The Project Gutenberg EBook of The Animal Parasites of Man, by
H. B. Fantham and J. W. W. Stephens and F. V. Theobald
This eBook is for the use of anyone anywhere in the United States and most
other parts of the world at no cost and with almost no restrictions
whatsoever. You may copy it, give it away or re-use it under the terms of
the Project Gutenberg License included with this eBook or online at
www.gutenberg.org. If you are not located in the United States, you'll have
to check the laws of the country where you are located before using this ebook.
Title: The Animal Parasites of Man
Author: H. B. Fantham
J. W. W. Stephens
F. V. Theobald
Contributor: Max Braun
Otto Seifert
Release Date: August 17, 2018 [EBook #57713]
Language: English
Character set encoding: UTF-8
*** START OF THIS PROJECT GUTENBERG EBOOK THE ANIMAL PARASITES OF MAN ***
Produced by Thiers Halliwell, Chris Curnow and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive)
Transcriber’s notes:
Small viewing devices might not display this e-book correctly
because of the complexity of the text, its length, and the presence
within it of some uncommon typographic characters. It is best viewed on
a large screen with character encoding set to Unicode (UTF-8).
The text has mostly been preserved in its original form, including
inconsistent italics, capitalisation, hyphenation and diacritics;
however, some infelicities have been corrected silently, e.g.
missing/inconsistent punctuation, parentheses and spacing. Several
footnote markers have been inserted where they were missing from the
text. The very long table of contents has somewhat inconsistent
levels of indentation that don’t correspond accurately with headings
used in the text. Some illustrations have been moved nearer to the
relevant text and their locations may differ from those specified
in the list of illustrations. A few illustrations that lacked an
identifying number have been correctly numbered. The book contains
extensive sections of quoted text but these are identified by square
brackets rather than by traditional quotation marks. Several short
passages of text were inserted by the printer at the last minute, and
these are enclosed in lightly shaded text boxes. Two different sized
fonts were used to print the original body text, a feature that is
replicated in this e-transcription.
A black underline indicates a hyperlink to a page, illustration
or footnote. A red dashed underline indicates the presence
of a concealed comment that can be viewed by hovering the
mouse pointer over the underlined
text. Page numbers are shown in the
right margin and footnotes are listed at the end. Footnotes are listed at the end.
A
list of spelling corrections and
inconsistencies is appended at the end of the book.
The cover image of the book was created by the
transcriber and is placed in the public domain.
THE ANIMAL PARASITES OF MAN
BY
H. B. FANTHAM, M.A.Cantab., D.Sc.Lond.
Lecturer on Parasitology, Liverpool School of Tropical Medicine; Sectional Editor in
Protozoology, “Tropical Diseases Bulletin,” London, etc.
J. W. W. STEPHENS, M.D.Cantab., D.P.H.
Sir Alfred Jones Professor of Tropical Medicine, Liverpool University, etc.
AND
F. V. THEOBALD, M.A.Cantab., F.E.S., Hon. F.R.H.S.
Professor of Agricultural Zoology, London University; Vice-Principal and Zoologist of the
South-eastern Agricultural College; Mary Kingsley Medallist; Grande Médaille Geoffroy
St. Hilaire, Soc. Nat. d’Acclim. de France, etc.
PARTLY ADAPTED FROM
Dr. Max Braun’s “Die Tierischen Parasiten des Menschen” (4th Edition, 1908) and an
Appendix by Dr. Otto Seifert.
NEW YORK
WILLIAM WOOD AND COMPANY
MCMXX.
PREFACE.
The English edition of Braun’s “Die Tierischen Parasiten des
Menschen,” produced in 1906, being out of print, the publishers
decided to issue another edition based on the translation of Braun’s
fourth German edition, which appeared in 1908, to which had been
added an appendix, by Dr. Otto Seifert on Treatment, etc.
When the work was considered with a view to a new edition,
it was found that a vast amount of new matter had to be incorporated,
numerous alterations essential for bringing it up to date were necessitated,
and many omissions were inevitable. The result is that parts of
the book have been rewritten, and, apart from early historical references,
the work of Braun has disappeared. This is more particularly the case
with the Protozoa section of the present work. The numerous additions,
due to the great output of scientific literature and other delays in
publication, have led to the book being somewhat less homogeneous
than we desired, and have necessitated the use of appendices to
allow of the presentation of new facts only recently ascertained.
Many new illustrations have been added or substituted for older, less
detailed ones. Some of these new figures were drawn specially for
this book.
The first section, on the Protozoa, has been written by Dr. Fantham,
there being little of the original text left except parts of the historical
portions, and thus the section on Protozoa must be considered as
new. The second section, on Worms (except the Acanthocephala,
Gordiidæ and Hirudinea), has been remodelled by Professor Stephens
to such an extent that this, too, must not be looked upon as a
translation of Braun’s book. With regard to the Arthropoda,
much remains as in the last English edition, but some new matter
added by Braun in his fourth German edition is included, and much
new matter by Mr. Theobald has been incorporated. As regards
the Appendix by Dr. Seifert, the first section has been remodelled,
but the sections on the Helminthes and the Arthropoda are practically
translations of the original.
The authors desire to express their thanks to Miss A. Porter, D.Sc.,
J. P. Sharples, Esq., B.A., M.R.C.S., and H. F. Carter, Esq., F.E.S.,
for valuable help. They also wish to thank the authors, editors, and
publishers of several manuals and journals for their courtesy in
allowing the reproduction of certain of their illustrations. In this
connection mention must be made more particularly of Professor
Castellani, Dr. Chalmers, Professor Doflein, Dr. Leiper, the late
Professor Minchin, Professor Nuttall, Dr. Wenyon, Mr. Edw. Arnold,
Messrs. Baillière, Tindall and Cox, Messrs. Black, Messrs. Cassell,
Dr. Gustav Fischer, Messrs. Heinemann, the Cambridge University
Press, the Editors of the Annals of Tropical Medicine and Parasitology,
the Editors of the Journal of Experimental Medicine, and the Editor
of the Tropical Diseases Bulletin.
H. B. F.
J. W. W. S.
F. V. T.
December, 1915.
| PAGE |
| PREFACE | iii |
| ERRATA | xxxii |
| ON PARASITES IN GENERAL | |
| Occasional and Permanent Parasitism | |
| Entozoa, Endoparasites, Helminthes, Turbellaria | |
| Hermaphroditism | |
| Fertility of Parasites | |
| Transmigrations | |
| Commensals, Mutualists | |
| Incidental and Pseudo-parasites | |
| The Influence of Parasites on the Host | |
| Origin of Parasites | |
| Derivation of Parasites | |
| Change of Host | |
| Literature | |
|
| THE ANIMAL PARASITES OF MAN | |
| A. Protozoa | |
| Classification of the Protozoa. |
| Class I. Sarcodina | |
| Order. Amœbina | |
| Foraminifera | |
| Heliozoa | |
| Radiolaria | |
| Class II. Mastigophora | |
| III. Sporozoa | |
| Sub-class 1. Telosporidia | |
| Order. Gregarinida | |
| Coccidiidea | |
| Hæmosporidia | |
| Sub-class 2. Neosporidia | |
| Order. Myxosporidia | |
| Microsporidia | |
| Sarcosporidia | |
| Haplosporidia | |
| Class IV. Infusoria | |
| V. Suctoria | |
| Class I. Sarcodina, Bütschli, 1882 | |
| Order. Amœbina, Ehrenberg | |
| A. Human Intestinal Amœbæ | |
| Entamœba coli, Lösch, 1875, emend. Schaudinn, 1903 | |
| Entamœba histolytica, Schaudinn, 1903 | |
| Entamœba tetragena, Viereck, 1907 | |
| Noc’s Entamœba, 1909 | |
| Entamœba buccalis, Prowazek, 1904 | |
| Entamœba undulans, Castellani, 1905 | |
| Entamœba kartulisi, Doflein, 1901 | |
| Amœba gingivalis, A. buccalis, A. dentalis | |
| Genus. Paramœba, Schaudinn, 1896 | |
| Paramœba (Craigia) hominis, Craig, 1906 | |
| B. Amœbæ from other Organs | |
| Entamœba pulmonalis, Artault, 1898 | |
| Amœba urogenitalis, Baelz, 1883 | |
| Amœba miurai, Ijima, 1898 | |
| Appendix: “Rhizopods in Poliomyelitis Acuta” | |
| Order. Foraminifera, d’Orbigny | |
| Sub-order. Monothalamia (Testaceous Amœbæ) | |
| Genus. Chlamydophrys, Cienkowski, 1876 | |
| Chlamydophrys enchelys, Ehrenberg | |
| Leydenia gemmipara, Schaudinn, 1896 | |
| Class II. Mastigophora, Diesing | |
| Sub-class. Flagellata, Cohn emend. Bütschli | |
| Order. Polymastigina, Blochmann | |
| Genus. Trichomonas, Donné, 1837 | |
| Trichomonas vaginalis, Donné | |
| Trichomonas intestinalis, R. Leuckart, 1879 = Trichomonas hominis, Davaine, 1854 | |
| Genus. Tetramitus, Perty, 1852 | |
| Tetramitus mesnili, Wenyon, 1910 | |
| Genus. Lamblia, R. Blanchard, 1888 | |
| Lamblia intestinalis, Lambl, 1859 | |
| Order. Protomonadina, Blochmann | |
| Family. Cercomonadidæ, Kent emend. Bütschli | |
| Genus. Cercomonas, Dujardin emend. Bütschli | |
| Cercomonas hominis, Davaine, 1854 | |
| Monas pyophila, R. Blanchard, 1895 | |
| Family. Bodonidæ, Bütschli | |
| Genus. Prowazekia, Hartmann and Chagas, 1910 | |
| Prowazekia urinaria, Hassall, 1859 | |
| Prowazekia asiatica, Castellani and Chalmers, 1910 | |
| Prowazekia javanensis, Flu, 1912 | |
| Prowazekia cruzi, Hartmann and Chagas, 1910 | |
| Prowazekia weinbergi, Mathis and Léger, 1910 | |
| Prowazekia parva, Nägler, 1910 | |
| Family. Trypanosomidæ, Doflein | |
| Genus. Trypanosoma, Gruby, 1843 | |
| Historical | |
| General | |
| Morphology | |
| Trypanosoma gambiense, Dutton, 1902 | |
| Trypanosoma nigeriense, Macfie, 1913 | |
| Trypanosoma rhodesiense, Stephens and Fantham, 1910 | |
| General Note on Trypanosomes with Posterior Nuclei | |
| Trypanosoma cruzi, Chagas, 1909 | |
| Trypanosoma lewisi, Kent, 1881 | |
| Trypanosoma brucei, Plimmer and Bradford, 1899 | |
| Trypanosoma evansi, Steel, 1885 | |
| Trypanosoma equinum, Voges, 1901 | |
| Trypanosoma equiperdum, Doflein, 1901 | |
| Trypanosoma theileri, Bruce, 1902 | |
| Trypanosoma hippicum, Darling, 1910 | |
| Endotrypanum schaudinni, Mesnil and Brimont, 1908 | |
| Trypanosoma boylei, Lafont, 1912 | |
| Monomorphic Trypanosomes | |
| Trypanosoma vivax, Ziemann, 1905 | |
| Trypanosoma capræ, Kleine, 1910 | 100 |
| Trypanosoma congolense, Broden, 1904 | 100 |
| Trypanosoma simiæ, Bruce, 1912 | 100 |
| Trypanosoma uniforme, Bruce, 1910 | 101 |
| General Note on Development of Trypanosomes in Glossina | 101 |
| Adaptation of Trypanosomes | 101 |
| Genus. Herpetomonas, Saville Kent, 1881 | 102 |
| Genus. Crithidia, Léger, 1902, emend. Patton, 1908 | 104 |
| Genus. Leishmania, Ross, 1903 | 104 |
| Leishmania donovani, Laveran and Mesnil, 1903 | 105 |
| Leishmania tropica, Wright, 1903 | 107 |
| Leishmania infantum, Nicolle, 1908 | 109 |
| Genus. Histoplasma, Darling, 1906 | 112 |
| Genus. Toxoplasma, Nicolle and Manceaux, 1908 | 112 |
| The Spirochætes | 114 |
| The Spirochætes of the Blood | 116 |
| Spirochæta duttoni, Novy and Knapp, 1906 | 116 |
| Spirochæta gallinarum, Stephens and Christophers, 1905 (= Spirochæta marchouxi, Nuttall, 1905) | 119 |
| Spirochæta recurrentis, Lebert, 1874 | 120 |
| Spirochæta rossii, Nuttall, 1908 | 122 |
| Spirochæta novyi, Schellack, 1907 | 122 |
| Spirochæta carteri, Mackie and Manson, 1907 | 122 |
| Spirochæta berbera, Sergent and Foley, 1910 | 122 |
| Other Human Spirochætes | 122 |
| Some Animal Spirochætes | 122 |
| Treponemata | 124 |
| Treponema pallidum, Schaudinn, 1905 | 124 |
| Treponema pertenue, Castellani, 1905 | 127 |
| Class III. Sporozoa, Leuckart, 1879 | 128 |
| Sub-class. Telosporidia, Schaudinn | 129 |
| Order. Gregarinida, Aimé Schneider emend. Doflein | 129 |
| Order. Coccidiidea | 135 |
| Genus. Eimeria, Aimé Schneider, 1875 | 142 |
| Eimeria avium, Silvestrini and Rivolta | 142 |
| Eimeria stiedæ, Lindemann, 1865 | 145 |
| (a) Human Hepatic Coccidiosis | 148 |
| (b) Human Intestinal Coccidiosis | 148 |
| (c) Doubtful Cases | 149 |
| Genus. Isospora, Aimé Schneider, 1881 | 149 |
| Isospora bigemina, Stiles, 1891 | 149 |
| Doubtful Species | 150 |
| Order. Hæmosporidia, Danilewsky emend. Schaudinn | 151 |
| The Malarial Parasites of Man | 155 |
| Development of the Malarial Parasites of Man | 159 |
| The Species of the Malarial Parasites of Man | 164 |
| Plasmodium vivax, Grassi and Feletti, 1890 | 164 |
| Plasmodium malariæ, Laveran | 166 |
| Laverania malariæ, Grassi and Feletti, 1890 (= Plasmodium falciparum, Welch, 1897) | 167 |
| Plasmodium relictum, Sergent, 1907 (in birds) | 170 |
| Cultivation of Malarial Parasites | 170 |
| Differential Characters of the Human Malarial Parasites | 171 |
| Family. Piroplasmidæ, França, 1909 | 172 |
| Genus. Babesia, Starcovici, 1893 | 174 |
| Genus. Theileria, Bettencourt, França and Borges, 1907 | 178 |
| Theileria parva, Theiler, 1903 | 178 |
| Theileria mutans, Theiler, 1907 | 180 |
| Genus. Anaplasma, Theiler, 1910 | 180 |
| Genus. Paraplasma, Seidelin, 1911 | 180 |
| Sub-class. Neosporidia, Schaudinn | 181 |
| Order. Myxosporidia, Bütschli | 181 |
| Order. Microsporidia, Balbiani | 184 |
| Order. Actinomyxidia, Stolč. | 187 |
| Order. Sarcosporidia, Balbiani | 187 |
| Sarcosporidia observed in Man | 193 |
| Order. Haplosporidia, Caullery and Mesnil, 1899 | 194 |
| Rhinosporidium kinealyi, Minchin and Fantham, 1905 | 195 |
| Class IV. Infusoria, Ledermüller, 1763 | 198 |
| Genus. Balantidium, Claparède et Lachmann | 200 |
| Balantidium coli, Malmsten, 1857 | 200 |
| Balantidium minutum, Schaudinn, 1899 | 204 |
| Genus. Nyctotherus, Leidy, 1849 | 204 |
| Nyctotherus faba, Schaudinn, 1899 | 205 |
| Nyctotherus giganteus, P. Krause, 1906 | 205 |
| [Nyctotherus] africanus, Castellani, 1905 | 206 |
| The Chlamydozoa | 207 |
| Protozoa Incertæ Sedis | 210 |
| Sergentella hominis, Brumpt, 1910 | 210 |
|
| B. Platyhelminthes (or Flat Worms) | 211 |
| Classification of the Platyhelminthes. |
| Class I. Turbellaria (or Eddy Worms) | 212 |
| Order 1. Rhabdocœlida | 212 |
| 2. Tricladida | 212 |
| 3. Polycladida | 212 |
| Class II. Trematoda (Sucking Worms) | 212 |
| III. Cestoda (Tapeworms) | 212 |
| Class II. Trematoda, Rud. | 212 |
| Development of the Trematodes | 222 |
| Biology | 229 |
| Classification of the Trematodes of Man. |
| Order. Digenea, v. Beneden, 1858 | 230 |
| Sub-order. Prostomata, Odhner, 1905 | 230 |
| Group. Amphistomata, Rudolphi, 1801, ep., Nitzsch, 1819 | 230 |
| Family. Paramphistomidæ, Fischoeder, 1901 | 231 |
| Sub-family. Paramphistominæ, Fischoeder, 1901 | 231 |
| Cladorchiinæ, Fischoeder, 1901 | 231 |
| Family. Gastrodisciidæ, Stiles and Goldberger, 1910 | 231 |
| Group. Distomata, Retzius, 1782 | 231 |
| Family. Fasciolidæ, Railliet, 1895 | 231 |
| Sub-family. Fasciolinæ, Odhner, 1910 | 231 |
| Fasciolopsinæ, Odhner, 1910 | 231 |
| Family. Opisthorchiidæ, Braun, 1901, emend, auctor. | 232 |
| Sub-family. Opisthorchiinæ, Looss, 1899, emend, auctor. | 232 |
| Metorchiinæ, Lühe, 1909 | 232 |
| Family. Dicrocœliidæ, Odhner, 1910 | 232 |
| Heterophyiidæ, Odhner, 1914 | 232 |
| Troglotremidæ, Odhner, 1914 | 232 |
| Echinostomidæ, Looss, 1902 | 233 |
| Sub-family. Echinostominæ, Looss, 1899 | 233 |
| Himasthlinæ, Odhner, 1910 | 233 |
| Family. Schistosomidæ, Looss, 1899 | 233 |
| The Trematodes observed in Man | 234 |
| Family. Paramphistomidæ, Stiles and Goldberger, emend. 1910 | 234 |
| Sub-family. Cladorchiinæ, Fischoeder, 1901 | 234 |
| Genus. Watsonius, Stiles and Goldberger, 1910 | 234 |
| Watsonius watsoni, Stiles and Goldberger, 1910 | 234 |
| Family. Gastrodisciidæ | 236 |
| Genus. Gastrodiscus, Lkt., 1877 | 236 |
| Gastrodiscus hominis, Lewis and McConnell, 1876 | 236 |
| Family. Fasciolidæ, Railliet, 1895 | 237 |
| Sub-family. Fasciolinæ, Odhner, 1910 | 237 |
| Genus. Fasciola, L., 1758 | 237 |
| Fasciola hepatica, L., 1758 | 237 |
| Halzoun | 242 |
| Fasciola gigantica, Cobbold, 1856 | 244 |
| Sub-family. Fasciolopsinæ, Odhner, 1910 | 245 |
| Genus. Fasciolopsis, Looss, 1898 | 245 |
| Fasciolopsis buski, Lank., 1857 | 245 |
| Fasciolopsis rathouisi, Ward, 1903 | 246 |
| Fasciolopsis goddardi, Ward, 1910 | 247 |
| Fasciolopsis fülleborni, Rodenwaldt, 1909 | 247 |
| Family. Troglotremidæ, Odhner, 1914 | 249 |
| Genus. Paragonimus, Braun, 1899 | 249 |
| Paragonimus ringeri, Cobb., 1880 | 249 |
| Family. Opisthorchiidæ, Braun, 1901 | 252 |
| Sub-family. Opisthorchiinæ, Looss, 1899 | 252 |
| Genus. Opisthorchis, R. Blanch., 1845 | 252 |
| Opisthorchis felineus, Riv., 1885 | 252 |
| Genus. Paropisthorchis, Stephens, 1912 | 255 |
| Paropisthorchis caninus, Barker, 1912 | 255 |
| Genus. Amphimerus, Barker, 1912 (?) | 257 |
| Amphimerus noverca, Barker, 1912 (?) | 258 |
| Genus. Clonorchis, Looss, 1907 | 258 |
| Clonorchis sinensis, Cobbold, 1875 | 258 |
| Clonorchis endemicus, Baelz, 1883 | 259 |
| Sub-family. Metorchiinæ, Lühe, 1909 | 261 |
| Genus. Metorchis, Looss, 1899, emend. auctor. | 261 |
| Metorchis truncatus, Rud., 1819 | 261 |
| Family. Heterophyiidæ, Odhner, 1914 | 262 |
| Genus. Heterophyes, Cobbold, 1866 | 262 |
| Heterophyes heterophyes, v. Sieb., 1852 | 262 |
| Metagonimus, Katsurada, 1913; Yokogawa, Leiper, 1913 | 264 |
| Metagonimus yokogawai, Katsurada, 1913 | 264 |
| Family. Dicrocœliidæ, Odhner, 1910 | 265 |
| Genus. Dicrocœlium, Dujardin | 265 |
| Dicrocœlium dendriticum, Rud., 1819 | 266 |
| Family. Echinostomidæ, Looss, 1902 | 267 |
| Sub-family. Echinostominæ, Looss, 1899 | 267 |
| Genus. Echinostoma, Rud., 1809; Dietz, 1910 | 267 |
| Echinostoma ilocanum, Garrison, 1908 | 267 |
| Echinostoma malayanum, Leiper, 1911 | 268 |
| Sub-family. Himasthlinæ, Odhner, 1910 | 269 |
| Genus. Artyfechinostomum, Clayton-Lane, 1915 | 269 |
| Artyfechinostomum sufrartyfex, Clayton-Lane, 1915 | 269 |
| Family. Schistosomidæ, Looss, 1899 | 269 |
| Genus. Schistosoma, Weinl., 1858 | 269 |
| Schistosoma hæmatobium, Bilharz, 1852 | 270 |
| Schistosoma mansoni, Sambon, 1907 | 277 |
| Schistosoma japonicum, Katsurada, 1904 | 277 |
| Class III. Cestoda, Rud., 1808 | 282 |
| Anatomy of the Cestoda | 284 |
| Development of the Tapeworms | 297 |
| Biology | 306 |
| Classification of the Cestoda of Man. |
| Order. Pseudophyllidea, Carus, 1863 | 308 |
| Family. Dibothriocephalidæ, Lühe, 1902 | 308 |
| Sub-family. Dibothriocephalinæ, Lühe, 1899 | 308 |
| Order. Cyclophyllidea, v. Beneden | 308 |
| Family. Dipylidiidæ, Lühe, 1910 | 309 |
| Hymenolepididæ, Railliet and Henry, 1909 | 309 |
| Davaineidæ, Fuhrmann, 1907 | 309 |
| Sub-family. Davaineinæ, Braun, 1900 | 309 |
| Family. Tæniidæ, Ludwig, 1886 | 309 |
| The Cestodes of Man | 309 |
| Family. Dibothriocephalidæ | 309 |
| Sub-family. Dibothriocephalinæ | 309 |
| Genus. Dibothriocephalus, Lühe, 1899 | 309 |
| Dibothriocephalus latus, L., 1748 | 310 |
| Dibothriocephalus cordatus, R. Lkt., 1863 | 315 |
| Dibothriocephalus parvus, Stephens, 1908 | 316 |
| Genus. Diplogonoporus, Lönnbrg., 1892 | 316 |
| Diplogonoporus grandis, R. Blanch., 1894 | 316 |
| Sparganum, Diesing, 1854 | 317 |
| Sparganum mansoni, Cobb., 1883 | 317 |
| Sparganum proliferum, Ijima, 1905 | 318 |
| Family. Dipylidiidæ, Lühe, 1910 | 320 |
| Genus. Dipylidium, R. Lkt., 1863 | 320 |
| Dipylidium caninum, L. 1758 | 320 |
| Family. Hymenolepididæ, Railliet and Henry, 1909 | 323 |
| Genus. Hymenolepis, Weinland, 1858 | 323 |
| Hymenolepis nana, v. Sieb., 1852 | 323 |
| Hymenolepis diminuta, Rud., 1819 | 326 |
| Hymenolepis lanceolata, Bloch, 1782 | 328 |
| Family. Davaineidæ, Fuhrmann, 1907 | 329 |
| Sub-family. Davaineinæ, Braun, 1900 | 329 |
| Genus. Davainea, R. Blanch., 1891 | 329 |
| Davainea madagascariensis, Davaine, 1869 | 329 |
| Davainea (?) asiatica, v. Linst., 1901 | 330 |
| Family. Tæniidæ, Ludwig, 1886 | 331 |
| Genus. Tænia, L., 1758 | 331 |
| Tænia solium, L., p. p., 1767 | 331 |
| Cysticercus acanthotrias, Weinland, 1858 | 336 |
| Tænia bremneri, Stephens, 1908 | 337 |
| Tænia marginata, Batsch., 1786 | 338 |
| Tænia serrata, Goeze, 1782 | 338 |
| Tænia crassicollis, Rud., 1810 | 338 |
| Tænia saginata, Goeze, 1782 | 338 |
| Tænia africana, v. Linst., 1900 | 342 |
| Tænia confusa, Ward, 1896 | 343 |
| Tænia echinococcus, v. Sieb., 1853 | 344 |
| Structure and Development of Echinococcus(Hydatid) | 347 |
| Echinococcus multilocularis (Alveolar Colloid) | 356 |
| Serum Diagnosis of Echinococcus | 359 |
|
| C. Nemathelminthes | 360 |
| Class. Nematoda | 360 |
| Anatomy of the Nematodes | 360 |
| Development of the Nematodes | 371 |
| Classification of the Nematoda. |
| Family. Anguillulidæ, Gervais and van Beneden, 1859 | 374 |
| Angiostomidæ, Braun, 1895 | 374 |
| Gnathostomidæ | 374 |
| Dracunculidæ, Leiper, 1912 | 374 |
| Filariidæ, Claus, 1885 | 374 |
| Trichinellidæ, Stiles and Crane, 1910 | 375 |
| Dioctophymidæ | 375 |
| Strongylidæ, Cobbold, 1864 | 375 |
| Physalopteridæ | 375 |
| Ascaridæ, Cobbold, 1864 | 375 |
| Oxyuridæ | 375 |
| The Nematodes observed in Man | 376 |
| Family. Anguillulidæ | 377 |
| Genus. Rhabditis, Dujardin, 1845 | 377 |
| Rhabditis pellio, Schneider, 1866 | 377 |
| Rhabditis niellyi, Blanchard, 1885 | 378 |
| Rhabditis, sp. | 378 |
| Genus. Anguillula, Ehrenberg, 1826 | 379 |
| Anguillula aceti, Müller, 1783 | 379 |
| Genus. Anguillulina, Gervais and Beneden, 1859 | 379 |
| Anguillulina putrefaciens, Kühn, 1879 | 379 |
| Family. Angiostomidæ, Braun, 1895 | 379 |
| Genus. Strongyloides, Grassi, 1879 | 379 |
| Strongyloides stercoralis, Bavay, 1877 | 380 |
| Family. Gnathostomidæ | 384 |
| Genus. Gnathostoma, Owen, 1836 | 384 |
| Gnathostoma siamense, Levinson, 1889 | 384 |
| Gnathostoma spinigerum, Owen, 1836 | 385 |
| Family. Dracunculidæ, Leiper, 1912 | 385 |
| Genus. Dracunculus, Kniphoff, 1759 | 385 |
| Dracunculus medinensis, Velsch, 1674 | 386 |
| Genus (of Crustacea). Cyclops, Müller, 1776 | 390 |
| Family. Filariidæ | 390 |
| Sub-family. Filariinæ | 390 |
| Genus. Filaria, O. Fr. Müller, 1787 | 390 |
| Filaria bancrofti, Cobbold, 1877 | 390 |
| Filaria demarquayi, Manson, 1895 | 403 |
| Filaria taniguchi, Penel, 1905 | 404 |
| Filaria (?) conjunctivæ, Addario, 1885 | 404 |
| Group. Agamofilaria, Stiles, 1906 | 406 |
| Agamofilaria georgiana | 406 |
| Agamofilaria palpebralis, Pace, 1867 (nec Wilson, 1844) | 406 |
| Agamofilaria oculi humani, v. Nordmann, 1832 | 406 |
| Agamofilaria labialis, Pane, 1864 | 407 |
| Filaria (?) romanorum-orientalis, Sarcani, 1888 | 407 |
| Filaria (?) kilimaræ, Kolb, 1898 | 407 |
| Filaria (?) sp. ? | 407 |
| Genus. Setaria, Viborg, 1795 | 407 |
| Setaria equina, Abildg., 1789 | 408 |
| Genus. Loa, Stiles, 1905 | 409 |
| Loa loa, Guyot, 1778 | 409 |
| Genus. Acanthocheilonema, Cobbold, 1870 | 414 |
| Acanthocheilonema perstans, Manson, 1891 | 414 |
| Genus. Dirofilaria, Railliet and Henry, 1911 | 416 |
| Dirofilaria magalhãesi, R. Blanchard, 1895 | 417 |
| Sub-family. Onchocercinæ, Leiper, 1911 | 417 |
| Genus. Onchocerca, Diesing, 1841 | 417 |
| Onchocerca volvulus, R. Leuckart, 1893 | 417 |
| Family. Trichinellidæ, Stiles and Crane, 1910 | 419 |
| Sub-family. Trichurinæ, Ransom, 1911 | 419 |
| Genus. Trichuris, Röderer and Wagler, 1761 | 419 |
| Trichuris trichiura, Linnæus, 1761 | 419 |
| Sub-family. Trichinellinæ, Ransom, 1911 | 421 |
| Genus. Trichinella, Railliet, 1895 | 421 |
| Trichinella spiralis, Owen, 1835 | 421 |
| History of the Development of Trichinella spiralis | 423 |
| Family. Dioctophymidæ | 431 |
| Genus. Dioctophyme, Collet-Megret, 1802 | 431 |
| Dioctophyme gigas, Rudolphi, 1802 | 431 |
| Family. Strongylidæ | 432 |
| Sub-family. Metastrongylinæ, Leiper, 1908 | 432 |
| Genus. Metastrongylus, Molin, 1861 | 432 |
| Metastrongylus apri, Gmelin, 1789 | 432 |
| Sub-family. Trichostrongylinæ, Leiper, 1908 | 433 |
| Genus. Trichostrongylus, Looss, 1905 | 434 |
| Trichostrongylus instabilis, Railliet, 1893 | 434 |
| Trichostrongylus probolurus, Railliet, 1896 | 435 |
| Trichostrongylus vitrinus, Looss, 1905 | 435 |
| Genus. Hæmonchus, Cobb., 1898 | 436 |
| Hæmonchus contortus, Rudolphi, 1803; Cobb., 1898 | 436 |
| Genus. Nematodirus, Ransom, 1907, emend. Railliet, 1912 | 438 |
| Sub-genus. Mecistocirrus, Railliet, 1912 | 438 |
| Mecistocirrus fordi, Daniels, 1908 | 438 |
| Sub-family. Ancylostominæ, Railliet, 1909 | 438 |
| Group. Œsophagostomeæ, Railliet and Henry, 1909 | 439 |
| Genus. Ternidens, Railliet, 1909 | 439 |
| Ternidens deminutus, Railliet and Henry, 1905 | 440 |
| Genus. Œsophagostomum, Molin, 1861 | 441 |
| Œsophagostomum brumpti, Railliet and Henry, 1905 | 441 |
| Œsophagostomum stephanostomum var. thomasi, Railliet and Henry, 1909 | 443 |
| Œsophagostomum apiostomum, Willach, 1891 | 444 |
| Group. Ancylostomeæ, Railliet and Henry, 1909 | 445 |
| Genus. Ancylostoma, Dubini, 1843, emend. Looss, 1905 | 445 |
| Ancylostoma duodenale, Dubini, 1843 | 445 |
| Ancylostoma ceylanicum, Looss, 1911 | 456 |
| Ancylostoma braziliense, Gomez de Faria, 1910 | 456 |
| Group. Bunostomeæ, Railliet and Henry, 1909 | 456 |
| Genus. Necator, Stiles, 1903 | 457 |
| Necator americanus, Stiles, 1902 | 457 |
| Necator exilidens, Cummins, 1912 | 459 |
| Ancylostomiasis | 459 |
| Group. Syngameæ, Railliet and Henry, 1909 | 459 |
| Genus. Syngamus, von Siebold, 1836 | 459 |
| Syngamus kingi, Leiper, 1913 | 459 |
| Family. Physalopteridæ | 460 |
| Genus. Physaloptera, Rudolphi, 1819 | 460 |
| Physaloptera caucasica, v. Linstow, 1902 | 461 |
| Physaloptera mordens, Leiper, 1907 | 461 |
| Family. Ascaridæ, Cobbold, 1864 | 461 |
| Sub-family. Ascarinæ | 461 |
| Genus. Ascaris, L., 1758 | 461 |
| Ascaris lumbricoides, L., 1758 | 463 |
| Ascaris sp. | 465 |
| Ascaris texana, Smith et Goeth, 1914 | 465 |
| Ascaris maritima, Leuckart, 1876 | 465 |
| Genus. Toxascaris, Leiper, 1907 | 465 |
| Toxascaris limbata, Railliet and Henry, 1911 | 466 |
| Genus. Belascaris, Leiper, 1907 | 466 |
| Belascaris cati, Schrank, 1788 | 466 |
| Belascaris marginata, Rudolphi, 1802 | 466 |
| Genus. Lagocheilascaris, Leiper, 1909 | 466 |
| Lagocheilascaris minor, Leiper, 1909 | 467 |
| Family. Oxyuridæ | 467 |
| Genus. Oxyuris, Rudolphi, 1803 | 467 |
| Oxyuris vermicularis, Linnæus, 1767 | 467 |
| Family. Mermithidæ | 469 |
| Genus. Mermis, Dujardin, 1845 | 469 |
| Mermis hominis oris, Leidy, 1850 | 469 |
| Agamomermis, Stiles, 1903 | 470 |
| Agamomermis restiformis, Leidy, 1880 | 470 |
| Technique | 471 |
|
| D. Acanthocephala, Rud | 475 |
| Echinorhynchus gigas, Goeze, 1782 | 477 |
| Echinorhynchus hominis, Lambl, 1859 | 478 |
| Echinorhynchus moniliformis, Bremser, 1819 | 478 |
|
| E. Gordiidæ | 479 |
|
| F. Hirudinea s. Discophora (Leech) | 480 |
| Family. Gnathobdellidæ (Leeches with Jaws) | 481 |
| Genus. Hirudo, L., 1758 | 481 |
| Hirudo medicinalis, L., 1758 | 481 |
| Hirudo troctina, Johnston, 1816 | 482 |
| Genus. Limnatis, Moq.-Tandon, 1826 | 482 |
| Limnatis nilotica, Savigny, 1820 | 482 |
| Genus. Hæmadipsa, Tennent, 1861 | 482 |
| Family. Rhynchobdellidæ (Leeches with Rostrum) | 482 |
| Genus. Hæmentaria, de Filippi, 1849 | 482 |
| Hæmentaria officinalis, de Filippi | 482 |
| Genus. Placobdella, R. Blanchard | 482 |
| Placobdella catenigera, Moq.-Tandon | 482 |
|
| G. Arthropoda (Jointed-limbed Animals) | 483 |
| A. Arachnoidea (Spiders, Mites, etc.) | 483 |
| Order. Acarina (Mites) | 484 |
| Family. Trombidiidæ (Running Mites) | 485 |
| Genus. Trombidium, Latreille (and Leptus) | 485 |
| Leptus autumnalis, Shaw, 1790 | 485 |
| Trombidium tlalsahuate, Lemaire, 1867 | 486 |
| Akamushi or Kedani | 487 |
| Family. Tetranychidæ (Spinning Mites) | 488 |
| Genus. Tetranychus, Dufour | 488 |
| Tetranychus molestissimus, Weyenbergh, 1886 | 488 |
| Tetranychus telarius, L., 1758, var. russeolus, Koch | 488 |
| Family. Tarsonemidæ | 488 |
| Genus. Pediculoides | 489 |
| Pediculoides ventricosus, Newport, 1850 | 489 |
| Genus. Nephrophages | 490 |
| Nephrophages sanguinarius, Miyake and Scriba, 1893 | 490 |
| Family. Eupodidæ | 491 |
| Genus. Tydeus, Koch | 491 |
| Tydeus molestus, Moniez, 1889 | 491 |
| Family. Gamasidæ (Coleopterous or Insect Mites) | 491 |
| Genus. Dermanyssus, Dugès | 492 |
| Dermanyssus gallinæ, de Geer, 1778 | 492 |
| Dermanyssus hirundinis, Hermann, 1804 | 492 |
| Genus. Holothyrus | 493 |
| Holothyrus coccinella, Gervais, 1842 | 493 |
| Family. Ixodidæ (Ticks) | 493 |
| Classification of Ixodidæ | 496 |
| Synopsis of Genera | 496 |
| Genus. Ixodes, Latreille | 497 |
| Ixodes reduvius, L., 1758 | 497 |
| Ixodes holocyclus, Neumann, 1899 | 499 |
| Ixodes hexagonus, Leach, 1815 | 500 |
| Genus. Amblyomma, Koch | 500 |
| Amblyomma cayennense, Koch, 1844 | 500 |
| Amblyomma americana, Linnæus | 501 |
| Amblyomma maculatum, Koch | 501 |
| Genus. Hyalomma, Koch | 501 |
| Hyalomma ægyptium, L., 1758 | 501 |
| Genus. Hæmaphysalis, Koch | 502 |
| Hæmaphysalis punctata, Canestrini and Fanzago, 1877–1878 | 502 |
| Genus. Dermacentor, Koch | 502 |
| Dermacentor reticulatus, Fabricius, 1794 | 502 |
| Dermacentor venustus, Banks | 503 |
| Dermacentor occidentalis, Neumann | 504 |
| Dermacentor variabilis, Say | 505 |
| Genus. Margaropus, Karsch | 505 |
| Margaropus annulatus australis, Fuller | 505 |
| Margaropus microplus, Canestrini | 505 |
| Genus. Rhipicephalus, Koch | 505 |
| Rhipicephalus sanguineus, Latreille, 1804 | 505 |
| Neumann’s Table of Species of Argas | 505 |
| Genus. Argas, Latreille | 506 |
| Argas reflexus, Fabricius, 1794 | 506 |
| Argas persicus, Fischer de Waldheim, 1824 | 506 |
| Argas brumpti, Neumann | 507 |
| Argas chinche, Gervais, 1844 | 508 |
| Genus. Ornithodorus, Koch | 508 |
| Ornithodorus moubata, Murray, 1877 | 508 |
| Ornithodorus savignyi, Audouin, 1827 | 509 |
| Ornithodorus coriaceus, Koch | 509 |
| Ornithodorus talaje, Guerin, 1849 | 509 |
| Ornithodorus turicata, Dugès, 1876 | 509 |
| Ornithodorus tholozani, Laboulbène and Mégnin, 1882 | 510 |
| Ornithodorus mégnini, Dugès, 1883 | 510 |
| Family. Tyroglyphidæ | 511 |
| Sub-family. Tyroglyphinæ | 511 |
| Genus. Aleurobius, Canestrini | 511 |
| Aleurobius (Tyroglyphus) farinæ, de Geer (part), Koch | 511 |
| Genus. Tyroglyphus, Latreille | 511 |
| Tyroglyphus siro, L., 1756 | 511 |
| Tyroglyphus longior, Gervais, 1844 | 512 |
| Tyroglyphus minor var. castellani, Hirst | 513 |
| Genus. Glyciphagus, Hering, 1838 | 513 |
| Glyciphagus prunorum, Hering, and G. domesticus, de Geer | 513 |
| Glyciphagus cursor, Gervais | 513 |
| Glyciphagus buski, Murray | 513 |
| Genus. Rhizoglyphus, Claparède, 1869 | 514 |
| Rhizoglyphus parasiticus, Dalgetty, 1901 | 514 |
| Genus. Histiogaster, Berlese, 1883 | 515 |
| Histiogaster (entomophagus ?) spermaticus, Trouessart, 1900 | 515 |
| Genus. Cheyletus | 516 |
| Cheyletus mericourti, Lab. | 516 |
| Family. Sarcoptidæ (Itch Mites) | 516 |
| Sub-family. Sarcoptinæ | 518 |
| Genus. Sarcoptes, Latreille | 518 |
| Sarcoptes scabiei, de Geer, 1778 | 518 |
| Sarcoptes minor, Fürstenberg, 1861 | 520 |
| Family. Demodicidæ (Mites of the Hair-follicles) | 522 |
| Genus. Demodex, Owen | 522 |
| Demodex folliculorum, Simon, 1842 | 522 |
| Order. Pentastomida | 523 |
| Family. Linguatulidæ | 523 |
| Genus. Linguatula, Fröhlich | 524 |
| Linguatula rhinaria, Pilger, 1802 | 524 |
| Genus. Porocephalus | 526 |
| Porocephalus constrictus, v. Siebold, 1852 | 526 |
| B. Insecta (Hexapoda) | 529 |
| Classification of the Hexapoda. |
| (1) Aptera | 531 |
| (2) Neuroptera | 531 |
| (3) Orthoptera | 531 |
| (4) Thysanoptera | 531 |
| (5) Hemiptera | 531 |
| (6) Diptera | 532 |
| (7) Lepidoptera | 532 |
| (8) Hymenoptera | 532 |
| (9) Coleoptera | 532 |
| Order. Rhyncota | 532 |
| (a) Rhyncota aptera parasitica | 532 |
| Family. Pediculidæ (Lice) | 532 |
| Genus. Pediculus, Linnæus | 532 |
| Pediculus capitis, de Geer, 1778 | 532 |
| Pediculus vestimenti, Nitzsch, 1818 | 533 |
| Genus. Phthirius, Leach | 534 |
| Phthirius inguinalis, Redi, 1668 | 534 |
| (b) Rhyncota hemiptera | 534 |
| Family. Acanthiadæ | 534 |
| Genus. Cimex , Linnæus | 534 |
| Cimex lectularius , Linnæus | 534 |
| Cimex rotundatus, Signoret, 1852 | 536 |
| Cimex columbarius, Jenyns | 536 |
| Cimex ciliatus, Eversmann, 1841 | 537 |
| Family. Reduviidæ | 537 |
| Genus. Conorhinus, Lap. | 537 |
| Conorhinus megistus, Burm. | 537 |
| Conorhinus sanguisuga, Lec. (Blood-sucking Cone-nose) | 537 |
| Conorhinus, sp. novum (Monster Bug) | 538 |
| Conorhinus rubrofasciatus, de Geer (Malay Bug) | 538 |
| Conorhinus renggeri, Herr-Schäff (Great Black Bug of Pampas) | 539 |
| Conorhinus variegatus (Variegated Cone-nose) | 539 |
| Conorhinus nigrovarius | 539 |
| Conorhinus protractus | 539 |
| Genus. Reduvius, etc. | 539 |
| Reduvius personatus, Linné | 539 |
| Coriscus subcoleoptratus, Kirby, 1837 | 540 |
| Rasahus biguttatus, Say, 1831 | 540 |
| Melanolestes morio, Erichson, 1848 (non-Walker) | 540 |
| Melanolestes abdominalis, Herrich-Schäffer, 1848 | 540 |
| Phonergates bicoloripes | 541 |
| Family. Aradidæ | 541 |
| Dysodius lunatus, Fabr. (Pito Bug) | 541 |
| The Ochindundu | 541 |
| Family. Lygæidæ | 541 |
| Lyctocoris campestris, Fabricius | 541 |
| Rhodinus prolixus, Stål, 1859 | 541 |
| Order. Orthoptera | 542 |
| Locusts Injurious to Man | 542 |
| Order. Coleoptera | 542 |
| Silvanus surinamensis, Linnæus (Saw-toothed Grain Beetle) | 542 |
| Order. Diptera | 543 |
| Aphaniptera or Siphonaptera (Fleas) | 543 |
| Family. Sarcopsyllidæ (Jiggers) | 543 |
| Genus. Dermatophilus, Guérin | 544 |
| Dermatophilus cæcata, Enderl. | 544 |
| Dermatophilus penetrans, L., 1758 (Jigger, Chigoe) | 544 |
| Genus. Echidnophaga, Olliff | 544 |
| Echidnophaga gallinacea, Westwood (Chigoe of Fowls) | 544 |
| Family. Pulicidæ (True Fleas) | 545 |
| Genus. Pulex, Linn. | 545 |
| Pulex irritans, L., 1758 | 545 |
| Genus. Xenopsylla, Glink | 546 |
| Xenopsylla cheopis, Rothschild | 546 |
| Xenopsylla brasiliensis, Baker | 547 |
| Genus. Ctenocephalus, Kolenati | 547 |
| Genus. Hoplopsyllus, Baker | 547 |
| Hoplopsyllus anomalus, Baker | 547 |
| Genus. Ceratophyllus, Centis | 547 |
| Ceratophyllus fasciatus, Bosc | 547 |
| Genus. Ctenopsylla, Kolenati | 548 |
| Genus. Hystrichopsylla, Taschenberg | 548 |
| Pulex pallipes | 548 |
| Systematic Anatomical and Biological Remarks on Mosquitoes | 548 |
| Culicidæ or Mosquitoes | 555 |
| The Classification of Culicidæ | 561 |
| Notes on the Different Genera | 566 |
| Sub-family. Anophelina | 566 |
| Genus. Anopheles, Meigen | 566 |
| Genus. Myzomyia, Blanchard; Grassia, Theobald | 567 |
| Genus. Neomyzomyia, Theobald | 567 |
| Genus. Cycloleppteron, Theobald | 567 |
| Genus. Feltinella, Theobald | 567 |
| Genus. Stethomyia, Theobald | 567 |
| Genus. Pyretophorus, Blanchard; Howardia, Theobald | 567 |
| Genus. Myzorhynchella, Theobald | 568 |
| Genus. Manguinhosia, Cruz (in Peryassu) | 568 |
| Genus. Chrystya, Theobald | 568 |
| Genus. Lophoscelomyia, Theobald | 568 |
| Genus. Arribalzagia, Theobald | 568 |
| Genus. Myzorhynchus, Blanchard; Rossia, Theobald | 568 |
| Genus. Nyssorhynchus, Blanchard; Laverania, Theobald | 569 |
| Genus. Cellia, Theobald | 569 |
| Genus. Neocellia, Theobald | 569 |
| Genus. Kertészia, Theobald | 569 |
| Genus. Manguinhosia, Cruz | 569 |
| Genus. Chagasia, Cruz | 570 |
| Genus. Calvertina, Ludlow | 570 |
| Genus. Birónella, Theobald | 570 |
| Sub-family. Megarhininæ | 570 |
| Genus. Megarhinus, Robineau Desvoidy | 570 |
| Genus. Toxorhynchites, Theobald | 570 |
| Sub-family. Culicinæ | 571 |
| Genus. Mucidus, Theobald | 571 |
| Genus. Psorophora, Robineau Desvoidy | 571 |
| Genus. Janthinosoma, Arribalzaga | 571 |
| Genus. Stegomyia, Theobald | 571 |
| Stegomyia fasciata, Fabricius (Yellow Fever Mosquito) | 574 |
| Stegomyia scutellaris, Walker | 575 |
| Genus. Theobaldia, Neveu-Lemaire | 575 |
| Theobaldinella, Blanchard | 575 |
| Theobaldia annulata, Meigen | 575 |
| Genus. Culex, Linnæus | 575 |
| Genus. Melanoconion, Theobald | 576 |
| Genus. Grabhamia, Theobald | 576 |
| Genus. Pseudotæniorhynchus, Theobald; Tæniorhynchus, Theobald, non-Arribalzaga | 576 |
| Genus. Tæniorhynchus, Arribalzaga; Mansonia, Blanchard; Panoplites, Theobald | 577 |
| Genus. Chrysoconops, Goeldi | 577 |
| Other Nematocera | 577 |
| Family. Simulidæ | 577 |
| Family. Chironomidæ (Midges) | 579 |
| Sub-family. Ceratopogoninæ | 580 |
| Family. Psychodidæ (Owl Midges) | 581 |
| Brachycera (Flies) | 582 |
| Family. Phoridæ | 582 |
| Aphiochæta ferruginea, Brun | 583 |
| Phora rufipes, Meig. | 583 |
| Family. Sepsidæ | 583 |
| Piophila casei, L. | 583 |
| Family. Syrphidæ (Hover and Drone Flies) | 583 |
| Family. Drosophilidæ | 584 |
| Drosophila melanogaster, Br. | 584 |
| Family. Muscidæ | 584 |
| Teichomyza fusca, Macq. | 584 |
| Homalomyia canicularis, L., etc. | 584 |
| Homalomyia scalaris, Fabr. | 585 |
| Anthomyra desjardensii, Macq. | 585 |
| Hydrotæa meteorica, L. | 585 |
| Cyrtoneura stabulans | 585 |
| Musca domestica, Linn. (Common House-fly) | 585 |
| Genus. Chrysomyia, Rob. Desv. | 587 |
| Chrysomyia (Compsomyia) macellaria, Fabr.; Lucilia macellaria, Fabr. | 587 |
| Chrysomyia viridula, Rob. Desv. | 588 |
| Genus. Lucilia, Rob. Desv. | 588 |
| Lucilia nobilis, Meig. | 588 |
| Genus. Pycnosoma, Brauer and v. Bergenstamm | 588 |
| Genus. Sarcophaga, Mg. | 589 |
| Sarcophaga carnaria, L., 1758 | 589 |
| Sarcophaga magnifica, Schiner, 1862 | 589 |
| Sarcophaga chrysotoma, Wied | 590 |
| Sarcophaga plinthopyga, Wied | 590 |
| Ochromyia anthropophaga, E. Blanch.; Cordylobia arthrophaga, Grünberg | 590 |
| Auchmeromyia (Bengalia) depressa (Walker) | 591 |
| Genus. Cordylobia, Grünberg, 1903 | 591 |
| Cordylobia grünbergi, Dönitz | 591 |
| Cordylobia anthropophaga, Grünberg | 592 |
| Lund’s Larva | 593 |
| Auchmeromyia luteola, Fabricius | 593 |
| Family. Oestridæ | 594 |
| Cutaneous Oestridæ | 595 |
| Genus. Hypoderma, Latreille | 595 |
| Hypoderma bovis, de Geer | 595 |
| Hypoderma lineata, de Villers | 596 |
| Hypoderma diana, Brauer | 596 |
| Genus. Dermatobia, Brauer | 596 |
| Dermatobia cyaniventris, Macq. | 596 |
| Cavicolous Oestridæ | 598 |
| Genus. Oestrus, Linnæus | 598 |
| Oestrus (Cephalomyia) ovis, L. | 598 |
| Gastricolous Oestridæ | 599 |
| Genus. Gastrophilus, Leach | 599 |
| Biting-mouthed and other Noxious Diptera which may be Disease Carriers | 600 |
| Family. Tabanidæ (Gad Flies) | 600 |
| Family. Asilidæ (Wolf Flies) | 602 |
| Family. Leptidæ | 603 |
| Blood-sucking Muscidæ | 603 |
| Genus. Glossina, Westwood | 603 |
| Glossina palpalis, Rob. Desv. | 607 |
| Glossina morsitans, Westwood | 608 |
| Genus. Stomoxys, Geoffroy | 609 |
| Genus. Lyperosia, Rondani | 610 |
| Pupipara or Eproboscidæ | 611 |
| Insects and Epidemic Poliomyelitis | 612 |
| Addenda | 613 |
| Akamushi or Kedani Sickness | 613 |
| Ticks.—African Tick Fever | 613 |
| Tick Paralysis | 613 |
| Diptera.—Psychodidæ | 613 |
| Pulicidæ.—Dermatophilus (Sarcopsylla) penetrans, or the “Jigger” | 613 |
| Brachycera.—Leptidæ | 613 |
| Myiasis | 615 |
| Auricular Myiasis | 615 |
| Body, Head, and Clothes Lice | 615 |
|
| SUPPLEMENT: CLINICAL AND THERAPEUTICAL NOTES | 617 |
| Protozoa | 617 |
| Introduction | 617 |
| I.—Amœbic Dysentery | 618 |
| II.—Trypanosomiases | 620 |
| African Sleeping Sickness | 620 |
| South American Trypanosomiasis | 623 |
| III.—Flagellate Diarrhœa and Dysentery | 623 |
| IV.—Leishmaniases | 626 |
| A. Kala-azar | 626 |
| Indian | 626 |
| Infantile | 627 |
| B. Oriental Sore, due to Leishmania tropica | 627 |
| Naso-oral (Espundia) | 628 |
| V.—Spirochætoses | 629 |
| A. Relapsing Fevers | 629 |
| B. Yaws or Frambœsia tropica | 632 |
| C. Syphilis | 632 |
| D. Bronchial Spirochætosis | 632 |
| VI.—Malaria | 633 |
| VII.—Balantidian Dysentery | 637 |
| Plathelminthes (Flat Worms) | 638 |
| Fascioliasis | 638 |
| Fasciola hepatica | 638 |
| Fasciolopsis buski | 638 |
| Paragonimiasis | 639 |
| Paragonimus ringeri | 639 |
| Clonorchis sinensis | 640 |
| Bilharziasis | 641 |
| Schistosoma hæmatobium | 641 |
| Cestodes | 644 |
| General | 644 |
| Dibothriocephalus latus | 658 |
| Sparganum mansoni | 659 |
| Dipylidium caninum (Tænia cucumerina) | 659 |
| Hymenolepis nana | 661 |
| Tænia solium | 662 |
| Tænia saginata | 667 |
| Nematodes | 674 |
| Strongyloides stercoralis | 674 |
| Dracunculus medinensis (Dracontiasis) | 675 |
| Filaria bancrofti | 676 |
| Loa loa | 678 |
| Trichuris trichiura | 678 |
| Trichinella spiralis | 680 |
| Eustrongylus gigas | 681 |
| Ancylostoma duodenale (Ancylostomiasis) | 682 |
| Ascaris lumbricoides (Ascariasis) | 687 |
| Oxyuris vermicularis (Oxyuriasis) | 694 |
| Hirudinea (Leeches) | 699 |
| Arthropoda | 702 |
| Arachnoidea | 702 |
| Leptus autumnalis (Grass, Harvest, or Gooseberry Mite) | 702 |
| Kedani, Akaneesch (The Japanese River or Inundation Disease) | 703 |
| Dermanyssus gallinæ (avium) | 703 |
| Ixodes reduvius (ricinus ) | 704 |
| Sarcoptes scabiei (Scabies) | 704 |
| Demodex folliculorum | 708 |
| Demodex folliculorum canis | 709 |
| Insecta | 709 |
| Pediculus capitis (Head Louse) | 709 |
| Pediculus vestimenti (Clothes Louse) | 710 |
| Phthirius inguinalis (Pediculus pubis) (Crab Louse) | 711 |
| Cimex (Acanthia) lectularia (Cimex lectularius) (Bed Bug) | 713 |
| Pulex irritans (Human Flea) | 714 |
| Dermatophilus (Sarcopsylla) penetrans (Sand Flea) | 714 |
| Myiasis | 715 |
| Myiasis externa | 715 |
| Gastricolous Oestridæ (Creeping Disease) | 729 |
|
| APPENDIX ON PROTOZOOLOGY | 733 |
| I.—Notes on Recent Researches | 733 |
| Differences between Entamœba histolytica and E. coli | 733 |
| Phagedænic Amœbæ | 733 |
| Endamœba gingivalis | 733 |
| Entamœba kartulisi | 734 |
| Craigia and Craigiasis | 734 |
| Human Trichomoniasis | 734 |
| Chilomastix (Tetramitus) mesnili | 735 |
| Giardia (Lamblia) intestinalis | 736 |
| Cercomonas hominis | 736 |
| Transmissive Phase of Trypanosomes in Vertebrates | 737 |
| Trypanosoma lewisi | 737 |
| Blepharoplastless Trypanosomes | 737 |
| The Experimental Introduction of certain Insect Flagellates into various Vertebrates, and its bearing on the Evolution of Leishmaniasis | 737 |
| The Transmission of Spirochæta duttoni | 739 |
| Spirochæta bronchialis | 739 |
| The Spirochætes of the Human Mouth | 740 |
| Coccidia in Cattle | 741 |
| The Hæmosporidia | 742 |
| The Leucocytozoa of Birds | 742 |
| II.—Formulæ of some Culture Media | 742 |
| Culture Media for growing Amœbæ | 742 |
| Culture Media for the growth of Protozoa parasitic in the Blood | 744 |
| III.—Brief Notes on General Protozoological Technique | 745 |
| Fresh Material | 745 |
| Stained Material | 747 |
| Fixatives | 748 |
| Stains | 749 |
| APPENDIX ON TREMATODA AND NEMATODA | 753 |
| Trematoda | 753 |
| Artyfechinostomum sufrartyfex | 753 |
| Metagonimus (Yokogawa) yokogawai | 753 |
| Opisthorchis sp. | 753 |
| Schistosome cercariæ | 753 |
| Distomata cercariæ | 753 |
| Group. Ferrocercous cercariæ | 753 |
| Family. Schistosomidæ | 753 |
| Cercaria bilharzia, Leiper, 1915 | 754 |
| Cercaria bilharziella, Leiper, 1915 | 754 |
| Schistosoma mansoni, Sambon, 1907 | 754 |
| Nematoda | 754 |
| Ancylostomiasis | 754 |
| Ground Itch | 754 |
| Ascaris lumbricoides | 754 |
| Filariasis | 755 |
| Onchocerca volvulus | 755 |
| Strongyloides stercoralis | 755 |
|
| BIBLIOGRAPHY | 756 |
| INDEX | 836 |
LIST OF ILLUSTRATIONS.
| Fig. | | | PAGE |
| 1 | | Amœba coli. (After Loesch) | |
| 2 | | Encysted intestinal amœbæ. (After Grassi) | |
| 3 | | Entamœba coli, life-cycle. (After Castellani and Chalmers) | |
| 4 | | Entamœba coli, so-called autogamy. (From Minchin) | |
| 5 | | Entamœba histolytica (tetragena form). (After Hartmann) | |
| 6 | | Entamœba histolytica, ingestion of red blood corpuscles. (After Hartmann) | |
| 7 | | Entamœba histolytica, section through infected intestinal ulcer. (After Harris) | |
| 8 | | Entamœba histolytica (tetragena), trophozoite and nuclei. (After Hartmann) | |
| 9 | | Entamœba histolytica (tetragena), cysts. (After Hartmann) | |
| 10 | | Entamœba buccalis. (After Leyden and Löwenthal) | |
| 11 | | Entamœba kartulisi. (After Kartulis) | |
| 12 | | Amœba miurai. (After Ijima) | |
| 13 | | Chlamydophrys enchelys. (After Cienkowski) | |
| 14 | | Chlamydophrys enchelys, encysted. (After Cienkowski) | |
| 15 | | Leydenia gemmipara, Schaudinn | |
| 16 | | Trichomonas vaginalis. (After Künstler) | |
| 17 | | Trichomonas intestinalis. (After Grassi) | |
| 18 | | Trichomonas intestinalis. (Original, Fantham) | |
| 19 | | Lamblia intestinalis. (After Wenyon, from Minchin) | |
| 20 | | Lamblia intestinalis. (After Grassi and Schewiakoff) | |
| 21 | | Cercomonas hominis. (After Davaine) | |
| 22 | | Cercomonas hominis, from an echinococcus cyst. (After Lambl) | |
| 23 | | Monas pyophila. (After Grimm) | |
| 24 | | Prowazekia urinaria. (After Sinton) | |
| 25 | | Prowazekia urinaria, excystation. (After Sinton) | |
| 26 | | Trypanosoma brucei in division. (After Laveran and Mesnil) | |
| 27 | | Trypanosoma lewisi, rosettes. (After Laveran and Mesnil) | |
| 28 | | Trypanosoma gambiense. (After Dutton) | |
| 29 | | Trypanosoma gambiense, development in vertebrate host. (Original, Fantham) | |
| 30 | | Trypanosoma gambiense, development in Glossina palpalis. (After Robertson) | |
| 31 | | Trypanosoma rhodesiense. (After Stephens and Fantham) | |
| 32 | | Chart showing daily counts of number of Trypanosomes per cubic millimetre of peripheral blood from a case of Rhodesian sleeping sickness. (After Ross and Thomson) | |
| 33 | | Trypanosoma cruzi, schizogony. (After Chagas, from Castellani and Chalmers) | |
| 34 | | Trypanosoma cruzi in muscle. (After Vianna, from Castellani and Chalmers) | |
| 35 | | Trypanosoma cruzi, development in Triatoma megista. (After Chagas, from Castellani and Chalmers) | |
| 36 | | Trypanosoma cruzi, forms found in salivary glands of Triatoma. (After Chagas, from Castellani and Chalmers) | |
| 37 | | Trypanosoma lewisi, from rat’s blood. (After Minchin) | |
| 38 | | Trypanosoma lewisi, from stomach of rat-flea. (After Minchin) | |
| 39 | | Trypanosoma lewisi, from rectum of rat-flea. (After Minchin) | |
| 40 | | Trypanosoma brucei. (After Laveran and Mesnil) | |
| 41 | | Trypanosoma evansi. (Original, Fantham) | |
| 42 | | Trypanosoma equinum. (After Laveran and Mesnil) | |
| 43 | | Trypanosoma equiperdum. (Original, Fantham) | |
| 44 | | Trypanosoma theileri. (After Laveran and Mesnil) | |
| 45 | | Trypanosoma vivax. (Original, Fantham) | 100 |
| 46 | | Trypanosoma congolense. (Original, Fantham) | 100 |
| 47 | | Trypanosoma uniforme. (Original, Fantham) | 100 |
| 48 | | Trypanosoma rotatorium. (After Laveran and Mesnil) | 101 |
| 49 | | Herpetomonas, Crithidia, Trypanosoma. (After Porter) | 103 |
| 50 | | Leishmania donovani. (After Christophers, Patton, Leishman; from Castellani and Chalmers) | 106 |
| 51 | | Toxoplasma gondii. (After Laveran and Marullaz, from Trop. Dis. Bulletin) | 113 |
| 52 | | Toxoplasma pyrogenes. (After Castellani, from Trop. Dis. Bulletin) | 113 |
| 53 | | Spirochæta balbianii. (After Fantham and Porter) | 114 |
| 54 | | Spirochæta duttoni. (After Fantham) | 117 |
| 55 | | Spirochæta duttoni and its coccoid bodies in the tick. (After Fantham) | 118 |
| 56 | | Treponema pallidum. (After Bell, from Castellani and Chalmers) | 124 |
| 57 | | Treponema pallidum, apparatus for cultivation of. (After Noguchi) | 125 |
| 58 | | Treponema pertenue. (After Castellani and Chalmers) | 127 |
| 59 | | Monocystis agilis. (After Stein) | 130 |
| 60 | | Gregarina longa, stages of growth of trophozoite | 130 |
| 61 | | Xyphorhynchus firmus. (After Léger) | 131 |
| 62 | | Gregarina munieri. (After Schewiakoff) | 131 |
| 63 | | Monocystis agilis, spores. (After Bütschli) | 132 |
| 64 | | Gregarines, conjugation and spore formation. (After Calkins and Siedlecki, modified) | 133 |
| 65 | | Stylorhynchus oblongatus, cyst and gametes. (After Léger) | 133 |
| 66 | | Gregarines, various spores. (After Léger) | 134 |
| 67 | | Eimeria (Coccidium) schubergi, life-cycle diagram of. (After Schaudinn) | 139 |
| 68 | | Eimeria avium in gut epithelium of grouse chick. (After Fantham) | 143 |
| 69 | | Eimeria avium, life-cycle, diagram of. (After Fantham) | 144 |
| 70 | | Eimeria stiedæ in section of rabbit’s intestine | 145 |
| 71 | | Eimeria stiedæ, oöcysts from rabbit’s liver. (After Leuckart) | 146 |
| 72 | | Eimeria stiedæ, spores. (After Balbiani) | 146 |
| 73 | | Eimeria stiedæ, schizogony. (After R. Pfeiffer) | 146 |
| 74 | | Eimeria stiedæ, section through infected nodule of liver | 147 |
| 75 | | Isospora bigemina. (After Stiles) | 150 |
| 76 | | Hæmoproteus (Halteridium) columbæ, life-cycle. (After Aragão, from Castellani and Chalmers) | 152 |
| 77 | | Leucocytozoön lovati. (After Fantham) | 153 |
| 78 | | Hæmogregarines from lizards. (After França) | 154 |
| 79 | | Leucocytogregarina canis, life-cycle. (After Christophers, from Castellani and Chalmers) | 155 |
| 80 | | Plasmodium vivax, life-cycle. (After Schaudinn and Grassi) | 160 |
| 81 | | Malignant tertian malarial parasite in intestine of Anopheles. (After Grassi) | 162 |
| 82 | | Oökinete of malignant tertian malaria in stomach of Anopheles. (After Grassi) | 162 |
| 83 | | Section of stomach of Anopheles with malarial oöcysts. (After Grassi) | 163 |
| 84 | | Sporulation of malarial parasites in Anopheles. (After Grassi) | 163 |
| 85 | | Tertian malarial parasite in human red blood corpuscles. (After Mannaberg) | 165 |
| 86 | | Quartan malarial parasite in human red corpuscles. (After Manson) | 166 |
| 87 | | Malignant malarial parasite in human red corpuscles. (After Manson) | 168 |
| 88 | | Malarial crescents. (After Mannaberg) | 168 |
| 89 | | Section through tubule of salivary gland of Anopheles infected with malarial sporozoites. (After Grassi) | 169 |
| 90 | | Nuttallia equi, life-cycle in red blood corpuscles. (After Nuttall and Strickland) | 173 |
| 91 | | Babesia (Piroplasma) canis, life-cycle in blood of dog. (After Nuttall and Graham-Smith) | 175 |
| 92 | | Theileria parva. (After Nuttall and Fantham) | 179 |
| 93 | | Myxosporidian spores and infected gill of fish. (After J. Müller) | 181 |
| 94 | | Myxobolus mülleri, spore. (After Bütschli) | 181 |
| 95 | | Myxobolus, schema of spore. (After Doflein) | 182 |
| 96 | | Chloromyxum leydigi. (After Thélohan) | 182 |
| 97 | | Myxobolus pfeifferi, spore formation. (After Keysselitz, from Minchin) | 183 |
| 98 | | Nosema apis. (After Fantham and Porter) | 185 |
| 99 | | Nosema bombycis from silkworm. (After Balbiani) | 186 |
| 100 | | Nosema bombycis, spores. (After Thélohan) | 186 |
| 101 | | Hexactinomyxon psammoryctis, spore. (After Stolč) | 187 |
| 102 |  | Sarcocystis miescheriana in muscle of pig. (After Kühn) | 188 |
| 103 |
| 104 | | Sarcocystis miescheriana, mature trophozoite | 189 |
| 105 | | Sarcocystis tenella in section, as seen in œsophagus of sheep | 190 |
| 106 | | Sarcocystis tenella, young trophozoite. (After Bertram) | 190 |
| 107 | | Sarcocystis miescheriana, end portion of trophozoite. (After Bertram) | 190 |
| 108 | | Sarcocystis blanchardi from ox. (From Wasielewski, after van Eecke) | 190 |
| 109 | | Sarcocystis tenella. (After Laveran and Mesnil) | 191 |
| 110 | | Haplosporidium heterocirri. (After Caullery and Mesnil) | 195 |
| 111 | | Haplosporidian spores. (After Caullery and Mesnil) | 195 |
| 112 | | Rhinosporidium kinealyi, portion of ripe cyst. (After Minchin and Fantham) | 197 |
| 113 | | Balantidium coli. (After Leuckart) | 200 |
| 114 | | Balantidium coli, free and encysted. (After Casagrandi and Barbagallo) | 200 |
| 115 | | Balantidium minutum. (After Schaudinn) | 204 |
| 116 | | Nyctotherus faba. (After Schaudinn) | 205 |
| 117 | | Nyctotherus giganteus. (After Krause) | 206 |
| 118 | | Nyctotherus africanus. (After Castellani) | 206 |
| 119 | | Trachoma bodies in conjunctival cells. (Original, Fantham) | 209 |
| 120 | | Half of a transverse section through Fasciola hepatica, L. | 214 |
| 121 | | Harmostomum leptostomum, Olss. | 215 |
| 122 | | Median section through the anterior part of Fasciola hepatica | 217 |
| 123 | | Polystomum integerrimum. (After Zeller) | 218 |
| 124 | | Allocreadium isoporum, Looss. (After Looss) | 218 |
| 125 | | Terminal flame cell of the excretory system. (Stephens) | 219 |
| 126 | | Diagram of female genitalia. (Stephens) | 220 |
| 127 | | Diagram of male and part of female genitalia. (Stephens) | 220 |
| 128 | | Ovum of Fasciola hepatica, L. | 223 |
| 129 | | Miracidium of Fasciola hepatica. (After Leuckart) | 223 |
| 130 | | A group of cercariæ of Echinostoma sp. | 225 |
| 131 | | Development of Fasciola hepatica, L. (After Leuckart) | 226 |
| 132 | | Young redia of Fasciola hepatica. (From Leuckart) | 227 |
| 133 | | Older redia of Distoma echinatum | 227 |
| 134 | | Cercaria of Fasciola hepatica. (After Leuckart) | 228 |
| 135 | | Encysted cercaria of Fasciola hepatica. (After Leuckart) | 228 |
| 136 | | Watsonius watsoni. (After Shipley) | 234 |
| 137 | | Watsonius watsoni: ventral projection composed from a series of transverse sections. (After Stiles and Goldberger) | 235 |
| 138 | | Gastrodiscus hominis. (After Leuckart) | 236 |
| 139 | | Fasciola hepatica, L. | 238 |
| 140 | | Fasciola hepatica, showing the gut and its branches | 239 |
| 141 | | Fasciola hepatica, L. (After Claus) | 239 |
| 142 | | Fasciola hepatica: egg from liver of sheep. (After Thomas) | 240 |
| 143 | | Limnæus truncatulus, Müll. (From Leuckart) | 240 |
| 144 | | Young Fasciola hepatica. (From Leuckart) | 242 |
| 145 | | Fasciola gigantica. (After Looss) | 243 |
| 146 | | Fasciolopsis buski, Lank. (After Odhner) | 245 |
| 147 | | Fasciolopsis rathouisi, Poir. (After Claus) | 246 |
| 148 | | Fasciolopsis fülleborni. (After Fülleborn) | 248 |
| 149 | | Paragonimus ringeri, Cobb. (After Katsurada) | 250 |
| 150 | | Paragonimus ringeri, Cobb. (After Kubo) | 250 |
| 150A | | Paragonimus westermanii, Kerb. (After Leuckart) | 250 |
| 151 | | Egg of Paragonimus ringeri, Cobb. (After Katsurada) | 251 |
| 152 | | Egg of Opisthorchis felineus | 253 |
| 153 | | Opisthorchis felineus. (After Stiles and Hassall) | 253 |
| 154 | | Opisthorchis pseudofelineus. (After Stiles) | 254 |
| 155 | | Parapisthorchis caninus. (After Stephens) | 256 |
| 156 | | Amphimerus noverca, Braun. (After McConnell) | 257 |
| 157 | | Metorchis conjunctus. (After Cobbold) | 258 |
| 158 | | Clonorchis sinensis. (After Looss) | 259 |
| 159 | | Ova of Clonorchis sinensis. (After Looss) | 259 |
| 160 | | Clonorchis endemicus. (After Looss) | 260 |
| 161 | | Clonorchis endemicus: eggs. (After Looss) | 260 |
| 162 | | Metorchis truncatus | 262 |
| 163 | | Heterophyes heterophyes. (After Looss) | 263 |
| 164 | | Metagonimus yokogawai. (After Leiper) | 264 |
| 165 | | Dicrocœlium dendriticum | 265 |
| 166 | | Eggs of Dicrocœlium dendriticum | 266 |
| 167 | | Miracidia of Dicrocœlium dendriticum. (After Leuckart) | 266 |
| 168 | | Echinostoma ilocanum. (After Brumpt) | 268 |
| 169 | | Echinostoma ilocanum. (After Leiper) | 268 |
| 170 | | Echinostoma malayanum, Leiper. (After Leiper) | 269 |
| 171 | | Schistosoma hæmatobium. (After Looss) | 270 |
| 172 | | Transverse section through a pair of Schistosoma hæmatobium in copulâ. (After Leuckart) | 271 |
| 173 | | Anterior end of the male Schistosoma hæmatobium. (After Looss) | 271 |
| 174 | | Schistosoma hæmatobium. (After Leuckart) | 276 |
| 175 | | Schistosoma hæmatobium, ovum of. (After Looss) | 277 |
| 176 | | Schistosoma japonicum. (After Katsurada) | 278 |
| 177 | | Schistosoma japonicum. (After Katsurada) | 279 |
| 178 | | Schistosoma japonicum. (After Looss) | 279 |
| 179 |  | Schistosoma japonicum from dog. (After Katsurada) | 280 |
| 180 | |
| 181 | |
| 182 | | Schistosoma japonicum. (After Catto) | 281 |
| 183 | | Schistosoma japonicum. (After Katsurada) | 282 |
| 184 | | Schematic representation of a small part of a transverse section of Ligula sp. (After Blochmann) | 287 |
| 185 | | Half of a transverse section through a proglottis of Tænia crassicollis | 288 |
| 186 | | Dipylidium caninum. (After Benham) | 289 |
| 187 | | Longitudinal section of the head and neck of Tænia crassicollis | 290 |
| 188 | | Tænia cœnurus. (After Niemisec) | 291 |
| 189 | | Young Acanthobothrium coronatum. (After Pintner) | 292 |
| 190 | | Scolex of a cysticercoid from Arion sp. (After Pintner) | 292 |
| 191 | | Proglottis of Tænia saginata, Goeze, showing genitalia | 293 |
| 192 | | Dibothriocephalus latus. (After Benham and Sommer and Landois) | 294 |
| 193 | | Diagram of genitalia of a Cestode. (Stephens) | 295 |
| 194 | | Part of a transverse section through a proglottis of Dibothriocephalus latus | 296 |
| 195 | | Egg of Diplogonoporus grandis. (After Kurimoto) | 298 |
| 196 | | Uterine egg of Tænia saginata. (After Leuckart) | 298 |
| 197 | | Oncosphere of Tænia africana (after v. Linstow) and oncosphere of Dipylidium caninum. (After Grassi and Rovelli) | 299 |
| 198 | | Diagram of a cysticercoid. (Stephens) | 301 |
| 199 | | Diagram of a cysticercus. (Stephens) | 301 |
| 200 | | Diagram of development of a cysticercus. (Stephens) | 303 |
| 201 | | Section through a piece of a Cœnurus cerebralis | 304 |
| 202 | | Median section through a cysticercus. (After Leuckart) | 304 |
| 203 | | Cysticercus pisiformis in an evaginated condition | 304 |
| 204 | | Various chains of segments of Dibothriocephalus latus | 311 |
| 205 | | Transverse section of the head of Dibothriocephalus latus | 311 |
| 206 | | Fairly mature proglottis of Dibothriocephalus latus | 311 |
| 207 | | Dibothriocephalus latus. (After Benham and Schauinsland) | 312 |
| 208 | | Plerocercoid of Dibothriocephalus latus | 313 |
| 209 | | A piece of the body wall of the Burbot, Lota vulgaris | 313 |
| 210 | | Cephalic end of Dibothriocephalus cordatus. (After Leuckart) | 315 |
| 211 | | Diplogonoporus grandis, Lühe, 1899. (After Ijima and Kurimoto) | 317 |
| 212 | | Diplogonoporus grandis. (After Ijima and Kurimoto) | 317 |
| 213 | | Cephalic end of Sparganum mansoni, Cobb. (After Leuckart) | 318 |
| 214 | | Sparganum mansoni. (After Ijima and Murata) | 318 |
| 215 | | Sparganum prolifer. (After Ijima) | 319 |
| 216 | | Sparganum proliferum. (After Stiles) | 319 |
| 217 | | Dipylidium caninum. (After Diamare) | 320 |
| 218 | | Dipylidium caninum. (After Benham and Moniez) | 320 |
| 219 | | Dipylidium caninum: central portion of a proglottis. (After Neumann and Railliet) | 321 |
| 220 | | Dipylidium caninum: development of embryo. (After Benham, Grassi, and Rovelli) | 321 |
| 221 | | Larva (cysticercoid) of Dipylidium caninum. (After Grassi and Rovelli) | 322 |
| 222 | | Hymenolepis nana, v. Sieb. (After Leuckart) | 324 |
| 223 | | Hymenolepis nana: head. (After Mertens) | 324 |
| 224 | | Hymenolepis nana: an egg. (After Grassi) | 324 |
| 225 | | Longitudinal section through the intestinal villus of a rat. (After Grassi and Rovelli) | 324 |
| 226 | | Hymenolepis nana (murina): cross-section of proglottis from a rat. (After v. Linstow) | 325 |
| 227 | | Hymenolepis nana: longitudinal section of an embryo. (After Grassi and Rovelli) | 325 |
| 228 | | Hymenolepis diminuta. (After Zschokke) | 326 |
| 229 | | Hymenolepis diminuta. (After Grassi) | 326 |
| 230 | | Hymenolepis diminuta. (After Bizzozero) | 326 |
| 231 | | Hymenolepis diminuta. (Stephens, after Nicoll and Minchin) | 327 |
| 232 | | Hymenolepis lanceolata. (After Krabbe) | 328 |
| 233 | | Hymenolepis lanceolata. (After Wolffhügel) | 328 |
| 234 | | Scolex of Davainea madagascariensis. (After Blanchard) | 330 |
| 235 | | Two fairly mature proglottids of Tænia solium | 332 |
| 236 | | Head of Tænia solium | 332 |
| 237 | | Large and small hooks of Tænia solium. (After Leuckart) | 333 |
| 238 | | Tænia solium. (After Leuckart) | 333 |
| 239 | | Two mature proglottids of Tænia solium | 333 |
| 240 | | Large and small hooklets of Tænia marginata. (After Leuckart) | 338 |
| 241 | | Mature segment of Tænia saginata | 339 |
| 242 | | Cephalic end of Tænia saginata | 339 |
| 243 | | Tænia saginata. (After Leuckart) | 339 |
| 244 | | A piece of the muscle of the ox, with three specimens of Cysticercus bovis. (After Ostertag) | 340 |
| 245 | | Mature segment of Tænia africana. (After v. Linstow) | 342 |
| 246 | | Proglottis of Tænia africana. (After v. Linstow) | 343 |
| 247 | | Head of Tænia africana. (After v. Linstow) | 343 |
| 248 | | Tania confusa. (After Guyer) | 344 |
| 249 | | Tania confusa. (After Ward) | 344 |
| 250 | | Tania echinococcus | 345 |
| 251 | | Echinococcus veterinorum. (After Leuckart) | 347 |
| 252 | | Diagrams of mode of formation of brood capsule and scolices (Stephens) | 348 |
| 252A |
| 253 | | Section through an invaginated echinococcus scolex. (After Dévé) | 350 |
| 254 | | A piece of the wall of an Echinococcus veterinorum stretched out and seen from the internal surface | 350 |
| 255 | | Echinococcus hominis in the liver. (After Ostertag, from Thomas) | 351 |
| 256 | | Section through an echinococcus scolex in process of vesicular metamorphosis. (After Dévé) | 351 |
| 257 | | Diagram of transformation of a scolex into a daughter cyst. (Stephens) | 352 |
| 257A |
| 258 | | Hooklets of echinococcus. (After Leuckart) | 355 |
| 259 | | Echinococcus multilocularis in the liver of the ox. (After Ostertag) | 357 |
| 260 | | Diagram of a transverse section of Ascaris lumbricoides. (After Brandes) | 362 |
| 261 | | Anterior end of an Ascaris megalocephala. (After Nassonow) | 362 |
| 262 | | Transverse section through Ascaris lumbricoides at the level of the œsophagus behind the nerve ring. (After Goldschmidt) | 364 |
| 263 | | Schematic representation of the nervous system of a male Ascaris megalocephala. (After Brandes) | 365 |
| 264 | | Diagram of female genitalia | 368 |
| 264A | | Diagram of male genitalia of a strongylid | 368 |
| 265 | | Transverse section through the ovarian tube of Belascaris cati of the cat | 369 |
| 266 | | Male of the rhabditic form of Angiostomum nigrovenosum | 370 |
| 267 | | Transverse section through the posterior extremity of the body of Ascaris lumbricoides (male) | 370 |
| 268 | | Hind end of a male Ascaris lumbricoides cut across at the level of the dilator cells of the gut. (After Goldschmidt) | 371 |
| 269 | | A piece of the trunk muscle of the pig with encapsuled embryonic Trichinæ | 373 |
| 270 | | Strongyloides stercoralis, female. (After Looss) | 380 |
| 271 | | Strongyloides stercoralis, male. (After Looss) | 380 |
| 272 | | Strongyloides stercoralis, female. (After Looss) | 382 |
| 273 | | Strongyloides stercoralis. (After Looss) | 382 |
| 274 | | Strongyloides stercoralis. (After Looss) | 383 |
| 275 | | Gnathostoma siamense. (After Levinsen) | 385 |
| 276 | | Guinea worm (Dracunculus medinensis). (After Leuckart) | 387 |
| 277 | | Anterior extremity of Guinea worm. (After Leuckart) | 387 |
| 278 | | Dracunculus medinensis. (After Claus) | 387 |
| 279 | | Transverse section of female Guinea worm. (After Leuckart) | 388 |
| 280 | | Cyclops virescens, female | 389 |
| 281 | | Filaria bancrofti. (After Leiper) | 391 |
| 282 | | Mf. bancrofti in thick film, dried and stained with hæmatoxylin. (After Fülleborn) | 397 |
| 283 | | Schematic drawings of the anatomy of Ml. loa and Mf. bancrofti. (After Fülleborn) | 399 |
| 284 | | F. demarquayi. (After Leiper) | 403 |
| 285 | | Mf. demarquayi in thick film, dried and stained with hæmatoxylin. (After Fülleborn) | 404 |
| 286 | | Filaria (?) conjunctivæ. (After Addario) | 405 |
| 287 | | Filaria (?) conjunctivæ. (After Grassi) | 405 |
| 288 | | Setaria equina. (After Railliet) | 408 |
| 289 | | Setaria equina: anterior end. (After Railliet) | 408 |
| 290 | | Loa loa: the anterior end of the male. (After R. Blanchard) | 410 |
| 291 | | Loa loa: anterior portion of the female. (After Looss) | 410 |
| 292 | | Loa loa in situ. (After Fülleborn and Rodenwaldt) | 410 |
| 293 | | Loa loa: male and female. (After Looss) | 410 |
| 294 | | Loa loa: the hind end of a male and of a female. (After Looss) | 411 |
| 295 | | Loa loa: lateral view of tail of male showing papillæ. (After Lane and Leiper) | 411 |
| 296 | | Loa loa. (After Leiper) | 411 |
| 297 | | Mf. loa: in thick film, dried and stained with hæmatoxylin. (After Fülleborn) | 413 |
| 298 | | Acanthocheilonema perstans. (After Leiper) | 414 |
| 299 | | Mf. perstans. (After Fülleborn) | 415 |
| 300 | | Dirofilaria magalhãesi. (After v. Linstow) | 417 |
| 301 | | Trichuris trichiura | 420 |
| 302 | | Trichinella spiralis. (After Claus) | 422 |
| 303 | | Isolated muscular fibre of a rat, invaded by Trichinella. (After Hertwig-Graham) | 425 |
| 304 | | Calcified Trichinella in the muscular system of a pig. (After Ostertag) | 426 |
| 305 | | Various phases of the calcification of Trichinella of the muscles | 426 |
| 306 | | Dioctophyme gigas. (After Railliet) | 432 |
| 307 | | Eggs of Dioctophyme gigas. (After Railliet) | 432 |
| 308 | | Metastrongylus apri. (Stephens) | 433 |
| 309 | | Trichostrongylus instabilis. (After Looss) | 434 |
| 310 |
| 311 | | Trichostrongylus probolurus. (After Looss) | 435 |
| 312 |
| 313 | | Trichostrongylus vitrinus. (After Looss) | 436 |
| 314 |
| 315 | | Hæmonchus contortus. (After Ransom) | 437 |
| 316 |
| 316 | | Mecistocirrus fordi. (After Stephens) | 439 |
| 317 |
| 318 | | Ternidens deminutus. (After Railliet and Henry) | 440 |
| 319 | | Œsophagostomum stephanostomum var. thomasi. (After Thomas) | 442 |
| 320 |
| 321 | | Œsophagostomum stephanostomum var. thomasi. (After Thomas) | 444 |
| 322 |
| 323 | | Ancylostoma duodenale, male and female. (After Looss) | 446 |
| 324 |
| 325 | | Ancylostoma duodenale, showing ventral teeth. (After Looss) | 447 |
| 326 | | Ancylostoma duodenale: diagrammatic representation of excretory system. (After a drawing by Looss) | 448 |
| 327 | | Ancylostoma duodenale. (After Railliet) | 449 |
| 328 | | Ancylostoma duodenale: bursa of male. (After Looss) | 450 |
| 329 | | Ancylostoma duodenale: eggs in different stages of development. (After Looss) | 451 |
| 330 | | Ancylostoma duodenale: larva. (After Leichtenstern) | 452 |
| 331 | | Ancylostoma duodenale. (After Looss) | 453 |
| 332 | | Ancylostoma ceylanicum. (After Looss) | 456 |
| 333 | | Ancylostoma braziliense. (After Gomez de Faria) | 456 |
| 334 | | Necator americanus. (After Looss) | 457 |
| 335 | | Necator americanus: lateral view. (After Looss) | 458 |
| 336 | | Necator americanus: bursa of male. (After Looss) | 458 |
| 337 | | Syngamus kingi: anterior end of male. (After Leiper) | 460 |
| 338 | | Syngamus kingi: anterior end of female. (After Leiper) | 460 |
| 339 | | Bursa of Syngamus trachealis. (Stephens) | 461 |
| 340 | | Physaloptera mordens, Leiper, 1907. (After Leiper) | 462 |
| 341 | | Ascaris lumbricoides. (From Claus) | 463 |
| 342 | | Ovum of Ascaris lumbricoides | 463 |
| 343 | | Ovum of Toxascaris limbata | 466 |
| 344 | | Transverse section through the head part of Belascaris cati from the cat. (After Leuckart) | 466 |
| 345 | | Male female of Oxyuris vermicularis | 468 |
| 346 |
| 347 | | Oxyuris vermicularis: egg freshly deposited | 468 |
| 348 | | Oxyuris vermicularis: egg twelve hours after deposition | 468 |
| 348A | | The male of Echinorhynchus augustatus | 476 |
| 348B | | Anterior portion of the female apparatus of Echinorhynchus acus. (After Wagener) | 476 |
| 348C | | Egg of Echinorhynchus gigas. (After Leuckart) | 477 |
| 348D | | The internal organs of the leech. (After Kennel) | 480 |
| 348E | | Hirudo medicinalis. (After Claus) | 481 |
| 349 | | Leptus autumnalis. (After Gudden) | 485 |
| 350 | | Leptus autumnalis. (After Trouessart) | 485 |
| 351 | | The kedani mite. (After Tanaka) | 487 |
| 352 | | Tetranychus telarius var. russeolus, Koch. (After Artault) | 488 |
| 353 | | Pediculoides ventricosus. (After Laboulbène and Mégnin) | 489 |
| 354 | | Nephrophages sanguinarius: male, ventral surface. (After Miyake and Scriba) | 490 |
| 355 | | Nephrophages sanguinarius: female, dorsal aspect. (After Miyake and Scriba) | 490 |
| 356 | | Tydeus molestus. (After Moniez) | 491 |
| 357 | | Dermanyssus gallinæ. (After Berlese) | 492 |
| 358 | | Dermanyssus hirundinis. (After Delafond) | 492 |
| 359 | | Ixodes ricinus, male. (After Pagenstecher) | 498 |
| 360 | | Female of Ixodes ricinus. (After Pagenstecher) | 498 |
| 361 | | Argas reflexus. (After Pagenstecher) | 506 |
| 362 | | Argas persicus. (After Mégnin) | 507 |
| 363 | | Tyroglyphus farinæ: male. (After Berlese) | 512 |
| 364 | | Tyroglyphus longior, Gerv. (After Fum. and Robin) | 512 |
| 365 | | Rhizoglyphus parasiticus: male and female. (After Dalgetty) | 514 |
| 366 | | Histiogaster (entomophagus ?) spermaticus. (After E. Trouessart) | 515 |
| 367 | | Sarcoptes scabiei. (After Fürstenberg) | 518 |
| 368 | | Sarcoptes scabiei: male, ventral aspect. (After Fürstenberg) | 519 |
| 369 | | Sarcoptes minor var. cati. (After Railliet) | 521 |
| 370 | | Demodex folliculorum of the dog. (After Mégnin) | 522 |
| 371 | | Linguatula rhinaria: female | 524 |
| 372 | | Larva of Linguatula rhinaria (Pentastoma denticulatum). (After Leuckart) | 524 |
| 373 | | Linguatula rhinaria. (After M. Koch) | 525 |
| 374 | | Mouth-parts of Pediculus vestimenti. (After Denny) | 533 |
| 375 | | Ovum of the head louse | 533 |
| 376 | | Head louse, male | 533 |
| 377 | | Pediculus vestimenti, Burm.: adult female | 533 |
| 378 | | Phthirius inguinalis, Leach | 534 |
| 379 | | Head of the bed bug from the ventral surface | 535 |
| 380 | | Dermatophilus penetrans: young female. (After Moniez) | 544 |
| 381 | | Dermatophilus penetrans: older female. (After Moniez) | 544 |
| 382 | | Pulex irritans | 546 |
| 383 | | Larva of flea. (After Railliet) | 546 |
| 384 | | Pulex serraticeps | 546 |
| 385 | | Head of a male and of a female Anopheles. (After Giles) | 549 |
| 386 | | Head of a male and of a female Culex. (After Giles) | 549 |
| 387 | | Mouth-parts of Anopheles claviger. (After Grassi) | 550 |
| 388 | | Anopheles maculipennis. (After Nuttall and Shipley) | 550 |
| 389 | | Longitudinal section of an Anopheles, showing alimentary canal. (After Grassi) | 551 |
| 390 | | Anopheles maculipennis, Meigen. (After Grassi) | 552 |
| 391 | | Larva of Anopheles maculipennis, Fabr. (After Grassi) | 553 |
| 392 | | Larva of Culex. (After Grassi) | 553 |
| 393 | | Pupa of Anopheles maculipennis, Meig. (After Grassi) | 554 |
| 394 | | Heads of Culex and Anopheles. (After Daniels) | 556 |
| 395 | | Eggs of Culex, of Anopheles, of Stegomyia, of Tæniorhynchus, and of Psorophora | 557 |
| 396 | | Diagram showing the structure of a typical mosquito. (Theobald) | 558 |
| 397 | | Types of scales, head and scutellar ornamentation, forms of clypeus. (Theobald, etc., etc.) | 559 |
| 398 | | Neuration of wing. Explanation of wing veins and cells. (Theobald) | 560 |
| 399 | | Wing of Anopheles maculipennis, Meigen | 566 |
| 400 | | Wing of a Culex | 575 |
| 401 | | Wing of Simulium | 579 |
| 402 | | Wing of Chironomus | 579 |
| 403 | | A Ceratopogon, or midge | 580 |
| 404 | | An owl midge, Phlebotomus sp. (From Giles’s “Gnats or Mosquitoes”) | 581 |
| 405 | | Larva of Homalomyia canicularis | 585 |
| 406 | | Larvæ of Calliphora vomitoria | 585 |
| 407 | | Larva of Chrysomyia macellaria. (After Conil) | 585 |
| 408 | | The screw-worm fly (Chrysomyia macellaria) | 587 |
| 409 | | Ochromyia larva on the skin of man, South Africa. (After Blanchard) | 590 |
| 410 | | Head end of “larva of Natal.” (After Gedoelst) | 591 |
| 411 | | Lund’s larva. (After Gedoelst) | 593 |
| 412 | | Dermatobia noxialis, Goudot | 597 |
| 413 | | Larva of Dermatobia cyaniventris. (After Blanchard) | 597 |
| 414 | | Larva of Dermatobia cyaniventris. (After Blanchard) | 597 |
| 415 | | The ox gad fly (Tabanus bovinus, Linn.) | 601 |
| 416 | | The brimp (Hæmatopota pluvialis, Linn.) | 602 |
| 417 | | Head of Glossina longipalpis. (After Grünberg) | 604 |
| 418 | | Antenna of Glossina pallidipes, male. (After Austen) | 604 |
| 419 | | Glossina palpalis and puparium. (After Brumpt) | 607 |
| 420 | | The tsetse-fly (Glossina morsitans, Westwood) | 608 |
| 421 | | The stinging fly (Stomoxys calcitrans, Linn.) | 609 |
| 422 | | Trichomonas from cæcum and gut of rat. (Original, Fantham) | 735 |
| 423 | | Chilomastix (Tetramitus) mesnili. (Original, Fantham) | 736 |
We regret to have taken without permission from the
“Transactions of The Society of Tropical Medicine and
Hygiene,” London, the following diagrams:—
| Pages | Figures |
| 268 | No. 169 |
| 269 | " 170 |
| 391 | " 281 |
| 411 | " 295 and 296 |
| 414 | " 298 |
| 460 | " 337 and 338 |
and tender our regret to the Society in question for having
done so.
P. 31, line 6 from bottom: delete “human,” as Leidy really worked with Endamœba
blattæ, parasitic in the gut of the cockroach.
P. 43, line 12 from bottom: for “John’s” read “Johns.”
P. 44, line 13 from bottom: for “Amœba buccalis, Sternberg,” read “Amœba buccalis,
Steinberg.”
P. 46, line 13 from top: for “breath” read “breadth.”
P. 53, In footnote 1, line 6 from bottom: insert “see” before Arch. f. Protistenk.
P. 75: To paragraph regarding development of the parasite in the fly’s salivary glands,
add that the crithidial phase takes two to five days.
P. 111, line 8 from top: the date of Sangiorgi should be 1911.
P. 142, line 7 from top: insert “Genus.” before Eimeria.
P. 252, Insert heading “Family. Opisthorchiidæ, Braun, 1901,” above “Sub-family.
Opisthorchiinæ, Looss, 1899.”
P. 351, description of fig. 255, line 3: for “Thoma” read “Thomas.”
P. 471, line 15 from bottom: for “alcohol 100 parts” read “alcohol 100 c.c.”
P. 472, line 11 from bottom: for “Or (2) 10 per cent. formalin,” read “Or (2) fix in hot
10 per cent. formalin.”
P. 493, line 21 from top: for “Conoy” read “Couvy.”
P. 589, line 2 from top: for “carnosa” read “carnaria.”
P. 620, line 15 from top: for “fo” read “of.”
P. 622, line 12 from bottom: delete comma after quantity.
P. 626, line 6 from bottom: delete comma after Mackie (1915).
P. 638: insert title “TREMATODES” above that of “Fascioliasis.”
P. 709, line 9 from bottom: omit second Pediculus capitis.
P. 748, line 8 from top: for “cytologica” read “cytological.”
P. 753, line 4 from bottom: for “Fercocercous” read “Furcocercous.”
P. 755 line 7: for “Oncocerca” read “Onchocerca.”
ON PARASITES IN GENERAL.
By the term PARASITES is understood living organisms which,
for the purpose of procuring food, take up their abode, temporarily
or permanently, on or within other living organisms. There are
both plants and animals (Phytoparasites and Zoöparasites) which
lead a parasitic life in or upon other plants and other animals.
Phytoparasites are not included in the following descriptions of
the forms of parasitism, but a very large number of animal parasites
(zoöparasites) are described. The number of the latter, as a rule,
is very much underrated. How great a number of animal parasites
exists may be gathered from the fact that all classes of animals are
subject to them. Some of the larger groups, such as Sporozoa, Cestoda,
Trematoda and Acanthocephala, consist entirely of parasitic species,
and parasitism even occurs among the vertebrates (Myxine). It
therefore follows that the characteristics of parasites lie, not in their
structure, but in the manner of their existence.
Parasitism itself occurs in various ways and degrees. According
to R. Leuckart, we should distinguish between OCCASIONAL (temporary)
and PERMANENT (stationary) PARASITISM. Occasional parasites, such
as the flea (Pulex irritans), the bed-bug (Cimex lectularius), the leech
(Hirudo medicinalis), and others, only seek their “host” to obtain
nourishment and find shelter while thus occupied. Without being
bound to the host, they usually abandon the latter soon after the
attainment of their object (Cimex, Hirudo), or they may remain on
the body of their host throughout their entire development from the
hatching of the egg (Pediculus). It follows from this mode of living
that the occasional parasites become sometimes distinguishable from
their free-living relatives, though only to a slight extent. It is, therefore,
seldom difficult to determine the systematic position of temporary
parasites from their structure.
In consequence of their mode of life, all these temporary parasites
live on the external surface of the body of their host, though more
rarely they take up their abode in cavities easily accessible from the
exterior, such as the mouth, nose and gills. They are therefore
frequently called Epizoa or Ectoparasites; but these designations
do not cover only the temporary parasites, because numerous epizoa
(as for instance the louse) are parasitic during their entire life.
In contradistinction to these temporary parasites, the permanent
parasites obtain shelter as well as food from their host for a long
period, sometimes during the entire course of their life. They do not
seek their host only when requiring nourishment, but always remain
with it, thus acquiring substantial protection. The permanent parasites,
as a rule, live within the internal organs, preferably in those
which are easily accessible from the exterior, such as the intestine, with
its appendages. Nevertheless, permanent parasites are also found in
separate organs and systems, such as the muscular and vascular
systems, hollow bones and brain, while some live on the outer skin.
Here again, the terms Entozoa and Endoparasites do not include all
stationary parasites; to the latter, for instance, the lice belong, which
pass all their life on the surface of the body of their host, where they
find shelter and food and go through their entire development. The
ectoparasitic trematodes, numerous insects, crustacea, and other
animals live in the same manner.
All “Helminthes,” however, belong to the group of permanent
parasites. This term is now applied to designate certain lowly worms
which lead a parasitic life (intestinal worms); but they are not all
so termed. For instance, the few parasitic Turbellaria are never
classed with the helminthes, although closely related to them. The
turbellarians, in fact, belong to a group of animals of which only
a few members are parasitic, whereas the helminthes comprise those
groups of worms of which all species (Cestoda, Trematoda, Acanthocephala),
or at least the majority of species (Nematoda), are parasitic.
Formerly the Linguatulidæ (Pentastoma) were classed with the
helminthes because their existence is also endoparasitic, and because
the shape of their body exhibits a great similarity to that of the true
helminthes. Since the study of the development of the Linguatulidæ
(P. J. van Beneden, 1848, and R. Leuckart, 1858) has demonstrated
that they are really degenerate arachnoids, they have been separated
from the helminthes.
It is hardly necessary to emphasize the fact that the helminthes
or intestinal worms do not represent a systematic group of animals,
but only a biological one, and that the helminthes can only be discussed
in the same sense as land and water animals are mentioned,
i.e., without conveying the idea of a classification in such a grouping.
It is true that formerly this was universally done, but very soon the
error of such a classification was recognized. Still, until the middle
of last century, the helminthes were regarded as a systematic group,
although C. E. v. Baer (1827) and F. S. Leuckart (1827) strenuously
opposed this view. Under the active leadership of J. A. E. Goeze,
J. G. H. Zeder, J. G. Bremser, K. A. Rudolphi and F. Dujardin, the
knowledge of the helminthes (helminthology) developed into a special
study, but unfortunately it lost all connection with zoology. It
required the intervention of Carl Vogt to disestablish the helminthes
as one class of animals, by uniting the various groups with those of
the free-living animals most closely related to them (Platyhelminthes,
Nemathelminthes).
Permanent parasitism in the course of time has caused
animals adopting this mode of life to undergo considerable, sometimes
even striking, bodily changes, permanent ectoparasites having
as yet undergone least alteration. The latter sometimes bear so
unmistakably the likeness to the group to which they belong, that
even a superficial knowledge of their structure and appearance often
suffices for the recognition of their systematic position. For instance,
though the louse, like many decidedly temporary parasites, has lost
its wings—a characteristic of insects—in consequence of parasitism,
yet nobody would deny its insect nature; such also occurs in other
temporary parasites (Cimex, Pulex). On the other hand, the changes
in a number of permanent ectoparasites (such as parasitic Crustacea)
are far more considerable, and correspond with those that have
occurred in permanent endoparasites.
These alterations depend partly on retrogression and partly on
the acquisition of new peculiarities. In the former case, the change
consists in the loss of those organs which have become useless in a
permanent parasitic condition of existence, such as wings in the
louse, and the articulated extremities seen in the larval stage of
parasitic Crustacea. The loss of these organs goes hand in hand
with the cohesion of segments of the body that were originally
separate, and alterations in the muscular and nervous systems. In
the same manner another means of locomotion is lost—the ciliated
coat—which is possessed by many permanent parasites during their
larval period. To all appearances, this character is not secondary
and recently acquired, but represents a primary character inherited
from free-living progenitors, and still transmitted to the altered
descendants, because of its use during the larval stage (e.g., the larvæ
of a great many Trematodes, the oncospheres of some Cestodes).
Amongst the retrogressions, the loss of the organs of sense may
be mentioned, particularly the eyes, which are still present, not only
in the nearest free-living forms but also in the free-living larvæ of
true parasites. It is only quite exceptionally that the eyes are
subsequently retained, as a rule they are lost. Lastly, in a great
many cases the digestive system also disappears, as in parasitic
Crustacea, in a few nematodes and trematodes, in all cestodes and
Acanthocephala. There remain at most the rudiments of the muscles
of the fore-gut, but these are adapted to entirely different uses.
The new characters which permanent parasites may acquire are,
first of all, the remarkably manifold CLASPING and CLINGING ORGANS,
which are seldom (as in parasitic Crustacea) directly joined on to
already existing structures. In those instances in which organs
for the conveyance of food are retained, these likewise frequently
undergo transformation, in consequence of the altered food and
manner of feeding. Such alterations consist, for instance, in the
transformation of a masticating mouth apparatus into the piercing
and sucking organs of parasitic insects.
Hermaphroditism (as in Trematodes, Cestodes, and a few Nematodes)
is a further peculiarity of many permanent parasites; moreover,
the association in couples that occurs, especially in trematodes, may
lead to complete cohesion and, exceptionally, also to re-separation
of the sexes. In many cases the females only are parasitic, while
the males live a free life, or there may be in addition the so-called
complementary males. Occasionally the male alone is parasitic, and
in that case lives within the female of the same species, which may
live free, like certain Gephyrea (Bonellia); or the female also may
be parasitic, as Trichosoma crassicaudum, which lives in the bladder
of the sewer rat (Mus decumanus).
We have numerous proofs that demonstrate how considerably
the original features of many parasites have become changed. We
need only draw attention to the aforementioned Linguatulidæ, also
to many of the parasitic Crustacea belonging to various orders. In
all of these a knowledge of the larval stages—in which there is no
alteration, or at most only a slight degree of change—serves to
determine their systematic position, i.e., the nearest conditions of
relationship.
The most remarkable changes are observed in those groups that
contain only a few parasitic members, the majority leading a free
life. A striking instance is afforded by a snail, the well-known Entoconcha
mirabilis, Müller. This mollusc consists merely of an elongated
sac living in a Holothurian (Synapta digitata). It possesses none
of the characteristics of either the Gastropoda or any molluscs,
and in its interior there is nothing to be observed but the organs
of generation and the embryos. Nevertheless, the Entoconcha is
decidedly a parasitic snail, as is clearly proved by its larvæ, but
it is a snail which, in consequence of parasitism, has lost all the
characteristics of molluscs in its mature condition, but still exhibits
them in the early stages of development.
Certain nematodes show very clearly to what devious courses
parasitism may lead. The Atractonema gibbosum, the life-history of
which has been described by R. Leuckart, and which lives in the
larvæ and pupæ of a dipterous insect (Cecidomyia), exhibits, in its
early stage, the ordinary characteristics of other threadworms. A
few weeks later—the males having died off immediately after copulation—the
females are transformed into spindle-shaped bodies, the
mouth and anus of which are closed. They carry with them an irregularly
shaped appendage, in which the segmenting ova are situated,
and in which the further conditions of life of the Atractonema are
accomplished. A minute examination has demonstrated that this
appendage is the prolapsed and enlarged vagina of the animal which
has become merely a supplementary attachment. The conditions
present in the Sphærularia, the nematoid nature of which was long
undiscovered, are still more remarkable. It was only when Siebold
proved that typical nematodes were hatched from their eggs
that their nature was recognized. The nematodes thus produced
have not the slightest resemblance to the parent.
The researches of Lubbock, A. Schneider, and more particularly
of R. Leuckart, have shown that what we call Sphærularia bombi is not
an animal but merely an organ—the vagina—of a nematode worm.
This vagina at first grows, sac-like, from the body of the tiny nematode;
it gradually assumes enormous dimensions (2 cm. in length);
it contains the sexual organs and parts of the intestine. The remaining
portion of the actual animal then becomes small and shrivelled;
it may be easily overlooked, being but an appendage to the vagina
with its independent existence, and it finally disappears altogether.
The GREAT FERTILITY of parasites is another of their peculiarities,
though this may be also the case to a certain degree with some of
the free-living animals, the progeny of which are likewise exposed
to enormous destruction.
More remarkable, however, is the fact that the young of the
endoparasites only very exceptionally grow to maturity by the side
of their parents. Sooner or later they leave the organ inhabited by
the parents, frequently reach the open, and after a shorter or longer
period of free existence seek new hosts. During their free period,
moreover, a considerable growth may be attained, or metamorphosis
may take place, or even multiplication. In the exceptional cases in
which the young remain within the same host, they nevertheless
usually quit the organ inhabited by the parents. They likewise rarely
attain maturity within the host inhabited by the parents, but only,
as in other cases, after having gained access to fresh hosts.
These transmigrations play a very important rôle in the natural
history of the internal parasites, but they frequently conceal the cycle
of development, for sometimes there are INTERMEDIATE GENERATIONS,
which themselves invade intermediate hosts. Even when there are no
intermediate generations, THE SYSTEM OF INTERMEDIATE HOSTS is
frequently maintained by the endoparasites.
According to the kind of food ingested by parasites, it has
recently become usual to separate the true parasites from those
animals that feed on the superfluity of the food of the host, or on
products which are no longer necessary to him, and to call the latter
MESSMATES or COMMENSALS. As examples, the Ricinidæ are thus
designated, because, like actual lice, they dwell among the fur of
mammals or the plumage of birds. They do not, however, suck
blood, for which their mouth apparatus is unsuited, but subsist
on useless epidermic scales. These epizoa, according to J. P. van
Beneden, are, to a certain extent, useful to their hosts by removing
deciduous materials which under certain circumstances might become
harmful to them.1 This investigator, who has contributed so greatly
to our knowledge of parasites, assigns the Ricines to the MUTUALISTS,
under which term he comprises animals of various species which
live in common, and confer certain benefits on one another. The
mutualists are usually intimately connected in a mutually advantageous
association known as “symbiosis.”2
Incidental and Pseudo Parasites.—In many cases the parasites are
confined to certain hosts, and may therefore be designated as specific
to such hosts. Thus, hitherto, Tænia solium and Tænia saginata
in their adult condition have only been found in man; Tænia crassicollis
only in the cat; Brandesia (Distoma) turgida and Halipegus
(Distoma) ovocaudatas only in Rana esculenta, and so forth. In many
other cases, however, certain species of parasites are common to several,
and sometimes many, species of hosts; Dipylidium caninum is found
in the domestic cat as well as in the dog; Fasciola hepatica is found in
a large number of herbivorous mammals (nineteen species), Diplodiscus
(Amphistomum) subclavatus in numerous urodele and ecaudate amphibia,
Holostomum variabile in about twenty-four species of birds,
and so on. In these cases the hosts are almost invariably closely related,
belonging, as a rule, to the same family or order, or at any rate to the
same class. Trichinella spiralis, which is found in man, and in the pig,
bear, rat, mouse, cat, fox, badger, polecat and marten, and is capable
of being artificially cultivated in the dog, rabbit, sheep, horse, in
other mammals, and even in birds, is one of the most striking
exceptions.
Some parasites are so strictly confined to one species of host that,
even when artificially introduced into animals very closely related
to their normal host, they do not thrive, but sooner or later, often
very quickly, die off, and very rarely establish themselves. For
example, repeated attempts have been made to rear the adult Tænia
solium in the dog, or to rear Cysticercus cellulosæ in the ox, or the
Cysticercus of Tænia saginata in the pig, but they have always proved
unsuccessful. Only exceptionally has it been possible to transfer
Cœnurus cerebralis, the larval stage of a tapeworm (Tænia cœnurus) of
the dog from the brain of the sheep to that of the domestic goat.
On the other hand, in the case of the Trichinellæ transference to
a different host is easily accomplished.
Under natural conditions, it is not uncommon for certain kinds
of specific parasites to occur occasionally in unusual hosts. Their
relationship to the latter is that of INCIDENTAL PARASITES. Thus
Echinorhynchus gigas, a specific parasite of the pig, is only an
incidental parasite of man; Fasciola hepatica and Dicrocœlium
lanceatum are specific to numerous kinds of mammals, but may
be found incidentally in man. On the other hand, Dibothriocephalus
latus, a specific parasite of man, may occasionally take up its abode
in the dog, cat and fox. As a rule, all those parasites of man that
are only rarely met with, notwithstanding that human beings are
constantly being observed and examined by medical men, are
termed INCIDENTAL PARASITES OF MAN. In many cases we are
acquainted with the normal or specific host of these parasites.
Thus we know the specific host of Balantidium coli, Eimeria stiedæ,
Fasciola hepatica, Dipylidium caninum, etc.; in others the host is as
yet unknown. In the latter case the question partly relates to such
forms as have been so deficiently described that their recognition
is impossible, partly to parasites of man in various regions of the
earth, the Helminthes and parasites of which are totally unknown
or only slightly known, or finally to early developmental stages that
are difficult to identify. Animals that usually live free, and exceptionally
become parasitic, may likewise be called incidental parasites.
In this category are included a few Anguillulidæ that have been
observed in man; also Leptodera appendiculata, which usually lives
free, but may occasionally become parasitic in black slugs (Arion
empiricorum): when parasitic it attains a larger size, and produces
far more eggs than when living a free life. In order to avoid errors,
the term “incidental parasites” should be confined to true parasites
which, besides living in their normal host, may also live in other
hosts. Leuckart speaks of FACULTATIVE PARASITISM in such forms
as Leptodera. L. Oerley3 succeeded in artificially causing Leptodera
(Rhabditis) pellio to assume facultative parasitism by introducing these
worms into the vagina of mice, where the parasites remained alive and
multiplied. Leptodera pellio dies in the intestines of mammals and
man; it remains alive in frogs, but always escapes into the open
with the fæces.
Recently the incidental parasites of man have also been called
“Pseudo-parasites” or “Pseudo-helminthes.” Formerly, however,
these terms were applied not only to living organisms that do
not and cannot live parasitically, and that only exceptionally and
incidentally get into man, but also to any foreign bodies, portions
of animals and plants, or even pathological formations that left the
human system through the natural channels, and the true nature
of which was misunderstood. Frequently these bodies were described
as living or dead parasites and labelled with scientific names, as if
they were true parasites. A study of these errors, which formerly
occurred very frequently, would be as interesting as it would be
instructive. It is better not to use the expression pseudo-parasites for
incidental parasites, but to keep to the original meaning, for it is not
at all certain that pseudo-parasites are not described, even nowadays.
The Influence of Parasites on the Host.—In a great many cases,
we are not in a position to state anything regarding any marked
influence exercised by the parasite on the organism, and on the conditions
of life, of the host. Most animals and many persons exhibit
few signs of such influence, an exception being infestation with
helminthes and certain other parasites which produce eosinophilia
in the blood. As a general rule, the parasite, which is always smaller
and weaker than its host, does not attempt to endanger the life
of the latter, as simultaneously its own existence would be threatened.
The parasite, of course, robs its host, but usually in a scanty and
sparing manner, and the injuries it inflicts can hardly be taken into
account. There are, however, numerous cases4 in which the situation
of the parasites or the nature of their food, added to their number and
movements, may cause more or less injury, and even threaten the life
of the host. It stands to reason that a Cysticercus cellulosæ situated in
the skin is of but slight importance, whereas one that has penetrated
the eye or the brain must give rise to serious disorders. A cuticular
or intestinal parasite is, as a rule, less harmful than a blood parasite.
A helminth, such as an Ascaris lumbricoides or a tapeworm, that
feeds on the residues of foodstuffs within the intestine, will hardly
affect its host by depriving it of this material. The case is different
when the parasites are very numerous, especially when the heavily
infested host happens to be a young individual needing all it ingests
for its own requirements, and therefore unable to sustain the drain of
numerous intruders in the intestine. Disturbances also set in more
rapidly when the intestinal helminthes are blood-suckers, the injury
to the host resulting from the kind of food taken by the parasite.
Generally, the disorders caused by loss of chyle are insignificant
when compared with those induced by the GROWTH and agglomeration
of the helminthes. The latter may cause chiefly obstructions of
small vessels or symptoms of pressure in affected or contiguous organs,
with all those complications which may arise secondarily, or they may
even lead to the complete obliteration of the organ invaded. Of course
the symptoms will vary according to the nature of the organ attacked.
In consequence also of the MOVEMENTS of the parasites, disorders
are set up that may tend to serious pathological changes of the
affected organs. The collective migrations, undertaken chiefly by
the embryos of certain parasites (as in trichinosis, acute cestode
tuberculosis), are still more harmful, as are also the unusual migrations
of other parasites, which, incidentally, may lead to the formation
of so-called worm abscesses or to abnormal communications (fistulæ)
between organs that are contiguous but possess no direct connection.
Recently, several authors have called attention to the fact that
the helminthes produce substances that are TOXIC to their host; and
the effects of such poisons explain the pathology of helminthiasis
far more satisfactorily than the theory of reflex action.
In a number of cases these toxic materials (leucomaines) have been
isolated and their effects on living organisms demonstrated by actual
experiments. It also appears that the absorption of materials formed
by the decomposition of dead helminthes may likewise cause toxic
effects. However, our knowledge of these conditions is as yet in
its initial stage.5
Nearly all the symptoms caused directly or indirectly by parasites
are of such a nature that the presence of the parasites cannot be
diagnosed with any certainty, or only very rarely. The most that
can be done is to deduce the presence of parasites by the exclusion
of other causes. Fortunately, however, there are sufficient means
by which we may confirm the diagnosis in a great many cases. Such
means consist not only in a minute examination of the patient by
palpation, percussion and local inspection, but also in the microscopical
examination of the natural secretions and excretions of the
body, such as sputum, nasal mucus, urine and fæces. Though such
examinations may entail loss of time, they are necessary in the interest
of the patient. It appears, moreover, that quackery, which has
gained considerable ground even in the treatment of the helminthic
diseases of man, can thus be considerably limited.
Origin of Parasites.6—In former times, when the only correct views
that existed related to the origin of the higher animals, the mode
of multiplication of parasites as well as of other lowly animals
was ascribed to SPONTANEOUS GENERATION (generatio æquivoca), and
this opinion prevailed throughout the middle ages. The writers on
natural science merely devoted their time to the interpretation of the
views of the old authors, and perpetuated the opinions of the ancients
on questions, which, even in those days, could have been correctly
explained merely by observation.
It was only when observations were again recommenced, and the
microscope was invented, that the idea of spontaneous generation
became limited. Not only did the microscope reveal the organs of
generation or their products (eggs) in numerous animals, but Redi
succeeded in proving that the so-called Helcophagi (flesh maggots) are
only the progeny of flies, and never appear in the flesh of slaughtered
animals when fully developed flies are prevented from approaching
and depositing their eggs on it. Swammerdam likewise knew that
the “worms” living in the caterpillars of butterflies were the larvæ
of other insects (ichneumon flies) which had laid their eggs in their
bodies; he also discovered the ova of lice. The two authors mentioned
were, however, unwilling to see that the experience they had
gained regarding insects applied to the helminthes. Leeuwenhoek
also vehemently opposed the theory of a spontaneous generation,
maintaining that, on a basis of common-sense, eggs, or at all events
germs, must exist, even though they could not be seen.
The use of the microscope also revealed a large number of very
small organisms in the water and moist soil, some of which undoubtedly
resembled helminthes. Considering the wide dissemination
of these minute organisms, it was natural to conjecture that after their
almost unavoidable introduction into the human system they should
grow into helminthes (Boerhave, Hoffmann). Linnæus went even
further, for he traced the descent of the liver-fluke of sheep from
a free-living planaria (Dendrocœlum lacteum), the Oxyuris vermicularis
from free-living nematodes, and the Tænia lata (i.e., Dibothriocephalus
latus) from a tapeworm (Schistocephalus solidus) found free in the
water. Linnæus’ statements met with general approval. However,
we must bear in mind that at that time the number of helminthes
known was very small, and many of the forms that we have long
ago learned to differentiate as specific were then regarded as belonging
to one species. Linnæus’ statements were partly supported by similar
discoveries by other investigators, such as Unzer, and partly also by
the discovery of eggs in many helminthes. It was believed that the
eggs hatched in the outside world gave rise to free-living creatures,
and that these, after their introduction into the intestine, were transformed
into helminthes. By means of these eggs the old investigators
tried to explain the HEREDITARY TRANSMISSION of the intestinal
worms, which was universally believed until the commencement of
the last century. Some authors went so far as to regard the intestinal
worms as congenital or inherited; they maintained the possibility
of direct transmission, as in suckling, and denied that the eggs
reaching the external world had anything to do with the propagation
of the parasites.
The more minute comparison between the supposed free-living
stages of the helminthes and their adult forms, and the impossibility
of finding corresponding free forms for the ever-increasing number
of parasitic species, revealed the improbability of Linnæus’ statements
(O. Fr. Müller). It was the latter author also who recognized the
origin of the tapeworms (Schistocephalus, Ligula) found free in the
water. They originate from fishes which they quit spontaneously.
However, in spite of the fact that van Doeveren and Pallas
correctly recognized the significance of the eggs in the transmission
of intestinal worms, these statements remained disregarded,
as did Abildgaard’s observation, experimentally confirmed, that the
(immature) cestodes from the abdominal cavity of sticklebacks became
mature in the intestines of aquatic birds. Moreover, at the end of
the eighteenth and the commencement of the nineteenth centuries,
after helminthology had been raised to a special branch of study
by the successful results of the investigations of numerous authors
(Goeze, Bloch, Pallas, Müller, Batsch, Rudolphi, Bremser), many
of whom experienced a “divine joy” in searching the intestines of
animals for helminthes, some authors reverted to generatio æquivoca,
without, however, entirely denying the existence of organs of generation
and eggs. The fact that a few nematodes bore living progeny—a
fact of which Goeze was already aware—had no influence on the
erroneous opinion, as in such cases it was considered that the young
continued to develop beside the old forms. There were also
many helminthes known that never developed sexual organs and
never produced eggs, and which therefore were referred to generatio
æquivoca. People were convinced that the intestinal mucous membrane
or an intestinal villus could transform itself into a worm,
either in a general morbid condition of the body, or in pathological
changes of a more local character. The appearance of helminthes
was even regarded as useful and as a means for the expulsion of
injurious matter.
These views, firmly rooted and supported by such eminent
authorities as Rudolphi and Bremser, could not easily be overthrown.
First, a change took place in the knowledge of the trematodes.
In 1773, O. Fr. Müller discovered Cercariæ living free in water.
He regarded them as independent creatures and gave them the name
that is still used at the present time. Nitzsch, who also minutely
studied these organisms and who recognized the resemblance of
the anterior part of their bodies to a Fasciola, did not, however, arrive
at a correct conclusion. He regarded the combination rather as
that of a Fasciola with a Vibrio, for which he mistook the characteristic
tail of the cercaria. He also noticed the encystment (transformation
into the “pupa”) on foreign bodies of many species of
these animals, but was of opinion that this process signified only the
termination of life.
Considerable attention was attracted to the matter when Bojanus
first published a paper entitled “A Short Note on Cercaria and their
Place of Origin.” He pointed out that the cercariæ creep out of the
“royal yellow worms,” which occur in freshwater snails (Limnæa,
Paludina), and are probably generated in these worms.
Oken, in whose journal, Isis (1818, p. 729), Bojanus published his
discovery, remarks in an annotation, “One might lay a wager that
these Cercariæ are the embryos of Distomes.” Soon after (1827),
C. E. v. Baer was able to confirm Bojanus’ hypothesis that the
cercariæ as a “heterogeneous brood” originated from spores in parasitic
tubes in snails (germinating tubes). Moreover, Mehlis (Isis, 1831,
p. 190) not only discovered the opercula of the ova of Distoma, but
likewise saw the infusorian-like embryo emerge from the eggs of
Typhlocœlum (Monostomum) flavum and Cathæmasia (Distoma) hians.
A few years later (1835) v. Siebold observed the embryos (miracidia) of
the Cyclocœlum (Monostomum) mutabile, and discovered in their interior
a cylindrical body that behaved like an independent being (“necessary
parasite”), and was so similar in appearance to the “royal yellow
worms” (Bojanus) that Siebold considered the origin of the latter from
the embryos of trematodes as, at all events, possible. Meanwhile,
v. Nordmann of Helsingfors had in 1832 seen the miracidia of flukes
provided with eyes swimming in water; v. Siebold (1835) had observed
the embryos, or oncospheres, of tapeworms furnished with six
hooklets in the so-called eggs of the Tænia; while Creplin (1837) had
discovered the “infusorial” young of the Diphyllobothrium (Bothriocephalus)
ditremum, and conjectured that similar embryos were to be
found in other cestodes with operculated eggs. At all events, the fact
was established that the progeny of the helminthes appeared in various
forms and was partly free living. The researches of Eschricht (1841)
were likewise of influence, as they elucidated the structure of the
Bothriocephali, and proved that the encysted and sexless helminthes
were merely immature stages.
J. I. Steenstrup (1842) was, however, the first to furnish explanations
for the numerous isolated and uncomprehended discoveries.
Commencing with the remarkable development of the Cœlenterata,
he established the fact that the Helminthes, especially the endoparasitic
trematodes, multiply by means of alternating and differently
formed generations. Just as the polyp originating from the egg of
a medusa represents a generation of medusæ, so does the germinal
tube (“royal yellow worm”) originating from the ciliated embryo of
a Distoma, etc., represent the cercaria. These were consequently
regarded as the progeny of trematodes, and Steenstrup, guided by
his observations, conjectured that the cercaria, whose entrance into
the snails he had observed accompanied by the simultaneous loss of
the propelling tail, finally penetrated into other animals, in which they
became flukes.
Part of this hypothetical cycle of development was erroneous, and
in other particulars positive observation was lacking, but the path
pursued was in the right direction. Immediately after the appearance
of Steenstrup’s celebrated work, v. Siebold expressed his opinion
that the encapsuled flukes certainly had to travel, i.e., to be transmitted
with their bearers into other hosts, before becoming mature.
This view was experimentally confirmed by de Filippi, La Valette
St. George (1855), as well as by Pagenstecher (1857), while the metamorphosis
of the ciliated embryo of Distoma into a germinal tube
was first seen by G. Wagener (1857) in Gorgodera (Distoma) cygnoides
of frogs. All that we have subsequently learned from the works of
numerous investigators about the development of endoparasitic trematodes
has certainly increased our knowledge in various directions, and,
apart from the deviating development of the Holostomidæ has, as a
whole, confirmed the briefly sketched cycle of development.
Steenstrup’s work on the cestodes did not attract the same attention
as his work on trematodes. Steenstrup always insisted on the
“nurse” nature of the cysticerci and other bladder-worms. Abildgaard
(1790), as well as Creplin (1829 and 1839), had already furnished
the information that certain sexless cestodes (Schistocephalus and
Ligula) from the abdomen of fishes only become mature after their
transference to the intestine of aquatic birds. These passive
migrations were confirmed in an entire series of other cestodes,
particularly by v. Siebold (1844, 1848, 1850) and E. J. van Beneden
(1849), not by actual experiment, but by undoubted observation.
It was correctly believed that the ova or oncospheres penetrate
into certain intermediate hosts, in which they develop into unsegmented
larvæ. Here they remain until, with their host, they are
swallowed by some predacious animal. They then reach the intestine,
being freed from the surrounding membranes through the process
of digestion, and settle themselves there to form the adult chain
of proglottides. Though some few scientists, such as P. J. van
Beneden and Em. Blanchard, deduced from these observations that
the bladder-worms (Cysticerci), which had hitherto been regarded as
a separate class of helminthes, were only larval Tæniæ, this correct
view was not at first universally accepted. The foundation was too
slight, and van Beneden was of opinion that the Cysticerci were not
necessary, but only appeared incidentally.
v. Siebold was a strenuous opponent to this theory, notwithstanding
his experiences on the change of hosts of the Tetrarhynchus.
Together with Dujardin (1850) he conjectured that the
Tæniæ underwent a deviating cycle of development. He was of
opinion that the six-hooked oncospheres left the intestine, in
which the older generation lived, and were scattered about with
the fæces, and finally re-entered per os (i.e., with water and food)
a host similar to the one they had left, in the intestine of which
they were directly transformed into tapeworms. A change of host
such as occurred in other cestodes was not supposed to take place
(the history of the cestodes was at this time not entirely established).
As the oncospheres of the Tænia are enveloped in one
calcareous or several softer coverings which they cannot leave
actively, and as, in consequence of this condition, innumerable
oncospheres cannot penetrate into an animal, and others cannot
reach the proper animal, v. Siebold conceded, at least for the
latter, the possibility of a further development. But this was only
supposed to occur because they had either invaded wrong hosts,
or, having reached the right hosts, had penetrated organs unsuitable
to their development, and had thus gone astray in their
travels, and had become hydropically degenerated tæniæ. This
was v. Siebold’s explanation of bladder-worms. Naturally, v. Siebold
himself conjectured that a recovery of the diseased tapeworm might
occur, in a few exceptional cases, after transmission into the correct
host, as, for instance, in the Cysticercus fasciolaris of mice, the host of
which is the domestic cat, and in which there is a seemingly normally
developed piece of tapeworm situated between the caudal vesicle and
the cysticercus head.
Guided by correct views, F. Küchenmeister undertook in Zittau
the task of confirming the metamorphosis of Cysticercus pisiformis
of hares and rabbits, into tapeworms in the intestine of the dog
by means of feeding experiments. The first reports on the subject,
published in 1851, were not likely to meet with universal approval,
because Küchenmeister first diagnosed the actual tapeworm he had
been rearing as Tænia crassiceps, afterwards as Tænia serrata, and
finally as Tænia pisiformis n. sp. However, in any case, Küchenmeister,
by means of the reintroduction of experimental investigation, rendered
a great service to helminthology.
The publication of Küchenmeister’s works induced v. Siebold
to undertake similar experiments (1852 and 1853), which were
partly published by his pupil Lewald in 1852. But the positive
results obtained hardly changed Siebold’s opinion, for although he
no longer considered the bladder-worms as hydropically degenerated
tapeworms, he still regarded them as tæniæ that had strayed. The
change of opinion was partly due to an important work of the
Prague zoologist, v. Stein (1853). He was able to examine the
development of a small bladder-worm in the larvæ of the well-known
meal-worm (Tenebrio molitor) and to demonstrate that, as Goeze had
already proved in the case of Cysticercus fasciolaris of mice, first the
caudal vesicle is formed and then the scolex, whereas Siebold believed
that in bladder-worms the posterior end of the scolex was formed
first, and that this posterior end underwent a secondary hydropic
degeneration.
In opposition to v. Siebold, Küchenmeister successfully proved
the necessity of the bladder-worm stage by rearing tapeworms
in dogs from the Cysticercus tenuicollis of domestic mammals and
from the Cœnurus cerebralis of sheep. He, and simultaneously
several other investigators independently, succeeded, with material
provided by Küchenmeister, in rearing the Cœnurus cerebralis in
sheep from the oncospheres of the Tænia cœnurus of the dog (1854).
R. Leuckart obtained similar results in mice by feeding them with the
mature proglottides of the Tænia crassicollis of cats (1854).
Küchenmeister also repeatedly reared the Tænia solium of man
from the Cysticercus cellulosæ of pigs (1855), and from the embryos
of this parasite P. J. van Beneden succeeded in obtaining the same
Cysticercus in the pig (1854). As Küchenmeister distinguished the
Tænia mediocanellata, known to Goeze as Tænia saginata, amongst
the large tæniæ of man (1851), so it was not long before R. Leuckart
(1862) succeeded in rearing the cysticercus of the hookless tapeworm
in the ox. It is particularly to this last-named investigator that
helminthology is indebted more than to any other author. He
followed the gradual metamorphosis from oncospheres to cystic
worms in all its details.
In view of all the researches that were made, and which are too
numerous to mention individually, the idea that bladder-worms are
abnormal or only incidental forms had to be abandoned. Everything
pointed to the fact that in all cestodes the development is
divided between two kinds of animals; in one—the host, the adult
tapeworm is found; while in the other, the intermediate host, we
find some form or other of an intermediate stage (cysticercus in
the broadest sense). The practical application of this knowledge is
self-evident. If no infected pork or beef is ingested, no tapeworm
can be acquired, and also the rearing of cysticerci in the human body
is prevented by avoiding the introduction of the eggs of tapeworms.
Though these results were definitely proved by numerous
researches, yet they have been repeatedly challenged, notably by
J. Knoch (1862) in Petrograd, who, on the basis of experiments,
sought to confirm a direct development without an intermediate host
and ciliated stage, at all events as regards Dibothriocephalus latus.
However, the repeated communications of this author met with
but little favour from competent persons, partly because the
experiments were conducted very carelessly, and partly because
their repetition on dog and man (R. Leuckart) had no results
(1863). It was only in 1883 that Braun was able to prove that the
developmental cycle of Dibothriocephalus latus is similar to that of
other Cestodes. The results obtained in other places by Parona,
Grassi, Ijima and Zschokke render any discussion of Küchenmeister’s
conclusions unnecessary.7 Long after Knoch, a French author,
P. Mégnin, also pleaded for the direct development of some
cestodes, and especially some tæniæ. He (1879) also sought to
prove a genetic connection between the hookless and armed tapeworms
of mammals, but the arguments he adduced, so far as they
rest on observations, can be easily refuted or attributed to misinterpretation.
Only one of these arguments is correct, namely,
that the number of the species of tæniæ with which we are acquainted
is far larger than that of the corresponding cystic forms; but this
disparity alone cannot be taken as a proof of direct development.
It can only be said that our knowledge in this respect is deficient.
As a matter of fact, we have during recent years become acquainted
with a large number of cystic forms, hitherto unknown, belonging
to tæniæ which have long been familiar. It must also be borne in
mind that no man in his lifetime can complete an examination for
bladder-worms of the large number of insects, for instance, which
may destroy an entire generation of an insectivorous species of bird
within a small district.
Naturally it does not follow that direct development in the
cestodes is altogether lacking. The researches of Grassi (1889) have
furnished an example in Hymenolepis (Tænia) murina, which shows
that development may sometimes take place without an intermediate
host, notwithstanding the retention of the cystic stage. It was found
that the oncospheres of this species, introduced into rats of a certain
age, after a time grow into tapeworms without leaving the intestine,
but not directly, for they bore into the intestinal wall, where they
pass the cystic stage, the cysts afterwards falling into the intestinal
lumen, where they develop into tapeworms. The recent experiments
of Nicoll (1911) show that the larval stages of Hymenolepis murina
also occur in the rat-flea, Ceratophyllus fasciatus.
Important observations were soon made on the remaining
groups of helminthes. The discussion on the origin of parasites
soon became confined to the helminthes. Amongst the Nematoda,
it had long been known that encapsuled forms existed that had
at first been regarded as independent species, but very soon they were
pronounced to be immature forms, in consequence of their lack
of sexual organs. Though Dujardin and also v. Siebold regarded
them as “strayed” animals, v. Stein (1853) very promptly demonstrated
that the progeny of the nematodes were destined to travel
by discovering a perforating organ in the larval nematodes of the
mealworm. This was first experimentally confirmed (1860) by
R. Leuckart, R. Virchow and Zenker, all of whom succeeded not
only in bringing to maturity the muscle Trichinæ (known since 1830)
in the intestine of the animals experimented upon, but were likewise
able to follow the migrations of the progeny. Of course, the
encapsulating brood remained in the same organism, and in this
respect deviated from the broods of other helminthes which escape
into the outer world and find their way into other animals, but
the encapsuled nematodes could no longer be regarded as the
result of straying. Subsequently, R. Leuckart worked out, more
or less completely, the history of the development of numerous
nematodes, or pointed out the way in which further investigations
should be made. It has been found that in nematodes far more
frequently than in other helminthes, the typical course of development
is subject partly to curtailment and partly to complications,
which sometimes considerably increase the difficulties of investigation
and have hitherto prevented the attainment of a definite conclusion,
though the way to it is now clear.
In a similar manner the works of R. Leuckart have cleared
up the development of the Acanthocephala and Linguatulida. Of
course, much still remains to be done. So far, we do not even
know all the helminthes of man and of the domestic animals
in all their phases of life, and still less is known of those of
other animals. We are indebted to the discoveries of the
last fifty years for the knowledge arrived at, though comparatively
few names are connected with it. The gross framework
is revealed, but the gaps have only been filled up here and
there. However, we may trustfully leave the completion of the
whole to the future, without fear that any essential alterations will
take place.
The deductions to be drawn are as follows: That the helminthes
like the ectoparasites multiply by sexual processes, that the entire course
of development of the helminthes is rarely or never gone through
in the same host as is the case with several ectoparasites, that the
progeny at an earlier or later stage of development, as eggs, embryos,
or larvæ, quit the host inhabited by the older generation, and almost
always attain the outer world: only in Trichinella does the development
take place directly in the definite host. Where the eggs have
not yet developed they go through the embryonic evolution in the
outer world. The young larvæ are transmitted, either still enclosed
within the egg or embryonic covering, to the intermediate host or
more rarely they are transferred straight to the final host. In other
cases they may hatch out from their envelopes, and after a longer or
shorter period of free life, during which they may partake of food and
grow, they, as before, penetrate, usually in an active way, into an
intermediate host, or at once invade the final host. Exceptionally
(e.g., Rhabdonema), during the free life there may be a propagation of
the parasitic generation, and in this case only the succeeding generation
again becomes parasitic, and then at once reaches its final host.
The young forms which have invaded the final host become mature in
the latter, or after a longer or shorter period of parasitism again wander
forth (as the Œstridæ, Ichneumonidæ, etc.), and reach the adult stage
in the outer world. The young stages, during which the parasites
undergo metamorphoses or are even capable of producing one or
several intermediate generations, are passed in the intermediate hosts
until, as a rule, they are passively carried into the final host and there
complete their cycle of development by the formation of the organs
of generation. This mode of development, the spending of life in
two different kinds of animals (intermediate and final host), is typical
of the helminthes. This is manifested in the Acanthocephala, the
Cestoda, the majority of the endoparasitic Trematoda, a number of
the Nematoda, and the Linguatulidæ. There are now and then
exceptions, however, in which, for instance, the host and intermediate
host change order (Trichinella, Hymenolepis murina).
Parasites are hardly ever inherited amongst animals.8 According
to a few statements, however, Trichinella and Cœnurus are supposed
to be transmissible from the infected mother to the fœtus. Otherwise
most animals acquire their parasites, especially the Entozoa, from
without, the parasites penetrating either actively, as in animals living
in the water, or passively with food and drink. A particular predisposition
to worms is not more likely than a spontaneous origin
of parasites.
Derivation of Parasites.—Doubt now no longer exists as to the
derivation of the temporary and of many of the stationary ectoparasites
from free-living forms. This conclusion is founded on the
circumstance that not only are there numerous intermediate degrees
in the manner of living and feeding between predacious and parasitic
animals, but that there is more or less uniformity in their structure.
The differences that exist are easily explained as consequences of
altered conditions of life. The case is more difficult in regard to
groups that are exclusively parasitic (Cestoda, Trematoda, Acanthocephala,
Linguatulidæ, and Sporozoa), or groups that are chiefly
parasitic (Nematoda), because in these cases the gulf that divides
these forms from free-living animals is wider. It is true that we know
that the nearest relatives of the Linguatulidæ are found amongst the
Arachnoidea, and indeed in the Acarina; that, moreover, the structure
and development of the Sporozoa refers them to the Protozoa, and
allows some of them to be regarded as the descendants of the lowest
Rhizopoda. We know that the Trematoda, and through these the
Cestoda, are closely related to the Turbellaria, from which they may
be traced. The Nematoda, and still more the Acanthocephala, stand
apart. This is less evident, however, in the Nematoda, for there
are numerous free-living members of these from which it is possible
that the parasitic species may be descended. Indeed, this seems
more than probable if such examples as Leptodera, Rhabdonema and
Strongyloides are taken into consideration, as well as the conditions
of life of free-living nematodes. These mostly, if not exclusively,
spend their lives in places where decomposing organic substances
are present in quantities; some species attain maturity only in such
localities, and there propagate very rapidly. Should the favourable
conditions for feeding be changed, the animals seek out other
localities, or they remain in the larval form for some time until more
favourable conditions set in. It is comprehensible that such forms
are very likely to adopt a parasitic manner of life which at first is
facultative (Leptodera, Anguillula), but may be regarded as the transition
to true parasitism. The great advantages attached to a parasitic
life consist not only in protection, but also in the supply of suitable
food, and consequently in the easier and greater production of eggs,
and thus fully account for the gradual passage of facultative parasitism
into true parasitism. In many forms the young stages live free for
some time (Strongylidæ), in others, as is the case in Rhabdonema,
parasitic and free-living generations alternate; in others, again, the
free period is limited to the egg stage or entirely suppressed.
Though it is possible thus to connect the parasitic with the free-living
nematodes, by taking their manner of life into account, this
matter presents greater difficulties in regard to other helminthes. It
is true that the segmented Cestoda may be connected with and traced
from the less known and interesting single-jointed Cestoda (Amphilina,
Archigetes, Caryophyllæus, Gyrocotyle). Trematodes are all parasites,
with the exception of one group, Temnocephalidæ, several genera and
species of which live on the surface of the bodies of Crustacea and
turtles of tropical and sub-tropical freshwaters. Temnocephalidæ are,
nevertheless, predacious. They feed on Infusoria, the larvæ of small
insects and Crustacea. So far as is known they do not nourish themselves
on part of the host. They belong to the group of commensals,
or more correctly, to that of the SPACE PARASITES, which simply dwell
with their host and do not even take a portion of the superfluity of
its food. However, space parasitism may still be regarded as the
first stage of commensalism, which is again to be regarded as a sort
of transition to true parasitism.
It is possible that parasitism came about in this way in the
trematodes, in which connection we must first consider the turbellaria-like
ancestors of the trematodes. Much can be said in favour of
such a genetic relationship between turbellaria and trematodes, and
hardly anything against it. It should also be remembered that
amongst the few parasitic turbellaria there are some that possess
clinging discs or suctorial pores, and these are only differentiated
from ectoparasitic trematodes by the possession of a ciliated integument,
which is found only in the larval stages of the latter.
The Acanthocephala occupy an isolated position. Most authors
certainly regard them as related to the nematodes; in any case,
the connection is not a close one, and the far-reaching alterations
which must have occurred prevent a clear view. Perhaps the free
original forms of Acanthocephala are no longer in existence, but
that such must have existed is a foregone conclusion.
An explanation of the CHANGE OF HOST so frequent in parasites
is more difficult than that of their descent. R. Leuckart is of opinion
that the present intermediate hosts, which belong principally to the
lower animals, were the original hosts of the parasites, and fostered
both their larval and adult stages. It was only in course of time that
the original hosts sank to the position of intermediate hosts, the cause
for this alteration being that the development of parasites, especially
of the helminthes, through further development and differentiation
extended over a larger number of stages. The earlier stages remained
in their original hosts, but the later stages sought out other hosts
(higher animals). To prove this, Leuckart points out that the mature
stages of the helminthes, with but few exceptions, occur only in the
vertebrates which appeared later in the development of the animal
kingdom, while the great majority of intestinal worms of the lower
animals only represent young stages, which require transmission into
a vertebrate animal before they can become mature. The few
helminthes that attain maturity in the lower animals (Aspidogaster,
Archigetes) are therefore regarded by Leuckart as primitive forms,
and he compares them with the developmental stages of helminthes,
Aspidogaster with rediæ, Archigetes with cysticercoids. He classes
the nematodes that become mature in the invertebrates with Anguillulidæ,
i.e., with saprophagous nematodes from which the parasitic
species descend.
Leuckart therefore regards the change of hosts as secondary,
so does Sabatier. The latter, however, adduces other reasons for
this (lack of clinging organs and the necessity to develop them in an
intermediary stage); but in this connection he only considers the
Cestoda. In opposition to Leuckart, R. Moniez, however, is convinced
that the migrations of the helminthes, as well as the system
of intermediate hosts, represent the original order of things. Moniez
traces all Entozoa from saprophytes, but only a few of these were
able to settle directly in the intestine and there continue their
development. These are forms that at the present day still lack
an intermediate host, such as Trichocephalus, Ascaris, and Oxyuris.
In most other cases the embryos, however, consisted of such saprophytes
as were, in other respects, suitable to become parasites, but
were incapable of resisting the mechanical and chemical influences
of the intestinal contents. They were therefore obliged to leave the
intestine at once, and accomplished this by penetrating the intestinal
walls and burrowing in the tissues of their carriers. In this position,
assisted by the favourable conditions of nutrition, they could attain
a relatively high degree of development. Mechanical reasons prevented
a return to the intestines, where the eggs could be deposited.
Most of them doubtless died off as parasites, as also their young
stages do at present when they penetrate wrong hosts. Some of
them, nevertheless, passively reached the intestine of beasts of prey.
Many were destroyed in the process of mastication; for a small part,
however, there was the chance of reaching the intestine of a beast
of prey undamaged, and there, having become larger and more
capable of resistance, maturity was attained. By means of this incidental
coincidence of various favourable circumstances, these processes,
according to Moniez, have been established by heredity and
have become normal.
This is not the place to express an opinion either for or against
the various hypotheses advanced, but the existence of these diametrically
opposed views alone will show the great difficulty of the
question. Independently, however, it appears more natural to come
to the conclusion that parasitism, as well as change of hosts, were
gradual transitions.
As a conclusion to this introductory chapter, a list of some of
the most important works on the parasitology of man and animals
is appended.
LITERATURE.
Goeze, J. A. E. Versuch einer Naturgeschichte der Eingeweidewürmer thierischer
Körper. Blankenburg, 1782. 4to, 471 pp., with 44 plates.
Zeder, J. G. H. Erster Nachtrag zur Naturgeschichte der Eingeweidewürmer.
von J. A. E. Goeze. Leipzig, 1800. 4to, with 6 tables.
Rudolphi, C. A. Entozoorum sive vermium intestinalium historia naturalis.
I, Amstelod., 1808; ii, 1809. 8vo, with 18 plates.
Rudolphi, C. A. Entozoorum synopsis. Berol., 1819. 8vo, with 3 plates.
Bremser, J. G. Ueber lebende Würmer im lebenden Menschen. Wien, 1819.
8vo, with 4 plates.
Bremser, J. G. Icones helminthum, systema Rudolphii entozoologicum illustrantes.
Viennae, 1824. Fol. (Paris, 1837).
Dujardin, F. Histoire naturelle des helminthes ou vers intestinaux. Paris,
1845. 8vo, with 12 plates.
Diesing, C. M. Systema helminthum. 2 vols. Vindobonnae, 1850, 1851. 8vo.
Supplements by the same author: Revision der Myzhelminthen (Report
of the Session of the Imp. Acad. of Science. Wien, xxxii, 1858); with
addendum (ibid., xxxv, 1859); Revision der Cephalocotyleen (ibid., xlix,
1864, and xlviii, 1864); Revision der Nematoden (ibid., xlii, 1861); Supplements
(ibid., xliii, 1862).
Beneden, P. J. VAN. Mémoire sur les Vers intestinaux. Paris, 1858. 4to, with
12 plates.
Küchenmeister, F. Die in und an dem Körper des lebenden Menschen vorkommenden
Parasiten. Leipzig, 1855. 8vo, with 14 plates.
Leuckart, R. Die menschlichen Parasiten und die von ihnen herrührenden
Krankheiten. I, Leipzig, 1863; II, Leipzig, 1876. 8vo.
Cobbold, T. Sp. Entozoa; an Introduction to the Study of Helminthology.
London, 1864. 8vo. Supplement, London, 1869.
Davaine, C. Traité des entozoaires et des maladies vermineuses de l’homme et
des animaux domestiques. 2nd edit. Paris, 1877. 8vo.
Linstow, O. V. Compendium der Helminthologie, ein Verzeichniss der
bekannten Helminthen, die frei oder in thierischen Körpern leben,
geordnet nach ihren Wohnthieren, unter Angabe der Organe, in denen
sie gefunden sind, und mit Beifügung der Litteraturquellen. Hanov.,
1878. 8vo. Supplement, including the years 1878–1888, Hanov., 1888.
Cobbold, T. Sp. Parasites; a Treatise on the Entozoa of Man and Animals,
including some Account of the Entozoa. London, 1879. 8vo.
Leuckart, R. Die Parasiten des Menschen und die von ihnen herrührenden
Krankheiten. 2nd edit. Leipzig, 1879–1886. The Protozoa, Cestodes,
Trematodes and Hirudinea have hitherto appeared (continued by
Brandes).
BÜtschli, O. Protozoa in Bronn’s Klass. u. Ordn. d. Thierreichs. Vol. i,
Leipzig, 1880–1889. 8vo, with 79 plates.
Braun, M. Trematodes in Bronn’s Klass. u. Ordn. d. Thierreichs. Vol. iv, 1,
Leipzig, 1879–1893. 8vo, with 33 tables. (The first thirteen sheets, comprising
the history of the worms up to 1830, were compiled by H.
Pagenstecher.)
Zürn, F. A. Die thierischen Parasiten auf und in dem Körper unserer Haussäugethiere,
sowie die durch erstere veranlassten Krankheiten, deren
Behandlung und Verhütung. 2nd edit. Weimar, 1882. 8vo, with 4
plates.
Cobbold, T. Sp. Human Parasites; a Manual of Reference to all the Known
Species of Entozoa and Ectozoa. London, 1882. 8vo.
Küchenmeister, F., and F. A. Zürn. Die Parasiten des Menschen. 2nd edit.
Leipzig, 1888., 8vo, with 15 plates.
Blanchard, R. Traité de zoologie médicale. I, Paris, 1889; II, 1890. 8vo.
Neumann, L. G. Traité des maladies parasitaires non microbiennes des
animaux domestiques. 2nd edit. Paris, 1892. 8vo. English edit., translated
by G. Fleming. 2nd edit., revised by J. Macqueen. 1905. London:
Baillière, Tindall and Cox.
Looss, A. Schmarotzerthum in der Thierwelt. Leipzig, 1892. 8vo.
Railliet, A. Traité de zoologie médicale et agricole. 2nd edit. I, Paris,
1895. 8vo.
Parona, C. L’elmintologia italiana da’ suoi primi tempi all’ anno 1890.
Genova, 1894. 8vo.
Braun, M. Cestoda in Bronn’s Klass. u. Ordn. d. Thierreichs. Vol. iv, 2,
Leipzig, 1894–1900. 8vo, with 24 plates.
Mosler, F., and E. Peiper. Thier Parasit. (Spec. Path. u. Ther. v. H.
Nothnagel. Vol. vi.) Wien, 1894. 8vo, with 124 illustrations.
Laveran, A., et R. Blanchard. Les hématozoaires de l’homme et des anim.
Paris, 1895. 12mo, with 30 figs.
Sluiter, C. R. De dierl. paras. v. d. mensch en van onze huisdier. Haag,
1895. 8vo.
Blanchard, R. Malad. parasit., paras. animaux, paras. végét. à l’exclus. des
bacter. (Traité de pathol. gén. de Ch. Bouchard, vol. ii.) Paris, 1895.
8vo, with 70 figs.
Huber, J. Ch. Bibliographie der klin. Helminthol. München, 1895. 8vo.
With Supplement, 1898, and continued as Bibl. d. klin. Entomol.
München, 1899–1900.
Moniez, R. Traité de parasitol. anim. et veget. appl. à la médecine. Paris,
1896. 8vo, with 116 figs.
Weichselbaum. Parasitologie (Weil’s Handb. d. Hyg.). Jena, 1898. 8vo,
with 78 illustrations.
Kraemer, A. Die thierischen Schmarotzer des Auges (Gräfe and Sämische’s
Handb. d. ges Augenheilk.). Leipzig, 1899. 8vo, with 16 illustrations.
Cholodkowsky, N. A. Icones helm. hominis. St. Petersburg, 1898–99. Fol.
(atlas with 15 plates).
Perroncito, E. I parassiti dell’ uomo e degli animali utili e le più comuni
malattie da essi prodotti. II_{a} ed. Milano 1902. 8o. con 276 fig. e 25 tav.
Stiles, Ch. W. and A. Hassall. Index Catalogue of Medicine and Veterinary
Zoology. Washington, 1902 (U.S. Dept. of Agric., Bur. of Anim. Ind.,
Bull. No. 39).
Neveu-Lemaire, M. Précis de parasitologie humaine, parasites végétaux et
animaux. 4e édit. Paris, 1911.
Hofer, B. Handbuch der Fischkrankheiten. München, 1904. 8o. 18 Taf.
222 Abb.
Guiart, J., and L. Grimbert. Précis de Diagnostic chimique, microscopique et
parasitologique. Paris, 1906. With 500 figs.
Ostertag, R. Handbuch der Fleischbeschau. V. Aufl. mit 265 Abb. Stuttgart,
1904.
Stiles, Ch. W. The International Code of Zoological Nomenclature as applied
to Medicine (Hygienic Lab., Bull. No. 24, Washington, 1905).
Stiles, C. W., and Hassall, A. Trematoda and Trematode Diseases. (Index
Catalogue of Med. and Vet. Zoology.) Hygienic Lab., Bull. No. 37,
Washington, 1908.
Stiles, C. W., and Hassall, A. Cestoda and Cestodaria. Hygienic Lab.,
Bull. No. 85, Washington, 1912.
Laloy, L. Parasitisme et mutualisme dans la nature. Paris, 1906. 8vo,
284 pp., 82 figs.
Theobald, F. V. A Monograph of the Culicidæ of the World. 5 vols. and
plates. 1901–1910. London: Brit. Museum, Nat. Hist.
James, S. P., and Liston, W. G. The Anopheline Mosquitoes of India.
2nd edit. 1911. Calcutta: Thacker, Spink and Co.
Howard, L. O., Dyar, H. G., and Knab, F. The Mosquitoes of North and
Central America and the West Indies. 2 vols. 1912. Washington:
Carnegie Institution.
Austen, E. E. African Blood-sucking Flies. 1909. London: Brit. Museum,
Nat. History.
Austen, E. E. A Handbook of Tsetse-flies. 1911. London: Brit. Museum,
Nat. History.
Castellani, A., and Chalmers, A. J. Manual of Tropical Medicine. 2nd edit.
1,747 pp. 1913. London: Baillière, Tindall and Cox.
Kolle and Wassermann. Handbuch der pathogenen mikroorganismen. Jena:
Gustav Fischer.
Minchin, E. A. An Introduction to the Study of the Protozoa. 1912.
London: Arnold.
Laveran, A., et Mesnil, F. Trypanosomes et Trypanosomiases. 2nd edit.
1912. Paris: Masson and Co.
Doflein, F. Lehrbuch der Protozoenkunde. 3rd edit. 1911. Jena: Gustav
Fischer.
Nuttall, G. H. F., Warburton, C., Cooper, W. F., and Robinson, L. E.
Ticks—a Monograph of the Ixodoidea. Pt. I (1908). Pt. II. (1911).
University Press, Cambridge, England.
Brumpt, E. Précis de Parasitologie. 2nd edit. 1913. Paris: Masson and Co.
Patton, W. S., and Cragg, F. W. A Text-book of Medical Entomology. 1913.
Christian Literature Society of India: London, Madras, and Calcutta.
JOURNALS.
For current researches the following, among others, should be consulted:—
Annals of Tropical Medicine and Parasitology, Liverpool.
Annales de l’Institut Pasteur, Paris.
Archives de Parasitologie, Paris.
Archives de Zoologie Expérimentale et Générale, Paris.
Archiv für Protistenkunde, Jena.
Archiv für Schiffs- und Tropen-Hygiene, Leipzig.
Bulletin of Entomological Research, London.
Bulletin de l’Institut Pasteur, Paris.
Bulletin de la Société de Pathologie Exotique, Paris.
Bulletins of the Bureau of Animal Industry, Washington.
Centralblatt für Bakteriologie und Parasitenkunde, Jena.
Compt. Rend. Acad. Sci., Paris.
Compt. Rend. Soc. Biol., Paris.
Indian Journal of Medical Research, Calcutta.
Journal of Experimental Medicine, New York.
Journal of Medical Research, Boston.
Memorias do Instituto Oswaldo Cruz, Rio de Janeiro.
Parasitology, Cambridge.
Proceedings of the Royal Society, London.
Quarterly Journal of Microscopical Science, London.
Review of Applied Entomology, London.
Tropical Diseases Bulletin (London: Tropical Diseases Bureau).
Zeitschrift für Infektionskrankheiten, Berlin.
THE ANIMAL PARASITES OF MAN.
Man is one of those organisms in on on which a whole host of parasites find
conditions suitable for their existence: Protozoa, Platyhelminthes, Nematoda,
Acanthocephala, Hirudinea, and a large number of Arthropoda (Arachnida as well
as Insects) all include members which are parasites of man. These animals either
live on the external surface of the body or within the intestine and its appendages.
Other organs and systems are not quite free from foreign organisms—we are
acquainted with parasites in the skeletal system, in the circulatory system, in the
brain, in the muscles, in the excretory and genital organs, and even in the organs of
sense.
It is possible, and perhaps might be advantageous, to arrange and describe the
parasites of man according to the situations in which they are found (parasites of the
skin, intestinal parasites, etc.). Their description in the various stages of development
would, however, be disturbed when, as is generally the case, the different stages
are passed in different organs, and a work which treats more fully of the natural
history of the parasites than of the local disorders to which they give rise would
suffer thereby. It is, therefore, preferable to describe the parasites of man in their
systematic order, and to mention their different situations in man in describing each
species.
A. PROTOZOA,
BY
H. B. FANTHAM, M.A., D.Sc.
All those animal organisms which throughout their entire life never rise above
the unicellular stage, or merely form simple, loose colonies of similar unicellular
animals, are grouped under the term Protozoa (Goldfuss, 1820), as the simplest types
of animal life. All the vital functions of these, the lowest forms of animals, are
carried out by their body substance, the protoplasm (sarcode). Often particular
parts possess special functions, but the limits of a cell are never over-stepped
thereby. These special parts of the cell are called “cell-organs”; recently they
have been termed “organellæ.”
The living protoplasm has the appearance of a finely granular, viscid substance
which, as a rule, when not surrounded by dense investing membranes or skeletons,
exhibits a distinct kind of movement, which has been termed amœboid. According
to the species, processes of different forms and varying numbers called pseudopodia
are protruded and withdrawn, and with their assistance these tiny organisms glide
along—it might almost be said flow along—over the surface. In most Protozoa two
layers of cytoplasm may be recognised, and distinguished by their appearance and
structure, namely, the superficially situated, viscid, and quite hyaline ectosarc or
ectoplasm, and the more fluid and always granular endosarc or endoplasm, which
is entirely enveloped by the ectoplasm. The two layers have different functions; the
movements originate from the ectoplasm, which also undoubtedly fulfils the functions
of breathing, introduction of food and excretion. The endoplasm, which in some forms
(Radiolaria) is separated from the ectoplasm by a membrane, undertakes the digestion
of the food. To this distribution of functions between the various layers of cytoplasm
is due the development of particular cellular organs, such as the appearance
of cilia, flagella, suctorial tubules (in the Suctoria) and the myophan striations, which
are contractile parts of the ectoplasm in Infusoria and Gregarines. In many cases
(Flagellata, Ciliata), an area is differentiated for the ingestion of food (oral part,
cytostome) to which there is often added a straight or curved opening (cytopharynx),
through which the food reaches the endoplasm. The indigestible residue is either cast
off through the oral part or excreted by a special anal part (cytopyge). In rare
cases, structures sensitive to light, the so-called pigment or eye spots are developed,
e.g., Euglena. In the case of Infusoria the endoplasm circulates slowly, and
agglomerations of fluids (food vacuoles) sometimes appear around each bolus of
food; in these vacuoles the food is digested under the action of certain materials
(ferments). Even in the lowliest Protozoa fluids to be excreted are, as a rule,
gathered into one, or, more rarely, several contractile vacuoles, which regularly
discharge their contents. This action, however, is to a certain extent governed
by the temperature of the surrounding medium. In some Infusoria a tube-like
channel in the cytoplasm is joined to the contractile vacuole which usually occupies
a certain position; this forms a sort of excretory duct, and there are also supply-canals
leading to these organellæ.
Very frequently various substances are deposited in the endoplasm, such as fatty
granules, drops of oil, pigment granules, bubbles of gas or crystals. More solid
skeletal substances are secreted in or on the ectoplasm. To the latter belong the
cuticle of the Sporozoa and Infusoria, the chalky shells containing one or several
chambers of the Foraminifera, the siliceous and very ornamental framework of the
Radiolaria, and the chitinous coat of many Flagellata, Infusoria, etc. Some forms
make use of foreign bodies found in their surroundings, such as grains of sand, to
construct their protective coverings.
The food often consists of small animal or vegetable organisms and of organic
waste; it is usually introduced in toto into the endoplasm. On the other hand, the
Suctoria extract nourishment from their prey by means of their tentacles. Many
parasitic species also ingest solid food, others feed by endosmosis.
In all cases one nucleus at least is present. It is true that the existence of non-nucleated
Protozoa, the so-called Monera, is still insisted upon, but some of these
have already proved to be nucleated, and the presence of nuclei in the others will no
doubt be established. Very often the number of nuclei increases considerably, but
these multinucleate stages are always preceded by uninucleate stages. In the
Infusoria, in addition to the larger or principal nucleus (macronucleus) there is
usually a smaller reproductive nucleus (micronucleus). This dualism of the nuclear
apparatus is considered by some to be general, and usually to appear first at the
onset of reproduction.
The form and structure of the nucleus vary greatly in different species. There
are elongate, kidney-shaped, or even branched nuclei as well as spherical or oval
ones. In addition to vesicular nuclei with a distinct karyosome and incidentally
also with a nuclear membrane, homogeneous and more solid formations are
frequently encountered. The nuclei are always differentiated from the protoplasm
by their reactions, particularly in regard to certain stains.
In many Protozoa an extra-nuclear mass, sometimes compact, sometimes diffuse,
arises from or near the nucleus. This mass, whose staining reactions resemble those
of the nucleus, is termed the chromidial apparatus. On the dualistic hypothesis, two
varieties of chromidia occur, one originating from the vegetative nucleus (macronucleus),
being chromidia in the restricted sense, the other derived from the
reproductive or micronucleus being termed sporetia. Chromidia consist of altered
(? katabolic) nuclear material.
The nucleus plays the same part in the life of the single celled organisms as it
does in the cells of the Metazoa and Metaphyta. It appears to influence in a certain
manner all, or at least most, of the processes of life, such as motility, regeneration,
growth, and generally also digestion. Its principal influence, however, is exercised
in the propagation of the cells, as this is always brought about by the nucleus.
The PROPAGATION of the Protozoa is effected either by division or by means of
direct budding. In division, which is preceded by direct or indirect (mitotic)
division of the nucleus, the body separates into two, several, or even a great many
segments. In this process the entire substance of the body is involved, or a small
residual fragment may be left, which does not undergo further division and finally
perishes. In the budding method of multiplication a large number of buds are
formed, either on the surface or in the interior of the organism. Where divisions or
buddings follow one another rapidly, without the segments separating immediately
after their production, numerous forms develop, which are often unlike the parental
forms, and these are termed swarm spores or spores. Divisions imperfectly
accomplished lead to the formation of protozoal colonies.
Sometimes encystment9 takes place previous to division. Frequently, also,
sexual processes appear, such as the union of two similar (isogamous) or dissimilar
(anisogamous) individuals. In the latter case sexual dimorphism occurs, with the
formation of males (microgametes) and of females (macrogametes). The union may
be permanent (copulation), the process being comparable with the fertilisation of the
ovum by a spermatozoon. On the other hand, attachment may be transient (conjugation)
when, after the exchange of portions of the nucleus, the couple separate,
to multiply independently of each other. Sometimes there is an ALTERNATION OF
GENERATIONS, as there may be several methods of propagation combined in the
same species, either direct multiplication, conjugation, or copulation being practised;
the different generations may thus, in certain cases, be unlike morphologically.
Protozoa inhabit salt water as well as fresh water; they are also found on land
in very damp places, and invade animals as parasites.
Classification of the Protozoa.
Class I.—Sarcodina (Rhizopoda). Protozoa, the body substance of which forms
pseudopodia; many of them are capable of developing chitinous, chalky, or siliceous
coverings or skeletal structures, which, however, permit the protrusion of the
pseudopodia either over the entire periphery or at certain points. They possess one
nucleus or several.
Order 1.—Amœbina (Lobosa) naked or with a simple shell, sometimes formed
of a foreign substance; the pseudopodia may be lobose or finger-shaped;
there may be a contractile vacuole; generally only one nucleus. They
live in fresh or salt water, in the soil, and also parasitically.
Order 2.—Foraminifera (Reticularia). Mostly provided with a calcareous
shell, usually consisting of several chambers, and allowing the protrusion
of the pseudopodia either at the periphery or only at the opening. The
pseudopodia are filamentous and frequently anastomosed; there is no
contractile vacuole; there are usually several nuclei. Mostly marine.
Order 3.—Heliozoa. Naked, or with a chitinous or simple radial siliceous
skeleton; the pseudopodia are filamentous, and are frequently supported
by firmer axes, which exhibit no tendency to anastomosis; there is a
contractile vacuole; one or several nuclei. Live in fresh water.
Order 4.—Radiolaria. The body has radially-disposed filamentous pseudopodia,
and the nucleus is hidden in the central capsule; there is almost
always a siliceous framework, consisting of pieces arranged radially,
tangentially, or lattice-like; there is no contractile vacuole, but fluid-containing
hydrostatic vacuoles are present in the peripheral protoplasm.
Marine.
Class II.—Mastigophora (Flagellata). Protozoa with one or several long
flagella used for locomotion and for acquiring food; in stationary forms their only
function is to take in food. Cytostome and contractile vacuole may be present.
May be either naked or provided with protective coverings; one or more nuclei.
They live either in fresh or salt water, or may be parasitic.
This class is again divided into several sub-classes and orders, of which only the
Euflagellata, with the Protomonadina and Polymastigoda are of interest here.
Class III.—Sporozoa. Protozoa that only live parasitically in the cells, tissues,
or organs of other animals. They ingest liquid food by osmosis; the surface of the
body is covered with an ectoplasmic layer, or cuticle; they have no cilia in the adult
state, but may form pseudopodia. Flagella occur, but only on the male propagating
individuals. There may be one or numerous nuclei, but no contractile vacuole.
Propagation by means of spores, mostly provided with sporocysts, is characteristic.
Sub-class 1.—Telosporidia. These are usually of constant form, rarely amœboid;
they are uninucleate in the mature state; they live within host cells in the
first stage. Spore-formation occurs at the end of the life-cycle.
Order 1.—Gregarinida. Body of a constant, usually elongate form, surrounded
by a cuticle. In the early stage they lead an intracellular existence; in
the mature stage they live within the intestine or body cavity of invertebrate
animals, especially the Arthropoda, and, like intestinal parasites,
are provided with clinging organs. Copulation usually isogamous;
the spores have coats (chlamydospores) and usually contain several
minute germs (sporozoites).
Order 2.—Coccidiidea. Body of uniform spherical or oval shape: they lead
an intracellular life, but are not freely motile in cavities of the body.
Fertilization is anisogamous; the spores have coats or shells (sporocysts),
and usually contain several sporozoites. Exhibit alternation of generations.
Order 3.—Hæmosporidia. Parasites of the blood corpuscles of vertebrate
animals; they exhibit amœboid movement; fertilization is anisogamous;
many present alternation of generations and hosts; spores naked.
Sub-class 2.—Neosporidia. They are multinucleate when adult, and the form of
the body varies exceedingly (often amœboid); spore-formation commences
before the completion of growth.
Order 1.—Myxosporidia. The spores have valvular coats, with or without
caudal appendages, with two, rarely four, polar capsules. They live free
in such organs as the gall or urinary bladder, but are chiefly found in
connective tissue. They occur especially in fishes.
Order 2.—Microsporidia. Spores with coats or sporocysts; no caudal
appendage, with one polar capsule. They usually live in the tissues of
Arthropoda.
Order 3.—Sarcosporidia. Elongate parasites of the muscular fibres of
amniotic vertebrates, on rare occasions they occur also in the connective
tissue; the spores, which are kidney or sickle-shaped, are naked and
apparently have no obvious polar capsule.
Order 4.—Haplosporidia. Simple organisms, forming simple spores; they
occur in Rotifers, Polychætes, Fish and Man.
Class IV.—Infusoria (Ciliata). The body is generally uniform in shape, with
cilia and contractile vacuole, frequently also with cytostome; usually has macro-
and micro-nucleus; live free in water and also parasitically.
The orders Holotricha, Heterotricha, Oligotricha, Hypotricha and Peritricha
are classified according to the arrangement of the cilia.
Class V.—Suctoria. Bodies with suctorial tubes, contractile vacuoles, macro-
and micro-nucleus, no cytostome. They generally invade aquatic animals as cavity
parasites, yet also attack plants; early stage ciliated. Live sometimes as parasites
on Infusoria. [The Suctoria are frequently regarded as a sub-class of the Infusoria.]
The Protozoa and Protophyta are sometimes united under the term Protista
(Haeckel, 1866). The Spirochætes are Protists (see pp. 114–128).
Class I. SARCODINA, Bütschli, 1882.
Order. Amœbina, Ehrenberg.
A. Human Intestinal Amœbæ.
The first record of the occurrence of amœba-like organisms in the human intestine,
that is, in intestinal evacuations, was that of Lambl (1859); nevertheless, the case
was not quite conclusive, as the occurrence of testaceous amœbæ of fresh water
(Arcella, Difflugia) was also reported. In 1870 Lewis found amœbæ associated with
disorders of the large intestine in patients in Calcutta. A year later Cunningham
reported from the same locality that he had observed on eighteen occasions, in one
hundred examinations of dejecta from cholera patients, colourless bodies with amœboid
movements, which became encysted and multiplied by fission. The daughter forms
were said to be capable of dividing again, but they might also remain in contact.
Contractile vacuoles were not noticed. The same bodies were observed also in
simple diarrhœa (twenty-eight cases out of one hundred.)
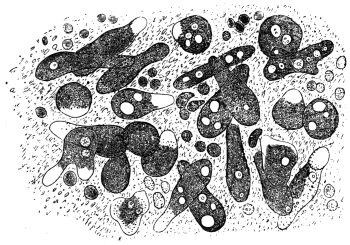
Fig. 1.—Amœba coli, Lösch, in the intestinal mucus. (After Lösch.)
The case reported by Lösch in 1875 attracted more attention. It was that of
a peasant, aged 24, who came from the province of Archangel. He was admitted
into Eichwald’s clinic at Petrograd with symptoms of dysentery. In the discharges
containing blood and pus, Lösch found amœbæ in large numbers. When
at rest these amœbæ measured from 20 µ to 35 µ; in a state of movement their length
might extend up to 60 µ (fig. 1). The pseudopodia appeared only singly, and,
since they were hyaline (ectoplasmic), were thus distinguished from the markedly
granular endoplasm that enclosed a spherical nucleus of from 5 µ to 7 µ in diameter.
One or more non-contractile vacuoles were present. Quinine enemata had the
effect of making the amœbæ disappear from the fæces and thus causing the diarrhœa
to abate. Four months after admission the patient died from the results of
intercurrent pneumonia. At the autopsy ulceration of the large intestine was found,
especially in the lower parts. Lösch connected the amœbæ with the ulcerations by
experiments made on four dogs by injecting them with recently passed stools
(per os et anum). Eight days after the last injection numerous amœbæ were found
in the fæces of one of these dogs; eighteen days after the injection the animal
was killed. The mucosa of the rectum was inflamed, covered with blood-stained
mucus and ulcerated in three places. Numbers of amœbæ were found both in
the pus of the ulcers and in the mucus. The three other dogs remained healthy.
From these observations Lösch concluded that the species of amœba described by him
as Amœba coli could not be regarded as the primary cause of the disease, but that it
was certainly capable of increasing a lesion of the large intestine already present,
or at least of preventing its healing.
B. Grassi (1879) found in the stools of healthy as well as in those of diarrhœic
patients from various localities in Northern Italy, amœbæ similar to those discovered
by Lösch. As this was of frequent occurrence, the pathogenicity could not be
definitely established. Normand, formerly naval surgeon at Hong-Kong, observed
numerous amœbæ in the dejecta of two patients suffering from colitis.
Many further investigations, which cannot be quoted in detail, showed not only
that intestinal amœbæ were widely distributed in man, but indicated with greater
certainty their rôle as agents of dysentery. The Commission sent out by the German
Government in the year 1883 to investigate cholera in India and Egypt—whose
members discovered the cholera bacillus—also collected information with regard to
dysentery. In five cases of dysentery examined post mortem at Alexandria, with
the exception of one case in which ulceration of the colon had already cicatrized or
was approaching cicatrization, R. Koch found amœbæ as well as bacteria in sections
from the base of the ulcers, although such had previously escaped notice in examination
of the dejecta. Encouraged by these results, Kartulis (1885), who had discovered
amœba-like bodies in the stools of patients suffering from intestinal complaints at
Alexandria, continued his investigations. The results, obtained from more than 500
cases, gave rise to the theory that typical dysentery was caused by amœbæ as were
also the liver-abscesses that often accompany it. Kartulis supported his theory not
only by the regular occurrence of amœbæ in the stools of dysenteric patients and
their absence in other diseases, and by the occurrence of the parasites in ulcers of the
large intestine and in the pus from liver-abscesses, but also by experiments which
he performed on cats. These were infected by injection per anum of stool material
rich in amœbæ from subjects of dysentery. The infection took place also when
amœba-containing, but bacteria-free, pus from liver-abscesses was used. It has been
objected that the infection of man with Amœba coli, as the dysenteric amœbæ were
then generally designated, does not take place per anum but per os. This difficulty,
however, diminished in proportion as the encysted states of amœbæ (fig. 2), long
known in the case of other Protozoa, became understood. The infection of man
(Calandruccio, 1890) and of cats (Quincke and Roos) succeeded solely when material
containing such stages was used. Amœbæ introduced into the intestine multiply
there by fission (Harris, 1894). However, this theory, to which various other authors
gave support on the grounds of their own observations, encountered opposition. Thus
it was established that amœbæ were not found in patients in every place where
dysentery was endemic, or else they were much rarer than was expected. Further,
amœbæ were present in the most varied kinds of intestinal diseases, both of infective
and non-infective characters. Also they were present in quite healthy persons.
Moreover, for various reasons, infection experiments on animals failed to supply
proof, and finally a bacterium was discovered (Shiga, 1898) to be the excitant of one
form of dysentery. Agglutination attested the specific part played by this organism,
as it was produced by the blood serum of a person suffering from or recovered from
dysentery, but not by the serum of one who was uninfected. Bacillary dysentery
consequently was a distinct entity. The final step to be taken was to decide whether
there was a specific amœbic enteritis (amœbic dysentery or amœbiasis, according to
Musgrave).

Fig. 2.—Encysted intestinal amœbæ showing nuclear multiplication.
(After B. Grassi.)
This question should decidedly be regarded from the positive point of view.
It is intimately connected with another, namely, whether there are not several species
of intestinal amœbæ. The possibility of this had already been recognized. In
addition to the Amœba coli Lösch, R. Blanchard distinguished yet another, Amœba
intestinalis, and designated thereby the large amœbæ described in the first communication
made by Kartulis; later on he stated the distinction between the species.
Councilman and Lafleur10 (1891) considered the amœba of dysentery to be Amœba coli
Lösch and so re-named the species Amœba dysenteriæ. Kruse and Pasquale (1893)
employed the same nomenclature, but retained the old name Amœba coli Lösch for
the non-infectious species. Quincke and Roos (1893) set forth three species: a
smaller species (25 µ) finely granular, pathogenic for men and cats (Amœba coli
Lösch); a larger species (40 µ) coarsely granular, pathogenic for men but not for
cats (A. coli mitis); and a similar species non-pathogenic either for man or cat
(A. intestini vulgaris). Celli and Fiocca (1894–6) went still further, they distinguished:
(1) Amœba lobosa variety guttula (= A. guttula Duj), variety oblonga (= A. oblonga
Schm.) and variety coli (= A. coli Lösch).
(2) Amœba spinosa n. sp. occurring in the vagina as well as in the intestine of
human patients suffering from diarrhœa and dysentery.
(3) Amœba diaphana n. sp. found in the human intestine in cases of dysentery.
(4) Amœba vermicularis Weisse, present in the vagina and in dysentery; and
(5) Amœba reticularis n. sp. in dysentery.
Shiga distinguished two species; a larger pathogenic species with a somewhat
active movement, and a small harmless species with a somewhat sluggish movement.
Bowman mentions two varieties, Strong and Musgrave (1900) two species—the pathogenic
Amœba dysenteriæ and the non-pathogenic Amœba coli; Jäger (1902) and
Jürgens (1902) mention at least two species. In the following year (1903) a work by
Schaudinn was published which marked a real advance. This, in conjunction with
the establishing of a special genus (Endamœba or Entamœba) for human intestinal
amœbæ first by Leidy11 and then by Casagrandi and Barbagallo,12 for the time cleared
up the confused nomenclature, the old name Amœba coli being retained for the
harmless intestinal amœbæ of man, whereas the pathogenic species was designated
Entamœba histolytica. The history of more recent work is incorporated in the
accounts of the entamœbæ given below.
Entamœba coli, Lösch, 1875, emend. Schaudinn, 1903.
Syn.: Amœba coli, Lösch, 1875. Entamœba hominis, Casagr. et Barbag. 1897.
The amœboid trophozoite, according to Lösch, measures 26 µ to
30 µ and upwards; according to Grassi 8 µ to 22 µ; according to
Schuberg 12 µ to 26 µ. A separation of the body substance into
ectoplasm and endoplasm is only perceived during movement. The
pseudopodia, which are generally only protruded singly, are broad
and rounded at the end (lobopodia) and are hyaline, while the
remainder of the body is granular. The ectoplasm is less refractile
than the rest of the cytoplasm; it also stains less intensely (fig. 1),
and is best seen on protrusion of a pseudopodium. Red blood
corpuscles are rarely, if ever, found ingested in the cytoplasm.

Fig. 3.—Entamœba coli: life-cycle, a-e, stages in binary fission; A-D, schizogony, with
formation of eight merozoites; 2–10, cyst formation or sporogony, with formation of eight
nucleate cysts. (After Castellani and Chalmers)
The nucleus is vesicular, and is spherical when inactive, measuring
5 µ to 7 µ, with a thick nuclear membrane. In the centre of the
nucleus is a chromatinic body or karyosome or sometimes several
small nuclear bodies formed of plastin and chromatin; the remaining
chromatin is arranged on the achromatic network in the form of fine
granules, especially thickly deposited on the nuclear membrane.
Entamœba coli lives as a commensal in the upper portion of the
large intestine, where the fæces still possess a pulpy consistency.
With their concentration and change in reaction lower in the
bowel, the parasites either die or else if they are at a suitable stage of
development form resistant cysts. These cysts (fig. 2) can be found
in great abundance in normal fæces, as Grassi first observed. Slight
laxantia or intestinal diseases of any kind producing increased
peristalsis, however, show amœbæ even in the unencysted condition,
provided that the person harbours intestinal amœbæ generally. The
intensity of infection varies according to the locality; thus Schaudinn
found that 50 per cent. of the persons examined were infected with
harmless amœbæ in East Prussia, 20 per cent. in Berlin and about 66
per cent. on the Austrian littoral.
The life-history (fig. 3) of the parasite exhibits two phases:
(a) asexual multiplication in the intestine, either by binary fission or
by schizogony with formation of eight merozoites, and (b) sporogony
leading to the production of eight-nucleate cysts. Infection results
from ingestion of cysts. Only cysts with eight nuclei are infective.
The diameter of such cysts is about 15 µ to 20 µ.
There are varying accounts of the details of the life-cycle of Entamœba coli in its
different stages. Thus, regarding schizogony or multiple fission it was formerly
stated that the nucleus of the parent amœba divided into eight portions, which after
dissolution of the nuclear membrane, passed outwards into the cytoplasm, which
segregated around each. Eight merozoites were thus produced. More recently the
process of schizogony has been considered to consist in the repeated division of the
nucleus into two, four, and finally eight nuclei (fig. 3, A-D), and the formation of
eight merozoites or amœbulæ.
The process of encystment is initiated by the extrusion of all liquid and foreign
bodies from the protoplasm, which assumes a spherical form (fig. 4, A). The rounded
uninucleate amœba then secretes a soft gelatinous coat, which finally differentiates
into a double contoured cyst wall in older cysts. According to Casagrandi and
Barbagallo, the size of the cyst varies from 8 µ to 30 µ, and averages about 15 µ.
According to Schaudinn (1903) the cytological changes during cyst formation are as
follows. The nucleus of a rounded uninucleate form divides into two (fig. 4, B).
Each of these nuclei fragments into chromidia (fig. 4, C), some of which are absorbed,
while others reunite so that the cell becomes binucleate again. Each of these
nuclei, by a twice repeated division, produces three nuclei (fig. 4, D), the smaller
two of which degenerate and were regarded as reduction nuclei. There is a clear
zone or vacuole in the middle of the cyst during these maturation processes, dividing
the cyst into two halves. After the nuclear reduction the clear space disappears,
and each nucleus (termed by some a gamete nucleus) divides into two pronuclei
(fig. 4, E). The pronuclei of the pairs were said by Schaudinn to differ slightly.
Copulation occurs between pairs of unlike pronuclei, and is an example of autogamy
(fig. 4, F). When complete, each of the fusion nuclei (synkarya) divides twice, giving
rise first to four and finally to eight nuclei. Eight amœbulæ are thus formed within
the cyst.
According to Hartmann and Whitmore (1911)13, however, autogamy does not
occur within the cysts of E. coli. They consider that eight small amœbulæ are
formed (fig. 3, 2-10) which escape from the cyst and then conjugate in pairs
(fig. 3, 10-12), afterwards growing into a new generation of trophozoites.
Only some 10 to 20 per cent. of the cysts evacuated with the fæces undergo the
full course of development, the majority perish previously. In old dry fæces, only
cysts with eight nuclei are found, and it is these alone that cause the infection.
Entamœba williamsi, E. bütschlii, E. hartmanni and E. poleki (Prowazek) are
probably only varieties of E. coli.
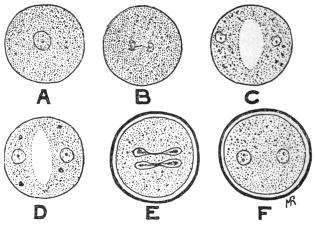
Fig. 4.—So-called autogamy of Entamœba coli. A, rounded amœba; B, nucleus dividing;
C, the two daughter-nuclei giving off chromidia; D, each nucleus has formed two reduction
nuclei; E, cyst membrane formed, and gamete nuclei are dividing; F, cyst with two synkarya.
The principal feature distinguishing Entamœba coli from E.
histolytica is the formation of eight-nucleate cysts by the former as
contrasted with the tetra-nucleate cysts of the latter. The cyst-wall of
E. coli is thicker than that of E. histolytica (tetragena). Further, E. coli
does not usually ingest red blood corpuscles, nor are “chromidial
blocks” present inside its cyst (see p. 40).
According to Chatton and Lalung-Bonnaire14 (1912) the entamœbæ
of vertebrates should be placed in a separate genus Löschia, as they
differ in their life-history from E. blattæ, the type species of Entamœba.
Leidy (1879), however, named the genus Endamœba, but further
researches are necessary on biological variation among these
organisms.
Entamœba histolytica, Schaudinn, 1903.
Syn.: Amœba coli, autt. p. p. Amœba dysenteriæ, autt. p. p.
The average size of the amœboid trophozoite is 25 µ to 30 µ.
In fæces diluted with salt solution the amœbæ swell to 40 µ and more.
There is sometimes separation of the body substance into a strongly
refractile vitreous ectoplasm and a corneous endoplasm, pronounced
even in repose, although the former is not equally thick at all parts of
the periphery. In the endoplasm generally there are numerous foreign
bodies (bacteria, epithelial cells, colourless and red blood corpuscles
(fig. 6), and occasionally living flagellates of the intestine). The
nucleus is 4 µ to 6 µ in diameter, and may be difficult to recognize
because it is sometimes weakly refractile and poor in chromatin. Its
shape is slightly variable; it is usually excentric, sometimes wholly
peripheral at the limit of the two parts of the body. Vacuoles are not
present in quite fresh specimens, but appear later. In the study of
E. histolytica, the morphological characters of the trophozoite or
vegetative stage of the organism formerly separated as E. tetragena
(figs. 5, 6, 8a) must be considered (see p. 38).
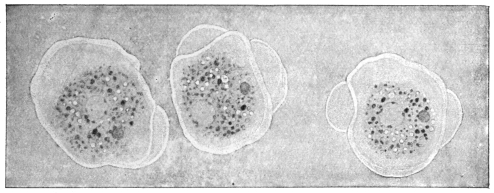
Fig. 5.—Entamœba histolytica (tetragena form), showing three successive changes of form
due to movement. × 1100. (After Hartmann.)
The history of the development of these species, which give rise to amœbic
enteritis as distinguished from bacillary dysentery, was formerly not so well known
as that of E. coli. Upon being introduced into cats (per anum) dysenteric amœbæ
provoke symptoms similar to those in man. In the
latter, besides metastatic liver abscesses, abscesses
of the lungs, and, according to Kartulis, cerebral
abscesses are occasionally produced. Marchoux
(1899) states that when the disease has lasted for
some time liver abscesses are produced in cats also.
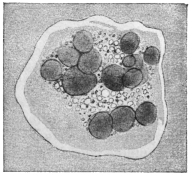
Fig. 6.—Entamœba histolytica
which has ingested many red blood
corpuscles. × 1100. (After Hartmann.)
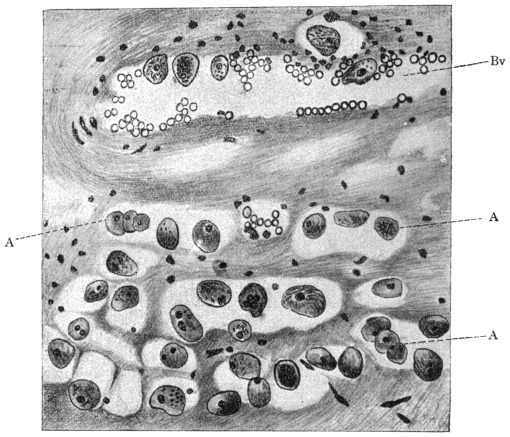
Fig. 7.—Section through wall of large intestine (of a man) close under an ulcer caused by
Entamœba histolytica. A, amœbæ that have penetrated partly in blood-vessels (Bv), partly
in tissue of submucosa to the muscularis. Magnified. (After Harris.)
In the large intestine of infected cats the
amœbæ creep over the epithelium, and here and
there they force the epithelial cells apart, as well
as removing them or pushing them in front of
them; the amœbæ thus insert themselves into the
narrowest fissures. They penetrate also into the
glands through the epithelium, and thence into
the connective tissue of the mucosa. Intestinal
and glandular epithelia perish under the influence
of these parasites: the cells are pushed aside,
fall to pieces or are absorbed by the amœbæ. In the connective tissue of the
mucosa the amœbæ migrate further, and often accumulate above the muscles.
Finally they rupture this and force their way into the submucosa. In cats,
apparently, the penetration is not so great as in men, according to Kruse and
Pasquale. During their migration the parasites also gain access to the lymph-follicles
of the wall of the intestine, which become swollen and commence
to suppurate; follicular abscesses arise and after their rupture follicular ulcers.
The diseased patches in the mucosa are markedly hyperæmic and numerous hæmorrhages
are set up. Roos and Harris state that the amœbæ also penetrate into
the blood-vessels (fig. 7) and this explains the occurrence of metastatic abscesses.15
The whole submucosa is severely swollen at the diseased spot and undergoes small-celled
infiltration in the neighbourhood of the colonies of amœbæ. From these
findings Jürgens (1902) draws the conclusion16 which is followed here, that the amœbæ
are causative agents of the enteritis of cats, which disease is well defined, both
pathologically and anatomically. Subsequent researches confirm the experience of
earlier authors; great precautions were taken to exclude errors, hence, as with
Gross and Harris, no exception can be taken to their results. The inoculation
material was derived from soldiers who suffered from amœbic enteritis in China
and who were admitted into the garrison hospital at Berlin. In order to be
independent of the patients themselves, transmission experiments from cat to cat
were performed, after the first experiments on cats yielded positive results. This
was also effected by rectal feeding as employed by earlier workers. Such appeared
necessary in order to prevent the evacuation of the inoculation material per anum,
as well as to avoid the employment of morphia and ether narcosis. Forty-six
cats were used for the experiments. Ten cats received tested stools containing
motile amœbæ from soldiers suffering from amœbic enteritis contracted in China.
Sixteen other cats received stools from cats infected by inoculation. All the animals
sickened and suffered from the disease. Five cats received dejecta from human
amœbic enteritis in which, however, no motile amœbæ were present. Thirteen cats
received stools from soldiers who suffered from bacillary dysentery. None of the
latter cats took the complaint and none showed changes in the large intestine upon
sectioning. The injection of various bacteria, obtained from a stool of amœbic
enteritis pathogenic to cats, remained without result in both the cats employed for
this experiment. Lastly, two cats, which had been kept with those artificially
infected, were taken ill spontaneously and suffered from the disease. In the opinion
of Harris, who ascertained the harmless nature of bacteria derived from the
intestinal flora containing dysenteric amœbæ, young dogs are capable of being
infected.
Within the large intestine an active increase of Entamœba histolytica must occur.
Nevertheless, Jürgens did not definitely find changes that might be interpreted in this
sense. Schaudinn (1903) observed division and gemmation in vivo. Both processes,
in which the nucleus divides by amitosis, can only be distinguished by the fact that
the daughter individuals are similar in binary fission but dissimilar in gemmation,
whether they make their appearance singly or in greater numbers. Schizogony,
resulting in the formation of eight individuals, which is so characteristic for Entamœba
coli, was not observed. (But schizogony, into four merozoites, is now known
to occur. Gemmation processes are apparently degenerative.)
Resistant stages, which serve for transmission to other hosts, are according to
Schaudinn17 first formed when the diseased portions commence to heal, or more
accurately, the recovery commences when the vegetative increase of the amœbæ in the
intestine discontinues. The so-called spores of E. histolytica were distinguished very
definitely from those of E. coli; they were said to consist of spheres of only 3 to
7 µ in diameter, which were surrounded by a double membrane, at first colourless, but
becoming a light brownish yellow colour after a few hours, and possessing a protoplasmic
content containing chromidia. They were said to arise by fragments of chromatin
passing outwards from the nucleus of the amœba into the surrounding cytoplasm
(fig. 9, a) and undergoing so marked an increase that finally the whole cytoplasm
became filled with chromidia. The remainder of the nucleus underwent degeneration
and became extruded. On the surface of the cytoplasm there then arose small
protuberances containing chromidia. These processes had been observed in the
living organisms. They gradually divided and separated from membranes which later
became yellow. The remainder of the amœba perished. Craig18 had also seen phases
of this process of development. It must be remarked that, according to recent
researches, these processes of exogenous sporulation are degenerative in character (see
p. 41). The small spores may be fungi. The “sporulation” processes are only mentioned
here as a warning. They are now only of historic interest. By means of an
experiment made on a cat, Schaudinn ascertained that ingestion of permanent cysts,
which resist desiccation, is the cause of the infection. The animal took food containing
dry fæces with amœba cysts; these fæces came from a patient suffering from amœbic
enteritis in China. On the evening of the third day the cat evacuated blood-stained
mucous fæces which contained large numbers of typical Entamœba histolytica. On
the fourth day after the infection the animal experimented upon died, and the large
intestine showed the changes previously stated.
E. histolytica also is found in the large intestine. This was originally shown
to be the case by Kartulis, and the fact has recently been confirmed from many
quarters. It is also present in the metastatic abscesses of which it is the cause
(cf. among other authors, Rogers, Brit. Med. Journ., 1902, ii, No. 2,177, p. 844; and
1903, i, No. 2,214, p. 1315).
It should lastly be pointed out in this connection that mixed infections also take
place. For instance, in addition to E. histolytica, E. coli, and, under certain circumstances,
flagellates may be found together. In the same way E. coli may come
under observation even in bacillary dysentery. On the other hand, Schaudinn
stated that in cases of dysentery endemic in Istria, Entamœba coli, if it had hitherto
been present, disappeared, to return again after recovery from the illness.
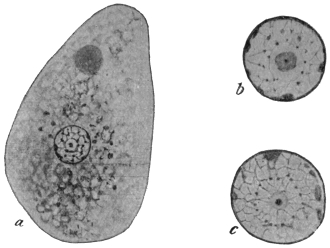
Fig. 8.—Entamœba histolytica. a, trophozoite (tetragena type) containing red blood
corpuscles, × 1,300; b and c, two isolated nuclei showing different appearances of karyosome,
centriole and nuclear membrane, × 2,600. (After Hartmann.)
(Entamœba tetragena, Viereck, 1907.)
This amœba must now be considered to be a part of the lifecycle
of Entamœba histolytica, in fact a very important part of that
cycle, especially in its tetranucleate cystic stages.
This organism, the so-called Entamœba tetragena, may occur in
the human intestine in cases of amœbic dysentery, especially in
mild or chronic cases. It was discovered by Viereck in 1907 in
patients suffering from dysentery contracted in Africa. Soon afterwards
an independent description was published by Hartmann, who
called the amœba E. africana. It was also studied by Bensen and
Werner. Recently (1912–13) much work has been published on
this amœba by Darling and others; in this way its relationship to
Schaudinn’s E. histolytica has been made known.
In general morphology it somewhat resembles Entamœba coli, and
its discoverer at first mistook it for a variety of that species. According
to Hartmann, a distinct ectoplasm is only clearly visible when
a pseudopodium is protruded (fig. 5). The granular endoplasm may
contain ingested red blood corpuscles (fig. 6). The large, round
nucleus is visible in the fresh state (fig. 8, a). So-called chromidial
masses (? crystalloidal substances) may occur in the cytoplasm.
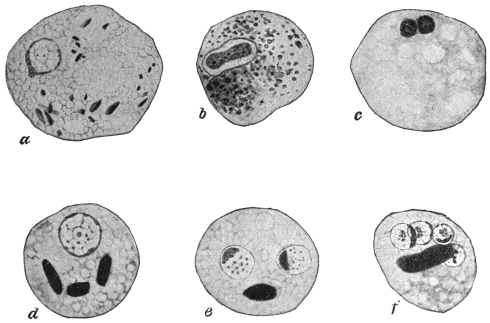
Fig. 9.—Entamœba histolytica (tetragena form). a, emission of chromatin from nucleus;
b, nuclear division; c, degenerating form with two nuclei; d, e, f, cysts containing one, two
and four nuclei respectively, and showing chromidial blocks. × 2,000. (After Hartmann.)
Some investigators, as Hartmann,19 lay stress on the internal
structure of the nucleus (fig. 8, b, c), best seen in preparations fixed wet
and stained with iron-hæmatoxylin. The nucleus is limited by a well-marked
nuclear membrane, on the inside of which granules or nodules
of chromatin may occur. There is a karyosome, which, in successfully
stained specimens, shows, at times, a central dot called a
centriole. (The nucleus of Entamœba coli does not contain such a
centriole.) However, the structure of the nucleus varies at different
periods during the life-cycle.
The diameter of the trophozoites or vegetative forms (fig. 8, a) is
variously given as from 20 µ to 40 µ. Multiplication proceeds by
binary fission and also by schizogony into four merozoites.20
Reproduction takes place by endogenous encystment (fig. 9, d-f),
which is preceded by nuclear division into two, reduction and then
autogamy. The interpretation of the latter phenomenon as autogamy
is disputed by some authors. The round cysts, which may measure
12 µ to 15 µ in diameter, contain four nuclei, together with darkly
staining masses of various shapes, the so-called “chromidial blocks”
(fig. 9, f). The cyst-wall of E. histolytica (tetragena) is thinner than
that of E. coli, and the diameter of the cyst is rather less. E. histolytica
has not yet been cultivated.
Infection in man occurs by way of the mouth by the ingestion
of cysts. A patient showing acute symptoms of dysentery is not
usually infective, for he is merely harbouring the large trophozoites,
which, by experiment, have been shown not to be infective to animals
(kittens) when administered by the mouth. The stools of recovered
patients may still contain cysts, and they may thus act as cyst-carriers
or reservoirs of disease by infecting water and soil. The stools of such
cyst-carriers are often solid, and so cysts of E. histolytica (tetragena)
are easily overlooked. Mathis (1913)21 points out that healthy carriers
of E. histolytica may be found; 8 per cent. of the natives of Tonkin
examined by him were healthy carriers of cysts.
In return cases, or prolonged untreated cases of entamœbic
dysentery, a generation of smaller trophozoites is associated with, or
replaces the larger ones. In stools they are frequently refractile and
consequently stain slowly intra vitam. These trophozoites are the
“smaller, senile, or pre-cyst generation” of Darling. This pre-cyst
generation is characterized by the presence of blocks of crystalloidal
substance in the cytoplasm, and by the possession of a prominent,
densely stainable karyosome. Darling believes this generation to
be the same as that described by Elmassian as Entamœba minuta.22
Walker,23 Darling,24 Wenyon25 and others believe that Entamœba
histolytica, which was only seen by Schaudinn in a single case, that of
a Chinaman, is really E. tetragena. Darling states that if the published
illustrations of E. histolytica and of E. tetragena are collected from the
literature and compared, it will be seen that the writers have been
calling E. histolytica the large trophozoites seen in dysenteric stools.
These large trophozoites frequently display no karyosome, but they
can be demonstrated as E. tetragena by animal inoculation, or by the
history of the case. On the other hand, the illustrations of E. tetragena
show that the authors have been dealing with the small generation or
reduced forms (“E. minuta”), which are the direct descendants of the
large trophozoites. If kittens are inoculated rectally with dysenteric
material containing large trophozoites, the strain may be carried in
successive kittens for four to six transfers. If, on the other hand,
kittens are inoculated rectally with small trophozoites of the pre-cyst
generation, the transmission cannot be carried through more than one
or two kittens. Wenyon has succeeded in maintaining E. tetragena in
kittens for several generations.
In some of the preparations from the last remove, pathological
forms of the trophozoites may be seen. These show abnormal forms
of budding, especially peripherally, such as have been described by
Schaudinn and by Craig as characteristic of E. histolytica. Schaudinn’s
small peripheral, exogenous buds and cysts are thus explained. Craig
has latterly changed his views.
Further, Darling states that tetragena cysts fed by the mouth to
kittens produce bowel lesions in which trophozoites having the characters
of E. tetragena, E. histolytica and E. nipponica (Koidzumi) occur.
In view of the work of recent observers, the peculiar exogenous
encystment which Schaudinn made characteristic of Entamœba
histolytica has been shown to be due to degenerative changes in senile
races of the amœba. E. histolytica and E. tetragena are one and the
same species, and its trophozoite is subject to variation. According
to some observers the histolytica type of nucleus—described by
Schaudinn as being poor in chromatin and not easily seen in the
fresh state—occurs frequently in patients with severe symptoms of
dysentery; on the other hand, the tetragena type of nucleus—round
and easily seen in the fresh state—may occur in cases presenting slight
dysenteric symptoms. Intermediate types of nuclei are seen. The
name of this species, the principal pathogenic amœba of man, must
then be E. histolytica by priority. The cystic stages of E. histolytica are
those first recorded by Viereck and formerly described as E. tetragena.
The geographical distribution of E. histolytica is wide.
Noc’s Entamœba (1909).
A species of Entamœba was cultivated by Noc26 in 1909 from cysts
derived from liver abscesses, from dysenteric stools and from the
water supply of Saigon, Cochin China. He cultivated it in association
with bacteria. It is pathogenic. It has been considered allied
to E. histolytica, and shows internal segmentation or schizogony. It
exhibits polymorphism. This amœba has been found by Greig and
Wells (1911) in cases of dysentery in India. It is an important
organism and requires further investigation.
Certain other Entamœbæ27 have been described at various times from
the intestinal tract of man. Probably most, if not all, of these are not
good species and in some cases much more information is needed.
Entamœba tropicalis (Lesage, 1908). This parasite is said to be
non-pathogenic, and to occur in the intestine of man in the tropics.
It has a general resemblance to E. coli, but forms small cysts (6 µ to
10 µ in diameter). The nucleus of the cyst is said to break up into a
variable number of daughter nuclei, from three to thirteen having
been noted. Lesage states that it is culturable in symbiosis with
bacteria. It is probably a variety of E. coli, if not a cultural amœba.
Entamœba hominis (Walker, 1908) has a diameter of 6 µ to 15 µ. A
contractile vacuole is present. Encystment is total, and small cysts are
formed. It is culturable. The original strain, now lost, was obtained
from an autopsy in Boston Hospital. This organism is probably
a cultural amœba.
Entamœba phagocytoides (Gauducheau, 1908). This parasite was
discovered in a case of dysentery at Hanoi, Indo-China. The amœba
is small, 2 µ to 15 µ in diameter. It is active. It ingests bacteria and
red blood corpuscles, while peculiar spirilla-like bodies are found in
its cytoplasm. It multiplies by binary and multiple fission. It can
be cultivated. More recently (1912) the author appears to consider
the amœba to be a stage of a Trichomonas, but abandons the view
later (1914). Further researches on this organism are needed.
Entamœba minuta (Elmassian, 1909)28 was found, in association
with E. coli, in a case of chronic dysentery in Paraguay. It resembles
E. tetragena but is smaller, rarely exceeding 14 µ in diameter. Schizogony
occurs, four merozoites being produced. The encystment is
total and endogenous, giving rise to cysts containing four nuclei.
This amœba is considered by Darling and others to be the pre-cyst
trophozoite stage of E. histolytica (tetragena).
Entamœba nipponica (Koidzumi, 1909) was found in the motions
of Japanese suffering from dysentery or from diarrhœa, in the former
case in company with Entamœba histolytica. Its diameter is 15 µ to
30 µ. The endoplasm is phagocytic for red blood corpuscles. The
nucleus is well defined, resembling that of E. coli and of E. tetragena.
Multiplication occurs by binary fission and by schizogony. Encystment
is total, but has not been completely followed. Darling and
others consider that this is an abnormal form of E. histolytica,
while Akashi (1913) doubts if it is an amœba at all, but rather is
to be regarded as shed epithelial cells.
General Remark.—It is now considered by some workers that
true Entamœbæ cannot be cultivated on artificial media. Quite
recently Williams and Calkins (1913)29 have somewhat doubted this
opinion, and state that certain cultural amœbæ, originally obtained
from Musgrave in Manila, exhibit the various morphological variations
associated with true entamœbæ of the human digestive tract.
Entamœba buccalis, Prowazek, 1904.
The size varies from 6 µ to 32 µ. Ectoplasm is always present;
the endoplasm contains numerous food-vacuoles. The nucleus is
vesicular, with a greenish tinted membrane which is poor in
chromatin. The size of the nucleus is from 1·5 µ to 4·5 µ. A contractile
vacuole is not visible. The pseudopodium is broad. It was
discovered in the mouths of persons with dental caries at Rovigno and
also at Trieste, being most easily found in dense masses of leucocytes,
also among leptothrix and spirochæte clusters. It can be easily distinguished
from leucocytes by more intense staining with neutral red.
Multiplication proceeds by fission. Transmission may take place
through the small spherical cysts. This species (fig. 10) has since been
observed in Berlin, and is also occasionally found in carcinoma of
various regions of the oral cavity. (Leyden and Löwenthal, 1905).
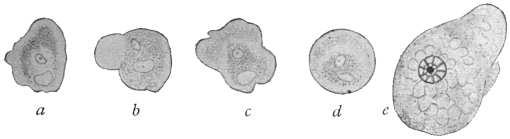
Fig. 10.—Entamœba buccalis, Prow. a-d, the same specimen observed during five minutes.
× 1,000. e, amœba fixed and stained with iron-hæmatoxylin. × 1,500. (After Leyden
and Löwenthal.)
Entamœba buccalis, Prow., is said to be allied to a protozoön which
A. Tietze has found either encysted or free in the lumen of the orifice
of the parotid gland of an infant aged 4 months. The gland had
undergone pathological change, and had therefore been extirpated.
The organisms, which were roundish and three to four times the size
of the normal epithelial cells of the gland, were without a membrane
and possessed a nucleus in which the chromatic substance appeared
to be contained in a karyosome. Bass and John’s30 (Feb. 1915) and
Smith, Middleton and Barrett (1914) state that E. buccalis is the cause
of pyorrhœa alveolaris.
Entamœba undulans, Aldo Castellani, 1905.
Under this name a protozoön is described which A. Castellani found in addition
to Entamœba histolytica and Trichomonas intestinalis in the fæces of an European
planter living in Ceylon, who had suffered from amœbic enteritis and liver abscess.
The shape of the body was roundish or oval, 25 µ to 30 µ in the greatest diameter.
It was without a flagellum, but with an undulating membrane, and capable of protruding
a long pseudopodium from different parts of its body at short intervals. The
nucleus could not always be recognized in life; it was, however, always demonstrable
by staining. One or two contractile vacuoles were present. The protoplasm was
finely granular, showing no differentiation into ecto- and endo-plasm. According to
Braun, in spite of the author declaring himself expressly against the flagellate nature
of the parasite, such a nature may be assumed to be tolerably certain in view of the
description and illustration.
It is now considered that Entamœba undulans is a portion of a flagellate,
namely, Trichomonas.
Entamœba kartulisi, Doflein, 1901.
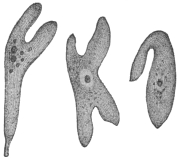
Fig. 11.—Entamœba kartulisi,
Dofl., from the pus of an
abscess in the lower jaw, showing
different stages of movement.
(After Kartulis.)
Doflein gave this name to amœbæ, from 30 µ to 38 µ in diameter,
which Kartulis (1893) found on examining the pus of an abscess in the
right lower jaw of an Arab, aged 43, and in a portion of bone that had been
extracted. The movements of the amœbæ
(fig. 11) were more active than those of
“dysenteric amœbæ.” Their coarsely
granular cytoplasm contained blood and
pus corpuscles, and a nucleus was generally
only recognizable after staining. Vacuoles
were not seen with certainty. Flexner reported
upon a similar case, and Kartulis
published five additional cases. As in these
cases dental caries was present the infection
is likely to have proceeded from the oral
cavity as a result of the carious teeth.
Craig31 (1911) considers that this parasite is
probably identical with Entamœba histolytica.
In the literature the following species have been reported as
occurring in the oral cavity of man:—
Amœba gingivalis, Gros, 1849. [? identical with Entamœba buccalis.]
Amœba buccalis, Sternberg, 1862.
Amœba dentalis, Grassi, 1879.
Far too little, however, is known concerning these to regard them as definite
species, that is, independent organisms; Grassi thinks it even possible there may
have been a confusion in their case with salivary corpuscles. If they really are
amœbæ they are all of them probably identical with Entamœba buccalis.
Genus Paramœba, Schaudinn, 1896.
Schaudinn established the genus Paramœba for a marine rhizopod which multiplied
by division, became encysted at the end of its vegetative life and then
segmented into swarm bodies with two flagella. These multiplied by longitudinal
fission, and finally passed into the condition of Amœbæ. Whether the human parasite
described by C. F. Craig (1906) as
Paramœba hominis.
belonged to this genus was for a time uncertain. It is now placed in a new genus
Craigia, Calkins, 1912, since it possesses only one flagellum.32
In the amœbic stage it is 15 µ to 25 µ in diameter; ecto- and endo-plasm during
rest are indistinguishable. The body substance is granular, with a spherical, sharply
contoured nucleus and an accessory nuclear body. No vacuoles are present, but
occasionally the endoplasm contains red blood corpuscles. The pseudopodia are
hyaline, finger- or lobe-shaped, and are protruded either singly or in twos.
Multiplication is by binary fission and by the formation of spherical cysts (15 µ to
20 µ in diameter) in which occurs successive division of the nuclei, ultimately forming
ten to twelve roundish bodies each of which soon develops a flagellum. The flagellate
stages have similarly a spherical shape and attain a diameter of 10 µ to 15 µ.
They also occasionally contain red blood corpuscles and pass either directly or after
longitudinal division into the amœboid phase.
Craig found these Amœbæ and the flagellate stage belonging to them in six
patients in the military hospital at Manila (Philippine Islands), five of whom were
suffering from simple diarrhœa whilst the sixth exhibited an amœbic enteritis and
contained also Paramœba hominis, with Entamœba histolytica, Schaudinn. In one
of the other cases, Trichomonas intestinalis was present.
B. Amœbæ from other Organs.
Entamœba pulmonalis, Artault, 1898.
Artault33 discovered a few amœboid forms with nucleus and
vacuole in the contents of a lung cavity. In the fresh condition they
were distinguishable from leucocytes by their remarkable capacity of
light refraction. They were also much slower than the latter in
staining with methylene blue or fuchsine. Their movements became
more lively in a strong light. Water and other reagents killed them,
and then, even when stained, they could not be distinguished from
leucocytes. They have also been seen by Brumpt. R. Blanchard found
amœbæ which may belong here in the lungs of sheep. A. pulmonalis
is perhaps the same as Entamœba buccalis. Smith and Weidman34
(1910, 1914) described an entamœba, E. mortinatalium, from the
lungs and other organs of infants in America.
Amœba urogenitalis, Baelz, 1883.
This species was found in masses in the sanguineous urine as well
as in the vagina of a patient in Japan, aged 23. Shortly before the
death of the patient, which was caused by pulmonary tuberculosis,
hæmaturia with severe tenesmus of the bladder had set in. The
amœba, which showed great motility, and had a diameter of about
50 µ when quiescent, exhibited a granular cytoplasm and a vesicular
nucleus. Baelz is of opinion that these parasites were introduced into
the vulva with the water used for washing the parts, and thence had
penetrated into the bladder and vagina. Doflein places the organism
in the genus Entamœba, and it is perhaps identical with E. histolytica.
Similar cases are also reported (1892–3) by other authors: Jürgens, Kartulis, Posner,
and Wijnhoff. Jürgens found small mucous cysts, filled with amœboid bodies, in the
bladder of an old woman suffering from chronic cystitis; they were also found
in the vagina. The amœba observed by Kartulis in the sanguineous urine of
a woman, aged 58, suffering from a tumour of the bladder, measured 12 µ to 20 µ,
and exhibited slow movements by protruding short pseudopodia. The vacuoles and
nucleus became visible only after staining with methylene blue.
Posner’s case related to a man, aged 37, who had hitherto been quite healthy and
had never been out of Berlin. Suddenly, after a rigor, he passed urine tinged with
blood. This contained, besides red and white blood corpuscles and hyaline and
granular casts, large granular bodies (about 50 µ in length and 28 µ in breadth), which
slowly altered their shape, and contained red blood corpuscles in addition to other
foreign matter. These bodies exhibited one or several nuclei and some vacuoles.
From the course of the disease, which extended over a year, and during which
similar attacks recurred, Posner came to the conclusion that the amœbæ which had
originally invaded the bladder had penetrated into the pelvis of the kidney, where
they probably had settled in a cyst, and thence induced the repeated attacks.
Wijnhoff observed four cases of amœburia in Utrecht.
Amœba miurai, Ijima, 1898.
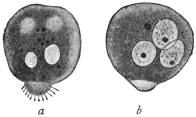
Fig. 12.—Amœba miurai, Ij.
× 500. a, fresh; b, after treatment
with dilute acetic acid.
(After Ijima.)
Under this term the author describes protoplasmic bodies which Miura, in
Tokyo, found in the serous fluid of a woman, aged 26, who had died from pleuritis
and peritonitis endotheliomatosa. Two days before death these same forms had
also appeared in the hæmorrhagic fæces of the
patient. The bodies were usually spherical or
ellipsoidal, and at one pole carried a small protuberance
(fig. 12) beset with filamentous short
“pseudopodia” (really a pseudopodium covered
with cilia). Their size varied between 15 µ and
38 µ. The cytoplasm was finely granular, and
no difference was observable in the ecto- and
endo-plasm, only the villous appendage
was clearer. The cytoplasm contained vacuoles
more or less numerous, none of which was
contractile. After the addition of acetic acid one
to three nuclei could be distinguished, 8 µ to 15 µ in size. Actual movements were
not observed. Taking everything into consideration, the independent nature of these
bodies is, to say the least, doubtful, although it cannot be denied that they possess a
certain similarity to the marine Amœba fluida, Grüber or Greeff, and to a few other
species. (It is likely that cells present in serous exudation were mistaken for amœbæ.)
Appendix.
“Rhizopods in Poliomyelitis acuta.”
In three cases of poliomyelitis acuta which were investigated by Ellermann, the
spinal fluid obtained by puncture of the cord contained bodies, from 10 µ to 15 µ in
size, which had amœboid movements and exhibited variously shaped pseudopodia in
large numbers. After staining, a usually excentric nucleus, about 1·5 µ in size, was
demonstrated in them.
Order. Foraminifera, d’Orbigny.
The order is divided by Max Schultze into Monothalamia and Polythalamia. Only
a few of the former can be considered here.
Sub-Order. Monothalamia. (Testaceous Amœbæ).
These forms occur frequently in fresh water, rarely in sea water.
They possess a shell which is either pseudo-chitinous in character, or
consists of foreign particles, or in a few cases is composed of siliceous
lamellæ. There is usually an orifice for the protrusion of pseudopodia.
The only representative of the order of interest here is:—
Genus. Chlamydophrys, Cienkowski, 1876.
The genus is based on a form which A. Schneider carefully investigated and
considered to be the Difflugia enchelys of Ehrenberg. L. Cienkowski rediscovered
this same form and created for it the genus Chlamydophrys. We agree
with this view, but not with the renaming of the organism (so common at the time).
If the parasite in dung, Chlamydophrys stercorea Cienk. is identical with Difflugia
enchelys of Ehrenberg, the old specific name should be retained.
The genus is characterized by the possession of a hyaline, structureless,
slightly flexible shell which is ovoid or reniform. At the more
pointed pole there is an orifice situated terminally or somewhat laterally,
serving for the emergence of the filiform pseudopodia (fig. 13, a).
The protoplasm does not entirely fill the interior of the shell. An
equatorial zone bearing excretory granules divides the shell internally
into two almost equal portions. The anterior portion is rich in
vacuoles and serves for the reception of nutriment and for digestion.
The posterior part is vitreous, and contains the nucleus. One to three
contractile vacuoles are situated in the equatorial zone.
Chlamydophrys enchelys, Ehrbg.
Syn.: Chlamydophrys stercorea, L. Cienkowski.
This species (fig. 13) is found in the fæces of various animals (cattle,
rabbits, mice, and lizards), and also in quite fresh human fæces.
According to Schaudinn, the parasite occurs so frequently in the
human fæces that it must be considered of wide distribution. The
species must traverse the intestine of man and animals during one
stage of its life cycle, as Schaudinn showed by experiments on himself
and on mice. He infected himself with cysts (fig. 14) by swallowing
them, and evacuated the first Chlamydophrys as early as the following
day. After the evacuation of numerous specimens on one of the
following days the infection ceased.
The nucleus of a living specimen is surrounded by a hyaline,
strongly refractile chromidial mass, arranged in the form of a ring.
Chromatin stains colour it darkly.
Asexual multiplication (fig. 13, b), which takes place in fæces, follows
a similar course to that of allied forms (e.g., Euglypha, Centropyxis). It
commences by the cytoplasm issuing from the orifice of the shell and
assuming the shape characteristic of the mother organism, but in
a reverse position. The nucleus then divides by mitosis, when the
daughter nuclei move apart from one another. The chromidial ring
also divides into two portions by a process of dumb-bell like constriction.
The one daughter nucleus remains in the mother organism,
the other moves towards the daughter individual, which then separates
from the parent.
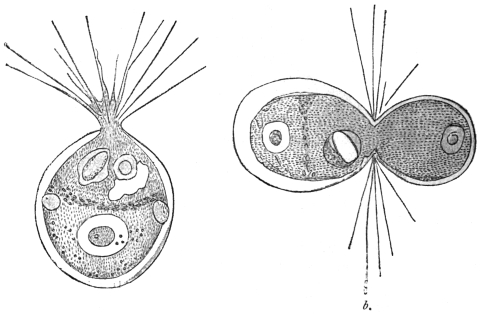
Fig. 13.—Chlamydophrys enchelys. a, free, motile form, showing nucleus, equatorial
granules, vacuoles and pseudopodia; b, dividing organism. × 760. (After Cienkowski.)
In this species plasmogamic union of two or more individuals (up to twenty) is
frequently observed. Such colonies may similarly divide, and in this way monstrosities
frequently arise. When drying of the fæces, or deficiency of food occurs,
encystment takes place apparently spontaneously. The whole body, as stated by
Cienkowski, issues from the shell, assumes a spherical shape (probably with discharge
of water) and becomes surrounded with a thick membrane (fig. 14). After the addition
of water and the escape of the encysted Chlamydophrys, a new shell must be formed.
Schaudinn, who has not given a more detailed description of the process of encystment
in this species, but refers to Cienkowski and to similar observations made on
Centropyxis, states of the latter that the encystment takes place within the shell.
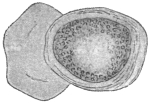
Fig. 14.—Chlamydophrys
enchelys, encysted;
on the left the old capsule.
× 760. (After Cienkowski.)
The sexual multiplication is accompanied by shedding of all the
foreign bodies and of the degenerating nucleus. The protoplasm,
now contracting into a sphere, remains behind in the shell with the
chromidial mass. From the latter several new nuclei arise (sexual
nuclei) often eight in number. The cytoplasmic sphere then segregates
into as many spherical portions as there are nuclei present.
When they have assumed an oval form, two flagella develop at one
pole and the flagellispores swarm out of the shell.35 The biflagellate
swarm-spores, or gametes, copulate in pairs and apparently the individuals
of the pairs of gametes arise from different mother organisms.
The zygote secretes a thick covering which soon
becomes brown and rough. These zygote cysts
or resistant spores must now pass from the intestine
of an animal in order to complete their
development. The escape of the cyst contents
does not always take place in the intestine;
often it does not occur until after defæcation.
These shell-less individuals (amœbulæ) soon
become invested with a shell. But in the
alkaline intestinal contents, shell formation may
proceed even while the organism is in the intestine,
and multiplication may take place.
Schaudinn’s further communication was of special interest; it was
to the effect that Chlamydophrys was related to
Leydenia gemmipara, Schaudinn, 1896.
In the fluid removed by puncture from two patients suffering
from ascites in the first medical clinic in Berlin, cellular bodies
with spontaneous movement were found, which Leyden and
Schaudinn regard as distinct organisms. They remained alive
without the use of the warm stage for four or five hours, the
external temperature being 24° to 25° C. In a quiescent condition
they were of a spherical or irregular polygonal form. Their surface was
rarely smooth, being beset with protuberances and excrescences (fig. 15).
The substance of the body was thickly permeated with light refractile
granules with a yellowish shimmer. The hyaline ectoplasm was
rarely seen distinctly. All sizes from 3 µ to 36 µ in diameter were
observed. The movements were rather sluggish, the ectoplasm in
the meantime appearing in the form of one or several lamellæ, in
which also strings of the granular endoplasm occurred, and frequently
protruded over the border of the hyaline pseudopodia. The tendency
for the joining of several individuals by means of their pseudopodia
was so marked that associations ensued similar to those known in
free-living Rhizopoda.
The cytoplasm enclosed blood corpuscles as well as numerous
vacuoles, one of which pulsated slowly about every quarter of an
hour. A vesicular nucleus the diameter of which was about equal
to one-fifth of the body was present.
Multiplication took place by means of division and budding (fig. 15, c),
after previous direct division of the nucleus. The buds were supposed
to divide repeatedly soon after their appearance, thus giving rise to
minute forms of 3 µ.
There was a suspicion in both cases that the ascites was associated
with malignant neoplasms in the abdomen, and autopsy confirmed
this view in one case.
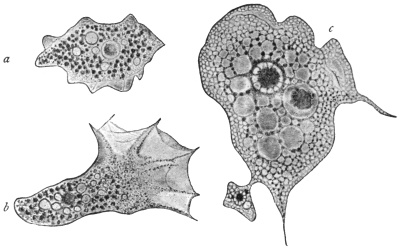
Fig. 15.—Leydenia gemmipara, Schaud. a, in a quiescent condition, × 1000; b, in the act of
moving, × 1000; c, from a fixed preparation, showing a bud, × 1500.
The parasite, which has seldom been observed, has been variously
interpreted; for example, it has been regarded merely as altered tissue
cells. It is now known, from Schaudinn’s researches, that Leydenia
gemmipara is connected with abnormal conditions of Chlamydophrys,
occasionally occurring as a commensal in the ascitic fluid. The form
is produced when pathological conditions of the large intestine create
an alkaline reaction of its whole contents. The formation of shells
then often ceases, and these naked Chlamydophrys are enabled to
multiply atypically by division and gemmation. Such stages, which
are no longer capable of a normal development, are the Leydenia, as
Schaudinn has demonstrated.
Class II. MASTIGOPHORA, Diesing.
Sub-Class. FLAGELLATA, Cohn emend. Bütschli.
During the motile part of their life the Flagellata possess one or more flagella
which serve for locomotion, and in many cases also for the capture of food. A few
groups (Euglenoidinæ, Choanoflagellata) have only one flagellum, others two or
several of about equal length (Isomastigoda), or of various lengths (Monadina,
Heteromastigoda, Dinoflagellata). The long flagellum is the principal one; the
smaller ones on the same organism are accessory flagella. The flagella directed
backwards, which occur in the Heteromastigoda and are used for clinging, are
termed trailing flagella or tractella. At the base of the flagellum, which is almost
always at the anterior end, a Choanoflagellate possesses a cytoplasmic funnel-shaped
neck or collar. In the parasitic forms an undulating membrane is often present.
The body of the Flagellata is usually small, generally elongate and of unchangeable
form. It is frequently covered by a distinct cuticle, and, in certain groups,
by a hard envelope, or it may be more or less loosely enveloped by a gelatinous or
membranous covering. An ectoplasmic layer is thin and not always obvious. The
granular cytoplasm contains a varying number of vacuoles, one of which may be
contractile, and is generally situated near the area from which the flagella arise,
that is, at the anterior extremity. The cytoplasm, moreover, contains the nucleus,
which is nearly always single; and in many species there are also yellow, brown,
or green chromatophores of various shapes, such as occur in plants. Some
species feed after the manner of green plants (holophytic), or of plants devoid of
chlorophyll (saprophytic); others, again, ingest solid food, and for this purpose
usually possess a cytostome; the latter, however, in a few forms is not used for its
original function, but is connected with the contractile vacuole. Many parasitic
forms feed by endosmosis. A few species possess eye-spots with or without light-refracting
bodies.
Variation in the form of the nuclear apparatus occurs. One nucleus only, which
may be compact or vesicular, is known in many species. This nucleus is situated
either centrally or sometimes near the flagellar end of the body, but its position is
subject to variation. The flagella may arise near the nucleus. Other structures,
such as an axial filament and a rhizoplast, may be present. Some flagellates are
binucleate, the two nuclei—which often differ in size and shape—being separated
from each other. One of these nuclei is the principal, vegetative or trophic nucleus;
the other is an accessory nucleus, frequently termed the blepharoplast, flagellar
or kinetic nucleus. One or more small basal granules are often present at or very
near the origin of the flagella.
Multiplication is by fission, usually longitudinal, which may occur in either the
free or encysted forms. Division is initiated by that of the nucleus or nuclei
(especially the kinetic nucleus). The basal granule divides also. Collars and
chromatophores, if present, likewise separate into two. Variation in the method of
doubling the original number of flagella occurs. In most organisms, especially
uniflagellate forms, the flagellum splits lengthwise, after division of the basal granule,
blepharoplast and nucleus. The daughter flagella may be of the same or different
lengths and thicknesses. Other flagellates at division are said to produce new
flagella in the neighbourhood of the original ones. The daughter organisms in such
cases are provided with one or more parental flagella in addition to newly formed
ones. It has been stated that in certain cases the parent flagellate retains all its
flagella, while new ones arise ab initio in the cytoplasm of the daughter forms.
Multiplication by longitudinal fission may be interrupted sooner or later by the
production of gametes, which form zygotes, from which new generations of individuals
arise. In many flagellates gamete formation and sporogony are unknown,
and asexual reproduction by fission alone prevails.
Incomplete division results in the formation of colonies of individuals. These
colonies must not be confused with the aggregation rosettes of flagellates found
among the parasitic Mastigophora. The individuals of aggregation rosettes are
capable of immediate separation from the rosette at will.
A number of parasitic Flagellata produce non-flagellate stages which are very
resistant to external conditions, the assumption of which forms serves to protect the
organisms during their transference from one host to another. Such non-flagellate
forms possess one or more nuclei, are usually of an oval or rounded contour, and
are capable of developing into the full flagellate on the return of more favourable
conditions. These forms are often known as the post-flagellate stage of the
organism. When ingested by a new host, the post-flagellate coat becomes more
flexible, and the phase of the organism which now recommences growth is known as
the pre-flagellate stage; it gradually develops into the typical flagellate organism.
Many Flagellata live free in fresh and salt water. They prefer stagnant water,
rich in organic products of decomposition, such as puddles, swamps and pools.
Those forms developing shells and colonies are, as a rule, adherent. A number of
species are parasitic in man and animals, living mostly within the intestine or in the
blood.
It is usual to classify the Flagellata in four orders: Euflagellata, Dinoflagellata,
Choanoflagellata, and Cystoflagellata, of which only the Euflagellata are of interest
to us. This is a group comprising numerous species, for the further classification
of which the number and position of the flagella are utilised.
The Euflagellata observed in man belong to the Protomonadina as well as to the
Polymastigina. The former possess either only one or two similar flagella, or one
principal and one or two accessory flagella. The Polymastigina possess at least
three flagella of equal size, or four to eight of unequal size, inserted at different
points. An undulating membrane may be present in members of both groups.
It must also be pointed out that unicellular organisms with one or several
flagella are not always classified with flagellates, for such forms occur in Rhizopods
as well as temporarily in the lower plants. In addition, the examination of the
flagellates, especially the parasitic species, is very difficult on account of their
diminutive size and great activity; thus it happens that certain forms cannot with
certainty be included in the group because their description is insufficient.
Order. Polymastigina, Blochmann.
The Polymastigina contains flagellates with three to eight flagella.
Some of the Flagellata parasitic in man belong to the Polymastigina,
and to two or three genera that are easily distinguishable.
Genus. Trichomonas, Donné, 1837.
The body is generally pyriform, the anterior part usually rounded, the posterior
part pointed. There are at the anterior extremity three (? four) equally long flagella
that are sometimes matted together. A blepharoplast (kinetic nucleus) and basal
granule are present, together with a supporting structure known as an axial
filament or axostyle. In addition there is an undulating membrane, bordered by a
trailing flagellum, that commences at the anterior extremity and proceeds obliquely
backwards. The nucleus, which is vesicular, is situated near the anterior extremity,
and behind it are one or more vacuoles, none of which seems to be contractile.
These flagellates are parasitic in vertebrate animals, and live chiefly in the intestine.
Trichomonas vaginalis, Donné.
The form of the body is very variable, and is elongate, fusiform
or pear-shaped, also amœboid. The length varies between 15 µ and
25 µ, and the breadth between 7 µ and 12 µ. The posterior extremity
is drawn out to a point and is about half the length of the remainder
of the body. The cuticle is very thin and the body substance finely
granular. At the anterior extremity there are three—some say four36—flagella
of equal length which are frequently united together, at least
at the base, and are easily detached.
There is an undulating membrane (fig. 16)
which runs spirally across the body, arising
from the place of insertion of the flagella, and
terminating at the base of the caudal process.
A cytostome seldom is recognizable in fresh
specimens, but is apparently present. The
nucleus is vesicular, elliptical and situated near
the anterior extremity.37
Multiplication takes place by division
(Marchand). Encysted forms are almost unknown.
Trichomonas vaginalis lives in the vaginal mucus of
women of various ages, not in normal mucus, but in
mucus of acid reaction. It is found in menstruating
females as well as in females who have passed the
menopause. It occurs in pregnant and non-pregnant
women, even in very young girls, provided always that
they have a vaginal catarrh with acid reaction of the
secretion. Should the acid reaction change, as, for
instance, during menstruation, the parasites disappear,
as they do likewise on injection of any alkaline fluid into
the vagina. A low temperature (below +15° C.) is also
fatal to the parasites. These flagellates can pass from
the vagina through the urethra into the bladder, and
produce severe catarrh, and are not easily removed.
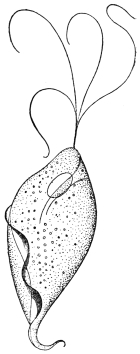
Fig. 16.—Trichomonas
vaginalis, Donné. × 2,000
approx. (After Künstler.)
Four flagella are represented,
but usually only
three are present.
T. vaginalis appeared to be a parasite specific to the female organs
and not transmissible to man. However, several observations have
since been made that confirm the occurrence of this species in the
urethra of the male. The infection apparently takes place through
coitus when changes are present in the urethral mucous membrane.
At any rate, three cases observed point to this circumstance.
Attempts at experimental transmission to rabbits, guinea-pigs and
dogs failed (Blochmann, Dock). So far, the manner in which women
become infected is unknown.
Trichomonas intestinalis, R. Leuckart, 1879 = Trichomonas
hominis, Davaine, 1854.
Some authors believe that a second trichomonad inhabiting man,
Trichomonas intestinalis, R. Lkt., is identical with Trichomonas vaginalis,
Donné. Leuckart’s species was based on the discoveries of Marchand
(1875) and Zunker (1878), who stated that according to all appearances,
and in their opinion, it was the same as Cercomonas intestinalis, Lambl,
1875 (nec 1859), which they found in
the fæces of patients suffering from
intestinal disorders. The organism
is described by them as being pear-shaped
and 10 µ to 15 µ in length
and 3 µ to 4 µ in breadth. The
posterior extremity terminated in a
point (fig. 17).
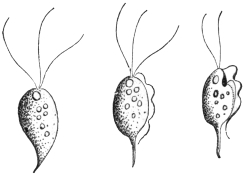
Fig. 17.—Trichomonas intestinalis, Lkt.
(After Grassi.)
A row of twelve or more cilia was said to
commence at the anterior end and extend
over the body. Leuckart stated that this
parasite, placed by the two authors in the
genus Cercomonas, was a Trichomonas, and
that they mistook the undulating membrane for cilia, and overlooked the flagella.
Notwithstanding its striking similarity with T. vaginalis, it was said to be distinguishable
from that species by differences in the undulating membrane. Lambl’s
C. intestinalis38 (of 1875) which corresponds with C. hominis, Davaine39 (1854), is
regarded by Leuckart as a true Cercomonad (characterized by a flagellum and the
absence of an undulating membrane, see p. 61), and is thus generically distinct
from Trichomonas.
The correctness of Leuckart’s judgment in regard to Marchand-Zunker’s flagellate
was demonstrated by Grassi’s researches, accounts of which were published soon
after. In about 100 cases of bowel complaints in North Italy and Sicily, Grassi
found Flagellata in the stools, which he first named Monocercomonas and Cimænomonas,
but later termed Trichomonas. However, in opposition to Leuckart, Grassi
has also classified Davaine’s C. hominis (= C. intestinalis, Lambl, 1875) as
Trichomonas, and most authors have followed his example. Hence arose the use
of the name Trichomonas hominis. It was through Janowski (1896) that the
former view was again taken up. After a review of the literature, the occurrence
of Cercomonads in the intestine of human beings in addition to Trichomonads
was considered by the author to have been proved, and he added a
description of the Trichomonads. According to this, all morphological distinction
between T. vaginalis, Donné, and T. intestinalis, Leuckart, disappeared. On the
other hand, it is worthy of note that the smaller size, the more pear-shaped form, and
the longer flagella differentiate T. intestinalis (= T. hominis) from T. vaginalis.40
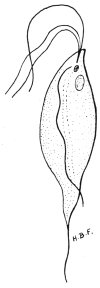
Fig. 18.—Trichomonas
intestinalis
from man,
showing anterior
flagella, cytostomic
depression anteriorly,
undulating
membrane, nuclei,
and axostyle.
×2,500. Original.
The easily deformed pear-shaped body has three free flagella
anteriorly, and an undulating membrane with its flagellar border
terminating in a short free flagellum posteriorly (figs.
17, 18). The undulating membrane may coil itself
spirally round the body. A supporting rod or axostyle
projects as a posterior spine. It appears to
begin near the nucleus and blepharoplast, which are
situated near the more rounded, anterior end of the
body. There may be a chromatoid basal supporting
line along the body for the undulating membrane.
Rows of chromatoid granules are sometimes situated
along one side of the axostyle. A cytostome may
sometimes be seen. In mice, Wenyon (1907) found
these parasites to vary in length from 3 µ to 20 µ.
They occur in the cæcum and intestine of mice,
where their internal structure seems more obvious
than in man. The flagellates divide by longitudinal
fission.
T. intestinalis, R. Leuckart, appears to be capable
of settling in all parts of the human intestine in which
the contents have an alkaline reaction. Trichomonads
have been cited as occurring in the oral cavity by
Steinberg, Zunker, Rappin and Prowazek; in the
œsophagus by Cohnheim, and in the stomach by
Strube, Cohnheim, Zabel, Hensen and Rosenfeld.
The normal situation seems to be the small intestine.
The parasites then appear in the dejecta, especially in various intestinal
diseases the course of which is connected with an increased peristalsis.
They are also found in healthy persons, from whom they are obtained
after the administration of laxatives. They have been regarded by some
workers as commensals, which, however, have the power of accelerating
the onset of intestinal complaints, or at least of adding to them. They
have been found in cases of carcinoma of the stomach, and in other
diseases of that organ in which the acid reaction ceased.
Naturally, whether all the reports relate to the same species of Trichomonas
must remain undecided. Certain authors (Steinberg, Cohnheim, van Emden)
accept several species. Prowazek speaks of a variety of T. intestinalis inhabiting
the oral cavity. This was distinguished by a posterior process exceeding the
length of the body fourfold, and by a somewhat unusual course of the undulating
membrane. The food of this form, which was found in the whitish deposit present,
especially in the cavities of carious teeth, consisted almost exclusively of micrococci.
Schmidt and St. Artault named the Trichomonads found in pathological products
(e.g., gangrene, putrid bronchitis, phthisis) of the lungs of man, as Trichomonas
pulmonalis. Trichomonads have also been found by Wieting in lobular pneumonia
in the lungs of pigs.
It is still uncertain in what way the infection takes place. Experiments in the
transmission of free trichomonads to mammals (per os), in which the same or allied
species occur (guinea-pigs, rats, apes), have been without result. Probably encystment
is necessary. Such conditions are mentioned by May, Künstler, Roos,
Schurmayer, van Emden, Prowazek, Galli-Valerio and Schaudinn. According to
Prowazek, intestinal trichomonads of rats become encysted for conjugation. In the
cyst an accumulation of reserve food material occurs, causing distension. The nuclei
of the conjugants each give off a reduction body and, after fusion, produce the
nuclei for the daughter individuals. According to Schaudinn the intestinal trichomonads
lose their flagella before conjugation, become amœboid and encyst in twos,
the formation of a large agglomeration of reserve substance accompanying this.
Galli-Valerio found double-contoured cysts in the fæces of trichomonad-infected
guinea-pigs, after the fæces had been kept for a month in a damp chamber. When
exposed to heat small flagellates escaped from them. Administration of such material
containing cysts resulted in severe infection with trichomonads, and death of the
experimental guinea-pigs followed. The cyst wall is clearly a protection against the
deleterious acid reaction of the stomach contents. Alexeieff (1911) and Brumpt (1912)
think that the trichomonad cysts of man are really fungi, while other workers also
doubt encystment among trichomonads. Wenyon (1907) states that T. intestinalis in
mice produces spherical contracted forms which escape from the body in the fæces.
Air, water, and under certain circumstances even food may be regarded as
vectors for the trichomonads. The occurrence of the organisms in the oral cavity,
and still more so in the lungs, is in favour of the air being the transmitting agent.
An observation made by Epstein supports the idea of water transmission. Multiplication
of the trichomonads, once they have gained access to the body, is effected by
longitudinal division commencing at the anterior end (Künstler). “Cercomonads”
with several flagella and an undulating membrane, as well as trichomonads, have
been observed by Ross in some cases of cutaneous ulcers.
Mello-Leitao (1913)41 has described flagellate dysentery in children
in Rio de Janeiro. He states that it is due to T. intestinalis
and Lamblia intestinalis either separately or together. Flagellate
dysentery, he thinks, is benign and is the most frequent form of
dysentery in infants. The flagellates are pathogenic to infants under
three years of age. Escomel (1913)42 found 152 cases of dysentery in
Peru due solely to Trichomonas. Such cases are probably widespread.
Genus. Tetramitus, Perty, 1852.
Tetramitus mesnili, Wenyon, 1910.
Syn.: Macrostoma mesnili, Chilomastix mesnili, Fanapapea intestinalis.
The genus Tetramitus differs from Trichomonas in possessing an
undulating membrane inserted in a deep groove or cytostome. There
are three anterior flagella. The pear-shaped organism measures 14 µ
by 7 µ, but smaller examples occur. T. mesnili occurs in the human
intestine, having been described by Wenyon43 (1910) from a man from
the Bahamas in the Seamen’s Hospital, London. Its occurrence
is widespread. Alexeieff considers that Macrostoma and Tetramitus
are synonymous. The parasite is the same as Fanapapea intestinalis,
Prowazek, 1911, from Samoa. Brumpt (1912) found T. mesnili to be
the causal agent of colitis in a Frenchwoman. Nattan-Larrier (1912)
considers it of little pathological importance.
Gäbel44 (1914) described an interesting case of seasonal diarrhœa
acquired in Tunis, in which a new Tetramitid was the causal agent.
The organism was pear-shaped, without an undulating membrane,
and measured 6·5 µ to 8 µ by 5 µ to 6 µ. The cytostome was large,
and there was no skeletal support. Encystment occurred. Gäbel
named the organism Difämus tunensis and considered that it was
pathogenic.
Genus. Lamblia, R. Blanchard, 1888.
Syn., Dimorphus, Grassi, 1879, nec Haller, 1878; Megastoma, Grassi, 1881, nec
de Blainville.
The body is pear-shaped, with a hollow on the under surface anteriorly. It has
four pairs of flagella directed backwards, of which three pairs lie on the borders of
the hollow disc, and the fourth arises from the pointed posterior extremity.
Lamblia intestinalis, Lambl, 1859.
Syn.: Cercomonas intestinalis, Lambl, 1859 (nec 1875); Hexamitus duodenalis,
Davaine, 1875; Dimorphus muris, Grassi, 1879; Megastoma entericum, Grassi,
1881; Megastoma intestinale, R. Blanch., 1886; Lamblia duodenalis, Stiles, 1902.
The organism is pear-shaped and bilaterally symmetrical. It is
from 10 µ to 21 µ long and 5 µ to 12 µ broad and possesses a thin
cuticle. Anteriorly an oblique depression is present, which functions
as a sucking disc (fig. 19, s). Its edges are raised above the general
surface and are contractile. It corresponds to a peristome and acts as
an adhesive organ (fig. 20, b, c). No true cytostome is present. A
double longitudinal ridge, representing axostyles, extends from the
sucking disc to the tapering posterior extremity, which is prolonged
as two flagella from 9 µ to 14 µ long.
Lamblia intestinalis possesses eight flagella (fig. 19). The first pair
of flagella, which cross one another, arise in a groove formed by the
anterior edge of the sucking disc. Two pairs of flagella (lateral and
median) are inserted on the posterior edge of the disc, while the
posterior flagella occur at the tapering posterior extremity of the body.
Basal granules are found at the bases of the flagella. The median
flagella are most active in movement, the anterior and lateral flagella
being less motile, as they are partially united to the body for part of
their length.
The nuclear apparatus is situated in the thin, anterior, hollowed
part of the body. It is at first dumb-bell shaped, the “handle” of the
dumb-bell being formed by a very slight connecting strand, which
eventually separates, so that the flagellate becomes binucleate, and thus
completes the general bisymmetry of the organism.
There is a karyosome in each nucleus. Other bodies of unknown
function, and possibly composed of chromatin, occur on or near
the axostyles.
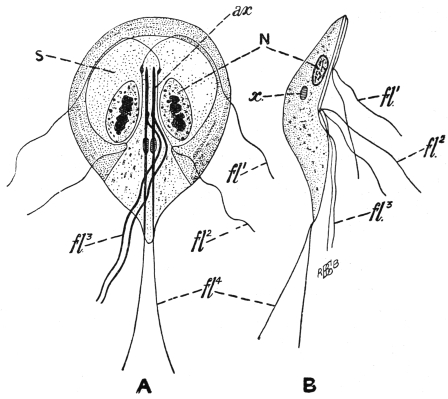
Fig. 19.—Lamblia intestinalis. A, ventral view; B, side view; N, one of the two nuclei;
ax., axostyles; fl1, fl2, fl3, fl4, the four pairs of flagella; s, sucker-like depressed area on the
ventral surface; x, bodies of unknown function. (After Wenyon.)
Division has not been observed in the flagellate stages of the
Lamblia, but it occurs within the cysts. The resistant cysts (fig. 20, e)
are oval and are surrounded by a fairly thick, hyaline cyst wall.
They measure 10 µ to 15 µ by 7 µ to 9 µ, and may be tetranucleate.
According to Schaudinn, the cysts arise from the conjugation of two
individuals, and nuclear rearrangement occurs.
L. intestinalis occurs in its flagellate stage in the duodenum and
jejunum, and rarely as such in the other parts of the intestine.
Normally it is found in the large intestine as cysts, which are voided
with the fæces. The hosts of Lamblia include Mus musculus, M.
rattus, M. decumanus, M. silvestris, Arvicola arvensis and A. amphibius,
the dog and cat, rabbit, sheep and man. Cysts voided with the fæces
of infected animals reach plants or drinking water, and thence are
transferred to man.
The flagellate in these different hosts exhibits some variation in
size and in the problematic chromatic bodies. Bensen has suggested
the species L. intestinalis from man, L. muris from the mouse and
L. cuniculi from the rabbit. It is not certain whether these different
species are necessary, as the variation may be due to differences of
environment.
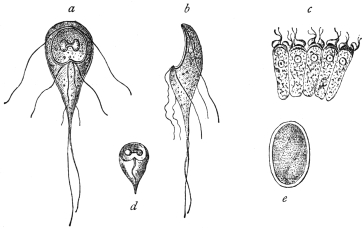
Fig. 20.—Lamblia intestinalis. a, from the surface; b, from the side; c, on intestinal
epithelium cells; d, dead and e, encysted. (After Grassi and Schewiakoff.)
Like Trichomonas, Lamblia can multiply under inflammatory
conditions of the alimentary tract. Thus they are found in cases
of diarrhœa, carcinoma of the stomach, etc. The parasites attach
themselves by their sucking discs to the epithelial cells of the gut
(fig. 20, c), and though their numbers may be very great, their direct
pathological significance is not fully known. Their occurrence in
cases of diarrhœa has been explained as being due to the increased
peristalsis, which has detached the parasites from the epithelium.
Free flagellate forms perish in stools if kept, more especially if the
temperature falls below 0° C. or rises above 40° C. Lamblia has
often been found in dysenteric diseases, especially in the East, and
is said to be the causal agent of certain diarrhœas in India. Mathis
(1914)45 found Lamblia in cases of diarrhœa with dysenteriform stools
in Tonkin. He also discovered healthy carriers of Lamblia cysts.
The parasite under discussion was first observed by Lambl (1859) in the mucous
evacuations of children. He regarded the parasite as a Cercomonad and termed
it Cercomonas intestinalis, which name as a rule is applied to Cercomonas hominis,
Davaine, although Stein had already pointed out the difference between the two species.
Grassi (1879) observed this species first in mice (calling it Dimorphus muris), and
subsequently in human beings in Upper Italy and named it Megastoma entericum.
Bütschli and Blanchard then laid stress on the identity of this species with Lambl’s
C. intestinalis (1859), and consequently called it Megastoma intestinale. Later,
Blanchard drew attention to the circumstance that the generic name Megastoma
chosen by Grassi had already been used four times for various kinds of animals,
and established the genus Lamblia. Accordingly, L. intestinalis is the valid name,
and should be generally adopted.
In Upper Italy the parasite in the encysted condition has also been seen by
Perroncito in man. At the same time, Grassi and Schewiakoff began a new
investigation of specimens from mice and rats. In Germany, L. intestinalis
was found by Moritz and Hölzl, Roos, Schuberg and Salomon. Moritz and Hölzl
confirmed the relative frequency of the species. In Königsberg, Prussia, a student
found encysted Lamblia in his fæces. One case was reported from Finland by
Sievers, another case from Scandinavia by Müller. Frshezjesski and Ucke reported
cases from Russia. Jaksch announced the occurrence of the parasite in Austria;
Piccardi mentioned their presence again in Italy. They were reported from Egypt
by Kruse and Pasquale, and from North America (Baltimore) by Stiles. Noc stated
that 50 per cent. of the population of Tonkin harboured Lamblia. Finally, the
structure of L. intestinalis has been described by Metzner (1901), and by Wenyon46
(1907) in mice.
In all these cases L. intestinalis has been observed in the small
intestine, or in the evacuations of patients with intestinal diseases.
It has also been found in the intestine of healthy subjects. Just as
Trichomonas intestinalis may be found inhabiting the stomach in
diseases of that organ, in which an alkaline reaction is present
(carcinoma), so has L. intestinalis been found to occur under
similar circumstances (Cohnheim, Zabel). However, in Schmidt’s
case, 1 per cent. hydrochloric acid was certainly stated to be present.
Infection takes place by the ingestion of cysts (fig. 20, e), as was
established by Grassi, experimentally on himself. Cereal food-stuffs,
contaminated with Lamblia cysts from vermin of the locality, such as
rats and mice, serve to convey the infection to man. Such cysts may
probably be found in street-dust, etc. Stiles induced infection in
guinea-pigs, and Perroncito in mice and rabbits, by means of cysts
of Lamblia from human beings. Stiles suspected that flies could
transport Lamblia cysts. Mathis (1914) found that L. intestinalis
was not amenable to emetine, at any rate in its cystic stage.
Order. Protomonadina, Blochmann.
The smallness of the Protomonadines and their less superficial
situation than the Polymastigines, may be the cause that so far as
the species occurring in man are concerned, they were formerly less
well known. As regards parasitic species, this group may be divided
as follows, according to the number of flagella and the presence or
absence of an undulating membrane:—
(1) Cercomonadidæ, with one flagellum at the anterior extremity,
without an undulating membrane.
(2) Bodonidæ, with two flagella, without an undulating membrane,
except in Trypanoplasma.
(3) Trypanosomidæ, with one flagellum, and an undulating membrane
along the length of the body in some genera.
Family. Cercomonadidæ, Kent emend. Bütschli.
Small uniflagellate forms, without cytostome.
Genus. Cercomonas, Dujardin emend. Bütschli.
Oval or rounded organisms, with the aflagellar end often drawn
out into a tail-like process.
Cercomonas hominis, Davaine, 1854.
Davaine found flagellates in the dejecta of cholera patients. They
had pear-shaped bodies, lengthening to a point posteriorly. Their
length was from 10 µ to 12 µ, and a flagellum about twice as long as
the body projected from one extremity (fig. 21). A nucleus was
hardly recognizable. Occasionally a somewhat long structure (cytostome?)
appeared at the anterior extremity. The animals moved with
remarkable activity. They also attached themselves by means of their
posterior extremities and swung about around the point of attachment.
Davaine found a smaller variety, only about 8 µ long, in the dejecta
of a typhoid patient (fig. 21, b).

Fig. 21.—Cercomonas hominis, Dav. a, larger,
b, smaller variety. Enlarged. (After Davaine.)
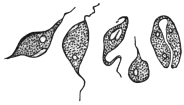
Fig. 22.—Cercomonas hominis,
Dav. From an Echinococcus
cyst. (After Lambl.)
The Flagellata observed by Ekeckrantz (1869) in the intestine of man belong to
this form—at least to the larger variety—and Tham (1870) reported fresh cases soon
after. Lambl’s publication of 1875, which was written in Russian, and became known
through Leuckart’s work on parasites, also alludes to apparently typical Cercomonads,
which, however, were discovered, not in the intestine, but in an Echinococcus cyst in
the liver (fig. 22). The elliptical, fusiform, rarely pear-shaped or cylindrical bodies
of the parasites measured 5 µ to 14 µ in length, and were provided with a flagellum
at one end, while the other extremity usually terminated in a long point. An oral
aperture occurred at the base of the flagellum, and there were one or two vacuoles
near the posterior extremity. Longitudinal division was also observed (fig. 22).
As already mentioned, this form, which Lambl termed Cercomonas
intestinalis, differs considerably from the form found by the same
author in 1859, which received the same designation (cf. Lamblia intestinalis,
p. 60), but it corresponds with Cercomonas hominis, Davaine.
The latter, as well as C. intestinalis, Lambl, 1875, is usually classed
with the Trichomonads, but, as has already been remarked (cf.
Trichomonas intestinalis, p. 54), this cannot be considered correct, as
only one flagellum is present.
Cercomonas vaginalis (Castellani and Chalmers, 1909) was found
in the vagina of native women in Ceylon.
Other species of Cercomonas have, at various times, been recorded
from man. However, the parasitic species of the genus Cercomonas
require further investigation.
According to Janowski (1896–7), typical Cercomonads have also been observed in
the intestine of man by Escherich, also by Cahen, Massiutin, Fenoglio, Councilman and
Lafleur, Dock, Kruse and Pasquale, Zunker, Quincke and Roos, and others. However,
it is an open question whether the Flagellata observed by Roos in one of his cases
belonged to Davaine’s species, the size showing some deviation (14 µ to 16 µ). In
his, as in many other cases, doubts have been raised as to whether the flagellates
found in the stools had actually lived in the intestine, or had subsequently appeared
in the fæces: for this a surprisingly short time only is necessary. Salomon also
appears to have observed Cercomonads (Berl. klin. Wochenschr., 1899, No. 46).
As with T. intestinalis so with C. hominis, it appears
that the parasite settles not only in the intestine but also in
the air-passages. This is demonstrated by the statements of
Kannenberg and Streng of the occurrence of Monads and
Cercomonads in the sputum and putrid expectoration in
gangrene of the lungs, which no doubt apply to C. hominis
(cf. also Artault). Possibly also the Flagellata observed in the
pleural exudation by Litten and Roos may be included here;
this is the more probable in Roos’s case as the process ensued
in the pleura after the breaking through of a vomica.
Perroncito and Piccardi have described encysted stages of
Cercomonads.

Fig. 23.—Monas
pyophila, R. Blanch.
(After Grimm.)
Monas pyophila, R. Blanch., 1895.
R. Blanchard thus designates a Flagellate that Grimm
found in the sputum, as well as in the pus of a pulmonary and
hepatic abscess, in the case of a Japanese woman living in
Sapporo. The parasites resemble large spermatozoa (fig. 23).
The body, 30 µ to 60 µ, has the shape of a heart or a myrtle leaf, and is surrounded by a
thick cuticle which is supposed to extend into the interior of the body, dividing it into
three parts. A long appendix at the rounded pole is covered for the greater part of
its length by the cuticle; the extremity, however, is free and resembles a flagellum.
The parasites were very active, frequently changed their shape, and were able to
retract the long appendix within the body, which then assumed a round form.
[This organism requires further investigation.]
Family. Bodonidæ, Bütschli.
Protomonadina which are either free-living or parasitic, with two
dissimilar flagella, while the possession of an undulating membrane
and of a kinetic nucleus or blepharoplast is variable.
There are three genera:—
1. Bodo, Stein, 1878, without a kinetic nucleus and undulating
membrane.
2. Prowazekia, Hartmann and Chagas, 1910, with a kinetic
nucleus and without an undulating membrane.
3. Trypanoplasma, Laveran and Mesnil, 1901, with a kinetic
nucleus and undulating membrane.
Of these genera Prowazekia must be discussed. Bodo does not
occur in man. Species of Trypanoplasma occur in the blood and in
the gut of various fishes, in the seminal receptacle of certain snails,
in the gut and genitalia of a flatworm (Dendrocœlum lacteum) and in
the vagina of a leech. Closely allied to Trypanoplasma is the genus
Trypanophis, parasitic in the cœlenteric cavity of Siphonophores.
Genus. Prowazekia, Hartmann and Chagas, 1910.
The genus was founded for a flagellate parasite, Prowazekia cruzi,
discovered in a culture of human fæces in Brazil. Various other
species have been referred thereto. The genus is separated from
Bodo by the possession of a second nucleus, the so-called kinetonucleus
or blepharoplast. It differs from Trypanoplasma in the absence of an
undulating membrane. It is heteromastigote, that is, it possesses two
dissimilar flagella, one anteriorly directed and the other lateral and
trailing.
The principal species are:
Prowazekia urinaria, Hassall, 1859.
Syn.: Bodo urinarius, Hassall, 1859; Trichomonas irregularis, Salisbury, 1868;
Cystomonas urinaria, Blanchard, 1885; Plagiomonas urinaria, Braun, 1895.
Hassall47 in 1859 first found Bodo-like flagellates in human urine.
He examined fifty samples of urine from patients suffering from
albuminuria and from cholera. The reaction of the urine was alkaline
or sometimes only feebly acid. The flagellates were only seen after
the urine had been standing for several days. Hassall named the
organism Bodo urinarius, and gave a very good description of it with
illustrations. The flagellate, which was round or oval, measured 14 µ
by 8 µ. The organism had “one, usually two, and sometimes three
lashes or cilia.” In 1868 Salisbury described a similar flagellate in
the urine under the name Trichomonas irregularis. Künstler in 1883
described the latter parasite under the name B. urinarius. In 1885
Blanchard, considering Künstler’s organism a different parasite from
Hassall’s, called it Cystomonas urinaria. Braun, in 1895, gave the
name Plagiomonas urinaria. Barrois (1894) considered Künstler’s
and Hassall’s organisms to be identical and not to be true parasites
of man. Sinton,48 in 1912, found the flagellate in the deposit, after
centrifuging, of a 24-hour old specimen of alkaline urine from a
Mexican sailor in the Royal Southern Hospital, Liverpool. Sinton
found a kinetic nucleus or blepharoplast in the organism, and
therefore placed it in the genus Prowazekia.
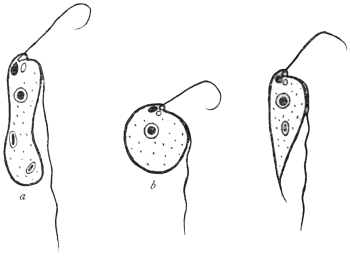
Fig. 24.—Types of Prowazekia urinaria. (a) sausage-shaped; (b) round; (c) carrot-shaped
form. (After Sinton.)
The flagellate stage (fig. 24) of the organism is polymorphic, and may
be either (a) sausage-shaped, 10 µ to 25 µ in length by 2·5 µ to 6 µ in
breadth; (b) round or oval, varying from 4 µ in diameter to oval
forms 15 µ by 10 µ; (c) a carrot-shaped form, of varying size up to
25 µ by 4 µ. The kinetic nucleus is large and pear-shaped. Near it
are basal granules, closely applied to one another, from which the
flagella arise. There is a small cytostome near the roots of the
flagella. There is a well-marked karyosome in the nucleus. The
movement is jerky. The shorter, anterior flagellum may be used in
food-capture. In life, bacteria have been seen to be ingested. Food-vacuoles
tend to accumulate at the posterior (aflagellar) end. A
contractile vacuole may be present, near the base of the cytostome,
and may really be the dilated fundus of the latter. Division occurs by
binary fission. The organism can encyst (fig. 25, a), when the flagella are
lost, and round or oval cysts are found, 5 µ to 7 µ in diameter. After
a time flagella are formed inside the cyst, and the organism emerges
therefrom in its typical flagellate form (fig. 25, b-f).
Sinton’s case is interesting. He obtained the flagellate only twice
from the same patient, a Mexican then in hospital in Liverpool. The
flagellate was not found in the patient’s fæces, nor was it found in
the urine on later occasions when taken aseptically.
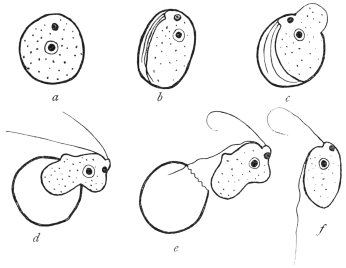
Fig. 25.—Prowazekia urinaria. Flagellate emerging from cyst. (After Sinton.)
In cultures Prowazekia urinaria was always found in association
with bacteria. The cultures died at a temperature of 37° C., but grew
well at 20° C. Various media were useful at the lower temperature,
such as urine, salt agar, nutrient agar, serum agar, blood agar, peptone
salt solution, and diluted blood serum. The flagellate was, then,
considered to be an accidental contamination and not a true parasite
of human urine.
Prowazekia asiatica, Castellani and Chalmers, 1910.
The flagellate was found by the discoverers in the stools of patients
suffering from ankylostomiasis and diarrhœa in Ceylon. It was referred
by them to the genus Bodo, but in 1911 Whitmore49 further studied
it and placed it in the genus Prowazekia. In the stools the flagellate
is found either as a long, slender form measuring 10 µ to 16 µ by 5 µ
to 8 µ or as a rounded form 8 µ to 10 µ in diameter. Its cytoplasm is
alveolar. A rhizoplast connects the basal granules to the kinetic
nucleus. There is multiplication and cyst formation as before. The
organism is easily cultivated, especially in the condensation water of
nutrose agar and maltose agar. The pathogenicity is stated to be nil.
Prowazekia javanensis, Flu, 1912.
Found in agar cultures from the motions of patients at Weltevreden,
Dutch East Indies.50 The flagellates are 12 µ long and 5 µ broad.
The lateral flagellum is stated to be attached to the cell body for a
short distance. Regarding the karyosome in the nucleus, the author
states that the smaller the karyosome the more chromatin is deposited
on the nuclear membrane. Flu mentions that the specific name
javanensis is a temporary one, as in the course of time it may be
shown that there is only one species of Prowazekia.
Prowazekia cruzi, Hartmann and Chagas, 1910.
Found in a culture from human fæces on an agar plate in Brazil,
and considered to be a free-living form.51 The organism is oval or
pear-shaped, 8 µ to 12 µ long and 5 µ to 6 µ broad. In human stools
at Tsingtau, China, a Prowazekia has been found by Martini which
he thinks is the same as Prowazekia cruzi. He considers it to be a
cause of human diarrhœa and intestinal catarrh.
Prowazekia weinbergi, Mathis and Léger, 1910.
This species was found in the fæces of men, both healthy and
diarrhœic, in Tonkin.52 It is pear-shaped, 8 µ to 15 µ long by 4 µ to
6·5 µ broad. The flagella occur at the broad end.
The discoverers think that Prowazekia weinbergi is an intestinal
inhabitant, but non-pathogenic, since it was found to occur in the
fæces even when obtained with aseptic precautions.
Prowazekia parva, Nägler, 1910.
A free-living form found in the slime on the stones at the biological
station at Lunz. Another Prowazekia was found in 1914 in tap-water
in Calcutta.
Family. Trypanosomidæ, Doflein.
The Trypanosomidæ, broadly considered, are uniflagellate
organisms, the flagellum being at the anterior end. The flagellum
arises near the blepharoplast (kinetic nucleus), which lies anterior,
near or posterior to the nucleus.
The following genera will be considered:—
Trypanosoma—with an undulating membrane along the length
of the body.
Crithidia—with a less well-developed undulating membrane
anteriorly (see fig. 49).
Herpetomonas—including the so-called Leptomonas, with anterior
free flagellum only, and no undulating membrane.
Leishmania—non-flagellate forms in mammalian blood, flagellate
herpetomonad stages in culture, probably occurring naturally in Arthropods.
Genus. Trypanosoma, Gruby, 1843.
The members of the genus possess a single flagellum, which
arises posteriorly, adjacent to a blepharoplast or kinetic nucleus. The
flagellum forms a margin to an undulating membrane, and may
or may not be continued beyond the body as a free flagellum.
Many species are parasitic in vertebrate blood and in the digestive
tracts of insects.
Historical.
The history of blood flagellates goes back to the year 1841, in which Valentin
discovered in the blood of a brook-trout (Salmo fario L.) minute bodies, from 7 µ
to 13 µ in length, with active movements and presenting marked changes in form.
Valentin considered the parasite a new species of the old genus Proteus or
Amœba, Ehrbg. This announcement led Gluge (1842) to publish a similar discovery
he had made in frog’s blood. The latter forms were called by Mayer (1843) Amœba
rotatoria, Paramœcium loricatum and P. costatum, while Gruby (1843) called them
Trypanosoma sanguinis.53 Later it was discovered that similar organisms occurred
also in the blood of birds (Wedl (1850), Danilewsky) and of mammals. Gros (1845)
found them in the mouse and mole, Chaussat (1850) in the house rat, Lewis (1879) in
the Indian rat, Wittich (1881) in the hamster. Danilewsky (1886–89) and Chalachnikow
(1888) investigated the structure and division of trypanosomes.
In the case of all these forms, there was no discussion as to a pathogenic
influence on the host. Opinion, however, as to the action of trypanosomes
changed when, in 1880, Evans found flagellates in the blood of horses in India
that suffered from a disease endemic there called “surra,” and associated
the parasites with the disease. Steel and Evans were successful in transmitting
the parasites—first known as Spirochæta evansi, Steel, then as Trichomonas
evansi, Crookshank, and finally as Trypanosoma evansi—to dogs, mules and horses.
They recognized that the above mentioned flagellates in the blood of the experimental
animals were the causal agents of the disease.
From that time there was a considerable increase in the literature, the
contents of which have been summarized by Laveran and Blanchard. In 1894
Rouget discovered trypanosomes in the blood of African horses that suffer
from “stallion’s disease” (dourine). In 1894 Bruce found similar forms (T. brucei) in
the blood of South African mammals suffering from “nagana,” and in consequence
attention was drawn to the part which the much dreaded tsetse-fly played in
the transmission of “nagana.” In 1901 Elmassian discovered trypanosomes in
the blood of horses that were stricken with “mal de caderas,” which is very
common in the Argentine. The disease in cattle named “galziekte” (gall-sickness),
occurring in the Transvaal, was also at one time attributed to
a trypanosome remarkable for its great size, and like some other species, bearing
the name of its discoverer (T. theileri).
The study of the species hitherto known has been carried on partly by the
above mentioned authors and in part by others, e.g., Rabinowitsch and Kempner,
Laveran and Mesnil, Wasiliewski, Senn. It was greatly advanced by the method
of double staining (with alkaline methylene blue and eosin) introduced by
Romanowsky (1891) and elaborated by Ziemann, Leishman, Giemsa and others.
By this means the presence of a terminal flagellum and of an undulating membrane
at the side of the flattened and extended body was demonstrated. Laveran and
Mesnil (1901) discovered allied flagellates in the blood of the fish, Scardinius
erythrophthalmus. These flagellates, now placed in the genus Trypanoplasma, had
a second free flagellum in addition to the one bordering the undulating membrane.
Trypanoplasms have since been found in both freshwater and marine fishes. The
transmission of trypanoplasms of freshwater fishes is effected by leeches. Trypanoplasma
varium from Cobitis is transmitted by Hemiclepsis marginata according to
Léger, while the Trypanoplasmata of Cyprinus carpio and Abramis brama reach
new hosts by the agency of Piscicola according to Keysselitz.
Another ally of the Trypanosomidæ, Trypanophis, lives in the cœlenteric
cavity of Siphonophores. It has also an extra terminal flagellum (Poche,
Keysselitz). [Trypanoplasma and Trypanophis belong to the Bodonidæ, see p. 63].
Finally it was shown that Trypanosomes occurred in human beings. Although
Nepveu’s early report of trypanosomes in the blood of malarial patients may be
doubtful, subsequent researches by Forde and Dutton demonstrated trypanosomes
(fig. 28) in the blood of a European, apparently suffering from malaria, living in the
Gambia. Dutton (1902) called the human trypanosome, T. gambiense. The expedition
despatched by the Liverpool School of Tropical Medicine (1902) to Senegambia
found trypanosome infections in six cases among a thousand inhabitants examined.
About the same time attention was devoted to the disease of West African
negroes known for a century as “sleeping sickness.” Castellani (1903) was the
first to succeed in demonstrating the presence of trypanosomes (at first called
T. ugandense) in centrifugalized cerebro-spinal fluid obtained by puncture from cases
of sleeping sickness in Uganda. Similar discoveries were made by Bruce, who also
found trypanosomes in the blood of those attacked with sleeping sickness. Sambon
regarded a species of Glossina as the transmitter. From consideration of the
geographical distribution of the disease Christy regarded Glossina palpalis as the
transmitter. Brumpt first thought it was G. morsitans, but, later, supported the
view of G. palpalis. Bruce, Nabarro and Greig also named the same insect as
the transmitter, not only for geographical reasons but also because healthy apes
became infected by the bite of certain G. palpalis. The inoculation of cerebro-spinal
fluid from subjects of sleeping sickness into the spinal canal of apes (Macacus)
had the same result.
Just as the discovery of the malarial parasites called forth a whole flood
of research memoirs which were followed by a second series on the relation of
the mosquitoes to malaria, so a similar outpouring occurred after the discovery
of the pathogenic trypanosomes of mammals and men. In both cases the inquiry
was not limited to the stages in man and other vertebrate hosts, but the fate
of the parasites in the intermediate (invertebrate) hosts was investigated, and
allied species were obtained from many different hosts.
Novy and MacNeal (1903) were the first to cultivate trypanosomes in artificial
media (blood-agar).
In 1910 Stephens and Fantham recorded the presence of another human
trypanosome, T. rhodesiense, from a case of sleeping sickness in Rhodesia, where
G. palpalis was absent. Kinghorn has since demonstrated that T. rhodesiense is
transmitted by G. morsitans. Kinghorn and Yorke believe that big game (e.g.,
antelope) is the reservoir of T. rhodesiense.
The output of literature on trypanosomiasis in men and animals is enormous.
To cope with it the Sleeping Sickness Bureau Bulletin was founded in 1908,
and it is now (since November 1912) continued as a section of the Tropical Diseases
Bulletin, wherein current literature is reviewed.
General.
Trypanosomes occur in the blood of representatives of all the
vertebrate classes. Often the trypanosomes occur so scantily in the
blood that they are overlooked on examination. A useful aid in
detecting the flagellates in such cases consists in the use of cultures
of the blood of the host on artificial media. Stimulated by the
medium multiplication occurs, and hence the parasites are more
easily detected. [For the composition of such culture media see
Appendix.]
There is a periodicity in the appearance of the trypanosomes in
the peripheral blood of the host, due to alternating phases of multiplication
and of rest on the part of the parasites. Such periodicity
has been established both by biological and enumerative methods.
Again, a seasonal variation has been observed in the occurrence
of certain trypanosomes in the peripheral circulation of the hosts; for
example, some trypanosomes (e.g., T. noctuæ in birds) are found only
in the summer in the blood, while in the winter they occur in the
internal organs.
Recent cultural researches have established that trypanosomes, e.g.,
T. americanum, may be present in very small numbers in hosts, such as
cattle, which are quite unharmed by them, and in which the presence
of these flagellates formerly was never suspected (“cryptic trypanosomiasis.”)
However, the majority of the trypanosomes occurring in
domestic animals are usually deleterious or even lethal to their hosts.
Many wild animals, such as various species of antelope, harbour
trypanosomes without being injured thereby. In such cases it is
probable that the vertebrate hosts have been so long parasitized in the
past, that they have become tolerant and immune to the effects of the
flagellates. Should such trypanosomes of wild animals be transmitted
to domesticated stock or man, they may re-acquire their initial virulence
and become pathogenic to the new host. As a general statement,
the newer a parasite is to its host the greater is its virulence.
For example, T. gambiense, T. rhodesiense and T. brucei are innocuous
to big game in Africa, but are pathogenic to man and domestic animals
respectively. Pathogenic trypanosomes appear to have a wider range
of hosts, that is, to be less limited to one specific host than non-pathogenic
forms. Thus, T. rhodesiense is pathogenic to man and all
laboratory animals, while it is non-pathogenic to antelopes and their
kind.
Morphology.
The general structure of the various trypanosomes shows much
uniformity, though variations in size and shape occur. Typically the
body is elongate and sinuous. The flagellar end tapers gradually to
a point, the aflagellar extremity usually being rounded or more blunt.
In some trypanosomes there is much diversity in size, the organisms
varying from long, slender forms to short, stumpy ones; in other
species relative constancy of size is maintained. The former are
known as polymorphic trypanosomes, the latter as monomorphic
forms.
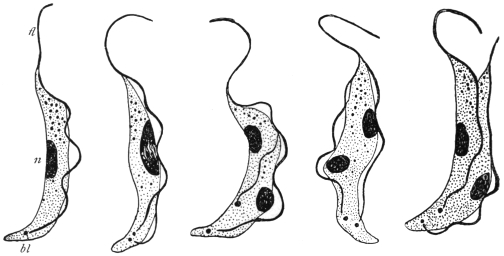
Fig. 26.—Trypanosoma brucei in division. n, nucleus; bl, blepharoplast; fl, flagellum.
× 2,000. (After Laveran and Mesnil.)
Two nuclei are present. The main or principal nucleus, sometimes
termed the trophic nucleus, is often situated towards the centre
of the body; it is frequently of the vesicular type, containing a
karyosome. The blepharoplast or kinetic nucleus is posterior to the
nucleus, and usually is rod-like. The flagellum arises close to the
blepharoplast, and forms an edge to the undulating membrane.
It may or may not extend beyond the limits of the undulating
membrane. If it does so, the unattached part is known as the free
flagellum. Sometimes a small granule is found at the origin of the
flagellum. This is the basal granule, and is considered by some to
function as the centriole of the kinetic nucleus.
The undulating membrane is a lateral extension of the ectoplasm
or periplast, and is the main agent in locomotion. It is edged by the
flagellum, which forms a deeply stainable border to it. Within the
membrane substance, often arranged parallel with its edge, are a
number of fine contractile elements, the myonemes. These contractile
elements may also occur on the body of the trypanosome.
They are easily seen in some large trypanosomes, but are difficult of
demonstration in others, owing to their great fineness.

Fig. 27.—Trypanosoma lewisi. Multiplication rosettes. × 1,000. (After Laveran and Mesnil.)
Multiplication of trypanosomes in the blood is brought about by
binary longitudinal fission (fig. 26). Division is initiated by that of
the blepharoplast and nucleus. The division may be equal or subequal,
whereby differences in size of individuals partly arise. Multiple
division by repeated binary fission, without complete separation of
the daughter forms, is known in some trypanosomes (e.g., T. lewisi),
and rosettes of parasites thereby are produced (fig. 27).
The classification of trypanosomes is very difficult. Laveran
(1911)54 has suggested the examination of the relative length of the
flagellum as a diagnostic character, and so arranged these flagellates
in mammals in three groups. The first group included those trypanosomes
always having part of the flagellum free (e.g., T. evansi,
T. vivax); the second group comprised forms without a part of
the flagellum free (e.g., T. congolense), while the third group included
forms some members of which have free flagella, while others have
not (e.g., T. gambiense). Bruce55 (1914) and Yorke and Blacklock56
(1914) have also devised classifications.
Resting stages of some trypanosomes have been found in the
internal organs of their vertebrate hosts. The formation of these
oval, Leishmania-like bodies will be noted in individual cases later.
Similar small oval bodies form an important phase in the life-history
of T. cruzi, which multiplies normally by multiple fission or schizogony
into these oval, daughter elements, and not by binary longitudinal
fission in the circulating blood.
Polymorphism in trypanosomes (e.g., T. gambiense, T. rhodesiense)
is now interpreted as a phenomenon resulting from growth and
division.57 Long, thin forms are those about to divide. Fully mature
forms are shorter and broader. Various intermediate types occur and
represent growth forms. Formerly, polymorphism was interpreted in
terms of sex, thin forms being regarded as males, broad forms as
females, while the intermediate types were termed indifferent. Conjugation
was not observed, and there is no evidence in support of
the sexual interpretation.
The transmission of trypanosomes from one vertebrate host to
another is usually accomplished by the intermediation of some biting
arthropod in the case of terrestrial animals, while leeches are usually
considered to act as transmitters in the case of the trypanosomes
occurring in aquatic animals. Developmental phases of the life-histories
of trypanosomes occur in the invertebrate transmitters, and
will be considered in individual cases.
Trypanosoma gambiense, Dutton, 1902.
Syn.: Trypanosoma hominis, Manson, 1903. Trypanosoma nepveui, Sambon, 1903.
Trypanosoma castellanii, Kruse, 1903. Trypanosoma ugandense, Castellani,
1903. Trypanosoma fordii, Maxwell Adams.
In vertebrate blood Trypanosoma gambiense is polymorphic, for
long, thin forms may be seen in contrast with short, stumpy forms,
as well as intermediate forms (fig. 29, a—c). This polymorphism has
been interpreted in terms of sex, especially by German investigators,
following Schaudinn (see above). However, there is no evidence of
conjugation, and the polymorphic forms are more easily interpreted
in terms of growth and division, for the long thin forms are potential
dividing organisms, and the stumpy or short parasites, with little or
no free flagellum, are the adult individuals.
Morphology of T. gambiense in the Circulating Blood.
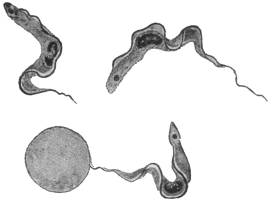
Fig. 28.—Trypanosoma gambiense. × 1,700.
(After Dutton.)
T. gambiense varies from 13 µ to 36 µ in length, its average length
being 24·8 µ, as was determined in 1913 by exact biometrical methods
by Stephens and Fantham.58 Three forms of parasite occur. According
to Miss Robertson,59 the relatively
short forms from 13 µ to 21 µ
long may be regarded as the
mature or “adult” type of parasite
in the blood. They carry on the
cycle in the vertebrate. From
them intermediate forms, which
are longer than the “adult” but
at first have the same breadth,
arise by growth. They possess
a free flagellum. The intermediate
forms grow into long individuals,
which are those about to
divide. The products of division give rise, directly or indirectly, to
the adult forms.
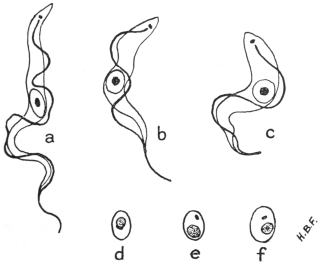
Fig. 29.—Trypanosoma gambiense. Development in vertebrate host. a, long, slender,
b, intermediate and c, short, stumpy forms, found in the blood; d, e, f, non-flagellate, latent
forms from internal organs. × 2,000. (Original. From preparations by Fantham.)
The organism has an elongate body with an anterior or flagellar
end and a blunter posterior or non-flagellar end. The protoplasm
is finely granular, large inclusions being rare. The central nucleus is
oval and large, often containing most of its chromatin concentrated as
a karyosome, with small granules only scattered near or on the fine
nuclear membrane. The blepharoplast is either rounded or rod-shaped.
The undulating membrane is thrown into folds and is
bordered by the flagellum. A small basal granule may be present
near, or at the actual origin of the flagellum.
Multiplication in the vertebrate is brought about by longitudinal
division. According to the recent account of division by Miss
Robertson, the blepharoplast doubles, then the flagellum splits for the
greater part of its length, and the daughter flagella separate, one being
shorter than the parent flagellum. The nucleus often shows two well
marked dark granules on the membrane at opposite poles, and these
appear to act as centrosomes. Nuclear constriction occurs and the
halves gradually separate. Finally the two daughter organisms become
free, the aflagellar end splitting last. The products of division may be
equal or unequal. Repeated division goes on in the general circulation
until the blood swarms with parasites. Then the trypanosomes gradually
disappear, and a period occurs when it is practically impossible to
demonstrate the parasite in the blood. At such a period, trypanosomes
can be obtained by puncture of the enlarged lymphatic glands or of
the spinal canal, or can be found in the internal organs, more particularly
in the spleen, lungs, liver and bone-marrow. In the latter
organs, latent bodies are produced (fig. 29, d—f) which are capable of
again becoming flagellates and entering the general circulation.
Their formation was described by Fantham (1911).60 The parasite
contracts, the blepharoplast migrates towards the nucleus, a very thin
coat differentiates around the two nuclei and a certain amount of cytoplasm,
and the parts exterior to the coat disintegrate, leaving a small,
oval body behind. Fuller details are given in connection with
T. rhodesiense. Laveran (1911)61 considers that latent bodies are
“involution” forms, but acknowledges that they can flagellate and
become infective in fresh blood.
No multiplication of the trypanosomes within the cells of the
lung, liver or spleen of infected monkeys was found by Miss
Robertson in her recent researches.
There appear to be negative periods in infected monkeys, since,
although trypanosomes may occur in their blood at such times, they
are not infective to Glossina.
Development in Glossina palpalis.—The principal accounts are
those by Sir D. Bruce and his colleagues (1911),62 and by Miss
Robertson63 (1912), whose results will be followed. According to the
latter investigator T. gambiense never enters the body cells of the fly
(G. palpalis), nor does it penetrate the gut wall into the body cavity.
Practically no crithidial stage occurs in the fly’s main gut, but a
trypanosome facies is retained therein.
After the trypanosomes are ingested by the fly during a meal of
infected blood, sooner or later multiplication occurs. This development
usually begins in the middle or posterior part of the mid gut,
and trypanosomes of varying sizes are produced. After the tenth
or twelfth day, many long, slender trypanosomes (fig. 30, a) are
found, which gradually move forwards into the proventriculus. Such
long, slender forms represent the limit of development in the lumen
of the main gut. The proventricular type, developed about the
eighth to the eighteenth or twentieth day, is not infective; it may
occur in the crop, but is not to be found permanently there.
Between the tenth and the fifteenth days multinucleate forms of
trypanosomes are found, and may be styled multiple forms (fig. 30, b).
Some of these latter may be degenerative.
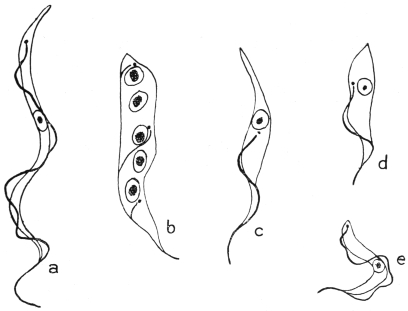
Fig. 30.—Trypanosoma gambiense. Development in the fly, Glossina palpalis. a, slender,
proventricular form; b, multinucleate form; c, d, crithidial forms; e, infective type of
trypanosome found in salivary gland. × 2,500. (After Robertson.)
Invasion of the Salivary Glands of the Fly.—Long, slender trypanosomes
from the proventriculus pass forward into the hypopharynx.
They then pass back along the salivary ducts, about sixteen to thirty
days after the fly’s feed. The trypanosomes reach the salivary glands
as long, slender forms. In the glands they become shorter and
broader, attach themselves to the surrounding structures, and assume
the crithidial facies (fig. 30, c, d). As crithidial forms they remain
attached to the wall and multiply in the glands. These crithidial
stages differentiate into the short, broad trypanosome forms, capable
of swimming freely (fig. 30, e).
Miss Robertson considers the development in the main gut to be
indifferent multiplication, and that salivary fluid seems necessary to
stimulate trypanosomes to the apparently essential reversion to the
crithidial type. The second development in the salivary gland is the
essential feature. The short, stumpy forms of trypanosomes (fig. 30, e)
finally produced in the salivary glands are alone infective. No
conjugation of trypanosomes occurs in the fly. Only about 5 per
cent. of captive tsetse flies fed on trypanosome-infected blood become
infective, but they probably remain infective for the rest of their lives.
J. G. Thomson and Sinton (1912)64 have obtained in cultures the
various trypanosome forms of T. gambiense seen in the fly’s main gut.
Duke (1912)65 found T. gambiense in a species of antelope, the
situtunga (Tragelaphus spekei), on Damba Island in Victoria Nyanza.
Wild G. palpalis could be infected therefrom. The antelope may
then act as a sleeping sickness reservoir in that district, but men are
apparently the chief reservoir.
Trypanosoma nigeriense, Macfie, 1913.66
Macfie has recently (August, 1913) described a human trypanosome
from the Eket district of Southern Nigeria. It is common in
young people. The disease produced does not seem to be of a
virulent type in Nigeria, and does not occur in epidemic form. In
the early stages the glands of the neck are enlarged. In the later
stages—cases of which are rarer—lethargy appears. The parasite
is a polymorphic trypanosome, morphologically almost indistinguishable
from T. gambiense, though it may be slightly shorter. Macfie
recorded the occurrence in his preparations of a few trypanosomes
appearing to have a flagellum free during their whole length.
Some of the parasites, as seen in a sub-inoculated guinea-pig, are very
small (8 µ long). Other trypanosomes have their nuclei displaced
somewhat anteriorly. This parasite may only be a variety of
T. gambiense. The parasite is perhaps spread by Glossina tachinoides.
Trypanosoma rhodesiense, Stephens and Fantham, 1910.
The parasite was found in the blood of a young Englishman who
had contracted sleeping sickness in the Luangwa Valley, North-eastern
Rhodesia, in the autumn of 1909. The patient had never
been in an area infested with Glossina palpalis.
(1) Morphology.—The morphology of the parasite in man and
sub-inoculated rats was studied by Stephens and Fantham in 1910.67
They pointed out a morphological peculiarity in the presence of
certain trypanosomes with posterior nuclei in sub-inoculated animals,
that is, parasites in which the nucleus (trophonucleus) was situated
towards the posterior or aflagellar end, close up to or even beyond the
blepharoplast or kinetic nucleus (fig. 31, 4, 5). When the nucleus was
beside the blepharoplast, the former was seen to be kidney-shaped
(fig. 31, 4). The posterior nuclear forms were of the stout and stumpy
variety, and about 6 per cent. of the stumpy forms were found to have
their nuclei displaced from the centre. The anterior or flagellar end
of these trypanosomes often contained chromatoid granules. T.
rhodesiense varies in length from 12 µ to 39 µ68; short stumpy forms
vary from 13 µ to 21 µ, intermediate forms from 21 µ to 24 µ, and long,
slender forms from 25 µ onwards. The average length is 24·1 µ.
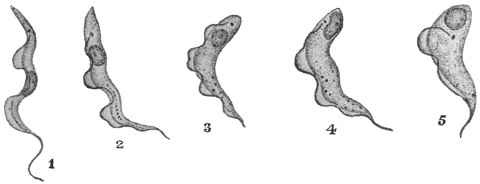
Fig. 31.—Trypanosoma rhodesiense. 1, Long narrow form; 2–4, nucleus passing to
posterior (aflagellar) end; 5, nucleus quite posterior. × 1,800. (After Stephens and Fantham.)
Certain regular periods occur in the course of the trypanosomiasis
when few or no flagellate trypanosomes are found in the peripheral
blood of the patient or of the sub-inoculated animal. These periods
can be explained in terms of morphology, for the trypanosomes are
capable of assuming a non-flagellate form in the internal organs of the
host, particularly in the lungs and in the spleen. Such forms are
known as “latent” or “resting” forms. The term “latent body”
was first used by Moore and Breinl in 190769 in connection with
T. gambiense. Fantham70 (1911) has described the process of formation
of latent from motile forms and the reconversion of the latent
bodies into active flagellates. Fresh preparations of splenic blood
or lung blood containing trypanosomes were made. A trypanosome
gradually withdrew or cast off its flagellum, concentrated its
cytoplasm, and became more or less elongate oval. Nucleus and
blepharoplast approached one another and came to lie more or less
side by side. Then an opaque line often made its appearance around
the nuclear area and differentiated as a slight envelope or covering,
the cytoplasm external to this merely degenerating. The small, oval,
refractile body (fig. 29, d—f) thus formed was a non-flagellate latent
body, 2 µ to 4 µ in diameter, like Leishmania or the non-flagellate,
multiplicative forms of T. cruzi (fig. 34), and remains temporarily
inactive in the internal organs of the host. After this period of
inactivity, the non-flagellate body, recuperated by its rest, begins
to elongate again. The nuclei separate. From a small vacuole-like
portion the flagellum differentiates and forces out the ectoplasm,
which assumes the form of the undulating membrane with its
flagellar border. Subsequent growth results in the production of the
typical trypanosome form, which re-enters the circulating blood and
multiplies by longitudinal binary fission. Division of the parasite
prior to the formation of a latent body may occur and division
of the latent forms themselves is known, though less common.
Consequently latent bodies, like the flagellate forms themselves,
show diversity in size. The blepharoplast of the latent bodies is
sometimes less well marked than in Leishmania (see fig. 29, d-f).
Laveran’s views on these bodies have already been given on p. 74.
(2) Animal Reactions.—The posterior nuclear trypanosomes were
found in all sub-inoculated animals, such as rats, guinea-pigs, dogs,
mice, Macacus, rabbits and horses, but were not seen in the human
patient, as few trypanosomes occurred in his peripheral blood.
R. Ross and D. Thomson71 found a periodic, cyclical variation in the
number of the parasites in the patient’s blood from day to day, the
cyclical period being about a week (fig. 32). Fantham and J. G.
Thomson72 (1911) found a similar periodic, cyclical variation in the trypanosomes
in the blood of sub-inoculated rats, guinea-pigs and rabbits.
On counting the parasites in the blood of similar animals inoculated
with T. gambiense, they established, by enumerative methods, that
T. rhodesiense was more virulent than T. gambiense, while Yorke also
showed this marked virulence of T. rhodesiense in practically all
laboratory animals. In other words the duration of infection in the
case of T. rhodesiense was shorter. It was also found that T. rhodesiense
was resistant to atoxyl. The patient, from whom the original
strain was obtained, died about nine months after the probable date
of infection. Some patients infected with T. rhodesiense have died in
an even shorter period, such as four or five months.
In sheep and goats T. rhodesiense causes an acute disease, marked
by high fever, œdema of the face, and keratitis, as shown by Bevan
and others, death resulting after a relatively short period. T. gambiense
gives rise, in these animals, to no symptoms except fever, which
may be overlooked. T. rhodesiense produces keratitis in dogs.
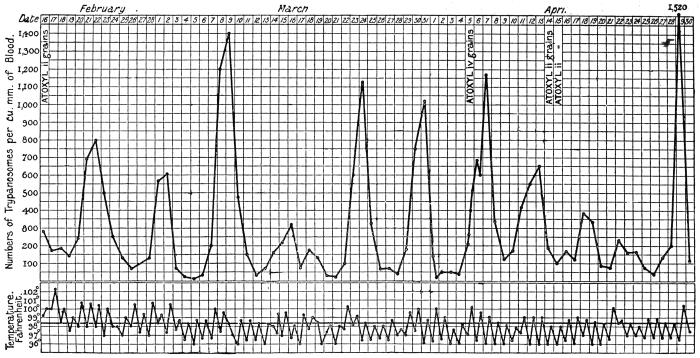
Fig. 32.—Chart showing daily counts of number of trypanosomes per cubic millimetre of peripheral blood from a case of
Rhodesian sleeping sickness. (After R. Ross and D. Thomson.)
Stannus and Yorke (1911) observed T. rhodesiense in animals
inoculated from a case of sleeping sickness in Nyasaland. Sir D. Bruce
and his colleagues73 have shown (1912) that T. rhodesiense is the
parasite usually found in man and in animals sub-inoculated from
cases of sleeping sickness in Nyasaland. It has since been found in
German East Africa and Portuguese East Africa, while Ellacombe
has described a case from North-western Rhodesia.
(3) Serum Reactions.—Interesting experiments on this subject were
performed during 1911 and 1912 by various French investigators.
(a) Action of Immune Serum (Mesnil and Ringenbach)74: (1) A goat
was infected with T. rhodesiense. Twenty-two days later its serum
mixed with T. rhodesiense was injected into a mouse. Result: Protection.
(2) The serum mixed with T. gambiense was injected into
a mouse. Result: Infection.
(b) Action of Baboon Serum.—Contrary to T. gambiense, T. rhodesiense
is very susceptible to human and baboon sera. Mesnil and
Ringenbach75 showed that a dose of 1 c.c. of baboon (Papio anubis)
serum cured mice infected with T. rhodesiense. In the same dose it
acted very feebly on T. gambiense.
(c) Action of Human Serum.—1 c.c. of human serum cured T.
rhodesiense mice in three out of four cases; on T. gambiense mice
there was no appreciable effect.
Laveran and Nattan-Larrier76 have shown the same, namely, that
human sera act on T. rhodesiense, but are quite without action on
T. gambiense.
(d) Trypanolytic Reactions.—Mesnil and Ringenbach77 have also
shown that the sera of animals (man, monkey and guinea-pig)
infected with T. gambiense are trypanolytic for the homologous
trypanosome, that is, T. gambiense, but have no action on the heterologous
trypanosome, that is, T. rhodesiense.
(4) Cross Immunity Experiments.—(a) Mesnil and Ringenbach78
immunized a monkey (Macacus rhesus) against T. gambiense. It was
inoculated with T. rhodesiense on June 7, 1911; on June 27 trypanosomes
appeared, the infection being slight; on July 4 it died. A
control died in ten and a half days.
(b) Laveran79 immunized a goat and mice against T. gambiense.
When they had acquired a solid immunity, they were inoculated with
T. rhodesiense. They became infected like the controls.
(c) Laveran and Nattan-Larrier80 immunized a ram against T. brucei,
it subsequently became infected with T. rhodesiense.
(d) Laveran81 immunized a ram and a sheep against different strains
of T. brucei. Inoculated with T. rhodesiense they both acquired acute
infections and died. Conclusion: T. rhodesiense is not T. brucei.
When the converse set of experiments is tried, namely, immunizing
an animal against T. rhodesiense, and then inoculating with
T. gambiense, the difficulty immediately arises that it is impossible to
immunize an animal against T. rhodesiense, owing to its virulence.
But a partial and transitory immunity to T. rhodesiense can be obtained
by treating the infected animal with drugs, such as arsenophenylglycin.
The results, so far as they go, seem to show that an animal immunized
against T. rhodesiense is immune not only to T. rhodesiense, but also
to T. gambiense, a fact which, according to Mesnil and Léger, does
not invalidate the specificity of T. rhodesiense, but tends to show that
the two trypanosomes are closely related.
(5) Mode of Transmission and Reservoir.—Kinghorn has shown that
T. rhodesiense is transmitted by Glossina morsitans in which it undergoes
development. Kinghorn and Yorke82 found that about 16 per
cent. of the wild game examined in Northern Rhodesia was naturally
infected with T. rhodesiense. The wild game examined included waterbuck,
hartebeest, mpala, bushbuck and warthogs. One native dog near
the Nyasaland border was found infected, but not domestic stock.
Taute doubts whether T. rhodesiense really occurs in wild game.
Approximately 3·5 per cent. of the tsetse flies fed on infected animals
may become permanently infected with T. rhodesiense, and capable
of infecting clean animals. Furthermore, a tsetse fly when once
infective probably remains infective for the rest of its life.
Kinghorn and Yorke, however, have shown that climatic conditions,
namely, those of temperature, also affect the infectivity of the
tsetse fly, as the ratio of flies capable of transmitting T. rhodesiense to
those incapable of transmitting the virus is 1 : 534 in hot valley
districts (e.g., Nawalia, Luangwa Valley, temperature 75° to 85° F.),
while on elevated plateaux (e.g., Ngoa, on the Congo-Zambesi watershed,
temperature 60° to 70° F.) the ratio falls to 1 : 1312.
Mechanical transmission by the tsetse fly does not occur, if
a period of twenty-four hours has elapsed since the infecting meal.
Developmental Cycle in the Fly.—The period which elapses between
the infecting feed of the flies and the date on which they become
infective varies from eleven to twenty-five days in the Luangwa
Valley, according to Kinghorn and Yorke. Attempts carried out at
laboratory temperature on the Congo-Zambesi plateau, during the
cold season, to transmit T. rhodesiense by means of G. morsitans
were always unsuccessful. The developmental cycle of the trypanosome
in the fly is influenced by the temperature to which the flies
are subjected (as stated above). The first portion of the developmental
cycle proceeds at the lower temperatures (60° to 70° F.), but
higher temperatures are necessary for the completion of the development
of the trypanosome. Kinghorn and Yorke found that the
trypanosomes may persist in the fly, at an incomplete stage of their
development, for at least sixty days when the climatic conditions
were unfavourable.
The first portion of the developmental cycle of the trypanosome
takes place in the gut of the fly. Invasion of the salivary glands of
the tsetse is secondary to that of the intestine, but is necessary for the
infectivity of the fly. A relatively high mean temperature, 75° to
85° F., is essential for the passage of the trypanosomes into the
salivary glands and the completion of their development therein.
Kinghorn and Yorke83 state that the predominant type of trypanosome
in the intestine of infected G. morsitans was a large broad form,
quite different from that which is most common in the salivary
glands. The trypanosome in the glands resembles the short form
seen in the blood of the vertebrate host. The authors quoted state
that both the intestinal and salivary gland forms of infective G.
morsitans are virulent when inoculated into healthy animals.
Bruce and colleagues84 have quite recently (June, 1914) published
an account of their investigations of T. rhodesiense in G. morsitans
in Nyasaland. (Incidentally it may be remarked that Bruce considers
T. rhodesiense to be identical with a polymorphic strain of T. brucei—see
pp. 83, 94). The development of T. rhodesiense takes place in the
alimentary canal and salivary glands, not in the proboscis, of the
tsetse fly. In feeding experiments with laboratory bred flies, as
well as with a few wild flies, fed on infected dogs or monkeys, only
8 per cent. of the flies were found to be infected on dissection. Of
such infected flies, however, only some allow of the complete development
of the trypanosomes within them, in other words only about
1 per cent of the flies become infective. The length of time which
elapses before a fly becomes infective varies from fourteen to thirty-one
days, averaging twenty-three days, when kept at 84° F. (29° C.).
The dominant intestinal type of flagellate in the fly is that seen in
the proventriculus, which contains many long, slender trypanosomes.
These proventricular forms find their way to the salivary glands,
wherein crithidial and encysted forms are seen. They change into
“blood forms,” which are short, stumpy trypanosomes and are
infective. “The infective type of trypanosome in the salivary glands—corresponding
to the final stage of the cycle of development—is
similar to the short and stumpy form found in the blood of the vertebrate
host.” The cycle is thus very similar to that of T. gambiense in
G. palpalis (fig. 30).
Culture.—J. G. Thomson (1912),85 and subsequently Thomson
and Sinton, succeeded in cultivating T. rhodesiense in a modified
Novy-MacNeal medium. The development obtained resembled that
of the trypanosome in the intestine of Glossina.
General Note on Trypanosomes with Posterior Nuclei.
Posteriorly placed nuclei have been found to occur not only in
T. rhodesiense by Stephens and Fantham (1910), but also in T. pecaudi
by Wenyon (1912), in T. brucei by Blacklock (1912), and in T. equiperdum
by Yorke and Blacklock (1912).
Recently Stephens and Blacklock (1913)86 have shown that two
trypanosomes, different morphologically, have been confused under
the name T. brucei. One of these is polymorphic (i.e., it exhibits
long and slender as well as short and stumpy forms) and came from
Uganda, while the other is monomorphic and is the original Zululand
strain described by Bruce from cattle suffering from “nagana.” Bruce
(1914) considers that morphological change has occurred in T. brucei
in its passage through laboratory animals, and thus explains the
diversity of views. The posterior nuclear forms described by
Blacklock occurred in the Uganda strain of T. brucei. (See p. 95.)
Similarly, a posterior nuclear form, T. equi, has been separated from
T. equiperdum. (See p. 98.)
Again, Bruce and his colleagues on the Royal Society Commission
investigating sleeping sickness in Nyasaland, have stated (April,
1913) that “evidence is accumulating that T. rhodesiense and T. brucei
(Plimmer and Bradford) are identical.” The exact identity of trypanosomes
showing posterior nuclei is, then, far from settled, although
Laveran by cross immunity tests has declared that T. brucei is distinct
from T. rhodesiense. No one has yet seen posterior nuclei in T. gambiense.
Trypanosoma cruzi, Chagas, 1909.
Syn.: Schizotrypanum cruzi, Chagas, 1909.
The trypanosome was discovered by Chagas87 in the intestine of
the bug, Triatoma (Conorhinus) megista, in Brazil, and then in the
blood of a small monkey bitten by the bug. A little later it was found
in the blood of a child, aged two years, suffering from irregular fever,
extreme anæmia and enlarged glands in the State of Minas Geraes,
Brazil. Chagas found that he was able to infect many of the usual
laboratory animals with the trypanosome, by allowing the bug to bite
them. He was also able to culture the parasite on blood agar.
Chagas found the Reduviid bug, Triatoma megista, in the houses
of the poorer inhabitants of the Brazilian mining State, and that it
attacked the people, more especially the children, at night, biting the
face. On this account the insect is called “barbeiro” by the
inhabitants. The bite is somewhat painful. The disease has since
been found in other parts of Brazil, e.g., Matta de São João in Bahia
province, Goyaz, Matto Grosso and São Paulo provinces, as well as
in Minas Geraes.
Morphology.—The trypanosome has a large blepharoplast or kinetic
nucleus. It is stated to occur both free and in the red blood corpuscles
in the peripheral blood. It is about 20 µ long, on an average.
Two forms of the parasite (fig. 33, 6, 7) are described in the human
blood. In one free form there is a large egg-shaped blepharoplast
and the posterior (aflagellar) end of the parasite is drawn out. The
blepharoplast (kinetic nucleus) may have a chromatin appendage.
The nucleus is oval or band-like, containing a karyosome. The
flagellum, starting close to the blepharoplast or its appendage, has a
free portion of variable length. The other free form in the blood has
a more or less round, terminal blepharoplast, smaller than in the first
form, without a chromatin appendage as a rule. The body of this
second form is decidedly broader than that of the first mentioned.
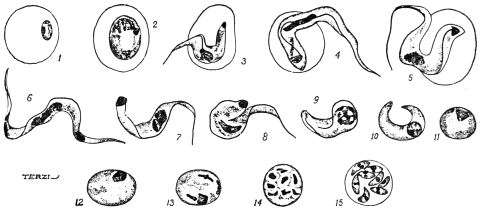
Fig. 33.—Trypanosoma cruzi. Schizogony. 1, merozoite in red blood corpuscle; 2,
parasite totally enclosed in red cell, no flagellum or undulating membrane; 3-5, parasites
partially enclosed in red cell; 6, 7, parasites in human blood; 8-11, parasites in lungs of the
monkey, Callithrix; 12, 13, initial forms of schizogony; 14, 15, schizogony in the lungs of
Callithrix. (After Chagas.)
The dimorphism has been interpreted sexually, the first mentioned
forms being termed males, the second ones females. The correctness
of this interpretation is very doubtful.
No sign of longitudinal division was ever seen in the peripheral
blood or in the internal organs. The “endocorpuscular” forms may
be completely or partially enclosed in the red cell or only attached
thereto (fig. 33, 1-5). At the beginning of infection the endocorpuscular
forms are the more numerous. Some authorities, however,
doubt these stages.
Life-history in the Vertebrate Host.—Chagas found fluctuations in
the number of the parasites in the peripheral blood. He believes the
increase of the parasites to be periodic.
The investigations of Chagas and of Hartmann have revealed two
types of multiplication which take place in the internal organs of the
vertebrate host.
(a) The first type—which possibly belongs to another organism,
Pneumocystis carinii, see p. 90—occurs in the capillaries of the lungs.
The flagellate parasite entering the lung capillaries loses its flagellum
and undulating membrane. Its body becomes curved, and the two
ends fuse, and so an oval mass is formed (fig. 33, 8-11). In some
cases the blepharoplast disappears, in other cases it blends or fuses
with the nucleus. The nucleus of the rounded parasite then divides
into eight by successive divisions (fig. 33, 12-15). Next the body,
which is surrounded by its own periplast, also divides, giving rise
to eight tiny daughter individuals or merozoites (fig. 33, 15). The
merozoites lie inside the periplast, which acts as a sort of “cyst
wall.” The merozoites are said to exhibit dimorphism, and Chagas
has interpreted the dimorphism in terms of sex. The daughter forms,
produced by the parent trypanosomes which kept their blepharoplasts,
themselves have blepharoplasts as well as nuclei, and have been termed
“males” or “microgametes.” The merozoites, arising from parent
trypanosomes which lost their blepharoplasts, have themselves only
nuclei, and have been called “females” or “macrogametes.” In the
case of the so-called “female” forms the single nucleus divides into
two unequal parts, of which the smaller becomes the blepharoplast,
and a flagellum is formed later. The so-called “males” possess early
a rudiment of a flagellum. Both kinds of merozoites escape from the
parent periplast wall, and enter red blood corpuscles. They grow into
flagellates within the corpuscles, and then become
free as adult trypanosomes in the blood-stream.
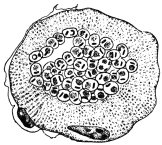
Fig. 34.—Trypanosoma
cruzi. Transverse section
of a striated muscle containing
rounded forms of
the parasite in the central
portion. × 1,000 approx.
(After Vianna.)
(b) The second mode of multiplication
is one of asexual reproduction (schizogony
or agamogony). It was first described by
Hartmann from hypertrophied endothelial cells
of the lungs. It has since been found in the
cardiac muscle, in the neuroglia of the central
nervous system, and in striped muscle (fig. 34).
In laboratory animals it has also been found
in the testicle and suprarenal capsules. In
these tissues the parasite is intracellular, appearing
as a small rounded body with nucleus
and blepharoplast, without flagellum or undulating
membrane. In other words the parasite is Leishmania-like in
the body tissues, and recalls the organism of kala-azar.
Chagas considers this second mode of multiplication to be strictly
asexual. By this means the number of parasites in the vertebrate
host is increased, and symptoms are produced. On the other hand
the first mode of multiplication, seen in the lung capillaries, is
considered by Chagas to be a process of gametogony, in which sexual
forms are differentiated. He finds that (1) the adult trypanosomes
exhibit a dimorphism in human blood rarely seen in artificially
infected guinea-pigs. In these guinea-pigs (infected from guinea-pigs)
the so-called gametogony in the lungs is seldom seen. (2) The
intermediate host, Triatoma (Conorhinus), becomes infective if fed
directly on infected human blood, but very rarely so if fed on guinea-pigs.
Chagas is led to believe that the occurrence of sexual forms
constantly in the blood of man implies a greater resistance to infection
on the part of man than on the part of guinea-pigs or other animals,
assuming the general hypothesis that the formation of gametes
represents a reaction of the Protozoön to unfavourable conditions.
In human infection the number of parasites is always less than in
laboratory animals, and their presence in the blood is transitory,
lasting from fifteen to thirty days in acute cases. In many cases
examination of the tissues at death has shown the presence of parasites
in patients who did not exhibit them in the general circulation.
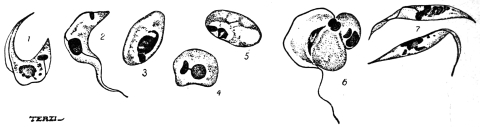
Fig. 35.—Trypanosoma cruzi. Development in Triatoma megista. 1-6, forms found in
the mid gut of Triatoma; 7 flagellate forms found in the posterior part of the gut of Triatoma.
(After Chagas.)
Life History in the Invertebrate Host.—About six hours after the
ingestion of infected blood by the bug (Triatoma megista), the kinetic
nucleus of the trypanosome moves towards the nucleus, and the
flagellum is usually lost (fig. 35, 1-5). The parasite becomes rounded
and Leishmania-like (fig. 35, 3-5), and multiplies rapidly by division.
After a time, multiplication having ceased, the rounded forms become
pear-shaped and develop a flagellum at the more pointed end.
Crithidial forms (fig. 35, 7) are thus produced and pass into the intestine,
where they multiply and may be seen in about twenty-five hours
after the ingestion of blood. The crithidial forms may also be
found in the rectum and fæces. The last stage in the invertebrate
is a small, trypanosome-like type, long and thin with a band-like
nucleus and conspicuous kinetic nucleus. These parasites are found
in the hind gut and in the body cavity. They find their way into
the salivary glands, and are the forms (fig. 36) which are transmissible
to a new vertebrate host. The development in the bug takes about
eight days altogether, after which time the bugs are infective.
There are thus three principal
phases in the development
of T. cruzi in Triatoma
megista: (1) A multiplicative
phase (Leishmania-like) in the
stomach of the bug, (2) a
crithidial phase, which is also
multiplicative, in the hind-gut,
and (3) a trypanosome phase, which is “propagative,” and apparently
passes through the wall of the alimentary canal into the body cavity
and so into the salivary glands.

