Mount Rainier from Indian Henry's Hunting Ground, July, 1934. (Fish and Wildlife Service photo by Victor B. Scheffer. No. 864.)
BY
WALTER W. DALQUEST
University of Kansas Publications
Museum of Natural History
Vol. 2, pp. 1-444, 140 figures in text
April 9, 1948
UNIVERSITY OF KANSAS
LAWRENCE
1948
MAMMALS OF WASHINGTON
MAMMALS OF WASHINGTON
BY
WALTER W. DALQUEST
(Contribution from the Museum of Natural History, University of Kansas)
University of Kansas Publications
Museum of Natural History
Volume 2, pp. 1-444, 140 figures in text
April 9, 1948
UNIVERSITY OF KANSAS
Lawrence
1948
(7)
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall and Donald F. Hoffmeister
Volume 2, pp. 1-444. 140 figures in text
April 9, 1948
University of Kansas
Lawrence, Kansas
Printed by
Ferd Voiland Jr., State Printer
Topeka, Kansas
1948

21-1993
| PAGE | |
| Introduction | 13 |
| Physiographic Provinces of the State | 16 |
| Distributional Areas | 20 |
| Climate and Vegetation | 25 |
| Life-zones and Ecology | 32 |
| Geologic History of Washington | 46 |
| The Faunas | 52 |
| Speculation as to Emigrational History of the Mammals | 54 |
| Speculation as to the Later Distributional History of the Mammals | 68 |
| Explanation of Treatment | 110 |
| Check List of Mammals | 112 |
| Accounts of Species and Subspecies | 121 |
| Addenda | 416 |
| Bibliography | 417 |
| Index | 430 |
|
Topography |
||
| FIGURE | PAGE | |
| Mount Rainier from Indian Henrys | frontispiece | |
| 1. | Cascade Mountains at Canadian Boundary | 17 |
| 2. | Columbia River one mile south of Kellers Ferry | 19 |
| 4. | Cascade Mountains in Chelan National Forest | 21 |
| 5. | Mount Rainier, Yakima Park | 22 |
| 6. | Columbia River in Stevens County | 23 |
| 7. | Blue Mountains, Washington | 24 |
| 8. | North Side of Mount Rainier | 26 |
| 9. | Mount Rainier: Cowlitz Chimneys | 28 |
| 10. | A "pothole" crowded by drifting sand | 30 |
| 12. | Arctic-Alpine Life-zone on Mount Rainier | 33 |
| 13. | Humid subdivision of the Transition Life-zone | 34 |
| 14. | Timbered, arid subdivision of the Transition Life-zone | 35 |
| 15. | Upper Sonoran Life-zone | 37 |
| 16. | Canadian Life-zone | 38 |
| 18. | McDowell Lake. Little Pend Oreille Wildlife Refuge | 50 |
| 19. | Pend Oreille River near Newport | 59 |
| 20. | Rocky bluff along north bank of the Columbia River | 65 |
|
Mammals |
||
| FIGURE | PAGE | |
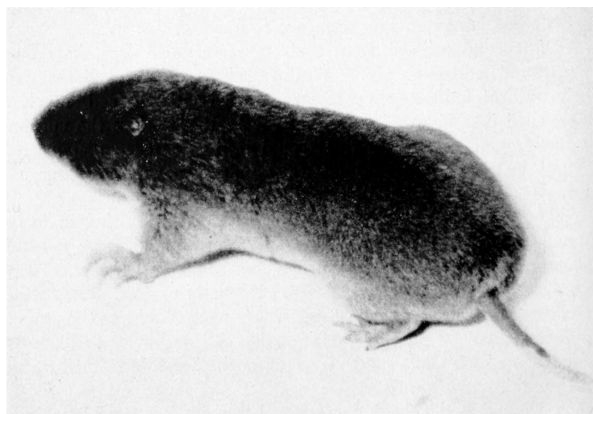
| 21. | Gibbs shrew-mole | 122 |
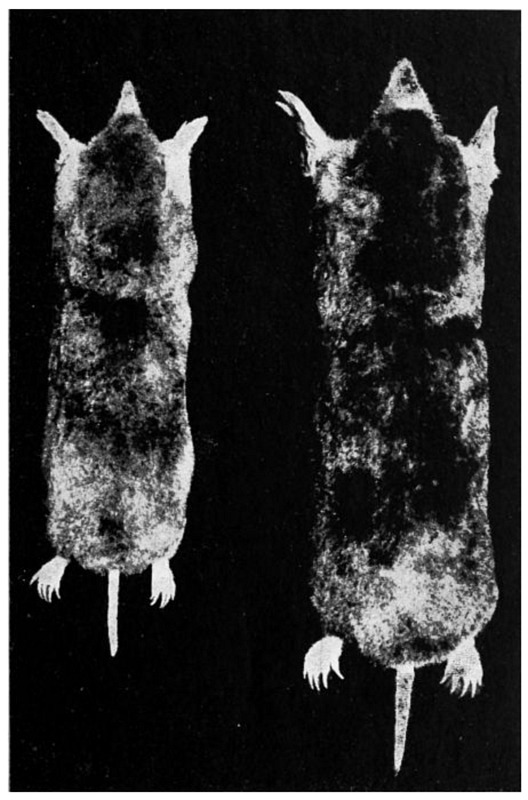
| 23. | Coast mole and Townsend mole | 125 |
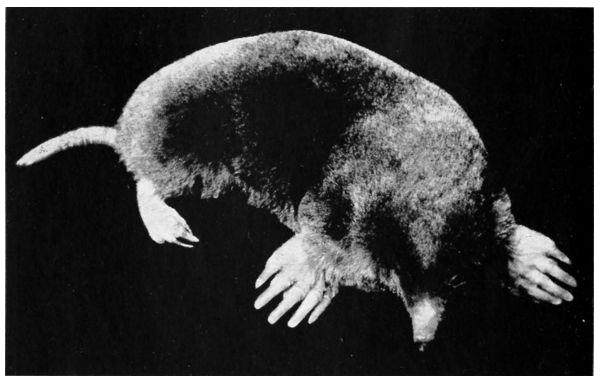
| 25. | Coast mole | 128 |
| 41. | Long-eared bat: female with young | 161 |
| 42. | Boulder Cave: habitat of long-eared bats | 162 |
| 48. | Female black bear and two cubs | 171 |
| 49. | Black bear in "hibernation" | 173 |
| 54. | Fisher | 187 |
| 59. | Wolverine | 202 |
| 60. | Wolverine: dried pelt | 203 |
| 62. | River otter | 208 |
| 66. | Badger | 220 |
| 69. | Coyote | 226 |
| 72. | Cougar or mountain lion: skin | 234 |
| 73. | Cougar or mountain lion: pelts | 235 |
| 75. | Canadian lynx | 238 |
| 76. | Trapper's catch of nine Canadian lynx | 239 |
| 78. | Bobcat | 241 |
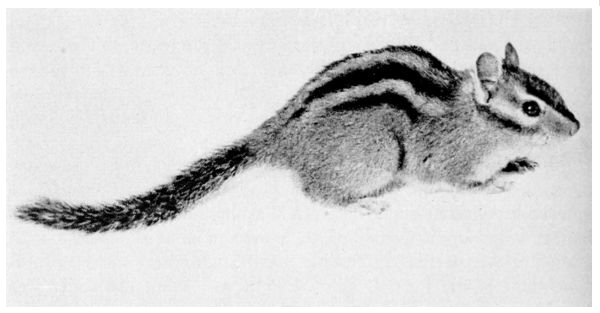
| 82. | Townsend chipmunk | 260 |
| 87. | Golden-mantled ground squirrel | 282 |
| 89. | Feeding station of Douglas squirrel | 287 |
| 91. | Northern flying squirrel | 292 |
| 95. | Northern pocket gopher | 302 |
| 96. | Giant mounds formed by pocket gophers | 304 |
| [Pg 11] 97. | Food cache of northern pocket gopher | 306 |
| 99. | Beaver | 315 |
| 100. | Beaver lodge and pond | 316 |
| 101. | Cottonwood pole carved by beaver | 317 |
| 102. | Road flooded by beavers | 320 |
| 106. | Deer mouse | 327 |
| 112. | Pennsylvania meadow mouse | 345 |
| 115. | Runways of Townsend meadow mice | 350 |
| 119. | Muskrat | 360 |
| 121. | Mountain beaver | 366 |
| 123. | Big jumping mouse in hibernation | 370 |
| 130. | Elk | 392 |
| 131. | Group of elk | 393 |
| 132. | White-tailed deer | 395 |
| 133. | White-tailed deer: fawn | 396 |
| 134. | Left antler of white-tailed deer | 397 |
| 135. | Antlers of white-tailed deer | 398 |
| 136. | Mule deer | 400 |
| 137. | Black-tailed deer | 401 |
| 138. | Mountain goat | 407 |
| 139. | Group of mountain goats | 408 |
|
Distribution Maps |
||
| 3. | Mammalian distributional areas | 20 |
| 11. | Life-zones of Washington | 32 |
| 17. | Extent of Vashon-Wisconsin ice | 48 |
| 22. | Gibbs shrew-mole | 123 |
| 24. | Townsend mole | 127 |
| 26. | Coast mole | 129 |
| 27. | Cinereous shrew | 133 |
| 28. | Merriam shrew and Trowbridge shrew | 134 |
| 29. | Wandering shrew | 137 |
| 30. | Dusky shrew | 140 |
| 31. | Mountain water shrew | 141 |
| 32. | Bendire water shrew and pigmy shrew | 143 |
| 33. | Big myotis | 147 |
| 34. | Yuma myotis | 149 |
| 35. | Fringe-tailed myotis and Keen myotis | 150 |
| 36. | Long-eared myotis | 152 |
| 37. | Hairy-winged myotis | 155 |
| 38. | California myotis | 156 |
| 39. | Small-footed myotis | 157 |
| 40. | Silver-haired bat | 160 |
| 43. | Long-eared bat | 163 |
| 44. | Western pipistrelle | 164 |
| 45. | Big-brown bat | 166 |
| 46. | Hoary bat | 168 |
| 47. | Pallid bat | 170 |
| 50. | Black bear | 175 |
| 51. | Grizzly bears | 177 |
| [Pg 12] 52. | Raccoon | 181 |
| 53. | Western marten | 185 |
| 55. | Fisher | 188 |
| 56. | Ermine | 192 |
| 57. | Long-tailed weasel | 196 |
| 58. | Mink | 199 |
| 61. | Wolverine | 204 |
| 63. | River otter | 209 |
| 64. | Civet cat | 214 |
| 65. | Striped skunk | 218 |
| 67. | Badger | 221 |
| 68. | Red fox | 225 |
| 70. | Coyote | 230 |
| 71. | Wolf | 232 |
| 74. | Cougar | 236 |
| 77. | Canadian lynx | 240 |
| 79. | Bobcat | 243 |
| 80. | Least chipmunk | 252 |
| 81. | Yellow-pine chipmunk | 256 |
| 83. | Townsend chipmunk | 261 |
| 84A. | Marmots | 266 |
| 84B. | Townsend and Washington ground squirrels | 268 |
| 85. | Columbian and Beechey ground squirrels | 274 |
| 86. | Golden-mantled ground squirrels | 280 |
| 88. | Western gray squirrel | 284 |
| 90. | Red and Douglas squirrels | 289 |
| 92. | Northern flying squirrel | 294 |
| 93. | Great Basin pocket mouse | 298 |
| 94. | Ord kangaroo rat | 300 |
| 98. | Northern pocket gopher | 308 |
| 103. | Beaver | 321 |
| 104. | Northern grasshopper mouse | 323 |
| 105. | Western harvest mouse | 325 |
| 107. | Deer mouse | 329 |
| 108. | Bushy-tailed wood rat | 335 |
| 109. | Northern lemming mouse | 337 |
| 110. | Heather vole | 339 |
| 111. | Gapper and California red-backed mice | 342 |
| 113. | Pennsylvania meadow mouse | 346 |
| 114. | Montane and Townsend meadow mice | 348 |
| 116. | Long-tailed meadow mouse | 352 |
| 117. | Water rat | 356 |
| 118. | Creeping mouse | 357 |
| 120. | Muskrat | 362 |
| 122. | Mountain beaver | 368 |
| 124. | Big jumping mouse | 372 |
| 125. | Porcupine | 375 |
| 126. | Pika | 378 |
| 127. | Snowshoe rabbit | 383 |
| 128. | Black-tailed jack rabbit | 386 |
| 129. | Nuttall cottontail | 388 |
Mammals of Washington are of especial interest to the naturalist because many of them are recent immigrants; much of Washington was buried under thick glacial ice until relatively recently and many of the mammals, therefore, have inhabited the area only since the ice disappeared. The evolution or development of certain subspecies, in Washington, has certainly occurred within the last few thousand years. To be able thus to date such evolutionary changes as have occurred is of course a matter both of importance and interest to zoölogists. The evolutionary changes in several species are relatively great. In color, for example, the bobcat in the humid coastal area of western Washington is notable for its dark coloration, whereas in the more arid area of southeastern Washington it is remarkably pale and of a different subspecies. Within the limits of the state of Washington, elevations ranging from sea level to more than 14,000 feet occur. Since different elevations have their characteristic mammals, more kinds are found in Washington than in other areas of corresponding size that lack such topographic diversity. Expressed in terms of the life-zone concept, Washington includes faunas ranging from the Upper Sonoran Life-zone to those of the Arctic-Alpine Life-zone.
The basis for a study of the mammals of Washington was laid in 1929 by W. P. Taylor and W. T. Shaw in the "Provisional List of the Land Mammals of the state of Washington." Bailey's "Mammals and Life Zones of Oregon" and "The Recent Mammals of Idaho" by W. B. Davis deal with the habits and distribution of mammals in the areas bordering Washington on the south and east, and were very useful in the organization of the present report.
The study was first planned from the taxonomic and ecologic point of view. Such a study, of necessity, involves the classification and distribution of the forms concerned. Classification has required more work than any other part of the study and has been, in a sense, the nucleus of the study. Nevertheless, as the report began to take form it was recognized that the part dealing with classification and other purely technical aspects of the paper probably would be uninteresting to the average reader. Therefore it was felt that a greater impetus to the study of the mammals of Washington would be given by reducing the taxonomic accounts to the minimum and dealing principally with the problems of distribution.
As originally planned, the present report was to be of joint authorship by Dr. Victor B. Scheffer of the United States Fish and Wildlife Service, Seattle, and the writer. The press of other work prevented Dr. Scheffer from devoting as much time as he had planned to the project. He has, however, contributed his field notes, specimens, and photographs, and in many other ways assisted in the project.
Field work on mammals of Washington was carried out by the writer from 1936 to 1940 but a decision to prepare a complete report was not reached until 1938. Intensive field work was done between 1938 and 1940. In August, 1941, the author took up residence at Berkeley, California. Drafting of the manuscript was begun at that time as a student under Professor E. Raymond Hall. War conditions and the press of other work delayed completion. Subsequently, the manuscript was put in final form at the University of Kansas.
Many persons in addition to Dr. Scheffer have given assistance in the course of this work. Dr. E. Raymond Hall, in particular, encouraged the project and gave assistance in various ways including critical attention to the manuscript. I am indebted also to Dr. Trevor Kincaid and Mrs. Martha Flahaut of the University of Washington, Dr. Seth B. Benson and Dr. Alden H. Miller of the University of California, Dr. H. H. T. Jackson of the U. S. Fish and Wildlife Service, Dr. George E. Hudson of the Charles R. Conner Museum, Mr. Burton Lauckhart of the State of Washington Department of Game, and Mr. Ernest Booth of Walla Walla College. Mrs. Peggy B. Dalquest typed and edited the several preliminary drafts of the manuscript and aided in the laboratory and field work. Thanks are due also to many others, including game protectors, hunters and trappers, who have given assistance. The names of some of them are mentioned in the following pages.
Approximately ten thousand specimens of mammals were used. In decreasing order, according to the number of specimens studied from Washington, the following collections are to be mentioned: materials obtained principally from southern Washington in the years 1939 to 1942 as a result of the interest of Miss Annie M. Alexander and Dr. E. Raymond Hall; these materials are in the University of California Museum of Vertebrate Zoölogy. The writer's own collection which at one time numbered 2,500 specimens was the second source. The residue, the part not destroyed by fire at the writer's home in the spring of 1942, in Seattle, now is in the [Pg 15] Museum of Vertebrate Zoölogy at the University of California and the Museum of Natural History at the University of Kansas. The other collections are those of the Biological Surveys of the United States Fish and Wildlife Service, the Washington State Museum at Seattle, the Charles R. Conner Museum at Washington State College, and the Museum of Natural History at the University of Kansas. In the latter collection are some materials obtained nearly half a century ago by the late L. L. Dyche, some recently taken specimens added by reason of the provision for work of this kind by the University of Kansas Endowment Association and, as noted above, a part of the author's original collection. Selected specimens from several other collections have been used and these are indicated in the text when particular reference is made to the specimens. Most of the specimens studied were conventional study skins with skulls. In some instances skeletons, skins alone, skulls alone, or entire animals preserved in alcohol have been used.
The state of Washington was divided into seven physiographic provinces by Culver (1936). Culver points out that the physiography, though complicated in detail, is basically simple.
The state, including Puget Sound and other inland waters, is nearly rectangular in shape and is 69,127 square miles in area. Its western boundary is the Pacific Ocean. Politically, it is bounded on the north by the United States-Canadian Boundary (49° north lat.), on the east by the state of Idaho, and on the south by the state of Oregon.
The Cascade Mountain Range, or Cascade Mountains Province, runs from the northern to the southern boundary and divides the state into two sections, of which the eastern is slightly the larger. The mountain range trends approximately 10° east of north and continues uninterruptedly into British Columbia, but on the south the Columbia River separates the Washington Cascades from the Cascades of Oregon. Near the northern border of the state the range is wide, extending from the Mount Baker Range on the west to Mount Chopaka, 100 miles to the east. In the central part of the state it is more compact, being some 60 miles wide in the vicinity of Mount Rainier. Farther south it expands to approximately 100 miles.
The Cascades of Washington possess five great volcanic cones. These are Mount Baker (10,750 feet elevation) on the north, Glacier Peak (10,436 feet) in the north-central part, Mount Rainier (14,408 feet) in the central area, and Mount Adams (12,326 feet) and Mount St. Helens (9,697 feet) on the south. Excluding these volcanic peaks, the crests of the Cascades have a relatively uniform level descending from an average of 8,000 feet at the north to 5,000 feet at the south. Ranges jutting southeastwardly from the north-central Cascades parallel elongate intermontane valleys. These include the Entiat and Wenatchee mountains. The latter range reaches the Columbia River and forms an important barrier to mammalian movements.
There is an extensive area of anticlinal ridges extending from the southern Cascades to the Columbia River. This area includes the Horse Heaven and other hills. Most of it is drained by the Yakima River and is termed, in this report, the Yakima Valley Area.
Fig. 1. Cascade Mountains at Canadian boundary, looking west along boundary trail. Monument 100 in foreground, headwaters of Ashnola River in broad valley at right. (Forest Service photo, No. 4328.)
The Puget Sound Trough, or Puget Sound Province, is immediately west of and parallel to the Cascade Mountains. It is part of a structural downwarp that extends southward into Oregon. Most of the area is below 1,000 feet elevation, and much of the northern part is below sea level and therefore flooded by the marine waters of Puget Sound. The most prominent feature of this area is Puget Sound. This is a glacially-carved and drowned river valley, studded with islands, peninsulas, fjords and bays that all possess a general north-south orientation resulting from the direction of ice movement. Puget Sound is connected with the Pacific Ocean by the Strait of Juan De Fuca, a wide channel separating the state of Washington and Vancouver Island.
The San Juan Islands represent the glaciated remnants of mountains that, in preglacial time, may have connected the mountains on Vancouver Island with the Cascades of Washington. The San Juan Islands lie at the junction of Puget Sound, the Strait of Georgia, and the Strait of Juan De Fuca. As a result of a boundary dispute and subsequent arbitration, the islands were apportioned, on the basis of the deepest channel separating them, between Canada and the United States. The American portion includes more than 400 islands. These vary in size from mere rocks above high tide to Orcas Island, 60 square miles in area.
The Olympic Peninsula, or Olympic Province, lies between Puget Sound and the Pacific Ocean. The Strait of Juan De Fuca separates this peninsula from Vancouver Island on the north. In the south the valley of the Chehalis River is a convenient boundary for the province. The central portion of the peninsula is occupied by the Olympic Mountain Range. This range is nearly oblong in shape, measuring some 70 miles east to west by 45 miles north to south. The mountains are extremely rough and jagged. They rise from sea level to above 6,000 feet. The highest peak, Mount Olympus, is 8,150 feet in elevation.
South of the Olympic Province and west of the Puget Sound Trough is an area of low, rough hills. Culver called it the Willapa Hills Province.
The northern third of the land east of the Cascade Mountains, or northeastern Washington, is termed the Okanogan Highland Province by Culver. Its southern boundary is set at the east-west flow of the Spokane and Columbia rivers. The outstanding physiographic feature of this area is its division into north-south trending areas of lowland with intervening highlands and mountain ranges. The rivers are, from east to west, the Clark Fork, Colville, Columbia, Kettle, San Poil and Okanogan. Not all intervening highlands are separately designated as mountains. Among these named are the Pend Oreille, Huckleberry, Kettle River, and Okanogan ranges.
The part of eastern Washington south of the Okanogan Highland Province, save the extreme southeastern corner of the state, constitutes the Columbia Lava Province. This is an extensive, relatively level plateau that lies mainly below 2,000 feet elevation. The plateau consists of gently folded lava flows that reach a depth of 4,000 feet in some places (Russell, 1893) and slope inward from the [Pg 19] east, north, and, in part, the west (Flint, 1938). These horizontal layers of basalt are extremely resistant to erosion by other than large rivers. Two great gashes cross the Plateau diagonally from the northeast to the southwest; these are Moses Coulee and the Grand Coulee. These old coulees are the former valleys of the Columbia River, and were formed at the time when the course of the river was successively blocked by the advance of Pleistocene ice. The Snake River crosses the southern edge of the Columbia Lava Province and separates the plateau proper from an area of similar land to the southward.
Fig. 2. Columbia River one mile west of Kellers Ferry, Washington, elevation 1,060 feet, April 16, 1940. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 933.)
The Blue Mountains Province is an area of relatively small extent in the extreme southeastern corner of the state of Washington. There, the province concerned constitutes, as it were, a northward extension of the Blue Mountains of Oregon. The mountains rise to only 5,000 feet elevation in the Washington part of the Blue Mountains Province.
The physiographic provinces are areas of land form. The form of the land has a considerable effect on the temperature, humidity, drainage, weathering, soil, and other non-organic features that combine to produce the various life-zones and influence the distribution of mammals. One might therefore expect a close correlation of mammalian distributional areas with physiographic provinces. Although there is a correlation, it is not exact because the distribution of mammals is influenced also by certain other factors. Among these are historical factors and isolation by geographic barriers.
