be termed membranous; but they vary in this respect, some being much thicker than others, either partially or totally: in spotted wings, as in those of many Libellulina, Tettigoniæ F., &c., the dark opaque parts are denser than those that are transparent: in several Orthopterous insects, as in Phasma, some Mantes, &c., the Costal Area or covering part of the wing is of a substance equally firm with that of the tegmen. This is a compensating contrivance, that where the latter is shorter and smaller than the former, its membranous part, when folded, may be protected from injury. Another similar contrivance of Divine Wisdom is exhibited by those Pterophyllæ K. (Locusta F.) whose tegmina resemble the leaves of plants (Pt. laurifolia, &c.); in these the tip of the wings when folded being longer, is not covered by the tegmina, and therefore exposed to injury; to prevent which this small piece, while the whole wing, as far as covered by those organs, is membranous, is of the same substance with them[1854]. The wings of most Coleoptera, Orthoptera, Hemiptera, and Thereva coleoptrata, in the Diptera, &c., are of a firmer substance than those of the other Orders; in many Locustæ Leach, Fulgoræ, &c., they are nearly as firm as the tegmina; and in Ascalaphus italicus, except at their base, the secondary wings are less membranous than the primary. M. Chabrier has observed[1855] that the wings of insects in general diminish in thickness from their[Pg 622] base to their apex, and from their anterior to their posterior margin.
3. I should have had, it is probable, but little original matter to communicate under the head of the composition and neuration of wings, had M. Jurine, who has written so ably on those of Hymenoptera, undertaken a survey of the organs of flight in every Order of insects: but as his views were confined to only two of the Linnean Orders, it is not wonderful that his system and set of terms should fail where a generalization is necessary; and I may stand acquitted of presumption and conceit if I attempt to substitute a system and body of terms more universally applicable. Had the plan of this able Entomologist led him to pay attention to tegmina and hemelytra, their division into three longitudinal areas would have immediately struck him; and having acquired this outline of the greater natural divisions, he would have applied it to the Orders that have wings only, and having discovered that it is to be traced in all, the result would have probably superseded my labors. Had his life been longer spared, perhaps something of this kind would have been effected by him; but as he, alas! is gone, and no abler hand seems to have undertaken the task, I will do what I can to give you satisfaction on this subject[1856]. You[Pg 623] have already got a tolerably good idea of these areas from what has been said upon the subject under tegmina and hemelytra; but I shall now more particularly state to you how they are circumstanced in wings. I shall first explain the general law as to their limits. The Costal Area[1857] is all that longitudinal portion of the wing that lies between the anterior margin and the postcostal nervure; the Intermediate Area[1858] is all that longitudinal portion of the wing that lies between the postcostal and the anal nervures; and the Anal Area[1859] is all that longitudinal portion of the wing that lies between the anal nervure and the posterior margin. But there are other helps to enable you to distinguish the areas in the different Orders. The Anal Area in all Orders forms the posterior fold of the wing; in Coleoptera turned under when in repose; in Orthoptera folded like a fan; in Lepidoptera, in some Papilionidæ, forming an arch over the abdomen. Again, in Blatta, the Costal Area is distinguished chiefly by longitudinal nervures; the Intermediate by oblique ones; and the Anal by radiating ones; and in both this tribe and the Mantidæ this last Area is marked out from the Intermediate by a marginal notch, which is not present in Phasma, but is found in both sections of the Hemiptera. In Locusta Leach the notch is between the Costal and Intermediate Areas: in Phasma the nervures of the Intermediate Area are branches of the externo-medial, while those of the Anal, as they do in all the Orthoptera, diverge[Pg 624] from the base of the wing: in many, as in Pterophylla K., the part of the wing lately alluded to, that is longer than the tegmen, and of the same substance, points out the limit of the Costal Area; and in others this part terminates in a segment of a circle and is differently reticulated at the apex from the Intermediate: in the Homopterous Hemiptera and the Libellulina, in which the areas at first seem indistinct, they may generally be easily traced by following them from the axes. The separation of the Costal from the Intermediate in the remaining Orders seems less easy on account of the branching of the nervures: in the rest of the Neuroptera and the Lepidoptera, if the posterior branches of the postcostal nervure are not included, you will have a narrow Postcostal Area, which in most cases forms an angle more or less prominent, in Corydalis almost a right angle, with the Intermediate: in Hemerobius and affinities this part is distinguished by areolets formed by transverse nervures, while those of the rest of the wing are longitudinal[1860]: but if the posterior branches are included, the Costal Area will be more ample: a similar observation applies to the Hymenoptera and Diptera; in these, in all cases, the areolets adjoining the anterior margin, which follow the stigma, should be regarded as belonging to the Area in question[1861]. In those tribes of the former Order, whose wings are without nervures, the areas are often marked by folds.
M. Chabrier has observed that in Coleoptera the specific weight of the margin of the wing, and its means of resistance, are augmented by a liquid which is introduced, at the will of the animal, into a long pocket under the[Pg 625] brachial, here called the costal and mediastinal nervures, covered by a supple membrane, which in a state of repose becomes flaccid[1862]: it is easily detected, being of a paler colour than the nervures between which it lies; this is what I call the Phialum; we have before seen that it exists also in Elytra and some Hemelytra[1863]; but I have not detected it in any other wings.
I have before given you a sufficiently full account of the alulæ or winglets of Diptera[1864]; and shall here only observe that they are not confined to one particular tribe, as has been usually imagined; but though sometimes extremely minute, simple, and not easily detected, are an universal distinction of the Order.
Having thus endeavoured to elucidate the larger Areas into which wings appear to be divided; I shall next say something on the smaller ones produced by the intersection or ramification of the nervures; these had been named areolets (areolæ) several years before M. Jurine's work, in which he calls them, I think improperly, cellules (cellulæ), was published; I therefore retain the prior term. The general structure of the nervures of the wings of insects having been before explained[1865], I shall not here repeat what I then said; but there is a curious circumstance connected with it, particularly visible in the wings of certain Hymenoptera, that I must not pass without notice. If you examine attentively with a microscope against the light the wing of any Nomada or Andrena, you will discover little transparent points in some of the smaller transverse nervures that form the middle areolets, in which the nervure becomes white and looks as if it[Pg 626] was interrupted, though in substance it seems continued: these little points, somewhat resembling minute air bubbles detained in the tubes, are what M. Jurine, who first discovered them, has, on that account, named bullæ, which he thus further describes:—"When the tube (of the nervure) arrives at the spot where a bulla is to be formed, it extends itself on all sides in minute threads in the upper membrane of the wing, losing its colour and tubular structure, which it resumes immediately after the formation of the bulla[1866]." But if you look closely at them you will find that there is always a slight fold of the wing that cuts the nervure exactly at the bullæ, and if the fold changes its direction they accompany it; their object, therefore, is clearly to relax the tension so as to admit a little motion where the fold is; consequently, rather than bullæ (bubbles), they should be denominated articulations. A similar construction, but on a larger scale, may be observed in the wings of Coleoptera[1867] and some others, as Psocus, where the folds traverse the nervures. I shall next make a few observations on the principal nervures; and first a word upon their names. M. Jurine, being of opinion that a striking analogy exists between the wings of insects and those of birds, in which M. Chabrier seems to agree with him, has named the nervures in the anterior margin of the wings of the former, radius and cubitus, as corresponding with the bones so named in the fore-arm of the latter, and the plate which often terminates these nervures in Hymenoptera, he names the carpus; it may look like presumption to differ from two such weighty authorities, but as their observations seem to have been too[Pg 627] limited, in one case to the Hymenoptera and Diptera only; and in various Orders there is nothing analogous to the stigma or carpus, and all the other nervures of an insect's wing have no analogue in that of a bird, but more especially as M. Latreille seems to think with me on this subject[1868], I have retained Linné's term for the marginal nervure, and for most of the others have adopted those of the great French Entomologist just mentioned. I shall here only further observe,—and it seems to me an observation of prime importance, in the determination of the question of the analogy of the wings of insects,—that they are not, as in birds, the fore-leg converted into an organ of flight, but, like the wing of the Draco, an organ superadded to the legs; and, further, that the connection is not with the fore-legs, but, as has been before observed[1869], with the two posterior pairs.
The Costa[1870] is usually the strongest of the nervures, and that upon which the wing seems to be built; but in some cases, as in Blatta, Scutellera, Cynips, &c., it is represented by the mere membrane of the anterior margin; in some Coleoptera, as in Geotrupes, Dytiscus, &c., its structure, except at the base, appears to be annular or nearly so, at least a vast number of corrugations, running transversely, are observable on its upper and lower surfaces; it is thus capable of greater tension and relaxation, and more flexile. The stigma or carpus[1871], though most conspicuous in the Hymenoptera Order, may be traced in some Coleoptera, Heteropterous Hemiptera, the Libellulina, &c.; but it has no representative in the Orthoptera, Lepidoptera, Trichoptera, &c. The mediastinal is usually a[Pg 628] very slender nervure, placed between the costa and postcosta, sometimes terminating in the former[1872], and at others in the latter[1873]: in the Orthoptera, Lepidoptera, &c., however, and some others, it is a very conspicuous and principal one[1873]; in the Hymenoptera it is obsolete, merging in those nervures[1874]. The Postcosta is the principal nervure of the wing in Scutellera, but in Staphylinus it is wanting; in Chalcis sispes it is the only true nervure of that organ, the others being represented by spurious ones[1875]. The externomedial and internomedial are sometimes distinct at their origin, but more frequently are branches from a common stem.
Having made these general remarks, I shall now consider particularly the neuration of the wings in the different Orders, beginning with the Coleoptera. The first thing that strikes the physiologist in surveying a wing belonging to an insect of this Order, is the general arrangement of the nervures[1876]; which are so placed that the required degree of tension may be given to every part of this organ: thus some are nearly straight[1877]; others run in a serpentine direction[1878]; others are forked with one branch recurrent and another proceeding onwards[1879]; others again are insulated, or do not originate from the base of the wing, or from other nervures, but are merely placed to strengthen an open space of it[1880]: these nervures are also usually broader and more substantial than those of the wings of the subsequent Orders. Another striking circumstance with regard to them is that the nervures form few or no[Pg 629] closed areolets, except in the Costal Area, where they are inconspicuous; in Dytiscus marginalis, indeed, and Tenebrio Molitor one or two may be found, but in general there are none. In many of this tribe the postcosta, which terminates at the joint of the wing, becomes recurrent, so as to form a hook, which perhaps represents the stigma, as in Dynastes[1881]; in Creophilus K., a rove-beetle, there is no hook but a broad plate adjacent to the costa. In the Strepsiptera Order the neuration is extremely simple, the nervures, except one insulated one, diverging from the base of the wing[1882]: in this respect, as well as in the form of that organ, an approach is made to the Orthoptera. In the Dermaptera this approach is still more evident; in the common earwig[1883], the diverging nervures become numerous; between each is an insulated one, taking its origin in the middle of the wing, and running to the margin; a little nearer to the latter all the nervures are dilated into a plate; those of the anal area are angular[1884], and the exposed part of the costal is as hard as the elytra. The neuration in the Orthoptera Order may be called radiate, the longitudinal nervures for the most part diverging from the base of the wing like rays: in some few instances[1885], but not often, I believe, an insulated nervure intervenes between each; traversing or connecting nervures, cutting the longitudinal ones in various directions, ornament these wings with an infinity of areolets, causing them to resemble fine gauze or beautiful lace or net-work; very often these areolets are quadrangular, sometimes rhomboidal,[Pg 630] frequently nearly circular, and differing occasionally, as has been before observed[1886], in the different areas: it sometimes occurs that there are no traversing nervures[1887], when the wing of course is without areolets. In the Heteropterous Hemiptera the type of neuration, as to the wing, seems borrowed from the Coleoptera, a further proof that these are the analogues of that Order amongst the Haustellata Clairv. In these the nervures usually are few and dispersed, and seldom form any closed areolets. If you examine any Scutellera, Pentatoma, or Lygæus, you may trace the uncinated, forked, serpentine, and insulated nervures of Coleopterous insects; in Gerris and Velia there is an approach to the neuration of some Homopterous species, and in Belostoma &c. the wing is reticulated by spurious nervures. In the Homopterous section there are several types of neuration; thus the Fulgoræ resemble the Orthoptera in this respect; while the Tettigoniæ F., &c., approach nearer to the Hymenoptera and Diptera, and have their apical areolets circumscribed within the margin by a traversing nervure; in Flata, &c., the areolets are mostly formed, not by traversing nervures, but by the branching of the longitudinal ones; in this respect they are not unlike the Lepidoptera. In this last-named Order there are some variations with regard to their neuration—thus, amongst the butterflies in Urania, &c., there is no closed areolet in any of the wings, and almost all the nervures diverge from the base[1888]; in Morpho, &c., there is only one in the primary wing[1889]; in Heliconia, &c., there is one in both wings; amongst[Pg 631] the moths, in the Bombyces L., this is divided into two, and in Cossus labyrinthicus Don. into three areolets: in some butterflies (Lycæna) there is one insulated nervure[1890], and in others (Hesperia) there are two[1891]; in these two last, and Heliconia, Urania, &c., the end of the Costal Area is divided into several areolets by oblique nervures[1892], which gives them some analogy to the wings of many Neuroptera; and at the base of this Area, in Morpho, is a roundish areolet[1893]. In this Order the externo-medial and interno-medial nervures coalesce into one, and are only represented separately by their first and third branches[1894]. In the Neuroptera Order the general type of neuration is borrowed from the Orthoptera; but in Osmylus, Termes, &c., there is an approach to that of Flata in the Homopterous Hemiptera, and in Psocus to others of that section; in the second of these genera the nervures, except those of the costal margin, are spurious.
I now come to the Order in which M. Jurine has laboured with so much success, I mean the Hymenoptera; and I only regret that his labours were directed to so small a portion of the Class Insecta, and in that portion only to a part of the upper wing; I say only a part, because all those areolets of the posterior part of the wing, in some cases amounting to five[1895], that lie behind his cubital cellules, are not employed by him as diagnostics, and are left without a name. By dividing the areolets of the[Pg 632] Intermediate Area of these wings into three portions, the basal, medial, and apical[1896], I have endeavoured to remedy this defect, and by naming each set of areolets in the middle portion, as you will see in the Orismological Definitions, under the term Areolets, you will find it easy to describe any given areolet and its place in the wing; those of the base may be called the anterior, intermediate, and posterior, where three occur; and the first and last of these terms will suffice where there are only two; the apical areolets, or those that are open to the margin, may be called, first, second, and third in the order of their occurrence, reckoning from the anterior or costal margin.
In this Order it is curious to trace the progress of neuration in the wings of different genera. Thus in Psilus only the costal nervure and the stigma are to be traced[1897]; in Chalcis the postcostal and stigma[1898]; in Codrus and Leucospis the costal, postcostal, stigma, and a nervure representing the externo-medial and interno-medial coalescing into one[1899]; in Omalus the basilar areolets appear[1900]; in Crabro both basilar and medial[1901]; in Cynips basilar, medial, and apical[1902]; and in Hylotoma the wing is filled with its greatest complement of areolets[1903]. The medial areolets of the Intermediate Area, as you will see in the definitions, form three distinct series; these may be called the protomesal, deuteromesal, and tritomesal, reckoning from the postcostal areolets; the first of these corresponds with the cubital cellules of Jurine. These series may be[Pg 633] expressed, according to the number of their areolets; by figures, the protomesal standing first. They vary much in this respect in the different genera. Thus in Cyclostoma K.[1904], reckoning the didymous areolet as two, the numbers will stand 4:2:1; in Hylotoma, &c., 3:2:1[1905]; in Aulacus, &c., 2:2:1[1906]; in Bracon, &c., 2:1:1[1907]; in Chelonus, 2:0:1[1908]; in Cynips erythrocephalus Jur., 2:0:0[1909]; in Formica, 1:1:1[1910]; in Oxybelus, 1:0:1[1911]; in Chrysis, 0:1:1[1912]; and in Cynips Rubi K., 1:0:0[1913]. The most natural number is 3:2:1. The next in importance to the medial areolets of the Intermediate Area are the apical, or those open to the margin; the most usual number of them, excluding the postcostal areolets which belong to the Costal Area, is three; but in Sirex there is an approach to four[1914]; in Evania there are only two[1915]; and in Philanthus there are none[1916]; in many, as Prosopis, Nomada, Andrena[1917], though there is the usual number, they are incomplete and do not reach the margin. The basal areas are of little importance in assisting to determine genera; they are most commonly two in number, but in Cynips, &c., there is only one[1918]. The shape and other circumstances of the areolets vary considerably[Pg 634] in different genera and species: upon these however I shall not enlarge further, but proceed in the next place to consider very briefly the wings of the Diptera Order as to their neuration. These are not so easily made subservient to a general plan. The basilar areolets are now reduced considerably in length, occupying merely the base of the wing[1919]; the medial are become less numerous and important[1920]; and the apical, in a variety of instances, are the most conspicuous[1921]; in some wings, as in those of Penthetria, the Intermediate Area has no nervures or areolets, or only spurious ones; in Psychoda the nervures diverge from the base almost without branching, so as to form no closed areolets[1922]; in many, the lower medial areolets are very long, resembling the basilar in Hymenoptera[1923]; these are often crowned by a single small one, as in the Stratyomidæ, Tipula, &c., from which numerous branches proceed to the margin[1924]; but in Musca two large ones approach the margin, the anterior one having an angle open to it[1925]; in the Hippoboscidæ almost the whole of the wing is occupied by the apical areolets[1926]; though in some cases they are incomplete[1927].
4. I am next to consider the position of wings in repose and their folding. The most important object of this is that when unemployed they may occupy less space, be less in the way of the insect, and be most effectually protected from injury. Another end is also served by this structure,—that wings can thus be very ample, and present[Pg 635] a large surface to the action of the atmosphere without incommoding the insect when it has not occasion to use them.
With respect to this head, insects may be divided into two classes—namely, those whose wings in repose are covered by wing-cases harder than the wings themselves, and those that have no such protection. In the former the wings, though the rule admits several exceptions, have more folds than in the latter. As the different mode of folding the wings has been assumed for a characteristic of the earlier Orders, I shall explain to you with as much brevity as possible how each is circumstanced in this respect, beginning as usual with the Coleoptera.
There are two principal folds of the wing in this Order, which may be named the anal and the apical: the former is when the Anal Area or part of it is folded on the under surface of the base of the wing; this fold is always more or less longitudinal: the latter, the apical fold, is by means of the commissura or joint of the postcosta lately mentioned: which in Hister, Staphylinus, &c., for obvious reasons[1928] is nearer the base of the wing; in Necrophorus in the middle; in Dynastes Aloeus beyond the middle[1929]; in Tenebrio Molitor near the apex; and in Dytiscus marginalis there appears to be no joint at all; but the fact is, that in this insect the postcosta,—the termination of which really forms the joint, the costa itself being only flexible at that point,—stands at a greater distance from the latter at its end. Well, at this joint the above fold is made, the apex of the wing, being first folded longitudinally, turning under and inwards, and forming an[Pg 636] angle, more or less acute, with the joint or costal margin, so that the fold is not quite but nearly transverse: this at least is the case in Geotrupes stercorarius and other Lamellicorns: in Staphylinus, &c., there are several transverse and longitudinal folds, and thus the wing is more easily packed under the short elytra; in Molorchus, Necydalis, &c., in which it is left uncovered, except at its base, the anal fold takes place, and the apical in some degree; a short portion near the apex forming an obtuse angle with the margin; in Atractocerus the wing appears to be only longitudinally folded; and in Buprestis vittata only the anal fold is to be detected. Besides these transverse and longitudinal folds these organs, in many beetles, have an infinity of fine corrugations, which ramify like the nervures of the tegmina of Flata[1930], &c., proceeding from the Costal Area or the disk of the wing to the posterior margin; the object of these plicatures is doubtless to present a more ample surface to the action of the atmosphere in flight[1931]. When all these folds have been made in a Coleopterous wing, the apex of the one at its posterior margin crosses or rests upon that of the other[1932].
In the Dermaptera[1933], at least the common earwig, there is a triple transverse fold of the wing, and besides this it has numerous longitudinal ones like those of a fan, each of the diverging nervures representing one of the sticks. In the Strepsiptera the folds are only longitudinal; a circumstance which, besides the form and neuration of the wing, sufficiently attests that its station is more near the Orthoptera and Coleoptera than the Diptera.[Pg 637] We next come to the Orthoptera[1934]; in these the folds in general are longitudinal; and those of the Anal Area in particular, either in whole or in part, exact counterparts of a fan: wherever there is a straight nervure, there is usually a fold or a tendency to it; this is the case even with the short oblique ones observable in the Intermediate Area of Blatta: in this tribe the Anal Area, or a considerable portion of it, is folded under the rest of the wing, and the whole lies on the back of the animal, so that in this wing there are only two primary folds; but in those with a narrower body, as Phasma, &c., there are more, and the Anal Area, folded like a fan, lies horizontally on the back; the Costal is vertically applied to the sides, and the Intermediate is between both, as in the tegmina[1935]. In Gryllus Latr., Gryllotalpa, &c., when the wings are folded, the end of the Anal Area projects so as to present the appearance of two tails[1936]; and in that remarkable Chinese animal Gryllus monstrosus, in which these tails are very long, they are convolute like those of some quadrupeds[1937]. It is to be observed that in the secondary folds of these wings the angles of the folds are surmounted by a nervure.
In both sections of the Hemiptera Order, as in the Coleoptera, the Anal Area is turned under the wing and lies over the back of the insect; this is the only primary fold, but besides there are several longitudinal semifolds or secondary ones, in which one part of the surface forms an obtuse angle with another; and in Tettigonia, &c., these folds ramify in the wings as well as in the tegmina at the[Pg 638] margin: a number of semifolds also, sometimes transverse and sometimes oblique, run in pairs from each side of every nervure of the disk of both tegmina and wings in the genus last named, the use of which has been before mentioned[1938].
We now come to those Orders that have four membranous wings: first, I shall consider the Lepidoptera. With respect to the position of their wings in repose some variations take place. In the majority of the day-fliers (Papilio L.), when the animal reposes the wings are applied to each other by their upper surface so as to be vertical; but in the skippers (Hesperia), the secondary wings assume a horizontal position, while the primary are vertical but applied to each other. In the Crepuscular tribes (Sphinx L.) the upper wings are incumbent on the lower, and deflexed. In the night-fliers (Phalæna L.) the types of position are various. In some Attacus, Saturnia, Noctua, &c., the wings cover each other, and are a little inclined from a horizontal position; in Gastropacha, Odenesis, and some other Bombycidæ, they are deflexed, and the anterior margin of the under wing projects beyond that of the upper: in some of the Tineæ L., as Crambus, the wings are convoluted, and in others, Galleria, they are applied close to the sides of the body, and being elevated at the apex, terminate, to use a French term—en queue de coq: in Noctua, Geometra, &c., the wings usually cover the abdomen, and are nearly horizontal. With regard to the folds of their wings, the Anal Area of the secondary is the only part that has any striking one; in Papilio Hector and affinities it turns[Pg 639] up so as to defend the sides and part of the back of the abdomen; in Morpho Teucer it turns down, and meeting that of the opposite wing, forms a semitube which receives and shelters that part below. In the Crepuscular and Nocturnal Lepidoptera this fold, especially in the former, is very slight. With respect to semifolds in the Diurnal, there is one originating in the disk, between each of the nervures, that goes to the margin of the wing; likewise the under wings, particularly of many Noctuæ, Arctiæ, &c., have many longitudinal semifolds.
In the Neuroptera Order several variations take place with regard to the position of these organs in repose: thus, in Æshna, Libellula, &c., they continue expanded; in Argion they are applied to the body; in Myrmeleon the upper are horizontally incumbent on the lower; in Hemerobius they incline to the horizon. With regard to their folds in Æshna, &c., the longitudinal nervures alternately form the summit or the bottom of a semifold, as do those branches that terminate in the posterior margin; this kind of plicature may be observed, but in a less degree, in Ascalaphus, Myrmeleon, &c.; in Panorpa every nervure is the ridge of a slight fold; in Termes, on the contrary, it forms its bottom. In the Trichoptera, the under wing being much more ample than the upper, the Anal Area forms a fold under the wing, and there seem longitudinal secondary folds besides.
We now come to the Hymenoptera. In this Order the wings, as to their position in repose, are usually incumbent upon each other, and cover the abdomen; in the Vespidæ, however, they are placed parallel to the body, but do not cover it. Before I notice the plicature of these wings, I must recall your attention to what I lately[Pg 640] observed[1939] with regard to Jurine's bullæ (bubbles), but which are really the joints of the nervures, as they are to be found only where the folds pass; and where they exist they are an index by which the folds, or rather semifolds, may be traced. I counted eleven of these little joints in the upper wing of Andrena cineraria; sometimes, however, instead of a bulla, a nervure stops short to admit the fold. Wings in this Order have often three longitudinal semifolds more or less conspicuous; these you may trace in the saw-flies (Tenthredo L.), whose wings Linné terms tumidæ, by which term he would indicate the elevation of the whole surface produced by this structure; in the under wings of these, and Scolia, Bembex, &c., the Anal Area is turned under the wing, as in many preceding tribes[1940]: in Sirex, &c., that Area of the upper wing turns upwards, forming an acute angle with the rest of the organ; the same circumstance distinguishes the under wing in the Ichneumonidæ. Several apical semifolds, marked by a pellucid streak, distinguish Tiphia F., and in Bombus, Bembex, &c., an infinity of branching ones, like those before described in Coleoptera, corrugate the apical margin. In the Vespidæ the upper wings are folded longitudinally into three nearly equal portions, but in the under ones the Anal Area only forms the fold.
In the Diptera Order, as to their position when at rest, the wings are mostly incumbent one on the other; but in Psychoda they are deflexed, so as to form a kind of penthouse. With regard to their plication, in some, Tipula oleracea, &c., a slight oblique semifold runs from the[Pg 641] stigma to the apical margin, and the Anal Area has two, as it has in many Muscidæ, itself forming nearly a right angle with the rest of the wing; besides these it is corrugated with minute transverse semifolds, which are observable also in several other Dipterous insects; in many Stratyomidæ they are oblique, and run from the disk to the posterior margin; and in Asilus, Bombylius, &c., they are wavy.
5. We are next to say something upon the shape of wings: this, though apparently extremely various in the different Orders and tribes, may I think be traced in every wing to one original prototype, a triangle with the largest angle rounded and subtended by the anterior or costal margin: in some, as the Coleoptera, Orthoptera, &c., this type of formation is a right-angled triangle[1941]; and in others, as in the Hymenoptera, Diptera, &c., the majority of the Neuroptera, &c., it is an obtusangled one[1942]; it may be further observed, that in receding from these forms wings very often assume that of the half or quadrant of some regular figure, as we shall see when we consider those of the different Orders. Another general observation I shall first mention,—that these organs are universally narrowest at their base and widest at the apex, provided we consider as the apex the termination outwards of the three Areas; otherwise we might say that wings in the Coleoptera, Orthoptera, &c., were wider at the base than at the apex[1943]. The wings in the former Order, and in several of the Heteropterous Hemiptera, as Gerris, Velia, &c., may in general, as to their shape, be termed semicordate or semiovate[1944]; in the Dermaptera they incline to an oval figure[1945]: in the[Pg 642] Strepsiptera, Orthoptera, most Homopterous and many Heteropterous Hemiptera, they approach to the quadrant of a circle; in a considerable portion of the Lepidoptera the two under wings, if united at their posterior margin, approach a circular form; the upper ones vary a little from the prototype of the under ones, forming an obtusangled triangle[1946]; in many Neuroptera the primary wings may be called oblong or linear-oblong, while the secondary betray more evidently the right-angled or obtusangled triangle; in the Hymenoptera this latter form is every where conspicuous, with little deviation, except in the rounding of the angles[1947]; and, finally, in the Diptera this form shades off again into an oblong, ovate, or linear shape, the wing being most commonly attenuated at the base into a kind of footstalk[1948]. Some singular variations with respect to the termination or marginal processes of the wings are exhibited by many Lepidoptera; thus in Attacus Atlas, &c., the primary wings are falcated or hooked at their apex[1949]; and in great numbers both wings are there scolloped into alternate bays and capes, if I may so speak, varying in depth and length[1950]. There is usually a sinus between every pair of nervures, each of which terminates in the adjoining prominence, as a fold does in the sinus[1951]. Where present, in the primary wings there are eight of these sinuses, and in the secondary, where they are most usual, seven; some are remarkable for the long tails which distinguish their secondary wings; those in Papilio are usually an elongation of the fifth, from[Pg 643] the anterior margin, of the prominences before mentioned, into a spathula-shaped diverging process, varying in length and width[1952]: but in P. Ulysses it does not diverge; and in P. Podalirius it is linear. They are found also in other subgenera; thus in Urania Patroclus there are two; in U. Riphæus three; in Erycina Cupido five; and in E. Endymion six of these tails; in some, as in E. Dorylas, the whole wing seems to form the tail; in others again, as in Hesperia Proteus and Bombyx Luna, it is an elongation of the anal angle. Other wings in this Order are divided into lobes resembling feathers, as you may see in Pterophorus hexadactylus, &c.[1953]
6. We are next to consider the clothing of wings: these, in the Orders in which they are covered by elytra, tegmina, or hemelytra, are generally naked, except that the spots in those of Fulgora laternaria, serrata, &c., and the whole wing in Flata, Aleyrodes, and others, are covered with a kind of farinaceous powder; but in all the remaining Orders, hairs or scales are more or less implanted in these organs: as the Lepidoptera are the most remarkable for the clothing of their wings, I shall leave them till the last, and begin with the Neuroptera. If you lightly pass your finger over the wing of any dragon-fly (Libellula F., Æshna F.), from the apex towards the base, you will find that the longitudinal nervures are, as it were, serrulated with very minute bristles, which point towards the extremity; if you next move the finger across the wing, from the posterior to the anterior margin, a similar circumstance will strike you. M. Chabrier conjectures that, amongst other uses[1954], these hairs may contribute[Pg 644] to fix the atmospheric fluid when the wings are depressed in flight, while it glides over them as they rise[1955]; in Ascalaphus, Myrmeleon, Nemoptera, Hemerobius, &c., the nervures are more visibly bristled; the bristles diverging on each side from the longitudinal ones, but all pointing towards the apex from the connecting or transverse ones; in Panorpa, besides these bristles, short hairs, pointing the same way, are thickly planted in the membrane of the wing; and in Hemerobius the margins of the wing are fringed; in the Ephemerina, Corydalis, &c., the wings are naked. In the Trichoptera Order, as their name imports, they are covered with minute decumbent hairs, less easily seen but still existing in the secondary pair. In the Hymenoptera in general the wings are covered with minute hairs or bristles; but in Tiphia, Scolia—with the exception of S. Radula and affinities in which they are hairy—and others, the wings are nearly naked; in Pompilus, Pepsis, &c., the hairs are infinitely numerous and very short; in the Sphecidæ, Mutilla, &c., they are more distinct, longer, and less numerous; in the humble-bee (Bombus) and many others the apex of the wing is darkened by a large number of more conspicuous hairs, each of which seems to spring from a minute tubercle: as these tubercles are in a part of the wing that is strengthened by few nervures, they may probably be intended to supply their place, in giving firmness and tension to this part. The wings of Diptera, under the present head, may be viewed with regard to the hairs that are implanted in the membrane of the wing, in its nervures, and in its margin. In the first view, in[Pg 645] Stratyomis and immediate affinities the wing is nearly naked; but in Xylophagus, Beris, and the great majority of the Order, the membrane of the wings is thickly planted with innumerable very minute bristles, not to be seen but under a powerful lens, often black, and seemingly crowning a little prominence, and giving the wing an appearance of the finest net-work. As to the clothing of the nervures, the costal, in Anthrax, Bombylius, &c., is often remarkably bristly at the base, with hairs intermixed; in Œstrus Ovis, in the inner margin or edge of this nervure, is a single series of bristles, or rather short spines, like so many black points; in Œ. Equi the whole costa is covered with short decumbent hairs or bristles; in Musca pagana F., just at the apex of the costal areolet, that nervure is armed with a spur or diverging bristle larger than the rest, which is also to be found in many others of the Muscidæ, some of which have two and others more of these spurs. The little moth-like midges (Psychoda Latr., Hirtæa F.) at first appear to have the whole surface of their wings covered with hairs; but upon a closer examination it will be seen that they are planted in the nervures, from each of which they diverge, so as under a lens to give it a very elegant appearance[1956]. This fly has its wings beautifully fringed with fine hairs, the third circumstance to be attended to under this head; in the Tipulidans, and many others of this Order, the apex and posterior margin are also finely fringed with short hairs. Some Dipterous insects make a near approach to the Lepidoptera in the covering of their wings: in the common gnat, when the wings are not rubbed, the nervures are adorned by a double series of scales, and the[Pg 646] marginal fringe also consists of them[1957]; and in a Georgian genus, which appears in some degree to connect Culex with Anthrax &c., there are scales scattered upon the membrane as well as upon the nervures; besides, its antennæ[1958] and abdomen are also covered with them.
The Order, the clothing of whose organs of flight excites the admiration of the most incurious beholder, is that to which the excursive butterfly belongs, the Lepidoptera. The gorgeous wings of these universal favourites, as well as those of the hawk-moths and moths, owe all their beauty, not to the substance of which they are composed, but to an infinite number of little plumes or scales so thickly planted in their upper and under surface, as in the great majority entirely to conceal that substance. Whether these are really most analogous to plumes or scales has been thought doubtful. De Geer is inclined to think, from their terminating at their lower end in little quills and other circumstances, that they resemble feathers as much as scales[1959]; Reaumur on the contrary suspects that they come nearer to scales[1960]. Their substance, approaching to membrane, seems to make further for the former opinion, and their shape and the indentations that often occur in their extremity, furnish an additional argument for the latter. Their numbers are infinite; Leeuwenhoek found more than 400,000 on the wings of the silk-worm moth (Bombyx Mori)[1961]; and in those of some of the larger moths and butterflies the number must greatly exceed this. You will observe[Pg 647] however that in many Lepidoptera the wings are partially, and in some instances generally, transparent: thus in Hesperia Proteus, a butterfly before noticed for the long tail that distinguishes its secondary wings, there are many transparent spots; in Attacus Atlas, one of the largest of moths, and its affinities, there is as it were a window in each wing formed by a transparent triangular space; in A. Polyphemus, Paphia, &c., the pupil of the ocellus is transparent, which in the former is divided by a nervure. In several of the Heliconian butterflies, and in Zygæna F., &c., the greater part of both wings is transparent, with scales only upon their nervures, round their margin, or forming certain bands or spots upon them; in Parnassius Apollo, Mnemosyne, &c., the scales are so arranged as not wholly to cover the wings, which renders them semidiaphanous; and in some (Nudaria) the wings are intirely denuded. With regard to size, the scales vary often considerably in different tribes; in Heliconia they appear to be more minute than in the rest; and in Castnia they are the largest and coarsest; the extremity of the wings of Lepidopterous insects in general is fringed with longer scales than their surfaces, and even those of the last in the same wing; sometimes vary in magnitude. The little seeming tooth that projects from the middle of the posterior margin in the upper wings of Notodonta, a subgenus of Bombyx L., is merely produced by some longer diverging hairs. The shape and figure also of scales are very various—some being long and slender; others short and broad; some nearly round; others oval, ovate, or oblong; others spathulate; others panduriform or parabolical; some again almost square or rhomboidal; many triangular; some representing an isosceles triangle, and[Pg 648] others an equilateral one; lastly, some are lanceolate and others linear; again, some have a very short pedicle and others a very long one: with regard to their extremity; some are intire, without projecting points or incisions, while others are furnished with them: of these some terminate in a single long mucro, others have several shorter ones; some are armed with teeth, varying in number from two to thirteen in different species[1962]. Many other forms might be enumerated, but these are sufficient to give you a general notion of the infinite variety of this part of the works of the Creator. I must next say a word or two upon their arrangement on the wing. In most instances this is in transverse lines, which sometimes vary a little from a rectilinear course, and the extremity of the scales of one row reposes on the base of those of the succeeding one, so that in this respect their arrangement is like that of tiles in a roof: in some cases it is not so regular: thus the minute scales on the wings of Parnassius Apollo, and others with subdiaphanous wings, are arranged without order; in Pieris and other Diurnal Lepidoptera, and many of the Crepuscular and Nocturnal, there appears to be a double layer of scales on both sides of the wing; the under layer usually consisting of white ones. If you denude the wings of any butterfly, which you may easily do by scraping it lightly on both sides with a penknife, you will be amused to trace the lines in which the scales were planted, consisting of innumerable minute dots: the lines of the under side, in some cases, so cut those of the upper side, as by[Pg 649] their intersection to form lozenges. With regard to the position of the scales on the wing, they usually lie flat, but sometimes their extremity is incurved: in the beautiful Argynnis Vanillæ a very singular appearance of numerous transverse ridges is produced by the extremity of those scales that cover the longitudinal nervures of the primary wings, except at the base, being recurved.
But though the general clothing of the wings of Lepidoptera consists of these little scales, yet in some cases they are either replaced by hairs or mixed with them. Thus, in the clear parts of the wings of Heliconians, Attaci, &c., short inconspicuous hairs are planted; in a large number of the Orders the upper side of the Anal Area of the secondary wings is hairy; in several Crepusculars (Sphinx Phœnix, &c.), where there is a double layer as before mentioned, the upper one consists of dense hairs, except at the apex, and the lower one of scales; and in most of them the scales of the primary wings are piliform, and the secondary are covered by what approach very near to real hairs; many of the Attaci are similarly circumstanced: the four wings of A. Cytherea are also covered externally with hair.
7. Before I conclude this long diatribe on the organs of flight of insects, I must not omit some notice of the infinite diversity of colours with which their wings are often variegated and adorned by the Creator, who loves to delight us by the beauty, as well as to astonish and awe us by the immensity and grandeur of his works. Though the wings in every Order exhibit instances of brilliant and beautiful colouring, yet those of the Lepidoptera in this respect infinitely excel them all, and to these, under this head, after noticing a few in the less privileged Orders,[Pg 650] my observations will be confined. Although in the Coleoptera the wings are seldom distinguished by their splendour; yet those of some Cetoniadæ, as Cetonia africana, are extremely brilliant, and resemble those of many Xylocopæ in the lovely violet hue that adorns them: amongst the Orthoptera some Pterophyllæ, and in the Homopterous Hemiptera some Fulgoræ, emulate the Lepidoptera in the ocelli that give a kind of life to these organs[1963]; and a vast number of the destructive tribe of locusts (Locusta Leach) are remarkable for the fine colours and gaiety of their wings[1964]; in the Neuroptera numerous Libellulinæ emulate the Heliconian butterflies by their maculation; and in the genus Ascalaphus, which represents the Lepidoptera by its clubbed antennæ[1965], many also have the resemblance increased by the painting of their wings, so that some Entomologists have actually considered some of them as belonging to that Order[1966]; the wings of the Xylocopæ, before alluded to, sometimes add to the deep tints of the violet—which also prevail in the wings of several Diptera—towards their extremity the most brilliant metallic green or copper varying,
and even those wings that consist of clear colourless membrane are often rendered extremely beautiful from the reflection of the prismatic colours. I should undertake an endless task did I attempt to specify all the modes of marking, clouding, and spotting, that variegate a wing, and all the shades of colour that paint it, amongst the Lepidopterous[Pg 651] tribes; I shall therefore confine myself to a few of the principal, especially those that distinguish particular tribes and families. Of whole coloured wings—I know none that dazzle the eye of the beholder so much as the upper surface of those of Morpho Menelaus and Telemachus: Linné justly observes that there is scarcely any thing in nature that for brightness and splendour can be paralleled with this colour; it is a kind of rich ultramarine that vies with the deepest and purest azure of the sky; and what must cause a striking contrast in flight, the prone surface of the wings is as dull and dark as the supine is brilliant, so that one can conceive this animal to appear like a planet in full radiance, and under eclipse, as its wings open and shut in the blaze of a tropical sun: another butterfly, Papilio Ulysses, by its radiating cerulean disk, surrounded on every side by a margin intensely black, gives the idea of light first emerging from primeval obscurity; it was probably this idea of light shining in darkness that induced Linné to give it the name of the wisest of the Greeks in a dark and barbarous age. I know no insect upon which the sight rests with such untired pleasure, as upon the lovely butterfly that bears the name of the unhappy Trojan king (P. Priamus); the contrast of the rich green and black of the velvet of its wings with each other, and with the orange of its abdomen, is beyond expression regal and magnificent. But peculiar beauties of colour sometimes distinguish whole tribes as well as individuals. What can be more lovely than that tribe of little butterflies that flit around us every where in our summer rambles, which are called blues, and which exhibit the various tints of the sky? Lycæna Adonis of this tribe scarcely yields to any exotic butterfly in the[Pg 652] celestial purity of its azure wings: our native coppers also, Lycæna dispar[1967], Virgaureæ, &c., are remarkable for the fulgid colour of these organs; in Argynnis the upper side of their wings is tawny, spotted with black, while the under side of the secondary ones is very often adorned by the appearance of silver spots. How this remarkable effect of metallic lustre, so often reflected by spots in the wings of butterflies, is produced, seems not to have occupied the attention of Entomologists. M. Audebert is of opinion that the similar lustre of the plumes of the humming birds (Trochilus) is owing to their density, to the polish of their surface, and to the great number of little minute concave mirrors which are observable on their little beards[1968]. But these observations will not apply to the scales of the wings of butterflies, which are always very thin and generally very flat: in some instances, as in Morpho Menelaus, there appears more than one very slight channel upon a scale; but this takes place also in others that reflect no lustre. Their metallic hues must therefore principally be occasioned by the high polish of their surface and the richness of their tints. It is the purity of the white, in conjunction with their shining surface, contrasted with the dull opaque colour of the under side of the secondary wings, that causes the spots that decorate those of the Fritillaries (Argynnis) to emulate the lustre of silver. In Papilio the Trojans are distinguished by the black wings with sanguine spots, and the Greeks by the same with yellow spots; but these have proved in some instances only sexual distinctions[1969]. In the Danai candidi L. the colour of the tribe may be described as[Pg 653] sacred to the day, since every shade, from white or the palest yellow to full orange, is exhibited by them. The yellows prevail also in those Noctuæ, the trivial names of which Linné made to end in ago, as N. Fulvago, Citrago, &c. I must not conclude this part of my subject without noticing one of the most striking ornaments of the wings of Lepidoptera, the many-coloured eyes which decorate so large a number of them. Some few birds, as the Peacock and Argus Pheasant, have been decked by their Creator very conspicuously with this almost dazzling glory; but in the insects just named it meets us every where. Some, as one of our most beautiful butterflies, Vanessa Io[1970], have them both on the primary and secondary wings; others, as Noctua Bubo[1971], only on the primary; others again, as Smerinthus ocellata[1972], only on the secondary: in some also they are on both sides of the wing, as in Hipparchia Ægeria[1973], and in others only on the upper side, as in Vanessa Io; in others again only on the under side, as in Morpho Teucer[1974]: in some likewise they are very large, as in the secondary wings of the same butterfly: and in others very small, as in those in the wings of the blues (Lycæna). Once more, in some they consist only of iris and pupil, as in Hipparchia Semele, and in others of many concentric circles besides, as in Morpho Teucer, &c.
v. Legs[1975]. We are next to consider those organs of motion affixed to the trunk, by which insects transport themselves from one place to another on the earth or in[Pg 654] the water, and by which also they perform various operations connected with their economy[1976]. In treating of them we should consider their number; kind; substance; articulation with the trunk; position; proportions; clothing; composition; folding; and motions.