Fig. 36.—Trypanosoma cruzi. Forms found
in the salivary glands of Triatoma megista.
(After Chagas.)
Brumpt found that T. cruzi could live in Cimex lectularius, C. boueti,
and Ornithodorus moubata. The Cimex fæces may be infective.
Blacklock found multiplication of the parasite in C. lectularius.
Culture.—The trypanosome can be cultivated on Novy-MacNeal’s
blood agar, and the cultural forms resemble those described in
the bug.
Possible Reservoir.—Chagas thinks that probably the armadillo or
“tatu” (Dasypus novemcinctus) may be the reservoir of T. cruzi. He
also thinks that Triatoma geniculata is a transmitter; it lives in the
burrows of the armadillo. Other carriers may be Triatoma infestans
and T. sordida.
Clinical Features.—The trypanosomiasis of Brazil, produced by
T. cruzi and spread by Triatoma spp. has received various names,
such as oppilação, canguary, parasitic thyroiditis, and coreotrypanosis.
It is also known as the human trypanosomiasis of Brazil, South
American trypanosomiasis, and Chagas’ disease.
Chagas88 reports two principal forms—acute and chronic. The
acute infection is rare, and is characterized by increase in the volume
of the thyroid gland, pyrexia, a sensation of crackling in the skin,
enlarged lymphatic glands in the neck, axilla, etc., while the liver and
spleen are increased in volume. Sclerosis of the thyroid gland is
found at autopsy and fatty degeneration of the liver. During an
attack of fever, trypanosomes are found in the blood. The acute
form was only observed in children.
In the chronic form Chagas reports several varieties: (a) A
pseudo-myxœdematous form, occurring in most cases, especially up
to the age of 15. There is hypertrophy of the thyroid gland or
at least signs of hypothyroidism, general hypertrophy of glands,
disturbance of heart rhythm, and nervous symptoms. (b) The myxœdematous
form is characterized by similar symptoms, especially by
considerable swelling of the thyroid body, and myxœdema of the
subcutaneous cellular tissue; sometimes there is a true pachydermic
cachexia. (c) In the nervous form there are motor disturbances,
aphasia, disturbances of intelligence or signs of infantilism, athetosis
of the extremities and idiocy. There are also paralytic symptoms of
bulbar origin, disturbances of mastication, phonation and deglutition,
and in some cases convulsive attacks. (d) The cardiac form, characterized
by disturbance of the heart rhythm. In all these forms the
parasite is found at autopsy in the nervous substance, brain, bulb and
heart.
Vianna (1911)89 has studied the histopathology of the disease.
Some of the chief points are: in the heart muscle destruction of
the sarcoplasm, followed by interstitial myocarditis; in the central
nervous system invasion of the neuroglia cells and inflammatory
reaction; in the suprarenal capsule invasion of medulla or cortex;
inflammatory reaction can also be seen in the kidneys, the hypophysis
and thyroid gland.
Recently Chagas states90 that “schizotrypanosomiasis” has been
found in a child 15 to 20 days old, and that Trypanosoma cruzi
has also been found in a fœtus—the mother being infected with
the trypanosome. The trypanosomiasis can, then, be transmitted
hereditarily.
Trypanosoma lewisi, Kent, 1881.
The trypanosome has a nucleus somewhat displaced anteriorly, about
one-third of the way from the anterior (flagellar) end of the body, a
relatively straight edge to the undulating membrane, and a rod-shaped
blepharoplast (fig. 37, A). It averages about 25 µ long and 1·5 µ broad.
Much attention has been devoted in recent years to the elucidation
of the life history of the rat parasite, Trypanosoma lewisi. It is
usually non-pathogenic to its host. It has been shown that the
trypanosome can be transmitted from rat to rat by the rat-flea,
Ceratophyllus fasciatus, and by Ctenocephalus canis (the so-called dog-flea).
(See also p. 92). The flagellate may also persist, but doubtfully
develop, in the rat-louse, Hæmatopinus spinulosus. These researches
may now be summarized.
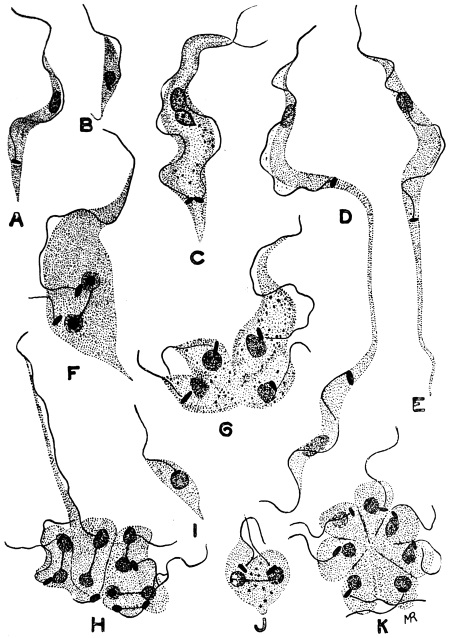
Fig. 37.—Trypanosoma lewisi, from rat’s blood. a, ordinary form; b, small form;
c, d, stages in equal binary fission; e, elongate form (longocaudense type), resulting from
division as seen in d; f, unequal binary fission; g, h, multiple fission into four and eight;
i, small form; j, binary fission of small form; K, division rosette. × 2,000. (After
Minchin and Thomson.)
Life Cycle in the Vertebrate Host.—After infection of a rat, the
trypanosomes usually appear in the animal’s blood in five to seven
days. This incubation period applies either to a natural or an artificial
infection. The trypanosomes first observed in the rat’s blood are
diverse in form (fig. 37), being small, medium and large in size. This
diversity is explained by the rapid multiplication taking place. A
trypanosome may divide by equal longitudinal fission (fig. 37, C, D),
but more commonly multiple fission occurs (fig. 37, G, H), and is
unequal. Rosette forms are produced, in which the parent form can
be recognized by its long flagellum (fig. 37, H) and attached to it are
daughter individuals, smaller in size, from which flagella are growing.
Minchin and J. D. Thomson (1912) find that the daughter forms may
be set free sometimes with a crithidia-like facies (fig. 37, I), the
blepharoplast being anterior but near to the nucleus. The daughter
forms, when set free, may themselves divide by binary or multiple
fission, in the latter case forming rosettes (fig. 37, K). Rosette
forms were described by Moore, Breinl and Hindle in 1908.
Lingard, some years ago, described as a distinct species, T. longocaudense,
certain forms with markedly elongate posterior ends
(fig. 37, E). According to Minchin, “these forms appear to arise by
binary fission” (fig. 37, D). These long drawn-out forms “are of
constant occurrence and very numerous at a certain stage of the
multiplication period.” It is about the eighth or tenth day after
infection that the multiplication of T. lewisi is at its maximum in the
rat’s blood. About the twelfth or thirteenth day the trypanosomes
seen in the blood appear uniform. According to Minchin (1912)91
the rat “gets rid of its infection entirely sooner or later, without
having suffered, apparently, any marked inconvenience from it, and
is then immune against a fresh infection with this species of trypanosome.”
There is, then, a cycle of development in the vertebrate host.
Minchin notes that the records of the pathogenicity of T. lewisi in
rats, causing their death, need further investigation.
T. lewisi inoculated into dormice (Myoxus nitela) and jerboas may
become pathogenic thereto.
Carini found cysts in the lungs of rats infected with T. lewisi.
He thought the cysts were schizogonic stages of the trypanosome,
comparable with those found in the lungs of animals sub-inoculated
with T. cruzi. Delanoë (1912)92 has found, however, that such
cysts, containing eight vermicules, occurred in rats uninfected with
T. lewisi. Delanoë concludes that the pneumocysts are independent
of T. lewisi, and represent a new parasite, Pneumocystis carinii. The
pneumocysts may be allied to the Coccidia, and must be considered
when investigating the life-cycle of a trypanosome in a vertebrate
host. Some of the stages of T. cruzi may possibly be of this nature.
Life-cycle in the Invertebrate Host.—This occurs in fleas, and has
been investigated in considerable detail by Minchin and Thomson
in Ceratophyllus fasciatus, and by Nöller in Ctenocephalus canis and
Ctenopsylla musculi.
When infected rat’s blood is taken up by the flea, the parasites pass
with the ingested blood direct to the mid-gut of the Siphonapteran.
In the flea’s stomach they multiply in a somewhat remarkable manner,
namely, by penetration of the cells of the lining epithelium, and
division inside the epithelial cells. Inside these lining cells the
trypanosomes first grow to a large size and then form large spherical
bodies, within which nuclear multiplication occurs (fig. 38, A-F).
Any one of these large spherical bodies contains at first a number of
nuclei, blepharoplasts and developing flagella, the original flagellum
still remaining attached for a time. The cytoplasm then divides into
daughter trypanosomes which are contained within an envelope,
formed by the periplast of the parent parasite. Inside the periplast
envelope are a number of daughter trypanosomes “wriggling very
actively; the envelope becomes more and more tense, and finally
bursts with explosive suddenness, setting free the flagellates, usually
about eight in number, within the host-cell” (fig. 38, F). The daughter
forms escaping from the host cell into the stomach lumen of the flea
are fully formed, long trypanosomes.
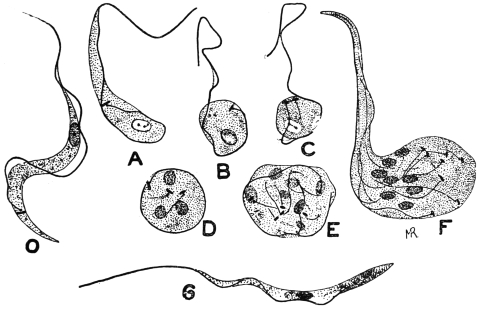
Fig. 38.—Trypanosoma lewisi. Developmental stages from stomach of rat flea. O,
ordinary blood type; A-F, stages occurring in gut-epithelium of flea, when the trypanosome
becomes rounded and undergoes multiplication, forming in F eight daughter trypanosomes;
G, type of trypanosome resulting from such division which passes back to the rectum.
× 2,000. (After Minchin.)
The trypanosomes (fig. 38, G) pass into the flea’s rectum. The
next phase is a crithidial one. The parasites become pear-shaped,
in which the blepharoplast (kinetic nucleus) has travelled anteriorly
past the nucleus towards the flagellum (fig. 39). The crithidial forms
attach themselves to the wall of the rectum, and multiply by binary
fission (fig. 39, D). A stock of parasites is thus formed which,
according to Minchin and Thomson, “persist for a long time in
the flea—probably under favourable conditions, for the whole life
of the insect” (fig. 39, A–I).
From the crithidial forms of the rectum, according to Minchin,
small infective trypanosomes arise by modification morphologically
(fig. 39, J–M). The flagellum grows longer and draws out more the
anterior part of the body, the blepharoplast migrates posteriorly,
behind the nucleus, and carries with it the flagellar origin. These
trypanosomes are small, but broad and stumpy (fig. 39, N), and can
infect a rat. Minchin and Thomson formerly considered that the
small, stumpy, infective trypanosomes pass forwards from the rectum
into the stomach, and “appear to be regurgitated into the rat’s blood
when the flea feeds.” However, the small infective trypanosomes
were previously described by Swellengrebel and Strickland.93 They
may be found in the flea’s fæces. Nöller (1912)94 has found that the
development of T. lewisi proceeds quite well in the dog flea (Ctenocephalus
canis) in Germany. Wenyon confirms this, and states that
the human flea, Pulex irritans, and the Indian rat-flea, Xenopsylla
cheopis, are also able to serve as true hosts for T. lewisi.
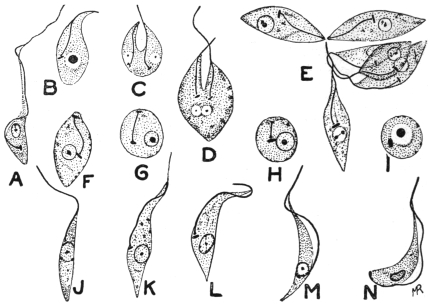
Fig. 39.—Trypanosoma lewisi. Developmental stages from rectum of rat-flea. A, early
rectal form; C, D, division of crithidial form; E, group of crithidial forms; F–I, crithidial
forms without free flagella, some becoming rounded; J–M, transitional forms to trypanosome
type seen in N, which represents the final form in the flea. × 2,000. (After Minchin.)
Nöller stated that rats were not infected with T. lewisi by infective
fleas biting them, but by the rats licking up the fæces passed by the
fleas while feeding. This is not in agreement with Minchin and
Thomson’s earlier views of regurgitation, which, apparently, they
have now abandoned.95 Wenyon (1912) confirms Nöller’s experiments.
He took a dog flea, containing infective trypanosomes in
its fæces, and allowed it to feed on a clean rat. The fæces of the
flea, passed while feeding, were carefully “collected on a cover glass
and taken up in culture fluid with a fine glass pipette.” The contents
of the pipette were discharged into the mouth of a second clean rat.
Injury to the rat’s mouth was carefully avoided. The first rat, on
which the infective flea was fed, did not become infected, while the
second rat, in whose mouth infective flea fæces were placed, became
infected in six days.
When infective forms of T. lewisi have been developed within the
gut of a rat flea, they may enter and infect the vertebrate host
by96 (a) being crushed and eaten by the rodent; (b) the rat may lick
its fur on which an infected flea has just passed infective excrement;
or (c) the rat may lick, and infect with flea excrement, the wound
produced by the bite of the flea.
The time taken for the full development of T. lewisi in the flea
is about six days. The intracellular phase is at its height about
the end of the first day; the crithidial phase, in the flea’s rectum,
begins during the second day; the stumpy, infective trypanosomes
are developed in the rectum about the end of the fifth day.
Wenyon97 writes that, “the fleas, when once infected with T. lewisi,
remain infected for long periods, for though many small infective
trypanosomes are washed out of the gut at each feed, those that
remain behind multiply to re-establish the infection of the hind gut.
Further, the infection is still maintained even if the flea is nourished
on a human being, so that fresh human blood does not appear to be
destructive to the infective forms in the flea.”
The best method of controlling fleas during experiments is that
due to Nöller. He adopted the method of showmen who exhibit
performing fleas, and secure them on very fine silver wire.
Of fleas fed on an infected rat only about 20 per cent. become
infective. About 80 per cent. are immune. If fleas are examined
twenty-four hours after feeding, trypanosomes will be found in all, so
that many of the parasites are destined to degenerate.
It may be of interest to note that Gonder98 (1911) has shown that
a strain of T. lewisi resistant to arsenophenylglycin loses its resistance
after passage through the rat-louse, Hæmatopinus spinulosus. These
experiments suggest that physiological “acquired characters” may
be lost by passage through an invertebrate host.
Trypanosoma brucei, Plimmer and Bradford, 1899.
Trypanosoma brucei was discovered by Sir D. Bruce in 1894 in
cattle in Zululand and was named T. brucei by Plimmer and Bradford
in 1899 in honour of its discoverer. This trypanosome is of considerable
economic importance, as it is responsible for the fatal
tsetse fly disease, or “nagana,” in cattle, horses and dogs. The
disease is widely distributed in Africa and is transmitted from host to
host by the tsetse, Glossina morsitans, and other species of Glossina.
The virus is maintained in nature in certain big game, such as wildebeest,
bushbuck and koodoo, which thus act as living reservoirs of
disease from which the tsetse may become infected. These reservoir
hosts are not injured, apparently, by the presence of the parasites.
T. brucei is rapidly fatal to the small laboratory animals, such as
rats and mice. Horses, asses and dogs practically always succumb
to its attacks, while a very small number of cattle recover from
“nagana.” The disease is characterized by fever, destruction of red
blood corpuscles, severe emaciation and by an infiltration of coagulated
lymph in the subcutaneous tissue of the
neck, abdomen and extremities giving a swollen
appearance thereto. The natural reservoirs in
which T. brucei has been long acclimatized are
unaffected by the trypanosomes, while the newer
hosts, such as imported cattle in Africa, are
rapidly destroyed by their action.

Fig. 40.—Trypanosoma
brucei. × 2,000. (After
Laveran and Mesnil.)
The general morphology and life history in
the vertebrate host is that of a typical trypanosome
(fig. 40). Its length is from 12 µ to 35 µ,
its breadth from 1·5 µ to 4 µ. Multiplication by
longitudinal division proceeds in the peripheral
blood (fig. 26), while latent, leishmaniform
bodies are produced in the internal organs.
Bruce and colleagues99 have quite recently
(June, 1914) described the development of a
Zululand strain of T. brucei in G. morsitans. The tsetse flies were
bred out in Nyasaland. In vertebrate blood the brucei strain
was polymorphic. The development was like that found for
T. gambiense in G. palpalis (fig. 30), and by Bruce and colleagues for
T. rhodesiense in G. morsitans in Nyasaland. Long trypanosomes
were found in the proventriculus of the tsetse. Crithidial, rounded
or encysted, and immature “blood forms” occurred in the salivary
glands; and finally infective, stumpy, “blood forms” were differentiated
in the salivary glands. The period of development of T. brucei
in G. morsitans takes about three weeks, and then the fly becomes
infective. Bruce believes that T. rhodesiense of Nyasaland and
T. brucei of Zululand are the same, their cycles of development
in G. morsitans being “marvellously alike.” (But see Laveran, p. 80.)
T. brucei has been cultivated with difficulty by Novy and MacNeal,
using blood agar. The best treatment for nagana is arsenic in some form.
It is probable that more than one trypanosome has been confused
under the name T. brucei, more especially as the occurrence of
many species of trypanosomes in various animals in Africa was not
suspected until comparatively recent times. It has been shown by
Stephens and Blacklock (1913) that the original Zululand strain of
T. brucei was monomorphic, while the organism sent from Uganda,
and at the time believed by Bruce to be the same as the Zululand
trypanosome, has been found to be polymorphic, with morphological
resemblances to T. rhodesiense. Stephens and Blacklock100 have
suggested the name T. ugandæ for the polymorphic trypanosome,
which, however, has marked resemblances with Trypanosoma
pecaudi, and they are, perhaps, identical. T. pecaudi was the name
given by Laveran101 in 1907 to the causal agent of “baleri” in equines
and sheep in the French Sudan. T. pecaudi, which is dimorphic,
is widely distributed in Africa. An extremely small number of both
T. pecaudi and T. ugandæ have been shown to possess posterior
nuclei. T. pecaudi is transmitted by various species of Glossina,
and is said to develop in the gut and proboscis of the fly.
On the other hand, Bruce and colleagues (1914), examining a
strain sent from Zululand in 1913, state that T. brucei is polymorphic.
Bruce (1914) suggests that passage through laboratory hosts has
influenced and altered the morphology of the parasite.
Trypanosoma evansi, Steel, 1885.
Syn.: Spirochæta evansi, Steel, 1885; Hæmatomonas evansi, Crookshank, 1886;
Trichomonas evansi, Crookshank, 1886.
Trypanosoma evansi, first found by Evans in 1880, in India, is the
causal agent of the disease known as “surra.” The malady affects
more particularly horses, mules, camels and cattle in India and neighbouring
countries, such as Burma and Indo-China. It occurs also in
Java, the Philippines, Mauritius and North Africa. Elephants may
be affected. A serious outbreak among cattle in Mauritius occurred
in 1902, the disease being imported into the island. The symptoms
are fever, emaciation, œdema, great muscular weakness and paralysis
culminating in death.
T. evansi varies from 18 µ to 34 µ in length and 1·5 µ to 2 µ in
breadth. It has a pointed posterior extremity, and, anteriorly, there
is a free portion to the flagellum (fig. 41). It is possibly monomorphic,
but a few broad forms occur. The trypanosome multiplies
by longitudinal fission in the blood. Rounded leishmaniform stages
occur in the spleen of the vertebrate host, which stages Walker102 (1912)
considers to be phases of schizogony.
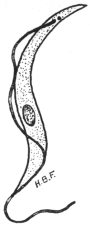
Fig. 41.—Trypanosoma
evansi. × 2,000. (Original.
From preparation by Fantham.)
The parasite is transmitted in nature by various species of Tabanus
and Stomoxys, though at present little is known of the life-history
within these invertebrate hosts.
Dogs are said to contract the disease by feeding on animals
dead of surra.
A variety of T. evansi is the cause of “mbori” in dromedaries
in Africa (Sahara and Sudan). Another possible variety, or closely
allied form, is T. soudanense, the causal agent
of “el debab” in camels and horses in North
Africa, especially Algeria and Egypt.
An extraordinary example of the possible
infection of a human being with an animal
trypanosome is recorded in the case of Professor
Lanfranchi, of the Veterinary School,
Parma. The Professor became infected with
trypanosomes, although only nagana and surra
were maintained in his laboratory, and he
himself had never visited the tropics. He
suffered from irregular attacks of fever and
was œdematous, but his mind remained clear.
The identification of the trypanosome from
Lanfranchi’s blood has been a matter of great
difficulty. Apparently Mesnil and Blanchard
(1914)103 consider the strain found in the patient is almost indistinguishable
in its reactions from T. gambiense, though the parasite is monomorphic.
Lanfranchi considers that he was infected with T. evansi.
Trypanosoma equinum, Voges, 1901.
Syn.: Trypanosoma elmassiani, Lignières.
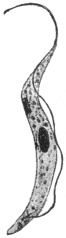
Fig. 42.—Trypanosoma
equinum. × 2,000. (After
Laveran and Mesnil.)
Trypanosoma equinum was found by Elmassian to be the cause of
the fatal disease, “mal de caderas,” of horses
and dogs, in South America (Paraguay, Argentine,
Bolivia). The name refers to the fact that
in the disease, as in other trypanosomiases,
the hind quarters become paralysed. Cattle
are refractory to inoculation.
T. equinum is about 22 µ to 24 µ long
and about 1·5 µ broad (fig. 42). Although this
trypanosome is very active, yet it is characterized
by the blepharoplast (kinetic nucleus)
being very minute or even absent, as the
granule sometimes seen may be the basal
granule of the flagellum.
The mode of transmission of T. equinum
is not known with absolute certainty. Migone
has shown that the parasite causes a fatal disease in the large South
American rodent, the capybara (Hydrochœrus capybara). This animal
appears to be a reservoir of the parasite. Dogs may become infected
by eating diseased capybaras, and it is suggested that the infection is
spread from the dogs to horses by the agency of fleas. Some authorities
consider that T. equinum may be spread by various Tabanidæ and by
Stomoxys. Neiva (1913)104 doubts all these modes of transmission in
Brazil, and suggests Chrysops or Triatoma as vectors.
Trypanosoma equiperdum, Doflein, 1901.
Syn.: Trypanosoma rougeti, Laveran and Mesnil.
The malady of horses known as “dourine” or “mal du coït” is
due to a trypanosome, T. equiperdum, discovered by Rouget in 1894.
“Dourine”—also known as “stallion disease” or “covering disease”—is
found among horses and asses in Europe,
India, North Africa and North America. The
trypanosome is transmitted by coitus, and so
far as is known not by insect agency.
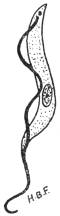
Fig. 43.—Trypanosoma
equiperdum. × 2000 approximately.
(Original. From
preparation by Fantham.)
The progress of the disease may be considered
under three periods. The period of
œdema, when signs of œdema of the genitalia
are seen. The œdema is generally painless
and non-inflammatory. This period lasts about
a month. It is succeeded by the period of
eruption, which sets in about two months
after infection. Circular œdematous areas
(“plaques”), often about the size of a two-shilling
piece, appear under the skin of the sides
and hind quarters, and also, at times, under the
skin of the neck, thighs and shoulders. The
eruption is variable, but usually lasts about a week and leaves the
animal in an enfeebled condition. Gland enlargement and swelling
of the joints and synovia also may occur. The third period of the
disease is described as that of anæmia and paralysis. The animal
becomes very anæmic, emaciation is marked, superficial non-healing
abscesses often form, and conjunctivitis and ulcerative keratitis can
occur. Paralysis ensues, and in from two to eighteen months the
animal dies. In the acute form of the disease the animal may die
after the first period from acute paralysis.
It is difficult to find the trypanosomes in naturally infected
animals, and they are best obtained from the plaques of the
eruption. Apparently the parasite occurs more in the lymph than
in the blood.
Ruminants are said to be refractory to this trypanosome.
T. equiperdum is about 25 µ to 28 µ in length on an average, but
varies from 16 µ to 35 µ. Its cytoplasm is relatively clear, and does not
show chromatic granules (fig. 43). It is stated to be monomorphic.
It has been shown recently by Blacklock and Yorke (1913)105 that
there is another trypanosome giving rise to dourine in horses. This
trypanosome is dimorphic (resembling T. pecaudi and T. ugandæ),
and is named T. equi. Previously T. equiperdum and T. equi had
been confused.
Uhlenhuth, Hübner and Worthe have demonstrated the presence
of endotoxins in T. equiperdum. These endotoxins may be set free
by trypanolysis.
Trypanosoma theileri, Bruce, 1902.
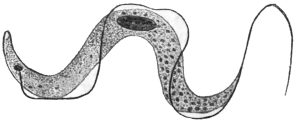
Fig. 44.—Trypanosoma theileri. × 2,000.
(After Laveran and Mesnil.)
This parasite, 60 µ to 70 µ long, and 4 µ to 5 µ broad, is distinguished
for its large size, though it is not so large as T. ingens
from Uganda oxen, whose length may be 72 µ to 122 µ, and breadth
7 µ to 10 µ. The posterior
end of T. theileri is drawn
out. Small forms of the flagellate
are known, 25 µ to 53 µ
in length. Probably other
forms of the parasite have the
nucleus posterior, and these
flagellates were formerly separated
as T. transvaaliense
(Laveran, 1902). Myoneme fibrils may be seen on its body. The
pathogenicity of this organism is doubtful, it was formerly thought
to be the causal agent of “gall-sickness” in cattle in South Africa.
T. theileri also occurs in Togoland, German East Africa, and Transcaucasia.
Allied or identical parasites occur in cattle in India.
Trypanosoma theileri, specific to cattle, is perhaps transmitted by
the fly Hippobosca rufipes in South Africa.
Trypanosoma hippicum, Darling, 1910.
Trypanosoma hippicum causes the disease of mules known as
“murrina.”106 It was found in mules imported to Panama from the
United States. It can live in other equines. The parasite varies from
18 µ to 28 µ in length, and is from 1·5 µ to 3 µ broad. Its undulating
membrane is little folded. The trypanosome has a noticeable
blepharoplast. It can penetrate mucous membranes, and it is thought
that the trypanosome may be transmitted during coitus. It may also
be spread mechanically by species of Musca, Sarcophaga and Compsomyia,
sucking the wounds of infected animals and carrying over the
trypanosomes to wounds on healthy ones.
Endotrypanum schaudinni, Mesnil and Brimont, 1908.
This organism was discovered in the blood of a sloth (Cholœpus
didactylus), in South America (French Guiana).107 It possesses special
interest, in that the best known form of the organism is endoglobular,
inhabiting the erythrocytes of the sloth. A free trypanosome in the
same animal was considered to be different from the endoglobular
form, which was somewhat like a peg-top, and possessed a short
flagellum. Darling108 (November, 1914) has seen the organism in
Panama. He describes free crithidial forms in shed blood, but not
in the blood-stream of the sloth.
Trypanosoma boylei, Lafont, 1912.
This is a parasite of the Reduviid bug, Conorhinus rubrofasciatus.
The insect attacks man in Mauritius, Réunion and other places.
Lafont infected rats and mice by intraperitoneal injection with the
gut-contents of infected bugs. Trypanosomes appeared in the mice.
Other flagellate types were assumed by the parasites in the bug.
Monomorphic Trypanosomes.
A number of trypanosomes, characterized by relative uniformity
in size and structure, may be considered under this heading. They
occur in cattle, sheep, goats and horses in Africa, especially West
Africa. Morphologically, they are characterized by the posterior
(aflagellar) part of the body being swollen, while the anterior part
narrows. The nucleus is central and situated at the commencement
of the narrowing of the body. The blepharoplast is almost
terminal, the undulating membrane is narrow and not markedly
folded, so that the flagellar border lies close to or along the body.
The flagellum may or may not possess a free portion.
Some recent workers have considered that T. brucei (Zululand
strain) and T. evansi are also monomorphic, but they do not exhibit
the general characteristics outlined above. T. brucei and T. evansi
have already been considered separately.
The monomorphic trypanosomes, as defined above, include:—
Trypanosoma vivax, Ziemann, 1905.
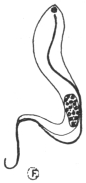
Fig. 45.—Trypanosoma
vivax. × 2,000. (Original.
From preparation by Fantham.)
This trypanosome109 occurs in cattle, sheep and goats, and was first
found in the Cameroons. It is fatal to cattle. Equines are also
affected. Antelopes are the possible reservoirs of the trypanosome.
It is probably transmitted by Glossina palpalis and other tsetse flies.
Its movement is very active. It possesses a free flagellum (fig. 45) and
it averages 23 µ to 24 µ in length. T. cazalboui (Laveran, 1906)—the
causal agent of “souma” in bovines and equines in the French Sudan—is
probably synonymous with T. vivax.
Trypanosoma capræ (Kleine, 1910) is allied,
but is somewhat broader and more massive.
It was found in goats in Tanganyika.
Trypanosoma congolense, Broden, 1904.
Probable synonyms.—Trypanosoma dimorphon, Laveran
and Mesnil, 1904; Trypanosoma nanum, Laveran,
1905; Trypanosoma pecorum, Bruce, 1910; Trypanosoma
confusum, Montgomery, 1909.
This trypanosome causes disease among
horses (e.g., Gambia horse sickness), cattle,
sheep, goats, pigs, and dogs. It is widely distributed
in Central Africa (e.g., Gambia, Congo,
Uganda, Nyasaland), the strain probably being
maintained naturally in big game. It is transmitted
by various Glossinæ, and perhaps by Tabanus and Stomoxys. It
is said to develop in the gut and proboscis of Glossina palpalis and
G. morsitans. The trypanosome averages 13 µ to 14 µ in length and
has no free flagellum (fig. 46). It is about 2 µ broad. Formerly
T. nanum and T. pecorum were said to differ in their pathogenicity, the
former being said not to infect the smaller laboratory animals. Yorke
and Blacklock (1913), however, consider
that the virulence varies and that these
trypanosomes are probably the same.
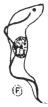
Fig. 46.—Trypanosoma
congolense. × 2,000. (Original.
From preparation by
Fantham.)
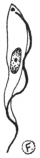
Fig. 47.—Trypanosoma
uniforme. × 2,000. (Original.
From preparation by
Fantham.)
The T. dimorphon originally obtained by Dutton and Todd (1903)
in Gambian horse sickness has been shown to be a mixture of T. vivax
and T. congolense.
Trypanosoma simiae (T. ignotum) is like T. congolense. It averages
17·5 µ long. It is virulent to monkeys and pigs.
Trypanosoma uniforme, Bruce, 1910.
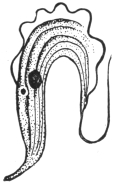
Fig. 48.—Trypanosoma
rotatorium, from blood of
a frog. × 1,400. (After
Laveran and Mesnil.)
This trypanosome was found in oxen in Uganda.110 It can be inoculated
to oxen, goats and sheep, but is refractory to dogs, rats and
guinea-pigs. It has been found in antelopes. It resembles T. vivax,
but is smaller (fig. 47), averaging 16 µ in length. A free flagellum
is present. It is transmitted by Glossinæ.
Many other trypanosomes occur in mammals,
while birds, reptiles, amphibia (fig. 48)
and fish also harbour them. The discussion of
these forms does not come within the scope
of the present work. They are dealt with
in Laveran and Mesnil’s “Trypanosomes et
Trypanosomiases,” 2nd edit., 1912.
General Note on Development of
Trypanosomes in Glossina.
Before concluding the account of trypanosomes,
it may be of interest to remark that
several African trypanosomes develop in various species of Glossina,
and are found in different parts of the alimentary tract and in the
proboscis. Thus (a) T. vivax, T. uniforme and T. capræ develop in the
fly’s proboscis (labial cavity and hypopharynx) only; (b) T. congolense,
T. simiæ and T. pecaudi develop first in the gut of the fly and then
pass forward to its proboscis; and (c) T. gambiense and T. rhodesiense
develop first in the gut and later invade the salivary glands of the
tsetse. The proboscis or the salivary glands in such cases are termed
by Duke111 the anterior station of the trypanosome, wherein it completes
its development.
Adaptation of Trypanosomes.
These flagellates may exhibit power of adaptation to changes of
environment, such as those due to the administration of drugs, change
of host, etc. A few examples of such mutations may be briefly
considered:—
(1) Blepharoplastless Trypanosomes.—T. brucei may become resistant
to pyronin and oxazine. Accompanying this drug resistance is a
change in morphology, namely, the loss of the blepharoplast
(Werbitzki).112 A race or strain of blepharoplastless trypanosomes may
be thus produced which retains its characteristic feature after as many
as 130 passages (Laveran).113 Oxazine is the more powerful drug, and
it acts directly on the blepharoplast. (Compare the natural blepharoplastless
character of T. equinum.)
(2) Reference has been made on p. 93 to the experiments of
Gonder, who showed that a strain of T. lewisi rendered resistant to
arsenophenylglycin lost its resistance after passage through the rat
louse. This is in marked contrast with the retention of drug resistance
during passage by inoculation from rat to rat.
(3) T. lewisi from the blood of a rat when transferred to a snake
seems largely to disappear, as very few flagellates are seen. When
blood from the snake is inoculated into a clean rat, then trypanosomes
reappear in the rat, but they are not all like those originally
inoculated. It seems certain that, in such a case, changes in form
and virulence of the trypanosome have occurred. Similar experiments
were made with T. brucei from rats to adders and other animals and
back to rats. Changes in the form and virulence of T. brucei occurred.
These interesting experiments were performed by Wendelstadt and
Fellmer.114
Genus. Herpetomonas, Saville Kent, 1881.
Herpetomonas is a generic name for certain flagellates possessing
a vermiform or snake-like body, a nucleus placed approximately centrally,
and a blepharoplast (kinetic nucleus) near the flagellar end.
There is no undulating membrane (fig. 49, a). The organisms included
in this genus certainly possess one flagellum, while according
to Prowazek (1904) Herpetomonas muscæ-domesticæ, the type species,
possesses two flagella united by a membrane. Patton,115 Porter116 and
others affirm, however, that the biflagellate character of H. muscæ-domesticæ
(from the gut of the house-fly) is merely due to precocious
division. The matter is further complicated by the generic name
Leptomonas, given by Kent in 1881, to an uniflagellate organism found
by Bütschli in the intestine of the Nematode worm, Trilobus gracilis.
This parasite, Leptomonas bütschlii, has not yet been completely
studied. Until these controversial points relating to the identity or
separation of Herpetomonas and Leptomonas have been satisfactorily
settled, we may retain the better known name Herpetomonas for such
uniflagellate, vermiform organisms. However, the name Leptomonas,
having been used by Kent two pages earlier in his book (“Manual of
the Infusoria”) than Herpetomonas, would have priority if the two
generic names were ultimately shown to be synonymous.
A full discussion of these interesting and important flagellates
hardly comes within the purview of the present work; brief mention
can only be given here to certain species.
The Herpetomonads occur principally in the digestive tracts of
insects, such as Diptera and Hemiptera. They are also known in the
guts of fleas and lice, but are not confined to blood-sucking insects.
One example, H. ctenocephali (Fantham, 1912)117 occurs in the digestive
tracts of dog fleas, Ctenocephalus canis, in England, France, Germany,
Italy, India, Tunis, etc. It is a natural flagellate of the flea, and might
easily be confused with stages of blood parasites in the gut of the dog
flea. Dog fleas are stated by Basile to transmit canine kala-azar, which
is believed to be the same as human infantile kala-azar. Confusion is
further likely to arise since herpetomonads pass through pre-flagellate,
flagellate and post-flagellate or encysted stages; pre- and post-flagellate
stages being oval or rounded and Leishmania-like. The post-flagellate
stages are shed in the fæces, and are the cross-infective stages by
means of which new hosts are infected by the mouth. The possible
presence of such natural flagellates must always be considered when
experimenting with fleas,
lice, mosquitoes, etc., as
possible vectors of pathogenic
flagellates like Leishmania
and Trypanosoma.
H. pediculi (Fantham,
1912) occurs in human
body lice.118 See further
remarks on pp. 107, 112.
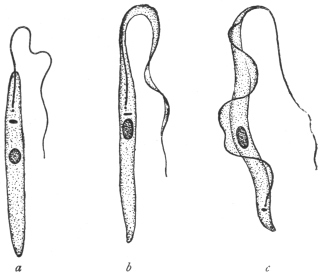
Fig. 49.—a, Herpetomonas; b, Crithidia;
c, Trypanosoma. (After Porter.)
Laveran and Franchini
(1913–14)119 have recently
succeeded in inoculating
Herpetomonas ctenocephali,
from the gut of the dog
flea, intraperitoneally into
white mice, and producing
an experimental leishmaniasis
in the mice. A
dog was also infected. They have also succeeded in infecting mice
with H. pattoni—a natural flagellate of the rat flea—by mixing infected
rat fleas with the food of the mice, and by causing them to
ingest infected fæces of rat fleas. Further, they have shown that
infection with the herpetomonas occurs naturally by this method,
that is, by the rodents eating the fleas and not by the insects inoculating
the flagellates into the vertebrates when sucking blood. These
experiments shed an interesting light on the probable origin of
Leishmania and its cultural herpetomonad stage, which were very
probably once parasitic flagellates in the gut of an insect.
Fantham and Porter120 (1914–15) have shown that young mice may
be inoculated or fed with Herpetomonas jaculum, from the gut of the
Hemipteran, Nepa cinerea (the so-called “water-scorpion”), with fatal
results. The pathogenic effects are like those of kala-azar. They also
showed that the post-flagellate stages of the herpetomonads seemed
most capable of developing in the vertebrate.
A herpetomonad, H. davidi, has been found in the latex of species of the plant-genus
Euphorbia in Mauritius, India, Portugal, etc. It is apparently transmitted to
the plants by Hemiptera. The plants sometimes suffer from “flagellosis.”
Franchini (1913)121 has described a new parasite, Hæmocystozoon
brasiliense, from the blood of a man who had lived in Brazil for many
years. It possesses flagellate and rounded stages, and is closely allied
to the herpetomonads.
Genus. Crithidia, Léger, 1902, emend. Patton, 1908.
Crithidia is the generic name of vermiform flagellates with a
central nucleus, a blepharoplast or kinetic nucleus in the neighbourhood
of the principal nucleus, and a rudimentary undulating membrane
bordered by a flagellum arising from a basal granule, which is the
centrosome of the kinetic nucleus (fig. 49b). The anterior or flagellar
end of the body is attenuated and fades off as the undulating membrane.
Crithidia fasciculata, the type species, was found by Léger in the
alimentary canal of Anopheles maculipennis. Crithidia occur in bugs,
flies, fleas,122 and ticks. Some of them are found in the body-fluid
of the invertebrate host as well as in the gut. Others may be restricted
to the body cavity or intestine respectively. C. melophagia from the
sheep-ked, Melophagus ovinus, and C. hyalommæ from the hæmocœlic
fluid of the tick, Hyalomma ægyptium, pass into the ovaries and eggs
of their hosts, and the young keds or ticks are born infected.
C. fasciculata has been shown by Laveran and Franchini to be
inoculable into white mice, producing a sort of experimental leishmaniasis
therein. In one case cutaneous lesions were produced like
those of Oriental sore.
Crithidia are natural flagellates of Arthropoda, with their own
pre-flagellate, flagellate and post-flagellate stages, and must not be
confused with transitory crithidial stages of trypanosomes.
Genus. Leishmania, Ross, 1903.
With an oval body containing nucleus and blepharoplast (kinetic nucleus)
but no flagellum. An intracellular parasite in the vertebrate host.
Included in the genus Leishmania are three species, namely:—
(1) Leishmania donovani, Laveran and Mesnil, 1903, the parasite
of Indian kala-azar, a generalized systemic disease, usually
fatal, occurring in subjects of all ages.
(2) Leishmania tropica, Wright, 1903, the parasite of Delhi boil,
Oriental sore, Aleppo button—a localized, cutaneous disease,
usually benign.
(3) Leishmania infantum, Nicolle, 1908, the parasite of infantile
kala-azar, occurring in children (and a few adults) around
the shores of the Mediterranean. The disease is perhaps a
form of Indian kala-azar, and the parasite is probably identical
with L. donovani.
These diseases may be termed collectively leishmaniases. The
morphology of the various species is practically identical.
Leishmania donovani, Laveran and Mesnil, 1903.
Syn.: Piroplasma donovani, Laveran and Mesnil.
The parasite of Indian kala-azar was demonstrated in 1900 by
Leishman from a post-mortem examination of a case of “Dum-Dum
fever,” but details were not published till May, 1903. In July, 1903,
Donovan found similar bodies from cases in Madras. Rogers
succeeded in cultivating the parasite in July, 1904.123 The original
centre of the disease was probably Assam; it occurs also in Madras,
Ceylon, Burma, Indo-China, China and Syria. A variety of this leishmaniasis
is found in the Sudan. The patient becomes emaciated,
with a greatly enlarged spleen. There is anæmia and leucopenia.
The parasite, commonly known as the Leishman-Donovan body,
is intracellular (fig. 50, 2, 3). It is found in the endothelial cells of the
capillaries of the liver, spleen, bone-marrow, lymphatic glands and
intestinal mucosa, and in the macrophages of the spleen and bone-marrow.
Some host cells may contain many parasites. It is rather
rare in the circulating blood, but may be found in the blood from
the femoral, portal and hepatic veins. It does not occur in the red
blood corpuscles as was formerly thought. The parasites liberated
from the endothelial cells are taken up by the mononuclear and
polymorphonuclear leucocytes. The Leishman-Donovan body is the
resting stage of a flagellate. As found in man it is a small, oval
organism, about 2·5 µ to 3·5 µ in length by 2 µ in breadth, and containing
two chromatinic bodies, corresponding to the nucleus and
kinetic nucleus (blepharoplast) of a flagellate. The latter element is
the smaller and more deeply staining, and is usually placed at the
periphery, transversely to the longer axis of the oval organism.
There is sometimes a very short, slightly curved filament to be seen,
which may be a rhizoplast. Multiplication takes place by binary or
multiple fission. The presence of the parasite used to be demonstrated
by splenic or hepatic puncture; nowadays it can be demonstrated in
peripheral blood, e.g., of the finger, or by culture of infected blood.
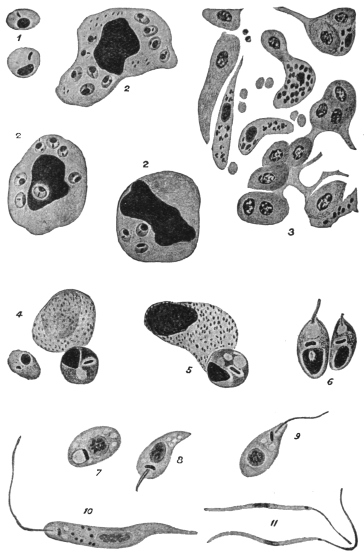
Fig. 50.—Leishmania donovani. 1, Free forms, each with nucleus and rod-shaped blepharoplast
(after Christophers); 2, endothelial cell and leucocytes containing parasites (after
Christophers); 3, capillary in the liver showing endothelial cells containing parasites (after
Christophers); 4, two parasites escaping from a leucocyte in the alimentary canal of the bug
(after Patton); 5, further development in bug (after Patton); 6, young flagellate forms in bug
(after Patton); 7-11, culture forms (after Leishman); 7, 8, 9, show development of flagellum.
L. donovani can be cultivated in citrated splenic blood, under
aerobic conditions, at 22° to 25° C. This was first accomplished
by Rogers (1904). It is not so easily culturable as L. infantum on
the Novy-MacNeal-Nicolle medium.124 L. donovani is inoculable with
some difficulty into experimental animals—in India, white rats, white
mice, dogs and monkeys (Macacus spp.), have been inoculated. The
Sudan variety, somewhat less virulent, is inoculable to monkeys. Row
also produced a local lesion in Macacus sinicus by subcutaneous inoculation
of L. donovani. Parasites taken from such a local lesion were
found to be capable of producing a generalised infection in Macacus
sinicus and white mice.
In cultures the various species of Leishmania all grow into
herpetomonad, uniflagellate organisms (fig. 50, 10), about 12 µ to 20 µ
in body length. On this account Rogers125 and Patton place the
Leishman-Donovan body within the genus Herpetomonas. The
method of culture may be used in diagnosing leishmaniases.
Kala-azar is very probably an insect-borne disease. Patton126 suspects
the bed-bug to be the transmitter and finds (fig. 50, 4-6) that the
Leishman-Donovan body can develop into the flagellate stage in the
digestive tract of the bed-bug. Feeding experiments are unsatisfactory,
since there are very few cases in which the parasites occur in sufficient
numbers in the peripheral blood to make the infection of the insect
possible, or at any rate easy. In examining the alimentary tracts of
insects for possible flagellate stages of Leishmania, it must be remembered
that in many insects natural flagellate parasites, belonging to the
genus Herpetomonas, may occur therein; such natural insect flagellates
may be harmless, and have no connection with the life-cycle of
L. donovani. Natural herpetomonads are known to occur in the
alimentary tracts of flies, mosquitoes, sand-flies, fleas and lice, but
not in bed-bugs. Further, if such flagellates are able to be inoculated
into and live within vertebrate hosts, producing symptoms like those
of leishmaniasis, the origin of kala-azar is indicated (see pp. 104, 112).
Leishmania tropica, Wright, 1903.
Syn.: Helcosoma tropicum, Wright, 1903; L. wrighti, Nicolle, 1908; Ovoplasma
orientale, Marzinowsky and Bogrow.
It is believed by some that the parasite was first described by Cunningham in
1885, and studied by Firth in 1891, being called by him Sporozoon furunculosum.
If these earlier studies were of the parasite, then its correct name is L. furunculosa,
Firth, 1891.
The benign disease produced by this parasite has received many
names, among the best known being Oriental sore, Tropical sore,
Delhi boil and Aleppo button. These names, however, are not happy
ones, as cutaneous leishmaniasis (e.g., on the ear) is now known to
occur in the New World, for example in Mexico, Venezuela, Brazil
and neighbouring States. However, it may be necessary to subdivide
cutaneous leishmaniases later.
In the Old World the disease occurs in India, Persia, Arabia and
Transcaucasia. It is also known in Algeria, Northern Nigeria, Egypt,
Sudan, Crete, Calabria, Sicily and Greece.
The boils often occur on the face, and before ulceration the
parasites may be found in the cells at the margin and floor of the
“button.” In searching for parasites the scab should be removed and
scrapings made from the floor and edges. Where lesions occur
atrophy of the epidermis takes place, and infiltration of mononuclear
cells (e.g., plasma cells, lymphoid and endothelial cells) follows.
The parasites are intracellular, being found inside mononuclear cells.
In non-ulcerating sores, Cardamitis found some free parasites. Non-ulcerating
forms are said to occur in the Sudan. In the Old World
the sores are often limited to exposed surfaces of the body. Infection
of mucous membranes (such as the lip, palate, buccal and nasal
membranes) may occur, especially in South America, and are often
known there as “Espundia.” Christopherson (1914) has recorded a
case in Khartoum.
Leishmania tropica is equally well cultivated on Novy-MacNeal-Nicolle
medium or on citrated blood. The usual temperature for
cultivation is 22° to 28° C., though Marzinowski claims to have
cultivated the parasite at 37° C. L. tropica can be inoculated
into monkeys and dogs, with the production of local lesions.
Material from a human sore or flagellates from a culture may be thus
successfully inoculated. Also infected material may be rubbed
directly into a scarified surface. The incubation period is long,
extending over several months. The duration of the disease may be
from twelve to eighteen months. Recovery from one attack of tropical
sore confers immunity, and the Jews in Bagdad inoculate their children
with the disease on a part of the body which will be covered, and
so secure immunity in adult life.
The mode of transmission of L. tropica is unknown. Wenyon
(1911)127 has found that the parasite develops into the flagellate stage
in the digestive tract of Stegomyia fasciata in Bagdad. Patton (1912)128
has found similar development in the bed-bug in Cambay. The
house-fly, Phlebotomus and Simulium have been suspected as transmitters
in different parts of the world.
An interesting announcement has been made recently (May, 1913),
that Neligan has found that L. tropica occurs in dogs in Teheran,
Persia, producing ulcers on the dogs’ faces (cf. natural occurrence of
L. infantum in dogs—see p. 110). Yakimoff and Schokhor (1914),129
have found the disease in dogs in Tashkent.
Gonder130 (1913) has performed some interesting experiments
showing the relation of infantile kala-azar to Oriental sore. Gonder
infected mice with L. infantum and with L. tropica. He used culture
material and injected intraperitoneally or intravenously. In each a
general infection resulted, with enlargement of the liver and spleen.
Later, however, mice injected with Oriental sore (North African
variety) developed peripheral lesions on the feet, tail and head, and
the lesions contained Leishmania. No such peripheral lesions
developed in the case of the mice infected with the kala-azar virus.
Gonder suggested that Oriental sore, like kala-azar, is really a general
infection overlooked in its earlier stages, and that it is in the later
stages that peripheral lesions on the skin are developed. Row (1914)131
also obtained a general infection in a mouse by the injection of
cultures of L. tropica from Oriental sore of Cambay.
Leishmania infantum, Nicolle, 1908.132
Infantile splenic anæmia has been long known in Italy. It also
occurs in Algeria, Tunis, Tripoli, Syria, Greece, Turkey, Crete, Sicily,
Malta,133 Spain and Portugal. This leishmaniasis is, then, distributed along
the Mediterranean littoral; also in Russia. Cathoire (1904) in Tunis
and Pianese (1905) in Italy were among the first to see the parasite.
Nicolle then found the parasite in patients in Tunis, and further found
spontaneous infection in dogs. The patients are usually children
between the ages of 2 and 5 years. There are a few cases known in
which the infantile type of leishmaniasis occurred in youths and adults
of the ages of 17 to 19, while one patient in Calabria was 38 years old.
The symptoms are like those of Indian kala-azar. Several Italian
investigators and others consider that L. infantum is the same as
L. donovani, and that the latter name should be used for the parasite
of Mediterranean leishmaniasis. This view, as to the identity of
L. donovani and L. infantum, seems coming into general favour.
There are, however, differences between the Indian and infantile
kala-azars, in addition to the ages of the patients affected, thus:
(a) As regards cultures, it is found that L. infantum is readily grown
on the Novy-MacNeal-Nicolle (“N.N.N.”) medium (saline blood-agar),
and that sub-cultures are easily obtained; in citrated blood
L. infantum grows with difficulty. The reverse is the case with
regard to culture media for L. donovani, which grows with difficulty
on the N.N.N. medium, but relatively easily in citrated splenic blood.
(b) Considering inoculability into experimental animals, it is found that
L. donovani is inoculated generally with some difficulty into white rats,
white mice and monkeys, and with greater difficulty into dogs, while
L. infantum can be inoculated into several experimental animals,
especially into dogs and monkeys, with ease. (c) At present L. donovani
is not known to occur spontaneously in animals, but L. infantum is
found naturally in dogs in the Mediterranean region, and the disease in
dogs is often referred to as canine kala-azar. Kittens have occasionally
been found infected. However, these differences must not be emphasized
too much.
The material for cultivation is obtained from punctures of spleen,
liver or bone-marrow of cases infected with L. infantum. It is not
always easy, however, to infect from cultures, as the cultural flagellates
inoculated into the body are often phagocytosed.
Similarly, the material for animal inoculation is obtained from
emulsions of infected spleen, liver or bone-marrow. Dogs and
monkeys are easily inoculated with such material; Nicolle inoculates
into the liver or the peritoneal cavity. Mice, white rats, guinea-pigs
and rabbits only show slight infections after such inoculations.
Dogs infected experimentally with infantile leishmaniasis may
show either acute or chronic symptoms. The acute course occurs
more often in young dogs, and is usually fatal in three to five months.
The chronic course is found more commonly in older dogs, and may
last seventeen to eighteen months. In acute forms there is irregular
fever, progressive wasting, diarrhœa occasionally, motor disturbances
involving the hind quarters, and the animal dies in a comatose condition.
In the chronic form the animal may appear well, except for
loss of weight. The parasites may be found in the internal organs of
these experimental dogs, but are not numerous in the peripheral
blood except at times of high fever. Experimental monkeys live about
three months.
It may be interesting to record the number of dogs found to be
infected naturally with leishmaniasis in various countries. In Tunis,
Nicolle and Yakimoff found about 2 per cent. infected out of about
500 dogs examined. Sergent in Algiers found 9 infected out of 125
dogs examined. In Italy and Sicily, Basile found about 40 per cent.
of the dogs to be infected out of 93 examined at Rome and
Bordonaro. Cardamitis found 15 infected out of 184 examined in
Athens. In Malta, Critien found 3 infected out of 30 dogs examined.
Alvares found 1 infected dog out of 19 examined in Lisbon. Pringault
has recently (December, 1913) found an infected dog in Marseilles.134
Yakimoff and Schokhor found 24 per cent. infected out of 647 dogs
examined in Turkestan.
The distribution of the parasites in the body of the human patient
is much the same as in the case of Indian kala-azar. Critien records
the finding of parasites in the mucous flakes of the stools of a three-year-old
Maltese child.135 Intestinal lesions rarely occur in infantile
leishmaniasis.
Ætiology.—Infantile leishmaniasis is stated to be transmitted by
fleas, especially dog fleas, Ctenocephalus canis (= Pulex serraticeps), and
by Pulex irritans. Children living in contact with infected dogs may be
bitten by infected dog fleas, and so contract the disease. Basile (1910–11)
and Sangiorgi (1910) state that they found L. infantum parasites in the
digestive tract of the dog flea. After searching they found infected
dog fleas on the beds, mattresses, and pillows used by children
suffering from the disease. Franchini (1912) thinks that Anopheles
maculipennis may be concerned in the transmission.
Basile136 tried a number of experiments to show that infantile
leishmaniasis is transmitted by fleas, thus:—
(1) Fleas were taken from a healthy dog. They were placed in
vessels containing infected spleen-pulp and allowed to feed thereon.
The fleas were then killed and dissected, and portions of the gut-contents
examined for parasites. The remainder of the gut was
emulsified and injected into a young puppy, whose bone-marrow had
been shown previously to be uninfected. Basile states that the puppy
became infected. The parasites are said to increase in number in
the flea’s gut.
(2) Two healthy pups, each a month old, and born in the laboratory,
were placed in a disinfected, flea-proof cage. A few days after,
an infected dog was placed in the cage, so that fleas from the
infected dog could pass on to the puppies. A month later the two
pups became infected, parasites being found in them after liver
puncture. A number of control puppies from the same litter
remained uninfected and in good health.
(3) Basile next used other laboratory-born puppies, a month old.
Four of the litter were placed in a disinfected, flea-proof gauze cage
in Rome. The cage was isolated from other dogs. Fleas obtained
from an infected area in Sicily were placed in the cage. The puppies
were examined by hepatic puncture, but were found to be negative
for two months. Then two of the puppies showed infection, and
six days later the remaining two puppies were found to be infected,
and all four died. They showed irregular temperatures, and were
getting thin. Control puppies remained healthy.
From these experiments Basile concludes that fleas transmit
leishmaniasis. However, Basile did not exclude the possible occurrence
of natural herpetomonads in the gut of the fleas.137 Herpetomonas
ctenocephali is known to occur in the gut of Ctenocephalus canis.
A natural Herpetomonas is also known in the gut of Pulex irritans,
as well as a Crithidia (C. pulicis, Porter). These natural flagellates of
the fleas pass through non-flagellate stages, like the Leishman-Donovan
body. In consequence Wenyon and Patton, among others, have
criticized Basile’s results. Further, other investigators, such as
Wenyon and Da Silva (1913), have repeated Basile’s flea experiments
and been unable to confirm them.
In feeding and inoculation experiments the incubation period of
the parasite may be long, and so it is necessary to wait a long time to
see whether the parasite will develop.
Immunity.—Nicolle has tried some experiments with L. infantum
and L. tropica. He finds that in animals recovery from an attack of the
former confers immunity against infection by the latter and vice-versâ.
Laveran138 records that a monkey having an immunity against L.
infantum was also immune to L. donovani.
As mentioned on p. 103, Laveran and Franchini (1913), working in
Paris, have succeeded in inoculating Herpetomonas ctenocephali, a
natural flagellate in the gut of the flea, Ctenocephalus canis, into
white mice. Leishmaniform stages of the flea flagellate were recovered
from the peritoneal exudate, blood and organs of the mice
some weeks after inoculation. The parasites may also be conveyed by
way of the digestive tract of the vertebrate. Similar experiments have
succeeded with H. pattoni. These experiments go to show, together
with those of Fantham and Porter with H. jaculum (see p. 104), that, in
the words of the latter authors, “it may be expected that the various
leishmaniases, occurring in different parts of the world, will prove to
be insect-borne herpetomoniases.”
Genus. Histoplasma, Darling, 1906.
Under the name Histoplasma capsulatum,139 Darling described small
round or oval parasites, enclosed in a refractile capsule, and each containing
a single nucleus. The bodies were found in cases of splenomegaly
in Panama. They occurred in the endothelial cells of the
small blood-vessels of the liver, spleen, lungs, intestine and lymphatic
glands, and also within the leucocytes. A few flagellates were stated to
occur in the lungs. The parasite has usually been placed near Leishmania,
but recently Rocha-Lima has stated that Histoplasma is a yeast.
Genus. Toxoplasma, Nicolle and Manceaux, 1908.
The genus was created for crescentic, oval or reniform parasites, 2·5 µ to 6 µ by
2 µ to 3 µ, possessing a single nucleus and multiplying by binary fission. They occur
in mononuclear and polymorphonuclear cells in the blood, spleen, liver, peritoneum
etc. (fig. 51). The parasites have been found in the gondi, dog, rabbit, mole, mouse,
pigeon and other birds. Although various species names have been given to the
parasites in these hosts, it seems probable, from cross infection experiments, that
there is but one species with several physiological races. Splendore140 (1913) has
described a flagellate stage.
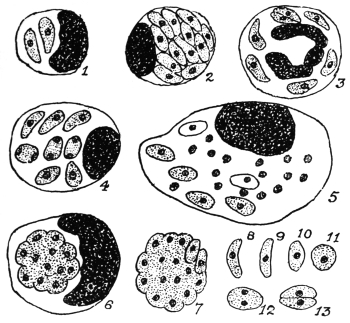
Fig. 51.—Toxoplasma gondii, endocellular or free in the peritoneal exudate of infected
mice. 1, 2, mononuclear leucocytes containing toxoplasms. 3, polynuclear, containing
parasites. 4, 5, 6, endothelial cells containing toxoplasms, agglomerated in 6. 7, agglomeration
forms. 8–11, free forms. 12–13, division stages. × 1,600. (After Laveran and Marullaz.)
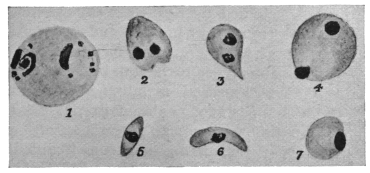
Fig. 52.—Toxoplasma pyrogenes. 1, body found in blood. 2–7, bodies found
in spleen.
[1 is about the size of a red blood corpuscle, as drawn in the figures]. Magnification not
stated. (After Castellani.)]
Castellani (1913–14)141 has described similar parasites from a case of
splenomegaly, with fever of long standing, in a Sinhalese boy. The
bodies were found in the spleen and more rarely in the blood (fig. 52).
Castellani has named them Toxoplasma pyrogenes. Further researches
are needed.
THE SPIROCHÆTES.
The Spirochætes are long, narrow, wavy, thread-like organisms,
with a firm yet flexible outer covering or periplast. There is a diffuse
nucleus internally in the form of bars or rodlets of chromatin
distributed along the body. In some forms there is a membrane or
crista present (fig. 53), which in the past was compared with the
undulating membrane of a trypanosome, but the membrane of a
spirochæte does not undulate. Progression is very rapid, corkscrew-like
and undulatory movements occurring simultaneously.
The genus Spirochæta was founded by
Ehrenberg in 1833 for an organism which
he discovered in stagnant water in Berlin.
Ehrenberg named the organism Spirochæta
plicatilis. According to Zuelzer (1912) S.
plicatilis does not possess a membrane or
crista, but an axial filament. S. gigantea has
been described by Warming from sea-water.
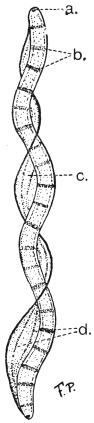
Fig. 53.—Spirochæta balbianii.
a, basal granule or
polar cap. b, chromatin rodlets.
c, membrane (“crista”).
d, myonemes in membrane.
(After Fantham and Porter.)
Spirochætes occur in the crystalline style
and digestive tract of many bivalve molluscs.
The first molluscan spirochæte to be studied
was that of the oyster, named by Certes (1882)
“Trypanosoma” balbianii (fig. 53). Similar
spirochætes, probably belonging to the same
species, occur in various species of Tapes and
in Pecten (the scallop). S. balbianii has rounded
ends (fig. 53). Other spirochætes occur in freshwater
mussels (Anodonta spp). S. anodontæ,
studied by Keysselitz (1906) and by Fantham
(1907), has pointed ends. Gross (1911) suggested
the generic name Cristispira for molluscan
spirochætes, because they possess a
well-marked membrane or “crista,” which
appears to be absent from S. plicatilis, according
to Zuelzer’s researches.
Schaudinn in 1905 founded the genus
Treponema for the parasite of syphilis (T.
pallidum), discovered by him and by Hoffmann.
According to Schaudinn the Treponemata have no membrane or
crista. The pathogenic agent of yaws or frambœsia, discovered by
Castellani, is also placed in the genus Treponema, as T. pertenue.
There remain the blood spirochætes. It is somewhat disputed as
to whether these organisms possess a membrane. The present writer
considers that they have a slight membrane or crista. The name of
the genus in which to place the blood-inhabiting forms is somewhat
uncertain and disputed. Various generic names given to them are
Spirochæta, Treponema, Spiroschaudinnia (Sambon) and Borrelia
(Swellengrebel). Included in this division are the causal agents of
relapsing or recurrent fever. These Protists will be named, for
description, Spirochætes without prejudice as to the ultimate correct
generic name.
It is sometimes made a matter of argument as to whether the
spirochætes are Protozoa or Bacteria. Such arguments are somewhat
unprofitable. Morphologically the spirochætes are like the Bacteria
in possessing a diffuse nucleus. They differ from Spirillum, an
undoubted bacterial genus, in being flexible and not possessing
flagella. Molluscan spirochætes, however, may appear to have
flagella if their membrane becomes frayed or ruptured, when the
myonemes therein (fig. 53), becoming separated, form apparent
threads or flagella (Fantham, 1907–08).142
Again, the mode of division of spirochætes has been used as a
criterion of their bacterial or protozoal affinity. They have been
stated to divide transversely, longitudinally, and by “incurvation,”
or bending on themselves in the form of a U, “a form of transverse
fission.” The present writer believes that they divide both transversely
and longitudinally, and that there is a periodicity in their mode of
division at first longitudinal (when there are few spirochætes in, say,
the blood) and then transversely (when spirochætes are numerous in
the blood).143 Some authors consider that longitudinal division is explained
by “incurvation.”
The spirochætes of relapsing fever show a remarkable periodic
increase and decrease in numbers in the blood. They are transmitted
by ticks or by lice. They react to drugs (e.g., salvarsan or “606”)
rather like trypanosomes, and—like Protozoa, but unlike Bacteria—they
are cultivated with difficulty. These and other criteria have been
used to endeavour to determine whether they are Protozoa or Bacteria.
The present writer believes that they are intermediate in character,
showing morphological affinities with the Bacteria and physiological
and therapeutical affinities with the Protozoa. The group Spirochætacea,
as an appendix to the Protozoa, has been created for them
by the present writer (Jan., 1908). Others have placed them in the
Spirochætoidea of the Bacteria or with the Spirillacea. Doflein (1909)
called them Proflagellata. Further discussion is unnecessary, as they
are undoubtedly Protista (see p. 29).
There is no true conjugation, sex or encystment in spirochætes,
but morphological variation may occur.144 They may agglomerate.
The Spirochætes form an interesting chapter in the evolution of
parasites. There are free living forms, parasitic forms in the guts
of both vertebrates and invertebrates, and blood-inhabiting forms.
These probably represent the order of evolution of parasitism. The
blood-inhabiting forms are pathogenic to warm-blooded hosts.
We must now consider the blood Spirochætes and the Treponemata
(organisms of syphilis and of yaws).
THE SPIROCHÆTES OF THE BLOOD.
There are at least two important human parasites included
hereunder:—
(a) Spirochæta recurrentis (=S. obermeieri), (b) Spirochæta duttoni.
More is known of the life-cycle of Spirochæta duttoni, and it will
be convenient to consider that first.
Spirochæta duttoni, Novy and Knapp, 1906.
The specific name duttoni was also given, independently, to this parasite in 1906
by Breinl and Kinghorn.
S. duttoni is the pathogenic agent of African tick fever in man,
prevalent in the Congo State and other parts of Africa. The full-grown
organism is about 16 µ to 24 µ long, and has pointed ends. It
is 0·25 µ to 0·5 µ broad. P. H. Ross and Nabarro were among the
earliest to see a spirochæte in the blood of patients in Uganda. It is
transmitted by the tick, Ornithodorus moubata.
In the blood of the patient some of the spirochætes may
show, after staining, lighter and darker portions (chromatin dots) and
evidence of the possession of a very narrow membrane (fig. 54).
The mode of division has already been discussed. Periodicity in
the direction of division was first described by Fantham and Porter,145
(1909). Just before the crisis in African tick fever, Breinl has stated
that S. duttoni becomes thinner in the spleen and bone-marrow
and rolls up into skein-like forms, which are surrounded by a thin
“cyst” wall (probably the periplast). Such occur in apyrexial
periods. Inside the cyst the spirochæte breaks up into granules.
Balfour and Sambon have described somewhat similar rolled up
forms, breaking into granules, inside the red blood cells of Sudanese
fowls in the case of S. granulosa (possibly only a variety of S. gallinarum).
The intracorpuscular stage is not definitely established.
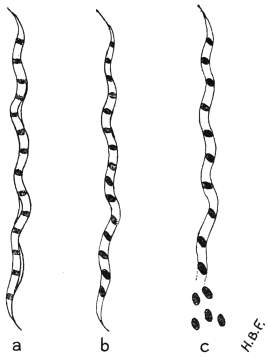
Fig. 54.—Spirochæta duttoni. a, blood
form showing slight membrane; b, granules or
coccoid bodies clearly formed within the organism;
c, beginning of extrusion of coccoid bodies in the
tick. (After Fantham.)
The granule phase, however, is an essential one in the invertebrate
transmitter (fig. 54c). In 1905,146 Dutton and Todd proved experimentally
that O. moubata transmitted S. duttoni. They fed ticks, obtained
from Congo native huts in which infected persons lived, on monkeys
and the latter became infected. Dutton and Todd also found the
offspring of infected ticks to be capable of transmitting the infection
to experimental animals. They concluded that O. moubata was a
true intermediate host.
A little later in 1905, Koch stated that spirochætes from the gut
of the tick penetrated the gut wall and tissues and found their
way into the eggs in the ovary. Koch figured tangled masses of
spirochætes as occurring in the tick eggs. He found ticks infective
to the third generation. He thought that the infection was spread
by the salivary fluid of the tick, in the act of biting. (This is now
known to be incorrect.) Markham Carter (1907) corroborated Koch’s
work on the spirochætes in the
tick eggs, and they have been
seen since by Kleine and Eckard
(1913).
Sir William Leishman,147 in
1909–10, found that at ordinary
temperatures the salivary glands
of infected ticks (O. moubata)
were not themselves infective,
and that the infection occurred
by way of the ticks’ excretion.
The spirochætes (contained in
the ticks’ excrement) found their
way into the vertebrate host
through the wound made by
biting. While feeding, ticks
pass large quantities of clear
fluid from the coxal glands;
in this fluid an anticoagulin
occurs. Some of the ticks also
pass thick, white Malpighian
secretion, that is, excrement,
towards the end of the feed.
Leishman, using experimental
monkeys, showed that if infected ticks were interrupted while
feeding, then no infection resulted in the monkeys. If, however, the
ticks were allowed to finish their feed, and the Malpighian secretions
were passed, then the experimental monkeys became infected.
Fantham148 and Hindle149 (1911), independently, have repeated the
experiments with mice.
Leishman’s methods and results may be summarized thus:
Saline emulsions of the organs of infected ticks were made, after
the organs had been most carefully dissected out. The ticks were
first kept for several days at certain constant temperatures, such as
24° to 25° C. or blood heat, 37° C. The saline emulsions of the
organs were inoculated, separately, into experimental animals, and the
results recorded:—
| At 24° C. | At 37° C. |
| Salivary glands | Negative | Positive |
| Malpighian tubules | Positive | Positive |
| Gut and contents | Positive | Positive |
| Excrement | Positive | Positive |
| Genital organs | Positive | Positive |
Coxal fluid is usually negative; thick, white excrement from
Malpighian tubes is positive.
When the ticks were
incubated at 21° to 24° C.
no spirochætes, as such,
were seen in the organs,
except perhaps in the gut,
where they often disappeared
in a few days.
When the ticks were previously
incubated at 35° to
37° C. for two to three days,
spirochætes, as such, reappear
in the gut, organs
and hæmocœlic fluid.
The infection proceeds,
not from the salivary gland, but from the infective excrement,
that is, from the thick, white material voided by the tick while
feeding, usually towards the end of the meal. This Malpighian
excrement passes into the wound caused by the bite, being greatly
aided by the clear and more limpid coxa fluid, which bathes the under
surface of the tick’s body, and mixes with and carries the infective
excrement into the wound. Ticks remain infective for a long time.
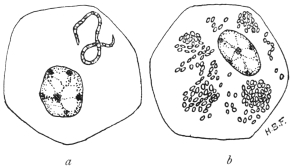
Fig. 55.—Spirochæta duttoni and its coccoid bodies
in the tick (O. moubata).—Mononuclear cells of the tick
(O. moubata) containing (a) Spirochæte breaking up into
coccoid bodies; (b) Similar tick-cell containing coccoid
bodies or granules. Such mononuclear cells occur in
various organs of ticks and in developing Malpighian
tubules. (Original. From preparations by Fantham.)
The spirochætes in the gut of infected ticks divide by a process
of multiple transverse fission into granules, which are composed of
chromatin (fig. 54). These granules—sometimes known as coccoid
bodies—are capable of multiplication. Leishman first found them in
clumps inside the cells of the Malpighian tubules (cf. fig. 55).
To summarize, when spirochætes are ingested by a tick, some of
them pass through the gut-wall into the hæmocœlic (body) fluid.
They then bore their way into the cells of various organs (fig. 55a)
and break up into coccoid bodies. In this manner the granules find
their way into the ovaries and ova, thus explaining how the young
ticks are born infected. Inoculation of these chromatinic granules
usually produces infection. Infective granules are also seen in the
rudiments of the Malpighian tubules of embryo ticks. Bosanquet
and Fantham (1911), independently, have shown that molluscan
spirochætes also break up into similar granules or coccoid bodies.
Gross has also demonstrated multiple transverse fission in molluscan
forms. Marchoux and Couvy (1913) and Wolbach (1914) consider
the granules or coccoid bodies to be degeneration products. This is
unlikely (see below).
Schuberg and Manteufel have found that certain O. moubata,
perhaps 30 per cent. of the specimens of a given neighbourhood, may
acquire a natural active immunity against infection with S. duttoni.
S. duttoni, or a closely allied form (by some termed S. novyi), occurs
in Colombia, and is spread by the tick Ornithodorus turicata. In
Panama a similar spirochæte is probably spread by O. talaje.
Spirochæta gallinarum, Stephens and Christophers, 1905
(= Spirochæta marchouxi, Nuttall, 1905).
This Spirochæte, which occurs in fowls and is pathogenic, is
transmitted by the tick Argas persicus. It is about 10 µ to 20 µ long.
There is a pathogenic spirochæte known to occur in geese, named by
Sakharoff (1891) S. anserina, and found in Caucasia. This may be
the same as S. gallinarum, in which case the name S. anserina will
have priority. These organisms cause fever, diarrhœa, anæmia and
death. The life history of the avian pathogenic spirochætes has been
studied by Balfour, by Hindle150 and by Fantham.151 It is essentially
similar to that of S. duttoni.
Marchoux and Couvy152 (1913) consider that the “fragmentation of
the chromatin” in spirochætes is a process of degeneration. Working
with A. persicus and S. gallinarum, they state that a large number of
the spirochætes ingested by the Argas almost immediately pass
through the wall of the alimentary canal and appear in the hæmocœlic
fluid. Marchoux and Couvy consider that Leishman’s granules
may be found in the Malpighian tubules of various Arachnids. They
found spirochætes in the cephalic glands of infected Argas. They
consider that spirochætes remain as wavy spirochætes within the tick,
if they are to be infective, though the spirochætes may become so thin
as to be invisible! The latter argument is obviously weak, and it
was never asserted that all granules in the Malpighian tubules of
infected ticks were derived from spirochætes. With dark-ground
illumination small, refractile spirochætal granules may be seen to grow
into spirochætes. The granule phase of spirochætes has recently
been discussed by Fantham153 (1914).
Spirochæta recurrentis, Lebert, 1874.
Syn.: Spirochæta obermeieri, Cohn, 1875.
This organism was discovered by Obermeier (1873) in cases of
relapsing fever in Berlin. Short forms 7 µ to 9 µ long, and longer
(probably adult) forms, 16 µ to 19 µ, are found in the blood. The
width is 0·25 µ. Parasites 12 µ or 13 µ long are often observed.
The spirochæte is found in the blood during febrile attacks and
relapses, but not during intervening periods. It can be inoculated
into monkeys, rats and mice. It can live in the bed-bug, Cimex
lectularius, and Nuttall has succeeded in transmitting S. recurrentis
from mouse to mouse by the bites of the same bug. The French
investigators Sergent and Foley (1908–9) in Algeria, and Nicolle,
Blaizot and Conseil (1912) in Tunis, have shown experimentally that
S. recurrentis (var. berbera) is transmitted by lice. The latter workers
also demonstrated the method of infection that commonly occurs,
namely, by the scratching of the skin and crushing of lice containing
spirochætes on the excoriated surface of the body.
Lice as transmitting agents for relapsing fever were indicated by Mackie154 in
1907. An epidemic among Indian school children furnished the materials.155 It was
noted that out of 170 boys, 137 were infected, and the boys were very verminous.
Among the girls, 35 out of 114 suffered, and few lice were found on them. Twenty-four
per cent. of the lice taken from the boys contained spirochætes as compared
with 3 per cent. of those from the girls. As the epidemic died out among the boys,
the lice also became fewer, and an increase in the number of cases among the girls
coincided with an increase in the number of lice. Spirochætes were found in the
gut, Malpighian tubules and genital organs of the lice. Mackie thought that infection
of the patients was brought about by the regurgitation of the spirochætes when
the lice fed, but proof of this was lacking.
In 1912, Nicolle, Blaizot and Conseil,156 working in Tunis and
using chiefly an Algerian strain of relapsing fever spirochætes (sometimes
called S. berbera), showed by direct experiments that infection
by means of the bites of Pediculus vestimenti and P. capitis was
untenable. As many as 4,707 infected lice were fed on one man, and
6,515 on another occasion were allowed to bite a man after they
had fed on a monkey heavily infected with spirochætes, yet no
infection of the man followed. Examination of the lice showed
that the spirochætes left the gut soon after they were ingested, and
passed into the body cavity, which swarmed with spirochætes. The
contents of the alimentary tract and the fæces of the lice alike were
uninfective. The spirochætes did not reappear in the gut till eight
days after an infective feed, but some persisted as late as the nineteenth
day when kept at 28° C.
It was noted that the irritation due to the lice caused scratching,
and that thereby lice became crushed on to the skin. An emulsion
was made of two infected lice and rubbed on to the slightly excoriated
skin of one of the above workers. Infection followed five days
later. A drop of emulsion placed on the conjunctiva of the human
eye produced spirochætosis after an incubation of seven days. The
body contents of such lice, then, produce infection when they reach
the blood by any excoriated or penetrable surface. The stages leading
up to infection in nature briefly are: The irritation due to the louse
bites causes scratching, and the lice are crushed on to the skin. The
slight abrasion is quite sufficient to permit the entry of the parasite.
The louse bite alone is harmless. Infection by way of the eye is
quite probable in Africa, remembering the constant trouble due to
sand, dust, insects, etc., resulting in frequent touching of the eyes.
The spirochætes occur in the body fluid of the lice and can pass
in it to the adjacent organs. Thus they probably find their way into
the genital organs, and into the eggs of the lice. Eggs laid twenty
to thirty days after the parent became infected have retained the
infection, and the larvæ issuing from such eggs must have contained
some form of spirochætes, for an emulsion of either the eggs or
the larvæ produced spirochætosis when inoculated into monkeys.
Further details regarding the spirochætosis in the eggs of the lice and
in the larvæ are needed. Hereditary infection, however, has been
demonstrated, but is not very common. Sergent and Foley (1914)
state that the spirochæte possesses a very small and virulent form
which it assumes during apyrexial periods in man and during a
period following an infecting meal in the louse. Nicolle and Blanc
(1914) find that the organisms are infective in the louse just before
they reappear as spirochætes. Nicolle and Blaizot found that female
lice were more susceptible to spirochætes than males, four times
as many females as males being infected.
Tictin (1897) found S. recurrentis in bugs recently fed on patients, and infected a
monkey with the fluids of crushed bugs. Karlinski (1902) found the spirochætes
in bed-bugs in infected houses. There is some other evidence to show that bugs
may transmit the spirochæte in Nature. Further researches are needed regarding
the relationship of bed-bugs and human spirochætosis.
Multiplication of S. recurrentis is by longitudinal and transverse
division (including so-called “incurvation”), and the organism forms
small, ovoid bodies (“coccoid” bodies) in the same way as S. duttoni.
S. recurrentis is the cause of European relapsing fever, and a
number of possible varieties of it are associated with relapsing fevers
in other parts of the world. Such spirochætes only differ by biological
reactions, such as acquired immunity tests. They include:—
S. rossii, the agent of East African relapsing fever; S. novyi, the
agent of North American relapsing fever; S. carteri, the agent of
Indian relapsing fever; S. berbera, the agent of North African and
Egyptian relapsing fever.
Other Human Spirochætes are:—
S. schaudinni. This organism, according to Prowazek, is the
agent of ulcus tropicum. It varies in length from 10 µ to 20 µ.
S. aboriginalis has been found in cases of granuloma inguinale
in British New Guinea and Western Australia. It also occurs in
dogs, and may not be truly parasitic.
S. vincenti. This spirochæte is 12 µ to 25 µ in length, tapers at
both ends and has few coils. It has been associated with angina
vincenti. It often occurs in company with fusiform bacilli.
S. bronchialis, found by Castellani in 1907 in cases of bronchitis
in Ceylon. The parasites are delicate, but show morphological
variation. This organism is important and has since been found
in the West Indies, India, Philippine Islands and various parts of
Africa, such as the Anglo-Egyptian Sudan, Uganda and West Africa.
It has recently been the subject of research by Chalmers and
O’Farrell, Taylor, and Fantham.
S. phagedenis was found by Noguchi in a ten days old ulcerated
swelling of the labium. The organism shows much variation in size,
being 4 µ to 30 µ in length.
S. refringens (Schaudinn, 1905) occurs in association with
Treponema pallidum in syphilitic lesions, but is non-pathogenic. It
is 20 µ to 35 µ long and 0·5 µ to 0·75 µ broad, being larger than
T. pallidum and more easily stained.
Various spirochætes have also been notified in vomits, chiefly in
Australia; others from the human intestinal tract, e.g., S. eurygyrata;
S. stenogyrata (Werner); S. hachaizæ (Kowalski), in cholera motions;
S. buccalis (Cohn, 1875) and S. dentium occurring in the human
mouth and in carious teeth (S. dentium, Koch, 1877, being the
smaller); S. acuminata and S. obtusa found by Castellani in open
sores in cases of yaws.
Animal spirochætes of economic importance include:—
S. anserina, highly pathogenic to geese.
S. gallinarum (= S. marchouxi) in fowls. (See p. 119.)
S. theileri in cattle and S. ovina in sheep also occur in Africa;
their pathogenicity is not clear.
S. laverani (= S. muris), occurring in the blood of and pathogenic
to mice, is probably the smallest spirochæte from the blood,
being only 3 µ to 6 µ long.
Numerous spirochætes have been recorded from the guts of
various mammals, birds, fishes, amphibia and insects.
Cultivation of Spirochætes.—Cultures of spirochætes have
been made with little success or with great difficulty until comparatively
recently, when Noguchi (1912) devised a means whereby he
has cultivated most of the pathogenic spirochætes as well as some
Treponemata.
Noguchi has now cultivated S. duttoni, S. recurrentis, S. rossii,
S. novyi and S. gallinarum from the blood; S. phagedenis157 from human
phagedænic lesions; S. refringens158 and spirochætes from the teeth.
His method is as follows:—
A piece of fresh, sterile tissue, usually rabbit kidney, is placed in a
sterile test-tube. A few drops of citrated blood from the heart of an
infected animal, e.g., rat or mouse, is added, and about 15 c.c. of sterile
ascitic or hydrocœle fluid is poured quickly into the tube. Some of
the tubes are covered with a layer of sterile paraffin oil, others are
left uncovered. The tubes are incubated at 37° C. The best results
are obtained if the blood is taken from an animal forty-eight to seventy-two
hours after it has been inoculated, that is, before the spirochætes
reach their maximum multiplicative period in the blood. The presence
of some oxygen seems indispensable for these blood spirochætes, and
they fail to develop in vacuo or in an atmosphere of hydrogen.
For subcultures, 0·5 c.c. of a culture is added to the medium
instead of citrated blood, and it is useful to add a little fresh, normal
blood, either human or from an animal, such as a rat.
Noguchi found that the events in cultures were:—
S. duttoni,159 maximum multiplication on the eighth to ninth day;
disintegration beginning on the tenth day, spirochætes disappeared
after about the fifteenth day. No diminution of virulence was found
at the ninth day.
S. rossii (= S. kochi).160 Maximum development on the ninth day,
after which the virulence diminishes. The incubation period is also
prolonged.
S. recurrentis161 (= S. obermeieri). Maximum growth on the seventh day.
S. novyi.162—Maximum development on the seventh day. It is
more difficult to grow than the preceding forms.
All the above spirochætes showed undoubted longitudinal division
and transverse division was observed in part.
S. gallinarum163 can be cultivated as above, but transverse division
was usual here. Maximum growth occurred in the culture about the
fifth day.
Treponemata.
The genus Treponema (Schaudinn, 1905), includes minute, thread-like
organisms, with spirally coiled bodies, the spirals being preformed
or fixed. No membrane or crista is present, according to Schaudinn,
though a slight one is said by Blanchard to be present in the organism
of yaws. The ends of the organisms are tapering and pointed.
Multiplication is by longitudinal and transverse division. The most
important members of the genus are T. pallidum, the agent of syphilis,
and T. pertenue, which is responsible for frambœsia or yaws.
Treponema pallidum, Schaudinn, 1905.
Syn.: Spirochæta pallida.
Treponema pallidum was first described by Schaudinn and Hoffmann
in 1905 under the name of Spirochæta pallida. It has also been
described under the names
of Spironema pallida, Microspironema
pallida and
Trypanosoma luis. Siegel
in 1905 described an organism
which he called Cytorhyctes
luis and considered
to be the agent of syphilis.
Schaudinn reinvestigated
Siegel’s work and found
T. pallidum, which he considered
to be the causal
agent of the disease, and
pronounced against Cytorhyctes
luis. It is probable
now that both workers
were correct, for Balfour
(1911) has seen the emission
of minute granules
or “coccoid” bodies from T. pallidum and these granules probably
correspond to the C. luis of Siegel. Recently E. H. Ross, having
observed a spirochæte stage in the development of Kurloff bodies,
thinks that T. pallidum is a stage in the life-history of a Lymphocytozoon.
MacDonagh has also described a complicated and somewhat
similar cycle, but these observations require further study and
confirmation.

Fig. 56.—Treponema pallidum. (After Bell, from
Castellani and Chalmers.)
T. pallidum varies from 4 µ to 10 µ in length, its average length
being 7 µ, while its width is usually about 0·25 µ. Longer individuals
of 16 µ to 20 µ have been recorded. The body has from eight to ten
spiral turns and forms a tapering process at each end (fig. 56). The
organism is most difficult to stain, and its internal structure is little
known. It is possibly like that of Spirochæta duttoni or S. balbianii,
as the “granule shedding” observed by Balfour is strongly suggestive
of the formation of resistant bodies by those spirochætes. Hoffmann
(1912) has seen the formation of spores in T. pallidum.
The Treponemata occur in the primary
and secondary sores, but are difficult to
find in the tertiary eruptions of syphilis.
Noguchi and Moore (1913) and Mott164
(1913) have demonstrated T. pallidum in
the brain in cases of general paralysis of
the insane. Marie and Levaditi (1914),
however, consider that the treponeme
found in the brain in such cases is
different from T. pallidum.
Cultivation of T. pallidum.—This has
been accomplished successfully by Noguchi,165
using a modification of his
method for spirochæte cultivation, for
T. pallidum is much more difficult to
grow than spirochætes, being a strict
anaerobe.
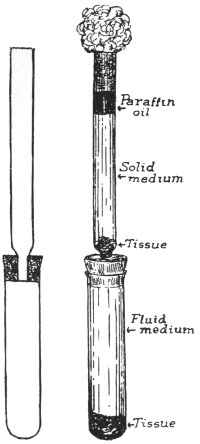
Fig. 57.—Diagram of apparatus
for cultivation of Treponema pallidum
by Noguchi’s method. (After
Noguchi.)
The apparatus consists of two glass
tubes, the upper being connected to the
lower by a narrower tube passing through
a rubber cork (fig. 57). Both tubes are
carefully sterilized.
A piece of fresh, sterile rabbit’s kidney
is placed in the lower tube, which is
filled with ascitic fluid, or ascitic fluid
and bouillon mixture. The tube is inoculated
with syphilitic material and
corked by inserting the upper tube. In the bottom of the upper tube
a piece of sterile rabbit’s kidney is placed and syphilitic material
poured over it. A mixture of one part ascitic fluid and two parts of
slightly alkaline agar is then poured over the tissue and allowed to
solidify. When solid, a layer of sterile paraffin oil is poured on top
of it, and the top plugged with cotton wool (fig. 57). The whole is then
incubated at 37° C. for two or three weeks. The tissue removes traces
of oxygen from the lower levels of the medium and also probably
provides a special form of nourishment. At first T. pallidum grows in
the solid medium, and then when the cultural conditions in the lower
fluid portion become favourable, the organisms migrate thither and
multiply abundantly. At first the culture is impure, but after several
transferences a pure culture is obtained readily.
The syphilitic material for culture is prepared by cutting off
pieces of tissue from the lesions, washing in sterile salt solution
containing 1 per cent. sodium citrate, and then emulsifying the
tissue in a mortar with sodium citrate.
Good cultures show rapid multiplication, which is invariably
by longitudinal division.
In his various cultivation experiments Noguchi166 found morphological
and pathogenic variations in T. pallidum. Three forms of the
organism were found, namely, thicker, average and thinner types.
The lesions caused in the testicle of the rabbit differ according to
the variety inoculated, but more work is necessary on the subject.
Noguchi167 has cultivated a separate organism, T. calligyrum,
from the surface of human genital or anal lesions, either syphilitic
or non-syphilitic. It is apparently non-pathogenic, and is 6 µ to
14 µ long.
Hata (1913)168 has modified the Noguchi technique for the cultivation
of spirochætes and treponemes, with a view to simplification and
convenience. Hata substitutes normal horse serum for ascitic fluid
and the “buffy coat” of the clot of horse blood in place of the small
pieces of rabbit’s kidney. It is unnecessary to place sterile paraffin on
the surface of the medium.
The horse serum is mixed with twice its volume of physiological
saline solution. The mixture is placed in tubes which are heated on a
water-bath at 58° C., the temperature being raised gradually until it
reaches 70° or 71° C. in three hours. The tubes are then heated at
71° C. for half an hour. After cooling, the contents will consist of an
opaque semi-coagulated mass. This semi-coagulated serum and saline
mixture may be substituted for Noguchi’s ascitic fluid.
The buff coagulum is cut into small pieces, about 1 c.c. in volume.
They must be forced with a sterile glass rod to the bottom of the semi-coagulated
serum and saline mixture. The medium is inoculated with
a small quantity of infected blood and kept at 37° C. In the case of
S. recurrentis, growth of spirochætes is observed on the second day,
reaching a maximum in five to seven days. The growth of the
organisms proceeds rather more slowly, they live for a longer period
and maintain their virulence better than in Noguchi’s medium.
Treponema pertenue, Castellani, 1905.
Syn.: Spirochæta pertenuis; S. pallidula, Castellani, 1905.
Castellani discovered the organism in 1905, in scrapings of yaws
pustules. He first described it under the name of Spirochæta pertenuis.
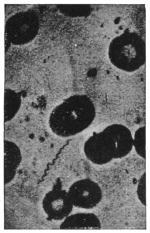
Fig. 58.—Treponema pertenue.
(After Castellani and Chalmers.)
Treponema pertenue (fig. 58), though delicate and slender, shows
great morphological variation both in length and thickness. It
may be short, e.g., 7 µ, but can attain 18 µ to 20 µ in length and may
be even larger. In cultures made by Noguchi, thick, medium and
thin forms were found, each giving rise to a different type of frambœsial
lesion when inoculated into the testicles of rabbits, thus
suggesting the possibility of the occurrence of varieties of T. pertenue.
The organism is difficult to stain, but occasionally deeper
staining granules are found along its body. They may represent
a diffuse nucleus. Granule formation
similar to that of T. pallidum has been
observed by Ranken, using dark-ground
illumination.
Many experiments have been made
with a view to establishing the identity
of the organism of yaws and also of
differentiating between the causative
agents of yaws and syphilis. Both
monkeys and the human subject have
been experimentally inoculated with yaws
material and have developed the disease.
In an early experiment, negroes were
inoculated with the secretion from lesions
of yaws. All of them developed the
disease, nodules appearing, chiefly at the
seat of inoculation, in from twelve to twenty days, followed by the
usual eruption. Similar results were obtained with thirty-two Chinese
prisoners, who were inoculated with yaws, twenty-eight becoming
infected.
A naturally infected yaws patient when inoculated with syphilis,
contracted that infection, thus showing that yaws does not confer
immunity to syphilis. This has also been observed naturally, when
yaws patients have contracted syphilis.
Experiments with monkeys have been successfully performed.
The incubation period varies from sixteen to ninety-two days.
Lesions appear first at the seat of inoculation, and in some
monkeys the eruption is localized to this spot, though the infection is
general, T. pertenue occurring in the spleen, lymphatics, etc. Monkeys
inoculated with splenic blood of a yaws patient, and also sometimes
with blood from the general circulation, have become infected.
Castellani and others have shown that monkeys successfully
inoculated with syphilis do not become immune to yaws, and vice-versâ.
Craig and Ashburn, using the monkey Cynomolgus philippinensis,
found these animals susceptible to yaws but not to syphilis.
The ulcerated lesions of frambœsia are rapidly invaded by
numerous bacteria as well as by different spirochætes, of which
Castellani has described three distinct species. One is identical
with Spirochæta refringens, Schaudinn, the other two are thin and
delicate. One, S. obtusa, has blunt ends; the other S. acuminata, has
pointed ends. T. pertenue is also present.
The reasons for considering T. pertenue to be the specific cause of
frambœsia are:—
(1) T. pertenue is the only organism present in non-ulcerated
papules, in the spleen and in the lymphatics of yaws patients, or
of monkeys artificially infected with the disease. By no method
has any other organism been obtained.
(2) Extract of frambœsia material, free from all organisms other
than T. pertenue, reproduces the disease if inoculated.
(3) Extract of frambœsia material deprived by filtration of
T. pertenue is no longer infective on inoculation.
The method of infection is contaminative, by direct contact.
Women in Ceylon are frequently infected by their children. Any
slight skin abrasion is sufficient to admit the parasite. In some
cases, insects may carry the disease from person to person, and even
in hospitals, when dressings are removed, it has been noticed that
flies greedily suck the secretion from the ulcers. T. pertenue has
been recovered from flies that have fed on yaws, and monkeys have
contracted the disease when flies were placed and retained on them
for a short time, after the insects had fed on yaws material.
Cultivation.—T. pertenue has been cultivated by Noguchi, who
finds three types of parasites in his cultures, as before mentioned. Its
multiplication is by longitudinal division.
Noguchi169 (1912), has cultivated species of Treponema from the
human mouth, e.g., T. macrodentium, T. microdentium and T. mucosum,
the latter from pyorrhea alveolaris. These parasites in the past may
have been confused under the name Spirochæta dentium.
Class III. SPOROZOA, Leuckart, 1879.
The third group of the Protozoa consists entirely of parasitic
organisms forming the class known as the Sporozoa or spore-producing
animals. The members of this class are characterized
by possessing very great powers of multiplication, coupled with a
capacity for producing forms that serve for the transference of the
organisms to other hosts. These reproductive bodies, whether for
increase of numbers within one host or for transmission to another
host, are called spores. But, strictly, the term spore should be used
only in the latter connection, when a protective or resistant coat
known as a sporocyst envelops the body of the spore.
The Sporozoa are widely distributed, occurring in various tissues
and organs of Annelids, Molluscs, Arthropods, and Vertebrates.
Their food, which is fluid, is absorbed osmotically. The life-cycle
of a Sporozoön may be completed within one host or may be distributed
between two different hosts.
The Sporozoa were divided by Schaudinn into two groups or
sub-classes, called (1) the Telosporidia, and (2) the Neosporidia.
The Telosporidia are Sporozoa in which the reproductive phase
of the parasites is distinct from the growing or trophic phase, and
follows after it. The Neosporidia include Sporozoa in which growth
and spore-formation go on simultaneously. This classification is not
final, for certain exceptions and difficulties are already known with
regard to it. It is possible that the class Sporozoa is not a natural
entity, but should be replaced by two classes of equal rank, corresponding
in most respects with the Telosporidia and Neosporidia.
The Telosporidia comprise the Gregarinida, the Coccidiidea, and
the Hæmosporidia. Doflein combines the two latter orders into one
known as the Coccidiomorpha.
The Neosporidia comprise the Myxosporidia, the Microsporidia,
the Actinomyxidia, the Sarcosporidia, and the Haplosporidia.
Doflein combines the first three orders into one, the Cnidosporidia.
Sub-Class. TELOSPORIDIA, Schaudinn.
Sporozoa in which the reproductive phases follow completion of
growth.
Order. Gregarinida, Aimé Schneider emend. Doflein.
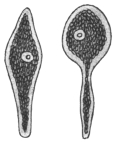
Fig. 59.—Monocystis
agilis from seminal
vesicles of Lumbricus
× 250. (After Stein.)
Knowledge of the Gregarinida probably goes back as far as the year 1684, when
Redi observed gregarines in the crab, Cancer pagurus. Von Cavolini (1787) found
them in Cancer depressus. The name Gregarina was created by L. Dufour (1828),
who observed masses of these organisms in the gut of insects of different orders.
Hammerschmidt (1838) and von Siebold found rich infestations in insects, while
Dujardin (1835) and Henle described various genera from segmented worms.
Henle (1835) also observed cysts containing “navicellæ” in the sperm-sacs of
segmented worms, and attention was drawn to his researches by the discovery by
von Siebold (1839) of “pseudonavicellæ” in the gut of Sciara nitidicollis. Up to this
time many workers considered the gregarines to be worms, but Kölliker (1845)
investigated many of them and maintained their unicellular nature, while Stein’s
work (1848) showed the interrelation of the pseudonavicellæ and the gregarines.
The discovery of amœboid germs in the pseudonavicellæ by Lieberkühn (1855) and
the demonstration of myonemes further aided in the elucidation of their true
systematic position. The entire process of conjugation, of which Dufour had seen
one phase, was followed by Giard under the microscope.
From 1873 onwards Aimé Schneider made important additions to the knowledge
of the morphology, life-history, and systematic position of numerous gregarines.
Bütschli (1881) and L. Léger (1892) also contributed much work on the subject. The
discoveries of Schaudinn with regard to the life-cycle of Coccidia gave a fresh
stimulus to the study of the Gregarines, whereby the life-cycles of numerous forms
and the phases thereof have been elucidated.
Asexual multiplication is not common among the Gregarines, but is known to
occur in the sub-order Schizogregarinea, formerly known as the Amœbosporidia.
Although the Gregarinida are not known to be parasitic in man or other vertebrates,
they are of great interest, inasmuch as they are among the earliest known
Sporozoa, and therefore will be briefly described here.
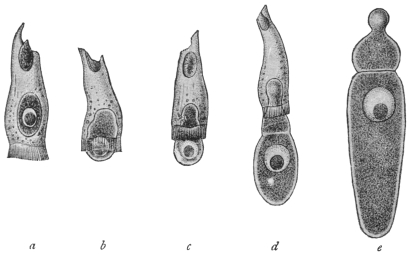
Fig. 60.—Gregarina longa from larva of crane-fly (Tipula). a, in epithelial cell of host;
b, c, gradually leaving host-cell; d, adhering to host-cell; e, fully developed free trophozoite.

Fig. 61.—Xyphorhynchus
firmus with epimerite in
intestinal epithelial cell of
host. (After Léger.)
The Gregarines are usually elongate, somewhat
flattened organisms (figs. 59, 60), whose bodies are
enclosed in an elastic and often thick cuticle. The
enclosed living substance shows a separation into
ectoplasm and endoplasm, as is common among
Protozoa. The cuticle is sometimes regarded as the
outer portion or epicyte of the ectoplasm. A single,
vesicular, spherical, or elliptical, large nucleus, with
its chromatin concentrated to form a spherical
karyosome, is present. The body of some gregarines
may be divided by ingrowing ectoplasmic
partitions or septa, and are then said to be “septate” or
“polycystid” (fig. 61). Other gregarines remain simple and non-septate,
and are termed “monocystid” (fig. 59). The monocystid gregarines
occur especially in the body cavity of Chætopoda and Insecta, more
rarely in Echinodermata, in the parenchyma of Platyhelminthes,
also in the gut of Tunicata and Insecta (fig. 60) and in the seminal
vesicles of Annelida. In the polycystid gregarines a single septum only
is present as a rule, and thus the body presents two portions: (1) an
anterior portion termed the protomerite; (2) a
posterior, larger portion, known as the deutomerite,
which generally contains the nucleus.
The protomerite is often modified anteriorly
to form an organ of attachment, termed the
epimerite (fig. 61), which is developed from the
pointed rostrum of the sporozoite or primary
infecting young gregarine. The structure of
the epimerite may be complicated, being provided
with hooks, spines, knobs, and other
appendages. An extension of the polycystid
condition is seen in Tæniocystis mira Léger
(from the dipteran larva, Ceratopogon solstitialis),
whose body shows a number of partitions,
giving the organism a superficial resemblance
to a tapeworm.
The ectoplasm of a gregarine exhibits three
layers: (1) An epicyte (cuticle) externally of
which the epimerite is composed; (2) a
sarcocyte which forms the septa if present;
(3) the deeper myocyte layer containing contractile
elements in the form of fibrils or threads
termed myonemes (fig. 62).
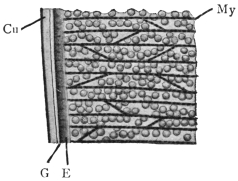
Fig. 62.—Gregarina munieri (from
the beetle, Chrysomela hæmoptera). Section
through surface layers. Cu, cuticle;
E, ectoplasm proper; G, gelatinous
layer; My, myonemes in myocyte layer.
× 1500. (After Schewiakoff.)
The endoplasm is fluid and granular, containing
many enclosures, which are of the nature of reserve food
materials. They consist of fat droplets
or of paraglycogen, and give
the organisms an opaque appearance.
Lithocystis contains crystals
of calcium oxalate in its endoplasm.
Many gregarines are capable of
active movements, though they do not
possess obvious locomotor organs.
The movement is of a smooth,
gliding character and two suggestions
have been put forward to
explain it. According to Schewiakoff,
a gelatinous substance is secreted between
the layers of the ectoplasm.
This is extruded posteriorly and thus
the animal is pushed forward. On the other hand, Crawley considers
that the movements are produced by contractions of the myonemes.
These two explanations are probably correct as far as each goes, and
are to be regarded as supplementary to one another.
Occasionally, temporary associations of gregarines are formed
by a number of individuals adhering to
one another end to end. Such temporary
associations are examples of syzygy. Such
syzygies must not be confused with true
associations which form an essential part of
the life-cycle.
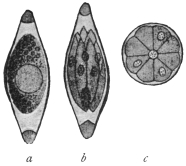
Fig. 63.—Monocystis agilis.
Spores from vesicula seminalis
of the Earthworm. a, Sporoblast
with single nucleus, enclosed in
sporocyst; b, mature spore containing
sporozoites; c, diagrammatic
cross-section of spore,
showing eight sporozoites round
residual protoplasm. (After
Bütschli.)
The life-cycle of a relatively simple gregarine,
such as Monocystis agilis (fig. 59),
parasitic in earthworms, may now be considered.
The gregarines, being members of
the Sporozoa, produce spores at one phase
of the life-cycle. Each gregarine spore
(fig. 63) develops within itself a number of
minute, sickle-shaped or vermicular bodies,
known as sporozoites or primary infecting
germs. Eight sporozoites are often formed
within each spore. When absorbed by a new
host, the spore softens and the sporozoites issue from it. They are
capable of active movement and may or may not enter a cell, such as
one of those of the digestive tract, or, as in Monocystis, a cell lining the
vesicula seminalis which becomes a sperm-cell aggregate (sperm morula).
When the sporozoite has reached the place of its choice in the host
it ceases active movements and proceeds to feed passively on the fluid
substances around it, whether they be those of tissues or body fluids.
This passive, growing and feeding form is known as the trophozoite.
After a trophic existence of longer or shorter duration, the trophozoite
ceases to feed and prepares for reproduction. Two trophozoites
associate together, each of them first becoming somewhat rounded.
The two trophozoites, now known as sporonts or gametocytes, become
invested in a single common envelope or cyst (fig. 64, a). The nucleus
of each gametocyte then divides by a series of binary fissions (fig. 64, b),
and the daughter nuclei thus produced arrange themselves at the
periphery of the parent cells (fig. 64, c). Cytoplasm collects around
each of these nuclei, and thus a number of gametes are formed within
each gametocyte. The gametes for a time exhibit active movements,
and ultimately ripe gametes of different parentage fuse in pairs, that
is, conjugation occurs (fig. 64, d). In this way zygotes are produced,
the nucleus of each zygote being formed by the fusion of two gamete
nuclei.
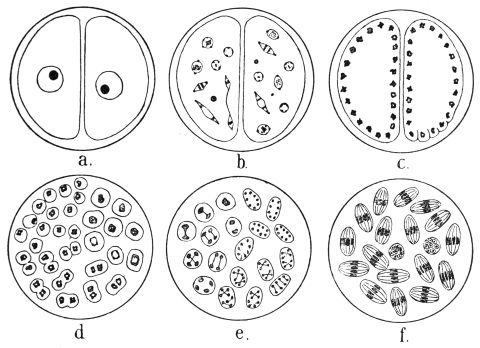
Fig. 64.—Schematic figures of conjugation and spore formation in Gregarines. For details
see text. (After Calkins and Siedlecki, modified.)
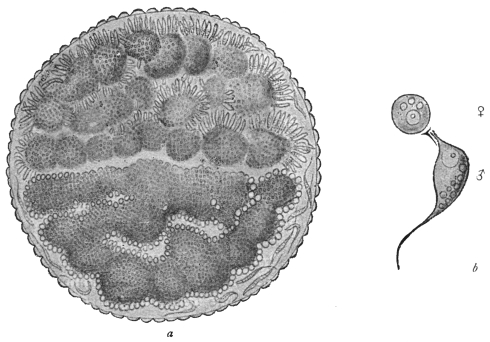
Fig. 65.—Stylorhynchus oblongatus. a, cyst containing two sporonts or gametocytes, each
full of gametes, those in the upper one being male. b, ripe male and female gametes. × 1,600.
(After L. Léger.)
The zygote grows slightly and becomes oval or elongate, and at
this period is often called the sporoblast. It then secretes an external
membrane, the sporocyst. Nuclear division occurs inside the sporocyst
by a series of three binary fissions (fig. 64, e), so that each sporocyst,
now usually referred to as a spore, contains eight nuclei. The
cytoplasm collects around each nucleus and eight vermicular sporozoites
are produced within each spore (fig. 64, f), thus completing
the life-cycle.
It will be noticed that in the above life-cycle no asexual multiplication
occurs. These organisms, such as Monocystis, are known as
the Eugregarines, and include the majority of the gregarines. The
remainder, which have introduced schizogony into their life-cycle, are
known as the Schizogregarines.
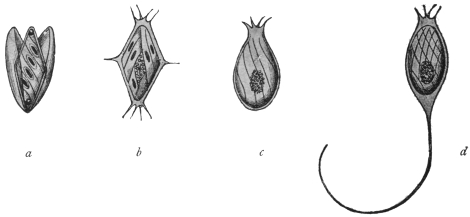
Fig. 66.—Spores of various Gregarines. a, Xiphorhynchus.
b, Ancyrophora. c, Gonospora. d, Ceratospora.
(After Léger.)
There are variations in the morphology and life-cycle of gregarines
besides those that have been mentioned. It is not within the scope
of this book to discuss them in detail, but the following may be
noted:—
Morphological differentiation of gametes may occur as in Stylorhynchus
oblongatus (fig. 65), which differentiation is probably of a
sexual nature.
The sporocyst really consists of two layers, an epispore and an
endospore. Externally the spores of different gregarines show great
variety in shape and markings, and spines, or long processes may
be present (fig. 66).
The resistant spores serve for the transmission of the gregarines
from host to host. The mode of infection is contaminative, the
spores expelled with the dejecta of one host being absorbed with the
food of a new host.
The Gregarinida may be classified as follows:—
Sub-order I.—Eugregarinea, without schizogony.
Tribe 1.—Acephalina.—Without an epimerite and non-septate;
often “cœlomic” (body-cavity) parasites. E.g.: Monocystis, with
several species parasitic in the seminal vesicles of earthworms. Many
other genera parasitic in Echinodermata, Tunicata, Arthropoda, etc.
Tribe 2.—Cephalina.—With an epimerite, either temporarily or
permanently, in the trophic phase. Usually septate (except Doliocystidæ).
Many families, genera and species. Common in the digestive
tracts of insects. E.g.: Gregarina, with several species, Gregarina
ovata in the earwig, Gregarina blattarum in the cockroach, Stylorhynchus
in beetles, Pterocephalus in centipedes, etc.
Sub-order II.—Schizogregarinea, with schizogony.
Tribe 1.—Endoschiza.170—With schizogony occurring in the intracellular
phase, e.g., Selenidium (from Annelida and Gephyrea), Merogregarina
(from an Ascidian).
Tribe 2.—Ectoschiza.—In which the schizont is free, and schizogony
is extracellular, e.g., Ophryocystis (from Blaps, a beetle), and Schizocystis
(from Ceratopogon larva).
Order. Coccidiidea.
Hake (1839) first saw the organisms now termed Coccidia during his investigations
on the so-called coccidial nodules of rabbits. The opinions as to the nature of these
peculiar formations were very diverse. The discoverer considered them to be a sort
of pus corpuscle; Nasse (1843) took them for epithelial cells of the biliary passages,
others for helminthes, especially the ova of trematodes (Dujardin, Küchenmeister,
Gubler, etc). Remak (1845) was the first to draw attention to their relation to the
Psorospermia (Myxosporidia), and this investigator found them also in the small
intestine and vermiform appendix of rabbits. Lieberkühn (1854), who examined not
only the coccidia of rabbits, but found similar forms in the kidneys of frogs, likewise
called them definitely psorosperms. To differentiate Müller’s psorosperms, which are
found in fishes, from those of rabbits, etc., the latter were called egg-shaped psorosperms
(Eimer), until R. Leuckart (1879) named them Coccidia and placed them in
a group of the Sporozoa analogous to that of the Gregarinida, Myxosporidia, etc.
Numerous works confirmed the occurrence of coccidia, not only in all classes of
vertebrate animals, but also in invertebrates (Mollusca, Myriapoda, Annelida, etc.).
A large number of genera and species have in the course of time been described which
inhabit the epithelium of the intestine and its appendages for choice, but are also
found in other organs (kidneys, spleen, ovaries, vas deferens, testicles). Some also
live in the connective tissue of various organs, more particularly of the intestine.
The knowledge of the development of the coccidia was of particular importance
in determining their classification. By means of encysted coccidia from the liver of
rabbits, Kauffmann (1847) first confirmed the fact that the cyst, which was partly or
entirely filled with granular contents, divided into three or four pale bodies (fig. 71)
after a long sojourn in water. Lieberkühn observed the same process in the host in
the case of the coccidia of the kidney of the frog. Stieda (1865) studied more minutely
the changes that occur within the encysted coccidia of the liver of rabbits after the
death of the host. He discovered that the bodies now known as “spores” were
oval formations pointed at one pole, and surrounded by a delicate membrane, which
exhibited a certain thickness at the pointed extremity and enclosed a slightly bent
rodlet, expanding at either end into a strongly light-refracting globule; a finely
granular globule was present in the middle of the spore. Waldenburg (1862) saw the
same phenomenon in coccidia from the epithelium of the villi and Lieberkühn’s
glands of the intestine of the rabbit; but the process in this case took place in a
much shorter time.
According to the discovery of Kloss (1855), the spores of the coccidia of the
urinary organ of the garden snail were formed in far greater numbers: the round
spores also harboured several (five to six) rodlets, which after the bursting of the
spore-envelope became free. Eimer’s researches (1870) afforded information
regarding a Coccidium from the intestine of the mouse, which was transformed in
toto into a “spore,” containing small sickle-shaped bodies. The fact was, moreover,
established that the little bodies left the delicate envelope when in the intestine,
made movements of flexion and extension, and were finally transformed into
amœboid organisms, which apparently penetrated the epithelial cells; at all events,
similar bodies of various sizes were seen in these cells. Taking the immense
number of these parasites into account and the lack of any other cause, Eimer
attributed the sudden death of his mice to the Gregarina falciformis, as the parasite
was then called, just in the same way as a few years previously Reincke ascribed the
acute and fatal intestinal catarrh of rabbits to the invasion of intestinal coccidia.
All that had become known about coccidia up to 1879 was then compiled by
Leuckart, and completed by his own observations on the coccidia of the liver of the
rabbit. Experimental infections had already been conducted by Waldenburg (1862)
with intestinal coccidia of rabbits, and by Rivolta (1869–73) with the coccidia of fowls,
which experiments confirmed the importance of the spores, or bodies enclosed in them,
in the transmission of the parasites to other animals. Accordingly, it was assumed that
after the entry of the spores into the intestine the sporozoites were set free, actively
penetrated into the intestinal cells, where they grew into coccidia, and finally became
encysted. The further development, i.e., the formation of spores, took place outside
the host’s body in these cases; in other cases (Kloss, Eimer) it took place within
the host. Although much regarding the cycle of development was still hypothetical,
the ideas coincided with the observations, and were therefore universally regarded as
established. Further research confirmed this view in numerous new forms.
L. Pfeiffer’s statements (1891) on the part that certain coccidia or their sporozoites
played, or seemed to play, as causes of disease gave a renewed impetus to the
investigation of the coccidia. The ingestion of even very numerous spores did not
appear to account for the mass infection so frequently observed, even after Balbiani
had confirmed the fact that there were two, and not one, sporozoites contained in every
spore of the coccidia of rabbits (fig. 72). The hypothesis was therefore advanced
that the sporozoites or young coccidia were able to divide once again by sporulating.
The question was finally solved quite differently. R. Pfeiffer (1892) first confirmed
the fact that in addition to the well-known method of sporulation in the coccidia of
the rabbit that causes the infection of fresh hosts (“exogenous sporulation”), an
enormous increase may follow in the already infected host in a manner that Eimer
first observed in the coccidia of the intestine of the mouse (“endogenous sporulation”).
It had hitherto been believed that some of the species of coccidia increased
like the rabbit parasite, then known as Coccidium oviforme, and others like Eimeria
falciformis, and this difference had been made the foundation of a classification.
R. Pfeiffer was successful in observing the occurrence of both kinds of development
in the same species, and expressed the opinion that endogenous sporulation (fig. 73),
which takes place within the host, was the cause of the mass-infection that is mostly
accompanied by serious consequences (fig. 74). L. Pfeiffer sought, especially, to
demonstrate the correctness of this view as regards other species of coccidia
and for this purpose he utilized the experiences already published. Coccidia were
known to exist in a number of different hosts, and to propagate in some according
to the Coccidium type, in others according to the Eimeria type. It therefore was
reasoned that in this case it was not a question of two species belonging to different
genera living side by side, with a different mode of development, but of one species,
in the life of which both forms of development occurred alternately.
This interpretation of facts was combated especially by A. Schneider (1892) and by
Labbé, but has, nevertheless, proved true, apart from the circumstance that Schuberg
succeeded in discovering the hitherto unknown Coccidium form in the intestine of
the mouse; and that, moreover, Léger confirmed the fact that there are no Arthropoda
in which Eimeria are not found together with coccidia. The question was
finally settled by experiments made by Léger with the coccidia of Scolopendra
cingulata, by Schaudinn and Siedlecki with those of Lithobius forficatus, and by
Simond with the coccidia of the rabbit. On Simond’s suggestion the sickle-shaped
germs corresponding to the sporozoites, which are formed by endogenous sporulation,
are generally termed merozoites; and in accordance with Schaudinn’s
suggestion, those individuals which form merozoites are termed schizonts, and
those which produce spores are called sporonts. In contradistinction to sporogony
(exogenous sporulation), the term schizogony (endogenous sporulation) is used.
The more minute examination of these processes at last led to the discovery of
sexual dimorphism, of copulation and of alternation of generations in the coccidia.
Schuberg was the first to consider the possibility of copulation in coccidia; in
addition to the formations which now are termed merozoites, he observed very
diminutive bodies (“microsporozoites”) in the coccidia of the intestine of the mouse,
which were able eventually to copulate. Labbé confirmed this observation in some
of the species, and Simond expressed the opinion that bodies termed “chromatozoites”
occurred in all coccidia. Copulation itself was then observed by Schaudinn
and Siedlecki (1897). The copulating bodies were termed gametes. As, however, they
differed considerably one from the other, the males were called microgametes (i.e.,
the microsporozoites of Labbé and the chromatozoites of Simond) and the females
macrogametes. After copulation was completed sporogony took place, and in the
cycle of development of one species this regularly alternated with schizogony (asexual
multiplication). Schaudinn in 1900 described in detail the life-cycle of Eimeria
(Coccidium) schubergi.
The recognition of this unsuspected complicated process was bound to effect
reforms in the classification of the coccidia; and all the forms that had been
regarded as developmental stages (Eimeria, etc.) had to be reconsidered.
Occurrence.—The Coccidiidea in their mature condition usually
live within the epithelial cells of various organs, and by choice inhabit
those of the intestine and of its associated organs. They also occur
frequently in the excretory organs of mammals, birds, amphibia,
molluscs, arthropods, and may also be found in the testes and vas
deferens, but the statement that they live in hen’s eggs, as well as
in the oviducts of fowls, has not been confirmed.171 Some species
inhabit the nuclei of cells, others live in the connective tissue, but
their presence in the latter situation is probably only secondary,
that is, they have only reached it from the epithelium of the affected
organs.
The size of the Coccidiidea, corresponding as a rule to the capacity
of their habitat, is usually small, but there are said to be species
that attain a diameter of 1 mm. Their form172 is globular, oval, or
elliptical. External appendages are lacking, at least during the trophic
or vegetative period of their life, which is spent in epithelial cells,
within which they increase in size. Frequently one only is present
in each cell, but more can occur. The body substance is composed
of a more or less finely granular or distinctly alveolar protoplasm
which exhibits no differentiation into ecto- and endoplasm. All
species possess a nucleus that enlarges with their growth; sometimes
it only shows through the cytoplasm as a lighter spot, or may
even be quite concealed. It is vesicular, and besides containing very
delicate threads of chromatin in the clear nucleoplasm, it contains
generally only one large karyosome.
The infected epithelial cells degenerate sooner or later as the
parasite feeds on them (fig. 67, II-IV). After their form has been
changed by the action of the growing parasite, they finally perish.
The cell membrane then alone surrounds the coccidia, which, at
least in the species sufficiently known, begin to propagate by an
asexual process (schizogony), the parasites themselves becoming
schizonts, as the initial stage is usually called. They differ from later
stages (sporonts or gametocytes), which resemble them in form, by
the absence of granulations in the cytoplasm, as well as by the
vesicular nucleus. The form is not always the same, for in some
cases, at least, many schizonts assume a globular form.
Schizogony (fig. 67, V-VII) commences with a division of the
nucleus, which takes place in different ways in the various species.
This finally leads to the formation of numerous daughter nuclei
which become smaller and smaller, and which collect beneath
the surface of the schizonts. In some species the daughter nuclei
collect only in one half of the schizont. A part of the protoplasm
of the periphery now divides around each daughter nucleus, the
remaining part (residual body) being left in the centre or on one
side. The segments of the divided cytoplasm, each containing
a nucleus, assume a fusiform shape and become merozoites, which
very soon become free (fig. 67, VIII) and leave the residual body.
They are distinguishable from the very similar sporozoites (fig. 67, I),
as the merozoites possess a karyosome.
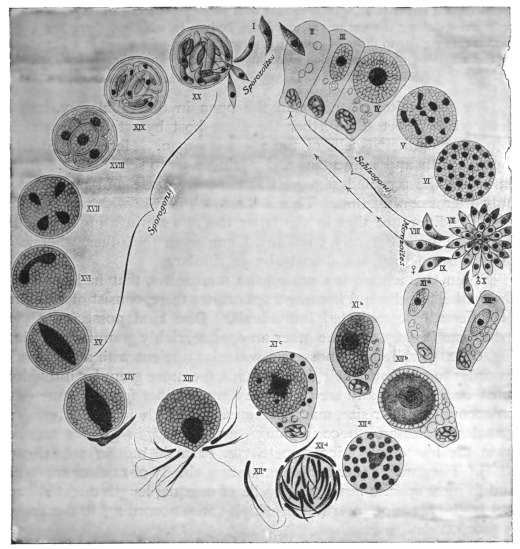
Fig. 67.—Life-cycle of Eimeria (Coccidium) schubergi, Schaud., from the intestine of
Lithobius. (After Schaudinn.) The infection is caused by a cyst (XX), containing spores,
which reaches the intestine of a Lithobius, where it discharges the sporozoites (I). II, A
sporozoite invading an intestinal epithelial cell; III, intestinal epithelial cell with young
trophozoite; IV, intestinal epithelial cell with a globular schizont; V, nuclear segmentation
within the schizont; VI, the daughter nuclei arranging themselves superficially; VII, formation
of the merozoites; VIII, merozoites that have become free, and which, penetrating into
other epithelial cells of the same intestine, repeat the schizogony (II-VIII); IX and X,
merozoites which, likewise invading the epithelial cells of the same intestine, become
sexually differentiated; xia, young macrogametocyte; XIb, older macrogametocyte; XIc,
mature macrogametocyte (discharging particles of chromatin); XIIa, young microgametocyte;
XIIb, older microgametocyte; XIIc, increase of nuclei in the microgametocyte; XIId, the
globular residual body around which numerous microgametes have formed; XIIe, an isolated
microgamete; XIII, the mature macrogamete surrounded by numerous microgametes and
forming a cone of reception or fertilization prominence; XIV, shows the nucleus of a
microgamete that has penetrated and fused with the nucleus of the macrogamete (fertilization)—the
latter forms a membrane and becomes an oöcyst; XV, XVI, XVII, nuclear segmentation
in the oöcyst; XVIII, oöcyst with four sporoblasts; XIX, the sporoblasts transformed into
spores, each containing two sporozoites; XX, the cyst introduced into the intestine and
liberating the sporozoites by bursting.
The merozoites move in a manner similar to that of the sporozoites. The movements
consist either of slow incurvations with subsequent straightenings, or annular
contractions along the entire extent of the body. In addition, there are gliding
movements similar to those of many gregarines, and brought about in a like manner
by the secretion at the posterior extremity of a gelatinous substance that hardens
rapidly.
The merozoites do not gain the open in the usual way, but are
destined to infect still further the same host by actively penetrating
into other epithelial cells of the affected organ. Here they continue
their growth and may again and again undergo schizogony. In the
Infusoria the repeated segmentations finally cease and are renewed
only after a conjugation. This is likewise the case with the Coccidia,
with the difference that in the latter the two conjugating individuals
(gametes) are differently constituted one from the other, whereas in
the Infusoria they are almost always similar.
When the schizogony ceases, the merozoites, that had penetrated
the epithelial cells and become trophozoites there, consist of two kinds
of differently constituted individuals. One kind possesses a clear
cytoplasm (fig. 67, XII), the other an opaque, richly granular cytoplasm
(fig. 67, XI), while both possess a vesicular nucleus with a karyosome.
In order to continue their development, the more granular individuals
must be fertilized, and are therefore termed either female gametes or,
on account of their size, macrogametes. The male individuals (microgametes)
necessary to conjugation, are formed in greater numbers
from the less dense microgametocytes or male mother-cells (fig. 67,
XIId). They are slender bodies consisting chiefly of nuclear substance,
and in most species bear two flagella of unequal length directed backwards,
the place of insertion of which varies according to the species
(fig. 67, XIIe).
While the development of the microgametes is rapidly advancing
a change occurs in the nucleus of the female parent forms or macrogametocytes.
Parts of the karyosome are extruded (fig. 67, XIc), and
the nucleus loses at the same time its vesicular form. One macrogamete
results, after nuclear maturation, from one macrogametocyte.
By this time the macrogametes are capable of conjugation, and
the process takes place within the host, generally, however, outside
the affected and degenerated host cells. The microgametes that
have now become free from the very large residual body, crowd
around the mature macrogametes, which often send out a small
prominence (“cone of reception” or fertilization protuberance) for
their reception (fig. 67, XIII). As soon as a microgamete comes in
contact with this and penetrates into the cytoplasm of the macrogamete,
the latter surrounds itself with a membrane which prevents
the intrusion of other microgametes. The nucleus of the microgamete
that has gained entry unites with the nucleus of the macrogamete,
which latter is afterwards capable of forming the well-known
spores. The parasite is now called an encysted zygote or oöcyst. The
oöcysts of some other members of the Coccidiidea, e.g., Eimeria avium,
can form their walls prior to fertilization. In such cases, a weak spot
is left at one place in the cyst wall, forming a micropyle, that permits of
the entry of the male, immediately after which the micropyle is closed.
The reduced nucleus of the macrogamete elongates on the entry of the microgamete,
and becomes a fertilization spindle to which the male pronucleus (from the
microgamete) becomes attached (fig. 67, XIV and XV). Thereupon the spindle
divides into two daughter nuclei (fig. 67, XVI) which assume a round shape. The
protoplasm at this stage may at once divide, or another segmentation of the daughter
nuclei may first occur. In the former case the two halves divide again, so that
finally four nucleated segments, the sporoblasts, are formed, whereas in the latter
case the four sporoblasts appear simultaneously (fig. 67, XVII). In both cases a
residual body of varying size is separated from the protoplasm of the oöcyst. As
a rule the oöcysts have already been discharged from the body of the host, and in
the manner described above, form the sporoblasts after a longer or shorter period
of incubation.
The sporoblasts are originally naked, but each soon secretes a
homogeneous membrane, the sporocyst, in which it becomes enveloped
(fig. 67, XVIII). After the segmentation of the nucleus the contents
divide into two sickle-shaped sporozoites, in addition to which there
is generally also a residual body (fig. 67, XIX).
This terminates the development. The spores are intended for
the infection of other hosts. If they reach the intestine of suitable
hosts, either free or enclosed in the oöcyst wall, the action of the
intestinal juices causes them to open and permits the escape of the
sporozoites (fig. 67, XX). The latter move exactly like the merozoites
and soon make their way into epithelial cells (fig. 67, I), where they
become schizonts, and thus repeat the life cycle.
Although our knowledge of the development of the coccidia is but of recent date,
yet it already extends to a large number of species, which exhibit various deviations
from the cycle of development described above. For instance, in addition to
differences in the gametocytes, the schizonts of Adelea and Cyclospora also show
differentiation and give rise to macromerozoites and micromerozoites, whilst in
Adelea and Klossia a precocious association of the gametocytes precedes the true
copulation of the ripe gametes.
The classification of the Coccidiidea is based chiefly on the
number of sporozoites found in each spore, and the number of
sporocysts (spores) found in one oöcyst. Léger173 recognises two
great legions, the Eimeridea and the Adeleidea, the former comprising
the greater number of genera, including the genus of most economic
importance, Eimeria. It must be noted that, though a member of
this genus may be frequently referred to as Coccidium, strictly it
should be termed Eimeria, that name having priority. The name
of the disease resulting from the action of such parasites is, however,
established and remains as coccidiosis.
Certain of the more important of the Coccidiidea may now be
considered.
Genus. Eimeria, Aimé Schneider, 1875.
Syn.: Psorospermium, Rivolta, 1878; Cytospermium, Rivolta, 1878; Coccidium,
R. Leuckart, 1879; Pfeifferia, Labbé, 1894; Pfeifferella, Labbé, 1899.
The Eimeria belong to Léger’s old family, the Tetrasporocystidæ,
which comprises forms producing oöcysts with four sporocysts, each
containing two sporozoites. The cysts are spherical or oval, as are
also usually the schizonts. The members of the genus are confined
chiefly to vertebrate hosts, the more important economically occurring
in mammals and birds. From the mammalian hosts very rarely the
parasites may reach man. Eimeria (Coccidium) avium of wild birds
and poultry, and Eimeria stiedæ parasitic in rabbits, may be considered.
There is a general similarity in their life-cycles and each is
of great practical importance.
Eimeria avium, Silvestrini and Rivolta.
Eimeria avium is responsible for fatal epizoötics among game birds such as
grouse, pheasants and partridges, and domestic poultry such as fowls, ducks, pigeons
and turkeys, and can pass from any one of these hosts to any of the others with the
same effect. The organism is parasitic in the alimentary tract of the host, affecting
more especially the small intestine (duodenum) and the cæca, but in some cases
penetrating to the liver and multiplying there (as in turkeys), producing necrotic
cheesy patches, that ultimately become full of oöcysts. The gut is rendered very
frail by the action of the parasites, its mucous membrane is greatly injured, and
is often reduced to an almost structureless pulp, riddled with parasites (fig. 68).
Infection is conveyed from host to host by the ingestion of food or drink contaminated
with the oöcysts voided in the fæces of infected birds. Oval oöcysts from 24 µ to
35 µ long and from 14 µ to 20 µ broad are the means of infection. The oöcysts
develop internally four sporocysts or spores, from each of which two sporozoites are
produced. The life-history174 presents two phases: (1) The asexual multiplicative
phase, schizogony, for the increase in numbers of the parasites within the same host;
(2) the reproductive phase, following the formation of gametes (gametogony), leading
to the production of resistant oöcysts, destined for the transference of the parasite
to new hosts (sporogony).
The oöcysts usually reach the duodenum unharmed, with food or drink. Under
the influence of the powerful digestive juices (especially the pancreatic) now encountered,
the oöcysts soften, as do the sporocysts, and ultimately two sporozoites
emerge from each sporocyst. The sporozoites are from 7 µ to 10 µ long, and each is
vermicular with a uniform nucleus (fig. 69, A). After a short period of active movement
in the gut, each sporozoite penetrates an epithelial cell (figs. 68 spz, 69, B), and
once within, gradually becomes rounded (fig. 69, B, C). It grows rapidly, feeding on
the contents of the host cell and living as a trophozoite (fig. 69, D). When the parasite
is from 10 µ to 12 µ in diameter, usually multiplication by schizogony (fig. 69, E-H)
begins. The nucleus of the parent cell, now called a schizont, divides into a number
of portions that become arranged at the periphery (fig. 69, E). Cytoplasm collects
around each nucleus (fig. 69, E, F) and gradually a group of daughter individuals
(merozoites) is produced (fig. 69, G), the nucleus of each merozoite showing a
karyosome.
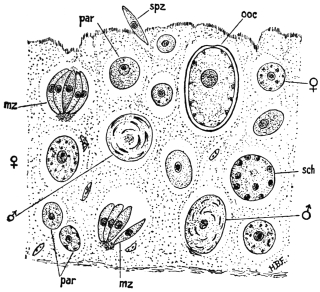
Fig. 68.—Small piece of epithelial lining of gut of heavily infected Grouse chick, showing
various stages in life history of the parasite Eimeria avium; par, parasite (trophozoite);
mz, merozoite; sch, schizont; spz, sporozoite; ooc, oöcyst; ♂, male gametocyte; ♀, female
gametocyte. × 750. (After Fantham.)
The merozoites of Eimeria avium are arranged “en barillet,” like the segments of
an orange (figs. 68 mz, 69, G), therein differing from those of E. schubergi, which are
arranged “en rosace.” They separate from one another (fig. 69, H), penetrate other
epithelial cells, where they may, in turn, become schizonts. Eight to fourteen merozoites
are usually formed by each schizont, twenty have been found, while in cases
of intense infection when space has become limited, the number may be only four.
After a number of generations of merozoites have been formed, a limit is reached
both to the multiplicative capacity of the parasite and to the power of the bird
to provide the invader with food. Consequently, resistant forms of the parasite
are necessary, and the trophozoites begin to show sexual differentiation instead
of forming schizonts, that is, gametogony commences.
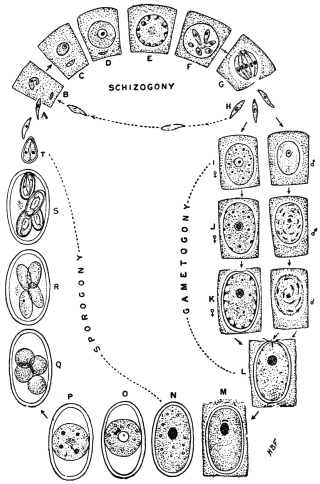
Fig. 69.—Eimeria avium. Diagram of life-cycle. For explanation see text.
(After Fantham.)
Certain trophozoites store food and become large and granular. These are
macrogametocytes (fig. 69, I, ♀). The microgametocytes (fig. 69, I, ♂) are
smaller and far less granular. The macrogametocyte continues to grow, and
becomes loaded with chromatoid and plastinoid granules (fig. 69, J, ♀), while
the microgametocyte has its nucleus divide to form a number of bent, rod-like
portions (fig. 69, J, ♂). The macrogametocyte gives rise to a single macrogamete,
which forms a cyst wall for itself, leaving a thin spot (micropyle) for the entry
of the male (fig 69, K, ♀). The microgametocyte gives rise to numerous small,
biflagellate microgametes (fig. 69, K, ♂) around a large, central residual mass,
from which they ultimately break free, and swim away. When a macrogamete
is reached, the microgamete enters through the micropyle (fig. 69, L)—which
then closes, thus excluding the other males—and applies itself to the female
nucleus (fig. 69, M). Nuclear fusion occurs, the oöcyst (encysted zygote) being thus
produced. Sporogony then ensues. The oöcyst (fig. 69, N) at first has its contents
completely filling it. They then concentrate into a central spherical mass (fig. 69, O)
which gradually becomes tetranucleate (fig. 69, P). Cytoplasm collects around each
nucleus, and four sporoblasts are thus formed (fig. 69, Q). Each sporoblast becomes
oval (fig. 69, R) and produces a sporocyst. Ultimately two sporozoites are formed in
each sporocyst or spore, at first lying tête-bêche (fig. 69, S), but finally twisting
to assume the position most convenient for emergence (fig. 69, T) when they reach a
new host. The period of the life-cycle of Eimeria avium (as well as the details
of the life-cycle) was determined by Fantham to be from eight to ten days, of which
period schizogony occupies four to five days.
The method of infection175 is contaminative, by way of food or drink. Young
birds are especially susceptible to infection. Certain birds, particularly older ones,
may act as reservoirs of oöcysts, being continuously infected themselves, without
showing any marked ill effects from the parasite, but being highly infectious
to other birds. Much moisture retards the development of sporocysts considerably.
The duration of vitality of the infective oöcysts has been determined
experimentally to extend well over two years, and in certain cases longer.
Eimeria avium is the causal agent of “white diarrhœa” or “white scour” in
fowls, and of “blackhead” in turkeys.
Eimeria avium of birds and E. stiedæ of rabbits closely resemble
one another, but are not the same parasite, for E. avium is not infective
to rabbits, nor E. stiedæ to poultry.
Eimeria stiedæ, Lindemann, 1865.
Syn.: Monocystis stiedæ, Lindemann, 1865; Psorospermium cuniculi, Rivolta,
1878; Cytospermium hominis, Rivolta, 1878; Coccidium oviforme, Leuckart, 1879;
Coccidium perforans, Leuckart, 1879; Coccidium cuniculi.
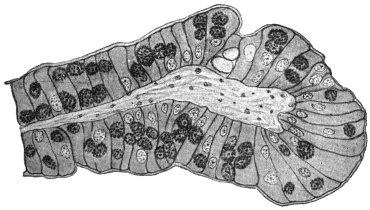
Fig. 70.—Eimeria stiedæ. Section through an infected villus of rabbit’s intestine. × 260.
Eimeria stiedæ is parasitic in the gut epithelium (fig. 70), liver, and
epithelium of the bile ducts of rabbits, and is usually considered to
be the parasite very occasionally found in man. The life-cycle
resembles that of Eimeria avium in its general outlines (see fig. 69)
and therefore will not be detailed in full here. The oöcysts (fig. 71)
are large, elongate-oval, greenish in fresh preparations and vary in
size from 24 µ to 49 µ long and 12·8 µ to 28 µ broad, the gut forms
being usually smaller than those occurring in the liver, owing to the
more confined space in which they are formed. Formerly, the parasites
in the liver were described under the name of Coccidium oviforme,
while those from the intestine were termed Coccidium perforans. This
distinction has now broken down.
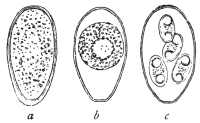
Fig. 71.—Eimeria stiedæ, from
the liver of the rabbit, oöcysts
in various stages of development.
(After Leuckart.)
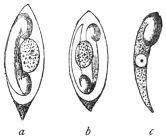
Fig. 72.—a, b, spores of
Eimeria stiedæ (Riv.), with
two sporozoites and residual
bodies; c represents a free
sporozoite. (After Balbiani.)
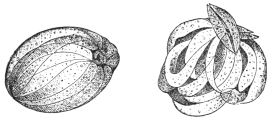
Fig. 73.—So-called swarm cysts (endogenous
sporulation or schizogony) of the Coccidium of
the rabbit. The daughter forms are called
merozoites. (After R. Pfeiffer.)
The oöcysts176 are thick-walled, somewhat flattened at one pole,
where a large micropyle is present. Four egg-shaped spores (sporocysts)
are formed within, each about 12 µ to 15 µ long and 7 µ broad
(fig. 72). The oöcysts are voided with the fæces. Sporogony takes,
in nature, about three days in the excrement. Fæcal contamination
of the food of rabbits results, and coccidian oöcysts are swallowed.
Under the influence of the pancreatic juice of a new host, the
sporozoites (fig. 72, a—c) are liberated from the spores and proceed
to attack the epithelium and multiply within it, as in the case of
Eimeria avium. From the gut,
infection spreads to the liver,
where multiplication of the parasite
goes on actively, resulting
in the formation of the whitish
coccidial nodules, which may be
very conspicuous (fig. 74). Proliferation
of the connective tissue
may occur around the coccidial
nodules, which then contain large
numbers of oöcysts in various
stages of development. It is said that the oöcysts in the older nodules
do not seem to be capable of further development. Schizogony (fig. 73)
and gametogony in all stages can be found in both liver and gut.
Young rabbits often die of intestinal coccidiosis before infection
of the liver occurs. The repeated schizogony of Eimeria stiedæ in the
gut is sufficient to cause death.
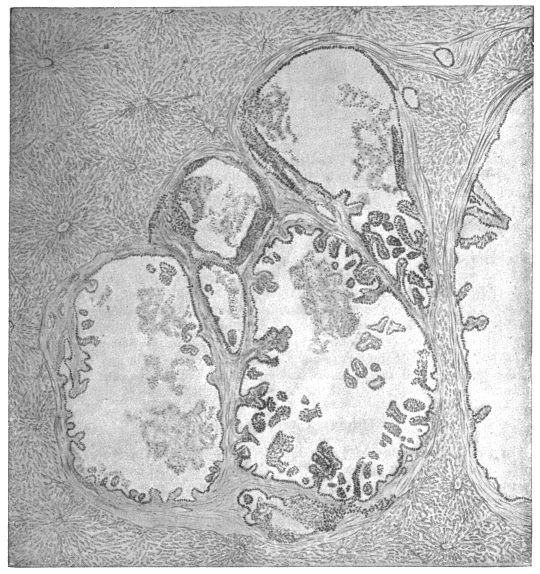
Fig. 74.—Eimeria stiedæ. Section through coccidian nodule in infected rabbit’s liver. × 55.
The disease of cattle popularly known as “red dysentery” is also ascribed to the
action of Eimeria stiedæ. The fæces of infected cattle show blood clots of various
sizes and in severe cases watery diarrhœa is present. Acute cases end fatally in
about two days. Numerous oöcysts, considered to be those of Eimeria stiedæ, occur
in the fæces, and there is a heavy infection of the gut, especially the large intestine
and rectum, all stages of the parasite being found in the epithelium. It is
suspected that cattle contract the disease by feeding on fresh grass contaminated
with oöcysts. The disease is recorded from Switzerland and from East Africa.
As before mentioned, Eimeria stiedæ is considered to be the
organism found in a few cases in man, possibly acquired by eating
the insufficiently cooked livers of diseased rabbits. These cases may
now be described.
(a) Human Hepatic Coccidiosis.
(1) Gubler’s Case. A stone-breaker, aged 45, was admitted to a Paris hospital
suffering from digestive disturbances and severe anæmia. On examination the liver
was found to be enlarged and presented a prominent swelling, which was regarded as
being due to Echinococcus. At the autopsy of the man, who succumbed to intercurrent
peritonitis, twenty cysts were found averaging 2 to 3 cm. in diameter, and one
measuring 12 to 15 cm. The caseous contents consisted of detritus, pus corpuscles,
and oval-shelled formations, which were considered to be Distoma eggs, but which,
in accordance with Leuckart’s conjecture, proved to be Coccidia.177
(2) Dressler’s Case (Prague). Relates to three cysts, varying from the size of
a hemp-seed to that of a pea, and containing Coccidia, found in a man’s liver.178
(3) Sattler’s Case (Vienna). Coccidia were in this case observed in the dilated
biliary duct of a human liver.179
(4) Perls’ Case (Giessen). Perls discovered Coccidia in an old preparation of
Sömmering’s agglomerations.180
(5) Silcock’s Case (London).181 The patient, aged 50, who had fallen ill with
serious symptoms, exhibited fever, enlarged liver and spleen, and had a dry, coated
tongue. At the autopsy numerous caseous centres, mostly immediately beneath the
surface, were found, while the contiguous parts of the liver were inflamed. Microscopical
examination demonstrated numerous Coccidia in the hepatic cells as well
as in the epithelium of the biliary ducts. A deposit of Coccidia was likewise found
in the spleen, which the parasites had probably reached by means of the blood-stream.182
(b) Human Intestinal Coccidiosis.
In two cadavers at the Pathological Institute in Berlin, Eimer183 found the
epithelium of the intestine permeated by Coccidia. Railliet and Lucet’s case may
be traced back to intestinal Coccidia, which were found in the fæces of a woman
and her child, who had both suffered for some time from chronic diarrhœa.184 In
other cases (Grassi, Rivolta), where only the existence of Coccidia in the fæces was
known, it is doubtful whether the parasites originated in the intestine or in the liver.
(c) Doubtful Cases.
To these belong Virchow’s case185 where, in the liver of an elderly woman, a thick
walled tumour measuring 9 to 11 mm. was found. Among the contents of this
tumour there were oval formations 56 µ long, surrounded by two membranes and
enclosing a number of round substances. Virchow considered these foreign bodies
to be eggs of pentastomes in various stages of development, others consider them
to be Coccidia.
The Coccidia which Podwyssotzki claims to have seen in the liver of a man, not
only in the liver cells, but also in the nuclei, are also problematic.186 The parasite
was called Caryophagus hominis.
Again, other explanations can be given to an observation by Thomas, on the
occurrence of Coccidium oviforme in a cerebral tumour of a woman aged 40. The
growth was as large as a pea and surrounded by a bony substance.187
Genus. Isospora, Aimé Schneider, 1881.
Syn.: Diplospora, Labbé, 1893.
Belonging to the section Disporea, that is, forming only two spores,
each with four sporozoites.
Isospora bigemina, Stiles, 1891.
Syn.: “Cytospermium villorum intestinalium canis et felis,” Rivolta, 1874;
“Coccidium Rivolta,” Grassi, 1882; Coccidium bigeminum, Stiles, 1891.
This parasite lives in the intestinal villi of dogs, cats, and the
polecat (Mustela putorius, L.). According to Stiles,188 the oöcyst
divides into two equal ellipsoidal portions or sporoblasts which
become spores and then each forms four sporozoites. The oöcysts
of this species vary from 22 µ to 40 µ in length and from 19 µ to
28 µ in breadth. Each spore is 10 µ to 18 µ long and contains
four sporozoites. The parasites live and multiply, not only in the
gut epithelium, but also in the connective tissue of the intestinal
submucosa. Wasielewski has seen merozoites in the gut of the cat.
Isospora bigemina (fig. 75) appears to occur also in man, for
Virchow published a case which was communicated to him by
Kjellberg, and attributed the illness to this parasite.189 Possibly also it
would be more correct to ascribe the observation of Railliet and
Lucet, which is mentioned under “Human Intestinal Coccidiosis,”
p. 148, to this species, as the Coccidia in that case were distinguished
by their diminutive size (length 15 µ, breadth 10 µ). The case communicated
by Grunow may also possibly refer to Isospora bigemina.190
Roundish or oval structures of 6 µ to 13 µ in diameter occurred in the
mucous membrane of the gut and in the fæces of a case of enteritis.
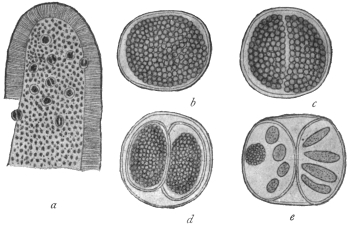
Fig. 75.—Isospora bigemina, Stiles (from the intestine of a dog). a, Piece of an intestinal
villus beset with Isospora, slightly enlarged; b, Isospora bigemina (15 µ in diameter), shortly
before division; c, divided; d, each portion encysted forming two spores; e, four sporozoites
in each part, on the left seen in optical section, together with a residual body—highly magnified.
(After Stiles.)
Doubtful Species.
In literature many other statements are found as to the occurrence of
Coccidia-like organisms in different diseases of man. In some of the cases the
parasites proved to be fungi. This was the case with the parasites of a severe
skin disease of man, formerly called Coccidioides immitis and Coccidioides pyogenes.
Other statements are founded on misapprehensions, or are still much disputed.
If reference is here made to “Eimeria hominis,” R. Blanchard, 1895, this
is done on the authority of the investigator mentioned. The structures in
question are nucleated spindle-shaped bodies of very different lengths (18 µ
to 100 µ), which either occurred isolated or were enclosed in large globular or
oval cysts, alone or with a larger tuberculated body (“residual body”). These
formations were found by J. Künstler and A. Pitres in the pleural exudation removed
from a man by tapping. The man was employed on the ships plying between
Bordeaux and the Senegal River.
Blanchard looks upon the fusiform bodies as merozoites and the cysts as
schizonts of a Coccidium. On the other hand, Moniez declares the spindle bodies
to be the ova and the supposed residual bodies to be “floating ovaries” of an
Echinorhynchus.
Severi’s “monocystid Gregarines,” which were taken from the lung tissue of a
still-born child, are also quite problematical.
No less doubtful are the bodies which Perroncito calls Coccidium jalinum, and
which he found in severe diseases of the intestine in human beings, pigs, and guinea-pigs;
Borini also reported another case.
Order. Hæmosporidia, Danilewsky emend. Schaudinn.
The Hæmosporidia are a group of blood parasites, comprising
forms differing greatly among themselves. Some of the forms need
much further investigation. However, there are certain true Hæmosporidia
which present close affinities with the Coccidia, leading
Doflein to use the term Coccidiomorpha for the two orders conjoined.
The Hæmosporidia present the following general characteristics:—
(1) They are parasites of either red or white blood corpuscles of
vertebrates during one period of their life-history.
(2) They exhibit alternation of generations—asexual phases or
schizogony alternating with sexual phases or sporogony—as do the
Coccidia.
(3) There is also an alternation of hosts in those cases which have
so far been completely investigated. The schizogony occurs in the
blood or internal organs of some vertebrates while the sporogony
occurs in an invertebrate, such as a blood-sucking arthropod or leech.
(4) Unlike the Coccidia, resistant spores in sporocysts are not
generally produced, such protective phases in the life-cycle being
unnecessary, as the Hæmosporidia are contained within either the
vertebrate or invertebrate host during the whole of their life.
The Hæmosporidia may be considered for convenience under
five main types:—
(1) The Plasmodium or Hæmamœba type. This includes the
malarial parasites of man and of birds. The asexual multiplicative
or schizogonic phases occur inside red blood corpuscles and are
amœboid. They produce distinctive, darkish pigment termed melanin
or hæmozoin. Infected blood drawn and cooled on a slide may
exhibit “exflagellation” of the male gametocytes, i.e., the formation
of filamentous male gametes. The invertebrate host is a mosquito.
The malarial parasites of man are discussed at length on p. 155.
Similar pigmented hæmamœboid parasites have been described in
antelopes, dogs, and other mammals, and even reptiles.
(2) The Halteridium type. The trophozoite stage inside the red
blood corpuscle is halter-shaped. Pigment is produced, especially near
the ends of the organism. The parasites occur in the blood of birds.
The invertebrate host of H. columbæ of pigeons in Europe, Africa,
Brazil and India, is a hippoboscid fly, belonging to the genus Lynchia.
Halteridium parasites are common in the blood of passerine birds,
such as pigeons, finches, stone owls, Java sparrows, parrots, etc.
The Halteridium embraces or grows around the nucleus of the host
red cell without displacing the nucleus. Young forms and multiplicative
stages of H. columbæ have been found in leucocytes in the
lungs of the pigeon (fig. 76, 8-12). Male and female forms (gametocytes)
are seen in the blood (fig. 76, 3a, 3b). The cytoplasm of the
male gametocytes is pale-staining and the nucleus is elongate, while
the cytoplasm of the females is darker and the nucleus is smaller
and round. Formation of male gametes from male gametocytes
(the so-called process of “exflagellation”) may occur on a slide
of drawn infected blood, also fertilization, and formation of the
oökinete, as first seen by MacCallum. The correct generic name for
Halteridia is, apparently, Hæmoproteus. Wasielewski (1913), working
on H. danilewskyi (var. falconis), in kestrels, finds that the halteridium
may be pathogenic to nestlings. The cycle of H. noctuæ
described by Schaudinn (1904) lacks confirmation. The account of
the life-cycle of H. columbæ given by Aragão (1908) is illustrated in
fig. 76. It agrees with the work of Sergent (1906–7) and Gonder
(1915). Mrs. Adie (1915) states that the cycle in Lynchia is like that
of a Plasmodium.
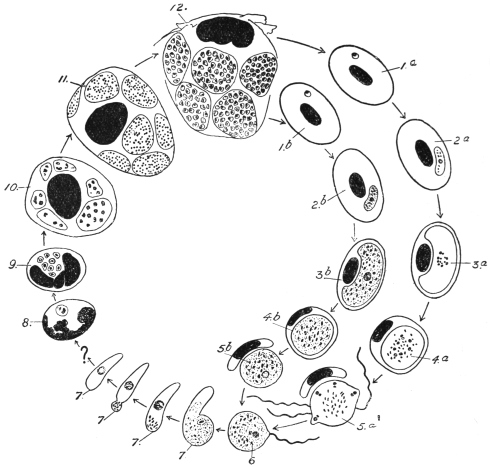
Fig. 76.—Hæmoproteus (Halteridium) columbæ. Life-cycle diagram: 1, 2, stages in red
blood corpuscle of bird; 3, 4, gametocytes (3a ♂, 3b ♀); 5a, formation of microgametes;
6, fertilization (in fly’s gut); 7, oökinete; 8–12, stages in mononuclear leucocytes in lungs.
(After Aragão.)
(3) The Leucocytozoön type. The trophozoites and gametocytes
occur within mononuclear leucocytes and young red cells (erythroblasts)
in the blood of birds. Laveran and França consider that
the Leucocytozoa occur in erythrocytes. The host cells are often
greatly altered by the parasites, becoming hypertrophied and the ends
usually drawn into horn-like processes (fig. 77), though some remain
rounded. Leucocytozoa are limited to birds, and very rarely produce
pigment. Male and female forms (gametocytes) are distinguishable
in the blood (fig. 77), and the formation of male gametes (“exflagellation”)
may occur in drawn blood.
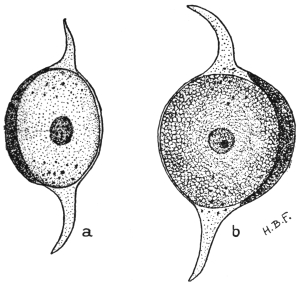
Fig. 77.—Leucocytozoön lovati. a, Male parasite
(microgametocyte), within host cell, whose ends are
drawn out; b, female parasite (macrogametocyte)
from blood of grouse. × 1,800. (After Fantham.)
The Leucocytozoa were first seen by Danilewsky in 1884. They
are usually oval or spherical. It is not easy sometimes to distinguish
the altered host cell from the parasite, as the nucleus of the former
is pushed to one side by the
leucocytozoön. The cytoplasm
of the female parasite
stains deeply, and the nucleus
is rather small, containing a
karyosome. In the male the
cytoplasm stains lightly and
the nucleus is larger, with a
loose, granular structure.
Many species of Leucocytozoa
are recorded, but
schizogony has only been
described by Fantham (1910)191
in L. lovati in the spleen of
the grouse (Lagopus scoticus),
and by Moldovan192 (1913) in
L. ziemanni in the internal
organs of screech-owls.
M. and A. Leger193 (1914)
propose to classify Leucocytozoa,
provisionally, according as the host cells are fusiform or
rounded.
(4) The Hæmogregarina type. Included herein are many parasites
of red blood corpuscles, with a few (the leucocytogregarines) parasitic
in the white cells of certain mammals and a few birds. They are not
amœboid but gregarine-like, vermicular or sausage-shaped (fig. 78).
They do not produce pigment. They are widely distributed among
the vertebrata, but are most numerous in cold-blooded vertebrates
(fishes, amphibia and reptiles). The hæmogregarines of aquatic hosts
are transmitted by leeches, those of terrestrial hosts by arthropods.
The nucleus of hæmogregarines is usually near the middle of the
parasite, but may be situated nearer one end. The body of the
parasite may be lodged in a capsule (“cytocyst”). There is much
variation in size and appearance among hæmogregarines. Some are
small (Lankesterella); some attack the nucleus of the host cell (Karyolysus);
others have full grown vermicules larger than the containing
host corpuscle, and so the hæmogregarines bend on themselves in
the form of U (fig. 78, b). Schizogony often occurs in the internal
organs of the host, sometimes in the circulating blood.
The hæmogregarines occurring in the white cells (mononuclears
or polymorphonuclears) of mammals have been referred to a separate
genus, Leucocytogregarina (Porter) or Hepatozoön (Miller). Such
leucocytogregarines are known in the dog (fig. 79), rat, mouse, palm-squirrel,
rabbit, cat, etc. Schizogony of these forms occurs in the
internal organs, such as the liver, lung and bone-marrow of the hosts.
They are apparently transmitted by ectoparasitic arthropods, such as
ticks, mites, and lice.
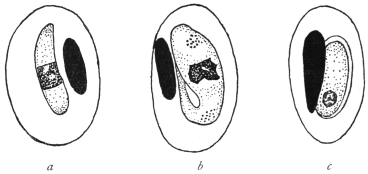
Fig. 78.—Hæmogregarines from lizards, a, H. schaudinni, var. africana, from Lacerta
ocellata; b, H. nobrei from Lacerta muralis; c, H. marceaui in cytocyst, from Lacerta
muralis. (After França.)
A few hæmogregarines are known to be parasitic in the red blood
corpuscles of mammals. Such are H. gerbilli in the Indian field rat,
Gerbillus indicus; H. balfouri (jaculi) in the jerboa, Jaculus jaculus,
and a few species briefly described from marsupials. These parasites
do not form pigment.
Strict leucocytic gregarines have been described from a few birds
by Aragão and by Todd.
The sporogony of hæmogregarines is only known in a few cases,
and in those affinity with the Coccidia is exhibited. In fact, the
Hæmogregarines are now classified by some authors with the
Coccidia.
(5) The Babesia or Piroplasma type. These are small parasites of
red blood corpuscles of mammals. They do not produce pigment.
They are pear-shaped, round or amœboid in Babesia, bacilliform
and oval in other forms referred to this group. Piroplasms are
transmitted by ticks. These parasites are described at length on
p. 172.
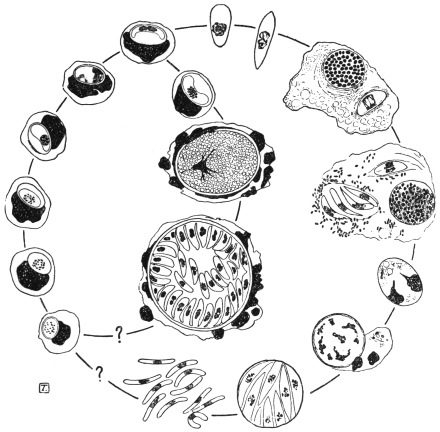
Fig. 79.—Leucocytogregarina canis. Life-cycle diagram. Constructed from drawings by
Christophers. (After Castellani and Chalmers.) Schizogony occurs in the bone-marrow. The
parasite is transmitted from dog to dog by the tick, Rhipicephalus sanguineus, development
in which, so far as known, is shown on the right.
THE MALARIAL PARASITES OF MAN.
Malaria, otherwise known as febris intermittens, chill-fever, ague,
marsh fever, paludism, etc., is the name given to a disease of man,
which begins with fever. It has been known since ancient times and
is distributed over almost all the world, although very unevenly, but
does not occur in waterless deserts and the Polar regions. In many
places, especially in the civilized countries of Central Europe, the
disease is extinct or occurs only sporadically, and large tracts of land
have become free from malaria.
The rhythmical course of the fever is characteristic. It begins
apparently suddenly with chilliness or typical shivering, whilst the
temperature of the body rises, the pulse becomes low and tense and
the number of beats of the pulse increases considerably. After half
to two hours the heat stage begins. The patient himself feels the rise
of his temperature (shown by feeling of heat, dry tongue, headache,
thirst). The temperature may reach 41°C or more. At the same time
there is sensitiveness in the region of the spleen and enlargement of
that organ. After four to six hours an improvement takes place, and
with profuse perspiration the body temperature falls rapidly, not
often below normal. After the attack the patient feels languid, but
otherwise well until certain prodromal symptoms (heaviness in the
body, headache) which were not noticed at first, denote the approach
of another attack of fever, which proceeds in the same way.
The intervals between the attacks are of varying length which
permit of a distinction in the kinds of fever. If the attacks intermit
one day, occurring on the first, third and fifth days of the illness
and always at the same time of day, it is termed febris tertiana; if
two days occur between fever days, it is called febris quartana. In
the case of the fever recurring daily, later writers speak of typical
febris quotidiana. But a quotidian fever may arise when two tertian
fevers differing by about twenty-four hours exist at the same time
(febris tertiana duplex). The patient has a daily attack, but the fever
of the first, third and fifth days differs in some point (hour of
occurrence, height of temperature, duration of cold or hot stage)
from the fever of the second, fourth and sixth days. Similarly,
two or three quartan fevers which differ by about twenty-four hours
each may be observed together (febris quartana duplex or triplex);
in the latter case the result is also a quotidian fever.
Two kinds of tertian fever are differentiated—a milder form
occurring especially in the spring (spring tertian fever), and a more
severe form appearing in the summer and autumn in warmer
districts, especially in the tropics (summer or autumn fever, febris
æstivo-autumnalis, febris tropica, febris perniciosa). The latter often
becomes a quotidian fever.
All the afore-mentioned infections are termed acute. They are
distinguished from the very different chronic malarial infection by the
frequent occurrence of relapses, which finally lead to changes of
some organs and particularly of the blood. The relapses are then
generally marked by an irregular course of fever.
The term masked malaria is used when any disturbance of the state
of health of a periodic character shows itself and disappears after
treatment with quinine.194 Generally it is a question of neuralgia.
That intermittent fever was an infectious disease, although not one which was
transmitted direct from man to man, had been assumed for a long time. Therefore
it was natural, at a time when bacteriology was triumphing, to look for a living agent
causing infection in malaria, which search was, seemingly, successful (Klebs,
Tomasi-Crudeli, 1879). Hence it was not surprising that the discovery of the
real malarial parasites in November, 1880, by the military doctor A. Laveran195 in
Constantine (Algeria), at first met with violent opposition, even after Richard
(1882) had confirmed it and Marchiafava, Celli, Grassi and others, had
further extended it. Not that the existence of structures found in the blood of
malaria patients by Laveran and Richard was denied; on the contrary, the investigations
of the opponents furnished many valuable discoveries, but the organisms were
differently interpreted and considered to be degeneration products of red blood corpuscles.
Only when Marchiafava and Celli (1885) saw movements in the parasites,
which Laveran called Oscillaria malariæ and later Hæmatozoön malariæ, was their
animal nature admitted and the parasites were named Plasmodium malariæ. Shortly
before this, Gerhardt (1884) had stated that the disease could be transmitted by the
injection of the blood of a malarial patient to a healthy person.
This supplied the starting point for further investigations, which were made not
exclusively, but principally, by Italian investigators (Golgi, Marchiafava and Celli,
Bignami and Bastianelli, Grassi and Feletti, Mannaberg, Romanowsky, Osier,
Thayer and others). In 1885 Golgi described the asexual cycle in the blood,
in the case of the quartan parasite. These investigations, after attention had been
drawn by Danilewsky (1890) to the occurrence of similar endoglobular parasites in
birds, were extended to the latter (Grassi and Feletti, Celli and Sanfelice, Kruse,
Labbé and others).
The result was as follows: Malaria in man (and birds) is the result of peculiar
parasites included in the Sporozoa by Metchnikoff, which parasites live in the erythrocytes,
grow in size and finally “sporulate,” that is, separate into a number of “spores”
which leave the erythrocytes and infect other blood corpuscles. Morphologically and
biologically several species (and respectively several varieties) of malarial parasites
may be distinguished, on which the different intermittent forms depend. Transmission
of the blood of patients to healthy people produces a malarial affection which
corresponds in character to the fever of the patient from whom the inoculation was
made. The combined types of fever (tertiana duplex, quartana duplex or triplex)
are explained by the fact that the patient harbours two or three groups of parasites
which differ in their development by about twenty-four hours, whilst the irregular fevers
depend on deviation from the typical course of development of the parasites. In
addition to stages of the parasites which could easily be arranged in a developmental
series concurrent with the course of the disease, other phases of the parasites also
became known, such as spheres, crescents, polymitus forms, which seemed not to be
included in the series and, therefore, were very differently interpreted.
The decision reached at the beginning of the last decade of the last century, which
found expression in comprehensive statements (Mannaberg, Ziemann and others),
only concerned a part of the complete development of the malarial parasites. No one
could with any degree of certainty demonstrate how man became infected, nor were
there reliable hypotheses based on analogy with other parasites concerning the exit
of the excitants of malaria from the infected person and their further behaviour.
Numerous hypotheses had been advanced, but none was able to elucidate the
various observations made from time to time in dealing with malaria. One hypothesis
only seemed to have a better foundation. Manson (1894), who knew from his
own experience the part played by mosquitoes in the development of Filaria from
the blood of man, applied this also to the malarial parasites living in the blood,
whereby at least the way was indicated by which the Hæmosporidia could leave man.
The parasites were said finally to get into water through mosquitoes which had
sucked the blood of malarial patients, and the germ spread thence to men who drank
the water. In some cases the parasites were supposed to reach man by the inhaling of
the dust of dried marshes. On the other hand, Bignami believed that the mosquitoes
were infected in the open air by malarial parasites which occurred there in an unknown
stage and the insects transmitted the germs to man when biting. R. Koch combined
both hypotheses, without, however, producing positive proof. R. Ross, then (1897–8)
an English military doctor in India, was the first to succeed in this. He had been
encouraged by Manson to study the fate of malarial Plasmodia which had entered
the intestine of mosquitoes with malaria-infected blood, especially in the case of the
Plasmodium (Proteosoma) living in the blood of birds. He showed that the Proteosoma
penetrate the intestinal wall of the mosquitoes, grow and develop into large
cysts which produce innumerable rod-like germs, which burst into the body cavity
and penetrate the salivary glands. Ross allowed mosquitoes to suck the blood of
birds affected by malaria, and some nine days later, let the infected mosquitoes
which had been isolated suck healthy birds. After five to nine days Proteosoma
were found to occur in the blood of the birds used. The Proteosoma and Halteridium
of birds were also further investigated by MacCallum (1897–8), Koch and others, and
important results followed.
In any case Ross (1898) had clearly established the importance of mosquitoes in
the spread of malaria among birds. It was now only a question of proving whether,
and how far, mosquitoes were concerned with human malaria. Ross himself worked
to this end. Here the experiments of Italian investigators (Bignami, Bastianelli,
Grassi)196 were of importance. These investigators studied the fate of malarial
parasites in man, produced malaria in men experimentally by the bites of infected
mosquitoes, and established that only mosquitoes belonging to the genus Anopheles
were concerned, and not species of Culex. These latter are only able to transmit
Proteosoma to birds. It is true that Culex can ingest the human malarial parasites,
but the latter do not develop in them. Development only occurs in species of
Anopheles. In Anopheles (and similarly for Proteosoma in Culex) sexual reproduction
takes place; crescents, spheres and polymitus forms are necessary stages
of development in the mosquito.
With these discoveries the campaign against malaria became more definite. It
was directed partly against the transmitters, whose biology and life-cycle were more
thoroughly investigated, instead of merely against the infection of the adult Anopheles.
The latter do not, as was believed for some time, transmit the malarial germs to their
offspring. They always infect themselves from human beings, whereby the relapses
appearing in early summer, and the latent infection, especially of children of natives,
play a principal part (Stephens and Christophers, Koch). Further, the crusade was
directed against the infection of man by the bites of Anopheles. Important results
have been obtained in these directions. Low and Sambon in 1900 lived in a
mosquito-screened hut in a malarial part of the Roman Campagna for three of the
most malarious months and did not contract the disease. In the same year Dr. P. T.
Manson was infected with malaria by infected mosquitoes sent from Italy. The rôle
of mosquitoes having been proved, it may be hoped that ultimately the eradication
of malaria, or at least a considerable restriction of it, will be achieved.
It is of importance to record that, although malarial parasites occur in mammals
(monkeys, bats, etc.) the human ones are not transmissible to mammals, not even to
monkeys. The species, therefore, are specific to the different hosts (Dionisi, Kossel,
Ziemann, Vassall).
An important work dealing with the modern applications of the mosquito-malaria
theory in all parts of the Tropics was published by Sir Ronald Ross in 1911. It is
entitled “The Prevention of Malaria” (John Murray, London, 21s.).
DEVELOPMENT OF THE MALARIAL PARASITES OF MAN.
The commencement of the developmental cycle and of the infection
of man, is the sporozoites (fig. 80, 1) which are passed into the blood
of a person by the bite of an infected mosquito. Prior to this the parasites
collect in the excretory ducts of the salivary glands (fig. 80, 27)
of the Anopheles. The sporozoites are elongate and spindle-shaped,
10 µ to 20 µ long and 1 µ to 2 µ broad, with an oval nucleus situated
in the middle. They are able to glide, perform peristaltic contractions,
or curve laterally. Schaudinn has studied the penetration of the red
blood corpuscles (fig. 80, 2) by the sporozoites in the case of the
living tertian parasite. The process takes forty to sixty minutes in
drawn blood. After its entrance the parasite, which is now called a
trophozoite, contracts, and becomes an active amœbula (fig. 80, 3).
It develops a food vacuole and grows at the expense of the invaded
blood corpuscle (fig. 80, 4), which is shown by the appearance of
pigment granules (transformed hæmoglobin) in it. When the maximum
size is attained, multiplication by schizogony (fig. 80, 5-7) begins
with a division of the nucleus, which is followed by further divisions of
the daughter nuclei, the number of which varies up to 16 or even 32,
depending on the species of the parasite. Then the cytoplasm divides
into as many portions as there are nuclei, the result being a structure
suggestive of the spokes of a wheel or of a daisy, the centre of the
resulting rosette being occupied by dark pigment. Finally, the parts
separate from one another, leaving behind a residual body containing
the pigment, and the daughter forms issue into the blood plasma
as merozoites (fig. 80, 7). They are actively amœboid (fig. 80, 8) and
soon begin to enter other blood corpuscles of their host, for the entry
into which thirty to sixty minutes are necessary, according to
Schaudinn’s observations.197
Here they behave like sporozoites which previously entered and
again produce merozoites. This process is repeated until the number
of parasites is so large that, at the next migration of the merozoites, the
body of the person infected reacts with an attack of fever,198 which is
repeated with the occurrence of the next or following generations.
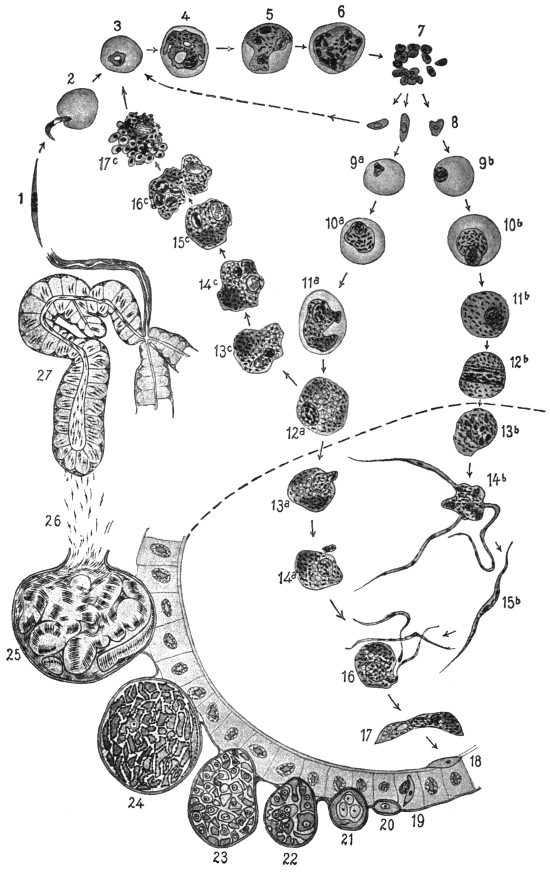
Fig. 80.—Life-cycle of the tertian parasite (Plasmodium vivax). Figs. 1 to 17, × 1,200;
figs. 18 to 27, × 600. (After Lühe, based on figures by Schaudinn and Grassi.) 1, sporozoite;
2, entrance of the sporozoite into a red blood corpuscle; 3, 4, growth of the parasite, now
sometimes called a trophozoite; 5, 6, nuclear division in schizont; 7, free merozoites; 8, the
merozoites which have developed making their way into blood corpuscles, (arrow pointing to the
left) and increase by schizogony (3–7); after some duration of disease the sexual individuals
appear; 9a-12a, macrogametocytes; 9b-12b, microgametocytes, both still in the blood-vessels
of man. If macrogametocytes (12a) do not get into the intestine of Anopheles they may perhaps
increase parthenogenetically according to Schaudinn (12a; 13c-17c). The merozoites which
have arisen (17c) become schizonts 3–7. The phases shown underneath the dotted line
(13–17) proceed in the stomach of Anopheles. 13b and 14b, formation of microgametes; 13a
and 14a, maturation of the macrogametes; 15b, microgamete; 16, fertilization; 17, oökinete;
18, oökinete in the walls of the stomach; 19, penetration of the epithelium of the stomach;
20–25, stages of sporogony on the outer surface of the intestinal wall; 26, migration of the
sporozoites to the salivary gland; 27, salivary gland with sporozoites.
The growth and schizogony last different times, according to the
species of the parasite, about forty-eight hours in the case of the
parasite of febris tertiana or tropica, and seventy-two hours for the
quartan parasite. The various intermittent forms produced by them
depend on this specific difference in the malarial parasites.
The schizogony can, however, only be repeated a certain number
of times, supposing that the disease has not been checked prematurely
by the administration of quinine, which is able to kill the parasites.
It appears that after a number of attacks of fever the conditions of
existence in man are unfavourable for the malarial parasites, and this
brings about the production of other forms which have long been
known, but also long misunderstood (spheres, crescents, polymitus).
The merozoites in this case no longer grow into schizonts, or at least
not all of them, but become sexual individuals called gametocytes
(fig. 80, 9-12), which only start their further development when they
have reached the intestine of Anopheles. This does not take place
in every case, nor with all the gametocytes which exist in the blood of
patients with intermittent fever. Of those parasites which remain
in the human blood the male ones (microgametocytes) soon perish,
the females (macrogametocytes) persist for some long time, and perhaps
at last acquire the capacity of increasing by schizogony. They
might thus form merozoites which behave in the body as if they had
proceeded from ordinary schizonts (fig. 80, 13c-17c). If their
number increases sufficiently, in course of time the patient, who
was apparently recovering, has a new series of fever attacks, or
relapses, without there having been a new infection. This is the
view of Schaudinn, who from researches of his own concluded that
relapses were brought about by a sort of parthenogenetic reproduction
of macrogametocytes. R. Ross, on the contrary, believes that in the
relatively healthy periods the number of parasites in the blood falls
below that necessary to provoke febrile symptoms; relapses then
result merely from increase in the numbers of the parasites present in
the individual. Ross’s view is now generally accepted.

Fig. 81.—Stages of development of pernicious or malignant tertian parasites in the intestine
of Anopheles macultpennis. (After Grassi.) a, macrogametocyte (crescent) still attached to
human blood corpuscles; b, macrogametocyte (sphere) half an hour after ingestion by the
mosquito; c, microgametocyte (crescent) attached to the blood corpuscle; d, microgametocyte
(sphere) half an hour after ingestion; the nucleus has divided several times; e, microgametes
attached to the residual body (polymitus stage).
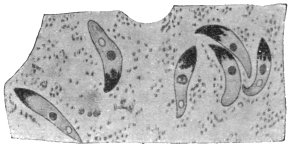
Fig. 82.—Oökinete of the malignant tertian parasite in the stomach of Anopheles maculipennis,
thirty-two hours after ingestion of blood. (After Grassi.)
If the gametocytes, which are globular, or in the pernicious
or malignant tertian parasite crescentic (fig. 81), gain access to the
intestine of an Anopheline,199 they mature. The macrogametocytes
extrude a part of their nuclear substance (fig. 80, 13a, 14a) and
thereby become females or macrogametes. The microgametocytes, on
the other hand, undergo repeated nuclear division, preparation for
this being made apparently whilst in the blood of man. This results
in the formation of threadlike bodies which move like flagella and
finally detach themselves from the residual body (fig. 80, 13b, 14b).
These are the males or microgametes200 (fig. 80, 15b).

Fig. 83.—Section of the
stomach of an Anopheles, with
cysts (oöcysts) of the malignant
tertian parasite. (After Grassi).
Copulation takes place in the stomach of the Anopheline (fig. 80, 16).
A microgamete penetrates a macrogamete and coalesces with it. The
fertilized females elongate very soon and are called oökinetes or
“vermicules” (figs. 80, 17; 82). They penetrate the walls of the
stomach, pierce the epithelium (fig. 80, 18, 19), and remain lying
between it and the superficial stratum
(tunica elastico-muscularis). Then they
become rounded and gradually develop
into cysts which grow larger and are
finally visible to the naked eye, being
called oöcysts (figs. 80, 20-24; 83). Their
size at the beginning is about 5 µ, the
maximum that they attain is 60 µ, only
exceptionally are they larger.
The sporulation (figs. 80, 21-25; 84),
which now follows, begins with repeated
multiple fission of the nucleus. Long
before the definitive number of nuclei,
which varies with the individual, is attained
the protoplasm, according to Grassi, begins to segment around the
individual large nuclei but without separating completely into cell
areas. According to Schaudinn,
however, there is a condensation
of the outstanding protoplasmic
strands. It is certain that the number
of nuclei increases with simultaneous
decrease in size. They
soon appear on the surface of the
strands or sporoblasts, surround
themselves with some cytoplasm
and then elongate (fig. 84). In
this manner the sporozoites are
formed and break away from the
unused remains of the cytoplasmic
strands of the sporoblasts
(fig. 80, 26). The number of the
sporozoites in an oöcyst varies from
several hundreds to ten thousand.
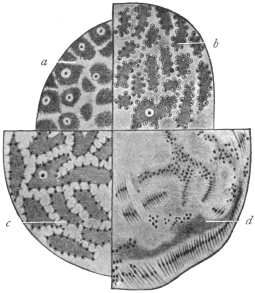
Fig. 84.—Four different sporulation stages
of malarial parasites from Anopheles maculipennis,
much magnified. a-c, of the
malignant tertian parasite; a, four to four
and a half days after sucking; b and c, five
to six days after sucking; d, of the tertian
parasite, eight days after sucking. (After
Grassi.)
The sporulation is influenced in its
duration by the external temperature
(Grassi, Jansci, Schoo). In the tertian
parasite it takes place quickest at a
temperature of 25° to 30° C. and takes
eight to nine days. A temperature a
few degrees lower has a retarding effect (eighteen to nineteen days at 18° to 20° C).
A still lower one has a restraining or even destructive effect. Temperatures over
35° C. also exercise a harmful effect. The malignant tertian parasite seems to need
a somewhat higher temperature and the quartan parasite a lower one.
The sporozoites of the various malarial parasites show no specific
differences. They were stated by Schaudinn to occur in three forms,
and these were described as indifferent (neuter), female and male.
There is, however, little or no evidence for this hypothetical differentiation.
The last were said to perish prematurely, that is, in the
oöcyst. The others after the rupture of the oöcysts enter the body
cavity of the Anophelines, whence they are carried along in the course
of the blood. Finally they penetrate the salivary glands (fig. 80, 27)
probably by their own activity, break through their epithelia and
accumulate in the salivary duct (fig. 80, 27). At the next bite by the
mosquito they are transmitted to the blood-vessels of man.
The Species of Malarial Parasites of Man.
In view of the differences in opinion regarding “species” and “varieties,” the dispute
whether the malarial parasites of man represent one species with several varieties, or several
species is almost superfluous. If necessary two genera may be distinguished.
The parasites of the tertian and quartan fever are alike in that
their gametocytes have a rounded shape (figs. 80, 12, 13), whilst the
corresponding stages of the pernicious or malignant tertian parasites
are crescentic (figs. 81, 88). These differences are used by some writers
as the distinguishing characteristic of two genera: Plasmodium, Marchiafava
and Celli, 1885, for the first mentioned species; Laverania,
Grassi and Feletti, 1889, for the pernicious or malignant tertian parasite.
Whether there is a genuine quotidian fever and accordingly a special
quotidian parasite is still disputed.
These parasites are treated in practical detail in Stephens and Christophers’
“Practical Study of Malaria,” 3rd edition, 1908.
Plasmodium vivax, Grassi and Feletti, 1890.
Syn.: Hæmamœba vivax, Grassi and Feletti, 1890; Plasmodium malariæ var.
tertianæ, Celli and Sanfelice, 1891; Hæmamœba laverani var. tertiana, Labbé,
1894; Hæmosporidium tertianum, Lewkowitz, 1897; Plasmodium malariæ tertianum,
Labbé, 1899: Hæmamœba malariæ var. magna, Laveran, 1900, p.p.; Hæmamœba
malariæ var. tertianæ, Laveran, 1901.
This species, P. vivax,201 is the causal agent of the simple or spring
tertian fever and is, therefore, named directly the tertian or benign
tertian parasite (figs. 80, 3-8; 85). During the afebrile period in the
patient, the young trophozoites or amœbulæ appear on or in the red
blood corpuscles as pale bodies of 1·5 µ to 2 µ diameter which at
first show only slow amœboid movements. Their nucleus is difficult
to recognize in the early stage. Soon the food vacuole is formed and
this grows concomitantly with the trophozoite and the parasite has a
ring-like appearance. Afterwards the vacuole diminishes, and at this
period the first brownish melanin granule appears. From this time
the activity and number of the pigment granules increase with continuous
growth. When the parasite has grown to about one-third
the diameter of the erythrocyte the latter shows characteristic red
Schüffner’s dots or “fine stippling,” after staining with Romanowsky’s
solution. Later, after about twenty-four hours, the blood corpuscles
begin to grow pale, then to increase in size, and after thirty-six hours,
that is, about twelve hours before the next attack of fever, schizogony
of the parasite is initiated by the division of the nucleus. The parasite
at this time occupies half to two-thirds of the enlarged blood corpuscle.
The daughter nuclei continue dividing until sixteen, and
occasionally twenty-four, daughter nuclei are produced. The pigment
which, up till now lies nearer the periphery, moves to the middle,
while the nuclei lie nearer the surface.
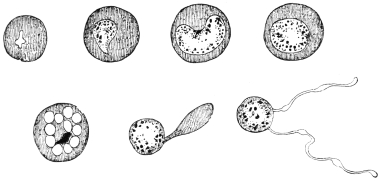
Fig. 85.—Development of the tertian parasite in the red blood corpuscles of man;
on the right a “Polymitus.” (After Mannaberg.) See also fig. 80, 3—7.
Around each nucleus a portion of cytoplasm collects and thus
young merozoites are produced. These separate from each other and
from the little residual masses202 which contain the melanin and pass
from the blood corpuscles, which now can hardly be recognized, to
the blood plasma, where they soon attack new erythrocytes.
The migration of the merozoites initiates a new attack of fever
and two groups of tertian parasites in the blood, differing in development
by about twenty-four hours, are the conditions for febris tertiana
duplex.
After a lengthy duration of fever the gametocytes (figs. 80, 9—12)
appear. They are uninucleate. The microgametocytes are about the
size of fully developed schizonts, the macrogametocytes are somewhat
larger. Their further development takes place in Anophelines.
The chief distinctive characteristics of the simple tertian parasite,
as seen in infected blood, are:—(1) The infected red-cell is usually
enlarged; (2) the presence of fine red granules known as Schüffner’s
dots in the red blood corpuscles, after Romanowsky staining; (3)
the fragile appearance of the parasite compared with other species.
Large forms are pigmented, irregular and “flimsy-looking,” sometimes
appearing to consist of separate parts. Irregularity of contour
is common.
Ahmed Emin203 (1914) has described a small variety of P. vivax.
Plasmodium malariæ, Laveran.
Syn.: Oscillaria malariæ, Laveran, p.p., 1883; Hæmamœba malariæ, Gr. et Fel.,
1890; Plasmodium malariæ var. quartanæ, Celli et Sanfel., 1891; Hæmamœba
laverani var. quartana, Labbé, 1894; Hæmosporidium quartanæ, Lewkowitz, 1897;
Plasmodium malariæ quartanum, Labbé, 1899; Plasmodium golgii, Sambon, 1902;
Laverania malariæ, Jancso, 1905 nec Grassi et Fel. 1890; Hæmomœba malariæ
var. quartanæ; Lav., 1901.
Plasmodium malariæ is the parasite of quartan malaria (fig. 86).
The trophozoites of the quartan parasite differ from the corresponding
stages of the tertian parasite in that their motility is less and soon
ceases. They differ also in their slower growth, by the early
disappearance of the food vacuole, by the more marked formation
of the dark brown pigment, and by the fact that the red blood
corpuscles attacked are not altered either in colour or size.
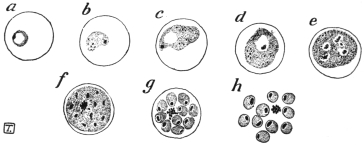
Fig. 86.—Development of the quartan parasite in the red corpuscles of man—asexual
stages. (After Manson.)
When the parasites have grown almost to the size of the erythrocytes
schizogony occurs. The pigment granules arrange themselves in lines
radiating towards the centre and the merozoites are also radially
disposed in groups of 6, 8, 10 or even 12, but are often arranged
less regularly. The whole development, growth and schizogony,
occupies seventy-two hours.
The appearance of quartana duplex or triplex is conditional on the
presence in the blood of the patient of two or three groups of
Plasmodia differing in their development by twenty-four hours.
The chief distinctive characters of the quartan parasite are: (1)
The erythrocyte is unchanged in size; (2) the rings are compact and
show pigment early; in the larger forms the chromatin is dense and
relatively plentiful; (3) the pigment, which is relatively well-marked,
may be arranged at the periphery.
Laverania malariæ, Grassi and Feletti, 1890 = Plasmodium
falciparum, Welch, 1897.
Syn.: Plasmodium malariæ var. quotidianæ, Celli et Sanf., 1891; Hæmamœba
malariæ præcox, Gr. et Fel., 1892 (nec H. præcox, Gr. et Fel., 1890); Hæmamœba
laverani, Labbé, 1894; Hæmatozoön falciparum, Welch, 1897; Hæmosporidium
undecimanæ and H. sedecimanæ and H. vigesimo-tertianæ, Lewkowitz, 1897; Hæmamœba
malariæ parva, Lav., 1900; Plasmodium præcox, Dofl., 1901; Plasmodium
immaculatum, Schaud., 1902; Plasmodium falciparum, Blanch., 1905.
The names most commonly used for the parasite of malignant
tertian malaria are Plasmodium falciparum and Laverania malariæ.
The summer and autumn fever (febris æstivo-autumnalis), also
called malignant tertian or sub-tertian, is caused by a malarial parasite
which is distinguished by the small size of its schizont, while the
gametocytes are crescentic (figs. 81, 88).
Most authors identify this kind of fever or the parasites which cause it (Laverania
malariæ) with the pernicious malaria of the tropics. Ziemann, however, repeatedly
has drawn attention to certain small but definite differences between the usual malignant
tertian or pernicious parasites which occur in the tropics and the tropical parasites
of some malarial districts, particularly of West Africa, and insists that at least two
varieties or sub-species occur. Other investigators distinguish from this or these
forms a quotidian parasite. On the other hand, the assertion is made that there are
no specific differences, but that the malignant or pernicious tertian parasite which
normally needs forty-eight hours for its development in the blood of man, can
also develop in twenty-four hours. The establishment of the duration of the
development is a matter of especial difficulty, because the stages of schizogony
are far less numerous in the peripheral blood than in that of the internal organs. It
is also stated that the tropical parasite very seldom forms crescentic but rather
rounded gametocytes. According to such an observation the organism would belong
to Plasmodium and not to Laverania. The question whether the tropical fevers are
caused by two different parasites does not seem to be definitely settled.
The young trophozoite of the malignant, pernicious tertian, or sub-tertian
parasite (fig. 87) are but slightly active and are very small, even
after the formation of the comparatively large food vacuole, which
makes the body appear annular (“signet ring” stage). Often two and
even more parasites are found in one blood corpuscle.
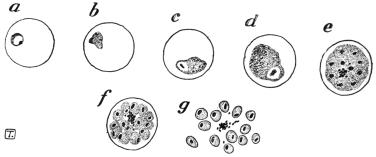
Fig. 87.—The pernicious malignant or sub-tertian parasite in the red corpuscles of man,
asexual stages. (After Manson.)
Fully grown they only attain two-thirds or less of the diameter of
the erythrocytes, which display an inclination to shrink and then appear
darker than the normal (brass-coloured). In the early stage dots or
stippling—sometimes called Maurer’s dots—appear on the blood corpuscles
as in those attacked by the ordinary tertian parasite (Plasmodium
vivax), but the Maurer’s dots are relatively coarse and few, and
are not easily stained. These dots were first described by Stephens
and Christophers in 1900, and subsequently by Maurer in 1902.
About thirty hours after the entrance into the blood corpuscles,
the parasites are rarely found in the peripheral blood, but they are
present in the internal organs, and especially in the spleen. The
schizogony, which now begins in the internal organs, proceeds on the
same lines as that of the quartan parasite, that is, usually with the
merozoites radially arranged around a central agglomeration of dark
brown pigment.
The number of merozoites formed is quoted differently, e.g., 8 to 24,
on an average 12 to 16. However, according to the recent cultural
researches of J. G. and D. Thomson204 (1913) the number of merozoites
of P. falciparum is 32. D. Thomson, from examination of spleen
smears at autopsy, also concludes that the number of merozoites may
reach 32. During their formation the blood corpuscle which is
attacked gets paler and disintegrates.

Fig. 88.—The crescents of the malignant tertian parasite. (After Mannaberg.)
See also fig. 81.
The gametocytes which finally appear are attenuated, curved
bodies, rounded at each end and known as crescents (figs. 81, 88), and
are provided with a nucleus and with coarse pigment masses. In the
males the pigment is more scattered than in the females, where it is
around the nucleus. Their length is 9 µ to 14 µ, and their breadth is
2 µ to 3 µ. At first they are still in the pale blood corpuscles, later
they free themselves and are found in numbers in the peripheral
blood in cases of pernicious malaria of Southern Europe and the
tropics, while, on the other hand, they occur much more rarely in
the peripheral blood in West African malignant tertian. Their further
development takes place under the same conditions as in the other
malarial parasites.
D. Thomson (1914),205 from studies of autopsy smears, has shown
that crescents develop chiefly in the bone-marrow and spleen, and
take about ten days to grow into the adult state in the internal organs.
He believes that crescents are produced from ordinary asexual spores.
Quinine, he states, has no direct destructive action on crescents, but
it destroys the asexual source of supply.
The sporozoites of Laverania malariæ (P. falciparum) are represented
in fig. 89.

Fig. 89.—Section through a tubule of the salivary gland of an Anopheles with sporozoites
of the malignant tertian parasites; on the left at the top a single sporozoite greatly magnified.
(After Grassi).
The principal distinctive characters of the malignant tertian parasite
are: (1) The ring forms are very small, occasionally bacilliform,
and may be marginal (“accolé” of Laveran); (2) the larger trophozoites
are often ovoid, and about one-third or one-half of the
erythrocyte in size; (3) the infected red cells sometimes show coarse
stippling (Maurer’s dots); (4) the gametocytes, or sexual forms, are
crescentic in shape.
J. W. W. Stephens (1914) has described a new malarial parasite of
man; it is called Plasmodium tenue. It is very amœboid, with scanty
cytoplasm and much chromatin, sometimes rod-like or irregular.
The parasite was described from a blood-smear of an Indian child.
The creation of a new species for this parasite has been criticized by
Balfour and Wenyon, and by Craig.
Plasmodium relictum, Sergent, 1907.
Syn.: Plasmodium præcox, Grassi and Feletti, 1890; Plasmodium danilewskyi,
Gr. et Fel., 1890; Hæmamœba relicta, Gr. et Fel., 1891; Proteosoma grassii,
Labbé, 1894.
Hæmamœboid, pigment-producing, malarial parasites are often found in birds.
Like the human malarial parasites they have been variously named. Labbé created
the genus Proteosoma for them, and this name is still often used as a distinctive one
unofficially. The correct name is stated to be either Plasmodium relictum or
P. præcox, or possibly even P. danilewskyi, assuming that there is only one species.
The nomenclature of the malarial parasites is most confused. The avian malarial
parasites are transmitted by Culicine mosquitoes.
The organism was discovered by Grassi in the blood of birds in Italy, and causes
a fatal disease in partridges in Hungary. Sparrows are affected in India, and it
was this Plasmodium in which Ross first traced the development of a malarial
parasite in a mosquito. The parasite may be transmitted from bird to bird by
blood-inoculation, canaries being very susceptible.
The principal stages of the avian plasmodium closely resemble those of the
malarial parasites of man. In its earliest stage P. relictum is unpigmented,
but soon the trophozoite grows and becomes pigmented, meanwhile displacing
the nucleus of the avian red-blood corpuscle, a characteristic feature, distinguishing
it from Halteridium. Schizonts are formed, each of which gives rise to
about nine merozoites in the circulating blood. Sexual forms or gametocytes also
occur in the blood. These develop in Culex fatigans, C. pipiens and C. nemorosus.
Oökinetes or vermicules are formed in twelve to fifteen hours in the
stomach of the mosquito, and in one to two days well-developed round oöcysts may
be seen. In three to four days sporoblasts have formed within the oöcysts and
young sporozoites begin to develop. In nine to ten days the oöcysts are mature,
being filled with sporozoites. The oöcysts then burst and the sporozoites travel
through the thoracic muscles to the salivary glands of the Culicine.
Neumann, experimenting with canaries, found that Stegomyia fasciata could
transmit the infection, but less efficiently than species of Culex.
The Cultivation of Malarial Parasites.
The successful cultivation of malarial parasites in vitro was first
recorded by C. C. Bass and by Bass and Johns (1912).206 Since then,
J. G. and D. Thomson,207 and McLellan (1912–13), Ziemann208 and others
have repeated the experiments.
DIFFERENTIAL CHARACTERS OF THE HUMAN MALARIAL PARASITES.
| Character | Plasmodium malariæ
(Quartan) | Plasmodium vivax
(Benign tertian) | Laverania malariæ =
Plasmodium falciparum
(Malignant tertian) |
|---|
| Schizogony | Complete in seventy-two hours | Complete in forty-eight hours | Complete in forty-eight hours or less |
| Trophozoite | Smaller than P. vivax, larger than L. malariæ | Young trophozoite large. | Young trophozoite small |
| Pseudopodia not marked or long | Long pseudopodia | |
| Movements | Rather slow in immature forms | Active amœboid movements | Sometimes actively motile |
| Pigment | Coarse granules, peripherally arranged, little movement | Fine granules, with active movement | Granules fine and scanty, movement oscillatory |
| Schizont | Smaller than red corpuscle | Larger than red blood corpuscle | Smaller than red corpuscle |
| Merozoites | 6 to 12 forming rosette | 15 to 20 regularly arranged | 8 to 32 (according to different authors) arranged irregularly |
| Gametocytes | Spherical | Spherical | Crescentic |
| Distribution of parasites in vertebrate host | About equal number in peripheral and visceral blood | Larger numbers in visceral blood | Scanty in peripheral blood compared with the enormous numbers in the internal organs. The latter part of the cycle (schizogony) may occur in the internal organs only |
| Alterations in erythrocytes | Almost normal | Pale and hypertrophied. | Corpuscle may be shrunken and dark, or may be colourless. Maurer’s coarse dots sometimes seen |
| Schüffner’s dots seen in deeply stained specimens |
Essentially the method of cultivation, as used by Thomson,
is as follows: 10 c.c. of infected blood are drawn from a vein and
transferred to a sterile test tube, in which is a thick wire leading to
the bottom of the tube. One-tenth of a cubic centimetre of a
50 per cent. aqueous solution of glucose or dextrose is placed
in the test tube, preferably before adding the blood. The blood
is defibrinated by stirring gently with the wire. When defibrination
is complete the wire and the clot are removed, and the glucose-blood
is transferred, in portions, to several smaller sterile tubes, each containing
a column of blood about one inch in height. The tubes
are plugged and capped and then transferred, standing upright, to an
incubator kept at a temperature of 37° C. to 41° C. The blood corpuscles
soon settle, leaving a column of serum at the top, to the extent
of about half an inch in each tube. The leucocytes need not be
removed by centrifugalization. J. G. Thomson (1913) and his
collaborators did not find it necessary to destroy the complement in
the serum, and they found that the malarial parasites developed at all
levels in the column of corpuscles, and not merely on the surface
layer of the corpuscles as first stated by Bass and Johns.
So far only the asexual generation of the malarial parasites has
been grown in vitro. Thomson rarely observed hæmolysis in the
cultures. Clumping of the malignant tertian parasites occurred.
In cultures of the benign tertian parasite (Plasmodium vivax) clumping
was not observed. J. G. and D. Thomson consider that this
difference as regards clumping explains why only young forms of
malignant tertian are found in peripheral blood, as the clumping
tendency of the larger forms causes them to be arrested in the finer
capillaries of the internal organs. It also explains the tendency to
pernicious symptoms, such as coma, in malignant tertian malaria.
Further it was found from cultures that P. falciparum was capable
of producing thirty-two spores (merozoites) in maximum segmentation,
while P. vivax produced sixteen spores (merozoites) as a rule,
though the number might be greater than sixteen. (Quartan parasites
produce eight spores or merozoites in schizogony.)
It may also be mentioned here that Babesia (Piroplasma) canis has
been successfully cultivated in vitro by Bass’s method. This has been
accomplished by Thomson and Fantham,209 Ziemann, and Toyoda in
1913. J. G. Thomson and Fantham used the simplified Bass technique
recorded above, namely, infected blood and glucose, incubating at
37° C. In one of the B. canis cultures, starting with heart blood of a
dog containing corpuscles infected with one, two, or, exceptionally,
four piroplasmata, Thomson and Fantham succeeded in obtaining a
maximum of thirty-two merozoites in a corpuscle. The cultures are
infective to dogs and sub-cultures have been obtained.
Family. Piroplasmidæ, França.
The parasites included in this provisional family or group belong
to the Hæmosporidia. They are minute organisms, sometimes
amœboid, but usually possessing a definite form. They are endoglobular,
being contained within mammalian red blood corpuscles,
but they produce no pigment. The true Piroplasmata, belonging
to the genus Babesia, destroy the host corpuscles, setting free the
hæmoglobin, which is excreted by the kidneys of the cow, sheep,
horse, dog, etc., acting as host. The disease produced, variously called
piroplasmosis or babesiasis, is consequently characterized by a red
coloration of the urine known as hæmoglobinuria, or popularly as
“red-water.” One of the best known piroplasms is Piroplasma
bigeminum or Babesia bovis (probably the latter name is correct),
which is the causal agent of “Texas fever” or “red-water” in cattle
and is spread by ticks.
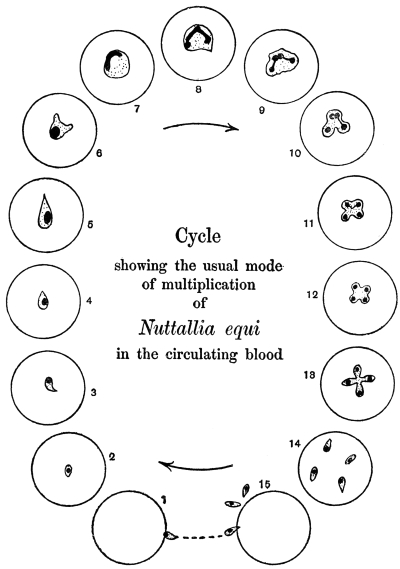
Fig. 90.—Nuttallia equi, life-cycle as seen in red blood corpuscles in stained preparations
of peripheral blood. (After Nuttall and Strickland.)
Of recent years, researches on the morphology of these blood
parasites has led to their separation into various genera and species.
However, our knowledge is still very far from complete. The various
genera recognized by França210 (1909), and placed in a provisional
family, Piroplasmidæ, may be listed, though further research may
lead to emendations:—
(1) Babesia (Starcovici) or Piroplasma (Patton). Pyriform
parasites, dividing by a special form of budding or gemmation with
chromatin forking, as well as by direct binary fission. Parasitic in
oxen, dogs, sheep, horses, etc.
(2) Theileria (Bettencourt, França and Borges). Rod-shaped and
oval parasites occurring in cattle and deer. T. parva is the pathogenic
agent of African East Coast fever in cattle.
(3) Nuttallia (França). Oval or pear-shaped parasites, with multiplication
in the form of a cross. N. equi211 (fig. 90) of equine “piroplasmosis”
(nuttalliosis). N. herpestidis in a mongoose.
(4) Nicollia (Nuttall). Oval or pear-shaped parasites with
characteristic nuclear dimorphism, and with quadruple division at
first fan-like, then like a four-leaved clover. N. quadrigemina from
the gondi.
(5) Smithia (França). Pear-shaped, single forms stretching across
the blood corpuscle. Multiplication into four in the form of a cross.
S. microti from Microtus arvalis, S. talpæ from the mole.
(6) Rossiella (Nuttall). This belongs to the family Piroplasmidæ
of França. It is intracorpuscular and non-pigment forming, occurring
singly, in pairs, or occasionally in fours. It is usually round and
larger than Babesia. The parasite multiplies by binary fission.
R. rossi in the jackal.
The genus Babesia is the best known and most important, and
will be considered next.
Genus. Babesia, Starcovici, 1893.
Syn.: Pyrosoma, Smith and Kilborne, 1893; Apiosoma, Wandolleck, 1895; Piroplasma,
W. H. Patton, 1895; Amœbosporidium, Bonome, 1895.
The organisms belonging to this genus are pyriform, round or
amœboid. The characteristic mode of division is as follows: Just
before division the parasite becomes amœboid and irregular in shape,
(fig. 91, 1–5) with a compact nucleus. The latter gives off a nuclear
bud. This nuclear bud divides into two by forking (fig. 91, 6, 7).
The chromatin forks grow towards the surface of the body of the
rounded parasite, and then two cytoplasmic buds grow out. The
forking nuclear buds, which are Y-shaped, pass into the cytoplasmic
outgrowths212 (fig. 91, 8, 9). The buds gradually increase in size at
the expense of the parent form until they become two pear-shaped
parasites joined at their pointed ends. The connecting strand shrinks
and the two daughter forms separate (fig. 91, 10–14). The pyriform
parasites after having exhausted the blood corpuscle escape from it
(fig. 91, 15), and seek out fresh host corpuscles, entering by the rounded,
blunt end (fig. 91, 1). It is the pyriform phase of the parasite which
penetrates red blood corpuscles, not rounded forms, which die if set
free. The pyriform parasite, however, becomes rounded (fig. 91, 2, 3),
soon after its entry into a fresh host cell. This interesting mode of
division by gemmation and chromatin forking has been made diagnostic
of the genus Babesia by Nuttall.213 Rounded forms of Babesia
divide by binary fission, and this direct method can also be adopted
by the other forms of Babesia.
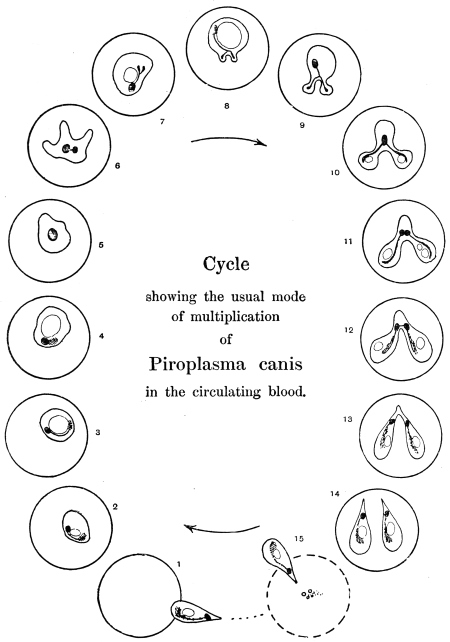
Fig. 91.—Babesia (Piroplasma) canis, life-cycle in stained preparations of infected blood
of dog. (After Nuttall and Graham-Smith.)
The distribution of the chromatin in the pear-shaped Babesia, as seen in B. canis
and B. bovis, is interesting. The main nuclear body consists of a karyosome
surrounded by a clear area. There is also a loose (chromidial) mass of chromatin
representing the remains of the chromatin forks seen during the formation of the
parasite as a daughter form by gemmation. Occasionally there is a small dot or
point, the so-called “blepharoplast” of Schaudinn and Lühe. This minute dot is
not a flagellate blepharoplast, for there is no flagellate stage in the life-history of
Babesia. These nuclear phenomena have been described by Nuttall and Graham-Smith
and Christophers (1907)214 for B. canis, by Fantham (1907)215 for B. bovis, and
by Thomson and Fantham (1913) from glucose-blood cultures of B. canis.
Babesia are tick borne, as was first shown by Smith and Kilborne
(1893). The developmental cycle in the tick is incompletely known.
The best accounts are those of Christophers (1907)216 for B. canis and
Koch (1906) for B. bovis, and these accounts are supplementary.
The principal stages, so far as known, may be summarized thus:—
(1) The piroplasms taken by the tick in feeding on blood pass into the tick’s
stomach. The pyriform parasites, which alone are capable of further development,
are set free from the blood corpuscles. In about twelve to eighteen hours they
become amœboid, sending out long, stiff, slender, pointed pseudopodia. The
nucleus of each parasite divides unequally into two. Similar forms have been
obtained in cultures. These stellate forms may be gametes, and according to Koch
fuse in pairs.
(2) A spherical stage follows, possibly representing the zygote. This grows, and
a uninucleate globular mass results. This form is found in large numbers on the
third day, according to the observations of Koch.
(3) A club-shaped organism is next formed. This may represent an oökinete
stage. The club-shaped bodies are motile and gregarine-like, and are about four
times the size of the blood forms. These club-shaped bodies and subsequent stages
were described by Christophers in the development of B. canis in the dog-tick,
Rhipicephalus sanguineus.
(4) The club-shaped bodies pass from the gut of the tick into the ovary, and so
get into the ova. There they become globular, and later are found in the cells of
the developing tick-embryo. The parasites are, then, transmitted hereditarily.
Similar globular bodies are found in the tissue cells of the body of tick nymphs
which have taken up piroplasms. The globular stage was called the “zygote” by
Christophers, but it may correspond to the oöcyst of Plasmodia.
(5) The globular body divides into a number of “sporoblasts,” which become
scattered through the tissues of the larval or nymphal tick, as the case may be.
(6) The sporoblasts themselves divide into a large number of sporozoites, which
are small uninucleate bodies, somewhat resembling blood piroplasms. The sporozoites
collect in the salivary glands of the tick. They are inoculated into the
vertebrate when the tick next feeds.
The chief species of Babesia and their pathogenic importance
may be listed thus:—
(1) Babesia bovis (Babes) produces infectious hæmoglobinuria
of cattle in Europe and North Africa. It is transmitted by Ixodes
ricinus. A similar parasite also occurs in deer.
(2) Babesia bigemina (Smith and Kilborne) produces Texas
fever, tristeza, or red-water in cattle in North and South America,
South Africa and Australia. It is transmitted by Boöphilus annulatus
in North America, by B. australis in Australia, South America, and
the Philippines, and by B. decoloratus in South Africa.
The parasite is from 2 µ to 4 µ long, and from 1·5 µ to 2 µ broad.
Babesia bigemina may be the same parasite as B. bovis.
(3) Babesia divergens (MacFadyean and Stockman) is a small
parasite. It is found in cattle suffering from red-water in Norway,
Germany, Russia, Hungary, Ireland, Finland, and France, and is
transmitted by Ixodes ricinus.
(4) Babesia canis (Piana and Galli-Valerio) gives rise to malignant
jaundice or infectious icterus in dogs in Southern Europe, India, and
other parts of Asia and North Africa, where it is transmitted by
Rhipicephalus sanguineus. In Africa generally, especially South
Africa, the disease is transmitted by Hæmaphysalis leachi. Babesia
canis varies from 0·7 µ to 5 µ, the size depending partly on the
number of parasites within the corpuscle. It averages about 3 µ. It
has been cultivated in Bass’ medium (glucose and infected blood),
see p. 172.
In India Piroplasma gibsoni (Patton) infects hunt dogs and jackals.
It is annular or oval in shape.
(5) Babesia ovis (Babes) produces “Carceag,” a disease of sheep
in Roumania, the Balkan Peninsula, Italy, and Transcaucasia. It
varies in size from 1 µ to 3 µ. It is transmitted by Rhipicephalus
bursa. The parasite has recently been recorded from Rhodesia.
(6) Babesia caballi (Nuttall and Strickland) causes “biliary
fever” in equines. The parasite occurs in Russia, Roumania, and
Transcaucasia. It varies in size from 1 µ to 2 µ. It is transmitted by
Dermacentor reticulatus.
It should be mentioned that Nuttallia equi also causes “piroplasmosis” in
equines, with symptoms of hæmoglobinuria and jaundice in Italy, Sardinia, many
parts of Africa, Transcaucasia, India, and Brazil. In Africa it is transmitted by
Rhipicephalus evertsi. It has been shown experimentally that a horse recovered
from Babesia caballi was susceptible to the inoculation of Nuttallia equi blood.
(7) Babesia pitheci (P. H. Ross) was found in a monkey,
Cercopithecus sp., in Uganda. The pear-shaped forms measure 1·5 µ
by 2·5 µ.
(8) Babesia muris (Fantham)217 was found in white rats. The
pyriform parasites are 2 µ to 3 µ long and 1 µ to 1·5 µ broad; oval
forms are 0·5 to 1·5 µ diameter.
The usual symptoms of babesiasis (piroplasmosis) are high fever,
loss of appetite, hæmoglobinuria, icterus, anæmia, paralysis, and
death in about a week in acute cases. In chronic cases there is
anæmia, and hæmoglobinuria is less marked. When animals recover,
there are still some piroplasms left in the blood. “Recovered” or
“salted” animals are not susceptible to reinfection, but ticks feeding
on them acquire piroplasms, and are a source of danger to freshly
imported animals.
Treatment.—Trypan-blue is the best drug, as shown by Nuttall and Hadwen218
(1909). It should be administered intravenously in 1 to 1·5 per cent. aqueous solution.
A dose of 5 to 10 c.c. is curative for dogs, one of 100 to 150 c.c. for horses and
cattle. Unfortunately, the tissues are coloured blue by the drug. The “salted”
animals, after trypan-blue treatment, still harbour the parasites in their blood for
years.
Genus. Theileria, Bettencourt, França and Borges, 1907.
The organisms belonging to this genus are rod-like or bacilliform, and coccoid
or round.
The best known of the species of Theileria is T. parva, the pathogenic agent of
East Coast fever or Rhodesian fever in cattle in Africa.
Theileria parva, Theiler, 1903.
Syn.: Piroplasma parvum.
In the blood corpuscles of infected cattle minute rod-like and
oval parasites are seen. Some are comma shaped and others are
clubbed (fig. 92, 1–12). The rod-like forms measure 1 µ to 3 µ in
length by 0·5 µ in breadth; the oval forms are 0·7 µ to 1·5 µ in
diameter. The intracorpuscular parasites are said by R. Gonder
(1910) to be gametocytes, the rod-like forms being thought to
be males, the oval forms to be females. Free parasites are practically
never seen in the blood. It is known that it is impossible to
produce the disease in a healthy animal by blood inoculation, but
only by intraperitoneal transplantation of large pieces of infected
spleen (Meyer). There may be as many as eight parasites in a corpuscle.
The chromatin is usually at one end of the organism. In
some parasites the appearance of the chromatin suggests division, but
such division, if it takes place, must be very slow, as it has not been
actually seen in progress. The red blood corpuscles appear merely to
act as vehicles for the parasites (Nuttall, Fantham, and Porter).219
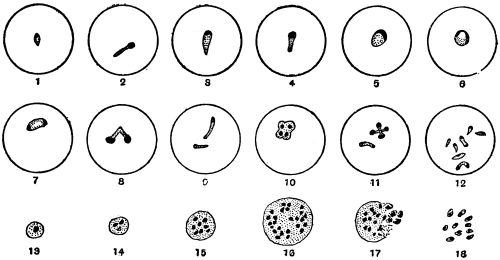
Fig. 92.—Theileria parva. 1–12, intracorpuscular parasites, stained. (After Nuttall and
Fantham); 13–18, Koch’s blue bodies, from stained spleen smear; 17–18, breaking up of
Koch’s body. (After Nuttall.)
In the internal organs, especially the lymphatic glands, spleen and bone-marrow,
are found multinucleate bodies known as Koch’s blue bodies (fig. 92, 13–18). These
are schizonts, according to Gonder.220 The actual Koch’s blue bodies are said to be
extracellular, but similar multinucleate bodies, schizonts, occur in lymphocytes. The
schizonts divide and the merozoites resulting probably invade the red blood corpuscles
in the internal organs. Gonder considers that the sporozoites injected by
the tick collect in the spleen and lymphatic glands, penetrate the lymphocytes and
give rise to the schizonts.
Gonder has studied the cycle of T. parva in the tick. He states that the
gametocytes leave the host corpuscles and give rise to gametes, then conjugation
occurs producing zygotes. The zygotes are then said to become active to form
ookinetes, and to enter the salivary glands of the tick. Multiplication is said to
occur therein, producing a swarm of sporozoites. This work needs confirmation.
T. parva is transmitted by Rhipicephalus appendiculatus, R. simus, R. evertsi,
R. nitens, and R. capensis. The parasites are not hereditarily transmitted in
Rhipicephalus, but when taken by the transmitter at one stage of its development
the tick is infective in its next stage (e.g., if the larva becomes infected, then the
nymph is infective; if the nymph becomes infected, then the adult is infective).
An animal recovered from Theileria parva is incapable of infecting ticks, but few
animals recover from East Coast fever. Animals suffering therefrom do not show
hæmoglobinuria.
Theileria mutans, Theiler, 1907·
Syn.: Piroplasma mutans.
This is transmissible experimentally by blood inoculation. It occurs in cattle in
South Africa and Madagascar and is apparently non-pathogenic. No Koch’s blue
bodies are formed. It is transmitted by ticks.
Theileria annulata (Dschunkowsky and Luhs) occurs in cattle in Transcaucasia.
A Theileria (T. stordii) has been found in a gazelle (França, 1912).
Genus. Anaplasma, Theiler, 1910.
This genus221 may be mentioned here. The organisms included therein are,
according to Theiler, coccus-like, consisting of chromatin, and are devoid of cytoplasm.
They occur in the red blood corpuscles of cattle, causing a disease characterized
by destruction of red cells, fever and anæmia, but with yellow urine. The
disease is tick transmitted. The bodies now called Anaplasma marginale were
formerly described as marginal points. They multiply by simple fission. They are
said by Theiler to cause gall-sickness in cattle in South Africa. Some authors doubt
whether these bodies are organismal.
Genus. Paraplasma, Seidelin, 1911.
Under this generic name Seidelin described certain bodies found
by him in cases of yellow fever in 1909. The type species is
P. flavigenum,222 and is claimed by Seidelin to be the causal agent of
yellow fever.
Paraplasma flavigenum occurs in the early days of the disease as
small chromatin granules with or without a faint trace of cytoplasm.
The bodies are usually intracorpuscular. Also, somewhat larger forms,
with distinct cytoplasm, are seen in small numbers. During the later
days of the disease still larger forms are found, and these occur also
in sections of organs (e.g., kidney) made post-mortem. Some of these
larger forms are perhaps schizonts. In the second period of the
disease possible micro- and macro-gametes may be found, some of
which are extracorpuscular. Some small free bodies have been seen.
Recently schizogony has been stated to occur in the lungs, and it is
said that guinea-pigs can be inoculated with Paraplasma flavigenum,
and show yellow pigment in the spleen.
Seidelin places Paraplasma in the Babesiidæ, with resemblances
more particularly to Theileria. V. Schilling-Torgau and Agramonte
have criticized these findings; the former considers them to be the
resultant of certain blood conditions.
P. subflavigenum was found by Seidelin in 1912 in a man suffering
from an unclassified fever in Mexico.
Further, it is now known that a Paraplasma occurs naturally in
guinea-pigs. More researches are needed on these matters, as some
writers (e.g., Wenyon and Low) claim that the bodies are not
organismal.
Paraplasma flavigenum.—The Yellow Fever Commission (West
Africa) in their third report, dated 1915, have come to the conclusion
that there is no evidence that the bodies termed Paraplasma flavigenum
are of protozoal nature or that they are the causal agents of
yellow fever.
Sub-class. NEOSPORIDIA, Schaudinn.
Sporozoa in which growth and spore formation usually go on
together.
Order. Myxosporidia, Bütschli.
These parasites, which were discovered by Johannes Müller (1841), live principally
in fishes, and occasionally cause destructive epizoötics amongst their hosts. Müller
first observed them in the form of whitish-yellow pustules on the skin or on the gills
of various fishes. These pustules contained masses of small shell-covered bodies with
or without tails (“psorosperms,” see fig. 93). Similar bodies were also found in the
air bladders of certain fish. Creplin (1842) demonstrated the resemblance of the cysts
(“psorosperm tubes”) harbouring the psorosperms to the “pseudonavicella-cysts” of
a gregarine, as described by v. Siebold. Dujardin (1845) considered that there was
possibly some connection between the protoplasmic “psorosperm tubes” and the
spores they contained, and the developmental stages of monocystid gregarines from the
vesiculæ seminales of earth-worms. The relationship of the “fish psorosperms” was
placed on a firmer basis by Leydig (1851) and Lieberkühn. The former found numerous
forms in marine fish, and he discovered in species which live free in the gall bladder
of cartilaginous fishes that the psorosperms originated in a manner similar to the
gregarines. Lieberkühn (1854) studied the Myxosporidia in the bladder of the pike
(fig. 93, a, b, d), and observed their amœboid movements, as well as the formation of
the spores, from each of which a small amœboid body escaped, a discovery that
was confirmed by Balbiani. The same author also found that spiral filaments were
enclosed in the so-called polar body, i.e., the polar capsule of the psorosperm spores,
and that these could be protruded (fig. 93, d, and fig. 95).

Fig. 93.—Upper figure, part of
a gill of a roach, Leuciscus rutilus
(natural size), with two myxosporidia.
Lower figures, a, b, d, spores
of myxosporidia from a pike, Esox
lucius. c, Spore from Platystoma
fasciatum. (After J. Müller.)
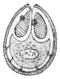
Fig. 94.—The tailless spore of
Myxobolus mülleri, with the polar
bodies and their nuclei and the
sporozoite. (After Bütschli.)
The term Myxosporidia, which at the present day is universally applied to the
“psorosperm tubes,” was introduced by Bütschli in 1881, who studied not only the
structure and development of the spores, but also the protoplasmic body of the
parasites (fig. 96), and confirmed the occurrence of numerous nuclei. Many authors
have made important additions to our knowledge of the Myxosporidia: Perugia,
Thélohan, Mingazzini, L. Pfeiffer, L. Cohn, Doflein, Mercier, Schröder and Auerbach;
while the presence of this parasite outside the class of fishes has become known
through Lutz, Laveran, and others. The species causing disease in fishes have been
described by Ludwig, Railliet, Weltner, L. Pfeiffer, Zschokke, Hofer, Doflein, Gurley,
Plehn, Schuberg, Fantham and Porter. With regard to classification the works of
Thélohan (1895) and Gurley (1894) may be mentioned.
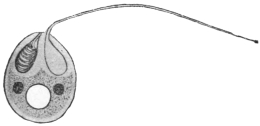
Fig. 95.—Schematic representation of a
spore of Myxobolus. One polar capsule has
protruded its filament; two nuclei and a
“vacuole” in the sporozoite. (After Doflein.)
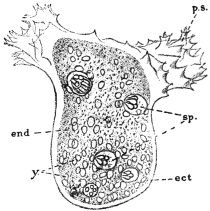
Fig. 96.—Chloromyxum leydigi. Active
trophozoite (parasitic in gall-bladder of skates,
rays, dog-fish). Ect, ectoplasm; ps, pseudopodia;
end, endoplasm; y, yellow globules
in endoplasm; sp, spores, each with four
polar capsules. × 525. (After Thélohan.)
The Myxosporidia live either free on the epithelial surface of hollow organs
(gall or urinary bladder, renal tubules, but never in the intestine), or are enclosed in
the tissues of their host. The gills and muscular system are their favourite
habitat, but other tissues or organs may be attacked. Species of Myxosporidia
are also known from Amphibia, Reptilia, and a few invertebrates.
The free forms, which are often amœboid (fig. 96), move by the aid of variously
shaped pseudopodia, have a constant form, or may exhibit contractions of the body.
The tissue parasites often reach a considerable size, so that the integument of the
host forms protuberances over them. They are of a roundish or irregular shape.
Frequently they are enveloped in a connective tissue covering formed by the host.
The protoplasmic body in the trophic phase (fig. 96) shows a distinct ectoplasm
which is finely granular or sometimes striated, and an endoplasm which is coarsely
granular and contains many nuclei as well as cell inclusions, such as crystals, pigment
grains and fat globules. The nuclei originate by division from the primitive
nucleus of the amœboid germ that issues from the spore. This amœbula may or
may not live intra-cellularly during the early stages of its existence.
The multinucleate trophozoite of a Myxosporidian forms spores in its endoplasm
practically throughout its whole period of growth (fig. 96). Vegetative reproduction
by a process of external budding or plasmotomy may also occur, as in Myxidium
lieberkühni from the urinary bladder of the pike.
The myxosporidian trophozoite may produce two spores within itself, when
it is placed in the sub-order Disporea, or it may produce numerous spores, which
is characteristic of the sub-order, Polysporea. The phenomenon of spore formation
is not simple (fig. 97), and the spore itself is surrounded by a bivalved shell or sporocyst
and contains polar capsules in addition to the amœboid germ (fig. 97, G, H). The
valves of the sporocyst and the polar capsules are really differentiated nucleate cells,
so that each spore is an aggregate of cells rather than one cell, though only a single
amœbula issues from a spore. The accounts of spore formation vary somewhat
according to the different workers.
Spore formation is usually very complicated and there are differences of opinion
as to the interpretation of various stages, particularly as to whether conjugation
occurs therein. The process is initiated by the concentration of cytoplasm around
one of the nuclei of the endoplasm, so that a small spherical mass or initial corpuscle
is produced, the pansporoblast (Gurley) or primitive sphere (Thélohan). Some
authors state that a pansporoblast really results from a conjugation of two initial
corpuscles (fig. 97, A-D). Nuclear multiplication occurs within the pansporoblast
(fig. 97, E), and sooner or later two multinucleate sporoblasts are formed within it
(fig. 97, F). Each sporoblast gives rise to a single spore, which consists of a sporocyst
or envelope composed of two valves each secreted by a cell, two polar capsules
each secreted by a cell, and the sporoplasm or amœbula which becomes binucleate
(fig. 97, G). During the process of spore formation (fig. 97) various vegetative and
reduction nuclei may be produced, in addition to those which are essentially involved
in spore formation, and the sporocyst cells may be developed early.
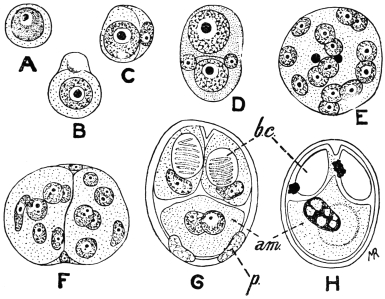
Fig. 97.—Myxobolus pfeifferi. Spore formation. A, reproductive cell from plasmodial
trophozoite; B, cell divided unequally into two; C, smaller cell forming envelope to larger
one; D, pansporoblast formed by union of two forms like C; E, multinucleate pansporoblast,
two of the nuclei being those of the envelope; F, pansporoblast divided into two multinucleate
sporoblasts; G, spore differentiation; p, two parietal cells forming sporocyst; bc, polar capsules;
am, binucleate amœbula; H, ripe spore in which the two nuclei of the amœbula have
fused. (After Keysselitz.)
Each spore contains two (figs. 94, 95) or more polar capsules which are clearly
visible in the fresh condition. Each polar capsule is a hollow, more or less pear-shaped
body, secreted by a cell and having a well defined contour. Within it, a
long, delicate, elastic filament, the polar filament, is formed, and lies spirally coiled
in the polar capsule until just before the emergence of the amœbula from the spore
(fig. 95). The polar filament is ejected, probably under the influence of the digestive
juice, when the spore reaches a new host, and serves to anchor the spore to the
tissue with which it is in contact, and thus allow of the emergence of the amœbula
in a situation suitable for its development. The polar capsule with its contained
polar filament has been compared with the stinging cells or nematocysts of the
Cœlentera, but it has a totally different function.
The spores fulfil the purpose of effecting transmission to other hosts. Infection
occurs by the ingestion of the parasites per os after their escape by some
means from their host. Thélohan and others have demonstrated that the valves of
the spores soon open under the influence of the digestive juices, thus allowing the
young myxosporidia to escape. Their further history is unknown; but it may be
surmised that they either travel direct to the organs usually affected (gall bladder,
urinary bladder), or are distributed in the body by means of the circulatory or
lymphatic systems.
The Myxosporidia that invade tissues are often deadly to their hosts. They may
be present in a state of “diffuse infiltration” when practically every organ of the
body may be infected, as in barbel disease (due to Myxobolus pfeifferi). On the
other hand, the parasites may be concentrated at one spot, when cysts, either large
or small, are produced. Such cysts occur on the gills of many fishes. A few additional
important pathogenic forms are Myxobolus cyprini, the excitant of “pockenkrankheit”
of carp, and Lentospora cerebralis, parasitic in the skeleton of Salmonidæ
and Gadidæ. The skeletons of the tail, fins and skull particularly are seats of infection,
and from the skull the Lentospora can spread to the semicircular canals, resulting
in loss of power to maintain its balance on the part of the fish. On this account
the malady is termed “drehkrankheit.” Young fish are more particularly infected.
Myxobolus neurobius infects the spinal cord and nerves of trout.
Myxosporidia are divided into two sub-orders—Disporea and Polysporea—according
to whether they form only two or several spores during their growth.
The former include two genera limited to fishes, which are easily distinguishable by
the shape of the spores: Leptotheca, Thél., with a rounded spore, and Ceratomyxa,
Thél., with a very elongate spore. The larger number of genera belong to the
Polysporea, which are divided into three families:
| (1) Amœboid germ with a vacuole the contents of which do not stain with iodine. | 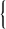 | (a) With two polar capsules.—Myxidiidæ. |
| (b) With four polar capsules.—Chloromyxidæ. |
| (2) Amœboid germ with a vacuole stainable with iodine. Spores with two polar capsules.—Myxobolidæ. |
| For further subdivisions the differences in the spores are principally utilized. |
Order. Microsporidia, Balbiani.
These are the organisms discovered in the stickleback by Gluge in 1834, and
in Coccus hesperidum by Leydig in 1853. They have since been found in numerous
other arthropods, especially insects. They acquired particular importance when it
was discovered that they were the cause of the “pébrine” disease (“gattina” of
the Italians) which caused so much destruction amongst silkworms (Bombyx mori).
Pasteur (1867–70) and especially Balbiani (1866) participated in the researches on
Nosema bombycis, and it was the latter who classed the “pébrine bodies” or “psorospermia
of the arthropoda” amongst the Sporozoa as Microsporidia (1882).223 The complete
life cycle of N. bombycis was described in 1909 by Stempell. The Microsporidia
are not confined to insects and arachnoids, they are now known to occur also in
crustacea, worms, bryozoa, fishes, amphibians and reptiles. Certain tumours in
fishes, similar to those formed by many Myxosporidia, are produced by Microsporidia.
Fantham and Porter found that Nosema apis was pathogenic to bees
and other insects, and was the causal agent of the so-called “Isle of Wight”
disease in bees224 in Great Britain.
The Microsporidia, as their name implies, form minute spores which usually are
oval or pear-shaped. Each spore contains a single polar capsule which is not easily
visible in the fresh state (fig. 98, f) and a single amœboid germ issues from the spore
(fig. 99, b).
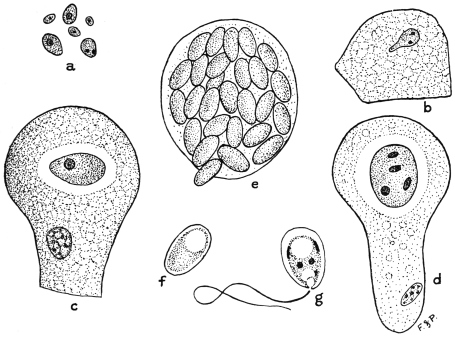
Fig. 98.—Nosema apis. Various stages in life-cycle. a, planonts or amœbulæ from chyle
stomach of bee; b, amœboid planont creeping over surface of gut epithelial cell; c, uninucleate
trophozoite within epithelial cell; d, meront with nucleus divided into four, about to form
four spores; e, epithelial cell crowded with spores; f, young spore; g, spore showing five
nuclei, polar filament ejected, and amœbula, about to issue. × 1,500, a-e; × 2,150, f-g.
(After Fantham and Porter.)
The life cycle of Nosema apis, parasitic in bees, may be taken as an example of
that of a microsporidian. The infection of the host is initiated by the ingestion of
spores of N. apis in food or drink contaminated with the excrement of other infected
bees. Under the influence of the digestive juice of the bee the spore-coat (sporocyst)
softens, the polar filament is ejected and anchors the spore to the gut epithelium,
and the minute amœbula contained in the spore emerges. The amœbula is capable
of active amœboid movements (fig. 98, b) and so is termed the planont or wandering
form (fig. 98, a). After a short time each planont penetrates between or into the cells
of the epithelium of the gut, a few only passing through into the body cavity. Within
the cells the amœbulæ become more or less rounded, lose their power of movement,
and after a period of growth of the trophozoite (fig. 98, c) commence to divide actively,
these dividing forms being known as meronts (fig. 98, d). Various forms of fission
occur, and during this phase, termed merogony, the numbers of the parasite within
the host are greatly increased, with concomitant destruction of the epithelium
(fig. 98, e). After a time sporogony commences. The full-grown meront becomes
successively the pansporoblast and sporoblast. Nuclear multiplication and differentiation
ensue and five nuclei are ultimately produced. At the same time a sporocyst
is secreted, and two vacuoles are produced within. One is the polar capsule, and
within it the polar filament is differentiated; the other forms the posterior vacuole
(fig. 98, g). Between the two vacuoles the body cytoplasm or sporoplasm forms a
girdle-like mass. Of the nuclei, one regulates the polar capsule, two control the
secretion of the sporocyst, and two remain in the sporoplasm. The polar capsule and
polar filament are not usually visible in the fresh condition, but can be demonstrated
by the use of various chemical reagents (fig. 100). The sporoplasm ultimately becomes
the amœbula (fig. 98, g) which issues from the spore after the ejection of the polar
filament.
Fig. 99.—a, section through the abdominal
wall of a silkworm, whose epithelial cells
contain Microsporidia (Nosema bombycis); b,
a spore, the contents of which are escaping.
(After Balbiani.)
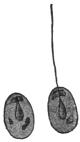
Fig. 100.—Nosema bombycis, Naeg.
Spores treated with nitric acid, thus
rendering the polar capsule perceptible,
and the filament has protruded from one
of the spores. (After Thélohan.)
A trophozoite (meront) of N. apis becomes a single pansporoblast which gives
rise to one sporoblast producing one spore, and this procedure is characteristic of
the genus Nosema. In other genera the trophozoite may form more than one pansporoblast
and each pansporoblast may form a variable number of spores in different
cases. Various attempts at classification have been based on these characteristics.
It must suffice here to note that in the cases where the trophozoite becomes one
pansporoblast, the latter can produce four spores in the genus Gurleya, eight spores
in Thélohania and many spores in Pleistophora. In other cases, where the trophozoites
give rise to many pansporoblasts, each of the latter may form many spores,
as in the genus Glugea.
A few pathogenic microsporidian parasites other than N. apis may be mentioned.
N. bombycis, causing pébrine in silkworms, may infect any or all the tissues of the
host (fig. 99). The larvæ of the host, i.e., the “silkworms,” may become infected by
eating food contaminated with spore-containing excrement of already infected silkworms.
In cases of heavy infection the silkworm dies, but should the infection be
less intense the larva becomes a pupa in which the parasite persists, so that the moth
emerges from the cocoon already infected. Not only is the moth parasitized itself,
but the Nosema reaches the generative organs of both sexes and penetrates the
ovaries of the female, with the result that the ova are deposited infected. Such
infected eggs are capable of developing, so that infection may be transmitted
hereditarily as well as by the contaminative method. Infected eggs can be recognized
by microscopic examination, as Pasteur showed, and thus preventive measures
may be adopted.
A microsporidian parasite is known to occur on the roots of the spinal and cranial
nerves of Lophius piscatorius, the angler fish. This parasite is variously referred to
the genera Nosema and Glugea.
Thélohania contejeani, parasitic in the muscles of crayfish, is believed by some
to be the causal agent of recent epizoötics among them, though others believe the
disease to be really due to a bacillus. It may be that the one organism aids in the
entry of the other into the host.
Order. Actinomyxidia, Stolč.
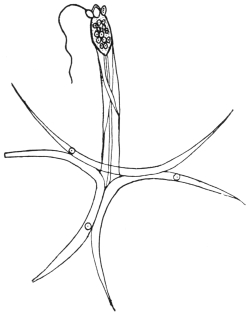
Fig. 101.—Spore of Hexactinomyxon
psammoryctis. At top of figure three polar
capsules, one with polar filament extended.
× 450. (After Stolč.)
A brief mention may be made of the Actinomyxidia (fig. 101), which were first
described by Stolč in 1899 as parasites of Oligochætes. They have also been
investigated by Mrazek, and a detailed study of certain species was made by
Caullery and Mesnil (1905). The trophozoite
is small and amœboid. The spores
are large, and exhibit tri-radiate symmetry.
Spore formation is complicated and sexual
processes occur therein. Many amœbulæ
are set free from each spore.
Order. Sarcosporidia, Balbiani.
The first member of this group was
discovered by Miescher in 1843. This
author found white filaments running
parallel with the direction of the fibres
in the voluntary muscles of mice. They
were visible to the naked eye, and proved
to be cylindrical tubes tapering at each
end. They were as long as the muscular
fibres, were enveloped in a membrane, and
contained innumerable elongate or kidney-shaped
bodies and a smaller number
of little spherical forms. Th. v. Hessling
confirmed (1853) the occurrence of these
“Miescher’s tubes” within the muscular
fibres, this author having discovered the
same structures in the heart muscles of
deer, cattle, and sheep. Both investigators considered them to be pathological transformations
of the muscles. v. Siebold, from his own experiences, regarded them
as fungus-like entophytes.
Rainey (1858) discovered similar structures in the muscular system of pigs, and
considered them to be early stages of Cysticercus cellulosæ, which error Leuckart
rectified, simultaneously emphasizing their relationship with Myxosporidia. Both
these authors found them in the muscular fibres, and both observed that they
possessed a thick striated membrane. Manz (1867) published the results of more
minute investigations on the structure and contents of the cylinders. This observer
also recognized the disease in rabbits and attempted to cultivate the parasites. He
also tried to induce experimental infection in guinea-pigs, rats, and mice, but the
result was negative.
However, domestic and wild mammals are not the only hosts of Sarcosporidia;
these parasites are also harboured by birds. Thus, according to Kühn, they are
found in the domestic fowl; according to Rivolta in Turdus, Corvus, and other
birds; according to Stiles in North American birds; while Fantham found Sarcosporidia
in the African mouse-bird, Colius. Reptiles also are parasitized occasionally.
Bertram found them in the gecko, Lühe in the wall-lizard. It was found also that
the Sarcosporidia could develop not only in the muscles but also in the connective
tissue. This led to the foundation of a new, but provisional, classification by Blanchard,
using the generic name Miescheria for the parasites in the muscles and
Balbiania for those in the connective tissue. Finally, Sarcosporidia have also been
observed in man.
The relation of these parasites to certain diseases of domestic animals has been
studied by veterinary surgeons. Sarcosporidia may cause fatal epizoötics among
sheep.
There is still a wide field open for research in regard to the structure and development
of these parasites, and the manner in which the hosts become infected.

Fig. 102.—Longitudinal section of a
muscle of the domestic pig, with Sarcocystis
miescheriana. × 30. (After
Kühn.)
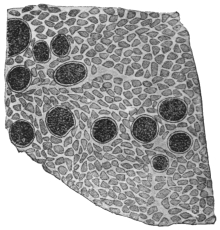
Fig. 103.—Transverse section of the
muscle of a pig, with Sarcocystis
miescheriana. × 38. (After Kühn.)
The Sarcosporidia usually appear as elongate, cylindrical, or
fusiform bodies, rounded at both extremities and of various lengths
and breadths (fig. 102). In some species they may be from 16 mm.
to 50 mm. long, as in the sheep and roebuck. These bodies are the
so-called sarcocysts or Miescher’s tubes. They lie in transversely
striated muscular fibres which they distend more or less. The forms
found in the connective tissue are apparently parasites which originally
inhabited the muscular fibres, and only on disintegration of the fibres
reached the connective tissue, where they grow to large oval or globular
bodies (fig. 105). The mammalian muscles usually infected are those
of the œsophagus, larynx, diaphragm, body-wall, and the psoas muscles.
The skeletal muscles may be affected in acute cases, as well as those
of the tongue and eye. The heart muscles are sometimes parasitized.
In fresh material cut into thin slices the parasites are frequently recognizable, even
with the naked eye, because of their yellowish-white colour. Under the microscope
they appear to be coarsely granular (fig. 103). Beginners may find some difficulty
in distinguishing them from other foreign bodies, such as dead and calcified encapsuled
Trichinæ, or from Cysticerci that have died and become calcified in the early
stages, more particularly as the Sarcosporidia also occasionally may become calcified.
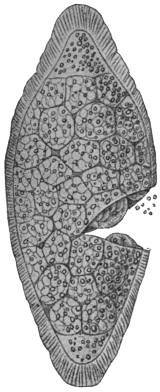
Fig. 104.—Sarcocystis miescheriana
from pig. Late
stage in which body is divided
into numerous chambers or
alveoli, each containing many
spores. (From Wasielewski,
after Manz.)
The Sarcosporidia are always enveloped in a membrane, which
is probably formed at an early stage. In a few cases it remains thin
and simple, in other cases a radially striated ectoplasmic layer is
present (figs. 104, 108), which has been variously described. From the
inner integument, which may be homogeneous or fibrous, thick or
thin, membranes or trabeculæ pass into the
interior of the body, forming anastomosing
partitions, and so producing a system of
chambers of various sizes that do not communicate
with one another (figs. 104, 108).
These chambers are occupied by sickle- or
bean-shaped bodies (spores or sporozoites),
or various developmental stages of them.
The oldest spores are found in the centre of
the Miescher’s tubes or trophozoites. If they
are not liberated they die there, so that the
central chambers of the tube are empty and
hollow.
In the youngest Sarcosporidia (40 µ in
length) from the muscles of the sheep there
occur, according to Bertram, small roundish
or oval cells (4 µ to 5 µ), the nuclei of which
are half their size, and are embedded in a
granular protoplasmic mass. In somewhat
larger, and therefore older, cylinders, the
investing membrane of which already shows
both layers, the cells have become larger (to
7 µ) and are more sharply outlined from each
other (fig. 106). These uninucleate cells may
be considered as pansporoblasts. In each
pansporoblast division of the nucleus occurs
(fig. 107), and meanwhile the pansporoblasts
become isolated within the chambers, the dividing partitions
of which originate from the granular protoplasm which is present
between the pansporoblasts. The numerous uninucleate daughter
forms produced within the chambers become spores direct (fig. 108).
The process commences in the centre of the cylinders or sarcocysts,
and then progresses towards the extremities, the parasites meanwhile
increasing in size, and new pansporoblasts being continually
formed at the extremities (fig. 107).
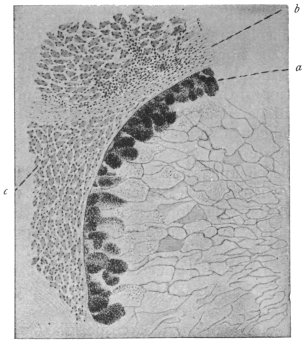
Fig. 105.—Transverse section of Sarcocystis tenella,
Raill. From the œsophagus of the sheep, Ovis aries.
× 38. a, marginal chambers filled with spores; b, connective
tissue of the œsophagus; c, muscles of the
œsophagus.
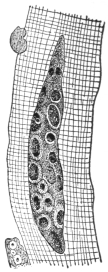
Fig. 106.—Young
Sarcocystis tenella of
the sheep, 47 µ
in length. (After
Bertram.)
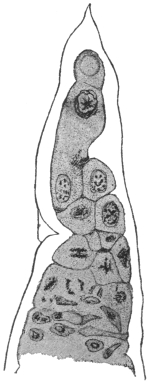
Fig. 107.—End of a
trophozoite of Sarcocystis
miescheriana from the
diaphragm of the pig,
showing division in pansporoblasts. × 800.
(After Bertram.)
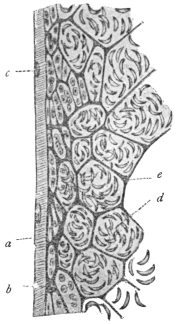
Fig. 108.—Sarcocystis
blanchardi of the ox. Longitudinal
section of sarcocyst or
Miescher’s tube. a, substance
of muscle fibre; b,
envelope of sarcocyst; c, muscle
nuclei; d, spores in chambers;
e, ground substance.
× 400. (From Wasielewski,
after van Eecke.)
The spores (sometimes called Rainey’s corpuscles), vary in shape
according to the species, but are also of different form individually.
They are mostly kidney-, bean- or sickle-shaped (fig. 109), and of
small size, sometimes reaching 14 µ by 3 µ to 5 µ. They are
apparently surrounded by a thin membrane, and at one extremity
(according to the discovery of L. Pfeiffer, confirmed by van Eecke,
Laveran and Mesnil) contain an obliquely striated body (fig. 109) often
homologized with the polar capsule, while the greater part of the spore
is taken up by the nucleate sporozoite. Several authors state that they
have also observed filamentous appendages (polar filaments) at one
end of the spores, and have seen two kinds of spores in the same
Sarcosporidium. Spores of various species of Sarcosporidia may
contain metachromatic granules, often centrally placed (fig. 109).
These granules may be metabolic or possibly may contain toxin
(see below).
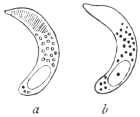
Fig. 109.—Spores of Sarcocystis
tenella, Raill. a, fresh,
showing the polar capsule;
b, stained, showing metachromatic
granules and nucleus.
× 1,000. (After
Laveran and Mesnil.)
The gymnospores of Sarcocystis muris, from the mouse, show active
boring movements when kept in saline solution warmed to 35° or
37° C. S. muris is very deadly to its host. From their structure the
spores do not appear to have great powers of resistance to external
conditions. They measure 12 µ by 3 µ to 4 µ or less.
Laveran and Mesnil (1899) isolated a toxin from S. tenella of
the sheep and called it sarcocystin. This substance is especially
pathogenic to experimental rabbits.
The duration of life of the Sarcosporidia is a comparatively long
one. The affected muscular fibres may remain intact and capable of
performing their functions for a long time,
but at last they perish, if the host lives long
enough. Thus the Sarcosporidia of the
muscles are then enveloped only by sarcolemma,
and finally, when this likewise disappears,
they fall into the intra-muscular
connective tissue. In many cases the Sarcosporidia
die off within their hosts, this,
according to Bertram, being brought about
by a disintegration of the spores in the central
chambers. In other cases the leucocytes
play a part in the destruction of the Sarcosporidia,
and sometimes it happens that lime
salts are deposited in and around the vacant cylinders.
In some places pigs, sheep, mice and rats are infected with
sarcosporidiosis to a remarkable extent, in certain cases almost reaching
100 per cent. Young animals also are infected, and perhaps
infection only takes place during youth.
Although the natural mode of transmission of the Sarcosporidia
remains to be determined, yet various experimental researches on
the problem are of interest and importance. Theobald Smith (1901)
found that mice could be experimentally infected with S. muris
by feeding them with the flesh of other infected mice. The incubation
period was a long one, namely forty to fifty days. Thus, on the
forty-fifth day after feeding young Sarcosporidia were found, and
seventy days after feeding spore formation began. Ripe spores were
found two and a half to three months after the commencement of
these experiments. This mode of infection—a cannibalistic one—hardly
seems likely to be the natural method for the infection of sheep
and ruminants generally. Smith’s researches have been confirmed.
Nègre225 (1910) found that the fæces of mice fed on infected muscular
tissue were infective to other mice when ingested by them. Negri226
infected guinea-pigs with S. muris by feeding them on infected mouse
flesh, and found that the parasite in guinea-pigs showed different
characters from those exhibited by it in mice. Darling227 also succeeded
in infecting guinea-pigs with S. muris, and Erdmann infected mice
with S. tenella (from the sheep).
According to Erdmann228 (1910) the Sarcosporidian spore germinates
in the intestine of the host, which has recently ingested infected
material. The spore liberates its contained toxin—sarcocystin—which
acts upon the adjacent intestinal epithelium, whereby the latter
is shed, and an amœbula creeps out of the spore. The amœbula is
able to penetrate the denuded area and get directly into the lymph-spaces
of the submucous coat of the intestine. The first period of
development, lasting some twenty-eight to thirty days, is said to be
passed in the lymph-spaces of the intestine. Later the amœbula
reaches a muscle fibre. Writing in May, 1914, Erdmann229 records the
appearance of small amœboid and schizogony forms six days after
infection of the host. Crawley230 (1913) controverts some of these
statements and considers that the Sarcosporidian spore, still sickle-shaped,
bores its way into the epithelial cells of the intestine and
comes to rest there. The spore then becomes round or elliptical, and
peripheral masses of chromatin appear within it, suggesting schizogony.
This happens about twelve hours after feeding, and in twenty-four
hours the spores appear to have left the intestine. More recently (May,
1914), Crawley231 considers that there is sexual differentiation among the
Sarcosporidian spores, a few hours after their ingestion by the host.
Interesting discussions have occurred as to the site of the toxic
sarcocystin within the spore. Metachromatic granules occur in the
middle of the Sarcosporidian spore (fig. 109), and the toxin may be
contained in these grains, as they disappear, according to Erdmann,
before the amœbula penetrates the denuded intestinal wall. However,
a polar capsule, containing a polar filament, may be present at one end
of a Sarcosporidian spore. Laveran and Mesnil described a striated
area at the more pointed end of the spore of S. tenella, which
area they consider to represent a polar capsule. Fantham232 (1913)
found a vacuole-like, polar capsule area in the spores of S. colii
from the African mouse-bird. The sarcocystin may be contained
in the polar capsule. The nucleus of the spore is generally at the
opposite, blunter end.
Again, various authors have stated that Sarcosporidian spores may
occur in the blood of the host at times. If so, then an intermediate
host may be concerned in their transmission. Perrin suggested that
Sarcosporidia might be spread by blow-flies and flesh-flies.
The classification of the Sarcosporidia as proposed by R.
Blanchard, which was based on their various habitats, can no longer
hold, because the same species may occur in the muscles as well as
in the connective tissue. For the present, the few species that are
known may be placed in one genus, Sarcocystis, Ray Lankester, 1882.
The following species of Sarcocystis are of interest:—
S. miescheriana, Kühn, 1865, in the pig.
S. bertrami, Doflein, 1901, in the horse.
S. tenella, Railliet, 1886, in sheep. S. tenella bubali in buffaloes in
Ceylon and Egypt.
S. blanchardi, Doflein, 1901, in cattle.
S. muris, Blanchard, 1885, in the mouse, to which it is lethal.
S. hueti, Blanchard, 1885, in the seal.
S. colii, Fantham, 1913, in the African mouse-bird, Colius
erythromelon.
Also various Sarcosporidia from antelopes, monkeys, opossum,
birds, the gecko and wall-lizard are known.
The spores of S. muris, S. bertrami, S. tenella, and S. colii can
multiply by longitudinal fission.
Sarcosporidia observed in Man.
(1) Lindemann233 found on the valves and in the myocardium of
a person who had died of dropsy certain brownish masses, 3 mm. in
length and 1·5 mm. in breadth which he regarded as gregarines. If
these were actually independent animal organisms it may be suggested
that they were Sarcosporidia. Rivolta (1878) named the species
S. lindemanni.
(2) Rosenberg234 found a cyst 5 mm. in length and 2 mm. in breadth
in a papillary muscle of the mitral valve of a woman, aged 40,
who had died from pleuritis and endocarditis. The cyst contained
no scolex nor hooklets of tænia. Numerous small refracting bodies,
round, oval or kidney-shaped, were found in a daughter cyst, as well
as sickle-shaped bodies. The description hardly appears to indicate
Sarcosporidia.
(3) Kartulis235 observed Miescher’s cylinders of various sizes in
the liver (?) and in the muscular system, of a Sudanese who had
succumbed to multiple abscesses of the liver and abdominal muscles.
This may be considered as the first actual case of the occurrence of
Sarcosporidia in man. Koch in 1887 described a case in Egypt.
(4) The case reported by Baraban and St. Remy236 was at once
demonstrated as certain. It related to a man who had been executed,
and in the laryngeal muscles of whom Sarcosporidia were found; the
length of the parasites varied between 150 µ and 1,600 µ, their
breadth between 77 µ and 168 µ. The affected muscular fibres were
distended to four times their normal thickness. This species was
described by Blanchard as “Miescheria” muris, but according to
Vuillemin, it was more probably Sarcocystis tenella of the sheep.
(5) Vuillemin has also described a case of Sarcosporidia found
in the muscles of a man who died from tubercle at Nancy. The
author considered that the parasite corresponded to S. tenella.
(6) Darling237 (1909) found Sarcosporidia in the biceps of a negro
from Barbados.
The Myxosporidia, Microsporidia, Actinomyxidia and possibly the
Sarcosporidia may be included within the section Cnidosporidia
(Doflein), since they possess spores containing polar capsules.
Order. Haplosporidia, Caullery and Mesnil.
The Haplosporidia are a group of organisms having both a simple structure and
life-history. The simplicity may represent a primitive condition or may be due to
degradation resultant on parasitism, and thus it is possible that the group is not
a homogeneous one. The order Haplosporidia was created by Caullery and Mesnil
in 1899, and includes parasites of rotifers, annelids (fig. 110), crustacea, fish, prochordates
and man. They may be present in the body cavity or alimentary tract,
and can also occur in the septum nasi of man, in the nervous system of Cephalodiscus,
and in tumours of fish.
As the name implies, the spores of the Haplosporidia are simple, without polar
capsules, and are uninucleate. In some genera, e.g., Haplosporidium, Urosporidium
(fig. 111) there is a spore-coat or sporocyst which may be elongate or spiny.
The developmental cycle of a Haplosporidian, such as Haplosporidium or Bertramia,
begins with a small, uninucleate cell, often rounded, possessing a cell membrane that
may be prolonged into processes. Growth takes place, coupled with an increase in
the number of nuclei, so that a multinucleate trophozoite is produced. Later, this
multinucleate trophozoite becomes segmented into a number of ovoid or spherical
pansporoblasts, which give rise to few (one to four) spores. Such a spore, when set
free, begins the life cycle over again.
More recently (1905–1907) two important organisms have been described and
included in this group, namely, Neurosporidium cephalodisci238 (Ridewood and
Fantham) from the nervous system of the prochordate, Cephalodiscus nigrescens,
and Rhinosporidium kinealyi (or seeberi) from the septum nasi of man. In the case
of Rhinosporidium and Neurosporidium, after the uninucleate spore has grown into
a multinucleate trophozoite, the latter segments into uninucleate pansporoblasts,
as in the preceding cases. A difference then occurs, for each pansporoblast
enlarges, its nucleus divides and a “spore-morula” is formed. Thus a multinucleate
pansporoblast or spore-morula, divided into many uninucleate sporoblasts (spore
mother cells) is produced, and each sporoblast without further change becomes
a uninucleate spore.
The Haplosporidia have therefore been divided by Ridewood and Fantham (1907)239
into two sections:—
(1) The Polysporulea, wherein the pansporoblast gives rise to a number of
spores (nine or more), e.g., Rhinosporidium, Neurosporidium.
(2) The Oligosporulea, wherein the pansporoblasts give rise each to a few (four)
spores or to only a single spore, e.g., Haplosporidium, Bertramia, Cœlosporidium,
Ichthyosporidium.
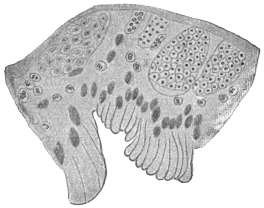
Fig. 110.—Haplosporidium heterocirri.
Section throughout wall of the Polychæte
worm, Heterocirrus viridis, showing various
developmental stages of the Haplosporidium.
× 550. (After Caullery and Mesnil.)
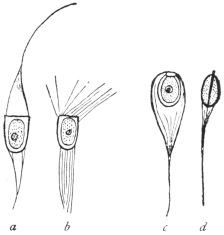
Fig. 111.—Haplosporidian spores.
a, b, Haplosporidium heterocirri. a,
fresh; b, after immersion in sea water;
c, d, Urosporidium fuliginosum.
× 1000. (After Caullery and Mesnil.)
Rhinosporidium kinealyi, Minchin and Fantham, 1905.
Rhinosporidium kinealyi, parasitic in man, must now be considered
in greater detail. This organism was found in nasal polypus
in India, and has since been recorded from the ear as small nodules
in the external auditory meatus. The Indian cases came from the
neighbourhoods of Calcutta and Madras, and the parasite has been
seen in Ceylon. Similar structures have since been described from
the United States and South America.
The Rhinosporidium polypus is said not to be particularly painful,
though nasal forms must interfere with breathing to some extent. The
first nasal polyp reported from India formed a vascular pedunculated
growth on the septum nasi and was about the size of a large pea or
raspberry. It was compared with a raspberry, being red in colour
with a number of small whitish dots upon its surface. When the
tumour was cut, a number of similar whitish dots were seen within.
These were the cysts of Rhinosporidium. According to Minchin
and Fantham240 (1905), they vary considerably in size and measure up
to 200 µ or 250 µ in diameter. Each possesses a cyst wall which varies
in thickness in different cysts. Its outer wall is always firm and
distinct, the inner limit being less definite at times. Each large cyst
is filled with numbers of spherical or oval bodies, showing every
gradation between small ones at the periphery and large ones at the
centre (fig. 112). Roughly, three zones of parasites can be distinguished
in a large cyst, a peripheral set consisting of the youngest
parasites, an intermediate group and a central, oldest zone. A large
cyst may possess a pore for the egress of its contents. Some of the
cysts show polar distribution of the zones.
The youngest forms of Rhinosporidium are difficult to detect.
They are small, granular masses, round, ovoid or irregular and at times
even amœboid in appearance. These are young trophozoites. They
increase in size, but encystment occurs early, the outer layer becoming
firm so that the organisms have a definite contour. Each is soon
multinucleate and the cytoplasm segments around the nuclei. The
cyst thus becomes full of uninucleate pansporoblasts or sporonts,
with a peripheral layer of undifferentiated protoplasm. The pansporoblasts
grow in size. In the larger cysts the formation of
pansporoblasts progresses at the expense of the peripheral layer of
protoplasm, which, however, continues to grow, so that the cyst as
a whole increases in size. The pansporoblasts at first are uninucleate
(fig. 112, a), and then undergo nuclear multiplication. This is well seen
in the intermediate zone of parasites, where the pansporoblasts show
first one, then two, then four or more spores (fig. 112, b), while in the
oldest centrally placed pansporoblasts, about a dozen or sixteen
closely packed spores (fig. 112, c), can be seen. The spore is small
and rounded, and its nucleus is clear and distinct. The fully formed
pansporoblast or spore morula becomes surrounded by a membrane.
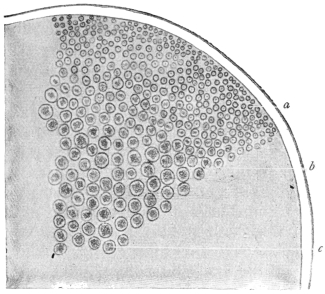
Fig. 112.—Rhinosporidium kinealyi. Portion of ripe
cyst containing pansporoblasts of various ages. × 480.
(After Minchin and Fantham.)
Certain of the cysts have been found in a ruptured condition,
whereby the spores have been liberated into the surrounding tissue.
It is almost certain that the spores serve for the auto-infection of the
host, for though the tumours of Rhinosporidium seemed to have
been removed entirely, it has been found that they recur, some
minute fragment of the parasite having probably been left behind.
The method whereby the parasite reaches new hosts has not yet been
determined, and it would be of interest if its life-history could be
more fully investigated.
The Asiatic specimens of R. kinealyi were first described in detail
by Minchin and Fantham (1905) from material briefly reported to the
Laryngological Society of London in 1903, by O’Kinealy. Material
obtained by Dr. Nair, of Madras, was described by Beattie241 in 1906.
This material came from Cochin. Castellani and Chalmers have
found similar polypi in Ceylon.
Wright242 has described the parasite from Memphis, Tennessee.
Seeber243 in 1896 described nasal polypi in Buenos Ayres, and in 1900
Wernicke named the parasite therein Coccidium seeberi. Seeber’s
parasite is a Rhinosporidium, R. seeberi, and may ultimately be found
to be the same as R. kinealyi. Ingram244 reports Rhinosporidium cysts,
with pores in the cyst walls, in conjunctival polypus and in papilloma
of the penis in India. Zschokke has reported the presence of Rhinosporidium
in horses in South Africa.
Class IV. INFUSORIA, Ledermüller, 1763.
The Infusoria (or Heterokaryota, Hickson, or Ciliophora, Doflein)
include the Ciliata and the Suctoria. A few authorities, including
Braun, raise the Suctoria (or Acinetaria) to separate rank as a class,
but this is not widely followed.
The body of the Ciliata usually is bilaterally symmetrical and is
enveloped in a cuticle which has numerous openings for the
protrusion of the cilia. Most kinds have a fixed shape, whilst
changes in the form of others are brought about by the contractions
of the body substance. The latter exhibits hyaline ectoplasm, in
which myonemes, and occasionally also trichocysts (minute spindle-shaped
bodies) appear, and granular endoplasm which may contain
numerous vacuoles. The cilia, on whose various arrangements
the classification is based, are always processes of the ectoplasm.
Their form varies; they may be hair-like, or more rarely thorn-like,
spur-like, or hook-shaped; undulatory membranes also may occur,
which are probably composed of fused cilia.
With the exception of some of the parasitic species, an oral cavity,
peristome or cytostome, is always present. It is frequently beset
with cilia or provided with undulatory membranes, which help to
waft the food inwards; sometimes there is an anal aperture (cytopyge)
generally placed at the opposite pole of the organism. A cytopharynx
fringed with cilia or sometimes with a specialized supporting
apparatus is connected with the peristome. Vacuoles form round
the ingested food, and in many species a constant rotation goes on
in the endoplasm. Often one, and sometimes two contractile vacuoles
are present, the frequency of the pulsations of which depends on the
surrounding temperature. Sometimes special conducting channels
lead to the vacuoles, or there are outlet channels leading to the exterior.
There is in almost every case a large nucleus (macronucleus),
and lying close up to it a small nucleus (micronucleus). The form
of the large nucleus varies according to the species. Numerous
nuclei are not very common, but these occur in Opalina, which lives
in the hind-gut of amphibia, and is also distinguished by the absence
of an oral aperture.
Reproduction is effected by binary fission; less commonly, after
encystment, by multiple division, or by budding. The divisions can
be repeated many times, but finally cease, and then the conjugation
of two specimens brings about a regeneration, particularly of the
nuclei. Numerous examinations (Bütschli, Hertwig, Maupas, Calkins)
have demonstrated that after two individuals have associated by
homologous parts of the body, the micronucleus separates from
the macronucleus, becomes larger and divides twice by mitosis, so
that four micronuclei are present in each one of the two individuals
forming the couple. Three of these nuclei perish and become
absorbed, the fourth gradually passes to the portion of protoplasm
connecting the two conjugants, which has originated by absorption
of the cuticle at the point of contact of the conjugants. After a
further division one micronucleus of each conjugant passes over
into the other conjugant, and fusion ensues between the two micronuclei
of each individual. Complicated changes and divisions may
occur, but only the main principles can be noted here. A new nuclear
body is thus formed in each conjugant, and soon divides into two.
Of the segments thus produced one becomes a micronucleus, and
one or several of the others, as the case may be, form or amalgamate
into a new macronucleus, the old macronucleus usually perishing or
becoming absorbed during the conjugation. Usually, sooner or later,
the two conjugants separate, or may have separated already, and again
multiply independently by fission until a series of divisions by simple
fission is again followed by conjugation. The theoretical significance
of conjugation cannot be dealt with fully here. It may be
remarked, however, that the macronucleus plays no part in it, but
governs entirely the metabolism of the Infusorian, whereas the
micronucleus is essentially a generative nucleus from which macro-
and micro-nuclei are again and again produced.
Encystment amongst the Infusoria is very general, and is essentially
a means of protection when the surrounding medium dries up.
Doubtless these cysts are frequently carried long distances by the wind,
which explains the wide geographical distribution of most species.
Also, multiplication often takes place in the encysted condition.
Some Infusoria live a free life, others are sedentary; the latter
form colonies in fresh as well as in salt water. Numerous species
are parasites of various lower and higher animals,245 and a few also are
parasitic in man.
The Prague zoologist, v. Stein, introduced a classification of the
Infusoria that has been almost universally adopted. It is founded
on the different position of the cilia on the body. Though, no
doubt, artificial, it is a convenient system. Bütschli has compiled a
better one.246 But for our purpose Stein’s system is sufficient:—
Order 1. Holotricha, Infusoria with cilia that are evenly distributed over the
entire body.
Order 2. Heterotricha, ciliated all over like the Holotricha, but having stouter
cilia about the peristome.
Order 3. Hypotricha, only ciliated on the ventral surface.
Order 4. Peritricha, with only a ring of spiral cilia, mostly sedentary.
The Infusoria observed in man belong to the order Heterotricha,
with few exceptions.
Genus. Balantidium, Claparède et Lachmann.
Heterotrichous Infusoria of oval or bag-like form and almost circular on transverse
section; the anterior extremity narrowed, the posterior end broad and
rounded off, or also narrowed; the peristome starting at the anterior end is
broadest there and becomes narrower as it gradually obliquely approaches towards
the posterior extremity. There are coarse cilia along the entire left border and the
anterior part of the right border. Longitudinal striation is distinct and regular.
There are two contractile vacuoles on the right, and occasionally also two or more to
the left. The anus (cytopyge) is terminal. The macronucleus is usually horse-shoe
or kidney-shaped, sometimes oval; the micronucleus contiguous. Reproduction is
by binary fission and conjugation, and encystment occurs. The cysts are spherical
or oval. These ciliates are parasitic in the large intestine of human beings and
pigs, in Amphibia, and in the body cavity of polychæte Annelida.
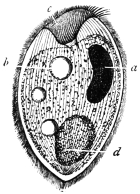
Fig. 113.—Balantidium
coli. a, nucleus;
b, vacuole;
c, peristome; d, bolus
of food. (After
Leuckart.)
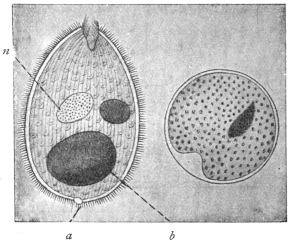
Fig. 114.—Balantidium coli, free and encysted;
a, anus or cytopyge; n, macronucleus; b, bolus of
food. (After Casagrandi and Barbagallo.)
Balantidium coli, Malmsten, 1857.
Syn.: Paramæcium coli, Malmsten, 1857.
The body is oval, 60 µ to 100 µ in length (up to 200 µ according
to Janowski), and 50 µ to 70 µ in breadth. The peristome is funnel-shaped
or contracted, the anterior end being then broadened or
pointed according to the degree of contraction (figs. 113, 114). The
ecto- and endo-plasm are distinct, the latter is granular, containing
drops of fat and mucus, granules of starch, bacteria, and occasionally
also red and white blood corpuscles. There are usually two contractile
vacuoles, seldom more. The anus (cytopyge) opens at the
posterior extremity. The macronucleus is bean- or kidney-shaped,
rarely oval; the micronucleus is spherical.
Balantidium coli lives in the large intestine of man, in the rectum of
the domestic pig, and has been found in monkeys. It propagates by
transverse division, but conjugation and encystment are known to
take place.247 Transmission to other hosts is effected by the cysts of
the parasite (fig. 114).
Balantidium coli, first seen by Leeuwenhoek, was described
by Malmsten in 1857 in a man aged 35 years, who had two
years previously suffered from cholera, and since then had been
subject to diarrhœa. The examination showed an ulcer in the rectum
above the mid sphincter ani, in the sanguineous purulent secretion of
which numerous Balantidia were swimming about. Although the
ulcer was made to heal, the diarrhœa did not cease and the stools
contained numerous Balantidia, the number of which could only be
decreased by extensive enemas of hydrochloric acid.
The second case related to a woman who was suffering from
severe colitis, and who died ten days after admission. The malodorous,
watery evacuations contained innumerable Balantidia, in
addition to pus, and at the autopsy the anterior portion of the large
intestine was found to be infested with them.
Subsequently this parasite has often been observed in human
beings, and various cases have been recorded. These occurred in
Russia, Scandinavia, Finland, Cochin China, Italy, Germany, Serbia,
Sunda Islands, Philippine Islands, China, and in other parts of Asia
and in America. Other cases were reported by Askanazy, Ehrnroth,
Klimenko, Nagel, Koslowsky, Kossler, Waljeff, Strong and Musgrave,
Glaessner248 and others. Sievers found B. coli very common in Finland.
In the majority of the cases described by Sievers from Finland,
and in other cases from Central Europe, the patients suffered from
obstinate intestinal catarrh, which did not always cease even after
the Balantidia had disappeared. On the other hand, Balantidia have
occasionally still been found to persist, though in small numbers, after
the catarrh has been cured. Some authors, nevertheless, do not
regard Balantidia as the primary cause of the various diseases of the
large intestine, which often commence with the development of
ulcers, but they consider that they may aggravate these diseases and
render them obstinate. According to Solowjew, Askanazy, Klimenko
and Strong and Musgrave, however, the parasites penetrate the intestinal
wall, and give rise to ulcerations which may extend deeply into
the submucosa, and even be found in the blood and lymphatic vessels
of the intestinal wall. According to Stokvis, B. coli occurs also in
the lung; at all events this author states that he found one living and
several dead paramæcia (?) in the sputum of a soldier, returned from
the Sunda Islands, who was suffering from a pulmonary abscess.
Sievers has shown that B. coli might occur in persons not suffering
from intestinal complaints, but E. L. Walker249 (1913) states that every
person parasitised with B. coli is liable sooner or later to develop
balantidian dysentery.
Since Leuckart confirmed the frequent presence of B. coli in the
rectum of pigs, and corresponding observations were made in other
countries, the pig is universally considered to be the means of the
transmission of Balantidium to man. The encysted stages only serve
for transmission, because, according to all observations, the free
parasites have a very small power of resistance. They perish when
the fæces have become cool; they cannot live in ordinary, slimy, or
salt water. As they are killed by acids even when much diluted, they
cannot pass through the normal stomach alive except under the most
unusual circumstances. The pigs, in whose intestines the Balantidium
appears to cause little or no disturbance, evacuate numerous encysted
Balantidia with the fæces, and their occasional transference to man
brings about their colonization there, but perhaps only when a disease
of the colon already exists.
Experimental transmission of the free parasites to animals (per os
or per anum) yielded negative results, even in the case of pigs.
Casagrandi and Barbagallo (1896), however, had positive, as well as
negative, results. They employed healthy young cats, or cats in which
catarrhal entero-colitis had been artificially induced (which in other
experiments is apt to cause the death of the animals experimented upon
in about six or seven days), or finally cats that had dilatation of the
rectum with alkaline reaction of the fæces. An attempt to infect three
healthy cats by injecting human fæces containing Balantidium into the
rectum proved negative, in so far as the fæces of the experimental
animals had an acid reaction and contained no Balantidia, but at the
autopsy performed eight days after infection a few encysted parasites
were found in the mucus of the ileum. In the case of four cats
suffering from entero-colitis, into which human fæces containing
Balantidia were introduced per os, Balantidium cysts were found in
the fæces three days after the last ingestion. Great numbers, moreover,
were found in the cæcum and the posterior part of the small
intestine at the autopsy of the animals, which died about eight days
after the commencement of the experiments. Actual colonization,
therefore, was not effected in either series of experiments. Free or
encysted Balantidia of pigs were used for further experiments. The
experiments proved negative when fæces containing cysts were injected
into the rectum of healthy cats (three experiments), or cats (two)
suffering from spontaneous intestinal catarrh, or when such material
was introduced per os into three healthy cats. In the case of two
cats with intestinal catarrh artificially produced, a small number of the
active Balantidia injected into the rectum remained alive. Larger
quantities of fæces containing encysted Balantidia were introduced
into two other cats affected with the same complaint. These, certainly,
did not appear in the fæces, but small numbers, free and alive, were
found in the cæcum. Similarly, encysted Balantidia were introduced
into two cats with dilated rectum, and whose fæces had an alkaline
reaction. In these cases no parasites appeared in the fæces, but three
and five days later, when the two animals were examined, a very small
number were discovered free in the large intestine. Klimenko did
not succeed in infection experiments with B. coli on young dogs,
whose intestines had been artificially affected by disease.
More recent experiments by Brumpt have shown that young
sucking pigs can be infected with Balantidium from infected monkeys
(Macacus cynomolgus) and suffer heavily from the same, whereas the
Balantidium of the pig is rarely harmful to its host. This and previous
experiments may be thought to suggest that there are perhaps
several pathogenic species, and also that harmless strains of Balantidium
may occur. At the same time, it must be remembered that
a large proportion of the cases recorded of Balantidian colitis occur
among swineherds and butchers, that is, among people in frequent
contact with pigs. Morphologically, there are practically no differences
between the Balantidia found in man, monkeys and pigs, and
it is probable that one species only, under slightly different environmental
conditions, may be responsible for the colitis observed. In
any case, efficient prophylactic measures should be taken against
balantidiasis in countries where it may occur, by confining the pigs
and not allowing them to run in yards and dwellings.
E. L. Walker (1913) has given a good summary of work on
balantidiasis. His own researches in the Philippines showed that
monkeys could be infected by Balantidia both from pigs and men.
Parasites may appear in the stools only at infrequent intervals. He
believes that the ciliates are the primary etiologic factor in the
symptoms and lesions of balantidian dysentery.
Behrenroth (1913) has given an interesting account of Balantidium
coli and its pathogenic significance.
Balantidium minutum, Schaudinn, 1899.
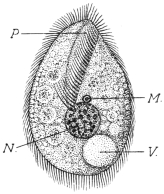
Fig. 115.—Balantidium minutum.
P, peristome; N, nucleus;
M, micronucleus; V, contractile
vacuole. Food vacuoles are represented
in the endoplasm. (After
Schaudinn.)
The body is of oval form, with the anterior extremity pointed, and
posterior extremity broad and rounded (fig. 115). The length is 20 µ
to 32 µ, and the breadth is 14 µ to 20 µ. The peristome, which is
fissure-like, extends to the centre of the body (fig. 115). The right
lateral border of the peristome is beset with cilia the same length as
those of the body, the left side terminates in a thin hyaline membrane
that extends towards the back, and can
pass over to the right side. A row of
longer and stronger cilia (cirri) are on the
left border of the peristome. The cuticle
is refractile, the ectoplasm hyaline and
the endoplasm granular, with numerous
food vacuoles.
A single contractile vacuole lies dorsally
and to one side at the posterior
extremity. The macronucleus, which is
always spherical, is central and is 6 µ to
7 µ in diameter. The micronucleus, close
in front of it, only measures 1 µ (fig. 115).
The cysts are oval.
These parasites were found in numbers
in the evacuations of a man aged 30,
who was born in Germany and had repeatedly travelled between
Hamburg and North America, where he made long stays. The patient
came to the Charité in Berlin to seek advice for constipation alternating
with diarrhœa accompanied by abdominal pain.
A second case (the parasite of which was described as Colpoda
cucullus by Schulz) was observed in a patient in the same institution.
As, in both cases, the parasites only appeared during the diarrhœa,
and disappeared as soon as the fæces had assumed a normal consistency,
or could only be demonstrated in a few encysted specimens,
it may be assumed that the small intestine or the duodenum is
their habitat.
Genus. Nyctotherus, Leidy, 1849.
Flat, heterotrichous Infusoria, kidney- or bean-shaped. The peristome commences
at the anterior pole of the body and extends along the concave side to the
middle, where the oral aperture is situated. The cytopharynx is oblique and is more
or less curved. The cytopyge is at the posterior extremity, where a single contractile
vacuole is also situated. The macronucleus is almost in the centre of the parasite.
The members of this genus live parasitically in the intestine of amphibia, insects
and myriapods, and at least one species is found in man.
Nyctotherus faba, Schaudinn, 1899.
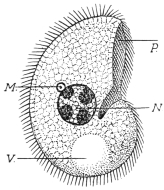
Fig. 116.—Nyctotherus faba. P,
peristome; N, nucleus; M, micronucleus;
V, contractile vacuole.
(After Schaudinn.)
The body is bean-shaped, and a little flattened dorso-ventrally.
It is 26 µ to 28 µ long, 16 µ to 18 µ broad, and 10 µ to 12 µ thick
(fig. 116). The peristome is on the right
border and extends to the middle; at
the left there are large adoral cilia, the
cilia on the right border not being larger
than those on the body. The cytopharynx
is short, slightly curved and
turned backwards. The contractile vacuole
is large, spherical, situated at the
posterior extremity, and its contents are
voided through the anus at its left. The
macronucleus is in the centre of the body;
it is globular (6 µ to 7 µ in size), and
contains four or five chromatin masses.
The micronucleus lies close to it, and is
spherical or somewhat elongate measuring
1 µ to 1·5 µ (fig. 116). The cysts are oval.
This species has hitherto only been seen once in the same patient
in whom Balantidium minutum was discovered.
Nyctotherus giganteus, P. Krause, 1906.
Under the name Balantidium giganteum n. sp., P. Krause described
an Infusorian which was repeatedly observed with Trichomonas intestinalis
in the alkaline evacuations of a typhoid patient in Breslau.
The body is ovoid, narrower and rounded anteriorly and broader
and stunted posteriorly. The peristome lies to one side; the macronucleus
is bean-shaped, the micronucleus small and globular; one
or two vacuoles are present. The anus is at the farther end. The
organism is 90 µ to 400 µ long, 60 µ to 150 µ broad (fig. 117). After a
prolonged stay outside the body, it becomes rounded and encystment
occurs. In the thermostat the Infusoria remain alive at 37° C. for five
weeks.
The species, however, hardly belongs to Balantidium, but to all
appearances is a Nyctotherus and is distinguished from N. faba by the
difference in size.
[Nyctotherus] africanus, Castellani, 1905.
In the fæces of a native of Uganda who suffered from sleeping
sickness and diarrhœa and had in his intestine Ascaris lumbricoides,
Trichocephalus trichiurus and Ancylostoma duodenale, Castellani found
a curiously shaped Infusorian, 40 µ to 50 µ long, and 35 µ to 40 µ
broad, with spherical macro- and micronucleus and a contractile
vacuole (fig. 118). He included the organism in the genus Nyctotherus,
perhaps wrongly, or the parasite may have been deformed. After the
patient’s death the same parasite was found in the intestine and
especially in the cæcum.
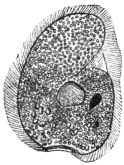
Fig. 117.—Nyctotherus
giganteus.
(After Krause.)
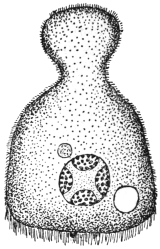
Fig. 118.—Nyctotherus
africanus. (After Castellani.)
G. Lindner, in Cassel, studied certain peritrichal Infusoria (stalkless Vorticella),
and connected them, probably incorrectly, with the most varied diseases of man and
domestic animals, even with Sarcosporidia of pigs. It may be mentioned that
according to a communication by letter from Schaudinn, Vorticella may be found
in freshly evacuated fæces, but always only after the administration of a water
enema. In spite of this, several other investigators mention Vorticellæ as
intestinal parasites of man.
The Chilodon dentatus (Ehrenberg) recorded in 1903 by J. Guiart as a parasite
of man, which may be found in all infusions, can hardly have lived in the man
from whose fæces it was cultivated, but may represent a chance admixture both in
the fæces and the cultivations. C. uncinatus was also found as a chance parasite
of man by Manson and Sambon. According to Doflein250 (1911) certain Chilodon-like
organisms have been found by Selenew in prostate secretions in gonorrhœa.
Other species of the genus Chilodon are known, but only as ectoparasites (e.g.,
Chilodon cyprini, Moroff, 1902, from the skin and gills of diseased carp).
A number of other parasitic Ciliates are known, among which Ichthyophthirius
multifiliis, destructive to fish, is important. It lives in the skin and the layers
immediately below it, forming small whitish pustules which may become confluent.
The pustules are most common on the head and fins, but occur also on the eyes and
gills of the host. The young parasite, which is one of many formed in a cyst, is very
small. At first it is free swimming, but soon attaches itself to the skin of a fish. It
bores inwards and becomes surrounded by the irritated skin. There it attains a
relatively large size, being 500 µ to 750 µ and occasionally more in diameter. The
body has a rounded terminal mouth, short cytopharynx and a number of minute
contractile vacuoles. The macronucleus is large and horseshoe-shaped; the small
micronucleus is only seen in the very young animal. When full grown, the organism
encysts and forces its way to the surface and bursts through, leaving a small, gaping
wound behind. The cyst sinks to the bottom of the water, nuclear multiplication
occurs and a number of young parasites are produced, which leave the cyst and
either attack new hosts or else perish.
Opalina ranarum, parasitic in the rectum and urinary bladder of frogs and toads,
shows great degradation and simplification due to parasitism, possessing no separate
micronuclei, no cytostome, cytopharynx or cytopyge. It has many macronuclei, and
is a large parasite. During summer and autumn nuclear multiplication followed by
division of the body occurs, the process being repeated after the daughter forms
have grown to the size of their parent. In spring, the Opalina divide rapidly, but
do not grow much before dividing again. Finally, tiny forms, containing three to
six nuclei, encyst and pass from the host with the fæces. As these latter are greedily
devoured by tadpoles, the Opalina gain new hosts in which they develop.
THE CHLAMYDOZOA.
The name Chlamydozoa was proposed by Prowazek in 1907 for a
number of minute, problematic organisms (fig. 119) believed to be
the causal agents of certain diseases in man and animals, such as
vaccinia and variola, trachoma, inclusion blenorrhœa in infants,
molluscum contagiosum, and bird epithelioma contagiosum. Other
diseases possibly due to Chlamydozoa251 are hydrophobia, measles,
scarlet fever, foot-and-mouth disease, the “Gelbsucht” disease of silkworms,
and perhaps even typhus (Prowazek, 1913). The subject is
difficult and controversial and can only be briefly discussed here.
It is known that the viruses in all these diseases can pass through
ordinary bacterial filters, that is, they belong to the group of “filterable
viruses.” At such periods the organisms are extracellular or free. It
is also known that in many of these cases the virus produces definite
and characteristic reaction-products or cell-inclusions in the infected
cells, during the intracellular phase of the life-history of the organism.
As the organisms to be considered are problematic, it will be convenient
to summarize their history:—
(1) Cell-inclusions, usually named after their discoverers, have
been found in certain diseases, thus: In vaccinia Guarnieri’s bodies,
in scarlet fever Mallory’s bodies, in hydrophobia Negri’s bodies, in
trachoma Prowazek’s bodies occur.
(2) At first these characteristic cell-inclusions were considered to
be actual parasitic organisms causing the diseases in question. The
bodies received zoological names and attempts were made to work
out their supposed development cycles. The supposed parasites of
vaccinia and variola were referred to a so-called genus Cytoryctes,
those of hydrophobia to Neuroryctes, of scarlet fever to Cyclasterium,
while those of molluscum contagiosum were referred to the Coccidia.
Calkins in 1904 studied in detail the cell-inclusions of vaccinia and
small-pox, calling them Cytoryctes variolæ, Guarnieri. Calkins considered
that in the stratified cells of the epidermis they passed through
two cycles, the one cytoplasmic, the other intranuclear. The first is
the vaccinia cycle, the second the pathogenic (intranuclear) variola
cycle. It is hardly necessary to follow all Calkins’ stages here.
Negri (1909) described a cycle for Neuroryctes hydrophobiæ. Calkins
refers both Cytoryctes variolæ and Neuroryctes hydrophobiæ to the
Rhizopoda.
Siegel (1905) described quite different organisms under the name
Cytorhyctes. He listed several species: C. vacciniæ; of vaccinia and
small-pox, C. scarlatinæ of scarlet fever, C. luis of syphilis (this is
probably the granule stage of Treponema pallidum), and C. aphtharum
of foot-and-mouth disease.
(3) The afore-mentioned views were criticized, and the bodies
were not considered to be living organisms but merely reaction
products or cell-inclusions due to the effects of the virus on the host
cells. Thus Guarnieri’s bodies were stated to consist of extruded
nucleolar or plastin material, having no developmental cycle. It was
further asserted that infection could be produced by lymph in which
Guarnieri’s bodies had been destroyed. Similar assertions have been
made regarding the Negri bodies, and others. The Cytoryctes,
Neuroryctes, etc., are considered, according to these views, to be
degeneration products of the nucleus or to be of a mucoid nature.
(4) More recently a positive belief has gained ground that there
are true parasitic organisms causing these diseases, and that the
parasites are very minute, being termed Chlamydozoa by Prowazek
and Strongyloplasmata by Lipschütz.
The Chlamydozoa are characterized by (a) their very minute size,
smaller than any bacteria, so that they can pass through bacterial
filters; (b) they pass through intracellular stages, in the cytoplasm
or the nucleus of the host cell, producing therein the reaction
products or inclusions in the cell already recorded as characteristic
or diagnostic of the diseases produced; (c) they pass through definite
developmental cycles. Such a cycle consists essentially of growth
and division. The mode of division of the Chlamydozoa resembles
that of the centriole of a cell, by the formation of a dumb-bell-shaped
figure. Two dots are observed connected by a fine line or strand
which becomes drawn out and finally snaps across the middle.
Prowazek and Aragão (1909) working on smallpox in Rio de Janeiro
found that the chlamydozoal granules passed through a Berkefeld
filter and that the filtrate was virulent. But if an “ultra-filter” were
used, i.e., one coated with agar, then the granules were retained and
the filtrate was no longer virulent. The surface of the ultra-filter was
found to contain many granules.
The Chlamydozoa are parasites of epiblastic tissues (e.g., epidermal
cells, nerve cells, conjunctival cells).
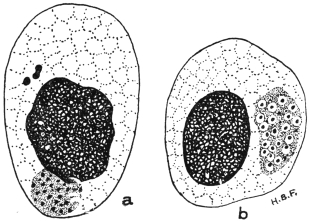
Fig. 119.—Chlamydozoa. Trachoma bodies in infected
epithelial cells of the conjunctiva. (a) initial bodies (above)
and cluster of elementary bodies (touching the nucleus);
(b) cluster of granules surrounded by mantles. × 2,000
approx. (Original. From preparation by Fantham.)
The life-history of a Chlamydozoön (fig. 119), such as that of
vaccinia, is, according to Prowazek, Hartmann and their school, as
follows:—
1. The infection begins with elementary bodies or elementary
corpuscles which live at first extracellularly. An elementary body is
a minute speck of chromatin, apparently devoid of cytoplasm, which
can pass through a bacterial filter. It can enter a host cell, but the
entry is not a process of phagocytosis.
2. Inside the host cell the elementary body grows in size, and
becomes an initial body (fig. 119, a).
3. A reaction on the part of the host cell results, for nucleolar,
plastin substance is extruded from the cell-nucleus and surrounds
the parasitic initial body. The latter is thus enveloped in a mantle
(hence the name Chlamydozoa, from χλαμὑς, a mantle), and the
characteristic cell-inclusion (Guarnieri’s body, Negri’s body, etc.) is
produced. The nucleolar, mantle substance probably represents the
“cytoplasm” of Cytoryctes, described by Calkins.
4. The body next breaks up into a number of smaller bodies
known as initial corpuscles. These, in their turn, divide by simple
division (in the manner already described) into numerous elementary
bodies (fig. 119). Thus, the life-cycle is completed.
The Chlamydozoa are, then, the minute granules inside the body of
the Cytoryctes variolæ or the Neuroryctes hydrophobiæ, so that the whole
body of the Cytoryctes or Neuroryctes corresponds to the mantle and
parasite of the Chlamydozoön. The Cytoryctes group is said to
cause destruction of the host cell. The Cytoöikon group (e.g.,
trachoma bodies) causes proliferation of the host cell.
In September, 1913, Noguchi252 described the cultivation of the
parasite of rabies in an artificial medium, similar to that used by him
for the cultivation of Spirochæta recurrentis. The cultures were stated
to be infective to dogs, rabbits and guinea-pigs. Levaditi, in
December, 1913, stated that he had succeeded in cultivating spinal
ganglia of rabid monkeys in monkey plasma.
Noguchi and Cohen (November, 1913)253 have succeeded in
cultivating the so-called trachoma bodies, or at any rate bodies
very closely resembling them morphologically. The medium
employed was Noguchi’s ascitic fluid and rabbit kidney medium, as
used for spirochætes. The coarser cultural forms stained blue with
Giemsa’s solution, the finer ones stained red. Attempts to infect
monkeys from the culture tubes failed.
From their behaviour on treatment with such reagents as saponin,
bile and sodium taurocholate, Prowazek considers that the Chlamydozoa
approach the Protozoa.
PROTOZOA INCERTÆ SEDIS.
Sergentella hominis, Brumpt, 1910.
Et. and Ed. Sergent in 1908 found vermiform bodies about 40 µ
long by 1 µ to 1·5 µ broad in the blood of an Algerian suffering from
nausea and cold sweats, without other symptoms. The bodies were
pointed at each end, with a somewhat ill-defined nucleus in the
middle. Their systematic position is doubtful.
Note.—An Appendix on Protozoology will be
found on pp. 733–752. This has been prepared in
order to incorporate a number of new additions to
knowledge made since the body of the book was
printed off.
B. PLATYHELMINTHES, or Flat Worms.
BY
J. W. W. STEPHENS, M.D., B.C., D.P.H.
Definition: Bilaterally symmetrical animals without limbs, the form of which
is leaf or tape-like, rarely cylindrical, and whose primary body cavity (segmentation
cavity) is absent, the cavity being filled by a mesenchymatous tissue (parenchyma).
The mouth is either situated at the anterior end of the body, or is shifted more
or less backwards on to the flat ventral surface. The alimentary canal consists of a
short fore-gut, which is frequently provided with a muscular pharynx, and of a simple
forked or branched mid-gut; there is neither a hind-gut nor an anus; in one class,
the Cestodes, the alimentary canal has entirely disappeared except for muscular
remnants in the scolex.
The INTEGUMENT OF THE BODY consists either of a ciliated epithelium of only
one layer (Turbellaria), or of a cuticle and gland-like cells embedded in the
parenchyma, or subcuticular layer (Cestodes, Trematodes). The dermo-muscular
layer consists of annular, longitudinal, and even diagonal fibres, while the
parenchyma is traversed by dorso-ventral fibres.
The central NERVOUS SYSTEM, which is embedded in the parenchyma of the
body, consists of cerebral ganglia, united together in the shape of dumb-bells, and
of two or more longitudinal MEDULLARY FASCICLES, often forming transverse
anastomoses. Organs of sense usually occur only in the free-living species, more
rarely during the free-living stages of a few parasitic species and in a few ectoparasitic
forms.
[In Platyhelminthes simple eye-spots frequently occur, and in a few an auditory
vesicle.]
Blood-vessels and definite RESPIRATORY ORGANS are lacking [except in
Nemertinea]; the EXCRETORY APPARATUS (formerly termed water-vascular system)
is typical of the entire class. It commences in the interstices of the parenchyma,
with peculiar terminal cells (ciliated funnels), which will be described later (p. 219),
the capillary processes of which go on uniting into larger branches, and finally form
two large collecting vessels, which, sometimes separately and sometimes united,
open to the exterior through one, two, or numerous pores.
Nearly all the Platyhelminthes are HERMAPHRODITIC, and in nearly all there are,
in addition to the ovaries producing ova, other glands attached to the female genital
apparatus, namely, the vitellaria or yolk glands, which provide a substance termed
yolk, which serves as nourishment for the embryo. The fully formed eggs have
shells and are “compound,” i.e., composed of the egg or ovarian cell, which is
surrounded by numerous yolk cells or their products of disintegration. The two
sexual openings usually lie close together, frequently in the fundus of a genital
atrium; they are rarely separated from one another. Shell glands also usually
occur (p. 221).
Reproduction is sexual, often, however, combined with asexual methods of
propagation (segmentation, budding). The Platyhelminthes live partly free in fresh
or salt water, exceptionally also on land. The greater part, however, live as
parasites on or in animals.
Classification of the Platyhelminthes.
Class I.—Turbellaria (or Eddy Worms). Flat worms for the most part, free
living, and always covered with a ciliated epithelium.
Order 1.—Rhabdocœlida, gut unbranched.
Order 2.—Tricladida, gut with three main branches.
Order 3.—Polycladida, a central gut with lateral cæca. Development direct
or through metamorphosis. They live in fresh and salt water or on land;
very seldom as parasites.
Class II.—Trematoda (Sucking Worms254). [Usually known as Flukes.—F. V. T.]
Flat worms, living as ecto- or endoparasites, that are only ciliated in the larval
condition, and in their adult state are covered with a cuticle, the matrix cells of
which lie in the parenchyma. They have either one, a few, or several suckers,255
and frequently also possess chitinous fixation and adhesive organs. The intestine
is single, but generally bifurcated, and not uncommonly there are transverse anastomoses
between the forks or diverticula on them. Excretory organs double, with
two orifices at the anterior extremity or a single one at the posterior end.
Development takes place by a metamorphosis or alternation of generations (p. 283).
These worms are almost always hermaphroditic, with two or more female and one
male sexual orifice. They live, almost without exception, as parasites on vertebrate
animals, but the intermediate generations are passed in molluscs.
Class III.—Cestoda (Tapeworms). Endoparasitic flat worms without an alimentary
canal. The larval stages are rarely ciliated, but are usually provided with six
spines; the adult worm is covered with a cuticle, the matrix cells of which are embedded
in the parenchyma. The body consists of a single segment (Cestodaria) or a
chain of segments, in which case it consists of the scolex and the segments containing
the sexual organs (proglottides) (Cestodes s. str.). The scolex is provided with
various adhesive and fixation organs, and there are calcareous corpuscles in the
parenchyma. Excretory organs symmetrical, opening at the posterior end. These
worms are always hermaphroditic, and then possess one or two female and one
male sexual orifice. During development a larval intermediate stage (“measle”)
occurs and almost always in a different host to that in which the adult sexual worm
lives. The adult stage is parasitic in vertebrate animals; but the larval stage may
occur in invertebrates.
Class II. TREMATODA, Rud.
These worms are usually leaf- or tongue-shaped, but also barrel-shaped
or conical; they vary from 0·1 mm. to almost 1 m.256 in
length; most of them, however, are small (5 mm. to 15 mm.). The surface
on which the orifice of the uterus and the male sexual opening
are situated is termed the ventral surface; the oral aperture, which
also acts as anus, is always at the anterior end in the sub-order
Prostomata (p. 230), but in the sub-order Gasterostomata it is ventral.
Suckers are always present and occur in varying numbers and
positions at the anterior extremities as well as on the ventral surface,
and occasionally on the lateral margin and on the dorsum; the
beginning of the intestine (mouth) is always surrounded by a sucker
in the Prostomata.
In or near the suckers there may be chitinous hooks, claws or
claspers, or the surface of the body is more or less covered with spines,
scales or prickles; in one genus (Rhopalias) there are projectile tentacles
beset with spines on the sides of the anterior part of the body.
The body of adult Trematodes is covered by a homogeneous layer
of varying thickness, which either lies directly over the external
layer (basement membrane) of the parenchyma, or over the muscles
embedded in the parenchyma. This investing membrane (cuticle)
arises from pear-shaped or spindle-shaped cells arranged singly or in
groups (which lie between or internal to the diagonal muscles), and
is connected with them by processes; these cells one may regard as
epithelial cells which have sunk down, or possibly as parenchymatous
cells. An epithelium of one layer is also found on the body of
young stages, but it disappears during growth, and only occasionally
do its nuclei persist until adult life. In its place we then find the
cuticle, which, moreover, extends into all the body openings more or
less deeply.
It is thus a debatable point whether the “investing layer” of flukes
is a cuticle—that is, consists of modified epithelial cells—or whether it is
a basement membrane, i.e., compressed and modified connective tissue
cells; in this latter case the true epidermis and cuticle have been cast
off. In the former case the epidermal cells are the pear-shaped cells
referred to above. According to recent authors it consists of two
parts, an outer true cuticle and an inner basement membrane. There
are also unicellular cuticular glands, lying isolated or in groups, which
are termed cephalic, abdominal, or dorsal glands according to the
position of their orifice.
The PARENCHYMA is a connective substance, the structure of
which is still a matter of dispute. It consists, according to some
authors, of multipolar cells, the offshoots from which anastomose with
each other so that a network, permeating the entire body and encompassing
all the organs, is produced. There exists also, as part of it, a
homogeneous matrix, in the form of lamellæ and trabeculæ that border
small cavities communicating with each other and filled with fluid.
According to other authors, the parenchyma of the Trematodes consisted
originally of cells, of which, however, only the cell membranes
remain, while the protoplasm has been liquefied except for small residua
around the nucleus. Between these cells an intercellular mass has
appeared. By partial absorption of the walls, adjoining spaces unite,
and the originally flat cell walls become transformed into trabeculæ.
According to this view the cavities filled with fluid are intra-cellular,
according to the former view inter-cellular. Pigment cells occur only
in a few species.
The MUSCULAR SYSTEM of the Trematodes is composed of (1) a
dermo-muscular tube, (2) the dorso-ventral or parenchymal muscles,
(3) the suckers, and (4) the special muscles of certain organs.
The dermo-muscular tube, which lies fairly close to the cuticle,
consists of annular, diagonal, and longitudinal fibres which surround
the entire body in one or several layers, and as a rule are more strongly
developed on the ventral surface as well as in the anterior part of the
body. The MUSCLES OF THE PARENCHYMA are found chiefly in the
lateral parts of the body and pass through the parenchyma in a
dorso-ventral direction; their diverging brush-like ends are inserted on
the inner surface of the cuticle (fig. 120).
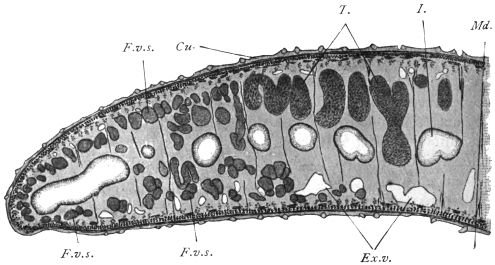
Fig. 120.—Half of a transverse section through Fasciola hepatica, L. 25/1. Cu., Cuticle
with scales; under the cuticle are circular muscles, and adjoining them the longitudinal and
diagonal muscles; internal to the latter are the matrix cells of the cuticle; I., gut; the
other similarly contoured cavities are gut diverticula that have been transversely or obliquely
sectioned; F.v.s., vitellaria; Ex.v., excretory vessels; T., testes; Md., median plane; the
fibres passing from the ventral to the dorsal surface are the muscles of the parenchyma. The
parenchyma itself is omitted.
The suckers are specially differentiated parts of the dermo-muscular
tube. Their concave inner surface is lined by the continuation
of the cuticle and their convex external surface is covered
by a more dense tissue that frequently takes the form of a refractive
membrane, thus separating them from the parenchymal muscles.
The principal mass of the suckers consists of muscular fibres which
run in three directions—equatorial, meridional and radial. The
equatorial fibres correspond to the annular muscles, the meridional
fibres to the longitudinal muscles, and the radial fibres to the muscles
of the parenchyma; the radial fibres are always the most strongly
developed. The function of these muscles is evident from their
position; the meridional fibres flatten the suctorial disc and diminish
the depth of its cavity, so that the internal surface may adhere to
the object to be held; if the equatorial fibres now contract, the
sucker rises by elongating longitudinally, and its inner surface is
drawn in by the contraction of the radial muscles. Thus the
sucking disc becomes adherent. Usually also there is a sphincter
at the border of the suckers, which plays its part during the act of
adhesion by constricting in a circular manner that part of the mucous
membrane to which it is attached. The loosening of the fixed sucker
is effected by relaxation chiefly of the radial fibres, by the contraction
of the meridional fibres and certain bundles of muscles situated at
the base and at the periphery of the suckers. The connective and
elastic tissues between the muscles of the suckers probably also take
part in the process.
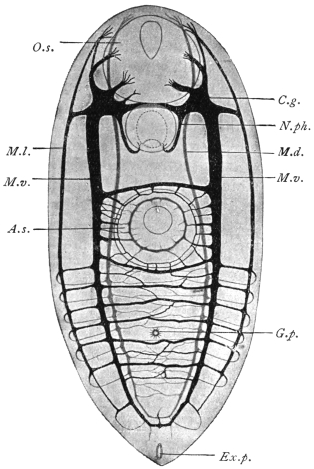
Fig. 121.—Harmostomum leptostomum, Olss., an immature
specimen from Helix hortensis. Nervous system, according to
Bettendorf. A.s., ventral sucker; C.g., cerebral ganglion;
Ex.p., excretory pore; G.p., genital pore; O.s., oral sucker;
M.d., dorsal medullary nerve; M.l., lateral medullary nerve;
N.ph., pharyngeal nerve; M.v., ventral medullary nerve.
Magnified.
Of the muscles of the organs which have developed from the
parenchyma muscles we may briefly mention those bundles that are
attached to certain parts of the genital apparatus, to the suckers,
to the hooks and claws, and also, at all events in Fasciola hepatica,
to the spines. The sheaths used for the projection of the tentacles
of the Rhopaliadæ are also muscular.
The contractile elements consist of fibres of various lengths
that are mostly parallel to one another, and frequently anastomose;
a cortical substance finely fibrillated can usually be distinguished
from an internal homogeneous mass; large nucleated cells of uniform
size are always connected with them; these have been variously
interpreted, but have been proved to be myoblasts, one or more
of their processes constituting the muscular fibres.
The MOVEMENTS of the Trematodes consist in alterations of form
and position of the body, as well as in creeping movements.
In the NERVOUS SYSTEM (fig. 121) can be distinguished a cerebral
portion as well as strands (medullary strands) running from it,
and peripheral nerves. The cerebral portion always consists of
two large ganglia situated in the anterior end of the body which
pass dorsally over the œsophagus and are connected by means
of a broad and thick commissure composed of fibres only. From
each ganglion three nerves run anteriorly—the inner and dorsal
nerve for supplying the anterior dorsal part of the body; the
median and ventral for the oral sucker; and the exterior and lateral
likewise for the supply of the sucker.
In a similar manner three strands run backwards from each
ganglion—one dorsal, one lateral and one ventral. The dorsal and
ventral strands become united and curve backwards; the symmetrical
lateral strands are connected by means of transverse commissures,
the number of which vary according to the species. Such commissures
also exist between the lateral and the two other strands on each
side. There are ganglion cells along the entire course of the posterior
cords, more particularly at the points of origin of the commissures.
There also appears to be in addition a fourth anterior and posterior
pair of nerves, the front pair for the oral sucker and the hind pair
for the pharynx.
The peripheral nerves, which spring from the posterior strands
as well as from the commissures, either pass directly to the muscular
fibres or to the sensory cells that are situated at the level of the
subcuticular cells, or they reach these after the formation of a plexus
situated immediately beneath the dermo-muscular layer; the processes
directed outwards terminate in small vesicles in the cuticle.
As to other ORGANS OF SENSE, simple eyes, two or four in
number, are known in several ectoparasitic species as well as in a
few free-living larval stages (Cercariæ) of endoparasitic forms. In the
adult stage, however, they usually undergo complete atrophy.
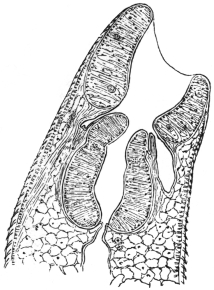
Fig. 122.—Median section through
the anterior part of Fasciola hepatica:
the oral sucker, pharyngeal pouches,
pharynx, œsophagus, cuticle with
spines, and the body parenchyma.
The ALIMENTARY CANAL commences with an oral aperture,
generally terminal or sub-terminal (ventral) at the anterior extremity,
which leads into an oral cavity usually surrounded by a sucker;
the œsophagus, of various lengths, is directed backwards and is
generally surrounded by a muscular pharynx (fig. 122). In some
cases there exists between the sucker and pharynx, pharyngeal pouches
(præpharynx). Sooner or later the
intestine divides into two lateral
branches directed backwards, both of
which end blindly (cæca) at the same
level.257 In many ectoparasites (Monogenea
[p. 222]) a connection exists
between the genital glands and one of
the intestinal branches (ductus vitello-intestinalis
[fig. 123]).
The oral cavity, pharyngeal pouches,
pharynx, and œsophagus are lined with
a continuation of the cuticle of the
body; the gut cæca are lined with tall
cylindrical epithelium (fig. 120). The
œsophagus and intestinal branches
often have also one layer of annular
and longitudinal muscles; the pharynx
has essentially the structure of a sucker
(fig. 122).
The accessory organs of the alimentary
canal consist of groups of unicellular SALIVARY GLANDS that
discharge into the œsophagus in front of or behind the pharynx, or
even into the pharynx itself.
The food of the Trematodes consists of mucus, epithelial cells, the
intestinal contents of the hosts, and often also of blood, and this not
only in those species living in the vascular system, but also in species
living as ectoparasites or in the intestine or biliary passages of their
hosts.
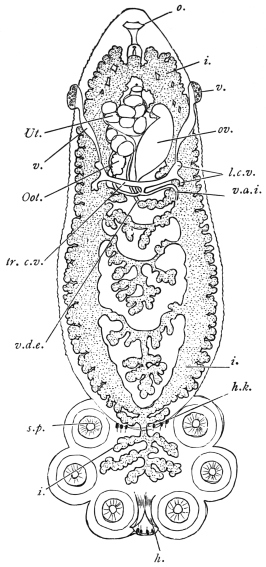
Fig. 123.—Polystomum integerrimum, a
monogenetic fluke from the urinary bladder
of the frog. i., intestine; h., large hooks
of the sucking disc; h.k., smaller hooklets;
l.c.v., longitudinal vitelline ducts; o., oral
orifice; Oot., oötype; ov., ovary; s.p., suckers
of the disc; tr.c.v., transverse vitelline ducts;
Ut., uterus with ova; v., entrance to the
vagina; v.d.e., vas deferens; v.d.i., ductus
vitello-intestinalis; the vitellaria and testes
are not shown. Magnified. (After Zeller.)
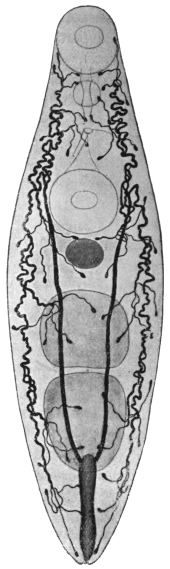
Fig. 124.—Allocreadium isoporum,
Looss. Excretory apparatus.
Of the other organs, the
oral sucker, pharynx, genital
pore, ventral sucker, ovary and
testes are shown; the cylindrical
excretory bladder is in
the posterior end. 38/1. (After
Looss.)
The final products of assimilation dissolved in the fluids of the
body are distributed throughout the parenchyma and are thence
expelled by a definite tubular system (excretory apparatus, proto-nephridia,
formerly also termed the water-vascular system). This
system, which is distributed throughout the entire body (fig. 124), is
symmetrically developed, and, in the ectoparasitic Trematodes, it
opens, right and left, at the anterior end on the dorsal surface; in all
other flukes, however, it opens singly into the excretory pore (foramen
caudale) at the centre of the posterior border; in those cases, however,
where a sucker is present at the posterior end, as in the Amphistomata,
the excretory pore is situated on the dorsal surface close in front of
the sucker.
The EXCRETORY SYSTEM258 consists of several parts: (1) of the more
or less numerous terminal “flame” cells or funnel cells (figs. 124, 125);
(2) of the capillaries ending in them; (3) of
larger vessels receiving the capillaries; and (4)
of the excretory bladder. Terminal cells and
capillaries may be compared to unicellular
glands with long excretory ducts; the cellular
body (fig. 125) is comparatively large, stretched
longitudinally, more rarely transversely, and
provided with numerous processes, that are
lost in the parenchyma; within is a conical
cavity (analogous to the secretory cavity of
unicellular glands) which is continued directly
into the structureless capillary; at its blind
end is a bunch of cilia projecting into the
cavity, and which, during life, shows a flickering
motion (ciliary flame). The nucleus is
situated in the protoplasm of the terminal cell
at its blind end.
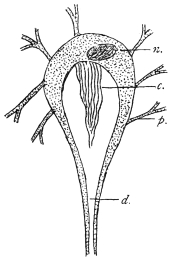
Fig. 125.—Terminal flame
cell of the excretory system.
n., nucleus of cell; c., bundle
of cilia forming the “flame”;
p., processes of cell extending
into parenchyma; d., excretory
capillary. (Stephens.)
The entire apparatus thus begins blindly—i.e.,
within the terminal cells, to which must
be ascribed the capacity of taking up from the fluid that permeates
the parenchyma the products which are first collected into their own
cavities and thence excreted by means of the capillaries and vessels.
The vessels possess definite walls, consisting of a membrane and
a nucleated protoplasmic layer. They unite at many points on either
side, and again pass into other canals (COLLECTING TUBES), which
finally, travelling towards the posterior end, discharge into the
excretory bladder (fig. 124).
The form and size of the bladder vary much according to the
different species, but it always possesses its own flattened epithelium,
surrounded by circular and longitudinal muscles, the circular muscles
forming a sphincter around the opening. Frequently also the
structure of the bladder extends to the tubules discharging into
it, which therefore are not to be regarded as separate “vessels,”
but rather as tubular diverticula of the bladder, directed anteriorly.
In some few species the diverticula also branch and the branches
anastomose, so that a network of tubules ensues which receives the
vessels or capillaries. In such cases there are also ciliary tracts in
the tubules.
The contents of the entire apparatus
usually consist of a clear or sometimes
reddish fluid; in some species
there are larger or smaller granules, and
occasionally also concretions occur.
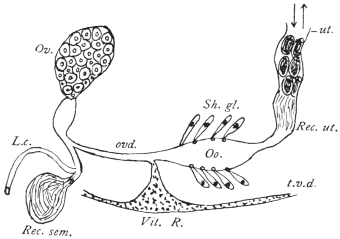
Fig. 126.—Diagram of female genitalia. Ov.,
ovary; ovd., oviduct; L.c., Laurer’s canal;
Rec. sem., receptaculum seminis; Vit. R., vitellarian
reservoir; t.v.d., transverse vitelline duct;
Oo., oötype; Sh. gl., shell gland; Rec. ut.,
receptaculum uterinum; ut., uterus. (The various
parts are not to the same scale.) (Stephens.)
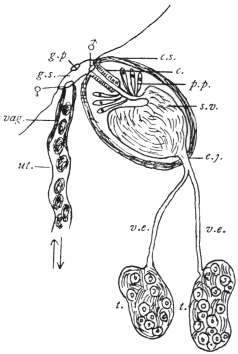
Fig. 127.—Diagram of male and part
of female genitalia. ut., uterus; vag.,
vagina; ♀, opening of vagina; g.s., genital
sinus; g.p., genital pore; ♂, opening
of ejaculatory duct or vas deferens; c.s.,
cirrus sac; c., cirrus; p.p., pars prostatica;
s.v., seminal vesicle; e.j., ejaculatory duct
or vas deferens; v.e., vas efferens; t., testis.
(Stephens.)
Sexual Organs.—Nearly all the Trematodes are hermaphrodites,
and only a few (Schistosomidæ, Koellikeria) are sexually differentiated.
The sexual organs usually lie in the “central field” limited
by the gut cæca; the vitellaria, on the other hand, are, as a rule,
external to the gut cæca in the “lateral fields.”
The male apparatus259 is composed of two variously formed
testes (fig. 127) (globular, oval, indented, lobed, or ramified), which
may lie side by side or one behind the other; from each testicle
a tube (vas efferens) originates; sooner or later, both tubes as a rule
unite to form the ejaculatory duct or vas deferens, which is frequently
enclosed in a muscular CIRRUS SAC, or more rarely passes directly into
the genital pore. The cirrus, which is the thick muscular terminal
portion of the vas deferens, can be everted and protruded from the
cirrus sac and serves as an organ of copulation. The walls of the
muscular portion of the tube (the cirrus) are attached to the walls of
the cirrus sac, and hence when the sac contracts the cirrus cannot
be protruded except by evagination of its lumen. Opening into the
middle portion of the vas deferens, and as a rule enclosed in the cirrus
sac, is found a mass of unicellular glands (prostate), the vesicula
seminalis (which is likewise within, or may also be outside the sac)
being the dilated first portion of the vas.
The female genitalia (fig. 126) consist of an ovary, usually situated
in front of the testes, the form of which varies according to the
species, the usually double vitellaria, the ducts and a number of
auxiliary organs; the short oviduct directed towards the centre
arises from the ovary, and is connected in the median line with
the excretory duct of the vitelline glands. These grape-like glands
possess longitudinal excretory ducts, which assume a transverse
direction behind the ovary, unite together at the median line and
form a single duct, often dilated into a vitelline receptacle, that unites
with the oviduct. Near this point, moreover, there frequently opens
a canal (Laurer’s canal) which begins on the dorsal surface, and
on the inner end of which a vesicle filled with sperm (receptaculum
seminis) usually occurs (fig. 126). Moreover, there are also numerous
radial unicellular glands (shell glands) at or beyond the point of
junction of the oviduct, vitelline ducts and Laurer’s canal. In this
portion of the duct (oötype), which is usually dilated, the ovarian
cells are fertilized, surrounded with yolk cells and shell material,
and as ova with shells they pass into the uterus (a direct continuation
of the oviduct), which, with its many convolutions, occupies a
larger or smaller portion of the central field, and runs either direct
to the genital pore or, forming convolutions, first runs posteriorly
and then bends forward (descending and ascending limbs). In both
cases the terminal part lies beside the cirrus pouch and discharges
beside the male orifice either on the surface of the body or into a
genital atrium. The terminal portion of the uterus, which is often
of a particular structure, serves as a vagina (METRATERM).
The cirrus sac may include (1) the genital atrium (i.e., the
common sinus, into which the vas deferens and vagina may open),
or (2) a variable extent of the vas from cirrus to seminal vesicle. Thus
the latter may be outside the sac. In the absence of a sac, the genital
sinus may be surrounded by a pseudo-sucker, as in Heterophyes (in
some cases the ventral sucker itself, from its close proximity to the
genital pore, serves as an accessory copulatory organ). In other cases
copulatory organs are formed by hooks projecting into the lumen
of the terminal portion of the vas.
The GENITAL PORE, which is the opening from the genital sinus on
to the surface, is generally situated at or near to the median line on
the ventral surface and in the anterior region of the body; in most
of the Distomata it is in front of the ventral sucker, in other cases,
e.g., in the Cryptocotylinæ, it is behind.260
The spermatozoa do not differ essentially in their structure from
those of other animals; the ovarian or egg cells are cells without
integument and contain a large nucleus and a little protoplasm; the
vitellaria also produce nucleated cells, in the plasm of which there
are numerous yellow yolk granules; the yolk cells detach themselves,
like the ovarian cells, from the ovarium, and pass into the oviduct
to surround each ovarian cell in the oötype. They disintegrate sooner
or later in the completely formed egg and are utilized as food by the
developing embryo.
Development of the Trematodes.
(1) Copulation.—Observation has demonstrated that the one or
two vaginæ occurring in the ectoparasitic Trematodes are utilized
as female organs of copulation, and that the copulation is cross;
it is also known that Laurer’s canal, which was formerly generally
regarded as the vagina, has only quite exceptionally, if at all, served
the digenetic Trematodes as such—it appears to be homologous with
the canalis vitello-intestinalis of the Monogenea261—but the terminal
portion of the uterus, termed the metraterm, is used for copulation.
Cross-copulation occurs as well as auto-copulation and auto-fecundation.
The spermatozoa subsequently pass through the entire uterus,
which is still quite short at the time the male organs are matured; the
maturation of which, as usually is the case in hermaphrodites, precedes
that of the female organs. It is only later with the onset of egg formation
that the uterus is fully developed. Copulation, however, takes place
also in the case of fully grown forms with completely developed uteri.
Fig. 128.—Ovum of Fasciola
hepatica, L., cut longitudinally.
The lid has been lifted in the
process. Within the egg are
numerous yolk cells, and at the
lid end there is the still unsegmented
ovum (dark). 240/1.
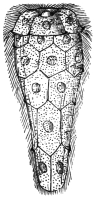
Fig. 129.—Miracidium of
Fasciola hepatica that has just
hatched from the egg, with a distinct
cuticular ciliated epithelium.
Magnified. (From Leuckart.)
(2) Formation of the Ova.—The ovarian cells arising from the ovary
first become mature after their entry into the oötype by the formation
of three polar bodies, fertilization then taking place. At the same
time as the ovarian cell a number of yolk cells from the vitellarium
and secretion, drop by drop, from the shell gland reach the oötype.262
The shell is then formed during the generally active contractions of
the oötype walls and then passes on into the uterus. In the uterus of
the endoparasitic trematodes the eggs accumulate more and more,
often in large quantities, while in ectoparasitic species generally only
one or some few eggs can be found. The completed ova are of
various forms and sizes. They are mostly oval, at all events in the
digenetic trematodes, and the yellowish or brown shell is provided
with an opening at one pole which is closed by a watch-glass-shaped
lid (operculum). Appendages (filaments) on the shell—at one or both
poles—are uncommon, but are the rule in the ova of the Monogenea
(ectoparasitic species).
(3) Deposition of the Ova.—Soon after their formation, the
Monogenea (ectoparasitic trematodes) deposit round the place of
their attachment on the skin or the gills or other organs of their
hosts, eggs which attach themselves by means of their filaments.
The embryonic development thus takes place outside the parent.
This also holds good for the eggs of many endoparasitic species,
although as a rule in these the eggs are always retained for a
longer time in the uterus. Moreover, they usually here undergo a
part or a whole of their development, and are eventually deposited
in those organs in which the adult forms are parasitic, but this is
not always the case, as the egg, e.g., of F. hepatica appears in bile (and
fæces) quite unchanged. By the natural passages they eventually get
out of the body, and in cases where such do not exist, as in the case
of the blood-vessels, the eggs pass out by means of the kidneys.
(4) The embryonic development, after irregular segmentation of the
ovum into a number of blastomeres, leads to the formation of a solid
blastosphere or morula, which is surrounded by a cellular investing
membrane (yolk envelope), while the principal mass of the cells forms
the embryo, which uses for its nourishment the yolk cells, which have
in the meantime disintegrated (cf. footnote, p. 223). Usually, after
the ova have reached water the embryos hatch out, leaving the yolk
envelope in the egg-shell; in other cases, however, the embryos only
hatch out after having been subjected to the influence of the intestinal
juices, that is to say, in the intestine of an intermediate host which has
ingested with its food the ova that have escaped from the primary host.
(5) The post-embryonic development of the Trematodes is accomplished
in various ways; the process is the most simple in the ectoparasitic
species (Monogenea), the young of which should certainly
be regarded as larvæ, because they possess characteristics (cilia,
simple gut, etc.) that are lacking in the adult worms, but which,
nevertheless, pass into the adult state direct after a relatively simple
metamorphosis. In the Holostomata,263 a group found chiefly in the
intestine of aquatic birds, and which rarely occur in other vertebrates,
the ova develop in water. The young are ciliated all over, and, after
having entered an intermediate host (leeches, molluscs, arthropods,
amphibians, fishes) living in the water, they undergo a metamorphosis
into a second larval stage; they then encyst and await transmission
into the final host, where they become adult Metastatic
trematodes, i.e., trematodes without asexually produced generations
(p. 229).
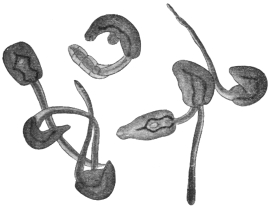
Fig. 130.—A group of cercariæ of Echinostoma sp. (from
fresh water). 25/1.
In the remaining so-called digenetic trematodes (p. 230) one or two
asexual generations interpose between the miracidium and terminal
stage, so that quite a number of adult worms may originate from
one egg. Usually the young, which are termed MIRACIDIA264 (fig. 129),
hatch in water, where they move with the aid of their cilia. Sooner
or later they penetrate into an intermediate host, which is always
a snail or a mussel, and while certain of their organs disappear,
they grow into a gutless germinal tube (SPOROCYST, fig. 131). These
are simple elongated sacs with a central body cavity. They may or
may not have excretory tubules. In these, according to the species,
the larval stages (CERCARIÆ) that will ultimately become adult worms
are produced, or another intermediate generation is first formed, viz.,
that of the REDIÆ265 (figs. 132, 133), which are always provided with an
intestine, and these then give rise to cercariæ (figs. 130, 134). The
cercariæ, as a rule, leave their host and move about in the water
with the assistance of their rudder-like tails. After a little time, however,
they usually again invade an aquatic animal (worms, molluscs,
arthropods, fishes, amphibians), then they lose their tails and become
encysted (fig. 135); here they wait until they attain, together with
their host, the suitable terminal host, and in this new situation they
establish themselves and reach maturity. Or, again, the cercariæ may
themselves encyst in water or on foreign bodies (plants) and wait until
they are taken up directly by the terminal host, e.g., sheep.
Accordingly the following conditions are necessary for the completion
of the entire development: (1) The terminal host in which
the adult stage lives; (2) an intermediate host into which the
miracidia penetrate and in which they become sporocysts; (3) a
second intermediate host in which the cercariæ become encysted.
In certain species, as in Fasciola hepatica, this second host is omitted,
as the cercariæ spontaneously encyst on plants, or again (in other
species) encystment may occur within the first intermediate host, when,
in fact, the cercariæ (which in this case do not acquire an oar-like tail)
do not swarm out of, but encyst themselves within their sporocysts.
The development, moreover, may be further complicated by rediæ
appearing in addition to the sporocysts, though this occurs in the
first intermediate host and not in a second one.
Animals that harbour adult digenetic Trematodes thus become
infected by ingesting encysted cercariæ, which either occur (1) in
certain animals (second intermediate hosts) on which they feed, or
(2) in water, or (3) on plants, or finally (4) in the first intermediate
host; whereas animals harbouring encysted cercariæ have been directly
infected by the corresponding tailed stage, and animals harbouring
germinal tubes (sporocysts or rediæ) have been infected by the
miracidia.
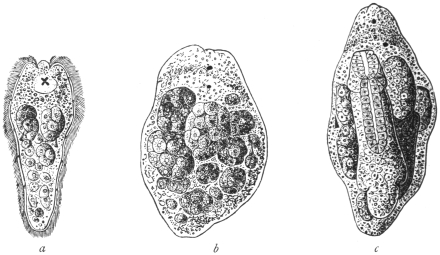
Fig. 131.—Development of Fasciola hepatica, L. a, the miracidium in optical section
showing cephalic lobe, x-shaped eye-spot resting on the cerebral ganglion, two germ balls;
below each of these a flame cell, and still lower germ cells lying in a cavity (primitive body
cavity). b, young sporocyst with two eye-spots, and germ balls; the cells lining the cavity
are not shown. c, older sporocyst with a young redia. Magnified. (After Leuckart.)
Thus certain species of ducks and geese become infected with
Echinostoma echinatum by devouring certain water-snails (Limnæus,
Paludina) in which the encysted cercariæ occur. Oxen become infected
with Paramphistomum cervi (= Amphistomum conicum) by swallowing
with water, cysts of this species which occur at the bottom of puddles
and pits. Sheep are infected with Fasciola hepatica by eating grass
to which the encysted cercariæ of the liver-fluke are attached; our
song-birds infect themselves or their young with Urogonimus macrostomus
by tearing off pieces containing the corresponding sporocysts
which are full of encysted cercariæ from snails (Succinea amphibia),
which act as the first intermediate hosts, and eating, or offering their
young these pieces.
(1) The MIRACIDIA of the digenetic Trematodes are comparatively
highly organized, and the mode of their formation from the
segmentation cells of the ovum is only imperfectly known. They have
a cuticular epithelium (fig. 129) entirely or partly covered with cilia,
beneath this a dermo-muscular tube composed of circular and longitudinal
muscles; also, a simple gut sac with an œsophagus, occasionally
also with pharynx, salivary glands and boring spine, also
a cerebral ganglion on which, in some species, there are eyes
(fig. 131, a). As to the excretory organs, they are represented by two
symmetrically placed terminal flame cells, with excretory vessels
opening separately; there is a
more or less ample (primary) body
cavity between the parietes of the
body and the gut; from the cellular
parietal lining of this cavity
single cells (germ cells) become
free (fig. 131, a, b), and become
rediæ or cercariæ.
[The germ cells of the miracidium
and the germ balls of the
sporocyst arise, according to some
observers, by further division of
undifferentiated blastomeres; according
to others from the cells of
the lining wall of its body cavity.
It is from these free germ balls
that the redia stage is developed.
[In the germ ball or morula
appears an invagination, giving
rise to the cup-shaped gastrula
stage. This elongates and forms
the REDIA (fig. 131, c).
[In the interior of the redia
cells are budded off and develop
into gastrulæ, as in the case of the
sporocyst. These become a fresh
generation of rediæ or give rise
to the third stage (CERCARIA).]
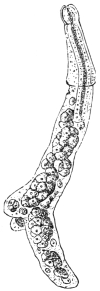
Fig. 132.—Young
redia of Fasciola hepatica,
with pharynx
and intestine, with a
circular ridge anteriorly
and a pair
of processes posteriorly
and masses of
cells (germ balls) in
the interior. Magnified. (From
Leuckart.)
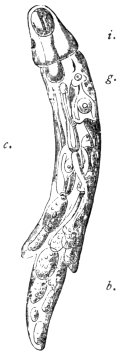
Fig. 133.—Older
redia of Distoma
echinatum, with rudimentary
intestine
i.; cercariæ, c.; germ
balls, b.; and birth
pore, g. Magnified.
(2) The SPOROCYSTS, on the contrary, which are produced direct
from the miracidia, are very simple, as all the organs of the latter
disappear, even to the muscles and excretory organs, during or after
penetration into the intermediate host, whereas the budded and still
budding cells of the wall of the (primary) body cavity continue to
develop rapidly and form germ balls. The sporocysts when fully
developed have the appearance of tubes or fusiform bodies with
rounded edge; they are frequently of a yellow colour. Their length
rarely exceeds a few millimetres; in some species their size increases
exceedingly through proliferation, and they then occupy a large portion
of the body of the intermediate host.
(3) The REDIÆ (figs. 132, 133), on the other
hand, are more cylindrical and always have a
simple intestine of varying length, provided
with a pharynx; they likewise possess, situated
near the circular ridge, a “birth pore” which
serves for the exit of the cercariæ originating
within them.
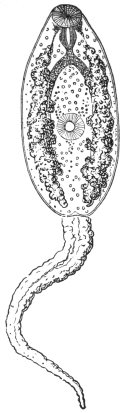
Fig. 134.—Cercaria of
Fasciola hepatica; the cutaneous
glands at the side of
the anterior body. Magnified.
(After Leuckart.)
(4) The CERCARIÆ266 are very different; typically
they consist of the anterior body and the
oar-like tail at the posterior end (fig. 134). The
former, even to the genitalia, has the organization
of the adult digenetic Trematodes, and
thus allows the easy recognition of at least the
characters of that large group to which the
species in question belongs. On the other
hand, however, there are also organs that are
lacking in the adult form, such as, in many, the
boring spine in the oral sucker, or the eyes
situated on the cerebral ganglion; moreover,
also, cutaneous glands (fig. 134), the secretion of
which forms the cyst membrane. The oar-like
tail may be long or short (stumpy-tailed cercaria)
or entirely absent; its free end may be
partly split (furcate cercaria), or split to its base
(bucephalus); in various forms also the anterior
end of the tail is hollow, and has enclosed
within it the anterior body, which is otherwise
free. The size also of the cercaria belonging
to the different species is very diverse; in
addition to forms swimming in the water that
have the appearance of minute milky-white
bodies, there are forms which measure as much
as 6 mm. in length.
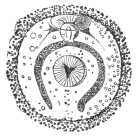
Fig. 135.—Encysted cercaria
of Fasciola hepatica.
Magnified. (After Leuckart.)
The encysted cercariæ (fig. 135) are globular
or oval, and are surrounded by a homogeneous
membrane, which may be striated or contain granules. The tail is
always cast off when encystment occurs, and organs peculiar to the
cercaria stage (boring papilla, eyes) almost entirely disappear. On
the other hand, the genitalia appear or become more or less highly
developed, in extreme cases to such an extent that they become
functional, and after autocopulation the creatures produce ova within
the cysts.
The cycle of development of the digenetic Trematodes has hitherto
been generally explained as a typical ALTERNATION OF GENERATIONS,
one sexual generation regularly alternating with one or two asexually
reproducing generations. Recent authors, however, regard the cells
in the sporocysts from which rediæ or eventually cercariæ arise as
parthenogenetically developing ova, and the sporocysts as well as the
rediæ as generations propagating parthenogenetically. In this case,
however, it is an alternation of a sexual not with an asexual but with
firstly a parthenogenetic generation (the sporocyst), the central cells
of which are regarded as ova which develop parthenogenetically into
the redia, and this the second parthenogenetic generation finally
produces larvæ (cercariæ) capable of developing into the sexually
mature form.
Other authors, again, regard the development of the Digenea as
only a complicated metamorphosis (p. 283), which is distributed over
several generations before it is concluded.
Biology.
Endoparasitic Trematodes, as fully developed organisms, occur in
vertebrate animals only, with very few exceptions; they inhabit almost
all the organs (with the exception of the nervous and osseous systems
and the male genitalia), but by preference the intestine in all its
extent from the oral cavity to the anus; and, further, certain species
or groups inhabit only quite restricted parts of the intestine. Besides
in the intestine other species live in the liver, or in the bile-ducts,
or in the gall-bladder; other accessory organs of the intestine, such
as the pancreas, bursa Fabricii (of birds), are only infected by a few
species. Many inhabit the lungs, or the air sacs in fowls, a few
the trachea. Trematodes have also been known to occur in the
urinary bladder, the urethra and the kidneys of all classes of
vertebrates; they are also present in the vascular system of a few
tortoises, birds and mammals; in birds they even penetrate from
the cloaca into the oviducts, and are occasionally found enclosed in
the laid eggs; one species is known to occur in the cavum
tympani and in the Eustachian tube of a mammal (Dugong),
another in the frontal sinus of the polecat; several species infest the
conjunctival sac under the membrana nictitans of birds, one species
even lives in cysts in the skin of song-birds. In an analogous manner
the ectoparasitic Trematodes are not entirely confined to the surface of
the body or the trachea of the lower vertebrate animals; a few species
appear exclusively in the urinary bladder, in the œsophagus, and in the
case of sharks in an accessory gland of the rectum.
Trematodes live free and active within the organs attacked,
though they may attach themselves by suction for a longer or shorter
period; in other cases, however, they bore more or less deeply
into the intestinal wall with their anterior end, or lie in cysts
of the intestinal wall which only communicate with the lumen
through a small opening; in those species living in the lungs of
mammals the host likewise produces a cyst, which usually encloses
two specimens; such association of a pair is also observed in
other situations, and, though this is the rule in species sexually
distinct, it is not entirely confined to these.
As regards the AGE attained by endoparasitic Trematodes, there
are but few reliable records, and these differ considerably; the
overwhelming majority of species certainly live about a year, or
perhaps a little longer, but there are some whose term of life
extends to several or many years.
Trematodes are but rarely found encysted in the higher vertebrate
animals; the condition, however, is more frequent in amphibians,
and especially in fishes, as well as in numerous invertebrate
animals.
Classification of the Trematodes of Man.
The following classification, partly artificial, partly natural,
embraces only the flukes found in man:—
Order. Digenea, v. Beneden, 1858.
Anterior sucker single and median, present. Eggs few. The (specialized)
terminal portion of the uterus serves as a vagina. Development indirect, i.e., an
intermediate host is required.
Sub-order. Prostomata, Odhner, 1905.
Mouth surrounded by the anterior sucker.
Group. Amphistomata, Rudolphi, 1801, ep., Nitzsch, 1819.
Gut forked, two suckers, the posterior sucker (acetabulum) terminal or ventro-terminal
behind the genitalia, or at most embraced by the vitellaria. Skin with no
spines. Excretory bladder a simple sac opening dorsally near hind end. Testes in
front of ovary. Genital pore, median in anterior third of body. Thick flukes, almost
circular in cross section.
Family. Paramphistomidæ, Fischoeder, 1901.
Amphistomata: Body not divided into a conical anterior portion and disc-like
caudal portion. Ventral pouch absent.
Sub-family. Paramphistominæ, Fisch., 1901.
Paramphistomidæ: Oral sucker without evaginations. Not in man.
Sub-family. Cladorchiinæ, Fisch., 1901.
Paramphistomidæ: Oral sucker with evaginations; testes, two, deeply cleft
(fig. 137). Genera: Watsonius, Cladorchis, etc.
Family. Gastrodisciidæ, Stiles and Goldberger, 1910.
Amphistomata: With body divided into a conical cephalic and disc-like caudal
portion (fig. 138). Posterior sucker ventro-terminal. Oral sucker with evaginations.
Genera: Gastrodiscus and Homalogaster.
Group. Distomata, Retzius, 1782.
Gut forked, two suckers, the posterior sucker (acetabulum) ventral. It is
always separated from the hind end by at least a part of the genitalia.
Family. Fasciolidæ, Railliet, 1895.
Large flat forms, genital pore in front of ventral sucker, the latter powerful.
Vitellariæ of numerous follicles, united by branching vitellarian ducts, at the sides of
the body meeting posteriorly and extending ventrally and dorsally. Cirrus and
vagina without spines. No crown of strong spines around sucker. Testes much
branched. Uterus not well developed. Excretory bladder much branched. Eggs
large.
Sub-family. Fasciolinæ, Odhner, 1910.
Large or median forms, gut much branched. Body has a shoulder separating
head from body. Receptaculum seminis absent. Ovary branched, ventral sucker
in anterior part of body. Genus: Fasciola.
Sub-family. Fasciolopsinæ, Odhner, 1910.
Shoulder absent. Receptaculum seminis present. Ovary branched, gut takes
a zig-zag course with kinks on it, ventral sucker in anterior part of body.
Genus: Fasciolopsis.
Family. Opisthorchiidæ, Braun, 1901, emend. auctor.
Ovary in front of testes. Small to medium flukes, very transparent, tapering
anteriorly. Vitellaria moderately developed not extending in front of sucker.
Cirrus absent. Seminal vesicle a twisted tube free in parenchyma. Testes near
hind end one behind the other, lobed or branched, but not dendritically. Excretory
bladder Y-shaped, the two limbs short, the stem S-shaped passing between the
testes. Receptaculum seminis well developed. Laurer’s canal present. Uterine
coils transverse, numerous. Eggs small.
Sub-family. Opisthorchiinæ, Looss, 1899, emend. auctor.
Opisthorchiidæ in which the excretory pore is terminal. Excretory bladder long,
dorsal to testes. Uterine coils not overlapping gut forks. Genera: Opisthorchis,
Paropisthorchis, Clonorchis, Amphimerus, etc.
Sub-family. Metorchiinæ, Lühe, 1909.
Opisthorchiidæ in which the excretory pore is ventral. Excretory bladder short,
ventral to testes. Uterine coils partly overlapping gut forks and extend anteriorly
beyond the sucker. Vitellaria compressed on the sides of the body. Genus: Metorchis.
Family. Dicrocœliidæ, Odhner, 1910.
Ovary behind testes. Testes behind the ventral sucker, between it and the ovary.
Body thin and transparent. Cirrus sac encloses the pars prostatica and seminal
vesicle. Skin smooth. Gut forks do not reach posterior end. Receptaculum seminis
and Laurer’s canal present. Vitellaria, moderate, lateral in mid-body slightly overlapping
the gut. Uterus with an ascending and descending branch and numerous
transverse coils extending to hind end. Eggs dark brown, 25 µ to 60 µ. Excretory
bladder tubular in posterior third or half of body. Parasitic in bile-ducts of mammals
and birds. Genus: Dicrocœlium.
Family. Heterophyiidæ, Odhner, 1914.
Ovary in front of testes. Genital pore behind or on a level with ventral sucker.
Genital pore surrounded by a pseudo-sucker (i.e., its muscle is not sharply separated
from but blends with the body muscles). Cirrus sac absent, consequently vesicula
seminalis and pars prostatica lie free. Vagina and ejaculatory duct unite into a
common duct before opening. Small and very small forms. Body covered with
scales. Genera: Heterophyes, Metagonimus, etc.
Family. Troglotremidæ, Odhner, 1914.
More or less flattened Distomes of compact form, 2 to 13 mm. long. Ventral
surface flat or somewhat hollowed, dorsal surface arched. Skin completely covered
with pointed spines. Musculature weakly developed also in the suckers in those
forms that inhabit cysts. Gut with pharynx and a not very long œsophagus and
cæca, which end more or less shortly before the hind end. Excretory bladder
Y-shaped or tubular. Pars prostatica and seminal vesicle always distinct. Testes
elongated, symmetrically placed in or behind the middle of the body. Ovary directly
in front of the testes, right-sided, generally much lobed. Receptaculum seminis
and Laurer’s canal present. Vitellaria generally well developed, exclusively or for
the most part confined to the dorsal surface, leaving only a median band unoccupied.
Uterus either very long, coiling here and there, or shorter and more convoluted.
Eggs in first case small 17 µ to 25 µ, in the second much larger 63 µ to 85 µ or
even 120 µ (?) long. Parasitic in carnivora or birds, generally occurring in pairs
in cyst-like cavities. Genera: Paragonimus, Pholeter, Collyriclum, Troglotrema.
Family. Echinostomidæ, Looss, 1902.
More or less elongated flukes, small or very large, much flattened anteriorly,
less so posteriorly, or even round. Suckers near one another, the anterior small and
weak, the posterior large and powerful directed obliquely backwards. Surrounding
the oral sucker dorsally and laterally but not ventrally is a fold or “collar” bearing
a row or rows of pointed spines which are continued round laterally on to the ventral
corners, the number being constant for each species, the corner spines large or
specialized, skin anteriorly scaled or spiny. Alimentary canal consists of a pharynx,
epithelial “pseudo-œsophagus” and gut cæca reaching to posterior end. Testes behind
one another in hind body. Ovary on right side or median directly in front of the
testes. Vitellaria lateral, usually extending to the hind end and not beyond the
ventral sucker anteriorly. Genital pore just in front of ventral sucker. Uterus in
transverse loops. Genital sinus absent or present. Receptaculum seminis and
Laurer’s canal present. Eggs thin shelled and large, bright yellow, 65 µ to 120 µ long.
Excretory bladder Y-shaped. Parasitic in gut of vertebrates, especially birds.
Sub-family. Echinostominæ, Looss, 1899.
Cirrus sac usually reaching to centre of ventral sucker, but not beyond. Cirrus
long, usually without spines, coiled when retracted. Seminal vesicle tubular, twisted.
On the head a ventral uniting ridge between the angles of the collar. Dorsal circlet
of spines, single or double, not interrupted unless the collar itself is dorsally divided.
Genera: Echinostoma, etc.
Sub-family. Himasthlinæ, Odhner, 1910.
Cirrus sac reaching far beyond ventral sucker. Cirrus armed with strong
rose-thorn-shaped hooks. Vesicula seminalis tubular not coiled. Cervical collar
not continued across ventral aspect. Spines on collar in one row. Body armed
with fine needle-shaped spines.
Family. Schistosomidæ, Looss, 1899.
Sexes separate. Genital pore behind the ventral sucker. Ventral sucker
elevated above the surface. Pharynx absent. Gut forks reunite to form a single
stem. In ♂ four or more testicular follicles. In ♀ a single ovary, just in front
of the union of the gut forks. Vitellaria on either side of the united gut stem.
The Trematodes Observed in Man.
Family. Paramphistomidæ, Stiles and Goldberger, emend. 1910.
Sub-family. Cladorchiinæ, Fisch., 1901.
Genus. Watsonius, Stiles and Goldberger, 1910.
Cladorchinæ.—Body pyriform. Ventral pouch absent. Acetabulum ventral or
(?) ventro-subterminal, very large, margins projecting, aperture small. Genital
pore in front of bifurcation of gut, not surrounded by a sucker; ductus hermaphroditicus
apparently absent. Excretory pore at posterior end of excretory vesicle,
behind Laurer’s canal. Oral sucker with a pair of irregularly globular suctorial
pouches; œsophagus thickened distally; cæca long, not wavy; end in acetabular
region.
Male Organs.—Testes two lobed, smaller than acetabulum; longitudinally,
nearly or quite coinciding; transversely they abut or slightly overlap; preovarial
in equatorial and caudal thirds. Pars musculosa not largely developed; cirrus
pouch absent.
Female Organs.—Ovary and shell gland post-testicular. Vitellaria extend from
gut fork to slightly beyond gut ending; uterus intercæcal, partly post-testicular.
Laurer’s canal in front of excretory vesicle.
Type Species.—Watsonius watsoni, Conyngham, 1904.
Watsonius watsoni, Stiles and Goldberger, 1910.
Syn.: Amphistomum watsoni, Conyngham, 1904; Cladorchis watsoni, Shipley, 1905.
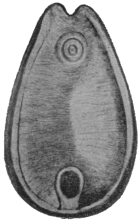
Fig. 136.—Watsonius
watsoni: ventral view.
4/1. (After Shipley.)
Body, 8 to 10 mm. long, by 4 to 5 mm. broad, by 4 mm. thick; tapers
anteriorly to 2·5 mm. Caudal extremity bluntly rounded, venter
surrounded by an elevated ridge, surface with
transverse ridges best defined ventrally. Genital
pore median about one-quarter of body length
from anterior end at level of suctorial pouches.
Acetabulum 1 mm. in diameter, margin projecting,
aperture small. Mouth in a groove with
digitate papillæ. Oral sucker very large, one-fifth
of length of body, with a pair of irregularly
globular pouches. Œsophagus somewhat longer
than sucker. Excretory pore at the level of
the acetabular aperture. The vesicle extends
from the plane of the transverse vitelline ducts
to centre of acetabulum.
Male Organs.—Testes deeply notched adjoining
one another. Vesicula seminalis much coiled
and dilated, pars musculosa not coiled. Pars prostatica (?) dilated,
ejaculatory duct long and narrow, opening on a papilla; genital
atrium papillated.
Female Organs.—Ovary dorso-posterior of posterior testis. Shell
gland dorsal to ovary. Vitellaria ventral and lateral to gut cæca
extending from gut fork to equator of acetabulum. Uterus dorsal
to testes, ductus hermaphroditicus absent. Laurer’s canal opens in
dorso-median line slightly behind anterior border of sucker.