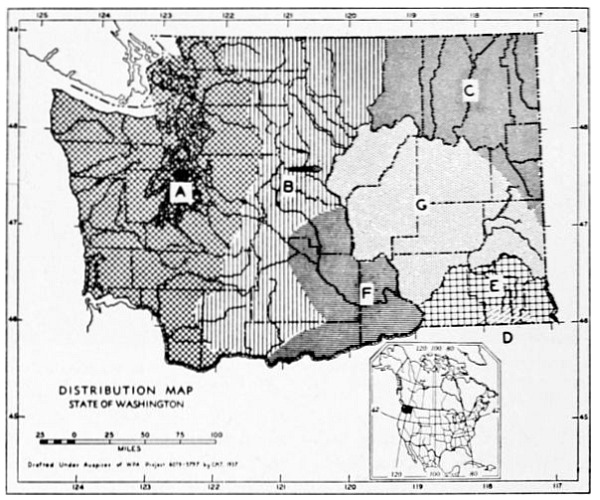
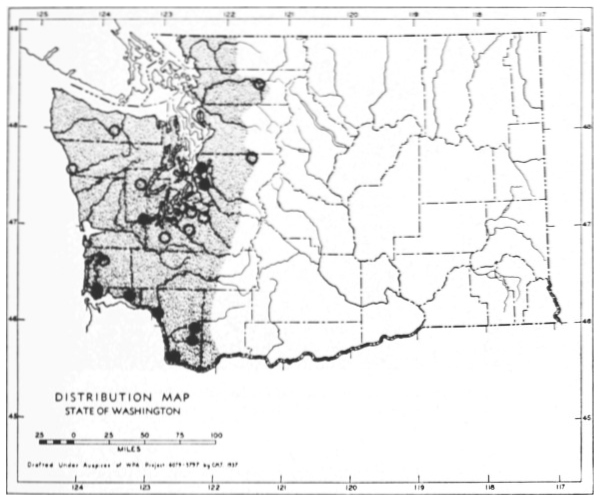
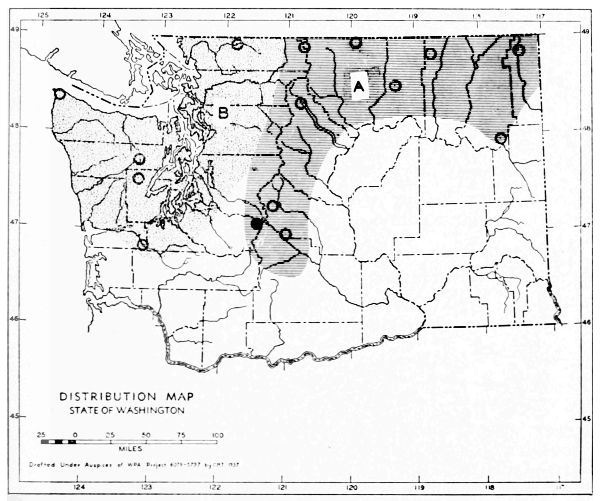
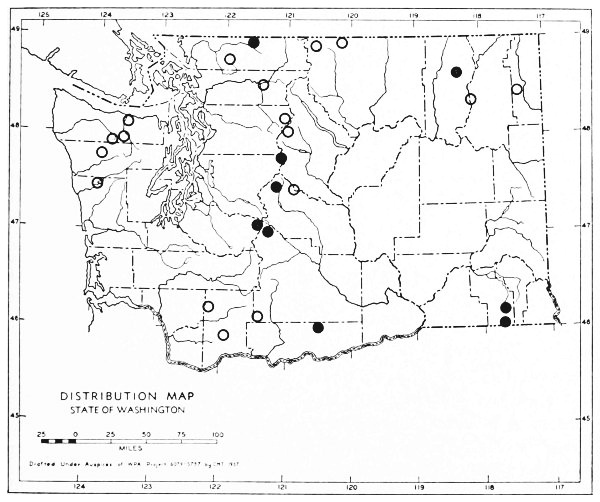
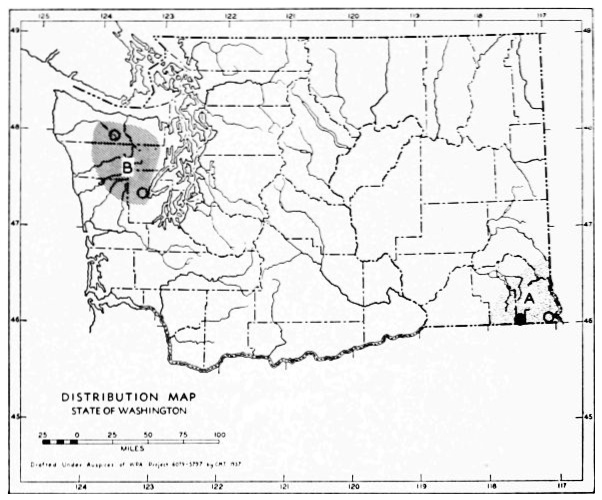
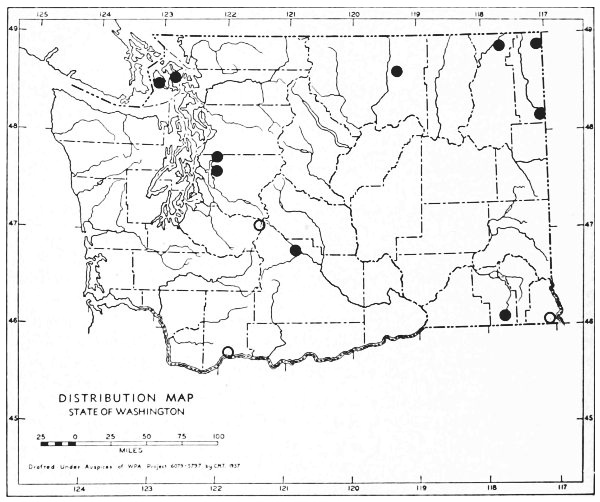
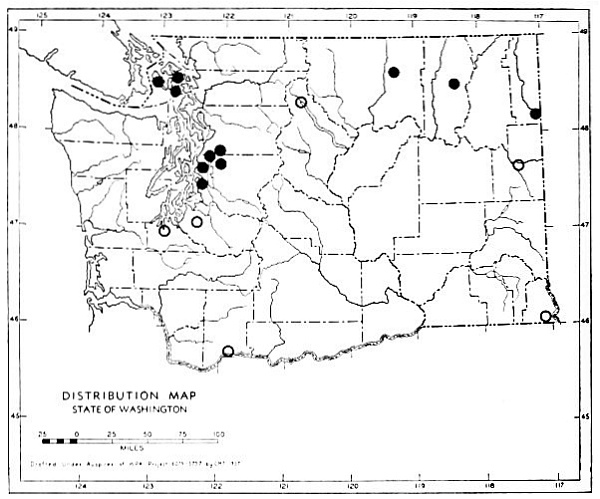
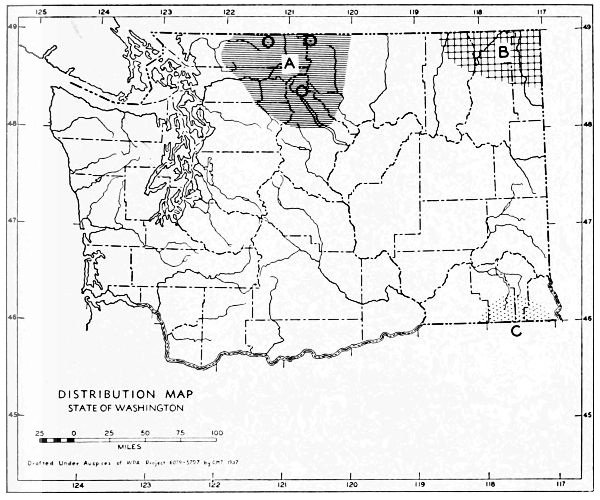
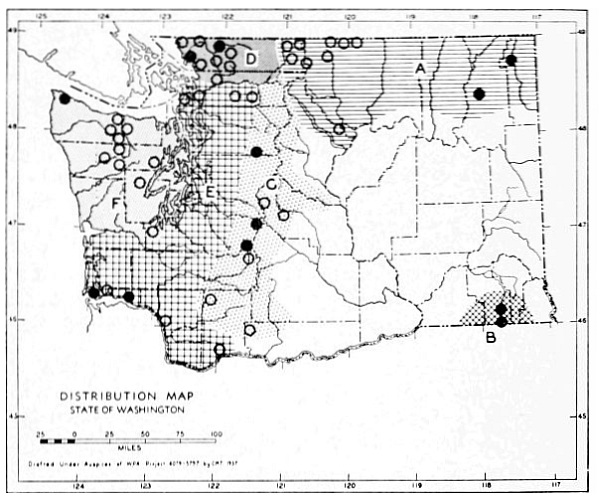
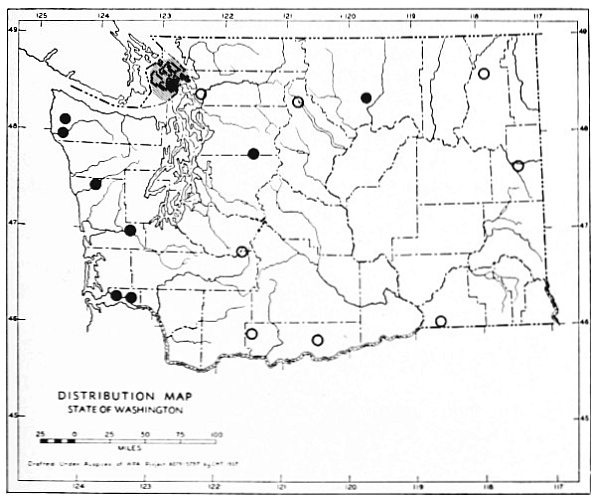
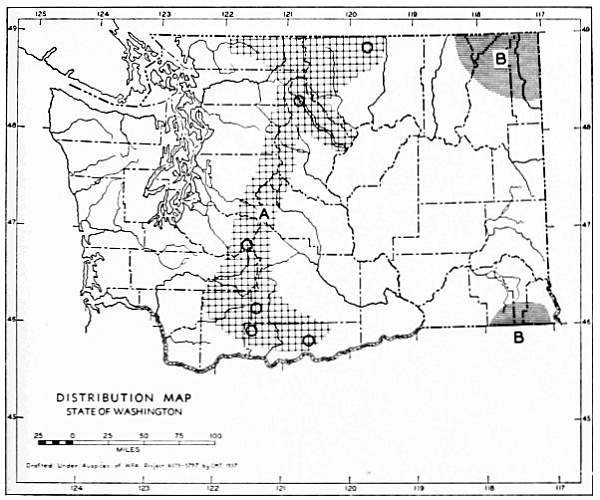
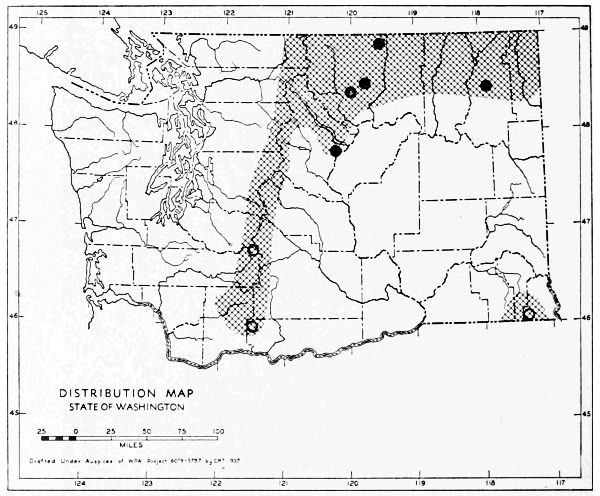
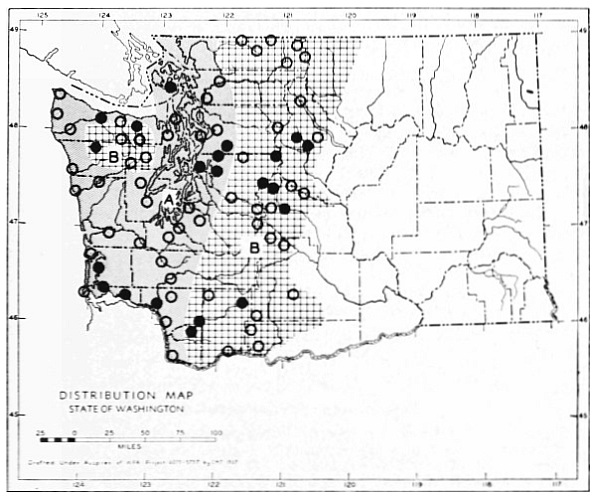
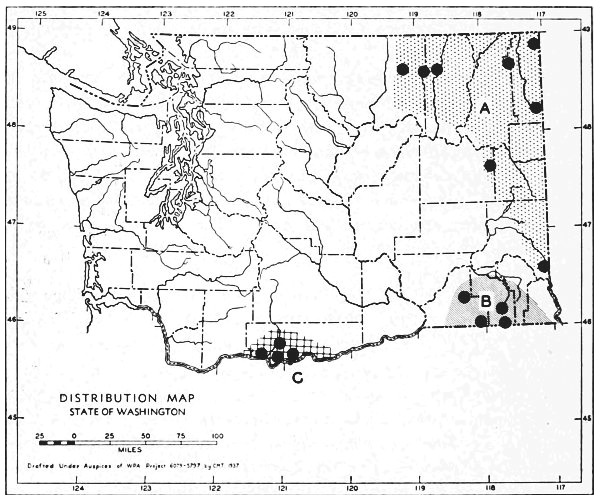
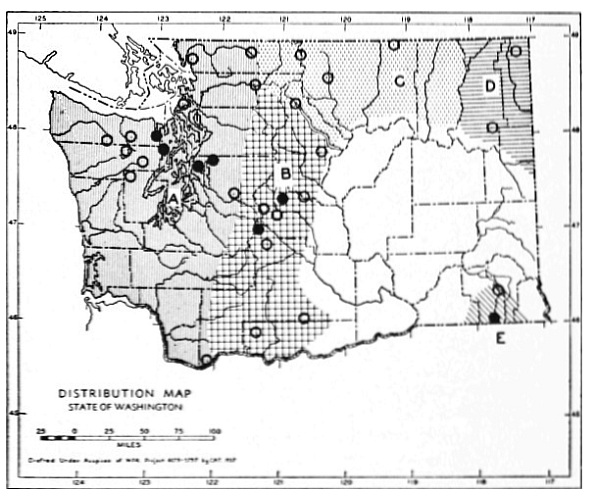
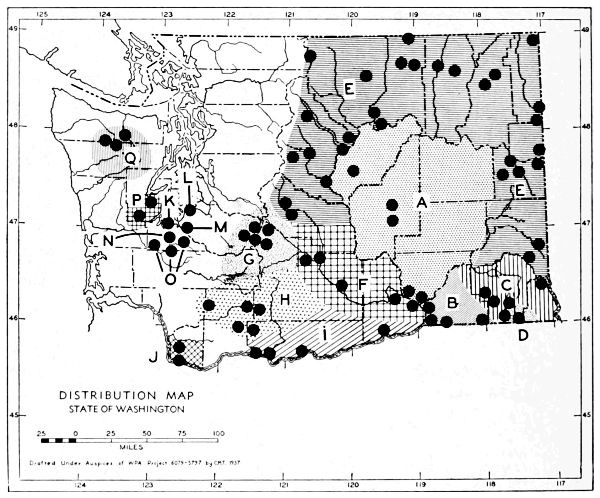
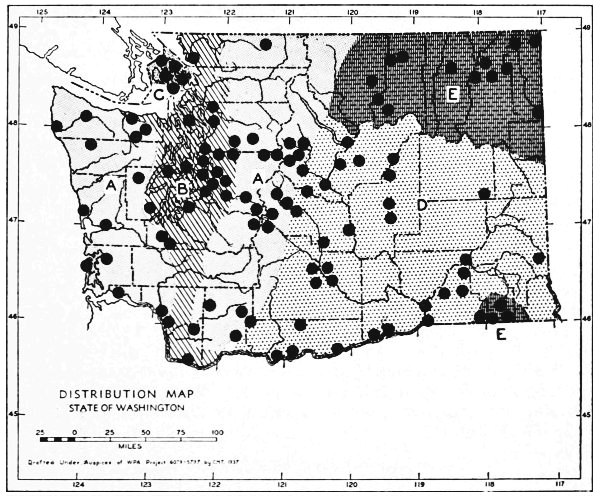
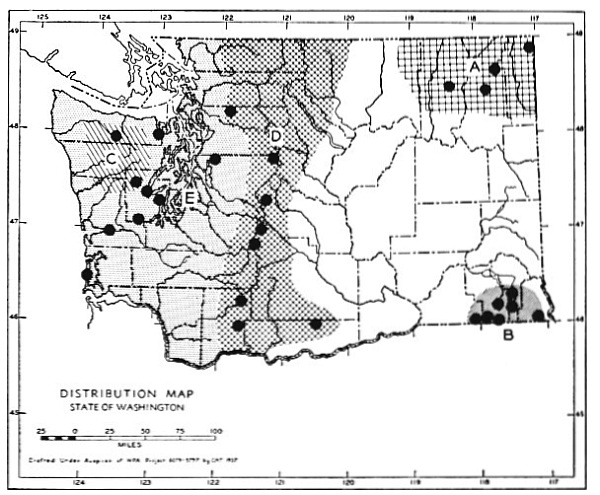
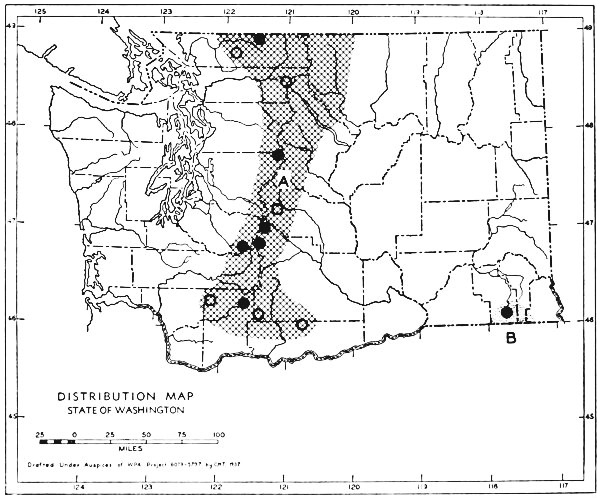
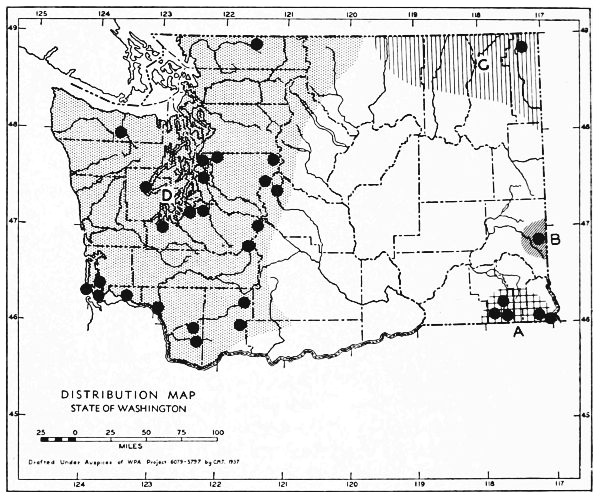
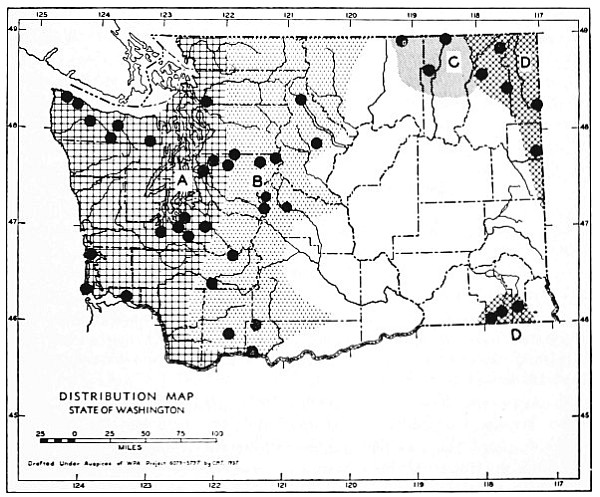
Fig. 3. Mammalian distributional areas of Washington. A. Western Washington. B. Cascade Mountains. C. Northeastern Washington. D. Blue Mountains. E. Southeastern Washington. F. Yakima Valley. G. Columbian Plateau.
The Cascade Mountains Province of Culver includes the Yakima Valley Area. This province contains two completely different mammalian distributional areas. The higher mountains possess a boreal, alpine fauna; the Cascade Range itself is called the Cascade Area [Pg 21] in this report. The Yakima Valley Area possesses a desert fauna derived from the desert of eastern Oregon.
The land west of the Cascades is separated into three physiographic provinces, the Puget Sound, Willapa Hills, and Olympic Mountains Province. The differences between the mammalian faunas of the Puget Sound and Willapa Hills provinces are slight. The Olympic Mountains possess a few species not found in the lower areas. The similarities of the faunas of the three provinces far outnumber their differences, and it seems best to consider them subdivisions of one distributional area.
Fig. 4. Cascade Mountains in Chelan National Forest, looking southwest at Straight Ridge; Cataract Creek (Methow watershed) at left. (Forest Service photo. No. 4260.)
The Okanogan Highland Province extends, from a physiographic point of view, west of the Okanogan River Valley. This valley, however, is a fairly efficient barrier to mammals. Thus the part of the state east of the Okanogan Valley and north of the east-west flow of the Spokane and Columbia rivers may be called the Northeastern Washington Distributional Area.
The Columbia Lava Province includes the land both north and south of the Snake River. Since the Snake River serves as a barrier to some species, it seems better to term the area north of the Snake River the Columbian Plateau Area and that to the south the Southeastern Washington Distributional Area.
The Blue Mountains Province and the Blue Mountains Distributional Area are the same.
Fig. 5. Yakima Park (or Sunrise Park), elevation 6,000 feet, Mount Rainier, August 29, 1932. (Photo by 116th Photo Section, Washington National Guard, No. 014-36A-116.)
Fig. 6. Columbia River at Hunters Ferry, Stevens County, Washington, April 15, 1940. (Fish and Wildlife Service photo by Victor B. Scheffer. No. 932.)
Fig. 7. Blue Mountains, Umatilla National Forest, Washington, looking north-northeast across Al Williams Ridge to Tucannon River; 1933. (Forest Service photo, No. 4437.)
|
Physiographic Provinces
|
Distributional Areas
|
||||
| Cascade Mountains | { { |
Cascade Mountains Yakima Valley |
|||
|
Puget Sound Willapa Hills Olympic Mountains |
} } } |
Western Washington | |||
| Okanogan Highlands | Northeastern Washington | ||||
| Columbia Lava | { { |
Columbian Plateau Southeastern Washington |
|||
| Blue Mountains | Blue Mountains | ||||
The life-zone theory of plant and animal distribution was proposed by Merriam (1892). Merriam's life-zones have been severely criticized by many authors, especially because an error was made in computing some of the data on temperature. However, zonation of vegetation and animals is obvious in Washington, and the life-zone concept has been employed in Washington by numerous botanists and zoölogists. Among them are: Piper (1906), Taylor and Shaw (1927), Jones (1936, 1938) and St. John (1937).
The higher parts of the Cascade Mountains are in the Arctic-alpine Life-zone. This is the area of wind-swept ridges, living glaciers, and permanent snow fields.
Trees are absent but a few shrubs are present; these include: Juniperus sibirica, Salix cascadensis, Salix nivalis, Gaultheria humifusa, Empetrum nigrum, and the heathers, Phyllodoce glanduliflora, Cassiope mertensiana and Cassiope stelleriana. Jones (1938) lists a total of 98 species of plants from the Arctic-alpine Life-zone of Mount Rainier. Many of these plants are most abundant in the next life-zone lower, and are of but incidental occurrence in the Arctic-alpine Life-zone. No mammalian species is resident but individuals of several species regularly visit and occasionally breed there.
Below the Arctic-alpine the Hudsonian Life-zone stretches the entire length of the Cascades. Temperatures are low, especially in winter; then the thermometer does not rise above zero for weeks at a time. The average annual temperature at Paradise, 5500 feet, Mt. Rainier, is 38.6° (all temperatures given here are in degrees Fahrenheit). Snowfall is heavy. The average yearly snowfall, for four years, at Mt. Baker Lodge, at 4200 feet elevation, Whatcom County, was 478 inches; at Goat Lake, 2900 feet, Snohomish County, 261 inches; Tye, Stevens Pass, 3010 feet, King County, 398 inches; Paradise, 5500 feet, Mt. Rainier, 587 inches. The deepest snow recorded at Paradise was 27 feet, 2 inches on April 2, 1917. Following the spring thaws the mountain passes are opened to travel, usually in April or May, although nightly temperatures in April and May are still below zero. Spring precipitation is heavy, the monthly average for a twelve-year period at Paradise being 6.78 inches in April and 5.5 inches in May. Summer temperatures are high in the daytime, when the sun beats down through the rarefied atmosphere, but cool at night when accumulated heat is lost through the thin atmospheric blanket. In summer precipitation is light, averaging, at Paradise, 3.46 inches in June, .9 inches in July, and 3.44 inches in August. In the autumn the temperature, both daily and nightly, drops somewhat, and rain and cloudiness are the rule. At Paradise the average precipitation in September is 8.29 inches and in October 10.02 inches. The winter snows usually arrive by the middle of November.
Fig. 8. North side of Mount Rainier, 14,408 feet, with Mount Adams at left and Mount St. Helens at right. June 19, 1932. (Photo by 116th Photo Section, Washington National Guard, No. 011-36A-116.)
Trees that are characteristic of parts of the Hudsonian Life-zone include the alpine fir (Abies lasiocarpa), mountain hemlock (Tsuga mertensiana), Alaska cedar (Chamaecyparis nootkatensis) and white-barked pine (Pinus albicaulis). The following shrubs are listed by Jones (1938) as common in the Hudsonian Life-zone on Mt. Rainier: Salix barclayi, Salix commutata, Juniperus sibirica, Alnus sinuata, Ribes howellii, Lutkea pectinata, Potentilla fruticosa, Sorbus occidentalis, Spiraea densiflora, Pachistima myrsinites, Arctostaphylos nevadensis, Arctostaphylos uva-ursi, Cassiope mertensiana, Cassiope stelleriana, Phyllodoce empetriformis, Rhododendron albiflorum and Gaultheria ovalifolia.
There are extensive coniferous forests in the Canadian Life-zone, still lower on the mountain slopes. This is an area of lesser temperature extremes than is the Hudsonian Life-zone. The average annual temperature at Longmire, 2761 feet, Mt. Rainier, is 43.8°. The average temperature for the winter months, however, is below freezing. In July and August the temperatures are high, especially in the daytime. The eighteen year average for Longmire during these months is 60.6°. Snow is regular but the fall is lighter than in the Hudsonian Life-zone. The annual average, over a period of nineteen years, is 184.4 inches at Longmire. Precipitation is similar to that in the Hudsonian Life-zone, averaging perhaps slightly less.
In the Cascades the typical feature of the Canadian Life-zone is the extensive coniferous forest that extends, almost without a break, the entire length of the Cascades on both sides of the main crest. In addition to Douglas fir, the following trees occur in this forest: western hemlock (Tsuga heterophylla), amabalis fir (Abies amabalis), white pine (Pinus monticola) and noble fir (Abies nobilis). Other plants include Vaccinnium ovalifolium, Vaccinnium membranaceum, Menziesia ferruginea, Alnus sinuata, Acer circinatum, Sorbus cascadensis, Cornus canadensis, Clintonia uniflora, Stenauthium occidentale, Galium oreganum, and Prenanthes lessingii. Saprophytes abundant in, if not confined to, this zone are listed by Jones as: Monotropa uniflora, Monotropa hypopitys, Allotropa virguta, Newberrya congesta, Pterospora andromedea, Corallorrhiza maculata, Corallorrhiza mertensiana and Corallorrhiza striata.
West of the Canadian Life-zone in the western Cascades, the coniferous forests merge with the lowland forests of western Washington. To the east of the Canadian Life-zone in the eastern Cascades, there is a distinct change to a more arid climate and flora. At Leavenworth, 1167 feet, Chelan County, the annual average precipitation is but 19.5 inches and at Cle Elum, 1930 feet, but 23.23 inches. Temperatures are higher, the annual average of the above two localities being 47.2° and 45.4° respectively. The winter months are cold, with the average temperature in January and February below freezing. In summer the averages in July and August at Leavenworth are 68.8° and 68.0°, with the average maximum being 87.5° and 86.1°. Snowfall is heavy, the yearly average at Leavenworth being 98.5 inches and at Cle Elum 86.3 inches. The effect of this more arid climate is seen in the vegetation. The dense Douglas fir forest, is replaced by more open forests of yellow pine (Pinus ponderosus). Groves of oak (Quercus garryana) are found near streams. The open forests give way to the extensive grasslands bordering the desert. The transition of vegetation is similar to that occurring in the Yakima Valley Area.
In the Yakima Valley Area, arid conditions prevail. The average yearly precipitation at Yakima is 6.67 inches. Only in November, December and January may more than one inch of precipitation be expected monthly. Snow may be expected in the winter months and the yearly average snowfall is 22.1 inches. Winter temperatures are low, the average for December and [Pg 28] January being but slightly above freezing. Summer temperatures are extreme; the July average is 73.1° and the average maximum for the same month is 89°. The highest temperature recorded is 111°.
The open pine forests of the eastern Cascades give way to grasslands. Grasses of several species are common but the bunchgrass (Agropyron spicatum) is most important. Other plants include the primrose (Oenothera pallida), lupines (Lupinus), and Mertensia. In ravines and near watercourses such shrubs as hawthorn (Crataegus douglasii), service-berry (Amelanchier cusickii, Amelanchier utahensis), aspen (Populus tremuloides), syringa (Philadelphus lewisii), snowberry (Symphoricarpos albus), choke-berry (Prunus melanocarpa) and elderberry (Sambucus caerulae) form thickets. Lower in the valley the vegetation is xerophytic, similar to that of the Columbian Plateau. Sagebrush (Artemisiae tridentata) is dominant. Other shrubs include rabbit brush (Chrysothamnus nauseosus, Chrysothamnus viscidiflorus), hop sage (Grayia spinosa), black sage (Purshia tridentata) and greasewood (Sarcobatus vermiculatus).
Fig. 9. Mount Rainier. Washington: Cowlitz Chimneys from base of Dege Peak, July 19, 1933. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 859.)
Climatic conditions in the Olympic Mountains are, in general, similar to those of the higher Cascade Mountains. There is a limited area of Arctic-alpine Life-zone on Mount Olympus. The principal life-zone is the Hudsonian. The Canadian merges with the lowland Humid-Transition and is difficult to ascertain as a separate zone. In the Hudsonian Life-zone the average temperatures are low. Winter climate is bitter and the snow lies deep. In the [Pg 29] absence of government weather stations in the Olympics, detailed descriptions of climatic conditions can not be given. Vegetation of the Hudsonian Life-zone of the Olympic Mountains is, in general, similar to that of the same life-zone of the Cascades (see Jones, 1936, Botanical Survey of the Olympic Peninsula).
The lowlands of western Washington have a cool, humid climate. The average annual temperature of the area varies little from 50°. In winter the temperature, especially in January and February, commonly drops below the freezing point at night. Summer temperatures are moderate, rarely reaching 90°. Snowfall is light, averaging about 10 inches. The prevailing winds are from the west and are moisture-laden. They rise over the Olympic Mountains and loose heavy rains along the coastal area of the lowlands. The average annual precipitation at La Push, Clallam County, is 97.9 inches; at Clearwater, Jefferson County, 124.98 inches; at Aberdeen, Grays Harbor County, 81.58 inches; at South Bend, Pacific County, 83.35 inches. In contrast, the more inland areas receive less than half as much precipitation. The yearly average at Bellingham is 31.09 inches; at Seattle, 30.07 inches; at Tacoma, 39.53 inches; at Vancouver, Clark County, 37.24 inches.
The outstanding feature of the vegetation of western Washington is the coniferous forest. Previous to the logging activities a dense cover of Douglas fir, western hemlock and red cedar spread almost unbroken over the area. The openings in the forest and the marshy ravines and river valleys supported growths of underbrush and deciduous trees so dense and luxuriant as to compare with a tropical jungle. In the dense rain forests along the coast, mosses and lichens develop an understory vegetation many inches deep and clothe the branches of the forest trees. The mild temperature and excessive rainfall cause some species that usually are of bush or shrub size to reach the proportions of small trees. In some places one can climb twenty feet from the ground in a huckleberry tree, the trunk of which is five inches in diameter. The coniferous forest is made up of several species of trees. Most important among these are the western hemlock (Tsuga heterophylla), Douglas fir (Pseudotsuga taxifolia), and red cedar (Thuja plicata). Locally the western yew (Taxus brevifolia), lodgepole pine (Pinus contorta) and spruce (Picea sitchensis) may be common. Deciduous trees are numerous and include several willows (Salix sp.), aspen (Populus tremuloides), hazel (Corylus californica), alder (Alnus oregona), oak (Quercus garryana), broadleaf maple (Acer macrophyllum), vine maple (Acer circinatum), and flowering dogwood (Cornus nuttallii). Mosses and ferns are abundant. The sword fern (Polystichum munitum) and bracken (Pteridium aquilinum) are especially common. Space prevents listing all but a fraction of the typical shrubs but these include huckleberry (Vaccinium parvifolium, Vaccinium ovatum), Oregon grape (Berberis nervosa), salal (Gaultheria shallon), rose (Rosa gymnocarpa), thimbleberry (Rubus parviflorus), salmonberry (Rubus spectabilis), blackcap raspberry (Rubus leucodermis) and wild blackberry (Rubus macropetalus).
The higher parts of some of the ranges of northeastern Washington are in the Hudsonian Life-zone, but most of the mountains are in the Canadian Life-zone. The valleys are in the Transition Life-zone. Climatic conditions are similar to those of the eastern slopes of the Cascades. Winter temperatures are low, the average for December, January and February being below freezing. Summer temperatures are high, the July average for Colville being 67.2° [Pg 30] and the July average maximum being 87.4°. Vegetation consists principally of coniferous forests in the mountains and deciduous woods in the valleys. Among the interesting features of the vegetation are the extensive stands of almost pure larch (Larix occidentalis). In most respects the flora closely resembles that of the Blue Mountains.
The Blue Mountains of southeastern Washington differ from other ranges in Washington in their relative aridity. There are few streams and a single river drains the area. There are no government weather stations in the Blue Mountains. The winter temperatures are low and the snow deep and lasting. Summer temperatures are high and humidity and precipitation low. Coniferous forests of the type of arid regions form the principal tree cover. Typical plant species include the white fir (Abies grandis), alpine fir (Abies lasiocarpa), larch (Larix occidentalis), spruce (Picea columbiana), and such shrubs as fool huckleberry (Menziesia ferruginea), Pachystima myrsinites, dogwood (Cornus canadensis), wild current (Ribes petiolare), mountain mahogany (Cercocarpus ledifolius), spirea (Spiraea sp.), lupines (Lupinus) of several species, maple (Acer douglasii), buckbrush (Ceanothus sanguineus), sticky brush (Ceanothus velutinus), and huckleberry (Vaccinium membranaceum).
Fig. 10. A "pothole" being crowded by drifting sand, ten miles south of Moses Lake, Washington, March 23, 1940. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 925.)
The Columbian Plateau and southeastern Washington present desert conditions. At Odessa, 1590 feet, Lincoln County, the average annual precipitation is only 9.38 inches, and only in the winter may more than one inch of precipitation per month be expected. The average temperature is 48.5°. In the winter the average is below freezing but in July it is 71.3°. The average maximum for July is 90° and an extreme of 111° is recorded. Walla Walla, 991 feet, has a higher annual precipitation (16.66 inches) but higher temperature (yearly average 53.5°, July average 75.0°, average July maximum 88.6°, extreme 113°). Winter temperatures on the Columbian Plateau are low. The January average at Odessa is 25.3° and at Walla Walla 32.4° The average [Pg 31] yearly snowfall at Odessa is 19.4 inches and at Walla Walla 23.5 inches. Vegetation of the Columbian Plateau and southeastern Washington is of the desert type. A few pines and junipers grow in favored places. Along streams the cottonwood (Populus hastata) and willow (Salix) of several species are common. Most typical are grasses and shrubs such as the bunch grass (Agropyron inerme, Agropyron spictatum), foxtail (Alopecurus aequalis), cheat grass (Bromus tectorum), saltbrush (Atriplex truncata), greasewood (Sarcobatus vermiculatus) mustard (Arabis sp., Brassica sp.), sagebrush (Artemisia rigida, Artemisia tridentata), rabbit brush (Chrysothamnus nauseosus, Chrysothamnus viscidiflorus) and cactus (Opuntia polyacantha). The arid climate of the Columbian Plateau affects, to some extent, surrounding areas. Thus the Yakima Valley Area, the Columbia Valley, where it borders the Plateau, and the Okanogan Valley possess vegetation typical of the Columbia Plateau.
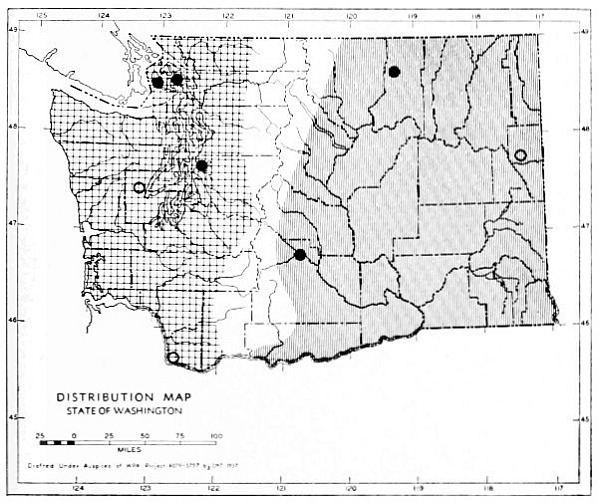
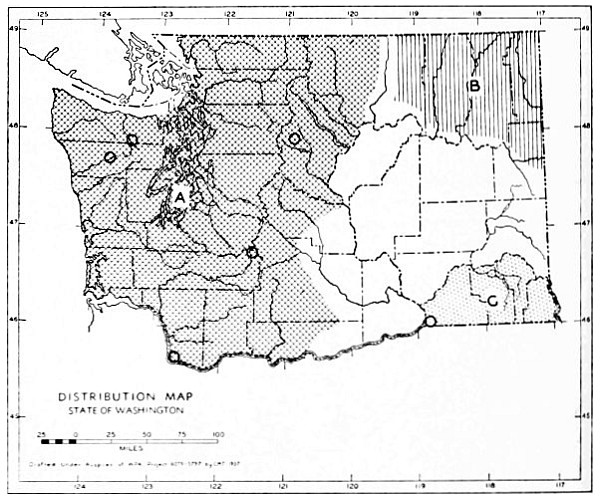
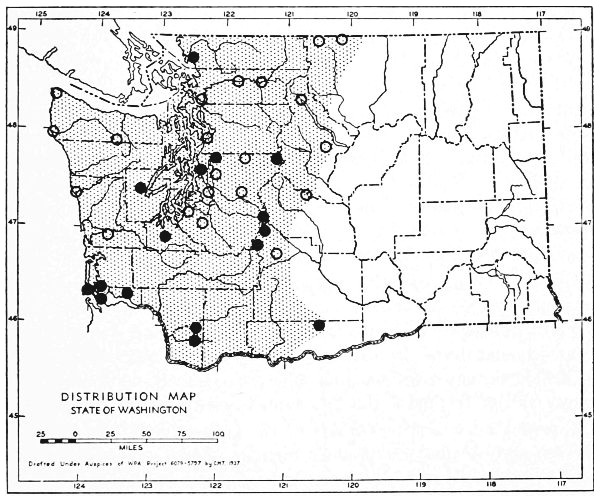
The Transition Life-zone is the principal life-zone in Washington. It is divisible into three subdivisions: Humid, Arid-timbered and Arid-grasslands (Fig. 11) subdivisions. The Humid and Arid-timbered subdivisions of the Transition life-zone are closely related in some respects but different in others. They are separated by the Cascade Mountains. All of the Transition Life-zone west of the Cascades belongs to the Humid subdivision and the timbered Transition Life-zone east of the Cascades belongs to the Arid-timbered subdivision.