1. Number. Having before very fully explained to you the number and kind of the legs of insects in their preparatory states[1977], I shall now confine myself to the consideration of these organs in their perfect or last state; beginning with their number. Insects, properly so called, as I formerly observed[1978], in this state, including the anterior pair or arms, have only six legs, none exceeding or falling short of this number; but in several of the Diurnal Lepidoptera (Vanessa, &c.) the anterior pair are spurious, or at least not used as legs, the tarsi having neither joints nor claws[1979]; this in some cases is said to be only a sexual distinction[1980]. In Onitis, Phanæus, and some other Scarabæidæ McL., the arm has either none or a spurious tarsus or manus[1981]; which in the first of these genera is also a sexual character. From both these instances we see that walking is only a secondary use of forelegs in the insect tribes. Besides insects proper, a whole tribe of mites (Caris Latr., Leptus Latr., Astoma Latr., Ocypete Leach) have only six legs; the rest, and the Arachnida in general, have eight; in the Myriapods, Pollyxenus has twelve pairs; Scutigera has fifteen; the terrestrial Glomerides (G. Armadillo, &c.) sixteen; and the oceanic (G. ovalis) twenty; the oriental Scolopendræ Leach, twenty-one; Polydesmus has usually about thirty[Pg 655] pairs; Craspedosoma, fifty; Geophilus electricus at least sixty; in Iulus terrestris there are more than seventy; in I. sabulosus nearly one hundred; in I. fuscus, 124; and in I. maximus 134 pairs or 268 single legs. But with respect to the Geophili, Iuli, &c., it is to be observed, that the number of pairs varies in different individuals; and the circumstance that has been before mentioned[1982], that these animals keep acquiring legs in their progress to the perfect state, instead of losing them, renders it difficult to ascertain what is the natural number of pairs in any species.
2. Kinds. Upon a former occasion I gave you a sufficiently full account of the kinds of legs[1982], and I have also assigned my reasons for giving a different denomination to the anterior legs under certain circumstances[1983]; I shall not therefore enlarge further upon this head.
3. Substance. The substance of the legs is generally regulated more or less by that of the rest of the body, only in soft-bodied insects they seem usually more firm and unbending. Each joint is a tube, including the moving muscles, nerves, and air vessels.
4. Articulation with the Trunk. M. Cuvier has observed that the hip (coxa), which is the joint that unites the leg with the body, rather inosculates, in its acetabulum, than articulates in any precise manner[1984]; but this observation, though true of a great many, will not apply universally, for the legs of Orthopterous insects, and of most of the subsequent Orders, are suspended rather than inosculating. Even in many Coleoptera a difference is observable in this respect. I have before mentioned that[Pg 656] what are called the puncta ordinaria, which distinguish the sides of the prothorax of many Scarabæidæ and Geotrupidæ, form a base for an elevation of the interior surface with which the extremity of the base of the clavicle, which plunges deep into the breast, ginglymates[1985]; this structure may also be found in other Lamellicorns, as the stag-beetle (Lucanus) and Dynastes, that have not those excavations; in these last it is an elevated ridge forming a segment of a circle with, it should seem, a posterior channel, receiving a corresponding cavity and protuberance of the clavicle. With regard to the mid-leg, in Copris, the coxa is emboxed in a nearly longitudinal cavity of the medipectus, and the coxa of the hind-leg anteriorly is suspended to a transverse cavity of the postpectus, but posteriorly it is received by a cavity of the first segment of the abdomen; so that it may be regarded as suspended anteriorly, and inosculating posteriorly.
In some tribes of this Order, as the Weevils (Curculio L.) and Capricorns (Cerambyx), the coxæ of the four anterior legs are subglobose[1986] and extremely lubricous, and are received each by a socket that fits it, and is equally lubricous. In the bottom of this externally, and in the head of the coxa, is an orifice for the transmission of muscles, nerves, and bronchiæ; but the coxa is suspended by ligament in the socket. This structure approaches as near the ball and socket as the nature of the insect skeleton will permit; the high polish of the articulations acts the part of synovia, and the motion is in some degree rotatory or versatile, whereas in Copris, &c., lately mentioned, it seems to be more limited, and is probably,[Pg 657] at least in the mid- and hind-legs, only in two directions; in the middle pair, probably, from the coxæ being in a position parallel with the breast, opposite to that of the hind pair. In Dytiscus L., Carabus L., and some other beetles, the coxæ, especially the posterior pair, appear to be fixed and incapable of motion. In many insects these coxæ seem to belong as much to the abdomen as to the trunk. We have just seen this to be the case in Copris, &c.; and in the Lepidoptera, if the former be separated from the latter, the legs will be detached with it.
4. Location. We are now to consider the location and position of the legs, both in general and with respect to each other. And first, as has been before stated, we may observe that, in the hexapods with wings, the arms belong to the manitrunk, and are attached to the antepectus on each side the prosternum; and the two pair of legs to the alitrunk, the mid-legs being attached to the medipectus, between the scapularia and mesosternum; and the hind-legs to the postpectus, between the parapleura and the posternum; and further, that the arms are opposed to the prothorax: the mid-legs to the mesothorax and the primary organs of flight; and the hind-legs to the metathorax and the secondary organs of flight; though in some cases the wings appear to be behind the legs and in others before them: thus, in Panorpa, the former are nearer the head than the latter; but in the Libellulina the reverse of this takes place, the legs being much nearer the head than the wings: in both cases, however, the scapularia and parapleuræ run from the legs to the wings, but in an oblique direction; and in Panorpa these pieces assume the appearance of articulations of the legs.[Pg 658] In most of the apterous hexapods they appear to be attached laterally between the thorax and the pectus[1987]; but in the flea (Pulex) they are ventral. In this tribe the arms are usually stated to be inserted in the head[1988]: but I once succeeded in separating the head of a flea from the trunk, and these organs remained attached to the latter[1989]. As to the Octopods and Arachnidæ, in the mites (Acarus L.) they are lateral, and in their analogues, the spiders (Aranea L.), they emerge between the thorax and the breast, which last they nearly surround; in the Phalangidæ the bases of the coxæ approach near to each other, being separated only by a narrow sternum; in their antagonists, Chelifer and Scorpio, they apply to each other, the anterior ones acting as maxillæ. In the myriapods the legs of the Chilopoda Latr., and some Chilognatha, as Glomeris, are inserted laterally, a single pair in a segment; but in Iulus L. their attachment is ventral, the coxæ seem to spring from a common base, and there are two pair to each segment[1990], except the three first, which bear each a single pair.
I shall next consider how the legs are located with respect to each other. To render this clear to you I shall represent each of the variations, which amount in all to[Pg 659] twelve in the hexapods that have fallen under my notice, by six dots.
1.  In this arrangement the legs are all planted
near to each other, there being little or no interval between
the pairs, and between the legs of each pair. It
is exemplified in the Lepidoptera, Blatta, and many
Diptera.
In this arrangement the legs are all planted
near to each other, there being little or no interval between
the pairs, and between the legs of each pair. It
is exemplified in the Lepidoptera, Blatta, and many
Diptera.
2.  Similar to the preceding, but the anterior pair
are distant from the two posterior; exemplified in the
bees (Apis) and most Hymenoptera; Chironomus; Scutellera;
Pachysoma K.[1991]
Similar to the preceding, but the anterior pair
are distant from the two posterior; exemplified in the
bees (Apis) and most Hymenoptera; Chironomus; Scutellera;
Pachysoma K.[1991]
3.  Like the last, but the posterior pair is distant
from the two anterior. Examples: Silpha, Necrophorus,
Telephorus, &c.
Like the last, but the posterior pair is distant
from the two anterior. Examples: Silpha, Necrophorus,
Telephorus, &c.
4.  Similar to the last, but the legs of the posterior
pair are more distant from each other than the four anterior.
Ex. Curculio L.
Similar to the last, but the legs of the posterior
pair are more distant from each other than the four anterior.
Ex. Curculio L.
5.  The legs of each pair near each other, but the
pairs distant. Ex. Gibbium.
The legs of each pair near each other, but the
pairs distant. Ex. Gibbium.
6.  Both the legs of each pair and the pairs distant.
Ex. Blaps, &c.
Both the legs of each pair and the pairs distant.
Ex. Blaps, &c.
7.  Anterior pair distant from the two posterior,
and the legs of the middle pair rather more distant
from each other than those of the other pairs. Ex. Scarabæus
McL.
Anterior pair distant from the two posterior,
and the legs of the middle pair rather more distant
from each other than those of the other pairs. Ex. Scarabæus
McL.
8.  Like the preceding, only the legs of the middle
pair are at a much greater distance from each other.
Ex. Copris McL.
Like the preceding, only the legs of the middle
pair are at a much greater distance from each other.
Ex. Copris McL.
9.  Legs of the two posterior pairs distant. Ex.
Hister, Scaphidium.
Legs of the two posterior pairs distant. Ex.
Hister, Scaphidium.
10.  Like the preceding, but the posterior legs
more distant than those of the middle pair. Ex. Lygæus.
Like the preceding, but the posterior legs
more distant than those of the middle pair. Ex. Lygæus.
11. Like the last, but the legs of the anterior
pair also distant. Ex. Velia.
12. The arms distant, intermediate legs more distant,
posterior legs close together. Ex. Byrrhus L.
5. Proportions. In general the legs of some insects are disproportionally long and slender, as in Phalangium Opilio and some species of Gonyleptes[1992]: those of others are disproportionally short, as in Elater, &c. With regard to their relative proportions, the most general rule is, in Hexapods, that the anterior pair shall be the shortest and most slender, and the posterior the longest and thickest; but there are many exceptions: thus, in Macropus longimanus, Clytra longimana, &c., in the male the arms are the longest; again, a thing that very rarely occurs, in the same sex of Podalirius retusa the intermediate legs are the longest[1993]; but in Rhina barbirostris and many weevils they are the shortest: in Saperda hirtipes Oliv.[1994] the hind-legs are disproportionally long: with regard to thickness, they are in general extremely slender in Cicindela, and in the Scarabæidæ very thick. In Goliathus Cacicus the arms are more robust than the four legs[1995]; in Gyrinus the latter are more dilated than the former; in many Rutelidæ, and particularly in the celebrated Kanguroo beetle (Scarabæus Macropus Franc.) the hind-legs are much the thickest; in a new genus of weevils from Brazil (Plectropus K.), the intermediate pair are more slender than either the arms or the posterior pair.
6. Clothing. The hairs on the legs of insects, though at first sight they may seem unimportant, in many cases are of great use to them, both in their ordinary avocations and motions: but as most of these were sufficiently noticed when I treated of the sexes of insects[1996], I shall not here repeat my observations, but confine myself to cases not then adverted to. Some insects have all their legs very hairy, as many spiders, the diamond beetle (Entimus imperialis), or at least a species very near it and common in Brazil[1997], &c.: in others they are nearly naked, as in the stag-beetle. In the Crepuscular Lepidoptera (Sphinx L.) and some of the Nocturnal ones (Bombyx L.) the thighs are much more hairy than the rest of the legs: and in Lucanus, Geotrupes, and many other Lamellicorns, &c., the anterior ones have a yellow or golden spot at their base, composed of decumbent hairs, which prevent them from suffering by the violent friction to which they are exposed in burrowing. In most Petalocerous beetles the tibiæ are set with scattered bristles, and sometimes the thighs. The Tiger beetles (Cicindela) are similarly circumstanced: but the bristles, which are white, are generally arranged in rows. In Dytiscus, Hydrophilus, &c., the four posterior tarsi; and in Notonecta the posterior pair, and also the tibiæ—are fringed on each side with a dense series of hairs, which structure assists them in swimming[1998]. The tarsi, especially the anterior pair, in a certain family of Lamia F. (L. papulosa, &c.[1999]), are similarly fringed, only the hairs curl inwards; and the hand[Pg 662] in Sphex and Ammophila, but not in Pelopæus and Chlorion, is fringed externally with long bristles.
7. Composition. With regard to their composition, both arms and legs generally consist of five pieces, which Entomologists have denominated—the coxa or hip—the trochanter—the femur or thigh—the tibia or shank—and the tarsus or foot. Where the structure and use of the fore-leg is different from that of the four hind-legs, I propose calling these pieces by names corresponding with those which anatomists have appropriated to the arm in the higher vertebrate animals: thus, as you will see in the table, I call the whole fore-leg the brachium or arm; and the coxa becomes the clavicula or collar-bone; the trochanter, the scapula or shoulder-blade; the femur, the humerus or shoulder; the tibia, the cubitus or arm; the tarsus, the manus or hand. But let me not lead you to suppose that the pieces, either in the arms or legs of insects, which are there named after certain others in vertebrate animals, precisely correspond with them—by no means—since that is a very doubtful point; and some of them, as the trochanter, clearly do not. Many gentlemen skilled in anatomy, as I have before observed[2000], have thought that what is regarded as the coxa in insects really represents the femur: but there are considerable difficulties in the way of this supposition, several of which I then stated. I shall not however enter further into the subject, and take the above names; since this application of them is so general and so well understood, except with[Pg 663] regard to the fore-leg, under certain circumstances, as I find them. I shall now consider them in the order in which I have named them.
a. Coxa or Clavicula[2001]. The coxa is the joint that connects the leg with the trunk of the insect. With regard to their shape, the most general form of the four anterior is more or less that of a truncated cone: in the Staphylinidæ, however, they tend to a pyramidal or four-sided figure; as do the whole six in the Trichoptera: in numbers of the weevils and capricorns they are subglobose; in the Lamellicorns they are mostly oblong, and not prominent: the posterior pair in the Coleoptera are generally flat and placed in a transverse position, and more or less oblong and quadrangular: in Elater, &c., they are cuneiform: in Haliplus Latr. they are dilated, and cover the thigh[2002]: in Buprestis, Copris, &c., they have a cavity that partly receives it: the corresponding part, the clavicle, in the arm of Gryllotalpa, is very large and remarkable; viewed underneath it is triangular, and trifid where the trochanter articulates with it: in that of Megachile Willughbiella the clavicle is armed with a spine[2003]. As to their proportions, the most general law seems to be, that the anterior pair shall be the shortest and smallest, and the posterior the longest and largest. In some instances, as in Buprestis, the two anterior pair are nearly equal; in others (Mantis, Eurhinus K.), the anterior are the longest, in the former as long as the thigh, and the four posterior the shortest: in the Trichoptera, Lepidoptera, &c., all are nearly equal; in Mantis the two posterior, and in Phengodes the intermediate pair are the largest; but[Pg 664] in Necrophorus they are the smallest:—though almost universally without articulations, in Galeodes the clavicle consists of two and the coxa of three[2004].
b. Trochanter or Scapula[2005]. This is the second joint of the leg: and if the coxa is regarded as the analogue of the thigh in vertebrate animals, this should seem to represent the patella or rotula, vulgarly called the knee-pan. Latreille and Dr. Virey consider this articulation as merely a joint of the coxa[2006]; but if closely examined, especially in Coleopterous insects, you will find it so fixed to the thigh as scarcely to have separate motion from it, and in many cases it seems to be merely its fulcrum; but I am not aware that any instance occurs in which it has not motion separate from that of the former joint.
As to its articulation with the coxa,—in the Coleoptera it appears to be of a mixed kind; for it inosculates in that joint, is suspended by ligament to its orifice, and its protuberances are received by corresponding cavities in it; and its cavities receive protuberances, which belongs to a ginglymous articulation. I have observed two variations in this Order, in one of which the motion of the thigh and trochanter is only in two directions, and in the other it is nearly versatile or rotatory. The Lamellicorns afford an example of the first, and the Rhyncophorous beetles or weevils of the second. If you extract from the coxa the thigh with the trochanter of the larger species of Dynastes McL., you will find that the head of the latter is divided into two obtuse incurving[Pg 665] lobes or condyles: that on the inner side being the smallest and shortest, and constricted just below its apex: and that under this is a shallow or glenoid cavity, terminating posteriorly in a lubricous flat curvilinear ridge. If you next examine the trochanter in articulation with the coxa, you will perceive that the head of the former inosculates in it, that the lower condyle is received by a sinus of the coxa, which also has a lubricous very shallow cavity corresponding with the ridge, in which it turns; and in the head of the coxa, on the lower side, is an external condyle, which is received by a sinus common to both, of the head of the thigh and of the exterior side of the trochanter[2007], in which it likewise turns: this last condyle has also an internal protuberance, which appears to ginglymate with a cavity of the trochanter: from this structure the leg is limited chiefly to a motion up and down upon two pivots, or to fold and extend itself. You will find an articulation very near this, but on a smaller scale, in the stag-beetle. In the other kind of articulation, which admits of freer motion, the head of the trochanter is prolonged, and the process terminates in a short interior condyle, which appears to work in a corresponding cavity of the interior of the coxa; and the base of the process is encompassed by a ridge with a cavity behind it, which is received by another of the lower part of that piece, and admits a corresponding ridge—a structure that allows a rotatory motion. In the hind-legs of this tribe the motion is chiefly limited to folding and extending; in Carabus, &c., also the head of the trochanter is nearly hemispherical, and the articulation[Pg 666] approaches ball and socket. In most of the other Orders, the Hymenoptera excepted, there is little or no inosculation, the trochanter being simply suspended by ligament to the coxa as well as to the thigh; its connection with the latter is similar in Coleoptera; but in Cicindela, &c., it inosculates in it. The part we are considering varies in its position with respect to the thigh: in the hind-legs of Carabus, &c., it forms a lateral fulcrum on the inner side of that part, and does not intervene between its base and the coxa; the muscles from the latter entering the former, not at the bottom of the base, but at its side: but in the four anterior legs it forms their base, as it does in all the legs in Apion, and in all the Orders except the Coleoptera, cutting them entirely off from contact with the coxa: in the Lamellicorns they cut off part of the base obliquely, but so as to permit their coming in contact with the condyle of the coxa, as before mentioned. In the Ichneumonidæ and some other Hymenoptera the trochanter appears to consist of two joints particularly visible in the posterior legs[2008].
As to size in general,—the part in question is smaller than the coxa; but in Notonecta it is larger, and in the dog-tick (Ixodes Ricinus) longer than that joint. It exhibits few variations in its shape or appendages worthy of particular notice. In general, in the Coleoptera it is triangular or trigonal; but in Carabus L., in the hind-leg it is oblong or rather kidney-shaped; in that of Necrophorus[2009] it terminates in one or two teeth or spines, varying in length in the different species: in the other Orders it is not remarkable in this respect.
c. Femur or Humerus[2010]. The femur or thigh is the third, and usually the largest and most conspicuous joint of the leg. In the hypothesis before alluded to[2011] it is considered as the analogue of the tibia of vertebrate animals. With regard to the articulation of this part with the trochanter, it has been sufficiently explained under that head, and that with the tibia I shall treat of when I come to that joint. As to the size of the thighs, and their relative proportions to each other and to the remaining joints of the leg, the most general law is, that the anterior pair shall be the shortest and smallest, and the posterior the longest and largest. With respect to the remaining articulations, most commonly the thigh is longer and larger than the tibia, and the tibia than the tarsus. But there are numerous exceptions to both these rules. With respect to the first, we may begin by observing that the increase of the magnitude of the thigh, from the anterior to the posterior pair, is usually gradual: but in many jumping insects, and likewise many that do not jump, the posterior pair are suddenly and disproportionally thicker than the rest[2012]. Again, in many insects the anterior pair are the longest and thickest, as in Macropus longimanus, Bibio, Nabis, &c.: in others, the intermediate exceed the rest in magnitude, as in Onitis Aygulus, cupreus; Sicus flavipes, &c.; in many Lamellicorns all the thighs are incrassated and nearly equal in size: but in some, as Ryssonotus nebulosus McL.[2013], the intermediate pair are rather smaller than the rest. With respect to the second rule—in some, as in the male of Macropus longimanus, the anterior tibia, though more[Pg 668] slender, is longer than the thigh; in Hololepta maxillosa it is longer and more dilated; in Lamia marmorata, or one related to it from Brazil, the intermediate pair are longer; in Ateuchus gibbus and others of that tribe the posterior thighs are smaller than the tibiæ: and, to mention no more; in Callichroma latipes the posterior tibia is wider than the part last named. Again, the tarsi are as long as either tibia or thigh in many of the larger Dynastidæ, as Megasoma Actæon, &c.; longer than either in Melolontha subspinosa F.; and in Tiphia, Scolia and affinities, often as long, or longer than both together.
As to shape,—the thigh, especially in the fore-leg, varies considerably: most generally it is flat, linear, and a little thicker where it is united to the tibia, on the outer side convex, and concave next the body; but in many it is gradually thicker from the base to the apex: in some Cerambyces (C. thoracicus) it is clavate; in others of this genus and Molorchus they may be called capitate; in Pterostichus they are rather lanceolate; in Onitis Sphinx the humerus is triangular, and the intermediate thigh rhomboidal; in Bruchus Bactris it is bent like a bow; and in some Brazilian Halticæ it is nearly semicircular. The humerus in Phasma is attenuated at the base; in Empusa gongyloides it is at first ovato-lanceolate, and terminates below in a kind of footstalk[2014]; in Phasma flabelliforme it is dolabriform[2015]; in Mantis often semioval or semielliptical, and thickest at the inner edge, which affords space for two rows of spines with which it is planted. In Phyllium siccifolium all the thighs are furnished on both sides with a foliaceous appendage nearly[Pg 669] from base to apex[2016]: in a species of Empusa (E. macroptera), the four posterior ones are so distinguished only on their posterior side[2017]: others of this last genus, as E. gongyloides, have an alary appendage on both sides at the apex of these thighs[2018]; and another family, as E. pauperata, have only one on the posterior side[2019]. The thighs of no insect are more remarkable for their elegant shape,—tapering gradually from the base to the apex, where they swell again into a kind of knee,—than the posterior ones of the locusts (Locusta Leach); each side of these thighs is strengthened with three longitudinal nearly parallel ridges, and the upper and under sides are adorned by a double series, in some coalescing as they approach the tibia, of oblique quadrangular elevations resembling scales[2020].
I shall next say a few words upon the spines and other processes which arm the thigh. Those moveable ones of Mantis which help to form a fearful instrument of destruction, have just been mentioned, and similar ones, but less conspicuous, arm the intermediate thighs of Sicus flavipes: other appendages of this kind are for a less destructive purpose—to keep the tibia when folded in its place. This seems to be the use of the serratures and spine that arm the thigh of Bruchus Bactris, or the Hymenopterous genera Leucospis, Chalcis, &c.; in Onitis Aygulus a short filiform horn arms the humerus, and a longer crooked one that of many species of Scaurus[2021]. In many Stenocori the thighs terminate in two spines, and[Pg 670] in Gonyleptes K. the posterior ones are armed internally with very strong ones; with which, as the legs converge at their knee[2022], they may probably detain their prey. The knee-pan (Gonytheca) of the thigh, or the cavity at its end, which receives the head of the tibia, is very conspicuous in the weevils; but in no insects more than in Locusta[2023], in which tribe it deserves your particular attention.
d. Tibia or Cubitus[2024]. The tibia or shank is the fourth joint of the leg, which according to the hypothesis lately alluded to is the analogue, in the anterior leg of the carpus or carpal bones, and in the four posterior ones of the tarsus or tarsal bones of vertebrate animals. This may be called the most conspicuous of the articulations of the leg; for though it is generally more slender and often shorter than the thigh, it falls more under the eye of the observer, that joint being more or less concealed by the body: it consists in general of a single joint; but in the Araneidæ and Phalangidæ it has an accessory one, often incrassated at its base, which I have named the Epicnemis[2025].
With respect to the articulation of the tibia with the thigh—we may observe that in general it is by means of three processes or condyles, two lateral and one intermediate, of the head of the former joint[2026]: the lateral ones are usually received by a cavity or sinus of the gonytheca[Pg 671] of the thigh[2027]; and upon these the tibia turns, with a semirotatory motion up and down as upon a pair of pivots: at the same time the mola or head of the latter joint, which has often a flexure so as to form an elbow with the rest of it, inosculates in the gonytheca, and is also suspended by ligament to the orifice through which the muscles, nerves, and bronchiæ are transmitted: so that in fact the articulation, strictly speaking, belongs exclusively to none of the kinds observable in vertebrate animals, but partakes of several, and may properly be denominated a mixed articulation,—a term applicable in numerous instances also to the other articulations of the legs of insects. In the different Orders some variations in this respect take place,—I will notice some of the most remarkable. In no Coleopterous insects is the structure more distinctly visible than in the larger Lamellicorns. In Copris bucephalus, for instance, if you divide the thigh longitudinally, you will find on each side, at the head, that it is furnished with a nearly hemispherical protuberance, perforated in the centre for the transmission of muscles, and surrounded externally by a ridge, leaving a semicircular cavity between them[2028]: if you next examine the tibia, after having extracted it, you will find on each side, at the base, a cavity corresponding with the protuberance of the thigh which it receives, having likewise a central orifice, and surrounded by a semicircular ridge corresponding with the cavity in the thigh in which it acts: below this ridge another cavity, forming a small segment of a circle, receives the ridge of the thigh[2029]. You will observe that the ridge of the tibia represents the[Pg 672] lateral condyle lately noticed: in the Dynastidæ this is more prominent, and often forms a smaller segment of a circle. In these also the protuberance of the thigh is more minute, and its ridge is received by a cavity of the tibia nearly semicircular[2030]; in Geotrupes Latr. the articulation is not very different, though on a reduced scale; in Calandra Palmarum the lateral condyles of the tibiæ are flatter and broader[2031]; and the articulation not being quite so complex, this joint is kept steady by an intermediate process observable in the gonytheca[2032]. From the above description it appears that the dislocation of the tibia is effectually prevented in the Lamellicorns by the protuberance and ridge of the thigh working in their corresponding cavities, while the condyle of that part turns with a rotatory motion in the cavity of the thigh. In the Orthoptera Order the tibia is suspended by a ligament, in the gonytheca the lateral condyles, which are very prominent, working in a sinus of that part[2033]. The subsequent Orders exhibit no very striking variations from these types of articulation, I shall therefore not detain you longer upon this head.
With regard to the proportions and magnitude of the joint we are considering,—the most general law is, that the anterior pair should be shorter and more slender than the intermediate; and the intermediate than the posterior; and that all the tibiæ should be shorter and more slender[Pg 673] than the thighs, and longer and thicker than the tarsi. Various exceptions, however, to this rule in all these cases might be produced; but I shall only observe that in all those insects in which the fore-legs are calculated for digging or seizing their prey, as in the Petalocerous beetles, the Gryllotalpa, Mantis, &c., this joint of the leg is usually much enlarged and more conspicuous than the others.
As to its figure and shape—most commonly the tibia grows thicker from the base to the apex, as in the majority of Coleoptera, Hymenoptera, &c.; in the Orthoptera, Neuroptera, &c., it is generally equally thick every where. Another peculiarity relating to this head observable in it, is its tendency to a trigonal figure: this, however, though very general, is not universal;—thus, in some Orthoptera, as Pterophylla K., its horizontal section is quadrangular; in others, as Locusta Leach and many other insects, it is nearly a circle; in some scorpions it is almost a hexagon. The superficial shape also of this joint in numerous instances is more or less triangular, but it sometimes recedes from this form:—thus, in Callichroma latipes it is a segment of a circle; in some Empides it is clavate; in Onitis Sphinx, dolabriform; in the Orthoptera, Neuroptera, &c., it is usually linear; in some Lygæi it is angular[2034]: but the most remarkable tibiæ in this respect are those of such species of this last genus as have the posterior ones winged or foliaceous, so that they resemble the leaf of some plant—the tibia being the rachis, and the wing (which in some species is veined) representing the leaf itself. This structure is[Pg 674] exemplified in Lygæus compressipes, phyllopus, foliaceus, &c.[2035] Under this head I must say a few words upon the flexure of this joint, which in some cases merits notice. I have before mentioned its bend at the knee[2036] or base: the apex also is sometimes incurved—in the anterior one of the male of Macropus longimanus so as almost to form a hook[2037]: in Lygæus Pharaonis the posterior pair are flexuose[2038]; in Bruchus Bactris, Leucospis, and several species of Chalcis, these tibiæ curve so as to adapt themselves to the bend of the thigh when folded. The notch on the inside of the anterior pair, in a large majority of Carabus L., armed above by a spur[2039], a structure which probably assists them in seizing and detaining their prey, may also here be introduced: in the generality it is a little removed from the apex of the joint in question; but in Pamborus it is very near to it, and in Cychrus, Carabus, &c., it becomes obsolete. I may mention here also a singular character which distinguishes the cubit of both sexes of Gryllus campestris, domesticus, &c. At the base there is an aperture which passes through the joint—anteriorly it is oval, and posteriorly elliptical and much larger, and on both sides is closed by a tense membrane.
The most striking peculiarities as to the clothing of his joint have been chiefly noticed under the sexual characters of insects[2040], but some appear not to be of that description. In Sphæridium Leach, while the thighs and tarsi are naked, the posterior tibiæ are remarkably beset[Pg 675] with stiff bristles; in Empis pennipes they are thickly fringed on both sides; in Scarabæus McL. only externally, and in Dytiscus serricornis internally; in Necydalis barpipes K. this fringe is longer at the apex; and in Saperda hirtipes Ol. the same tibiæ at that part are adorned with a large brush, like that observable in the antennæ of some Lamiæ[2041].
I must next call your attention to the teeth, spines, and spurs with which the tibiæ of insects are sometimes armed. With regard to teeth, you have doubtless often observed those that distinguish the cubitus of the arm of most Lamellicorn beetles: these vary in number from one, as in Trox suberosus, to seven, as in Geotrupes autumnalis; but the most universal number is three: in some species of Geotrupes, as G. stercorarius, &c., the third tooth from the apex, and those that follow it, may be called double. These teeth, in their cubit or anterior shank, doubtless assist these insects in burrowing. The four posterior tibiæ in this tribe are also distinguished by a kind of teeth which occupy their whole diameter, and resemble so many steps. I have before noticed the remarkable cubit of the Gryllotalpa, and likewise that of Scarites, Pasimachus, &c., in which some of the teeth are prolonged into spines[2042], which are the next description of tibial arms that I mentioned. Spines are of two kinds—those which are merely processes of the crust of the tibia, and those that are implanted in it, and seem to have a gomphosis or perhaps an amphiarthrosis articulation[2043]. An instance of the first kind may be seen in the hind-legs of some[Pg 676] grasshoppers[2044] (Locusta Leach), the Rutelidæ, &c. though in others they are implanted:—of the second, in the cubitus of the Mantidæ, and of all the tibiæ of the dragon-flies (Libellulina McL.)[2045];—and of both kinds in the hind-legs of Acrida K., those which arm the upper angles of the tibiæ being processes, and those of the lower being implanted. The term spine I think ought to be restricted to the first kind; the second ought rather to be denominated spurs (calcaria), and may perhaps be regarded as in some degree synonymous with those most important appendages of the joint in question, that are implanted in or near their apex, which have been hitherto distinguished by this last denomination, and which I am next to consider. But though I have not altered a term generally adopted, I must here express my opinion that they ought rather to be considered as minute toes or fingers, and that the denomination best agreeing with their functions, as accessories to the main toe, would be digituli: this is proved particularly by a character peculiar to those of many species of the genus Cimbex amongst the saw-flies, in which these organs are furnished with a sucker or pulvillus (as they are also in Œnas a kind of blister beetle), as well as the joints of the tarsi[2046]; which makes it evident that they are applied by the animal to surfaces, and assist it in walking or climbing; and in general it may be observed that in most insects their principal use is connected with these motions, and with burrowing. This circumstance tends to prove[Pg 677] that the generality of insects (for all have not these organs) have really a didactyle or tridactyle hand or foot; and the hypothesis so often alluded to—that the cubitus or tibia, &c., is really analogous to the carpus or tarsus in vertebrate animals[2047]—seems to receive no small confirmation from it; since, if the spurs be really analogous to fingers or toes, the part they articulate with cannot be the tibia, &c. Though the parts in question did not escape the notice of Reaumur, Linné, De Geer, Latreille, &c., yet they have not been employed in the determination of tribes, genera, &c., except by the author last named, but perhaps adopted from Bonelli[2048], in the subgenera Zabrus and Pelorus: in many instances, however, they afford excellent subsidiary characters, sometimes common to a whole Order, and at others distinguishing its various subdivisions. With regard to their number—I have noticed many variations which I will now state to you, first observing that I shall express them by three figures, the first representing the number of spurs on the anterior leg, the second that of those on the intermediate, and the third on the posterior; and where there are spurs, as in the Trichoptera and Lepidoptera, on the middle as well as at the end of the tibia, I shall express it by one figure over another, the upper one representing the number of the middle spurs. If you make an examination yourself, it will be proper to remind you that these little organs are extremely liable to be broken off, but the socket in which they were planted is usually very visible. The most natural number is represented by 2:2:2; this[Pg 678] you will find very prevalent in the Coleoptera Order, as in the Predaceous and numerous other beetles: in the Orthoptera and Hemiptera Orders, however, I have not discovered an instance of it; but in all the rest it more or less occurs: next to this number—tibiæ with obsolete or no spurs seem most prevalent, particularly in the Hemiptera; not a single instance of an insect furnished with them occurring to me in the Heteropterous section; and it is doubtful whether there are any in the Homopterous.—Having stated the most universal structure in this respect, I will next consider the Orders seriatim. Amongst the Coleoptera though the numbers 2:2:2 are most frequent in occurrence, yet there are numerous exceptions. Thus, in the Lamellicorns, 1:1:1 represents the calcaria of one tribe of the Scarabæidæ McL. formed of the genus Scarabæus McL.; 1:2:1 represents those of another tribe of that family, including the subgenera Ateuchus, Copris, Phanæus, &c.; 1:2:2 again forms the character in this respect of Aphodius and the great majority of the Lamellicorns; while 2:2:2 is confined in this section to Æsalus F. and Melolontha chrysomeloides Schranck (Psephus McL. MS.). In the other tribes of Coleoptera other numbers occur. Thus, 0:1:1 characterizes Hylœcetus; 0:1:2 Mordella; 0:2:2 Macropus; 1⁄1:2:2 Harpalus, and all those Carabi L., except Zabrus, that have a notch in their anterior tibiæ; ½:2:2 Zabrus. In the Orthoptera Order it is not easy to distinguish the real spurs from the implanted spines that frequently arm the legs: these in Blatta are extremely numerous, even at the apex of the tibiæ; but I cannot distinguish any that can be regarded as true analogues of the former: the most natural number of spurs[Pg 679] in this Order is represented by 0:0:4; this you will see in all the Locusts; in Acrida, Conocephala, Pterophylla; and in Truxalis, Pneumora, &c.; in Phasma there are none. In Mantis, if the terminal process of the cubitus is excluded, it will be 0:2:2; in Gryllotalpa, admitting the terminal teeth of that part[2049] as analogues of spurs, the number is 4:4:4; in Tridactylus Latr. 0:0:5[2050]; in Gryllus Latr. 3:3:5; in Gryllus monstrosus, 4:4:6. In the whole Hemiptera Order I have discovered no instance of an insect furnished with the real spurs: for though in Tettigonia F., Cercopis, &c., there are implanted spines in the posterior tibia, and several at the apex, there are none of them clearly analogous to real spurs. In the Lepidoptera the most general arrangement appears to be 1⁄0:2:2/2; and next to this, 1⁄0:2:2. In this Order most commonly there is no spur at the end of the cubit, but one resembling a thumb[2051] arms its middle; in Pieris, &c., this thumb is not present, so that the number is 0:2:2; in Agarista Leach, Erebus, &c., you will find 1⁄0:2:4, the posterior calcaria being all terminal; and in Attacus Atlas, all these organs are obsolete except the thumb. In the Neuroptera the most general arrangement is 2:2:2; but in the Libellulina, although the legs are very spinose, there are no spurs. In the Trichoptera K., in Phryganea rhombica and affinities, the number of them is expressed by ½:½:½; and in those with long antennæ, P. atra, &c., by 2⁄2:2⁄2:2⁄2. In the Hymenoptera the number 1:2:2 is most prevalent; and next to this, as in Apis L., 1:1:2. In the Ichneumones minuti L. the spurs are[Pg 680] 1:1:1; in Atta Latreille, a kind of ant[2052], 1:0:0. In the Diptera it is often difficult to distinguish the spurs from the spines; but the number most universal is, I think, 2:2:2; in Tipula it is 1:2:2; in the Tabanidæ 0:2:0; and in Culex, Limonia, &c., there are none. Amongst the insects with more than six legs, most commonly the tibiæ have no spurs; but in the Araneidæ each is armed with two, a circumstance which also distinguishes the corresponding joint of the pedipalpi.
These little organs inosculate each in an appropriate socket of the end, or in many cases of the middle of the tibia; and that part of their head or base that is received by it, is often constricted for the purpose: from hence it follows that they are capable of some degree of motion, but in some insects, as those on the four posterior legs of Scarabæus sacer and its more immediate affinities, and those at the end of the cubit of Gryllotalpa, they are immoveable, and appear almost processes of the joint to which they belong. They are commonly sharp, of a subtriquetrous figure, with the lower side flat: where there are two, the outer one is usually the longest; and in general the spurs on the hind legs are longer than those on the four anterior: but there are exceptions—thus, in Acanthopus Latr. the intermediate spurs are the longest; and in Cicindela the anterior are longer than the former; in Blaps mortisaga those on all the legs are nearly equal in length. They vary sometimes in shape—those on the middle of the cubit of many Lepidoptera,[Pg 681] which may be regarded as a kind of thumb[2053], are of a lanceolate shape; in Meloe the external posterior one is flat and obtuse; in Œnas Latr. it is obconical, concave at the extremity, and apparently furnished with a sucker; in Ateuchus smaragdulus the anterior, and in Copris Carolina the posterior is forked and emarginate; in Sirex the former is hooked and winged; in Lamprima it is triangular and dilated; in Aphodius analis it is dolabriform; in Dynastes retusus and Juvencus the spurs are bent like a bow. In many Hymenoptera, as the Sphecidæ, they are pectinated[2054], with a series of minute parallel spines—a structure which assists the animal in burrowing[2055]; in Acanthopus Latr. they are armed with little teeth or spines[2056]; in the hive bee the spur of the cubit is furnished with a membranous appendage which I have called the velum[2057]; and in a subgenus related to Saropoda Latr. (Ctenoplectra K. MS.), the interior spur of the posterior leg is crescent-shaped, fixed transversely, and fitted on the inner side with a membrane, the edge of which is finally pectinated.
e. Tarsus or Manus[2058]. This is the last portion of the leg, usually supposed to be analogous to the hand or foot of vertebrate animals; but, according to the hypothesis so often alluded to, rather the representative of their jointed finger or toe. In treating of this part I shall consider its articulation with the tibia, and of its joints inter se; the number of those joints; their proportion and shape; their parts and appendages.
I seem to have observed three kinds of tarsal articulation. The first is a species of enarthrosis or ball and socket, the joints terminating in a globular head, perforated indeed for the transmission of muscles, &c., and which is received by a corresponding cavity of the tibia or preceding joint, as may be seen in many weevils (Curculio L.[2059]). This admits of some rotatory motion.—The second is a mixed articulation between enarthrosis and ginglymus, when at the base of the ball a deep transverse channel receives a corresponding ridge of the tibiæ or preceding joint: this may be found in Rutela and probably many other Lamellicorn beetles; and something very similar in the Predaceous ones.—The third kind is where there is little or no inosculation, and the joints are scarcely more than suspended: this takes place in the Orthoptera, Neuroptera, &c.; but in Blatta and the hind legs of Mantis there is some approach to the foregoing kinds.
We are now to consider the number of joints of the tarsus, which varies considerably in the different Orders, and in one has been assumed as a clue for a subdivision of it into sections[2060], which, though not perfectly natural, is very convenient, and has been adopted by most modern Entomologists. In treating of this head, I shall use those denominations that have been employed by M. Latreille and others to express the variations of the number of the tarsal joints in the Coleoptera, but shall apply them to insects in general. Insects in this view, therefore, may be called pentamerous; heteromerous; tetramerous; trimerous; dimerous; or monomerous.
Pentamerous insects are those which have five joints in all their tarsi. This is the most universal, and may be called the natural number of these joints. More than half the Coleoptera belong to this section; in the Orthoptera—the Blattidæ, Mantidæ, and Phasmidæ; all the Lepidoptera except those butterflies called tetrapi (Vanessa, &c.); all the Trichoptera, Hymenoptera, and Diptera; in the Neuroptera—Ascalaphus, Myrmeleon, Hemerobius, Corydalis, &c.; and in the Aptera—Pulex[2061].