Fig. 137.—Watsonius watsoni: ventral projection composed
from a series of transverse sections. o.s., oral sucker; s.p., suctorial
pouch; ga., genital atrium; d.e., ejaculatory duct;
es., œsophagus; e.g., œsophageal ganglion; p.p., pars prostatica;
p.m., pars musculosa; i., gut; ut., uterus; v.e., vas efferens;
v.e.s., left vas efferens; v.e.d., right vas efferens; v.g., vitellarium;
t., testes; ov., ovary; s.g., shell gland; t.vd., transverse vitelline
duct. (After Stiles and Goldberger.)
Eggs.—123 µ to 133 µ long by 75 µ to 80 µ broad.
Habitat.—Jejunum and duodenum of man, German West Africa.
The parasite has only been found once in man. The patient, a negro
from German West Africa, died at Zola, Northern Nigeria. The
symptoms were persistent watery diarrhœa without blood or mucus.
The parasites were also passed in the stools. It occurs also in
monkeys.
Family. Gastrodisciidæ.
Genus. Gastrodiscus, Lkt., 1877.
Acetabulum small, caudal and ventral margin raised, aperture relatively large.
Genital pore without sucker. Excretory pore post-vesicular, posterior to opening
of Laurer’s canal. Œsophagus with muscular thickening; cæca not wavy, long,
end post-equatorial and post-testicular.
Male Genitalia.—Testes two, branched pre-ovarial.
Female genitalia.—Ovary and shell gland post-testicular. Vitellaria extracæcal;
uterus intercæcal; Laurer’s canal entirely prevesicular.
Type.—Gastrodiscus ægyptiacus, Cobbold, 1876.
Gastrodiscus hominis, Lewis and McConnell, 1876.267
Syn.: Amphistomum hominis, Lew. and McConn.
Body, reddish in the fresh, 5 to 8 mm. long; posteriorly, 3 to
4 mm. broad. The disc has incurved edges which are interrupted in
front where it joins the anterior cylindrical portion and posteriorly
behind the ventral sucker. The disc itself and ventral surface are
covered with a number of (microscopic) papillæ.
Pharynx provided with two diverticula or pouches.
The bifurcation of the gut lies sometimes above,
sometimes below the level of the genital pore. The
gut cæca end about the level of the centre of the
acetabulum.

Fig. 138.—Gastrodiscus hominis.
Slightly magnified. (After Lerckart.)
Genital Pore.—About the middle of the conical
anterior portion. (It appears to be surrounded by a
muscular sucker.) Leiper (1913) describes the ducts
as discharging at the tip of a large fleshy papilla, the
surface of which bears cuticular bosses.
Testes much lobed, the anterior is smaller than the posterior and
lies at about the level where the anterior conical portion joins the
disc. The posterior testis just in front of the anterior margin of the
acetabulum separated from it by the ovary. The ovary, somewhat
oval in shape or slightly constricted in the middle, lies slightly to the
right of the median line. Dorsal to it lies the well-developed shell
gland, Laurer’s canal opening in front of the excretory bladder.
The excretory bladder is a long sac with its opening at its posterior
extremity about the level of the middle of the acetabulum. The
vitellaria are restricted in extent. They do not extend forward
beyond the anterior border of the posterior testis. They are best
developed in the area between the acetabulum and the termination
of the gut cæca.
The eggs are oval and measure 150 µ in length by 72 µ in
breadth.
Habitat.—Cæcum and large intestine of man. Also in the pig
(5 per cent.) in Annam.
Distribution.—This parasite has been recorded from Assam (not
uncommon), British Guiana (Indian immigrants), and Cochin China.
Gastrodiscus ægyptiacus, Cobbold, 1876, and G. secundus, Looss,
1907, occur in the horse; G. minor, Leiper, 1913, in the pig in
Nigeria and Uganda.
Family. Fasciolidæ, Raill., 1895.
Sub-family. Fasciolinæ, Odhner, 1910.
Genus. Fasciola, L., 1758.
The ventral sucker is situated at the level of the junction of the cone with the
body, viz., at the level of the “shoulder,” and is large and powerful. The cuticle
is covered with strong spines; the gut cæca run in the mid-line to the hind end,
and are provided with numerous long lateral and fewer and shorter median branches.
The ovary lies on one side in front of the transverse vitelline duct; the testes
lie obliquely one behind the other. The uterus, in the shape of a rosette, lies
in front of the genitalia. Laurer’s canal is present; the vesicula seminalis lies
in the cirrus pouch; the ova are large, not very numerous, and only develop after
they have been deposited. Parasites of the biliary ducts of herbivorous animals.
Fasciola hepatica, L., 1758.
Syn.: Distomum hepaticum, Retz., 1786; Fasciola Humana, Gmel., 1789;
Distomum caviæ, Sons., 1890; Cladocœlium hepaticum, Stoss., 1892.
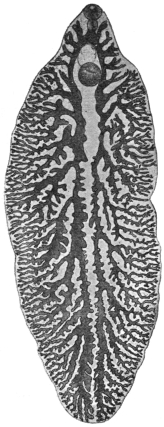
Fig. 139.—Fasciola hepatica,
L. From a specimen
that is not yet mature,
showing the gut and its
branches. 5/1.
Length 20 to 30 mm., breadth 8 to 13 mm., cephalic cone 4 to 5 mm.
in length and sharply differentiated from the body by a shoulder on
each side. Spines in alternating transverse rows and extending on
the ventral surface to the posterior border of the testes, and on the
dorsal surface not quite so far. The spines are smaller on the cephalic
cone than on the posterior part of the body, where they are discernible
with the naked eye. The suckers are hemispherical, and near each
other; the oral sucker is about 1 mm. and the ventral sucker about
1·6 mm. in diameter. The pharynx, which includes almost the entire
œsophagus, measures 0·7 mm. in length and 0·4 mm. in breadth.
The intestine bifurcates at the limit of the cephalic cone and the
branches are even here furnished with diverticula directed outwardly.
The ovary is ramified and situated in front of the transverse vitelline
duct, usually on the right side; the shell gland lies near the ovary in
the median line; posterior to the transverse vitelline ducts are the
greatly ramified testes, which occupy the greater portion of the
posterior part of the body, with the exception
of the lateral and posterior border; the long
vasa efferentia only unite as they enter the
cirrus pouch. The vitellaria occupy the sides
of the posterior part of the body, commencing
at the level of the ventral sucker and uniting
behind the testes. The ova are yellowish-brown,
oval, operculated, 130 µ to 145 µ in length, 70 µ
to 90 µ in breadth (average size 132 µ by 70 µ).
The Liver Fluke inhabits the bile-ducts of
numerous herbivorous mammals (sheep, ox,
goat, horse, ass, rabbit,268 guinea-pig, squirrel,
beaver, deer, roe, antelope, camel, kangaroo,
and others), and is distributed over the whole
of Europe, though not to an equal extent. It
is further known in North Africa, in North and
South America, as well as in Australia; it is
also found in Asia, as it has been reported
from Japan, China, and Tonkin (Gaide, two
cases in man). In some districts of Germany
it is very frequent, and the slaughter-house
statistics of various places show that it is of
daily occurrence. Fasciola magna occurs in
herbivora in America.
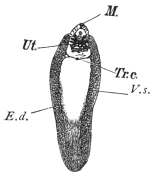
Fig. 140.—Fasciola hepatica. M., mouth;
Ut., uterine rosette; Tr.c., transverse vitelline
ducts uniting to form a vitelline receptacle in
the mid-line; E.d., longitudinal vitelline ducts;
V.s., vitellaria. The clear space in the centre
represents the position of the ramifying testes
and part of the gut. Natural size. (Mull. fluid,
alcohol, creosote, Canada balsam.)
The liver fluke, however, is by no means a
harmless parasite, for it produces in domestic
animals, more especially in sheep, a disease of the liver that appears
epidemically in certain years and districts, and commits great ravages
amongst the flocks.
[The following records show the enormous loss caused in sheep
by this parasite. In 1812, in the Midi, principally in the Departments
of the Rhône, Herault, and Gard, the disease was rampant; 300,000
sheep perished in the Arles territory, and 90,000 in the Arrondissements
of Nîmes and Montpellier. In 1829 and 1830, in the Department
of the Meuse and near localities, not only sheep but oxen died in
enormous numbers; for instance, in the Arrondissement of Verdun
out of 50,000 sheep 20,000 died, and out of 20,000 cattle 2,200 died.
In England, in 1830, 2,000,000 sheep were carried off; whilst in 1862
60 per cent. of the sheep died in Ireland; and in 1879 over 300,000
were lost in England; whilst as late
as 1891 one owner in the same
country lost over 10,000 sheep (Live
Stock Journal, October 30, 1891).—F.
V. T.]
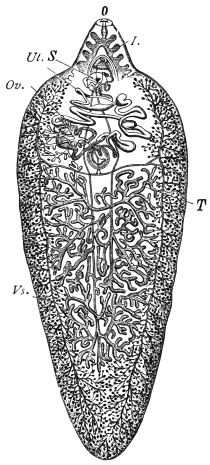
Fig. 141.—Fasciola hepatica, L. I.,
intestine; Vs., vitellaria; Ov., ovary;
O., oral aperture; Ut., uterus; S., ventral
sucker; T., testes. In front of the
testes are seen the transverse vitelline
ducts uniting to form the pyriform vitelline
receptacle. Immediately in front of
this the spherical shell gland. The two
vasa efferentia can also be seen running
up in the mid-line. The branches of the
gut are only shown in the cephalic cone.
(After Claus.)
The disease usually commences
towards the end of summer with an
enlargement of the liver, induced by
the invasion of numerous young
flukes; in the autumn and winter the
animals suffer from the consequences
of disordered biliary secretion; they
become feverish, emaciated, and
anæmic, and lose their appetite.
In consequence of the consecutive
atrophy of the liver, œdema and
ascites set in, and many animals succumb to this “liver rot.” On
examination the liver is found to be shrunken, the bile-ducts are
enormously dilated and in parts saccular and full of flukes. Should
the animals survive this stage, spontaneous recovery ensues in
consequence of the flukes commencing to leave the liver in the
spring, but the liver remains changed and its sale is prohibited269 when
the changes are extensive.270
[The following stages may be noticed in sheep suffering from
fascioliasis. Gerlach recognized four stages, based on the varied
relations that the flukes contract with the liver of their host. These
periods are sometimes very marked, but at others, owing to subsequent
infections, the features become merged and so obliterated. But
when a single infestation occurs they are very marked.
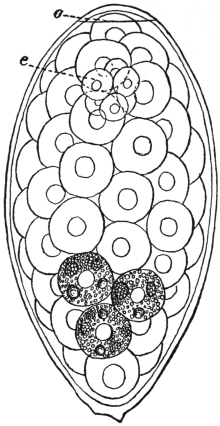
Fig. 142.—Fasciola hepatica: egg from liver
of sheep. o, operculum, e, segmenting ovum.
The rest of the space is occupied by yolk cells,
the granules in three only being shown. × 680.
(After Thomas.)
[The first period is called the
PERIOD OF IMMIGRATION. This
occurs at the fall of the year and
generally passes unperceived, as
the young flukes do little harm
to the liver. It varies from four
to thirteen weeks. Gerlach has
remarked upon cases of death
from apoplexy at this period.
[The second period is the
PERIOD OF ANÆMIA. This occurs
in November and December.
The sheep at first fatten rapidly,
but later the mucous membranes
become pale and of a yellowish
hue, and the sheep become sluggish and cease to feed. The fæces
are normal, but may contain fluke ova.
[The third period is the PERIOD OF WASTING. This corresponds
with the beginning of January—about three months after the
entry of the larvæ. Emaciation now becomes very marked, the
skin and mucous membranes blanched, temperature variable and
marked by an irregular curve; respiration laboured and quick;
appetite regular; abortion frequently occurs in pregnant ewes;
pressure on the back causes the animals to fall; local œdemas
occur, the most perceptible in the submaxillary space, extending
below the larynx and over the cheeks and parotids (called
“bourse,” “boule” in France; “watery poke” or “cockered” in
England). Death usually occurs at this period, but a fourth
stage may occur.
[The fourth period is the PERIOD OF MIGRATION OF THE
FLUKES. This is a period of convalescence and recovery, generally
in May and June.—F. V. T.]
Oxen suffer less in general, but even in these animals “stray”
hepatic flukes are occasionally found in the lungs, enclosed in thick-walled
cysts.
Pathological Anatomy.—The bile-ducts are conspicuous on the
surface of the liver. They are thickened and much dilated and in
parts saccular, and considerable atrophy of the liver cells accompanies
the condition. Histologically there is immense proliferation
of the epithelium of the bile-ducts leading to “adenomata.”

Fig. 143.—Limnæus truncatulus, Müll.,
the intermediate host of Fasciola hepatica.
a., natural size; b., magnified. (From
Leuckart.)
The LIFE-HISTORY of the liver fluke was discovered by
R. Leuckart and P. Thomas. According to these investigators the
elongated miracidium (fig. 131, a) ciliated all over develops from the
eggs a few weeks after the latter (fig. 142) have reached the water, and
after it has become free the embryo penetrates and becomes a sporocyst
(fig. 131, b) in a water-snail (Limnæus truncatulus, Müll. =
L. minutus, Drap.) that is common in fresh water, and can live in the
smallest collection of water as well as in fields that have been flooded.
The sporocyst first of all produces rediæ, which remain in the same
host (and under certain circumstances, e.g. in summer, these develop a
second generation of rediæ), and these finally form cercariæ (fig. 134).
The latter become encysted on blades of grass and are taken up by
the respective hosts with their food; this takes place towards the
end of summer, while the sheep feeding on the pasture land in the
spring spread the eggs of the fluke, and sometimes the fluke itself,
by passing them with their fæces.
In districts where Limnæus truncatulus is absent, analogous
species act as the intermediary hosts, of which one example
according to Lutz is Limnæus oahuensis in the Sandwich Islands.
[The host in Europe is Limnæus truncatulus. This snail
extends from Siberia to Sicily and Algeria, and according to
Captain Hutton is a native of Afghanistan. It also occurs in
Thibet, Amoor, Morocco, Tunis, Canary Islands and the Faroe
Islands. It deposits its eggs or spawn upon the mud around ponds,
ditches and streams. The eggs are laid in batches of thirty to a
hundred, each snail laying as many as 1,500 eggs; they are united
into strips of a gelatinous substance. In about two weeks young
snails appear. It is amphibious, being more frequently met with out
of the water than in it. It occurs in elevated spots as well as in
low-lying districts. Moquin-Tandon found it at 4,000 feet in the
Pyrenees. In the allied species, L. peregra, the fluke will develop
up to a certain stage, but never completes all its varied phases.
[In South America the host is probably Limnæus viator, Orb.,
and in North America Limnæus humilis, Say.—F. V. T.]
In human beings as well as in some of the mammals quoted
above, the liver fluke is only a casual parasite, and hitherto only
twenty-eight cases have been observed in man; the infection was
mostly a mild one and there were no symptoms, or only very
trifling ones; a few isolated cases were only discovered post mortem.
Occasionally, however, even when the infection was inconsiderable,
severe symptoms were set up, which in isolated cases led to death.
The symptoms (enlargement and painfulness of the liver, icterus)
merely pointed to a disease of the liver.
Diagnosis can only be established by finding eggs in the fæces.
Care should be taken not to confuse them with those of Dibothriocephalus
latus.
Halzoun.
Fig. 144.—Young Fasciola
hepatica, soon after
entry into the liver. The
intestinal cæca have lateral
diverticula. Magnified.
(From Leuckart.)
In North Lebanon, the liver fluke is, according to A. Khouri,
a frequent parasite of man, not in the liver, however, but in the
pharynx. The occurrence in this unusual site
is effected by the eating of raw infected livers,
especially those of goats (Capra hircus). The
flukes thus taken in do not all reach the
stomach, where they would be soon killed, but
some of them attach themselves to the pharyngeal
mucosa and to the adjoining parts, and
there cause inflammation and swelling, which
lead to dyspnœa, dysphagia, dysphonia and
congestion of the head, sometimes even to
still more severe symptoms, and even death.
The affection termed “Halzoun” lasts some
hours or several days, and after vomiting
recovery sets in. In other cases man becomes
infected in the usual way by ingesting cysts attached to grass or
the underside of leaves of plants (e.g., Rumex sp.), where they
are overlooked from
their scanty size (0·2 to
0·3 mm.).
As the liver fluke
feeds on blood it is
possible that it also
reaches, particularly
when young, the circulatory
system, and cases
have been known in
which it has been carried
by the blood into organs
far from its original situation.
Such cases also
have been repeatedly
observed in men. Probably
the parasite described
by Treutler,
1793, as Hexathyridium
venarum, which protruded
from the ruptured
anterior tibial vein
of a man, was a young
liver fluke. A few adult
specimens were found
by Duval in the portal
and other veins post
mortem at Rennes (1842)
in a man, aged 49, and
a similar statement is
reported by Vital from
Constantine (1874).
Giesker, in 1850, found
two hepatic flukes in a
swelling on the sole of
the foot of a woman.
Penn Harris states that
he observed six specimens
in Liverpool in a
spontaneously ruptured
abscess of the occiput
of a two months old
infant. Another case
which, like the previous
one, is reported by Lankester,271 relates to a sailor who suffered from
an abscess behind the ear, and from which a liver fluke was expelled.
Finally, Dionis de Carrières reports the case of a man, aged 35, in
whose right hypochondriac region a tumour the size of a pigeon’s egg
had formed, and from which a young liver fluke was extracted.
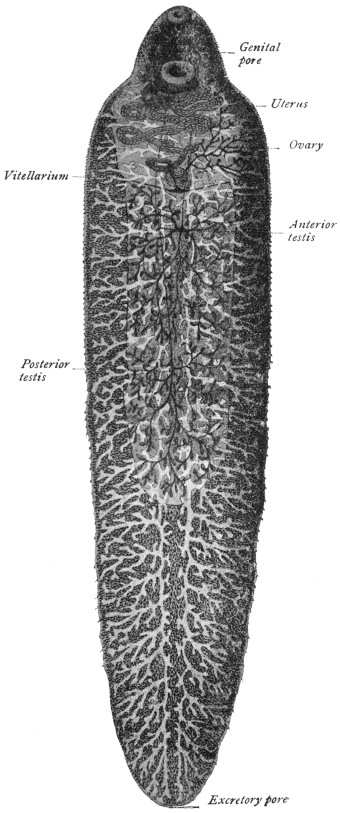
Fig. 145.—Fasciola gigantica. × 6 1/2
(After Looss.)
From such records it is not impossible that Distomum oculi
humani, Ammon, 1833, as well as Monostomum lentis, v. Nordm.,
1832, may have been very young hepatic flukes that had strayed.
Ammon found four specimens (length 0·5 to 1 mm.) of his species
(named Distomum ophthalmobium by Diesing in 1850) between the
opaque lens and the capsule of a five months old child in Dresden,
and von Nordmann discovered his Monostomum lentis to the number
of eight specimens (only 0·3 mm. in length) in the opaque lens of
an old woman. Minute white bodies which Greef found in the cortex
of the lens of a fisherman, aged 55, removed on account of cataract,
were with some reserve regarded as Trematode larvæ. The fact that
Ammon found that the intestinal cæca of the worm discovered by
him had no lateral branches does not negative the above opinion, as
in the liver fluke the intestinal cæca are originally unbranched, and
according to Lutz they only develop lateral ramifications later, between
the twelfth and twenty-second day of infection (fig. 144).
Fasciola gigantica, Cobbold, 1856.
Syn.: Distomum giganteum, Diesing, 1858; Fasciola gigantea, Cobbold, 1858;
Cladocœlium giganteum, Stoss., 1892; Fasciola hepatica var. angusta, Raill., 1895;
Fasciola hepatica var. ægyptiaca, Looss, 1896.
This species is closely allied to Fasciola hepatica, but is distinguished
by its elongated body, short cephalic cone, almost parallel
sides, larger ventral sucker, which is also closer to the oral sucker,
and by its larger eggs. Length up to 75 mm., width up to 12 mm.
Oral sucker 1 to 1·2 mm., ventral sucker up to 1·7 mm. in diameter.
Eggs 150 µ to 190 µ long by 75 µ to 90 µ broad.
Habitat.—Bile-ducts of Camelopardalis giraffa, Bos taurus, Bos
indicus, Bos bubalis, Ovis aries and Capra hircus.
Distribution.—Africa.
This species has once been observed in man by Gouvea, in Rio de
Janeiro, in a French naval officer who became ill with fever, cough
and slight blood-spitting. The lungs were normal except for a
sharply circumscribed spot at the base of the left lung. Twenty
days later during a fit of coughing the patient spat up a fluke 25 mm.
long, characterized by its slender aspect and by the size of its ventral
sucker, and its close proximity to the oral sucker. Considering the
fact that Gouvea’s patient had spent many weeks in July of the same
year in Dakar (Senegambia), where according to Railliet Fasciola
gigantica is common in slaughtered animals, and considering also
the characters of the fluke, Railliet rightly assumes that one had to
do with the African giant fluke and that the patient had infected
himself in Dakar.
Sub-family. Fasciolopsinæ, Odhner, 1910.
Genus. Fasciolopsis, Looss, 1898.
Ventral sucker large, and elongated
posteriorly into a sac. Cirrus pouch
long and cylindrical, its greatest length
being occupied by the sinuous tubular
seminal vesicle, on which exists a peculiar
cæcal appendage. Laurer’s canal present.
Fasciolopsis buski, Lank., 1857.
Syn.: Distomum buski, Lank., 1857;
Dist. crassum, Cobbold, 1860, nec v. Sieb.,
1836.
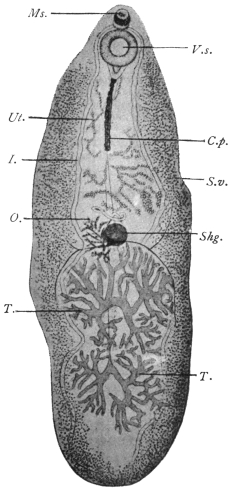
Fig. 146.—Fasciolopsis buski, Lank. V.s.,
ventral sucker; C.p., cirrus pouch; I., intestinal
fork; S.v., vitellaria; T., testes; O.,
ovary; Ms., sucker; Shg., shell gland;
Ut., uterus. Magnified. (After Odhner.)
The length of the body varies;
it may measure 24 to 37 or even
attain 70 mm.; the breadth is from
5·5 to 12 to 14 mm. In the pig the
fresh parasites measure, smallest,
12 to 8 mm.; largest, 35 to 16 mm.
(Mathis and Léger). Skin without
spines, but according to Heanly
always present in man and pig
specimens. The oral sucker
measures 0·5 mm. in diameter;
the ventral sucker is three to four
times as large; the pharynx is
globular, 0·7 mm. in diameter;
the prepharynx is provided with a
sphincter; the intestinal cæca extend
to the posterior border with
two characteristic curves, one at the anterior border of the anterior
testis, the other between the two testes. The genital pore is at the
anterior border of the ventral sucker; the cylindrical cirrus pouch
extends from behind the ventral sucker to half-way to the shell gland.
The seminal vesicle extends forwards within the cirrus pouch as a
convoluted tube. From its anterior portion is given off the cæcal
appendage, which has itself short lateral diverticula.
It runs backwards, ending blindly
about 0·5 mm. from the posterior end of the
cirrus sac. The seminal vesicle is continued as
the pars prostatica (?) 0·5 mm. long, and this
by the very short ejaculatory duct (13 µ), and
finally by the fairly long cirrus, which is beset
with very fine spines except at either extremity.
The ovary and shell gland are situated at about
the middle of the body with the testes behind
them, and the uterus in front. The vitellaria
extend from the ventral sucker to the posterior
border. The eggs measure 120 µ to 130 µ in
length and 77 µ to 80 µ in breadth, and resemble
those of Echinochasma sp. in dogs. The larval
stages are said to occur in shrimps.
Habitat.—Intestine of pig and man.
Distribution.—In man: India, Siam, China,
Assam, Sumatra. It is common in Cochin
China (16 out of 133 Annamites, Noc.), in
Tonkin very rare. Dr. J. Bell has sent me
[J. W. W. S.] human specimens from Hong Kong. In pigs: very
common in South China (Heanly). Common in pigs in Hong Kong.
Sixteen out of 248 pigs (i.e., 6 per cent.) infected in Hanoi.
Fasciolopsis rathouisi, Ward, 1903.
Syn.: Distomum rathouisi, Poirier, 1887.

Fig. 147.—Fasciolopsis
rathouisi, Poir.: the
mouth at the top, and under
it the genital pore and ventral
sucker, behind which
again is the uterus. The
vitellaria are at the sides,
and posteriorly in the central
field the ramified testes; the
ovary is in front of the right
testis. (After Claus.)
Fifteen to 19 mm. long by 8·5 to 10·5 mm. broad by about 3 mm.
thick. Skin with spines (Leiper). Bluntly oval or elliptical with
short cephalic cone which is absent in Fasciolopsis buski. Oral sucker
subterminal, 0·25 to 0·29 mm. broad by 0·2 mm. in antero-posterior
diameter. Distant from ventral sucker by about twice its diameter.
Ventral sucker 1·32 to 1·38 mm. broad by 0·68 to 0·7 mm. in antero-posterior
diameter. Œsophagus extremely short. Cirrus sac not
conspicuous and straight as in Fasciolopsis buski, but is convoluted.
Testes one behind the other (according to Poirier they lie beside one
another), more compactly branched, broader and denser than in
Fasciolopsis buski. Ovary on right side, small, coarsely branched.
Uterus in broad, closely grouped coils, packed with ova anterior to
ovary. Vitellarian acini more numerous and somewhat differently
distributed. Eggs 150 µ by 80 µ, thin shelled. [H. B. Ward, who has
examined this species, and from whose account the above is mainly
taken, considers that it is a good species, although the differences
between it and Fasciolopsis buski are slight, while Odhner, who examined
the original species, is of the opposite opinion.—J. W. W. S.] The
parasite appears to cause diarrhœa, wasting and occasionally jaundice.
Habitat.—Intestine of man.
Distribution.—China, common in some parts (Goddard).
Fasciolopsis goddardi, Ward, 1910.
Twenty-one to 22 mm. long, 9 mm. broad. Skin with spines
(Leiper). Uterus very closely coiled, most striking character is the
large size of the vitelline acini. Imperfectly known.
Distribution.—China (Shanghai).
Fasciolopsis fülleborni, Rodenwaldt, 1909.
The fully extended fluke is tongue-shaped, 50 by 14 mm.; two
contracted specimens measured 40 by 15 mm. and 30 by 16 mm.
respectively. Skin without spines, with according to Leiper cephalic
cone not clearly defined. Oral sucker circular, 0·75 mm. in
diameter, slightly larger than that of Fasciolopsis buski. Ventral sucker
2·6 mm. in diameter (that of Fasciolopsis buski 1·6 to 2 mm.). Length
2·9 mm. (as in Fasciolopsis rathouisi), the excess of length over breadth
being due to the posterior elongated sac-like prolongation of the
sucker. Prepharyngeal sphincter present. Pharynx 0·7 mm. in
diameter. Œsophagus practically absent. Gut cæca similar to those
of Fasciolopsis buski.
Testes—regularly branched, separated by an incurving of the cæca,
the anterior occupying a smaller area than the posterior.
Ovary—very small, as in Fasciolopsis buski, on the right side.
Shell Gland—almond-shaped, 2·3 by 1·2 mm. In Fasciolopsis buski
it is round and smaller, 1 to 1·5 mm. in diameter.
Vitellaria—similar in distribution to those of Fasciolopsis buski, but
the acini are strikingly small.
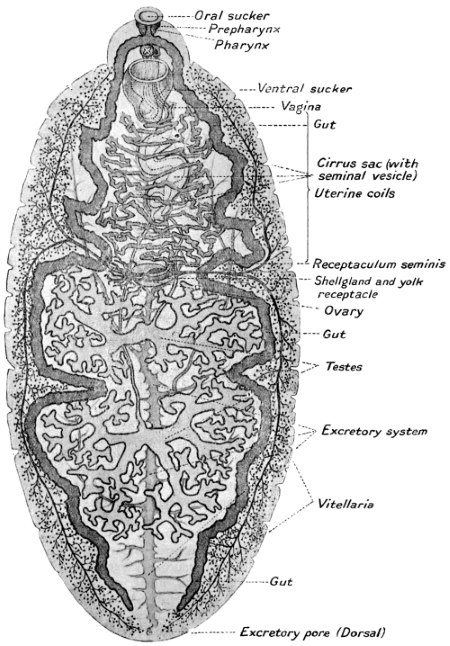
Fig. 148.—Fasciolopsis fülleborni, ventral aspect. (After Fülleborn.)
Cirrus Sac—is the most characteristic feature of this species. It is
a powerfully built, convoluted sac standing out clearly on the body.
It is not a uniform, straight cylinder 0·25 to 0·33 mm. in diameter, as
in Fasciolopsis buski, but even in fully extended flukes is typically
convoluted. It is 1 mm. thick in the middle, but in other parts varies
much from this. The posterior end of the cirrus sac is at two-thirds
or more of the distance from ventral sucker to shell gland. In the
case of Fasciolopsis buski the posterior end of the sac only extends
half-way.
Seminal Vesicle—has a peculiar convoluted, saccular and angular
course, but the cæcal appendage characteristic of the genus appears
to be absent!
Excretory System.—The main stem gives off very regular transverse
branches which are well seen posteriorly.
Eggs.—100 µ by 73 µ. Thin shelled.
Habitat.—Intestine. Mahommedan from Calcutta.
[It is evident that a re-examination of fresh material is required
before the validity of all these species can be accepted.—J. W. W. S.]
Family. Troglotremidæ, Odhner, 1914.
Genus. Paragonimus, Braun, 1899.
Body egg-shaped or somewhat elongated, generally more broadly rounded in
front than behind. Covered all over with spear-shaped spines arranged in groups.
Gut cæca winding with dilatations or constrictions in parts. Ventral sucker in or
in front of the middle of the body. Excretory bladder cylindrical, very long and
broad, reaching in front to the bifurcation of the gut. The lateral excretory canals
join the bladder only a little in front of the excretory pore. Genital pore median
just behind the ventral sucker. Genital sinus duct-like. Cirrus sac absent. Male
terminal organs very small. Ejaculatory duct present. Testes and ovary deeply
lobed, the testes in or just behind the middle, the ovary somewhat laterally placed
just behind the ventral sucker. Uterus forms a coil behind the ventral sucker.
Eggs rather large, thin shelled, the ovarian cell still unsegmented on deposition.
Receptaculum seminis, small.
Parasitic in the lungs of mammals, enclosed in cyst-like cavities, generally in
pairs.
Type Species.—P. westermanii in the tiger.
Paragonimus ringeri, Cobb., 1880.
Syn.: Distoma ringeri, Cobb., 1880; Distoma pulmonale, Baelz, 1883;
Distoma pulmonis, Suga, 1883.

Fig. 149.—Paragonimus
ringeri, Cobb.: to the right,
dorsal aspect; to the left, ventral
aspect. Natural size. (After
Katsurada.)
The body is of a faint reddish-brown colour and plump oval
shape. The ventral surface a little flattened; 7·5 to 12 mm. in length,
4 to 6 mm. in breadth, and 3·5 to 5 mm. thick (in man). The oral
sucker (0·75 mm.) is subterminal; the ventral sucker (0·8 mm.)
somewhat in front of the middle of the body. Pharynx spherical,
0·3 mm. in diameter, or 0·4 by 0·3 mm.; œsophagus, 0·02 mm.;
intestinal cæca convoluted, asymmetrical, the first part having the
same structure as the œsophagus. The
cuticle is covered with spines in groups;
the excretory pore opens at the posterior
end rather on the ventral surface, the excretory
ducts open into the elongated bladder
at the hind end near the pore. Genital
pore behind the ventral sucker and
median. Genital sinus 0·2 mm. long
with thick wall, ejaculatory duct 0·13 mm.,
pars prostatica 0·2 mm., seminal vesicle duct-like of irregular outline.
Behind the sucker the ovary on the left, and the closely packed
uterine coil on the right (though amphitypy of these two organs is
common); the two irregularly lobed testes lie side by side posteriorly.
Vitellaria extensive, leaving only a median dorsal and ventral space
free. Seminal receptacle probably absent; Laurer’s canal present.
The eggs are oval, brownish-yellow, fairly thin shelled, and measure
on an average 81·2 µ by 49·2 µ.
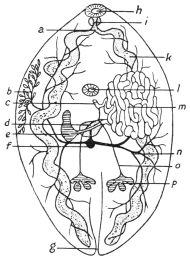
Fig. 150.—Paragonimus ringeri, Cobb.:
diagram of the internal organs. a, œsophagus;
b, vitellaria (a portion only shown);
c, common genital duct; d, shell gland with
oviduct, Laurer’s canal and vitelline duct; e,
ovary; f, vitelline receptacle; g, excretory
pore; h, oral sucker; i, pharynx; k, gut; l,
ventral sucker; m, uterine coils; n, vitellarian
ducts; o, vas efferens; p, testis. (After Kubo.)
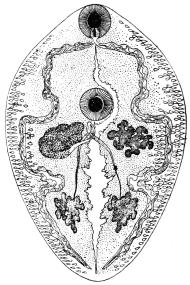
Fig. 150a.—Paragonimus westermanii,
Kerb.: seen from the ventral surface.
Mouth, pharynx, intestinal cæca, at the sides
of which the vitellaria are observed. The
genital pore is behind the ventral sucker, and
next to it, on the left, the ovary; on the right,
the uterus; the two testes posteriorly; the
excretory vessel in the middle, 10/1. (After
Leuckart.)
The following species are also known:—P. westermanii, Kerb.,
1878, in the tiger, and P. kellicotti, Ward, 1908, in the pig, dog, and
cat (N. America). Ward and Hirsch give the following differences
between the spines of the three forms:—
| P. ringeri. | P. westermanii. | P. kellicotti. |
| Shape | Chisel-shaped, mod-
erately heavy. | Lancet-shaped,
very slender. | Chisel-shaped,
heavy. |
| Distribution | Circular rows, in
groups. | Circular rows,
in groups. | Circular rows,
singly. |
Two other species, P. rudis, Diesing, 1850, in a Brazilian otter
(Lutra brasiliensis) and P. compactus, Cobbold, 1859, in the Indian
ichneumon, are but little known.
Habitat.—Lungs, pleuræ, and especially the bronchi of man and
dog. The alleged occurrence (of eggs) in other organs may be due
to confusion with those of Schistosoma japonicum.
Fig. 151.—Egg of Paragonimus
ringeri, Cobb.,
from the sputum. Showing
the ovarian cell and vitelline
cells and granules.
1,000/1. (After Katsurada.)
Distribution.—China, Korea, and especially in Japan, where, according
to Katsurada, there are no districts that are entirely free from pulmonary
flukes. The mountainous provinces of Okayama, Kumamoto,
Nagano and Tokushima are the principal
centres.
Pathology.—The number present in the
lung varies from two to twenty, about.
Usually one cyst contains one worm, but in
the dog each cyst contains two. The cysts
admit the tip of the finger, and have a fibrous
wall 1 mm. thick. They originate partly from
dilatation of bronchi and bronchioles. Others
arise from the inflammatory reaction of lung
tissue into which the worms have wandered.
The worms and their eggs cause bronchitis
and peribronchitis, catarrhal, hæmorrhagic, or
purulent, and areas of consolidation. Areas
containing eggs in their centre resembling
tubercle nodules are not uncommon, and
extensive cirrhosis of the lung may be found.
As a result of these changes, emphysema and bronchiectasis also
occur.
As to the development, only the following details are known:
that the eggs, which before segmentation of the ovum reach the open
in the sputum and through being swallowed also in the fæces,
develop in water into a miracidium ciliated all over, which hatches
and swims about freely. According to Manson this takes place in
four to six weeks.
Sub-family. Opisthorchiinæ, Looss, 1899.
Genus. Opisthorchis, R. Blanch., 1845.
Opisthorchiinæ with lobed testes. Laurer’s canal present. Parasitic in the bile-ducts
of mammals and birds.
Opisthorchis felineus, Riv., 1885.
Syn.: Distoma conus, Gurlt, 1831 (nec Creplin, 1825); Distoma lanceolatum,
v. Sieb., 1836, v. Tright, 1889 (nec Mehlis, 1825 = Fasciolo lanceolata, Rud., 1803);
Distoma sibiricum, Winogr., 1892; Distoma tenuicolle, Mühl., 1896.

Fig. 152.—Egg
of Opisthorchis
felineus,
Riv.
830/1.
This parasite is yellowish-red in the fresh condition, and almost
transparent. The body is flat, with a conical neck at the level of the
ventral sucker marked by a shallow constriction; this, however,
is only noticeable in fresh and somewhat contracted specimens.
Posteriorly to the ventral sucker the lateral borders run fairly parallel;
the posterior end is either pointed or rounded off. The length and
breadth vary according to the contraction, being usually 8 to 11 mm.
by 1·5 to 2 mm. The suckers are about one-fifth to one-sixth of
the length of the body distant from each other, and of about
equal size (0·23 to 0·25 mm.). The œsophagus is hardly any longer
than the pharynx, which lies close behind the oral sucker; the
intestinal cæca reach almost to the posterior border and are often
filled with blood. The excretory pore is at the posterior extremity,
and the excretory bladder forks in front of the anterior testis. The
testes in the posterior fourth of the body lie obliquely one behind
the other; the anterior one has four lobes, the posterior one five
lobes; the ovary is in the median line transversely, simple or
slightly lobed; behind it lies the large pear- or retort-shaped receptaculum
seminis and Laurer’s canal. The uterus is in the median field.
The vitellaria occupy the fairly broad lateral areas, in about the
central third of the body, beginning behind the ventral sucker and
terminating at about the level of the ovary; the acini are small and
arranged in groups of seven to eight, separated by interstices. The
genital pore is close in front of the ventral sucker. The eggs are
oval with sharply defined operculum at the pointed pole, 30 µ, by 11 µ.
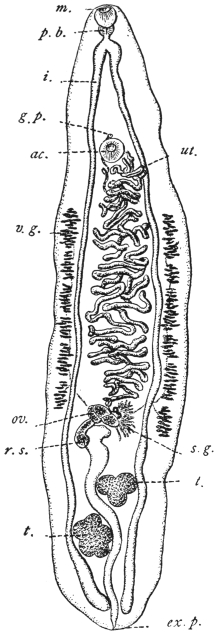
Fig. 153.—Opisthorchis felineus:
from the cat. m., mouth; p.b.,
pharynx; i., gut; g.p., genital pore;
ac., ventral sucker; ut., uterus; v.g.,
vitellarium; ov., ovary; s.g., shell
gland; r.s., receptaculum seminis;
t. testes; ex. p., excretory pore.
(After Stiles and Hassall.)
This species, which is frequently confused with others, inhabits the gall-bladder
and bile-ducts of the domestic cat especially; but is also found in the
dog, in the fox, and in the glutton (Gulo borealis). It has been observed in
France, Holland, North Germany (being particularly frequent in East Prussia), in
Russia, Scandinavia, Siberia, Japan, Tonkin, Hungary, and Italy. The North
American form (from cats and Canis latrans) is a distinct species (Opisthorchis
pseudofelineus).
In man this species was first found by Winogradoff in Tomsk (nine
cases), then by Kholodkowsky in a peasant from the neighbourhood
of Petrograd who had travelled a
great deal in Siberia, and finally by
Askanazy in five persons who were
natives of the East Prussian district of
Heydekrug. In Tomsk, Opisthorchis
felineus is the most frequent parasite
of man that comes under observation
at post mortem (6·45 per cent.), whereas
Tænia saginata has only been found
in 3·2 per cent., Echinococcus in 2·4 per
cent., Ascaris lumbricoides in 1·6 per
cent., and Oxyuris vermicularis in 0·8
per cent. of the autopsies. In the district
of Heydekrug, however, the species in
question is also frequent, as in a few
years five cases came to our knowledge
(of which three were diagnosed by the
discovery of the eggs in the fæces).
In none of Winogradoff’s nine cases
had the death of the patient been caused
direct by the parasites, yet more or less
extensive changes in the liver were
found in all of them; such as dilatation
of the bile-ducts with inflammation and
thickening of their walls, and foci of
inflammation or atrophy in the liver
substance; icterus was present five
times and atrophy of the liver an equal
number of times; ascites
was observed three times,
and in two cases, probably
of recent date, the organ
was enlarged. The number
of parasites found
fluctuated between a few
and several hundreds.
In two of Askanazy’s
cases, which he examined
more closely, carcinoma
which had developed at
the places most invaded by flukes was found at the post-mortem, so
that perhaps there may be grounds for the connection which the author
seeks to establish between cancer of the liver and the changes induced
by the parasites; these changes consist of numerous and even ramified
proliferations of the epithelium of the
biliary duct into the connective
tissue, which is likewise proliferated.
The number of worms found in one
case amounted to over 100; in a
second case, in which the parasites
had also invaded the pancreatic duct,
their number was even larger.
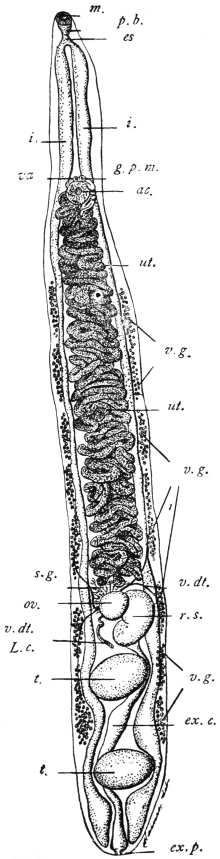
Fig. 154.—Opisthorchis pseudofelineus: from the
bile-duct of the cat (Iowa), m., oral sucker; p.b.,
pharyngeal bulb; es., œsophagus; i., intestine; va.,
vagina; g.p.m., male orifice; ac., ventral sucker;
ut., uterus; v.g. vitellarium; s.g., shell gland; v.dt.,
vitelline duct; ov., ovary; r.s., receptaculum seminis;
L.c., Laurer’s canal; t., testis; ex.c., excretory
bladder; ex.p., excretory pore. (After Stiles.)
Winogradoff as well as Askanazy
found isolated flukes in the intestine
also.
Unfortunately, nothing much is
known of the history of the development
of Opisthorchis felineus; we only
know that when deposited the eggs
already contain a ciliated miracidium,
which, however, according
to my experience, does not hatch out
in water, but only after the entry of
the eggs into the intestine of young
Limnæus stagnalis; no further development,
however, occurs. Winogradoff
states that he has seen the
miracidia hatch after the eggs had
been kept in water for a month at
37° C.; and has even observed free
miracidia in the bile of man and of
a dog respectively. Although the
whole post-embryonal development
of the cat fluke remains yet to be
investigated, Askanazy by a series of
experiments on cats and dogs has discovered
the mode of infection. The
intermediate hosts are fish, and
mainly the ide, in this country called
Tapar (Idus melanotus, H. and Kr.),
and of subsidiary importance the
roach (Leuciscus rutilus). Both species of fish as well as others are
readily eaten raw by man on the Courland lagoon (Baltic). It is, moreover,
significant that those persons whom Askanazy found infected
with the cat fluke were also infected with Dibothriocephalus latus, the
intermediate host of which is also fish (Lota sp., Esox sp., Perca sp.).
In one of his nine cases Winogradoff also saw a small fluke
covered all over with spines, which he conjectured to be the young
stage of Opisthorchis felineus; as, however, according to my experience,
this species, even in smaller specimens, is always without spines,
the above hypothesis cannot be accepted. It is much more probable
that one of the other species that also invade the liver of cats may
accidentally be introduced into man; we know, in fact, that Metorchis
albidus, Braun, and Metorchis truncatus, Rud., are both covered with
spines. As, however, the spines of the first-named species are rather
apt to fall off, and also as it possesses a different shape (spatula-shaped),
it may be assumed that probably Winogradoff had found Metorchis
truncatus, Rud., 1819, in his patient.
Genus. Paropisthorchis, Stephens, 1912.
Structure as in Opisthorchis, except that the ventral sucker and genital pore occur
on the apex of a process or pedicle projecting from the anterior portion of the body.
This process is about 1/2 mm. long, and is retractile.
Paropisthorchis caninus, Barker, 1912.
Syn.: Distoma conjunctum, Lewis and Cunningham, 1872; Opisthorchis noverca,
M. Braun, 1903 (pro parte); Opisthorchis caninus, Barker, 1912 (?).
Length varies from 2·75 to 5·75 mm. in preserved specimens, average
3·6 to 5·2 mm. Body uniformly spinose, though as a rule spines are
not present on the pedicle. Body slightly concavo-convex, the concavity
being ventral. Oral sucker 0·28 mm. Pharynx 0·224 by
0·184 mm. Œsophagus 0·04 mm. Ventral sucker 0·176 mm. in
diameter. Pedicle about 1/2 mm. long, may be completely retracted.
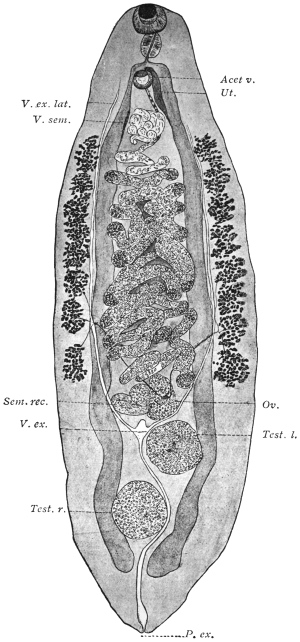
Fig. 155.—Paropisthorchis caninus: from the bile-ducts of the pariah dog, India. Acet. v.,
ventral sucker; Ut., uterus; V. ex. lat., longitudinal excretory duct; V. sem., seminal vesicle;
Sem. rec., seminal receptacle; Ov., ovary; V. ex., excretory bladder; Test. l., left testis;
Test. r., right testis; P. ex., excretory pore. × 40. (After Stephens.)
Genital Pore—opens on the apex of the pedicle in front of the
ventral sucker. Its exact position varies with the state of contraction
of the parts. In certain cases it actually opens within the cuticular
border of the sucker, in other cases it opens externally to the sucker
and anterior to it. The opening is covered with scales. The vas
deferens and uterus run alongside one another until they merge near
the apex of the pedicle into a common sinus.
Vitellaria—consist of eight acini on each side, extending from
slightly behind the base of the pedicle to the anterior border of the
ovary, or as far back as a line separating the posterior border of the
ovary from the anterior border of the anterior testis.
Testes.—Anterior testis 0·496 by 0·44 mm.; posterior testis 0·52 by
0·48 mm., usually ovoid, though both may be regularly lobed. The
anterior testis is usually on the left side.
Ovary—multilobular, the lobes 6 to 8 being irregular in size and
shape.
Shell Gland—extensive and diffuse, occupying an area which approximately
corresponds with the loop of the transverse vitelline ducts.
Seminal Receptacle—globular, to the right of and dorsal to the
posterior lobe of the ovary.
Laurer’s Canal—generally
runs from the end of the receptacle
with a single curve
medially and backwards.
Uterine Coils—form loosely
packed transverse coils terminating
slightly in front of
the level of the first vitelline
acini. From here the uterus
passes forwards into the pedicle
to the left and ventral to the
seminal vesicle.
Seminal Vesicle—commences
about the level of the
first vitelline acini. The coils
displace the uterus ventrally and
to the left. In the pedicle the
vesicle diminishes in extent and
lies in its dorsal (anterior) side.
Habitat.—Liver of pariah
dogs, India. In North-Western
Provinces about 40 per cent.
are infected. This fluke appears
to be different from
Amphimerus (Opisthorchis)
noverca in man, as the latter has
not the pedicle on the summit
of which lie the sucker and
common genital pore.
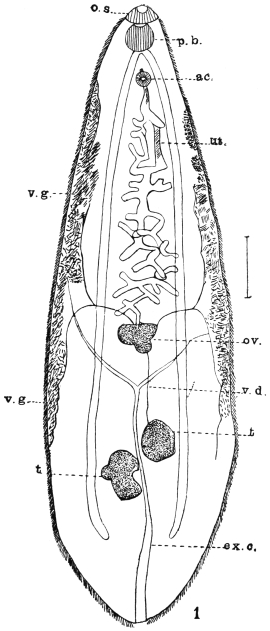
Fig. 156.—Amphimerus noverca, Braun. o.s.,
oral sucker; p.b., pharynx; ac., ventral sucker;
ut., uterus; v.g., vitellarium; ov., ovary; v.d., vas
efferens; ex.c., excretory canal; t., testis. (After
McConnell.)
Genus. Amphimerus, Barker,
1912 (?).
Structure as in Opisthorchis, except
that the vitellaria are separated
into two portions, an ant-ovarial and
a post-ovarial.
Amphimerus noverca, Barker, 1912 (?).
Syn.: Distomum conjunctum, McConnell, 1876 (nec Cobbold, 1859); Opisthorchis
noverca, M. Braun, 1903 pro parte.
At the autopsy of two Mahommedans who died in Calcutta,
McConnell found a large number of Distomata in the thickened and
dilated bile-ducts. The worms were lancet-shaped, covered with
spines, and measured 9·5 to 12·7 mm. in length and 2·5 mm. in breadth.
The two suckers lie very close to one another, the anterior one being
larger than the ventral; the genital pore opens immediately in front
of the ventral sucker; pharynx spherical; intestinal cæca extending far
back. At the commencement of the posterior third of the body the
two testes, somewhat apart, the anterior one roundish, the posterior one
distinctly lobed. The transverse
and slightly lobed ovary in front
of the bifurcation of the Y-shaped
excretory bladder, whence the
uterus, in convolutions barely
spreading beyond the central
field, extends to the pore; the
vitellaria in the lateral areas
commence behind the ventral
sucker and extend to the testes.
Cirrus pouch absent. Eggs oval,
34 µ by 21 µ.
Genus. Clonorchis, Looss, 1907.
Structure as in Opisthorchis, distinguished,
however, by the branched
testes situated one behind the other,
the branches of which ventrally encroach
upon the gut forks; dorsal
to the testes the S-shaped excretory
bladder, the main branches of which,
arising at the level of the bifurcation
of the gut, open into the bladder below
its anterior end. Parasitic in the bile-ducts
of mammals and man.
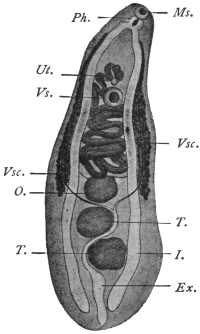
Fig. 157.—Metorchis conjunctus,272 (Syn.:
Distomum conjunctum, Cobb., nec Lew. and
Cunn., nec McConn.): from Canis fulvus.
Vs., ventral sucker; I., intestine; Vsc. vitellaria;
Ex., excretory bladder; T., testes;
O., ovary; Ms., oral sucker; Ph., pharynx;
Ut., uterus. (After Cobbold.)
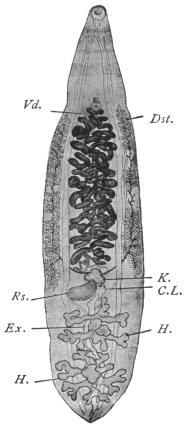
Fig. 158.—Clonorchis sinensis.
C.L., Laurer’s canal; Dst., vitellaria;
Ex., excretory bladder;
H., testes; K., ovary; R.s., receptaculum
seminis; Vd., terminal
section of vas deferens.
Magnified 4 1/2 times. (After
Looss.)
Clonorchis sinensis, Cobbold, 1875.
Syn.: Distoma sinense, Cobbold, 1875; Distoma spathulatum, R. Leuckart, 1876
(nec Rudolphi, 1819); Distoma hepatis innocuum, Baelz, 1883.
In shape resembles Opisthorchis felineus, 13 to 19 mm. long, 3 to
4 mm. broad, at the beginning of sexual maturity 12 to 13 mm. long, 2·5
to 3 mm. broad. Oral sticker 0·58 to 0·62 mm., ventral sucker 0·45 to
0·49 mm. in transverse diameter. In the parenchyma numerous
yellowish or brownish granules, especially behind the oral sucker and
at the posterior end. Testicular branches very long, in the anterior
testis often four, in the posterior testis five branches. Ovary generally
with three large lobes and a smaller lobe. Vitellaria not always symmetrical,
generally extending laterally from the ventral sucker to the
ovary, interrupted in parts.
Eggs 26 µ to 30 µ by 15 µ to 17 µ.
Average 29 µ by 16 µ.
Fig. 159.—Ova of Clonorchis sinensis. The knobs on the ends of
the eggs are not shown. 900/1. (After Looss.)
This (?) species was discovered in 1874
by McConnell, in Calcutta, in the bile-ducts
of a Chinaman who died shortly after being
admitted into hospital.
Habitat.—Bile-ducts of man, dog and cat.
Distribution.—Especially in China, apparently
rare in Japan.
Clonorchis endemicus, Baelz, 1883.
Syn.: Distoma sinense s. spathulatum p.p.;
Distoma hepatis endemicum s. perniciosum, Baelz,
1883; Distoma japonicum, R. Blanchard, 1886.
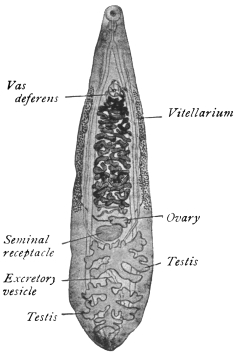
Fig. 160.—Clonorchis endemicus. × 6
about. (After Looss.)
Very similar to the previous species and
consequently generally confused with it.
Length between 6 and 13 mm., width
varying between 1·8 and 2·6 mm. Oral
sucker 0·37 to 0·5 mm., usually 0·43 to
0·45 mm. in transverse diameter; ventral
sucker 0·33 to 0·45 mm., usually 0·37 to
0·40 mm. No pigment in parenchyma;
anterior testis with four, posterior testis with
five branches. Vitellaria continuous, ova
26 µ by 13 µ to 16 µ.
Habitat.—Bile-ducts of man, dog, cat
and pig.
Distribution.—This species occurs very frequently in man, in
certain districts of Japan, especially in the province of Okayama,
Central Japan, in particular localities of which above 60 per cent. of
the population are infected. The worms are sometimes found in
enormous numbers in the liver (upwards of 4,000), also in the
pancreas and rarely in the duodenum.
It is common in Tonkin
and Indo-China. Léger in Tonkin
found 50 per cent. of people apparently
in normal health infected,
so that probably symptoms only
arise when the infection is intense.
[The exact distribution of these two
species is, however, not precisely
defined at present, as commonly no
distinction is made between them.—J.
W. W. S.]
Fig. 161.—Clonorchis endemicus: eggs. The knobs
on the eggs are not shown. × 900. (After Looss.)
Verdun and Bruyant deny, in
opposition to Looss, the possibility
of being able to distinguish within
the genus Clonorchis the two species
described, but they admit the justification
for the new genus. They
also report the occurrence of Opisthorchis
felineus in man in Tonkin
(Compt. Rend. Soc. de Biol., lxii, 1907).
Pathology.—Both species of Clonorchis
give rise to grave symptoms. The liver is generally enlarged,
though when the infection has lasted some time it begins to contract.
The surface of the organ is studded with white vesicles, and on cutting
into it one sees numerous
cavities with thickened
walls (distended
bile-ducts) filled with a
brownish fluid containing
innumerable eggs,
which cause its colour.
Microscopically, the epithelium
of the bile-ducts
is either (1) entirely destroyed,
or (2) actively
proliferates, forming an adenomatous outgrowth. Occasionally this
proliferation is not limited by the wall of the bile-duct but penetrates
it and leads to a growth of numerous new ducts, forming a
malignant biliary adenoma. The bile-ducts have their connective
tissue wall greatly sclerosed. These fuse with one another, forming
areas of sclerosis devoid of liver tissue. As a result of these changes
the liver cells atrophy and undergo fatty pigmentary and granular
degeneration. Besides these changes, due probably to the toxic action
of the flukes, mechanical obstruction due to the actual plugging of the
ducts by the flukes causes retention of bile and icterus, and through
pressure on veins, ascites and hypertrophy of the spleen.
To what extent blood or bile respectively forms the food of the
flukes is uncertain.
Life-history.—(Kobayashi, 1911, Mitteilungen aus dem kaiserlichen
Institut für Infektions-Krankheiten zu Tokio, pp. 58–62.)
It results from the work of Kobayashi in Japan that fresh-water fish
form the second intermediate host for Clonorchis endemicus. He fed
cats with encysted flukes (cercariæ) from various fish and easily
succeeded in infecting them, e.g. a kitten, proved to be uninfected by
repeated examination of its fæces, was fed on infected fish; a month
later innumerable flukes were found in the bile-ducts, gall-bladder,
pancreas and even in the duodenum. The fish infected were Leucogobis
güntheri, Pseudorasbora parva, and to a less extent Acheclognathus
lanceolata, Acheclognathus limbata, Paracheclognathus rhombea, Pseudoperilampus
typus, Abbottina psegma, Biwia zezera and Sarcocheilichthys
variegatus. The cysts occur throughout the muscles and subcutaneous
tissue of the fish. Length 0·13 mm., breadth 0·1 mm. The cercaria
lies folded in the cyst, length 0·5 mm. breadth 0·1 mm. It tapers
posteriorly. Skin at first covered with fine spines, disappearing as they
grow older. Body dotted with fine pigment.
The first intermediate host is still unknown.
Sub-family. Metorchiinæ, Lühe, 1909.
Genus. Metorchis, Looss, 1899, emend. auctor.
Hind end rounded. Gut forks reach extreme end. Testes only slightly lobed,
filling the hind end.
Metorchis truncatus, Rud., 1819.
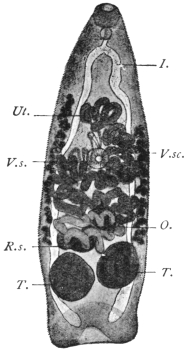
Fig. 162.—Metorchis truncatus,
Rud.: from the biliary ducts of the
domestic cat. V.s., ventral sucker;
I., gut; V.sc., vitellaria; T., testes;
O., ovary; R.s., receptaculum seminis;
Ut., uterus. 25/1.
This species, which attains a length of 2 mm., is slender and conical,
the anterior end is pointed and the posterior truncated, and provided
with a muscular tuberosity that resembles a terminal sucker; for this
reason the discoverer of the species (Rudolphi) classed it with the
Amphistomes. The cuticle in the young, as well as in the adult
specimens, is entirely and closely covered with spines. Suckers about
equal in size (0·134 to 0·172 mm.); the ventral sticker lies somewhat
in front of the middle of the body. The pharynx is small (0·09 mm.),
the œsophagus minute, the intestinal cæca reach to the posterior
extremity. Between them, and in front of
their blind ends, lie the two elliptical testes,
one generally a little in front of the other.
In front of them, either in the median
line or somewhat laterally, the spheroidal
ovary is situated; in front, again, is the
uterus, the coils of which usually extend
beyond the median field. The vitellaria
are at the sides of the central third of the
body, thus commencing in front of the
ventral sucker; cirrus pouch absent; the
genital pore is close in front of the
acetabulum. The excretory pore is terminal
(?). Eggs 29 µ by 11 µ.
Metorchis truncatus lives in the bile-ducts
of the seal, cat, dog, fox, and
glutton (Gulo borealis). The source of
infection is unknown, although one would
suspect fish. Askanazy did not succeed
in getting this fluke in his feeding experiments,
but another species, Metorchis
albidus, not uncommon in cats by feeding
them on roach (Leuciscus rutilus).
Family. Heterophyiidæ, Odhner, 1914.
Genus. Heterophyes, Cobbold, 1866.
Syn.: Cotylogonimus, Lühe, 1899; Cænogonimus, Looss, 1899.
No crown of spines on head. Body divided into a narrow, movable, anterior
part (neck), and a broader, less movable, posterior portion, which contains the
genitalia. The suckers separated from one another by a space equal to half the
length of the body or more; the pharynx is close behind the oral sucker; the
œsophagus is long; the intestinal cæca extend to the posterior border; the genital
pore is placed laterally, and behind the ventral sucker. Genital sucker provided
with a circlet of chitinous rodlets, shaped like stags’ horns. The testes are at the
posterior end, the ovary in a median position in front of them. Laurer’s canal with
receptaculum seminis present; the small vitellaria are at the sides of the posterior
part of the body. Parasitic in the intestine of mammals and birds.
Heterophyes heterophyes, v. Sieb., 1852.
Syn.: Distomum heterophyes, v. Siebold, 1852; Heterophyes ægyptica, Cobbold,
1866; Mesogonimus heterophyes, Railliet, 1890; Cœnogonimus heterophyes, Looss
1900; Cotylogonimus heterophyes, Braun, 1901.
Length up to 2 mm., breadth 0·4 mm.; the neck not sharply defined;
in life it stretches to double the length of the hind body. The scales
are rectangular, 5 µ to 6 µ by 4 µ, their posterior margin serrate with
seven to nine teeth. Cuticular glands are numerous on the ventral
surface, especially in the fore part of the body, and partly discharge
at the anterior border of the oral sucker. The oral sucker is 0·09 mm.,
the ventral sucker 0·23 mm. in diameter; the pharynx measures
0·05 to 0·07 mm. in length;
the œsophagus is about
three times as long; posteriorly
the intestinal cæca
are directed one towards
the other and terminate
beside the excretory
bladder. Close in front
of the posterior ends of
the intestinal branches are
the two elliptical testes,
which are not exactly on
the same level. In the
middle in front of them is
the receptaculum seminis,
and in front of the latter
lies the spherical or elliptical
ovary. The two vasa
efferentia unite to form the
vas deferens, which after
a short course passes over
into the angularly bent
seminal vesicle; after the
entry of the prostatic
glands it becomes united
with the metraterm
(vagina), and the common
duct opens into the genital
sucker. The latter is somewhat
smaller than the ventral
sucker, lateral to and
close (0·15 mm.) behind it, and bears a not entirely closed ring of
from seventy-five to eighty chitinous rods (20 µ in length). The vitellaria
on either side consist of about fourteen acini. The uterus is
spread almost throughout the entire posterior part of the body. The
eggs have thick shells with a knob resembling that of Clonorchis eggs
but not so prominent, and measure 30 µ by 17 µ; they contain a
completely ciliated miracidium with a rudimentary intestinal sac.
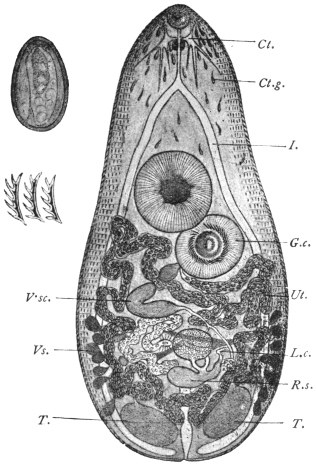
Fig. 163.—Heterophyes heterophyes, v. Sieb.
C., cerebral ganglion; I., intestinal cæca; Ct.g.,
cuticular glands; V.sc., vitellaria; Ut., genital sucker;
T., testes—the excretory bladder between them; L.c.,
Laurer’s canal; R.s., receptaculum seminis, with the
ovary in front of it; G.c., ventral sucker; Vs., vesicula
seminalis, 53/1. On the left side above, an egg, 700/1,
is depicted, and below it three chitinous rodlets from the
genital sucker. 700/1. (After Looss.)
This species was discovered in 1851 by Bilharz in the intestine
of a boy who died in Cairo; a second case was only found in 1891
and published by R. Blanchard, so that it appeared as if the species
were very scarce. According to Looss, this is, however, not the case,
but the species easily escapes notice on account of its small size.
Looss found it in Alexandria twice in nine autopsies, and once in
Cairo, and has recently stated that in man “it is not at all uncommon
to meet with the parasite in cadavers, and the eggs of the worm in
the stools of the patients.” Leiper records one case from Japan
and one from China. The parasites occupy the middle third of the
small intestine, and even when present in large numbers appear to be
harmless.
This small species, according to
Looss, frequently occurs in Egyptian
dogs, less so in cats, and has also been
found in the fox, as well as once in
Milvus parasiticus; Janson also reports
this species from the intestine of the
dog in Japan.
Metagonimus, Katsurada, 1913;
Yokogawa, Leiper, 1913.
Resembles in general structure
Heterophyes. In the arrangement of
its ventral genital suckers resembles
but differs from that of Tocotrema,273
Looss. The ventral and genital suckers
lie laterally and on the right.
Metagonimus yokogawai. Katsurada,
1913.
Syn.: Yokogawa yokogawai, Leiper, 1913.
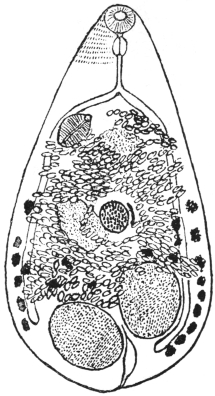
Fig. 164.—Metagonimus yokogawai,
Katsurada, 1913: the spines are only
shown over a small part of the skin.
(After Leiper.)
One to 1·5 mm. long, seldom 2·5 mm.,
and 0·4 to 0·7 mm. broad; elliptical in
shape. The body is thickly covered
with nail-shaped spines about 10 µ
long. Oral sucker 77 µ, to 85 µ in diameter. Ventral sucker characteristic
and peculiar 0·12 to 0·14 mm. by 0·08 to 1 mm. It is a sac-like
organ placed deeply in the body, but does not open as in other flukes on
the ventral surface. Testes elliptical, not quite symmetrically placed at
the hind end of the body. Vesicula seminalis retort-shaped, situated
transversely, internal to the ventral sucker. Pars prostatica present.
Ejaculatory duct opens with the uterus into a genital sinus, which,
together with the internal opening of the ventral sucker, opens into a
pit at the front of the ventral sucker. The opening of the genital sinus
and that of the ventral sucker are furnished with a complex muscular apparatus.
Ovary spherical, 0·12 to 0·13 mm. in diameter, lies in the middle
of the hind body. Receptaculum seminis and Laurer’s canal present.
Vitellaria in the hind half of the body, consisting of about ten acini on
each side. Shell gland to the left of the
ovary. Uterus forms three to four transverse
coils. Eggs elliptical, double
contoured, yellowish-brown in colour.
There is no shoulder below the operculum
as in the eggs of Cl. sinensis.
At the rounder end there is a thickening
or knob different from the spine-like
or hook-like process seen in Cl.
sinensis. Dimensions 28 µ by 16 µ.
Habitat.—Mainly in upper or
middle portion of jejunum, rarely in
cæcum. They penetrate deep into the
mucosa, but not into the submucosa,
and post mortem appear as a number
of small brown points. They frequently
occur in the solitary glands,
which they destroy. They cause
chronic catarrh of the gut. Parasitic
in man and mammals.
Geographical Distribution.—Japan.
Life-history.—The cercarial stage
occurs in a trout (Plecoglossus altivelis)
and seldom in Crassius sp. and Cyprinus
sp. Infection takes place through
the eating of the fish raw. Seven to
sixteen days later eggs appear in the
fæces (of dog).
Family. Dicrocœliidæ, Odhner, 1910.
Genus. Dicrocœlium, Dujardin.
Dicrocœliidæ, with leaf-shaped bodies,
pointed posteriorly and anteriorly. Greatest
width behind the mid-line. Vitellaria double.
The testes smooth or indented, lying symmetrically or obliquely beside or behind
the ventral sucker. The ovary approaches the median line behind one testis.
Parasitic in the liver and gall-bladder (rarely in the intestine) of members of all
classes of vertebrate animals—by preference in birds and mammals.
Dicrocœlium dendriticum, Rud., 1819.
Syn.: Dicrocœlium lanceatum, Stil. and Hass., 1896; Fasciola lanceolata,
Rud., 1803 (nec Schrank, 1790); Distomum lanceolatum, Mehlis, 1825; Dicrocœlium
lanceolatum, Dujardin, 1845.
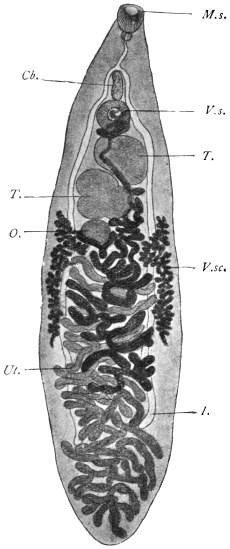
Fig. 165.—Dicrocœlium dendriticum,
Rud. V.s., ventral sucker; Cb., cirrus
pouch; I., intestinal cæca; V.sc., vitellaria;
T., testicles; O., ovary; M.s.,
oral sucker; Ut., uterus. 15/1.
Body lancet-shaped, narrowing especially at the anterior extremity;
length 8 to 10 mm., breadth 1·5 to 2·5 mm., the greatest breadth usually
behind the middle of the body. Suckers distant from each other by
about one-fifth the length of the body; oral sucker about 0·5 mm., ventral
sucker about 0·6 mm. Pharynx globular, adjoining the oral sucker;
œsophagus 0·6 mm. in length; intestinal cæca reach to four-fifths of
the body length. Genital pore at the level of the bifurcation of the
intestine; cirrus pouch small and slender. The large, slightly lobed
testes lie obliquely one behind the other behind the ventral sucker;
the ovary, which is considerably smaller, is placed behind the posterior
one; the vitellaria, commencing at the level of the posterior
testis, terminate far before the cæca. The uterus, situated behind the
ovary, extends throughout the posterior end, not confined to the central
field, but overlapping the lateral fields with its transverse coils; at
the posterior edge of the body it turns back again and winds forwards
to the ovary in transverse loops, then between the testes, and finally,
dorsal to the ventral sucker, terminates in the genital pore. The
thick-shelled eggs when young are yellowish, when older dark brown.
They measure 38 µ to 45 µ by 22 µ to 30 µ. They contain an oval or
roundish miracidium, only the anterior part of which is ciliated, and
which possesses a rudimentary intestinal sac with a boring spine.
The miracidia do not hatch out in water spontaneously, but, according
to Leuckart, in the intestines of slugs (Limax, Avion), but they
do not develop either in these (slugs) or in water-snails.
The lancet fluke inhabits the biliary duct of herbivorous and
omnivorous mammals (sheep, ox, goat, ass, horse, deer, hare, rabbit,
pig), and is often found associated with the liver fluke; it is not,
however, so common nor so widely disseminated, nevertheless, it has
been met with outside of Europe, namely, in Algeria, Egypt, Siberia,
Turkestan, and North and South America.
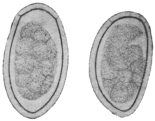
Fig. 166.—Eggs of Dicrocœlium
dendriticum, Rud.
To the left seen flat, to right
lying on one side. 600/1.
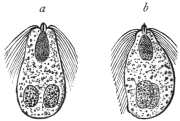
Fig. 167.—Miracidia of
Dicrocœlium dendriticum. a,
from the dorsum; b, from the
side. (After Leuckart.)
In man it is still more uncommon than the liver fluke, and has
hitherto only been observed seven times (Germany, Bohemia, Italy,
France, and Egypt); it may, however, have occurred more frequently,
and have been overlooked, as in slight infections it produces no
special symptoms.
The intermediate host is still unknown. Leuckart for some time
held the opinion that small species of Planorbis from fresh water,
which contain encysted Distomata, were to blame, and he supported
his views by a feeding experiment which seemingly yielded positive
results; this, however, is not definitely proved. Piana’s statement
that small land snails are the intermediate hosts has also not been
proved.
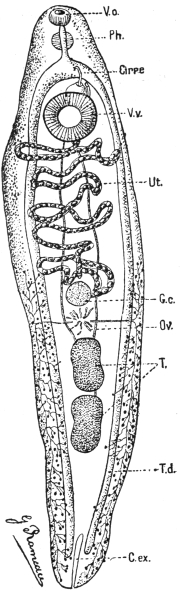
Fig. 168.—Echinostoma ilocanum. Vo.,
oral sucker; Ph., pharynx; Cirre, cirrus;
V.v., ventral sucker; Ut., uterus; G.c.,
ovary; Ov., shell gland; T., testes;
T.d., vitellarium; C.ex., excretory vesicle.
(After Brumpt.)
Family. Echinostomidæ, Looss, 1902.
Sub-family. Echinostominæ, Looss, 1899.
Genus. Echinostoma, Rud. 1809; Dietz, 1910.
Fore-body not bulging. Greatest width at or behind the ventral
sucker. Oral sucker not atrophied. Collar kidney-shaped with a double
dorsally unbroken row of spines, terminating in four to five angle spines.
The border spines of the aboral series not larger than the oral. Skin
spined or smooth. Body elongated. Uterus long with numerous
transverse coils. Ventral sucker in the anterior quarter of body. Cirrus
sac small, almost completely in front of the ventral sucker. Testes round
or oval, smooth incurved or lobed, in the hinder half of body. Ovary
median or lateral in front of testes. Vitellaria from hinder margin of
ventral sucker to end of body. Eggs oval, 84 µ to 126 µ by 48 µ to 82 µ.
The spines placed most ventrally, or those placed most medially
on ventral surface, are from differences of position or form termed
“angle” spines, the rest “border” spines.
Type.—Echinostoma echinatum, Rud.
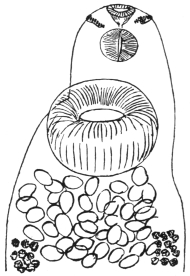
Fig. 169.—Echinostoma ilocanum,
Garrison, 1908: head end showing collar
of spines, ventral view. (After Leiper.)
Echinostoma ilocanum, Garrison, 1908.
Length 4 to 5 mm., breadth 1 to 1·35 mm., thickness 0·5 to
0·6 mm. The circum-oral disc 0·3 mm. broad, separated by a shallow
groove from the body. Crown of forty-nine spines and five to six
angle spines on each side continuous with an irregularly alternating
series of fourteen spines on the dorsum. Largest spines are
34 µ long, 8 µ thick at the base. The remainder of the dorsal spines
are 24 µ by 6 µ. Skin thickly covered with scales on the margins
of the body as far back as the level of the hind testis. Oral sucker,
0·18 mm.; ventral sucker, 0·4 to 0·46 mm. Its anterior border about
0·07 mm. from the anterior end. Pharynx 0·17 mm. long, 0·11 mm.
broad. Testes about mid-line of the body, much lobed; the lobes of
the anterior testis run transversely, while the axis of the posterior testis
is longitudinal, as often occurs in the Echinostomidæ. Cirrus sac
reaches to the centre of the ventral sucker. Ovary transversely oval
in front of the testes. Vitellaria commence about half-way between
the ventral sucker and ovary and extend to the posterior end. Eggs
numerous, 92 µ to 114 µ by 53 µ to 82 µ.
Average.—99·5 µ by 56 µ.
Habitat.—Gut of man (Filipinos),
Philippine Islands.
Echinostoma malayanum,
Leiper, 1911.
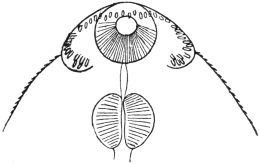
Fig. 170.—Echinostoma malayanum, Leiper,
1912: anterior end showing collar of spines,
ventral view. (After Leiper.)
Twelve millimetres long, 3 mm.
broad, 1·3 mm. thick. Ends bluntly
rounded. At the anterior end a
ventral furrow on either side, one-third
the width of the body,
marking off the circum-oral collar.
Along its edge is a row of forty-three
spines extending across the
middle line dorsally but not ventrally.
The spines vary in size from
0·07 mm. in length (ventrally) to 0·05 to 0·016 mm. (dorsally). Cuticular
spines also exist on the ventral side as far back as posterior end of
body, but dorsally limited to a triangular area ending in front of the
ventral sucker. Oral sucker 0·07 mm. thick, occupying the middle
third of the circum-oral disc; pharynx 0·25 mm. in diameter; œsophagus
0·04 mm. long; gut cæca simple, extending to end of body;
ventral sucker 0·9 mm. long by 0·75 mm. broad by 0·7 mm. deep; wall
about 0·25 mm. thick. The
sucker is inclined at an angle
of 40° to the ventral surface.
Testes lobed, one behind the
other, behind the ventral sucker.
Cirrus pouch well developed,
reaching to the posterior edge
of the sucker. Genital pore in
the angle between neck and
anterior lip of ventral sucker.
Ovary smooth, 0·3 mm. in
diameter, 0·85 mm. behind
ventral sucker. Vitellaria very
numerous, extending from posterior margin of sucker to posterior end
of body, where they intermingle. Eggs few in number, brown and large.
Habitat.—Gut of man (Tamils), Malay States.
Sub-family. Himasthlinæ, Odhner, 1910.
Genus. Artyfechinostomum, Clayton-Lane, 1915.
Crown of thirty-nine spines, continuous over dorsum. Two corner
spines long. Vitellaria extend from posterior margin of sucker to posterior
end of fluke. Eggs without filament. [Although the possession
of strong rose-thorn hooks is given by Odhner as a sub-family characteristic,
yet in this genus assigned to this sub-family they have not
been seen.—J. W. W. S.]
Artyfechinostomum sufrartyfex, Clayton-Lane, 1915.
Spirit specimens: 9 by 2·5 by 0·8 mm. thick. Ventral sucker
conspicuous, 1 mm. in diameter. Cirrus sac 2 mm. long. Testes
lobed, about 1·5 mm. in diameter. Posterior border of posterior
testes 1 mm. from posterior end. Vitellaria meet posteriorly behind
the posterior testis.
Family. Schistosomidæ, Looss, 1899.
Genus. Schistosoma, Weinl, 1858.
Syn.: Gynæcophorus, Dies., 1858; Bilharzia, Cobb., 1859; Thecosoma,
Moq. Tandon, 1860.

Fig. 171.—Schistosoma hæmatobium, Bil.:
male carrying the female in the canalis gynæcophorus.
12/1. (After Looss.)
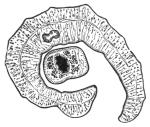
Fig. 172.—Transverse section
through a pair of Schistosoma
hæmatobium in copulâ. In the male
the point of reunion of the intestinal
forks has been cut across. (After
Leuckart.)
The males have bodies that widen out considerably behind the ventral sucker,
the lateral parts of which in-roll ventrally, forming the almost completely closed
canalis gynæcophorus, within which the female is enclosed. There is no cirrus
pouch. The male has five or six testes, the females are filiform; the uterus is long.
There is no Laurer’s canal. The ova almost equally attenuated at either extremity;
they have a small terminal spine, and are not provided with a lid. They contain
a miracidium, ciliated on all sides, which is characterized by the possession of two
large glandular cells, which discharge anteriorly beside the gastric sac. They live
in the vascular system of mammals. (An allied genus [Bilharziella] lives in the
blood-vessels of birds.)
Schistosoma hæmatobium, Bilharz, 1852.
Syn.: Distoma hæmatobium, Bilh.; Distoma capense, Harley, 1864.
The Male is whitish, 12 to 14 mm. in length, but is already mature
when 4 mm. long. The anterior end is 0·6 mm. or a little over in
length. The suckers are near
each other, the oral sucker is infundibular,
and the dorsal lip is
longer than the ventral one. The
ventral sucker is a little larger,
0·28 mm., and is pedunculated.
A little behind the ventral sucker
the body broadens to a width of
1 mm., decreasing, however, in
thickness; the lateral edges in-roll
ventrally, so that the posterior
part of the body appears almost
cylindrical, 0·4 to 0·5 mm. in
diameter; the posterior extremity
is somewhat more attenuated.
The dorsal surface of the posterior
part of the body is covered
with spinous papillæ. There are
delicate spines on the suckers, and
larger ones invest the entire internal
surface of the gynæcophoric canal,
as well as a longitudinal zone at
the edge of that side of the external
surface that is covered by the other
side rolling over it. The œsophagus
is beset with numerous
glandular cells (fig. 173), and presents
two dilatations; the intestinal
bifurcation is close in front of
the ventral sucker, the two
branches uniting sooner or later behind the testes into a median
trunk, which may again divide at short intervals. The excretory
pore is at the posterior end, but placed somewhat dorsally; the
genital pore is at the beginning of the gynæcophoric canal, thus
behind the ventral sucker; into it opens the vas deferens which,
posteriorly, broadens into the seminal vesicle and then continues
as the vasa efferentia of the four or five testes (fig. 173).
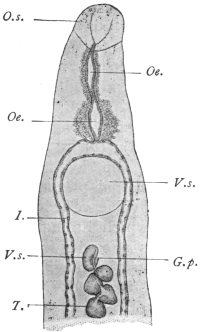
Fig. 173.—Anterior end of the
male Schistosoma hæmatobium, Bilh.
V.s., ventral sucker; I., gut cæca;
G.p., genital pore; T., testes;
O.s., oral sucker; Oe., œsophagus
with glandular cells; V.s., vesicula
seminalis. 40/1. (After Looss.)
The Female—filiform, about 20 mm. in length, pointed at each end,
and measuring 0·25 mm. in diameter in
the middle. Their colour varies according
to the condition of the contents of
the intestine. (Posteriorly they are dark
brown or blackish.) The cuticle is smooth
except in the sucker, where there are very
delicate spines, and at the posterior end,
where there are other larger spines. The
oral sucker is a little larger than the
pedunculated ventral sucker (0·07 and
0·059 mm. respectively). The anterior
part of the body, 0·2 to 0·3 mm. in length;
the œsophagus is as in the male. The
intestinal bifurcation is in front of the
ventral sucker, the two branches uniting
behind the ovary and the trunk running
in a zigzag manner to the posterior border.
There are indications of diverticula at the
flexures. The ovary is median. In young
females it is of an elongated oval shape;
in older females the posterior end becomes
club-shaped, whereas the anterior end
becomes attenuated; the oviduct originates
at the posterior end, but immediately
turns forwards and joins the parallel
vitelline duct in front of the ovary (fig. 174),
where the shell gland cells open; the
common canal becomes dilated to form
the oötype, and then proceeds as the
uterus, with only slight convolutions,
along the central field to the genital pore,
which lies in the middle line immediately
behind the ventral sucker. The single
vitellarium starts behind the ovary and
extends to the posterior end. The acini
are situated at the sides of the excretory
duct, which runs a median course. The eggs are compact spindles,
much dilated in the middle; they have no lid, and are provided with a
terminal spine (rudimentary filament) at the posterior end, measuring
120 µ to 150 µ in length and 40 µ to 60 µ in breadth, but vary in size
and shape (fig. 175).
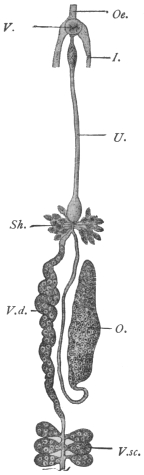
Fig. 174.—Schistosoma
hæmatobium, Bilh.: genitalia
of the female. V.s.,
ventral sucker; I., gut cæca;
V.d., vitelline duct; V.sc.,
vitellarium; O., ovary; Oe.,
œsophagus; Sh., shell
gland; U., uterus. Magnified.
(After Leuckart.)
Distribution.—In order to understand the distribution of the worms
and eggs in the body, it may be well to recall the blood supply of the
abdominal and pelvic organs. It is generally assumed that the early
life (? cercarial stage) of the worms occurs in the liver, and that the
young worms travel from here, where they are invariably found, to
their various sites along the portal vein and its tributaries and so
against the blood stream. The tributaries of the portal vein are:—
(1) Superior mesenteric, the tributaries of which are: (a) the veins
of the small intestine; (b) ileo-colic; (c) right colic; (d) middle colic;
(e) right gastro-epiploic; and (f) inferior pancreatic. By these paths
infection of the small intestine, ascending and transverse colon and
pancreas would occur.
(2) Splenic. (Ova have been recorded by Symmers in the spleen.)
(3) Inferior mesenteric, the tributaries of which are (a) superior
hæmorrhoidal veins from the upper part of the hæmorrhoidal plexus;
(b) sigmoid veins from sigmoid flexure and lower portion of descending
colon; (c) left colic vein draining descending colon.
The superior hæmorrhoidal veins form a rich plexus in the rectum,
and below this level in the upper and middle parts of the anal canal.
The plexus forms two networks, an internal plexus in the submucosa
and an external on the outer surface. The internal plexus opens at
the anal orifice into: (a) branches of the inferior hæmorrhoidal vein
(from the pudic); (b) the external plexus. The external plexus gives
off: (a) inferior hæmorrhoidal opening into internal pudic (of internal
iliac vein); (b) mid-hæmorrhoidal into internal iliac or its branches;
and (c) superior hæmorrhoidal opening into inferior mesenteric.
The external plexus further communicates with the vesico-prostatic
plexus. The vesico-prostatic (vaginal) plexus opens into the vesical
veins, which drain into the interior iliac vein. This plexus also
receives afferents from the pudendal plexus, the chief tributary of
which is the dorsal vein of the penis. The pudendal plexus also
receives branches from the inferior pudic and the anterior surface of
the bladder.
There is thus a communication between the portal vein and the
vena cava by means of these plexuses, viz., through the inferior and
middle hæmorrhoidals, and by the inferior hæmorrhoidals to the
bladder and thence by the vesical veins or the pudic to the caval
system (interior iliac).
It is thus by the inferior mesenteric and its tributaries that the
worms reach the descending colon, rectum, anal canal, and eventually
the bladder, and in some cases the caval system.
Before considering what is actually found post mortem in these
veins and the organs drained by them, we may further recall the fact
that the calibre of “medium” veins is 4 to 8 mm., “small” veins
less than 40 µ in diameter and capillaries 8 µ to 20 µ. Further, the
maximum diameter of the male worm is 1 mm., that of the female
280 µ and eggs in utero 80 µ to 90 µ long by 30 µ to 40 µ.
Liver and Portal Vein.—Here worms are most easily found post
mortem. Often only males are found and these of the same size,
and if females occur only a few worms are found in copulâ. The
worms are frequently not full size and the males may contain no free
spermatozoa in their testes, and as regards the females some may
be fertilized, others not, as shown by the presence or absence of
spermatozoa in the seminal receptacle or uterus. In either case they
may contain eggs—lateral-spined—usually one, less often two, but
there may be as many as five or six. These eggs may also show
some abnormality, which takes the form of: (1) abnormal contents,
viz., disintegrating yolk cells with or without an ovarian cell;
(2) abnormal shape but with normal contents and probably represented
by the collapsed and empty egg-shells which are found in the
tissues.
As to the interpretation of these facts, Looss believes that these
lateral-spined eggs are products of young females whose egg-laying
is not at first properly regulated. The shape that the eggs take, viz.,
with a lateral spine, is determined by an excess of material—ovarian
and yolk cells—being present in the oötype. The shape of eggs
depends upon the position they have in the oötype during their
formation. In young females an excess of cells—yolk cells especially—accumulates,
distending not only the dorsal wall but a portion also
of the short duct joining the oötype to the uterus. The result of this
is that the axis of the oötype and egg is almost transverse to the body,
and the posterior funnel-shaped portion of the oötype, instead of
being terminal, has now a lateral or rather a ventral position, so that
the spine which occupies this portion, instead of being terminal, is
now lateral. It is noteworthy that these lateral-spined eggs are
thicker, owing to the excess of material present, and not uncommonly
have a curved anterior border, due to a projection of the anterior end
into the anterior opening of the oötype.
As these eggs are being laid by females in the portal vein they are
carried back to the liver by the blood stream. The liver is one of
the commonest sites for these eggs; also terminal-spined eggs may be
found here for the same reason.
Hæmorrhoidal Veins.—Mature worms, generally in copulâ, are
usually found here, though young not fully grown females may also
occur. The tissues of the rectal wall (or colon) show, as a rule, large
quantities of lateral-spined eggs, though less often only terminal-spined
eggs may be found.
Vesico-prostatic Plexus.—Worms in copulâ are found in the veins
of the submucosa in the bladder, and the eggs in the mucosa, and
those voided are usually terminal-spined, though lateral-spined eggs
are not so rare as generally thought. The problem next arises as to
how the eggs get to the lumen of the gut or bladder.
The female worm is 280 µ in diameter. Veins in the submucosa
of the rectum less than 178 µ in diameter are not affected with endophlebitis.
It is probable that the female even by stretching could not
penetrate much beyond this. Eggs are probably then laid in the
submucosa as near the muscularis mucosa as possible. Now if the
eggs are laid in a vein of larger calibre than the worm fills, the eggs
would be carried back to the inferior mesenteric vein, so that presumably
the worm must succeed in blocking the vein already narrowed
by endophlebitis, so that by the stasis which ensues the eggs may
escape from the veins. How this occurs is not exactly known; it is
not necessarily due to the spine, as the same escape into the tissues
occurs in spineless eggs, such as those of Schistosoma japonicum. The
eggs, then, pass as foreign bodies through the tissues. Another hypothesis
is that the worms leave the veins in order to lay their eggs,
but the evidence is against this.
Caval System.—Occasionally worms that have passed through the
vesical plexus may be found in the iliac vein, inferior vena cava,
and even the lungs. If the worms are young they contain a lateral-spined
egg; if adult, numerous (50 to 100) terminal-spined eggs.
Lungs.—When the liver is strongly infected with (terminal-spined)
eggs it is possible that by passive movements some may pass into the
intralobular veins, and thence by the inferior vena cava to the lungs.
Gall-bladder.—Similarly terminal-spined eggs pass into the bile-capillaries
and gall-bladder (where they may be abundant), and so into
the fæces.
Detection of Eggs.—Occasionally eggs may be found in various other
parts of the body. They are best detected by macerating pieces of the
tissue in question in about 1/4 per cent. hydrochloric acid at 50 to 60°C.
(Looss).
Pathological changes:—
Rectum.—These have been studied thoroughly by Letulle in the
case of an apparently pure infection of the rectum.274 They take the
form of a chronic diffuse inflammation, which may result in—(1) ulceration,
or (2) hyperplasia of the mucosa, producing adenomata.
Ulcerative Form.—The mucosa is transformed into a mass of
vascular connective tissue. The connective tissue spaces next become
invaded by numerous mononuclear cells. The tissue itself
undergoes diffuse sclerosis, becoming hard and fibroid. Eventually
ulcerative necrosis sets in. During these changes the Lieberkühn
glands are destroyed. The process does not extend to the submucosa,
in this respect differing from that in chronic dysentery.
Hyperplastic Form.—The Lieberkühn glands of the mucosa at first
hypertrophy; then there is an actual hyperplasia resulting in adenomata.
The interstitial tissue of the glands is also greatly hypertrophied,
giving rise to very vascular granulations. These growths
are often hollow and contain worms. Many eggs are found in the
mucosa on their way to the lumen of the gut.
The muscularis mucosa is thickened up to twice or even ten times
the normal. Its vessels are dilated (36 µ to 80 µ), but they do not
allow of the passage of worms.
The submucosa is profoundly changed; rigid and hard instead of
supple. It is here that the greatest number of eggs occur. A remarkable
condition of endophlebitis exists in the veins of the
submucosa, not only in the smaller ones but also in the larger ones
(370 µ by 270 µ). This endophlebitis results in a more or less complete
occlusion of the vessels of the lumen.
The muscular coats are free from change, also their veins.
The Serous Coats.—The veins about 1,900 µ, also show endophlebitis.
Besides the rectum, in extreme cases even the transverse colon, the
cæcum and small intestine may be affected.
Bladder.—In the early stages the mucosa is deep red and swollen
like velvet, or there may be localized patches of hyperæmia or
extravasation. The subsequent changes take two chief forms:—
(1) Sandy Patches.—The mucosa looks as if it were impregnated
with a fine brownish or yellowish powder (myriads of ova). This is
accompanied by a gradual hypertrophy and new formation of connective
tissue, so that dry, hard or plate-like patches with this sandy
appearance arise; the thickening eventually affects all the coats of
the bladder. In the older patches many of the eggs are calcified.
These patches sooner or later break down, ulcerate and necrose.
Phosphatic deposits are abundant and stone is common. These
patches are not found in the rectum.
(2) Papillomata.—Where the inflammatory change produced by
the eggs gives rise to hypertrophy and hyperplasia of the mucosa,
papillomata result, the axis of which is formed by connective tissue
of the submucosa. These are most variable in shape and form and
bleed readily, and sometimes contain cavities of extravasated blood.
As in the rectum, it is in the submucosa that eggs are most
abundant, and worms in copulâ occur in the veins of this layer, but
endophlebitis is not as general as described in the rectum. Malignant
disease of the bladder is not an uncommon sequela of bilharziasis.
Besides the bladder, the ureters and kidneys may in advanced cases
be involved. The prostate and vesiculæ seminales are commonly
diseased. Eggs have been recorded in the semen. The urethra is
frequently attacked; the vagina in the female.
Eggs also occur in the lymphatic glands of the gut.
Geographical Distribution.—East Africa:
Nile Valley, Red Sea Coast, Zanzibar, Portuguese
East Africa, Delagoa Bay, Natal, Port
Elizabeth.
South Africa: Cape Colony, Orange Free
State, Transvaal, Mauritius, Bourbon, Madagascar.
West Africa: Angola, Cameroons, Gold
Coast, Gambia, Senegal, Sierra Leone, Lagos,
Nigeria.
North Africa: Tripoli, Tunis, Algeria, parts
of the Sahara.
Central Africa: Sudan, various portions.
Uganda, Nyasaland.
It occurs with varying frequency in these
regions. It is probably more widely spread
than this list implies, as undoubtedly many
cases are seen which are not recorded.
Isolated cases have been recorded from
Arabia, India,275 Greece, Cyprus.
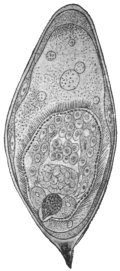
Fig. 175.—Ovum of
Schistosoma hæmatobium,
Bilh., with miracidium,
which has turned its anterior
end towards the
posterior end of the egg.
275/1. (After Looss.)
The means by which infection is brought
about are still uncertain; we only know that
the miracidia (fig. 175) enclosed in the discharged
eggs do not hatch if the eggs remain
in the urine, but after cooling perish. As
soon, however, as the urine is diluted with
water the shell swells, generally bursting lengthways,
and releases the miracidium from its
investing membrane, so that it can swim about
with the aid of its cilia. In its structure it
differs but little from the miracidium of Fasciola
hepatica, as, for instance, in the lack of eyes;
the two large gland cells situated on either side of the intestinal
sac are also present in the miracidia of Fasciola hepatica.
Sarcode Globules.—This is a term applied to certain globules which at
times appear in the miracidium and are later ejected. Some authors
consider them as indicative that the miracidium has developed into
a sporocyst, but Looss considers them to be degeneration products.
The Bilharzia mission, under R. T. Leiper, sent to Egypt by the
War Office early in 1915, reports that cercariæ of bilharzia type were
recognized in four of the commonest fresh-water molluscs around
Cairo.
With material obtained from naturally infected Planorbis boissyi
acute bilharziosis was experimentally produced in rats, mice, and
monkeys. Infection takes place experimentally through the skin and
also through the mucous membrane of the mouth and œsophagus.
The miracidium, after entering the mollusc, develops into a sporocyst.
This gives rise not to rediæ, but to secondary sporocysts, which, in
turn, produce cercariæ. These, like the adult worm, differ from other
distomes in lacking a muscular pharynx.
Schistosoma mansoni, Sambon, 1907.
According to Manson, Sambon and others, the eggs with lateral
spines belong to a species different from Schistosoma hæmatobium.
Infections with this species only are said to
occur in the Congo, Southern States of North
America, West Indies (Guadeloupe) and Brazil
(Bahia). The following characters, according to
Flu, differentiate this species: (1) In the male
the transition from the anterior portion of the
worm to the lateral fields (the infolded portions
which form the gynæcophoric canal) is not a
gradual one as in Schistosoma hæmatobium,
but in this case the lateral fields rise suddenly,
almost at right angles to the anterior portion.
(2) The ovaries have a well-marked convoluted
course as in no other schistosome. (3) The
oötype is symmetrical in reference to the long
axis of the body, its duct being lateral on the
ventral side (Looss’ explanation of this we have
already given). (4) The worms live exclusively
in portal vein and tract. (As lateral-spined eggs
occur also in the bladder, this is not exactly true.)
Schistosoma hæmatobium, Bilharz, 1852.
Male, four or five large testes. Gut forks unite late, so that the
single gut stem is short. Female, ovary in posterior half of body.
Uterus very long, voluminous, with many terminal-spined eggs, some
lying in pairs. Vitellaria in posterior fourth of body. Cercariæ in
Bullinus contortus and Bullinus dybowski (syn.: Physa alexandrina) in
Egypt.
Schistosoma mansoni, Sambon, 1907.
Male, eight small testes. Gut forks unite early, so that the single
gut stem is very long. Females, ovary in anterior half of body. Uterus
very short; usually only one lateral-spined egg at a time in utero.
Vitellaria occupy posterior two-thirds of body. Cercariæ in Planorbis
boissyi in Egypt.
The above morphological descriptions are founded on worms of
each species, derived from experimentally infected mice (Leiper, R. T.,
Brit. Med. Journ., March 18, 1916, p. 411).
Schistosoma japonicum, Katsurada, 1904.
Syn.: S. cattoi, Blanchard, 1905.

Fig. 176.—Schistosoma japonicum:
anterior end with testes;
posterior end with point of union
of cæca. Length of worm about
10 mm. (After Katsurada.)
Male.—Eight to 19 mm., but extreme limits are 5 to 22·5 mm. Consists
of a short fore-body, separated by the ventral sucker from the
hind-body. The ventral sucker is stalked and somewhat larger than
the oral sucker. Both suckers are larger than the corresponding ones
in S. hæmatobium. Body usually smooth, but in the fresh state
numerous fairly evident spines along the margin of the canal. Œsophagus:
two bulbs. The junction of the gut forks more posterior than
in S. hæmatobium, the median united gut
stem occupying a quarter to one-fifth to
one-sixth of the body length. An excretory
canal runs along each side of the
body, opening into the dorsal excretory
pore. Testes irregularly elliptical, six to
eight in number, in the anterior part of
hind-body. The vasa efferentia unite
into a common vas deferens which opens
directly behind the ventral sucker. The
seminal vesicle lies just behind this.
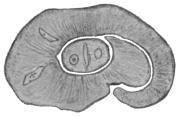
Fig. 177.—Schistosoma japonicum,
male and female in copulâ.
× 60. (After Katsurada.)
Female.—Up to 26 mm., generally
thinner than the male. Surface smooth.
Suckers armed with fine spines. Ventral
sucker larger than oral. Body thicker
behind the region of the ovary. The
gut forks unite immediately behind the
ovary. The united gut much thicker
than in S. hæmatobium. Ovary elliptical,
almost in the mid-body, its hinder portion
dilated. The oviduct arises from
its posterior end and then runs sinuously
forward, where it is joined by the vitellarian
duct; the vitellarium well developed,
extending from behind the ovary
almost but not quite to the posterior
end as in S. hæmatobium. Shell gland
ducts enter at the junction point of
oviduct and vitelline duct. The canal
here forms an oötype and then proceeds
as the uterus to open directly behind
the ventral sucker. The uterus occupies
almost half the hind-body. In S. hæmatobium
this is not so. The uterine canal
is cleft-like, i.e., its dorso-ventral diameter
is much greater than its lateral diameter.
The number of eggs varies from about
50 to 300 from observations made in various hosts.
Eggs.—In utero assume various shapes, as they are soft; the lumen
of the uterus is narrow. Outside they are oval, faint yellow, double
contoured. In fæces the eggs measure 83·5 µ, by 62·5 µ (man); 85 µ
by 61·5 µ (cattle); 98·2 µ by 73·8 µ (dog). The eggs have either
small lateral spines or thickenings, and Looss at the opposite
side has described cap-like thickenings.
The eggs in the tissues undergo various deformities,
and may contain a miracidium, as
also the eggs in fæces do; or the contents
may consist of granular matter or amorphous
masses or they may be calcified. Lymphocytes
and giant cells may also invade the
eggs.
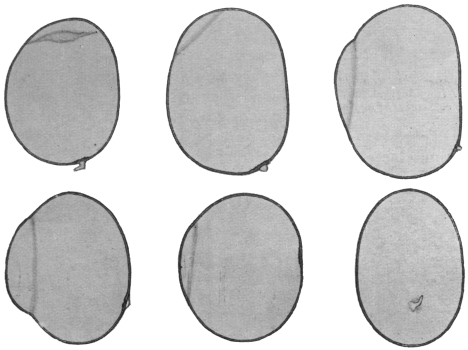
Fig. 178.—Schistosoma japonicum: eggs from human liver, showing “spines” and
“hoods” at opposite pole. (After Looss.)
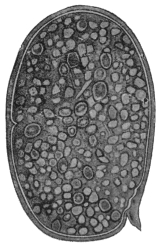
Fig. 179.—Schistosoma japonicum:
from dog. Uterine
egg. × c. 800. (After Katsurada.)
Mode of Infection.—The miracidia hatch
in water in as little as fifteen minutes, but
the majority in one to three hours. They will live in water for about
twenty-four hours. In water they undergo a transformation into
“larvæ,” which then penetrate the skin, as has been shown by
Japanese writers to hold good for man, cattle, dog and cat. The
penetration of the skin is attended with an eruption on the legs,
“Kabure.” The exact route by which the worms reach the portal vein
is uncertain. Infection in Japan takes place from spring to autumn,
especially May to July, when the soil is contaminated with manure
of cattle infected with S. japonicum. They also appear to develop
in molluscs. Leiper and Atkinson found cercariæ (in sporocysts)
in the liver of a mollusc, Katayama nosophora. They infected mice
by immersing them in water containing liver emulsion and so free
cercariæ, thus confirming the similar results of Miyairi and Suzuki.
Habitat.—The worm occurs in Japan, China, and the Philippines.
The normal host is man and mammals. Cattle, dog and cat are
often found naturally infected. Mice can also be experimentally
infected. Their seat of election is the portal vein and its branches,
especially the mesenteric veins. They either
swim free in the blood or remain fixed by
their suckers to the intima of the vessels.
They have also been found in the vena cava
and right heart of a cat, but not so far in the
vesical plexus.
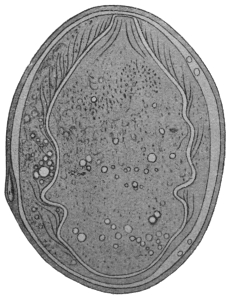
Fig. 180.—Schistosoma japonicum: from
dog. × c. 800. (After Katsurada.)

Fig. 181.—Schistosoma japonicum:
from dog. Egg from fæces. × c. 800.
(After Katsurada.)
Eggs are found in the submucosa and
mucosa of the gut, especially the colon, and
at times in the serosa and subserosa of the
small intestine, where they give rise to new
growths. Occasionally eggs are found in
the brain. The life of the worms is at least
two years.
Pathogenic Effects.—Anæmia through loss
of blood due to worms; enlarged spleen,
toxic in origin (?); phlebitis, thrombosis, due
to portal stasis; the eggs, however, cause the
greatest mischief. They are carried by the circulation to various
organs where they produce inflammation, granulation tissue, and later
connective tissue.
Liver.—The eggs reaching this organ give rise to granulomata and
hence enlarged liver, and later, when connective tissue is formed, to
contraction. The surface is rough and irregularly granular, “parasitic
embolic cirrhosis” of Yamagiwa.
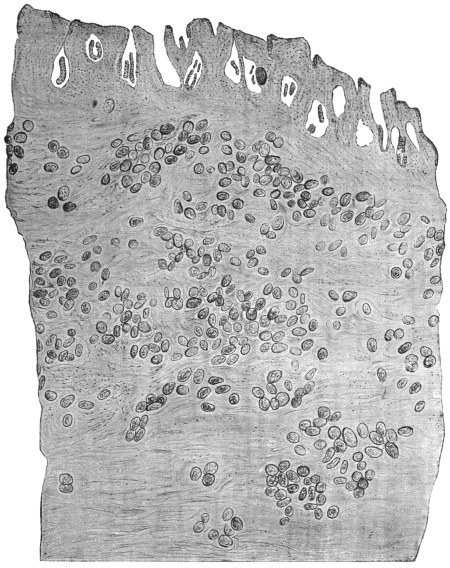
Fig. 182.—Schistosoma japonicum: section through the gut of a Chinaman showing eggs.
× 58. (After Catto.)
Gut.—The eggs in the mucosa and submucosa cause catarrh and
destruction of tissue or new growth. In the small intestine the eggs
are mainly in the serosa and subserosa, where they give rise to polypoid
or branched growths.
Spleen.—Enlarged, at first due to toxin (?) and later due to portal
stasis. Eggs in the spleen are uncommon.
Ascites also arises from the portal stasis, and is generally present
in advanced cases.
Eggs may be found in many other situations: glands (numerous),
mesentery, stomach, pancreas, kidney, etc. The bladder remains free.
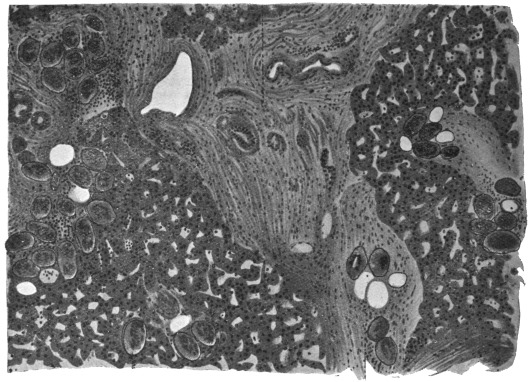
Fig. 183.—Schistosoma japonicum: liver showing eggs in the intra- and interlobular
connective tissue. × c. 80. (After Katsurada.)
Class III. CESTODA, Rud., 1808.
Tapeworms have been known from ancient times—at all events, the large
species inhabiting the intestines of man—and there has never been a doubt as
to their animal nature. The large cysticerci of the domestic animals (occasionally
of man also) have been known for an equally long period, but they were generally
regarded as growths, or “hydatids,” until almost simultaneously Redi in Italy, and
Hartmann and Wepfer in Germany, concluded from their movements and organization
that they were of animal nature. From that time the cysticerci have been
included amongst the other intestinal worms, and Zeder (1800) established a special
class (Cystici, Rud., 1808) for the bladder worms. Things remained in this condition
until the middle of the last century, when Küchenmeister, by means of successful
feeding experiments, demonstrated that the cysticerci were definite stages of
development of certain tapeworms. Before Küchenmeister, E. Blanchard, van
Beneden, and v. Siebold had held the same opinion in regard to other asexual
Cestodes.
Since the most remote period another question has again and again occupied
the attention of naturalists, the question of the morphological nature—that of the
INDIVIDUALITY OF THE TAPEWORM. The ancients, who were well acquainted
with the proglottids (Vermes cucurbitani) that are frequently evacuated, were
of the opinion that the tapeworm originated through the union of these separate
proglottids, and this view was maintained until the end of the seventeenth
century. In 1683 Tyson discovered the head with the double circlet of hooks in
a large tapeworm of the dog; Redi (1684) was also acquainted with the head and
the suckers of several Tæniæ. Andry (1700) found the head of Tænia saginata, and
Bonnet (1777) and Gleichen-Rusworm (1779) found the head of Dibothriocephalus
latus. Consequently most authors, on the ground of this discovery, considered the
tapeworm as a single animal, that maintains its hold in the intestine by means of the
head, and likewise feeds itself through it. The fact was recognized that there were
longitudinal canals running through the entire length of the worm, and it was thought
that these originated in the suckers, and that the entire apparatus was an intestine.
As, moreover, the segments form at the neck, and are cast off from the opposite
extremity, the tapeworm was also compared with the polyps, which were formerly
regarded as independent beings.
Steenstrup, in his celebrated work on the alternation of generations (1841), was
the first to give another explanation. This has been elaborated still further by
van Beneden, v. Siebold and Leuckart, and until a few years ago all authorities
adopted his views. According to this view, the tapeworm is composed of numerous
individuals, something like a polyp colony, and, in addition to the proglottids—the
sexual individuals which are usually present in large numbers—there is ONE
individual of different structure, the scolex, which not only fastens the entire colony
to the intestine, but actually produces this colony from itself, and therefore is
present earlier than the proglottids. The scolex is a “nurse,” which, though itself
produced by sexual means, increases asexually like a Scyphistoma polyp; the
tapeworm chain has therefore been termed a strobila. Consequently the development
of the tapeworms was explained by an alternation of generations. In support
of this opinion it was demonstrated not only that the adult sexual creatures, the
proglottids, can separate from the colony and live independently for a time, but
that in certain Tæniæ, and especially in many Cestodes of the shark, the proglottids
detach themselves long before they have attained their ultimate size, and thus
separated continue to develop, grow and finally multiply; the scolex also exhibits a
certain independence in so far as, though not, as a rule, capable of a free life, yet
it in some cases lives as a free being, partly on the surface of the body of marine
fishes and partly in the sea. With the more intimate knowledge of the development
of the cysticerci, the independent nature of the scolex was recognized. It is
formed by a budding of the bladder that has developed from the oncosphere, in
some cases (Cœnurus) in large numbers, in other cases (Echinococcus) only after
the parent cyst has developed several daughter cysts. Released from its mother
cyst and placed in suitable conditions, it goes on living, and gives rise at its
posterior end by budding to the strobila, the proglottids of which eventually become
sexual individuals.
In order to make this clearer we will briefly summarize what takes place in
the jelly-fishes.
By metamorphosis is meant a developmental change in the same individual, while
alternation of generations, or metagenesis, implies a stage in which reproduction of
individuals takes place by a process of budding or fission. This asexual reproductive
stage alternates with the sexual mode of reproduction. Thus in the development
of the Scyphozoa (jelly-fishes) we have:—
(1) The fertilized egg cell divides regularly and forms a morula.
(2) By accumulation of fluid in the interior this becomes a closed sac with a wall
formed of a single layer of cells, forming the blastosphere or blastula.
(3) One end of the sac is invaginated, forming a gastrula.
(4) The gastrula pore or mouth closes, forming again a sac, the walls of which
have two layers, forming a planula.
(5) This becomes fixed to a rock, an invagination forms at one end, a depression—the
stomodæum—communicating with the enteric cavity. Tentacles grow out
and we have a Scyphozoön polype, Scyphistoma or Scyphula. It is to this stage that
Steenstrup gave the name “nurse” (“wet-nurse”), because it nourished or produced
asexually the succeeding forms.
(6) Asexual reproduction by transverse fission occurs in this, forming a pile of
saucer- or pine-cone-like animals which before this time had been considered to
be a distinct animal, which was called strobila from its resemblance to a pine-cone.
This is the alternate generation.
(7) The individuals of the strobila become free and are called Ephyrulæ.
(8) These develop finally into adult sexual jelly-fish, Scyphozoa, so that comparing
a tapeworm with this we have (a) egg, (b) scolex (= Scyphula or “nurse”),
(c) asexual reproduction of the tapeworm chain (= strobila), (d) development of the
individuals of the chain (proglottids) into sexual adults.
Van Beneden’s terminology for these stages is the following: Ciliated embryo
= protoscolex; scyphistoma = deutoscolex (or scolex); free Ephyrula = proglottis.
According to this view, as is the case in many endoparasitic Trematodes, asexual
reproduction by budding occurs at two stages of the whole cycle of development,
viz. (1) in the formation of the scolex by budding from the bladder (“nurse”),
(2) in the formation of the strobila by budding from the scolex (“nurse”).
But in cysticercal larval forms it appears that the scolex does not arise in this
way but is simply a part of the proscolex (hexacanth embryo), becoming invaginated
into it for protection, so that there is no asexual gemmation here. It has been
questioned also whether the strobila also arises by gemmation. If it does, the
tapeworm is a colony of zoöids produced by budding from the asexual scolex; if it
is not produced in this way, then the tapeworm is to be regarded as an individual
in which growth is accompanied by segmentation. Against the “colony” view are
the facts that the muscular, nervous, and excretory systems are continuous throughout
the worm, and that some tapeworms, such as Ligula, are unsegmented.
Finally, if the tapeworm is an individual the question arises which is the head
end. As new segments are formed at the neck, and as this point in annelids is
the antepenultimate segment, the scolex must be the last or posterior segment. The
caudal vesicle or bladder of larval forms is consequently anterior. According to
this view, in tapeworms as among many endoparasitic flukes, an asexual multiplication
occurs at two points of the whole cycle of development, which is as follows:
(1) egg, (2) oncosphere or hexacanth embryo, (3) bladder (cysticercus or hydatid),
(4) (after digestion of the bladder) by budding, the scolex, (5) by budding from the
scolex the sexual proglottids, (6) the egg; (4) and (5) being the two asexual stages.
Anatomy of the Cestoda.
If we except the tapeworms with only one proglottis, the
Cestoidea Monozoa, Lang = Cestodaria, Monticelli, we can always
distinguish in the Cestodes, in the narrower sense, one scolex or head
and a large or small number of segments (proglottids). The SCOLEX
serves the entire tapeworm for fastening it to the internal surface of the
intestinal wall, and therefore carries at its end various organs which
assist in this function, and which are as follows: (i) Suctorial
organs, i.e., the four suckers (acetabula), which are placed crosswise
at the circumference of the thickened end of the scolex; further, the
double or quadruple groove-like suckers (bothridia), which are
diversely shaped in the various genera and families.276 (2) Fixation
organs (hooklets)277 that likewise occur in varying numbers and
different positions; they may be in the suckers, or outside them on the
apex of the scolex; for instance, in many of the Tæniidæ they appear
in a circle around a single protractile organ, the rostellum, or the latter
may be rudimentary, and is then replaced by a terminal sucker.
(3) Proboscis. One family of the Cestodes, the Rhynchobothriidæ,
carries four proboscides, moved by their own muscular apparatus,
on the scolex, and they are beset with the most diverse hooks.
(4) Tentacle-like formations are only known in one genus
(Polypocephalus).
The thickened part of the scolex that carries the suckers is usually
called the head; the following flat (unsegmented) part connecting it
with the proglottids is called the neck, and is sometimes quite small.
In a few cases the entire scolex (or head) disappears, and its function is
then undertaken by the contiguous portion of the chain of proglottids,
which is transformed into a variously shaped PSEUDO-SCOLEX.
The proglottids are joined to the scolex in a longitudinal row,
and are arranged according to age in such a manner that the oldest
proglottis is farthest from the scolex, and the youngest nearest to it.
The number of segments varies, according to the species, from
only a few to several thousands; they are either quadrangular or
rectangular; in the latter case their longitudinal axis falls either
longitudinal or transverse to that of the entire chain, according as
the segments are longer than broad or broader than long. When the
number of segments is very large, the youngest ones are, as a rule,
transversely oblong, the middle ones are squarish, and the mature
ones longitudinally oblong. The posterior border of the segments,
as a rule, carries a longitudinal groove for the reception of the shorter
anterior border of the following proglottis. The two lateral borders
of the segment are rectilinear, but converge more or less towards the
front, or they are bent outwards. In most of the Cestodes the
segments, just as the neck, are very flat; in rare cases their
transverse diameter is equal to their dorso-ventral diameter. As a
rule the segments, singly or several united together, detach themselves
from the posterior end, in many cases only after complete
maturity is attained, and in others much earlier; they then continue
to live near their parent colony, to still call it by that name, in the
same intestine and continue their development. Even when evacuated
from the intestine the proglottids under favourable circumstances
can continue to live and creep about, until sooner or later they
perish.
The first proglottis formed, and which in a complete tapeworm
[i.e., sexually complete] is the most posterior, is as a rule smaller and
of different shape, it also frequently remains sterile, as likewise happens
in the next (younger) segments in a few species; otherwise, however,
sooner or later the generative organs develop in all the segments,
mostly singly, sometimes in pairs; in the latter case they may be quite
distinct from each other or possess some parts in common. The
term “mature” is used for a proglottid that has the sexual organs
fully developed, while “gravid” is used for one containing eggs.
Most of the species combine male and female genitalia in the same
segment, only a few are sexually distinct (Diœcocestus). In the
hermaphrodite species one male and one female sexual orifice are
always present, and, in addition, there may be a second female orifice,
the uterine opening; as a rule, however, this is lacking, and in one
sub-family, the Acoleinæ, to which also the genus Diœcocestus belongs,
the other sexual orifice, the opening of the vagina, is also absent.
The position of these orifices varies; the cirrus and vagina usually
open into a common atrium on one lateral border or on a surface
of the segments; the orifice of the uterus may be on the same
surface or on the opposite one.
The surface on which the uterus opens is termed the VENTRAL
SURFACE; if this orifice is absent, one must depend on the ovary,
which almost always approaches one of the two surfaces; this surface
is then called the ventral.
The length of the Cestodes—independently of their age—depends
on the number and size of the segments, as well as on their
contraction; the smallest species (Davainea proglottina) is 0·5 to
1·0 mm. in length; the largest may attain a length of 10 m., and
even more.
The entire superficial surface of the tapeworms is covered with
a fairly resistant and elastic layer, which exhibits several indistinctly
limited layers and which is usually called a cuticle, which also covers
the suckers, and is reflected inwardly at the sexual orifices. In some
species fine hairs appear, either on the entire body or only in the
region of the neck, on the external surface. In the cuticle there can be
recognized, besides the pores, which no doubt are concerned with
nutrition, spaces in which lie the ends of sensory cells. Close under
the cuticle lies the external layer of the parenchyma (basal membrane),
and below this the circular and longitudinal muscles forming the
dermo-muscular coat. The matrix cells of the cuticle occur as in
the Trematodes, only on the
inner side of the peripheral
muscles in the external zone
of the parenchyma; they are
fusiform cells, forming one
or two layers, but are not
arranged in the manner of
epithelial cells (fig. 184, Sc.c.).
They have fine branching
processes which run between
the dermal muscles, pass
through the basal membrane
and penetrate the internal surface
of the cuticle with small
pistil-like enlargements, expanding
on the internal
surface of the cuticle into
a thin plasma layer.
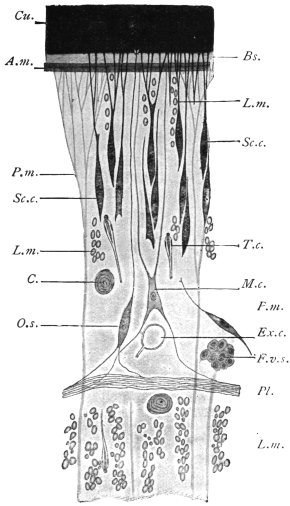
Fig. 184.—Schematic representation of a small
part of a transverse section of Ligula sp. Bs., basal
membrane; Cu., cuticle; at its base are the endplates
of the subcuticular (epithelial) cells; in the
centre a cuticular sense organ, O.s.; F.v.s., vitelline
follicle; Exc., excretory vessel; C., calcareous corpuscle;
L.m., longitudinal muscles; M.c., myoblast;
P.m., parenchymatous or dorso-ventral muscles;
Pl., plexus of nerve fibres; A.m., circular muscles;
Sc.c., subcuticular or matrix cell; T.c., terminal
flame cell. 500/1. (After Blochmann.)
In addition to the above
mentioned, there are other
cuticular formations occurring
on the cuticle of some
Cestodes, such as immobile
hairs and variously formed
hooks, such as are seen principally
on the scolex. Their
development is only roughly
known in a few species; they
are usually already present in
the larval stage, and of the
same arrangement and shape
as in the fully developed
tapeworms; a matter of
importance, because by these
structures larvæ can be recognized
as being those of a certain species of tapeworm.
The CUTICULAR GLANDS in Cestodes are scarce.
The PARENCHYMA forms the chief tissue of the entire body, and in
all essentials its structure is similar to that of the Trematodes.
The same doubt exists here also as to the nature of the parenchyma.
Recent authors consider that it consists of highly branched cells, the
processes of which ramify in all directions. These cells lie in a non-cellular
matrix containing fluid vacuoles. This matrix spreads in
between and so breaks the continuity of the epidermal cells.
In the parenchyma of almost all the Cestodes there are found in
adult specimens, as well as in larvæ, light-refracting concentrically
striated structures, of a spherical or broad elliptical shape, which, on
account of their containing carbonate of lime, are termed CALCAREOUS
CORPUSCLES (fig. 184, C.). Their size, between 3 µ and 30 µ, varies
according to the species; their frequency and distribution in the parenchyma
also varies, but they are chiefly found in the cortical layer.
They are the product of certain parenchymatous cells, in the interior
of which they lie like a fat globule in a fat cell, but according to
others they are intercellular in origin.
The MUSCULAR SYSTEM of the proglottids is composed of—(1) the
subcuticular muscles (figs. 184 and 185), as a rule consisting of a
single layer of annular muscles; (2) longitudinal muscles; (3) dorso-ventral
fibres extending singly from one surface to the other, and
at both ends expanding in a brush-like manner, and inserted into
the basal membrane, consisting of an outer, more numerous, and an
inner, less numerous but more powerful layer (the number of bundles
in this layer being in certain cases of specific importance); (4) transverse
fibres, the elements of which penetrate to the borders of the segments,
thus passing through the longitudinal muscles and reaching
the cuticle. In the region of the septa the transverse and dorso-ventral
muscles form a kind of plate.
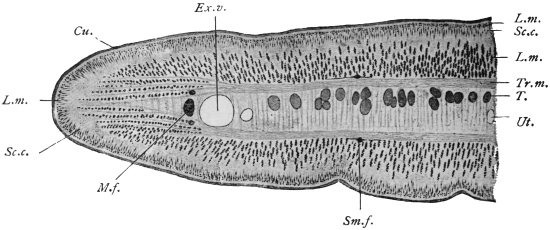
Fig. 185.—Half of a transverse section through a proglottis of Tænia crassicollis.
Cu., cuticle; Ex.v., external excretory vessel, to the right of which there is the smaller
internal one; T., testicular vesicles; L.m., longitudinal muscles (outer and inner); M.f., lateral
nerve with the two accessory nerves; Sc.c., subcuticular matrix cells; Sm.f., submedian
nerve; Tr.m., transverse muscles; Ut., the uterus, and the middle of the entire transverse
section. 44/1.
The mass of parenchyma bounded by the transverse muscles is
termed the MEDULLARY layer, while the mass lying outside them
is termed the CORTICAL LAYER.
It was known long ago that the myoblasts adhere to the dorso-ventral
fibres as thickenings, but it is only recently that large star-shaped
cells (fig. 184), separated from but connected with them by
processes, have been recognized as
the myoblasts of other fibres (Blochmann,
Zernecke).
Within the scolex the direction
and course of the muscular layers
change.
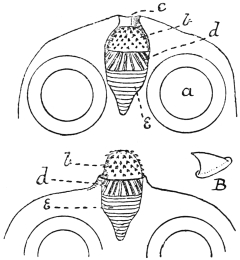
Fig. 186.—Dipylidium caninum: from
the cat. In the upper figure the rostellum is
retracted, in the lower protruded, a, sucker;
b, hooks of rostellum; B, enlarged hook;
c, apical aperture on scolex; d, longitudinal
muscles; e, circular muscles. (After
Benham.)
The SUCKERS are parts of the
musculature, locally transformed,
with a powerful development of
the dorso-ventral muscles, now
become radial fibres.
The ROSTELLUM of the armed
Tæniæ, like the proboscis of the
Rhynchobothriidæ, also belongs to
the same category of organs.
In the simplest form, the rostellum,
or top of the head (as
in Dipylidium caninum), appears
as a hollow oval sac, the anterior
part of which, projecting beyond
the upper surface of the head, carries several rows of hooks (fig. 186).
The entire internal space of the sac is occupied by an elastic, slightly
fibrous mass, while the anterior half of the surface of the rostellum is
covered by longitudinal fibres and the posterior half by circular fibres.
On contraction of the latter the entire mass is protruded through the
apical aperture, the surface of the rostellum becomes more arched, and
the position of the hooks is, in consequence, altered. The rostellum
of the large-hooked Tæniidæ, which inhabit the intestine of man and
beasts of prey, is of a far more complicated structure, for, in addition
to the somewhat lens-shaped rostellum carrying the hooks on its outer
surface, there are secondary muscles grouped in a cup-like manner
(fig. 187). Every change in the curvature of the surface of the rostellum
induces an alteration in the position of the hooks. In the hookless
Tæniidæ the muscular system of the rostellum is altered in a very
different manner; in a few forms a typical sucker appears in its place.
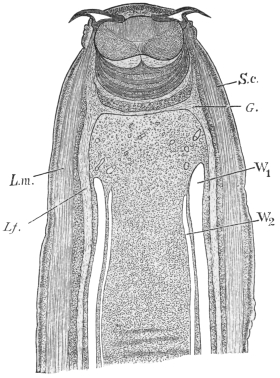
Fig. 187.—Longitudinal section of the head
and neck of Tænia crassicollis, showing the
lens-shaped muscular rostellum, with two hooks
lying in the concentric cup-like mass of muscles.
L.m., longitudinal muscles of the neck; L.f., left
lateral nerve; G., ganglion; S.c., subcuticular
layer; W1, external, W2, internal excretory
vessel. 30/1.
The NERVOUS SYSTEM commences in the scolex and runs through
the neck and the entire series of proglottids. Within the proglottids
it consists of a number of longitudinal nerve fibres of which those at
each lateral border are usually the largest. In the Tæniæ the lateral
nerves are accompanied both
dorsally and ventrally by a
thinner nerve (accessory nerve)
(fig. 185); on each surface,
moreover, between the lateral
nerve and the median plane,
there are two somewhat
stronger bundles (sub-median),
so that there is a total of ten
longitudinal nerve bundles.
They lie externally to the transverse
muscle plates, and the
lateral and accessory bundles
lie externally to the principal
excretory vessels, and are everywhere
connected by numerous
anastomoses and secondary
anastomoses; one typical ring
commissure is usually found
at the posterior border of the
segments. In the Bothriocephalidæ
the distribution of
the nerve bundles is different
(for instance, two lie in the
medullary layer), or they are
split up into a larger number
of branches. In the scolex the nerve bundles are connected in a very
remarkable manner by commissures with that which is generally
termed the central part of the entire nervous system. There occurs
normally a commissure between the two lateral nerves; at the same
level, the dorsal and ventral median nerves are also connected at each
surface as well with each other as with the lateral nerves, so that a
hexagonal or octagonal figure is formed. The so-called apical nerves
pass from this commissural system anteriorly, embrace the secondary
muscular system of the rostellum semicircularly, and form an annular
commissure (rostellar ring) at the inner part of the rostellum.
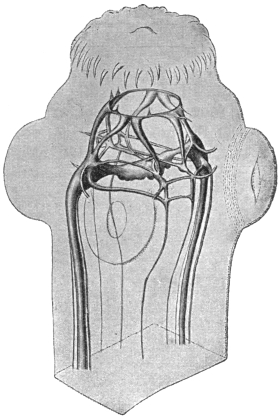
Fig. 188.—Tænia cœnurus, head and part of
neck showing nervous system. Enlarged. (After
Niemiec.)
The peripheral nerves arise from the nerve bundles as well as
from the commissures situated in the scolex; some go direct to the
muscles, while others form a close plexus of nerves external to the
inner longitudinal muscles, which plexus likewise sends out fibres to
the muscles, but principally to numerous fusiform sense organs
(fig. 184, Pl.); they lie internal to the subcuticular cells and, piercing the
cuticle with their peripheral processes, end as projecting “receptor”
hairs. Higher organs of sense are not known.
The EXCRETORY APPARATUS of the Cestodes is similar to that of
other flat worms. The terminal
(flame) cells, which hardly differ
in appearance from those of
the Trematodes, are distributed
throughout the parenchyma, but
are more common in the cortical
than in the medullary layer
(fig. 184, T.c.). Before opening
into a collecting tube, the capillaries
run straight, tortuously,
or in convolutions, anastomosing
frequently with one another
or forming a rete mirabile. The
collecting tubes, which have
their own epithelial and cuticular
wall, and which also
appear to be provided with
muscular fibres, occur typically
as four canals passing through
the entire length of the worm
(fig. 189); they lie side by side,
two (a wider thin-walled ventral,
and a narrower thick-walled
dorsal one) in either lateral field;
in the head the two vessels on
each side unite by means of a loop, at the posterior extremity they
open into a short pyriform or fusiform terminal bladder which discharges
in the middle of the posterior edge of the original terminal
proglottis.
This primitive type (fig. 189) of arrangement of collective tubes is
subject to variation in most Cestodes, in the scolex as well as in the
segments. Indeed, even the lumen of the four longitudinal tubes
does not remain equal, as the dorsal or external tubes are more fully
developed and become thicker, whereas the ventral or internal ones
remain thin, and in some species quite disappear in the older segments
(figs. 185, 187). Moreover, very frequently connections are
established between the right and left longitudinal branches, as in the
head, where a “frontal anastomosis” develops, which in the Tæniidæ
usually takes the form of a ring encircling the rostellum (fig. 190), and
in the segments of a transverse anastomosis at each posterior border,
especially between the larger branches, and more rarely between the
smaller collecting tubes also (fig. 191).
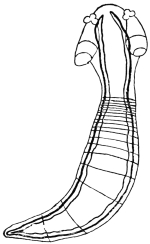
Fig. 189.—Young Acanthobothrium
coronatum, v. Ben., with the excretory
vessels outlined. Slightly enlarged. (After
Pintner.)
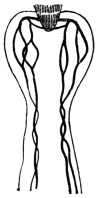
Fig. 190.—Scolex of a cysticercoid
from Arion sp., with the excretory
vessels outlined. (After Pintner.)
The so-called “island” formation is another modification, i.e.,
at any spot a vessel may divide and after a longer or shorter course
the two branches reunite, and this may appear in the collecting tubes
themselves as well as in their anastomoses. The above-mentioned ring
in the frontal commissure of the Tæniidæ is such an island; similar
rings also frequently encircle the suckers (fig. 190). In extreme cases
(Triænophorus, Ligula, Dibothriocephalus, etc.) this island formation
extends to all the collecting tubes and their anastomoses. Instead of
two or four longitudinal canals only, connected by transverse anastomoses
at the posterior border of the segments, there is an irregular
network of vessels, situated in the cortical layer, from which the
longitudinal branches, having again subdivided, can only be distinguished
at intervals, and even then not in their usual number.
The opening of the longitudinal branches at the posterior end
requires more accurate investigation; it is true that a single terminal
bladder is mentioned as being present in many species, but this is
also disputed; when the original end proglottis has been cast off, the
longitudinal branches discharge separately. Some species possess the
so-called foramina secundaria, which serve as outlets for the collecting
tubes; they are generally at the neck, but may be situated on the
segments.
The contents of the excretory vessels is a clear fluid, the regurgitation
of which is prevented by the valves present at the points of
origin of the transverse anastomoses. The fluid contains in solution
a substance similar to guanine and xanthine.
Genital Organs.—With the exception of one genus (Diœcocestus,
Fuhrm.), in which the species are sexually differentiated, all the
Cestodes are hermaphroditic; the genitalia develop gradually in
the segments (never in the scolex), the male organs, as is usual
in hermaphroditic animals, forming earlier than the female. The
youngest proglottids generally do not exhibit even traces of genitalia:
these, as a rule, develop first in the older segments, and the development
proceeds onwards from segment to segment. In a few exceptional
cases (Ligula) the sexual organs are already developed in
the larval stage, but are only functional after the entry of the parasite
into the final host.
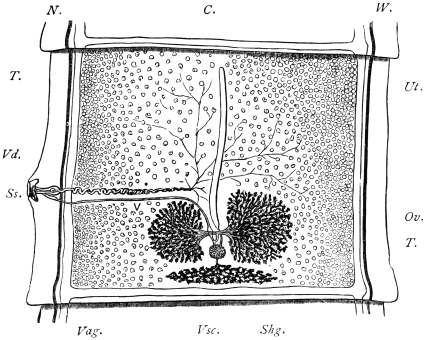
Fig. 191.—Proglottis of Tænia saginata, Goeze, showing genitalia. C., transverse
excretory canal; N., lateral longitudinal nerve; W., longitudinal excretory
canal; T., testicles scattered throughout the proglottis; Ut., opposite the central
uterine stem (a closed sac); Ss., genital pore leading into the genital sinus; above
the cirrus and coiled vas deferens (V.d.), below the vagina (Vag.), bearing near
its termination a dilatation, the seminal receptacle; Vsc., the triangular vitellarium,
and above it (Shg.) the shell gland; leading from this to the uterus is seen
the short uterine canal, on either side of this the two lobes of the ovary (Ov.). 10/1.
With the exception of the end portions of the vagina, cirrus and
uterus, all the parts of the genital apparatus lie in the medullary layer,
except only the vitellaria, which in many species are in the cortical
layer. The male apparatus consists of the testes, of which, as a rule,
there are a large number,278 and which lie dorsal to the median plane
(fig. 185, T.); a vas efferens arises from each testis, unites with
contiguous vasa, and finally discharges into the muscular vas deferens
that is situated in about the middle of the segment. According to
the position of the genital pore, the vas deferens opens on the lateral
margin or in the middle line in the front of the segment; it is much
convoluted or twisted, and frequently possesses a dilatation termed
the vesicula seminalis. It finally enters the cirrus pouch, which is
usually elongated; within the cirrus pouch lies the protrusible cirrus,
which is not uncommonly provided with hooklets.

Fig. 192.—Dibothriocephalus latus. Upper figure: female genitalia, ventral
view. Lower figure: male genitalia, dorsal view. The central portion only of
the proglottis is shown. a, cirrus sac; b, partly everted cirrus; c, genital atrium
and pore; d, vaginal pore; e, uterus; f, uterine pore; g, vagina; h, ovary;
i, shell gland; j, vitelline duct; k, lateral nerve; l, vitellarium; n, vas deferens
(muscular portion); p, vas deferens; q, seminal vesicle; r and x, vasa efferentia;
s, lateral excretory canal; t, testicular follicles. (After Benham and Sommer
and Landois.)
The male sexual orifice almost always opens with that of the
vagina into a genital atrium, the raised border of which rises above
the edge of the segment and forms the genital papilla (fig. 191).
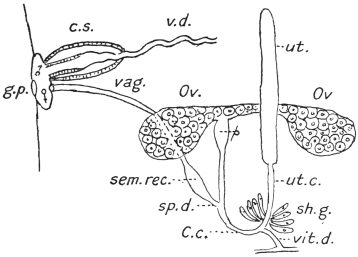
Fig. 193.—Diagram of genitalia of a Cestode. g.p., genital pore; ♀ ♂, male
and female ducts opening into genital sinus; c.s., cirrus sac; v.d., coiled vas
deferens (“outer seminal vesicle”); vag., vagina; sem. rec., seminal receptacle;
sp. d., spermatic duct; C.c., fertilization canal; vit. d., vitelline duct; sh. g., shell
gland; ut. c., uterine canal; ut., uterus; Ov., ovary; p, pumping organ.
Cf. figs. 191 and 233. (Stephens.)
The vagina, like the vas deferens, usually runs inwardly and
posteriorly, where it forms a spindle-shaped dilatation (receptaculum
seminis); its continuation, the spermatic duct, unites with the
oviduct, the common duct of the ovaries (fig. 191). The ovaries,
usually two in number, are compound tubular glands in the posterior
half of the proglottis, which extend into the medullary layer, but
ventral to the median plane.
At the origin of the oviduct there is frequently a dilatation provided
with circular muscles (suction apparatus), which receives the
ovarian cells and propels them forward. After the oviduct has
received the spermatic duct the canal proceeds as the fertilization
canal, and after a very short course receives the vitelline duct or
ducts, and then the numerous ducts of the shell glands (oötype).
[Although the nomenclature of these parts varies, we may consider
the oviduct as extending from the ovary to the shell gland and
as receiving the spermatic duct and then the vitelline duct and the
ducts of the shell gland. The short piece into which the shell gland
ducts open corresponds to the oötype in the flukes, but in the tapeworms
this portion of the canal is seldom dilated. From this point
the oviduct is continued as a shorter or longer tube, the uterine canal
or true oviduct opening into the uterus proper.—J. W. W. S.]
The vitellarium may be single, but often exhibits its primitive
duplication more or less distinctly, in which case it is situated at the
posterior border of the segments in the medullary layer (fig. 191).
The original position of the double organ is, moreover, the same
as in the Trematodes, i.e., at the sides of the proglottids, and thence
eventually extending more or less on both surfaces (figs. 192 and 194);
the gland is then distinctly grape-like and the follicles lie mostly in
the cortical layer.
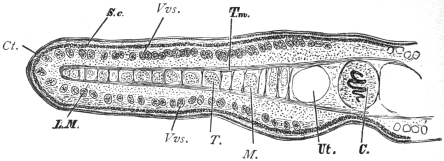
Fig. 194.—Part of a transverse section through a proglottis of Dibothriocephalus latus. Ct.,
cuticle; C., cirrus; Vvs., vitelline follicles; L.M., longitudinal muscles; T., testicles;
M., medullary nerve; S.c., subcuticle; T.m., transverse muscles; Ut., uterus. 20/1.
The egg cell that has been fertilized and supplied with yolk cells
receives the shell material at the point of entry of the shell gland ducts,
and, as a complete egg, then moves onward to the uterus. In those
cases in which the uterus in its further course presents a convoluted
canal, and may form a rosette (pseudo-phyllidea), there is an external
opening which is usually separate from the genital pore, and lies on
the same or the opposite surface. In all other cases, however, the
uterus terminates blindly and is represented by a longer or shorter sac
lying in the longitudinal axis (fig. 191), but in many forms transversely.
With the accumulation of eggs it becomes modified in
various ways: (1) it sends out lateral branches (fig. 241), or (2) forms
numerous isolated sacs (PARENCHYMAL CAPSULES) containing single
eggs or groups of eggs (fig. 217); further, (3) in some cases at the
blind end one or more special thick-walled cavities are formed
(PARUTERINE ORGANS or UTERINE CAPSULES), in which all or most of
the eggs are collected, the uterus then undergoing atrophy.
In species in which the uterus lacks an opening, simultaneously
with the growth of this organ an atrophy of the male apparatus, at
least of the testes and their excretory ducts, takes place; this atrophy
also frequently occurs in the female glands, so that the entire mature
segments have besides the uterus only traces of the genitalia left.
In the Acoleïnæ the vagina is more or less extensively atrophied,
and in any case has no external opening.
A number of genera are distinguished by the duplication of
the genitalia in every segment; the genital apparatus in its entirety,
or with the exception of the uterus, is double, or the genital glands
and the uterus are single, but the cirrus, vas deferens and vagina
are double.
On comparing the genitalia of the Trematodes and Cestodes the
parts will be found to agree, but the vagina of the Cestodes
corresponds with the uterus of the Trematodes, and the uterus of the
tapeworms to Laurer’s canal of the Trematodes, which in most of the
Cestodes has lost its external orifice.
Development of the Tapeworms.
Copulation.—As each proglottis possesses its own genital apparatus,
and male as well as female organs are present, the following processes
may occur: (1) self- or auto-fecundation (without immissio cirri);
(2) self- or auto-copulation (with immissio cirri); (3) cross-copulation
between proglottids of the same or different chains (of the same
species); and (4) cross-copulation in the same proglottis in species
with double genital pores. These various modes have actually been
observed.
In those species which lack the vagina (Acoleïnæ) it appears that
the cirri, which are always furnished with hooks, are driven into
the tissues and for the most part reach the receptaculum seminis.
The eggs of all Cestodes are provided with shells, but the shells,
like their contents, vary. In genera that possess a uterine pore the
mature eggs frequently do not differ from those of the Distomata;
they have a brown or yellow shell of oval form provided with an
operculum, and contain a number of yolk cells in addition to the
fertilized ovarian cell (fig. 128), but in other genera (with a uterine
pore) the lid is absent and the egg-shell is very thin, the eggs of these
genera resembling those of Cestodes in which the secretion of the
vitellarium is a light albumin-like substance that contains only a
few granules, and in which the egg-shell is very delicate and without
operculum.
The eggs of Tæniidæ, for example, at first consist of egg-shell
(oötype), ovum and yolk cells. The egg-shell is as a rule soft, colourless
and frequently deciduous, and the yolk is scanty in amount and
contains few granules. The eggs are, moreover, more complicated
than this. They enlarge and change their shape and various envelopes
are developed around the embryo. The egg-shell proper often disappears,
and one or more embryonal envelopes, or protoplasmic
layers, arise, so that eventually it is difficult to say whether the whole
egg is present, and, if not, what the layers that remain really are.
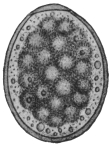
Fig. 195.—Egg of Diplogonoporus
grandis, showing the
morula surrounded by yolk cells
and granules. 440/1. (After
Kurimoto.)
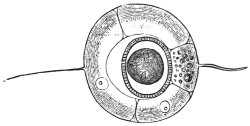
Fig. 196.—Uterine egg of Tænia saginata, G.
Uterine shell with filaments; the oncosphere with
embryonal shell (embryophore) in the centre.
500/1. (After Leuckart.)
The embryonal development in most species takes place during the
stay of the eggs in the uterus; in other species it takes place after the
eggs have been deposited and are in water. Separate cells or a layer of
cells always separate from the segmentation cells, as well as from the
cells of the developing embryo, and form one or more envelopes round
the embryo; usually two such envelopes are formed, the inner one
of which stands in intimate relationship with the embryo itself and
is often erroneously termed the egg-shell, but more correctly the
embryonal shell or embryophore. In some species it carries long cilia,
as in Dibothriocephalus latus, by aid of which the young swim about
when released from the egg-shell; as a general rule, however, there
are no cilia and this envelope is homogeneous, or is composed of
numerous rods and is calcified, as in Tænia spp. (fig. 197). The
second outer envelope (“yolk envelope”) (fig. 207, 3) lies close within
the true (oötype) egg-shell, and remains within it when the embryo
hatches out, and in many species, as in Tænia spp., it perishes at the
end of the embryonal development with the delicate egg-shell which
was formed in the oötype, so that one observes not the entire egg
with egg-shell but only the embryo in its embryonal shell, viz., the
embryophore (fig. 197, a.).
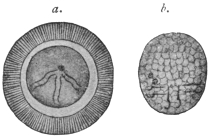
Fig. 197.—a., oncosphere, in its
radially striated embryophore (erroneously
termed egg-shell) of Tænia
africana. Greatly magnified. (After
von Linstow.) b., freed oncosphere of
Dipylidium caninum. (After Grassi
and Rovelli.) Both oncospheres show
six spines.
The embryo (the ONCOSPHERE) enclosed within the embryonal
shell (embryophore) is of spheroidal or ovoid form (fig. 197, b.), and is
distinguished by the possession of three pairs of spines, a few terminal
(flame) cells of the excretory system, and muscles to move the spines.
No further development of the oncosphere takes place, either
in the parent organism or in the open; in fact, in all cases in which
the oncospheres are already formed within the proglottids they do
not become free, but remain in their shell; it is only when the
oncospheres are provided with a ciliated embryophore that they leave
the egg-shell, and they even cast this ciliated envelope after having
swum about in water by its means for a week or so. Sooner or later,
however, all the oncospheres leave the host that harbours the parental
tapeworm and reach the open, either still enclosed in the uterus of
the evacuated proglottids, after the disintegration of which they
then become free, or after being deposited as eggs in the intestine
of the host; they then leave it with the fæces. In the former case
also, the slightest injury to the mature proglottids while still in the
intestine suffices to allow a part of the oncospheres in their embryophores
to be released and mingled with the fæces. Here they are the
generally, but falsely, so-called Tæniæ “eggs.” For, as stated above,
the “yolk” envelope and the true shell deposited in the oötype have
before this disintegrated.
In other cases, e.g., Hymenolepis spp., the uterine (oötype) shell
persists in fæces (fig. 230).
In any case the oncospheres
must be transmitted into suitable
animals to effect their further development;
in only very rare cases
might an active invasion be possible,
as, for instance, takes place with the
miracidia of many Trematodes. The
entry into an animal is, as a rule,
entirely passive, that is to say, the
oncospheres are swallowed with the
food or water. Many animals are
coprophagous and ingest the oncospheres
direct with the fæces; others
swallow them with water, mud, or
food contaminated by such fæces. Infection is easily produced
artificially by feeding suitable animals with mature proglottids of
certain Cestodes or introducing the oncospheres with the food. As
the mature tapeworm frequently finds the conditions suitable for its
development in only one species of host, or in species nearly related,
and perishes when artificially introduced into other hosts, experiment
has taught us that to succeed in cultivating the oncospheres certain
species of animals are necessary. Thus we are aware that the oncospheres
of Tænia solium, which lives in the intestine of man, develop
only in the pig, and only quite exceptionally develop into the stage
characteristic of all Cestodes—the cysticercus in the wide sense of the
word—in a few other mammals. The oncospheres of T. saginata
develop further only in the ox; those of T. marginata (of the dog) in
the pig, goat, and sheep; those of T. serrata (of the dog) in hares and
rabbits; those of Dipylidium caninum (of the dog and cat) in parasitic
insects of the dog and cat, etc. It is not unusual that young animals
only appear to be capable of infection, while older animals of the
same species are not so.
Once introduced into a suitable animal, which is only exceptionally
the same individual or belongs to the same species as the
one which harbours the adult tapeworm, the oncosphere passes
into the larval stage common to all Cestodes, but varying in structure
according to the species. In the simplest case—as, e.g., in Dibothriocephalus—such
a larva resembles the scolex of the corresponding
tapeworm, only that the head, provided with suckers, is retracted
within the fore-part of the neck. Such a larval form is known as a
plerocercoid (πλήερης, full; κέρκος, tail). They differ from the cysticercoids
in being solid larval forms, elongated, tape-like or oval, with the
head invaginated. The conditions appear to be similar in Ligula,
Schistocephalus, Triænophorus, but here the larvæ are very large, indeed
as large in the first-mentioned genera as the tapeworms originating
from them, and the sexual organs are already outlined; doubtless,
however, this stage is preceded by one that corresponds to the scolex
of the genus in question, and which represents the actual larval stage.
In such cases the development of the body of the tapeworm from the
scolex has already begun within the first or intermediate host; in
other cases, except in the single-jointed (monozootic) Cestodes, this
only takes place in the definitive host. The direct metamorphosis
of the oncosphere into the larval forms termed PLEROCERCOID has
hitherto not been investigated, although Ligula, Schistocephalus and
Bothriocephalus are very common parasites, but many circumstances
point to the conclusions arrived at by us and by other observers. In
the larval stages of other tapeworms we can always distinguish the
scolex and a caudal-like appendage, vesicular in the cysticerci (fig. 200),
compact in the cysticercoids (fig. 231). The scolex alone forms the
future tapeworm, the variously formed appendage perishing.
It has now been proved that the appendage, the caudal vesicle,
originates direct from the body of the oncosphere, and therefore is
primary, and that the scolex only subsequently forms through proliferation
on the surface of this appendage. On account of this
origin the scolex is generally regarded as the daughter, and the part
usually designated as the appendage as the mother, originating from
the oncosphere.
Accordingly, two modes of development of the larval stage may
be distinguished; in the one case, plerocerci and plerocercoids, the
oncosphere changes directly into the scolex, thus forming the body
of the tapeworm within the primary host; in the other case, cysticerci
and cysticercoids, the scolex only forms secondarily in the transformed
body of the oncosphere, which later on perishes, the scolex
alone remaining as the originator of the tapeworm colony.
We may summarize briefly what has been said regarding these
larval forms. We have, firstly, solid larval forms without any bladder.
These arise directly from the oncosphere and are of two kinds,
plerocercus and plerocercoid. Plerocercus is a solid globular larva with
the head invaginated into the posterior portion. Plerocercoid (fig. 208)
is a solid elongated larva also with the head invaginated into the
posterior portion, which is sometimes very long. Secondly, we
have larval forms with bladders from which the scolices arise thus
indirectly from the oncosphere. They are of two kinds, cysticercoid
and cysticercus.
Cysticercoid.—The bladder is but slightly developed and is usually
absorbed again. The anterior portion is, moreover, retracted into
the posterior, and in some cases there is a long or a stumpy tail
(figs. 220, 231).
Cysticercus, or true bladder worms. (These may be divided into
(1) cysticercus proper, consisting of a bladder and one scolex;
(2) cœnurus, a bladder and many scolices; (3) echinococcus, a
bladder in which daughter bladders or cysts are developed, and then
in these multiple scolices.)
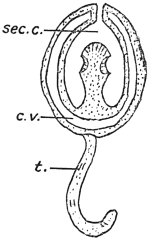
Fig. 198.—Diagram of a cysticercoid. Cf.
figs. 220, 227. c.v., caudal vesicle or bladder
(small); sec. c., secondary cavity caused by the
growth forward of the hind-body; t., tail bearing
six spines. (Stephens.)
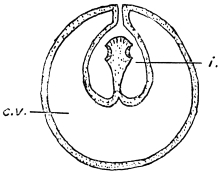
Fig. 199.—Diagram of a cysticercus.
c.v., caudal vesicle or bladder; i., invagination
of wall of bladder. (Stephens.)
In the case of cysticerci a papilliform invagination forms, projecting
into the interior of the bladder (fig. 201). The layer of cells
forming the papilla becomes divided into two laminæ, the outer279 of
which forms a kind of investing membrane (receptaculum capitis) for
the papilla. The head and suckers are now developed on the walls
bounding the axial lumen of the papilla. The papilla eventually
evaginates, so that the receptaculum capitis now forms the inner
surface of the hollow head, which eventually becomes solid.
Our knowledge of the development of cysticerci in the wide
sense of the word is limited almost exclusively to that of a few true
“bladder worms” (cysticerci); in other cases we know either only
the terminal stage, i.e., the complete larva, or, exceptionally, one of
the intermediate stages, but we are not acquainted with a complete
series; the description must therefore be incomplete.
We know from feeding experiments that, after the introduction
of mature proglottids or of the fully developed ova of Tænia
crassicollis (of the cat) into the stomach of mice, the oncospheres
escape from the shell in the middle portion of the small intestine,
and a few hours later penetrate into the intestinal wall by means
of a boring movement; they have been found in this position twenty-seven
to thirty hours after the infection. By means of this migration,
for which purpose they employ their spines, they attain the blood-vessels
of the intestine; indeed, already nine hours after the infection
and later they are found in the blood of the portal vein, and in the
course of the second day after infection they are found in the
capillaries of the liver, which these larvæ do not leave.
Leuckart, in experimental feeding of rabbits with oncospheres
of Tænia serrata (of the dog), found free oncospheres in the stomach
of the experimental animal, but not in the intestine: however,
he came across them again in the blood of the portal vein. The
passage through the blood-vessels to the liver is the normal one
for those species of Tænia the eggs of which become larvae in
mammals; even in those cases in which the oncospheres develop
further in the omentum or in the abdominal cavity (Cysticercus
tenuicollis, C. pisiformis), there are distinct changes observable in
the liver that lead one to the conclusion that there has been a
secondary migration out of the liver into the abdominal cavity.
Indeed, one must not imagine that the young stages of the Cestodes
are absolutely passive; once they have invaded an organ they travel
actively, and leave distinct traces of their passage.
In other cases the oncospheres leave the liver with the circulation,
and are thus distributed further in the body; they may settle and
develop in one or more organs or tissues. Many oncospheres may,
by travelling through the intestinal wall, penetrate through it and
attain the abdominal cavity direct; some, perhaps, pass also into the
lymph stream. Where there are no blood and lymphatic vessels in
the intestinal wall, as in insects, the oncospheres attain the body
cavity or its organs direct; in short, they never remain in the
intestinal lumen itself, and only rarely—as in Hymenolepis murina
of the rat—do they remain in the intestinal wall.
When the infection has been intense, and the body is crowded with numerous
oncospheres, acute feverish symptoms, are induced, to which the infected animals
usually succumb (“acute cestode tuberculosis”); while in other cases the alterations
in the organs attacked—as the liver in mice and the brain in sheep—may cause death.
Sooner or later the oncospheres of tapeworms come to rest,
and are first transformed into a bladder, which may be round or
oval according to the species. The embryonal spines disappear sooner
or later, or remain close together or spread over some part of the
bladder wall (fig. 200). Their discovery by V. Stein in the bladder
worm of the “meal worm” (the larva of a beetle, Tenebrio molitor)
first led to the conclusion that bladder worms (cysticerci) actually
originate from the oncospheres of Tæniidæ.
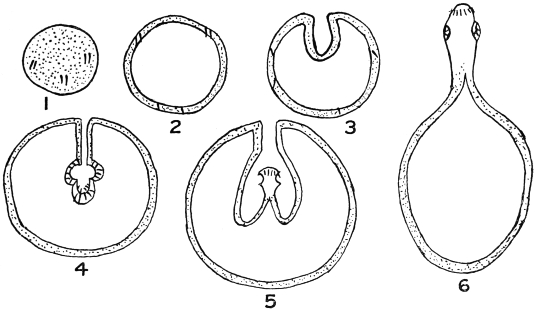
Fig. 200.—Diagram of development of a cysticercus. 1, solid oncosphere with six spines;
2, bladder formed by liquefaction of contents; 3, invagination of bladder wall; 4, formation
of rostellum (with hooklets) and suckers at the bottom of the invagination; 5, evagination of
head; 6, complete evagination effected by pressure. (Stephens.)
The bladder may remain as a bladder, and then by proliferation
the scolex forms on its wall (fig. 202), or it may divide into an
anterior so-called “cystic” portion and a solid tail-like appendage of
various lengths, on which the embryonal hooks are to be found,
and this is particularly the case in those larval forms (cysticercoids),
e.g., those of Dipylidium caninum, that develop in invertebrate animals,
such as Arthropoda.
As mentioned above one may regard the scolex as an individual
that originates through proliferation of the wall of the parent cyst,
mostly singly, but in those cysticerci that are termed cœnurus
(fig. 201) many scolices occur, whereas in those called echinococcus
the parent cyst originating from the oncosphere of Tænia echinococcus
(of the dog) first produces a number of daughter cysts, which in their
turn form numerous scolices. Echinococcus-like conditions also
occur in cysticercoids, as, for instance, in those peculiar to earthworms;
and similar conditions prevail in a larval form known as
Staphylocystis, found in the wood-louse (Glomeris). Thus it happens in
these cases that finally one tapeworm egg produces not one, but
numerous tapeworms, for, under
favourable conditions, each scolex
can form a tapeworm.
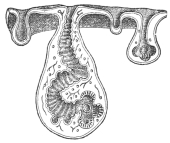
Fig. 201.—Section through a
piece of a Cœnurus cerebralis,
with four cephalic invaginations
in different stages of development.
At the bottom of the
invaginations the rostellum, hooks
and suckers develop. (From a
wax model.)
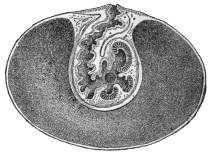
Fig. 202.—Median section through
a cysticercus, with developed scolex
at the bottom of the invagination.
(After Leuckart.)
The rudiment of the scolex appears as a hollow bud, the cephalic invagination
usually directed towards the interior of the bladder cavity; on its invaginated
surface arise the four suckers, and the rostellum with the
hook apparatus is formed in its blind end; we thus get a
Tænia head, but with the position of the parts reversed
(fig. 201). In many cysticerci the head rises up from the
base of the cephalic invagination and is then surrounded by
the latter. A more or less elongated piece of neck also
develops, and even proglottids may appear, as in Cysticercus
fasciolaris (the larva of Tænia crassicollis of the cat) of the
Muridæ, a process somewhat analogous to that of Ligula, etc.
The period that elapses from the time of
infection till the cysticercus is fully developed
varies according to the species; the cysticercus
of Tænia saginata requires twenty-eight weeks,
that of T. marginata seven to eight weeks, that
of T. solium three to four months, and that of
T. echinococcus longer still.
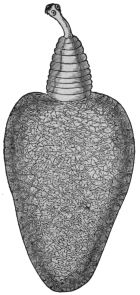
Fig. 203.—Cysticercus
pisiformis in an evaginated
condition, with
neck, fore-body and
bladder, with excretory
network in its wall. 18/1.
With one single exception (Archigetes) the
larvæ do not become sexually mature in the organ
where they have developed; they must enter the
terminal host, a matter that is usually purely
passive, the carriers of the larvæ or infected parts
of them being usually devoured by other animals. In this manner, for
instance, the larvæ (Cysticercus fasciolaris) found in mice and rats reach
the intestine of cats; those of the hare and rabbit (C. pisiformis) reach
the intestine of dogs; those of the pig (C. cellulosæ) are introduced into
man; those of insects are swallowed by insectivorous birds; those of
crustaceans are ingested by ducks and other water fowl; perhaps,
also, the infection of herbivorous mammals is caused by their
accidentally swallowing smaller creatures infected by larvæ. Indeed,
the researches of Grassi and Rovelli have taught us that such an
intermediate host is not always necessary; Hymenolepis murina of
rats and mice in its larval stage lives in the intestinal wall of these
rodents, and as a larva it passes into the intestinal lumen and develops
into a tapeworm in exactly the same way as the larvæ of other species
that reach the intestine of the terminal host by means of an intermediate
carrier. Probably this curtailed manner of transmission also
occurs in many other species. In some cases the larvæ actively quit
the body of the intermediate host, as in the case of Ligula and Schistocephalus,
which travel out of the body cavity of infected fish and
reach the water, where they may be observed in hundreds in summer,
at all events in some localities. The larval stage of Calliobothrium—wrongly
termed Scolex—has been observed swimming free in the sea,
and the scolices of Rhynchobothrium, without their mother cysts, have
been observed free within the tissues of several marine animals. In
any case there is almost always a change of hosts, even in the single-jointed
Cestodes, for the larva of Caryophyllæus, which lives in fishes
of the carp family, is found in limicoline Oligochætes, that of Gyrocotyle
(Chimæra) in shell-fish (Mactra), and different conditions can
hardly be possible for Amphilina. Archigetes alone becomes sexually
mature in the larval stage, but the life-history of this creature is not
well known, so that it is not impossible that the attainment of sexual
maturity as a larva in invertebrates (Oligochætes) is perhaps abnormal,
and somewhat analogous to the maturity of some encysted
Trematodes.
The METAMORPHOSIS OF THE LARVA into the tapeworm is rarely
accomplished in a simple manner; the transformation, however, is
not complex in the single-jointed Cestodes, nor in Ligula and Schistocephalus;
the latter is swallowed by birds (Mergus, Anas, etc.), produces
eggs after only a few days, and very soon quits the intestine of its
terminal host. In all other cases it is the scolex only which, by
proliferating at its posterior extremity, forms the proglottids, after
having invaded as a larva the intestine of a suitable host. The mother
cysts, or what corresponds to them, die, are digested, absorbed, or
perhaps even eliminated; on the contrary, segments found on the
scolex during the larval stage, also in the case of Cysticercus fasciolaris,
are retained. It is not certain whether the larvæ of Dibothriocephalus
lose any part.
The time required by the scolex to complete the entire chain
of proglottids does not depend only on the number it has to
produce, for Tænia echinococcus, which, as a rule, only possesses
three or four segments, takes quite as long a time for their growth
(eleven to twelve weeks) as T. solium with its numerous segments;
T. cœnurus is fully developed in three to four weeks, and
the same holds good for Dibothriocephalus latus, which possesses
many more segments than the above-mentioned Tænia of the dog.
In a number of species it has been possible to determine fairly
accurately the average daily growth; for instance, in Dibothriocephalus
latus the daily growth is 8 cm., in Tænia saginata 7 cm., etc.
The history of the development of the Cestodes demonstrates that
persons and beasts harbouring larval tapeworms have become
infected by having swallowed the oncospheres of the species of
tapeworm to which they belong. In regard to Hymenolepis murina
alone, it is known that the introduction of the oncospheres into those
species of animals which harbour the adult tapeworm leads to the
formation of the latter after the development of a larval stage in the
intestinal wall; nevertheless, only young animals (rats) are capable of
infection, for a previous infection, or the presence of mature tapeworms
in the intestine, appears to produce a kind of immunity.
Biology.
In their adult stage, the tapeworms inhabit almost exclusively
the alimentary canal of vertebrate animals, with but few exceptions
the small intestine, and a few species select definite parts of it. A
small number of Rhynchobothriidæ of marine fishes live apparently
always in the stomach, while in rays and sharks the spiral intestine is
their exclusive site. Bothriocephali generally attach themselves with
their head on to the appendices of the pylorus of fishes; other
species (Hymenolepis diminuta) occasionally fix their head in the
ductus choledochus, and this is more frequent still in the tapeworms
of the rock badger (Hyrax), which occasionally penetrate entirely
into the biliary ducts. Stilesia hepatica, Wolffh., has so far only been
found in the bile-ducts of its host (sheep and goat, East Africa).
In the disease of sheep induced by Cestodes, the worms have
been observed also in the pancreas. Specimens found in the large
intestines were probably being evacuated.
The Cestodes are looked upon as fairly inert creatures, this
opinion having been formed by observing their condition in the cold
cadavers of warm-blooded animals. Actually, however, they are
exceedingly active, and accomplish local movements within the
intestine, for they have been found in the ducts communicating with
the bowel, or in the stomach, and may even make their way forward
into the œsophagus.
They also invade other abdominal organs through abnormal communications,
or through any that may be temporarily open between
the intestine and such organs; they thus reach the abdominal cavity or
the urinary bladder, or they work their way through the peritoneum.
They produce changes in the intestinal mucous membrane at the
place of their attachment, the alterations varying in intensity according
to the structure of the fixation organs. The mucous membrane
is elevated in knob-like areas by the suckers; the epithelial cells
become atrophied or may be entirely obliterated. Dipylidium
caninum bores into the openings of Lieberkühn’s glands with its
rostellum, dilating the lumen to two or three times its normal size,
while the suckers remain fixed between the basal parts of the cells.
Species with powerful armatures penetrate deeper into the submucosa,
and some that are not provided with exceptionally strong armatures,
or are even unarmed, may be actually found with the scolex embedded
in the muscles of the intestinal walls or even protruding beyond
(Tænia tetragona, Mol., in fowls, etc.). Other species, again, even
cause perforation of the walls of the intestine of their hosts.
It is generally assumed that tapeworms, which almost without
exception live in the gut of vertebrates, get their nutriment from the
gut contents, which apparently they absorb through the whole body
surface (cuticular trophopores). In favour of this view is the
existence of fat drops in the proglottids, the identity in colour in
certain forms between that of the fresh worm and the gut contents
and the passage of certain substances derived from medicines (iron
and mercury preparation) into the worms in the gut, etc. Whether
the suckers are concerned in the absorption of nutriment and to
what extent is still questionable.
The length of life of the adult tapeworm certainly varies;
as a rule it appears to last only about a year; in other cases (Ligula)
it averages only a few days, but we are likewise aware that certain
species of Cestodes of man attain an age of several or many years
(thirty-five). The natural death of Cestodes often appears to be
brought about by alterations in the scolex, such as loss of the hooks,
atrophy of the suckers and rostellum, finally the dropping off of the
scolex; it is unknown whether a chain of segments deprived of its
scolex then perishes or whether it first attains maturity. It has already
been mentioned that in a few species the foremost proglottids are
transformed into organs of fixation on the normal loss of the scolex.
Abnormalities and malformations are encountered relatively frequently in the
Cestodes—such as abnormally short or long segments; the so-called triangular
tapeworms, which—if belonging to the Tæniidæ—always possess six suckers; often
also club-shaped segments occur between normal ones, or there may be a defect
in one segment or in the centre of a number following one another (fenestrated
segments); bifurcated chains of segments have likewise been observed, as well as
incomplete or complete union of the proglottids, abnormal increase of the genital
pores, reversion of the genitalia. Besides the above-mentioned increase of the
number of suckers on the scolex (in Tæniæ), there may be a decrease in the
number; in other cases the crown of hooks may be absent, or abnormally shaped
hooks may be formed.
Classification of the Cestoda of Man.
Order. Pseudophyllidea, Carus, 1863.
Scolex without proboscis or rostellum. Head “stalk” absent.
Scolex never with four, generally with two (or one terminal) bothria.280 Vitellaria
numerous. Uterine opening present. Genitalia do not atrophy when uterus is
developed. In large majority of proglottids eggs (or, if formed, their contents) are
at the same stage of development.
Family. Dibothriocephalidæ, Lühe, 1902.
Syn.: Diphyllobothriidæ, Lühe, 1910.
Genitalia repeated in each proglottid (polyzootic Cestodes). Ventral and dorsal
surfaces flat. Cirrus unarmed. Cirrus and vagina if non-marginal open on the
same surface as the uterus. Uterus long, convoluted, often forming a “rosette,”
never dilates into a uterine cavity. Eggs thick shelled, operculated, constantly
being formed in mature proglottids.
Sub-family. Dibothriocephalinæ, Lühe, 1899.
Syn.: Diphyllobothriinæ, Lühe, 1910.
Segmentation distinct. Scolex unarmed, elongated, sharply separated (generally
by a neck) from the first proglottis. Cirrus and vagina open ventrally. Genital
pores non-alternating. Vas deferens surrounded by a muscular bulb. Receptaculum
seminis large, sharply separated from the spermatic duct.
Order. Cyclophyllidea, v. Beneden.
Four suckers always present. Uterine opening absent. Vitellarium single.
Genitalia atrophy when uterus is fully developed.
Family. Dipylidiidæ, Lühe, 1910.
Rostellum if present armed. Suckers unarmed. Uterus breaks up into egg
capsules. Paruterine organs absent.
Family. Hymenolepididæ, Railliet and Henry, 1909.
Segment always broader than long. Genitalia single. Longitudinal muscles in
two layers. Genital pores unilateral. Testes one to four. Uterus persistent, sac-like.
Eggs with three shells.
Family. Davaineidæ, Fuhrmann, 1907.
Rostellum cushion-shaped. Armed with numerous (sixty to several thousand)
hammer-shaped hooks in two (rarely one) rows.
Sub-family. Davaineinæ, Braun, 1900.
Suckers armed. Uterus breaks up into egg capsules. Paruterine organs absent.
Family. Tæniidæ, Ludwig, 1886.
Suckers unarmed. Uterus with median longitudinal stem and lateral branches.
Female genitalia at the hind end of the proglottis. Genital pore irregularly alternating.
Testes numerous in front of female genitalia. Ovary with two lobes (wings).
Vitellarium behind the ovary. Embryophore radially striated.
The Cestodes of Man.
Most of the species to be mentioned live in man in their adult
stage and occupy the small intestine; man is the definite host of these
parasites, but is not the specific host for all the species; some of these
species, as well as others (of mammals), may occur in man also in the
larval stage.
Family. Dibothriocephalidæ.
Sub-family. Dibothriocephalinæ.
Genus. Dibothriocephalus, Lühe, 1899.
Syn.: Diphyllobothrium, Cobbold, 1858; Bothriocephalus, p. p. Rud., 1819;
Dibothrius, p. p. Rud., 1819; Dibothrium, p. p. Dies., 1850.
Scolex egg-shaped; dorsal and ventral bothria elongated, moderately strong,
cutting rather deeply into the head; genitalia single in each proglottis; papillæ in
the vicinity of the genital atrium; the testes and vitellaria are in the lateral fields, the
former in the medullary layer, the latter in the cortical layer on both surfaces, and
occasionally extending to the median line; the ovary ventral, the shell gland dorsal.
The uterus is in the central field, taking a zigzag course, and frequently forms a
rosette.
Dibothriocephalus latus, L., 1748.
Syn.: Tænia lata, L., 1748; Tænia vulgaris, L., 1748; Tænia grisea, Pallas,
1796; Tænia membranacea, Pall., 1781; Tænia tenella, Pall., 1781; Tænia dentata,
Batsch, 1786; Bothriocephalus latus, Bremser, 1819; Dibothrium latum, Dies.,
1850; Bothriocephalus cristatus, Davaine, 1874281; Bothriocephalus balticus,
Kchnmstr., 1855; Bothriocephalus latissimus, Bugn., 1886.
Length 2 to 9 m. or more; colour yellowish-grey; after lying in
water the lateral areas become brownish and the uterine rosette
brown. The head is almond-shaped, 2 to 3 mm. in length, the dorso-ventral
axis is longer than the transverse diameter; the head, therefore,
generally lying flat, conceals the suctorial grooves at the borders;
these suckers are deep and have sharp edges (fig. 205). The neck
varies in length according to the degree of contraction and is very
thin; there are 3,000 to 4,200 proglottids and there may be more;
their breadth is usually greater than their length, but in the posterior
third of the body they are almost square, and the very oldest are not
uncommonly longer than they are broad. There are numerous testes
situated dorsally in the medullary layer of the lateral fields; the vas
deferens (fig. 192) passes dorsally in transverse loops in the central
field anteriorly and forms a seminal vesicle before its entry into the
large cirrus pouch.
The orifice of the vagina is close behind the orifice of the cirrus;
the former passes almost straight along the median line posteriorly,
and widens into a receptaculum seminis shortly before its junction
with the oviduct; the ovary is bilobed, in shape like the wings
of a butterfly, ventrally in the medullary layer; the shell glands lie
in the posterior recess of the ovary; the uterus, forming numerous
transverse convolutions, passes ventral to the vas deferens forwards.
Eggs (fig. 207) large, with brownish shells and small lids, 68 µ to 71 µ
by 45 µ; the ovarian cell, which is already, as a rule, in process of
segmentation, is surrounded by numerous large yolk cells; the
proglottids nearest the posterior extremity are frequently eggless.
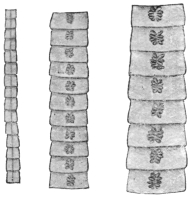
Fig. 204.—Various chains of
segments of Dibothriocephalus
latus, showing the central
uterine rosette. (Natural size.)
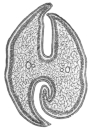
Fig. 205.—Transverse section
of the head of Dibothriocephalus
latus. 30/1.
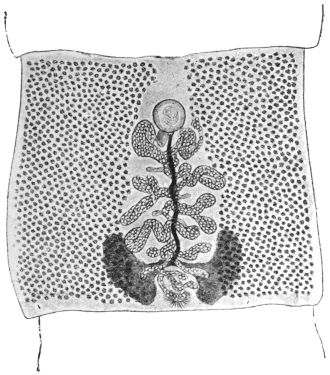
Fig. 206.—Fairly mature proglottis of Dibothriocephalus
latus. The vitellaria are at the sides; the uterus, filled
with eggs, is in the middle, also the vagina (the dark stripe
passing almost straight from the front to the back), and the
vas deferens (almost hidden by the uterus). Above in the
centre is the cirrus sac, and below the shell gland and ovary
are seen. 15/1. (From a stained preparation.)
The eggs, which are deposited in the intestine and evacuated
with the fæces, hatch in water after a fortnight or more; the embryonal
integument (embryophore) of the oncosphere is provided with cilia;
after bursting open the lid of the egg the oncosphere in its embryophore
(fig. 207) reaches the water and swims slowly about; often it
slips out of its ciliated embryophore, sinks to the bottom and is capable
of a creeping motion; sooner or later it dies in the water. The
manner and means of its invasion of an intermediate host are still
unknown; yet we are aware that the larval stage (plerocercoid,
fig. 208), which resembles the scolex and may reach a length of 30 mm.,
lives in the intestine, in the intestinal wall, in the liver, spleen, genital
glands and muscular system (fig. 209) of various fresh-water fish,
the pike (Esox lucius), the miller’s thumb (Lota vulgaris), the perch
(Perea fluviatilis), Salmo umbla, Trutta vulgaris, Tr. lacustris, Thymallis
vulgaris (grayling), Coregonus lavaretus, C. albula (in Europe) and
Onchorhynchus perryi (in Japan). The transmission of the plerocercoids
from these fish to the dog, cat and man (Braun, Parona, Grassi
and Ferrara, Grassi and Rovelli, Ijima, Zschokke, Schroeder) leads to
the development of the broad tapeworm, the growth of which is rapid.
In my experiments on human beings the average number of proglottids
formed per diem averaged thirty-one to thirty-two for five
weeks, with a length of 8 to 9 cm. According to Parona the eggs
appear twenty-four days after man has been infected. Zschokke found
the average growth in the experimental infection of man between 5·2
and 8·2 cm. per diem, and the person experimented upon by Ijima
evacuated a piece of a Dibothriocephalus latus, 22·5 cm. in length,
only twenty-one days after the infection.
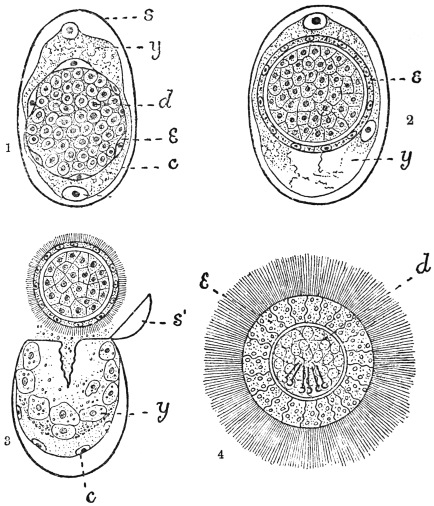
Fig. 207.—Dibothriocephalus latus: development of egg. 1, segmentation complete; some
cells of the blastosphere have migrated through the yolk and have flattened to form c, the
yolk envelope; others form a layer of flattened cells (e) forming the embryophore; the
remaining cells (d) of the blastosphere form the hexacanth embryo. 2, embryophore (e) is
becoming thicker. 3, the ciliated embryo has been pressed out of the shell; s′, the operculum;
c, the yolk envelope remaining in the shell (s); y, the yolk consisting of separate cells. 4,
a free-swimming larva much swollen by the water. (After Benham and Schauinsland.)

Fig. 208.—Plerocercoid
of Dibothricephalus
latus. A., with the head
evaginated; B., with
the head invaginated.
From the muscle of
the pike.
The “broad tapeworm” is a frequent parasite
of man in some districts, but it also occurs in the
domestic dog, and on rare occasions is found in
the domestic cat (together with Dibothriocephalus
felis, Crepl.) and fox. French Switzerland and the
Baltic Provinces of Russia are the centres of distribution;
from the former districts the distribution
radiates to France and Italy (Lombardy, Piedmont);
from the Baltic Provinces over Ingermanland
to Petrograd, over Finland to Sweden (on the
shore of the Gulf of Bothnia), in a southerly direction
to Poland, and into the Russian Empire and
across it to Roumania, and towards the west along
the coast of the Baltic Sea to the North Sea, where, however, its
frequency considerably diminishes (Holland, Belgium, and the
North of France).
In Turkestan and Japan the “broad tapeworm” is the most
frequent parasite of man; it has been reported in Africa from the
vicinity of Lake N’gami as well as from Madagascar; cases, in
part at least imported, have also come under observation in North
America.
In Germany Dibothriocephalus latus—apart from the fact that
it is undoubtedly imported from Switzerland, Russia or Italy—is
particularly frequent in East Prussia amongst the inhabitants of
the Courland Lagoon district, on the Baltic; it is, moreover, also
found in the Province and even in the City of Königsberg. In West
Prussia and Pomerania it is very much scarcer.
It is also found in Munich and in the vicinity of the Lake of
Starnberg (Bollinger).
Krabbe found it in 10 per cent. of the sufferers from tapeworms
in Denmark; Szydlowski found the ova of this worm in
Dorpat in 10 per cent. of the fæces examined; Kruse found the
worm in 6 per cent. of post-mortems; Kessler, in Petrograd,
found the eggs in the fæces in 7·8 per cent.; at post-mortems he
found the worms in 1·17 per cent., though Winogradoff only found
it in 0·8 per cent. In Moscow, according to Baranovsky, 8·9
per cent. of the fæces examined contained the ova of Dibothriocephalus.
In the interior and southern provinces of Sweden the
worm, according to Lönnberg, is only found sporadically, but, on
the other hand, in Angermanland about 10 per cent. of the population
is affected; while again in Norbotten the majority of persons
are affected, and in Haparanda the entire population (with the
exception of infants) harbour this parasite. In Switzerland
D. latus is very frequent in close proximity to the lakes of Bieler,
Neuchatel, Morat and Geneva (according to Zaeslin 10 to 15 to
20 per cent. of the population are affected); the parasite is less
frequent in districts one to four hours removed from these lakes.
Of the fish from Swiss lakes examined by Schor those from
Lake Geneva were most commonly infected, and especially Lota sp.
and Perea sp.
The frequency and distribution have, nevertheless, decreased
perceptibly in places; at the commencement of the eighteenth
century the broad tapeworm was very common in Paris, at the
present date it only occurs when imported (Blanchard); in
Geneva, also, according to Zschokke, it has become rarer (formerly
10 per cent., now only 1 per cent.).
The disturbances produced in man by the presence of broad
tapeworms are, as a rule, very trifling; in other cases they produce
partly gastric disorders and partly nervous symptoms; in a number
of cases, again, they set up severe anæmia, apparently caused by
toxins produced by the worms and absorbed by the host. There
is no danger of auto-infection, as the larval stage lives only in
fishes, not in warm-blooded animals. The case reported by
Meschede (ova like those of Dibothriocephalus latus in the brain of
a man who had suffered from epilepsy for six years) must be otherwise
explained.

Fig. 209.—A piece of the body wall of the Burbot, Lota vulgaris. The
tangential section has exposed the muscles of the trunk, with a plerocercoid of
Dibothriocephalus latus. Natural size.
Human beings, like other hosts, can only acquire the broad
tapeworm by ingesting its plerocercoids with the previously mentioned
fresh-water fishes; the opportunity for such infection is
afforded the more readily by the fact that not only do the lower
classes not pay sufficient attention to the cooking of fish, so that
all the larvæ that are present may be killed, but also in certain
localities the custom exists of eating some parts of these fishes in a
raw condition; even the mere handling of the usually severely
infected intermediary hosts may occasionally cause infection. The
plerocercoids are as well known as, but differ materially in appearance
from, the cysticerci (Cysticercus cellulosæ) of pig’s flesh. In
Germany the occurrence of the plerocercoids of Dibothriocephalus
latus has been confirmed in the pike, miller’s thumb and perch of
East Prussia, and more particularly in those taken from the Courland
Lagoon.
The life of D. latus is a very long one (six to fourteen years),
as is deduced from persons who have left D. latus regions after they
have been infected.
According to the experiments of M. Schor, plerocercoids of
D. latus placed in slowly warmed water completely lose their movement
at 54° to 55° C.; they survive the death of their host for several
days; they are killed by low temperatures -3° to +1° C. in two
days; strong acids and salt solutions kill them at once, also high
temperatures, but all the same at least ten minutes is required in
boiling or frying fish in order to kill the plerocercoids with
certainty.
Dibothriocephalus cordatus, R. Lkt.,
1863.
Syn.: Bothriocephalus cordatus, R. Lkt.
Fig. 210.—Cephalic end
of Dibothriocephalus cordatus;
on the left viewed
sideways, on the right from
the dorsal surface, showing
a suctorial groove. (After
Leuckart.)
Length, 80 to 115 cm.; the head is heart-shaped
and measures 2 by 2 mm. The
suctorial grooves are on the flat surface; the
segments commence close behind the head and
increase rapidly in breadth. At only 3 cm.
behind the head they are already mature; the
greatest breadth attained by them averages 7 to 8 mm., the length 3 to
4 mm.; the number of proglottids averages 600; the most posterior ones
are usually square. The uterine rosette is generally formed of six to
eight lateral loops. The eggs are operculated and measure 75 µ by 50 µ.
Dibothriocephalus cordatus is a common parasite of the seal, the
walrus and the dog in Greenland and Iceland, occasionally of man
also. No doubt its larva lives in fishes.
The statement that D. cordatus also occurs in Dorpat in human beings
has been proved erroneous (Zool. Anzeiger, 1882, v, p. 46), as also has the
report that this worm lives in hares in the neighbourhood of Berlin, whither it
was supposed to have been carried by Esquimaux dogs (Rosenkranz in Deutsch.
med. Wochenschr., 1877, iii, p. 620). The parasite stated by the author to be
D. cordatus is Tænia pectinata, Goeze, which has been known since 1766.
Dibothriocephalus parvus, Stephens, 1908.
Largest gravid segments 5 by 3 mm. Uterus forms a central
rosette with four to five loops on each side of median line. In a proglottid
measuring 3·5 by 2·25 mm. the genital atrium is situated 0·4
to 0·5 mm. behind the anterior margin and the uterine opening the
same distance behind the genital atrium. Calcareous corpuscles absent
in the preserved specimens. Eggs operculated, 59·2 µ by 40·7 µ.
Distinguished from Dibothriocephalus latus—(1) by the size of gravid
segments (the minimum width of gravid segments of D. latus is 10
to 12 mm., so that D. parvus is a much smaller worm); (2) quadrate
segments of D. latus measure 6 by 6 mm., those of D. parvus 4 by
4 mm.; (3) by the eggs.
From D. cordatus it is distinguished by—(1) D. cordatus has only
fifty immature segments, D. parvus has at least 200, possibly more;
(2) mature segments of D. cordatus measure 7 to 8 mm., maximum
width of D. parvus is 5 mm.; (3) quadrate segments of D. cordatus
measure 5 to 6 mm.; (4) D. cordatus has six to eight uterine loops;
(5) D. cordatus measures 75 µ to 80 µ by 50 µ.
Habitat.—Intestine of man (Syrian, in Tasmania).
Genus. Diplogonoporus, Lönnbrg., 1892.
Syn.: Krabbea, R. Blanch., 1894.
The scolex is short and has powerful suctorial grooves; no neck;
the proglottids are short and broad; there are two sets of genital
organs side by side in each segment, which in all essentials resemble
the single one of Dibothriocephalus.
Parasitic in whales and seals, occasionally in man.
Diplogonoporus grandis, R. Blanch., 1894.
Syn.: Bothriocephalus sp., Ijima et Kurimoto, 1894; Krabbea grandis, R. Blanch.
Scolex unknown; chain of proglottids over 10 m. in length,
1·5 mm. broad anteriorly, 25 mm. broad posteriorly. The proglottids
are very short (0·45 mm.), but 14 to 16 mm. broad. On either side to
the right and left of the worm, along the entire ventral surface, there
is a longitudinal groove; these grooves are nearer to each other than
to the lateral margin; in them lie the genital pores, and they are in
the same sequence as in Dibothriocephalus; corresponding to the
scanty length (0·45 mm.) of the proglottids, the ovary is only developed
transversely; the uterus only makes a few loops. Eggs (fig. 195)
thick shelled, brown, 63 µ by 48 µ to 50 µ. This parasite has hitherto
been observed twice in Japanese. Similar species are known in Cetacea
and seals.
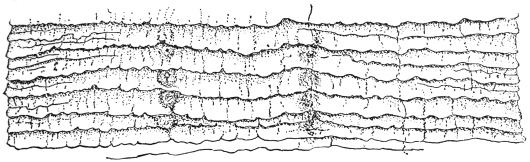
Fig. 211.—Diplogonoporus grandis, Lühe, 1899: ventral view of a portion of the
strobila, showing two rows of genital pores and partially extruded cirri. (After Ijima and
Kurimoto.)
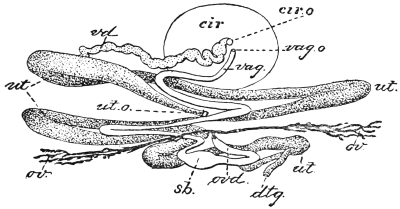
Fig. 212.—Diplogonoporus grandis: ventral view (diagrammatic) of genitalia
of left side; cir, cirrus; cir.o, cirrus opening; dtg., vitelline duct; ov., ovary;
ovd., oviduct; sb., receptaculum seminis; ut., uterus; ut.o., uterine pore; vag.,
vagina; vag.o., vaginal pore; vd, vas deferens. × 150. (After Ijima and
Kurimoto.)
Sparganum, Diesing, 1854.
The term Sparganum, invented by Diesing, is used as a group name
of larval bothriocephalid Cestodes whose development is not sufficiently
advanced to enable them to be assigned to any particular genus.
Sparganum mansoni, Cobb., 1883.
Syn.: Ligula mansoni, Cobbold, 1883; Bothriocephalus linguloides, R. Lkt.,
1886; Bothriocephalus mansoni, R. Blanch., 1886.
Fig. 213.—Cephalic
end of Sparganum
mansoni, Cobb.
(After Leuckart.)
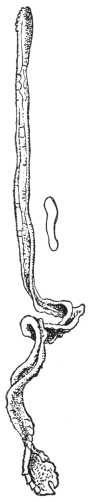
Fig. 214.—Sparganum
mansoni: on
the right in transverse
section. Natural size.
(After Ijima and
Murata.)
These plerocercoids were discovered in 1882 by P. Manson
during the post-mortem on a Chinaman who had died in Amoy,
twelve specimens being found beneath the peritoneum
and one free in the abdominal cavity.
Cobbold described them as Ligula mansoni, and
Leuckart, who contemporaneously reported a case
in Japan, termed them Bothriocephalus liguloides.
Ijima and Murata reported eight further cases, and
Miyake records nine further cases, seven of which
are recorded in Japanese literature.
The plerocercoid, which hitherto alone is known
to us, attains a length of 30 cm. and a breadth of
3 to 6 to 12 mm. The ribbon-shaped body is
wrinkled, the lateral borders are often somewhat
thickened, so that the transverse section has the
form of a biscuit; the anterior end is usually wider
and has the head provided with two weak suctorial
grooves, either retracted or protracted.
The parasite makes migrations within the body,
and thus may reach the urinary passages; then it is
either evacuated with the urine or has to be removed
from the urethra; not rarely it causes non-inflammatory
tumours on various parts of the skin, which
are at times painful and at times vary in size.
Nothing is known of its development and origin.
Sparganum proliferum, Ijima, 1905.
Syn.: Plerocercoides prolifer, Ijima, 1905; Sparganum
prolifer, Verdun, Manson, 1907.
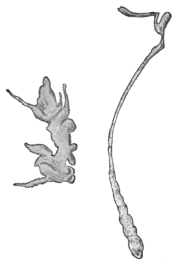
Fig. 215.—Sparganum prolifer:
left with buds, right extended.
× 4. (After Ijima.)
These plerocercoids produce an acne-like condition
of the skin. The condition is really one of
capsules in great abundance in the subcutaneous
tissue and less so in the corium and in the intermuscular
connective tissue. The encapsuled worms
in the corium feel like embedded rice grains and
raise the epidermis, giving rise to an acne-like condition.
Many thousands occur scattered over the
body; in Ijima’s Japanese case there were over
10,000 in the left thigh. The worms when they first
appear in the skin cause itching. The capsules are
ovoid, generally about 1 to 2 mm. in diameter, but
they may be smaller and also much larger. The
larger ones occur in the subcutaneous tissue. The
capsules consist of dense tough connective tissue.
Each capsule, as a rule, contains one worm, but as many as seven
may occur. The skin of areas that have been long infected is swollen
and indurated or adherent, giving a somewhat elephantoid appearance.
The subcutaneous tissue is thick and filled with slimy fluid or in
other parts sclerosed.
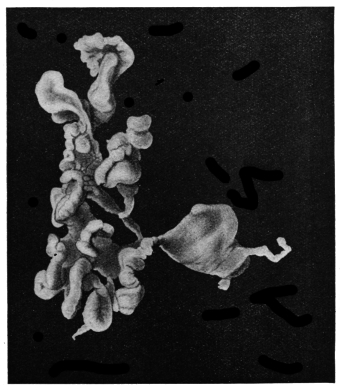
Fig. 216.—Sparganum proliferum. × 10. (After Stiles.)
The Worm.—The chief peculiarity is its
irregular shape and its reproduction in the
larval stage by forming supernumerary heads,
which are supposed to wander about the
body.
The simplest forms are thread-like bodies,
flat or round, 3 mm. long and 0·3 mm. in
diameter, but they may be 12 mm. long by
2·5 mm. broad. The narrow end is the head,
which in life invaginates and evaginates, but
there is no indication of any suckers, except
an inconstant terminal depression. In addition
to these simple forms the most complicated and
irregular forms occur, due to the
formation of buds (heads) at various parts.
The detachment and growth of a head account for the presence of
more than one worm in a cyst. The irregularity in form is also
increased by the presence in the subcuticular tissue of the worm of
reserve food bodies. These bodies are supposed to be of this nature
and are spherical, 100 µ to 300 µ in diameter, but also much elongated.
Calcareous bodies in the Japanese worms were 7·5 µ to 12 µ; in the
Florida worms 8·8 µ to 17·6 µ.
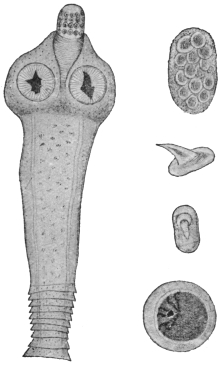
Fig. 217.—Dipylidium caninum: on the
left, the scolex, neck and the first proglottids;
on the right, at the top, a packet of
ova; below, hooks of the rostellum, side
and front views; below, an ovum. Various
magnifications. (After Diamare.)
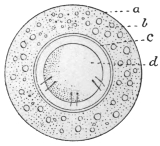
Fig. 218.—Dipylidium caninum; egg showing
a, egg-shell (vitelline membrane of Moniez);
b, albuminous coat; c, internal shell formed of
or secreted by an outer layer of blastomeres
(Moniez); d, hexacanth embryo. (After Benham
and Moniez.)
Mode of Infection.—Probably from eating uncooked fish.
Distribution.—Japan, Florida.
Family. Dipylidiidæ, Lühe, 1910.
Genus. Dipylidium, R. Lkt., 1863.
Rostellum retractile, with several rings of alternating hooks; the latter with a
disc-like base, having the shape of the thorns of a rose. Genital pores opposite;
genitalia double. Testes very numerous in the central field; ovary with two lobes;
the vitellaria, which are smaller, behind them; the uterus forms a reticulum, in the
network of which the testicular vesicles lie; later on it breaks up into sacs enclosing
one or several eggs. The eggs have a double shell.
Dipylidium caninum, L., 1758.
Syn.: Tænia canina, L., 1758, p. p.; Tænia
moniliformis, Pallas, 1781; Tænia cucumerina,
Bloch, 1782; Tænia elliptica, Batsch, 1786;
Dipylidium cucumerinum, Lkt., 1863.
This worm measures 15 to 35 cm.
in length and 1·5 to 3 mm. in breadth.
The scolex is small, rhomboidal, and
has a club-shaped rostellum on which
there are, in three to four rings, forty-eight to sixty hooks resembling
rose thorns, the size of those in the foremost being 11 µ to 15 µ and those
in the hindmost ring 6 µ. The neck is very short, the most anterior
segments broad and short, the middle as long as they are broad; the
mature segments are longer than wide (6 to 7 mm. by 2 to 3 mm.), fairly
thick, are frequently of a reddish colour, and when cast off resemble
cucumber seeds. The genital pores lie symmetrically at the lateral
margins; the roundish egg sacs, arising from the uterine reticulum,
contain eight to fifteen eggs embedded in a reddish cement substance
(in life). The eggs are globular
(43 µ to 50 µ,); the embryonal
shell (embryophore) is
thin, the oncosphere measures
32 µ to 36 µ. Surrounding the
embryophore is an albuminous
coating, and outside this
the thin vitelline envelope
(fig. 218).
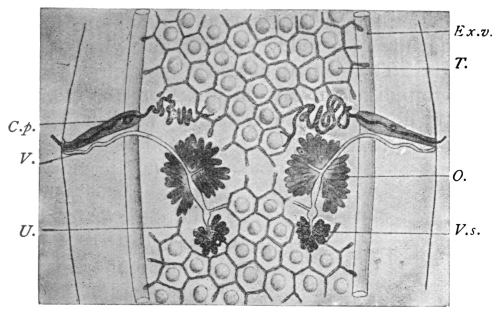
Fig. 219.—Dipylidium caninum: central portion of a proglottis. C.p., cirrus
sac; V.s., vitellaria; Ex.v., excretory vessels; T., testicles lying in the meshes
of the uterine reticulum which laterally forms pouches; O., ovary; U., reticulum
of uterus; V., vagina and seminal receptacle (below ovary). Magnified. (After
Neumann and Railliet.)
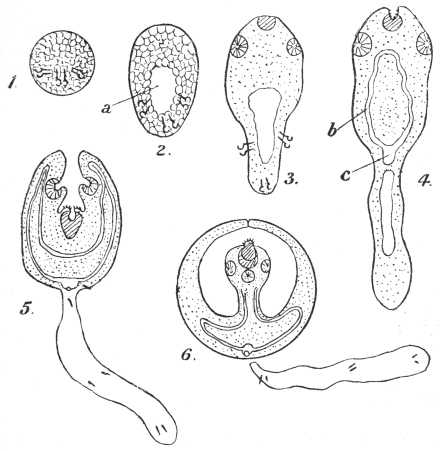
Fig. 220.—Dipylidium caninum: development of embryo. 1, solid hexacanth embryo;
2, primitive lacuna (a) in the embryo; 3, elongation of hinder part, rudiments of sucker and
rostellum appearing; 4, “body” and “tail” distinct, (b) and (c) excretory system; 5, fore-body
invaginates into hind-body, excretory bladder has a pore; 6, tail has dropped off; scolex
growing up into secondary cavity formed by fore-body; the primitive cavity has been absorbed
at stage 4. (After Benham, Grassi and Rovelli.)
Dipylidium caninum is a
common intestinal parasite of
dogs, in which it grows larger
(Tænia cucumerina, Bloch)
than in cats (T. elliptica,
Batsch); it has, however, also
been found in jackals, as well
as in human beings, though
in the latter it is of comparatively
rare occurrence (twenty-four
cases), and almost always
affects children, generally of
tender age. One-third of all
the cases in children were
sucklings, about a quarter of
all the cases recorded were
adults, and these occurred
throughout all Europe with
the exception of Spain and
Italy.
The proglottids, which
leave the intestine spontaneously,
are recognizable by the
naked eye on account of their
form and reddish colour, as
well as their two genital pores. As a rule, the presence of this parasite
sets up no marked symptom in the patient.
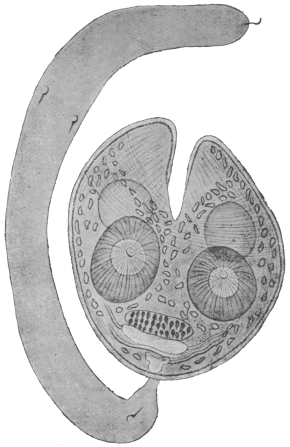
Fig. 221.—Larva (cysticercoid) of Dipylidium
caninum, consisting of body and tail. The latter
is solid and bears on it the embryonal spines. The
bladder, which was only slightly developed, has
disappeared, and the fore-part of the body bearing
the rostellum is now seen invaginated into the
hind portion. The hooklets are shown in front
of the excretory system which has now developed.
At a further stage the tail drops off; the head now
evaginates, but is still enclosed in a double-walled
sac formed by the prolongation upwards on each
side of the topmost parts of the body shown in
the figure. Cf. fig. 220, 6. Enlarged. (After
Grassi and Rovelli.)
The corresponding larval form (cysticercoid) lives in the louse
of the dog (Trichodectes canis), a fact that was first established by
Melnikow and Leuckart; according to Grassi and Rovelli, as well
as Sonsino, it also lives in the flea of the dog (Ctenocephalus canis)
and in the flea of man (Pulex irritans), but not in its larva. The
adult segments, which also leave the rectum of dogs and cats
spontaneously, creep about around the anus and get into the hair,
and are thus partly dried and disintegrated. Part of the segments, or
the oncospheres released by disintegration, are then taken up by lice
and fleas, within which they develop into larvæ (cysticercoids). Dogs
and cats are thus infected by their own skin parasites, which they bite
and swallow whilst gnawing at their fur. The infection of human
beings must occur in an analogous manner, by transmission of the
cysticercoids present on the lips or tongue of dogs when the latter lick
them, or it may be that the vermin of cats and dogs harbouring cysticercoids
are accidentally and directly swallowed by human beings.
Family. Hymenolepididæ, Railliet and Henry, 1909.
Genus. Hymenolepis,282 Weinland, 1858.
Accessory sac (opening into genital atrium) usually absent. Vas deferens with
an external (outside cirrus sac) and an internal (inside cirrus sac) “seminal vesicle.”
Three testes in each proglottis. The eggs are round or oval with two to four distinct
envelopes. In mammals and birds.

Fig. 222.—Hymenolepis
nana, v.
Sieb. About 12/1.
(After Leuckart.)
Hymenolepis nana, v. Sieb., 1852.
Syn.: Tænia nana, v. Sieb., 1852, nec van Beneden, 1867; Tænia ægyptiaca,
Bil., 1852; Diplacanthus-nanus, Weinld., 1858; Tænia (Hymenolepis) nana, Lkt.,
1863.
The worm is 10 to 45 mm. in length and 0·5 to 0·7 mm. in breadth;
the head is globular, 0·25 to 0·30 mm. in diameter. The rostellum
has a single circlet consisting of twenty-four or twenty-eight to thirty
hooks, which are only 14 µ to 18 µ in length. The neck is moderately
long; the proglottids are very narrow, up to 200 in number,
0·4 to 0·9 mm. in breadth, and 0·014 to 0·030 mm. in length. The
eggs are globular or oval, 30 µ to 37 µ to 48 µ; the oncospheres measure
16 µ to 19 µ in diameter, with two coats, separated by an intervening
semi-fluid substance (fig. 224).
This species was discovered by Bilharz in Cairo in 1851; it was
found by him in great numbers in the intestine of a boy who had
died of meningitis. For several years this was the only case, until
1885, since when numerous cases have come to light. Spooner (1873)
even reported a case from North America, which may, however, have
related to Hymenolepis diminuta. In Europe the worm is particularly
frequent in Sicily, but it has also been repeatedly observed in
North Italy; it has, moreover, been reported from Russia, Servia,
England, France, Germany, North and South America, the Philippines,
Siam and Japan, in all over 100 cases. Notwithstanding its
small size this worm causes considerable disorders in its hosts—mostly
children—as it sets up loss of appetite, diarrhœa, various
nervous disturbances, and even epilepsy; all these symptoms, however,
disappear after the expulsion of the parasites, which are generally
present in large numbers.
Fig. 223.—Hymenolepis
nana: head. Enlarged.
(After Mertens.)

Fig. 224.—Hymenolepis
nana: an egg. Highly magnified.
(After Grassi.)

Fig. 225.—Longitudinal
section through the intestinal
villus of a rat, with the
larva (cysticercoid) of Hymenolepis
murina. Magnified.
(After Grassi and
Rovelli.)
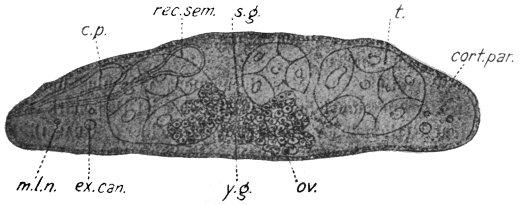
Fig. 226.—Hymenolepis nana (murina): cross section of proglottis from a rat. c.p.,
cirrus sac; rec. sem., receptaculum seminis; s.g., shell gland; ov., ovary; t., testis; cort.
par., cortical parenchyma; m.l.n., main lateral nerve; ex. can., excretory canal; y.g.,
vitellarium. (After v. Linstow.)
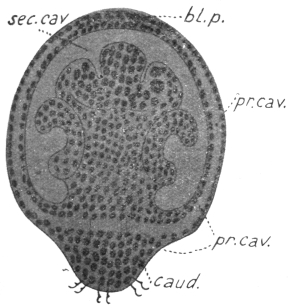
Fig. 227.—Hymenolepis nana: longitudinal section of an embryo. bl.p.,
anterior opening of secondary cavity; caud., caudal appendage; pr. cav., primary
cavity; sec. cav., secondary cavity. Enlarged. (After Grassi and Rovelli.)

Fig. 228.—Hymenolepis
diminuta:
scolex. Magnified.
(After Zschokke.)
The development as well as the manner of infection is still
unknown; Grassi is of opinion that Hymenolepis nana is indeed
merely a variety of Hymenolepis murina, Duj., which lives in rats.
According to Grassi direct development takes place with omission
of the intermediate host, but with the retention of the larval stage;
that is to say, rats infect themselves directly with Hymenolepis murina,
by ingesting the mature segments or oncospheres of this species, from
which subsequently the small larvæ originate in the intestinal wall
(fig. 225); when fully developed they fall into the intestinal lumen
and become tapeworms. The identity of the two forms has nevertheless
been disputed (Moniez, R. Blanchard, v. Linstow), though their
near relationship cannot be denied. Grassi gave mature segments of
Hymenolepis murina to six persons, but only one person evacuated
a tapeworm. This, however, proves nothing in a district where
Hymenolepis nana frequently occurs in man; it was, moreover, not
possible to infect rats with segments of Hymenolepis nana (of man).
Accordingly this form may represent an independent species, which,
however, like Hymenolepis murina, also omits an intermediate host.
Hymenolepis diminuta, Rud., 1819.
Syn.: Tænia diminuta, Rud., 1819; Tænia leptocephala, Crepl., 1825; Tænia
flavopunctata, Weinld., 1858; Tænia varesina, E. Parona, 1884; Tænia minima,
Grassi, 1886.
This species measures 20 to 60 cm. in length, and up to 3·5 mm.
in breadth; there are from 600 to 1,000 segments. The head is very
small (0·2 to 0·5 mm.), it is club-shaped and has
a rudimentary unarmed rostellum; the neck is short;
the mature segments are 3·5 mm. in breadth, 0·66 mm.
in length; the eggs are round or oval. The outer
egg-shell is yellowish and thickened, with indistinct
radial stripes; the inner embryonal shell (embryophore)
double, thin; the outer layer is somewhat
pointed at the poles; oncosphere 28 µ by 36 µ.
Between the inner and outer shells is a middle
granular layer.
Hymenolepis diminuta lives in the intestine of
rats—Mus decumanus (the sewer rat), Mus rattus (the
black rat), and Mus alexandrinus, rarely in mice; it
is occasionally also found in human beings.
Weinland described it from specimens collected
by Dr. E. Palmer in 1842, in Boston, from a child
aged 19 months, as T. flavopunctata. A second case
relating to a three year old child, from Philadelphia,
was only reported in 1889 by Leidy; a third case
was simultaneously reported of a two year old girl
in Varese (T. varesina); and Grassi
described another case relating to a
twelve year old girl from Catania
(Sicily). Sonsino and Previtera reported
the same species in Italy, Zschokke in France, Lutz and Magalhães
in South America, and Packard in North America: a total of twelve
cases, five from America, the rest from Europe (Ransom).
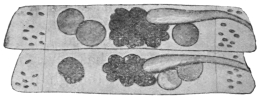
Fig. 229.—Hymenolepis diminuta: two proglottids
showing testes (3), ovary and vagina.
Slightly enlarged. (After Grassi.)

Fig. 230.—Hymenolepis diminuta:
egg from man. (After Bizzozero.)
According to Grassi and Rovelli the larval stage lives in a small
moth (Asopia farinalis), as well as in its larva, in an orthopteron
(Anisolabis annulipes), and in coleoptera (Acis spinosa and Scaurus
striatus). Experimental infections have been successful on rats as
well as on human beings. In America other species of insects may
be the intermediary hosts.
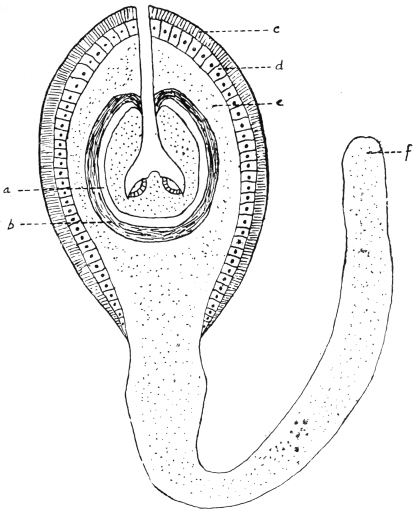
Fig. 231.—Hymenolepis diminuta: cysticercoid from the rat flea (Ceratophyllus fasciatus).
a, remains of primary vesicle; b, fibrous layer; c, radially striated layer resembling cuticle;
d, layer of columnar cells; e, parenchymatous layer of irregularly disposed cells; f, parenchymatous
layer. (Stephens, after Nicoll and Minchin.)
Nicoll and Minchin283 found in the body cavity of 4 per cent.
of rat fleas (Ceratophyllus fasciatus) the cysticercoid of Hymenolepis
diminuta. That it belonged to this species was shown by its unarmed
rostellum and by feeding; 340 fleas were fed to white rats and fourteen
worms obtained, i.e., about 4 per cent., thus corresponding to the infection
of the fleas. The development in the flea probably begins in the
pupal stage, the eggs being ingested by the older flea larvæ. The larva
is 0·31 by 0·25 mm.; tail 0·8 mm., scolex 0·075 by 0·09 mm., suckers,
0·055 mm. in diameter. Microscopically it shows—(1) externally
a radially striated layer resembling cuticle, (2) a layer of columnar cells,
(3) parenchymatous layer continuous with the tail, (4) fibrous layer
around the small caudal vesicle, then the parenchymatous scolex at
the bottom of the secondary cavity.
Nicoll and Minchin (loc. cit.) found a cysticercoid284 in the rat flea
Ceratophyllus fasciatus which was probably that of Hymenolepis murina.
Body 0·16 mm., tail 0·19 mm., scolex 0·096 mm. in diameter. Rostellum
has twenty-three spines in a single row. Length 0·017 mm.,
handle 0·01 mm., guard 0·007 mm., prong 0·007 mm. Sucker 0·042 mm.
Although this cycle, then, for H. murina also exists, it is not probable
that rats (or man in the case of H. nana if this be considered distinct)
infect themselves in this way, as they hardly
ingest all the necessary fleas to account for the
massive infection which frequently exists in
rats (and man), so that Grassi’s cycle holds
good as the predominant method. Xenopsylla
cheopis has also been found by Johnston to
harbour both cysticercoids in Australia.

Fig. 232.—Hymenolepis
lanceolata. Natural size.
(After Goeze.) To the
right above, two hooks.
120/1. (After Krabbe.)
Hymenolepis lanceolata, Bloch, 1782.
Syn.: Tænia lanceolata, Bloch, 1782; Drepanidotænia
lanceolata, Railliet, 1892.
The parasite measures 30 to 130 mm. in
length and 5 to 18 mm. in breadth; the
head is globular and very small; the
rostellum is cylindrical, with a circlet composed of eight hooks (31 µ
to 35 µ in length). The neck is very short. The short segments
increase gradually and equally in breadth, but only a little in
length; the female glands lie on the side opposite to that on which
the genital pore is situated; the three elliptical testes are on the
same side as the pores; the cirrus is armed and slender. The
eggs have three envelopes and are oval (50 µ by 35 µ), the external
envelope is thin, the middle intermediate layer or envelope is not
so marked as in H. diminuta, and the internal one is very thin and
sometimes has polar papillæ, as in Hymenolepis diminuta and
H. nana.
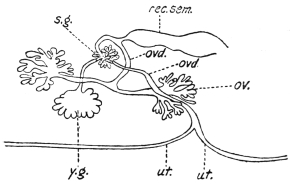
Fig. 233.—Hymenolepis lanceolata: diagram of
female genitalia. ov., ovary; ovd., oviduct; rec. sem.,
receptaculum seminis; s.g., shell gland; ut., uterus;
y.g., vitellarium. (After Wolffhügel.)
It inhabits the intestine of the following birds: Domesticated
ducks and geese, the Muscovy duck (Cairina moschata), white-headed
duck (Erismatura leucocephala), the pochard (Nyroca rufina), and
the flamingo (Phœnicopterus antiquorum). It has been recorded from
Great Britain, France, Denmark, Austria and Germany.
Zschokke reports the receipt of two specimens which a twelve
year old boy in Breslau evacuated spontaneously at two different
times.
The corresponding larva, according to Mrázek, lives in fresh water
Cyclops; according to Dadai it is likewise found in another copepod,
Diaptomus spinosus, but the hooks of Dadai’s larva differed in size.
Family. Davaineidæ, Fuhrmann, 1907.
Sub-family. Davaineinæ, Braun, 1900.
Genus. Davainea, R. Blanch., 1891.
The large scolex is more or less globular, much wider than the rostellum, which is
furnished with two rings of very small and numerous hooks. Neck absent, proglottids
few, genitalia single. Parasitic chiefly in birds.285

Fig. 234.—Scolex
of Davainea madagascariensis.
The
hooks have fallen
off. 14/1. (After
Blanchard.)
Davainea madagascariensis, Davaine, 1869.
Syn.: Tænia madagascariensis, Dav.; Tænia demerariensis, Daniels, 1895.
This worm measures 25 to 30 cm. in length; the head has four
large round suckers; the rostellum has ninety hooks (18 µ in length);
there are 500 to 700 segments, of which the last 100 are filled
with eggs and form half of the entire worm. The segments, when
mature, measure 2 mm. in length by 1·4 mm. in breadth; genital
pores unilateral; about fifty testes; the uterus consists of a number of
loops, which at each side are rolled up into an almost spherical ball;
when filled with eggs the convolutions unwind, permeate the segment
and then lose their wall; the eggs lying free in the parenchyma
become finally surrounded, one, or several together, by proliferating
parenchymatous cells; this is how the 300 to 400 egg masses, taking
up the entire mature segment, are formed. The globular oncosphere
(8 µ) is surrounded by two perfectly transparent shells, the outer of
which terminates in two pointed processes.
Davainea madagascariensis has hitherto been found in man only
(eight times). Davaine described this species from fragments sent
to him from Mayotta (Comoro Islands), which were found in two
Creole children. Chevreau observed four cases in Port Louis
(Mauritius), likewise in children; Leuckart received the first perfect
specimen—it was obtained from a three year old boy, the son
of a Danish captain, in Bangkok; Daniels, at the post-mortem of
an adult native of George Town, Guiana, found
two specimens (Tænia demerariensis); and finally
Blanchard describes another perfect specimen
which was in Davaine’s collection of helminthes
in Paris, and which was obtained from a little girl
3 years old, of Nossi-Bé (Madagascar). The intermediate
host is unknown.
Davainea (?) asiatica, v. Linst., 1901.
Syn.: Tænia asiatica, v. Linstow.
There exists only one headless specimen of this
species, which is not quite adult, and which is preserved
in the Zoological Museum of the Imperial
Academy of Science in Petrograd. It came
from a human being and was found by Anger in
Aschabad (Asiatic Russia, near the northern frontier
of Persia). The specimen measures 298 mm. in
length. The breadth anteriorly is only 0·16 mm.,
the posterior part measures 1·78 mm. across. The
number of segments is about 750. The genital
pores are unilateral; the testes are globular and
lie in a dorsal and ventral layer in the medullary layer; the cirrus
pouch is pyriform, 0·079 mm. in length and 0·049 mm. in breadth;
the female glands lie in the fore-part of the segments, the ovary
reaching to the excretory vessels; the vitellarium is small and round.
The vagina has a large fusiform receptaculum seminis; the uterus
breaks up into sixty to seventy large, irregularly polyhedric eggsacs.
Family. Tæniidæ, Ludwig, 1886.
Genus. Tænia, L., 1758.286
With the characters of the family. In the genus Cladotænia recognized by
some authors, the testes encroach on the mid field and the uterine stem reaches the
anterior end of the segment.
Tænia solium, L., p. p., 1767.
Syn.: Tænia cucurbitina, Pall., 1781; Tænia pellucida, Goeze, 1782; Tænia
vulgaris, Werner, 1782; Tænia dentata, Gmel., 1790; Halysis solium, Zeder, 1800;
Tænia humana armata, Brera, 1802; Tænia (Cystotænia) solium, Lkt., 1862.
The average length of the entire tapeworm is about 2 to 3 m.,
but may be even more; the head is globular, 0·6 to 0·8 to 1·0 mm.
in diameter. The rostellum is short with a double circlet of hooks,
twenty-two to thirty-two in number, usually twenty-six to twenty-eight;
large and small hooks alternate regularly; the length of the large hooks
is 0·16 to 0·18 mm., of the small ones 0·11 to 0·14 mm. The rostellum
is sometimes pigmented. The suckers are hemispherical, 0·4 to
0·5 mm. in diameter. The neck is fairly thin and long (5 to 10 mm.).
The proglottids, the number of which averages from 800 to 900, increase
in size very gradually; at about 1 m. behind the head they are square
and have the genitalia fully developed. Segments sufficiently mature for
detachment measure 10 to 12 mm. in length by 5 to 6 mm. in breadth.
The genital pores alternate fairly evenly at the lateral margin a little
behind the middle of each segment. The fully developed uterus consists
of a median trunk, with seven to ten lateral branches at either side,
some of which are again ramified. The eggs are oval, the egg-shell
very thin and delicate; the embryonal shell (embryophore) is thick,
with radial stripes; it is of a pale yellowish colour, globular, and
measures 31 µ to 36 µ in diameter; the oncospheres, with six hooks,
are likewise globular, and measure 20 µ in diameter (fig. 238).
Malformations are not so common as in T. saginata; they consist in two or
several proglottids being partly or entirely fused, formation of single club-shaped
segments, fenestration of long or short series of segments and so-called double formation,
in which the head has six suckers and the segments exhibit a Y-shaped transverse
section. The oncospheres occasionally also possess more than six hooklets.
Very slender specimens have led to the description of a particular species or variety
(T. tenella).
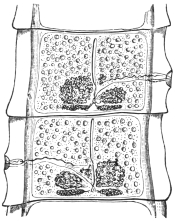
Fig. 235.—Two fairly mature proglottids
of Tænia solium, showing ovaries (one
bi-lobed), vitellaria, central uterine stem,
cirrus and vas deferens (above), vagina
(below), testes (scattered), longitudinal and
transverse excretory vessels.
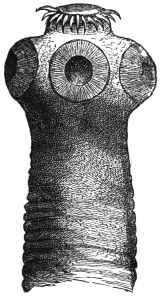
Fig. 236.—Head of Tænia solium.
45/1.
In its fully developed condition T. solium is found exclusively
in man; the head is usually attached in the anterior third of the
small intestine and the chain, in numerous convolutions, extends
backwards; a few mature detached proglottids usually lie at the
most posterior part, and these are usually evacuated during defæcation.
In exceptional cases single proglottids or whole worms may
reach contiguous organs if abnormal communications with them
exist; thus they may reach the abdominal cavity and the urinary
bladder, or they may be found in a so-called worm abscess of the
peritoneum; occasionally, in vomiting, single segments or several
together may be brought up. Exceptionally it induces severe anæmia.
The larval stage (Cysticercus cellulosæ) that gives rise to Tænia
solium lives normally in the intramuscular connective tissue and
other organs of the domestic pig, but it is known to exist also in
a few other mammals, such as the wild boar, the sheep,287 the stag,
dog, cat, brown bear and monkey, as well as in man. The cysticercus
of the pig is an elliptical vesicle with a longitudinal diameter of
6 to 20 mm., and a transverse diameter of 5 to 10 mm.
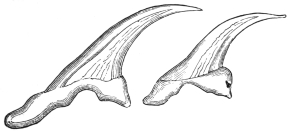
Fig. 237.—Large and small hooks of Tænia solium. 280/1.
(After Leuckart.)
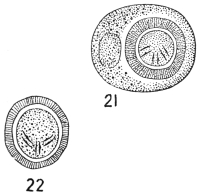
Fig. 238.—Tænia solium. 21, Egg with external membrane;
22, without (embryophore). (After Leuckart.)
Even with the naked eye a white spot may be observed in the
centre of the long equator, this being the invaginated head; it is easy
to make it project by pressing on the vesicle (after tearing off with the
finger-nail the investing connective tissue), and on examining it under
the microscope one can convince oneself that it corresponds with the
head of Tænia solium.
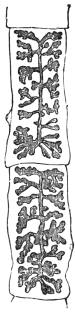
Fig. 239.—Two
mature proglottids
of Tænia solium with
fully developed
uterus. 2/1.
Numerous experiments have proved that the Cysticercus cellulosæ
of the pig, if introduced into the intestine of man, grows to a Tænia
solium (Küchenmeister, 1855; Humbert, 1856; Leuckart, 1856; Hollenbach,
1859; Heller, 1876); the cysticercus has frequently also been
cultivated purposely by feeding pigs with mature proglottids of
T. solium (P. J. van Beneden, 1853; Haubner and Küchenmeister,
1855; Leuckart, 1856; Mosler, 1865; Gerlach, 1870; etc.), but success
did not attend the efforts to make Cysticercus cellulosæ establish themselves
in the intestines of pigs, dogs, guinea-pigs, rabbits and monkeys
(Macacus cynomolgus), and so become adult Tæniæ; the attempts, also,
to infect dogs with cysticerci by means of ova were likewise, as a
rule, abortive.288
The development of Cysticercus cellulosæ takes two and a half to three or four
months; it is not known how long the cysticerci remain alive in animals; not uncommonly
they perish at earlier or later stages, and become calcified or caseated.
Extracted cysticerci die in water at a temperature of 47° to 48° C., in flesh at normal
temperature they remain alive for twenty-nine days or more. On account of the
present rapid means of pickling and smoking meat, the cysticerci as a rule are not
killed, also the effect of cold on them for some time in cold chambers of slaughterhouses
is not lethal, but freezing is fatal (Ostertag).
There is not the least doubt that human beings are almost exclusively
infected with Tænia solium by eating pork containing cysticerci
in a condition that does not endanger the life of the cysticerci. The
infection may likewise be caused in man by eating the infected meat
of other animals subject to this species of bladder worm, mainly, as a
matter of fact, deer and wild boar.
The frequency of cysticerci in pigs’ flesh has considerably decreased since the
introduction of meat inspection; in the Kingdom of Prussia there was on an average
1 infected pig to every 305 slaughtered between 1876 to 1882; from 1886 to 1889, there
was 1 to 551; from 1890 to 1892, there was 1 to 817; in 1896, 1 to 1,470; and in 1899,
1 to 2,102; in the Kingdom of Saxony in 1894 there was 1 infected pig to every 636;
in 1895 there was 1 to every 2,049, and in 1896 only 1 infected pig was found of
5,886 slaughtered. In South Germany pigs with cysticerci are very rare, but are
more frequent in the eastern provinces of Prussia; in 1892 the number of infected
pigs compared with the total slaughtered was as follows:—
| In the district of Marienwerder | 1 : 28 |
| " " Oppeln | 1 : 80 |
| " " Königsberg | 1 : 108 |
| " " Stralsund and Posen | 1 : 187 |
| " " Danzig, Frankfort a. O. and Bromberg | 1 : 250 |
| As compared with the district of Arnsberg | 1 : 865 |
| " " " Coblenz | 1 : 975 |
| " " " Düsseldorf | 1 : 1,070 |
| " " " Münster and Wiesbaden | 1 : 1,900 |
The average for the whole of Prussia in the same year was 1 : 1,290; for the
eastern provinces, on the other hand, 1 : 604. Even more unfavourable are the proportions
in Russian Poland (over 1 per cent. of pigs measly), in Prague (over 3 per
cent.), in Bosnia and Herzegovina (6 to 7 per cent.). The cause for this is most
likely attributable to the manner in which the pigs are kept. When allowed to be
in the farmyards of the small farmers for the whole day, or allowed to wander in the
village streets and pasture lands, they are more liable to take up the oncospheres of
the T. solium than when shut up in good pig-styes.
The geographical distribution of T. solium generally corresponds
with that of the domestic pig and the custom of eating pork in any
form insufficiently cooked or raw. There are, or were, some isolated
districts in Germany, France, Italy and England where the “armed
tapeworm” was frequent (for instance, Thuringia, Brunswick, Saxony,
Hesse, Westphalia, whereas it is and was very scarce in South Germany);
it is thus easily understood why it occurs very rarely in the
East, in Asia and in Africa, in consequence of the Mahommedans,
Jews, etc., not eating pork. In North America, also, T. solium is very
rare; the tapeworm which is known there by this name is generally
T. saginata, Stiles. During the last decade T. solium infection has,
however, very markedly decreased in North and East Germany in
consequence of the precautions exercised by the public in the choice
of pork to avoid trichinosis, especially, however, because measly meat
must be sold as such and must be thoroughly cooked before being
placed on the market; indeed, if badly infected it may not be sold
for food, but can be turned to account for industrial purposes.
The occurrence of Cysticercus cellulosæ in man has been known
since 1558 (Rumler, Obs. med., liii, p. 32); there is hardly an organ
in man in which cysticerci have not been observed at some time;
they are most frequently found in the brain,289 where they grow to a
variety known as Cysticercus racemosus; next in frequency they are
found in the eye, in the muscular system, in the heart, in the subcutaneous
connective tissue, the liver, lungs, abdominal cavity, etc.
The number of cysticerci observed in one man varies between a few
and several thousands. Of the sexes, men are most subject (60 to
66 per cent. of the number attacked). The disturbances caused in
man by cysticerci vary according to the nature or position of the
organs attacked; when situated in the cerebral meninges they have
the same effect as tumours.
During the last decades, however, these cases have also become
less common. In Rudolphi’s time 2 per cent. of post-mortems in
Berlin showed cysticerci; in the ’sixties, according to Virchow, about
the same; in 1875 the number fell to 1·6 per cent.; in 1881 to 0·5 per
cent.; in 1882 to 0·2 per cent.; in 1900 to 0·15 per cent., and in 1903
to 0·16 per cent. Hirschberg between 1869 and 1885 discovered
cysticerci in the eye seventy times in 60,000 ophthalmic cases, but
during the following six years the parasite was only present twice
amongst a total of 46,000 cases of ophthalmic diseases, and since
1895 no ophthalmic case has been met with.
The infection of human beings with the cysticerci can only take
place by the introduction of the oncospheres of Tænia solium into
the stomach with vegetable foods, salads that have been washed
in impure water containing oncospheres, also by drinking contaminated
water; the carriers of T. solium, moreover, infect themselves
still more frequently through uncleanliness in defæcation, the privies
in public localities and many private houses affording striking testimony
of this. The minute oncospheres can thus easily reach the
fingers and thence the mouth (as in twirling the moustache, biting the
nail). More rarely a third manner of transmission or internal auto-infection
may possibly take place, as when, in the act of vomiting,
mature proglottids near the stomach are drawn into it; the oncospheres
or segments there retained are then in the same position as
if they had been introduced through the mouth.
On account of these dangers of internal or external auto-infection, it is therefore
the duty of the medical attendant, after recognizing the presence of tapeworms, to
expel them,290 and in doing so to employ every possible means to prevent vomiting
setting in; it is, however, equally important to take the necessary steps to destroy
the parasites evacuated. It may be incidentally mentioned that in using certain
remedies the scolex not rarely remains in the intestine; the cure in such cases has
not been accomplished, as the scolex again produces new proglottids, and after
about eleven weeks the first formed ones are again mature and appear in the fæces.
Amongst the cysticerci also many malformations appear; thus absence of the
rostellum and the hooks, or double formation with six suckers, or abnormalities of
growth on account of the surroundings, which have had a special name given to
them, viz., Cysticercus racemosus, Zenk. (= C. botryoides, Hell.; C. multilocularis,
Kchnmstr.); these forms are more especially found at the base of the brain, are
irregularly ramified and often without the head.
A certain interest is attached to those forms that have led to the
establishment of a distinct species:—
Cysticercus acanthotrias, Weinld., 1858.
In making the autopsy of a white Virginian woman who had died of phthisis, a
cysticercus was found in the dura mater, and eleven or twelve specimens in the
muscles and subcutaneous tissue. Weinland and Leuckart, who examined the
specimens, found that they resembled Cysticercus cellulosæ in form and size, but that
they carried on the rostellum a triple crown, each consisting of fourteen to sixteen
hooks, which differed from the hooks of C. cellulosæ or of Tænia solium by the
greater length of the posterior root process and the more slender form of the
hooks; the large hooks measured 0·153 to 0·196 mm., the medium-sized hooks,
0·114 to 0·14 mm., and the small ones 0·063 to 0·07 mm.
On account of these differences a distinct species of cysticercus
was established, and this naturally presupposed a corresponding
species of Tænia (T. acanthotrias, Lkt.); this could be done with
justice so long as the case remained isolated, i.e., in America, as
there was the possibility of the corresponding Tænia being found. In
this respect, however, the position has changed; Delore first described
a cysticercus the size of a nut from the biceps muscle of the arm of
a silk-worker in Lyons; according to Bertolis this specimen possessed
hooks of three different sizes, the dimensions of which corresponded
with the figures given by Weinland and Leuckart; the correctness of
the diagnosis could hardly be doubted, as Bertolis was known to be
a very exact observer. A second case has become known through
Cobbold, who regards a specimen of a cysticercus in Dallinger’s
collection as likewise belonging to Cysticercus acanthotrias; this
specimen also came from a man’s brain; finally a third case, also
from France, has been published by Redon. This author, amongst
numerous C. cellulosæ of a man, found one that had forty-one hooks in
three rows, and he was the first to express the opinion that C. acanthotrias
does not represent a distinct species, but is only an abnormality
of C. cellulosæ. This view was also taken by Blanchard and Railliet, and
is probably correct, as the discovery of the large corresponding Tænia
furnished with three rows of hooks is not to be expected in European
beasts of prey, and in Redon’s case C. acanthotrias as well as C. cellulosæ
occurred simultaneously.
The duration of life of C. cellulosæ in man is very long; cysticerci
of the eye have been known to persist for twenty years, and in cysticercus
of the brain ten to nineteen years may elapse from the first
appearance of cerebral symptoms until death. Dead cysticerci may
shrivel up or become calcified, perhaps also undergo fatty degeneration
and then absorption. Finally, it may be mentioned that if particular
proof is required that C. cellulosæ of man belongs to the cycle of
development of the Tænia solium, such proof has been furnished by
Redon.
Note.—Tænia tenella, mentioned on p. 332, was ascribed by Cobbold to
cysticerci of the muscular system of sheep. It has, however, been demonstrated
that these cysticerci belong to the cycle of development of Tænia marginata
(dog) (Cysticercus tenuicollis, from the omentum of sheep); but as already stated
C. cellulosæ also occurs in sheep. Chatin himself swallowed the cysticercus, which
Cobbold termed C. ovis, without causing a Tænia to develop in his intestine. Müller
also vainly sought to induce infection with C. tenuicollis in his own person. On the
other hand, the feeding of a dog with Cysticercus ovis resulted in the production of
Tænia marginata.
Tænia bremneri, Stephens, 1908.
Characterized by the large size of the gravid segments. The
largest was 32 by 9 mm. Smallest 21 by 6 mm. Average 28·6 by
8·5 mm. Mode 21 by 6 mm. Uterine branches twenty-two to
twenty-four in number. Calcareous bodies numerous, 15·2 µ in
diameter. Eggs maximum 45·6 µ by 41·8 µ. Smallest 34·2 µ by
30·4 µ. Mode 38 µ by 30·4 µ.
Tænia marginata, Batsch, 1786.
Syn.: T. e. Cysticerco tenuicolli, Küchenmeister, 1853.
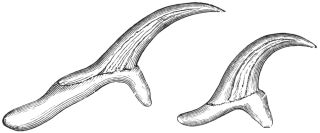
Fig. 240.—Large and small hooklets of Tænia
marginata. 280/1. (After Leuckart.)
This species, which in structure resembles Tænia solium, lives in the intestine
of the dog and the wolf. It attains 1·5 to 4 m. in length, possesses a double crown
of thirty to forty hooks, on
an average thirty-six to thirty-eight
hooks, and in its larval
stage (Cysticercus tenuicollis)
lives in the peritoneal cavity of
ruminants and the pig, occasionally
in the monkey and squirrel.
It is included in this work
because, according to one statement,
C. tenuicollis is supposed
to have been observed in man
in North America; but the case
is not quite certain, as the number of hooks was less than in C. tenuicollis and
coincided with C. cellulosæ, although the size of the cysticercus appeared to point to
C. tenuicollis. A yet earlier statement of Eschricht, that Cysticercus tenuicollis had
been observed in Iceland in the liver of a man, is undoubtedly due to an error.
Tænia serrata, Goeze, 1782.
This parasite attains a length of from 0·5 to 2 m., possesses a double crown of
thirty-four to forty-eight (mostly forty) hooks. It lives exclusively in the intestine
of the dog, the corresponding cysticercus (Cysticercus pisiformis) living in the
mesentery of the hare and rabbit. We mention this species with all reserve
amongst the parasites of man, because Vital states that he has observed it twice in
Constantine (Algeria) in human beings. The data, however, are not sufficient to
characterize the species. It is highly probable that they relate to Tænia solium.
Galli-Valerio even swallowed five specimens of Cysticercus pisiformis, but without
result.
Tænia crassicollis, Rud., 1810.
I only mention this species from the intestine of the domestic cat because
Krabbe regards its occurrence in man as possible. It attains a length of 60 cm.
and is armed; its cysticercus (Cysticercus fasciolaris) lives in the liver of mice and
rats. In Jutland, according to Krabbe, chopped-up mice (spread on bread) are
eaten raw, being a national remedy for retention of urine, and this custom affords
the possibility of the introduction of C. fasciolaris into the intestine of man (Nord.
med. Arkiv, 1880, xii).
Tænia saginata, Goeze, 1782.
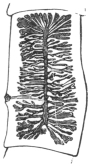
Fig. 241.—Mature
segment of Tænia
saginata, G., with
distended uterus.
2/1.
Syn.: Tænia solium, L., 1767 (pro parte); Tænia cucurbitina, Pallas, 1781 (p.p.);
Tænia inermis, Brera, 1802. Moquin-Tandon, 1860; Tænia dentata, Nicolai, 1830;
Tænia lata, Pruner, 1847; Bothriocephalis tropicus, Schmidtmuller, 1847; Tænia
mediocanellata, Küchenmeister, 1855; Tænia zittavensis, Küchenmeister, 1855; Tænia
tropica, Moquin-Tandon, 1860; Tænia (Cystotænia) mediocanellata, Leuckart, 1863.

Fig. 242.—Cephalic
end of Tænia
saginata in the contracted
condition.
8/1.
The length of the entire tapeworm averages 4 to 8 to 10 m. and
more, even up to 36 m. According to Bérenger-Feraud it attains
a length of 74 m. (?) The head is cubical in shape, 1·5 to 2 mm.
in diameter; the suckers are hemispherical (0·8 mm.) and are
frequently pigmented; there is a sucker-like organ in place of the
rostellum, and this also is frequently pigmented. The neck is
moderately long and about half the breadth of the head; the
proglottids, the number of which averages more than 1,000,
gradually increase in size; the mature detached segments are shaped
exactly like pumpkin-seeds, and are about 16 to 20 mm. in length
and 4 to 7 mm. in breadth. The genital pores alternate irregularly
and are situated somewhat behind the middle of the lateral margin.
There are twenty to thirty-five
lateral branches at each side of the
median trunk of the uterus, and
these again ramify. The eggs are
more or less globular, the egg-shell
frequently remains intact and
carries one or two filaments; the
embryonal shell (embryophore) is thick, radially striated, is transparent
and oval; it is 30 µ to 40 µ in length, and 20 µ to 30 µ in breadth.
Several segments simultaneously are usually passed spontaneously
with defæcation.
Malformations are not uncommon, and resemble those of Tænia solium;
a triangular form has been termed T. capensis by Küchenmeister, and T. lophosoma
by Cobbold, names that naturally possess as little value as does the term
T. fenestrata for fenestrated specimens. Moreover, T. solium, var. abietina, Weinld.,
1858, which was evacuated by an Indian, was probably a T. saginata with somewhat
close uterine branches. It is regarded by Stiles and Goldberger as a doubtful subspecies.
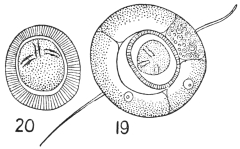
Fig. 243.—Tænia saginata. 19, egg with
external shell. 20, without (embryophore).
(After Leuckart.)
T. saginata in its adult condition lives exclusively in the intestinal
canal of man.291 The corresponding cysticercus is Cysticercus bovis, and
is found almost exclusively in the ox; it is small, 7·5 to 9 mm.
in length and 5·5 mm. in breadth, may easily escape notice, and
requires from three to six months for its development. Numerous
experiments have confirmed the connection of Cysticercus bovis with
Tænia saginata; indeed, the cysticercus was only discovered by feeding
experiments after attention had been called to the ox as the probable
intermediary host of this Tænia.

Fig. 244.—A piece of
the muscle of the ox, with
three specimens of Cysticercus
bovis. Natural size.
(After Ostertag.)
Medical men observed that weakly children who were ordered to eat raw scraped
beef to strengthen them contracted T. saginata. It was found, moreover, that
Jews, who are prohibited from eating pork from religious motives, suffered especially
from T. saginata; when T. solium was found to occur in a Jew he often confessed
to having eaten pork; and finally it was found that certain nations—for
instance, the Abyssinians—frequently harbour T. saginata, and only eat beef—raw
by preference.
These observations led Leuckart, in 1861, to feed
young calves with the proglottids of T. saginata in
order to discover the corresponding cysticercus, which
was then not known. This experiment was successful.
Similar experiments, with similar results, were then conducted
by Mosler (1863), Cobbold and Simonds (1864
and 1872), Röll (1865), Gerlach (1870), Zürn (1872),
Saint Cyr, Jolicœur (1873), Masse and Pourquier
(1876), and Perroncito, in 1876. The attempts to
infect goats, sheep, dogs, pigs, rabbits and monkeys
were unsuccessful. Only Zenker and Heller were able
to infect kids, and Heller infected one sheep, but these
are exceptions.
Artificial infections of human beings with
Cysticercus bovis to obtain the tapeworm were
less numerous, and indeed quite superfluous,
yet this was also done by Oliver (1869) in India,
and Perroncito (1877) in Italy. The experiments
of the latter prove that the extracted
cysticerci of the ox certainly perish in water at
47° to 48° C.
It is a remarkable circumstance that, at least as
regards Central Europe, C. bovis in the ox, after natural
infection, was so seldom found that almost every case
was published as a rarity; whereas the Tænia is very frequent in man. The
reason for this is that in Germany cattle are not severely infected, and that the
small, easily dried-up cysticerci easily escape notice in the large body of the host.
Hertwig, the late director of the town cattle market in Berlin, in 1888, pointed out
that the cysticercus of the ox is found chiefly in the musculi pterygoide externi
and interni, and since that time a far greater number of infected oxen have been
found in Berlin.
Year | Number of oxen
slaughtered | Infected | Proportion |
1888–89 | 141,814 | 113 | 1 : 1,255 |
1889–90 | 154,218 | 390 | 1 : 395 |
1890–91 | 124,593 | 263 | 1 : 474 |
1891–92 | 136,368 | 252 | 1 : 541 |
1892–93 | 142,874 | 214 | 1 : 672 |
Since 1892 an increase has taken place in the number of oxen infected with
cysticercus, but this is probably attributable to the more general and searching
examinations. In the slaughter-houses of Prussia the number of infected beasts was
as follows:—
| 1892 | 567 |
| 1893 | 686 |
| 1894 | 748 |
| 1895 | 1,143 |
| 1896 | 1,981 |
| 1897 | 2,629 |
The condition was most frequent in Neisse (3·2 to 4 per cent.), Eisenach
(1·91 per cent.), Ohlau (1·57 per cent.), Oels i. Schles. (1 per cent.), Marienwerder
(0·34 to 1·2 per cent.). The flesh of oxen only slightly infected (containing not more
than ten living cysticerci) is sold in pieces of not more than 5 lb. to consumers
after having been rendered innocuous by cooking, or by pickling for twenty-one days
in 25 per cent. salt brine, or hanging for twenty-one days in suitable refrigerators;
oxen in which only one cysticercus is found are allowed free commerce, and those
strongly infected (i.e., containing more than ten living cysticerci) may only be used
for industrial purposes.
It is a striking fact that more bulls than cows are infected (according to Reissmann,
in Berlin, from 1895 to 1902, 0·446 per cent. bulls, 0·439 per cent. oxen, and
0·262 per cent. cows), the explanation of which, according to Ostertag, is that most
oxen are killed when young, when also infection most readily takes place, and,
further, that the larva later on in life can be completely atrophied.
The cysticercus of the ox has hitherto been found in man on very
rare occasions. Arndt (Zeitschr. f. Psychiat., xxiv) mentions a case in
the brain, Heller in the eye, and Nabiers and Dubreith also in the
brain (Journ. méd. Bordeaux, 1889–1890, p. 209); but the diagnoses
are not quite certain, as absence of hooks occasionally occurs in
Cysticercus cellulosæ.
Tænia saginata is the most frequent tapeworm of man (with the
exception of Dibothriocephalus latus in a few districts), and the parasite
is widely distributed over the surface of the globe; it has been known
in the East for ages, so far as data are available; it is frequent in
Africa, America, and Europe. Its frequency has perceptibly increased
during the last few years, but a decrease should soon take place in
consequence of the extent and improvement of the official inspection
of meat.
The following table shows the relative frequency of the Cestodes
of man:—
Author | Year | | Number of
cases | T. saginata | T.
solium | Dibr.
latus | Dipyl.
canin. | Unde-
termined |
| Parona (Milan) | 1899 | | 150 | 121 | | 11 | 4 | — | 14 |
| Parona (Italy) | 1868–99 | | 513 | 397 | | 71 | 26 | — | 19 |
| Krabbe (Denmark) | 1869 | | 100 | 37 | | 53 | 9 | 1 | — |
| " " | 1869–86 | | 200 | 153 | | 24 | 16 | 8 | — |
| " " | 1887–95 | | 100 | 89 | | — | 5 | 6 | — |
| " " | 1896–1904 | | 50 | 41 | | 1 | 5 | 3 | — |
| Blanchard (Paris) | 1895 | | ? | 1,000 | | 21 | — | — | — |
| Stiles (United States) | 1895 |  | more than
300 | more than
300 |  | — | 3 | — | — |
| Schoch (Switzerland) | 1869 | | 19 | 16 | | 1 | 2 | — | — |
| Zaeslein (Switzerland) | 1881 | | ? | 180 | | 19 | ? | — | — |
| Kessler (Petrograd) | 1888 | | ? | 22 | | 16 | 47 | — | — |
| Mosler (Greifswald) | 1894 | | 181 | 112 | | 64 | 5 | — | — |
| Bollinger (Munich) | 1885 | | 25 | 16 | | 1 | 8 | — | — |
| Vierordt (Tübingen) | 1885 | | 121 | 113 | | 8 | — | — | — |
| Mangold (Tübingen) | 1885–94 | | 128 | 120 | 6 | 8 | — | — |
Tænia africana, v. Linst., 1900.
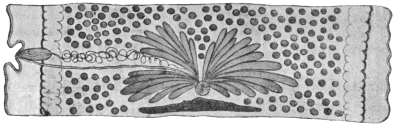
Fig. 245.—Mature segment of Tænia africana. The ovary is in the
middle, and behind it are the shell gland and vitellarium; at the sides are
the testicles, and externally the excretory canals; the cirrus pouch, the vas
deferens and the vagina are on the left. Magnified. (After v. Linstow.)
This worm measures over 1·3 m. in length. The segments are
all broader than they are long. The scolex is unarmed and is
provided with an apical sucker (0·16 mm.). The parasite measures
1·38 mm. in breadth, 1·03 mm. in width; the suckers measure
0·63 mm. in diameter. The neck is very short and somewhat broader
than the scolex; number of segments about 600; the hindmost
segments measure 7 mm. in length and 12 to 15 mm. in breadth.
The genital pores alternate irregularly in the middle of the lateral
margin; the testes are very numerous and occupy the entire medullary
layer; the vas deferens is much convoluted; the cirrus pouch
is pyriform and thick walled; the cirrus and vagina are beset with
bristles directed outwards; the receptaculum seminis is fusiform; the
ovary is large and double, and consists of radially placed club-shaped
tubes that do not anastomose and do not branch; the vitellarium is
at the posterior border of the proglottids, the round shell gland in
front of it; the uterus consists of a median trunk and fifteen to
twenty-four non-ramified lateral branches
on each side; the embryonal shell is thick
and has radial stripes—it may be round
(31·2 µ to 33·8 µ) or oval (39 µ, by 33·8 µ);
the spines of the oncospheres measure
7 µ to 8 µ in length (fig. 197).

Fig. 246.—Proglottis of Tænia africana, with uterus. Magnified.
(After v. Linstow.)
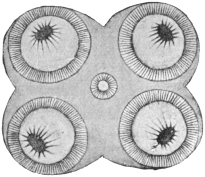
Fig. 247.—Head of Tænia africana;
apical surface. Magnified.
(After v. Linstow.)
At present only two specimens are
known; they came from a black soldier
from the vicinity of Lake Nyasa. The
cysticercus is unknown; perhaps it lives
in the zebu, the flesh of which the Askaris
are in the habit of devouring uncooked.
Tænia confusa, Ward, 1896.
Length 8·5 m., breadth about 5 mm. The scolex is unknown;
there is no neck; number of proglottids 700 to 800, always longer
than they are broad; the hindmost measure 35 mm. in length, 4 to
5 mm. in breadth; the genital pores alternate irregularly behind the
middle of the lateral margin; testicles numerous; vas deferens not
much coiled; the cirrus pouch thick walled, elongated and club-shaped,
with globular vesicula seminalis; the cirrus is beset with little
hairs; the receptaculum seminis is globular; ovary small, double;
each half is bean-shaped; vitellarium narrow, triangular; shell
gland globular; uterus with median trunk and fourteen to eighteen
short ramified lateral branches on either side. The embryophores are
oval (39 µ by 30 µ), thick and radially striated.
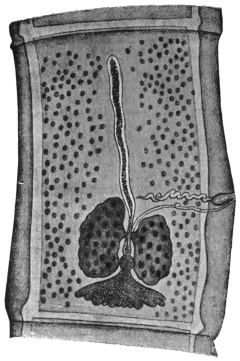
Fig. 248.—Tænia confusa: mature
segment showing central uterine stem,
bilobed ovary, globular shell gland, triangular
vitellarium, scattered testes, cirrus,
vas deferens, and vagina. 15/1. (After
Guyer.)

Fig. 249.—Tænia confusa:
gravid segment.
25/1. (After Ward.)
Of this species only two specimens have been recorded; they
occurred in human beings and were sent at different times to the
first describer of them by a doctor in Lincoln (Nebr.). Perhaps
Tænia solium, var. abietina, Weinld., which was found in a Chipeway
Indian, is of the same species in spite of the shorter segments.
Tænia echinococcus, v. Sieb., 1853.
Syn.: Tænia nana, v. Ben., 1861 (nec v. Sieb., 1853); Echinococcifer
echinococcus, Weinld., 1861.