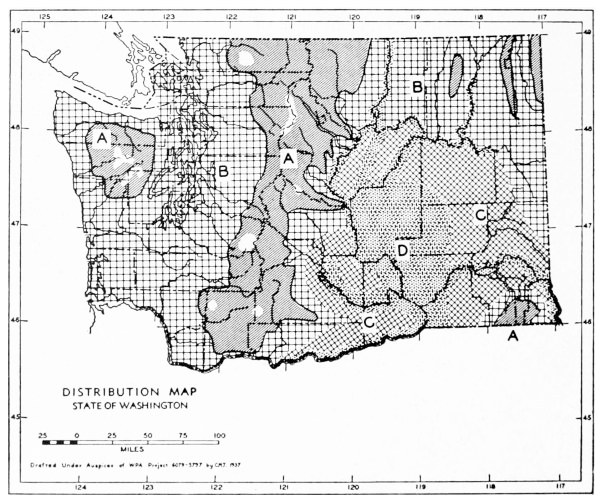
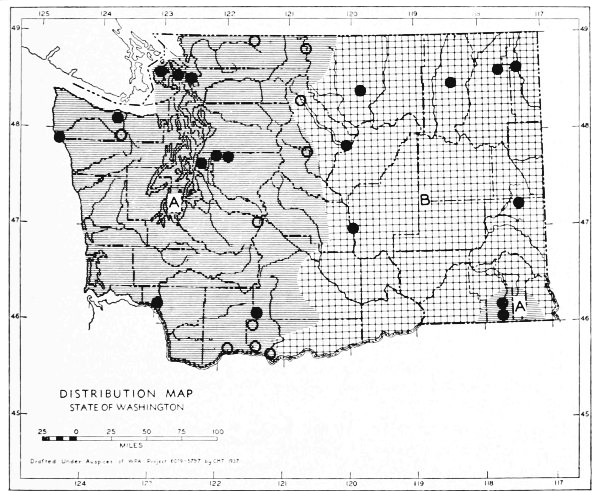
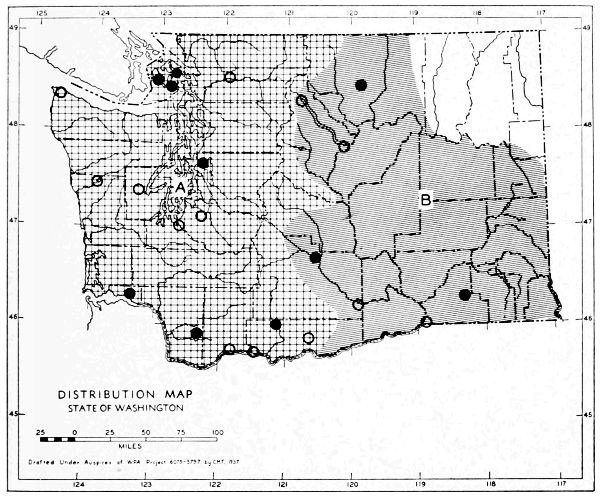
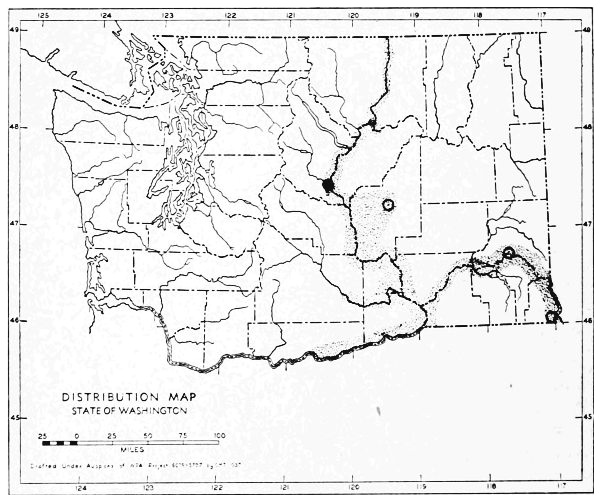
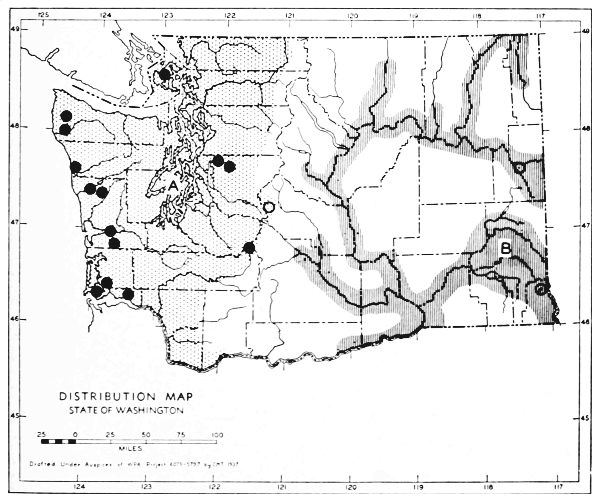
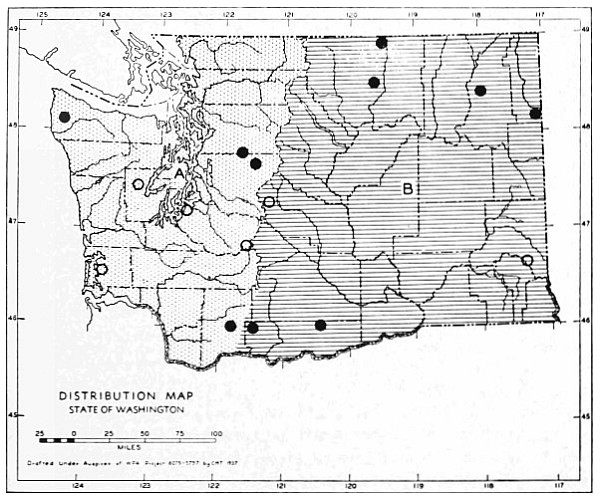
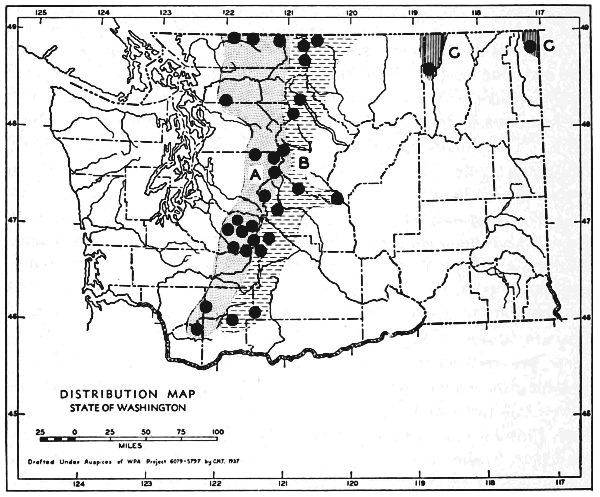
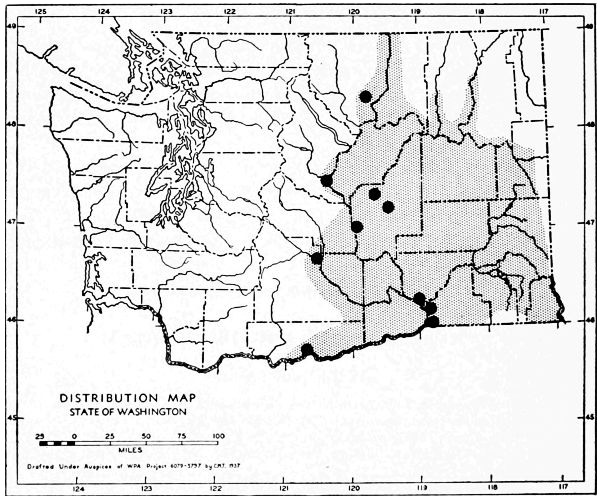
Fig. 11. Life-zones of Washington. Arctic-alpine not shaded. A. Hudsonian and Canadian (mapped together). B. Forested Transition (Humid and Arid subdivisions). C. Arid-grasslands of the Transition. D. Upper Sonoran.
The Arid-grasslands are of minor geographic extent. Although this subdivision is relatively distinct as concerns the distribution of plants, insects and birds, it is of little importance as concerns the distribution of mammals. For the most part, the mammals [Pg 33] occupying it are more representative of surrounding areas. Large parts of the Arid-grasslands have been taken over for agriculture, especially wheat raising. Perhaps the greatest extent of the Arid-grasslands existing in a natural state is along the eastern Cascade Mountains and along the eastern side of the Columbian Plateau. These are truly transition areas, situated where the arid pine forests are replaced by open, sagebrush desert.
Fig. 12. Arctic-alpine Life-zone, Mount Rainier, Washington: Cowlitz Glacier from elevation of 9,500 feet. (Fish and Wildlife Service photo by Victor B. Scheffer. No. 900.)
The Upper Sonoran Life-zone includes the desert areas of Washington (figure 11 D). Its principal extent is the central Columbian Plateau. From the central Columbian Plateau, fingerlike projections of desert extend along the principal valleys.
The Arctic-alpine Life-zone occurs in the high Olympic Mountains and on the higher peaks of the Cascades. This is shown on the life-zone map, Fig. 11, as white, unshaded areas.
The mammalian faunas of the Hudsonian and Canadian life-zones resemble each other closely. The boundary between them is too complex to permit separating them on a small-scale map. Consequently they are mapped together on Fig. 11 as A.
This subdivision is remarkably uniform in composition over western Washington. The greatest difference is in precipitation. Rainfall along the coast is heavier than that in the interior. Consequently vegetation is more dense and luxuriant along the coast.
Four habitats may be distinguished in the Humid subdivision and further subdivision is possible. The dominant and most extensive habitat is the forest. Dominant mammalian species include: Peromyscus maniculatus, Sorex trowbridgii, Sorex obscurus, Tamiasciurus douglasii, Clethrionomys californicus, Aplodontia rufa, Glaucomys sabrinus and Odocoileus hemionus. Mammals are scarce and nocturnal forms prevail. As a rule, a line of mouse traps set in a forest habitat will take principally Peromyscus maniculatus with a few Sorex trowbridgii and Sorex obscurus and rarely a Clethrionomys californicus. In some places, especially where the moss is deep, a line of mouse traps will catch only shrews.
Fig. 13. Humid subdivision of the Transition Life-zone, Headley's Marsh, five miles east of Granite Falls, Washington, June 4, 1938. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 60.)
A second important habitat is the deciduous jungle. This differs from the forest habitat in that the dominant trees are of the deciduous type and in that understory vegetation, such as shrubs and annuals, is dense. The jungle habitat occurs in ravines and in valleys [Pg 35] of streams and rivers and, in general, covers the lower, poorly drained portions of the Humid subdivision of the Transition Life-zone. Mammals are abundant and varied in the jungle habitat. The deer mouse (Peromyscus maniculatus) is the most common mammal but a line of mouse traps might also catch: Neurotrichus gibbsii, Scapanus orarius, Sorex vagrans, Microtus oregoni, or Zapus p. trinotatus. The mountain beaver, snowshoe rabbit, and Townsend chipmunk also occur there.
The prairies form a third habitat. These areas of native grasslands are of minor extent but are the principal home of several races of gophers and the Townsend mole (Scapanus townsendii). Deer and elk also browse on the prairies.
Fig. 14. Timbered arid subdivision of the Transition Life-zone, Kettle Falls on the Columbia River (now beneath Coulee Dam backwater), Stevens County, Washington, June 15, 1938. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 72.)
A fourth habitat is the marsh. In it there is here included the sphagnum bogs and marshy shores of lakes and streams. This habitat is characterized by damp ground, standing water, and dense vegetation. Typical mammalian species include the water shrew (Sorex bendirii), Townsend meadow mouse, muskrat and mink.
Other habitats, such as aerial for the bats and aquatic for the beaver and otter, might be listed.
In Washington the Arid timbered subdivision of the Transition Life-zone is the open, pine forest. Because of the aridity of this [Pg 36] habitat, marshes and streamside thickets are uncommon, but where habitats of this kind do occur they have a fauna distinct from that of other habitats.
The pine forest habitat includes many diurnal species, such as the red squirrel, yellow-pine chipmunk, and Columbian ground squirrel. The white-tailed deer occurs here and, for most of the year, the mule deer. Snowshoe rabbits are usually present. Near rocks the bushy-tailed wood rat is common. Mice are scarce, probably because of the open nature of the surface of the ground. A night's trapping usually yields only a few Peromyscus maniculatus.
The mammalian fauna of the marshes and streamside thickets is similar. Shrews including Sorex vagrans and Sorex obscurus are uncommon. Meadow mice, including Microtus pennsylvanicus, Microtus longicaudus, and more rarely Microtus montanus, are taken.
This subdivision is so much utilized by man where it occupies any considerable areas, and is of such a transitional nature elsewhere, that it is important for only a few native wild mammals. The sagebrush vole (Lagurus curtatus) seems to be confined to the arid grasslands. The white-tailed jack rabbit is now found principally in the arid grasslands, but its confinement there has resulted probably from competition with the black-tailed jack rabbit. The montane meadow mouse (Microtus montanus) is the only common, representative species. Many species from the Upper Sonoran Life-zone extend into the arid grasslands where conditions are suitable. These include Reithrodontomys megalotis, Perognathus parvus, Citellus washingtoni and Marmota flaviventris. A few species more typical of the Arid timbered subdivision of the Transition Life-zone stray onto the arid grasslands. Citellus columbianus and Microtus longicaudus may be included here.
The sagebrush desert in Washington is relatively uniform in nature. Several different habitats may be distinguished, such as sandy areas, open sage, dense sage, stony ground, and talus. Qualitatively, however, the mammalian fauna of these areas is surprisingly similar. Quantitatively, there are great differences. For example, the grasshopper mouse is rare in the open sage areas with hard, claylike soil but common on drifted sand. The harvest mouse is common in dense sage but rare in open sage or in open, sandy areas. Mammals are abundant on the sagebrush desert and typical species include: [Pg 37] the black-tailed jack rabbit, Nuttall cottontail, Ord kangaroo rat, Great Basin pocket mouse, Townsend ground squirrel, Washington ground squirrel.
Marshes are not uncommon on the Columbian Plateau and elsewhere in the Upper Sonoran Life-zone in Washington. They do not possess a fauna that is strictly Upper Sonoran but instead contain species more typical of the Arid-timbered subdivision of the Transition Life-zone. Meadow mice found in desert marshes include Microtus montanus and Microtus pennsylvanicus. The only shrew we have found is Sorex vagrans. The harvest mouse (Reithrodontomys megalotis) is often abundant in marshes.
Fig. 15. Upper Sonoran Life-zone, sand and basalt cliffs along the east bank of the Columbia River, at Vantage, Washington, 1930. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 848.)
The Canadian Life-zone is a relatively uniform area in which the forest habitat is most important. Talus and swift, cold streams bring some typical mammalian species into the Canadian Life-zone from the Hudsonian. Mammals are usually common; they are abundant only in the upper portion of the Life-zone. Arboreal species and forms adapted to life beneath the forest cover are dominant. The Douglas squirrel, red squirrel, northern flying squirrel and Townsend chipmunk are typical arboreal species. Traps set beneath the trees might catch Peromyscus maniculatus, Clethrionomys gapperi, Neotoma cinerea, Sorex obscurus, or Sorex trowbridgii.
Fig. 16. Canadian Life-zone forest on Mount Rainier, Washington, elevation 2,800 feet, September 14, 1934. Western hemlock, Douglas fir, western red cedar, and grand fir. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 325.)
In Washington the Hudsonian Life-zone is of greater diversity than any other. Frost and steep slopes have formed great masses of talus and, in this talus, certain species of mammals, such as Ochotona princeps, Marmota caligata and Marmota olympus are found. Other species, such as the golden-mantled ground squirrels, mountain chipmunk, bushy-tailed wood rat, red-backed mouse and long-tailed meadow mouse find the talus an ideal home. Glacial action has produced, in the Hudsonian Life-zone, numerous level or concave areas that contain small lakes and slow-moving streams. Dense, herbaceous vegetation is abundant nearby. Small mammals abound and a line of mouse traps will almost certainly catch a few such typical species as: Sorex palustris, Sorex obscurus, Microtus oregoni, Microtus richardsoni, Microtus longicaudus and Zapus princeps as well as the ever present Peromyscus maniculatus. The shrew-mole or heather vole might also be taken, though the latter is more apt to be found in nearby heather meadows.
| Species and Subspecies. | Humid Transition |
Arid-timbered Transition |
Arid- grasslands Transition |
Upper Sonoran |
Canadian | Hudsonian |
| Neurotrichus gibbsii gibbsii | ? | C | ||||
gibbsii minor |
A | |||||
| Scapanus townsendii | A | ? | C | |||
orarius orarius |
A | |||||
orarius schefferi |
C | |||||
orarius yakimensis |
A | |||||
| Sorex cinereus cinereus | R | R | ||||
cinereus streatori |
R | |||||
merriami merriami |
R | |||||
trowbridgii trowbridgii |
A | C | ||||
trowbridgii destructioni |
A | |||||
vagrans vagrans |
A | R | ||||
vagrans monticola |
C | C | A | C | ||
obscurus obscurus |
R | |||||
obscurus setosus |
C | C | A | |||
palustris navigator |
C | A | ||||
bendirii bendirii |
A | C | ||||
bendirii albiventer |
C | |||||
| Microsorex hoyi washingtoni | R | |||||
| Myotis lucifugus carissima | C | C | ||||
lucifugus alascensis |
A | C | ||||
yumanensis sociabilis |
C | C | ||||
yumanensis saturatus |
A | |||||
keenii keenii |
R | R | ||||
evotis evotis |
R | |||||
evotis pacificus |
R | |||||
thysanodes thysanodes |
R | |||||
volans longicrus |
C | |||||
volans interior |
C; | |||||
californicus californicus |
R | R | ||||
californicus caurinus |
A | C | ||||
subulatus melanorhinus |
R | A | ||||
| Lasionycteris noctivagans | A | A | C | |||
| Corynorhinus rafinesquii townsendii | R | |||||
rafinesquii intermedius |
R | |||||
| [Pg 41] Pipistrellus hesperus hesperus | R | |||||
| Eptesicus fuscus bernardinus | A | A | ||||
| Lasiurus cinereus cinereus | R | R | R | R | ||
| Antrozous pallidus cantwelli | R | R | ||||
| Ursus americanus altifrontalis | A | C | A | A | ||
americanus cinnamomum |
A | C | ||||
chelan |
R | |||||
| Procyon lotor psora | A | C | ||||
lotor excelsus |
R | R | C | |||
| Martes caurina caurina | A | C | ||||
caurina origenes |
A | C | ||||
pennanti |
R | R | ||||
| Mustela erminea invicta | C | C | ||||
erminea gulosa |
C | C | ||||
erminea murica |
R | |||||
erminea fallenda |
R | |||||
erminea streatori |
R | C | ||||
erminea olympica |
R | |||||
frenata nevadensis |
C | C | C | C | ||
frenata effera |
A | C | C | C | ||
frenata washingtoni |
C | C | ||||
frenata altifrontalis |
A | C | C | |||
vison energumenos |
C | C | C | C | C | |
| Gulo luscus luteus | R | R | R | |||
| Lutra canadensis pacifica | C | R | R | R | C | |
| Spilogale gracilis saxatilis | R | R | ||||
gracilis latifrons |
A | |||||
| Mephitis mephitis hudsonica | A | |||||
mephitis major |
R | R | ||||
mephitis notata |
C | R | ||||
mephitis spissigrada |
A | |||||
| Taxidea taxus taxus | C | C | C | R | ||
| Vulpes fulva cascadensis | R | |||||
| [Pg 42] Canis latrans lestes | C | A | A | A | C | R |
latrans incolatus |
A | C | C | A | ||
lupus fuscus |
R | R | R? | R | ||
| Felis concolor missoulensis | C | C | ||||
concolor oregonensis |
C | C | C | |||
| Lynx canadensis | R | |||||
rufus fasciatus |
A | C | ||||
rufus pallescens |
A | C | C | C | ||
| Tamias minimus scrutator | C | |||||
minimus grisescens |
R | |||||
amoenus caurinus |
A | A | ||||
amoenus felix |
A | A | ||||
amoenus ludibundus |
A | A | ||||
amoenus affinis |
A | |||||
amoenus canicaudus |
A; | |||||
amoenus luteiventris |
C | A | ||||
ruficaudus simulans |
C | A | ||||
townsendii townsendii |
A | |||||
townsendii cooperi |
C | A | R | |||
| Marmota monax petrensis | R | |||||
flaviventris avara |
R | C | A | |||
caligata cascadensis |
R | A | ||||
olympus |
C | A | ||||
| Citellus townsendii townsendii | A | |||||
washingtoni |
C | A | ||||
columbianus columbianus |
A | C | ||||
columbianus ruficaudus |
A | |||||
beecheyi douglasii |
A | |||||
lateralis tescorum |
C | |||||
lateralis connectens |
C | |||||
saturatus |
C | A | ||||
| Tamiasciurus hudsonicus richardsoni | A | A | ||||
hudsonicus streatori |
A | A | R | |||
douglasii douglasii |
A | A | A | C | ||
| Sciurus griseus griseus | C | C | ||||
| [Pg 43] Glaucomys sabrinus oregonensis | C | |||||
sabrinus fuliginosus |
A | |||||
sabrinus columbiensis |
A | |||||
sabrinus latipes |
A | A | ||||
sabrinus bangsi |
R | C | ||||
| Perognathus parvus parvus | C | A | ||||
parvus lordi |
C | A | ||||
parvus columbianus |
A | |||||
| Dipodomys ordii columbianus | A | |||||
| Thomomys talpoides devexus | A | |||||
talpoides columbianus |
A | |||||
talpoides aequalidens |
A | |||||
talpoides wallowa |
A | |||||
talpoides fuscus |
A | C | R | C | C | |
talpoides yakimensis |
C | A | ||||
talpoides shawi |
C | A | ||||
talpoides immunis |
A | A | ||||
talpoides limosus |
C | A | ||||
talpoides douglasii |
A | |||||
talpoides glacialis |
A | |||||
talpoides tacomensis |
A | |||||
talpoides pugetensis |
A | |||||
talpoides tumuli |
A | |||||
talpoides yelmensis |
A | |||||
talpoides couchi |
A | |||||
talpoides melanops |
C | A | ||||
| Castor canadensis leucodonta | A | A | C | C | ||
canadensis idoneus |
A | |||||
| Onychomys leucogaster fuscogriseus | A | |||||
| Reithrodontomys megalotis megalotis | C | A | ||||
| Peromyscus maniculatus oreas | A | A | A | |||
maniculatus hollisteri |
A | |||||
maniculatus austerus |
A | |||||
maniculatus rubidus |
A | |||||
maniculatus gambelii |
C | R | A | |||
maniculatus artemisiae |
A | R | R | C | C | |
| [Pg 44] Neotoma cinerea occidentalis | C | R | A | C | A | |
cinerea alticola |
C | A | C | |||
| Synaptomys borealis wrangeli | A | |||||
| Phenacomys intermedius intermedius | C | A | ||||
intermedius oramontis |
C | A | ||||
| Clethrionomys gapperi saturatus | R | A | A | |||
gapperi idahoensis |
R | A | A | |||
gapperi nivarius |
A | A | ||||
californicus occidentalis |
A | |||||
| Microtus pennsylvanicus funebris | A | R | C | |||
pennsylvanicus kincaidi |
A | |||||
montanus nanus |
A | C | ||||
montanus canescens |
C | A | A | |||
townsendii townsendii |
A | |||||
townsendii pugeti |
A | |||||
longicaudus halli |
A | C | C | C | ||
longicaudus macrurus |
R | C | A | |||
richardsoni arvicoloides |
C | A | ||||
richardsoni macropus |
C | A | ||||
oregoni oregoni |
A | A | ||||
| Ondatra zibethicus osoyoosensis | A | A | C | |||
zibethicus occipitalis |
A | |||||
| Zapus princeps oregonus | A | |||||
princeps kootenayensis |
A | |||||
princeps idahoensis |
A | |||||
princeps trinotatus |
A | C | A | |||
| Aplodontia rufa rufa | A | |||||
rufa rainieri |
A | |||||
| Erethizon dorsatum epixanthum | A | R | C | A | R | |
dorsatum nigrescens |
A | R | C | A | R | |
| Ochotona princeps cuppes | A | |||||
princeps fenisex |
A | |||||
orinceps brunnescens |
C | A | ||||
| Lepus townsendii townsendii | C | A | ||||
californicus deserticola |
C | A | ||||
americanus washingtonii |
A | A | ||||
| [Pg 45] americanus cascadensis | C | A | C | |||
americanus pineus |
C | A | C | |||
americanus columbiensis |
A | |||||
| Sylvilagus nuttallii nuttallii | A | |||||
idahoensis |
A | |||||
| Cervus canadensis roosevelti | A | A | R | |||
canadensis nelsoni |
C | A | ||||
| Odocoileus virginianus leucurus | A | |||||
virginianus ochrourus |
A | |||||
hemionus hemionus |
A | A | C | |||
hemionus columbianus |
A | C | ||||
| Ovis canadensis canadensis | A | A | A | A | A | |
canadensis californiana |
A | A | A | A | ||
| Oreamnos americanus americanus | A |
The composition of the mammalian fauna of any area is dependent on several factors. These include the composition of the original fauna, species which have since invaded the area, and quantitative and qualitative changes that have occurred in the area. The latter two factors refer to changes in relative numbers or extermination of species through environmental changes or competition with other forms and evolutionary changes that have occurred in the species making up the mammalian fauna.
Our knowledge and understanding of the distribution and history of the species of mammals occurring in Washington decreases rapidly as we go back in time. The distribution of the modern fauna at the present time is fairly well known. The distribution of species 100 years ago is less well understood. This is especially true of certain game species and carnivores whose distribution has been altered by man. Our knowledge of the distribution of mammals in the Pleistocene and earlier times is based on fossil skeletons. Such knowledge must necessarily be meager, for conditions favorable to fossilization and the preservation of fossils until their subsequent discovery by man, were not of common occurrence.
In the Cascades and in eastern Washington, the Miocene was a time of orogeny and great volcanism. Great flows of lava, 4,000 feet thick in the Snake River area (Russell, 1893), emerging from fissures in the Snake River area, formed the Columbian Plateau. The Columbian basalt slopes inward centripetally from the eastern, northern, and western margins of the Columbian Plateau with an average descent of 25 feet to the mile (Flint, 1938). The dip of the lava flows results in the basalt-marginal course of the Spokane and Columbia rivers today, along the northern edge of the Columbian Plateau. The earlier part of the Pliocene was a period of erosion and deformation. In the early Pleistocene the five great volcanic cones of the Cascades, Mount Baker, Glacier Peak, Mount Rainier, Mount Adams and Mount St. Helens were formed. In eastern Washington a gentle folding of the Miocene lava flows occurred. The folding took place slowly and the Columbia River in its course along the eastern edge of the Cascades cut through the folds as they formed, making a series of water gaps. Farther south, the Simcoe-Frenchman Hills anticline seems to have arisen [Pg 47] more rapidly and the Columbia River was forced eastward before it became impounded and rose over the barrier and plunged down, tearing out the great Wallula Water Gap (Flint, 1938). This gap is a mile wide, eight miles long and, in places, a thousand feet deep. The impounding of the Columbia by the Simcoe-Frenchman Hills anticline resulted in a lake several hundred miles in area. Sediments deposited in this lake form the Ringold formation. The Ringold formation possesses a very early Pleistocene mammalian fauna.
The Pleistocene was a time of great change in the mammalian fauna of the world. Unfortunately the beautiful glacial sequence revealed in Europe and the Mississippi Valley cannot be detected in Washington. In western Washington the deposits of the last continental glaciation and fluvial deposits of the last interglacial period almost everywhere obscure evidence of earlier glaciations. Deposits of an earlier glaciation, named Admiralty by Bretz (1913), have been detected in places. Deposits of greater age, that may represent a still earlier glaciation, have been noted. In eastern Washington the only definite proof of multiple glaciation is of one glaciation preceding the last. This is the Spokane glaciation of Bretz (1923). That multiple glaciation in the sequence reported from the Mississippi Valley affected Washington seems probable. The lack of evidence of a complete sequence is negative evidence. In western Washington the earliest glacial deposits might be beneath the later deposits or they may have been removed or reworked by subsequent glaciations, whereas in eastern Washington they may have been removed by subsequent glaciation and erosion.
The time interval between the two known glaciations appears to have been of greater duration than the Recent. The drift of the earlier period is sometimes found covered by the till of the later glaciation, and preserved by it. The early material is deeply weathered and all save the hardest pebbles and quartzites, for example, are rotten and disintegrate at the touch. In contrast, the later deposits are almost unweathered. Pebbles are hard, and ring when struck. A zone of leaching and oxidation of the finer materials reaches a depth of some 30 inches, below which the till is fresh.
Two names are currently applied to the last continental glaciation of the state of Washington. That west of the Cascade Mountains, studied and described by Bretz (1913), was termed "Vashon." The interglacial cycle preceding it was called "Puyallup." The glaciation of eastern Washington has been called "Wisconsin," after the Mississippi [Pg 48] Valley terminology, by several writers. Papers by Flint (1935, 1937) describe and map it.
The Vashon and Wisconsin glaciations probably occupied the same time interval, although this has not certainly been established. In the present report I have used the term "Vashon-Wisconsin" in speaking of the entire period, or the glaciers both east and west of the Cascades together. Vashon, alone, is restricted to western Washington and Wisconsin to eastern Washington.