Heteromerous insects are those in which the number of these joints varies in the different pairs of legs[2062]. These variations, like the spurs, may be expressed by three figures, the first representing the anterior tarsus, the second the intermediate, and the third the posterior. I begin with 5:5:4. This number represents those beetles that have been exclusively regarded as heteromerous by modern Entomologists—of this description is the Linnean Tenebrio, Meloe, &c., now subdivided into numerous genera; they have five joints in the two anterior pair, and four in the posterior. The tarsal joints of the aquatic genus Hydroporus (a singular anomaly in the Order to which they belong) are expressed by 4:4:5,[Pg 684] thus reversing the number in the preceding tribe: other Heteromerous genera are to be found amongst the Hemiptera. Thus, in Ranatra the numbers are 2:1:1; in Sigara and Nauceris 1:2:2; in a new subgenus between Belostoma and Naucoris (Xiphostoma K. MS.), brought by Dr. Bigsby from Canada, 3:2:2; in the Lepidoptera the butterflies called tetrapi (Vanessa, &c.) may be expressed by 1:5:5. Amongst the Aptera and Arachnida there are three remarkable genera, which if their pedipalps are included may be deemed Heteromerous. I mean Phrynus, Thelyphena, and Galeodes;—in the former the numbers will be *:4:4:4, the asterisk denoting more than ten; in the second, 8:4:4:4.; and in Galeodes (in which the first pair of pedipalps are not chelate, the mandibles performing their office) the numbers are 1:1:3:3:3.[2063]
Tetramerous insects are those in which all the tarsi consist of four joints; these in the Coleoptera are next in number to the pentamerous—indeed a very large proportion of them strictly speaking are really of the latter description, since in Linné's four great genera, Curculio, Cerambyx, Chrysomela, and Cassida and some others, the claw-joint (ungula) consists of two articulations, one very short, forming merely the ball at its base[2064], which inosculates in the socket of the preceding joint, and the other constituting the remainder: if you carefully separate these two pieces, you will find that the last inosculates[Pg 685] in the summit of the ball, and is moved by appropriate muscles[2065]. This structure probably permits the readier elevation and depression of this joint. In the Orthoptera the tetramerous genera are those which Linné called Tettigonia amongst his Grylli (Locusta F.); Acheta monstrosa also, and in the Neuroptera, Raphidia belong to this section.
Trimerous insects are those whose tarsi consist of only
three joints. Amongst beetles the Lady-birds (Coccinella
L.) are remarkable for this structure, but in them
the claw-joint is also biarticulate, so that strictly speaking
they are tetramerous; in the Orthopterous Order the
migratory locusts (Locusta Leach) belong to this section,
as likewise Gryllus Latr. and Gryllotalpa Latr.:
in the first of these genera is an appearance of there being
more joints in the tarsus, because there is more than
one cushion below the first[2066]. To this section also belong
the great majority of the Hemiptera, excluding only those
tribes that connect the two sections of the Order constituting
the two Linnean genera Nepa and Notonecta; the
Libellulina likewise belong here, as do also the Scorpionidæ
and Scolopendridæ.
Dimerous insects are those that have two joints in all their tarsi. Such are the Pselaphidæ in the Coleoptera Order[2067]; in the Hemiptera—Belostoma and Notonecta; in the hexapod Aptera—Pediculus; in the octopod—the Acari of Linné; in the myriapod—Iulus; and in the Arachnida—the Araneidæ.
Monomerous insects are those which have only a single tarsal joint. Only one Coleopterous and also one Hemipterous genus is so distinguished: the first is Dermestes Armadillus De Geer[2068], and the second the common water-scorpion, Nepa Latr. Among the Aptera we find Nirmus, Podura, Sminthurus, &c., that belong to this section.
To the above sections another may be added for those insects whose tarsi have more than five joints, which may be denominated Polymerous. Here belong the genera Gonyleptes K., Phalangium and Scutigera Latr. In the first the number of joints varies from six to eleven, and in the two last they far exceed that number, amounting in some species of Phalangium to more than fifty, and becoming convolute like the antennæ of Ichneumons[2069].
I am next to notice the proportions and shape of the tarsus and its joints. The most general law is, that it shall be shorter and more slender than the tibia; but it admits of several exceptions—thus, in Megasoma K.[2070], in all the legs; in Agrostiphila McL. MS.[2071] in the intermediate, and in Amphicoma lineata in the posterior pair the tarsi are the longest; in Trichius Delta these last are longer than the thigh and tibia together. In some insects the tarsi are disproportionally short, as in Cassida, the Pselaphidæ, Locusta Leach, &c. Though generally more slender than the tibia, in several instances[Pg 687] they are as thick or thicker, or more dilated, as in most of the tetramerous beetles, which being climbers require a dilated tarsus. Again, comparing the three pairs of this joint with each other, the most general rule is, that the anterior should be the shortest, and the posterior the longest: but in some, as the Capricorn beetles, &c., they are nearly equal in length; in others, as Lytta marginata, the anterior pair, and in Rhipiphorus the intermediate, are the longest; in Trichius Delta these last are the shortest. With respect to thickness, the anterior tarsi, except in many males[2072], are not very strikingly different from the rest.
With regard to the proportion of the joints of the tarsus to each other,—according to the most general law, the first is the longest, the last next in length, then the second and third, and the fourth is the shortest. In Gonyleptes K. and other Phalangidæ the first is almost thrice the length of all the rest taken together; but there are numerous exceptions to the rule. In the female Carabi the first joint is not longer than the last, and in the males not so long; and in Hydrophilus, &c., it is the shortest of all. Again, the second joint is longer than the three following ones in Dasytes ater[2073]; and than the last in Cicindela sylvatica: the third joint is shorter than the fourth in Lampyris ignita: it is longer than the first in Donacia, many Melolonthidæ, &c. Once more, the fourth joint, usually the shortest of all, is longer than the second and third in Anthia, &c. Lastly, the claw-joint, usually the second in length, in the Eproboscidea Latr. (Hippobosca L.) is very long and large, while the four first joints are so[Pg 688] extremely short as to be scarcely distinguishable from each other: it is the shortest of all in Colymbetes, &c.; it is of the length of the third in Cicindela sylvatica, of the fourth in C. sexguttata. Though commonly the slenderest joint of all, particularly so in Raphidia, in many Heteromerous and Lamellicorn beetles it is the largest, conspicuously so in Mellinus tricinctus. Sometimes, as in Buprestis chrysis, &c., all the tarsal joints are nearly equal in length and thickness.
We are next to say something upon the shape of the tarsi and their joints. In general we may first observe that their upper surface is commonly more or less convex, and the lower flat or concave: in insects that are swift runners, as the terrestrial Predaceous beetles, they are usually slender and filiform[2074]; in those that swim, as Dytiscus, the two posterior pair taper nearly to a point from the base to the apex[2075]; in some that climb, as Buprestis, they are rather flat and linear; and in others (the Weevils, Curculio L.) they grow gradually wider towards the claw-joint[2076]; sometimes, as in Mordella Latr., the four anterior tarsi are of this shape, and the posterior pair setaceous. In Gyrinus the four posterior are flat and triangular; and in that extraordinary insect Gryllus monstrosus the tarsi are foliaceous and lobed[2077]. In many males and some others the anterior pair or hands are of a different shape from the two posterior: thus, in several Carabi they are lanceolate; in Staphylinus, Creophilus, &c. in both sexes they are often nearly circular, like those of male Dytisci[2078]. With regard to the shape of[Pg 689] individual joints it may be said in general that they are rather triangular, with an anterior sinus for the reception of the succeeding joint: the first joint usually departs most from this form; in the bees it is commonly much larger than the rest, especially in the last pair of legs, and nearly forming a parallelogram[2079]; in Euglossa it is trapezoidal; in the majority nearly linear or filiform. With regard to their termination—in Brachycerus and some ants (Ponera, Myrmica, &c., Latr.) the three first joints; in Dascillus, Lycus reticulatus and affinities, the third and fourth; and in the great majority of the Tetramerous insects the penultimate joint is bilobed; although in most Predaceous beetles this joint is entire or simply emarginate, yet in Colliuris it terminates in a single oblique lobe; and in Lebia, Drypta, &c., it is nearly bipartite. I must now advert to the Ungula or claw-joint: it is usually clavate or thickest at the end and curved; but in the Asilidæ it is shaped like a vase or cup; in Phanæus, in the four posterior tarsi, in which the claws are obsolete, it is thickest at the base and sharpest at the extremity[2080]; it usually forms an angle with the rest of the tarsus, rising upwards, which enables the insect to move more easily without hindrance from the claws, and also more readily to lay hold of any object it meets with; but in the Lamellicorn beetles and many other insects it is in the same line with it. As in the beetles last mentioned this joint is often inserted in the extremity of the preceding one; but in Œdemera it articulates with the middle of its upper surface; and in Lycus and a numerous host of[Pg 690] Tetramerous beetles it springs from its base, just behind where it diverges into two lobes.
I shall next call your attention to the different kinds of appendages with which the tarsi are furnished. They are seldom armed, like the tibiæ, with teeth, or spines, or horns; but something of the kind occasionally distinguishes them. In Phileurus, Oryctes, and several other Dynastidæ, the first joint is armed at the apex externally with a considerable mucro; in the fore-leg of Dasytes ater a similar process is prolonged into a crooked horn[2081]. But the most important appendages of the tarsi are the claws which almost universally arm their extremity, and which appear clearly analogous to those of birds, quadrupeds, &c., though probably differing as to their substance[2082]. Some few, however, are without them; this, as I lately observed, is the case with Phanæus with respect to the four posterior legs; the anterior ones of Vanessa amongst the Lepidoptera, and all those of Stylops and many Acari L., are also without them: this is likewise the case with the first pair of legs, or the second of the pedipalps of Galeodes. In this genus these organs consist of two joints[2083]. With respect to number they vary in different tribes, but not so much as the calcaria: these variations may likewise be represented by three numbers. The most natural is two in all the tarsi, exhibited by the Predaceous beetles and the great majority; 2.2.1. are to be found in Hoplia, Anisonyx, &c.[2084]; 1.2.2. in Belostoma; three in all the legs in the[Pg 691] Araneidæ[2085]; in Meloe[2086], Elater, &c., each claw is double or consisting of two, which makes four in each leg; and in many Hippoboscidæ there are six[2087]; in Nepa and the Myriapods there is only one. In most insects, perhaps, the claws are simple or undivided[2088]; but in Galeruca, Melolontha subspinosa[2089], &c., they are bifid at the apex; as is the exterior claw of the four posterior legs in Chasmodia and Macraspis[2090] McL., and of all in Melolontha horticola; in Serica brunnea McL. the claws are all cleft at the extremity, but the internal tooth is broad, flat, and obtuse[2091]; in Melolontha vulgaris and Pelidnota punctata McL.[2092], the claws are armed with an internal tooth near the base[2093]. In the Araneidæ, which have three claws, the two external ones are furnished with several parallel teeth, which the animal uses to keep separate the threads of its web, and probably for other purposes[2094]; and some Predaceous beetles, as Lebia and Cymindis, have both their claws similarly furnished[2095]. These organs vary in their relative proportions: thus, in Anoplognathus the inner claw is much smaller than the other[2096]; and in Elater sulcatus, fuscipes, &c., it is represented by a mere bristle; in Hoplia, in the anterior tarsus it is not half the length of the outer one[2097]; in Areoda and Pelidnota McL. this last is the smallest. They vary also in length—in Rynchænus, Ascalaphus, &c., they are very short; in the Lamellicorns, Galeodes, &c., very long; and in Myrmeleon[Pg 692] longer than the claw-joint. With regard to their curvature they generally form the segment of a circle; in many Asilidæ they are crooked like the claws of the eagle[2098], and the posterior one of the Hopliæ is bent like a hook[2099]; they most commonly diverge from each other; but in the Rutelidæ, Anoplognathidæ, &c., they are perfectly parallel, and in the former often inflexed[2100]. With regard to other appendages of the part we are treating of, if you examine the stag-beetle and many other Lamellicorns, you will find between the claws a minute but conspicuous joint terminated by two bristles which seem to mimic the ungula and its claws; these parts are what are denominated in the table the palmula, plantula, and pseudonychia: in the stag-beetle these are long[2101]; in the Melolonthidæ short[2102]; and in many Cetoniadæ they resemble an intermediate claw.
The most remarkable of the appendages of the tarsi are to be looked for on their under side or sole (solea), and are the means by which numbers of insects can overcome atmospheric pressure and walk against gravity. Many of these have been fully described in a former letter[2103]; but much that relates to them was there omitted, which I shall now detail to you. Four kinds of pulvilli, as I would call these appendages, are found in the sole of insects, upon each of which I shall make a few remarks.
The first is a cushion or brush composed of very thickly set hairs or short bristles: examples of this you will find in the majority of Tetramerous and Trimerous beetles. In Chrysomela, Timarcha, &c., there is one of these cushions[Pg 693] on each of the three first joints; in Prionus, Liparus, &c., there is a pair; and in Coccinella on the two first; in others (Balaninus Nucum, &c.) a pair only on the penultimate joint; in Calandra Palmarum, Rhina barbirostris, &c., that joint has an intire cushion; in Eurynotus muricatus K.[2104] the three first joints of the four anterior tarsi are similarly circumstanced, but the cushions resemble sponge[2105].
The second kind of cushion is a vesicular membrane capable of being inflated. This distinguishes the tarsi of Thrips[2106], and many Acari L.[2107]; likewise those of Xenos[2108]; and also of many Orthoptera fully described on a former occasion[2109], though the fact of their capacity of inflation has not been ascertained, belong to this section.
The third kind of covering of the sole is when the three or four first joints of the tarsus each terminate in one or two membranous lobes or appendages: of the first description is Priocera K., in which the lobes are involute[2110]; and of the second Rhipicera Latr.[2111], in which there is a pair on each joint, in the Brazil species set with very fine hairs.
The fourth and last kind are what may with the utmost propriety be denominated suckers, since their use as such is clearly ascertained. These are not only to be found in a large proportion of the Diptera, in some of which there are two of them, as in the Asilidæ[2112]; and in others[Pg 694] three, as the Tabanidæ[2113]; but also in many of the subsequent Orders: thus, in the Heteropterous Hemiptera, in Scutellera and Pentatoma, but not the Reduviadæ, and in the Neuropterous genus Nymphes Leach there is a minute one under each claw. It is discoverable between the claws in many Hymenoptera, as Apis[2114], Vespa, &c. But the genus that exhibits to the curious Entomologist the most singular and elaborate apparatus of this kind is Dytiscus Latr.; and the examination of the under side of the hand of any male of this genus will almost compel the most inattentive observer to glorify the wisdom and skill of the Allfather so conspicuously manifested in the structure of these complex organs. For this part in these, instead of two or three pedunculate cups as in the insects just mentioned, is composed of a vast number, some large and some small. If you take a male specimen of the common D. marginalis, you will find that the three first joints of the hand are very much dilated, so as to form a plate or shield nearly circular, fringed all round with stiffish hairs; if you next examine the under side of this plate with a good magnifier, you will discover at the base, where it is united to the cubit, two circular cups, the external one more than three times the size of the other, with an umbilicated centre[2115]; besides these two larger cups the rest of the shield is covered by a vast number of minute ones of a similar construction[2116]: the larger cups are nearly sessile, but the smaller are elevated upon a tubular footstalk[2117]; the three first joints of the intermediate tarsi are also dilated, but not into an[Pg 695] orbicular shield, and thickly set with minute pedunculated suckers[2118]. The structure varies however in different species. Thus in D. limbatus the shield is triangular with the smaller suckers at the base, and two rows of larger oblong ones, concave but not umbilicated, at the apex; in another Brazilian undescribed species (D. obovatus K. MS.) the shield is oblong and quite covered with suckers like those last mentioned; in D. sulcatus (Acilius Leach) almost the whole plate is occupied by a very large sucker, above which, at some distance in the inner side, are two smaller ones, while the extremity of the shield is covered by minute ones elevated on long footstalks: the central umbilicated elevation of the large one, which nearly fills its cavity, is in this species beautifully radiated. The male of Colymbetes transversalis has also an orbicular shield, but the suckers are much less strongly marked. The use of this organ has been before sufficiently explained[2119].
A few words will be necessary upon the folding of the legs in repose. When insects walk, the thigh is usually in an ascending position, rising above the horizontal line, the tibia forming with it rather an obtuse angle, and the tarsus nearly a right one with the tibia; but in the Myriapods, as far as I can unravel their swift many-footed motions, these angles in walking do not take place; in repose however, in many insects, the coxa forms an angle with the thigh below the horizontal line and with the tibia above it, and the tibia and tarsus continue in the same line, and point downwards nearly vertically; in others, as in the Tetramerous beetles, the last-mentioned joints[Pg 696] form an angle with each other and turn upwards, the tibia having an external oblique cavity to permit this; but the insects most remarkable for packing close their legs are those carnivorous genera Dermestes, Anthrenus, Byrrhus, &c. In the last-mentioned genus there are cavities in the under side of the trunk, in each division of the breast, and at the base of the abdomen, to receive the legs when folded; the coxæ have also a cavity to receive the base of the thigh. In the anterior legs this last part has a longitudinal one on its upper side, and in the four posterior on the under, which receives the tibiæ, which at the inner edge are straight, and at the outer curvilinear, and the tarsi are turned up and received by the concave part, on the anterior side of the first pair and the posterior side of the two last of the tibiæ, so as to lie between it and the body: when the legs are close packed, the animal looks almost as if it had none. I have observed that when Dytisci repose on the water, the posterior legs are turned up and laid over the elytra, and curved towards the head.
vi. Pectines. I must next say a few words upon a remarkable organ, which seems in some degree supplementary to the legs, by which the Creator has distinguished the genus Scorpio, called from its parallel teeth, set in a back, their pecten or comb[2120]. This back consists of two or more articulations, is attached by its anterior extremity to the sides of the posterior piece of the mesostethium, and is marked by a longitudinal furrow or channel. The teeth, which vary in number in the different species, and[Pg 697] in the same species at different periods of its growth, are usually ovato-lanceolate, or obtusangular, furnished on their exterior edge with what appears to be a longitudinal sucker, and supported between their bases, or at the base, both within and without, by triangular, conical, or subglobose props. With regard to the use of these organs, it has not been clearly ascertained. Amouroux states that he has seen the animals use them as feet, and he conjectures that by them they may fix themselves and turn upon them as on a pivot, when they have to make a retrograde movement[2121]. M. Latreille, from their having branchial pouches immediately under them, seems to think that they are connected with respiration[2122]. This may be true; but from the suckers just described, I am inclined to think with Amouroux, that they are useful to the animal in its motions, and that like the suckers of the Gecko, flies, &c., they enable it to support itself against gravity and to climb perpendicular surfaces.
Whether the five obtriangular plates, elevated on a pedicle, which are found arranged in a series on the under side of each of the jointed coxæ of the posterior legs in Galeodes, are at all analogous to the pectens of scorpions, has not been ascertained[2123]. M. Leon Dufour watched them very attentively in one species (G. intrepidus), but he could observe no motion in them[2124].
The abdomen of insects, which we are next to consider, is the third great section of the body, and is the seat of the organs of generation, as well as of a principal part of those connected with respiration. My remarks upon it will be under the following heads: Its substance; articulation with the trunk; composition; shape and proportions; its appendages; and its clothing.
i. Substance. Under this head I may observe in general, that where the abdomen is protected by hard elytra or tegmina, as in most Coleoptera, and many Heteropterous Hemiptera, the upper side is generally of a softer and more flexible substance than the under, which from its exposure requires a greater degree of hardness and firmness to prevent its being injured. In some,—as the Dynastidæ and those beetles whose elytra are connate, or as it were soldered together, the former is scarcely more than membrane. In others of the above tribes, nearly the whole of the back of whose abdomen, as in Staphylinus; or only its anal extremity, as in Melolontha; or its sides, as in Lygæus, &c., is not covered by the elytra[Pg 699] or tegmina, that part, as was requisite for its protection, is harder than the covered portion.
ii. Articulation with the trunk. Two distinct modes of this articulation take place:—in the first the abdomen is united to the trunk by the whole diameter of its base, without any appearance of incision; in the other only a small part of that diameter, with a very visible incision. All the Orders, except the majority of the Hymenoptera and Diptera, and the Araneidæ, belong to the first of these sections; for in all these the aperture by which the abdomen is suspended to the trunk, occupies the whole of the base; I say suspended, because, though in many cases it inosculates in the posterior cavity of the latter part, it does not in all, and the margins of the orifice are united by ligament to those of that cavity. Indeed, in the Coleoptera and others that have a somewhat prominent metaphragm[2125], the trunk may with more propriety be said to inosculate in the abdomen. With regard to the second section,—those in which the orifice is of less diameter than the base, occupying only a portion of it,—it may be further subdivided into those whose abdomen is sessile, and those in which it is united to the trunk by the intervention of a long or short pedicle or footstalk: to the first of these subdivisions belong all those Diptera that have an incision between the trunk and abdomen—for many tribes of this Order, as the Tipulidæ, Asilidæ, &c., belong rather to the first section—and the Araneidæ; the abdomen, however, in all is merely suspended, without any inosculation. To the second subdivision belong all the Hymenoptera, except the[Pg 700] Tenthredinidæ and Siricidæ, the abdomen of which is united to the trunk by the whole diameter of its base; these may be further subdivided into those that have a very short pedicle and those that have a long one; but as the mode of articulation in both these is the same, there will be no necessity to consider them separately. M. Cuvier has included the Diptera and Araneidæ in the same tribe with such Hymenoptera as have a petiolate abdomen[2126]; but as the manner in which the latter articulates with the trunk is widely different from that of the Diptera &c., I thought it best to consider them as distinct; especially as in the Diptera there is no tendency to a pedicle, while only the above two tribes of Hymenoptera are wholly without it. This learned author thus describes the articulation where the abdomen is connected by a pedicle. "They have," says he, "a real solid articulation, a kind of hinge in which the first segment is emarginate above, and receives a saliant portion of the trunk upon which it moves; this articulation is rendered solid by elastic and powerful ligaments; muscles which have their attachment in the interior of the trunk are inserted in this first segment, and determine the extent of its movement[2126]." But this passage by no means conveys an adequate idea of the singular mechanism by which the Divine Artificer has enabled these little creatures to impart the necessary movements to an organ so bulky compared with its very diminutive point of attachment. As no author that has fallen in my way has examined the articulation of the abdomen with the trunk in these Hymenoptera with the attention which it merits[2127],[Pg 701] I shall enlarge a little upon it. You would be surprised, and not without reason incredulous, were I seriously to assert that these insects lift their weighty posteriors by means of a rope and pulley; yet something like this really does take place, though not with all in a manner equally striking. The point of articulation in the insects in question, except in Evania, is at the base of the metathorax just above the posterior pair of legs: here you see a small orifice, either insulated or connected by a narrow opening with the larger one, when the abdomen is removed, which in many instances, as in the common wasp, is surmounted by another still smaller, through which, if you examine it attentively, you will find there is transmitted a flat and sometimes broadish ligament or rather tendon, in which the levator muscles of the abdomen, attached by their other end to the metaphragm[2128], terminate: another minute orifice above the base of the pedicle affords a point of attachment to the tendon, so as to give it prize upon the abdomen. Here the upper orifice in the trunk is the pulley (trochlea)[2129], the tendon is the rope (funiculus)[2129], and the abdomen is the weight to be lifted. When the muscles contract, the tendon, running over the edge of the aperture, is pulled in, and the part just named is elevated; and when they are relaxed the tendon is let out, and it falls. Some little variation in the structure takes place in different tribes: thus, in the[Pg 702] Formicidæ, Scoliadæ, &c., instead of a separate orifice, the part I call the pulley is merely an upper sinus of the large orifice that receives the pedicle of the abdomen. The shape of these orifices, both of the trunk and abdomen, varies in different genera: thus, in the bee it is triangular, with the vertex reversed; and in the wasp the upper one is circular, and the lower one transversely oblong; but in all, the apertures of the trunk correspond with those of the abdomen. In Evania, in which the minute abdomen is inserted in the upper side of the metathorax, there is scarcely any trace of this structure. With regard to the articulation of the pedicle itself with the lower orifice of the trunk, it appears simply suspended, with little or no inosculation. I may observe under this head, that though the abdomen in almost all insects is wholly clear of the cavity of the trunk, yet in some Phalangidæ (Gonyleptes K.) it appears almost retracted within it[2130].
iii. Composition. I shall next consider the segments into which the abdomen is usually divided, their number, and other circumstances connected with them. In the Hippoboscidæ, Acaridæ, Phalangidæ, and Araneidæ, the part we are considering is not divided into segments, though in some instances, as in Gonyleptes and the cancriform Epeiræ[2131], they are represented by folds; but in the great majority of insects it consists of several dorsal and ventral pieces or segments, forming by their union the annuli or rings into which it appears divided. The number of these abdominal segments varies in different[Pg 703] insects; I have noticed more than twenty such variations, and probably there are many more. Before I give you them in detail, I must first observe that the dorsal and ventral segments, though sometimes they correspond in number, yet very often do not, the dorsal most commonly exceeding the ventral by a segment; in a few cases however the reverse takes place. In the sexes also there is frequently a difference in the number of segments, as has been before observed[2132]. I shall express the variations in question by two figures, the first representing the number of dorsal segments, and the second the ventral—they usually only express the apparent segments: perhaps a very general examination and dissection might bring many of them nearer to a common type.
| 1:1. | Chelonus. |
| 3:3. | Chrysidæ[2133]. |
| 4:2. | Leucospis[2134]. |
| 5:5. | Syrphus. |
| 5:6. | Halictus ♀. |
| 6:5. | Nepa. |
| 6:6. | Halictus ♂. |
| Belostoma. | |
| 7:5. | Curculio L. |
| Cerambyx L.[Pg 704] | |
| 7:6. | Dytiscus ♀. |
| 7:7. | Ammophila, &c. |
| 8:6. | Dytiscus ♂. |
| Lucanus, &c. | |
| 8:7. | Geotrupes Latr. |
| 8:8. | Pimpla. |
| 8:10. | Euchlora McL.[2135]? |
| 8:13. | Scutigera. |
| 9:5. | Carabus Latr. |
| 9:6. | Gymnopleurus Ill. |
| 9:7. | Perga Leach ♀. |
| 9:8. | Perga ♂. |
| 10:7. | Locusta Leach ♀. |
| 10:8. | —————– ♂. |
| 10:10. | Æshna. |
| 11:7. | Phasma. |
| 11:9. | Chelifer. |
| 12:11. | Thelyphonus. |
| Many: | Myriapoda. |
I shall next explain the articulation of the segments with each other, both that of the rings formed by the union of the dorsal and ventral pieces, and that of those pieces themselves. In general it may be stated with respect to the former, that each ring is suspended by ligament to that which precedes it; but this takes place in three ways—in some the margins of the suspended rings touch each other only, with little or no inosculation; in others the dorsal segments only touch, and the base of each ventral is covered more or less by the apex of the preceding one; and in others again the base of the whole ring, both above and below, is so covered, or inosculates. The first kind here mentioned you will find exemplified in Melolontha, Geotrupes, Musca, &c.; the second in Scorpio; and the third in Staphylinus, the Hymenoptera, and many others. In the Coleoptera, says M. Cuvier, speaking of the movements of the abdomen, the rings only touch each other at the margin, and the[Pg 705] movement is very limited; whilst in the Hymenoptera they are so many little hoops, which inosculate in each other as the tubes of a telescope, one third only of their extent often appearing uncovered[2136]. We see the reason of this structure when we consider the calls they have for greater powers of movement in this part in laying their eggs, and annoying their enemies and assailants; and also in the Staphylinidæ to enable them to turn up their abdomen like a scorpion, both as a posture of attack, and to fold their wings: in all cases, however, as far as my observation goes, these animals, when they want to lengthen this part, can disengage the rings from almost all inosculation, so that no impediment remains to any movement.
The articulation of the dorsal and ventral segments with each other is next to be considered. In Iulus and some Centroti the ring appears to be formed of a single piece, with scarcely any trace of the existence of any such division; it is however almost universal, and is of three descriptions; in the first the dorsal segments are united to the ventral at the lateral margin or edge of the abdomen; in the second it is above this margin, and in the third below it. You will find that in Fulgora and many other Homopterous Hemiptera these segments unite at the margin, as they do likewise in Cimex lectularius belonging to the other Hemipterous section; but in the rest of the Heteropterous tribes, the ventral segments turn upwards, and their union with the dorsal is in the back of the abdomen; in these the Hemelytra and wings only cover the dorsal segments, leaving the edge, formed[Pg 706] of the ends of the ventral, uncovered. The Lamellicorn beetles also, and many other Coleoptera, exhibit the same structure. To the last description, in which the dorsal segments turn down to meet the ventral, belong the Lepidoptera, Locusta Leach; likewise Sirex, Chrysis, and many other Hymenoptera. The articulation between these segments is by means of an elastic membranous ligament, which usually is not externally visible; but in many instances, in which the connecting ligament is of a firmer substance, as in Scorpio, Thelyphonus, and Phrynus, it is very conspicuous, and in the latter genus exhibits many longitudinal folds, as it does likewise in Gryllotalpa, which must permit a vast extension of the abdomen. In this membrane, in some cases, as in Dynastes McL., Melolontha, &c., the two or three first spiracles are fixed[2137]. In the Hymenoptera and many other insects the dorsal segments do not unite by their margin with the ventral, but the end of each dorsal laps over that of the corresponding ventral.
Dorsal segments[2138]. I shall next notice the segments seriatim, in the order of their occurrence, beginning with the dorsal ones. The most remarkable circumstance with respect to these that occurs to my recollection takes place in the Cancroid spiders (Epeira cancriformis, aculeata, &c.), in which the back of the abdomen is formed by a plate, in some extended in a transverse direction (E. cancriformis), in others in a longitudinal one (E. aculeata), of a much harder substance than the under side and quite flat, set with strong sharp spines, in the former species apparently moveable, and terminating behind[Pg 707] in a piece resembling in some measure the scutellum of the Stratyomidæ and similarly armed with a pair of spines[2139]: in E. aculeata the sides of the abdomen, under the plate, have a number of longitudinal folds like those of Phrynus. In Cryptocerus, a genus of ants peculiar to South America, the first segment, not reckoning the pedicle, forms almost the whole back of the abdomen, and the three last are so minute as scarcely to be distinguishable. Nothing very remarkable is exhibited by the other segments, except that in Trichius the penultimate is the largest; in some Staphylinidæ (S. splendens) and Brachini (B. melanocephalus) it is emarginate, and in the former tribe also often terminating in a white membrane. The dorsal segment most worthy of notice is the last, which is called the podex; for though in general it is a minute piece, often retracted within the abdomen and invisible, as in many Diptera, yet sometimes it is the most conspicuous of the dorsal segments. It is most commonly triangular, and usually deflexed and forming an angle with a horizontal line; but in Clytra, Chlamys, and Oryctes, it is inflexed; in many Lamellicorns it is nearly vertical. In Tettigonia F., many other Homopterous Hemiptera, and some Hymenoptera (Cimbex), its sides turn down and become ventral; on its lower side it has in these a longitudinal cavity which receives the ovipositor in repose[2140]. In many other insects it unites with the last ventral segment, the hypopygium, to form a tube for that organ, as you will find in Callidium violaceum[2141], many Muscidæ, and Thelyphonus. As to its termination the podex is sometimes bifid, Blatta; bipartite, Ranatra;[Pg 708] mucronate, Sirex; acuminate, Melolontha vulgaris, Trichius hemipterus. Generally this part is flat; but the disk is elevated or gibbous in Oryctes and some other Lamellicorns. In the majority of the Coleoptera Order it is quite covered by the wings and elytra; but in many of the last-mentioned tribe, and sometimes the penultimate segment also, it is not covered by them[2142]. In some insects the piece we are considering appears to consist of two segments; in the male of Locusta morbillosa the whole podex is rhomboidal, but it is formed by two triangular pieces which articulate with each other; this structure permits the more easy elevation of the terminal one for the extrusion of the feces.
Ventral Segments[2143]. We are now to turn our attention to the ventral segments of the abdomen. The first of them is what is called the epigastrium[2144] in the table. This part, according to M. Chabrier, is of considerable importance to the animal in flight, as, by its pressure against the trunk, not only regulating the movements of the abdomen, but as, in his opinion, contributing to push forward the trunk[2145] in the descent of the animal. It is remarkable only in the Coleoptera and Heteropterous Hemiptera, to which my observations upon it will be confined. It may be stated as usually consisting of two articulations, that nearest the trunk being narrow, and in the Predaceous beetles[2146], as also in Scutellera, Pentatoma,[Pg 709] &c., interrupted in the middle[2147]. In many Lamellicorns this joint is concealed under the posterior coxæ, and with the anterior part of the second forms a hollow cavity for their reception; this last joint is what is properly the Epigastrium, the former, especially when distinct, being called in the table the Hypochondria. In Sagra and Brentus the epigastrium is particularly conspicuous for its size, in the former occupying half, and in the latter nearly two-thirds of the under side of the abdomen; but in general it is distinguished from the remaining segments only by the central mucro or point that terminates it towards the trunk[2148], and which is received by a sinus of the metasternum; this point is generally minute and triangular, but in Sagra it is large and rounded at the extremity, and in Calandra it terminates nearly in a transverse line somewhat waving. It is most remarkable, however, in some species of the Heteropterous genus Edessa F.; for in E. nigripes and affinities it is a sharp sterniform conical horn, which passing between the four posterior legs covers the end of the promuscis. In fact, this part appears a kind of abdominal sternum. In the Cetoniadæ, &c., the Hypochondria unite before this mucro, and form a ridge which articulates with it, and dips towards the abdominal cavity; in Scolytus the epigastrium is much elevated from the rest of the ventral segments, so that the under side of the abdomen appears as if it were suddenly cut off, whence Herbst's awkward though not inexpressive name, Ekkoptogaster; this part in this genus has something of a posterior mucro.
The intermediate ventral segments exhibiting no very[Pg 710] remarkable peculiarities, I shall pass them without further notice, and call your attention to the last, which is opposed to the podex, and which I have named the hypopygium[2149]. Though usually a single small piece, in Edessa and many Pentatomæ it consists of several plates; and in Trichius it is very large: it is mostly intire, but in the male Dytisci it is cleft; in Lamia ocellata trilobed; in Edessa tripartite; in Centrotus Taurus it is boat-shaped and hollowed out to receive the stalk of the ovipositor. It is also generally in the same line with the body, but in Xenos it is turned up and bent inwards[2150].
iv. Shape. With regard to shape, in some Orders the abdomen varies considerably; but the most general form is one that approaches to trigonal, so that a transverse section will be a triangle, with the vertex more or less obtuse, and the base more or less convex; some tendency to this form will often be found even in those insects whose abdomen appears almost as flat as a leaf, as in many Aradi. In the hive-bee the transverse section is almost an equilateral triangle; in Belostoma grandis the disk of the under side of the part in question is longitudinally elevated into a trigonal ridge, the section of which is an equilateral triangle, the sides being quite flat. In general, in the vertical section of an abdomen, the vertex of the triangle points downwards, but in Libellula F. it points upwards. In Blatta this section is nearly lanceolate; in Staphylinus olens it is a segment of a circle with the convex side downwards; in Æshna F. with that side upwards; and in Agrion the section is circular. In Copris, Ateuchus, &c., the abdomen is very short and[Pg 711] thick; in Staphylinus slender and long; in Aradus, Nepa, &c., depressed and flat; compressed in Ophion and Evania; conical in Cœlyoxis; rhomboidal in many Mantes; boat-shaped in many Lygæi; fusiform in various Papilionidæ; lanceolate in some Ichneumonidæ, falcate in others; nearly round in Diapria pupurascens; ovate in Lyrops; elliptical in Andrena; oblong in many Xylocopæ; heart-shaped in the naked Euglossæ; triangular in Dytiscus; gibbous in Flata; and vaulted in Chrysis. At its base it is truncated in Sirex; retuse in most bees; forming the segment of a circle in Andrena; in general sessile, but in the majority of Hymenoptera, as has been already observed, terminating in a pedicle. The pedicle is very short in the Andrenidæ and Apidæ; long in the Sphecidæ; thick in the Formicidæ; slender in Evania; fusiform in Pelecinus; clavate in Ammophila; campanulate in many Vespidæ; nodose in Myrmica[2151]; squamigerous in Formica[2152]: it sometimes also consists of two joints, as in Ammophila and many Vespidæ. As to margin, some have none, as Centrotus; in others, as Dytiscus, it is very narrow; in others again, wide and flat, as in the Nepidæ; in Staphylinus, &c., it is distinguishable only on the upper side of the abdomen; in Locusta Leach only on the under side, though mostly intire; it is serrated in Blatta, sinuated in Acanthia paradoxa, and crenated in Cerceris.
v. Proportions. These vary greatly in the different tribes; in some the abdomen is long and slender, as in Locusta, and Staphylinus; disproportionably so in a remarkable degree in some Agrionidæ from South America,[Pg 712] as A. lineare, &c.[2153]; in others it is extremely short and thick, as in Copris, &c.; a mere appendage in Evania; it is shorter than the elytra in Trox; of the same length in most beetles; longer in Melolontha, Hister, &c.; disproportionably so in Staphylinus: though usually of the same width with the trunk, in many Mantidæ it is much wider[2154]; and more slender in the Libellulina, Myrmeleon, &c.
vi. Arms and Appendages[2155]. These are various; and may be considered under the following heads: processes; organs of respiration, motion and prehension; weapons; and other anal appendages the use of which is unknown.
1. Processes. Under this term I include all prominences of whatever kind, whether tubercles, teeth, spines, or horns, that arm any part of the abdomen. Many of these are sexual characters, and have been sufficiently described in a former letter[2156]; I need not therefore detain you long on this head. Of the first kind is a remarkable elevation that distinguishes the second ventral segment of Scolytus Destructor (Ips Scolytus Marsh.) or of a species allied to it[2157]; in S. pygmæus (I. multistriatus Marsh.) the same segment is armed by a flat horizontal tooth or horn; in an Aradus from Brazil, before alluded to[2158] (A. laminatus K. MS.), the margin of the abdomen is surrounded by eight flat subquadrangular laminæ; in another species figured by Stoll[2159], it is cut out into bays by a number of[Pg 713] denticulated teeth; and in Acanthia paradoxa by long spinose lobes[2160]. In Edessa F., another genus of bugs, the abdomen usually terminates in four strong sharp dentiform spines, the intermediate ones being the shortest, and in some the margin is also armed with spines[2161]; occasionally the anal spines are very long[2162]. In addition to the ventral horns before mentioned that distinguish the sexes of some insects[2163]; the males of the genus Conops, a two-winged fly, have, on the antepenultimate ventral segment, a singular process, varying in length and shape in the different species, standing nearly at right angles with the belly, convex towards the trunk, and concave towards the anus. De Geer supposes that with the anal extremity this forms a forceps with which this fly seizes the other sex[2164].
2. Organs of respiration[2165]. I shall defer my account of the spiracles, and other external respiratory organs, till I come to treat of the system of respiration in insects, when every thing connected with that subject will be most properly discussed; but there are certain appearances in some insects, which at first sight seem to partake of the same character, but which being really independent of that vital function, may here have their place. If you examine the abdomen of the mole-cricket (Gryllotalpa vulgaris), you will easily discover the true spiracles in the folds of the pulmonarium, which separates the back of that part from the belly; if you next inspect the five intermediate segments of the latter, you will discover on each nearer the base a pair of oblique little channels,[Pg 714] which precisely resemble closed spiracles. These may be denominated false or blind spiracles. Again, if you examine the pupa of any Scutellera or Pentatoma, in which tribe the true spiracles are ventral, you will discover, placed in a square on the two or three intermediate dorsal segments, four or six elevated points resembling spiracles, but not perforated, connected often by corrugations in the skin or crust[2166]; in the larvæ also of some Reduvii the first minute dorsal segment, at each lateral extremity, has a similar elevation with a central umbilicus precisely resembling a spiracle, but still not perforated: another instance of false spiracles in this section of the Hemiptera, is furnished by Aradus laminatus before mentioned, in the perfect insect; between the spiracle and the margin of each ventral segment is a white round callus, with a dark point resembling a perforation on its exterior side, and terminating internally in a channel covered by membrane leading to the disk of the segment, so that the whole in shape resembles a tobacco-pipe[2167]. A number of similar callosities with a central impression, but without any channel, variously disposed, are also to be found in another bug, Rhinuchus compressipes K.[2168] In the Homopterous section of this Order, a series of impressed points, which may be easily mistaken for spiracles, are to be discovered on both sides of the abdomen, at the margin in Centrotus, in which the real spiracles are quite concealed.
In spiders, as we learn from Treviranus, the open ventral spiracles of the scorpion are replaced by pseudo-spiracles;[Pg 715] these in Epeira Diadema are three pair of small black points: on the back of the abdomen also are four pair, but in some species there are only two[2169]: the most remarkable, however, are exhibited by the cancriform spiders before noticed[2170]: in Epeira cancriformis, in the plate which covers the abdomen, they are dark red spots with an elevated rim and centre[2171] exactly resembling spiracles, except that they are not perforated; there are twenty-four of them, twenty arranged round the margin, and four in a square in the disk.
3. Organs of motion. In a former letter you were told that several insects are enabled to leap by means of organs in their abdomen; I shall now describe such of them as require further elucidation. I then said that Podura and Sminthurus, two apterous genera, take their leaps by means of an anal fork[2172]. In the former genus the fork consists of a single piece attached to the under side of the anus, and terminating in a pair of long slender sharp processes which articulate with it and form the fork or saltatorious instrument[2173]. In Sminthurus the tines, as they may be called, of the fork do not articulate with the base, but are of the same piece and consist of two joints, the terminal one being flat and obtuse[2174]. Machilis to the anal fork adds eight pair of ventral linear springs (Elastes), which are covered with hair or scales, and terminate in a bristle or two. I have on a former occasion mentioned the natatorious laminæ with which the anus[Pg 716] of the larva of Agrion and of some Diptera is furnished[2175]; the same part in that of Dytiscus ends in a pair of tapering organs, fringed on each side like the hind-legs of the imago[2176], which doubtless assist it in swimming; those respiratory foliaceous laminæ which so singularly distinguish the abdomen of the larvæ of Ephemera, like the legs of the Branchiopod Crustacea, are probably used in some degree as fins, and aid their motions in the water[2177]. Under this head may also be mentioned the many-jointed bristles that form the long tails[2178] of the fly that proceeds from these larvæ, whose interesting history I long since enlarged upon; for when they fly the two lateral ones diverge from the central one, and perhaps perform the same office as the tail feathers (rectrices) of birds. These bristles are also to be found in Machilis[2179], and probably, as its leaps are almost as long as flights, for a similar purpose, to steady their motion. I may here lastly state that I once saw a Cryptophagus (Corticaria Marsh.), but I forgot to note the species, walking upon my window, which when it wanted to turn fixed itself to the glass by an inflated anal vesicle, and so accomplished its purpose.