Fig. 250.—Tænia echinococcus:
the cirrus sac, the
vagina, uterus, ovary, shell gland
and vitellarium, and the testicles
at the sides are recognizable in
the second proglottis; the posterior
proglottis shows the uterus
partly filled with eggs, as well
as the cirrus sac and the vagina.
50/1.
Tænia echinococcus measures 2·5 to 5 or 6 mm. in length; the head
is 0·3 mm. in breadth, and has a double row of twenty-eight to fifty
hooklets (on an average thirty-six to thirty-eight) on the rostellum.
The size and form of these hooklets vary (the larger ones are
0·040 to 0·045 mm. in length, the smaller ones are 0·030 to
0·038 mm. in length). The suckers measure 0·13 mm. in diameter;
the neck is short; there are only three or four segments, the posterior
segment being about 2 mm. in length and 0·5 mm. in breadth. The
genital pores alternate; there are forty to fifty testicles; the vas deferens
is spirally coiled; the cirrus pouch is pyriform. The ovary is horseshoe-shaped
with the concavity directed backwards; the vitellarium
double, each half almost bean-shaped, at
right angles to the plane of the segment;
the shell gland is round. The median trunk
of the uterus is dilated when filled with eggs
and (instead of lateral branches) has lateral
diverticula. It is not unusual for the eggs
to form local heaps. The embryonal shell
(embryophore) is moderately thin, with radial
striæ, almost globular, 30 µ to 36 µ in
diameter.
When mature this parasite lives in the
small intestine of the domestic dog, the
jackal, and the wolf, and apparently also in
Felis concolor, and is usually present in great
numbers; it can also be transmitted experimentally
to the domestic cat, one successful
result out of seven (Dévé).292 The
larval stage (Echinococcus polymorphus) lives
in various organs—chiefly in the liver and
lungs—of numerous species of mammals
(twenty-seven), especially in sheep, ox and
pig, and it is even not uncommon in man,
though the Tænia itself has never been found
in a human being; accordingly man can only
acquire the echinococcus by ingesting the
eggs of the “dog worm.” The dogs disseminate
the eggs of Tænia echinococcus
wherever they go, or carry them to their
mouths and coats by biting up the
evacuated segments, and are thus able to
transmit them directly to human beings (by
licking them or making use of the same
crockery, etc.). In other cases the oncospheres,
enclosed in the embryophores, must
withstand desiccation for a time and then
(as when the dogs are “kissed” or otherwise
caressed) are transmitted into or on to
man. As echinococcus disease in man is always very dangerous, it
would be a matter of general interest to prevent dogs being infected
by destroying the echinococci,293 and all measures would be justifiable
which would diminish the superfluous number of house-dogs (for
instance, high taxes); measures should also be adopted to limit the
association of men with dogs, particularly in such frequented places
as restaurants, railway carriages and tram-cars.
Echinococcus is very common in slaughtered animals; in Germany, however,
the figures in the reports of the abattoirs present an erroneous view in so far as,
besides the total number of animals slaughtered, only the numbers of those organs
(liver and lungs) are published that were so severely infected with echinococci that,
even when the parasites were “shelled” out, the flesh could not be placed upon the
market and was therefore “condemned.”
In Berlin the following animals were slaughtered:—
Year | 1889–90 | 1890–91 | 1891–92 | 1892–93 | 1896–97 | 1902 |
| Oxen | 154,218 | 124,593 | 136,368 | 142,874 | 146,612 | 153,748 |
| Sheep | 430,362 | 371,943 | 367,933 | 355,949 | 395,769 | 434,155 |
| Pigs | 442,115 | 472,859 | 530,551 | 518,073 | 694,170 | 778,538 |
During the same years the following were condemned in consequence of being
infected with echinococci:—
| Lung | Liver | Lung | Liver | Lung | Liver | Lung | Liver | Lung | Liver | Lung | Liver |
| Oxen | 7,266 | 2,418 | 5,792 | 1,938 | 4,497 | 1,721 | 2,563 | 739 | 3,284 | 1,156 | 2,507 | 791 |
| Sheep | 5,479 | 2,742 | 4,595 | 2,059 | 4,435 | 1,669 | 3,331 | 1,161 | 4,561 | 1,939 | 11,138 | 4,437 |
| Pigs | 6,523 | 5,078 | 5,083 | 3,735 | 6,037 | 4,374 | 6,785 | 4,312 | 7,888 | 5,398 | 9,544 | 9,233 |
Nevertheless there are statistics that give the total number of animals infected
with echinococcus:—
Author | Place | Oxen | Sheep | Pigs |
| Längrich | Rostock i. M. | 26 | ·2 per cent. | 37 | ·0 per cent. | 5 | ·4 per cent. |
| Olt | Stettin | 7 | ·1 " | 25 | ·8 " | 7 | ·3 " |
| Steuding | Gotha | 24 | ·6 " | 35 | ·4 " | 21 | ·4 " |
| Prettner | Prague | 23 | ·2 " | 5 | ·5 " | ? |
In Güstrow, in Mecklenburg, half of the animals slaughtered are said to be
infected with echinococcus; in Wismar 25 per cent. of the oxen, 15 per cent. of the
sheep and 5 per cent. of the pigs are infected; according to Mayer, in Leipzig,
3·79 per cent. native pigs, 24·47 per cent. Hungarian pigs, and 13·09 per cent. of sheep
were infected with echinococcus; at the same time it was stated that in regard to
the native pigs the liver was more frequently affected than the lungs (3·81 per cent.
as compared with 0·26 per cent.); in sheep the lungs were more frequently infected
(12·71 per cent. to 3·73 per cent.), whereas in the Hungarian pigs both organs were
almost equally infected (14·78 per cent. to 12·03 per cent.).
The data of Lichtenheld, in Leipzig, give the frequency with which various
organs were affected, as shown in the following table:—
| Cattle | Pigs | Sheep | Horses |
| | | ♂ | ♀ | | |
| Lungs | 69 | ·3 per cent. | 16 | ·2 per cent. | 21 | ·4 per cent. | 52 | ·2 per cent. | 5 | ·5 per cent. |
| Liver | 27 | ·0 " | 74 | ·2 " | 72 | ·0 " | 44 | ·9 " | 94 | ·5 " |
| Spleen | 2 | ·2 " | 3 | ·2 " | 2 | ·7 " | 2 | ·9 " | — |
| Heart | 0 | ·75 " | 3 | ·2 " | 1 | ·3 " | — | — |
| Kidneys | 0 | ·75 " | 3 | ·2 " | 1 | ·3 " | — | — |
| Subperitoneal tissue | — | — | 1 | ·3 " | — | — |
Structure and Development of Echinococcus (Hydatid).
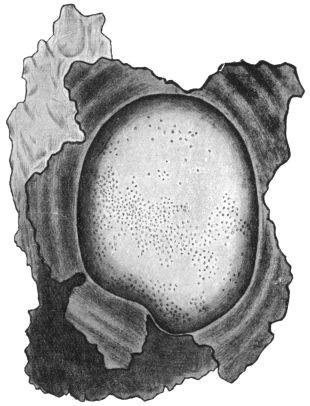
Fig. 251.—Echinococcus veterinorum: the fibrous sac enclosing the
echinococcus has been opened and laid back in five parts, so that the
surface of the bladder worm may be seen, with the brood capsules, visible
to the naked eye, showing through it. Natural size. (After Leuckart.)
An echinococcus is a spherical or roundish bladder full of a
watery liquid, which originates by liquefaction of the oncosphere, and
in man may attain the size of a child’s head, but remains smaller in
cattle (the size of an orange or apple). The thin wall of the bladder
is composed of an external laminated cuticle (ectocyst) and an internal
germinal or parenchymatous layer (endocyst). The latter again
exhibits two layers: an outer layer of small cells that are less sharply
defined, and an inner layer of larger cells. It contains, in addition,
calcareous corpuscles, muscular fibres and excretory vessels. It is
rich in glycogen.
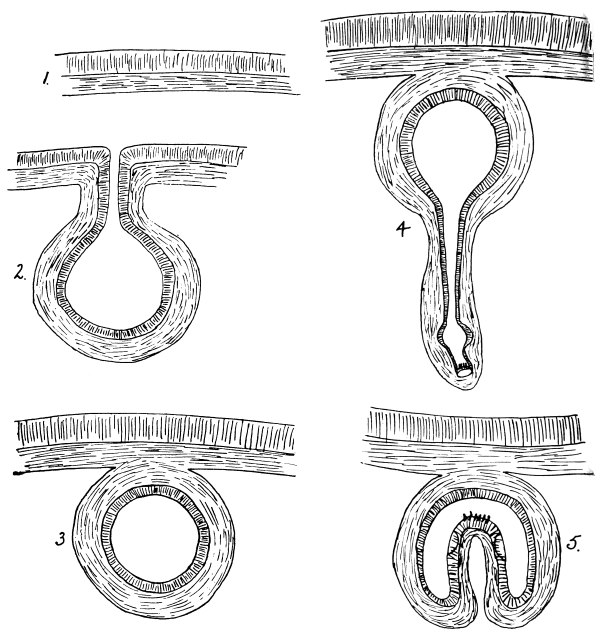
Fig. 252.
Figs. 252 and 252A.—Diagrams of mode of formation of brood capsule and scolices.
(1) Wall of mother cyst, consisting of ectocyst and endocyst; (2) theoretical stage of invagination
of wall; (3) a brood capsule with the layers of the wall in the reverse position to that in the
mother cyst; (4) evagination of wall; (5) invagination; (6) fusion to form the solid scolex;
(7) invagination of fore-part of scolex into hind-part. (Note.—The size of the scolex is much
out of proportion to the brood capsule.) (Stephens.)
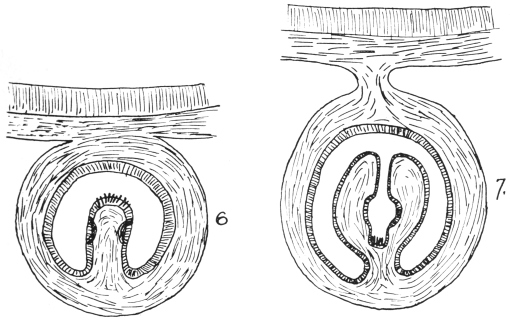
The development in cattle often remains stationary at the bladder
stage, and they are then called “acephalocysts,” or Echinococcus cysticus
sterilis. According to Lichtenheld, sterile cysts occur in 80 per cent.
of cases in cattle, in 20 per cent. in pigs, and in 7·5 per cent. in sheep.
In other cases large numbers of small, hollow BROOD CAPSULES are
formed in the germ layer, but are not arranged in any particular order.
The order of the layers is just the reverse in them to what it is in the
parent cyst, that is to say, they have inside a thin non-laminated cuticle
and the parenchymatous layer on their external surface. These,
theoretically at least, may be regarded as invaginations of the bladder
wall giving rise to a cavity with the cuticle internal and the
parenchymatous layer external. If we suppose the orifice to close,
we should then get an isolated
cavity with cuticle internal and
parenchymatous layer external, as
in the brood capsule (fig. 252).
If we next suppose an evagination of the wall of the brood capsule to
occur at one point we should get a hollow process lined with cuticle;
at the bottom of this we get the scolex and hooklets formed, and a
little higher up the tube the suckers (fig. 252, 4). If this hollow scolex
is now pictured as being invaginated we get a hollow scolex covered
with cuticle and lined by a parenchymatous layer projecting into the
cavity of the brood capsule. The two sides of this hollow scolex now
fuse and we get a solid scolex projecting into the cavity. Finally, if
we imagine once more the rostellum and suckers invaginated into the
posterior part of the scolex we get the condition as frequently found
in the brood capsules, i.e., a scolex covered with cuticle projecting into
the cavity, with the rostellum and suckers invaginated into the posterior
portion of the scolex (fig. 252A, 7).
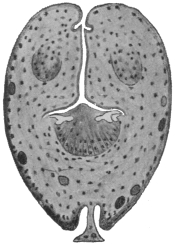
Fig. 253.—Section through
an invaginated echinococcus
scolex. Cf. fig. 252A, 7. × 300.
(After Dévé.)
A large hydatid may contain many thousands of brood capsules.
Each brood capsule is about as big as a pin’s head, and may contain
ten to thirty or more scolices. The delicate wall of the brood capsules
may rupture, so that the scolices are now free in the mother cyst. These
free scolices and also free brood capsules constitute what is known as
“hydatid sand,” which settles at the bottom of a glass when hydatid
fluid is poured into it. This form occurs chiefly in domesticated
animals and is termed E. veterinorum, Rud., or E. cysticus fertilis.
In man, and only rarely in cattle, the mother cyst first forms
“daughter cysts” (E. hominis, Rud. [fig. 255]), which, though smaller
than the “mother cyst,” resemble
it in the structure of
their walls; thus they are
covered externally by a laminated
cuticle and internally
by the parenchymatous layer.
They originate:
(1) Between the laminæ of the cuticle of the mother cyst from
small, detached portions of the parenchymatous layer; during their
growth they bulge inwardly or outwardly and may separate themselves
entirely from their parent cyst. In the latter case they lie
between the mother cyst and the capsule of connective tissue
formed by the host (E. granulosus or E. hydatidosus exogenus);
when growing inwardly they reach the interior of the mother cyst
(E. hydatidosus endogenus). Their number is very variable and does
not depend on the size of the mother cyst. They are as big as, or
bigger than, gooseberries.
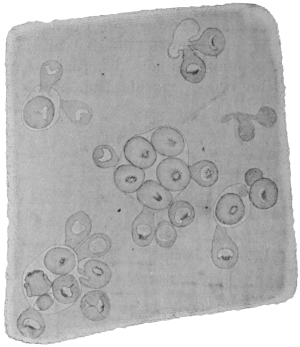
Fig. 254.—A piece of the wall of an Echinococcus
veterinorum stretched out and seen from the internal
surface. A few brood capsules (the outline of which
is only faintly shown), with scolices directed towards
their interior and exterior. 50/1.
(2) According to some authors, endogenous daughter cysts arise also
from a metamorphosis of scolices that have separated off from the brood
capsule. This takes place in the following way: Fluid accumulates
in the interior of the scolex, so that eventually nothing remains except
a sac consisting of cuticle lined by parenchyma. The cuticle gradually
thickens and several layers form (fig. 257).
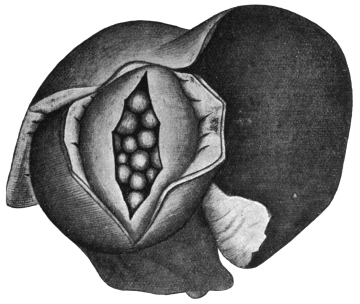
Fig. 255.—Echinococcus hominis in the liver. The fibrous capsule and
the wall of the echinococcus have been incised, so that the endogenous
daughter cysts may be seen. Reduced. (After Ostertag, from Thoma.)
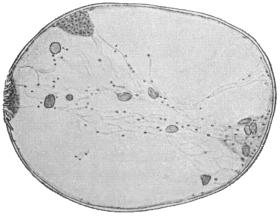
Fig. 256.—Section through an echinococcus
scolex in process of vesicular metamorphosis,
twenty-six days after insertion in the pleural
cavity. × 250. (After Dévé.)
(3) Transformation of Brood Capsules into Daughter Cysts.—This is
also held to be possible by various observers. New epithelial layers are
deposited between the cuticle
which lines the brood capsule
and the outer parenchymatous
layer. This parenchymatous
layer gradually disappears and a
new parenchymatous layer forms
in the interior from the parenchyma
of the scolex or scolices.
Although it appears strange that
a completely formed scolex with
specifically differentiated tissues
and organs should retrogress
to more primitively organized
matter, and again become a
proliferating bladder, yet we can
hardly doubt that the older observations, regarding such a vesicular
metamorphosis, of Bremser (1819), v. Siebold (1837), Naunyn (1862),
Rasmusser (1866), Leuckart (1881), Alexinsky (1898), Riemann (1899),
Dévé (1901), and Perroncito (1902) are correct.
(4) Further, a fourth method of formation of daughter cysts is
described by Naunyn as occurring in sterile hydatids, i.e., those containing
no brood capsules. In this case a portion of the mother wall
of the hydatid gets constricted off.
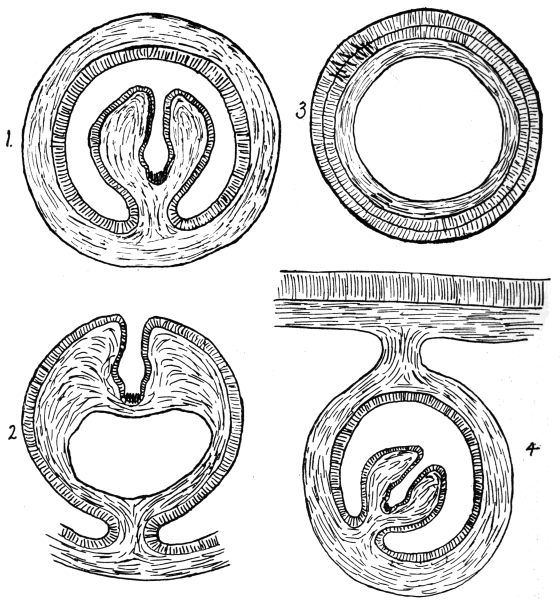
Fig. 257.
Figs. 257 and 257a.—Diagram of transformation of a scolex into a daughter cyst (1 to 3):
1, scolex in brood capsule; 2, liquefaction of scolex; 3, daughter cyst; and (4 to 6) of a brood
capsule into a daughter cyst; 4, brood capsule with scolex; 5, deposition of new epithelial
layers on the inner layer of the parenchyma; 6, disappearance of outer parenchyma and formation
of inner parenchyma from the parenchyma of scolex, which has now disappeared. (Note.—The
scolices are out of proportion to the brood capsules and to the daughter cysts. Stephens.)
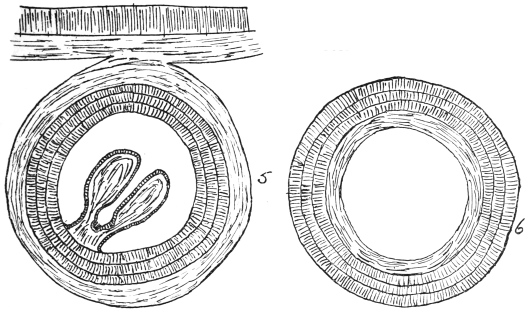
It has also been established that not only daughter cysts transplanted
into animals develop further (Lebedeff, Andrejew, Stadnitzky, Alexinsky,
Riemann), but that this also holds good if only hydatid scolices from
man or animals are transplanted into animals (rabbits). They develop
into echinococci and can then give rise to brood capsules and scolices.
As Dévé further established, hydatid scolices are not capable of
developing in guinea-pigs, while corresponding experiments with
rabbits are in the large majority of cases successful where the scolices
are introduced subcutaneously or into the pleural or peritoneal cavities.
It is only in the case of daughter cysts that further growth is obtained
in the case of guinea-pigs. Finally it appears, as has been already
stated, that brood capsules can transform themselves into daughter
cysts, but according to Dévé only within the mother cyst, not after
transplantation. Daughter cysts that have been formed in the mother
cyst of man and animals behave themselves just as the mother cyst
does, i.e., they can remain sterile,
or give rise to brood capsules
and scolices, or even again
to fresh cysts—granddaughter
cysts. The mother cyst can also die, so that the daughter cysts then
lie in the cavity of the connective tissue capsule. The number of the
daughter cysts in either case may attain several thousands.
The echinococcus fluid, which originally is formed from the blood of the host, is
light yellow, with a neutral or slightly acid reaction; its specific gravity averages 1009
to 1015. It contains about 1·5 per cent. of inorganic salts, half of which is common
salt; in addition (besides water) it contains sugar, inosite, leucine, tyrosin, succinic
acid (associated with lime or soda) and albumens which are not coagulated by heat;
occasionally also the fluid has been found to contain hæmatoidin and uric acid salts
(in echinococcus of the kidneys), which doubtless demonstrates that the echinococcus
liquid originates from the host. It has been generally assumed that echinococcus
fluid contains a toxic substance the escape of which into the body cavity (at operation
or by bursting of a hydatid cyst) produces more or less severe symptoms (fever,
peritonitis, urticaria), so much so that one speaks of hydatid intoxication. The
investigations of Kobert, Joest, etc., have, however, shown the harmlessness of fresh
undecomposed hydatid and cysticercus fluid for rabbits, mice and guinea-pigs, whether
inoculated intraperitoneally, subcutaneously or intravenously. Contrary data or
clinical experience must accordingly depend on other factors.
According to the researches of Leuckart, the growth of the
echinococcus is very slow; four weeks after infection the average
size is only 0·25 to 0·35 mm., at the age of eight weeks it is 1 to
2·5 mm., and at this period the formation of the central cavity commences;
at the age of five months, and with a size of 15 to 20 mm.,
the first brood capsules with scolices are formed. The consequence
of this gradual increase of size is that the organ attacked
can maintain its functions by vicarious hypertrophy, and that many
echinococci induce no special symptoms and cannot even be
diagnosed, the latter circumstance being due to their hidden position.
The echinococcus cannot be said to be scarce in man, as is
shown by the following table for Central Europe:—
Place | Period | No. of
post-mortems | No. of cases
of echino. | Percentage |
| Rostock | 1861–83 | 1,026 | 25 | 2·43 |
| Greifswald | 1862–93 | | | 1·48 |
| Jena | 1866–87 | 4,998 | 42 | 0·84 |
| Breslau | 1866–76 | 5,128 | 39 | 0·761 |
| Berlin | 1859–68 | 4,770 | 33 | 0·69 |
| Würzburg | — | 2,280 | 11 | 0·48 |
| Göttingen | — | 639 | 3 | 0·469 |
| Dresden | 1852–62 | 1,939 | 7 | 0·36 |
| Münich | 1854–87 | 14,183 | 35 | 0·25 |
| Vienna | 1860 | 1,229 | 3 | 0·24 |
| Prague | — | 1,287 | 3 | 0·23 |
| Kiel | 1872–87 | 3,581 | 7 | 0·19 |
| Zürich, Basle, Berne | — | 7,982 | 11 | 0·13 |
| Erlangen | 1862–73 | 1,755 | 2 | 0·11 |
These, however, are only cases that have become known by post-mortem;
in addition, there are cases that have been treated medically,
of which there are a few statements, at all events relating to the
principal districts of Germany. According to Madelung, one case
of echinococcus occurs in every 1,056 inhabitants in the town of
Rostock, in the district of Rostock one to every 1,283, in Schwerin
one to every 5,887, and in Ludwigsort one to every 23,685; according
to Peiper, in Upper Pomerania one case occurs to every 3,336,
in the district of Greifswald one to every 1,535 inhabitants. The
northern districts of Pomerania are more affected than the southern
ones.
Accordingly, echinococcus is also considerably more frequent in cattle in
Pomerania. On an average in Germany 10·39 per cent. oxen, 9·83 per cent. sheep,
and 6·47 per cent. pigs are infected, whereas in Upper Pomerania 37·73 per cent.
oxen, 27·1 per cent. sheep, and 12·8 per cent. pigs are infected; in Greifswald, indeed,
64·58 per cent. oxen, 51·02 per cent. sheep, but only 4·93 per cent. pigs are infected.
In accordance with these figures Tænia echinococcus must be frequent in dogs in
Pomerania, especially in Upper Pomerania; on the other hand, the conjecture that
the frequency of echinococcus in Mecklenburg is explained by the occurrence of
Tænia echinococcus in foxes has not been confirmed, as the fox does not harbour this
worm in Mecklenburg.
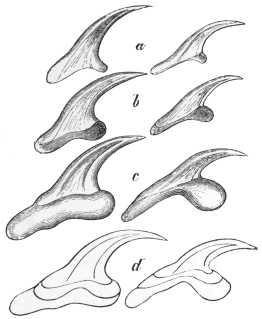
Fig. 258.—Hooklets of echinococcus. a, of
Echinococcus veterinorum; b, of Tænia echinococcus,
three weeks after infection; c, of the
adult Tænia echinococcus; d, the three forms
of hooklets outlined one within the other.
600/1. (After Leuckart.)
Beyond the European continent, echinococcus is frequent in
the inhabitants of Iceland, Argentine, Paraguay and Australia. In
Iceland, according to Finsen, 1 in every 43 inhabitants is affected
with echinococcus; according to Jonassen the proportion is 1 to 63;
this is due to the habits of the people of Iceland or, in fact,
to the frequency of Tænia echinococcus
in dogs, and the prevalence
of the hydatid in cattle. In certain
districts of Australia it is
just as frequent. In Cape Colony,
Egypt and Algeria echinococcus
is not rare, but it is scarce in
America and in Asia, with the
exception of the nomadic tribes
of Lake Baikal.
Echinococcus attacks persons
of every age, though it is rare in
children up to 10 years of age
and in old people. It occurs
most frequently between the ages
of 21 and 40 years. According
to all statistics it preponderates
in women (about two-thirds of the
cases). The liver is its favourite
seat (57·1 per cent. of the cases);
next in order come the lungs
(8 per cent.), kidneys (6 per cent.),
cranial cavity, genitalia, organs of circulation, spleen (3·8 per cent.), etc.
As a rule one organ only is invaded; multiple occurrence may originate
from one infection, or eventually from a later infection (?), or it may
come to pass that from some cause (through the spontaneous rupture of
an echinococcus, or the rupture of one caused by an injury or surgical
operation) daughter cysts, brood capsules or scolices escape into the
abdominal cavity,294 where they settle or become transformed and go
on growing. In the distribution of this secondary echinococcus the
great powers of motility of the free scolices must be taken into
account (Sabrazès, Muratet, and Husnot).
Human echinococci may also die at various stages of development, become
caseous or calcified, or may be absorbed, the cause for this being either disease of
the hydatid itself or inflammation of its connective tissue capsule; the discovery of
the laminated cuticle, which has great powers of resistance, or the finding of the
hooklets of the scolices is sufficient to form a conclusion as to the nature of such
formations.
Siebold (1853) was the first to rear Tænia echinococcus in the dog
by feeding it with the echinococcus of cattle and especially of sheep.
Küchenmeister, van Beneden, Leuckart, Railliet and others obtained
similar results, and Thomas, Naunyn, Krabbe and Finsen succeeded
in rearing T. echinococcus in dogs from the bladder worms of human
beings; these grow comparatively slowly (one to three months295) and
only during the process of growth develop their hooklets in their
definite form (fig. 258). It lies in the nature of things that dogs,
whether experimentally or naturally infected, almost always harbour
T. echinococcus in large quantities. That cats exceptionally harbour
these worms has been already mentioned (Dévé). Finally, Leuckart
infected young pigs by feeding them with mature segments.
Echinococcus multilocularis (alveolar colloid).
In addition to the form of echinococcus already described, and
which is also frequently termed Echinococcus unilocularis, there is a
second form which occurs in man as well as in animals, and which
is termed E. multilocularis, s. alveolaris (alveolar colloid).
It was originally regarded as a tumour; its animal nature was first
established by Zeller and R. Virchow. The parasite, which varies
in size from that of a fist to a child’s head, presents a collection of
numerous cysts, measuring between 0·1 and 3 to 4 mm. to 5 mm.
in diameter, which are embedded at first in a soft, connective tissue
stroma; the cut surface has therefore a honeycomb appearance. The
cysts are surrounded by a pellucid and laminated cuticle, and each
according to its size encloses either a small-celled tissue or a cavity
lined by a parenchymatous layer; the fluid contained in such a cavity
may be transparent, or is rendered opaque by globules of fat, bile-pigment,
hæmatoidin and fat crystals. According to some authors
all or most of these cysts intercommunicate; others state that this is
the case at least as regards the cuticle. The scolices are by no means
found in all the cysts, and when present only a few, rarely half, of the
cysts contain scolices (one or more); it is supposed that at least some
of these scolices are formed in brood capsules, and that the former
are capable of undergoing a cystic metamorphosis.
One circumstance is peculiar to the multilocular echinococcus
of man, namely, the disintegration that sets in at certain stages; in
the centre of the parasite a cavity forms that frequently becomes very
large and is filled with a purulent or brownish or brownish-green
viscid fluid; in this fluid one finds shreds of the wall of the cavity,
calcareous bodies, echinococcus cysts, also scolices and hooklets, as
well as fat globules and crystals of hæmatoidin, margarine and
cholesterin and concretions of lime. Such ulcerative processes,
according to Ostertag, are never present in the multilocular echinococcus
of oxen,296 in which the separate cysts are larger and the
connective tissue integument less powerfully developed.
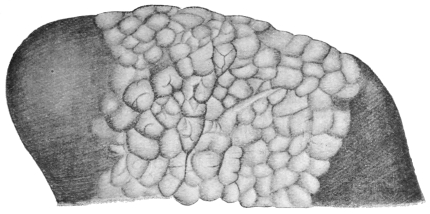
Fig. 259.—Echinococcus multilocularis in the liver of the ox. Natural size.
(After Ostertag.)
Hardly anything positive is known with regard to the development
of the alveolar echinococcus; its peculiar conformation is attributed
by some to enormous infection of oncospheres, by others to the
abnormal situation of one oncosphere; a few authors ascribe it to
infection of lymphatic vessels, others to infection of the biliary ducts
or to peculiarities of the surrounding hepatic tissue; Leuckart ascribes
it to a grape-like variety of form which continues budding; a few
more recent authors consider multilocular echinococcus to be specifically
different from unilocular echinococcus, and therefore also
different the species of Tænia arising from them. Melnikow-Raswedenkow
is also of this opinion. According to this author the
oncospheres infect the lumen of a branch of the portal vein in
Glisson’s capsule of the liver and grow into an irregularly shaped
formation (chitinous coil), which breaks through the vascular walls
and thus forms the alveoli. So far the data coincide well with
Leuckart’s opinion of the original grape-like form of the Echinococcus
multilocularis; according to Melnikow-Raswedenkow the “granular
protoplasmic substance” (parenchymatous layer) is not only present
in the interior of the loculi but also outside, and, moreover, “ovoid
embryos” are supposed to develop in the chitinous coils, which,
“thanks to their amœboid movements, reach the lumen of a vessel,
where, under favourable circumstances, they begin to develop
further,” that is to say, they become “chitinous cysts with fantastic
outlines,” or also “single-chambered chitinous cysts”; scolices may
develop in both. Dévé, however, considers that these embryos are
only prolongations of the protoplasmic layer which secondarily
cuticularize.
The multilocular echinococcus, which in man produces a severe disease and
almost always leads to premature death, infects most frequently the liver, but may
also be found primarily in the brain, the spleen and the suprarenal capsule; from
the liver by means of metastasis it may reach the most various organs, especially
those of the abdomen, but also the lungs, the heart, etc. Up to 1902, 235 cases have
been described and up to 1906, 265, being 70 from Russia, 56 from Bavaria, 32 from
Switzerland, 30 from the Austrian Alps, 25 from Würtemberg; the remaining cases
are distributed over Central Germany, Baden, Alsace, France, Upper Italy, North
America. In some the origin is doubtful; in any case after Russia, the mountainous
South of Europe is the principal region of distribution. As to the domesticated
animals, the same parasite is found principally in the ox (according to Meyer, in
Leipzig, in 7 per cent. of the oxen affected with echinococcus); it is rarer in the
sheep and very scarce in the pig.
It has already been mentioned above that recently the multilocular
echinococcus has been stated to be specifically different from hydatid
or unilocular echinococcus. To this may be added the fact that
Mangold, who fed a young pig with oncospheres of a Tænia reared
from the multilocular echinococcus, found two growths in the liver
four months later, which he took to be E. multilocularis, and consequently
one has to assume the existence of two different worms.
The chief defender of this view, already put forward by Vogler,
Mangold, and Müller, is Possett. He bases his opinions on (1) the
more restricted distribution of the multilocular hydatid, the former
occurring in districts where only cattle are raised, the latter where
sheep-breeding is established; (2) that those engaged in looking after
sheep are attacked by multilocular, whereas those looking after cattle
are attacked by unilocular hydatid; (3) that among the cases of
unilocular hydatid occurring in the distribution areas of multilocular
hydatid no transitions between the two forms are observed; (4) on
the difference in the hooks both in the hydatid as well as in the Tænia
stage; the hooks of Tænia echinococcus are plump, sharply curved,
and have a short posterior root process the length of which is to that
of the total length as 1 to 4·7, whereas on the contrary the hooks of the
alveolar echinococcus are more slender, slightly bent, and have a
long posterior root process (1 to 2·5); and (5) on the form of the
uterus, which in the alveolar Tænia has the form of a spherically
distended sac anteriorly.
Serum Diagnosis of Echinococcus.
(1) Precipitin Reaction.—Mix equal parts of hydatid fluid (of the
sheep) and serum of patient. Keep at 37° C. The reaction is not
decisive as it may be given by normal sera.
(2) Complement Deviation.—Required: (1) Hydatid fluid of sheep
(antigen), (2) guinea-pig complement, (3) patient’s serum, (4) red
cells of sheep, (5) hæmolytic serum (of rabbit) against sheep’s red
cells, (6) 0·8 per cent. salt solution. Mix the antigen + patient’s
serum (heated) + complement + salt solution at 37° C. for one hour.
Add red cells of sheep + hæmolytic serum. Allow to stand for
half an hour at 37° C. It is imperative to make adequate control
observations. An example will indicate the method. Salt solution
1·3 c.c. + patient’s serum (heated) 0·2 c.c. + hydatid fluid 0·4 c.c. +
complement 0·1 c.c. of serum diluted to a quarter strength + hæmolytic
serum and red cell emulsion 1 c.c. Result: no hæmolysis, i.e.,
the patient’s serum contains specific (echinococcus) antibodies.
C. NEMATHELMINTHES.
BY
J. W. W. STEPHENS, M.D., B.C., D.P.H.
Bilaterally symmetrical animals, without limbs and with a body cavity in
which the gut or other organs float. They are generally cylindrical.
Class. NEMATODA.
Nemathelminthes with an alimentary canal.
Nematodes are as a rule elongated round worms of a filiform or fusiform shape;
their length varies according to the species from about 1 mm. to 40 to 80 cm.
The outer surface of the body is smooth or annulated, and at certain points provided
with papillæ, occasionally also with bristles and alar appendages. The anterior end
carrying the oral aperture is usually rather slender, occasionally quite thin; the
posterior end is pointed or rounded; the anus, as a rule, lies somewhat in front of
the posterior extremity. The sexes are almost always separate, and the male can
as a rule be easily distinguished from the female because the former is smaller and
more slender, its posterior extremity is often spiral or incurved, or carries an alar
appendage, whereas the female is larger and thicker, and its posterior extremity
is straight. In the male the genitalia open into the anus; the sexual orifice of the
female opens ventrally along the median line in the anterior half of the body, in the
middle, or a little further back. Both sexes, moreover, have an orifice, the excretory
pore, which is situated ventrally in the median line and about the level of the
œsophageal nerve ring.
In large species, even with the naked eye, two lighter transparent bands—the
lateral lines—may be distinguished; they run along the sides of the body from the
anterior to the posterior end, while two other bands, the median lines, running
along the ventral and dorsal mid-lines, are less evident; in exceptional cases there
are also four sub-median lines. These bands or lines are inward projections of the
ectoderm, and in them lie the nerves and excretory vessels (fig. 260).
Some Nematodes live free in fresh or salt water, in soil, mud or decaying
vegetable matter, others parasitically in the most various organs of animals,
frequently also in plants.
Anatomy of the Nematodes.
All the Nematodes are covered by (1) a CUTICLE, which in the
small species is thin and delicate, while in the larger species it is
thickened, and may consist of several layers of complicated structure.
Canalicular pores do not occur. According to general opinion,
which is confirmed by the history of development, the cuticle is a
product of (2) the EPITHELIUM or ectoderm that had formerly existed
or is still found beneath it; in young specimens and small species it
is perceptible, but in older worms it frequently alters so considerably
that not only do the borders of the cells disappear,297 but a fine fibrous
differentiation appears in their cytoplasm. The matrix or ectoderm
then has the appearance of an ectodermal syncytium permeated by
fibres and strewn with nuclei, so that it is hardly distinguishable from
the tissue of (3) the CUTIS, which is always present, though developed
to a varying degree. Both layers, matrix and cutis, project internally
as ridges and form the lateral lines, while the less marked median
lines are produced apparently only by the ectoderm (fig. 260).

Fig. 260.—Diagram of a transverse section of Ascaris
lumbricoides, showing thick cuticle, and beneath it the
matrix or syncytial ectoderm. The flat intestine is in
the middle, and to the right and left near it in the body
wall the lateral lines with excretory vessel and lateral
nerves; above and below in the centre the dorsal or
ventral median lines with the nerves radiating to the
muscles, also the muscle cells with their striated outer
contractile portion and inner nucleated vesicular protoplasmic
portion. About 50/1. (After Brandes.)
Unicellular cutaneous glands are known in parasitic as well as in
free-living species; they vary in number and arrangement, and are
found discharging some at the anterior extremity and others in the
vicinity of the genital orifices. In other cases large numbers of them
are present along the lateral lines; they are strongly developed in
most of the Trichotrachelidæ, where they discharge either along a
part of the ventral surface or along the lateral and median lines; they
are placed so closely together that the ridges of the cuticle perforated
by the orifices have long been known, and have been described, as
“rodlet borders,” or “fields of rods.”
As the cutis is immediately adjacent to (4) the DERMO-MUSCULAR
TUBE the simple layer of the muscular cells is divided into four quadrants
by the longitudinal lines—two dorsal and two ventral (fig. 260).
The MUSCLES are in the simplest cases large rhomboid cells that lie
two by two in each quadrant, so that on transverse section of the
entire worm only eight cells are perceptible. The outer border of
the cells is converted into contractile fibrils, while the contiguous
inner portion has remained protoplasmic, and contains the nucleus.
In large species the muscular cells do not only increase in length
(up to 3 mm.) and in number in every quadrant, but their contractile
portion curves up to form a groove (like that of a dead leaf) thereby
even becoming thicker; simultaneously space is gained for more
cells, the protoplasmic parts of these cells (on transverse section) project
out of the grooves like vesicles. In all cases there is only one
layer of longitudinal muscular cells, which, by contracting, can shorten
the body or, by contracting one side, can bend it. In the latter case
the muscles of the opposite side have an antagonistic effect, or when
all the muscles are contracted, the elasticity of the cuticle acts in the
same way. Special muscles exist at the beginning of the gut and at
sections of the genital apparatus.
The existence of a cavity between the body and the gut wall has
hitherto been generally assumed, and has been referred to the cleavage
cavity, and consequently designated as a primary body cavity. More
recent investigators, however, state that such a cavity does not exist,
but that the space between the longitudinal muscles or their protoplasmic
portions and the gut epithelium is filled by a complicated
“isolation tissue.” This in the main proceeds from a large cell (Is.,
fig. 262) which lies directly behind the nerve ring dorsal to the
œsophagus, and consists of a system of lamellæ which sheathe the
muscles and penetrate through them to the cutis and also cover the
gut in a thin layer.

Fig. 261.—Anterior
end of an Ascaris megalocephala
cut open and
showing the four tuft-like
organs lying on the
lateral lines. Natural size.
(After Nassonow.)
We may now consider the “tuft-like” or “phagocytic” organs,
which attain 1 cm. in size, and consist of four, six, or even more
ramified cells, which lie close to the walls of the body (fig. 261).
They are found either only in the anterior part of the body (Ascaris),
or throughout the whole length of the body (Strongylus, syn., Sclerostomum),
and their position usually corresponds to the lateral lines.
In some species there are small protoplasmic cells on the processes
of these organs. In consequence of their size they can be recognized
with the naked eye, especially when they are loaded with granules of
stain (carmine, Indian ink) injected into the body cavity.
Intestinal canal.—The oral aperture, which is situated at the
tip of the anterior extremity, is frequently surrounded by thick lips,
or small bristles, or papillæ; it leads to a more or less strongly
developed buccal cavity, which is lined by a continuation of the body
cuticle, and which in some species is provided with “teeth,” representing
differentiated portions of the cuticle.
The œsophagus (fig. 262), which arises from the base of the oral
cavity, is as a rule a short, bottle-shaped tube with triradiate lumen;
its wall is chiefly composed of radiating muscular fibres, which give
it the appearance of being transversely striped when viewed from the
surface. There exist also in its wall three large gland cells (œsophageal
glands) and nerves arising from the lateral lines and running
forward. The radial fibres cause a dilatation of the lumen, and
exercise an effect antagonistic to the elasticity of the cuticle lining
the inner surface. The latter has its own particular layer, which is
not in direct connection with that of the oral cavity. Special dilator
muscles, arising from the dermo-muscular tube and situated at the
commencement of the œsophagus, are only known in a few species.
The posterior end of the œsophagus presents a bulb-like dilatation,
and is frequently provided with small chitinous movable valves. In
a few forms, which belong to the Trichotrachelidæ (Trichocephalus,
Trichinella), the œsophagus is a very long cuticular tube, beset on its
dorsal surface with a series of large nucleated cells. In others (Cucullanus,
Ascaris, etc.), a tube, the so-called glandular stomach, lined
only by epithelial cells, follows behind the muscular œsophagus.
This glandular stomach is, from its structure, easily distinguished
from the mid-gut, or chyle intestine, which is likewise cellular. The
so-called mid-gut is a tube lined by flat, cubical, or cylindrical cells
(fig. 260) surrounded by “isolation tissue”; its transverse section is
circular or flattened dorso-ventrally; the lumen may run in a straight
line, or it runs a sinuous course through the alternating prominences
of the then flat epithelial cells.
The ectodermal hind gut is, as a rule, very short. At the anal
opening the cuticle and the subcuticular layers are reflected inwards,
forming the lining of the hind gut. In large species the subcuticular
tissue forms large cells on which anteriorly lie in addition large
“glandular cells.”298 In the male the ejaculatory duct opens at this
point. Around the end part of the gut, either on the chyle intestine
or at the beginning of the end gut, there exists a sphincter muscle
arising from a muscle cell which acts antagonistically to the two
diaphragm-like dilator muscle cells which stretch from the gut to the
body wall. In many species large stretches of the gut are provided
with dilator muscles. There is sometimes a retrogressive absorption
of the gut in the adult stage of a few parasitic species.
Intestinal cæca and œsophageal glands sometimes exist as
intestinal appendages;
the former are tubular
appendages of various
size, running backwards
or forwards, and arising
from the posterior
extremity of the œsophagus.
They are lacking
in many species.
The œsophageal glands
are unicellular; a dorsal
and two subventral
glands may be distinguished
according to
their position; as a rule
they open into the
œsophagus at a distance
from one another. The
body of the gland lies
in the bulb of the œsophagus,
or in the dorsal
cul-de-sac arising
from it.
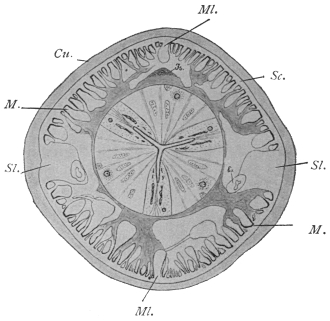
Fig. 262.—Transverse section through Ascaris lumbricoides
at the level of the œsophagus behind the nerve ring. Cu.,
cuticle; Sc., subcuticular layer; Ex., excretory vessel; Is.,
isolation cell and the system of lamellæ proceeding from it;
M., muscles; Ml., median line; Sl., lateral line. Magnified.
(After Goldschmidt.)
The nervous system is sufficiently known in a few species only;
it consists of a ring containing fifty to sixty fibres closely surrounding
the œsophagus, various groups of ganglion cells, and a certain
number of nerves extending anteriorly as well as posteriorly. The
remarkably small number of fibres, as well as ganglion cells, is
characteristic of the nervous system of all Nematodes. Immediately
behind the œsophageal ring (fig. 263, Lg.) an agglomeration of ganglion
cells lies at either side (lateral ganglia); part of their off-shoots
form the œsophageal ring, and part are directed posteriorly and
ventrally, and unite partly in front of and partly at the back of the
excretory pore, with fibres originating direct from the œsophageal
ring, and passing along the ventral median line to the back; these
fibres then together form the ventral median nerve (fig. 263, V.m.n.).
This nerve, originally consisting of thirty to fifty fibres, becomes
in the female attenuated quite evenly in its further course. There is
also an agglomeration of ganglion cells close in front of the anus (anal
ganglia), and then the median nerve divides in order to combine with
the lateral nerves on either side. In the male the median nerve
enlarges to nearly the original number of fibres in front of the anal
ganglion, which contains seven
cells; there is also an anal ring
embracing the terminal gut, and
there are two ganglion cells in it on
each side. In the dorsal median
line the dorsal median nerve is alike
in both sexes; arising in front with
a single root from the œsophageal
ring, it gathers its fibres from the
lateral ganglia; in the anterior part
of the body it consists of thirteen
to twenty fibres; in the posterior
part of the body the fibres are reduced
to four or six; behind the
anus it divides and combines with
the lateral nerves; the latter
consists of two fascicles at either
side right up to their most posterior
extent—one dorsal and one
ventral—which in the greater part
of the body do not run in, but
beside the lateral lines, and exhibit
a different origin anteriorly. The
ventral fascicle at each side branches
off from the ventral median nerve
in front of the excretory pore,
whereas the dorsal fascicles originate
from the œsophageal ring close
to the lateral ganglia. Each of the
four fascicles contains only two or
three fibres, which run backwards
parallel to the lateral lines; a few
centimetres in front of the caudal
extremity they enter the lateral
lines and remain separate from one
another up to the level of the anal
ganglion; here they amalgamate on
either side, after each interpolating
one ganglion cell, with the single
short lateral nerve which first takes
up the forked ends of the ventral and then of the dorsal median nerve;
finally, both lateral nerves unite with each other at the back in an
arch-like manner.
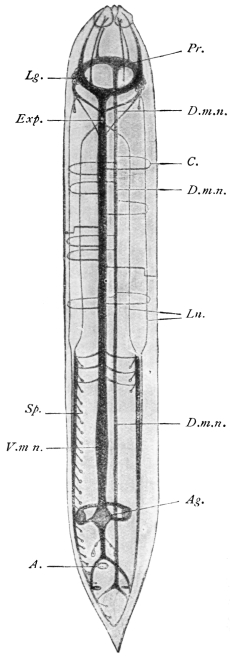
Fig. 263.—Schematic representation of
the nervous system of a male Ascaris megalocephala.
A., anus; Ag., anal ganglion;
C., commissures; D.m.n., dorsal median
nerve; Exp., excretory pore; Pr., œsophageal
sensory ring; Lg., lateral ganglia;
Ln., lateral nerve; Sp., papilla; V.m.n.,
ventral median nerve. (After Brandes.)
In the male each ventral part of the lateral nerves becomes
thickened by taking up fibres from the ventral nerves, which become
thickened posteriorly to the nervus bursalis, which towards the
middle gives off a mass of fibres to the “genital papillæ” situated
in front of and behind the anus; the number of these fibres averages
eighty to 100; in its further course the bursal nerve resembles the
corresponding ventral part of the lateral nerves of the female.
The ventral and dorsal nerves are connected by a number of
semicircular commissures, which originate from the ventral nerves
and serve to supply the dorsal nerve, which is always being
decreased by fibres departing from it. It is remarkable that these
commissures are not placed symmetrically, and their position also
is different in the two sexes; in the female there are thirty-one on
the right side and only thirteen on the left side. In the male there
are thirty-three commissures on the right side and fourteen on the
left, which run into the subcuticular layer, generally in pairs, and
usually cross at the level of the lateral lines.
The fibres of the two median nerves are chiefly motor; fascicular
processes run from each protoplasmic part of the muscular cells to
the median nerves; from these they take up bundles of primitive
fibrils, which separate, pass through the protoplasmic part and enter
the contractile part (fig. 260). One part of the fibrils, however,
penetrates beyond the muscles into the subcuticular layer, where
they form a network, probably of a sensory nature, with contiguous
fibrils. Nerves directed anteriorly finally originate from the
œsophageal ring; they consist each of three fibres, carry three
ganglion cells at their point of origin, and enter the sensory organs
of the three papillæ surrounding the oral aperture. Two of these
little trunks lie in the lateral lines, the remaining four are situated
in the middle of the four quadrants (Nn. sub-mediani anteriores).
Parasitic species lack higher ORGANS OF SENSE; free-living worms
occasionally have two rust-red eyes, sometimes with lenses, at the
anterior part of the body. In addition to the above-mentioned
sensory papillæ surrounding the oral aperture and the genital papillæ
of the male at the end of the body, another pair exist in the vicinity
of the lateral ganglia, the “cervical papillæ,” and two dorsal papillæ
in the central region of the body and two lateral ones near the tip of
the tail (Ascaridæ). The differences in the distribution and number
of the sensory papillæ serve for characterizing the larger and smaller
groups of Nematodes.
The excretory organs of the Nematodes are variable. In a
great many cases the apparatus is symmetrical, and consists of a vessel
commencing in the posterior extremity in each lateral line (fig. 260),
and passing anteriorly. In the vicinity of the anterior extremity both
tubes pass out of the lateral lines, bend ventrally, and, in the median
ventral line, unite into a short vesicle formed by an ectodermal cell—the
cavity of which is lined by a continuation of the cuticle of the
body—which opens into the excretory pore (fig. 263, Exp.). Asymmetry
is occasioned through the excretory duct proceeding from the
ventral pore to the lateral line, and it here proceeds as (or takes up)
the left excretory canal, which anteriorly is a broader tube and runs
along the left lateral line; shortly before its union with the excretory
duct it throws out a branch to the right towards the lateral line,
which, however, always remains weak, and runs posteriorly in the
right lateral line; a few smaller branches in addition spring from
the left main stem. In other species the right branch is completely
suppressed; the entire organ thus lies in the left lateral line, and
consists of the excretory duct, which occasionally opens quite in front
near the lips, as well as the excretory canal, which throws out a
number of lateral branches.
This excretory vesicle is a single elongated or horse-shoe-shaped
cell, with a large nucleus and an intracellular tubular system, which is
connected with the excretory duct arising from the excretory pore on
the outer surface (fig. 326). The so-called ventral gland is the only excretory
organ of marine Nematodes, and probably represents a primitive
form. Goldschmidt, who has investigated the excretory apparatus
of Ascaris lumbricoides, considers that the vessels running in the lateral
lines are only ducts to which belong a glandular system hitherto
overlooked or otherwise interpreted. This system also lies in the
lateral lines, and takes the form of two glandular tracts, forming
a syncytial tissue in which lie the ducts, one dorsal, one ventral.
In parts these tracts are connected by commissures, although their
junction with the excretory vessels cannot be clearly made out. These
statements, however, require confirmation. The author has further
found that the anterior ends of the lateral canals, directly before they
bend ventrally, anastomose with one another and give off anteriorly
a small blind process, which can be interpreted as a rudiment
of a canal coming from the head end, and as a matter of fact,
according to Golowin, such anterior excretory canals exist in a
number of genera.
In a number of Nematodes (Cheiracanthus, Capillaria, Trichocephalus,
Trichinella, etc.), however, special excretory organs are
lacking; possibly the cutaneous glands, which are in these species
generally powerfully developed, replace these organs.
Sexual organs.—With the exception of a few species, the
Nematodes are sexually differentiated.
(a) Female Sexual Organs.—The sexual orifice (vulva), surrounded
by thick labia, is, as a rule, ventral and varies in position from near the
head to near the anus. It leads into a short or long vagina (ectodermic),
bifurcating into the two uteri, which may be long or short; the long
filiform ovaries are continuations of them (fig. 264). Further there is
often, e.g., in Ankylostoma, a differentiation into the following parts:
(1) Ovejector: the specialized portion of the uterus before it joins the
vagina; there may be a separate one for each uterus, or a common
one for both uteri. (2) Seminal receptacle: at the other extremity of
the uterus. (3) Oviduct: a narrow tube connecting the ovary with
the uterus proper. (For the explanation of the terms convergent and
divergent uteri vide footnote p. 432.) Uterus and ovaries, which arise
in the first place from a single cell, lie between the body wall and the
gut and are surrounded by connective tissue. In some species (for
instance, Trichinella) the ovary is single.
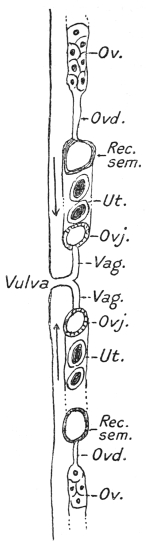
Fig. 264.—Diagram of
female genitalia. Ov., ovary (in
part); Ovd., oviduct; Rec. sem.,
seminal receptacle; Ut., uterus
(in part); Ovj., ovejector; Vag.,
vagina.
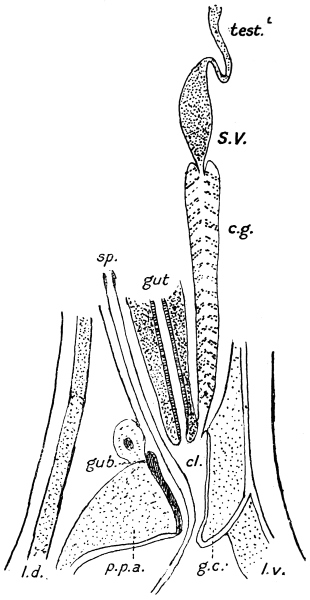
Fig. 264a.—Diagram of male genitalia of a strongylid.
Test., testis (in part); S.V., seminal vesicle; c.g.,
cement gland surrounding ejaculatory duct; sp., spicules;
cl., cloaca; gub., gubernaculum; p.p.a., pulvillus
post-analis; g.c., genital cone; l.d., dorsal lateral
line; l.v., ventral lateral line (the bursa is not shown).
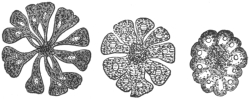
Fig. 265.—Transverse section through the
ovarian tube of Belascaris cati of the cat
at various levels. To demonstrate the
development (right to left) of the ova and
of the rhachis. Magnified.
At the blind end of the ovary there is a mass of protoplasm with numerous nuclei
that multiply continuously. Gradually the nuclei arrange themselves in longitudinal
rows (fig. 265) and the protoplasm commences to leave the periphery and surround
each nucleus. The nearer to the uterus the more progressive is this loosening
process, until club-shaped cells each containing a nucleus are developed. The most
pointed end of each, however, is still attached to an axial fibre of protoplasm, the
rhachis; probably this has some connection with the nutrition of the ova. Finally
the ova fall off and reach the uterus, where they are fertilized and enclosed in shells.
(b) Male Sexual Organs.—There is never more than one testis
(fig. 266), which is a straight or sinuous tube of the same construction
as an ovary, and in which the mother cells originate in the same
manner as the ova. In the same way as the ovary passes into the
uterus, so does the testis pass into the spermatic duct; the latter is
often divided into the somewhat dilated seminal vesicle and into
the muscular ductus ejaculatorius, which, running ventral to the
intestine backward (fig. 267), finally opens into the cloaca. In
many species, e.g., A. duodenale, the ejaculatory duct is surrounded for
a greater or less portion of its extent by the cement gland, the secretion
of which (brownish or blackish in colour) serves for copulation. The
ejaculatory duct of the large Ascaridæ is for the most of its course
surrounded by a muscular network which takes its origin from the
two dilator cells of the gut (fig. 268
F.). The spermatozoa of the
Nematodes, it may be noted, only
attain their full development after
the sperm mother cells have been
conveyed by copulation into the
uteri of the female genitalia. In
their form (sheathless, capable of
amœboid motion) they differ from
those of most other animals.
Spicules.—The male genital apparatus is also provided with one
or two sacs, situated on the dorsal side of the intestine, and opening
into the cloaca. In each sac there is a chitinous rod-like body, the
spicule. Further, in many cases there exists, more or less fixed in the
dorsal wall of the cloaca, a chitinous structure, the accessory piece or
gubernaculum, the latter name implying its function of guiding the
spicules during copulation (fig. 264A). A special muscular apparatus,
consisting of protractors and retractors, moves the spicules. The
protractors or exsertors in the large Ascaridæ consist of four flat
band-like muscles which surround the spicule sac. Two long muscle
cells which arise proportionally far forward on the dorsal side of the
lateral line and are inserted into the base of the spicules serve as
retractors. The spicules can be projected
from the cloacal orifice (anus) during copulation,
and when they are introduced into
the vagina they serve as prehensile organs,
perhaps also as stimulatory organs.
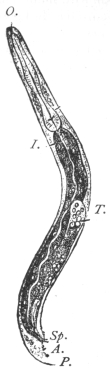
Fig. 266.—Male of the
rhabditic form of Angiostomum
nigrovenosum.
A., anus; I., mid-gut;
T., testicular tube; O.,
oral orifice; P., papillæ;
Sp., spicule. Magnified.

Fig. 267.—Transverse section through the
posterior extremity of the body of Ascaris
lumbricoides (male). The intestine is in the
middle, and the lateral lines are subjoined
thereto; above the intestine the two spicule
sacs are seen; below is the ductus ejaculatorius.
The muscular fibres are between the lateral
and median lines. Magnified.
Bursa copulatrix.—The males in many genera possess epidermal
wing-like appendages at their posterior extremity. These are supported
by elongated tactile papillæ called ribs. In the most highly developed
bursæ, e.g., in the Strongylidæ, the ribs are called rays, as they consist
not only of nerve fibres but mainly of “pulp,” i.e., prolongations of
the subcuticular layer. Bursæ are either open, i.e., bilaterally
symmetrical, or closed, when the posterior border is continuous all
round. A pseudo-bursa is one unsupported by ribs or rays, e.g., in
Trichuris. The bursa serves as an organ of prehension during
copulation. Some forms, moreover, carry a sucker at the posterior
extremity (e.g., Heterakis); in others the spicules and other prehensile
organs are absent; they are then replaced by an evertible
cloaca, e.g., Trichinella.
Development of the Nematodes.
After impregnation, the ovum develops around itself a delicate
membrane (vitelline membrane), and subsequently an egg-shell is
formed. This is derived either as a secretion from the uterine wall
or it is a further differentiation of the vitelline membrane, the original
single membrane splitting into two, the outer becoming the egg-shell.
Further the uterus often secretes a special albuminous covering
around the egg-shell. The “yolk” granules of the ovum are secretions
of the protoplasm of the ovum itself and first appear when the
rhachis is formed. In certain cases ova lie in follicles or capsules
formed of epithelium cells derived from the ovarian tubes. These
cells subsequently fuse and form a membrane—the CHORION.
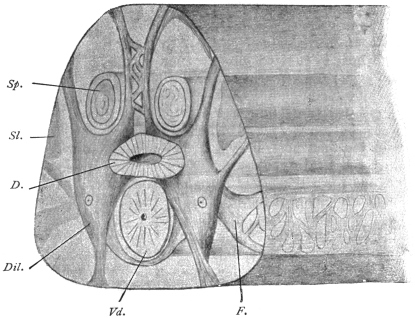
Fig. 268.—Hind end of a male Ascaris lumbricoides cut across at the level of the dilator
cells of the gut. D., gut; Dil., dilator cells of the gut; F., a process of the dilator cells
forming a network over the vas deferens; Sl., lateral line; Sp., spicule; Vd., vas deferens.
The anterior end of the worm lies to the right. Magnified. (After Goldschmidt.)
The shape of the completed eggs is characteristic of the different
species, and therefore a single egg often suffices to diagnose the
species. According to the species, the eggs may be deposited sooner
or later, either before or during segmentation, or with the embryo
perfectly developed. Only a few species are viviparous, e.g., Dracunculus
mediensis, Trichinella spiralis; in the other Nematodes the
further development of the extruded eggs takes place after various
lengths of time in the open, in moist earth, or in water. Thick-shelled
eggs can maintain their developmental capacity for a long time, even
after prolonged desiccation.
Finally, a nematode-like embryo develops, which usually lies
somewhat coiled up within the shell, and varies in its further
development according to the species to which it belongs.
In the simplest forms, as in the free-living Nematodes, the
embryos, apart from their size, resemble their parents, and grow up
into these after leaving the egg-shell. In many parasitical Nematodes,
however, the young must be called larvæ, as they present
characters which are subsequently lost.
The manner of conveyance of the eggs or the embryos contained
in them after they have left the body into the definite host is very
different in the various species.
(1) Without Intermediate Host.—(a) In many the conveyance into
the definite host is effected directly after the larvæ have developed
within the eggs; thus, for instance, the feeding of suitable animals
with the embryo-containing eggs of species of Trichocephalus and
Ascaris leads to an infection of the gut, for the young Trichocephali
or Ascarides only leave the egg-shell when they have attained the
intestine of the final host, in which they become adult.
In other cases (b) Ancylostoma, Necator, the larvæ hatch in the
open, and live for a time free, changing their form; they grow, cast
their skin, and finally gain the intestine of the host by means of water
or through the skin, when they lose their larval characters and
assume the structure of the adult worm.
(c) In a number of Nematodes, however, HETEROGONY occurs.
This terms signifies a mode of development in which two structurally
different sexual generations of the same species alternate with each
other. To these appertains, for instance, Angiostomum (syn.:
Rhabdonema) nigrovenosum, which lives in the lungs of frogs and
toads; this Nematode measures about 1 cm. in length and is
hermaphrodite (protandric). The eggs are deposited in the pulmonary
cavity, and through the cilia of the same reach the oral cavity, where
they are swallowed and thus conveyed into the intestine. They pass
through the entire gut, and are finally evacuated with the fæces; often,
indeed, the young themselves emerge from the egg-shell within the
hind-gut of the frogs. These young forms become sexually differentiated,
remain much smaller than the parent, their œsophagus is
differently constructed (rhabditis form), and they are non-parasitic
(fig. 266). After having grown in the open they copulate; the males
die soon after copulation, and the females in their own bodies develop
a few young, which, given the opportunity to get into frogs, infect
them, and are transformed into the hermaphroditic Angiostomum.
The same manner of development occurs in other species of the same
genus, and also in the case of Strongyloides stercoralis.

Fig. 269.—A piece of the trunk muscle of the
pig with encapsuled embryonic Trichinæ. Magnified.
(2) With Intermediate Host.—(a)
Frequently, however, the
larvæ of Nematodes make use
of one or even two intermediate
hosts; their condition then
resembles that of Cestodes or
Trematodes, excepting that
there is never a multiplication
within the intermediate hosts.
The larvæ become encapsuled
amongst the tissues of the intermediate
host, and wait till
they are introduced with the
latter into the final host. For
instance, Ollulanus tricuspis, the
adult form of which is found in
cats, previously lives encysted
in the muscular system of mice.
Cucullanus elegans, which attains
the adult stage in fishes
(perch, etc.), is found encysted
in species of Cyclops. Other
examples of species that require an intermediate host are Filaria
bancrofti and Dracunculus medinensis.
Peculiar conditions prevail in the case of (b) Trichinella spiralis.
This species, which in its adult state lives in the intestine of man and
of various mammals, is viviparous; the young Trichinæ, however, do
not leave the intestine, but reach the intestinal wall (Cerfontaine,
Askanazy) in the following way: the female intestinal Trichinæ bore
into the intestinal wall, where they are found in the submucosa, or
in the lumen of the dilated lacteal vessels. Here the young are born,
in the intestinal wall, and leave this position with the lymph stream.
Some of them, no doubt, actively bore through the intestinal wall,
reaching the lymph or blood-stream, or even pass into the body
cavity. What occurs during their further migrations is difficult to
say at present. It has hitherto been maintained that the wandering
is entirely active; for instance, the ligaturing of an artery would be
no protection against the part of the body supplied by such artery
being invaded by Trichinella. This observation cannot be otherwise
explained than by the active progress of the young Trichinella. The
question, however, may be mooted as to where and when the worms
quit the blood-vessels, which they for the most part reach through
the thoracic duct, the natural connection between the vascular system
and the lymphatic system, to wander further independently, and
ultimately reach the muscular system, in which they become encysted
(fig. 269). Thus the progeny does not leave the body of the host
inhabited by the parents, as is generally the case amongst helminthes,
but uses it as an intermediate carrier to reach another host, which
is then the final host. The latter may belong to another species,
or may be another individual of the same species. This second
migration is, of course, purely passive.
Classification of the Nematoda.
The Nematodes are usually divided into a number of families, some of which
it is at present impossible to define accurately; moreover, the definition of many
genera is also in an unsatisfactory state.
Family. Anguillulidæ, Gervais and van Beneden, 1859.
A “family” name not definable. They comprise a vast number of small forms,
most of which live free in fresh water, in soil, or in macerating substances; amongst
them there are some which live parasitically on plants, more rarely on animals.
They do not exceed 8 mm. in length. The large majority are only 1 to 2 mm., or
even 0·5 mm. The uterus is straight. Eggs in the uterus at one time, one to
four. Genera very numerous, but many of them insufficiently defined (Anguillula,
Anguillulina, Rhabditis, Heterodera, etc.).
Family. Angiostomidæ, Braun, 1895.
Small Nematodes undefined morphologically, but characterized by heterogony,
i.e., there is a free-living “rhabditic” generation and a parasitic “filariform” generation
which succeed one another (e.g., Angiostomum, Strongyloides, Probstmayria).
Family. Gnathostomidæ.
Cuticle covered totally or partly with cuticular laminæ fringed posteriorly with
multiple points. Head subglobular, covered with simple spines. Two spicules.
Vulva behind middle of body, parasitic in vertebrates, especially mammals (e.g.,
Gnathostoma, Tanqua, Rictularia).
Family. Dracunculidæ, Leiper, 1912.
Males very small in proportion to females. Anus absent. Vulva absent (?).
Genera: Dracunculus, Icthyonema (in body cavity of eel and other fish).
Family. Filariidæ, Claus, 1885.
Long thread-like Nematodes. Anus present. Œsophagus without bulb.
Vulva usually in anterior half of body. Two ovaries. Generally ovoviviparous.
Development often requires an intermediate host. This family is at present
ill-defined, but has been already subdivided into several sub-families, Filariinæ,
Onchocercinæ, Arduenninæ.
Family. Trichinellidæ, Stiles and Crane, 1910.
Œsophagus consisting of a chain of single cells, the lumen of the œsophagus
passing through their centre. Ovary single. Vulva at junction of anterior and
posterior portions. Sub-families: (1) Trichurinæ, (2) Trichinellinæ.
Family. Dioctophymidæ.
Body anteriorly armed with spines or unarmed; mouth without lips, with
six, twelve, or eighteen papillæ in one or two circles; œsophagus very long
without a bulb; anus terminal in female; one ovary; vagina very long; spicule
in male very long; bursa cup-shaped without rays (Dioctophyme, Hystrichis,
Eustrongylides).
Family. Strongylidæ, Cobbold, 1864.
Bursa, supported by rays, always present. Oviparous.
Family. Physalopteridæ.
Mouth with two large lips. Bursa with supporting papillæ in form of a
lanceolate cuticular expansion, with genus Physaloptera.
Family. Ascaridæ, Cobbold, 1864.
Rather thick Nematodes. Mouth with three lips—one dorsal, two latero-ventral.
Sub-families: (1) Ascarinæ, (2) Heterakinæ, etc.
Family. Oxyuridæ.
Smallish forms, 4 to 45 mm., with cuticle thickened on each side for the
whole length of body in the form of a lateral flange or wing. Œsophagus long
with a well-marked bulb containing a valvular apparatus. Tail end of female
drawn out into a long point. Eggs asymmetrical. Males very small (about
2 mm.). One spicule. Genera: Oxyuris, Passalurus, Ozolaimus, Atractis, etc.
Mermithidæ, greatly elongated “Nematodes,” which, in the larval stage, are
parasitic in insects, but in their adult condition are free living. Cuticle with
diagonal striation. Without an open mouth or anus. Oral papillæ present.
Characteristic eggs with two processes, ending in a tuft of filaments. Larvæ
with a movable boring spine at the head end.
Gordiidæ.—Long, thread-like “Nematodes.” Mouth and anterior portion of
gut atrophied in adult. Oral papillæ absent.
The Nematodes Observed in Man.
| Family | Sub-family | Genus | Species |
| Anguillulidæ | — | Rhabditis | R. pellio. |
| | | R. niellyi. |
| | | Rhabditis sp. |
| | Anguillula | A. aceti. |
| | Anguillulina | A. putrefaciens. |
| Angiostomidæ | — | Strongyloides | St. stercoralis. |
| Gnathostomidæ | — | Gnathostoma | Gn. siamense. |
| | | Gn. spinigerum. |
| Dracunculidæ | — | Dracunculus | D. medinensis. |
| Filariidæ | Filariinæ | Filaria | F. bancrofti. |
| | | F. demarquayi. |
| | | F. taniguchi. |
| | | F. (?) conjunctivæ. |
Group. | Agamofilaria | — | Ag. georgiana. |
| | | Ag. palpebralis. |
| | | Ag. oculi humani. |
| | | Ag. labialis. |
| | | F. (?) romanorum-
orientalis. |
| | | F. (?) kilimaræ. |
| | | F. (?) sp. ? |
| | (Mikrofilaria) | Mf. powelli. |
| | | Mf. philippinensis. |
| | Setaria | S. equina. |
| | Loa | L. loa. |
| | Acanthocheilonema | Ac. perstans. |
| | Dirofilaria | Di. magalhãesi. |
| Onchocercinæ | Onchocerca | O. volvulus. |
| Trichinellidæ | Trichurinæ | Trichuris | T. trichiura. |
| Trichinellinæ | Trichinella | T. spiralis. |
| Dioctophymidæ | — | Dioctophyme | D. gigas. |
| Strongylidæ | Metastrongylinæ | Metastrongylus | M. apri. |
| Trichostrongylinæ | Trichostrongylus | T. instabilis. |
| | | T. probolurus. |
| | | T. vitrinus. |
| | Hæmonchus | H. contortus. |
| | Mecistocirrus
(Nematodirus) | M. fordi. |
| Ancylostominæ | | |
Group. | Œsophagostomeæ | Ternidens | T. deminutus. |
| | Œsophagostomum | Œ. brumpti. |
| | | Œ. stephanostomum
var. thomasi. |
| | | Œ. apiostomum. |
Group. | Ancylostomeæ | Ancylostoma | A. duodenale. |
| | | A. ceylanicum. |
| | | A. braziliense. |
Group. | Bunostomeæ | Necator | N. americanus. |
| | | N. exilidens. |
Group. | Syngameæ | Syngamus | S. kingi. |
| Physalopteridæ | — | Physaloptera | P. caucasica. |
| | | P. mordens. |
| Ascaridæ | Ascarinæ | Ascaris | A. lumbricoides. |
| | | A. sp. |
| | | A. texana. |
| | | A. maritima. |
| | Toxascaris | T. limbata. |
| | Belascaris | B. cati. |
| | | B. marginata. |
| | Lagocheilascaris | L. minor. |
| Oxyuridæ | — | Oxyuris | O. vermicularis. |
| Mermithidæ | — | Mermis | M. hominis oris. |
| | (Agamomermis) | Ag. restiformis. |
Family. Anguillulidæ.
Genus. Rhabditis, Dujardin, 1845.
Buccal cavity elongated, with lips. Its chitinous wall uniformly thick. Lateral
lines absent. Males with bursa.
Rhabditis pellio, Schneider, 1866.
Syn.: Pelodera pellio, Schn., 1866; Rhabditis genitalis, Scheiber, 1880;
Rhabditis pellio, Schn., 1866.
Males 0·8 to 1·05 mm. in length; females, 0·9 to 1·3 mm. in length.
The posterior extremity of the body of the male has a heart-shaped
bursa, and seven to ten ribs on each side; the bursa may, however,
be lacking. The spicules measure 0·027 to 0·033 mm. in length, but
are never quite alike. The posterior extremity of the female is long
and pointed; the vulva lies somewhat behind the middle of the body,
the ovary is single, the eggs are oval, 60 µ by 35 µ.
This species was found in Stuhlweissenburg by Scheiber in the acid urine
(containing albumin, pus and blood) of a woman suffering from pyelonephritis,
pneumonia and acute intestinal catarrh; the observer was able to convince himself
that the Nematodes which were found during the whole period of the illness lived
in the vagina, and were evacuated with the urine.
Oerley proved that this species had long been known; during
its larval stage (Anguillula mucronata, Grube, 1849) it lives in earthworms;
in its adult stage it lives in decomposing matter in the soil.
By introducing individuals of this species into the vagina of mice,
Oerley succeeded in obtaining infection and multiplication (facultative
parasitism). These Nematodes must in some such way have got into
the vagina of Scheiber’s patient.
Two other cases described by Baginsky and Peiper probably
belonged to the same or a nearly related species.
Rhabditis niellyi, Blanchard, 1885.
Syn.: Leptodera niellyi, Blanchard, 1885.
In 1882 Nielly had a cabin-boy, aged 14, under observation
in Brest. The lad had never left the neighbourhood of Brest,
and had suffered from itching papules on the skin for five or
six weeks; in the papules the observer found one or several rhabdites,
measuring 0·33 mm. in length by 0·30 mm. in breadth.
Their cuticle presented a delicate transverse striation; the intestine
was the only internal organ recognizable, and it opened somewhat
in front of the posterior extremity. Therefore, it must have belonged
to the rhabditis-like larva of a Nematode, the adult stage of which
is unknown.
The manner of infection was established almost certainly by a further observation
of Nielly’s: at the commencement of the illness small Nematodes were
found in the blood of the patient; later on, however, they disappeared, neither were
Nematodes found in the fæces, urine or sputum. Therefore it must be concluded
that the cabin-boy, who was in the habit of drinking water from brooks, had thus
ingested embryo-containing eggs of a Nematode; the young hatched out in the
intestine, perforated it, reached the blood and then settled in the skin; but, on the
other hand, the entry may have been direct through the skin.
In connection with the foregoing, reference should be made to a communication
by Whittles, insufficient from a zoological point of view. In a case of hypertrophic
gingivitis occurring in a female patient, aged 19, who had never left Birmingham,
he found Nematode larvæ in the periosteum of the upper jaw, which was excised
after extraction of the right premolar; the genital rudiment could be recognized in
them. Similar larvæ were found in the same patient in abscesses in various regions
of the skin, and in the case of her mother in the blood. The author considers that
the infection took place through a dog, and refers to the case of O’Neil (1875), who
found Filariæ in the skin (in the condition known as “craw-craw”), referred by
Manson to Filaria perstans. O’Neil’s case was quoted, and attributed to Filaria
sanguinis hominis. In conclusion, the author states that he has repeatedly found
Nematode larvæ in the blood of persons who suffered from pruritus; in his opinion
the parasite had been imported through the agency of troops returned from South
Africa. Glatzel found true Filaria larvæ in a pustule of a cutaneous eruption of the
trunk and extremities in a patient at Dar-es-Salam.
Skin diseases which are caused by young Nematodes are also observed in dogs
(Siedamgrotzky, Möller, J. G. Schneider, Künnemann), foxes (Leuckart), and horses
(Semmer). Zürn found young Nematodes (Anguillulidæ) also in pig’s flesh. In
Künnemann’s case it was shown that the adult Rhabdites lived in the straw upon
which the dog lay.
Rhabditis, sp.
In the fluid obtained by lavage from the stomach of a female patient, aged 16,
suffering from ozæna, O. Frese found during two consecutive months Rhabdites of
various ages, 0·275 to 0·64 mm. in length, the adults all with eggs; males were
not found; transmission into rabbit’s stomach failed, but they could be kept alive in
much diluted hydrochloric acid (2 : 1,000) for several weeks. Neither eggs nor larvæ
appeared in the fæces of the patient. The nature of the infection, which was perhaps
of unique occurrence, remained doubtful.
Genus. Anguillula, Ehrenberg, 1826.
Buccal cavity very small, without lips. Males without bursa, but with a series of
papillæ. Lateral lines absent.
Anguillula aceti, Müller, 1783.
Cuticle unstriped, body cylindrical, anterior end tapering but little,
posterior end long, pointed. Male up to 1·45 mm. long, 0·024 to
0·028 mm. wide; two pre-anal papillæ, one post-anal; spicules equal,
curved, 0·038 mm. long; gubernaculum present; testis extending in
front of mid-line of body. Female up to 2·4 mm. long, 0·040 to
0·072 mm. wide; anterior uterus reaching to near the œsophagus,
posterior to hind gut. Viviparous; embryos in both or only in one
uterine horn, 0·22 mm. long, 0·012 mm. broad.
The species is a frequent inhabitant of vinegar (prepared by older methods), and
was once observed for some time by Stiles and Frankland in the urine of a woman;
the urine had an acid reaction, and once had a distinct odour of vinegar. It was
assumed that the patient, who was hysterical and suffered from chronic nephritis,
employed vaginal douches with diluted vinegar, perhaps to deceive her physician or
to protect herself against conception. According to Ward, Billings and Miller are
said to have reported on two other cases. Ill-effects which might be connected with
the vinegar eel (Anguillula aceti) were not present.
Genus. Anguillulina, Gervais and Beneden, 1859.
Syn.: Tylenchus, Bastian, 1864.
Characterized by the possession in the buccal cavity of a spine knobbed posteriorly;
bursa present; uterus asymmetrical. Numerous species parasitic in
plants.
Anguillulina putrefaciens, Kühn, 1879.
Syn.: Tylenchus putrefaciens, Kühn; Trichina contorta, Botkin, 1883.
In 1883 Botkin (Pet. klin. Wochenschr., 1883) found a small Nematode, which was,
however, entirely mistaken, in the material vomited by a Russian; this was not a
species of Trichinella, but an Anguillulina living in onions which had already, in
1879, been described by Kühn as Tylenchus putrefaciens; the Nematodes got into
the stomach with the onions, causing nausea and vomiting.
Family. Angiostomidæ, Braun, 1895.
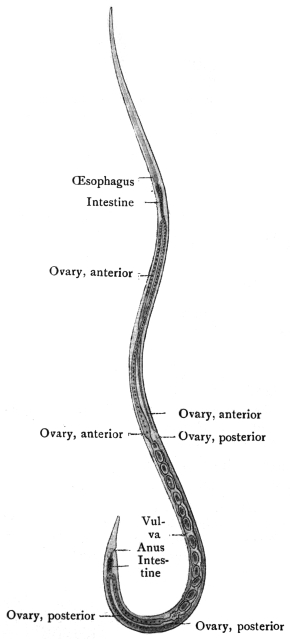
Fig. 270.—Strongyloides stercoralis, female: parasitic
generation from gut of man. × 70. (After Looss.)
Genus. Strongyloides, Grassi, 1879.
Syn.: Pseudorhabditis, Perroncito, 1881; Rhabdonema, Leuckart, 1882, p.p.
The genus is insufficiently defined. The parasitic form possesses a simple mouth
opening directly into the long cylindrical œsophagus which occupies the anterior
third of the body. The free-living forms possess a small buccal cavity; the
œsophagus is short, with a double bulb, in the hinder one there is a Y-shaped
chitinous valve; two spicules of equal size.
Strongyloides stercoralis, Bavay, 1877.
Syn.: Anguillula intestinalis et stercoralis, Bavay, 1877; Leptodera intestinalis
et stercoralis, Cobb.; Pseudorhabditis stercoralis, Perroncito, 1881; Rhabdonema
strongyloides, Leuckart, 1883; Strongyloides intestinalis, Grassi, 1883; Rhabdonema
intestinale, Blanchard, 1886.
In 1876, a number of French
soldiers returned to Toulon from
Cochin China suffering from
severe diarrhœa. Dr. Normand,
under whose treatment they
were, discovered a large number
of Nematodes in the evacuated
fæces, and Bavay described
them as Anguillula stercoralis. Soon after Normand, at the post-mortem of five
patients who had died of Cochin China diarrhœa, found numerous other
Nematodes in the intestine, from the stomach to the rectum, in the bile-ducts and
in the pancreas, and these he handed over to Bavay. The latter diagnosed another
species, and described them as A. intestinalis. Both forms were then regarded
as the cause of Cochin China diarrhœa until, in 1882, Leuckart was able to
demonstrate that the two forms are only two succeeding generations of the same
species, of which the one (A. intestinalis) lives parasitically in the intestine,
whereas its young (A. stercoralis) attain the open, where they come to maturity
and propagate. The young of these again live parasitically. There thus exists
the same heterogony as was discovered by Leuckart in Angiostomum nigrovenosum
of frogs, which heterogony, indeed, according to v. Linstow, appertains
to the entire family of the Angiostomidæ.
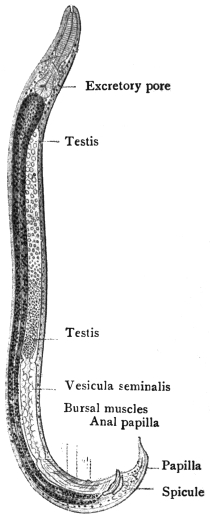
Fig. 271.—Strongyloides stercoralis,
male: free-living generation.
× 170. (After Looss.)
(1) The parasitic generation (strongyloid or filariform ♀) is quite
colourless and cannot be seen in situ even with a lens. To detect
them it is necessary to scrape the mucosa of the jejunum and examine
the scrapings microscopically. It measures 2·2 mm. in length, and
34 µ to 70 µ in breadth; the cuticle is finely transversely striated;
the mouth is surrounded by four lips; the œsophagus is almost cylindrical
and a third the length of the entire body. The anus opens
shortly in front of the pointed posterior extremity; the vulva is
situated at junction of middle and posterior thirds of the body; the
uterus has no special ovejector; the eggs measure 50 µ to 58 µ in
length, and 30 µ to 34 µ in breadth, and lie in a chain one behind
the other (fig. 270). As in the case of Angiostomum nigrovenosum,
Leuckart considers this stage to be hermaphroditic, the testes
degenerating after having functioned; other authors (Rovelli) regard
it as a female reproducing by parthenogenesis.
(2) The free-living generation (♂ and ♀) has a smooth body,
cylindrical, somewhat more slender at the anterior extremity and
pointed at the tail end. The mouth has four indistinct lips; the
œsophagus is short with a double (rhabditis-like) bulb; there is a
Y-shaped valve in the posterior bulb; the anus opens in front of the
tail end. The males measure 0·7 mm. in length, 0·035 mm. in breadth.
Their posterior end is rolled up; the two brown spicules are small
(38 µ) and much curved. There is also a gubernaculum. The
females measure 1 mm. in length or a little over; 0·05 mm. in breadth.
The tail end is straight and pointed; the vulva lies somewhat behind
the middle of the body. The yellowish, thin-shelled ova measure
70 µ in length and 45 µ in breadth.
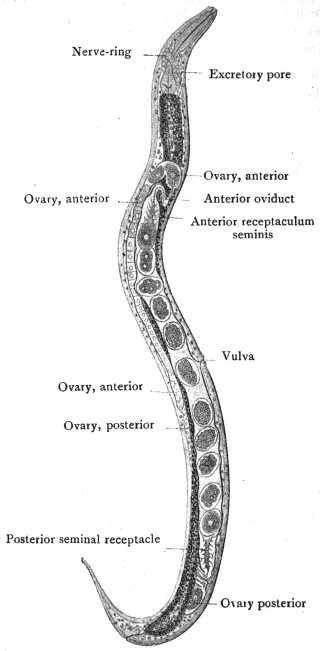
Fig. 272.—Strongyloides stercoralis, female; free-living
generation, × 170. (After Looss.)
As Askanazy has shown, the parasitic form bores deeply into the
mucous membrane of the intestine, and frequently into the epithelium
of Lieberkühn’s glands, both for nourishment and oviposition. The
eggs then develop in the intestinal wall. The eggs which are found in
scrapings from the mucosa occur, at least in the case of Strongyloides
of the sheep, in chains enclosed in a thin tube or sheath, the origin
of which is doubtful; possibly it is the uterus. The eggs themselves
are only rarely found in stools, e.g., after a strong purge. The
larvæ, which are hatched out, and measure 0·2 to 0·25 mm. long by
0·016 mm. broad, again reach the lumen of the intestine,299 and grow to
double or three times that size, until they are passed out with the
fæces. They already differ from the parent (♀) in the shape (rhabditiform)
of the œsophagus. When the external temperature is
sufficiently high (26° to
35° C.), they become
sexually mature after
moulting. In about
thirty hours they are completely
developed and
copulate, now forming
the free-living rhabditiform
generation. At
lower temperatures the
larvæ only moult, but
do not escape from the
old cuticle and do not
develop further. At a temperature of about 25° C. only some of the
larvæ attain maturity.
Fig. 273.—Strongyloides
stercoralis: larva from fresh
human fæces. × 310. (After
Looss.)
The females of the free-living
generation (rhabditiform) deposit from
thirty to forty eggs, which develop
rapidly, sometimes even within the
uterus in the case of old females.
After the larvæ have emerged from the
egg-shell, they measure 0·22 mm. in
length, and possess the characteristics
of the parents (rhabditiform larvæ).
When they have grown to 0·55 mm.
they moult, and while losing their
own characteristics they acquire the
characteristics of their parasitic grandparents
(strongyloid or filariform).
After about eight days the free-living
adult generation in the cultures have
disappeared, and all the rhabditiform
larvæ have been transformed into
strongyloid or filariform larvæ; they
then die off unless they reach the
intestine.
Fig. 274.—Strongyloides stercoralis:
mature filariform larva showing long
transparent œsophagus, slender granular
intestine and characteristic tip to
the tail ending in two small points.
× 620. (After Looss).
This cycle of development holds
good for Strongyloides stercoralis of
tropical origin (Bavay, Leuckart, Leichtenstern,
Zinn). In the European
Strongyloides the free-living generation,
as a rule, is absent (Grassi,
Sonsino, Leichtenstern, Braun); the
rhabditis-like larvæ evacuated with the
fæces are transformed into the strongyloid
or filariform type of larva (in
cultures which are easily made) which
will only become adult if introduced
into man.
So that we have these two cycles:
(A) (1) ♀ parasitic, (2) eggs, the rhabditiform
larvæ in fæces, (3) free-living
strongyloid or filariform larva,
(4) ♀ parasitic. (B) (1) (2) (3) as before,
then (4) adult ♀ and ♂, free living,
(5) eggs, (6) rhabditiform larva,
(7) strongyloid or filariform larva,
(8) ♀ parasitic.
Infection of man results not only
from direct entry into the stomach but also, according to van Durme
and Looss, through the skin.
Occurrence in Man.—As already mentioned, Strongyloides stercoralis was
first observed in persons suffering from so-called Cochin China diarrhœa.
From the enormous numbers of parasites evacuated with the fæces, the cause
of the disease was apparently evident. It appeared, however, that only some of
the soldiers returning from Cochin China and Martinique, and suffering from
diarrhœa, harboured Strongyloides (Chauvin). Breton made the same observations
in Cochin China and found that only 10·4 per cent. of cases of chronic dysentery,
and 8·8 per cent. of chronic diarrhœa, show Strongyloides. Normand, moreover,
found that only a few of the Europeans residing in Cochin China are exempt
from S. intestinalis, yet the people exhibit no intestinal symptoms; if, however,
from any cause a catarrhal condition of the intestine supervenes the condition
is changed, the parasites appear in larger numbers, and the disorder is
considerably intensified.
S. intestinalis, besides being present in the Indo-China region, also occurs
in the Antilles, in Brazil, Africa, and Europe; in 1878 it was discovered
in Italy by Grassi and C. and E. Parona; in 1880 it was also found in the
labourers working at the St. Gothard tunnel. It was imported into Germany,
Belgium, and the Netherlands by Italian labourers. One sporadic case has been
observed in East Prussia, and the worm has also been reported from Siberia.
In mammals the following species are found: Probstmayria (Strongyloides)
vivipara, Ransom, 1907, in Equus caballus; Strongyloides fülleborni, v. Linst., in
Anthropopithecus troglodytes and Cynocephalus babuin.
Their development is, so far as is known, the same as that of Strongyloides
stercoralis (v. Linstow, Centralbl. f. Bakt., Path. u. Infektionsk., 1905, Orig. xxxviii,
p. 532).
Family. Gnathostomidæ.
Genus. Gnathostoma, Owen, 1836.
Syn.: Cheiracanthus, Diesing, 1839.
Easily recognizable by the numerous spines which cover the entire body or
only the anterior extremity, and terminate in several points; head globular and
beset with bristles; mouth with two lips; two spicules; vulva situated behind the
middle of the body.
Gnathostoma siamense, Levinsen, 1889.
Syn.: Cheiracanthus siamense, Lev., 1889.
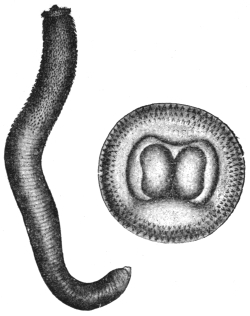
Fig. 275.—Gnathostoma siamense: to the
left, the entire worm (8/1); to the right the
head seen from above, with two fleshy lips
(about 40/1). (After Levinsen.)
Female measures 9 mm. in length, 1 mm. in breadth. There are
eight rows of simple spines on the head; the armature of spines extends
over the anterior third of the body only; each spine on the anterior
region of the body spreads into three points, of which the middle one
is the longest; the posterior spines are simple; they gradually become
smaller and then disappear entirely. The vulva is situated behind
the middle of the body.
Male.—10·5 mm. long by 0·6 mm. broad. Head terminates in a
globular swelling with two large lips. Neck 3 mm. broad. In front of
neck eight rows of simple spines directed backwards. Anterior half of
body with cuticular laminæ, posterior unarmed. Two pre-anal and
two post-anal papillæ. Bursa wanting.
Spicules 1·1 and 0·4 mm. respectively.
Leiper considers Gnathostoma siamense to be identical with
Gnathostoma spinigerum.
The single specimen described by Levinsen was found by Deuntzer in
Bangkok (Siam), and was obtained from a young Siamese woman who suffered from
a small tumour of the breast which had developed in the course of a few days.
After the disappearance of the tumour, nodules the size of beans were found in the
skin; out of one of these the worm was obtained. The same observer saw this
affection in two other persons.
A closely related species, Gnathostoma
spinigerum, Ow., lives in the stomach of
wild cat (Felis catus), puma (Felis concolor),
tiger (Felis tigris), and domestic cat
(India); another species, Gnathostoma
hispidum, Fedsch., 1839, in the stomach
of pigs in Turkestan, Annam, Hungary,
Congo, and by Collin in the stomach of an
ox (Berlin).
Gnathostoma sp. in pariah dogs,
Calcutta. Gnathostoma sp. in monkeys,
French Guiana. They produce large
fibrous thickenings in the stomach wall.
Gnathostoma spinigerum,
Owen, 1836.
Cuticle of bulb with eight rows
of chitinous laminæ with their
posterior edges notched into spines.
The laminæ on the anterior portion
of the body are similar trident
laminæ. In the middle of the
body, the laminæ are simple and conical, cuticle posteriorly is unarmed.
Mouth with two fleshy lips.
Male 5 mm. long by 0·5 mm. broad; tail spiral, four pairs of papillæ.
Female about twice as long; tail straight, trilobed.
Family. Dracunculidæ, Leiper, 1912.
Genus. Dracunculus, Kniphoff, 1759.
Anterior end rounded with a cuticular thickening or shield.
Mouth triangular with two lips. Alimentary canal atrophied.
Dracunculus medinensis, Velsch, 1674.
Syn.: Vena medinensis, Velsch, 1674; Dracunculus persarum, Kämpfer, 1694;
Gordius medinensis, Linné, 1758; Filaria dracunculus, Bremser, 1819; Filaria
æthiopica, Valenciennes, 1856; Dracunculus medinensis, Cobbold, 1864; Guinea
worm, Medina worm.
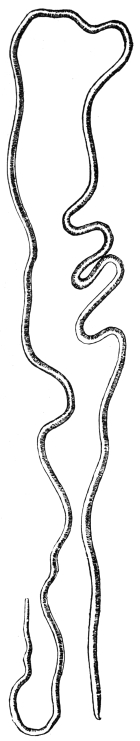
Fig. 276.—Guinea
worm (Dracunculus medinensis).
(After Leuckart.)
The females attain a length of 50 to 80 cm., or even more, and
average 1·5 to 1·7 mm. in diameter. They are whitish or yellowish
in colour. The anterior extremity is roundish and bears a cuticular
thickening or shield. The triangular mouth opening is surrounded
by two projections or lips, behind which on the shield there are two
lateral and four sub-median papillæ; the posterior end terminates in
a spine, ventrally directed, and about 1 mm. in length; the alimentary
canal below the œsophagus is atrophied, but not entirely obliterated;
anus absent; the lateral lines are very flat. The greater part of the
body is occupied by the long uterus, in which a great number of
young larvæ are always found. The ovaries probably lie at the ends
of the uterus; the vulva lies just behind the cephalic shield. During
parturition the uterus is prolapsed through this opening.
Fig. 277.—Anterior extremity
of Guinea worm,
showing dorsal and ventral
lips, one lateral and two submedian
papillæ and the lateral
line. (After Leuckart.)
The male is almost unknown. Leiper in an experimentally infected
monkey found two males 22 mm. long, one from the psoas
muscle, the other from the connective tissue behind the œsophagus.
Occurrence.—Filaria medinensis has been known since the most remote
period. The “fiery serpents” that molested the Israelites by the Red Sea, and
which Moses mentioned, were probably filariæ. The term Δρακὁντιον occurs in
Agatharchides (140 B.C.). Galen called the disorder dracontiasis; the Arabian
authors were well acquainted with the worm. It is found not only in Medina or
Arabia, but also in Persia, Turkestan, Hindustan. The Guinea worm is also widely
distributed in Africa, on the coasts as well as in the interior. It occurs in the Fiji
Islands. It was carried to South America by negro slaves, but is said at the present
time to exist in only quite a few places (British Guiana, Brazil [Bahia]); it is also
observed in mammals (ox, horse, dog, leopard, jackal [Canis lapuster], etc.).
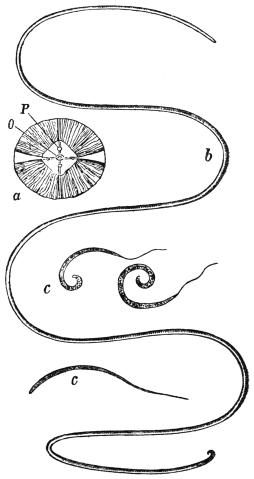
Fig. 278.—Dracunculus medinensis. a,
anterior extremity seen end on; O, mouth;
P, papillæ; b, female reduced more than
half; c, larvæ enlarged. (After Claus.)
Dracunculus medinensis in its adult stage lives in superficial ulcers
on the body surface; it is seen most frequently on the lower extremities,
more especially in the region of the ankle, but it also occurs in
other parts of the body—on the trunk, scrotum, perineum, on the
upper extremities, and in the eyelids and tongue. Sometimes there
is only one ulcer and one worm, but more commonly several. It
attacks man without distinction of race, age or sex. It is observed
most frequently during the months of June to August.
Life history.300—When about a year old the worm seeks the surface
of the body and produces there a thickening as big as a florin. Over
this a vesicle forms which eventually ruptures, and at the bottom
of the ulcer can be seen a hole from which a part of the worm may
project. On bathing the sides of the ulcer with water, a drop of
fluid, at first clear then milky, exudes. This contains numerous larvæ.
In other cases a thin tube an inch long is prolapsed (through the
vulva). This is probably the uterus, but the mechanism of parturition
is not clearly known. It lasts for about a fortnight. An abundant
supply of larvæ can be got by placing wet compresses on a fresh
ulcer. In a few hours a mass of larvæ is obtained.
The larvæ are 500 µ to 750 µ by 15 µ to 25 µ, with a long slender
tail about one-third of the total length. The cuticle is transversely
striated. The body is flattened. They possess an œsophagus and
gut. At the anus there are apparently glandular structures.
The larvæ live
and move actively in
water for about two
days, the majority
dying on the third
(Leiper). If a number
of Cyclops sp.
have been collected
and isolated in clean
water, and the larvæ
are now added, the
further development
can be traced.
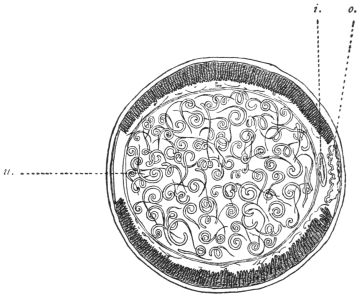
Fig. 279.—Transverse section of female Guinea worm;
u., uterus containing embryos; i., intestinal canal; o., ovary.
(After Leuckart.)
The larvæ enter
the Cyclops, according
to most authorities,
by penetrating
the exoskeleton, but
according to Leiper this is impossible; they must enter by the mouth
and penetrate the gut in order to reach the body cavity. In eight days
moult 1 takes place, the striated cuticle being cast off. In ten days
moult 2 takes place. In five weeks the larva is mature. If now the
infected Cyclops is placed in 0·2 per cent. HCl solution the Cyclops is
killed immediately, but the larvæ are stirred into activity, escape from
the body, and swim about in the acid. This suggests that infection
in nature probably takes place by the swallowing of infected Cyclops;
Leiper, by feeding Cyclops containing mature larvæ to a monkey, found
in it, post mortem six months later, two immature females 30 cm. long
and two males 22 mm. long.
In certain areas the new cases occur principally in June. Five
weeks later the larvæ will become mature in Cyclops, so that infection
of Cyclops is taking place in July or August, and from then to June
about ten months elapse, giving the period of development in man.
Pathology.—The initial induration is accompanied by itching.
Urticarial eruptions are described in Dahomey and Mauretania
accompanied by fever, rigors, blood-shot conjunctiva, and prostration
resembling fungus poisoning. Symptoms last for one to two days,
later the worms appear on the surface.
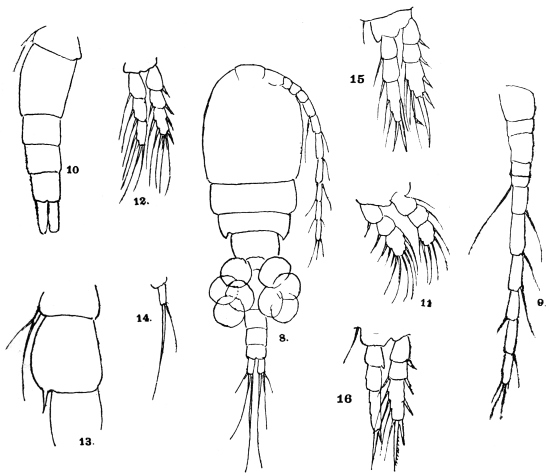
Fig. 280.—Cyclops virescens, ♀. 8, Female, ventral view, × 120; 9, anterior antennæ
× 240; 10, urosome and last thoracic segment, × 240; 11, foot of first pair, × 320;
12, 15, 16, foot of second, third and fourth pairs, × 240; 14, foot of fifth pair, × 440; 13,
last thoracic segment and first segment of urosome of male, × 240.
If the worm is ruptured in an attempt to extract it, disastrous
results may occur through the escape of the larvæ into the tissues:
fever, inflammation, abscess, sloughing, ankylosis, even death from
sepsis. Eosinophilia is often marked, 11 to 13 or even 50 per cent.
Extraction.—(1) The native method consists in rolling the worm
round a stick; 1 in. to 2 in. are extracted each day, the process
taking about a fortnight; (2) Emily used injections of 1 in 1,000
sublimate into the swelling or into the worm itself fixed by a ligature.
(3) Béclère chloroforms the worm; (4) the worm can be more
easily removed when all the embryos have been deposited (two to
three weeks).
Cyclopidæ.—Cephalothorax ovate, clearly separated from abdomen.
Anterior antennæ of female when bent back scarcely ever stretch
beyond the cephalothorax. The second antennæ are unbranched.
First four pairs of feet two-branched, outer branches three-jointed.
The fifth pair of limbs are rudimentary alike in both sexes, usually
one-jointed. There is no heart. The female has two egg sacs
containing about fifty eggs.
Genus. Cyclops, Müller, 1776.
Mandible palp rudimentary, reduced to a tubercle bearing two
branchial filaments. Maxillary palp rudimentary (obsolete). Lower
foot-jaw non-prehensile. Head ankylosed to first thoracic segment.
Family. Filariidæ.
Sub-family. Filariinæ.
The residue after exclusion of the Arduenninæ and Onchocercinæ.
Genus. Filaria, O. Fr. Müller, 1787.
Very long, slender Nematodes, without excretory vessels or
excretory pore, the males of which are usually considerably smaller
than the females. Mouth round, without lips, unarmed. The lateral
lines occupy one-sixth of the circumference of body. The tails of the
males are bent or spirally rolled, and bear little wing-like appendages.
The two spicules are unequal; almost always there are four pre-anal
papillæ, but the number of post-anal papillæ varies. The vulva is
always situated at the anterior extremity. Parasitic chiefly in the serous
cavities and in the subcutaneous connective tissue. Insufficiently
defined.
Filaria bancrofti, Cobbold, 1877.
Syn.: Trichina cystica, Salisbury,301 1868 (nec Filaria cystica, Rud., 1819);
Filaria sanguinis hominis, Lewis, 1872; Filaria sanguinis hominis ægyptiaca,
Sonsino, 1875; Filaria wüchereri, da Silva Lima; Filaria sanguinis hominum,
Hall, 1885; Filaria sanguinis hominis nocturna, Manson, 1891; Filaria nocturna,
Manson, 1891.
These parasites of man were for a long time only known in their larval
stage. They were discovered in 1863 in Paris by Demarquay, in the hydrocele
fluid of a Havanese emptied by puncture; they were next observed by Wücherer,
in Bahia, in the urine of twenty-eight cases of tropical chyluria; they were likewise
observed in North America by Salisbury, who gave them the name of Trichina
cystica. The next discoveries (in Calcutta, Guadeloupe, and Port Natal) related to
chyluria patients, until Lewis discovered the larvæ in the blood of man (India), and
found they were almost always present in persons suffering from chyluria, elephantiasis,
and lymphatic enlargements; he also, in exceptional cases, found them in
apparently healthy persons (Filaria sanguinis hominis). Lewis and Manson studied
the disease and the filariæ of the blood very minutely, and became aware that the
filariæ were sucked up by mosquitoes with the blood. Manson described the
metamorphoses that take place within the body of the mosquito. The adult
female was discovered in Queensland by Bancroft, and soon after Lewis found it
in Calcutta; it was described by Cobbold as F. bancrofti. The male was first
seen by Bourne in 1888.
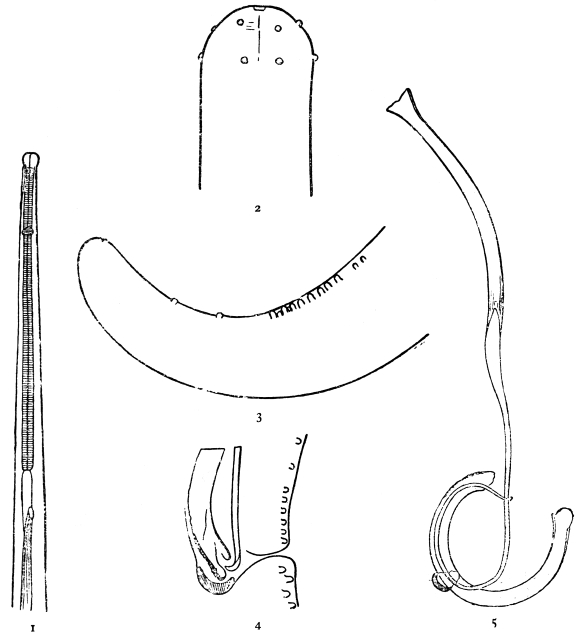
Fig. 281.—Filaria bancrofti. 1, Anterior portion of male; 2, two rows of papillæ on
head; 3, papillæ of tail of male; 4, cloaca of male showing tips of spicules and gubernaculum;
5, the spicules and gubernaculum of male. (After Leiper.)
Head bougie-like, i.e., separated by a narrowing from the neck,
having two rows of minute papillæ. Cuticle has extremely fine
striations.
Female.—50 to 65 mm. long by 1·5 to 2 mm. broad. Vulva
0·4 to 0·7 mm. behind the head. Anus about 1/4 mm. from the tip
of the tail (vulva 1 to 1·3 mm. from head, and anus 0·17 to 28 mm. from
tail according to other authors). The vagina is a muscular tube forming
three bold loops, and has terminally a pyriform enlargement.
Uterus double (or single). Ovoviviparous.
Male.—25 to 30 mm. long by 0·1 mm. thick (40 by 0·1 mm. according
to various authors). Probably two pairs of pre-anal papillæ,
eight pairs of peri-anal, two pairs of post-anal papillæ, and one pair
terminal. Tail curved. Two spicules, 0·2 and 0·6 mm. respectively,
and a cup-like gubernaculum. The long spicule is cylindrical,
expanded proximally and tapering distally to a filament with wings.
At the tip it is spoon-like. The short spicule is of the same diameter
throughout. It is gutter-like, coarsely marked. Testis uncoiled,
terminating in a snowdrop-like process (Leiper).
Eggs.—40 µ by 25 µ. They do not appear to possess a true shell,
but only an embryonal or vitelline membrane secreted by the ovum.
Embryos.—In the posterior part of the uterus eggs occur, in the
anterior part embryos; the larvæ at birth measure 127 µ to 200 µ by
8µ to 10 µ. In the blood they measure in the fresh 260 µ by 7·5 µ to 8 µ.
In stained films, owing to shrinkage, there is great variation in size, from
154 µ to 311 µ. Probably 260 µ to 285 µ is the average in stained
films.
Geographical Distribution.—Europe: Two cases recorded, one
from near Barcelona. The patient suffered from hæmato-chyluria
and enlarged scrotum with mikrofilariæ in the blood. A second
case from Siena. Africa: The filarial index has not been estimated
for various parts. In Nigeria it is about 10 per cent.
Habitat.—Lymphatic glands: e.g., inguinal, femoral, iliac, lumbar,
mesenteric, bronchial, superficial cervical, epitrochlear.
Lymphatic vessels: e.g., those draining into the receptaculum chyli
of the spermatic cord, in the thoracic duct and in various different parts.
Organs, etc.: Testis, epididymis, spermatic cord, tunica vaginalis,
mammary cyst, and in abscesses.
They may occur in masses, but usually only a few (one to eight).
Females are commoner than males. Dead and calcified worms are
common in the various sites.
Distribution of Larvæ in Body.—These are by no means uniformly
distributed, but occur in greater number in the capillaries of the
lungs. Besides the lungs they occur in the capillaries of other organs,
as the following data of Rodenwaldt show:—
| Mikrofilariæ | | Mikrofilariæ |
| Lungs | 134,821† | | Spleen | 1,666 |
| Liver | 4,884 | | Brain | 3,833 |
| Kidneys | 15,253 | | Glands | 0 |
| Glomeruli | 8,008 | | Marrow | 0 |
| Parenchyma | 7,245 | | Blood | 3,000 |
† These figures refer to 1 c.c. of each organ, and were estimated by cutting sections of
definite thickness (30 µ to 40 µ) and counting the filariæ in a definite area of section, e.g.,
1/4 cm.2 The organs before removal from the body have their vessels tied, and are then fixed in
hot alcohol.
The following data of Rodenwaldt refer to the larvæ of Filaria
immitis in the dog. They are commoner in organs than in vessels,
and especially in the capillaries of the organs, but in the lungs they
appear to be equally distributed in capillaries, arteries and veins.
The length of life of larvæ is unknown, but they appear to be
destroyed in the kidneys, as dead calcified specimens are fairly
numerous in the capillaries of the vasa recta of the medullary
substance.
Kidneys: mainly in the glomerular capillaries and those of the
vasa recta.
Liver: in the capillaries of the portal system, especially in those
between the interlobular and the central intralobular veins.
Periodicity of Larvæ.302—Roughly speaking, the larvæ of Filaria
bancrofti are found in the peripheral blood only during the night,
disappearing (but not entirely) during the daytime. Their periodicity
and that of Loa loa larvæ is shown by the table on p. 394, based on
that of Smith and Rivas (Amer. Journ. Trop. Dis. and Prev. Med.,
1914, vol. iii, p. 361).
It was discovered by Mackenzie that this periodicity could be
reversed by making the patient sleep during the daytime, showing
that the phenomenon was in some way dependent on sleep or its
attendant phenomena. Rodenwaldt gives the following explanation
of the phenomenon of periodicity:—
Mikrofilariæ come to rest in capillaries. After passing up the
thoracic duct they would reach the capillaries of the lungs by the
superior vena cava. Here they occur in immense numbers. In the
case of Loa loa larvæ (which have a diurnal periodicity) some of
these are forced out by the increased force and rapidity of the
pulmonary circulation during the day, but are able to rest (owing
to their sticky sheath?) in the peripheral capillaries on their way to
the capillaries of the organs. During the night the force of the
current through the lungs is relaxed and consequently they are able
to remain in the pulmonary capillaries and do not appear in the
capillaries of the systemic circulation. If it is true that the periodicity
of Loa loa cannot be reversed by changing the hours of sleep, then
the explanation is incomplete. In the case of the larvæ of Filaria
bancrofti (which have a nocturnal periodicity), in order to apply the
same explanation we must further assume that the mikrofilariæ have
less power of resisting the force of the capillary current (i.e., are less
sticky). They are washed out of the pulmonary capillaries by day
and by night, but it is only at night, when the blood stream in systemic
capillaries is less rapid, that they are able to rest there. In the daytime
they are washed on until they reach the capillaries of the organs
(possibly again the lungs). The reversal of the periodicity by sleeping
during the daytime admits of a similar explanation. If this explanation
be true, then a prolongation of the day conditions, e.g., by
continued exercise, should result in still keeping the larvæ out of the
circulation, but this does not appear to be the case.
| Larvæ of L.
loa in equal
quantities
of blood | Average 132.
Deviations
from average | Case 1.
F. bancrofti
larvæ in 1 c.c.
of blood | Average 1,000
(about).
Deviations
from average | Case 2.
F. bancrofti
larvæ in 1 c.c.
of blood | Average 1,570
(about).
Deviations
from average |
2 | a.m. | 9 | – 123 | 3,500 | + 2,500 | 6,500 | + 3,930 |
4 | a.m. | 11 | – 121 | 3,200 | + 2,200 | 5,200 | + 3,630 |
6 | a.m. | 41 | – 91 | 2,800 | + 1,800 | 2,000 | + 430 |
8 | a.m. | 168 | + 36 | 900 | – 100 | 1,100 | – 470 |
10 | a.m. | 298 | + 166 | 210 | – 790 | 350 | – 1,220 |
12 | noon | 531 | + 389 | 30 | – 970 | 50 | – 1,520 |
2 | p.m. | 252 | + 120 | 20 | – 980 | 40 | – 1,530 |
4 | p.m. | 146 | + 14 | 10 | – 990 | 30 | – 1,540 |
6 | p.m. | 91 | – 41 | 40 | – 960 | 40 | – 1,530 |
8 | p.m. | 23 | – 99 | 60 | – 940 | 100 | – 1,470 |
10 | p.m. | 5 | – 127 | 600 | – 400 | 800 | – 770 |
| 12 | midnight | 5 | – 127 | 750 | – 250 | 2,600 | + 1,030 |
Total | 1,580 | — | 12,120 | — | 18,810 | — |
In certain countries, e.g., Fiji, Samoa, Philippines, West Africa,
larvæ, apparently those of Filaria bancrofti, show no periodicity. In
Fiji the usual intermediate host is Stegomyia pseudoscutellaris, a day-biting
mosquito, so that possibly, as Bahr suggests, the mikrofilariæ
have partly adapted themselves to the habits of their intermediate
host, as the nocturnal mikrofilariæ are adapted for transmission by a
nocturnal feeding mosquito, e.g., Culex fatigans, but how this could
come about is a mystery. It is not certain in all cases whether
the non-periodic mikrofilariæ really belong to Filaria bancrofti;
some may be L. loa larvæ, or possibly unknown larvæ. An exact
morphological description of these larvæ is therefore always necessary.
Preservation of Living Larvæ.—Blood from the vein (or finger
puncture) is shaken up with twenty times its volume of sterile 0·9 per
cent. salt solution, and kept in an ice cupboard (Fülleborn).
Concentration of Larvæ.—(a) The above mixture is hæmolysed with
water and then sufficient salt solution added to make up to 0·9 per
cent. The solution is allowed to stand or can be centrifugalized.
(b) The blood is mixed with sodium citrate and centrifugalized; the
larvæ are found in the leucocytic layer (Bahr). (c) Allow blood to
clot in a small tube; the larvæ appear on the surface of the clot and
are so got in pure serum. A drop of blood may also be allowed to
clot on the slide; the larvæ are found in the clear areas of serum.
(d) Hæmolyse blood with water or acetic acid. Centrifugalize, make
smears from, or examine the sediment.
Removal of Red Corpuscles.—The blood film is allowed to stand for
some minutes in a moist atmosphere. The staining solution is sucked
through with blotting paper: the larvæ stick to the slide, while the
corpuscles are washed out.
Morphology of Larvæ.—Wet staining: Azur II one part, 0·9 per
cent., salt solution 3,000, or very dilute Giemsa or ripened methylene
blue or neutral red solutions. Place a drop on the slide and add a
drop of blood to this. The larvæ remain alive for one or more days;
it sometimes takes twenty-four hours to stain some particular structure.
Differentiation by drawing through weak eosin solution is often
useful. This method is the best for finest details. The excretory
pore, anal pore, excretory cell, and chief “genital” cell stain first,
then the matrix cells and finally the column of nuclei.
Wet fixation and staining: The blood is spread on a large cover-glass—floated
on the surface of 70 per cent. alcohol heated to about
70° C. Wash in water, (1) overstain with 1 in 1,000 azur II solution,
warming slightly; (2) differentiate with (a) absolute alcohol (containing,
if necessary, a trace of HCl), or (b) with absolute alcohol
96 per cent. ninety parts, anilin oil ten parts; (3) clear in origanum,
bergamot or cajeput oil; (4) mount in balsam. Or stain with hæmatoxylin,
e.g., Mayer’s glycerine alumhæmatein, heating till slightly
steaming. Differentiate with acid (2 per cent. HCl) alcohol if overstained.
Clear and mount as above.
Dry fixation and staining: (1) With azur II as above, or (2) with
hæmatein (warm). Examine the dried films in the usual way without
a cover-glass. The azur stains the excretory and genital cells clearly.
Thick films: (1) The blood is smeared out fairly thickly over an
area as big as a sixpence.
(2) Dry quickly to prevent shrinking, using carefully a spirit lamp
in a moist climate.
(3) Place films downwards in water for a few minutes.
(4) Fix in alcohol.
(5) Stain with azur II, 1 in 1,000. Differentiate as above. Examine
as a dry film. This method suffices for showing the excretory cell and
the G1 cell; or
(6) Stain with hæmatein (slightly steaming), especially for the
column of nuclei and the sheath. The fixation in alcohol in this
case may be omitted.
(7) The removal of the hæmoglobin and the fixation may be
combined by using Ruge’s mixture (formalin 2 per cent., containing
1 per cent. acetic acid) or acetic alcohol (glacial acetic 1, alcohol 3).303
Structure of Larvæ.—(1) Subcuticular cells: By vital staining, at
intervals underneath the cuticle are seen a series of spindle-shaped
cells—the subcuticular matrix cells of Rodenwaldt, the muscle cells
of Fülleborn. There are thirty or forty or more of these.
(2) Nerve ring: Appears as a break in the nuclear column about
20 per cent. of total length from the head.
(3) Excretory system: Consists of a lateral spherical hollow
excretory pore which shows a radial striation. Connected with the
pore is an excretory cell which appears to be canalized. Excretory
pore, 29·6 per cent. of length from head. Excretory cell, 30·6 per cent.
of length from head.
(4) “Genital” cells and anal pore: Consists of a pore opening
ventrally on a very fine papilla with which are connected four
other cells in series, the chief “genital” cell (G1) being some distance
from the three others, which lie close to the pore. G1, 70·6 per cent.,
anal pore, 82·4 per cent. of length from head.
(5) Internal body, viscus, or reserve material: Best shown by vital
staining with neutral red. This is a granular strand-like body extending
from 52·7 per cent. to 65 per cent. of length from head.
(6) Tail end: (i) Rod-like structures resembling those in the head,
90 per cent. of length. (ii) The column of nuclei extends to 95 per cent.
of length, so that the terminal portion is free from nuclei.
(7) Mouth: Terminal according to some authors, lateral according
to others. Some describe a fang on the head, others not. By vital
staining and eosin differentiation two rod-like structures with mushroom-like
caps can be seen behind the head.
(8) Cuticle: Transversely striated. There is a longitudinal break in
the striation on each side corresponding to the lateral lines. The striation
is best shown by vital staining with azur II and eosin differentiation.
(9) Column of nuclei: These nuclei of the gut cells form the
main feature in ordinary dry films stained with hæmatoxylin. They
are separated by a space from the subcuticular cells.
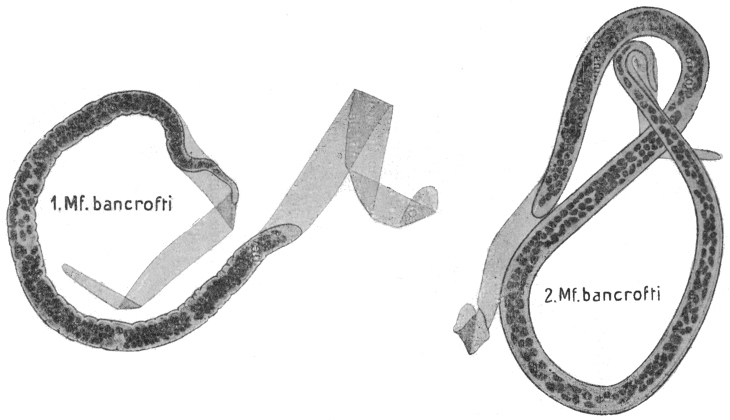
Fig. 282.—Mf. bancrofti in thick film, dried and stained with hæmatoxylin: 1, shrunken; 2, unshrunken.
× 1,000. (After Fülleborn)
Distinction between Mikrofilaria bancrofti AND Mikroloa loa.
| Dry Films, Hæmatoxylin Staining:— |
| Mf. bancrofti. | Ml. loa. |
| (1) In graceful curves (but only if quickly dried). | (1) Kinked. |
| (2) Tip of tail free from nuclei. | (2) Nuclei extend to tip. |
| (3) Column of nuclei separated by a space from the cuticle. | (3) Not so distinctly. |
| Azur Staining:— |
| (4) G1 cell small, easily overlooked. | (4) G1 cell large, stains deep blue, cell protoplasm = twice width of larva, easily seen. |
| (5) Excretory cell close to excretory pore, 2 per cent. of length. | (5) Excretory cell farther from pore, 4 per cent. of length. |
| Vital Staining with Neutral Red:— |
| (6) Internal body or reserve material clearly shown. | (6) Not shown. |
Life History.—In the stomach of the mosquito the larvæ cast
their sheath in the thickened blood in one to two hours. In twenty-four
hours the majority have reached the thoracic muscles, where
development proceeds. They are at first immobile and of a “sausage”
form (110µ by 13 µ), with a short spiky tail. In three to five days the
œsophagus is formed, the larva now being 0·5 mm. long. The larva
appears to moult at this time. After the gut is formed papillæ, three
or four in number, appear at the tail end. In two to three weeks
the larvæ are 1·5 mm. long. They now leave the thorax and reach the
labium, but they may be found in various parts of the body, e.g., the
legs. They bore through Dutton’s membrane and so arrive on the
surface of the skin, which they rapidly enter. Their development in
man is unknown, but it may be very long, as children are not infected
till 4 to 5, or even 10 years old, but this may be due to unknown
causes.
Development takes place in numerous mosquitoes. Anophelines:
Myzomyia rossii, Pyretophorus costalis, Myzorhynchus sinensis, Myzorhynchus
barbirostris, Myzorhynchus peditæniatus.
Culicines: Culex pipiens, Culex fatigans, Culex skusei, Culex
gelidus, Culex sitiens, Culex albopictus, Stegomyia fasciata, Stegomyia
pseudoscutellaris, Stegomyia gracilis, Stegomyia perplexa, Mansonioides
uniformis, Mansonioides annulipes, Scutomyia albolineata, Tæniorhynchus
domesticus.
Partial development takes place in other species.
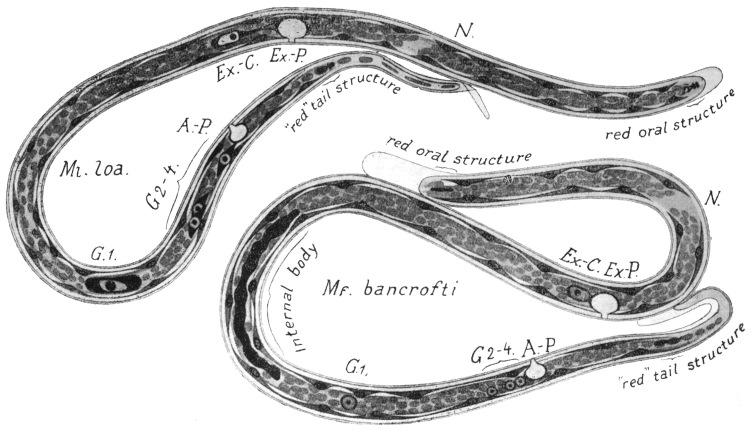
Fig. 283.—Schematic drawings of the anatomy of Ml. loa and Mf. bancrofti combined from specimens stained in different ways.
The position of the organs has not been based on the average values of a large series of specimens, but on that of a single specimen.
G1, chief genital cell; G2–4, other genital cells; Ex.-C., excretory cell; Ex.-P., excretory pore; A-P., anal pore; N., nerve ring.
Magnification circa 1,000. (After Fülleborn).
Pathology.—Among the conditions which Filaria bancrofti is
believed to produce are lymphangitis, varicose glands, especially
inguinal and epitrochlear, chyluria, chylocele, lymph scrotum,
orchitis, abscess, and elephantiasis. The evidence that these
so-called “filarial diseases” are produced by F. bancrofti is (1) geographical
and statistical; (2) pathological. Bahr has contributed
evidence of the former kind from his researches in Fiji, on which
we may base the following statements:—
(1) The prevalence of filarial diseases is proportional to the
prevalence of Mikrofilaria bancrofti in the blood. Thus in four
villages examined by him he got the following figures:—
| Village A | Village B | Village C | Village D |
| Mf. bancrofti | 12 | ·5 | per cent. | 25 | per cent. | 31 | per cent. | 33 | per cent. |
| Filarial diseases | 29 | | " | 39 | " | 58 | " | 34 | " |
| Total population | 168 | | | 114 | | 425 | | 222 |
(2) Out of 257 people with Mf. bancrofti in the blood, 153 were
suffering from filarial diseases, i.e., 59 per cent.
(3) Whereas of 672 people without Mf. bancrofti in the blood,
only 263 were suffering from filarial diseases, i.e., 37·6 per cent.
(4) Again out of 416 people suffering from filarial disease, 153
showed Mf. bancrofti in their blood, i.e., 36·7 per cent.
It is generally assumed that all people suffering from filarial
disease show at some (presumably early) stage larvæ in the blood;
but we do not consider that this must necessarily be so. It appears
to us quite possible that living adult filariæ may be present in the
body, producing disease, without their larvæ appearing in the blood.
The absence of larvæ from the blood in 63·3 per cent. of persons
suffering from filarial disease is, however, generally explained otherwise.
The adults which occur in enlarged glands, etc., get eventually
destroyed by inflammatory reaction, so that larvæ are no longer
being produced, while the enlarged gland, etc., which the adults have
produced remains. This explanation assumes that the larvæ of the
original worm die in the circulation or elsewhere, e.g., kidney, but
we have no evidence as to the duration of life of larvæ in the human
body; but also it assumes that a person cannot be reinfected with
filaria, for otherwise there is no reason why the diseased should not be
infected in the same proportion as the non-diseased. But assuming
the explanation to be true, it would explain why a diseased population
show larvæ in only about one-third of the cases. It must be borne
in mind also that the figures are rather small.
Pathology.—In order to explain the effects which do or may be
expected to occur from obstruction of lymphatics, it is necessary to
have an accurate knowledge of the distribution and connections of
lymphatic vessels (and glands) and the anastomoses of these vessels.
We can only briefly summarize our knowledge here.
We should recall also that considerable destruction or obstruction
of lymphatics or glands may occur without necessarily producing any
lymphatic obstruction, at least, of a permanent nature, e.g., when a
mass of lymphatic glands is destroyed by a bubo in the groin or,
again, when a carcinomatous mass of glands is removed from the
axilla. Again, to take the case of chyluria—where it is generally
assumed that obstruction must occur higher up than the point at
which the intestinal lacteals enter the juxta-aortic glands—this disease
may occur, e.g., in temperate regions, quite apart from such obstruction.
It is true that some of these cases of chyluria are not cases of
chyle in the urine, but, as little or no fat is present, lymphuria.
These do not require the above assumption, but seeing that true
chyluria may apparently occur without such obstruction, we should
be cautious about explaining this and other symptoms on the basis
of obstructions which theory may demand, for only too often there
are no post-mortem facts at our disposal.
Lymphangitis: What this is due to is unknown. There is no
actual evidence of the occurrence of adults in the inflamed vessel.
Complete disappearance, not to reappear, of (non-periodic) mikrofilariæ
from the blood has been shown by Bahr and others to occur
within twenty-four hours after an attack of lymphangitis, orchitis
adenitis or simply a high temperature. This mysterious phenomenon
requires explanation. If the mikrofilariæ were being killed by the
attack, their dead bodies should still be found in the blood; or if the
adults were being killed, for all we know to the contrary, the larvæ
might well survive. We consider there is no evidence that either
are affected, but that for some reason, as little understood as in
periodicity, the larvæ now remain in the organs.
Abscess: In Fiji, by Bahr, they have been found in the substance
of various muscles, e.g., quadriceps extensor, latissimus dorsi, serratus
magnus, in the popliteal space, groin, axilla, and over the internal condyle
of the humerus, and in the upper extremity they are frequently
infected with cocci. They not infrequently contain fragments of
dead adult filariæ. Their mode of origin is not clear. They form
nearly 30 per cent. of cases of filariasis in Fiji. Of 95 cases, 41
showed mikrofilariæ in blood, 54 did not.
Hydrocele and enlarged testis: In Fiji they form about 10 per
cent. (36 out of 343) of cases of filariasis. The fluid is usually sterile;
mikrofilariæ were present in the fluid in 1 out of 11 cases. In the wall
numerous calcified adult filariæ may be found. The walls consist
chiefly of hypertrophied muscle with fibrous tissue, dilated blood-vessels
and lymphatics, the lining epithelium of which appears to be
absent; of 38 cases 14 had mikrofilariæ in the blood, 24 had not.
Most of the cases are associated with elephantiasis of the scrotum
(11 out of 12 cases).
Enlarged glands form over 40 per cent. (153 out of 343) of cases
of filariasis, so that they are the commonest expression of filariasis
met with in Fiji. The glands are enlarged, fibrotic, and the trabeculæ
are thickened. The lymphatics are thickened or represented merely
by fibrous tissue. The gland also shows dilated blood-vessels and
numerous spaces filled with lymph. Giant-cells are common in
those glands which contain remnants of filariæ. Masses of lymphocytes
enclosed by inflammatory or fibrous tissue are common.
Eosinophile cells are also extremely common, not only in the fibrous
tissue of the glands, but in other inflammatory or fibrotic conditions:
in other organs living or calcified filariæ are “usually” present.
Only about 33 per cent. show mikrofilariæ in the blood. The
epitrochlear gland is frequently enlarged in Fiji.
Breinl has examined enlarged glands and finds loose vascular
fibrous tissue with lymphocytic invasion. In parts, the lymphocytes
collect into areas 200 µ to 800 µ in diameter. The lymph tissue
surrounding the spermatic cord showed abundance of vessels—(1)
large, (2) small. The large had thick walls and wide lumina. In
other cases the lumina were nearly filled by a thrombus of newly
formed, fine, loose connective tissue.
Varicose glands: In about 7 per cent. (24 out of 343 cases) of
filariasis, mikrofilariæ are found in the blood in 50 per cent. (12 out
of 24).
Elephantiasis.—Elephantiasis scroti is associated with hydrocele
in 50 per cent. of cases (12 out of 23); in 65 per cent. of cases (15 out
of 23) there are associated enlarged glands in one or both groins,
though also hydrocele and enlarged glands occur without elephantiasis
scroti. In 13 out of 27, i.e., about 50 per cent., cases of elephantiasis
in various regions, no associated enlargement of glands is found.
Elephantiasis forms in Fiji less than 10 per cent. of cases of filariasis.
Mikrofilariæ are present in the blood in 36 per cent. (12 out of 33)
of cases.
Chyluria.—Exceedingly rare in Fiji. Theory would demand an
obstruction above the point of entry of the lacteals, viz., the pre-aortic
lymphatic glands, but in cases in temperate regions it may occur
without any such lesion. In some of these cases the fluid is not
chyle (fat absent), but presumably lymph. A discussion of the mode
of production of chyluria, lymph scrotum, elephantiasis, etc., is at
present premature; theory has far outrun fact. Too much stress had
been laid on the mechanical action of the worms to the almost total
exclusion of their (or possibly their larval) toxic action. The above
analysis has been made in the hope of acquiring more extended
observations similar to those made by Bahr.
Geographical Distribution.—Filaria bancrofti is known in nearly all
tropical countries. It occurs in India, China, Indo-China, Japan,
Australia, Queensland, the Islands of Polynesia (with the exception
of the Sandwich Islands), Egypt, Algeria, Tunis, Madagascar, Zanzibar,
Sudan, etc., the south of the United States of America, Brazil,
the Antilles, etc. Whether it is the same species in all cases is
questionable.
Filaria demarquayi, Manson, 1895.
Syn.: F. ozzardi, Manson, 1897.
Fig. 284.—F. demarquayi:
tail, showing
paired large fleshy
papillæ. (After Leiper.)
The adult female F. demarquayi measures from 65 to 80 mm.
in length by 0·21 to 0·25 mm. in breadth. The head has a diameter
of from 0·09 to 0·1 mm. The mouth is terminal. The genital
pore opens at 0·76 mm. from the head. The alimentary canal is
nearly straight and terminates in an anus, which
is subterminal. The opening of the anus is
marked by a slight papilla. The tail is curved.
It rapidly diminishes in size just below the anal
papilla. A characteristic pair of fleshy papillæ
project from the tip of the tail. The diameter near
the tip of the tail before its termination is 0·03 mm.
F. demarquayi is a thicker worm than Ac. perstans.
It differs from F. bancrofti in the greater size of
the head, in the smaller tail, and particularly in the
marked fleshy papillæ at the tip of the tail. These
papillæ are knobby, and not simply cuticular as
in Ac. perstans.
The male of Filaria demarquayi has still to be found.
The adult female form of F. demarquayi was found by Dr. Galgey
in the body of a native of St. Lucia in whose blood the larvæ had
been found during life. Five adult females were found in the connective
tissue of the mesentery.
The larva measures 200 µ in length by 5 µ in breadth; it is sharp-tailed,
and has no sheath. Its movements are very active, and the
absence of a sheath enables it to glide along freely all over the slide.
It observes no periodicity, being present in the peripheral circulation
both by day and by night. As a rule, some eight or ten parasites are
found in an ordinary preparation. Sometimes hundreds of these
larval filariæ may be counted on every slide.
The intermediate host has not been discovered.
Geographical Distribution.—St. Vincent, Dominica, Trinidad, and
St. Lucia (West Indies), British Guiana, New Guinea (?).

Fig. 285.—Mf. demarquayi in thick film, dried and stained with hæmatoxylin.
6, unshrunken; 7, shrunken. × 1,000. (After Fülleborn.)
Filaria taniguchi, Penel, 1905.
Female 68 by 0·2 mm. in breadth. Cuticle non-striated. Mouth
two pairs of papillæ. Anus 23 mm. from extremity. Vulva 1·3 mm.
from mouth. Larva 164 µ by 8 µ, sheathed. Tail truncated.
Periodicity nocturnal.
Habitat.—Lymphatic glands of man. Japan.
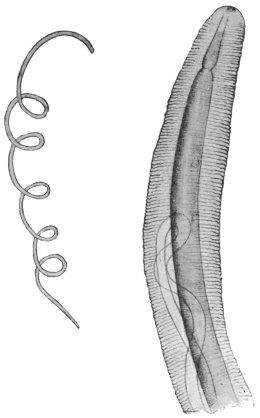
Fig. 286.—Filaria (?) conjunctivæ: to the left, life size; to the right, the
anterior extremity magnified. (After Addario.)
Filaria (?) conjunctivæ, Addario, 1885.
Syn.: Filaria peritonei hominis, Babes, 1880; Filaria inermis, Grassi, 1887;
Filaria apapillocephata, Condorelli-Francaviglia, 1892.
The female only of this species is known. It measures 16 to
20 cm. in length and 0·5 mm. in breadth, and is of a whitish or
brownish tint. The cuticle is striated with fine transverse and more
marked longitudinal striæ with the exception of a small field surrounding
the mouth, which is terminal and has neither papillæ nor
lips. The œsophagus measures 0·6 mm. in length. The anus is 3 mm.
in front of the rounded posterior extremity, and behind it there are
two (glandular?) sacs. The vulva is close behind the oral aperture;
the vagina soon divides into two convoluted uteri, which are filled
with eggs and embryos. Embryos 350 µ by 5·5 µ.
This species (115 mm. long) was first observed in Milan by Dubini in the eye of
a man; subsequently it was observed, encysted and calcified (190 mm. long), by
Babes in the gastro-splenic omentum of a woman in Budapest, and finally one
(95 mm. long) was extracted by Vadela from a tumour the size of a pea in the ocular
conjunctiva of a woman in Catania (Sicily), which case has been described by
Addario. Possibly Agamofilaria palpebralis, Pace, 1867 (nec Wilson, 1844), and
A. oculi humani, v. Nordm., 1832, are the same species.
Fig. 287.—Filaria (?) conjunctivæ: anterior end greatly magnified;
the mouth with the pharynx in the middle; in the cuticle on the right
side the opening of the vagina, and behind it the excretory pore.
(After Grassi.)
Filaria (?) conjunctivæ is certainly only an incidental parasite of
man; the horse and ass are its normal hosts, but it is not common
in these animals, or is frequently confused with Hamularia equi,
Gmelin, 1789.
Group. Agamofilaria, Stiles, 1906.
Not a generic but a group name for immature Filariidæ the development
of which does not admit of generic determination.
Agamofilaria georgiana.
Adult unknown, length from 32 to 53 mm. Maximum diameter
560 µ to 640 µ. Head no cephalic cone. Mouth small, circular,
surrounded by six papillæ (two small latero-median and four
sub-median). The larger papillæ are 24 µ from base to tip. Excretory
pore about 0·5 mm. from head. Anus 64 µ to 128 µ from
tip. Cuticle fine striæ near anus, occasionally elsewhere. Lateral
lines clearly marked. Œsophagus 2·5 to 2·9 mm. Rectum 200 µ
long.
Habitat.—Superficial sores on the ankle of a negress, Georgia,
U.S.A.
Agamofilaria palpebralis, Pace, 1867 (nec Wilson, 1844).
100 by 1·5 mm., removed from a cyst in the left upper eyelid of
a boy by Pace, in Palermo.
Agamofilaria oculi humani, v. Nordmann, 1832.
Syn.: Filaria lentis, Diesing, 1851.
The sexless Nematodes observed in the lens of the human eye
were termed Filaria oculi humani. Only three cases are known.
v. Nordmann observed very small round worms in the lens of a
man and woman with cataract, and Gescheidt once found three
specimens in the lens of a woman similarly affected.
The demonstration of nematode-like formations in the vitreous
remains uncertain even when movements are observed, and when
they cannot be extracted and examined microscopically the doubt
may occur that one may have mistaken the remains of the hyaloid
artery for a worm, which it resembles in form, size and colour; the
slightest movement of the eye also causes it to move so that it
simulates a living organism.
Accordingly it would be more correct to exclude all the cases known only
ophthalmoscopically (Quadri, 1857; Fano, 1868; Schoeler, 1875; Eversbusch, 1891).
There then remains only one positive case, described by Kühnt in 1891. In this
case it was possible to follow the gradual growth of the parasite for some time, and
the worm, which measured only 0·38 mm. in length, was finally extracted.
Agamofilaria labialis, Pane, 1864.
The parasite measures 30 mm. in length; the anterior extremity
is pointed; the terminal oral aperture is surrounded by four papillæ;
the anus opens 0·5 mm. in front of the posterior extremity; the vulva
is 2·5 mm. in front of the anus; the uterus is double; the anterior one
passes with convolutions forward to the cephalic end; the posterior
one is directed backwards and remains rudimentary.
Extracted from a small pustule on the inner surface of the upper lip. Also
found in Naples by Pierantoni in 1908.
The position of many of these worms is doubtful, and still more so
is that of many other imperfectly described “Filariæ,” which are
hardly more than useless and confusing names. These include the
following:—
Filaria (?) romanorum-orientalis, Sarcani, 1888.
Observed in the blood of a Roumanian woman; 1 mm. in length,
0·03 mm. in breadth; tail end pointed, a tongue-like appendage on
the head. Eggs the size of a red cell with developed embryo,
apparently viviparous.
Filaria (?) kilimaræ, Kolb, 1898.
Several female specimens, 10 to 20 cm. long by 0·5 to 1 mm. broad,
were once found free in the abdomen of a fallen Kitú warrior;
according to Spengel, who examined them, the oral papillæ of these
worms were similar to those of Dracunculus medinensis. Moreover,
Kolb classifies together Nematodes that probably have no connection
with each other.
Filaria (?) sp.?
Cholodkowsky calls attention to Filariæ that are still unknown
which cause tumours resembling whitlows on the fingers of peasants
of the Twer Government.
Mikrofilaria powelli, Penel, 1905. In Bombay.
Mikrofilaria philippinensis, Ashburn and Craig, 1906. In the
Philippines.
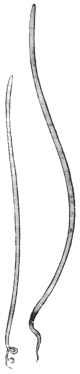
Fig. 288.—Setaria
equina: left, male;
right, female. Natural
size. (After Railliet.)
Genus. Setaria, Viborg, 1795.
Syn.: Hamularia, Treutler, 1793; Tentacularia, Zeder, 1800 (nec Bosc, 1797).
Mouth with projecting peribuccal armature deeply notched on the
lateral margins, less so dorsally and ventrally. Tail in both sexes
with peculiar caudal appendages.
Parasitic in serous cavities, especially of ruminants.
Setaria equina, Abildg., 1789.
Syn.: Gordius equinus, Abildg., 1789; Filaria equi, Gmelin, 1789; Hamularia
lymphatica, Treutler, 1793; Tentacularia subcompressa, Zedder, 1800; Filaria
papillosa, Rud., 1802; Filaria hominis bronchialis, Rud., 1819; Filaria hominis,
Dies., 1851; Strongylus bronchialis, Cobb., 1879.
The body is whitish, filiform, pointed posteriorly. The cuticle
presents a delicate transverse striation. The mouth is small, round,
and surrounded by a chitinous ring, the border of which carries, at the
sides, two semilunar lips, and there is on the dorsal as well as on the
ventral surface a papilliform process; on the
tail, corresponding with each sub-median
line, is a conical papilla. The male measures
6 to 8 cm. in length; the posterior
extremity ends in a corkscrew spiral; there
are on each side four pairs of pre-anal and
four or five post-anal papillæ; the spicules
are unequal. The female measures 9 to
12 cm. in length and is viviparous; the
embryos measure 0·28 mm. in length and
0·007 mm. in breadth.
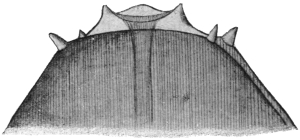
Fig. 289.—Setaria equina: anterior end, magnified.
(After Railliet.)
Setaria equina is a frequent parasite of horses and asses; it inhabits the
peritoneal cavity, and from there occasionally invades the female genitalia or even
the liver; it is found more rarely in the pleural cavity or in the cranium. The
statement that it also occurs in the subcutaneous connective tissue is probably due
to confusion with Setaria (Filaria) hæmorrhagica, Raill., 1885 (Filaria multipapillosa,
Cond. et Drouilly, 1878). Setaria labiata papillosa (immature form)
occurs in the eye of the horse, adults in the peritoneal cavity.
Treutler, in 1790, found a filaria in the enlarged bronchial lymphatic
gland of a patient suffering from phthisis. It measured 26 mm. in
length and had two spicules, which Treutler mistook for mouth hooks,
hence the name Hamularia. Blanchard mentions another case from
Geneva, Brera a third and v. Linstow a fourth. As shown by the
synonyms, a few authors consider this form to be a distinct species,
which is hardly probable.
Genus. Loa, Stiles, 1905.
Characterized by the possession of cuticular bosses in both sexes
(fig. 294).
Loa loa, Guyot, 1778.
Syn.: Filaria oculi, Gerv. et v. Ben., 1859; Dracunculus oculi, Diesing, 1860;
Dracunculus loa, Cobbold, 1864; Filaria subconjunctivalis, Guyon, 1864.

Fig. 290.—Loa loa: the
anterior end of the male, magnified.
(After R. Blanchard.)
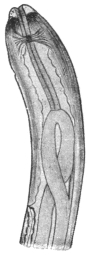
Fig. 291.—Loa loa: anterior
portion of the female as far as
vulva. (After Looss.)
The male measures 25 to 35 mm. in length, and 0·3 to 0·4 mm. in
breadth; the cuticle is not striated, but, with the exception of the
anterior and posterior extremities (1·5 mm.), is beset with numerous
irregularly distributed bosses (4 µ to 12 µ high by 12 µ to 27 µ
broad). The anterior extremity is somewhat attenuated, and in front
is conical and transversely truncated. At the anterior limit of the
conical part is a small papilla corresponding with the dorsal and ventral
median lines, and a little in front six non-projecting sensory papillæ
(two lateral, four sub-median). Excretory pore 0·65 mm. from the
anterior end. The posterior extremity is attenuated and somewhat
curved ventrally; the anus is 0·082 mm. distant from the rounded
posterior border. In front of the anus on each side are three globular
and pedunculated papillæ of different sizes, set close one behind the
other but asymmetrically; behind the anus on either side are two
smaller papillæ of a different shape; the anterior one resembles
the pre-anal papillæ in form, but is smaller; the posterior one is
conical, and rests with a broad base on the cuticle. The spicules
are 0·113 and 0·176 mm. long.
The female measures 45 to 63 mm. in length by 0·5 mm. in
breadth. It is also beset with irregularly distributed bosses, which in
places lie close to each other, and extend to the anterior extremity;
posteriorly they become less frequent, but are not entirely absent.
The anterior extremity is conical, the posterior one straight, attenuated,
rounded off, 0·17 mm. from the anus. The uteri contain eggs in the
most various stages of development, as well as hatched-out larvæ,
253 µ to 262 µ in length and 4·7 µ to 5 µ in breadth. The vulva lies
about 2 mm. from the head end. The vagina, 9 mm. long, divides
into two branches, which at first run posteriorly and parallel to one
another for about 18 mm. One then bends forward, runs as far as
the œsophagus, bends here again and runs backward to end at the
point of its first bending. The other branch at first runs straight
backward and then bends forward, but before reaching the point of
the first bend of the anterior tube bends backward again, forms
again a loop and ends at the level of the anus. The tubes consist in
the main of the uterus, then a club-shaped swelling, the receptaculum
seminis, then the oviduct 2 mm. long, and finally the ovary.

Fig. 292.—Loa loa in situ.
Natural size. (After Fülleborn
and Rodenwaldt.)
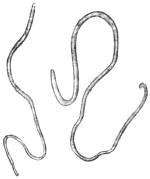
Fig. 293.—Loa loa: male
on the left, female on the
right. × 2. (After Looss.)
Unsegmented eggs measure 32 µ by 17 µ, in the morula stage
40 µ by 25 µ, and when containing embryos 50 µ by 25 µ. The
vitelline “shell” of the egg is, according to most authors, stretched by
the embryo and becomes the sheath of the hatched larva. While
still in the vulva, the larva measures 217 µ to 274 µ (average 246 µ) in
fresh, 146 to 226 µ (average 192 µ) stained.

Fig. 294.—Loa loa: on the left, the hind end of a male; on the right, of a female.
Note the cuticular bosses shown in the figure of the female. × 285. (After Looss.)
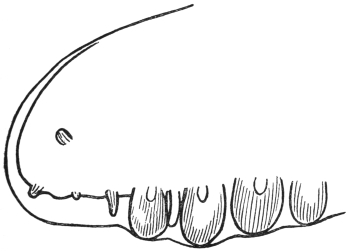
Fig. 295.—Loa loa: lateral view of tail of male showing
papillæ. (After Lane and Leiper.)
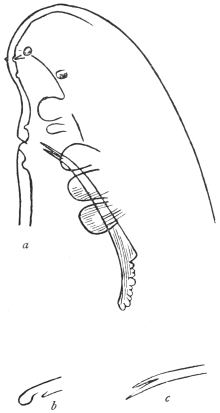
Fig. 296.—Loa loa. a, ventro-lateral
aspect of tail showing papillæ and one
spicule; b and c, terminations of the
two spicules. (After Leiper.)
Site of Worms.—In various localities; under the muscular aponeuroses
on extensor surfaces of arms and legs, fingers, trunk, eyelid,
conjunctiva, frænum linguæ, penis, pericardium, anterior chamber of
eye, and, according to some authorities, in lymphatic vessels, e.g.,
those of spermatic cord. As many as thirty adults may be found.
The worms appear to be frequently immature, and it has been stated
that worms in superficial parts are immature, those situated deeply
are mature, but the data are few.
The first accounts of Loa loa—long since forgotten—were reported by
Pigafetta, and are contained in a book of travels on the Congo printed in 1598.
In an accompanying illustration is depicted, not only the ancient method of
extraction of the Medina worm, but also the operative removal of the filaria from
the conjunctiva. Subsequently the presence of the worm in negroes was confirmed
by Bajon in Guiana (1768) and by Mongin in Mariborou (San Domingo), likewise in
a negro (1770). At about this time a French ship’s doctor, Guyot, was cruising on
the West Coast of Africa; he observed the parasite termed “loa” by the natives,
and learned that it was frequent in the negroes of the Congo district. Since that
time numerous observations have been reported. It was formerly common in South
America, where the parasite was imported by slaves, but it disappeared when the
traffic ceased; it was particularly prevalent in the Congo, where it occurs not only
in natives, but also in Europeans. During recent times it has repeatedly been
observed in Europe in negroes as well as in white men who have lived on the West
Coast of Africa.
Nematodes of different size have been repeatedly observed in the
eye of man, in the anterior chamber, lens and vitreous. For example,
Mercier, in 1771 and 1774, removed a filaria out of the anterior
chamber of two negroes in St. Domingo. One was 36 mm. long.
Barkan, in 1876, in San Francisco, removed one from the eye of an
Australian. Again, Cappez and Lacompte, in Brussels, in 1894,
observed for some weeks immature Nematodes in the eye of a negro
girl, aged 2 1/2 years, and then removed them. What these Nematodes
actually were in these cases it is impossible to say.
Structure of Larvæ.—In dried films the larva varies in size from
140·5 µ to 166·5 µ, average, 152·5 µ; while another set of measurements
gave the values 131 µ, to 150 µ, average, 143·6. In films fixed
with hot alcohol the dimensions were 208 µ to 254 µ, average, 231 µ.
The nerve ring 21·4 to 21·8 per cent. Excretory pore 30·4 to 31·8
per cent. Excretory cell 34·8 to 37·3 per cent. G1 cell 68·2 to 68·5
per cent. Anal pore 81·6 to 82·4 per cent. of total length. For other
details cf. Filaria bancrofti.

Fig. 297.—Mf. loa: in thick film, dried
and stained with hæmatoxylin. × 1,000.
(After Fülleborn.)
Larvæ in Blood.—These from their diurnal periodicity are known
as Mikrofilaria diurna. The evidence that these larvæ are the young
of the adult worm Loa loa is: (1) They are identical in structure with
larvæ taken from the uterus of L. loa; (2) their geographical distribution
is the same as that of L. loa; (3) they eventually occur in
the blood of patients suffering from Calabar swellings, a condition
due to L. loa. Their occurrence in the blood in this latter condition
and in L. loa infections we shall consider later.
Periodicity.—Here, as in the case of the larvæ of Filaria bancrofti,
the larvæ that appear in the blood
are probably the overflow simply of
the larvæ which we assume, on
analogy, to have their principal site
in the lungs. They appear in the
blood about the time of getting
up, 6 to 8 a.m. (10 in 20 mm.3),
at 12 noon there are twenty-four,
at 8 p.m. the number has fallen to
eighteen, and at midnight to one,
while from 2 a.m. to 6 a.m. none,
or one only, may be found. This
periodicity is, as a rule, a very
constant one, but there are exceptions,
and in certain cases more
have been found at midnight than
at 9 a.m. The periodicity is also
lost in pathological conditions,
e.g., sleeping sickness (vide also
under Filaria bancrofti). The
possibility of non-periodic Loa loa
larvæ should also be considered.
Pathology.—The parasite wanders
about the body, and may be seen
under the skin in thin parts. Their
advance is in some cases at the rate
of an inch in two minutes. During
their progress they give rise to
creeping sensations and to a condition
of transient œdematous areas
known as Calabar swellings on
various parts of the body, e.g., arm.
These vary in diameter from 1 to
10 cm., and often shift their position an inch or so a day. They give
rise to a certain amount of redness, tension and heat, and their development
is promoted by muscular action of the part. They disappear to
reappear elsewhere. The condition is associated with a high eosinophilia,
50 per cent. being not uncommon. Patients known to harbour
L. loa, e.g., native children, frequently show no larvæ in their blood,
but they may do so after years of infection. Again, in patients having
an infection of Mikrofilaria diurna, there is frequently at the time no
evidence of the presence of Loa loa adults. Here again they may
appear later, but the conditions which determine whether persons
infected with L. loa show larvæ in the blood, or persons infected
with Mikrofilaria diurna also show L. loa, are unknown, though
explanations unsupported by facts
abound. Likewise also the mode
of production of the swellings is
unknown.
Not uncommonly Mikrofilaria
perstans occurs in the blood together
with M. diurna.
Duration of Life.—This is long,
as some cases have been observed
five to six years after leaving
Africa. The incubation period
is about a year.
Life-history.—Development of
the larvæ takes place in the salivary
glands of Chrysops sp. as shown
by Leiper.
Geographical Distribution.— West
Africa, especially in Congo.
Genus. Acanthocheilonema, Cobbold, 1870.
Cuticle striated longitudinally.
Œsophagus divided into
two portions. Tail in both sexes
with short lateral conical cuticular
appendages. Spicules unequal,
the larger membranous distally,
the smaller hooked. Vulva in
œsophageal region.
Acanthocheilonema perstans, Manson, 1891.
Syn.: Filaria perstans, P. Manson, 1891; Filaria sanguinis hominis var. minor,
Manson, 1891.
The adult female Ac. perstans measures 70 to 80 mm. in length
by 120 µ to 140 µ in breadth. The head is club-shaped and measures
0·07 mm. in diameter. The vulva opens at 0·6 to 1·0 mm. from
the head. The tail is curved and presents a cuticular thickening which
forms two triangular appendages. The anus opens at the apex of a
papilla situated in the concavity of the curve formed by the tail 150 µ
from the end. The diameter of the tail just before termination is
0·02 mm.
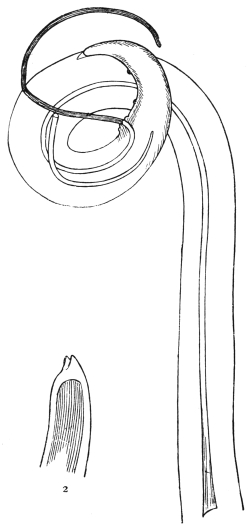
Fig. 298.—Acanthocheilonema perstans.
1, tail of male; 2, tail showing cuticular
flaps devoid of fleshy contents. (After
Leiper.)

Fig. 299.—Mf. perstans in thick film, dried and stained with hæmatoxylin;
4, unshrunken; 5, shrunken. × 1,000. (After Fülleborn.)
The adult male measures 45 mm. in length by 60 µ to 80 µ in
breadth. The diameter of the head is 0·04 mm. The tail is much
curved. There are four pairs of pre-anal papillæ and two pairs of
post-anal papillæ. Spicules very unequal in size. Cloaca 121 µ from
the tail end. At the tail end two triangular cuticular appendages.
The adult worms inhabit the connective tissue at the base of
the mesentery, especially in the region of the pancreas, abdominal
aorta and suprarenals. To find them the mesentery should be
removed, placed in a 2 per cent. solution of formalin, and then
carefully examined at leisure.
Mikrofilaria perstans.—160 µ to 210 µ by 5 µ to 6 µ broad. Has no
sheath. Cuticle transversely striated. Tail rounded off, not pointed.
Nerve ring at 34 µ. Excretory pore 49 µ, genital pore 125 µ from
head. Smaller larvæ 90 µ to 110 µ by 4 µ broad. A “fang” is also
described on the head.
Mf. perstans. | Mf. demarquayi. |
| (1) Tail stumpy. | (1) Tail pointed. |
| (2) Column of nuclei extends to tip of tail. | (2) Does not extend to tip. |
Periodicity.—None.
Life-history.—Unknown.
Geographical Distribution.—Very common in many parts of Africa:
Sierra Leone, Dahomey, Northern Nigeria, Southern Nigeria, Cameroons,
Ivory Coast, Gold Coast, Old Calabar, Congo, Uganda. Absent
from Zululand, Basutoland. On the East Coast of Africa it is not
found in the towns of Zanzibar and Mombasa, neither is it found in
the country of the Masi, nor amongst the Kavirondo, who dwell
along the north-east shores of Lake Victoria.
In South America, Ac. perstans is very common amongst the
aboriginal Indians in the interior of British Guiana. However,
it is not found in Georgetown and in New Amsterdam, neither is
it found in the cultivated strip of coast lying between these two
towns, but it is common on the coast farther north near the Venezuelan
boundary, where the forests stretch to the sea. The Waran
Indians, who live at the mouth of the Waini river, harbour this
parasite. It is absent in the West Indies.
Topographically, Ac. perstans is found only in areas covered by
dense forest growth and abounding in swamps. In Kavirondo,
where the forest disappears and the land is covered with scrub and
short grass, it is not found; likewise it is not found on the grassy
plains of the highlands of British East Africa. Towns and cultivated
areas are free from it.
Genus. Dirofilaria. Railliet and Henry, 1911.
Body very long, thread-like, cuticle transversely striated. Mouth
with six papillæ. Male tail spiral with voluminous pre-anal and some
large post-anal papillæ; spicules unequal. Vulva near the anterior
hundredth of body; viviparous. Parasitic in heart or blood-vessels
and subcutaneous tissue.
Dirofilaria magalhãesi, R. Blanchard, 1895.
Syn.: Filaria bancrofti, v. Linstow, 1892; Filaria bancrofti, P. S. de Magalhães,
1892 (nec Cobbold, 1877).
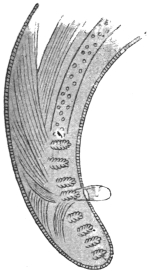
Fig. 300.—Dirofilaria
magalhãesi: posterior extremity.
(After v. Linstow.)
The male measures 83 mm. in length by 0·28 to 0·40 mm. in
breadth. The anterior extremity is rounded, and has no papillæ (?6);
the posterior extremity exhibits a double curve,
with four pre-anal and four post-anal papillæ on
each side. These are large and have a villous
appearance. The mouth is round and unarmed,
the pharynx measures 1 mm. in length, is cylindrical,
very muscular, and its hinder part is
dilated. The anus is situated 0·11 mm. in front
of the hind end. There are probably two
unequal spicules; one only, however, is known—apparently
the shorter one—the length of
which is given as 0·17 to 0·23 mm.
The female measures 155 mm. in length
and 0·6 to 0·8 mm. in breadth; the rings of
the cuticle are 0·005 mm. apart (in the male
0·003 mm. apart); the anterior extremity is
slightly thickened and club-like, the posterior
extremity is slender, and terminates obtusely;
the lateral line is 0·127 mm. in breadth (that of
the male 0·007 to 0·008 mm.); the anus opens 0·13 mm. in front of
the hind end, the vulva is 2·5 mm. distant from the mouth, the ovaries
are two much convoluted tubes. The eggs measure 38 µ by 11 µ.
This species was first discovered at a post-mortem, in the left ventricle, by J. P.
Figueira de Saboia in Rio de Janeiro, and has been described by P. S. de Magalhães.
D. immitis occurs in the right ventricle of the heart of the dog in Europe and
the Tropics.
D. repens is also a common subcutaneous Nematode in dogs in Annam.
Sub-family. Onchocercinæ, Leiper, 1911.
Cuticle with spiral thickenings.
Genus. Onchocerca, Diesing, 1841.
Male with four pre-anal papillæ. Female with vulva situated
anteriorly.
Onchocerca volvulus, R. Leuckart, 1893.
Syn.: Filaria volvulus, R. Leuckart, 1893.
The adult male measures 30 to 35 mm. in length by 0·14 mm. in
breadth. The body is white, filiform, attenuated at both ends. The
head is rounded and has a diameter of 0·048 mm. The cuticle
is distinctly transversely striated. The mouth is unarmed. The
alimentary canal is straight, the anus opening 0·07 mm. from the tip
of the tail. The tail is strongly curved and somewhat flattened on
the concave surface. There are three papillæ, one large and two
small, on each side of the cloaca and one large and two post-anal
small papillæ. Two curved spicules, 0·166 and 0·08 mm. respectively.
The adult female is of uncertain length, but much longer than
the male, probably about 10 to 12 cm. The head is rounded and
truncated; it measures 0·065 mm. in diameter. The tail is curved.
The vulva opens 0·55 mm. from the head. The hand-like cuticular
thickenings are well marked. Eggs ovoid with a prolongation at
each pole “like an orange wrapped in tissue paper.” The larva
measures about 300 µ by 7 µ to 8 µ; it has no “sheath.” The body
tapers from about the last fifth of its length, and terminates in a
sharply pointed tail. At about the anterior fifth of the body there
is a V spot.
O. volvulus is found in peculiar subcutaneous tumours, the size of
a pea to that of a pigeon’s egg. The same patient may present one
or several of these tumours. The regions of the body most frequently
affected are those in which the peripheral lymphatics converge.
Thus they are usually found in the axilla, in the popliteal
space, about the elbow, in the sub-occipital region and in the intercostal
spaces. The tumours are never adherent to the surrounding
structures, and can be easily enucleated. They are formed of a dense
connective tissue wall and internally a looser fibrous meshwork.
This is traversed by a series of canals in which the worms lie, but
they are also partly embedded in the denser wall. The canals
apparently dilate into cavities filled with slimy pus-like fluid consisting
largely of larvæ. According to Brumpt the posterior extremity
of the male, and the anterior extremity of the female with its vaginal
opening, are free in one of the spaces for the purpose of copulation
and parturition. If a tumour be cut into and placed in salt solution,
Rodenwaldt states that the undamaged males wander out into the
solution.
The formation of the tumours is elucidated by Labadie-Lagrave
and Deguy’s case. The authors found an immature female Onchocerca
volvulus in a lymphatic vessel partly obstructed by an infiltration of
fibrin and leucocytes. It appears, therefore, that the presence of the
parasites within the lymphatics gives rise to an inflammatory process,
and that the consequent fibrinous deposit envelops the parasites,
obliterates the lumen of the vessel, and ultimately isolates the affected
tract. At any rate, in young tumours the worms appear to lie in
a structureless substance permeated by leucocytes in which connective
tissue is gradually organized from the periphery, thus isolating the
worms.
In cases of infection with O. volvulus larvæ have been found by
Ouizilleau, Fülleborn, and Simon in lymph glands, and in the finger
blood if considerable pressure is used so as to squeeze lymph out
of the tissues. They are sheathless, and the following are the
dimensions in ordinary dried films: Length, 274 µ; nerve ring,
23·7 per cent.; G1 cell, 69·6 per cent.; end of last tail cell, 96·3 per
cent. The dimensions of larvæ of O. volvulus taken from the uterus
and prepared in the same way are: Length, 224·5 µ; nerve ring,
24·3 per cent.; G1 cell, 68·9 per cent.; end of the last tail cell,
95·5 per cent. In all probability the larvæ in the glands and blood
are those of O. volvulus.
According to the natives, the tumours may last indefinitely and
never ulcerate. Some old patients told Brumpt that their tumours
had been present since childhood. Probably Onchocerca volvulus,
like some other Filariidæ, may live for many years.
O. volvulus occurs in various parts of West Africa: Gold Coast,
Sierra Leone, Dahomey, Lagos, Cameroons. Brumpt, on the banks
of the Welle between Dongon and M’Binia (Belgian Congo), found
about 5 per cent. of the riverine population affected.
Family. Trichinellidæ, Stiles and Crane, 1910.
Sub-family. Trichurinæ, Ransom, 1911.
Male with a single long spicule, with sleeve-like sheath. One
ovary. Eggs with an opening at each pole closed by a plug-like
operculum. Eggs hatch on being swallowed by a new host. Genera:
Trichuris, Capillaria.
Genus. Trichuris, Röderer and Wagler, 1761.
Syn.: Trichocephalus, Goeze, 1782 (nec Trichiurus, L., 1758); Mastigodes,
Zeder, 1803.
The anterior part of the body is very long and thread-like; the posterior, much
shorter part, is thicker, rounded posteriorly, and the anus is terminal. The males
have the posterior extremity spirally rolled; the vulva is situated at the commencement
of the posterior part of the body. The Trichocephali live in the large intestine
of mammals, the cæcum by predilection; their development is direct, infection occurs
through the ingestion of embryo-containing eggs.
Trichuris trichiura, Linnæus, 1761.
Syn.: Trichocephalus trichiurus, L., 1771; Ascaris trichiura, L., 1771;
Trichocephalus hominis, Schrank, 1788; Trichocephalus dispar, Rud., 1801.
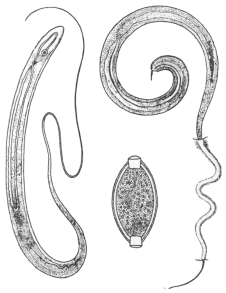
Fig. 301.—Trichuris trichiura: on
the left, male; on the right, female with
the anterior extremity embedded in the
mucous membrane of the intestine;
below, egg.
The male measures 40 to 45 mm. in length, the spicule is
2·5 mm. long, its retractile sheath is beset with spines. The female
measures 45 to 50 mm. in length, of which two-fifths appertain
to the posterior part of the body. The ova are barrel-shaped and
have a thick brownish shell which is perforated at the poles. Each
opening is closed by a light-coloured plug. The eggs measure
50 µ to 54 µ in length and 23 µ in breadth; they are deposited
before segmentation. Trichuris trichiura usually lives in the cæcum
of man, and is also occasionally found in the vermiform appendix
and in the colon, exceptionally also in the small intestine; usually
only a few specimens are present, and these do not cause any particular
disturbance, although, as Askanazy found, they feed on blood;
in other cases cerebral symptoms of more or less severity are observed
when Trichocephali are present in large numbers. At post-mortems
performed soon after death the filiform anterior extremity of the worm
is frequently found embedded in the
mucous membrane (Askanazy).
The whip worm is one of the most common
parasites of man and appears to be distributed
over the entire surface of the globe; it is, however,
more frequent in the warmer regions. It
is found in persons of both sexes and all ages
with the exception of infants. In autopsies it
is found in the following numbers: In Dresden
in 2·5 per cent., in Erlangen in 11·1 per cent.,
in Kiel in 31·8 per cent., in Munich in 9·3 per
cent., in Petrograd in 0·18 per cent., in
Göttingen in 46·1 per cent., in Basle in 23·7 per
cent., in Greenwich in 68 per cent., in Dublin
in 89 per cent., in Paris in about 50 per cent.,
and in Southern Italy in almost 100 per cent.
On examining the fæces the eggs of the whip
worm were found as follows: In Munich in 8·26
per cent., in Kiel in 45·2 per cent., in Greifswald
in 45 per cent., in North Holland in 7 per cent.,
in Novgorod in 26·4 per cent., in Petrograd
in 5 per cent., in Moscow in 5·3 per cent.
The development of the eggs is completed in water or in moist
soil, and occupies a longer or shorter time according to the season;
the eggs possess great powers of resistance, as do the larvæ, which,
according to Davaine, may remain as long as five years in the eggshell
without losing their vitality. Leuckart proved by experiment
that direct infection with Trichuris ovis (Ovis aries) and T. crenata
(Sus scrofa dom.) was produced by embryo-containing eggs; Railliet
obtained the same results with T. depressiuscula of dogs, and Grassi
subsequently, by means of two experiments, demonstrated the direct
development of Trichuris trichiura. In one case embryo-containing
eggs were swallowed on June 27, 1884, and on July 24 the ova of
Trichocephali were found in the fæces for the first time.
Trichuris trichiura is found not only in man, but also in various
monkeys (T. palæformis, Rud.), as well as in lemurs (T. lemuris, Rud.).
Other species are T. crenata in pig; T. ovis in cattle, sheep, goat, and
pig (?); T. depressiuscula in dog; T. campanula in cat; T. unguiculata
in rabbit and hare; T. cameli in camel; T. discolor in humped cattle;
T. nodosus in mouse; T. alcocki in the thamin (India); T. globulosa in
camel; T. giraffæ in giraffe.
Sub-family. Trichinellinæ, Ransom, 1911.
Male without spicule; females ovoviviparous. Larvæ penetrate
muscles of host and become encysted. Genus: Trichinella.
Genus. Trichinella, Railliet, 1895.
Syn.: Trichina, Owen, 1835 (nec Meigen, 1830).
Very small Trichinellinæ, the males of which have two conical appendages at the
caudal extremity; the vulva is situated at the border of the anterior fifth of the
body. There is only one species.
Trichinella spiralis, Owen, 1835.
Syn.: Trichina spiralis, Owen, 1835.
The male measures 1·4 to 1·6 mm. in length and 0·04 mm. in
diameter. The anterior part of the body is narrowed, the orifice of
the cloaca is terminal and lies between the two caudal appendages;
internal to these are two pairs of papillæ, dorsal one behind the other.
The cloaca is evertible for copulation. The females measure 3 to
4 mm. in length and 0·06 mm. in diameter; anus terminal.
Trichinella spiralis in its adult stage inhabits the small intestine
of man, pig, wild boar, rat. The young do not leave the body of the
host but become encysted in the muscles. Experimentally it develops
in the black rat (Mus rattus), the sewer rat (M. decumanus), the
domestic pig (Sus scrofa dom.), the wild boar (Sus scrofa ferox), the
domestic dog (Canis familiaris), the fox (C. vulpes) the badger (Meles
taxus), the polecat (Putorius fœtidus), the marten (Mustela foina), the
raccoon (Procyon lotor), the hippopotamus and the cat, and many
other mammals (rodents and carnivora); Trichinellæ have been artificially
introduced, by administering the encysted stage, into the dog,
the mole (Talpa europæa), the mouse (Mus musculus), the hare (Lepus
timidus), the rabbit (L. cuniculus), the hedgehog (Erinaceus europæus),
the marmot (Cricetus vulgaris), the vole, the dormouse, the sheep, the
calf, the horse, etc. Human beings and the pig, rat, mouse, guinea-pig
and rabbit are most easily infected; less easily the sheep, calf and
horse; with difficulty the cat, dog and badger. Trichinella can also
be reared in birds (fowl, pigeon and duck), but the young do not
encyst in the muscular system, but are expelled with the fæces. By
cold-blooded animals as well as by insects (Calliphora vomitaria),
encysted Trichinellæ are evacuated without undergoing any change,
but they will still develop if subsequently ingested, say, by rabbits.
According to Gujon, however, Trichinella can develop in salamanders,
because he has found Trichinella of the muscles in these animals
after they had been fed on encysted specimens. A high temperature
(30°C.) must be provided in which to keep the experimental animals
to ensure the success of the infection.
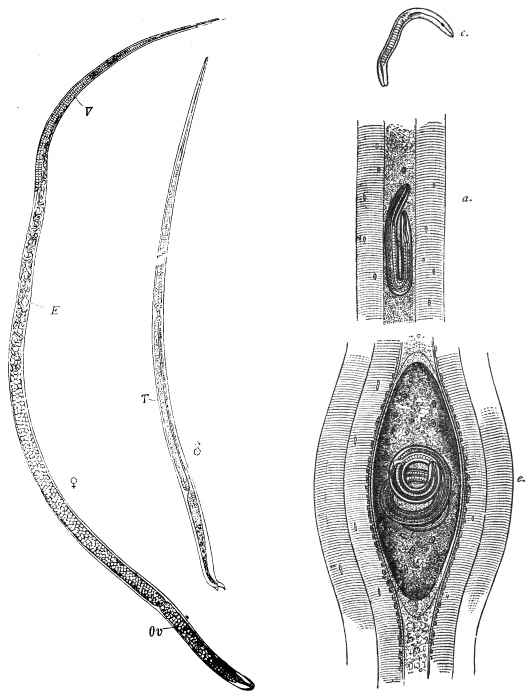
Fig. 302.—Trichinella spiralis. ♀, mature female: E, embryos; V, vulva; Ov, ovary.
♂, mature male: T, testes. c., newly born larva. d., larva in the muscles. e., encapsuled larva
in the muscles. Magnified. (After Claus.)
History.—Encapsuled Trichinellæ had been observed in London by Peacock
(1828) and by J. Hilton (1833) in the muscular system of man; soon after
(1835), Paget found them in London in an Italian who had died of tuberculosis,
and recognized them to be encysted entozoa, which R. Owen described as Trichina
spiralis. Soon after, some further observations were reported on the occurrence of
encysted Trichinellæ, in man, in England, Berlin, Heidelberg, Denmark, North
America; they were also found in the pig (Leidy, Philadelphia) and the cat (Herbst,
Göttingen, and Gurlt, Berlin). Herbst even succeeded in infecting a badger with
encysted Trichinellæ, and subsequently infected two dogs with the flesh of this
badger (1850). In 1855 R. Leuckart (Giessen) also commenced feeding experiments,
and, like Küchenmeister and Virchow (1859), first went on the wrong track because
it was believed at that time that Trichinellæ were the larvæ either of Trichocephalus
or Strongylus. Nevertheless, these experiments yielded some important results;
they showed that Trichinellæ become adult in the intestine within a few days, and
that the females are viviparous (Leuckart). Until that time Trichinellæ had been
regarded as fairly harmless guests of man, but opinions soon changed when Zenker
in Dresden (January, 1860), in performing the autopsy of a girl, aged 10, who had
entered the hospital with typhoid symptoms and there died, found Trichinellæ (not
yet encysted) in the muscles; the intestinal lesions characteristic of typhoid were
lacking, but numerous adult Trichinellæ were found in the intestine. Inquiries
elicited the fact that at about Christmas time the girl had been taken ill after eating
pork, and at the same time the butcher from whom the meat was bought as well
as several of his customers fell sick: the pickled pieces of the same meat were full
of Trichinellæ. In the face of this information it was not difficult to ascertain the
cause of the disease and the manner of infection in Zenker’s case, and it was not
long before Leuckart, Virchow and Zenker were able by renewed experiments to
demonstrate the cycle of development of Trichinella spiralis. Similar investigations
followed by Claus in Würzburg, Davaine in Paris, Fuchs and Pagenstecher in
Heidelberg, etc.
Hardly had Zenker’s case been published than numerous observations on
trichinosis in man appeared, some referring to isolated cases, others to small or
great epidemics, and nearly all from North Germany. The worst epidemic was
that of Hadersleben (1865), in which place, numbering hardly 2,000 inhabitants,
337 persons were taken ill within a short time, and of these 101 died. The source
of infection proved to be a single pig, the flesh of which had been mixed with that
of three other pigs; 200 of the badly infected persons had exclusively eaten raw
pork.
Moreover, it soon became clear that epidemics of trichinosis had been observed
in Germany prior to 1860, but that their nature had not been recognized, although
in a few cases Trichinellæ had been found in the muscles of those who had
succumbed.
History of the Development of Trichinella spiralis.
Shortly after their introduction into the intestine of experimental
animals the encysted Trichinellæ escape from their capsules, which
are destroyed by the gastric juices, and they then enter the duodenum
and jejunum, where they become adult. During this period they do
not grow much, the males from 0·8 to 1·0 to 1·2 to 1·5 mm.; the
females to 1·5 to 1·8 mm. Soon after copulation, which takes place
in the course of two days, the males die; the females, which during
the following days attain a length of 3 to 3·5 mm., either bore more
or less deeply into the villi or, by means of Lieberkühn’s glands, into
the mucous membrane (Askanazy, Cerfontaine, Geisse), and thus
usually attain the lymph spaces. A few also pierce the intestinal wall
and are then found in the mesentery and glands. The females deposit
their young, the number of which, according to Leuckart, averages at
least 1,500, in the lymph spaces; the newly born larvæ measure 90 µ to
100 µ in length, 6 µ in diameter, and they do not appear to increase in
size during their migrations. The migrations are mostly passive, that
is to say, the larvæ are carried along mainly by the lymph stream to
the heart, but sometimes they are active, as may be inferred from the
fact that young Trichinellæ are found in various parts of the intestinal
wall beyond the chyle and lymph spaces, as well as in abundance
in the abdominal cavity. Trichinellæ occur in the heart’s
blood of artificially infected animals seven to twenty-three days
after infection. If scanty, dilute the blood with about ten times the
amount of 3 per cent. acetic acid and centrifugalize.
The young brood is distributed from the heart throughout the
entire body, but the conditions necessary to its further development
are found only in striated muscle; the young Nematodes penetrate
the capillaries, attain the intramuscular connective tissue and then
invade the fibres (Virchow, Leuckart, Graham304). On the ninth or
tenth day after infection the first Trichinellæ have reached their
destination; but further invasions are constantly taking place because
the intestinal Trichinellæ live from five to seven weeks, and continue
to produce their young.
Symptoms.—(1) Period of invasion: Gastro-intestinal symptoms—nausea,
vomiting, watery diarrhœa, colic. Muscular pains may occur
even at this period. Recurrent abdominal pains about the eighth day,
a temporary œdema. Embryos are abundant in the serous cavities.
(2) Period of dissemination: Second week. Myositis, variable in
amount, is the predominant symptom. The biceps and calf may be
hard and tender. Mastication, speech, respiration, etc., may be
difficult and painful. Dyspnœa may be intense. Temperature
104° to 105° F.
(3) Period of encystment: Symptoms of marked cachexia. Third
week: Second period of œdema, especially of face. Delirium,
somnolence, lung affections. Death or gradual subsidence of
symptoms in mild cases.
Eosinophilia (50 per cent. or more) is present.
In consequence of the new batches of young produced during several weeks,
the above-mentioned symptoms of disease are often considerably aggravated; the
fever increases, delirium may arise, and infiltration of the lungs, fatty degeneration
of the liver and inflammation of the kidneys may ensue; the initial slight œdema
may extend, the strength dwindles, and in many cases the patients succumb to the
trichinosis. In severe cases improvement of the condition is only apt to occur
in the fourth or fifth week; the convalescence is always protracted.
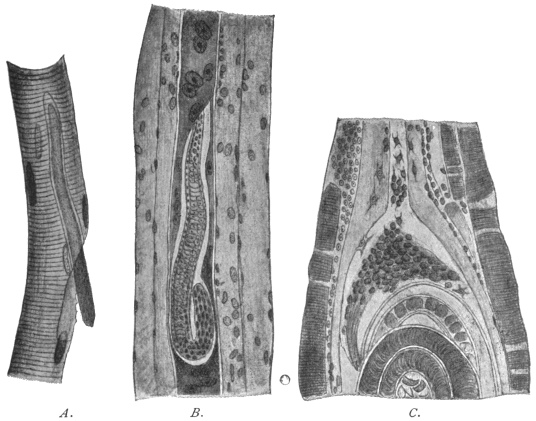
Fig. 303.—A., isolated muscular fibre of a rat, invaded by Trichinella. 510/1. B.,
section through the muscle of a rat; the infected fibre has lost its transverse striation; its
nuclei are enlarged and multiplied. 310/1. C., portion of a Trichinella capsule, at the pole
of which connective tissue cells are penetrating the thickened sarcolemma. (After Hertwig-Graham.)
The muscular fibres attacked degenerate, the transverse striation
at first disappearing; the fibres then assume a granular appearance,
the nuclei multiply and become enlarged, and are surrounded by an
area of granular material, which stains more deeply than the remaining
contents of the sarcolemma. Two or three weeks after infection, the
spirally rolled-up Trichinellæ have grown to 0·8 to 1·0 mm., and in
their vicinity the muscular fibre is swollen, spindle-shaped, and the
sarcolemma is glassy and thickened. The inflammation also extends
to contiguous fibres, especially to the intramuscular tissue, which
proliferates greatly, especially in the vicinity of the degenerated fibres.
While the latter become more and more absorbed, the capsule is
formed by the inflamed connective tissue, which, penetrating into the
glassy and thickened sarcolemma from the poles of the spindle, forms
the cystic membrane. According to other authorities, the larvæ settle
in the intermuscular connective tissue which forms the cyst and not
in the muscular fibres within the sarcolemma. The cysts are lemon-shaped
and usually lie with their longitudinal axis in the direction of
the muscular fibres; on an average they measure 400 µ in length by
250 µ in breadth.
Later on fat cells appear at their poles, and after about six or
nine months they commence to calcify, the process starting at the
poles (fig. 305). Finally, sometimes after the lapse of years, the
captive Trichinellæ themselves become calcified.
Fig. 304.—Calcified Trichinella in the
muscular system of a pig; the capsules are
not calcified. (After Ostertag.)

Fig. 305.—Various phases of the calcification of
Trichinella of the muscles, which starts at the poles
of the capsule.
According to experience, Trichinellæ are not evenly distributed
in the muscular system of pigs; the diaphragm, the muscles of the
larynx, tongue, abdomen and intercostal spaces are their favourite
positions; this predilection for the respiratory muscles is explained
by their regular contractions, owing to which regular narrowings
of the capillaries take place, thus favouring the settling of the circulating
Trichinellæ. The same circumstance probably explains the
frequency of the parasites in the tongue.
Possibly also the Trichinellæ that bore direct through the intestine
may, from the abdominal cavity, penetrate the muscles in the vicinity.
Frequently also encysted Trichinellæ are found in remarkable numbers
in the vicinity of the points of insertion of the tendons, this proclivity
being probably connected with the fact that the Trichinellæ
first of all wander into the muscular fibres and find a natural barrier
at the points of insertion of the tendons.
The Trichinellæ, in their encysted condition, may remain alive
and capable of development for many years—in the pig eleven years
and in man as much as twenty-five to thirty-one years. Encystment,
however, is not a necessary condition for the development of the
brood, that is to say, Trichinellæ which reach the gut of suitable
animals become sexually mature and multiply provided that they have
developed so far as to possess a rudimentary genital spot, which
occurs when the body is 0·5 to 0·75 mm. long, but all the same
a great part of non-encapsuled Trichinæ perish on their passage
through the stomach.
The black rat (Mus rattus), and more particularly the sewer rat
(Mus decumanus305), are the normal hosts of Trichinella spiralis. These
animals, especially the last-named species, infect themselves very
easily, as they are cannibalistic, and they also transmit trichinosis
to other species by which they are devoured, such as pigs, dogs,
cats, foxes, bears and martens. Rats are infected also by the ingestion
of fæcal matter from infected animals which contains
trichinæ (Höyberg). Man becomes infected with Trichinella by
eating the flesh, insufficiently cooked, of infected pigs, also, but
more rarely, by eating the infected flesh of wild boars, dogs, cats,
bears and foxes.
The infection of pigs may likewise take place by their having access to the offal
of trichinous pigs, or being actually fed on it. These are, however, exceptions,
which, as a matter of course, are of great importance in certain places. As
a matter of fact, the rats examined for Trichinella were always found to be severely
infected. Thus Billings, in the knackers’ yard at Boston, found that 76 per cent.
of the rats were infected, and in an export slaughterhouse 100 per cent. were found
to harbour the parasite; in the city of Boston 10 per cent. of the rats had trichinosis.
Heller found that of 704 rats, from twenty-nine different places in Saxony, Bavaria,
Würtemberg and Austria, 8·3 per cent. were infected with Trichinellæ; of the rats
caught in the knackers’ yards, 22·1 were diseased; of those taken in slaughterhouses,
2·3 were infected, and of rats from other localities only 0·3 per cent.
harboured the parasite. Leisering found almost the same figures, but in rats from
slaughterhouses 5·3 per cent. were infected.
The geographical distribution of T. spiralis does not correspond
with the occurrence of trichinosis in man; local customs are
an important factor; for instance, the custom of eating pork in
a condition that does not affect the life of the enclosed trichinella.
In places where such customs do not prevail, epidemics do not
occur—at the most there are isolated cases of the disease, although
there be a great number of infected pigs. The following conditions
prevail in North America: In Boston, Billings found that 4 to 5·7 per
cent. of the pigs examined were trichinous; Belfield and Atwood
found that 8 per cent. were infected in Chicago; Salmon found
on an average that 2·7 per cent. were infected (but at various places
the percentage fluctuated between 0·28 to 16·3 per cent.), yet epidemics
of trichinosis hardly ever occur in North America, and only isolated
cases of the disease are met with in German immigrants, who keep
to their native customs.
This report, according to the researches of H. U. Williams, must be considerably
modified. This author has examined the muscular system of human cadavers
according to the method employed by inspectors of meat for pigs. The investigations
were conducted in the Pathological Institute of the University of Buffalo, and
the observer has examined 505 bodies since 1894, of which 27 (= 5·34 per cent.)
were invaded by Trichinella. The cases, according to the nationality, are divided
as follows:—
| Examined | Trichinella | Percentage
of positive
results |
Absent | Present |
| Americans: | | | |
| (a) Whites | 207 | 201 | 6 | 2 | ·89 |
| (b) Negroes | 70 | 65 | 5 | 7 | ·14 |
| British and Irish | 62 | 57 | 5 | 8 | ·06 |
| Canadians | 12 | 10 | 2 | 16 | ·66 |
| Germans | 49 | 43 | 6 | 12 | ·24 |
| Italians | 12 | 10 | 2 | 16 | ·66 |
| Other nationalities | 27 | 27 | 0 | 0 | |
| Nationality unknown | 66 | 65 | 1 | 1 | ·51 |
Total | 505 | 478 | 27 | 5 | ·34 |
It is worthy of remark that half of all the positive cases were mental patients,
who were found to be affected with Trichinella to well-nigh 12 per cent. Trichinosis
was not, however, the cause of death in any case. Very frequently the Trichinellæ
were found calcified and dead.
Conditions are similar in most countries of Europe, where, of
course, the number of infected pigs is considerably smaller, but
the disease depends less on this than on the way in which the
pork is prepared.
Cases of trichinosis have been known to occur in nearly all
the countries of Europe; further, in Egypt, Algeria, East Africa,
Syria, India, Australia, and America. North Germany, more
especially the Saxe-Thüringian states, is the classical land for
epidemics of trichinosis; the mortality varies, but it may be
very high.306
Prophylaxis.—The grave nature of the disease and the comparatively high
mortality relating to trichinosis led the authorities to adopt certain preventive
measures, which are the more necessary as national customs cannot be altered in
a short time. As the usual process of pickling and smoking, even when long
continued, does not certainly ensure the death of the Trichinellæ contained in the
meat, and also because in roasting and boiling large pieces of pork a considerable
time is necessary to permit the temperature required to kill off the parasites
(62° to 70° C.) to penetrate to the middle of the joint, it appeared to be most
practical to have all pigs microscopically examined for Trichinellæ before they, or
parts of them, were placed on the market, and all infected meat condemned, no
matter whether the Trichinellæ were present in large or small numbers, still
undeveloped or calcified. Since 1877 obligatory examination of pork has been
introduced in Prussia, though as yet it is not thoroughly carried out; other states
of North Germany as well as the larger towns of South Germany soon followed;
a complete army of trichina inspectors, officially examined and periodically controlled
by experts, and whose number in Prussia amounted to 27,602 in 1896,
this being even increased to 28,224 in 1899, have the charge of examining
pork on certain lines laid down. These are at the present time uniformly
administered. The proceeding is usually that the trichina inspector himself goes to
the slaughterhouses, or special samplers take pieces of the muscles that are known
to be the favourite seats of the parasite (pillars of the diaphragm, the costal part of
the diaphragm, muscles of the tongue and larynx, intercostal and abdominal muscles);
six small portions are separated from each piece, pressed between slides or special
compressors, and carefully gone through by examining them with a low power of
the microscope. The pigs free from Trichinellæ are passed for commerce; trichinous
pigs, on the other hand, in Prussia, are only allowed to be used for industrial
purposes, i.e., the hide and bristles are used, the fat is allowed to be melted down,
or certain parts are used for the manufacture of soap or glue. In Saxony, however,
it is still permitted to place trichinous flesh on the market, fully declaring its nature,
and after having been heated to its deepest strata at a temperature of 100° C. in a
suitable apparatus, and under the supervision of a veterinary surgeon.
As to the proportion of trichinous pigs to healthy ones, the following
tables give the figures for Prussia:—
Year | Number of
pigs examined | Number of
trichinous pigs | Proportion |
1878 | 2,524,105 | 1,222 | 1 : 2,065 |
1879 | 3,164,656 | 1,938 | 1 : 1,632 |
1881 | 3,118,780 | 1,695 | 1 : 1,839 |
1882 | 3,808,142 | 1,852 | 1 : 2,056 |
1883 | 4,248,767 | 2,199 | 1 : 1,932 |
1884 | 4,611,689 | 2,624 | 1 : 1,741 |
1885 | 4,421,208 | 2,387 | 1 : 1,852 |
1886 | 4,834,898 | 2,114 | 1 : 2,287 |
1887 | 5,486,416 | 2,776 | 1 : 1,988 |
1888 | 6,051,249 | 3,111 | 1 : 1,945 |
1889 | 5,500,678 | 3,026 | 1 : 1,818 |
1890 | 5,590,510 | 1,756 | 1 : 3,183 |
1891 | 6,550,182 | 2,187 | 1 : 2,996 |
1892 | 6,234,559 | 2,085 | 1 : 2,992 |
1896 | 8,759,490 | 1,877 | 1 : 4,666 |
1899 | 9,230,353 | 1,021 | 1 : 9,040 |
1902 | 9,093,210 | 725 | 1 : 12,397 |
The proportion, however, is not only subject to variation in separate years, but
differs according to the district; thus, in 1884, in the state district of Minden there
was one trichinous pig to 30,146 healthy animals, in Erfurt 1 to 14,563, in the district
of Gnesen 1 to 101, in Schrimm 1 to 86, and in Schroda 1 to 68.
In Germany Trichinella is becoming LESS COMMON in pigs (Ostertag):—
(a) Prussia. |
Year | | Pigs found
to be trichinous |
| 1878–1885 | | 0· | 061– | 0·048 | per cent. |
| 1886–1892 | | 0· | 033– | 0·043 | " |
| 1896 | | | | 0·021 | " |
| 1899 | | | | 0·014 | " |
| 1902 | | | | 0·011 | " |
(b) Saxony. |
Year | | Number of pigs
found to be trichinous |
| 1891 | | 0· | 014 | per cent. |
| 1892 | | 0· | 011 | " |
| 1893 | | 0· | 008 | " |
| 1894 | | 0· | 007 | " |
| 1895 | | 0· | 012 | " |
| 1896 | | 0· | 0102 | " |
| 1899 | | 0· | 004 | " |
| 1902 | | 0· | 0056 | " |
(c) City of Berlin. |
Year | | Number of pigs
found to be trichinous |
| 1883–1893 | | 0· | 035– | 0·064 | per cent. |
| 1893–1897 | | 0· | 022– | 0·015 | " |
| 1902 | | 0· | 006 | | " |
There is no doubt that the excellent preventive measure of
official inspection for Trichinella has led to the avoidance of grave
disasters; its introduction has not yet caused an entire cessation
of trichinosis in man, because inspection of pork is not obligatory
everywhere, so that human beings may become infected by
unexamined trichinous pigs from their own country or from abroad,
and also because an infection may occasionally escape notice. For
these reasons the meat imported into Berlin from abroad as free
from Trichinæ is examined again and not always in vain; finally, also,
gross negligence may at times occur, or fatal errors may be made.
In addition private prophylaxis must not be neglected, and its
chief aim should be directed to the suitable preparation of pork.
Family. Dioctophymidæ.
Genus. Dioctophyme, Collet-Megret, 1802.
Syn.: Eustrongylus, Dies., 1851.
Large worms. Anterior extremity unarmed; the mouth is surrounded by six
papillæ. One ovary. The vulva is in the anterior region of the body.
Dioctophyme gigas, Rudolphi, 1802.
Syn.: Dioctophyme renale, Goeze, 1782; Ascaris canis et martis, Schrank, 1788;
Ascaris visceralis et renalis, Gmelin, 1789; Strongylus gigas, Rud., 1802; Eustrongylus
gigas, Dies., 1851; Strongylus renalis, Moq. Tand., 1860; Eustrongylus visceralis,
Raill., 1885.
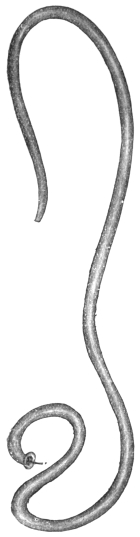
Fig. 306.—Dioctophyme
gigas, male.
Natural size. (After
Railliet.)
Colour blood-red; the anterior extremity somewhat slender;
there is a series of about 150 papillæ along the lateral lines; the
sub-median lines are strongly developed, and from them spring the
radial muscles for the intestine.
The males attain a length of 40 cm. and a diameter of 4 to
6 mm.; the posterior extremity is transversely truncated; the anal
orifice is within the base of the collar-like bursa, the thickened edges
of which are beset with papillæ; the spicule measures 5 to 6 mm.
in length.

Fig. 307.—Eggs of
Dioctophyme gigas;
above seen from the
flat, below in optical
section. 400/1. (After
Railliet.)
The females attain a length of 100 cm. and a breadth of 12 mm.
The anus is crescent-shaped and terminal. The vulva is 50 to
70 mm. distant from the anterior extremity. The eggs are oval and
have a thick shell presenting numerous depressions; the shell itself
is brownish, but it is colourless at the somewhat thickened poles;
it measures 60 µ in length by 40 µ in breadth. The larva measures
240 µ by 14 µ.
Dioctophyme gigas lives in the pelvis of the kidney, more rarely in
the abdominal cavity of the seal, otter, dog, wolf, fox, horse, marten
and polecat, exceptionally also in human beings. It also occurs in
tumours of the mamma and perinæum. Most of the cases in which
this parasite has been reported as occurring in man may be traced
back to unrecognized Ascaris lumbricoides or to clots of fibrin; seven
certain cases, eight more or less doubtful, however, remain.
The source of infection is unknown, but according to Balbiani the
eggs develop an embryo in water or moist soil,
and this embryo may remain alive several years
without hatching; the infection of dogs with
embryo-containing eggs did not succeed; an
intermediate stage in fishes is conjectured,
but still the infection of cattle and horses is
unintelligible.
Family. Strongylidæ.
Sub-family. Metastrongylinæ, Leiper, 1908.
Buccal capsule absent or
slightly developed, vagina elongate,
uteri convergent307 and have
a simple musculature. Parasitic
in the respiratory or circulatory
system. Genera: Metastrongylus,
Synthetocaulus.
Genus. Metastrongylus, Molin, 1861.
Mouth with six lips, of
which the two lateral are the
largest. Postero- and postero-external
rays308 of bursa thin, the
rest thick. Only the median
ray double. Spicules very long
and slender, striated. Vulva
immediately in front of anus.
Eggs contain an embryo when
laid.
Metastrongylus apri, Gmelin, 1789.
Syn.: Gordius pulmonalis apri, Ebel, 1777; Ascaris apri, Gmelin, 1789;
Strongylus suis, Rud., 1809; Strongylus paradoxus, Mehlis, 1831; Strongylus
elongatus, Duj., 1845; Strongylus longevaginatus, Dies., 1851.
The male measures 12 to 25 mm. in length; the bursa is bilobed;
there are five rays in each lobe; the spicules are thin and up to
4 mm. in length. The females measure 20 to 50 mm. in length,
the anus is close in front of the posterior extremity, which
has a recurved, hook-like process; the vulva is close in front of the
anus. The eggs are elliptical, 57 µ to 100 µ in length, 39 µ to 72 µ in
breadth; when the eggs are deposited the embryo is already formed,
220 µ to 350 µ by 10 µ to 12 µ.
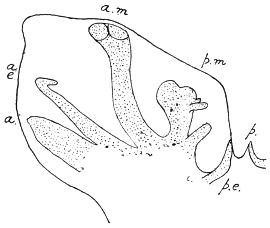
Fig. 308.—Metastrongylus apri: one side
of bursa. a., anterior; a.e., antero-external;
a.m., antero-median; p.m., postero-median;
p.e., postero-external; p., one division of
posterior ray. (Stephens.)
Metastrongylus apri frequently lives in the bronchial tubes—usually
the smaller ones—of the pig309 and wild boar; it is also found occasionally
in sheep and in man; in young pigs it is apt to set up
a bronchitis, which frequently causes death.
The first communication as to the occurrence of this species in
man was that of Diesing, who, in 1845, in Klausenburg, had the
opportunity of examining Strongylidæ found by Jortsits in the lung
of a little boy, aged 6, in Transylvania; probably also the Nematodes
found in the trachea and
larynx of man, and described by
Rainey and Bristowe as specimens
of Filaria trachealis, belong to
this group; according to Chatin,
Metastrongylus apri may also occur
in the intestine of man; this occurrence,
however, may in all
probability have been due to an
accidental introduction of adult
worms into the intestine, and
should not be attributed to an
infection by the larval stage.
No experiments to induce infection
have been made; it is probable,
however, that infection is
direct and without the aid of an
intermediate host.
Sub-family. Trichostrongylinæ, Leiper, 1908.
Strongylidæ with buccal capsule absent or slightly developed,
vagina short, uteri divergent (i.e., anterior and posterior), ovejectors
differentiated. Parasitic in the alimentary canal. Contains the
genera Trichostrongylus, Hæmonchus, Ostertagia, Nematodirus,
Cooperia, Dictyocaulus.310
Genus. Trichostrongylus, Looss, 1905.
Very small Strongylidæ. Mouth with three small lips and nodular
or punctiform papillæ. Cervical papillæ absent. Bursa entirely
closed, with large lateral lobes, and median lobe not distinctly defined.
Anterior311 rays double, the branches widely divergent, one thin,
the other thick, and close to the antero-median. The postero-median
ray is thin and close to the postero-external. Posterior ray
bifurcate, each branch bifid at the tip (fig. 311). Spicules short, spoon
or spatula-like, with on the broad anterior end a lateral knob or disc
and in front of the point an angular projection. Gubernaculum of a
peculiar canoe or shoe shape in profile. Vulva in the hinder half of
the body. Tail with two minute papillæ just in front of tip. Eggs
thin shelled; when laid they show eight to thirty-two segments.
Parasitic in duodenum, seldom in the stomach of herbivora.
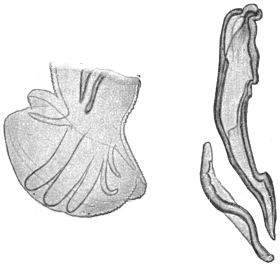
Fig. 309.—Trichostrongylus instabilis: left,
posterior end of male; right, spicule and
gubernaculum, side view. Cf. fig. 311. Magnified.
(After Looss.)
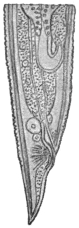
Fig. 310.—Trichostrongylus instabilis:
posterior end of female. Magnified.
(After Looss.)
Trichostrongylus instabilis,312 Railliet, 1893.
Syn.: Strongylus instabilis, Railliet, 1893; Strongylus subtilis, Looss, 1895.
Male 4 to 5·5 mm. long, 0·08 mm. thick in front of bursa.
Spicule 0·135 to 0·145 mm. long, accessory piece (gubernaculum)
0·07 mm. thick. Antero-external ray usually thickest of all, occasionally
only as thick as the
antero-median; postero-median
far more slender than the antero-external
and antero-median and
nearer to the postero-external than to the antero-median. Female
5 to 6 mm. long, vulva 1·05 to 1·2 mm. distant from the tip of the
tail, placed longitudinally, 50 µ to 55 µ long, always shorter than
the unpaired portion of the canal formed by the union of the two
ovejectors; anus 0·055 to 0·07 mm. distant from tip of the tail;
ova 73 µ to 80 µ by 40 µ to 43 µ.
This species lives in the duodenum, exceptionally also in the
stomach of Ovis aries, O. laticauda, Antilope dorcas, Camelus dromedarius
(Egypt), Cynocephalus hamadryas (North Africa), sheep and
goats (France), and has been found by Looss in bodies of fellaheen
at Alexandria and in the stomach of a Japanese female by Ijima.
Trichostrongylus probolurus, Railliet, 1896.
Syn.: Strongylus probolurus, Railliet, 1896.
Male 4·5 to 5·5 mm. long, in front of bursa 0·08 mm. thick;
spicule 0·126 to 0·134 mm. long, gubernaculum 0·075 to 0·08 mm.
long. Bursa: latero-anterior rib thickest; antero-external thicker
than antero-median, postero-median and postero-external very short
and close together. Female 4·5 to 6 mm. long, vulval opening
1·08 to 1·25 mm. from tip
of tail, placed longitudinally,
and slightly curved, 76 µ long,
always longer than the unpaired portion of the ovejector; anus 0·040
to 0·05 mm. distant from tip of tail. Posterior end thick, point of tail
short. Ova 76 µ to 80 µ by 43 µ to 46 µ.

Fig. 311.—Trichostrongylus probolurus: tail of
male from left side. d., posterior; e.d., postero-external;
p.l., postero-median; m.l., antero-median;
e.l., antero-external; l.v., latero-anterior;
v.v., antero-anterior; gub., portion of gubernaculum;
sp., portion of spicules. × c. 300.
(After Looss.)
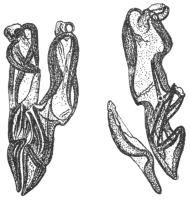
Fig. 312.—Trichostrongylus probolurus:
spicules and gubernaculum of male; on
left, ventral view; on right, lateral view.
× c. 300. (After Looss.)
Habitat.—In the duodenum of Ovis aries, O. laticauda, Antilope
dorcas, Camelus dromedarius (Egypt) and occasionally also in man
(Egypt).
Trichostrongylus vitrinus, Looss, 1905.
Male 4 to 5·5 mm. long, in front of bursa 0·085 mm. thick.
Bursa larger than in the other two species, antero-external rib thickest,
antero-anterior and postero-median equally thick, straight. Spicule
0·16 to 0·17 mm. long, gubernaculum 0·085 to 0·095 mm. long.
Female 5 to 6·5 mm. long, vulval opening 1·15 to 1·25 mm. distant
from tip of tail, crescent shaped, oblique to body axis, and around
it irregular thickenings. Ova 84 µ to 90 µ by 46 µ to 50 µ.
In duodenum of Ovis
aries, O. laticauda, occasionally
in Camelus dromedarius
and in man (Egypt).
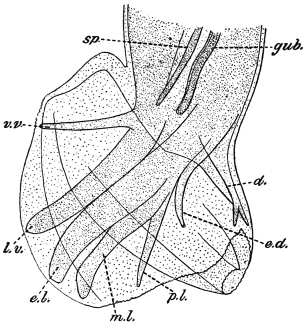
Fig. 313.—Trichostrongylus vitrinus: tail of male
from left side. d., posterior; e.d., postero-external;
p.l., postero-median; m.l., antero-median; e.l., antero-external;
l.v., latero-anterior; v.v., antero-anterior;
gub., portion of gubernaculum; sp., portion of spicule.
× c. 300. (After Looss.)
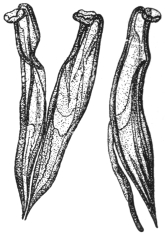
Fig. 314.—Trichostrongylus
vitrinus: spicules and gubernaculum;
on left, ventral
view; on right, lateral view.
× c. 300. (After Looss.)
Genus. Hæmonchus, Cobb., 1898.
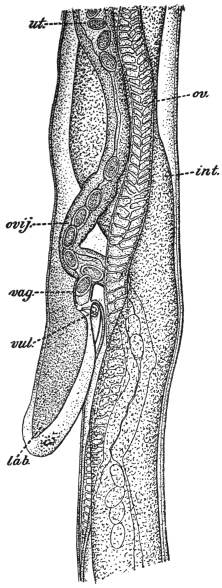
Fig. 315.—Hæmonchus contortus:
vulval region of female viewed from
left side. int., intestine; lab., linguiform
process covering vulva; ov., ovary;
ovij., ovejector; ut., uterus; vag.,
vagina; vul., vulva. × 75. (After
Ransom.)
Small mouth cavity contains a “tooth” or “lancet” arising from
the dorsal side. Cuticle of head and neck not inflated. Cervical
papillæ well marked. Bursa bilateral, with large lateral lobes and a
small dorsal lobe not median, but lateral, attached to the base of one
of the lateral lobes (fig. 316). Posterior ray bifurcate, each branch
bifid apically. Each lateral lobe six rays. Anterior rays separated
distally, curving forward. Antero-median and postero-median rays
distally curve away from the antero-external. Postero-external ray
long and slender. Spicules less than 1 mm. Gubernaculum present.
Vulva in posterior part of body covered by a prominent tongue-like
flap. Eggs ellipsoidal.
Hæmonchus contortus, Rudolphi, 1803; Cobb., 1898.
Dorsal “tooth” or “lancet” 10 µ to 15 µ long. Cervical papillæ
0·3 mm. from head.
Male 20 mm. long by 400 µ thick (maximum). Asymmetrical lobe
of bursa 150 µ by 125 µ attached to left lateral lobe. Posterior
ray bifurcate; each branch bifid. Stem of ray less than twice as long as
its branches. Spicules 300 µ to 500 µ with knobbed tips, and the left
spicule with a barb 20 µ from the tip, right spicule with a barb 40 µ
from tip. Gubernaculum 200 µ by
25 µ to 35 µ, fusiform with thickened
edges.
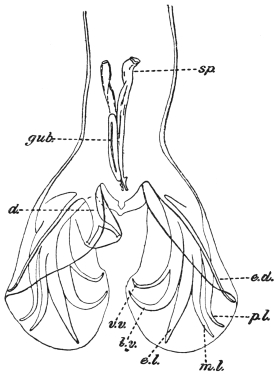
Fig. 316.—Hæmonchus contortus: tail of
male, dorsal view, d., posterior ray of the asymmetrically
placed posterior lobe; e.d., postero-external;
p.l., postero-median; m.l., antero-median;
e.l., antero-external; l.v., latero-anterior;
v.v., antero-anterior; gub., gubernaculum;
sp., spicule. × 75. (After Ransom.)
Female 18 to 30 mm. by 500 µ
(maximum). Vulva 3 to 4·5 mm.
from tip. Linguiform flap 0·5 mm.
(a second one exists, according to
Brumpt). Anus 400 µ to 630 µ from
tip. Tail acutely pointed. Eggs 75 µ
to 95 µ by 40 µ to 50 µ.
Habitat.—Fourth stomach of cattle, sheep, antelope.
Distribution.—Europe, America, Africa, Asia, Australia, New
Zealand. Once in man in South America by de Magalhães.
Pathology.—Produces anæmia, emaciation, dropsy in sheep; and
in the human case the symptoms were mistaken for those of
ancylostomiasis.
Life-history.—Rhabditic embryos easily hatch in water, then
moult several times, becoming eventually “filariform” larvæ enclosed
in the moulted skin. These crawl up blades of grass and are
swallowed by sheep, etc.
Genus. Nematodirus, Ransom, 1907, emend. Railliet, 1912.
Head over 50 µ in diameter. Cuticle may be slightly inflated and
often transversely striated. Cuticle with eighteen distinct longitudinal
ridges. Cervical papillæ absent (?). Posterior lobe of bursa
reduced to short lobules each with a dorsal ray. Antero-anterior
+ latero-anterior (= anterior double) rays close together, parallel;
antero-external ray diverges widely from antero- and postero-median,
which are close together and parallel. Postero-external ray slender.
Spicules more than 0·5 mm. long, at most one-twelfth of body, united
by a membrane throughout their length or only distally. Gubernaculum
absent. Vulva behind middle of body. Eggs ellipsoidal,
shell rather thick.
Habitat.—Duodenum of ruminants.
Sub-genus. Mecistocirrus, Railliet, 1912.
Head slightly inflated, with transverse striations. Skin with
eighteen longitudinal ridges, but little apparent; cervical papillæ
distinct. Bursa bilobed; median ray double (= postero-median +
antero-median); very large antero-external at the edge, close to the
anterior. Spicules very long, slender, one-sixth length of body
(3·5 mm.); tail pointed. Vulva immediately in front of anus.
Habitat.—Stomach of ruminants.
Mecistocirrus fordi, Daniels, 1908.
Syn.: Strongylus fordi, Daniels, 1908; Strongylus gibsoni, Stephens, 1909;
Nematodirus fordi, Leiper, 1911.
Male 21 mm. long by 0·4 mm. thick. Cervical papillæ 0·45 mm.
behind the head. Spicules about 7 mm. long, i.e., one-third of the
body length. At the level of the postero-external rays of the bursa,
the bursa has a projecting lobule.
Fig. 317.—Mecistocirrus fordi: bursa of male, dorsal view. The rays
are (1) postero-external, (2) median (= postero-median + antero-median), (3)
antero-external, (4) latero-anterior, (5) antero-anterior. These two latter are
parallel. The posterior ray is absent. (After Stephens.)
Female 25 mm. long. Anus 0·2 mm., vulva 0·5 mm. from the tip
of tail. Eggs 100 µ by 53 µ.
Sub-family. Ancylostominæ, Railliet, 1909.
Strongylidæ with buccal capsule, well developed. Uteri divergent.
Parasitic in the alimentary canal, exceptionally in the respiratory
system.
Group. Œsophagostomeæ, Railliet and Henry, 1909.
Bursa with anterior and median ray cleft (not double), postero-
and postero-external arising from a common trunk, posterior bifurcated,
each limb bidigitate.
Contains at present four genera: (1) Ternidens, (2) Chabertia,
(3) Œsophagostomum, (4) Agriostomum.
Genus. Ternidens, Railliet, 1909.
Buccal capsule sub-globular, opening obliquely in the dorsal surface,
and having at the bottom three complex teeth resembling those of
Triodontophorus.313 Two crowns of leaflets; peristomic collar moderate,
edge of bursa slightly toothed.
Type.—T. deminutus, Railliet and Henry.
Ternidens deminutus, Raill. and Henry, 1905.
Syn.: Triodontophorus deminutus, Raill. and Henry, 1905.
Body relatively thick. Cervical papillæ 0·5 mm. behind the head.
Buccal capsule 40 µ deep. Teeth 40 µ long.
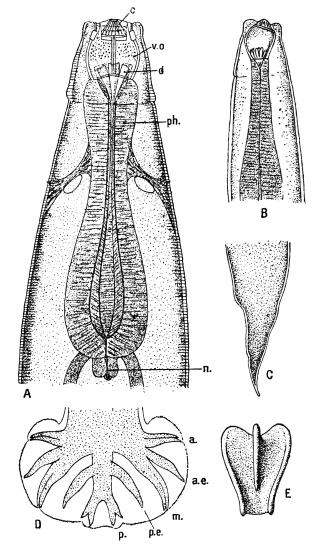
Fig. 318.—Ternidens deminutus. A, head end, ventral view: c, crown of
leaflets; v.o., buccal cavity; d, pharyngeal plates; ph., pharynx; n., valve.
B, lateral view. C, tail of female. D, bursa of male: a., anterior ray; a.e.,
antero-external; m., median; p.e., postero-external; p., posterior. E, pharyngeal
plate. Enlarged. (After Railliet and Henry.)
Male 9·5 mm. long by 560 µ thick. Œsophagus 660 µ long.
Bursa broader than long, the lateral lobes united by a small posterior
lobe with slightly sinuous margin; edge of bursa finely toothed.
Spicules about 900 µ long.
Female 12 to 16 mm. long by 650 µ to 730 µ thick. Œsophagus
860 µ long. Vulva forms a distinct projection 480 µ from tip of tail.
Anus 240 µ to 270 µ from tip. Eggs 60 µ to 80 µ by 40 µ.
Habitat.—Large intestine of a negro (Comoro Islands) and in the
natives of Nyasaland and Portuguese East Africa. Also in large
intestine of Macacus sinensis and Macacus cynomolgus.
Genus. Œsophagostomum, Molin, 1861.
No teeth. Cuticle around the mouth dilated to form a narrow
cuticular “peristomic collar.” Separated by a constriction from this
is a much more extensive inflation, the “cephalic vesicle,” bounded
abruptly behind on the ventral side by a transverse groove, the
“ventral cleft,” which is always present even in absence of the vesicle.
Buccal cavity of slight depth with a short dorsal tunnel. Internal
margin of the mouth armed with chitinous leaflets (“external crown”);
internal border of the buccal capsule armed with short tongue-like
leaflets (internal crown). Lateral membranous wings may extend
backwards from the ventral cleft. Cervical papillæ present. Bursa
with two lateral lobes united by a smaller median lobe. Spicules
over 5 mm. long, slender; gubernaculum inconspicuous. Vulva
in front of anus. Adults usually in large intestine of ruminants,
suidæ, tapirs, edentates and apes. Larvæ sometimes in nodules in
intestinal wall.
Œsophagostomum brumpti, Railliet and Henry, 1905.
Female immature, 8·5 to 10·2 mm. long, 0·295 to 0·325 mm.
thick. Cuticle transversely striated. The cephalic vesicle immediately
behind the vestibulum oris, embracing the anterior two-fifths
of the œsophagus, extending ventrally, however, towards its posterior
end. Vestibulum oris formed by a cuticular band provided with a
crown of twelve apical leaflets directed forward and inwards; six
cephalic papillæ (two lateral, four submedian); buccal capsule in front
of cervical swelling not delineated circularly behind, but provided
with three wide incisions (one dorsal, two sub-ventral). Œsophagus,
0·470 to 0·500 mm. long, two cervical papillæ at five-eighths of
its length. Vulva 0·350 to 0·475 mm., anus 0·170 to 0·200 mm.,
before tip of tail.
Habitat.—Found by Brumpt in tumours of the cæcum and colon
of a native of the River Omo (Lake Rudolph), East Africa. Immature
forms only were present. Adults have been found in similar tumours
in monkeys.
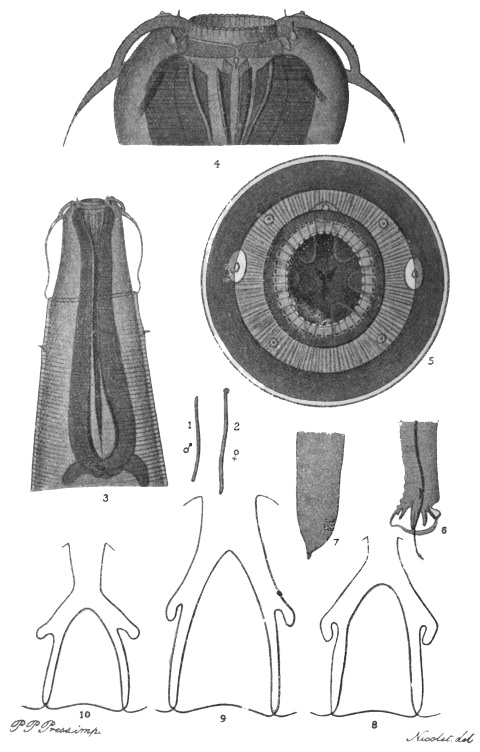
Fig. 319.—Œsophagostomum stephanostomum var. thomasi. 1, male, natural size; 2,
female, natural size; 3, head of female, ventral view, showing cephalic vesicle and ventral cleft
limiting it behind, × 55; 4, head of female, dorsal view, × 225; 5, head of male, end view,
showing external and internal leaf crowns, × 225; 6, tail of male, lateral view (cf. fig. 318, D),
× 20; 7, tail of female, lateral view, × 20; 8, Œs. thomasi, posterior ray of bursa, × 150;
9, Œs. dentigerum, from chimpanzee, posterior ray of bursa, × 150; 10, Œs. stephanostomum,
from gorilla, posterior ray of bursa, × 150.
Pathology.—They occur in hæmorrhagic cysts in the submucosa or
muscularis mucosæ of the gut wall. The cysts project internally and
externally, and contain immature adults, which eventually escape into
the lumen of the gut.
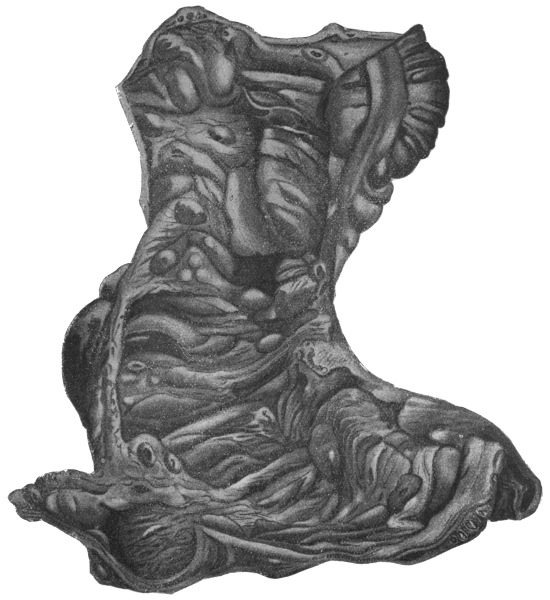
Fig. 320.—Œsophagostomum stephanostomum var. thomasi: cæcum and ascending colon.
Subperitoneal cysts are seen on the top right hand, and in the lumen of the gut numerous
cysts arranged transversely. The small roundish patches are areas of necrosis in the cyst walls.
(After Thomas.)
Œsophagostomum stephanostomum var. thomasi,
Raill. and Henry, 1909.
Body thick, pointed only at the ends. Buccal capsule much
reduced. External crown of thirty-eight leaflets (the “crown” nearest
the centre of fig. 319, 5). Male 17 to 22 mm. long by 750 µ thick.
Spicule 1·380 to 1·475 mm., slightly curved at the tip. Female,
immature, 16 to 20 mm. long by 900 µ thick, tail ending in a little
conical appendage. Anus 230 µ, vulva 500 µ to 525 µ from tip.
Ovejectors close together. Uteri very short in form of oblong pouch.
Œs. stephanostomum, Stossich, 1904, in the large intestine of gorilla.
Œs. stephanostomum var. dentigera, Raill. and Henry, 1909, in the
chimpanzee.
Habitat.—In large and small intestine of man, Brazil.
Pathology.—Nodules occur in the gut wall; 187 were found by
Thomas in his, the sole case. The tumours contain each a single worm.

Fig. 321.—Œsophagostomum stephanostomum
var. thomasi: portion of the ileum, showing a cyst
with protruding worm. × 8. (After Thomas.)

Fig. 322.—Œsophagostomum stephanostomum
var. thomasi: colon with
œsophagostome withdrawn from its cyst
cavity. × 20. (After Thomas.)
Œsophagostomum apiostomum, Willach, 1891.
According to Leiper, Œs. brumpti is identical with, and hence a
synonym of, this species. Parasitic in large intestines of monkeys,
producing dysentery, and in man (Northern Nigeria).
According to Walker this species is common in Philippine monkeys.
Ova are scanty in the fæces. They measure 73 µ to 84 µ by 44 µ to
57 µ and are in the morula stage. They are easily cultivated. The
rhabditiform larva is 340 µ by 16 µ and has a long filiform tail. It
moults twice, and at the second moult becomes a filariform larva
retaining the skin of this moult, this stage being that of the mature
larva. It now measures 9 mm. long by 30 µ thick. Walker suggests
that the mode of infection is similar to that of ancylostomes.
Group. Ancylostomeæ, Railliet and Henry, 1909.
Bursa with anterior ray cleft, median double,314 postero- and postero-external
arising from a common trunk, posterior bifurcate, each limb
being tridigitate. Vulva in posterior third of body. Uteri divergent.
Contains the following genera: (1) Strongylus,315 (2) Ancylostoma,
(3) Uncinaria, (4) Characostomum, etc.
Genus. Ancylostoma, Dubini, 1843, emend. Looss, 1905.
Ventral margin of mouth capsule armed with teeth, the “roots”
of which are continued backwards and appear on the external surface
of capsule as rib-like thickenings. Terminal third of dorsal ray
cleft. Genital tubes very long, with short, closely packed diagonal
convolutions.
Ancylostoma duodenale, Dubini, 1843.
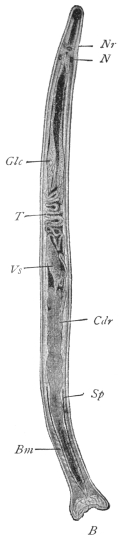
Fig. 323.—Ancylostoma duodenale,
male. B, bursa; Bm, bursal muscles;
Cdr, cement gland surrounding the ejaculatory
duct; Glc, cervical glands; N,
nucleus of cephalic gland; Nr, nerve ring;
T, testes; Sp, spicule; Vs, vesicula seminalis.
× 15. (After Looss.)
Buccal Capsule.—The buccal capsule is bent dorsally, 0·21 mm.
long, 0·19 mm. broad. If a worm is rolled under the cover-glass so
that the dorsal side is upwards, we observe the following features
(fig. 325): In the dorsal edge of the chitinous capsule there is a gap
as if a U-piece had been punched out. This is the “dorsal gap or
incision.” The so-called “dorsal teeth” are simply the rounded
edges of the tips of this gap. They project beyond the skin which
covers the capsule externally. Below this gap is seen a curved line
which, if followed along the sides of the capsule on each side, merges
into the base of the most posterior ventral tooth. This line is the
optical expression of a very shallow groove on the inside of the
capsule. The skin on the outside of the capsule, which is reflected
over the edge of and into the capsule, dips into this groove, which
gives it a firm attachment. Below the middle (dorsally) of this curved
line there is a thickening in the capsule wall, which is perforated by the
opening of the dorsal œsophageal gland. This is the “dorsal ridge”; in
optical section it has a conical appearance with a lumen (of the duct).
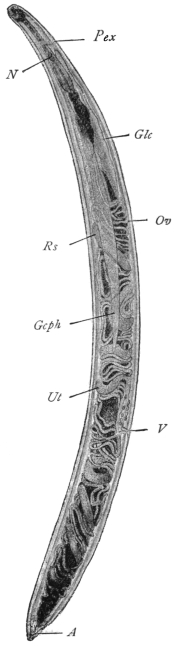
Fig. 324.—Ancylostoma duodenale,
female. A, anus; Gcph, cephalic gland;
N, nucleus of cephalic gland; Glc, cervical
gland; Ov, ovary; Pex, excretory pore;
Rs, receptaculum seminis; Ut, uterus;
V, vagina. × 15. (After Looss.)
Male 9 mm. long by 0·45 mm. thick, female 12 mm. long by
0·6 mm. thick. Pale flesh colour, or an intense red in posterior third.
Anteriorly may be more or less black due to (blood) pigment in the
cells of the chyle intestine (= stomach + small intestine). The worm
is about the same thickness all through and is plump and rigid.
Cuticle striated. The body has a marked torsion, so that if the ventral
side of the head is upwards the anus appears to open laterally and
vice versâ. The dorsal curve of the head end is only slight and
the œsophagus is roughly cylindrical.
On the ventral wall one sees the two pairs of strong teeth, their
points being directed somewhat backwards. They are covered by
cuticle above and below, but their points are free, piercing the cuticle.
The “roots” of these teeth followed backwards appear as two
thickenings or ribs on the outside of the capsule wall, so that the
outside wall is not smooth—a characteristic of the genus Ancylostoma.
In the space between these ribs lies the ventral nerve papilla,
and lying against the outside of the outer root the lateral nerve
papilla. The nerve papillæ are thus, as it were, concealed by these
roots, and not conspicuous as they are in Necator. Following the
ventral curve of the capsule on the inside, posteriorly we next find
two triangular ventral lancets.316 These stand straight up into the
capsule on either side of the longitudinal axis, converging at their
summits. So that to sum up, the cutting apparatus is entirely
ventral, consisting of two pairs of cutting teeth and a pair of lancets.
Cervical Papillæ.—Two, one on each side behind the head at the
level of the excretory pore. They consist of “pulp,” i.e., extensions
of the substance of the lateral
bands covered by cuticle and
supplied with a nerve (fig. 326).
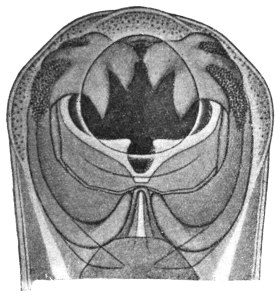
Fig. 325.—Ancylostoma duodenale: showing
ventral teeth, dorsal cleft, and behind it the dorsal
ridge with duct of dorsal œsophageal gland.
× c. 200. (After Looss.)
Œsophageal Glands (3).—The
chitin of the triradiate
œsophagus is continuous with
that of the buccal capsule. In
its muscular walls are three
glands—one dorsal, two sub-ventral.
The dorsal gland opens
into the buccal cavity through
the dorsal ridge; the two others
into the lumen of the œsophagus
at the nerve ring. They
branch freely amidst the muscles.
They are probably digestive in
function.
Cephalic Glands (2).—Lie in
the lateral lines or bands on
either side. They begin about
the middle line of the body, and their ducts open at the base of the outer
ventral tooth on the surface of the skin on each side. Each is 0·15 mm.
thick in the middle, and has a single nucleus about as big as an ancylostome
egg (N, fig. 323). They probably function as poison glands.
Excretory System and Cervical Glands (2).—The excretory pore lies
in the mid line ventrally behind the œsophageal nerve ring (figs. 324
and 326). It opens into the excretory vesicle, a cavity in a large
cell with lateral appendages which fuse with the lateral lines, this
cell thus forming the “bridge” of the excretory system. Adhering
to this (bridge) cell are the spindle-shaped cervical glands (Glc,
fig. 324), and branches from the excretory vesicle enter the glands,
which are excretory in function; the vesicle also receives branches
from the lateral excretory canals (fig. 326) running in the lateral
lines or bands. The cervical glands are swollen anteriorly, forming
the so-called ampullæ just in front of the bridge. They extend
backwards a little beyond the anterior loop of the testis.
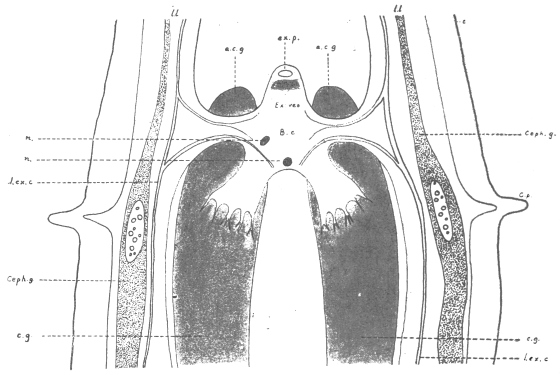
Fig. 326.—Ancylostoma duodenale: diagrammatic representation of excretory system.
ex.p., excretory pore; e.c.g., excretory cervical gland; Ex. ves., excretory vesicle in B.c.,
bridge cell, which is connected with c.g., cervical gland, and l.l., lateral lines; ceph.g.,
cephalic gland; l.ex.c., lateral excretory canal passing into the bridge cell; l.l., lateral line
containing excretory canal and cephalic gland; c.p., cervical papilla; n., nuclei of bridge
cell. (After a drawing of Looss.)
Lateral Lines.—(1) Are broad elevations of the subcuticle, in
which, here and there, a nucleus occurs. (2) Near the bursa in the
male they increase in volume, and finally divide into branches which
form the “pulp” of the different rays. (3) In addition to the lateral
lines or bands, there is also a dorsal and ventral band. (4) The
ventral band is well developed caudally, forming a large pad dorsal
to the cloaca, “pulvillus post-analis.”
The bursal rays are outgrowths of the lateral lines. Beside this
“pulp” they contain a nerve, and at their bases complex muscles.
The Bursa is closed on all sides with a short median (ventral) lobe,
which may be tucked inwards. It is an outgrowth of the inner layer
of the skin pushing the outer layer before it, so that it consists of three
layers, not four, as it would be if it were a fold. The bursa is twice
as broad as long. It is supported by a variety of rays, the arrangement
of which is best followed from the figure (fig. 327). The
different terminology for these rays as used by various authors
should be noted: Ventral = anterior; externo-lateral = antero-external;
medio-lateral + postero-lateral or antero-median + postero-median
= median (doubled); externo-dorsal = postero-external;
dorsal = posterior. All the rays end in tactile papillæ, seven, on each
side; the postero-external and antero-external on the outer surface of
the bursa, the five others on the inner surface.317 Of the six terminal
digitations of the dorsal ray, only the external two contain tactile
papillæ.
In the male there are prebursal papillæ and minute caudal papillæ
in the female.
In the female the inner layer of the cuticle projects at the posterior
end as a sharp spike, 20 µ long, which may sometimes be broken off.
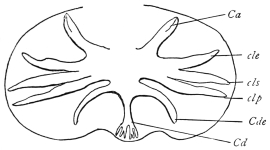
Fig. 327.—Ancylostoma duodenale: bursa enlarged. Ca, anterior ray
cleft; cle, antero-external; cls, antero-median; clp, postero-median; Cde,
postero external; Cd, posterior bifurcated, each bifurcation tridigitate.
(After Railliet.)
Ovaries.—The anterior tube runs from the cephalic to the posterior
end and back again. The posterior tube begins anteriorly, runs to the
posterior end of the body, and then back to the cephalic end, forming
a vulval loop before ending. The ovaries on the whole run in
oblique coils. The uterus is the thicker portion of the tube, 5 mm.
long. A short tube connecting the ovary and uterus is the oviduct.
The two uteri unite to form a single duct, the vagina, opening 1 mm.
behind the middle line. The portion of the uterus next to the oviduct
functions as a seminal receptacle, whereas the part next the vagina
functions as an ovejector.
Testis.—The blind end begins a little behind the beginning of
the cement gland. The transverse coils occupy the middle third of
the body. About the middle of the body it passes into the spindle-shaped
seminal vesicle, which, with the spicular canal and rectum,
opens into the cloaca. An anterior longitudinal coil pushing in
between the cervical glands is characteristic of Ancylostoma. The
cement gland surrounds the ejaculatory duct for practically its whole
course, and it occupies nearly the posterior half of the body and
secretes a brown or black cement. The spermatozoa are curved
rods about 2 µ long.
Spicules are 2 mm. long, ending in a fine point. They are moved
by exsertor and retractor muscles. At first they lie free in the body
cavity; next in a groove in the dorsal wall of the cloaca; then in an
isolated canal, and finally in two canals. Anteriorly each has two
longitudinal crests on its inner surface. These meet the corresponding
crests of the other spicule, and so form a canal along which the
sperm passes into the female. The gubernaculum is a thickening
of the dorsal wall of the cloaca. It is not a free piece, but is moved
by various muscles.
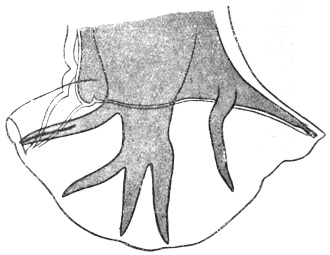
Fig. 328.—Ancylostoma duodenale: bursa of male. The rays
from left to right are: (1) anterior cleft; (2) antero-external; (3)
and (4) median doubled, i.e., antero-median and postero-median;
(5) postero-external arising from a common trunk with the posterior.
× c. 120. (After Looss.)
Genital Cone is a prominence on the floor of the bursa on the
ventral side of the body, on which the genito-anal orifice opens.
The cone is only slightly marked in Ancylostoma duodenale, but is
much more prominent in Necator americanus.
Distribution.—Africa, Egypt, Europe, Japan, China (mainly), but in
association with Necator americanus in Southern States of America,
British India, Assam, Burma, Hongkong, Liberia, Jamaica, Martinique,
Costa Rica, Colombia, Antigua, Guadeloupe.
Habitat.—The worms live in the jejunum, less frequently in the
duodenum, of man only.
Food.—The worms feed on the mucous membrane of the gut,
attaching themselves to the base of the villi, sucking these in; and
when these are destroyed they attack further the submucosa. As
a rule the worms have no blood in the gut, but in their attack on
the submucosa a blood-vessel may be eroded, and so the gut of the
worm filled with blood.
Development.—The eggs are oval with broadly rounded poles,
56 µ to 61 µ by 34 µ to 38 µ. In fresh fæces they contain four
granular nucleated segmentation masses of the ovum (fig. 329)
separated by a clear space from the shell.
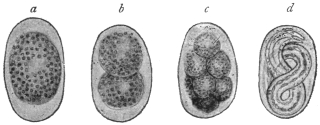
Fig. 329.—Ancylostoma duodenale: eggs in different stages of development.
a to c, in fresh fæces; d, containing a larva, only in old fæces.
× 336. (After Looss.)
Egg of Ancylostome appears to have a single contour. Under high
powers this appears double, but they are the outer and inner surface
of the true (chitinous) egg-shell. Internal to this is the extremely
delicate yolk-envelope, a kind of skin secreted by the egg cell around
itself for protection. The function of this is probably to absorb
water to swell and burst the outer chitinous shell. The embryos
when hatched are termed larvæ.
Embryos which are ready to hatch have their bodies almost free
from granules; others, though apparently mature, that have granules
will not hatch.318
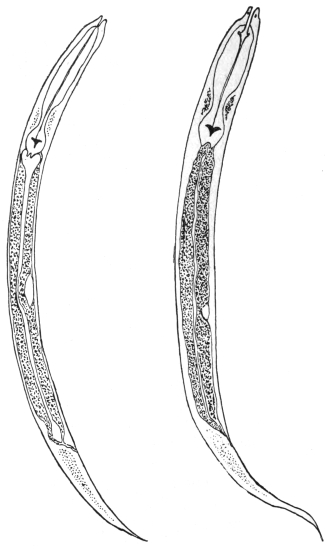
Fig. 330.—Ancylostoma duodenale larva on fourth day of culture on right;
Strongyloides stercoralis larva on left. (After Leichtenstern.)
Larva.—Stage I: Average length, 25 mm. Maximum thickness
in œsophageal region, 17 µ. Head end fairly blunt, from behind the
anus (the tail) tapering in an uniform manner. Buccal cavity is
characteristic, 10 µ to 12 µ by 1 µ to 8 µ, longer and narrower than the
corresponding larvæ of Strongyloides stercoralis. Œsophagus “rhabditic”
in character, i.e., it has three sections, but they are not so clearly
marked off as in larvæ of the genus Rhabditis. The posterior bulb has
a Y-shaped valve, the function of which, according to Looss, is to
prevent regurgitation of food. The granules of the gut serve as a
reserve of food, and are used up if the larvæ are starved. The genital
rudiment consists of two cells half-way between the end of the
œsophagus and the anus in the mid-ventral line. The larva lives
on fæcal matter and grows to about 0·4 mm., then moult319 I takes
place in two days or more, the skin being ruptured by the activity of
the larva.
Stage II: The larva is now in this stage, which does not differ
much from the previous one. It grows to 0·5 mm. The mouth
opening closes. The œsophagus elongates, becoming cylindrical or
“filariform”; a new skin is formed underneath the old one, and in
about a week moult II takes place.
Stage III: The mature larva remains enclosed in the old skin.
Its movements are now much more active and of a boring character.
Length is now 0·6 mm. This mature stage has been erroneously
called the encysted larva, because there is no cyst secreted from its
surface by the larva, but it is simply the old skin, which is not cast off,
but is retained for purposes of protection, as the larva is free living,
but casts it as soon as it assumes parasitic life again. From the egg
to this mature stage is thus six to ten days.
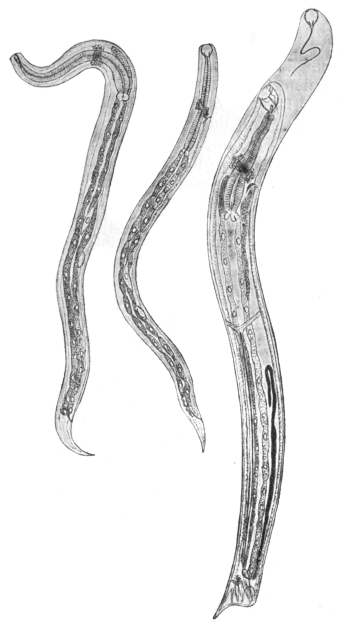
Fig. 331.—Ancylostoma duodenale: left, four days after transmission into
dog, 190/1; in the centre, at the commencement of the second stage of development
(five to six days), 105/1; on the right, fourteen to fifteen days after transmission.
42/1. (After Looss.)
Bionomics of Development.—Air: Eggs can develop when shut
off from the air for a “comparatively long” time.
Temperature: Hatching takes from eight hours upwards. Eggs
develop best at 25° to 30° C., but will not develop below 8° to 10° C.
The larvæ, however, will stand freezing.
Moisture: Eggs and larvæ do not live long under water, because
they suffocate or starve, but mature larvæ will live for months (six to
twelve) in water; they require no food—in fact, can take none in—but
live on their reserve granules, and in course of time become as
clear as glass.
Thigmotropism: The mature larvæ, after casting their skin, will
penetrate pith, climb up stems, stalks, etc., and creep into any pore.
It is important to recognize that this third stage of the mature
larva is the only infective one.
Mode of Entry into the Body.—Infection is effected through the
mouth (Leichtenstern and others), and also through the skin, as was
first discovered by Looss and afterwards confirmed from the most
diverse quarters, partly in the case of Ancylostoma duodenale,
partly in that of A. caninum in dog, man, and monkey. The
larvæ that gain access to the intestine partly through contaminated
food, or through unwashed hands, or under some circumstances
through water, first throw off their “sheath”—that is, they complete
moult II. Moult III takes place four to five days after they have
reached the gut, and they now have a mouth capsule supplied with
four small teeth arranged crosswise, enabling them to fasten on to
the intestinal epithelium, upon which they feed. On this food the
worms grow in four to six days to 3 to 5 mm. in length, and now
moult IV. takes place, thus attaining their definite shape and
distinctive character. About eight days later the sexual organs
commence to function; at this time the first copulation should be
taking place—it will later be frequently repeated—and a few days
later the first ova are laid, first in less and later in larger numbers, so
that they appear in the fæces about four to five weeks after the
infection.320
Infection by the Skin.—Mature larvæ, which are placed on the
skin of man or suitable animals, cast their “sheath” and bore their
way through delicate fissures either horizontal in the superficial
scales of the epidermis, or through vertical fissures into hair follicles
where these exist, and then they invade the cutis. Now according
as they migrate further into the lymphatic vessels or the small vesicles,
the final path to the gut differs to some extent. The blood path
leads to the right heart, and from there into the lungs; here the
larvæ leave the blood stream and enter the air passages, over the
mucosa of which they travel further headwards, through the bronchi
into the trachea and larynx, and from hence through the œsophagus
to the stomach; in some cases also they are swallowed. The
lymphatic path leads finally also into the blood stream, but the
lymphatic glands must first be passed, and in these many larvæ are
retained and perish. In the cutaneous infection seven to ten weeks
elapse till the time of appearance of the first ova in the fæces.
The penetration of the skin by the larvæ also in man causes reddening and
burning at the affected points, and this is followed in a few days by transitory swelling
in the subcutaneous connective tissue. Skin affections can also be set up by
such Ancylostoma (and Strongyloides) larvæ as do not gain access to the blood or
lymphatic vessels or gut; such larvæ apparently wander further in the connective
tissue, and, as Looss has in his own person observed, gain access to the cutis at
different points, thus causing progressive swellings (accompanied by intense itching),
which cease when the worm again penetrates into the deep tissues. Skin affections
such as “ground-itch” or “pani-ghao” occurring in the tropics and only attacking
the feet, or other affections (e.g., sump bunches) are now well recognized as being
due to the invasion of Ancylostoma larvæ.
Other names for these skin affections are water-sore, sore feet of coolies, maza-morra,
bunches, botches, quaddeln, krätze, ampoules, gourmes, taons, pitirr.
Whether oral or dermal infection is the more important one further observation
must decide.
The duration of life of Ancylostoma duodenale, which is a specific
parasite of man and has not been observed in other mammals,
amounts to about five years, as strayed larvæ according to Looss
wander for this extent of time in the body.
Cultivation of Larvæ.—(1) Mix the fæces (free from drugs such as
salines or thymol) with animal charcoal, adding water if necessary
till a consistence of porridge is obtained. If the stools are very fluid,
allow to sediment first and pour off the fluid. The best charcoal is
that made from bones, and should not have an acid reaction. Charcoal
is necessary in order to prevent fermentation, which kills the
larvæ. Spread in layers 2 to 3 mm. thick in Petri dishes. Incubate
at room temperature. To extract the larvæ from the culture allow
the surface thoroughly to dry, then pour on water; the larvæ
wander out and are poured off and subsequently further purified by
sedimentation or filtering through blotting paper, the larvæ passing
through.
(2) A funnel is plugged with cotton wool, then filled with washed
sand to within a centimetre or two of the rim. Stand this in a jar of
water so that the level of the water is slightly below that of the sand.
On the surface of the wet sand now place layers of blotting paper,
and spread the fæces, diluted if necessary, on this in layers of a few
millimetres thick (vide p. 474).
Detection of Eggs.—Vide p. 473.
Dermal Infection of Dogs.—Infection with larvæ of A. caninum.
In two hours most of the larvæ are free in the cutis and in four hours
in the subcutaneous tissue. By scraping a few days later the mucosa
of the trachea large numbers of larvæ are found there.
Ancylostoma ceylanicum, Looss, 1911.
Fig. 332.—Ancylostoma ceylanicum:
head end, two teeth on each side, the
inner almost concealed by the outer.
× c. 200. (After Looss.)
At the anterior edge of mouth capsule one large tooth; below or
behind this towards the middle line a very small tooth, the tip only
of which is seen. Male 5 mm. average. Lobes of bursa almost as
long as broad, strongly projecting
towards the ventral side. Rays short
and relatively thick. Female 7 mm.
Habitat.—Intestine civet cat
(Viverricula malacensis), Ceylon, and man in
Bengal according to Clayton-Lane.
Other species are: A. caninum
(Ercolani), in cat and dog, Europe
and Africa; A. malayanum (Alessandrini),
1905, in the Malay bear
(Helaretos malayanus); A. pluridentatum
(Alessandrini), 1905, in Felis
mitis, Brazil.
Ancylostoma braziliense, Gomez de Faria, 1910.
In cats (and dog), Brazil. Female 8·5 mm., male 7·5 mm. long.
Eggs 65 µ by 32 µ. Leiper considers it to be identical with A. ceylanicum.
Fig. 333.—Ancylostoma braziliense: bursa of male. (After Gomez de Faria.)
Group. Bunostomeæ, Railliet and Henry, 1909.
Bursa with median double, postero- and postero-external arising
from a common trunk, posterior bifurcated, each limb bidigitate
(fig. 336). Vulva in middle of body or a little in front. Uteri divergent.
Contains the following genera: (1) Bunostomum (= Monodontus);
(2) Necator; (3) Bathmostomum; (4) Gaigeria.
Genus. Necator, Stiles, 1903.
Mouth capsule small, narrowed anteriorly (ventrally) by chitinous
plates, as in Uncinaria. On each side of the base of the dorsal cone
a lateral chitinous plate or lancet with smooth edge (not serrated),
ventral lancets as in Ancylostoma. No ridges on outside of ventral
wall. Aperture of dorsal œsophageal gland on tip of a cone projecting
freely into the buccal capsule. Bursa closed. Posterior ray
cleft to its root.
Necator americanus, Stiles, 1902.
Syn.: N. africanus, Harrison, 1910.
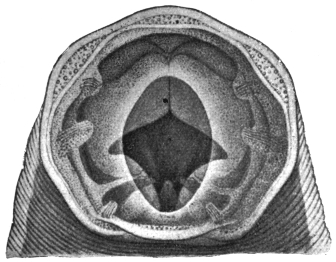
Fig. 334.—Necator americanus. Showing cutting plates and the projecting
dorsal ridge, and deep in the cavity the edges of the ventral lancets. × c. 475.
(After Looss.)
Male 8 mm. long, female 10 mm. The head is strongly bent dorsalwards
so that almost by this character alone it can be distinguished
from Ancylostoma duodenale. The buccal capsule is markedly small—in
the male, 0·093 by 0·084 mm., in the female 0·11 by 0·097 mm.
There are no teeth anteriorly on the ventral side of the capsule, but
instead there are two cutting chitinous plates, the anterior portions
of which are prominent and angular, and meet in the middle line
in front. Posteriorly on each side the plate projects less, while
between the anterior and posterior parts there is a deep angle. The
inner (posterior) ventral lancets which also occur in A. duodenale are
large, and project far into the lumen, the tips of these, of the lateral
lancets, and of the dorsal cone almost meeting in the centre of the
lumen. As already stated in the definition of the genus Necator, there
are also lateral lancets which start from the base of the dorsal cone.
This dorsal ridge, or rather in this case cone, is a striking object in
the mouth, and projects right out into the cavity, and on its summit
opens the dorsal œsophageal gland.
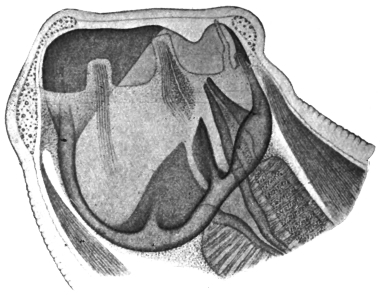
Fig. 335.—Necator americanus: lateral view, showing the dorsal ridge perforated
by the duct of the dorsal œsophageal gland, the lateral lancet and ventral
lancet and the nerve papillæ. × c. 475. (After Looss.)
The bursa is about as long as broad, but has the lateral lobes
strikingly lengthened, giving a trilobed appearance (fig. 336), but as
in Ancylostoma duodenale it
is closed on the ventral side.
The distribution of the rays
is best understood from the
figure. The genital aperture
lies on a marked conical protuberance;
the cement gland
is bilobed in transverse section.
In the female the opening of
the vulva is in front of the
middle line, in A. duodenale it
is behind.
The spicules, 0·92 mm. long
are hooked at the extremity.
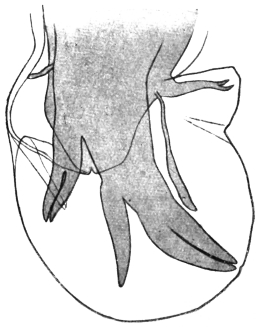
Fig. 336.—Necator americanus: bursa of male.
The rays from right (top) to left are: (1) posterior,
(2) postero-external, (3) and (4) median
doubled, i.e., postero-median and antero-median,
(5) antero-external, (6) anterior (cleft), and above
it on left a pre-bursal ray. × c. 120. (After
Looss.)
Eggs more pointed at the
poles than those of A. duodenale,
64 µ to 72 µ by 36 µ, so that
it may not be possible to distinguish
single eggs owing to
individual variations, yet on
comparing a number they can
be distinguished.
Geographical Distribution.—Brazil, Porto Rico, Cuba, Central
Africa, East Africa, Victoria Nyanza, Gold Coast, Uganda, North-Western
Rhodesia, Ceylon, Mysore. For other localities where
A. duodenale is also found see p. 450.
Habitat.—In small intestine of man and gorilla (Troglodytes gorilla).
Necator exilidens, Cummins, 1912.
Syn.: N. africanus, Looss, 1911.
Male 7 mm., female 9 mm. long. The edges of the cutting plates
are rounded, not angular, and do not meet in the middle line. Inner
(posterior) ventral lancets very small. Lateral lobes of bursa broader
than long. Rays thick and plump.
Habitat.—In the chimpanzee (Anthropopithecus troglodytes).
Ancylostomiasis.
Morbid Anatomy.—Organs pale and bloodless. Abdominal
organs sodden, and there is fluid in the serous cavities. Lungs:
œdema. Kidneys: fatty changes, especially large pale kidney.
Liver and heart also show fatty changes—there is much hæmosiderin
in the liver cells. Blood: early stages, a leucocytosis 20,000
upwards, and eosinophilia 50 per cent. Later, anæmia (hydræmia).
The number of worms found varies from ten to 1,000. They are rare
in the duodenum, but occur as far as 6 ft. from the pylorus.
Group. Syngameæ, Railliet and Henry, 1909.
Bursa with anterior and median ray cleft; antero-external, close
to median; postero-external, arising separately from posterior; posterior
bifurcate to base, each branch bifurcate or trifurcate. Vulva in
the anterior fourth of body. Uteri divergent.
Genus. Syngamus, von Siebold, 1836.
Head thickened, not tapering; broad mouth with gaping buccal
capsule.
Male and female often in permanent copulâ.
Parasitic in respiratory passages of birds and mammals.
Habitat.—S. trachealis in poultry; S. bronchialis in goose; S. laryngeus
in cattle; S. vasicola in goats, etc.
Syngamus kingi, Leiper, 1913.
Buccal capsules of male and female on same level. In S. trachealis
and S. laryngeus, that of male in front of that of female. In S. dispar,
that of male behind that of female. Œsophagus of male one-sixth,
that of female one-ninth of total length. Mouth capsule in male
and female terminal; it is dorsal in S. trachealis and in mammalian
species. Tail of female bluntly pointed. Ovary reaches to anus.
Excretory pore opposite the middle of the bulb of œsophagus. In
S. trachealis it is opposite the œsophageal valves.
Habitat.—Found in sputum of patient by King in St. Lucia.
Normal host probably a carnivore.
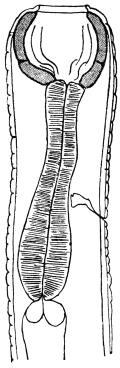
Fig. 337.—Syngamus kingi: anterior
end of male. (After Leiper.)
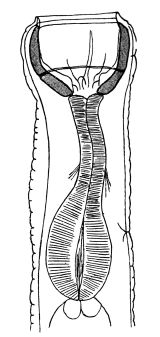
Fig. 338.—Syngamus kingi: anterior
end of female. (After Leiper.)
Family. Physalopteridæ.
Genus. Physaloptera, Rudolphi, 1819.
Mouth surrounded by two large lateral lips bounded posteriorly
by a cuticular band projecting anteriorly, forming a collar. Each lip
bears anteriorly and inwardly a cuticular appendage, the external tooth.
Immediately below and internal to the external teeth the internal teeth,
one on each lip. Each lip bears two large submedian papillæ. Tail
of male with four pairs of pedunculated papillæ in a row on each
side external to the six pairs of unpedunculated papillæ. Spicules
unequal. Vulva in the anterior region of the body. Eggs with a
characteristic thick smooth shell.
Parasitic in the intestine, more especially the stomach, of mammals
(twenty species), birds (twelve species), reptiles (fourteen species).
Physaloptera caucasica, v. Linstow, 1902.
The male measures 14·2 mm. in length and 0·71 mm. in breadth;
the bursa is broad, rounded off in front and narrower at the back;
the right spicule measures 0·62 mm. in length, the left spicule
1·76 mm.; there are two papillæ in
front of the orifice of the cloaca, four
behind it and six unpedunculated on
the tail. The female measures 27 mm.
in length, 1·14 mm. in breadth; the
caudal extremity is rounded off; the
vulva is on the border of the first
and second sixth of the length of the
body; the eggs have thick shells,
and measure 57 µ by 39 µ. It has
hitherto only been observed once,
by Ménétriés in the intestine of man
(Caucasus).
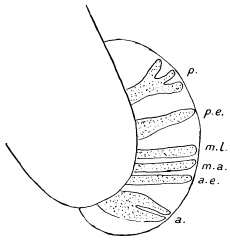
Fig. 339.—Bursa of Syngamus trachealis.
a., anterior ray cleft; a.e., antero-external;
m.a., antero-median; m.l., postero-median;
p.e., postero-external; p., one branch of
posterior (trifurcate). (Stephens.)
Physaloptera mordens, Leiper, 1907.
Large worms resembling an immature
Ascaris lumbricoides.
The inner lancet-shaped teeth have a sharp cutting edge towards
the lumen. Below each is a cuticular boss projecting into the mouth
(fig. 340).
Male 30 to 50 mm., bursa with four pairs of pedunculated papillæ,
the second and third lying external to the first and fourth on each side.
Spicules unequal, one slender (4·6 mm.), the other stouter (6 mm.).
Female 40 to 55 mm. Tail sharp. Vulva opens between the
anterior fourth and fifth of the body. Eggs 43·6 µ by 35·3 µ with a
thick smooth shell.
Habitat.—Œsophagus, stomach, small intestine of man (several
cases). Nyasaland and Portuguese East Africa.
Family. Ascaridæ, Cobbold, 1864.
Sub-family. Ascarinæ.
Without œsophageal or intestinal diverticula; spicules without
flanges.
Genus. Ascaris, L., 1758.
Intermediate lips and auricles absent. Lips edged with fine teeth. Lips triangular
in cross section. Not grooved on internal surface.
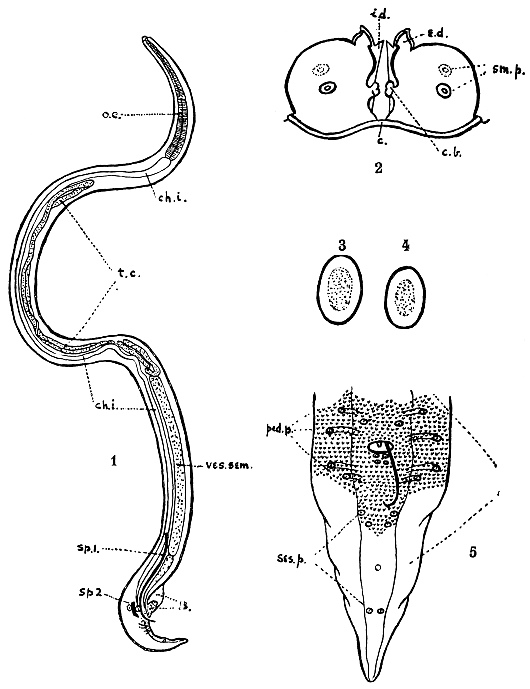
Fig. 340.—Physaloptera mordens, Leiper, 1907. (1) adult male: o.e., œsophagus; ch.i.,
chyle intestine; t.c., testicular coils; ves. sem., vesicula seminalis; sp. 1, long spicule; sp. 2,
short spicule; B., bursa. (2) Mouth parts: c., cuticular collar embracing the two lips
posteriorly; c.b., cuticular bosses guarding the mouth laterally; e.d., external tooth;
i.d., internal tooth; sm.p., submedian papillæ. (3) egg of P. caucasica. (4) egg of
P. mordens. (5) bursa enlarged: ped.p., pedunculated papillæ; ses.p., sessile papillæ.
(After Leiper.)
Ascaris lumbricoides, L., 1758.
The colouring, in the fresh condition, is reddish-yellow or greyish-yellow;
the body is of an elongated spindle shape. The oral papillæ
are finely toothed. The dorsal papilla carries two sensory papillæ,
the two ventral papillæ each one sensory papilla. The male measures
from 15 to 17 to 25 cm. in length, and about 3 mm. in diameter;
the posterior extremity is conical and bent hook-like ventrally; the
spicules measure 2 mm. in length, are curved, and somewhat broadened
at their free end; on each side around the orifice of the cloaca there
are seventy to seventy-five papillæ, of which seven pairs are post-anal.
The testicular tube is much folded, showing through the body integument,
and is about eight times the length of the body. The female
measures 20 to 25 to 40 cm. in length and about 5 mm. in diameter;
the posterior extremity is conical and straight. The vulva is at the
junction of the anterior and middle thirds of the body, which, at
this point, has a slight ring-like constriction; the convoluted ovaries
measure ten times the length of the body.
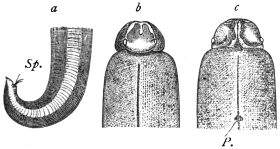
Fig. 341.—Ascaris lumbricoides. a, posterior
extremity of the male with the spicules protruding
from the orifice of the cloaca (Sp.); b,
anterior extremity from the dorsal surface, the
two lobes of the pulp of the lip separated by the
“saddle”; c, anterior extremity from the ventral
surface; P., excretory pore. (From Claus.)
Fig. 342.—Ovum of Ascaris
lumbricoides, with shell and
albuminous envelope. 400/1.
The ova are elliptical with a thick (4 µ) transparent shell (fig. 342)
and an external albuminous coating which forms protuberances; the
ova measure 50 µ to 70 µ in length, 40 µ to 50 µ in breadth; they are
deposited before segmentation; the albuminous coating is stained
yellow by the colouring matter of the fæces, but is sometimes absent.
The egg cell is unsegmented, it almost completely fills the shell, and
its nucleus is concealed by the large amount of coarse yolk granules.
Abnormal or unfertilized eggs also occur in fæces. They are
distinguished by their elongated form (80 µ by 45 µ), irregularly
cylindrical, its contents consisting of refractive granules.
Ascaris lumbricoides is one of the most frequent parasites of man;
it is distributed all over the inhabited parts of the world, and though
it is particularly frequent in the warmer regions, yet it also occurs
in Finland, Greenland, etc. In temperate climates A. lumbricoides
occurs most frequently in young children; it is, moreover, more
common amongst country dwellers than amongst the inhabitants of
towns, but is not lacking in infants, adults and aged persons. As a
rule only a few specimens are present in the intestine, but many cases
are known in temperate zones in which several hundreds of worms
have been found in the same patient. This species is particularly
numerous in the negroes of Africa and America. It occurs also in
the monkey, dog and pig (? A. suilla).
The parasite was known in ancient times; the Greeks called
it ἐλμινς στρογγύλη, Plinius termed it Tinea rotunda, later on
it was named Lumbricus teres. The ἄσκαρις of the Greeks is our
Oxyuris.
The small intestine is the normal habitat of Ascaris lumbricoides; the worms,
however, often leave this part of the intestine and wander into the stomach, whence
they are frequently evacuated by vomiting, or they may creep through the œsophagus
into the pharynx and crawl out through the nose or mouth; very rarely they
may find their way into the Eustachian tube or into the naso-lachrymal duct, or into
the excretory ducts of the liver and pancreas; exceptionally they may gain the
trachea, and they have also been found in the abdominal cavity. They may bore
through adhesions between the intestinal wall and the omentum (worm abscess);
they occasionally penetrate the urinary apparatus and are passed with the urine;
in febrile diseases A. lumbricoides usually leaves the intestine spontaneously. It is
obvious that these wanderings may be accompanied by the most serious symptoms,
but in sensitive persons the invasion of even only a few intestinal Ascarides gives
rise to a series of almost inexplicable symptoms (hysterical, epileptiform attacks,
cerebral congestion, aphonia, etc.), which cease with the expulsion of the worms, so
that many authors are driven to the conclusion that these Ascarides secrete a toxin.
Fortunately, the presence of A. lumbricoides in the intestine is easily demonstrated
by the microscopical examination of the fæces.
Development.—Several authors (Gros, Schubart, Richter, Leuckart
and Davaine) have demonstrated that the ova of Ascaris develop
in water or moist earth after a long period of incubation. Freezing
and desiccation (if not too long) do not injure their powers of
development; the duration of the development depends on the
degree of the surrounding temperature. At a medium temperature,
after a varying period of incubation, it takes from thirty to forty
days for the embryo to become formed. The spirally rolled up
embryo, with its so-called “tooth,” formed by three papillæ close
together, never leaves the egg-shell in the open, even if the eggs are
kept for years under favourable conditions. Davaine proved that the
larvæ hatch out in the intestine of the rat, but are again expelled with
the fæces; he therefore concluded that the hatching likewise takes
place within the intestine of man, but is followed by the invasion
of the larvæ. In the meantime Leuckart had sought to infect himself
by swallowing embryo-containing eggs, but without results; he
therefore conjectured that there must be an intermediary host, and
v. Linstow thought he had found it in myriapods (Julus guttulatus).
Subsequently, Davaine’s opinion proved correct. First of all Grassi
succeeded in infecting himself by swallowing 100 embryo-containing
eggs of Ascaris lumbricoides; five weeks after ingestion the worms had
attained maturity and their ova appeared in the fæces. Calandruccio
also sought to infect himself, but failed, yet he succeeded in infecting
a little boy aged 7. Lutz also reports a successful experiment which
must have been positive, as young worms 5·5 to 18 mm. long were
expelled. Lutz proved that the eggs lost their albuminous shell by
long lying in water and then died when introduced into the stomach;
this would explain the failure of Leuckart’s experiment; in moist earth
the albuminous shell is retained. Finally, Epstein conducted unimpeachable
experiments on three children which place direct infection
with embryo-containing eggs beyond doubt; he, moreover,
proved that the development of the eggs takes place more rapidly in
the fæces when there is free admission of air, sun, and a sufficiency
of moisture.
Accordingly, infection occurs partly through water, but principally
direct from the soil.
Ascaris, sp.
Wellmann states that yet another species of Ascaris in man occurs
in the highlands of Angola: up to the present nothing certain
is known about it (Welland, “Critical Notes on Tropical Diseases
of the Angola Highlands,” New York Med. Journ. and Philadelphia
Med. Journ., August 12 to September 2, 1905.)
Ascaris texana, Smith et Goeth, 1914.
Female alone known; 58 to 60 mm. and upwards in length;
characterized by the serration of the anterior border of the lip and by
the appearance of interlabia. Evacuated by a white settler in Texas.
Position of this worm doubtful.
Ascaris maritima, Leuckart, 1876.
Only one immature specimen, a female (43 mm. in length and
1 mm. in breadth), has hitherto been described, and it was vomited by
a child in North Greenland in 1865. (R. Leuckart, “Die menschlichen
Parasiten,” 1876, edition 2, i, p. 877.)
Genus. Toxascaris (τόξον, an arrow), Leiper, 1907.
Body anteriorly bent dorsally, cuticle finely striated. Œsophagus
without a distinct bulb. Tail of male tapers to a point. Testis in
anterior portion of posterior half of body. Vulva about middle of
body. Eggs oval and smooth.
Toxascaris limbata, Railliet and Henry, 1911.
Syn.: Lumbricus canis, Werner, 1782; Ascaris teres, Goeze, 1782; Ascaris cati
et caniculæ, Schrank, 1788; Ascaris canis et felis, Gmelin, 1789; Ascaris tricuspidata
et felis, Bruguiere, 1791; Ascaris werneri, Rud., 1793; Fusaria mystax, Zeder,
1800; Ascaris marginata et mystax, Rud., 1802; Ascaris alata, Bellingham, 1839.
Striations 6 µ to 12 µ apart. Cephalic wings long, narrow, semi-lanceolate.
Male, 4 to 6 cm. Spicules, 1,002 µ and 1,005 µ.
Female, 0·5 to 10 cm. Eggs, 75 µ to 85 µ, shell thick and smooth.
Host: dog, occasionally man.

Fig. 343.—Ovum
of Toxascaris limbata,
with thin albuminous
envelope.
Magnified.
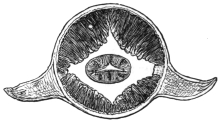
Fig. 344.—Transverse section through the head
part of Belascaris cati from the cat, with the lateral
wings. In addition, one may note the four fields of
muscles, the longitudinal lines with the œsophagus
in the centre. Magnified. (After Leuckart.)
Genus. Belascaris (Βέλος, an arrow), Leiper, 1907.
Body anteriorly bent ventrally, cuticle coarsely striated.
Œsophagus with a distinct bulb. Tail of male conical. A papillæ-bearing
protuberance behind the anus. Testis in anterior half of body.
Vulva in anterior part of body. Eggs corrugated.
Belascaris cati, Schrank, 1788.
Syn.: Belascaris mystax, Leiper, 1907; Ascaris mystax.
Striations 12 µ to 16 µ apart. Cephalic wings lanceolate. Male
3 to 6 cm. Spicules 17 to 1·9 mm. Female 4 to 10 cm. Eggs, 65 µ
to 75 µ in diameter, surface finely honeycombed. Host: domestic
cat, and man, eight or nine cases.
Belascaris marginata, Rudolphi, 1802.
Striations 16 µ to 22 µ apart. Cephalic wings long, narrow, semi-lanceolate.
Male, 5 to 10 cm. Spicules, 750 µ and 950 µ. Female,
9 to 18 cm. Eggs, 75 µ to 80 µ. Shell finely honeycombed. Host:
dog.
Genus. Lagocheilascaris, Leiper, 1909.
Thick lips separated by a furrow from the body; between the lips
small intermediate lips without “pulp.” The cutting angle of each
lip bifurcated. Along each lateral line a cuticular wing extending the
whole length of the body. Eggs, thick shell with a mosaic pattern.
Lagocheilascaris minor, Leiper, 1909.
Male, 9 mm., tail sharply curved. Spicules colourless, 3·5 and
4 mm. long. More than twenty-four pairs of pre-anal papillæ, at least
five pairs of post-anal. Female, 15 mm. Straight posteriorly. Vulva
6 mm. from head with two lips. Eggs, 65 µ in diameter. Host:
man, cutaneous abscesses. Trinidad.
Family. Oxyuridæ.
Genus. Oxyuris, Rudolphi, 1803.
Mouth unarmed. The three labial papillæ are only slightly protuberant, the
œsophagus is long and presents two well-marked bulbs. The vulva is in the anterior
part of the body.
Oxyuris vermicularis, Linnæus, 1767.
Syn.: Ascaris vermicularis, L.; Fusaria, Zeder, 1803.

Fig. 345.—A,
male, and B, female,
of Oxyuris
vermicularis. 5/1.
Colour white, the striated cuticle forms projections at the anterior
end which extend some distance back along the middle of the ventral
and dorsal surfaces; the longitudinal lateral flanges of the skin corresponding
to the lateral lines are well seen in transverse sections; there
are three small retractile labial papillæ around the mouth. The
male measures 3 to 5 mm. in length, and shortens on death; the
posterior extremity of the body is curved ventrally and presents six
papillæ. Spicule 70 µ long, hook-like. The female is 10 mm. in length
and 0·6 mm. in diameter; the anus is about 2 mm. in front of the tip of
the tail; the vulva is in the anterior third of the body; the eggs are
oval, asymmetrical, with double-contoured shells, and measure 50 µ to
55 µ by 16 µ to 25 µ; they are deposited with clear, non-granular
tadpole-like embryos already developed.
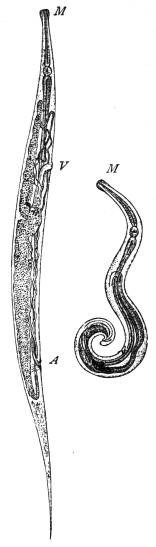
Fig. 346.—On the left,
female; on the right, male.
A, anus; M, mouth; V,
vulva. Greatly enlarged.
(After Claus.)
Habitat.—Adults in large intestine of man. Young forms in small
intestine and often in the appendix.
The worm lives in the lower part of the small intestine, cæcum and vermiform
appendix, and before becoming adult undergoes two or three moults (Heller).
According to Wagener the worms at times live in the gut wall, giving rise to calcareous
nodules. When the uterus of the fertilized females begins to fill with eggs they leave
the cæcum and travel through the colon to the rectum. The uterus is now packed
with eggs which contain a tadpole-shaped embryo. Egg-laying now takes place,
partly in the rectum, partly outside, the mode of exit being not only passive through
defecation but also an active one on the part of the worms when the patient is in bed.
In this case the worms crawl out of the anus, producing a most intolerable itching as
they scatter their eggs between the nates and the perinæum. From here in the case
of girls they may get occasionally into the vulva and vagina, and even into the
oviducts and so into the body cavity. The worms also may wander through the
alimentary canal in the opposite direction, getting out occasionally through the
mouth. Recently a rôle has been assigned to them, as to other gut parasites, in
appendicitis and typhlitis.
It is stated that the males die after fertilizing the females, thus explaining why
they are so rarely met with in fæces [but it is probable that they often escape notice
from their small size.—J. W. W. S.].
Development.—The eggs, which often adhere together, contain a
tadpole-like embryo, the thin tail of which is bent upwards ventrally;
the embryo in a short time, given a sufficiently high temperature, passes
into a second folded nematode-like embryonal stage, lying in the egg-shell,
either in the fæces, with which also numerous females pass out,
or in the moisture of the groove between the buttocks, and they there
await the opportunity of being reintroduced into man per os. It is
very improbable that infection takes place directly in the large intestine,
as is occasionally stated, because although the harbourers of Oxyuris
are frequently liable to auto-infection, this takes place exclusively
through the mouth, and is conveyed by the fingers, on which the ova
of Oxyuris, and occasionally the female worms, have clung.

Fig. 347.—Oxyuris
vermicularis:
egg freshly deposited,
with tadpole-like
embryo. × 640.

Fig. 348.—Oxyuris
vermicularis:
egg twelve hours after
deposition, with nematode-like
embryo.
× 640.
The opportunity for this is afforded every evening, as naturally the troublesome
itching caused by the wandering of the worms is met by scratching and rubbing with
the fingers. It is therefore possible that the eggs may even thus be introduced into
the nose, where the young Oxyuris are perhaps hatched out, if they get high enough
up on the moist pituitary mucous membrane. As a matter of fact, the larvæ of
Oxyuris have been found in the nose. Moreover, one can understand that the eggs
of Oxyuris are transferred from person to person by the hand, directly or indirectly.
This again explains the wholesale infections which occur in collective dwellings,
after a person harbouring Oxyuris has been admitted into boarding-houses, etc.
The primary infection may be also caused in other ways—by foods, fruits, vegetables
and other articles that are eaten raw, and are polluted with the ova. Perhaps also
flies or their excrement play a part in the distribution of the parasite, similar to that
demonstrated by Grassi as taking place in the spread of the ova of Trichocephalus
and Tænia.
The assumption of a direct development without an intermediary
host was first substantiated by Leuckart by experiments on himself
and three of his students; about fourteen days after swallowing the
eggs the Oxyuris has attained 6 to 7 mm. in length; Grassi, and
later on Calandruccio, infected themselves by swallowing adult female
Oxyuris, with the same results. Heller found worms in the gut
(appendix vermiformis) of a male child five weeks old.
Other species are: O. compar in the cat; O. curvula and O. mastigodes in horse,
ass, mule; O. ambigua in the rabbit; O. poculum in the horse; O. tenuicauda in the
horse. Many species occur in insects, especially in Blattidæ and Hydrophilidæ
(aquatic beetles).
Family. Mermithidæ.
Genus. Mermis, Dujardin, 1845.
With characters of the family.
Mermis hominis oris, Leidy, 1850.
Fourteen centimetres in length, 0·16 mm. in breadth; mouth
terminal; posterior extremity obtuse and provided with a recurved
hook 50 µ long.
The parasite was “obtained from the mouth of a child.” Stiles considers it to
be probably a Mermis, possibly swallowed in an apple.
Agamomermis, Stiles, 1903.
Group name for immature Mermithidæ.
Agamomermis restiformis, Leidy, 1880.
This worm measures 65 cm. in length, pointed anteriorly, the
posterior extremity broadened and rounded off (1·5 mm. in breadth);
the mouth is terminal, without lips. Behind the mouth six papillæ;
the œsophagus measures 1·125 mm. in length; the intestine appears
to terminate blindly.
This parasite was obtained in West Virginia from the urethra of a young man,
aged 20, who for a few days previous to expelling the worm passed turbid and bloody
urine.
TECHNIQUE.
Preservation and Examination of Flukes.
Fixation.—(Method A.) (1) Place the flukes in a test tube or small bottle
a quarter full of normal saline. Shake the contents as hard as possible (the
object of this is to extend the flukes) for half a minute.
(2) Add immediately an equal bulk of saturated aqueous solution of corrosive
sublimate and shake again as vigorously as possible for a few minutes.
(3) Transfer when convenient to 70 per cent. alcohol. (Before staining
and mounting remove the sublimate with tincture of iodine.)
(Method B.) In case of large flukes, e.g., Fasciola hepatica, Fasciolopsis
buski, compress the flukes between two glass slides with rubber bands or
thread. Fix in sublimate or in absolute alcohol, or in 10 per cent. formalin.
(Method C.) Place the flukes in 10 per cent. formalin solution.
Staining is successfully effected by using quite dilute solutions of carmine
or hæmatein overnight. This is far preferable to using strong solutions,
as it may be almost impossible to remove a too intense stain. Almost any
dilute carmine solution suffices. One of the best is acetic-alum carmine
(boil excess of carmine in a saturated aqueous solution of potash-alum for
about fifteen minutes; add glacial acetic acid to the extent of 10 per cent.;
let it stand for a week; filter). For use, dilute about thirty times with
water. Place the flukes directly in the stain. Stain overnight or longer.
Differentiation.—In order to get the sharpest picture, it is best now to
differentiate (but this may often be omitted) with acid alcohol (70 per cent.
alcohol 100 parts, HCl 5 drops). Allow to act from one to twenty-four
hours, according to the appearance of the flukes. Similarly, in staining with
hæmatoxylin solution, dilute twenty to thirty times so that the water is merely
tinged with the stain. Differentiate as before. After staining, dehydrate,
clear, and mount in balsam if required.
Clearing and Mounting.—(1) Carbolic acid (carbolic acid 94, water 6) is
a very convenient clearing agent. It may be used for stained or unstained
specimens. It will clear rapidly without previous dehydration. If it is
required to mount a specimen permanently, transfer from carbolic to
alcohol, then cedar-wood oil (or xylol, etc.), then balsam.
(2) Creasote.—Dehydrate the specimen, stained or unstained, transfer to
creasote. If it is desired to mount permanently, transfer back to alcohol,
then cedar-wood oil, then balsam.
(3) Cedar-wood Oil.—Preferable to xylol or oil of cloves. Dehydrate
the specimen in alcohol. To mount permanently, transfer to balsam.
(4) Glycerine.—Vide under methods of preservation of ova; to mount
permanently, transfer to glycerine jelly; subsequently to harden the jelly,
expose to formalin vapour.
Of these media, carbolic acid has the greatest refractive index excepting
that of balsam. The latter may, in some cases, render structures too
transparent, and it may be advisable to use only glycerine or glycerine jelly.
Preservation of Ova in Fæces, Urine, Bile, etc.
Heat some 70 per cent. alcohol in a basin to about 60 to 70° C. (until
bubbles begin to appear). Add the fæces, etc., in the proportion of one
part to about nine of fixative; keep stirring. Allow the sediment to settle.
Transfer to a bottle with some fresh 70 per cent. alcohol.
Transference to Glycerine.—Prepare 5 per cent., 10 per cent., 20 per cent.
solutions of glycerine in 70 per cent. alcohol. Pour off the alcohol in the
bottle of fæces, etc., and replace by 5 per cent. glycerine solution. Allow
to stand an hour or so. Then in the same way replace the 5 per cent. by
a 10 per cent. glycerine, and finally by a 20 per cent. glycerine solution.
When in this latter expose freely to the air (protecting from dust), so
as to allow the alcohol and water to evaporate. Add a few drops of
glycerine from time to time till eventually the ova are in pure glycerine.
(In a very moist climate it may be necessary to use lime or calcium
chloride to dry the air.) To mount permanently transfer some of the
sediment to glycerine jelly.
Preservation and Examination of Cestodes.
Fixation.—(1) Saturated aqueous corrosive sublimate.—Add to this glacial
acetic acid to the extent of 1 per cent. (Note this fixative will dissolve
the “calcareous corpuscles”; 10 grammes of sublimate to 160 c.c. of
water will give a saturated solution.) Warm the fixative to 70° to 80° C.
(Avoid the use of needles.) Use plenty of fixative. Allow to act for a
quarter of an hour or so. (a) Transfer to 70 per cent. alcohol. (It is
advisable to remove the sublimate by the use of Lugol’s solution, or a
solution containing tincture of iodine, adding this until the iodine colour
is permanent.) Or (b) transfer for preservation to 10 per cent. formalin.
Or (2) 10 per cent. formalin.—In order to prevent contraction it is
advisable to extend the tapeworm and keep it fixed by glass plates, or
wind the worm around a wide glass tube or bottle, and then fix it.
Or (3) fix in hot alcohol.
Staining.—As under flukes. It is necessary to sacrifice portions of the
tapeworm for this purpose, cutting out, e.g., mature segments, so as to
study the topography of the genitalia.
Clearing.—As under flukes. To examine the hooks satisfactorily it is
best to cut off the head with a sharp knife and mount. A certain amount
of pressure is then advisable in order to view the hooks completely so as
to measure them.
Preservation of Ova in Fæces, etc.
As under flukes.
Preservation and Examination of Nematodes.
Fixation.—(1) Thoroughly wash the worms to get rid of mucus, etc., by
shaking up in warm saline (or water) till the washings are clean. Then
transfer to 70 per cent. alcohol heated to about 70° C. It is absolutely
necessary to use hot fixatives in order to extend the worms. If no alcohol
or spirit is immediately available, drop the worms into hot water, or saline,
and transfer later to 70 per cent. alcohol.
(2) Drop into hot 10 per cent. formalin.
Clearing.—(1) Carbolic acid, vide p. 471.
(2) Creasote, vide p. 471.
(3) Glycerine, vide p. 472.
Staining.—In case of quite small Nematodes, e.g., Anguillulidæ, carmine
may be used, but as a rule staining is not advantageous.
Rolling.—In order to study the mouth parts, or bursa, etc., it is necessary
to place the worm in any desired position. This is done as one would
roll a penholder along the table by one’s finger placed on top of it. In the
case of a worm, one edge of the cover-glass is placed over the worm, the
other is supported by a strip of cardboard. By tapping the cover-glass
the worm will now revolve as much as required provided it is round and
straight. In certain cases it may be necessary for this purpose to cut off
the head or tail. Roll these separately.
When a suitable position is got, the worm may be fixed in this position
by pressure on the cover-glass, so as slightly to flatten it.
Mounting the Head.—If it is required to get an end view, it is necessary to
cut off the head transversely as near the end as possible, and then mount.
Detection of Eggs (Bass and Hall).—Mix the fæces thoroughly with ten
times the volume of water. Filter through gauze. Centrifugalize the
filtrate. Wash the sediment and centrifugalize. Repeat twice. To sediment
add CaCl2 solution, sp. gr. 1250. The eggs float to surface. Pour
off surface fluid. Dilute to sp. gr. 1050. Centrifugalize. Examine sediment,
which contains practically all the eggs in the stool.
Detection of Small Nematodes.—Mix the fæces thoroughly with water.
Allow to settle for five minutes. Carefully decant off, or better, syphon off
the fluid. Mix the sediment again with water. Allow to settle. Remove
the fluid. Repeat several times. Examine the sediment in a Petri dish.
As the fluid is poured off, the worms will be seen collected in the backwater.
Remove them with a brush. Fix in hot 70 per cent. alcohol.
Cultivation of Larval Forms of Ancylostoma and Strongyloides.
A modification of the second method of Looss (p. 455) is that of
Fülleborn. A glass filter funnel is lined with linen or with cotton wool
dyed black with iron-tannin. On this is placed a layer of sterile sand, and
on top of this the fæces. The whole is moistened. The larvæ hatch out
and wander through the meshes of the wool, appearing on the edges of the
same as white threads visible to the naked eye. With a platinum needle
these can be easily removed. The glass filter can be placed on a glass
cylinder, and this in another large stoppered cylinder containing caustic
potash solution at the bottom, so that any larvæ escaping from the funnel
are killed.
D. ACANTHOCEPHALA, Rud.
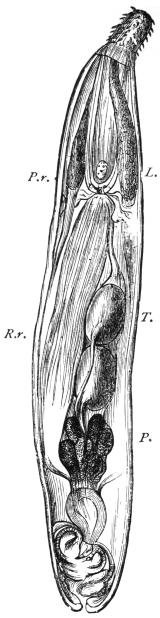
Fig. 348a.—The male of
Echinorhynchus augustatus.
L., lemnisci; T., testicles; P.,
prostatic glands; P.r., sheath of
proboscis, with ganglion; R.r.,
retractor of sheath of proboscis.
25/1.
Gutless, nematode-like worms that carry at their anterior end a retractile
rostrum beset with hooks. In their adult condition they only live in vertebrate
animals. During their larval stage they are often parasitic in invertebrate animals.
The Acanthocephala are elongated cylindrical worms, with a rounded posterior
end. In some species an annulation is distinctly recognizable; they are, however,
not segmented. The size varies according to the species, between about 5 to 10 mm.
and 40 to 50 cm.; in general, however, there is a preponderance of the small species.
The sexes are separate, and the males can easily be distinguished from the females
without examination of the genitalia, as the females are both larger and thicker.
The body wall of Echinorhynchus is limited by a thin cuticle, which is attached
inwardly to the hypodermis. In only exceptional cases a syncytium with large
nuclei, even in the adult condition, is represented by the hypodermis; and in it fibre
systems, the elements of which run in layers in various directions, appear, and it is
only towards the interior from these strata of fibres that the nuclei of the hypodermis
are found. As a rule, these fibres, at all events the radiary fibres, are regarded as
muscles. Hamann describes them as elastic fibres, which lie in a viscid gelatinous
connective substance (transformed protoplasm?); a lacune system filled with a
granular fluid, the central part of which are two longitudinal lacunes lying at the
sides, also belongs to the cutaneous strata, as do the so-called lemnisci, two short,
flat organs suspended in the body cavity, and the pedicles of which are attached
anteriorly at the border between the rostrum and body; their structure as well as
their origin permit them to be traced to the skin (fig. 348A).
Finally, inwardly below the skin there follows a layer of annular, and after these
a layer of longitudinal muscles, the structure cells of which remain present in the
residues, carrying nuclei. The motor apparatus of the rostrum, the sheath of the
rostrum, and the lemnisci also belong to the muscular system. The rostrum
represents a finger-shaped hollow process of the cutaneous layer; but, according
to Hamann, it originates from the entoderm and passes through the skin secondarily.
It is covered by a thin cuticle, and as a rule contains a large number of regularly
placed chitinous hooks that adjoin a granular formation tissue. From the base of
the rostrum springs a tubular hollow muscle extending into the body cavity; this
is the RECEPTACULUM PROBOSCIDIS, from the base of which again bundles of
longitudinal muscles originate, which pass along its axis and that of the rostrum
itself, and are inserted at the inner surface of its anterior end (RETRACTOR
PROBOSCIDIS). These muscles when they contract invaginate the proboscis and
draw it into the receptaculum; when reversed they act again as PROTRUSOR
PROBOSCIDIS. The whole of the anterior body, however, can be invaginated, and
for this purpose there is a muscle that originates from the body wall at a variable
distance back, and which is joined to the receptaculum (RETRACTOR RECEPTACULI);
there is also a bell-shaped muscle which springs from the body wall behind the
lemnisci in rings, and passes forward to the spot of attachment of the lemnisci.
The nervous system consists of a cluster of ganglia situated at the base of the
rostrum, from which three nerves pass towards the front and two towards the back.
No sensory organs are known.
The excretory organs, according to Kaiser, lie at the upper border of the ductus
ejaculatorius in the male and at the so-called bell in the female. Here they
represent the long-known villous tufts, placed on disc-like cushions. In each of the
cylindrical villi—which terminate blindly towards the body cavity—there lies a
cilium, which springs from the membrane lining the villus, and which lies in a space
cavity of the villus, which ultimately proceeds as a
little canal. There are three canals discharging into
the uterus that serve to conduct the excretory materials
from the body cavity; special glandular cells corresponding
to the terminal cells of the Platyhelminths,
at the commencement of the system, are not present
in the Acanthocephala.
Sexual Organs.
(a) Male Organs.—The greatest part of the male
genital apparatus is contained in a muscular sheath—the
ligament—which originates at the posterior end
of the receptaculum proboscidis, passes along longitudinally
through the body cavity, and is inserted at
the posterior end of the worm. The two oval testicles
usually lie one behind the other; their vasa efferentia
unite sooner or later into a vas deferens which passes
backwards, and finally terminates in the penis; the
terminal portion of the conducting apparatus is surrounded
by six large glandular cells (prostatic glands)
the excretory ducts of which open into the vas deferens. The penis itself is placed
at the base of a bell-shaped invagination of the posterior end, the bursa, which is
everted during copulation.
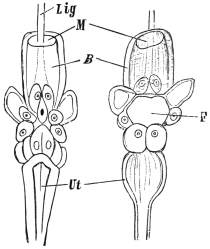
Fig. 348b.—Anterior portion of the female
apparatus of Echinorhynchus acus. On the
left seen from behind, on the right seen from
the front. F, inferior orifice of the bell; B,
bell; Lig, ligament; M, mouth of bell; Ut,
uterus. Magnified. (After Wagener.)
(b) Female Organs.—There are only two ovaries present in the ligament during
the larval stage. During the course of growth they divide into accumulations of
cells (placentulæ, loose or floating ovaries), which finally cause the ligament to burst
and they thus attain the body cavity. Thence a peculiarly constructed apparatus
finally conveys the eggs out. This apparatus consists of the uterine bell and vagina,
the latter discharging at the posterior extremity of the body. The bell is a muscular
canal provided with apertures at both the anterior and posterior extremities. Its
interior space is in direct communication with the body cavity, and the anterior
orifice takes up all materials floating in the cavity—egg-balls, mature and immature
eggs—and pushes these further backwards. The continuation of the bell lumen is
now narrowed by a number of large cells in such a manner that only bodies of a
certain form can pass through this tract and attain the uterus; everything else is
conveyed back into the body cavity through the posterior opening of the bell.
The eggs are already fertilized in the body cavity, and in this position go through
their development to the formation of the embryo. Completely developed eggs are
surrounded by three shells, and are generally fusiform. The eggs agglomerate
in masses in the uterus until they are finally deposited through the vagina and vulva.
For the further development, the transmission of the eggs into an intermediary host—usually
a crustacean or an insect—is necessary; the metamorphosis is very complicated;
but this transmission may be very easily effected artificially by feeding suitable
crustaceans (Asellus, Gammarus, etc.) with the eggs of Acanthocephala; this being
the only method of inducing the larva to hatch out so that its structure may be
studied. The larva appears in the form of an elongated, somewhat bent body, at the
stumpy anterior end of which there is a crown of hooks or spines, whereas the
posterior end is pointed. Especial retractors draw in the hook-beset anterior surface,
and an elastic cushion beneath them jerks them forward again when required. In
the middle of the body a roundish heap of small cells is seen, from which the entire
body of the Echinorhynchus originates, even to the cutaneous layer; the latter is
also the larval skin in which the small Echinorhynchus gradually grows. The
development of all the organs takes place within the intermediary host, and the
parasite only needs to be imported into the terminal host to attain the adult stage after
a certain growth. In some cases, however, a second intermediary host is utilized.
Species of Acanthocephala only occur exceptionally in human beings.
Echinorhynchus gigas, Goeze, 1782.
Syn.: Tænia hirudinacea, Pallas, 1781.
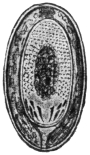
Fig. 348c.—Egg
of Echinorhynchus
gigas. 300/1. (After
Leuckart.)
The body is elongated, gradually decreasing in thickness towards the back.
The rostrum is almost spherical, and is beset with five or six rows of recurved
hooks. The males measure 10 to 15 cm. in length, the females 30 to 50 cm.;
the eggs are provided with three shells, of which the middle one is the thickest.
The eggs measure 0·08 to 0.1 mm. in length. The giant
Echinorhynchus occurs especially in the intestinal canal
of the domestic pig; it is less common in other mammals.
It bores deep into the mucous membrane with its rostrum,
and causes an annular proliferation around the perforated
spot; occasionally also it causes perforation of the intestine.
It is doubtful whether the giant Echinorhynchus occurs
in man. Leuckart admitted that there were a few positive
cases. According to Lindemann, Ech. gigas occurs in
human beings in South Russia, and its presence is not rare.
This statement, however, has not been confirmed. Its
presence in man is by no means impossible, as its intermediary
host, the cankerworm, or cock-chafer (Melolontha),
is, according to Schneider, occasionally eaten raw by human
beings. According to Kaiser, the golden beetle (Cetonia aurata) and, according
to Stiles, another beetle in America (Lachnosterna arcuata) are also intermediary
hosts.
Echinorhynchus hominis, Lambl, 1859.
This term is applied to an Echinorhynchus found by Lambl in the intestine
of a boy who had died of leucæmia; the worm was 5·6 mm. in length, and the
almost spherical head was beset with twelve transverse rows of hooks.
Echinorhynchus moniliformis, Bremser, 1819.
The male is 4 cm. in length, the female 8 cm. long. This species lives in the
intestine of field-mice, rats, marmots and Myoxus quercinius. A beetle (Blaps
mucronata) is the intermediary host.
This species has also once been artificially cultivated in man (Grassi and
Calandruccio).
E. GORDIIDAE.
Very long thin worms similar to Filariæ, which, in their adult condition, live
free in brooks, pools and springs; the mouth and the commencement of the
intestine are obliterated; there are no lateral ridges, and the muscular system
presents a structure different to that of the Nematoda. The posterior end of the
male is split, and spicules are lacking; there are two testicles. In both sexes the
genitalia discharge through the terminal gut.
The larvæ, which carry a rostrum beset with hooks, force themselves into the
larvæ of water-insects; more rarely they invade molluscs, and they then become
encysted within the body of the host. According to Villot, at least a part of them
attain the intestine of fishes, where they again become encysted, and after a period
of rest they travel into the tissues of their hosts, and finally again reach the exterior
by way of the intestine, where they then become adult. In most cases, however,
the gordius larvæ are taken up by predacious water insects; they live for a while
in the body cavity of these insects, undergo a metamorphosis, and finally wander
into the water.
A few species invade man accidentally with water, in which case they are
usually vomited up:—
Gordius aquaticus, Dujardin, 30 to 90 cm. in length (Aldrovandi, Degland,
Siebold, Patruban).
Gordius tolosanus, Duj., 11 to 13 cm. in length (Fiori).
Gordius varius, Leidy, 10 to 16 cm., female, up to 30 cm. in length (Diesing).
Gordius chilensis, Blanch. (Guy). Gordius villoti, Rosa (Bercutti, Camerano);
Gordius tricuspidatus, L. Def. (R. Blanchard), Gordius violaceus, Baird (Topsent),
and Gordius pustulosus, Baird (Parona).
F. HIRUDINEA s. DISCOPHORA (Leech).