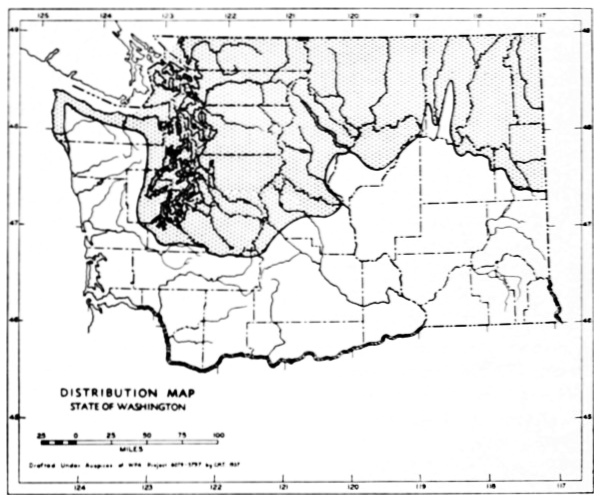
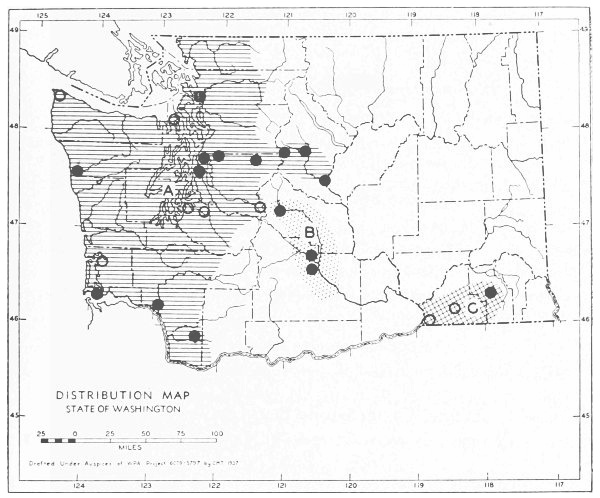
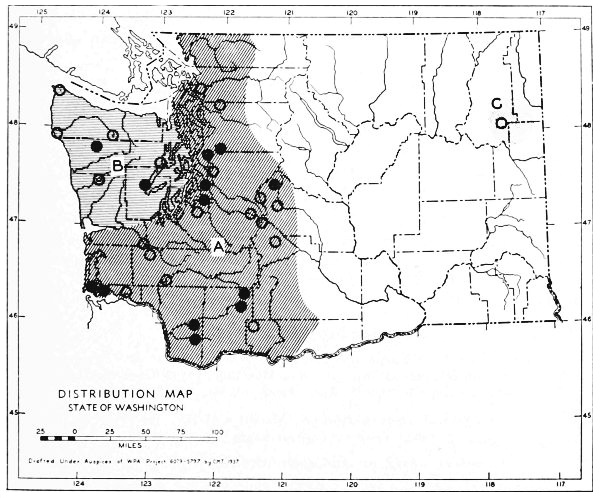
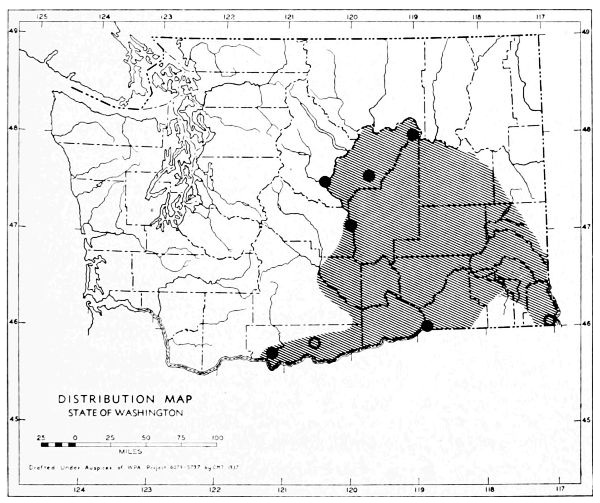
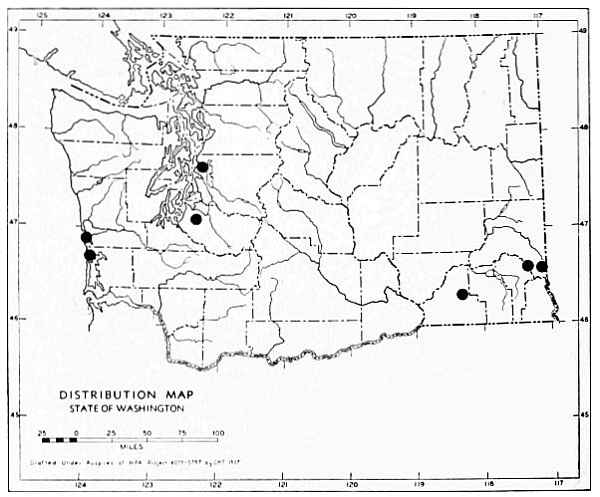
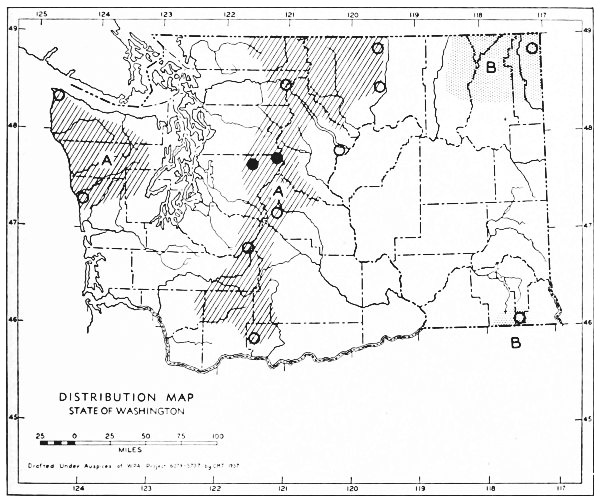
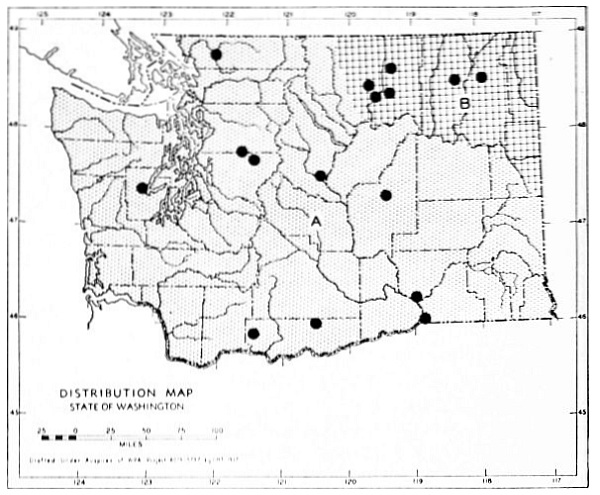
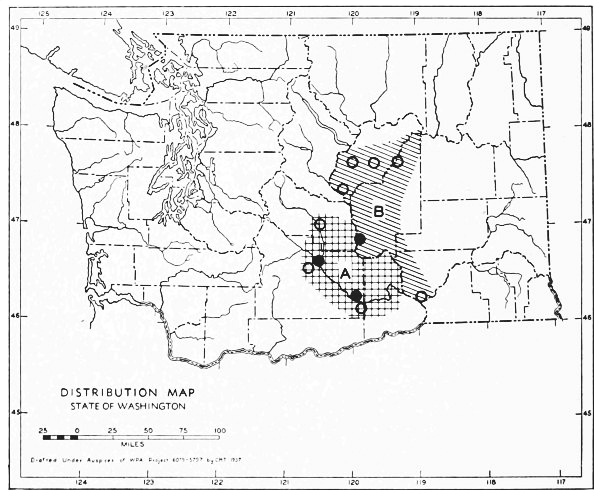
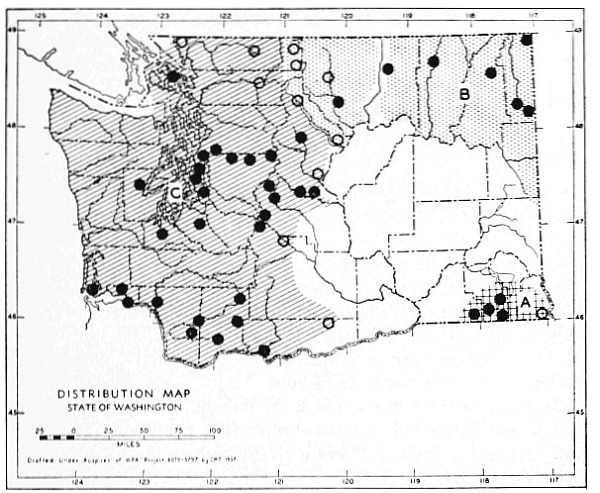
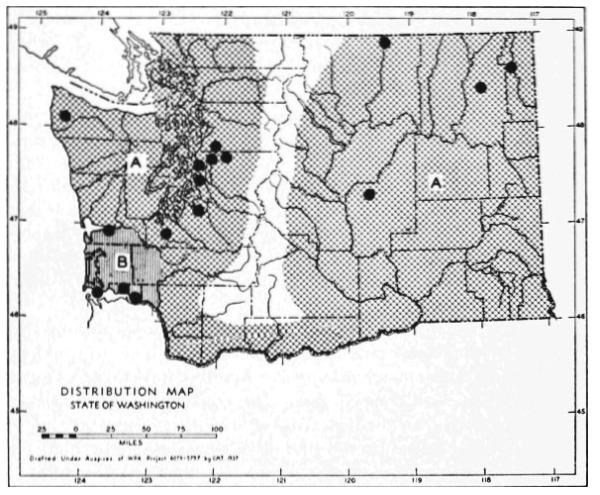
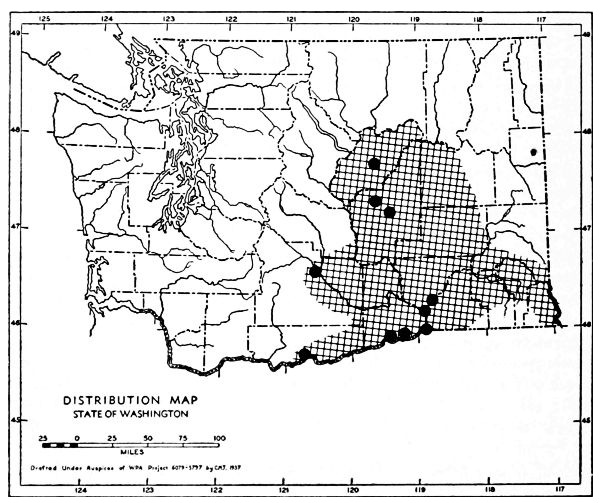
Fig. 17. Extent of Vashon-Wisconsin ice over Washington. The Wisconsin and Vashon glaciers have been connected through the Mount Rainier section of the Cascades because it is thought that few or no mammalian species lived in the Cascades north of Mount Rainier while the ice was in place. Data generalized from Flint (1937), Bretz (1913), Culver (1936) and other sources.
The Vashon glaciation seems to have consisted of an ice dome centering in Puget Sound (the Puget Glacier of Bretz, 1913) and flooding the lowlands from the Olympic Mountains to the Cascade Mountains. The southern edge of the Puget Glacier was slightly south of the present terminus of Puget Sound. Fingerlike projections of ice were forced up valleys of the western Cascades and the northern and eastern Olympics. Some of these upward moving fingers of ice met and coalesced with valley glaciers descending from the mountains. At the southern edge of the glacier, the Black Hills [Pg 49] and Porcupine Hills remained above the ice although partially surrounded by it.
The Wisconsin glacier, according to Flint (1935), was a great piedmont glacier, fed by valley glaciers from the Cascades and Coast Ranges to the west and the Rockies to the east. It extended from the Idaho boundary to the Cascade Mountains. From the Canadian Boundary it sloped down to an approximate elevation of some 6500 feet at Republic and to 2500 feet on the northern edge of the Columbian Plateau which was the southern edge of the glacier. The Kettle River Mountains, in almost the center of the glacier, remained a peninsula or driftless area that divided the glacier into two lobes. The Pend Oreille, Huckleberry and other mountain ranges, formed nunataks, or islands above the ice, at the southern part of the glacier.
The behavior of valley glaciers in the northern Cascade Mountains during Vashon-Wisconsin time, seems to have been variable. Some depression of the snow line, at least in the north, seems probable.
The Vashon Glacier impinged on the eastern, northern and to some extent the western, slopes of the Olympic Mountains. Late Pleistocene valley glaciers in the Olympics, however, seem to have been inconsequential.
The time of the retreat of the Vashon-Wisconsin glaciers is a subject of special interest to the mammalogist in that it represents time for invasion and dispersal of species and in that it represents generations of individuals upon which natural selection might act. It is generally agreed that a period of approximately ten thousand years has elapsed since the retreat of the Vashon-Wisconsin glaciers from Washington.
Information on the climate of the state of Washington previous to the period of the last continental glaciation is understandably meager. Bretz (1913) considers the Puyallup period a time of excessive precipitation and erosion. Bits of lignite from Puyallup sediments seem to be of Douglas fir. Presumably the climate was slightly warmer and more humid than it is today. Vegetation possibly consisted of coniferous forests.
With the advance of the Vashon ice, mammals north of the ice border were all or mostly eliminated. Climatic conditions south of the border of the ice probably were strongly affected by it. Remains of mammoths have been found in Vashon till. The presence of many non-boreal species of mammals in southwestern Washington [Pg 50] indicates their persistence there and that conditions therefore were not intolerable for them. Probably the climate of southwestern Washington was cool and dry. Fir, spruce, and Douglas fir may have been the dominant trees. Hansen (1941 A: 209) found evidence from studies of pollen that coniferous forests were growing in west-central Oregon in late glacial time. These pollen studies of postglacial peat bogs by Henry P. Hansen give evidence of postglacial climatic changes. Hansen points out (1941 B, 1941 C) that climatic changes west of the Cascades were probably slight because of the influence of the Pacific Ocean. Pollen profiles indicate an early, cool, dry climate followed by a warmer one and increasing humidity. The present climate may be considered cool and humid. Most of western Washington lies in the Humid subdivision of the Transition Life-zone.
Fig. 18. McDowell Lake, Little Pend Oreille Wildlife Refuge, Stevens County, Washington, September 29, 1939. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 730.)
The pre-Wisconsin climate of eastern Washington is unknown. [Pg 51] From the loessial nature of the Palouse Soil (Bryan, 1927), a preglacial deposit, the area would seem to have been arid, probably a grassland or a sagebrush desert. If the fossil fauna discovered at Washtuckna Lake, Adams County, is of this period, the forest conditions of the Blue Mountains were slightly more extensive than at present. The glacial climate of the Columbian Plateau in Wisconsin time was probably cool and arid. Pollen studies by Hansen (1939, 1940) indicate increasing dryness and warmth since the retreat of the Wisconsin ice. Present-day climate on the Columbian Plateau is warm and dry. The plateau is principally a sagebrush desert. The glaciated area to the north is cooler and more humid, supporting extensive forests of yellow pine and other conifers.
We have mentioned elsewhere that three different mammalian faunas occur in Washington. These may be described as follows.
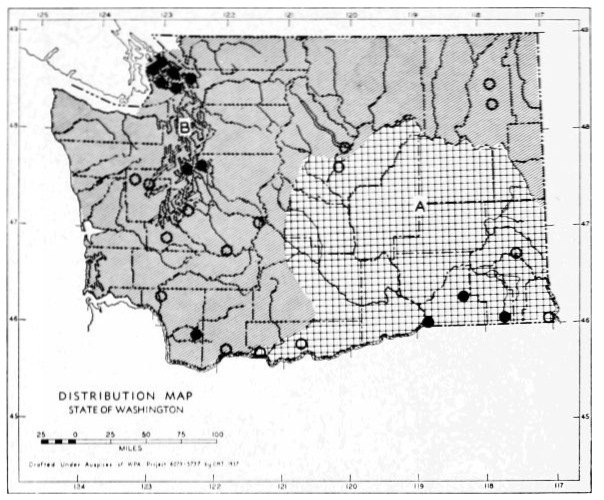
1. The Great Basin Fauna. This fauna is best typified by such genera as Perognathus and Dipodomys. Species that, at least in Washington, are confined to this fauna are:
| Sorex merriami | Tamias minimus | |
| Myotis thysanodes | Perognathus parvus | |
| Myotis subulatus | Dipodomys ordii | |
| Pipistrellus hesperus | Onychomys leucogaster | |
| Antrozous pallidus | Reithrodontomys megalotis | |
| Taxidea taxus | Lagurus curtatus | |
| Marmota flaviventris | Lepus californicus | |
| Citellus townsendii | Sylvilagus nuttallii | |
| Citellus washingtoni | Sylvilagus idahoensis |
The break between the Great Basin Fauna and the other two faunas is extremely sharp, probably as a consequence of a sharp break in the flora.
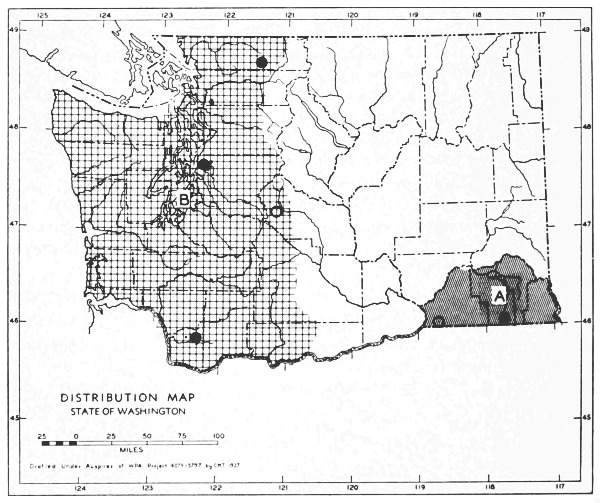
2. The Pacific Coastal Fauna. The mammals of the humid coastal district range from the Fraser River, British Columbia, southward to the vicinity of Monterey Bay, California. In Washington typical genera are Aplodontia, Neurotrichus and Scapanus. The following species are typical of the Pacific Coastal Fauna in Washington:
| Neurotrichus gibbsii | Tamiasciurus douglasii | |
| Scapanus townsendii | Sciurus griseus | |
| Sorex trowbridgii | Clethrionomys californicus | |
| Sorex bendirii | Microtus townsendii | |
| Marmota olympus | Microtus oregoni | |
| Tamias townsendii | Aplodontia rufa |
Some species which range outside this faunal area have strongly marked races confined to it. Glaucomys sabrinus oregonensis and Lepus americanus washingtonii are examples. The Pacific Coastal Fauna is a forest fauna. Like the Great Basin Fauna, it reaches its northern limit of distribution in Washington and is better represented farther south. Unlike the Great Basin Fauna, the break between the Pacific Coastal and the surrounding fauna is not sharp, because forests continue into the more boreal faunal areas to the north and east. There, some mingling of coastal and Rocky Mountain faunas occurs.
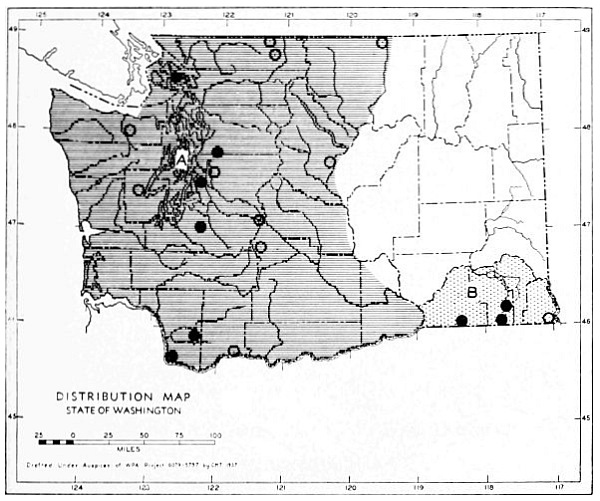
3. Rocky Mountain Fauna. If this fauna be thought of as including mammals of the Rocky Mountains of the United States, and also those of the subarctic faunal area to the east of these mountains in Canada, the species in Washington are as follows:
| Sorex palustris | Synaptomys borealis | |
| Microsorex hoyi | Phenacomys intermedius | |
| Lynx canadensis | Clethrionomys gapperi | |
| Marmota caligata | Microtus richardsoni | |
| Citellus lateralis | Ochotona princeps | |
| Citellus columbianus | Lepus americanus | |
| Tamias amoenus | Rangifer montanus | |
| Tamiasciurus hudsonicus | Oreamnos americanus | |
| Glaucomys sabrinus |
Of the three, the Great Basin Fauna is the most distinct. Only about twenty species which occur within the Great Basin Faunal Area, occur also outside of it in one or both of the two other faunal areas. Most of these twenty are subspecifically different in the Great Basin Faunal Area as contrasted with one or both of the other areas. Each of the other two areas has no less than 32 species that are not restricted to it.
The present fauna of the state of Washington was derived in part from Asia and in part from native forms. Great changes occurred in early Pleistocene through emigration. By the late Pleistocene most of the mammals now occurring in the state of Washington were as they are today. The greatest changes that seem to have occurred in the late Pleistocene are the extinctions of numerous groups, locally or totally. Among the carnivores, Matthew (1902: 321) reports remains of the great lion, Felis atrox, associated with such familiar species as the badger, cougar, lynx and mountain goat. The great lion was very similar to the modern African lion but was fully a fourth larger. Associated with the great lion in the California tar pits are the carnivorous short-faced bears (Tremarctotherium), as large as the Alaskan brown bears; dire wolves (Aenocyon), larger than timber wolves; and saber-tooth tigers (Smilodon). These forms were probably also present in Washington in the late Pleistocene. Matthew (loc. cit.) reports remains of the giant beaver, Castoroides, from the Silver Lake deposit of Oregon. This great beaver, as large as a black bear, was doubtless a resident of Washington also. Peccaries, camels, bison, horses and giant ground sloths have been recorded from Pleistocene deposits of Washington and nearby areas. Of the elephant tribe, the mastodon and several species of mammoths were present.
These extinct forms have doubtless exerted some influence on the past distribution of mammals in Washington and possibly have had an effect on the distribution of members of the living fauna. One species of the mammoth, at least, existed in Washington in postglacial time. Remains of this form, Elephas columbi, have been found in Vashon till.
The three faunas of Washington can be placed in two categories. One is Sonoran, essentially a desert type, and occupies the Columbian Plateau. The other two are forest faunas, predominantly boreal in complexion, and are closely related. The Rocky Mountain Fauna is found in the Blue Mountains and in northeastern Washington. The Pacific Coastal Fauna is found in western Washington.
The ice sheets of Vashon-Wisconsin time descended southward to southern Puget Sound and to the northern edge of the Columbian [Pg 55] Plateau. If the area of the ice sheet be superimposed on a map of distributional areas of Washington, it is seen that the area occupied by the Rocky Mountain Fauna in northeastern Washington is eliminated. Thus, at the maximum descent of Wisconsin ice, the Rocky Mountain type of mammalian fauna was found only in extreme southeastern Washington. No point of contact between the forest fauna of the Rocky Mountains and the fauna of the Pacific coast exists, anywhere, because desert areas, or at least barren plains, lie between them from the border of the glaciers south to Mexico. For the entire period, perhaps thousands of years long, while the glaciers were in place, the two forest faunas were separated. Repeated separation of the faunas by successive glaciations is thought to be responsible for many of the differences now existing between them.
Following the retreat of the ice, the Pacific Coastal Fauna extended its range northward to the Fraser River and, in part, into the Cascade Mountains. The Rocky Mountain Fauna invaded northeastern Washington and boreal Canada, including the Pacific Coast north of the Fraser River. Certain parts of the Rocky Mountain Fauna also invaded the Cascade Mountains.
Inasmuch as the Cascades were invaded by species from both faunas, a detailed analysis of the mammals existing there now seems justified. Several significant features of the composition of the mammal fauna of the Cascades are apparent. First, several species typical of the Pacific Coastal Fauna are present, such as Neurotrichus gibbsii, Sorex trowbridgii, Sorex bendirii, Tamias townsendii, Microtus oregoni and Aplodontia rufa. Each of these species has no close relatives in the Rocky Mountain Fauna and, save perhaps Sorex trowbridgii, occupies a unique ecological niche and has no counterpart in the Rocky Mountain Fauna.
A second group includes species with close relatives in both the Rocky Mountain and Pacific Coastal faunas. This group is remarkable in that it is composed of either very closely related species or very strongly differentiated subspecies in each fauna. For example, the golden-mantled ground squirrel (Citellus saturatus) of the Cascade Mountains is specifically distinct from Citellus lateralis. Supposedly the Cascade form was isolated in the southern Cascades during Vashon-Wisconsin time. The Douglas squirrel (Tamiasciurus douglasii) of the Cascades, which has a red belly, is the same as the squirrel of the lowlands of western Washington but is specifically distinct from the red squirrel (Tamiasciurus hudsonicus) of the Rocky Mountain Fauna, which has a white belly. In the extreme [Pg 56] northeastern Cascades the two species come together. They do not interbreed but seem to compete, for they do not occur together. The flying squirrel (Glaucomys sabrinus fuliginosus) of the Cascades is only slightly differentiated from other races of the Rocky Mountain Fauna but is much different, as are all Rocky Mountain races, from the western Washington subspecies (Glaucomys s. oregonensis). The red-backed mouse of the Cascades is Clethrionomys gapperi, a species distinct from Clethrionomys californicus of western Washington. The jumping mouse of the Cascades is Zapus princeps trinotatus, the same race that occurs in western Washington. It is quite distinct from, and has previously been considered a species separate from, the races of the Rocky Mountain Fauna. The snowshoe rabbit of the Cascades is closely related to other races of the Rocky Mountain Fauna but is distinct from L. a. washingtonii of western Washington. The pika (Ochotona princeps) of the Cascades was apparently isolated in the southern part of the range during the glaciation. After the retreat of the glaciers it extended its range northward. Competition between two subspecies has resulted in parallel distributions due to relative body size. The two races freely intergrade and the differences between them are not so great as in the other forms mentioned.
The third group of mammalian species of the Cascades is composed of species typical of the Rocky Mountain Fauna such as: Marmota caligata, Synaptomys borealis, and Orcamnos americanus. Each has no ecological counterpart in the Pacific Coastal Fauna. Each is absent from the Cascades of Oregon.
We interpret the mixture of faunas in the Cascades as follows: The Vashon-Wisconsin ice sheet was in place for a long period of time, longer, probably, than the Recent. During this time, forest mammals of the Pacific Coast were isolated from forest mammals farther east by glaciers to the north and desert to the east. Changes took place in both of the separated forest faunas. Certain species, perhaps, such as the mammoth, became extinct. Other forms were exterminated then or at an earlier time in one fauna or the other. If Aplodontia, Neurotrichus or Scapanus occurred in the Rocky Mountain faunal area, it lived in an inland area of rigorous climate, and disappeared there because it was unable to adapt itself to the cold. In the mild climate caused by proximity of the ocean, mild even in Vashon-Wisconsin time to judge from evidence yielded by study of fossil pollens, primitive forms such as moles, the Bendire shrew, and mountain beaver persisted along the coast, where there [Pg 57] were no boreal conditions. Some alpine forms, such as Marmota olympus, Ochotona princeps brunnescens and Citellus saturatus persisted in the Olympic or Cascade mountains as relic species. On the whole, however, the glacial divergence resulted in a boreal forest fauna and a temperate forest fauna.
In addition to change in component species, there were evolutionary changes in the species themselves. In some these were considerable, as shown by the differences between related forms of the two faunas. In most species, however, evolutionary changes have resulted in only subspecific differences.
Following the retreat of the glaciers and the establishment of vegetation on the deglaciated areas, movements of the faunas occurred. The Rocky Mountain Fauna spread northward and westward, to northeastern Washington and, in Canada to the Pacific, occupying most of the land exposed by the glaciers. The Pacific Coastal Fauna spread northward only as far as the relatively slight barrier of the Fraser River. The Cascade Mountains became a "no-man's land." The pika and golden-mantled ground squirrel of the southern Cascades spread northward. Boreal Rocky Mountain forms with no ecologic competitors from the Pacific Coastal Fauna occupied the Cascades. Also, coastal species with no Rocky Mountain competitors occupied the Cascades. Nevertheless, some competition between members of the two faunas ultimately occurred, and in instances where closely related forms occurred in the two faunas, one or the other prevailed in the Cascade Range. For example, the Douglas squirrel and big jumping mouse are now established in that range, but the relative of each occurring in the Rocky Mountains is present in the extreme northeastern Cascades. It is possible that in these two cases, the related form occurring in the Rocky Mountains has just entered the area and that competition has just begun. With regard to the flying squirrel, red-backed mouse and snowshoe rabbit, the more boreal Rocky Mountain representatives have definitely displaced the coastal forms.
Certain mass movements of mammals are popularly believed to have occurred with the advance of the ice sheets of the Pleistocene. The boreal birds and plants on higher peaks of the Cascades and the Sierra Nevada of California are thought to represent relics of faunas that moved northward. Such mass movements probably did occur and there is some evidence of their occurrence in Washington. Probably the pre-Wisconsin flora of coastal British Columbia [Pg 58] consisted of coniferous forest similar to that of western Washington today. If this were the case, the mammalian species in British Columbia corresponded closely to those of western Washington. An influx of such a fauna into coastal Washington would scarcely be evident today if, indeed, it was noticeable even then. In eastern Washington, forest species forced southward would come upon the barren, inhospitable plains and deserts of the Columbian Plateau.
The greater part of the southward moving forms found refuge in the Cascade Mountains where, for most of Wisconsin time, they were isolated in the southern Cascades. Examples are Sorex palustris, Martes caurina, Martes pennanti, Gulo luscus, Vulpes fulva, Lynx canadensis, Tamias amoenus, Thomomys talpoides (douglasii group), Phenacomys intermedius, Microtus richardsoni and Ochotona princeps. In each of these species little or no subspecific variation has occurred between the populations in the Cascades of Washington and the Cascades of Oregon.
While the ice sheet existed in Washington there may have been relatively little movement of the mammalian fauna. There is definite evidence of a brief contact between the Rocky Mountain Fauna of the Blue Mountains and the fauna of the southern Cascades. For example, the pocket gopher of southeastern Washington (Thomomys talpoides aequalidens) is most closely related to the gopher of the Simcoe Anticline, and the long-tailed meadow mouse (Microtus longicaudus halli) of the Blue Mountains closely resembles the meadow mouse of the Yakima Valley.
Mammals of the Blue Mountains and those of the southern Cascades may have come into contact on the Simcoe-Horseheaven Hills Anticline, which now stretches 150 miles from the Cascades to the Wallula Water Gap. Excepting the easternmost 40 miles, it is timbered. East of the Columbia, a continuation of the anticline and other hills reaches to the Blue Mountains. Supposedly, in Wisconsin Time, this anticline possessed a more humid climate and the habitat was essentially the same as that of an alpine meadow today. The forms on the two ends of the anticline that are closely related inhabit humid, meadow habitat. In an earlier paper, Dalquest and Scheffer (1944: 316) named this connection the Simcoe Bridge. Its existence was so strongly indicated by the distribution of pocket gophers in Washington that we supposed that the study of many other species would show that they crossed this bridge. However, study of additional species shows that for them the Simcoe [Pg 59] Bridge was of only slight importance; there appears to have been but little mingling of the fauna of the Blue Mountains and the Cascades by way of the bridge. The Columbia River probably acted as an effective barrier to many forms that might otherwise have utilized it. The forms that did cross on this bridge are species known to be active in winter and to emigrate over considerable areas through tunnels under the snow (Davis, 1939: 257). The pocket gopher and long-tailed meadow mouse may have crossed the Columbia, under a cover of snow, when the river was frozen over. The Columbia has frozen over at the Wallula Water Gap in historic times.
Fig. 19. Pend Oreille River (or Clark Fork of the Columbia) from a point near Newport, Washington, looking south, June 13, 1938. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 67.)
Great movements of mammal species came after the retreat of the Vashon-Wisconsin ice. The greatest of these was the spread of the Rocky Mountain Fauna northward and eastward to the Pacific. In this process, northeastern Washington was inhabited by animals that probably came from Idaho and Montana. Some of the species from farther north, as for example the caribou, may first have been forced into Idaho and Montana by the glaciers. The invasion of northeastern Washington probably was not a mass movement of an entire fauna, because invasion seems to be still going on. As yet the woodchuck and striped ground squirrel of the Pend Oreille Mountains have not crossed the Columbia River, a [Pg 60] relatively minor barrier in northeastern Washington. The mountains west of the Columbia are occupied instead by the yellow-bellied marmot, a member of the Great Basin Fauna, and there is no ecologic counterpart in these mountains of the golden-mantled ground squirrel.
Also the invasion of the Cascade Mountains by a number of Rocky Mountain species may have been an intermittant or gradual movement. The red squirrel and Rocky Mountain subspecies of jumping mouse now are present in the extreme northeastern Cascades, where they possibly arrived relatively recently. The squirrel is competing with the coastal species already present and may eventually supplant it. The same may be true of the two forms of jumping mouse.
The invasion by the Rocky Mountain Fauna was rapid as compared with that of the Pacific Coastal Fauna. Evidence of this was presented in an earlier paper (Dalquest and Scheffer, 1944: 310), where it was shown that the gophers of the douglasii group, isolated during the Vashon time in the southern Cascades, made only a few feeble postglacial movements and then only when conditions were ideal. In this same time the fuscus group of gophers moved from Idaho and virtually surrounded the range of the douglasii group. This tendency to immobility seems to have been characteristic of every member of the Pacific Coastal Fauna. If the retreat of the Vashon and Wisconsin glaciers occurred at the same time, both faunas had an equal opportunity to invade the deglaciated area. Yet, the Pacific Coastal Fauna actually moved northward along the coast only to the Fraser River area, and slightly farther in the Cascades.
Possibly the two glaciers retreated at different times. The Wisconsin Glacier was a piedmont glacier, fed by valley glaciers to the west and east, and may have disappeared when the feeder glaciers dried up. The Vashon Glacier was instead an ice cap, supposedly self-supporting much in the same manner as is the Greenland Ice Cap, and may have persisted longer than the Wisconsin glacier. If it did persist longer it formed a barrier to the northward emigration of coastal species of mammals.
Possibly, also, the whole of the Pacific Coastal Fauna possessed an inherent sluggishness resulting from their long residence in the uniform climate and habitat of the Pacific Coast. Certainly the species show today great habitat specialization as compared with species of the Rocky Mountain Fauna. Also, there are fewer individual mammals per unit of area in western Washington than [Pg 61] in northeastern Washington. The persistence of the Vashon Glacier, an inherent lack of incentive to emigrate, or retention of a favorable environment, may account for the relatively small area invaded by the Pacific Coastal Fauna.