4. Organs of Prehension[2180]. The abdominal organs of prehension are various; but as the great body of them are connected with the sexual intercourse of insects, I shall not consider them till I come to treat on that subject. The only remarkable one that is common to both sexes is that of the earwig, which is too well known to every child to call for any long description. The external organs[Pg 717] of oviposition I shall also describe hereafter, and likewise those of secretion that have not already been noticed.
5. Weapons. As the stings of some Hymenoptera are analogous to the ovipositors of the majority of that Order, I shall consider them both together when I treat of the sexual organs of insects; but there is one, and that a tremendous one, not connected with those organs, which may be noticed here. I mean the sting of the scorpion. There appears to be some analogy between the poisonous fangs of one tribe of the Ophidian reptiles[2181], the mandibulæ of spiders[2182], the second pair of pedipalps, or the fangs of the Scolopendridæ[2183], and the organ in question[2184]; but the last possesses this peculiarity, that it is placed at the opposite extremity of the body, where it is preceded by a long jointed tail, which properly speaking is merely a continuation of the abdomen, since the spinal marrow, the intestinal canal, and the pseudocardia, are extended into it[2185]. Providence might have a double view in thus contracting the dimensions of this part of the abdomen; in the first place, the animal is by this enabled to turn its tail over its back preparatory to its inflicting a wound, and in the second, perhaps, this formation favours the sublimation of the venom, the long tail acting as an alembic for that purpose. This machine consists of six angular joints including the sting, the last but one being the longest, and the last inflated, as it were, at the base, and terminating in a sharp subulato-conical point which curves downwards,[Pg 718] and has an orifice in a channel at the end on each side. Treviranus could not discover these orifices in the sting of Scorpio europæus[2186]; they may however be readily seen if viewed with a sufficiently high power, though not under a common pocket microscope. Whether the very slender, many-jointed, real tail of the remarkable genus Thelyphonus is used in any respect as a weapon, has not been ascertained: it is a filiform hairy organ consisting in some specimens of more than twenty joints, the first being very much larger than the rest[2187].
6. Appendages[2188]. We are lastly to advert to those appendages of the abdomen of which the use is not at present discovered. These are the styles (styli) of the Staphylinidæ; the leaflets (foliola) of the Libellulina; the floret (flosculus) of the Fulgoræ; the cerci of the Blattidæ and Gryllina; and the threads (fila) of Machilis: but having nothing important to add concerning them, the definitions of those terms will give you a sufficiently clear idea of them[2189]. As they are common to both sexes, if their use is connected with the sexual intercourse, it must be similar to that which Treviranus ascribes to the pectens of scorpions, they must be instruments of excitement.
And now, after this long discourse on the External Anatomy and structure of these little beings, you may think perhaps at first that the subject is exhausted; and[Pg 719] that I must have discovered and described every part and every variation of every part of the crust of an insect. But when you go on to reflect what a comparatively small number of these creatures have fallen under my examination, and in those, after all my laborious and painful researches, from my limited faculties and other imperfections of our common nature, how much will probably have eluded my notice, you may conclude that thousands of facts still remain concealed to reward the patient assiduity of future investigators. Such are the immensity and variety of the works of the Creator in this department, that it would require a long life, and fill volumes upon volumes, to discover and give a description of all the peculiarities of structure of the insects that are already known; and could all that exist[2190] be so studied and explained in full detail, the library that the Calif Omar ordered to be burned at Alexandria could scarcely have contained more books than would be required to receive the results. But "who is sufficient for these things[2191]?" This is a question that the most able and most experienced physiologist must often feel disposed to put to himself when, lost in the intricate labyrinth of the works of his Maker, he sees all things arranged, "wheel within wheel," in an order that he can only partially unravel, instead of tracing the "regular confusion" through all its windings. But glimpses of light, and points of irradiation, here and there discover to fragments of the truth of things, and such vestiges[Pg 720] of the grand system of the Deity, as enable him in some degree to appreciate, and dispose him humbly to adore that Wisdom, Power, and Goodness, that at first created and now sustains in its full beauty and harmony the wondrous whole.
I am, &c.
END OF THE THIRD VOLUME.
Printed by Richard Taylor,
Shoe-Lane, London.
PLATE VI.[2193]
| FIG. | |
| 1. | Head of Mylabris. Upper side, or face. |
| 2. | –——————— Under side, or subface. |
| 3. | Trophi of Dytiscus. Six pieces. |
| 4. | Head of Locusta. Face. |
| 5. | ——————— Front view, to exhibit the mouth. |
| 6. | Trophi of ditto. Seven pieces. |
| 7. | Head of Cicada. Face. |
| 8. | ——— Scutellera. Subface. |
| 9. | Trophi, or promuscis of Hemiptera. Three pieces. |
| 10. | Head of Æshna. Face. |
| 11. | ——————– Front view. |
| 12. | Trophi of ditto. Seven pieces. |
| 13. | ——– or antlia of Lepidoptera. Four pieces. |
| 14. | ——– of Panorpa. Three pieces. |
PLATE VII.[2194]
| FIG. | |
| 1. | Head and trophi of Phryganea L. Face. |
| 2. | –——————— Vespa Crabro. Ditto. |
| 3. | Trophi of Bombus. |
| 4. | Head of Tabanus L. Face. |
| 5. | Trophi or proboscis of ditto. (Reaum.) |
| 6. | ————————— Bombylius[2195]. |
| 7. | Head of Oxypterum. Face. |
| 8.[Pg 722] | Head of Pulex, with its antennæ and trophi, or rostrulum. Side view. |
| 9. | ——— Araneidæ, with the trunk. |
| 10. | Trophi of ditto. |
| 11. | Head of Scolopendra morsitans. Subface. |
| 12. | ————————————— Front view, to show the mouth. |
| 13. | Trophi of ditto. |
| 14. | Pharynx of Pentatoma. (Savigny.) |
PLATE VIII.[2196]
| FIG. | |
| 1. | Prothorax of Lucanus. a. Apex. b. Base. c d. Sides. a. Disk. |
| 2. | Antepectus of ditto. |
| 3. | Alitrunk of ditto. Upper side. a. A piece between the metathorax and metaphragm. |
| 4. | ——————— Under side. |
| 5. | Abdomen. Upper side, or tergum. |
| 6. | ———— Under side, or venter. |
| 7. | Antepectus of Hydrophilus piceus. |
| 8. | Alitrunk of ditto. Under side, to show the metasternum. |
| 9. | Abdomen of Dynastes Aloeus, to show the dorsal and ventral spiracles. |
| 10. | Prothorax of Locusta. a. Apex. b. Base, c c. Sides. |
| 11. | Antepectus of ditto, to show the prosternum. |
| 12. | Alitrunk of ditto. Upper side. |
| 13. | ——————— Under side. |
| 14. | ——————— Lateral view. Vol. III. p. 48. |
| 15. | Abdomen of ditto. Lateral view. |
| 16. | Alitrunk of Cicada Latr. Upper side. |
| 17. | —————————— Under side. a. The piece in the ♀ corresponding to the drum-covers of the ♂. |
| 18. | Abdomen, and part of postpectus of ♂ ditto. Under side, to show the drums. Vol. II. p. 405—. |
| 19.[Pg 723] | Abdomen, and part of postpectus of ♂. Lateral view, with the covers removed to show the machinery. |
| 20. | Alitrunk. Upper side. Pentatoma. |
PLATE IX.[2197]
| FIG. | |
| 1. | Alitrunk of Cossus ligniperda. Upper side. |
| 2. | Part of ditto, to show the mesophragm. |
| 3. | ————— Under side. |
| 4. | Patagia of Lepidoptera. Upper and under sides. Vol. III. p. 368, 539. |
| 5. | Tegulæ of ditto. Two species. Vol. III. p. 378. |
| 6. | Prothorax of Æshna. a. The base elevated and forming an obtuse angle with the rest. |
| 7. | Alitrunk of ditto. Upper side. a. Two elevated areas of the posterior parts of the collar, strengthened by a marginal ridge and denticles, internally connected by an elastic ligament, apparently to aid and sustain the powerful action of the wing-muscles. |
| 8. | ——————— Lateral view. a. A piece by which the mid-leg is connected with the scapular. Vol. III. p. 48, 565. |
| 9. | Part of the abdomen of Libellula. |
| 10. | Trunk of Semblis F. Upper side. |
| 11. | Alitrunk of Vespa Crabro. Upper side. a. Aperture in the trunk for the passage of the ligament that elevates the abdomen. |
| 12. | Lateral view of ditto. |
| 13. | —————— posterior part of ditto, and of the base of the abdomen, to show the above apparatus. a. The aperture. Vol. III. p. 701. |
| 14. | Head and part of the manitrunk of Tenthredo L. to show the membrane a. representing the prothorax. Vol. III. p. 550—. |
| 15.[Pg 724] | Alitrunk of Xiphydria. Upper side. |
| 16. | ————————— Lateral view. |
| 17. | Part of trunk and abdomen of Formica, to show the squama. Vol. III. p. 389. 3. |
| 18. | ——————————– of Myrmica, to show the nodus. Vol. III. p. 389. 4. |
| 19. | Alitrunk of Musca. Upper side. a. Alula or winglet. |
| 20. | Metathorax of ditto. |
| 21. | Alitrunk of ditto. Lateral view. |
| 22. | Abdomen of ditto. Venter. |
PLATE. X.[2198]
N.B. In this plate the red points out the costal, and the yellow the anal areas, the intermediate being uncoloured.
| FIG. | |
| 1. | Elytra, a. Base. b. Shoulder. c. Lateral margin. d. Apex. |
| 2. | Tegmina. Blatta. |
| 3. | Hemelytra. Pentatoma. |
| 4. | Wing. Coleoptera. a. An insulated nervure. Vol. III. p. 625. |
| 5. | ——– Dermaptera. |
| 6. | ——– Lepidoptera. |
| 7. | ——– Neuroptera. |
| 8. | ——– Hymenoptera. Tenthredo L. |
| 9. | ————————– Bombus. |
| 10. | Under wing. Hymenoptera. |
| 11. | ——————————— Proctotrupes. |
| 12. | ————— Diptera. Tipula. |
| 13. | ————————— Psychoda. Vol. III. p. 645. |
| 14. | ————————— Musca. a b. Two areolets between the costal and mediastinal nervures. c. Areolet between the mediastinal and postcostal nervures.[Pg 725] d. Areolet between the postcostal and subcostal nervures, e. Open areolet. Vol. III. p. 634. |
| 15. | Under wing. Diptera. Stratyomis. a b. The two areolets between the costal and postcostal nervures; the mediastinal being nearly obsolete. c. Middle areolets crowned by a small one, d. |
PLATE XI.[2199] Antennæ.
| FIG. | FIG. | ||
| 1. | Setaceous. | 13. | Distichous. |
| 2. | Capillary. | 14. | Pectinate. |
| 3. | Filiform. | 15. | Duplicato-pectinate. |
| 4. | Incrassate. | 16. | Ciliate. |
| 5. | Fusiform. | 17. | Flabellate. |
| 6. | Prismatic. | 18. | Ramose. |
| 7. | Ensiform. | 19. | Furcate. |
| 8. | Falciform. | 20. | Auriculate. a. The auricle. |
| 9. | Moniliform. | 21. | Palmate. |
| 10. | Dentate. | 22. | Irregular. |
| 11. | Serrate. | 23. | Perfoliate. |
| 12. | Imbricate. |
PLATE XII. Antennæ.
| FIG. | FIG. | ||
| 1. | Capillaceous. | 16. | Subulate. |
| 2. | Mucronate. | 17. | Filate, simple.. |
| 3. | Uncinate. | 18. | |
| 4. | Clavate. | 19. | Filate, compound. a. Joints. |
| 5. | Nodose, or Biclavate. | 20. | Filate. |
| 6. | Convolute. | 21. | Aristate. Setarious a. Bristle. |
| 7. | Geniculate. | 22. | Aristate. Plumate a. Bristle. |
| 8.[Pg 726] | Capitate with a tunicate knob. | 23. | Stupeous. Vol. III. p. 646. |
| 9. | Capitate with a solid knob. | 24. | Plumose. |
| 10. | Capitate with a perfoliate knob. | 25. | Scopiferous. a. Brush. |
| 11. | Filiform. | 26. | Barbate. |
| 12. | Globiferous. | 27. | Verticillate. |
| 13. | Connate. | 28. | Inflated. |
| 14. | Setigerous. | 29. | Auriculate. a. Auricle. |
| 15. |
PLATE XIII.[2200]
| FIG. | |
| 1. | Unguiculate feeler. Gonyleptes. a. Claw. |
| 2. | Securiform ditto. Cychrus. a. Terminal joint. |
| 3. | Inflated ditto. Araneidæ ♂. a. ditto. |
| 4. | Lunulate ditto. Oxyporus. a. ditto. |
| 5. | Dentate mandible. Megachile. |
| 6. | Suctorious ditto. Larva of Dytiscus. a. Aperture. |
| 7. | Prosthecate ditto. Staphylinus. Vol. III. pp. 356, 439. |
| 8. | Trophi of Curculio L. |
| 9. | Pedunculate eyes. Diopsis. a. Footstalk. |
| 10. | Compound ditto. Muscidæ. Vol. III. p. 494. 3. |
| 11. | Conglomerate ditto. Iulus. Ibid. p. 494. 2. |
| 12. | Rostrate head. Balaninus. |
| 13. | Capistrate ditto. Nitidula. |
| 14. | Clypeate ditto. Copris. |
| 15. | Lychnidiate ditto. Fulgora. |
| 16. | Buccate ditto. Myops. a. The inflated part. |
| 17. | Cruciate prothorax. Locusta. |
| 18. | Cucullate and alate ditto. Tingis. |
| 19. | Subulate elytra. Sitaris. |
| 20. | Ampliate ditto. Lycus. |
PLATE XIV.[2201]
| FIG. | |
| 1. | Ideal wing, to exemplify painting. Vol. IV. p. 286—. a. Anterior or exterior margin. b. Interior ditto. c. Posterior ditto. d. Humeral angle. e. Scutellar ditto. f. Posterior ditto. g. Anal ditto. a. Articulate fascia, or band. b. Macular ditto. c d. Sesquialterous ditto. d e. Sesquitertious ditto. f. Dimidiate ditto. g. Abbreviate ditto. h. Pyramidate ditto. i. Supercilium. k. Hastate pupil. l. Compound eyelet or ocellus. m. Nictitant ditto. n. Simple ditto. o. Annulet. p. Bipupillate eyelet. q. Sesquialterous ditto. r. Double ditto. s. Caudate wing. t. Pupil. u. Iris. v. Atmosphere. |
| 2. | Reversed wings. Gastrophaca. |
| 3. | Digitate ditto. Pterodactylus. |
| 4. | Falcate ditto. Attacus. |
| 5. | Saltatorious leg, with loricate thigh. Locusta. |
| 6. | Natatorious ditto. Dytiscus. |
| 7. | Ambulatorious ditto. Lucanus. |
| 8. | Prehensorious ditto. Gonyleptes. |
PLATE XV.[2202]
| FIG. | |
| 1. | Laminate coxa. Haliplus. |
| 2. | Alate tibia. Lygæus phyllopus, a. The appendage. |
| 3. | Clypeate ditto. Crabro ♂. a. The clypeus. Vol. III. p. 334. |
| 4. | Dolabriform ditto. Curculio maritimus E.B. |
| 5. | Fossorious leg, with palmate tibia. Clivina. Vol. II. p. 365. |
| 6. | —————— with digitate ditto. Gryllotalpa. Ibid. p. 366. |
| 7. | Chelate feeler. Scorpio. |
| 8. | Scutate tarsus. Hydrophilus piceus ♂. Vol. III. p. 336. |
| 9. | Patellate ditto. Dytiscus marginalis ♂. a. Cups. Ibid. p. 336, 694—. |
| 10.[Pg 728] | Obumbrate abdomen. Epeira cancriformis. |
| 11. | Retracted ditto. Gonyleptes. |
| 12. | Cheliferous tail. Panorpa ♂. |
| 13. | Flosculiferous ditto. Fulgora. |
| 14. | Saltatorious ditto. Podura. |
| 15. | Folioliferous ditto. Æshna. |
| 16. | Cauduliferous, and filiferous ditto. Machilis. |
| 17. | Styliferous ditto. Staphylinus. |
| 18. | Unciferous ovipositor. Locusta. |
| 19. | Ensate ditto. Acrida. |
| 20. | Navicular ditto. Cicada. |
| 21. | Serrulate ditto. Tenthredo L. |
| 22. | Telescopiform ditto. Chrysis. |
| 23. | Anal apparatus of Blatta. |
PLATE XVI.[2203]
| FIG. | |
| 1. | Extricated ovipositor. Pimpla. Two pieces. |
| 2. | Telescopiform ditto. Stomoxys calcitrans? (Reaum.) |
| 3. | ———————— Œstrus. (Ibid.) Vol. I. p. 150. |
| 4. | Semicomplete pupa. Cicada. |
| 5. | Subsemicomplete ditto. Libellula. a. Mask. Vol. III. p. 125—. |
| 6. | Incomplete ditto. Hydrophilus. (Lyonnet.) |
| 7. | –—————— Myrmeleon emerging from its cocoon. (Reaum.) |
| 8. | –—————— Vespa vulgaris. |
| 9. | –—————— Chironomus plumosus. (Reaum.) a b. Respiratory plumes. |
| 10. | Obtected pupa. Apatura Iris. |
| 11. | —————— Vanessa Urticæ. a. Head-case with two points. |
| 12. | —————— Gonepteryx Rhamni. a. Head-case with one point. |
| 13.[Pg 729] | Obtected pupa. Sphinx Ligustri. a. The tongue-case. b. The eye-case. c. The trunk-case. d. First segment of the abdomen. e. The adminicula. f. The mucro, or point of the tail. Vol. III. p. 249—. |
| 14. | Hairy obtected pupa of Laria fascelina. |
PLATE XVII.[2204]
| FIG. | |
| 1. | Coarctate pupa. Œstrus hæmorrhoidalis. (Reaum.) |
| 2. | —————— Stratyomis chamæleon. (Ibid.) a. The pupa as formed within the skin of the larva. |
| 4. | Oviform body which many pupæ of Diptera at first assume under the skin of the larva. (Ibid.). Vol. III. p. 235. |
| 3. | The same when the parts begin to show themselves. (Ibid.) |
| 5. | Cocoon of Saturnia pavonia. a. Pupa. b. Threads that close the orifice. Vol. III. p. 217, 279. |
| 6. | Loose and irregular ditto, of Arctia villica. Ibid. p. 220. |
| 7. | Boatshaped ditto, of Tortrix prasinana. Ibid. p. 221. |
| 8. | Network ditto, attached to the stalk of a plant. |
| 9. | Ditto, imitating the scales of fish. (Reaum.) Vol. I. p. 462. |
| 10. | Spiral case of Trichopterous larva, formed of pieces of leaf. (De Geer.) |
| 11. | Grate spun by these larvæ to prevent ingress. (Ibid.) Vol. II. p. 264. |
| 12. | Chilopodimorphous larva of Melolontha vulgaris. Vol. III. p. 163. |
| 13. | Araneidiform? ditto of Cicindela campestris. Ibid. 152, 163. |
PLATE XVIII.[2205]
| FIG. | |
| 1. | Anoplurimorphous larva. Chrysomela Populi. a. Osmateria, or scent organs. Vol. II. p. 245. III. p. 163, 166. |
| 2.[Pg 730] | Anoplurimorphous larva. Cassida. a. The fecifork covered with excrement. Vol. IV. p. 353. 5. |
| 3. | Helminthimorphous or vermiform ditto of Balaninus Nucum. Vol. III. p. 163. |
| 4. | Chilognathimorphous ditto of Elater Segetum. a. a. Spiracles. |
| 5. | Decapodimorphous ditto of Dytiscus marginalis. Vol. III. p. 165. |
| 6. | Chilopodimorphous ditto of Staphylinus? a. Anal proleg. |
| 7. | Amphipodimorphous ditto of Acrida. Vol. III. p. 165. |
| 8. | Larva of Zelus. |
| 9. | Helminthimorphous ditto. Apis mellifica. (Reaum.) |
| 10. | Larva of Sirex. |
| 11. | –——— Tenthredo L. (Reaum.) a. 6 legs. b. 16 prolegs. |
| 12. | –——— Sphinx. a. 6 legs. b. 10 prolegs. c. Anal horn. |
| 13. | Spinose ditto of Vanessa Io. |
PLATE XIX.[2206]
| FIG. | |
| 1. | Larva of Papilio Machaon, a. Its retractile osmaterium emerging from its neck. Vol. II. p. 244—. III. p. 148. |
| 2. | Larva of Cerura Vinula. a. Its anal mastigia. Vol. III. p. 151. |
| 3. | Onisciform ditto of Thecla Rubi. |
| 4. | Larva of Stauropus Fagi. (Rösel.) Vol. III. p. 133. noted. |
| 5. | –——— Notodonta ziczac. (Reaum.) |
| 6. | Laria fascelina. a. Pencil of hairs, b. Verricules of ditto. c. Fascicule of ditto. Vol. IV. p. 277. 3, 5, 7. |
| 7. | –——— of one of the Geometers in their attitude of surveying. |
| 8.[Pg 731] | Araneidiform larva of Myrmeleon. (Reaum.) |
| 9. | Larva of Culex pipiens. (Reaum.) a. Tail. b. Respiratory apparatus. |
| 10. | –—— of Chironomus plumosus. (Reaum.) a. Respiratory organs. |
| 11. | –—— of a Volucella inhabiting the nests of humble-bees. (Reaum.) a. Anal radii. |
| 12. | –—— of Elophilus pendulus. (Reaum.) a. Respiratory tubes. |
| 13. | –—— of Stratyomis Chamæleon. (Swamm.) a. Plumes of respiratory orifice. |
PLATE XX.[2207]
| FIG. | |
| 1. | Larva of a Musca. |
| 2. | ———– an Œstrus. |
| 3. | Egg of Vanessa Urticæ. (Sepp.) |
| 4. | –—— Hipparchia Pilosellæ. (Ibid.) |
| 5. | –——————— Hyperanthus. (Ibid.) |
| 6. | –—— Geometra Cratægata. (Ibid.) |
| 7. | –—— Pieris Brassicæ. (Ibid.) |
| 8. | –—— Hipparchia Ægeria. (Ibid.) |
| 9. | –—— Ourapteryx Sambucaria. (Ibid.) |
| 10. | –—— Noctua nupta. (Ibid.) |
| 11. | –————— Fraxini. (Ibid.) |
| 12. | –—— Geometra prunaria. (Ibid.) |
| 13. | –—————— armillata. (Ibid.) |
| 14. | –—— Lasiocampa neustria. (Reaum.) |
| 15. | –—— Hipparchia Jurtina. (Sepp.) |
| 16. | –—— Pentatoma. a. Bow-shaped spring, by which the operculum is thrown off. Vol. III. p. 104. |
| 17. | –—— Apis mellifica. (Reaum.) |
| 18. | –—— Culex pipiens. (Ibid.) a. Summit. |
| 19. | –—— Scatophaga. (Ibid.) a a. Auricles. |
| 20.[Pg 732] | Necklace of eggs. Vol. III. p. 67. |
| 21. | Egg of Tipula oleracea. (Reaum.) |
| 22. | –—— Ophion luteum. (De Geer.) Vol. IV. p. 213—. |
| 23. | –—— Nepa cinerea. (Swamm.) |
| 24. | Jelly, with a necklace of eggs running in a spiral direction from end to end, taken out of the water. |
| 25. | Jelly of more consistence, enveloping the eggs of Phryganea atrata. Vol. III. p. 68. |
Printed by Richard Taylor,
Shoe-Lane, London.
[1] Εναιμα, Αναιμα. Hist. Animal. l. i. c. 6.
[2] Hist. Animal. l. i. c. 5, 6: compare 1. v. c. 3 and 33, and De Partibus Animal. l. iv. c. 1 and 11.
[3] Το δε σκληρον αυτων ου σθραυσον αλλα φλασον.
[4] Hist. Animal. l. iv. c. 1.
[5] Εντομα πολυποδα μεν γαρεσι παντα. De Part. Animal. l. iv. c. 6.
[6] Hist. Animal. l. iv. c. 19.
[7] The insection that distinguishes these parts, the abdomen especially, is most visible in the majority of the Hymenoptera and Diptera orders; next in some Coleoptera, as the Lamellicorn tribes, &c. and the Lepidoptera. Latreille is of opinion, that the two last segments of the thorax in some insects are represented by the first of the abdomen, and that the upper half segment of this part in Coleoptera also represents the same. Latr. De quelques Appendices, &c. Annales Générales des Sciences Physiques. A Bruxelles, vi. livrais, xviii. 14. In fact, in the Lepidoptera, when the abdomen is separated from the trunk, this segment usually remains attached to the latter. In the Myriapods, the trunk is to be distinguished from the abdomen only by its bearing the three first pair of legs.
[8] There is no general rule without exceptions, and no character is so universal as to be distinctly exhibited by every member of a class or other natural group. Thus, in the majority of the mites (Acarus L.) the body is marked by no segments, and the only articulation or incision is in the legs, palpi, &c. But as the exception does not make void the rule, so neither does the extenuation or absence of some primary character at its points of junction with others, in some individuals, annihilate the class or group.
[9] Hist. Nat. l. xi. c. 1.
[10] Animalcula polypoda, spiraculis lateralibus respirantia, cute ossea cataphracta; antennis mobilibus sensoriis instruuntur. Syst. Nat. ed. 12. i. 533.
[11] Quoted by Mr. Wm. MacLeay in his very remarkable and learned work Horæ Entomologicæ, in which he inclines to the same opinion. 383.
[12] Treviranus (Ueber den innern Bau der Arachniden, &c. 22.) always calls the palpi of spiders "Fülhörner." In Scorpio he regards them as palpi (Palpen).
[13] N. Dict. d'Hist. Nat. xvi. 181.
[14] Treviranus, ut supra, 48. For the nervous system of scorpions, see t. i. f. 13; and for that of spiders, t. v. f. 45.
[15] Plate XXIX. Fig. 2. Treviranus, t. i. f. 1.
[16] Cuvier Anat. Comp. iv. 407.
[17] N. Dict. d'Hist. Nat. ix. 190.
[18] The females of Dorthesia, however, a genus related to Coccus, are said to survive laying their eggs. N. Dict. d'Hist. Nat. ix. 553.
[19] Anim. sans Vertebr. i. 381.
[20] Anim. sans Vertebr. i. 457.
[21] The number five, which Mr. MacLeay assumes for one basis of his system as consecrated in Nature, seems to me to yield to the number seven, which is consecrated both in Nature and Scripture. Metaphysicians reckon seven principal operations of the mind; musicians seven principal musical tones; and opticians seven primary colours. In Scripture the abstract idea of this number is—completion—fullness—perfection. I have a notion, but not yet sufficiently matured, that Mr. MacLeay's quinaries are resolvable into septenaries.
[22] Anim. sans Vertebr. i. 381.
[23] See on this point MacLeay, Hor. Entomolog. 209—.
[24] Anim. sans Vertebr. iii. 243.
[25] Ibid. iii. 245.
[26] Anim. sans Vertebr. iii. 245.
[27] Ibid.
[28] Des Rapports généraux, &c. des Anim. invertebr. artic., Ann. du Mus.
[29] Ibid. Hor. Entomolog. 383.
[30] Leach in Entomologist's Useful Compendium, by Samouelle, 75.
[31] Hor. Entomolog. 348.
[32] Ibid. 354.
[33] Ibid. 373.
[34] Ibid. 381.
[35] Ibid. 389.
[36] There is some reason for thinking, though the octopod and myriapod insects breathe by tracheæ, that there is no small difference in the distribution of these organs. The Trachean Arachnida have only a pair of spiracles, from which the tracheæ must radiate, if I may so apply the term, in order to convey the necessary supply of air to every part of the body. Scutigera, as far as I can discover, has only a single series of dorsal spiracles (see Plate XXIX. Fig. 20)—an unusual situation for them: in these also, to attain the above end, each trachea must also radiate, so as to supply each part of the segment it is in. Those of Iulus, according to the observations of Savi (Osservaz. per servire alla Storia di una Specie de Iulus, &c. 15—), consist of bundles of parallel tracheæ. Perhaps these circumstances would warrant the considering of these Arachnida and the Myriapoda as primary classes? The genus Galeodes is said to breathe by gills similar to those of the Araneidæ, which structure, probably, carries with it a system of circulation, and exhibits a third type in the Arachnida, with four palpi, six legs, and a distinct thorax. This genus, then, is the corresponding point in the Arachnida to the Hexapod Aptera, as the Scorpions are to the Cheliferidæ or Pseudo-Scorpions, and the Araneidæ to the other Octopods; and these analogies furnish a strong proof, that the Tracheans belong rather to Insecta than Arachnida. Comp. N. Dict. d'Hist. Nat. xxvi. 445; and Description de six Arachnid. nouv. &c. par Leon Dufour, 16.
[37] Mr. MacLeay observes with regard to the Tardigrade, described by Spallanzani and Dutrochet, that "it proves that an animal may exist without antennæ or distinct annular segments to the body, but having two eyes and six articulate legs." (Hor. Entomolog. 350—.) Many Acari prove the same thing. De Geer, vii. t. vii. f. 14.
[38] De Geer, vii. t. iii. f. 8.
[39] Hor. Entomolog. 351.
[40] De Geer, Ibid. 571, 583. t. xxxvi. f. 20, 21.
[41] Dufour ubi supra. Hor. Entomolog. 382.
[42] Male Insecta in some instances engender more than once. Mr. MacLeay sen. has observed this with regard to Chrysomela Polygoni, and I have noticed it in Bombyx Mori.
[43] Hor. Entomolog. 134. 200.
[44] Zoolog. Miscell. iii. t. 146. In this figure the segments are made much more distinct than they are in my specimen.
[45] Hor. Entomolog. 422—.
[46] See above, Vol. I. 4th Ed. p. 66. Note[90].
[47] Surely the denomination ought to have been Arachnido-Crustacea, since the learned author considers them as belonging to the Crustacea class.
[48] It may not be without use to give here a short definition of the Annulosa; I mean excluding the Vermes, which Mr. W. MacLeay has included; and the Annelida, which Latreille has made the fifth of his Annulose classes. Ann. du Mus. 1821.
Annulosa. Animal invertebrate, oviparous; external integument of a firmer consistence than the internal substance, serving as a general point of attachment to the muscles; eyes immoveable; legs more than four, jointed.
Classes.
1. Crustacea. Gills external; more than eight legs.
2. Arachnida. Gills internal; spiracles; eight legs.
3. Insecta. Tracheæ; spiracles; six to eight thoracic legs.
[49] What L. Dufour regards as the liver in Scorpio (N. Dict. d'Hist. Nat. xxx. 421.) Treviranus looks upon as an Epiploon (Fettkörper) both in Scorpio and Aranea. 6. t. i. f. 6. A A. t. ii. f. 24. dd. Hepatic ducts: t. i. f. 6. ii. t. ii. f. 24. β. β. β. β.
[50] S. minutissima of Marsham is synonymous with Dermestes atomarius De Geer, Scaphidium atomarium Gyllenh., and Latridius fascicularis Herbst., but surely arranging with none of these genera, being sufficiently distinguished from them and every other insect by its singular capillary wings. In my cabinet it stands under the name of Trichopteryx K.
[51] Panz. Fn. Germ. Init. lxii. 24. Comp. Hor. Entomolog. Addenda, &c. 523.
[52] The Annelida have, however, sometimes jointed organs, which facilitate their progressive motion whether vermicular or undulatory; but they cannot be deemed legs, since they neither support the body nor enable it to walk, &c. Latreille Anim. invertebr. Artic. 126. Ann. du Mus. 1821.
[53] De Antennis Insect. ii. 65.
[54] Nouv. Obs. sur les Abeilles, ii. 376—. It appears from M. Huber's experiment, that it was only when the hair-pencil, impregnated with the oil of turpentine, was presented "près de la cavité, au dessus de l'insertion de la trompe," that the bee was sensible of the odour.
[55] Anim. sans Vertebr. I. i. Mem. i.
[56] Plate VIII. Fig. 10-14; IX. Fig. 6-8.
[57] Coquebert Illust. Ic. iii. t. xxi. f. 3.
[58] Hor. Entomolog. 413—.
[59] N. Dict. d'Hist. Nat. xxviii.; compare 104 and 110.
[60] See above, Vol. I. Ed. 4. p. 63—.
[61] The word μεταμορφοω, and its derivative μεταμορφωσις, are not extant in any Greek writer before the date of the New Testament. They are used to express any external change of form or colour, and metaphorically an inward change and progressive improvement of the mind. Comp. Matth. xvii. 2. Ælian. Var. Hist. l. i. c. 1. Rom. xiii. 2. 2 Cor. iii. 18. They are, therefore, not improperly applied, as some have supposed, to the changes of insects.
[62] Entwickelungsgeschichte der Schmetterlinge 12-27. 105—.
[63] Dr. Virey's observations under the article Embryo (N. Dict. d'Hist. Nat. x. 195.) deserve here to be considered. "Il y a donc quelque chose au dessus de l'intelligence humaine dans cette formation des êtres; en vain on veut l'approfondir, c'est un abime dans lequel on ne voit que la main de Dieu. A quoi bon s'appesantir sur le mystère de la formation des êtres, sans esperance de l'expliquer? Ne vaut-il pas mieux observer les opérations de la nature autant qu'il est permis à l'œil humain de les appercevoir?"
[64] § xiv.
[65] N. Dict. d'Hist. Nat. x. 193.
[66] Œuv. v. 279. "Il n'est pas exact de dire que le cœur, la tête, et la moelle épinière, sont formés les premiers dans les fœtus des animaux à sang rouge et vertébrés," says Dr. Virey; "mais il faut dire seulement que tel est l'ordre dans lequel ces organes commencent à devenir visibles." N. Dict. d'Hist. Nat. x. 196.
[67] Ibid. 193.
[68] Œuvr. viii. 315.
[69] Hor. Entomolog. 446.
[70] See on this subject N. Dict. d'Hist. Nat. xx. article Metamorphosis.
[71] N. Dict. d'Hist. Nat. xx. 349—.
[72] N. Dict. d'Hist. Nat. xx. 348.
[73] Bibl. Nat. Ed. Hill. ii. 138.
[74] Œuvr. v. 283—.
[75] N. Dict. d'Hist. Nat. xx. 355.
[76] Leeuwenhoek discovered in the incipient fœtus of a sheep, not larger than the eighth part of a pea, all the principal parts of the future animal. Arc. Nat. I. ii. 165, 173.
[77] Bonnet, Œuvr. v. 284.
[78] N. Dict. d'Hist. Nat. xx. 352.
[79] Select Works by Hoole, i. 132. The fact is confirmed by M. L. Dufour, who, having opened the abdomen of a female scorpion, found in the midst of some eggs nearly mature a little scorpion a quarter of an inch long; it lay without motion, with its tail folded under the body. N. Dict. d'Hist. Nat. xxx. 426.
[80] Reaum. iv. 425—.
[81] Ibid. 428—. t. xxix. f. 10, 11.
[82] Busch, a German author, affirms that many Cimicidæ are subject to this law. Schneid. i. 206.
[83] Quoted in Huber Fourmis, 208. Some reptiles also are at one time oviparous, and at another ovo-viviparous. N. Dict. d'Hist. Nat. xii. 568.
[84] I say almost all insects, because the larvæ of Hymenoptera and Diptera are supposed not to undergo this change. N. Dict. d'Hist. Nat. xx. 365.
[85] Reaum. vi. Mém. xiv. De Geer, vi. 280.
[88] De Geer i. 494—.
[89] Called by M. l'Abbé Preaux, who observed it near Lisieux in Normandy, Mouche Baliste. N. Dict. d'Hist. Nat. xxi. 442.
[91] Reaum. vi. 509. t. xlv. f. 11, 12.
[92] Reaum. vi. 434.
[93] Ibid. vi. 494.
[94] The vesicles, which Reaumur thinks may be pulmonary vesicles, as well as assisting in the extrusion of the masses of eggs, he has figured t. xliv. f. 10. u u.
[95] De Geer ii. 534. t. xiii. f. 13.
[96] Coquebert Illustr. Ic. t. i. f. A. B.
[98] Ent. Carniol. 269. n. 705.
[99] Reaum. ii. 401.
[100] In Raii Hist. Ins. 264.
[102] Goeze Naturf. xvii. 183—. t. iv. f. 16-19. Comp. N. Dict. d' Hist. Nat. iii. 475. and xix. 239. De Geer iii. 533.
[103] Second Journey through Persia, 100—.
[106] Latr. Hist. Nat. des Fourmis, 334. N. Dict. d'Hist. Nat. ii. 284.
[107] Lister De Aran. Tit. 13, 14. N. Dict. d'Hist. Nat. ii. 284.
[108] Lister Ibid. 56. Tit. 15.
[109] N. Dict. d'Hist. Nat. xxvi. 447.
[110] Ins. Surinam, t. i.
[111] A striking instance of this may be seen in her forty-ninth plate, in which she has clapped the rostrated head of Fulgora laternaria upon the body of a Cicada Latr., affirming it to be the former fly in its previous state! This might be a trick upon her.
[112] N. Dict. d'Hist. Nat. xv. 489.
[113] Lesser L. i. 300.
[114] Annales du Muséum, xiv. 441.
[115] Lesser L. i. t. ii. f. xvi.
[116] Miger Ann. du Mus. ubi supr. Comp. N. Dict. d'Hist. Nat. xv. 482—.
[117] De Geer i. 192.
[118] Ibid. ii. 982.
[119] Reaum. ii. 97. 159.
[120] Ibid. 107—. t. iii. f. 15.
[121] De Geer iii. 48. 51.
[122] Reaum. v. 122.
[123] See above, Vol. I. p. 196. 202.
[124] Journ. de Phys. Philos. Mag. ix. 244.
[125] Reaum. iv. Mem. i.
[126] Rösel, ix. 157. t. 265?
[127] Ibid. iii. 197.
[128] See above, Vol. I. p. 195.
[130] Reaum. i. 95— f. 1-13.
[131] Reaum. iv. 615—. t. xliv. f. 2-7.
[132] N. Dict. d'Hist. Nat. xv. 445. Rös. iii. 156.
[133] Epist. 1687.
[135] See above, Vol. I. p. 344—.
[136] Reaum. iii. 8—.
[138] N. Dict. d'Hist. Nat. xxviii. 271.
[139] De Geer vii. 194.
[140] Compare N. Dict. d'Hist. Nat. xvi. 246. with xx. 352-; but as the Amnios immediately envelops the fœtus, the pellicle seems most analogous to it, and the shell to the Chorion.
[141] Swamm. Bibl. Nat. ed. Hill. l. 133. a. Comp. N. Dict. d'Hist. Nat. xvi. 246.
[142] Swamm. Ibid.
[143] Sepp. iv. t. iii. f. i. c. v. t. iv. f. 2.
[144] See above, Vol. I. p. 208: it is there called an Aphis, but it is a distinct genus.
[145] De Geer iii. 245. t. xiii. f. 20-22.
[146] Sepp. iv. t. xiii. f. 2. 3.
[147] The sturgeon is said to lay 1,500,000 eggs, and the cod-fish 9,000,000.
[148] Reaum. iv. 392.
[149] See above, Vol. I. p. 350.
[150] De Geer vii. 159.
[151] See above, Vol. II. p. 109.
[154] iv. t. xviii. f. 4. 5.
[155] De Geer ii. 638.
[156] Bibl. Nat. i. 132. b.
[157] Gould 36.
[158] Reaum. v. 477.
[159] Ibid. iii. 579. v. 121.
[160] Fourmis, 69—.
[161] Rösel iii. 152.
[162] De Geer vii. 145.
[163] Ibid. 123—. See above, Vol. I. p. 393.
[164] Raii Hist. Ins. 265.
[165] Eggs of various shapes are given Plate XX. Fig. 3-23. See also Brunnich. Entomologia 4. N. Dict. d'Hist. Nat. xvi. 245. Reaum. ii. t. iii. iv. xiv. xxvi. xxvii. &c.
[167] Plate XX. Fig. 23. Swamm. Bibl. Nat. t. iii. f. 7, 8. In a specimen I opened of this insect the bristles converged so as to form a kind of tail to the egg.
[168] Darwin Phytolog. 512.
[169] Geoffr. Ins. Par. i. 480. t. x. f. 1. b. c.
[170] See above, Vol. I. p. 261.
[171] Reaum. iii. 386—. t. xxxii. f. 1. t. xxxiii. f. 5.
[172] I allude to Ophion luteum F. (Ichneumon L.) Vol. i. Ed. 3. p. 269, figured Plate XX. Fig. 22; and the Hydrachnæ or Trombidia. See above, and De Geer vii. 145.
[173] From this circumstance called πολυποικιλος σοφια by the Apostle, Ephes. iii. 10.
[174] Nat. Theol. 11th Ed. 375.
[176] Reaum. iv. 376—. t. xxvii. f. 9, 10.
[177] Hist. Nat. gen. et partic. des Crust. et Ins. xii. 282.
[178] Reaum. iv. 381. t. xxvi. f. 19, 20.
[179] Roxburgh in Linn. Trans. vii. 34.
[180] Some of the Noctuæ have similar eggs, as N. Lappa. Sepp iv. t. iii. f. 1. c.
[181] Reaum. ubi supr. f. 22, 23.
[184] Ibid. Fig. 3. 4. 7. 9. &c.
[186] Bonnet Œuvr. ii. 9.
[187] Naturf. xiii. 229.
[188] N. Dict. d'Hist. Nat. xvi. 245.
[189] Reaum. ii. 286.
[190] Plate XX. Fig. 11. Sepp t. iv. f. 2.
[191] Reaum. iv. 617.
[192] Philos. Ent. 76.
[193] See above, Vol. I. 358—.
[194] See above, Vol. I. Ibid.
[195] Young's France, ii. 34. This author asserts, that no art will hatch the eggs of the common silk-worms the first year, or that in which they are laid; but that there is a sort brought from Persia, which are hatched three times a year, and which will hatch in fifteen days in the proper heat. In 1765, it is said, the common sort hatched in the first year. Ibid. 226—.
[196] In the N. Dict. d'Hist. Nat. xii. 564. the eggs of the flesh-fly are said to hatch in two hours. This is true I believe in very warm weather.
[197] Brahm. 310.
[198] Rimrod Naturf. xvi. 131.
[199] Fourmis. 69.
[200] De Geer vii. 195.
[201] Ibid. 196.
[202] Reaum. ii. 167.