Fig. 348d.—The
internal organs of
the leech. The
creature has been
opened from the
dorsal surface, and
part of the intestine
has been removed.
The testicles, with
vas deferens, may be
seen between the
blind ducts of the
intestine; beyond
these on either side
the segmental organs.
The female genital
organs are in front
of the most anterior
pair of testicles.
(After Kennel.)
The Hirudinea, which have been appropriately included amongst the Annelida,
differ in many respects from the typical members of the group; their body is long
and flat, it lacks the parapodia that are characteristic to all forms of Annelida; but,
on the other hand, possesses a terminal posterior sucker, and in many species there
is also an anterior sucker. The mouth is terminal at the anterior end, the anus lies
dorsally above the posterior sucker (fig. 348D). The body is
segmented, but this is less manifest in the body covering than
it is in the arrangement of the internal organs; the segmentation,
nevertheless, is also indicated exteriorly by the appearance
of the cutaneous sensory organs which correspond to the
segments. This shows what the condition of the ganglia in the
abdominal ganglion chain has taught us, that the anterior and
the most posterior segments are considerably abbreviated—a
part of the latter taking part in the formation of the suctorial
organs. In a great many species the skin is distinctly annulated,
four or five of such rings, at least in the central region
of the body, appearing on one segment of the body. The
condition of their body cavity is another peculiarity of the
Hirudinea; it is narrowed by the powerful development of the
connective tissue and the muscular system into four tubular
sinuses, which have the appearance of blood-vessels. There
are usually one dorsal and one ventral median trunks, as well
as two lateral trunks; in addition, a particular blood-vessel
system exists.
The skin consists of a very thin cuticle that is cast off from
time to time; it is secreted by the underlying cylindrical epithelium,
which contains numerous goblet cells. The muscular
system is strongly developed; it consists of long tubular fibres,
which run circularly, longitudinally and in the dorso-ventral
direction; the muscular system is subject to a particular
expansion in the clinging organs and at the commencement
of the intestine. On the whole, the alimentary canal represents
a tube running straight from the mouth to the anus, which
possesses a number of blind sac-like protuberances at the
sides varying according to the species. The most anterior
section, the pharynx, in the leeches with maxillæ carries three
chitinous, semicircular plates furnished with teeth—the jaws—which
serve to tear up the epidermis in order to open the
blood-vessels; in the leeches with rostra a long protractile
proboscis rises from the base of the elongated pharynx.
Numerous salivary glands, the secretion from which possesses
toxic properties, discharge into the pharynx. The œsophagus,
which follows the pharynx, and to the exterior of which
numerous radiary muscles are fixed, is a suctorial organ in
its entire structure. The nutriment in the larger species consists
of the blood of vertebrate animals, in smaller species and
in the young stages the food consists of small invertebrate
animals.
The NERVOUS SYSTEM exhibits the typical structure of other segmented worms;
the sensory organs consist of the previously mentioned goblet-shaped cutaneous
sensory organs, of the organs of taste, and of eyes, the latter frequently being
present in large numbers.
The EXCRETORY or segmental organs exhibit many peculiarities, which cannot,
however, be detailed here. They commence with funnels in the lacunes of the body
cavity, and usually discharge on the ventral surface.
Almost all the Hirudinea are hermaphrodite and copulate reciprocally. The
two ovaries are very small, and the oviducts that proceed from them soon unite into
a common duct, which then passes into the uterus and discharges through the
short vagina in the median line of the ventral surface behind the male organs into
the so-called clitellar region. The male sexual apparatus consists of symmetrically
arranged testicles, varying in number according to the species, the short vasa
efferentia of which, one by one, run into the vas deferens, passing towards the front
on each side. In front, at about the level, or a little in front, of the female genitalia,
the two vessels pass into a convoluted mass of tubes to the so-called epididymis, and
then discharge into the single protractile penis (fig. 348D).
All leeches deposit so-called COCOONS. These are small barrel-shaped or pouch-like
bodies, which are surrounded by a thicker shell and contain a number of eggs
in a large mass of albumen; the albumen originates from glands of the generative
organs, the shell substance from cutaneous glands of the clitellar region.
Family. Gnathobdellidæ (Leeches with Jaws).
These are distinguished by the possession of usually three jaws in the pharynx;
the body consists of twenty-six segments. The posterior sucker is large and flat;
the anterior sucker is smaller. The Hirudinea have five pairs of eyes, the Nephelinæ
have four pairs.
Genus. Hirudo, L., 1758.
The entire body consists of 102 annulations, five appearing on one segment in
the central region of the body. The pharynx has three semicircular jaws, the arched
border of which is beset with numerous teeth (50 to 100). The male sexual orifice
lies between the thirtieth and thirty-first rings, the female orifice between the thirty-fifth
and thirty-sixth. There are numerous species, some of which are utilized for
medicinal purposes.
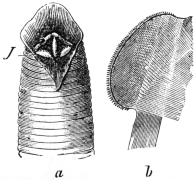
Fig. 348e.—Hirudo medicinalis.
a, anterior end, with
open buccal cavity, with the jaws,
J, at the; b, one jaw isolated.
(After Claus.)
Hirudo medicinalis, L., 1758.
It occurs in numerous colour varieties, one of which has been designated
Hirudo officinalis, Moq.-Tandon. Usually the dorsal surface is greyish-green
and is marked with six rusty-red longitudinal
stripes. The ventral surface is olive-green, more
or less spotted with black, and marked at the
sides with a black longitudinal line. The length
averages 8 to 12 to 20 cm. This leech lives in
swamps, ponds and brooks, overgrown with plants
and having a muddy bed. The cocoons are
deposited in the soil at the sides. Europe, as
well as North Africa, is its home. At the present
day it has been exterminated from most parts of
Central Europe, but it is still very common in
Hungary. Its use for medicinal purposes is well
known. A large leech can suck about 15 grs. of
blood, and about the same amount is lost through
secondary hæmorrhage.
Hirudo troctina, Johnston, 1816.
Syn.: Hirudo interrupta, Moq.-Tandon, 1826.
This species measures 8 to 10 cm. in length. The back is greenish, with six
rows of black spots surrounded by red; the lateral borders are orange-coloured;
the abdomen spotted or unspotted. Its habitat is in North Africa and Sardinia. It
is applied medicinally in England, Spain, France, Algeria, etc.
Genus. Limnatis, Moq.-Tandon, 1826.
Nearly related to Hirudo, but is differentiated by a longitudinal groove on the
inner surface of the upper lip of the anterior sucker. The jaws are furnished with
over 100 very sharp toothlets.
Limnatis nilotica, Savigny, 1820.
Syn.: Bdella nilotica, Sav.; L. nilotica, Moq.-Tandon; Hæmopis (vorax),
Moq.-Tandon, 1826, p. p.; Hæmopis sanguisuga, Moq.-Tandon, 1846 (nec Hir.
sanguis, Bergm., 1757).
This species measures 8 to 10 cm. in length, and becomes gradually more
pointed towards the front; the body is always soft. The back is brown or greenish,
and has usually six longitudinal rows (rarely only two or four) of black dots. The
abdomen is dark; but numerous colour variations occur.
The native place is North Africa, especially the coastal regions; it is also found
in the Canaries, the Azores, Syria, Armenia, Turkestan, perhaps also Southern
Europe. It is taken into the mouth with drinking water, and may settle in the
pharynx, larynx, œsophagus, and nasal cavities of human beings. This species has
also been observed in the vagina and on the conjunctiva. It is equally fond of
attacking domestic animals.
Hirudo mysomelas (Senegambia) and Hirudo granulosa (India) are placed with
this genus, and, like our leech, are also used for medicinal purposes.
Genus. Hæmadipsa, Tennent, 1861.
These leeches live on land, and measure 2 to 3 cm. in length. About a dozen
species are known. They are a veritable scourge to persons in the tropics (Asia,
South America), as they attack them to suck their blood. They are able to force
their way even under close-fitting garments, so that it is difficult to protect oneself
from their assaults (Hæmadipsa ceylonica, Bl., and other species).
Family. Rhynchobdellidæ (Leeches with Rostrum).
These are furnished with a proboscis in lieu of the jaws; the segment consists
of three annulations.
Genus. Hæmentaria, de Filippi, 1849.
Hæmentaria officinalis, de Fil.
Inhabit Mexico, where they are used for medicinal purposes.
Genus. Placobdella, R. Blanch.
Placobdella catenigera, Moq.-Tandon.
Indigenous to South Russia, Hungary, Italy and South France. It is a
parasite of the swamp turtle, but frequently attacks human beings.
G. ARTHROPODA (Jointed-limbed Animals).
BY
FRED. V. THEOBALD, M.A.
Bilaterally symmetrical segmented animals which are covered with a thick
cuticle that is frequently calcareous (Crustacea), but always thinner between the
segments; they carry (primitively) a pair of jointed appendages on every segment.321
The segments of the body are uniform in certain regions, but differ from those of
contiguous regions, so that it is easy to distinguish three parts (head, thorax and
abdomen), each composed of segments. The cephalic segments are always formed
into a uniform head, the segmentation being scarcely recognizable at either end;
the thoracic segments may also fuse, or part or all of them may coalesce with the
head; the abdomen, as a rule, retains its segmentation, but this may possibly also
be lost, in which case it is [sometimes] united to the cephalothorax. The structure
of the three regions depends mostly on the varying form and function of the
appendages: those on the head are primitively locomotive organs (and frequently
are still so in the early stages), but they become transformed into feelers and mouth-parts
(mandibles, maxillæ); the limbs of the thorax, however, usually retain their
ambulatory functions, as frequently do those of the abdomen; sometimes, however,
the abdominal limbs disappear, entirely or partly; in the latter case they are then
utilized for other purposes.
In their organization the Arthropoda approach the segmented worms.
The Arthropoda are generally divided into five groups (Crustacea,322 Protracheata,
Arachnoidea, Myriapoda,323 and Insecta or Hexapoda), of which only the Arachnoidea
and the Hexapoda interest us here.
A. ARACHNOIDEA (Spiders, Mites, etc.).
The head and thorax are always united together; the abdomen is
either segmented or without exterior segmentation, in which case it is
united with the cephalothorax.324 The number of pairs of appendages
amount to six, of which the two front pairs, the cheliceræ and the
pedipalpi, are attached to the head region and the four remaining
pairs to the thoracic region.325 The abdomen in the adult condition
has no appendages. The Arachnoids are air-breathers, and for this
purpose are either provided with tracheæ or with so-called lung-sacs,
or they breathe through the surface of the body. Some aquatic forms
breathe by gills.
There are eight or ten orders of Arachnoids,326 of which, however,
only two, the Acarina and the Linguatulida, have to be considered
here.327
Order. Acarina (Mites).
Small Arachnoids, the three parts of the body of which are, as a rule, coalesced;
it is only rarely that a faint line indicates the division between a cephalothorax and
abdomen. The two appendages on the head are designed for biting or puncturing
and sucking, and vary according to their use. The cheliceræ328 are fang-like jaws
or puncturing bristles forming a kind of rostrum, the pedipalpi are claw-like or shear-shaped,
or form a suctorial proboscis.329 The four pairs of legs are usually well
developed, more rarely they are rudimentary or have partly vanished; many
parasitic forms are provided with pedunculated suckers [ambulacra—F. V. T.].
Respiratory organs (tracheal tufts) may be present or absent. The nervous system
is reduced to a minimum, eyes are usually lacking. The intestine, situated in the
central part, generally has three blind appendages; the anus is situated on the venter
above the posterior end. Sexes separated; nearly all the species deposit eggs, from
which six-legged larvæ hatch. The Acarina live either free in the water or in moist
soil, or they are parasitic on plants and animals.330
Family. Trombidiidæ (Running Mites).
Soft-skinned Acarina with tracheæ and with two eyes, usually pedunculated; they
are often brightly coloured; cheliceræ lancet- or claw-shaped; pedipalpi claw-like;
legs composed of six segments, with suctorial discs between the terminal ungues.331
Larvæ six-legged. To the latter belong the larvæ of several species of Trombidium
such as:—
Genus. Trombidium, Latreille (and Leptus).
Leptus autumnalis, Shaw, 1790.
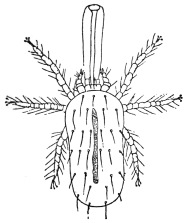
Fig. 349.—Leptus autumnalis, with so-called
sucking proboscis. Enlarged. (After Gudden.)
Leptus occur as parasites in the human skin and cause a cutaneous
disease known as autumn erythema, and produce a very unpleasant
sensation on account of the troublesome
itching; in children it is very
often accompanied by fever.332
Formerly these mites were considered
adult forms, but when they were recognized
as mite larvæ they were taken
for those of the spider-mite (Tetranychus
telarius); the investigations of Hanstein,
however, showed this to be a mistake.
When Henking first investigated the development of Trombidium fuliginosum,
parasitic in the larval stage on vine-fretters, he demonstrated the occurrence of a
form very similar to Leptus autumnalis, and the “autumn, grass, or gooseberry”
louse was commonly designated the Trombidium larva. Even before Henking’s
work it had been described by Mégnin as the larva of Trombidium holosericeum, a
red-coloured species, frequently occurring in spring and summer on the ground, trees,
etc. This assumption, however, as Moniez was the first to explain, is not correct;
indeed, as many as three species come under consideration: T. gymnopterosum,
L., T. fuliginosum, Herm. (according to Brucker), and two species known hitherto
only in the early stage, T. striaticeps, Helm. et Oudem., and T. poriceps, Helm. et
Oudem., which are not only parasitic on mammals, but on birds, on Arthropods and
especially on insects. Arthropods appear to be the normal hosts for the larvæ.
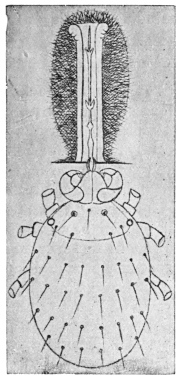
Fig. 350.—Leptus autumnalis: the
so-called proboscis is formed around
the hypopharynx sunk into the skin.
100/1. (After Trouessart.)
The above-mentioned forms invade the skin of man by means
of their oral apparatus, by preference invading the orifices of the
sebaceous glands so as to suck the blood; around the point attacked
there arises a wheal about the size of a lentil, and around the inserted
hypopharynx a fibrinous secretion, the “proboscis,” which, however,
is a product of the host, just as chitinous secretions are provoked by
Trombidia parasitic on Arthropods.
Further species, analogous in habit to Leptus autumnalis, are described by Riley
from Central and South America as L. americanus and L. irritans.
[L. autumnalis attacks small mammals by preference, such as moles
and hares, which are often literally covered with them. Dogs are
also subject to their attack, and cats suffer similarly. This mite also
frequently appears in colonies on cows; cavalry horses after autumn
manœuvres often suffer from an erythematous affection about the
hocks and knees due to this pest.
[A number of Leptus, so far undescribed, occur abroad which
attack man in the same way as L. autumnalis in Europe. Dr. Durham
has brought me specimens from British Guiana called bête rouge;
this species works under the skin much as does our European species,
but it is very distinct, being considerably larger.—F. V. T.]
Trombidium tlalsahuate, Lemaire, 1867.
T. tlalsahuate occurs in Mexico under conditions similar to those
of Leptus here. It also frequently attacks men, and especially fastens
itself on to the eyelids, in the axillae, navel, or on the prepuce; it
induces itching and swelling of the parts affected, and sometimes
even causes suppuration; the symptoms, however, generally disappear
after a week and remain localized.333
Other species of mites which attack man are reported, mostly
by travellers, from various other places; zoologically, however,
there is little known about them. The pou d’agouti in Guiana,
niaibi in New Granada, colorada in Cuba, mouqui in Para, and the
buschmucker in New Guinea represent a few of these.
Akamushi or Kedani.
In a few districts of Japan there occurs a serious illness, with
a mortality of 40 to 70 per cent. It is called river or flood
fever, and the Japanese doctors have connected it with a small
mite (akamushi, kedani). Baelz has opposed this opinion on the
grounds that he has repeatedly observed the same species of mite
in his dwelling without any subsequent illness occurring. According
to Keïsuke Tanaka, however, a connection certainly does exist,
inasmuch as the akamushi, like Leptus, attacks persons to suck
blood. If the mite is not removed, or if the spot attacked is
injured by scratching, etc., a papule surrounded by a red area
forms, and a pustule ensues; and finally a black scab covers the
seat of injury. The lesion becomes the point of entrance of bacteria,
especially a species of proteus which produces river fever. If the
mites are carefully removed no general illness takes place.
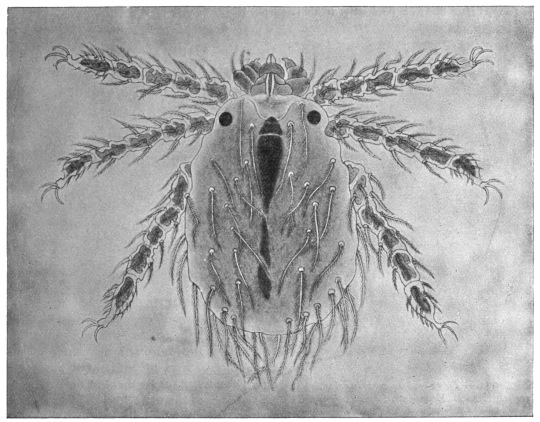
Fig. 351.—The kedani mite. Enlarged. (After Tanaka.)
The orange-red mites, which we only know in their larval
condition, measure 0·16 to 0·38 mm. in length by 0·10 to 0·24 mm.
in breadth. They have leg-like palpi with three joints, hirsute
bodies, and very hairy legs composed of five segments, terminating
with three ungues.
Family. Tetranychidæ (Spinning Mites).
These have tracheæ and eyes; the palpi are composed of four segments, of
which the last but one has a powerful claw. The legs have six segments with sucker
discs between the claws.
[The red spiders or spinning mites (Tetranychi) are usually placed
in the family Trombidiidæ.—F. V. T.]
Genus. Tetranychus, Dufour.
Tetranychus molestissimus, Weyenbergh, 1886.334
Found in Argentine and Uruguay on the under surface of the
leaves of Xanthium macrocarpum; it attacks mammals and men,
producing severe itching, accompanied by fever in the latter.
It has been asserted by Haller
that the Cape ailment (Port Natal
sickness) is caused by mites, but this
statement has been contested.
Tetranychus telarius, L., 1758,335 var. russeolus, Koch.
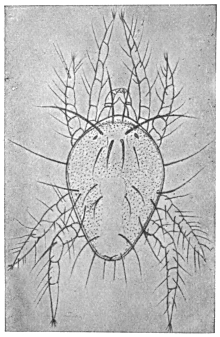
Fig. 352.—Tetranychus telarius var. russeolus,
Koch. Enlarged. (After Artault.)
This common spinning mite likewise
attacks human beings, but the
papules produced by it very soon
disappear.
Family. Tarsonemidæ.
A family distinguished by complete
sexual dimorphism, the species of which are
provided with tracheæ; the legs have five
segments; the terminal segments of the
front pair of legs of both sexes possess a
claw; the terminal segment of the posterior
pair of legs of the male likewise has a claw.
In the female this pair of legs, like the
second and third pairs of both sexes, is provided with two hooklets and a sucking
disc. The cuticle of the body on the back is “annulated.”
[This family of small transparent mites live normally as plant
parasites. The last two pairs of legs are widely separated from the
two front pairs.—F. V. T.]
Genus. Pediculoides.
Pediculoides ventricosus, Newport, 1850.
Syn.: Heteropus ventricosus, Newport, 1850; Acarus tritici, Lagrèze-Fossot,
1851; Physogaster larvarum, Lichtenstein, 1868; Sphærogyna ventricosa, Laboulbène
and Mégnin, 1885.
Males are oval in shape, 0·12 mm. in length and 0·08 mm. in
breadth, flattened. There are six pairs of chitinous hairs on the
dorsal surface and a lyre-shaped lamella on the posterior part. The
female in the non-gravid state is cylindrical in form, 0·2 mm. in
length and 0·07 mm. in breadth; when gravid the posterior part of
the body becomes enlarged into a ball, which may attain 1·5 mm. in
size, as in the case of Pulex penetrans and of the female Termites.
On emerging the young are already provided with four pairs of legs
and copulate soon after birth.
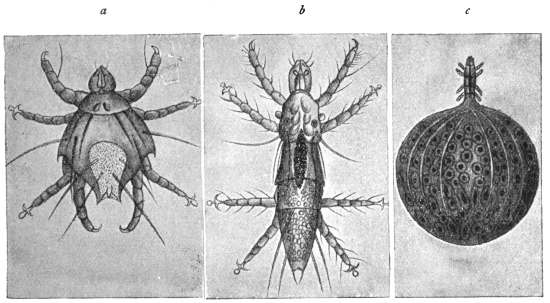
Fig. 353.—Pediculoides ventricosus. a, male; b, young female; c, gravid female.
Enlarged. (After Laboulbène and Mégnin.)
These animals live on the stalks of cereals, and feed on vegetable and animal
juices; they are also found on corn-infesting insects. They invade the barns and
seek out the insects living in the dry grains of corn, or wait for an opportunity of
obtaining food. They have been repeatedly observed on human beings, particularly
labourers occupied in handling grain; their bite causes severe irritation, local
elevation and reddening of the epidermis, as well as fever. It cannot be positively
asserted that all cases of the occurrence of cereal mites on man relate to P. ventricosus,
as the descriptions are often insufficient. Geber states that one form is Chrithoptes
monunguiculosus, or Acarus hordei; Flemming mentions Tarsonemus uncinatus;
Koller Oribates sp.; and Karpelles Tarsonemus intectus.
[The pregnant female Pediculoides has a large round inflated abdomen, in
which the ova hatch and the young mature. Later they escape from the parent as
adults.—F. V. T.]
Genus. Nephrophages.
Nephrophages sanguinarius, Miyake and Scriba, 1893.
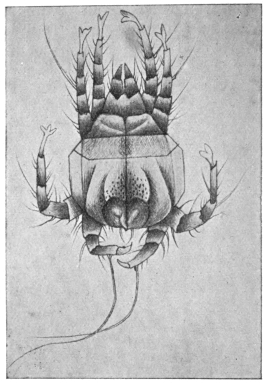
Fig. 354.—Nephrophages sanguinarius: male,
ventral surface. Enlarged. (After Miyake and
Scriba.)
Males measure 0·117 mm. in length and 0·079 mm. in breadth;
females up to 0·360 mm. in length by 0·120 mm. in breadth. The head
is provided with two very large scissors-like jaws and two large round
eyes. The legs are composed of five segments and are all of equal
length; the three anterior pairs of legs have pedunculated ambulacra,
the posterior ones terminate in a claw. The cuticle on the back is
thickened in three places, shield-like; the abdominal surface without
scutellum is longitudinally striped and is beset with chitinous hairs.
Colour greenish to brownish-yellow.
Eggs 0·046 to
0·040 mm.
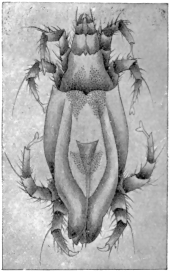
Fig. 355.—Nephrophages sanguinarius:
female, dorsal aspect. Enlarged.
(After Miyake and Scriba.)
The authors discovered these mites, but always dead, in the urine of a Japanese
suffering from fibrinuria complicated with chyluria and hæmaturia. They surmised
that they were endoparasites, probably situated in the kidney; but this view is not
convincing, though they also report that for a week, day after day, the mites were
found in the patient’s urine, as well as in urine drawn off by means of a catheter,
and in the water used to wash out the bladder (one or two specimens and an egg).
The statement that these mites have large eyes makes the discovery suspicious, to
say the least. The significance of the discovery is not supported by the further
statement that Disse is supposed to have found an encapsuled mite closely related
to the Tyroglyphides on the wall of the vena cava.
In the case of Marpmann, who found a dead Acarid in the urine of a man
suffering from chronic nephritis, and in whom later examinations proved negative,
the author himself was of opinion that the mite had reached the urine from
outside.
We are certainly acquainted with mites living endoparasitically, namely, the
Cysticolæ, Analgesinæ, of which Laminosioptes gallinarum live in the intramuscular
and subcutaneous connective tissue of fowls, and Cytoleichus sarcoptoïdes in their
air sacs. Another kind of mite (Halarachne halichœri) is occasionally found in the
nasal mucous membrane of the seal (Halichœrus grypus), and, quite recently,
Pneumonyssus simicola, which is more nearly related to Halarachne, has been found
in the lung of Cynocephalus sp. It is therefore not improbable that endoparasitic
mites are found in man; but no definite discovery has yet been made.
Family. Eupodidæ.
Small tracheate mites, with moderately long or short pedipalpi, composed of
four segments, of which the last segments bend; cheliceræ forceps-shaped, with
serrated edges; legs with two claws, more rarely with one, and terminating in
a tuft ornamented with fine hairs; genital orifices on the abdomen, surrounded
by a circle of little hairs. Most species live free—one lives parasitically on the
bodies of slugs.
Genus. Tydeus, Koch.
Tydeus molestus, Moniez, 1889.
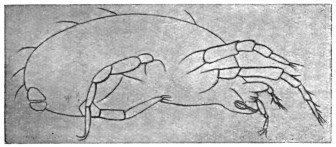
Fig. 356.—Tydeus molestus: seen in profile. Enlarged.
(After Moniez.)
Male, 0·2 mm. in length, 0·125 mm. in breadth. Females, 0·225 mm.
in length, 0·135 mm. in breadth; gravid female 0·315 to 0·360 mm.
in length and 0·180 mm.
in breadth. They were
observed by Moniez on
an estate in Belgium,
whither the creature
had apparently been imported
twenty-five years
previously with Peruvian
guano; they appeared
regularly in the
summer and remained
until the first frost set
in; they were found on grass-plots, on trees and bushes in masses;
they regularly attacked human beings, mammals and birds, tormenting
their hosts in a terrible manner.
Family. Gamasidæ (Coleopterous or Insect Mites).
Cheliceræ chelate or styliform; pedipalpi filiform; the legs are composed of six
segments with two terminal ungues and a bladder-like sucking disc [caruncle—F. V. T.].
Stigmata situated between the third and fourth pairs of legs; the
cuticle thickened, leather-like; no eyes; the larvæ have six legs.
The Gamasidæ are predaceous on small insects and other mites; some are
parasitic on insects, and one is noticeable as a pest on birds, etc.
Genus. Dermanyssus, Dugés.
Dermanyssus gallinæ, de Geer, 1778.
Syn.: Pulex gallinæ, Redi, 1674; Atarus gallinæ, de Geer, 1778;
Dermanyssus avium, Dugés, 1834.
The male measures 0·6 mm. in length by 0·32 mm. in breadth; the
female 0·7 to 0·75 mm. in length by 0·4 mm. in breadth. The body
is somewhat pear-shaped; the colour whitish, reddish, or reddish-black,
according to the contents of the intestine. The legs are
fairly short and strong. During the day they live concealed in the
nests, cracks, etc., of the hen-house, and at night attack the inmates
in order to suck their blood; they rarely remain long on the birds.
They have been repeatedly found on persons, on whose skin they
produce an itching eruption.
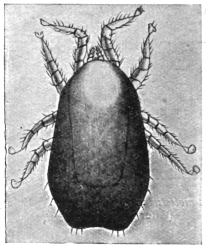
Fig. 357.—Dermanyssus gallinæ. Enlarged.
(After Berlese.)

Fig. 358.—Dermanyssus hirundinis.
40/1. (After Delafond.)
Dermanyssus hirundinis, Hermann, 1804.
Syn.: Acarus hirundinis, Herm., 1804.
Of a brownish colour, 1·2 or 1·4 mm. in length; lives in the nests
of swallows and is occasionally found on man.
[The red hen mite (Dermanyssus gallinæ) not only attacks
poultry and man, as stated above, but is found on all birds and
many mammals. The D. gallinæ is the same as D. avium. The
species found in swallows’ nests is also said to be the same. This
mite can remain for weeks without any food from its normal
host. They only attack man when entering or cleaning dirty and
neglected fowl-houses; they do not produce a true dermatosis. They
chiefly attack the backs of the hands and forearms of those who constantly
attend poultry and give rise to symptoms similar to the
papular eczema of scabies. That they may remain some time upon
the human body we know from the following cases out of many
recorded: Geber observed that the Dermanyssus had caused a diffused
eczema on a woman, which lasted four weeks and then disappeared.
The tique of F. V. Raspail is the bird Dermanyssus; he records
children and adults being attacked not only when handling pigeons,
but even when walking in a garden manured with pigeons’ dung.
The affection soon disappeared when the pigeons were destroyed and
the excreta buried. I have frequently heard of poultrymen being
seriously attacked by this pest.—F. V. T.]
Genus. Holothyrus.
Holothyrus coccinella, Gervais, 1842.
Measures 5 mm. in size; lives on birds in the Island of Mauritius;
ducks and geese frequently fall victims to its bite; it also attacks
human beings, on whose skin it causes severe burning and swelling,
but no reddening; it may be dangerous to children, especially by
settling in the oral cavity.
Other Gamasides occasionally occur in man, for instance, according to Moniez,
Leignathus sylviarum, Canestr. et Fanzago; according to Neumann Lælaps stabularis.
The former live normally in the nests of various species of Sylvia, Lælaps on dried
vegetable substances, also in houses.
[Marchoux and Conoy (Bull. Soc. Path. exot., 1912, v, No. 10,
pp. 796–798) found Leishman granules in Lælaps echidninus. It is
assumed that Leishman granules may be found in most Arachnoids,
and have no connection with Spirochæta.—F. V. T.]
Family. Ixodidæ (Ticks).
Comparatively large Acarines with a leathery skin; they are flattened in form,
but after sucking blood the abdomen becomes spherical; the cheliceræ are rod-like
and possess a serrated terminal joint, bent hook-like; the median parts of the
pedipalpi (maxillæ) form a rostrum furnished with barbed hooks (fig. 359); the
maxillary palpi themselves are club-like or rounded; the legs are composed of six
segments with two terminal ungues, often also with “sucking discs”; the stigmata
are at the sides of the body, posterior to the fourth or third pair of legs. The larvæ
are six-legged.
[The true ticks (Ixodidæ) are all blood-suckers, and as far as is
known they do not take vegetable food at all. Not only are the
Ixodidæ important as actual parasites, but they are most so on
account of the fact that they are the active agents in carrying various
diseases in animals and apparently in man. It has been conclusively
proved that the bont tick (Amblyomma hebræum) is the carrier
of the fatal “heart-water fever” so rife amongst sheep in South
Africa, that the dog tick (Hæmaphysalis leachi) is the agent by which the
protozoa that cause malignant jaundice in dogs are distributed, that
Texas fever in cattle is spread by Rhipicephalus annulatus, and Coast
or Rhodesian fever by R. appendiculatus and R. simus. Their importance
as disease carriers amongst mammals is therefore considerable,
and it may prove to be so for man.336 They frequently attack
man, but chiefly, according to my observations, in their early stages
in Europe; this is not so, however, abroad. The life-history of
a number of ticks has been clearly demonstrated. Mr. Wheler has
shown that in Ixodes reduvius it is as follows: the female deposits
her eggs in masses upon the ground, gradually reducing in size
as the eggs pass out, until she finally remains a mere shrivelled
empty bag and then dies. The eggs are oval, golden brown in
colour and smooth; in length they are 0·59 mm.; as in all Ixodidæ
they are covered with a glutinous secretion, by means of which
they adhere together in masses. These egg masses may be deposited
anywhere on the ground, but amongst rough, coarse herbage seems
to be the favourite place. The egg stage may last as long as
twenty-two weeks, or it may only take eight weeks. In the case
of the bont tick a single female may deposit 15,000 or more eggs.
The process of egg-laying is described as follows by Mr. Wheler:
“When egg-laying is about to take place, the head is further
depressed till it rests close against the under side of the body.
In this attitude the end of the rostrum actually touches the genital
orifice, the palpi being at the same time widely opened out. Behind
the head and from beneath the shield, at what for the purposes
of explanation may be described as the back of the neck, a white,
perfectly transparent, delicate gelatinous membrane is brought down
through inflation, either with air or with a transparent fluid, above
the head, which it temporarily conceals. The end of this membrane
terminates in two conical points which appear to be covered
with a glutinous secretion, and at the same time an ovipositor of a
somewhat similar character, but only semi-transparent, is pushed
forward from the genital orifice. This latter is a tube, within which
is the egg. As the ovipositor projects it turns itself inside out, like
the finger of a glove, leaving the egg protruded at the end and lying
between the two finger-like points of the membrane. The membrane
and the ovipositor are then withdrawn each from the other. The
egg adheres to the former, which collapses through the withdrawal
of its contents, dragging the head forward and depositing it on the
top of the head. Neither legs, palpi, nor the organs of the mouth
take any part in oviposition, but after the collapse of the membrane
the palpi are closed and the head is raised, by which action the
egg is pushed forward to the front edge of the shield, forming
in time an adherent mass of eggs, which are deposited in front of
the tick.”
[The egg gives rise to the larval form, the so-called “seed-tick”
stage. At first these minute specks are pallid and soft, but they
soon harden and darken in colour. These larvæ are six-legged and
crawl up grasses and various plants, and there await a passing host,
waving their two front legs in the air and becoming attached by this
means. The larval ticks feed upon the blood of the host, and when
replete fall to the ground, the body becoming inflated in the
meanwhile. These larvæ may remain on the host only two days, or
they may remain much longer. Eventually they moult on the ground
and change to the nymph or pupal stage, which has four pairs of legs.
This pupa acts just as the larva, crawls up plants and waits to regain
the host. After a time the nymphs, having gorged themselves with
blood, fall off and remain on the ground for nearly three months;
they then moult and become adult males and females. In about ten
days they assume their normal colour and regain the host afresh;
the female gradually swells until she attains that large inflated form
so characteristic of ticks. The male does not swell, but nevertheless
feeds upon the host and fertilizes the female.
[The act of coitus is strange: the male tick inserts its rostrum and
other mouth organs into the sexual orifice of the female, between the
base of the posterior pair of legs. The males then die and the
females fall to the ground and deposit the ova. There are variations
in the different species, of course, from those given above, which
apply solely to Ixodes reduvius. The larvæ and nymphs seem to
attack most animals, but the adults mainly keep to the same host.
The periods in the life-cycle of ticks not only vary in the different
species, but in each species according to climatic conditions; for
instance, in the bont tick (Amblyomma hebræum, Koch), Lounsbury
has shown that the development is rapid in summer, slow in winter.
The period from the time that the female drops to the time she
commences to lay eggs varied in specimens observed by him from
twelve days in summer to twelve weeks in winter, and the complete
period from the dropping of the female to the hatching of the eggs,
from eleven weeks in summer to thirty-six weeks through the winter.
Other stages vary in a similar manner.
[Ticks may live a long time away from the host provided they are
supplied with a certain amount of moisture. Mr. Wheler kept dog
ticks (Ixodes plumbeus) in the larval stage for ten months; the pupæ,
male and female, of I. reduvius for six months.
[I have kept Ornithodorus moubata alive for eighteen months
without food.
[In many species moulting takes place off the host, but in I. bovis,
now known as Rhipicephalus annulatus, Say (the carrier of Texas
fever), moulting takes place on the host, and in many other species
also.337 Some species of ticks leave their host on its death (as the dog
tick, Hæmaphysalis leachi), but others die with the host (bont tick,
Amblyomma hebræum).
[Two species are of special importance, namely Ornithodorus
moubata, Murray, which may infect human beings with the spirillum
of African tick fever, and Dermatocentor reticulatus var. occidentalis,
which is said to be the carrier of Rocky Mountain spotted fever.
Classification of Ixodidæ.
[The ticks, or Ixodidæ, are divided into two groups, known as
(1) Argantinæ, (2) Ixodinæ. The Argantinæ are told from the
Ixodinæ by the absence of dorsal or ventral shields in both sexes,
and also by the rostrum being placed beneath the cephalothorax,
which covers it over: except in the larval stage, in which it is subterminal,
and in the pupal, when it partly projects. Legs nearly
equal in length. The sexual orifice is situated between the two first
pairs of legs. The males usually smaller than the females.
[The Ixodinæ have the legs unequal, of six segments with two
false segments, making them look as if composed of eight segments.
The rostrum is terminal and never hidden beneath the body.
The sexual orifice is situated between the bases of the first three
pairs of legs. In the males the orifice is obsolete or very rudimentary,
sexual intercourse being effected by the rostrum. The
males are smaller than the females. The shield in the females never
covers so much as one-half of the body even when fasting, also
in the larvæ and nymphs; but in the males, which do not distend,
the shield covers the body entirely, or all but a narrow margin.
The Ixodinæ are divided into two groups: (i) the Ixodæ, and (ii) the
Rhipicephalæ. The former have a long proboscis reaching nearly to
the end of the palpi or even a little longer than the palpi. The
palpi are longer than broad. The Rhipicephalæ have short palpi,
nearly or quite as broad as long, more or less conical or subtriangular.
They were called Conipalpi by Canestrini.
Synopsis of Genera.
| [A. Argantinæ: Rostrum concealed in adult, partly exposed in larvæ and nymphs. No dorsal and ventral shields. |
| Body flat with thin edges, finely shagreened and punctate Argas. |
| Body with numerous small round granules and with thick sides | Ornithodorus. |
| [B. Ixodinæ: Rostrum terminal. Body with dorsal shield over some part of it. |
I. | Rostrum and palpi longer than broad (Ixodæ). |
α. | A groove around anus in front. |
| Palpi caniculated in both sexes | Ixodes. |
| Palpi claviform, not caniculated in the male; legs very long | Eschatocephalus. |
| Palpi claviform, not caniculated in the male; anal groove absent in the female | Ceratixodes. |
β. | A groove around the anus behind. |
| No eyes; adanal shields | Aponomma. |
| Eyes present. |
| Males with no adanal shields | Amblyomma. |
| Males with adanal shields | Hyalomma. |
II. | Labium and palpi short; palpi not, or very little, longer than broad (Rhipicephalæ). |
α. | No eyes. |
| Rostrum rectangular; no ventral shields in the male | Hæmaphysalis. |
β. | Eyes present. |
| No adanal shields, but usually with greatly developed coxæ on fourth pair of legs. Capitulum quadrangular | Dermacentor. |
| Capitulum hexagonal | Rhipicentor. |
| Adanal shields in male. Stigmata comma-shaped | Rhipicephalus. |
| Stigmata oval or round; legs normal | Boophilus. |
| Segments of legs expanded | Margaropus. |
| [The genus Ceratixodes of Neumann, 1902, occurs on birds. |
| [The genus Eschatocephalus of Frauenfeld, 1853, of which seven species are known, is mostly parasitic on bats, and is found in holes, caves, and church towers. |
| [The genus Aponomma of Neumann, 1899, is exotic, and almost entirely confined to snakes and saurians. |
| [The following are synonyms of the different genera:— |
| Argas, Latreille, 1796 (Rhynchoprion, Hermann, 1804). |
| Ixodes, Latreille, 1795 (Acarus, Linnæus, 1758; Cynorhæstes, Hermann, 1804; Crotonus, Dumeril, 1822). |
| Ceratixodes, Neumann, 1902 (Ixodes, Cambridge, 1879; Hyalomma, Cambridge, 1879). |
| Eschatocephalus, Frauenfeld, 1853 (Sarconyssus, Kolenati, 1857). |
| Amblyomma, Koch, 1844 (Ixodes, Latreille, 1795). |
| Hæmaphysalis, Koch, 1844 (Rhipistoma, Koch, 1844; Gonixodes, Dugès, 1888; Opitodon, Canestrini, 1897). |
| Rhipicephalus, Koch, 1844 (Acarus, Linnæus, 1758; Ixodes, Latreille, 1795; Phanloixodes, Berlese, 1889; Boophilus, Curtice, 1890). |
| Dermacentor, Koch, 1844 (Ixodes, Latreille, 1795; Pseudixodes, Haller, 1882).—F. V. T.] |
Genus. Ixodes, Latreille.
Ixodes reduvius, L., 1758.338
Syn.: Acarus reduvius and ricinus, L.; Ixodes ricinus, Latreille, 1806.
The males are oval; their length 1·2 to 2 mm.; they are brownish-red
or black in colour; the females are yellowish-red, 4 mm. long;
when gorged they are lead-coloured, and may attain 12 mm. in
length by 6 to 7 mm. in breadth.
The dog tick (fig. 360) lives in thickets on leaves, etc., and attacks sheep and
oxen, and more rarely dogs, horses, and human beings, into the skin of which the
female bores with the rostrum in order to suck blood; the bite is not dangerous, and
sometimes is not even felt. Inflammation, however, is set up if the creatures are
forcibly removed from the wound, as the rostrum as a rule is torn off in the process.
If left alone or smeared over with some grease—vaseline, oil, butter, etc.—the
creatures drop off spontaneously. Sometimes the entire tick bores itself into the
skin; they also appear to be permanent inmates of kennels.
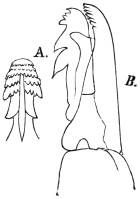
Fig. 359.—A., the rostrum of Ixodes
ricinus (male); B>., the terminal joint of the
maxillary palpi of the female. Enlarged.
(After Pagenstecher.)
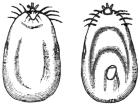
Fig. 360.—Female of Ixodes ricinus,
gorged full, dorsal and ventral surfaces.
2/1. (After Pagenstecher.)
[The species I. reduvius is the same as I. ricinus, Latreille.
The male is 2·35 to 2·80 mm. long; the body is dark brown, almost
black, with a pale, almost white, margin; there are also traces
of reddish mottling. Coxæ of the first pair of legs with a short
spine. Rostrum much shorter than that of the female; shield oval;
anal shield small, about one-third the length of ventral shield. The
adult female varies from 2·80
to 3·5 mm. when not distended,
but when gorged may reach
10 mm. long. The shield and
legs are dark blackish-brown, body deep orange-red with four dark
longitudinal lines, paler beneath and light grey in front. When distending
it is pale red to grey or white; when fully gorged olive-green,
or dark red to black, with irregular yellow streaks on the back and sides
just before egg-laying. Sexual orifice opposite fourth pair of legs.
The nymph varies from 1·60 to 1·70 mm. long when fasting; the
body is olive-white, opaque, with four distinct brown posterior
markings and similar anterior ones, leaving a pale centre to the
shield. When fully gorged it is 3 mm. long. As the nymph distends,
it changes from opaque white to blue-black, and finally black. The
little larva is 0·80 to 1·50 mm. long, transparent with olive-green
intestinal markings; as it becomes inflated it changes to blue-black,
and then black. There are no eyes. It is widely distributed, and
chiefly attacks sheep; sometimes it occurs on dogs and also attacks
man. Mégnin records it from horses in the nymph stage. Amongst
its other numerous hosts are goats, cattle, deer, hedgehogs, moles, bats,
birds, and lizards. It is usually known as the grass tick and bottle-nosed
tick. This species occurs in Europe, Asia, North Africa, and
North America.
[Synonyms.—Considerable confusion exists over the name of this
and other common ticks, owing to the same species having been
described under a great many names. Observers have taken the
same species on different animals and in various stages to be distinct,
and have described them accordingly.
[The name Ixodes reduvius, Leach, does not stand, as Leach was
describing quite a different parasite. The name I. ricinus, Latreille,
1806, is now substituted by Neumann and Wheler.
[The synonyms given by Wheler are as follow: Reduvius,
Charleton, 1668; Ricinus caninus, Ray, 1710; Acarus ricinoides,
de Geer, 1778; Acarus ricinus, Linnæus, 1788; Cynorhæstes reduvius,
Hermann, 1804; Cynorhæstes ricinus, Hermann, 1804; Ixodes
megathyreus, Leach, 1815; Ixodes bipunctatus, Risso, 1826; Cynorhæstes
hermanni, Risso, 1826; Crotonus ricinus, Dumeril, 1829; Ixodes
trabeatus, Audouin, 1832; Ixodes plumbeus, Dugés, 1834; Ixodes
reduvius, Hahn, 1834; Ixodes fuscus, Koch, 1835 (?); Ixodes lacertæ,
Koch, 1835 (?); Ixodes pustularum, Lucas, 1866; Ixodes fodiens, Murray,
1877; Ixodes rufus, Ixodes sulcatus, and Ixodes sciuri, Koch.—F. V. T.]
Ixodes holocyclus, Neumann, 1899.
[Under the name I. holocyclus, Cleland (Journ. Trop. Med. and Hyg.,
1913, xvi, No. 3, pp. 43–45) says that: “This tick is common in man
where there is dense scrub and tropical jungle along the east coast of
Australia at certain times of the year. It may cause severe symptoms
in children resulting in death.” He records a child being attacked in
1884 which died, and another case from which 200 ticks were
removed, the symptoms being weak heart, collapse, syncope, but the
patient recovered under treatment; again, in the same journal
(pp. 188, 189), the case of a 4 1/2-year-old girl who was bitten showed
widespread muscular paralysis, and other cases resembling conium
poison.
[Taylor (Rep. Ent. Aust. Inst. Trop. Med., 1911, p. 21, 1913)
refers to this species as the scrub tick of New South Wales.
The partially fed female has a dark reddish-yellow scutum and is
almost as broad as long, punctations very numerous, equal and
confluent in places, long white hairs on the lower half of each coxa.
He records it as attacking man commonly, mentioning Kamerunga,
Cairns district, Queensland, and Sydney, N.S.W., as localities.—F. V. T.]
Ixodes hexagonus, Leach, 1815.
Syn.: Ixodes sexpunctatus, Koch, 1897; I. vulpis, Pagenstecher, 1861.
Lives in the same manner as the foregoing; especially attacks
hounds, but also other mammals and even birds. The difference
consists in the shape of the legs, the shorter rostrum, and the larger
size of the male. It also occasionally attacks man, but is usually
confused with the previously mentioned species.
[The synonyms of this species are as follow:339 Ixodes autumnalis,
Leach, 1815; I. erinacei, Audouin, 1832; I. reduvius, Audouin, 1832;
I. crenulatus, Koch; I. erinaceus, Murray, 1877; I. ricinus, Mégnin,
1880. Two other synonyms are given above by Braun.
[The female when fully replete is 11 mm. long, when fasting
3·86 mm.; the shield is heart-shaped and punctate, body finely hairy;
palpi short and broad; labium shorter, and tarsi of all the legs more
truncate than in I. ricinus. The colour of the distended body is
drab and somewhat waxy; rostrum, shield and legs light testaceous.
The male varies from 3·5 to 4·0 mm. long, and is reddish-brown
in colour with lighter legs; the shield is punctate and leaves a
narrow margin around the body; the body is elliptical, almost as
large in front as behind. There is a spine on the coxæ of the first
pair of legs, which is shorter than in the male I. ricinus and longer
than in the female. The genital orifice is opposite the interval
between the second and third pair of legs. The fasting nymph is
1·76 mm. long, light bluish-grey, margined and transparent, with
four large posterior intestinal marks joined together behind the
shield and smaller ones extending to the front and sides. When
fully distended it is uniformly brownish-white; shield, legs and
rostrum pale testaceous. The larva varies from 0·88 mm. when
fasting to 1·76 mm. when gorged. Its body is light, but gradually
becomes darker, with similar intestinal marks to ricinus.
[This tick is very common, especially on ferrets, stoats and hedgehogs.
It is also found on sheep, cattle, etc. The males do not
generally occur in company with the females on the host. Pairing
probably takes place on the ground.—F. V. T.]
Genus. Amblyomma, Koch.
Amblyomma cayennense, Koch, 1844.
Syn.: Amblyomma mixtum, Koch, 1844; Ixodes herreræ, Dugés, 1887;
Amblyomma sculptum, Berlese, 1888.
Characterized by the possession of eyes. The male measures
3·8 mm. in length by 3 mm. in breadth; the female 4 mm. in length
by 3 mm. in breadth, but when full of blood may become 13 mm.
in length and 11 mm. in breadth. They are common in the whole
of Central America (Carrapatas), and attack mammals, amphibious
animals and man.340
[This species was described by Fabricius. It occurs in Cayenne,
Guiana, in Southern Texas, Florida, California, Mexico, Guatemala,
Honduras, Nicaragua, Costa Rica, Panama, Bermuda, Cuba, Jamaica,
Trinidad, Colombia, Venezuela, French Guiana, Brazil, Paraguay
and the Argentine. It is called the silver tick. It frequently attacks
man. Schwarz and Bishopp (Bull. 105, U.S. Dept. Agric., p. 158)
heard of one man whose legs were well covered with suppurating
sores and who was ill from the attack of these ticks and the wounds
produced by scratching, and records other cases of their swarming
on man. Newstead (Ann. Trop. Med. and Par., 1909, iii, No. 4, p. 442)
records it as the worst pest to man in Jamaica.—F. V. T.J
Amblyomma americana, Linnæus.
The so-called long star tick, from the silvery spot on the apex of
the scutum of the female. It will attack any mammal and even birds
and also man. It occurs in North America, and also in Brazil,
Guiana and Guatemala. Its punctures frequently end in suppuration.
In the Eastern and Southern States man is more frequently attacked
by this species than any other. Moss-gatherers in Louisiana are
badly attacked by it.341 It also attacks the milkers in dairies. Attempts
to transmit Texas fever failed with this species.
Amblyomma maculatum, Koch.
The so-called Gulf Coast tick, of the Gulf Coast, occurs on birds,
mammals and man, especially cattle, and attacks the ears.
Genus. Hyalomma, Koch.
Hyalomma ægyptium, L., 1758.
Syn.: Acarus ægyptius, L., 1758; Ixodes camelinus, Fischer, 1823.
A species frequently found in Africa, particularly in Egypt and
Algeria, and which also occurs in France and Italy, as well as in
Asia. Male 8 mm. in length, 4·5 mm. in breadth. Female up to
24 mm. in length and 15 mm. in breadth. It infests large and small
animals as well as human beings.342
[This is one of the largest ticks, nearly reaching the size of the
bont tick. It is known in Africa as the bont leg-tick; all farm stock
is attacked, but sheep and goats suffer most. Only one generation
appears to occur each year. The male is almost black with a pale
marginal stripe; the replete female brown with irregular light blue
stripes. It is abundant in parts of South Africa.]
Genus. Hæmaphysalis, Koch.
Hæmaphysalis punctata, Canestrini and Fanzago, 1877–1878.
Syn.: Hæmaphysalis sulcata, Canestrini and Fanzago, 1877–1878; Rhicocephalus
expositicius, Koch, 1877; Hæmaphysalis peregrinus, Cambridge, 1889;
Herpetobia sulcata, Canestrini, 1890.
[This species does not appear to be common. It occurs on sheep,
goats, horses and cattle. I have seen a female taken from man in
Britain. The female when fasting is 3·44 mm. long, when gorged
12 mm. long. Colour, reddish-brown, leaden-grey when gorged;
dorsal shield deeply indented in front; rostrum, shield and legs
brownish; body finely punctate, both above and below; sexual
opening opposite the coxæ of the second pair of legs in both sexes.
Palpi a little longer than the labium; first segment short and narrow,
second and third widened on the dorsal face. Coxæ with a short,
broad blunt spine; tarsi short, terminated with a spur on the first
pair. The male is 3·10 mm. long. Body rather narrow, yellowish
to reddish-brown; dorsal shield nearly covers the whole body;
numerous punctures over the whole surface. Eleven indentations on
the posterior margin of the body; peritremes lighter in colour, large
and comma-shaped. The three anterior pairs of legs with a short
spine on the haunches, the fourth with a very long one directed
backwards. The nymph varies from 2·5 to 3·0 mm., is oval, and
light yellow to dark red in colour. Dorsal shield rounded with few
punctations. No spur on tarsi, and sexual orifice nearly obsolete.
Larva short and oval. Length 1·20 mm.—F. V. T.]
Genus. Dermacentor, Koch.
Dermacentor reticulatus, Fabricius, 1794.
Syn.: Acarus reticulatus, Fabr., 1794; Ixodes reticulatus, Latreille, 1806;
I. marmoratus, Risso, 1826.
This tick is provided with eyes, but it is distinguished from Ixodes
and analogous genera by the lack of the abdominal plastron in the
male, which measures 5 to 6 mm. in length by 3·5 mm. in breadth.
The female may attain 16 mm. in length and 10 mm. in breadth.
It is found in the South of Europe, in Asia, and in America;
it attacks chiefly oxen, sheep and goats, and occasionally man.343
[This tick sometimes causes much annoyance to human beings.
It was once most troublesome at Revelstoke. Specimens have
recently been found on fowls, turkeys and pheasants in Kent.
[Other synonyms are as follows: Cynorhæstes pictus, Hermann,
1804; Crotonus variegatus, Dumeril, 1829; I. pictus, Gervais, 1844;
Dermacentor albicollis, Koch, 1844–1847; D. pardalinus, Koch, 1844–1847;
D. ferrugineus, Koch, 1844–1847; Ixodes holsatus, Kolenati,
1857; Pseudixodes-holsatus, Haller, 1882; Hæmaphysalis marmorata,
Berlese, 1887.
[The female when fasting is 3·86 mm. long by 2 mm. wide.
The body is depressed, larger behind and reddish-brown in colour.
The shield is very large and extends to the level of the third pair
of legs, with a few large and many small punctations, milky white,
variegated with reddish-brown. Sexual orifice opposite the coxæ
of the second pair of legs. Coxæ of the front legs are deeply
bifid, the others with a moderate spine. When gorged light
brown, and may reach 16 mm. When depositing eggs the female
is mottled with dark brown above and below. The male is like
the female. The shield is reddish-brown, variegated with a milky
white pattern. Coxæ of the fourth pair of legs three times the
size of the third. There is a sharp backwardly pointing spine on
the second palpal segment, also seen (but smaller) in the female.
Length 4·20 mm.
[According to Mr. Wheler this is a very variable species both
in size and colour. It occurs in England on sheep, but not commonly.
It has probably been introduced into Britain. Besides
the animals mentioned above it is also found on deer.—F. V. T.]
Dermacentor venustus, Banks.
[The Rocky Mountain tick fever tick. This species has been
wrongly called Dermatocentor reticulatus var. occidentalis. The correct
name of the carrier of Rocky Mountain tick fever is Dermacentor
venustus, Banks (Hooker, Bishopp and Wood, Bull. 106, U.S. Dept.
Agric., Div. Ent., p. 165).
[The female is from 13·8 by 10 by 6·4 mm. to 16·5 by 11·4 by
6·9 mm. when gorged; the male from 2·1 by 1·5 mm. to 6 by
3·7 by 1·4 mm. The male reddish-brown; scutum with an extensive
pattern of white lines, usually but little white on the mid-posterior
region, legs slightly lighter than scutum, joints tipped with
white. Female with scutum mostly covered with white, abdomen
reddish-brown, legs as in male. The nymph when unengorged
reddish-brown, when gorged dark bluish-grey; the larva is yellowish-brown
when unengorged, slate blue when engorged. The ova light
brown, shiny and smooth.
[The chief wild hosts are the brown bear, coyote, woodchuck,
rabbit, wild cat, badger and mountain goat for the larvæ; practically
all small mammals act as hosts for larvæ and nymphæ, whilst the
adults are seldom found on other than large domestic animals; horses
and cattle are preferred. It occurs in British Columbia, southward
to Northern New Mexico, and from the foothills of the Rocky
Mountains in Colorado to the base of the Cascade Range in Oregon
and California; abundant in Western Montana, Idaho, Eastern
Washington, Oregon, North Utah, West Wyoming and North-west
Colorado.
[Of great importance in the Bitter Root Valley of Montana, where
a number of cases of fever occur each year, with a mortality of about
70 per cent. In British Columbia this tick causes tick paralysis in
man and sheep. Only the adults seem to attack man and animals
there (Hadwen and Nuttall, Parasitology, 1913, vi, No. 3, pp. 288–297
and 298–301) according to the Canadian Medical Association
Journal, December, 1912. The symptoms are unlike spotted
fever. For full details of this tick vide Bulls. 105 and 106, U.S.
Dept. Agric.]
Dermacentor occidentalis, Neumann.
This tick only occurs in the Pacific Coast region of the United
States. Owing to the fact that it frequently attacks man as well as
occurring in great abundance in Oregon and California, it is of considerable
economic importance. It is spoken of as the wood tick, and
in the regions where found is the most common tick to attack man.
Hooker, Bishopp and Wood (Bull. 106, U.S. Dept. Agric., Div. Ent.,
1912, p. 189) state that a number of cases have been brought to their
notice where the bite of this tick has caused considerable local inflammation,
which in some cases required physicians’ attention. It has
been supposed to be connected with Rocky Mountain spotted fever,
but it is doubtful if it is concerned in its transmission. The engorged
female is steel grey, the dorsum with an olive-green surface colour,
which covers the grey except in small spots, giving a mottled
appearance. The unengorged males and females are reddish-brown,
scutum covered with a whitish bloom, interrupted by many red
punctures. The female is 9 by 6·1 by 3·3 mm. to 11·8 by 7·6 by
5·6 mm.; the male 2·8 by 1·6 mm. to 4·2 by 2·3 mm. The larvæ
are bluish-grey when engorged, reddish-brown when unengorged.
The nymph is light brown, sides of scutum darker, and the
intestines dark brown. It is confined to the Coast Range and
Sierra Nevada Mountains in California and Oregon and the small
mountain range to the south-west.
Dermacentor variabilis, Say.
The American dog tick has also been found on man, but it is of
little economic importance as it is easily removed from its host.
Genus. Margaropus, Karsch.
Margaropus annulatus australis, Fuller.
The so-called Australian cattle tick. Newstead344 reports this as a
great pest to man in Jamaica in its larval stage. Its chief hosts are
cattle, horses, goats, sheep, dogs and rabbits.
Margaropus microplus, Canestrini.
Recorded by Aragão (Mem. Inst. Oswaldo Cruz, 1911, iii, fasc. 2,
p. 163) as occurring in larval stage on man in Brazil.
Genus. Rhipicephalus, Koch.
Rhipicephalus sanguineus, Latreille, 1804.
Syn.: Ixodes sanguineus, Latr., 1804; I. rufus, Koch, 1844; Rhipicephalus limbatus,
Koch, 1844; Rh. siculus, Koch, 1844; Rh. stigmaticus, Gerstäcker, 1873.
Spread over almost the entire tropical and sub-tropical regions,
occurring in Europe in the South of France and in Italy; it infests
dogs and more rarely sheep; oxen, cats, foxes and human beings
are also attacked.345
Neumann’s Table of Species of Argas.
| 1 | | Body elliptical (sides curved) | 2. |
| Body oblong (sides straight), ending anteriorly in a point | 7. |
| 2 | | Body transversely oval | vespertilionis. |
| Body elongate oval | 3. |
| 3 | | Margin of body striated | 4. |
| Margin of body formed by quadrangular areolæ | PERSICUS. |
| 4 |  | Body flat, integument plainly wrinkled | 5. |
| Body tumid, elongate; integument finely wrinkled; coxæ of fourth pair of legs near anterior third of body | hermanii. |
| 5 | | Body oval, narrowed anteriorly | REFLEXUS. |
| Body elliptical, blunt, hardly narrowed anteriorly | 6. |
| 6 | | Body twice as long as broad | cucumerinus. |
| Body hardly longer than broad | transgariepinus. |
| 7 |  | Dorsal integument with large polygonal depressions; tarsi appearing bifid | BRUMPTII. |
| Dorsal integument almost smooth; tarsi not appearing bifid | æqualis. |
The Argantinæ are distinguished from the Ixodinæ by the head, which in
the former is situated on the inferior aspect of the cephalothorax, while
in the Ixodinæ it projects freely; also by the very short proboscis, the small
club-like palpi, the lack of suckers on the legs, as well as by the scutellum, which
covers the entire back and is bent up round the borders. Two genera are distinguished:
Argas, Latreille, 1796 (Rhynchoprion, Hermann, 1804), and Ornithodorus,
Koch, 1844. The species live on mammals, but more especially on birds.
Genus. Argas, Latreille.
Argas reflexus, Fabricius, 1794.
Syn.: Acarus reflexus, Fabricius, 1794; A. marginatus, Fabricius, 1794;
Rhynchoprion columbæ, Hermann, 1804.
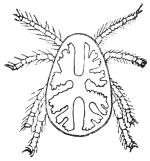
Fig. 361.—Argas reflexus:
from the dorsal surface, the
intestine showing through
the integuments. (After
Pagenstecher.)
The European marginated tick, Argas reflexus (length of male
4 mm., breadth 3 mm., length of female 6 to 8 mm., breadth
4 mm.), is of a yellowish colour and has yellowish-white legs. The
ingested blood shows red or brown through the intestine, which is
provided with blind sacs. It lives in dovecots. It remains hidden
during the day and at night crawls on to the sleeping pigeons to suck
their blood. It has been observed in France, England, Italy, Germany,
and Russia. Persons sleeping near infected dovecots, or in apartments
formed from pigeon-lofts, are also attacked, even when the
room in question has not been used for sheltering pigeons for years,
as “marginated ticks” can live in a fasting condition for a very long
time. The bite sometimes gives rise to serious symptoms, such as
general erythema and sudden œdema.
[This pest more often feeds on the blood
of man than is imagined. Blanchard states
that he has received them from men’s clothes
in Strasburg. Boschulte, of Westphalia, records
these parasites in a bedroom inhabited
by children and connected with a pigeon-house.
The children were bitten during sleep
on the hands and feet. The result of the bite
was intense itching along the nerves, the bite
only being marked by a red spot. In a girl
of 14 or 15, vesicles were formed similar to
those produced by burns, and in an old man
an ulcer formed. Others record painful punctures
and persistent œdema produced by this
pigeon pest. It was once abundant in Canterbury Cathedral, and
often caused much annoyance, I am told, to the worshippers; the
ticks falling down from the roof, where they were living, derived
from the numerous pigeons that breed in the towers. This Acarus
has enormous powers of vitality, living without food for months at
a time.—F. V. T.]
Argas persicus, Fischer de Waldheim, 1824.
Of oval form and brownish-red colour. The male measures 4 to
5 mm. in length by 3 mm. in breadth; the female 7 to 10 mm. in
length by 5 to 6 mm. in breadth. It frequents the entire north-west
and north-east of Persia (the gerib-gez or malleh of the Persians, the
miana bug of travellers). It lives concealed in houses and attacks
man at night to suck his blood. Its bite is much dreaded, but the
serious results may probably be attributed to unsuitable treatment of
the wound or its invasion by bacteria.

Fig. 362.—Argas persicus:
ventral aspect. 7/1. (After
Mégnin.)
[This tick, sometimes called the tampan and wandlius in South
Africa, is mainly a fowl parasite. Fowls and ducks frequently die
under its attack, particularly young ones, death being due to loss
of blood. This tick remains attached to its host during its larval
stage for about five days; it then leaves and moults in concealment.
In its subsequent stages it visits its host by night and remains for
about an hour only, during which time it distends itself fully with
blood. As a nymph it moults twice, not once as do the cattle ticks.
This tick and other Argas become larger with each moult, but retain
their same general appearance. The female visits the host every now
and then, and, between, deposits eggs in sheltered crevices. About
fifty to 120 are deposited at once. Four weeks seems a necessary
period to intervene between visits to the host, and the interval may
be extended to upwards of a year according to Lounsbury.346
[It is found in the Sudan, where Balfour
has found granules derived from the segmentation
of spirilla in their digestive tract.
Fantham and Hindl have confirmed this. It
has been assumed that these granules carry
infection.
[This so-called Persian tick, the miana,
which is such a scourge to travellers in Persia,
appears to infest the huts of natives in that
country. It has been sent me from Quetta,
where it has invaded houses to such an extent
the natives cannot live in them. The virulence
of its bite is probably due to the tick transmitting
fever germs from natives, probably inured, to strangers, who
would be susceptible.—F. V. T.]
Argas brumpti, Neumann.
[Found in Somaliland, by Brumpt, and in the Sudan. This tick
attacks man as well as wild animals and produces a painful swelling
according to King,347 but as pointed out by that naturalist it probably
relies on other than human food.—F. V. T.]
Argas chinche, Gervais, 1844.
This Acarus, a native of the temperate parts of Colombia, is
very troublesome to man. It is probably identical with A. americanus,
Packard, which infests domestic fowls and turkeys, and occasionally
also cattle, and is differentiated from A. reflexus by the sculpturing
of the cuticle.
Genus. Ornithodorus, Koch.
Neumann’s Synopsis of the Genus Ornithodorus is as
follows:—
| 1 | | Hypostome unarmed; integument in nymph stage and partly in adult spinulose | MÉGNINI. |
| Hypostome armed with recurved teeth; integument not spinulose | 2. |
| 2 | | Camerostome with movable lateral flaps | TALAJE. |
| Camerostome without movable lateral flaps | 3. |
| 3 | | Anterior border of distal segments of legs with tubercles or festoons | 4. |
| Anterior border of segments of legs without tubercles or festoons | 8. |
| 4 | | Body not much contracted anteriorly | 5. |
| Body pointed anteriorly | 7. |
| 5 | | Tubercles of distal segments of legs higher than broad, distant | 6. |
| Festoons of distal segments of legs as broad as high, contiguous | pavimentosus. |
| 6 | | Eyes present | SAVIGNYI. |
| No eyes | MOUBATA. |
| 7 | | Eyes present | coriaceus. |
| No eyes | TURICATA. |
| 8 | | Integument with fine radiating wrinkles | lahorensis. |
| Integument granular | 9. |
| 9 | | Tarsi appearing bifid at apex | furcosus. |
| Tarsi not appearing bifid at apex | 10. |
| 10 | | Tarsi of first pair of legs with three dorsal tubercles, of other legs with one | canestrinii. |
| Tarsi without dorsal tubercles or with only one | 11. |
| 11 | | Tarsi of last three pairs of legs with pronounced dorsal protuberance | tholozanii. |
| Tarsi of legs with indistinct dorsal protuberance | erraticus. |
Ornithodorus moubata, Murray, 1877.
An abundant African tick which is one of the carriers of the
spirillum of African relapsing fever and can also carry Filaria perstans
(Christy). Its body is oval, yellowish-brown when young, greenish-brown
when mature. The integument is covered with mamillose
tubercles. No eyes and the stout legs granular above, the tibiæ and
tarsi fringed with tubercles on the upper side. Pocock348 records it from
Uganda and German East Africa, Congo and Angola, to Namaqualand
and the Transvaal in the south. It is called bibo in Uganda, moubata
in Angola, and tampan on the Lower Zambesi. It feeds on animals
and birds as well as man. Its bite is very painful. This tick is found
in native huts, living in cracks and crevices and in the thatch roofs.
The female tick infected with the spirillum transmits the infection
to the eggs and the next generation. They appear to be able to live
without food a long time, and probably live for years. They lay their
eggs in masses on the ground or in crevices, and when they hatch
they are in the nymph stage with four pairs of legs. O. moubata also
occurs in Madagascar with recurrent fever (Lamoureux, Bull. Soc.
Path, exot., 1913, vi, No. 3, pp. 146–149).
Ornithodorus savignyi, Audouin, 1827.
At one time considered the same as the preceding species, but can
be easily separated by the presence of two pairs of eyes. It is widely
spread over Africa and has been found in South India and at Aden.
In the Sudan it occurs in large numbers. King349 records that a few
miles N.N.E. of Khartoum 370 specimens were collected in two
hours under a single tree by a well. It is found in Somaliland,
where relapsing fever occurs and no O. moubata, which it probably
replaces as a transmitter (Drake-Brockman, “Rep. Col. Office,” April 6
and April 16, 1913). It also occurs in Tunis, where the natives call it
“tobbiah” (Weiss, Arch. de l’Inst. Pasteur de Tunis, 1912, pt. 4,
p. 226).
Ornithodorus coriaceus, Koch.
Found in Mexico, Paraguay and California. Attacks man.
Ornithodorus talaje, Guerin, 1849.
An eyeless species with somewhat elongate pentagonal body found
in Mexico and South America, called the “chinche.” A variety of
it (coniceps) is found at Venice, etc., and another variety on various
islands in the Indian Ocean and South Atlantic. Its bite is very
painful to man.
Ornithodorus turicata, Dugès, 1876.
Without eyes. Indigenous in Central America; attacks human
beings and pigs. The bite is painful and is often followed by
serious consequences.
[So virulent is this species that pigs put in an infested sty often
die in a night. This “turicatas” of Mexico often reaches 6 mm.
in length.—F. V. T.]
Ornithodorus tholozani, Laboulbène and Mégnin, 1882.
Syn.: Argas tholozani, Lab. and Még., 1882.
Without eyes. Males 4 to 6 mm. in length and 2 to 4 mm. in
breadth; females 8 to 10 mm. in length and 4 to 5 mm. in breadth.
It especially attacks sheep. Native of Persia and Asia Minor.
[This species is reputed as being very dangerous to man. It is
locally know as the kéné, or sheep-bug. In its fully gorged state it is
deep violet.—F. V. T.]
Ornithodorus mégnini, Dugès, 1883.
Syn.: Argas mégnini, Dugès, 1883.
Length 8·5 mm., breadth 5·5 mm. Native of Mexico.
[Another synonym for this species is Rhynchoprion spinosum,
Marx. The adult males and females are grey to dark brown, the
male somewhat the smaller; female 5 by 3·5 by 2·5 mm. to 10 by
6 by 3·5 mm. The larvæ at the seed tick stage are dark grey,
turning to pink, then to a whitish grey when engorged. The nymph
when young is blood-red in front, rest pearly white; later they turn
reddish-brown.
[Intense pain may be caused by its presence in and around
the ears.
[Two specimens in the nymphal state were taken from the ears of
a visitor to Cambridge by Dr. J. Christian Simpson. They were
supposed to have entered the ears when the gentleman was camping
out in Arizona (Lancet, 1901, i, No. 4,052, p. 1198, fig. 3).
[This species attacks the horse, ass, dog, cats and oxen, generally
around the ears, and also attacks man. It is well known in the
United States as infesting the ears of children (New York Ent. Soc.
Journ., 1893, pp. 49–52).
[It occurs in Texas, Arizona, New Mexico and California as well
as Mexico, Brazil, and possibly many parts of South America; and
recently Bedford (“Sec. Report Div. Vet. Res., S. African Union,”
1912, pp. 343, 344) has shown it to occur at Vryburg and Fauresmith,
in the Transvaal, on stock. It also occurs in the Sudan.—F. V. T.]
Other Literature on Ixodidæ.
(1) “Pénétration de l’Ixodes ricinus sous la peau de l’homme,” Compt. rend. Soc. de Biol.,
1891, xliii, ser. 9, iii, pp. 689–691, R. Blanchard.
(2) “Notas sobre Ixodidas brazileiros,” Mem. Inst. Oswaldo Cruz, 1911, iii, fasc. 2,
pp. 145–195, pls. 11 and 12, Dr. H. de Beaurepaire Aragão. Table of Brazilian Species.
(3) “Contribuicão para a sistematica e biolojia dos Ixodidas,” Mem. Inst. Oswaldo Cruz,
1912, iv, fasc. 1, pp. 96–120, pls. 2 and 3, Dr. H. de Beaurepaire Aragão.
Family. Tyroglyphidæ.
Very small mites without eyes and without tracheæ, with smooth skin.
The males usually have a suctorial pore on either side of the anus, which is used
during copulation, or suckers may be found in both sexes near the sexual orifice.
The mouth parts form a cone with chelate cheliceræ, and three-jointed pedipalpi;
the legs are usually short, have five segments with a terminal claw and suckers, or
either one or other of these organs. The numerous species and genera live free and
from choice in slowly decomposing vegetable and animal matter (cheese, cereals, flour,
sugar, preserves, dried anatomical preparations, bacon, dried fruits and fungi), also in
the corners of dwellings, etc.; they incidentally get into or on to man, or are found in
chamber utensils and in spittoons; actual parasites are rarely found amongst them.
[The chief genera are Tyroglyphus, Rhizoglyphus, Glyciphagus, Aleurobius
and Histiogaster. The first three have typical characters referred to, but are
distinguished from each other by the two former having the hairs on the dorsum
smooth, whilst in Glyciphagus they are hairy, plumose, or feathered. Rhizoglyphus
can be told from Tyroglyphus by having claws on the tarsi without any suckers;
Tyroglyphus has both claws and suckers. The larvæ are hexapod and may become
adult in the usual way by repeated moults, or they enter the so-called hypopial
stage. In this the eight-legged nymph becomes quiescent, and during this stage
it fixes itself to some insect or other animal by a patch of suckers on the lower
surface of the hind end of the body, and is so carried from place to place. The
hypopus does not feed and has a hard shell and short legs. When it has reached
a new home it moults and development proceeds in the normal way. Canestrini
and Kramer treat the Tyroglyphidæ as a sub-family of the Sarcoptidæ, calling them
sub-family Tyroglyphinæ, the other sub-families being Sarcoptinæ, Canestriniinæ and
Analsinæ.—F. V. T.]
Sub-family. Tyroglyphinæ.
Genus. Aleurobius, Canestrini.
Aleurobius (Tyroglyphus) farinæ, de Geer (part), Koch.
The male measures 0·33 mm. in length by 0·16 mm. in breadth;
the female 0·6 mm. in length by 0·3 mm. in breadth. These mites
possess five pairs of suctorial organs of a light colour; the legs are
reddish. Moniez observed them in Lille on the skin of labourers
who had been unloading Russian corn. A few of the species
generally mentioned under the designation of Tyroglyphus siro are
probably the common flour-mite, which also occurs on dry cheese.
[The farinæ of de Geer is an Aleurobius described by him in 1778
(“Mém. Hist. Ins.,” vii, t. 5, f. 15, p. 97) as Acarus farinæ.—F. V. T.]
Genus. Tyroglyphus, Latreille.
Tyroglyphus siro, L., 1756.
(Defined by Gervais, 1844.)
Male 0·5 mm. in length by 0·25 mm. in breadth; female 0·53 mm.
in length by 0·28 mm. in breadth; the males have two suckers on the
tarsi of the fourth pair of legs. Penis straight, colour whitish or
reddish.
Tyroglyphus longior, Gervais, 1844.
White or yellowish, with two black spots on the abdomen.
Male 0·55 mm. in length, 0·28 mm. in breadth; penis bent.
Female 0·61 mm. in length and 0·28 mm. in breadth.
T. siro and T. longior live on dry cheese, in flour, on dried fruits,
etc., and have been occasionally observed in the stools, urine, or pus
of human beings, and also on their skin. The so-called vanillismus
is to be attributed to these species.
[T. siro and T. farinæ of Schrank (non Geer) are the same. They
are described under other names, such as Acarus lactis, Linn.;
A. favorum, Herm., etc.; A. lactis in milk, farinæ in flour, and siro
in cheese; and as A. dysenteriæ, Linnæus (“Syst. Nat.,” ed. 12,
pp. 1024–1767).]
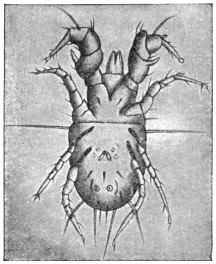
Fig. 363.—Tyroglyphus farinæ: male.
Enlarged. (After Berlese.)
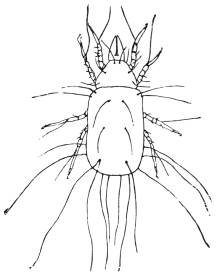
Fig. 364.—Tyroglyphus longior,
Gerv. (After Fum. and Robin.)
It is to these species that a case of dysentery was referred.
Rolander, who studied under Linnæus, was attacked by what was
called dysentery. The complaint soon gave way to treatment,
but eight days after it returned, soon disappeared, but again
came a third time. All the time Rolander had been living like
the other inmates of the house, who all escaped. Linnæus, aware
that Bartholemy had attributed dysentery to insects which he
said he had seen, advised his student to examine his stool. The
result was that innumerable mites were found to be present. Their
presence was easily accounted for by the fact that they were found
in numbers in a cup made of juniper wood from which the student
alone drank of a night, and they were found to be of the same species.
What this species is we do not know. Linnæus called it Acarus
dysenteriæ, but it was the same as his Acarus siro. No records have
occurred since. It cannot be, as Latreille supposed, the cheese mite,
for they have been eaten by millions since, and it is strange no such
case has occurred again.
[Tyroglyphus minor var. Castellani, Hirst,
causes the copra itch in persons employed in the copra mills in
Ceylon. The skin of the hands, arms, legs and even body
becomes covered with pruriginous papules, papulo-pustules and
pustules near the head. The eruption begins as a rule on the
hands. The mites live in the copra dust. They produce dermatitis.
Castellani produced the disease experimentally by rubbing copra
dust containing mites on the skin of healthy people. Beta-naphthol
ointment (5 to 10 per cent.) proved useful in treatment (Journ. Trop.
Med. and Hyg., December 16, 1912, Castellani and Hirst).—F. V. T.]
Genus. Glyciphagus, Hering, 1838.
Glyciphagus prunorum, Her., and G. domesticus, de Geer.
The Glyciphagi are differentiated from the Tyroglyphi in that
the chitinous hairs on the body are fringed or feathered, and that
they lack a furrow dividing the cephalothorax from the abdomen.
They live under similar conditions to the Tyroglyphi and are
occasionally found on man or in fæces.
[Sugar merchants and grocers are frequently troubled by swarms
of G. domesticus, which leave the stores when being handled, and
especially shopmen, who handle sugar kept in small stores for some
time. These are the Acari that cause that irritating temporary affection
known as “grocer’s itch.”—F. V. T.]
Glyciphagus cursor, Gervais.
Under this name Signor Moriggia figures a horny excrescence
of great length growing from a woman’s hand, and containing in
its cavities quantities of Acarus. This species is really G. domesticus,
de Geer. G. domesticus has also been described by Gervais (Ann. Sci.
Nat., 1841, ser. 2, xv, p. 8) as G. hippopodes.
Glyciphagus buski, Murray.350
[This is a mite found by Busk and named after him by Murray.
It was taken from beneath the cuticle of the sole of the foot of
a negro in the Seamen’s Hospital Ship on the Thames in 1841,
in large sores of a peculiar character confined to the soles of the
feet. It appeared that the disease was caused by its burrowing
beneath the thick cuticle. The disease was attributed to the
wearing of a pair of shoes which had been lent to another negro
whose feet had been similarly affected for nearly a year. The negro
to whom the shoes were lent came from Sierra Leone. Mr. Busk
stated that some water brought by Dr. Stranger from the River
Sinoe, on the coast of Africa, contained one nearly perfect specimen,
and fragments of others very similar to if not identical with this
Acarus. Mr. Busk adds that he had been informed by Staff-Assistant
Surgeon P. D. Murray that at Sierra Leone there is a
native pustular disease called craw-craw—a species of itch breaking
into open sores.
[From Busk’s original figure I see no reason to doubt that this
is a Glyciphagus.—F. V. T.]
Genus. Rhizoglyphus, Claparède, 1869.
Rhizoglyphus parasiticus, Dalgetty, 1901.
The Rhizoglyphii are to be recognized by their short legs, which are beset
with spines, and by the tarsi, which terminate in a claw. They live on plants,
roots and bulbs, especially the bulbs of lilies.
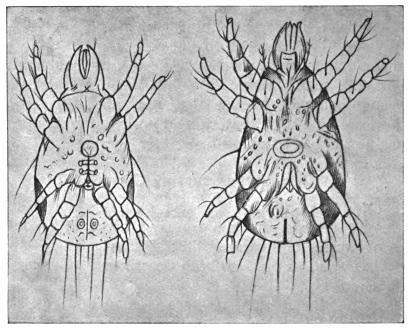
Fig. 365.—Rhizoglyphus parasiticus. a., male; b., female. Enlarged. (After Dalgetty.)
This species has been observed on the feet of Indian coolies
working in the tea plantations; they produce a skin disease
which always commences with blebs between the toes, and which
almost always extends to the malleoli, but not beyond. The
Acari have an elliptical body, which is grey, but varies from
greenish-yellow to greenish-brown when the stomach is full. Eyes
are absent. The legs are composed of five segments and terminate
with a claw. The males measure 0·18 mm. in length by 0·08 mm.
in breadth, and possess genital and anal pores; the females measure
0·2 mm. in length by 0·09 mm. in breadth.351 [This is also known as
coolie itch and is common in Indian tea plantations.—F. V. T.]
Genus. Histiogaster, Berlese, 1883.
Histiogaster (entomophagus?) spermaticus, Trouessart, 1900.
The genus Histiogaster, which also approaches the Tyroglyphinæ, is
characterized by the circumstance that the males possess suctorial pores
used in copulation, as well as leaf-shaped appendages at the posterior end
of the body. They feed on vegetables, especially on small fungi.
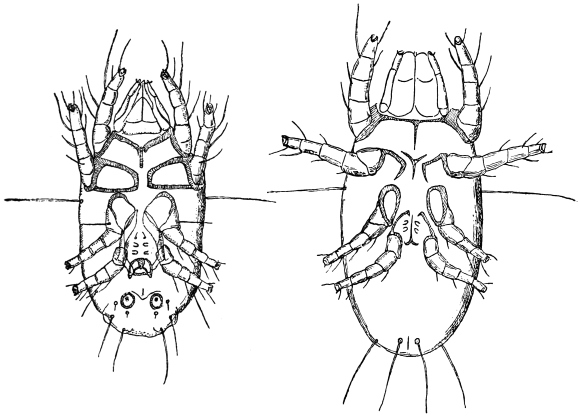
Fig. 366.—Histiogaster (entomophagus?) spermaticus: on left, male; on right, female—both
from the abdominal aspect. 200/1. (After E. Trouessart.)
This species has been described by Trouessart,352 who found
numerous specimens, some adult, others in the developmental stage
(larvæ, nymphs), and ova, in the fluid removed by puncture from
a cyst of the right testis. The males measure 0·25 mm., the females
0·32 mm., and the larvæ 0·1 mm. in length. The author is of opinion
that the animal—perhaps a fertilized female—was introduced by a
catheter, and, as a matter of fact, it was afterwards found that the
patient had once had the catheter passed in India while suffering from
pernicious fever.
It would here rather appear to be the case of a facultative parasitism
of an otherwise free-living species. Histiogaster entomophagus,
Laboulbène, is found occasionally in collections of insects feeding
on larger species containing much fat; the species also occurs on dry
cantharides; it appears to belong to the region of South Europe,
where, however, it is widely spread.
[Entomophagus occurs all over Europe and in America. It has
been described under the following names: Acarus malus, Shimer, 1868
(Trans. Illinois Hort. Soc.); Dermaleichus mali, Riley, 1873 (Rep. Ins.
Missouri, v, p. 87); Tyroglyphus mali, Murray, 1877 (“Eco. Ent. Apt.,”
p. 275); T. corticalis, Michael, 1885 (Trans. Roy. Micros. Soc., ser. 2, v,
3, p. 27, figs. 1 to 14); Histiogaster corticalis, Canestrini, 1888
(Prosp. Acarof., iii, p. 397); H. aleurophagus, Sicherin, 1894, Canestrini,
Prosp. Acarof., vi, p. 815. Trouessart’s species is evidently distinct.—F. V. T.]
Genus. Cheyletus.
Cheyletus mericourti, Lab.
Acaropsis mericourti, Moq. Tand.
[This mite has been described from three specimens found in pus
which flowed from an abscess in the ear of a naval officer, produced
by inflammation of the auditory passage. Where the mites came
from we do not know, as they were found near the Bank of Newfoundland.
This genus of Acari has enormous mandibles and a
peculiar tracheal system; two ungues and appendages to the tarsi.—F. V. T.]
Family. Sarcoptidæ (Itch Mites).
Small mites without eyes and tracheæ, and with delicate, transversely striated
cuticle. The mouth parts form a cone, over which the shield-shaped upper lip
protrudes; the cheliceræ are chelate; the pedipalpi (or maxillary palpi) have three
joints; the legs are short and compact, and composed of five segments; the terminal
joints have pedunculated suckers (ambulacra) or a long bristle. The larvæ are six-legged.
They live on or under the skin of birds and mammals, on which they
produce the skin disease known as scabies, or itch.
[The Sarcoptidæ attack the hairs, feathers or epidermis of birds, animals and
man, living as permanent parasites. The punctures they produce are followed by
the formation of more or less thick crusts or scabs, beneath which the mites live and
breed (so called scab, mange and itch). Most are oviparous, some ovoviviparous.
The eggs are minute, ovoid, with a thin semi-transparent shell. They incubate in
a few days, varying from two to ten or eleven, as a rule. Generally sarcoptic
diseases lie dormant in winter and revive in spring and summer in man; but in
animals with long wool, such as sheep, they are most active during winter, although
revival of active reproduction takes place in spring.
[Speaking generally, for the Sarcoptidæ there are three distinct stages in the
development of the male, four in the female, as follows:—
[(1) The larva. In this stage only three pairs of legs occur.
[(2) The nymph, in which a fourth pair of legs appear, and which thus approaches
the adult; but so far no sexual organs occur. Nymphs are of two sizes—the
smaller being future males, the larger females.
[(3) The next stage in the female is the age of puberty, the female now being
provided with a vulvo-anal slit; this so-called pubescent female is fertilized by the male.
The male then dies. But the female again casts her skin and enters another stage—
[(4) The ovigerous female—the egg-laying female—which has differently modified
legs.
[The rate at which these Acari breed is very great. Gerlach has found that
roughly, in each Sarcopt gallery, a female produces fifteen individuals—ten females
and five males—and that the progeny reproduce again in fifteen days. The table
given below thus shows that one pair may produce the enormous number of
1,500,000 descendants in three months:—
| First generation after 15 days | | 10 | females | | 5 | males |
| Second | " " 30 " | | 100 | " | | 50 | " |
| Third | " " 45 " | | 1,000 | " | | 500 | " |
| Fourth | " " 60 " | | 10,000 | " | | 5,000 | " |
| Fifth | " " 75 " | | 100,000 | " | | 50,000 | " |
| Sixth | " " 90 " | | 1,000,000 | " | | 500,000 | " |
= 1,500,000 individuals. |
[These Acarinæ are divided into three distinct sub-families, namely the Cytolichinæ,
Sarcoptinæ, Canestriniinæ.
[The Sarcoptinæ alone interest us here, and of the nine genera the three following
are the most important:—
[(1) Sarcoptes, Latreille; Eusarcoptes.
[(2) Psoroptes, Gerv.; Dermatodectes, Gerlach; Dermatocoptes, Fürstenberg.
[(3) Chorioptes, Gerv.; Symbiotes, Gerlach; Dermatophagus, Fürst.; Sarco-dermatocedes,
Del.
[The following are the main characters of these three genera:—
[Sarcoptes—round or slightly oval; the two posterior pairs of legs being nearly or
quite concealed beneath the body; the tarsi end in simple long pedicles, with
ambulatory suckers.
[Psoroptes—oval; the legs are all visible outside the margin of the body; the
ambulatory suckers are carried on long triangulated stalks; the male has copulatory
suckers and abdominal prolongations.
[Chorioptes—oval; legs long, thick, all visible; ambulatory suckers very wide,
carried at the end of simple, short pedicles.
[Sarcoptes make channels or furrows beneath the epidermis, and in these the
female lays her eggs. This form of acariasis is thus difficult to cure. It is the cause
of human itch (vide Sarcoptes scabiei).
[Psoroptes do not make sub-epidermic galleries; they live and breed in colonies
beneath crusts or scabs formed by the changes they produce in their host’s skin.
Sheep scab is a common type of disease produced by Psoroptes. This genus is of
little importance as a parasite to man.
[Chorioptes live as Psoroptes; they also do not affect man. Otodectes, Can.,
affecting cats and dogs, and others occur, but do not affect man as far as we know at
present (“Demodicidae und Sarcoptidae,” von Professor G. Canestrini und
P. Kramer, Das Tierreich, 1899).—F. V. T.]
Sub-family. Sarcoptinæ.
Genus. Sarcoptes, Latreille.
Sarcoptes scabiei, de Geer, 1778.
Syn.: Acarus scabiei, de Geer, 1778; A. psoricus, Pallas, 1760; A. siro, L., 1778;
Sarcoptes exulcerans,? Linn., 1758, Nitsch, 1818; S. hominis, Raspail, 1834, and
Hering, 1838; S. galei, Owen, 1853; S. communis, Delaf. et Bourg., 1862; S. scabiei
var. hominis, Mégnin, 1880.
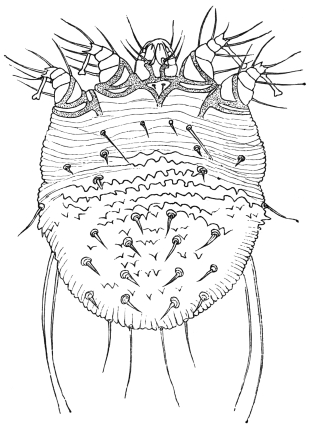
Fig. 367.—Sarcoptes scabiei: female, dorsal aspect.
200/1. (After Fürstenberg.)
The body is oval or nearly circular and whitish in colour, with transverse
rows of striæ partly interrupted on the back. There are transverse
rows of small bristles
on the dorsal surface, and
groups of trichomæ on
the front, sides and back.
There are chitinous hairs at
the base of the legs; the two first
pairs are provided with
pedunculated ambulacra in
both sexes, the two posterior
pairs terminate each with a
long bristle in the female;
in the male the third pair
of legs terminate in a bristle,
the fourth pair with a pedunculated
ambulacrum. The
anus is situated at the posterior
border of the dorsal
surface.
At one time numerous species
were differentiated, according to
the form of the Acarus, the number,
position and size of the hairs
and spines, even according to the
hosts, etc. All these characteristics,
however, fluctuate so considerably
that absolute differentiation is impossible; the supposed species may be
regarded in the same light as Mégnin did, as varieties. It is also hardly possible
to distinguish the mite of human scabies (S. hominis) from that of a number of
domestic animals (S. squamiferus). It is best, therefore, to accept one single species
(S. scabiei), which may give rise to different races or castes by living in the skin
of man and mammals, but can pass from one host to the other.
[Canestrini and Kramer, in their monograph of the Sarcoptidæ,
enumerate eighteen distinct species of this genus, from the dog, goat,
camel, horse, ferret, lion, wolf, sheep, pig, etc., and two species
parasites of man (scabiei and scabiei-crustosæ). There is no doubt
that they are distinct species.—F. V. T.]
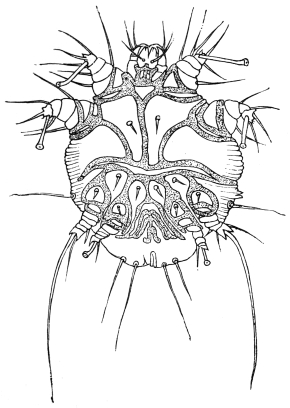
Fig. 368.—Sarcoptes scabiei: male, ventral aspect.
200/1. (After Fürstenberg.)
The S. scabiei of man (S. scabiei var. hominis) (length of male
0·2 to 0·3 mm., and breadth 0·145 to 0·190 mm.; length of female
0·33 to 0·45 mm., and breadth 0·25 to 0·35 mm.) lives in the tunnels
that it excavates in the epidermis, and attacks by preference places
with thin skin, such as between the fingers, in the bend of the
elbows and knees, in the inguinal region, on the penis, on the
mammæ, but may also affect other parts. The tunnels, which
vary from a few millimetres to a centimetre and more long, do not
run straight, but are somewhat
tortuous; the female is found
at the terminal end. The tunnels
contain the excrement and
oval eggs (0·14 mm. in length)
of the parasite; the males are
rarely met with, as they die off
after copulation; the females
die after depositing their eggs.
The six-legged larvæ hatch
out after four to eight days,
and after about a fortnight,
during which time they change
their skins three times and
undergo metamorphosis, they
begin themselves to burrow.
Transmission from person to
person rarely is effected
through linen, but by direct
contact (as in coitus); transmission
can be artificially
effected on horses, dogs and
monkeys, but not on cats.
The smaller S. scabiei-crustosæ,
Fürstenberg, is the cause of the itch that occurs chiefly in
Norway; it is not certain whether this is a distinct species of
itch mite.
[This is quite a distinct species, which is recorded from Germany and
France. Mégnin (Parasitology, 1880, p. 165) described this as S. scabiei
var. lupi. The female is 140 µ long, 340 µ broad; the male is 170 µ long
by 150 µ broad. In Science (March 3, 1893, p. 125) is recorded that
at the Indiana Academy of Science Dr. Robert Hessler referred to
“a case of that extremely rare and almost extinct form of itch
known as ‘Norway itch,’ the scabies norvegica of Hebra, 1852.”
The afflicted man was covered with thick, creamy white, leathery
scales; some of these scales measured over an inch in diameter and
1/10 in. thick. A constant shedding of scales went on, a handful
being gathered daily. They were found full of mites and eggs and
riddled with passages. Under treatment the mites were killed and
the skin became normal. Dr. Hessler made a calculation of the
number of eggs and mites, amounting to ova and shells 7,004,000,
mites in all stages 2,009,000.—F. V. T.]
The following forms may be transmitted from DOMESTIC ANIMALS
to MAN:—
(1) S. scabiei var. equi. Male, 0·2 to 0·23 mm. long, 0·16 to 0·17 mm. broad.
Female, 0·40 to 0·42 mm. long, 0·28 to 0·32 mm. broad. The horse is the normal host.
(2) S. scabiei var. ovis. Male, 0·22 mm. long, 0·16 mm. broad. Females,
0·32 to 0·44 mm. long, 0·24 to 0·36 mm. broad. This mite lives on sheep, and passes
over to goats and human beings; it may also be artificially transferred to horses,
oxen and dogs.353
(3) S. scabiei var. capræ. Male, 0·24 mm. long, 0·188 mm. broad. Female,
0·345 mm. long, 0·342 mm. broad. On goats, passing from them to horse, ox, sheep,
pig and man. On the latter, in contradistinction to the varieties (1) and (2), it produces
a severe affection.
(4) S. scabiei var. cameli. Frequently observed in man, chiefly in Africa. A
few cases have been observed in Europe; the affection induced by it is severe.
(5) S. scabiei var. aucheniæ. Male, 0·245 mm. long, 0·182 mm. broad. Female,
0·34 mm. long, 0·264 mm. broad. It lives on the llama, and may be transmitted
to man.
(6) S. scabiei var. suis. Male, 0·25 to 0·35 mm. long, 0·19 to 0·3 mm. broad.
Female, 0·4 to 0·5 mm. long, 0·3 to 0·39 mm. broad. In the domestic pig and wild
boar; occasionally also in man. The settlement, however, is usually of short
duration.
(7) S. scabiei var. canis. Male, 0·19 to 0·23 mm. long, 0·14 to 0·17 mm. broad.
Female, 0·29 to 0·38 mm. long, 0·23 to 0·28 mm. broad. In the house-dog, and also,
not unusually, in human beings.
(8) and (9) S. scabiei var. vulpis and S. scabiei var. leonis of the fox and lion
have likewise been observed on man.
These are all distinct species and should read as follows: S. canis, Gerl.; S. ovis,
Mégn.; S. equi, Gerl.; S. dromedarii, Gerv. (cameli, Mégn.); S. aucheniæ, Raill.;
S. suis, Gerl.; S. vulpis, Fürst.; S. leonis, Can.
Sarcoptes minor, Fürstenberg, 1861.
Anus situated on the back, legs short, pedunculated ambulacra
broad; living on cats (S. minor var. cati) and rabbits (S. minor var.
cuniculi). In cats this mite usually lives in the cervical region, and
thence spreads to the ears and head; it usually causes the death of
the infected animals; it is easily transferable from cat to cat, is
difficult to transmit to rabbits, but once settled on them can easily
infect other rabbits. On the other hand, the transmission of the
itch mite of the rabbit to the cat does not succeed. In man S. minor
induces an eruption that disappears after about a fortnight.
[S. minor, Fürstenberg, 1861 (“Krätzm.,” viii, p. 218), comes in
Railliet’s sub-genus Notoedres, 1893 (“Zool.,” ed. 2, p. 660). Canestrini
raised this to generic rank in 1894 (Prosp. Acarof., vi, p. 724).
[There are three species: (1) N. notoedres, Mégnin = Sarcoptes
alepis, Railliet and Lucet (Compt. rend. Soc. de Biol., 1893, xlv, p. 404),
and Sarcoptes notoedres var. muris, Mégnin (Parasitology, 1880,
pp. 172–174). This occurs on the black and brown rats and the
water-vole.
[(2) N. cati, Hering, 1838 (N. acta. ac. Leop., ii, 18, xliv, p. 605,
figs. 9 and 10), = Sarcoptes minor, Fürstenberg (“Krätzm.,” 1861, viii,
p. 215). Found on the cat in Germany, France, Italy, and Britain.
[(3) N. cuniculi, Gerlach, 1857, “Krätzm.,” iii, figs. 20, 21. It
lives on the rabbit and is found in Germany and France.—F. V. T.]
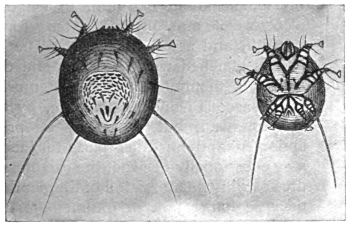
Fig. 369.—Sarcoptes minor var. cati: on the left, female (lying on its
abdomen); on the right, male (lying on its back). (After Railliet.)
The itch mites of domestic animals, which belong to the genera Psoroptes
(= Dermatodectes = Dermatocoptes) and Chorioptes (Symbiotes = Dermatophagus),
as a rule do not infest and live on man, even when artificially transmitted. It is,
however, possible for this to occur. Moniez (“Traité de par.,” 1896, p. 559)
mentions that a species of Chorioptes—probably Ch. bovis—had been found on man,
as had also Demodex folliculorum. This author also includes Dermatophagoides
scheremetewskyi, Bogdanoff (Bull. soc. imp. d. natural., Moscou, 1864, xxxvii,
p. 341), which has repeatedly been found on man in Moscow and Leipzig (Zürn,
Ber. d. med. Ges., Leipzig, 1877, p. 38), as Chorioptes bovis.
Other References to Scabies crustosæ AND norvegica, ETC.
(1) “Ein Fall von Scabies crustosa norvegica,” Würzb. med. Zeitschr., l, pp. 134–139,
pl. 3, H. Bamberger.
(2) “Ueber die Krätzmilbe (Acarus scabiei),” Notiz. a. d. Geb. d. Nat. u. Heilk., Weimar
(1913), xlii (11), Oct., pp. 161–166 (1834), de Blainville.
(3) “Rapport sur le ciron de la gale (Acarus scabiei),” Ann. de Mus. d’Hist. nat.,
1831; Parasitology, iv, pp. 213–232, de Blainville.
Family. Demodicidæ (Mites of the Hair-follicles).
Small Acarina, elongated in worm-like fashion, with annulated abdomen, and
without eyes or tracheæ. The mouth parts consist of a suctorial proboscis and
three-jointed palpi; the legs are short, and have three segments with small terminal
ungues. The anus is situated on the anterior border of the abdomen; oviparous;
the larvæ have six stumpy legs. These mites live in the hair-follicles of mammals.
Genus. Demodex, Owen.
Demodex folliculorum, Simon, 1842.
Syn.: Acarus folliculorum, Sim., 1843; Demodex folliculorum, Owen, 1843;
Macrogaster platypus, Miescher, 1843; Simonea folliculorum, P. Gervais, 1844;
Steatozoon folliculorum, Wilson, 1847.
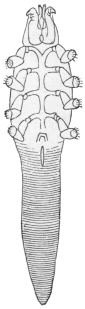
Fig. 370.—Demodex
folliculorum of
the dog. (After
Mégnin.)
As in Sarcoptes scabiei, numerous varieties of this species are
known; the form parasitic on man lives in the hair-follicles, the
meibomian and sebaceous glands, and hardly ever causes inconvenience;
the male measures 0·3 mm. in length and
the female about 0·4 mm. in length. The eggs
0·06 to 0·08 mm. in length, 0·04 to 0·05 mm. in
breadth, and are thin-shelled. The creatures are
always attached with the head end downwards in
the parts mentioned; they are most frequent in
the sebaceous glands of the face, by the nose, lips
and forehead, but they may be present on the
abdomen and on other parts of the body. They
may occasionally obstruct the excretory gland ducts,
thus causing inflammation of the gland (comedones);
their agglomeration in the meibomian glands sets
up inflammation of the margins of the eyelids.
There are generally only a few specimens in a gland.
According to some statements Demodex occurs in
50 per cent. of mankind and even in children; they
survive the death of their hosts by several days.
The variety living in the dog (D. folliculorum var. canis)
is smaller than the variety living in man, and produces a skin
disease resembling scabies in these animals. According to
Zürn they may also live on man; nevertheless, no other investigator has recorded
a similar observation, and attempts at artificial infection have proved negative.354
[Ten distinct species of Demodex are given by Canestrini and
Kramer (“Demodicidae und Sarcoptidae,” Das Tierreich, 1899, vii).
The species are certainly distinct.
[The species living on the dog (D. canis, Leydig, 1844) is cosmopolitan.
According to the British Medical Journal (February 22, 1913,
p. 407), dog mange may be caught by humans. Whitfield and
Hobday describe in the Veterinary Journal seventeen cases which
have come under their observation.—F. V. T.]
Order. Pentastomida.
Family. Linguatulidæ.
Arachnida greatly altered in consequence of their parasitic manner of life;
for a long time they were regarded as helminthes. The body is elongated,
vermiform, flattened or cylindrical, and more or less distinctly annulated. The
head, thorax, and abdomen are not defined from each other (fig. 371). The elliptical
mouth, surrounded by a chitinous ring, is situated at the anterior end, on the
ventral surface, and the intestine leading straight through the body opens at the
posterior end. Two retractile hooks are at the sides of the mouth (fig. 372); these
are usually considered to be the terminal joints of two pairs of legs, but it appears
to be more correct to regard them as the remains of the antennæ and palpi (Stiles).
According to this opinion, the legs in the adult state are completely degenerated.
The nervous system is reduced to an œsophageal ring. No organs of sense are
recognizable except the papillæ at the anterior end. There are neither organs of
circulation nor of respiration.355
The sexes are distinct. In the small male the sexual orifice is situated ventrally
in the anterior part of the body; in the female it is placed near the anus. The
Linguatulidæ lay eggs, and from each egg, after being conveyed into an intermediate
host, a four-legged larva, with rudimentary mouth parts, hatches out. It
goes through a series of metamorphoses, and passes through a second larval
condition, which, however, possesses the essential characteristics of the fully
developed form. Sooner or later it migrates during this stage, and reaches its final
host, mammal or reptile, in the nostrils or lungs of which the adult Linguatulidæ
live.
[As adults they live as internal blood feeders in various birds,
reptiles and mammals, especially in the nasal and respiratory
passages. The larval stage occurs in another host in an encysted
condition; this host is usually an animal preyed upon by the species
in which the sexual forms are found. The larvæ bore through the
walls of the host’s stomach and enter liver and spleen or brain, where
they encyst; here they grow until they assume almost the appearance
of the adult. These encysted larvæ on being eaten later make their
way into the nasal passages and lungs, where they mature. Both
adults and larvæ occur in man, as mentioned later.
[Three genera are recognized in this family:—
[(1) Linguatula.—Body flat, annulated. Adults live in the nasal sinus.
[(2) Porocephalus.—Body cylindrical, elongate, with often deeply cut rings. Adult
in respiratory organs of snakes, larvæ in animals and man.
[(3) Reighardia.—Cylindrical, but not ringed. Not found in humans.—F. V. T.]
Genus. Linguatula, Fröhlich.
Linguatula rhinaria, Pilger, 1802.
Syn.: Tænia rhinaria, Pilger, 1802; Polystoma tænioides, Rud., 1810;
Linguatula tænioides, Lam., 1816; Pentastoma tænioides, Rud., 1819.

Fig. 371.—Linguatula
rhinaria: female.
Natural size.
The male is white in colour, 18 to 20 mm. in length, anterior
portion 3 to 4 mm. in breadth, posterior part 0·5 mm. in breadth.
The female is of a yellowish colour, 8, 10, or 13 cm. long,
anterior part 8 to 10 mm. and posterior part 2 mm. wide. The
brownish eggs can be seen in the median line. The body is
elongated, rather flat, and exhibits about ninety rings
or segments with crenellated borders. The hooks
round the mouth are strongly curved and are articulated
to a basilar support. Eggs oval, 0·09 µ in length,
0·07 µ in breadth.
L. rhinaria, in the adult condition,
lives in the nasal cavity
and frontal sinus of the dog,
wolf, fox, horse, goat, and
occasionally of man; it causes
severe catarrh, epistaxis and
suppuration.

Fig. 372.—Larva of
Linguatula rhinaria
(Pentastoma denticulatum).
Enlarged. (After
Leuckart.)
Development.—The ova,
which are found in masses in
the nasal mucus, already possess
an embryo; they are expelled
with the nasal secretion, and
are swallowed by herbivorous
mammals with their food,
mostly by hares and rabbits,
but also by sheep, goats, oxen,
horses, antelopes, fallow deer,
pigs, cats, and occasionally also
by human beings. The young
larvæ hatch out in the stomach;
they possess a thickened anterior
body with rudimentary
mouth parts and two pairs of
limbs; the body gradually tapers to a short tail.
The larvæ of the Linguatulidæ bore through the intestinal wall
and reach the liver, more rarely the mesenteric glands, etc.; they
here become encysted and enter a sort of pupal stage in which they
lose their limbs; after several moultings and gradual growth the
second larval stage, having the appearance of the adult Linguatula,
sets in. About five to six months after infection the creatures have
become 4 to 6 mm. long, possess eighty to ninety rings, which have
a series of fine points on their posterior border; the mouth and
intestine are formed, the sexual organs mature and the two pairs of
hooks are near the mouth. This larval stage (fig. 372) has been
known for a long time, but it was regarded as an independent species
of animal, and therefore had a separate name (Linguatula serrata,
Fr.; Pentastoma denticalatum, Rud., etc.).
Later these Linguatula larvæ make an attempt to escape from their
hosts, and this, of course, can only be effected by means of an active
migration; they leave the cysts, and according to their respective
positions in the abdominal or pleural cavities they reach the bronchi
or the intestine, and finally pass out; they may be again sniffed up
by dogs and settle in their nasal cavities. Still this outward migration
does not appear to be necessary for further development. A
portion of the larvæ gain access to the nasal cavities directly through
the trachea, and thus herbivorous mammals certainly become directly
infected. In most cases the infection of dogs, wolves and foxes, that
is, of carnivorous mammals, takes place through consuming the
bodies of mammals, or parts of them, such as the liver and
lungs, which are affected with the second larval form; in any case
most larvæ obtain access first to the stomach of their host, from
here they make an active migration through the œsophagus to the
oral and nasal cavities, in which they settle. It is possible also that
the same larvæ which are free in the oral cavity when the food is
being eaten migrate into the nasal cavities. After being stationary
a fresh skin is formed and the spine-bearing cuticula are thrown off.
The male attains its full size in the fourth, and the female in the sixth
month. The duration of life is stated to be from fifteen months to
several years.
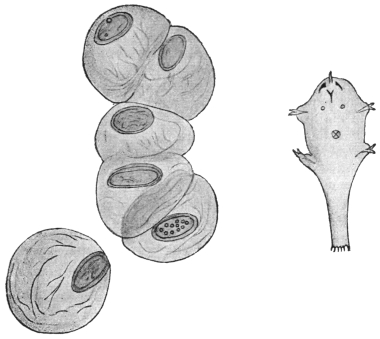
Fig. 373.—Linguatula rhinaria: on left, eggs in gelatinous covering, 110/1.
On right, first larval stage. 300/1. (After M. Koch.)
L. rhinaria has been observed in man in the adult as well as in the larval
condition (Pentastoma denticulatum). Zenker first called attention to the occurrence
of the larva in man, having found it nine times in the liver in 168 autopsies. Heschl
found it twice in Vienna in twenty autopsies, Virchow found it in Würzburg and Berlin,
Wagner in Leipzig (10 per cent.), and Frerichs in Breslau five times in forty-seven
autopsies. The parasite is much less frequent in Switzerland. According to Klebs,
one case occurs in 900 autopsies, and according to Zaeslin two cases occurred in Basle
to 1,914 autopsies. In the Seamen’s Hospital in Kronstadt P. denticulatum has been
found six times in 659 autopsies. It was almost always the liver that contained one
or a few specimens. The parasite was very rarely found in the kidney or spleen, or
encysted in the intestinal wall. The adult L. rhinaria is far more rarely observed
in man.
A case reported by Landon that related to a blacksmith of Elbing is particularly
interesting. This man accompanied the campaign of 1870; he soon, however, fell
ill with pains in the liver, accompanied by icterus and intestinal disorders. Soon
after the war, and after the symptoms were reduced to icterus and weakness, bleeding
of the nose set in and continued with slight intermissions for seven years; an
unpleasant sensation of pressure in the left nasal cavity set in, with inflammatory
swelling of the mucous membrane. At last, in the summer of 1878, when the
pressure in the nose had considerably increased, a Linguatula was expelled from the
nose with a violent attack of sneezing, and lived for three days longer in water. The
bleeding of the nose then ceased and the patient soon recovered. There can be no
doubt that the first illness was connected with the invasion in the liver of numerous
larvæ of Pentastoma, and disappeared after their encystment; one or a few of these
must subsequently have found its way to the nose and settled there.
Genus. Porocephalus.
Porocephalus constrictus, v. Siebold, 1852.
Syn.: Nematoideum hominis, Diesing, 1851; Pentastomum constrictum,
v. Sieb., 1852; Porocephalus constrictus, Stiles, 1893.
Porocephalus is distinguished from Linguatula by its cylindrical
body and by certain internal structures. Porocephalus constrictus
is at present only known in its larval stage. It is milk white in
colour with golden-yellow hooklets. Number of rings, twenty-three.
Length 13 mm., breadth 2·2 mm. There are no prickles on the
posterior border of the annulations of the body.
This species was first discovered by Pruner encysted in the livers of two
negroes in Cairo. Bilharz reported two further cases in which the parasites
were encysted in the liver and in the mucosa of the intestine; a few other observations
have been made by Fenger, Aitken, Giard and Chalmas. Aitken’s report deals
with soldiers of the British Colonies in Africa. The parasites were discovered in
the liver as well as in the lung, and appear to have been the cause of death in one
case (pneumonia, peritonitis).
Pruner has found the same parasite also in the liver of the giraffe.
It has recently been assumed that Porocephalus constrictus is the larva of
Pentastoma moniliforme, Diesing, 1835, that attains a length of 70 mm. and lives
in the lungs of African Pythonides. The larva is known to have been ejected
from monkeys (Cercopithecus albogularis, Cynocephalus maimon), from the giraffe
(Camelopardalis giraffa), from a species of hyæna (Proteles cristatus), and should
be expected to occur frequently in smaller mammals which have been swallowed
by African serpents of enormous size.
[The three species of Pentastomidæ, or tongue worms, found in
man are Linguatula serrata, Frölich; Porocephalus armillatus, Wyman;
and Pentastoma moniliformis, Diesing.
[(1) Linguatula serrata has been referred to under a great number
of names.356 It is a frequent parasite in dogs, oxen and sheep; as an
adult in the dog and also in the fox and wolf. The nymphal stage is
found in rats, hares, rabbits, the horse, oxen, sheep, goats, pigs, camels,
deer, the African and long-eared hedgehogs, porcupine, guinea-pig
and peccary. In man it is found in both adult and nymphal stages.
Sambon says the nymphal stage is of frequent occurrence, but is
usually overlooked. Zenker, who first found it in man, obtained it in
nine out of 160 post-mortems, usually encysted in the liver. It is then
said to be harmless. Landon, in 1878, found the adult in man, the
patient suffering from epistaxis for about seven years; in the end
during a fit of sneezing the living parasite was ejected through the
nostril. This case is of particular interest as it appears to suggest
that this Acarid may now and then pass its entire development in the
same host, or at any rate may actively migrate from the liver to the
nasal cavities after a period of encystment in the liver or elsewhere,
which has also been observed in herbivorous animals (vide also p. 526).
[It is recorded from man in Central America (Darling, Bull.
Soc. Path. exot., 1912, v, p. 118; and again Arch. Int. Med., 1912, v,
p. 401), also from Rio de Janeiro (Mem. Inst. Oswaldo Cruz, 1913,
fasc. ii, p. 125) by Faria and Travassos.
[(2) Porocephalus armillatus, Wyman, is also known under a variety
of names.357 This species is widely spread over tropical Africa. The
adult stage is found in pythons and puff-adders, the nymphal in the
chimpanzee, Sykes monkey, mandrill and other monkeys, the lion,
leopard, banded ichneumon, Aard wolf, dog, black rat, South African
reedbuck and the giraffe. The adult has never been found in man
or any mammal. No fewer than sixteen cases of the nymphal form,
Sambon tells us, have been found in man, and it is probably much
more widespread than at present known. So far it has only been
found in the African natives. This species has sixteen to seventeen
body rings in the male, eighteen to twenty-two in the female, and the
body does not taper as much as in the next species.
[(3) Pentastoma moniliformis, Diesing,358 is an Oriental species, found
in India, Indo-China and South China, and the Malay Archipelago.
The adult occurs in both the Indian and reticulated pythons.
The nymphal stage has been found in monkeys, the tiger, the civet and
the Indian otter.
[The nymph has twice been found in man; in one case in the liver
of a Filipino, the other in the serous coat of the small intestine of a
native of Sumatra.
[This species can be told by the female having twenty-nine to
thirty-three body rings, the male twenty-six, and the annulations are
more bead-like and less prominent than in the African species.
[In addition to these three, Sambon thinks it probable that others
occur in man.—F. V. T.]
Other References to Pentastomidæ.
(1) “Die Wanderung des Pentastomum denticulatum beim Rinde,” Centralbl. f. Bakt.
u. Parasitenk., Jan. 2, 1889, v (1), pp. 1–5, V. Bates.
(2) “Il Pentastoma moniliforme, Dies., nella pantera,” Med.-vet. Torino, 1877, 4 s.,
vi (12), pp. 529–532, R. Bassi.
(3) “On the Organization and Development of Linguatula (Pentastoma), accompanied
with the description of a new species from the abdominal cavity of the mandrill,” Ann. and
Mag. Nat. Hist., 1848, 2 s. ii (7), 2, pp. 69–70, v. Beneden.
(4) “De la Linguatula ferox (Pentastoma denticulatum aut serratum),” Bull. Acad. roy.
d. Sci. d. Belg., 1855, xxii, pt. 1 (1), pp. 4–10, v. Beneden.
(5) “Note sur quelques pentastomes,” Bull. Acad. roy. d. Sci. de Belg., 1857, 26,
2 s., ii (5), pp. 29–30, v. Beneden.
(6) “Ueber das Pentastoma in de gekrösdrusen den Schafe,” Repert. d. Thierh. Stuttg.,
1861, xxii, pp. 37–38, Collin.
(7) “Eine Linguatula aus der Mesenterialdrüse des Schafes und Dromedars als zweites
ungesche. Stadium von Pent. taenioides,” Notiz. u. Tagsber. u. d. Geb. d. Nat. u. Heilk.
Jena, 1862, v, pp. 127, 128, Colin.
B. INSECTA (Hexapoda).
Three separate regions can always be distinguished in the body of insects,
namely, the head, thorax and abdomen. The HEAD is a roundish unsegmented
capsule and possesses four pairs of appendages. The first pair are the various
shaped feelers (antennæ), which are placed on the superior surface of the head next
to the eyes; then more ventrally placed a pair of upper jaws (mandibles) without
palpi and without articulations; they are powerful masticatory organs.359 The first
pair of lower jaws (maxillæ) are jointed and bear a palpus (palpus maxillaris); the
second pair of maxillæ are soldered together and form the lower lip (labium), and
likewise carry a palpus labialis on each side. The upper lip (labrum), as well as
the other parts (which, however, are only appendages), belong to the mouth, which
is really formed of a number of closely united pieces. The mouth parts are modified
according to the functions required of them. Coleoptera, Neuroptera, and Orthoptera
have biting or masticatory mouth parts which conform with the scheme
described above. In the licking mouth parts of the Hymenoptera the maxillæ and
under lip are considerably elongated, while the mandibles retain their form
and are used for triturating the food; in the Lepidoptera nearly all the mouth
parts are shortened except the maxillæ, which form a long and sometimes spirally
rolled suctorial proboscis; the Diptera and Rhynchota have piercing and sucking
mouth parts. The mandibles and maxillæ are metamorphosed into needle-like
structures, while the suctorial apparatus is formed by the labrum.
The thorax consists of three segments, which are frequently united; ventrally it
carries three pairs of legs, which consist of a definite number of articulated pieces
joined together. Their form also changes according to their function, so that legs
for running, walking, digging, swimming, jumping, and preying are seen. A pair
of wings are respectively attached to the last and last but one thoracic rings, and
these may be traced back, not to metamorphosed appendages, but to tracheal
branchia. They are composed of chitinous membranes supported by branched
structures (veins or ribs). Their size and formation vary; they are seldom of equal
size and form (Neuroptera); often the posterior wings are larger than the anterior
wings, the former then only serving as protective coverings for the latter
(Coleoptera), or the anterior wings are larger (Lepidoptera), or the posterior wings
are shortened or are entirely absent (Diptera); and finally there are insects in
which both pairs of wings are lacking.360
The abdomen retains its segmentation, but, with the exception of a few groups
related to the primitive forms of insects, has no appendages in the imago condition;
the abdomen usually consists of ten segments, on the last of which the anus
is situated.
We need only observe the following characters in considering the anatomy of
insects:—
The EPIDERMIS consists of the chitinous cuticle, which is separate from the
cellular layer beneath (hypodermis); the various appendages are supported by the
chitinous layer.
The INTESTINAL CANAL usually consists of the anterior, median and terminal
intestine, and as a rule passes straight through the body; salivary glands discharge
into the anterior part, and, in some cases, yield a stiffening secretion which serves
for spinning webs; numerous or scanty hepatic tubes are appended to the median
intestine, while on the border between the median and terminal intestine open four
to six long tubes (vasa malpighiana), which act as urinary organs. Finally the end
portion of the intestine carries various glands (anal and rectal glands, etc.).
The CENTRAL NERVOUS SYSTEM agrees in structure with that of the Annelids,
but is more highly developed. The pharyngeal ring surrounds the front part of the
intestine; the sensory nerves originate from its SUPERIOR PHARYNGEAL GANGLIA
and are the seat of the higher psychical functions; the INFERIOR PHARYNGEAL
GANGLIA govern the mouth parts, and in addition appear to regulate the movements
(cerebellum) of the vertebrates.
The chain of GANGLIA lying on the ventral side of the abdomen consists primitively
of pairs of ganglia corresponding with the twelve segments, which are connected
by longitudinal and transverse commissures. But many changes in the ganglia may
be seen in insects caused by partial or entire amalgamation of single ganglia, so that
in a few cases only one abdominal ganglion is present. In conclusion, a definite
INTESTINAL NERVOUS SYSTEM is always present.
Of the organs of sense the FACETTED EYES, situated at the sides of the head,
deserve special mention, as do also the ORGANS OF TOUCH and SMELL, situated on
the antennæ, and the ORGANS OF HEARING and taste, or finer sensations, situated at
the mouth and in the buccal cavity.
The sounds emitted by insects are, as a rule, produced by the friction or beating
of certain chitinous parts, but sounds are also produced in breathing (flies).
The ORGANS OF RESPIRATION, the so-called tracheæ, are highly developed; there
are openings (stigmata) at the sides of the body which draw in air by means of the
active participation of the muscles of the body. The number of stigmata varies
between two and ten pairs; the tracheæ themselves branch off from the trunks in
the most varied manner, and carry air to the internal organs.
The colourless BLOOD circulates between the tissues and organs, and is kept
circulating by the contraction of a chambered dorsal vessel provided with ostia,
and which terminates in a short aorta opening at the anterior end.
Insects are SEXUALLY DISTINCT; their sexual glands are in pairs and have a
tubular structure, but the testicular tubules are united together by a capsule into an
oval testicle; exceptionally, also, the excretory canals are double, as also the sexual
orifices; usually the paired canals unite into a single oviduct or spermatic duct
which terminates at the posterior end of the body after receiving the products of
various glands.
As to the HISTORY OF THE DEVELOPMENT of insects, all that is necessary to
mention here is that the young hatched from eggs only exceptionally (as in
Apterygota) resemble the adult parent (insecta ametabola); as a rule they differ
from them not only in the shape of the body, but also more or less by their manner
of life, and only attain the form of the parent through METAMORPHOSIS. This is a
gradual process (insecta hemimetabola) in the Rhynchota and Orthoptera, or a
sudden one with a stage of inanition (insecta metabola) in the other orders. This
stage of rest or inanition, the PUPA, concludes the larval life (caterpillar, maggot,
etc.); during the pupal stage no nourishment at all is taken, but the internal
organs undergo changes; in some forms the rest is not absolute, as voluntary
local movements may take place (pupæ of gnats).
The insects are divided into numerous orders according to the form of the
mouth parts, the structure of the wings, as well as the manner of the development;
with the exception of the lowest group (Apterygota), which is most nearly related to
the ancestors of the insects, and which has no wings and undergoes no metamorphosis,
all the remaining orders, which are termed Pterygota, have wings on the thorax,
though there are, of course, a few species and families of this group which have lost
their wings.
The Pterygota include—
(1) Orthoptera.—Biting mouth parts, anterior wings leathery, posterior wings
thin, folded longitudinally; metamorphosis incomplete (grasshoppers,
crickets, cockroaches).
(2) Pseudoneuroptera.—Biting mouth parts, wings of equal size, thin, not folded
up (dragon-flies, hair and feather lice, termites).
(3) Rhynchota or Hemiptera.—Mouth parts formed for puncturing and sucking;
wings alike, or the anterior wings may be thickened, parchment-like at
their base (plant lice, cicadæ, bugs and true lice).
(4) Neuroptera.—Biting mouth parts; wings alike, thin; metamorphosis
complete (ant-lions, lace-wing flies, etc.).
(5) Trichoptera.—Licking mouth parts; anterior wings narrow, posterior wings
longitudinally folded, both ornamented with little hairs; the larvæ are
worm-like in form, live in water, and breathe through tracheal gills
(may flies, etc.).
(6) Lepidoptera.—Suctorial mouth parts; wings covered with scales (butterflies).
(7) Coleoptera.—Biting mouth parts; anterior wings thickened and differ in
colour, appearance and function from the thin, folded posterior wings
(beetles).
(8) Hymenoptera.—Mouth parts for licking and biting; the wings alike,
membranous (ichneumon flies, ants, wasps, bees, humble bees).
(9) Diptera.—Mouth parts formed for puncturing, sucking or licking; posterior
wings degenerated (gnats, flies, gadflies, fleas).
(10) Strepsiptera.—Anterior wings shortened; the female without wings and
living parasitically (fan-wings).
The parasites of man occur amongst the Rhynchota, Coleoptera,
and amongst the Diptera.
[The most usual and recent classification of the Hexapoda is the following:—
(1) Aptera.—Wingless insects; scarcely any metamorphosis.
(2) Neuroptera.—Four membranous wings, frequently with much network;
the front pair not much, if at all, harder than the under pair; the latter
with but little or no fan-like action in closing; mandibulate; metamorphosis
variable, but rarely complete.
(3) Orthoptera.—Four wings; front pair coriaceous or leather-like, usually
smaller than the other pair, which are of more delicate texture and
contract in repose like a fan; mandibulate; metamorphosis complete.
(4) Thysanoptera.—Four very narrow fringed wings; mouth imperfectly
suctorial; metamorphosis slight.
(5) Hemiptera.—Four wings; the front pair either all transparent or with the
basal half leathery; mouth suctorial; metamorphosis slight.
(6) Diptera.—Two membranous wings only; mouth suctorial, very varied;
metamorphosis complete.
(7) Lepidoptera.—Four large wings covered with scales; mouth suctorial,
metamorphosis great.
(8) Hymenoptera.—Four membranous wings; front pair larger than hind, which
do not fold up in repose; mandibulate, sometimes with a tubular
proboscis; metamorphosis complete.
(9) Coleoptera.—Four wings, the front pair hard and horny (elytra), meeting in
a line over the back and covering the delicate hind pair; mandibulate;
metamorphosis complete.
[There are two other well-known arrangements, namely, Packard’s and Brauer’s,
of recent date, but the one given here, which is based on Linnaeus’
grouping by Dr. Sharp, is by far the simplest.—F. V. T.]
Order. Rhyncota.361
The lower lip forms a long thin tube that can be turned back (rostrum), and
within which lie the setaceous mandibles and maxillæ; the first thoracic segment is
not united with the two posterior ones; the anterior wings are usually leathery as
far as the centre.
(a) Rhyncota aptera parasitica.
Family. Pediculidæ (Lice).
The lower lip is transformed into a projecting rostrum provided with barbed
hooklets in which the hollow extensile sucker (maxillæ and mandibles) is situated;
no wings; no metamorphosis; only simple eyes; the antennæ are five-jointed,
the feet possess hook-like terminal structures; the barrel-shaped eggs (nits) are
deposited on the hair of the host.
[The lice or Pediculidæ are also known as Anoplura and Siphunculata.
[They have been split up into a number of families and sub-families and a
number of genera, but as far as this work is concerned it is best to retain the single
family Pediculidæ.
[Only the three species mentioned here are common parasites of man, but now
and then horse and cattle and sheep lice, Hæmatopinus, may cause transitory
annoyance.—F. V. T.]
Genus. Pediculus, Linnæus.
Pediculus capitis, de Geer, 1778.
Fig. 374.—Mouth
parts of Pediculus
vestimenti. Enlarged.
(After Denny.)

Fig. 375.—Ovum
of the head louse.
70/1.
Male 1 to 1·5 mm. in length, female 1·8 to 2·0 mm. in length.
The colour varies from light grey to black according to the colour of
the hair of the human race upon which they are parasitic. The
abdomen has eight segments, of which the six central ones are
each provided with a pair of stigmata. The thorax is as broad as the
abdomen. Eggs 0·6 mm. in length; about fifty are deposited by a
female head louse. The young can propagate when eighteen days old.
The head louse lives especially in the hairy parts of the head
of human beings; more rarely it is found on other hairy parts
of the body. It is spread over the entire surface of the globe,
and was present in America before the arrival of Europeans.
Quite exceptionally it is said that it bores itself deep into the
epidermis and can live in ulcers.
[The eggs are pear-shaped and are attached to the hairs
near the roots by means of a clasping collar. They hatch
in about seven days. The young are like the adults and
mature in a month. Its general colour varies with that of
its host. In West Africans nearly black, in Hindoos dark and smoky, on Chinese
and Japanese yellow, on Hottentots orange, on South American Indians dark brown
(Murray).—F. V. T.]
Pediculus vestimenti, Nitzsch, 1818.
The head in front is somewhat rounded. Antennæ longer than
in the head louse; 2 to 3·5 to 4 mm. in length; whitish-grey; the
abdomen is broader than the thorax; stigmata as in P. capitis.
Eggs 0·7 to 0·9 mm. in length; about seventy are deposited.
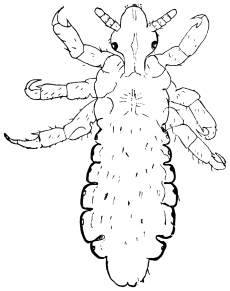
Fig. 376.—Head louse, male. 15/1.
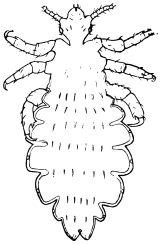
Fig. 377.—Pediculus vestimenti,
Burm.: adult female.
15/1.
P. vestimenti lives on the neck, throat and trunk of persons, and the clothing next
the body, in which also the eggs are deposited. The louse of so-called pedicular
disease (P. tabescentium) is, according to Landois’ researches, only the usual
P. vestimenti; moreover, many cases of phthiriasis are attributable to mites
or fly maggots.
[This parasite has often been a great pest amongst soldiers during long
campaigns, especially amongst the Russians during the Crimean War. Vide also
notes in Addenda (p. 615) under “Body, Head and Clothes Lice.”—F. V. T.]
Genus. Phthirius, Leach.
Phthirius inguinalis, Redi, 1668.
Syn.: Pediculus pubis, L.
Male 0·8 to 1·0 mm. in length; female 1·12 mm. in length; colour
greyish-white; form subquadrate; the two posterior pairs of legs are
strong; the abdomen has nine segments and six pairs of stigmata;
and still another pair of stigmata is situated between the two anterior
limbs. Eggs pear-shaped, 0·8 to 0·9 mm. in length, 0·4 to 0·5 mm.
in breadth, and are deposited in rows of about ten on the hairs.
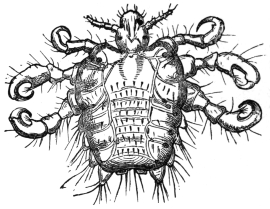
Fig. 378.—Phthirius inguinalis, Leach: they are distinguished by the larger
tracheal trunks originating from the stigmata. Enlarged.
Pediculus pubis, which is found almost exclusively in the Caucasian race, lives on
hairy parts of the body, but hardly ever on the skin of the head; the pubic
region is its favourite place of abode.
[This species reproduces more rapidly than other lice, and is communicated
much more freely. The eggs are often laid singly attached to the hairs near their
apex. It is known as the “crab louse.”—F. V. T.]
(b) Rhyncota hemiptera.
Family. Acanthiadæ.
Body flattened, antennæ four-jointed, rostrum three-jointed, wings atrophied.
[This family, the Cimicidæ, includes the bed bugs; the proboscis, which lies in a
groove, is of three segments; the front wings are shown by two small elytra, there is
no trace of hind wings. Two species are known commonly to attack man.—F. V. T.]
Genus. Cimex, Linnæus.
Cimex lectularius, Linnæus.
Syn.: Acanthia lectularia, Fabricius, 1794.
It measures 4 to 5 mm. in length, 3 mm. in breadth; brownish-red;
eight abdominal segments. The female deposits fifty whitish
eggs at a time (1·12 mm. in length) three or four times a year; the
entire development up to complete maturity takes about eleven
months. [They will breed all the year round, but less so in cold
weather.—F. V. T.]
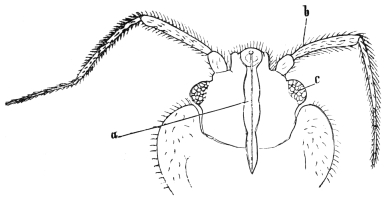
Fig. 379.—Head of the bed bug from the ventral surface. a, the rostrum;
b, the antenna; and c, the eye. 70/1.
The bed bugs live in the cracks and fissures of human habitations, under carpets,
behind pictures, in furniture, bedsteads, etc.; hidden during the day, they attack
persons at night to suck their blood. The alkaline secretion of the salivary glands
dropped into the wound around the separate bites causes the so-called “wheals.”
The bed bugs were known in bygone days by the Greeks (κάριο) and the Romans
(cimex). They were first mentioned from Strasburg in the eleventh century, and
in England about 1500.
[This is the common bed bug of northern latitudes and must not be confused with
the tropical bed bug (C. rotundatus). The bed bug can migrate from one house to
another; this especially takes place when a house is uninhabited. They escape
from windows and pass along walls, water-pipes and gutters, and so reach adjoining
houses. This noxious pest accompanies man wherever he goes; ships and trains
become infested, especially the former.
[A characteristic feature in this animal is the peculiar odour it produces, like
many others in the same group of insects. This odour comes from a clear, oily
volatile liquid secreted by glands in various parts of the body. Although the normal
food is man’s blood, the bed bug can subsist upon moist wood, dust and dirt that
collects in crevices in floors, walls, furniture, etc. The puncturing mouth consists of
a fleshy under lip, within which lie four thread-like hard filaments which pierce the
flesh, the blood being drawn up through the beak.
[The eggs are oval, white, with a projecting rim around one end, with a lid which
is pushed off when the young hatch; they are laid in cracks and crevices in batches
of from twelve to fifty. The egg stage lasts from seven to ten days. The larval
stage so gradually passes into the adult that one scarcely notices the change;
during its growth the skin is cast five times, and at the change the little wing-pads
are seen, showing that the adult stage is reached. The young larva is at first pale
yellowish-white. It resembles the parent, but has no trace of elytra. Although
eleven weeks is said to be necessary for their development, the stages may be gone
through much more rapidly; Howard and Marlatt362 give seven weeks in some
instances. It seems pretty certain that these Cimex only take one meal of blood
between each moult and another preceding egg laying.—F. V. T.]
Cimex rotundatus, Signoret, 1852.
[This bug is common in warm climates; it is an abundant insect
in India, and King has found it in the Sudan, where C. lectularius is,
however, the common species. It is usually known as the tropical
bed bug. Signoret’s bug can be told from the other common species
by the shape of the pronotum. In C. rotundatus it is uniformly
convex, whilst in C. lectularius the lateral edges are flat and sometimes
even concave. The abdomen of rotundatus is also rather more
elongate.
[This species is of considerable importance, as according to Patton
it may act partly as the intermediary host of the piroplasma of
kala-azar.
[Wenyon found at Bagdad that Cimex sp. would take up Leishmania
from Oriental sore, and that the parasite developed into flagellate
form. Patton came to the conclusion that the bed bug transmitted
Oriental sore in Cambay, India, but Wenyon contests this view
(vide Journ. Lond. School Trop. Med., 1912, ii, pt. 1, pp. 13–26).
Franchini (Bull. Soc. Path, exot., 1912, v, No. 10, pp. 817–819)
was unable to connect Cimex with this disease. At present nothing
seems proved. Besides their possible connection with kala-azar, it
has been shown by Howard and Clark (Journ. Exp. Med., 1912, xvi,
No. 6, pp. 850–859) that they can carry the virus of poliomyelitis.
[This bed bug was originally described from the Island of Réunion
in 1852 by Signoret. A similar insect was described from Burma by
Fieber, in 1861, as C. macrocephalus. This is the same as Signoret’s
species.
[The distribution given by Patton363 is as follows: India, Burma,
Assam, Malay, Aden, Islands of Mauritius and Réunion. Patton in
this paper refers to an erroneous statement made in a recent edition
of this book (the last English edition). As I have personally kept
lectularius in moist dirt, wood and refuse for over two years, the
statement as far as I am concerned is not erroneous. Moreover, since
his doubting this fact the same experiment has been twice repeated
with the same results. What they did and do persist on I cannot
say.—F. V. T.]
whilst collecting them. It is rounder and has shorter antennæ than
Cimex columbarius, Jenyns.
[This is common in parts of Europe in pigeon nests, and also
amongst poultry (vide Report Econ. Zool. for year ending
September 30, 1913, pp. 142–144, Theobald). It occurs in Britain on
the latter and will attack man. I have personally been badly bitten
C. lectularius. Jenyns also described a more pubescent species from
swallows as C. hirundinis. I have recently received an account of the
swallow bug invading a house in Kent and causing much annoyance.—F. V. T.]
Cimex ciliatus, Eversmann, 1841.
3·3 mm. in length, yellowish-red, thickly covered with hair;
indigenous in Russia (Kasan).
[From a single specimen seen it is evidently distinct.—F. V. T.]
Family. Reduviidæ.
Head long, narrowed behind into a neck; eyes large, prominent; rostrum
thick and curved; antennæ moderately long, slender at the tip; legs long and
stiff; carnivorous.
Amongst the Reduviidæ one genus is of particular importance,
namely the genus Conorhinus, which has a long head and the first
segment of the beak very much shorter than the second, and the
posterior tibiæ longer than the femora.
These large bugs have a wide distribution, the Oriental region,
North and South America, and the West Indies, Madagascar and
West and Central Africa.
These large bugs may cause very nasty wounds by their bites, but
beyond that it has recently been shown that one interposes in the life-cycle
of a trypanosome, namely—
Genus. Conorhinus, Lap.
Conorhinus megistus, Burm.
This large bug has recently been shown by Chagas to be the agent
in the development of the trypanosome (T. cruzii) which is the cause
of the well-known disease in many parts of Brazil called Barbeiro
(Barbier). This insect is about 1 in. long, black, with four red spots
on the pronotum, and six red lateral lines on the abdomen, black legs,
head and beak. The insect is figured in a coloured plate (No. 9) in
Mem. Inst. Oswaldo Cruz, 1909, i, fasc. 2, pp. 158–218.
A further account is given by Neiva.364
Conorhinus sanguisuga, Lec. (Blood-sucking Cone-nose).
This bug is also known as the Texas or Mexican bed bug, also as
the big bed bug. It is particularly troublesome in the Mississippi
Valley in bedrooms. The bite is very severe and results in more
pronounced swelling and inflammation than that of the Cimex.
Normally this genus feeds upon the blood of mammals and insects.
Its fondness for human blood appears to be quite a new habit, and
appears limited to the mature insect only. It is nearly an inch long,
flat, head very narrow and long, the rostrum short and thick. In
colour it is dark brown with pink markings. They are fully winged
when adult, and they fly with ease, entering houses on the wing,
especially being attracted by lights in windows; they also run swiftly.
Like the bed bug they conceal themselves during the day and come
out at night and bite the sleeper. The effect of the bite is very varied,
but as a rule a sore, itching wound, accompanied by burning pain
and swellings, which may extend over a good deal of the body, occur.
A specific poison is undoubtedly injected into the puncture; but no
doubt serious results are also due to the beak being contaminated
through the insects feeding upon foul carrion. Mr. Lembert, when
bitten by a Conorhinus sp. (?) on the Pacific slope, exhibited the
following symptoms: an itching sensation extending up the leg,
large blotches manifesting themselves on the upper part of the
limb and extending up to the hands and arms; his lips swelled
and the itching and swelling extended over the head; there was
also much nausea. Similar results are recorded from other regions.365
The eggs of the C. sanguisuga are at first white, then become
yellow, then pink; the young hatch in twenty days. There are two
larval and two pupal stages, the latter showing wing-pads. The eggs
are laid and the young feed out of doors, chiefly upon insects. It is
particularly abundant in April and May indoors.
Conorhinus, sp. novum (Monster Bug).
Another species; acts in a very similar way in California, the
bite being very poisonous.
Conorhinus rubrofasciatus, de Geer366 (Malay Bug).
This large bug attacks man in Malaysia and elsewhere. It is recorded
as inflicting “a very nasty sting, which is done by the huge proboscis.”
Acute pain and inflammation follow in a few minutes. In one case
the whole leg became swollen. This species occurs over the whole
Oriental region, in Madagascar and Sierra Leone. It is dark brown
in colour with dusky yellow or brick-red markings on the pronotum
and elytra. Donovan suggests that it may be connected with the
kala-azar piroplasma.
Conorhinus renggeri, Herr-Schäff
(Great Black Bug of Pampas).
This large black bug is mentioned by Darwin,367 who states as
follows: “At night I experienced an attack (for it deserves no less
a name) of the benchuca, a species of Reduvius, the great black
bug of the Pampas. It is most disgusting to feel soft wingless
insects, about an inch long, crawling over one’s body. Before sucking
they are quite thin, but afterwards they become round and
bloated with blood, and in this state are easily crushed. One which
I caught at Iquique (for they are found in Chili and Peru) was very
empty. When placed on a table, and though surrounded by people,
if a finger was presented the bold insect would immediately protrude
its sucker, make a charge and, if allowed, draw blood. No pain was
caused by the wound. It was curious to watch its body during the
action of sucking, as in less than ten minutes it changed from being
flat as a wafer to a globular form. This one feast, for which the
benchuca was indebted to one of the officers, kept it fat during four
whole months, but after the first fortnight it was quite ready to have
another suck.” Mr. Kirby368 also refers to this species.
Conorhinus variegatus (Variegated Cone-nose).
Occurs in Florida in houses, and chases bugs (Cimex) and flies;
not definitely known to bite man.
Conorhinus nigrovarius.
This species occurs in South America. It is one of the forms
known as bichuque. Its bite makes a troublesome swelling.
Conorhinus protractus.
also attacks man in Utah.369 It has been called the “big bed bug.”
Genus. Reduvius, etc.
Reduvius personatus, Linné.
Syn.: Reduvius personatus, Leconte, 1855.
European, but also found in the United States. The bite causes
intense pain. It bites when caught or handled, but does not seem
to do so voluntarily. Swelling and irritation result which may last
a week, and may even cause death.370 In 1899 it was very abundant
at Washington and elsewhere; other species occurred, and so no
definite opinion existed as to the actual biter, but some people took
R. personatus actually biting. It was first described as a parasite of
man in America by Walsh and Riley.371
A popular name for this bug is the wheel or masked bug—a
black insect, three-fourths of an inch long. The larva of this
bug is carnivorous and covers its body with dust so as to conceal
itself from its prey. The adult is active on the wing.
Coriscus subcoleoptratus, Kirby, 1837.
Syn.: Nabicula subcoleoptrata, Kirby, 1837; Nabis subcoleoptratus,
Reuter, 1872; Coriscus subcoleoptratus, Stål, 1873.
Northern United States. Howard was bitten by one between the
fingers—the pain was intense, like a needle prick, but the swelling
was small.372 No other case known.
Rasahus biguttatus, Say, 1831.
Syn.: Pirates biguttatus, Stål, 1862; Callisphodrus biguttatus, Stål, 1866;
Rasahus biguttatus, Stål, 1872.
Common in southern United States, and found in Cuba, Panama
and Pará, etc. Known as the two-spotted corsair on account of the
great spot on the hemielytra. Frequently found in houses, where
it chases the bed bug. It also bites man frequently. From
1869 Walsh and Riley placed it amongst the parasites of man.
In the United States Davidson373 is of opinion that all cases attributed
to spider bites are due to this insect.
Melanolestes morio, Erichson, 1848 (Non-walker).
Syn.: Pirates morio, Erichson, 1848; Melanolestes morio, Stål, 1866;
Pirates picipes, Herrich-Schäffer, 1848; Melanolestes picipes, Howard, 1900.
Guiana and Mexico and eastern and southern United States.
Length 20 mm., hides under stones and logs during daylight, and flies
at night. Attracted by lights into houses. Very abundant in 1899
at Washington. Howard cites cases where it was proved to bite man.
Melanolestes abdominalis, Herrich-Schäffer, 1848.
Syn.: Pirates abdominalis, Herrich-Schäffer; Melanolestes abdominalis, Uhler, 1875.
Allied to the former; some say similar, but can be told by the
shorter wings on the female. It occurs in the same localities as M. morio.
Phonergates bicoloripes.
This reduvid attacks man in Africa.
Family. Aradidæ.
Broad and very flat bugs, with antennæ of four segments and the beak of three;
scutellum short, no cuneus to elytra and the tarsi of two segments. They normally
live under the bark of trees, etc., and are found in most parts of the world.
Dysodius lunatus, Fabr. (Pito Bug).
A large species which is found in South America, frequenting
houses, and bites very severely.
The Ochindundu.
The bug is described by Wellman (Journ. Trop. Med., April 2,
1906, p. 97) as not only feeding on ticks, such as Ornithodorus
moubata, but as also attacking man. It is called by the Angola Bantus
the ochindundu. It is black in colour; the first two pairs of legs
are of a bright red hue. It has curious paddle-like structures on the
front four legs, which seem to be designed for securely holding the
ticks. It infects native kraals for the sake of preying on ticks. The
natives also state that it inflicts a bite which far exceeds in painfulness
that of the tick. They compare the bite with that of a poisonous
snake.
Family. Lygæidæ.
Scutellum short; antennæ four-jointed; ocelli present; membranous part
of hemielytra with never more than five nervures. Nearly all vegetable feeders.
A few are recorded here as biting man.
Lyctocoris campestris, Fabricius.
Syn.: Acanthia campestris, Fabr. (Lyctocoris domesticus).
Rare in habitations, lives on human blood. Found by Blanchard
in a bed at an hotel at Liverpool. The bite is undoubtedly worse
than that of Cimex; cosmopolitan. In colour it is ferruginous,
shining, legs testaceous; hemielytra slightly shorter and narrower
than the abdomen; membranous portion transparent, the apex
broadly fuscous. Length 3·8 to 4·8 mm.
Rhodinus prolixus, Stål, 1859.
Sometimes attacks man, and the bite is very painful. It is
25 mm. long and 8 mm. broad, and occurs in Colombia. It is
found also in Cayenne and Venezuela. This like other species
is known in South America as bichuque or benchuca.
[A few other unimportant species are also recorded as biting man,
such as Harpactor cruentas, in the South of France; Eulyes amœna,
from Borneo and Java; Arilus carinatus, Forster, from Brazil.
The latter appears to be the same as the Acanthia serratus,
Fabricius.—F. V. T.]
Order. Orthoptera.
[The only Orthoptera recorded as doing actual harm to man are certain wingless
locusts found in Africa. The cysticercus stage of a small tapeworm found in
rats and man has been found in an earwig (Alcock).
[The strange Hemimeridæ found in West Africa, resembling wingless cockroaches,
are parasitic on rats (Cricetomys). Phasmidæ, or stick insects, are said to be
able to eject a fluid which may cause blindness if it comes in contact with the eyes.
Locusts Injurious to Man.
[A wingless locust—Enyaliopsis durandi, Luc—is recorded by Wiggins374 as
injurious to man in Uganda. “The bite of this insect,” it is said, “gives rise to a
very nasty eruption, which may extend over the whole body, with high temperature
and general malaise. The skin at the site of the bite sloughs away, and generally
leaves a large deep cavity, which heals very slowly.”
[An allied species—E. petersi, Schaum—emits a clear yellow fluid, but according
to Marshall this does no harm.375 Stannus writes that “for some years I have been
cognizant of the fact that among the natives of Nyasaland an allied if not the
same species is held to cause skin lesions by the emission of a fluid on the bare skin
surface of the body. I have seen cases of ulcers on various parts of the body, for
which the ‘nantundua’ was assigned as the cause.” He then describes the destruction
of the superficial layers of the skin which he observed after the yellow fluid had
been on the skin twelve hours.—F. V. T.]
Order. Coleoptera.
The larvæ of beetles, similarly to those of some other Arthropoda (myriapods
and the larvæ of gnats), have sometimes been observed in man as purely accidental
guests. In one case or another, such accounts may have originated through a
mistake of the observer. Thus English doctors report the presence of the larvæ of
Blaps mortisaga in the stools of human beings, Sandberg of the larva of Agrypnus
murinus in his ten year old son, and Blanchard mentions the larva of a beetle that
was vomited by a child. All these cases, however, do not represent actual parasitism,
although there are beetles living parasitically.376
Silvanus surinamensis, Linnæus (Saw-toothed Grain Beetle).
[Taschenberg records this beetle as having invaded some sleeping
apartments adjoining a brewery where stores were kept, and
annoying the sleepers at night by nipping them when in their beds.
[This beetle is common in many parts of the world amongst
groceries, corn, meal, seeds, dried fruits, etc. It is about 1/10 in. long,
much flattened and chocolate-brown in colour. The thorax has two
shallow grooves and bears six minute teeth on each side. The jaws
are strong, but the bite cannot be very serious.—F. V. T.]
Order. Diptera or Siphonaptera.
Aphaniptera (Fleas).
Wingless, the thoracic rings distinct and free; antennæ of three segments; legs
very powerful; abdomen with nine segments. [Ten segments are present, but only
nine are visible.—F. V. T.] The mandibles transformed into serrated puncturing
organs, which are situated in the split sheath of the rostrum; the maxillæ are
laminated and have palpi, and more or less conceal the other parts.
The importance of fleas lies mainly in the fact that they act as plague carriers.
About 150 species have already been described. The only ones of importance for
this work are those found on man and those on rats and mice. The two families
in which these are found are known as Pulicidæ and Sarcopsyllidæ.
The eggs of fleas are laid on the ground, on rugs, etc., and in birds’ and rodents’
nests. They hatch rapidly in warm weather and in warm climates, varying from
two to five days; in cold countries they may take two or three weeks to incubate.
The larva is a footless creature, pearly white in colour, the head sometimes
being darkened, composed of fourteen segments including the head, and although
apodal can move with considerable agility. It lives amongst dust and dirt, and
feeds upon any organic matter it can find. In about two weeks it is said to become
mature, and then spins a cocoon in which pupation takes place.
The cocoons of the common human flea and the fowl flea become covered with
dust and dirt. The period of pupal life seems varied, for I have had the fowl flea
hatch out in ten days, and others in three weeks at the same time of year.
The adults are blood suckers and cause considerable irritation as well as
acting as disease carriers, and in the Sarcopsyllidæ the females attach themselves
permanently to their hosts, embedding themselves under the skin, where they
become pregnant. Some kinds harbour the cystic stage of tapeworms, and the rat
trypanosome passes certain stages in the rat flea. Most fleas have definite hosts,
but some, like the rat and fowl fleas, attack man.
The fleas which can carry the bacillus of plague are Xenopsylla cheopis, Pulex
irritans, Ceratophyllus fasciatus and Hoplopsyllus anomalus.
The two families, Pulicidæ and Sarcopsyllidæ, can be distinguished as follows:
| Thoracic segments much foreshortened, coxæ and femora of hind legs very slightly enlarged | Sarcopsyllidæ. |
| Thoracic segments normal, coxæ and femora of all the legs much enlarged | Pulicidæ. |
Family. Sarcopsyllidæ (Jiggers).
| The members of this family are not confined to one host. |
| Three genera are known and tabulate as follows:— |
| α. Hind coxa without a patch of spines on the inside. |
| α1. Hind femur simple | 1. Dermatophilus. |
| α2. Hind femur with a large tooth-like projection near the base | 2. Hectopsylla. |
| β. Hind coxa with a patch of short spines on the inside | 3. Echidnophaga. |
Genus. Dermatophilus, Guérin.
Dermatophilus cæcata, Enderl.
The eyes of the female vestigial. Taken on and behind the ears of Mus rattus
in Brazil.
Dermatophilus penetrans, L., 1758 (Jigger, Chigoe).
Syn.: Sarcopsylla penetrans.
About 1 to 1·2 mm. in length; brown in colour. Eyes distinct.
The males only occasionally visit man to bite; the fertilized female,
on the other hand, bores into the skin with her head, particularly
about the toes of the host, and then attains considerable dimensions.
The eggs develop on the
soil with a metamorphosis
similar to that of the
common flea.
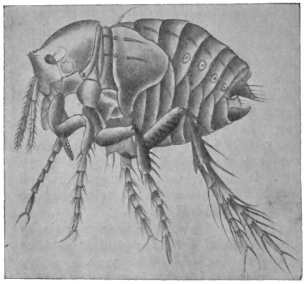
Fig. 380.—Dermatophilus penetrans: young
female. Highly magnified. (After Moniez.)

Fig. 381.—Dermatophilus penetrans:
older female. Enlarged. (After Moniez.)
The sand flea (nigua) particularly infests Central and South America, and,
in 1873, was carried by ships from Brazil to the West Coast of Africa. In a comparatively
short time it has become disseminated throughout Africa and has also
appeared in Madagascar; recently also it has been reported from China.
Besides attacking man, it also settles on mammals, for instance, on dogs, pigs,
etc. According to Jullien the wound or little swelling caused by the female has no
particular significance, as children infested with ten or eleven sand fleas quietly
proceeded with their games. It will be understood, however, that the wound easily
affords the opportunity for the setting up of inflammation or even septic processes,
as is the case in any kind of wound.
[The jigger is also well known in the West Indies.—F. V. T.]
Genus. Echidnophaga, Olliff.
Four species found on rats, etc.
Echidnophaga gallinacea, Westwood (Chigoe of Fowls).
[This flea is a native of tropical Asia and Africa. It lives on the
fowl chiefly, attacking the neck and around the eyes. Specimens
were sent me from Texas, where they not only attack poultry but also
children, the latter somewhat severely. It also occurs on cats, and is
found on rats in Africa. It has been introduced into North America.
[Three other species are found on rats, viz.: E. myrmecobii,
Rothsch., from Australia; E. murina, Tirah, from Southern Europe;
and E. liopus, Rothsch., India and Western Australia.—F. V. T.]
Family. Pulicidæ (True Fleas).
Rothschild’s classification is as follows:—
| Section I.—Club of antennæ distinctly segmented only on the hind side. |
Key to Genera. |
| a. No comb on head and thorax. |
| a1. The internal incrassation, which extends from the insertion of the mid coxa into the thorax, joins the anterior edge of the mesosternite | Pulex. |
| b1. This incrassation joins the upper edge of the mesosternite | Xenopsylla. |
| b. With a comb on the pronotum only | Hoplopsyllus. |
| c. With a comb on the pronotum and at the lower edge of the head | Ctenocephalus. |
| Section II.—Club of antennæ distinctly segmented all round. |
Key to Genera. |
| a. Eye developed. |
| a1. No comb on head. |
| a2. Pygidium not projecting backwards; frons with tubercle | Ceratophyllus. |
| b2. Pygidium strongly convex, projecting backwards; frons without tubercle | Pygiopsylla. |
| b1. Two spines at angle of genæ | Chiastopsylla. |
| b. Eye vestigial or absent. |
| a1. Abdomen without comb. |
| a2. Hind edge of tibiæ with about eight short and several long bristles, which do not form a comb. |
| a3. Fifth segment in fore and mid tarsi with five, and in hind tarsus with four bristles | Neopsylla. |
| b3. Fifth segment in fore and mid tarsi with four, and in hind tarsus with three lateral bristles, there being an additional pair of bristles in all the tarsi on the ventral surface in between the first pair | Ctenophthalmus. |
| b2. Hind edge of tibiæ with about twelve short and three long bristles, the short ones forming a kind of comb | Ctenopsylla. |
| b1. Abdomen with at least one comb | Hystricopsylla. |
Genus. Pulex, Linn.
Pulex irritans, L., 1758.
Male 2 to 2·5 mm. in length, females about 4 mm.; reddish or
dark brown; head without bristles; thoracic and abdominal rings of
bristles on the dorsal aspect, and small hairs directed backwards at
the posterior margin. The barrel-shaped white eggs are deposited
in cracks in the boards, sweepings, spittoons, etc.; they produce legless
larvæ consisting of fourteen segments, which, after about eleven days,
are transformed into pupæ; after another eleven days the flea emerges.
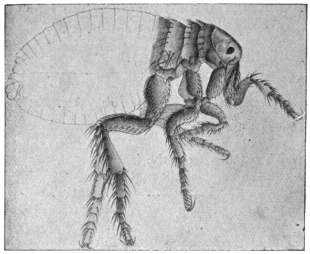
Fig. 382.—Pulex irritans. 14/1.

Fig. 383.—Larva of flea.
Enlarged. (After Railliet.)
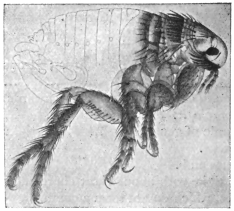
Fig. 384.—Pulex serraticeps. 22/1.
Fleas live in human dwellings all over the world, and periodically pass on to
persons to suck their blood. They may deposit their eggs on very uncleanly
individuals, and even undergo development, therefore it is possible to find larvæ
and pupæ on such persons.
The dog flea, Pulex serraticeps, is easily distinguished from the flea of man by
the large thick bristles on the posterior margin of the first thoracic ring (fig. 384).
Genus. Xenopsylla, Glink.
Xenopsylla cheopis, Rothschild.
This is the common rat flea of tropical countries. Rothschild377
says: “Although practically cosmopolitan, it cannot apparently flourish
in temperate and cold climates.”
In the male the bristles of the flap-like process of the clasper all
slender; in the female the narrow portion of the receptaculum
seminis long. Originally discovered in Egypt.
This is apparently the chief plague flea. The Indian Plague
Committee have proved that this flea is easily infected when fed on
plague rats, and that the bacillus multiplies rapidly in the flea’s
stomach and that the fleas may remain infective for fifteen days. How
the flea infects man does not apparently seem to have been proved, as
it does not do so through its bite, but the excrement is highly infective.
It is probable that this poisoned fæcal matter gets to the wound
caused by the piercing mouth.
Xenopsylla brasiliensis, Baker,
occurs on rats in West Africa and has been introduced into Brazil.
Genus. Ctenocephalus, Kolen.
Includes the cat and dog fleas. The dog flea, C. canis, Dugès,
is found on the dog all over the world, but especially in temperate
climates. It also occurs on rats. Man is often badly bitten by this
insect and it overruns houses. The eggs are laid on rugs, carpets and
dust and dirt and amongst dogs’ hair, but are not fastened to it and
fall anywhere. The ova may hatch in about fifty hours and the larvæ
live for seven days and then spin their cocoons amongst dust and dirt.
The pupal stage lasts about eight days.
The cat flea (C. felis) is widely distributed over the world, and
occurs on many mammals beyond the cat, and is also found on rats.
Genus. Hoplopsyllus, Baker.
A genus found in North America related to Pulex, but at once
recognized by the prothorax bearing a comb.
Hoplopsyllus anomalus, Baker,
which is found on the ground squirrel (Citellus beecheyi) in California,
and according to Rothschild once found on the rat, has been proved
to carry the plague bacillus and to play an important part in plague
infection in California.378
Genus. Ceratophyllus, Centis.
Ceratophyllus fasciatus, Bosc.
This flea is also found on the rat in Europe and will attack man.
It is a plague carrier. It has eighteen to twenty teeth on the
prothoracic comb and no black spines on the head. The genus
Ceratophyllus of Centis has a pronotal comb and three ante-pygidial
chætæ on each side. Two other specimens recorded: C. londiniensis,
Rothsch., and C. anisus, Rothsch. The former on rats and mice in
London, the latter on Felis sp. from Japan and Mus norvegicus in
California.
Genus. Ctenopsylla, Kolen.
This genus contains a very abundant rat and mouse species,
C. musculi, Dugès, which is widely distributed over the globe.
Genus. Hystrichopsylla, Tasch.
Large hairy fleas, with no eyes and one or more combs on the
abdomen. In the Mediterranean area one species, H. tripectinata,
Tirah, is common on rats and mice, and also in the Azores. Several
others occur on rats and mice. For information concerning these the
reader is referred to Rothschild’s papers.
Pulex pallipes is another species found on the rat and man.
Systematic, Anatomical, and Biological Remarks on Mosquitoes.
Mosquitoes come in the Nematocera, one of the sub-orders of the Diptera,
and are divided into numerous families, of which, however, the Culicidæ are of most
interest to us here. Other families as the Psychodidæ and Chironomidæ are also
of considerable importance, vide following pages. The head is small, the facetted
eyes are placed laterally, but there are no accessory eyes (ocelli). In front of the
eyes are situated the comparatively long antennæ, the differences of which strongly
mark the distinction of sex.379
The antennæ are composed of fifteen or sixteen segments. In the male they
are covered with long whorl-like hairs, while in the female the antennal hairs are
short—differences that are perceptible even with the naked eye.380 The proboscis,
which is longer than the antennæ, protrudes from the inferior aspect of the head and
is composed of the following parts (figs. 387 and 388): Two grooved half tubes,
facing one another, of which the upper one is the upper lip (labrum) and the lower
one the lower lip (labium), which represents a pair of coalesced maxillæ. Within
the tube formed by the labrum and labium are the mandibles and maxillæ, transformed
into instruments for piercing, and a single puncturing organ, the hypopharynx.
On the right and left, next to the proboscis, are placed the straight five-jointed
palpi, the final joint of which is thickened in the male.381 In biting, the
labrum, which is swollen at its free end, is not introduced into the wound like the
other mouth parts, but is bent backwards. The labium and hypopharynx push
direct into the skin; the maxillæ and mandibles, however, which are needle-like and
serrated at the tips, penetrate with a saw-like movement. [The swollen free end of
the labrum really means the labellæ, two articulated pieces, supposed by some to be
the labial palpi. In most species the mandibles are not serrated at their ends.—F. V. T.]
The saliva is introduced into the wound through the lumen of the
hypopharynx, while the blood is sucked up by the mosquito in the groove of the
labium.
The three thoracic segments are soldered together. The central one carries the
membranous wings on the sides of the dorsal surface; the posterior somite carries
the small halteres (rudimentary posterior wings). There are three pairs of long
slender legs on the lower side.

Fig. 385.—Head of a male (a) and of a female (b) Anopheles. Slightly enlarged.
(After Giles.)
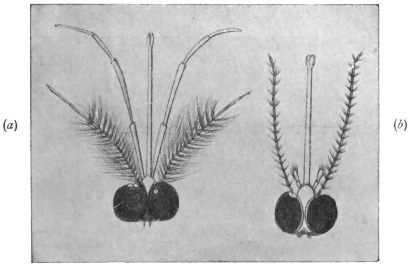
Fig. 386.—Head of a male (a) and of a female (b) Culex. (After Giles.)
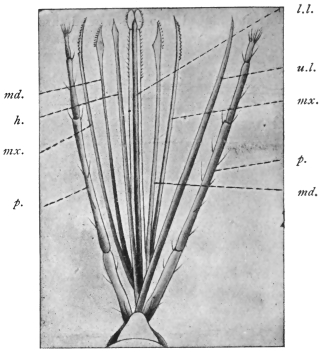
Fig. 387.—Mouth parts of Anopheles claviger.† h., hypopharynx;
md., mandible; mx., maxilla; u.l., upper lip; l.l.,
lower lip; p., palpi. (After Grassi.)
† [This should read Anopheles maculipennis, Meig.; there was no type of A. claviger.—F. V. T.]
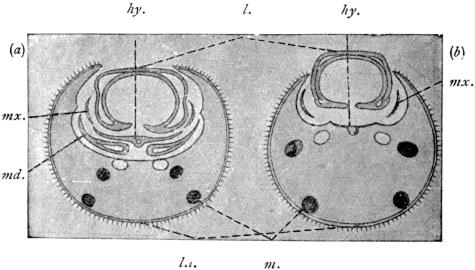
Fig. 388.—Anopheles maculipennis: transverse section through the
proboscis of a female (a) and a male (b). hy., hypopharynx, with duct
of the salivary gland; m., muscles; md., mandibles; mx., maxillæ;
l., labium; l.l., labrum. (After Nuttall and Shipley.)
The abdomen has no limbs, is composed of eight (rarely nine) distinct segments;
the sexual and anal orifices are at the posterior end, the stigmata on the sides. The
intestinal canal (fig. 389) is composed of three principal divisions; the anterior
part reaches as far as the front pair of legs, and consists of the œsophagus, which
is provided with two small lateral diverticula. [At the commencement of the
œsophagus are one or more diverticula, which vary in size; they contain air, food
and bacteria.—F. V. T.] The mid gut reaches as far as the fifth and sixth abdominal
ring; in front it is thin, and has numerous small supra-œsophageal ganglia; the
posterior part is, however, more dilated. Four or five Malpighian tubes, the excretory
organs, discharge at the place where the mid gut passes into the terminal gut.
The pair of salivary glands have one common excretory duct leading into the
hypopharynx.
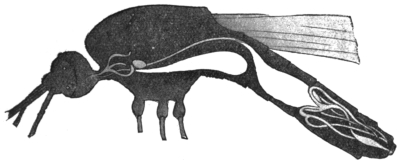
Fig. 389.—Longitudinal section of an Anopheles, showing alimentary canal. In the forepart
of the thorax is the salivary gland consisting of three tubules; ventrally, the suctorial stomach
extending into the abdominal cavity; the stomach, and at the posterior end of the abdomen
the Malpighian vessels. (After Grassi.)
These glandular bodies are situated in the thorax; each consists of three slightly
serpentine tubules, the dorsal and ventral tubes being long, the central one shorter.
The above-named characteristics apply to both genera Culex and Anopheles, but in
the genus Culex is smaller, Anopheles larger. [In Anopheles the ends of the ducts
in the lobules are dilated, whilst in most of the genera the ducts are the same
size all along. The lobules may bifurcate, and in Psorophora there are five lobules.—F. V. T.]
The legs of the genus Culex are about the same length as the whole body;
in Anopheles they are double that length.382 In Anopheles the palpi and proboscis are
of equal length; in Culex the condition is different, according to sex. In the male
the palpi are longer than the proboscis; in the female considerably shorter and the
number of segments diminished. The venation of the wings exhibits further points of
differentiation, as also their adornment, though this last sign is not by any means
always conclusive; most species of the genus Culex have unspotted wings, whilst
those of Anopheles are usually spotted. More important is the fact that in Culex the
abdomen is decorated with small scales, similar to those on butterflies, whereas
there are small bristles on the abdomen of Anopheles. [This cannot be said to be a
character by which an Anopheline may be told from a Culicine, for in such common
Anopheline genera as Cellia and Neocellia we get plenty of scales on the abdomen.—F. V. T.]
An experienced observer can, however, separate the two genera by the
difference in size and their manner of resting. When settled they either touch the
resting place with all the legs or only with the four anterior legs. In consequence
of the different length of the legs, the body of Culex approaches the resting place
more closely; moreover, Culex holds the abdomen parallel or at an acute angle to
the resting surface, whereas Anopheles carries the abdomen directed upwards (at an
angle of about 145°) and holds the head down. Both genera, however, usually only
rest on the four anterior legs, and then, as has long been known, Culex carries the
third pair directed towards the dorsum, while those of Anopheles hang down.
In regard to the differentiation of the species, I must refer you to the special
literature, and content myself by observing that about 150 species of Culex and
about fifty species of Anopheles have been described, of which fifty about four are
found in Europe. [The number of known Anophelines now is more—100 species—of
other Culicidæ over 700.—F. V. T.] According to our present knowledge it
appears that the entire genus Anopheles can transmit malaria to man; this observation
has been confirmed in Anopheles claviger, Fabr.; A. maculipennis, Meig.;
A. bifurcatus, L.; A. superpictus, Grassi; A. pseudopictus, Gr., all of which are
found in Italy,383 Germany, etc., as well as in the tropics. Moreover, in A. costalis,
Loew; A. funestus, Giles (Africa); A. quadrimaculatus, Say (North America), and
A. rossii, Giles; the latter is perhaps identical with A. superpictus, Gr., as well as
with A. culicifacies (India). [Anopheles maculipennis and A. claviger are the same.
Certainly neither maculipennis nor bifurcatus has been found in the tropics.
Anopheles quadrimaculatus, Say, is the same as A. maculipennis. There is no
evidence that all Anophelines carry malaria, but there is much to show that certain
species only are capable of so doing. A list of known carriers is given later.—F. V. T.]

Fig. 390.—Anopheles maculipennis, Meigen. Enlarged.
(After Grassi.)
Everyone is aware that mosquitoes swarm at sunset in fine weather, and then
seek out human beings and other warm-blooded animals to take food. In this
regard, however, the sexes differ, for it is almost without exception that the females
only suck blood, while the males subsist on the juices of plants (blossoms or fruits).384
After sucking, and when night has fallen, the mosquitoes find a place of refuge, for
which purpose they utilize the grasses or foliage of trees and bushes, or inhabited or
uninhabited rooms of houses, also cellars, stables, verandahs, etc., where they also
pass the day.
[Some mosquitoes bite in the daytime—Stegoymia and some Anophelines; some
bite right into the night, as Culex fatigans and C. pipiens.—F. V. T.]
The period required for digestion varies according to the temperature. It takes
two days in summer, and may take up to ten days or more in cool weather. After
digestion is complete more food is taken up, this being necessary [in some species
only—F. V. T.] for the maturing of the sexual products in the female.

Fig. 391.—Larva of Anopheles maculipennis,
Fabr. Enlarged. (After Grassi.)
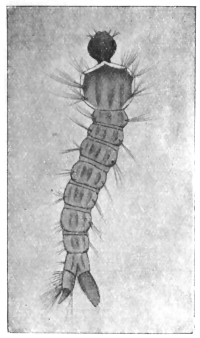
Fig. 392.—Larva of Culex. Enlarged.
(After Grassi.)
It is still unknown under what circumstances copulation takes place;385 in any
case, sooner or later the females are fecundated, and when the ova have become
mature, and the season is not too far advanced, they seek a suitable place in which
to deposit them.386 These are larger or smaller, permanent or temporary, collections
of standing water, pools, puddles, lakes, pits, water in rain-water barrels, basins, etc.
Nevertheless, certain kinds prefer certain waters; thus Anopheles (claviger) maculipennis
and several of the Culices seek stagnant water overgrown with swamp vegetation
and decomposing vegetable matter; A. bifurcatus and certain Culices, clear
water with some vegetation (such as fountains and the lakes in gardens and parks);
Culex pipiens has a preference for rain-water barrels, even though the water be dirty
and evil-smelling. [I have found the larvæ of Anopheles bifurcatus living in great
numbers in ponds and lakes completely overgrown with floating water-weeds, and
those of Culex pipiens in liquid manure.
Sexual Organs of the Mosquito.—The female has a pair of ovaries, opening into
a single tube by the ovarian tubes; into the single tube opens a duct coming from
the spermathecæ, and also a mucous gland. The spermathecæ store up the male
cells. The male organs consist of two testes joined by ducts (vasa deferentia) to the
ejaculatory duct formed by their union. Each vas deferens is joined by a short tube
with the sac-like vesicula seminalis.—F. V. T.]
There is also a difference in the manner in which Culex and Anopheles deposit
their ova. Culex deposits two to three hundred eggs in compact heaps that float
on the water, and in which the eggs stand perpendicularly one next the other;
whereas Anopheles maculipennis deposits only three or four up to twenty eggs,
united in groups that float horizontally on the water; the eggs of A. bifurcatus,
again, are arranged in star-like groups. The eggs are about 0·75 mm. in length,
and assume a dark hue soon after being laid. The development only occupies a
few days. The young larvæ grow rapidly, changing their integument several times;
the larvæ also differ in the various genera, though they have a general resemblance
(figs. 391 and 392).
The long legless larva has a flattened head, a fairly broad, rectangular, or
trapeziform thorax, on which there are bristles, and an abdomen distinctly segmented,
and on the segments of which there are also lateral bristles. The situation
of the stigmata marks the difference between the two genera. Though in both
genera the stigmata are at the posterior end and on the dorsal surface, they are in
Anopheles close to the surface of the body; in Culex, however, they are on the free
end of a long tube (siphon).
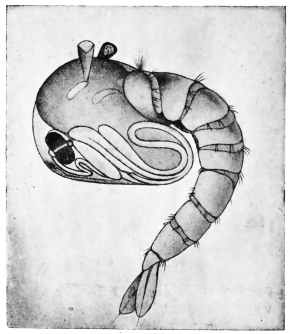
Fig. 393.—Pupa of Anopheles maculipennis, Meig.
Enlarged. (After Grassi.)
The position of the larva in the water also differs. The larva of Anopheles lies
almost horizontally beneath the surface of the water, the posterior border of the
penultimate abdominal segment, upon which the stigmata are situated, being on the
surface; whereas the larva of Culex hangs head downwards perpendicularly in the
water, the point of the siphon only touching the surface.
In about a fortnight the larva is fully grown and becomes a pupa. The pupa
(fig. 393), which moves in jerky movements, remains in the water, but partakes of
no food. In shape it somewhat resembles a tadpole, that is to say, it consists of a
bulky anterior portion, on the surface of which the head, with its appendages, is
recognizable, and a more slender segmented abdomen. Above, on the thorax, there
are two small trumpet-shaped breathing tubes for the conveyance of air to the
tracheal system. After three or four days the perfect mosquito hatches out, remains
a short time on the surface of the water until its chitinous integument is hardened,
and then flies away.
The females that are fertilized in the autumn hibernate in sheltered spots in the
open air, or in houses, cellars, under stairs, in stables, barns, etc., and are the
progenitors of the first generation of the following year.
In accordance with the climate of a country, or the kind of weather of a year,
the conditions in regard to the manner of life and the duration of the development
of the mosquito vary. At all events, the life-history of the mosquito elucidates
many points relating to malaria which were hitherto not understood.
[The length of the egg, larval and pupal life varies so much that it is not
possible to give an account of any value here. Frequently the eggs may incubate
in two days, whilst I have had Stegomyia fasciata eggs from Cuba that have hatched
out under abnormal circumstances more than two months after they were laid
(“Mono. Culicid.,” iii, p. 6). Some larvæ, as Anopheles bifurcatus, live for
months during the winter. Some mosquitoes therefore hibernate as larvæ. The
larvæ and pupæ of the different genera present very marked characters, mainly in
regard to the structure of the siphons. Specific differences may be found in the
frontal hairs of Anopheline larvæ and in the number and arrangement of a group
of spines at the base of the siphon in Culicines.—F. V. T.]
Culicidæ or Mosquitoes.
The importance of these insects to man is very great. They not only produce
painful bites, which may become inflamed and give rise to a considerable amount
of œdema, but they are more important on account of the part they play in the
distribution of various diseases. Culicidæ may not only carry disease germs, but
act as intermediate hosts for certain parasites, such as some of the Anophelina for
malarial parasites, Culex for Filariæ, and Stegomyia for yellow fever, etc.; the last-named
is in any case the distributor of that fatal disease. It is therefore very
necessary to know the life-history, habits and characters of these pests.
Mosquitoes exist in almost all parts of the world from the Arctic circle to the
tropics; temperate regions suffer from them less than the two extremes, but even
there they form not only a source of great annoyance but of danger as malaria and
possibly now and again yellow fever carriers. A few years ago comparatively few
species were known, now some 800 odd have been described. Their number will
probably not stop far short of 1,000, in spite of the fact that many have been
described under different names, yet really the same species. Some are purely
domestic, others entirely sylvan; the former, as we might expect, often have a very
wide distribution, having been taken from place to place in boats and trains. The
more rapid transport becomes, the greater becomes the possibility of this wide
distribution of many species increasing, and the spread of other species from their
natural home to foreign parts by sea and then by trains further inland.
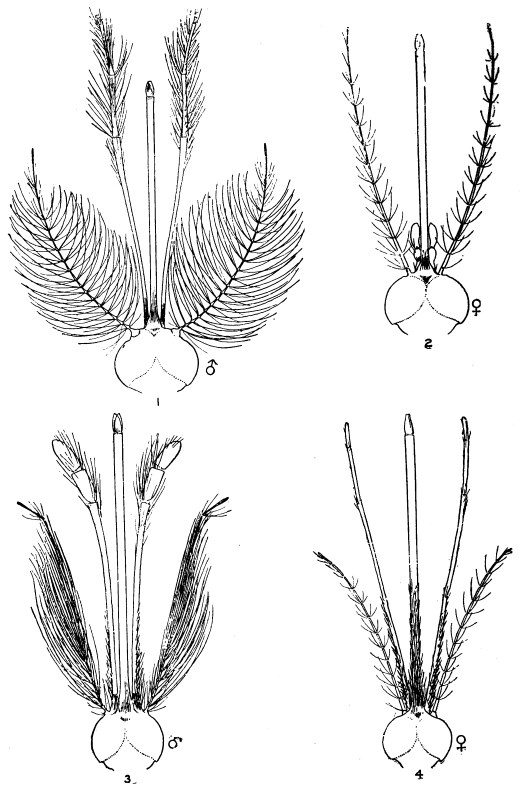
Fig. 394.—Heads of Culex and Anopheles: (1) Culex male; (2) Culex female;
(3) Anopheles male; (4) Anopheles female. (After Daniels.)
All Culicidæ are aquatic in their larval and pupal stages. Almost all small
collections of water, both natural and artificial, may form breeding grounds for
these pests. Some even breed in pitcher plants and many in bromelias. The
favourite resorts for the larvæ of Anophelina are small natural collections of water,
such as puddles, ditches and small pools around swamps; certain species (A. maculipennis,
etc.) live in rain barrels as well. They may also occur in the sluggish water
at the edges of rivers or even in mid river, where the flow is checked by masses
of water weeds (Myzomyia funesta, etc.). The Stegomyias prefer artificial collections
of water, but also occur in natural pools. The yellow fever species (S. fasciata)
prefers small collections, such as in barrels, pots, jars, etc. Culex occur in all
manner of places—rain barrels, tanks, cisterns, ponds and ditches. Some of the
South American species of Culex, Wyeomyia, Joblotia, etc., breed in the collections
of water at the base of bromelia leaves.387 Very few Culicid larvæ live in salt water
except in Australia, where Dr. Bancroft has found them in salt water of specific
gravity 1·040 (Mucidus alternans and Culex annulirostris). Other salt water
mosquitoes are known in America. The food of the larvæ is very varied; the
majority appear to feed upon confervæ, small crustacea and insects; some are
cannibals, readily devouring others of their own kind. The larger larvæ of Megarhinus,
Psorophora, Toxorhynchites and Mucidus are extremely ravenous and devour
one another.
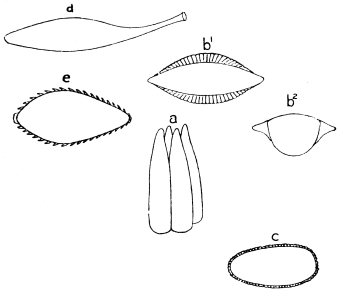
Fig. 395.—a, eggs of Culex; b1 b2, eggs of Anopheles; c, egg of Stegomyia;
d, egg of Tæniorhynchus; e, egg of Psorophora.
There are two main types of larvæ, the Anopheline and Culicine; in the former
there is no respiratory siphon, in the latter the siphon is long or moderately long.
The head offers certain marked peculiarities which are of specific value; this
especially applies to the Anophelina, in which the frontal hairs are of great service
in distinguishing the larvæ,388 whilst in Culex the number and position of the spines
at the base of and on the siphon are characteristic. The position assumed by the
larvæ in the water also varies in the different groups; most of the Anophelines lie
horizontally, most of the Culicina and Ædeomyina hang head downwards. The
pupæ also vary, but not to the same extent; the chief differences to be noticed are
in the form of the two respiratory trumpets.
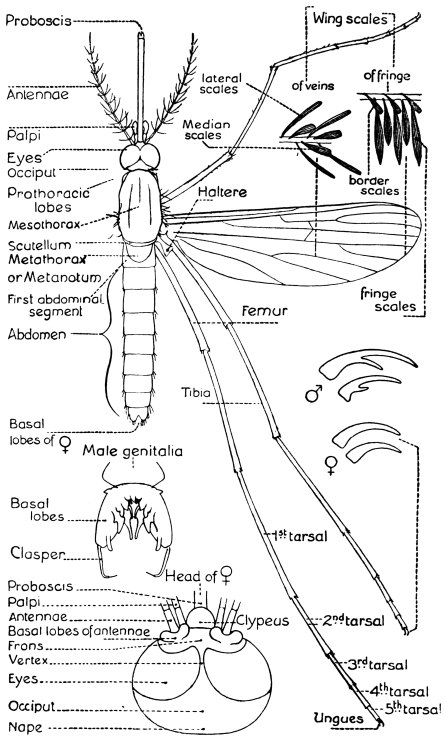
Fig. 396.—Diagram showing the structure of a typical mosquito. (Theobald.)
The eggs, which may be laid separately (Anopheles maculipennis, Stegomyia
fasciata, Joblotia nivipes, etc.), or in rafts (Culex pipiens, C. fatigans) or in chains
(Pseudotæniorhynchus fasciolatus), present a great variety of forms. The most
peculiar are shown in fig. 395 (Tæniorhynchus, Culex, Stegomyia, Anopheles,
Psorophora).
As in all insects, they differ very materially in each species of one genus. Those
best known are the Anopheline eggs.
The eggs always float on the surface of the water; immersion soon destroys
them, but many may occur in mud and can resist desiccation.
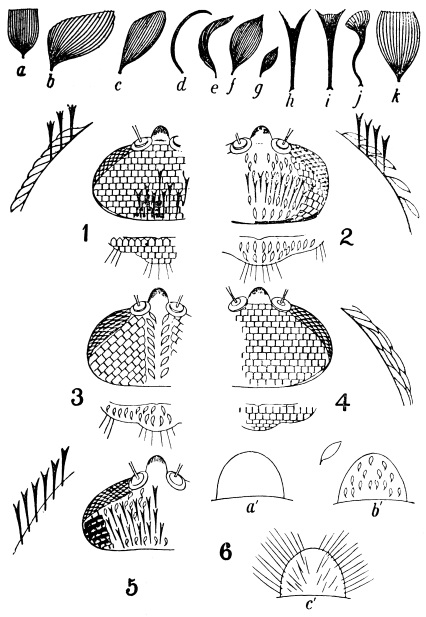
Fig. 397.—Types of scales, a to k; head and scutellar ornamentation, 1 to 5; forms of
clypeus, 6. (Theobald.) 1, head and scutellum of Stegomyia, etc.; 2, of Culex and
Mansonia; 3, of Howardina, Ædes, etc.; 4, of Megarhinus and Toxorhynchites, etc.;
5, of Cellia and some other Anophelines; 6, a′, clypeus of Culex; b′ of Stegomyia;
c′, of Joblotia.
Characters of Adult Culicidæ.—The chief characters by which true mosquitoes,
or Culicidæ, are known are the following:—
(1) Wings always with the veins covered with scales; the longitudinal veins,
usually six in number (in one genus seven); the costal vein carried round the border
of the wing.
(2) Head, thorax and abdomen usually, but not always (Anopheles, etc.), covered
with scales.
(3) Mouth parts formed into a long piercing proboscis.
As a rule the males may be told from the females by their antennæ being
plumose, whilst in the females they are pilose (vide fig. 394), but this does not
invariably hold good, for in Deinocerites, Theobald, and Sabethes, Desvoidy, and
others, they are pilose in both sexes. The labial palpi are very variable in regard
to their form and the number of segments; in the Anophelina they are long in both
sexes, as long or nearly so as the proboscis, more or less clubbed in the males; in
Culicina, Joblotina and Heptaphlebomyia, they are long in the males, short in the
females; in Ædeomyina, short in both sexes.
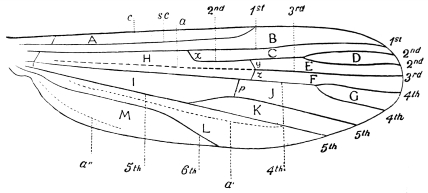
Fig. 398.—Neuration of Wing. Explanation of Wing, Veins and Cells.—A, costal cell;
B, sub-costal cell; C, marginal cell; D, first sub-marginal cell (= first fork cell); E, second
sub-marginal cell; F, first posterior cell; G, second posterior cell (= second fork cell); H, first
basal cell; I, second basal cell; J, third posterior cell; K, anal cell; L, auxiliary cell;
M, spurious cell; c, costal vein; 1st—6th, first to sixth longitudinal veins; a, a′ and a′′, incrassations
(a′ called by Austen the sixth vein, a′′ the eighth vein); y, supernumerary cross vein;
z, mid cross vein; p, posterior cross vein; s.c., sub-costal. (Theobald.)
Scales.—The most important structural peculiarities in Culicidæ are the scales,
which form the chief and most readily observed characters for separating genera
and species. The importance of scale structure has been recently ignored by some
workers, who are probably right academically, but as a means of separating groups,
and so more easily running down a species, the practical man is strongly advised to
follow this method. As to what a genus is, is purely a matter of personal opinion.
If one examines any recent standard work on entomology one will find a species
being placed in varied genera by the varied authorities.
The head, thorax, abdomen and wings are in nearly all cases clothed with
squamæ of varied form, of which the following are the main types (fig. 397):—
(1) Flat, spade-shaped scales (a).
(2) Narrow curved scales (e).
(3) Hair-like curved scales (d).
(4) Spindle-shaped scales (f).
(5) Small spindle-shaped scales (g).
(6) Upright forked scales (h) and (i).
(7) Twisted upright scales (j).
(8) Inflated or pyriform scales (k).
(9) Mansonia scales (b).
(10) Small broad asymmetrical scales (c).
Various other varieties are found on the wings, such as:—
(1) Narrow linear lateral scales.
(2) Narrow lanceolate scales.
(3) Broad lanceolate scales.
(4) Elongated, broad, truncated scales (= Pseudotæniorhynchus-like scales).
(5) Pyriform scales.
(6) Asymmetrical broad or Tæniorhynchus scales.
(7) Flat spade-like scales.389
The wings have a series of scales along the middle line of the veins, and also
lateral scales to all or nearly all the veins. The wing is also fringed by a series of
scales (fig. 396), which, however, are of little systematic importance; the so-called
“border scales” (b.s.) vary, however, to some extent, and are useful characters in
separating some of the Tæniorhynchus.
The Classification of Culicidæ.
Section A.—Proboscis formed for piercing; metanotum nude.
Scutellum simple.
I. | Wings with six-scaled longitudinal veins. |
A. | Palpi long in the male. |
α. | Palpi long in both sexes, clavate in ♂ Anophelina. |
I. | First submarginal cell as long or longer than the second posterior cell. |
| Antennal segments without dense lateral scale tufts. |
| Thorax and abdomen with hair-like scales |  | Prothoracic lobes simple; no flat head scales |  | Wing scales lanceolate | Anopheles, Meigen. |
| Wing scales mostly long and narrow | Myzomyia, Blanchard. |
| Wing scales as above, but fourth long vein near base of third and outstanding scales on prothoracic lobe | Neomyzomyia, Theobald. |
| Wing scales partly large and inflated | Cycloleppteron, Theobald. |
| Prothoracic lobes mammillated; some flat head scales. Basal lobe of ♂ genitalia of two segments | Stethomyia, Theobald. |
| Prothoracic lobes with dense outstanding scales | Feltinella, Theobald. |
| Thorax with some narrow curved scales; abdomen hairy | Pyretophorus, Blanchard. |
| Wing scales small and lanceolate. Wing scales broad and lanceolate | Myzorhynchella, Theobald. |
| Thorax with hair-like curved scales, some narrow curved ones in front; abdomen with apical lateral scale tufts, scaly venter; no ventral tuft | Arribalzagia, Theobald. |
| Thorax with hair-like curved scales; abdominal scales on venter only, with a distinct ventral apical tuft | Myzorhynchus, Blanchard. |
| Much as above, but abdomen with long spine-like dense lateral tufts | Chrystia, Theobald. |
| Thorax with very long hair-like curved scales; abdomen pilose, except last two segments which are scaly; dense scale tufts on third femora; wings with broadish, blunt, lanceolate scales | Lophoscelomyia, Theobald. |
| Thorax and abdomen with scales |  | Abdominal scales as lateral dorsal patches of small flat scales; thoracic scales narrow and curved, or spindle-shaped | Nyssorhynchus, Blanchard. |
| Abdomen nearly completely covered with irregular scales and with lateral tufts | Cellia, Theobald. |
| No lateral scale tufts | Neocellia, Theobald. |
| Thoracic scales hair-like except a few narrow curved ones in front; abdominal scales long, broad and irregular | Kerteszia, Theobald. |
| Thorax with hair-like curved scales and some broad straight scales, others spatulate on sides. Abdomen covered with fine hairs except last three segments, which are scaly. Tufts of scales on hind femora. Wing scales lanceolate | Manguinhosia, Cruz. |
| Antennal segments with many dense scaly tufts | Chagasia, Cruz. |
| Antennæ with outstanding scales on second segment, more appressed ones on the first. At least one segment of abdomen with long flat more or less spatulate scales | Calvertina, Ludlow. |
II. | First submarginal cell very small | Bironella, Theobald. |
| With a distinct cylindrical tubercle projecting obliquely from the prothoracic region | Dactylomyia, Newstead and Carter.390 |
| Scutellum trilobed. |
| First submarginal cell much smaller than the second posterior cell; proboscis long and bent | Megarhininæ. |
| Palpi long in both sexes | Megarhinus, Rob. Desvoidy. |
| Last segment of ♂ palpi blunt. Last segment of ♂ palpi long and pointed | Ankylorhynchus, Lutz. |
β. | Palpi short in the female | Toxorhynchites, Theobald. |
| First submarginal cell longer than the second posterior cell | Culicinæ. |
| Legs more or less densely scaly; head not entirely clothed with flat scales; all the legs densely scaly. |
| Wings with large pyriform scales | Mucidus, Theobald. |
| Wings with narrow scales | Psorophora, Rob. Desvoidy. |
| Hind legs only densely scaled | Janthinosoma, Arribalzaga. |
| Head entirely clothed with flat scales. Legs uniformly scaled with flat scales. Head and scutellar scales all flat and broad. Palpi of ♀ short, of ♂ thickened apically and tufted | Stegomyia, Theobald. |
| Palpi of ♀ longer than in Stegomyia and in ♂ long and thin, acuminate, simple | Desvoidea, Blanchard. |
| Head scales mostly flat, but a median line of narrow curved ones; scutellar scales flat on mid lobe, narrow curved on lateral lobes and palpi longer than proboscis | Macleayia, Theobald. |
| Head scales mostly flat, irregular, narrow curved ones behind; mid lobe scutellum with flat scales, lateral with narrow curved; ♂ palpi shorter than proboscis | Catageiomyia, Theobald. |
| Head scales mostly flat, but a few narrow curved ones in middle in front; scutellar scales all flat | Scutomyia, Theobald. |
| Head scales all flat; scutellar scales all narrow curved | Skusea, Theobald. |
| Head with flat scales, except a small median area of narrow curved ones; scutellar scales all narrow curved | Howardina, Theobald. |
| Head with all flat scales except a thin line of narrow curved ones behind; scutellar scales all narrow curved | Danielsia, Theobald. |
| Head with small flat scales over most of surface, with median line and line around eyes of narrow curved ones; scutellar scales bluntly spindle or club-shaped | Hulecoetomyia, Theobald. |
| Head and scutellar scales narrow curved. Wing scales long, narrowly lanceolate, collected in spots; palpi clubbed in ♂; five-jointed and rather long in ♀ | Theobaldia, Neveu-Lemaire. |
| Wing scales (lateral) long and narrow, and ♀ palpi three-jointed, ♂ not clubbed and hairy | Culex, Linnæus. |
| Wing scales at apex of veins dense and rather broad, femora swollen; small dark species | Melanoconion, Theobald. |
| Wings with short, thick, median scales and short, broadish lateral ones on some of the veins; scales mottled; fork-cells rather short | Grabhamia, Theobald. |
| Wings with dense, broadish, elongated, truncated scales | Pseudotæniorhynchus, Theobald. |
| Wings with broad, short, asymmetrical scales | Tæniorhynchus, Arribalzaga. |
| Head covered with rather broad, flat, spindle-shaped scales; scutellum with small flat scales to mid lobe | Gilesia, Theobald. |
| Head clothed with flat, irregularly disposed scales all over, with patches of narrow curved ones; ♂ palpi clubbed | Acartomyia, Theobald. |
| Abdomen with projecting flat lateral scales with deeply dentate apices; wings not ornamented | Lasioconops, Theobald. |
| Wings ornamented; scutellum with flat and narrow curved scales | Finlaya, Theobald. |
γ. | Palpi short in ♂ and ♀ | Ædeomyina. |
| Wings unornamented. |
| Antennæ pilose in ♂ and ♀; second joint very long | Deinocerites, Theobald. |
| Antennæ plumose in the ♂. |
| Head clothed with narrow curved and flat scales. |
| Mid-lobe of scutellum with six border-bristles. |
| Scutellum with narrow curved scales. |
| Palpi in ♀ four-jointed, in ♂ two-jointed | Ædes, Meigen. |
| Mid-lobe of scutellum with four border bristles. |
| Scutellum with flat scales. |
| Head clothed with flat scales only. |
| Fork-cells normal length. |
| Mid-lobe of scutellum with four border-bristles. |
| Palpi of ♀ two-jointed | Verallina, Theobald. |
| Palpi of ♀ five-jointed, metallic | Hæmagogus, Williston. |
| Fork cells very small or small. |
| Scutellar scales flat. |
| First submarginal cell longer than the second posterior cell; no flat scales on mesothorax | Ficalbia, Theobald. |
| First submarginal cell smaller than the second posterior cell; flat scales on mesothorax | Uranotænia, Arribalzaga. |
| Scutellar scales narrow curved. |
| First submarginal cell as in Uranotænia | Mimomyia, Theobald. |
| Wings ornamented with Mansonia-like scales | Ædeomyia, Theobald. |
Section B.—Metanotum ornamented with chætæ, squamæ or both.
α. | With chætæ only. |
| Proboscis longer than whole body; lateral wing scales Tæniorhynchus-like | Phoniomyia, Theobald. |
| Proboscis as long as whole body in ♀ frons drawn out into a prominence; wing scales rather broad and long | Binotia, Blanchard =
Runchiomyia, Theobald. |
| Proboscis not as long as the whole body; lateral vein scales narrow | Wyeomyia, Theobald. |
| Proboscis not as long as whole body, swollen apically; wing scales long and broad | Dendriomyia, Theobald. |
β. | Metanotum with squamæ and chætæ. |
| Palpi short in ♂ and ♀. |
| Proboscis straight in ♀ and ♂; legs with scaly paddles | Sabethes, Rob. Desvoidy. |
| Venation like Sabethes. |
| Legs simple | Sabethoides, Theobald. |
| Venation like Culex | Goeldia, Theobald. |
| Proboscis in ♂ elbowed, with two scaly tufts | Limatus, Theobald. |
| Palpi long in ♂, short in ♀ | Joblotina, Blanchard. |
II. | Wings with seven-scaled longitudinal veins: Culex type | Heptaphlebomyia, Theobald. |
Section C.—Proboscis short, not formed for
| piercing | Corethrina. |
| Metatarsus longer than first tarsal joint | Corethra, Linnæus. |
| Metatarsus shorter than first tarsal joint | Mochlonyx, Ruthe.391 |
Notes on the Different Genera.
Sub-family. Anophelina.
The following Anophelines have been recorded as malaria
carriers:—
*Anopheles maculipennis, Meigen.
Anopheles bifurcatus, Linnæus.
*Myzomyia funesta, Giles.
Myzomyia lutzii, Theobald.
*Myzomyia rossii, Giles.
Myzomyia listonii, Liston.
Myzomyia culicifacies, Giles.
Pyretophorus superpictus, Grassi.
*Pyretophorus costalis, Loew.
Pyretophorus chaudoyei, Theobald.
*Cellia argyrotarsis, Robineau Desvoidy.
Myzorhynchus pseudopictus, Grassi.
Myzorhynchus barbirostris, Van der Wulp.
Myzorhynchus sinensis, Wiedemann.
Myzorhynchus paludis, Theobald.
Myzorhynchus mauritianus, Grandpré.
Neocellia stephensii, Liston.
Neocellia willmori, James.
Nyssorhynchus theobaldii, Giles.
Nyssorhynchus fuliginosus, Giles.
Nyssorhynchus annulipes, Walker.
Those marked with an asterisk (*) also carry the larvæ of Filaria bancrofti, as
also do Myzorhynchus minutus, Theobald, and Myzorhynchus nigerrimus, Giles.
Genus. Anopheles, Meigen.
“Syst. Beschr. Europ. zwei. Ins. I,” 1818, ii, p. 2, Meigen; “Mono. Culicid.,”
1903, i, p. 191; iii, p. 17; and 1910, v, p. 3, Theobald.
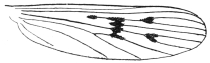
Fig. 399.—Wing of Anopheles maculipennis, Meigen.
This genus contains a few large species found either in temperate climates or
in hills and mountains of warm climates. The type is the European and North
American A. maculipennis.
A. maculipennis, Meigen. This species and A. bifurcatus are malaria carriers.
True Anopheles only occur in Europe, North America, the North of Africa and in
the mountains of India, and one has been found by Bancroft similar to A. bifurcatus
in Queensland. They are easily told by the absence of scales on thorax
and abdomen, and by the rather densely scaled wings with lanceolate scales.
Genus. Myzomyia, Blanchard; Grassia, Theobald.
Comp. rend. heb. Soc. Biol., No. 23, p. 795, Blanchard; “Mono. Culicid.,”
1910, iii, p. 24; v, p. 16, Theobald.
This genus occurs in Asia, Africa and South America, Europe and East Indies.
The type is M. funesta, Giles, found in Central and West Africa. Although structurally
there is not much difference between this genus and Anopheles, they differ
greatly in appearance, and there are usually a few narrow curved thoracic scales
projecting over the head, whilst the wing scales are much smaller in proportion, and
the wings more uniformly spotted, always so along the costa. Funesta and lutzii
are undoubtedly malaria bearers and also rossii.
Genus. Neomyzomyia, Theobald.
“Mono. Culicid.,” 1910, v, p. 29.
A single species only occurs in this genus, N. elegans, James, from India. In
this genus, which is near to Myzomyia, the fourth long vein is very near the base
of the third, and there are outstanding scales on the prothoracic lobes, and there is
a marked tuft of dense scales at the posterior angles of the head.
Genus. Cycloleppteron, Theobald.
“Mono. Culicid.,” 1903, ii, p. 312; 1903, iii, p. 58; 1910, v, p. 33.
Two common species only occur in this genus, C. grabhamii, Theob., from
Jamaica, and C. mediopunctatus, Theob. (Lutz., ms.), from South America. The
chief character is the presence of large black inflated pyriform scales on the wings.
The palpi are densely scaled. Neither have been shown to be malaria bearers.
Genus. Feltinella, Theobald.
“Mono. Culicid.,” 1907, iv, p. 56.
A single species, so far only found in this genus. The basal lobes of the male
genitalia of two segments, the prothoracic lobes with dense outstanding scales.
The species, F. pallidopalpi, Theob., occurs in Sierra Leone.
Genus. Stethomyia, Theobald.
“Mono. Culicid.,” 1903, iii, p. 13; 1907, iv, p. 59; 1910, v, p. 35.
Four species occur in this marked genus—one S. nimba, Theob., from British
Guiana and Para, another S. fragalis, Theob., from the Malay States, S. culiciformis,
James and Liston, from India, and S. pallida, Ludlow, from India.
The former may be a malaria carrier, for Dr. Low says: “Malarial fever is got
amongst the Indians and often of a severe type. In that connection it is interesting
that in the interior, at a place called Corato, I got an entirely new Anopheles in
large numbers.” The genus is easily told by its unornamented wings, flat head
scales, mammillated prothoracic lobes and long thin legs.
Genus. Pyretophorus, Blanchard; Howardia, Theobald.
Compt. rend. heb. Soc. Biol., No. 23, p. 705, Blanchard; Journ. Trop. Med., v, p. 181;
and “Mono. Culicid.,” 1903, iii, p. 13; 1910, v, p. 36, Theobald.
Forty-four species come in this genus, of which Anopheles costalis, Loew, is the
type.
This genus is found in Africa, India, Europe and in Australia. Three species
are proved malaria bearers, namely, P. costalis, Loew, P. chaudoyei, Theob., and
P. superpictus, Grassi. Members of this genus can be told by having narrow
curved thoracic scales, hairy abdomen, and much-spotted wings.
Genus. Myzorhynchella, Theobald.
“Mono. Culicid.,” 1907, iv, p. 78.
In this genus the thorax has distinct, narrow curved scales, and the abdomen
is hairy, the wing scales broad and lanceolate, and the head with broad scales not
closely appressed, but not forked or fimbriated.
Five species are known: lutzi, Cruz; parva, Chagas; nigritarsis, Chagas;
tibiomaculata, Neiva; gilesi, Neiva; and nigra, Theobald. They are all
recorded from Brazil, and nigra also from Mexico.
Genus. Manguinhosia, Cruz, in Peryassu.
“Os Culicideos do Brazil,” 1908, p. 112.
A single marked species from the Brazils. The thorax has piliform curved
scales, and some narrow curved and flattened ones on the sides. Abdomen pilose,
except the last three segments which are scaled. No tufts of scales on posterior
femora.
Allied to Lophoscelomyia, but at once told by the absence of scale tufts on the
hind femora. M. lutzi, Cruz, Brazil.
Genus. Chrystya, Theobald.
“Rep. Sleeping Sickness, Roy. Soc. Eng.,” 1903, vii, p. 34.
A very marked genus in which the hairy abdomen has very long, dense, hair-like,
apical, scaly tufts to the segments. A single species only so far known, C. implexa,
Theobald, from Africa (Uganda, Sudan, etc.).
Genus. Lophoscelomyia, Theobald.
Entomologist, 1904, xxxvi, p. 12.
A single species only, from the Federated Malay States. The hind femora have
dense, apical scale tufts; the thorax long, hair-like curved scales; abdomen pilose,
except the last two segments which are scaly; wings with broad, blunt, lanceolate
scales.
Genus. Arribalzagia, Theobald.
“Mono. Culicid.,” 1903, iii, pp. 13 and 81; and 1910, v, p. 48.
Two species only occur, found in South America. The thorax and abdomen
have scales and hairs respectively, as in Pyretophorus, but the abdomen has in
addition prominent lateral apical scale tufts to the segments and a scaly venter.
Wings with membrane tinged in patches and wing scales bluntly lanceolate and
very dense. The type is A. maculipes, Theob. found in Trinidad and Brazil;
A. pseudomaculipes, Cruz, also in Brazil.
Genus. Myzorhynchus, Blanchard; Rossia, Theobald.
Compt. rend. heb. Soc. Biol., 1902, No. 23, p. 795, Blanchard; Journ. Trop.
Med., 1902, p. 181, Theobald; “Mono. Culicid.,” 1903, iii, p. 84; 1907, iv, p. 80;
1910, v, p. 49.
A very marked genus of large, dark, densely scaled species, found in Europe,
Asia, Africa and Australia. The thorax with hair-like curved scales; the abdomen
with ventral and apical scales, and a median ventral apical tuft, and with very
densely scaled palpi in the female, and densely scaled proboscis. It seems to be
mainly an Asiatic and East Indian genus, but three species occur in Africa and
one in Australia. They are mostly sylvan species and bite severely.
Fourteen species are known. Five are malaria carriers (vide list, p. 566).
Genus. Nyssorhynchus, Blanchard; Laverania, Theobald.
“Mono. Culicid.,” 1910, iii, p. 14; v, p. 55, Theobald; Compt. rend. heb. Soc. Biol.,
No. 23, p. 795, Blanchard.
A group of small, closely allied species found in Asia, Africa and Australia,
twelve out of the twenty species coming from India.
The thorax is covered with narrow curved and spindle-shaped scales, abdomen
with small, flat or narrow curved dorsal scales, especially on the apical segments or
in patches; the legs are always banded or spotted with white, and the tarsi have as
a rule one or more pure white segments. (This banding and spotting is of no
generic value, however.)
The species show considerable seasonal variation. The type of the genus is
N. maculatus, Theobald.
Three are malaria carriers (vide list, p. 566).
Genus. Cellia, Theobald.
“Mono. Culicid.,” 1903, iii, p. 107; 1910, v, p. 67.
Very marked Anophelines, with densely scaly abdomens, the scales irregularly
disposed on the dorsum and forming dense lateral tufts; thorax with flat spindle-shaped
scales; palpi densely scaled and also the wings.
The type of the genus is the African C. pharoensis, Theob. It is represented in
Asia by C. kochii, Dönitz; in West Indies and South America by C. argyrotarsis,
Desvoidy, and C. bigotii, Theob.; in Africa by C. squamosa, Theob., etc.
C. argyrotarsis, Desvoidy, and C. albimana, Wiedemann, are undoubtedly
malaria bearers.
Genus. Neocellia, Theobald.
“Mono. Culicid.,” 1907, iv, p. 111.
Allied to Cellia, but has no lateral scale tufts. Three species recorded from
India.
Genus. Kertészia, Theobald.
“Ann. Mus. Nat., Hung.,” 1905, iii, p. 66.
This genus has the thoracic scales hair-like, except a few narrow curved ones
in front; abdominal scales long, broad and irregular.
A single species, K. boliviensis, Theob. from Bolivia.
Genus. Manguinhosia, Cruz.
The thorax has narrow hair-like curved scales and some broad straight scales;
others spatulate on the sides. Abdomen with fine hairs, except the last three
segments which are scaly. Tufts of scales on the hind femora. Wing scales
lanceolate.
The type is M. lutzi, Cruz, from Brazil.
Genus. Chagasia, Cruz.
“Brazil-Medico,” 1906, xx, pp. 20, 199.
This genus can at once be told by the antennal segments having many dense
scaly tufts. Type, C. fajardoi, Lutz, from Brazil.
Genus. Calvertina, Ludlow.
Canadian Entomologist, 1909, xli, pp. 22, 234.
The antennæ in this genus have outstanding scales on the second segment, more
appressed ones on the first. At least one abdominal segment with long, flat, more
or less spatulate scales. Type, C. lineata, Ludlow, from Philippine Islands.
Genus. Birónella, Theobald.
“Ann. Mus. Nat. Hung.,” 1905, iii, p. 69.
At once told by the first submarginal cell being very small. Type, B. gracilis,
Theob. from New Guinea.
Sub-family. Megarhininæ.
Three genera occur in this marked sub-family; they are the largest of all mosquitoes,
and are very brilliantly coloured, and many have tail fans. They occur
in North and South America, Asia, Africa, and Australia. The long curved
proboscis is very marked. They are usually spoken of as elephant mosquitoes;
some are vicious blood-suckers at times.
The three genera tabulate as follows:—
| α. | Palpi long in both sexes. |
| β. | Last segment of ♀ palp round or blunt as if broken | Genus Megarhinus, R. Desvoidy. |
| ββ. | Last segment of ♀ palp long and pointed | Genus Ankylorhynchus, Lutz. |
| αα. | Palpi of female short of male long.
Palpi of female not more than one-third length of proboscis | Genus Toxorhynchites, Theobald. |
Genus. Megarhinus, Robineau Desvoidy.
“Mém. Soc. d’Hist. nat. de Paris,” 1827, iii, p. 412; “Mono. Culicid.,”
1901, i, p. 215; 1903, iv, p. 163; 1907, iv, p. 128; 1910, v, p. 89.
All large brilliant mosquitoes with long palpi in both sexes and, as a rule, with
a caudal fan of scales; the proboscis is long and bent. They are all sylvan species,
and are not so far recorded as biting man.
Genus. Toxorhynchites, Theobald.
“Mono. Culicid.,” 1901, i, p. 244; 1903, iii, p. 119; 1907, iv, p. 140; 1910, v, p. 95.
Differs from the former genus in that the female palpi are short. The palpi may
have one, two or three minute terminal segments. Banks’s genus Worcesteria has
three.
The elephant mosquito of India (T. immisericors), Walker, bites very severely.
They are sylvan species.
Sub-family. Culicinæ.
Genus. Mucidus, Theobald.
“Mono. Culicid.,” 1901, i, p. 268; 1910, v, p. 125.
This genus is so far confined to Australia, West and Central Africa, India, East
Indies and Malay Peninsula. They are all large mosquitoes, easily told by the
whole body being more or less covered with long twisted scales, giving them a
mouldy appearance, and the legs densely scaled with outstanding scales; the wings
with large parti-coloured scales. The Australian M. alternans, Walker, occurs in
larval form both in fresh and salt water. The adults bite man.
Genus. Psorophora, Robineau Desvoidy.
“Mém. de la Soc. d’Hist. nat. de Paris,” 1827, iii, p. 412, R. Desvoidy; “Mono.
Culicid.,” 1901, i, p. 259; 1903, iii, p. 130; 1907, iv, p. 158; 1910, v, p. 123, Theobald.
This genus is confined to the Americas and the West Indies. Several species
exist which can easily be told from Mucidus by the absence of long twisted scales
and the narrower wing scales. The legs are densely scaled and the thorax ornamented
with flat spindle-shaped scales.
P. ciliata, Robineau Desvoidy, occurs in both North and South America, and
bites man.
Genus. Janthinosoma, Arribalzaga.
“Dipt. Arg.,” 1891, p. 52, Arribalzaga; “Mono. Culicid.,” 1901, i, p. 253;
1903, iii, p. 124; 1907, iv, p. 152; and 1910, v, p. 118, Theobald.
Hind legs only densely scaled; some of the hind tarsi are always white. The
venation is as in Culex. The abdomen is metallic and iridescent. They all bite
man and occur only in the Americas and West Indies.
Genus. Stegomyia, Theobald.
“Mono. Culicid.,” 1901, i, p. 283; 1903, iii, p. 130; 1907, iv, p. 170; 1910, v, p. 151.
This, the most important genus in the Culicinæ, can be told by the head and
scutellum being clothed with flat scales and the thorax with narrow curved ones.
About forty species are known in this genus, occurring in Southern Europe, Asia,
Africa, Australia, the Americas, East and West Indies, and on most oceanic islands.
Many of them seem to be vicious blood-suckers. They are mostly black and white
mosquitoes, and several seem to go by the name of tiger mosquitoes. The genus
contains the yellow fever mosquito (S. fasciata, Fabricius), the only one that need
be dealt with in detail here. The chief known species tabulate as follows:—
A. | Proboscis banded. |
α. | Legs basally banded. |
| Thorax brown, with scattered creamy-white scales | annulirostris, Theobald. |
| Thorax black, with narrow, curved golden scales | periskelta, Giles. |
αα. | Legs with basal and apical banding. Fore legs with no bands; mid with apical and basal bands on first and second tarsals, hind with basal bands. |
| Thorax white in front, with a brown eye-like spot on each side | thomsoni, Theobald. |
AA. | Proboscis unbanded. |
β. | Legs basally banded. |
γ. | Abdomen basally banded. |
| Thorax with one median silvery-white line | scutellaris, Walker. |
| Thorax as above, but pleuræ with white lines | pseudoscutellaris, Theobald. |
| Thorax similar, but two white spots near where line ends | gelebinensis, Theobald. |
| Thorax with two median yellow lines and lateral curved silvery lines | fasciata, Fabr. |
| Thorax with two short median lines and a white patch on each side | nigeria, Theobald. |
| Thorax with large lateral white spots in front, smaller ones by wings, two narrow median lines and two posterior sub-median white lines | lilii, Theobald. |
| Thorax with a white W-shaped area in front, a prolongation curved on each side enclosing a brown eye-like spot | W-alba, Theobald. |
| Thorax with white frontal median spot, two large lateral spots, a small one in front of the wings, a narrow median white line and narrow sub-median ones on posterior half. Last two hind tarsi white | wellmannii, Theobald. |
| Thorax brown, with broad white line in front extending laterally towards wings, where they swell into a large patch, a white line on each side just past wing roots. Last two hind tarsi white | albipes, Theobald. |
| Thorax with silvery white spot on each side in front, small one over roots of wings and white over their base. Last two hind tarsi white | pseudonigeria, Theobald. |
| Thorax with two lateral white spots, front ones the largest, a small median one near head, two yellow median lines, a short silvery one on each side before the scutellum | simpsoni, Theobald. |
| Thorax with silvery-white scaled area in front and another on each side in front of wings | argenteomaculata, Theobald. |
| Thorax with median yellowish-white line, a silvery patch on each side in front of wings extending as a fine yellow line to the scutellum, and another silvery spot before base of each wing | poweri, Theobald. |
| Thorax with small grey-scaled area in front of wing roots and three short creamy lines behind | minutissima, Theobald. |
| Thorax (?) denuded; abdomen black; fifth segment with yellow basal band; sixth unbanded; seventh, two median lateral white spots; eighth, two basal lateral white spots; second hind tarsal nearly all white | dubia, Theobald. |
γγ. | Abdomen unbanded. |
| First hind tarsal all white, second basally white, last two dark. Thorax chestnut brown, with a broad patch of white scales on each side in front and a median pale line | terreus, Walker. |
ββ. | Legs with white lines as well as basal bands. |
| Thorax brown, with white lines; abdomen with basal bands | grantii, Theobald. |
βββ. | Fore and mid legs with apical bands, hind basal. |
| Fourth tarsal of hind legs nearly all white | mediopunctata, Theobald. |
| Mid metatarsi with basal pale banding, base and apex of hind, also base of first tarsal pale | assamensis, Theobald. |
ββββ. | Legs unbanded. |
δ. | Abdomen basally banded. |
| Thorax with front half white, rest bronzy-brown | pseudonivea, Ludlow. |
| Thorax deep brown, with scattered golden scales, showing two dark eye-like spots; head white, dark on each side and behind | albocephala, Theobald. |
| Thorax brown with golden stripes; abdomen with narrow basal bands on fifth and sixth segments only | auriostriata, Banks. |
δδ. | Abdominal banding indistinct. |
| Thorax with broad silvery white patch on each side in front | albolateralis, Theobald. |
δδδ. | Abdomen unbanded. |
| Thorax with six silvery spots | argenteopunctata, Theobald. |
δδδδ. | Abdomen with apical white lateral spots. |
| Thorax unadorned, except for pale scaled lines laterally | punctolateralis, Theobald. |
δδδδδ. | Abdomen with basal white lateral spots. |
| Thorax with two pale median parallel lines and two silvery lateral spots | ininuta, Theobald. |
| Thorax unadorned. |
| A white spot middle of head | tripunctata, Theobald. |
| No white spot | amesii, Ludlow. |
AAA. | Proboscis yellow basally, dark apically. |
| Abdomen with apical pale bands | crassipes, Van der Wulp. |
AAAA. | Proboscis with median interrupted white line on basal half. |
| Head black, anterior margin grey | albomarginata, Newstead. |
Stegomyia fasciata, Fabricius (Yellow Fever Mosquito).
This insect, which is the proven carrier of yellow fever, is commonly called the
tiger, brindled, spotted day or striped mosquito. It is also referred to by some
writers as S. calopus, Meigen. It is subject to considerable variation in colour,
but the thoracic markings are generally very constant. The general colour is
almost black to deep brown, the head with a median white area, white at the sides
and in front around the eyes; the thorax has two median parallel yellow lines,
a broad curved silvery one on each side and white spots at the sides; the scales on
the intervening spaces of the thorax are brown. The dark abdomen has basal white
bands and basal white lateral spots. The dark legs have basal white bands, the
last segment of the hind legs being all white except in a variety from South America
and the West Indies (luciensis), which has the tip of the last hind tarsal dark. The
abdomen may also vary in colour, some having pale scales over most of the surface
(queenslandensis).
The food of the adult female consists mainly of man’s blood, but she will also
feed on dogs and other animals. The male has been said to bite, but such is very
unusual. This mosquito bites mainly in the daytime up till about 5 p.m.
The adults breed the first day after emergence. They may live a considerable
time, Bancroft having kept females for two months in confinement. The
ova are laid separately, often in chains; they are black, oval, with a reticulated
membrane outside, some of the reticulated cells containing air. They may hatch
in from six to twenty hours, the larval stage nine days, the pupal stage three; thus
the whole cycle may be completed in from twelve to thirteen days. The ova when
dry can remain undeveloped for a considerable time. The larvæ are greyish-white,
with short, thick siphon, and feed at the bottom of the water, only coming to
the surface now and again to breathe. This is almost entirely a domesticated
gnat, seldom being found far from man’s habitations. Its larvæ occur in such
small collections of water as old sardine tins, jam-pots, calabashes, puddles, barrels,
wells—in fact, wherever water is held up, even to the gutters of houses. Not
only are they found breeding on land, but also on board ship, although they
prefer artificial collections of water. They may also breed in larger natural
collections.
This insect is easily transported by steam and sailing ships and by train, and
this doubtless explains its very wide distribution. The adults may live for fifty
days, and it is on this account and their frequent occurrence on ships that danger
lies in regard to the Panama Canal. An infected insect may leave that endemic
centre of yellow fever and live until the vessel arrived at the Philippine Islands and
fly ashore, and so introduce the disease for the native fasciata possibly to spread.
Roughly the distribution of this pest is as follows: Africa from South to North,
but especially along the coast and up the Nile. In Asia, in India, Ceylon, Burma,
Siam, along the ports of the Malay Peninsula, in French Cochin China, Philippine
Islands, the Andaman and Nicobar Islands, Japan, Malay Archipelago, and East
Indies, Turkey in Asia, Arabia and Palestine.
In Australia it occurs in Queensland, New South Wales, Victoria and South
Australia.
In Europe in Italy, Spain, Portugal, Greece, in the Mediterranean Islands.
In South America, Central America, Mexico, North America, and the West
Indies it is very abundant, and it also is found in the Bahama Islands, Fiji, Sandwich
Islands, Samoa, the Azores, Teneriffe and Santa Cruz, Pitcairn Islands and Bermuda.
For a full account of its distribution the reader is referred to the following:
“The Distribution of the Yellow Fever Mosquito (Stegomyia fasciata, Fabricius)
and General Notes on its Bionomics;” “Mém. 1er Congrès international
d’Entomologie, 1911, ii, pp. 145–170, F. V. Theobald.” In addition to being the
yellow fever carrier, it is supposed by Wenyon to be the intermediate host of the
parasite of Bagdad sore.
Stegomyia scutellaris, Walker.
A vicious biter, found in India, China, Malay, East Indies, and Ceylon. The
thorax has one median silvery stripe, and so can easily be told from S. fasciata.
A very similar species occurs in Fiji, but can be told by the pleuræ having white
lines, not spots (S. pseudoscutellaris, Theobald). It is the intermediate host of
filaria in Fiji (Bahr).
A number of nearly allied genera occur here (vide synoptic table).
Genus. Theobaldia, Neveu-Lemaire.
Theobaldinella, Blanchard.
Includes several large Culicines, of which T. annulata, Meigen, is the type. The
wings are usually spotted (annulata, incidens, etc.), but may be nearly plain (spathipalpis).
The males have the palpi swollen apically, and the females have long
five-jointed palps.
Several of these are vicious biters.
Theobaldia annulata, Meigen.
This large gnat (6 mm. long) can be told by its wings having five large spots of
dark scales and by its legs having broad basal white bands to the tarsi. The larvæ
occur in rain barrels and small pools. It is essentially a domestic form, occurring in
houses and privies. Its distribution is Europe generally and North America. The
bite is very severe, and in some districts gives rise to painful œdema.392
Theobaldia spathipalpis, Rondani, occurs in Italy, Mediterranean Islands,
Palestine, the Himalayas, Khartoum, and in South Africa. It is about the same size
as T. annulata, but is yellowish-brown in colour, with striped thorax and mottled and
banded legs. It occurs in privies and bites very severely.
Genus. Culex, Linnæus.
“Syst. Nat. Ed.,” 1758, x, Linnæus; “Mono. Culicid.,” 1901, i, p. 326;
1910, v, p. 322, Theobald.
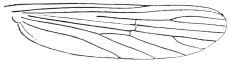
Fig. 400.—Wing of a Culex.
This large genus still contains many forms which should be excluded. The
species normally have narrow curved median head-scales, and similar ones on the
scutellum; the female palpi are shorter than in the former genus and the male
palpi are pointed; the lateral vein-scales are narrow and linear.
The type is Culex pipiens, Linn., the common gnat of Europe. The thorax
is covered with narrow curved golden-brown scales, the abdomen has basal pale
bands to the segments and the legs and proboscis are unbanded. The stem of the
first submarginal cell is always less than one-fifth the length of the cell. It lays its
eggs in rafts in water-butts, etc., and even in the foulest water. They are first
deposited in England in June and July, and again soon after hatching in August. In
some districts this gnat bites man viciously, in others not at all.
The common tropical gnat (Culex fatigans, Wied). This resembles the
European Culex pipiens, but can always be told by the stem of the first submarginal
cell always being much longer than it is in C. pipiens. This is one of the species
that has been proved to transmit filariæ to man, etc. Varieties of it occur in almost
every country between 40° N. and S., having a very similar range to S. fasciata. In
all countries it appears to be connected with the transmission of Filaria bancrofti,
and it is also said to carry the micrococcus of dengue fever.
Genus. Melanoconion, Theobald.
“Mono. Culicid.,” 1903, iii, p. 238; 1907, iv, p. 507; 1910, v, p. 455.
This genus is composed of eight species, most of which are small black gnats
which bite viciously and which occur in swamps and jungles. They can at once
be told from Culex by the veins of the wings having dense broadened scales on their
apical areas and along the upper costal border. The femora and apices of the tibiæ
are swollen.
The black mosquito, Melanoconion atratus, Theob. This small gnat is a very
troublesome pest in swamps in the West Indies. The female bites both by day and
by night, and the bite causes severe irritation. The larvæ live in permanent ponds.
It is almost black in colour, but sometimes presents a dull coppery sheen; each
segment has small lateral basal white spots. Length 2·5 to 3 mm.
It occurs in Para and British Guiana as well as in the West Indies.
Ordinary mosquito netting is no use for keeping off this pest.
Genus. Grabhamia, Theobald.
“Mono. Culicid.,” 1903, iii, p. 243; 1907, iv, p. 284; and 1910, v, p. 277.
Allied to Culex, but separated by the wings having short fork-cells, mottled scales,
the median ones thick and also some of the lateral ones short and broad; the last
two joints of the male palps are very slightly swollen. The eggs are laid singly, not
in rafts, and the larvæ have short, thick siphons. Ten species occur and are found
in Europe, North America, West Indies and Natal. G. dorsalis, Meigen, bites
severely in Europe. G. sollicitans, Walker, is a great scourge along the New Jersey
Coast and at Virginia summer resorts and in Florida. It breeds in brackish water
and is the most common mosquito of the Atlantic seaboard.
Genus. Pseudotæniorhynchus, Theobald;
Tæniorhynchus, Theobald, non-Arribalzaga.
Differs from the former in having the whole wing veins clothed with dense,
broadish elongated scales. They occur in South America (T. fasciolatus, Arri.), in
Africa (T. tenax, Theob.), in Europe (T. richardii, Ficalbi). The latter bites very
severely.
Genus. Tæniorhynchus, Arribalzaga; Mansonia, Blanchard;
Panoplites, Theobald.
Compt. rend. heb. Soc. Biol., 1901, iii, 37, p. 1046; “Mono. Culicid.,”
1901, ii, p. 173; and 1910, v, p. 446, Theobald.
A very marked genus, easily told by the broad asymmetrical wing scales. It
occurs in Africa (T. africana and T. major, Theob.); in Asia (T. uniformis, Theob.;
T. annulipes, Walker, etc.) and in Australia (T. australiensis); in the Americas and
West Indies (T. titillans, Walker). The eggs (fig. 395, d) are peculiar in form and
are laid separately; the larva has not been described; the pupa has long curved
siphons. They mostly occur along rivers, in swamps and forests, and bite very
severely. They also enter houses (T. titillans). T. uniformis is most troublesome
during the rains. The saliva is strongly acid. Both these species carry the larvæ of
Filaria bancrofti.
Genus. Chrysoconops, Goeldi.
“Os Mosq. no Para,” 1905, p. 114, Goeldi; “Mono. Culicid.,” 1910,
v, p. 433, Theobald.
Bright yellow or yellow and purple mosquitoes, with rather dense wing scales.
Numerous species occur in Africa (aurites, annettii, fuscopennatus, etc.), others in
India, Australia and South America.
Low found filariæ in the thoracic muscles of fuscopennatus in Uganda.
Several of the Ædeomyina bite, especially the small Uranotænias. They are all
sylvan species, seldom entering houses. They need not, therefore, be referred to here.
For full details of the Culicid genera and species the reader is referred to my
monograph393 and other works mentioned below.
Other Nematocera.
Other nematocerous flies are midges, daddy-long-legs and sand-flies. The ones
which cause annoyance to man besides Culicidæ are the following:—
Sand-flies (Simulidæ), certain midges (Chironomidæ), and a few owl midges
(Psychodidæ).
The Nematocera have long thread-like jointed antennæ and their pupæ are, as a
rule, naked; the larvæ have a distinct head and can thus be told from the next
section (Brachycera).
Family. Simulidæ.
This family consists of a single genus, Simulium, Latreille, which Roubaud has
recently divided into two sub-genera called Pro-Simulium and Eu-Simulium. These
insects, which are frequently spoken of as sand-flies, are found in all parts of the
world; they are all small insects varying from 1·5 to 3 mm. The females are very
bloodthirsty, but the males appear to be incapable of sucking blood.
The head sunk under the humped thorax; antennæ short, straight; palpi short
and broad, of four segments, bent; wings broad and in some iridescent, legs stout.
The male has holoptic eyes, whilst in the female they are small and widely separate.
The sucking proboscis is short. The thorax and abdomen are clothed with short
hairs which may form spots and markings; these are golden, silvery, grey, or brownish.
In the sub-genus Pro-Simulium the second segment of the hind tarsi in both
sexes is elongate, linear, and without a basal notch; in Eu-Simulium it is short,
curved, and dorsally notched at the base.
Simulidæ often occur in swarms, and attack not only man but cattle, horses,
and poultry. In some districts they are more annoying than mosquitoes.
Their life-cycle has been most completely worked out by King, in Africa.
The larvæ and pupæ occur in swiftly flowing water, by waterfalls, in rapids, etc.
The ova are laid in gelatinous masses on plants or rocks close to or overhanging the
water. The larva is cylindrical, enlarged posteriorly, where it is provided with a
sucker, by means of which it attaches itself to a rock, water weeds, debris, etc.;
anteriorly it has a proleg close behind the head on the lower surface. The head is
dark and chitinous. The respiration takes place by means of branched tracheal gills
which protrude from the dorsal surface of the last body segment; they are retractile.
The colour varies from deep green to yellow or almost black. Their food consists of
algæ and other organisms in the water brought to their mouth by two fan-like organs
placed on the head. The larvæ can crawl from place to place by means of the
thoracic proleg; they occur in masses, usually in a more or less erect attitude. A network
of threads is spun on their support, by means of which King tells us “they are
enabled to maintain their position against the strongest current; frequently they will
leave their support and let themselves out into the stream anchored by threads of
silk and enabled by them to return.”
When full fed the larva spins a pocket-shaped cocoon on the support, within
which it pupates. The pupa is motionless and has a pair of branched spiracles projecting
from behind the head. When the adult emerges, a bubble of air collects
around it, and in this it floats to the surface and at once takes wing. The European
species take a month to complete larval life, a week being spent in the pupal stage.
The flies are most restless, and even when stationary continually move their legs
about like feelers. Sometimes the swarms consist entirely of females, sometimes
early in the season mostly of males.
The females pierce the skin of humans on tender spots, such as ears, the forehead,
around the eyes and nose, and crawl into the cavities. They are quite harmless
at night, mainly attacking about sunrise and sunset. Some crawl up the arms and
legs and down the neck, and leave behind little red weals which itch intensely
(S. damnosum, Theob.), and blood may flow freely from the wounds.
The following are some of the worst species:—
Simulium columbaschensis, the “Kolumbatz fly,” which abounds in the damp
marshy lands along the Danube, and is a great plague to man and beasts in Hungary,
and is also abundant in Austria and Moravia, and is most numerous after inundations
from the Danube. They sometimes appear in such swarms that it is impossible
to breathe without getting them into one’s mouth. There are instances of children
being killed by these flies when left on the ground by their mothers when working
in the fields.
S. damnosum, Theob. This occurs throughout Equatorial Africa and is known
as the “jinja fly” in Uganda, the “fouron” in the French Congo, the “kilteb”
in the Sudan. It is a most vicious biter, and in some parts occurs in “belts”;
Dr. Christy found one such extending from the shores of the Victoria Nyanza
northwards along the right bank of the Nile for twelve or fifteen miles or more, and
perhaps three or four miles wide. In this area the flies swarm in millions at certain
seasons, so much so that the natives have to leave their plantations. The bite
causes a weal, marked by a drop of blood.
S. griseicollis, Becker. The so-called “nimitti” occurs in Upper Egypt and
the Anglo-Egyptian Sudan. It lives near the river and is not found more than
half a mile from it. Human beings are bitten on the face and hands, animals in
the region of the pudenda.
S. latipes, Meigen. This is a European species, also found in Natal.
S. wellmanni, Roubaud. The “ohomono” of Angola, where it bites viciously
and is dreaded by the naked porters.
S. buissoni, Roubaud. Occurs in abundance in the Marquesas Islands. It has
been suggested that this species may help to propagate leprosy.394
A large number of these insects have been described by Lutz in Brazil.395
A Simulium sp. (?) is very harmful to poultry in Cape Colony.396
In America, Simulidæ are most annoying. One, S. meridionale, Riley, also
known as the turkey gnat in the Mississippi Valley, has been supposed to be the
carrier of chicken cholera; anyhow, it has caused the death of thousands of chickens
and turkeys in Virginia annually.397
In Mexico Townsend found a Simulium which was named S. occidentalis, which
caused great annoyance to man, many people being so susceptible to them as to
preserve through the gnat season a chronic inflammation of the exposed parts of
face and neck, resulting from the repeated bites giving rise to sores.398
Men and horses have been partially incapacitated by the bites of sand-flies or
Simulium in a Hampshire wood (Cantlie, Brit. Med. Journ., April 28, 1900,
v, No. 2,052, p. 1023).

Fig. 401.—Wing of Simulium.

Fig. 402.—Wing of Chironomus.
Family. Chironomidæ (Midges).
The Chironomidæ or midges are not only frequently mistaken for mosquitoes,
but some are very annoying to man by biting him as mosquitoes do. They are
easily distinguished from true mosquitoes (Culicidæ) by the following characters:
(1) head small, often retracted under the cowl-like thorax; (2) no scales to the wings
or body; and (3) the different arrangement of veins on the wings (fig. 402).
Two genera are important as annoying man, namely, Culicoides, Latreille, and
Johannseniella, Williston. The larvæ of Chironomidæ are either aquatic, both
fresh water and marine, and help to make the former foul,399 according to Slater, or
may, as in Ceratopogoninæ, live beneath the bark of trees, etc. The pupæ are very
varied and also the life-histories of the different genera.400 The blood-sucking habit
is confined to the sub-family Ceratopogoninæ.
Sub-family. Ceratopogoninæ.
This sub-family of midges consists of very small species varying from 1 to
2 mm. in length; the wings have darkened areas, and the second longitudinal vein
is wanting, and the first and third veins are stouter than the others and placed close
to the anterior margin, the fourth and fifth are forked; the antennæ in both male
and female are composed of fourteen segments, six or eight in the males bearing
long hairs.
The chief blood-sucking species belong to the genera Culicoides, Latreille,
and Johannseniella, Williston. The latter genus differs from the former in the
absence of an empodium or median appendage on the last segment of the tarsi.
The genus Ceratopogon, as restricted by Kieffer, is not supposed to take vertebrate
blood, but Austen has recently noticed that the type specimen of C. castaneus,
Walker, and a new species described by him, apparently have their bodies distended
with blood. The wings in the Ceratopogoninæ are carried flat when at rest.
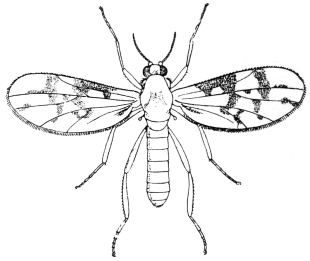
Fig. 403.—A Ceratopogon, or midge. Greatly enlarged.
In spite of their small size the females are the most bloodthirsty and annoying of
all insects. The Culicoides, which are often called “sand-flies,” bite during the day
and rarely at night. Usually they are most troublesome between 3 and 6 p.m.
They frequently attack in swarms, especially in the open, and owing to their minute
size can get through fine mosquito netting. Some of them produce a distinct “buzz”
when on the wing. These insects are found in all parts of the world. No species
has been definitely connected with any disease, but Culicoides has been suspected of
carrying the germs of Delhi boil. The larvæ of Culicoides are elongate in form and
have smooth bodies composed of thirteen segments including the head, which is
horny; there is no proleg on the first segment as seen in Chironomus, and on the
anal segment are retractile gills. They are very active and live in the sap of various
trees which saturates diseased bark.
The pupæ are smooth, but the abdominal segments bear a transverse row of
small spines. Austen describes a number of Culicoides and one Johannseniella and
three Ceratopogons from Africa,401 and Lutz402 a number of this sub-family from Brazil,
including a new genus, Centrorhyncus. Another genus, Tersesthes, Townsend
(Centrotypus, Grassi; Mycterotypus, Noe), also occurs in Brazil.
Culicoides ornatus, Taylor, is described from Townsville, Australia, found in
mangrove swamps. It is a very vicious biter and causes considerable irritation,
settling on hands and wrists (Taylor, Rep. Ent. Aust. Inst. Trop. Med. (1912),
1913, p. 24).
Family. Psychodidæ (Owl Midges).
This family of diptera is of considerable importance, not only on account of the
blood-sucking habits of some species, but especially on account of one at least having
been proved to be the carrying agent of “papataci” fever, a three-day fever very
prevalent in Malta and several parts of Southern Europe in the autumn.
It is also possible that these small flies are connected with the formation of
“Delhi boil,” caused by a protozoan parasite.
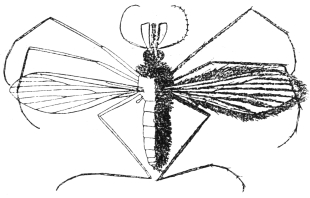
Fig. 404.—An owl midge, Phlebotomus sp. Greatly enlarged.
(From Giles’s “Gnats or Mosquitoes.”)
Psychodidæ are all very small flies, many of which have a moth-like appearance,
and owing to their fluffy nature are spoken of in Britain as “owl flies,” sometimes
also as “window flies.” Their bodies and wings are covered with hairs, densely in
some (sub-family Psychodinæ), and in a few with patches of flat squamæ. In the non-blood-sucking
Psychodinæ the wings are carried in a peculiar manner downwards
over the body, to a slight extent resembling the Hepialidæ, or swift moths. The
wings may be ovoid or lanceolate, and have a marked venation as seen in the figure.
The proboscis is short and non-suctorial in the majority of genera, but in the sub-family
Phlebotominæ it is elongated and hard. The antennæ are long and of sixteen
segments, and bear whorls of fine hair.
There are two sub-families, Psychodinæ and Phlebotominæ; in the former the
mouth is not suctorial; the female has a horny ovipositor and the second longitudinal
vein is branched at the root of the wing; in the second sub-family the proboscis
may be formed for sucking, the female has no horny ovipositor, and the second long
vein has its first fork near the middle of the wing.
The sub-family Phlebotominæ contains the genus Phlebotomus, which occurs in
South Europe, South Asia, Africa, North and South tropical America. They are
all small grey, brown, or dull yellow-coloured flies, and carry their wings when at rest
upwards like a butterfly. The proboscis is moderately long and the legs long and thin.
The females are most vicious blood-suckers, but in some species anyhow the
males also bite (P. duboscii). They are mainly nocturnal feeders and hide away
during the day in any dark corners or crevices.
The life-cycle has been worked out by Newstead403 and Grassi404 in Europe, and
by Howlett405 in India.
The larvæ have been found in crevices in rocks and caves, in dirty cellars, and
dark damp places containing rubbish, and are also said to live in crevices in the
walls of privies and cesspits.
The minute larva is very marked; as figured by Newstead it has two long chætæ
projecting upwards, in some stages branched, in others simple, and on the segments
a few blunt spine-like processes. The pupæ are found in similar situations.
The ova are very minute, elongate, translucent white, and covered with a thin coating
of viscous matter when first laid; soon after they become dark brown, shiny, with
long black wavy lines. Newstead found the incubation period in Malta to last for
about nine days in P. papatacii. Five species are known in Europe, five in Africa,406
two in North America, and eight are described by Annandale407 in the Oriental region.
Lutz and Neiva have described three species from Brazil408 (P. longipalpis, intermedius
and squamiventris).
Brachycera (Flies).
The antennæ as a rule have three segments, and are usually shorter than the head.
The first segment of the antennæ is frequently very small, and the third one is generally
the largest, and sometimes possesses a terminal annulated bristle. The palpi have
from one to three segments; the mandibles are covered by the labium. The three
thoracic rings are coalesced; wings are almost always present, the posterior ones
being rudimentary and covered with a little scale. From the ova legless maggots
are hatched, which as a rule have not a distinct head, but occasionally possess
two claw-like hooklets. These maggots live in decomposing organic matter; they
rarely live in water and some of them are parasitic. They either become barrel-shaped
pupæ within the last larval integument or, after casting it, are transformed
into naked pupæ. The larvæ of numerous Brachycera have been observed
in man, some in ulcers or on mucous membranes, others in the skin or in the
intestine, etc. In many cases the report only mentions the presence of the larvæ of
flies; in other cases the species has been determined; whilst in still other cases the
corresponding adult creature is unknown. We must therefore confine ourselves to
describing the most common varieties.
Family. Phoridæ.
These flies belong to the same division of Diptera, the Aschiza, as the family
Syrphidæ or “hover flies.” They are all small insects with marked antennæ and
wings; the former have the third segment globular and enlarged, and thus hiding
the first two; the wings are short and broad, the venation shows two short, thick,
long veins with four thin ones running out from them. The larvæ normally live in
decaying animal and vegetable matter, but one species, Aphiochæta ferruginea,
Brun., has been found as an intestinal parasite of man.
Aphiochæta ferruginea, Brun.
This small fly belonging to this family is of an orange-ochreous colour, the
upper part of the thorax tawny, and with dark bands on the abdomen, legs pale
yellow, the hind femora tipped with dark brown. It measures only 2 to 3 mm. in
length. This insect is shown by Austen to be widely distributed in the tropics,
being found in India, Burma, West Africa, and Central America. The larvæ breed
in decaying animal matter, such as putrid meat, decomposing shell-fish, etc.
Heusner bred out sixty-three flies from larvæ taken from an Indian’s foot.
Baker (Proc. Burma Branch Brit. Med. Assoc., 1891, p. 11–16) found that
the maggots of this fly were passed per anum by a European at intervals during
a period of ten months. Baker found that the larvæ fed on human fæces; from
the egg stage to the deposition of eggs from the resultant brood of flies occupied
twenty-two days. He concludes that they are capable of propagating, and do
so while living within the human intestines. He also records the larvæ in two
girls.
The larva does not seem to have been described, but Austen describes the pupa
(Trans. Soc. Trop. Med. and Hyg., iii, No. 5, p. 229).
Phora rufipes, Meig.
The larvæ of the “hump-backed fly” live in rotting potatoes, mushrooms,
radishes, etc., and when accidentally introduced into the intestine of man can,
like other larvæ, live there twenty-four hours and even more, and may set up serious
gastric disturbances.
P. rufipes is the same as P. pallipes, Latr.
Family. Sepsidæ.
Small blackish flies, elongate, with abdomen narrowed at the base, thickened
and curved downwards towards the extremity. Larvæ often found in decaying
vegetables, ham, cheese, etc. The larvae have the power of skipping; conical in
form, pointed in front, truncated behind, about 5 mm. long, shiny and smooth, the
anal segment with fleshy protuberances. The genus Piophila has a short proboscis
and the cross-veins of the wings approximate.
Piophila casei, L.
Cheese flies. The larvæ live in ripe cheese, with which they are sometimes
introduced into human beings (Meschede).
The larvæ of the cheese flies (Piophila casei) may pass through the alimentary
canal of human beings alive, and have been occasionally referred to in cases of
internal myiasis. It also breeds in dead bodies in adipose tissue. Howard records
it on human excrement. It is thus possible that some of the recorded cases of this
pest being passed alive may be due to eggs deposited on human fæces.
Family. Syrphidæ (Hover and Drone Flies).
Amongst the large family of Syrphidæ is found a section known as the Eristalinæ
or drone flies, whose curious long-tailed larvæ are popularly called “rat-tail
larvæ,” on account of the end of the body being drawn out into a long telescopic tail
of two segments, at the end of which are placed the breathing pores. These larvæ
live in water, no matter how foul, and in liquid manure. They have occasionally
been obtained in foul drinking water by human beings and from eating watercress
improperly washed or from badly kept beds. Austen (Trans. Soc. Trop. Med. and
Hyg., iii, No. 6, p. 221) records that in the autumn of 1907 a number of the larvæ
of the common drone fly (Eristalis tenax) were passed per rectum by a woman in
Hampshire who had recently arrived from France. The patient had eaten a considerable
quantity of watercress before leaving France. I have twice found small
Eristalis larvæ clinging by their long tails on watercress served at table.
Family. Drosophilidæ.
Small, rather plump flies, with short, broad abdomen, with bristles on the head
and legs. Often abundant in decomposing fruit, and may occur in dense masses.
Drosophila melanogaster, Br.
The larvæ of this fly occur in over-ripe fruit and in fungi, often also in human
habitations, and live in substances undergoing acid fermentation (vinegar, decaying
fungi, rotting fruit, in damaged spots in diseased trees), much more rarely in animal
substances, and they occasionally gain access to the human intestine (for example, by
the medium of sour milk). When introduced in any quantity, they cause vomiting
or attacks resembling colic; when taken in the pupal stage no unpleasant results
are produced.
Family. Muscidæ.
Teichomyza fusca, Macq.
Syn.: Scatella urinaria, Rob. Desv.; Ephydra longipennis, Meigen.
The larvæ live in the urine in privies. Several authors state they have found
them in fresh fæces or in vomited matter. Pruvot states that they continue for three
days in the stomach of rats into which they have been intentionally introduced.
(Pruvot, G., “Contrib. à l’étude des larves de dipt, trouv. dans le corps humain,”
Thèse de Par., 1882; Chatin, J., in Comp. rend. Soc. de Biol., Paris, 1888 (8), v,
p. 396; Roger, H., ibid., 1851 (1), iii, pp. 88, etc.)
Homalomyia canicularis, L., etc.
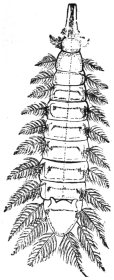
Fig. 405.—Larva of
Homalomyia canicularis.
Enlarged.
Homalomyia manicata, Meig., live as larvæ in decomposing vegetable matter or
in cultivated vegetables (cabbage); they are easily recognizable by their plumed
bristles, which are situated laterally on the body segments. They obtain access
fairly often to the human intestine and give rise to very uncomfortable symptoms.
Cases have been recorded from Germany, Austria, France, England, North
America (Wacker, in Artzl. Intelligenzbl., 1883, xxx, p. 109; Florentin, in Compt.
rend. Soc. de Biol., Paris, 1904, lvi, p. 525; and other authors).
The larvæ of an allied genus (Anthomyia), which, however, does not possess
plumed bristles, has been found in the external auditory meatus of a man (A. pluvialis,
according to Danthon).
[H. canicularis is common to Europe and North America, and is an abundant
house-fly. It is the small house-fly so often seen on windows. Besides living on
vegetable matter, they have also been found in the nests of the humble bee. Larvæ
of this species (fig. 405) were sent to the British Museum, taken from the fæces of
a woman suffering from cancer.409 They were found at Shrewsbury. Hagen410 reports
the larvæ of this fly as occurring alive in the urethra of a patient.—F. V. T.]
Homalomyia scalaris, Fabr.
[This is not a synonym of the above, but a distinct species.
[H. manicata, Meigen, is also distinct.—F. V. T.]
Anthomyia desjardensii, Macq.
This fly, allied to Homalomyia, is the cause of both intestinal and cutaneous
myiasis at Bihé, Angola (Wellman, Journ. Trop. Med. and Hyg., June, 1907,
x, p. 186).
Hydrotæa meteorica, L.
The larvæ live in decaying vegetable substances, also in dung, and have been
evacuated in some cases by man (Zetterstedt, Joseph).
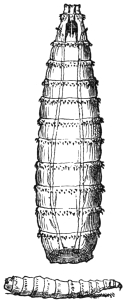
Fig. 406.—Larvæ of
Calliphora vomitoria.
Enlarged.
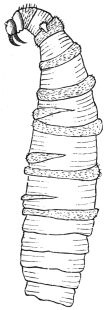
Fig. 407.—Larva of
Chrysomyia macellaria.
4/1. (After Conil.)
Cyrtoneura stabulans.
Larvæ in fungi, but occasionally also on larvæ of butterflies and Hymenoptera;
occasionally introduced into the human intestine (Joseph).
Musca domestica, L.,
and M. (Calliphora) vomitoria, L., and allied species; larvæ of these have been
repeatedly found in the intestine and nose of man (Mankiewicz, etc.).411
Musca domestica, Linn. (Common House-fly).
It is not only on account of a few larvæ of the common house-fly (Musca
domestica) being found in the intestines of man that it is of importance medically.
It is far more important on account of the part it plays in the spread of diseases
of the intestines, such as typhoid fever and cholera, infantile diarrhœa and
dysentery.
Howard and Clark (Journ. Exp. Med., 1912, xvi, No. 6, pp. 850–859) have shown
that the house-fly is capable of carrying the virus of poliomyelitis for several days
on the surface of the body and for several hours in the gastro-intestinal tract.
The house-fly may also distribute the ova of Tænia solium and the white worms
(Oxyuris and Ascaris). It has also been proved that they may carry the germs
of tuberculosis, and it is said that they play an important part in the spread
of infectious ophthalmia in Egypt.
This insect is found in all parts of the world. In warm countries it breeds
all the year round, and it may do so even in temperate climates in warm places,
such as stove houses. Most, however, die off in the autumn; but some survive the
winter as adults, in such places as kitchens, restaurants, and warm houses. I have
never failed to find a few Musca domestica in houses during the winter. The
majority, however, hibernate as puparia.
The females deposit from 120 to 150 eggs in a batch in stable manure, rotting
vegetation, house refuse, spent hops, old soiled bedding, etc. A single female
may lay as many as six batches of ova during her life. The eggs are shiny white,
and hatch in from eight to twenty-four hours in warm weather to three or four
days in cool weather. The white footless maggots are cylindrical, tapering to
a point at the head end, truncated posteriorly. The head consists of two dark
mandibular hooks and two short antennæ. On the tail end are two plates, the
stigmata, in which the main tracheal trunks open; in the second segment are
a small pair of projecting stigmata. The larval stage lasts from seven to five
days in hot weather; but in cold weather in temperate climes it may last six or
eight weeks.
The larva on reaching maturity becomes a barrel-shaped puparium of a dark
brown to black colour, and in this case changes to the pupa. This stage lasts from
three days in the tropics to four or five weeks in cold weather, the life cycle thus
varying from ten days in the tropics to fourteen in warm weather in Europe up to
three or four months under unfavourable conditions.
All breeding grounds should be burnt or otherwise done away with, such
as stable manure, house and kitchen refuse, human excrement and soiled substances,
also decaying vegetation as soon as possible, certainly by every sixth day. Stable
manure should be kept in closed receptacles and should be removed by every sixth
day to at least one mile from habitations and sprinkled with chloride of lime.
All kitchen and household refuse should be burnt at once or buried in pits and
covered with soil. Latrines should be as far as possible from hospitals, mess rooms
and tents. Food—especially milk, sugar and fruit—should be kept screened with
muslin when house-flies are about. Mess rooms and tents and hospitals should
have doors and windows screened with fine wire gauze during the fly season.
All possible steps should be taken to prevent them contaminating man’s food and
from breeding in human excrement and from entering hospitals. When present
in dwelling-houses in numbers they may be killed by fumigation with pyrethrum
or sulphur.
Genus. Chrysomyia, Rob. Desv.
Chrysomyia (Compsomyia) macellaria, Fabr.;
Lucilia macellaria, Fabr.
Syn.: Lucilia hominivorax, Coq.; Calliphora infesta, Phil.; Calliphora
anthropophaga, Conil.
A species distributed from the Argentine to the south of the United States
which deposits its ova on ulcers, in the aural meatus or in the nasal cavities of
persons who sleep in the open air. The larvæ are yellowish white, 16 mm. long, are
armed with two strong mouth hooks, and provided with spinous rings (screw-worm);
they lie hid in the nasal and frontal sinuses, in the pharynx, larynx, etc.; they perforate
the mucous membranes, even cartilage, migrate into the eyes, the cranial
cavity, middle ear, and cause severe disturbances; after the mature stage, in which
the larvæ leave the host to enter the pupal state, these symptoms often spontaneously
abate after a lapse of eight days, leaving behind greater or less cicatrices, and consequently
also defects in function of the organs attacked. Very often, however,
sepsis sets in, usually with a fatal termination.
(Coquerel in: Arch. gén. de méd., 1858 (5), p. 513; 1859, xiii, p. 685; Ann. Soc. ent.
France, 1858 (3), vi, p. 171; 1859, vii, p. 234. Weber in: Rec. de mém. de méd. milit., 1867
(3), xviii, p. 159. Francius, A., in: Arch. f. path. Anat., 1868, xliii, p. 98. Conil in:
Bol. Acad. nac. cienc. Cordoba, 1881, iii, p. 296. Humbert, Fr., in: Proc. U.S. Nat. Mus.,
1883, vi, p. 103; Amer. Nat., 1884, xviii, p. 540. Lindsay in: Journ. Trop. Med., 1902;
v, p. 220, and other authors.)
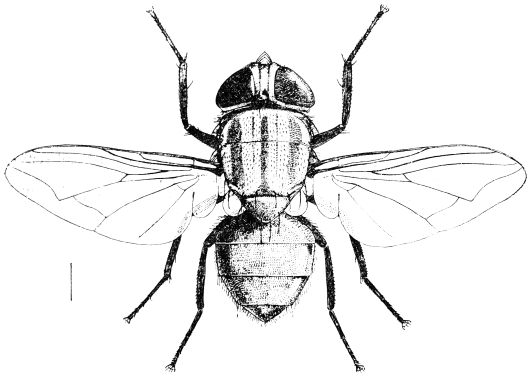
Fig. 408.—The screw-worm fly (Chrysomyia macellaria).
[This species is known as the screw-worm fly. It attacks animals as well as
man, especially laying its eggs on wounds formed by barbed wire. It may also be
found on dead flesh. Dr. St. George Gray sent me specimens from St. Lucia, from
the nose and mouth of a patient in Victoria Hospital. Others were found in the
vagina of another patient. Out of the four patients attacked, two occupied the
same bed, one after the other, and a third the next bed to it. The other case was
in a more remote part of the hospital. There are numerous records of this fly
attacking man. It occurs from the Argentine to Texas.—F. V. T.]
Chrysomyia viridula, Rob. Desv.
[This species is somewhat larger than the former; the body is metallic bluish-green,
the dorsum of the thorax with three blackish, longitudinal stripes, and the
face ochraceous; about 10 mm. long. Austen records this species from man,
Dr. Daniels having bred it from larvæ from a sore on a human being in New
Amsterdam, British Guiana. Dr. Laurence also bred it in Trinidad. In the latter
case between 100 and 150 maggots were discharged from the nose of a woman
suffering from facial myiasis (Brit. Med. Journ., January 9, 1909, p. 88 + fig.).—F. V. T.]
Genus. Lucilia, Rob. Desv.
Lucilia nobilis, Meig.
The larvæ were observed by Meinert in Copenhagen in the auditory meatus
of a person who, after taking a bath, fell asleep in the open air, and on waking
felt singing in the ears, and had a sensation as if there were water in the auditory
canal. During the next days severe pains set in, and there was a discharge of
blood and pus from both ears, as well as from the nose. On washing out the
meatus the maggots made their appearance.
Lucilia cæsar and L. sericata have also been observed in the larval state in
man (Thompson, Hope, Henneberg and Calendoli, Napoli, 1907).
[This golden-green fly usually lays its eggs on decomposing organic matter;
now and again it lays its eggs in wounds on man.—F. V. T.]
Genus. Pycnosoma, Brauer and v. Bergenstamm.
The species of this genus have a general resemblance to the Lucilias and
Chrysomyias, but the body is stouter and the abdomen banded. The genus can be
distinguished from Chrysomyia by the absence of the three thoracic stripes and
by the eyes of the male, in which the facets forming the upper portion are much
enlarged, whereas in Chrysomyia they are not noticeably larger. Austen also points
out that the sterno-pleural bristles in Pycnosoma are 1 : 1, in Chrysomyia 2 : 1.
The genus is found in tropical Asia and Africa only. All records of Chrysomyia
(Compsomyia) in India must be referred to this genus. Bezzi and Stein (“Katalog
der Palăarktischen Dipteren,” 1907, iii, p. 543), however, regard the two as
synonymous.
The larvæ are frequently found in the nostrils of man and burrow into the sinus,
but normally they live on decaying animal matter.
Pycnosoma forms the so-called Indian screw-worm. Patterson (Ind. Med. Gaz.,
October, 1909, xliv, No. 10) records the case of a woman at Tezpin, Assam, from
whom as many as 100 larvæ were removed at one time, and later the left orbital
cavity was found packed with hundreds of maggots; eventually the patient died.
It is possible that this, however, was due to a species of Sarcophaga. Austen
undoubtedly records this genus causing nasal myiasis in India (Trans. Soc. Trop.
Med. and Hyg., iii, p. 235). At Dehra Doon, U.P., a woman discharged 100
larvæ from her nose, with great pain in the nasal region and frontal sinuses.
The so-called “peenash,” a common malady in Rajputana, is a true nasal myiasis.
Genus. Sarcophaga, Mg.
Sarcophaga carnosa, L., 1758.
Larvæ of flesh-flies provided with two claws at the anterior end, which settle
on raw or cooked meat, and in the open on carcases of animals; they are often
observed in man, both in the intestine (introduced with food) and in the nasal
cavities, frontal sinus, conjunctiva, aural meatus, anus, vulva, vagina, prepuce,
and open ulcers, often migrating further from the regions first attacked. (Gayot
in Compt. rend. Acad. Sci., Paris, 1838, vii, p. 125. Grube in Arch. f. Naturg., 1853,
xix, 1, p. 282. Legrand du Saulle in Compt. rend. Acad. Sci., Paris, 1857, xlv, p. 600,
and other authors.)
[This fly is viviparous. The fly varies from 10 to 30 mm. in length, and is
of a general ash-grey colour; the thorax with three dark stripes, the abdomen
light grey with three black spots on each segment; legs black; base of wings
yellow. It also attacks animals and birds, especially geese. The genus Sarcophaga
is universally distributed. The maggots are whitish or yellowish footless larvæ
of twelve segments, tapering to a point in front, broadened posteriorly. There
are two mouth hooks, by means of which they rasp their food. The breathing
pores are at the end and consist of two groups of three slits, each surrounded by
a hardened area. They pupate in their old skin, which turns brown.—F. V. T.]
Sarcophaga magnifica, Schiner, 1862.412
Syn.: Sarcophaga wohlfahrti, Portschinsky, 1875.
A species widely distributed over the whole of Europe, occurring especially in
Russia (Mohilew); the presence of the larvæ in man was first observed by
Wohlfahrt (1768). The larvæ settle in the pharynx, in the nose, the aural meatus,
the conjunctiva, and in other regions of the human body; they also attack domestic
animals and birds. As Portschinsky has shown, they cause severe inflammations,
hæmorrhages and suppurations in the organs in which they occur; children are
especially attacked. A number of cases have been observed also in Central and
Western Europe. [The fly has a light grey abdomen with shiny black spots which
do not change their shape and appearance according to the angle in which the fly
is viewed.—F. V. T.]
(Wohlfahrt: “Observ. de vermibus per nares excretis,” Halæ, 1768; Nov. Act.
Acad. Caes. Nat. curios., 1770, iv, p. 277. Gerstäcker in: Sitzungsb. Ges. nat. Frde.
Berl., 1875, p. 108. Portschinsky in: Horæ soc. entom. ross., 1875, 1884, p. 123.
Laboulbène in: Ann. Soc. ent. France, 1883 (6), iii; Bull., p. xcii. Leon in: Bull.
Soc. des Méd. et Nat. de Jassy, 1905, xix, p. i. Freund, L., in: Verh. Ges. deutsch.
Naturf. u. Ärzte, Homburg (1902), 1902, ii, 2, p. 450, and other authors.) [Probably
most cases of attack in Europe are due to this species.—F. V. T.]
The above cited do not exhaust the number of observations of diptera larvæ
parasitic in man; there are yet to be mentioned the larvæ of S. hæmorrhoidalis,
S. hæmatodes (of G. Joseph), those of S. ruficornis (excitants of a cutaneous myiasis
in the East Indies), those of species of Eristalis (of Hanby and others), and those of
Phora rufipes (of Kahl, of Warsaw, and others). In many cases the determination
of the diptera larvæ has been omitted (or must be omitted); such is the case with
diptera larvæ in the eye (Schultz-Zehden in: Berl. klin. Wochenschr., 1906, p. 286.
Ollendorf in: Med. Korrespondenzbl. d. würt. ärtzl. Landesver., 1904, p. 1017.
Kayser in: Klin. Monatsbl. f. Augenheilkunde, 1905, xliii, i, p. 205. Ewetzky and
v. Kennel in: Zeitschr. f. Augenheilkunde, 1904, xii, p. 337, and other cases). Austen
records several cases of myiasis due to Sarcophaga (vide Trans. Soc. Trop. Med.
and Hyg., 1910, iii, No. 6).
The larvæ of African Muscidæ have now become of greater
interest; like several Oestrid larvæ they live normally in the skin of
mammals, but also attack man. The knowledge of these species is
certainly very insufficient, but this is not likely to be the case much
longer, as medical men practising in the Colonies are giving their
attention to these parasites. At the present time four distinct forms
are recognized according to Gedoelst.413
Sarcophaga chrysotoma, Wied.
[This species is recorded as attacking human beings at New Amsterdam, British
Guiana. The fly is 15 mm. long, has a golden-coloured face, three broad black
thoracic stripes and ochraceous buff anal segments. It was bred from larvæ
obtained by Dr. Roland from a sore on a girl’s foot. It is known to occur in the
Brazils and the West Indies. Another species was also bred which Austen was
unable to identify.—F. V. T.]
Sarcophaga plinthopyga, Wied.
[This and other species of Sarcophaga are called “yaw flies” in Dominica, as
they are believed to be concerned in the dissemination of frambœsia or yaws
(Nicholls) (vide Austen, Trans. Soc. Trop. Med. and Hyg.,
1910, iii, p. 239).—F. V. T.]

Fig. 409.—Ochromyia
larva on the skin of
man, South Africa.
3/1. (After Blanchard.)
Ochromyia anthropophaga, E. Blanch.; Cordylobia arthrophaga, Grünberg.
Indigenous to the Senegal and neighbouring districts;
in the district of Cayor (between the mouth of the Senegal
and Cape Verde) the larva is known as the “ver de Cayor.”
It lives under the skin, especially at the lower extremities
and the lower region of the trunk, producing small boils,
which cause pain, but after about eight days, when the
larva leaves the body to enter the pupal stage, the pain
discontinues. Besides man the larva occurs in dogs, goats,
cats, and in the jackal. It is still questionable whether
the fly deposits its eggs direct or on the ground, from
whence the larvæ as they emerge gain access to animals
and man. Larvæ yellowish-white, 14 mm. long, 4 mm. wide,
eleven segments414; head with two globular antennæ-like
appendages, two black curved mouth hooks, and two wart-shaped,
finely spinous structures at their base. Body evenly
covered to the seventh segment with small black prickles,
which are stronger at the sides and the anterior borders of the segments; from the
seventh they increase in size, on the two hindermost they are wanting; on the last
segment two deep yellow spiracles, each with three markedly curved fissures; in
addition two stigmata on the posterior border of the first segment. Duration of the
larval stage about eight days. Upon the construction of roads in Guinea the larva
is spread by dogs far into the interior.
Auchmeromyia (Bengalia) depressa (Walker).415
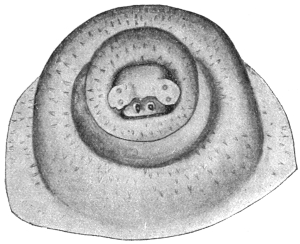
Fig. 410.—Head end of “larva of Natal.”
Magnified. (After Gedoelst.)
Distributed in the region of Natal and apparently over the whole of South
Africa. The “larva of Natal,” as one may still term the species provisionally, as
its identity is not certain, possesses on its head (besides the mouth hooks) lateral
protuberances beset with a row of chitinous spines. The cuticle of the body is
spinose. The spines are difficult to recognize on account of their transparency and
want of colour; they are longest over the anterior segments, from the fifth they
become smaller, and over the
hindermost they are very small.
Apart from the foremost segment,
the position they take is that
of rows running transversely or
obliquely, two to four generally
in juxtaposition; the number of
spines in the groups gradually
increases posteriorly, attaining the
number of eight to twelve on the
sixth segment, and this number is
maintained to the end of the body.
Isolated spines are found over the
head; over the second, third and
fourth segments single ones are
still found adjoining the groups of
spines, from the fifth onward they
are wanting. From here the spines
cover the whole free surface of the segments; over the fourth the anterior three-quarters,
over the third two-thirds and over the first and second only the anterior
half. The stigmata found at the anterior end also serve as distinguishing characters.
The parasitic stage appears to last about fourteen days. [Fuller (Agric. Journ.,
Dept. Agric. and Mines, Natal, 1901, iv, p. 606) refers to this as Bengalia depressa
also.—F. V. T.]
Genus. Cordylobia, Grünberg, 1903.
Cordylobia grünbergi, Dönitz.
Syn.: Ochromyia anthropophaga, Grünberg, nec Blanch.; Cordylobia
anthropophaga, Grünberg.
Endemic in German East Africa and neighbouring regions. Larva up to
14 mm. long, 4 to 5·5 mm. wide, of cylindrical shape, slightly narrowed behind,
truncated, gradually tapering in front; antennæ-like processes, cone-shaped, blunt.
Smaller cylindrical formations at the base of the mouth hooks surrounded by a
circle of chitinous hooks. Body from the first segment covered with small brown
squamous spines which are disposed in numerous irregular transverse rows. The
spines are small over the two first segments, the two posterior thirds of all the
segments, as well as from the eighth; over the third to the seventh they are larger,
but between these there are very small spines. The breathing pores of the stigmata
at the anterior end are kidney-shaped; the orifices are elongated and very tortuous,
each divided into three. The larval period appears to last several weeks.
Cordylobia anthropophaga, Grünberg.
This well-known cutaneous African parasite seems to have been the cause
of much confusion in regard to names. It belongs to the genus Cordylobia
of Grünberg, and is one of the family Muscidæ, and differs from Auchmeromyia
in that the second abdominal segment of the female is of normal size, whilst
in Auchmeromyia it is more than half the length of the whole abdomen, and in the
male the eyes are holoptic or close together, whilst in Auchmeromyia they are wide
apart. The flies of this genus (three so far described) attack man in their larval
stage (anyway two of the three), and also dogs and other animals, by burrowing into
the skin and producing painful boils.
[C. anthropophaga, Grünberg, is widely distributed in Africa, extending from
Senegal, where its maggot is known as the “ver de Cayor,” and is referred
to on p. 590 as Ochromyia anthropophaga, E. Blanchard, to Natal, where it is
known as the “Natal worm,” and referred to erroneously on p. 591 as Bengalia
depressa, Walker.
[It is a thick-set Muscid of a general straw-yellow colour, with blackish markings
on the dorsum of both thorax and abdomen, about 9·5 mm. long. The larva is fat
and when mature about 12 mm. long, bluntly pointed in front, truncate behind;
from the third to eleventh segments it is thickly covered with minute recurved
spines of a brownish colour, arranged in transverse series of groups of two or more,
which form more or less distinct irregular transverse rows. On each of the two
posterior stigmatic plates, the respiratory slit on either side of the median one
is characteristically curved, resembling an inverted note of interrogation. The
puparium is brown to ferruginous or black and about 10 mm. long. The maggots
are found in both natives and white men, and occur as a severe pest in dogs, also
in monkeys, rats, and other mammals. In Sierra Leone it is called the “tumba
fly.” The larvæ have been frequently found as true subcutaneous parasites, each
larva living singly and forming a boil or warble in the skin, with an opening just as in
an ox-warble, through which the maggot breathes and eventually escapes. Although
they more usually occur as isolated specimens, Marshall found in Salisbury,
South Rhodesia, that sixty were extracted from one lady, and Bérenger-Féraud, in
Senegal, that more than 300 occurred in a single spaniel puppy.
[Neave (Bull. Ent. Res., 1912, iii, p. 217) records it from ulcers in a native
at Lourenço Marques in 1908, and at the same time from ulcers in a dog, and that
it is a severe pest to man in Mozambique and parts of the Transvaal. It seems
to be more abundant in North Rhodesia and Nyasaland than to the north (Neave,
Bull. Ent. Res., 1912, iii, p. 310). It is also recorded in Zanzibar, German East
Africa, Uganda, East Tropical Africa (Neave).
[Simpson (Bull. Ent. Res., iii, p. 170) records a Muscid larva taken from the
breast of a European in South Nigeria that was probably Cordylobia.
[It is not known how infection takes place. Neave (Bull. Ent. Res., iii, p. 310)
says: “Many instances in human beings would preclude the possibility of eggs
having been laid direct on the skin: in these cases they have probably been laid
on the clothing put out to dry.”
[Gedoelst has described another species, C. rodhani, and Austen a third species,
C. prægrandis, from Nyasaland, Cape Colony, Transvaal, Natal, North-west
Rhodesia, and German East Africa.
[The following are some papers dealing with this subject: Proc. Ent. Soc.,
London, for year 1907, p. xlvii; Journ. R.A.M.C., 1908, pp. 5–11, figs. 1 and 2,
by Austen; Journ. R.A.M.C., 1908, pp. 1 and 2, by Major F. Smith; Trans. Soc.
Trop. Med. and Hyg., 1910, iii, pp. 223–225, by Austen.—F. V. T.]
Lund’s Larva.
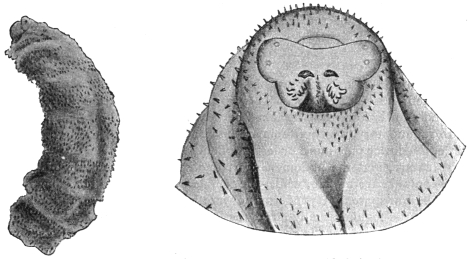
Fig. 411.—Lund’s larva: on the left, the whole larva, magnified six times. On the right,
the head end, much enlarged. (After Gedoelst.)
Endemic in the region of the Congo State; called after Commander Lund, from
the skin of whose arm it was extracted; 12·5 mm. long, 4·5 mm. broad; colour
yellowish, with brown rings, on account of the division of the brown spines; head
cone-shaped, with two hemispherical smooth antennæ, two thick black mouth hooks
and wart-shaped bodies, between which are situate two to three longitudinal rows
of dark brown chitinous laminæ. The body segments are covered over their whole
surface with irregularly distributed triangular yellow spines, the points of which are
coloured dark brown. Its size increases from the second to the sixth segment,
diminishes from the seventh to the ninth, at the tenth it is reduced, and at the
eleventh quite small. The posterior stigmata are bean-shaped, each with three
markedly tortuous openings. Duration of the larval stage unknown; the same
applies to the pupal and imago stages.
Auchmeromyia luteola, Fabricius.
[This fly, the parent of the so-called Congo floor maggot,416 belongs to a nearly
allied Muscid genus to Cordylobia, but which can at once be told by the great
length of the second abdominal segment. The maggot occurs in numbers in the
native huts in the Congo region and is fairly common in central and northern parts
of Mozambique; it is also recorded from the Zambesi River and the vicinity of
Barberton in the Eastern Transvaal (Bull. Ent. Res., 1912, iii, p. 216), in German
East Africa, in Nyasaland, and British East Africa. It is also recorded from
Bara, Kordofan,417 where they occurred on the floor of the men’s prison and bit the
prisoners. They were destroyed by sprinkling Jeyes’ fluid on the floor. Neave
states (ibid., p. 310) that it occurs in the more neglected huts in native villages
throughout tropical Africa, and frequently enters a tent when pitched near a village.
It is also found in West Africa. The fly is thick-set and about the size and build of
a bluebottle fly; length 10 to 12 mm.; tawny in colour to dirty yellowish-brown,
with dusky hairs, giving it a smoky appearance; the flattened thorax has long dark
stripes and the abdomen a dusky line in the centre of the second segment, which
meets a dark line on its posterior border; the dusky third segment has a narrow
yellowish anterior line; the fourth segment is also dusky; legs buff with black hairs;
the fifth tarsal segment black. The larvæ are whitish, becoming reddish after a feast
of blood, with much wrinkled skin and rather flat and broad. They live in crevices
of the mud floor, under sleeping mats during the daytime, and come out at night and
suck the blood of sleepers and then retire to shelter again. Dutton, Todd, and
Christy noticed that where people slept on beds or platforms raised above the floor
the maggots were not so numerous as under the sleeping mats laid on the ground.
They turned up many of the maggots from a depth of three inches or more.418—F. V. T.]
Family. Oestridæ.
[The family of Oestridæ or warble flies are all parasitic in their larval stage,
usually termed the “bot” stage. They are found as parasites in warm-blooded
animals, and man is frequently attacked by them. The members of this family have
the mouth rudimentary, many of them are hairy and bee-like, with large eyes and the
head large, the lower part more or less swollen. The thorax is large with a distinct
transverse suture, and the abdomen short and stumpy or very slightly elongated.
The male genitalia are hidden, whilst the female ovipositor is often elongated. The
wings may be transparent (Hypoderma) or mottled (Gastrophilus), and have muscid-like
venation; the tegulæ usually large, the legs moderately long.
[As a rule each species is confined to a particular host, but as we see recorded
here those that attack animals may also attack man. The flies occur in warm weather
and usually during the warmest part of the day, and have a strong dislike to shade
and water. The genus Hypoderma attack oxen, sheep, goats, antelope and musk
deer; Oestrus, sheep, antelope and horses; Gastrophilus, the horse and ass;
Cephenomyia, the deer; Cepholomyia, the camel and buffalo; Dermatobia, dogs,
cats, oxen, deer, apes and man; Cuterebra and Rogenhofera, rodents and opossums.
[Some live as parasites in the stomach and intestines (Gastrophilus); others
infest the skin (Hypoderma, Dermatobia and Oestromyia, the latter on Lagomys and
Hypodæus); Œdemagena tarandi also infests the skin of the reindeer in Siberia and
boreal America. Oestrus lives in the nasal sinus, and Cephalomyia in the throat as
well, Cuterebra and Rogenhofera, the skin or scrotum, so that we have really three
groups of parasitic oestride larvæ: (i) cutaneous, (ii) intestinal, and (iii) facial.
[No species seems confined to man, but the so-called “creeping disease,” caused
by Hypodermæ, and the attack of sheep nasal fly are comparatively common, as also
is the Dermatobia attack.—F. V. T.].
Cutaneous Oestridæ.
The eggs are deposited on the surface of the body; the larvæ burrow in the
skin, which they reach after somewhat long peregrination.
Genus. Hypoderma, Latreille.
Hypoderma bovis, de Geer.
The cattle fly or warble fly, which swarms during the hot season, settles on the
head or on the hair of grazing cattle: through the young being licked off they gain
access to the mouth and are swallowed.419 The larvæ appear first in the commencing
portion of the stomach, to escape, as some state, into the preceding sections of the
alimentary canal; at any rate, they are found from July onward regularly in the
submucous tissue of the pharynx, in which they travel about for several months
(up to November, and in isolated cases up to February); they then penetrate the
muscularis and migrate by way of the subserosa along the mediastinum, the crura
of the diaphragm, the renal capsules, and the intermuscular connective tissue of the
psoas muscle in the direction of the spinal canal, into which they penetrate by way
of the muscles and nerves, through the intervertebral foramina. Here they stay for
about two to three months, then they leave the spinal canal again through the
vertebral foramina and make their way (from January to March) through the intermuscular
connective tissue of the muscles of the back to the skin of the back, where
sooner or later (from January to June) they arrive and enter a resting stage, which
commences with penetration of the skin and terminates with outward migration
from the boils due to the wound set up by the maggot. At the commencement
of this period the larvæ cast their skin, and their form, hitherto cylindrical, becomes
oval. After about a month, a second moulting of the skin takes place—the third
larval stage, which lasts about two and a half months (up to June). The approaching
end of the same is indicated by a change of colour on the part of the larva from the
hitherto yellowish-white to brown and finally to blackish-brown. When they have
become mature the larvæ leave the warbles, drop on to the ground and pass into
the pupal stage in the superficial layers of the soil within twelve to thirty-six hours.
After about a month the flies emerge. Irregularities with regard to the time and
direction of the migrations of the larvæ take place (Jost, H., in Zeitschr. f. wiss.
Zool., 1907, xxxvi, p. 644).
In a number of cases the larva of the cattle fly has been observed
in the human integument, usually in the winter months, that is,
during the migration period; consequently, it is not surprising that
the larvæ before they enter on the resting stage and produce a warble
undergo migrations. But that this takes place subcutaneously—which
does not appear to be so in the case of cattle—is perhaps
explained by the fact that in man, on account of the short space that
has to be traversed, the larvæ are not sufficiently developed to enter
on the resting stage simultaneously upon having obtained access
to the integument. Whether the Oestrid larvæ in Bulgaria that
similarly migrate beneath the skin in man belong to the cattle fly
or to another species, or even another genus, has not yet been
ascertained. (Doctorow, in Arch. de Par., 1906, x, p. 309; Spring, A.,
in Bull. Acad. sci. Belg., 1861 (2), iv, p. 172; Walker, R., in Brit. Med.
Journ., 1870, i, p. 151; Kjelgaard, in Ugeskr. f. Laeger, 1904, p. 535;
Condorelli, M., in Bull. Soc. Zool. Ital., 1904, xiii, p. 171.)
Hypoderma lineata, de Villers.
The larvæ of this species, that occurs not only in Europe but in North America,
live under similar conditions in the skin, very rarely in man; also migrating
subcutaneously (Topsent in Arch. de Par., 1901, iv, p. 609).
[In Sweden, the ox warble fly (H. bovis) is well known to attack man.
Schoyen states “that over 100 years ago up to the present time cases of travelling
grubs under the human skin in some districts of Sweden were well known.”
The species appeared to be H. bovis, many of which he had examined. They
accomplished long ramblings under the skin, always in an upward direction,
previous to their appearance through an opening in a tumour on the upper part of
the body, on the head, neck, or shoulders. An interesting case is recorded in
Insect Life, ii, pp. 238–239. A bot similar to H. diana was taken from the eye
and cheek of a child at Kane, McKean County, Pa., U.S.A. It was said
to have travelled in five months from the elbow to the eye. Riley later (Insect
Life, iv, p. 310) was inclined to think the maggot was that of H. lineata,
the common American ox warble, which is also found in Europe in great
numbers. I have recorded another case in England (Rept. Econ. Zool. for
year ending September 30, 1910, p. 128), where Dr. Menzies removed the larva
of H. bovis from the upper eyelid of a patient. It caused considerable swelling
of the face, much pain and distress; but the case did well, and the wound healed
at once. The larva was nearly mature. Numerous other references to this
so-called creeping disease will be found in the Supplement.
[It is quite probable that bovis and lineata are confused in the latter accounts.
The larvæ are, however, easily distinguished if carefully examined.—F. V. T.]
Hypoderma diana, Brauer.
In its larval stage it lives like other species of Hypoderma, attacking the red
deer (Cervas elaphas) and roe deer (Cervas capreolus); it is occasionally also found
in man (Joseph, in “Myiasis externa dermatosa,” Hamburg, 1800; Völkel, in
Berl. klin. Wochenschr., 1883, xx, p. 209).
Genus. Dermatobia, Brauer.
Dermatobia cyaniventris, Macq.
Syn.: Dermatobia noxialis, J. Goudot.
The genus Dermatobia represents the subcutaneous Oestridæ of Europe in
warmer parts of America. Both domesticated and wild mammals are attacked,
according to one statement birds also (Ramphastus), and man with fair frequency.420
It is assumed that in all cases one and the same species is concerned, for which
recently a name originating from C. Linné, jun. (Oestrus hominis), has been
employed. Three larval stages are recognized in the skin; the two first appear to
resemble one another in the club-shaped or tadpole-like appearance (called macaque
in Cayenne, mayacuil [mayoquil] in Mexico), the third is swollen spindle-shaped
(Berne, called torcel). Segments 2 to 4 in the club-shaped larvæ are closely beset
with small black spines, segments 5 to 7 bear at the anterior border a complete ring
of strong black hooks, segments 4 to 6 a similar ring, which, however, is interrupted at
the ventral surface. The four last segments forming the tail are smooth, only at the
posterior end are there small spines. The arrangement of spines of the third stage
differs from this. Italian workmen that have been employed in Brazil show the
presence of Dermatobia larvæ on their return (Blanchard, in Bull. Soc. Ent.
France, 1893, p. 24; Bull. Soc. centr. de Méd. vet., 1896; Ann. Soc. Ent.
France, 1894, lxiii, p. 142; Ward, H. B., in Mark Annivers. Vol., Article 25,
p. 483, New York, 1903).
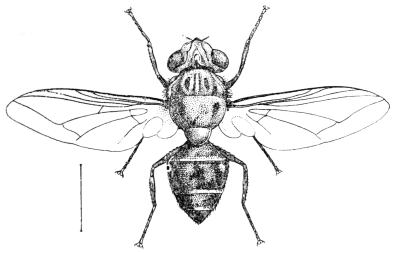
Fig. 412.—Dermatobia noxialis, Goudot.
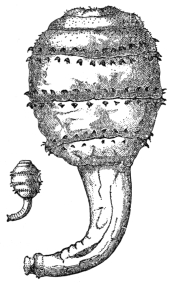
Fig. 413.—Larva of Dermatobia
cyaniventris in its
natural size and magnified.
(After Blanchard.)

Fig. 414.—Larva of Dermatobia
cyaniventris. Enlarged.
(After Blanchard.)
[Dermatobia cyaniventris, Macquart, 1843, is said not to be the same as noxialis
(vide Brauer, “Mono. Oestriden,” 1863, p. 266). It is known by various other names,
as nuche or gusano in New Granada, the ura in Brazil, and the macaw fly in
Cayenne. It occurs in Central and South America and the West Indies. According
to Goudot the fly is found in great numbers on the borders of large woods and
lands covered with underwood.
[It is seldom that more than one larva is found in each individual. It is generally
found in the arm and leg, but now and then the face. The perfect insect has never
been bred from a larva removed from a human being, so that there is still uncertainty
as to the actual species. D. cyaniventris is 11 to 12 mm. long, has an ochraceous
buff-coloured face, dark grey thorax, metallic dark blue to purple abdomen, and
brownish wings. D. noxialis is somewhat larger.
[In the Journal of Tropical Medicine and Hygiene, January 15, 1905, viii, p. 23,
reference is made to this Oestrid in Trinidad, where it is called the “mosquito
worm.” One case here recorded showed no fewer than four worms on the chin and
one on the hand. It is here stated that the fly never attacks man or animals directly,
as it is said to do by Scheube, but that the eggs are deposited on leaves and branches
in wooded lands and forests, and thus man, hunting dogs and wild animals in
passing through get the larvæ deposited on them accidentally. The affection is
common in Trinidad. Mention is made that a little 1 in 40 carbolic lotion syringed
into the aperture in the skin over the worm quickly killed it.
[The cattle worm, or founzaia ngómbe, is the name given to a larva which
develops beneath the skin of oxen and men in Central Africa, especially amongst the
natives and stock of Unyamonezi. According to P. Dutrieux, the egg is laid by a
large fly that accompanies cattle. It is unknown between the central plateau or the
Ugogo and the East Coast.—F. V. T.]
Cavicolous Oestridæ.
The forms belonging to this group inhabit as larvæ the nasal and
frontal sinuses of ruminants, Equidæ and Proboscidæ, which they leave
for the pupal stage. The larva of—
Genus. Oestrus, Linnæus,
Oestrus (Cephalomyia) ovis, L.,
occurring in sheep, has also been observed in man in six cases in the
nose and larynx (Saitta in Gaz. d. Osp. d. Clinic, 1903, No. 128). So
far as is known, the eggs are deposited in the nasal cavity.
[Oestrus ovis frequently occurs in man. MM. Sergent (Ann. de l’Inst. Pasteur,
1907, pp. 392–399) mention that they lay their ova on the noses, eyes and mouth of
humans in Algeria whilst flying, but that they disappear after three to ten days or
the inflammation produced by them. Portschinsky (Mem. Bur. Ent. Sci. Com. Cent.
Bd. Land Adm. and Agric., 1913, x, No. 3, p. 63) also gives cases. He doubts
that ova are laid on the nose; evidently the Russian habit is anomalous, for the
Sergents, Collings and myself find ova laid as a common occurrence. I have often
seen them on the nose of sheep. This fly also occurs in the Argentine (Serres, in
Gaceta Rural, April, 1913, vi, pp. 759–761).
[The tamné or thimni of the Kabyles, a human myiasis of the Tuareg mountains
in the Sahara, is caused by Oestrus ovis. Here the larvæ are said to be ejected on
to the conjunctival and nasal mucous membrane of humans.
[Ed. and Lt. Sergent (Bull. Soc. Path. exot., 1913, vi, No. 7, pp. 487–488) report
their attack from the Ahaggar mountains, in Central Sahara. The Tuareg name for
the fly, tamné, is the Targui form of the word thimni used by the Kabyles.—F. V. T.]
Gastricolous Oestridæ.
The eggs are deposited on the hairs of Equidæ, and the larvæ
escaping from them are licked up and swallowed. They pass their
larval stage, according to the species, in various parts of the
intestine and stomach, and when mature, pass out per anum in
order to undergo the pupal stage.
Genus. Gastrophilus, Leach.
One of the most frequent species is Gastrophilus equi, Fabr.; the eggs are laid
on the hairs; the larvæ live some ten months in the stomach, living attached
to the inner surface. The eggs of G. hæmorrhoidalis, L., are deposited on
the lips or the long hairs on them. The larvæ adhere to the cardiac end of
the stomach, to the stomach itself, and finally to the terminal portion of the
intestine. Here, however, and elsewhere in the intestine, the larvæ of G. pecorum,
Fabr., are also met with, whilst the larvæ of G. nasalis (so called because the
eggs are deposited in the nasal orifices) almost exclusively inhabit the anterior
section of the duodenum.
Cholodkowsky attributes the “wormlet” observed by Samson and Sokolew
(Wratsch, 1895, Nos. 48 and 57) and others (ibid., 1896–98) to Gastrophilus larvæ.
It burrows into the epidermis of man by minute passages. This observation
should, however, be verified. The phenomenon is designated as skin-mole, larva
migrans, and creeping eruption.
Other Papers on Dipterous Larvæ, etc., in Man.
(1) “Ein Fall von lebenden Fliegenlarven im menschlichen Magen,” Deutsch med.
Wochenschr., Leipz. and Berl., xxiv (12), pp. 193–194. Bachmann, and review of same,
“Living Fly Larvæ in the Human Stomach,” Philadelphia Med. Journ., 1898, i, 18, p. 773.
(2) “Sudi una larva di dittero parassita della congiuntiva umana,” Ann. di ottal., Paira,
1895, xxiv (4), pp. 329–336, 1 fig., E. Baquis.
(3) “Sur quelques diptères suceurs de sang, observé à Terre-Neuve,” Arch. de Par.,
Paris, 1900, iii (1), pp. 202–204, E. Barret.
(4) “An Account of the Larvæ of two Species of Insects discharged from the Human
Body,” Edin. Med. and Surg. Journ., January 1, 1811, vii (25), pp. 41–48, 1 pl., figs. 1 to 8,
T. Bateman.
(5) “Un cas de myiase par la Sarcophaga magnifica en Roumanie,” Bull. Soc. Zool.
de France, Par., 1891, xvi (2), pp. 25–26, R. Blanchard.
(6) “Sur les oestrides américains dont la larve vit dans la peau de l’homme,” Ann. Soc.
ent. de France, 1892, v, pp. 109–154, figs. 1–12, R. Blanchard.
(7) “Note additionnelle sur les oestrides américans dont la larve vit dans la peau de
l’homme,” Bull. Soc. ent. de France, Paris, 1894, xiv, pp. 209–211, R. Blanchard.
(8) “Note sur des larves de Dermatobia provenant de Brésil,” Bull. Soc. ent. de France,
Paris, 1893 (2), pp. 24–27, R. Blanchard.
(9) “Larven der Wohlfahrtfliege (Sarcophila wolfahrtii) im Zahnfleische eines Menchen,”
Wratsch., St. Petersburg, 1888, 5–6, E. K. Brandt.
(10) “Ueber den sogenannten Oestrus hominis und die oftmals besichteten Verirrungen von
Oestriden der Säugetheiere zum Menchen,” Verhandl. d. k. zool.-bot. Gesellsch., 1860, x
Abhandl., pp. 57–72, Brauer.
(11) “Ueber die Larven der Gattung Cuterebra, Clk.,” Verhandl. d. k. zool.-bot.
Gesellsch., 1860, x Abhand., pp. 777–786, Brauer.
(12) “Des désordres produits chez l’homme par les larves de la Lucilia hominivorax,”
Thèse, Paris, 1864, 43 pp., V. Audouit.
(13) “Note on the ‘Flesh Worm,’ ” Med. Press and Circ., London, April 12, 1882,
lxxxii (N.S. xxxiii), p. 314, P. S. Abraham.
(14) “Larvas de la Calliphora limensis en fosas nasalis,” 1855, 18 pp., F. Aguirre.
(15) “Raro caso di parasitismo nell ’uomo dovuto alla larva di una mosca (Sarcophaga
affinis, Meigen),” Boll. d. Soc. Rom. per gli Stud. Zool., Roma, 1893, iv (5–6), pp. 278–289,
1 pl., 3 figs., Giulo Alessandrini.
(16) “Observations sur l’espèce de ver nommé Macaque (Oestrus),” Mém. Acad. Sci.
par Hist., 1753, p. 72, F. Artur.
(17) “Contribuição ao estudo da biologia da Dermatobia cyaniventris,” Trav. do Inst. de
Manguinhos, 1908.
Biting-mouthed and other Noxious Diptera which may be
Disease Carriers.
[Amongst the division Brachycera (as meant in this work) we get several groups
of flies which, like the fleas and mosquitoes, are partially parasitic on man, the adults,
mainly in the female sex, being provided with a piercing mouth with which they
extract the blood of man and animals. The importance of these parasites is not the
mere fact that they feed upon our blood, but that they often carry germs from man
to man (tsetse-flies and trypanosomiasis, Tabanidæ and anthrax). Amongst the
most important biting-mouthed Diptera in this section are the following: Tabanidæ,
or gad-flies; Glossinæ, or tsetse-flies; and certain other Muscidæ. Some of the
exotic Asilidæ and a few Leptidæ also bite man.
Family. Tabanidæ (Gad-flies).
[The Tabanidæ have a broad, rather flattened body and a large head; eyes united
in the male (except in some Chrysops). The antennæ are composed of three segments,
have the third joint composed of five to eight annuli—in Chrysops they are
fairly long. The proboscis is projecting, and sometimes much elongated. The legs
are moderately stout. The venation of the wings is shown in fig. 415.
[This family of gad or horse flies contains a great number of genera, all of
which may bite animals and man more or less severely. The female alone is
blood-sucking, the males feed upon the juices of flowers. The females deposit their
spindle-shaped white, black, or brown eggs on leaves, stems of plants that either
overhang or stand in water, and amongst rushes; they are at first white, but become
brown or black. The eggs are laid in rounded, flattened or conical masses composed
of layers one upon the other. The larvæ are carnivorous, feeding upon snails,
worms, other larvæ, etc., and have a distinct head; they are cylindrical, composed
of eleven segments, the last with a vertical breathing pore, or the last two
segments may form a breathing tube. The majority taper to a point at each end, in
colour shining white or dull grey to yellowish, many of the larger specimens mottled
or banded with dark brown or black. The first seven abdominal segments are
encircled near the anterior margin with a ring of fleshy protuberances consisting of
a transverse dorsal ridge which may be divided by a depression into two. The
young larvæ burrow into any soft vegetable substance; they live both in the water
and under damp soil surrounding water, also in damp earth generally. The larvæ
are not only carnivorous, but they are cannibals, frequently devouring their own
species. They may take more than a year to mature.
[The pupæ are found close to the surface of mud and earth, and are mostly dull
yellowish to brown in colour, with rows of spines on the distal third of each
abdominal segment; the thorax bears a pair of ear-shaped spiracular structures, and
there are also six denticles at the apex of the abdomen.
[A habit common to the adults of most of the Tabanidæ of considerable economic
importance is that of the adults coming to water to drink. Portschinsky421 has found
that by applying kerosene to the pool they frequent the adults are killed, and
Hine422 that the same oil kills the larvæ that fall into the water from eggs laid on
plants above.
[Tabanidæ are not only of importance as purely biting insects, for they may
often convey pathogenic organisms from one animal to another, such as the
bacillus of anthrax, which they are known to carry, and possibly also trypanosomes
in regard to man. Chrysops also acts as a host of Filaria loa in South
Nigeria (Leiper, Brit. Med. Journ., January, 1912, pp. 39–40). Two species are
incriminated, viz., C. silacea and C. dimidiata. With animals these flies play
a more important part, for MM. Sergent, in Algeria, have proved that species of
Tabanus are able to transmit three forms of animal trypanosomes by biting a
healthy animal as long as twenty-two hours after having bitten an unhealthy
one. In India they have also been shown to transmit the parasite of “surra”
in dogs and rabbits by Rogers. Other observers have since corroborated these
results, and Mitzmain, who has recently performed valuable work in this connection,
states that T. striatus is undoubtedly the carrier of this disease in the Philippine
Islands. Certain members of the genus Hæmatopota have also been shown to be
capable of the direct transmission of Trypanosoma evansi. Martoglio (Ann.
d’Ig. sper., 1913, xxiii, N.S., No. 3, pp. 363–366) states that the trypanosome disease
of dromedaries known as salaf is transmitted by Tabanidæ, especially Pangonia
(P. magretti and P. beckeri) in Italian Somaliland. It is quite likely that these
flies play a much greater part in the spread of such diseases than is imagined
at the present time.