The Olympic Mountains, on the Olympic Peninsula, rise above the timber-line and are surrounded by forested lowlands which in a sense isolates this mountain range. Early workers, notably Elliot, obtained specimens of mammals from the Olympics and described numerous races, principally, it appears, on the supposition that because the range was somewhat isolated it should possess a unique fauna. Subsequent revisions of groups of mammals have indicated that most of the names proposed, on the basis of specimens from the Olympics, were either invalid or pertained to mammals found also in the Cascades.
The mammals of the Olympic Peninsula appear to be divisible into three groups. A majority of them fall within the first group, namely coastal races possessing wide ranges in the lowlands of western Washington. The second group consists of species of the Rocky Mountain Fauna but with close relatives in the Cascades. The third group includes but two forms, both unique and found only on the Olympic Peninsula.
The first group includes nonalpine forms of the lowlands surrounding the Olympic Mountains. For the most part these are identical with races of the Puget Sound area. A few are slightly differentiated from the mammals of the Puget Sound area but are the same as mammals from southwestern Washington. As will be shown later, some differentiation in the Pacific Coastal Fauna has occurred. This is thought to be evolution in situ, rather than the result of mass movements. Many nonalpine Coastal mammals occur in alpine habitat in the Olympics.
The second group consists of species of the Rocky Mountain Fauna. Their relationship to the mammals of the Cascades is indicated in the two parallel columns below.
| Olympics | Cascades | |
| Sorex palustris navigator | Sorex palustris navigator | |
| Martes caurina caurina | Martes caurina caurina | |
| Martes pennanti | Martes pennanti | |
| Tamias amoenus caurinus | Tamias amoenus ludibundus | |
| Phenacomys intermedius oramontis | Phenacomys intermedius oramontis | |
| Clethrionomys gapperi nivarius | Clethrionomys gapperi saturatus |
Only two of these are racially distinct from their relatives in the Cascades. Of these the chipmunk is a plastic species and breaks down into many races in Washington. The chipmunks of the [Pg 62] Olympics and of Mt. Rainier are so similar that Howell (1929: 77) considered them as identical and mapped Mount Rainier as an isolated part of the range of the Olympic form (see account of T. a. caurinus). The relationship of the red-backed mice, also, is close, but has been obscured by the usual assumption of relationship between californicus (occidentalis) and gapperi. The principal difference between the alpine forms is the pallor of nivarius. This pallor of mammals in general from the Olympic Mountains is noteworthy, but in the red-backed mouse is exceptionally noticeable. This pallor is discussed beyond in the paragraphs dealing with differentiation. Mention should be made here of Myotis keenii. This is a species which seems to have extended its range to Washington from the north. The power of flight, of course, removes it from consideration in attempting to reconstruct routes followed by terrestrial mammals.
The route of the pocket gopher (Thomomys) in emigrating from the Cascades to the Olympics (Dalquest and Scheffer, 1944: 310), was over the outwash train of the Mount Rainier Glaciers, especially the Nisqualli Glacier, to the extensive outwash aprons of the Vashon Glacier around southern Puget Sound, and thence into the Olympic Mountains. Under the conditions in early postglacial time this invasion route, hereinafter termed the Puget Bridge, around the Pleistocene Lake Russell (present Puget Sound), is thought to have been mainly an alpine meadow. Indeed, the isolated prairies remaining today are the unforested remnants of the outwash aprons (see Dalquest and Scheffer, 1942: 69) and possess several species of alpine plants, notably the shooting star, camas, and bear grass.
If the Vashon Glacier remained in place considerably longer than the Wisconsin Glacier, these Rocky Mountain species may have invaded the Cascades from northeastern Washington and travelled around the southern edge of the Puget Glacier or of Lake Russell. The close relationship of the races involved, however, suggests that the emigration took place much more recently. The barriers to such movement even today are slight, consisting principally of narrow areas of forest. For the water shrew, an almost continuous water habitat still exists, by way of the Nisqualli River, streams in the Puget Sound area, and the Satsop River in the Olympics. Tree-living forms such as the fisher and marten might easily travel the intervening distance today, and, by going along the forests north of the Chehalis River, reach the Olympics without crossing more than small streams and virtually without descending to the ground. [Pg 63] Chipmunks and mice probably utilized the prairie or meadow area of the Puget Bridge, as did the gophers.
Considering the long existence of the Puget Bridge, it is surprising that such forms as the pika, water rat and golden-mantled ground squirrel did not cross to the Olympics. These forms are, however, species of the higher or eastern slopes of the Cascades.
The third group of Olympic mammals includes the white-bellied water shrew and the Olympic marmot, both indigenous forms.
The Bendire water shrew, Sorex bendirii albiventer, is not restricted to alpine habitat but occurs throughout the Olympic Peninsula. Its nearest relative is S. b. bendirii of the rest of western Washington. S. b. albiventer differs from bendirii only in possessing a partially white ventral surface. We can only conclude that the white belly of albiventer is a mutation that the local environment has favored and that the characters have, therefore, spread through the population on the Olympic Peninsula. Occasional specimens are taken with dark bellies characteristic of bendirii (Jackson, 1928: 199).
The Olympic marmot, Marmota olympus, specifically distinct, and apparently the only preglacial relic species of alpine mammal in the Olympics, is most nearly related to Marmota vancouverensis of the unglaciated mountains of Vancouver Island, British Columbia. Both olympus and vancouverensis are close relatives of Marmota caligata which ranges southward into the Cascades of Washington.
The Columbia River in its course westward through the Cascade Mountains, might be expected to act as a highway for the movement of mammals, but the extent to which it has done so seems to be slight, at least in postglacial time. The pocket gopher of southwestern Washington reached the area about Vancouver from the southern Cascades by way of meadows on the gravel terraces of Wisconsin glacial drift. No other mammal seems to have extended this far. Several Great Basin species, such as the cottontail, extend westward in the valley of the Columbia to the vicinity of Bingen.
The mammals of western Oregon and southwestern Washington are closely similar as are the plants and climate, despite the fact that the broad Columbia River courses through the area and did so all through Recent and Pleistocene times. Many species would be expected to have crossed this barrier by swimming and rafting, and that they did so is indicated by the large number of mammals which are identical or very closely related on the two sides of the river.
Mammals which seem not to differ on the two sides of the river include:
| Scapanus townsendii | Citellus beecheyi | |
| Scapanus orarius | Tamias townsendii | |
| Sorex trowbridgii | Sciurus griseus | |
| Sorex vagrans | Glaucomys sabrinus | |
| Canis lupus | Castor canadensis | |
| Felis concolor | Microtus townsendii | |
| Lynx rufus | Microtus oregoni | |
| Mephitis mephitis | Ondatra zibethicus | |
| Spilogale gracilis | Zapus princeps | |
| Procyon lotor | Odocoileus hemionus | |
| Ursus americanus |
The following mammals are subspecifically distinct in western Washington and western Oregon:
| Washington | Oregon | |
| Sorex bendirii bendirii | Sorex bendirii palmeri | |
| Sorex obscurus setosus | Sorex obscurus bairdi | |
| Neotoma cinerea occidentalis | Neotoma cinerea fusca | |
| Peromyscus maniculatus austerus | Peromyscus maniculatus rubidus | |
| Clethrionomys californicus occidentalis | Clethrionomys californicus californicus | |
| Microtus longicaudus macrurus | Microtus longicaudus abditus | |
| Aplodontia rufa rufa | Aplodontia rufa pacifica |
The following species are found in western Oregon but do not occur in western Washington:
| Vulpes fulva | Phenacomys albipes | |
| Urocyon cinereoargenteus | Microtus canicaudus | |
| Neotoma fuscipes | Thomomys bulbivorus | |
| Phenacomys silvicola | Lepus californicus | |
| Phenacomys longicaudus | Sylvilagus bachmani |
Several of these mammals which occur south of the river but not north of it are common on the south bank, a few miles from favorable but uninhabited territory on the north.
Seemingly the pre-Vashon faunas of western Oregon and Washington were similar. Some species became extinct in Washington in the course of Vashon isolation. Others persisted. The very close relationship of the mammals of the first group indicates some crossing of the river. The best known of such crossings was that of the Beechey ground squirrel which, previous to 1915, was unknown in Washington. In 1915, when there was no man-made bridge at White Salmon, it crossed the river and since has spread over an area of at least 50 square miles. The distribution of the mountain beavers is unusual in that the form in the lowlands of Washington is indistinguishable from the subspecies in the Cascades of Oregon.
The mammals that are racially distinct on the two sides of the Columbia River merit careful scrutiny. The Peromyscus of the two sides more closely resemble one another than those of southern Oregon resemble those of northern Oregon or than those of southern Washington resemble those of northern Washington. For Peromyscus maniculatus, the Columbia River is simply a convenient boundary for the separation of two slightly different races. The Oregon race of the bushy-tailed wood rat is a coastal type but the Washington form is the same as that of eastern Washington. Seemingly the more eastern race spread to an unoccupied habitat in western Washington. Other races that differ on the two sides of the Columbia probably developed while separated by the river.
Fig. 20. Rocky bluff along north bank of the Columbia River near Lyle. Washington. March 20. 1939. Habitat of Beechey ground squirrel and yellow-bellied marmot. (Fish and Wildlife Service photo by Victor B. Scheffer. No. 640.)
The San Juan Islands now possess a limited mammalian fauna. Unfortunately the activities of man have somewhat changed the native populations, especially by the introduction of the domestic rabbit which is now a serious pest in the islands. The Douglas squirrel, present on Blakeley Island, is said to have been introduced [Pg 66] and one resident claims to have first brought it to the island. Two different persons claim credit for introducing the Townsend chipmunk on Orcas Island but do not account for its presence on Lopez Island. The three mammals most abundant and widely distributed in the islands are Sorex vagrans, Peromyscus maniculatus and Microtus townsendii. These species, at least, probably reached the islands at an early time. The two last named are now subspecifically distinct from their mainland relatives. Other mammals which probably were established before the arrival of the white man include the mink, otter, beaver, muskrat, raccoon and black-tailed deer.
The Great Basin Fauna of eastern Washington exists as three units, one on the Columbian Plateau, another in southeastern Washington and the third in the Yakima Valley area. The desert species of the Yakima Valley are more closely related to the species of eastern Oregon than they are to those of the Columbian Plateau. In a number of respects the Columbian Plateau gives indications of age. The ground squirrel, Citellus washingtoni, is related to, but specifically distinct from Citellus townsendii of the Yakima Valley and eastern Oregon. Perognathus parvus lordi is a well-marked race, as is Microtus pennsylvanicus kincaidi and Thomomys talpoides devexus. We suppose that these species were present on the Columbian Plateau at least through the Recent and probably through all of Wisconsin Time. The loess deposits of eastern Washington seem to have been laid down in Wisconsin and Recent times. These indicate an arid climate which, although probably cool, was probably not so cold as to exterminate these species. On the other hand, some species that are now abundant on the Columbian Plateau seem to have arrived there relatively recently. The black-tailed jack rabbit, for example, was unknown in eastern Washington before 1870 when it appeared in Walla Walla County. In 1905 it crossed the Snake River on ice and invaded the Columbian Plateau where it rapidly spread over the whole area. In January, 1920, it crossed the Columbia in two places and spread over the Yakima Valley.
The known facts of this movement were sufficiently impressive to cause the author to study rather closely the distribution of mammals in this area. The collection of bones from a cave along the Columbia River near Vantage, Grant County, on the Columbian Plateau, is especially helpful in this respect. This cave was first visited in 1938. It had been the habitat of owls, bats, and primitive man. The floor of the cave was buried under from one to three [Pg 67] feet of bat guano, much of which had been hauled away for fertilizer. Here and there we found traces of fire and occasional piles of mussel shells. Some arrowheads and one beautiful obsidian spear head were found, all buried in guano and about midway between the floor and the top of the deposit. Remains of mammals were abundant through the bat guano, and apparently had been brought to the cave both by man and owls. The jaw of a mountain sheep was found. This species was known to be present when the first settlers reached the area (Cowan, 1940: 558). The remains of smaller mammals included gopher, pocket mouse, muskrat, meadow mouse, deer mouse, coyote and white-tailed jack rabbit. No remains of cottontail, black-tailed jack rabbit or harvest mouse were found. The absence of the cottontail was especially surprising, in that fully thirty skulls of white-tailed jack rabbits were noted. The grasshopper mouse (Onychomys) was also absent, but this species is not common. The two rabbits and the harvest mouse, however, are abundant in the area today. The cottontail and harvest mouse have only recently been recorded from the Okanogan Valley of British Columbia (Cowan and Hatter, 1940: 9). The black-tailed jack rabbit has never been taken there.
Apparently then, some species have only recently entered the Upper Sonoran Life-zone of eastern Washington. They have, of course, reached the state from Oregon. The first step in the invasion probably was the occupation of southeastern Washington. No barrier prevents mammals from reaching southeastern Washington from eastern Oregon but the Columbia to the north and west prevents them from occupying the Yakima Valley, and the Snake River prevents them from reaching the Columbian Plateau. The kangaroo rat, Great Basin striped skunk and Great Basin spotted skunk now are at this stage of invasion. The second stage was the crossing of the Columbia River to the Yakima Valley. This has been accomplished by the black-tailed jack rabbit and, earlier, by the pocket mouse, Perognathus parvus parvus, and ground squirrel, Citellus townsendii. The third stage was the crossing of the Snake River and occupation of the Columbian Plateau. The final stage is the crossing of the northern Columbia River and occupation of the Okanogan Valley.
Whereas it is probable that a few of the species now occurring in Washington evolved there, most are immigrants from other areas. The success of a given species in any area is dependent on numerous factors which may be classified under food, shelter from the elements, protection from enemies and safe breeding places. The factors may be of an inorganic nature, such as climate, soil and exposure or they may be organic, such as vegetation, competition for food and response to enemies. Abundance results in population pressure and a tendency for the range of a species to expand.
Mammalian populations are dynamic and change in accordance with alterations in environment. Because the later geologic history of the state of Washington was violent, with resultant changes in climate and geography, the mammalian populations and the distribution of the species have changed much. With changes in environment, rare species may become common; common and widespread species may become rare or extinct; species foreign to the area may enter, become established and affect the distribution of other forms.
Subspecies are groups of individuals with similar genetic components or are groups of microgeographic races. In instances where the phenotypic expression of these similar genetic factors, or the "characters," are, as a unit, uniformly different from those of animals of the same species in another geographic area, it is convenient to give recognition to the two kinds by separate subspecific name. Intergradation between two geographically adjacent subspecies occurs, directly or where impassable barriers separate them, indirectly by way of one or more other subspecies. Subspecies of mammals are geographic races, which means that to warrant recognition by subspecific name, there must be a logical geographic range in addition to morphological characters.
Timofeef-Ressovsky (1932, 1940) advances the theory of harmoniously stabilized gene-complexes to account for the persistence of subspecies. The persistence of subspecies as genetic units has been best explained, I feel, by Sumner (1932: 84-86) who theorizes as follows:
1. The number of young produced by a subspecies is greater than the carrying capacity of the land they occupy, at least at certain times or in some years.
2. Population pressure results, with a tendency of individuals to emigrate outwards, to the border of the range of the subspecies, where the population pressure is less.
3. The outward moving tendency keeps the center of the range of the subspecies genetically "pure."
4. The peripheral wave continues, as long as favorable habitat is encountered, until an oppositely directed wave of another race is encountered.
5. Areas of intergradation represent local mingling of genetic factors and do not affect the "pure" individuals of the central part of the range of the subspecies.
Certain aspects of this hypothesis are strongly supported by the distribution of mammals in Washington. Witness the rapid invasion of Citellus beecheyi and Lepus californicus in Washington, and the eastern cottontail in western Washington and the domestic rabbit in the San Juan Islands. The volume of the "wave of population pressure" where no opposing force is met, is scarcely believable. In seven years the eastern cottontails released in southwestern Washington multiplied from a maximum of 12 individuals to a minimum of 40,000.
Competition between subspecies where their ranges come into contact seems to be exceptional. Peromyscus maniculatus oreas and P. m. austerus seem to afford an example of this. However, in a few cases subspecies seem to be determined in part by adaptation to restricted environments; each race lives only where local conditions favor its respective adaptations.
In the pocket gophers, where restricted habitat and fossorial habits cause numerous microgeographic races, these microgeographic races may be potential subspecies. This is especially true in the Puget Sound area, where six races occur in a small area. These races meet all the requirements of subspecies and are recognized as such. It should be pointed out, however, that these races and probably many other races produced by isolation, may represent degenerative mutations of the type mentioned by Wright (in Huxley, 1940). The principal differences of such races seem to have resulted from the loss of factors of original multiple factor series, with resultant homogeneity of the race. Inherent variability is another thing that has to be taken into account when considering the differentiation of the mammals of Washington into subspecies. The pocket gopher is an extremely plastic species, especially in Washington, whereas the Douglas squirrel is less so. The flying squirrels, the yellow-pine chipmunk and the snowshoe rabbit are the other plastic species. These species are not so likely to break up into numerous subspecies over all of their ranges as they are in Washington where [Pg 70] in a small area the topography is highly varied. The range of the one subspecies, Tamias amoenus amoenus, to the southeast of Washington is larger than the combined ranges of all six races occurring in Washington but, so far as I can see, the topography and environment are no more varied in Washington than in the mentioned area to the southeast of it. The range of one subspecies, Lepus americanus americanus, in Canada is several times larger than the entire state of Washington, in which four races are found.
The shrews are poor subjects for a study of differentiation, principally because their small size makes it difficult to see morphological variations that may be present. The difficulty is increased because cranial sutures become ossified at an early age. Although it is difficult to evaluate the differentiation in them, there is some. The bats, especially the Myotis, are less restricted by geographic barriers than are terrestial mammals. Nevertheless, obvious differentiation exists. The larger predatory mammals and the artiodactyls are able to move over large areas, at least in the breeding season, but in these animals also, some differentiation has occurred.
The greatest changes, other than the extinctions, to occur in the mammalian fauna of Washington since the late Pleistocene, are changes in distribution. The interglacial cycle preceding the Vashon-Wisconsin glaciation was of far greater duration than the Recent. Presumably the mammalian fauna had, from a distributional standpoint, reached a relatively stable condition. The descent of the Vashon-Wisconsin ice destroyed the stability and set parts of the fauna in motion. Probably no stability was reached before the ice began to recede, and when it did so the previous movements of the various species were, at least in part, reversed. Stability has not yet been reached by the mammalian fauna of Washington. Great changes have occurred in historic times and other changes probably are under way at present.
In the following pages an attempt has been made to interpret the probable late Pleistocene and Recent distributional history of the species of mammals occurring in Washington. The interpretations are made in the light of what is known of the physical history of the state and are to be accepted as such rather than as evidence for the conclusions made concerning the physical history of the state of Washington and adjacent areas.
Scapanus townsendii.—Probably this animal was confined to the Humid Transition Life-zone of the Pacific Coast since the Pliocene.
Scapanus orarius.—This species probably had a history similar to that of townsendii up to the late Pleistocene. It seems slightly more adaptable than townsendii, and to be able to extend higher into the mountains. The distribution of the subspecies orarius is almost exactly that of S. townsendii. In Oregon, orarius extended eastward over the Cascades where the subspecies schefferi developed. Perhaps this subspecies developed since the Pleistocene and since that time extended along the Columbia River Valley to southeastern Washington. The race yakimensis, in the Yakima Valley area, is closely related to schefferi, and seemingly could have been developed from a stock of schefferi that migrated westward across the Simcoe Bridge.
Neurotrichus gibbsii.—The history of Neurotrichus in North America was probably similar to that of the two species of Scapanus. It tolerates environmental differences to about the same degree that Scapanus orarius does but occurs much farther south (Monterey County, California) than S. orarius. This may be because Neurotrichus has no counterpart to compete with it in the south, whereas Scapanus orarius must compete in northern California with the morphologically similar Scapanus latimanus. S. orarius stops short at this place and S. latimanus occupies all the territory to the south.
The shrew-mole of the lowland of Washington (N. g. minor) probably became distinct from the mountain subspecies (gibbsii) in Vashon-Wisconsin time.
Sorex cinereus.—It is reasonable to suppose that the cinereous shrew had a continuous range across the forested area of British Columbia in pre-Wisconsin time. Without having been isolated, the dark coastal race (streatori) may have developed from the wider-ranging inland cinereus, as a response to the denser, humid, coastal forest-habitat, after having been forced southward to Washington by the Vashon Glaciation. Since that time it is presumed to have reoccupied the coast of British Columbia and southern Alaska. This coastal race might have developed in Vashon time, while isolated in southwestern Washington. The Cascades are populated by a race of the Rocky Mountain Fauna, S. c. cinereus, which probably entered the Cascades from northeastern Washington or British Columbia in Recent Time. The absence of the species in western Oregon, its rarity in western Washington, and its abundance farther north suggest a northern origin and northward rather than southward postglacial movement. Had the full species cinereus been a [Pg 72] preglacial resident of western Washington we would expect streatori or a race related to it to occur in the Cascades.
Sorex merriami.—The periphery of the range of this member of the Great Basin Fauna may have been in southeastern Washington since pre-Wisconsin time.
Sorex trowbridgii.—This shrew is a typical Pacific coastal species with an extensive range along the Pacific Coast south of Washington. The Washington population may have been isolated in southwestern Washington during Vashon time or may have crossed the Columbia into Washington from western Oregon early in the Recent. Since the retreat of the ice it has extended northward to southern British Columbia and eastward to the eastern side of the Cascades. Save for crossing the Cascades its postglacial movements have been slight, as is typical of Pacific Coastal species. The race destructioni probably has been isolated on Destruction Island for several thousand years.
Sorex vagrans.—This species probably has had a continuous range over the western United States since the late Pleistocene. The dark coastal race (vagrans) probably was differentiated from the paler races of the Great Basin in response to the more humid climate along the coast.
Sorex obscurus.—The history of this shrew of alpine predilection probably corresponded closely to that of Sorex cinereus. The derivation of the dark, long-tailed, coastal race (S. o. setosus) from the smaller, paler, inland race (obscurus) probably occurred before Vashon-Wisconsin Time. Sorex o. setosus is one of a complex of races distributed along the Pacific Coast from Alaska to California.
Sorex palustris.—This species has a wide range in North America and extends southward in the Cascade-Sierra Nevada Chain to southern California. Its extensive range at present in this mountain chain suggests that it was resident in the Cascades previous to Wisconsin time. Mountain water shrews probably reached the Olympic Mountains from the Cascades by way of the Puget Bridge in early Recent Time.
Sorex bendirii.—This Pacific Coastal species probably had a history very similar to that of Neurotrichus and Scapanus orarius.
The difference between the Bendire water shrews of western Washington and western Oregon indicates that the Washington population was separated from the shrews of western Oregon during Vashon Time. The white-bellied race of the Olympic Peninsula is probably of local origin.
Microsorex hoyi.—The Washington record of this shrew at Loon Lake, Stevens County, is in an area where mammals typical of the Rocky Mountain Fauna occur.
Myotis lucifugus.—The dark race of this species (alascensis) may have persisted through the glacial period in southwestern Washington. The race carissima, of the Great Basin Fauna, may have entered the state since the glacial period, from the south, of course. Habitat selection determines their range at present.
Myotis yumanensis.—The dark, coastal race (saturatus) seems to be an established member of the Pacific Coastal Fauna. Unlike lucifugus, the coastal race is not found east of the Cascades. The race sociabilis, of the Great Basin, has doubtless entered the desert of eastern Washington from eastern Oregon.
Myotis keenii.—The southernmost record station for this north coastal species is on the Olympic Peninsula of Washington. It probably developed in the humid, northern part of the Pacific Coastal area previous to the last Pleistocene glaciation and extended its range to the south in Vashon-Wisconsin Time. The range of tolerance in M. k. keenii seems to be more restricted than that of M. lucifugus alascensis.
Myotis evotis.—In Washington, the distribution of this bat is similar to that of Myotis lucifugus. The dark, forest race probably originated in the north-coastal region. The paler race, that developed in the southwest, entered eastern Washington from Oregon.
Myotis thysanodes.—In Washington this species has been recorded only in the southeastern part where the Great Basin Fauna occurs. It probably originated in the southwestern United States, and a point in British Columbia a little way north of Washington marks the northern edge of its natural range.
Myotis volans and Myotis californicus.—Remarks made about Myotis lucifugus apply also to these two species.
Myotis subulatus.—The northwestern periphery of the range of this species seems to be in eastern Washington.
Lasionycteris noctivagans.—Undifferentiated subspecifically from coast to coast, no basis is provided for judging the route by which this species entered the state.
Pipistrellus hesperus.—The northwestern periphery of the range of this bat, also, lies in eastern Washington.
Eptesicus fuscus.—Big brown bats from both eastern and western Washington seem to have been derived from the Pacific [Pg 74] Coastal race of the species. Presumably it extended its range westward across the Cascades in early post-Pleistocene Time.
Lasiurus cinereus.—No speculation as to the distributional history of the hoary bat seems justified at present.
Corynorhinus rafinesquii.—The dark, coastal race of this bat probably persisted in southwestern Washington and western Oregon through Vashon Time and moved northward in the Recent. The paler intermedius probably invaded eastern Washington from eastern Oregon in the Recent.
Antrozous pallidus.—This species strays into eastern Washington from Oregon as part of the Great Basin Fauna.
Ursus americanus.—The dark, western race of the black bear (altifrontalis) and the paler, inland race (cinnamomum) were probably separated by a glacial divergence. The inland race has entered northeastern Washington in the Recent with other members of the Rocky Mountain Fauna.
Ursus chelan, etc.—The apparent past distribution of chelan indicates it to have invaded Washington from British Columbia since the Pleistocene.
The apparent absence of grizzly bears from the southern Cascades and western Washington may indicate their absence from these areas immediately before pre-Vashon time, or their extermination in or shortly after that period.
Procyon lotor.—The raccoon of western Washington seems to be the Pacific Coastal race which occurs also in western Oregon and northwestern California. This indicates that the coastal race (psora) was confined to the coastal area south of Washington during Vashon Time and has only recently reinvaded western Washington. It is possible, though less likely, that raccoons existed in southwestern Washington during Vashon Time but did not develop racial characters, or that the Columbia was crossed so frequently that genetic differences were dispersed throughout the entire population.
Reasons why the second hypothesis is inadequate are: (1) Raccoons range but little north of the state of Washington, both east and west of the Cascades. (2) Raccoons of western Washington and the area about San Francisco Bay, California, are as much alike as are raccoons from southwestern Washington and northwestern Oregon. It is thought that raccoons, if resident in western Washington since interglacial time, would have developed strong racial characters, and the fact that they have not indicates that they have entered the state at a relatively recent date.
The raccoon of eastern Washington (excelsus) is a member of the Great Basin Fauna and has probably included southeastern Washington in its natural range for a long period of time. The raccoon has not extended its normal range into northeastern Washington, although it is seemingly ideal raccoon habitat; only an occasional vagrant occurs there. A stock of raccoons from which emigrants might come has existed in southeastern Washington and the Yakima Valley for some time. The Columbia River might serve as a highway by which emigrants could reach northeastern Washington.
Martes caurina.—The earlier distributional history of the western marten has been postulated by Davis (1939: 131-132), who stated: "When the ancestral stock split into the two groups, the one that gave rise to americana may have pushed eastward across Canada to the Atlantic Coast; the other, giving rise to caurina, may have migrated southward along the Sierra Nevada-Cascade and Rocky mountains. Perhaps the great ice sheet was instrumental in pushing americana eastward and separating it geographically from caurina." The present occurrence of americana in Alaska and British Columbia is thought to have been by invasion from the east in postglacial time.
Davis' theory seems basically correct but subject to correction in detail. The presence of caurina in the southern Rocky Mountains suggests that it is not a Pacific Coastal species in the common sense. Had americana occupied northern British Columbia in pre-Wisconsin Time, it and not caurina would be expected to occur in the southern Rocky Mountains today, for the form found in British Columbia almost certainly would have been forced into the Rockies. The range now occupied by caurina in the Rocky Mountains is so extensive as to suggest that martens could not have migrated into all of it from the Pacific Coast since Vashon Time, even had the region been unoccupied by any species of marten. The presence of americana in Alaska and British Columbia suggests that it arrived in those areas before caurina and that had the Rocky Mountains been unoccupied by martens in pre-Wisconsin time, americana and not caurina would have reached the Rockies first. It appears that caurina occupied much of western North America in pre-Wisconsin Time and was forced southward into the southern Rocky Mountains and along the Pacific Coast by Vashon-Wisconsin ice.
The separation of americana and caurina may be supposed to have occurred before the pre-Vashon-Wisconsin interglacial interval, perhaps by a glacier similar to but antedating the Vashon-Wisconsin glaciation.
The martens of western Washington (Martes caurina caurina) are a coastal race. Those of northeastern Washington belong to a race of the Rocky Mountain Fauna, and are referred to M. c. origenes. Davis (1939: 132) refers the martens of Idaho to Martes caurina caurina. I have compared specimens from Idaho with animals trapped for fur from the Pacific Coast proper and feel that the animals from northeastern Washington and those from Idaho are more like origenes than caurina, although perhaps not typical. The animals from the Pacific Coast proper are caurina and have darker heads and brown instead of yellow patches on the throat.
Martes pennanti.—Fishers are found throughout the Cascade Mountains and probably were widely distributed over western North America in pre-Wisconsin Time.
Mustela erminea.—The distribution of ermines along the coast of northern California and in the Cascade-Sierra Nevada of Oregon and California indicates, as does their differentiation there, that they ranged southward to these areas before and during Vashon-Wisconsin Time.
In immediate pre-Vashon-Wisconsin Time, the dark race streatori probably occurred in western Washington. The race murica probably occurred in the Blue Mountains then, as it does today, but probably occurred also in the Cascades of Washington. The descent of the Vashon glaciers probably displaced streatori from the northern part of its range, at least temporarily. In the Cascades, murica was likewise forced southward. Ermines related to the northern richardsonii were forced into northern Washington and Idaho by the Wisconsin ice. They probably were unable to live on the barren, unglaciated plains of eastern Washington but persisted in Idaho.
The ranges of the three forms at the maximum extent of the Vashon-Wisconsin may be reconstructed as follows: streatori in southwestern Washington; murica in the southern Cascades and the Blue Mountains; invicta stock in northern Idaho. While so isolated, the ermines of the southern Cascades probably mingled, to a certain extent, with streatori and developed the characters that now separate gulosa from both murica and streatori. The intermediate nature of gulosa has been mentioned by Hall (1945: 85).
The retreat of the ice allowed streatori to move north and invicta to move north and east into Washington and the northeastern Cascades. To a lesser extent, gulosa may have moved north. The poorly-marked race olympica probably evolved from streatori in the Recent. It is difficult to account for the dark race fallenda.
It must have evolved from streatori in the Recent but the origin of such a strongly marked race in such a short time is surprising. It might be mentioned that a similarly differentiated race of chipmunk, Eutamias amoenus felix, occupies much the same range.
Mustela frenata.—The long-tailed weasels of the Pacific Coast behave as a plastic group and clearly show the effect of the Vashon-Wisconsin Divergence. The range of the coastal race, altifrontalis, indicates that it was isolated in southwestern Washington during Vashon Time. In that period, or shortly after, it extended its range southward but only along the extreme, coastal area of Oregon (see Hall, 1936: 101). Following the retreat of the ice it extended its range northward to the deglaciated area of western Washington.
Also following the retreat of the ice, a Great Basin subspecies (nevadensis) extended its range northward. This race seems to have been more adaptable and successful than other kinds of Great Basin mammals, for it extended its range farther northward, eastward and westward than most.
A third race, washingtoni, was isolated in the southern Cascade Mountains during Vashon Time and became differentiated from both altifrontalis and nevadensis. It is now found in the Cascades from central Oregon north to Mount Rainier. It is difficult to see why it did not extend its range to include the northern Cascades when the glacial ice left, but it did not. Instead altifrontalis entered the northern Cascades from the west and nevadensis did the same from the east. Weasels obtained in habitats north of Mt. Rainier are intergrades between altifrontalis and nevadensis.