[203] Brahm. 249. Rösel. iv. 130. Swamm. Bibl. Nat. t. i. f. 2.
[204] By Mr. White, jun. cordwainer at Ipswich.
[206] De Geer vii. 197.
[207] De Geer vii. 197.
[208] Ibid. 85.
[209] Epist. lxvii. 1694. 390.
[210] Enum. Ins. Austr. 575.
[211] N. Dict. d'Hist. Nat. i. 74.
[212] De Geer vii. 576.
[213] Ibid. 584.
[214] Considerat. Géner. 21. Horæ Entomolog. 353.
[215] De Geer, Ibid. Mr. W. MacLeay observes of the Chilopoda, or Centipedes, that they moult in the manner of Crustacea. ubi supr. 352.
[216] De Geer iii. 549. The figure of the forceps in De Geer (Ibid. t. xxv. f. 21) is not quite correct. The styles do not taper to a point, but are filiform and acute.
[217] Compare De Geer iii. t. xviii. f. 2 and 12. q.
[218] See above, Vol. II. p. 401.
[219] Plate XVI. Fig. 4. c. Reaum. v. t. xix. f. 16. De Geer ubi supr. t. xxxii. f. 26. According to Reaumur, the larva as well as the pupa of Chermes Ficus has wing-cases (iii. 353).
[220] These are in the female sex of some Coleoptera, as Lampyris, &c. which retain in the perfect state nearly the same form which they had when larvæ. The larvæ of some Staphylini are not very dissimilar in form to the perfect insect.
[221] The larvæ described in the first Section, which resemble the imago, are usually covered with a skin not materially different from that of the insect in that state.
[222] Huber Fourmis. 73; N. Dict. d'Hist. Nat. vi. 250.
[223] Reaum. v. 40. t. vi. f. 4-15.
[224] Müller, the Danish zoologist, relates, that he once met with a papilio which, with the true wings of the genus, had a head without antennæ or tongue, furnished with mandibles; and, in short, that of a true caterpillar. It was a female, which deposited eggs that proved barren. If this solitary instance was not a mistake, is it possible that some parasitic larva had devoured only the inclosed head of the butterfly, or so injured it that it could not reject the hard skin of the larva, and yet not be destroyed?
[225] The only larvæ which have a visible distinct neck are those of some Dytisci, Staphylini, and a few others, in which this part is quite distinct: proving the erroneousness of the opinion of those German entomologists, who consider the thorax as analogous to the neck of other animals, and hence call it Halsschild. In some lepidopterous larvæ, however, as in that of Pieris Brassicæ, though no visible neck presents itself, one is very perceptible when the insect stretches the head forward considerably. Reaum. i. 460.
[227] Reaum. v. t. vi. f. 7. i. c.
[228] In fact, in almost all Lepidopterous larvæ the head may be regarded as divided into two lobes or eye-shaped portions, which include in the angle formed by their recession anteriorly from each other, the nasus (clypeus F.), the labrum, and other instruments of manducation. Posteriorly these lobes generally come into contact; but I have a specimen in which there is a narrow space between them.
[229] Ins. Surinam. t. xvii.
[230] Ins. Surinam. t. liii.
[231] Ibid. t. xxxii.
[232] Ibid. t. viii.
[233] Ibid. t. xxiii.
[234] Ibid. t. xiv.
[235] I purchased this singular caterpillar from the collection of the late Mr. Francillon, with his other exotic larvæ; but without any indication of the fly to which it belonged.
[236] De Geer vi. 352.
[237] De Geer iv. 66. ii. 922.
[238] De Geer v. 170.
[239] De Geer says, he could not make out the number of eyes of the larva of the whirlwig (Gyrinus): probably, as in that of Dytiscus, there are six. iv. 362. 385.
[240] Pez. 188.
[241] ii. 923, t. xxxvi. f. 1, b b. Fabr. Philos. Ent. 60.
[242] Lyonnet 41. t. ii. f. 1. c.
[243] De Geer vi. 307.
[244] Ibid, ii. t. xvi. Comp. f. 2 a a with f. 14 a a.
[245] In the larva of Cicindela there are six palpi, as in the perfect insect.
[246] Lyonnet, t. i. f. 7. e. In the larva of Callidium violaceum, however, this part is of a singular shape, being orbicular. Kirby Linn. Trans. v. t. xii. f. 12. a.
[247] It is affirmed (N. Dict. d'Hist. Nat. vii. 333) that the larvæ of those Coleoptera that live in carcases have mandibles almost membranous: those, however, of that of Silpha rugosa are horny and hard.
[248] Lyonnet, t. ii. f. 1. d d, and f. 2, 3, 4.
[249] Kirby in Linn. Trans. v. t. xii. f. 7 b.
[250] Cuvier Anat. Comp. iii. 322.
[251] Reaum. vi. 340.
[252] The larva of Cicindela campestris has mandibles of this description. Plate XVII. Fig. 13. c´.
[253] See above, Vol. II. 275—.
[254] Reaum. v. 9. t. i. f. 4. c c. l l.
[255] Traité Anatom. t. ii. f. 1. h h.
[256] Reaum. ii. t. 40. f. 4.
[257] De Geer v. 229.
[258] Ibid. iv. t. xi. f. 16. p p.
[259] Linn. Trans. v. t. xii. f. 10.
[260] Cuvier Anat. Comp. iii. 323.
[261] De Geer iv. t. xv. f. 9. b b. The exterior and interior palpi are both represented in this figure.
[262] Reaum. vi. t. xxxvii. f. 5. e e.
[263] Ibid. i. 125.
[264] Plate XXI. Fig. 9. The organ with which the larvæ of Hemerobius, Myrmeleon, and Hydrophilus, spin their cocoons, is situated in the anus. The spinneret of the Cossus is figured by Lyonnet Anatom. t. ii. f. 1. i. and fig. 9.
[265] De Geer vi. 370. This species (Tipula Agarici seticornis De Geer) has two separate spinnerets. t. xx. f. 8. m m.
[266] Lyonnet 55—.
[267] Reaum. iv. 166.
[268] Reaum. v. 155.
[269] Ibid. vi. t. xxxvii. f. 7. b p.
[270] Ibid. m e e.
[271] Ibid. f. 6. p.
[272] Ibid. Compare f. 4 with f. 6, 7.
[273] Ibid. t. xxxvi. f. 12. s u e.
[274] Ibid. n e, and xxxviii. f. 7, d c.; De Geer ii. t. xix. f. 17. d g.
[275] Reaum. vi. t. xxxvii. f. 4-6. 8.
[276] Ibid. t. xxxviii. First joint f. 8. b f p.; jaws f. 7. c d.; opening o, Ligula, f. 6. l.
[277] De Geer ii. f. 17. Jaws g g; claw d; tooth h.
[278] Ibid. 674.
[279] Ibid. ii. 674.
[280] Reaum. iv. 376.
[281] N. Dict. d'Hist. Nat. xii. 64.
[282] Anat. Comp. iii. 322.
[283] At first in the Dytisci they appear to have five joints; but, as I before observed, the first joint must be regarded as representing the maxilla.
[284] Lyonnet Anatom. 55, 58.
[285] De Geer v. 203.
[286] De Geer iv. 5. Legs of this kind are figured Plate XXIII. Fig. 7.
[287] In the larva, however, of Sialis, or some kindred genus, in which, like those of Scolopendra, the prolegs are jointed, a pair distinguishes each abdominal segment. See Reaum. iv. t. xv. f. 1, 2. Compare De Geer ii. t. xxiii. f. 11.
[288] See above, Vol. II 286—.
[290] Lyonnet Anatom. t. iii. f. 8. Coxa b. Trochanter c. Femur d. Tibia e. Tarsus f. Claw g.
[291] De Geer iv. t. xiii. f. 20; and t. xv. f. 16.
[292] Ibid. ii. t. xvi. f. 5, 6, 7. d e: and t. xix. f. 4. c f g h.
[293] The larva of a scarce moth (Stauropus Fagi. See Plate XIX. Fig. 4) is an exception to this. The first pair of its legs are of the ordinary stature, but the two next are remarkably long, and so thin and weak as to be unable to bear the body. Pezold. 119. Another minute caterpillar described by Reaumur has the third pair of the legs apparently fleshy and singularly incrassated at the apex into a pyriform figure, terminated by a pair of claws. This conformation is for some particular purpose in the economy of the animal, since they are the most busily employed of all in arranging the threads of her web. Reaum. ii. 258. In the larva of a geometer (Geometra lunaria) the third pair are remarkably long. Illig. Mag. 402. In that of another moth, according to Kuhn (Naturf. xvi. 78. t. iv. f. 3), the third pair of the fore-legs is remarkably incrassated, being twice as thick and long as the other pair, though consisting of the same number of joints, the last of which has claws.
[294] On the legs and prolegs see also what is said above, Vol. II. p. 286—.
[295] In some few instances these legs are dorsal. Ibid. 281.
[296] The claws or crotchets, though general, are not universal, in Lepidopterous larvæ. An exception is furnished to the rule by the singular limaciform ones of Hepialus Testudo and Asellus of Fabricius, two moths forming Haworth's genus Apoda, which have no distinct prolegs, but in their stead a number of small transparent shining tubercles without claws. The larva also of one of the subcutaneous moths first discovered by De Geer in the leaves of the rose (i. 446), but whose history is fully given by Goeze, Naturf. xv. 37-48, (who has satisfactorily ascertained that it is the true larva of a Tinea of Linné, but of a different habit from that of most subcutaneous ones), has no true legs, and eighteen prolegs without any claws. Another subcutaneous larva, for the history of which we are indebted to M. Godeheu de Riville, is according to him entirely deprived of legs of any kind (Bonnet ix. 196—.); as is another of the same tribe that feeds on the poplar, an account of which is given by Goeze Naturf. xiv. 105.
[297] Plate XXIII. Fig. 7. See also below, p. 137.
[298] Lyonnet Anatom. 84. t. iii. f. 11, 12.
[299] Hist. Vermium, 130.
[302] Account of Locust-tree Insects, 69.
[303] Reaum. iv. 443. t. xxx. f. 6. l l. t. xxii. f. 6. l l.
[304] De Geer vi. 383. and 137. t. viii. f. 8, 9.
[305] See above, Vol. II. p. 278. De Geer ubi supr. 376.
[306] Reaum. iv. 184. t. xv. f. 12. c c.
[307] De Geer v. 203.
[309] Some few subcutaneous larvæ have more, as that, before mentioned, observed by De Geer in the leaves of the rose; which has eighteen prolegs, and no true ones.
[310] De Geer ii. t. xl. f. 15, 16. Bergman has added to these four classes of the larvæ of saw-flies, a fifth; the insects belonging to which, he affirms, though they have sixteen prolegs, are without the anal pair. Ibid. 931. But as neither De Geer nor Reaumur ever met with one of this description, it is probable he was mistaken. Reaumur thought he had seen one with eighteen prolegs upon Erysimum alliaria (v. 91), but he does not speak positively.
[311] De Geer v. 288.
[312] De Geer iv. 157.
[313] Ibid. v. 36. t. ii. f. 11.
[314] See above, Vol. I. p. 171.
[315] De Geer v. 228.
[316] Ibid. 233.
[317] See above, Vol. II. p. 281.
[318] De Geer vi. 388.
[319] Ibid. 389.
[320] Reaum. v. t. v. f. 10.
[321] Ibid. 31. This larva has also a pair of pediform processes at the anus, surrounded at the end with claws (t. v. f. 4, 5, s s), which he saw the animal use in locomotion; but which he suspects to be respiratory organs (Ibid. 33), which Latreille asserts they are. Gen. Crust. et Ins. iv. 249.
[322] De Geer Ibid. t. xxiv. f. 15-17.
[323] Ibid. 383.
[324] Ibid. 111. t. vi. f. 14-16.
[325] Merian Ins. Sur. t. xx.
[326] Ibid. t. xxxiv.
[327] I have a caterpillar, I believe from Georgia, in which this horn is nearly an inch long, filiform, slender, and tortuous.
[328] Plate XVIII. Fig. 12. c.
[329] That of Sphinx Iatrophæ L. appears to be jointed, at least it is moniliform. Merian Surinam. t. xxxviii. Compare also t. iii.
[330] N. Dict. d'Hist. Nat. vi. 252.
[331] Schellenberg Entomolog. Beytr. t. 1.
[332] Smith's Abbott's Insects of Georgia, t. xiii.
[333] De Geer ii. 507. t. xi. f. 16. m n. t. xiv. f. 7.
[334] N. Dict. d'Hist. Nat. vi. 256.
[335] See above, Vol. II. p. 244—.
[337] Reaum. i. t. xxx. f. 2. N. Dict. d'Hist. Nat. xxiv. 490, 497—.
[338] Ray says he found it feeding on common fennel, about Middleton in Yorkshire: Lett. 69. The indefatigable Mr. Dale recently found many in the neighbourhood of Whittlesea-mere, feeding on Selinum palustre. It will also eat the wild carrot.
[339] This gentleman was remarkable for the admirable manner in which he prepared caterpillars, so as scarcely to differ from life.
[340] Reaum. i. 92.
[341] Bonnet ii. 84—. iii. 1.
[342] See above, Vol. II. 251—.
[343] Bonnet ii. 88.
[344] De Geer ii. 507. t. xi. f. 16. c.
[345] Rös. iv. 162.
[346] De Geer i. 322—. See Plate XIX. Fig. 2. a a.
[347] Reaum. ii. 275. t. xxii. f. 3.
[348] Ibid. 276. t. xxii. f. 4, 5.
[349] Ins. Surinam. t. vii. Nymphalis Amphinome xxiii. Morpho Teucer t. xxxii. Papilio Cassiæ.
[350] This is not, however, universally the case, for the caterpillar of a Geometer described by Reaumur (ii. 363. t. xxix. f. 8.) (G. amatoria) has a pair of fleshy anal horns, terminating, it should seem from his figure, in a minute hook that the animal uses as a forceps; which has at the same time the anal legs, of which indeed these horns seem to be appendages.
[351] Sepp. iv. t. l. f. 6-8.
[352] Plate XIX. Fig. 5. a b. Sepp. iv. t. xxi. f. 4-7.
[353] Rös. iii. 69.
[355] N. Dict. d'Hist. Nat. vii. 95.
[356] De Geer v. 170— t. v. f. 19-23. Compare Reaum. iii. 235—.
[357] Plate XIX. Fig. 11. a. De Geer vi. 137. Reaum. iv. 482.
[358] Reaum. iv. t. xiv. f. 9, 10.
[359] Reaum. v. 32. t. v. f. 3-5. Latr. Gen. Crust. et Ins. iv. 249.
[360] De Geer ii. 1031. t. xl. f. 13, 14. k k.
[361] N. Dict. d'Hist. Nat. x. 430.
[362] De Geer ii. 697. t. xxi. f. 4, 5. b b b.
[363] Reaum. v. t. vi. f. 7. n.
[365] Reaum. ii. t. xxv. f. 20.
[366] See above, Vol. II. p. 245—.
[367] Reaum. iii. 384. vi. 366. t. xxxii. f. 7, 8.
[368] Rös. iii. t. lxviii. f. 1. Meinecken Naturf. vi. 120.
[369] Ibid. xiii. 175.
[370] In the larva of Tenthredo Cerasi L., and some others, no traces of segments are to be seen; and in many coleopterous and dipterous ones the folds of the skin prevent the segments from being distinctly perceptible.
[371] Reaum. ii. 361. In the larva of a small common moth often met with in houses (Aglossa pinguinalis), every segment is divided into two parts, and underneath has two deep folds, by means of which these two parts can separate to a certain point, or approach again, according to circumstances. Thus Providence has enabled them to prevent their spiracles from being stopped by the greasy substances on which they often feed. N. Dict. d'Hist. Nat. i. 208.
[373] N. Dict. d'Hist. Nat. xvii. 329.
[374] Hor. Entomolog. 285. 397—. 422. 462—. &c.
[375] Ibid. 399-401.
[376] Hor. Entomolog. 423.
[378] The Intestinaux cavitaires of Cuvier, and the Epizoaria of Lamarck. See Hor. Entomolog. 286—.
[379] Hor. Entomolog. 422. comp. 463. Mr. MacLeay's idea of the larva of Meloe is taken from the animal which Frisch, Goedart, and De Geer imagined to be such; but upon this opinion there rest great doubts. (See Kirby Mon. Ap. Angl. ii. 168, and Latreille N. Dict. d'Hist. Nat. xx. 109.) At p. 464 he gives also Mordella and many Heteromera as having Thysanuriform larvæ. He thinks, that probably that of Clerus is of the same description; to which he suspects that many of Latreille's Malacoderma likewise belong.
[380] Plate XVIII. Fig. 1. as to the thoracic shield.
[381] May 27, 1822. This day, T. Allen, Esq. F.L.S. brought me in a phial a vast number of the little insect which Goedart, Frisch, and De Geer took for the larva of Meloe Proscarabæus, which he found on the leaves of Achillea Millefolium. These little animals were coursing each other with wonderful velocity over the sides of the phial. To assist them in their motions, they applied to the surface of the glass the end of their abdomen, using it, like many larvæ of Coleoptera, as a seventh leg. This circumstance excited a suspicion in the minds of both Mr. MacLeay sen., then visiting me, and myself, that after all they might be coleopterous larvæ. One, amongst other circumstances, however, seemed to militate strongly against this opinion; namely, that in this infinite number none appeared to differ in size.
[383] Ibid. Fig. 12.; Plate XVIII. Fig. 4, 11, 13, &c.
[386] Ibid. Fig. 3. Reaum. v. 97. t. xii. f. 17, 18.; De Geer ii. 1004. t. xxviii. f. 12.
[387] See above, p. 110, 114, 138, 142.
[392] Hor. Entomolog. 465.
[393] De Geer iv. 66. t. ii. f. 5-8.
[394] Ibid. t. xiii. f. 16-19. A very singular larva, which preys upon that of Aleyrodes proletella Latr., if Reaumur's figure be correct (ii. t. xxv. f. 18-20), is of a perfect Chilopodiform type, the abdominal legs being represented by a tubercle crowned by a bristle: yet even this, which turns to a minute beetle (Ibid. f. 21), has some tendency to the Anopluriform type.
[395] "Squilla insectum a squilla pisce parum differt." Mouffet, 319.
[396] A remarkable difference obtains between the larva of the wire-worm and that of Elater undulatus. In the former, the last segment is longer than the preceding one, terminating in a small acute mucro at the apex, with a deep cavity, perhaps a spiracle, on each side, at the base. In the latter, this segment is shorter than the preceding one, forming above a nearly circular plate; the margin of which is a little elevated, and armed on each side with three teeth, and at the apex with a pair of furcate recurved horns, and without any basal spiracle. De Geer iv. 156. t. v. f. 25. I have a similar larva, but not the same species.
[397] Hor. Entomolog. 397.
[398] Ibid. 399.
[399] Ibid. 438. Note *.
[400] Traité Element. ii. 35. n. 577.
[401] Trans. Linn. Soc. vii. 66. t. vi. f. 3.
[402] Compare De Geer iii. t. xi. f. 3. and t. xvii. f. 14. &c.
[403] Ibid. t. i. f. 4, 9. t. ii. f. 15. t. ix. f. 4.
[405] Compare Plate VI. Fig. 6. with Fig. 12 c, d, d.
[406] De Geer ii. t. xxi. f. 4, 5.
[407] Swamm. Bibl. Nat. t. xiii. f. 1.
[408] Hor. Entomolog. 438.
[409] See above, Vol. II. p. 256.
[410] De Geer ii. t. xxiii. f. 9-14. Comp. Reaum. iv. t. xv. f. 1, 2.
[411] De Geer ii. t. xiv. f. 7. &c. The caterpillar of P. G. Scratiotata L. like those of Phryganeæ, has these respiratory threads. Ibid. i. t. xxxvii. f. 2-6. De Geer has described the larva of a Phryganea L. which is without any respiratory threads, ii. 569. t. xv. f. 10.
[412] Hor. Entomolog. 401. Montagu in Linn. Trans. vii. 67.
[413] Ins. Surinam. t. xxviii. Compare Ibid. t. xix. right-hand figure.
[415] Swamm. Bibl. Nat. t. xxxix. Plate XIX. Fig. 13.
[416] Lyonnet 69—.
[417] Surinam, t. lvii. right-hand figure.
[418] Sepp iv. t. ii. f. 3. t. xvi. f. 2, 3.
[419] N. Dict. d'Hist. Nat. vi. 254.
[420] Plate XIX. Fig. 6. One of these larvæ was taken at Melville Island. See Parry's Voyage, Appendix No. x. 37.
[421] Sepp. iv. t. viii. f. 4. Some species have three, others four, and others even five of these brushes. N. Dict. d'Hist. Nat. vi. 255.
[422] Ibid. Merian Eruc. xxxiv. upper left hand figure.
[423] Merian Ins. Surinam. t. lx.
[424] Ibid. t. xl.
[425] See above, Vol. I. p. 238.
[426] De Geer iv. 207. t. viii. f. 4-6.
[427] Ins. Sur. t. xix. right hand caterpillar.
[428] Ibid. xli.
[430] Ins. Sur. t. xxix.
[431] Ibid. t. vii. liii.
[432] Smith's Abbott's Ins. of Georg. Pref. vi.
[433] Prodromus Entomology.
[434] Ins. Sur. t. xliii. The figure represents only the two spines near the head as thus circumstanced.
[435] Reaum. v. t. xii. f. 8, 14. Plate XVIII. Fig. 11.
[436] See above, Vol. II. p. 238. This, with B. imperatoria, &c. in the modern system, should form a genus.
[437] Ins. Sur. t. xlviii. right hand figure.
[438] Ibid. t. xi.
[439] Ibid. t. xxiii.
[440] Ibid. t. xxix.
[441] Reaum. v. 95.
[442] Huber Mœurs des Fourmis, 79.
[443] See above, Vol. II. p. 276—.
[444] Reaum. v. 72. t. ix. f. 2-4.
[445] Rös. t. 211.
[446] See above, Vol. I. p. 29, 198—.
[447] De Geer iii. 111. Comp. 121. It would be as well to adopt the French word flocon, instead of locks or flocks, which strictly mean very different things.
[448] vii. 604. t. xliv. f. 26.
[449] Fn. Germ. Init. xxxvi. 21.
[450] Syst. Rhyng. 311. 29.
[451] N. Dict. d'Hist. Nat. ix. 554.
[452] Natural History of the Slug-worm, 7.
[453] Ins. Surinam. t. xv. xvii.
[454] The larvæ of Carabus L. form one, being generally black.
[455] Annales de Chimie ii.
[456] Wien. Verz. 219.
[457] Wien. Verz. 4.
[458] Reaum. v. 92.
[459] Ins. Surinam. t. xi.
[460] ii. 1017.
[461] De Geer i. 57.
[462] Ibid. 58. Reaum. i. t. xxxix. f. 13, 14.
[463] De Geer ii. 400.
[464] See above, Vol. I. Letters xii. xiii.
[465] Bonnet (ii. 18) mentions, that the young larvæ of a butterfly (Pieris Cratægi), after devouring the exuviæ of the eggs from which they were hatched, gnawed those which were not so: not, however, so as to destroy the included animal, but rather to facilitate its egress. Those also of Coccinella bipunctata which I lately bred from the egg, as soon as hatched began to devour the unhatched ones around them, which they seemed to relish highly. I am inclined to believe, however, that this unnatural procedure was to be attributed to the circumstance of the female not having had it in her power to place her eggs in the midst of Aphides, their proper food.
[466] N. Dict. d'Hist. Nat. xx. 359.
[467] In the human species, after certain fevers a simultaneous and total moult, if the term may be so applied, takes place. I experienced this myself in my boyhood; when convalescent from Scarlatina, the skin of my whole body, or nearly so, peeled off.
[468] The translator, more ignorant of natural history than his author, has turned the "linguis micat ore trisulcis" of Virgil, into "darts his forky sting."
[471] Cuvier Anat. Comp. ii. 596. N. Dict. d'Hist. Nat. xxvi. 165.
[472] Cuvier Ibid. 624.
[473] Reaum. i. 182.
[474] N. Dict. d'Hist. Nat. vi. 290.
[475] Those Diptera whose metamorphosis is coarctate (Vol. I. p. 67), bees, the female Cocci, &c. do not cast their skin in the larva state. Reaum. iv. 364. N. Dict. d'Hist. Nat. xx. 365.
[476] N. Dict. d'Hist. Nat. vi. 289. xx. 372. Cuvier Anat. Comp. ii. 548. M. Cuvier (Ibid. 547.) asserts, that most Papiliones and Bombyces moult seven times.
[477] Œuvr. ii. 71.
[478] Reaum. ii. 75.
[479] Bibl. Nat. E. Trans. i. 135. col. b. t. xxvii. f. 6.
[480] Œuvres, viii. 303.
[481] Entwickelungsgeschichte, &c. 34, 88. Swammerdam on the contrary affirms, that "on the hinder part of the cast skin where it is twisted and complicated, whoever accurately examines the skin itself may still observe the coat that was cast by the intestinum rectum." Ubi supr. 136. col. a.
[482] N. Dict. d'Hist. Nat. vi. 290
[483] Reaum. iv. 604.
[484] Ibid. 364. N. Dict. d'Hist. Nat. xx. 365. Huber Fourmis 78. M. Huber does not say expressly that the grubs of ants do not change their skin; but his account seems to imply that they change it only previously to their metamorphosis.
[485] Lyonnet 11.
[486] N. Dict. d'Hist. Nat. vi. 290.
[487] De Bombycibus, 68.
[488] Opusc. i. 27.
[489] Linn. Trans. x. 399.
[490] N. Dict. d'Hist. Nat. vii. 129.
[491] As the larvæ of Ephemeræ usually live in the submerged part of the banks of rivers, perhaps they may be regarded as following the economy of subterranean terrestrial larvæ.
[492] A caterpillar nearly answering to the description of that of Bombyx camelina, which I found upon the hazel, after a few days produced sixteen grubs of some Ichneumon. At first these grubs were green, but they became gradually paler; and after a day or two became pupæ. But I mention this circumstance here for another reason: upon examining them after this last occurrence, I observed that they adhered to the lid of the box in which I kept the larva, arranged somewhat circularly; and at a little distance from the anus of each was a pea-green mass, consisting of about eight oval granules, which appeared like so many minute eggs. These were the excrement evacuated by each grub previously to its becoming a pupa. The appearance of this little group, with their verdant appendage, formed a curious spectacle: they are still pupæ, July 30, 1822.
[493] Except some species of Polyammatus Latr. (Thecla, Argynnis F.), P. Argiolus, Corydon, &c., and Hesperia Rubi, Betulæ F., &c. Some of the larvæ of the former become pupæ within the stalk of some plant, or partly under the earth: those of the latter usually in a leaf to which the abdomen is fastened by various threads. These last are the rouleuses of the butterfly-tribe, living, like some moths, in leaves that they have rolled up. N. Dict. d'Hist. Nat. xxiv. 499.
[496] Bonnet is of opinion that this twirling process is not with any view to get rid of the exuviæ, but is caused only by the irritation occasioned by the spines of the skin of the caterpillar when they touch that of the pupa. Œuv. ii. 109.
[497] For the above account see Reaum. i. Mem. x. xi.
[498] N. Dict. d'Hist. Nat. vi. 291—.
[499] De Bombyc. 24.
[500] i. 498.
[501] De Bombyc. 43.
[502] N. Dict. d'Hist. Nat. vi. 294.
[503] Lesser. L. ii. 150, note 22. Boyle says an English lady found that the silk of a single cocoon would extend 300 English leagues or 900 miles. But this must be a mistake.
[504] Reaum. i. 555—.
[506] De Geer i. t. xxxii. f. 3-6.
[507] De Geer i. 463—.
[508] Reaum. ii. Mem. xi. Comp. De Geer ii. 162. Reaum. ii. 424.
[509] B. Catax—Pupa arcte folliculata. Fab.
[510] Travels in Greece, 285.
[511] See above, Vol. I. p. 476—.
[512] Merian Surinam. t. xv.
[513] Reaum. ii. t. xxiii. f. 5.
[514] Sepp. iv. t. viii. f. 5.
[515] Reaum. i. t. xliv. f. 2.
[517] I have a black one from Mr. Francillon's cabinet.
[518] N. Dict. d'Hist. Nat. vi. 294.
[519] See above, Vol. II. p. 298—.
[520] Reaum. ii. 436.
[521] Reaum. i. 503.
[522] Peck on Locust-tree Insects, 69.
[523] Bonnet ii. 260.
[524] Sepp. iv. t. ii. f. 4.
[525] Brahm. Ins. Kal. 289.
[527] The thick cocoons of Attacus Paphia, Polyphemus, &c. are also thus fastened between leaves.
[528] Merian Europ. ii. t. ix.
[529] Reaum. ii. 284.
[530] Ibid. i. 524.
[531] Bonnet ii. 297.
[532] Ibid. ix. 181.
[533] Reaum. v. 102.
[534] Ibid. iv. 269.
[535] De Geer ii. 1084. Comp. Ray Hist. Ins. Præf. xi. It is the opinion of M. P. Huber, that in this case the naked pupæ are deprived of their cocoons by the neuters: he states, indeed, that he has often seen them pulled off by them, and also by those of F. canicularia; and he seems to think that these larvæ are never developed. Mœurs des Fourmis, 84. note 1.
[536] II. viii. 16.
[537] Linn. Trans. vii. t. ii. f. 5, 6.
[538] Wien. Verz. I possess a cocoon of this kind from New Holland, even now quite solid, and retaining its form. No silk appears to have been used in its composition.
[539] Reaum. i. 579.
[540] Ibid. vi. 368.
[541] Ibid. i. 542.
[542] Ibid. 543.
[543] Linn. Trans. i. 196.
[544] Reaum. i. 545—.
[545] Pyral. 8. 3. t. iii. f. 16.
[546] See above, Vol. I. p. 172—.
[547] Reaum. ii. 491.
[548] Reaum i. 540.
[549] See above, Vol. I. 167—. II. 264.
[550] See above, Vol. I. p. 67.
[551] N. Dict. d'Hist. Nat. xvi. 269—. xxii. 76.
[552] Reaum. iv. 32. The author here quoted asserts that the grub of Ichneumon Larvarum L. retains its skin, which, he says, is so transparent that the form of the nymph can be seen through it. Ibid. ii. 447. De Geer, however, found that this really did cast its skin, which is so transparent as to be scarcely visible, by pushing it gradually towards the anus, where it soon dries up and cannot then be discovered. De Geer ii. 893—. According to Rösel the same circumstance attends the transformation of Coccinella renipustulata Illig. (C. Cacti Ent. Brit.), which at first perplexed him not a little. It is probable that in this case the retention of the skin was accidental; for some of the grubs of a Mycetophila, the transformation of which I observed, became pupæ within their last skin, while others wholly disengaged themselves from it. The cause of this variation, I conjectured, arose from the former being too weak to extricate themselves from the skin.
[553] See above, Vol. I. p. 238. Byrrhus Musæorum belongs to this genus.
[554] N. Dict. d'Hist. Nat. ii. 161.
[555] Pezold. 102.
[556] De Geer i. 339—.
[557] Reaum. ii. 423, and iii. 497.
[558] Ibid. i. 605.
[559] De Geer ii. 941.
[560] Brahm Insek. 72.
[561] Reaum. ubi supra.
[562] In the Hemiptera the male Cocci (Reaum. iv. 32.) and Aleyrodes (Ibid. ii. 311.) belong to the second division.
[563] The terms larva and pupa, applied to the insects of this subdivision, are perhaps not strictly proper.
[564] The larvæ and pupæ of many of the homopterous section of Hemiptera differ often from the imago, not only in their fore-legs (Plate XVI. Fig. 4.), but also in other respects. I have the larva of a Centrotus from Canada, given me by Dr. Bigsby, which has a long anal process or tail.
[566] iii. 135.
[567] The pupæ of Cassida, Imatidium, &c. seem to vary somewhat from this type, the upper part being neither membranous nor exhibiting distinctly the form of the inclosed imago.
[568] The following arrangement of pupæ is perhaps in some respects better than that above given. But it is scarcely possible to propose one free from objections.
Lamarck divides the pupæ of insects that undergo a metamorphosis into three kinds, which he names—Chrysalis, Mumia, and Nympha.
i. Chrysalis. Under this denomination he includes all inactive pupæ inclosed in an opaque puparium which entirely conceals them. These he further subdivides into two kinds.
1. Chrysalis signata. This term is synonymous with the Pupa obtecta of Linné, or the Chrysalis of Lepidoptera and some Diptera.
2. Chrysalis dolioloides. Equivalent to the Pupa coarctata Linn. peculiar to those Diptera that assume this state in the skin of the larva.
ii. Mumia. All inactive pupæ which are covered by a transparent skin, through which all the parts of the inclosed imago may be seen, subdivided also into two.
1. Mumia coarctata. Corresponding with the Pupa incompleta Linn., which includes the Coleoptera and most of the Hymenoptera.
2. Mumia pseudonympha, confined to the Pupa of Phryganea and some others. This might be named Pupa subincompleta.
iii. Nympha. Under this denomination are included all insects that undergo only a partial metamorphosis, and are active in their pupa state, corresponding with the Pupa semicompleta Linn. and also subsemicompleta MacLeay. See Anim. sans Vertebr. iii. 285—.
M. Latreille has started an ingenious idea on this subject with regard to these kinds of metamorphosis, which comprehends both larva and pupa under a distinct denomination: as thus—
[569] N. Dict. d'Hist. Nat. vii. 57.
[570] De Geer ii. 105.
[571] Reaum. ii. 428—.
[572] Swamm. Bibl. Nat. Engl. Tr. ii. 32. t. xli. f. 2. Comp. Reaum. iv. t. xxv. f. 1.
[573] Ibid. i. 144.
[574] Reaum. i. 355.
[575] N. Dict. d'Hist. Nat. ubi sup. 59.
[576] Plate XVI. Fig. 14. N. Dict. d'Hist. Nat. vii. 59.
[577] Ins. Surinam. t. xliv.
[578] De Geer v. 47. t. ii. f. 29-31.
[579] In the pupa of Hydrophilus piccus (Lesser L. t. ii. f. 13, 14), the arrangement of the parts is nearly the same, but the tarsi are not reflexed.
[580] Ibid. f. 9, 10. De Geer ii. t. xxxii. f. 5. Reaum. v. t. xxxvi. f. 14.
[581] Reaum. Ibid. t. ii. f. 9.
[582] The legs of Tipula replicata L. are placed in a similar way. De Geer vi. t. xx. f. 12. l.
[583] Rös t. 81.
[584] Ibid. t. 95.
[585] De Geer vi. 237. t. xiv. f. 8.
[586] Reaum. v. t. ii. f. 7. The anal and ventral spines of Tipula replicata are also remarkable. De Geer vi. t. xx. f. 14.
[587] De Geer Ibid. 377. t. xxiii. f. 8, 9. n. Reaum. v. 42. t. vi. f. 9. m n.
[588] The caterpillar consists of twelve segments (Lyonnet t. i. f. 4, 5), excluding the head; on each of which, except the 2d, 3d, and 12th, there is a pair of spiracles. The chrysalis usually exhibits an analogy to this structure, though the first, second, and last pair of spiracles are more or less obsolete in most.
[589] De Geer ii. 847. t. xxix. f. 7. a b.
[590] Animaux sans Vertebres, iii. 287.
[591] N. Dict. d'Hist. Nat. vii. 57.
[592] Sepp ii. t. i. f. 4. t. ii. f. 4. t. iv. f. 5.
[595] Ins. Surinam. t. lx. It is singular that the chrysalis of its congener, Morpho Teucer, which she figures t. xxiii., exhibits no such process. The larvæ also widely differ.
[597] Sepp ii. t. iii. f. 5.
[598] Sepp i. t. vii. f. 5.
[599] De Insectis, ed. Lister. t. 1.
[600] Ins. Surinam. t. liii.
[601] Sepp i. t. ii. f. 6.
[602] N. Dict. d'Hist. Nat. vii. 60.
[603] Ibid. 57.
[604] See above, Vol. I. p. 131.
[605] Reaum. ii. 158. t. viii. f. 4, 5.
[606] Lesser L. i. 160. note. t. ii. f. 19.
[607] N. Dict. d'Hist. Nat. xxvi. 165. Reaum. i. 347. Rösel says this is present only in some individuals. I. ii. 47.
[609] Ins. Surinam. t. iii.
[610] De Geer ii. 433. t. viii. f. 4. t.
[611] See above, Vol. II. p. 300.
[612] This description was taken from a puparium in my own cabinet; it is similarly described by De Geer i. 490. t. vii. f. 2.
[614] Kliemann Beitrage, 304.
[616] Von Scheven in Naturf. stk. xx. 64. t. ii. f. 4.
[617] Plate XVII. Fig. 2. Lesser L. t. ii. f. 26.
[618] Plate XVII. Fig. 1. Lesser L. t. ii. f. 24, 25.
[619] Whether M. Meigen has separated this fly generically from others, I am not aware: in my catalogue it stands under the name of Tyrophaga.
[620] Reaum. iii. 376. t. xxxi. f. 7.
[621] Ibid. iv. 318. t. xxiii. f. 1-4. xxv. f. 1.
[622] Ins. Surinam. t. xxix.
[623] Ins. Surinam. t. xxxii. Lister imitated the gilding of Chrysalises by putting a small piece of a black gall in a strong decoction of nettles: this produced a scum, which when left on cup-paper, he says, will exquisitely gild it.—Ray's Letters, 87. 90.
[624] Beitrage, 181.
[625] Sepp. pt. ii. t. ii. f. 4.
[626] Rösel. I. i. 61. ii. 5.
[627] Reaum. i. 383.
[628] Lister's Goedart. 122.
[629] Reaum. ii. 10—.
[630] Ibid. 24.
[631] This is a legend of Virgil, of which an account is given in The Lay of the Last Minstrel, Note xv. 12mo ed. 1822, p. 257.
[632] Haworth Lepidopt. Britann. i. 125. An instance is recorded in Scriba's Journal, in which a pupa was not disclosed until the fourth year. B. i. st. iii. 222. Pezold. 170.
[633] Marsham in Linn. Trans. x. 402.
[634] Meinecken found, that of several pupæ of Saturnia pavonia, some kept all winter in a room heated daily by a stove, and others in a cold chamber, some of both parcels appeared in March (none earlier), and some of both had not appeared in July, though evidently healthy. Naturf. viii. 143.
[635] The exclusion of certain moths, &c. from the pupa is probably regulated by the time their eggs require to be hatched, and the appearance of the leaves that constitute their appropriate food.
[636] Mr. Marsham makes a similar observation in Linn. Trans., ubi supr.
[638] The appearance of them sometimes continues to near the end of the month: it began on the 19th, when Reaumur observed them. vi. 480. 488.
[639] Bibl. Nat. E. Transl. i. 103—.
[640] Reaum. vi. 486.
[641] Brahm. 423. 421.
[642] Naturf. xxi. 75.
[643] Reaum. ii. 423.
[644] De Geer ii. 370. It is not certain, however, that De Geer did not, in this instance, mistake the winter habitation of a larva for a cocoon intended to shelter the future chrysalis; since Lyonnet informs us that they spin a habitation to pass the winter in. Traité Anatomique, &c. 9.
[645] De Geer i. 490. t. vii. f. 3, 4.
[646] Œuv. ii. 1.
[647] De Geer v. 229.
[650] I. iv. 101.
[651] Reaum. vi. 407.
[652] Haworth Lepidopt. Britann. i. 127.
[653] De Geer ii. 566.
[654] Reaum. iii. t. xlv. f. 12-14.
[655] For this whole account, see Reaum. iv. Mem. viii.
[656] Ibid. 472.
[657] See above, p. 255—. and Vol. II. p. 301—.
[658] Bonnet, Œuv. ii. 169.
[659] Bonnet, Œuvr. ii. 207.
[660] Rös. I. iv. 209. t. lxiii. ccxii.
[662] Bonnet, Œuvr. ii. 229.
[663] De Geer ii. 477.
[664] Sepp. iv. t. xi. f. 8.
[665] Plate XVII. Fig. 5. N.B. Sepp's figure represents the exterior funnel; and this, which exhibits the cocoon divided longitudinally, the interior one, or dome.
[666] Rös. I. iv. 31.
[667] Naturf. viii. 133.
[668] De Bombyc. 29.
[669] Reaum. i. 624.
[670] Trans. of the Society of Arts, vii. 131.
[671] Reaum. ubi supr.
[672] ii. 359.
[673] Linn. Trans. vii. 35.
[674] Pezold. 171.
[675] Lyonnet 16.
[676] Some Notice of the Insect which destroys the Locust-trees, 70. This Memoir is in some American periodical work, of which I have not the title.
[677] Huber Fourmis 82.
[680] De Geer ii. 519.
[681] Reaum. iv. 610—.
[682] Ibid. v. 30—. t. v. f. 1-10. See above, p. 153—.
[683] See above, Vol. II. p. 346.
[684] Reaum. i. Mem. ult. De Geer i. 73. Swamm. Bibl. Nat. i. 184.
[685] Swamm. Ibid.
[686] Jurine Hymenopt. 16.
[687] iv. 342. Herold also attributes the rapid expansion of the wing to the flow of an aqueous fluid, which he calls blood, into the nervures, the orifices of which open into the breast. Entwickelungs. der Schmetterl. 101. sect. 106.—M. Chabrier, in his admirable Essai sur le Vol des Insectes (Mém. du Mus. 4ieme, ann. 325), having observed a fluid in the interior of the nervures of the wings of insects, thinks it probable that they can introduce it into them and withdraw it at their pleasure: the object of which, he conjectures, is either to strengthen them and facilitate their unfolding, or to vary the centre of gravity in flight, and increase the intensity of the centrifugal force.
[688] Ibid. 340.
[689] Brahm. Insek. ii. 423.
[690] Reaum. vi. 505—. t. xlvi. f. 9. Comp. De Geer ii. 627—.
[691] Reaum. iii. 378.
[692] Ibid. 385.
[693] Insects of the beetle tribe, especially such as undergo their metamorphosis under ground, in the trunks of trees, &c., are often a considerable time after quitting the puparium before their organs acquire the requisite hardness to enable them to make their way to the surface. Thus, the newly-disclosed imago of Cetonia aurata remains a fortnight under the earth, and that of Lucanus Cervus, according to Rösel, not less than three weeks.
[694] See above, Vol. I. p. 34—.