Fig. 415.—The ox gad fly (Tabanus bovinus, Linn.).
[The Tabanidæ are divided into two groups or subfamilies: (1) The Pangoninæ,
and (2) the Tabaninæ; the former have spurs on the hind tibiæ and usually ocelli;
the latter have neither tibial spurs nor ocelli.
[The Pangoninæ contain two main genera, Pangonia and Chrysops. In the
former the proboscis is much elongated, and the third antennal segment is composed
of eight rings, and is never angulated or ungulated at the base. The proboscis
is often very long.
[In Chrysops, the so-called blinding storm flies, all the three segments of the
antennæ are long, the third having only five annulations, and the proboscis short
but very strong.
[There are many genera in the Tabaninæ, which are found in all parts of the
world, of which two only are shown here—viz., Tabanus and Hæmatopota. The
former has the first two segments of the antennæ short, the third angulated
at the base, sometimes spurred and composed of five annulations; the second
has the second segment short, and the third composed of four annulations—never
angulated nor spurred at the base—and the wings are adorned with grey or
brown markings. These latter are usually called “brimps” and “clegs” in Britain,
the former gad or horse flies, the seruts and mangrove flies of tropical countries.
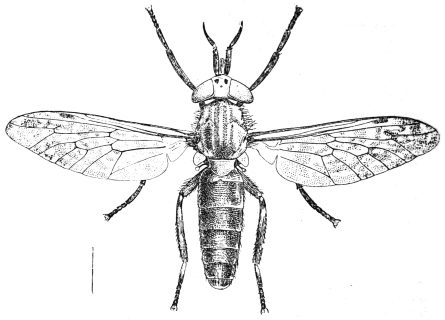
Fig. 416.—The brimp (Hæmatopota pluvialis, Linn.).
Family. Asilidæ (Wolf Flies).
[These flies are of little importance in regard to the subject dealt with in this
book; but I have notes sent concerning the biting habits of one or more species
belonging to this family from the Malay States and Africa.
[Asilidæ, or wolf flies, are easily told by the following characters: Large or
moderate-sized flies, thickly hairy; head separated from thorax by a narrow neck;
eyes separated in both sexes; proboscis firm and horny, adapted for piercing;
abdomen long, pointed, and composed of eight segments. Legs strong and bristly,
of moderate length. Wings sometimes mottled, lying parallel over the abdomen
when at rest. There are nearly 3,000 species. They live mostly upon insects, but
some are said to bite animals and man. They are, however, of little importance in
this respect.
Family. Leptidæ.
[This widely distributed family of flies has a few species which suck the blood of
man, and the writer has been personally badly bitten in Norway by a Leptis which
was apparently Leptis scolopacea.
[The Leptidæ have usually blotched wings and similar venation to Tabanus; they
are elongated flies of moderate or large size, and of dull colours. The antennæ are
varied and consist of three segments, either with or without a terminal bristle or with
the third segment compound, and in a few they may be almost nematocerous. The
wing veins are distinct, very crowded anteriorly, the third long vein is furcate, basal
cells large, and there are usually five posterior cells, the anal cell being open in some;
the squamæ are always small, sometimes only rudimentary.
[Four are known to be blood-suckers, namely the American Symphoromyia,
Trichopalpus obscurus in Chili, and Leptis strigosa and L. scolopacea in Europe.
The genus Symphoromyia has a single spur on the hind tibiæ, none on the fore or
mid tibiæ, the third segment of the three-ringed antennæ kidney-shaped, and a short
proboscis. In the genus Leptis the hind tibiæ have two spurs, and the third antennal
segment is not reniform.
[The other biting genus Trichopalpus can be told at once by the elongated
proboscis. Most of this family live upon other insects. The larvæ live in earth,
decaying wood, sand, stagnant waters, and the nests of wood-boring beetles; they
are usually cylindrical and may have fleshy abdominal legs; the anal segment has
a transverse cleft, and often two posteriorly directed processes and two stigmata
between them. They are all predaceous, and in one genus (Vermileo) make pitfalls
in sand like the ant lions (Myrmeleon).
Blood-sucking Muscidæ.
[The blood-sucking Muscidæ are mainly contained in the following genera:
Glossina, Stomoxys, Hæmatobia, Lyperosia, Stygeromyia, Philæmatomyia and
Bdellolarynx.
[The first is the most important genus on account of the part it plays in the
spread of trypanosome diseases. Stomoxys may also serve as a disease carrier.
The remainder and a few more genera cause considerable annoyance by their
bites, and may also act as occasional carriers of pathogenic organisms. All these
flies have their mouth parts elongated to some extent, forming a distinct proboscis,
which becomes more or less strongly chitinized; the labella are usually serrated
or spiny, and thus form a structure easily capable of piercing the skin. Unlike
the Culicidæ, the blood-sucking Muscidæ have the sanguinary habit common to both
sexes.
Genus. Glossina, Westwood.
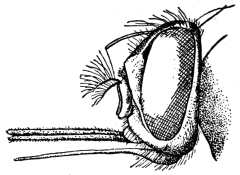
Fig. 417.—Head of Glossina longipalpis,
Wied. (After Grünberg.)
[This genus contains sixteen species,423 all of which are confined to the Ethiopian
region. Glossina may be distinguished from other allied genera by the proboscis,
the antennæ, wings, and male genitalia. The proboscis projects forwards and has
a swollen bulb-like base to the slender labium which holds the two structures, the
needle-like epipharynx and the thread-like hypopharynx; the whole proboscis is
ensheathed in the maxillary palpi. The antennæ have the first two segments small,
the third large with a marked pore, the orifice of the sense organ near the base;
from the base of the third segment also arises the three-jointed arista, the first two
segments being, however, minute; the
third bears a series of from seventeen
to twenty-one fine branched hairs on one
side. The male genitalia or hypopygium
is more or less oval and tumid, its long
axis lying in the antero-posterior direction,
with a vulviform median groove (the
anus) running from the anterior margin
to beyond the middle.
[Newstead has shown the importance
of the study of the genitalia in separating
species (vide Bull. Ent. Res., ii, pp. 9–36
and 107–110, and iii, pp. 355–360; and
Ann. Trop. Med. and Par., vii, No. 2,
pp. 331–334).
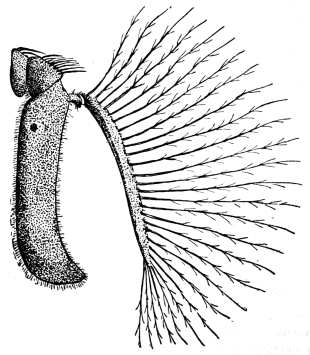
Fig. 418.—Antenna of Glossina pallidipes, male. (After Austen.)
[The tsetse-flies reproduce differently from all other Muscidæ. The female produces
at each birth a single full-grown larva, which is retained within the oviduct and
there nourished by the secretion of special glands, and on being born crawls to some
hiding place and at once becomes a puparium.
[The larva is a yellowish footless maggot nearly as large as the mother’s body,
the skin shagreened and the anal extremity having a pair of large, black, granular
prominences separated by a depression containing the breathing pores.
[The puparium is brown of various shades, the tumid lips of the larva being conspicuous,
the size and shape of the lips enabling the puparia to be identified.
[These puparia are often found in masses at the base of trees, in hollows in trees
and rocks just buried under vegetal debris. These insects are generally confined
to definite tracts known as “fly-belts.” They usually occur in damp, hot places on
the borders of rivers and lakes, and never far from water in the case of the palpalis
group, although others of the morsitans group may be found a considerable distance
from water. They are usually absent on grass plains, but may now and then occur
there (Kinghorn, vide Hindles’ “Flies and Disease, Blood-sucking Flies,” 1914,
p. 274); cover of trees, shrubs, or thick reeds is essential to them.
[Their range in Africa extends roughly from 18° N. to 31° S.
[Glossina palpalis is the chief carrier of the more prevalent type of sleeping
sickness. Two distinct types of parasites can produce this disease, viz., Trypanosoma
gambiense, which produces the ordinary sleeping sickness, transmitted by
G. palpalis, and Trypanosoma rhodesiense the Rhodesian or Nyasaland sleeping
sickness, transmitted by G. morsitans, and possibly identical with T. brucei, the
parasite of N’agana. Koch has also shown that G. pallidipes, Austen, and G. fusca,
Walker, can be artificially infected with the human trypanosome. It appears
probable that Koch used G. brevipalpis, not G. fusca, in his transmission experiments,
as at that time fusca included nearly all the large tsetses, but brevipalpis
is its Eastern representative.
[A Table of Species (modified after Austen) is appended here:—
I. |
Glossina palpalis Group. |
| 1. | Dorsum of abdomen ochraceous buff or buff; third
and following segments exhibiting sharply defined,
dark brown or clove brown, interrupted transverse bands | tachinoides, Westwood. |
| Dorsum of abdomen not so marked | 2. |
| 2. | Third joint of antennæ pale (cream buff to ochraceous
buff), clothed with long and fine hair, forming
a conspicuous fringe on front and hind margins | pallicera, Bigot. |
| Third joint of antennæ entirely dark (mouse-grey)
except at extreme base on outer side, and without
a conspicuous fringe of long and fine hair | 3. |
| 3. | Dorsal surface of abdomen dark sepia brown;
median paler area on second segment broad, and
more or less quadrate or irregular in outline;
hypopygium of ♂ buff or ochraceous buff | caliginea, Austen. |
| Dorsal surface of abdomen blackish-brown; median
paler area cuneate (i.e., triangular in outline);
hypopygium of ♂ grey | palpalis, Rob. Desv. |
II. |
Glossina morsitans Group. |
| 1. | Hind tarsi entirely dark; small slender species;
abdomen bright ochreous or reddish ochreous
with dark lateral markings | austenii, Newstead. |
| Hind tarsi not entirely dark; abdomen drab-grey,
buff or ochreous buff with conspicuous dark
interrupted transverse bands | 2. |
| 2. | Last two joints of front and middle tarsi with
sharply defined clove brown or black tips | 3. |
| Last two joints of front and middle tarsi without
sharply defined clove brown or black tips (front
and middle tarsi either entirely pale or, at most,
two joints of front tarsi faintly brownish at
the tips), and last joint and distal half of penultimate
joint of middle tarsi light brown, never so
dark as to form a sharp contrast with the
remaining joints | pallidipes, Austen. |
| 3. | Third joint of antennæ with a distinct fringe of fine
hair on front margin; dark brown or clove-brown
bands on abdominal segments extending close to
hind margins (i.e., pale ground colour, apart from
the median interspace, confined to a very narrow
hind border) | longipalpis, Wiedeman. |
| Third joint of antennæ without a distinct fringe of
fine hair on front margin; dark brown or clove-brown
bands on abdominal segments not extending
close to hind margins | morsitans, Westwood. |
III. |
Glossina fusca Group. |
| 1. | Third joint of antennæ fringed with fine hair on
anterior and posterior margins; fringe on anterior
margin conspicuous under a hand lens magnifying
15 diameters (nominal) when head is viewed in profile | 2. |
| Third joint of antennæ with fringe of fine hair on
anterior margin so short as to be scarcely noticeable
under a hand lens magnifying 15 diameters
(nominal) when head is viewed in profile (longest
hairs in fringe in length not exceeding one-sixth
of width of third joint); palpi long and slender | 3. |
| 2. | Longest hairs in fringe on front margin of third
joint of antennæ, in length equal to from one-fourth
to one-third (not exceeding one-third) of width of
third joint; palpi of moderate length | tabaniformis, Westwood. |
| Longest hairs in fringe on front margin of third joint
of antennæ in length equal to from one-half to
three-fourths of width of third joint; palpi
noticeably long and slender | nigrofusca, Newstead. |
| 3. | Pleuræ drab-grey or isabella-coloured, hind coxæ
buff or greyish-buff | fusca, Walker. |
| Pleuræ dark grey; hind coxæ mouse-grey | fuscipleuris, Austen. |
IV. |
Glossina brevipalpis Group. |
| 1. | Dorsum of thorax with four sharply defined brown,
more or less oval or elongate spots, arranged in
a parallelogram, two in front and two behind the
transverse suture; proboscis bulb with a sharply
defined brown or dark brown tip | longipennis, Corti. |
| Dorsum of thorax without such spots; proboscis
bulb not brown or dark brown at tip | 2. |
| Wings with upper thickened portion of anterior
transverse vein much darker in colour than
adjacent veins and thus standing out conspicuously
against the rest of the wing | brevipalpis, Newstead. |
| Wings with upper, thickened portion of anterior
transverse vein not much darker in colour than
adjacent veins, and thus not standing out conspicuously
against the rest of the wings (wings practically unicolorous) | medicorum, Austen.424 |
Glossina palpalis, Rob. Desv.
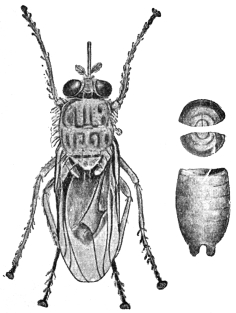
Fig. 419.—Glossina palpalis and puparium.
(After Brumpt.)
[This is the chief carrier of sleeping sickness in Nature. It is found in places
over the whole of West Africa from the mouth of the Senegal River to Angola,
and extends eastwards into the Bahr-el-Ghazal. The eastern boundary follows the
valley of the Nile and includes the eastern
shores of Lakes Victoria and Tanganyika;
from the southern end of the lake the
boundary tends south-west, approximately
following the frontier between North-eastern
Rhodesia and the Congo Free State, and
passing through the Katanga district of
the latter country into Angola (Austen).
It may occur up to 3,000 ft.; but, according
to Bagshawe, it has not been recorded
above 4,000 ft. It feeds on the blood of
many animals, including reptiles, amphibia,
birds, and even amphibious fishes, as well
as all the wild mammals. It seems, however,
to possess a decided predilection for
man, and undoubtedly thrives better upon
mammals and birds than upon cold-blooded
animals.
[It is not usually found far from water,
requiring a humid atmosphere and temperature
of about 85° F. (shade). But a
marked seasonal distribution is shown, the
flies considerably extending their range
during the rainy season, and thus visiting districts which are dry for the greater
part of the year; as the rains diminish the fly gradually leaves the temporary
haunts and returns to the more permanent ones. It bites only by day, and then
only in sunny weather, and usually lives in shade.
[Roubaud has shown that the first larva produced is about three weeks after
copulation, and that others are produced at an interval of nine or ten days. The
puparium stage is rapidly produced after the expulsion of the larva, often in
three-quarters of an hour. The puparium stage lasts from thirty-two to thirty-five
days. The puparia occur in well-drained humus close to water, sheltered by trees
or bushes, in crevices in rocks, and between the exposed roots of trees, sometimes
in sand.
[Bruce has shown that only a very small percentage of flies fed experimentally
on infected animals ultimately become infective, and that the infectivity of this
small percentage depends upon a delayed infection of the salivary glands.
[A variety, wellmani of Austen, is found in Angola, Gambia, the Katanga district
of the Congo Free State, the Matondwi Islands of Tanganyika, etc.
Glossina morsitans, Westwood.
[This species has been shown by Kinghorn and Yorke, and also by Bruce, to be
responsible for the transmission of Trypanosoma rhodesiense, the micro-organism
producing sleeping sickness in man in Rhodesia and Nyasaland and also in parts
of German and Portuguese East Africa. Fisher and Taute have demonstrated
experimentally that Trypanosoma gambiense—the sleeping sickness parasite of other
parts of Africa—may also be transmitted by this fly, and in addition it is known to
be capable of disseminating several species of trypanosomes pathogenic to animals.
Of these, T. brucei (=? T. rhodesiense), the parasite of tsetse disease, first
incriminated by Bruce, is perhaps the most important.
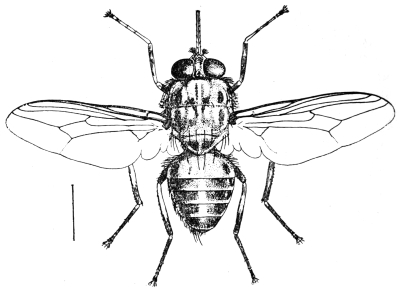
Fig. 420.—The tsetse-fly (Glossina morsitans, Westwood).
[It is the most widely spread of all tsetse-flies; its range extends from Senegambia
in the north-west to Southern Kordofan and Southern Abyssinia in the
north-east, and then southwards to the Bechuanaland Protectorate, North-eastern
Transvaal and Zululand. The actual localities given by Austen are Gambia, French
Guinea, Gold Coast, Togoland, Dahomey, Northern Nigeria, Congo Free State, the
Bahr-el-Ghazal, the Uganda Protectorate, German East Africa, and Portuguese East
Africa.
[This species is confined to “belts,” often of very limited extent, and appears
to prefer regions where there is sufficient vegetation for moderate but not excessive
cover and a hot, moderately dry climate. It is not nearly so dependent upon
water as is G. palpalis, and generally is most active in a dry atmosphere; some
observers, however, state that in certain districts it is more common along the
banks and edges of rivers. This tsetse-fly has been taken as high as 5,500 ft.
altitude. It infests native villages as well as the bush. Like other tsetse-flies
it bites not only during the hottest part of the day, but also on bright warm moonlight
nights, and it feeds on the blood of all mammals.
[The structure of the male genitalia of those representatives of G. morsitans
occurring on the West Coast of Africa and in parts of the Soudan presents certain
constant differences from that of the typical form of this species; this form is
known as G. morsitans, race submorsitans, Newst.
Genus. Stomoxys, Geoffroy.
[The members of this genus which occur in temperate and tropical countries
are provided with a hard, slender, shiny black proboscis which projects horizontally
from beneath the head; by means of this structure they can bite severely.
In general appearance they resemble house flies, but the proboscis at once
distinguishes them. In many parts of Britain they are known as storm flies
on account of their frequent appearance indoors previous to a storm of rain or
wind, which I have invariably found to be correct; they are also called stinging
flies. In colour they are greyish, dusky or brownish-grey or black, varying from
5 to 7 mm. in length; the thorax has dark longitudinal stripes and the abdomen
dark spots or bands. In the male the eyes are closer together than in the female.
These flies usually occur in stables and farmyards, along woods and in lanes, and
mainly attack mammals.
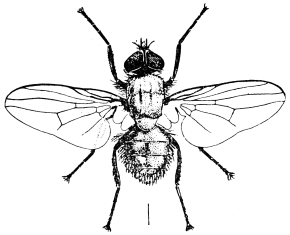
Fig. 421.—The stinging fly (Stomoxys calcitrans, Linn.).
[One species (Stomoxys calcitrans, Linnæus) occurs practically all over the world.
The female lays her eggs in moist, warm, decaying vegetation; as many as eighty
may be laid by a single female. The ova are white, banana-shaped, with a broad
groove on the shorter curvature; they may hatch in two or three days. The
creamy-white larva tapers to a point at the head end, and is truncated at the tail
end. Two black mouth hooks are plainly visible at the cephalic extremity. There
are two plates on the posterior surface of the last segment which bear the respiratory
pores, nearly circular in outline. It reaches maturity in fourteen to twenty-one
days; when mature it is 11 mm. long. The pupal stage is passed in the old larva
skin and lasts from nine to thirteen days; it is barrel-shaped, 5 to 8 mm. long, and
of a bright reddish-brown to dark chestnut-brown colour.
[This insect may act as a carrier of anthrax, and has been proved to be the agent
of an extensive epidemic of malignant pustule in the Isle of Pines, New Caledonia.425
[Noè’s426 experiments tend to show that it is an intermediate host and transmitter
of Filaria labiato-papillosa of the ox.
[Surra is generally stated to be transmitted by Stomoxys as well as Tabanus,
and yet Nitzman in the Philippines obtained uniformly negative results in exhaustive
experiments. Others have also been unsuccessful. Certainly Stomoxys can transmit
the disease in French West Africa (Bonet and Roubaud), and mechanically has
been proved to be capable of disseminating other trypanosomes (experimentally):
sleeping sickness (T. gambiense); nagana (T. brucei); souma (T. cazalboui); and
el debat (T. soudanense).
[S. calcitrans may also be a carrier of poliomyelitis (Rosenau and Brues,
Harvard Alumni Bulletin, 1912, xv, No. 9, pp. 140–142). Several species are
now known (S. brunnipes, Grünb.; S. inornata, Grünb.; S. nigra, Macq.;
S. omega, Newst.; S. ochrosoma, Speiser, etc.).
Genus. Lyperosia, Rondani.
[A genus of small flies which bite man and animals, but are not so far connected
with the transmission of any disease in man, but in Java it appears to carry surra
(P. Schat, Meeledeel Praefstation Oost-Java, 1903, 3e ser., No. 44), the species being
Lyperosia exigua, Meijere. These flies can be told from Stomoxys by the palpi
being broader, flattened laterally, and as long, or nearly so, as the proboscis.
When not feeding the palpi enclose the proboscis, as in Glossina. They are usually
about half the size of Stomoxys, and are the smallest blood-sucking Muscidæ.
They frequently swarm around and upon domesticated animals.
[The life-history of the horn fly in America (L. irritans, Linn.) is well known.
It lays its ova singly in freshly dropped cow-dung, and there the maggots feed,
pupating in the soil beneath.
[Patton and Cragg also give some details as to the life-history of Liperosia
exigua (“Medical Entomology,” p. 375) as follows: “L. exigua, whose habits
have been observed in Madras, usually lays twelve eggs at a time. The flies
immediately return to the cow and the process is repeated when the dung is again
dropped. The larvæ migrate from the dung when about to pupate, and the puparia
are always found in the earth at some distance away or under the sides of the patch
of dung. The fly usually hatches out in five days, though sometimes as late as the
eighth. Weiss has studied the life-history of irritans var. weisii from Algeria;
its larval stage lasts five days, and the flies hatch out of the puparia in another
five days.”
[The other biting genera of Muscidæ, Hæmatobia, Hæmatobosca, Bdellolarynx,
Stygeromyia, and Philæmatomyia, although sometimes annoying to man, have
not in any way been connected with any disease.
[The horse fly (Hæmatobia irritans, L.427) attacks cattle chiefly, but now and then
man is bitten. The different species can be told from Stomoxys by the palpi being
nearly as long as the proboscis.
[The genus Philæmatomyia, Austen, is intermediate between Stomoxys and
Musca in structure, and between the non-blood-sucking Musca, as M. domestica,
and the blood-sucking Musca pattoni, Austen, which feeds on the blood exuding
from the bites of true blood-suckers. They occur in Central Africa and India,
Ceylon and Cyprus (vide “The Life-history of Philæmatomyia insignis, Austen,”
Ann. Trop. Med. and Par., 1912, v, p. 515).
[Two flies belonging to the family Anthomyidæ also attack man, namely:—
[Hydrotæa meteorica, L. (the meteoric fly). This fly attacks man as well as
animals. They especially bite around the eyes and nostrils of animals, but are not
so particular with man; the head, however, is usually chosen. Linnæus called it
the meteoric fly because it often forms clouds around horses’ heads at the approach
of rain. The Hydrotæas are usually black or blue-black in colour with bare eyes
and simple abdomen, the front femora peculiarly constructed. H. meteorica, L.,
occurs in Britain.
[The members of the genus Hydrophoria, Desvoidy, also bite man.
Pupipara or Eproboscidæ.
[The Pupipara are all blood-suckers, the majority occurring as parasites on
mammals and birds, where they are more or less permanent parasites. Occasionally
some may attack man. They all produce their young fully formed, and they assume
the pupal stage immediately after extrusion. The puparia are large. They are
mostly flat, louse-like flies which may or may not be winged. In the case of Melophagus
I have found the puparia are often passed by the female. The winged forms
have a short quick flight, and when disturbed will seek shelter in man’s hair or
beard. Two main families occur: (1) the Hippoboscidæ, and (2) the Nycteribiidæ.
The former occur on animals and birds, the latter on bats only, but may invade
man. Two other families are known—the Braulidæ (bee parasites) and the Streblidæ
(bat parasites).
[The mouth of the Hippoboscidæ is long and sharp, forming a proboscis. The
thorax and abdomen are flat and leathery. The legs are stout and strong, and
terminate in large dentate claws and other structures of use in holding on to the
hair or feathers of their host when blood-sucking.
[Austen says it is probable that the Hippoboscidæ are descended from ancestors
belonging to the Muscidæ, which underwent modification in bodily structure as the
consequence of the adoption of a parasitic mode of life.
[Two wings are present in the true Hippoboscæ, Hippobosca equina (of the horse),
H. camelina (of the camel), H. maculata (of oxen), and H. capensis (of dogs), but
are absent in Melophagus, the sheep tick or ked fly (M. ovinus).
[In two genera, Lipoptena and Echestypus, wings are at first present, but are
lost as soon as the fly finds its permanent host.
[With regard to their biting man, such is only occasional. I have known sheep
shearers to be badly bitten by Melophagus ovinus, and have more than once been
attacked myself when standing where shearing is taking place. Sharp records the
grouse parasite, Ornithomyia lagopodis, as once biting severely a gamekeeper in
Scotland. There are also records of H. maculata biting man in Africa and India.
[Although so far not connected with any human disease, it is interesting to note
Theiler has shown that Hippobosca rufipes, v. Olfers, and H. maculata, Leach, are
capable of transmitting Trypanosoma theileri, Laveran, the cause of gall sickness
amongst cattle in the Transvaal. It is now considered, however, that Trypanosoma
theileri is non-pathogenic, and that the cause of gall sickness is a piroplasma-like
organism known as Anaplasma marginale. Theiler, Laveran and
Mesnil all hold this view (vide Laveran and Mesnil, “Trypanosomes and Trypanosomiases,”
second edition, 1912, p. 330).
[Lynchia.—Three members of this genus have been shown to transmit the
non-pathogenic (?) organism, Hæmoproteus columbæ amongst pigeons in Algeria
and S. America.
Insects and Epidemic Poliomyelitis.
[In a recent number of the Journal of Economic Entomology,428 Brues and
Sheppard point out the possibility of acute epidemic poliomyelitis (infantile paralysis)
being an insect-borne disease. They summarize as follows:—
[Many facts connected with the distribution of cases and the spread of epidemics
of this disease with histories of insects bites, suggest at least that the disease may
be insect-borne. Field work during the past summer, together with a consideration
of the epidemiology of the disease so far known, points strongly towards biting
flies as possible carriers of the virus. It seems probable that the common stable
fly (Stomoxys calcitrans, L.) may be responsible to a certain extent for the spread
of acute epidemic poliomyelitis, possibly aided by other biting flies such as
Tabanus lineola. No facts which disprove such a hypothesis have as yet been
adduced, and experiments based upon it are now in progress.
[If the disease should prove to be common to any species of domestic animals,
as is now strongly suspected, a secondary connection of ticks in spreading the
disease among such animals seems probable, as has been mentioned.
[The following is some of the more important literature on Diptera in general: Meigen,
J. W., “Syst. Besch. d. bek. europ. zweiflügligen insecten,” 1818–1838, 7 vols.; Brauer, F.,
“Monographie der Oestriden,” Wien, 1863; Idem, “Nachtr. hiersu,” Wien. ent. Zeit.,
1887, vi, pp. 4, 71; Schiner, J. R., “Fauna austriaca: die Fliegen,” Wien, 1860–64; Löw, Fr.,
“Ueber Myiasis und ihre Erzeuger,” Wien. med. Wochenschr., 1882, xxii, p. 247; 1883,
xxxiii, p. 972; Joseph, G., “Ueb. Fliegen als Schädlinge und Parasiten des Menschen,”
Deutsch. med. Zeit., 1885, i, p. 37; 1887, iii, pp. 713 and 725; Peiper, E., “Fliegenlarven als
gelegentl. Paras. d. Mensch.,” Berlin, 1900; Theobald, F. V., “Monograph of the Culicidæ
of the World,” 1901–1911, 5 vols. and 1 atlas, plates; Austen, E., “A Monograph of Glossina
Tsetse-flies,” 1903, 1 vol.; Van der Wulp, “Diptera neerlandica,” 1877; Walker, “Insecta
Britannica: Diptera,” 1851–53 and 1856; Lundbeck, “Diptera danica,” 1907–12; Zetterstedt,
“Diptera scandinaviæ,” 1850; Theobald, “British Flies,” 1892; Aldrich, “N. American
Diptera,” 1905; Loew and Osten Sacken, “Monographs of the N. American Diptera,”
1862–63 and 1869; Macquart, “Diptera exotique,” 1830–47; Rondani, “Diptera exotica et
Italica,” 1863–68; Williston, “Manual of Families and Genera of N. American Diptera,”
second edition; Verrall, “British Flies.” A fuller literature will be found in Peiper, as well as
in Huber’s “Bibliographie d. klin. Ent.,” 1899, iii, Jena, in the Bibliography at the end of
this work and in the Rev. of App. Ent. (Dulau and Co., London), where all references to
modern research can be found.—F. V. T.]
Akamushi or Kedani Sickness (vide also p. 487).—Schuffner (Far
East. Assoc. Trop. Med., Compt. rend. Trois. Cong. Biennial, 1913,
Saigon, 1914, pp. 309–315) states he observed a peculiar fever in Deli,
Sumatra, somewhat resembling typhoid. This he traced either to a
mite or tick. He figures the possible carriers, namely, a Trombidium
and Cheyletidæ. He calls this disease pseudo-typhus—a variant of
Japanese kedani sickness, which, he says, also occurs in the Philippines.
Ticks.—African Tick Fever: Marzinovsky (Proc. of Conference of
Bacteriologists and Representatives of Medical Sanitary Authorities on
the Campaign against Infectious Diseases in connection with the War,
Soc. Russ. Physicians in mem. Pirosov, Moscow, 1915, pp. 56–68), states
that African tick fever has been imported into Persia, and that it is
there carried by Ornithodorus tholosani.
Tick Paralysis: Todd (“Paralysis and Tick-bite,” Can. Med.
Assoc. Journ., 1914, iv, No. 9, pp. 825–826) refers to paralysis ascribed
to the bites of ticks in children, and possibly adults, in America,
British Columbia and Australia. He states that a young child,
perfectly well one day, has more or less complete paresis or paralysis
on the next, fever, a rapid pulse, and other constitutional symptoms.
The child may be dull and stupid, and may have convulsions. If
the tick is not found and removed the child may die, but if it is
removed, the symptoms disappear and recovery is complete in a
few hours. The tick must be entirely removed.
Diptera.—Psychodidæ: Bolt (China Med. Journ., Shanghai,
xxix, No. 2, pp. 78–86) states that sand-flies (Plebotomus) and the
fever due to them are common in North China, May and June being
the worst months. The natives of the region appear to be immune,
but all others suffer. Old ruined buildings are the favourite haunts
of the Phlebotomus. The species of Phlebotomus has not been
determined.
Pulicidæ.—Dermatophilus (Sarcopsylla) penetrans, or the
“Jigger.”—This flea (vide p. 544) is believed by Lama (Giorn. Ital.
Mal. Ven., Milan, 1914, xlix, pp. 465–472) frequently to carry leprosy
and he points out that the early lesions of leprosy usually appear
on the uncovered parts of the body. This flea also attacks rats.
Brachycera.—Leptidæ (vide p. 603): White, A. (“The
Diptera-Brachycera of Tasmania,” part I, Papers and Proc. Roy. Soc. of
Tasmania for 1914, 1915, pp. 35–74), describes a new blood-sucking
Leptid, Spaniopsis tabaniformis, which resembles a small gad fly
(Tabanus) in appearance.
Pycnosoma putorium: This is believed by Roubaud (“Les Producteurs
de Myiases et Agents similaires chez l’homme et les animaux,”
Paris, 1914, part I) to be largely concerned in the spread of
amœbic dysentery in French West Africa.
Lucilia argyrocephala, Macquart: This green-bottle fly is described
by Roubaud as producing myiasis in Africa (“Les Producteurs de
Myiases et Agents similaires chez l’homme et les animaux,” 1914,
Paris, part I). It attacks ulcers and sores in man and animals.
Auchmeromyia luteola, Fabr.: Schwetz (Ann. Trop. Med. and Par.,
1914, viii, No. 3, pp. 497–507), collected a large quantity of this
insect at Kabinda. He placed them in flasks with sand and a few
days later they pupated, and in fifteen days several flies hatched
out. The larval period varies from an unknown minimum up to
several months. The larva may live for at least two months without
food. A female oviposited on the 17th, and on the 18th one larva
hatched. The pupal stage seems to last eight to fifteen days. The
larvæ appear to bite by day as well as night according to native
information.
Cordylobia anthropophaga, Grünb.: Roubaud (“Etudes sur la
Faune parasitaire de l’Afrique occidentale française,” part I, “Les
Producteurs des Myiases et Agents similaires chez l’homme et les
animaux,” Paris, 1914) gives the life-history of this species. One
fly laid 150 ova in a glass vessel, on the sides, and on some rotten
fruit, and died the following day. He found that fifteen larvæ just
hatched placed on sand in a glass vessel with a guinea-pig gave rise
to characteristic tumours on the ventral surface of the body and
the anus. Other experiments failed. It thus seems that infection
takes place from larvæ which have hatched apart from the host.
Infection of man is regarded as accidental; no positive infection of
horses, oxen, sheep or pigs is known—it is rare in goats, and poultry
never seem to be attacked. The result of experiments tends to show
that the apparent choice of a host is mainly a question of body
temperature. The larva, whether freshly emerged or eight to ten days
old, penetrates the skin immediately, boring obliquely between the
epidermis and dermis. Once removed from the tumour the maggot
cannot bore again. The first moult takes place about three days
after penetration, and the total period of residence in the host is
seven to eight days. Upon emerging the larva falls to the ground
and buries itself. In two or three days it pupates and this stage lasts
no longer than twenty days. High temperatures, such as 95° F.,
appear to be fatal.
Myiasis.—Coates, G. M., “A Case of Myiasis Aurium accompanying
the Radical Mastoid Operation,” Journ. Amer. Med. Assoc.,
Chicago, Ill., 1914, lxiii, pp. 479–480: Apparently C. macellaria, forty
to fifty coming away with the gauze after the operation.
Huber, G. U., and Flack, F. L., “An Unusual Case of Screw-worms
in the Nose and Nasal Accessory Sinuses,” Journ. Amer. Med.
Assoc., Chicago, 1914, lxiii, pp. 2288–2289.
Auricular Myiasis.—Francaviglia, M. C., “An cora sulla myiasi
auricolare,” Boll. Sedute Accad. Gioenia, Catania, 1914, No. 31, pp.
15–23. This writer mentions the following parasites in the human ear:
Sarcophaga carnaría, L.; Wohlfartia magnifica, Schiner; Chrysomyia
macellaria, F.; Calliphora vomitoria, L.; and Anthomyia pluvialis, L.
He refers to a severe myiasis in Russia, due to a fly variously
recorded as Sarcophaga wohlfarti, Rond.; S. ruralis, Meig.; or Sarcophila
meigeni, Portsch. These are all probably synonyms of
W. magnifica. Chrysomyia macellaria, in Central America and
South America, is quite as harmful as S. carnaria, causing perforation
of the tympanum and meningitis. Lucilia nobilis and L. cæsar
have also been incriminated. Of the sub-family Anthomyinæ, the
larvæ of Fannia scalaris, Meig., F. canicularis, Meig., F. incisurata,
Zett, and Hydrotæa meteorica, L., are chiefly associated with
myiasis. He recommends, if the larvæ are outside the tympanum,
an injection of chloroform vapour by a few drops of water saturated
with chloroform, by an emulsion of 5 per cent. carbon bisulphide
or with benzine. When detached they may be removed with forceps
or a solution of boric acid. If the tympanum has been perforated,
the larvæ must be removed at once.
Francaviglia also records the larva of Oestrus ovis in the human
ear (Boll. Sedute Accad. Gioenia, Catania, 1914, No. 31, pp. 23–27).
Body, Head, and Clothes Lice.—Lobaczewski (Wien. klin.
Wochenschr., Vienna, 1915, xxviii, pp. 373–374) recommends the
impregnation of body linen with a 30 per cent. solution of oleum betæ
in 96 per cent. alcohol as an efficient method of keeping the body
free of lice. But the process must be renewed each time the linen is
washed and it takes fifteen minutes to carry out. On adding the
oil to the alcohol, a portion of the former is precipitated, the supernatant
fluid is decanted and poured over the linen, which is wrung
out in it and dried. The garments retain their lice-proof properties
until washed. Three days after wearing the clothes thus treated no
lice remain on the body.
Portnikov, Proc. of Conference of Bacteriologists and Representatives
of Medical Sanitary Authorities on the Campaign against Infectious
Diseases in connection with the War, Soc. Russ. Physicians in mem.
Pirosov, Moscow, 1915, p. 131.
Pediculus capitis and Phthirus pubis are shown to be successfully
controlled by applying spirit extract of sabadilla and both white and
grey mercury ointment, solution of corrosive sublimate of a strength
of 1 in 250 to 1 in 100, amyl and ethyl alcohol, benzine, chloroform,
carbon tetrachloride, methane, birch tar, liquid of malinin, etc. The
control of Pediculus vestimenti by the mixture of tartaric acid and
sodium sulphite slightly moistened with water is advised. It is placed
in small linen bags underneath the shirt; the heat of the body produces
a reaction which continues for two days, giving off a large amount of
SO2, which spreads beneath the shirt and kills all the parasites but does
not affect the skin. Marzinovsky, in the same Proceedings (pp. 56–68),
gives a number of remedies for Pediculus vestimenti (called humanus),
and mentions quinine or mercury, which latter the natives in Turkestan
carry on their hands and legs in bracelets soaked in mercury
compounds. He also mentions ethereal oils, the most effective being
clove oil, eucalyptus, oil of anise and camphor. He recommends for
disinfecting clothing for army purposes the chamber used by the
Japanese on a large scale. Kummerfelds’ wash is advised, and is prepared
as follows: 20 parts of precipitated sulphur are incorporated in
a mortar with 50 parts of glycerine; 2 parts of camphor are separately
ground with 50 of eau-de-Cologne and 20 of borax, and 870 parts of
distilled water are added; the whole is mixed together and 3 drops of
an extract of musk are added; shake in order to prevent the sulphur
settling down; 50 parts of ether are added to the mixture. This
sounds an expensive and troublesome preparation to make.
Shipley A. E., “Flowers of Sulphur and Lice,” Brit. Med. Journ.,
1915, p. 295. It is here stated by Dr. Lounsbury that the South
African troops were supplied by the Government with bags of flowers
of sulphur sewn in small calico bags and secured to the underclothing
next the skin as a preventive of lice. The bags were 2 in. square,
one on the trunk and one against each leg. This is a generally
accepted preventive, but is best mixed with equal parts of creosote
and naphthalene.
Shipley, A. E., “Insects and War,” Brit. Med. Journ., September
19 to November 14, 1914. General advice given re lice.
SUPPLEMENT:
CLINICAL AND THERAPEUTICAL NOTES.
PROTOZOA.
INTRODUCTION.
The aim of the present volume is to give an account of the animal
parasites of man, the number of which is very large. The Protozoa
that infest man are very important, and the literature relating to them
and to the treatment of the diseases that they produce is very extensive.
All that can be done in this Appendix is to give a very brief
outline of some of the more recent and approved methods of treatment,
for further details of which the reader should refer to standard
medical works, among which the following are noteworthy:—
Allbutt and Rolleston (1907): “System of Medicine,” vol. ii,
part 2, “Tropical Diseases and Animal Parasites,” London.
Castellani and Chalmers (1913): “Manual of Tropical Medicine”
(second edition), London.
Laveran and Mesnil (1912): “Trypanosomes et Trypanosomiases”
(second edition), Paris.
Manson (1914): “Tropical Diseases” (fifth edition), London.
Mense (1905): “Handbuch der Tropenkrankheiten,” Leipzig.
Ross (1911): “The Prevention of Malaria,” London.
Scheube (1910): “Die Krankheiten der Warmen Länder,” Jena.
References to the treatments tried in many parasitic diseases can
be found in the Sleeping Sickness Bulletin and Kala-azar Bulletin, both
now superseded and greatly extended in scope in the Tropical Diseases
Bulletin, published by the Tropical Diseases Bureau, Imperial Institute,
London, S.W.
The following diseases, due to protozoa and allied forms, are
discussed:—
| I. | Amœbic Dysentery. |
| II. | Trypanosomiases. |
| III. | Flagellate Diarrhœa and Dysentery. |
| IV. | Leishmaniases—Kala-azar and Oriental Sore. |
| V. | Spirochætoses—Relapsing Fevers, Yaws, Syphilis and Bronchial. |
| VI. | Malaria. |
| VII. | Balantidian or Ciliate Dysentery. |
I.—AMŒBIC DYSENTERY.
Amœbic dysentery, due to Entamœba histolytica (see pp. 34–41),
is present throughout the tropical world and also occurs in temperate
zones.
Walker and Sellards429 (1913) conducted important experiments
with amœbæ on prisoners in the Philippine Islands. They showed
experimentally that cultural amœbæ are non-pathogenic. As regards
experiments with Entamœba coli, after feeding to twenty individuals
they concluded that E. coli is a parasite of the human intestine but
non-pathogenic and non-culturable. In a third series of experiments,
after feeding with motile Entamœba histolytica, tetragena cysts were
found in the stools later; when tetragena cysts were administered,
motile E. histolytica were present in the subsequent stools. Some of
the histolytica cases developed dysentery after a time. They lay stress
on the necessity for the frequent examination of stools in order to
detect carriers. The incubation period of entamœbic dysentery is
usually long.
With regard to the symptomatology of amœbic dysentery, Castellani
and Chalmers distinguish four types—the acute, chronic, latent, and
mixed types.
The acute type has an abrupt onset; pain is felt in the lower part
of the abdomen, and the motions, rarely exceeding thirty daily, are
accompanied by much griping and straining. Blood and mucus are
present in the motions, and occasionally greyish material, consisting
of leucocytes, mucus, Charcot-Leyden crystals, amœbæ, and bacteria,
sometimes with particles of tissue. Nausea and vomiting may occur.
Digestion is usually deranged. The abdomen is sunken, the liver and
spleen are normal, but tenderness is felt along the course of the large
intestine. The urine may be diminished in quantity.
The chronic type may succeed the acute, or appear like diarrhœa,
the motions being fæculent and containing mucus. Between exacerbations,
constipation may occur. The number of motions may only be
twelve to fourteen per diem. Gangrenous complications may occur
at any time, and chronic dysentery may persist for many years.
The latent type is important, as the patients, though free from
dysenteric symptoms, harbour amœbæ and act as parasite carriers.
The latent condition may lead to acute attacks or to liver abscess.
The mixed type occurs where amœbic and bacillary dysentery are
combined. There is much fever, nausea, and vomiting. The motions
are numerous and often very offensive.
Treatment.—The most modern method of treatment, due to
Leonard Rogers, is by emetine. According to Castellani and
Chalmers, it is well to relieve griping and straining by either a hypodermic
injection of morphia or by small enemata of 40 minims of
laudanum in 1 oz. of mucilage of starch or by using 1/4 gr. morphia
or 1/4 gr. codeine suppository. A dose of castor oil (ʒiv to ʒvi) with
or without a few minims of liquor opii sedativus or a few doses of
saline may be given during the first twenty-four hours. After the
castor oil has acted or simultaneously, emetine treatment should be
commenced; 1/3 to 1/2 gr. of emetine hydrochloride, dissolved in sterile
normal salt solution, is injected hypodermically three times a day for
two or three days.
If emetine cannot be obtained, 5 gr. doses of ipecacuanha every
three to six hours in the form of membroids, or as pills coated with
salol or keratin, can be substituted.
After acute symptoms have disappeared, intestinal irrigations once
or twice daily, on alternate days, are useful. A solution of tannic acid
(3 to 5 per 1,000) or of quinine bihydrochloride varying in strength
from 1 in 5,000 to 1 in 750 is very slowly injected in quantities of
1/2 to 3 pints by means of a long, soft, rectal tube.
For gangrenous dysentery Castellani and Chalmers state that
appendicostomy, with irrigation of the whole lower bowel with
quinine lotion (1 in 1,000) or collargol (1 in 500), is the only chance.
The use of emetine should be continued in smaller doses after the
dysenteric symptoms have ceased, in order to prevent relapses and as
a possible safeguard against the development of a liver abscess.
Recently (July, 1914), Dr. W. E. Deeks430 has given an account of
his successful procedure in dealing with the dysenteries in the Ancon
Hospital, Panama Canal Zone, of which medical clinic he is the chief.
With regard to amœbic dysentery he advocates: (1) Rest, to increase
the patient’s resistance; (2) a generous milk diet, which is practically
all absorbed before it reaches the large bowel; (3) saline or plain
water irrigations, one to three daily; (4) the administration of
bismuth sub-nitrate in heroic doses; 180 gr. is given mechanically
suspended in about a tumbler of plain or effervescent water every
three hours, day and night in severe cases, only lessening the amount
when improvement takes place. Mechanical suspension in a large
quantity of water is essential. When the stools begin to decrease in
number and the tongue becomes clean, the number of doses is reduced
to three or four daily. In very chronic cases one or two doses daily
for a month after convalescence are recommended.
In exceptional cases of extreme emaciation and exhaustion,
showing marked toxic symptoms, surgical treatment is necessary,
and at Ancon a wide, open cæcostomy is performed.
The treatment of dysentery with bismuth sub-nitrate has been in
use for some years at Ancon. Latterly, a combined treatment by
hypodermic injections of emetine and bismuth sub-nitrate by the
mouth has been used, and the authorities there consider that it is
better to combine the two drugs rather than use each singly. Emetine
probably acts as a direct poison to the amœbæ, while the bismuth
probably acts by destroying the symbiotic organisms necessary for
their growth.
With regard to preventive measures, all drinking water should be
filtered and boiled, and uncooked vegetables and salads avoided.
Scrupulous care with regard to personal cleanliness, and avoidance
of touching the mouth or lips after contact with dysenteric patients,
are essential. Isolation of parasite carriers is of great use in combating
and controlling outbreaks of amœbic dysentery. The pollution
of soil and water must be rigorously prevented.
Liver abscess due to amœbæ must be localized by exploratory
punctures, and then opened and drained. Intramuscular injections
of emetine hydrochloride, 1/6 gr. to 1/2 gr. every day, will reduce the
temperature and afford relief.
Oral endamœbiasis has been recently investigated by Bass and
Johns, Smith and Barrett and colleagues (see pp. 43, 733). It responds
to treatment with emetine, and 1/2 gr. of emetine hydrochloride
administered hypodermically each day is of service. Rinsing the
mouth with a solution of fluid extract of ipecacuanha is also useful.
Rogers431 (1915) recommends a combined treatment of emetine
and streptococcal vaccines for pyorrhœa alveolaris.
II.—TRYPANOSOMIASES.
The human trypanosomiases are those occurring in Africa, due to
Trypanosoma gambiense and T. rhodesiense and spread by Glossinæ,
and that due to T. cruzi, occurring in South America and spread
by the Reduviid bugs, Triatoma spp. These trypanosomiases present
different clinical features and are best dealt with separately.
African Sleeping Sickness.
Sleeping sickness, due to Trypanosoma gambiense or varieties
thereof, was first reported from West Africa and is now present, not
only along the West Coast and in Nigeria, but throughout the Congo
basin into Uganda, north of which it exists in the Bahr-el-Ghazal
province of the Sudan. In Nyasaland and Rhodesia a more virulent but
less widely distributed disease is produced by Trypanosoma rhodesiense.
There is a general similarity between the two diseases, and the
symptoms as described by the leading authorities agree in the main.
The malady due to T. rhodesiense has been known only since 1910
and the differences between the malady due to it and to T. gambiense
will be indicated.
The course of the disease may be roughly divided into three
stages, the incubation, the febrile or glandular, and the cerebral stage.
The exact incubation period is not known with certainty in man.
Probably, in most cases, it does not exceed two to three weeks, but
disease signs may not appear for months. The bite of the Glossina
gives rise to local irritation, which may be overlooked. The irritation
usually subsides in the course of a few days.
The febrile, or glandular stage, is marked by attacks of fever of an
intermittent type. An erythematous eruption is often found on Europeans.
This rash begins as irregularly shaped pinkish patches which
clear in the centre until a ring is produced. It may occur on any
part of the body but is more frequent on the trunk. A typical
symptom is the enlargement of one or more of the lymphatic glands,
especially those of the neck. A general, deep hyperæsthesia, known
as Kerandel’s sign, may be present, and if the patient strikes a limb
against any hard object, a feeling of acute pain is felt, the sensation
being slightly delayed. As repeated attacks of fever increase, the
patient may become anæmic. The febrile stage may last for years,
and cure may be brought about at this phase, but frequently, after the
febrile stage has lasted some time, the cerebral stage is reached.
Tachycardia is also a symptom. Auto-agglutination of the red blood
corpuscles is another useful characteristic, as it is said to occur rarely
in other tropical diseases, but some workers doubt its value.
The cerebral, or true sleeping sickness stage is marked by a great
change in the habits of the victim, who becomes apathetic and dull,
careless and dirty in habits, and begins to experience difficulty in
walking. Tremors of varying degrees of severity are common and
the gait is peculiar. There is usually fever with rise of temperature
from 100° F. to 104° F. in the evening, becoming subnormal in the
morning. For some days before death, it often becomes permanently
subnormal. Congestion and œdema of the lungs, with patches of
pneumonia, are not infrequently observed before death. The torpor
gradually deepens, and the patient loses flesh. Frequently the lips
swell and saliva dribbles. The patient usually becomes comatose and
death ensues. Mania and delusions, and psychical and physical
symptoms resembling those found in general paralysis of the insane,
sometimes occur, and death may arise from secondary complications
such as pneumonia or dysentery.
Pathologically, the disease seems to consist of a chronic inflammation
of the lymphatic system. The trypanosomes reach the
lymphatic glands which become inflamed, and gradually invade the
blood and the cerebrospinal fluid. Sooner or later, as a result of
the lymphatic disease, changes occur in the membranes and
substances of the brain and spinal cord. There is round-celled
perivascular infiltration of the pia-arachnoid of the brain and spinal
cord. These changes cause compression of the blood-vessels, and
so lessen the supply of blood to the brain and spinal cord. Further
changes in the latter organs result in the production of the symptoms
that have given the disease the name of “sleeping sickness.”
The disease due to Trypanosoma rhodesiense generally runs a more
rapid course than that due to T. gambiense. The torpor and sleepiness
may not be obvious or be very slight, and the enlargement of
the lymphatic glands of the neck also may not be marked or may
appear to be absent. The duration of the disease often appears
to be from three to six months.
Treatment is only of use if commenced in the earlier stages of
the disease. The substances of most value so far are arsenic in
the form of atoxyl (introduced by Wolferstan Thomas in 1905) and
antimony in the form of tartar emetic. Castellani and Chalmers and
Manson recommend treatment by combining the use of both substances.
The combined treatment is recommended not only because both substances
have been proved of service independently, but also because
certain strains of trypanosomes resistant to arsenic are known, and
trypanosomes can develop a resistance to arsenic. Such forms, that
would not be affected by the atoxyl, are left open to attack by the
antimony salt. Daniels also recommends combined arsenic and
antimony treatment, and (1915) uses atoxyl and antiluetin.
Atoxyl is best given intramuscularly in 10 per cent. solution in sterile
normal saline solution. Galyl is also said to have given good results.
Castellani and Chalmers recommend: (1) Manson’s method of
administration of atoxyl, viz., 2 to 3 gr. of atoxyl are given by
intramuscular injection every third day for at least two years; or
(2) Broden and Rodhain’s method, 7 1/2 gr. of atoxyl by intramuscular
injection every fifth day. For the combined therapy by atoxyl and
antimony they recommend the following:—“An atoxyl injection
(3 gr.) is given every third day or 7 1/2 gr. every fifth day, and sodiotartrate
of antimony (Plimmer’s salt) is administered daily, 2 gr.
dissolved in a large quantity of water (2 pints) by the mouth or
by the rectum. Tartar emetic, however, is best given by intravenous
injections, using solutions of 1 in 100 or 1 in 1,000. The dose of
the drug to be given is 5 to 10 cg. per injection. It is important
that none of the fluid of the injection should escape into the
surrounding tissues, as a violent inflammation may result. These
injections should be administered monthly on ten consecutive days
for a long period.”
Macfie and Gallagher (1914) injected 6 gr. of atoxyl intramuscularly
every week in cases infected with T. nigeriense in the Eket
district of Southern Nigeria.
Large doses of atoxyl were often said to cause distressing results
such as optic atrophy, and when the onset of such occurred the drug was
usually discontinued. However, Daniels432 (July, 1915) points out that
eye troubles, such as iridocyclitis, are symptoms of trypanosomiasis.
Other arsenical preparations such as soamin and arsenophenylglycin
have been used, but less successfully than atoxyl. Fowler’s solution,
well diluted, has been given by the mouth when treatment by injection
was not possible, the doses commencing with 5 minims and increasing
to 15 minims.
Salvarsan and neo-salvarsan have also been tried for sleeping
sickness. Plimmer recommended powdered antimony suspended
in sterile olive oil. Ranken used precipitated metallic antimony in
normal saline solution injected intravenously.
Laveran and Thiroux have recommended a combined treatment
of atoxyl and an inorganic salt of arsenic such as orpiment. The
orpiment is given as pills, in doses of 2 gr. of orpiment two or three
times daily. Opium is added to the orpiment to prevent diarrhœa.
This treatment is said to have been used in man with good results.
Trypanosoma rhodesiense seems less amenable to treatment than
T. gambiense.
The main preventive measures seem to lie in segregation of the
sick in areas not infested with Glossinæ, and in measures against these
flies, such as bush clearing and destruction, to some extent, of proved
reservoirs in big game.
South American Trypanosomiasis.
The chief clinical features of the trypanosomiasis occurring in
Brazil have already been indicated (see p. 87). With regard to
treatment, according to Castellani and Chalmers the indications are
the same as those for African trypanosomiasis, together with treatment
for hypothyroidism. Preventive measures are directed against
the Reduviid bug, Triatoma megista, that transmits the disease. The
bugs occur in numbers in the cracks of the houses of the poor of
Minas Geraes, and may be destroyed by sulphur fumigation, lime-washing
or whitewashing.
III.—FLAGELLATE DIARRHŒA AND DYSENTERY.
The chief causal agents are Trichomonas hominis (T. intestinalis),
Chilomastix (Tetramitus) mesnili and allied organisms (see pp. 54 to 57),
and Lamblia intestinalis (see pp. 57 to 60 and Appendix pp. 734 to 736).
These parasites and the associated diarrhœas occur in temperate
as well as in warm climates. Probably some of the diarrhœas in India
are thus caused. The same, or similar parasites occur in various
Muridæ, especially rats and mice, which may act as reservoirs.
(i) Mello-Leitao433 (1913), writing from Rio de Janeiro, states that
there is a primary flagellate dysentery, due to Trichomonas intestinalis
(Leuckart) and to Lamblia intestinalis (Lambl), either separately or in
combination. He considers it a benign disease, and the most frequent
form of dysentery in young children. Trichomonas and Lamblia
were found to be pathogenic to children under 3 years of age.
Escomel434 (1913) collected 152 cases of dysentery in Peru due
solely to Trichomonas. Examination of the reservoirs containing
the water used for drinking purposes showed the presence of Trichomonas.
After the reservoirs were cleaned no more Trichomonas was
found and the cases of dysentery ceased.
Brumpt435 (1912) described a colitis due to Trichomonas intestinalis
in a patient returned from Tonkin.
Cases of infection by Chilomastix (Tetramitus) mesnili, with colitis
or dysenteric symptoms, are recorded by Brumpt (1912) from France,
and by Nattan-Larrier436 (1912) from the Ivory Coast respectively.
Marques da Cunha and Torres437 (1914) describe five cases of
chronic diarrhœa in Brazilian children due to the Chilomastix
(Tetramitus).
Gäbel438 (1914) described a case of seasonal diarrhœa contracted in
Tunis and caused by a Tetramitid parasite which he named Difämus
tunensis, as the discoverer considered that it lacked an undulating
membrane in its large cytostome.
Derrieu and Raynaud439 (1914) record a case of chronic dysentery
in Algeria due to a Trichomonad possessing an undulating membrane
and five free flagella. The parasite was named Hexamastix ardindelteili,
but the generic name Hexamastix is pre-occupied. Chatterjee’s
Pentatrichomonas bengalensis (1915) is possibly the same organism.
Treatment.—Escomel (1913), finding ipecacuanha and calomel
useless, recommends turpentine for Trichomonad dysentery. Two to
4 gr of essence of turpentine in an emulsion are given by the mouth,
and enemata containing 15 to 20 drops of turpentine emulsified in
the yolk of an egg to which is added a little water and tincture of
opium. Derrieu and Raynaud found this treatment effective in Algeria.
Smithies440 (1912) reports two cures of cases of severe dyspepsia, in
which Trichomonads were found in the stomach contents, after
administration of a single dose of 50 to 60 gr. of thymol, given at
bed-time, together with 2 gr. of calomel, and followed by an ounce
of Carlsbad salts in the morning. The patients came from the
Southern United States, and had been in the habit of drinking
unfiltered surface water in the localities in which they lived. Mello-Leitao441
used magnesium sulphate and water or milk diet. Sometimes
enemata of collargol (1 per cent.) or electrargol were required.
Rosenfeld recommended calomel. Methylene blue has also been
tried. Recently, Escomel442 (1914) recommends enemata of an aqueous
solution of iodine (1 per 1,000) and farinaceous diet. Lynch443 (1915),
working in South Carolina, recommends a mouth wash of saturated
solution of bicarbonate of soda three times daily in oral infections.
A similar solution was used as a douche in vaginal trichomoniasis.
Stiles (1913) points out that when amœbæ or flagellates are found
in a large percentage (10 to 40, or even 60) of the members of a
community, means should be taken to improve the methods for the
disposal of the dejecta, so that the food supply may be carefully
protected against fæcal contamination. Cysts of the parasites may be
air-borne or conveyed to food on the bodies of house-flies.
(ii) Lamblia intestinalis in man may cause diarrhœa with dysenteriform
stools. The diarrhœa may be of a chronic recurrent character.
The flagellate, or a variety of it, is fairly common in the digestive tract
of rats and mice.
Mathis444 (1914) gives an interesting account of cases in Tonkin.
In a child, aged 3, the stools were at first glairy and blood-stained,
containing many encysted Lamblia. The child’s home was infested
with mice. In another case, the house of the patient harboured
numerous rats.
According to Mathis, prognosis is favourable, but emetine hydrochloride
is without action on Lamblia. Prowazek and Werner445
(1914), however, state that emetine will act upon the flagellates, but
not upon the cysts. They recommend uzara (two tablets, three times
daily) and extract of male fern as useful in certain cases. Martin
Mayer (1914) found emetine hydrochloride successful in a case in the
Hamburg Seamen’s Hospital, but Assmy (1914) points out that a
suitable diet and daily doses of magnesium sulphate are sufficient,
in his experience, to effect an improvement, and he doubts the specific
action of emetine. Escomel (1914) recommends milk diet, then
calomel succeeded by castor oil.
According to Noc, Lamblia may also be water-borne. Healthy
carriers of Lamblia cysts are known. Food should be protected
from being soiled by rats and mice.
IV.—LEISHMANIASES.
A. Kala-azar.
(i) “Indian” Kala-azar due to Leishmania donovani.
Indian kala-azar due to Leishmania donovani is a very fatal disease
with a rate of mortality varying from 70 to 98 per cent. of the cases.
The incubation period is very variable and the early symptoms
not well defined. The incubation period seems to range from three
weeks to several months after exposure to infection. The onset seems
to commence with a rigor and attack of irregular, remittent fever, which
may show two remissions per day in a four-hourly temperature chart.
Rogers considers the daily double remission almost diagnostic. The
duration of this first attack is from two to six weeks. The spleen
and liver enlarge, especially the former, and are painful and tender.
Towards the end of the time the temperature declines and the first
period of the disease ends. After this period an apyrexial interval
occurs, which, after some weeks, ends in an attack of fever resembling
the first. Periods of pyrexia and apyrexia alternate. Anæmia commences
and asthenia appears and deepens steadily. The patient is
now thin and wasted, the abdomen much swollen and protuberant,
the ribs show clearly, the limbs are wasted and skin and tongue
darker than normal. In Europeans the skin is of a remarkable earthy
hue, and in natives of India darker than normal, approaching black.
Intestinal disturbances, often in the form of very obstinate and
intractable diarrhœa or dysenteric attacks, are common. Papular
eruptions often appear, particularly on the thighs; hæmorrhages also
may occur. The disease lasts for periods varying from seven months
to two years, and usually ends fatally.
Treatment, unfortunately, has not been very successful up to 1915.
Manson has reported two cases of cure by intramuscular injections of
atoxyl daily or every other day in doses of 3 gr. Rogers has advocated
large doses of quinine, 60 to 90 gr. daily until the temperature falls and
then 20 gr. daily. Castellani and Chalmers consider the best results
are obtained by large doses of quinine given intramuscularly, supplemented
by a course of quinine cacodylate injections or atoxyl injections.
Tartar emetic should be tried (see pp. 627, 629), especially
as L. Rogers (July, 1915) has had promising results in ten cases.
Castellani (1914) and Mackie (1915), have also had successful results.
Leishman states that the administration of red bone-marrow, either
raw or in the form of tablets, may be beneficial. Good nursing and
careful diet are essential, and diarrhœa or dysentery must receive the
appropriate treatment.
With regard to preventive measures, the extermination of bugs
and other biting insects seems to be of most service. Domestic and
personal cleanliness is of great importance. Patients should be
segregated. It would probably be as well if houses in which many
cases of kala-azar occurred were destroyed. Dodds Price, in Assam
tea gardens, moves the coolie lines 300 to 800 yards from old infected
ones, with satisfactory results.
(ii) Infantile Kala-azar due to Leishmania infantum.
This malady is found among children, rarely in adults, along the
Mediterranean littoral.
The disease commences insidiously and is often unrecognized until
some intestinal disturbance occurs. The spleen is then found to be
somewhat enlarged, and the case has often been regarded as one of
malaria. The child becomes anæmic, suffers from diarrhœa, alternating
with constipation, and has attacks of irregular fever. The spleen
continues to enlarge and protrudes from under the cover of the ribs.
Hæmorrhages from the nose and gums and into the skin occur.
Anæmia and wasting set in. The abdomen then becomes very
enlarged. The child becomes much less active both physically
and mentally, and looks prematurely old. Death often occurs from
exhaustion, though some cases of spontaneous recovery are known.
Treatment up till recently has been unsatisfactory. Some of the
remedies tried, as quoted by Castellani and Chalmers, are 15 cg. doses
of atoxyl, benzoate of mercury (2 to 4 mg. as a daily injection),
thiarsol (5 to 15 mg. by subcutaneous injection), salvarsan, etc.
Recently Cristina and Caronia (1915)446 have given repeated intravenous
injections of 1 per cent. aqueous solution of tartar emetic, the dose
varying from 2 to 10 cg. The treatment in various cases has lasted
from 15 to 40 days.
Prophylactic measures seem to lie in the destruction of infected
dogs and diminishing the breeding of fleas (see p. 111).
B. Oriental Sore, due to Leishmania tropica.
Oriental sore, known under many other names (see p. 107), is a
local infection of the skin due to Leishmania tropica. The incubation
period varies from a few days to some weeks, or even months, and
then one or several small itching papules appear. Each spot becomes
red and shotty, the papules increase slowly in size and the surface
becomes covered with papery scales. After a variable time, usually
not exceeding three to four months, ulceration occurs and a yellowish
secretion is exuded that soon dries into a scab. Under the scab
ulceration continues by erosion of the edges, and subsidiary sores
arise around the parent ulcer and usually fuse with it. Healing commences
after six to twelve months. Granulation begins at the centre
and spreads outwards, and when healing is complete, a depressed,
whitish or pinkish scar remains.
Many treatments for Oriental sore have been devised but do not
seem particularly satisfactory. Castellani and Chalmers state that
the scabs should be removed by boracic acid fomentations, and the
ulcers thoroughly disinfected once or twice daily with a 1 per 1,000
solution of perchloride of mercury, after which an ordinary antiseptic
ointment is applied.
The use of permanganate of potash has been advocated both by
French and English doctors. Both large and small sores can be
treated. The patient’s skin around the sore is protected by a thick
layer of vaseline, and the surface of the ulcer powdered with potassium
permanganate, which is kept in position by a pad of gauze and a
bandage. The treatment is said to cause great pain for six to eight
hours, but at the most, three treatments are necessary before the sore
becomes a simple ulcer, well on the way to healing. The permanganate
may also be used in ointment. Excision of the ulcer when
small is advisable when the site of the ulcer permits of this. According
to Manson, reports on treatment by radium, salvarsan and carbon
dioxide snow are decidedly promising. Mitchell (1914)447 reports
favourably on the use of carbon dioxide snow in the form of a pencil,
in India. In Brazil several workers (1914) record successful results
from the intravenous injection of a 1 per cent. solution of tartar
emetic in distilled water. Low (1915) has successfully treated a case by
direct local application of tartar emetic. Row (1912) has treated cases
of Oriental sore by inoculation of killed cultures of the causal organism.
As the disease is very contagious, the slightest wound, and any
insect bite, should be thoroughly disinfected with 5 per cent. carbolic
acid or iodine. Destruction of bugs, lice, and other biting insects
should be enforced. As dogs may contract the disease (see p. 108),
it is well not to allow them in the house and not to encourage undue
contact with them.
Naso-oral Leishmaniasis (Espundia) due to Leishmania tropica.
This form of Leishmaniasis has been reported from South America
and recently by Christopherson448 (1914) from the Sudan. In South
America it is often called Espundia, also Buba and Forestal Leishmaniasis.
The primary lesion is found usually on the forearms,
legs, chest or trunk. This ulcer is of the Oriental sore type, and
after some months, or even as long as two years, heals up, leaving a
thick scar. While the ulcer is open, or more often after it has healed,
lesions appear on the mucosa of the mouth and nose. The hard and
soft palate, gums and lips all may be attacked. The mucosa of the
nose is usually attacked and the cartilages become destroyed, producing
great deformity. In bad cases the pharynx and larynx may
become infected.
Till recently it was believed that treatment was of little use unless
the case could be investigated early. Escomel considered that if the
primary cutaneous lesion was excised or destroyed, further progress
of the disease was prevented. When lesions have appeared on the
mucosa of the mouth or nose, little could be done. The ulcers might
be cauterized and mild antiseptic mouth washes used.
In 1913 Vianna, working in Brazil, introduced treatment by tartar
emetic, which is now becoming more widely known and proving
efficacious. Carini449 (1914) applies it thus. Tartar emetic (that is,
potassium antimonyl tartrate) in 1 per cent. aqueous solution is introduced
slowly into a vein, such as the vein at the bend of the elbow,
in doses of 5 to 10 c.c. daily or on alternate days according to the
tolerance of the patient to the drug. Eighteen to forty injections
have been used. In some of the memoirs on the subject, the drug is
referred to as antimony tartrate.
The course of the disease is chronic and may last for twenty to
thirty years, death usually resulting from some intercurrent disease.
At present the actual transmitter of Espundia is not known with
certainty. Various sand-flies (Simulidæ) have been suspected of
transmitting the disease, though so far proof is wanting. It has also
been suggested that the natural food sources of some Simulidæ known
to bite man, namely, certain snakes450 and lizards,451 are possible
reservoirs of the disease.
Prophylactic measures would seem to consist in the immediate
disinfection of insect bites by tincture of iodine, and by avoidance of
areas known to be infested with snakes and lizards, and insects that
prey on them and man indifferently. The destruction of the primary
lesion as soon as detected is essential, and the isolation of advanced
cases of the disease seems advisable.
V.—SPIROCHÆTOSES.
A. Relapsing Fevers.
The relapsing fevers of Europe and of America, due to Spirochæta
recurrentis and S. novyi (probably a race of S. recurrentis), present
much the same symptoms, which differ in some respects from those
due to S. duttoni, the excitant of “tick” or “relapsing” fever in
Africa (see pp. 116–122).
The incubation period of S. recurrentis varies from two to
twelve days, during which time a very slight indisposition may be
noticed. The onset is usually sudden, with severe headache, pains
in the back, limbs and stomach and a feeling of weakness. There
is a rise of temperature to 103° F. or 104° F., and the temperature
continues high till about the sixth or seventh day. The skin is
yellowish, hot and damp; a rash, disappearing on pressure, may occur
on the trunk and legs, nausea is always present and thirst is usual.
The liver and spleen both enlarge. The number of respirations and
pulse-rate become increased. On the sixth or seventh day a crisis
occurs. There is violent perspiration, with a rapid fall of temperature,
pulse and respiration become normal and the patient sleeps and
awakes better. Improvement continues for some days, and recovery
may ensue, but usually about the fourteenth day relapse occurs,
lasting usually three or four days. A second relapse is unusual.
Numerous complications are known, e.g., bronchitis, pneumonia,
diarrhœa and dysentery.
With regard to treatment, the specific appears to be salvarsan.
Castellani and Chalmers recommend salvarsan administered intravenously.
Intramuscular inoculations (for example, into the buttock)
of a suspension of “606” in oil can also be given. The drug is very
efficacious, but large doses should not be given. An intravenous
injection of 4 or 5 gr. does not give rise to unpleasant symptoms
but is sufficient to effect a cure.
The incubation period for the American form of the disease is at
least five to seven days, and the first attack lasts about five to six days.
The treatment is by salvarsan as detailed previously.
As relapsing fever is spread by body lice and possibly by bugs,
preventive measures are directed against these insects. Strict
cleanliness of person, clothing, bedding and dwellings is essential.
Furniture, e.g., wooden bedsteads, liable to harbour such insects
should not be used.
The principal and best-known relapsing fever of Africa is that
excited by Spirochæta duttoni, and transmitted to man by ticks, chiefly
Ornithodorus moubata. The incubation period is usually about
seven days but may be longer. The patient is dull and lethargic,
perspires freely and is often constipated. The temperature rises to
103° F. or 105° F., there is headache, pains in the back and limbs,
general chilliness and great pain in the region of the spleen, which
often enlarges. The symptoms become worse, there is a fall of
temperature with improvement in the morning, and a rise, with
increase of pain, in the evening. Spirochætes are now found in
the blood in greater numbers. The symptoms last three to four days
and end in a crisis with profuse sweating and fall of temperature
below normal. The day before the crisis there is a pseudo-crisis,
when the temperature falls but there is no improvement. The
patient is left weak and tired. Recovery may follow, but more usually
a relapse occurs. The intermission period varies; five to eight days
is common. The symptoms of the relapses are like those of the first
attack. The number of relapses varies, five to eleven may occur.
The treatment recommended is by salvarsan, as for the European
relapsing fever.
With regard to prophylaxis, localities where ticks abound must
be avoided and the parasites themselves destroyed. Native huts
should be avoided. Mosquito nets, a bed well off the ground and
the use of night lights are advised by Manson to avoid attacks by
ticks, which are often nocturnal in their habits.
In North Africa (Algeria, Tunis, Tripoli, Egypt), and sometimes
in the Anglo-Egyptian Sudan, a spirochætosis due to S. berbera
occurs. According to Castellani and Chalmers, the incubation period
varies somewhat. The fever reaches its height during the first twenty-four
hours, and afterwards shows a morning remission. Jaundice
is often absent, but there may be hepatic tenderness and splenic
enlargement. One or two relapses usually occur. The treatment is
on the same lines as for the other spirochætal fevers. Sergent and
Gillot452 (1911), working at the Institut Pasteur of Algeria, have had
good results by using injections of salvarsan in doses of 0·75 to
1·0 cg. per kilogramme weight of the patient. The prophylactic
measures are directed against lice and other biting insects. Personal
cleanliness is most necessary.
In Asia, a relapsing fever, due to the spirochæte named S. carteri
by Manson in 1907, producing a mortality of about 18 per cent.,
occurs. The symptoms have a general resemblance to those produced
by S. recurrentis, but on the fall of temperature to subnormal on the
sixth or seventh day, when profuse perspiration and polyuria occur,
instead of improvement following, the patient often becomes collapsed,
with a clammy skin and feeble pulse. Improvement is slow. The
first relapse occurs about the fourteenth day of the attack, when the
temperature may be higher than for the first attack. There are
seldom more than four relapses. The treatment is by salvarsan, of
which doses of not more than 5 gr. intravenously should be given.
Sudden heart failure being common, Castellani and Chalmers state
that cardiac stimulants should be given. Prophylaxis is the same as
for European relapsing fever.
B. Yaws or Frambœsia tropica.
Yaws is essentially a tropical disease, though it is found in the
tropical and subtropical zones in all parts of the world, except in the
mountains and cold districts. In 1905, Castellani found the causal
organism, Treponema pertenue (sometimes called Spirochæta pertennis)
(see p. 127). The disease shows three periods: (1) The primary stage,
consisting of the development of the primary lesion or papule, which
is usually extragenital. The papule dries into a crust beneath which
an ulcer lies. (2) The secondary or granulomatous stage, which commences
from one to three months after the primary lesion is first seen.
It consists of a general eruption of small papules, some of which enlarge
and become granulomatous nodules covered with a yellowish crust.
They are common on the limbs and face. (3) The tertiary stage, in
which deep ulcerations and gummatous nodules appear. Any of the
tissues may be involved. Osseous lesions may occur. The disease
does not appear to be hereditary; it is usually spread by contact.
The best treatment appears to be by salvarsan or neo-salvarsan.
Castellani and Chalmers recommend intramuscular and intravenous
injections. For intramuscular injection an alkaline or neutral solution
of the drug is preferable, or a suspension of the drug in oil
may be used. The dose varies from 0·3 to 0·5 gr according to
the age and sex of the patient. For use intravenously, a slightly
smaller dose is required. Galyl is also being used.
In countries where frambœsia is endemic, slight skin abrasions
should be carefully treated with antiseptics. Yaws patients should be
isolated till cured, and their dwellings and personal possessions
disinfected.
C. Syphilis.
Syphilis, due to Treponema pallidum (sometimes called Spirochæta
pallida), is prevalent throughout the tropics as well as in temperate
zones. The disease is amenable to treatment by salvarsan and neo-salvarsan,
for administration of which see relapsing fever and yaws.
Galyl is also being used with favourable results. Lambkin’s mercury
cream has been found useful in treating numerous cases in Uganda.
The life-history of the parasite is given on p. 124, and further medical
details hardly come within the purview of this book.
D. Bronchial Spirochætosis.
Bronchial spirochætosis, due to Spirochæta bronchialis (see
pp. 122, 739) is probably of wide distribution in the tropics. The
spirochætes have been found in cases of chest complaints, especially
those with bronchitic symptoms. The disease may be suspected in
atypical cases of pneumonia and bronchitis, and may be mistaken for
incipient phthisis.
Chalmers and O’Farrell453 (1913), writing from Khartoum, recommended
rest in bed, good food and ventilation, coupled with
treatment by arsenic in some form, preferably associated with glycerophosphates.
These may be given by the mouth, or intramuscularly
as an injection of:—
| Sodium cinnamate | 0· | 05 grm. |
| Sodium cacodylate | 0· | 10 " |
| Sodium glycerophosphate | 0· | 10 " |
Taylor454 (1913–14), writing from Entebbe, Uganda, prescribes
arsenious acid by the mouth in increasing doses. Creosote has been
used in West Africa.
VI.—MALARIA.
Malaria, known also under the names of ague, paludism, marsh
fever, remittent fever, intermittent fever and climatic fever, among
others, is a very widely spread disease. It is most prevalent in the
equatorial regions and gradually diminishes north and south of the
equator. The various malarial parasites (see pp. 155 to 172) are spread
by species of Anophelines, and hence malaria is present in districts
favourable to these intermediate hosts, that is, in places where there
is a considerable amount of atmospheric moisture and rain, as well
as heat.
The principal malarial parasites are: Plasmodium vivax, the agent
of simple tertian fever; Plasmodium malariæ, the parasite of quartan
malaria, and Laverania malariæ or Plasmodium falciparum, producing
malignant tertian or sub-tertian malaria (and quotidian, see p. 167).
These various malarial fevers present certain clinical features in
common, which will be stated here (see also pp. 155 to 157).
For further particulars regarding malaria in all its aspects the reader
is referred to the book by Sir Ronald Ross on “The Prevention of
Malaria,” to the “Manual of Tropical Medicine,” by Drs. Castellani
and Chalmers, and to the “Tropical Diseases” of Sir Patrick Manson.
Typical malarial fevers consist of a series of pyrexial attacks which
recur at definite intervals of twenty-four (quotidian), forty-eight or
seventy-two hours, according to the parasite present in the patient’s
blood. Each attack shows three stages, a stage of rigor, a heat stage
and a stage of profuse perspiration. Following on these three stages,
there is an interval relatively or actually without pyrexia. Then the
fever returns again. A rise of temperature, often accompanied by a
general feeling of malaise, may precede the initial stage of rigor. When
the latter sets in, the patient feels intensely cold, shivers violently, the
skin becomes cold and the features pinched. There may be violent
vomiting and convulsive attacks in young children. The temperature,
however, is really above the normal, and continues to rise. After
about an hour, the shivering abates and the heat stage succeeds it.
The temperature rises rapidly, even to 106° F. The patient becomes
very flushed, the pulse is rapid, headache may be intense and the skin
dry and burning. This stage, that causes acute distress to the patient,
may last for one or often three to four hours, and then the patient
commences to perspire profusely, the clothing and bedding often
being saturated with sweat. After this, the fever rapidly declines, and
when the sweating ceases, the patient may feel almost well although
somewhat languid. The sweating stage persists from two to four
hours, so that the attack lasts as a rule from six to ten hours. After
an interval of one, two or three days, a recurrence takes place.
During the early part of the attack, especially at the stage of rigor,
there is great splenic enlargement. At first the enlargement disappears
in the interval, but in the case of repeated attacks the spleen tends to
become permanently enlarged. During malarial attacks and during
the intermission period, there is a great increase in the amount of
nitrogen excreted by the kidneys, while the excretion of iron and bile
in the fæces is increased.
Stitt455 (1914) points out that it is characteristic of malignant tertian
paroxysms that they set in with chilly sensations rather than a frank,
definite chill, and that the fever is of the remittent type.
Plasmodium malariæ and P. vivax rarely produce marked lesions
in the bodies of their hosts, as they sporulate in the circulating blood
and so do not accumulate in any one organ. On the other hand,
Laverania malariæ (Plasmodium falciparum) multiplies within the
internal organs of its host, and consequently aggregates or clusters
of the parasites occur therein. The organ in which most sporulation
occurs suffers most. The liver is generally enlarged, soft and congested.
The capsule of the spleen is tense, but the splenic consistency
is less than normal. The bone-marrow is often dark and congested
in the spongy bones and brownish-red in long bones. The blood-capillaries
of the brain and spinal cord are often filled or blocked
with sporulating parasites and large quantities of pigment are found
in these organs. Even if the parasites are absent, the pigment is
present in the endothelial cells. Pigment is found in most organs of
the body.
Atypical forms of malaria may occur in which some or all of
the symptoms are much modified. Irregular fevers also may be
produced by successive infections by the same parasite, or by the
presence of two different malarial parasites.
As regards the diagnosis of malaria, according to Manson the
three pathognomonic signs are—periodicity, the effect of quinine, and
the presence of the malarial parasite.
Treatment.—The great specific for malaria is quinine. It attacks
the merozoites or asexual generation. The drug can be administered
by the mouth, by the rectum, by intramuscular injections or
by intravenous injections, the two latter methods being adopted in
serious infections or where gastric complications are present. When
quinine is taken by the mouth, the more soluble acid salts, e.g.,
quinine bihydrochloride and bisulphate, are better than the sulphate,
the form in which quinine is usually sold. Tablets, pills and capsules
are convenient means of taking quinine but must not be old
or hard, or they may pass unchanged through the body. In the
case of mild tertian or quartan malaria, Castellani and Chalmers
recommend the administration of a dose of quinine four hours
before the sporulation of the parasite is due. Another modification
is to give 10 gr. of quinine by the mouth in the morning and a second
dose of 10 gr. as above. In many cases they give 5 to 10 gr.
of the drug three times a day. Administration of quinine per
rectum may be useful but they recommend intramuscular inoculation.
The solutions used must be sterile, and the “sterilettes,”
small, hermetically sealed vials, containing 1 grm.) or 1/2 grm.
(7 1/2 grm.) of quinine in solution, are recommended. A deep injection
into the deltoid or gluteus muscle is usual.
For pernicious infections, intravenous inoculation with not less
than 1 grm. at a time is recommended.
After the fever has subsided, the administration of quinine in
smaller doses must be continued for some time, in order to avoid
relapses.
Stitt (1914) writes that “there now seems to be a tendency to use
the alkaloid itself instead of its salts, it having been found that the
alkaloid and its very insoluble tannate are absorbed from the digestive
tract equally as well as the soluble salts.” Euquinine or ethylcarbonate
of quinine contains 81 per cent. of quinine, but is expensive.
During malarial attacks, constipation must not be allowed.
Headache can be relieved by cold applications, and perspiration must
be encouraged in the early stage by hot tea, warm lime drinks, etc.
After bad attacks, a change to a cooler climate is desirable, but the
quinine treatment must not be discontinued.
Preventive measures take two main forms, directed respectively
against the malarial parasites in man, and against the mosquitoes
that convey the parasite from man to man.
With regard to man, houses should be built away from low-lying
marshy ground, and kept free from vegetation such as grass or
brush which furnishes shelter to the mosquitoes. In the tropics, the
chief reservoirs of the malarial parasites are the native children, hence
European quarters should be away from native dwellings as far as
possible. Mosquito nets, having twenty to twenty-four meshes per
square inch, should be used invariably, and houses should be screened.
Malaria-conveying mosquitoes bite chiefly towards evening. Quinine
treatment for preventive purposes is important. A dose of 5 gr. of
quinine daily, with a dose of 10 gr. on the seventh day (Castellani), is
efficacious. Some workers, however, recommend a large dose (15 gr.)
on two consecutive days every eight or ten days for three months,
while others recommend 10 gr. twice a week. Celli administered 3 gr.
of quinine morning and evening.
The second line of attack is directed against mosquitoes, especially
Anophelines, on the lines so well set forth by Sir Ronald Ross.456 The
accumulation of small quantities of water in various vessels, many of
them unnecessary, should be prevented, as Stegomyia (Culicines) breed
in such receptacles. Anophelines breed in small pools. All drinking
water and household vessels, water-butts and cisterns must be
effectively screened with wire gauze. Cesspools, etc., must also be
screened, and they, and all collections of water, should be oiled with
crude petroleum sprays every week or ten days, or fortnight according
to some workers. The petroleum is a good larvicide and suffocates
the Anopheline larvæ, while its presence renders the site obnoxious
to the adult mosquitoes. The amount of crude petroleum or kerosene
will vary according to the locality concerned, due regard being paid
to its powers of spreading on the surface treated. Different authorities
have used different quantities, such as 1 oz. of oil to 1 square yard
or to 15 square feet. Others have used 1 pint of the petroleum to
a circle of 20 feet in diameter, while 1/2 pint for every 100 square feet
of surface has also been recommended. The larvicide used so
successfully in Panama consisted of:—
| Average mixture |
| Crude carbolic acid (containing 15 per cent. phenol) | 300 | gallons |
| Caustic soda | 30 | lb. |
| Resin | 200 | lb. |
One part of this mixture in 5,000 parts of water containing
mosquito larvæ destroys them within five minutes; 1 part in 8,000
of water kills larvæ in thirty minutes. Small fish, such as the
“millions” fish, that feed on the larvæ, can be introduced into
collections of water and are of local service. Ducks may also act as
destroyers of larvæ. The growth of water-weeds and rank vegetation,
that affords shelter to the larvae, must be prevented as far as possible.
Wherever possible hollows should be filled up, swamps and roads
should be well drained. Much good has followed the use of such
measures in Panama, Egypt, British Guiana and other places. The
ideal conditions for malaria reduction appear to consist in a combination
of general quinine prophylaxis with anti-mosquito measures.
VII.—BALANTIDIAN DYSENTERY.
This disease is also known as ciliate or ciliary dysentery. The
chief causal agent is Balantidium coli. Others are Balantidium
minutum, Nyctotherus faba, etc. (see pp. 200–206).
Balantidiasis is insidious and is marked by alternate attacks of
diarrhœa and constipation with vomiting, while mucus is passed in
the motions, which are foul smelling. There may be chronic ulceration
of the colon. Œdema of the face and limbs and anæmia may
occur.
Treatment is at present rather unsatisfactory. Castellani and
Chalmers state that “the symptomatic treatment for entamœbic
dysentery may be tried.” Various treatments, more or less empirical,
by calomel, quinine, carbolic acid in pill form, salicylic acid,
extract of male fern, methylene blue, iodine solution, rice water and
tannin enemata are mentioned by Prowazek457 (1913) and by Seifert.
E. L. Walker458 (1913) found, from experimental work, that organic
compounds of silver, e.g., protargol, were most effective. Local treatment
by large enemata of collargol or protargol seems to be indicated.
Behrenroth459 (1913) successfully treated a Prussian case with thymol,
given in 4 grm. doses every two days, followed at the end of a
fortnight by de-emetinized ipecacuanha, given in pills containing
6 cg. each, to the number of thirty a day. In about another fortnight
the symptoms had subsided. The thymol checked the
diarrhœa, but it was necessary to give the de-emetinized ipecacuanha
to kill off the balantidia still present. Phillips (1915) also recommends
thymol. Ardin-Delteil, Raynaud, Coudray and Derrieu (1914)
found neither emetine hydrochloride nor protargol of use.
As regards prophylaxis Walker states that pigs “should be
confined and not allowed to run in yards and dwellings.” Behrenroth
considers that dirty hands, for example, those of farm workers
brought into contact with pigs, are probably the medium of infection.
The personal cleanliness of such persons is, then, of the
greatest importance.
This section, except for minor corrections, is
practically a translation of the original.
PLATHELMINTHES (Flat Worms).
BY
J. W. W. STEPHENS, M.D., B.C., D.P.H.
FASCIOLIASIS.
Fasciola hepatica.
The symptoms of disease evoked by Fasciola hepatica are rarely
observed in our part of the world, whereas Kermogant460 states them
to be of frequent occurrence in Tonkin461; the parasites are there
called “Douves.” In our experience they are only accidentally
found post mortem in a certain number of cases, as no changes are
manifested during life which would permit of any conclusion being
drawn as to the presence of these parasites. In three cases (Bierner,462
Bostroem463 and Sagarra464) icterus was present; in a fourth case, recorded
by Duffek,465 the parasites had led to a severe and acute distomiasis of
the liver, combined with chronic purulent and ulcerative cholecystitis,
with purulent cholangitis and dilation of the bile-ducts and numerous
small abscesses of the liver. The total number of flukes found in
these cases amounted to about fifty. The parasites passed from the
duodenum into the bile-ducts, and first obstructed the flow of bile
and then set up icterus, followed by cholecystitis and cholangitis.
As regards localization of the liver fluke in the pharynx, see p. 242.
The treatment must be directed to the principal symptoms;
prophylaxis is especially important in districts where distomiasis is
of frequent occurrence. As the embryos live in water, only boiled
or filtered water should be drunk. The attempts of Tappeiner466 to
discover an effective remedy against liver-fluke disease (liver rot), so
prevalent among sheep, were unsuccessful.
Fasciolopsis buski.
This parasite lives in the intestine, not in the liver of man; it
produces bloody stools and typical symptoms—high fever and a
condition of apathy (Odhner).467
PARAGONIMIASIS.
Paragonimus ringeri.
The disease produced by the lung fluke is specially endemic in
Japan, also in isolated parts of China, Formosa and Korea. The
fact that the lung-fluke disease is most frequently found in mountainous
districts (Katsurada468) is worthy of special attention. The onset
of pulmonary paragonimiasis is generally insidious (Looss469); generally
the only symptom is a slight cough, occurring at first at longer, and
later at shorter intervals; it is accompanied by the expectoration of
discoloured sputum, frequently blood-stained. Though now and
then severe hæmorrhages result, up to the present no case has been
established in which they have been the direct cause of death.
Examination of the thorax frequently fails to reveal anything
abnormal. Inouye470 states that the most frequently observed changes
consist in retraction of the thorax and in a contraction of its infrascapular
portion. Scheube471 repeatedly observed that the one side,
presumably that which harboured the worm, moved less freely than
the other. The physical changes are not uniformly spread over the
whole lung, but are localized. The disease may come to a standstill
for long intervals and then set in again, lasting on the whole from ten
to twenty years. In addition to paragonimiasis of the lungs, cysts are
frequently found on the eyelids, which occasionally extend deeply into
the orbit and hinder the movements of the eyes. Post mortem, cysts
the size of hazel nuts containing one, two, or three adult worms
are found in the lungs, and in addition, not uncommonly there exist
pulmonary emphysema and bronchiectasis. Besides being present in
the lungs and in the eyelids, the parasites have also been found in the
pleura, the liver, the intestinal wall, the peritoneum, the cervical
glands, and in the scrotum, without actually occasioning any actual
symptoms in these tracts.
The most dangerous locality is in the brain. Otani,472 Inouye,473
Yamagiva,474 and recently also Taniguchi,475 have found post mortem
the worms and their ova in tumours of the brain, or, in areas of
softening in cases of Jacksonian epilepsy; in Taniguchi’s case the
eggs were found in masses in the inflammatory areas of softening.
In the nineteen cases of paragonimiasis of the brain collected by
Inouye, the following symptoms were observed: general convulsions
on eight occasions, unilateral convulsions on six occasions, convulsions
with paralysis on the same side and hemiplegia, five times each;
in Taniguchi’s case, attacks of cortical epilepsy, choreiform twitchings
in the right extremities, which gradually become athetotic. The
following were symptoms of rarer occurrence: paresis of the right
upper extremity, vertigo, dementia, and amnesic aphasia, disturbances
of vision. Paragonimiasis of the brain appears to arise by
embolism from a primary pulmonary lesion.
The diagnosis depends upon the finding of ova in the sputa; if
together with ova in the sputa, cerebral disturbances make their
appearance, in all probability the cause is the presence of worms or
ova in the brain.
The prognosis of pulmonary paragonimiasis is favourable; on
the other hand, that of cerebral paragonimiasis is very doubtful.
The treatment of the pulmonary lesion consists only in paying
attention to the general condition (good food, rest, cough remedies),
as all attempts to destroy the worms in the lungs by means of vermicidal
drugs administered internally or by way of inhalation have so
far been without result. The treatment of the cerebral lesion is
entirely hopeless. Trephining has been proposed for cases the condition
of which is more favourable, but it has not reached the stage
of performance.
Prophylaxis consists in general management: cleansing and if
need be boiling of everything that is eaten or drunk.
Clonorchis sinensis.
According to our present knowledge Clonorchis sinensis is only
found in China and Japan; even the post-mortem case reported by
Laspeyres476 was that of an Asiatic sailor who was admitted into the
General Hospital St. George, Hamburg, in a moribund condition
with the clinical diagnosis of beri-beri. The bile-ducts are the usual
site of the parasite, though Katsurada477 has found them also in the
pancreatic ducts. In addition, it is found not uncommonly in the
upper portion of the small intestine, especially in the duodenum,
also, though decidedly rarely, in the stomach. As these sites, however,
do not afford the conditions necessary to life, they are only found
here on their way out of the body of the host.
The initial stage of infection with this fluke generally runs a
symptomless course; in proportion as the worms multiply the
following symptoms are manifested: First there is a morbid sense of
hunger and irregularity in defæcation; at the same time the patient
experiences a feeling of pressure and pain in the epigastrium and right
hypochondrium, or just a dull pain. Pressure increases the pain
considerably. The liver appears to be enlarged, sometimes the
enlargement is specially perceptible over the left lobe of the liver.
The patients maintain a proportionately good general state of health in
this state for a long time and may hope to recover. In severe cases
there occurs copious and generally bloody diarrhœa, also icterus.
The next stages are anæmia, emaciation, epistaxis, ascites, enlarged
spleen, and cachexia, to which the patient finally succumbs. In
general the course of the disease is very chronic and irregular; in
winter and spring there is generally improvement, in the summer and
autumn the patient gets worse. At post-mortem the bile-ducts are
enlarged and thickened, there is interstitial hepatitis with enlargement
of the liver, but not to such an extent as in hypertrophic cirrhosis.
After the initial enlargement contraction of the liver sets in, the
peritoneal coat and capsule proper of the liver become more or less
thickened in places. In the pancreas also dilatation and thickening
of the ducts occur, as well as interstitial inflammatory processes.
Obstructions in the portal circulation may lead to catarrhal changes
in the stomach.
The diagnosis is based on the demonstration of ova in the fæces.
As a radical treatment is still unknown, consequently it can only be
purely symptomatic. Prophylaxis consists in the prohibition of
drinking unboiled water or eating uncooked molluscs, fish, etc., of
canal water. Leaving the epidemic region may bring about gradual
recovery.
BILHARZIASIS.
Schistosoma hæmatobium.
The symptoms of bilharziasis are manifested chiefly in the urinary
apparatus, and above all as hæmaturia, at the outset without any
special troubles. Later, however, it is accompanied by subjective
symptoms in the shape of feelings of pain, and of vague pains in the
perinæum and lumbar region, and of burning in the urethra during
the passing of urine. All the symptoms are usually aggravated after
excesses in eating and drinking, and after considerable bodily exertion.
Another condition found, but not often mentioned, is lipuria (Stock478);
the highest amount has been 2 per cent. fat in the urine. Stock found
6 to 20 per cent. of eosinophile cells in ten cases examined by him.
They appear to be increased, especially in the early cases; Kautsky479
also called attention to the excessive degree of eosinophilia, whilst
Goebel480 expresses the opinion that a specific toxic action on the
organism generally is not developed in bilharziasis. Kautsky481 assumes
a toxic anæmia as in the case of ancylostomiasis. English authors
also have called attention to the eosinophilia and to a considerable
amount of leucocytosis (Balfour,482 Douglas and Hardy483). The
severe forms occur almost exclusively in men; symptoms of catarrh
of the bladder make their appearance, vesical calculi are frequently
found, whilst the formation of stone in the kidneys and ureters is
rare. Urethral fistula occurs in bilharziasis, often without stricture,
and if granulations occur the fistula is distal to them. Goebel484
regards the bilharzia fistula as a chronic burrowing of pus, caused by
the irritation set up by the ova as foreign bodies and consecutive
restricted suppuration; and secondly as due to the passage of urine
through the defect in the epithelium or the wall of the urethra. The
fistulæ, which are generally situated at the neck of the bladder and at
the membranous portion, are very tortuous and frequently very
numerous; they often lie embedded in well-marked tumours—in fact,
in granulation tumours with marked inclination to excessive formation
of cicatricial tissue. The opening generally is in the perineal and
scrotal regions. In the case of a patient, aged 21, from the Transvaal,
Kutner485 found by cystoscopic examinations the whole summit and
walls of the bladder covered with large and small tumours. In
addition to smooth glistening tumours, others were more or less disintegrated,
and scattered large and small cauliflower-like growths
occurred. Like malignant growths, the tumours were inclined to
break down, the process extending from within outwards towards the
surface. Whether the hydrocele so frequent in Egypt has any
connection with bilharzia is not known. A frequent sequela of
bilharziasis is complete sexual impotence (Petrie486).
Bilharziasis of the rectum is manifested by symptoms of
dysentery; the repeated violent attempts at defæcation lead in time to
prolapse of the rectum, which sooner or later induces septic infection
and so death. In the mucosa of the rectum, polypoid growths similar
to those in the bladder are met with, due to the ova of the parasites
in the mucosa and submucosa. In the case of a man, aged 36, who
had lived for a long time in South Africa, Burfield487 found in the
excised vermiform appendix ova of Schistosoma hæmatobium; he
assumed this to be a gradual secondary infection of the appendix,
whilst Kelly488 mentions a case of primary bilharziasis of the appendix;
the eggs lay in the submucosa directly above the muscularis.
Tumours containing numerous ova are frequently found in the region
of the genitalia, thighs and scrotum. In one case Symmers489 found
numerous male schistosomes in the portal blood and a copulating
pair in the left lung. Though schistosome eggs have been found by
some observers in the lung tissue, this is nevertheless the first case in
which living parasites have been found in the lesser circulation.
Perhaps they got there by way of the external iliac vein from the
veins of the bladder and rectum.
In the female sex bilharziasis is incomparably rarer than in the
male and is generally limited to hæmaturia. Bilharziasis of the
vagina, which takes the form of an acute vaginitis, is frequent
according to Milton.490 Horwood491 found in one case a polypoid
tumour of the cervix uteri, and in the connective tissue of the tumour
Schistosoma ova, both in masses and singly. It could not be
established whether the ova reached the vagina and thence the cervix
directly, or through the urine from the bladder.
The course of the disease is chronic, and in slight cases, provided
fresh infections do not occur, is not unfavourable; in severe cases
the cachexia caused by loss of blood, or intercurrent diseases to
which the patients easily succumb—e.g., pyelitis, pyelonephritis,
pyæmia, or uræmia—lead to a fatal issue.
In regions in which Schistosoma hæmatobium is endemic, or in
patients from such regions, the diagnosis is easy by microscopically
finding the eggs in the urine.
As regards the treatment of the affection this much must be said,
that so far there is in existence no certain remedy. In countries
where bilharziasis is endemic copaiva balsam is considered a specific.
Kutner (loc. cit.), however, in the case of his patient who for a long
time had taken no inconsiderable amounts of copaiva, had no success
worth speaking of to record. Urotropin (three times daily, 1 grm.)
has similarly failed, salol (0·75 grm. several times daily) perhaps affords
relief in affection of the bladder (Milton). Methylene blue, oil of
turpentine with extract of male fern (Brock492), or the latter alone and
santonin given in small doses for a week at a time, in the morning, are
said by Petrie493 to be of value. Sandwith494 and Harley495 were not
very successful. By way of experiment Kutner for some time used
collargol per rectum, proceeding on the assumption that this preparation,
which has proved of such remarkable service in bacterial infection,
would perhaps render a continuance of life difficult for the
bilharzia worms. But this hope proved illusory. In order so far
as possible to limit the loss of blood, Kutner regularly employed
stypticin for long periods (three times daily, two tabloids of 0·01 grm.)
with undoubted success, in so far that the hæmorrhages became
considerably less in amount. As two patients in the course of enteric
fever lost their hæmaturia, Stock accordingly recommends subcutaneous
injections of Wright’s typhoid vaccine. In the early stages
of the rectal lesion suppositories of iodoform, ichthyol, or narcotics
might possibly be of use. In the case of urethral fistulæ, division,
excision and scraping out of the granulation tissue are recommended;
in cystitis with formation of tumours high resection with curetting
of the tumours or their destruction with the cautery; in the case of
vesical calculi, high resection, curetting the bladder, and then
drainage. Tumours of the rectum must also be removed by
operation.
Prophylaxis is important; it should be extended to all modes of
using water, only filtered water being drunk, and only boiled water
being used for washing. This advice should be given to tourists who
travel through the infected districts, and is also recommended to
soldiers and officials who are despatched to the Colonies. The favourable
influence of change of climate can only show itself where fresh
infections are avoided.
CESTODES.
GENERAL.
It seems advisable to preface the section on the Cestodes with
some general observations on the symptoms of disease provoked by
tapeworms, especially so far as they relate to the question of toxic
effects, and to include the Nematodes in this discussion. After this
will follow a brief exposition of the most important intestinal lesions
causally connected with intestinal parasites.
It is known to every experienced practitioner that the different
intestinal parasites can give rise to a series of nervous symptoms,
slight or severe, and produce, above all, blood changes—anæmia of
the most varied nature, to the extent of severe progressive anæmia.
These symptoms are regarded by many authors as reflex, or, as in
the case of ancylostomiasis, the main feature from the loss of blood
caused by the habit of life of the intestinal parasites. More frequently,
however, they are regarded as toxic conditions produced by
the parasites. In view of this divergence of opinion there appears
to be some advantage in defining clearly the present position as to
the toxic action of parasites. Most interesting in this respect are
Dibothriocephalus latus and Ancylostoma duodenale.
We are indebted to the clinic at Helsingfors for our most detailed
knowledge of bothriocephalus anæmia. Reyher496 was the first to
demonstrate that this parasite under certain circumstances can produce
a severe, progressive and sometimes fatal anæmia, which can be
cured, generally in a surprisingly short time, by expulsion of the
worm. Among the various hypotheses which have been advanced
as to the mode of origin of bothriocephalus anæmia, the greatest
importance has been attached to the assumption already mentioned
by Reyher, but definitely expressed by von Shapiro,497 to the effect that
Bothriocephalus latus produces a poison which is absorbed by the
intestine and exercises a deleterious influence on the composition of
the blood, especially on the erythrocytes, perhaps also on the blood-forming
organs. This assumption is supported by no slight number
of clinical and experimental investigations. Podwissotsky498 observed
severe blood changes in a child, aged 4 1/2, affected with B. latus. In
the case reported by Pariser499 the severe anæmia in a girl disappeared
fairly soon after expulsion of the worm. In that reported
by Schaumann500 high fever accompanied the bothriocephalus anæmia;
he also proved the hæmolytic properties of the broad tapeworm.
The case reported by F. Müller501 was one of severe anæmia. Also, in
the first of the cases described by Kurimoto502 of Diplogonoporus
grandis there were present the same symptoms of anæmia as in the
case of B. latus. Meyer503 observed severe anæmia in two youths
caused by B. latus. Rosenquist504 has discussed the proteid metabolism
in anæmia. The presence of B. latus produces in the majority of
cases an increased proteid consumption, to which the blood change
generally corresponds—toxic anæmia; in a further communication
he reports on twenty cases of bothriocephalus anæmia, nineteen of
which were cured by expulsion of the worms, while one case proved
fatal, and he again emphasizes the toxic properties of the intestinal
parasites. In the case reported by Bendix,505 that of a girl, aged 4 1/2,
the anæmia was moderate, whilst in the case of Zinn506 (a woman,
aged 30) the anæmia was so excessive that the patient succumbed
five days after expulsion of six bothriocephalus heads. Isaac and
van den Velden507 have established that in the serum of patients who
suffer from anæmia due to B. latus, parasitic products are dissolved,
as shown by a distinct precipitin reaction. Galli-Valerio508 considers
it likely that toxic substances are secreted by the living helminthes
which produce a lowering or raising of the body temperature, nervous
disturbances and hæmolysis. Tallqvist509 succeeded in extracting
from B. latus a lipoid-like body which had a strong hæmolytic action.
The experimental anæmia thereby produced differed in no respect
from the severe chronic bothriocephalus anæmia of man. The
question as to under what special conditions severe, and sometimes
fatal bothriocephalus anæmia is developed is answered by Leichtenstern510
and by Lenhartz,511 by the assumption that among the Bothriocephali
some are toxic, that is, manufacture a poison which, when
absorbed by the host, produces a severe anæmia.
Certain factors lead him to conclude that an accumulation of
poison, dependent on time and place, occurs in the Bothriocephali.
In the case of ancylostome anæmia, experience so far, according
to Leichtenstern,512 by no means supports the hypothesis of a difference
in virulence of the worms according to time and locality, ancylostome
anæmia being rather, so far as is known at present, in all races of
man, everywhere and at all times, simply and solely dependent on
the number of ancylostomes, the duration of the disease and—within
certain narrow limits—on the individual capability of resisting
the loss of blood and the toxic effect of the parasites. As is shown
by a short historical résumé of the toxic action that has to be considered
in ancylostome anæmia, we must admit that doubtless here,
as in the case of bothriocephalus anæmia, the toxins secreted by the
parasites exercise a hæmolytic action, even while admitting Leichtenstern’s
contention that the significance of the loss of blood due to
ancylostomes must not be underrated. The toxic hypothesis acquired
a definite standing through a series of experiments of Lussana513 on
rabbits, where he succeeded in producing anæmia by injecting urinary
extracts of ancylostome patients. Arslan514 extracted toxins from the
urine of two ancylostome patients and injected them into rabbits,
which thereupon sickened and showed the same blood changes as
the ancylostome patients. Retinal hæmorrhages, so frequent in
ancylostome anæmia, which, according to Fischer515 and Samelsohn,516
are not due to direct loss of blood, must also be ascribed to a parasitic
toxin. A further argument in favour of the toxic hypothesis
is furnished by the blood changes recorded by Zappert,517 Müller and
Rieder,518 Bücklers,519 and Neusser,520 which must be regarded as the
expression of toxic action, especially with reference to eosinophilia.
The striking increase in proteid destruction in ancylostomiasis
observed by Bohland,521 and which ceased after the parasites had been
expelled, also gives additional support to the assumption of toxic
action. The observation of Daniels522 also deserves consideration in
this connection, according to which the presence of yellow pigment
in the liver and kidney cells is to be attributed to blood destruction
by a verminous toxin absorbed from the gut. Looss523 considers it
not at all improbable—in fact, almost certain—that Ancylostoma, in
addition to withdrawing blood, exert a kind of toxic action on their
host.
Scheube524 attributes almost equal importance to the loss of blood,
the digestive disturbances, and the intoxication induced by certain
metabolic products of the parasites. According to v. Jaksch525 ancylostome
anæmia is not induced solely by loss of blood, but by the
fact that the parasites produce a ferment which has a toxic action
and produces stimulation in those organs in which the eosinophile
cells arise. The hæmolytic action of ancylostomes has frequently
been observed by Galvagno526 in men employed in sulphur mines.
According to Loeb and Smith527 the anterior half of the body of
ancylostomes contains a substance which probably causes anæmia.
Bauer528 found in the urine of ancylostome patients glycuronic acid,
which he considers to be a sign of metabolic disturbance due to
parasitic toxins. As has been demonstrated by Allessandrini,529 the
secretion of glands in the anterior part of the body has a distinct
hæmolytic effect on the erythrocytes. While the worm attaches
itself to the mucosa by means of its teeth, these glands discharge
their secretion, producing hyperæmia. The extravasated blood is
acted on by this secretion, so that it can serve as food for the
parasites. Hynek530 attributes eosinophilia (up to 20 per cent.) to
a toxic action. Goldmann531 expresses a similar opinion, though he
assumes that the anæmia is secondary, as the toxin of the cephalic
glands, as the parasites bite, penetrates the mucosa and thence into
the blood, where it dissolves the red blood corpuscles. Romani532
discusses the agglutinating hæmolytic action of the serum of ancylostome
patients. Whether Ancylostoma produce toxins and what
is their nature, or whether the loss of blood causes the anæmia,
Liefmann533 was unable definitely to determine; hæmolytic substances
do not appear to take any part in it.
Berti534 also is inclined to attribute the anæmia to metabolic
products of the ancylostomes; he found, in fact, that a serum obtained
from a sheep (after subcutaneous injections of the culture fluid of
ancylostome larvæ) was efficacious in the treatment of ancylostome
anæmia. Peiper535 likewise assumes that the parasite secretes a cell
toxin. Löbker536 at the present day still maintains that the cause of
the disease must be looked for really, if not perhaps entirely, in the
continued withdrawal of blood by the parasites; the secretion of
toxins by ancylostomes has not yet, in his opinion, been conclusively
proved. Except in the case of Bothriocephalus latus, referred to
previously, toxic action appears to be of quite subordinate importance
for the other Cestodes occurring in man—especially Tænia solium and
T. saginata, which are most frequently found; thus Cao537 flatly denies
the presence of toxins in the body of Tæniæ, while others, such as
Messineo and Calmida,538 Jammes and Mandoul,539 consider they are
justified from their investigations in concluding that Tæniæ contain
a specific toxin. Messineo540 injected, with all bacteriological precautions,
extracts of Tænia, dissolved in physiological salt solution.
He invariably obtained severe motor disturbances and frequently
death. The observation by Pereira541 of a case of chorea in which
rheumatic and cardiac symptoms were absent and which after expulsion
of a Tænia was quickly cured, also favours the view of a toxic
action. Barnabo,542 however, was unable to obtain a toxin from Tænia
saginata. Gagnoni,543 on account of a marked eosinophilia which,
after expulsion of a Tænia saginata, fell within fourteen days to
1 per cent., assumes the formation of a Tænia toxin. Dirksen’s544
observation has reference to a sailor affected with serious anæmia,
who, after expulsion of twelve pieces of Tænia solium, was rapidly
cured. A portion of the worm was already breaking down, the
absorption introducing into the body highly toxic hæmolytic products,
to which the anæmia must be ascribed. How far the serious
disturbances of the nervous system, frequently to be observed in
cases of Hymenolepis nana, are to be considered as of purely reflex
nature or toxic must remain an open question; the same applies to
Dipylidium caninum, in which case Brandt545 observed serious central
nervous symptoms. Caution is necessary in judging as to any connection
between worm stimulus and nervous symptoms in cases of
Ascaris infection. Peiper546 is inclined to regard such nervous
symptoms not as reflex, but rather as due to a toxin contained in
the helminthes, or metabolic in origin.
In cases of pernicious anæmia when the symptoms disappear after
expulsion of Ascaridæ a toxic action must be assumed (Demme547).
Additional clinical observations do not, indeed, lead to any definite
conclusion as to the question whether Ascaridæ produce a toxin which
is capable of causing more or less injury either to the nervous system
or to the blood, yet it may be worth while to give a brief review
of this question. In a case of Kutner’s,548 that of a girl, aged 12,
there was a hæmolysis which was cured after expulsion of twenty-four
Ascaridæ. Attacks of opisthotonos in a girl, aged 16, ceased
after seventy-eight Ascaridæ had been expelled (Lutz549). Unusually
serious disturbances were observed in a man, aged 26, who was rapidly
cured by Drouillard550 by the removal of a great number of Ascaridæ.
The observations on pseudomeningitis are of especial interest; they
are evidently toxic in origin as in the case of Annaratone,551 of a man
who was taken ill with gastro-intestinal symptoms and who died with
meningitic symptoms. Post mortem the brain was normal, but the
stomach contained a great coil of Ascaridæ. The cases of Delille,552
Mériel,553 Papi554 (the occurrence of Cheyne-Stokes respiration has been
ascribed to the action upon the centre in the medulla oblongata of
the products of the Ascaridæ), and Taillens555 related to children in
which the meningitic symptoms (meningismus), partly serious, disappeared
with the removal of the Ascaridæ. Máreo556 designates this
disease helminthiasis meningitiformis, which exhibits all the symptoms
of meningitis, but which is caused by the metabolic products of
Ascaridæ.
Schupfer,557 Duprey558 (observations in the West Indies, where such
symptoms are said to be of very frequent occurrence), Naab559 (the flow
of water from the mouth at night is mentioned as a remarkable fact),
and Hammiss560 assume the action of an Ascaris toxin in the clinical
observations made by them, mostly children with fever and intestinal
symptoms. Schupfer assumes in such cases, as he observed it once
in a man, aged 23, that the disease termed Lombricoise à forme typhoïde
by Chauffard was due to B. coli of marked virulence due to the
action of the Ascaridæ. The Widal reaction was negative. Koneff561
reports a case in which acute attacks of cramp, trismus, and rigidity
of the pupil disappeared after expulsion of seven Ascaridæ. Tetanus,
as observed by Buchholz562 in a girl, aged 17, and rapidly cured after
expulsion of sixteen Ascaridæ, is manifestly rare, since only Rose563
mentions this as a cause in his article on Tetanus. Only a few
experimental data exist. Cattaneo564 could detect only a very weak
toxin in Ascaris, while Messineo,565 by injecting into animals extracts
in physiological salt solution, invariably succeeded in producing serious
motor disturbances and frequently death. Interesting also are the
observations of Huber,566 who, after working with Ascaridæ, suffered
from itching of the head and neck, blisters, swelling of the ear, conjunctivitis,
ecchymosis and troublesome palpitation in the head. He
consequently assumes that Ascaridæ can induce irritation by chemical
(toxic) means.
In the case of Trichocephalus dispar no more than in the case of
Ascaris lumbricoides can we speak with certainty of a toxic effect, even
though a number of observations are available which might justify
such an assumption as regards these intestinal parasites. Barth567 found
the brain normal in a man who had died with meningitic symptoms,
but the intestines were full of Trichocephalus dispar; Gibson568 records
the rapid cure of serious cerebral symptoms after expulsion of Trichocephalus,
so also Pascal,569 Burchhardt570 and Rippe.571 Moosbrugger572
was the first to draw attention to grave anæmic conditions induced by
Trichocephalus, Morsasca573 and Becker574 to progressive grave anæmia
(trichocephalus anæmia is accompanied by marked reduction of the
number of red blood corpuscles, of the specific gravity and of the
hæmoglobin, well-marked morphological changes of the red cell,
micro-, macro-, and poikilocytosis and nucleated red cells). Sandler,575
in his case of a boy, aged 11, who died of anæmia, assumes a trichocephalus
toxin to be the cause of the disease, and Kahane also reports
on anæmic conditions induced by Trichocephalus. Girard,576 in addition
to symptoms in the gastro-intestinal tract, calls attention to those
arising in the blood—anæmia and its sequelæ—and also to nervous
symptoms: cerebral phenomena, headache, giddiness, aphonia, symptoms
of meningitis. In a case of Schiller’s577 high fever was present,
which probably set in when the Trichocephali present in the gut in
great numbers commenced their parasitic activity. Hausmann,578 in
order to explain the adaptability of Trichocephalus, assumes that
according to the locus minoris resistentiæ, at one time the reflex at
another the toxic action is effective, now on one organ, then on
another; anæmia being present in most cases, frequently general
and local neuroses and cerebral symptoms of various kinds.
With regard to the toxic action of Oxyuris there is only the single
record of Hartmann,579 who noticed the disappearance of epileptic
fits and psychic disturbances in a girl, aged 13, after the removal of
Oxyuris. Nervous disturbances and blood changes can but rarely be
attributed to Strongyloides. Silvester580 and Valdes581 report on giddiness,
headache and anuria in cases observed by them; whether the
eosinophilia recorded by Bücklers582 and Bruns583 is due to the toxin
of Strongyloides must remain an open question.
Reference has already been made to the possibility that intestinal
ciliates (Balantidium coli) can also produce toxins.
The contents of echinococcus cysts appear to contain a substance
only moderately toxic, giving rise to urticaria, in a series of cases
where the fluid has escaped into the abdominal cavity (during
puncture). D. Müller584 has collected nine such cases out of the
literature, to which may be added six cases of Finsen585 in which the
escape of fluid into the peritoneal cavity led to severely itching
urticaria, which usually disappeared again after one or two days.
On one occasion, indeed, urticaria occurred after rupture into the
pleural cavity. In the case recorded by Caffarena586 of echinococcus
of the right lobe of the liver, widespread urticaria developed as the
result of the exploratory puncture. In the case of an echinococcus
of the liver rupturing into the abdominal cavity La Spada587 ascribed
the symptoms leading to death to toxic influence while the peritoneal
symptoms were less marked. Eosinophilia in hydatid disease is slight
according to the investigations of Bindi588 and Santucci,589 and is,
according to Welsh and Barling,590 no certain sign of echinococcus;
it is independent of the age, sex and temperature of the patient, but
upon rupture of the cyst eosinophilia invariably sets in.
The question as to the importance of helminthes in relation to
certain diseases of the gut requires special discussion, but it concerns
only Ascaris lumbricoides, Oxyuris vermicularis, and Trichocephalus
dispar, and the question of appendicitis first of all. The entrance of
intestinal parasites into the vermiform appendix was already known
to medical men in the fifties of last century, as is shown by the works
of Merling591 (1836), Zebert592 (1859), Platonor593 (1853), and Schachtinger594
(1861). Most of these authors have considered intestinal worms,
together with other foreign bodies, to be the cause of appendicitis.
As regards the part played by these intestinal parasites in the etiology
of appendicitis, so much discussion has taken place during the last
few years that it is worth while to give a résumé of the later views on
this question, even though at the outset it must be admitted that the
matter is not cleared up. Bergmann595 records a case in which an
Ascaris perforated the appendix and got into the peritoneal cavity.
Strümpell596 reckons among the symptoms of Trichocephalus the
possibility of a “typhlitis.” On account of the marked sensitiveness
of the ileo-cæcal region, Boas597 mentions the possibility of confusing
it with appendicitis. Still598 regards Oxyuris as a principal cause of
catarrhal affections of the appendix. Arboré-Rally599 regarded severe
symptoms of appendicitis in a boy, aged 10, as due to Ascarides.
In all cases of appendicitis Metschnikoff600 requires a microscopical
examination to be made for eggs, and considers treatment for worms
carried out otherwise as a cause of the frequency of perityphlitis.
Matignon601 does not agree with this opinion, as in spite of the extraordinary
frequency of intestinal worms in China, he has only seen
one case of appendicitis in four and a half years, and Des Barres602
expresses himself in similar fashion. Out of twenty-one cases of
appendicitis Kirmisson603 discovered the ova of Trichocephalus eighteen
times and the ova of Ascarides in three of these cases; in twelve cases
of enteric fever the examination for eggs was negative nine times.
Moty604 considers Oxyuris to be the sole cause in his three cases of
appendicitis. Girard605 ascribes to Trichocephali the rôle of more
or less septic foreign bodies which may bring about the entry of
intestinal bacteria into the appendix, and Triboulet606 describes a
case of appendicitis which he considers was due to Ascaris. In
Morkowitin’s607 case numerous Oxyuris had clearly caused the
appendicitis. von Genser608 records the case of a boy, aged 5, who
was operated on for appendicitis, and who passed through the operation
wound a living Ascaris on the eighteenth day after the operation.
In the first case communicated by Schiller609 the disappearance of the
typhlitic swelling after the discharge of the Ascarides pointed to the
etiological significance of the parasites, and the same obtained in a
further case published at an earlier date by Czerny and Heddäus.610
In a case abstracted by Kaposi611 Trichocephali appear to have been
a contributory cause in the production of the appendicitis. In a
further case reported by Schiller, where the appendix was removed,
it was shown that Oxyuris had given rise to a pronounced appendicular
colic. In a girl, aged 13, who died from diffuse peritonitis,
Schwankhaus612 found that an Ascaris had perforated the appendix.
Ramstedt613 found in an extirpated appendix a whole “tangle” of
Oxyuris, and believes in the possibility of their having provoked the
inflammation; he recommends an examination for entozoa before
the operation, without, however, after Metschnikoff’s example, substituting
worm treatment for the operation. Rostowzeff614 ascribes
only a minimal direct etiological significance to intestinal worms
in the origin of appendicitis; in 163 cases he found worms in three
instances. Wirsaladze615 expresses himself in a similar fashion.
Oppe616 observed Oxyuris six times in excised appendices, and
emphasizes the opinion that in appendicitis the question of a worm
cure ought to be taken into consideration. Ascaris and Oxyuris, if
no contra-indication exists, may be expelled, but in the case of
Trichocephalus, which frequently defies all expulsive treatment, no
attempt should be made, but operation proceeded to forthwith.
In a case briefly reported by Hanau617 Oxyuris was undoubtedly the
etiological starting-point; in a case of Galli-Vallerio618 Oxyuris and
Trichocephalus. In the opinion of Ssaweljews619 in some cases of
appendicitis, in addition to other causes, intestinal parasites play a
prominent part. The case recorded by Nason620 is an interesting one;
in this an Ascaris in the appendix became twisted with it round a coil
of gut, causing obstruction. Spieler621 argues against the underestimation
by many authors as to the part played by intestinal worms
in producing appendicitis, although he also does not regard them
as a frequent, to say nothing of an exclusive, cause of the disease.
In a case recorded by Bégonin622 fifteen Oxyuris were found in the
excised appendix (the mucosa showed some ulceration), and in another
recorded by Putnam623 twenty Oxyuris were present in the appendix,
in which there was no evidence of any change. The standpoint
Schilling624 takes is to the effect that entozoa irritate the mucosa and
can increase an already existing inflammation, but he considers it
very questionable whether they can produce appendicitis. Blanchard625
assumes the possibility of a secondary infection arising from lesions
of the mucosa produced by helminthes (Ascaris and Oxyuris).
Moore626 considers Trichocephalus the excitant of the appendicitis
in his case. In a second case of appendicitis recorded by Auley627
operation became unnecessary owing to the passage of the Ascaridæ.
Page’s628 case is an interesting one; it was that of a man who came
up for operation with a diagnosis of appendicitis. On incising the
abdominal wall numerous Ascarides were found at the base of the
wound, lying in cavities; even after eight days Ascarides escaped
from the wound. The author assumes there was a perforation of the
gut wall; it is strange that the worms were able to exist a proportionately
long time in the muscular tissue. Schoeppler629 states that there
is the danger of an appendicitis even after the death of an Oxyuris
that has found its way into the appendix. Oui630 met with two
specimens of Trichocephalus which had become embedded by their
thin ends deep in the mucosa. Frangenheim631 is not in a position to
pronounce any opinion as to what part intestinal parasites play in the
etiology of appendicitis. In a case recorded by Kahane632 many
Trichocephali were found partly free in the appendix and partly
embedded in the mucosa; microscopically appendicitis was diagnosed.
At a laparotomy for salpingitis Heekes633 found the appendix
elongated, thickened, and containing about eleven Oxyuris without
the mucosa being in any way changed. In one case Andrews634 claims
Ascarides to have been the direct cause of the appendicitis. The
literature dealing with this question, so important in our time, has
been collected almost without any omissions, but, unfortunately,
no decisive opinion as to the significance of parasites in appendicitis
can be inferred from it. The vexed question whether intestinal
parasites, especially Ascaris, are able to penetrate the intestinal wall is
just as little finally decided. Leuckart,635 Heller,636 Mosler and Peiper,637
Henoch,638 Davaine,639 Küchenmeister,640 and Bremser641 are opposed
to the idea that the healthy intestinal wall can be penetrated by
intestinal worms, especially Ascarides, whilst a whole series of other
authors are of the opinion that even the healthy intestinal mucosa can
be perforated. Among these is numbered Mondière,642 who is of the
opinion that Ascaris, by violent pressure against the mucosa, forces it
so much apart that it is enabled to escape through the gap thus
formed into the peritoneal cavity; this opinion is shared by
v. Siebold.643 Rokitansky644 considers perforation of the gut by Ascaris
as at least a rare occurrence. Gerhardt645 does not doubt that the
worms can actively perforate the intestine. Cases like those of
Abrault,646 Apostolides,647 Marcus648 (recorded by Perls as a valid
example of “ascaridophagous” gut perforation), Wischnewsky,649
Galvagno,650 Salieri651 certainly show that perforation of the healthy
gut wall cannot be denied, but at the same time that this occurrence,
compared with the frequency of Ascaridæ, should be regarded as
exceedingly rare. It is another matter as to whether it is possible
for the worms to penetrate an intestinal wall already diseased,
especially when ulcerated; a whole series of observations are in
favour of this. In Lini’s652 case (fifty-six Ascarides escaped from the
umbilicus of a girl, aged 7), in Gräffe’s653 (eighty Ascarides escaped
from an inguinal tumour), in Nicolino’s654 (perforation of the intestinal
wall with strangulated hernia), in Liesen’s655 (a living Ascaris in the
peritoneal cavity in a woman suffering from a peritoneal abscess)—in
these it is clear that disease processes in the intestine preceded the
exit of the worms. In a case described by Boloff656 the Ascarides
appear to have produced, by forming a tight coil, necrosis of the
gut with perforative peritonitis. In a case recorded by Lutz657 the
perforative peritonitis was without doubt provoked by Ascaris, and
in one by Schiller658 the Ascaris had clearly gained access to the
peritoneal cavity through a gunshot wound opening. In a case
observed by Rehn659 the worm probably entered through a gangrenous
portion of the intestine in a hernial sac. Broca660 is unable to determine
whether in his case the intestinal perforation was primary
(a worm escaped from the abdominal wound about two months after
a laparotomy for suppurative peritonitis). The case reported by
Lutz661 is of special interest: it was that of a young man who had
shot himself in the region of the abdomen, and who died after fifteen
days. At the post-mortem two Ascarides were found in the pulmonary
artery; they had probably escaped from the intestine, and had gained
access to the inferior vena cava. Froelich662 assumes that in his case
(a boy, aged 11) the Oxyuris were able to penetrate the whole
intestinal wall, but Vuillemin663 considers this improbable, and is
more inclined to think that the Oxyurides penetrated the rectum at
small ulcerated points, and thus gained access to the perirectal
connective tissue. In females Oxyuris not only have the power
of penetrating far into the sexual organs (Marro664), and perhaps causing
a parasitic endometritis (Simons665), but also clearly of gaining access
to the peritoneal cavity by way of the tubes, as is to be assumed in
the case recorded by Kolb666 (that of a woman, aged 42, in whom post
mortem nodules were found over the peritoneum of Douglas’s pouch,
in which the pressure of encapsuled Oxyuris could be demonstrated),
in that reported by Chiari667 (adult Oxyuris in Douglas’s pouch) and
by Schneider668 (an Oxyuris encapsuled in the pelvic peritoneum).
Sehrt’s669 case is worthy of attention; in this an abscess was found in
the omentum with numerous Ascaris ova in the pus and a nodular
lesion of the peritoneum, with Ascaris ova encapsuled in the nodules.
Massive accumulation of Ascarides may give rise to a complete
occlusion of the gut. Such an occurrence is not so surprising as
might be thought when one reflects that the number of Ascarides in
one individual may amount to several hundreds. For instance, one
boy evacuated within a single day 600 Ascarides (Fauconneau-Dufresne670)
and within three years 5,126 worms. In the case
recorded by Tschernomikow671 a boy, aged 2 1/2, evacuated during a
day 208 worms, partly through the stomach, partly through the
intestine. Coil-formation of such masses of Ascarides renders
possible not only constipation, but also complete obstruction with
symptoms of ileus, as shown by the five cases quoted by Mosler
and Peiper,672 as well as from observations made by Raie,673 Schulhof,674
Rehberg,675 Rocheblave,676 Heller,677 Leichtenstern,678 Huber,679 and
Wilms.680 In two cases of Black681 and Parkinson682 the intestinal
obstruction was caused by a coil of tapeworms.
In the earlier history of medicine the helminthes played a great
part as the excitants of many intestinal diseases and of enteric
as well. Even if to-day they no longer be regarded as such, the
conception that they represent the predisposing factor in typhoid
infection through the injury they inflict on the mucosa (Guiart,683
Blanchard,684 Vivaldi and Tonello685) must not be summarily rejected.
Vivaldi and Tonello found helminthes in 80 per cent. of their
typhoid patients, numbering among these Trichocephalus dispar,
Oxyuris vermicularis, Ancylostoma duodenale, and Ascaridæ. The
report of Leuckart686 is here worth citing, to the effect that Thiebault
never failed to find Trichocephalus in his cholera patients at Naples.
Blanchard687 goes so far as to express the desire that in every febrile
affection of the intestine an anthelmintic treatment with thymol
should be undertaken as early as possible, even before learning the
results of serum diagnosis.
The lesions of the liver and pancreas due to Ascaridæ are briefly
discussed in the chapter on Ascariasis (p. 687).
A discussion of the intestinal helminthes from the clinical and
therapeutical point of view follows these general considerations.
Dibothriocephalus latus.
From what is known as to the development of Dibothriocephalus
latus, the way by which man is infected is self-evident: infection
can only take place through the ingestion of insufficiently cooked
fresh-water fish (pike, burbot, perch, grayling and vendace); what
degree of temperature is necessary to kill the larval forms is still
unknown. Dibothriocephalus latus lives in the small intestine of man,
alone or in some numbers, frequently also together with Tænia solium.
The proglottides are passed always united in large pieces, the ova are
deposited through the uterine pore, while the worm is still in the
intestine, so that they are easily found in the fæces. The proglottides
are so characteristic that they cannot be confused with those of other
species. In reference to whether age or sex is spared by D. latus,
it is not possible to make any definite statement, especially so far
as the endemic area is concerned, whether a person resides in it
continuously or visits it, so long as his habit of life is in accordance
with those of the country. Bendix688 certainly emphasizes the fact
that early childhood is as a rule immune: his case was that of
a child, aged 4 1/2 years.
Sparganum mansoni.
According to our present knowledge (Miyake689) the disease occurs
almost exclusively in China and Japan. On the main island it occurs in
all districts, though rarely under observation. It is especially frequent
in the neighbourhood of Kioto and Osaka; these places are very
near together, and between them there is mutually active intercourse,
so that taken together they may be regarded as one district infested
by this worm disease. As regards localization in the body, there
appears to exist a certain predisposition for definite regions, for
instance, the eye and genito-urinary tract. In some cases the parasite
manifested the peculiarity of wandering about the body and of
appearing at certain favourite points (musc. quadriceps femoris)
(Hashimoto690). Most patients complain more or less of the onset of
attacks of pain and of sensitiveness to pressure. In those cases in
which the patients evacuated the worm during micturition, the
symptoms were variable; sometimes there was tenesmus of the
bladder, sometimes pains in the inguinal region, sometimes
hæmaturia. None of these troubles is characteristic of the disease,
and does no more than represent the symptoms that follow a
mechanical irritation that any kind of foreign body may produce.
Besides the onset of attacks of pain, swelling of the regions affected,
if superficial, may often be recognized, when a superficial diffuse
soft tumour can be felt which often gives pseudo-fluctuation. Sometimes
a peculiar crackling can be detected internally, as in the making
of a snowball. During the further course an abscess not infrequently
forms around the worm. When the situation of the worm is superficial,
“an inflammatory tumour with a tendency to migrate” is
stated by Omi691 to be an important diagnostic sign. That, however,
is not always the case, as the observation made by Inoye692 shows. It
would be better to add to this sign the onset of paroxysmal pain and
the temporary change in volume of the tumour. When once the
parasite is removed, the wound heals just as satisfactorily as any
other fresh wounds made at operation.
Dipylidium caninum (Tænia cucumerina).
This species belongs to parasites of rare occurrence. Up to the
year 1905 Bollinger693 collected thirty-six cases from the literature,
twenty-nine of which were children and seven adults. Since then
some further cases have come to light, so that the number now
observed amounts to ninety, and among them only eight adults.
The youngest child was 6 weeks old (Köhl694), in which the first proglottides
were passed when the child was 40 days old. This preponderating
occurrence in children is clearly connected with the
close intercourse between children and dogs, and also cats. Bollinger
believes that D. caninum in reality occurs more frequently in adults
than has hitherto been supposed. In addition, it must be mentioned
that this species is quite unknown to many physicians, and is
occasionally confused with Tænia solium. One notices almost daily
a large quantity of cucumber-seed-like bodies, reddish or whitish-grey,
about 1 cm. long and 2 mm. broad, discharged with the stools.
Lindblad695 remarks that these bodies have lively movements, that they
perish rapidly in fresh water, and become white and smooth. These
Cestodes, in isolated cases, are parasitic in the intestine in large
numbers. Sonnenschein696 expelled four fragments in the case of a
boy, aged 4 months; Asam697 three fragments in the case of a child, aged
19 months; and Zschokke698 as many as five or six in that of a boy,
aged 4. They do not always produce such striking symptoms as
occurred in Pollak’s case.699 In other cases gastro-intestinal disturbances
with or without fever (Krüger700), emaciation (Zschokke),
or even nervous symptoms of central origin in the form of convulsions
(Brandt701) have been observed. From the nature and mode
of infection children must be kept from close contact with dogs and
cats as much as possible to ensure prophylaxis. The appropriate
treatment, as it mainly affects children, deserves special mention,
whilst the expulsion of the remaining Cestodes may be described in
this connection. Among the drugs one may mention flor. kousso
1·0 grm., pulpa tamar. depur. 2 grm., syrup of sugar 50·0 grm., one-third
to be taken every hour (Lindblad). Kamala appears to have no
effect, although Huber702 recommends it in small doses according to
age from 0·5 to 3·0 grm. He warns against Filix mas preparations,
which otherwise, even in early childhood, under careful dosage gives the
best results. Young children are given 1·0 to 2·0 grm. extr. fil. maris,
with mint syrup or raspberry syrup 30·0 grm., in the morning twice an
hour by the mouth, or 1·0 grm. extr. fil. maris is mixed with syrup
of mint, and given by means of a stomach tube (Rosenberg703). A few
hours afterwards a mild laxative may be taken—one to two tablespoonfuls
of aqueous tincture of rhubarb (Asam)—or an enema may
be given. In a case reported by Sonnenschein decoction of pomegranate
root had no effect, as it was vomited up.
Hymenolepis nana.
This species, very rare in Central and Northern Europe, inhabits
the small intestine, especially of children; it burrows very deeply into
the mucosa. Not uncommonly several thousand have been found in
one case (Nicolo,704 E. Stoerk and Haendel705). It is remarkable that
these Cestodes have been found so frequently post mortem and after
vermifuges given for other reasons. Thus the clinical symptoms
must often be very indefinite (Stoerk and Haendel), so that one
may assume that only a slight percentage of cases of Hymenolepis
nana come under observation and are published as such. On the
other hand, it is certainly conceivable that with the large number of
parasites that frequently occur in one individual a whole series of
symptoms, in part quite severe, are capable of being produced.
These are partly symptoms of intestinal catarrh, consisting of
abdominal pains, constipation, alternating with attacks of diarrhœa,
perverse appetite, and boulimia, abdominal pains of a cramp-like
nature, followed by emaciation, headache, sleeplessness, pallor,
lassitude, and in part nervous symptoms—epileptiform attacks
without loss of consciousness, weakness of memory, melancholia,
irregular febrile attacks (Lutz706). Possibly, too, Hymenolepis nana
infects the urinary organs, producing true chyluria (Predtetschensky707).
Stoerk and Haendel are inclined to think that this species, unlike
other Cestodes parasitic in man and domestic animals, needs no
intermediate host for its development, and that the larval forms
(cysticercoid) live in the same host as the adults. The diagnosis
is based on the demonstration of ova in the stools. As far as
expulsion of this Cestode is concerned, santonin, kamala, kousso
flowers and thymol appear to have no effect of importance; whilst
extract of male fern, recommended by Grassi708 as a result of his
considerable and successful experience, has been given, with the
result that the worms really are expelled, and that after the treatment
neither worms nor ova are any longer demonstrable in the stools of
patients. In his cases of chyluria Predtetschensky prescribed ol.
terebinth. 20 drops three times daily for a fortnight, then acid. gallic.
0·5 grm. three times a day for two days, then 1·0 grm. three times a
day; the urine became clear, but whether permanent cure resulted
remained doubtful.
Hymenolepis diminuta, H. lanceolata, Davainea asiatica, and D.
madagascarensis possess no actual clinical interest; with regard to
the latter it need only be pointed out that Bordier709 in studying a
case of chyluria found this species in the kidneys of a person in
Madagascar.
Tænia solium.
Tænia solium inhabits the small intestine of man; single
proglottides or whole worms may get into the abdominal cavity and
the bladder through fistulæ, and penetrating the abdominal wall
escape outwards or become discharged with the urine. Symptoms of
intestinal stenosis are certainly very rare, as in the case recorded by
Steinhaus710 of a child, aged 9, the stenosis ceasing after the expulsion
of the segments. The usual position of the worm in the small
intestine is with the head closely adherent to the mucosa and the
proglottides lying along the intestine; from time to time portions are
discharged with the fæces per rectum. Its position can also be
reversed, and the proglottides in the gut become thus discharged by
vomiting.
The diagnosis depends upon the proglottides being generally
discharged in pieces in the stools, or eventually an examination for
eggs. Larval infection (Cysticercus cellulosæ) occurs also in man
through auto-infection or through food.
Cysticercus cellulosæ of the skin and subcutaneous tissue occurs
very seldom singly; as a rule they are found in hundreds and
thousands in the same individual. They occur in different parts of
the body, especially on the flexor surfaces of the extremities (generally
symmetrically), small globular swellings, the size of a pea or a hazel
nut, smooth, of a tough cartilaginous consistence, fairly movable
under the skin, in the muscles less so. They never degenerate or
cause the surrounding skin to lose its colour. It is an interesting
fact that in the case described by Posselt711 nodules on the face,
namely in the neighbourhood of the left cheek and behind the left
ear, reformed. The following are, according to Posselt, characteristic
for cutaneous tumours due to cysticerci: (1) the position in the
subcutaneous connective tissue (and almost always simultaneously
in the muscles); (2) the approximately equal size and regularly
rounded oval form; (3) the peculiar density, almost reminding one
of cartilage in its hardness and the sensation of tightly distended
thick-walled bladders; (4) proportionately slight mobility; (5) with
painlessness, absence of any cutaneous reaction (hyperæmia or
swelling of the skin or pigmentation). The very gradual appearance
generally of the tumours supports the diagnosis, and in
addition to this evidence we may emphasize the preponderating
liability of the upper part of the body to attack and the symmetrical
arrangement of the nodules. Cutaneous and muscular cysticerci
cause the most varied symptoms, sensory disturbances, abnormal
sensations, depression and a feeling of weariness whenever the diseased
parts are moved, weakness in the lower extremities, pains in the course
of the sciatic nerve, in addition to those which simulate cramp in the
calves, numbness in the hands, pains upon their being moved. In
the case of a cysticercus situated in the elbow-joint, painful dragging
sensation in the course of the ulnar nerve persisted. In other cases
the arm was almost paralysed, or it could not be completely extended;
stiffness and bending of the little finger were noticed. Cysticerci of
the gluteal muscle cause trouble upon sitting and upon defæcation.
Remittent unilateral headaches were present in the case of a cysticercus
of the region of the right eyebrow; pains of a neuralgic
character radiated from the diseased temporal region. The cysts may
be inflamed and may suppurate; this especially happens in the case
of solitary cutaneous and muscle cysticerci. The best treatment
consists in puncture of the cysts with a Pravaz syringe and subsequent
injection of a drop of 1 per cent. sublimate solution. Tincture of
iodine has similarly been proposed (Wolff712). Frangenheim713 recommends
early extirpation (this, however, only in the case of
solitary cysts). Pelagutti714 believes that in his case diminution in
the size of the cysts was obtained by the use of anthelminthic remedies
continued over a long period combined with potassium iodide and
calcium salts (internally). Cysticercus is very rarely found in the
tongue; there the worms generally lie in front of the sulcus terminalis,
corresponding to the middle of the tongue, according to Glas.715 In
the case recorded by Gaetano716 (a boy, aged 10) there was a nodule
on the left side of the tongue which grew very rapidly till it reached
the size of a nut; it was embedded in the muscle and covered over
by normal mucosa. Cysticerci are just as rare in the pleuræ, in the
lungs, in the intestinal submucosa, in the submucosa of the small
intestine, in the mesenteric glands, in the liver, pancreas, spleen and
kidneys, in the mamma, in the heart, in the bones and in the great
vessels (Huber717). Cysticercus of the eye deserves special mention;
in rare cases the cysticercus has been met with in the subcutaneous
cellular tissue of the eyelid, once in the muscle bundles of the
musculus orbicularis. Subconjunctival cysts are found chiefly in
youthful individuals. Their position is most varied, generally in the
neighbourhood of the inner angle of the eye. Dilated vessels pass
right over the cysts, which are generally movable, together with the base
they rest upon, producing a spherical protrusion. The head of the
worm can sometimes be seen shining through as a whitish speck.
The only symptoms are those of a slight irritation of the connective
tissue and some difficulty in closing the lid; larger cysts dislocate the
globe. The diagnosis has the rapid growth of the cystic tumour to
support it; there is the possibility of its being mistaken for a foreign
body (Kaldrovils718). After division of the connective tissue capsule
extraction is easily performed. It is most rare for the cysticercus to
occur in the orbit. Suppuration of the cyst may have serious consequences
for the eye. It is only exceptionally that the cysticerci
gain access to the anterior chamber of the eye.
Subretinal cysticerci or those localized in the vitreous are more
frequent. Upon examination with the ophthalmoscope there is seen
in the vitreous a bluish bladder with a smooth surface. The head is
seen as a white patch, and the circle of hooks and the suckers
also come into view, also the frequent movements which the head
and neck make in the vitreous. Operation generally yields good
results; in rare instances the globe is atrophied and must be
enucleated.
Formerly cysticerci in the brain were met with in fair frequency,
but the number of such cases has generally decreased of late years in
a remarkable way, in correspondence with the diminution of cysticerci,
which is to be attributed to compulsory meat inspection. Whilst,
for example, the post-mortem records of the Pathological Institute in
Berlin before the year 1875 showed 20 per cent. cysticerci affecting
the brain, this number declined later to 16·3 per cent., and of late
years has fallen to 1 per cent. (Orth719). Nevertheless even now
cysticercus still plays no inconsiderable part in the etiology of
cerebral diseases. For example, in the clinic of de Amicis at Naples,
among seven cases of cysticerci of the skin, they were found four
times also in the brain (Sipari720). Cysticerci may occur in the dura
mater, arachnoid, pia mater, choroid plexus, the surface of the cerebral
hemisphere, the medullary substance, the ventricles, the aqueduct,
the corpus striatum, corpora quadrigemina, the pineal gland, the pons,
the cerebellum, the olfactory trigone, the bulb, the medulla oblongata,
and the olive. They are most frequently found in the cortical substance
and in the ventricles; the frequency of the latter situation may
be explained by the flow of the fluid (Henneberg721). The severity of
the symptoms is not always in proportion to the number of cysticerci.
Cases have been known in which ten, twenty and forty cysticerci have
been found (Hagen-Thorn722), and yet the clinical symptoms have
been remarkably slight. On the other hand, solitary cysts may
both run a course completely without symptoms and also cause the
severest symptoms when located in specially important parts of
the brain (crus, pons, central convolutions). In the case mentioned
by Jacobson723 the invasion of the brain by cysticerci was immense;
the largest cyst was found in the cerebral cortex. The chief
symptoms of cysticercus of the brain substance consist in the
onset of cortical epilepsy, which sometimes runs a very pernicious
course, frequently with psychical disturbances, whilst paralyses are
absent. Perhaps, too, the localization of pain, spontaneous and on
pressure, corresponding with the points observed on the cranium,
is of importance. Cysticerci may also change their position
in the brain; patients who had earlier suffered from epileptiform
convulsions later showed intra-ocular cysticerci after the cerebral
symptoms had completely disappeared. Treatment can only be
surgical; v. Bergmann724 operated in two cases with well-marked
improvement. Parasites in the ventricles are especially dangerous,
more especially so when free in the ventricles, and so capable of
giving rise to the danger of sudden closure of the foramen of Majendie
(Simmonds,725 Versé726). Stern727 states the symptoms of cysticercus
in the fourth ventricle to be the following: general cerebral pressure
symptoms (headache, vertigo, vomiting, somnolence, congested disc
caused by internal hydrocephalus); in addition, there are symptoms
which point to disease of the hind-brain—pain and stiffness in the
neck, vertigo and cerebellar ataxy, violent and persistent vomiting,
slowness of pulse; and lastly those rare but certain symptoms of a
lesion of the bulb, such as diabetes, respiratory disturbances and
paralysis of cerebral nerves, especially of the abducens. These are
far less marked than the general symptoms of cerebral pressure. One
characteristic is the remarkable alternation between severe general
symptoms and periods of complete sense of well-being; in this way
a functional nervous affection may be simulated (Jolasse728). Brun’s
symptom (in the widest sense, sudden onset of violent cerebral
symptoms upon change of head-posture) is a specially characteristic
sign of free cysticercus in the fourth ventricle; the disease generally
terminates with sudden death from cessation of the heart’s action.
Defects in motor power, convulsions, implication of other nerves,
are rare and unessential complications (Hartmann729). Carefully
carried out, lumbar puncture may possess some diagnostic and
therapeutic value. Treatment is purely symptomatic, or eventually
Neisser’s ventricle puncture may be considered.
At the base of the brain the cysticerci, as a rule, assume that
form which is designated as C. racemosus, and consists of rows of
delicate grape-like bladders in groups, sometimes also markedly
branched, but generally sterile, which develop in the meshes of the
soft meninges and may envelop the nerves and vessels of the base of
the brain. Such tumours bring about hydrocephalus and chronic
leptomeningitis, which must be regarded as the causes of the clinical
disturbances (cysticercus meningitis), attacks of loss of consciousness,
dementia and apathy, dulness and confusion and headaches.
In the case recorded by Meyer730 symptoms which resembled paralysis
agitans were noteworthy, and defects in speech in the case recorded
by Durst731 (C. racemosus in the region of the left Sylvian fossa).
According to Markwald732 C. racemosus of the fourth ventricle is said
to represent a characteristic clinical picture: violent headaches,
attacks of vertigo followed very soon by deep coma and death
in a few days. Treatment in Cysticercus racemosus is ineffectual.
In the diagnosis of cerebral cysticerci in general the recognition of
multiple cysticerci in the skin and muscle and of the tapeworm is
of importance. In cases of cerebral diseases in which cysticerci may
be a possible cause, Remmert733 recommends that the skin of the
whole body should be palpated.
Cysticercus in the spinal cord and in the vertebral column is
occasionally observed; as a rule, other organs, above all the brain
and its membranes, are simultaneously affected. Here, too, the
cysticercus occurs in two forms—sometimes the cysts are roundish
or oval, solitary or multiple, and at other times Cysticercus racemosus
occurs.
Tænia saginata.
Occurs in the small intestine of man. It is characteristic of the
habit of life of this parasite that once it has become mature its
proglottides are dropped off daily in increasing numbers because
its growth is extraordinarily rapid. The joints are discharged
generally spontaneously during the whole day without a stool. An
extraordinarily unpleasant sensation is produced by the damp, cool
joints slipping down into one’s lower garments and over one’s legs
when walking; women especially, in whom the proglottides slip
through their petticoats on to their legs, complain bitterly of this
troublesome symptom. Another unpleasant symptom is superadded
in the shape of the proglottides tickling the rectum, and this excites
irritable people to the last degree. Different species of tapeworms are
not mutually exclusive. B. latus and T. solium frequently occur side
by side, so also T. solium and T. saginata—for instance, in a butcher’s
assistant we once expelled twelve T. solium and one T. saginata at
the same time. The greatest number of Tæniæ which have been
observed at one time amounted to forty T. solium (Kleefeld734).
Even though the cysticercus of T. saginata is not, as in the case of
T. solium, particularly dangerous to man, a parasite, nevertheless,
which requires so much nutrient material during its rapid growth,
and thereby sets up manifold disturbances in the general condition
of health, ought to be expelled as rapidly and thoroughly as possible.
Tapeworms are found not uncommonly with other intestinal
parasites, such as Ascaris, Oxyuris, Trichocephalus or Ancylostoma.
Prunac735 described a case in which a woman passed a Tænia through
the anus while she vomited a Fasciola hepatica.
The symptomatology of these three large species of Cestodes,
Dibothriocephalus latus, Tænia solium, and T. saginata, may very well
be summarized together, as, apart from some peculiarities, the clinical
symptoms, especially so far as their localization in the intestine is
concerned, are practically the same for all three species. In a large
number of cases the hosts have no suspicion whatever that they are
harbouring a tapeworm; they feel quite well and free from any
disquieting symptoms whatever, and only become aware of the fact
that they are the carriers of a tapeworm when the discharge of the
segments takes place; on the other hand, it is often difficult to rid
people of the idea that they are harbouring a Tænia (Küchenmeister
calls such Tænia imaginata); usually it is undigested fibrous shreds
of beefsteak which are regarded by the patients as proglottides of
tæniæ.
In a large number of cases, disturbances of the intestinal tract
set in, e.g., sense of pressure in the abdomen, which sometimes
becomes constant on one and the same side, or sometimes
changes, now at the umbilicus and again at the epigastrium; here
and there colicky pains are present. Derangements of appetite and
digestion are frequently complained of; the most frequent are the
sensations of morbid hunger or irregular appetite, nausea and
vomiting. Thus, at the Third Congress of Internal Medicine,
Senator recorded a case in which there were symptoms of nervous
dyspepsia, cured after a successful vermifuge. There is either
constipation or diarrhœa, so that many of such patients are brought
for treatment with the diagnosis of “chronic intestinal catarrh” and
correspondingly treated. As to the treatment of toxic action of the
Tæniæ when such arises, see the special section on the subject
(bothriocephalus anæmia, p. 644). The frequent disturbances of the
general condition, so-called reflex phenomena, so far as the action of
toxic substances is not in question, may be explained by the fact of
their occurrence in specially sensitive individuals who are affected by
such phenomena. The proof that a diseased condition is produced
by a tapeworm will be forthcoming with some degree of certainty
if the symptoms cease immediately after the removal of the parasites.
As a whole series of troubles, which certainly have nothing to do
with them, are erroneously ascribed to the tapeworm, as is frequently
assumed, one will do well to be somewhat critical in this respect.
The treatment is of a threefold nature: prophylactic, symptomatic
and radical.
Under any circumstances, the best prophylaxis is that which
consists in only eating the flesh of those animals in which any of the
three larval forms occur (pig, cattle, salmon, pike, burbot, etc.)
so prepared that the larval forms have been destroyed and the food
thus rendered innocuous. For domestic and public use the rule
prescribed by Küchenmeister is under all circumstances most easily
understood, namely to roast or boil till the flesh appear greyish-white
and sufficiently done by reason of the coagulation of the
albumen and decolorization of the blood. The general prophylaxis
simply concerns the tapeworm carriers trying to limit as far as
possible the further extension of the parasites in the animal world
by carefully rendering the expelled segments and worms harmless
(pouring sulphuric acid over the fæces and burning the worms) and
also by strictly adhering to official regulations. The official system
of meat inspection in this respect has been of immense service, and
much can still be done by means of thorough official control over
cleanliness in abattoirs and butchers’ shops. Galli-Valerio736 very
rightly desires the abolition of the custom of manuring fruit-plants
such as strawberries, vegetables and salad with the contents of privies,
and would extend the use of privies in the country.
Symptomatic treatment consists, in the case of those Tæniæ which
resist radical attempts at expulsion, of repeated use of drugs injurious
to the worm as soon as ever new proglottides are formed, or in
special cases, as in the case of persons weakened by diseases or
operations, or frail old people, or patients with severe heart failure,
gastric or intestinal carcinoma, or in pregnancy, in effecting the
expulsion of a large chain of proglottides by the mildest measures
possible.
Radical treatment of the Tænia is not always equally easy in all
three species, even when the means used are the same; the easiest to
expel is T. solium, then D. latus, and the most difficult T. saginata.
That as yet no certain cure exists for Cestodes is clear from the large
number of drugs recommended from time to time, and the increase
of bungling treatment in this respect; in addition, there is no department
in which there is so much quackery as in vermifuges. The
treatment proper should always be preceded by thorough preparatory
treatment, the purpose of which is to render the gut as empty as
possible once for all, and on the other hand to put the worms themselves
into a diseased condition. How far the host himself has been
made ill by such preliminary cures (herring, pickle, garlic, onions,
preserved strawberries), many a person who has had to do with such
things can recount. In the opinion of Fischer737 strict preparatory
treatment appears to favour the development of toxic substances, or
else it disposes to vomiting; as a rule it causes the patient far more
discomfort than the treatment itself. In recent times far less weight
is attached to these preparatory treatments than to carefully prepared
and correctly dosed drugs; the preparation is generally limited to
relieving the intestine in a simple way, the day before the treatment,
of the densest fæcal masses, by a simple aperient or water enema.
We recommend the following, which has always proved itself to
be the best and simplest remedy against T. saginata. The patient
takes early in the evening before the treatment nothing but a plate of
soup or a glass of milk, and then takes a laxative (electuar. lenit or
infus. sennæ compos. or an enema), so that later in the evening one
to two stools are passed. In this connection we fail to agree with
Grawitz738 and Boas,739 who consider that at least preliminary evacuation
of the intestines can be dispensed with. On the following
morning the patient should take a cup of black coffee or tea without
anything else, and half an hour later the vermifuge.
The best drug is extract. filicis maris æther., which also forms the
main constituent of most of the secret remedies recommended for
tapeworms. Earlier mishaps with this preparation had their origin
principally in insufficient dosage. Also, in addition to correct dosage,
extract. filic. maris needs very careful preparation if satisfactory results
are to be attained. If preparations with the trade mark “Helfenberg”
or “Wohnar” are not used, but the male fern extract has been
prepared by a chemist, one must make certain that the roots of the
Aspidium filix-mas have been collected in May or October, and only
green sappy specimens selected, and that the attached paleæ have
been separated, that they have been broken up small and ether
poured over them with a little spirits of wine while quite fresh. The
whole mass is to be kept in a cool place, but not too closely covered.
If at any time a certain quantity is to be used, it is taken out, the
ether carefully distilled in a retort till the extract has a suitable fluid
consistency. Fischer attaches great importance to the direction in
the Pharmacopœia being exactly followed, to the effect that the
extract is to be carefully stirred before prescribing, as the active substances
undergo partial crystallization if kept for any length of time
and sink to the bottom, so that the preparation has a different strength
and toxicity in different layers. Of this extract 10 to 12 to 15 grm. are
to be taken in gelatine capsules within half an hour. We consider it
unjustifiable to give greater doses than 15 grm. to adults, as many
cases are known in which to some extent severe toxic symptoms have
followed, such as headache, sensation of giddiness, dyspnœa and
cyanosis, yellow vision (xanthopsia), delirium, stupor, the most severe
cramps in the extremities, rapidly fatal trismus and tetanus. The
most serious are defects of vision of various kinds, which may end
in amblyopia and amaurosis, with permanent blindness. A complete
collection of toxicological literature up to the year 1903 is to be
found in Marx’s740 Dissertation. Since that time further instances
of such intoxications have been made known. Nagel741 observed
them only in severe cases. O. Meyer742 lays special stress on the bad
prognosis of the disturbances of vision evoked by poisoning with
extract. filicis maris. Studt743 has seen two cases of optic neuritis, one
with circumscribed, the other with diffuse retinal œdema. Uhthoff744
has only seen one case; in that reported by Noiszewski745 the toxic
retinitis was cured; in Viereck’s746 case bilateral concentric limitation
of the field of vision followed three days after taking 8·0 grm. extract.
filicis maris. Stuelp747 attributes the amaurosis occurring after taking
filix mas to a toxic action on the muscularis of the central retinal
artery; there followed paralysis of the vessel, vascular engorgement,
and thereby nutritional defects of the nervous elements followed.
In children one has to diminish the dose correspondingly, as with
them, still more so than with adults, severe disturbances arise. Huber748
claims that this drug should not be given to children indiscriminately.
The view is frequently expressed that a combination of extractum
filicis maris with fatty oils in which the active constituents are soluble
favours intoxication. Marx749 also argues from this standpoint and
assumes that the ideal preparation, free from objection, would be got
if from filix-mas extract a preparation free from fatty oils could be
made, and he considers it advisable to limit the use of castor oil as an
aperient before and after taking the “cure” and to prescribe instead
a saline laxative, such as Epsom salts or Glauber’s salts. Sonnenschein750
also advises against the simultaneous exhibition of extractum
filicis maris with oleum ricini, as is the case with Helfenberg’s
capsules, and Boas751 is likewise anxious that ol. ricini should be
avoided. Lenhartz752 appears to consider the warning against the
simultaneous combination of the extract with fats or ethereal oils, and
especially against the employment of castor oil as an after-treatment,
as without justification, and we, too, in the course of our many
filix treatments, have never yet witnessed any unfavourable effect from
the use of castor oil in the after-treatment. The surest way of
obviating the toxic effects of extractum filicis is to give a laxative
(ol. ricini) as soon as the extract has left the stomach, say, about half
an hour, so that it need not stay longer than necessary in the gut and
become absorbed. Perhaps in most cases of poisoning, transgressions
against this rule have been the cause of the toxic action. The nausea
that sets in the day after taking the drug and the inclination to vomit
are best resisted by giving iced coffee, iced tea, iced pills, peppermint
tea, cognac, one to two wafer powders of menthol and sacch. lactis
āā 0·2 grm. (Apolant753) half an hour before the drug is taken.
Fischer754 considers that lying still in the horizontal position is the
best remedy. Boas755 recommends the injection of the drug into the
stomachs of patients who tolerate extractum filicis badly, in the form of
a thin emulsion (with gi. arab.). In the case of children the extract
is prescribed with honey as an electuary. The method recommended
by Fowler756 is without doubt too detailed; he prescribes before the
treatment two to three to four days’ rest in bed; special diet, tablets of
cascara sagrada three times daily, on the fourth day senna infusion,
and then to give the extractum filicis maris in capsules in four doses,
to be taken every quarter of an hour.
Under Jaquet’s757 direction, Kraft has prepared an amorphous acid
from the fern root extract which is designated filmaron. As a vermifuge
the drug is prescribed for children of 2 to 5 years of age in
doses up to 0·2 to 0·3 grm., for children of from 8 to 12 years in doses
up to 0·5 to 0·7 grm., and for adults up to 0·7 to 1·0 grm., so as to expel
the parasites. Bodenstein758 gives the filmaron oil introduced into
commerce by the firm of Boehringer (one part filmaron and nine parts
castor oil) in still greater dosage, either fasting or, in the case of
sensitive patients, one hour after a cup of tea; he gives peppermint
tablets against possible nausea. Brieger759 tested the preparation in
twenty-three cases; in twenty-one of these he prescribed it as an ether-castor
oil mixture, and in two as capsules. The action always took
effect in from two to five hours, and only in three cases were
unpleasant after-effects in the shape of colic observed; in sixteen
cases the result was positive, in seven negative.
The attempts made by Goldmann760 to prepare from the bark of
Musenna abyssinica, a plant of the order Myrsinaceæ, indigenous to
Persia, the active substance, namely sebirol, have shown that when
this is given alone it certainly acts as a vermicide, but not as a
vermifuge; on the other hand, the results of a combination of sebirol
with thymol and salicylates were surprisingly good; this mixture has
been introduced into commerce as tæniol, in the shape of pastilles
prepared with chocolate for children. The method of giving tæniol
is as follows: On the day before the administration a light diet and
thorough purging with calomel are ordered; and then on the day of
the treatment itself, after a breakfast consisting of a cup of tea, in the
case of adults, thirteen to fifteen tæniol pastilles are taken in some red
wine at intervals of ten minutes respectively. In the middle of this
treatment an interval of some hours is interposed. After the pastilles
have been taken a calomel purge is again given. The results obtained
by Liermberger761 are sufficiently encouraging to be put to further test.
Fischer762 has tested in some of his cases extracts of some new
species of fern root; he employed the extract from the rhizomes of
Aspidium spinulosum and A. dilatatum, two fern roots indigenous to
Sweden, and obtained remarkable results (doses of 4 grm.). Laurén763
had previously recorded similar results, and recently Friedjung,764
using extr. aspid. spinulos.
Cortex radicis granati as fresh bark is a very good drug, and is
usually given as a decoction: 180·0 bark to 1,000·0 water, boiled for
forty hours to 240·0, and a small cupful to be given every half an hour;
colic, vomiting and diarrhœa, are, however, easily induced. The chief
constituent of the granate root, pelletierinum, possesses vermicidal
properties, and is much recommended, especially in France. Sequelæ
easily arise (vertigo, hazy vision, malaise, vomiting, quickened heart’s
action, muscular tremors, cramps in the calves), especially in delicate
persons and children, so that one should refrain from giving it to the
latter especially (Drivon765). Sometimes, judging by the experience of
Sobotta766 and Boas,767 the action is problematical. Where it is desired
to employ it in the case of adults, the following is prescribed: pellet.
sulfur. 0·3 to 0·4 grm., acid. tannic. 0·5 grm., sir. rub. jd. 30·0 grm., to
be taken at one time, and a quarter to half an hour after a purgative
(senna infusion). In the case of children it is better to employ
semina cucurbitæ maximæ instead of extractum filicis maris. Sixty
to 100 pumpkin seeds are pounded up with sugar, which yield a
pleasant-tasting electuary, and which are taken all at once; half an
hour afterwards a laxative is taken (Storch,768 Pick769), Jungklauss’s
preparation is nothing else than a pumpkin extract; its action is
favourable; it is, however, too expensive (Ritter770). Flores kousso
up to 15 to 20 grm. in compressed form or in sugar or honey in the
form of electuaries (children 2·0 to 10·0 grm. according to age) is not to
be relied upon; kussin, prepared from kousso flowers (Bedall, Munich),
is not a pure body; when taken it is divided into four parts up to
1·0 to 2·0 grm. with elæosaccharum menthæ, at half-hourly intervals;
it is said to be less unpleasant than treatment with flores kousso (Liebreich
and Langgard771). Kosinum crystallisatum (dose 1·5 to 2·0 grm.)
is prepared by the firm of Merck. Kamala is the least potent of the
tapeworm drugs in use, and is principally to be recommended in the
treatment of children: 1·5 to 3·0 grm. in electuaries. According to
Leichtenstern772 and White773 chloroform, even in toxic doses, cannot
do any harm to the tapeworm, nevertheless it has been recently
recommended by Carratú774; chloroform 6·0, sirup. 60·0, one teaspoonful
to be taken every hour (fasting). Salol is recommended by
Galli-Valerio775 as an absolutely harmless tapeworm drug; thymotal
(a derivative of thymol) by Pool,776 3 grm. to be given up to three to
four times on four consecutive days.
The drug well known long ago, cuprum oxyd. nigr., has been
recently brought into fresh notice by Dörr.777 It is also the chief constituent
of the tapeworm drug introduced into commerce by the firm
of Dehlsen (Itzehoe) (Koch778). The coconut is absolutely ineffectual,
also naphthalin, croton-chloral, ether, gallanol, strontium lactate,
glycerine and bromide of potash.
Where possible one should endeavour to discover the head or the
heads of the tapeworm in the stools, so as to make certain whether
the treatment has been successful; this search is best carried out by
immediately and carefully pouring water over the total quantity of
evacuations collected in the night stool, without stirring them up, till
only the tapeworm is found lying at the bottom of the vessel.
NEMATODES.
Strongyloides stercoralis.
The pathological significance of this intestinal parasite is not yet
fully demonstrated. In Seifert’s779 observation, on what Leichtenstern780
called the celebrated Würzburg case, the patient had suffered many
times from attacks of blood-stained diarrhœa with tenesmus, as in
Zinn’s781 case of a three year old boy who had bloody purulent diarrhœa.
Schlüter782 speaks of a hæmorrhagic enteritis produced by Strongyloides.
In other cases besides diarrhœa (either with or without blood) there
were noted: pains in the body (Schlüter), tenderness of the abdomen,
loss of appetite, gastric troubles of a general kind, headache, giddiness,
fainting attacks, anæmia (Silvestri,783 Valdes,784 and Trappe785), so
that even if in isolated cases (Fülleborn786) symptoms are absent, some
significance cannot be denied these parasites as a matter of course
(Bruns,787 Leichtenstern788). According to Kurlow,789 in Siberia there is a
form of sporadic bloody diarrhœa which has its origin in the presence
of Strongyloides stercoralis. The parasite does not live only in the
intestinal lumen, but also in the intestinal wall, where it causes
abscesses, fistulæ and effusions of blood.
Diagnosis is easily made by the detection of the actively moving
larvæ in the stools.
Treatment is rather difficult, as it is not always successful in
getting rid of the parasites. Authors differ as to the effectiveness of
extr. fil. maris. Goldmann790 still considers this preparation as the most
effective; he recommends preliminary treatment with calomel 0·2 grm.
and tuber. jalapæ 0·5 grm. a day before the special treatment, which
consists of gelatine capsules of 15·0 grm. extr. fil. maris (to be taken
in the course of four hours); afterwards rectified oil of turpentine
in gelatine capsules. The thymol treatment (vide Ancylostomiasis,
p. 682), thymol alone or in combination with calomel (Schlüter,791 Valdes,792
Soussino,793 Goldmann794), has often caused diminution of the number
of larvæ, but also often remains resultless. Teissier795 maintains that by
degrees he procured complete cure by the administration of mercury
in the form of blue pill. In our case neither thymol nor calomel,
santonin, extr. fil. maris, decoct, rad. granat., had any result whatever.
Davaine796 believes he attained decrease and final disappearance of
the larvæ by protracted milk-cure. Santonin, tannalbin and other
preparations seem ineffectual. Tannin enemata (Mildner797), high
injections with starch enemata (Schlüter798), may alleviate in persistent
diarrhœa. Travellers who are visiting regions the native home of
Strongyloides must exercise the most extreme care and scrupulous
cleanliness, and these are also necessary in patients already suffering
from Strongyloides, to prevent auto-reinfection (Trappe799).
Dracunculus medinensis (Dracontiasis).
The guinea worm develops in the dermis of human beings without
any symptoms; only when it is completely grown does it form
boil-like, extremely painful abscesses, in the greater majority of cases
in the legs, in the region of the ankle, and is accompanied by general
disturbance and a feeling of heaviness, dragging and pricking of the
affected part; it occurs more rarely in the arms, certain parts of the
back, the head, neck, scrotum and penis; in a superficial position
the worm can occasionally be felt through the skin. In most cases
there is only one worm and one abscess, but here and there one finds
patients with three, four or even up to eight worms, and very exceptionally
still more, as in the cases described by Poupée-Desportes800
(fifty worms) and by Harington801 (seventeen worms).
Diagnosis offers no difficulty when the worms are presenting or
can be felt under the skin.
The inhabitants of the native home of the guinea worm, as a rule,
quietly wait till it has got so far out that it can be conveniently
grasped; it is then bound round with thread and fastened between
the tips of a split piece of wood and slowly wound out. In ten to
twelve days it can be wound out in this way. Emily802 makes injections
of a 1 in 1,000 solution of sublimate either in the neighbourhood
of the worm or directly into its body. Mense803 managed to
remove the worm in one sitting by laying a wad of cotton wool
soaked in chloroform on the exposed portion, thus stupefying it.
Our therapeutic observations (Frangenheim804) favour the free laying
open of the existing abscess and the consequent complete extraction
of the worm.
Prophylaxis depends on care in the use of water in the guinea
worm countries, especially dangerous being permanent waters infested
by Cyclops sp.
Filaria bancrofti.
The parasitism of this filaria leads to the formation of lymphangitis,
elephantiasis, chyluria, orchitis, chylocele, abscesses, lymphatic varices,
perhaps also to chylous ascites and chylous diarrhœa.
Lymphangitis usually attacks the extremities, beginning generally
with a rigor and swelling of the lymphatic vessels with adjoining
lymph glands. The lymphatics become hard, knotty and extremely
painful, the overlying skin red and swollen in longitudinal lines
(Looss), high fever sets in with, to some extent, severe general disturbance.
After some days the attack subsides, the swelling then
partially disappears, but not completely, and often abscesses develop
in consequence of the lymphangitis. Children, as a rule, suffer from
such lymphangitic attacks (Finucane805).
Diagnosis is not easy, for many other causes frequently produce
lymphangitis.
Treatment consists in rest, raising the affected limb, applications
of vinegar and alum or liquor plumbi, in some cases incisions into the
swollen part under antiseptic precautions.
Elephantiasis (Arabian) is usually situated in the lower extremities,
in men in the scrotum and penis, in women in the labium pudendi,
mons veneris, and the mammæ; more rarely it attacks the upper
extremities or, indeed, the head. The disease develops during repeated
attacks, which occur at irregular intervals of weeks, months or years,
of fever accompanied by symptoms of lymphangitis and erysipelas
(elephantoid fever), and especially as the result of different accidental
occurrences such as chills, bodily exertions, external irritation. The
extremities become shapeless, heavy cylinders, the scrotum occasionally
a colossal tumour, the female genitalia and the mammæ smaller or
larger tumours; the penis often shares in the general thickening, the
inguinal glands form large hard prominent masses, and enormous
deformity is caused. The cause is more often seen in men than
women, rarely in children over 10, never in younger children.
Treatment of elephantiasis of the extremities consists in raising the
affected part, massage, bandaging, vapour baths; the large elephantoid
tumours of the genitalia and mammæ can only be treated by operative
removal.
Chyluria (hæmato-chyluria), as a rule, begins by a series of attacks
and often ceases for weeks or months, the attacks being accompanied
by fever, pain in the back and lumbar region, about the kidneys and
in the perinæum. The attacks are separated by intervals of months’
or even years’ duration, a continuous chyluria being quite rare. The
disease may last many years without the constitution being markedly
weakened, but in other cases anæmia and debility ensue and result
in death from marasmus. In chyluria the urine becomes completely
opaque like milk; but sometimes, from the presence of blood, is of a
peach-like redness: the sediment contains clotted blood, and microscopically
one finds fine dust-like fat granules and red cells and
leucocytes, and usually, but not always, filaria larvæ. Sclerodermia
may possibly be caused by Filaria (Bancroft806).
Treatment, consists in administration of ol. santali, methylene blue
(0·12 grm. dose several times daily), ichthyol (in pills from 0·5 to
1·5 grm. per day), ol. terebinthinæ (0·5 to 1·5 gr. per day), thymol
(Ziemann807 had no result from either thymol or methylene blue),
together with absolute rest in bed, diminution of all fatty nourishment
and administration of light purgatives.
Orchitis is in acute attacks a relatively frequent symptom in
the East; the chylocele is rarely marked; the fluid usually shows
numerous larvæ; in the case of abscesses they are generally caused
directly by the adult parasites, as they have often been found in them;
varices of the lymphatic vessels are either superficial or deep;
lymphorrhagia arises from rupture of the dilated vessels; chylous
ascites and chylous diarrhœa may also be produced by Filariæ.
Loa loa.
Loa loa, according to modern investigations, is a parasite of the
subcutaneous connective tissue of man, and its appearance in the
conjunctiva somewhat accidental; in earlier times it seems to have
been less common (Ziemann808). A number of cases are seen in
Europe of patients who have lived in filaria regions, and on return
have been found to have this Nematode in the subconjunctival tissue.
Pick,809 in the case of a man who had lived in the Cameroons,
found the parasites in active motion under the connective tissue of
the eyeball right over the cornea; extraction was easy. Ziemann810
noted three cases of Loa loa in the eye accompanied by temporary
migratory swellings in different parts of the body. In one case,
observed by Wurtz and Cleri811 (a woman from the French Congo),
Loa loa was the cause of intermittent elastic swellings in the
subcutaneous and subconjunctival tissue (marked eosinophilia). In
the case recorded by Pollack812 (for thirty years police commissioner
in the Cameroons) the worm under the connective tissue of the left
eye by its snake-like movements caused an unpleasant itching. With
cocaine and adrenalin the worm can be made visible, and by means
of a strabismus hook can be drawn out of a small wound in the
connective tissue. Martens813 exhibited a Filaria extracted from the
eyelid under local anæsthesia.
Trichuris trichiura.
Whilst many authors consider the whip-worm as a harmless
parasite of the large intestine (Leichtenstern,814 Eichhorst,815
Askanazy816), the number of severe and even fatal cases of diseases
caused by it (trichocephaliasis) increase so much that the Trichuris
trichiura must be excluded from the group of harmless intestinal
parasites. (For disturbances of the nervous system and of the blood
[anæmia] from trichocephaliasis, see p. 650). Infection in human
beings results from the eggs that have developed outside the body,
which probably reach the digestive tract on the hands soiled with dirt
or earth, or possibly through drinking water. (Moosbrugger817 and
Kahane818 mention in their cases that the children had an absolute
passion for earth-eating.) Possibly, too, patients reinfect themselves
anew, as an intermediate host is not necessary.
The anterior part of the body of the parasite is usually fixed in the
mucous membrane, and according to Askanazy feeds on the blood
of its host. Moosbrugger,817 Schulze,819 Kahane,818 Vix,820 Girard821 and
Blanchard822 all found changes in the mucous membrane of the gut,
showing that the parasites had been in the gut for a considerable
time. Kahane818 had an opportunity of seeing at the Pasteur
Institute Trichocephali with the anterior part of the body penetrating
not only the mucosa but also deep into the muscularis of the gut
wall. From this mode of attachment to the wall it is easily understood
how Trichocephali, especially when they are numerous in the
gut, cause local irritation and inflammatory conditions consisting of
frequent attacks of diarrhœa, sometimes twenty times a day, lasting
for months, resisting all remedies, and often accompanied by colicky
pains and symptoms of peritonitis. The stools often have blood
mixed with the fluid, very glassy, jelly-like mucus, more or less
abundantly as in the cases of Moesasca, Moosbrugger,817 Kahane,818
Girard,821 Poledne,823 and Rippe.824 Nausea and vomiting are rarer
symptoms.
Diagnosis as a rule can only be made by microscopical examination
of the stools; together with the eggs, regular and beautifully
formed Charcot-Leyden crystals occur.
The prognosis is unfavourable in severe infections, in slighter
cases, where only a few worms are present, the danger of important
symptoms is less. Treatment consists in administration per os of
vermicides and in local treatment of the large gut. A remedy which
was once much used was calomel, which is much lauded by Gibson
and given as follows: calomel 0·06 grm., rheum. 0·3 grm., tinct. ferri
sesquichlor. 1·2 c.c., aq. dest. 90·0 grm., six dessert-spoonfuls three times
daily. Rippe appears to have got no result from the use of this prescription.
Thymol, especially in conjunction with local treatment of
the large intestine, had unquestionably some effect in certain cases,
such as those of Girard, Poledne, Hausmann, Kahane and Schiller.
The local treatment of the large bowel is most effectual when high injections
of water and benzine are given. Becker825 obviously used too
much benzine (1 dessert-spoonful to 1 litre of water), for severe
irritation was set up, whilst Peiper826 used only a few drops of benzine,
5 drops to 1 litre of water being enough (Schiller). Instead of
benzine enemata, garlic, 1 per cent. thymol solution, and physiological
saline injections have been used, but the benzine enemata seem to be
far and away the most effective. In Schiller’s case 2,000 worms came
away on the first day as the result of such a combined treatment
(thymol internally and benzine enemata).
Trichinella spiralis.
Trichinosis is, happily, becoming so much rarer that many
doctors get no opportunity, either in their student days or in private
practice, of seeing this severe disease; we ourselves remember having
observed one typical case of a peasant, aged 17, from Metz in Med.-Rat
Merkel’s clinic in Nuremberg in the year 1879. In the description
of the disease we follow Merkel’s827 observations.
The eating of flesh containing Trichinæ is often followed, if not
invariably so, by gastric disturbances of different kinds, especially by
vomiting and diarrhœa, with colic, great muscular fatigue, œdema
of the eyelids, muscular swellings with hardness and extreme painfulness,
disturbance of ocular movements, of deglutition and of breathing,
hoarseness, aphonia, intestinal hæmorrhage, bleeding of the nose,
ecchymosis of the skin and mucosæ, prurigo, herpes, miliaria, pustules,
boils, severe sweating, œdema of the extremities, and, finally,
desquamation of the skin; more rarely there is considerable
decubitus, bronchial catarrh, hypostatic and catarrhal pneumonia,
with dry and purulent pleurisy, and in severe cases symptoms of
collapse with delirium close the scene. Slight cases last from three
to six weeks, severe ones for several months, and in the latter
convalescence is very slow. It is remarkable that in cases of
trichinosis of long duration, cancer of the breast was observed at
the same time (Klopsch,828 Langenbeck,829 Babes830). Death during
epidemics occurred in 30 per cent. of all cases. The disease begins
generally from one to ten days after eating trichinous flesh, yet there
have been cases noted in which the disease began several weeks after.
Diagnosis in the presence of several cases, or in epidemics, is not
difficult, but in isolated cases, on the other hand, it is not easy.
If there is a suspicion of trichinosis, from the muscular fatigue and
the œdema of the eyelids, the diagnosis can be made by excision of
a piece of muscle and by finding the Trichinæ in the tissue, taken
with the results of the examination of the previously eaten sausage or
meat. In contradistinction to this circumstantial process, there is
the examination of the blood, which, according to Schleip831
(Homburg trichinosis epidemic, August 19 to 26, 1903, 130 cases), is
the most valuable method of diagnosing trichinosis when the Trichinæ
have not yet penetrated the muscles, for a blood examination shows
a large increase in the numbers of the eosinophile cells; Stäubli
detected his seven cases in this way, four of the severe ones showing
a marked hyperleucocytosis, and a combination of Kernig’s sign with
absence of the patellar reflex. On account of the rarity of these two
signs in combination in other infective diseases, they have a certain
diagnostic value. Stäubli832 also observed in trichinosis the constant
appearance of a remarkably strong positive diazo-reaction of the
urine.
Prophylaxis in trichinosis is fully considered under Trichinella
spiralis (p. 429).
Treatment consists in those cases where it is known that trichinous
flesh has been swallowed in the first place of washing out the
stomach, but still more in a thorough evacuation of the bowels, for
which calomel (0·5 grm.), ol. ricini (a dessert-spoonful till the action
becomes marked), infusion of senna with sulphate of magnesia and
large enemata are employed, and should be repeated at intervals
during the first few weeks. Alcohol (cognac up to 250 c.c. a
day) is recommended by some, also glycerine (150 grm. at a dose)
and large doses of dilute hydrochloric acid. Beside these, a large
number of other remedies are recommended, of which, perhaps,
benzine and thymol, especially in the form of enemata, are worthy
of notice.
When the disease is fully developed the treatment should be
symptomatic; a protracted practically continuous luke-warm bath is
especially useful.
Eustrongylus gigas.
Eustrongylus gigas is most frequently found in the pelvis of the
kidney. Infection in the majority of cases leads to pyelitis. The
inflammation extends to the capsule from the pelvis, resulting in a
purulent nephritis. In infections of longer duration, the affected
kidneys become changed into so-called kidney sacs, while the kidney
itself continuously shrinks. Owing to the worm fixing its posterior end
in the ureter, and owing to an inflammatory swelling of the mucosa
of the ureter, the passage of urine becomes very difficult.
The symptoms resemble those caused by a foreign body, e.g.,
kidney pain, suppression of urine, dysuria, discharge of blood and
pus with the urine. But these symptoms are not sufficient for
a diagnosis; this can only be established by finding eggs or the
parasite itself in the urine.
Moscato833 records a case with chyluria, pain in the region of the
right kidney, and hysterical symptoms. During an hysterical attack
a specimen of Eustrongylus gigas was discharged in the urine, and the
chyluria and nervous affections disappeared. In a case described
by Stuertz834 of an Australian with chyluria due to Eustrongylus gigas
the chyluria had existed for seven years. In the urine the eggs of
Eustrongylus gigas were found. The cystoscopic examination showed
that turbid urine was discharging from the left ureter. Nephrectomy
was considered.
Ancylostoma duodenale (Ancylostomiasis).
Whilst up to quite modern times it has been generally maintained
that the great majority of worm diseases cause more or less marked
symptoms, the exact investigations of the last few years have made
it plain that the great majority of people with worms are not only
perfectly healthy, but the most careful clinical observations show no
single sign of any ill-effect of the intestinal parasites on the health
of the host (Löbker and Bruns835). If infection has led to the
development of only a few ancylostomes, then injury to the general
health is, as a rule, scarcely noticeable. In order to produce severe
illness the presence of several hundred worms in the intestine is
necessary, and in general the intensity of illness varies in exact
proportion to the number of worms. Then the duration of the infection
comes into play: the longer the human organism is submitted
to the injurious effect of the parasite, the clearer is the effect on the
host. Besides, the resistance of the individual has to be considered.
Whilst a more robust person can harbour without ill-effect for a
longer time a larger number of ancylostomes, the symptoms of the
disease become more markedly and much sooner apparent in weakly
persons or in those weakened by other diseases.
The first symptom is disturbance of the digestive system; more
often there is a feeling of pain in the epigastrium, more severe upon
pressure, heartburn, nausea, vomiting of mucus or food at different
times of the day (occasionally ancylostome ova have been found in
the vomit). Whether the eggs which reach the frontal sinus with the
vomit can develop into larvæ there is questionable, but the records
of v. Ziemssen836 and Huppertz,837 to the effect that in some instances
ancylostomes have been discharged from the frontal sinus, are of
interest. The five cases recorded by the latter had a fatal termination
from œdematous swellings of the face with severe inflammation of
the meninges. The tongue is furred, and extensive catarrhal stomatitis
and ptyalism are recorded. The appetite is variable, increasing
or diminishing, there is loathing of nourishment or a marked longing
for acid food and unripe fruit, whilst ordinary meals are rejected.
At first there is often constipation, later diarrhœa with abundant
mucus, and often blood in the stools; microscopically eggs and
Charcot-Leyden crystals were found.
In the further course of the disease symptoms due to increasing
anæmia predominate; the hæmoglobin of the blood diminishes from
one-fourth to one-fifth of the normal (Baravalle838), the eosinophile cells
increase considerably (Boycott,839 Lohr840), yet in regard to diagnosis
eosinophilia cannot be regarded as of equal value to a microscopical
examination of the fæces (Bruns, Liefmann, and Meckel841). The disturbances
of the circulatory system take the form of more or less
severe palpitation, pain in the region of the heart, quick pulse, œdema
of the eyelids, of the face, of the lower limbs, and even of the whole
body. Disturbance of the sexual functions (impotence, irregular
menstruation, delayed onset of puberty) are not infrequently observed.
Infection in human beings takes place by the mouth, if uncleansed
vegetables are eaten—in Japan especially, where human fæces are used—and
articles of food are not sufficiently carefully cleaned (Inouye842),
or from putting food into the mouth with dirty hands. Looss843 does
not think that drinking water is dangerous as a rule, for the larvæ
sink to the bottom in standing water, and are only brought to the
top by shaking. Looss has done most valuable service by discovering
that infection can arise also through the skin. During the last few
years so many authors have confirmed this at first doubted source
of infection, that one must accept this source of infection now, even
though it is undecided which mode of infection is the more prevalent,
by the mouth or through the skin. Some authors have described
the changes induced in the skin by the penetration of the larvæ; for
instance, Looss and Schaudinn,844 itching papules in their own skin,
and Dieminger845 a skin affection in the Graf Schwerin mine which
was called the “Schweriner itch,” and a skin affection not unlike
scabies in the tea plantations of Assam and South America; pani-ghao
(water itch) (Dubreuilh846); the penetration of the larvæ through
the skin also explains the frequent appearance of boils and itching
purulent eczema in miners in infected pits (Goldmann847).
The absolute diagnosis of ancylostomiasis depends on the detection
of the ancylostome eggs in the fæces, and presents no difficulties.
Prophylaxis is of the greatest importance, especially to miners.
The spread of ancylostomiasis seems to depend only on fæces
deposited in damp places, so that on the one hand the deposition
of fæces must be prevented, and on the other the fæces must be
rendered as far as possible harmless; in addition, there is the
individual prophylaxis.
General prophylaxis requires:—
(1) Examination immediately for ancylostomes of miners seeking
work and of those newly taken on five to six weeks after.
(2) Indentured workers who are infected with worms are not
allowed to work underground until a medical certificate in writing
is brought to the effect that they are no more infected with eggs (the
same procedure applies to workmen in brick kilns) (Goldmann848).
(3) Indentured workers infected with worms must submit themselves
to the prescribed treatment, and after its completion further
submit their stools to three examinations at intervals of about four
weeks.
(4) Special supervision of miners and brick-makers coming from
the Italian frontier.
(5) Workmen must be given instructions, both by word of mouth
and in writing in their mother tongue, as to the infectivity and danger
of ancylostomiasis both to themselves and others.
(6) Orders are to be given as to washing, baths, and changing
of clothes at the end of the work.
(7) During the hours of working in the pits, taking of food is
strictly forbidden without thorough and entire washing.
(8) All privies must be so arranged that the vessels used for the
reception of the excreta must not leak, must be protected by a cover,
and easily transportable. The emptying of these vessels must be
carried out in specially constructed impenetrable pits.
(9) Defæcation in any other place than a privy is forbidden (alike
for miners and brick-makers).
(10) The manure of horses used in the mines is to be regularly
removed; possibly infection takes place in this way also. [This is
impossible.—J. W. W. S.]
How far it is possible to disinfect a mine already severely infected
is a matter of question; Tenholt,849 Goldmann,850 and Dieminger851
recommend washing out with freshly prepared lime water with the
addition of caustic soda; Calmette852 and Manouriez853 spraying with
salt water. Theoretically spraying with hot water or steam should be
done every now and again for the destruction of the larvæ (Looss854).
Personal prophylaxis is partially included in the general prophylaxis
in so far as it is a case of oral infection, but something more can be
done for the individual to avert the danger of cutaneous infection.
According to Manson855 it is advisable in the tropics to cover the
naked hands and feet with green Barbados tar, and the tarred parts
thickly with flour; Fabre856 recommends that miners who might come
in contact with infected water should anoint the unprotected parts
(hands and feet), as then the larvæ cannot penetrate the skin; this last
procedure can easily be carried out on account of its simplicity and
cheapness.
Among the usual remedies for the expulsion of ancylostomes
thymol certainly comes first, introduced by Bozzolo857 and since
used by many other authors, partly with good and partly with less
good results. The day before the beginning of treatment one should
endeavour to procure a thorough evacuation of the bowels by means
of calomel (Lutz,858 Grünberger,859 Smith860) or cascara sagrada (Mann861),
only fluid food should be taken the evening before, and on the day
of treatment thymol is given in a quantity of 6, 8, 10 or 15 grm., in
single doses of 2 grm. with one or two hours’ interval, and some hours
after an aperient. As a rule, one day of this treatment is not enough.
(Prowe862), but one is compelled to repeat it on two consecutive days,
or even oftener, with subsequent intervals of many days. Thymol is
either given in wafers, gelatine capsules or mixed with sugar. Caution
should be used in giving brandy at the same time or[sic] bodies which
dissolve thymol (oil, fat) and thereby considerably favour its absorption.
It has been shown in many cases from toxic phenomena
that thymol is by no means an indifferent drug; violent burning in
the stomach and alimentary canal, lowering of the temperature,
shortness of breath and feeble pulse, giddiness, delirium and fainting
have all been observed. Sandwith863 and Thornhill,864 as well as
Leichtenstern,865 even record cases of death after the use of thymol;
4 grm. thymol caused severe symptoms of poisoning in Grünberger’s866
case. The black colour of the urine (thymoluria) which so often sets
in after the first dose is quite harmless, and is no contra-indication
to the continuance of the cure. Now and again there are traces of
albumin in the urine, but it is very seldom there is any severe acute
inflammation of the kidneys. Thymol is contra-indicated in advanced
old age and in debility, also in cases with a tendency to vomiting, in
gastritis, dysentery, heart or kidney affections.
The combination recommended by Goldmann867 under the name
of taeniol, already mentioned under the treatment of tapeworms, and
which consists of thymol, sebirol and salicylate, appears also to render
good service in the treatment of ancylostomiasis (Goldmann868 and
Liermberger869).
A carbonate of thymol, thymotal, from which thymol separates off
in the intestine, is given three to four times a day, in doses of
3 grm. per diem (children up to 1·0 grm.) on four consecutive days,
and at the end of the treatment a purge (Pool,870 Bauer871); Leonardi872
speaks well of thymol essence (4·0 c.c. per diem) in an emulsion with
plenty of water.
The next drug for the expulsion of ancylostomes is extractum
filicis maris, which is to be employed as in tapeworm treatment, but
has not always had the desired result, whilst in such cases as resist
the fern extract, thymol attains the desired effect (Mann873), whilst the
reverse is frequently observed (Grünberger874). Nagel875 prescribes
extr. fil. 8 to 10 grm., chloroform 10 to 15 drops, syr. sennæ 16 grm.;
before taking, the glass must be placed in hot water, otherwise the
contents will not pour freely. Zinn876 prefers extract. filicis maris
(freshly prepared) to all other drugs. Warburg877 considers the treatment
with extr. fil. to be all the more certain the more thoroughly the
preliminary treatment is carried out. Filmaron 0·7 grm., thymol
5·0 grm., chloroform 1·5 grm., ol. ricini 20·0 grm. gave good results
after being given two to three times (Nagel878). Opinions are divided
as to the combination of thymol and extractum filicis maris (Hynek,879
Stockman,880 Boycott and Haldane,881 Adams882). As regards other remedies,
eucalyptus oil is well spoken of by Philips883 and Hermann884: ol. eucalypti
2·0 grm., chloroform 3·0 grm., ol. ricini 30·0 grm., to be taken at one
time or in three separate doses in the morning (on the previous evening
a saline purgative). Neumann885 recommends podophyllin, to be taken
twice on three consecutive days in doses of 0·035 grm. Podophyllin
appears to produce quite a peculiar condition of the intestinal mucosa
which is very prejudicial to the Ancylostoma adhering to it. Bentley886
regards β-naphthol as the best drug; after previous examination of the
bowels he gives it two or three times at two-hourly intervals, in doses
up to 1·0 grm. (Vide also the Appendix, p. 754, for other drugs.)
For the treatment of the anæmia, which often persists very obstinately,
good and abundant food, iron and arsenic preparations, Levico water
(Goldmann,887 Liermberger888) are suitable.
Ascaris lumbricoides (Ascariasis).
Ascaris lumbricoides is one of the most frequent parasites that
occur in man, both in adults as well as in children; as a rule, indeed,
it most frequently infects children of medium age. The normal
situation is the small intestine; this, however, is frequently left, and
the Ascarides travel into the stomach, œsophagus, pharynx, bronchi,
the nasal cavities and still other regions. It is a peculiarity of the
Ascarides that they are prone to glide into narrow canals; for example,
Clason889 records that in the case of an idiot whose custom it was to
swallow glass beads, the Ascarides showed a predilection for sticking
in the beads and were passed in the fæces. The disturbances which
Ascarides occasion in the intestine itself vary; isolated species do not
give rise to any symptoms at all, whereas a large number may
eventually give rise to severe local symptoms, or those of a toxic or
reflex nature which have been discussed in the General Section.
Among the local symptoms are the following: loss of appetite,
excessive appetite, perverted sense of taste, fœtid breath, sensitiveness
to pressure over the abdomen, colicky pains and irregularity of the
bowels. The appearance and state of health suffer; the patients,
children in especial frequency, become remarkably pale; their complexions
undergo rapid change, and rings of grey or bluish-brown are
seen about the eyes. Children may become so reduced by this rare
condition, enteritis verminosa, due to Ascarides in large numbers, that
suspicion of the existence of intestinal tuberculosis arises. Emaciation
to a skeleton, excessive meteorism, and evacuations of thin gruel-like
stools, sometimes blood-stained, are observed in these cases. Even in
the case of adults, chronic uncontrollable vomiting with severe
inanition due to the Ascarides has been observed. When the
Ascarides escape spontaneously per anum, they frequently cause an
exceedingly troublesome irritation in the anal region (pruritus ani).
The most disagreeable symptoms and those most dangerous to
life arise from the migrations of Ascarides when they invade the bile-ducts;
no inconsiderable number of cases of this kind are recorded
in the literature (summarized, up to the year 1901, in Sick’s890 Dissertation).
Penetration post mortem (or shortly before death) of the
worms into the bile-ducts cannot be considered as a rarity; the laxity
of the muscular orifices easily allows of this invasion also in other
directions on the part of the parasite in its escape from the body of its
dead host. The occurrence of the worm in the biliary passages in the
living is to be regarded as still less frequent, but nevertheless often
enough according to the records in literature. Sick891 was able to
collect as many as sixty-one such cases, to which he added two further
fresh cases from the Tübingen clinic, that is, from the material provided
by his father. In the year 1891 Borger892 collected fifty-nine
cases relating to the invasion by Ascaridæ of the bile-ducts and
passages, and Dauernheim’s893 Dissertation treats of this question as
well. A further case of Ascaris in the ductus choledochus (choledochotomy)
is recorded by Neugebauer.894 In the case of Schupper895
(woman, aged 52), all the biliary passages were distended and filled
with fourteen living Ascaridæ (perhaps as they were living they had
not led to a septic infection of the biliary passages); in the case
communicated by Schiller,896 an Ascaris had gained access to the
biliary passages after an operation for cholelithiasis (with distension
of the gall-bladder and formation of a fistula); it had kept itself alive
here eighteen days and was extracted from the fistulous opening.
Epstein897 confirms the correctness of the explanation of the mark of
strangulation in an Ascaris in Mertens’898 case (in a woman, aged 30,
there was first icterus, later ascites, anasarca, swelling of the liver,
then the discharge of two dead Ascaridæ, one of which exhibited a
constriction somewhat behind its centre; after that there was rapid
improvement in all the symptoms); in his case there was icterus in
consequence of closure of the ductus choledochus by an Ascaris.
After the discharge of the worm the symptoms persisted; one of the
Ascaridæ had a typical strangulation mark. From the observation
recorded by Vierordt899 it follows that, without doubt, mature females
can penetrate into the liver and there deposit eggs; in addition, that
such eggs appear exceptionally to undergo segmentation. A unique
feature in this case consisted in the exclusive discharge of immature
worms almost regularly throughout an interval of nine weeks; this
cannot be explained from our present knowledge of the biology and
pathology of the Ascaridæ. These worms clearly make their way from
the intestine outwards, through the opening into the duodenum of
the common bile-duct, and unquestionably the fully developed
Ascarides, with the aid of their conical head end, are enabled gradually
to penetrate the wall of the ductus choledochus (Quincke900), and gain
access to the gall-bladder, the hepatic duct and its branches.
The changes in the biliary passages and the liver are, on the one
hand, the mechanical results of a partial or total obstruction to the
flow of the bile, and, on the other, of inflammatory processes. The
blocking of the common bile-duct and of the trunk of the hepatic
duct leads to the well-known symptoms of biliary engorgement;
protracted continuance of this condition has, as its sequela, general
distension of the whole biliary system and degenerative destruction
of the liver-cells. If the Ascaris is situated at some other part of the
biliary system, its presence causes a partial arrest of the flow of bile,
with the corresponding sequelæ. Many Ascarides perish in the ductus
choledochus, and here and in the gall-bladder they may supply the
nucleus of a gall-stone; deeper in the liver this does not appear to
happen; the dead Ascaridæ here undergo a kind of maceration, disintegrate,
and may be completely absorbed; in many cases the worms
continue to live for a very long time in the biliary passages. When
the worms infect the biliary passages through the invasion of intestinal
bacteria, liver abscesses arise (Dauernheim,901 Saltykow902).
Leer903 goes so far as to maintain that Ascaridæ may be the second
most frequent cause of liver abscesses. That Ascaris in the pancreas
may simulate liver abscess in a remarkable fashion is shown by
Vierordt’s904 observation, which is quite unique, while Ascaridæ have
been found to occur in isolated instances in the excretory ducts of
the pancreas and in its branches, where they have remained living for
a long time.
It is no rare occurrence for Ascaridæ, in consequence of their
migration into the stomach, to be ejected by the act of vomiting, and
in such way to gain access into the upper air passages, or to find
their way during sleep into the nose or accessory sinuses (Mosler and
Peiper905) without giving rise to special symptoms. For example,
Troja906 found in the frontal sinus of a cadaver a large coiled-up
Ascaris which occupied the whole cavity. Wrisberg907 made the same
observation in the cadaver of a boy. Deschamps908 and Fortessin909
mention an Ascaris being met with in the antrum of Highmore.
Observations of the discharge of living or dead Ascarides from the
nose are frequently recorded. To this class belongs the case
mentioned by Albrecht,910 in which an Ascaris was removed from the
nose of a girl, aged 7; also the case recorded by Benievini,911 from
the nose of one of whose friends a worm escaped; he had suffered
from the most violent headaches, fainting fits, dimness of vision and
vomiting; after the escape those untoward symptoms disappeared.
Similar records have been made by Forest,912 Lanzoni,913 Langelott,914
Tulpe,915 Reisel,916 Fehr,917 Bruckmann,918 Bahr,919 Slabber,920 Lange,921
and Chiari.922 A rarer case is that recorded by Haffner,923 that of a
child, aged 4, in whom an Ascaris reached the nasal cavity through
the act of vomiting, and from there it gained access through the
naso-lachrymal duct and the inferior lachrymal sac into the lower
punctum lachrymale, from which half of it protruded.
Among the rarer causes of the occurrence of strange bodies in the
pharynx and naso-pharyngeal cavity, Jurasz924 mentions in the first
place vomiting, which may afford opportunity for the more solid
bodies of the stomach contents, and even parasites of the digestive
tract, especially Ascaridæ, to become firmly lodged in the pharyngeal
or naso-pharyngeal cavity. Ascaridæ may obtain access from the
naso-pharyngeal cavity to the middle ear by way of the Eustachian
tube, as has been observed by Reynolds925 and Wagenhäuser926; in the
case recorded by Turnbull927 (girl, aged 8, with pains in her ear) the
Ascaris apparently reached the external auditory meatus by the same
route.
The irritation of the larynx and air passages by Ascaridæ is far
more dangerous than their penetration into the nose and naso-pharyngeal
cavity, because not only are attacks of suffocation, but
sudden suffocation thereby induced. Oesterlein928 records a fatal
attack of choking from Ascaridæ in the trachea. In a case recorded
by Smyly929 of a boy, aged 3 1/2, tracheotomy for extreme asphyxia was
performed without relief. At the post-mortem the cause of the asphyxia
was found to be an Ascaris in the trachea. Fürst930 collected twenty-five
observations of invasion of the larynx and trachea by Ascaris.
Mosler931 reports the case of a patient with aphonia and dyspnœa
from whose larynx an Ascaris was removed. Donati932 reports a case
of four Ascarides in the larynx, and Cerchez933 of asphyxia from
Ascarides in the larynx or trachea. Wagner934 records the case of
a boy, aged 8, in whom a coil of worms was ejected from the stomach
by vomiting; the mass blocked the entrance to the larynx and led to
death from suffocation. A case similar to that recorded by Smyly is
communicated by Rabot935; it was that of a child who underwent
tracheotomy for diphtheria, and who was not relieved by the operation;
when, however, an Ascaris appeared in the cannula and the
parasite was removed the child breathed well. In Negresco’s936 case,
that of a boy, aged 3, an Ascaris gained access to the larynx and from
there into the trachea, and a fatal issue from asphyxia resulted.
The route by which Ascaridæ obtain access to the urinary passages
must remain undecided. Schlüter937 treated a woman, aged 60, with
retention of urine. Upon catheterization the hinder end of an
Ascaris hung out from the catheter opening; the anterior end was
fixed in the tube and the lumen was obstructed. Perhaps in the
female sex Ascaridæ travel from the gut into the vulva and from there
into the bladder, as they have already been observed in the vagina,
where they cause troublesome symptoms (pruritus pudendi).
The diagnosis of ascariasis is not in general difficult; now and
then the worms are discharged spontaneously; if not, the ova, which
cannot be mistaken, can easily be detected in the fæces upon microscopical
examination. Epstein’s938 method—namely, on every occasion
to obtain fresh material for examination—is much to be recommended.
This consists in introducing a Nelaton’s catheter into the rectum with
a rotatory motion and then drawing it out. A small portion of fæces
forced into the catheter opening is more than sufficient to demonstrate
the presence of ova of the parasites upon microscopical
examination of a preparation.
In spite of all pressure on the part of relatives, treatment directly
against Ascaridæ should not be carried out until the diagnosis is
certain.
As regards prophylaxis, much can be done by not throwing the
worms, when expelled, on to the dung-hill or into the privy, but
straightway into the fire. Metschnikoff939 has issued a warning
against the consumption of unboiled or badly washed vegetables,
salad, strawberries, etc., and also against drinking polluted water.
For the expulsion of the worms flores cinæ were formerly considered
the most useful means; now, however, santonic lactone—santonin—which
is prepared from them, is almost universally
preferred. By many, especially in practising among children, flores
cinæ are still recommended in the form of Störk’s worm electuary
(consisting of flores cinæ, rad. jalapæ, valerian and oxymel simplex).
Guermonprez940 recommends them because he thinks that santonin
only excites the worms and consequently causes unpleasant symptoms.
Besides, in the form of the above-mentioned electuary, flores
cinæ can also be given several times daily with raspberry jelly up to
0·5 grm. to 2 grm. (children and adults).
Santonin is prescribed either in single doses from 0·03 to 0·05 to
0·1 grm. with sugar in the form of powder, or else in oily solution.
When given in the latter form the absorption of the santonin in the
stomach is excluded and the whole quantity introduced is thus
enabled to reach the worms in the intestinal canal. Küchenmeister941
has already recommended combination of santonin with ol. ricini.
Lewin,942 however, states that ol. morrhuæ, ol. olivarum, ol. cocos
and ol. cinæ can also be taken. In prescribing santonin in oily solution
Henoch943 also prefers the combination with ol. ricini. According
to Lewin’s direction the prescription would run as follows:—
| ℞ | Santonin | 0· | 2 grm. |
| Ol. ricini. | 20· | 0 grm. |
| Ol. cinæ æth. | gtt. | iv. |
| M., d.s. |
S., one tablespoonful to be taken two to three times. |
If the patients should manifest a repugnance to castor oil,
Starke’s ricinus paste may be selected:—
| ℞ | Santonin | 0· | 2 grm. |
| Ol. ricini | 20· | 0 grm. |
| Ol. cinæ æth. | gtt. | iv. |
| Sacch. albi. | q.s. | |
| Pasta moliis. |
S., to be used for two days. |
If necessary the first-mentioned mixture might be given in gelatine
capsules. Small children should be given 0·025 grm. santonin in
warm olive oil slightly sweetened with sugar (a teaspoonful) in the
morning; if in the course of the forenoon specimens of Ascaris
escape, a second dose should follow in the afternoon about two
hours after the meal. Older children should be given santonin in
combination with castor oil or calomel:—
| ℞ | Santonini | 0· | 01 to 0·02 to 0·03 grm. |
| Calomelan | 0· | 025 | grm. |
| Sacch. albi. | 0· | 5 | grm. |
| M.f.p. D. tal. dos. x. |
| S., one powder about six, seven, and eight o’clock on three consecutive days. |
As santonin causes slight toxic symptoms such as urticaria,
vomiting, retention of urine, headache, vertigo, yellow vision
(xanthopsia), it is in every case advisable to follow with a laxative to
expel the drug from the body as speedily as possible. The urine
is coloured yellow from one to two days and assumes a scarlet
red colour upon the addition of alkalis; this, however, soon
disappears, while it persists in the case of rhubarb and senna.
In the place of santonin iodoform in the form of a powder mixed
with bicarbonate of soda is given by Schidlowsky944 in doses up to
0·01 to 0·06 grm. three times daily, and a dose of castor oil on the day
after the iodoform is given. Thymol in addition to thymol enemas
may be tried, in doses up to 0·5 to 2·0 grm. per diem (Calderone,945
Hausmann946), also β-naphthol up to 0·45 grm. three times daily (Du
Bois947), and—
| ℞ | Benzo-naphthol | 2·0 | grm. |
| Semin cinæ | 1·0 | grm. |
| Sacch. albi. | 0·5 | grm. |
| M., f.p. Divide in part. æq. xxii.
S., three to five powders daily. |
(Ferran948), filmaron oil 1·0 to 2·0 to 3·0 grm. in gelatine capsules,
according to age (Bodenstein949). Brüning950,951 recommends the so-called
American worm-seed oil, derived from a plant native to the United
States, Chenopodium anthelminticum, Gray. It is given in emulsion
(ol. chenopodii anthelm. 10·0 grm., vitelli ovi unius, ol. amygd., gi.
arab. pulver. āā 10·0 grm., aq. destill. 200 grm.; f. emulsio) up to 0·25
to 0·5 grm. three times daily at one to two-hourly intervals, or as a pure
oil from 8 to 15 drops in sugar and water; to be followed an hour
after the last dose by oleum ricini or pulvis curellæ. If no action
takes place by the afternoon, a laxative should again be given. The
treatment frequently must be repeated the next day. Thelen952 appears
to have had good results from this drug.
Corsican moss (mousse de Corse), kamala, Artemisia absinthium,
valerian, semen sabadillæ, have all been supplanted by santonin and
at most are used as adjuvants for the latter.
Oxyuris vermicularis (Oxyuriasis).
Oxyuridæ do not remain at rest in the gut, but leave it, generally
at night time, to migrate around the anus, into the gluteal folds, and
in females into the vulva and vagina and still higher up, giving rise
in these different sites to a whole series of irritative symptoms. In the
rectum, also, Oxyuridæ give rise to such symptoms, which are manifested
in the form of catarrhal inflammation; numerous chronic
intestinal catarrhs are thus explained. The frequent coincidence of
hæmorrhoidal troubles with Oxyuridæ may be attributed to the fact
that the veins of the rectum participate in those changes which have
been described as occurring in the intestinal mucosa. Oxyuridæ
may also give rise to prolapse of the anus, either by the tenesmus
they bring about having such a prolapse as its direct sequel, or the
proctitis that supervenes constituting a further etiological factor for
its occurrence (Ungar953). Anal fistulæ which still further increase
the trouble, and even rectal fistulæ, appear to be capable of onset in
consequence of the irritation of the mucosa brought about by Oxyuridæ
(Trendelenburg954). The conditions recorded by von Wagener955
and Ruffer956 appear to be of interest. At the post-mortem on a child,
aged 5, the former found fifteen to twenty quite minute nodules on
some Peyer’s patches, and in several of these Oxyuridæ were found
upon microscopical examination between the calcareous concretions
within the patches. He presumes that the parasites penetrated the
follicular ulcers, and after healing of the latter that they died and
became calcified. In the case of a man who died from cirrhosis
of the liver, Ruffer found in the rectum, at a distance of about 6 in.
from the anal orifice, several tumours covered by the intestinal
mucosa, the smallest of which was the size of a pin’s head and the
largest that of a walnut. The tumours looked like calculi overgrown
by connective tissue; under the microscope, countless Oxyuridæ ova
were found in their interior.
The symptoms of irritation set up by these migrations from the
intestine are troublesome to the last degree; the pruritus thereby
induced is often unendurable; as this irritation from itching comes
on with especial severity during the night, the night’s rest is grievously
interfered with; many attacks of night terrors appear to be occasioned
by these worms. But the general condition suffers as well; the
children become pallid and affected with nervous excitability.
Through the act of scratching the irritated parts the ova of the
parasites may be conveyed by contaminated fingers directly into the
oral or nasal cavities, certainly also into the oral cavity by the
contamination of food (auto-infection). In the case of boys the
sexual organs may be excited sympathetically through irritation of
the sacral nerves of the rectum; girls may be induced to practise
onanism in consequence of the entrance of the worms into the vulva.
As a result of the itching irritation which the scratching gives rise
to, and of the irritation due to the parasites migrating to the area
surrounding the anus, congestion and inflammatory symptoms may
arise in the peri-anal and perineal regions (weeping eczema, Seifert),957
and these do not abate till after the removal of the oxyuriasis. Some
authors speak of an oxyuriasis cutanea (Majochi958), in the more
limited sense of a dermatitis intertriginoides. So far five such cases
have been recorded, one each by Szerlecky,959 Michelson,960 Majochi,961
Barbagallo962 and Vignolo-Lutati.963 Szerlecky’s case was that of
a young woman with intertrigo over the thighs (the skin was covered
as if with leather); Michelson’s case was that of a boy, aged 13,
with intertrigo on the skin of the genito-crural fold, of the scrotum
and of the thigh; Majochi’s was that of a man, aged 38, with the
same localization; Barbagallo’s case was that of a boy, aged 14,
in whom the dermatitis extended to the hypogastrium (rhagades on
the scrotum); and Vignolo-Lutati’s case was that of a man, aged 24,
with intertrigo of the peri-anal and perineal region, of the scrotum
and the inner side of the thigh.
On leaving the gut, Oxyuridæ frequently migrate to the stomach,
to the œsophagus, to the mouth, to the nasopharyngeal cavity, and
into the nose (Zarniko964) (the localization in the nose has been
referred to as associated with the possibility of auto-infection—see
p. 695 as to the development of embryos from the ova in the
moist nasal mucosa). Still the occurrence of Oxyuridæ in the nose
is among the greatest of rarities. Chiari965 records the case of a girl,
aged 14, who suffered from pains at the root of the nose and in
the left side of the forehead; female specimens of Oxyuris vermicularis
were evacuated from her nose on several occasions. A similar case
is recorded by Hartmann966; it was that of a girl, aged 13, with
epileptiform convulsions and psychic disturbances; numerous
Oxyurides frequently escaped from her nose. With their departure
the symptoms of irritation of the central nervous system also disappeared.
Rheins967 records a case, that of a woman, in which a
specimen of Oxyuris vermicularis was discharged from the right
nostril during the act of sneezing. Proskauer968 found in the nose
of a woman, aged 30, a conglomerate of from fifteen to twenty very
small worms which proved to be Oxyuris embryos.
The diagnosis of oxyuriasis is not difficult to make, as the troublesome
sensations in the anus and about the genitals necessarily suggest
the presence of Oxyuridæ. As a rule the small white worms are
seen crawling about over recently evacuated fæces, or the ova are
found upon microscopical examination of soiled matter adhering to
the anus, or in scrapings removed with the spatula from the surface
of the skin (in the case of oxyuriasis cutanea).
Prophylaxis has to be directed to infection with Oxyurides
generally, on the one hand, and, on the other, to the possibility of
auto-infection. With reference to the first-mentioned point, Metschnikoff’s969
directions should be borne in mind, to the effect that badly
washed vegetables, salad, etc., ought not to be eaten (vegetables to be
rinsed with boiling water), and also that the members of the family
of the diseased individual should be examined for Oxyuridæ and
eventually be treated (Heller970). With regard to the second point,
one has to observe strict cleanliness in general (Barbagallo971 found
ova of the parasites in the layer of dirt under the finger-nails).
Treatment of oxyuriasis must be of a twofold nature; first,
medicinal, the administration per os of vermicidal drugs in combination
with purgatives; and secondly, local treatment of the gut by
means of enemata, suppositories and high injections. Following
the method prescribed by Ungar,972 pulv. glycyrrhizæ co. is first given
in the case of smaller children, castor oil or calomel in that of those
older, in order to evacuate the intestine, and four times daily on two
days following one another a dose of naphthalin, not directly after
meal-time, but as far as possible in the interval between two meals,
and at the same time the ingestion of fatty or oily nutriment is as
far as possible to be avoided. After eight days this treatment should
be repeated, and under certain circumstances once again after
a further interval of a fortnight. The dose varies between 0·05 and
0·1 grm. (children of 1 year old), 0·1 to 0·2 grm. (children of 2 to 3 years
old) and 0·2 to 0·4 grm. (children of 4 to 10 years old). Dornblüth973
employs the same medicament in a form only slightly modified from
Ungar’s method, Barbagallo974 gives internally only a purgative (decoct.
sennæ cum natr. sulfur). Thymol, santonin, kousso, kamala or valerian
may be tried instead of naphthalin. For enemata the following are
employed: naphthalin in a solution of 1 in 50, ol. olivar. or thymol
0·1 in 200 aq. destill., diluted solutions of lysol, menthol in 1/2 per cent.
oily solution, salicylate of soda in watery solution, decoctum
tannaceti with santonin, with the addition of some drops of ol.
terebinth. (Barbagallo). Decoctions of garlic, infusion of valerian,
sulphur water (sublimate is to be avoided), aq. calcariæ, ol. olivarum
camphoratum (Vignolo-Lutati). Santonin 0·1 grm. is the best to
employ for suppositories.
For high injections, large quantities of plain water are employed
(2 to 4 litres), or soapy water (0·2 to 0·5 per cent. solution of sapo
medicatus, Heller,975 Still976), 1/2 per cent. salicylic acid solution or liq.
alum. acet. (one tablespoonful to a litre of water, Dornblüth977), or
gujanosol (2 to 3 to 4 to 5 per cent. solution, Rahn978). The employment
of benzine for such high injections is not advisable according to
the experience of Senger,979 owing to the symptoms of poisoning after
the external application of benzine, at least not in the case of young
children.
That diseases of the intestine which are accompanied by frequent
thin fluid evacuations may lead to recovery from oxyuriasis has
frequently been observed by us in the case of young children who
have suffered from dysentery (Seifert980). Inunctions of cod-liver oil
appear to be very valuable in the treatment of oxyuriasis (Szerlecky,
Vignolo-Lutati), whilst those with mercurial ointment may easily
increase the inflammatory symptoms. The luxury recommended
by Esser,981 that patients every evening before going to sleep should
have the female Oxyuridæ picked from the anal fold in the knee-elbow
position is one which is certainly only in the power of a few
people to carry into execution.
An essay has been published by Hippius and Lewinson (Deutsch. med. Wochenschr.,
1907, xliii.) in which the relationship of Oxyuridæ to appendicitis is considered and the treatment
of oxyuriasis is discussed. The instructive case recorded appears to show that germs
through Oxyuridæ gain access to the tissue of the appendix, and, indeed, are carried in by
them. In view of this more recent communication as to the part which intestinal parasites
play in the etiology of appendicitis, it seemed to me [O. S.] to be worth while to interrogate my
surgical colleagues as to this point. About 2,000 appendicectomies have been jointly performed
by Drs. Burkhardt, Enderlen, Pretzfelder, Riedinger, Rosenberger and Siber, and in
not one of these cases could entozoa be found to be a possible cause of the appendicitis.
Such figures without doubt speak in favour of the fact that even if in individual cases entozoa
might come into reckoning as a possible cause, such an etiological factor must be classed
among the greatest of rarities. My colleague, Dr. Ries, who practised for ten years in Mexico,
informed me that there practically speaking every Indian without exception harboured parasites
of the most varied kind, and that in spite of the very extensive professional standing he
enjoyed among these people he never had under observation among them a single case of
appendicitis. As far as the observation of the authors in question as to the treatment of
oxyuriasis is concerned, it must be energetically directed to the employment of local measures
for the intestine; they maintain that the use of enemata would be irrational, and that it is
astonishing that this method has been able to maintain its standing down to the present day.
HIRUDINEI (Leeches).
The only one of the leeches that comes under consideration from
the clinical point of view is Limnatis nilotica (Hæmopsis sanguisuga),
which obtains access to the mouth with drinking water, and becomes
lodged, even in the case of man, in the pharynx, larynx, trachea,
œsophagus and nose.
Amongst the causes of severe hæmorrhage from the pharynx
Jurasz982 mentions the occurrence of leeches in that region: in
Northern Europe this must be accounted one of the greatest of rarities,
whilst at all times in southern countries, such as South Italy, Spain,
Greece, Algiers, Tunis and Egypt, it appears to have been more
frequent. Even the physicians of antiquity had much to say about
it. Upon the occurrence of blood-stained expectoration, Hippocrates
recommends the oral cavity to be examined to see whether a leech
is not present in it. Galen speaks of hæmatemesis due to the presence
of leeches in the pharynx and stomach. Similar mention is found
in the writings of Celsus, Asclepiades, Scribonius Largus, Dioscorides,
Aëtius, Oribasius, Paulus Aegineta and others. In recent times,
Cortial983 has published observations relating to this subject which
he had the opportunity of making in Constantine. Palazzolo984
also in Sicily found leeches in two cases in the pharynx, in one case
on the posterior wall, in the other in the crypt over the left tonsil.
According to Roset,985 leeches adhere by preference behind the uvula,
simulating hæmatemesis and hæmoptysis, and the persistent hæmorrhages
they give rise to may lead to severe anæmia. Leeches are found
in still greater frequency in the larynx than in the pharyngeal cavity.
Huber986 records several observations of this kind in his historical
and therapeutical study. In the case of a man, aged 64, Ramon
de la Sota y Lastra987 observed a leech on the nodulus epiglottidis;
this was removed with the forceps. In the case recorded by
Photiades,988 a leech had remained adherent to the vocal cord for
more than twenty-two days. Maissurianz989 records two such cases:
in one the leech had remained in the sinus morgagni for three weeks,
in the other in the same place for ten days. The case recorded by
Schmolitschew990 is an interesting one; it was that of a woman who
for four days had suffered from violent hæmoptysis, the cause of
which was a leech that was fixed on the laryngeal wall of the epiglottis
close above the vocal cords. In his case (that of a soldier), Godet991
was forced to perform thyrotomy to remove the leech from the larynx.
Ficano992 removed a live leech with the forceps from the lower
laryngeal cavity in a man, aged 30. Massei993 reports a similar case.
The case reported by Winternitz and Karbinski994 was that of a
peasant girl, aged 16, who suffered from coughing, hoarseness, and
blood-stained expectoration; a leech had lodged on the root of the
epiglottis. Aubert995 removed a leech from the larynx of a woman
after the performance of tracheotomy. Seifert996 reports three cases:
in the first the leech had become fixed to the left vocal cord, in the
second it was found in the lower laryngeal cavity, and in the third
on the border of the left ligamentum aryepiglotticum. Leone997 has
published the case of a leech in the larynx, Martin998 two cases with
the leech lodged in the lower laryngeal cavity, Berthoud999 a similar
case, Palazzolo1000 two such cases, Panzat1001 one case (lower laryngeal
cavity). Moucharinski1002 reports a case in which the leech had stayed
more than twenty days in the larynx. Martin1003 easily removed a
leech from the posterior portion of the vocal cord with the forceps.
Vieus and Nepeon1004 record a case of a leech in the larynx. It is
quite exceptional for leeches to gain access to the trachea; cases of
this kind have been recorded by Aubert,1005 Vicano,1006 Ridola1007 and
Tapin1008 (the leech was firmly fixed to the bifurcation and caused
coughing, hæmoptysis and attacks of asphyxia; it was easily removed
by the aid of a tracheal tube). Now and then leeches are found in
the nose.
Lusitanus1009 relates the case of a man who suffered from severe
headaches. A medical man ordered the application of a leech to the
anterior portion of the nostril. Owing to the carelessness of the
surgeon the leech crawled right into the nose; it was impossible to
extract the leech or to kill it, and it produced a severe hæmorrhage
which led to the death of the patient within two days. In a case recorded
by Sinclair,1010 a leech, Hæmopsis sanguisuga, gained access to the nose
of a boy, aged 3; it remained there a fortnight; it caused frequent
attacks of epistaxis and in the end it was removed by means of
forceps. Condorelli-Francaviglia1011 records a case in which severe
epistaxis was caused by a leech which had probably entered the
anterior portion of the left nostril by way of the pharynx and become
tightly fixed there. It was seen by posterior rhinoscopy, and
was removed from in front by means of slightly curved forceps.
Sota y Lastra1012 mentions the occurrence of leeches in the nose, and
Keng1013 reports the case of nasal obstruction from a leech. The
removal of leeches is effected by means of injections or by the direct
sprinkling of salt or acid solutions on their bodies, which brings about
their detachment. When possible a previous attempt should be made
to seize them with forceps so as to make their immediate extraction
possible. The species of Hæmadipsa (Looss1014) live in tropical regions
in moist places on the ground or in the jungle. They climb bushes
and even trees with astonishing rapidity upon the approach of larger
animals and also of man (whom they clearly recognize from the
vibration of the ground caused by footsteps). From thence they let
themselves fall on their victims to suck their blood. Their bites are
generally painless, and of themselves not dangerous, but if they are
unusually numerous they rapidly accumulate on the body in large
numbers and give rise to marked debility and, if the wound become
infected, to severe complications and even death. On the other hand,
under careful treatment the wounds heal easily and fairly rapidly.
Firm leather and firmly adhering clothes afford no certain protection
against the attacks of these leeches, as they know how to
force themselves with extraordinary rapidity through the narrowest
interstices between the clothes and thus gain access to the skin.
When they have sucked their fill—and this may take several hours to
accomplish—they fall off of themselves. To effect an earlier removal
drops of irritative or corrosive fluids are employed (salt solutions,
acids, etc.). Tearing away the leech by force should be avoided,
as in this way portions of the leech’s body may be left behind in the
wound and inflammation be set up.
ARTHROPODA.
Leptus autumnalis (Grass, Harvest, or Gooseberry Mite1015).
In the hot season of the year, that is, during the months of July
and August, it is noticed that those people who stray amongst
syringa bushes or who pick gooseberries or kidney beans are
attacked by the Leptus autumnalis. On the uncovered parts of the
body there appear numerous red spots and papules, which itch and
burn smartly. The itching does not commence diffusely, as in the
case of scabies (MacLennars1016), but is limited to the particular
points where the parasite is situated. There are especial outbreaks
of itching in the morning, arising perhaps from the hatching of ova
in the host after lying in the warmth of the bed.1017 Leptus frequently
provokes general erythema, eczematization or severe feverish
urticaria, which in France is known by the name of fièvre de grain
(Mégnieu, Besnier1018). If the individual efflorescences be carefully
examined, there will be noticed almost without exception a minute
boss towards the centre, noticeable by its yellowish-red colour. If
an attempt is made to remove it with the point of a needle or to
scrape it off the surface, one can often perceive, even with the naked
eye, a small reddish creature moving actively about. The treatment
of these very troublesome symptoms consists in warm baths with
soapy lavages, also lavages with alcohol, spirit salmiac (G. P.), 5 per
cent. carbol or creolin solution, diluted vinegar, benzine, emulsions of
balsam of Peru, rubbing in sulphur ointment (Sandwith1019); ointments
of creosote or eucalyptus are recommended. Other grass and grain
mites also occasionally penetrate the skin of man and produce transitory
but sometimes very severe eruptions, urticaria and eczema
papulosum, as Geber1020 and subsequent to him Josai1021 have reported
of the barley mite. In sensitive individuals the skin becomes bright
red, to a greater or less extent their temperature is raised and frequently
slight febrile affections are present. If the inflammatory
skin symptoms have reached their culminating point after three or
four days and no fresh complications arise, they only remain for
a short while, the effects of scratching and pigment spots being
left.
Kedani, Akaneesch (The Japanese River or Inundation Disease).
This disease is only known in Japan, and is limited to the
neighbourhood of some great rivers on the west coast. The people
mostly attacked are those who cut the hemp harvest in the infected
localities, occasionally those who transport it or come into contact
with it (Looss1022). The disease is frequently manifested in the form
of indefinite disturbances of the general condition; it commences
generally on the sixth day after the presumed infection with rigors,
headaches, feeling of weakness, swelling of the lymphatic glands in
the loin or in the arm-pits; in the periphery a black dry scab is
formed. In addition there is an intense conjunctivitis, and added
to symptoms of fever an exanthema resembling measles that lasts
from four to seven days. There is frequent delirium and difficulty
of hearing which persist for a long while. Obstinate constipation is
a striking symptom. At the end of a fortnight, earlier in slighter
cases, the fever commences to abate and a rapid convalescence sets
in. In pregnant women abortion with fatal issue is frequent. With
regard to prophylaxis, Baelz1023 recommends as rapid a cultivation of
the soil as possible, which has led to a speedy disappearance of
the disease in districts where it was once dreaded. Treatment is
symptomatic. Japanese do not tolerate antipyretic drugs as well
as Europeans.
Dermanyssus gallinæ (avium).
During the day the resort of bird mites is in the droppings and
in the woodwork, etc., of cages in which canaries, crossbills and
parrots are kept; in the crevices of doors, in the chinks between
the board planks of bedsteads, so that at night they may seek some
domestic animal to suck the blood and so satisfy their hunger. It
is by no means rare for young animals, chickens and unfledged
pigeons, etc., to perish in consequence of the great loss of blood.
This nocturnal habit of life explains why no mites can be found
during the day in spite of the most careful examination of the human
body, to which they may be transmitted. On the uncovered parts
of the body they not only cause severe irritation, but also severe
diffuse itching erythema and eczema. Thorough disinfection of the
cages by hot solution of caustic potash, in addition, sprinkling over
with tar, red carbolic acid or petroleum, thoroughly powdering over
the birds with flores pyrethræ, washing with water containing oleum
anisi, washing the walls, doors and bedsteads with soap, disinfection
of the mattresses, linen and clothes, will protect against further
infection. In the case of man the disease needs no special treatment,
as the eruptions generally disappear after some days. Heinecke1024
recommends lavages with 1 per cent. carbolic acid solution. [Vide
also p. 492 in body of this work.—F. V. T.]
[Dermanyssus hirundinis, Hermann, is identical with this species.
By far the best treatment is with paraffin or kerosene oil applied to
the places where they pass the day.—F. V. T.]
Ixodes reduvius (ricinus).
The female is occasionally transmitted to the human skin, and
bores its proboscis deep into it and sucks itself full of blood. At
sensitive points of the cutaneous surface—for example over the skin
of the penis—a feeling of severe pain is produced. Buy’s1025 observations
as to the geographical distribution of the Ixodinæ show that in all
lands in which cattle, horses, sheep and dogs exist, Ixodinæ are to be
found. Recent observations show that the Ixodinæ play an important
part in the transmission of Hæmosporidia (vide body of work,
pp. 493, 494). Sprinkling with oil, vaseline, benzine, ether, petroleum,
naphtha, turpentine (Jelgenum1026), will easily lead to the removal of
the parasite; if the body is torn away with violence and the proboscis
is left sticking in the skin, the presence of the latter will give rise to
inflammation and suppuration.
Sarcoptes scabiei (Scabies).
The disease produced by Sarcoptes scabiei shows itself in polymorphous
areas, such as accompany eczema, and are produced on
the one hand by the Sarcoptes alone and on the other hand by
the scratching with the nails. The localization of both kinds of
efflorescences is different from those which are produced by the
Sarcoptes; they occur as papules, vesicles, pustules and mite-tracks,
and their usual situation is between the fingers, on the ulnar border
of the hand, on the wrist, on the palm of the hand, on the anterior
border of the axilla, on the penis and at the base of the thorax. The
excoriations are situated on the forearm, over the thigh, over the
abdomen, and may be distributed in greater or less degree over the
whole body; the back and the face only remain free. The symptoms
consist in violent itching, the onset of which specially takes place
at night.
The mite-tracks are fine curving lines, curved like a, u, c, or s,
which appear as if they had been scratched with a fine needle. Upon
closer examination with the magnifying glass one sees in their course
small openings. These openings, in persons who keep themselves
clean, are scarcely coloured; but in patients whose occupations
necessitate their being associated with coloured or dirty substances,
they are dark. The length of the tracks varies from some millimetres to
1 1/2 to 2 cm. They are at the one end, where the Sarcoptes is embedded
in the epidermis, widened like a funnel and slightly exfoliated. The
track at this point is sharply defined; the mite shows through the
epidermis as a yellowish round point. In the course of the track there
develop papulæ, vesicles or pustules, which raise the level of the track.
The intensity of these inflammatory appearances depends upon the
susceptibility of the human individual and upon the capability of
the reaction of the skin. There are people in whom scarcely
any inflammatory symptoms make their appearance; on the other
hand there are some, especially children and lymphatic individuals,
in whom severe impetiginous ecthymatous pustules, together with
their sequelæ, are set up.
The results produced by scratching consist in papules, which
usually bear a small scab of blood, and are arranged in the form of
striæ, in eczematous surfaces, weeping or sanguineous scabs, vesicles,
pustules, etc. The complications that set in are frequently urticaria
and even furuncles, lymphangitis and inflammation of the glands,
which now and then is followed by the formation of abscesses in the
glands.
The duration of the disease is unlimited; when untreated it leads
to a form of rare occurrence, that of scabies norvegica1027; in this the
collection of crusts and scales, in which a quantity of dead mites,
larvæ and ova are present, may become colossal.
The symptoms of scabies abate in the presence of intercurrent
acute diseases and reappear after the malady is over. The fact has
for long contributed to the idea of scabies being regarded as a disease
capable of being “driven in” upon the internal organs and forming
metastases.
The diagnosis is rendered certain upon the discovery of a track.
Traces of scratching on the extremities and on the abdomen, papular
or pustular efflorescences between the fingers, toes, in the neighbourhood
of the wrist, of the elbow, on the anterior border of the arm-pit,
on the tuber ischii, in the girdle region, and especially the presence of
disintegrated tracts over the penis (prepuce and glans), will allow of
the diagnosis being made. Certain occupational eczemas (grocers,
lime-workers, maltsters, bakers and others), also prurigo, must be
borne in mind when diagnosing this disease.
The prognosis is always a favourable one. Even after such a
long duration and after such severe symptoms the disease may
completely clear up. There are, however, frequently left behind post-scabious
inflammatous and pruriginous conditions which only yield
after protracted treatment. Scabiophilia, which persists in certain
patients for a long time after the scabies has been cured, must here
be mentioned.
In the treatment of scabies four points must be kept in view.
(1) The mites and the ova must be killed by the treatment; (2) the
treatment must have regard to the intensity of the inflammatory
symptoms; (3) the clothes (body-linen) of the patients must be
disinfected; the bed-linen, the beds and the bedsteads must be
cleansed; (4) when a person suffers from scabies his entourage
must be examined, and all diseased conditions treated in the same
way as under (3).
The treatment (1) should be preceded by a bath with thorough
soap ablution, and when the inflammatory symptoms are not too
severe, with green soap. After the bath the skin is dried and the
scabies remedy proper applied in warmth. Sulphur preparations
receive first consideration; among such Vlemingkz’s mixture occupies
a prominent position; this is rubbed in for half an hour by means of
a strong camel-hair brush, to be followed by another bath and
powder applications after drying. Repeat this method for three days
one after the other, or for two days, and a third time eight days later.
The latter method is worthy of recommendation as the ova, which
perhaps resist the parasiticide action, have by this time developed into
larvæ, and the latter can then be destroyed with certainty. The
remaining sulphur preparations, which are specially employed in the
form of ointments, are more complex, as the ointment should remain
on the skin. Helmerisch’s and Wilkinson’s ointments are the kinds
specially employed. Nagelschmidt1028 recommends thiopinol as a very
suitable sulphur preparation in the form of baths or as a 10 or
5 per cent. ointment in the following way: Upon his reception the
patient is given a thiopinol bath, in which he remains for thirty
minutes. Immediately afterwards 30 to 40 grm. 10 per cent. thiopinol
vaseline is carefully rubbed in. The rubbing is repeated daily, and
the treatment is concluded on the second to fourth day with a second
thiopinol bath. Thiopinol produces no more irritation than the
ordinary sulphur ointments; it is, however, much more penetrative
and more capable of absorption.
We frequently make use of Kaposi’s naphthol ointment, as it
renders the skin supple, causes proportionately little irritation, and
has but little smell. Treatment with balsam of Peru is certainly
expensive, but in the slighter attacks it is relatively the simplest. We
give the patient a bath, have him thoroughly dried and rub in 30 to
40 to 50 grm. balsam of Peru carefully and evenly all over, wrap
him in a covering of wool, and make him rest in bed for twelve to
fifteen hours, to be followed by a bath with careful cleansing with
soap; this treatment need rarely be repeated. The balsam of Peru
can be applied undiluted for the rubbings or mixed with ung.
glycerini, or resorbin or glycerine in equal parts. [Norman Walker
uses balsam of Peru 1/2 oz. dissolved in rectified spirit; to be painted
on with a brush.]—J. P. S. The manufacturers name the undiluted
product of the active constituent of balsam of Peru, benzoic acid
benzyl-ester, Peruscabin. For the treatment of scabies it is recommended
by Sachs1029 that it should only be administered when mixed
with ricinus oil, under the name of Peru oil, in applications repeated
three times within thirty-six hours.
Sack1030 also considers Peru oil a non-irritant, effectual, pleasant,
inodorous and non-staining drug. But he only allows the applications
to be used every twelve hours for three to four consecutive days
(altogether 200 to 300 grm. of Peru oil are requisite), and after the
sixth or seventh rubbing a bath should be taken with the use of
Dutch soap. Juliusberg1031 considers this treatment specially suited
for private practice. Another modern drug is epicarin ([Beta]-oxy-naphthyl-ortho-oxy-meta-tolyol
acid); this is applied in 10 to 20 per
cent. ointments (Pfeiffenberger1032), epicarin 7·0 grm., cretæ alb. 2·0 grm.,
vasel. flavi 30·0 grm., lanolin 15·0 grm., axungia poric. 45·0 grm.
(Rille1033); epicarin 15·0 grm., sapon. virid. 5·0 grm., axung. poric. 100·0 grm.,
cretæ alb. 10·0 grm. (Kraus1034); for children, epicarin 5·0 grm.,
lanolin 90·0 grm., ol. olivar. 10·0 grm. (Kaposi1035). Siebert1036 lays stress
upon the odourlessness and colourlessness of epicarin ointment as a
strong reason for its use, and points out that it is a harmless drug, the
action of which is certain. Endermol (salicylic acid ointment) has a
destructive action on the mites even in a 0·1 per cent. ointment
(Wolters,1037 Demitsch1038); it is, however, very expensive and not wholly
free from danger; and the same applies to nicotiana soap (Taenzer,1039
Schumann1040).
To give an account in detail of the drugs and methods—old and
new—used in the treatment of scabies would far outrun the limits of
this work.
Demodex folliculorum.
It is not yet certain whether the Demodex folliculorum is capable
of developing pathological conditions in man. Veiel1041 assumes that
the hair follicle mite has no connection either with the formation
of comedones or even with sebaceous gland disease. Kaposi1042 considers
that they cause no disease in man and cannot be regarded as
a cause of acne. Saalfeld1043 clearly adheres to the same standpoint,
similarly so Jessner,1044 who, when discussing comedones, makes no
mention of acne of hair follicle mites. Weyl1045 and Geber1046 adhere to
the opinion that the presence of a Demodex in man in contradistinction
to its presence in animals possesses absolutely no pathogenic
influence. On the other hand de Amicis,1047 Majochi,1048 and Dubreuilh1049
report single cases of pronounced circumscribed clear brown
pigmentations which they attribute to Demodex folliculorum. In all
these cases, moreover, as regards localization the affection had a
certain resemblance to pityriasis versicolor; nevertheless, in the scales
separated off with the scalpel no fungi were found, but on the other
hand Demodices in moderate quantity. In his earlier cases Majochi
has seen the Demodex in the secretion from meibomian glands and
had claimed it to be the excitant of chalazion and, as Mibelli1050 did,
considered it to be the cause of some diseases of the eyelids. Ivers1051
found the parasite in 69 per cent. of normal borders of the eyelids,
and attributes a pathological signification to it. Hünsche1052 and
Mulder1053 arrive at the same conclusions; in the light of their investigations
the Demodex is found as a constant accessory—certainly not in
the meibomian glands, as it is limited only to the internal part of the
hair follicle. Lewandowsky1054 considers that it can hardly be demonstrated
at present that the same parasite which in individual specimens
causes no symptoms is capable of producing pathological conditions
when markedly increased in numbers.
Treatment is by the removal of the comedones, above all, by their
mechanical removal by pressure with a watch-key and with the
various comedo-compressors, and by subsequent cleansing of the
skin with ether, benzine or spirit. If the eyelids should be affected
with blepharitis due to the presence of Demodex in large numbers,
epilation and administration of a parasiticide is recommended.
Demodex folliculorum canis.
Transmission from dog to man is in any case very rare, and by
many its occurrence is generally doubted. Nevertheless Gruby1055 and
Remak1056 claim that it is transmissible—an opinion which has also
been shared by Neumann1057 and Zürn.1058 The latter saw in the case
of a married couple who had the care of mangy dogs the onset of
diseased areas on their hands and feet, which were like those on the
dogs and contained the same parasites.
A. Babes1059 also reports several observations which go to show
that persons who, to some extent, have been shown to have been in
contact with mange-stricken dogs have been attacked by a scabies-like
eruption localized over the thorax, abdomen, back and extremities;
large numbers of Demodices were found in the follicular
pustules. Lewandowsky1060 reports one case—that of an Italian
workman, who suffered from an outbreak on the face, like impetigo;
there was crust formation and at the edge of the crusts the epidermis
appeared like a narrow row or border of vesicles. A small portion
of the covering of the row of vesicles was lifted off, and after slight
warming examined in 40 per cent. liquor potassæ. In this a large
number of animal parasites of the Demodex group were found, and
without doubt Demodex folliculorum canis alone. Hünsche1061 assumes
that Demodex folliculorum penetrates into the tissues and produces
abscesses.
Treatment first consisted in dusting with zinc amyl powder, but
after four days there was no change. After the regular use of xeroform
as a powder application, the affection cleared up within
fourteen days.
INSECTA.
Pediculus capitis (Pediculus capitis) (Head Louse).
We find Pediculus capitis in very young children and in others
more grown up to be the incessant and frequent cause of impetiginous
crust-forming eczemas. It is more frequent in girls than in
boys. In families it is endemic, in schools epidemic, but it also
occurs in fair frequency in female adults (servant maids, waitresses)
who may pay little attention to bodily cleanliness. The puncture
of the parasites sets up a severe irritation, which leads to violent
scratching. The consequences of this are the formation of nodules
and pustules, crusts and “weeping” patches; the hairs become felted
and the final clinical picture is that of plica polonica. The conditions
of irritation which are produced by these parasites and then by the
scratchings of the impetiginous, and frequently the very severe
suppurative processes of the hair-bed, lead to swellings in the neck
and sometimes even to glandular suppurations. The eczematous
processes not infrequently extend over the face, the neck and the
thorax. Blepharitis and conjunctivitis may be due to Pediculus
capitis.
The means of infection are often very remarkable. Transmission
from one individual to another certainly often occurs, but infection
may take place in railway carriages and in other ways. A case under
the observation of a colleague in Frankfort is a most remarkable one:
he diagnosed pediculosis as the cause of a head eczema occurring
among the children of one of the best families there. The infection
took place through dolls adorned with human hair, in which the
presence of nits could be demonstrated.
The diagnosis of Pediculus capitis is not difficult to make when
the hairs and hairy scalp are carefully examined for nits and living
parasites. In better families it is a good plan to point out the
corpora delicti to their possessors and to make them aware of the
possible sources of infection.
As regards treatment, lotions of sabadill vinegar are recommended;
in slighter cases these are quite sufficient. In severe cases
cure will not result unless dressings of petroleum, naphthol ointment
(5 to 10 per cent.) and balsam of Peru be applied. In the case of
plica polonica, the hair must be cut quite short (even in adults) so
as to control matting of the hair. To get rid of nits from hair that
is not matted, careful combing and washing with strongly alkaline
fluids or with hot vinegar is suitable.
Pediculus vestimenti (Clothes Louse).
The clothes louse attacks adults by preference, and with especial
frequency old and emaciated persons. It lives in the clothes, but
derives its nourishment from the body. At the moment at which
the clothes louse inserts its proboscis into the skin the person experiences
a slight sting, which, however, at once ceases to hurt. If the
body of the louse is sucked full of blood it falls off and the individual
has rest from it for a time. A wheal develops around the hæmorrhagic
area of the bitten spot and itches severely. The itching goes
on until the eruption is scratched all over. This is followed by crust
formation. When many parasites are present the itching reflexes
become more severe, and the patients scratch themselves considerably
and make long marks at those places where the Pediculi have been.
The localization of the scratching effects is characteristic, corresponding
with folds between portions of clothing (regions between the
shoulder-blades, wrist and neck). If the condition lasts for a month,
the scratching effects extend over the whole body, and secondary
efflorescences become associated with it, such as pustules, ulcers and
eczemas. Intermediate between this we find cicatrices and pigmentation,
the latter under certain circumstances extending over the whole
body. Sulla, Herod, Cardinal Dupet, Philip II, and others are said
to have died from louse disease. That even at present many human
beings are exposed to the danger of being devoured by lice is a fact
that we have had the opportunity of observing on several occasions.
Only to record one instance, a man, aged 65, was received into our
clinic some time ago in an absolutely neglected condition (he had
been staying for some weeks in a stable, lying on a wretched bed).
The whole of the surface of his body was covered with countless
furuncles, of greater and less size, which had partly become changed
into undermined ulcers. Over the ulcers and beneath their undermined
edges Pediculi were swarming.
Phthirius inguinalis (Pediculus pubis) (Crab Louse).
The transmission of these parasites generally takes place during
coitus, and therefore they especially occur in the pubes. It is possible
also that transmission is effected through dirty clothes and bed-linen
and privy seats.1062 Starting from the pubes the animals crawl out over
the other parts of the body provided with hairs to the abdominal
wall and the thorax (so far as these parts are furnished with thick
hair) to the arm-pits, the beard, the eyebrows; not, however, to the
hair of the head, or rarely so; among our numerous cases we have
never met with an example of the crab louse attacking the hair of
the head.
The irritation produced by the crab louse is extraordinarily severe,
especially during the night, as the warmth of the bed incites the lice
to active sucking. In consequence of the violent scratching indulged
in, eczemas are set up at the points attacked, and these often spread
to the neighbouring parts not covered with hair.
Of special interest is the onset of maculæ cæruleæ (tâches bleues)
in some persons affected with crab lice (people disposed to sweating
seem to be peculiarly liable to these). They consist in pale blue
patches of various size and shape, varying from that of a hemp-seed
to that of a lentil, and again to that of a nail in size and form. These
are found over the cutaneous surface of the abdomen, thorax and
thigh, and are often only seen by a good lateral illumination. Duguet1063
considers that the condition is a toxic erythema, that it is set up, on
the occasion of the bite of the parasite penetrating the skin, by the
poisonous substance derived from it. Oppenheim1064 considers that it
is a colouring substance that is formed in the salivary glands of the
parasites, and which penetrates the skin when the insects bite, and
thus forms the maculæ cæruleæ. We have on several occasions
emulated the experiment of Duguet (trituration in a mortar of crab
lice freshly taken from the human body and inoculating the mass
thus obtained beneath the skin), and have similarly been enabled to
produce the maculæ cæruleæ experimentally, but we have certainly
been unable to determine which of the hypotheses is the correct
one, the toxic erythema or the colouring substance inhibition
theory.
The diagnosis of phthirasis is very easy, for either the sexually
mature parasites or the nits are found on the hairs.
As regards treatment, grey ointment is regarded as a generally
useful application; it gives rise, however, to a slight eczema of the
genitals, especially in males, when injudiciously used. Geber1065
recommends petroleum or balsam of Peru, Oppenheim1066 a 1 per cent.
sublimate solution for lotions, or a mixture of equal parts of petroleum
and benzine when the sublimate cannot be borne. The use of
a 5 per cent. ointment with hydrarg. oxid. flavum is worth considering
in treatment of pediculosis of the eyebrows and eyelashes. The
simplest method of treatment, and one with a radical effect, is that by
sulphuric ether recommended by Thomer.1067 It certainly produces
a sharp burning sensation, but the living parasites and nits are
destroyed in one sitting. We prefer ether lotions as a rule, and we
thoroughly rub the affected parts with a pad of wadding well soaked
with the ether. The dead parasites and the nits fall on to what lies
beneath when the rubbing is done thoroughly, and the burning sensation
caused by the ether only lasts a few minutes.
Cimex (Acanthia) lectularia1068 (Cimex lectularius) (Bed Bug).
The puncture in the skin made by the bed bug gives rise to an
extraordinary amount of severe itching and a burning sensation, and
when the skin is sensitive wheals of remarkable size (urticaria ex
cimicibus). These eruptions that cause such severe itching are
scratched by those attacked, till very soon blood begins to flow, and
this generally leads to the formation of a dried crust of blood at the
point of eruption.
The diagnosis is not always easy, as urticaria arising in other ways
frequently leads to similar vigorous scratching and formation of crusts
of dried blood. Men who have some experience in this matter (for
example, commercial travellers), when they are attacked by severe
itching at night, are in the habit of striking a light and searching in
their bed and body-linen for the bugs, in order to be able to hand
over the corpora delicti to the landlord if need be. The assumption
that the bugs in the East play an actual part in the propagation of
tuberculosis and bubonic plague has been proved by investigations
made by Nuttall1069 to be at least very exaggerated if not wholly without
foundation. Further investigations may decide how far the bugs
participate in the transmission of kala-azar, as is believed by Rogers
to take place.
The bed bugs must be exterminated by spraying the chinks and
joints in the boards with petroleum and benzine, pulling up the
carpets and cleansing the bedsteads. For the treatment of the bite
itself the methods recommended as an antidote against insects’ stings
in general are suitable: 2 per cent. carbol vaseline (Rosenbach1070),
thymol dissolved in spirit (1 in 501071), æthrol or deci-æthrol, form-æthrol
(manufactured by Dr. Nordlinger, Flörsheim a. /M.), formol1072
(formol 15 parts, xylol 5 parts, acetone 44 parts, Canada balsam 1 part),
with the aid of a pad of wadding placed over the part bitten, lavages
with vinegar, citron juice and spirit of salmiac.
Pulex irritans (Human Flea).
The bite of the flea produces a slight discharge of blood about the
size of a pin’s head, which rapidly becomes surrounded with a circular
area similar to a patch of roseola. The redness fades away after
a longer or shorter while (several hours), whilst the discharge of blood
is to be seen for one or two days longer. In dirty people the whole
body may be covered with such discharges of blood. Individuals
with very delicate, sensitive skin, especially small children, show true
wheal formation at the site of the bite. In certain cases there
develops from one such single bite an urticaria that extends over
a large part of the body. The manner by which an irritating substance
is introduced into the skin upon biting by the bed bug and
also by the flea is clear. The bite is followed by a feeling of itching,
which is liable to rob nervous persons of their sleep. Sensitive
individuals are upset even by the fleas moving over the surface of the
skin during their rest at night.
Treatment consists in extreme cleanliness, capture of the parasites,
sprinkling the body and bed-linen with insect powders. The fleas
are difficult to remove from barracks, schools and hospitals.
Dermatophilus (Sarcopsylla) penetrans (Sand Flea).
The fertilized females penetrate into the skin with their heads, and
here they swell, in consequence of the numerous and growing eggs
and larvæ, to a white ball the size of a small pea, on which the head
is recognizable only as a small brown point.
In this way a small brown tumour arises, over which, at the
commencement, the skin is not reddened; after some days, however,
it becomes inflamed; in the centre of it a small opening is seen.
If the parasite is not extracted the skin that lies over it becomes
destroyed by suppuration, and thus becomes removed. At the
commencement the part affected itches, with increasing inflammation;
the symptoms of irritation become more severe and may amount
to actual pain. If the small suppurative processes be neglected,
inflammation and gangrenous and septic processes may arise. The
region of the body sought out by preference by the sand flea is