One is reminded here of the douglasii group of Thomomys talpoides in which subspecies did not move north of Mt. Rainier in postglacial time. The area north of Mt. Rainier was populated instead by gophers of the fuscus group, subspecies of which invaded the area from the east. Perhaps Mt. Rainier itself served as a barrier to alpine mammals in the immediate post-Pleistocene. Perhaps Mustela f. washingtoni will eventually extend its range northward, displacing the altifrontalis-nevadensis intergrades from the habitats to which washingtoni may be better adapted.
The Blue Mountains of southeastern Washington are occupied by a weasel (effera) that has a more extensive range in eastern Oregon. The range of this race has probably not changed materially for a long period of time.
One would expect the weasels from extreme northeastern Washington to be referable to the race oribasa, of the Rocky Mountain [Pg 78] Fauna. Instead they are intermediate between that race and the Great Basin race, nevadensis. Apparently nevadensis was so dynamic and adaptable that it actually entered the geographic ranges of surrounding races for some distance. In view of Sumner's theory for the retention of subspecies, one might say that the population pressure of nevadensis on the periphery of its range is stronger than the opposing pressure of some surrounding races.
Mustela vison.—Pending a review of the minks of North America, little can be said concerning their historical distribution in the state of Washington. From the general range of the species in western North America, one would expect some effect of the Vashon-Wisconsin Divergence to be apparent. There is some evidence for this. Minks from Idaho and adjacent parts of British Columbia are distinctly less reddish than minks from the area about Puget Sound, as noted by Davis (1939: 138).
Gulo luscus.—The range of the subspecies luteus, peculiar to the Cascades and Sierra Nevada suggest that the wolverine may have been forced southward in the Cascades and there isolated during Vashon Time. The differences separating the southern race from the northern may have been developed while the two populations were isolated. The range of the wolverine was probably more extensive in glacial and immediate postglacial time than at present.
Lutra canadensis.—The otter of western Washington seems to be a member of the Pacific Coastal Fauna. Little can be said regarding the distributional history of the species in the state, for specimens from eastern Washington are not numerous enough to permit of a person certainly establishing their systematic position.
Spilogale gracilis.—The western race of the civet cat (latifrons) seems to be a coastal race, isolated in southwestern Washington and western Oregon during Vashon time. The eastern race, saxatilis, is a race of the Great Basin Fauna, that has entered the state from Oregon and that will probably extend its range to the north.
Mephitis mephitis.—Of the four subspecies of skunks occurring in Washington, two seem to have been resident in the state during Vashon Time. The western race, spissigrada, was probably isolated in southwestern Washington and extended its range northward, in the deglaciated area of western Washington, after the retreat of the ice. Another race (notata) was probably isolated in the southeastern Cascades and adjacent Oregon. M. m. hudsonica of the Rocky Mountain Fauna entered the northeastern part of Washington after the ice retreated from there. A race of [Pg 79] the Great Basin Fauna, major, entered southeastern Washington from Oregon and may eventually extend its range farther north.
It is interesting to note that both of the western races, spissigrada and notata, both of which probably developed in Washington during Vashon Time, occupy limited ranges in adjacent Oregon (Bailey, 1936: 308).
Taxidea taxus.—This species has probably long been resident on the Columbian Plateau and in southeastern Washington. For the early distributional history of the species see Hall (1944: 17). Pleistocene remains, referable to this race, have been found in Franklin County.
Vulpes fulva.—The red fox of the Cascades was probably isolated there during Vashon Time by glacial ice. Its range extends southward in the Cascades to Oregon. The fox of eastern Washington is probably a member of the Rocky Mountain Fauna that lived in the Blue Mountains of southeastern Washington in Wisconsin Time and that emigrated to northeastern Washington in Recent Time.
Canis latrans.—The distributional history of the coyote in Washington is not clear.
Canis lupus.—The dark wolf (fuscus) of western Washington is probably a coastal race. The race that may have occurred in northeastern Washington probably was an invader from the Rocky Mountain Fauna, and the race that possibly occurred in southeastern Washington would be assumed to have long been a resident of the area.
Felis concolor.—The cougar of western Washington is a coastal race, probably developed while isolated in southwestern Washington and western Oregon. The cougar of northeastern Washington probably entered the state with other Rocky Mountain species, early in the Recent. The cougar of the Blue Mountains of southeastern Washington has probably long been resident there.
Lynx rufus.—The bobcat of western Washington seems to be a coastal race that was isolated in either southwestern Washington or western Oregon by Vashon ice. It has since extended its range into southern British Columbia. The bobcat of eastern Washington seems to be a member of the Great Basin Fauna that has spread to some forested areas on the periphery of the more arid life-zones.
Lynx canadensis.—The lynx is an element of a northern fauna that was probably forced southward into the Cascades and Rocky [Pg 80] Mountains. Its range was probably more extensive, as is indicated by the scattered records of its occurrence in Oregon (Bailey, 1936:271).
Marmota monax.—The woodchuck invaded northeastern Washington in the early Recent with the Rocky Mountain Fauna.
Marmota flaviventris.—The yellow-bellied marmot is a typical member of the faunas of the Great Basin and the southern Rocky Mountains. It has doubtless entered southeastern Washington from eastern Oregon at an early time. In northeastern Washington, west of the Columbia River, it occupies alpine habitat, but it does not occur farther east, where Marmota monax is found, or in the Cascades where Marmota caligata lives.
The yellow-bellied marmots are great wanderers, and commonly are found in scattered outcrops far out on the Columbian Plateau. There is even one record for western Washington, near Bellingham, Whatcom County. This individual must have crossed some low pass in the Cascades from the area about Lake Chelan. There are records of eastern Washington birds occurring in this same area, so it seems likely that the marmot was a natural stray and not an animal that escaped from captivity.
Marmota caligata.—The absence of the hoary marmot from the Cascades of Oregon, and the presence there of Marmota flaviventris, indicates that the species did not occur in the southern Cascades of Washington during Vashon Time. Presumably the hoary marmot is a member of the fauna of the northern Rocky Mountains and entered the Cascades of Washington in the Recent, after which it spread widely and rapidly owing to lack of competition with any established species of marmot.
Marmota olympus.—This species has probably lived in the Olympic Mountains since pre-Vashon Time.
Citellus washingtoni.—This ground squirrel has probably lived on the Columbian Plateau since before Wisconsin Time.
Citellus townsendii.—The Townsend ground squirrel probably entered the Yakima Valley area from Oregon. The differences between it and its relatives in Oregon indicate a considerable period of isolation but one far shorter than the period during which washingtoni is presumed to have been isolated from townsendii.
Citellus columbianus.—The Columbian ground squirrel might have been forced southward in the Rocky Mountain area by the Wisconsin glaciation, might have lived in southeastern Washington since [Pg 81] then, and might have invaded northeastern Washington in the Recent with other species of the Rocky Mountain Fauna.
Citellus beecheyi.—This ground squirrel is known to have entered Washington about 1915 from Oregon.
Citellus saturatus.—The mantled ground squirrel of the Cascades probably evolved, from the lateralis stock, as a separate species while isolated in the southern Cascades during Vashon Time. It is a poorly differentiated species and may actually be instead a strongly marked subspecies.
Citellus lateralis.—The golden-mantled ground squirrels of northeastern and southeastern Washington are closely similar. It is deduced that connectens of southeastern Washington developed the differences that characterize it while isolated, from the main stock, in the Blue Mountains area of Washington and Oregon.
The race found in extreme northeastern Washington (tescorum) probably reached that area in relatively recent times. Its range in Washington is more restricted than that of several other members of the Rocky Mountain Fauna; areas of suitable habitat west of the Columbia River are not inhabited by these ground squirrels. Its range in Washington is almost exactly that of (Marmota monax).
Tamias minimus.—The least chipmunk of the Yakima Valley is the same race (scrutator) as that occupying the Great Basin area of Oregon and Nevada. It must have crossed the Columbia in relatively recent times. Had it been resident in the isolated Yakima Valley area for any considerable period of time, the development of distinctive racial characters there would be expected. Perhaps, then, it has not been resident there as long as has the Townsend ground squirrel which, though closely related to the ground squirrel of eastern Oregon, is racially distinct.
The least chipmunk of the Columbian Plateau is thought to be racially distinct from its relatives in the Yakima Valley and eastern Oregon. Probably it reached the Plateau very early in the Recent. It has probably not been separated from the parent stock as long as has the ground squirrel (Citellus washingtoni) of the plateau. The ground squirrel is specifically rather than racially distinct.
Tamias amoenus.—The distributional picture of the yellow pine chipmunks in Washington is complex. (Fig. 81.) Certain habits of these mammals doubtless have modified what was probably the original postglacial distribution of the species. Chipmunks are [Pg 82] diurnal and natural selective factors for color possibly operate more strongly on animals active by day than on nocturnal animals. Yellow pine chipmunks are neither forest nor desert inhabitants. Indeed, dense forests or open deserts serve as barriers to their distribution. They prefer brush lands, open woods, and other habitats where there is food and cover but abundant sunlight. In such habitats they are almost independent of altitude, temperature and humidity. They live in the Olympic Mountains where rainfall is heavy and humidity high. They live and breed at considerable altitudes in the Cascades, even in the crater of Mount Rainier, where snow, ice and freezing conditions exist the year around. On the other extreme, they occupy the low, open pine forests and brush lands at the lower edge of the Arid Transition Life-zone where temperatures, in summer, are high and rainfall scarce.
We find in the present distribution of the species in the Cascade-Sierra Nevada chain and the Rocky Mountains, indication that the species had a wide geographic range over western North America previous to the Vashon-Wisconsin glacial interval. Probably the range of the species extended in an arc, from the Rocky Mountains across northern Washington to the Cascades, around the basaltic plateau desert in eastern Washington and Oregon. Presumably the descent of the Vashon-Wisconsin glaciers broke this arc into two parallel geographic ranges, the Rocky Mountains and the Cascade-Sierra Nevada chain, with a desert area between.
Almost every species of forest-dwelling animal had its range separated into two parts by the southward movement of the glaciers. Most of these forest-dwelling species were composed of relatively homogeneous stocks, although the yellow pine chipmunk probably was not. The extensive range of tolerance of the yellow pine chipmunk to altitude and climate and its unique habitat requirements cause it to meet radically different natural selective factors. The predators of the chipmunks near Wenatchee, Chelan County, would include: rattlesnake, gopher snake, badger, striped skunk, prairie falcon, red-tailed hawk and other predominantly desert-dwelling species. The chipmunks at Stevens Pass, in the mountains to the west, would have to contend with: marten, black bear, goshawk, bald eagle and other alpine predators. At the present time, the chipmunk of the eastern Cascades is racially distinct from that of the higher Cascades. Geologic and botanical evidence indicates that the Columbian Plateau was a desert in pre-Wisconsin Time. We suppose that a transition from alpine conditions in the Cascades [Pg 83] to desert conditions on the Columbian Plateau existed even in pre-Wisconsin Times. We suppose also that the chipmunk existed in this transition area and in the Cascades before Wisconsin Time and in the southern and southeastern Cascades during Wisconsin Time. We further suppose that the differences separating the transition area race (Tamias amoenus affinis) from the mountain race (T. a. ludibundus) came about through natural selection and not as a result of geographic isolation. The principal difference between the two is the paler color of the race in the transition area.
The descent of the Vashon-Wisconsin glaciers, then, found two races of the yellow pine chipmunk in the Cascades. Chipmunks living north of the Columbian Plateau, in northern Washington and British Columbia, were probably forced southward onto the inhospitable plains of the plateau and exterminated. Farther east, north of northeastern Washington, chipmunks from the north were probably forced southward to compete with resident chipmunks.
The range of Tamias amoenus luteiventris in Washington, Idaho and Montana is most unusual (See Howell, 1929; Davis, 1939). From a compact range in Montana, two long fingers reach northward and westward. The western finger crosses Idaho to end in the Blue Mountains of Oregon and Washington. The northern finger crosses northern Idaho, northeastern Washington and extends on into southern British Columbia. Between these two fingers of the range of luteiventris another race (canicaudus) is found. This race occupies a more lowland area than does luteiventris. The range of luteiventris in the northern Rocky Mountains is extensive. Presumably this race occupied an area farther north in pre-Wisconsin Time and was forced southward to its present range by the Wisconsin glaciers. The original population occupying extreme eastern Washington and adjacent Idaho was Tamias amoenus canicaudus. The pre-Wisconsin range of this race might have been more extensive. At any rate, luteiventris which was driven southward displaced canicaudus, or some other race of chipmunk, from much of the Rocky Mountains south of the glacier. The northern chipmunks were adapted to more boreal conditions and perhaps otherwise better suited to environmental conditions of the northern Rocky Mountains. A small population of the older established race (canicaudus) persisted in lowland areas of eastern Washington and adjacent Idaho.
Regarding the range of canicaudus, surrounded by the range of luteiventris on three sides and faced by desert on the west, Davis (1939: 220) writes, "It may be that, of these two races, luteiventris [Pg 84] has a greater range of tolerance to environmental conditions and, thus, is able to succeed in areas to which canicaudus is not adapted. This inference is supported by the fact that luteiventris occupies a large range which is diversified geographically and climatically, whereas canicaudus seems to be limited to a much smaller, more nearly uniform area." Seemingly canicaudus now exists only in an area ideally suited to it, and one where it can successfully compete with the generally more adaptable and successful luteiventris. The maximum extent of the glacial ice, then, found luteiventris the dominant chipmunk in the northern Rocky Mountains, with an isolated population of canicaudus in eastern Washington and adjacent Idaho.
The topography of the ground moraine exposed by the retreat of the Vashon-Wisconsin glaciers was a barrier to many species of mammals. The rough, rocky surface with thin soil probably first supported mosses and grasses, then brush, and later trees. The earlier stages of plant succession on the deglaciated ground probably presented ideal habitat for yellow pine chipmunks. Certainly the races immediately adjacent to the glaciers extended their ranges farther north than many species. In eastern Washington, T. a. luteiventris spread to the northeastern corner of the state and on into British Columbia. In the northern Cascades, T. a. affinis spread northward and eastward, across the Okanogan River, into northeastern Washington as far as the range of luteiventris. The chipmunk of the higher Cascades (ludibundus) likewise extended its range northward into British Columbia. In the northwestern Cascades of northern Washington and southern British Columbia, a richly-colored race, T. a. felix, now occupies a limited geographic range. This race doubtless originated from ludibundus stock but the method of its development is unknown. Perhaps in early postglacial time, selective factors developed in chipmunks of the western slopes of the Cascade Mountains the rich, dark color of felix. The ancestral ludibundus may have given rise to a pale race, affinis, in the arid eastern Cascades and a dark race, felix, on the humid western slope of the Cascades. This seems improbable for there is no trend to darker color on the western border of the range of ludibundus south of the range of felix, and instead, affinis may have given rise to ludibundus. A more appealing hypothesis is that a local mutation in some ludibundus stock so changed the range of tolerance of a portion of the population that it was allowed to enter the more dense habitat along the coast north of the Fraser River and, there, isolated by habitat selection, it developed the characters of [Pg 85] felix. Population pressure later forced it eastward until the eastern border of its range again met the range of the ancestral race, ludibundus.
The chipmunks of the Olympic Mountains probably reached their present range from the Cascades. Their probable path of emigration was westward from Mt. Rainier, along the glacial outwash train of Nisqualli Glacier, to the moraine and outwash apron of the Vashon Glacier and thence to the Olympics. So similar are the chipmunks of Mt. Rainier and the Olympic Mountains that Howell (1929) included Mt. Rainier in the range of caurinus.
Briefly summarized, the probable pre-Vashon-Wisconsin distribution of chipmunks of the species Tamias amoenus in Washington was: ludibundus in the higher Cascades; affinis in the eastern Cascades; canicaudus in eastern Washington and adjacent Idaho; and luteiventris in the area north of the range of canicaudus. The descent of the Vashon-Wisconsin ice restricted but did not materially alter the ranges of ludibundus or affinis. On the east, luteiventris was forced southward to compete with canicaudus and displaced it over a large region, especially in mountainous areas. Following the retreat of the ice, luteiventris, affinis, and ludibundus extended their ranges northward over the deglaciated territory. A stock of ludibundus that moved westward from Mt. Rainier became isolated and gave rise to caurinus. In some less obvious development, ludibundus stock gave rise to felix north of the Fraser River in the Cascades.
Tamias ruficaudus.—Until a better understanding of the range of this chipmunk and its relation to other Tamias is gained, uncertainty will remain concerning its distribution in the past.
Tamias townsendii.—This is a typical coastal species that ranges southward, along the coast, to California. The lowland race of western Oregon and Washington (townsendii) probably occurred no farther north than southwestern Washington when the Vashon Glacier was in place. Chipmunks of this species in the Cascades and in the southern Olympic Mountains probably developed independently the slightly paler color that separates cooperi from townsendii. The tendency for species of the Pacific Coastal Fauna of the Cascades and the Olympic Mountains to be paler than their lowland relatives is widespread.
After the retreat of the ice, both races probably moved northward. Perhaps because of its alpine adaptations, cooperi has moved farther than townsendii. Also, townsendii, in the lowlands, ranges to the Fraser River, a barrier not encountered by cooperi.
Sciurus griseus.—This species of the Pacific Coastal Fauna probably entered Washington from Oregon since the retreat of the Vashon Glacier. It has probably entered the state in relatively recent times.
Tamiasciurus hudsonicus.—The two species of red squirrels, T. hudsonicus and T. douglasii, are specifically distinct and probably became differentiated in the Pleistocene when southward moving glaciers cut in two the range of the ancestral stock. The morphological differences are too great, comparatively, to have occurred during the Vashon-Wisconsin Divergence. T. hudsonicus probably occupied a range in pre-Wisconsin Time that included the Rocky Mountains and areas to the north. Glacial ice probably restricted the range of hudsonicus in Wisconsin Time but after the retreat of the ice hudsonicus moved northward to reoccupy its former range. It also moved westward across northern Washington to the Cascades, where it met the range of douglasii. Farther north, it moved westward to the Pacific, thus occupying an area that, in pre-Vashon time, probably was occupied by douglasii.
Tamiasciurus douglasii.—The Douglas squirrel probably occupied the coastal region of Oregon, Washington and British Columbia in pre-Vashon Time. The descent of the ice restricted its range to southwestern Washington and areas to the south. After the retreat of the ice it moved northward somewhat but, like other coastal species, the movement was slow. Meanwhile hudsonicus from the Rocky Mountain Fauna, had spread to the coast of British Columbia.
Glaucomys sabrinus.—This flying squirrel is a plastic species. It inhabits all of the forested parts of Washington. The distributional picture presented by the 5 races (Fig. 92) which occur in Washington is complicated. The ranges of 3 of these lie principally outside the state of Washington.
The race oregonensis occupies Washington and Oregon west of the Cascades; fuliginosus occupies the Cascades of Washington, Oregon and southern British Columbia; columbiensis occupies the interior valleys of British Columbia and adjoining Washington; latipes occupies the northern Rocky Mountains of British Columbia, northern Idaho and extreme northeastern Washington; and bangsi occupies the Blue Mountains of Washington and Oregon and a wide range in Idaho and eastward.
The differences separating the race oregonensis from other subspecies found in Washington are relatively great. This lowland [Pg 87] race is smaller and richer in color. The other races exhibit slight but relatively constant differences. The relatively great difference between oregonensis and the other races indicates that oregonensis was isolated from the remainder of the species for a considerable time. Presumably oregonensis was a strongly differentiated coastal race in pre-Vashon Time and occupied most of western Washington and Oregon. The descent of the Vashon ice restricted the range of oregonensis to southwestern Washington and western Oregon. The descent of the ice forced a northern race, fuliginosus, southward into the range of oregonensis. The northern race, adapted to boreal conditions, was able to compete successfully with the established oregonensis only in mountainous areas. In the Cascade Mountains, fuliginosus extended its range southward to southern Oregon.
The descent of the Wisconsin ice in eastern Washington forced the flying squirrels of adjacent British Columbia southward into the Rocky Mountains. These squirrels were probably closely related to fuliginosus, or to bangsi, which latter race already may have been established farther south in the Rocky Mountains. The Blue Mountains of southeastern Washington were probably inhabited by bangsi in Wisconsin times, or even earlier. The retreat of the Vashon-Wisconsin glaciers allowed the flying squirrels to extend their ranges northward. In western Washington oregonensis moved to southern British Columbia. In the Cascade Mountains the more boreal fuliginosus moved much farther northward and, north of the Okanogan Valley, spread eastward to the arid, interior valleys of British Columbia. Subsequent differentiation in the population of the arid, interior valleys developed the slightly differentiated race columbiensis. Farther east, flying squirrels from the northern Rocky Mountains moved northward. Northeastern Washington and adjacent British Columbia were occupied by latipes, derived from bangsi.
Thomomys talpoides.—Views as to the probable historical distribution of this plastic group have been presented in an earlier report (Dalquest and Scheffer, 1944: 308-333). This may be briefly summarized as follows.
Previous to Vashon-Wisconsin Times, pocket gophers occupied at least the Cascade Mountains and the Columbian Plateau of Washington. The race occupying the Columbian Plateau, devexus, was probably racially distinct in pre-Wisconsin time. The descent of the Vashon-Wisconsin glaciers isolated gophers in the southern [Pg 88] Cascades. Three racial stocks developed there: shawi in the Mount Rainier area; limosus in the Columbia River Valley south of the Cascades; and immunis in the mountainous area between the other two races. At the maximum extent of the Wisconsin ice, gophers from the Columbia River Valley (limosus) were able to cross the Simcoe Bridge and reach the Blue Mountains. With the retreat of the Wisconsin ice, the Simcoe Bridge was closed. Gophers isolated in the Blue Mountains developed the racial characters of acqualidens and those between the Blue Mountains and the Columbia River intergraded with the desert race, devexus. These intergrades, which have, also, some characters of their own, bear the name columbianus. Gophers in the southern Cascades (limosus) moved westward on prairie-like river terraces to Clark County where, isolated, they became racially distinct (douglasii).
Gophers from the Mount Rainier area (shawi) moved westward on glacial outwash trains to the extensive outwash aprons of the Vashon glaciers in the area about Puget Sound. Here they multiplied and spread to the Olympic Mountains. Growth of forest on the original outwash apron broke the area into numerous isolated prairies. Gophers in the Olympic Mountains (melanops) were isolated from those in the area about Puget Sound. Six distinct races originated on the isolated prairie (glacialis, tacomensis, pugetensis, yelmensis, tumuli, couchi).
Following the retreat of the glacial ice from eastern Washington, pocket gophers from the Blue Mountains of Oregon (wallowa) moved northward into Washington and gophers from the Rocky Mountain Fauna of Idaho moved onto the deglaciated part of northeastern Washington. From northeastern Washington they spread westward to the Cascades and thence southward to meet the native gophers of the Cascades in the Yakima Valley Area. No racial differentiation in these gophers occurred; all are referable to fuscus. Where fuscus and the native gophers came together in the Yakima Valley, a new race, yakimensis, developed.
Perognathus parvus.—Three races of the pocket mouse occur in Washington. Two of these (lordi and columbianus) occur on the Columbian Plateau. Like many desert species that occur on the Columbian Plateau, the pocket mice are rather different than their relatives in eastern Oregon. Presumably they have been isolated on the plateau since before Vashon-Wisconsin Times.
The range of the pocket mouse of southeastern Washington, Perognathus parvus parvus, is continuous with the range of the race [Pg 89] in Oregon. This same race occurs in the Yakima Valley, whence it probably arrived from Oregon in relatively recent time.
The distribution of pocket mice on the Columbian Plateau, in eastern Oregon and in the Yakima Valley resembles that of the least chipmunk in those areas. It is also similar to, but of more recent origin than, that of the ground squirrels, Citellus washingtoni, and townsendii.
Dipodomys ordii.—This kangaroo rat enters the desert area of southeastern Washington from Oregon. It may be expected eventually to cross the Columbia River to the Yakima Valley and the Snake River to the Columbian Plateau.
Castor canadensis.—Two races of beavers occur in Washington. One, found in southwestern Washington and northwestern Oregon, is dark with a short, wide skull. The other, ranging over most of the state, is paler with a longer, narrower skull.
The form now found in southwestern Washington and adjacent Oregon (idoneus) was probably isolated there by the Vashon glaciation and developed its characters while isolated. The other race, leucodonta, was probably widely spread in Wisconsin Time. Beavers are present in Moses Lake, in almost the center of the Columbian Plateau. Beavers might well have lived in the streams of melt water that emerged from the Wisconsin Glacier. The beavers of western Washington, save those in the extreme southwest, are like the beavers of eastern Washington. It seems likely that the race leucodonta originated north of the state of Washington and was forced southward by the Vashon-Wisconsin glaciers. This northern race, adapted to boreal conditions, competed with the resident coastal race, idoneus, and occupied much of its range. The distribution of the races of muskrat in Washington closely resembles that of the beavers.
Onychomys leucogaster.—The desert-dwelling grasshopper mouse has doubtless entered eastern Washington and the Yakima Valley from eastern Oregon at a relatively recent time.
Reithrodontomys megalotis.—The harvest mouse, like the grasshopper mouse, seems to have entered Washington from Oregon at a relatively recent date. Within the last ten years it has extended its range into the Okanogan Valley in British Columbia.
Peromyscus maniculatus.—Six subspecies of Peromyscus maniculatus occur in the state of Washington. The geographic range of one of these (rubidus) lies mainly in the states of California and Oregon and includes, so far as is known, a single small island in [Pg 90] the Columbia River that is politically within the state of Washington. Another (hollisteri) is restricted to certain islands in northern Puget Sound and obviously has become subspecifically differentiated in postglacial time. The remaining four subspecies, namely oreas, austerus, artemisiae and gambelii, have extensive geographic ranges. These subspecies are not confined to their geographic ranges by geographic barriers. Deer mice occur in the deep forests and the open desert, on high mountains and in low valleys, and are almost everywhere the commonest species of mammal present.
The study of several populations of deer mice from any general area usually shows small but constant differences between them. Dice (1939: 21) studied stocks of deer mice from nine localities in southeastern Washington and found significant differences between several of them. A statistical study of mice from the San Juan islands shows that the population of almost every island is different in one or more respects from the mice of any other island. Geographically separated populations of "wild caught" mice of the subspecies austerus, of the mainland, were statistically compared and significant differences were found between these populations, too. Small, differentiated populations are to be found in many parts of the state, and each subspecies appears to be an assemblage of such tiny genetic units.
These genetic units probably are the microgeographic races of Debzhansky (1937). They have been intensively studied by Sumner (1917 A, B) and Dice. An especially important paper by the latter author (1940) summarizes much of the available information on speciation in Peromyscus and clearly discusses the microgeographic races of Peromyscus.
The numerous microgeographic races of Peromyscus maniculatus in Washington present diverse combinations of characters which could result from the random fixation and elimination of genes (Wright. 1932: 360-362). Such a hypothesis, however, requires at least partial isolation of the populations involved. The mechanism of such isolation, in such populations of deer mice as we have studied, is not readily apparent. Some microgeographic races are not separated by noticeable geographic or ecologic barriers and the distance between their ranges is not too great to be traversed by a deer mouse. The tendency to remain on a home range may have the same effect as isolation would be supposed to have.
The work of Murie and Murie (1931: 200-209, 1932: 79) is enlightening in this respect. These authors found that mice residing [Pg 91] in a locality tended to remain in that locality; individuals trapped and marked were retaken in the same locality a year later. Individuals released some distance from the point of capture remained where released or returned to the point of capture. Transported individuals did not spread at random. The home instinct was developed in young as well as in old mice. Two mice in the gray pelage, four to eight weeks old, returned to their home ranges from distances one and two miles away. The authors fix the home range of an individual Peromyscus m. artemisiae in Teton County, Wyoming, at approximately one hundred yards in diameter.
This home-range instinct is essentially a lack of incentive for individual mice to emigrate to new localities where mice of the same species are already established. This may partly account for the microgeographic races of deer mice in Washington.
Dice (1939: 21) pointed out that, except in color, the differences in nine stocks of mice from southeastern Washington could not be correlated with environmental factors. We have found this to be true of microgeographic races throughout the state of Washington.
Of the four subspecies of deer mice that occupy extensive geographic ranges in Washington, one, oreas, is a long-tailed form that seems not to intergrade with austerus, a neighbor in western Washington that has a tail of moderate length. These two and gambelii, a short-tailed form with which oreas intergrades, are easily distinguished. In eastern Washington two short-tailed subspecies, gambelii and artemisiae, are currently recognized. The taxonomic relationships of these two subspecies are complex. The subspecies gambelii has an extensive geographic range in Oregon and California. These mice, with short tails, occur in the Wallula Water Gap of southeastern Washington and on the Columbian Plateau. To the west the desert conditions of the Columbian Plateau fade into the Transition Life-zone forests of the eastern Cascade Mountains. The pale, short-tailed desert mice (gambelii) gradually change to the dark, long-tailed subspecies, oreas, that occupies the Cascade Mountains.
North of the Columbian Plateau, in northeastern Washington, the deer mice are darker and relatively longer-tailed than on the Columbian Plateau. Some populations are distinctly reddish, almost as reddish as oreas. Although assigned to artemisiae, they are almost identical with populations of deer mice from the eastern Cascade Mountains, known to be intergrades between oreas and gambelii. This fact, and the presence of surprisingly oreas-like characters in [Pg 92] some microgeographic races in extreme northeastern Washington, may indicate that the race called artemisiae is a group of intergrades between gambelii and an oreas-like mouse that has become extinct.
Intergradation between gambelii and artemisiae is normal and takes place gradually where the ranges of the two subspecies meet. The same is true of intergradation between oreas and gambelii in the eastern Cascade Mountains. West of the Cascade Mountains the range of oreas meets the range of austerus. These two subspecies appear not to interbreed in nature, since no intergrades were taken at any of the numerous localities in western Washington where the mice were trapped. Pure populations of the two subspecies exist within a few miles of each other. In the valley of the Skykomish River, in the western Cascades Mountains, from the town of Skykomish, King County, to the lowlands to the west, only austerus was found. In the coniferous forests of the mountains above Skykomish, only oreas was taken. Several pairs, each an oreas and an austerus of the other sex, were kept from four to six months, and one pair was kept for a year, but they did not reproduce. The oreas were from the upper Skykomish Valley and the austerus were from Seattle, King County. Along the border of the ranges of the two subspecies in the Skykomish River Valley, a definite habitat preference was noted. The coniferous forests were occupied by oreas and brush or deciduous forests by austerus. Within the range of austerus and within the range of oreas only one subspecies is found whether the habitat be coniferous forest or thickets of alder and willow, but where the ranges of the two subspecies meet austerus occurs only in the thickets of aspen and willow and oreas occurs only in the coniferous forest.
The morphological characters that separate oreas from austerus, namely, larger size, richer color, and longer tail, are all features that might be associated with a more arboreal existence in dark, coniferous forests. Our observations show that oreas is, to a large extent, arboreal. Traps nailed to tree trunks six to eight feet from the ground and set for flying squirrels often caught oreas in the Skykomish River Valley. On one occasion I walked up a leaning tree trunk to set a trap, fifty feet from the ground on the trunk of another tree that was upright. An oreas was found in this trap the next morning. Svihla (1933: 13) relates how a specimen of oreas that escaped from a live trap took refuge in a tree when pursued. We have set numerous traps for flying squirrels in the area about Puget Sound. As far as memory and field notes serve, we [Pg 93] have never taken austerus in these traps. Our observations on the habits of austerus are much more abundant than those on oreas, but for austerus no evidence of arboreal activities has been obtained.
There are, then, two subspecies that do not interbreed, each capable of existing in any ecologic niche that will support deer mice. Where the ranges of the two subspecies come together, they compete. The large size, richer color, longer tail, and perhaps arboreal habits of oreas give it an advantage in coniferous forests. The small size and dark color of austerus give it an advantage in other habitats, especially, perhaps, in winter, when the deciduous trees have shed their leaves.