[695] Jurine Hymenopt. 9. Note 1.
[696] Oliv. N. i. t. i. f. 1. c. f. N. 3. t. iii. f. 22. a b c. t. v. f. 33. t. vi. f. 5. t. xiii. f. 124. a b.
[697] Reaum. vi. 423.
[698] Kirby Mon. Ap. Angl. ii. t. xvi. f. 12, 13. t. xvii. f. 10-12.
[699] Reaum. iv. 393.
[700] See above, Vol. I. 473—.
[701] De Geer vii. 304.
[702] Reaum. iv. 30.
[703] Ibid. t. iv. f. 15.
[705] De Geer iii. 25.
[706] Linn. Trans. iv. 54—.
[707] ix. 65. n. 110.
[708] vi. 423.
[709] Entomologische, &c. 224.
[710] De Geer ii. 847. 850. Jurine Hymenopt. 100.
[711] Kirby Mon. Ap. Angl. ii. 296. 264.
[712] Ibid. ii. 142—. 144, 147, 148, &c.
[713] A remarkable anomalous exception to this rule sometimes occurs in the female of D. marginalis, which has smooth elytra like the male (Gyll. Ins. Suec. i. 467-). I have this variety from the Rev. Mr. Dalton, of Copgrove, Yorkshire.
[714] De Geer i. t. vii. f. 11.
[715] See above, Vol. II. 125, Note [135].
[716] Melitta ** c. Kirby Mon. Ap. Angl. i. 140.
[717] Ibid. t. iv. f. 10. a. b. f. 14.
[718] Ibid. t. xiii. f. 20. a.
[719] Kirby Mon. Ap. Angl. i. t. xi. Apis **. d. 2. α. β. f. 18 a. b. c. d.
[720] Coquebert Illustr. Icon. i. t. vi. f. 6.
[721] Kirby Mon. Ap. Angl. i. Apis **. c. 1. α. **. c. 1. β. **. c. 2. α. **. c. 2. β. **. c. 2. γ. **. c. 2. δ.
[722] Ibid. t. viii. f. 28. f. g.
[723] Christ. Hymenopt. t. iv. f. 3. b.
[724] Kirby Mon. Ap. Angl. i. t. iv. Melitta **. c. f. 1. a.
[725] Scheven Naturfors. stk. xx. 65. t. ii. f. 4. Compare Ibid. x. 101.
[726] Reaum. iii. t. xv. f. 18, 19.
[727] Oliv. no. 84. Brentus, t. i. f. 1. b. c. t. ii. f. 17. a. b.
[728] Oliv. no. 3. Scarabæus, t. xviii. f. 169.
[729] Oliv. Scarabæus, t. xii. f. 114. t. xv. f. 138. a.
[730] Ibid. t. v. f. 33.
[731] Ibid. t. xii. f. 112.
[732] Linn. Trans. vi. t. xix. f. 12. t. xx. f. 2.
[733] Oliv. no. 57. Tenebrio, t. i. f. 2.
[734] Oliv. ubi supr. No. 3. t. i. f. 1.
[735] Oliv. no. 3. t. iii. f. 20. a.
[736] Ibid. no. 55. Diaperis, t. i. f. 3.
[737] Oliv. Scarabæus, t. xx. f. 185.
[738] As Dynastes Actæon, Elephas, Typhon, &c. differ from D. Hercules, &c., not only in their general habits, horns, &c., but also in their maxillæ and labium,—the former in D. Actæon being simple, and in D. Hercules toothed, and the labium of the first bilobed at the apex, and in the last entire and acute,—according to the modern system they ought, therefore, to be considered as distinct genera. I would restrict the name Dynastes to D. Hercules and its affinities: D. Actæon, &c. I would call Megasoma.
[739] Oliv. Scarabæus, t. xvii. f. 156.
[740] Ibid. t. viii. f. 63.
[741] This insect is beautifully figured in M. Latreille's Insectes sacres des Egyptiens, f. 11. See Luke xi. 15. Heb. בעלןבול Dominus stercoris.
[742] Oliv. no. 83. 160. t. vi. f. 60. ♂. t. v. f. 45. ♀?
[743] Ibid. no. 36. t. ii. f. 12.
[744] Ibid. no. 6. t. vii. f. 61.
[745] See above, Vol. II. 224—.
[746] Coquebert Illustr. Icon. iii. t. xxi. f. 2.
[747] Stoll Cigales, t. xviii. f. a b c. Grillons t. iv. f. 16-18. This singular animal, which was found by Mr. Patterson at the Cape of Good Hope, is stated to be an aquatic; and affords the only known instance of an Orthopterous insect inhabiting the waters. The Gryllotalpa loves the vicinity of water.
[748] Mon. Ap. Angl. i. Melitta **. b. 139. t. ii. f. 4-6.
[749] Ibid. **. a. f. 4, 5.
[750] Ibid. Apis *. b. 190—. t. v. f. 18 b.
[751] By Rösel, by a friend of De Geer's, and by M. Marechal. De Geer iv. 331—. Nouv. Dict. d'Hist. Nat. xviii. 225.
[752] Oliv. no. i. Lucanus, t. ii. f. 3.
[753] Ibid. t. iii. f. 7.
[754] Linn. Trans. xii. 410. t. xxi. f. 12.
[755] Ibid. vi. 185. t. xx. f. 1.
[756] Oliv. ubi supr. t. ii. f. 4.
[757] Regne Animal, iii. t. xiii. f. 3.
[758] See Vol. I. Plate I. Fig. 3.
[759] Christ. Hymenopt. t. xviii. f. 2.
[760] Mon. Ap. Angl. i. Melitta *. a. t. i. f. 5. ♀. 7. ♂.
[761] Ibid. Melitta **. a. t. ii. f. 6. ♀. 7. ♂. and **. b. t. iii. f. 3. ♀. 4. ♂.
[762] Ibid. t. viii. f. 11. ♀. 12. ♂.
[763] Ibid. i. t. viii. f. 9. ♀. 10. ♂.
[764] Ibid. Apis **. c. 2. β. t. ix. f. 6. ♀. 7. ♂.
[765] See above, Vol. II. 125. Note[135].
[766] Mon. Ap. Angl. ubi supr. t. xiii. f. 13. ♀. 14. ♂.
[767] MacLeay Hor. Entomolog. 4—.
[768] N. Dict. d'Hist. Nat. xxii. 488.
[769] Linn. Trans. xii. 425—. t. xxii. f. 6.
[772] Oliv. no. xxv. Lymexylon, t. 1. f. 1.
[773] De Geer vii. 249—. t. xiv. f. 20, 21. Treviranus Arachnid. 36—. t. ii. f. 16. a b c. t. iv. f. 35-37.
[774] Mon. Ap. Angl. i. Apis **. c. 2. γ. t. ix. f. 7. ♀. 9. ♂.
[775] Ibid. Melitta **. a. t. ii. f. 8. ♀. 9. ♂. and **. b. t. iii. f. 6. ♀. 7. ♂. **. c. t. iv. f. 11. ♀. 12. ♂.
[776] Jurine Hymenopt. t. 11. f. 2.
[777] Mon. Ap. Angl. i. Apis **. d. 1. t. x. f. 7.
[778] Oliv. no. 80. Macrocephalus, t. i. f. 2.
[779] Latr. Gen. Crust. et Ins. iv. 156.
[780] Jurine Hymenopt. 61. t. vi. f. 8.
[781] Ibid. 289.
[782] Mon. Ap. Angl. i. t. ix. Apis **. c. 2. γ. f. 9.
[787] Plate XXV. Fig. 11. Linn. Trans. xii. t. xxi. f. 4. a.
[788] Ibid. f. 3.
[790] De Geer i. t. xix. f. 11, 12.
[791] Jurine Hymenopt. t. vi. f. 8.
[794] Reaum. iv. t. xl. f. 2. a a. ♂. t. xxxix. f. 3. ♀. In the last the hairs are too conspicuous.
[796] Jurine Hymenopt. t. vi. f. 3.
[797] Plate XII. Fig. 25, 26. XXV. Fig. 17, 32.
[800] N. Dict. d'Hist. Nat. xiv. 395.
[804] Linn. Trans. xii. t. xxii. f. 8. e. ♂. f. ♀.
[805] Hist. Nat. des Fourmis, 195—. 270—.
[806] De Geer ii. 1094.
[807] Ibid. 650. Mon. Ap. Angl. i. t. xi. Apis xx. c. 1. f. 2. ♂. t. xii. f. 3. ♀.
[808] Plate XXVI. Fig. 39. De Geer ii. 651. 659.
[809] Voet Coleopt. i. t. xxxix. f. 47, 48. ♂. 46. ♀.
[810] Oliv. no. 3. t. vi. f. 46. a. ♂. b. ♀.
[811] Ibid. t. i. f. 1. iv. x. f. 31. xi. f. 102. xii. f. 114.
[812] Ibid. t. xxvi. f. 219.
[813] Ibid. t. i. f. 2.
[814] Ibid. t. xxiii. f. 35.
[815] Ibid. t. ii. f. 7.
[816] Ibid. t. v. f. 40.
[817] Ibid. xix. f. 175.
[818] Ibid. t. xii. f. 115.
[819] Copris floriger Kirby in Linn. Trans. xii. 396.
[820] Oliv. no. 3. t. ii. f. 18.
[821] Ibid. t. xxii. f. 32.
[822] Ibid. t. ix. f. 85.
[823] Ibid. t. iii. f. 22.
[824] Ibid. t. xiii. f. 124. a.
[825] Ibid. t. v. f. 38.
[826] Ibid. t. xxviii. f. 242. t. xviii. f. 169.
[827] Ibid. t. xvi. f. 152.
[828] Ibid. t. xxviii. f. 247.
[829] Ibid. t. xv. f. 138. a. t. v. f. 33.
[830] Samouelle's Compend. t. i. f. 1.
[831] Oliv. no. 3. t. v. f. 36. a.
[832] Schon. Synon. i. t. 1.
[833] Oliv. no. 3. t. xxiv. f. 208.
[834] Ibid t. x. f. 88.
[835] Ibid. f. 87.
[836] Ibid. t. xx. f. 185.
[837] Ibid. t. vi. f. 42. a.
[838] Ibid. n. 83. Curculio t. xxii. f. 295.
[839] Oliv. no. 81. Attelabus t. ii. f. 27. b. 28.
[840] De Geer ii. t. xxxi. f. 18-22.
[841] Ibid. iii. 21.
[842] Lesser L. i. 185.
[843] De Geer iii. 308.
[844] See above, Vol. II. 394—.
[845] Linn. Trans. i. 145. 135—.
[846] Ibid. t. xiii. f. 1. 2. ♂. 3. ♀.
[847] Mon. Ap. Angl. i. t. xi. Apis **, a. 2. α. β. f. 18.
[848] Oliv. no. 68. Saperda t. i. f. 8.
[849] Mon. Ap. Angl. i. t. viii. f. 28. c.
[850] Ibid. t. ix. Apis **. c. 2. β. f. 12.
[851] Clairv. Ent. Helv. ii. t. xii. f. B.
[852] Oliv. Ins. no. 66. t. iii. iv. f. 12.
[853] Ibid. no. 3. t. iv. f. 27.
[854] Punaises, t. iii. f. 20.
[855] Mr. Marsham has made two species of this from this circumstance, viz. Necydalis Podagrariæ and simplex.
[856] Oliv. n. 3. t. xxvii. f. 27. ♀. and t. iv. f. 27. ♂.
[857] Ibid. t. vii. f. 58. ♂. f. 57. ♀.
[858] Ibid. n. 95. Hispa t. i. f. 4. Plate XXVII. Fig. 24.
[860] De Geer i. t. vii. f. 14, 15.
[861] Coquebert Illust. Icon. i. t. vi. f. 6. Plate XXVII. Fig. 32.
[862] Illig. Mag. iv. 214. Gyllenhal. Insect. Suec. i. 168.
[865] Christ. Hymenopt. 118. t. iv. f. 3.
[866] Mon. Ap. Angl. i. t. xi. Apis **. e. 1. ♂. f. 8. e. and t. xii. **. e. 1. neut. f. 19. c.
[867] Hor. Entomolog. 144.
[868] Plate XXVII. Fig. 45. a.
[869] De Geer ii. t. xxviii. f. 2.
[870] Mon. Ap. Angl. i. t. v. Apis **. a. f. 10. ♂. 11. ♀.
[871] Ibid. t. vii. Apis **. c. 1. α. 17. ♀. 18. ♂.
[872] Ibid. t. viii. f. 30. ♂. 31. ♀.
[873] Ibid. t. xi. Apis **. e. 1. mas. f. 9. t. xii. Apis **. e. 1. fem. f. 9. and neut. f. 22.
[874] De Geer vi. t. xviii. f. 12, 13.
[875] De Geer v. 151—.
[876] Mon. Ap. Angl. i. 177. t. ix. Apis **. c. 2.γ. f. 11. a, d.
[877] Ibid. f. 13. a.
[878] See above, Vol. II. 395.
[879] Plate XXIX. Fig. 13. Stoll. Spectres, &c. t. xxv. f. 99.
[880] Sparrman. Voyage, i. 312—.
[881] Coleopt. Micropt. 16.
[882] Mon. Ap. Angl. i. t. viii. f. 25. De Geer iii. 255. t. xiv. f. 8.
[883] Mon. Ap. Angl. i. t. viii. f. 24.
[884] Ibid. t. ix. Apis xx. c. 2. γ. f. 12.
[885] Ibid. Apis **. c. 2. β. f. 11.
[886] Ibid. t. vii. Apis **. c. 1. α. f. 11, 12. ♀. 13, 14. ♂.
[887] Plate XV. Fig. 12. De Geer ii. t. xxiv. f. 9, 10. ♀. t. xxv. f. 2, 3. ♂.
[888] Reaum. vi. 494. t. xliv. f. 3-11.
[889] De Geer ii. t. xvii. f. 5-7.
[890] N. Dict. d'Hist. Nat. xxxii. 443.
[891] See above, Vol. II. 110, 118.
[893] II. i. 6.
[894] Clairville Ent. Helvet. ii. 214—. I have seen it asserted in some popular work on Natural History, (the title of which I do not recollect,) that Mantis religiosa has been known to live ten years; and a flea, when fed and taken care of, six. But this is so contrary to experience in other cases, that the statement seems quite incredible.
[895] Rösel III. 379. N. Dict. d'Hist. Nat. ii. 285.
[896] Dumeril Traité Elément. ii. 87. n. 683.
[897] De Geer ii. 288.
[898] Huber i. 106.
[899] N. Dict. d'Hist. Nat. ix. 553.
[901] Morier's Second Journey through Persia, 100.
[903] Lamarck Hist. Nat. des Anim. sans Vertèbr. i. 311, 214.
[904] Ibid. 162. Compare the Systême des Anim. sans Vertèbr. of the same author, p. 12—.
[905] The doctrine of Epicurus—that the Deity concerns not himself with the affairs of the world or its inhabitants, which, as Cicero has judiciously observed (De Nat. Deor. 1. 1. ad calcem), while it acknowledges a God in words, denies him in reality; has furnished the original stock upon which most of these bitter fruits of modern infidelity are grafted. Nature, in the eyes of a large proportion of the enemies of Revelation, occupies the place and does the work of its Great Author. Thus Hume, when he writes against miracles, appears to think that the Deity has delegated some or all of his powers to nature, and will not interfere with that trust. Essays, ii. 75—. And to name no more, Lamarck, treading in some measure in the steps of Robinet (who supposes that all the links of the animal kingdom, in which nature gradually ascends from low to high, were experiments in her progress towards her great and ultimate aim—the formation of man. Barclay On Organization, &c. 263), thus states his opinion: "La nature, dans toutes ses opérations, ne pouvant procéder que graduellement, n'a pu produire tous les animaux à-la-fois: elle n'a d'abord formé que les plus simples; et passant de ceux-ci jusques aux plus composés, elle a établi successivement en eux différens systêmes d'organes particuliers, les a multipliés, en a augmenté de plus en plus l'énergie, et, les cumulant dans les plus parfaits, elle a fait exister tous les animaux connus avec l'organisation et les facultés que nous leur observons." (Anim. sans Vertèbr. i. 123.) Thus denying to the Creator the glory of forming those works of creation, the animal and vegetable kingdom (for he assigns to both the same origin, Ibid. 83), in which his glorious attributes are most conspicuously manifested; and ascribing them to nature, or a certain order of things, as he defines it (214)—a blind power, that operates necessarily (311); which he admits, however, to be the product of the will of the Supreme Being (216). It is remarkable, that in his earlier works, in which he broaches a similar opinion, we find no mention of a Supreme Being. (See his Systême des Animaux sans Vertèbres, Discours d'Ouverture.) Thus we may say that, like his forerunner Epicurus, Re tollit, dum oratione relinquit Deum. But though he ascribes all to nature; yet as the immediate cause of all the animal forms, he refers to the local circumstances, wants, and habits of individual animals themselves; these he regards as the modifiers of their organization and structure (162). To show the absurd nonplus to which this his favourite theory has reduced him, it will only be necessary to mention the individual instances which in different works he adduces to exemplify it. In his Systême, he supposes that the web-footed birds (Anseres) acquired their natatory feet by frequently separating their toes as far as possible from each other in their efforts to swim. Thus the skin that unites these toes at their base contracted a habit of stretching itself; and thus in time the web-foot of the duck and the goose were produced. The waders (Grallæ), which, in order to procure their food, must stand in the water, but do not love to swim, from their constant efforts to keep their bodies from submersion, were in the habit of always stretching their legs with this view, till they grew long enough to save them the trouble!!! (13—). How the poor birds escaped drowning before they had got their web feet and long legs, the author does not inform us. In another work, which I have not now by me, I recollect he attributes the long neck of the camelopard to its efforts to reach the boughs of the mimosa, which, after the lapse of a few thousand years, it at length accomplished!!! In his last work, he selects as an example one of the Molluscæ, which, as it moved along, felt an inclination to explore by means of touch the bodies in its path: for this purpose it caused the nervous and other fluids to move in masses successively to certain points of its head, and thus in process of time it acquired its horns or tentacula!! Anim. sans Vertèbr. i. 188. It is grievous that this eminent zoologist, who in other respects stands at the head of his science, should patronize notions so confessedly absurd and childish.
[906] Lyonnet Traité, &c. Pref. xxii. Want of due encouragement, it is to be feared, caused the abortion of these valuable treatises. The MSS. are, I believe, still in existence. It would probably answer now to publish them.
[909] There are certain processes which are a continuation of the internal surface of the crust; and serve, as well as the rest of it, for points of attachment to the muscles: these, though completely internal, must be considered as parts of the external skeleton.
[911] The crust which covers the body of insects is lined internally with a kind of fibrous cuticle. Query, Whether in any degree analogous to the Periosteum of Vertebrate animals?
[912] We employ this term instead of Instrumenta Cibaria F., to avoid circumlocution.
[913] Plates VI. VII. &c. a´, and XXVI. Fig. 30-33.
[914] Ibid. Fig. 30. Mon. Ap. Angl. i. 139. Melitta **. b. t. ii. f. 4, 5.
[915] Plates VI. VII. &c. and XXVI. Fig. 23-29. b´.
[916] Plates VI. and VII. a´´, and XXVI. Fig. 34, 35.
The part in this work regarded as the mentum, does not in all cases accord with what MM. Latreille, Savigny, &c. have regarded as entitled to that denomination. Thus in Hymenoptera, their Mentum is what we term the Labium, while our Mentum is the small piece upon which that part sits (Plate VII. Fig. 3. a´´). This is called the Fulcrum in Mon. Ap. Angl. (See i. Explan. of the Plates.) Our Mentum may generally be known by its situation between the hinges and base of the Maxillæ.
[917] Plates VI., VII., and XXVI. b´´.
[919] Plate XIII. Fig. 7. c´´.
[920] Marcel de Serres Comparaison des Organes de la Mastication des Orthoptères. 7. Ann. du Mus. 11.
[921] Plate VI. Fig. 6. c´, a´´´. and XIII. Fig. 5. a´´´.
[922] Plate VI. Fig. 12. b´´´. and XIII. Fig. 5. b´´´.
[923] Plate XXVI. Fig. 16. c´´´.
[925] Cuv. Anat. Comp. iii. 322—.
[926] Plates VI. VII. d´. and XXVI. Fig. 9-15.
[929] Ibid. and XXVI. Fig. 13-15.
[931] Plate XXVI. Fig. 11. d´´´.
[932] Plate VI. Fig. 6, 12. d´´´.
[933] Ibid. Fig. 3, 6, 12. and XXVI. Fig. 9, 10. e´´´.
[934] Ibid. VI. Fig. 3, 12. f´´´.
[935] Plates VI. VII. h´´. XIII. Fig. 1-4, 8. h´´. and XXVI. Fig. 1-8.
[936] Plate VI. Fig. 6, 12. e´.
[937] Plate XXVI. Fig. 26, 29. e´.
[938] Plate VII. Fig. 2, 3, e´.—What is here called the Lingua in Hymenoptera has been usually regarded as the Labium; but surely that organ which collects, and as it were laps the honey, and passes it down to the Pharynx, is properly to be considered as the tongue. The Labium itself appears to be represented by what has been called the Mentum, and the true Mentum, as was lately observed, is at the base of the part last mentioned, in the usual situation of that piece. This, though long since noticed (Kirby Mon. Ap. Angl. i. 103—), has not been much attended to by modern entomologists.
[939] Huber Fourmis, 4—.
[940] Plate VII. Fig. 2, 3. and XXVI. Fig. 28. i´´.
[942] Ibid. Fig. 2. k´´. This is M. Savigny's name for this part. It has also been called Epiglossa. Latreille Organisation Extérieure des Insectes, 185.
[943] Vide Savigny Mém. sur les Anim. sans Vertèbr. I. i. 12—.
[944] The majority of Hymenopterous insects, though they have the ordinary Trophi, are not masticators, using their mandibulæ only for purposes connected with their economy.
[945] See his Mémoires sur les Animaux sans Vertèbres, I. i.
[946] I have used this word here and on a former occasion (see above, p. 29), perhaps not with strict propriety, in the sense of the French word remplacer, for which we seem to have no single corresponding word in our language.
[948] Plate VI. Fig. 7, 9. b´.
[954] Ibid. a. The Labella have been usually thought confined, or nearly so, to the genus Musca L.; but they may be traced in all genuine Diptera, i. e. excluding Hippobosca L.
[955] Plate VII. Fig. 5. a´, c´, d´.
[956] Plate VII. Fig. 5, 6. a´.
[958] Ibid. d´. It has not yet been satisfactorily ascertained, whether all the ordinary Trophi are represented in every Dipterous mouth, the number of the lancets seeming in some cases to vary.
[962] Ibid. Labrum a´; Mandibulæ c´; Maxillary Palpus h´´.
[965] Ibid. d´. Maxillary Palpi h´´.
[967] Plate VII. Fig. 11, 13. f´.
[971] Plate VI. Fig. 1, 4, 10. â.
[979] Plates VI. VII. and XXVI. h.
[980] Plate VI. Fig. 1. and VII. Fig. 2. h´.
[981] Plate VI. Fig. 4, 10. VII. Fig. 1, 2, 4. and XXVI. Fig. 39-41. i.
[982] Plates XI. XII. and XXV.
[983] Plate VI. Fig. 1, 2. and VII. Fig. 1. i´.
[984] Ibid. XII. Fig. 6. 9. k´.
[988] Ibid. Fig. 6, 8-10. m´´.
[990] Plate VII. Fig. 2. l. Mon. Ap. Angl. i. t. xiii. f. 1. a, e.
[994] Plate XXVII. Fig. 1, 3, 4. n.
[996] Plate IX. Fig. 7, 10, 11, &c. and XVI. Fig. 4, 8. B.
[997] M. Chabrier, in his admirable Mémoires sur le Vol des Insectes. uses the term Tronc Alifère, which suggested the terms here employed.
[998] Plate IX. Fig. 3, 12, 16, &c.
[999] Ibid. Fig. 1, 2, 10, 11, &c.
[1002] Ibid. VIII. Fig. 3, 11.
[1003] Plate XXIX. Fig. 12. c´.
[1004] Plate VIII. Fig. 2, 11. d´.
[1005] Plate XXII. Fig. 7. e´.
[1006] M. Latreille, in his Organisation Extérieure des Insectes (Mem. du Mus. viii. 198.) proposes calling the fore-legs of Hexapods Propedes; but having long ago applied this term to the false legs of caterpillars (see above, Vol. II. p. 288. &c.), we shall not adopt it.
[1007] Plate XXVII. Fig. 36. a´.
[1009] Plate XXVII. Fig. 59. a.
[1010] Plate VIII. Fig. 3, 4, 12-14, 16, 17. IX. Fig. 1, 3, 7, 8, 10-12, 15.
[1012] Plate IX. Fig. 7, 11, 12, 15, 19. g´. The Collare of Hymenoptera and Diptera has usually been regarded as representing the Prothorax of Coleoptera, Orthoptera, &c. But this difference obtains between them—the latter evidently belongs to the Manitrunk, and its muscles do not appertain at all to the Alitrunk; whereas the Collare as evidently is a part of the latter, its muscles belong to it, and its functions in assisting in flight are important. These reasons, and others we shall state hereafter, induced us long ago to consider this part as not representing the Prothorax; and they seem to have induced M. Chabrier almost to adopt a similar opinion. Sur le Vol des Insectes. Ann. du Mus. 3eme Ann. 414. et 4eme Ann. 54—.
[1013] Plate XXII. Fig. 8, 11. h´.
[1014] Ibid. Fig. 8. Plate VIII. Fig. 3, 12, 14, 16. IX. Fig. 1, 7, 8, 10-12, 15, 19, 21. i´.
[1015] Plate VIII. Fig. 14, 20. IX. Fig. 11, 12. and XXII. Fig. 8. b´´.
[1016] Plate X. Fig. 1.; and XXVIII. Fig. 1-8, 10.
[1017] Plate XXVIII. Fig. 3-5. b´´´.
[1019] Plate XXVIII. Fig. 6-8. d´´´.
[1020] Plate XXIII. Fig. 6. e´´´.
[1021] Plate XXVIII. Fig. 2. a´´´.
[1022] Ibid. Fig. 19. and Plate X. Fig. 2.
[1023] The upper organs of flight of many of the homopterous section of the Hemiptera seem altogether membranous, and may almost be included under the term Alæ Superiores.
[1027] Ibid. Fig. 5-9, 11-15. and Plate XXVIII. Fig. 18.
[1029] N.B. In the Plate the Costal Area is red, the Intermediate white, and the Anal yellow. When the Hemelytra are considered as divided into Areas, the Membrana might be denominated the Apical Area.
[1030] Plate X. Fig. 2, 3, b..
[1032] Plate X. Fig. 2, 3. d..
[1037] Chabrier Sur le Vol des Insectes, Ann. du Mus. 3eme ann. 428, 4eme ann. 325—. 3d Cahier 78.
[1038] Linn. Trans. i. t. xiii. f. 2. 3. d.
[1039] Plate X. Fig. 5-15. i..
[1043] Ibid. Fig. 5, 6, 13. b*.
[1045] Ibid. Fig. 5, 6, 13. c*.
[1046] Plate X. Fig. 5-15. n..
[1050] Ibid. Fig. 14, 15. n´´´.
[1051] Plate IX. Fig. 5, 11. g´´.
[1052] Plates VIII. IX. XXVIII. k´.
[1055] Plate VIII. Fig. 4, 13, 17.; and IX. Fig. 3, 8, 12. d.
[1058] Plate VIII. Fig. 4, 8, 13, 17. p´.
[1059] Plate XXII. Fig. 6. q´.
[1060] Plate XVI. Fig. 4-6. r´.
[1061] Plate VIII. Fig. 3, 12.; and IX. Fig. 1, 7, 10, 11, 12, 15. c.
[1062] Plate XXII. Fig. 9, 11. s´.
[1063] Plate VIII. Fig. 3, 12, 20. and IX. Fig. 7, 10-12, 15, 20. t´.
[1065] Plates VIII. IX. XXVIII. v´.
[1066] Plates VIII. and IX. w´.
[1069] Linn. Trans. i. t. xiii. f. 1. b. 3. a.
[1070] Kirby Mon. Ap. Angl. i. t. xiii. f. 19.
[1071] De Geer ii. t. ix. f. 9. d.
[1072] Ibid. vi. t. ii. f. 23. a a.
[1073] Plate IX. Fig. 19. p´´´.
[1074] Ibid. Fig. 7. and Plate XXII. Fig. 14. k´´.
[1075] Plate XXII. Fig. 10, 11. x´.
[1077] Plates VIII. and IX. f.
[1080] Plate XXIX. Fig. 15. m´´.
[1083] Plate XXII. Fig. 3. b†.
[1084] Plate VIII. Fig. 18; and XXII. Fig. 13. c†.
[1086] Plate VIII. Fig. 2, 4, 11, 13, &c. o´´.
[1088] Plate XIV. Fig. 6-8; and XXVII. Fig. 12. p´´.
[1090] Plate XIV. Fig. 5-8; and XXVII. Fig. 6-8. r´´.
[1091] Plate XXVII. Fig. 7, 8, 15. r´´´.
[1092] Plate XIV. Fig. 5-8. s´´.
[1093] Plate XXVII. Fig. 21. s´´´.
[1094] Ibid. Fig. 9, 10, 16, 17. t´´´.
[1095] Ibid. Fig. 34-36. u´´´.
[1096] Plate XIV. Fig. 6; and XXVII. Fig. 29-36. v´´´.
[1097] Plate XIV. Fig. 5-8; and XXVII. Fig. 44, 45, 62, 63. t´´.
[1098] Plate XXVII. Fig. 25, 26, 41. w´´´.
[1099] Plate XXVII. Fig. 25, 26. x´´´.
[1100] Plate XXVI. Fig. 47, 48; and XXVII. Fig. 43. r..
[1102] Plate XXVI. Fig. 47, 48. d*.
[1103] Plate XXVII. Fig. 37-57. e*.
[1107] Plate XV. Fig. 9; and XXVII. Fig. 35, 59-61. t..
[1108] Plate VIII. Fig. 5, 6, 9, 15, 18, 19.
[1112] Ibid. Fig. 5, 9, 15. A´´.
[1113] Plate VIII. Fig. 6, 9, 15. B.
[1114] The scientific reader must recollect that these terms are employed, not because these parts are thought to be true representatives of the Epigastrium and Hypochondria of vertebrate animals, but merely on account of some analogy between them.
[1115] Plate VIII. Fig. 6. C´.
[1118] Ibid. Fig. 18, 19. C´´.
[1119] Ibid. Fig. 6, 9, 15. E´.
[1120] Plate VIII. Fig. 9. D´.
[1121] Plate IX. Fig. 17, 18. C.
[1127] Plate VIII. Fig. 5, 15. K´.
[1128] Ibid. Fig. 6, 15, 18. L´.
[1129] Plate XV. Fig. 18-22; and XVI. Fig. 1-3.
[1131] Plate XV. Fig. 22; and XVI. Fig. 2, 3.
[1132] Ibid. Fig. 20, 21; and XVI. Fig. 1. F´´.
[1133] Plate XV. Fig. 20. G´´.
[1134] Ibid. Fig. 20, 21; and XVI. Fig. 1. II´´.
[1135] Kirby Mon. Ap. Angl. i. t. xii. Apis **. e. 1. neut. f. 23-25; and t. xiii. f. 27, 28.
[1136] Ibid. t. xiii. f. 30, 31.
[1137] Ibid. a.
[1138] Plate XV. Fig. 12. L´´.
[1139] Plate XV. Fig. 14. M´´.
[1144] De Geer iii. t. xxii. f. 10. a a.
[1145] Ibid. t. xxiv. f. 2. c; and f. 11.
[1146] Plate XV. Fig. 16. R´´.
[1148] Ibid. Fig. 10; and Plate XXIII. Fig. 16, 17. T*´´.
[1149] Ibid. Fig. 12. B´´´. 15.
[1150] De Geer ubi supr. t. iii. f. 5, 20, 21. c.
[1151] See above, p. 86, 110, 243—.
[1152] Many species of Hister, Curculio L., Doryphora Illig. are extremely hard, while Cantharis Geoffr., Meloe F., and Telephorus Geoffr., are very soft.
[1153] Thenard Traité de Chimie Elémentaire, iii. 637. n. 2005. The other products he mentions are—a green oil, a yellow substance, a black ditto, acetic acid, uric acid, phosphate of magnesia. The vesicant matter consists of little micaceous laminæ soluble in boiling alcohol and oil, but insoluble in water.
[1154] Coquebert Illustr. Icon. ii. t. xviii. f. 14, 15.
[1155] Linn. Trans. xii. t. xxii. f. 16.
[1156] This name I would give to Locusta F., reserving, with Dr. Leach, the latter name to the true locust (Gryllus F.). The name Conocephalus, by which Locusta F. has been distinguished, is better restricted to those with a conical head.
[1157] Plate XXVIII. Fig. 1, 2.
[1158] Huber Nouv. Obs. ii. 317.
[1160] Hair, in the Holy Scriptures, is used as the symbol of strength or power. Judges xvi. 17—. 1 Cor. xi. 10.
[1161] Anat. Compar. ii. 624.
[1162] Anat. Compar. i. 119.
[1163] Ibid. ii. 540.
[1164] Ibid. 547.
[1165] Ibid. 553.
[1166] Anat. Compar. ii. 553.
[1167] Ibid. 557.
[1168] Ibid. 560.
[1169] Plate XXVIII. Fig. 2. a´´´.
[1170] Anat. Compar. ii. 557.
[1171] A harmonic suture is when the margins of two flat bones simply touch each other, without any intermediate substance; and a squamose, when the thin margin of one covers that of the other. Anat. Compar. i. 124. With regard to the flat portions of the integument of insects, they have some motion; whereas a suture is an articulation without movement. Ibid.
[1172] Their connexion by means of a ligament classes them under Synneurosis (Monro On the Bones, Dr. Kirby's edit. 29), but even this not strictly, since a common ligament connects them all. Those of the trunk, as admitting a slight degree of motion, belong to Amphiarthrosis (Anat. Compar. i. 126), and those of the abdomen, which are capable of larger movements, to Diarthrosis (Ibid. 127).
[1174] In the hornet and other wasps, this line on the inside of the head furnishes a foundation for a septum, which in the sides of the nose is very high, and connects also with the hind part of the head.
[1175] Anat. Compar. i. 445—.
[1176] Ibid. 447.
[1177] Mém. sur les Anim. sans Vertèbr. I. i, 11—. Comp. Anat. Compar. iii. 314—.
[1178] It is probable that M. Cuvier took his idea of this first kind of articulation, by contact of solid parts, from this individual insect; since, besides its very prominent throat, there is on each side of the lower part of the occiput a small elevation, or approach to a tubercle.
[1179] Gen. des Crustac. et Ins. ii. 246. Regne Anim. iii. 325.
[1180] This was written directly after the experiment recommended in the text had been tried, with the result there stated.
[1181] Reaum. iv. 40. Latreille Fourmis, 328—.
[1182] Plate VII. Fig. 2. k´´.
[1183] Clairville (Ent. Helvet. i. 44) appears to have been the first who classed insects according to their mode of taking their food.
[1184] Plate VI. VII. XXVI. a´.
[1185] In Lucanus, Lamprina, &c. the labrum seems to form the under-side of the nose, and to be connate with it.
[1186] Kirby Mon. Ap. Angl. i. t. v. Apis *. b. f. 18. b.
[1187] Ibid. t. ii. Melitta **. b. f. 4, 5. Plate XXVI. Fig. 30.
[1188] Plate XXVI. Fig. 31. Mon. Ap. Angl. i. t. x. Apis **. c. 2. δ. f. 13. c.
[1189] Plates VI. VII. and XXVI. b.
[1190] De Geer iv. 124. t. iv. f. 12. iii. 415. t. xxi. f. 4.
[1191] Ibid. iv. 281—. t. xi. f. 7.
[1192] Ibid. 329. t. xii. f. 3.
[1193] Ibid. ii. 775—. t. xxvi. f. 10. b c, b c.
[1194] Philos. Entom. 18.
[1195] Syst. Eleuth. i. Præf. iv.
[1196] Gen. Crustac. et Ins. i. 180.
[1197] N. Dict. d'Hist. Nat. iv. 246.
[1198] Plate VI. Fig. 6. b´. a´´.
[1199] Plate VII. Fig. 3. b´. a´´.
[1200] Kirby Linn. Trans. xii. t. xxi. f. 8. f.
[1201] Ibid. t. xxi. f. 10. d. MacLeay Hor. Entomol. i. t. iii. f. 26, 27.
[1205] Plate XXVI. Fig. 24. b´.
[1206] Plate VII. Fig. 3. a´´.
[1207] N. Dict. d'Hist. Nat. xxiv. 171.
[1209] Plate VII. Fig. 3. i´´.
[1213] Plates VI. VII. XXVI. b´´.
[1215] Hor. Entomolog. i. t. i. f. 1. g.
[1216] Ibid. t. ii. f. 18. g.
[1217] Kirby Mon. Ap. Angl. i. t. xii. neut. f. 1. g. c.
[1218] Ibid. 93. 103—. t. vi. Apis **. b. f. 3. b. c.
[1219] Ibid. t. i. *. a. f. 3. b.
[1220] Ibid. t. ix. Apis **. c. 2. γ. f. 3. b.
[1221] Clairv. Ent. Helvet. ii. t. xxiv. f. 1. c.
[1222] Plate XXVI. Fig. 24, 28. b´´.
[1223] Mon. Ap. Angl. i. t. ii. Melitta **. b. f. 2. c.
[1224] Plate XIII. Fig. 2. Linn. Trans. xii. t. xxi. f. 6. b.
[1225] This is the case with Oxyporus F. Plate XIII. Fig. 4.
[1226] Plate VI. Fig. 12. b´´. Latreille, N. Dict. d'Hist. Nat. xvii. 545, seems not to regard these as palpi; but from their tubular form, and insertion in the socket of the labium, it is clear that they ought to be so considered.
[1227] Plates VI. VII. XXVI. c´.
[1228] A corresponding articulation takes place between the tibia and thigh of some of the Scarabæidæ, which will be hereafter described. See Plate XXVII. Fig. 8-11.
[1229] Comparaison des Organes de la Mastication des Orthoptères, 2.
[1230] Comparaison des Organes de la Mastication des Orthoptères, 2.
[1231] See above, p. 407. note1174.
[1232] Ubi supra, 4.
[1233] Ibid.
[1234] Ibid. 5.
[1235] Oliv. Ins. no. 66. Prionus. t. xiii. f. 54.
[1236] Ibid. no. 83. Curculio. t. iv. f. 37.
[1237] Linn. Trans. xii. t. xxi. f. 12.
[1238] For Mandibles of Locusta see Plate VI. Fig. 6. c´. of Lampyris Oliv. Ins. no. 28. t. i. f. 1. of Buprestis, Ibid. no. 32. t. iii. f. 17. of Lucanus, Ibid. no. 1. t. i-v. and of Prionus, Ibid. no. 66. t. ii. f. 8.
[1239] Gomphosis is, when one bone is immoveably fixed in another as a nail in a board.
[1240] Marcel de Serres ubi supra. 7.
[1241] See Plate XIII. Fig. 7. Kirby Mon. Ap. Angl. i. t. xiii. f. 13. and t. xii. neut. f. 10.
[1243] Oliv. Ins. no. 42. t. i. f. 1. and no. 2. t. i. f. 1. b.
[1244] Plate XXVI. Fig. 16, 18.
[1246] Mon. Ap. Angl. i. t. iv. Melitta. f. 5-8.
[1247] Drury Ins. ii. t. xlviii. f. 3. See above, p. 315.
[1248] Oliv. no. 1. t. v. f. 16. &c. t. iii. f. 7.
[1249] Comparaison des Organes, &c. 7—.
[1250] Plate VI. Fig. 6. and XIII. Fig. 5. a´´´.
[1251] Plate VI. Fig. 12. and XIII. Fig. 5. b´´´.
[1253] I was not aware that Knoch had observed this part, till some time after the publication of my paper On Mr. William MacLeay's Doctrine of Affinity and Analogy (see Linn. Trans. xiv. 105—), when I happened to meet with it in a letter from a friend, received more than thirteen years ago; but without any reference to the work of Knoch, in which it was stated. It was doubtless taken from his Beiträge zur Insektengeschichte.
[1254] Anat. Comp. iii. 321—.
[1255] One of those mandibles is represented in Plate XXVI. Fig. 20. a´´´. incisive teeth d´´´. molary plate. Comp. Linn. Trans. ubi supr. t. iii. f. 4. c a b.
[1257] In the Myrmeleon, or ant-lion, the suction is promoted by the action of a piston, that pumps up the juices. Reaum. vi. 369.
[1258] De Geer iv. 386—. t. xv. f. 10. See above, p. 121.
[1259] Plate XIII. Fig. 7. c´´.
[1260] Oliv. Ins. no. 42. Staphylinus. t. i. f. 1. b.
[1261] Plates VI. VII. XXVI. d´.
[1262] Plate VI. Fig. 3, 6, 12. VII. Fig. 3. c´´.
[1263] Plate VII. Fig. 3. a´´. e´´. Mon. Ap. Angl. i. t. xiii. f. 1. e.
[1264] Ibid. f. 3. a.
[1265] Clairv. Ent. Helvet. ii. 146. t. xxiii. f. super. b.
[1266] N. Dict. d'Hist. Nat. iv. 243.
[1267] In Anoplognathus, however, though it has neither teeth nor spines, it is as hard as the mandibles.
[1269] Plate XXVI. Fig. 10, 11. d´´´. e´´´.
[1270] Ibid. Fig. 9. d´´´. e´´´.
[1271] Plate VI. Fig. 6, 12. d´´´. e´´´. Oliv. Ins. no. 45. Meloe. t. i. f. 1. c. These are what Fabricius calls galeate maxillæ, on which he founded his class Ulonata.
[1272] Plate XXVI. Fig. 11. d´´´. e´´´.
[1273] Plate VI. Fig. 3. d´´´.
[1274] Clairv. Ent. Helvet. t. i. t. xviii. f. super. b.
[1275] Ibid. t. xix. b. This genus may be the analogue of some heteromerous one yet undiscovered, as Calosoma is of Adelium (Kirby Linn. Trans. xii. t. xxii. f. 2.)
[1276] Clairv. Ent. Helvet. ii. t. xxiv. f. super. b.