the sole of the foot, the toes, under the free ends of the nails and
the digito-plantoid folds—more rarely the scrotum, thigh and other
parts are attacked (Scheube1073). The number of parasites found on
one person may amount to several hundreds.
Treatment consists in the removal of the parasites from the skin
with a needle or a small sharp knife and the application of a bandage.
Rubbing the feet with copaiba or Peru balsam, sprinkling them with
insect powder, or washing them with bay rum (Berger1074) acts as a
prophylactic or removes the irritation of the skin produced by the
parasites.
Myiasis.
Under the name of myiasis we designate the complex symptoms
which parasitic dipterous larvæ give rise to in man (Braun), and we
conceive under the term myiasis externa (dermatosa s. cutanea) all
lesions of the human integument caused by fly larvæ and of the
cavities covered with mucosa therewith connected, such as the
external auditory meatus, the oro-nasal cavity, the urethra and vagina.
The occurrence of dipterous larvæ in the digestive tract is named
myiasis intestinalia or interna.
Myiasis externa.
The larvæ of a species of fly belonging to the Muscidæ, Lucilia
macellaria,1075 are found in relative frequency in the nose, especially
in America and India.1076 Riley1077 has stated that the screw-worm of
Central America and of the United States is nothing else than the
larva of Lucilia macellaria, and also that the Brazilian fly named
“berna” may be no other than Lucilia macellaria. Their offspring
may set up inflammatory disturbances in the soft tissues of man. This
fly has a wide distribution, from the Argentine Republic to Canada,
also in the British portions of the East Indies, where the disease is
named “peenash.” This word is derived from the Sanskrit, and is
said to be a collective name for all diseases of the nose. Lahory1078
states that within a period of nine years ninety-one cases of “peenash”
occurred in Allyghar, two of these ending fatally. Lucilia macellaria
is not at all timid but bold, like the house-flies and blue-bottles,
its relatives. It not only lives at no great distance from human
dwellings, and forces its way into villas and country houses, but even
attacks its victims without awaking them from their sleep. Although
this species shows a certain preference for nasal cavities affected with
catarrh or pus (v. Frantzius1079), and also the external auditory meatus,
as well as ulcerated or wounded parts of the body, and even badly
ulcerated skin carcinoma (Lutz1080), it is not a rare thing for it to
penetrate into one of the above-mentioned cavities rapidly to deposit
its eggs, without these parts having been previously affected. The
report also of Conil,1081 in which these flies bear the name of Calliphora
anthropophaga1082 is an interesting one. Probably it was the same species
of Muscid in the cases of myiasis nasi observed by von Tengemann,
Delasiauve,1083 Weber,1084 Mankiewicz,1085 and Kirschmann.1086 In the case
recorded by Prima,1087 and in that recorded by Britton,1088 the issue was
a fatal one; in the latter the larvæ escaped through the pharynx
and nose; the hyoid bone and the soft parts of the palate were
destroyed, the speech and power of swallowing were hindered. At
the post-mortem extensive destruction of the internal nose was found,
so that the nasal bones could only be kept in their position by the
aid of the external skin. Even during life 227 larvæ escaped.
Similar destructive processes were found in the case communicated
by Richardson.1089 In two cases reported by Schmidt1090 300 and 350
larvæ were respectively removed from the nose, and the patients
recovered. Wolinz1091 found his patient had lost consciousness, and
that in the pus filling up the entrances to the nose numerous larvæ
were moving; recovery followed. In the case communicated by
Adler,1092 more than 150 larvæ escaped from the nose of an old man.
Curran1093 states that people suffering from “peenash” frequently die
from meningitis. The cases reported by Pierre1094 related to the forms
of severe myiasis frequently to be observed in Guiana. In a patient
who was suffering from typhus (? typhoid), Douglas1095 found the
conjunctival sacs full of larvæ; in two other individuals the nasal
cavities were attacked.
The case observed by Summa1096 was that of a man, aged 28, who
suffered from nasal obstruction, fœtor, epistaxis and pain in the nose.
Out of seven of the cases occurring at Fort Clark, U.S.A., and in its
neighbourhood, six ended fatally; in all these cases Kimball1097 diagnosed
ozæna; attracted by the strong odour the flies forced their
way into the noses of the patients when asleep and there deposited
their ova. In a case reported by Carrière1098 an abscess of the nasal
septum was produced by the larvæ of flies; Chiodi1099 reports seven
cases of myiasis due to Lucilia macellaria; among these was a case
of rhinitis myiatica, in which a cerebral abscess leading to a fatal
termination developed, being produced by the migration of a larva
into the brain. Among the three cases of Lesbini1100 was that of a girl,
aged 16, with 250 larvæ in the diseased nasal cavity. Quintano1101
observed larvæ beneath the eyelids in one case. It is possible that
the cases of Cesare1102 and Calamida1103 were those of myiasis nasi due to
Lucilia macellaria. The larvæ are also found in the nasal accessory
sinuses, as is seen from the cases reported by De Saulle1104 (frontal
sinus), Delasiauve1105 (frontal sinus), MacGregor1106 (antrum of Highmore),
and Bordenave1107 (antrum of Highmore).
If a survey is made of the literature of the cases described of
myiasis nasi produced by Lucilia macellaria1108 the following information
is forthcoming: In Europe this form of the disease is of very rare
occurrence, whilst in America and India1109 it is frequent. Persons
suffering from ozæna are rendered the most liable to danger as the
penetrating odour entices the flies in tropical countries with intense
frequency, so much so that v. Frantzius does not consider this
myiasis as an independent disease, but as a complication of ozæna of
frequent occurrence in warm countries. The infection is so far
of interest in its nature, in that it only takes place during the day.
The fly is on the wing only by day when the sun is shining, and
consequently only deposits its eggs at this time. Therefore persons
suffering from ozæna are principally exposed to the danger of being
pursued by the flies when they succumb to sleep during the mid-day
hours in the open or in dwellings that are not closed up.
Headache is the symptom which most troubles the patients.
It extends over the whole cranium and persists uninterruptedly, with
more or less severe periods. Violent headaches in the frontal and
buccal regions are almost always present in this complaint; they are
experienced either only on one side or on both simultaneously;
sometimes the pain is extended to the lower jaw and region of the
neck, following the whole extent of the trigeminal nerve. The
inflammation of the nasal mucosa produced by the penetration into
it of the larvæ extends right into the frontal sinus and antrum.
Simultaneously the patients, at the height of their trouble, suffer
from persistent sleeplessness and severe vertigo, so that they reel and
cannot walk straight; excessive sneezing always sets in at the commencement.
The larvæ immediately spread over the nasal mucosa to
seek a place suitable to feed, and irritate the nasal mucous membrane
by the tickling sensation they produce. Later the patients frequently
sneeze when the maggots move to and fro.
One very characteristic symptom consists in the peculiar swelling
of the face, which is extended either over the whole or only one half
of it, and may alternate with attacks of erysipelas (Brokaw1110).
The discharge from the nose is of special diagnostic value. It
consists of a blood-stained serous matter or blood-stained fluid,
which is perpetually trickling from one or both nostrils. The larvæ
especially choose the anterior portions of the nasal cavity, where they
can be seen lying in groups together at the base of the choanæ.
The consequence of this is that the soft palate becomes intensely
swollen, and this in turn makes swallowing very difficult; speech is
impeded, and the voice acquires a nasal intonation. Symptoms of
fever become more or less pronounced according to the number
of larvæ present, and according to the nature and constitution of the
individual. The appetite is in abeyance throughout the whole duration
of the illness, and sometimes there is the onset of slight attacks
of diarrhœa.
If the larvæ are not removed in good time there follows excessive
destruction of the interior of the nose and of the turbinals; and the
whole nasal framework undergoes disintegration, frequently, too, the
velum palati, so that the larvæ come into sight in the oral cavity.
Individuals thus severely attacked succumb through exhaustion,
symptoms of meningitis (cerebral abscess) or septicæmia (Prima1111).
Twenty-one out of thirty-eight cases recorded (collected) by Maillard1112
died.
The method of prophylaxis is self-evident from what has been
stated. On bright summer days neither the healthy nor those suffering
from diseases of the nose should sleep during the day-time in the
open or in public habitations; sufferers from nasal diseases should
pay special attention to this.
Treatment consists in the removal of the larvæ; this, however, is
not always easy.
With regard to the methods which have proved to be effectual in
the destruction of living larvæ and their expulsion from the nose,
strongly smelling and easily diluted fluids come first, such as alcohol,
eau-de-Cologne, and ether, which should kill the creatures when
injected into the nostrils. The earlier physicians, such as Salzmann,1113
Honold,1114 and Henkel,1115 have seen good results from the use of these
methods, whilst Mankiewicz1116 and Goldstein1117 obtained no results
whatever. Kimball’s1118 careful investigations have shown that a decoction
of bitter herbs recommended by Behrends1119 (tansy, wormwood)
have just as little effect as the tobacco decoction employed by
Boerhave1120 and Kilgour.1121 The sternutatories employed by the older
physicians are entirely neglected. Delasiauve1122 experienced good
results from the inhalation of the smoke of paper cigarettes, which
were soaked with a solution of 2·0 pot. arsenic in 30·0 distilled water.
Whilst, according to Kimball, balsam of Peru had no effect on the
larvæ, Mankiewicz succeeded in removing the larvæ from the nose
with the help of that drug. Turpentine steam or mixtures of turpentine
employed by Indian physicians have not been very effectual according
to Moore,1123 Kimball and Goldstein. Success has been attained in
some cases by the use of insufflations of calomel (Roura,1124 Cerna,1125
Schmidt1126) or of iodoform (Pascal1127). Joseph1128 recommends concentrated
alum solution being sniffed up into the nose as very effectual.
Sublimate and carbol solutions do not appear to be very successful
(Kimball, Moore, Goldstein), whilst benzine inhalations (Pierre1129)
have shown better results. Scheppegrell1130 strongly recommends
injections of oil which kill the larvæ, while it is perfectly harmless
to the nasal mucosa. Cesare1131 employed nasal lavages with solutions
of salicylate of soda with good results, and Calamida1132 lavages with
physiological saline solution. Bresgen1133 recommends the nose being
cocainized and the larvæ being removed with a pincette. Roorda-Smit1134
cocainized the nose, then insufflated calomel and plugged the
nose with a gauze tampon dusted with calomel. After two hours
fifty-six larvæ crawled out along the plug. Continuation of the
treatment resulted in a complete cure.
Injections of chloroform water (Jourdran1135) or chloroform inhalations,
or injections of pure chloroform into the nose, have proved
the most effectual (Goldstein,1136 Osborn,1137 Jourdran, Durham,1138
Jennings,1139 Kimball,1140 Mackenzie,1141 Oatmann,1142 Zarniko,1143 Antony,1144
Folkes1145). Camphorated carbolic solutions are very well spoken of:
Grayson1146 states that these kill the larvæ immediately. Some authors
have removed the larvæ with forceps (Goldstein1147), others with
pincettes; thus Brokaw extracted 200 fragments with the forceps,
Pascal eighty fragments with the pincettes, and Wolinz1148 also appears
to have removed the larvæ with forceps.
Greater operative measures than these do not appear to have been
undertaken in latter days; yet Morgagni1149 states that the army
surgeon, Cæsar Mogatus, at Bologna, first trephined the frontal sinus
and then extracted a “worm” from it.
Larvæ of other Muscidæ have come under observation much more
rarely (Cheval1150 [larvæ of Galleria mellonella1151], Bond,1152 Dumesnil1153
[larvæ of Piophila casei]). Species of the genus Scolopendra
(Myriapoda), which all shun the light and seek their food during the
night—which consists of animal and vegetable substances—frequently
make their way into the nasal cavities of people when asleep. They
are found not only in the nose, but in the accessory cavities. In the
chapter on the “Parasites of the Nose”1154 we have collected striking
instances, but we have omitted to mention the observation made by
Bertrand1155 (Scolopendra in sinus maxillaris) and that made by Bergmann1156
(Scolopendra in sinus frontalis). In the same chapter some
remarks are made as to the occurrence in the nose of earwigs, caterpillars,
scorpions and termites, as well as of animals which have not
been identified.
The larvæ that develop in the auditory meatus penetrate the
membrana tympani, destroy the middle ear and may produce meningitis
and intracranial suppurations. In one case Vesescu1157 extracted
seven living larvæ from the ear with the aid of a thin pair of pincettes.
Köhler1158 recommends the infusion of drops of ol. terebinth. to destroy
the larvæ, Quintano1159 the insufflation of the following powder: Oxid.
hydrarg. rubr., sulfur., āā 1·0 grm., pulv. gi. arab. 8·0 grm.; Lesbini1160
recommends tincture of iodine. In the case reported by Henneberg1161
the larvæ were those of Lucilia cæsar.
Eye affections due to Lucilia macellaria are very uncommon; the
literature relating to the lesions of the eye produced by the larvæ of
flies has been collected in Kayser’s1162 work. In the cases under the
observation of Schultz-Zeyden1163 both the eyes of a female tramp were
destroyed, and quantities of larvæ were also found in the nasal fossæ
and in the ears.
The Lucilia is found relatively seldom on the cutaneous surface.
Henneberg’s1164 case was that of a neglected girl, aged 20, in whom
countless larvæ (L. cæsar) were found in a plica polonica; after the
plica polonica had been removed the scalp was found to be covered
with a large quantity of ulcers which swarmed with larvæ, large and
small. The skin of the trunk was also much macerated and covered
with larvæ. Death resulted from sepsis; Westenhöffer1165 remarks on
this case that a lesion of the head from which the patient had suffered
previously and the perpetual state of intoxication in which she was
had probably given rise to the lodgment of the fly larvæ. Whether
the communications made by Munk1166 of maggots in the mouth
relate to Lucilia I do not know. Vesescu,1167 in one case with extensive
ulceration and deep fistulæ in the skin, removed 176 larvæ with the
pincette. In Roorda-Smit’s1168 case there were two ulcers in the neck
of a girl, aged 17, and larvæ appeared at their base. After dusting
with calomel and the application of a bandage the next day fifty-two
dead or half-dead larvæ came to light. Recovery took place.
Lesbini,1169 in the case of an old lady, saw numerous larvæ in an ulcer
of the leg she was suffering from. Hector’s1170 case appears to have
been one of myiasis cutanea provoked by Lucilia.
The first exact observations of myiasis cutanea from Sarcophaga
magnifica are due to Wohlfahrt,1171 in whose honour Portschinsky1172
named this species of fly S. wohlfahrti. Portschinsky ascertained that
S. wohlfahrti was not confined to man as its sole host, but that several
of our domestic animals, such as cattle, horses, pigs, dogs and geese,
were visited. In these animals small wounds serve to entice the flies
and to supply them with a suitable site for the deposition of their
eggs. The oral armature of the young larvæ renders it easy for them
to penetrate not only the mucosa and cutaneous surface but also
intact places in the submucous connective tissue. In many localities
more than half the herds have proved to be infected by the flies.
The fly only frequents open spaces and never enters human dwellings,
and is so timid that it approaches man only during sleep; infection,
therefore, takes place only out of doors, in summer, in clear, warm
weather, and only in such individuals as sleep in the open air.
Individuals are most exposed to risk who suffer from catarrhs or
inflammations, combined with purulent secretions of the nasal cavity
(ozæna), or otorrhœa, or ulcers in any parts of the body accessible to
the female fly.
The frequency and intensity of the infection will be in inverse
proportion to the advance in civilization of the inhabitants, their idea
of cleanliness, their having timely medical aid and the chances of their
being rapidly attended to. On that account the majority of cases of
myiasis (Sarcophaga) are reported from Russia. The literature of this
kind of myiasis nasalis is not very extensive; in addition to Wohlfahrt,
Portschinsky and Joseph,1173 there is a communication by Gerstäcker,1174
who found fifteen adult larvæ of S. wohlfahrti in the nasal cavity of
one man. The larvæ transmitted from Ordruf by Dr. Thomas to
Löw,1175 in Vienna, which were discharged from the nose of a woman,
aged 71, suffering from ozæna, were recognized by the well-known
dipterologist Braun as belonging to S. wohlfahrti. Among the cases
reported by Joseph, one only affected the nose; it was that of a
peasant girl, aged 11, who had suffered from ozæna; she had
travelled on the open road and had there gone to sleep. Severe
symptoms set in and death followed under delirium. In making the
post-mortem it was found that the interior of the nose was extensively
destroyed by larvæ of S. wohlfahrti. Powell found Sarcophaga
larvæ in two persons who had slept in the open air; the larvæ were
killed by injections of chloroform and sublimate. Destruction of the
eyes by S. wohlfahrti has only been observed in a few cases; it is
reported by Cloquet1176 that, in the case of a ragman who had lain
some time in the fields, both eyes were pierced by larvæ. On the outer
skin the larvæ of S. wohlfahrti have been found more than once in
inflammatory or festering areas. Freund1177 demonstrated that from
a five year old child, which had suffered for some time from an
impetiginous eczema of the skin of the head, from two suppurating
abscess cavities which extended to the periosteum, which was already
affected, twenty-one living larvæ were taken; rapid healing took place
under antiseptic bandaging.
The small treatise by Balzer and Schimpff1178 contains two new
observations on myiasis externa; in the one case an ulcer on a man’s
foot was full of larvæ, in the other case the head of a woman showed
numerous larvæ without the skin of the head being destroyed.
Brandt’s1179 observation is interesting, for he found such larvæ in the
gums of a sick person.
The impression which one obtains of the active movement of
larvæ on wounds is a strange and at the same time uncanny one.
One finds that the larvæ to obtain protection against the drying of
the surface of the abscess almost incessantly burrow with their heads,
first contracting and then expanding the body, which rises and falls,
and keeping the tail upwards. Owing to these movements producing
irritation, increase of inflammation may ultimately arise, causing
erysipelas and cellulitis.
The treatment of myiasis nasalis caused by Sarcophaga is the same
as in myiasis caused by Lucilia, and in the other places where found it is
merely a question of the removal of the larvæ and the subsequent
proper treatment of the surface of the abscess. In Northern Nigeria
Lelean1180 found Auchmeromyia depressa to be the cause of myiasis
externa.1181
The occurrence of Oestrid larvæ in a human being is very rare,
at least up till now myiasis oestrosa has been very seldom observed
in man in Europe. Whilst the hosts of the Muscidæ comprise a
considerable number of warm-blooded animals, on which the larvæ
develop, each species of the Oestridæ appears, on the other hand, to
have a definite host or some definite hosts of the class Mammalia.
No species of Oestrid is peculiar to man. Although in America, as
well as in Europe, Oestrus hominis was spoken of up to the middle of
the last century, no such species exists.
But in both hemispheres, in America much more often than in
Europe, Oestrid larvæ have been found in man. In Florida, Mexico,
New Granada, Argentina, Brazil, Costa Rica and other districts,
and especially where large herds of cattle are kept, myiasis oestrosa
has been observed in shepherds, huntsmen and amongst the rural
population. The larvæ of Hypoderma bovis, according to the observations
of Goudot,1182 occur as a parasite in man. Poilroux1183 found larvæ
of cavicolous Oestridæ in the nose of a man, aged 55. Amongst
the species of warble flies, whose larvæ are parasites in domestic
animals and game in Europe, reliable observers have found larvæ of
two kinds, Hypoderma bovis and Hypoderma diana, also in man.1184
The larvæ of H. bovis have very seldom been observed in the nose.
The case quoted by Kirschmann,1185 which was that of a peasant
woman, aged 50, who was suffering from ozæna, and in which
violent attacks of sneezing, epistaxis, pain in the forehead, and swelling
of the face were observed, is, according to Löw1186 and Joseph,1187 not
an Oestrid; Muscid larvæ were evidently the cause. By the injection
of diluted iron chloride solution seventy-nine larvæ were removed
from the nose. In the case reported by Razoux1188 the species
of larva is not definitely known—at least, v. Frantzius1189 did not
consider them Oestrid larvæ. Joseph does not definitely say that
Oestrid larvæ were the cause of a case which he quotes. He was sent
a number of uninjured larvæ of Oestrus ovis ready to pupate, which
were said to have been expelled, during violent sneezing, from the
nose of a peasant woman who had suffered for six months from
continuous frontal headache and chronic nasal catarrh.
The Oestrides prefer to use the surfaces of wounds on the skin of
man to lay their eggs, which develop into larvæ; but they often use
their ovipositors1190 to make a fresh wound. In this case there arise in
the skin, and particularly in the subcutaneous connective tissue of the
neck, in the region of the shoulder, as well as in other parts of the
body painful, furuncle-like inflammations which are known under
the name of gad-fly boils. These boils may become the size of pigeons’
eggs; if several are together, they appear to form a connected tumour.
Each tumour is elastic and somewhat movable, and has an orifice
through which the larva breathes and discharges its excreta. At
times these turn to festers and gangrenous disintegrations, which may
even cause the loss of a limb. Wilms1191 had the opportunity a few
years ago of observing a case of myiasis dermatosa oestrosa in Leipzig.
The fistula which led to the larva was slit open and the larva extracted.
As a notable characteristic of myiasis oestrosa Joseph states that the
larvæ grow very slowly. The flight time of the Oestridæ is the hot
summer months.
Adams1192 observed on the Isthmus of Panama a number of cases
of a skin disease which is caused by the larvæ of Dermatobia noxialis
(Gusano-peludo-Muche). The larvæ penetrate not only the skin but
also the mucous membrane of the pharynx and larynx, and from there
proceed through the tissue to the subcutaneous cellular tissue. The
infection seems to result from bathing.
The study of “thimni,” a human myiasis caused by Oestrus ovis,
by Ed. and Et. Sergent,1193 deals more with the zoology and with the
geographical distribution of this insect in North Africa than with the
clinical appearances of myiasis. [This paper deals with matters of
great interest, with important facts.—F. V. T.]
The treatment consists in the removal of the larvæ (from the
nose); in Brazil it is the custom to drop tobacco juice into the
boil in order to kill the larvæ (Strauch1194).
One is only justified in speaking of myiasis intestinalis when there
is no doubt that living fly maggots or flies themselves can be proved
to have been found in the fresh contents of the stomach or intestine
(Schlesinger and Weichselbaum1195). In the discussion of myiasis
intestinalis we give the evidence of Schlesinger and Weichselbaum,
as well as that of Wirsing,1196 to which must be added a number
of other investigations.
In a great number of acute cases apparently only the stomach was
affected, there being no signs in the intestine. In these cases sudden
illness is noticed, colic, sometimes unbearable pains in the region of
the stomach, pyrosis, vomiting or continuous intense inclination to
vomit, occasionally even with the mixture of blood. Frequently a
general feeling of malaise, twinges of pain in the muscles, and attacks
of giddiness were notified, very rarely fever. Generally all the
symptoms disappeared in a short time when the larvæ had been
removed by an act of vomiting or by washing out the stomach.
It is well to note that in the history of many cases the pains
preceding the expulsion of the larvæ are stated to be extremely
violent.
Acute myiasis of the intestinal canal frequently runs a course
without special symptoms and is only an accidental condition; one
has, however, in such cases to guard against errors. The fæces may
be deposited in vessels or places where fly larvæ are in great numbers,
or a subsequent infection of the fæces with the eggs or larvæ of flies
may have taken place. Only when the inspection of the excrement
immediately following defæcation proves the presence of living larvæ,
and when there were certainly no fly larvæ in the vessel previously,
can one speak of the passing of fly larvæ from the intestine. More
frequent than the cases showing no special symptoms are those with
pronounced disturbances in the intestinal passage, obstruction or
diarrhœa (also constipation and diarrhœa alternately), violent and
sometimes agonizing abdominal pains (Pottiez1197), which preceded
the evacuation of the larvæ and subsided after their removal. General
symptoms, like weakness, languor, transitory vague pains, loss of
appetite, sickness, rarely fever, giddiness, attacks of faintness, epileptic
attacks (Krause1198) are observed. In a few cases blood and pus have
been noticed in the evacuation of the bowels.
In the cases of chronic myiasis of the intestine the aspect of the
disease is dominated by the complex symptom of colitis mucosa.
The following features are noticeable, namely, the intermittent
passing of blood, the influence over the expulsion of the larvæ of
mechanical procedure (massaging of the abdomen), the duration of
the process for several years, the sometimes enormous number of
insects contained in the dejecta. Another clinically important factor
is the passing of the larvæ in batches. While for some time no larvæ
may appear in the stools, they may suddenly be ejected in great
numbers, either because the conditions of feeding are not suitable,
or because medicaments remove them from the intestine. The
hæmorrhage is ascribed by Schlesinger and Weichselbaum directly
to lesions of the mucous membrane caused by the larvæ; in the case
reported by these writers there were found shreds of tissue as well as
pus in the stool. The pains occurring spontaneously in the abdomen
are at times influenced by position and attitude of the body, often
they were more violent after rest and after evacuation of the bowels;
often they were continuous, but in that case less intense; pressure on
the abdomen is generally little felt. The condition of the blood was
in two cases (Pasquale1199 and Schlesinger and Weichselbaum) a marked
chlorotic one. The state of nutrition seems almost always to suffer
with prolongation of the disease, but in Peiper’s1200 cases this was not
so. The condition of the appetite was in some instances good, in
others very bad. A frequent symptom is headache of a migraine-like
character and neuralgic pains in different parts.
Schlesinger and Weichselbaum’s case shows that there are forms
of myiasis intestinalis which, after prolonged sickness, lead to death,
and that in consequence of the formation of intestinal abscesses
stricture of the intestine may arise from the subsequent formation of
a scar.
The question of the mode of infection is interesting; in this
mouth, nose and anus must be considered. The most frequent way
is certainly by means of food on which flies have laid their eggs, or
which is permeated with young maggots. This may be raw (especially
grated) meat, cheese, fruit, salad, milk, cabbage, cold farinaceous
foods, raspberries. When the stomach is affected, when the gastric
juice has lost acidity and power of digestion, the larvæ will be able
to stay and develop more easily. According to Csokor,1201 if the eggs
get into the gastro-intestinal canal of man with the food, the delicate
stages of the young larvæ would certainly not survive the action of
the gastric juice. Salzmann1202 assumed that the invasion occasionally
occurred through the rectum, the larvæ creeping into the anus while
the person is asleep. Wirsing accepts this method of infection for
two of his cases, where it was a question of the infection of an infant.
Salzmann1202 reports a case where the maggots of Anthomyia1203 scalaris
were passed in great numbers from the urethra of an old man. The
patient had been catheterized on account of urethral stricture and was
probably infected with eggs or larvæ at the same time.
The diagnosis of the affection is easy and sure, if living larvæ are
found in the contents of the stomach or in the stools, and if contamination
is out of the question.
The number of different species of flies whose larvæ are found in
myiasis intestinalis is considerable. The larvæ of species of Anthomyia
(A. canicularis,1204 A. scalaris, etc.), of Sarcophaga carnaria and
S. magnifica and of Musca vomitoria1205 are especially observed.
The prognosis is certainly generally favourable, but must be made
with some reserve in chronic cases, in view of the observations of
Schlesinger and Weichselbaum (intestinal stenosis).
The treatment must aim at removing the larvæ as soon as possible
from the digestive canal.
In cases of myiasis of the stomach, a thorough washing out of the
stomach (Joseph,1206 Staniek1207) is to be preferred to emetics used with
success in individual instances; perhaps it would be advisable to add
menthol or thymol to the mixture.
In myiasis of the intestine internal remedies and local treatment
of the intestine must be considered.
So far santonin seems to have proved to be the best remedy.
In some cases extract. filicis maris, calomel, semina cucurbitæ,
naphthalene 0·1 to 0·5 (Peiper1208), infus. of Persian insect powder
(5 in 200), mineral waters, Carlsbad water, seem to have had good
results.
For irrigation of the rectum, weak solutions of argentum nitricum,
tannin, thymol, gelatine, ol. ricini, naphthalene may be used. Wirsing
administered an aperient (Rurella compound liquorice powder) and
a soap enema after the passing of the first larvæ.
The principal thing is the prophylaxis, which must include the
careful protection of articles of food, on which flies may lay their
eggs (protection by glass dishes, tulle or fine wire nets). Fruit
should not be eaten before being washed or rubbed with a cloth.
Gastricolous Oestridæ (Creeping Disease).
Syn.: Creeping eruption; Larva migrans; Hautmaulwurf; Dermatomyiasis
linearis migrans oestrosa; Hyponomoderma; Dermatitis linearis migrans; Linea
migrans; Epidermiditis linearis migrans Wolossatik; Kriechkrankheit; Hautkratzschorf;
Myiase hypodermique.
Under the name “creeping disease,” R. J. Lee1209 has recorded
a peculiar affection of the skin in a three year old girl, which appeared
first in the form of pale red, thread-like irregular protuberances,
which seemed partly to become entwined on the right malleolus
and had spread without causing special disturbances to the abdomen.
Dickinson, Fox and Duckworth1210 reported, in connection with this,
that they observed a growth of this red line of about 1 in. per
diem. Since then a number of similar cases have been reported
which, without doubt, were cases of larvæ creeping under the
skin. Crocker1211 saw such a case in a two year old girl, the progress
of the red line varying in one night between 4 and 7 1/2 in. In Europe
the first case was observed in Vienna, by v. Neumann and Rille,1212
also in a two year old girl.
v. Samson-Himmelstjerna,1213 Sokoloff,1214 Rawnitzky1215 found larvæ
at the end of the tract, which had been recorded as larvæ of Gastrophilus
by Cholodowsky.1216 According to Blanchard (Arch. f. Par.,
1901) the larvæ were those of Hypoderma bovis.
How these larvæ get into the skin has not yet been definitely
ascertained; v. Samson is of the opinion that they usually obtain
access to man as larvæ, Stelwagon1217 believes that the infection
generally occurs in a seaside watering place; a patient of
Ehrmann’s1218 fell ill when he returned from the manœuvres, where
he had lain for some time on the ground. Here and there it is
reported that the eruption was preceded for a longer or shorter time
by lesions of the skin (incised wounds, furuncles, slight excoriations,
v. Harlingen1219).
Twice it has been suggested that perhaps the parasites might come
from vineyard snails (Crocker, Lenglet and Delaunay1220), and it is
pointed out by v. Samson that in Russia the infection of peasants
who work in the fields was specially frequent. It is noticeable how
frequently the affection begins on uncovered parts of the body (face,
hands, arms); but that fact, on the whole, is not in conflict with the
statement (Kengsep1221) that the disease makes its first appearance
over the nates, because children often sit on the ground and play
with that part of their body uncovered. A case observed by us was
that of an elderly lady who did not do this and was properly clothed,
yet showed the typical lines of creeping disease on the nates, and
asserted again and again that she had the feeling as if a worm were
creeping under her skin.
The disease occurs in children as well as adults, so that age, sex
and calling offer no determining point etiologically.
The clinical symptoms of the disease consist in the sudden
appearance of itching and burning; if the cause is looked for one
perceives a red line, raised but little above the surface of the skin,
with irregular curves, never branched, but often entwined, broadening
more or less rapidly at one end (1 to 15 cm. in twenty-four hours).
The larva can be seen sometimes with a lens under pressure of the
skin as a dark spot; formations of pus, such as other larvæ produce,
are not noticed; now and again there is a formation of little vesicles
(Hamburger,1222 v. Harlingen,1223 Bruno,1224 Ehrmann,1225 Brodier and
Fouquet,1226 Rawnitzky1227). It may happen that the parasite burrows
through a small region of the skin with many close curves for some
time; on the other hand, observations exist where it covered large
tracts in a short time. The itching and smarting cease in the place
left by the larva, so that the patients even in the shortest tract can
point out at which end the larva is, even if they have not watched
the lengthening of the tract. Very rarely the larva invades the
mucous membrane of the mouth, the nose, and the conjunctiva,
proceeding from thence to the external cutaneous area.
The localization of the affection is very varied; the primary seat
has been observed on the glutei muscles (Lee, Kengsep, Morris,1228
Rille, Seifert) and their surroundings (Stelwagon, Hamburger,
Bruno), on the lower extremities (Stelwagon, Lenglet and Delaunay,
Hutchins, Moorhead, Lee, Crocker, Schmid,1229 v. Harlingen), on the
upper extremities (Samson, Meade and Freeman, Hutchins, Sokoloff,
v. Harlingen, Brodier and Fouquet, Shelmire,1230 Stelwagon), on the
face (Sokoloff, Moorhead, Kumberg,1231 Rawnitzky, Crocker, Boas1232),
on the neck (Sokoloff), and on the body (Ehrmann, Brodier and
Fouquet, Kaposi,1233 Topsent1234).
The duration of the affection varies very much; it varies between
a few hours and some years1235; several times a spontaneous recovery
has been reported.
The diagnosis of the disease is not at all difficult owing to its
peculiar appearance.
The treatment can only consist in the removal or killing of the
larvæ, since one cannot rely on spontaneous recovery, even if it has
occurred in some cases. If one should succeed in locating the larva
as a black spot at the end of the tract, its removal by means
of a needle is the simplest method (Quortrup and Boas1236). In
some instances a cure has been successfully accomplished by excision
of the active end of the tract (v. Neumann and Rille, Schmid). In
opposition to this method, which not all patients will allow, the
method practised by Arab women (Rille and Riecke1237) of killing the
worm with red hot needles is quite rational. Shelmire1238 used the
electrolytic needle for the destruction of the maggots, Stelwagon1239
made use of cataphoresis, by means of which he applied a sublimate
solution, afterwards cauterizing with a drop of nitric acid,
as excision was refused. Crocker1240 and v. Harlingen1241 injected
small quantities of carbolic acid; Moorhead1242 by a single freezing
of the skin with ethyl chloride, attained a definite cessation of the
attack at the active end. Hutchins1243 in one case made use of hypodermic
injection of a few drops of solution of cocaine and afterwards
of 1 to 2 drops of chloroform; in a second case of repeated applications
of tincture of iodide, as Lenglet and Delaunay1244 did.
v. Harlingen1245 allayed the affection in his first case by rubbing in
sapo viridis and tar, in Kensep’s1246 case the cure seems to have been
accomplished by an ointment containing resorcin, in Meade and
Freeman’s1247 case by a 20 per cent. ichthyol paste. In our case we
made exclusive use of Lassar’s paste; within four weeks a cure
resulted, probably spontaneously, since one cannot ascribe any
essential effect to this paste.
APPENDIX ON PROTOZOOLOGY,
Comprising Notes on Recent Researches, Formulæ of some
Culture Media, and Brief Notes on General Protozoological
Technique.
BY
H. B. FANTHAM, M.A., D.Sc.
I.—NOTES ON RECENT RESEARCHES.
Since the foregoing section on Protozoology was sent to press,
certain interesting observations and results have been published.
Brief notes on such, and some references thereto, are now added.
It is necessary, however, to remark that sometimes it is impossible
to give a precise or rigid definition to a genus of Protozoa, owing to
differences of opinion, to differences regarding nomenclature or to
incompleteness of knowledge. Such a lack of definition, while inconvenient
for the time being, is not unhopeful, as it directs attention to
the necessity for further work, which is inevitable in such a relatively
new and wide subject as protozoology. Thus, it may be noted in
illustration that Minchin, in 1912, in his text-book regarding the
genus Entamœba writes: “The entozoic amœbæ are commonly
placed in a distinct genus, Entamœba, distinguished from the free-living
forms by little, however, except their habitat and the general
(but not invariable) absence of a contractile vacuole.”
Differences between Entamœba histolytica and E. coli.—In
continuation of the remarks on pp. 34 and 40, it may be added that
Lugol’s solution (iodine in aqueous potassium iodide solution) in
fresh specimens shows by brownish staining the presence of glycogen
in the vacuoles of Entamœba coli. Such a reaction is rarely or never
given by E. histolytica.
Phagedænic Amœbæ.—Carini and others record cases in which
the skin around an operation wound in connection with liver abscess
became gangrenous. Amœbæ, possibly Entamœba histolytica, were
found therein and may have been responsible for the gangreno-phagedænic
action.
Endamœba gingivalis (see pp. 43, 44).—Smith and Barrett,1248 after
analysing the early literature, state (June, 1915) that Endamœba
gingivalis, Gros, 1849, is the correct name for the following organisms:
E. buccalis, Prowazek, 1904 (see p. 43); Amœba gingivalis, Gros, 1849;
Amœba buccalis, Steinberg, 1862, and Amœba dentalis, Grassi, 1879.
They conclude that E. gingivalis is the causal agent of pyorrhœa
alveolaris, and that this disease responds to treatment with emetine.
Entamœba kartulisi (see p. 44), synonym E. maxillaris, Kartulis,
is considered to be E. gingivalis.
Smith and Barrett adopt the generic name Endamœba, Leidy,
1879 (see footnote on p. 31, also p. 34). Leidy worked on Endamœba
blattæ.
Craigia and Craigiasis (see p. 45).—Barlow1249 (May, 1915) found
Craigia (Paramœba) hominis in cases of chronic diarrhœa and mild
dysentery in Honduras. He also described a new species of Craigia
under the name of C. migrans. Fifty-six cases were studied, five of
which were due to Craigia hominis, the remainder to C. migrans.
In C. migrans, each flagellate, on attaining full development, becomes
an amœba without dividing. Each amœba encysts and produces
a number of flagellates which are somewhat like cercomonads. On
the other hand, in C. hominis the flagellate form produces, by longitudinal
fission, several generations of flagellates before entering upon
the amœbic stage. The cysts of C. migrans contain fewer “swarmers”
(flagellulæ) than those of C. hominis, but the “swarmers” are somewhat
larger, namely, 5 µ instead of 3 µ in diameter. Further, there is
no accessory nuclear body in C. migrans, but its flagellum stains
more deeply than that of C. hominis and has a peculiar banded
appearance.
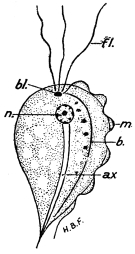
Fig. 422.—Trichomonas from cæcum and gut of rat: n, nucleus;
bl, blepharoplast; fl, flagella; ax, axostyle; m, undulating membrane;
b, line of attachment of undulating membrane to the body. × 2,000
approx. (Original.)
Human Trichomoniasis (see pp. 52–56).—Lynch1250 (April and
May, 1915), working in Charleston, seems to favour the view that
the trichomonads found in the vagina, urethra, mouth, lungs and
alimentary tract are one and the same organism, and that these
flagellates may further excite already existing inflammatory conditions.
He gives detailed histories of cases of (a) infection of the
vagina and gums, and (b) intestinal infection manifested as intermittent
attacks of diarrhœa. The flagellates were found in catarrhal
vaginal discharge, in blood-stained scrapings from the gums (together
with Endamœba buccalis), and in stools after a purge of magnesium
sulphate. The parasites were tetratrichomonads (see footnote, p. 53),
that is, each possessed four flagella anteriorly as well as an undulating
membrane. Lynch successfully infected rabbits from the cases and
from cultures of the parasite. Encysted trichomonads were seen in a
patient’s stools, in rabbits infected therefrom and in cultures. The
culture medium used was bouillon acidified with about 0·05 per cent.
acetic acid and the cultures were maintained at 30° C.
Trichomonads occur in the digestive tracts, for example, the cæca
of rats and mice (fig. 422). In man allied flagellates can occur in
similar situations, as well as in other parts of the intestine.
Other trichomonad-like organisms have been recently described
from the fæces of man, more particularly from cases of chronic
dysentery in the tropics. Derrieu and Raynaud1251 (July, 1914), working
in Algeria, found a flagellate possessing five free flagella anteriorly
and an undulating membrane apparently lateral. They named the
parasite Hexamastix ardin-delteili, but the generic name Hexamastix
is pre-occupied. Chatterjee1252 (January, 1915), working in India, found
probably the same flagellate and called it Pentatrichomonas bengalensis.
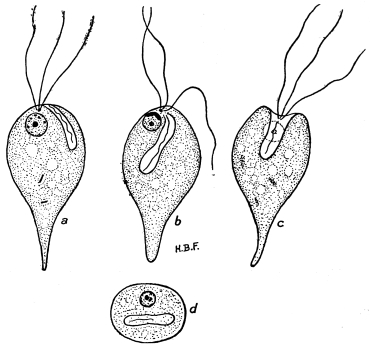
Fig. 423.—Chilomastix (Tetramitus) mesnili. a, b, c, flagellate forms;
d, rounded or encysted form. × 2,500. (Original.)
Chilomastix (Tetramitus) mesnili (see p. 57).—Alexeieff1253 (1914)
now places the parasite originally called Macrostoma mesnili, by
Wenyon (1910), in the genus Chilomastix, Alexeieff. The differential
characters of the genera Tetramitus and Chilomastix are not especially
well marked. According to Alexeieff, Tetramitus is characterized by
four unequal flagella (which he figures anteriorly), a ventral cytostome
in the form of a linear cleft and a pulsatile vacuole in front of the
anterior nucleus. Chilomastix, according to the same author, has
three forwardly directed flagella and a fourth backwardly directed one
in the cytostome, which is well developed (fig. 423). Some authors
consider that the fourth flagellum forms the edge of an undulating
membrane in the cytostome.
Diagrams of Chilomastix mesnili are given in fig. 423.
Giardia (Lamblia) intestinalis (see p. 57).—Alexeieff1254 (1914) considers
that Lamblia intestinalis, Lambl, should be placed in the genus
Giardia, Kunstler, 1882. Bipartition occurs in the encysted state.
The axostyles persist in the quadrinucleate cyst.
Cercomonas hominis (see p. 61).—This parasite is considered by
some authors to be of a doubtful nature, as it is thought to have been
mistaken for deformed or incompletely observed Trichomonas or Chilomastix
or even Lamblia.
Wenyon1255 (1910) described Cercomonas longicauda from cultures
of human fæces. It is considered that the genus is very confused,
and the author points out that the tail flagellum has been overlooked.
He considers that the genus Cercomonas should include flagellates
with an anterior blunt end from which arises a single long flagellum,
and a posterior tapering end also with a flagellum, which can be traced
over the surface of the body towards the insertion of the anterior
flagellum.
Another species, Cercomonas parva, has been found in cultures of
human fæces by Hartmann and Chagas1256 (1910). It has a somewhat
different structure.
Further researches are necessary on the organisms variously
referred to the genus Cercomonas.
Transmissive Phase of Trypanosomes in Vertebrates.—In
addition to the general remarks on the morphology of trypanosomes
set forth on pp. 70 to 72, it may be noted that Woodcock1257 (November,
1914) states that, in certain cases, there is a definite transmissive phase of
a trypanosome in its vertebrate host. He quotes the work of Minchin
and himself on T. noctuæ of the little owl, in which the transmissive
form is spindle-shaped and occurs in the bird’s peripheral blood
during the early summer months (see p. 69). A similar phase occurs
in T. fringillarum, and Robertson1258 has found that the short, stumpy
form of T. gambiense is its transmissive phase in vertebrates.
Trypanosoma lewisi (see p. 88).—Brown (1914–15) has published
some interesting results on the potential pathogenicity of T. lewisi.
Blepharoplastless Trypanosomes (see p. 101).—Laveran1259 (April,
1915) suggested a practical use of strains of blepharoplastless trypanosomes
produced by the action of drugs. He finds that tryposafrol
will also produce such strains, and remarks on blepharoplastless
strains of T. evansi and T. brucei, which in the former case can
undergo 450 passages without reversion, and in the latter 273
passages. He states that if it is desired to inoculate surra or nagana
to Capridæ or Bovidæ in order to produce immunity, use should be
made of the blepharoplastless races of the respective trypanosomes,
which races are a little less virulent than the corresponding normal
ones. Also, the immunity which follows from an infection due to
blepharoplastless T. evansi or T. brucei is only a little less complete
than that following infections from either of the respective normal
strains.
The Experimental Introduction of certain Insect Flagellates
into various Vertebrates, and its bearing on the Evolution of
Leishmaniasis.—In continuation of the remarks on pp. 103, 104,
and 112, further researches have been conducted on the introduction
into vertebrates of flagellates normally parasitic in insects. The
vertebrates became infected by inoculation with the flagellates or by
being fed on insects containing the protozoa. Fantham and Porter1260
(June, 1915) published the following results. Flagellates from
sanguivorous and non-sanguivorous insects were used, and cold-blooded
as well as warm-blooded vertebrates as hosts. The introduced
protozoa were pathogenic to the mammals, but not markedly
so to the cold-blooded vertebrates. Herpetomonas jaculum, H. stratiomyiæ,
H. pediculi, and Crithidia gerridis (parasitic in certain waterbugs)
proved pathogenic to mice. A puppy was infected by way
of the digestive tract with H. ctenocephali. Frogs became infected
with H. jaculum and with C. gerridis, toads and grass snakes with
H. jaculum, lizards with C. gerridis, and sticklebacks with H. jaculum.
Second and third passages of some of the parasites were obtained.
The protozoa, whether Herpetomonas or Crithidia, were present in
the vertebrate hosts in either the non-flagellate or the flagellate form,
or usually both. They were more abundant in the internal organs
of the hosts, more particularly in the liver, spleen and bone-marrow.
In all experiments in which C. gerridis was used the parasite
invariably retained the crithidial facies in the vertebrate host. No
transition to a trypanosome was ever seen. Infections in adult
animals were not so heavy as in the young ones, and the parasites
were more virulent in young hosts, as is the case with Mediterranean
kala-azar in children.
The mode of infection of the vertebrate in Nature seems to be
contaminative, either by its food or through an already existing
abrasion or puncture on the surface of its body. Cases in which the
flagellate-infected insects have been allowed to suck the blood of
vertebrates have proved negative up to the present. In areas where
leishmaniases are endemic, an examination should be made of all
insects and other invertebrates likely to come into contact with men
or dogs, or rats and mice (see below), in order to ascertain if these
invertebrates harbour herpetomonads. Preventive measures should
be directed against such invertebrates, especially arthropods. Further,
it is likely that certain vertebrates, such as reptiles and amphibia
(especially those that are insectivorous), may serve as reservoirs
of leishmaniases, or, as they should preferably be termed, herpetomoniases.
From such reservoirs the herpetomonads may reach
man by the agency of ectoparasites or flies, especially such as are
sanguivorous.
That vertebrates in Nature can harbour herpetomonads in their
blood has been shown by the work of Dutton and Todd (1903) on
the herpetomonads of Gambian mice, while the recently published
investigations of Fantham and Porter1261 (June, 1915) on natural herpetomonads
in the blood of mice in England have shown that these
rodents may be a natural reservoir of herpetomoniasis. The origin
of the infection of mice is to be sought in a flagellate of an ectoparasite
of the mouse, very probably Herpetomonas pattoni parasitic in
various fleas, which protozoön can adapt itself to life in the blood
of mice. Herpetomonads were also found naturally in the blood
of birds by Sergent (1907). Recently, Fantham and Porter have
successfully infected birds with herpetomonads experimentally.
The significance of insect flagellates in relation to the evolution of
disease has recently been set forth by Fantham1262 (June, 1915). The
deductions to be made from the occurrence of a herpetomonad stage
in Leishmania, especially in L. tropica, in man himself, and of flagellate
stages of the so-called Histoplasma capsulatum in man are fully
discussed and correlated. It is pointed out that flagellosis of plants
(see p. 104) may possibly be connected with leishmaniasis. The
evolution of Leishmania from flagellates of invertebrates is thus
traced and the way again indicated for preventive measures against
leishmaniasis, as first set forth by Dodds Price and Rogers.
Franchini and Mantovani (March, 1915) have successfully
infected rats and mice by inoculation or by feeding with Herpetomonas
muscæ domesticæ obtained from flies and from cultures.
It is of great interest to note that the recent observations of
Ed. and Et. Sergent, Lemaire and Senevet1263 (1914) have demonstrated
the presence of a herpetomonad flagellate in cultures of the blood
and organs of geckos obtained from areas in Algeria in which Oriental
sore, due to L. tropica, is prevalent. Phlebotomus flies, which may
harbour a natural herpetomonad, feed on the geckos and on men.
Hence animals like geckos may possibly act as reservoirs of
leishmaniasis. Lindsay1264 (1914) writes that the parasite of dermo-mucosal
leishmaniasis in Paraguay is believed by native sufferers to
be conserved in rattlesnakes, and spread by ticks or flies (Simulium)
feeding on the reptiles and transferring the parasite to man.
The Transmission of Spirochæta duttoni (see p. 116).—It is
probable that Ornithodorus savignyi acts as the transmitting agent of
S. duttoni in places like Somaliland (Drake-Brockman, 1915).1265
Spirochæta bronchialis (see p. 122).—The morphology and life-history
of S. bronchialis have been investigated by Fantham1266 (July,
1915). From researches conducted in the Anglo-Egyptian Sudan, he
found that S. bronchialis is an organism presenting marked polymorphism,
a feature that has only been determined by the examination
of numerous preparations from the deeper bronchial regions of various
patients.
S. bronchialis varies in length from 5 µ to 27 µ, and its breadth is
about 0·2 µ to 0·6 µ. These variations are due to the processes of
growth and division. Many of the parasites measure either 14 µ to
16 µ long, or 7 µ, to 9 µ, the latter resulting from transverse division
of the former. The ends show much variation in form, but approach
the acuminate type on the whole. The discrepancies in dimensions
given by the very few previous workers on the subject are probably
the result of the measurement of a limited number of parasites. All
such sizes can be found on some occasion during the progress of the
disease, when a larger number of spirochætes is examined.
The movements of S. bronchialis are active, but of relatively short
duration, when it is removed from the body. The number of coils
of the spirochæte is rather an index of its rapidity of motion than a
fixed characteristic of the species.
The motile phase of S. bronchialis is succeeded by one of granule
formation, the granules or coccoid bodies serving as a resting stage
from which new spirochætes are produced. The formation of
coccoid bodies and reproduction of spirochætes from them can be
observed in life.
S. bronchialis is a species distinct from the spirochætes occurring
in the mouth. It differs from them in morphology, pathogenicity
and in staining reactions. It is not a developmental form of any
bacterium, and is an entity in itself.
The passage from man to man is effected most probably by means
of spirochætes, and especially coccoid bodies, that leave the body in
the spray with expired air and by way of the nasal secretions. Owing
to the fragility and short life of S. bronchialis extracorporeally, the
resistant coccoid bodies in air, in dried sputum and dust, and possibly
also on the bodies of flies and other insects, are probably instrumental
in inducing attacks of bronchial spirochætosis in human beings, especially
those having a lowered bodily resistance, such as occurs after
a chill. Lurie (December, 1915), has described a case from Serbia.
The Spirochætes of the Human Mouth (see p. 122).—Two
species of spirochætes were recorded as occurring in the human
mouth about forty or fifty years ago. These are Spirochæta buccalis,
Steinberg (often ascribed to Cohn, 1875), and S. dentium, Miller
(often attributed to Koch, 1877).
The most recent work on S. dentium and S. buccalis is that of
Fantham1267 (July, 1915), who observed the parasites ascribed to Cohn
and to Koch, these being the two common spirochætes seen in the
mouths of natives of the Sudan and of Europeans in England, as
well as the forms described and cultivated by recent investigators.
Some of the mouth spirochætes are not very active, but there is
marked corkscrew and boring movement, and they are flexible.
Tangles or tomenta of these mouth spirochætes are common.
Internal structure is seen with some difficulty, but in some specimens
it can be determined, and chromatin granules are then seen. Mühlens
(1907) figured stained specimens of S. buccalis and S. dentium, in
which chromatin-coloured granules were distributed along the bodies
of the organisms.
S. dentium has tapering ends, and varies in length from 4 µ to 10 µ.
S. dentium is rather like Treponema pallidum, and has been placed by
some workers—for example, Dobell—in the genus Treponema. It has
already been mentioned, on p. 128, that Noguchi cultivated three
species of Treponema from the human mouth—namely, T. macrodentium,
T. microdentium, and T. mucosum, but they cannot be easily
distinguished morphologically, and so may appear to be biological
varieties of S. dentium.
S. buccalis has somewhat rounded or bluntly acuminate ends
and varies in length from 9 µ to 22 µ. A slight membrane or crest
may sometimes be observed. S. buccalis was found to be the predominant
spirochæte in the mouths of eight natives examined by
Fantham in the Anglo-Egyptian Sudan.
S. buccalis and S. dentium take up stains well and with relative
ease. Intracellular stages of the parasites are uncommon. Multiplication
by binary fission has also been observed. Coccoid bodies or
granule stages of the mouth spirochætes are formed, but appear to be
relatively few in number.
J. G. and D. Thomson1268 (1914) have written an interesting paper
on various spirochætes occurring in the alimentary tract of man and
of some of the lower animals. They have also given a useful list of
references, and the work of some of the earlier authors is discussed in
the paper.
With regard to the general morphology of spirochætes, it may be
noted that the so-called axial fibre of Zuelzer is acknowledged to be
homologous with the membrane or crista of molluscan spirochætes.
Coccidia in Cattle.—Regarding the remarks on coccidiosis or
“red dysentery” in cattle on p. 147, it may be added that Schultz1269
(July, 1915) has found the malady among cattle in the Philippine
Islands. He states that some irregular or atypical cases of apparent
rinderpest are really due to coccidia. As has been pointed out by
Montgomery, rinderpest can be transmitted by blood inoculation,
while coccidiosis cannot be so transmitted, but may be diagnosed by
the microscope. These differences should be remembered as the two
diseases are often found to be associated and are difficult to separate
clinically. Coccidia have also been found in Australian cattle.
The Hæmosporidia.—It is likely that this order (see p. 151) may
be soon abolished. Mesnil1270 (April, 1915) considers that the grouping
of the three families, Plasmodiidæ (or Hæmamœbidæ), Hæmogregarinidæ
and Piroplasmidæ in the order Hæmosporidia is no longer
possible, because of the coccidian nature of the Hæmogregarines
(see p. 154). The Coccidia are divisible into the Adeleidea and the
Eimeridea (see p. 141). The Hæmogregarinidæ are allied to the
former, and the Plasmodiidæ to the latter. The Piroplasmidæ, until
more is known of their life-cycle in the invertebrate host, cannot be
more definitely placed.
The Leucocytozoa of Birds.—Regarding the statement, on p. 153,
that Laveran and França consider that avian leucocytozoa may inhabit
red blood cells, it may be added that França1271 (April, 1915) remarks
that the action of the parasites on the red cells is very rapid and
very intense. The host cells become so altered that it is difficult
to recognize their true nature. He used very young birds in his
researches. Two shapes of host cell are considered, namely, those
with fusiform prolongations, and those which are rounded and without
such prolongations (see p. 153). The movements and form of
the Leucocytozoa determine the shape of the host cell, as was pointed
out by Fantham1272 in 1910.
Schizogony of these parasites has been seen by França (1915) and
by Coles (1914), in addition to Fantham (1910), and to Moldovan
(1913), mentioned on p. 153. Schizogony may also take place in the
lungs of the host. The genus Leucocytozoön, established by Ziemann
in 1898, belongs to the family Hæmamœbidæ.
II.—FORMULÆ OF SOME CULTURE MEDIA.
(1) Culture Media for growing Amœbæ.—There has been much
discussion as to whether the true parasitic Entamœbæ or Endamœbæ
can be grown on culture media (see p. 42). Undoubtedly certain
free-living amœbæ can be so grown, and it is considered that some
of the earlier researches on the so-called artificial growth of the
dysenteric amœbæ were really due to contaminations with free-living
forms. The following media are worthy of note:—
Musgrave and Clegg in 1904 devised a culture medium for
amœbæ. The organisms grown by them were probably not dysenteric
amœbæ, as was thought, but free-living forms. Phillips1273 (1915)
gives a slightly modified formula of Musgrave and Clegg’s medium,
thus:—
| Agar-agar | 2· | 5 | grm. |
| Sodium chloride | 0· | 05 | " |
| Liebig’s beef extract | 0· | 05 | " |
| Normal sodium hydroxide | 2· | 0 | c.c. |
| Distilled water | 100· | 0 | " |
Without clarifying, sterilize at 7 kilograms pressure per square centimetre
for about three-quarters of an hour. It should be neutral to
phenolphthalein.
Anna W. Williams1274 (1911) described a medium consisting of fresh
tissue spread on agar plates for the culture of amœbæ. There are
three stages in the procedure: (1) obtaining living amœbæ free from
other living organisms; (2) obtaining sterile tissue; and (3) making
successive transplants of amœbæ and tissue, and showing that every
transplant is free from other living organisms. Each step requires
many controls. The essentials of the method may now be given.
Remove aseptically and rapidly the tissue required, such as brain,
liver, kidney, or spleen, from a freshly killed animal (guinea-pig,
rabbit, or dog). Put each tissue on a separate agar plate. Cut the
selected tissue into tiny pieces, and spread them over freshly made
agar plates. Place these plates in a thermostat at 36° C. for twenty-four
hours to insure sterility. Add the broken up tissue to the
amœbæ, free from bacteria, and maintain the cultures in thermostats,
some at 36° C., and some at 20° C. to 24° C. Emulsions of liver and
brain in sterile neutral glycerine may also be used. The freshly
removed tissue serves as food for the amœbæ.
The cultural amœbæ mentioned on p. 42 were grown on such
media or modifications thereof. One modified medium actually used
was brain tissue, to which blood was added from day to day, and an
easily assimilable bacterium (one of the influenza group of bacilli)
was present, which did not overgrow the medium at a temperature of
38° C. Different conditions of food and of temperature produced
morphological variations in the cultural amœbæ.
Couret and J. Walker1275 (1913) state that they have cultivated five
varieties of intestinal amœbæ, the associated bacteria having been
previously separated. They used a medium consisting of agar to
which sterile autolysed tissue had been added. The sterile tissue,
such as brain or liver, was kept in a sterile thermostat at a temperature
of 40° C. for ten to twenty days. The surface of the agar should
be broken up before use, and the medium must not be too acid (not
over 1·5 per cent.). They consider that autolysed tissue is necessary
for the growth of Entamœbæ, and that naturally associated bacteria
aid growth by autolysing the tissues.
(2) Culture Media for the growth of Protozoa parasitic in the
Blood.—MacNeal and Novy,1276 in 1903, used a mixture of blood and
agar for the cultivation of trypanosomes such as T. lewisi and T. brucei.
They employed varying proportions of the blood and agar, a medium
consisting of two parts of defibrinated rabbit’s blood mixed with one
part of agar being useful. The trypanosomes grew in the water of
condensation. Some of the authors’ earlier formulæ contained
different proportions of blood and agar with a little peptone, while
one of these media contained meat extract, agar, peptone, salt and
sodium carbonate. The temperature, like the proportion of blood
and agar, varied with the trypanosome investigated, but the optimum
was 25° C.
Mathis1277 (1906) somewhat simplified the technique of Novy and
MacNeal. He collected the blood of a suitable animal, such as rabbit,
cow or dog, strict asepsis not being essential. The blood was
defibrinated in the ordinary way. One part of blood was added
to two parts of agar at 50° C. The mixture was sterilized several
times by heating to 75° C. or 100 ° C. Slopes were made and the
water of condensation was inoculated with a little blood containing
the trypanosomes. Blood may be obtained from a superficial vein
or from the heart.
Novy-MacNeal-Nicolle or N. N. N. Medium.—In 1908 C. Nicolle1278
brought forward a modification of the Novy-MacNeal (N.N.)
medium. The formula is as follows:—
| Agar | 14 | grm. |
| Sea salt | 6 | " |
| Water | 900 | " |
Apparently pure sodium chloride can be substituted equally
well for sea salt. The mixture is placed in tubes and sterilized in
an autoclave. To each tube one-third of its volume of rabbit
blood, taken by aseptic puncture of the heart, is added. The salt
agar is kept liquid at 45° C. to 50° C. and the blood is added to
the mixture. The culture medium so prepared is maintained for
five days at 37° C., and then for a few days at room temperature.
This medium was devised for the cultivation of Leishmania (see
p. 106), but trypanosomes may also be grown thereon. Subsequently,
Nicolle recommended the use of citrated rat’s blood heated to 45° C.
for half an hour, instead of defibrinated rabbit’s blood. On such a
medium, J. G. Thomson and Sinton1279 (1912) succeeded in growing
Trypanosoma gambiense and T. rhodesiense (see pp. 76, 83).
Noguchi’s media for the cultivation of Spirochætes and Treponemata
are described on pp. 123, 125. Hata’s modification is
discussed on p. 126.
Bass’s glucose-blood medium for the cultivation of malarial
parasites is described on pp. 170–172. It has also been used
successfully for the cultivation of Piroplasma or Babesia (see p. 172).
III.—BRIEF NOTES ON GENERAL PROTOZOOLOGICAL TECHNIQUE.
The object of this book is to give accounts of the structure and
life-histories of the numerous parasitic organisms that affect man
more particularly. It is, therefore, inappropriate to devote much
space to a consideration of technique, regarding which many volumes
have already been written. Methods of procedure are largely matters
of opinion, and the technique that gives brilliant results when used
by one investigator may be a complete failure in the hands of
another. In the present appendix, brief notes regarding certain
relatively simple methods only can be given, because the number of
fixatives in use is very great; there are also large numbers of stains
as well as many modifications of them, while the methods of applying
both fixatives and stains are, perhaps, still more numerous.
There are so many, in fact, that confusion frequently arises from
the multiplicity of choice presented to the worker. Those desiring
more information on the subject of technique are advised to consult
the treatises of Bolles Lee1280 and of Langeron.1281
Fresh Material.
(a) Simple Examination.
Fluid Substances, such as Blood and Sputum.—A small quantity of
the substance to be examined is taken on a sterile platinum loop and
transferred to a perfectly clean glass slide. A clean cover-slip is gently
lowered on to the drop, air bubbles being avoided. The preparation
is luted with vaseline or paraffin and examined first with a low power
and then with a high power objective. The light is cut down by
partly closing the diaphragm of the substage of the microscope.
Skin Ulcers and Similar Sores.—Scrapings are made from the edge
of the sore, mixed with sterile physiological salt solution, and
prepared and examined as above.
Fæces.—A small portion of fæces, or flakes of mucus (which may
be blood-stained) from the same, is removed on a sterile platinum
loop, spread out thinly after dilution, if necessary, with physiological
salt solution on a slide, covered and examined as before.
Alternatively, hanging drop preparations of blood, ulcerative tissue,
or fæces, appropriately diluted if necessary with sodium citrate or
physiological salt solution, may be made on a cover-slip, which is
inverted over a slide with a well in it. The cover-slip is then luted and
examined.
For the elucidation of the developmental processes of such
organisms as trypanosomes, spirochætes and piroplasms, fresh
preparations may be often kept under observation longer by the
use of a thermostat, maintained at or near blood heat, in which the
microscope is inserted.
(b) Intra vitam Staining of fresh Preparations.
Intra vitam staining is of service on some occasions, more
particularly for the study of the nucleus and other chromatoid
substances of the living organism. Two methods are in common
use. In the first case, the stain, employed usually in very dilute
solution, is mixed with the medium containing the organism. The
latter takes up some of the stain, the amount of coloration depending
on the organism concerned and on the stain employed.
The commoner intra vitam stains are pure, medicinal (zinc-free)
methylene blue and neutral red, used in aqueous solutions. A solution
of methylene blue of 1 per 1,000 of water may be tried, while neutral
red in the proportion of 1 per 3,000 parts of water has proved of
service.
The second method of vital colouring consists in placing a drop of
1 per cent. solution of methylene blue on a slide or cover-slip, slightly
spreading it, and allowing it to dry. The living organism is then
placed in a drop of saline on the prepared slide or cover-slip, which
is then mounted and examined under the microscope. Progressive
staining of the organism occurs and its internal structure can be seen.
A similar procedure may be followed for neutral red. Intra vitam
staining is useful for relatively large and easily deformed protozoa
such as ciliates, as well as for amœbæ and flagellata of the gut.
When examining very actively motile organisms, it is sometimes
useful to endeavour to restrict their movements by adding a little gum
or gelatine to the medium.
(c) Examination by aid of the Paraboloid Condenser.
The use of one of the dark-ground illuminators (so-called ultra-microscopes)
is of service for the detection of minute living organisms
or of organisms present in small numbers only. The forms of
paraboloid condenser manufactured by the firms of Zeiss and Leitz
can be recommended. For details of their methods of employment,
reference should be made to the leaflets of the firms supplying the
said instruments. By the use of the paraboloid condenser, the finer
details of certain stages of life-cycles, such as the formation of
granules in spirochætes and treponemata, can be observed more
readily than by using the ordinary substage of the microscope.
The use of the paraboloid condenser for the detection of small
numbers of living organisms renders it of value for rapid diagnostic
purposes.
Stained Material.
Fuller accounts of the technique of fixed and stained material
will be found in Bolles Lee and in Langeron, already mentioned.
Thin Films.—For the examination of blood-inhabiting Protozoa,
it is necessary to make first thin films or smears of blood. There are
many ways of doing this, and opinions differ as to their respective
merits. A simple method is to take a straight surgical needle about
2 in. long, the eye of which has been removed, and a clean glass slide.
The patient’s skin is pricked, and when the bead of blood reaches
the size of a small pin’s head, the slide is applied to the surface of
the blood, about 1/3 in. from the far (left-hand) end of the slide.
The shaft of the needle is laid across the drop of blood, which
spreads between the slide and the needle. The latter is drawn evenly
along the slide towards the right. The film is dried by waving it in
the air. The film should possess a straight edge parallel with that of
the slide and should be as uniform and thin as possible. Another
glass slide may be used as a spreader, or a cover-slip or thin glass
rod may be employed.
Thick Films.—These are of service in detecting malarial parasites
or trypanosomes, especially when the parasites are few. The method
of Ross, or a modification thereof, has been much used. A small drop
of fresh blood is spread evenly and quickly with a needle-point over
a square area somewhat less than that of an ordinary square cover-glass.
The blood is allowed to dry. The film is then carefully
dehæmoglobinized in water in which there is a trace of acetic acid.
The dehæmoglobinizing fluid is then carefully drained off and the
film again dried. It is fixed in absolute alcohol and stained with
Romanowsky’s solution. A cubic millimetre of blood divided into
quarters may be thus dehæmoglobinized and stained. The parasites
in such a cubic millimetre of blood may be counted. Such a procedure
was followed by R. Ross and D. Thomson,1282 in determining the
periodic variation of the numbers of trypanosomes in the blood of
a patient, as mentioned and figured on pp. 78 and 79.
For cytological details of various Protozoa, thin film preparations
on cover-slips or slides are often useful. Cover-slip preparations are
preferable, unless the organisms under investigation are extremely
scanty. The medium containing the organisms, such as blood,
lymph, intestinal contents, sputum, scrapings of ulcers, and urine, is
spread thinly, either alone or diluted with a little physiological salt
solution, on the cover-slip. Fixation while still wet is necessary.
Various methods are employed.
Fixatives.—A useful procedure is to fix the wet film by exposure
to 4 per cent. osmic acid vapour for ten to thirty seconds, then place
in absolute alcohol for five minutes to harden. Grade down from
absolute alcohol through 90 per cent., 70 per cent., 50 per cent., and
30 per cent. alcohols to water. Stain wet with a suitable stain such
as hæmatoxylin, and gradually dehydrate by grading through the
necessary strengths of alcohol, clear in xylol or other oily clearing
medium and mount in Canada balsam.
Other fixatives may be employed, such as are also useful for fixing
pieces of tissue for sectioning. Films or smears on cover-slips while
still wet are floated on the surface of the fixative in a watch glass.
Some good fixatives of wide application are:—
Schaudinn’s Fluid.—This consists of a mixture of
| Saturated aqueous solution of corrosive sublimate | 2 | volumes |
| Absolute alcohol | 1 | volume |
Two modifications of Schaudinn’s formula may be found useful.
A saturated solution of corrosive sublimate in physiological salt
solution may be substituted for the aqueous one, and the addition of
a few drops of glacial acetic acid to either of the preceding mixtures
may be made.
Some workers prefer to use hot fixatives, raised to a temperature
of about 50° C.
Fixation by corrosive sublimate solutions must be followed by
thorough removal of the mercury salt by washing repeatedly in
30 per cent. alcohol or with iodine-alcohol.
Bouin’s Fluid, or modifications thereof, is also very useful for wet
fixation. Bouin’s picro-formol solution consists of:—
| Saturated aqueous solution of picric acid | 30 | volumes |
| Formalin, 40 per cent. | 10 | " |
| Acetic acid, glacial | 2 | " |
The best-known modification is one due to Duboscq and Brasil,
and often known as Bouin-Duboscq Fluid. Its formula is as follows:—
| Alcohol, 80 per cent. | 150 | c.c. |
| Formalin, 40 per cent. | 60 | " |
| Acetic acid, glacial | 15 | " |
| Picric acid | 1 | grm. |
Thorough washing of the smear or cover-slip preparation with
70 per cent. alcohol until the yellow colour disappears is necessary
to remove excess of fixative.
Other fixatives, which may be of use, more especially for fixing
small pieces of tissue for sectioning, are the solutions of Flemming
(chromo-aceto-osmic acids) and of Zenker (sublimate-bichromate-acetic,
with sodium sulphate).
Regarding the time of fixation, there is much difference of opinion.
Usually, exposure to or contact with the fixative for five minutes is
sufficient in the case of films or smears. Material for sections
should be cut into small cubic pieces, of a thickness of about
5 mm. ( 1/5 in.). One or two hours should be sufficient time for the
fixation of such pieces of tissue, though some, as Langeron, prefer
a longer time of fixation. On the other hand, Gustav Mann1283 recommends
a short fixation period. The excess of fixative should be
thoroughly washed out of the tissue in the manner appropriate to
the particular fixative used. If it is desired to keep the tissue for some
time before sectioning and staining, it should be transferred to 70 per
cent. alcohol.
When fluid fixatives are employed, large quantities of the fixing
media are necessary. The volume of the fixative should be at least
ten to twenty times that of the object, and the latter should be suspended
in the middle of the fixative. The tissue should be fixed as
soon as possible after the death of the host.
For sectioning tissue parasitized by Protozoa, embedding in
paraffin is generally recommended. Microtome sections should not,
if possible, exceed 5 µ in thickness. Details of special procedures
must be sought in larger works.
Staining.—Here, as with fixatives, much choice is presented. The
various modifications of the Romanowsky stain have aided greatly in
the detection of various Protozoa parasitic in the blood. Such
stains, however, leave something to be desired in the revealing of finer
cytological details. Other stains, more especially the hæmatoxylins,
must be employed for cytological purposes.
Formulæ of some of the principal Romanowsky and hæmatoxylin
stains may now be given.
The underlying principle of the Romanowsky Stain is the reaction
between alkaline methylene blue and eosin, forming the so-called
eosinate of methylene blue which stains chromatin purplish-red.
A solution of medicinal methylene blue after having been subjected
to the action of an alkali, such as sodium carbonate, becomes partly
converted into certain derivatives, the chief of which are methylene
azure and methylene violet. These substances are also present in
matured polychrome methylene blue.
The formula of a slightly modified Romanowsky Stain which gives
excellent results is given below:—
Two stock solutions are required—
| Solution A.— | Methylene blue, pure medicinal | 1·0 | grm. |
| Sodium carbonate | 0·5 | " |
| Water | 100·0 | c.c. |
Keep in a warm incubator for two or three days, until the solution is distinctly purple in
colour. It improves with age.
| Solution B.— | Eosin, water soluble, extra B.A. | 1·0 | grm. |
| Water | 1,000·0 | c.c. |
This solution must be kept in the dark, in dark-tinted (amber-coloured) bottles, as unfortunately
it is decolorized by light.
Before use each stock solution must be diluted. Thus, make up
5 c.c. of each stock solution to 100 c.c. by adding distilled water.
For staining, 1 volume of solution A is added to 2 or 3 volumes of
solution B. Mix thoroughly by shaking, pour the mixture over the
film, previously fixed in absolute alcohol, and stain for ten to fifteen
minutes. Wash carefully in running water, then dry. The cytoplasm
of a protozoan parasite will be stained blue, the chromatin purplish-red
and vacuoles or very tenuous protoplasm will remain colourless.
The exact proportions of solutions A and B, which must be
mixed together, should be determined by experiment. Freshly mixed
stain must be used on each occasion.
Leishman’s Stain is the precipitate resulting from the interaction
of alkaline methylene blue and eosin. The washed and dried
precipitate is collected and dissolved in pure methyl alcohol, which
acts as a fixative; 0·015 grm. of Leishman powder may be dissolved
in 10 c.c. of methyl alcohol for staining films. The film is covered
with the solution for one minute, twice the volume of water is then
added and mixed with the stain on the slide. The staining is then
continued for five to ten minutes, and the film is finally washed with
water.
Giemsa’s Stain.—This should be procured ready made. Azure II
is a mixture of methylene azure and methylene blue. (Methylene
azure is sometimes known as Giemsa’s Azure I.) The formula given
by Giemsa himself in 1912 is:—
| Azure II-eosin | 3·0 | grm. |
| Azure II | 0·8 | " |
| Glycerine, pure | 125·0 | " |
| Methyl alcohol, pure | 375·0 | " |
The film is first fixed in absolute alcohol. The proportion of
stain usually used is one drop of stain to 1 c.c. of water. Stain for
about ten minutes and then wash in water.
The details of the application of the Giemsa stain to films fixed
wet and to sections must be sought in larger works on technique.
These works should also be consulted for information regarding
the use of Pappenheim’s Panchrome mixture.
There are numerous formulae of stains containing ripened Hæmatoxylin
or its essential principle, Hæmatein. A mordant is necessary,
one of the alums being usually employed. The mordant may be
included as an ingredient in the staining mixture, or it may be used
separately as in the case of the so-called iron-hæmatoxylins, wherein
ferric ammonium alum is used separately and is followed by staining
with hæmatoxylin or hæmatein. A few of these stains of general
application may now be mentioned.
Delafield’s (or Grenachier’s) Hæmatoxylin.
| Hæmatoxylin crystals | 4 | grm. |
| Absolute alcohol | 25 | c.c. |
| Saturated aqueous solution of ammonia-alum | 400 | " |
Mix these ingredients, and leave exposed to light and air for three to four days. Filter
and add—
| Glycerine | 100 | c.c. |
| Methyl alcohol | 100 | " |
Allow the mixture to stand until the colour is sufficiently deep,
then filter and place in a stoppered bottle. The solution should be
allowed to ripen for at least two months before use. Dilute aqueous
solutions of the stain are of service for films and for sections. A
trace of acetic acid may be added at the moment of use, for sharp
differentiation.
Ehrlich’s acid hæmatoxylin, Mayer’s hæmalum, and Mayer’s
glychæmalum are also useful. Their formulæ will be found in larger
works.
The chief Iron-Hæmatoxylin Stain is that devised by Heidenhain.
Unfortunately the procedure involved is a long one, and various
modifications have been made to obviate this disadvantage. Hæmatein
may be used instead of ripened hæmatoxylin.
One efficacious modification of Heidenhain’s stain is that of
Rosenbusch. The smear or tissue, after fixation, must be graded
downwards through the alcohols to water. Mordant for one and a
half hours in a 3 1/2 per cent. aqueous solution of ferric ammonium
sulphate. Stain for about three minutes in 1 per cent. solution of
ripe hæmatoxylin or hæmatein in absolute or 96 per cent. alcohol,
to which a drop of saturated aqueous solution of lithium carbonate,
sufficient to produce a wine-red colour, has been added. Differentiate
under the microscope with a very dilute solution of the ferric
ammonium sulphate. Wash, gradually dehydrate, clear and mount
in balsam. It must be remarked that iron-hæmatoxylin is a regressive
stain, hence great care must be exercised in differentiating with the
iron alum.
Gentian Violet.—A 1 per cent. alcoholic solution of gentian violet,
or of methyl violet, or of crystal violet, will be found useful for staining
spirochætes.
Methyl Green.—This substance is considered to be a chromatin
stain, for either fresh or perhaps recently fixed tissues. A concentrated
aqueous solution contains about 1 per cent. of the stain. This
should be added to a 1 per cent. solution of acetic acid. It may
be used for demonstrating the nuclei of ciliates.
In conclusion it is essential to remember that the actual magnification
of figures of Protozoa should be given, and not merely the
combination of objective and ocular that has been used, for unless
the tube-length and distance of the drawing board from the ocular be
also given, it is not possible to compute the magnification from such
information. Drawings should always be made with the aid of
a camera lucida, drawing prism or other form of projection apparatus.
APPENDIX ON TREMATODA AND NEMATODA.
BY
J. W. W. STEPHENS, M.D., B.C., D.P.H.
TREMATODA.
Artyfechinostomum sufrartyfex, Clayton Lane, 1915.—Leiper
thinks this may be the same as Echinostoma malayanum, Leiper,
1911, which species Odhner assigns to the genus Euparyphium.
Metagonimus (Yokogawa) yokogawai occurs in dogs in Shanghai.
Encysted cercariæ probably in the perch.
Opisthorchis sp.—Skin covered with spines. Gut forks almost
reach end of body. Œsophagus two to three times length of
pharynx. Ovary multilobed. Ovary and testes in posterior fourth
of body. Vitellaria end opposite the ovary. Distinguished from
O. felineus by presence of spines and lobed ovary; from O. pseudofelineus
and O. noverca by the lobed ovary, and by the fact that the
yolk glands do not extend as far as the anterior testis. It agrees
with Poirier’s description of O. viverrini in the Indian civet cat, but
whether this species has spines on the cuticle is not known.
Habitat.—Man in Chiengmai (Malay States). Fifteen per cent.
of prisoners in the jail showed the ova of this species in their fæces.
Schistosome cercariæ.
Schistosome cercariæ belong to the furcocercous division of the
Distomata cercariæ.
Distomata cercariæ.
Body without a floating membrane. Tail absent, or if present
not cleft to the base. Mouth anterior, gut forked. Oral sucker
present. Ventral sucker near middle of body. Eyes generally absent.
Group Fercocercous cercariæ.
Cercariæ single (not in colonies). Tail forked at its end.
Family. Schistosomidæ.
Pharynx absent.
Cercaria bilharzia, Leiper, 1915.
Pigment spots (eyes) anterior to ventral sucker absent, cuticular
keel on forks of tail absent.
In Bullinus sp. and Planorbis boissyi in Egypt, (?) in Physopsis
africana, South Africa. Adult form, Schistosoma hæmatobium.
Cercaria bilharziella, Leiper, 1915.
Cuticular keel on tail forks present. Pigment spots (eyes) in front
of ventral sucker present.
In Planorbis boissyi and P. mareoticus, and in Melania sp. Adult
form (?).
For characters of numerous other cercariæ which occur in fresh
water molluscs see “Die Susswasserfauna Deutschlands,” Max Lühe,
H. 17 (Gustav Fischer, Jena, 1909).
The characters of Cercaria japonica of S. japonicum in the mollusc
Katayama nosophora and of C. mansoni have still to be defined.
Schistosoma mansoni, Sambon, 1907.
The evidence appears to be strong that terminal-spined eggs are
not found in the West Indies, and that therefore the lateral-spined
eggs found in fæces there belong probably to S. mansoni. If this
be true, then the egg described by Stephens and Christophers
in man in India probably also belongs to another species of
Schistosome.
NEMATODA.
Ancylostomiasis.—Treatment: (1) Oleum chenopodii (U.S.P.),
dose ♏ x to ♏ xv on a lump of sugar, three doses at two-hourly
intervals, preceded and followed by a purge. It is cheap, not
unpleasant to take, and non-toxic. Effective also against Ascaris
lumbricoides.
(2) Milk of the higueron Ficus laurifolia. A spoonful in milk,
three times daily for three days followed by a purge. Described as a
harmless but very successful form of treatment.
Ground-itch.—Completely cured in a few days by a 3 per cent.
solution of salicylic acid in ethyl alcohol. Apply for five minutes
twice daily.
Ascaris lumbricoides can be kept alive for twelve days in
Kronecker’s solution; NaOH 0·069 grammes, normal saline 1,000 c.c.
Eggs are laid and develop in about a fortnight at ordinary room
temperature. At 70° C. they are readily killed.
Filariasis.—Dutcher and Whitmarsh have cultivated from the
blood and from the exudation fluids of cases of filariasis (elephantiasis,
lymphangitis, etc.), in about sixteen cases, a bacillus resembling
B. subtilis. Controls were negative. They propose the name Bacillus
lymphangiticus for this organism, and they believe it to be the cause
of the diseases grouped under the designation “filariasis.”
Oncocerca volvulus.—Unsheathed embryos (indistinguishable from
those taken from the uterus of this worm) have been found in lymphatic
glands and in the blood (if considerable pressure is used so as
to squeeze out lymph at the time of taking the finger blood, otherwise
none occurs in the specimens). The measurements in dried films
are: Nerve ring 23·7 per cent. of length; G1 cell 69·6 per cent.;
end of last tail cell 96·3 per cent; total length 274·3 µ.
Strongyloides stercoralis.—Pathology: They occur in the wall
of the intestine and may be associated with ulceration. They also
occur in lymphatics and blood-vessels.
[In the following pages the letters C. f. B., P. u. Inf. are used to indicate
the Centralblatt für Bakteriologie, Pathologie und Infektions-Krankheiten.]
(A) PROTOZOA (pp. 25 to 210, 617 to 637, and 733 to 742).
[This list applies to the earlier literature only. More recent references are
given as footnotes in the text.]
(a) General.
Bütschli, O. Protozoa in Bronn’s Klass. u. Ordn. d. Tierreichs, Leipz., 1880–1889.
Calkins, G. N. The Protozoa, Columbia Univ. Biol. Ser., vi, New York, 1901.
Delage, Y., and E. Hérouard. Traité de Zool. Concr., i, La cellule et les
protozoaires, Paris, 1896.
Farmer, J. B., J. J. Lister, E. A. Minchin and S. J. Hickson. Protozoa, in
A Treatise on Zoology, edited by E. Ray Lankester, London, 1903, i, 2.
Lang, A. Lehrb. d. vergl. Anatomie d. wirbellos. Tiere, 2. Aufl., 2. Lief,
Protozoa, Jena, 1901.
(b) Pathogenic Protozoa in General.
Doflein, F. Die Protozoen als Parasiten u. Krankheitserreger, Jena, 1901;
Lehrbuch der Protozoenkunde, 1912.
Doflein, F., and S. v. Prowazek. Die pathog. Protoz. mit Ausnahme d.
Hämospor., in Handb. d. path. Mikroorganism.; issued by W. Kolle and
A. Wassermann, 11. and 12. Lief, Jena, 1903.
Kästner, P. Die tierpathogenen Protozoen, Berlin, 1906.
Kisskalt, K., and M. Hartmann. Praktikum der Bakteriologie und Protozoologie,
Jena, 1907.
Lühe, M. Die im Blute schmarotzenden Protozoen und ihre nächsten Verwandten,
in Handb. d. Tropenkrankh., issued by C. Mense, Leipz., 1906, iii.
Pfeiffer, L. Die Protozoen als Krankheitserreger, 2. Aufl., Jena, 1891; Supplement, Jena, 1895.
Roos, E. Die im menschl. Darm vork. Protozoen u. ihre Bedeutg., Med.
Klinik, 1905, i, p. 1328.
Schneidemühl, G. Die Protozoen als Krankheitserreger der Menschen und der
Haustiere, Leipz., 1898.
Sievers, R. Zur Kenntn. d. Verbreitg. d. Darmparas. d. Mensch, Helsingfors,
1905, Festschrift f. Palmèn.
Ward, H. B. Protozoa, Wood’s Ref. Handbook of the Med Sci., 1904, viii.
Class I.—Sarcodina (pp. 29 to 50).
Order. Amœbina (p. 29).
Entamœba coli; Entamœba histolytica (pp. 32 to 41, 618 to 620, and 733).
Blanchard, R. Traité de Zool. médic., 1885, i, Paris, p. 15.—Maladies
paras., 1895, p. 658.
Bowmann, M. H. Dysentery in the Philippines, Journ. Trop. Med., 1901, iv,
p. 420.
Bunting, C. H. Hæmatogenous Amœbic Abscess of the Lung, Arch. f. Schiffs-
u. Tropenhyg., 1906, x, p. 73.
Calandruccio. Anim. par. dell’ uomo in Sicilia, Atti Accad. Gioen., iv, 1890,
ii, p. 95.
Casagrandi, O., and P. Barbagallo. Sull’ amœba coli, Boll. Accad. Gioen. sci.
nat., Catania, 1895.
— — Entamœba hominis s. Amœba coli Lösch, Annal. d’Igiene sperim., 1897,
vii, 1.
Celli, A., and R. Fiocca. Beitr. z. Amoebenforsch., ii, C. f. B. u. Par., 1894,
xvi, p. 329; Ric. int. alla biol. d. Amœbe, Bull. Accad. med. Roma, 1894–95,
xxi, p. 285; abstracted in C. f. B., P. u. Inf., 1897, i, xxi, p. 290.
Councilman, W. P., and H. A. Lafleur. Amœbic Dysentery, Johns Hopkins
Hosp. Rep., 1891, ii, p. 395.
Craig, C. F. Etiology and Pathology of Amœbic Infection of the Intestines
and Liver, Intern. Clin., Philad., 1905 (14), iv, p. 242.
Cunningham, D. Seventh Ann. Rep. San. Comm. of India, Calcutta, 1871.
— Unters. üb. d. Verh. mikrosk. Organ. z. Cholera in Indien, Zeitschr. f. Biol.,
1872, viii, p. 251; Quart. Journ. Micros. Sci., 1881 (2), xxi, p. 234.
Grassi, B. Dei protozoi par. e spec. di quelli che sono nell’ uomo, Gazz. med.
ital.-lomb., 1879 (8), i, p. 445; Int. ad alc. prot. endop., Atti soc. ital. sci.
nat., 1882, xxiv, p. 1; Morf. e sist. di alc. prot. par., Atti Acc. Lincei.
Rendic. (4), iv, 1, p. 5; Signif. patol. d. prot. par. dell’ uomo, ibid., p. 83.
Gross, A. Beobacht. üb. Amoebenenteritis, Arch. f. klin. Med., 1903, lxxvi,
p. 429.
Harris, H. F. Amœbic Dysentery, Amer. Journ. of Med. Sci., April, 1898.
— Experimentell bei Hunden erzeugte Dysent., Arch. f. path. Anat., 1901,
clxvi, p. 66; On the Alterations Produced in the Large Intestine of
Dogs by the Amœba coli, Philadelphia, 1901.
Hoppe-Seyler, G. Üb. Erkrankung des Wurmfortsatzes bei chron. Amoebenenteritis,
Münch. med. Wochenschr., 1904, No. 15.
Jaeger, H. Die in Ostpreuss. heim. Ruhr eine Amoebendysent., C. f. B., P. u.
Inf., 1902, i Abt. Orig., xxxi, p. 551.
— Erwiderg. a. d. Bemerk. Shigas, ibid., 1902, xxxii, p. 865.
Janowski, W. Zur Ätiol. d. Dys., C. f. B., P. u. Inf., 1897, i, xxi, pp. 88, 151,
194, 234.
Jürgens. Zur Kenntn. d. Darm-Amoeb. u. d. Amoeben-Enteritis, Veröff. a. d.
Geb. d. Milit.-Sanitätswes., Berl., 1902, Heft 20, p. 111.
Kartulis. Über Riesenamoeben (?) bei chron. Darmentdg. d. Ägypt, Virch.
Arch. f. Path., 1885, xcix, p. 145; Zur Ätiol. d. Dysent. in Ägypt, C. f. B.
u. Par., 1887, v, p. 745; Üb. trop. Leberabsc. u. ihr Verh. z. Dysent.,
Virch. Arch. f. Path., 1889, cxviii, p. 97; Einiges üb. d. Path. d.
Dysenterie-Amœb., C. f. B. u. Par., 1891, ix, p. 365; Article: Dysenterie
in Spec. Path. u. Ther. v. H. Nothnagel, Wien, 1896, v, 3.
— Gehirnabscesse nach dysent. Leberabsc., C. f. B., P. u. Inf., 1904, i Orig.,
xxxvii, p. 527.
Koch, R., and G. Gaffky. Bericht üb. d. Tätigkeit d. z. Erforschg. d. Cholera
ents. Kommiss., Arb. a. d. kais. Gesundheitsamt, 1887, iii.
Kovacs, F. Beob. u. Vers. üb. d. sog. Amoeben-Dys., Zeitschr. f. Heilkde.,
1892, xiii, p. 509.
Kruse, W., and Pasquale. Eine Exped. nach Ägypt, Deutsch. med. Wochenschr.,
1893, No. 15, p. 354; No. 16, p. 368.
— — Untersuch. üb. Dys. u. Leberabsc., Zeitschr. f. Hyg., 1894, xvi, p. 1.
Lambl. Aus d. Franz-Joseph-Kinderspit. in Prag, 1860, i, p. 362.
Lesage, A. Culture de l’amibe de la dysenterie des pays chauds, Ann. Inst.
Pasteur, 1905, xviii, p. 9; 1905, xix, p. 8.
Lewis. Sixth Ann. Rep. San. Comm. of India, Calcutta, 1870.
Lösch, F. Massenh. Entw. v. Amoeben im Dickd., Virch. Arch. f. Path., 1875,
lxv, p. 196.
Marchoux. Note sur la dysentérie d. pays chauds, C. R. Soc. Biol., Paris, 1899
(11), i, p. 870.
Musgrave, W. E., and M. T. Clegg. Amœbæ, their Cultivation and Etiological
Significance. “Treatment of Intestinal Amœbiasis,” in the Trop. Manila,
1904, Bur. of Govern. Lab., Biol. Lab., No. 18.
Normand. Note sur deux cas de colite parasit., Arch. méd. nav., 1879, xxxii,
p. 211.
Quincke and Roos. Über Amoebenenteritis, Berl. klin. Wochenschr., 1893,
xxx, No. 45, p. 1089.
Roos, E. Zur Kenntn. d. Amoebenenteritis, Arch. f. exper. Path. u. Pharm.,
1894, xxxiii, p. 389.
Ruge, A. Amoebenruhr., Handb. d. Tropenkrankh., issued by C. Mense, 1906,
iii, p. 1.
Schaudinn, Fr. Unters. üb. d. Fortpflanz. einig. Rhizopod, Arb. a. d. kais.
Gesundheitsamt, 1903, xix, 3, p. 547.
Schuberg, A. Die paras. Amoeb. d. menschl. Darms, C. f. B. u. Par., 1893,
xiii, pp. 598, 654, 701.
Shiga, K. Bemerk. zu Jaegers Die in Ostpreuss. einh. Ruhr eine Amoebendys.,
C. f. B., P. u. Inf., 1902, i Abt. Orig., xxxii, p. 352.
Strong, R. P., and W. E. Musgrave. Report on the Etiology of the Dysentery
of Manila, Rept. Surg.-Gen. of the Army to the Secretary of War, Washington,
1900, p. 251.
Verdun. Sur quelq. caract. spécif. de l’amibe de la dysenterie et des abscès
trop. du foie, C. R. Soc. Biol., 1904, lvi, p. 183.
Woolley, P. G., and W. E. Musgrave. The Pathology of Intestinal Amœbiasis,
Dept. of the Int. Bureau of Government Lab., No. 32, Manila, 1905,
Journ. Amer. Med. Assoc., Chicago, 1905, xlv, p. 1371.
Entamœba buccalis (pp. 43 and 620).
Leyden E. v., and W. Loewenthal. Entam. bucc. Prow, bei einem Fall von
Carcinom d. Mundbod, Charité-Ann., Berl., 1905, xxix; Berl. klin.
Wochenschr., 1905, xlii, No. 7, p. 187.
Prowazek, S. Entamœba buccalis n. sp., Arb. a. d. kais. Gesundheitsamt,
Berl., 1904, xxi, 1, p. 42.
Tietze, Al. Ein Protozoenbef. i. ein. erkrankt. Parotis., Mitt. Grenzgeb. Med.
u. Chirurg., 1905, xiv, p. 302.
Entamœba undulans (p. 43).
Castellani, A. Dysentery in Ceylon, Journ. of the Ceylon Branch of the Brit.
Med. Assoc., 1904.
— Observations on some Protozoa found in Human Fæces, C. f. B., P. u. Inf.,
1905, i Abt. Orig., xxxviii, p. 67.
Entamœba kartulisi (pp. 44 and 734).
Doflein, F. Die Protoz. als Paras. u. Krankheitserreg., Jena, 1901, p. 30.
Flexner. Amœbæ in an Abscess of the Jaw, Johns Hopkins Hosp. Bull., 1892,
No. xxv; abstracted in C. f. B., P. u. Inf., 1893, xiv, p. 288.
Kartulis. Üb. pathog. Protoz. b. Mensch., Zeitschr. f. Hyg., 1893, xiii, p. 9.
— Über Amoebenosteomyelitis d. Unterkief, C. f. B., P. u. Inf., 1903, i Abt.,
Ref. xxxiii, p. 471.
Amœba gingivalis, A. buccalis and A. dentalis (pp. 44 and 733).
Grassi, B. Gazz. med. ital.-lomb., 1879 (8), i, No. 45, p. 445.
Gros, G. Fragm. d’helm. et de phys. micros., Bull. soc. Imp. d. natur. de
Moscou, 1849, i, 2, P. 555.
Steinberg. In Zeitschr. f. neuere Med. (Russ.), issued by Walter in Kiew,
1862, Nos. 21–24.
Craigia (Paramœba) hominis (pp. 45 and 734).
Craig, Ch. F. A new Intestinal Parasite of Man: Paramœba hominis, Amer.
Journ. Med. Sci., 1906, N.S. cxxxii, Philad. and New York, p. 214.
Schaudinn, Fr. Über den Zeugungskreis von Paramœba eilhardi n. g. n. sp.,
Sitzgsber. Kgl. Pr. Akad. d. Wiss., Berlin, Phys.-math. Cl., 1896, No. 2.
Entamœba pulmonalis (p. 45).
Artault, St. Flore et faune d. cav. pulm., Arch. de paras., 1898, i, p. 275.
Blanc, L. Sur une Amibe viv. accid. dans le poumon du mouton, Ann. Soc.
Linn. Lyon, 1899 (2), xlv, p. 529.
Amœba urogenitalis (pp. 45, 46).
Baelz, E. Üb. einige neue Paras, d. Mensch., Berl. klin. Wochenschr., 1883,
p. 237.
Jeffries. Present. of a Specimen of Urine containing Amœbæ. Med. Rec.,
New York, 1904, xlvi, p. 356.
Jürgens. In Deutsche med. Wochenschr., 1892, p. 454.
Kartulis. Pathog. Prot. b. Mensch, Zeitschr. f. Hyg., 1893, xiii, p. 2, Anm. 2.
Posner, C. Üb. Amoeben im Harn, Berl. klin. Wochenschr., 1893, xxx,
No. 28, p. 674.
Wijnhoff, J. A. Over amoeburie, Nederl. Tijdschr. v. Geneeskde., 1895,
p. 107.
Amœba miurai (p. 46).
Ijima, J. On a New Rhizopod Parasite of Man, Ann. zool. japon., 1898, ii, 3,
p. 85; abstracted in C. f. B., P. u. Inf., 1899 (i), xxv, p. 885.
Miura, K. Amoebenfund i. d. Punktionsflüss. bei Tumoren d. Peritonealh.,
Mitt. med. Facult. d. kais. Jap. Univ., Tokyo, 1901, v, p. 1.
Chlamydophrys and Leydenia (pp. 47 to 50).
Cienkowski, L. Üb. einige Rhizop. u. verwandte Organismen, Arch. f. mikr.
Anat., 1876, xii, p. 39.
Lauenstein, C. Üb. ein. Fund von Leyd. gemmip., Deutsche med. Wochenschr.,
1897, xxiii, p. 733.
Leyden, E. v., and F. Schaudinn. Leyd. gemmip., ein neuer i. d. Ascites-Flüssigk.
d. leb. Mensch. gefund. amoebenähnl. Rhizop., Sitzgsb. kgl.
Preuss. Akad. d. Wiss., Berlin, 1896, xxxix, p. 951.
Leyden, E. v. Zur Ätiol. d. Carcin., Zeitschr. f. klin. Med., 1901, xliii, p. 4.
Schaudinn, Fr. Untersuch. üb. d. Fortpflanz. einig. Rhizopod., Arb. a. d.
kais. Gesundheitsamt, 1903, xix, 3, p. 560.
Schneider, A. Beitr. z. Naturgesch. d. Infus., Müllers Arch. f. Anat., Phys.
u. wiss. Med., Jahrg. 1854, p. 191.
Class II.—Mastigophora (pp. 50 to 128, 620 to 633, and 734 to 741).
Biland, J. Beitr. z. Frage d. Pathog. d. Flagellat., Deutsch. Arch. f. klin.
Med., 1905, lxxxvi, p. 274.
Blochmann, F. Mikrosk. Tierw. d. Süsswassers, 2. Aufl., 1895.
Kent, W. S. Manual of the Infusoria, London, 1880–81.
Prowazek, S. Flagellatenstudien, Arch. f. Protistenkde., 1903, ii, p. 195.
Senn, G. Flagellata in Engler und Prantl, Die natürlich. Pflanzenfam., Lief
202, 203, Leipzig, 1900.
Stein, F. v. Der Organismus der Infus., iii, Der Org. d. Flagellaten, Leipzig,
1878.
Trichomonas vaginalis (pp. 52, 53, and 734).
Baatz, P. Trich. vag. in der weibl. Harnblase, Monatsber. f. Urol., 1902, vii, 8.
Blochmann, F. Bemerk. über einige Flagell., Z. f. wiss. Zool., 1884, xl, p. 42.
Dock, G. Flagellate Protozoa in the freshly passed Urine of a Man, Med.
News, 1894, lxv, p. 640.
— Trichomonas as a Parasite of Man, Amer. Journ. Med. Sci., 1896, p. 1.
Donné, A. Rech. sur la nature du mucus, Paris, 1837.
Hausmann. Die Paras, der weibl. Geschlechtsorg., Berlin, 1870.
Kunstler, J. Trichom. vag. D., Journ. de Micrographie, 1884, viii, p. 317.
Laveran, A., and F. Mesnil. Sur la morph. et la syst. d. Flag. à membr.
ondul., C. R. Acad. Sci., Paris, 1904, cxxxiii, p. 131.
Marchand, F. Über das Vorkomm. v. Trichom. im Harne eines Mannes, C. f.
B. u. Par., 1894, xv, p. 709.
— Remarks [in the paper by K. Miura], ibid., 1894, xvi, p. 74.
Miura, K. Trichom. vag. im frischgelass. Urin eines Mannes, ibid., 1894, xvi,
p. 67.
Scanzoni, F. W. Beitr. z. Geburtskde., 2, Würzb., 1855, p. 131.
Scanzoni, F. W., and A. Koelliker. Quelq. rem. sur le Trichom. vag., C. R.
Acad. Sci., Paris, 1868, xl, p. 1076.
Trichomonas intestinalis (pp. 54 to 56, 623, and 734).
Boas. In Deutsch. med. Wochenschr., 1896, p. 214.
Cohnheim, P. Über Infus. im Magen und im Darmkanal des Menschen,
Deutsch. med. Wochenschr., 1903, xxix, Nos. 12–14.
— Zur klinisch-mikrosk. Diagnose der nicht-pylor. Magencarcinome, Festschr.
f. Jul. Lazarus, Berlin, 1899, p. 65.
Davaine, C. Sur les anim. infus. trouv. dans les selles d. malad. atteints du
choléra et d’autr. malad., C. R. Soc. Biol., 1854 (2), i, p. 129.
Emden, J. E. G. van. Flagell. en hunne beteeknis voor de pathol., Handel 8,
Nederl. Natuur- en Geneesk. Cong., Rotterdam, 11–14 April, 1901, p. 186.
Epstein, A. Beob. üb. Monocercomonas hominis und Amœba coli, Prag. med.
Wochenschr., 1893, Nos. 38–40.
Galli-Valerio, B. Note de parasitol., C. f. B., P. u. Inf., i Abt., 1900, xxvi,
p. 305.
Grassi, B. Int. ad alc. prot. entopar., Atti soc. ital. sci. nat. Milano, 1882,
xxiv, p. 135.
— Sur quelq. protoz. endop., Arch. ital. de biol., 1882, ii, p. 402; 1883, iii,
p. 23.
— Signific. pathol. dei protoz. paras. dell’ uomo, Atti R. accad. d. Lincei.
Rendic., 1888, iv, p. 83.
Hensen, H. In Deutsch. Arch. f. klin. Med., 1897, lix, p. 450.
Janowski, W. Flagellat. i. d. mensch. Faeces, Zeitschr. f. klin. Med., 1897,
xxxi, p. 442.
Kunstler, J. Observ. sur le Trichom. intest., Bull. scientif. de la France et
de la Belg., 1898, xxxi.
Leuckart, R. Die Paras. d. Mensch., Leipzig, 1879–86, 2. Aufl., i, 1, p. 131.
Marchand. Ein Fall von Infus. im Typhusstuhl, Virch. Arch. f. path. Anat.,
1875, lxiv, p. 293.
Prowazek, S. Notiz über Trichom. hominis, Arch. f. Protistenkde., 1902, i,
p. 166.
— Unters. über einige paras. Flagell., Arbeit. aus dem kais. Gesundheitsamt,
1904, xxi, 1, p. 1.
Rappin, G. Contrib. à l’étude des bact. de la bouche, Thèse Paris, 1881.
Roos, E. Über Infusorien-Diarrhöe, Deutsch. Arch. f. klin. Med., 1893, li,
p. 505.
Rosenfeld, A. Die Bedeutung der Flagellaten im Magen und Darm des
Menschen, Deutsch. med. Wochenschr., 1904, No. 47.
Ross, R. Cercomonads in Ulcers, Indian Med. Gaz., 1902, xxxvii, 4, p. 157.
Schaudinn, Fr. Unters. Fortpfl. einig. Rhizop., Arbeit. kais. Gesundheitsamt,
xix, 3, p. 547.
Schuberg, A. Die paras. Amoeb. d. menschl. Darms, C. f. B. u. Par., 1893,
xiii, p. 598.
Schürmayer, B. Über das Vorkommen der Flagell. im Darmkanal des
Menschen, C. f. B. u. Par., 1895 (1), xviii, p. 324.
Strube, G. Trichom. hominis im Mageninh. bei Carcin. cardiae, Berl. klin.
Wochenschr., 1898, p. 708.
Zabel, E. Flagellaten im Magen, Wien. klin. Wochenschr., 1904, xvii, No. 38.
Zuncker. Über das Vorkommen des Cercom. intest. im Digestionskanal des
Menschen, Deutsch. Zeitschr. f. prakt. Med., 1878, p. 1.
Lamblia intestinalis (pp. 57 to 60, 625, and 736).
Blanchard, R. Traité de Zool. méd., 1885, i, Paris, p. 91.
— Rem. sur le Megast. ent., Bull. Soc. Zool. France, 1888, xiii, p. 18.
Bütschli, O. Protozoa in Bronn’s Kl. u. Ord. d. Tierr., 1884, p. 843.
Cohnheim, P. Über Infus. im Magen- und Darmkanal des Menschen und ihre
klin. Bedeutung, Deutsch. med. Wochenschr., 1903, xxix, Nos. 12–14.
Frshezjesski. Über die Rolle des Megast. ent. bei chron. Darmkatarrh, Russk.
Arch. Patol., klin. Med. i. Bakt., 1867, ii.
Grassi, B. Des protoz. par. e spec. di quelli che sono nell’ uomo, Gazz. med.
ital.-lomb., Milano, 1879, No. 45.
— Di un nuova paras. dell’ uomo (Megastoma entericum), Gazz. degli ospit.,
1881, ii, Nos. 13–15.
— Intorno ad alc. prot. endop., Atti soc. ital. sci. nat., 1882, xxiv, and Arch.
ital. de biol., 1882, ii, p. 421.
Grassi, B., and W. Schewiakoff. Beitr. zur Kenntnis des Megastoma
entericum, Zeitschr. f. wiss. Zool., 1888, xlvi, p. 143.
Jaksch, v. Über das Vorkommen von tier. Paras. in den Faeces der Kinder,
Wiener klin. Wochenschr., 1888, No. 25, p. 511.
Kruse, W., and A. Pasquale. Unters. über Dysent. und Leberabsc., Zeitschr.
f. Hyg., 1894, xvi, p. 19.
Lambl. Unters. der Darmexkrete, Vierteljahrsschr. f. prakt. Heilkunde, 1859,
lxi, Prag, p. 51.
— Aus dem Franz-Josef-Kinderspit. in Prag, 1860, i, Prag, p. 360.
Metzner, R. Unters. an Megast. ent. aus dem Kaninchendarm, Zeitschr.
f. w. Zool., 1901, lxx, p. 299.
Moritz, E., and Hölzl. Über Häufigk. und Bedeutung des Vorkommens des
Megastoma entericum im Darmkanal des Menschen, Münch. med.
Wochenschr., 1892, xxxix, No. 47; Sitzungsb. ärztl. Ver. in München,
1893, ii, p. 89.
Müller, E. Cercom. intest. i. jejunum fran männ, Nord. med. ark., 1889, xxi,
No. 21; Förhdlg. biol. Föreng. Stockh., 1890, ii, p. 42; abstracted in
C. f. B. u. Par., 1890, viii, p. 592.
Perroncito, E. Über die Einkaps. des Megast. intest., C. f. B. u. Par., 1887,
ii, p. 738, and Giorn. R. Accad. med., Torino, 1887.
— Une maladie mort. du lapin prod. par la Lamblia intest. de l’homme et du
rat, Bull. Soc. Zool. France, 1902, xxvii, p. 151.
— La Lamblia intest. di R. Blanchard nell’ uomo e nei topi causa di moria deiconigli,
Ann. R. Accad. d. Agric., Torino, 1902, xlv.
Piccardi. Alc. prot. delle feci dell’ uomo, Giorn. R. Accad. med., Torino, 1895,
lviii, 1; Progr. méd., 1895, No. 23, p. 377.
Quincke, H. Über Protozoen-Enteritis, Berl. klin. Wochenschr., 1899, Nos.
46, 47.
Roos. Über Infusorien-Diarrhöe, Deutsch. Arch. f. klin. Med., 1893, li, p. 505.
Salomon, H. Über einen Fall von Infus.-Diarrhöe, Berl. klin. Wochenschr.,
1899, No. 46.
Schmidt, R. Infus. im Mageninhalt bei Ulcus ventriculi, Wiener klin. Wochen
schr., 1904, xvii, 48, p. 1304.
Schuberg, A. Abstract of the work of Moritz and Hölzl, C. f. B. u. Par., 1893,
xiv, p. 85.
Sievers. Balantidium coli, Megastoma entericum und Bothriocephalus latus
bei derselben Person, Zeitschr. f. klin. Med., 1896, xxxiii, p. 25.
Stiles, C. W. First American Case of Infection with Lamblia duodenalis,
Washington Med. Ann., 1902, i, No. 1, p. 64.
Ucke, A. Beobacht. über Flagell. in den Faeces des Menschen, C. f. B., P. u.
Inf., 1904, xxxiv, p. 772.
Zabel, E. Megastoma intestinale und andere Paras. in den Zotten eines Magenkrebses,
Arch. f. Verdauungskrankh., 1901, vii, 6, p. 509.
— Flagellaten im Magen, Wiener klin. Wochenschr., 1904, xvii, 38, p. 1007.
Cercomonas hominis (pp. 61, 62, and 736).
Artault, St. Flore et faune d. cav. plum., Arch. de Paras., 1898, i, p. 278.
Cahen. Über Protoz. im kindlichen Stuhl, Deutsch. med. Wochenschr., 1891,
No. 27, p. 853.
Davaine, C. Sur des animalcules inf. trouv. dans les selles d. mal. att. du
choléra et d’autres mal., C. R. Soc. Biol., Paris, 1854 (2), i, p. 129.
Ekecrantz. Bidr. t. känned. om de i mennisk. tarmkan. förek. infus., Nord.
med. ark., 1869, i, No. 20; Virchow-Hirschs Jahresber., 1869, i, p. 202.
Fenoglio. Entérocolite par Amœba coli, Arch. ital. de méd., 1890, xiv, p. 62.
Janowski, W. Flagellaten in den menschlichen Faeces, Zeitschr. f. klin.
Med., 1897, xxxi, p. 442.
Kannenberg. Über Infus. im Sputum, Virch. Arch. f. path. Anat., 1879, lxxv,
p. 471; Zeitschr. f. kl. Med., 1880, i, p. 228.
Kruse, W., and Pasquale. Unters. über Dysent., Zeitschr. f. Hyg., 1894, xvi,
p. 1.
Lambl. Cercomonas et Echinococcus in hepate hominis, Med. Wjestnik, 1875,
No. 33.
Leuckart, R. Die thier. Paras. des Menschen, 2. Aufl., i, p. 308.
Litten. Über Hydropneumothorax, Verh. Congr. f. inn. Med., 1886, p. 417.
Massiutin. Amöben als Paras. des Dickdarms, Wratsch, 1889, No. 25; C. f.
B. u. Par., 1889, vi, p. 451.
Perroncito, E. Über die Art der Verbreitung der Cercom. intest., C. f. B. u.
Par., 1888, iv, p. 220; Arch. ital. de biol., 1888, x, p. 257; Ann. R. Acc.
d’agric. Torino, 1889, xxxii.
Piccardi. Alc. prot. d. feci dell’ uomo, Giorn. R. Accad. d. med. Torino, 1895,
lviii, 1; Progr. méd., 1895, p. 377.
Quincke and Roos. Amöb.-Enteritis, Berl. klin. Wochenschr., 1893, p. 1089.
Roos, E. Über Infus.-Diarrhöe, Deutsch. Arch. f. klin. Med., 1893, li, p. 505.
Streng, W. Infusor. im Sputum bei Lungengangrän, Fortschr. d. Med., 1892,
x, p. 757.
Tham, P. V. S. Tvänne fall of Cercom., Upsala läkarefören. förhdlg., 1870,
v, p. 691; Virchow-Hirschs Jahresber., 1870, i, p. 314.
Zunker. Über die Verbr. v. Cerc. int. im Dig. des Menschen, Deutsch.
Zeitschr. f. prakt. Med., 1878, p. 1.
Monas pyophila (p. 62).
Blanchard, R. Maladies parasit., etc., Paris, 1895, p. 690.
Grimm, F. Über einen Leber- und Lungenabsc. mit Protoz., Arch. f. Chir.,
1894, xlviii, p. 478.
Prowazekia urinaria (pp. 63 to 65).
Barrois, Th. Quelq obs. au sujet du Bodo urinarius, Rev. biol. nord France,
1894–95, vii, p. 165.
Blanchard, R. Traité Zool. méd., i, Paris, 1885, p. 78.
— Malad. paras., etc., Paris, 1895, p. 691.
Braun, M. Die thier. Paras. d. Menschen, 2. Aufl., 1895, p. 108.
Cohnheim, P. Über Infus. im Magen, Deutsch. med. Wochenschr., 1903, xxix,
Nos. 12–14.
Hassall, A. H. On the Development and Significance of Vibrio lineola, Bodo
urinarius, in Alkaline and Albuminous Urine, Lancet, 1859, ii, p. 503.
Kunstler. Analys. micr. des urines d’un mal. atteint de pyélite chron. conséc.
à une opér. de taille, Bull. Soc. d’Anat. et de Phys. norm. et path.,
Bordeaux, 1883, iv, p. 215.
Salisbury. On the Parasitic Forms developed in Epithelial Cells of the
Urinary and Genital Organs, Amer. Journ. Med. Sci., 1868.
Trypanosomes (pp. 67 to 102, 620 to 623, and 737).
Bouffard, M., and G. Schneider. Etude exp. de la dourine du cheval, Bull.
Acad. Méd., 1900, 3e sér., xliv, p. 154; Rev. vétér., 1900, xxv, p. 589;
C. f. B., P. u. Inf., 1900, i Abt., xxvii, p. 882.
Broden, A. Rapport sur les travaux du labor. méd. de Léopoldville de 1900–1905,
Bruxelles, 1906, ii.
Broden, A. Trypanosomiasis et maladies du sommeil, Bruxelles, 1904, Publ.
Soc. d’étud. colon. de Belg.
— La Trypanosomiase chez l’Européen, Bruxelles, 1905, ibid.
Bruce, D. Preliminary Report on the Tsetse-fly Disease or Nagana in Zululand,
Durban, 1895, C. f. B., P. u. Inf., 1896, i Abt., xix, p. 955.
— Note on the Discovery of a new Trypanosome (T. theileri), Lancet, 1902, i,
p. 664.
Bruce, D., and D. Nabarro. Progress Report on Sleeping Sickness in Uganda,
Royal Society Report of the Sleeping Sickness Commission, No. 1,
London, 1903, p. 11; No. 4, p. 1.
Bruce, D., D. Nabarro and E. D. W. Greig. Further Report on Sleeping
Sickness in Uganda, Roy. Soc. Lond., Report of the Sleeping Sickness
Commission, No. 4, 1903.
Brumpt, E. Maladie du sommeil expér. chez le singe, C. R. Soc. Biol., Paris,
1903, lv, p. 1494.
— Les trypanos. chez les vertébrés, Arch. Méd. exp. et d’Anat. path., 1905,
xvii, p. 743.
— Maladie du sommeil, Arch. de Paras., 1905, ix, 2, p. 205.
Castellani, A. On the Discovery of a Species of Trypanosoma in the Cerebrospinal
Fluid of Cases of Sleeping Sickness, Proc. Roy. Soc., London,
1903, lxxi, p. 501.
— Die Ätiologie der Schlafkrankh. der Neger, C. f. B., P. u. Inf., 1903, i Abt.
Orig., xxxv, p. 62; Arch. f. Schiffs- u. Tropen-Hyg., 1902, vii, p. 382.
— Researches on the Etiology of Sleeping Sickness, Journ. Trop. Med., 1903.
Chalachnikow. Rech. sur les paras. du sang chez les animaux à sang froid et
à sang chaud, Charkow, 1888.
Chaussat, J. B. Rech. micr. appl. à la pathol. d. hématozoaires, Thèse de
Paris, 1850.
Christy, C. The Distribution of Sleeping Sickness in the Victoria Nyanza,
Brit. Med. Journ., 1903, ii, p. 648.
Danilewsky, B. Zur Parasitol. des Blutes, Biol. Ctrbl., 1885, v, p. 529.
— Matér. pour servir à la parasit. du sang, Arch. slav. de biol., 1886, i, pp. 85,
364; 1887, iii, pp. 33, 157, 257, 370.
— Rech. sur la parasit. comp. du sang; Zooparas. du sang des oiseaux;
Charkow, 1888 et 1889.
— Nouv. rech. sur les paras. du sang des oiseaux, Charkow, 1889.
Durme, P. van. Contrib. à l’étude des Trypanosomoses, repartition des tryp.
dans les organes, Arch. de Paras., 1906, x, 2, p. 160.
Dutton, J. E. Preliminary Note upon a Trypanosome occurring in the Blood
of Man, Thompson Yates Laboratory Report, Liverpool, 1902, iv, 2,
p. 453; Brit. Med. Journ., 1902, ii, No. 2,177, p. 881; Journ. Trop. Med.,
1902, v, p. 363.
Dutton, J. E., and J. L. Todd, in Brit. Med. Journ., 1903, i, ii; First Report
of the Trypanosome Expedition to Senegambia, Liverpool School of
Trop. Med., Memoir XI, 1903; Memoir XVI, 1905.
Elmassian, M. Mal de caderas, Annales Soc. rur. Argent., 1901, xxxvi, p. 195;
Berl. tierärztl. Wochenschr., 1901, p. 606; Rec. Méd. vét., 1901, 8e sér.,
viii, p. 786.
— Sur le mal de caderas ou flagellose parésiante des équidés sud-amér., Annal.
Inst. Pasteur, 1903, xvii, p. 241.
Evans, Griffith. Report on Surra Disease in the Dera Ismail Khan District
Military Depôt, 1880; Report by Punjab Govt. Military Dept., No. 439,
1880.
— On a Horse Disease in India known as Surra, probably due to a Hæmatozoon,
Vet. Journ., 1881, xiii, pp. 1, 83, 180, 326; 1882, xiv, pp. 97, 181.
Forde, R. M. Some Clinical Notes on a European Patient, in whose Blood a
Trypanosoma was observed, Journ. Trop. Med., 1902, v, p. 261.
Gluge. Über ein eig. Entozoon im Blute des Frosches, Müllers Arch. f. Anat.,
Phys. u. wiss. Med., 1842, p. 148.
Grassi, B. Sur quelq. protistes endopar., Arch. ital. de biol., 1882–83.
Gros. Observ. et induct, microsc. sur quelq. paras., Bull. Soc. imp. d. Natural,
Moscow, 1845, xviii, 1, p. 380.
Gruby, D. Rech. et obs. s. une nouv. esp. d’hématozoaires, C. R. Acad. Sci.,
Paris, 1843, xvii, 2, p. 1134; Ann. sci. nat. (Zool.), 1844, 3e sér., i, p. 104.
Keysselitz, G. Über flagell. Blutparas. b. Süsswasserfisch, Sitzungsber. Ges.
nat. Frde., Berl., 1904, p. 285.
— Über Trypanophis grobbeni, Arch. f. Protistenkde., 1904, iii, 3, p. 367.
Koch, R. Über die Trypanosomenkrankh., Dtsche. med. Wochenschr., 1904,
xxx, p. 1706.
— Über die Unterscheidg. d. Trypanosomen, Sitzungsber. kgl. preuss. Akad.
d. Wiss., 1905, No. 46.
Krueger. Bericht über die Schlafkrankh. in Togo, Arch. f. Schiffs- u. Tropen-Hyg.,
1904, viii, p. 479.
Krzystalowitz, Fr., and M. Siedlecki. Contribut. à l’étude de la struct. et du
cycle évol. de Spirochaeta pallida Schaud., Bull. Acad. sci. de Cracow,
Cl. d. sci. math. et natur., 1905, p. 713.
Laveran, A. Des trypanosomes parasites du sang, Arch. Méd. exp. et d’Anat.
path., 1e sér., 1892, i, p. 257.
— Sur un nouv. tryp. des bovidés (Tr. theileri), C. R. Acad. Sci., Paris, 1912,
cxxxiv, p. 512.
Laveran and R. Blanchard. Les hématozoaires de l’homme et des animaux,
Paris, 1895, Part I, p. 129.
Laveran, A., and F. Mesnil. Sur les Flag. à membr. ondul. des poiss., C. R.
Acad. Sci., Paris, 1901, cxxxiii, p. 670.
— — Des trypan. des poissons, Arch. f. Protistenkde., 1902, i, p. 475.
— — Rech. morph. et expér. sur le Tryp. des rats, Annal. Inst. Pasteur, 1901,
xv, p. 673.
— — Rech. morph. et exp. sur le Tryp. du Nagana, ibid., 1902, xvi, p. 1.
— — Trypanosomes et Trypanosomiasis, Paris, 1904.
Lebailly, Ch. Rech. sur les hématozoaires paras. des téléostéens marins, Arch.
de Paras., 1906, x, 3, p. 348.
Léger, L. Tryp. varium n. sp., paras. du sang, de Cobitis barbatula L.,
C. R. Soc. Biol., Paris, 1904, lvii, p. 345.
Lewis, F. R. Flagellate Organisms in the Blood of Healthy Rats, Fourteenth
Ann. Rept. San. Comm., Govt. India (1878), Appendix; Quart. Journ.
Micros. Sci., 1879, N.S. xix, p. 109.
Lühe, M. Die im Blute schmarotzenden Protozoen und ihre nächsten Verwandten,
Handb. d. Tropenkrankh., issued by Mense, 1906, iii.
Luhs, F. Trypanosoma theileri in Transkaukasien, Arch. de Paras., 1906, x, 2,
p. 170.
Manson, P. Trypanosoma occurring in the Blood of Man, Lancet, 1902, clxiii,
2, p. 1391.
Manson, P., and C. W.Daniels. Remarks on Case of Trypanosomiasis, Brit.
Med. Journ., 1903, i, p. 1249.
Martini. Trypanosomenkrankheiten (Schlafkrankheit) und Kala-azar, Jena,
1907.
Mayer, A. F. J. C. Specileg. obs. anat. de org. electr. in Rajis anelectr. et
de haematozois, Bonn, 1843.
Mitrophanow, P. Beitr. z. Kenntn. d. Haematoz., Biol. Ctrbl., 1883–84, iii,
p. 35.
Musgrave, W. E., and M. T. Clegg. Trypanosoma and Trypanosomiasis with
special reference to Surra in the Philippine Islands, Dept. of the Int.
Bur. of Govt. Lab., Biol. lab., No. 5, Manila, 1903.
Nepveu, G. Sur un trypanosome dans le sang de l’homme, C. R. Soc. Biol.,
Paris, 1898, 10e sér., v, p. 1172.
Nocht, B., and M. Meyer. Trypanosomen als Krankheitserreger, Ergänzungsbd.
zu Kolle u. Wassermann, Handb. d. pathog. Mikroorg., Jena,
1906.
Novy, F. G., and W. J. McNeal. On the Cultivation of Trypanosoma lewisi.
Contribution to med. res. dedicated to V. Cl. Vaughan, Ann Arbor,
Mich., 1903, p. 549.
— — The Cultivation of Tr. brucei, Journ. Amer. Med. Assoc., 1903, xli,
p. 1266, and 1904, xlii; Journ. of Inf. Dis., 1904, i, p. 1.
Poche, Fr. Über zwei neue in Siphonoph. vork. Flagell., Arb. a. d. zool. Inst.,
Wien, 1903, xiv, 3, p. 307.
Rabinowitsch, L., and W. Kempner. Die Trypanos. in der Menschen- u. Tierpathol.,
C. f. B., P. u. Inf., 1903, i Abt., xxxiv.
— — Beitr. z. Kenntn. d. Blutpar., speziell der Rattentrypanos., Zschr. f. Hyg.,
1899, xxx, p. 251.
Reports of the Sleeping Sickness Commission, Roy. Soc. Lond., 1903–1905,
Nos. 1–6.
Rouget, J. Contrib. à l’étude du Tryp. des mammif., Annal. Inst. Pasteur,
1896, x, p. 716.
Salmon, D. E., and Ch. W. Stiles. Emergency Report on Surra, with a
Bibliography of Surra and Allied Trypanosomatic Diseases, by A.
Hassall, U.S. Dept, of Agric., Bureau of Animal Industry, Bull. No. 42,
Wash., 1902.
Sambon, L. W. Sleeping Sickness in the Light of Recent Knowledge, Journ.
Trop. Med., London, 1903.
Schaudinn, Fr. Generations- und Wirtswechsel bei Trypanosoma und Spirochaete,
Arb. a. d. kais. Gesundheitsamt, 1904, xx, p. 387.
Schilling, A. Über die Tsetsekrankh. oder Nagana, Arb. Reichsges.-Amt,
1904, xxi, p. 476.
Steel, J. H. An Investigation into an Obscure and Fatal Disease among
Transport Mules in British Burma, 1885.
— On Relapsing Fever of Equines, Vet. Journ., 1886, xxii, pp. 166, 248.
— Le surra, malad. contag. des anim. dom. dans l’Inde, Rec. Méd. vétér.,
1886, 7 sér., v, p. 298.
Thiroux. Rech. morph. et exp. sur Trypanosoma duttoni Thir., Annal. Inst.
Pasteur, 1905, xix, p. 504.
Valentin. Über ein Entozoon im Blut von Salmo fario, Müllers Arch. f. Anat.,
Phys. u. wiss. Med., 1841, p. 435.
Wasiliewski, v., and G. Senn. Beitr. z. Kenntn. d. Flagell. d. Rattenbl.,
Zeitschr. f. Hyg., 1900, xxxiii, p. 444.
Wedl, C. Beitr. z. Lehre v. d. Haematozoen, Denkschr. k. k. Akad. d. Wiss.,
Wien, 1850.
Wittich. Spirillen im Blut von Hamstern, Centralbl. f. d. med. Wiss., 1881,
xix, p. 65.
Ziemann, H. Eine Methode der Doppelfärbung bei Flagell., C. f. B., P. u.
Inf., 1898, i Abt., xxiv, p. 945.
Leishmania (pp. 104 to 112 and 626 to 629).
Blanchard, R. Note critique sur les corpuscles de Leishman, Rev. de Méd. et
d’Hyg. trop., 1904, i, p. 37.
Cunningham, D. A Peculiar Parasitic Organism in the Delhi Boil, Scient.
Mem. of Med. Off. of the Army of India, Calcutta, 1885, i.
Donovan, C. Human Piroplasmosis, Lancet, 1904, ii, p. 744; Brit. Med.
Journ., 1904, ii, p. 651.
Firth, R. H. Note on the Appearance of Certain Sporozoon Bodies in the
Protoplasma of the Oriental Sore, Brit. Med. Journ., 1891.
Laveran, A., and F. Mesnil. Sur un protozoaire nouveau (Piroplasma donovani),
paras. d’une fièvre de l’Inde, C. R. Acad. Sci., Paris, 1903, cxxxvii,
p. 957; 1904, cxxxviii, p. 187.
Leishman, W. B. On the Possibility of the Occurrence of Trypanosomiasis in
India, Brit. Med. Journ., 1903, i, p. 1253.
— The Nature of the Leishman-Donovan Body, ibid., 1904, ii, pp. 29, 642.
Leishman, W. B., and J. C. B. Statham. The Development of the Leishman
Body in Cultivation, Journ. Roy. Army Med. Corps, March, 1905.
Manson, P., and G. C. Low. The Leishman-Donovan Body and Tropical
Splenomegaly, Brit. Med. Journ., 1904, i, p. 183.
Marchand, F., and J. C. G. Ledingham. Über die Infektion mit Leishmanschen
Körperchen (Kala-Azar?) und ihr Verhältnis zur Trypanosomenkrankheit,
Zeitschr. f. Hyg. u. Infkrankh., 1904, xlvii, i, p. 1.
Marzinowski, J. E., and S. L. Bogrow. Zur Ätiologie der Orientbeule, Arch.
f. path. Anat., 1904, clxxviii, 1, p. 112.
Riehl, G. Zur Anatomie und Ätiologie der Orientbeule, Vierteljahrsschr. f.
Dermat. u. Syph., 1886, p. 805.
Rogers, L. On the Development of Flagellated Organisms from the Spleen;
Protozoic Parasites of Cachexial Fevers and Kala-Azar, Quart. Journ.
Micros. Sci., 1904, xlviii, p. 367.
— Further Work on the Development of the Hepatomonas [Herpetomonas] of
Kala-azar and Cachexial Fever from Leishman-Donovan Bodies, Proc.
Roy. Soc., London, 1906, lxxvii, B, p. 284.
Wright, J. H. Protozoa in a Case of Tropical Ulcer, Journ. Med. Research,
1903, x, 3, p. 472.
Spirochætes (pp. 114 to 128, 629 to 633, and 739 to 741).
Schaudinn, Fr., and E. Hoffmann. Vorl. Ber. über das Vork. von Spirochaeten
in syph. Krankheitsprod. und bei Papillomen, Arb. a. d. kais.
Gesundheitsamt, 1905, xxii, p. 527.
Class III.—Sporozoa (pp. 128 to 197).
Balbiani, G. Leçons sur les Sporozoaires, Paris, 1884.
Hagenmüller, P. Bibliographie générale et spéciale des travaux concernant
les Sporozoaires parus antérieurement au 1er janvier, 1899, Ann. Mus.
d. ’Hist. nat., Marseille, 2 sér., 1899, i.
Labbé, A. Sporozoa, Berlin, 1899, Das Tierreich, 5. Lief.
Lühe, M. Ergebnisse der neueren Sporozoenforschung, Jena, 1900. Reprinted
from C. f. B., P. u. Inf., 1900, xxvii and xxviii.
Wasiliewski, Th. v. Sporozoenkunde, Jena, 1896.
Telosporidia.
Order. Gregarinida (pp. 129 to 135).
Beneden, E. van. Sur une nouvelle espèce de Grégarine désignée sous le nom
de Gregarina gigantea, Bull. Acad, roy., Belg., 2e sér., 1869, xxviii,
p. 444.
— Recherches sur l’évolution des Grégarines, ibid., 1871, xxxi, p. 325.
Berndt, A. Beitrag zur Kenntnis der im Darme der Larve von Tenebrio molitor
lebenden Gregarinen, Arch. f. Protistenkde., 1902, i, p. 375.
Brasil, L. Recherches sur la reproduction des Grégarines monocystidies, Arch.
Zool. exp., 4e sér., 1905, iii, p. 17; 1905, iv, p. 69.
Bütschli, O. Kleine Beiträge zur Kenntnis der Gregarinen, Zeitschr. f. wiss.
Zool., 1881, xxxv, p. 384.
— Bemerkungen über einen dem Glycogen verwandten Körper in den
Gregarinen, Zeitschr. f. Biol., 1885, xxi, p. 603.
Caullery, M., and Mesnil, F. Sur une Grégarine coelomique présentant dans
son cycle évolutif une phase de multiplication asporulée, C. R. Soc. Biol.,
Paris, 10e sér., 1908, v, p. 65; C. R. Acad. Sci., Paris, 1898, cxxvi, p. 262.
— — Le parasitisme intracellulaire et la multiplication asexué des grégarines,
C. R. Soc. Biol., Paris, 1901, liii, p. 84.
Cavolini, F. Memoria sulla generazione dei pesci e dei granchi, Napoli, 1787,
p. 169. Translation into the German by E. A. W. v. Zimmermann,
Berlin, 1792, p. 169.
Crawley, H. The Progressive Movement of Gregarines, Proc. Acad. Nat. Sci.,
Philadelphia, 1902, p. 4.
Cuénot, L. Recherches sur l’évolution et la conjugaison des grégarines, Arch.
de Biol., 1901, xvii, p. 581.
Drzewecki, W. Über vegetative Vorgänge im Kern und Plasma der Gregarinen
des Regenwurmhodens, Arch. f. Protistenkde., 1904, iii, p. 107.
Dufour, L. Note sur la Grégarine, nouveau genre de ver qui vit en troupeau
dans les intestins de divers insects, Ann. Sci. nat., 1e sér., 1828, xiii,
p. 366.
Dujardin, F. Recherches sur les organismes inférieures, II, Sur les Infusoires
appelées Protées, Ann. Sci. nat., 2e sér., Zool., 1835, iv, p. 352.
Giard, A. Contributions à l’histoire naturelle des Synascidies, IV, Sur une
Grégarine parasite d’un Amaraecium, Arch. Zool. expér., 1873, ii, p. 495.
Greeff, R. Über die pelagische Fauna an den Küsten der Guinea-Inseln,
Zeitschr. f. wiss. Zool., 1885, xlii, p. 452.
Henle, J. Über die Gattung Gregarina, Müllers Arch. f. Anat. u. Phys., 1845,
p. 369.
Kölliker, A. Beiträge zur Kenntnis niederer Tiere, I, Über die Gattung
Gregarina, Zeitschr. f. wiss. Zool., 1848, i, p. 1.
Léger, L. Recherches sur les Grégarines, Tabl. zool., 1892, iii, p. 1.
— Nouvelles recherches sur les Polycystidées parasites des Arthropodes terrestres,
Ann. Fac. Sci., Marseille, 1896, vi, p. 3.
— Sur un nouveau Sporozoaire des larves de Diptères, C. R. Acad. Sci., Paris,
1900, cxxxi, p. 722.
— La réproduction sexuée chez les Stylorhynchus, Arch. f. Protistenkde., 1904,
iii, p. 303.
Léger, L. Etude sur Taeniocystis mira, Grégarine métamérique, Arch. f.
Protistenkde., 1906, vii, p. 307.
Léger, L., and O. Duboscq. Les Grégarines et l’épithélium intestinal chez les
Trachéates, Arch. de Parasitologie, 1902, vi, p. 377.
— — Nouvelles recherches sur les Grégarines et l’épithélium intestinal chez les
Trachéates, Arch. f. Protistenkde., 1904, iv, p. 335.
— — La reproduction sexuée chez Pterocephalus, Arch. Zool. exper., 4e sér.,
1903, i; Notes et revue p. cxli.
Lieberkühn, N. Evolution des Grégarines, Mém. Cour. et Mém. d. Sav.
étrang., Acad. Belg., 1855, xxvi.
Lühe, M. Bau und Entwicklung der Gregarinen, I, Arch. f. Protistenkde.,
1904, iv, p. 88.
Maupas, E. Sur les granules amylacés du cytosome des Grégarines, C. R.
Acad. Sci., Paris, 1886, cii, p. 120.
Nusbaum, J. Über die geschlechtliche heterogame Fortpflanzung einer im
Darmkanale von Henlea leptodora schmarotzenden Gregarine (Schaudinella
henleae), Zeitschr. f. wiss. Zool., 1903, lxxv, p. 280.
Paehler, Fr. Über die Morphologie, Fortpflanzung und Entwickelung von
Gregarina ovata, Arch. f. Protistenkde., 1904, iv, p. 64.
Prowazek, S. Zur Entwickelung der Gregarinen, Arch. f. Protistenkunde,
1902, i, p. 297.
Redi, Fr. De animalculis vivis, quae in corporibus animalium viventium
reperiuntur, Amstelod., 1708, p. 270. [An earlier account was published
in 1684.]
Schewiakoff, W. Über die Ursache der fortschreitenden Bewegung der
Gregarinen, Zeitschr. f. wiss. Zool., 1894, lviii, p. 340.
Schneider, Ai. Sur quelques points de l’histoire du genre Gregarina, Arch.
Zool. exper., 1873, ii, p. 515.
— Contributions à l’histoire des Grégarines des Invertébrés de Paris et de
Roscoff, ibid., 1875, iv, p. 493; also Thèse de Paris, 1876.
— Second contribution à l’étude des Grégarines, ibid., 1882, x, p. 432.
— Sur le développement du Stylorhynchus longicollis, ibid., 2e sér., 1884, ii,
p. 1.
— Etudes sur le développement des Grégarines, Tabl. zool., 1885, i, pp. 10, 81.
— Grégarines nouvelles ou peu connues, ibid., pp. 25, 90; 1887, ii, p. 67.
Schnitzler, H. Über die Fortpflanzung von Clepsidrina ovata, Arch. f. Protistenkunde,
1905, vi, p. 309.
Siebold, Th. v. Beiträge zur Naturgeschichte der wirbellosen Tiere, Über die
zur Gattung Gregarina gehörenden Helminthen, Neueste Schrift d. naturforsch.
Ges., Danzig, 1839, iii, p. 56.
Siedlecki, M. Über die geschlechtliche Vermehrung der Monocystis ascidiae,
R. Lank., Anz. Akad. d. Wiss., Cracow, 1899, p. 515.
— Contributions à l’étude des changements cellulaires provoqués par les
Grégarines, Arch. d’anat. micros., 1901, iv, p. 87.
Stein, Fr. Über die Natur der Gregarinen, Müllers Arch. f. Anat. u. Phys.,
1848, p. 182.
— Neue Beiträge zur Kenntnis der Entwickelungsgeschichte und des feineren
Baues der Infusionstiere: I, Die Entwickelungsgeschichte der Vorticella
microstoma Ehrb., nebst vergleichenden Bemerkungen über die Entwickelungsweise
der Gregarinen, Zeitschr. f. wiss. Zool., 1852, iii, p. 474.
Wolters, M. Die Conjugation u. Sporenbildung bei Gregarinen, Arch. f. mikr.
Anat., 1891, xxxvii, p. 99.
Schizogregarinea (p. 135).
Léger, L. La reproduction sexuée chez les Ophryocystis, C. R. Soc. Biol.,
Paris, 1900, lii, p. 927.
— Les Schizogrégarines des Trachéates: I, Le genre Ophryocystis, Arch. f.
Protistenkunde, 1907, viii, p. 160.
Léger, L., and Hagenmüller. Morphologie et évolution de l’Ophryocystis
schneideri, Arch. Zool. exp., 3e sér., 1900, viii; Notes et revue p. xl.
Schneider, Ai. Ophryocystis bütschlii, sporozoaire d’un nouveau type, Arch.
Zool. expér., 2e sér., 1884, ii, p. 111.
— Ophryocystis francisci, Tabl. zool., 1885, i, p. 1.
Order. Coccidiidea (pp. 135 to 150, 741 and 742).
Balbiani, G. Leçons sur les Sporozoaires, Paris, 1884, p. 104.
Blanchard, R. Les coccidies et leur rôle path., Causeries scientif. Soc. Zool.
France, No. 5, Paris, 1900.
Eimer, Th. Über die ei- und kugelförmigen Psorospermien der Wirbelthiere,
Würzburg, 1870.
Hake, A. A Treatise on Varicose Capillaries as constituting the Structure of
Carcinoma of the Hepatic Ducts, with an account of a New Form of the
Pus Globule, London, 1839.
Kauffmann, W. Anal. ad tubercul. et entoz. cognitionem, Diss. inaug., Berol,
1847.
Kloss, H. Über die Parasiten der Niere von Helix, Abh. Senckenb. nat. Ges.
Frankf. a. M., 1855, i, p. 189.
Labbé, A. Recherches zoologiques, cytol. et biol. sur les Coccidies, Arch. Zool.
exp. 1896 (3), iv, p. 516.
Laveran, A., and F. Mesnil. Sur la coccidie trouvée dans le rein de la Rana
esculenta et sur l’infection générale qu’elle produit, C. R. Acad. Sci.,
Paris, 1902, cxxxv, p. 82.
Léger, L. Le cycle evol. d. Coccidies chez les Arthrop., C. R. Soc. Biol.,
Paris, 1897 (10), iv, p. 382.
— Coccidies nouv. du tube digest. d. Myriap., C. R. Acad. Sci., Paris, 1897,
cxxiv, p. 901.
Leuckart, R. Die Parasiten des Menschen, 2. Aufl., 1879, i, p. 248.
Lieberkühn, N. Über die Psorospermien, Müllers Arch. f. Anat. u. Phys.,
1854, p. 1.
— Evolution des grégarines, Mem. cour. et Mém. d. say. étrang., Acad. roy.
de Belg., 1855, xxvi.
Milian, G. Les sporozooses humaines, Thèse, Paris, 1899.
Moussu, G., and G. Marotel. La coccidiose du mouton et son paras., Arch. de
Paras., 1902, vi, p. 82.
Nasse, H. Üb. d. eiförmigen Zellen der tuberkelähnlichen Ablagerungen in
den Gallengängen der Kaninchen, Müllers Arch. f. Anat. u. Phys., 1843,
p. 209.
Pfeiffer, L. Beiträge z. Kenntnis d. pathogenen Gregarinen: II, Über
Gregarinose, ansteckendes Epitheliom und Flagellaten-Diphtherie der
Vögel, Zeitschr. f. Hyg. u. Inf., 1889, v, p. 363.
— Die Protozoen als Krankheitserreger: 1. Aufl., Jena, 1890; 2. Aufl., 1892.
Pfeiffer, R. Beiträge zur Protozoenforschung: I, Die Coccidienkrankheit der
Kaninchen, Berlin, 1892.
Reincke. Nonnulla quaedam de psorosp. cuniculi, Diss. inaug., Kiliae, 1866.
Remak, H. Diagnost. und pathog. Untersuchungen, Berlin, 1845.
Rivolta, G. Psorospermi i psorospermosi negli anim. dom., Med. veter., 1869
(3), iv.
Schaudinn, F. Unters. über d. Generationswechsel d. Coccidien, Zool. Jahrb.
Anat., 1900, Abt. xiii, p. 197.
— Studien über krankheitserr. Protozoen: I, Cyclospora caryolytica, der
Erreger der perniciösen Enteritis des Maulwurfes, Arb. kais. Gesundheitsamt,
1902, xviii, p. 378.
Schaudinn, F., and M. Siedlecki. Beitr. z. Kenntnis d. Coccidien, Verh. d. D.
zool. Ges., 1897, vii, p. 192.
Schneider, Ai., and M. L. Pfeiffer. Le cycle évolutif des Coccidies, Tabl.
zool., ii, p. 105.
Schuberg, A. Die Coccidien aus dem Darm der Maus, Verh. nat.-med. Ver.
Heidelberg, N. F. V., 1895, p. 369.
Simond, P. L. L’évolution d. sporoz. du genre Coccidium, Ann. Inst. Pasteur,
1897, xi, p. 545.
— Note sur une Coccidie nouv. (Cocc. kermorganti) paras. de Gavialis gangeticus,
C. R. Soc. Biol., Paris, 1901, liii, p. 483.
Smith, Th., and H. P. Johnson. On a Coccidium (Klossiella muris) parasitic
in the Renal Epithelium of the Mouse, Journ. Exp. Med., 1902, vi, p. 303.
Stieda, L. Über die Psorospermien der Kaninchenleber, Virchows Arch. f.
pathol. Anat., 1865, xxxii, p. 132.
Waldenburg, L. De struct. et origine cystidum verminos, Diss. inaug., Berol,
1860; Virchows Arch. f. path. Anat., 1862, xxiv, p. 149.
— Zur Entwickelung der Psorospermien, ibid., 1867, xl, p. 435.
Eimeria stiedae (pp. 145 to 148).
Felsenthal and Stamm. Veränder. i. Leber und Darm bei d. Coccidienkrankh.
d. Kaninchen, Virchows Arch. f. path. Anat., 1893, cxxxii, p. 36.
Lindemann, Kr. Weiteres über Gregarinen, Bull. soc. Imp. Nat., Moscow,
1865, xxxviii, 2, p. 385.
Lühe, M. Über Geltung und Bedeutung. d. Gattungsnamen Eimeria und
Coccidium, C. f. B., P. u. Inf., 1902, i Abt. Orig., xxxi, p. 771.
Metzner. Untersuchungen an Coccidium cuniculi I, Arch. f. Protistenkde.,
1903, ii, p. 13.
Pianese, G. Le fasi di sviluppo del coccidio oviforme e le lesioni istologiche
che induce, Arch. d. Paras., 1899, ii, p. 387.
Podwyssotzki, W. Zur Entwickel. d. Coccidium oviforme als Zellschmarotzer,
Bibl. med., 1895, Abt. D., ii, p. 4, Cassel.
Rieck, M. Sporozoen als Krankheitserreger bei Haustieren, Deutsche Zeitschr.
f. Tiermedizin u. vergl. Path., 1889, xiv, p. 52.
Stiles, Ch. W. Eimeria stiedae. Correct Name of the Hepatic Coccidia of
Rabbits, Eleven Miscellaneous Papers on Animal Parasites, U.S. Dept.
Agric., Bur. An. Ind., Bull. No. 35, Washington, 1902.
Wasiliewski, Th. v. Studien u. Microphot. z. Kenntn. d. pathog. Protozoen:
I, Bau Entw. u. pathog. Bedeutung d. Coccidien, Leipzig, 1904.
Isospora bigemina (pp. 149, 150).
Finck. Sur la physiol. de l’épith. intest., Thèse, Strasb., 1854, p. 17.
Grassi, B. Intorno ad alc. prot. endopar., Atti Soc. Ital. Sci. Nat. Milano,
1882, xxiv.
Grunow. Ein Fall von Protozoen- (Coccidien?) Erkrankung des Darmes,
Arch. f. exp. Path. u. Pharm., 1901, xlv, p. 262.
Railliet, A., and A. Lucet. Notes s. quelq. esp d. Cocc. encore peu conn.,
Bull. Soc. Zool. France, 1891, xvi, p. 249.
Stiles, Ch. W. Notes on Parasitology, No. 11, Journ. Comp. Med. and Vet.
Arch., 1892, xiii, p. 517; Bull. Soc. Zool. France, 1891, xvi, p. 163.
Virchow, R. Helminth. Notiz. 3, Virchows Arch. f. path. Anat., 1860, xviii,
pp. 342 and 527.
Wasiliewski, Th. v. Studien u. Mikrophot. zur Kenntnis d. path. Prot., I,
Leipzig, 1904, pp. 88 ff.
Doubtful Species (p. 150).
Blanchard, R. Les cocc. et leur rôle path., Caus. scient. Soc. Zool. France,
1900, No. 5.
Borini, A. Assoc. paras. ed il nuovo prot. di Perroncito, Giorn. R. Accad.
Med., Torino, 1899, No. 7.
Kunstler, J., and A. Pitres. Sur une psorospermie trouv. dans une humeur
pleurét., Journ. Microgr., 1884, viii, p. 469.
Moniez, R. Traité de parasit., Paris, 1896, p. 52.
Perroncito, E. Di un nuovo protoz. dell’ uomo e di talune specie, Giorn. R.
Accad. Med., Torino, 1899, No. 1; Cinquantenn. de la Soc. Biol., Paris,
1899, p. 184.
— Il coccidio jalino ed il microspor. poliedrico nell’ uomo, ibid., ann. 65, 1902,
p. 378.
Severi, A. Gregarinosi polmonale in infante natomorto, Rif. med., 1892, ii,
p. 54; Boll. Accad. med., Genova, 1892, vii, No. 2.
Order. Hæmosporidia (pp. 151 to 155, and 742).
Balfour, A. A Hæmogregarine of Mammals, Journ. Trop. Med., 1905, viii,
p. 241.
Börner, C. Unters. über Haemosporidien, I, Zeitschr. f. wiss. Zool., 1901,
lxix, p. 398.
Bütschli, O. Einige Bemerk. über d. rot. Blutk. d. Frosches, Abh. Senckenb.
nat., Ges., Frankf. a. M., 1876, p. 49.
Castellani, A., and A. Willey. Hæmatozoa of Vertebrates in Ceylon, Spolia
zeylanica, Colombo, 1904, ii, p. 2.
Celli, A., and F. Sanfelice. Paras, d. rot. Blutk. d. Mensch. u. d. Thiere,
Fortschr. d. Med., 1891, Nos. 11–15; Ann. ist. d’igien. esp., Roma, N.S.I.,
1891.
Chaussat. Des hématozoaires, Thèse, Paris, 1850.
Danilewsky, B. Die Haematozoen der Kaltblüter, Arch. f. mikr. Anat., 1885,
xxiv, p. 588.
— Matér. pour servir à la paras. du sang, Arch. slav. de biol., 1886, i, pp. 89,
364; 1887, ii, pp. 33, 157, 370; Biol. Centralb., 1885, v, p. 529.
Durham, H. E. Drepanidium in the Toad, Liverpool School of Trop. Med.
Memoir VII, Liverpool, 1902, p. 78.
Gaule, J. Über Würmchen, welche a. d. Froschblutkörp. auswandern, Arch.
f. Anat. u. Phys., Phys. Abt., 1880, p. 57.
— Die Beziehungen der Cytozoen zu den Zellkernen, ibid., 1881, p. 297.
Grassi, B., and R. Feletti. Malariaparas. in den Vögeln, C. f. B. u. Par.,
1891, ix, p. 403; 1891, x, p. 449.
Hintze, R. Lebensweise u. Entw. v. Lankesterella minima (Chauss)., Zool.
Jahrb., Anat. Abt., 1902, xv, p. 693.
Kruse, W. Über Blutparasiten, Arch. f. path. Anat., 1890, cxx, p. 541, and
cxxi, p. 359.
Labbé, A. Rech. zool. et biol. sur les paras. endoglob. du sang. d. vertébrés,
Arch. Zool. exp., 3 sér., 1894, ii, p. 55.
Lankester, E. Ray. On Undulina, the Type of a New Group of Infusoria,
Quart. Journ. Micros. Sci., 1871, xi, p. 387.
— On Dreipanidium ranarum, ibid., 1882, xxii, p. 53.
Lebailly, Ch. Rech. sur les hémat. par. des téléastéens marins, Arch. de Paras.,
1906, x, 3, p. 370.
Osler, W. An Account of Certain Organisms occurring in the Liquor
Sanguinis, Proc. Roy. Soc. Lond., 1874, xxii, p. 391.
Ruge, R. Unters. über d. deutsche Proteosoma, C. f. B., P. u. Inf., 1901 (1),
xxix, p. 398.
Siegel. Die geschlechtliche Entw. v. Haemogregarina stepanovi im Rüsselegel
Placobdella catenigera, Archiv für Protistenkunde, 1903, ii, p. 339, with
additions by F. Schaudinn.
Simond, P. L. Contrib. à l’étude des hématoz. endoglob. d. reptiles, Ann. Inst.
Pasteur, 1901, xv, p. 319.
Wallerstein. Über Drepanidium ranarum, Inaug.-Diss., Bonn, 1882.
Malarial Parasites of Man (pp. 155 to 172).
(a) Comprehensive Works.
Celli, A. La malaria sec. le nuove ricerche, Roma, 1900.
Kerschbaumer, Fr. Malaria, ihr Wesen, ihre Entstehung und ihre Verhütung,
Wien, 1901.
Laveran, A. Traité des fièvres palustres, Paris, 1884.
Lühe, M. Cf. p. 765, Die im Blute schmar. Prot., 1906.
Mannaberg, J. Die Malariaparasiten, Wien, 1893.
Neveu-Lemaire, M. Les hématozoaires du paludisme, Paris, 1901.
Reinhardt, Ludw. Die Malaria und deren Bekämpfung nach den Ergebn. d.
neuesten Forschg., Würzb., 1905.
Ruge, R. Einführung in das Studium der Malariakrankh. mit bes. Berücksicht.
d. Technik, Jena, 1901.
Stephens, J. W. W., and S. R. Christophers. The Practical Study of
Malaria and other Blood Parasites, 3rd edition, London, 1908.
Ziemann, H. Über Malaria- und andere Blutparasiten, Jena, 1898.
— Malaria, Handb. d. Tropenkrankh., Lpzg., 1906, iii, p. 269.
(b) Special Studies.
Argutinsky, P. Malariastudien, Arch. f. mikros. Anat., 1901, lix, p. 315; 1902,
lxi, p. 331.
— Zur Kenntn. d. Tropica-Paras., C. f. B., P. u. Inf., i Abt. Orig., xxxiv,
p. 144.
Atti della società per gli studi della malaria, Roma, 1899–1906, i-vii.
Bignami, A. Das Tropenfieber u. die Sommer-Herbstfieber d. gemäss. Klimate,
C. f. B., P. u. Inf., 1898, (1), xxiv, p. 650.
Bignami and Bastanielli. Osserv. nelle febbre malar. estivo-autunn., Rif.
med., 1890, p. i334.
— — Studi sull’ inf. mal., Bull. R. Accad. med., Roma, 1893–94, xx.
— — Sulla strutt. dei par. mal. e in specie dei gameti d. par. est.-aut., Atti
Soc. stud. d. mal., 1899, i.
Celli, A., and F. Sanfelice. Über d. Paras. d. roth. Blutk. im Menschen u.
in Thieren, Fortschr. d. Med., 1891, pp. 499, 541; Ann. istit. d’igiene
sperim., Roma, 1891, N.S. i.
Danilewski, B. Zur Parasit. d. Blutes, Biol. Centralbl., 1885–86, v, p. 529.
— La parasitologie comp. du sang, Charkow, 1889.
— Sur les microb. d’ infect. malar. aiguë et chron. chez les oiseaux et chez
l’homme, Ann. Inst. Pasteur, 1890, p. 753; 1891, p. 758.
— Über den Polymitus malariae, C. f. B. u. Par., 1891, ix, p. 397.
Dionisi, A. La malaria di alcune pipistrelli, Ann. d’igiene sperim., 1899, ix, 4;
Atti soc. ital. p. stud. d. malaria, 1899, i.
Gerhardt. Über Intermittensimpfungen, Arch. f. klin. Med., 1884, vii.
Golgi, C. Sull’ infezione malarica, Arch. p. le sci. med., 1886, x; Arch. ital.
de biol., 1887, viii; Fortschr. d. Med., 1889, 3.
— Sul ciclo evolutivo dei paras. mal. nella feb. terz., Arch. p. le sci. med.,
1889, xiii.
GRASSI, B. Preliminary Communication in: Rend. R. Accad. d. Lincei, Roma,
1898, ser. 5, vii, pp. 163, 234, 314; 1899, viii, p. 165.
— Studi di un zoologo sulla malaria, Atti R. Accad. dei Lincei, Mem. cl. fis.,
1900, ser. 5, iii; Roma, 1900; Die Malaria, Studien eines Zoologen,
2. Aufl., Jena, 1901; also in Italian, Roma, 1901.
— Documenti rig. la storia della scop. del modo di trasmiss. della malaria
umana, Milano, 1903.
Grassi, B., and A. Dionisi. Il ciclo. evol. degli emosporidi, Rend. R. Accad.
d. Lincei, Roma, 1898, ser. 5, vii, 2 sem., p. 308.
Grassi, B., and R. Feletti. Über d. Paras, d. Malaria, C. f. B. u. P., 1890,
vii, pp. 396, 340; Malariapares. in d. Vögeln, ibid., 1891, iv, pp. 403, 429,
461; Weiteres zur Malariafrage, ibid., 1891, x, pp. 449, 481, 517.
Jancsó, N. Zur Frage d. Inf. d. Anopheles claviger mit Mal.-Paras. b. nied.
Temp., C. f. B., P. u. Inf., 1904, i Abt. Orig., xxxvi, p. 624; Der Einfl.
d. Temp. a. d. geschl. Generationsentw. der Mal.-Paras. u. auf d. exper.
Mal.-Erkr., ibid., 1905, xxxviii, p. 650.
Koch, R., and R. Pfeiffer. Beiträge z. Protozoenforsch.: I, Die Coccidienkrankh.
d. Kaninchen, Berlin, 1892.
— — Ärztl. Beob. i. d. Tropen, Verhandl. d. D. Kol.-Ges. Abt. Charlottenburg-Berlin,
1897–98, Heft 7, p. 280.
— — Die Malaria in Deutsch-Ostafrika, Arb. kais. Gesundheitsamt, 1898, xiv,
p. 292.
— — Reiseberichte über Rinderpest . . . trop. Malaria . . . Berlin, 1898.
— — Ergebn. d. wiss. Exped. nach Italien z. Erf. d. Malaria, Deutsche med.
Wochensch., 1899, p. 69.
— — [Zwei] Berichte üb. d. Tätigkeit d. Malaria-Exp., ibid., p. 601; 1900,
p. 88.
— — Über die Entwickelung d. Mal.-Paras., Zeitschr. f. Hyg. u. Inf., 1899,
xxxii, p. 1.
KOSSEL, H. Über einen malariaähnl. Blutparas. b. Affen, Zeitschr. f. Hyg.,
1899, xxxii, p. 25.
Kruse, W. Über Blutparasiten, Virchows Arch. f. path. Anat., 1890, cxx,
p. 451; 1891, cxxi, p. 1395.
Labbé, A. Rech. zool. et biol. sur les par. endoglob. du sang d. vertébr., Arch.
Zool. exp., 1894 (3), ii, p. 55.
Laveran, A. Note sur un nouveau paras. trouvée dans le sang de plus. malad.
att. de fièvre palustre, Bull. Acad. de Méd., Paris, 1880, November 23
and December 28.
— Communication sur la nat. paras. des accid. de l’impalud., C. R. Acad. Sci.,
Paris, 1881, xciii, p. 627; 1882, xcv, p. 737.
— Nature parasitaire des accidents de l’impaludisme, Paris, 1881, 8vo.
McCallum, W. G. On the Hæmatozoan Infection of Birds, Journ. Exper.
Med., Baltimore, 1898, iii, p. 117.
Marchiafava, E., and A. Celli. Les altér. des glob. roug. dans l’infect. par
malaria, Arch. ital. de biol., 1884, v.
— — Nuove ricerche sulla infez. malarica, Annali di agric., 1885, 1886; Fortschr.
d. Med., 1885, Nos. 11 and 24; Arch. p. le sci. med., 1886, ix; 1888,
xii; 1889–90, xiv; Arch. ital. de. biol., 1887, viii, p. 131; 1888, ix.
Maurer, G. Die Tüpfelung der Wirtszelle des Tertianparasiten, C. f. B., P.
u. Inf., 1900, i Abt., xxviii, p. 114.
— Die Malaria perniciosa, C. f. B., P. u. Inf., 1902, i Abt. Orig., xxxii, p. 695.
Nuttall, G. H. F. Die Mosquito-Malariatheorie, C. f. B., P. u. Inf., 1899,
xxv, pp. 161, 209, 245, 285, 337.
— Neuere Forsch. üb. d. Rolle d. Mosqu. bei d. Verbreit. d. Mal., ibid.,
pp. 877, 903; xxvi, p. 140; 1900, xxvii, pp. 193, 218, 260, 328.
Panichi, M. Sulla sede del par. mal. nell’ eritrocito dell’ uomo, Arch. Farmacol.
sper. e scienze aff., 1902, i, pp. 418, 450.
Pezopoulo, N., and J. P. Cardamatis. Die Malaria in Athen, eine biolog. u.
histol. Studie üb. d. Malariaplasmodien, C. f. B., P. u. Inf., 1906, i Orig.,
xl, p. 344.
Plehn, F. Beitr. z. Lehre d. Malariainf., Zeitschr. f. Hyg., 1890, p. 78.
Reports to the Malaria Committee, Roy. Soc. Lond., 1900–1903, i-viii.
Romanowsky, D. Zur Frage d. Paras. . . . d. Malaria, St. Petersb. med.
Wochenschr., 1891, Nos. 34, 35.
Ross, R. Untersuchungen über Malaria (translated by Schilling), Jena, 1905.
Ruge, R. Über d. Plasmod. bei Malariaerkr., Deutsche mil.-ärztl. Zeitschr.,
1892, xxi, pp. 49, 109.
— Ein Beitr. z. Chromatinfärb. d. Mal.-Par., Zeitschr. f. Hyg., 1900, xxxiii,
p. 178.
— Zur Tüpfelung der rot. Blutscheib. bei Febr. interm. tert., Arch. f. klin.
Med., 1902, lxxii, p. 208.
Schaudinn, F. Über d. Generationswechsel d. Coccid. u. die neueren Mal.-Forsch.,
Sitzungsb. Ges. nat. Frde., Berlin, 1899, p. 159.
— Stud. üb. Krankheitserreg. Prot.: II, Plasmodium vivax, d. Erreger d.
Tertianfieb. b. Mensch., Arb. Reichsges.-Amt., 1902, xix, p. 169.
Schoo, H. J. M. Over Malaria: I, Welke Temperatur ist noodig voor de
Amphigonie von Plasmodium vivax? Nederl. Tijdschr. v. Geneeskde.,
1901, ii, p. 1338.
Schüffner, W. Beitr. zur Kenntn. der Malaria, Deutsch. Arch. f. kl. Med.,
1899, lxiv, p. 428; Zur Tüpfelung der r. Blutsch. bei Febr. int. tert.,
ibid., 1901, lxxi.
Serra. Contrib. allo stud. d. posiz. del par. mal. in rapp. glob. rossi, Giorn.
Accad. di med., Torino, 1905, Nos. 5 and 6.
Thayer, W. S. Recent Investigations upon Malaria, Med. News, 1899, lxxiv,
p. 617.
Vassall, J. J. Sur un hématoz. endoglob. nouv. d’un mammifère, Annal. Inst.
Pasteur, 1905, xxi, p. 224.
Ziemann, H. Blutparas. bei heim. und trop. Malaria, C. f. B., P. u. Inf., 1896,
i Abt., xx, p. 653; Z. Morph. d. Malariaparas., ibid., 1897, xxi, pp. 641, 805.
Babesia (pp. 174 to 178).
Babes, V. Die Ätiologie der seuchenhaften Hämoglobinurie des Rindes, Virch.
Arch. f. path. Anat., 1889, cxv.
— Bemerk. über d. Paras. des “Carceag.” der Schafe und die paras. Ictero-Hämaturie
der Schafe, ibid., 1895, cxxxix.
— Bemerk. über d. Entdeckung der seuchenhaften Hämoglobinurie des Rindes
und des Carceag. des Schafes, C. f. B., Par. u. Inf., 1903, i Abt., xxxiii,
p. 449.
Bonome. Über paras. Ictero-Hämaturie der Schafe, Virch. Arch. f. path. An.,
1895, Bd. cxxxix, p. 1.
Celli, A., and F. G. Santori. Die Rindermalaria in der Campagna von Rom,
C. f. B., P. u. Inf., 1897, xix, p. 561.
Chauvelot, E. Les babésioses, Paris, 1904.
Dschunkowski, E., and J. Luhs. Die Piroplasmosen der Rinder, C. f. B.,
P. u. Inf., 1904, i Abt. Orig., xxxv, p. 486, 3 Taf.
Fantham, H. B. Piroplasma muris from the Blood of the White Rat, Quart.
Journ. Micros. Sci., 1906, 1, p. 483, 1 pl.
Kinoshita, K. Untersuchungen üb. Babesia canis, Arch. f. Protistenkde., 1907,
viii, p. 294.
Kleine, F. K. Kultivierungsversuche d. Hundepiroplasmen, Zeitschr. f. Hyg.
u. Inf., 1906, liv, p. 10.
Koch, R. Reiseberichte, über Rinderpest . . . Texas-Fieber, Berlin, 1898.
— Vorl. Mitt. über die Ergebn. einer Forschungsreise nach Ostafrika, Deutsche
med. Wochenschr., 1905, No. 45.
— Beitr. z. Entwicklungsgesch. der Piroplasmen, Zeitschr. f. Hyg. u. Inf.,
1906, liv, p. 1.
Kossel, H., Weber, Schütz and Miessner. Über die Hämoglobinurie der
Rinder in Deutschland, Arb. a. d. Kais. Gesundheitsamt, 1903, xx, 1,
3 Taf.
Laveran, A. Contrib. à l’étude de Piroplasma equi, C. R. Soc. Biol., Paris,
1901, liii (14), p. 385.
Leblanc, P. Piroplasma canis, C. R. Soc. Biol., Paris, 1900, lii, p. 168.
Lingard, A. Can the Piroplasma bigeminum find a Habitat in the Human
Subject? C. f. B., P. u. Inf., 1904, i Abt. Orig., xxxvi, p. 214.
Lingard, A., and E. Jennings. A Preliminary Note on a Piroplasmosis found
in Man and in Some of the Lower Animals, Ind. Med. Gaz., 1904, xxxix,
p. 161.
Lühe, M. Zur Kenntnis von Bau und Entw. d. Babesien, Zool. Anz., 1906, xxx,
p. 45.
Miyajima and Shibayama. Über das in Japan beobachtete Rinderpiroplasma,
Zeitschrift f. Hyg., 1906, lii, p. 189.
Nocard and Motas. Contrib. à l’étude de la piropl. canine, Ann. Inst. Pasteur,
1902, xvi, 1, p. 257, 2 pl.
Nuttall, G. H. F., and G. S. Graham-Smith, Canine Piroplasmosis V, Journ.
of Hyg., 1906, vi, p. 586.
Schmidt, A. Die Zeckenkrankh. der Rinder, Arch. f. wiss. u. prakt. Tierheilkunde,
1904, xxx, p. 42.
Smith, T., and F. Kilborne. Investigations into the Nature, Causation, and
Prevention of Texas or Southern Cattle Fever, Eighth and Ninth Annual
Reports, Bureau of Animal Industry, Washington, U.S.A., 1893, pp. 177–304,
10 pl.
Theiler, A. Die Piroplasmose des Maultieres und des Esels, Zeitschr. f.
Tiermed., 1904, viii, p. 383.
Wilson, L. B., and W. M. Chowning. Studies in Piroplasmosis hominis,
Journ. Infect. Dis., 1904, p. 31, 2 pl.
Neosporidia.
Order. Myxosporidia (pp. 181 to 184).
Balbiani, G. Sur l’organis. et la nature d. psorosp., C. R. Acad. Sci., Paris,
1863, lvii, p. 157.
Bütschli, O. Zur Kenntn. der Fischpsorosp., Zeitschr. f. wiss. Zool., 1881,
xxxv, p. 629.
Cohn, L. Über die Myxospor. von Esox lucius u. Perca fluviatilis, In.-Diss.,
Königsberg, 1895, u. Zool. Jahrb. Anat., 1895, ix.
— Zur Kenntnis der Myxospor., C. f. B., P. u. Inf., 1902, i Abt. Orig., xxxii, p. 628.
Creplin, J. C. H. Beschreibung der Psorosporm. des Kaulbarsch. nebst Bemerkung
über die der Plötze, Arch. f. Naturg., 1842, viii, 1, p. 61.
Doflein, F. Stud. z. Nat. d. Prot.: III, Über Myxospor., Zool. Jahrb., Anat.,
1898, xi, p. 281.
Dujardin, F. Hist. nat. des helm., Paris, 1845, p. 643.
Gurley, R. On the Classification of the Myxosporidia, Bull. U.S. Comm. of
Fish and Fishermen, 1891, Washington, 1893, p. 407.
— The Myxosporidia or Psorosperms of Fishes and the Epidemics produced
by them, Rep. U.S. Comm. of Fish and Fishermen, 1892, Washington,
1894, p. 65.
Hofer, B. Die sogen. Pockenkrankh. d. Karpfen, Allg. Fisch.-Ztg., 1896,
pp. 2, 28; 1902, p. 22.
Joseph, H. Chloromyxum protei n. sp., Arch. f. Protistenkde., 1907, viii,
P· 398.
Laveran, A., and F. Mesnil. Sur une myxosp. des voies biliaires de l’Hippocampe,
C. R. Soc. Biol., Paris, 1900, lii, p. 380.
— — Sur la multiplic. endog. d. Myxosporidies, C. R. Soc. Biol., Paris, 1902,
liv, p. 469.
Leydig, F. Über Psorosperm. u. Gregarinen, Arch. f. Anat. u. Phys., 1851,
p. 221.
Lieberkühn, N. Über d. Psorosp., Arch. f. Anat. u. Phys., 1854, p. 349.
Ludwig, H. Über d. Myxospor. d. Barben in d. Mosel, Jahresber. d. Rhein.
Fisch.-Ver., 1888–89, p. 27.
Lühe, M. Cystodiscus immersus Lutz., Verh. d. D. zool. Ges., 1899, p. 291.
Mercier, L. Phénomènes de sexualité chez le Myxobolus pfeifferi, C. R. Soc.
Biol., Paris, 1906, lx, p. 427.
Müller, J. Über eine eigent. krankh. paras. Bildung mit specif. organis.
Samenkörp., Arch. f. Anat. u. Phys., 1841, p. 477.
Müller, J., and A. Retzius. Über paras. Bildungen, ibid., 1842, p. 193.
Perugia, A. Sulle myxosp. d. pesci marini, Bull. scientif., Ann. xii-xiii,
1889–90, p. 10.
Pfeiffer, L. Die Protoz. als Krankheitserreger, 2. Aufl., Jena, 1891.
— Unters. über d. Krebs, Jena, 1892.
Plehn, M. Über die Drehkrankh. der Salmoniden, Arch. f. Protistenkde.,
1905, v, p. 145.
Railliet, A. La maladie d. barbeaux de la Marne, Bull. soc. centr. d’aquicult.
France pour 1890, ii, p. 117.
Schröder, Ol. Eine neue Myxosporidienart aus den Kiemen von Acerina
cernua, Arch. f. Protistenkde., 1906, vii, p. 186.
Schuberg, A., and O. Schröder. Myxosp. a. d. Nervensyst. u. d. Haut der
Bachforelle, Arch. f. Protistenkde., 1905, vi, p. 47.
Thélohan, P. Rech. sur les Myxospor., Bull. scientif. France et Belg., 1895,
xlvi, p. 100.
— Observ. sur les myxosp. et essai de classific. d. ces. org., Bull. soc. philom.,
Paris, 1892 (8), iv, p. 165.
Weltner, W. Über Myxospor. in d. Eiern v. Esox lucius, Sitzungsber. Ges.
naturf. Frde., Berl., 1892, p. 28.
Zschokke, F. Myxospor. d. Gattg. Coregonus, C. f. B., P. u. Inf., 1898 (i),
xxiii, p. 602; Mitt. nat. Ges., Lucerne, 1898, 2, p. 205.
Order. Microsporidia (pp. 184 to 186).
Balbiani, G. Rech. sur les corpusc. de la pébrine, Journ. de l’Anat. et de la
Phys., 1866, iii, p. 599.
— Etud. sur la maladie psorosperm. des vers à soie, ibid., 1867, iv, pp. 263, 329.
Bolle, J. Der Seidenbau in Japan, Budapest, Wien, Leipzig, 1898, p. 94.
Gluge. Tumeurs enkystées observées sur la peau des épinoches, Bull. Acad.
roy. de Belg., 1838, v, p. 772.
Henneguy. Note sur un paras. d. muscl. du Palaemon rectirostris, Mém. soc.
philom. à l’occas. du centenn. de sa fondat, Paris, 1888.
Henneguy and Thélohan. Myxospor. par d. muscl. chez quelq. crust. décap.,
Ann. microgr., Paris, 1892, iv, C. R. Soc. Biol., Paris, 1892 (9), iv, C. R.
Acad. Sci., Paris, 1892, cxiv.
Hesse, E. Sur une nouv. microsp. tétraspor. du genre Gurleya, C. R. Soc.
Biol., Paris, 1903, lv, p. 495.
— Sur la prés. d. microsp. du genre Thelohania chez les insectes, C. R. Acad.
Sci., Paris, 1903, cxxxvii, p. 418.
— Thelohania legeri n. sp., microsp. nouv. d. larves d’Anopheles maculipennis
Meig., C. R. Soc. Biol., Paris, 1904, lvii, p. 570.
— Sur Myxocystis mrázeki, microsp. par de Limnodrilus hoffmeisteri Clap.,
C. R. Soc. Biol., Paris, 1905, lviii, pp. 12–15.
Korotneff, A. Myxosporidium bryozoides, Z. f. w. Zool., 1892, liii, p. 591.
Kulagin, N. Zur Entw. v. Glugea bombycis Thél., Zool. Anz., 1898, xxi,
p. 469.
Lebert, H. Über die gegenwärtig herrsch. Krankh. des Insects d. Seide, Berl.
entom. Ztschr., 1858, ii, p. 149.
Leydig, Fr. Zur Anat. v. Coccus hesferidum, Z. f. w. Zool., 1854, v, p. 11.
— Zum fein. Bau d. Arthropoden, Arch. f. An. u. Phys., 1855, p. 397.
— Über Paras. nied. Tiere, Arch. f. path. Anat., 1858, p. 280.
— Der Parasit in der neuen Krankh. d. Seidenraupe, Arch. f. Anat. u. Phys.,
1863.
Lutz, A., and A. Splendore. Über Pebrine u. verw. Microspor., C. f. B.,
P. u. Inf., 1903, i Abt., xxxiii, p. 150.
Moniez, R. Note sur des paras. d. helm., Bull. scient. du Départ. du Nord,
1879 (2), ii, p. 304.
— Observ. pour la revis. d. microsporid., C. R. Acad. Sci., Paris, 1887, civ,
p. 1312.
Mrázek. Sporozoenstudien: II, Glugea lophii Dofl., Sitzungsber. k. böhm.
Ges. der Wiss., math.-nat. Kl., 1899, Prag, 1900.
Pasteur, L. Etude sur la maladie des vers à soie, Paris, 1870.
Pérez, Ch. Sur une nouv. microsp. paras. d. Carcinus maenas, C. R. Soc.
Biol., Paris, 1904, lvii, p. 214.
Perroncito, E. Il coccidio jalino ed il microsp. poliedrico nell’ uomo, Giorn.
Accad. Med., Torino, 1902, Ann. 65, p. 378.
Pfeiffer, L. Beitr. zur Kenntn. d. pathog. Gregar.: I, Die Microsporidien
und die Fleckenkrankh. (Pébrine) d. Seidenspinners, Zeitschr. f. Hyg.,
1888, iii.
Stempell, W. Über Thélohania mülleri (L. Pfr.), Zool. Jahrb. Anat., 1902,
xvi, p. 235.
— Über Nosema anomalum Moniez, Arch. f. Protistenkunde., 1904, iv, p. 1.
Vaney, C., and A. Conte. Sur une nouv. microsp., Pleistophora mirandellae,
paras. de l’ovaire d’Alburnus mirandella Blanch., C. R. Acad. Sci.,
Paris, 1901, cxxxiii, p. 644.
Order. Actinomyxidia (p. 187).
Some earlier literature will be found quoted in:—
Caullery, M., and F. Mesnil. Recherches sur les Actinomyxidies, I, Arch. f.
Protistenkde., 1905, vi, 3, p. 272.
Order. Sarcosporidia (pp. 187 to 194).
Bertram, A. Beitr. zur Kenntn. d. Sarcosp., Zool. Jahrb., 1892, v, p. 581.
Blanchard, R. Sur un nouv. type d. Sarcospor., C. R. Acad. Sci., Paris, 1885,
c, p. 1599.
— Note sur les Sarcospor. et sur un ess. d. classif. d. ces sporoz., Bull. Soc.
Zool. France, 1885, x, p. 244.
Dammann, C. Psorosp.-Krankh. beim Schaf., Arch. f. path. Anat., 1867, xli,
p. 283.
Eecke, J. van. Sarcosporidien, Geneesk. Tijdschr. v. Nederl.-Indie, 1892,
xxxii; Jaarsverl. Labor. path. An. en Bact. te Weltevreden (1892),
Batavia, 1893.
Foret, P. Observ. rel. au dével. de la cuticle chez le Sarcocystis tenella,
Arch. d’Anat. Micr., 1903, vi, p. 86; C. R. Soc. Biol., Paris, 1903, lv,
p. 1054.
Hessling, V. Histol. Mittheil., Zeitschr. f. wiss. Zool., 1854, v, p. 189.
Koch, M. Über Sarcosporidien, Verh. V. intern. Zool. Congr., Berlin, Jena,
1902, p. 674.
— Die experimentelle Übertrag. d. Miescherschen Schläuche, Berl. klin.
Wochenschr., 1904, li, p. 321.
Korté, W. E. de. On the Presence of Sarcosporidia in the Thigh Muscle of
Macacus rhesus, Journ. of Hyg., Cambridge, 1905, v, p. 451.
Laveran, A., and F. Mesnil. Morph. d. sarcospor., C. R. Soc. Biol., 1899 (x),
vi, p. 245.
Leisering and Winkler. Psorosp.-Krankh. beim Schaf., Ber. üb. Veterin.-Wesen,
Königr., Sachsen, 1865; Arch. f. path. Anat., 1865, xxxvii, p. 431.
Manz, W. Beitr. z. Kenntn. d. Miescherschen Schläuche, Arch. mikr. Anat.,
1867, iii, p. 345.
Miescher, F. Über eigent. Schläuche in d. Musk. einer Hausmaus, Ber. über
die Verh. d. naturf. Ges., Basel, 1843, v, p. 198; Reprinted in Verh. d.
V. internat. Zool.-Congr., Berlin, Jena, 1902, p. 679.
Piana, G. P. Fasi evol. d. Sarcosp., La clinica veter., 1896, p. 145; C. f. B.,
P. u. Inf. (1), xx, p. 39.
Pluymers, L. Des sarcosp. et de leur rôle dans la pathog. d. myositis, Arch.
Méd. exp. et d’Anat. pathol., 1896, p. 761; C. f. B., P. u. Inf. (1), xxii,
p. 245.
Rainey, G. Structure and Development of Cysticercus Cells as found in the
Muscles of the Pig, Phil. Trans. Roy. Soc., 1858, cxlvii, p. 111.
Rieck, V. Sporozoen als Krankheitserreger, Deutsche Zeitschr. f. Thiermed.
u. vergl. Path., 1889, xiv, p. 75.
Rievel and Behrens. Beitr. zur Kenntn. d. Sarcosp. und deren Enzyme, C. f.
B., P. u. Inf., 1903, i Abt. Orig., xxxv, p. 341.
Rivolta. Dei paras. veget., Torino, 1873; Giorn. an., fis. e pat. d. anim., 1874,
vi, p. 25.
Schneidemühl, G. Über Sarcosporidien, Thiermed. Vortr., Leipzig, 1897, iii,
p. 11.
Siebold, C. Th. v. Zusatz [zu Hessling. histol. Mittheil.], Zeitschr. f. wiss.
Zool., 1854, v, p. 199.
Siedamgrotzky, O. Psorosp. in d. Musk. d. Pferde, Wochenschr. f. Thierheilkde.
u. Viehz., 1872, xvi, p. 97.
Smith, Th. The Production of Sarcosporidia in the Mouse by Feeding Infected
Muscle Tissue, Journ. Exp. Med., Baltimore, 1902, vi, p. 1.
— Further Observations on the Transmission of Sarcocystis muris by Feeding,
Journ. Med. Res., 1905, xiii, p. 429.
Stiles, Ch. W. Notes on Parasites: 18, Presence of Sarcosporidia in Birds,
U.S. Dept. of Agric., Bur. of An. Ind., Bull. 3, 1893, p. 79.
Sarcosporidia observed in Man (pp. 193, 194).
Baraban and St. Remy. Sur un cas d. tub. psorosp. obs. chez l’homme, C. R.
Soc. Biol., Paris, 1894 (x), i, p. 201.
— — Le parasitisme d. sarcosp. chez l’homme, Bibliogr. anat., 1894, p. 79.
Braun, M. Zum Vork. d. Sarcosporid. b. Menschen, C. f. B. u. Par., 1895 (1),
xviii, p. 13.
Kartulis. Über pathog. Protoz. b. Menschen, Zeitschr. f. Hyg., 1893, xiii, p. 1.
Lindemann. Über d. hygien. Bedeutung d. Gregarinen, Deutsche Zeitschr. f.
Staatsarzneikde., 1868.
Rosenberg. Ein Befund von Psorosp. im Herzmuskel d. Mensch., Zeitschr. f.
Hyg., 1892, xi, p. 435.
Vuillemin, P. Le Sarcocystis tenella, paras. de l’homme, C. R. Acad. Sci.,
Paris, 1902, cxxxiv, p. 1152.
Order. Haplosporidia (pp. 194 to 197).
Caullery, M., and F. Mesnil. Rech. sur les Haplosporidies, Arch. de Zool.
exp., 1905, sér. iv, iv, p. 101, in which a good bibliography is given.
Minchin, E. A., and H. B. Fantham. Rhinosporidium kinealyi n. g., n. sp., a
New Sporozoon from the Mucous Membrane of the Septum Nasi of Man,
Quart. Journ. Micros. Sci., 1905, xlix, p. 521.
Class IV.—Infusoria (pp. 198 to 210).
Bütschli, O. Studien über . . . die Conjugation d. Infusorien, Abh. d.
Senckenb. naturf. Ges., 1876, x.
Ehrenberg, Ch. G. Die Infusionsthierchen als vollkommene Organismen,
Leipz., 1838.
Guiart, J. Sur un nouv. infus. paras. de l’homme, C. R. Soc. Biol., Paris,
1903, lv, p. 245.
Hertwig, R. Über die Conjugation d. Infusorien, Abh. kgl. bayer. Akad. d.
Wiss., 1889, ii, Kl., xvii.
Kent, Sav. A Manual of the Infusoria, London, 1880–1882.
Maupas, E. Rech. expér. sur la multipl. des Infusoires ciliés, Arch. Zool. exp.,
1888 (2), vi.
— Le rajeunissement karyogamique chez les Ciliés, ibid., 1889, vii.
Stein, Fr. v. Der Organismus der Infusionsthiere, Leipz., 1859–1867.
Balantidium coli (pp. 200 to 204 and 637).
Askanazy, M. Pathog. Bedtg. d. Bal. coli, Wien. med. Wochenschr., 1903,
liii, p. 127; Verh. d. D. path. Ges., v (1902), Berlin, 1903, p. 224.
Casagrandi, O., and P. Barbagallo. Bal. coli s. Param. coli, Catania, 1896,
8vo.
Collmann, B. Fünf Fälle von Bal. coli im Darm d. Mensch., In.-Diss.,
Kgsbg., Pr., 1900.
Ehrnroth, E. Z. Frage der Pathogenität d. Bal. coli, Zeitschr. f. klin. Med.,
1903, xlix, p. 321.
Grassi, B. Signif. patol. d. prot. par. dell’ uomo, Atti Accad. Lincei, Rendic.,
1888 (4), iv, Sem. 1, p. 86.
Janowski, W. Ein Fall von Bal. coli im Stuhl, Zeitschr. f. klin. Med., 1897,
xxxii, p. 415. (With copious literature compiled by Shegalow, Solowjew
and Klimenko.)
Klimenko, W. Beitr. z. Pathol. d. Bal. coli, Beitr. z. path. Anat. u. allg.
Path., 1903, xxxiii, p. 281.
Koslowski, J. J. Zur Lehre v. d. Infus., die als Paras. im Verdauungskan.
d. Mensch. vork., Arch. f. Verdauungskrankh., 1905, xi, p. 31.
Kossler, K. Ein Fall von Balantidium-Colitis, Wien. med. Wochenschr.,
1906, lvi, p. 522.
Maggiora, A. Microsk. u. bacter. Beob. während einer epid. dysent. Dickdarmentzdg.,
C. f. B. u. Par., 1892, xi, p. 181.
Malmsten, P. H. Infusorien als Intestinalthiere b. Mensch., Arch. f. path.
Anat., 1857, xii, p. 302.
Nagel. Üb. ein. Fall v. Infusorienenteritis, Münch. med. Wochenschr., 1905,
No. 44.
Shegalow, J. P. Ein Fall von Bal. coli bei einem 5 jähr. Mädchen, Jahrb. f.
Kinderhlkde., 1899, xlix, p. 425.
Sievers, R. Über Bal. coli im menschl. Darm u. dessen Vork. in Schwed. u.
Finland, Arch. f. Verdauungskrankh., 1900, v. Abstracted in C. f. B.,
P. u. Inf., 1900 (1), xxviii, p. 328.
Sievers, R. Zur Kenntn. d. Verbreit. v. Darmparas. d. Menschen in Finland,
Helsingfors, 1905; Festschr. f. Palmén, No. 10.
Solowjew. Bal. coli als Erreger chron. Durchfälle, C. f. B., P. u. Inf., 1901
(1), xxix, pp. 821, 849. [Solowjew’s additional communication that
appeared in “Wratsch,” 1901, Nos. 12 and 14, as well as in the “Russki
Wratsch,” 1902, No. 14, has been translated into German by Klimenko
(l. c.).]
Stokvis, B. J. Paramaecium in sputa, Nederl. Tijdschr. v. Geneeskde., 1884
(2), xx.
Strong, R. P., and W. E. Musgrave. Preliminary Note of a Case of Infection
with Balantidium coli, Bull. Johns Hopkins Hosp., Baltimore, 1901, xii,
p. 31.
— — The Clinical and Pathological Significance of Balantidium coli, Dept. of
Int. Bureau, Govt. Labor. Biol., Manila, No. 26, 1905, p. 1.
Wlajeff, G. Zur Frage d. Ätiol. u. Behandlg. d. Dysenterie, Wracebraja
Gaseta, Kemmern, 1905, xii, p. 913; abstracted in C. f. B., P. u. Inf.,
1906, i, Ref. xxxvii, p. 757.
Woit, O. Drei neue Fälle von Bal. coli i. menschl. Darm., Deutsch. Arch. f.
klin. Med., 1898, lx, p. 363.
Balantidium minutum (pp. 204 and 637).
Jakoby, M., and F. Schaudinn. Üb. zwei neue Infus. i. Darm. d. Mensch.,
C. f. B., P. u. Inf., 1899 (i), xxv, p. 487.
Schulz. Colpoda cucullus im Darm d. Mensch., Berl. klin. Wochenschr., 1899,
No. 16, p. 353.
Nyctotherus (pp. 204 to 206 and 637).
Castellani, A. Observations on some Protozoa found in Human Fæces, C. f.
B., P. u. Inf., 1905, i Abt. Orig., xxxviii, p. 66.
Jakoby, M., and F. Schaudinn. Über zwei Infus. i. Darm d. Mensch., ibid.,
1899 (1), xxv, p. 487.
Krause, P. Üb. Infus. im Typhusstuhle nebst Beschreibg. einer bisher noch
nicht beob. Art. (Balantidium giganteum), Deutsch. Arch. f. klin. Med.,
1906, lxxxvi, p. 442.
Chlamydozoa (pp. 207 to 210).
Bosc, F. J. Les malad. bryocytiques (malad. à protozoaires), II, La maladie
vaccinale (Plasmodium vaccinæ), C. f. B., P. u. Inf., i Orig., xxxvi,
p. 630; xxxvii, pp. 39, 195.
— Les malad. bryocyt., III, La variole et son parasite (Plasmodium variolæ),
ibid., xxxix, pp. 36, 129, 247, 389, 594.
Calkins, G. N. The Life-history of Cytoryctes variola, Journ. Med. Research,
Boston, 1904, xi, p. 136.
Councilman, Magrath, Brinckenhoff, Tyzzer, Southard, Thompson, Bancroft
and Calkins. Studies on the Pathology and on the Etiology of
Variola and of Vaccinæ, Journ. Med. Research, Boston, 1904, xi, 1,
1904.
Gorini, C. Über die bei der mit Vaccine ausgef. Hornhautimpf. vorkomm.
Zelleinschlüsse, C. f. B., P. u. Inf., 1900, i, Abt. xxviii, pp. 233, 589;
1902, i Orig., xxii, p. 111.
Guarnieri, G. Ric. sulla patogenesi ed etiol. dell’ inf. vacc. e variolosa, Arch.
sci. med., Torino, 1892, xvi.
— Ulteriori ric. sulla etiol. e sulla patog. della inf. vacc., Clinica moderna,
Firenze, 1897, iii.
Hückel. Die Vaccinekörperchen, Beitr. z. pathol. Anat. u. z. allg. Path.,
Supp. II, 1898.
Loeff, A. van der, in Weekbl. van het Nederl. Tijdschr. v. Geneeskde., 1886,
No. 46.
Mühlens, P., and M. Hartmann, Zur Kenntnis d. Vaccineerregers, C. f. B.,
P. u. Inf., 1906, i Orig., xxxxi, pp. 41, 203, 338, 435.
Pröscher, F. Über d. künstl. Züchtung eines “unsichtbaren” Mikroorgan.
aus der Vaccine, C. f. B., P. u. Inf., 1906, i Orig., xl, 3, p. 337.
Prowazek, S. Unters. üb. d. Vaccine, I, Arb. a. d. kais. Gesundheitsamt, 1905,
xxii, p. 535.
— Unters. üb. d. Vaccine, II, ibid., 1906, xxiii, p. 525.
Salmon, P. Rech. sur l’infect. dans la vaccine et la variole, Annal. Inst.
Pasteur, 1897, xi, No. 4.
Schulze, F. E. Cytorrhyctes luis Siegel, Berl. klin. Wochenschr., 1905, No. 21.
Schulze, W. Impfungen mit Luesmaterial an Kaninchenaugen, Klin. Monatsbl.
f. Augenheilkde., 1905, xliii.
— Das Verhalten der Cytorrhyctes luis in der mit Syphilis geimpften Kanin
cheniris, Beitr. z. path. Anat. u. z. allg. Path., 1906, xxxix, p. 180.
Siegel, J. Zur Kritik der bisherigen Cytorrhyctesarbeiten, C. f. B., P. u. Inf.,
1906, i Orig., xlii, pp. 128, 225, 321, 480.
Wasielewski, v. Beitr. z. Kenntnis d. Vaccineerregers, Zeitschr. f. Hyg.,
1901, xxxviii, p. 212.
(B) PLATYHELMINTHES (pp. 211 to 359, 638 to 698 and 753 to 755).
Class II.—Trematodes (pp. 212 to 282, 638 to 644, 753, and 754).
[N.B.—The literature, which is very comprehensive, has, up to the year
1892, been quoted and critically examined in Braun’s monograph on the Trematodes:
Bd. iv, Abth. i, of Bronn’s “Klass. u. Ord. d. Thierreichs,” Leipz. Of
works that have appeared later it is not possible to do more than enumerate the
following.]
Bettendorf, H. Musculatur u. Sinneszell. d. Tremat., Zool. Jahrb. Anat.,
1897, x, p. 307.
Blochmann, F. Die Epithelfrage bei Cestoden u. Trematoden, Hamburg, 1896.
Braun, M. Arten d. Gattg. Clinostomum, Zool. Jahrb., 1900, Syst. xiv, p. 1.
— Trematoden d. Chelonier, Mitt. zool. Mus. Berlin, 1901, ii, p. 1.
— Trematoden d. Chiroptera, Annal. K. k. naturh. Hofmus., Wien, 1900, xv,
p. 217.
— Zur Kenntn. d. Tremat. d. Säugeth., Zool. Jahrb., 1901, Syst. xiv, p. 311.
— Fascioliden d. Võgel, ibid., 1902, xvi, p. 1.
Brugge, G. Zur Kenntn. d. Excretionsgefässsyst. d. Cestoden u. Tremat.,
Zool. Jahrb. Anat., 1902, xvi, p. 208.
Fischoeder, F. Die Paramphistomiden d. Säugeth., Zool. Jahrb., 1903, Syst.
xvii, p. 485.
Gronkowski, C. v. Zum feineren Bau d. Tremat., Poln. Arch. f. biol. u. med.
Wiss., 1902, i.
Hein, W. Zur Epithelfrage d. Tremat., Zeitschr. f. wiss. Zool., 1904, lxxvii,
p. 546.
Looss, A. Die Distomen unserer Fische und Frösche, Stuttg., 1894; Bibl.
zool., xvi.
— Rech. faune paras. de l’Egypte, I, Mém. Inst. égypt., 1896, iii, p. 1.
— Weit. Beitr. z. Kenntn. d. Tremat.-Fauna Ägypt, Zool. Jahrb., 1900, Syst.
xii, p. 521.
— Über neue u. bekannte Tremat. aus Seeschildkröten, ibid., 1902, xvi, p. 411.
Maclaren, W. Beitr. z. Kenntn. einig. Tremat., Jen. Zeitschr. f. Naturw.,
1903, xxxviii, p. 573.
Monticelli, F. S. Stud. tremat. Endopar., I, Zool. Jahrb., 1893, Suppl. iii.
Roewer, C. F. Beitr. z. Histogenese v. Cercariaeum helicis, Jen. Zeitschr. f.
Naturw., 1906, xli, p. 185.
Schubmann, W. Eibildung u. Embryonalentw. v. Fasciola hepatica, Zool.
Jahrb. Anat., 1905, xxi, p. 571.
Ziegler, H. E. Das Ectoderm d. Plathelminthen, Verh. D. zool. Ges., 1905,
p. 35.
Watsonius watsoni (pp. 234, 235).
Conyngham, H. F. A New Trematode of Man, Brit. Med. Journ., 1904, ii,
p. 663; Lancet, 1904, ii, p. 464.
Shipley, A. E. Cladorchis watsoni (Conyngham), a Human Parasite from
Africa, Thompson, Yates and Johnston Lab. Report, Liverpool, 1905,
vi, 1, p. 129.
Gastrodiscus hominis (pp. 236, 237).
Giles, G. M. A Report of an Investigation into the Causes of the Disease
known in Assam as Kála-azár and Beriberi, Shillong, 1890, p. 125.
Leuckart, R. Die Paras. d. Mensch., 2. Aufl., ii, p. 450, where the first discovery
is reported in greater detail.
Lewis, T. R., and McConnel. A New Parasite Affecting Man, Proc. Asiatic
Soc., Bengal, 1876, p. 182.
Fasciola hepatica (pp. 237 to 244, and 638).
Ammon. Klin. Darst. d. Krankh. d. menschl. Auges, Dresden, 1838.
Bossuat, E. Les helminth. dans le foie, Arch. de Paras., 1902, vi, p. 186. [The
author is in error when he writes “The name Dist. sibiricum originated
from M. Braun”!]
Coe, W. R. Bau des Embryos v. Dist. hep., Zool. Jahrb. Anat., 1896, ix, p. 561.
Duffek, E. Dist. hep. beim Mensch., Wien. klin. Wochenschr., 1902, p. 772.
Gaide, cf. under Clonorchis sinensis (p. 787).
Gescheidt and Ammon. Die Entoz. d. Auges, Zeitschr. f. Ophth., 1833, iii,
P. 405.
Greeff, R. Über d. Vork. v. Würmern im Auge, Arch. f. Augenheilkde., 1907,
lvi, p. 334.
Havet, J. Contrib. à l’étud. d. syst. nerv. d. Trémat., La Cellule, 1900, xvii,
p. 351.
Henneguy, L. F. Rech. sur la mode de form, de l’œuf du Dist. hep., Arch.
d’anat. micr., 1906, ix, p. 47.
Khouri, A. Le Halzoun, Arch. de Paras., 1904, ix, 1, p. 78.
Küchenmeister, F. On Animal and Vegetable Parasites of the Human Body,
translated by E. Lankester, London, 1857.
Leuckart, R. Z. Entw. d. Lebereg., Arch. f. Naturg., 1882, i, p. 80.
Lutz, A. Lebensgesch. d. Dist. hep., C. f. B. u. P., xi, p. 783; xiii, p. 320.
Malherbe. Progr. méd., 1898, vii, No. 4.
Marcinowski, K. Das untere Schlundgangl. von. Dist. hep., Jen. Zeitschr. f.
Naturw., xxxvii, 1903, p. 544.
Nordmann, A. v. Mikrograph. Beitr. z. Naturgesch. d. wirbellos. Thiere,
Berlin, 1832, ii, p. 9.
Pallas, P. S. De infestis viventibus intra viventia, Diss. in., Lugd., Batavia,
1760.
Saito, S. Beitr. z. Kenntn. d. geogr. Verbr. d. Dist. hep., C. f. B., P. u. Inf.,
1906, i Orig., xli, p. 822.
Schaper. Die Leberegelkrankheit. d. Schafe, Deutsche Zeitschr. f. Tiermed.,
1890, xvi, p. 1.
Sommer, L. Anat. d. Leberegels, Z. f. w. Zool., 1880, xxxiv, p. 539.
Stieda, L. Beitr. z. Anat. d. Plattw.: I, Arch. f. Anat. u. Phys., 1867, p. 52.
— Über d. angebl. inneren Zusammenhang d. männl. u. weibl. Org. b. Tremat.,
ibid., 1871, p. 31.
Stiles, C. W. Frogs, Toads and Carp as Eradicators of Fluke Diseases,
Ann. Rep. Bur. of Anim. Ind., 1901, Wash., 1902, xviii, p. 220.
Thomas, P. The Life-history of the Liver Fluke, Quart. Journ. Micros. Sci.,
1883, xxiii, p. 99.
[N.B.—A bibliography of cases has been compiled by Davaine (1877),
Leuckart (1889–1894), Moniez (1896), Blanchard (1889), and Huber (1895), in
addition to Khouri (l. c.).]
Fasciola gigantica (pp. 244, 245).
Cobbold, Th. Sp. Description of a New Trematode Worm (Fasciola gigantica),
Edin. New Phil. Journ., 1855, N.S. ii, p. 262.
— Entozoa, an Introduction to the Study of Helminthes, London, 1864, pl. i.
Gouvea, H. de. La distomatose pulm. par la douve du foie, Thèse, Paris, 1895.
Looss, A. Rech. sur la faune de l’Egypte, Mém. Inst, égypt., 1896, iii, p. 33.
— Obs. à prop. d’une note . . . C. f. B., P. u. Inf. (1), 1898, xxiii, p. 459.
Railliet, A. Sur une forme partic. de douve hépat. prov. de Senegal, C. R.
Soc. Biol., Paris, 1895, 10e sér., ii, p. 338.
Fasciolopsis buski (pp. 245, 246, and 638).
Budd, G. On Diseases of the Liver, London, 1852.
Cobbold, T. Sp. On the Supposed Rarity of . . . Dist. crassum, Journ.
Linn. Soc., 1875, xii, p. 285; Obs. on the Large Human Fluke, Veterinarian,
1876.
Giles, G. M., cf. under Gastrodiscus hominis (p. 784).
Lankester, E. Manual of Animal and Vegetable Parasites (Küchenmeister),
London, 1857, App. i, B. p. 437.
Leidy, J. On Distomum hepaticum, Proc. Acad. Nat. Sci., Philadelphia, 1873,
P· 364.
Odhner, Th. Fasciolopsis buski, C. f. B., P. u. Inf., i Orig., xxxi, p. 573.
Fasciolopsis rathouisi (pp. 246, 247).
Poirier, P. Note sur une nouv. esp. de Dist. paras. de l’homme, Arch. Zool.
exp., 1887 (2), v, p. 203.
Paragonimus ringeri (pp. 249 to 251, 639 and 640).
Baelz, E. Über paras. Haemopt., Centralbl. f. med. Wiss., 1880, p. 721.
— Über einig. neue Paras. d. Mensch., Berl. klin. Wochenschr., 1883, p. 234.
Inouye, J. Über d. Dist. ringeri Cobb, Zeitschr. f. klin. Med., 1903, l, p. 120,
with list of Japanese literature.
Janson, J. Die bish. in Japan bei Schweinen gef. Paras., Mitt. d. Ges. f.
Natur- u. Völkerkde. Ostasiens, 1897, Heft 59–60.
Katsurada, F. Beitr. z. Kenntn. d. Dist. westerm., Beitr. z. path. Anat. u. z.
allg. Path., 1900, xxviii, p. 506.
Kerbert, C. Zur Trem.-Kenntn., Zool. Anz., 1878, i, p. 271.
— Beitr. z. Kenntn. d. Tremat., Arch. f. mikros. Anat., 1881, xix, p. 519.
Manson, P. Dist. ringeri, Med. Times and Gaz., 1881, ii, p. 8; 1882, ii, p. 42.
Miura, M. Fibr. Tuberkel verurs. durch Parasiteneier, Arch. f. path. Anat.,
1889, cxvi.
Montel, R. Distomiase pulm. en Cochinchine, Annal. d’ Hyg. et de Méd. Col.,
1906, ix, p. 258.
Railliet, A. Paras, des anim. domest. du Japon, Le Natural., 1891, xii, p. 143.
Stiles, C. W. Notes on Parasites, No. 26; Dist. (Mesogon.) westermanni,
Discovery of a Parasite of Man, new to the United States, Vet. Journ.,
1894, p. 107.
Stiles, C. W., and A. Hassall. Notes on Parasites, No. 50: A Muscle Fluke
in American Swine, XVI Ann. Rep., Bur. of Anim. Industry (1899),
Wash., 1900, p. 559.
— No. 51, The Lung Fluke in Swine, ibid., p. 560.
Taniguchi. Ein Fall von Distomum-Erkrankung des Gehirns mit dem Symptomenkomplex
von Jacksonscher Epilepsie, Arch. f. Psych, u. Nervenheilk.,
1904, xxxviii, No. 1.
Ward, H. B. Dist. westerm. in den Vereinigten Staaten, C. f. B. u. P., 1894,
xiv, p. 362; 1895, xvii, p. 304.
Yamagiva, K. Lungendistomenkrankh. in Japan, Arch. f. path. Anat., 1892,
cxxvii; Zur Ätiologie der Jacksonschen Epilepsie, ibid., 1890, cxix.
Ophisthorchis felineus and Metorchis truncatus (pp. 252 to 255, 261 and 262).
Askanazy, M. Über Inf. d. Mensch. mit Dist. felin. in Ostpreussen u. ihren
Zusammenhang mit Leberkrebs, C. f. B., P. u. Inf. (1), 1900, xxviii,
p. 491; Verh. d. Deutsch. path. Ges., 1900, iii, p. 72.
— Die Ätiologie u. Path. d. Katzenegelerkrankg. d. Mensch., Deutsche med.
Wochenschrift, 1904, xxx, p. 689; Verh. d. Ver. f. wiss. Heilkde. i.
Königsb. i. Pr., 1904, iii, p. 3.
— Weitere Mitteil. üb. d. Quellen d. Inf. mit Dist. felineum, Schrift d. Phys.-oek.
Ges., Königsberg i. Pr. (1905), 1906, xlvi, p. 127.
Braun, M. Die Leberdistomen d. Hauskatze u. verw. Arten, C. f. B. u. Par.,
1893, xiv, p. 381.
— Über ein für den Menschen neues Distomum, ibid., 1894, xv, p. 602.
Cholodkowsky, N. Icones helm. hom., II, St. Petersb., 1898, Taf. xi, 115.
Kamensky, G. Not. helm., I, Charkow, 1900.
Kholodkowsky, N. Sur quelq. rar. paras. de l’homme en Russie, Arch. de
Paras., 1898, i, p. 354.
Rivolta. Sopra una spec. di Distoma nel gatto e nel cane, Giorn. anat., fisiol.
e pat. d. animali, 1884, xvi, p. 20.
Ward, H. B. On Dist. fel. in the United States, Vet. Mag., 1895.
— Notes on Parasites of the Lake Fish: III, On the Structure and the Copulatory
Organ in Microphallus, Stud. Zool. Lab., Univ. Nebraska, May,
1901, p. 174.
Winogradoff, K. Ein neues Dist. a. d. menschl. Leber, Nachr. v. d. k.
Tomskischen Univ. (1891), 1892, iv, p. 116; Ein zweiter Fall von Dist.
sib., ibid., p. 131.
— Über Würmer, welche im menschl. Körper paras., ibid. (1892), 1893, v.
Zwaardemaker, H. Cirrhosis parasit., Arch. f. path. Anat., 1890, cxx, p. 197.
Amphimerus noverca (p. 258).
Cobbold, T. Sp. Synopsis of the Distomidæ, Journ. Linn. Soc., London, 1859,
Zool., v, p. 8; Further Observations on Entozoa, with Experiments,
Trans. Linn. Soc., London, 1862; xxiii, p. 349, pl. 33, figs. 1 and 2.
Lewis, T. R., and D. Cunningham. Micros. and Phys. Res., XI Ann. Rep.
San. Comm. Govt. India, Calcutta, 1872, Appendix C, p. 168.
McConnell, J. F. P. On the Dist. conj. as a Human Entozoon, Lancet, 1876,
i, p. 343; 1878, i, p. 476.
Clonorchis sinensis and Cl. endemicus (pp. 258 to 261, 640, and 641).
Baelz, E. Über einige neue Parasiten des Menschen, Berl. klin. Wochenschr.,
1883, p. 235.
Blanchard, R. Lésions du foie déterm. par la prés. des Douves, Arch. de
Paras., 1901, iv, p. 581.
Cobbold, T. Sp. The New Human Fluke, Lancet, 1875, ii, p. 423.
Gaide. De la distomatose hépatique au Tonkin, Ann. d’Hyg. et de Méd.
Colon., 1905, viii, p. 568; abstracted in Arch. f. Schiffs- u. Trop.-Hyg.,
1906, x, p. 256.
Ijima, J. Dist. endemicum, Journ. Coll. Sci., Imp. Univ., Japan, 1886, i, p. 47.
Inouye, Z. Über d. Distom. spathulatum, Arch. f. Verdauungskrankh., 1903,
ix, p. 107.
Katsurada, F. Beitr. z. Kenntn. d. Dist. spathulat., Beitr. z. path. Anat. u. z.
allg. Path., 1900, xxviii, p. 479.
Looss, A. On some Parasites in the Museum of the School of Trop. Med.,
Liverpool, Ann. Trop. Med. and Par., 1907, i, p. 123.
McConnell, J. F. P. Remarks on the Anatomical and Pathological Relations
of a New Species of Liver-fluke, Lancet, 1875, ii, p. 271; 1878, i, p. 406.
McGregor. A New Form of Paralytic Disease Associated with the Presence
of a New Species of Liver Parasite, Lancet, 1877, i, p. 775.
Moty. Lésions anat. prod. par le Dist. sinense, C. R. Soc. Biol., Paris, 1893,
p. 224.
Saito, S. Über den Eiinhalt d. Dist. spathul. u. d. morphol. Beschaffenh.
seines Embryos, C. f. B., P. u. Inf., i Orig., 1906, xlii, p. 133.
Heterophyes heterophyes (pp. 262 to 264).
Blanchard, R. Note prél. sur le Dist. heterophyes, C. R. Soc. Biol., Paris,
1891 (9), iii, p. 792.
Looss, A. Über d. Bau von Dist. heteroph. u. D. fraternum n. sp., Cassel,
1894.
Looss, A. Not. z. Helminth. Ägypt., I, C. f. B., P. u. Inf., 1896 (1), xx,
p. 836.
— Weitere Beitr. z. Kenntn. d. Tremat.-Fna. Ägypt., Zool. Jahrb., Syst. 1899,
xii, p. 699.
— Notz. z. Helminth. Ägypt, V, C. f. B., P. u. Inf., 1902, i Orig., xxxii, p. 886.
Sandwith, F. M. Dist. heterophyes in a Living Patient, Lancet, 1899, ii, p. 888.
Siebold, C. Th. v. Beitr. zur Helminth. hum., Z. f. wiss. Zool., 1852, iv, p. 52.
Dicrocœlium dendriticum (pp. 266, 267).
Anglas, J., and E. de Ribaucourt. Etude anat. et hist. du Dist. lanceolatum,
Ann. Sci. Nat., 8vo, sér. xv, 1902, p. 313; cf. Zool. Centralbl., 1902, ix,
p. 840.
Aschoff, L. Ein Fall von Dist. lanc. i. d. menschlichen Leber, Arch. f. path.
Anat., 1892, cxxx, p. 493.
Brera, V. L. Memor. fis.-med. sopra i princ. vermi di corpo, Roma, 1811.
Dubini, A. Entozoograf. umana, Milano, 1850.
Galli-Valerio, B. Notes de parasitol., B, Paras. anim. (4) Œufs de Dicr. lanc.
dans les fèces de l’homme, C. f. B., P. u. Inf., 1905, i Orig., xxxix, p. 239.
Hollack, J. Z. Kenntn. der sexuellen Amphitypie bei Dicrocoel., C. f. B., P.
u. Inf., 1902, i Orig., xxxii, p. 867.
Kamensky, S. Notes helm. No. 2, Sur la prés. réelle du Dicroc. lanceol. chez
le chien, Trav. Soc. Nat. Univ. Charkow, 1902, xxxvi, p. 63.
Mehlis, C. F. S. Observ. anat. de Dist. hep. et lanceol., Gotting., 1825.
Piana, G. P. Le cercarie d. moll. stud. in rapp. colla pres. del Dist. epat. e
D. lanc., La clinica veter., 1882, v.
Walter, G. Beitr. zur Anat. und Histol. einig. Tremat., Arch. f. Naturg., 1858,
xxiv (1), p. 269.
Zschokke, F. Selt. Paras. des Menschen, C. f. B. u. P., 1892, xii, p. 500.
Schistosoma hæmatobium (pp. 270 to 277 and 641 to 644).
Balfour, A. Eosinophilia in Bilharzia dis. and Dracontiasis, Lancet, 1903, ii,
p. 1649.
Beyer, H. G. A Second Chinese Case of Infection with the Asiatic Blood
Fluke, Amer. Med., 1905, x, p. 578.
Bilharz, Th. Beitr. zur Helminth. hum., Z. f. w. Zool., 1852, ii, pp. 53, 454.
Catto, J. Schistosoma cattoi, a New Blood Fluke of Man, Brit. Med. Journ.,
January 7, 1915; abstracted in C. f. B., P. u. Inf., 1906, i, Ref. xxxvii,
p. 617.
Chaker, M. Et. sur l’hématurie d’Egypt, Thèse, Paris, 1890.
Chatin, J. Obs. sur le dével. et l’org. du proscol. de la Bilh., Ann. sc. nat.
Zool., 1881 (6), xi.
Fritsch, G. Zur Anat. d. Bilh. haemat., Arch. f. mikr. Anat., 1888, xxxi,
p. 192.
Fujinami, A. Weitere Mitt. über die path. Anat. d. sog. Katayama-Krankh.
und der Krankheits-Er. ders., Kyoto Igaku Zassi, March 1, 1904. (In
Japanese, with German abstract.)
Fujinami, A., and J. Kon. Beitr. z. Kenntn. der pathol. Anat. der sog. Katayama-Krankheit,
ibid., 1904, i, 1; abstracted in C. f. B., P. u. Inf., 1905,
i, Ref. xxxvi, p. 499.
Gunn. Bilharzia Disease, Journ. Amer. Med. Assoc., 1906, No. 14; abstracted
in C. f. B., P. u. Inf., 1907, i, Ref. xxxix, p. 217.
Kartulis. Vorkommen d. Eier von Dist. haemat., Arch. f. path. Anat., 1885,
xcix, p. 139.
— Weitere Beitr. z. path. Anat. d. Bilharz., ibid., 1898, clii, p. 474.
Kasai, K. Unters. über die sog. Katayama-Krankh., Mitt. der med. Ges.,
Tokio, 1904, xviii, p. 4; abstracted in C. f. B., P. u. Inf., 1905, i, Ref.
xxxvi, p. 499.
Katsurada, F. Schistosomum japonicum, ein neuer menschl. Paras., durch
welchen eine endem. Krankh. in versch. Gegend. Japans verursacht wird,
Annot. zool. japon., 1904, v, 3, p. 147.
Lahille, A. La bilharziose intest. aux Antilles, Ann. d’Hyg. et de Méd. Colon.,
1906, ix, p. 262.
Letulle, M. Bilharziose intestinale, Arch. de Paras., 1905, ix, p. 329.
Looss, A. Beob. über Eier und Embr. v. Bilh., in Leuckart: Die Paras. d.
Menschen, 2. Aufl., i, p. 521.
— Bemerkungen zur Lebensgeschichte der Bilh. haem., C. f. B. u. P., 1894, xvi,
pp. 286, 340.
— Rech. faun. paras. de l’Egypt, Mém. Inst. égypt., 1895, iii, p. 158.
— Zur Anat. und Histol. d. Bilh. haem., Arch. f. mikr. Anat., 1895, xlvi, p. 1.
— Bilharziosis, Handb. d. Tropenkrankh., 1905, i, p. 93.
— Schistosomum japonicum, eine neue asiatische Bilharzia d. Menschen,
C. f. B., P. u. Inf., 1905, i Orig., xxxix, p. 280.
Lortet and Vialleton. Etud. sur la Bilh. hæm., Paris, 1895; Ann. de l’Univ.
de Lyon, 1894, ix.
Ogawa. Beitr. zur Kenntn. der Katayama-Krankh., Kyoto Igaku Zassi, 1904,
i, p. 3; with German abstract.
Railliet, A. Obs. sur l’embr. du Gynæcoph. hæm., Bull. Soc. Zool. France,
1892, xvii, p. 101.
Rütimeyer, L. Über Bilharzia-Krankheit, Mitt. klin. u. med. Inst. d. Schweiz.,
i, 1894, xii, p. 871.
Scheube, B. Ein neues Schistosomum beim Menschen, Arch. f. Schiffs- u.
Tropen-Hyg., 1905, ix, p. 150.
Sonsino, P. Ric. s. sviluppo d. Bilh., Giorn. R. Ac. med., Torino, 1889, xxxii,
p. 380.
Stiles, C. W. The New Asiatic Blood Fluke . . . of Man and Cats, Amer.
Med., 1905, ix, p. 821.
Williamson. Bilharzia hæmatobium in Cyprus, Brit. Med. Journ., 1902, p. 956.
Woolley, P. G. The Occurrence of Schistosoma japonicum vel cattoi in the
Philippine Islands, Philipp. Journ. Sci., 1906, i, p. 83.
Class III.—Cestodes (pp. 282 to 359 and 644 to 674).
[An almost complete collection of the literature relating to Cestodes up to
1895 is to be found in Braun’s monograph on the tapeworms in Bronn’s
“Klassen und Ordnung des Thierreiches,” iv, p. 2; of later books the following
may be mentioned.]
Bartels, E. Cysticercus fasciolaris, Anat., Beitr. zur Entw. und Umwandl. in
Taenia crassicollis, Zool. Jahrb. Anat., 1902, xvi, p. 511.
Blanchard, R. Sur quelq. Cest. monstr., Progr. méd., 1894 (2), xx.
Blochmann, F. Die Epithelfrage bei Cestoden und Tremat., Hambg., 1896.
Boas, J. E. V. Triplotaenia mirabilis, Zool. Jahrb., 1902, Syst. xvii, p. 329.
Brandes, G. Teratol. Cestoden, Ztschr. f. d. ges. Nat., Halle, 1899, p. 105.
Bugge, G. Zur Kenntn. der Excretionsgefäss-Syst. der Cestoden und Tremat.,
Zool. Jahrb. Anat., 1902, xvi, p. 177.
Child, C. M. Abnormality in Moniezia expansa, Biol. Bull. Woods Holl.,
1902, iii, pp. 95, 143.
Cohn, L. Zur Anat. und Syst. der Vogelcestoden, Nov. Act. Acad. Caes. Leop.-Carol.
Nat. Cur., Halle, 1901, lxxix, No. 3.
Drago, U. Azione sperim. dei succhi diger. sull’ involucro della ova di alc.
Tenie, Arch. de Paras., 1906, x, p. 321.
Fuhrmann, O. Ein getrenntgeschlechtlicher Cestode, Zool. Jahrb., 1904, Syst.
xx, p. 131.
Grohmann, W. Die Abnormitäten in den Progl. der Cestoden, Inaug.-Dissert.,
Giessen, 1906.
Kunsemüller, F. Zur Kenntnis der polycephalen Blasenwürmer, insbesondere
des Coenurus cerebralis Rud. und des C. serialis Gerv., Zool. Jahrb.
Anat., 1903, xviii, p. 507.
Lühe, M. Zur Anat. und Syst. der Bothrioceph., Verhandl. der Deutsch. zool.
Ges., 1899, p. 30.
— Review of Braun’s Bothr.-Syst., C. f. B., P. u. Inf., 1902, i Orig., xxi,
p. 318.
Messineo, G. Sul veleno conten. in alcune Tenie dell’ uomo, Atti Accad.
Gioenia sci. nat., Catania, 1901 (4), xiv, No. 6.
Mingazzini, P. Sul vario modo di fissaz. delle Tenie alia parete intest., Rich.
Labor. anat., Roma, 1904, x, p. 5.
Rössler, P. Über den fein. Bau der Cysticerken, Zool. Jahrb. Anat., 1902,
xvi, p. 423.
Saint-Rémy, G. Dévelop. embr. Taenia serrata, Arch. de Paras., 1901, iv,
p. 143.
Schaaf, H. Zur Kenntn. der Kopfanlage der Cysticerken, insbes. des Cysticercus
der Taenia solium L., Zool. Jahrb. Anat., 1906, xxii, p. 435.
Spengel, J. W. Die Monozootie der Cestoden, Ztschr. f. w. Zool., 1905, lxxxii,
p. 252.
Stiles, Ch. W. Revision of Ad. Tapeworm of Hares and Rabbits, Proc. U.S.
Nat. Mus., 1896, xix.
Stiles, C. W., and A. Hassall. Tapeworms of Poultry, U.S. Dept of Agric.,
Bur. of Anim. Ind., 1896, Bull. 12.
Vigener, J. Über dreikant. Bandwürmer a. d. Fam. d. Taeniiden, Jahrb. nass.
Ver. f. Naturke., Wiesb., 1903, p. 115.
Zernecke, F. Unters. über d. fein. Bau d. Cestod., Inaug.-Diss., Rostock, 1895.
Dibothriocephalus latus (pp. 310 to 315, and 658).
(a) Anatomy.
Böttcher, A. Studien über den Bau des Bothr. latus, Arch. f. path. Anat.,
1864, xxx, p. 97; 1869, xlvii, p. 370.
Eschricht, D. F. Anat.-phys. Untersuchung. über die Bothrioceph., Nov. Act.
Ac. Caes. Leop.-Carol. nat. curios., 1841, xix, Suppl. ii.
Schmidt, F. Beitr. z. Kenntn. d. Entwickl. d. Geschlechtsorg. d. Cestoden,
Z. f. w. Zool., 1888, xlvi, p. 155.
Sommer, F., and L. Landois. Beitr. z. Anat. d. Plattw.: I, Bothr. latus, Z. f.
w. Zool., 1872, xxii, p. 40.
Stieda, L. Zur Anat. d. Bothr. latus, Arch. f. Anat. u. Phys., 1864, p. 174.
(b) Development of Embryos.
Bertolus. Sur le dévelopm. du Bothrioceph. de l’homme, C. R. Acad. Sci.,
Paris, 1863, lvii, p. 569.
Knock, J. Die Naturg. d. breiten Bandw. mit bes. Berücks. sein. Entw., Mém.
Acad. d. Sci., St. Pétersbourg, 1862 (7), v, p. 5; Journ. de l’Anat., 1879,
vi, p. 140.
Schauinsland, H. Die embryon. Entw. d. Bothrioceph., Jen. Zeitschr. f.
Naturw., 1885, xix, p. 520.
(c) Infection.
Alessandrini, G. Bothr. latus Br. nella prov. di Roma, Boll. soc. zool. ital.,
1906, ser. 2, vii, p. 231.
Braun, M. Zur Frage d. Zwischenwirth. von Bothr. latus, Zool. Anzeiger,
1881, iv, p. 593; 1882, v, pp. 39, 42, 194; 1883, vi, p. 97.
— Bothriocephalus latus u. seine Herkunft., Arch. f. path. Anat., 1883, xcii,
p. 364.
— Zur Entwickel. d. breit. Bandw., Würzburg, 1883.
— Salm oder Hecht? Berl. klin. Wochenschr., 1885, xxii, p. 807.
— Über den Zwischenwirth des breit. Bandw., eine Entgegnung an Küchenmeister,
Würzburg, 1886.
— Bothriocephalus-Finnen im Hecht des St. Petersb. Fischmarktes, St. Petersb.
med. Wochenschr., 1892, xvii, No. 28.
— Helminth. Notizen, I, C. f. B. u. Par., 1893, xiv, p. 802.
Grassi, B., and Ferrara. Zur Bothriocephalusfrage, D. med. Wochenschr.,
1886, p. 699.
Grassi, B., and G. Rovelli. Contrib. allo studio d. svil. d. Bothr. latus, Giorn.
R. Accad. Med., 1887, No. 11.
— — Bandwürmerentwickelung, C. f. B. u. Par., 1888, iii, p. 173.
Ijima, J. The Source of Bothr. latus in Japan, Journ. Coll. Sci., Imp. Univ.,
Tokio, 1888, ii, 1, p. 49.
Küchenmeister, F. Wie steckt sich der Mensch mit Bothr. latus an? Berl.
klin. Wochenschr., 1885, xxii, pp. 505, 527.
— Die Finne des Bothrioceph. u. seine Übertrag. a. d. Menschen, Leipzig,
1886.
— Weit. Bestätigung meiner Behamptung, die Finne des Hechts hat nichts mit
Bothr. latus zu thun, D. med. Wochenschr., 1886, p. 551.
Leuckart, R. Zur Bothriocephalusfrage, C. f. B. u. Par., 1887, i, pp. 1, 33.
Lönnberg, E. Über das Vork. d. breit. Bandw. in Schweden, C. f. B. u. Par.,
1892, xi, p. 189.
Parona, E. The Bothr. latus in Lombard., Rend. R. Istit. Lomb., 1886 (2), xix,
fasc. 14.
— Sulla quest. d. Bothr. latus, Gazz. med.-ital.-lomb., 1887.
Schroeder, A. v. Wie bek. die Einwohner St. Petersb. d. breit. Bandw., St.
Petersb. med. Wochenschr., 1892, xvii, No. 22.
— im Wratsch, 1894, No. 12; 1895, No. 15; Jesched. journ. prakt. med., 1896,
Nos. 19 and 27; abstracted in C. f. B. u. Par., xvi, p. 314; xviii, p. 24;
xx, p. 621.
Zschokke, F. Weit. Zwischenwirthe des Bothr. latus, C. f. B. u. Par., 1888,
iv, p. 417; 1890, vii, pp. 393, 435.
(d) Geographical Distribution Statistics.
Bavay. Sur la prés. du Bothr. latus à Madagascar, Bull. Soc. Zool. France,
1890, xv, p. 134.
Beneden, E. van. Sur la prés. en Belgique du Bothr. latus, Bull. Acad. roy.
Belg., 1886, 3e sér., xi, 8, p. 265.
Berkeley, W. N. A Case of Bothriocephalus with Remarks on the Occurrence
of Bothriocephalus in America, Med. Rec., New York, 1903, lxiii, p. 355.
Bollinger, O. Über das autochth. Vork d. Bothr. latus in München nebst
Bemerk. über die geogr. Verbreitung der Bandw., Dtsch. Arch. f. klin.
Med., 1885, xxxvi, p. 277.
Grusdieff, S. S. Zur Frage der Verbreit. tier. Darmparas. bei der Schuljugend,
Wratsch, 1891, No. 13.
Hahn, L. Le Bothriocéph., son dével., ses migrat., sa distrib. géogr. et sa
prophyl., Gaz. hebd. méd. et chir., 1885 (2), xxii, p. 450.
Huber, J. Über die Verbreitung der Cestoden in Schwaben., Ber. d. naturhist.
Ver., Augsburg, f. 1886, p. 85.
Kerbert, E. Bothr. latus Br. (in Nederland), Nederl. Tijdschr. v. Geneeskde.,
1889, i, p. 424; Handel van het II Nederl. Nat. en Geneesk. Congr.,
Leiden, 1889.
Kessler, D. A. Beitr. z. Statistik der Eingeweidew. bei den Einwohnern
Petersburgs, Wratsch, 1888, pp. 109, 128.
Kiaer, F. C. Baendelorm hos mennesk. i Norge, Tidsskrift. f. pr. med.,
Kristiania, 1889, p. 1; abstracted in C. f. B. u. Par., 1889, v, p. 353.
Krabbe, H. 300 Tilfälde af bändelorm hos mennesket jagt. i Danmark, Nord.
med. Arkiv, 1887, xix, 1.
— Über das Vork. von Bandw. beim Menschen in Dänemark, ibid., 1905,
Abt. ii, i, No. 2.
Leon, N. Note sur la fréq. des Bothrioc. en Roumanie, Bull. Soc. d. Sci. de
Bucarest, 1904, xiii, p. 286.
Nickerson, W. S. The Broad Tapeworm in Minnesota, Journ. Amer. Med.
Assoc., 1906, p. 711.
Schor, M. Contrib. à l’ét. du Bothr. latus dans le canton de Vaud, Thèse,
Lausanne, 1902; abstracted in C. f. B., P. u. Inf., 1903, Abt. i, Ref. xxxiii,
p. 286.
Sievers, R. Zur Kenntnis der Verbreit. von Darmparas. des Menschen in
Finnland, Festschr. f. Palmén, No. 10, Helsingfors, 1905.
Szydlowski. Beitr. zur Mikroskopie der Faeces, Inaug.-Diss., Dorpat, 1879.
Vanlair, C. Un nouv. cas de bothriocéphalie en Belg., Bull. Acad. roy. Belg.,
1889, 3e sér., xviii, p. 379.
Wellman, C. Notes on Tropical Diseases of the Angola Highlands, New York
Med. Journ., August 12, 1905, and Phil. Med. Journ., September 2, 1905.
Willson, R. N. Bothr. latus . . . Amer. Journ. Med. Sci., Philadelphia,
1902, p. 262.
Zaeslin, Th. Über die geogr. Verbreit. u. Häufigkeit der Entozoen in der
Schweiz, Corresp.-Bl. f. schweiz. Ärzte, 1881, xi, p. 673.
Zschokke, F. Der Bothr. latus in Genf, C. f. B. u. Par., 1887, i, pp. 377, 409.
(e) Bothriocephalus Anæmia.
Askanazy, M. Über Bothrioceph.-Anämie und die progn. Bedeutung d.
Megaloblast, Ztschr. f. klin. Med., xxvii, p. 492.
Babes, V. Über den Bothr. latus und die Bothr.-Anämie, Arch. f. path. An.,
1895, cxli, p. 204.
Fédorov, N. L’anémie bothriocéph., Arch. de Paras., 1902, vi, p. 207.
Isaac, S., and van den Velden. Eine specif. Präcipitinreaktion bei Bothr.
latus beherb. Menschen, Dtsche. med. Wchschr., 1904, xxx, p. 982.
Meschede. In: Tagebl. d. 45. Vers. dsch. Naturf. u. Ärzte in Leipzig, 1872,
p. 186; Ztschr. f. Psych., 1874, xxx, p. 109.
Möller, W. Stud. öfv. de hist, förändr. i. dig.-Kan. . . . vid Bothrioc.
anaem., Helsingfors, 1897.
Neubecker, O. Bothrioc.-Anämie ohne Bothrioceph., Inaug.-Diss., Königsb.,
1898.
Reyher, G. Beitr. zur Ätiol. u. Heilbark. der pernic. Anämie, Dtsch. Arch. f.
klin. Med., xxix, p. 31.
Rosenquist, E. Über den Eiweissstoffwechsel bei der pernic. Anämie, mit
spezieller Berücks. der Bothrioc.-Anämie, Ztschr. f. klin. Med., 1903,
xlix, p. 193.
Runeberg, J. W. Bothr. latus und pernic. Anämie, ibid., 1886, xli, p. 304.
Schapiro, H. Heilung der Biermerschen pernic. Anämie durch Abtreibung von
Bothr. latus, Ztschr. f. klin. Med., 1889, xiii, p. 416.
Schaumann, O. Zur Kenntnis der sog. Bothr.-Anämie, Berlin, 1894.
Thomson, W. G. A Case of Dibothriocephalus latus Infection causing Pernicious
Anæmia with Complications; Recovery, Med. News, 1905, lxxxvi,
p. 635.
Zinn, W. Tödl. Anämie durch Bothr. latus, D. med. Wchschr., 1903, xxix,
p. 264.
(f) Duration of Life.
Bremser. Über lebende Würmer im leb. Menschen, Wien, 1819.
Leuckart, R. Die Paras. d. Mensch., &c., Leipz., 1881, 2. Aufl., i, 2; and
Arch. f. Naturg., 40. Jahrg., 1874, ii, p. 446.
Mosler, F. Über Lebensdauer u. Renitenz d. Bothr. latus, Arch. f. path.
Anat., 1873, lvii, p. 529.
Dibothriocephalus cordatus (pp. 315, 316).
Braun, M. Berichtig. betr. d. Vork. v. Bothr. cordatus in Dorpat, Zool. Anz.,
1882, v, p. 46.
Leuckart, R. Jahresb. üb. d. wiss. Leist. i. d. Naturgesch. d. nied. Thiere f.
1861–62, Arch. f. Naturgesch., 29. Jahr., 1863, p. 149.
— Die menschl. Paras., 1863, i, p. 437.
Diplogonoporus grandis (pp. 316, 317).
Blanchard, R. Not. sur les paras. de l’homme, IV, C. R. Soc. Biol., Paris,
1894 (10), i, p. 699.
Ijima and T. Kurimoto. On a New Human Tapeworm, Journ. Coll. Sci., Imp.
Univ., Tokio, 1894, vi, p. 371.
Kurimoto, T. Diplogonoporus grandis, Zeitschr. f. klin. Med., 1900, xl, p. 1.
Sparganum mansoni (pp. 317, 318, and 659).
Cobbold, P. Sp. Description of Ligula mansoni, Linn. Soc. Journ. Zool.,
Lond., 1883, xvii, p. 78.
Ijima, J., and Murata. Some New Cases of the Occurrence of Bothriocephalus
ligula, Journ. Coll. Sci., Imp. Univ., Tokio, 1888, ii, p. 149.
Leuckart, R. Demonstr. eines selt. menschl. Entoz., Tagebl. 57, Vers. d.
Naturf. u. Ärzte zu Magdeburg, 1884, p. 321.
— Die Paras. d. Mensch., 2. Aufl., i, p. 941.
Manson, P. Case of Lymph Scrotum associated with Filariæ and other Parasites,
Lancet, 1882, ii, p. 616.
Miyake, H. Beitr. z. Kenntn. d. Bothrioc. ligul., Mitt. Grenzgeb. Med. u.
Chir., 1904, xiii, p. 145.
Stiles, C. W., and L. Tayler. A Larval Cestode of Man . . . U.S. Dept.
of Agric., Bur. of Anim. Ind., Bull. No. 35, Wash., 1902, p. 47.
Sparganum proliferum (pp. 318 to 320).
Ijima, J. On a New Cestode Larva Parasitic in Man, Journ. Coll. Sci., Imp.
Univ., Tokio, 1905, xx, Article 7.
Dipylidium caninum (pp. 320 to 323 and 659 to 661).
(a) Anatomy and Development.
Diamare, V. Il genere Dipylidium, Atti R. Accad. sci. fis. e met. Napoli,
1893, 2 ser., ii, No. 7.
Grassi, B., and G. Rovelli. Embryol. Forsch. an Cestoden, C. f. B. u. Par.,
1889, v, p. 370.
— — Ric. embryol. sui Cestodi, Atti Gioen. sci. nat., Catania, 1892, 4 ser., iv.
Melnikow, W. Über d. Jugendzust. d. Taen. cucum., Arch. f. Naturg., 1869,
xxxv, i, p. 62.
Sonsino, P. Ric. s. ematoz. del cane e sul ciclo evol. d. T. cucum., Atti soc.
tosc. sci. nat., 1888, x, p. 1.
Steudener, F. Unters. über d. fein. Bau d. Cestod., Abh. nat. Ges., Halle,
1877, xiii, p. 295.
(b) Case Histories (New Cases Only).
Asam, W. Taenia cucumerina bei einem Kinde, Münch. med. Wochenschr.,
1903, l, p. 334.
Bollinger, O. Über T. cucum. beim Mensch., Deutsch. Arch. f. klin. Med.,
1905, lxxxiv, p. 50.
Brandt, E. Zwei Fälle von T. cucum. beim Mensch., Zool. Anz., 1888, xi,
p. 481.
Freriks, B., and C. W. Broers. Een T. cucum. bij een Kind, Weekbl.
Nederl. Tijdsch. v. Geneesknde., 1904, Deel. ii, p. 33.
Hoffmann, A. Taen. cucum. bei einem 4 Monate alten Kinde, Jahrb. f.
Kinderheilkde., 1887, N.F. xxvi.
Köhl, O. Taen. cucum. bei einem sechs Wochen alten Kinde, Münch. med.
Wochenschr., 1904, li, p. 157.
Krüger, F. In St. Petersb. med. Wochenschr., 1887, No. 41.
Rosenberg, L. Zehn Bandwürm. bei einem vierzehn Monate alten Kinde,
Wien. med. Wochenschr., 1904, liv, p. 427.
Sonnenschein, G. Taen. cucum. s. elliptica bei einem sechs Monate alten
Kinde, Münch. med. Wochenschr., 1903, No. 52.
Triis. In Nord. Med. Arkiv., 1884, xvi, No. 6.
Zschokke, F. Ein neuer Fall von Dipylidium caninum beim Mensch., C. f. B.,
P. u. Inf., 1903, i Abt., xxxiv, p. 42.
— Dipyl. canin. als Schmarotz. d. Mensch., ibid., 1905, i Orig., xxxviii, p. 534.
Hymenolepis nana (pp. 323 to 326, 661 and 662).
Blanchard, R. Hist. zool. et méd. d. Téniad. du genre Hymenolepis, Paris,
1891.
Capazzo, Z. Due casi di T. nana, Riv. clin. pediatr., 1904, ii, p. 829.
Carrer, C. Un caso di T. nana, Riv. venet. sci. med., 1905, xxii, T. xliii, 2,
p. 509.
Comini, E. Epilessia rifl. da T. nana, Gazz. d. osp., 1887, viii, p. 174.
— Due casi di T. nana, Gazz. med. ital.-lomb., 1888, p. 81.
Foster, Ch. L. Two Cases of Infection with T. nana in the Philippine
Islands, Journ. Amer. Med. Assoc., 1906, xlvii., p. 685.
Grassi, B. Die T. nana und ihre med. Bedeutung, C. f. B. u. Par., 1887, i,
p. 97.
— Einige weit. Nachr. üb. T. nana, ibid., 1887, ii, p. 282.
— Entw. d. T. nana, ibid., p. 305.
— Cenno prev. int. ad una nuov. mal. par. nell’ uomo, Gazz. d. osp., 1886,
viii, pp. 450, 619.
Grassi, B., and G. Rovelli. Ric. embr. sui Cestodi, Atti Accad. Gioen. sci.
nat., Catania, 1892, 4. ser., iv.
Hallock. Tænia nana, Report of Two Cases, Journ. Amer. Med. Assoc.,
April 2, 1904.
Linstow, v. Über Taenia nana u. T. murina, Zeitschr. f. d. ges. Naturw.,
1896, p. 571.
Lutz, A. Beobacht. üb. d. als Taenia nana u. T. flavopunct. bek. Bandw. d.
Mensch., C. f. B. u. P., 1894, xvi, p. 61.
Mertens, in Berl. klin. Wochenschr., 1892, Nos. 44, 45.
Miura, K., and Yamazaki. Über T. nana, Mitt. med. Facult., kais. Univ.,
Tokio, 1897, iii, p. 239.
Moniez, R. Sur la Tænia nana, C. R. Acad. Sci., Paris, 1888, cvi, p. 368.
Orsi, F. Sei casi d. T. nana, Gazz. med. ital.-lomb., 1889, 9 ser., ii, xlviii,
p. 235.
Perroncito, E. Caso di T. nana, Giorn. R. Accad. Med., Torino, 1887, xxxv,
P. 7.
Perroncito, E., and P. Airoldi. Caso di T. medioc. e di molte T. nana, ibid.,
1888, xxxvi, p. 312; Gazz. d. osp., 1888, p. 554.
Ransom, B. H. An Account of the Tapeworms of the Genus Hymenolepis,
Parasites of Man, U.S. Hygien. Lab., Bull. No. 18, 1904.
Ransom, W. H. Probable Existence of T. nana . . . in England, Lancet,
1888, ii.
Rasch, Chr. Taenia nana in Siam, D. med. Ztg., 1895, p. 143.
Roeder, H. Über ein. weit. Fall v. T. nana in Deutschland, Münch. med.
Wochenschr., 1899, p. 344.
Senna. Stor. clin. dei sei casi d. T. nana, Gaz. med. ital.-lomb., 1889, xlviii,
9 ser., ii, p. 245.
Siebold, C. Th. v. Ein Beitr. z. Helminthogr. hum., Z. f. w. Zool., 1852, iv,
p. 64.
Sonsino, P. Tre casi d. T. nana nei dint. di Pisa, Riv. ital. clin. med., 1891, iii.
— Nuov. oss. di T. nana, Boll. soc. med., Pisa, 1895, i, p. 4.
Spooner, E. A. Species of T. nana, Amer. Journ. Med. Sci., 1873 (2), lxv,
p. 163.
Stiles, C. W. The Dwarf Tapeworm (H. nana), etc., New York Med. Journ.
and Philad. Med. Journ., 1903, p. 1.
W. S. Wiener Bericht., Med. Klin., 1904, i, p. 71.
Wani, S. Über T. nana in Japan; abstracted in C. f. B., P. u. Inf., 1905,
i, Ref. xxxvi, p. 500.
Wernicke, O. T. nana, Anal. circ. med. Argent., 1890, xiii, p. 349; C. R. Soc.
Biol., 1891 (9), iii, p. 441.
Zograf, N. Note sur la myol. d. Cestod., Congr. int. Zool., IIe sess., Moscou,
2e part., p. 23.
Hymenolepis diminuta (pp. 326 to 328 and 662).
Creplin, F. C. H. Observ. de entozois., I, Gryphisw., 1825, p. 71.
Grassi, B. Bestimmung d. 4 v. Parona . . . gefundenen Taenien, C. f. B.
u. Par., 1887, i, p. 257.
— Taenia flavop., leptoceph., diminuta, Atti R. Accad. Sci., Torino, 1888, xxiii,
p. 492.
Grassi, B., and G. Rovelli. Ric. embr. s. Cestodi, Catania, 1892.
Leidy, J. Occurrence of a Rare Human Tapeworm, Amer. Journ. Med. Sci.,
1884 (2), lxxxiii, p. 110; Proc. Accad. nat. sci., Philadelphia, 1884, p. 137.
Lutz, A. Beob. über d. als T. nana u. flavop. bek. Bandw. d. Mensch., C. f. B.
u. Par., 1894, xiv, p. 61.
Magalhaes, P. G. de. Ein zweit. Fall von Hym. diminuta als menschl. Paras.
in Brasil, ibid., 1896 (1), xx, p. 673.
Packard, F. A. Tænia flavop., with Description of a New Species, Journ.
Amer. Med. Assoc., 1900, xxxv, p. 1551.
Parona, E. Di un caso di T. flavopunct., Giorn. R. Accad. Med., Torino,
1882, xxxii, p. 99.
Previtera, G. Due casi prob. di T. leptoceph. nei minat. d. zolfare, Boll. Acc.
Gioen. sci. nat., Catania, 1900, S.N., fasc. 63, p. 9.
Sonsino, P. Su paras. dell’ uomo con un nuovo caso di T. flavopunct.,
C. f. B., P. u. Inf., 1896 (1), xix, p. 937.
Zschokke, F. Seltene Paras. d. Mensch., ibid., 1892, xii, p. 497.
Hymenolepis lanceolata (pp. 328, 329 and 662).
Bloch, M. E. Abhandlung v. d. Erzeugung d. Eing.-Würmer, Berlin, 1782.
Daday, E. V. Helminth. Stud. Einige in Süsswasser-Entomostr. leb. Cercocystis-Form,
Zool. Jahrb., 1901, Syst. xiv, p. 161.
Feuereisen, J. Beitr. z. Kenntn. d. Taenien, Z. f. w. Zool., 1868, xviii, p. 161.
Mrázek, A. Zur Entwicklung einiger Taenien, Sitzungsber. kgl. böhm. Ges.
d. Wiss., math.-nat. Kl., Prag, 1896, Art. xxxviii.
Wolffhügel, F. Drepanidot. lanceol., C. f. B., P. u. Inf., 1900 (1), xxviii,
p. 49.
Zschokke, F. Hymenol. lanc. als Schmarotzer d. Menschen, ibid., 1902,
Orig., xxxi, p. 331.
Davainea madagascariensis (pp. 329, 330 and 662).
Blanchard, R. Note sur quelq. vers par. de l’homme, C. R. Soc. Biol., Paris,
1891 (9), iii, p. 604.
— Le Dav. madag. à Guyane, Bull. Acad. Med., 1897 (3), xxxvii, p. 34.
— Un cas inéd. de Dav. madag., consid. sur le genre Davainea, Arch, de Paras.,
1899, ii, p. 200.
Chevreau, P. Le Tænia madag., Bull. Soc. Méd. de l’île Maurice, ix, p. 134.
Daniels, C. W. Tænia demerariensis, Brit. Guiana Med. Ann. Hosp. Rep. for
1895, Lancet, 1896, ii, p. 1455.
Grenet and Davaine. Note sur une nouv. esp. de Taenia rec. à Mayotte, Mém.
Soc. Biol., Paris, 1869 (5), i, p. 233; Arch. Méd. nav., 1870, xiii, p. 134.
Leuckart, R. Über Taenia madagascariensis, Verh. d. D. zool. Ges., Leipzig,
1891, i, p. 68.
Davainea (?) asiatica (pp. 330 and 662).s
Linstow, v. Taenia asiatica, eine neue Taenie des Menschen, C. f. B., P. u.
Inf., 1901 (1), xxix, p. 982.
Tænia solium and T. saginata (pp. 331 to 336, 338 to 342, and 662 to 674).
Beneden, E. van. Rech. sur le dével. embryol. d. quelq. Tén., Arch. de Biol.,
1881, ii, p. 183.
Bérenger-Feraud, L. J. B. Leç. clin. sur les Ténias de l’homme, Paris, 1888.
Borchmann. Über die Häufigkeit von Cyst. cellul. beim Reh., Zeitschr. f.
Fleisch- u. Milchhyg., 1904, xv, p. 39.
Dirksen, E. Über schwere Anämie d. Taenia solium, Deutsche med. Wochenschr.,
1903, No. 29.
Gerlach, A. C. Fütterungsvers. bei Schweinen mit T. solium, Jahresber.
klg. Thierarzneisch., Hannover (1869), 1870, ii, pp. 66, 69.
Haubner, G. C. Über d. Entw. d. Band- u. Blasenw. . . . Mag. f. d. ges.
Thierheilk., 1854, xx, pp. 243, 366; 1855, xxi, p. 100.
Küchenmeister, E. Über Cysticerk. im allg. u. die des Mensch. insbes.
Zittau, 1853.
— Experimenteller Nachweis, dass Cyst. cellulosae sich in Taenia solium
umwandelt., Wiener med. Wochenschr., 1855, p. 1; 1856, p. 319; Deutsche
Klinik, 1860, xii, p. 187.
Leuckart, R. Die Blasenbandw. u. ihre Entw., Giessen, 1856.
— Finnenzustand d. Taenia mediocan., Gött. Nachr., 1862, pp. 13; 195.
Moniez, R. Ess. monogr. sur les Cysticerques, Trav. Institut. Zool., Lille,
1880, iii, p. 1.
— Mém. sur les cestodes, ibid., 1881, iii, p. 2.
Mosler, F. Helminthol. Studien u. Beobacht., Berlin, 1864.
Perroncito, E. Gli Abissini e la Tænia mediocanellata, Giorn. R. Accad. d.
Med., Torino, 1891, Nos. 3, 4.
— Esper. s. prod. del Cystic. della T. medioc. . . . Ann. R. Accad. Agric.,
Torino, 1877, xx; Zeitschr. f. Vet.-Wiss., 1877, v.
Schwarz. Zur Unterscheidung d. Cyst. cell. v. C. tenuicollis, Zeitschr. f.
Fleisch- u. Milchhyg., 1893, iii, p. 89.
Sommer, F. Bau u. Entwickl. d. Geschlechtsorg. von T. medioc. u. T. solium,
Z. f. w. Zool., 1874, xxiv, p. 499.
Steudener, F. Unters. über d. Bau d. Cestod., Halle, 1877.
Cysticercus cellulosæ and C. bovis in Man (pp. 332 to 337, and 340 to 342).
Gross, J. Über Cyst. racemosus d. Gehirns, Inaug.-Diss., Leipzig, 1903.
Henneberg, R. Über Gehirncysticerken, insbesondere d. basale Cysticerken-meningitis,
Charité-Annal., 1906, xxx, p. 202.
Hirschberg, J. Die Verminderung der Finnenkrankheiten, Berl. klin. Wochenschr.,
1904, xli, p. 661.
Kühn, H. Cystic. cellul. i. d. Lungen, Inaug.-Diss., Leipzig, 1905.
Lewin. Cystic. cellul. u. sein Vork. in der Haut d. Menschen, Charité-Annal.,
1877, ii, p. 609; Arch. f. Dermatol., 1894, xxvi, pp. 71, 217.
Marchand, F. Über Gehirncysticerken, Volkmanns Sammlung klin. Vortr.,
1904, No. 371.
Utz, F. Ein Fall von Cyst. racem. d. Gehirnbasis, Inaug.-Diss., München,
1902.
Volovatz, E. Ladrerie ou cysticercose de l’homme, Thèse, Paris, 1902.
Zenker, F. A. Cystic. racemosus d. Gehirns, Bonn, 1882.
Cysticercus acanthotrias (pp. 336, 337).
Braun, M. Helminth. Notiz. III: Cyst. tenuicollis u. C. acanthotrias beim
Mensch., C. f. B. u. Par., 1894, xv, p. 409.
Cobbold, T. Sp. On a Rare and Remarkable Parasite from the Collection of
the Rev. W. Dallinger, Rep. 40th Meeting Brit. Assoc. Adv. Sci., 1870–71,
Note, p. 135.
Delore, X. Cystic. acanthotrias obs. chez une jeune fille, Mém. et C. R. Soc.
Sci. Med., Lyon, 1863, ii, p. 202.
Leuckart, R. Die menschl. Paras., 1863, i, p. 310.
Redon. Exp. sur le dével. rubanaire de Cyst. de l’homme, C. R. Acad. Sci.,
Paris, 1877, lxxxv, p. 675; Gaz. méd., Paris, 1877, xlviii, p. 519; Arch.
vétér. publ. à l’École d’Alfort, 1877, ii, p. 910; Ann. Sci. Nat. Zool.,
1877 (6), vi, No. 4.
Weinland, D. F. An Essay on the Tapeworms of Man, Cambridge, U.S.,
1858.
— System. Catalog aller Helminth., die im Menschen gefunden werden, Arch.
f. Naturg., 1859, xxv, i, p. 276.
— Beschreibg. zweier neuer Taenien a. d. Mensch., Nov. Act. Acad. Leop.-Caes.
nat. cur., 1861, xxviii.
Tænia marginata (p. 338).
Eschricht, D. F. Afhdlg. om de Hydat. d. fremkalde den i Island endem.
Leversyge, Overs. K. Dansk. Selsk. Forhdlg., 1853, p. 211.
Hodges. Cysticercus tenuicollis in the Human Body, Boston Med. and Surg.
Journ., 1866, lxxv, p. 185.
Krabbe, H. Helminthol. Unders. in Danmark og paa Island, Rech. helm. en
Danemark et en Islande, Copenh., 1866, p. 43.
Tænia serrata (p. 338).
Galli-Valerio, B. Not. helminthol. et bactériol., C. f. B., P. u. Inf., 1898
(1), xxiii, p. 940.
Vital, A. Les entozoaires à l’hôpital de Constantine, Gaz. méd., Paris, 1874,
p. 285.
Tænia africana (pp. 342, 343).
Linstow, v. Taenia africana, eine neue Taenie d. Mensch. aus Afrika, C. f.
B., P. u. Inf., 1900 (1), xxviii, p. 485.
— Helminth. vom Ufer d. Nyassasees, Jen. Zeitschr. f. Naturw., 1900, xxxv,
p. 420.
Tænia confusa (pp. 343, 344).
Guyer, M. F. On the Structure of T. confusa, Zool. Jahrb., 1898, Syst. xi,
p. 469.
Ward, H. B. Report of the Zoologist, Ann. Rept. Nebraska State Board Ag.
for 1895 (1896), p. 257; for 1896 (1897), p. 173.
— A New Human Tapeworm, West. Med. Rev., 1896, i, p. 35; Proc. Nebraska
State Med. for 1896, p. 83.
— Note on T. confusa, Zool. Anz., 1897, xx, p. 321.
Tænia echinococcus (pp. 344 to 347).
Blanchard, R. Prophyl. de la maladie hydatique, Arch. de Paras., 1905, ix,
p. 451.
Brault, A., and M. Loeper. La glycogène dans la membr. germ. d. kystes
hydat., Journ. d. Phys. et Path. gén., 1904, vi, p. 295; abstracted in
C. f. B., P. u. Inf., 1905, i, Ref. xxxvi, p. 689.
Bremser, J. G. Üb. lebende Würmer im leb. Menschen, Wien, 1819.
Dévé, F. De l’echinococcose secondaire, Paris, 1901.
— Sur l’évolution kystique du scolex échinoc., Arch. de Paras., 1902, vi, p. 54.
— Inoculations échinoc. aux cobayes, C. R. Soc. Biol., Paris, 1903, lv, p. 122.
— Les kystes hydat. du foie, Paris, 1905; abstracted in C. f. B., P. u. Inf.,
1906, i, Ref. xxxix, p. 64.
— Les deux scolex échinoc., C. R. Soc. Biol., Paris, 1906, lx, p. 956.
Erlanger, R. v. Der Geschlechtsapparat d. Taenia echinococcus, Z. f. w.
Zool., 1890, l, p. 555.
Eschricht, F. F. Om the hydatider, Overs. K. Danske Vid.-Selsk. Forh.,
1853, p. 211; 1856, p. 127; Zeitschr. f. d. ges. Naturwiss., Halle, 1857,
x, p. 231.
Goellner, A. Die Verbreitung der Echinococcenkrankh. in Els.-Loth., Mitt.
a. d. Grenzgeb. Med. Chir., 1903, xi, p. 80.
Goldschmidt, R. Zur Entwickel. d. Echinococcusköpfchen, Zool. Jahrb.
Anat., 1900, xiii, p. 467.
Joest, E. Studien über Echinokokken- und Zystizerkenflüssigkeit [sic],
Zeitschr. f. Infekt.-Krankh., paras. Krankh. u. Hyg. d. Haust., 1906,
ii, p. 1, and the literature there referred to.
Krabbe, H. Die isländ. Echinococcen, Arch. f. path. Anat., 1863, xxvii, p. 225;
Deutsch. Zeitschr. f. Thiermed., xvii.
Lebedew, A. J., and N. J. Andrejew. Transplant. v. Echinoc. v. Menschen
auf Kaninchen, Arch. f. path. Anat., 1889, cxviii, p. 552.
Lendenfeld, R. Tænia echinococcus, Zool. Jahrb., 1886, i, p. 409.
Leuckart, R. Über Echinococcus, Götting. Nachr., 1862, p. 13; Die Paras, d.
Mensch., 1. and 2. Aufl.
Lichtenfeld. Über Fertilität u. Sterilität der Echinococcen, C. f. B., P. u.
Inf., 1904, i Orig., xxxvi, p. 546; 1904, xxxvii, p. 64.
Madelung, O. W. Beitr. mecklenb. Ärzte z. Lehre v. d. Echinococcenkrankheiten,
Stuttg., 1885.
Moniez, R. Essai monogr. sur les Cysticerques, Thèse, Paris, 1880; Trav. de
l’Inst. Zool., Lille, 1880, iii, 1.
Mosler, F. Über Mittel zur Bekämpf. endem. vork. Echinococcuskrankh.,
Deutsche Med.-Ztg., 1889, No. 72.
Naunyn, B. Entwickel. d. Echinococcus, Arch. f. Anat. u. Phys., 1862, p. 612;
1863, p. 412.
Neisser, A. Die Echinococcenkrankheit, Berlin, 1877.
Peiper. Verbreitg. d. Echinococcenkrankh. in Vorpommern, Stuttg., 1894.
Perroncito, E. Dévelop. de nouv. kyst. aux dépens des scolex cont. dans les
kyst. hydat., Bull. Soc. Zool., France, 1902, xxvii, p. 150.
— Osservaz. fatte sulla tenia ech. e sulla rapidità del suo svil. dai dentoscolici,
Giorn. R. Accad. di med., Torino, 1906, Nos. 1 and 2.
Rasmussen, V. Bidr. til kundsk. on Echinococcernes udvikl., Vidensk.
meddel. fra naturh. Foren., Kjobenhavn (aar 1865), 1866, p. 1; translation
in Australian Med. Journ., Melbourne, 1869, xiv, pp. 33, 65.
Riemann, H. Über die Keimzerstreuung der Echinoc. im Peritoneum, Beitr. z.
klin. Chirurgie, xxiv; In.-Diss., Rostock, 1899.
Sabrazès, J., L. Muratet and P. Husnot. Motiité due scol. echinoc., C. R.
Acad. Sci., Paris, 1906, i; abstracted in C. f. B., P. u. Inf., 1906, i,
Ref. xxxix, p. 145.
Siebold, C. Th. v. Zur Entwickelung der Helminthen, in K. F. Burdach, Die
Physiol. als Erfahrungswiss., Leipzig, 1837, 2. Aufl., ii, p. 183.
— Über die Verwandlung von Echinococcusbrut in Taenien, Z. f. w. Zool.,
1853, iv, p. 409.
Thomas, J. D. Hydatid Disease . . . in Australia, Adelaide, 1884.
Tschötschel, K. Zur Casuist. d. Echinoc.-Krankheit in Vorpommern, Inaug-Diss.,
Greifswald, 1900.
Wiedemann, C. Zur Statistik d. Ech.-Krankheit in Vorpommern, Inaug.-Diss.,
Greifswald, 1895.
Echinococcus multilocularis (pp. 356 to 359).
Beha, R. Zur Kenntnis der Echinoc. alveol. der Leber, Inaug.-Diss., Freiburg
i. B., 1904.
Dévé, F. Sur quelq. caract. zool. de l’Echinococcose alvéol. bavaro-tyrol,
C. R. Soc. Biol., Paris, 1905, lviii, p. 126.
Guillebeau, A. Zur Histol. d. multil. Echinoc., Arch. f. path. Anat., 1890,
cxix, p. 108.
Linstow, v. Echinoc. alveolaris und Plerocercus lachesis, Zool. Anz., 1902,
xxvi, p. 162.
Mangold. Über die multil. Echinoc. und seine Taenie, Berlin, klin. Wochenschr.,
1892, No. 2; In.-Diss., Tübingen, 1892.
Melnikow-Raswedenkow, N. Stud. über die Echin. alveol. s. multil., Beitr.
z. path. Anat. u. z. allgem. Path., Suppl., 1901, iv.
Müller, A. Beitr. zur Kenntnis der Taenia echinoc., Münch. med. Wochenschr.,
1893, No. 13.
Ostertag, R. Über die Echinoc. multiloc. bei Rind und Schwein, Deutsch.
Zeitschr. f. Thiermed., 1891, xvii, p. 172.
Perroncito, E. Particol. interess. relat. alla cisti di echinococco, Giorn. R.
Accad. di med. di Torino, 1906, Nos. 1 and 2.
Posselt, A. Die Stellung des Alveolarechinococcus, Münch. med. Wochenschr.,
1906, liii, Nos. 12 and 13.
Schmidt, W. Über die geogr. Verbr. der Echin. multil. u. Hydat. in Bayern,
In.-Diss. München, 1899.
(C) NEMATHELMINTHES (pp. 360 to 470, 674 to 698, 754, and 755).
Bastian, H. C. On the Anatomy and Physiology of the Nematodes, Phil.
Trans. Roy. Soc., London, 1866, clvi, p. 2.
Bömmel, A. v. Über d. Cuticularbld. bei Nemat., Arb. zool.-zoot. Inst.,
Würzburg, 1895, x, p. 189.
Brandes, G. Das Nervensyst. der als Nemathelm. zusammengef. Wurmtypen,
Abh. nat. Ges., Halle, 1899, xxi, p. 273.
Bütschli, O. Beitr. zur Kenntnis des Nervens. der Nemat., Arch. f. mikr.
Anat., 1874, x, p. 74.
Dewitz, J. Lebensfähigkeit der Nemat. ausserhalb des Wirtes, Zool. Anz.,
1899, xxii, p. 91.
Domaschko, A. Die Wandung der Gonade von Ascaris megalocephala, Arb.
zool.-zoot. Inst., Wien, 1905, xv, p. 257.
Goldschmidt, R. Histol. Unters. an Nematoden: I, Die Sinnesorg. v. Asc.
lumbricoides und A. megalocephala, Zool. Jahrb. Anat., 1903, xviii, p. 1.
— Histol. Unters. an Nematoden: II, Der Chromidialapparat lebhaft funkt.
Gewebszellen, ibid., 1905, xxi, p. 49.
— Über die Cuticula vom Ascaris, Zool. Anz., 1905, xxviii, p. 259.
— Mitteil. zur Histol. von Ascaris, ibid., 1906, xxix, p. 719.
Hamann, O. Die Nemathelminthen, Jena, 1895, ii.
Hesse, R. Über das Nervensystem von Ascaris megalocephala, Zeitschr. f.
w. Zool., 1892, liv, p. 548.
Jaegerskiold, L. A. Über die Oesoph. der Nematoden, Bih. kgl. Svensk.
Vet.-Akad. Handl., 1897, xxiii, p. 4.
— Beitr. zur Kenntnis der Nemat., Zool. Jahrb. Anat., 1894, vii, p. 449.
— Weitere Beiträge zur Kenntnis der Nemat., Svensk. Vet.-Akad. Handl.,
1901–02, xxxv.
Jammes, L. Contrib. à l’étud. de la couche sous-cutic. d. Némat., Ann. Sci.
Nat. Zool., 1892 (7), xiii, p. 321.
Looss, A. Bau des Ösophag. bei einigen Ascariden, C. f. B., P. u. Inf., 1896
(1), xix, p. 1.
— The Sclerostomidæ of Horses and Donkeys in Egypt, Rec. Egypt. Govt.
School of Med., 1901.
Marcus, H. Ein Rhachiskern bei Ascariden, Biol. Centralbl., 1905, xxv,
p. 479.
Mayer, A. Zur Kenntnis der Rhachis im Ovarium und Hoden der Nematoden,
Zool. Anz., 1906, xxx, p. 289.
Nassonow, N. Sur les org. phagocyt. d. Ascar., Arch, de Paras., 1898, i,
p. 170; Arch. f. mikr. Anat., 1900, lv, p. 488.
Nedkoff, P. Die Metamorph. d. Geschlechtsapp. b. Ascaris nigrovenosa,
Inaug.-Diss., Leipzig, 1897.
Rauther, M. Über den Bau des Oesophagus und die Lokalisation der Nierenfunktion
bei freilebenden Nematoden, Zool. Jahrb. Anat., 1907, xxiii,
p. 703.
Retzius, G. Zur Kenntnis der Hautschicht der Nematoden, Biol. Unters.,
1906, N.F. xiii, No. 12.
Rohde, E. Beiträge zur Kenntnis der Nematoden, Zool. Beiträge (A.
Schneider), 1885, i, p. 11.
Schneider, A. Monographie der Nematoden, Berlin, 1866.
Toldt, C. Über den feineren Bau der Cuticula von Ascaris megalocephala,
Arb. zool.-zoot. Inst., Wien, 1899, p. 289.
— Die Saftbahnen in der Cuticula von Asc. megalocephala, Zool. Anz., 1904,
xxvii, p. 728.
Voltzenlogel, E. Unters. über d. anat. u. histol. Bau d. Hinterendes v.
Ascaris megalocephala u. A. lumbricoides, Zool. Jahrb. Anat., 1902, xvi,
p. 481.
Rhabditis pellio (p. 377).
Baginsky. Haemoglobinurie mit Auftreten von Rhabditiden im Urin, Deutsche
med. Wochenschr., 1887, p. 604.
Oerley, L. Die Rhabditiden und ihre medic. Bedeutung, Berlin, 1886.
Peiper and Westphal. Über d. Vorkom. v. Rhabditiden im Harne bei Haematurie,
Centralbl. f. klin. Med., 1888, ix, p. 145.
Scheiber, S. H. Ein Fall von mikrosk. kleinen Rundwürm. i. Urin eines
Kranken, Arch. f. path. Anat., 1880, lxxxii, p. 161.
Rhabditis nieillyi (p. 378).
Glatzel. Drei Monate im Serra Hadji Hospital zu Dar-es-Salam, Arch. f.
Schiffs- u. Tropen-Hyg., 1906, p. 284.
Künnemann. Rundwürmer (Rhabditis strongyloides) als Ursache eines
Hautausschlages beim Hunde, Deutsche tierärztl. Wochenschr., 1905,
No. 24.
Nielly. Un cas de dermatose paras. obs. pour la première fois en France,
Arch. méd. nav., 1882, xxxvii, pp. 337, 488; Bull. Acad. Méd., 1882 (2),
xi, pp. 395, 581.
O’Neill, J. On the Presence of a Filaria in Craw-craw, Lancet, 1875, i.
Schneider, J. G. Nematodenembr. i. d. Haut d. Hundes, In.-Diss., Basel,
1891.
Whittles, J. D. A Case of General Infection by a Nematode accompanied
by Hypertrophic Gingivitis, Lancet, 1903, i, p. 1435; 1903, ii, p. 57.
Rhabditis sp. (pp. 378, 379).
Frese, O. Über mikroskop. Würmer (Rhabditiden) im Magen einer Ozaenakranken,
Münch. med. Wochenschr., 1907, No. 11.
Anguillula aceti (p. 379).
Henneberg, W. Zur Biologie des Essigaales. Berlin, 1900, Deutsche Essigind.
Institut für Gährungsgewerbe, 1899, Nos. 45–52; 1900, Nos. 1–5.
Pallechi, P. Sulla resistenza vitale dell’ Anguillula dell’ aceto, Boll. Mus.
Zool. ed Anat. comp., Genova, 1893, No. 17.
Stiles, Ch. W., and W. A. Frankland. A Case of Vinegar Eel (Anguillula
aceti) Infection in the Human Bladder (eleven miscellaneous papers on
animal parasites), U.S. Dept, of Agric., Bur. of Animal Ind., Bull.
No. 35, Washington, 1902, p. 35.
Strongyloides stercoralis (pp. 380 to 384, 674, 675, and 755).
Askanazy, M. Über Art und Zweck der Invasion der Anguillula intestinalis in
die Darmwand, C. f. B., P. u. Inf., 1900, i Abt., xxvii, p. 569.
Bavay. Sur l’Anguillule stercorale, C. R. Acad. Sci., Paris, 1876, lxxxiii, p. 694.
— Sur l’Anguillule intestinale, ibid., 1877, lxxxiv, p. 266.
Braun, M. Bemerk. üb. d. sporad. Fall von Anguill. intest. in Ostpreussen,
C. R. Acad. Sci., Paris, 1899, xxvi, p. 612.
Brown, Ph. K. Report of Three Cases in which Embryos of the Strong.
intest. were found in the Stool, Boston Med. and Surg. Journ., 1903,
cxlviii, p. 583.
Chauvin. L’Anguillule stercorale dans la dysenterie des Antilles, Arch. Méd.
Nav., 1878, xxix, p. 154.
Durme, P. van. Quelq. notes sur les embryons de Strongyloides intestin. et
leur pénétrat par la peau, Thompson Yates Lab., Liverpool, 1902 (2),
iv, p. 471.
Grassi, B. Sovra l’Anguillula intestinale, Rend. Istit. lomb. sc. e lett., 1879
(2), xii, p. 228.
Grassi, B., and C. Parona. Sovra Anguillula intest., Arch. sci. med., 1879,
iii, p. 10.
Grassi, B., and R. Segrè. Nuove osserv. sull’ eterogonia del Rhabdon. intest.,
Atti R. Accad. d. Lincei Rendic., 1887 (4), iii, p. 100.
Kurlow, M. v. Anguillula intestinal. als Ursache akuter, blut. Durchfälle
beim Menschen, C. f. B., P. u. Inf., 1902, i Orig., xxxi, p. 614.
Leichtenstern, O. Über Anguillula intestinalis, Deutsche med. Wochenschr.,
1898, No. 8, p. 118.
— Zur Lebensgeschichte d. Ang. int., C. f. B., P. u. Inf., 1899, xxv, p. 226.
— Stud. üb. Strongyl. intest. nebst Bemerk. üb. Ancylost. duodenale, Arb.
kais. Gesundheitsamt, 1905, xxii, p. 309.
Leuckart, R. Über d. Lebensgesch. d. sogen. Anguillula stercoralis, Ber. d.
math.-phys. Class. d. k. sächs. Ges. d. Wiss., 1883, p. 85.
Looss, A. Die Wanderung der Ancylostomum- u. Strongyloides-Larven von
d. Haut nach dem Darm, C.R. 6me Congr. intern. de Zool., Berne, 1904
(1905), p. 225.
Normand, A. Sur la maladie dite diarrhée de Cochinchine, C. R. Acad. Sci.,
Paris, 1876, lxxxiii, p. 316.
— Mém. sur la diarrh. de Cochinchine, Arch. Méd. Nav., 1877, xxvii, p. 35.
— Du rôle étiol. de l’anguillule dans la diarrh. de Cochinchine, ibid., 1878,
xxx, p. 214.
Pappenheim. Ein sporad. Fall von Ang. intest. in Ostpreussen, C. f. B., P. u.
Inf., 1899, i Abt., xxvi, p. 608.
Perroncito, E. Obser. sur le dévelop. de l’Anguillula intestinalis, Journ. de
l’Anat. et de la Physiol., 1887, xvii, p. 499.
— Il Rhabdonema strongyloides Leuck., Anguillula intestinalis e Pseudorhabditis
stercoralis, Giorn. R. Accad. di Med., Torino, 1906, p. 3.
Price, Marshall Langton. Occurrence of Strongyloides intestinalis in the
United States, Journ. Amer. Med. Assoc., 1903, xli, p. 713.
Rovelli, G. Ric. s. org. genit. d. Strongyloides, Como, 1888.
Schlüter, H. Zur Kenntn. d. Anguillula-Erkrank. b. Menschen, In.-Diss.,
Kiel, 1905.
Seifert. Über Anguillula stercoralis, Sitzungsber. phys.-med. Ges., Würzburg,
1883, p. 22.
Sonsino, P. Tre casi d. mal. du Rhabdonema intest. o Rhabdonemiasi, Suppl.
d. Riv. gen. ital. di Clinica med., July 20, 1891.
Strong, R. P. Cases of Infection with Strongyloides intestinalis; First
reported cases in North America, Johns Hopk. Hosp. Rept., 1901, x, p. 91.
Stursberg. Über Anguillula intestinalis, Sitzungsber. niederrh. Ges. f. Natur- u.
Heilkde., Bonn., 1905, p. 28.
Teissier, P. Nouv. contrib. à l’étude de l’Anguill. stercoralis; Anguillulose
experim. de la grenouille, Arch. Méd. expér., 1896, p. 358.
Testi, A. Contrib. allo studio dell’ anguillulosi intest., Riv. crit. di clin.
med., 1904, v, Nos. 6–8.
Thayer, W. G. On the Occurrence of Strongyloides intestinalis in the United
States, Journ. Exper. Med., 1901 (1), vi, p. 75.
Zinn, W. Über Anguillula intestinalis, C. f. B., P. u. Inf., 1899, i Abt., xxvi,
p. 696.
Gnathostoma siamense (pp. 384, 385).
Levinsen, G. M. R. Om en ny rundworm hos mennesket, Vidensk. meddel. fra.
naturh. Foren., Kjöbenhavn, f. 1889, p. 323; abstracted in C. f. B., P. u.
Inf., 1890, viii, p. 182.
Dracunculus medinensis (pp. 386 to 390, 675, and 676).
Bastian, A. On the Structure and Nature of the Dracunculus, Trans. Linn.
Soc. Lond., 1863, xxiv, p. 101.
Blanchard, R. Malad. paras., paras., paras. animaux, Paris, 1895, p. 768.
Charles, R. H. Hist. of the male Filaria medin., Sci. Mem. Med. Offi., Army
of India, Calcutta, 1892, vii.
Coppola, N. Filaria medin. nella Colon. Eritrea, Giorn. med. del R. esercito,
1906, liv, p. 92; abstracted in Arch. f. Schiffs- u. Tropen-Hyg., 1906, x,
p. 518.
Fedschenko. Bau u. Entw. d. Filaria, Ber. d. k. Ges. d. Frd. d. Nat.,
Anthrop. u. Ethnogr., 1879, viii (1), p. 71.
Leuckart, R. Die menschlichen Parasiten, 1876, 1. Aufl., ii, p. 644.
Manson, P. On the Guinea-worm, Brit. Med. Journ., London, 1895, ii, p. 1350;
Lancet, 1895, ii, p. 309.
— The Life-span of Filaria medinensis, Brit. Med. Journ., 1903, ii, p. 10.
Mosler, F. Über die medic. Bedeutung d. Medinawurmes, Wien, Leipzig, 1885.
Plehn, F. Die Kamerunküste, Berlin, 1898, p. 294.
Powell, S. A. The Life-span of the Guinea-worm, Lancet, 1904, i, p. 76; Brit.
Med. Journ., 1904, i, p. 73.
Railliet, A. De l’occurr. de la Filaire de Médine chez les animaux, Bull. Soc.
Zool. France, 1889, xiv, p. 73.
Vortisch, H. Erfahrungen über einige spezifische Krankheiten an der Goldküste,
Arch. f. Schiffs- u. Tropen-Hyg., 1906, x, p. 537.
Filaria bancrofti (pp. 390 to 403, and 676 to 678).
Annett, H. E., J. E. Dutton and J. H. Elliot. Report of the Malaria Expedition
to Nigeria, II, Filariasis, Liverpool, 1901.
Bancroft, Th. On the Metamorphosis of the Young Form of Filaria bancrofti
in the body of Culex ciliaris, Proc. Roy. Soc. N. S. Wales, 1898, xxiii,
p. 48; 1901, xxxv.
Biondi, D. Chiluria da Filaria sanguinis hominis nocturna in Europa, Atti
Accad. d. Lincei, January 12, 1903 (5), p. 538.
Blanchard, R. Transmiss. de la filariose par les moustiques, Arch. de Paras.,
1900, iii, p. 280.
Bourne, G. A. Note on Filaria sanguinis hominis with description of a Male
Specimen, Brit. Med. Journ., 1888, No. 1429.
Cobbold, T. S. The Life-history of Filaria bancrofti, Journ. Linn. Soc.,
London, 1879, xiv, p. 356.
Demarquay. Note sur une tumeur de bourse . . . renferm. . . . des
helm. némat., Gaz. méd., Paris, 1863 (3), xviii, p. 665.
Lewis, T. R. On a Hæmatozoon inhabiting Human Blood, Calcutta, 1872;
2nd. edit., 1874.
— The Pathological Significance of Nematode Hæmatozoa, Calcutta, 1874.
Linstow, v. Über die Arten d. Blutfil. d. Menschen, Zool. Anz. 1900, xxiii,
p. 76.
Low, G. C. A Recent Observation on Fil. nocturna in Culex, Brit. Med.
Journ., 1900, i, p. 1456.
— The Development of Filaria nocturna in Different Species of Mosquitoes,
Brit. Med. Journ., London, 1901, i, p. 1336.
— Malaria and Filaria Diseases in Barbados, West Indies, Brit. Med. Journ.,
1901, ii, p. 687.
Lynch, G. W. A. Note on the Occurrence of Filaria in Fijians, Lancet, 1905,
i, p. 21.
Maitland, J. Note on the Etiology of Filariosis, Brit. Med. Journ., 1900, ii,
p. 537.
Manson, P. The Filaria sanguinis hominis . . . London, 1883.
— The Metamorphosis of Fil. sanguinis hominis in the Mosquito, Trans. Linn.
Soc., Lond., 1884 (2), ii, pp. 10, 367.
— The Filaria sanguinis and Filariasis, Trop. Diseases, New Edit., London,
1903, p. 545.
Penel, R. Les filaires du sang de l’homme, 2e édit., Paris, 1905; abstracted in
Arch. de Paras., 1905, ix, p. 187.
Scheube. Die Filariakrankheit, Volkmanns Samml. klin. Vortr., 1883, No. 232.
Sibthorpe. On the Adult Male of Filaria sanguinis hominis, Brit. Med. Journ.,
1889, No. 1485.
Solieri, S. Chiluria da Filaria bancrofti in Europa, Atti Accad. fisiocrat.
sci., 1904 (4), xv, p. 429.
Taniguchi, N. Über Filaria bancrofti, C. f. B., P. u. Inf., 1904, i Abt. Orig.,
xxxv, p. 492.
Thiesing, H. Beitr. zur Anat. der Filaria sang. hom., Inaug.-Diss., Basel,
Leipzig, 1892.
Triboudeau. Note sur la Filaria aux îles de la Société, C. R. Soc. Biol., Paris,
1903, lv, p. 996.
Vincent, G. A. Observations on Human Filariasis in Trinidad, Brit. Med.
Journ., 1902, i, p. 189.
Ziemann, H. Beitr. z. Filariakrankh. d. Menschen u. d. Tiere in d. Tropen,
Dtsche. med. Wchschr., 1905, No. 11.
Filaria demarquayi (pp. 403 and 404).
Daniels, C. W. Filariæ and Filarial Disease in British Guiana, Brit. Med.
Journ., 1898, ii, p. 878.
— Adult Form of Filaria demarquayi, Journ. Trop. Med., 1902, v, p. 357.
Galgey, O. Filaria demarquayi in St. Lucia, Brit. Med. Journ., 1891, i, p. 145.
Low, G. S. Notes on Filaria demarquayi, ibid., 1902, i, p. 196.
Manson, P. On Certain New Species of Nematode Hæmatozoa occurring in
America, ibid., 1897, ii, p. 1837.
Ozzard, A. T. Description of a Female Form of the Filaria demarquayi,
Journ. Trop. Med., 1902, v, p. 259.
Filaria (?) conjunctivæ (pp. 404 to 406).
Addario, C. Si di un nemat. dell’ occhio umano, Ann. d. ottalmol., 1885, xv.
Alessandrini, G. Nuovo caso di Filaria conjunctivæ Add., Boll. soc. zool.
ital., 1906, 2a ser., vii, p. 233.
Babes, V. Über einen neuen Parasiten d. Menschen, Med.-chir. Ctrlbl., Wien,
1879, xiv, p. 554.
— Über einen im menschl. Peritoneum gefund. Nemat., Arch. f. path. Anat.,
1880, lxxxi, p. 158.
Grassi, B. Filaria inermis, ein Parasit d. Menschen, d. Pferdes u. d. Esels,
C. f. B. u. Par., 1887, i, p. 617.
Agamofilaria oculi humani (p. 406).
Gescheidt. Die Entozoen des Auges, Zeitschr. f. Ophthalmol., 1833, iii, p. 405.
Kraemer, A. Die thier. Schmarotzer des Auges, Graefe-Saemischs Handb. d.
ges. Augenheilkde., Leipz., 1899, x, 2, chap. xviii.
Kuhnt, H. Extract. ein. neuen Entozoon a. d. Glaskörper, Arch. f. Augenheilkde.,
1891, xxiv, p. 205.
Nordmann, A. v. Mikrograph. Beitr. z. Naturgesch. d. wirbellos. Thiere,
Berlin, 1831, Heft 1, p. 7; Heft 2, p. 9.
Agamofilaria labialis (p. 407).
Pane. Nota su di un elminte nematode, Annal. Accad. degli aspiranti
naturalisti, Napoli, 1864 (3), iv.
Filaria (?) romanorum-orientalis (p. 407).
Sarcani, A. Filaria romanorum orientalis, Wien. med. Presse, 1888, No. 7,
p. 222.
Filaria (?) kilimaræ (p. 407).
Kolb, G. Filaria kilimaræ in Britisch-Ostafrika, Arch. f. Schiffs- u. Tropen-Hyg.,
1898, ii, p. 28.
Filaria (?) sp.? (p. 407).
Cholodkowsky, N. A., in Wratsch, 1896, No. 3.
— Über einige selten beim Menschen vork. Parasit., Sitzgsber. St. Petersb.
naturf. Ges., 1897, p. 185.
Mikrofilaria powelli (p. 407).
Penel, R. Les filaires du sang de l’homme, 2e ed., Paris, 1905, p. 126.
Powell, A. A Species of Blood Filaria, probably hitherto undescribed, Brit.
Med. Journ., 1903, i, p. 1145.
Setaria equina (pp. 408, 409).
Blanchard, R. Traité de Zool. méd., Paris, 1890, ii, p. 16.
Brera. Mem. phys.-med. sopra i princip. vermi del corpo umano, Crema, 1811,
p. 31.
Deupser. Zur Entw. d. Filaria papillosa, Zool. Anz., 1892, xv, p. 129; In.-Diss.,
Breslau, 1894.
Linstow, v. Beobacht. an neuen u. bekannten Nemathelm., Arch. f. mikr.
Anat. u. Entw., 1902, lx, p. 222.·
Treutler, F. A. Observat. pathol.-anat. auct. ad helminthol. humani corporis
continentes, Lipsiae, 1793.
Loa loa (pp. 409 to 414, and 678).
Billet, A. Un nouv. cas de Filaria loa male, C. R. Soc. Biol., Paris, 1906, lxi,
p. 507.
Blanchard, R. La Filaire sous-conjonctivale, Progr. Méd., Paris, 1886 (2), iv,
pp. 591, 611.
— Nouveau cas de Filaria loa, Arch. de Paras., 1899, ii, p. 504.
Brumpt, E. La Filaria loa est la forme adulte de la microfilaire designée sous
le nom Filaria diurna Mans., C. R. Soc. Biol., Paris, 1904, lvi, p. 630.
Gauthier, C. Microfilaires du sang coincidant avec une filaire de l’œil, Semaine
méd., 1905, p. 176; C. R. Soc. Biol., Paris, 1905, lviii, p. 632.
Habershon, J. H. Calabar Swellings on the Upper Congo, Journ. Trop. Med.,
1904, vii, p. 3.
Linon, J., and Pénaud. Un cas de Filaria loa avec oedèmes intermitt.,
microfil. dans le sang, l’urine et la salive, C. R. Soc. Biol., 1906, lxi,
p. 510.
Looss, A. Zur Kenntnis des Baues der Filaria loa G., Zool. Jahrb., 1904,
Syst. xx, p. 549.
Ludwig, H., and Th. Saemisch. Über Filaria loa im Auge des Menschen,
Z. f. w. Zool., 1895, lx, p. 726.
Manson, P. The Filaria sanguinis hominis major and minor, Two New
Species of Hæmatozoa, Lancet, 1891, i, p. 4; Ctrlbl. f. allg. Path., ii,
p. 298.
— Geographical Distribution of Filaria sanguinis hominis diurna . . . Trans.
VII. Int. Cong. of Hyg. and Demography, London, 1891, i, p. 79.
Mongin. Observ. sur un ver trouvé sous la conjonctive à Maribaron, île
St.-Domingue, Journ. de Méd., Chirurg., Pharmacie, Paris, 1770, xxxii,
p. 338.
Penel, R. Les Filaires du sang de l’homme, 2e édit., Paris, 1905, p. 127.
Plehn, F. Die Kamerunküste, Berlin, 1898, p. 296.
Sambon, L. W. Remarks on the Individuality of Filaria diurna, Journ. Trop. Med., 1903, vi, p. 26.
Thompstone, S. W. Calabar Swellings, Journ. Trop. Med., 1899–1900, ii, p. 89.
Ward, H. B. The Earliest Record of Filaria loa, Zool. Ann., 1906, i, p. 376;
Med., London, 1902, v, p. 381; vi, p. 26.
— Studies on Human Parasites in North America: I, Filaria loa, Stud. Zool.
Lab., Univ. Nebraska, 1906, No. 63.
Wurtz, R., and A. Clerc. Nouv. observ. de Filaria loa, Arch. Méd. exp. et
d’Anat. path., 1905, xvii, p. 261.
Acanthocheilonema perstans (pp. 414 to 416).
Christy, C. The Distribution of Sleeping Sickness; Filaria perstans . . .
in East Equatorial Africa, Rep. Sleeping Sickness Comm., London, 1903,
ii, p. 1.
Daniels, C. W. The Filaria sanguinis hominis perstans found in the
Aboriginals of British Guiana, Brit. Guiana Med. Ann., 1897, p. 28.
— Discovery of the Parental Form of a British Guiana Bloodworm, Brit. Med.
Journ., 1898, i, p. 1011.
Daniels, C. W. The Filaria ozzardi and their Adult Form, Brit. Guiana Med.
Ann., 1898, x, p. 1.
— Filariæ and Filarial Disease in British Guiana, Brit. Med. Journ., 1898, ii,
p. 878.
— The Probable Parental Form of a Sharp-tailed Filaria found in the Blood
of the Aboriginals of British Guiana, ibid., 1899, i, p. 1459.
— The Sharp-tailed Filaria in British Guiana, Journ. of Trop. Med., 1899–1900,
ii, p. 11.
— Filariasis in British Central Africa, Journ. Trop. Med., 1901, p. 193.
Firket, C. M. Filariose du sang chez les nègres du Congo, Bull. Acad. Roy.
méd. Belg., 1895 (4) ix; abstracted in C. f. B., P. u. Inf. (1), xix, p. 791.
Henry. Remarks on Filaria, Proc. Acad. Nat. Sci., Philadelphia, 1896, p. 271.
Hodges, A. Sleeping Sickness and Filaria perstans in Busoga, Journ. Trop.
Med., 1902, v, p. 298.
Low, G. C. Filaria perstans, Brit. Med. Journ., London, 1903, i, p. 722.
Manson, P. Cf. under Loa loa and Filaria demarquayi (pp. 805, 806).
Ozzard, A. T. A Supposed New Species of Filaria sanguinis hominis found in
the Interior of British Guiana, Brit. Guiana Med. Ann., 1897, p. 24.
Penel, R. Cf. under Loa loa (p. 806).
Ziemann, H. Über das Vorkommen von Filaria perstans und von Trypanosomen
beim Schimpansen, Arch. f. Schiffs- u. Tropen-Hyg., 1902, vi, p. 362.
— Beitr. zur Filariakrankheit d. Menschen u. d. Tiere in d. Tropen, Dtsche.
med. Wchschr., 1905, No. 11.
Dirofilaria magalhāesi (p. 417).
Blanchard, R. Maladies parasitaires . . . Paris, 1895, p. 782.
Linstow, v. Über Filaria bancrofti Cobb., C. f. B. u. Par., 1892, xii, p. 88.
— Über die Arten d. Blutfil. d. Menschen, Zool. Anz., 1900, xxiii, p. 79.
Magalhaes, P. S. de. Die Filaria bancrofti Cobb. u. die Fil. immitis Leidy,
C. f. B. u. Par., 1892, xii, p. 511.
Filaria gigas.
Prout, W. T. Filariasis in Sierra Leone, Brit. Med. Journ., 1902, ii, p. 879;
Journ. Trop. Med., 1902, v, p. 317.
Oncocerca volvulus (pp. 417 to 419, and 755).
Brumpt, E. A propos de la Filaria volvulus Leuck., Rev. d. Méd. et d’Hyg.
trop., 1904, i, p. 43; abstracted C. f. B., P. u. Inf., 1905, i, Ref. xxxvi,
p. 148.
Labadie-Lagrave and Deguy. Un cas de Filaria volvulus, Arch. de Paras.,
1899, ii, p. 451.
Leuckart, R. (in Manson, P.). Diseases of the Skin in Tropical Climates;
Davidson: Hygiene and Diseases of Warm Climates, Edinb., London,
1893, p. 963.
Penel, R., cf. under Filaria bancrofti (p. 804).
Prout, W. T. A Filaria found in Sierra Leone, Brit. Med. Journ., 1901, i,
p. 209.
— Observations on Filaria volvulus, Arch. de Paras., 1901, iv, p. 301.
Trichuris trichiura (pp. 419 to 421, and 678 to 680).
Askanazy, M. Der Peitschenwurm ein blutsaug. Parasit., Deutsch. Arch. f.
klin. Med., 1896, lvii, p. 104.
Davaine. Rech. sur le dévelop. et la propag. de l’Asc. lombr. et du Trichoc.
de l’homme, C. R. Acad. Sci., Paris, 1858, xlvi, p. 1217.
Eberth, J. G. Beitr. z. Anat. u. Phys. d. Trichoc. dispar., Z. f. w. Zool., 1860,
x, pp. 233, 383; 1862, xi, p. 96.
French, H. S., and A. E. Boycott. The Prevalence of Trichocephalus dispar,
Journ. of Hyg., 1905, v, p. 274.
Galli-Valerio, B. Die Verbreitg. d. Helminthen d. Mensch., Therap. Monatshefte,
July, 1905.
Goeze, J. A. E. Vers. ein. Naturgesch. d. Eingeweidewürmer, 1782, p. 182.
Grassi, R. Trichocephalus- und Ascaris-Entwickelung, C. f. B. u. Par., 1887,
i, p. 131.
Leuckart, R. Die menschlichen Paras., 1876, ii, p. 492.
Mayer, F. J. C. Beiträge zur Anat. der Entozoen, Bonn, 1841.
Morgagni, J. B. Epist. anat. XVIII ad scripta pertinentium celeb. viri A. M.
Valsalvae, Venetiis, 1740, ii Ep., xiv, p. 45.
Railliet, A. Not. helminth., Bull. Soc. centr. méd. vétér., 1884, p. 449.
Roederer, J. G. Nachr. von den Trichiuriden, Göttinger gel. Anz., 1761,
No. 25.
Wichmann. Über das Verhalt. der Trichoceph. zur Darmschleimhaut, Inaug.-Diss.,
Kiel, 1889.
Trichinella spiralis (pp. 421 to 431, 680, and 681).
Askanazy, M. Zur Lehre von der Trichinosis, C. f. B. u. Par., 1894, xv, p. 225.
— Arch. f. path. Anat., 1895, cxli, p. 42.
Babes, V. Ein 21 Jahre alter Fall von Trichinose mit lebenden Trichinen,
C. f. B., P. u. Inf., 1906, i Abt., Orig. xlii, p. 541.
Brown, T. R. Studies on Trichinosis, Bull. Johns Hopkins Hosp., 1897, viii,
No. 73.
Cerfontaine, P. Contribution à l’étude de la trichinose, Arch. de Biol., 1893,
xiii, p. 125; Bull. Acad. roy. de Belg., 1893 (3), xxv, p. 454.
Chatin, J. La trichine et la trichinose, Paris, 1883.
Ehrhardt, O. Zur Kenntnis der Muskelveränderung bei der Trichinose der
Kaninchen, Beitr. z. pathol. Anat. und zur allg. Path., 1896, xx, p. 1.
— Zur Kenntnis der Muskelveränderung bei der Trichinose des Menschen,
ibid., p. 44.
Geisse, A. Zur Frage der Trichinenwanderung, In.-Diss., Kiel, 1894.
Goujon, L. Expér. sur la Trichina spir., Thèse, Paris, 1866.
Graham, J. Y. Beitr. zur Naturgesch. der Trichina spiralis, Arch. f. mikros.
Anat., 1897, 1, p. 219.
Herbst, G. Beobacht. über Trichina spiralis, Göttinger Nachr., 1851, p. 260;
1852, p. 183.
Hertwig, R. Entwickelung der Trichinen, Münch. med. Wochenschr., 1895,
No. 21.
Höyberg, H. M. Fütterungsvers. mit trichinös. Faekal., C. f. B., P. u. Inf.,
1906, i Abt., Orig. xli, p. 210.
Johne, A. Der Trichinenschauer, 4. Aufl., Berlin, 1893.
Leuckart, R. Untersuchungen über Trichina spiralis, 1. Aufl., 1860; 2. Aufl.,
1866.
Lübke. Über das Vork. der Trichinen beim Dachs, Zeitschr. f. Fleisch- u.
Milchhyg., 1903, xiii, p. 116.
Mark, E. L. Trichinæ in Swine, 20th Ann. Rep. Mass. State Board of Health
for 1888, p. 113.
Opalka, L. Beitr. zum Vork. der Trichinen beim Menschen mit Rücksicht auf
die Prophylaxe, In.-Diss., Giessen, 1904, Arb. a. d. hyg. Inst. d. kgl.
tierärztl. Hochschule, Berlin, No. 3, Berlin, 1904.
Ossipow, N. Über histol. Veränderungen in Spätstadien der Muskeltrichinose,
Beitr. z. path. Anat. u. z. allg. Path., 1903, xxxiv, p. 253.
Owen, R. Description of a Microscopical Entozoon infesting the Muscles of
the Human Body, Trans. Zool. Soc., Lond., 1835, i, p. 315.
Pagenstecher, H. A. Die Trichinen, Wiesbaden, 1865.
Pirl. Das Vorkommen von Trichinen im Hundefleisch, Zeitschr. f. Fleisch- u.
Milchhyg., 1899, x, p. 5.
Schleip, K. Die Homberger Trichinosisepidemie und die für Trichinosis
pathognomische Eosinophilie, Deutsch. Arch. f. klin. Med., 1904, lxxx,
p. 1.
Stäubli, C. Beitr. z. Kenntnis d. Verbreitungsart der Trichinenembryomen,
Vierteljahrsschr. der naturf. Ges., Zürich, 1905, l, p. 163.
— Klin. u. exp. Unters. über Trichinosis u. über die Eosinophilie im allg.,
Deutsch. Arch. f. klin. Med., 1905, lxxxv, p. 286.
Stiles, Ch. W. Trichinosis in Germany, U.S. Dep. of Agric., Bur. of Anim.
Ind., Bull. No. 30, Washington, 1901.
— Trichinella spiralis, Trichinosis and Trichina Inspection, Proc. Path. Soc.,
Philadelphia, 1901, N.S. iv, p. 137.
Virchow, R. Zur Trichinenlehre, Arch. f. path. Anat., 1865, xxxii, p. 332.
— Darstellung der Lehre von den Trichinen, Berlin, 1864 and 1866.
Williams, H. U. The Frequency of Trichinosis in the United States, Journ.
of Med. Res., 1901, vi, p. 64.
Zenker, F. A. Über die Trichinenkrankh. des Menschen, Arch. f. path. Anat.,
1860, xviii, p. 561.
— Beitr. zur Lehre von der Trichinenkrankh., Deutsch. Arch. f. klin. Med.,
1866, i, p. 90; 1871, viii, p. 387.
Dioctophyme (Eustrongylus) gigas (pp. 431, 432, 681 and 682).
Balbiani, G. Recherch. sur le dévelop. et le mode de propagation du Strongyle
géant, Journ. de l’Anat., 1870, vii, p. 180; C. R. Soc. Biol., Paris, 1874
(6), i, p. 125.
Blanchard, R. Nouv. observ. de Strongyle géant chez l’homme, C. R. Soc.
Biol., Paris, 1886 (8), iii, p. 379.
Magueur. Strongyle géant du rein expulsé en partie par le canal de l’urètre
chez un enfant de deux ans et demi, Journ. méd., Bordeaux, 1887–88,
p. 337.
Rothstadt, J. Über das Vork. v. Eustrongylus gigas in Hunden der Stadt
Warschau, Arb. zool. Labor. Kais. Univ., Warschau, 1897.
Stiles, Ch. W. Notes on Parasites, 49, Med. Rec., 1898, liii, p. 469; abstracted
in C. f. B., P. u. Inf., xxiv, p. 505.
Stuertz. Eustrongylus gigas im menschl. Harnapparat mit einseit. Chylurie,
Dtsch. Arch. f. klin. Med., 1903, lxxviii, p. 557.
Trumbull, J. A Case of Eustrongylus gigas, Med. Rec., 1897, lii, p. 256;
abstracted in C. f. B. u. Par., xxii, p. 619.
Metastrongylus apri (pp. 432, 433).
Chatin, J. Le strongyle paradoxal chez l’homme, Bull. Acad. Méd., Paris,
1888, p. 483.
Diesing, C. M. Systema helminthum, II, Vindob, 1851, p. 317.
— Revision der Nematoden, Sitzungsb. K. Akad. d. Wiss., Wien, Math.-anat.
Kl., 1860, xlii, p. 722.
Jelkmann, F. Über den fein. Bau v. Strong. pulm. apri, In.-Diss., Basel, 1895.
Rainey. Entozoon found in the Larynx, Trans. Path. Soc., Lond., 1855, vi,
p. 370.
Spemann, H. Zur Entw. des Strongylus paradoxus, Zool. Jahrb. An., 1895,
viii, p. 301.
Wandolleck, B. Zur Embryonalentw. d. Strong. paradoxus, In.-Diss., Berlin,
1891, Arch. f. Naturg., 1892, lviii, p. 123.
Trichostrongylinæ (p. 433).
Ijima, J. Strongylus subtilis in Japan, Zool. Mag., 1896, vii, p. 155.
Looss, A. Strongylus subtilis, ein bisher unbekannter Parasit des Menschen in
Ägypten, C. f. B. u. Par., 1895 (1), xviii, p. 161.
— Notizen zur Helminthol. Ägyptens, I, ibid., 1896, xx, p. 864.
— Notizen zur Helminthol. Ägyptens, VI, ibid., 1905, i Abt. Orig., xxxix, p. 409.
Railliet, A. Traité de Zool. méd. et agric., 2e édit., Paris, 1895, p. 442.
— Sur les variations des Strongyles de l’appar. digest. et sur un nouv. Strongyle
du Dromadaire, C. R. Soc. Biol., Paris, 1896 (10), iii, p. 540.
Ternidens deminutus (pp. 440, 441).
Looss, A. Notiz z. Helminthol. Ägyptens, III, C. f. B., P. u. Inf., 1900 (1),
xxvii, p. 190.
— The Sclerostomidæ of Horses and Donkeys in Egypt, Records Egypt. Govt.
School of Med., 1901.
Railliet, A., and A. Henry. Un nouv. Sclérostom., paras. de l’homme, C. R.
Soc. Biol., Paris, 1905, lviii, p. 569; Bull. Mus. d’Hist. nat., 1905, p. 269.
Œsophagostomum brumpti (pp. 441 to 443).
Railliet, A., and A. Henry. Encore un nouveau Sclérostomien parasite de
l’homme, C. R. Soc. Biol., Paris, 1905, lviii, p. 643.
Ancylostoma and Necator (pp. 445 to 459, 682 to 687, and 754).
Alessandrini, G. Brevi osserv. sull. svil. e ciclo evolut dell’ Anchyl. duod.,
Boll. soc. zool. ital., Roma, 1904, xiii, p. 147.
— Ulter. ric. sul ciclo del svil. dell’ Uncinaria duod., ibid., 1904, xiv, p. 173.
— Su di alcune Uncinariæ paras. dell’ uomo e di altri vertebrati, ibid., xiv,
p. 23.
Bentley, C. A. On the Causal Relationship between “Ground Itch” or “Pani
ghao” and the Presence of the Ancylostoma duodenale in the Soil, Brit.
Med. Journ., 1902, p. 1900.
Boycott, A. E. A Case of Skin Infection with Ancylostoma, Journ. of Hyg.,
1905, v, p. 280; abstracted in C. f. B., P. u. Inf., 1906, i, Ref. xxxvii,
p. 749.
Boycott, A. E., and J. S. Haldane. An Outbreak of Ancylostomiasis in
England, Journ. of Hyg., 1903, iii, p. 95; 1904, iv, p. 73.
— — Uncinariasis in the Southern States, Journ. Amer. Med. Assoc., 1903, xl,
p. 36.
Bruns and W. Müller. Die Durchwanderung d. Ancylostoma-Larven durch
die menschl. Haut, Münch. med. Wochenschr., 1905, No. 31.
Bugnion, E. L’Ankylost. duodén. et l’anémie du St. Gotthard, Rev. méd.
suisse romande, Genève, 1881, Nos. 5, 7.
Calmette, A., and M. Breton. L’Ancylostomiase, Paris, 1905.
Dubini, A. Nuovo verme dell’ intest, umano, Annal. Univ. di Medic. d’Omodei,
1843, cvi, p. 51.
Ferrier. L’uncinariose en Algérie, Arch. de Paras., 1905–06, x, pp. 77, 458.
Firket. Rech. sur le trajet des larves de l’Ancylost. à travers les org. après
pénétrat. par voie ent., Bull. Acad. Roy. méd. Belg., 1905.
Galli-Valerio, B. Notes de parasit.; B. Paras. anim., 6, 7, 8, C. f. B., P. u.
Inf., 1905, i Orig., xxxix, p. 241.
Grassi, B. Anchilostomi ed Anguillule, Gas. d. ospit., 1882, No. 41; Giorn. R.
Accad. Med., Torino, 1883, xxxi, p. 119.
Griesinger. Klin. u. anat. Beob. üb. d. Krankh. v. Ägypten, Arch. f. phys.
Heilkde., 1854, xiii, p. 55.
Hermann. Note sur la pénétrat. des larves de l’Anchyl. duod. à travers la
peau humaine, Bull. Acad. Roy. Méd. Belg., 1905, 4e sér., xix, p. 181.
Inoye, J. Über Ancylostoma duodenale in Japan, Arch. f. Verdauungskrankh.,
1905, xi, p. 58.
Isola, D. Esiste in Italia l’Uncinaria americana? Boll. Mus. zool. e anat.
comp., Univ. Genova, 1904, No. 129.
Karschin. Ein Fall von Ancylostomiasis (in Sibirien), Sibir. ärztl. Nachr.,
1904, ii, No. 23; C. f. B., P. u. Inf., 1905, i, Ref. xxxvii, p. 504.
Lambinet, J. Recherch. sur la mode d’infect. de l’organ. anim. par les larves
d’Anchylost., Bull. Acad. Roy. méd. Belg., 1905, 4e sér., xix, p. 56.
Leichtenstern, O. Über Ancylostoma duodenale bei d. Ziegelarbeitern in der
Umgebung Cölns, Deutsche med. Wochenschr., 1885, xi, Nos. 28–30.
— Weitere Beitr. z. Ancylostomenfrage, ibid., 1886, xii, Nos. 11–14.
— Fütterungsvers. mit Ancylost.-Larven Centralbl. klin. Med., 1886, No. 39.
— Einiges über Ancylostoma duodenale, D. med. Wochenschr., 1887, xiii,
Nos. 26–32.
Liefmann, H. Beitr. zum Studium der Ancylostomiasis, Zeitschr. f. Hyg.,
1905, l, p. 349.
Linstow, v. Zwei wenig bekannte Ankylostomen . . . C. f. B., P. u. Inf.,
1906, i Orig., xliii, p. 89.
— Helminth. Beob. Ancyl. americ. aus Simia troglodytes, C. f. B., P. u. Inf.,
1903, i Orig., xxxiv, p. 526.
Löbker and H. Bruns. Über das Wesen u. die Verbreitung der Wurmkrankh.
mit besond. Berücks. ihres Auftretens in deutschen Bergwerken, Arb.
kais. Gesundheitsamt, 1906, xxiii, p. 421.
Loeb, L., and A. J. Smith. Über eine die Blutgerinnung hemmende Substanz
in Ancylostoma caninum, C. f. B., P. u. Inf., 1904, i Orig., xxxvii, p. 93;
1906, xl, p. 738.
Looss, A. Notizen z. Helminthol. Ägypt., I, C. f. B., P. u. Inf., 1896 (1), xx,
p. 865.
— Notizen z. Helminthol. Ägypt., II, ibid., 1897, xxi, p. 913.
— Zur Lebensgesch. d. Ancyl. duod., ibid., 1898, xxiv, p. 484.
Looss, A. Die Ancylostomafrage, ibid., 1899, xxv, p. 662.
— Über d. Eindringen der Ancylostomalarven in die menschliche Haut, ibid.,
i Abt., xxix, p. 733.
— Weiteres über d. Einwanderung der Ancylostomen von der Haut aus, ibid.,
1903, i Orig., xxxiii, p. 330.
— Zum Bau d. erwachs. Ancylostomum duodenale Dub., ibid., 1904, xxxv,
p. 752.
— Die Wanderung d. Ancylostomum- u. Strongyloides-Larven von der Haut
nach dem Darm, C. R. 6e Congr. int. de Zool. (Berne, 1904), 1905, p. 225.
— Note on Intestinal Worms found in African Pygmies, Lancet, 1905, ii, p. 430.
— The Anatomy and Life-history of Ancyclostoma duodenale Dub., Records of
Egypt. Govt. School of Medicine, Cairo, 1905, iii.
— Einige Betracht. über die Inf. mit Ankylost. duod., Zeitschr. f. klin. Med.,
1905, lviii, p. 1.
Lutz, A. Über Ancylost. duodenale, Volkm. Samml. klin. Vortr., 1888,
Nos. 255, 256 and 265.
Menche. Anchyl. duoden. bei d. Ziegelbrenner-Anaemie i. Deutschland,
Centralbl. f. klin. Med., 1882, p. 161; Zeitschr. f. klin. Med., 1883, vi,
p. 161.
Parona, C., and B. Grassi. Sull. svilup. dell’ Anchilost. duod., Atti soc. ital.
sci. nat., 1878, xxi, p. 53.
Perroncito, E. Helminth. Beobacht., Moleschotts Unters. z. Naturl. d.
Mensch., xii, p. 532.
— Der Dochmius und verwandte Helminth. in ihren Beziehungen zur sogen.
Bergcachexie, Centralbl. f. d. med. Wiss., 1881, No. 24.
— Les ancylostomes en France et la maladie des mineurs, C. R. Acad. Sci.,
Paris, 1882, p. 29.
— L’anémie d. mineurs au point de vue parasitol., Arch. ital. de biol., 1882, ii,
p. 315; 1883, iii, p. 7.
Pieri, G. Sol modo di trasmissione dell’ Anchilost. duodenale, Rendic. R.
Accad. Lincei, Roma, Cl. fis., mat. e nat., 1902, xi, p. 217; Arch. ital. de
biol., 1902, xxxvii, p. 269.
— Nuove ricerche sul modo in cui avviene l’infezione da Anchylostoma, Rendic.
R. Accad. Lincei., Cl. fis., mat. et nat., 1903, xii, p. 393.
— Osservaz. sulla biol. dell’ Ancylostoma, ibid., 1905, No. 12.
Schaudinn, F. Über die Einwanderung d. Ancylostomenlarven von der Haut
aus, Deutsche med. Wochenschr., 1904, xxx, p. 1338.
Schüffner, W. Über den neuen Infektionsweg der Ancylostomalarven durch
die Haut, C. f. B., P. u. Inf., 1906, i Orig., xl, p. 683.
Schulthess, W. Beitr. z. Anat. von Ancylostoma duodenale, Z. f. w. Zool.,
1882, xxxvii, p. 163.
Siccardi, P. D. Osserv. su l’Anchilostomiasi. L’Ancylostoma americanum,
Atti R. Ist. Veneto di sci., lett. e arti, 1904–05, lxiv, p. 1473.
— Per lo studio dell’ Anchilostomiasi (da Ancyl. amer.), ibid., 1905–06, lxv,
p. 69.
Siebold, C. Th. v. Ein Beitrag zur Helminthograph. humana, Z. f. wiss.
Zool., 1852, iv, p. 55.
Smith, C. A. Uncinariasis in the South, Journ. Amer. Med. Assoc., 1903, xli,
p. 709; 1904, xlii.
Stiles, Ch. W. A New Species of Hookworm (Uncinaria americana), parasitic
in Man, Amer. Med., 1902, iii, p. 777.
— The Signification of the Recent American Cases of Hookworm Disease in
Man, Eighteenth Ann. Rept. of the Bur. of Anim. Ind. (1901), Washington,
1902.
— Report upon the Prevalence and Geographical Distribution of Hookworm
Disease, 2nd ed., Washington, 1903 (Hyg. Lab. Bull. No. 10).
— Address on Hookworm Disease, or Uncinariasis, Journ. Mississ. State Med.
Assoc., 1904, ix, p. 123.
Stiles, Ch. W., and J. Goldberger. A Young Stage of the American Hookworm,
Eight to Twelve Days after Skin Infection in Rabbits and Dogs,
Amer. Med., 1906, xi, p. 63.
Tenholt. Die Untersuch. auf Anchylostomiasis mit bes. Berücks. d. wurmbehafteten
Bergleute, 2. Aufl., Bochum, 1904.
— Über die Ancylostomiasis, Deutsche Vierteljahrsschr. f. öffentl. Ges., 1906,
xxxviii, p. 271.
Wucherer. Über Anchylostomenkrankheit, trop. Chlorose oder trop.
Hyperämie, Dtsch. Arch. f. klin. Med., 1872, x, p. 379.
Zinn, W., and M. Jacoby. Ancylostomum duodenale, Leipzig, 1898.
Physaloptera caucasica (p. 461).
Linstow, v. Zwei neue Parasiten d. Menschen, C. f. B., P. u. Inf., 1902 (1),
Orig. xxxi, p. 769.
Molin, R. Una monografia del genere Physaloptera, Sitzungsber. kais. Akad.
d. Wiss., Wien, Math-phys. Cl., 1860, xxxix, p. 637.
Stossich, M. Il genere Physaloptera, Boll. Soc. Adriat. Sc. nat., Trieste, 1889,
xi.
Ascaris lumbricoides (pp. 463 to 465, 687 to 694, and 754).
Davaine, E. Rech. sur le dévelop. . . . de l’Ascar. lumbr., C. R. Acad. Sci.,
Paris, 1858, xlvi, p. 1217; Mém. Soc. Biol., Paris, 1862 (3), iv, p. 261.
Epstein, A. Über die Übertrag. d. menschl. Spulwurms, Jahrbüch f. Kinderheilk.,
1892, N.F. xxxiii, p. 3.
Epstein, W. Die Strangulationsmarke beim Spulwurm in ihrer diagnostischen
Bedeutung, Deutsches Arch. f. klin. Med., 1904, lxxxi, p. 543; cf. Huber
in: C. f. B., P. u. Inf., 1907, i, Ref. xxxix, p. 481.
Grassi, B. Trichocephalus- und Ascarisentwickelung, C. f. B. u. Par., 1887,
i, p. 131; 1888, iii, p. 748.
Guiart, J. Rôle pathol. de l’Asc. lumbr., Arch. de Paras., 1900, iii, p. 70.
Hallez, P. Rech. sur l’embryol. et sur les condit. du dével. de quelq. némat.,
Paris, 1885.
Leuckart, R. Die Übergangsweise der Asc. lumbric., C. f. B. u. Par., 1887, ii,
p. 713.
Linstow. Über d. Zwischenwirt von Asc. lumb., Zool. Anz., 1886, ix, p. 525.
Lutz, A. Zur Frage der Invas. von . . . Asc. lumbric., C. f. B. u. Par.,
1887, ii, p. 713.
— Weiteres zur Übertrag. d. Spulwurms, ibid., 1888, iii, p. 265.
Maurizi, A. Un nuovo caso di ascaridi nel fegato, Boll. soc. zool. ital., 1903
(2), xi, p. 198; collection of 130 cases.
Miura, K., and N. Nishiuchi. Über befruchtete und unbefruchtete Ascari-deneier
im menschl. Kote, C. f. B., P. u. Inf., 1902, i Orig., xxxii, p. 637.
Sick, C. Über Spulwürmer i. d. Gallenwegen, Tübingen, 1901.
Ascaris texana (p. 465).
Smith, A. J., and R. A. Goeth. Ascaris texana, a Note on a Hitherto Undescribed
Ascaris Parasitic in the Human Intestine, Journ. Amer. Med.
Assoc., 1904, xliii, p. 542; abstracted in C. f. B., P. u. Inf., 1905, i, Ref.
xxxvi, p. 499.
Toxascaris limbata (p. 466).
Bellingham, O. B. Undescribed Species of Human Intestinal Worm, Dub.
Med. Press, 1839, i, p. 104; Gaz. des Hôp., 1839 (2), i, p. 97.
Cobbold, T. Sp. On the Occurrence of Ascaris mystax in the Human Body,
Lancet, 1863, i, p. 31.
Davaine, C. Sur la constitution de l’œuf des cert. entoz., Mém. Soc. Biol.,
Paris, 1862 (3), iv, p. 273.
Grassi, B. Intorno all’ Ascaris mystax, Gazz. med. it.-lomb., 1878, xxxix,
p. 276.
— Beitr. z. Kenntn. d. Entwickelungscyclus von 5 Paras, d. Hundes, C. f. B.
u. Par., 1888, iv, p. 609.
Heller, C. Über Ascaris lumbricoides, Sitzgsber. Erlang. phys.-med. Societät,
1872, iv, p. 71.
Hering. Beitr. z. Entwickel. einig. Eingeweide-Würmer, Württemb. nat.
Jahreshefte, 1873, p. 305.
Kelly, H. A. The Occurrence of the Ascaris mystax in the Human Body,
Amer. Journ. Med. Sci., 1884 (2), lxxxviii, p. 483.
Leuckart, R. Menschl. Paras., 1. Aufl., ii, p. 261.
Morton, F. Ascaris mystax, Lancet, 1865, i, p. 278.
Petit, G., and M. Motas. Pénétration de l’Ascaris mystax dans les canaux
hépatiques du chien, Bull. Soc. Centr. Méd. Vétér., Paris, 1902 (2), xix,
p. 146.
Ragazzi, V. Sulla presenza dell’ Ascaris mystax nell’ uomo, Ann. med. nav.
Ann., May 2, 1904, ix, p. 509; abstracted in C. f. B., P. u. Inf., 1906,
i, Ref. xxxvii, p. 613.
Oxyuris vermicularis (pp. 467 to 469, and 694 to 698).
Blanchard, R. L’appendicite et la typhlo-colite sont très fréq. des affections
vermineuses, Arch. de Paras., 1906, x, p. 404.
Edens. Über Oxyuris vermicularis in der Darmwand, C. f. B., P. u. Inf.,
1905, i Orig., xl, p. 499.
Flögel, J. H. L. Über die Lippen einiger Oxyuris-Arten, Z. f. w. Zool., 1869,
xix, p. 234.
Grassi, B. I malefizi delle mosche, Gaz. Ospit., 1883, No. 59.
Heller, A. Über Oxyuris vermicularis, Deutsch. Arch. f. klin. Med., 1903,
lxxvii, p. 21.
Kolb, R. Über d. Befund von auf dem Peritoneum des Cavum Douglasi
angewachs. Oxyuriden, C. f. B., P. u. Inf., 1902, i Orig., xxxi, p. 268.
Michelson. Die Oberhaut der Genitocruralfalte und ihre Umgebung als Brutstätte
von Oxyuris vermicularis, Berl. klin. Wochenschr., 1877, xiv,
No. 33.
Proskauer, Th. Embryonen von Oxyuris in der Nase, Zeitschr. f. Ohrenheilkde.,
1891, xxi, p. 310.
Schneider, P. Oxyuris vermicularis im Beckenperitoneum eingekapselt, C. f.
B., P. u. Inf., 1904, i Orig., xxxvi, p. 550.
Schöppler, H. Eier von Oxyuris vermicularis im Wurmfortsatz, C. f. B.,
P. u. Inf., 1906, i Orig., xli, p. 453.
Stricker, W. Phys.-path. Bemerkungen über Oxyuris vermicularis, Arch. f.
path. Anat., 1861, xxi, p. 360.
Vix, E. Über Entozoën bei Geisteskranken, insbes. über . . . Oxyuris
vermicularis, Allg. Zeitschr. f. Psych., 1860, xvii, p. 149.
Vuillemin, P. Sur la pénétration des femelles d’Ox. vermic. à travers les
parois de l’intestin, C. f. B., P. u. Inf., 1903, i Orig., xxxii, p. 358.
Wagener, O. Oxyuris vermicularis in der Darmwand, Deutsch. Arch. f. klin.
Med., 1904, lxxxi, p. 328.
— Weitere Unters. über Oxyuris vermicularis in der Darmwand des Menschen,
Arch. f. path. Anat., 1905, clxxxii, p. 145.
Mermis hominis oris (p. 469).
Leidy, J. Description of Three Filariæ, Proc. Acad. Nat. Sci., Philadelphia,
1850, v, p. 117.
Agamomermis restiformis (p. 470).
Leidy, J. On a Filaria reported to have come from a Man, Proc. Acad. Nat.
Sci., Philadelphia, 1880, p. 130.
(D) ACANTHOCEPHALA (pp. 475 to 478).
Grassi, B., and Calandruccio. Über einen Echinorhynchus, der auch im
Menschen parasitiert, C. f. B. u. Par., 1888, iii, p. 521.
Hamann, O. Die Nemathelminthen, I, Monogr. d. Acanthocephalen, Jena,
1891.
Kaiser, J. Über die Entwickelung d. Echinorhynchus gigas, Zool. Anz., 1887,
x, p. 414.
Kaiser, J. E. Die Acanthocephalen und ihre Entwickelung, Cassel, 1893, Bibl.
Zool., vii.
Lambl, W. Mikroskop. Untersuch. d. Darmexcrete, Prag. Vierteljahresschrift
f. prakt. Heilkde., 1859, lxi, p. 45.
Leuckart, R. Commentatio de statu embryonali et larv. Echinorhynch.,
Lipsiae, 1873.
Schneider, A. Entwickelung d. Echinorhynchus gigas, Sitzgsb. Oberhess.
Ges. für Natur- u. Heilkde., 1871, p. 1.
Stiles, Ch. W. Sur l’hôte interméd. de l’Echinorh. gigas en Amérique, C. R.
Soc. Biol., Paris, 1891 (9), iii, p. 764.
(E) CORDIIDÆ (p. 479).
Aldrovandi, Ul. De animalibus insectis, 1638, Lib. vii, cap. x, p. 720.
Blanchard, R. Pseudoparas. d’un Gord. chez l’homme, Bull. Acad. Méd.,
Paris, 1897, xxxvii, p. 614.
Camerano, L. Ricerche intorn. al. parasit. ed al polimorf. dei Gordii, Mem.
R. Accad. Sci., Torino, 1887 (2), xxxviii, p. 495.
— Monografia d. Gordii, ibid., 1897, xlvii.
Degland, C. D. Descript. d’un ver filiforme rendu par vomissem, Rec. trav.
soc. d’amat. d. sci., de l’agricult. et des arts de Lille, 1819–1822, p. 166.
Guégneu, F. Nouv. cas de pseudopar. d’un Gordius dans le tube digest. de
l’homme, C. R. Soc. Biol., Paris, 1905, lix, p. 398.
Montgomery, Th. H. The Adult Organism of Paragordius, Zool. Jahrb.
Anat., 1903, xviii.
Parona, C. Altro caso di pseudopar. d. Gordio nell’ uomo, Clinica med., 1901,
No. 10.
Patruban, v. Vorkommen von Gordius aquaticus beim Menschen, Wien. med.
Jahrb., 1875, p. 69.
Rauther, M. Beitr. z. Kenntn. d. Morphol. u. d. phylogen. Beziehung. d.
Gordiiden, Jen. Zeitschr. f. Naturwiss., 1905, xl.
Topsent, E. Sur un cas de pseudopar. chez l’homme du Gordius viol., Bull.
soc. sci. et méd. de l’Ouest, 1900, ix, No. 1.
Villot, A. Evolution des Gordiens, Ann. Sci. Nat. Zool., 1891 (7), xi.
(F) HIRUDINEI (pp. 480 to 482, and 699 to 701).
Apathy. Analyse d. äuss. Körperf. d. Hirudineen, Mitt. zool. Stat. Neapel,
1888, viii, p. 153.
— Süsswasserhirudineen, Zool. Jahrb., Syst. iii, 1888, p. 725.
Blanchard, R. Article Hirudinées, Dict. encycl. d. Sci. méd., 1888, xiv, p. 129.
— Many articles in Bull. Soc. Zool. France, 1890–1899.
Ebrard. Nouv. monogr. des sangsues méd., Paris, 1857.
Leuckart, R. Die Paras. d. Mensch., 2. Aufl., i, ii, Hirudineen, fortges.
von G. Brandes, Lpzg., 1886–1901.
Moquin-Tandon. Monogr. de la famille des Hirudinées, 2e éd., Paris, 1846.
Whitman, C. O. The External Morphology of the Leech, Proc. Amer. Acad. of
Arts and Sci., 1884, xx, p. 76.
— The Segmentary Sense Organs of the Leech, Amer. Natural., 1884, xviii,
p. 1104.
— The Leeches of Japan, Quart. Journ. Micros. Sci., 1886 (2), xxvi, p. 317.
(G) ARTHROPODA (pp. 483 to 616 and 702 to 732).
(A) Arachnoidea (pp. 483 to 529).
Arachnida in General.
Lankester, E. Ray. Structure and Classification of the Arachnida, Quart.
Journ. of Micros. Sci., 1905, xlviii, p. 165.
Order. Acarina (pp. 484 to 523 and 702 to 709).
Acarina in General.
Banks, N. A Treatise on the Acarina or Mites, Proc. U.S. Nat. Mus., xxviii.
Megnin, P. Les Acariens Parasites, 1892.
Oudemanns, A. C. A Short Survey of the more Important Families of Acari,
Bull. Ent. Res., 1910, i, pt. 2.
Leptus autumnalis (pp. 485, 486, 702 and 703).
Brandis, F. Über Leptus autumnalis, Festschr. z. 50 jähr. Best. d. Prov.
Irrenanst., Nietleben bei Halle a. S., 1897, p. 417.
Brucker. Sur le rouget de l’homme, C. R. Acad. Sci., Paris, 1897 (2), cxxv,
p. 879.
Flögel, J. H. L. Über eine merkw. durch Paras. hervorger. Gewebsneubild,
Arch. f. Naturgesch., 1876 (1), xlii, p. 106.
Gruby. Herbsterytheme, Allg. Wien. med. Ztg., 1861, p. 19.
Hanstein, R. v. Beitr. z. Kenntn. d. Gttg. Tetranychus Duf. nebst Bemerk.
1871, lii, p. 255.
Hanstein, R. v. Beitr. z. Kenntn. d. Gttg. Tetranychus Duf. nebst. Bemerk.
über Leptus autumnalis, Zeitschr. f. wiss. Zool., 1901, lxx, p. 58.
Helm, F., and A. Oudemans. Sur deux nouv. formes larv. de Thrombidium,
paras. de l’homme, C. R. Acad. Sci., Paris, 1904, cxxxviii, p. 704.
— — Deux nouv. espèc. de Thrombidium de France, Bull. Soc. Ent. de France,
1904, p. 91.
Henking, H. Beitr. z. Anat., Entw. u. Biol. von Trombidium fuliginosum,
Zeitschr. f. wiss. Zool., 1882, xxxvii, p. 553.
Kraemer. Beitr. z. Kenntn. d. Leptus autumnalis, Arch. f. path. Anat., 1872,
lv, p. 354.
Küchenmeister, F. Über die sog. Stachelbeer- und Erntemilbe, Varga’s
Ztschr. f. Med., Chir. u. Geburtsh., 1862, N.F. i, p. 289.
Mégnin, P. Mém. sur la metamorph. des Acariens, Ann. sci. nat. Zool., 1876 (6), iv, Art. No. 5.
— Les Acarid. paras., Encycl. scientif. des aide-mém., Paris.
— Observations sur le Rouget, C. R. Acad. Sci., Paris, 1897, cxxv, p. 967.
Moniez, R. Sur les differ. Acar., qui s’attaq. à l’homme et qui out reçu le nom
de Rouget, Rev. biol. du Nord de la France, 1894–95, vii, p. 301.
Thiele, J. Die Gras- oder Erntemilbe, eine Plage der Feldarbeiter, Dtsche.
landw. Presse, 1898, No. 98, p. 1016.
Trouessart, E. L. Sur la piqûre du Rouget, Arch. de Paras., 1899, ii, p. 286.
Other Leptus, including Genus Trombidium (p. 486).
Altamirano, F., and A. Dugès. El tlalsahuate, El estudio, 1892, iv, p. 196.
Bonnet, G. Contrib. à l’étude du parasit., Thèse de Montpellier, 1870, p. 53.
Lemaire. Import. en France du tlalsahuate, C. R. Acad. Sci., Paris, 1867, lxv,
p. 215.
Riley, C. V. The Mexican Jigger or Tlalsahuate, Insect Life, 1893, iv, p. 211;
American Naturalist, 1873; abstracted in Handbook of the Med. Sci.,
1887, v, p. 745.
Kedani Mite (pp. 487, 613, and 703).
Baelz, E., and Kawakaini. Das japanische Fluss- oder Überschwemmungsfieber,
Arch. f. path. Anat., 1879, lxxviii, p. 373.
Schüffner. Far East Assoc. Trop. Med., C. R. Trois. Congrès Biennal,
Saigon, 1914.
Tanaka, K. Über Ätiol. u. Pathol. der Kedani-Krankh., C. f. B., P. u. Inf, 1899 (1), xxvi, p. 432.
— Über meine japanische Kedani-Krankh., ibid., 1906 (1), Orig. xlii, pp. 16,
104, 205, 320.
Tetranychus (p. 488).
Artault, L. Le platane et ses méfauts; un nouv. Acar. par. accid. de l’homme,
Arch. de Paras., 1900, iii, p. 115.
Fritsch, G. Bemerkgn. zu Herrn Hallers Aufs., Zool. Anz., 1866, x, p. 229.
Haller, G. Vorl. Nachr. über einige noch wenig bekannte Milben, ibid., p. 52.
Pediculoides, &c. (pp. 489, 615 and 616).
Brucker, C. A. Monographie de Pediculoides ventricosus, Bull. scientif. de
la France et de la Belg., 1900, xxxv, p. 365.
Flemming, J. Über eine geschlechtsreife Form der als Tarsonemus beschriebenen
Thiere, Ztschr. f. d. ges. Naturwiss., Halle, 1884 (4), iii, p. 472.
Geber, E. Entzündliche Processe der Haut durch eine . . . Milbe veranlasst,
Wiener med. Presse, 1879, xx.
Karpelles, L. Eine auf dem Menschen und auf Getreide lebende Milbe,
Anzgr. d. k. Akad. d. Wiss., Wien, 1885, xxii, p. 160.
Koller, J. Eine Getreidemilbe als Krankheitserregerin, Biol. Centralbl.,
1885, iii, p. 127; Pester med.-chir. Presse, 1882, No. 36.
Kramer. Zu Tarsonemus uncinatus Fl., Ztschr. f. d. ges. Naturw., Halle,
1884 (4), iii, p. 671.
Laboulbène, A., and P. Mégnin. Mém. sur le Sphærogyna ventricosa, Journ.
de l’Anat., 1885, xxi, p. 1.
Moniez, R. Sur l’habit. norm. dans les tiges d. céréal. d’un paras. accid. de
l’homme, Rev. biol. du Nord de la France, 1895, vii, p. 148.
Robin, Ch., and Rouyer. Erupt. cut. due à l’Acaris du blé, C. R. Soc. Biol.,
Paris, 1867 (4), iv, p. 178.
Willcocks, F. C. The Predacious Mite Pediculoides ventricosus, Agric.
Journ., Egypt, Cairo, 1914, iv, No. 1, pp. 17–51.
Nephrophages, &c. (pp. 490 and 491).
Allman, G. Description of a New Genus of Tracheal Arachnidæ, Ann. Mag.
Nat. Hist., 1847, xx, p. 47.
Banks, N. A New Genus of Endoparasitic Acarina, Geneesk. Tijdsch. v.
Nederl. Indie, 1901, deel 41, i, p. 334.
Castellani, A. Note on an Acarid-like Parasite found in the Omentum of a
Negro, C. f. B., P. and Inf., 1907, i Orig., xliii, p. 372.
Grijns, G., and J. de Haan. Acarid. als Endoparas., Geneesk. Tijdsch. v.
Nederl. Indie, 1901, deel 41, i, p. 176.
Haan, J. de. Gibt es beim Menschen endoparasitär lebende Acariden? C. F.
B., P. u. Inf., 1906, i Orig., xl, p. 693 [Carpoglyphus alienus].
Haan, J. de, and G. Grijns. Eine neue endoparasitäre Acaride, Pneumonyssus
simicola, C. f. B., P. u. Inf., 1901 (1), xxx, p. 7.
Harst, van der. Mijten in urine, Pharm. Weekbl., 1903, No. 6.
Kramer, P. Über Halarachne halichoeri, Ztschr. f. d. ges. Naturw., Halle,
1885, lviii, p. 46.
Lambl, W. Mikrosk. Unters. d. Darm-Excrete, Prager Vierteljschr. f. prakt.
Heilkde., 1859, lxi, p. 45.
Magalhaes, P. S. de. Um novo Acariano, Progr. medico, 1877, No. 4.
Marpmann. Über das Vork. v. Milben im Harn, C. f. B., P. u. Inf., 1898 (1),
xxv, p. 304.
Mégnin, P. Mém. sur les Acar. paras. du tissu cellul. et des bourses aérienn.
chez les oiseaux, Journ. de l’Anat. et de la Phys., 1879.
Miyake, H., and J. Scriba. Vorl. Mitteil. über einen neuen Paras. des Menschen,
Berl. klin. Wochenschr., 1893, No. 16, p. 374.
— — Nephrophages sang., ein neuerer menschl. Parasit im Urogenitalapparat,
Mitteil. a. d. med. Fakult. d. k. Japan. Univ., iii, p. 1.
Newstead and Todd in Thompson Yates Lab. Reports, 1906.
Nyander, J. C. Exanthemata viva, Linnaei amoenitates acad., 1757, v, p. 92,
Diss. lxxxii.
Oudemans. Over mijten in de urine en in de nieren, Med. Weekbl., 1904,
No. 12; Pharm. Weekbl., 1904, p. 269.
Silva, Aranjo A. P. da, in Gaz. med. da Bahia, 1877 (2), ii, No. 11; 1878, iii,
No. 1.
Tydeus (p. 491).
Moniez, R. Hist. natur. du Tydeus molestus, Rev. biol. du Nord de la France,
1893–94, vi, p. 419.
Dermanyssus (pp. 492, 493, 703 and 704).
Alt, Ch. H. De phthiriasi, Diss. Inaug., Bonn, 1824.
Blanchard, R. Nouv. cas de Dermanyssus gallinæ dans l’esp. hum., C. R.
Soc. Biol., Paris, 1894 (10), i, p. 460.
Geber, E., in Ziemssens Handb. d. spec. Path. u. Ther., 1884 (2), xiv, p. 394.
Heinicke, W. Zwei Fälle v. Urticaria, hervorgerufen durch die Vogelmilbe,
Münch. med. Wochenschr., 1901, No. 53.
Itzigsohn, H. Path. Bagatellen. I. Psora dermanyssica, Arch. f. path. Anat.,
1858, xv, p. 166.
Judée. Sur un nouv. paras. de la peau chez l’homme, C. R. Soc. Biol., Paris,
1867 (4), iv, p. 73.
Simon, G. Die Hautkrankheiten durch anat. Untersuchung erl., 2. Aufl.,
Berlin, 1851, p. 320.
Theobald, F. V. The Parasitic Diseases of Poultry, 1896, p. 49.
Wagner, A. Über das Vork. v. Derman. avium beim Menschen, Inaug. Diss.,
Greifswald, 1873.
Holothyrus (p. 493).
Gervais, P. Quinze espèces d’insect. apt., Ann. Soc. Ent. France, 1842, xi,
Bull. p. xiv.
Mégnin, P. Un Acarien dangereux de l’île Maurice, C. R. Soc. Biol., Paris,
1897 (10), iv, p. 251.
Leiognathus, Laelaps (p. 493).
Moniez, R. Leiognathus sylviarum, Rev. biol. Nord de la France, 1893, v,
p. 408.
Neumann, G. Pseudoparasitisme du Laelaps stabularis sur une femme, C. R.
Soc. Biol., 1893 (9), v, p. 161.
Liponyssus.
Porta, A. Dermatosi occasionale nell’ uomo dovuta ad un acara (Liponyssus
lobatus), Zool. Anz., Berlin, 1914, xliv, No. 11, p. 481.
Ixodidæ (pp. 493 to 500, 613, and 704).
Allen, W. E. Internal Morphology of the American Cattle Tick, Stud. Zool.
Lab., Univ. Nebraska, 1905.
Aragao, H. d. B. Notas sobre ixodidas brazileiros, Mem. Inst. Oswaldo Cruz,
1911, iii, pp. 145–195.
— Contribuiçao para a sistematica e biolojia dos ixodidas, Mem. Inst. Oswaldo
Cruz, 1912, iv, p. 96.
— Nota sobre algumas Coleçoes de Carrapatos brazileiros, Mem. Inst. Oswaldo
Cruz, 1913, v, pp. 261–270.
Dönitz. Die Zecken des Rindes als Krankheitsüberträger, Sitzungsb. Ges. nat.
Frde., Berlin, 1905, p. 105; 1906, No. 5.
King, H. H. Ticks of the Sudan, Fourth Rept. Well. Trop. Res. Lab., 1911,
pp. 128–130.
Koch, C. L. Uebers. des Arachnidensystems, Nürnberg, 1847, Hft. iv.
Neumann, G. Revision de la famille des Ixodidés, Mém. Soc. Zool. France,
x-xii.
— Das Tierreich, Lieferung 26, 1911.
— Notes sur les Ixodidés, Arch. de Paras., vi, viii-xi.
Nuttall, G. H. F. The Harben Lectures, 1908, Journ. Roy. Inst. Public
Health, 1908.
Nuttall, Warburton, Cooper, and Robinson. A Monograph of the Ixodoidea.
Pagenstecher, H. Beitr. zur Anat. der Milben, Leipzig, 1861, ii.
Rohr, C. J. Estudo sobre Ixodidas do Brazil.
Salmon, D. E. and Ch. W. Stiles. Cattle Ticks (Ixodoidea) of the United
States, Washington, 1902.
Ixodes (pp. 497 to 500 and 704).
Bertkau, P. Bruchstücke aus der Lebensgeschichte unserer Zecke, Verh. nat.
Ver. preuss. Rheinl. u. Westph., 1881; Sitzungsb. p. 145.
Blanchard, R. Pénétration de l’Ixodes ricinus sous la peau de l’homme, C. R.
Soc. Biol., 1891 (3), ix, p. 689.
Johannessen, A. Acute Polyurie bei einem Kinde nach dem Stiche eines
Ixodes ricinus, Arch. f. Kinderheilkde., 1885, vi, p. 337.
Mégnin, P. Encore un mot sur la biol. des Ixodes, C. R. Soc. Biol., Paris,
1903, lv, p. 175.
— Sur la biol. des tiques ou Ixodes, Journ. de l’anat. et da la phys., 1904, xl,
p. 569.
Nordenskiöld, E. Zur Anat. u. Histol. v. Ixodes reduvius, Zool. Anz., 1906,
xxx, p. 118.
Hyalomma (pp. 501, 502).
Ronsisvalle. Sui fenomeni morb. prod. nel uomo da un Ixodide denomin.
Hyalomma aegyptium, Bull. Accad. Gioenia sci. nat., 1891, xvii.
Dermacentor (pp. 502 to 505).
Cooley, R. A. The Spotted Fever Tick (Dermacentor venustus Banks) and its
Control in the Bitter Valley, Montana, Journ. Eco. Ent., 1915, viii,
pp. 47–54.
Argas and Ornithodoros (pp. 505 to 510).
Ajutolo, G. de. Dell’ Argas reflexus paras, dell’ uomo, Mem. Roy. Accad.
sci. Istit. Bologna, 1899 (5), viii.
— Nuovi casi di Argas reflexus par. dell’ uomo, Rendic. Accad. sci., Bologna,
1898, N.S. ii, p. 222.
Alt, K. Die Taubenzecke als Paras. des Mensch., Münch. med. Wochenschr.,
1892, No. 3; C. f. Bakt., 1893, xiv, p. 468.
Audouin. Descript. de l’Egypte, 2e éd., xxii, Zool., p. 426.
Blacklock, B. The Resistance of Ornithodoros moubata to various Sheep Dips,
Ann. Trop. Med. and Par., 1912, vi, p. 429.
Boschulte. Argas reflexus als Paras. des Menschen, Arch. f. path. Anat.,
1860, xviii, p. 554; 1879, lxxv, p. 562.
Brandes, G. Argas reflexus als gelegentl. Paras. des Menschen, C. f. B., P. u.
Inf., 1897 (1), xxii, p. 747.
Dugès, A. Piqûre de Turicata, C. R. Soc. Biol., Paris, 1885 (8), ii, p. 216.
Fischer de Waldheim, G. Note sur l’Acarus de Perse, Mém. soc. nat., Moscow,
1823, vi, No. 30; Ann sci. nat., 1824, ii, p. 77.
Fritsch, G. Über die giftige Wirkung des Argas persicus, Sitzungsber. Ges.
nat. Frde., Berlin, 1875, p. 61.
Gerstäcker, A. Argas reflexus, ein neuer Paras. des Menschen, Arch. f. path.
Anat., 1860, xix, p. 547.
— Gliederth. Ostafrikas von C. v. d. Deckens Reise, 1873, p. 464.
Gibert, J. M. L’Argas reflexus et son paras. chez l’homme, Thèse, Bordeaux,
1896.
Guérin-Méneville. Descript. de l’Argas talaje, Rev. et Mag. de Zool., 1849,
p. 342.
Heller, C. Zur Anat. d. Argas persicus, Sitzungsber. kais. Akad. d. Wiss.,
Wien; math.-nat. Kl., 1858, xxx, p. 297.
Laboulbène, A., and P. Mégnin. Mém. sur les Argas de Perse, Journ. de
l’Anat., 1882, xviii, p. 317.
Mégnin, P. Expér. sur l’action novice des Argas de Perse, C. R. Soc. Biol.,
Paris, 1882, p. 305.
— Les Argas du Mexique, Journ. de l’Anat. et da la Phys., 1885, xxi, p. 463.
Oken, L. Über giftige Milben in Persien, Isis, 1818, p. 1567.
Simpson, J. C. Case of Parasite (Argas mégnini) in Each Ear, Lancet, 1901,
p. 1197.
Strickland, C. Note on a Case of Tick Paralysis in Australia, Parasitology,
1914, xii, No. 4, p. 37.
Tholozan, J. D. Des phénom. morb. prod, par la piqûre . . . Argas de
Perse, C. R. Soc. Biol., Paris, 1882, p. 15.
Todd, J. L. Tick Paralysis, Journ. of Parasitology, 1914, i, pp. 55–64.
Tyroglyphidæ (pp. 511 to 513).
Canestrini and Krama. Das Tierreich, Lieferung, 1899.
Dalgetty, A. B. Water Itch, or Sore Feet of Coolies, Journ. Trop. Med., 1901,
iv, p. 73.
Fürstenberg, O. Die Kratzmilben des Menschen und der Thiere, 1861.
Layot, A. Etude sur le vanillisme, Rev. d’hyg. et de police sanit., 1883, v,
p. 711.
Ludwig, F. Die Milbenplage der Wohnungen, Leipzig, 1904.
Michael, A. D. British Tyroglyphidæ, 1901 (Roy. Society), i; 1903, ii.
Moniez, R. Sur les Tyroglyphes, qui vivent aux dépens. d. mat. alim. ou d.
prod. pharm., Rev. biol. Nord France, 1899, vi.
— Parasitisme accid. sur l’homme du Tyroglyphus farinæ, C. R. Acad. Sci.,
Paris, 1899, cviii, p. 1026.
Perrier, E. Cas de paras. passager du Glyciphagus domesticus, C. R. Acad.
Sci., Paris, 1896, cxxii, p. 859.
Trouessart, E. Faux parasit. d’une espèce de Sarcopt. détriticole dans un
kyste du testicule chez l’homme, C. R. Soc. Biol., Paris, 1900, lii, p. 742.
— Deuxième note sur l’Histiogaster spermaticus et sa prés. dans un kyste du
testic. chez l’homme, ibid., p. 893.
Trouessart, E. Endoparasitisme accid. chez l’homme d’une espèce de Sarcoptide
détriticole, Arch. de Paras., 1902, v, p. 449.
— Note compl. sur un Sarcopt. détriticole, ibid., 1906, x, p. 314.
Sarcoptidæ (pp. 516 to 521 and 704 to 708).
Alexander, A. Übertrag. d. Tierkrätze auf Menschen, Arch. f. Dermatol. u.
Syphilis, 1900, lii, p. 185.
Bergh, R. Über Borkenkrätze, Arch. f. path. Anat., 1860, xix, p. 1; Vierteljahrschr.
f. Dermatol. u. Syphilis, 1874, vi, p. 491.
Bourguignon, H. Rech. sur la contagios. de la gale des anim. à l’homme,
Mém. Soc. Biol., Paris, 1851, iii, p. 109; Ann. Sci. Nat., 1855 (4), iii,
p. 114.
Canestrini, G., and P. Kramer. Demodicidæ und Sarcoptidæ, Das Tierreich,
Berlin, 1899, Lief. 7.
Fürstenberg, M. H. F. Die Krätzmilben des Menschen u. der Thiere, Leipzig,
1861.
Gurlt and Hertwig. Vergl. Unters. üb. d. Haut des Menschen u. üb. d.
Krätzmilben, Berlin, 1844.
Hertwig, C. Über Krätz- u. Räudemilben, Arch. f. Naturgesch., 1835, i, p. 398.
Mégnin, P. Mém. sur l’acclim. des acar. psoriques des anim. sur d’autres espèc.
anim. et sur l’homme, La France méd., 1876, xxiii, p. 166.
— Sur certains détails anat. que présent. l’espèce Sarcoptes scabiei et ses nomb.
variat., C. R. Acad. Sci., Paris, 1875, lxxxi, p. 1058.
— Les Acariens parasites, Paris, Encycl. scientif. d. aide-mémoires.
Weydemann. Sarcoptes vulpis beim Menschen, C. f. B. u. Par., 1897, xxii,
p. 442.
Demodicidæ (pp. 522, 523, 708 and 709).
Guiart, J. La fréq. du Demodex chez l’homme, Bull. Soc. Zool., France, 1902,
xxvii, p. 128.
Ivers, K. Demodex s. Acarus folliculorum u. seine Beziehungen zur Lidrandentzdg.,
Deutsche med. Wochenschr., 1899, p. 220.
Kraus, A. Über färbetechn. Method. z. Nachweis d. Acarus folliculorum,
Arch. f. Dermatol. u. Syphilis, 1901, lviii, p. 351.
Landois, L. Über d. Haarbalgparas. d. Menschen, Greifswalder med. Beitr.,
1863, i, p. 17.
Leydig, F. Über Haarsackmilben u. Krätzmilben, Arch. f. Naturg., 1859 (1),
xxv, p. 338.
Majochi, D. Int. al Demodex folliculorum nelle ghiand. meibom., Bologna,
1897; C. f. B., P. u. Inf., xxv, p. 784.
— Demodex folliculorum in qualche rara affezione cutanea e speciale reperto
del medesimo nei follic. delle ciglia e delle vibrisse, Rend. sess. R. Accad.
sci. istit. Bologna, 1899, N.S. iii, 3, p. 90; Bull. sci. med., Bologna (7),
x, 5, p. 346.
Raehlmann, E. Über Blepharitis acarica, Klin. Mtsbl. f. Augenheilkde., 1899,
xxxvii, p. 33.
Simon, G. Sur les acares vivant dans les follicules pileux de l’homme, Arch.
Méd. compar., 1843, i, p. 45.
Stcherbatchoff. Le Demodex folliculorum dans les follicules ciliaires de
l’homme, Thèse, Lausanne, 1903; C. f. B., P. u. Inf., 1904, i, Ref. xxxiv,
p. 622.
Stieda, L. Über d. Vorkom. d. Haarbalgparas. an den Augenlidern, Centralb.
f. prakt. Augenheilkde., July, 1890, p. 193.
Thudichum, J. S. W. On the Demodex folliculorum as the Parasite causing
the Mange of Dog and its Transference upon Man, Med. Press and Circ.,
1894, p. 103.
Order. Pentastomida (pp. 523 to 529).
Linguatula rhinaria (pp. 524 to 526).
Beneden, P. J. van. Rech. sur l’organ. et dével. des Linguatiles, Nouv. Mém.
Acad. Belg., 1849, xxiii; Ann. Sci. Nat., 1849 (3), Zool. xi, p. 313.
Colin, in Bull soc. méd. vét., 1861 (2), v, p. 125; 1863, vii, p. 22; 1864, viii,
p. 108.
Koch, M. Zur Kenntn. des Parasit. d. Pentastomen, Arb. a. d. pathol. Inst.,
Berlin, 1906.
Kulagin, N. Zur Naturgesch. d. Pentastoma denticulatum, C. f. B., P. u.
Inf., 1898 (1), xxiv, p. 489.
Laengner, H. Über Pentastomum denticulatum beim Menschen, C. f. B., P.
u. Inf., 1906, i Orig., xl, p. 368.
Laudon. Ein kasuistischer Beitrag z. Ätiol. d. Nasenblutens, Berl. klin.
Wochenschr., 1878, xv, p. 730.
Leuckart, R. Bau und Entwickelungsgesch. d. Pentast., Leipzig, 1860.
Lohrmann, E. Unters. üb. d. anat. Bau d. Pentastomen, Arch. f. Naturgesch.,
1889, lv, 1, p. 303.
Rátz. St. v. Von der act. Wanderg. d. Pentast. denticulatum, C. f. B. u. Par.,
1892, xii, p. 329.
Schubart, T. D. Entwickelg. v. Pentastomum taenioides, Zeitschr. f. w. Zool.,
1852, iv, p. 116.
Shipley, A. E. An Attempt to Revise the Family Linguatulidæ, Arch. de
Paras., 1898, i, p. 52.
Stiles, C. W. Bau u. Entwickl. von Pentastomum proboscideum u. P. subcylindricum,
Zeitschr. f. w. Zool., 1891, lii, p. 85.
Virchow, R. Helminth. Notiz., I, Arch. f. path. Anat., 1857, xi, p. 81.
Welch, F. H. The Presence of an Encysted Echinorynchus in Man, Lancet,
1872, ii, p. 703.
Zenker, F. A. Über einen neuen thier. Paras. d. Menschen, Zeitschr. f. rat.
Med., 1854 (2), v, p. 212.
Porocephalus constrictus (pp. 526 to 528).
Aitken, W. On the Occurrence of Pentastoma constrictum in the Human Body
as a cause of Painful Disease and Death, Sci. and Pract. of Med.,
4th ed., London, 1865.
Bilharz, Th. Ein Beitr. z. Helminthographia humana, Zeitschr. f. w. Zool.,
1852, iv, p. 65.
— Über Pentastomum constrictum, ibid., 1856, vii, p. 329.
— Übers. über d. i. Ägypten beobacht. Eingeweidewürmer, Zeitschr. d. Ges. d.
Ärzte, Wien, 1858, i, p. 447.
Borden and Rodhaim. Contribution à l’étude de Porocephalus moniliformis,
Anns. Trop. Med. and Par., 1908, ii, p. 303.
Chalmers. A Case of Pentastoma constrictum, Lancet, June 24, 1899; C. f. B.,
P. u. Inf. (1), xxvi, p. 518.
Diesing, C. M. Vers. einer Monogr. d. Gttg. Pentastoma, Annal. d. wien.
Mus., 1835, i, p. 1.
Faria, de Gomes, and L. Travassor. Nota sobre a presence de lava de
Linguatula serrata, Froelich no intestino do homan no Brazil, Mem. Inst.
Oswaldo Cruz, 1913, v, p. 123.
Giard, A. Pentastomum constrictum, paras. du foie des nègres, C. R. Soc.
Biol., Paris, 1896 (10), iii; C. f. B., P. u. Inf. (1), xxiii, p. 1098.
Kiewiet de Jonge. Nadere inlichingen over de Porocephalus moniliformis,
Geneesk. Tijdschr. Nederl. Indie, 1906, xlvi, p. 524.
Neumann, G. Sur les Porocéphales du chien et de quelq. mammif., Arch. de
Paras., 1899, ii, p. 356.
Ouwens, P. A. Porocephalus moniliformis niet alleen tot Afrika, Geneesk.
Tijdschr. v. Nederl. Indie, 1906, xlvi, p. 423.
Pruner. Krankh. d. Orients, Erlangen, 1847, p. 249.
Spencer, W. B. The Anatomy of Pentastomum teretiusculum, Quart. Journ.
Micros. Sci., 1890, N.S. xxxiv, pt. 1.
Stiles, C. W. Bau und Entwickelungsgeschichte v. Pentastomum, Zeit. für
wiss. Zool., 1891, lii, p. 85.
Thiroux, M. Un cas de Pentastomum constrictum observ. au Sénégal, C. R.
Soc. Biol., Paris, 1905, lviii, p. 79. [Firket, who abstracted this paper in
Arch. f. Schiffs- u. Tropen-Hyg., 1906, x, p. 295, Anm., mentions that at
a meeting in Liége he showed an example of Pent. constr. which he had
discovered in the liver of a Congo negro.]
(B) Insecta (pp. 529 to 612 and 709 to 732).
Pediculidæ (pp. 532 to 534, and 709 to 713).
Artault de Vevey, St. Deux cas de pediculose accid. et intermitt. chez
l’homme, C. R. Soc. Biol., Paris, 1895, No. 29, p. 684.
Biebel, C. G. Insecta epizoa, Leipzig, 1874.
Cholodkowsky, N. Zur Morphologie der Pediculiden, Zool. Anz., 1903, xxvii,
p. 120.
— Zur Kenntn. d. Mundwerkz. u. Syst. d. Pediculiden, ibid., 1904, xxviii,
p. 368.
Enderlein, G. Läuse-Studien, Zool. Anz., 1904, xxviii, p. 121.
Graber, V. Anat.-phys. Studien über Phthirius inguinalis, Zeitschr. f. wiss.
Zool., 1872, xxii, p. 157.
Landois, L. Untersuch. über die am Menschen schmarotz. Pediculinen, ibid.,
1864, xiv, p. 1; 1865, xv, pp. 32, 494.
Marzocchi, V. Sur le Phthirius inguinalis, Archiv. Parasitologie, 1903,
pp. 314–317.
Pawlowsky, E. Über den Stech- und Saugapparat der Pediculiden, Zeitschr.
f. wiss. Insectenbiol., 1906, ii, Heft. 5–7.
Piaget. Les pédiculines, Leide, 1880, Suppl. 1885.
Acanthiadæ (pp. 534 to 537, and 713).
Bacot, A. W. The Influence of Temperature, Submersion, and Burial on the
survival of Eggs and Larvæ of Cimex lectularius, Bull. Ent. Res., 1914,
v, pt. 2, pp. 111–152.
— Notes on the Development of Bacillus pestis in Bugs (Cimex lectularius),
&c., Journ. Hygiene, 1915, iv, p. 777.
Blacklock, B. On the Resistance of Cimex lectularius to various Reagents,
Powders, Liquids, and Gases, Ann. Trop. Med. and Par., 1912, No. 4, vi,
p. 415.
Brumpt, E. Réduvides de l’Amérique du Nord capable de transmettre le
Trypanosoma cruzi, Bull. Soc. Path. Exot., Paris, 1914, vii, No. 2,
pp. 132–133.
— Un Réduvide du Vénézuela, le Rhodnius prolixus, chez lequel évolue
Trypanosoma cruzi, Bull. Soc. Path. Exot., Paris, 1913, pp. 382–383.
Eversmann, E. Quædam insectorum species novæ, Bull. soc. Impér. de Nat.,