The differences separating austerus from oreas would be expected to have developed under complete isolation. That oreas developed from austerus or austerus from oreas seems impossible. A glance at the distribution map (Fig. 107) shows that the range of austerus clearly separates the range of oreas into two segments. The range of austerus tapers out to the north, east and west. On the south austerus intergrades with rubidus from Oregon, a subspecies from which it is but slightly differentiated.
The geographic behavior of the four subspecies of deer mice that occupy extensive areas in Washington may be summarized as follows: P. m. gambelii occupies southeastern Washington and intergrades normally with oreas in the eastern Cascade Mountains and with artemisiae at the northern edge of the Columbian Plateau; artemisiae, occupying northeastern Washington, closely resembles populations of mice that are known to be intergrades between oreas and gambelii and itself intergrades with both oreas and gambelii; oreas occupies most of western Washington, intergrades with its neighbors artemisiae and gambelii to the east, but does not intergrade with austerus, its morphologically more similar neighbor in the west; austerus occupies a range in western Washington that is almost surrounded by the range of oreas, a subspecies with which it apparently does not interbreed.
The relations of these four subspecies appear to be the result of certain great changes in the range of deer mice in the Pacific Northwest that occurred during and since the last Pleistocene glaciation. We reconstruct these changes as follows. In the Puyallup interglacial cycle, just previous to the last (Vashon-Wisconsin) continental glaciation, clines, or gradual transitions existed in the ranges of the deer mice along the Pacific Coast. The small, dark, short-tailed mice of the coastal redwood forests of California and Oregon [Pg 94] became gradually larger, redder and longer-tailed to the north, climaxing in a large, red, long-tailed form in the spruce forests of southern Alaska and northern British Columbia. A similar cline existed inland. Small, pale, short-tailed mice of the Great Basin became increasingly larger, darker, and longer-tailed to the north, reaching a maximum in the spruce forests of northern British Columbia.
The development and advance of the Vashon-Wisconsin ice sheet exterminated mice over much of British Columbia and the northern United States. Long-tailed northern mice were forced southward and contacted the southern, short-tailed forms. The preglacial clines were thus destroyed.
What might be the southern part of the western cline may be noted in the deer mice of western Oregon today. From the southern coast of Oregon the mice (P. m. rubidus) do become larger, redder and longer-tailed to the north. The climax of this cline is now reached in austerus, of the Puget Sound area of Washington. The cline is not continued farther northward because the range of oreas is encountered.
The advance of the Vashon-Wisconsin ice from the north probably forced species of mammals southward, ahead of it, including the long-tailed northern deer mice which generation by generation encountered progressively shorter-tailed forms of resident mice. Perhaps the unfamiliar, and certainly the extremely frigid, conditions in the fore of the glacier exterminated the short-tailed individuals but favored the long-tailed mice, since the latter originally were adapted to a boreal environment. The climax of the ice advance found the glaciers just within the political limits of the United States and found the long-tailed mice spread before the ice front.
In western Washington the Vashon glacier advanced as far south as the southern edge of the Puget Sound area. Long-tailed northern mice advancing before it reached the Columbia River. This glacially swollen stream served as a natural barrier and prevented their southward extension. At this time the northern mice had traversed more than half the length of the original western cline. The northern mice, originating in a boreal habitat a thousand miles away, were unable to interbreed with the southern mice and such individuals as might have crossed the Columbia River never became established in Oregon. During the existence of the glacier in western Washington, the long-tailed northern emigrants competed with the resident deer mice of western Washington to the total elimination [Pg 95] of the resident mice. The retreat of the Vashon Glacier from Washington found the northern mice in complete control of that part of the state from the Pacific Ocean to the Cascade Mountains.
In eastern Washington the Wisconsin Glacier advanced south to the northern edge of the Columbian Plateau. Northern mice advancing before it probably did not survive on the treeless plateau but existed in forested areas of northern Idaho and driftless areas of northern Washington. No natural barriers separated the northern mice from the pale, short-tailed forms. The nonuniform topography perhaps allowed more mingling of the two types where climatic conditions permitted. Intergradation in some places as well as competition and elimination of one form or another in other places occurred. Following the withdrawal of the Vashon ice and the establishment of soil and forests on the deglaciated land, the long-tailed mice of western Washington (P. m. oreas) apparently spread northward, unhindered by competition, until they reached southern Alaska. The deep coniferous forests of western Washington presented conditions acceptable to oreas and it persisted there despite postglacial changes in climate.
Postglacial changes in climate did, however, permit the dark, short-tailed mice (rubidus) to cross the Columbia River and to become established in suitable habitats, namely the deciduous jungles of the river valleys. From these points the mice spread northward through the lowlands of western Washington, infiltrating the range of oreas, competing with it, and driving it from the lowlands. This movement is in progress today. By logging and clearing of lands for agriculture man has considerably speeded the invasion of the southern mice. Slight differentiation of the short-tailed mice north of the Columbia River (austerus) separates them from their parent race, rubidus.
In the dense forests along the Pacific Coast of Washington, austerus did not become established. This area includes most of the land west of Puget Sound. There is a narrow band of austerus that extends along the eastern and part of the northern edge of the Olympic Peninsula, where they have probably invaded in relatively recent time.
On the Oregon side of the Columbia River the range of rubidus is continuous from the Pacific to the Cascades. On the Washington side of the river, oreas extends from the Pacific to eastern Wahkiakum County, where the range of austerus starts. At the border of [Pg 96] the ranges of the two subspecies, ecological conditions determine the ranges; oreas occurs in the Douglas fir and hemlock forests and austerus occurs in the jungles of alder and salmonberry in the stream valleys. The range of austerus extends from eastern Wahkiakum County east along the Columbia, to western Klickitat County. In a north-south strip across the Columbia, through the ranges of rubidus in Oregon and austerus in Washington, normal intergradation is apparent. By gradual degrees rubidus changes to austerus. In a north-south strip farther west, through the ranges of rubidus and oreas, the same transition of the Oregon rubidus is seen, namely an approach toward austerus. The cline is, of course, abruptly terminated by the range of the dissimilar oreas.
On Puget Island, a small island lying in the Columbia River in western Wahkiakum County and politically within the state of Washington (see Scheffer, 1942) a population of deer mice occurs that resembles rubidus more closely than austerus. This island lies in the river between the ranges of rubidus and oreas. The lack of intergradation between these two subspecies has probably kept the Puget Island population pure rubidus. Puget Island is a junglelike lowland locally known as "tideland."
The San Juan Islands of northern Puget Sound were thoroughly glaciated in Wisconsin Time and probably were under thousands of feet of ice when the Vashon Glacier was at its maximum extent. The subspecies of deer mouse occurring there now is hollisteri, a race similar to oreas in color, body size and cranial characters and probably derived from oreas. P. m. hollisteri differs from both oreas and austerus in its much shorter tail. Shortness of tail is apparently a product of insularity in northwest coastal mice. The transition from long-tailed mainland mice to short-tailed island mice is shown by Hall (1938A: 461). When oreas first occupied the area about Puget Sound (this area is occupied by austerus today) some individuals probably reached the San Juan Islands soon thereafter. Contact between oreas and hollisteri is now prevented by the presence of austerus between their ranges. Of course, a water barrier separates austerus from hollisteri but austerus does occur in pure form on some islands in Puget Sound (not on any of the San Juan Islands), for instance, on Whidby Island. One wonders why austerus has not established itself on some of the San Juan Islands but considering the degree of difference between hollisteri and austerus, I doubt that the two would interbreed if they did occur together.
On Vancouver Island, British Columbia, a short-tailed, austerus-type [Pg 97] of mouse occurs in the lowlands and a long-tailed, oreas-type of mouse in the mountains. Vancouver Island is a large island (16,400 square miles). Apparently a stock of austerus from the mainland reached Vancouver Island and were able to find suitable habitat and compete with and drive out the established oreas in the manner that they probably did in the western Washington lowlands. The large Vancouver Island offers far more variety of habitat and opportunity for establishment of emigrating mice than do the smaller San Juan Islands, the largest of which is Orcas (57 square miles).
Following the withdrawal of the Wisconsin ice from eastern Washington, numerous minor movements and adjustments of deer mice seem to have taken place. Long-tailed, oreas-type of mice were exterminated on the Columbian Plateau if, indeed, they ever became established there. Long-tailed mice did apparently become established in driftless areas and forested areas to the south of the drift border. With the establishment of soil and forests on the deglaciated land, the short-tailed gambelii and the long-tailed mice became thoroughly intermixed. In some areas, especially near the type locality of artemisiae, the gambelii characters of the mixture predominate at the present time. Deer mice from the Okanogan Valley differ from gambelii only in slightly larger size and darker color. In other areas, notably near Metaline, Pend Oreille County, the oreas characters of the mixture dominate at the present time. Mice from here are large and red and differ from typical oreas in having relatively short tails. Other less marked segregations of characters are numerous, in Idaho and British Columbia as well as in Washington. Cowan (1937) has described Peromyscus m. alpinus, isolated in a mountainous area of British Columbia. This seems to be a form derived from oreas-type stock.
With the reëstablishment of soil and forests on the deglaciated land of British Columbia north of Washington, a northward extension of the gambelii-oreas mixture occurred. Another invasion was taking place at this time, however. Peromyscus m. arcticus spread onto the deglaciated land from the north or east, ahead of artemisiae (the gambelii-oreas mixture). This new invader intergraded with artemisiae as well as with oreas. Further collecting and studies are required in this area before the relationships of the three subspecies can be completely understood.
If geographic ranges are assigned to the named subspecies of deer mice that occur in Washington, it must, in part, be done arbitrarily. [Pg 98] The deer mice of the San Juan Islands are all referred to the subspecies hollisteri. Only Puget Island, Wahkiakum County, is considered to be inhabitated by rubidus. The boundary between the ranges of austerus and oreas is sharply defined and further collecting by resident naturalists should result in detailed mapping of the boundary.
The ranges and distribution of the deer mice of eastern Washington are less clearly defined than those of western Washington. Peromyscus m. gambelii exists in its purest state on the Columbian Plateau. Here the Columbia River makes a convenient boundary to the west. Pure Peromyscus m. oreas exist in the Cascade Mountains. In the area between the Columbian Plateau and the Cascades the deer mice are variously intermediate between gambelii and oreas and, over most of the area, are nearest to gambelii. This might be considered a natural area of intergradation such as commonly occurs between the ranges of subspecies. These intergrades, however, are similar to Peromyscus m. artemisiae and the area occupied by these intermediates is connected on the north with the range of artemisiae. Thus Osgood (1909: 61) regards specimens from Easton and Wawawai, in the area of intergradation, as artemisiae or intergrades between gambelii and artemisiae. Taylor and Shaw (1929: 22) include the entire area of intergradation in the range of artemisiae. This treatment, although convenient to the student of geographic variation, is apt to conceal the evolutionary and historical influences. These influences may be appreciated better if the intergrades from each locality are referred to the subspecies they most closely resemble.
The specimens upon which the name artemisiae was based came from a narrow tongue of zonally lower country that extends northward from the range of the lighter-colored gambelii. As might be expected, topotypes are lighter in color than specimens from the north, west and east. Nevertheless, the type locality is within a geographic area that is occupied principally by a darker race, artemisiae, to the north of gambelii. The topotypes of artemisiae may be considered to be intermediate between gambelii and the darker, northern race. Thus the name artemisiae becomes available for the mice of the general area in question. The mice of the area immediately to the east of the type locality, in Washington and presumably in British Columbia and Idaho also, are essentially a mixture of the subspecies gambelii and a now mostly extinct and unnamed race that probably resembled oreas. Local variations [Pg 99] of populations from within this area are extreme but some segregation of color and length of tail has taken place. Mice from mountainous areas resemble oreas while mice from the lowlands resemble gambelii, or, more exactly, mice from coniferous forests resemble oreas while mice from other areas resemble gambelii.
South of the area of racial mixture in northeastern Washington, in the Blue Mountains of southeastern Washington, the deer mice are like gambelii except in slightly darker color. Mice from the Blue Mountains are darker and browner than gambelii, not more reddish. There is no indication of adulteration with oreas stock. Since the Blue Mountains are a forested area and are far south of the drift border, we suppose that deer mice existed there through the last glacial period and that their dark color is an adaptation to forest habitat.
Mice similar to those of the Blue Mountains have an extensive range in Idaho (Davis 1939: 290). These mice have relatively uniform racial characters and constitute a "good" subspecies.
At present the deer mice of northern Washington, southeastern British Columbia, northern Idaho, northeastern Oregon, western Montana and northwestern Wyoming are called artemisiae. The mice of this extensive area are, however, of two genetic types: that type with mixed racial characters that lives in northeastern Washington and probably also farther east along the Wisconsin drift border, and that type that occurs farther south in Idaho and seems to constitute a stable subspecies.
The separation of these two types may be desirable. Detailed study of the deer mice from the area now assigned to the range of artemisiae may show that the name subarcticus (Peromyscus texanus subarcticus Allen, 1899) is applicable to the southern form. The type with mixed racial characters must be called artemisiae. If the two types are eventually separated, the mice from the Blue Mountains of Washington will be referable to the southern form.
Neotoma cinerea.—The wood rat found over most of Washington. Neotoma c. occidentalis, probably entered the state from eastern Oregon early in the Recent and spread over most of the state. The wood rat of the Blue Mountains (alticola) probably developed from occidentalis.
A coastal race of the wood rat (fusca) occurs in western Oregon. This race, if it occurred in western Washington in pre-Vashon Time, was eliminated in Vashon Time or subsequently. In Oregon it lives in deep forests (Bailey, 1936: 174). In Washington occidentalis occupies [Pg 100] but a small part of the ecologic niche occupied by fusca in Oregon. Elimination of fusca from Washington through competition with occidentalis seems highly improbable. Should fusca ever cross the Columbia River and become established it would probably spread to a considerable part of western Washington.
Synaptomys borealis.—The lemming mouse seems now to be retreating northward and was an arctic species forced southward by the Vashon-Wisconsin glaciers. Unlike other alpine species, it seems to be unable to exist for long in isolated mountain areas.
Phenacomys intermedius.—The heather vole, like the lemming mouse, probably was forced southward by the Vashon-Wisconsin glaciers. In the Cascade and Rocky mountains it found suitable habitat and spread southward to almost the lower end of the Sierra Nevada in California. The Cascade race moved eastward on the Puget Bridge to the Olympic Mountains after the retreat of the Vashon ice, and northward in the Cascades. Northeastern Washington was reinvaded by the subspecies of the Rocky Mountain Fauna.
Clethrionomys gapperi and CALIFORNICUS.—The two species of red-backed mouse found in Washington were probably distinct in pre-Vashon Times. At the maximum extent of the Vashon ice, californicus was probably found in western Oregon and gapperi in the Blue Mountains, where idahoensis was developed, and in the southern Cascades (cascadensis). During or shortly after Vashon Time, gapperi crossed the Puget Bridge to become established in the Olympic Mountains. After the retreat of the ice, gapperi moved northward and eastward from the Cascades and californicus crossed to western Washington from Oregon and displaced gapperi from the lowlands.
Hinton (1926: 215) separated the American Clethrionomys into three groups, suggesting that Old World counterparts of each group exist. Davis (1939: 308) gives an excellent analysis of the emigrational history of the American species, and points out the close relationship of the Siberian and Arctic-American forms. He suggests that the rutilus group invaded Arctic-America from Siberia at the close of the last glaciation.
Two other groups are recognized by Davis, who assumes that both arose from a common ancestral stock and divided into two stems, one of which (gapperi group) followed the main Rocky Mountain course southward and worked eastward across Canada while the other (californicus group) followed down the Pacific Coast. [Pg 101] Davis states, "This southward extension of range in America probably took place in the Pleistocene, but almost certainly the present southern range of the genus in the West was not reached until late in, or after, that period."
Our studies of the Clethrionomys of Washington indicate the essential correctness of Davis' conclusions. The separation of the two groups, however, probably was caused by southward moving glaciers and the separation of the mice into two stocks closely paralleled the divergence of other groups, such as that of the snowshoe rabbits. The two groups of red-backed mice are more sharply separated than was thought by Davis. The intermediate color of the back of C. g. idahoensis, mentioned by Davis, seems to be merely coincidental. The essential difference in the two groups is the sharply marked red band of the gapperi group as opposed to the general red area on the dorsal surface of the californicus group. Clethrionomys gapperi nivarius has been considered a derivative of occidentalis because the range of the latter surrounds the Olympic Mountains, where nivarius is found. Considering the immediate post-Pleistocene movements of mammals from the Cascades to the Olympic Mountains, so clearly illustrated in Thomomys, Tamias amoenus, and Phenacomys, a means is indicated by which Clethrionomys of the gapperi group might have reached the Olympics. The apparent lack of intergradation between occidentalis and nivarius gives proof of their relatively distant relationship. C. nivarius seems not to have been derived from occidentalis, and apparently does not belong to the californicus group. It belongs instead to the gapperi group, and I consider it to be a subspecies of gapperi. In no sense is it intermediate between the two groups. The other form considered by Davis to be intermediate between the two groups is caurinus. This mouse has not yet been taken in Washington, although it may eventually be found on Point Roberts, on the Fraser River delta. Its distribution is paralleled by that of many other mammals that are definitely not of the Pacific Coastal Fauna.
The californicus group, I feel, contains only the races of Clethrionomys californicus, while the gapperi group contains C. gapperi and its races, including caurinus, and possible other species.
Possibly intergradation occurs between C. californicus occidentalis and C. gapperi. Nevertheless, I fail to find evidence of such intergradation. I have taken C. gapperi nivarius and C. californicus occidentalis within ten miles of each other, but each retained its [Pg 102] distinctive characters with no evidence of intergradation. In the case of C. gapperi saturatus and C. californicus occidentalis the proof is less conclusive. In spite of numerous attempts to trap Clethrionomys in the area geographically intermediate between their ranges, I have taken none. Though common along the coast, occidentalis becomes progressively scarcer to the east, being rare in the vicinity of Seattle and apparently absent from the western base of the Cascades. So far as is known, the ranges of caurinus and occidentalis are separated by the Fraser River.
Microtus pennsylvanicus and MONTANUS.—The Pennsylvania meadow mouse is closely related to Microtus montanus. Certain races of montanus, notably those from southern Oregon, California, and northern Nevada, closely resemble pennsylvanicus externally and cranially. From the central part of its range northward, montanus becomes progressively less like pennsylvanicus. The races nanus and canicaudus are quite different from pennsylvanicus both externally and cranially, and in addition the anterior loop of the second molar is less constricted; often it is not constricted at all. Microtus pennsylvanicus and Microtus montanus occur together over parts of the northern Rocky Mountains. Where the two species came together, pennsylvanicus occurs with races of montanus that are most unlike it.
Microtus pennsylvanicus kincaidi closely resembles races of Microtus montanus that occur in southern Oregon, California, and Nevada. It is larger, darker, and longer-furred than Microtus pennsylvanicus funebris from Washington. Seemingly pennsylvanicus and montanus diverged previous to Vashon-Wisconsin Time. The stock that gave rise to montanus spread over the Great Basin while pennsylvanicus ranged farther east. Some of the montanus stock worked northward in the Rocky Mountains. Microtus pennsylvanicus had meanwhile moved westward to the Rocky Mountains. The two stocks met and behaved as full species.
Microtus pennsylvanicus probably occupied northeastern Washington in the interglacial cycle preceding the Recent. The advance of the Wisconsin ice eliminated most of these mice. The glacier dammed the Columbia River and caused it to turn southward from its basalt-marginal course and take a path over the Columbian Plateau. Along this glacial river a population of Microtus persisted to become kincaidi.
While the Wisconsin ice was at its maximum extent, Microtus montanus from the Blue Mountains crossed the Simcoe Bridge to the [Pg 103] Yakima Valley and the eastern Cascade Mountains. The closing of the Simcoe Bridge isolated these mice, which subsequently became slightly differentiated, as canescens. Another stock moved westward along the Columbia River to western Oregon. This stock is now called Microtus canicaudus but is probably racially rather than specifically distinct from Microtus montanus nanus.
With the retreat of the Wisconsin ice, montanus extended its range northward from the Yakima Valley along the eastern Cascade Mountains to extreme southern British Columbia. Microtus pennsylvanicus funebris entered northeastern Washington with other elements of the Rocky Mountain Fauna.
At present, pennsylvanicus occurs in northeastern Washington while montanus is found in southeastern Washington and the eastern Cascade Mountains. The Okanogan River Valley separates their ranges. Eventually montanus may extend its range to northeastern Washington and pennsylvanicus to the Cascades, the two forms occurring together as they do in Montana, Idaho and Colorado. The isolation of kincaidi on the Columbian Plateau seems complete and the probability of its range reaching that of its related species seems slight.
Microtus longicaudus.—The later distributional history of the long-tailed meadow mouse is not yet clear.
Microtus townsendii.—This Pacific Coastal species probably lived west of the Sierra Nevada-Cascade Mountains since the early Pleistocene. It has extended its range northward since the retreat of the Vashon ice and has reached some of the islands in Puget Sound and the Strait of Georgia. This species seems to be related to Microtus richardsoni but the two species probably separated at an early time.
Microtus richardsoni.—The water rat of the Cascade Mountains seems to be have been forced southward into Washington and Oregon by the descent of the Vashon-Wisconsin glaciers where it became isolated from the water rat of the northern Rocky Mountains. After the retreat of the ice, both forms have moved northward. Bailey (1900) records macropus from Bonner County, Idaho, not far from northeastern Washington but in Washington up to this time, macropus has been recorded only from the Blue Mountains of the southeastern part of the state.
Microtus oregoni.—The later historical distribution of the creeping mouse was probably the same as that of townsendii.
Lagurus curtatus.—The distribution of this species of the Great Basin Fauna has probably changed little if any since the late Pleistocene.
Ondatra zibethicus.—The distribution of the two races of muskrat that occur in Washington is almost exactly that of the beavers. Probably one form was isolated in southwestern Washington and northwestern Oregon during Vashon Time and another, more adaptable, race occurred in eastern Washington. After the retreat of the ice the adaptable race spread widely but the muskrats of the lower Columbia River changed their range little or not at all.
Aplodontia rufa.—The race of Aplodontia found in the Cascades of Washington was probably confined to the southern Cascades in Vashon Time and has since spread northward to reoccupy the range as far north as southern British Columbia.
The presence of a mountain beaver in western Washington that is indistinguishable from the race rufa, found in the Cascades of Oregon, is most surprising. In Oregon, pacifica occurs in the western lowlands and rufa in the mountains to the east. In Washington rufa occurs in the western lowlands and rainieri in the mountains to the east.
The offset in range of rufa seems best explained by assuming that all of western Washington was once occupied by mountain beavers (rufa) that spread from the Cascades of Oregon to the Cascades of Washington and thence to the western Washington lowlands. This must have occurred early in the interglacial cycle preceding the Vashon Glaciation. While confined to the southern Cascades, rainieri developed the larger size that now separates it from the ancestral rufa that occurs to the west and south of it.
Zapus princeps.—The coastal race of the jumping mouse was probably distinct from the more inland population before Vashon-Wisconsin Times. The differences between them were probably accentuated while the Vashon-Wisconsin glaciers separated their ranges. The western race (trinotatus) was isolated in the southern Cascades and southwestern Washington. After the retreat of the glaciers, trinotatus moved northward through western Washington and the Cascades into British Columbia. The race kootenayensis of the adjacent Rocky Mountains moved eastward through northeastern Washington to the Cascades. The race oregonus, found in the Blue Mountains, has probably been resident there since it evolved from the populations to the east.
Erethizon dorsatum.—I have inadequate basis for speculation concerning the historical distribution of the porcupine.
Ochotona princeps.—The descent of the Wisconsin ice separated the western pikas into two populations. One was confined to the Cascade-Sierra Nevada system and another to the Rocky Mountains. Two races are now found in the Cascades of Washington. One, brunnescens, inhabits the higher Cascades and another, fenisex, the eastern edge of the Cascades. The range of brunnescens extends southward into Oregon while the range of fenisex extends farther north in British Columbia than does that of brunnescens. The principal difference between the two races is the smaller size and paler color of fenisex. In Washington, fenisex occupies a slightly more arid habitat than brunnescens. Further, fenisex lives in talus principally of basaltic rock while brunnescens lives in talus of granitic rock. The basaltic talus is more finely fractured, offering smaller crevices in which pikas can conceal themselves. The granitic rock, on the other hand, forms talus composed of fragments of large size. Freshly fractured granite is pale, whitish gray. After weathering, however, it becomes blackish as the more soluble, pale feldspars are removed, leaving the black hornblend and biotite exposed. Freshly fractured Columbian basalt is blackish but, after weathering, becomes rusty, reddish brown.
In Washington the two races maintain their distinctiveness because selective factors in the basalt talus of the eastern Cascades favor the smaller size and paler color of fenisex while in the higher Cascades, selective factors in the granitic batholith favor larger size and darker color.
The present range of brunnescens in Oregon indicates that this race was the pre-Wisconsin resident of the Cascades of at least southern Washington. Probably fenisex evolved in the arid interior of British Columbia. In color fenisex is intermediate between the dark brunnescens and the pale cuppes of the western spurs of the Rocky Mountains. Probably fenisex was forced southward into the eastern Cascades by the Wisconsin ice and inhabited the area east of the range of brunnescens. This eastern area was not occupied by brunnescens because the basaltic talus was of small size. After the retreat of the ice, fenisex (adapted to more arid habitat than brunnescens), spread northward to the relatively dry valleys of the interior of British Columbia.
The distinctness of brunnescens and fenisex is maintained by selective factors of the habitats they occupy but the zone of intergradation [Pg 106] between the two is broad. A tendency towards paleness is noticeable in specimens from as far west as the eastern side of Mount Rainier.
The pikas in the Rocky Mountain area have given rise to a number of races. The pale, small race of northern Idaho, cuppes, entered northeastern Washington with other members of the Great Basin Fauna after the retreat of the ice.
Lepus townsendii.—The white-tailed jack rabbit has probably lived on the Columbian Plateau ever since or even before the time when the Wisconsin ice bordered the plateau on the north.
Lepus americanus.—Snowshoe rabbits were probably spread over forested parts of Washington in pre-Wisconsin time. A coastal race, washingtonii, was probably distinct from the remainder of the species at an early time. It, and its kindred races klamathensis and tahoensis, are very different from races derived later from the Rocky Mountain Fauna. From this it is deduced that washingtonii was a member of the Pacific Coastal Fauna before the advance of the last continental glaciers.
The snowshoe rabbits now found in the Cascade Mountains of Washington are racially distinct from populations occurring to the north and east, but are more closely related to the northern and eastern rabbits than they are to washingtonii. This race, cascadensis, probably lived in the area north of the state of Washington in pre-Wisconsin interglacial time. The snowshoe rabbit of northeastern Washington was probably a race having an extensive range in the Rocky Mountains from which pineus in the Blue Mountains developed. The descent of the Vashon-Wisconsin glaciers found washingtonii restricted to western Oregon. It might have persisted also in southwestern Washington but, had this been the case, we would expect a broader zone of intergradation between washingtonii and cascadensis than actually exists.
The glaciers forced cascadensis southward into the Cascades of southern Washington. The difference between washingtonii and cascadensis and the narrow zone of intergradation between them, indicate that the two races were not in contact in pre-Vashon Time. The presence in the Cascades of Oregon of a race related to washingtonii rather than cascadensis also supports this view. The snowshoe rabbits of northeastern Washington were probably eliminated from areas where the ice was in place. The effect of the glaciers in nearby areas was probably less on snowshoe rabbits than upon other species.
Following the retreat of the glaciers, washingtonii reinvaded western Washington and spread northward through the lowlands to the Fraser River. In the Cascades, cascadensis moved even farther north. L. a. pineus of the Rocky Mountain Fauna invaded northeastern Washington from adjacent areas to the south. Another race, columbiensis, spread from the interior valleys of British Columbia to the western part of northeastern Washington in probably relatively recent times. The post-Wisconsin history of the distribution of this race is uncertain.
Lepus californicus.—The black-tailed jack rabbit occurs throughout the sagebrush areas of eastern Washington, except in the Okanogan Valley. It invaded Washington from the south recently (Couch, 1927: 313). The first blacktails were thought to have entered Walla Walla County about 1870. Here they were checked by the Snake River. About 1905 they crossed the Snake River, on ice, and by 1920 had spread over much of the Columbian Plateau. In January, 1920, they crossed the Columbia River in two places, on ice jams, and by 1927 had spread north and west over the sagebrush areas between the Columbia River and the Cascade Mountains. When Couch, in 1927, published his account, he predicted that the blacktail would eventually spread to the Okanogan Valley. This has not yet occurred although it is still to be expected.
It should be noted that the migrational history of the black-tailed jack rabbit is known only because the animal is large, spectacular, and diurnal, and therefore noticed by resident ranchers, and because the facts came to the notice of a competent biologist. Migration of a less conspicuous mammal, for example, a mouse, would rarely be detected.
Sylvilagus nuttallii.—The cottontail might have been resident on the Columbian Plateau during Wisconsin Time but probably it invaded, or perhaps reinvaded, the Plateau from eastern Oregon in the Recent.
Sylvilagus idahoensis.—Too little is known about this rabbit in Washington to hazard a guess as to its recent distributional history.
Cervus canadensis.—The elk emigrated from Siberia to North America in the late Pleistocene but previous, certainly, to Vashon-Wisconsin time. Probably the coastal race was distinct from the interior population in the preceding interglacial cycle. The descent of the last continental glaciers, in Washington, separated the elk [Pg 108] of the Pacific Coastal Fauna from the Rocky Mountain Fauna. After the retreat of the ice the coastal race moved northward to southern British Columbia and the Rocky Mountain race moved northward and westward through British Columbia. Northeastern Washington was reoccupied by the Rocky Mountain elk.
Remains of elk have been found associated with human artifacts on the northern part of the Columbian Plateau. The plateau is poorly suited to elk but a few may have persisted there until late historic time. The remains may have been brought by Indians from northeastern Washington. The remains included teeth and portions of the skull and it seems unlikely that these would have been carried any great distance.
Odocoileus hemionus.—The American deer probably evolved in North America. In this respect they are unlike the elk, moose and caribou, all of which emigrated from Asia to America in the Pleistocene. Odocoileus hemionus and virginianus probably diverged from a common stock in the Pliocene.
The black-tailed deer was probably a member of the Pacific Coastal Fauna at an early time and distinct from the mule deer before the last interglacial cycle. Probably the mule deer lived in the eastern Cascades and on part of the Columbian Plateau in Wisconsin Time. They occur on the plateau in limited numbers at present and their remains have been found associated with human artifacts in the Grand Coulee area.
Odocoileus virginianus.—The white-tailed deer ranges from the Atlantic to the Pacific and from Canada to Mexico. The species was probably abundant in the Pacific Northwest in the Pleistocene, perhaps in Vashon-Wisconsin time, but has since largely given way to the black-tailed and mule deer. The race O. v. leucurus now occupies an extremely small range. The accounts of early naturalists indicate that it was more abundant and had a wider range 100 years ago. Probably leucurus was once an important member of the Pacific Coastal Fauna.
The inland race, ochrourus, is a typical member of the Rocky Mountain Fauna. It and leucurus were probably derived from a common ancestor in the late Pleistocene.
Alce americanus.—The moose occurs in Washington as a casual wanderer from the Rocky Mountain Fauna to the east.
Rangifer montanus.—The caribou occurs in northeastern Washington as a winter migrant from the north. It was probably of more regular and extensive occurrence in the past.
Antilocapra americana.—The antelope probably was a casual wanderer to southeastern Washington and perhaps to the Columbian Plateau from eastern Oregon before white man reached Washington.
Bison bison.—The buffalo, like the antelope, probably occurred in southeastern Washington and on the Columbian Plateau only as a casual wanderer from Oregon in postglacial time.