[1278] Oliv. Ins. no. 19. Trogosita. t. 1. f. d. no. 41 bis. Dryops. t. i. f. 1. c.
[1279] See above, Vol. I. p. 238.
[1280] Oliv. Ins. no. 17. Ptinus. t. i. f. 1. c.
[1281] Ibid. no. 60. Blaps t. i. f. 2. c.
[1282] Ibid. no. 88. Languria. t. i. f. 2. c.
[1283] Ibid. no. 89. Erotylus. t. ii. f. 12. c.
[1284] Oliv. Ins. no. 41. Gyrinus. t. i. f. 1. e.
[1285] Plate XXVI. Fig. 13. Hor. Entomolog. i. t. iii. f. 29, 30. E.
[1286] Mon. Ap. Angl. i. t. ii. Melitta. **. a. f. 2. t. v. Apis. *. b. f. 4. &c.
[1287] Oliv. Ins. no. 48. Cerocoma. t. i. f. 1. c.
[1288] Plate XXVI. Fig. 10-12.
[1289] Clairv. Ent. Helvet. ii. Cicindela. t. xxiv. f. super. b. for Carabidæ, Dytiscidæ, his other plates.
[1290] Hor. Entomolog. i. t. ii. f. 13. E.
[1291] Plate VI. Fig. 6, 12. f´´´.
[1292] Hor. Entomolog. t. i. f. 3. E.
[1293] Ibid. f. 4. E.
[1294] Oliv. Ins. no. 7. Hexodon. t. i. f. 1. e.
[1295] Ibid. no. 3. Scarabæus. t. l. f. 1. f.
[1296] Kirby in Linn. Trans. xiv. 102. t. iii. f. 4. d.
[1297] De Geer v. 417. t. iv. f. 12.
[1300] Mon. Ap. Angl. i. t. ix. 2. c. 2. β. f. 2. d. g. 4. t. xii. neut. f. 6. d. t. xiii. f. 3. b.
[1301] Clairv. Ent. Helv. ii. t. xxiii. f. 1.
[1302] Plate VII. Fig. 3. b´´. h´´.
[1303] Plate VI. Fig. 12. b´´. f´´´.
[1304] Ibid. Fig. 13. h´´. Savigny Anim. sans Vertèbr. l. i. 29—. t. i.-iii. ȯ.
[1305] De Geer vi. t. xix. f. 4. d.
[1306] Ibid. t. ix. f. 8. b b. t. xii. f. 20. b. t. xiv. f. 15. i i.
[1309] Kirby Mon. Ap. Angl. i. t. x. Apis. **. c. 2. δ. f. 3. a. and **. d. 1. f. 1. d.
[1310] Clairv. Ent. Helvet. ii. t. xxxi. f. super. b. Mon. Ap. Angl. i. t. xiv. f. 1. h. f. 3, 5. c.
[1312] Plate XXVI. Fig. 1. As the very remarkable maxillary palpi of that extraordinary Coleopterous genus Atractocerus seem not to have been so fully described as they deserve, I shall give here a minute detail of their composition. They consist of four joints: the first is wide and short, and somewhat platter-shaped; the second is much smaller and shorter: the third is concavo-convex, or shaped like a shallow bowl: towards the breast this joint is elevated, and on the elevated edge sits the last joint, which is longer than all the rest taken together. In my specimen it points towards the breast; its under side is entire and slightly curved, but in the upper side are two rows of lamellæ (b), placed alternately nine on each side, with an odd one at the end: these lamellæ are full of minute papillæ, and furrowed on the side next the mouth. From between the first pair a slender exarticulate hairy branch or appendage emerges (a), which forms nearly a right angle with the main stem. The labial palpi appear to consist of three joints; the two first very short, and the last large, ovate, and acute. This description is not taken from A. necydaloides, but from a Brazilian species more than five times its size, which I have named A. grandis.
[1313] Plate VI. VII. XXVI. e´.
[1314] Plate VI. Fig. 6, 12. e´. Cuvier Anat. Compar. iii. 347.
[1315] Cuvier Ibid.
[1316] Huber Fourmis, 4—.
[1317] Anim. sans Vertèbr. iii. 304.
[1319] Reaum. v. 309—.
[1321] Clairv. Ent. Helvet. ii. Pref. xxii.
[1322] Kirby Mon. Ap. Angl. i. t. xiv. (1) 2. b.
[1323] Plate XXVI. Fig. 24. e´.
[1324] Clairv. ubi supr. t. xx. c.
[1325] Plate XXVI. Fig. 28. e´.
[1326] Kirby ubi supr. fig. (8) 1. c c. The lateral pieces in the tongue in Vespa (Ibid. c c.) have been regarded as lobes of it, but they are rather Paraglossæ.
[1327] Plate XXVI. Fig. 29. e´.
[1328] Clairv. ubi supr. t. xxx. c.
[1329] Kirby ubi supr. no. 2. f. 1, 3.
[1330] Plate XXVI. Fig. 24. b.
[1331] Kirby ubi supr. t. x. Apis. c. 2. δ. f. 5. t. xii. neut. f. 2, 3. t. xiii. f. 1. h.
[1332] Plate XXVI. Fig. 26, 29.
[1333] Plate XXVI. Fig. 28. i´´.
[1334] Kirby Mon. Ap. Angl. i. t. xii. neut. f. 1. h h. t. xiii. f. 1. f f.
[1335] Ibid. t. x. **. d, 1. f. 2. b b.
[1337] Plate VII. Fig. 14. f´.
[1338] Reaum. v. 317.
[1339] Organisation exterieur des Ins. 184.
[1340] Ubi supra.
[1341] Anim. sans Vertèbr. I. i. 12.
[1342] De Geer ii. 778—. t. xxvi. f. 11. m. Plate VII. Fig. 2. k´´.
[1343] Ubi supra.
[1345] Organisation &c. 182.
[1346] Under this name M. Savigny includes the Myriapoda, Arachnida, and Crustacea. Anim. sans Vertèbr. I. i. 40.
[1347] Ibid. 43.
[1348] Vol. II. p. 275—. Also see above, p. 121—.
[1349] Anim. sans Vertèbr. I. i. t. ii. f. 2. a. a´.
[1350] Plate VII. Fig. 13. c´.
[1352] Anim. sans Vertèbr. I. i. 106. Plate VII. Fig. 13. b´.
[1353] Ubi supr. 45—.
[1354] Plate VII. Fig. 11. f´, a´´.
[1355] Ubi supra, 45.
[1356] Ibid. 44—.
[1357] N. Dict. d'Hist. Nat. ii. 275—. Plate VII. Fig. 10. c´.
[1358] De Geer t. xl. f. 4. t. x. f. 7, 8.
[1360] Savigny Anim. sans Vertèbr. I. i. 62.
[1361] N. Dict. d'Hist. Nat. ii. 277.
[1362] N. Dict. d'Hist. Nat. ii. 276.
[1363] Ibid.
[1364] Ubi supr. 58.
[1366] Plate VI. Fig. 7-9. a´, b´, c´, d´.
[1367] Magaz. 1806.
[1369] Ibid. Labium b´. Mandibulæ c´. Maxillæ d´.
[1370] Savigny Anim. sans Vertèbr. I. i. 37.
[1371] De Geer iii. 137—. t. ix. f. 4.
[1372] Reaum. iii. 335. t. xxviii. f. 8-14.
[1373] De Geer iii. 117. t. viii. f. 22. b.
[1374] Plate VII. Fig. 5, 6. a´, b´, c´, d´.
[1375] N. Dict. d'Hist. Nat. iv. 253.
[1376] Plate VII. Fig. 5, 6. a´.
[1378] Reaum. iv. t. xvi. Fig. 13. z.
[1379] Authors are not agreed as to the precise number of lancets contained in a gnat's proboscis. Swammerdam affirms there are six, including the labrum. i. 156. b. t. xxxii. f. 3. Reaumur could find only five. iv. 597—. t. xlii. f. 10. And Leeuwenhoeck only four.
[1381] Plate VII. Fig. 5. This figure is copied from Reaumur, and was engraved before this discovery was made.
[1382] M. Savigny is of opinion that the central lancet or lancets represent the Epipharynx and Hypopharynx; for which he does not state his reasons: but as these are properly covers of the pharynx, the idea seems incorrect. Ubi supr. 15.
[1383] N. Dict. d'Hist. Nat. ix. 489. and iv. 253—.
[1384] Ubi supr. 36.
[1385] Ibid. t. iv. f. 1. o. o.
[1387] Plate VI. Fig. 13. a´, b´, c´, d´.
[1389] Plate VI. Fig. 13. a´. Savigny Anim. sans Vertèbr. I. i. 3—. t. i.-iii. a.
[1390] Ibid. i. Plate VI. Fig. 13. c´.
[1391] Ibid. Fig. 13. h´´. Savigny ubi supr. o.
[1392] Plate VI. Fig. 13. d´. Savigny ubi supr. t. 1-3. o.
[1393] Ibid. ö. Plate VI. Fig. 13. b´.
[1394] N. Dict. d'Hist. Nat. xvii. 467.
[1395] N. Dict. d'Hist. Nat. iv. 253.
[1396] Plate VII. Fig. 8. c´, d´, e´, h´´.
[1401] Rösel. ii. t. iii. f. 15. Latreille Gen. Crust. et Ins. iv. 365.
[1402] Swammerdam Bibl. Nat. t. ii. f. 4.
[1403] N. Dict. d'Hist. Nat. xxviii. 266.
[1404] Ibid. xvi. 432. De Geer vii. t. vi. f. 4. Not quite accurate.
[1405] Plates VI. VII. XXVII. a.
[1406] Organisat. Extér. des Ins. 196.
[1407] In the Transactions of the Royal Society, this part in Anobium tessellatum is so called. xxxiii. 159—.
[1408] Plate XXVII. Fig. 4. a.
[1410] Observ. Nouv. sur les Hyménoptères (Ann. du Mus.) 5.
[1411] Those beetles whose posterior pair of tarsi have only four joints, and the two anterior five, are so called.
[1412] Kirby in Linn. Trans. xii. 464. t. xxiii. f. 6.
[1413] Ibid. xiii. t. i. f. 1. b.
[1419] Kirby Mon. Ap. Angl. i. t. i. Melitta. *. b. f. 3.
[1423] Plate XXVI. Fig. 41. i.
[1424] Stoll Punaises, t. xxxix. f. 279, 280.
[1425] Plates VI. VII. XXVI. d.
[1428] Plate XXVII. Fig. 4. f.
[1429] Cuv. Regne Animal. iii. t. xiii. f. 4.
[1430] This insect was taken both at Matlock and Exmouth. The body and thighs are of a light-brown, wings testaceous, legs pale; antennæ between setaceous and filiform, two-thirds the length of the body; first joint not much thicker than the rest.
[1432] De Geer iii. 561. t. xxvii. f. 1.
[1433] Plates VI. VII. XXVI. h.
[1434] Plate VII. Fig. 8, 9. XXVI. Fig. 43. h.
[1435] Viz. one on each side above, and one below.
[1436] Walckenaer Aranéïdes, t. v. f. 50, 52. t. viii. f. 82.
[1437] Treviranus (Arachnid. 4.) says that Scorpio Europæus has only two eyes. He appears to have overlooked the two on the anterior side of a tubercle at each angle of the head, where they are large, but not conspicuous, at least in my specimen.
[1438] De Geer vii. t. iii. f. 8, 9, 12.
[1439] Plate XXVI. Fig. 43., h.
[1440] Walck. Aran. t. i. f. 3.
[1441] Ibid. t. v. f. 42-48.
[1442] Ibid. t. iv. f. 41.
[1443] Walck. Aran. t. i. f. 2.
[1444] Ibid. t. i. f. 7.
[1445] Ibid. t. ii. f. 18, 20.
[1447] De Geer vii. 138. t. viii. f. 15. y y.
[1448] Ibid. t. xl. f. 3. o o, y y.
[1449] Plate XXVI. Fig. 43. h.
[1450] Segestria perfida, Walck. Aran. t. v. f. 52. &c.
[1451] Tetragnatha and Latrodectes, Ibid. t. vii. f. 64. and t. ix. f.. 84.
[1452] Nyssus coloripes, Ibid. t. vi. f. 58.
[1453] Dolomeda, Ibid. t. ii. f. 18, 20.
[1454] Sphasus, Ibid. t. iii. f. 24.
[1455] Mygale avicularia, Ibid. t. i. f. 3.
[1456] Sparasus, Ibid. t. iv. f. 41. Plate XXVI. Fig. 37.
[1457] Eresus, Ibid. t. iii. f. 26.
[1458] Storena, Ibid. t. ix. f. 86.
[1459] Argyroneta, Ibid. f. 88.
[1460] Pholeus, Ibid. t. viii. f. 80.
[1462] Plate XXIX. Fig. 11. h.
[1466] Reaum. iv. 245.
[1467] Microgr. 176.
[1468] Epist. Mar. 6. 1717.
[1469] Amœn. Academ. vii. 141.
[1470] I possess a specimen in which the eye is partly black and partly white: the lenses are invisible in the black part, but very visible in the white.
[1471] Philos. Entomolog. 19.
[1473] Hooke Microgr. schem. xxiv.
[1475] Cuvier Anat. Compar. ii. 442—. Compare Swammerdam Bibl. Nat. i. 211. t. xx. f. 45.
[1476] Vol. II. p. 4, 364, &c.
[1477] Plate XXVI. Fig. 36. h. Fabricius, and after him Olivier, though both quote Forster, regard one of these eyes in Lamia Tornator as a spot; but they could not have examined it attentively. Saperda præusta F. has also four eyes.
[1478] Plate XXVI. Fig. 39. h.
[1479] Latreille Gen. Crust. et Ins. iii. 73.
[1480] N. Dict. d'Hist. Nat. i. 479.
[1482] De Geer vii. 562.
[1484] Ent. Helvet. i. t. xii.
[1486] Kirby Mon. Ap. Angl. i. t. xi. Apis. **. e. 1. f. 2.
[1487] De Geer iii. t. xxxiv. f. 17, 18, 24. o o.
[1488] Mon. Ap. Angl. i. t. xiv. no. 11. f. 1. f. Linn. Trans. xi. t. ix. f. 10. d.
[1489] Plate XIII. Fig. 9. Fuessly Archiv. t. vi.
[1490] Schellenberg Cimices t. xiii. ix. f. 1. a. De Geer ii. t. xviii. f. 10.
[1491] Stoll Spectres, &c. t. iv. f. 14. t. x. f. 38, &c.
[1493] Linn. Trans. ubi supr.
[1494] Schellenberg Mouches, t. xxvii. f. 1, 2. a, d.
[1495] Ibid. t. ix. f. 3. a.
[1496] Ibid. t. ii. f. 2. a.
[1497] Mon. Ap. Angl. i. 148.
[1498] Plate XXVII. Fig. 4. h´.
[1499] This circumstance proves that Mr. W. S. MacLeay is correct in considering this as a subgenus; but it militates against its being connected with Lamprima.
[1500] Plate VI. Fig. 4, 10. VII. Fig. 1, 2, 4. XXVI. Fig. 39-42. i.
[1501] Bibl. Nat. i. 214.
[1502] Reaum. iv. 245.
[1503] Ibid. v. 287—.
[1504] Magas. der Entomolog. iv. 410.
[1505] Latreille speaks of Phasma as having no stemmata; but it should seem that he examined only the apterous ones, all the winged individuals, at least so far as I have examined them, having three very visible ones. It may, I think, be laid down as a rule, that the larvæ and pupæ of Orthoptera have not these organs. Probably their use is principally in flying?
[1506] Flata phalænoides F. and affinities have no stemmata, while Flata reticulata and affinities have them: a proof that these tribes are distinct genera.
[1508] Reaum. iv. 243. He refers for this insect to plate xiv. without adding any number for the figure; but no such is in that plate.
[1509] Plate XXVI. Fig. 40. i.
[1510] Cercopis, Ibid. Fig. 42; and Fulgora, Fig. 41. i.
[1512] Plate XXVI. Fig. 40. i.
[1514] Plate XXVI. Fig. 41. i.
[1515] De Geer iii. t. xxvii. f. 1. Reaum. iii. t. xxxii. f. 3, 9.
[1516] Plate XXVI. Fig. 40. i.
[1518] Linn. Trans. xiii.
[1519] Mag. der Entomolog. iv. 5.
[1520] Palpi quatuor, subæquales, cylindrici, ad basin clypei. Germ.
[1522] Plate VI. Fig. 1, 4. i´.
[1523] See above, p. 321. Linn. Trans. xii. t. xxi. f. 3.
[1524] Ibid. f. 7.
[1525] Plate VI. Fig. 4. c. i´.
[1526] Plate XII. Fig. 9. l´´. This circumstance was very recently discovered; which will account for this plate not being quite correct in this respect, the bulb being represented as a distinct joint in Fig. 6, 10, 26.
[1529] Oliv. Ins. no. 80. Macrocephalus t. i. f. 1-4.; Anthribus f. 5-12; and no. 83. Curculio t. ii. Calandra f. 16.
[1530] Schellenberg Cimices t. xiv. f. 1. b.
[1531] De Geer vii. t. iv. f. 7. a a.
[1533] Plates XII. Fig. 28; and XXV. Fig. 9, 24.
[1535] Plates XII. Fig. 29; and XXV. Fig. 28. a.
[1536] Plates XI. XII. XXV. k´´.
[1537] Plates XI. XII. XXV. l´.
[1539] Plates XII. Fig. 28; and XXV. Fig. 13. m´´.
[1540] Plate XXV. Fig. 2, 5, 21. m´´.
[1541] Plate XII. Fig. 10. m´´.
[1543] Plate XXV. Fig. 1. m´´.
[1547] Jurine Hymenopt. t. vii. f. 3.
[1553] Linn. Trans. xii. t. xxi. f. 3.
[1554] Latreille says six, but only five are discernible; the three last form a kind of bristle.
[1555] Latr. Fourmis, 323.
[1556] Jurine Hymenopt. t. vi. f. 3.
[1557] Ibid. f. 2.
[1558] Ibid. f. 1. Plate XXV. Fig. 7.
[1559] Plate XXV. Fig. 25, 26.
[1562] Linn. Trans. xxii. t. xxi. f. 3, 4. Plate XXV. Fig. 11.
[1564] Kirby Mon. Ap. Angl. i. 184. t. x. **. d. 1. f. 8.
[1570] Linn. Trans. xii. t. xxiii. f. 5. f.
[1571] Ibid. t. xxi. f. 8 g. 9, 10. c.
[1572] De Geer iv. 219. t. viii. f. 20.
[1573] See Vol. II. p. 65, 201—.
[1574] De Geer iv. t. vii. f. 22.
[1576] De Geer vi. t. i. f. 5.
[1581] Plate XXVII. Fig. 1, 3-5. n´.
[1585] For the reason which induced the authors to use this word instead of Terminology, before employed, see Vol. I. Pref. p. xii—. They are gratified to see that M. Latreille has adopted this term in the work quoted on the other side, p. 194.
[1586] Organisation Extérieure des Insectes, Mém. du Mus. t. viii.
[1587] Ibid. 199—. I have never been able to procure M. Audoin's Mémoire on this subject.
[1588] Fundament. Entomolog. in Amœn. Acad. vii. 143.
[1589] Terminologie, 1578, &c. He afterwards called the trunk Stethidium: Terminologie der Insekten Magaz. 1806. 14.
[1590] Organisation, &c. 198.
[1591] In Nirmus Anseris, &c. however, in this Order, the same distinction is observable.
[1594] If the head of any individual of these two Orders be carefully taken off, it will be found that above there is a very short piece representing the prothorax, and quite unlike the collar of Hymenoptera.
[1603] Coquebert Illustr. Ic. ii. t. xviii. f. 2, 4.
[1604] Stoll Cigales t. xxi. f. 116.
[1605] Ibid. t. xi. f. 53.
[1607] Plate XIII. Fig. 18. a.
[1608] Stoll Cigales t. xxviii. f. 163.
[1609] Ibid. t. xxi. f. 115. Coquebert Illust. Ic. ii. t. xviii. f. 5.
[1610] Stoll Spectres t. xi. f. 42. t. xii. f. 45. t. xvi. f. 58, 59.
[1612] Sur le Vol des Ins. c. vii. 374. t. xviii. f. 9. i i.
[1614] Theatr. Ins. 98.
[1615] De Geer iv. 74.
[1616] Organisation Extérieure des Ins. 177.
[1617] A subgenus, related to Lebia (Hexagonia K. MS.) and some Cimicidæ, are so circumstanced.
[1618] Regne Animal iii. t. xiii. f. 6.
[1623] Linn. Trans. vi. t. xxi. f. 10.
[1624] Ibid. f. 8.
[1627] Something of the kind is observable at the base of the other legs in this tribe.
[1628] Monro On the Bones, 160.
[1631] Kirby in Linn. Trans. xiv. t. iii. f. 4. i.
[1632] It is doubtful whether M. Latreille's Harpalus megacephalus is synonymous with Carabus megacephalus Fab. Comp. Gen. Crust. et Ins. i. 206. with Syst. Eleuth. i. 187. 95.
[1634] Plate VIII. Fig. 11. b.
[1635] Linn. Trans. iv. 53.
[1636] Syst. Nat. i. Cancer. Scorpio.
[1637] Moses, when he describes insects as going upon four legs, evidently considers the anterior pair as arms; Bochart does the same. Levit. xi. 20—. Hierozoic. ii. 497.
[1640] Samouelle t. v. f. 4.
[1642] Latreille Organization &c., 199. Chabrier Sur le Vol des Ins. c. i. 412. c. iv. 54, &c.
[1643] In Chlorion, Ammophila, &c., this part separates more readily from the alitrunk.
[1644] Chabrier Sur le Vol des Ins. c. i. 413—. c. iv. 54. This author seems to regard the collar as something peculiar to Hymenoptera.
[1649] M. Chabrier (Vol. des Ins.) supposes that the humming of insects is produced by the exit of the superfluous air from their thoracic spiracles, &c.; in Melolontha he thinks they are in the metathorax under the wings (c. i. 457—. Plate XXII. Fig. 13. c. †. represents the operculum of one of those of Dytiscus marginalis): in the Hymenoptera, in the mesothorax, near the posterior lobes of the collar (Ibid. 459. c. iv. 50.); and in the Diptera, in the metathorax, near the poiser (c. i. 457). I observed myself lately, that Elophilus tenax, if held by the anterior part of the body, when it hummed, alternately opened and shut this spiracle. The wings during the sound vibrated intensely. The hum ceased and was renewed, as they were restrained from this motion or released from restraint; when the wing was moved towards the head, a different sound was emitted from that produced when it merely vibrated.
[1652] When the prothorax is separated from the elytra by a kind of isthmus, as in Scarites, Passalus, &c., the dorsolum is more or less uncovered.
[1653] Plate VIII. Fig. 16. 20. i´.
[1658] Plates VIII. IX. XXVIII. k´.
[1659] Audoin, Chabrier, &c.
[1660] Olivier. He seems also to have thought that neither the Orthoptera nor Homopterous Hemiptera have this part. N. Dict. d'Hist. Nat. x. 112.
[1661] Plate VIII. Fig. 3. k´.
[1662] In Macraspis MacLeay it is often half as big as an elytrum.
[1663] Plate VIII. Fig. 12. k´.
[1664] Mr. W. S. MacLeay opposes the Hemiptera to the Orthoptera, the Homoptera to the Neuroptera, and the Aptera to the Coleoptera: but if analogous structure be made the guide, I think my arrangement will be found most correct. Hor. Entomolog. 367.
[1665] Plate VIII. Fig. 20. k´.
[1667] Plate IX. Fig. 1. k´. N. B. This is from Cossus F.
[1670] Mon. Ap. Angl. i. t. vi. Apis. ** a. f. 2. a a.
[1671] Stoll Cigales t. xxviii. f. 164.
[1673] Plates VIII. IX. XXVIII. l'.
[1674] Plate XXIII. Fig. 6. e´´´.
[1675] Plate VIII. Fig. 12. l´.
[1677] Plate XXVIII. Fig. 11. l´.
[1678] Chabrier Sur le Vol des Ins. c. iii. t. viii-v. B. i.
[1680] Chabrier Sur le Vol des Ins. c. iii. 354.
[1681] From πνυμι to breathe and σέγω to cover.
[1683] At first I had named this piece the antecosta, and the mesostethium the postcosta; and there is certainly some analogy between the thorax of insects, consisting of several pieces that follow each other, and the vertebral column; between their three sternums and the sternum, and between their other pieces and the ribs of vertebrate animals. Comp. Chabrier, ubi supr. c. iv. 49. note 1.
[1685] Plate VIII. Fig. 12. 13. o´. z´.
[1687] Plate IX. Fig. 8. o´. z´.
[1691] Plate VIII. Fig. 3, 13. p´.
[1692] Linn. Trans. xii. t. xxii. f. 4.
[1693] A remarkable instance of analogy is afforded by this genus. In Erotylus there are two groups; one distinguished by gibbous elytra, and the other by flatter ones. The same distinction is observable in Spheniscus; for to this genus belongs Helops fasciatus Oliv., which represents the flat Erotylí, and even individually Erotylus trifasciatus Oliv., E. fasciatus F.
[1694] I would restrict this name to the conical-headed Locustæ F.
[1695] Plate VIII. IX. t´. Linn. Trans. xi. t. ix. f. 16. c.
[1696] Plate VIII. Fig. 3. t´.
[1697] Ibid. Fig. 12. Comp. i´, k´, l´, with t´, u´, v´.
[1700] Plate IX. Fig. 19, 20. t´.
[1703] Plate VIII. Fig. 3. u´.
[1704] Ibid. VIII. Fig. 12. u´. Plate IX. 7. u´.
[1706] Plate VIII. Fig. 3. v´.
[1707] Plate VIII. Fig. 12, 16.; and Plate IX. Fig. 7. v´.
[1708] Plate VIII. Fig. 3. w´.
[1709] Plate XXII. Fig. 14. w´.
[1710] Plate XXIX. Fig. 25. w´.
[1711] Ibid. and Plate VIII. Fig. 12.; and Plate IX. Fig. 7. k´´.
[1712] Sur le Vol des Ins. c. iii. 354.
[1714] Plate IX. Fig. 15. k´´.
[1715] Plate IX. Fig. 11. k´´.
[1718] Plate VIII. Fig. 4. z´.
[1721] Plate VIII. Fig. 13. y´. a´ †.
[1724] Plate VIII. Fig. 4. z´.
[1725] Plate XXIX. Fig. 15. z´.
[1729] Plate VIII. Fig. 8. a †.
[1730] De Geer iv. t. iv. f. 3. d d. c c.
[1731] Plate VIII. Fig. 13. a †.
[1732] The history of this parasite has been traced by Dr. Reid; but alas! this learned and acute observer of nature did not live to give his discoveries to the world: it is hoped, however, they will not be lost, being in most able hands.
[1733] Plate VIII. Fig. 18. and XXII. Fig. 13. c †.
[1735] Sur le Vol des Ins. c. i. 459.
[1736] Ibid. 457—.
[1737] Plate XXII. Fig. 13. c †.
[1738] Mem. sur les Anim. sans Vertèbr. 45—. Hor. Entomolog. 411—.
[1739] Plate VIII. Fig. 3. x´. IX. Fig. 2. s´. and XXII. Fig. 5-14.
[1742] Plate XXII. Fig. 8, 11. h´.
[1743] Plate XXII. Fig. 9, 11. s´.
[1748] Plate XXII. Fig. 10, 11. x´. Comp. Linn. Trans. xi. t. ix. f. 16. g.
[1749] Plate XXII. Fig. 10, 11. d.
[1757] See above, Vol. I. p. 191. and II. p. 257, 366.
[1758] This machine is described by Dr. Eschscholtz, Beiträge zur Naturkunde, &c. Heft. i. 24—. t. i. ii.
[1763] MacLeay, Horæ Entomolog. 9. Chabrier, Sur le Vol des Ins. c. i. 417.
[1764] Plate XXII. Fig. 5. b b b.
[1766] Luke xii. 7.
[1769] Chabrier Sur le Vol des Ins. c. i. 413—.
[1771] Chabrier Sur le Vol des Ins. c. i. 446, 448, 451—.
[1772] Ibid. 412.
[1773] Ubi supr. c. ii. 333.
[1774] According to M. Chabrier, who agrees with him, M. Latreille also is of opinion, that the parapleura is the analogue of the posterior coxæ. Ubi supra, c. ii. 312. Note 2.
[1775] M. Latreille has changed the denomination of this Order to Rhiphiptera, because at first he thought that these organs were not at all analogous to elytra or wings; but since, upon further investigation, he appears to admit that they assist in flight (Annales Génér. des Scienc. Phys. VI. xviii. 8. Compare MacLeay, Hor. Entom. 423. Note *), in common justice he is bound to restore the name originally given to the Order. In the same place of the work here quoted, M. Latreille also speaks of these pseudelytra, as I would call them, as appendages of the mesothorax: but whoever consults Mr. Bauer's admirable figures of Xenos Peckii (Linn. Trans. xi. t. ix.), and is aware of the unimpeached and minute accuracy of that admirable microscopic artist, will be convinced that they belong to the anterior legs, and consequently to the prothorax.
[1776] Plate X. and Plate XXVIII. Fig. 18-23.
[1777] Chabrier, Analyse, &c. 27.
[1778] N. Dict. d'Hist. Nat. ix. 568. We have seen above (p. 578.) that the wings of insects are connected with their legs by the scapula and parapleura.
[1779] MacLeay, Hor. Entomolog. 413—. Mr. MacLeay's opinion seems to receive some confirmation from a circumstance overlooked when the larvæ of insects were treated of above (p. 130—), and to which he alludes (411); namely, that in that state they consist of two segments more than in the imago; these follow the three pedigerous segments, have no pro-legs, and are supposed to belong to the trunk rather than to the abdomen. To make this circumstance bear upon the question, it must be proved that in the perfect insect these segments in some manner become the back of the trunk and bear the wings. This would not be more wonderful than many changes that are known to occur in insects.
[1780] Latreille, Organization extérieure des Ins. 173—.
[1781] For instance Meloe, the female glow-worm, Lygæus brevipennis, Ephemera diptera, Cynips aptera, neuter ants, &c. &c.
[1783] Chionea araneoïdes Dolm.
[1784] See above, p. 560, and Vol. II. 348, 352—.
[1785] See above, p. 591, Note1775.
[1786] De Geer, iii. t. xvii. f. 10, 11. f f. M. Savigny has noticed a part in some Annelides, which he regards as analogous to elytra. Système des Annelides, 4, 9, 11.
[1788] Plates X. and XXVIII. b..
[1791] Plate XXVIII. Fig. 3-5. b´´´.
[1792] Chabrier Sur le Vol des Ins. c. i. 439.
[1794] Chabrier ubi supr.
[1795] Plate XXVIII. Fig. 6-8. d´´´.
[1800] Plate XXVIII. Fig. 6-8. d´´´.
[1805] Sur le Vol des Ins. i. c. 428—. c. ii. 325.
[1806] Oliv. Ins. No. 97. Cassida, t. i. f. 10.
[1809] M. Chabrier says that the arc described by the wings of Melolontha vulgaris to that of the elytra, is as 200 to less than 50. Sur le Vol des Ins. c. i. 440.
[1810] Plate X. Fig. 2. and XXVIII. Fig. 18-20.
[1811] Magas. 1806. Terminologie der Insekt. 18. 1675.
[1812] Plate X. Fig. 2. is the tegmen of a Blatta divided into areas.
[1813] Sur le Vol des Ins. c. ii. 327—.
[1817] Stoll, Cigales, t. viii. f. 39.
[1819] Stoll Grillons t. i. c. f. 2.
[1820] Ibid. Spectres t. xxv. f. 95. and xi. f. 42.
[1821] Ibid. t. ii. f. 5. Grillons t. i. c. f. 1.
[1822] Ibid. Spectres t. xvi. f. 58.
[1823] Ibid. Sauterelles à Sabr. t. iii. f. 7. By this name (Pterophylla) I distinguish those Locustæ F. without a conical head that are veined like leaves.
[1824] Stoll Ibid. t. vi. a. f. 18. and Plate XXVIII. Fig. 19.
[1825] Stoll Sauterel. à Sabr. t. i-iii.
[1826] Ibid. Spectres t. iv. f. 14.
[1827] Ibid. Cigales t. i. f. 1, 3-5. and t. vi. f. 31.
[1828] Stoll Cigales t. iii. f. 12-15. and t. xvii. f. 92.
[1829] Linn. Trans. xii. 449, no. 96.
[1831] Linn. Trans. xiii. t. i. f. 14. Flata should come before this genus.
[1832] Of this kind is one of Stoll's Cigales, t. xxv. f. 141.
[1834] Stoll Cigales t. i. f. 1. t. x. f. 46. t. xxix. f. 170. t. v. f. 22. t. iv. f. 19. &c.
[1836] In Latreille's whole genus Pentatoma, including several Fabrician genera, the Hemelytra are more substantial than in the subsequent tribes.
[1839] My insect, which nearly resembles the Coleopterous genus Cerylon Latr., agrees with Latreille's description in all respects, except that it cannot be said to be membrana nulla apicali.
[1840] Chabrier Analyse, &c. 24.
[1844] Plate XXVIII. Fig. 23. f´´´ is the corium and g´´´ the membrana of a species of Reduvius F.
[1846] Chabrier Analyse, &c. 23.
[1847] Chabrier Sur le Vol des Ins. c. ii. 325—. and 326. Note 1.
[1849] Plate XXVIII. Fig. 9. a.
[1851] Ibid. p. 560. and Plate XXVIII. Fig. 11. a.
[1853] Ibid. Fig. 11. 12. g´´.
[1854] Stoll Sauterelles à Sabr. t. iv. f. 12. t. vi. f. 21. &c.
[1855] Sur le Vol. des Ins. c. i. 424.
[1856] The idea of dividing the wing of an insect into larger areas seems first to have been acted upon in Monogr. Apum Angl. (1801), in which those of Hymenoptera were stated to consist of three portions, viz. Basis, Medium, Apex (i. 211.); which mode of dividing them was at first adopted by M. Latreille (Gen. Crust. et Ins. iii. 226. Note 1.) The same learned author (Ibid. iv. 239.), with regard to the Diptera, made a near approximation to the plan of dividing wings into longitudinal areas, but by the addition of a basal area, which interrupts the attention to the communication of the areas with their axes, he has rendered his system less perfect. Two of his terms—Costal Area and Intermediate Area, are here adopted; but his Internal is changed to the Anal Area, for the term internal belongs rather to the base of the wing. M. Latreille afterwards relinquished both these plans, N. Dict. d'Hist. Nat. i. 248—.
[1857] Plate X. and XXVIII. b.. i.. is the Postcostal Nervure.
[1858] Ibid. c.. n.. is the Anal Nervure.
[1860] Plate X. Fig. 7. h.. i.. k..
[1861] Ibid. Fig. 8. 9. 12. a *.
[1862] Sur le Vol. des Ins. c. i. 428.
[1863] See above, p. 600. 616.
[1864] Vol. II. p. 358—. See above, p. 559.
[1866] Jurine Hymenopt. 19. and t. v.
[1868] N. Dict. d'Hist. Nat. i. 251.
[1869] See above, p. 564, 578, 591.
[1871] Ibid. Fig. 4, 11. m´´´.
[1875] Jurine Hymenopt. t. v. Gen. 47.
[1882] Plate II. Fig. 1. Comp. Linn. Trans. xi. t. ix. f. 1.
[1885] Stoll Spectres, t. xviii. f. 65.
[1887] Stoll figures Empusa as without them, t. ix. f. 35. but? I have a nondesc. Phasma? without them.
[1888] Jones in Linn. Trans. ii. t. viii. f. 2.
[1889] Ibid. f. 5.
[1890] Jones in Linn. Trans. ii. t. viii. f. 7.
[1891] Ibid. f. 9.
[1892] Ibid. f. 2, 3, 6-9.
[1893] I wonder Mr. Jones's plan of ascertaining the divisions or subgenera of butterflies by the neuration of their wings has never been followed up; it would I think furnish an easy clue for the extrication of the tribes of all the Lepidoptera. I mean as subsidiary to more important characters.
[1894] Plate X. Fig. 6. l., m..
[1896] Plate X. Fig. 8. basal e., medial f., apical g..
[1897] Jurine Hymenopt. t. v. Gen. 48.
[1898] Ibid. Gen. 47.
[1899] Ibid. Gen. 45, 46. Comp. Plate X. Fig. 11.
[1900] Jurine Ibid. Gen. 43.
[1901] Ibid. t. iv. Gen. 47.
[1902] Ibid. t. v. Gen. 40.
[1903] Ibid. t. ii. Gen. 2.
[1904] This is a remarkable insect belonging to Vespa L., related to the hornets (V. Crabro), distinguished by having a semicircular piece taken out of the internal margin of each mandible, so that when these organs are closed there is a circular orifice,—whence the name Cyclostoma.
[1905] Jurine Hymenopt. t. ii. Gen. 2.
[1906] Ibid. Ord. ii. Gen. 2.
[1907] Ibid. t. iii. Gen. 3.
[1908] Ibid. t. v. Gen. 41.
[1909] Ibid. t. xii. Gen. 40.
[1910] Ibid. t. v. Gen. 39.
[1911] Ibid. t. iv. Gen. 29.
[1912] Ibid. t. v. Gen. 42.
[1913] This Cynips inhabits a long polythalamous gall of the bramble.
[1914] Jurine Ibid. t. ii. Gen. 11.
[1915] Ibid. Ord. ii. Gen. 1.
[1916] Ibid. t. iv. Gen. 23.
[1917] Ibid. Gen. 30-32.
[1918] Ibid. t. v. Gen. 40.
[1919] Plate X. Fig. 12, 15. c..
[1926] Leach on Eproboscideous Insects, Mem. Wern. Soc. 1817. t. xxv. f. 3, 5, 8, &c.
[1927] Ibid. t. xxvi. f. 7, 10, 13.
[1928] Insects with short elytra of course must fold their wings nearer the base than those with long ones.
[1931] See above, p. 613, and Chabrier Analyse, &c. 24.
[1936] Stoll Grillons, t. iii. c. f. 11-13.
[1937] Ibid. t. i. c. f. 1, 2.
[1941] Plate X. Fig. 4, 5. and XXVIII. Fig. 21, 22.
[1943] Ibid. Fig. 4, 5. and XXVIII. Fig. 21, 22.
[1951] In Gastropacha quercifolia, &c., amongst the Nocturnal Lepidoptera, these sinuses exist, in the upper wing ten, and in the lower nine, but without the folds.
[1954] For some uses of hairs, see above. p. 399—.
[1955] Analyse, 24. He seems to think that certain crooked hairs, in some wings, supply the place of folds. Ibid.
[1957] Reaum. iv. t. xxxix. f. 4-11.
[1958] A portion of the antenna of the insect here mentioned is figured Plate XII. Fig. 23.
[1959] De Geer i. 63—.
[1960] Reaum. i. 200.
[1961] Hoole's Leeuwenhoek. i. 63—.
[1962] De Geer has given 34 figures of different scales (i. t. iii. f. 28); and in Plate XXII. Fig. 6. a-w. 22 others, collected from Reaumur, are given.
[1963] Stoll Sauterelles à Sabre. Pterophylla ocellata t. i. ii., Cigales, Fulgora laternaria t. i. f. i., and F. serrata t. xxix. f. 170.
[1964] Ibid. Sauter. de Passage, Locusta Dux t. i. 5. L. carinata t. v. b. f. 16. L. cristata t. ix. b. f. 30. &c. &c.
[1966] Scopoli, Hubner.
[1968] N. Dict. d'Hist. Nat. viii. 257.
[1970] Sepp. I. i. t. vii. f. 6.
[1971] Rœmer Genera t. xxii. f. 2.
[1972] Sepp I. iii. t. ii. f. 7.
[1973] Ibid. i. t. vi. f. 7.
[1974] Rœmer ubi supr. t. xiv. f. 1.
[1975] Plates f., r., d†.
[1979] De Geer i. t. xx. f. 11.
[1980] Règne Animal. iii. 546.
[1981] Plate XXVII. Fig. 44, 45.
[1982] Vol. II. p. 312, 363, 365.
[1984] Anatom. Compar. i. 453.
[1986] Plate XXVII. Fig. 18, 19.
[1987] Mr. Montague describes the legs of Nycteribia, as dorsal (Linn. Trans. xi. 13); but Dr. Leach calls them lateral (Samouelle, 303).
[1988] N. Dict. d'Hist. Nat. xxviii. 247.
[1990] Plate XXVII. Fig. 58. M. Savigny affirms that these insects cannot have, and really have not, but one pair to each segment; only that the segments are alternately membranous and shelly, and that the former are concealed under the latter (Anim. sans Vertebr. I. i. 44.): but, pace tanti viri, I cannot discover that any suture separates these portions from each other: so that, admitting his theory, they must be regarded as two segments soldered together.
[1991] It is by this arrangement of the legs that Pachysoma is principally distinguished, as a subgenus, from Scarabæus McL.
[1993] Monogr. Ap. Angl. i. t. xi. Apis **. d. 2. α. f. 18. ii. 296—.
[1994] Oliv. Ins. 68. t. i. f. 8.
[1995] Ibid. n. 6. t. iv. f. 22.
[1997] This variety appears to differ very little from the Curculio imperialis of Fabricius and Olivier, except in the remarkable hairiness of its legs.
[1999] Oliv. Ins. n. 67. t. xx. f. 156.
[2000] See above, p. 591. Some physiologists have been of opinion, that in birds, what is called the thigh should properly be denominated the tibia, and that this last is really the tarsus. Illiger, Terminologie, 184. § 185. n. 1246.
[2001] Plates XIV. XV. XXVII. p.
[2002] Plate XV. Fig. 1. p´´, r´´.
[2004] L. Dufour, Descr. des six Arachn. &c.: Annales Generales, &c. 1820. 19. t. lxix. f. 7. d.
[2005] Plate XIV. XV. XXVII. q´´.
[2006] N. Dict. d'Hist. Nat. xvi. 195. xxvi. 157.
[2007] Plate XXVII. Fig. 12. b.
[2008] Plate XXVII. Fig. 20. q´´.
[2010] Plate XIV. XV. XXVII. r´´.
[2011] See above, p. 591, 662.
[2013] Linn. Trans. xii. t. xxi. f. 12.
[2014] Stoll Spectres t. xvi. f. 58, 59.
[2015] Ibid. t. xviii. f. 65.
[2016] Stoll Spectres t. vii. f. 25.