Moscow, 1841, xiv, p. 351.
Girault, A. A. An Account of the Habits of the Common Bed-bug and Bibliography,
Psyche, 1905, xii, p. 61; 1906, xiii, p. 42.
— Preliminary Studies of the Biology of the Bed-bug, Cimex lectularius,
Journ. Econ. Biol., London, 1914, ix, No. 7, pp. 25–45.
Joyeux, Ch. Biologie de Cimex boneti (rotundatus), Archiv. de Paras., 1913,
pp. 140–146.
Landois, L. Anatomie von Cimex lectularius, Zeitschr. f. wiss. Zool., 1868,
xviii, p. 206.
Neiva, A. Informaçoes sobre a biologia do Conorhinus megistus, Burm., Mem.
d. Inst. Oswaldo Cruz, 1910, ii, fas. 11, p. 206.
— Informaçoes sobre a biologia da Vinchuca-Triatoma infestans, Klug., Mem.
Instit. Oswaldo Cruz, 1913, v, p. 24.
Patton, W. S. Notes on the Distribution of the Two Species of Bed-bugs,
Ind. Med. Gaz., February, 1907, xlii, No. 2.
Rothschild, N. C. The Tropical Bed-bug, Clinocoris hemiptera, Fabr., Bull.
Ent. Res., 1913, iv, pt. 4, p. 345.
Signoret, V. Notice sur quelq. hemipt. nouv., Ann. soc. entom., France, 1852
(2), x, p. 539.
Thomson, D. Attempts to find Disease Germs in the European Bed-bug (Cimex
lectularius) after Feeding Experiments, Ann. Trop. Med. and Par.,
1914, viii, p. 19.
Pulicidæ (pp. 545 to 548, 613, 714, and 715).
Bacot, A. W. Observations on the Length of Time Fleas (Ceratophyllus fasciatus)
carry Bacillus pestis, &c., Journ. Hygiene, 1915, iv, p. 770.
Baker, C. F. Revision of American Siphonaptera with a Bibliography of the
Group, Proc. U.S. Nat. Mus., xvii.
Bergh, R. Flohlarven als Pseudoparasiten d. Menschen, Monatshefte f. pr.
Dermatologie, 1885, iv, p. 209.
Blanchard, R. Quelq. mots sur la chique, Bull. Soc. Zool., France, 1889, xiv,
p. 95.
— Présence de la chique à Madagascar, Arch. de Paras., 1899, ii, p. 627.
Blandfoord, W. H. F. The Chigoe in Asia, Ent. Monthly Mag., 1894 (2), v,
p. 228.
Cunha, de Almeida. Contribuiçao para o conhecimento dos siphonapteros
brazileiros, Mem. Inst. Oswaldo Cruz, 1914, vi, fas. 2, pp. 125–136.
Henning, G. Zur Geschichte des Sandflohs in Afrika, Naturw. Wchschr.
(Potonié), 1904, N. F. iii, p. 310.
Hettner, A. Die Ausbreitung des Sandflohs in Afrika, Geograph. Zeitschr.,
1899, v, p. 522.
Hindle, E. A Chinese Flea-trap, Proc. Camb. Phil. Soc., 1913, xvii, pt. 3,
p. 284.
Hirst, L. F. Identification of Rat-fleas in Colombo, Brit. Med. Journ.,
January, 1914, p. 85.
Jordan, K., and N. C. Rothschild. A Revision of the Non-combed-eyed
Siphonaptera, Parasitology Supp. Journ. Hygiene, 1908, i, No. 1.
— A Revision of Sarcopsyllidæ, Thompson Yates and Johnston’s Lab. Report,
1906, vii, p. 15.
Jullien, J. La chique sur la côte occident. d’Afrique, Bull. soc. Zool. France,
1889, xiv, p. 93.
Lama. Dermatophilus penetrans, Giorn. Ital. Mal. Ven. Milan, 1914, xlix,
pp. 465–472.
Lass, M. Beitr. z. Kenntn. des histol.-anat. Baues des weibl. Hundeflohs,
Ztschr. f. wiss. Zool., 1905, lxxix, p. 73.
Rothschild, C. N. A Synopsis of the British Siphonaptera, Entom. Mon. Mag.,
1915, li, No. 610, pp. 42–112.
Rothschild, N. C. On Three Species of Xenopsylla occurring on Rats in
India, Bull. Ent. Res., 1914, v, pt. 1, p. 83.
— A Synopsis of Fleas found on Mus norvegicus (decumanus), Mus rattus
(alexandrinus), and Mus musculus, Bull. Ent. Res., 1910, i, pt. 2.
Russell, H. The Flea, Camb. Univ. Press, 1913.
Schwellengrebel, N. H. Versuche und Beobachtungen über die Biologie von
Xenopsylla cheopis in Ost Java, C. f. B., P. u. Inf., Jena, 1914, lxxiv,
Nos. 5 and 6, pp. 456–466.
Schwellengrebel, N. H., and L. Otten. Experimentelle Beitrage zur Kenntnis
der Uebertragung der Pest durch Flöhe und Läuse, C. f. B., P. u. Inf.,
1914, lxxiv, No. 7, p. 592.
Strickland, C. The Biology of Ceratophyllus fasciatus, the Common Rat-flea
of Great Britain, 42nd Ann. Rept. Loc. Gov. Board, Suppl., App. B,
1913, No. J, pp. 401–412.
Taschenberg, O. Die Flöhe . . . monographisch dargestellt, Halle, 1880.
Tiraboschi, C. Les Rats, les Souris et leurs parasites cutanés, Arch, de Paras.,
1903–04, viii.
Troussaint. Accid. grav. prod. par le Sarcopsylla penetrans, Arch. Méd. et
Pharm. mil., 1902, xxxix, p. 42.
Wellman, F. C. Notes from Angola, Journ. Trop. Med., 1905.
Mosquitoes and other Nematocera (pp. 548 to 603).
Adie, H. A. Note on the Sex of Mosquito Larvæ, Ann. Trop. Med. and Par.,
1912, vi, p. 463.
Annandale, N. Indian Species of Phlebotomus, Rec. Ind. Mus., 1910, iv,
pp. 2 and 3.
Austen, E. E. Notes on African Blood-sucking Midges (family Chironomidæ,
sub-family Ceratopogoninæ), Bull. Ent. Res., 1912, iii, pp. 99–112.
Ballou, H. A. Millions and Mosquitoes, Pamphlet No. 55, Imperial Dept.
Agri., West Indies, 1908.
Blanchard, R. Les moustiques, Hist. nat. et méd., Paris, 1905.
Bolt. Sand-flies and Fever in China, China Med. Journ., Shanghai, 1915, xxix,
No. 2, pp. 78–86.
Boyce, R. Prevalence, Distribution, and Significance of Stegomyia fasciata
in West Africa, Bull. Ent. Res., May, 1911, ii, pt. 1.
Carter, H. F. On certain Mosquitoes of the genera Banksinella, Theobald, and
Tæniorhynchus, Arribalzaga, Ann. Trop. Med. and Par., 1913, vii, p. 581.
Christophers, S. R. Contribution to the Study of Colour Markings and other
variable characters of Anophelinæ, &c., Ann. Trop. Med. and Par., 1913,
vii, p. 45.
Coquillett, D. W. A Classification of Mosquitoes of North and Middle
America, U.S. Dept. Agri., Div. Ent., 1906.
Dyar, H. G. Key to the known Larvæ of the Mosquitoes of the United States,
U.S. Dept. Agri., Bur. Ent. Circular 72, 1906.
Dyé, L. Les parasites des Culicides, Arch. de Paras., 1904, ix, p. 5.
Edwards, F. W. On the Culicid Genus Eretmapodites, Theobald, Ann. Mag.
Nat. Hist., July, 1911, viii, ser. 8, pp. 67–73.
— The African Species of Culex and Allied Genera, Bull. Ent. Res., 1911, ii,
pp. 241–268.
— Some New West African Species of Anopheles, &c., Bull. Ent. Res., 1911,
ii, pt. 2, p. 141.
Eyssell, A. Die Steckmücken, Handb. d. Tropenkrankh., issued by C. Mense,
Leipzig, 1905, ii, p. 44·
Fabricius, J. C. Systema antliatorum, Brunsv., 1805–06.
Franca, C. Phlebotomus papatasii (Scopoli) et fièvre à Pappataci au Portugal,
Bull. Soc. Path. Exot., 1913, vi, pp. 123–4.
Galli-Valerio, B., and J. Rochaz de Jongh. Beobachtungen über Culiciden,
Centralbl. f. Bakt., &c., 1913, xvii, p. 472.
— Beobachtungen über Culiciden und Mittelung über das Vorkommen var.
Phlebotomus papatasii Scop. im Kanton Waadt, Centralbl. f. Bakt., &c.,
1912, lxiii, pp. 222–227.
Giles, G. M. J. A Handbook of Gnats or Mosquitoes, 1st ed., London, 1900;
2nd ed., London, 1902, 17 pl.
— A Revision of the Anophelinæ, London, 1904.
GOUGH, L. H. Preliminary Notes on Egyptian Mosquitoes, Bull. Ent. Res.,
1914, v, pt. 2, p. 133.
Grassi, B. Ricerche sui Flebotomi, Memorie della societa italiana della
Scienze, 1907, se. 3, xiv.
Green, E. E. Insects, Rec. Ind. Mus., 1912, vii, pt. iii, p. 309.
Grünberg, K. Die blutsaug. Dipteren, Jena, 1907.
Guenther, K. Culiciden, Zeits. f. wiss. Insektenbiol., 1913, ix, pp. 26–40.
Headlee, T. J. The Control of Mosquitoes in a Limited Locality, Journ.
Eco. Ent., 1915, viii, No. 1, pp. 40–47.
Howard, L. O. Mosquitoes: How they Live, How they Carry Disease, &c.,
New York, 1901.
— The Yellow Fever Mosquito, U.S. Dept. Agric., Farmers’ Bull., 1913,
No. 547.
Howard, L. O., H. C. Dyar, and F. Knab. The Mosquitoes of North and
Central America and the West Indies, 1912, i and ii; 1915, iii.
Howlett, F. M. The Influence of Temperature upon the Biting of Mosquitoes,
Parasitology, 1910, iii, No. 4, p. 478.
James, S. P. The Protection of India from Yellow Fever, Indian Journ. of
Med. Res., 1913, i, No. 2, pp. 213–217.
— Note on the Practicability of Stegomyia Reduction in Indian Seaports, Ind.
Journ. Med. Res., 1913, i, No. 2, pp. 258–262.
James, S. P., and W. G. Liston. A Monograph of the Anopheles Mosquitoes
of India, Calcutta, 1904, 4to, 30 pl.; 2nd ed., 1911.
King, H. H. Animals Injurious to Man and Animals, Mosquitoes, Fourth
Rept. Well. Lab. Trop. Res., 1911, pp. 99–112.
— On the Bionomics of the Sandflies of Tokar, Bull. Ent. Res., 1913, iv, pt. 1,
p. 83.
Lahille, F. Sobre un Anopheles, una Stegomyia y la Notaçion, de las Nervaduras
alares de los Mosquitos, Ann. d. Mus. Naçion. d. Hist. Nat. de
Buenos Aires, 1912, xxiii, p. 253–263.
Leicester, G. F. The Culicidæ of Malaya, Studies from the Inst. of Med.
Res., Federated Malay States, 1908, iii.
Leon, N. Vorl. Mitt. über den Saugrüssel der Anopheliden, Zool. Anz., 1904,
xxvii, p. 730.
Liston, W. G., and T. G. Akula. A Stegomyia Survey of the City and Island
of Bombay, Proc. Gen. Mal. Comm., Madras, November, 1912, Simla,
1913, p. 187.
Ludlow, C. S. Disease-bearing Mosquitoes of North and Central America,
the West Indies, and the Philippine Islands, Bull. 4, War Dept., Office
Surgeon-General, U.S.A., 1914.
— The Philippine Mosquitoes, Psyche, 1911, xviii, No. 4, pp. 125–133.
Lutz, A. Brazilian Simulidæ, Mem. d. Inst. Oswaldo Cruz, 1910.
— Contribuiçao para o estudo das Ceratopogoninas hematofagas encontradas
Mem. Inst. Oswaldo Cruz, 1913, v, pp. 45–72.
— Contribuiçao para o estudo das Ceratopogoninas hematofagas encontradas
no Brazil, Mem. Inst. Oswaldo Cruz, 1912, pp. 1–33.
— Contribuiçao para o conhecimento das Ceratopogoninæ do Brazil, Mem.
Inst. Oswaldo Cruz, 1914, v, pp. 81–98.
— Segunda contribuiçao para o conhecimento das especies brazileiras do genus
Simulium, Mem. do. Inst. Oswaldo Cruz, 1910, ii, p. 213.
Lutz, A., and A. Neiva. Contribuiçoes para o biologia des Megarininas,
Mem. Inst. Oswaldo Cruz, 1913, v, pp. 129–141.
— Contribuiçoes para o estudo das Megarhininæ, Mem. Inst. Oswaldo Cruz,
1914, pp. 50–56.
— Contribuiçao para o conhecimento das especies do genero Phlebotomus
existentes no Brazil, Mem. Inst. Oswaldo Cruz, 1912, iv, pp. 84–95.
MacFie, J. W. S. A Note on the Action of Common Salt on the Larvæ of
Stegomyia fasciata, Bull. Ent. Res., 1914, iv, pt. 4, pp. 339–344.
Malloch, J. R. American Black Flies or Buffalo Gnats, U.S. Dept. Agri.,
Bur. Ent. Tech. Sci., 1914., No. 26.
Mansion, J. Les Phlebotomes en Corse, Bull. Soc. Path. Exot., Paris, 1913,
vi, No. 9, pp. 637–641.
Mhaskar, K. S. Stegomyia in Karachi, Proc. Gen. Malaria Com., Madras,
1912, Simla, 1913.
Mitchell, E. G. Mosquito Life, New York and London, 1907.
Newstead, R. Papataci Flies (Phlebotomus) of the Maltese Islands, Bull.
Ent. Res., 1911, ii, pt. i.
— The Papataci Flies (Phlebotomus) of the Maltese Islands, Bull. Ent. Res.,
1911–12, ii, p. 47.
— Notes on Phlebotomus with descriptions of New Species, Bull. Ent. Res.,
1912, iii, p. 361.
— Notes on Phlebotomus with descriptions of New Species, Bull. Ent. Res.,
1914, v, pt. 2, pp. 129–136.
Newstead, R., and H. F. Carter. On Some New Species of African Mosquitoes
(Culicidæ), Ann. Trop. Med. and Par., vii, p. 233.
— — Description of a New Genus and three New Species of Anopheline Mosquito,
Ann. Trop. Med. and Par., 1910, iv, p. 377.
— — On a New Genus of Culicinæ from the Amazon Region, Ann. Trop. Med.
and Par., 1910, iv, p. 553.
Newstead, R., and H. W. Thomas. The Mosquitoes of the Amazon Region,
Ann. Trop. Med. and Par., 1910, iv, p. 141.
Nuttall and Shipley. Structure and Biology of Anopheles, Journ. of Hygiene,
January, April, and October, 1901, and January and April, 1903.
— — Studies in Relation to Malaria; II, Structure and Biology of Anopheles,
Journ. of Hyg., 1901, i; 1902, ii; 1903, iii.
Peryassu, A. Os Culicideos do Brazil, Trabalho do Instituto de Manguinhos,
1908.
Ross, E. H. The Reduction of Domestic Mosquitoes, London, John Murray.
Roubaud, E. Quelques mots sur les Phlébotomes de l’Afrique occidentale
française, Bull. Soc. Path. Exot., 1913, vi, pp. 126–128.
Schiner, J. R. Fauna austriaca, Die Fliegen (Diptera), Wien, 1860–64.
Schwetz, J. Preliminary Notes on the Mosquitoes of Kabinda, Belgian
Congo, Ann. Trop. Med. and Par., 1914, viii, p. 163.
Stanton, A. T. The Anopheles of Malaya, Bull. Ent. Res., 1913, pt. 1, iv,
pp. 129–133.
— The Anopheles Mosquitoes of Malaya and their Larvæ, with some Notes on
Malaria-carrying Species, Journ. London School Trop. Med., 1912, ii,
pt. 1, pp. 3–11.
Summers, S. L. M. A New Species of Phlebotomus from South America, Bull.
Ent. Res., 1912, iii, p. 209.
Taylor, F. H. The Culicidæ of Australia, Trans. Ent. Soc. Lond., March 31,
1914, pp. 683–708.
— A Revision of the Culicidæ in the Macleay Museum, Sydney, Proc. Linn.
Soc., N.S. Wales, 1913, xxxviii, pt. 4, pp. 747–760.
— Report of Entomologist, Reprint from Report for Year 1911 of the Australian
Inst. Trop. Med., 1913, pts. xiii, xiv, pp. 24.
— Description of Mosquitoes collected in the Northern Territory during the
Expedition, 1911, Rept. on Health and Disease in the Northern Territory,
Bull. No. 1A, 1912, p. 25.
— Culicidæ of Papua, Trans. Ent. Soc. Lond., 1914, pt. 1, pp. 185–205.
Theobald, F. V. A Monograph of the Mosquitoes of the World, 1901–1911,
vols. i-v, and atlas and plates.
— New Culicidæ from the Sudan, Ann. Trop. Med. and Par., 1913, vii, p. 591.
— A New Genus and Two New Species of Culicidæ from the Sudan, Fourth
Rept. Wellcome Trop. Res. Lab., 1911, Vol. B, Gen. Science, pp. 151–156.
— Three New Culicidæ from the Transvaal, Entomol., March, 1912.
— A New Mosquito from North China, Entomol., June, 1913, p. 179.
— A New Mosquito from Samoa, Entomol., January, 1914, p. 36.
— Culicidæ of the R. Zool. Soc., “Natura Artis Magistra,” Amsterdam Overgt.
uit het Tijdsch. v. Ent., 1911, liv, pp. 233–240.
— The Distribution of the Yellow Fever Mosquito (Stegomyia fasciata, Fabricius)
and general notes on its Bionomics, First Int. Cong. d’Ent., 1911,
pp. 145–170.
Theobald, F. V. A New Species of Culicidæ, Rev. Zool. africaine, 1912, ii,
fas. 1.
— Novæ Culicidæ, 1911, pt. i, pp. 35.
— Culicidæ (Percy Sladen Trust Exp. to Indian Ocean in 1905), Trans. Linn.
Soc. Lond., 1912 (2nd ser., Zool.), xv, pt. 1, pp. 81–94.
— Culicidæ from New Caledonia and the Loyalty Islands, Nova Cal., Zool.,
1913, i-iii, No. 3, p. 163.
— A New Genus and Two New Species of Culicidæ from the Sudan, Fourth
Report Wellcome Trop. Lab., 1911, Vol. B, Gen. Sci., pp. 151–156.
Townsend, C. H. T. A Phlebotomus the practically certain Carrier of
Verruga, Science, 1913, xxxviii, pp. 194–195.
— The Vector of Verruga, Phlebotomus vermucarum, sp. nov., Insecuta
Inscitiæ Menstruus, Washington, D.C., 1913, i, No. 9, pp. 107–109.
Urich, F. W. Mosquitoes of Trinidad, Proc. Agri. Soc. Trini. and Tobago,
1913, xiii, No. 10, pp. 525–530.
Willcocks, F. C. A Preliminary Note on the Prevalence of Mosquitoes in
Cairo and its Environs, Ann. Trop. Med. and Par., 1909, v, p. 583.
The House-fly (p. 586).
Bayon, H. Leprosy and House-flies, Ann. Trop. Med. and Par., 1915, ix,
pp. 1–90.
Bishopp, Dore, and Parman. Notes on Certain Points of the Economic
Importance in the Biology of the House-fly, Journ. Eco. Ent., 1914, viii,
pp. 54–71.
Felt, E. P. Methods of Controlling the House-fly and thus Preventing the
Dissemination of Disease, New York Med. Journ., April 2, 1910.
Hewitt, C. G. The Biology of the House-fly in relation to Public Health,
Journ. Roy. Inst. Pub. Health, October, 1908.
— On the Bionomics of certain Calyptrate Muscidæ and their Economic Significance,
with especial reference to Flies inhabiting Houses, Journ. Eco.
Biol., 1907, ii, p. 3.
— The Structure, Development, and Bionomics of the House-fly, Musca
domestica, II and III, Bionomics, &c., and the Relations to Human
Disease, Quart. Journ. Micros. Sci., 1908, and December, 1909, pt. 3, liv.
— A Preliminary Account of the Life-history of the Common House-fly (Musca
domestica, L.), Mems. and Proc. Manchester Lit. and Phil. Soc., 1906–1907,
pt. 1, li.
— House-flies and the Public Health, Ottawa Naturalist, May, 1910, xxiv,
pp. 31–38.
— Notes on the Pupation of the House-fly (Musca domestica) and its mode of
Over-wintering, Can. Ent., March, 1915, xlvii, pp. 73–78.
Hindle, E. The Flight of the House-fly, Proc. Camb. Phil. Soc., 1914, xvii,
pt. 4, pp. 310–313.
Howard, L. O. House-flies, U.S. Dept. Agri., Div. Ent., Circ. No. 71, 1906.
— The House-fly, Disease Carrier, New York, 1911.
Hutchinson, R. H. The Migratory Habit of the House-fly Larvæ as indicating
a Favourable Remedial Measure, Bull. U.S. Dept. Agri., 1914,
No. 14.
Jepson, F. P. The Breeding of the Common House-fly (Musca domestica)
during the Winter Months, Journ. Eco. Biol., 1909, iv, pt. 3.
Morrell, A. W. House-fly Baits and Poisons, Journ. Eco. Ent., 1914, vii,
No. 3, pp. 268–273.
Newstead, R. Preliminary Report on the Habits, Life Cycle, and Breeding-places
of the Common House-fly (Musca domestica, L.) as observed
in the City of Liverpool, &c., Health Committee Rept., City Liverpool,
1907.
— Report on the Habits, Life Cycle, and Breeding-places of the Common
House-fly, Liverpool, 1907, Ann. Trop. Med. and Par., February, 1908.
Packard, A. S. On the Transformation of the Common House-fly, with Notes
on Allied Forms, Proc. Boston Soc. Nat. Hist., 1874, xvi, p. 136.
Reports of the Local Government Board on Public Health, &c. New Series,
Nos. 5 and 16, 1909. Preliminary Reports on Flies as Carriers of Infection,
and further Preliminary Reports (Paper by Copeman, Jepson,
Nuttall, Graham Smith, and Austen), containing Bibliography and
Abstracts up to date.
Surface, H. A. To keep down House-flies, Zool. Press Bull., Div. Zool.,
Dept. Agric., Pennsylvania, 1915, No. 313. Recommends ground phosphate
rock, scattered over manure heaps.
Brachycera, &c. (pp. 600 to 612, and 613 to 615).
Ashley-Emile, L. E. Zambesi ulcer, Journ. Trop. Med., 1905; Arch. f. Schiffs.- u.
Tropen-Hyg., 1906, x, p. 164.
Austen, E. E. African Blood-sucking Flies other than Mosquitoes and Tsetse-flies,
1909.
— A Monograph of the Tsetse-flies, 1903.
— Supplementary Notes on Tsetse-flies, Brit. Med. Journ., 1904.
— A Handbook of the Tsetse-flies, 1891.
— Some Dipterous Insects which cause Myiasis in Man, Trans. Soc. Trop.
Med. and Hyg., March, 1910, p. 215.
— New African Tabanidæ, Bull. Ent. Res., 1914, iv, pt. 4, pp. 283–300.
Bezzi, M. Etudes systématiques sur les Muscides hématophages du genre
Lyperosia, Archiv. Parasit., 1911, xv, pp. 110–143.
— Die Gattungen der blutsaugenden Musciden, Zeitsch. f. Hymenop. u. Dipt.,
1907, vii, p. 413.
Blanchard, C. Contrib. à l’Etude des Diptères paras., Ann. Soc. Ent. France,
1896, lxv, p. 641.
Carter, H. F. Descriptions of Three New African Species of the genus
Tabanus, Ann. Trop. Med. and Par., 1914, vi, p. 435.
Chevrel, R. Sur le Myiase des Voies urinaires, Arch. de Paras., 1909, xii,
p. 369.
Cluss, F. Myiasis interna and externa, Inaug.-Diss., Tubingen, 1902.
Cockerill, J. D. A. A Dermatobia in Guatemala, Entomologist, 1914, xlvii,
p. 131.
Davy, J. B. Notes on the Habits of Glossina fusca, Bull. Ent. Res., 1910–1911,
i, p. 143.
Dönitz. Eine neue afrik. Fliege, Cordylobia murium, Sitz. Ges. naturf. Frde.
Berl., 1905, p. 248.
Dubreuilh, W. Les Diptères cutic. chez l’Homme, Arch. Méd. exp. et d’Anat.
path., 1894, vi, p. 328.
Enderlein, G. Die Respirationsorgane der Gastriden, Sitz. k. Akad. d. Wiss.,
Wien, Math-nat., 1899, Kl. cviii.
Fell, T. E. Notes on Tsetse-flies and on Prophylactic Measures against
Sleeping Sickness, &c., Bull. Ent. Res., 1912, iii, p. 227.
Fiske, W. F. The Bionomics of Glossina, Bull. Ent. Res., 1913, iv, pt. 2,
p. 95.
Francaviglia, M. C. An cora sulla myiasi auricolare, Boll. Sedute Accad.
Gioenia, Catania, 1914, No. 31, pp. 15–23.
Fuller, C. The Skin-maggot of Man (Cordylobia anthropophaga), Agri.
Journ. Union of South Africa, Pretoria, 1914, vii, No. 6, pp. 866–874.
Gedoelst, L. Contrib. à l’Etude des larves cuticoles des muscides africaines.
Arch. de Paras., 1905, ix, p. 568.
Giles, G. M. The Anatomy of the Biting Flies of the genus Stomoxys and
Glossina, Journ. Trop. Med., 1906, ix.
Grünberg. Ueber Blutsaugende Musciden, Zool. Anz., 1906, xxx, p. 78.
Grünberg, K. Afrik. Musciden mit parasit. lebenden Larven, Sitz. Ges. Nat.
Frde. Berl., 1903, p. 400.
— Die Blutsaugenden Dipteren.
— Ein neuer Fall des Vorkommens der Larve der Rinderdasselfliege im
menschlichen Auge, Sitz. Ges. Nat. Frde. Berl., 1913, 5 and 6, pp. 298–304.
Hewitt, C. G. Observations on the Feeding Habits of the Stable Fly,
Stomoxys calcitrans, L., Trans. Roy. Soc. Canada, 1914, viii, pp. 37–42.
Huber, G., and F. L. Flack. An Unusual Case of Screw-worms in the Nose
and Nasal Accessory Sinuses, Journ. Amer. Assoc. Chicago, 1914, lxiii,
No. 26, p. 228.
Joseph, G. Über Fliegen als Schädlinge und Paras. d. Menschen., Dtsche.
Med. Ztg., 1885, i, p. 37, and 1887, iii, pp. 713–725.
Jack, R. W. Observations on the Breeding Haunts of Glossina morsitans,
Bull. Ent. Res., 1911–1912, ii, p. 357.
King, H. H. Blood-sucking Flies other than Mosquitoes, Fourth Rept. Well.
Trop. Res. Lab., 1911, pp. 112–126.
— Observations on the Occurrence of Glossina in the Mongolla Province of
the Anglo-Egyptian Sudan, Bull. Ent. Res., 1912, iii, p. 89.
Kingham, A. Notes on the Preliminary Stages of Glossina morsitans, Bull.
Ent. Res., 1911–1912, ii, p. 291.
Le Dantec and Boyé. Note sur une myiase observée chez l’homme en Guinée
franç. (Réun. biol. de Bordeaux), Le Caducée, 1905, v, p. 9; Arch. f.
Schiffs- u. Tropen-Hyg., 1906, x, p. 71.
Lloyd, L. Notes on Glossina morsitans in the Luangwa Valley, N. Rhodesia,
Bull. Ent. Res., 1912, iii, p. 233.
Löw, F. Über Myiasis und ihre Erzeuger, Wien. med. Wchschr., 1882, xxii,
p. 247, and xxiii, p. 972.
Lowne, B. T. Physiology, Morphology, and Development of the Blow-fly,
two vols.
Lutz, A. Tabanidas do Brazil e de alguno Estados visinhos, Mem. Inst.
Oswaldo Cruz, 1913, v, pp. 143–190.
— Novas contribuiçoes para o con hecmento das Pangoninas e Chrysopinas do
Brazil, Mem. Inst. Oswaldo Cruz, 1911, iii, fas. 1, pp. 65–84.
Lutz, A., and A. Neiva. Los Tabanidæ do Estado do Rio de Janeiro, Mem.
Inst. Oswaldo Cruz, 1914, vi, pp. 69–80.
McConnell, R. E. Some Observations on the Larva of Auchmeromyia luteola,
Fabr., Bull. Ent. Res., 1913, iv, pt. 1, p. 29.
— Notes on the Occurrence and Habits of Glossina fuscipes in Uganda, Bull.
Ent. Res., 1912, iii, p. 55.
MacFie, J. W. S. Experiments and Observations upon Glossina palpalis, Bull.
Ent. Res., 1912, iii, p. 61.
Magalhaes, P. de. Subsidio ao estudo das Myiases Rio de Janeiro, 1892.
Meigen, J. W. Systemat. Beschr. d. bek. Europ. zweiflügel. Insecten, 1818–1838.
Minchin, Gray and Tulloch. Glossina palpalis and its relation to Trypanosoma
gambiense and other Trypanosomes, Proc. Roy. Soc., London,
Ser. B., 1906, lxxviii, p. 4.
Moiser, B. Notes on the Haunts and Habits of Glossina tachinoides near
Geidam, Bornu Province, N. Nigeria, Bull. Ent. Res., 1912, iii, p. 195.
Neave, S. A. Notes on the Blood-sucking Insects of Eastern Tropical Africa,
Bull. Ent. Res., 1912, iii, p. 275.
Neiva, A., and de Faria Gomes. Notas sobre um caso de Miiase humana
ocasionada por larvas de Sarcophaga pyophila n. sp., Mem. Inst. Oswaldo
Cruz, 1913, v. pp. 16–23.
Neish, W. D. The Tabanidæ and Anophelinæ of Jamaica, Rept. Dist. Med.
Officer, Spanish Town, Jamaica, 1913.
Newstead, R. On the Life-history of Stomoxys calcitrans, Journ. Eco. Biol.,
1906.
— On Three New Species of the genus Glossina, Ann. Trop. Med. and Par.,
1910, iv, p. 369.
— A New Tsetse-fly from British East Africa, Ann. Trop. Med. and Par.,
1912, iv, pt. 4, p. 129.
— A New Tsetse-fly from the Congo Free State, Ann. Trop. Med., 1913, vii,
p. 331.
— On the Characteristics of the Newly Discovered Tsetse-fly (Glossina austenii),
&c., Bull. Ent. Res., 1912, iii, p. 355.
— A Revision of the Tsetse-flies (Glossinæ) based on the study of the Male
Genital Armature, Bull. Ent. Res., 1911–1912, ii, p. 9.
Paris, P. Un cas de myiase intestinale, C. R. 41me Session Assoc, française
pour l’Avancement des Sciences, 1913, p. 447.
Peiper, E. Fliegenlarven als gelegentl. Paras. d. Menschen, Berlin, 1900.
Pollard, J. Notes on the Tsetse-flies in the Muri Province, N. Nigeria,
Bull. Ent. Res., 1912, iii, p. 219.
Richardo, G. Notes on the Tabanidæ, Ann. and Mag. Nat. Hist., January to
May, 1910.
Rodhain, J., C. Pons, van den Branden, and G. Bequaert. Rapport s. l.
Travaux d. l. Mission scientifique du Katanga. Annexe, Carte du
Katanga, Répartition des Glossines, 1913, Tabanidæ et Glossinæ.
Rodhain, J., and J. Bequaert. Nouvelles observations sur Auchmeromyia
luteola, Fabr., et Cordylobia anthropophaga, Grünb., Revue Zool.
africaine, 1913, ii, pt. 2, pp. 145–154.
Roorda-Smit, J. A. Die Fliegenkrankheit u. ihre Behandl., Dtsch. Med.
Wochenschr., 1906, p. 763.
Roubaud, E. Recherches sur les Auchmeromyies, Calliphorines, à larves
suceuses de sang de l’Afrique tropicale, Bull. Scient. d. l. France et d. l.
Belg., 1913, xlvii, fas. 2, pp. 105–202.
Sander, L. Die Tsetsen, Leipzig, 1905 (Reprint from Arch. f. Schiffs- u.
Tropen-Hyg., 1905, ix).
Sawyer, W. A., and W. B. Herms. Attempts to Transmit Poliomyelitis by
means of the Stable Fly (Stomoxys calcitrans), Journ. Amer. Med.
Assoc., 1913, lxi, pp. 161–166.
Schiner, J. R. Fauna austriaca, Die Fliegen, 1860–1864.
Schwertz, J. Quelques observations préliminaires sur la morphologie et la
biologie de la larve, de la nymphe, et de l’image de l’Auchmeromyia
luteola, Fabr., Ann. Trop. Med. and Par., Se. T.M., 1914, viii, No. 3,
pp. 497–507.
Sergent, Ed. and Et. La “Tamne,” myiase humaine des montagnes sahariennes
touareg, identique à la “Thimni” des Kabyles, due à Œstrus oris,
Bull. Soc. Path. Exot. Paris, 1913, vi, No. 7, pp. 487–488.
Speiser, P. Hippoboscidæ, Wien. ent. Zeitg., 1899, xviii, p. 201.
— Neue Arbeiten über blutsaugende und Krankheiten übertragende Insekten,
Zeit. f. wiss. Insektenbiol., 1908, pp. 241, 301, 420, 437.
— Die geographische Verbreitung der Diptera pupipara und ihr Phylogenie,
Zeit. f. Wiss. Insektenbiol., 1908, pp. 241, 301, 420, 437.
Wellman, F. C. Experimental Myiasis in Goats, Journ. Trop. Med., 1906.
Wirsing. Über Myiasis intestinalis, Zeitschr. f. klin. Med., 1906, lx, p. 122.
Woosnam, R. B. Report on the Search for a Glossina on the Amala River,
S. Masai Reserve, E. Africa Protectorate, Bull. Ent. Res., 1911, iv,
pt. 4, pp. 271–351.
General Works.
Blanchard, R. Traité de Zoologie médicale, Paris, ii, 1886.
Boyce, R., and F. C. Lewis. The Effects of Mosquito Larvæ on Drinking
Water, Ann. Trop. Med. and Par., 1909, v, p. 591.
Butler, E. A. Our Household Insects, 1893.
Doane, R. W. Insects and Disease.
— Disease-bearing Insects in Samoa, Bull. Ent. Res., 1913, iv, pt. 4, p. 265.
Fantham, H. B., and A. Porter. Some Minute Animal Parasites. 319 pp.
and 56 text-figs. London: Methuen and Co., 1914.
Graham-Smith, G. S. Flies in Relation to Disease (non-Blood-sucking),
2nd ed., Camb. Press.
Hindle, E. Flies and Disease (Blood-sucking), 1914.
Howard, C. W. Insects directly or indirectly Injurious to Man and Animals
in Mozambique, East Africa, Bull. Ent. Res., 1912, iii, pp. 211–218.
King, H. H. Report of the Entomological Section of the Wellcome Tropical
Research Laboratories, Fourth Report, Gen. Sci., 1911, vol. B, pp. 95–130.
Mégnin, P. Les Parasites et les Maladies Parasitaires chez l’Homme, &c.,
Paris, 1880.
Neumann, G. The Parasites of Domesticated Animals, 1892.
Newstead, R. Reports on the Twenty-first Expedition of the Liverpool School
of Tropical Medicine, Sec. 1, Med. and Econ. Ent., Jamaica, 1908–1909,
pt. 1.
— Tick and other Blood-sucking Animals, Ann. Trop. Med. and Par., 1909,
iii, p. 421.
Newstead, R., J. E. Dutton, and J. L. Todd. Insects and other Arthropoda
collected in the Congo Free State, Ann. Trop. Med. and Par., 1907,
No. 1, i, pp. 3–114.
Nicholls, L. The Transmission of Pathogenic Micro-organisms by Flies in
St. Lucia, Bull. Ent. Res., 1912, iii, p. 81.
Nuttall, G. H. F. On the Rôle of Insects, Arachnids and Myriapoda, as
Carriers in the Spread of Bacterial and Parasitic Diseases, Johns
Hopkins Hospital Reports, 1899, viii.
Railliet, A. Traité de Zoologie médicale et agricole, Paris.
Ross, R. The Prevention of Malaria.
Ross, Sir R., and E. S. Edie. Some Experiments on Larvicides, Ann. Trop.
Med. and Par., 1911, v, No. 3, p. 385.
Sergent, E. Les Insectes piqueurs-suceurs, Paris.
Stephenson, J. Medical Zoology, 1832.
alpha-table
A.
Abdomen, malignant growth, in, with ascites, Leydenia gemmipara associated with, 49, 50
Abdominal and pelvic organs, blood-supply of, as illustrating distribution of Schistosoma hæmatobium in body, 272
— operation wound, escape of Ascarides from, 654, 655
Abscess cavities, larvæ of Sarcophaga wohlfahrti from, 723
— in filariasis, 401
Abscesses, cutaneous, due to Lagocheilascaris minor, 467
Abyssinians, infection with Tœnia saginata, 340
Acanthiadæ, characters, 534
Acanthia lectularia, see Cimex lectularius
Acanthobothrium coronatum, excretory vessels, 292
Acanthocephala, 475
— development of, 17
— isolated position of, 20
— life spent in intermediate and final host, 18
— loss of digestive system in, 3
— morphology, 475
Acanthocheilonema, morphology, 414
— perstans, geographical distribution, 416
— — — — in South America, 416
— — morphology, 414
— — topographical distribution, 416
Acarina, characters and morphology, 484
— hosts, habitat and food of (and footnote), 484
Acartomyia, characters, 564
Acarus dysenteriæ, 512
— hordei, cereal mite, 489
Accessory sinuses, nasal, larvæ in, 717
Acephalina often “cœlomic” parasites, 134
Acid alcohol in differentiation of flukes, 471
Acinetaria, 198
Acne, lesions resembling, set up by Sparganum proliferum, 318
Acoleïnæ, vagina atrophied in, 297
Actinomyxidia, 129, 187
Adams, skin disease caused by larvæ of Dermatobia noxialis, 725
Addario, Filaria (?) conjunctivæ in man, 405
Adelea, 141
Adeleidea, 141, 742
Adie, Mrs., life-cycle of Hæmoproteus (Halteridium) columbæ, 152
Ædeomyia, characters, 565
Ædeomyina, characters, 564
Ædes, characters, 564
Africa, larvæ of Muscidæ causing myiasis in man in (footnote), 590
— South, farm stock in, attacked by Hyalomma ægyptium, 501, 502
— West, French, cause of myiasis in, 614
— — geographical distribution of Onchocerca volvulus in, 419
Agamofilaria, 406
— georgiana, habitat, 406
— — morphology, 406
— labialis, morphology, 407
— oculi humani, 405, 406
— palpebralis, 405, 406
Agamomermis, 470
— restiformis, morphology, 470
Agglutinating hæmolytic action of serum of ancylostome patients, 648
Ahmed Emin, small variety of Plasmodium vivax, 166
Air-passages, invasion by larvæ of screw-worm fly (Chrysomyia macellaria), 587
— — by Metastrongylus apri, 433
— upper, Ascarides in, 690
— — — danger of, 691
— — — how introduced, 690
— — leeches in, 699, 700
Aitken, Porocephalus constrictus, 526
Akamushi, see Kedani mite
Akaneesch, see Kedani mite
Alcohol, application in nasal myiasis, 719
Aleppo button, see Oriental sore
Aleurobius (Tyroglyphus) farinæ, characters of, 511
Alexeieff, Chilomastix (Tetramitus) mesnili, 735
— on genus Giardia, 736
— views on trichomonad cysts, 56
Alimentary canal of Hirudinea, 480
— tract, spirochætes in, 741
Allessandri, hæmolytic action of ancylostomes, 647
Allocreadium isoporum, excretory apparatus, 218
Alum solution in nasal myiasis, 719
Amaurosis following male fern poisoning, 671
Amblyomma americana, geographical distribution, 501
— — objects of attack, 501
— — suppuration resulting from punctures of, 501
— cayennense, characters, 500
— — geographical distribution, 501
— — ill-effects from attacks of, 501
Amblyomma cayennense, synonyms, 500
— characters of, 497
— hebræum (bont tick) carrier of heart-water fever in sheep, 493
— — life-cycle of, 495
— maculatum, 501
America, North, amount of prevalence of trichinosis in, 428
— South, geographical distribution of Acanthocheilonema perstans in, 416
Amicis, de, infection with Demodex folliculorum, 708
Amœba buccalis, 44, 734
— coli, 31
— dentalis, 44, 734
— diaphana, 31
— dysenteriæ, 31
— fluida, 46
— gingivalis, 44, 734
— lobosa, var. guttula, var. oblonga, 31
— miurai, 46
— — characters, 46
— reticularis, 31
— spinosa, 31
— urogenitalis, 45
— vermicularis, 31
Amœbæ, bodies resembling, found in serous fluid, 46
— cerebral abscess set up by, 35
— cultural, 42, 618
— culture media for, 742, 743
— experimental injection of, in cats producing dysentery, 35
— — — — producing enteritis, 36, 37
— experiments with, 618
— found in fæces, 47, 48
— — in lung, 45
— — in urine, 45, 46
— intestinal, association with colitis, 30
— — causal agent in production of dysentery, 30
— — — — — — experiments made to prove, 30
— — culture medium for, 743
— — discovery in case of dysentery, 29, 30
— — — in stools, 30
— — encysted, 31
— — human, 29
— — — discovery of, 29
— — ingestion of red blood corpuscles by, 35, 39, 42
— invading bladder, 46
— liver abscess set up by, 35
— penetration of intestinal blood-vessels by, 36
— phagedænic, 733
— pulmonary abscess set up by, 35
— testaceous (Monothalamia), characters of, 47
Amœbic dysentery, 618
Amœbina, characters and habitat, 27
Amœboid germs in pseudonavicellæ, 130
Amœbosporidia, 130
Amœbulæ, 34, 49, 183, 185
— formation of, 34
Amphimerus, 257
Amphimerus noverca, habitat, 258
— — morphology, 257, 258
— — synonyms, 258
Amphistomata, morphology, 230
Anæmia, case of, effect of expulsion of Tænia solium on, 648
— in ancylostomiasis, treatment, 687
— of dourine, 97
— period of, in liver-fluke disease in sheep, 240
— pernicious, symptoms disappearing after expulsion of Ascaridæ, 649
— proteid metabolism in, 645
— splenic, infantile, see Kala-azar, infantile
— see also Ancylostome anæmia
— see also Bothriocephalus anæmia
— see also Trichocephalus anæmia
Anal canal, means of access of Schistosoma hæmatobium to, 272
Anaplasma, 180
— marginale, 180, 611
— — cause of gall-sickness in cattle, 180
Ancylostoma braziliense, 456
— caninum, 456
— ceylanicum, habitat, 456
— — morphology, 456
— characters, 445
— duodenale and Strongyloides stercoralis, larvæ of, differences between, 451
— — buccal capsule, 445
— — bursa, 448, 449, 450
— — cephalic glands, 447
— — cervical papillæ, 447
— — development, 451
— — diagrammatic representation of male and female, 446
— — embryos, 451
— — excretory system and cervical glands, 447
— — — — — — diagrammatic representation, 448
— — food, 450
— — genital cone, 450
— — geographical distribution, 450
— — habitat, in man only, 450
— — infection by, 682
— — — modes of, 683
— — — must be on large scale to produce illness, 682
— — — see also Ancylostomiasis
— — invading frontal sinus, 683
— — larvæ of, bionomics of development, 453, 454
— — — infection by skin, 454, 455
— — — — — in dogs, 455
— — — infective stage, 454
— — — method of cultivation, 455
— — — mode of entry into body, 454
— — — morphology, 451, 452, 453
— — — stages, 451, 452
— — — thigmotropism in mature stage, 454
— — lateral lines, 448
— — morphology, 445
— — number of females present in intestine, mode of reckoning (footnote), 454
Ancylostoma duodenale, œsophageal glands, 447
— — organs of Necator americanus compared with those of, 458
— — ova of, 451
— — ovaries, 449
— — spicules, 450
— — testis, 449
— — ventral teeth, 446, 447
— larvæ of, cultivation, 474
— malayanicum, 456
— pluridentatum, 456
Ancylostome anæmia, etiology, 647, 648
— — experimental, 646
— — retinal hæmorrhages in, 646
— — toxic hypothesis, 646, 647
— — treatment, 687
Ancylostomeæ, 445
Ancylostomes, expulsion of, drugs for, 685, 686
— toxic action on hosts, 647
Ancylostomiasis, agglutinating hæmolytic action of serum of patients, 648
— eosinophilia in, 647
— morbid anatomy, 459
— prophylaxis against, 684
— — — in miners, 684
— proteid destruction in, 647
— symptoms, 683
— — set up by invasion by Hæmonchus contortus mistaken for those of, 438
— treatment, 754
Ancylostominæ, 438
Andrews, Oxyuris in appendix, 655
Angiostomidæ, 374, 379
Angiostomum nigrovenosum, heterogony in, 381
— — male of rhabditic form, 370
— — mode of generation, 372
Angola, highlands of, uncertain species of Ascaris occurring in, 465
Anguillula, 379
— aceti found in vinegar, 379
— — morphology, 379
— — occurrence in urine, 379
— intestinalis and A. stercoralis, see Strongyloides stercoralis
— mucronata, 377
Anguillulidæ, 377
— hosts of, 374
— morphology, 374
Anguillulina, 379
— putrefaciens living in onions, 379
— — synonyms, 379
Animal matter, decomposing, Tyroglyphidæ in, 511
Animals, mites living endoparasitically in, 491
Ankylorhynchus, characters, 563
Annaratone, pseudomeningitis due to Ascaridæ infection, 649
Anopheles and Culex, larvæ of, position in water compared, 554
— — ova of, method of depositing compared, 554
— — points of difference between, 551
Anopheles bifurcatus, ova of, localities selected for deposition, 553
— characters, 561, 566
— claviger, mouth parts, 550
— development of human malarial parasite only takes place in, 158, 159
— entire genus capable of transmitting malaria to man, 552
— head of male and female, 549, 556
— only genus of mosquito transmitting malaria, 158
— ova of, 557, 558
— maculipennis, 552
— — breeding places of, 557
— — Crithidia inhabiting, 104
— — intestine of, stages of development of pernicious or malignant tertian parasites in, 162
— — larva of, 553
— — ova of, localities selected for deposition, 553
— — pupa of, 554
— — sporulation stages of malarial parasites from, 163
— — stomach of, oöcysts and oökinetes of malignant tertian parasite in, 162, 163
— — transverse section through proboscis, 550
Anophelines, genera of (footnotes), 562, 563
— larvæ of, destruction in prevention of malaria, 636
— number of species, 552
— ova of, best known, 559
Anoplura, 532
— see also Pediculidæ
Antelope tolerant to trypanosomes, 69
— Trypanosoma gambiense in, 76
— T. rhodesiense in, 69, 70
Anterior station in Glossina of a trypanosome, 101
Anthomyia desjardensii, cause of intestinal and cutaneous myiasis, 585
— pluvialis, larvæ of, 584
— scalaris, maggots of, passed from urethra, 728
Anthomyidæ, flies belonging to, attacking man, 611
Anthrax transmitted by Stomoxys, 610
Antimony, use in sleeping sickness, 623
— and atoxyl, combined, in sleeping sickness, 622
Anus, exit of Oxyuris vermicularis from, 467
— prolapse of, set up by migrations of Oxyuris vermicularis, 695
Aphaniptera (fleas), characters, 543
— see also Fleas
Aphides (or plant lice) (footnote), 532
— — — said to have been passed in human urine (footnote), 532
Aphiochæta ferruginea, 582, 583
— — characters, 583
— — geographical distribution, 583
— — larvæ (maggots) of, 583
Aponomma, characters of, 497
— hosts of, 497
Apoplexy cause of death in first period of liver-fluke disease in sheep, 240
Appendicitis, association of Oxyuris vermicularis with, 467
— in relation to intestinal parasites, views of authors regarding, 652, 653, 654, 655
— relationship of Oxyuridæ to, 698
Appendicostomy in gangrenous dysentery, 619
Appendix vermiformis, ascaris in, causing intestinal obstruction, 654
— — bilharziasis of, 642
— — intestinal parasites invading, authors recording cases of, 652
— — Oxyuris in, 654, 655
— — perforation by Ascaris, rarity of, 655, 656
— — trichocephali in, 655
Aptera, 531
Apterygota, 531
Arachnoidea, characters, 483, 484
— orders of (footnote), 484
— relation to Linguatulidæ, 19
Aradidæ, characters, 541
Aragao, on Chlamydozoa, 209
— on Hæmoproteus, 152
— on leucocytogregarines in birds, 155
Archigetes, attains maturity in lower animals, 21
— stage of sexual maturity, 305
Argantinæ and Ixodinæ, distinguishing features between, 505
— characters of, 496
Argas brumpti, 507
— chinche, 508
— persicus, 506
— — appearance of Spirochæta gallinarum in hæmocœlic fluid of, 119
— — bite of, serious effects, 507
— — blood-sucking, 507
— — granules in digestive tract of, 507
— — hosts of, 507
— — transmission of Spirochæta gallinarum by, 119
— reflexus, bite of, symptoms set up by, 506
— — blood-sucking habits of, 506
— — geographical distribution, 506
— — habitat, 506
— — vitality of, 506
— species of, Neumann’s table, 505
Arilus carinatus, 542
Arion sp., scolex of cysticercoid from, with excretory vessels outlined, 292
Armadillo, possible reservoir of Trypanosoma cruzi, 87
Arribalzagia, characters, 562, 568
Arsenic and glycero-phosphates in bronchial spirochætosis, 633
— in sleeping sickness, 622, 623
— in treatment of nagana, 94
Arsenious acid in bronchial spirochætosis, 633
Arseno-phenyl-glycin in sleeping sickness, 623
— resistance of Trypanosoma lewisi to, how lost, 93
Arslan, experimental ancylostome anæmia, 646
Artault, Entamœba pulmonalis, 45
Arthropoda, 483
— natural flagellates of, 104
— pébrine bodies or Microsporidia in, 184
— segmented structure of, 483
— skin of, how hardened (footnote), 483
Artyfechinostomum, morphology, 269
— sufrartyfex, 753
— — morphology, 269
Ascariasis, diagnosis, 692
— — Epstein’s method, 692
Ascaridæ, 375, 461
— chemically toxic effects of, 650
— epidermal cells, isolated, of (footnote), 361
— expulsion of, favourable effects of, 649, 650
— infection by, causing pseudomeningitis, 649, 650
Ascarides causing constipation, 657
— — intestinal obstruction, 657
— escape from abdominal operation wound, 655
— — from inguinal tumour, 656
— — from umbilicus, 656
— evacuation in enormous numbers, 657
— expulsion of, drugs for, 692
— female, depositing ova in liver, 689
— in bile-ducts, 688, 689
— in pulmonary artery, 656
— in upper air-passages, 690
— — — danger of, 691
— — — how introduced, 690
— infection by, prophylaxis against, 692
— invading urinary passages, 692
— invasion of, causing liver abscess, 690
— massive accumulation causing occlusion of intestine, 657
Ascarinæ, 461
Ascaris causing perforative peritonitis, 656
— characters, 461
— in appendix causing intestinal obstruction, 654
— in peritoneal cavity, 656
— lacks intermediate host, 21
— lumbricoides, distribution world-wide, 463
— — excretory apparatus, 367
— — expulsion of, 754
— — hosts of, 464
— — infection by, 687
— — — experimental, 464, 465
— — — mode of, 464, 465
— — — symptoms, 688
— — injury inflicted by, depends on number in host, 8, 9
— — male, hind end, 371
— — — transverse section through posterior extremity of body, 370
— — method of keeping alive, 754
— — migration from small intestine to other parts of body, 464
Ascaris lumbricoides, morphology, 463
— — names by which known in antiquity, 464
— — normal habitat, small intestine, 464
— — organs of, 463
— — ova of, 463
— — prevalence in young children in temperate climates, 464
— — self-infection with, on part of experimenter, 464, 465
— — sites of body invaded by, 687, 688
— — transverse section showing organs, diagram of, 362
— — — — through, diagram showing, 364
— — unrecognized Dioctophyme gigas in man traced to, 431
— maritima, 465
— megalocephala, nervous system, schematic representation, 365
— — “tuft-like” or “phagocytic” organs, 362
— ova of, in pus in case of abscess of omentum, 657
— perforation of intestine by, 656
— — — following diseased processes, 656
— sp., 465
— texana, 465
Ascites and abdominal malignant growth, Leydenia gemmipara associated with, 49, 50
— association of Leydenia gemmipara with, 49
— chylous, from Filaria bancrofti infection, 678
— set up by invasion of ova of Schistosoma japonicum, 282
Ascitic fluid in cultivation of Treponema pallidum, 125, 126
Askanazy, mode of infection by Opisthorchis felineus, 254
— post-mortem discoveries of Opisthorchis felineus, 253
— Trichuris trichiura, 420
Asphyxia following invasion of upper air-passages by Ascarides, 691
Aspidogaster attains maturity in lower animals, 21
Asses, nagana fatal to, 94
Atoxyl and antimony combined in sleeping sickness, 622
— in Indian kala-azar, 626
— in infantile kala-azar, 627
— in sleeping sickness, 622, 623
— Trypanosoma rhodesiense resistant to, 78
Atractonema gibbosum, habitat, 4
— — peculiar characters of females, 5
Auchmeromyia (Bengalia) depressa as cause of myiasis externa, 724
— — — “larva of Natal,” characters, 591
— luteola, 593, 594
— — characters, 594
— — geographical distribution, 594
— — larva of (Congo floor maggot), 593, 594
— — — how destroyed, 594
— — life-history, 614
Auditory meatus, larvæ penetrating, 721
— — external, infected with Rhinosporidium, 195, 196
Austen, Chironomidæ described by, 580
— description of larva of Ochromyia anthropophaga (footnote), 590, 591
— myiasis due to Sarcophaga, 590
Autopsies at Tomsk, human parasites most frequently found at, 253
— occurrence of Linguatula rhinaria at, 526
B.
Babes, A., cases of transmission of Demodex folliculorum canis infection to man, 709
— Filaria (?) conjunctivæ in man, 405
Babesia, 155, 172, 173, 174, 177
— bigemina, geographical distribution of, 177
— bovis, causal agent of “Texas fever” or “red-water fever” in cattle, 173
— — cause of infectious hæmoglobinuria in cattle, 177
— — transmission of, by Ixodes ricinus, 177
— caballi, 177, 178
— — cause of biliary fever in equines, 177
— — geographical distribution of, 177
— — transmitting agent, 178
— canis and B. bovis, life-cycle in tick, stages of, 176, 177
— — agents of transmission, 177
— — cause of malignant jaundice in dogs, 177
— — cultivation in vitro by Bass’s method, 172, 177
— — life-cycle in infected blood of dog, 175
— distribution of chromatin in, 176
— divergens, cause of European red-water fever in cattle, 177
— — geographical distribution of, 177
— morphology and hosts of, 174
— muris, morphology, 178
— nuclear phenomena in species of, 176
— ovis, agent of transmission of, 177
— — cause of “carceag” in sheep, 177
— — geographical distribution of, 177
— — transmitting agent, 177
— parasites of red blood corpuscles of mammals, 154
— pitheci, 178
— species of, 177
— synonyms (see Piroplasma), 174
— tick borne, 176
— — development in, 176
Babesiasis, symptoms of, 178
— treatment of, 178
Baboon serum, action on Trypanosoma rhodesiense, 80
Bacillus lymphangiticus, 755
Baelz, prophylaxis against kedani, 703
Baer, C. E. von, views as to origin of cercariæ, 12
Bagdad sore, parasite of, supposed intermediate host, 575
Bahr, filariasis in Fiji, 401, 403
Baker, larvæ of Aphiochæta ferruginea, 583
Balantidiasis, see Dysentery, balantidian or ciliate
Balantidium, 200
— coli, 200, 201
— — dysentery associated with, 202, 203, 637
— — geographical distribution, 201
— — habitat in body, 201
— — hosts of, 7, 202
— — in man, cases recorded, 201
— — morphology, 200
— — transmission, 202
— giganteum, see Nyctotherus giganteus
— minutum, 204
— — habitat in body, 204
— — in diarrhœa, 204
— — morphology, 204
— morphology, 200
— reproduction of, 200
Balbiani, infection by Dioctophyme gigas, 432
— researches on silkworm disease, 184
Baleri, causal agent of, 95
Balfour, A., coccoid bodies of Treponema pallidum, 124, 125
— — granules in digestive tract of Argas persicus, 507
Balfour, A., and Sambon, researches on Spirochæta granulosa, 116
Balsam of Peru, application in scabies, 707
— — — in crab louse infection, 712
— — — in head louse infection, 710
— — — in nasal myiasis, 719
— permanent mounting agent for flukes, 471
Balzer and Schimpff, unusual situation of Sarcophaga larvæ, 723
Barbagallo, case of dermatitis set up by Oxyuris vermicularis, 696
— method of evacuation of Oxyuridæ, 697
Barbeiro, parasite causing, 537
Barbel disease, cause of, 184
Barkan, nematode in human eye, 412
Barlow, Craigia hominis, 734
Barth, pseudomeningitis following infection by Trichocephalus dispar, 650
Basile, experiments showing that infantile leishmaniasis is transmitted by fleas, 111
— transmission of canine kala-azar by dog fleas, 103
Bass, C. C., cultivation of malarial parasites, 170
Bass and Hall, detection of ancylostome eggs, 473
Bass and Johns on Entamœba buccalis, 43
— — treatment of oral endamœbiasis, 620
Bass’s method, cultivation of Babesia (Piroplasma) canis by, in vitro, 172, 177
— — — of malarial parasites, 170, 171, 172
Bastianelli, mosquitoes in relation to human malaria, 158
Bat parasites (Streblidæ), 611
Baths, luke-warm, in trichinosis, 681
Beattie on Rhinosporidium from Madras, 197
Becker, trichocephalus anæmia, 651
Béclère, method of extraction of Dracunculus medinensis, 390
Bed bug, development of Leishmania tropica in, 108
— — probable agent of transmission of kala-azar, 107
— — Texas or Mexican, see Conorhinus sanguisuga
— — see also Cimex (Acanthia) lectularia
Bee parasites (Braulidæ), 611
Bees, microsporidiosis, due to Nosema apis, in, 185
Beetle, intermediate host of Echinorhynchus moniliformis, 478
Bégonin, Oxyuris in appendix, 654
Behrenroth, prevention and treatment of balantidian dysentery, 637
Belascaris, morphology, 466
— cati, morphology, 466
— — ovarian tube, transverse section through, 369
— — transverse section through head part of, 466
— marginata, morphology, 466
Beneden, van, on commensals, 6
Bentley, beta-naphthol in expulsion of ancylostomes, 687
Benzine and petroleum in crab louse infection, 712
— and water enemata in Trichuris trichiura infections, 680
— enemata in arrest of trichinosis, 681
— high injections of, in evacuation of Oxyuridæ to be avoided, 698
— inhalations in nasal myiasis, 719
Bergmann, operation results of cysticercus of brain, 665
— Scolopendra in frontal sinus, 721
Berlin, number of oxen, sheep, and pigs slaughtered in, infected, 346
— — of pigs found trichinous in, 430
— oxen infected with Cysticercus bovis in, 341
Berti, cause of ancylostome anæmia, 648
Bertramia, 194, 195
Bertrand, scolopendra in maxillary sinus, 721
Beta-naphthol in expulsion of ancylostomes, 687
— ointment in copra itch, 513
Bête rouge, undescribed species of Leptus, 486
Betten, Caligus curtus invading cornea (footnote), 483
Big game reservoirs of nagana, 94
— — — of Trypanosoma rhodesiense, 69
— — trypanosomes innocuous to, 70
Bignami, mosquitoes in relation to human malaria, 158
Bile, preservation of ova of flukes in, 472
Bile-ducts, Ascarides in, 688, 689
— — in results of, 689, 690
— habitat of Clonorchis sinensis, 259
— human, thickened and dilated, Amphimerus noverca found in, 258
— inhabited by Metorchis truncatus, 262
Bile-ducts, invasion by Clonorchis sinensis, 641
— pathological changes in, set up by Clonorchis endemicus, 260, 261
— and liver, habitat of Clonorchis endemicus, 259, 260
Bilharz, discovery of Hymenolepis nana, 323
— Porocephalus constrictus, 526
Bilharzia Mission, report of, 277
Bilharziasis, diagnosis, 643
— prognosis, 643
— prophylaxis against, 644
— regions of body affected by, 642, 643
— symptoms, 641
— — mainly urinary, 641
— treatment, 643
Biliary fever in equines, cause of, 177
Billings, percentage of rats infected with trichinella, 427
— proportion of trichinous pigs found by, 428
Binotia, characters, 565
Bird epithelioma contagiosum, 207
Birds, blood of, Halteridium parasites occur in, 151
— development of Plasmodium relictum in, 170
— endoglobular parasites, similar to malarial, in, discovery of, 157
— experimental infection with herpetomonads, 739
— herpetomonads in blood of, 739
— malaria in, spread by mosquito, 158
— mites living endoparasitically in, 491
— sarcosporidia in, 187
— species of, inhabited by Hymenolepis lanceolata, 329
Bironella, characters, 562, 570
Bismuth subnitrate in dysentery, 619
Bitter Root Valley of Montana, mortality of Rocky Mountain tick fever at, 504
Blackhead in turkeys, causal agent, 145
Blacklock, experimental host of Trypanosoma cruzi, 87
— and Yorke, Trypanosoma equi, 98
— see also Yorke and Blacklock
Bladder, amœbæ invading, 46
— means of access of Schistosoma hæmatobium to, 272
— pathological changes in, due to Schistosoma hæmatobium, 275
— worms, development of, 15
— — explanation of, 14
Blaizot, mode of transmission of relapsing fever, 120
Blanchard, Myriapoda parasitic in intestine and nose of man, 483
— on Lamblia, 57, 60
— on Monas pyophila, 62
— on nomenclature of amœbæ, 31
Blaps mortisaga, larvæ of, in stools, 542
Blenorrhœa, inclusion, in infants, 207
Blepharitis due to head louse infection, 710
Blood, changes in, in ancylostomiasis, 683
— circulating, morphology of Trypanosoma gambiense in, 73
— citrated, cultivation method for Leishmania tropica, 108
— colourless, of insecta, 530
— corpuscle, red, number of malignant tertian parasites found in one, 167
— corpuscles, red, action of leucocytozoa on, 742
— — — attacked by quartan malarial parasite, not altered in size or colour, 166, 167
— — — development of benign tertian parasite in, 160, 164, 165
— — — — — — — appearance of Schüffner’s dots, 165, 166, 168
— — — — malignant tertian parasite in, appearance of Maurer’s dots, 168
— — — hæmogregarines in, 153, 154
— — — ingestion by amœbæ, 35, 39, 42
— — — life-cycle of Nuttalia equi in, 173
— — — separation from filaria larvæ, 395
— examination of, for protozoa, 745
— — in diagnosis of trichinosis, 681
— films, thick, method of making, 747
— — thin, method of making, 747
— flagellates, history of, 67
— — hosts of, 67
— inoculation, rinderpest transmissible by, 742
— larvæ of Loa loa in, 412, 414
— multiplication of trypanosomes in, 71
— peripheral, periodicity of larvæ of Filaria bancrofti in, 393, 394
— prevalence of filarial disease proportionate to amount of Mikrofilaria bancrofti in, 400
— protozoa parasitic in, culture media for, 744
— spirochætes, 114
— supply of abdominal and pelvic organs as illustrating distribution of Schistosoma hæmatobium in body, 272
— — trypanosomes in, cultures of, 69
— — cyclical variation, 78
— — daily number from case of Rhodesian sleeping sickness, 79
— — method of determining number, 748
— — periodicity of, 69
— — seasonal variation, 69
Blood-sucking habit of Argas persicus, 507
— — of Argas reflexus, 506
— — of Cimex lectularius (bed bug), 535
— — of Conorhinus renggeri (great black bug of Pampas), 539
— — — sanguisuga, 537
— — of Culicoides, 580
— — of fleas, 543
— — of Glossina palpalis, 607
— — of leeches, 701
— — of Leptidæ, 603
— — of Linguatulidæ, 523
— — of mosquitoes confined to females, 552
— — of Muscidæ, 603
— — of Phlebotomus, 581
— — of Pupipara, 611
Blood-vessels, migration of oncospheres from intestine to liver through, 302
Blood-vessels, Strongyloides stercoralis in, 755
Boas, injection of emulsion of male fern, 671
Bodo, 63
Bodonidæ, 61
— characters of, 63
— genera of, 63
Body lice, human, inhabited by Herpetomonas pediculi, 103
— — prophylaxis against, 615, 616
— — see also Pediculus vestimenti
Bohland, proteid destruction in ancylostomiasis, 647
Boils and ulcers due to invasion by larvæ of Cordylobia anthropophaga, 592
— produced by Oestridæ, 725
Bojanus, views as to origin of cercariæ, 12
Bollinger, cases of Dipylidium caninum infection, 659
Bolt, sand flies in North China, 613
Bond, larvæ of Muscidæ in nose, 720
Bone-marrow, development of crescents of tertian malignant parasite in, 169
— red, administration in Indian kala-azar, 626
Bont tick, see Amblyomma hebræum
Boophilus annulatus, transmission of Babesia bigemina by, 177
— characters of, 497
— species of, transmission of Babesia bigemina by, 177
Boracic acid fomentations in Oriental sore, 628
Bordier, Davainea madagascarensis, 662
Börger, cases of Ascarides in bile-ducts, 688
Borrelia, 115
Bosanquet, molluscan spirochætes breaking up into granules, 119
Boschulte, effects of bite of Argas reflexus, 506
Bothriocephalus anæmia, 644, 645
— — cases of, 645
— — dissolution of parasitic products in serum of patients with, 645
— — experimental, 646
Bouin-Duboscq fluid, 749
Bouin’s fluid, 749
Brachycera (flies), characters, 582
— — larvæ (maggots) of, parasitic in man, 582
Bradford, Sir J. Rose, see Plimmer, H. G.
Brain, abscess of, set up by amœbæ, 35
— cysticerci in, 335, 664, 665
— — change of position, 665
— — operation for, 665
— — percentage of cases, 664
— — site, 664, 665
— — symptoms, 665
— fourth ventricle, cysticerci in, symptoms, 665
— — — — treatment, 666
— paragonimiasis of, 639
— — diagnosis, 640
— — prognosis, 640
Brandesia turgida, host of, 6
Brandt, effect of invasion by Dipylidium caninum on nervous system, 649
Braulidæ (bee parasites), 611
Braun, developmental cycle of Dibothriocephalus latus, 16
Brazil, Trypanosoma cruzi prevalent in, 83
Breast, tumour of, infection by Gnathostoma siamense associated with, 385
Breinl, enlarged glands in filariasis, 402
— researches on Spirochæta duttoni, 116
Bremser, origin of helminthes, 12
Brieger, effects of filmaron oil, 672
Britton, fatal case of myiasis externa, 716
Brock, treatment of bilharziasis, 643
Broden and Rodhain’s method of administering atoxyl in sleeping sickness, 622
Bronchi, invasion by Paragonimus ringeri, 251
Bronchitis due to invasion of air passages by Paragonimus ringeri, 251
— in pigs set up by Metastrongylus apri, 433
— spirochæte associated with, 122, 632
Bruce, Sir D., classification of trypanosomes, 72
— — — development of Trypanosoma gambiense in Glossina palpalis, 74
— — — discovery of trypanosomes in blood of horses with “nagana,” 68
— — — investigation of sleeping sickness, 68
— — — investigations of Trypanosoma rhodesiense in Glossina morsitans, 82
— — — proportion of Glossina palpalis becoming infected, 608
— — — question of distinction or identity of Trypanosoma brucei and T. rhodesiense, 83
— — — tsetse-fly transmitting Trypanosoma rhodesiense, 608
— — — Zululand strain of Trypanosoma brucei, 94
Brues and Sheppard, insects transmitting epidemic poliomyelitis, 612
Brumpt, experimental hosts of Trypanosoma cruzi, 87
— on Sergentella hominis, 210
— subcutaneous tumours associated with invasion by Onchocerca volvulus, 418
— Tetramitus mesnili, 57, 624
— trichomonad cysts, 56
Brun’s symptom of cysticerci in fourth ventricle, 666
Büchholz, tetanus disappearing after expulsion of Ascaridæ, 650
Buff coagulum in cultivation of Treponema pallidum, 126
Bulgaria, oestrid larvæ in, invading human integument, 595
Bunostomeæ, 456
Burbot (Lota vulgaris), muscles of trunk containing plerocercoid, 313
Burfield, bilharziasis of appendix, 642
Bursa copulatrix of male nematodes, 370
Buschmucker, mite attacking man, 486
Bütschli, O., on Gregarines, 130
— — on Myxosporidia, 181
C.
Cæca, intestinal, of nematodes, 364
Cæcum, cysts of, œsophagostomum contained in, 441, 443, 444
Caffarena, echinococcus cysts causing urticaria, 652
Cairo, fresh-water molluscs round, cercariæ of bilharzia type in, 277
Calandruccio, experimental infection with Ascaris lumbricoides, 465
— — self-infection with Oxyuris vermicularis, 469
— on experimental amœbic infection, 30
Calcium oxalate crystals in endoplasm of Lithocystis, 131
— salts, internal administration for cutaneous and muscular cysticerci, 663
Caligus curtus invading cornea (footnote), 483
Calkins on cultural amœbæ, 42
— on genus Craigia, 45
— on vaccinia and variola, 208
Calliobothrium, larval stage, 305
Calomel in evacuation of Oxyuridæ, 697
— in flagellate dysentery, 624, 625
— in intestinal myiasis, 728
— insufflations of, in nasal myiasis, 719
— — — — following cocainization of nose, 720
Calvertina, characters, 562, 570
Camels, “surra” in, 95
Canaries, susceptibility to infection by Plasmodium relictum, 170
Cancer, association with invasion by Opisthorchis felineus, 254
— of oral cavity, association of Entamœba buccalis with, 43
Canestrini and Kramer, species of Demodex, 522
— — — of Sarcoptes enumerated by, 518
Canguary, synonym of Brazilian trypanosomiasis, 87
Canterbury Cathedral, Argas reflexus formerly abundant in, 506
Cantlie, J., sand flies biting in Hampshire, 579
Cape ailment (Port Natal sickness), 488
Cappez, nematodes in human eye, 412
Carbolic acid clearing agent for flukes, 471
— — injection in creeping disease, 731
Carbon dioxide snow, application in Oriental sore, 628
Carceag in sheep, cause of, 177
Cardiac form of Brazilian trypanosomiasis, 88
— stimulants in Asiatic relapsing fever, 631
Caries, dental, association of Entamœba buccalis with, 43
— — — — kartulisi with, 44
Carini, cysts in lungs of rats, 90
— phagedænic amœbæ, 733
— treatment of espundia, 629
Carlsbad water in intestinal myiasis, 728
Carmine, acetic-alum, solution of, in staining flukes, 471
— solutions of, in staining flukes, 471
Caryophyllæus, larvæ of, 305
Casagrandi and Barbagallo, Entamœbæ, 31, 33
Castellani, A., demonstration of trypanosomes in cerebrospinal fluid from sleeping sickness, 68
— — experimental production of copra itch, 513
— — Nyctotherus africanus, 206
— — Toxoplasma pyrogenes, 113
— — Treponema pertenue, 127
— and Chalmers, on Rhinosporidium in Ceylon, 197
— — — treatment of relapsing fever, 630, 631
— — — of balantidian dysentery, 637
— — — of gangrenous dysentery, 619
— — — of Indian kala-azar, 626
— — — of infantile kala-azar, 627
— — — of malaria, 635
— — — of Oriental sore, 628
— — — of sleeping sickness, 622
— — types of amœbic dysentery, 618
Castor oil in evacuation of Oxyuridæ, 697
— — in flagellate dysentery, 625
— — preliminary administration in amœbic dysentery, 619
Cat, Dipylidium caninum parasitic in, 322
— flea, see Ctenocephalus felis
— host of Dibothriocephalus latus, 313
— — of Paragonimus kellecotti, 251
— infected with Dipylidium caninum through skin parasites, 323
— intestine of, Isospora bigemina parasitic in, 149
— parasites found only in, 6
— production of dysentery in, by infection with amœbæ, 30, 35
— — of enteritis in, by injection of amœbæ into, 36, 37
— Tænia echinococcus in, 356
— and dog, parasites common to, 6
Catageiomyia, characters, 563
Cataphoresis in creeping disease, 731
Cataract, association of Filaria oculi humani with, 406
Catarrh, intestinal, associated with Balantidium coli, 201
Caterpillars, larvæ living in, discovery of, 10
Cathæmasia (Distoma) hians, progeny of, discovery, 12
Cattle, disease in, caused by Anaplasma organisms, 180
— East Coast fever in, cause of, 178
— — — — pathogenic agent, 174
— European red-water fever in, cause of, 177
— gall-sickness in, cause of, 180
— — supposed causal agent, 98
— infectious hæmoglobinuria in, cause of, 177
— migrations of larvæ of Hypoderma bovis in body of, 595
— nagana prevalent among, and generally fatal to, 93, 94
— organs infected with echinococcus, percentage of frequency, 347
— red dysentery in, cause of, 147
— Rhodesian fever in, carriers of, 494
— “surra” in, 95
— Texas or red-water fever in, carrier of, 494
— — — — causal agent, 173
— Trypanosoma vivax fatal to, 99
Caullery and Mesnil, Actinomyxidia, 187
— — Haplosporidia, 194
Caval system, means of access of Schistosoma hæmatobium to, 272
— — Schistosoma hæmatobium in, 274
Cedar-wood oil, mounting agent for flukes, 471
Cell inclusions, 207, 208
Celli, discovery of movements in malarial parasites, 157
— and Fiocca, species of amœba distinguished by, 31
Cellia, characters, 562, 569
Centrorhyncus, 581
Cephalina, morphology and hosts of, 135
Cephalodiscus nigrescens, 195
Ceratixodes, characters of, 497
— hosts of, 497
Ceratomyxa, 184
Ceratophyllus anisus, 548
— distinctive characters, 545
— fasciatus, carrier of plague bacillus, 543, 547
— — development of crithidial forms of Trypanosoma lewisi in rectum of, 91, 93
— — transmission of Trypanosoma lewisi by, 88
— londiniensis, 548
Ceratopogoninæ (midges), characters of, 580
— geographical distribution, 580, 581
Cercaria bilharzia, characters, 754
— bilharziella, characters, 754
Cercariæ, bilharzia type, in fresh-water molluscs round Cairo, 277
— (larval stages) of Trematodes, 225, 227, 228
— origin of, early views as to, 12
Cercomonadidæ, 61
Cercomonads, encysted stages of, 62
— presence in intestine, 62
Cercomonas, characters, 61, 736
— hominis, 61, 736
— — characters, 61
— — flagellum of, 61, 62
— intestinalis, characters of, 54
— longicauda, 736
— parva, 737
— vaginalis, 62
Cereals, mites infesting, effects on man, 489
Cerebrospinal fluid from cases of sleeping sickness, trypanosomes in, 68
Cesspools, screening against mosquitoes, 636
Cestoda, animal nature of, always known, 282
— cirrus pouch, 295
— cortical layer, 289
— development of, 13
— developmental cycle of, discovery, 16
— — direct, 17
Cestoda, egg-shell substance, 297
— eggs of, 297
— excretory apparatus, 291
— — vessels, collecting tubes, 291
— — — — — frontal anastomosis, 291
— — — — — island formation, 292
— fixation, staining and clearing of, 472
— genital apparatus, 293
— — — female, 295
— — — male, 293, 294
— — papilla, 295
— infection by, 644
— life spent in intermediate and final host, 18
— loss of digestive system in, 3
— medullary layer, 289
— myoblasts, 289
— nervous system of, 289, 290
— of man, 309
— — classification, 308
— — relative frequency, 342
— ovaries, 295
— oviduct, 295
— — course and situation, 295, 296
— parauterine organs or uterine capsules, 297
— preservation and examination of, 472
— relation to Turbellaria, 19
— rostellum, 289
— segmented and single-jointed connection, 20
— sexual orifice, male, 295
— suckers, 289
— testes, 293
— uterus of, 296
— — parenchymal capsules, 296
— vagina, 295
— vas deferens, 294
Cestode tuberculosis, acute, 303
Cestodes, see Cestoda (above)
Chagas, morphology and life-history of Trypanosoma cruzi, 84–87
— multiplication of Trypanosoma cruzi in vertebrate host, 85
— on Cercomonas parva, 737
— researches on Brazilian trypanosomiasis, 87, 88
— reservoir of Trypanosoma cruzi, 87
— schizotrypanosomiasis, 88
— Trypanosoma cruzi, 83
Chagasia, characters, 562, 570
Chagas’s disease, see Trypanosomiasis, Brazilian
Chalazion due to infection by Demodex folliculorum, 708
Chalmers and O’Farrell, treatment of bronchial spirochætosis, 633
— — see also Castellani and Chalmers
Chatin, Metastrongylus apri in human intestine, 433
Chatterjee, Pentatrichomonas bengalensis, 624, 735
Chatton and Lalung-Bonnaire, entamœbæ of vertebrates, 34
Cheese-fly, 583
Chelifer cancroides, pseudoparasite in man (footnote), 484
Cheyletus, characters, 516
— méricourti, 517
Chiari, case of Oxyuridæ in nose, 696
Chiastopsylla, distinctive characters, 545
Chicken cholera, possible carrier of, 579
Chigoe, see Dermatophilus penetrans
Children, administration of male fern to, 671, 672
— Dipylidium caninum infection prevalent in, 322, 659, 660
— enteritis verminosa in, 688
— flagellate dysentery in, 56
— invasion by Hymenolepis nana, 324, 661
— lymphangitic attacks in, from Filaria bancrofti infection, 676
— native, carriers of malarial parasites, 636
— — latent malaria in, 158
— oxyuriasis in, 695
— — treatment, 697, 698
— young, prevalence of Ascaris lumbricoides in, in temperate climates, 464
Chilodon dentatus, 206
— uncinatus, 206
Chilodon-like organisms found in gonorrhœa, 206
Chilomastix and Tetramitus, differential characters, 735, 736
— (Tetramitus) mesnili, 57, 735
— — — diagrams of, 736
China, North, sand flies and fever due to them in, 613
Chironomidæ (midges), distinguishing characters from mosquitoes, 579
— — larvæ of, 579
Chironomus, wing of, 579
Chlamydophrys, 47, 759
— characters of, 47
— enchelys, asexual multiplication in fæces, 47
— — characters of, 47
— — encysted form, 48, 49
— — free motile form and dividing organisms, 47, 48
— — habitat, 47
— — plasmogamic union, 48
— —sexual multiplication, 48
— stercorea, 47
— — see also Chlamydophrys enchelys
Chlamydozoa, 207, 210
— characteristic features, 208
— granules of, filtration experiments with, 209
— mode of division, 209
— possible cultivation, 210
— relationship of, 210
— systematic position, 210
— tissues inhabited by, 209
Chlamydozoon, life-history of, 209
Chloroform, administration in expulsion of ancylostomes, 686, 687
— as tapeworm drug, 674
— hypodermic injection in creeping disease, 731
— in expulsion of guinea-worm, 676
— injections or inhalations in nasal myiasis, 720
Chloromyxidæ, 184
Chloromyxum leydigi, trophozoite of, 182
Choanoflagellata, 52
Cholera Investigation Commission, work of, with regard to dysentery, 30
— motions, spirochætes found in, 122
— spread by house-fly, 586
Cholodkowsky, “wormlet” burrowing into human epidermis, 599
Chorea, case of, cured after expulsion of Tænia, 648
Chorion enveloping ova of nematodes, 371
Chorioptes bovis, 521
— characters, 517
— species found on man, 521
Christophers, on Babesia canis, 176, 177
— on Leucocytogregarina canis, 155
— see Stephens and Christophers
Christopherson, J. B., case of espundia, 108, 628
Chrithoptes monunguiculosus, 489
Chromidial apparatus of protozoa, 26
Chrysoconops, 577
Chrysomela hæmoptera, gregarine from, 131
Chrysomyia and Pycnosoma, distinguishing features, 588
— (Compsomyia) macellaria (screw-worm fly), 587
— — — — — larvæ of, invasion by, fatal results from, 587
— — — — — — regions of body invaded by, 587
— — — — — references to, 587
— — — — — synonyms, 587
— viridula, characters of, 588
— — larvæ (maggots) of, discharged from nose, 588
Chrysops host of Filaria loa, 601
— dimidiata, 601
— silacea, 601
— transmission of surra by, 601
Chrystia, characters, 562, 568
Chyluria, association of Hymenolepis madagascarensis with, 662
— following infection by Eustrongylus gigas, 683
— — — by Hymenolepis nana, 661
— from Filaria bancrofti infection, 677
— — treatment, 677
— in filariasis, 402
— without lymphatic obstruction, 401
Cigarettes, paper, smoking of, in nasal myiasis, 719
Ciliata, 198
— classification, 199
— macro-nucleus and micro-nucleus of, 198
— morphology of, 198
— peristome of, 198
— reproduction of, 198
Ciliophora, 198
Cimex (Acanthia) lectularia (bed bug), see Cimex lectularius (following)
— boneti, host of Trypanosoma cruzi, 87
— ciliatus, 537
Cimex columbarius, 536
— — bite of, 536
— hirundinis (swallow bug), 537
— lectularius (bed bug), artificial host of Trypanosoma cruzi, 87
— — — — bite of, treatment, 713
— — — — blood-sucking, 535
— — — — characters and habits, 534
— — — — extermination of, 713
— — — — infection by, 713
— — — — — diagnosis, 713
— — — — larval stage, 535
— — — — ova of, 535
— — — — peculiar odour emitted by, 535
— — — — persistent accompaniment of man by, 535
— — — — possible transmission of kala-azar by, 107, 713
— — — — transmission of Spirochæta recurrentis by, 120, 121
— macrocephalus, 536
— rotundatus (tropical bed bug), 535, 536
— — — — — carrier of virus of poliomyelitis, 536
— — — — — geographical distribution, 536
— — — — — points of distinction from C. lectularius, 536
— — — — — possible connection with kala-azar, 536
— sp., connection with Oriental sore, 536
Cimicidæ, characters, 534
Circulatory system, disturbances of, in ancylostomiasis, 683
Cisterns, screening against mosquitoes, 636
Cladorchiinæ, 231, 234
— male and female organs, 234
— morphology, 234
Clark, see Howard and Clark
Clasping and clinging organs in permanent parasites, 4
Climates, temperate, prevalence of Ascaris lumbricoides in young children in, 464
Clonorchis endemicus, geographical distribution, 260
— — habitat and hosts of, 259
— — intermediate host of, first unknown, 261
— — — — second, 261
— — life-history, 261
— — morphology, 259
— — synonyms, 259
— morphology, 258
— sinensis, geographical distribution, 259
— — habitat and hosts of, 259
— — infection by, 640
— — — diagnosis, 641
— — — prophylaxis against, 641
— — — symptoms, 640, 641
— — — treatment, symptomatic, 641
— — morphology, 258, 259
— — organs of, diagram showing, 259
— — ova of, 259
— — sites of invasion in body, 640
Cloquet, destruction of eyes by Sarcophaga larvæ, 723
Clothes louse, see Pediculus vestimenti
Cnidosporidia, 129, 194
Cobbold, Ligula mansoni, 318
Cocaine, application, followed by calomel insufflations, in nasal myiasis, 720
— hypodermic injection in creeping disease, 731
Coccidæ (scale insects), 532
Coccidia, copulation in, 137
— experimental infection with, 136
— ova of helminthes mistaken for (footnote), 137
— pathogenicity of, 136
Coccidia-like organisms in various diseases of man, 150
Coccidiidea, 129, 135, 137
— characters and habitat, 28
— classification of, 141
— gametes of, 137, 139, 140
— hosts of, 137
— life-cycle of, 138–141
— macrogametes, 139, 140
— merozoites of, 138, 139, 140
— microgametes of, 137, 139, 140
— morphology of, 138
— occurrence, 137
— oöcysts of, 141
— schizogony in, 138
— sporoblasts and sporocyst of, 141
— sporozoites of, 138, 139, 140
Coccidioides immitis, 150
— pyogenes, 150
Coccidiomorpha, 129, 151
Coccidiosis, avian, 142–145
— diagnosis of, 742
— human, doubtful cases, 149
— — hepatic cases, 148
— — intestinal cases, 148
— in cattle, 147, 741
— in rabbits, intestinal and hepatic, 145, 147
— rinderpest mistaken for, 741
Coccidium jalinum, 150
— — hosts of, 150
Cochin-China diarrhœa, see Diarrhœa (Cochin-China)
Cockchafer, intermediate host of Echinorhynchus gigas, 477
Cocoons of Hirudinea, 481
Cod-liver oil, inunctions of, in evacuation of Oxyuridæ, 698
Cœlosporidium, 195
Cœnurus, definition of, 301
— cerebralis, experimental rearing of tapeworms from, 15
— — reared experimentally, 15
— section showing cephalic invaginations, 304
— scolices in, 303
Coleoptera, characters, 531, 532
— larvæ of, accidental parasites, 542
Colitis associated with Balantidium coli, 201
— — with intestinal amœbæ, 30
— mucous, complicating intestinal myiasis, 726, 727
Collargol in balantidian dysentery, 637
Collargol, irrigation of lower bowel with, in gangrenous dysentery, 619
— rectal administration in bilharziasis, 643
Colon, cysts of, Œsophagostomum contained in, 441, 443, 444
— descending, means of access of Schistostoma hæmatobium to, 272
Colorada, mite attacking man, 486
Colpoda cucullus, 204
Comedones, removal in infection by Demodex folliculorum, 708
Commensals, 6, 20
— nature of, 6
Congo floor maggot, 593, 594
Conjunctiva, dipterous larvæ in, 716
— trachoma bodies in infected epithelial cells of, 209
Conjunctivitis due to head louse infection, 710
Connective tissue, subcutaneous invasion by Loa loa, 678
Conorhinus megistus intermediate host of Trypanosoma cruzi, 83, 537
— — see Triatoma megista
— nigrovarius, bite of, 539
— protractus, 539
— renggeri (great black bug of Pampas), blood-sucking, 539
— rubrofasciatus, bite of, 538
— — trypanosome inhabiting, 99
— sanguisuga (blood-sucking cone nose), bite of, 537, 538
— — — — — ova of, 538
— sp. (?), bite of, symptoms following, 538
— variegatus, 539
Conseil, mode of transmission of relapsing fever, 120
Constipation, prevention during malarial attacks, 635
— set up by Ascarides, 657
Copaiva balsam in bilharziasis, 643
Copper, black oxide of, as tapeworm drug, 674
Copra itch, mite causing, 513
— — treatment, 513
Copulation, modes of, in tapeworms, 297
Cordylobia anthropophaga, characters, 592
— — geographical distribution, 593
— — hosts of, 592
— — larvæ of, characters, 592
— — — lesions set up by invasion, 592
— — life-history, 592
— — references to, 593
— grünbergi, larvæ, characters, 591, 592
— — synonyms, 591
— rodhani, 593
— — geographical distribution, 593
Coreotrypanosis, 87
Corethra, 565
Corethrina, 565
Coriscus subcoleoptratus, bite of, 540
— — geographical distribution, 540
— — synonyms, 540
Cornea, parasitic crustacean (Caligus curtus) invading, 483
Cortical layer of Cestoda, 289
Councilman and Lafleur on nomenclature of amœbæ, 31
Couret and Walker, J., culture medium for intestinal amœbæ, 743, 744
Couvy, Spirochæta gallinarum, 119
— see Marchoux and Couvy
Cover-slip preparations, 748
Cows attacked by Leptus autumnalis, 486
Crab louse, see Phthirius inguinalis
Craig, C. F., on Entamœba histolytica, 37, 41
— — on Paramœba (Craigia) hominis, 44, 45
Craigia hominis, 45, 734
— migrans, 734
Craigiasis, 734
— nature of, 734
Crane-fly, Gregarina longa from larva of, 130
Craw-craw (filaria infection of the skin), 378, 514
Crawley, experimental infection with Sarcocystis, 192
— movements of gregarines, 131
Creeping disease, cases of, various authors reporting, 729
— — clinical symptoms, 730
— — duration, 731
— — localization of, 730
— — mode of origin, 729, 730
— — synonyms, 729
— — treatment, 731
— — — methods reported by various authors, 731
— eruptions, 599
Creosote in bronchial spirochætosis, 633
— mounting agent for flukes, 471
Creplin, discovery of progeny of Diphyllobothrium (Bothriocephalus) ditremum, 13
— psorosperms, 181
Cristina and Caronia, treatment of infantile kala-azar, 627
Crithidia, 67, 103
— inoculation experiments with, 104, 738
— fasciculata, 104
— — host of, 104
— gerridis, 738
— hosts of, 104
— hyalommæ, 104
— melophagia, host of, 104
— morphology, 104
— natural flagellates of Arthropoda, 104
Crustacea, parasitic, change of original features in, 4
— — loss of digestive system in, 3
— — or free-living, invading man abnormally (footnote), 483
Csokor, mode of infection in intestinal myiasis, 727
Ctenocephalus canis, herpetomonad inhabiting, 103
— — transmission of Trypanosoma lewisi by, 88, 90, 92
— distinctive characters, 545
— felis (cat flea), 547
Ctenocephalus, hosts of, 547
Ctenophthalmus, distinctive characters, 545
Ctenopsylla, 548
— distinctive characters, 545
— musculi, transmission of Trypanosoma lewisi by, 90
Cucumber seeds in intestinal myiasis, 728
Culex and Anopheles, larvæ of, position in water compared, 554
— — ova of, method of depositing compared, 554
— — points of difference between, 551
— characters, 564
— fatigans (common tropical gnat), distinguishing character, 576
— — transmission of Filariæ by, 576
— head of male and female, 549, 556
— human malaria not spread by species of, 158
— larva of, 553
— ova of, 557, 558
— pipiens (common gnat), 575
— — characters, 576
— — ova of, localities selected for deposition, 553
— species of, development of Plasmodium relictum in, 170
Culicidæ, classification of, 561
— number of species, 552
— scales of, 560
Culicinæ, characters, 563, 571
Culicoides, blood-sucking habits of, 580
— larvæ of, 580
— ornatus, bite of, 581
— possible carrier of germ of Delhi boil, 580
— pupæ of, 580
Culture media for amœbæ, 742
— — for blood protozoa, 744
Cunningham, discovery of intestinal amœbæ, 29
Cutaneous glands, unicellular, of nematodes, 361
— tumours due to cysticerci, characteristics of, 662, 663
Cyclasterium, 208
Cyclocœlum mutabile, progeny of, discovery, 12
Cycloleppteron, characters, 561, 567
Cyclophyllidea, 308
Cyclopidæ, characters, 390
Cyclops, characters, 390
— intermediate host of Dracunculus medinensis, 388
— virescens, 389
Cyclospora, 141
Cyrtoneura stabulans, larvæ of, habitat, 585
Cysticerci, cutaneous and muscular, symptoms set up by, 663
— — — treatment, 663
— development from oncosphere of Tæniidæ, 303
— early researches on, 282
— experimentally reared from tapeworms, 15
— number of, in relation to species of Tæniæ, 16
— origin of, 14
— regions of body site of, 663, 664
— subretinal, 664
— tapeworms experimentally reared from, 15
Cysticercoid, morphology of, diagram showing, 301
Cysticercoids, echinococcus-like conditions in, 304
Cysticercus acanthotrias, 336
— — and C. cellulosæ, 336, 337
— bovis, 340
— — amount of prevalence in ox, 340, 341
— — — — — in Prussia and Berlin, 341
— — artificial infection of human beings with, 340
— — rarity in man, 341
— cellulosæ, amount of injury inflicted by, depends on situation in body, 8
— — development to Tænia solium, 340
— — infection of skin and subcutaneous tissues, 662
— — and C. acanthotrias, 336, 337
— — decrease of frequency in pork, how effected, 334
— — development, time taken for, 334
— — habitat, 332, 333
— — hosts of, 332
— — how conveyed to man, 334, 335
— — in man, 335
— — — long persistence of, 337
— — in sheep, 337
— — organs of body invaded by, 335
— — sex distribution of invasion by, 335
— — vitality of, 334
— development of, 301, 302
— — diagram showing, 303
— fasciolaris, host of, 338
— — possible means of spread to man, 338
— forms of, 301
— morphology of, diagram showing, 301
— ovis, 337
— papilliform invagination into bladder, 301
— pisiformis, hosts of, 338
— — in evaginated condition, 304
— racemosus, 335, 336
— tenuicollis, 337, 338
— — experimental rearing of tapeworms from, 15
— with developed scolex at bottom of invagination, 304
Cystoflagellata, 52
Cysts, intestinal, Œsophagostomum contained in, 441, 443, 444
Cytorhyctes, 208
— aphtharum, 208
— cytoplasm of, 209
— luis, 124, 208
— scarlatinæ, 208
— vacciniæ, 208
— variolæ, 208
— — minute granules in, 210
D.
Dactylomyia, characters, 562
Daniels, C. W., iridocyclitis in trypanosomiasis, 623
— — parasitic coleopterous larva (footnote), 542
— — treatment of trypanosomiasis, 622
— — yellow pigment in kidney and liver cells in ancylostomiasis, 647
Danielsia, characters, 564
Danilewsky, discovery of endoglobular parasites, similar to malarial, in birds, 157
— — of Leucocytozoa by, 153
Danube, banks of, Simulium columbaschensis plague on, 578
Darling, experimental infection with Sarcocystis muris, 192
— Endotrypanum schaudinni, 99
— Histoplasma capsulatum, 112
— researches on Entamœba tetragena, 38, 40, 41
Darwin, Charles, great black bug of Pampas (Conorhinus renggeri), 539
Dauernheim, Ascarides in bile-ducts, 688
Daughter cysts of echinococcus, 350, 351
— — — mode of origin, 350, 351, 352
Davaine, Cercomonas hominis, 61
— milk cure in expulsion of Strongyloides stercoralis, 675
— mode of development of Ascaris lumbricoides, 464, 465
Davainea asiatica, 662
— — morphology, 330
— madagascarensis, association with chyluria, 662
— — cases of human infection by, 330
— — morphology, 329
— morphology, 329
Davaineidæ, 309, 329
Davaineinæ, 309, 329
Davidson, bite of Rasahus biguttatus, 540
Deeks, W. E., treatment of amœbic dysentery, 619
Deguy, see Labadie-Lagrave and Deguy
Deinocerites, characters, 564
Delanoë, pneumocysts in rats, 90
Delhi boil, see Oriental sore
Demarquay, observation of Filaria bancrofti in man, 390
Demodex folliculorum, 521
— — affecting eyelids, 708
— — as cause of chalazion, 708
— — characters, 522
— — infection by, treatment, 708
— — — views of different authors respecting, 708
— — synonyms, 522
— — var. canis, 522
— — — infection by, transmission from dog to man, 709
— — — — treatment, 709
Demodicidæ (mites of hair follicles), characters, 522
Dendriomyia, characters, 565
Dengue fever, micrococcus supposed carrier, 576
Dermacentor, characters of, 497
— occidentalis, bite of, effects, 504
— — characters and morphology, 504
— — (wood tick), characters, 504
— reticulatus, hosts of, 502, 503
— — transmission of Babesia caballi by, 178
— variabilis, 505
— venustus, characters and morphology, 503
— — hosts of, wild and domestic, 504
Dermanyssus gallinæ, disinfection methods against, 704
— — effect on skin of host, 492
— — infection, symptoms set up by, 703
— — morphology, 492
— — synonyms, 492
— hirundinis, disinfection against, 704
— — skin affections due to, 492
Dermatitis intertriginoides set up by Oxyuris vermicularis, 696
Dermatobia cyaniventris, 596, 597
— — characters, 598
— — geographical distribution, 598
— — hosts of, 596
— — larvæ of, characters, 597
— — local or vernacular names for, 598
— — see also Mosquito worm
— noxialis, 597
— — larvæ of, skin disease caused by, 725
Dermatocentor reticulatus, var. occidentalis, carrier of Rocky Mountain spotted fever, 496
Dermatodectes, 521
Dermatophagoides scheremetewskyi, 521
Dermatophilus, 543
— cæcata, 544
— penetrans (jigger, chigoe), characters and morphology, 544
— — — — geographical distribution, 544
— (Sarcopsylla) penetrans (sand flea), entrance beneath skin, 714
— — — host of, 613
— — — lesions produced by, 714
— — — — — treatment, 715
— — — possible carrier of leprosy, 613
Dermo-muscular layer of nematodes, 361
Derrieu and Raynaud, chronic dysentery due to trichomonads, 624
— — trichomonad-like organism discovered by, 624, 735
Desvoidea, characters, 563
Dévé, experimental development of hydatid scolices, 353
Diarrhœa associated with Balantidium coli, 201
— — with invasion by Balantidium minutum, 204
— — with Lamblia intestinalis, 59, 625
— — with Trichomonas hominis, 54, 624
— — with presence of Watsonius watsoni, 235
— — with Prowazekia asiatica, 65
— blood-stained, in Strongyloides stercoralis infection, 674, 675
— caused by Difämus tunensis, 57
— chylous, from Filaria bancrofti infection, 678
— (Cochin-China) in cases of infection with Strongyloides stercoralis, 380, 381
— flagellate, 623
— — climatic distribution, 623
— infantile, spread by house-fly, 586
— white (or white scour), in fowls, causal agent, 145
Dibothriocephalidæ, 308, 309
— morphology, 308
Dibothriocephalinæ, 308, 309
— morphology, 308
Dibothriocephalus, 309
— cordatus, 315
— — cephalic end, 315
— — hosts of, 315
— excretory apparatus, collecting tubes, island formation, 292
— felis, 313
— latus, 310
— — chains of segments, 311
— — development, 311
— — developmental cycle of, 16
— — disturbances produced by, in man, 314
— — duration of life, 315
— — embryophore of, 298
— — experimental infection of man with, 312
— — geographical distribution, 313, 314
— — growth of, 306
— — habitat in man, 658
— — head of transverse section, 311
— — hosts of, 313
— — human parasite invading animals, 7
— — in muscles of trunk of burbot, 313
— — intermediate host of, 255
— — means of transmission to man and other hosts, 314
— — mode of infection, 658
— — morphology, 310
— — ova of Fasciola hepatica to be distinguished from, 242
— — ovum of, development, 312
— — parasitic association with Tænia solium, 658
— — percentage in sufferers from tapeworms in various localities, 314
— — plerocercoids of, 313
— — — habitat and host, 311
— — — how destroyed, 315
— — — inhabiting fish, 314
— — proglottids of, average number found daily, 312
— — proglottis, fairly mature, stained preparation, 311
— — prophylaxis against, 668
— — supposed origin of, 11
— — symptoms produced by infection by, 667, 668
— — synonyms, 310
— — topographical anatomy, transverse section through proglottis showing, 296
— morphology, 309
— parvus, habitat, 316
Dibothriocephalus parvus, how distinguished from D. latus and D. cordatus, 316
— — morphology, 316
— plerocercoid of, 300
— synonyms, 309
Dicrocœiidæ, morphology, 232, 265
Dicrocœlium dendriticum, intermediate host unknown, 267
— — morphology, 266
— — organs of, diagram showing, 265
— — ova and miracidia, 266
— — synonyms, 266
— lanceolatum, incidental human parasite, 7
Diesing, first record of case of Metastrongylus apri in man, 433
Difämus tunensis, cause of diarrhœa, 57
— — characters, 57
Difflugia enchelys, 47
Digenea, morphology, 230
Digestive system, loss of, in parasites, 3
Dinoflagellata, 52
Dioctophyme, 431
— gigas, hosts and habitat in body, 431
— — in man, source of, 431
— — infection by, 431
— — morphology, 431
— — ova of, 432
— — synonyms, 431
Dioctophymidæ, 431
— characters, 375
Dionis de Carrières, liver-fluke in right hypochondriac region, 244
Diphyllobothrium (Bothriocephalus) ditremum, discovery of progeny of, 13
Diplodiscus subclavatus, hosts of, 6
Diplogonoporus, morphology, 316
— grandis, egg of, 298
— — morphology, 316
— — ventral view of genitalia of left side, 317
— — — — of portion of strobila, 317
Diptera, bibliography, 612
— biting-mouthed and other noxious carriers of disease, 600
— characters, 531, 532
— digestive tract of, inhabited by Herpetomonads, 102
— larvæ of, in man, references to, 599
Dipylidiidæ, 309, 320
Dipylidium, morphology, 320
— synonyms, 320
— caninum, 320
— — confused with Tænia solium, 660
— — cysticercoids of, 322
— — — hosts of, 322
— — dogs and cats infected with, through skin parasites, 323
— — embryo of, development, 321
— — expulsion of, drugs suitable for, 660
— — hosts of, 6, 7, 322
— — invasion by, effect on central nervous system, 649
— — morphology, 320
— — oncosphere of, 299
— — oncospheres of, animals selected as hosts for development, 299
Dipylidium caninum, prevalence of infection by, in children, 659, 660
— — proglottids of, 322
— — proglottis of, central portion, 321
— — region of human body inhabited by, 660
— — rostellum of, 289
Dirksen, effect on anæmia of expulsion of Tænia solium, 648
Dirofilaria immitis, 417
— magalhaesi, morphology, 417
— morphology, 416
— repens, 417
Discophora, see Hirudinea
Disporea in Myxosporidia, 182, 184
Distoma, opercula of ova of, discovery, 12
— echinatum, redia of, in later stage, 227
Distomata, morphology, 231
— cercariæ, 753
Distomum ophthalmobium, 244
Doeveren, van, on transmission of intestinal worms, 11
Doflein, Coccidiomorpha, 129, 151
— Cnidosporidia, 129, 194
— Entamœba kartulisi, 44
— Trypanosoma equiperdum, 97
Dog and cat, parasites common to, 6
— blood of, life-cycle of Babesia (Piroplasma) canis in, 175
— Dipylidium caninum parasite in, 322
— fleas, natural flagellates in, 111, 112
— — transmission of canine kala-azar by, 103
— — see also Ctenocephalus canis and Pulex serraticeps
— host of Dibothriocephalus latus, 313, 315
— — of Paragonimus kellicotti, 250
— intestine of, Isospora bigemina parasitic in, 149
— mange, transmission to man, 523
— rearing of Tænia echinococcus in, 356
— to dog, transmission of leucocytogregarine from, by tick, 155
— transmission of infection with Demodex folliculorum canis to man, 709
Dogs, contraction of surra by, 96
— dermal infection with larvæ of Ancylostomum duodenale, 455
— echinococci in, 346
— experiments with, to prove transmission of infantile kala-azar by fleas, 111
---- infected with Dipylidium caninum through skin parasites, 323
— infection with infantile leishmaniasis, 110
— — — — experimental, 110
— — — — natural, 110
— Leishmania tropica in, 108
— malignant jaundice in, carrier of, 493
— — — cause of, 177
— nagana fatal to, 94
— percentage infected with Tænia echinococcus in various cities and countries (footnote), 345
— prevention of echinococcus infection by, 346
— segregation of, as preventive against Oriental sore, 628
— skin diseases in, due to young nematodes, 378
Doliocystidæ, 135
Domestic animals, species of Sarcoptes transmissible from, to man, 520
— — trypanosomes deleterious or lethal to, 69, 70
Dormouse, inoculation of Trypanosoma lewisi into, 90
Dornblüth, method of evacuation of Oxyuridæ, 697
Dörr, tapeworm drug recommended by, 674
Dourine, anæmia and paralysis in, 97
— periods or stages of, 97
— trypanosome causing, 97
Dovecots, habitat of Argas reflexus, 506
Dracontiasis, disorder named by Galen, 386
Dracunculidæ, 385
— characters, 374
Dracunculus medinensis (Guinea worm), anterior extremity, 387
— — expulsion by extraction, 676
— — — methods and drugs for, 676
— — female, transverse section of, 388
— — infection by, prophylaxis against, 676
— — — symptoms and lesions following, 389
— — intermediate host, 388
— — larvæ of, 388
— — life-history, 386, 388
— — methods of extraction from body, 389
— — morphology, 386
— — period of development in man, 389
— — regions of body inhabited by, 386
— — synonyms, 386
— — viviparous nematode, 371
— morphology, 385
Dromedaries, “mbori” in, 96
Drone fly, 583, 584
Drosophila melanogaster, larvæ of, characters and habitat, 584
— — — effects produced by ingestion of, 584
Drosophilidæ, characters and habitat, 584
Drouillard, favourable effects of expulsion of Ascaridæ, 649
Drugs, reaction of spirochætes to, 115
Dubini, Filaria (?) conjunctivæ in man, 405
Dubreuilh, infection with Demodex folliculorum, 708
Ducks, destruction of mosquito larvæ by, 636
— how infected by Echinostoma echinatum, 226
Dufour, creation of name Gregarina by, 129
Duguet, maculæ cæruleæ (tâches bleues) due to infection by crab louse, 712
Dujardin, development of Tæniæ, 14
— psorosperms, 181
Duke, anterior station of trypanosomes in Glossinæ, 101
— Trypanosoma gambiense in antelope, 76
Dum-dum fever, 105
Dumesnil, larvæ of Muscidæ in nose, 720
Duodenum, flagellate stage of Lamblia intestinalis found in, 59
— human, habitat of Ancylostoma duodenale, 450
— possible invasion by Balantidium minutum, 204
— species of Trichostrongylus inhabiting, 435, 436
Durham, specimens of Leptus (bête rouge), 486
Dutton, Trypanosoma gambiense, 68
— and Todd, herpetomonads in mice, 738
— — researches on Spirochæta duttoni, 116, 117
Duval, liver-flukes in veins, 243
Dysentery, amœba as causal agent, 30, 34
— — — — experiments made to prove, 30, 40, 618
— amœbic, acute, symptoms, 618
— — carriers, 618
— — chronic, 618
— — experimental production, 618
— — gangrenous, treatment, 619
— — incubation period long, 618
— — latent, 618
— — — leading to liver abscess, 618
— — preventive measures, 620
— — relief of griping and straining, 618, 619
— — treatment by emetine hydrochloride,618, 619
— — — by ipecacuanha, 619
— — — by preliminary administration of castor oil, 619
— — — of liver abscess, 620
— — — surgical, 619
— association of Lamblia intestinalis with, 59, 625
— — of Noc’s entamœba with, 41
— — of Trichomonas with, 56, 624
— bacillary, 31
— — Entamœba coli present in cases of, 38
— bacillus, discovery of, 31
— balantidian or ciliate, 201, 202, 637
— — — prophylaxis, 637
— — — symptoms, 637
— — — treatment, 637
— discoveries of Cholera Investigation Commission with regard to, 30
— discovery of intestinal amœbæ in cases of, 30
— flagellate, 623
— — diet in, 625
— — geographical distribution, 624
— — prognosis, 625
— — prophylaxis, 625
— — protection of food supply against fæcal contamination in, 625
— — treatment, 624, 625
— followed by recovery from oxyuriasis, 698
— production by injection of amœbæ into cats, 35
— red, geographical distribution, 147, 741
— — in cattle, cause of, 147
— so-called, due to mites, 512
— spread by house-fly, 586
— treatment by bismuth subnitrate, 619
Dysodius lunatus, bite of, 541
E.
Ear, abscess of, liver-fluke in, 244
— parasites in, 615
— — see also Myiasis, auricular
Ears of hosts infested by Ornithodorus mégnini rendered painful, 510
Earth-eating in connection with Trichuris trichiura infection, 679
East Coast fever in cattle, cause of, 178, 179
— — — — pathogenic agent, 174
Eau de Cologne, application in nasal myiasis, 719
Echidnophaga, 543
Echinococcus, brood capsules, 349
— — — and scolices, mode of formation, 348, 349
— — — transformation into daughter cysts, 351, 353
— cysticus fertilis, 350
— cysts causing urticaria, 651
— daughter cysts, 350
— definition of, 301
— fluid, chemistry of, 353
— frequency of infection by, of various organs in slaughtered animals, 347
— hominis in liver, incised fibrous capsule and wall showing daughter cysts, 351
— in dogs, oxen, sheep and pigs, 346
— in man, age incidence, 355
— — death at various stages of development, 356
— — geographical distribution, 354, 355
— — organs of body invaded by, 355
— — percentage of prevalence in Central Europe, 354
— — secondary, 356
— — sex incidence, 355
— infection, prevention of spread by dogs, 346
— multilocularis (alveolar colloid), development, 357, 358
— — — — feeding experiments with Tænia from, 358
— — — — hooklets of, 359
— — — — in liver of ox, 357
— — — — in man, early disintegration, 357
— — — — — geographical distribution, 358
— — — — — invasion, results of, 358
— — — — — — site of, 358
— — — — morphology of, 356
— — — — reasons for distinction from hydatid or unilocular, 357, 358
— of liver rupturing into abdominal cavity, 652
— rate of growth, 354
— rich in glycogen, 348
— scolex, 349
— — development in rabbits, 353
Echinococcus scolex in process of vesicular metamorphosis, section through, 351
— — invaginated, section through, 350
— — transformation into daughter cyst, 352
— scolices in, 303, 349
— serum diagnosis of, 359
— structure and development, 347
— veterinorum, 350
— — brood capsules and scolices, 350
— — hooklets of, 355
Echinorhynchus, anatomy, 475
— excretory organs, 475, 476
— “floating ovaries” of, 150
— gigas, hosts of, 477
— — incidental human parasite, 7
— — intermediate hosts, 477
— — morphology, 477
— hominis, 478
— moniliformis, hosts, habitat and intermediate host of, 478
— nervous system, 475
— ova of, 477
— protrusor proboscidis, 475
— receptaculum proboscidis, 475
— retractor proboscidis, 475
— — receptaculi, 475
— sexual organs, 476
Echinostoma, cercariæ of, 225
— echinatum, method of infection of ducks and geese by, 226
— ilocanum, habitat, 268
— — morphology, 267
— — organs of, diagram showing, 268
— malayanum, habitat, 269
— — morphology, 268, 269
— morphology, 267
Echinostomidæ, 267
— morphology, 233
Echinostominæ, 267
— morphology, 233
Ectoparasites, 1
— permanent, changes in, 3
Ectoplasm of protozoa, 25, 26
— — substances deposited in, 26
Ectoschiza, 135
Eczema due to head louse infection, 709, 710
— following infection by crab louse, 712
— occupational, diagnosis from scabies, 706
— peri-anal and perineal, set up by migrations of Oxyuris vermicularis, 695
— purulent, following infection by ancylostomes, 684
— resulting from clothes louse infection, 711
— set up by infection with Dermanyssus gallinæ, 703
— — by Leptus autumnalis, 702
Ehrenberg, Spirochæta plicatilis, 114
Ehrlich’s acid hæmatoxylin, 751
Eimer, researches on coccidia, 136
Eimeria, 142
— avium, causal agent of white diarrhœa or white scour in fowls, and blackhead in turkeys, 145
— — cause of fatal epizootics among game birds and poultry, 142
Eimeria avium, infection by, method of, 145
— — life-cycle of, 142–145
— — — period, 144, 145
— — — phases, 142–144
— — merozoites of, 143
— — microgametes and macrogametes of, 143, 144
— — relation to E. stiedæ, 145
— — sporozoites of, 143
— — trophozoites of, 143
— falciformis, 136
— (Coccidium) schubergi, life-cycle of, 138–141
— hominis, 150
— — bodies described as, 150
— stiedæ, ascribed cause of “red dysentery” in cattle, 147
— — host of, 7, 145
— — oöcysts of, 142, 146
— — parasitic in rabbit and occasionally in man, 145, 148
— — schizogony, 147
— — — — effects, 147
— — synonyms, 145
— — see also Coccidiosis
— synonyms, 142
Eimeridea, 141, 742
Electrolytic needle, application in creeping disease, 731
Elephantiasis arabum from Filaria bancrofti infection, sites of body affected by, 677
— — — — — symptoms, 677
— — — — — treatment, 677
— scroti in filariasis, 402
Ellermann, rhizopods in poliomyelitis acuta, 46
Elmassian, discovery of Entamœba minuta, 42
— — of trypanosomes in “mal de caderas,” 68
Embryophore of tapeworms, 298
Emetine and vaccine treatment combined in pyorrhœa alveolaris, 620
— hydrochloride in flagellate dysentery, 625
— — in treatment of amœbic dysentery, 618, 619
— in oral endamœbiasis, 620
Emily, expulsion of guinea worm, 676
— method of extraction of Dracunculus medinensis, 389
Endamœba, 31, 34, 734
— see also Entamœba
Endamœbiasis, oral, 620
— — treatment, 620
Endermol, application in scabies, 707
Endoparasites, 1
— intermediate generations invading intermediate hosts, 5
— — hosts of, 5
— young of, leaving host or organ of host inhabited by parents, 5
Endophlebitis set up by Schistosoma hæmatobium, 274, 275
Endoplasm of protozoa, 25, 26
— — substances deposited in, 26
Endoschiza, 135
Endotoxins in Trypanosoma equiperdum, 98
Endotrypanum schaudinni, 99
Enemata, in evacuation of Oxyuridæ, 697
Entamœba, 31, 734
— africana, see under Entamœba tetragena
— buccalis, 43, 734
— — association with cancer of oral cavity, 43
— — — with dental caries, 43
— — — with pyorrhœa alveolaris, 43, 734
— — characters, 43
— — possible relation of E. pulmonalis to, 40
— bütschlii, 34
— coli, 32, 618, 733
— — characters, 33
— — cysts of, 33
— — — in normal fæces, 33
— — encystment process, 33
— — — — cytological changes during, 33
— — how distinguished from E. histolytica, 34, 40, 733
— — life-cycle of, 32, 33
— — non-pathogenic and non-culturable, 618
— — parasite of human intestine, 618
— — may be present in bacillary dysentery, 38
— — schizogony of, 32, 33
— — so-called autogamy of, 34
— — sporogony of, 32, 33
— gingivalis, 733
— — synonyms, 734
— hartmanni, 34
— histolytica, 32, 34, 45, 618, 733
— — causal agent of amœbic dysentery, 35
— — changes in intestine produced by, 35
— — characters, 34, 35
— — cysts of, permanent, injection producing infection, 37
— — dysentery following experimental infection with, 618
— — how distinguished from E. coli, 34, 40, 733
— — present in large intestine, 38
— — producing liver abscess, 35, 620
— — sporulation of (so-called), 37
— — and E. coli, differences between, 34, 40, 733
— — — — mixed infection, 38
— — and E. tetragena, identity of, 38, 40, 41
— hominis, 42
— kartulisi, 44, 734
— — association with dental caries, 44
— maxillaris, 734
— minuta, 42
— — relation to E. histolytica, 40, 42
— mortinatalium, 45
— nipponica, 42
— Noc’s, 41
— — association with liver abscess and dysentery, 41
— phagocytoides, 42
— poleki, 34
Entamœba pulmonalis, 45
— — relation with E. buccalis, 45
— tetragena, 38
— — characters of, 39
— — described as E. africana by Hartmann, 38
— — found in amœbic dysentery, 38
— — infection by, 40
— — multiplication of, 39
— — nucleus of, 39
— — part of life-cycle of E. histolytica, 38
— — reproduction of, 39
— — trophozoites, 39, 40
— — and E. histolytica identical, 38, 40, 41
— tropicalis, 41
— undulans, 43, 44
— — characters of, 43
— — probable flagellate nature of, 44
— williamsi, 34
Entamœbæ of vertebrates, 34
— question of cultivation on artificial media, 42, 742
Enteritis, hæmorrhagic, in Strongyloides stercoralis infection, 674
— produced by injection of amœbæ into cats, 36, 37
— verminosa in children, 688
Entozoa, 1
— derivation of, 21
Enyaliopsis durandi, bite of, 542
— petersi, 542
— species of, producing ulcers, 542
Eosinophilia in ancylostomiasis, 647
— in bilharziasis, 641
— in hydatid disease, 652
Epicarin, application in scabies, 707
Epidermis, human, excavation of tunnels in, by Sarcoptes scabiei, var. hominis, 519
— “wormlet” burrowing into, 599
Epistaxis, association with presence of Linguatula in nasal cavity, 526, 527
— leeches in nose causing, 701
Epithelium of nematodes, 360
Epizoa, 1
Eproboscidæ, see Pupipara
Epstein, experimental infection with Ascaris lumbricoides, 465
— transmission of trichomonad infection, 56
Epstein’s method of diagnosis of ascariasis, 692
Equines, baleri in, causal agent, 95
— biliary fever in, cause of, 177
Erdmann, experimental infection with Sarcosporidia, 192
Erythema, autumn, set up by Leptus autumnalis, 485, 486
— following bite of Argas reflexus, 506
— set up by infection with Dermanyssus gallinæ, 703
— — by Leptus autumnalis, 702
Eschatocephalus, characters of, 497
— hosts and habitat of, 497
Escomel, dysentery due to Trichomonas, 56, 624
— treatment of espundia, 629
— — of lamblial diarrhœa, 625
Espundia, 108, 628
— course of, 629
— geographical distribution, 628
— pathology of, 628
— prophylactic measures against, 629
— transmission of, 629
— treatment, 629
Ether, application in nasal myiasis, 719
— lotions in crab louse infection, 712
— sulphuric, in crab louse infection, 712
Ethyl chloride, freezing by, in creeping disease, 731
Eucalyptus oil in expulsion of ancylostomes, 687
Euflagellata, 52
— characters of, 52
Eugregarinea, schizogony absent in, 134
Eulyes amœna, 542
Euphorbia, infection by Herpetomonas davidi, 104
Eupodidæ, characters, 491
Euquinine in malaria, 635
Europe, Central, percentage of prevalence of echinococcus in man in, 354
Eustrongylus gigas, infection by, site of, 681
— — — symptoms, 682
Evans, discovery of trypanosomes in blood of horses with “surra” disease, 67
— see also Steel and Evans
Excretory apparatus of cestodes, 291
— or segmental organs of Hirudinea, 481
— organs of Echinorhynchus, 475
— — of nematodes, 366, 367
Eye, cysticerci in, 335, 664
— cysticercus of, 664
— — diagnosis from foreign body, 664
— diseases due to Lucilia macellaria, 721
— human, infection with filaria, 406
— nematodes observed in, 412
— invasion by Loa loa, 678
— paragonimiasis of, 639
Eyeball, bodies found in, probably young liver-flukes, 244
Eyebrows and eyelashes, pediculosis of, treatment, 712
Eyelid, upper, extraction of larva of Hypoderma bovis from, 596
Eyelids, Demodex folliculorum affecting, 708
Eyes, destruction of, by Sarcophaga wohlfahrti, 723
— loss of, in parasites, 3
F.
Fabre, prophylaxis against ancylostomiasis, 685
Face, swelling of, in nasal myiasis, 718
Fæces, amœbæ found in, 30, 34, 47, 48
— asexual multiplication of Chlamydophrys enchelys in, 47
— examination for protozoa, 746
— human, fresh, Strongyloides stercoralis larva from, 382
— larvæ of Homalomyia canicularis found in, 584
— — of Piophila casei found in, 583
— males of Oxyuris vermicularis rarely met with in, 468
— normal, cysts of Entamœba coli present in, 33
— preservation of ova of flukes in, 472
— Vorticella in, 206
Fanapapea intestinalis identical with Tetramitus mesnili, 57
Fantham, H. B., appendix on protozoology (recent researches, formulæ of culture media, general protozoological technique), 733
— — avian coccidiosis, 145
— — — pathogenic spirochætes, 119
— — classification of Haplosporidia, 195
— — — of Schizogregarines, 135
— — experimental infection with Spirochæta duttoni, 117
— — granule phase of spirochætes, 120
— — Herpetomonas ctenocephali, 103
— — — pediculi, 103
— — latent forms of trypanosomes, 73, 74, 77
— — molluscan spirochætes breaking up into granules, 119
— — morphology and life-cycle of Eimeria avium, 142–145
— — — and life-history of Spirochæta bronchialis, 739
— — nuclear phenomena of Babesia bovis, 176
— — Protozoa, 25
— — recent work on spirochætes of human mouth, 740, 741
— — Rhinosporidium kinealyi, 195–197
— — schizogony in Leucocytozoon lovati, 153
— — significance of insect flagellates in relation to disease, 104, 739
— — Theileria parva, 179
— — Trypanosoma rhodesiense, 69, 76
— — and Porter, experimental introduction of insect flagellates into vertebrates, 104, 112, 738
— — — inoculation experiments with Herpetomonas jaculum, 104, 738
— — — natural herpetomonads in mice, 739
— — — researches on Nosema apis, 185
— — — — on Spirochæta duttoni, 116
— — and Thomson, J. G., cultivation of Babesia canis, 172
— — — — periodic cyclical variation of trypanosomes in blood, 78
— — see also Stephens and Fantham
Fasciola, 237
— gigantica, distribution, 244
— — habitat, 244
— — invading lung, 245
— — morphology, 244
— — — section illustrating, 243
— — synonyms, 244
— hepatica, 237, 238, 239, 638
— — cercaria of, 228
— — — encysted, 228
— — development of, 226
— — fixation method, 471
— — geographical distribution, 238
— — half transverse section through, 214
— — hosts of, 6, 7, 238
— — incidental human parasite, 7
— — intermediate host, 240, 241
— — invading and infecting pharynx, 242
— — — regions of body other than liver, 243
— — life-history, 241
— — method of infection of sheep by, 226
— — miracidium of, 223
— — morphology, 237
— — — sections illustrating, 238, 239
— — ova of, to be distinguished from those of Dibothriocephalus latus, 242
— — ovum of, 223
— — — from liver of sheep, 240
— — redia, in early stage, 227
— — synonyms, 237
— morphology, 237
Fascioliasis, prevention and treatment, 638
— symptoms of, 638
— see also Liver-fluke disease
Fasciolidæ, 237
— morphology, 231
Fasciolinæ, 237
— morphology, 231
Fasciolopsinæ, 245
— morphology, 231
Fasciolopsis buski, fixation method, 471
— — geographical distribution, 246
— — habitat, 246, 638
— — morphology, 245, 246
— — symptoms set up by invasion by, 638
— fülleborni, cirrus sac, 247
— — habitat, 249
— — morphology, 247
— — — ventral aspect showing, 248
— goddardi, morphology and geographical distribution, 247
— rathouisi, geographical distribution, 247
— — habitat, 247
— — morphology, 246
— morphology, 245
Feltinella, characters, 561, 567
Females, greater prevalence of head louse infection among, 709, 710
— rarity of bilharziasis in, 643
Fern root, new species, effects as vermifuge, 673
Fibrin, clots of, Dioctophyme gigas in man traced to, 431
Ficalbia, characters, 565
Fièvre de grain, 702
Fiji, intermediate host of filaria in, 575
— manifestations of filariasis in, 402
Filaria associated with phthisis, 408
— bancrofti, anatomy of, diagrams showing, 391
— — discoveries relating to, 390
— — diseases following infection by, 398, 400
— — embryos, 392
— — female, characters, 392
— — geographical distribution, 392, 403
— — habitat in body, 392
— — infection by, diseases resulting from, 676, 677
— — larvæ of, absence from blood in those suffering from filarial disease, 400
— — — distribution in body, 392, 393
— — — method of concentration, 395
— — — — of preservation, 395
— — — morphology, determination by fixation and staining methods, 395, 396
— — — periodicity of, in peripheral blood, 393, 394
— — — separation of red corpuscles from, 395
— — — species of Tæniorhynchus carrying, 577
— — — structure, 396
— — life-history, 398
— — male, characters, 392
— — mosquitoes acting as hosts of, 398
— — ova of, 392
— — synonyms, 390
— — transmission of, 576
— (?) conjunctivæ, 404, 405
— — morphology, 404
— — normal hosts of, 406
— — sites of infection in man, 405
— — synonyms, 404
— demarquayi, 403, 404
— — geographical distribution, 403
— — morphology, 403
— infection of skin by, 378
— intermediate host of, in Fiji, 575
— (?) kilimaræ, 407
— loa, host of, 601
— medinensis, antiquity of knowledge concerning, 386
— morphology, 390
— oculi humani, association with cataract, 406
— perstans, carrier of, 508
— (?) romanorum orientalis, morphology, 407
— (?) sp. (?), 407
— taniguchi, 404
Filariasis, cultivation of bacillus from cases of, 755
— prevalence proportionate to prevalence of Mikrofilaria bancrofti in blood, 400
Filariidæ, 390
— characters, 374
Filariinæ, 390
Filmaron, administration in expulsion of ancylostomes, 687
— as vermicide, 672
— oil, dosage of, 672
— — effects of, 672
— — in expulsion of Ascarides, 694
“Filterable viruses,” 207
Filtration experiments with Chlamydozoon granules, 209
Finlaya, characters, 564
Finsen, echinococcus cysts causing urticaria, 651
Finucane, lymphangitic symptoms in children from Filaria bancrofti infection, 676
Fischer, effects of new species of fern root as a vermifuge, 672, 673
— retinal hæmorrhages in ancylostome anæmia, 646
Fish, destruction of mosquito larvæ by, 636
— disease of, due to invasion by Myxosporidia, 182, 184
— eating of raw or badly cooked, favours transmission of Dibothriocephalus latus in man, 315
— fresh-water, second intermediate host of Clonorchis endemicus, 261
— — — — — — — proved by feeding experiments, 261
— intermediate host of Dibothriocephalus latus, 255
— — — of Opisthorchis felineus, 255
— parasitic ciliate destructive to, 206
— plerocercoids of Dibothriocephalus latus inhabiting, 314
— trypanoplasms in, 68
Fistulæ, anal and rectal, set up by migration of Oxyuris vermicularis, 695
— formation of, through migration of parasites, 9
— urethral, arising from bilharziasis, 642
— — — — treatment, 644
Fixation, time of, 749
Fixatives, 748, 749
— for wet films, 748
— hot, 748
Flagella may occur among rhizopods, 52
— of Flagellata, 50, 51
Flagellata, 50
— characters of, 28, 50, 51
— classification, 52
— formation of colonies of individuals, 51
— habitat, 28, 52
— multiplication of, 51
— non-flagellate stages, 51
— nuclear apparatus of, 51
— post-flagellate and pre-flagellate stages, 52
Flagellates, aggregation rosettes of, 51
— dysentery in children due to, 56, 624
— in blood of horses, diseases associated with, 67, 68
— natural, in dog fleas, 112
— of invertebrates, evolution of Leishmania from, 739
— parasitic, in insects, experimental introduction into vertebrates, 104, 112, 737, 738
— — — — — — relation to evolution of leishmaniasis, 112, 737, 738
— — in relation to evolution of disease, 739
Flagellosis of plants, possible connection with leishmaniasis, 104, 739
Flat worms, see Platyhelminthes
Flea, human, see Pulex irritans
Fleas acting as intermediate hosts, 543
— blood-suckers, 543
— carriers of plague, 543
— cocoons of, 543
— fed on infected rat, percentage infected with trypanosomes, 93
— herpetomonads in gut of, 103
— larvæ of, 543
— life-cycle of Trypanosoma lewisi in, 88, 90
— method of controlling, during experiments, 93
— ova of, 543
— possible transmission of kala-azar by, 111
Flemming’s solution, 749
Flesh of animals containing larvæ of tapeworms must be thoroughly cooked before eating, 668
Flesh-fly, see Sarcophaga carnosa
Flexner, Entamœba kartulisi, 44
Flies, larvæ of different species of, found in intestinal myiasis, 728
Flukes, clearing and mounting agents, 471
— differentiation methods, 471
— fixation methods, 471
— miracidia of, discovery, 12
— ova of, preservation in fæces, urine, bile, 472
— — transference to glycerine, 472
— preservation and examination of, 471
— staining methods, 471
— see also Trematoda
Fœtus, Trypanosoma cruzi in, 88
Foley, transmission of relapsing fever, 120, 121
Food, transmission of trichomonad infection by, 56
Foods, decomposing, inhabited by and nutriment of Tyroglyphidæ, 511
Foot, sole of, hepatic flukes found in swelling on, 243
Foot-and-mouth disease, 207, 208
Foraminifera, 27, 47
— characters and habitat, 27
Forde and Dutton, discovery of human trypanosomes, 68
Foreign body, diagnosis of cysticercus of eye from, 664
Formalin, fixation of cestodes by, 472
Fowler, method of administering male fern to children, 671, 672
Fowler’s solution in sleeping sickness, 623
Fowls, fatal effects of Spirochæta gallinarum on, 119
Fox, host of Dibothriocephalus latus, 313
Frambœsia tropica, see Yaws
França, action of leucocytozoa on red cells, 153, 742
— genera of Piroplasmidæ, 174
Francaviglia, auricular myiasis, 615
Franchini, experimental infection of vertebrates with herpetomonads, 103, 104, 112, 739
—Hæmocystozoon brasiliense, 104
Frese, O., rhabdites found in gastric fluid obtained by lavage, 378
Freund, sarcophaga larvæ from abscess cavities, 723
Frog, rectum and bladder of, ciliates parasitic in, 207
Frontal sinus, invasion by Ancylostoma duodenale, 683
— — scolopendra in, 721
Fruit pickers affected by Leptus autumnalis (footnote), 485
Fülleborn, cultivation of larval forms of Ancylostoma and Strongyloides, 474
Furcocercous cercariæ, 753
Fürst, cases of Ascarides invading larynx and trachea, 691
G.
Gabel, diarrhœa due to Difämus tunensis, 624
— Difämus tunensis, 57, 624
Gad-flies, 600
— see also Tabanidæ
Gaetano, cysticercus of tongue, 663
Galen, disorder named “dracontiasis” by, 386
Gall-bladder, Schistosoma hæmatobium, eggs of, in, 274
Gall sickness in cattle, cause of, 98, 180, 611
Galleria melonella, larvæ of, in nose, 720
Galli-Vallerio, bothriocephalus anæmia, 646
— infection by trichomonads, 56
— Oxyuris and Trichocephalus infection in relation to appendicitis, 654
Galyl in syphilis, 632
— in trypanosomiasis, 622
Gamasidæ (coleopterous or insect mites), characters, 491
— — — — hosts and prey of, 491
Gambia horse sickness, cause of, 100
Game infested by Dermacentor reticulatus, 503
Game-birds, fatal epizootics among, due to Eimeria avium, 142
Gametes of Coccidiidea, 137, 139, 140
— of gregarines, 132, 133
— of malarial parasites, 162
Gametocytes of Coccidia, 140
— of gregarines, 132, 133
— of malarial parasites, 162
Gametogony, in Eimeria avium, 143
Gammarus pulex occasionally parasitic in man, 483
Garlic and saline injections in Trichuris trichiura infection, 680
Gastrodisciidæ, 236
— morphology, 231
Gastrodiscoides, how distinguished from Gastrodiscus, 236
Gastrodiscus, 236
— ægyptiacus, hosts of, 237
— hominis, 236
— — genital pore, 236
— — geographical distribution, 237
— — habitat, 237
— — morphology, 236
— — ova, 237
— — testes, 236
— male and female genitalia, 236
— morphology, 236
Gastrophilus, 599
— equi, 599
— hæmorrhoidalis, 599
— larvæ of, in stomach and intestine, 599
— nasalis, 599
— pecorum, 599
Geber, treatment of crab louse infection, 712
Gecko, blood and organs of, herpetomonad flagellate in cultures from, 739
Gedoelst, Cordylobia rodhani, 593
Geese, how infected with Echinostoma echinatum, 226
Genital apparatus of Cestoda, 293–296
Genitalia as means of distinguishing species of Glossina, 604
Genitals, external female, invaded by Oxyuris vermicularis, 467
Genser, von, Ascaris infection in relation to appendicitis, 653
Gentian violet, 752
Gerbillus indicus, 154
Gerlach, stages of liver-fluke disease in sheep, 240
Germany, districts of, percentage of occurrence of echinococcus in man in, 354
— trichinosis in, epidemics of, 423, 429
— — prophylaxis against, 429
Giard, microscopical investigations of conjugation in gregarines, 130
Giardia (Lamblia) intestinalis, 736
Giemsa’s stain, 751
— — formula of, 751
Giesker, liver-fluke in sole of foot, 243
Gilesia, characters, 564
Gingivitis, nematode larvæ in periosteum of upper jaw associated with, 378
— see also Entamœba gingivalis, 733; and E. buccalis, 43
Girard, effects of Trichocephalus infection, 651
— Trichocephalus infection in relation to appendicitis, 653
Glas, cysticercus of tongue, 663
Glossina austenii, characters, 605
— brevipalpis group, characters, 606, 607
— caliginea, characters, 605
— characters, 603, 604
— development of trypanosomes in, 101
— fusca, characters, 606
— — group, characters, 606
— fuscipleuris, characters, 606
— habitat of species, 605
— larvæ of, 604
— longipalpis, characters, 606
— — head of, 664
— longipennis, characters, 607
— medicorum, characters, 607
— morsitans, characters, 606
— — development of Trypanosoma brucei in, 94
— — development of Trypanosoma rhodesiense in, percentage of fly becoming infective, 82
— — developmental cycle of Trypanosoma rhodesiense in, 81, 82
— — geographical distribution, 608
— — group, characters, 605
— — — method of reproduction, 604
— — race submorsitans, 609
— — transmission of nagana (tsetse-fly disease) by, 93
— — — of Trypanosoma rhodesiense by, 69, 81, 608
— nigrofusca, characters, 606
— pallicera, characters, 605
— pallidipes, antenna of, 604
— — characters, 606
— palpalis, 100
— — blood-sucking, 607
— — carrier of sleeping sickness, 605, 607
— — characters, 605
— — development of Trypanosoma gambiense in, 74, 75
— — geographical distribution, 607
— — group, characters, 605
— — larval and pupal stages, 608
— — proportion becoming infected, 608
— — salivary glands of, invasion by Trypanosoma gambiense, 75
— — transmission of sleeping sickness infection by, 68
— puparia of, 604, 605
— special means of distinguishing species, 604
— species of, artificial infection with human trypanosome, 605
— spread of trypanosome diseases by, 603
— tabaniformis, characters, 606
— tachinoides, characters, 605
Glycerine, mounting agent for flukes, 472
— transference of ova of flukes to, 472
Glycerophosphates and arsenic in bronchial spirochætosis, 633
Glychæmalum, Mayer’s, 751
Glyciphagi, differentiation from Tyroglyphi, 513
— buski, 513
— cursor, 513
— domesticus, cause of grocer’s itch, 513
— hippopodes, 513
— prunorum, 513
Glycogen, echinococcus rich in, 348
Gnathobdellidæ, 481
Gnathostoma hispidum, hosts of, 385
— morphology, 384
— siamense, infection by, associated with tumour of breast, 385
— — morphology, 384
— sp., hosts of, 385
— spinigerum, 385
— — hosts of, 385
— — morphology, 385
Gnathostomidæ, 384
— characters, 374
Gnats, see Culex
Goebel, bilharziasis, 641, 642
Goeldia, characters, 565
Golden beetle, intermediate host of Echinorhynchus gigas, 478
Goldmann, male fern extract in expulsion of Strongyloides stercoralis, 675
— sebirol as vermicide, 672
— tæniol in ancylostomiasis, 686
Goldschmidt, excretory apparatus of Ascaris lumbricoides, 367
— formation of ova of trematodes, 223
Golgi, description of asexual cycle in blood in case of quartan parasite, 157
Gonder, relation of infantile kala-azar to Oriental sore, 108, 109
— strain of Trypanosoma lewisi losing resistance to arseno-phenyl-glycin, 93
— Theileria parva, 178
Gordiidæ, 375
— characters of, 479
— larvæ of, 479
Gordius aquaticus, 479
— chilensis, 479
— pustulosus, 479
— species invading man, 479
— tolosanus, 479
— tricuspidatus, 479
— varius, 479
— villoti, 479
— violaceus, 479
Grabhamia, characters, 564, 576
— dorsalis, 576
— geographical distribution, 576
— sollicitans, geographical distribution, 576
Gräffe, escape of ascarides from inguinal tumour, 656
Granate root as vermifuge, 673
Granuloma inguinale, spirochæte associated with, 122
Grass, harvest or gooseberry mite, see Leptus autumnalis
Grassi, Cercomonas and Trichomonas, 54
— development of cestodes without intermediate host, 17
— — of Trichuris trichiura, 420
— discovery of amœbæ in stools, 30
— experimental self-infection with Oxyuris vermicularis, 469
— expulsion of Hymenolepis nana, 661
— Hymenolepis nana, 324
— larval stage of Hymenolepis diminuta, 327
— mosquitoes in relation to human malaria, 158
— on Entamœba coli, 32, 33
— self-infection with Ascaris lumbricoides, 465
Great black bug of Pampas, see Conorhinus renggeri
Gregarina blattarum, 135
— longa from larva of crane-fly, 130
— munieri, from Chrysomela hæmoptera, 131
— ovata, host of, 135
Gregarines, ectoplasm of, 131
— endoplasm of, 131
— gametes of, 132, 133
— gametocytes of, 132, 133
— mode of infection, 134
— monocystid, 130
— — hosts of, 130
— morphology of, 130
— movements of, 131
— myonemes of, 130, 131
— polycystid, 130, 131
— — protomerite, deutomerite and epimerite of, 131
— resistant spores of, purpose of, 134
— spore-production, 132, 133
— sporocyst of, 134
— sporozoites of, 132, 133
— syzygy of, 132
— trophozoites of, 132, 133
— zygotes of, 132, 133
Gregarinida, characters and habitat, 28, 130
— classification, 134
— history of discoveries relating to, 129
Grey ointment in crab louse infection, 712
Grocer’s itch, cause of, 513
Ground-itch, skin affection set up by invasion of larvæ of Ancylostomum duodenale, 455
— treatment, 754
Guarnieri’s bodies, 207, 209
Gubler, case of human hepatic coccidiosis, 148
Guermonprez, method of expulsion of ascarides, 692
Guinea-pigs, experimental infection with Sarcocystis muris, 192
— natural occurrence of Paraplasma in, 180
Guinea worm, see Dracunculus medinensis
Gurley on Myxosporidia, 182, 183
H.
Hæmadipsa, 482
— blood-sucking pest in tropics, 482
Hæmagogus, characters, 565
Hæmalum, Mayer’s, 751
Hæmamœba, 151, 742
Hæmaphysalis, characters of, 497
— leachi (dog tick), carrier of malignant jaundice in dogs, 493
— — transmitting agent of Babesia canis, 177
— punctata, characters and morphology, 502
— — hosts of, 503
— — synonyms, 502, 503
Hæmatein, essential principle of hæmatoxylin, 751
— solutions of, in staining flukes, 471
Hæmatobia irritans, 610
Hæmatopinus spinulosus, 88
— — effect on strain of Trypanosoma lewisi being passed through, 93
Hæmatoxylin, Delafield’s (or Grenacher’s), 751
— Ehrlich’s acid, 751
Hæmaturia in bilharziasis, 641
Hæmentaria, 482
— officinalis used medicinally, 482
Hæmocystozoon brasiliense, 104
Hæmoglobinuria, infectious, in cattle, cause of, 177
Hæmogregarina balfouri (jaculi), 154
— gerbilli, 154
— marceaui, 154
— nobrei, 154
— schaudinni, var. africana, 154
Hæmogregarines, characters of, 154, 742
— hosts of, 153
— in red blood corpuscles, 154
— leucocytic, 154
— transmission of, 153
— variation in size and appearance of, 154
Hæmolysis, cure, after expulsion of Ascaridæ, 649
Hæmonchus contortus, diseases due to invasion by, 437, 438
— — geographical distribution, 437
— — habitat and hosts of, 437
— — life-history, 438
— — morphology, 436, 437
— — symptoms caused by invasion mistaken for those of ancylostomiasis, 438
— morphology, 436
Hæmoproteus (Halteridium) columbæ, insects transmitting, 151, 612
— — — life-cycle of, 152
— — danilewskyi, 152
Hæmorrhoidal veins, Schistosoma hæmatobium in, 273
— — superior, plexus formed by, in rectum, 272
Hæmorrhoids set up by migrations of Oxyuris vermicularis, 694
Hæmosporidia, 151
— Babesia or Piroplasma type, 154
— characters and habitat, 28, 151
— Hæmogregarina type, 153
— Halteridium type, 151
— Leucocytozoön type, 152
— Plasmodium or Hæmamœba type, 151
— recent views regarding, 742
— transmission by Ixodinæ, 704
Hagen-Thorn, cysticerci in brain, 665
Hair, methods of getting rid of nits from, 710
— follicles, mites of, 522
Hake, discovery of Coccidia by, 135
Halipegus ovocaudatus, host of, 6
Halteridium parasites occur in blood of birds, 151
“Halzoun,” affection of pharynx, produced by Fasciola hepatica, 242
Hampshire, sand flies biting in, 579
Hanau, Oxyuris infection in relation to appendicitis, 654
Haplosporidia, 129, 194
— characters and habitat, 29, 194
— classification, 195
— life-cycle, 194
Haplosporidium, 194, 195
— heterocirri, 195
Harington, guinea worm infection, 676
Harley, treatment of bilharziasis, 643
Harmostomum leptostomum, immature specimen, 215
Harpactor cruentas, 542
Harris, Penn, liver-flukes in abscess of occiput, 243
Hartmann, case of Oxyuridæ in nose, 696
— independent description of Entamœba tetragena under name of E. africana, by, 38
— multiplication of Trypanosoma cruzi in vertebrate host, 85
— toxic action of Oxyuris, 651
— and Chagas, Cercomonas parva, 737
— and Whitmore, formation of amœbulæ, 34
Hata, modification of Noguchi technique for cultivation of spirochætes and treponemes, 126
Hausmann, effects of trichocephalus infection, 651
Head louse, see Pediculus capitis
Headache in nasal myiasis, 717, 718
Heart-water fever in sheep, carrier of, 493
Hectopsylla, 543
Heekes, Oxyuris in appendix, 655
Heidenhain-Rosenbusch, iron-hæmatoxylin, 752
Heliozoa, characters and habitat, 27
Heller, life-history of Oxyuris vermicularis, 467, 469
— percentage of rats infected with Trichinella, 427
Helmerisch’s ointment, application in scabies, 706
Helminthes, 2
— biological, not systematic group, 2, 3
— dead, decomposition of, 9
— growth and agglomeration in host, 9
— life spent in intermediate and final host, 18
— life-history of, 18
— multiplication of, discovery of method, 13
— origin of, discoveries as to, 10, 11
— ova of, mistaken for coccidia (footnote), 137
— permanent parasites, 2
— producing substances toxic to host, 9
— separation of Linguatulidæ from, 2
Helminthiasis, 9
— meningitiformis, 649
Helsingfors, clinic of, work done on bothriocephalus anæmia at, 644
Hemiptera, characters, 531, 532
— digestive tract of, inhabited by Herpetomonads, 102
— sub-orders (footnote), 532
Henle, investigation of Gregarinida, 129
Henneberg, site of cysticerci in brain, 665
Henoch, expulsion of ascarides, 693
Hepatozoön, 154
Heptaphlebomyia, characters, 565
Herbst, experimental infection with encysted Trichinellæ, 423
Hermann, eucalyptus oil in expulsion of ancylostomes, 687
Hermaphroditism in permanent parasites, 4
Herpetomonad flagellate in cultures of blood and organs of gecko, 739
Herpetomonads, experimental infection of birds with, 739
— hosts of, 102
— in blood of birds, 739
— in mice, 738, 739
— stages of, 103
Herpetomonas, 67, 102
— ctenocephali, 103, 111, 112, 738
— — inoculation experiments with, 103
— davidi infecting plant genus Euphorbia, 104
— jaculum, inoculation experiments with, 104, 738
— life-history, stages of, 103
— muscæ domesticæ, 102, 739
— pattoni, 103, 739
— — inoculation experiments with, 103
— pediculi, 103, 738
— species of, introduction into vertebrates, 103, 104, 112, 738, 739
— stratiomyiæ, 738
Herpetomoniases, 112, 738
Hessler, R., Norway itch (scabies norvegica), 519, 520
Heterogony in nematodes, 372
Heterokaryota, 198
Heterophyes heterophyes, geographical distribution, 264
— — habitat, 264
— — morphology, 263
— — organs of, diagram showing, 263
— — synonyms, 262
— morphology, 262
Heterophyiidæ, 262
— morphology, 232
Heterotricha, 29, 200
Hexactinomyxon psammoryctis, spore of, 187
Hexamastix ardin-delteili, 624, 735
Hexapoda, classification, 431
Higueron, milk of, in ancylostomiasis, 754
Hilton, J., observation of encapsuled Trichinellæ, 423
Himasthlinæ, 269
— morphology, 233
Hindle, avian pathogenic spirochætes, 119
— experimental infection with Spirochæta duttoni, 117, 118
Hippius and Lewinson, relationship of Oxyuridæ to appendicitis, 698
Hippobosca camelina, 611
— capensis, 611
— equina, 611
— maculata, 611
— — bite of, 611
— rufipes, 98
— wings of various species, 611
Hippoboscidæ, 611
Hirudinea, 480
— alimentary canal of, 480
— anatomy of, 480
— body cavity of, 480
— cocoons of, 481
— excretory or segmental organs of, 481
— muscular system of, 480
— nervous system of, 481
— œsophagus of, 480
— pharynx of, 480
— sexual organs of, 481
Hirudo granulosa, used medicinally, 482
— medicinalis, geographical distribution, 481
— — habitat, 481
— — morphology, 481
— morphology, 481
— mysomelas, used medicinally, 482
— troctina, characters, 482
— — geographical distribution, 482
Histiogaster, characters, 515
— (entomophagus ?) spermaticus, 515
— — — habitat, 516
— — — synonyms, 516
— food of, 515
Histoplasma, 112
— association with splenomegaly, 112
— capsulatum, 112, 739
Hoffmann, formation of spores in Treponema pallidum, 125
Holostomata, ova of, development, 224
Holostomum variabile, hosts of, 6
Holothyrus coccinella, effects of bite of, 493
Holotricha, 29, 199
Homalomyia canicularis, larvæ of, 584, 585
— — — characters and habitat, 584
— scalaris, 585
Homoptera (footnote), 532
Hoplopsyllus anomalus, carrier of plague bacillus, 543, 547
— distinctive characters, 545, 547
Horse, piroplasmosis in, cause of, 174
— serum in cultivation of Treponema pallidum, 126
— sickness (Gambia), cause of, 100
Horses attacked by Leptus autumnalis, 486
— nagana fatal to, 94
— organs infected with echinococcus, percentage of frequency, 347
— “surra” in, 95
— trypanosomes in blood of, diseases associated with, 67, 68
Horwood, polypoid tumour of cervix uteri with Schistosoma infection, 643
Host, influence of parasites on, 8
House-fly, diseases spread by, 586
— see also Musca domestica
Howard, larvæ of Piophila casei, 583
— Melanolestes morio, 540
— and Clark, bug carrier of virus of poliomyelitis, 536
Howardia, characters, 567, 568
Howardina, characters, 564
Huber, chemically toxic effects of Ascaridæ, 650
— limitations in male fern administration, 671
— sites of cysticercus in body, 664
Hulecoetomyia, characters, 564
Human parasites, medical works on, 617
Humble-bee, nests of, larvæ of Homalomyia canicularis found in, 584
Hungary, Simulium columbaschensis plague in, 578
Hünsche, infection with Demodex folliculorum, 708
Hyalomma ægyptium, characters and morphology, 501, 502
— — Crithidia parasitic in, 104
Hyalomma ægyptium, farm stock sufferers from, in South Africa, 502
— characters of, 497
Hydatid, see Echinococcus
— disease, eosinophilia in, 652
— intoxication, 353
— sand, 350
Hydrocele in filariasis, 401
Hydrophobia, cell inclusions in, 207, 208
Hydrotæa meteorica, characters, 611
— — larvæ of, habitat, 585
Hymenolepididæ, 309, 323
Hymenolepis diminuta, 324, 662
— — hosts of, 326
— — — intermediate, 327, 328
— — larva of, morphology, 328
— — morphology, 326
— — occurrence in man, cases reported, 326
— — synonyms, 326
— lanceolata, 662
— — hosts of, 329
— — larva, host of, 329
— — morphology, 328
— — synonyms, 328
— morphology, 323
— murina, larval stage, development into tapeworm, 305
— nana, development, 324
— — diagnosis of presence in body, 661
— — expulsion of, drugs used for, 661, 662
— — geographical distribution, 324
— — habitat in body, 661
— — infection with, symptoms following, 324
— — morphology, 323
— — occurrence in man, 326
— — prevalence in children, 661
— — question of identity with H. murina, 324, 325
— — symptoms following infection by, 661
— — synonyms, 323
— (Tænia) murina, development without intermediate host, 17
— — — larval stages occurring in rat flea, 17
— — — omission of intermediate host by, 326
Hymenoptera, characters, 531, 532
Hypochondriac region, right, liver-fluke in, 244
Hypoderma bovis (cattle or warble fly), 595
— — — — — invading human integument, 595, 596
— — — — — larvæ of, in nose, 724
— — — — — — invading upper eyelid, 596
— — — — — — migration in body of cattle, 595
— diana, 596
— lineata, 596
— — geographical distribution, 596
Hypotricha, 29, 200
Hystrichopsylla, 548
— distinctive characters, 545
I.
Ichneumon, Indian, host of Paragonimus compactus, 251
Ichthyol in chyluria from Filaria bancrofti infection, 677
— paste, application in creeping disease, 732
Ichthyophthirius multifiliis, 206
— — morphology and life-history, 207
Ichthyosporidium, 195
Ijima, experimental infection of man with Dibothriocephalus latus, 312
— on Amœba miurai, 46
— Tristrongylus instabilis in man, 435
India, nasal myiasis in, 588
— North-West Provinces, percentage of pariah dogs in, affected with Paropisthorchis caninus, 257
Indian Plague Committee, proof of infection with Xenopsylla cheopis, 547
Infusoria, 198
— characters and habitat, 29, 198, 199
— classification, 199
— digestive processes, 26
— encystment of, 199
— hosts of, 199
— macronucleus and micronucleus of, 198
— mode of life, 199
— morphology of, 198
— reproduction of, 198
Ingram, Rhinosporidium cysts, 197
Inguinal tumour, ascarides escaping from, 656
Inouye, lung fluke disease, 639
Insecta, abdomen of, 529
— anatomy, 529
— blood of, colourless, 530
— development of, 530
— epidermis of, 530
— faceted eyes of, 530
— head of, 529
— intestinal canal of, 530
— metamorphosis of, 530
— nervous system of, central, 530
— — — intestinal, 530
— orders of, 531
— organs of respiration, 530
— — of touch, smell and hearing, 530
— pharyngeal ganglia of, 530
— sexual organs of, 530
— sexually distinct, 530
— thorax of, 529
Insect flagellates, 102, 104, 737
— — experimentally introduced into vertebrates, 104, 112, 737, 738, 739
Intestinal canal of Insecta, 530
— — of nematodes, 363
— obstruction, Ascaris in appendix causing, 654
— — set up by ascarides, 657
— — tract, habitat of Oxyuris vermicularis, 467
Intestine, blood-vessels of, penetration by amœbæ, 36
— coccidiosis of, in man, cases, 148
— Dipylidium caninum parasitic in, 660
— human, Entamœba coli parasite of, 618
— — invasion by Metastrongylus apri, 433
— — larvæ of Homalomyia found in, 584
— — Myriapoda parasitic in, 483
— large, cystic stage of Lamblia intestinalis found in, 59
— — Entamœba histolytica present in, 38
— — high injections into, in evacuation of Oxyuridæ, 698
— — human, Trichuris trichiura parasitic in, 678
— — irrigation in gangrenous dysentery, 619
— larvæ of Gastrophilus inhabiting, 599
— migration of oncospheres from, to liver, 302
— — of Oxyuris vermicularis from, lesions and irritative symptoms set up by, 694, 695
— number of females of Ancylostoma duodenale present in, mode of reckoning (footnote), 454
— occlusion of, due to massive accumulation of ascarides, 657
— parasites of, in relation to appendicitis, views of authors regarding, 652, 653, 654, 655
— — invading vermiform appendix, authors recording cases of, 652
— pathological changes in, due to ova of Schistosoma japonicum, 281
— perforation by Ascaris, 655, 656
— — — following diseased processes, 656
— small, Ascaris lumbricoides parasitic in, 687
— — Dibothriocephalus latus parasitic in, 658
— — human, inhabited by Tænia solium, 662
— — — Tænia saginata parasitic in, 667
— — inhabited by Hymenolepis nana, 661
— — migration of Ascaris lumbricoides from, to other parts of body, 464
— — normal habitat of Ascaris lumbricoides, 464
— — — situation of Trichomonas intestinalis, 55
— — possible invasion by Balantidium minutum, 204
— stenosis of, following infection by Tænia solium, 662
— Strongyloides stercoralis in, 755
— ulceration of, associated with Balantidium coli, 202
— Vorticellæ in, 206
— see also Myiasis, intestinal
Intra vitam staining of fresh preparations of Protozoa, 746
Inundation disease, see Kedani
Iodide, tincture of, applications in creeping disease, 731
Iodine enemata in flagellate dysentery, 625
— tincture of, in treatment of cutaneous and muscular cysticerci, 663
Iodoform with bicarbonate of soda, administration of, in expulsion of ascarides, 694
Ipecacuanha in amœbic dysentery, 619
— de-emetinized, in balantidian dysentery, 637
Iridocyclitis in trypanosomiasis, 623
Iron-hæmatoxylin stain, Heidenhain’s, 752
Isaac and van Velden, dissolution of parasitic products in serum of patients with bothriocephalus anæmia, 645
Isospora, 149
— bigemina, hosts of, 149
— — morphology, 149
— — occurrence in man, 149
— — synonyms, 149
Israelites, “fiery serpents” molesting, probable nature of, 386
Itch mites, see Sarcoptidæ
Itching resulting from clothes louse infection, 711
— set up by Tetranychus molestissimus, 488
Ivers, infection with Demodex folliculorum, 708
Ixodæ, characters of, 496, 497
Ixodes, characters of, 497
— hexagonus, characters and morphology, 500
— — hosts of, 500
— — synonyms, 500
— holocyclus, characters, 499
— — symptoms resulting from attacks of, 499
— plumbeus (dog tick), length of life apart from host, 495
— reduvius, act of coitus in, 495
— — disinfection against, 704
— — infection by, symptoms, 704
— — larvæ of, 495
— — life-history of, 494
— — method of oviposition, 494
— — see also Ixodes ricinus
— ricinus (dog tick), bite of, effects, 498
— — — prophylaxis against, 498
— — characters and morphology, 497, 498
— — confusion in nomenclature, 499
— — geographical distribution, 499
— — habitat and hosts of, 498, 499
— — synonyms, 499
— — transmission of Babesia bovis by, 177
Ixodidæ (ticks), carriers of various diseases to animals and man, 493
— — characters of, 493
— — classification of, 496
— — genera of, synonyms, 497
— — — synopsis of, 497
Ixodinæ, characters of, 496, 497
— transmission of Hæmosporidia by, 704
— and Argantinæ, distinguishing features between, 505
J.
Jackal, Dipylidium caninum parasitic in, 322
Jaksch, von, causation of ancylostoma anæmia, 647
Jamaica, Amblyomma cayennense pest in, 501
— Margaropus annulatus australis pest to man in, 505
James, genera of Anophelines (footnotes),562, 563
Janowski on the Trichomonads, 55
— presence of Cercomonads in intestine, 62
Janthinosoma, characters, 563, 571
Japan, Central, percentage of population infected with Clonorchis endemicus, 260
— illness set up by kedani mite in, 487
Japanese river or inundation disease, see Kedani
Jaundice, malignant, in dogs, carrier of, 493
— — — cause of, 177
Jaw, upper, nematode larvæ in periosteum of, associated with gingivitis, 378
Jejunum, flagellate stage of Lamblia intestinalis found in, 59
— human, habitat of Ancylostoma duodenale, 450
Jerboa, hæmogregarine in, 154
— inoculation of Trypanosoma lewisi into, 90
Jews, infection with Tænia saginata, 340
— inoculation against Oriental sore, 108
Jigger, see Dermatophilus penetrans
Joblotina, characters, 565
Johannseniella, 579, 580
Johns, cultivation of malarial parasites, 170
Jungklauss’s preparation as vermifuge, 673
Jürgens, case of amœbæ in urine, 46
— intestinal amœbæ, 36, 37
K.
Kabyles, tamné or thimni of, 598
Kahane, earth-eating in connection with Trichuris trichiura infection, 679
— Trichocephali in appendix, 655
— trichocephalus anæmia, 651
Kala-azar, canine, similarity to human infantile form, 103
— — transmission by dog fleas, 103
— dissemination of bug possibly connected with, 107, 536
— Indian, agents of transmission, 107
— — geographical distribution, 105
— — incubation period, 626
— — mortality great, 626
— — parasite of, 105, 626
— — preventive measures, 626, 627
— — symptoms and course, 626
— — treatment, 626
— infantile, 109, 627
— — ætiology, 111, 627
— — geographical distribution, 109
— — in adolescents and adults, 109
— — natural infection of dogs with, 110
— — preventive measures, 627
— — relation to Indian, 109
— — — to Oriental sore shown experimentally, 109
— — symptoms, 627
— — transmission by fleas, experiments to prove, 111
— — treatment, 627
— possible transmission by bed bug, 107, 713
Kaldrovils, cysticercus of eye mistaken for foreign body, 664
Kamala as vermifuge, 673
— in evacuation of Oxyuridæ, 697
Kaposi’s naphthol ointment, application in scabies, 707
Kartulis, case of amœbæ in urine, 46
— cerebral abscesses in amœbiasis, 35
— discovery of amœbæ in stools of dysentery patients, 30
— Entamœba kartulisi, 44
— experiments proving connection of amœbæ with dysentery, 30
— Sarcosporidia in man, 194
Karyolysus, 154
Kautsky, bilharziasis, 641
Kayser, eye affections due to Lucilia macellaria, 721
Kedani (Japanese river or inundation disease), 487, 703
— — — — — prophylaxis against, 703
— — — — — symptoms, 703
— mite, 487
— — characters of, 487
Kelly, bilharziasis of appendix, 642
Kent, Herpetomonas and Leptomonas, 102
Kerosene, destruction of Tabanidæ by, 601
Kerteszia, characters, 562, 569
Kholodkowsky, post-mortem discovery of Opisthorchis felineus, 253
Kidney and liver cells, yellow pigment in, in ancylostomiasis, 647
Kinghorn, transmission of Trypanosoma rhodesiense, 69
— and Yorke, tsetse-fly transmitting Trypanosoma rhodesiense, 608
— — transmission of Trypanosoma rhodesiense, 81
Kirmisson, trichocephalus infection in relation to appendicitis, 653
Klebs, early researches on malaria, 156
Klencke, early mention and depiction of malarial parasites, 157
Kloss, researches on Coccidia, 136
Klossia, 141
Knackers’ yards, infection of rats with Trichinella in, 427
Knoch, J., views on development of cestodes, 16
Kobayashi, second intermediate host for Clonorchis endemicus, 261
Koch, R., artificial infection of species of Glossina with human trypanosome, 605
— — discovery of amœbæ in stools of dysentery patients, 30
— — investigations of Proteosoma and Halteridium in birds, 158
— — relapses and latent infection of malaria, 158
— — researches on malaria, 158
— — — on Spirochæta duttoni, 117
Koch’s blue bodies in Theileria parva, 179
Kölliker, investigation of gregarines, 129
Koneff, favourable effects of expulsion of Ascaridæ, 650
Kousso flowers as vermifuge, 673
— in evacuation of Oxyuridæ, 697
Kraft, filmaron as vermifuge, 672
Kruse and Pasquale, nomenclature of amœbæ, 31
Küchenmeister, F., experimental rearing of tapeworms, 15
— — experiments as to metamorphosis of tapeworms, 15
— — expulsion of Ascaridæ, 693
— — nature of cysticerci, 282
Kuhnt, infection of human eye with filaria, 406
Kummerfeld’s wash for clothes lice, 616
Kunstler, genus Giardia, 736
Kurlow, blood-stained diarrhœa from Strongyloides stercoralis infection, 675
Kütner, favourable effect of expulsion of Ascaridæ, 649
— treatment of bilharziasis, 643
L.
Labadie-Lagrave and Deguy, invasion of lymphatic vessels by Onchocerca volvulus, 418
Labbé, copulation in Coccidia, 137
Lacompte, nematodes in human eye, 412
Lælaps echidninus, Leishman granules in, 493
— stabularis, 493
Lafleur, see Councilman and Lafleur
Lagocheilascaris, characters, 466
— minor, host of, 467
— — lesions set up by, 467
— — morphology, 467
Lakes, mosquitoes depositing ova in, 553
Lama, possible carrier of leprosy, 613
Lambkin’s mercury cream in treatment of syphilis prevailing in Uganda, 632
Lambl, discovery of human intestinal amœbæ, 29
Lamblia intestinalis, 57, 625, 736
— — association with diarrhœa, 59, 60, 625
— — — — treatment, 625
— — characters, 57
— — flagella of, 57, 58
— — hosts of, 59
— — infection with, 60
— — nuclear apparatus, 58
— — site in intestine of flagellate and cystic stages, 59
— — synonyms, 57, 736
Lankester, liver-fluke in abscess of ear, 244
— Sarcocystis, 193
Lankesterella, 154
Larva migrans, 599
Larvæ, dipterous, in conjunctiva, 716
— — in nasal accessory sinuses, 717
— — in nose in enormous numbers, 716, 717
— in wounds, movement of, 723
— see also under Names of Parasites
Larvicides, use in campaign against mosquitoes, 636
Larynx, ascarides invading, 691
— leeches in, 699, 700
La Spada, echinococcus of liver rupturing into abdominal cavity, 652
Lasioconops, characters, 564
Lassar’s paste, application in creeping disease, 732
Laurer’s canal of trematodes, 221, 222
Laveran, A., classification of trypanosomes, 71
— — cross-immunity experiments with Trypanosoma rhodesiense and T. brucei, 80, 94
— — — — with trypanosomes, 80
— — discovery of true malarial parasites by, 157
— — latent forms of trypanosomes, 74
— — on Leucocytozoa, 153, 742
— — Trypanosoma pecaudi, 95
— — and Franchini, inoculation experiments with Crithidia fasciculata, 104
— — — — — with Herpetomonas ctenocephali, 103
— — — — — with H. pattoni, 103
— — and Mesnil, isolation of sarcocystin, 191
— — — on the spore of Sarcocystis tenella, 193
— — — “Trypanosomes et Trypanosomiases,” 617
— — and Thiroux, treatment of sleeping sickness, 623
Laverania, characters, 164, 569
— malariæ (Plasmodium falciparum), crescents of, 162, 167, 168
— — — — — sites of development, 169
— — — — cultivation of, clumping in, 172
— — — — cultures of, number of spores produced, 172
— — — — development, duration of, 167
— — — — distinctive characters of, 169
— — — — invasion of spleen by, 168
— — — — merozoites, number of, 168
— — — — number in one red blood corpuscle, 167
— — — — oöcysts of, in stomach of Anopheles, 163
— — — — oökinete of, in stomach of Anopheles maculipennis, 162
— — — — parasite of malignant tertian or sub-tertian fever, 167
— — — — — — — — and quotidian malaria, 167, 633
— — — — pathological effects, 634
— — — — question of varieties or subspecies, 167
— — — — “signet-ring” stage, 167
— — — — sporozoites, 169
— — — — stages of development in intestine of Anopheles maculipennis, 162
— — — — synonyms, 167
— — — — trophozoites of, 167, 168
Lee, R. J., creeping disease, 729
Leeches in upper air passages, 699, 700
— — — — cases reported by various authors, 699, 700, 701
— — — — mention among ancient writers, 699, 700
— invading body, means of riddance, 701
— see also Hirudinea, Rhyncobdellidæ
Leeuwenhoek, opposition to theory of spontaneous generation, 10
Léger, L., classification of Coccidiidea, 141, 142
— genus Crithidia, 104
— researches on Coccidia, 137
Léger, M., proportion of population in Tonkin infected with Clonorchis endemicus, 260
Léger, M. and A., proposed classification of Leucocytozoa, 153
Leichtenstern, bothriocephalus anæmia, 646
— toxic symptoms following thymol administration, 686
Leidy, genus Endamœba, 31, 34, 734
Leignathus sylviarum, 493
Leiper, R. T., Gastrodiscoides, 236
— — host of Filaria loa, 601
— — identity of Œsophagostomum brumpti with Œs. apiostomum, 444
— — report of Bilharzia Mission under, 277
Leipzig, frequency of infection of various organs of animals with echinococcus slaughtered at, 347
Leisering, percentage of rats infected with Trichinella, 427
Leishman, Sir W. B., experimental researches on infection with Spirochæta duttoni, 117, 118
— — — on parasite of Indian kala-azar, 105
— — — treatment of Indian kala-azar, 626
Leishman-Donovan body, see Leishmania donovani
Leishman granules in Lælaps echidninus, 493
Leishmania, 67, 104
— donovani, 105
— — cause of Indian kala-azar, 105, 626
— — cultivation methods, 106
— — inoculation experiments with, 107
— — localization of infection, 105
— — morphology, 105, 106
— — possible mode of transmission, 107
— evolution from flagellates of invertebrates, 739
— infantum, cause of infantile kala-azar, 105, 109, 627
— — cultivation methods, 109
— — immunity to, 112
— — in dogs, 110
— — inoculation, 110
— — — animals suitable for, 110
— — probable transmitter, 111
— probable origin of, 103, 739
— tropica, 105, 107
— — cause of Oriental sore, 105, 107, 627
— — cultivation methods, 108
— — hosts of, 108
— — inoculation, experimental, 108
— — possible transmitters, 108
— — synonyms, 107
Leishmaniasis, cutaneous, 107
— dermo-mucosal, supposed mode of transmission in Paraguay, 739
— evolution of, relation of experimental introduction of insect flagellates into vertebrates on, 737, 738, 739
— experimental production in white mice, 103
— geographical distribution, 105, 107, 109
— infantile, see Kala-azar, infantile
— naso-oral, see Espundia
— possible reservoirs, 738, 739
— treatment, 626–629
Leishman’s stain, 750
Lemaire, herpetomonad flagellate in cultures of blood and organs of gecko, 739
Lenhartz, bothriocephalus anæmia, 646
Lentospora cerebralis, 184
Lepidoptera, characters, 531, 532
Leprosy, possible carrier of, 579, 613
Leptidæ, 603
— blood-sucking species, 603
— characters, 603
Leptis scolopacea, 603
— strigosa, 603
Leptodera, life-history of, 19
— appendiculata, occasional parasite, 7
— pellio, facultative parasitism of, 8
Leptomonas, 102
— bütschlii, 102
Leptotheca, 184
Leptus autumnalis (grass, harvest or gooseberry mite), animals attacked by, 486
— — skin irritation set up by, 702
— — habitat of, 485
— — hosts of, 485, 486
— — nut and fruit pickers affected by, 485
— — skin affection set up by, 485, 486
— — so-called proboscis of, 485, 486
— geographical distribution of species, 486
— undescribed species of, 486
Lesbini, dipterous larvæ in nose in enormous numbers, 717
Letulle, pathological changes in rectum due to Schistosoma hæmatobium, 274
Leuckart, R., advances in helminthology due to, 15, 16
— — attempt at self-infection with Ascaris lumbricoides, 464, 465
— — change of host in parasites, 20, 21
— — classes of parasites, 1
— — development of Acanthocephala and Linguatulida, 17
— — — of alveolar echinococcus, 357, 358
— — — of nematodes, 17
— — — of Trichinella spiralis, 423
— — distinction between Cercomonas and Trichomonas, 54
— — experimental self-infection with Oxyuris vermicularis, 469
— — facultative parasitism, 7
— — feeding experiments with Tænia saginata, 340
— — — — with Trichinellæ, 423
— — growth of echinococcus, 354
— — heterogony in Strongyloides stercoralis, 381
— — method of infection with Trichuris ovis, 420
— — migration of oncospheres, 302
— — name of Coccidia first given by, 135
— — Trichocephalus in association with cholera, 658
— — and Thomas, P., life-history of liver-fluke, 241
Leucocytogregarina, 154
— canis, life-cycle diagram, 155
— — transmission from dog to dog by tick, 155
Leucocytogregarines, 154
Leucocytosis in bilharziasis, 642
Leucocytozoa, action of, on red blood cells, 153, 742
— classification proposed, 153
— hosts of, 153
— morphology of, 153
— schizogony of, 153, 742
Leucocytozoon type of Hæmosporidia, 152
— lovati, schizogony in, 153
— ziemanni, schizogony in, 153
Leucomaines, effects on living organisms, 9
Levaditi, cultivation of spinal ganglia of rabid monkeys, 210
Lewandowsky, infection with Demodex folliculorum, 708
Lewin, expulsion of Ascarides, 693
Lewis, finding of intestinal amœbæ, 29
— studies of filariasis, 391
Leydenia, 49
— gemmipara, 49
— — in ascites, 49
— — association with possible ascites and malignant growth in abdomen, 49, 50
— — characters of, 49
— — cytoplasm containing blood corpuscles, 50
— — pseudopodia of, joining several individuals, 49
Leydig, psorosperms, 181
Lice, Herpetomonads in gut of, 103
— transmission of relapsing fever by, 120
— wingless, owing to parasitic life, 3
— see also Pediculidæ
Lieberkühn, investigations of Coccidia, 135
— — of gregarines, 130
— psorosperms, 181
Liesen, Ascaris in peritoneal cavity, 656
Ligula, excretory apparatus, collecting tubes, island formation, 292
— plerocercoid of, 300
Limatus, characters, 565
Limnæus, species other than L. truncatulus intermediate hosts of Fasciola hepatica, 242
— truncatulus, amount of ova deposited by, 242
— — geographical distribution, 241
— — intermediate host of Fasciola hepatica (?), 240, 241
— — — hosts of liver-fluke, 240, 241
Limnatis, characters, 482
— nilotica, characters, 482
— — geographical distribution, 482
— — habitat, 482
— — only leech of clinical importance as parasite, 699, 701
— — synonyms, 482
Lindblad, Dipylidium caninum, 660
Lindner, G., peritrichal Infusoria (stalkless Vorticella), 206
Lindsay, possible mode of transmission of dermo-mucosal leishmaniasis to man in Paraguay, 739
Linguatula, 523, 524
— rhinaria, characters and morphology, 524
— — development and life-history of, 524, 525, 526
— — larvæ of, 524, 525
— — occurrence at autopsies, 526
— — organs of body invaded by, 524, 525, 526
— — ova of, 524, 525
— — parasitic in nasal cavity of animals and man, 523, 524
— — synonyms, 524
— serrata, hosts of, 527
— — synonyms (footnote), 527
Linguatulida, development of, 17
Linguatulidæ, blood-sucking, 523
— change of original features in, 4
— characters and morphology, 523
— hosts of, 523
— larvæ of, 523
— nature of, 2
— relation to Arachnoidea, 19
— separation from Helminthes, 2
Lini, escape of Ascarides from umbilicus, 656
Linnæus, discoveries as to origin of Helminthes, 10, 11
— so-called dysentery infection due to mites, 512
Lipari, cysticerci of brain, 664
Lipuria in bilharziasis, 641
Lithocystis, endoplasm of, contents, 131
Liver, abscess of, association of Entamœba histolytica with, 35
— — — of Noc’s entamœba with, 41
— — caused by invasion of Ascaridæ, 690
— — due to amœbic dysentery, treatment, 620
— — set up by amœbæ, 35
— and bile-ducts, habitat of Clonorchis endemicus, 259, 260
— and kidney cells, yellow pigment in, in ancylostomiasis, 647
— and portal vein, Schistosoma hæmatobium most easily found post mortem in, 273
— coccidiosis of, in man, cases, 148
— encystment of Porocephalus constrictus in, 526, 527
— female Ascarides depositing ova in, 689
— human, eggs of Schistosoma japonicum, showing “spines” and “hoods” at opposite pole, 279
— invasion by larvæ of Linguatula rhinaria, 525, 526
— migration of oncospheres from intestine to, 302
— pathological changes associated with invasion by Opisthorchis felineus, 253
— — — in, due to ova of Schistosoma japonicum, 281
— — — set up by Clonorchis endemicus, 260
Liver-fluke, supposed origin of, 10
— see also Fasciola hepatica
— disease, diagnosis, 242
— — in man, 242
— — in sheep, 238
— — — ravages caused by, 238, 239
— — — stages of, 240, 241
— — pathological anatomy, 241
— — symptoms, 239
Liverpool School of Tropical Medicine, expedition to investigate trypanosome infections, 68
Lizards, hæmogregarines from, 154
Loa, morphology, 409, 411
— loa, duration of life of, 414
— — early historical accounts of, 412
— — geographical distribution, 414
— — larvæ of, in blood, 412, 414
— — — periodicity, 413
— — — structure, 412
— — lesions produced through invasion by, 413, 414
— — life-history, 414
— — morphology, 409, 411
— — ova of, 410
— — sites of body invaded by, 412, 678
— — synonyms, 409
Lobaczewski, prophylaxis against body, head and clothes lice, 615
Löbker, cause of ancylostome anæmia, 648
Locusts injurious to man, 542
Looss, infection by Ancylostoma duodenale through skin, 683
— origin of lateral-spined eggs of Schistosoma hæmatobium, 273
— prevalence of Heterophyes heterophyes, 264
— skin affections set up by invasion of larvæ of Ancylostoma duodenale, 455
— symptoms of lymphangitis from Filaria bancrofti infection, 676
— toxic action of ancylostomes, 647
— Trichostrongylus instabilis in man, 435
Lophius piscatorius, 186
Lophoscelomyia, characters, 562, 568
Lösch, discovery of intestinal amœbæ in case of dysentery, 29, 30, 32
Löschia, 34
Lota vulgaris, see Burbot
Lounsbury, life-cycle of Amblyomma hebræum, 495
Louse disease, historical instances of death from, 711
Low, personal experiments with regard to malaria infection, 158
— treatment of Oriental sore, 628
Lucilia argyrocephala cause of myiasis in French West Africa, 614
— cæsar, 588
— macellaria, larvæ of, causing eye diseases, 721
— — — in nose, 715, 716
— — — — see also Myiasis, nasal
— — — on cutaneous surface, 721, 722
— — — penetrating auditory meatus, 721
— nobilis, larvæ (maggots) of, discharge from auditory meatus, 588
— sericata, 588
Lumbricosis, typhoid, 650
Lumbricus, Monocystis agilis from seminal vessels of, 130, 132
— teres, see Ascaris lumbricoides, 464
Lund’s larva, characters, 593
Lung, abscess of, set up by amœbæ, 35
— amœbæ found in, 45
— Balantidium coli occurring in, 202
— bilharziasis of, 642, 643
— gangrene of, possible occurrence of Cercomonads in, 62
— invasion by Fasciola gigantica, 245
— — by Paragonimus ringeri, 251
— — by Schistosoma hæmatobium, 274
— Trichomonads found in, 56
Lung-fluke disease, geographical distribution, 639
— — prognosis, 640
— — symptoms, 639
— — treatment, 640
Lussana, toxic theory of ancylostome anæmia, 646
Lütz, ascarides in pulmonary artery, 656
— Ceratopogoninæ described by, 580
— experimental infection with Ascaris lumbricoides, 465
— favourable effects of expulsion of Ascaridæ, 649
— perforative peritonitis due to Ascaris, 656
Lyctocoris campestris, bite of, 541
— — characters, 541
Lygæidæ, characters, 541
Lymphangitis from Filaria bancrofti infection, symptoms, 676
— in filariasis, 401
Lymphatic glands, enlarged, in filariasis, 402
— vessels and glands, destruction without lymphatic obstruction, 401
— — — distribution and connections of, 400, 401
— vessels, invasion by Onchocerca volvulus, 418, 419
Lymphatics, Strongyloides stercoralis in, 755
Lynch, human trichomoniasis, 734
Lynchia, transmitting Halteridium, 151
Lyperosia, differentiation from Stomoxys, 610
— exigua, life-history, 610
— irritans, var. weisii, 610
M.
Macacus sinicus, inoculation with Leishmania donovani, 107
MacCallum, “exflagellation,” 152
— investigations of Proteosoma and Halteridium in birds, 158
McDonagh, J. E. R., life-cycle of organism of syphilis, 124
MacFadyean and Stockman, Babesia divergens, 177
Macfie, Trypanosoma nigeriense, 76
— and Gallagher, treatment of sleeping sickness, 622
Mackenzie, periodicity of larvæ of Filaria bancrofti in peripheral blood, 393
Mackie, suggested transmission of relapsing fever by lice, 120
— treatment of Indian kala-azar, 628
Macleayia, characters, 563
MacNeal, see Novy and MacNeal
Macrostoma mesnili, 57, 735
Maculæ cærulæ (taches bleues) due to infection by crab louse, 712
Maggots, see under Names of Parasites and Regions of Body
— in nose, see Myiasis, nasal
Magnesium sulphate in flagellate dysentery, 625
Maillard, fatal cases of nasal myiasis, 718
Majochi, case of intertrigo set up by Oxyuris vermicularis, 696
— infection with Demodex folliculorum, 708
Mal de caderas in horses, trypanosomes associated with, 68
— — trypanosome causing, 96
Malaria, acute, 156
— atypical forms, 634
— campaign against, commencement and progress of, 158
— chronic, 156
— development of parasites of, 159
— diagnosis (pathognomonic signs), 635
— geographical distribution, 155
— historical, 157
— in birds spread by Culex, 158
— in man, 155
— latent, in children of natives, 158
— masked, 156
— parasites of, 164–170, 633
— — asexual generation, cultivation in vitro, 170
— — copulation, 160, 161, 162
— — exflagellation (footnote), 162
— — gametocytes of, 160, 161, 162
— — human, development, 159
— — — — occurs only in Anopheles, 158, 159
— — — differential characters, 171
— — — species of, 164, 633
— — — — see also Laverania malariæ, Plasmodium malariæ, Plasmodium relictum, Plasmodium vivax
— — macrogametes of, 160, 161, 162
— — merozoites of, 159, 160
— — methods of detecting, 747
— — microgametes of, 160, 161, 162
— — movements, discovery of, 157
— — not transmissible to mammals, 159
— — oökinetes, 160, 161, 163
— — schizogony of, 161, 172
— — sporozoites of, 159, 160, 164
— — — penetration of red blood corpuscles by, 159, 160
— sporulation, 160, 161, 163
— pigmentation of organs, 165 (footnote), 634
— prevention of constipation during, 635
— preventive measures against mosquitoes, 635, 636
— — — against parasite in man, 635, 636
— — — by quinine administration, 636
— prophylaxis, 636, 637
— relief of symptoms, 635
— symptoms, 156, 633
— synonyms, 155, 633
— tertian, malignant, paroxysms of, 634
— treatment by quinine, 635
Malarial fever, quartan, 156
— — — duplex or triplex, appearance of, 167
— — — parasite of, 166
— — malignant or sub-tertian, parasite of, 167
— — — pernicious symptoms, explanation, 172
— — quotidian, 156
— — rhythmical, course of, 155
— — symptoms, 155, 633
— — tertian, 156
— — — simple or spring, parasite of, 164
— — typical, clinical features, 633, 634
Male fern, administration to children, 671, 672
— — emulsion, injection of, 671
— — ethereal extract best vermifuge for Tænia saginata, 670
— — — — dosage and method of administration, 670, 671
— — extract of, expulsion of Hymenolepis nana by, 661
— — — in bilharziasis, 643
— — — in expulsion of ancylostomes, 686
— — —in intestinal myiasis, 728
— — poisoning, 670, 671
— — — antidotes to and remedies for, 671
— — — bad effects on vision, 670
Malignant malarial parasites, sporulation, influence of temperature on, 163
— — — — stages of, 163
Mallory’s bodies, 208
Mamma, tumours of, association of Dioctophyme gigas with, 431
Mammals, human malarial parasites not transmissible to, 159
— leucocytogregarines in, 154, 155
— red blood corpuscles of, Babesia parasitic in, 154
Man, incidental parasites of, 7
— infection with animal trypanosome, 96
— parasites found only in, 6
Mange, see Dog mange
Mangold, feeding experiments with Tænia from multilocular echinococcus, 358
Manguinhosia, characters, 562, 568, 569
Manson, Sir Patrick, development of Paragonimus ringeri, 251
— — — discovery of Sparganum mansoni, 317
— — — infection of skin by Filaria perstans, 378
— — — on Spirochæta carteri, 631
— — — pathognomonic signs of malaria, 635
— — — prophylaxis against ancylostomiasis, 685
— — — researches on malaria, 158, 635
— — — studies of filariasis, 391
— — — treatment of Indian kala-azar, 626
— — — — of Oriental sore, 628
Manson, P. T., infected with malaria by infected mosquitoes, 158
Mansonia, 577
Manson’s method of administration of atoxyl in sleeping sickness, 622
Manteufel, immunity of Ornithodorus moubata against infection with Spirochæta duttoni, 119
Marchiafava, discovery of movements in malarial parasites, 157
Marchoux, amœbic abscesses in liver of experimental cats, 35
— Spirochæta gallinarum, 119
— and Couvy, Leishman granules in Lælaps echidninus, 493
Mareo, Helminthiasis meningitiformis, 649
Margaropus annulatus australis, hosts of, 505
— — — pest to man in Jamaica in larval stage, 505
— characters of, 497
— microplus, 505
Marx, male fern administration, 671
— toxic action of male fern, 670
Marzinovsky, prophylaxis against Pediculus vestimenti, 616
Mastigophora, 28, 50, 760
— aggregation rosettes of, 51
— characters and habitat, 28
Mathis, carriers of Entamœba histolytica, 40
— diarrhœa due to Lamblia intestinalis, 625
— Lamblia intestinalis, 59, 60
— modification of Novy-MacNeal medium, 744
Maurer’s dots, 168, 170, 171
Maxillary sinus, Scolopendra in, 721
Mayer’s glychæmalum, 751
— hæmalum, 751
Mbori in dromedaries, 96
Measles, 207
Meat inspection, decrease of cysticerci in pork effected by, 334
Meatus, auditory, larvæ of Anthomyia pluvialis found in, 584
— — Rhinosporidium in, 196
— — maggots of Lucilia nobilis in, 588
— — synonyms, 438
Mecistocirrus, habitat, 438
— morphology, 438
— fordi, morphology, 438, 439
— — — synonyms, 438
Medullary layer of Cestoda, 289
Megarhininæ, characters, 563, 570
Megarhinus, characters, 563, 570
Mégnin, development of cestodes, 16
Mehlis, discovery of progeny of Distoma, Typhlocœlum flavum, and Cathæmasia hians, 12
Mehlis’ gland secretion in trematodes, 223
Melanoconion, distinguishing characters, 564, 576
— atratus, characters, 576
— — geographical distribution, 576
Melanolestes abdominalis, 540
— morio, geographical distribution, 540
— — synonyms, 540
Mello-Leitao, flagellate dysentery in children, 56, 624
Melnikow-Raswedenkow, development of alveolar echinococcus, 357, 358
Melophagus ovinus (sheep ked), bite of, 611
— — — — Crithidia inhabiting, 104
Meningitis, fatal, peenash terminating in, 716
— symptoms of, due to Ascaridæ infection, 649
— terminating nasal myiasis fatally, 718
Mense, expulsion of Guinea worm, 676
Mercier, nematodes in human eye, 412
Mercury, benzoate of, in infantile kala-azar, 627
— cream (Lambkin’s) in syphilis prevailing in Uganda, 632
— in expulsion of Strongyloides stercoralis, 675
Mermis, 469
— hominis oris, 469
Mermithidæ, 469
— characters, 375
Merogregarina, 135
Merogony, 185
Meront, 185
Merozoites of Coccidiidea, 138, 139, 140
— of malarial parasites, 161
Mesenteric vein, superior, tributary of portal vein, 272
Mesnil, on Actinomyxidia, 187
— on Hæmosporidia, 742
— on Haplosporidia, 194
— and Ringenbach, cross-immunity experiments with trypanosomes, 80
— — trypanolytic reactions, 80
— see also Laveran and Mesnil
Messineo, effects of experimental injection of extracts of Tænia, 648
Metagonimus, 264
— Yokogawa yokogawai, 264, 753
— — — geographical distribution, 265
— — — habitat, 265
— — — host and intermediate host, 265
— — — life-history, 265
— — — morphology, 264
Metastrongylinæ, characters, 432
Metastrongylus, morphology, 432
— apri, hosts of, 433
— — in man, cases recorded, 433
— — invasion of air-passages by, 433
— — morphology, 432
— — synonyms, 432
Methyl green, 752
Methylene blue in bilharziasis, 643
— — in flagellate diarrhœa, 625
Metorchiinæ, 261
— morphology, 232
Metorchis, 261
— conjunctus, organs of, diagram showing, 258
— truncatus, habitat and hosts of, 262
— — morphology, 261, 262
— organs of, diagram showing, 262
Metschnikoff, intestinal parasites in relation to appendicitis, 652, 653
— prophylaxis against oxyuriasis, 697
Meyer, disturbances of vision in male fern poisoning, 670
Mibelli, infection with Demodex folliculorum, 708
Mice, experimental infection with herpetomonads, 103, 104, 112, 737, 738, 739
— — — with Sarcocystis muris, 191
— — — with Spirochæta duttoni, 117
— natural herpetomonads in, 738, 739
— occasionally hosts of Hymenolepis diminuta, 326
— Sarcosporidia in, 187
— spherical contracted forms of Trichomonas intestinalis in, 56
Michelson, case of intertrigo set up by Oxyuris vermicularis, 696
Microgametes of Coccidiidea, 137, 139, 140
Microscope, use of, discoveries of parasites from, 10
Microsporidia, 129, 184
— characters and habitat, 28
— morphology of, 185
— various pathogenic members, 186
Midges, see Chironomidæ, Ceratopogoninæ, Psychodidæ
Miescher’s tubes, 187, 188
Mikrofilaria bancrofti, prevalence in blood, prevalence of filarial diseases proportionate to, 400
— — and Mikroloa loa, distinction between, 398
— diurna, larvæ of Loa loa, 412
— — presence in blood, 412, 414
— perstans, morphology, 416
— — and M. diurna, simultaneous presence in blood, 414
— philippinensis, 407
— powelli, 407
Mikrofilariæ, periodic, 393, 394
Milk cure in expulsion of Strongyloides stercoralis, 675
Milton, bilharzial vaginitis, 643
— treatment of bilharziasis, 643
Mimomyia, characters, 565
Minchin on genus Entamœba, 733
— researches on Trypanosoma lewisi, 89–92
— see also Nicoll and Minchin
Minchin and Fantham, on Rhinosporidium kinealyi, 195, 196, 197
Minchin and Woodcock on Trypanosoma noctuæ, 737
Mineral waters in intestinal myiasis, 728
Miners, prophylaxis against ancylostomiasis in, 684
Mines infected with ancylostomes, disinfection of, 685
Miracidia of digenetic trematodes, morphology of, 226, 227
Miracidium, germ cells of, 227
Mitchell, treatment of Oriental sore, 628
Mites attacking man, geographical distribution of species, 486
— case of so-called dysentery said to be due to, 512
— living endoparasitically in animals and birds, 491
— see also Acarina
— see also Arachnoidea
Mochlonyx, 565
Moiriez, species of Chorioptes found on man, 521
Moldovan, schizogony in Leucocytozoon ziemanni, 153
Molluscs, spirochætes in, 114
— fresh-water, round Cairo, cercariæ of bilharzia type in, 277
Molluscum contagiosum, 207, 208
Monas pyophila, 62
— — characters of, 62
Mondière, perforation of appendix by Ascaris, 655
Monera, 26
Moniez, Aleurobius (Tyroglyphus) farinæ, 511
— on derivation of entozoa, 21
Monkeys, dysentery in, associated with presence of Œsophagostomum apiostomum, 444
— experimental infection with Spirochæta duttoni, 117
— inoculation experiments with yaws upon, 128
— rabid, spinal ganglia of, cultivation, 210
— Trypanosoma simiæ virulent to, 100
Monocystis, hosts of, 134, 135
— agilis from seminal vessels of Lumbricus, 130, 132
— life-cycle of, 132, 133
Monogenea, canalis vitello-intestinalis, 222
— ova of, deposition, 223, 224
— post-embryonic development in, 224
Monostomum lentis, 244
Monothalamia (testaceous amœbæ), characters of, 47
Montgomery, transmission of rinderpest, 742
Moore and Breinl, latent bodies of Trypanosoma gambiense, 77
Moosbrugger, earth-eating in connection with Trichuris trichiura infection, 679
— trichocephalus anæmia, 651
Moriggia, Glyciphagus cursor, 513
Morkowitin, Oxyuris infection in relation to appendicitis, 653
Morphia, injection of, in relief of griping and straining in amœbic dysentery, 618, 619
Morsasca, trichocephalus anæmia, 651
Moscato, chyluria following infection by Eustrongylus gigas, 682
Mosquito nets, use of, 636
— worm in Trinidad, 598
— — — how destroyed, 598
Mosquitoes, abdomen, 550
— acting as hosts of Filaria bancrofti, 398
— alimentary canal, 550, 551
— anatomical remarks on, 548
— antennæ of, 548
— aquatic in larval and pupal stages, 555
— breeding places of, 553, 557
— campaign against, in prevention of malaria, 636
— copulation of, 553
— distinguishing features of Chironomidæ (midges) from, 579
— females alone blood-suckers, 552
— — fertilized in autumn, hibernation of, 555
— first development of malarial parasite in, traced in Plasmodium relictum, 170
— labrum, labium, and hypopharynx, 548, 549
— larvæ, food of, 557
— — living in salt water, 557
— — position assumed in water, 557
— length of egg, larval and pupal life, 555
— maxillæ and mandibles, 548, 549
— ova of, 558
— — float on water, 559
— — localities for deposition of, 553
— proboscis of, 548
— pupæ of, 558
— spread of malaria in birds by, 158
— systematic remarks on, 548
— typical structure of, diagram showing, 558
— ubiquitous existence of, 555
— see also Culicidæ
Moth-like appearance of Psychodidæ, 581
Mott, F. W., association of Treponema with general paralysis, 125
Moty, Oxyuris infection in relation to appendicitis, 653
Mouqui, mite attacking man, 486
Mouth, human, cultivation of species of Treponema from, 741
— — spirochætes in, 122, 740
— infection with Oxyuris vermicularis solely through, 469
— maggots in, 721
Mucidus, characters, 563, 571
Mulder, infection with Demodex folliculorum, 708
Mules, murrina in, trypanosome causing, 98
— “surra” in, 95
Müller, D., echinococcus cysts causing urticaria, 651
Müller, J., discovery of Myxosporidia, 181
— O. F., discovery of and views as to cercariæ, 12
— — — of origin of tapeworms by, 11
Müller’s psorosperms, 135
Murrina in mules, trypanosome causing, 98
Musca domestica (common house-fly), 586
— — breeding grounds, destruction of, 586
— — characters, 585, 586
— — destruction of, methods, 586
— — diseases spread by, 586
— — hibernation as puparia, 586
— — larvæ (maggots) of, characters, 586
— — life-cycle of, 586
— — ova of, places where deposited, 586
— — pupa of, 586
— pattoni, 611
Muscidæ, 584
— African, larvæ of, 590
— — — causing myiasis in man (footnote), 590
— blood-sucking, 603
— larvæ of, other than Lucilia, in nose, 720
Muscles, encystment of Trichinella spiralis in, 425
— invasion by Trichinella spiralis, 424, 425
— of nematodes, 361
— sarcosporidia in, 191
Muscular system of Hirudinea, 480
Musgrave, on human intestinal amœbæ, 31
— and Clegg’s culture media for amœbæ, 743
Mussels, fresh-water, spirochætes of, 114
Mutualists, nature of, 6
Myiasis, 715
— auricular, 615
— — treatment, 615
— dermatosa œstrosa, 725
— due to Sarcophaga, 589, 590
— externa, 715
— — methods of treatment recommended by various authors, 719, 720
— — rare situations of, 723
— gastric, treatment, 728
— human, occurring in mountains of Central Sahara, 598
— in French West Africa, cause of, 614
— intestinal, 725, 726
— — chronic, 726
— — — complicated by mucous colitis, 726, 727
— — diagnosis, 728
— — irrigation of rectum in, 728
— — larvæ of different species of flies found in, 728
— — modes of infection, 727
— — — — views of various authors on, 727
— — prognosis, 728
— — prophylaxis, 728
— — symptoms, 726
— — treatment, 728
— — and cutaneous, fly causing, 585
— larvæ of African Muscidæ causing (footnote), 590
— nasal, 715
— — cases of, authors reporting, 716, 717
— — connection with ozæna, 717, 722, 723
— — discharge from nose in, 718
— — due to Sarcophaga, treatment, 723
— — fatal termination of, 718
— — from Sarcophaga wohlfahrti, 722, 723
— — maggots of flies setting up, 588
— — prophylaxis against, 718
— — symptoms, 717, 718
— — treatment, 719
— — see also Peenash
— œstrosa, geographical distribution, 724
— — prevalent among rural population, 724
— — rare in man, 724
— — treatment, 725
Myriapoda parasitic in intestine and nose of man, 483
Myxidiidæ, 184
Myxidium lieberkühni, 182
Myxobolidæ, 184
Myxobolus cyprini, 184
— pfeifferi, 184
— — cause of barbel disease, 184
— — spore formation, 183
— neurobius, 184
— schematic representation of spore of, 182
Myxœdematous form of Brazilian trypanosomiasis, 88
Myxosporidia, 129, 181
— authors adding to knowledge of, 182
— — describing species causing diseases in fishes, 182
— characters and habitat, 28, 182
— free forms of, 182
— introduction of term of, by Bütschli, 181
— invasion by, causing disease in fishes, 182, 184
— mode of infection, 184
— multinucleate trophozoite of, 182
— plasmotomy, 182
— spore formation, 182, 183
— tissue parasites, 182
Myzomyia, characters, 561, 567
— funesta, breeding places of, 557
Myzorhynchella, characters, 561, 568
Myzorhynchus, characters, 562, 568
N.
Nabarro, on sleeping sickness, 68
— on Spirochæta duttoni, 116
Nagana (tsetse-fly disease), agent of transmission, 93
— fatal to horses, asses and dogs, 94
— prevalent among and generally fatal to cattle, 93, 94
— treatment by arsenic, 94
— trypanosomes in blood of horses suffering from, 68
Nagel, chloroform and syrup of senna in expulsion of ancylostomes, 686
— filmaron in expulsion of ancylostomes, 687
Nagelschmidt, treatment of scabies, 706
Naphthalene in evacuation of Oxyuridæ, 697
— in intestinal myiasis, 728
Naphthol ointment, dressings of, in head louse infection, 710
— — (Kaposi’s), application in scabies, 707
Nasal cavity, deposition of ova of Oestrus ovis in, 598
— — see also Myiasis, nasal
— — Linguatula parasitic in, 523, 524, 526, 527
— polypus, Rhinosporidium causing, 195–197
Nason, Ascaris in appendix, causing intestinal obstruction, 654
Nasse, investigations of Coccidia, 135
Natal, larva of, characters, 591
Nattan-Larrier, cross-immunity experiments with trypanosomes, 80
— Tetramitus mesnili, 57, 624
Natural flagellates of insects, 103, 104, 107, 112, 739
Naunyn, mode of formation of daughter cysts of echinococcus, 352
Neave, S., ulcers set up by invasion by larvæ of Cordylobia anthropophaga, 592
Necator, 447
— americanus, 450
— — geographical distribution, 459
— — habitat, 459
— — morphology, 457, 458
— — organs compared with those of Ancylostoma duodenale, 458
— characters, 457
— exilidens, characters, 459
— — habitat, 459
Negri, experimental infection with Sarcocystis muris, 192
— on Neuroryctes, 208
Negri’s bodies, 208, 209
Neligan, Leishmania tropica in dogs, 108
Nemathelminthes, 360
Nematoda, see Nematodes
Nematode larvæ in blood in cases of pruritus, 378
— — in periosteum of upper jaw in case of gingivitis, 378
Nematodes, anatomy of, 360
— bursa copulatrix of males, 370
— chorion enveloping ova, 371
— classification of, 374
— clearing of, 473
— cutaneous glands, unicellular, 361
— cuticle of, 360
— cutis of, 361
— dermo-muscular layer of, 361
— development of, 17, 371
— embryos, 372
— encapsuled forms of, 17
— epithelium of, 360
— excretory canals, anterior, 367
— — organs, 366, 367
— — — special, lacking in certain genera, 367
— — pore and duct, 367
— — vesicle, 367
— fixation of, 473
— glandular stomach of, 363
— gubernaculum of male genital apparatus, 369
— hatched from eggs of Sphærularia, 5
— heterogony in, 372
— hind gut, 363
— infection by, 644
— intestinal cæca, 364
— — canal, 363
— “isolation tissue,” 362, 363, 364
— life spent in intermediate and final host, 18
— marine, ventral gland of (so-called), 367
— mounting head of, 473
— muscles of, 361
— nervous system, 364–366
— observed in human eye, 412
— — in man, 376
— œsophageal glands, 364
— œsophagus of, 363
— organs of sense lacking in parasitic species, 366
— ova of, 371
— — conveyance to definite host with intermediate host, 373
— — — — — without intermediate host, 372
— — detection, 473
— — developmental capacity, 371, 372
— ovejector, 368
— oviduct, 368
— parasitic and free-living, connection, 20
— preservation and examination of, 473
— rolling of, 473
— seminal receptacle, 368
— sexual organs, 367
— — — female, 367, 368
— — — — diagram of, 368
— — — male, 369
— — — — diagram of, 368
— small, detection of, 473
— spicules of male genital apparatus, 369
— staining of, 473
— testis of, 369
— “tuft-like” or “phagocytic” organs, 362
— viviparous species, 371
— young, skin diseases due to, in dogs, 378
Nematodirus, habitat, 438
— morphology, 438
Neocellia, characters, 562, 569
Neomyzomyia, characters, 561, 567
Neopsylla, distinctive characters, 545
Neosalvarsan in syphilis, 632
— in yaws, 632
Neosporidia, 129, 181
— characters, 28, 129, 181
Nephrophages sanguinarius, characters and morphology, 490
— — presence in urine, 490
Nervous system of Cestoda, 289, 290
— — of Echinorhynchus, 475
— — of Hirudinea, 481
— — of Insecta, 530
— — of nematodes, 364–366
— — central, effect of Dipylidium caninum on, 649
Neumann, mosquitoes transmitting Plasmodium relictum, 170
— podophyllin in expulsion of ancylostomes, 687
— synopsis of genus Ornithodorus, 508
— table of species of Argas, 505
Neuritis, optic, following male fern poisoning, 670
Neuroptera, characters, 531
Neuroryctes, 208
— hydrophobiæ, 208
— — minute granules in, 210
Neurosporidium, 195
— cephalodisci, 195
Newstead, Amblyomma cayennense, 501
— life-cycle of Phlebotomus, 582
— Margaropus annulatus australis, 505
— means of separating species of Glossina, 604
Niaibi, mite attacking man, 486
Nicoll, development of cestodes without intermediate host, 17
— and Minchin, cysticercoids in rat fleas, 327, 328
Nicolle, immunity experiments with Leishmania infantum and L. tropica, 112
— and others, transmission of relapsing fever by lice, 120, 121
Nicollia, 174
— quadrigemina, 174
Nicotiana soap, application in scabies, 707
Nits, methods of getting rid of, from hair, 710
Nitzsch, views as to cercariæ, 12
Noc, cultivation of species of amœba by, 41
— on Lamblia intestinalis, 60, 625
Noguchi, cultivation of parasite of rabies, 210
— — — of Treponema from human mouth, 128, 741
— — of Treponema pallidum, 125
— method of cultivation of spirochætes, 123
— Spirochæta phagedenis, 122
— Treponema calligyrum, 126
— and Cohen, cultivation of so-called trachoma bodies, 210
— and Moore, association of Treponema with general paralysis, 125
Nöller, development of Trypanosoma lewisi in dog flea (Ctenocephalus canis), 90, 92
— method of controlling fleas during experiments, 93
Nordmann, von, discovery of miracidia of flukes, 12
Normand, association of amœbæ with colitis, 30
— — of Strongyloides stercoralis with diarrhœa, 380
Norway itch (scabies norvegica), 520
Nose, ascarides in, 690
— dipterous larvæ in, in enormous numbers, 716, 717
— discharge from, in nasal myiasis, 718
— human, Myriapoda parasitic in, 483
— larvæ of Hypoderma bovis in, 724
— — of Lucilia macellaria in, 715, 716
— — — — see also Myiasis, nasal
— — of Oxyuris vermicularis in, 469
— leeches in, 700, 701
— — causing epistaxis, 701
— maggots in, 588
— Oxyuridæ migrating into, 695, 696
Nosema apis, 184
— — life-cycle of, 185
— — pansporoblast and sporoblast of, 185
— — planont of, 185
— bombycis, 184
— — spores of, 186
Notoedres cati, 521
— cuniculi, 521
— notoedres, 521
Novy and MacNeal, artificial cultivation of trypanosomes, 69
Novy-MacNeal medium, 744
— — Mathis’s modification, 744
Novy-MacNeal-Nicolle medium, best for cultivation of Leishmania infantum, 109
— — for cultivation of Leishmania tropica, 108
— — formula, 744
Nut-pickers affected by Leptus autumnalis (footnote), 485
Nuttall, Spirochæta marchouxi, 119
— Piroplasmidæ, 174
Nuttall and Hadwen, trypan-blue in treatment of piroplasmosis, 178
— and others, nuclear phenomena of Babesia canis, 176
— — Theileria parva, 179
Nuttallia, characters, 174
— equi, cause of equine piroplasmosis, 174, 178
— — life-cycle in red blood corpuscles, 173
— herpestidis, 174
Nycteribiidæ, 611
Nyctotherus, 204
Nyctotherus africanus, 206
— faba, 205
— — morphology, 205
— giganteus, 205, 206
— — morphology, 205
Nyssorhynchus, characters, 562, 569
O.
Occiput, abscess of, liver-fluke in, 243
Ochindundu, bite of, 541
— characters of, 541
Ochromyia anthropophaga, larvæ of, characters (footnote), 590, 591
— — — hosts of, 590
Œdema following bite of Argas reflexus, 506
Oerley, induction of facultative parasitism of Rhabditis pellio, 377
Œsophageal glands of nematodes, 364
Œsophagostomeæ, characters, 439
Œsophagostomum, morphology, 441
— apiostomum, habitat and host of, 444
— — morphology, 444
— brumpti, habitat, 441
— — morphology, 441
— stephanostomum, habitat, 444
— — var. thomasi, morphology, 442, 443, 444
Œsophagus of Hirudinea, 480
— — nematodes, 363
— trichomonads in, 55
Oestridæ (warble flies), boils produced by, 725
— cavicolous, 598
— cutaneous, 595
— flight time of, 725
— gastricolous, 599
— — infection by, 729
— — — see also Creeping disease
— hosts of, 594
— larvæ of, occurrence in man rare, 724
— method of depositing ova on skin of man, 725
Oestrus (Cephalomyia) ovis, 598
— — — geographical distribution, 598
— — — ova of, deposition in nasal cavity, 598
Oil, injections of, in nasal myiasis, 719
Ointment, application in scabies, 706
Oken, views as to origin of cercariæ, 12
Oleum chenopodii in ancylostomiasis, 754
Oligosporulea, 195
Oligotricha, 29
Oliver, artificial infection of human beings with Cysticercus bovis, 340
Omentum, abscess of, with Ascaris ova in pus, 657
Omi, diagnostic sign of presence of Sparganum mansoni in body, 659
Onchocerca, 417
— volvulus, 417
— — distribution in West Africa, 419
— — invading lymphatic vessels, 418, 419
— — invasion in man associated with formation of tumours, 418
— — measurements, 755
— — morphology, 417, 418
Onchocercinæ, 417
Oncospheres (embryos) of tapeworms, 298, 299
— — — certain species of animals necessary for, 299
— — — development into plerocercoid, 300
— — — further development must take place in suitable animals, 299
— migration in body, 302
O’Neil, filaria infection of skin, 378
Onions, Anguillulina putrefaciens living in, 379
Oöcysts of Coccidiidea, 141
— of malarial parasites, 163
Opalina, 198, 207
— ranarum, 207
Ophryocystis, 135
Opisthorchiidæ, morphology, 232
Opisthorchiinæ, 252
Opisthorchis, 252
— felineus, development, 254
— — geographical distribution, 252
— — hosts of, 252
— — — intermediate, 254
— — mode of infection by, 254
— — morphology, 252
— — synonyms, 252
— pseudofelineus, anatomy of, diagram illustrating, 254
— sp., habitat, 753
— — morphology, 753
Opisthotonos, disappearance after expulsion of Ascaridæ, 649
Oppenheim, maculæ cærulæ (taches bleues) due to infection by crab louse, 712
— treatment of crab louse infection, 712
Oppilaçao, synonym of Brazilian trypanosomiasis, 87
Oral cavity, cancer of, association of Entamœba buccalis with, 43
— — trichomonads in, 55, 56
Orbit, cysticercus of, 664
Orbital cavity, Pycnosoma maggots invading, 588
Orchitis from Filaria bancrofti infection, 677
Oribates sp., 489
Oriental sore, cause of, 107, 627
— — experimental production, 109
— — geographical distribution, 108
— — germ of, possible carrier, 580
— — immunity to, procured by inoculation, 108
— — objection to name, 107
— — occurrence in dogs, 108
— — parasite producing, 107, 627
— — pathology of, 627
— — preventive measures, 628
— — relation of infantile kala-azar to, shown experimentally, 109
— — sites of occurrence on body, 108
— — transmission of, bugs possibly connected with, 108, 536
— — treatment, 628
Ornithodorus coriaceus, geographical distribution, 509
— mégnini, characters, 510
— — ears of hosts infested by, 510
— — geographical distribution, 510
— — hosts of, 510
— moubata, carrier of African tick fever, 116, 496
— — — of Filaria perstans, 508
— — — of spirochæte of relapsing fever, 508
— — geographical distribution, 509
— — immunity against infection with Spirochæta duttoni, 119
— — length of life apart from host, 495
— — Malpighian secretion passed by, significance, 117
— — transmission of Spirochæta duttoni by, 116
— savignyi, 509
— — geographical distribution, 509
— — transmitting Spirochæta duttoni, 739
— synopsis of genus, 508
— talaje, 119
— — bite of, 509
— — geographical distribution, 509
— tholozani, geographical distribution, 510
— turicata, 119
— — bite of, effects, 509
Ornithomyia lagopodis, bite of, 611
Orthoptera, characters, 531
Otter, Brazilian, host of Paragonimus rudis, 251
Ova, transmission of intestinal worms by, 11
— see also under Names of parasites
Owen, Trichina spiralis, 423
Ox, liver of, Echinococcus multilocularis in, 357
— Sarcocystis blanchardi from, 190
— gad fly (Tabanus bovinus), 601
Oxazine producing blepharoplastless trypanosomes, 101
Oxen, amount of prevalence of Cysticercus bovis in, 340, 341
— echinococci in, 346
— how infected with Paramphistomum cervi, 226
Oxygen necessary in cultivation of spirochætes, 123
Oxyuriasis, diagnosis, 696
— dysentery followed by recovery from, 698
— in children, 695
— — — treatment, 697, 698
— prophylaxis against, 697
— treatment by drugs and purgatives, 697
— — local, 697
Oxyuridæ, 467
— migrating into nose, 695, 696
— morphology, 375
— relationship to appendicitis, 698
Oxyuris, 467
— ambigua, 469
— compar, 469
— curvula, 469
— encapsuled in female pelvis, 657
— in appendix, 654, 655
— infection in relation to appendicitis, 653
— invading peritoneal cavity, 657
— lacks intermediate host, 21
— mastigodes, 469
— poculum, 469
— tenuicauda, 469
— toxic action of, 651
— vermicularis, association with appendicitis and typhlitis, 467
— — development, 468
— — — direct, 469
— — experimental self-infection with, 469
— — habitat, 467
— — infection by, 694
— — — with, mode of, 469
— — larvæ of, found in nose, 469
— — life-history of, 467, 468, 469
— — males rarely met with in fæces, 468
— — migration from intestine, lesions and symptoms of irritation set up by, 694, 695
— — — of, in and from intestinal tract, 467
— — morphology, 467
— — ova of, where deposited, 467
— — supposed origin of, 11
Oyster, spirochæte of, 114
Ozæna, connection of nasal myiasis with, 717, 722, 723
P.
Page, case of escape, of ascarides from abdominal operation wound, 654, 655
Paget, observation of encapsuled Trichinellæ, 423
Pallas, on transmission of intestinal worms, 11
Panama, larvicide used at, in campaign against mosquitoes, 636
— Canal, Stegomyia fasciata source of danger to, 574
Pani-ghao, skin affection set up by penetration of larvæ of Ancylostomum duodenale, 455
Panoplites, 577
Pansporoblast, 183, 186
Papataci fever, carrying agent of, 582
Pappenheim’s panchrome mixture, 751
Paraboloid condenser, 747
Paraffin, embedding in, for sectioning tissue parasitized by protozoa, 749
Paragonimiasis, 639
— affecting regions other than lung, 639
— prophylaxis, 640
— see also Lung-fluke disease
Paragonimus, morphology, 249
— compactus, host of, 251
— kellicotti, hosts of, 250
— — spines of, 251
— ringeri (lung-fluke), 639
— — development, 251
— — diseases caused by, 251
— — habitat, 251
— — internal organs, diagram illustrating, 250
— — morphology, 249, 250
— — sites of body in which found post mortem, 639
— — spines of, 251
— — synonyms, 249
— rudis, host of, 251
— westermannii, host of, 250
— — morphology, diagram illustrating, 250
— — spines of, 251
Paraguay, supposed mode of transmission of dermo-mucosal leishmaniasis in, 739
Paralysis due to tick bites, 613
— — — — geographical distribution, 613
— of dourine, 97
Paramœba, 44
— hominis, 45, 734
— — characters of, 45
— — now called Craigia hominis, 45, 734
Paramphistomidæ, 231, 234
Paramphistominæ, 231
Paramphistomum cervi, method of infection of oxen by, 226
Paraplasma, 180
— doubt as to organismal nature, 180
— occurs naturally in guinea-pigs, 180
— flavigenum possibly associated with yellow fever, 180
— — morphology, 180
— subflavigenum, 180
Parasites, definition, 1
— derivation of, 19
— diagnosis of presence of, 10
— discoveries from use of microscope, 10
— great fertility of, 5
— hereditary transmission of, 19
— human, Opisthorchis felineus most frequently found at autopsies at Tomsk, 253
— incidental, 6
— — human, 7
— influence on host, 8
— invading many hosts, 6
— limited to closely related hosts, 6
— — to one species of host, 6
— migrations in host, injuries set up by, 9
— movements of, disorders set up by, 9
— occasional (temporary), 1
— origin of, 10
— permanent, bodily changes in, 3
— — clasping and clinging organs in, 4
— — classes of, 2
— — hermaphroditism in, 4
— — loss of organs in, 3
— — (stationary), 1, 2
— transference from one host to another, 7
Parasitic life, advantages of, 20
Parasitism, facultative, 7
Pariah dogs, liver of, habitat of Paropisthorchis caninus, 257
— — North-west Provinces, India, percentage infected with Paropisthorchis caninus, 257
Paropisthorchis, 255
— caninus, genital pore, 255
— — habitat, 257
— — morphology, 255
— — seminal vesicle, 257
— — synonyms, 255
— — uterine coils, 257
— — vitellaria, 255
Partridges, Plasmodium relictum cause of fatal disease in, 170
Pasquale, see Kruse and Pasquale
Pasteur, L., researches on silkworm disease, 184
Patterson, maggots of Pycnosoma removed from orbital cavity, 588
Patton, genus Crithidia, 104
— Herpetomonas muscæ domesticæ, 102
— Piroplasma gibsoni, 177
— places Leishman-Donovan body in genus Herpetomonas, 107
— probable transmission of Leishmania, 107, 108
— and Cragg, life-history of Lyperosia exigua, 610
Peacock, observation of encapsuled Trichinellæ, 423
Pébrine bodies or Nosema bombycis of Arthropoda, 184
Pediculidæ (lice), characters, 532
Pediculoides ventricosus, effects on man, 489
— — morphology, 489
— — shape of pregnant female, 489
— — synonyms, 489
Pediculis capitis (head louse), characters and morphology, 532, 533
— — geographical distribution, 533
— — habitat, 533
— — infection by, 709
— — — causing eczema, 709, 710
— — — diagnosis, 710
— — — greater prevalence among females, 709, 710
— — — remarkable instances, 710
— — — resulting in blepharitis and conjunctivitis, 710
— — — — in plica polonica, 710
— — — treatment, 710
— — mouth parts of, 533
— — ova of, 533
— — prophylaxis against, 615, 616
— vestimenti (clothes louse), characters, 533
— — habitat, 533
— — infection by, 710
— — — lesions and symptoms following, 711
— — pest among soldiers during campaigns, 533
— — prophylaxis against, 615, 616
— — transmission of relapsing fever by, 120, 630
Peenash (nasal myiasis), 588, 715
— ending in fatal meningitis, 716
Peiper, cause of ancylostome anæmia, 648
Pelagutti, treatment of cutaneous and muscular cysticerci, 663
Pelletierinum as vermifuge, 673
Pelvic and abdominal organs, blood-supply of, as illustrating distribution of Schistosoma hæmatobium in body, 272
Pelvis, female, Oxyuris encapsuled in, 657
Pentastoma armillatus, hosts of, 528
— denticulatum, former name of larval stage of Linguatula rhinaria, 525, 526
— moniliformis, hosts and habitat of, 528
— — synonyms (footnote), 528
Pentastomidæ, references to, 528
Pentateuch, “fiery serpents” mentioned in, probable identification, 386
Pentatrichomonas bengalensis, 624, 735
Pereira, case of chorea cured after expulsion of Tænia, 648
Perinæum, tumours of, association of Dioctophyme gigas with, 431
Peritoneal cavity, Ascaris in, 656
— — Oxyuris invading, 657
Peritonitis, perforative, due to Ascaris, 656
Peritricha, 29, 200
Perroncito, artificial infection of human beings with Cysticercus bovis, 340
— infection with Lamblia intestinalis, 60
Persia, importation of African tick fever into, 613
Persian insect powder infusion in intestinal myiasis, 728
Peru oil, application in scabies, 707
Petrie, treatment of bilharziasis, 643
Petroleum as larvicide in campaign against mosquitoes, 636
— dressings of, in head louse infection, 710
— in crab louse infection, 712
— and benzine in crab louse infection, 712
Pfeiffer, L., pathogenicity of Coccidia, 136
Pfeiffer, R., Coccidia, 136
Pharynx, ascarides invading, 691
— invasion and infection in man by Fasciola hepatica, 242
— leeches in, 699
— of Hirudinea, 480
Philæmatomyia, position of genus, 611
— insignis, 611
Philippine Islands, experiments on amœbæ in, 618
Philips, eucalyptus oil in expulsion of ancylostomes, 687
Phillips, L. P., on Musgrave and Clegg’s medium, 743
— — treatment of balantidian dysentery, 637
Phlebotominæ, characters, 581
Phlebotomus, blood-sucking, 581
— characters, 581
— duboscii, 582
— intermedius, 582
— geographical distribution, 581
— — — of species, 582
— larvæ of, habitat, 582
— longipalpis, 582
— papatacii, 581, 582
— squamiventris, 582
Phonergates bicoloripes, 541
Phoniomyia, characters, 565
Phora rufipes, 589
— — larvæ (maggots) of, habitat, 583
Phoridæ, characters, 582
Phthiriasis, agents of, 533
Phthirius inguinalis (crab louse), characters, 534
— — habitat, 534
— — infection by, diagnosis, 712
— — — how effected, 711
— — — lesions and symptoms following, 711, 712
— — — sites of body affected, 711
— — — treatment, 712
— — rapid reproduction of, 534
— pubis, prophylaxis against, 616
Phthisis, filaria associated with, 408
Physaloptera, habitat and hosts of species, 460
— caucasica, morphology, 461
— mordens, geographical distribution, 461
— — habitat and host, 461
— — morphology, 461, 462
— morphology, 460
Physalopteridæ, 375, 460
Phytoparasites, 1
Pierantoni, Agamofilaria labialis, 407
Pig concerned in transmission of Balantidium coli, 202
— development of Trichinella spiralis in, 426, 427
— domestic, normal host of Cysticercus cellulosæ, 332
— echinococci in, 346
— geographical distribution of Tænia solium corresponds with that of, 334
— host of Paragonimus kellicotti, 250
— intestine of, Fasciolopsis buski in, 246
— Metastrongylus apri in, 433
— organs infected with echinococcus, percentage of frequency, 347
— rectum of, Balantidium coli present in, 202
— Sarcocystis miescheriana in, 190
— Sarcosporidia in, 187
— trichinous, proportion to healthy, in Prussia, 429, 430
— Trypanosoma simiæ virulent to, 100
Pigeon lofts inhabited by Argas reflexus, 506
Piophila casei, characters, 583
— — larvæ of, found in fæces, 583
— — — in nose, 720
Piroplasma, 172, 173, 174
— see Babesia
— gibsoni, 177
— hosts of, 173, 174
Piroplasmidæ, 172, 742
— genera of, 174
Piroplasmosis, treatment of, 178
— — by trypan-blue, 178
— — symptoms of, 178
— — transmission by ticks from recovered to uninfected animals, 178
Placobdella, 482
— catenigera, geographical distribution, 482
Plague, fleas carriers of, 543, 547
Planont, 185
Plants, flagellosis of, possible connection with leishmaniasis, 739
Plasmodium, 151, 742
— falciparum, see Laverania malariæ
— malariæ, development in red corpuscles of man, asexual stage, 166
— — distinctive characters, 167
— — lesions set up by, not marked, 634
— — parasite of quartan malaria, 166, 633
— — pigment granules of, 166, 167
— — schizogony of, 166
— — synonyms, 166
— — trophozoites of, differ from those of tertian parasite, 166
— or hæmamœba type of Hæmosporidia includes malarial parasites of man and birds, 151
— relictum, first development of malarial parasite in mosquito traced in, 170
— — hosts of, 170
— — mosquitoes transmitting, 170
— — stages in life-history, 170
— — synonyms, 170
— species, differential table of, 171
— tenue, 170
— vivax, agent of simple tertian malarial fever, 164, 633
— — cultivation of, clumping not observed in, 172
— — — number of spores produced, 172
— — development in red blood corpuscles of man, 160, 164, 165
— — — of “Polymitus,” 160, 165
— — — time occupied by, 165
— — distinctive characters of, 166
— — lesions set up by, not marked, 634
— — life-cycle of, 160, 164
— — merozoites of, 165
— — — migration, 165
— — micro- and macrogametocytes of, 165
— — pigment granules, 165
— — small variety, 166
— — “stippling,” 165
— — synonyms, 164
Platyhelminthes (flat worms), 211
— central nervous system of, 211
— classification, 212
— definition, 211
— diseases caused by, 638
— excretory apparatus, 211
— hermaphroditic, 211
— integument of body of, 211
— method of reproduction, 211
— morphology, 211
Plerocercoid, definition of, 301
Plerocercus, definition of, 301
Pleuræ, invasion by Paragonimus ringeri, 251
Plica polonica due to head louse infection, 709, 710
— — — — — — treatment, 710
Plimmer, H. G., treatment of sleeping sickness with antimony, 623
— — and Bradford, Sir J. Rose, Trypanosoma brucei, 93
Pliny, Ascaris lumbricoides known to, 464
Pneumocystis carinii, 90
Pneumocysts in rats, 90
Pocock, geographical distribution of Ornithodorus moubata, 508, 509
Podophyllin in expulsion of ancylostomes, 687
Polar capsule, 181, 183, 184, 186
— filament, 183, 184, 186
Polecat, intestine of, Isospora bigemina parasitic in, 149
Poliomyelitis acuta, possible rhizopods in, 46
— carrier of, 610
— epidemic, insects transmitting, 612
— virus of, 536
— — carried by house-fly, 586
Pollack, invasion by Loa loa, 678
Polymastigina, 52
Polymitus of Plasmodium vivax, 160, 165
— form of malarial parasites (footnote), 162
Polypus, nasal, caused by Rhinosporidium kinealyi, 195, 196
Polysporea, 182, 184
Polysporulea, 195
Polystomum integerrimum, organs of, 218
Ponds, mosquitoes depositing ova in, 553
Pork, cysticerci in, cause of decrease, 334
— eating of, cause of trichinosis, 423
— — means of infecting man with cysticerci, 334
— inspection of, in prophylaxis against trichinosis, 429
Porocephalus, 523
— armillatus, 527
— — synonyms (footnote), 528
— constrictus, characters, 526
— — hosts of, 526, 527
— — organs of body invaded by, 526, 527
— — synonyms, 526
Port Natal sickness (Cape ailment), 488
Portal vein and liver, Schistosoma hæmatobium most easily found post mortem in, 273
— — and vena cava, communication between, how formed, 272
— — tributaries of, as illustrating distribution of Schistosoma hæmatobium in body, 272
Porter, A., Crithidia pulicis, 111
— — generic differences among insect flagellates, 103 (fig. 49)
— — Herpetomonas muscæ domesticæ, 102
— — Leucocytogregarina, 154
— — Theileria parva, 179
— see also Fantham and Porter
Portschinsky, deposition of ova of Oestrus ovis, 598
— method of destroying Tabanidæ, 601
Posner, case of amœbæ in urine, 46
Posselt, cutaneous tumours due to cysticerci, 662
— reasons for distinction of multilocular from hydatid or unilocular echinococcus, 358
Post-flagellate stage in herpetomonads, 103
— — in Crithidia, 104
Potassium iodide in treatment of cutaneous and muscular cysticerci, 663
— permanganate, application in Oriental sore, 628
Pou d’agouti, mite attacking man, 486
Poultry, fatal epizoötics among, due to Eimeria avium, 142
Poultrymen attacked with Dermanyssus gallinæ, 493
Poupée-Desportes, Guinea worm infection, 676
Powell, method of destruction of Sarcophaga larvæ, 723
Predtetschensky, expulsion of Hymenolepis nana, 661, 662
Pre-flagellate stage in herpetomonads, 103
— — in Crithidia, 104
Price, Dodds, method of prevention of Indian kala-azar, 627, 739
Prima, fatal case of myiasis externa, 716
Privies, disinfection of, as prophylactic against ancylostomiasis, 685
Proflagellata, 115
Proskauer, case of Oxyuridæ in nose, 696
Prostomata, 230
Protargol in balantidian dysentery, 637
Proteid destruction in ancylostomiasis, 647
— metabolism in anæmia, 645
Proteosoma, spread of malaria in birds by, 158
Protista defined, 29
— spirochætes classed among, 115
Protomonadina, 52, 60
— classification, 60, 61
Protozoa, 25, 756
— alternation of generations in, 27
— blood-inhabiting, examination of, 747
— characters, 25
— chromodial apparatus of, 26
— classification, 27
— clinical and therapeutical notes relating to, 617
— cytological details, method of examining, 748
— definition of, 25
— digestive apparatus, 26
— ectoplasm and endoplasm of, 25, 26
— encystment of, 27
— examination, methods for, 745, 746
— food of, 26
— genera of, precise definition sometimes impossible, 733
— hereditary transmission of (footnote), 19
— intra vitam staining of fresh preparations, 746
— nucleus of, 26, 27
— organellæ, 29
— parasitic in blood, culture media for, 744
— propagation of, 27
— sectioning tissue parasitized by, 749
— or bacteria, question whether spirochætes to be classed among, 115
Protozoology, notes on technique, 745–752
— recent researches in, 733
Prowazek, balantidian dysentery, 637
— Chlamydozoa, 207
— Entamœba bütschlii, &c., 34
— — buccalis, 43
— Herpetomonas muscæ domesticæ, 102
— lamblial diarrhœa, 625
— variety of Trichomonas intestinalis inhabiting oral cavity, 56
— and Aragao, filtration experiments with chlamydozoal granules, 209
Prowazekia, characters of, 63
— asiatica, 65
— cruzi, characters, 66
— javanensis, characters, 66
— parva, 66
— urinaria, 63
— — characters, 63
— — flagellate stage, 64
— — in cultures associated with bacteria, 65
— — synonyms, 63, 64
— weinbergi, characters, 66
Prowazek’s bodies, 208
Pruner, Porocephalus constrictus, 526, 527
Pruritus ani due to escape of ascarides, 688
— — set up by migration of Oxyuris vermicularis, 695
— nematode larvæ in blood associated with, 378
Prussia, oxen infected with Cysticercus bovis in, 341
— percentage of pigs infected with cysticerci in, 334
— proportion of trichinous to healthy pigs in, 429, 430
Pseudo-helminthes, 8
Pseudomeningitis due to Ascaridæ infection, 649, 650
Pseudo-myxœdematous form of Brazilian trypanosomiasis, 88
Pseudonavicellæ, 129, 130
— amœboid germs in, 130
Pseudoneuroptera, characters, 531
Pseudo-parasites, 6, 8
Pseudophyllidea, morphology, 308
Pseudotæniorhynchus, 576
— characters, 564
Psorophora, characters, 563, 571
— ovum of, 557, 558
Psoroptes, characters, 517
Psorospermia of Arthropoda, 184
Psorosperms (Myxosporidia), discovery of, 181
— egg-shaped, former name for Coccidia, 135
Psychodidæ (owl midges), moth-like appearance of, 581
Psychodinæ, characters, 581
Pterocephalus, host of, 135
Pterygota, classification, 531
Pulex, distinctive characters, 545
— irritans (human flea), bite of, effects, 714
— — — treatment, 714
— — carrier of plague bacillus, 543
— — characters, 545
— — larva of, 546
— — may transmit Trypanosoma lewisi, 92
— pallipes, 548
— serraticeps (dog flea), 546
Pulicidæ (true fleas), characters, 543
— classification of genera, 545
Pulmonary artery, ascarides in, 656
Pumpkin seeds as vermifuge, 673
Pupipara or Eproboscidæ, blood-sucking, 611
Purgatives for expulsion of ascarides, 693
— in arrest of development of trichinosis, 681
Pustules arising from clothes louse infection, 711
Putnam, Oxyuris in appendix, 654
Pycnosoma, characters of, 588
— and Chrysomyia, distinguishing features, 588
— putorium, spread of amœbic dysentery by, 614
Pyelitis following invasion by Eustrongylus gigas, 682
Pygiopsylla, distinctive characters, 545
Pyorrhœa alveolaris, association of Entamœba buccalis with, 43, 734
— — — of species of Treponema with, 128
— — treatment, 620
Pyretophorus, characters, 561, 567
Pyronin producing blepharoplastless trypanosomes (T. brucei), 101
Q.
Quincke and Roos, species of amœbæ named by, 31
Quinine, administration as preventive against malaria, 636
— administration in malaria, 635
— — — dosage, 635
— — — methods of administration, 635
— — — time for, 635
— — — treatment by, 635
— in Indian kala-azar, 626
— lotion, irrigation of lower bowel with, in gangrenous dysentery, 619
R.
Rabbit, development of hydatid scolices in, 353
— host of Eimeria stiedæ, 145
— intestinal coccidiosis in, 145, 147
— intestine of, section infected by Eimeria stiedæ, 145
— kidney of, use in cultivation of Treponema pallidum, 126
— liver of, section through nodule infected by Eimeria stiedæ, 147
— Sarcosporidia in, 187
Rabies, parasite of, cultivation, 210
Radiolaria, characters and habitat, 28
Radium treatment of Oriental sore, 628
Railliet, method of infection with Trichuris depressiuscula, 420
Rainey’s corpuscles, 189
Rain-water barrels, mosquitoes depositing ova in, 553, 557
Ramstedt, Oxyuris infection in relation to appendicitis, 653
Ranken, treatment of sleeping sickness with antimony, 623
Rasahus biguttatus, bite of, 540
— — geographical distribution, 540
— — synonyms, 540
Rat attacked by Dermatophilus (Sarcopsylla) penetrans, 613
— blood of, transference of Trypanosoma brucei from, to blood of snake, 102
— blood parasite, see Trypanosoma lewisi
— gut and cæcum of, Trichomonas from, 735
— infection with Trichinella, method of, 427
— — with Trichinella spiralis in slaughterhouses and knackers’ yards, 427
— — with Trypanosoma lewisi, mode of, 92, 93
— muscles of, invaded by Trichinella spiralis, 425
— normal host of Trichinella spiralis, 427
— pneumocysts in, 90
— sewer and black, hosts of Hymenolepis diminuta, 326
— flea (Ceratophyllus fasciatus), cysticercoid of Hymenolepis diminuta found in, 327, 328
— — host of rat trypanosome, 88, 90, 543
— — larval stages of Hymenolepis murina occurring in, 17
— — see also Ceratophyllus fasciatus
— Trypanosoma lewisi in, 88
Rectum, administration of quinine by, in malaria, 635
— bilharziasis of, 642
— — treatment, 644
— irrigation of, in intestinal myiasis, 728
— means of access of Schistosoma hæmatobium to, 272
— pathological changes in, due to Schistosoma hæmatobium, 274, 275
— plexus formed in, by superior hæmorrhoidal veins, 272
Redi, origin of flesh maggots, 10
Rediæ of trematodes, 225, 226, 227, 228
Reduviidæ, bites of, 537
— characters of, 537
— geographical distribution, 537
Reduvius personatus, bite of, sometimes fatal, 539
— — geographical distribution, 539
Red-water fever, European, in cattle, cause of, 177
Reighardia, 523
Relapsing fever, 120, 629
— — African, cause of, 116, 630
— — — incubation period, 630
— — — prophylaxis, 631
— — — symptoms, 630, 631
— — — treatment, 631
— — American, 630
— — Asiatic, mortality from, 631
— — — prophylactic measures, 631
— — — symptoms, 631
— — — treatment, 631
— — complications, 630
— — East African, cause of, 122
— — European, agent of, 122, 629
— — — incubation period, 630
— — — prophylaxis, 630
— — — symptoms, 630
— — — treatment, 630
— — Indian, cause of, 122
— — North African, prophylactic measures, 631
— — — — symptoms, 631
— — — — treatment, 631
— — — — and Egyptian, cause of, 122, 631
— — prophylactic measures, 630
— — spirochætes causing, 115, 120, 122, 508
— — transmission by lice, 120, 630
— — — by ticks, 117, 630, 631
— — treatment, 630
Remak, investigations of Coccidia, 135
Reptiles, hæmogregarines in, 153, 154
— Sarcosporidia in, 187
Resorcin ointment, application in creeping disease, 732
Respiration, organs of, in Insecta, 530
Retinal hæmorrhages in ancylostome anæmia, 646
Reyher, bothriocephalus anæmia, 644, 645
Rhabdites in gastric fluid obtained by lavage, 378
— mellio, presence in vagina, 377
— niellyi, 378
— — mode of infection in man, 378
— pellio, induction of facultative parasitism, 377
— — morphology, 377
— — synonyms, 377
Rhabdonema, alternation of parasitic and free-living generations, 20
— life-history of, 19
— propagation of, parasitic generation during free life, 18
Rheins, case of Oxyuridæ in nose, 696
Rhinosporidium, 195
— hosts of, 197
— in conjunctival polypus, 197
— in external auditory meatus, 196
— in horses, 197
— in nasal polypus, 195
— in papilloma of penis, 197
— kinealyi (or seeberi), 195, 197
— — causal agent of a nasal polypus, 195, 196
— — cysts of, 196
— — geographical distribution, 195, 196
— — pansporoblasts of, 196
— — trophozoites of, 196
— — tumours produced by, 197
Rhipicentor, characters of, 497
Rhipicephalæ, characters of, 496, 497
Rhipicephalus, characters of, 497
— species of, transmitting Theileria parva, 179
— annulatus, carrier of Texas fever in cattle, 494
— — moulting of, 496
— appendiculatus and R. simus, carriers of Rhodesian fever in cattle, 494
— bursa, transmitting agent of Babesia bovis, 177
— sanguineus, geographical distribution, 505
— — hosts of, 505
— — synonyms, 505
— — transmission of leucocytogregarine from dog to dog by, 155
— — transmitting agent of Babesia canis, 177
Rhizoglyphii, characters and habitat, 514
Rhizoglyphus parasiticus, characters, 514, 515
— — skin disease produced by, 514
Rhizopods, flagella occurring among, 52
— possible association with poliomyelitis acuta, 47
Rhodesian fever in cattle, carriers of, 494
Rhodinus prolixus, bite of, 541
— — geographical distribution, 542
Rhyncobdellidæ, 482
Rhyncobothrium, scolices of, 305
Rhyncota, see Hemiptera
Ricinidæ classed among mutualists, 6
Riley, see Walsh and Riley
Rinderpest and coccidiosis, 741
— method of transmission, 742
River fever set up by kedani mite in Japan, 487
Rivolta, experimental infection with Coccidia, 136
— on Sarcocystis lindemanni, 193
Robertson, Miss, development of Trypanosoma gambiense, 74, 75
— — — — — in Glossina palpalis, 74, 75
— — forms of Trypanosoma gambiense, 73, 737
Rocky Mountain spotted fever, carrier of, 496
— — tick fever, carrier of, 503
— — — — mortality, 504
Rodenwaldt, distribution of larvæ of Filaria immitis in body, 393
— periodicity of larvæ of Filaria bancrofti in peripheral blood, 393
Rogers, Sir L., agent transmitting kala-azar, 713
— — — cultivation of Leishmania donovani, 105, 106
— — — places Leishmann-Donovan body in genus Herpetomonas, 107
— — — transmission of surra by Chrysops, 601
— — — treatment of amœbic dysentery, 618
— — — treatment of Indian kala-azar, 626
— — — — of pyorrhœa alveolaris, 620
Rokitansky, perforation of intestine by Ascaris, 655
Romani, agglutinating hæmolytic action of serum of ancylostome patients, 648
Romanowsky stain, 749
— — slightly modified, formula of, 750
— — underlying principle of, 750
Roos, presence of cercomonads in gangrenous lung, 62
— and Harris, penetration of intestinal blood-vessels by amœbæ, 36
Rosenquist, proteid metabolism in anæmia, 645
Ross, E. H., Treponema pallidum, 124
Ross, Sir Ronald, campaign against mosquitoes in prevention of malaria, 636
— — — development of malarial parasite in mosquito traced in Plasmodium relictum by, 171
— — — discovery of transmission of malarial parasites by mosquito, 158
— — — “Prevention of Malaria,” 617, 633
— — — relapses in malarial fever, 161, 162
— — — trichomonads and cercomonads, 56
— — — and Thomson, D., cyclical variation of trypanosomes in blood, 78
— — — — — — method of determining number of trypanosomes in blood, 747, 748
Rossia, characters, 568
Rossiella, morphology, 174
— rossi, 174
Rostellum of Cestoda, 289
Rothschild, classification of genera of Pulicidæ, 545
Roubaud, cause of myiasis in French West Africa, 614
— life-history of Cordylobia anthropophaga, 614
— Pycnosoma putorium, 614
Rovelli, larval stage of Hymenolepis diminuta, 327
Row, experimental production of Oriental sore, 109
— treatment of Oriental sore, 628
Rudolphi, origin of helminthes, 12
Ruffer, lesions produced by Oxyuris vermicularis, 695
Runchiomyia, characters, 565
S.
Sabadill vinegar, lotions of, in head louse infection, 710
Sabatier on change of hosts, 21
Sabethes, characters, 565
Sabethoides, characters, 565
Sachs, treatment of scabies, 707
Sack, treatment of scabies, 707
Sahara, Central, human myiasis occurring in mountains of, 598
St. Artault, Trichomonas pulmonalis, 56
Saline solution, physiological, lavages of, in myiasis, 719
Salol as tapeworm drug, 674
Salt water, mosquito larvæ living in, 557
Salvarsan in Asiatic relapsing fever, 631
— in North African relapsing fever, 631
— in Oriental sore, 628
— in relapsing fever, 630
— in tropical syphilis, 632
— in trypanosomiasis, 623
— in yaws, 632
— — dosage, 632
Salzmann, mode of infection in intestinal myiasis, 727
Sambon, L. W., Linguatula serrata, 527
— — personal experiments with regard to malarial infection, 158
Samelsohn, retinal hæmorrhages in ancylostome anæmia, 646
Sandal oil in chyluria from Filaria bancrofti infection, 677
Sand flea, see Dermatophilus (Sarcopsylla) penetrans
— flies, 577
— — and fever due to them in North China, 613
— — biting in Hampshire, 579
Sandflies, haunts of, 613
— — see also Simulium
Sandler, trichocephalus anæmia, 651
Sandwith, F. M., toxic symptoms following thymol administration, 686
— — treatment of bilharziasis, 643
Santonin in bilharziasis, 643
— in expulsion of ascarides, 692
— — of Oxyuridæ, 697
— in intestinal myiasis, 728
Sapo viridis and tar, application in creeping disease, 732
Sarcocystin, isolation of, 191
Sarcocystis bertrami, 193
— blanchardi, 193
— — from ox, 190
— colii, 193
— — spore, 193
— hueti, 193
— lindemanni, 193
— miescheriana, 188, 193
— — from pig, 190
— muris, 193
— — deadly to host, 191
— — experimental infection with, 191, 192
— — gymnospores of, 191
— — spore of, site of sarcocystin, 192
— of muscles, 191
— pansporoblasts of, 189
— recognition from other foreign bodies, 188
— spores of, 189
— tenella, 193
— — from sheep, 190
— — spores of, 191
— — toxin isolated from, 191
Sarcodina, 27, 29
— characters and habitat, 27
Sarcoid globules in miracidium of Schistosoma hæmatobium, 276
Sarcophaga carnosa (flesh fly), characters, 589
— — larvæ of, 589
— — — regions of human body invaded by, 589
— — viviparous, 589
— chrysostoma, 590
— hæmatodes, 589
— hæmorrhoidalis, 589
— magnifica, geographical distribution, 589
— — larvæ of, regions of human body invaded by, 589
— — references to, 589
— plinthopyga, probably concerned in dissemination of yaws, 590
— ruficornis, 589
— wohlfahrti, larvæ of, method of destroying, 723
— — — unusual situations of, 723
— — nasal myiasis from, 722, 723
Sarcopsyllidæ, characters, 543
Sarcoptes, characters, 517
— species transmissible from domestic animals to man, 520
— auchenii, 520
— canis, 520
— dromedarii, 520
— equi, 520
— leonis, 520
— minor, hosts of, 520
— ovis, 520
— scabiei, characters, 518
— — infection by, disease produced by, 704
— — — see also Scabies
— — synonyms, 518
— — — crustosæ, 519
— — var. hominis, 519
— — — — development of, 519
— — — — excavation of tunnels in human epidermis by, 517, 519
— — — — transmission of, natural and artificial, 519
— suis, 520
— vulpis, 520
Sarcoptidæ (itch mites), characters, 516
— development, stages in male and female, 517
— hosts of, 516
— rate of breeding, 517
Sarcoptinæ, 517, 518
Sarcosporidia, 129, 187, 193
— chambers of, 189
— characters and habitat, 28, 188
— experimental transmission, 191, 192
— fatal to sheep, 188
— hosts of, 187
— in man, 193
— morphology, 188
— muscles affected, 188
Sarcosporidiosis, possible percentage of animals affected by, 191
Scabies, 704
— diagnosis, 705
— — from occupational eczema, 706
— mite tracks of, 705
— prognosis, 706
— symptoms of, 705
— treatment, 706, 707
— norvegica (Norway itch), 520, 705
Scabiophilia, 706
Scarlet fever, cell inclusions in, 208
Schaudinn, classification of intestinal amœbæ, 31
— cytological changes during encystment process of Entamœba coli, 33
— infection by trichomonads, 56
— intensity of infection with Entamœba coli, 33
— on Leydenia gemmipara, 49
— on Paramœba hominis, 44
— penetration of red blood corpuscles by sporozoites of tertian parasite, 159
— relapses in malarial fever, 161
— researches on Coccidia, 137, 138, 139
— — on Entamœba histolytica, 34, 37
— Treponema pallidum, 114
Schaudinn’s fluid, 748
Scheube, lung-fluke disease, 639
Schewiakoff, movements of gregarines, 131
Schiller, Ascaris and Oxyuris infection in relation to appendicitis, 653
— effects of trichocephalus infection, 651
Schistocephalus, pleroceroid of, 300
Schistosoma, morphology, 269
— synonyms, 269
— cercariæ, 753
— hæmatobium, distribution in body, 272
— — endophlebitis set up by, 274, 275
— — female, diameter, 273, 274
— — — morphology, 271
— — — genitalia, 276
— — geographical distribution, 276
— — in caval system, 274
— — in gall-bladder, 274
— — in hæmorrhoidal veins, 273
— — in lungs, 274
— — in vesico-prostatic plexus, 273, 274
— — infection by, 641
— — — see also Bilharziasis
— — male, anterior end, diagram showing organs, 271
— — — carrying female in canalis gynæcophorus, 270
— — — diameter, 273
— — — morphology, 270
— — — and female in copulâ, transverse section through, 271
— — means of access to descending colon, rectum, anal canal, bladder and caval system, 272
— — miracidium of, sarcoid globules in, 276
— — most easily found post mortem in portal vein and liver, 273
— — ova in utero, diameter, 273
— — — lateral spined, 273
— — — — — origin of, 273
— — ovum of, 277
— — pathological changes in rectum and bladder due to, 274, 275
— — synonyms, 270
— japonicum, 277
— — anterior end with testes, posterior end with point of union of cæca, 278
— — female, morphology, 278
— — from dog, 280
— — — egg from fæces, 280
— — — uterine egg, 280
— — habitat, 280
— — liver showing eggs in the intra- and interlobular connective tissue, 282
— — male, morphology, 277
— — — and female in copulâ, 279
— — mode of infection by, 279
— — ova of, 278
— — — from human liver, showing “spines” and “hoods” at opposite pole, 279
— — — sites in which found in body, 282
— mansoni, 754
Schistosomidæ, 269, 753
— morphology, 233
Schizocystis, 135
Schizogony absent in Eugregarinea, 134
— in Coccidiidea, 138
— in Leucocytozoa, 153
— of malarial parasites, 161, 172
Schizogregarinea, 135
Schizotrypanum cruzi, 83
Schleip, blood examination in diagnosis of trichinosis, 681
Schlesinger, intestinal myiasis, 727
Schlüter, hæmorrhagic enteritis from Strongyloides infection, 674
Schmidt, larvæ in nose in enormous numbers, 716
— Trichomonas pulmonalis, 56
Schneider, A., on Coccidia, 137
— — on Eimeria, 142
— — on gregarines, 130
Schuberg, copulation in Coccidia, 137
— immunity of Ornithodorus moubata against infection with Spirochæta duttoni, 119
Schüffner, peculiar fever resembling typhoid, 613
Schüffner’s dots, 165, 166, 171
Schultz, on Coccidia in cattle, 741
Schupfer, typhoid lumbricosis, 650
Schwankhaus, Ascaris infection in relation to appendicitis, 653
Schweriner itch following infection by ancylostomes, 684
Schwetz, life-history of Auchmeromyia luteola, 614
Scolex of tapeworms, 300, 303, 304
— — morphology, 304
Scolopendra in maxillary and frontal sinuses, 721
Screw worm, Indian, see Pycnosoma
— — fly, see Chrysomyia (Compsomyia) macellaria
Scutomyia, characters, 563
Seal, host of Dibothriocephalus latus, 315
Sebirol as vermicide, 672
Seeber, Rhinosporidium described by, 197
Sehrt, abscess of omentum with Ascaris ova in pus, 657
Seidelin, association of Paraplasma bodies with yellow fever, 180
Seifert, blood-stained diarrhœa from Strongyloides stercoralis infection, 674
Selenidium, 135
Sellards, see Walker and Sellards
Senevet, herpetomonad flagellate in cultures of blood and organs of geckos, 739
Senna, syrup of, in expulsion of ancylostomes, 686
Sense, organs of, lacking in parasitic nematodes, 366
Sepsidæ, characters, 583
— larvæ (maggots) of, 583
Septicæmia terminating nasal myiasis fatally, 718
Sergent, transmission of relapsing fever, 120, 121
Sergent, Ed. and Et., herpetomonad flagellate in cultures of blood and organs of gecko, 739
— — — “thymni,” 725
Sergent, E. and L., deposition of ova of Oestrus ovis, 598
— — — transmission of trypanosomes by species of Tabanus, 601
Sergent and Gillot, treatment of North African relapsing fever, 631
Sergentella hominis, 210
Serous fluid, bodies resembling amœbæ found in, 46
Serum diagnosis of echinococcus, 359
— — — complement deviation, 359
— — — precipitin reaction, 359
— human, action on Trypanosoma rhodesiense, 80
— immune, action on Trypanosoma rhodesiense, 80
Setaria, habitat, 407
— morphology, 407
— equina, hosts and habitat of, 408
— — morphology, 408
— — synonyms, 408
— (Filaria) hæmorrhagica, 408
— labiata papillosa, 408
Sexual organs, irritative effects on, set up by migrations of Oxyuris vermicularis, 695
— — of Echinorhynchus, 476
— — of Hirudinea, 481
— — of Insecta, 530
— — of nematodes, 367, 368, 369
Sheep, baleri in, causal agent, 95
— “carceag” in, cause of, 177
— Cysticercus cellulosæ in, 337
— echinococci in, 346
— heart-water fever in, carrier of, 493
— how infected by Fasciola hepatica, 226
— liver-fluke disease in, death from apoplexy in first period, 240
— — — period of anæmia, 240
— — — — of immigration, 240
— — — — of migration of flukes, 241
— — — — of wasting, 240
— — — ravages caused by, 238
— organs infected with echinococcus, percentage of frequency, 347
— Sarcosporidia fatal to, 188
— section of Sarcocystis tenella from, 190
Sheep-ked, see Melophagus ovinus
Shell gland secretion in trematodes, 223
Shiga, discovery of dysentery bacillus, 31
— species of amœbæ distinguished by, 31
Shipley, A. E., prophylaxis against clothes lice, 616
Sick, cases of ascarides in bile-ducts, 688
Siebert, application of epicarin in scabies, 707
Siebold, v., development of Tæniæ, 14
— explanation of bladder worms, 14
— feeding experiments with Tænia echinococcus, 356
— investigations of Gregarinida, 129
— observation of Pseudonavicellæ, 129
— psorosperms, 181
— views as to development of Helminthes, 13
Siedlecki, researches on Coccidia, 137
Siegel, Cytorhyctes, 208
— Cytorhyctes luis, 124, 208
Silcock, case of human hepatic coccidiosis, 148
Silkworm disease, “gelbsucht,” 207
— — Nosema cause of, 184
Silvanus surinamensis, characters and habitat, 543
Silver tick, see Amblyomma cayennense
Simond, researches on Coccidia, 137
Simulidæ, 577
Simulium, bite of species of, 578, 579
— characters, 577
— larvæ of, 578
— life-cycle of, 578
— wing of, 579
— buissoni, possible connection with spread of leprosy, 579
— columoaschensis, geographical distribution, 578
— damnosum, geographical distribution, 578
— griseicollis, geographical distribution, 579
— latipes, 579
— meridionale, possible carrier of chicken cholera, 579
— occidentalis, 579
— wellmanni, 579
Sinton, culture of trypanosome forms of T. gambiense, 76
— — of T. rhodesiense, 83
— Prowazekia urinaria, 64, 65
Siphunculata, 532
— see also Pediculidæ
Skin affections caused by cereal mites, 489
— — due to Dermanyssus hirundinis, 492
— — set up by Leptus autumnalis, remedies against, 702
— — — by Trombidium tlalsahuate, 486
— disease caused by larvæ of Dermatobia noxialis, 725
— — due to young nematodes in dogs, 378
— — produced by Rhizoglyphus parasiticus, 514
— diseases set up by penetration of larvæ of Ancylostoma duodenale, 455
— — — — — — — various names for, 455
— filaria infection of, 378
— infection by Ancylostoma duodenale through, 683
— — by larvæ of Ancylostoma duodenale, 454, 455
— lesions due to Sparganum mansoni, 318
— mole, 599
— parasites of dogs and cats infecting them with Dipylidium caninum, 323
— surface of, larvæ on, 721, 722
Skusea, characters, 563
Slaughter-houses, infection of rats with Trichinella in, 427
Sleeping sickness, 68, 69, 72, 76, 620
— — association of trypanosomes with, 68
— — cerebral stage, 621
— — due to Trypanosoma gambiense, 68, 72, 620
— — — — rhodesiense, 69, 76, 620
— — — — — symptoms, 622
— — febrile or glandular stage, 621
— — incubation period, 621
— — investigation of, 68
— — parasites producing, 72, 76, 605
— — pathology of, 621
— — preventive measures, 623
— — Rhodesian, daily number of trypanosomes in blood from case of, 79
— — transmission of, 68, 605, 607, 608
— — — experimental (with apes), 68
— — treatment by arsenic and arsenical preparations, 622, 623
— — — by atoxyl, 622
— — — by tartar emetic, 622
— — — must be commenced in early stages to be effective, 622
Sleeping Sickness Bureau Bulletin, foundation of, 69
Sloth, blood of, inhabited by Endotryparium schaudinni, 99
Smith, Theobald, experimental infection of mice with Sarcocystis muris, 191
— and Barrett, Endamœba, 734
— — Endamœba gingivalis, 733
— — treatment of oral endamœbiasis, 620
Smith and Kilborne, 174, 176, 177
— and Weidman, Entamœba mortinatalium, 45
Smithia, morphology, 174
— microti, 174
— talpæ, 174
Snake, blood of, transference of Trypanosoma brucei to, from blood of rat, 102
Soamin in sleeping sickness, 623
Soda, bicarbonate, with iodoform in expulsion of ascarides, 694
— salicylate of, lavages of, in nasal myiasis, 719
Soldiers, Pediculus vestimenti pest among, during campaigns, 533
— prophylaxis against clothes lice among, 616
Solium, derivation of specific term (footnote), 331
Souma in bovines and equines, causal agent, 100
Space parasites, 20
Spaniopsis tabaniformis, 614
Sparganum, 317
— mansoni, 317
— — cephalic end, 318
— — diagnostic signs of presence, 659
— — discovery of, 317
— — geographical distribution, 659
— — habitat in body of man, 659
— — migration in body, 318
— — plerocercoid of, 318
— — skin lesions due to, 318
— — symptoms set up by invasion, 659
— — synonyms, 317
— — transverse section of, 318
— proliferum, 318
— — acne-like condition set up by, 318
— — geographical distribution, 320
— — mode of infection, 320
— — morphology, 319
— — synonyms, 318
Spengel, Filaria (?) kilimaræ, 407
Spermatozoa of Trematoda, no essential difference in structure from those of other animals, 222
Sphærularia, nematodes hatched from eggs of, 5
Spiders, see Arachnoidea, 483
Spinal ganglia of rabid monkeys, cultivation, 210
Spinning mites, see Tetranychidæ
Spirochæta, 115
— aboriginalis, association with granuloma inguinale, 122
— acuminata, 122, 128
— anodontæ, 114
— anserina, 119, 122
— balbianii, 114
— berbera, agent of North African and Egyptian relapsing fever, 122
— bronchialis, 122, 632, 739
— — mode of infection, 740
— — morphology and life-history, 739, 740
— buccalis, 122
— — morphology, 741
— carteri, agent of Indian relapsing fever, 122
— dentium, 122, 128
— — morphology, 741
— duttoni, 116
— — agent transmitting, 116
— — cause of African relapsing fever, 116, 630
— — cultivation of, 123
— — geographical distribution, 116, 119
— — infection by, experimental, 117
— — — — summary of methods and results, 118, 119
— — — immunity of Ornithodorus moubata against, 119
— — transmission of, 739
— eurygyrata, 122
— gallinarum, 119, 122
— — agent of transmission, 119
— — appearance in hæmocœlic fluid of Argas persicus, 119
— — cultivation of, 123
— — fatal to fowls, 119
— gigantea, 114
— granulosa, 116
— hachaizæ in cholera motions, 122
— laverani, small size of, 122
— marchouxi, see Spirochæta gallinarum
— muris, 122
— novyi, agent of North American relapsing fever, 122
— — cultivation of, 123
— obermeieri, see Spirochæta recurrentis
— obtusa, 122, 128
— ovina, 122
— phagedenis, 122
— plicatilis, 114
— recurrentis, 120
— — agents of transmission, 120
— — cause of European relapsing fever, 120, 122
— — cultivation of, 123
— — incubation period, 630
— — morphology, 120
— refringens, 122, 128
— — association with Treponema pallidum, 122
— rossii, agent of East African relapsing fever, 122
— — cultivation of, 123
— schaudinni, agent of ulcus tropicum, 122
— stenogyrata, 122
— theileri, 122
— vincenti, 122
Spirochætacea, 115
Spirochætes, 114
— blood inhabiting, 116
— classed among Protista, 29, 115
— cultivation of, 123
— — presence of oxygen necessary for, 123
— granule phase of, 120
— hosts of, 114
— in alimentary tract, 741
— in human mouth, 122, 740
— in vomited matter, 122
— mode of division, 115
— molluscan, breaking up into granules, 119
— morphology and morphological variation, 114, 115
— of human mouth, recent work on, 740, 741
— of relapsing fever, periodic increase and decrease in blood, 115
— reaction to drugs, 115
— systematic position, 115
Spirochætoidea, 115
Spirochætoses, 629
— bronchial, diseases for which mistaken, 632
— — treatment, 633
— relapsing fever, 629
— syphilis, 632
— yaws, 632
Spiroschaudinnia, 115
Spleen, development of crescents of tertian malignant parasite in, 169
— enlargement due to ova of Schistosoma japonicum, 282
— — in malaria, 634
— pigmentation of, following malaria (footnote), 165
Splenic blood, citrated, cultivation of Leishmania donovani in, 106
— vein, tributary of portal vein, 272
Splenomegaly, association of Histoplasma capsulatum with, 112
— — of Toxoplasma pyrogenes with, 113
— infantile (kala-azar), 109, 627
Spontaneous generation, theory of, 10
— — — early opposition to, 10
Sporoblasts of Coccidiidea, 141
— of malarial parasites, 163
— of Myxosporidia, 183
Sporocyst, germ balls of, 227
— of Coccidiidea, 141
— of gregarines, 134
— of trematodes, 225, 227
Sporogony, 144, 186
Sporozoa, 28, 128
— characters and habitat, 28
— classification, 129
— hosts of, 129
— relation to Protozoa, 19
Sporozoites of Coccidiidea, 138, 139, 140
— of gregarines, 132, 133
— of malarial parasites, 159
Stained material, examination of, 747
Staining, 749
Stallion’s disease (dourine), trypanosomes in blood of horses with, 68
Stannus, species of Enyaliopsis producing ulcers, 542
— and Yorke, observation of Trypanosoma rhodesiense in animals inoculated from case of sleeping sickness, 78
Staphylocystis, 304
Stäubli, blood examination in diagnosis of trichinosis, 681
Steel and Evans, experimental transmission of Trypanosoma evansi, 67
Steenstrup, discovery of method of multiplication of Helminthes, 13
Stegomyia, breeding of, prevention, 636
— characters, 563, 571
— ovum of, 557, 558
— transmission of yellow fever by, 555
— albipes, characters, 572
— albocephala, characters, 573
— albolateralis, characters, 573
— albomarginata, characters, 573
— amesii, characters, 573
— argenteomaculata, characters, 572
— argenteopunctata, characters, 573
— assamensis, characters, 573
— auriostriata, characters, 573
— crassipes, characters, 573
— dubia, characters, 573
— fasciata, biting hours of, 574
— — breeding of, 574
— — carrier of yellow fever, 574
— — characters, 572, 574
— — development of Plasmodium relictum in, 171
— — distinguishing characters of S. scutellaris from, 575
— — domesticated species, 574
— — food of, 574
— — geographical distribution, 574
— — larvæ of, habitat, 574
— — ova of, 574
— — possible host of Leishmania tropica, 108
— — source of danger to Panama Canal, 574
— — supposed intermediate host of parasite of Bagdad sore, 575
— — transportation of, 574
— gelebinensis, characters, 572
— grantii, characters, 573
— lilii, characters, 572
— mediopunctata, characters, 573
— minuta, characters, 573
— minutissima, characters, 572
— nigeria, characters, 572
— poweri, characters, 572
— pseudonigeria, characters, 572
— pseudonivea, characters, 573
— pseudoscutellaris, 394, 575
— — characters, 572
— — intermediate host of filaria in Fiji, 575
— punctolateralis, characters, 573
— scutellaris, characters, 572
— — distinguishing character from S. fasciata, 575
— simpsoni, characters, 572
— terreus, characters, 573
— tripunctata, characters, 573
— W -alba, characters, 572
— wellmannii, characters, 572
Stein, interrelation of pseudonavicellæ and gregarines, 129
Stein, v., classification of Infusoria, 199
— discovery of meal worm in bladder worm, 303
Steinhaus, intestinal stenosis following infection by Tænia solium, 662
Stempell, on Nosema bombycis, 184
Stephens, J. W. W., appendix on Trematoda and Nematoda, 753
— — Nemathelminthes, 360
— — Plasmodium tenue, 170
— — Platyhelminthes or flat worms, 211, 638
— — and Christophers, Maurer’s dots, 168
— — — relapses and latent infection of malaria, 158
— — and Fantham, length of Trypanosoma gambiense, 73
— — — Trypanosoma rhodesiense, 69, 76
Stern, symptoms of cysticercus in fourth ventricle, 665
Stethomyia, characters, 561, 567
Stiles, C. W., infection with Lamblia intestinalis, 60
— — prophylaxis against flagellate diarrhœa, 625
Stillborn child, problematical “monocystid gregarine” from lung tissue of, 150
Stitt, alkaloid of quinine in malaria, 635
— paroxysms of malignant tertian fever, 634
Stock, bilharziasis, 641
— treatment of bilharziasis, 644
Stokvis, Balantidium coli occurring in lung, 202
Stomach, cancer of, Lamblia intestinalis in, 59, 60
— fluid from, obtained by lavage, rhabdites found in, 378
— larvæ of Gastrophilus inhabiting, 599
— trichomonads in, 55
— Tristrongylus instabilis in, 435
— wall, fibrous thickenings in, produced by species of Gnathostoma, 385
Stomoxys, characters, 609
— differentiation of Lyperosia from, 610
— disease carrier, 603
— species of, 610
— calcitrans (stinging or stable fly), 609
— — diseases transmitted by, 610
— — ova, larval and pupal stages, 609
— — transmission of epidemic poliomyelitis by, 612
Stools, larvæ of Blaps mortisaga in, 542
— method of discovering head of tapeworms in, 674
Streblidæ (bat parasites), 611
Strepsiptera, characters, 531
Strong and Musgrave, species of amœbæ distinguished by, 31
Strongylidæ, 375, 432
— free life of young stages, 20
Strongyloides, European, free-living generation generally absent in, 383
— fulleborni, 384
— intestinalis, geographical distribution, 384
— larvæ of, cultivation, 474
— life-history of, 19
— morphology, 379
— stercoralis, free-living form, morphology of, 381
— — — generation, female, 382
— — habitat in body, 755
— — heterogony of, 381
— — infection by, diagnosis, 675, 676
— — — diarrhœa associated with, 381
— — — expulsive treatment, 675
— — — pathological significance, 674
— — — prophylaxis against, 675
— — — symptoms, 674, 675
— — larva from fresh human fæces, 382
— — — mature filariform, 383
— — mode of development, 373
— — occurrence in man, 384
— — parasitic generation, morphology, 381
— — — — ova, 381, 382
— — synonyms, 380
— — and Ancylostoma duodenale, larvæ of, differences between, 451
— synonyms, 379
— toxic action of, 651
— vivipara, 384
Strongyloplasmata, 208
Stuelp, amaurosis following male fern poisoning, 671
Stuertz, chyluria following infection by Eustrongylus gigas, 682
Stylorhynchus, host of, 135
— oblongatus, gametes of, morphological differentiation, 133, 134
Stypticin in bilharziasis, 643
Sublimate, corrosive, saturated aqueous, fixation of cestodes by, 472
— — solutions, fixation by, 748
— solution in crab louse infection, 712
— — injection in expulsion of Guinea worm, 676
— — — into cutaneous and muscular cysticerci, 663
Suckers of Cestoda, 289
Sucking worms, see Trematoda
Suctoria, 29, 198
— characters and habitat, 29
Sulphur, flowers of, prophylactic against clothes lice, 616
— preparations, application in scabies, 706
Sump bunches, skin affection set up by penetration of larvæ of Ancylostoma duodenale, 455
Surra, animals among which prevailing, 95
— causal agent of, 95
— geographical distribution, 95
— transmission by Chrysops, 601
— — by Stomoxys, 96, 610
— — by Tabanus sp., 96
— trypanosomes in blood of horses with, 67
Swallow bug, see Cimex hirundinis
Swammerdam, discoveries of origin of parasites, 10
Swamps, drainage of, in prevention of malaria, 636
Sweden, ox warble fly (Hypoderma bovis) attacking man in, 596
Swellengrebel and Strickland, on Trypanosoma lewisi, 92
Symbiosis, 6
Symmers, bilharziasis of lung, 642, 643
Symphoromyia, characters, 603
Syngameæ, characters, 459
Syngamus, 459
— habitat and hosts of species, 459
— kingi, habitat and host, 460
— — morphology, 459, 460
— trachealis, bursa of, 461
Syphilis, inoculation with, producing no immunity to yaws, 128
— non-immunity to, produced by inoculation with yaws, 128
— parasite of, 114, 124, 125, 632
— tertiary eruptions of, Treponemata difficult to find in, 125
— treatment, 632
Syrphidæ, rat-tailed larvæ of, characters and habitat, 583, 584
Syzygy of gregarines, 132
Szerlecky, case of intertrigo set up by Oxyuris vermicularis, 696
T.
Tabanidæ (gad flies), characters, 600, 601
— diseases transmitted by, 601
— larvæ, 600
— method of destruction, 601
— ova, 600
— pupæ, 600
Tabanus, species of, transmitting trypanosomes, 96, 601
— bovinus (ox gad fly), 601
Tænia, 331
— africana, mature segment of, 342
— — morphology, 342
— — oncosphere of, 299
— — proglottis and head of, 343
— bremneri, morphology, 337
— capensis, 339
— cœnurus, nervous system, head and part of neck showing, 291
— confusa, mature and gravid segments, 344
— — morphology, 343
— crassicollis, anatomy of, longitudinal section showing, 290
— — cysticercus of, 338
— — host of, 6
— derivation of name (footnote), 331
— echinococcus, hooklets of, 355, 359
— — hosts of, 345
— — morphology, 344
— — organs of, 345
— — percentage of dogs infected with, in various cities and countries (footnote), 345
— — rearing of, in dog, 356
— — synonyms, 344
— — see also Echinococcus
— expulsion of, resulting in cure of chorea, 648
— extracts of, experimental injection, effects, 648
— lata (Dibothriocephalus latus), supposed origin of, 11
— lophosoma, 339
— marginata, 337
— — cysticercus of, 338
— — hooks of, 338
— — hosts of, 338
— oncospheres of species of, animals selected as hosts for development, 299
— — — migration from intestine through blood-vessels to liver, 302
— saginata, cysticercus of, 340
— — expulsion of, best method for, 669, 670
— — frequency in man, 341
— — genitalia, proglottis showing, 293
— — geographical distribution, 341
— — habitat in man, 667
— — host of, 6
— — malformations, 339
— — morphology, 339
— — parasitic in man in association with other tapeworms, 667
— — proglottids of, feeding experiments with, 340
— — prophylaxis against, 668
— — race incidence of infection, 340
— — symptoms produced by infection by, 667, 668
— — synonyms, 338
— — uterine egg, 298
— serrata, cysticercus of, 338
— — hooks of, 338
— — host of, 338
— solium, 339
— — carriers of, 335
— — diagnosis of presence in body, 662
— — Dipylidium caninum confused with, 660
— — expulsion from body, effect on anæmia, 648
— — — — measures for, must be thorough, 336
— — geographical distribution, 334
— — — — corresponds with that of domestic pig, 334
— — habitat in body of man, 662
— — head of, 332
— — host of, 6
— — in man, mode of infection, 335
— — larval infection, 662
— — — see also Cysticercus cellulosæ
— — malformations of, 332
— — modes of transmission, 336
— — morphology, 331
— — parasitic association with Dibothriocephalus latus, 658
— — proglottids, 332
— — prophylaxis against, 668
— — symptoms produced by infection by, 667, 668
— — synonyms, 331
— species of, respective times required for development of cysticercus from date of infection, 304
— — various, respective time required for growth, 306
Tæniæ, development of, 14
— infection by, treatment, symptomatic, 669
— nervous system of, 290
— oncospheres of, 14
— species of, in relation to cystic forms, 16
Tæniidæ, 331
— egg-shell substance, 297
— eggs of, 297
— morphology, 309
— oncospheres of, development of cysticerci from, 303
— rostellum, 289
— — of, ring encircling, 291, 292
Tæniol, administration in ancylostomiasis, 686
— effects of, 672
Tæniorhynchus, 576, 577
— africana, 577
— annulipes, 577
— australiensis, 577
— characters, 564
— major, 577
— ova of, 557, 558, 577
— titillans, 577
— — carrier of larvæ of Filaria bancrofti, 577
— uniformis, 577
— — carrier of larvæ of Filaria bancrofti, 577
Tallqvist, experimental bothriocephalus anæmia, 646
Tamné or thimni of Kabyles, 598
Taniguchi, paragonimiasis of brain, 639
Tapeworms, adult, length of life, 307
— biology, 307
— caudal vesicle, 300
— cysticerci experimentally reared from, 15
— development of, 297
— — embryonal, 298
— embryophore, 298
— experimental rearing of, 15
— — from cysticerci, 15
— expulsion by preliminary aperients, 669
— — by vermifuges, 669
— found in association with other intestinal parasites, 667
— individuality of, early researches as to, 283
— infection by, symptomatic treatment, 669
— injury inflicted by, depends on number in host, 9
— larvæ of, sexual maturity must take place in terminal host, 304
— larval stages, development, 298–301
— — — — modes of, 300
— metamorphosis of, 15
— — of larva into, 305
— method of discovering head in stools, 674
— oncospheres (embryos) of, 298, 299
— — transformation into bladder worms, 303
— origin of, discovery, 11
— — early researches as to, 283
— ova of, 297
— — consistency, 297
— plerocercoid of, 300
— scolex of, 300, 303, 304
Tar and sapo viridis, application in creeping disease, 732
Tarsonemidæ, characters of, 488
Tarsonemus intectus, 489
— uncinatus, 489
Tartar emetic in espundia, 629
— — in Indian kala-azar, 626
— — in infantile kala-azar, 627
— — in Oriental sore, 628
— — in sleeping sickness, 622
Taschenberg, Silvanus surinamensis, 542
Taylor, treatment of bronchial spirochætosis, 633
Technique, protozoological, fixed and stained material, 747
— — fresh material, 745
— — notes on, 745–752
Teeth, carious, spirochæte associated with, 122
Teichomyza fusca, larvæ of, habitat, 584
Teissier, mercury in expulsion of Strongyloides stercoralis, 675
Telosporidia, 28, 129
— characters, 28, 129
Temnocephalidæ, habitat and habits of, 20
Terebinthine oil in chyluria from Filaria bancrofti infection, 677
Ternidens, characters, 439
— deminutus, 439, 440
— — habitat, 441
Tersesthes, 581
Testis, enlarged, in filariasis, 401
— of Ancylostoma duodenale, 449
Tetramitus, 57
— and Chilomastix, differential characters, 735, 736
— how differing from Trichomonas, 57
— mesnili, causal agent of colitis, 57
— — Fanapapea intestinalis identical with, 57
— — habitat, 57
— — synonyms, 57
Tetranychidæ (spinning mites), characters of, 488
Tetranychus, 488
— molestissimmus, geographical distribution, 488
— — itching produced by, 488
— telarius, var. russeolus, effects produced by, 488
Tetratrichomonas, 53 (footnote), 734
Texas fever in cattle, carriers of, 177, 494
— — — causal agent, 173, 177
Theiler, 178, 180
Theileria, 174, 178
— annulata, 180
— characters of, 174
— mutans, 180
— parva, 178, 179
— — agents of transmission, 179
— — Koch’s blue bodies in, 179
— — life-cycle in tick, 179
— — morphology of, 178
— — pathogenic agent of East Coast fever in cattle, 174, 178
— stordii, 180
Thélohan on Myxosporidia, 182, 183
Thelohania contejeani, 186
Theobald, F. V., Arthropoda (jointed limbed animals), 483
Theobaldia, 575
— annulata, bite of, 575
— — characters, 575
— — domestic form, 575
— — geographical distribution, 575
— — larvæ of, habitat, 575
— characters, 564
— spathipalpis, bite of, 575
— — characters, 575
— — geographical distribution, 575
Theobaldinella, 575
Thiarsol in infantile kala-azar, 627
Thiopinol, application in scabies, 706
Thomas, W., introduction of atoxyl in trypanosomiasis, 622
Thomer, treatment of crab louse infection, 712
Thomson, D., sites of development of crescents of tertian malignant parasite, 169
— — see also Ross, Sir R., and Thomson, D.
Thomson, J. D., researches on Trypanosoma lewisi, 89, 90, 92
Thomson, J. G., and Fantham, cultivation of Babesia (Piroplasma) canis by Bass’s method, 172
— — — nuclear phenomena of Babesia canis in cultures, 176
— — see also Fantham and Thomson, J. G.
Thomson, J. G., and Sinton, culture of Trypanosoma rhodesiense, 82, 83
— — — culture of trypanosome forms of T. gambiense, 76
— — — medium employed by, for growth of Trypanosoma gambiense and T. rhodesiense, 745
— — and Thomson, D., methods of cultivation of malarial parasites, 171, 172
— — — number of merozoites of malignant tertian parasite, 168
— — — spirochætes in alimentary tract, 741
Thornhill, toxic symptoms following thymol administration, 686
“Thymni” or tamné of Kabyles, 598, 725
Thymol, administration of, in expulsion of ancylostomes, 685, 686
— — — — mode of, 685, 686
— — — of ascarides, 694
— — — of Oxyuridæ, 697
— — — of Strongyloides stercoralis, 675
— — in flagellate dysentery, 624
— — in Trichuris trichiura infection, 679, 680
— — — — — followed by benzene enemata, 680
— — toxic symptoms following, 686
— enemata in arrest of trichinosis, 681
— — in expulsion of ascarides, 694
Thymoluria, 686
Thymotol, administration in ancylostomiasis, 686
Thyroiditis, parasitic, 87
— see also Trypanosomiasis, Brazilian
Thysanoptera, characters, 531
Tick, stages of life-cycle of Babesia canis and B. bovis in, 176, 177
— bites, paralysis due to, 613
— or relapsing fever, 116, 630
— — — African, carrier of, 116, 496, 630
— — — — importation into Persia, 613
— — — — pathogenic agent, 116
— paralysis, cause of, 504
Ticks, transmission of piroplasmosis by, from recovered to uninfected animals, 178
Tiger, host of Paragonimus westermannii, 250
Tinea rotunda, see Ascaris lumbricoides
Toad, rectum and urinary bladder of, Opalina parasitic in, 207
Todd, on leucocytogregarines in birds, 154
— tick paralysis, 613
— see also Dutton and Todd
Tommasi-Crudeli, early researches on malaria, 156
Tomsk, Opisthorchis felineus, human parasite most frequently found at autopsies at, 253
Tongue, cysticercus of, 663
Townsend, Simulium occidentalis, 579
Toxascaris, characters, 465
— limbata, morphology, 466
— — ovum of, 466
— — synonyms, 466
Toxoplasma, 112
— hosts of, 113
— pyrogenes, association with splenomegaly, 113
Toxorhynchites, characters, 563, 570
Trachea, ascarides invading, 691
Trachoma bodies in infected epithelial cells of conjunctiva, 209 (fig. 119)
— — so-called, cultivation, 210
Trematoda, endoparasitic life spent in intermediate and final host, 18
— relation to Turbellaria, 19
Trematodes (sucking worms or flukes), 212
— age attained by, 230
— alimentary canal, 217
— asexual generations, 224, 225
— cercariæ (larval stages), 225, 227, 228
— cirrus sac, 221
— copulation in, 222
— — cross, 222
— development, 12, 222
— — embryonic and post-embryonic, 224
— — final, conditions necessary for, 225
— developmental cycle, 229
— digenetic, adult, animals harbouring, how infected, 226
— — development, 224, 226
— — miracidia of, 226
— endoparasitic, biology of, 229
— — hosts and habitat of, 229
— excretory bladder, 219
— — system, 219
— — — terminal flame cell, 219
— food of, 218
— found in man, classification, 230
— genital pore, 222
— intestine of, variation in, 217
— investing layer of, 213
— Laurer’s canal of, 221, 222
— metraterm of, 221, 222
— miracidia of, 223, 224
— morphology of, 212
— movements of, 216
— muscular system of, 214
— nervous system of, 216
— organs of sense, 216
— origin of, 12
— — of parasitism in, 20
— ova of, deposition, 223
— — formation, 223
— parenchyma of, 213
— — muscles of, 214
— rediæ of, 225, 226, 227, 228
— salivary glands, 217
— sexual organs, 220
— — — deviation from typical position (footnote), 222
— — — female, 220, 221
— — — male, 220
— shell gland secretion in, 223
— sporocyst of, 225, 227
— suckers of, 213, 214
— and turbellaria, genetic relationship between, 20
Treponema, 114, 115, 123
— calligyrum, 126
— cultivation of species from human mouth, 128, 741
— macrodentium, 128
— microdentium, 128
— morphology, 124
— mucosum, 128
— pallidum, 114
— — causal agent of syphilis, 124
— — cultivation of, method, 125
— — difficult to find in tertiary eruptions of syphilis, 125
— — granule formation, 124, 125, 127
— — morphological and pathogenic variations, 126
— — morphology, 124, 125
— — Spirochæta refringens associated with, 122
— — synonyms, 124
— pertenue, cultivation, 128
— — granule formation, 127
— — mode of infection, 128
— — morphology, 127
— — pathogenic agent of yaws, 114, 127
— — reasons for considering specific cause of yaws, 128
— species of, association with pyorrhœa alveolaris, 128
Treutler, filaria associated with phthisis, 408
— parasite, probably liver-fluke, in vein, 243
Triænophorus, excretory vessels, island formation, 292
— plerocercoid of, 300
Triatoma megista, discovery of Trypanosoma cruzi in, 83, 84
— — phases of development of Trypanosoma cruzi in, 87
— — preventive measures against, 623
Triboulet, Ascaris infection in relation to appendicitis, 653
Trichina spiralis, 423
Trichinella, 421
— development in definite host, 18
— spiralis, 421
— — development of, 373
— — — history of, 423, 424
— — geographical distribution not in correspondence with occurrence of trichinosis in man, 427, 428
— — hosts of, 6
— — in man, percentage of invasion according to nationalities determined by post-mortem examination, 428
— — infection by, 680
— — — distribution in body after, 424
— — — see also Trichinosis
— — invasion and encystment in muscles, 424, 425
— — mammals in which developed experimentally, 421
— — — infected by, in order of frequency, 421
— — — inhabited by, 421
— — morphology, 421
— — normal hosts of, 427
— — symptoms produced by, in periods of invasion, dissemination and encystment, 424, 425
— — viviparous nematode, 371
Trichinellæ, development in encysted condition, 427
— encysted, in man and other mammals, early observations of, 423
— fatal case of infection by, 423
— feeding experiments with, 423
Trichinellidæ, 419
— characters, 375
Trichinellinæ, 421
Trichinosis, amount of prevalence in North America, 428
— diagnosis, 681
— — by blood examination, 681
— epidemics of, 423
— — in Germany, 423, 429
— geographical distribution, 428
— in man, geographical distribution of Trichinella spiralis not in correspondence with occurrence of, 427, 428
— prophylaxis against, 429, 431
— symptoms of, 424, 425, 680
— treatment, before and after development, 681
Trichocephali in appendix, 655
Trichocephalus anæmia, 651
— infection by, effects of, 651
— — in relation to appendicitis, 653
— lacks intermediate host, 21
Trichomonads, habitat in body, 55, 735
— question of cysts of, 56
Trichomonas, 52
— characters of, 52
— diarrhœa due to, 57, 624, 734
— from gut and cæcum of rat, 735
— hominis same as T. intestinalis, 54
— intestinalis, 45, 54
— — axostyle of, 55
— — characters of, 55
— — flagella of, 55
— — relation to T. vaginalis, 54
— — spherical contracted forms in mice, 56
— — transmission, modes of, 56
— points of difference of Tetramitus from, 57
— regions of body other than intestine in which found, 55, 56
— vaginalis, 52, 760
— — characters of, 52, 53
— — flagella of, 53
— — nucleus of, 53
— presence in urethra of male, 53
Trichomoniasis, human, recent researches in, 734
— oral, treatment, 625
— vaginal, treatment, 625
Trichopalpus, 603
— larvæ, characters and habitat, 603
— obscurus, 603
Trichoptera, characters, 531
Trichosoma crassicaudum, female parasitic, 4
— — habitat of, 4
Trichostrongylinæ, characters and habitat, 433
Trichostrongylus, morphology, 434
— instabilis, habitat, 435
— — hosts of, 435
— — in man, cases recorded, 435
— — morphology, 434
— probolurus, habitat, 435
— — hosts of, 435
— — morphology, 435
— vitrinus, hosts of, 436
— — morphology, 435, 436
Trichotrachelidæ, œsophagus of, 363
— unicellular cutaneous glands of, 361
Trichurinæ, 419
Trichuris, morphology, 419
— alcocki, 421
— cameli, 421
— campanula, 421
— crenata, 421
— — infection with, 420
— depressiuscula, 420, 421
— — infection with, 420
— discolor, 421
— giraffæ, 421
— globulosa, 421
— nodosus, 421
— ovis, 421
— — infection with, 420
— trichiura, habitat in man, 420
— — infection with, sources, 679
— — — symptoms, 679
— — — treatment, 679
— — mammals inhabited by, 421
— — mode of attachment to wall of intestine, 679
— — morphology, 419
— — ova, development of, 420
— — — embryo-containing, 420
— — — — infection by, 420
— — parasitic in large intestine, 678
— — percentage found at autopsies, 420
— — synonyms, 419
— unguiculata, 421
Trinidad, mosquito worm in, 598
Triodontophorus, bursal formula (footnote), 439
Troglotremidæ, 249
— morphology, 232
Trombidiidæ, characters, 485
Trombidium, 485
— fuliginosum, 486
— gymnopterosum, 486
— serraticeps, 486
— tlalsahuate, skin affections set up by, 486
Trophozoites of Coccidia, 140, 143
— of Entamœba tetragena, 39, 40
— of gregarines, 132
— of malarial parasites, 159
— of Microsporidia, 185
— of Myxosporidia, 182
Tropical Diseases Bureau Bulletin, foundation of, 69
Tropical sore, see Oriental sore
Trouessart, Histiogaster (entomophagus ?) spermaticus, 515
Trypan-blue treatment of piroplasmosis, 178
— — — dosage for dogs, horses and cattle, 178
Trypanophis, 63
Trypanoplasma, characters of, 63
— hosts of, 63
Trypanoplasms in fish, 68
Trypanosoma, 67
— americanum, 69
— boylei, 99
— — experimental infection with, 99
— — host of, 99
— brucei, 93, 94
— — and T. rhodesiense, question of distinction or identity, 80, 83, 94
— — blepharoplastless strains, 101, 737
— — cause of nagana (tsetse-fly disease), 93
— — development in Glossina morsitans, 94
— — drug resistance of, 101
— — innocuous to big game, 70
— — morphology and life-history in vertebrate host, 94
— — nucleus, blepharoplast and flagellum of, 70
— — posterior nuclei in, 83
— — strain from Uganda, 95
— — — from Zululand, 94, 95
— capræ, monomorphic, 100
— cazalboui, causal agent of “souma,” 100
— — monomorphic, 100
— characters, 67
— congolense, agents of transmission, 100
— — cause of Gambia horse sickness, 100
— — geographical distribution, 100
— — monomorphic, 100
— — probable synonyms, 100
— cruzi, 83
— — crithidial forms, 86
— — culture, 87
— — geographical distribution, 83, 84
— — hosts of, 85, 86, 87
— — in fœtus, 88
— — invertebrate host of, 83, 84, 537
— — life-history in invertebrate host, 86
— — — in vertebrate host, 84
— — — — — modes of multiplication (“sexual” and asexual), 85, 86
— — microgametes and macrogametes, 85
— — morphology, 84
— — possible reservoir of, 87
— — schizogony of, 84, 85, 86
— dimorphon, 100
— equi, 83, 98
— equinum, cause of “mal de caderas,” 96
— — morphology, 96
— — transmission of, 97
— equiperdum, 97
— — cause of “dourine” or stallion disease, 97
— — endotoxins in, 98
— — morphology, 98
— — posterior nuclei in, 83
— — progress of disease, 97
— evansi, blepharoplastless strains, 737
— — causal agent of surra, 95
— — morphology, 95, 96
— — possible case in man, 96
— — synonyms, 95
— — transmission of, 95
— — — experimental, 67
— — variety causing “mbori” in dromedaries, 96
— fringillarum, 737
— gambiense, 68, 72
— — cause of sleeping sickness, 68, 605
— — cultivation of, medium used for, 745
— — cultures of trypanosome forms of, 76
— — development in Glossina palpalis, 74, 75
— — effect of serum reactions on, 80
— — immunization against, does not protect against infection by T. rhodesiense, 80
— — in antelope, 76
— — innocuous to big game, 70
— — invasion of salivary glands of Glossina palpalis, 75
— — latent forms of, 77
— — morphology, 72
— — — in circulating blood, 73
— — serum from animals infected with, no effect on T. rhodesiense, 80
— — — — — trypanolytic for, 80
— — synonyms, 72
— hippicum, agents transmitting, 99
— — cause of “murrina” in mules, 98
— — morphology, 98
— lewisi, crithidial forms, 91
— — — — development in rectum of rat flea, 91, 93
— — inoculation experiments, 90
— — life-cycle in invertebrate host, 90, 91
— — — in vertebrate host, 88, 89
— — morphology, 88
— — multiplication rosettes, 71
— — potential pathogenicity, 737
— — rosette forms, 89, 90
— — strain of, losing resistance to arsenophenyl-glycin, how effected, 93
— — transference from blood of rat to blood of snake, 102
— — transmission of, 88
— nanum, 100
— nigeriense, 76
— noctuæ, 69, 737
— pecaudi, 95
— — causal agent of baleri in sheep and equines, 95
— — posterior nuclei in, 83
— pecorum, 100
— rhodesiense, 69, 76
— — and T. brucei, question of distinction or identity, 80, 83, 94
— — animal reactions, 78
— — cause of Rhodesian sleeping sickness, 69, 76, 605
— — cultivation of, 83
— — — medium used for, 745
— — developmental cycle in Glossina morsitans, 81
— — effect of serum reactions on, 80
— — Glossina morsitans transmitting, 608
— — immunization against T. gambiense does prevent infection by, experiments proving, 80
— — latent or resting forms of, 77, 78
— — morphology, 76, 77
— — non-pathogenic to antelopes, 70
— — partial immunity against, 81
— — pathogenic to man and laboratory animals, 70
— — posterior nuclei in, 83
— — reservoir of, 81
— — resistant to atoxyl, 78
— — serum from animals infected with T. gambiense has no effect on, 80
— — transmission of, 69, 81
— — — climatic factors affecting, 81
— — virulence of, compared with that of T. gambiense, 78
— simiæ, virulent to monkeys and pigs, 100
— theileri, 98, 611
— — geographical distribution, 98
— — morphology, 98
— ugandæ, 95
— uniforme, hosts of, 101
— — monomorphic, 101
— vivax, fatal to cattle, 99
— — monomorphic, 99
— — transmission of, 100
Trypanosome, animal, infection of human being with, 96
— diseases spread by Glossina, 603
— human, 68, 69
— — artificial infection of species of Glossina with, 605
— infections, Liverpool School of Tropical Medicine Expedition sent to investigate, 68
Trypanosomes, adaptation of, 101
— artificial cultivation, 69
— blepharoplastless, 101, 737
— classification, 71, 72
— deleterious or fatal to domestic animals, 69
— general note on development in Glossina, 101
— hosts of, 67, 68, 69
— immunity to, in antelope, 69
— in blood, cultures aid in detection of, 69
— — cyclical variation, 78
— — daily number from case of Rhodesian sleeping sickness, 79
— — method of determining number, 748
— — multiplication, 71
— — periodicity, 69
— — seasonal variation, 69
— in cerebrospinal fluid from cases of sleeping sickness, 68
— latent forms, non-flagellate, from internal organs of vertebrates, 73, 74, 77
— monomorphic, 99
— morphology of, 70
— nuclei of, 70
— pathogenic to man and domestic animals, 70
— percentage of fleas fed on infected rat becoming infected with, 93
— polymorphism, 72
— posterior nuclei in, 83
— resting stages, 72
— transmission from one vertebrate host to another, 72
— transmissive stage in vertebrates, 737
— transmitted experimentally by Stomoxys, 610
— undulating membrane, 71
Trypanosomiasis, African, see Sleeping sickness
— Brazilian, acute, 87
— — chronic, varieties of, 87, 88
— — clinical features, 87
— — hereditary transmission, 88
— — histopathology, 88
— — suggested treatment, 623
— — synonyms, 87
— cryptic, 69
Trypanosomidæ, 61
— characters, 66
— genera of, 67
Tryposafrol, producing blepharoplastless trypanosomes, 737
Tsetse-fly, see Glossina morsitans
— disease, see Nagana
Tuberculosis, see Cestode tuberculosis
“Tuft-like” or “phagocytic” organs of nematodes, 362
Tumours, subcutaneous, associated with invasion by Onchocerca volvulus, 418
Turbellaria, parasitic, 2
— relation of Trematoda and Cestoda to, 19
— and trematodes, genetic relationship between, 20
Turkeys, blackhead in, 145
Turpentine in flagellate diarrhœa, 624
— in nasal myiasis, 719
— oil of, in bilharziasis, 643
Tydeus molestus, habitat, 491
— — host-tormenting, 491
Tylenchus putrefaciens, 379
Typhlitis, association of Oxyuris vermicularis with, 467
Typhlocœlum flavum, progeny of, discovery, 12
Typhoid fever, helminthes as predisposing factor of, 657
— — peculiar fever resembling, 613
— — spread by house-fly, 586
— — symptoms of, in lumbricosis, 650
— vaccine in bilharziasis, 644
Typhus, possibly due to a chlamydozoön, 207
Tyroglyphi, differentiation of Glyciphagi from, 513
Tyroglyphidæ, characters, 511
— habitat and food of, 511
Tyroglyphus longior, 512
— — characters of, 512
— — habitat, 512
— minor, var. castellani, cause of copra itch, 513
— siro, 512
— — characters of, 511
U.
Uganda, strain of Trypanosoma brucei from, 95
— syphilis in, treatment, 632
Uhlenhuth (and others), endotoxins in Trypanosoma equiperdum, 98
Ulcers arising from clothes louse infection, 711
— and boils due to invasion by Cordylobia anthropophaga, 592
— examination for protozoa, 746
— production by species of Enyaliopsis, 542
Ulcus tropicum, agent of, 122
Umbilicus, ascarides escaping from, 656
Unger, treatment of oxyuriases, 697
Uranotænia, characters, 565
Urethra, fistulæ of, arising from bilharziasis, 642
— — treatment, 644
— larvæ of Homalomyia canicularis found in, 585
— maggots passed from, 728
— male, presence of Trichomonas vaginalis in, 53
Urinary apparatus, symptoms of bilharziasis mainly centred in, 641
— passages, invasion by ascarides, 692
Urine, amœbæ found in, 45, 46
— human, aphides said to have been passed in (footnote), 532
— occurrence of Anguillula aceti in, 379
— presence of Nephrophages sanguinarius in, 490
— preservation of ova of flukes in, 472
Urosporidium, 194
— fuliginosum, 195
Urotropine in bilharziasis, 643
Urticaria, echinococcus cysts causing, 651, 652
— set up by Leptus autumnalis, 702
Uterus, cervix, polypoid tumour of, with Schistosoma infection, 643
Uzara in flagellate diarrhœa, 625
V.
Vaccine and emetine treatment combined in pyorrhœa alveolaris, 620
Vaccinia, cell inclusions in, 207, 208
Vagina atrophied in Acoleïnæ, 297
— presence of Rhabditis pellio in, 377
Vaginitis, acute, due to Schistosoma infection, 643
Vanillismus, so-called, cause of, 512
Varicose glands in filariasis, 402
Variola, cell inclusions in, 207, 208
Vegetable food, raw, avoidance of, in prophylaxis against Oxyuriasis, 697
— matter, decomposing, Tyroglyphidæ in, 511
— — larvæ of Homalomyia canicularis found in, 585
Veins, liver-flukes found in, 243
Vena cava and portal vein, communication between, how formed, 272
Verallina, characters, 565
Vermifuges, 669–675
Vertebrates, entamœbæ of, 34
— experimental introduction of insect flagellates into, 104, 112, 737, 738
— internal organs of, latent forms of trypanosomes from, 73, 74
— multiplication of trypanosomes in blood of, 71
— spirochætes in, 116, 122
Vesicles, formation of, in creeping disease, 730
Vesico-prostatic plexus, Schistosoma hæmatobium in, 273, 274
Vianna, histopathology of Brazilian trypanosomiasis, 88
— treatment of espundia, 629
Viereck, discovery of Entamœba tetragena by, 38
Vignolo-Lutari, case of intertrigo set up by Oxyuris vermicularis, 696
Villot, larvæ of Gordiidæ, 479
Vinegar, Anguillula aceti found in, 379
— see also Sabadill vinegar
Virchow, R., development of Trichinella spiralis, 423
— — doubtful case of human coccidiosis,149
— — Echinococcus multilocularis, 356
Vital, liver-fluke in vein, 243
Vlemingkz’s mixture, application in scabies, 706
Vogt, C., on the Helminthes, 3
Vomited matter, spirochætes in, 122
Vorticella in fæces, 206
W.
Wagener, von, lesions produced by Oxyuris vermicularis, 695
— life-history of Oxyuris vermicularis, 467
Waldenburg, experimental infection with Coccidia, 136
Walker and Sellards, experiments with dysenteric amœbæ, 618
Walker, E. L., balantidiasis, 203
— — on Entamœba histolytica, 40
— — prevention and treatment of balantidian dysentery, 637
Walker, Norman, treatment of scabies, 707
Walrus, host of Dibothriocephalus cordatus, 315
Walsh and Riley, Rasahus biguttatus, 540
— — Reduvius personatus, 540
Warble flies (Oestridæ), hosts of, 594
Warburg, extract of male fern in expulsion of ancylostomes, 687
Wasielewski, Hæmoproteus (Halteridium) danilewskyi, var. falconis, 152
Water, eggs of mosquitoes float on, 559
— filtered and boiled, as prophylactic against bilharziasis, 644
— infected, avoidance of, in prophylaxis against Guinea worm infection, 676
— larvæ of Stegomyia fasciata occur in all collections of, 574
— mature larvæ of Ancylostoma duodenale capable of living in, 454
— receptacles, screening against mosquitoes, 636
— stagnant, mosquitoes depositing ova in, 553, 557
— transmission of trichomonad infection by, 56, 624
— weeds harbouring mosquito larvæ, destruction of, 636
Watercress, passage of larvæ of Syrphidæ into human beings through eating, 584
Watsonius, 234
— watsoni, 234, 235
— — diarrhœa in host associated with, 235
— — female organs, 235
— — habitat, 235
— — male organs, 234
— — morphology, 234
— — ova, 235
— — synonyms, 234
Weichselbaum, intestinal myiasis, 726, 727
Weidman, see Smith and Weidman
Welland, Ascaris sp., 465
Wellmann, the ochindundu, 541
Wendelstadt and Fellmer, trypanosomes, mutation experiments with, 102
Wenyon, C. M., connection of Cimex sp. with Oriental sore, 536
— — possible host of Leishmania tropica, 108
— — on Entamœba histolytica, 40, 41
— — on genus Cercomonas, 736
— spherical contracted forms of Trichomonas intestinalis, 56
— supposed intermediate host of parasite of Bagdad sore, 575
— Tetramitus mesnili, 57
— transmission experiments with Trypanosoma lewisi, 92, 93
Werbitzki, blepharoplastless trypanosomes, 101
Wheler, Dermacentor reticulatus, 502
— length of life of Ixodes plumbeus (dog tick) apart from host, 495
— life-history of Ixodes reduvius, 494
Whip worm, see Trichuris trichiura
White mice, experimental production of disease like leishmaniasis in, 103
— — infection with Herpetomonas ctenocephali and H. pattoni, 103
— scour in fowls, causal agent, 145
Whitfield, A., and Hobday, F., transmission of dog mange to man, 523
Whittles, nematode larvæ in periosteum of upper jaw in case of gingivitis, 378
Wiggins, locust injurious to man, 542
Wijnhoff, cases of amœbæ in urine, 46
Wild game, Trypanosoma rhodesiense present in, 81
Wilkinson’s ointment, application in scabies, 706
Williams, Anna W., culture media for amœbæ, 743
— — on cultural amœbæ, 42
Williams, H. U., invasion of human beings by Trichinella according to nationalities, 428
Wilms, myiasis œstrosa dermatosa, 725
Winogradoff, post-mortem discoveries of Opisthorchis felineus, 252, 253
Wirsing, mode of infection of intestinal myiasis, 727
Wohlfahrt, myiasis cutanea from Sarcophaga, 722
Wolff, treatment of cutaneous and muscular cysticerci, 663
Woodcock, transmissive phase of trypanosomes, 737
Wood tick, see Dermacentor occidentalis
Worm abscesses, formation of, 9
— electuary (Störk’s) in expulsion of ascarides, 692
— seed oil in expulsion of ascarides, 694
“Wormlet” burrowing into human epidermis, 599
Worms, intestinal, hereditary transmission of, former belief in, 11
— — of lower animals represent young stages, 21
— — spontaneous generation, belief in, 12
— — transmission by ova, discovery of, 11
Wounds, larvæ in, movements of, 723
Wright, Rhinosporidium kinealyi, 197
Wurtz and Cleri, invasion by Loa loa, 678
Wyeomyia, characters, 565
X.
Xenopsylla, distinctive characters, 545
— host of cysticercoids of Hymenolepis murina and H. nana, 328
— brasiliensis, 547
— cheopis, 546
— — carrier of plague bacillus, 543, 547
— — host of Trypanosoma lewisi, 92
Xeroform, application in Demodex folliculorum canis infection, 709
Xyphorhyncus firmus, 131
Y.
Yaws, climatic distribution, 632
— inoculation with, experimental, 127
— — producing no immunity to syphilis, 128
— non-immunity to, produced by inoculation with syphilis, 128
— pathogenic agent of, 114, 127, 128, 632
— prophylaxis, 632
— species of Sarcophaga concerned in dissemination of, 590
— stages of, 632
— treatment, 632
Yellow fever, mosquito carrier of, 574
— — Paraplasma flavigenum said to be associated with, 180
— — transmission by Stegomyia, 555
— pigment in kidney and liver cells in ancylostomiasis, 647
Yorke and Blacklock, classification of trypanosomes, 72
— see also Blacklock and Yorke
— see also Stannus and Yorke
Z.
Zarniko, case of Oxyuridæ in nose, 696
Zeder, special class of cysticerci established by, 282
Zeller, Echinococcus multilocularis, 356
Zenker, development of Trichinella spiralis, 423
— fatal case of infection by Trichinellæ, 423
— Linguatula serrata, 527
Zenker’s solution, 749
Ziemann, infection by Loa loa, 678
— varieties or sub-species of malignant tertian parasite, 167
Zinn, blood-stained diarrhœa from Strongyloides stercoralis infection, 674
— extract of male fern in expulsion of ancylostomes, 687
Zooparasites, 1
Zschokke, experimental infection of man with Dibothriocephalus latus, 312
— Rhinosporidium in horses, 197
Zuelzer, on spirochætes, 114, 741
Zululand, strain of Trypanosoma brucei from, 94
Zürn, case of transmission of infection by Demodex folliculorum canis to man, 709
Zygotes of Coccidia, 141, 144
— of gregarines, 132, 133
Return to transcriber’s notes
Spelling inconsistencies:
Ankylostoma/Ancylostoma/Anchylostoma
anthelminthic/anthelmintic
Endamœba/Entamœba
proglottids/proglottides
Bilharziasis/Bilharziosis
Spelling corrections:
Ater → After
breath → breadth
Schizotrvpanum → Schizotrypanum
cyle → cycle
vertebrate → vertebrates
the tickis in completely known → the tick is incompletely known
epthelial → epithelial
Protion → Portion
ooks → looks
succeded → succeeded
imes → times
Furthur → Further
tell → tells
o → of
ow → now
fo → of
cytologica → cytological
sucessfu → successful
Agchylostoma → Ancylostoma
Ancylostomalarve → Ancylostomalarven
lombr. → lumbr.
hyatid → hydatid
Szerlicky → Szerlecky
genita → genital
cystercerci → cysticerci
diagnoiss → diagnosis
cysticerus → cysticercus
s → is
n → In
protanrdic → protandric
Cuticule → Cuticle
cel → cell
fron → front
brought → bought
inmature → immature
ater → later
Acarides → Ascarides
artifically → artificially
cauity → cavity
an daccording → and according
he → the
synonomy → synonymy
follow → follows
Ecchinococcus → Echinococcus
Brachyera → Brachycera
NaHO → NaOH
End of the Project Gutenberg EBook of The Animal Parasites of Man, by
H. B. Fantham and J. W. W. Stephens and F. V. Theobald
*** END OF THIS PROJECT GUTENBERG EBOOK THE ANIMAL PARASITES OF MAN ***
***** This file should be named 57713-h.htm or 57713-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/5/7/7/1/57713/
Produced by Thiers Halliwell, Chris Curnow and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive)
Updated editions will replace the previous one--the old editions will
be renamed.
Creating the works from print editions not protected by U.S. copyright
law means that no one owns a United States copyright in these works,
so the Foundation (and you!) can copy and distribute it in the United
States without permission and without paying copyright
royalties. Special rules, set forth in the General Terms of Use part
of this license, apply to copying and distributing Project
Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm
concept and trademark. Project Gutenberg is a registered trademark,
and may not be used if you charge for the eBooks, unless you receive
specific permission. If you do not charge anything for copies of this
eBook, complying with the rules is very easy. You may use this eBook
for nearly any purpose such as creation of derivative works, reports,
performances and research. They may be modified and printed and given
away--you may do practically ANYTHING in the United States with eBooks
not protected by U.S. copyright law. Redistribution is subject to the
trademark license, especially commercial redistribution.
START: FULL LICENSE
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full
Project Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project
Gutenberg-tm electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or
destroy all copies of Project Gutenberg-tm electronic works in your
possession. If you paid a fee for obtaining a copy of or access to a
Project Gutenberg-tm electronic work and you do not agree to be bound
by the terms of this agreement, you may obtain a refund from the
person or entity to whom you paid the fee as set forth in paragraph
1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this
agreement and help preserve free future access to Project Gutenberg-tm
electronic works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the
Foundation" or PGLAF), owns a compilation copyright in the collection
of Project Gutenberg-tm electronic works. Nearly all the individual
works in the collection are in the public domain in the United
States. If an individual work is unprotected by copyright law in the
United States and you are located in the United States, we do not
claim a right to prevent you from copying, distributing, performing,
displaying or creating derivative works based on the work as long as
all references to Project Gutenberg are removed. Of course, we hope
that you will support the Project Gutenberg-tm mission of promoting
free access to electronic works by freely sharing Project Gutenberg-tm
works in compliance with the terms of this agreement for keeping the
Project Gutenberg-tm name associated with the work. You can easily
comply with the terms of this agreement by keeping this work in the
same format with its attached full Project Gutenberg-tm License when
you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are
in a constant state of change. If you are outside the United States,
check the laws of your country in addition to the terms of this
agreement before downloading, copying, displaying, performing,
distributing or creating derivative works based on this work or any
other Project Gutenberg-tm work. The Foundation makes no
representations concerning the copyright status of any work in any
country outside the United States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other
immediate access to, the full Project Gutenberg-tm License must appear
prominently whenever any copy of a Project Gutenberg-tm work (any work
on which the phrase "Project Gutenberg" appears, or with which the
phrase "Project Gutenberg" is associated) is accessed, displayed,
performed, viewed, copied or distributed:
This eBook is for the use of anyone anywhere in the United States and
most other parts of the world at no cost and with almost no
restrictions whatsoever. You may copy it, give it away or re-use it
under the terms of the Project Gutenberg License included with this
eBook or online at www.gutenberg.org. If you are not located in the
United States, you'll have to check the laws of the country where you
are located before using this ebook.
1.E.2. If an individual Project Gutenberg-tm electronic work is
derived from texts not protected by U.S. copyright law (does not
contain a notice indicating that it is posted with permission of the
copyright holder), the work can be copied and distributed to anyone in
the United States without paying any fees or charges. If you are
redistributing or providing access to a work with the phrase "Project
Gutenberg" associated with or appearing on the work, you must comply
either with the requirements of paragraphs 1.E.1 through 1.E.7 or
obtain permission for the use of the work and the Project Gutenberg-tm
trademark as set forth in paragraphs 1.E.8 or 1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any
additional terms imposed by the copyright holder. Additional terms
will be linked to the Project Gutenberg-tm License for all works
posted with the permission of the copyright holder found at the
beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including
any word processing or hypertext form. However, if you provide access
to or distribute copies of a Project Gutenberg-tm work in a format
other than "Plain Vanilla ASCII" or other format used in the official
version posted on the official Project Gutenberg-tm web site
(www.gutenberg.org), you must, at no additional cost, fee or expense
to the user, provide a copy, a means of exporting a copy, or a means
of obtaining a copy upon request, of the work in its original "Plain
Vanilla ASCII" or other form. Any alternate format must include the
full Project Gutenberg-tm License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works
provided that
* You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is owed
to the owner of the Project Gutenberg-tm trademark, but he has
agreed to donate royalties under this paragraph to the Project
Gutenberg Literary Archive Foundation. Royalty payments must be paid
within 60 days following each date on which you prepare (or are
legally required to prepare) your periodic tax returns. Royalty
payments should be clearly marked as such and sent to the Project
Gutenberg Literary Archive Foundation at the address specified in
Section 4, "Information about donations to the Project Gutenberg
Literary Archive Foundation."
* You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or destroy all
copies of the works possessed in a physical medium and discontinue
all use of and all access to other copies of Project Gutenberg-tm
works.
* You provide, in accordance with paragraph 1.F.3, a full refund of
any money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days of
receipt of the work.
* You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project
Gutenberg-tm electronic work or group of works on different terms than
are set forth in this agreement, you must obtain permission in writing
from both the Project Gutenberg Literary Archive Foundation and The
Project Gutenberg Trademark LLC, the owner of the Project Gutenberg-tm
trademark. Contact the Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
works not protected by U.S. copyright law in creating the Project
Gutenberg-tm collection. Despite these efforts, Project Gutenberg-tm
electronic works, and the medium on which they may be stored, may
contain "Defects," such as, but not limited to, incomplete, inaccurate
or corrupt data, transcription errors, a copyright or other
intellectual property infringement, a defective or damaged disk or
other medium, a computer virus, or computer codes that damage or
cannot be read by your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium
with your written explanation. The person or entity that provided you
with the defective work may elect to provide a replacement copy in
lieu of a refund. If you received the work electronically, the person
or entity providing it to you may choose to give you a second
opportunity to receive the work electronically in lieu of a refund. If
the second copy is also defective, you may demand a refund in writing
without further opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO
OTHER WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT
LIMITED TO WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of
damages. If any disclaimer or limitation set forth in this agreement
violates the law of the state applicable to this agreement, the
agreement shall be interpreted to make the maximum disclaimer or
limitation permitted by the applicable state law. The invalidity or
unenforceability of any provision of this agreement shall not void the
remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in
accordance with this agreement, and any volunteers associated with the
production, promotion and distribution of Project Gutenberg-tm
electronic works, harmless from all liability, costs and expenses,
including legal fees, that arise directly or indirectly from any of
the following which you do or cause to occur: (a) distribution of this
or any Project Gutenberg-tm work, (b) alteration, modification, or
additions or deletions to any Project Gutenberg-tm work, and (c) any
Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of
computers including obsolete, old, middle-aged and new computers. It
exists because of the efforts of hundreds of volunteers and donations
from people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future
generations. To learn more about the Project Gutenberg Literary
Archive Foundation and how your efforts and donations can help, see
Sections 3 and 4 and the Foundation information page at
www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg Literary
Archive Foundation are tax deductible to the full extent permitted by
U.S. federal laws and your state's laws.
The Foundation's principal office is in Fairbanks, Alaska, with the
mailing address: PO Box 750175, Fairbanks, AK 99775, but its
volunteers and employees are scattered throughout numerous
locations. Its business office is located at 809 North 1500 West, Salt
Lake City, UT 84116, (801) 596-1887. Email contact links and up to
date contact information can be found at the Foundation's web site and
official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To SEND
DONATIONS or determine the status of compliance for any particular
state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations. To
donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic works.
Professor Michael S. Hart was the originator of the Project
Gutenberg-tm concept of a library of electronic works that could be
freely shared with anyone. For forty years, he produced and
distributed Project Gutenberg-tm eBooks with only a loose network of
volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as not protected by copyright in
the U.S. unless a copyright notice is included. Thus, we do not
necessarily keep eBooks in compliance with any particular paper
edition.
Most people start at our Web site which has the main PG search
facility: www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.