[2017] Ibid. t. viii. f. 30.
[2018] Ibid. ubi supr.
[2019] Ibid. t. x. f. 40.
[2020] Plate XIV. Fig. 5. This appearance of scales on the thighs is principally confined to this tribe.
[2022] Linn. Trans. xii. t. xxii. f. 16.
[2023] Plate XIV. Fig. 5, and XXVII. Fig. 15. r´´´.
[2024] Plates XIV. XV. XXVII. s´´.
[2025] Plate XXVII. Fig. 21. s´´´. M. Savigny (Anim. sans Vertèbr. 1. i. 46. Note b.) seems to think that this structure obtains in all his Apiropods; viz. the Octopod Aptera, Arachnida, and Myriapoda: but it seems to me evident only in the two tribes mentioned in the text.
[2026] Plate XXVII. Fig. 6, 16, 17. t´´´.
[2027] Plate XXVII. Fig. 15. r´´´.
[2030] Plate XXVII. Fig. 8. Thigh. a. Protuberance. b. Semicircular cavity, c. Ridge. Fig. 9. Tibia. a. Central cavity. b. Ridge. c. Exterior cavity.
[2033] Ibid. Fig. 15. Thigh of Locusta Leach, a. Sinus in which the condyle of the tibia works. Fig. 16. Tibia of Do. aa. Lateral condyles. b. Intermediate one.
[2034] Stoll Punaises, t. x. f. 67. t. xvi. f. 114.
[2035] Stoll Punaises, t. ii. f. 14. t. viii. f. 54. t. xxviii. f. 201. Plate XV. Fig. 2.
[2037] Oliv. Ins. n. 66. t. iii. f. 12. Compare Scarabæus longimanus, Ibid n. 3. t. iv. f. 27.
[2038] Stoll Punaises, t. iii. f. 20.
[2041] Oliv. Ins. n. 68. t. i. f. 8. comp. n. 67. t. xii. f. 83. and Plate XII. Fig. 25. a.
[2042] Vol. II. p. 365. and Plate XV. Fig. 5. 6.
[2043] See above, p. 433, Note1239. and 404, Note1172.
[2045] It is remarkable that in this tribe all the legs may be called raptorious, though the thighs are not incrassated, for they are armed with a double series or more of fine long spurs, which enable them to catch and retain their prey.
[2046] Plate XXVII. Fig. 35. v´´´. Philos. Trans. 1816. t. xix. f. 8, 9. See above, Vol. II. p. 331.
[2047] See above, p. 591, 667, &c.
[2048] Règne Animal, iii. 191. I have never had an opportunity to consult Bonelli's Observ. Entomolog. on the genus Carabus L. in the Memoirs of the Turin Academy.
[2049] Plate XV. Fig. 6. v´´´.
[2050] Coquebert Illustr. Ic. iii. t. xxi. f. 3. D.
[2051] Plate XXVII. Fig. 29. v´´´.
[2052] Most of Latreille's genera of ants are confirmed by differences in their spurs. Thus Formica is 1:1:1; Ponera 1:2:2 with the internal intermediate one pectinated: Myrmica 1:2:2 with all symmetrical, &c.
[2053] Plate XXVII. Fig. 29. v´´´.
[2055] Linn. Trans. iv. 200. Note a.
[2056] Plate XXVII. Fig. 32. v´´´.
[2058] Plates XIV. XV. XXVI. XXVII. a´´, t´´.
[2059] Plate XXVI. Fig. 44, 46, 47. a.
[2060] By Geoffroy—Hist. Ins. i. 58.
[2061] The Cleridæ, which M. Latreille has placed in the pentamerous section, vary considerably in the number of their tarsal joints. Thus in general in Thanasimus the tarsi are pentamerous; but in T. formicarius they appear to be heteromerous; and in Enoplium, Opilo, Clerus and Necrobia they are tetramerous. M. Latreille's expression, (N. Dict. d'Hist. Nat. vii. 172.) "le premier article etant fort court et caché sous le second," seems to indicate that there is a fifth joint in some of these, the first being concealed under the second; but I have never been able to discover it. Perhaps he reckoned the pulvillus as a joint?
[2062] The term heteromerous properly belongs to all insects in which the different pairs of tarsi vary inter se in the number of their joints, and it is here used in that large sense.
[2063] These three genera appear really to have only six legs, since the pedipalps or maxillary legs are not armed with claws, while the real representatives of the legs, or three last pair, are so distinguished. In Phrynus and Thelyphona the anterior pair are chelate; but in Galeodes they are pediform, as in the Araneidæ, and the great chelæ are the mandibles.
[2064] Plate XXVI. Fig. 47, 48. d *.
[2065] Plate XXVI. Fig. 49. s.. a.
[2067] Dr. Leach says there are three joints in this tribe. Nat. Misc. iii. 80.
[2068] From De Geer's description this insect seems related to Agathidium (iv. 221—. t. viii. f. 21-23). M. Leclerck de Laval discovered it to be monomerous. Règne Animal, iii. 365.
[2070] See above, p. 311. Note738.
[2071] Melolontha sericea and aurulenta. Linn. Trans. xii. 463. 400. belong to this subgenus.
[2074] Plate XIV. Fig. 7. t´´.
[2079] Mon. Ap. Angl. i. t. xii. neut. f. 20.
[2080] Plate XXVII. Fig. 44. s..
[2081] Plate XXVII. Fig. 26. w´´´.
[2083] L. Dufour Descr. de six Arachnides. Annales, &c. 1820. 19.
[2084] Plate XXVII. Fig. 51. is the posterior claw of Hoplia.
[2092] This structure is not general in this genus.
[2104] Linn. Trans. xii. t. xxii. f. 1.
[2105] For other instances of this structure, see above, p. 336.
[2106] De Geer, iii. 7.
[2107] Ibid. vii. 84. Plate XXVII. Fig. 60, 63.
[2111] Linn. Trans. xii. t. xxi. f. 3.
[2113] Plate XXVII. Fig. 54. Philos. Trans. 1816. t. xviii. f. 9-11.
[2114] Plate XXVII. Fig. 55. t.
[2117] Philos. Trans. 1816. t. xx. f. 9, 12-15.
[2118] Philos. Trans. 1816. t. xx. f. 4, 11.
[2121] Amouroux Insectes Venimeux, 44.
[2122] Observations Nouvelles, &c. Mém. du Mus. viii. 177.
[2123] N. Dict. d'Hist. Nat. xii. 370.
[2124] Descr. de six Arachnides, &c. Annales Gen. des Scienc. Phys. 1820. 19. t. lxix. f. 7. d.
[2125] Anatom. Compar. i. 450.
[2126] Anatom. Compar. i. 451.
[2127] De Geer notices something of the kind in Cimbex femorata. ii. 947.
[2128] It was omitted to be observed, when the supposed pneumatic pouches in the genus Vespa were mentioned (see above, p. 585), that they have also a very conspicuous metaphragm, as probably have most Hymenoptera, to which the muscles that move the wings are attached.
[2129] Plate IX. Fig. 13. F´ is the tendon, G´ the aperture in the abdomen C, and a, the aperture in the trunk B.
[2130] Plate XV. Fig. 11. Linn. Trans. xii. t. xxii. f. 16.
[2131] Surely these Epeiræ, of so different a habit from the rest, form a distinct genus?
[2133] There is reason to suppose that in Chelonus and the Chrysidæ several segments are retracted within the abdomen, and if the cavity of its under side in the latter be examined, it will be discovered that the epigastrium is divided in the middle into two pieces, and that both the sides of this and the following segments are covered by three narrow accessory plates, one to each, the last being the shortest. De Geer (ii. 833.) describes Chrysis ignita as having four abdominal rings; but this is only in appearance, there being really only three. This appearance is produced by the apex of the last dorsal segment being more depressed and marked with several deep little excavations that look like holes. In some species of Stilbum this segment consists as it were of three ridges or steps.
[2134] In this genus the ventral segments are replaced by a long narrow central plate, succeeded by a minute one.
[2135] In this genus the bed of the posterior coxæ appears to consist of two segments, which are beautifully fringed with parallel short bristles.
[2136] Anatom. Compar. i. 451.
[2137] Plate VIII. Fig. 9. A´´, B´.
[2140] Reaum. v. t. xvii. f. 14. a. a.
[2141] Linn. Trans. v. t. xii. f. 15.
[2142] Daldorf (Asiatic Society's Trans. vii.) has divided Geotrupes into two families, one with the podex covered (G. vernalis, &c.) which he calls modesti, the other with it uncovered (G. stercorarius, &c.) which he calls obscæni.
[2145] Sur le Vol des Ins. c. i. Addend. 299.
[2146] In Dytiscus marginalis the upper side of the margin of the Hypochondria is curiously cut into transverse corrugations.
[2147] Plate VIII. Fig. 6. C´.
[2150] Linn. Trans. xi. t. ix. f. 15. b.
[2153] Rœmer. Genera, &c. t. xxiv. f. 4.
[2154] Stoll Spectr. t. vii.
[2157] This tubercle I find only in a specimen from Sweden, sent to me by Major Gyllenhal, but not in any British one I possess. In this specimen the declivity before mentioned (see above, p. 709.) is observable in the first segment, but in the others it is formed by the second.
[2159] Punaises, t. xiii. f. 84.
[2160] Stoll Punaises, t. xiii. f. 101.
[2161] Ibid. t. xvii. f. 117.
[2162] Ibid. t. xxxvi. f. 253.
[2164] De Geer, vi. 260. t. xv. f. 8. d.
[2166] Plate XXIX. Fig. 22. is part of the back of the abdomen of the pupa of a Pentatoma. a the pseudo-spiracle, b the connecting corrugations.
[2169] Treviranus. Arachnid. 23—.
[2170] See above, p. 702, 706.
[2171] Plate XXIX. Fig. 26. represents one of them.
[2173] Plate XV. Fig. 14. M´´. De Geer, vii. t. ii. f. 5, 10, 21.
[2174] Ibid. t. iii. f. 4, 14.
[2176] Plate XVIII. Fig. 5. a.
[2177] Plate XXIX. Fig. 3, 4. De Geer, ii. t. xvii. f. 12. and t. xviii. f. 2.
[2178] Ibid. t. xvi. f. 8-13.
[2179] Plate XV. Fig. 16. S´´.
[2181] Philos. Trans. 1818. t. xxii.
[2182] N. Dict. d'Hist. Nat. ii. 275—. Hoole's Leeuwenh. i. t. ii. f. 19. I.
[2183] Leeuwenh. Epist. 17. Octobr. 1687. f. 10. C.
[2184] Hoole's Leeuwenh. i. t. v. f. 12, 13.
[2185] Treviranus, Arach. 4.
[2186] Treviranus, ubi supr. 14.
[2187] In my specimen including the first joint there are twenty, and some seem to have been broken off. In Rœmer's figure (Genera, t. xxix. f. 11.) there are only ten. Perhaps they vary in number according to the age of the animal.
[2188] Plate XV. Fig. 13, 16, 17.
[2190] I have heard it stated upon good authority that 40,000 species of insects are already known, as preserved in collections. How great, then, must be the number existing in this whole globe!
[2191] 2 Cor. ii. 16.
[2192] Recourse must be had to the synoptical table of the nomenclature of the parts of the external crust of Insects (Vol. III. p. 354) for the explanation of the reference letters not here given.
[2193] Vol. I. p. 394—. III. p. 355—, 394—. IV. p. 305—.
[2195] Probably e´ is resolvable into two pieces.
[2196] Vol. III. p. 367—, 529—. IV. p. 326—.
[2197] Vol. III. p. 367—, 529—. IV. p. 326—.
[2198] Vol. II. p. 347—. III. p. 372—, 595—.
[2199] Vol. III. p. 366, 510—. IV. p. 316—.
[2200] Vol. IV. p. 307. iii, iv. 309. b. 310. d. 313. viii. 328, 334.
[2201] Vol. IV. p. 286—, 338, 345—.
[2203] Vol. IV. p. 351. ii. III. Letter XXXII. Vol. I. p. 65—.
[2204] Ubi supr.

















{kind=link}
{kind=link}
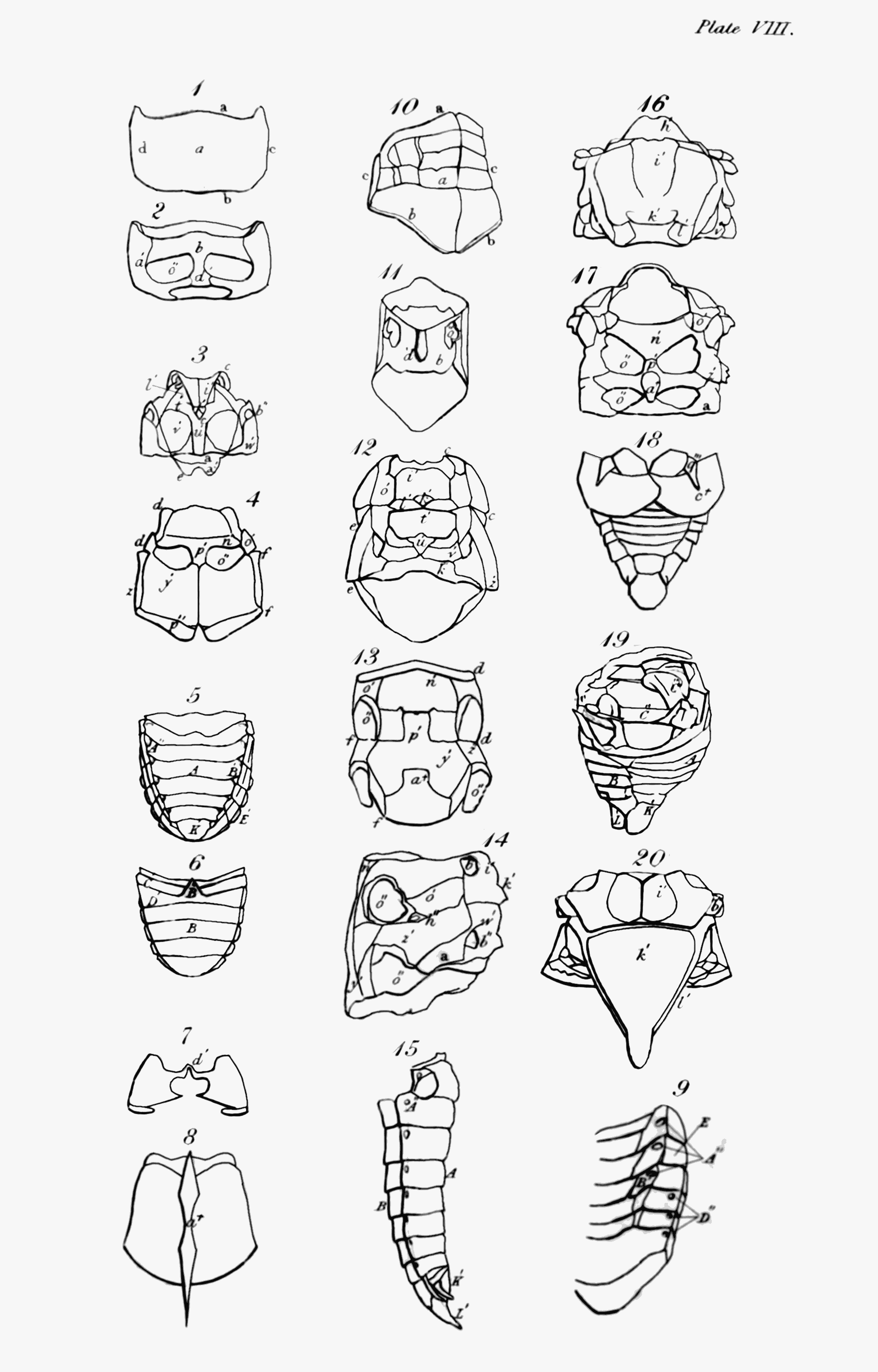{kind=link}
{kind=link}
{kind=link}
{kind=link}
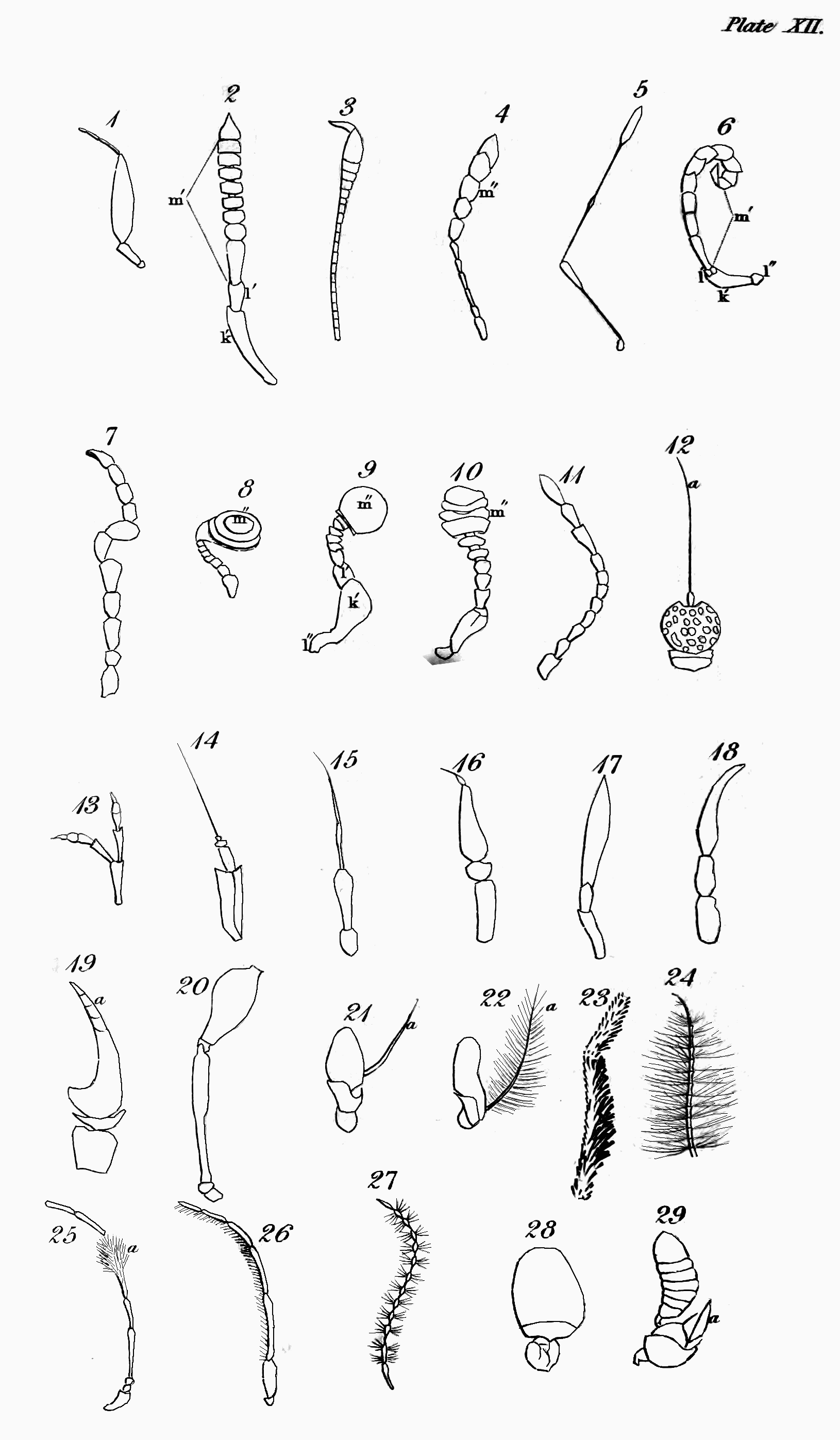{kind=link}
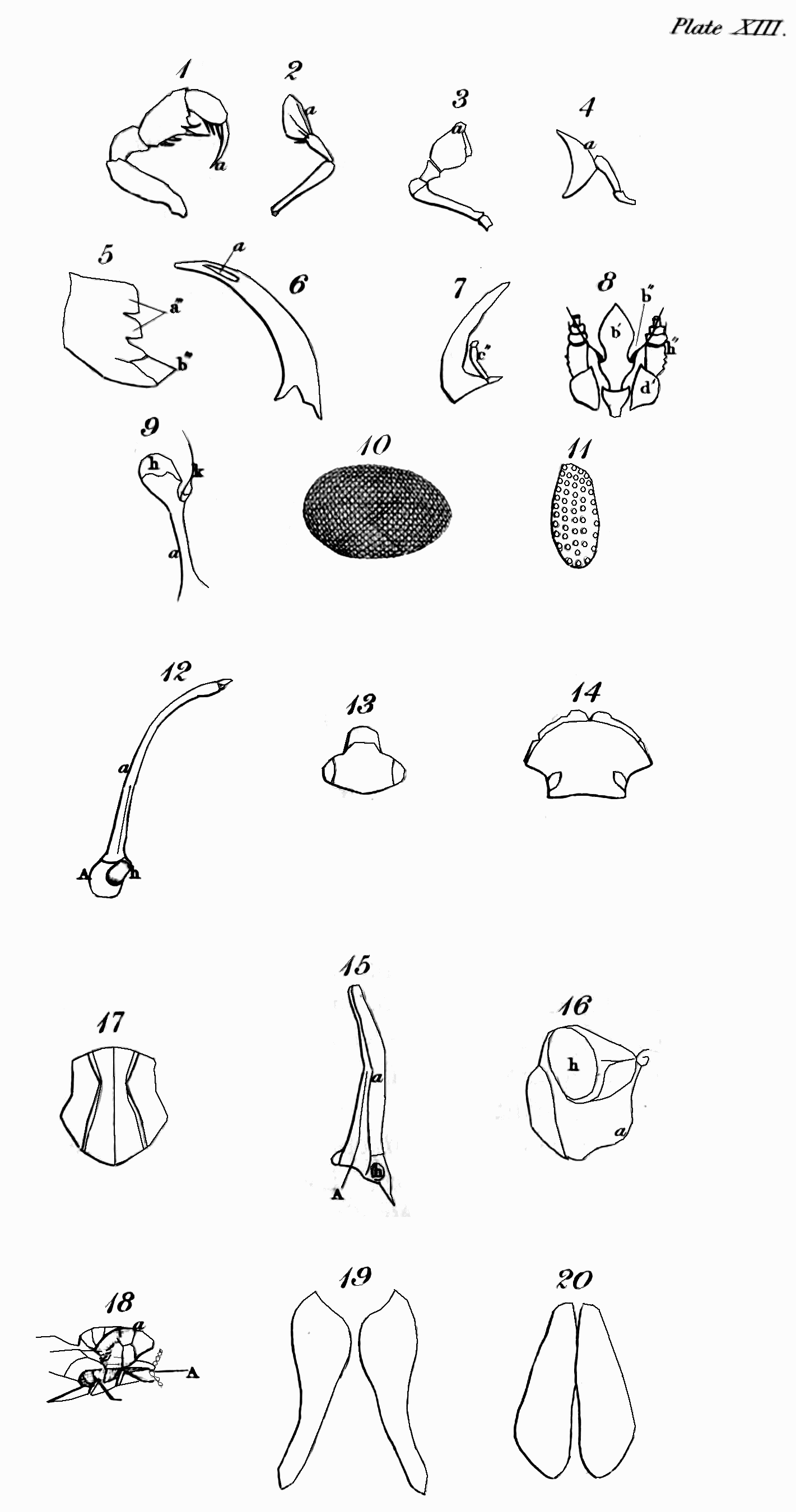{kind=link}
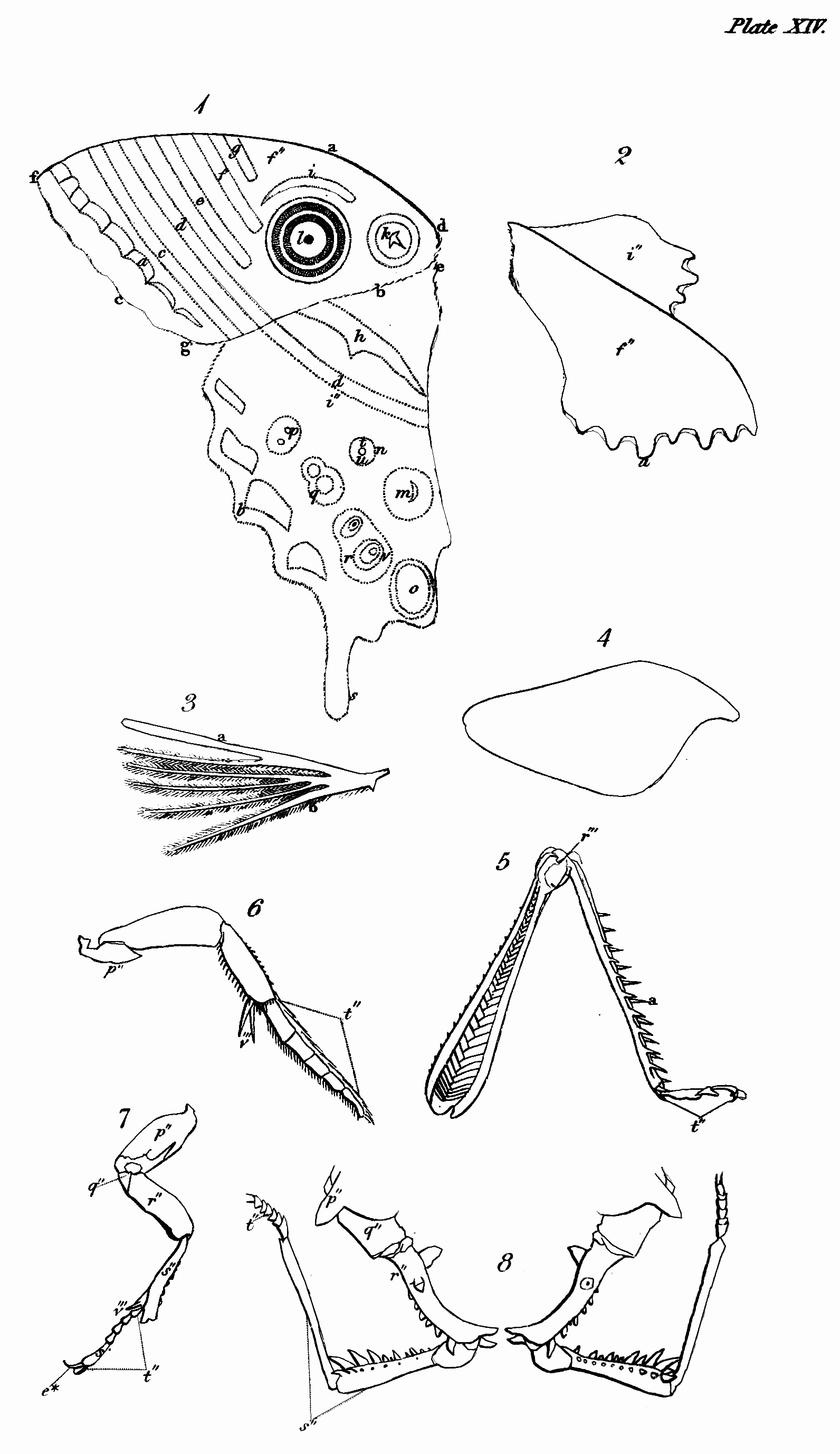{kind=link}
{kind=link}
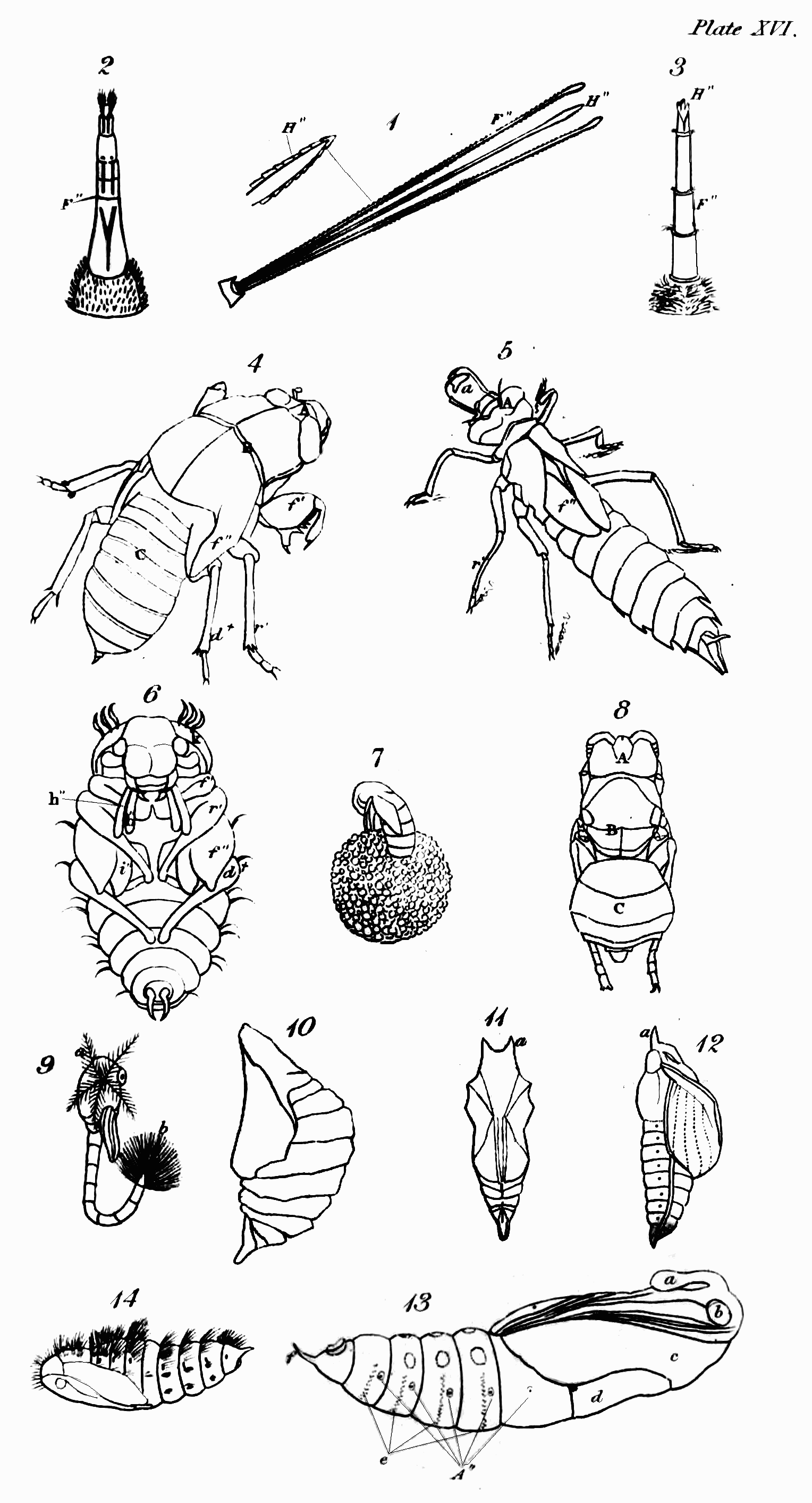{kind=link}
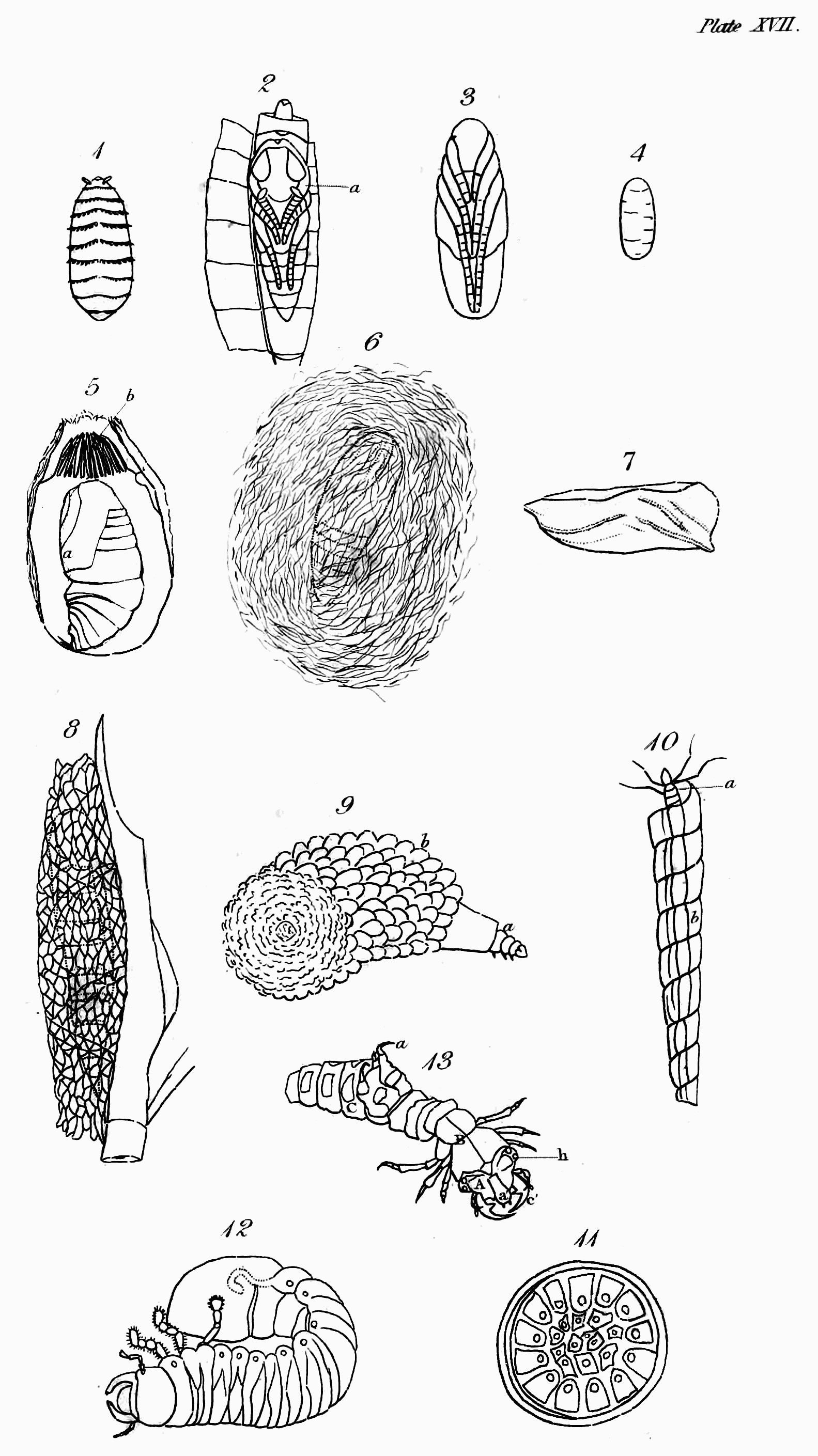{kind=link}
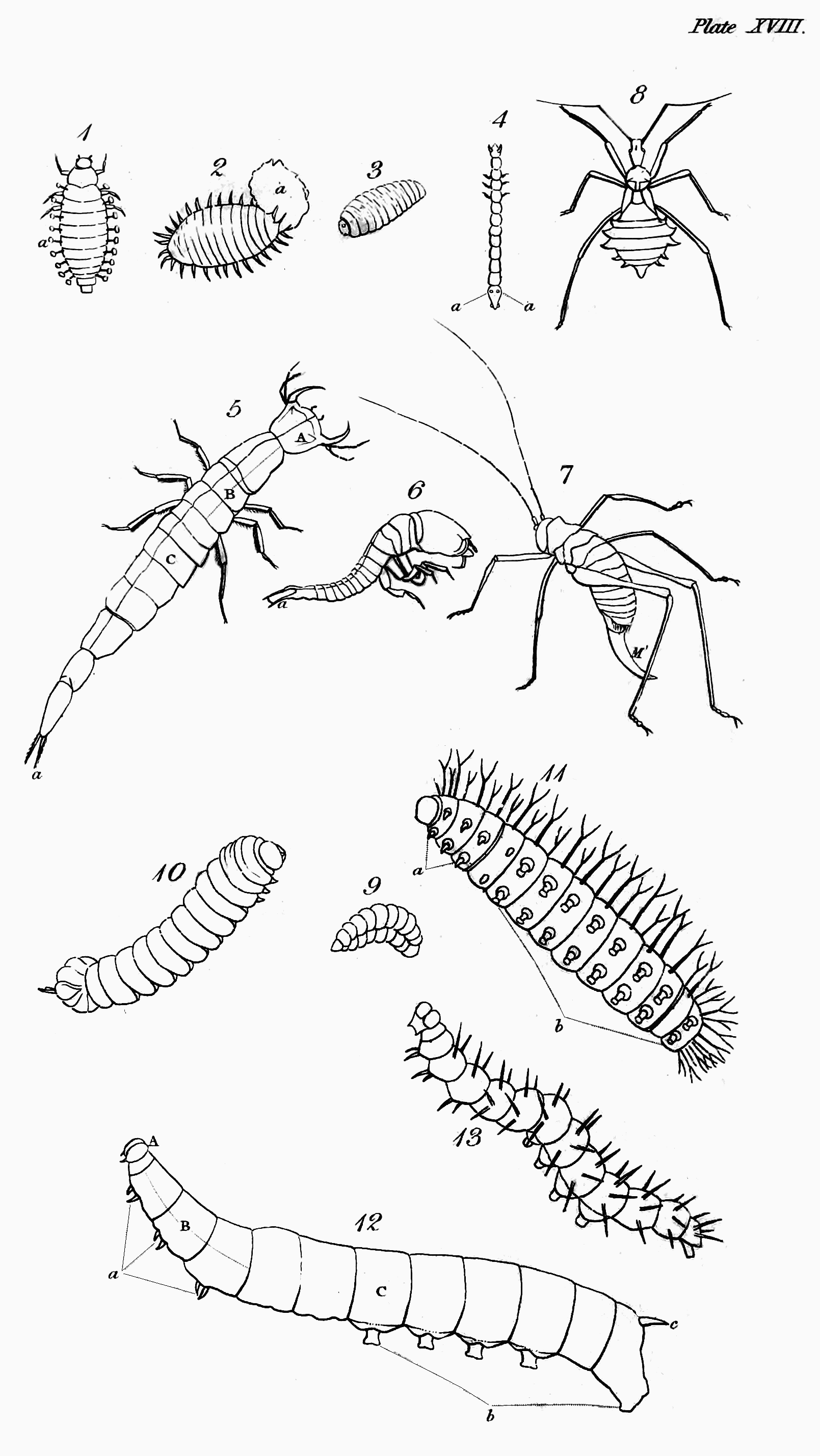{kind=link}
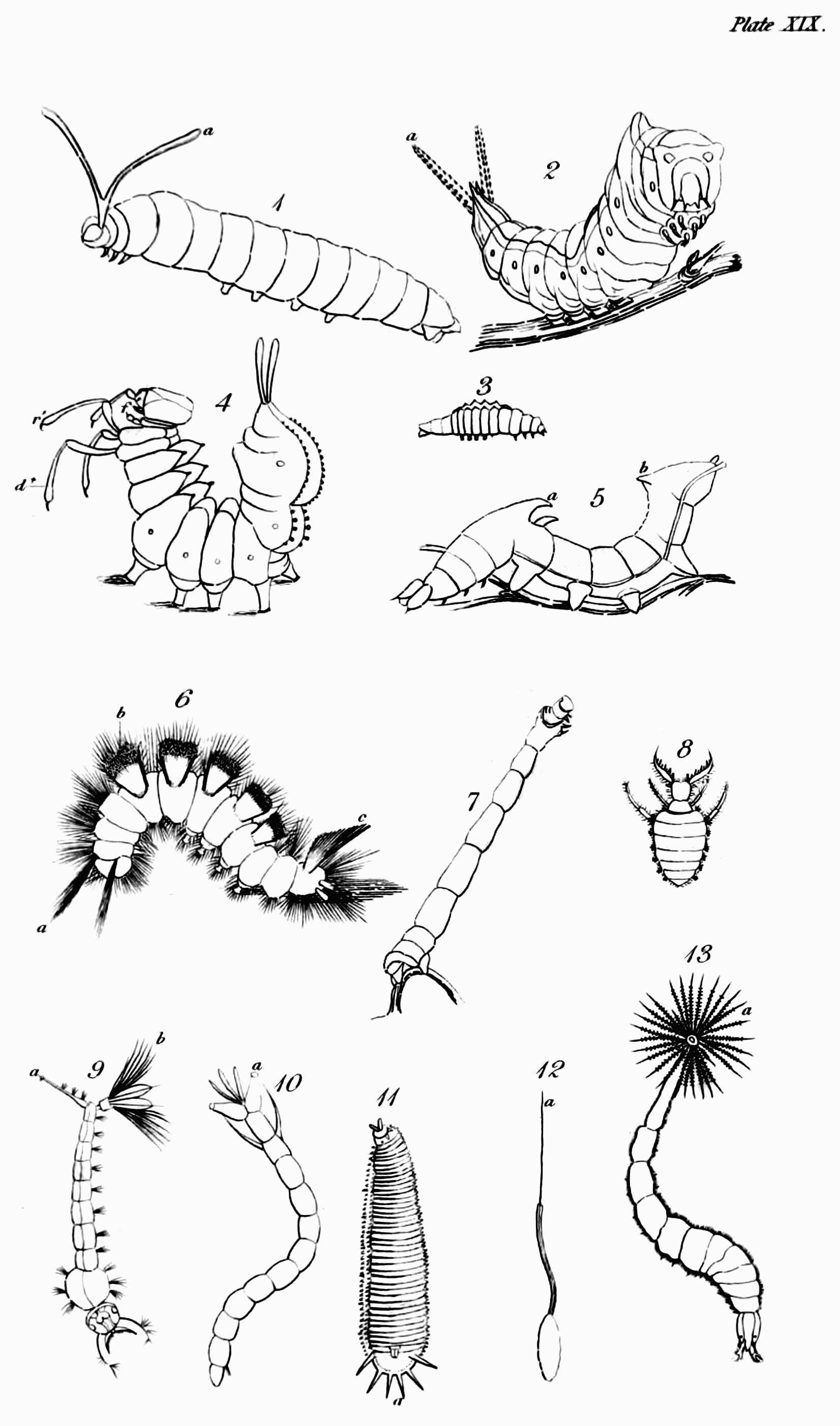{kind=link}
{kind=link}