Ovis canadensis.—Mountain sheep reached North America from Asia in the Pleistocene. By Vashon-Wisconsin Time they had spread southward, perhaps to Mexico. The descent of the Vashon-Wisconsin ice in Washington presumably separated the canadensis and californiana type of sheep, the former being confined to the Rocky Mountain Fauna and the latter to the Cascade-Sierra Nevada chain and adjacent parts of the Great Basin.
Mountain sheep probably persisted in the southern Cascades of Washington and on the Columbian Plateau during Vashon-Wisconsin times. After the retreat of the ice, these sheep (californiana) moved northward slightly, both in the Cascades and on the Columbian Plateau. Remains of sheep are to be found with human artifacts in the Grand Coulee area today. When white man first reached the state, sheep existed throughout the Cascades and on part of the Columbian Plateau. Only a pitiful remnant remains in the extreme northern Cascades.
Mountain sheep of the Rocky Mountain race probably existed in small numbers in the Blue Mountains of southeastern Washington until historic times. Sheep from the Blue Mountains or adjacent Idaho invaded northeastern Washington shortly after that region was freed from Wisconsin ice. These sheep were exterminated shortly after the coming of the white man.
Oreamnos americanus.—Fossil remains of the mountain goat have been discovered at Washtuckna Lake, associated with those of the lion, horse, and camel. Probably the mountain goat had an extensive range in Washington at the time of a glacial advance previous to the Wisconsin glaciation. In Wisconsin Time the mountain goat was confined to the southern Cascades. Strangely enough, it seems not to have crossed the Columbia River to become established in the Cascades of Oregon. After the retreat of the ice it moved northward through the Cascades.
A mountain goat recently taken in northeastern Washington seemingly wandered to the area from Idaho.
The order of arrangement of the following accounts is that of Miller (1924) with some modifications. For example, the Chiroptera are arranged according to Tate (1942), the Sciuridae according to Bryant (1945) and the Cetacea according to Scheffer (1942). A few other minor changes are included.
Although the principal purpose of this report is to describe and interpret the distribution of the various species and races of mammals that occur in Washington, a brief description and account of the habits of the animals is included. Each species account begins with a description, based principally on external characters. This is followed by pertinent information regarding the range of the species, its relationships within the genus and a brief account of its habits. The habits are dealt with in most detail for the species that are of greatest economic importance.
The accounts of subspecies are largely technical. The account of the type specimen is rather complete. Subspecific diagnoses are brief, stating often only the principal racial characters. Standard external measurements are usually given for each subspecies. All measurements, unless otherwise stated, are in millimeters and weights are in grams. Specimens are adult, unless otherwise stated.
No formal list of specimens examined is included. Localities from which specimens have been examined are usually shown on the distribution maps by solid circles. Open circles indicate localities from which specimens, not examined by me, have been recorded by other persons. Most of these records are published but a few are based on authentic trappers' reports, photographs, or other evidence. Unusual occurrences of animals outside their natural ranges are not shown on the maps. Not every record from well within the range of a subspecies has been plotted, but care has been taken to plot all records of occurrences from peripheral areas.
When more than one race of a species occurs in Washington, specimens from localities where the geographic range of one subspecies meets or approaches that of another are listed as "marginal occurrences" in the paragraph on the distribution of the race to which they are best referred. Marginal occurrences are listed from north to south and from west to east. The authority for the record, if published, is given. When the record is based on specimens examined by the author, the collection containing the specimen is indicated as follows:
(E.S.B.) Ernest S. Booth collection. College Place, Washington.
(J.M.E.) J. M. Edson collection, Bellingham, Washington.
(K.U.) University of Kansas, Museum of Natural History, Lawrence, Kansas.
(M.V.Z.) Museum of Vertebrate Zoölogy, University of California, Berkeley, California.
(U.S.N.M.) United States National Museum, Washington, D. C. Specimens from the Biological Surveys Collection are included here.
(V.B.S.) Victor B. Scheffer collection. United States National Museum, Washington, D. C.
(W.S.C.) Washington State College, Charles R. Conner Museum, Pullman, Washington.
(W.S.M.) Washington State Museum, University of Washington, Seattle, Washington.
(W.W.D.) Walter W. Dalquest collection, Seattle, Washington.
In all, 230 kinds (species and subspecies) of mammals are listed for the state of Washington. Of these, 9 kinds (opossum, eastern gray squirrel, fox squirrel, 3 kinds of Old World rats, house mouse, nutria, and eastern cottontail) have been introduced into Washington from elsewhere. The dog of the Indians might be listed as a tenth introduced species. The grizzly bear, the sea otter, the wolf, the moose, one race of mountain sheep, one race of mountain goat, and, if they ever occurred in the state, the pronghorn antelope and bison, are nearly or completely extinct in Washington. Mammals other than cetaceans possibly occurring in Washington, but of which satisfactory record is lacking, are mentioned in a hypothetical list at the end of the accounts of species. Of the 220 kinds of native mammals, known to occur in the state, 23 are marine (4 pinnipeds, 19 cetaceans). The 197 living, native, land mammals include 6 orders, 20 families, 58 genera and 101 full species.
Didelphis virginiana Kerr. Anim. Kingd., p. 193, 1792.
Type locality.—Virginia
Description.—Slightly smaller than a house cat; body in older animals heavy and fat; tail long, naked, scaled and prehensile. Ears large, naked and black with white tips; muzzle elongate and pointed; color of fur variable; overhair usually white and underfur white tipped with black; guard hairs long and coarse but underfur soft and dense; forefoot with opposable thumb; females with abdominal pouch.
Remarks.—The opossum has been introduced from the eastern United States into California and Oregon and has become well established in those states. Recent records from Clear Lake, Skagit County, and South Bend, Pacific County, indicate that the opossum is now resident in Washington and it may be expected to increase and spread (Scheffer, 1943). The animals may also enter the area about Walla Walla from Oregon. The source of the opossums which have appeared in Washington is not yet known.
Of this animal, Dr. Carl Hartman (1923: 347) has written:
"In the popular mind, the generation of no animal is so shrouded in mystery as that of the opossum. Throughout the country, among both whites and negroes, deeply rooted tradition has it that the opossum copulates through the nose and that the female blows the fruit of conception into the pouch. Other myths relating to details of the reproductive process in this species are current among the people.
"The growth of such legends need not surprise one, however, for the early birth of the embryos and the use of the pouch as an incubator certainly challenge the imagination. These phenomena attract the attention because they are unique, differing from the familiar method of rearing the young obtaining among the higher mammals, including man. Familiarity breeds contempt; the ordinary ceases to be marvelous. Thus on account of its rareness and its 'different' character the opossum, our only marsupial, figures in the folklore to a prominent degree."
Description.—The shrew-mole is tiny, possessing a head and body 2-1/2 to 3 inches long and a tail about 1-1/2 inches in length. The body is relatively stout but is less cylindrical than that of Scapanus. The eyes are nearly buried in the fur. The nose is long and pointed. The legs are short and the forefeet wide and powerful. The tail is thick, constricted at the base and clothed with short, stiff bristles. The fur is short and posteriorly directed. In color the shrew-mole is dark slate, almost black.
Fig. 21. Gibbs shrew-mole (Neurotrichus gibbsii minor), female in captivity; Seattle, Washington, September 12, 1939. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 719.)
Remarks.—Shrew-moles inhabit moist habitats from sea level to 8,000 feet. They are burrowing mammals and prefer to live in soft earth, free of sod. In the lowlands of western Washington, shrew-moles are most abundant in damp, shady ravines where the vegetation includes deciduous trees and dense underbrush with but little grass. In the mountains, shrew-moles are usually found near streams or rock slides, where the larger annuals grow densely on soil that is deep, soft, and free of turf. Vertically they range from the Humid [Pg 123] Transition Life-zone through the Canadian, and well into the Hudsonian Life-zone.
Fig. 22. Distribution of the Gibbs shrew-mole in Washington. A. Neurotrichus gibbsii gibbsii. B. Neurotrichus gibbsii minor.
Shrew-moles are both diurnal and nocturnal. They rest or sleep periodically, the length of their rest or sleep being longer when much food is eaten, and the intervals between their periods of rest or sleep is longer when less food is eaten. Owls and snakes appear to be their principal enemies. Predatory mammals eat some shrew-moles, and probably kill many that they do not eat. Shrew-moles are completely blind, and their long, prehensile nose guides all their activity. Their ordinary movements on the surface of the ground are slow and cautious. When frightened they break into a scuttling rush which ends beneath a leaf or bit of bark where the animal becomes motionless. Shrew-moles, on the surface of the ground, make considerable noise. They construct molelike burrows, but these are not as extensive or complicated as those of moles. The shrew-mole hunts for food in shallow trenches that it makes just under the layer of dead leaves and vegetable debris that covers the ground in their habitat. The food of the shrew-moles includes earthworms, isopods, [Pg 124] insect larvae, soft-bodied insects, and other animal matter. Some vegetable matter is eaten.
Breeding takes place at all seasons of the year, save perhaps in December and January. Embryos vary from one to four. The nest of a shrew-mole at Seattle, King County, consisted of a handful of damp leaves in a cavity of a rotten, punky, alder stump. The nest contained four half-grown young.
Urotrichus gibbsii Baird, Mamm. N. Amer., p. 76, 1857.
Neurotrichus [sic] gibbsii Günther, Proc. Zool. Soc. London, pl. 42, 1880.
Neurotrichus Gibbsii True, Proc. U. S. Nat. Mus., 7 (1884):607, 1885.
Neurotrichus gibbsii Bryant, Zoe. 1:359, February, 1891.
Neurotrichus gibbsii gibbsii Miller, U. S. Nat. Mus. Bull., 79:11, December 31, 1912.
Type.—Obtained at Naches Pass, 4,500 ft., Pierce County, Washington, by G. Gibbs on July 15, 1854 (see Dalquest and Burgner, 1941); type in United States National Museum.
Racial characters.—Size relatively large; tail relatively long; foreclaws straight on ventral surface.
Measurements.—Two females from Tye, 4,000 ft., Stevens Pass, King County, average: total length, 121.5; length of tail, 45; length of hind foot, 18.
Distribution.—The Cascade Mountains, from British Columbia south, and Destruction Island, Jefferson County. Records of occurrence are Baker Lake (J. M. E.), Tye (M. V. Z.), and Mount Rainier (Mount Rainier Ntl. Park Mus.)
Remarks.—Shrew-moles are present on Destruction Island, a small island in the Pacific off the coast of Jefferson County. These moles are large, and are like gibbsii. It is thought, however, that this resemblance is due to convergent evolution rather than a once-continuous range with gibbsii. It is significant that a shrew (Sorex trowbridgii destructioni), the only other native land mammal on the island, differs from its mainland counterpart in much the same way as does Neurotrichus g. gibbsii from Neurotrichus g. minor.
Type.—Obtained on the University of Washington Campus, Seattle, King County, Washington, by W. W. Dalquest on May 19, 1940; type in the Museum of Vertebrate Zoölogy.
Racial characters.—Size small; tail short; foreclaws light and weak as compared to those of gibbsii, with ventral surfaces curved.
Measurements.—Eighty-five specimens (males and females) from Seattle, King County, average: total length, 107.0; length of tail, 35.3; length of hind foot, 15.0.
Distribution.—The lowlands of western Washington. Marginal records are: Mt. Vernon (Jackson, 1915: 97), Cottage Lake (W.W.D.) and Yacolt (M. V.Z.).
Fig. 23. Coast mole (Scapanus orarius orarius) left and Townsend mole (Scapanus townsendii) right; Puyallup. Washington, May, 1914. (Fish and Wildlife Service photo by T. H. Scheffer, No. B-18637.)
Moles are of stocky build and have cylindrical, rounded bodies. The eyes are tiny, nearly concealed in the fur. They have no external ears. The legs are short; the forefeet wide, spadelike, and armed with powerful claws. The fur is erect, not posteriorly directed. The color of the fur is deep bluish or brownish slate. The short, nearly naked tail is pale pink or whitish. Moles of this genus are found only along the Pacific Coast of North America from southern Canada south into Baja California.
Scalops townsendii Bachman. Jour. Acad. Nat. Sci. Philadelphia, 8 (pt. 1):58, 1839.
Scapanus tow[n]sendii Pomel. Arch. Sci. Phys. Nat., Geneva, 9 (ser. 4):247, 1848.
Scapanus Townsendii True. Proc. U. S. Nat. Mus., 7 (1881):607, 1885.
Type.—Probably obtained at Fort Vancouver. Clark County. Washington, by J. K. Townsend. A cotype was obtained on May 9. 1835; type in Academy of Natural Sciences of Philadelphia.
Measurements.—Three males and 6 females from southwestern Washington average, respectively: total length 221, 214; length of tail 51, 46; hind foot 28.3, 26; weight 147, 117 grams.
Distribution.—The lowlands of western Washington. Marginal occurrences are: Sauk (Jackson. 1915: 61); Skykomish (Jackson, 1915: 61) and Yacolt (M.V.Z.).
Remarks.—Scapanus townsendii occurs only in a narrow belt extending from southwestern British Columbia to northwestern California. It seems to prefer a generally damper habitat than the smaller-sized coast mole, although both species are sometimes found in the same locality. The larger mole is abundant in the meadows on the flood plains of rivers at low elevations, and on the glacial outwash prairies. It is often numerous in the fir forests, although its workings and mounds are less conspicuous there. The bodies of nine drowned individuals were found in a well by an old cabin in dense fir forest near Duvall, King County. Townsend moles occasionally occur in the Canadian Life-zone, as at Staircase on the north side of Lake Cushman in Mason County where workings were observed, but most records are from the Humid division of the Transition Life-zone.
The Townsend mole is mainly nocturnal. If ridges of its runways are crushed down, they usually remain so throughout the day and are rebuilt the following night. Only about ten per cent of the ridges that were crushed were rebuilt in the daytime, and most of these were repaired in the early morning. The species is almost completely subterranean. Some individuals are crushed on highways by cars, showing that these moles occasionally travel on the surface of the ground.
Townsend moles throw up numerous mounds, each usually containing about a cubic foot of earth. The mounds commonly are built just about as far apart as a man can step. The general direction as well as the twists and turns of a mole's burrow can usually be determined from the mounds. Townsend moles also construct ridges on the surface of the ground by pushing up sod in building a [Pg 127] tunnel just below the grass roots. Smaller ridges are less commonly made by coast moles. More extensive tunnels, constructed deeper in the earth, serve as living quarters.
Fig. 24. Distribution of the Townsend mole, Scapanus townsendii, in Washington.
According to Wight (1928: 24), Scheffer (1922: 11) and Moore (1933: 39), the food of this large mole includes earthworms and ground-inhabiting insects, insect larvae, spiders, centipedes, flesh, and small amounts of soft vegetation. Scheffer (1922: 10) found that the large mole breeds in February and produces from two to four young at a litter, with an average of three.
Description.—The coast mole is almost identical with the larger mole in form of body but is smaller. Head and body are about 5-1/4 and tail about 1-1/2 inches in length.
The coast mole occupies all of the territory inhabited by the Townsend mole and ranges slightly farther northward, southward and eastward. However it does not range east of the boundaries of the three Pacific Coast states or British Columbia.
The mounds and workings of the coast mole are smaller than those of the Townsend mole and consequently are less noticed. It seems less prone to make numerous mounds, a pace apart, than the larger mole, and burrows tend to extend deeper in the ground. Upthrust ridges are less commonly built by orarius than by townsendii.
Fig. 25. Coast mole (Scapanus orarius orarius). Freshly killed; Seattle, Washington, June 9, 1938. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 64.)
Jackson (1915: 62) mentions the possibility of ecologic differences between orarius and townsendii. There are some differences in habitat and habits. The coast mole seems to live deeper in the ground, prefers better drained soil, and is less colonial than the Townsend mole. These are average differences, however, and the two species commonly occur together. Another difference is that the larger mole rarely enters the dense deciduous woods, such as the brush-grown alder and dogwood jungles along the stream valleys. I have taken the coast mole in such localities, and often have found their workings there. The surface of a nearby meadow may be dotted with mounds of both species, but the larger mole seems not to enter ground that is thickly grown with brush. The coast mole ascends to greater altitudes in the Cascade Mountains than does the Townsend mole.
Like the larger mole the coast mole feeds principally on earthworms and insects (Moore, 1933: 38). On September 29, 1939, in [Pg 129] a ravine at Seattle, in two baited traps set six inches apart, a creeping mouse (Microtus oregoni) and a coast mole were found. The mouse had come to the bait and been caught. The mole had seemingly emerged from the ground through a tunnel a foot away and had been attracted to the body of the mouse. It had eaten an area a half inch in diameter and three-quarters of an inch deep into the body of the mouse just behind the shoulder, when in shifting its position it had become caught in the unsprung trap behind it. When found the mole lay dead with its nose inside the body of the mouse and its back broken. This is the only case known to me of a coast mole appearing voluntarily on the surface of the ground. Never have I found coast moles crushed on the highway; several Townsend moles so killed have been found.
Fig. 26. Distribution of the coast mole in Washington. A. Scapanus orarius orarius. B. Scapanus orarius yakimensis. C. Scapanus orarius schefferi.
The coast mole seems to breed very early in the spring. Males with swollen testes are found late in January. The young usually number four and are born in late March or early April. The breeding season seems to be the same as that of townsendii but the number of young to a litter may average slightly greater.
Scapanus orarius True. Proc. U. S. Nat. Mus., 19:52, December 21, 1896.
Scapanus orarius orarius Jackson, N. Amer. Fauna, 38:61, September 30, 1915.
Type.—Obtained at Shoalwater (= Willapa) Bay, Pacific County, Washington, by J. G. Cooper on August 30, 1855; type in United States National Museum.
Racial Characters.—Color dark bluish; frontal region of skull not inflated.
Measurements.—Eight males and two females from Seattle, King County, average, respectively: total length 159, 155; length of tail 33, 31; hind foot 20.7, 20.5; weight 58.5, 55.8.
Distribution.—Western Washington. Marginal occurrences are: Skykomish (B.S.C.), Merritt (B.S.C.), Wenatchee (B.S.C.), Lester (Jackson, 1915: 64) and Yacolt (M.V.Z.).
Type.—Obtained 3/4 mile north of Union Gap, Yakima County, Washington, by J. A. Gray, Jr., on July 3, 1941; type in Museum of Vertebrate Zoölogy.
Racial Characters.—Color pale, grayish; skull narrow with inflated frontal region.
Measurements.—Six specimens (males and females) from Selah, Yakima County, average: total length 164; length of tail 37; length of hind foot 21.5. The type specimen weighed 58 grams.
Distribution.—The Yakima Valley area; recorded from the type locality northwestward to Easton (B.S.C.).
Scapanus orarius schefferi Jackson, N. Amer. Fauna, 38:63, September 30, 1915.
Type.—Obtained at Walla Walla, Walla Walla County, Washington, by T. H. Scheffer on August 8, 1914; type in United States National Museum.
Racial Characters.—Large size; pale color; large, wide skull with inflated frontal area.
Measurements.—Six topotypes average: total length 159; length of tail 35; hind foot 21.5.
Distribution.—Southeastern Washington; recorded from Fort Walla Walla (Jackson, 1915: 64), Walla Walla (Jackson, 1915: 64) and Dayton (M.V.Z.).
Shrews have tiny eyes, almost concealed in the fur. The body is slim; the nose elongate and pointed. The legs are short and the feet small and weak. The fur is short but soft and posteriorly directed. The cinereous shrew, for example, is about 4 inches in length, of which the tail comprises 1-3/4 inches. The upper parts are dark grayish brown and the underparts dull gray.
Shrews of the genus Sorex are cosmopolitan in distribution. In North America they range from the arctic south to Central America. Three subgenera are recognized by Jackson (1928: 27), all three of which are represented in the state of Washington. The twelve subspecies present occupy numerous habitats, and their ranges include almost all of the state.
Jackson pointed out (1928: 1) that "No other group of American mammals having a wide distribution, and in many localities an abundance of individuals, is so little known to the nonprofessional mammalogist as the long-tailed shrews." In Washington, especially in the coastal area of western Washington, shrews are widespread and abundant in many habitats. In some places they are the most common mammal present. In spite of this their presence is often unsuspected by persons that are otherwise alert to the animal life around them. This is even more surprising when one takes into account the facts that long-tailed shrews are diurnal as well as nocturnal and are less apt to detect the presence of man than are most other species of mammals.
An important factor making long-tailed shrews inconspicuous is their small size, and contributing factors include their rapid movements and dull colors. On several occasions the writer, after seeing a long-tailed shrew vanish soundlessly under a log or into a patch of dead leaves, was left wondering if one actually had been seen or if instead his imagination had conjured up an animal from a dust mote or wind-blown leaf.
The environment of the long-tailed shrews, except for the water shrews, is the zone at the very surface of the earth, just beneath the layer of moss, grass, dead leaves, and decaying vegetation. The removal of the covering vegetative layer reveals a maze of tiny tunnels, the branchings and complexities of which are infinite. Tiny [Pg 132] traps baited with oats or meat and set in these runways catch the long-tailed shrews that inhabit them.
The food of long-tailed shrews is varied. It is principally soft-bodied insects, insect pupae, and earthworms. At times a considerable quantity of soft vegetation and some seeds are eaten. Recent studies (Moore, 1940: 1942) have shown that by destroying seeds some shrews may adversely affect the reforestation of some coniferous trees. Shrews readily eat meat, and often destroy the small mammals, including other shrews, taken in the mammal collector's traps.
Hamilton (1940: 485) found that in one species of long-tailed shrew in the United States individuals rarely lived more than one year. This seems not to be true of at least some of the species found in Washington.
Parasites, internal or external, are not commonly found on long-tailed shrews. They are regularly eaten by owls and snakes, but most carnivorous mammals, though they readily kill them, rarely eat them.
The cinereous shrew ranges over most of Alaska, Canada and the northern half of the United States. A number of subspecies have been described, of which two have been reported from Washington. The cinereous shrew is of medium size and difficult to distinguish from the dusky and wandering shrews, especially in eastern Washington, without studying the skulls. In cinereus the fourth unicuspid tooth is smaller than the third; in vagrans and obscurus it is larger. The relatively narrow rostrum of cinereus also serves to separate it from the other two species.
The cinereous shrew seems to be less restricted to the vicinity of streams and marshes than Sorex vagrans, resembling Sorex obscurus and Sorex trowbridgii in this respect. In Washington it seems to be rare and has not been taken by the writer. Published records for Sorex c. cinereus indicate that this race is, in Washington, confined to mountainous areas. The coastal race, streatori, seems to be confined to the humid area.
Sorex arcticus cinereus Kerr. Anim. Kingd., p. 206, 1792.
Sorex cinereus cinereus Jackson. Jour. Mamm., 6: 56, February 9, 1925.
Type.—None. Name based on the account of a shrew seen at Fort Severn, Canada, by J. R. Forster in 1772.
Racial characters.—Small size and pale color.
Measurements.—Seven males and 8 females from Indian Point Lake, British Columbia, average, respectively: total length 99, 95; length of tail 43.5, 42.2; hind foot 12.7, 12.3.
Distribution.—Northeastern Washington (Metaline and Loon Lake, Jackson, 1928: 49) and the Cascades from Whatcom Pass (Jackson, 1928: 55) southward to Conrad Meadows (Jackson, 1928: 49). There is a specimen in the collection of the California Academy of Sciences, from Mt. Rainier.
Remarks.—This is the most wide-ranging subspecies of shrew, being recorded from Alaska, 10 Canadian provinces and 26 states (Jackson, 1928: 46-50). In the more northern and eastern parts of its range it is sometimes the commonest shrew.
Fig. 27. Distribution of the cinereous shrew in Washington. A. Sorex cinereus cinereus. B. Sorex cinereus streatori.
Sorex personatus streatori Merriam. N. Amer. Fauna, 10:62, December 31. 1895.
Sorex cinereus streatori Jackson. Jour. Mamm., 6:56. February 9, 1925.
Type.—Obtained at Yakutat, Alaska, by C. P. Streator on July 9, 1895; type in the United States National Museum.
Racial characters.—Larger and darker than Sorex cinereus cinereus.
Measurements.—Five females from Alta Lake, British Columbia, average: total length 103; length of tail 45; hind foot 12.5.
Distribution.—The western part of the northern Cascades (Glacier, Jackson, 1928: 55) and the Olympic Peninsula (Neah Bay south to Cedarville, Jackson, 1928: 55).
Remarks.—This shrew seems to occupy the same range as Sorex obscurus setosus but is rare where obscurus is common.
Sorex merriami Dobson. Monogr. Insectivora, pt. 3, fasc. 1, pl. 23, fig. 6, May, 1890.
Sorex merriami merriami Benson and Bond. Jour. Mamm., 20: 348, August 14, 1939.
Type.—Obtained on Little Bighorn River, about a mile and a half above Fort Custer, Crow Indian Reservation, Montana, by Charles E. Bendire on December 26, 1884; type in United States National Museum.
Measurements.—Of type: total length 90; length of tail 35; hind foot 11.5 (after Jackson, 1928: 80).
Distribution.—A single specimen of this rare shrew is known from Washington. Jackson (1928: 81) states that it "was collected by George G. Cantwell, November 18, 1919, at the entrance to an old badger digging on top of a 'high bunch grass hill' at Starbuck (altitude 645 feet), Columbia County, Wash."
Description.—Similar to Sorex cinereus but upper parts pale grayish and underparts white.
The Merriam shrew has been recorded from but a few localities in the western United States. It is one of the rarest of the small mammals known to occur in North America. All the known specimens have been found in desert areas.
Description.—The Trowbridge shrew closely resembles the cinereous shrew in body form but possesses a longer tail. The head and body of adults measure about 2-1/4 inches and the tail about 2 inches. The Trowbridge shrew may be separated from all other small shrews that occur in Washington by its dark bluish upper parts and bluish or slaty underparts. The tail is distinctly bicolor.
Trowbridge shrews occur from southern British Columbia south to central California. Their distribution, to the south, is more extensive than that of many members of the Pacific Coastal Fauna. They are forest animals, ranging widely over the dry ground beneath the fir forest, where they are usually the only shrews present. They are abundant in ravines and in some swampy woods when other shrews are absent, but they avoid open meadows or marshes. [Pg 135] Vertically, they occur from the humid division of the Transition Life-zone to the Hudsonian Life-zone.
These little shrews do not live well in captivity and I have learned relatively little concerning their habits from live specimens. They seem to be slower-moving and less aggressive than the wandering shrew. Population studies showed that the Trowbridge shrew is unable to compete with the wandering shrew in ravine habitats (Dalquest, 1941A: 173). The principal food of the Trowbridge shrew includes soft-bodied insects and insect pupae.
Fig. 28. Distribution of the Merriam and Trowbridge shrews in Washington. A. Sorex merriami merriami. B. Sorex trowbridgii trowbridgii. C. Sorex trowbridgii destructioni.
Moore (1942) has shown that shrews eat the seeds of the Douglas fir and may be a serious check on the reproduction of this important tree. The Trowbridge shrew is the most abundant shrew in the fir forests and probably constitutes the principal shrew that might be classed as a pest.
A specimen containing 4 embryos was taken near Shelton, Mason County, on April 23, 1937. Males with enlarged, greenish testes were taken in April of 1938, 1939 and 1940. Specimens obtained in other months showed no indications of breeding.
Sorex trowbridgii Baird, Rept. Pacific R. R. Survey, 8 (pt. 1):13, 1857.
Type.—Two cotypes were obtained at Astoria, Clatsop County, Oregon. The skin of one (the lectotype) was entered in the U. S. National Museum catalogue in July, 1855, and the skull in January, 1857. The other was obtained by J. Wayne on July 10, 1855.
Racial characters.—Small size and narrow skull.
Measurements.—The average measurements of 38 adults from King County, Washington, are: total length 115.3; length of tail 54.4; hind foot 13.4.
Distribution.—Forested areas from the Pacific Coast eastward through the Cascades to Stehekin (Jackson, 1928: 96), 2 mi. S. Blewitt Pass (Jackson, 1928: 96) and Satus Pass (M.V.Z.).
Type.—Obtained on Destruction Island, Jefferson County, Washington, by V. B. Scheffer on April 22, 1941; type in United States National Museum.
Racial characters.—Large size and wide skull.
Measurements.—Thirty topotypes average: total length 122.5; length of tail 56.7; hind foot 14.3; weight 7.5 grams.
Distribution.—Known only from Destruction Island, 35 acres in area, lying 4 miles off the Washington Coast.
Description.—This species closely resembles the cinereous shrew in body form. Its head and body measure about 2-1/4 inches; the tail slightly less than 2 inches. In summer the upper parts are reddish brown and the underparts gray tinged with brownish. In winter the upper parts are more dusky.
Several races of the wandering shrew range over western North America from southwestern British Columbia south to southern Mexico. Two races occur in the state of Washington.
Marshy areas and damp places are the habitat of the wandering shrew. Cattail and tule marshes, sphagnum bogs, and meadows are favored. They frequent streams through forests but rarely are taken in places away from water. On some of the San Juan Islands, wandering shrews were found along the beaches where they were feeding on the amphipods that live in the dead seaweed and litter at the high tide line. In a favorable habitat, wandering shrews may be the most abundant mammal present. Specimens are occasionally taken in 90 per cent or more of a mammal collector's traps.
The preference of the wandering shrew for damp areas makes it more or less independent of life-zones, for marshy areas, whether [Pg 137] in Transition, Canadian, or Upper Sonoran life-zones, present comparable ecological conditions.
Broadbrooks (1939: 65) found that captives taken at Seattle ate rolled oats, apple, fresh or cooked meat, sow bugs, centipedes, earthworms, frogs (Hyla regilla), a salamander (Plethodon vehiculum), and small, black slugs (Arean arean). Wandering shrews proved incapable of destroying snails (Helisoma occidentalis) and large slugs. The captive shrews kept by Broadbrooks ate an average of 1.3 times their own weight in food each day.
Fig. 29. Distribution of the wandering shrew in Washington. A. Sorex vagrans vagrans. B. Sorex vagrans monticola.
The wandering shrew changes from winter to summer pelage in a few days, seemingly in the second week of October (Dalquest, 1944: 147). The spring molt occurs rapidly but perhaps at a less regular date. Rarely a midsummer molt occurs.
The earliest evidence of breeding in a wandering shrew was discovered on January 27, 1937. Most adult females taken in February, March, April and May were pregnant. Embryos were less often found in summer and fall, but one pregnant female was taken in November. Embryos varied in number from 3 to 8 with an average of six.
Sorex vagrans Baird, Rept. Pacific R. R. Survey, 8 (pt. 1):15, 1857.
Sorex suckleyi Baird, Rept. Pacific R. R. Survey, 8 (pt. 1):18, 1857 (type from Steilacoom Pierce County, Washington).
Type.—Obtained at Willapa Bay [Shoalwater Bay], Pacific County, Washington, by J. G. Cooper; entered in U. S. Nat. Mus. catalogue on October 23, 1856.
Racial characters.—Dark color.
Measurements.—Twenty-five males and 25 females from Seattle, King County, average, respectively: total length 110.5, 107; length of tail 44.6, 45; hind foot 12.4, 12.4.
Distribution.—This is the common marsh shrew of western Washington and occurs from the Pacific Ocean east to the Cascades at Lake Keechelus (W.S.M.) and 15 mi. NW White Salmon (Jackson, 1928: 106).
Sorex monticolus Merriam, N. Amer. Fauna, 3:43, September 11, 1896.
Sorex vagrans monticola Merriam, N. Amer. Fauna, 10:69, December 31, 1895.
Type.—Obtained on San Francisco Mountain, 1150 ft. altitude, Coconino County, Arizona, by C. H. Merriam and V. Bailey on August 28, 1889; type in United States National Museum.
Racial characters.—Pale color.
Measurements.—Five males and 5 females from Selah, Yakima County, average, respectively: total length 100, 98.4; length of tail 40.6, 39.2; hind foot 12.2, 12.
Distribution.—Eastern Washington. The range of this shrew extends west to Bauerman Ridge (Jackson, 1928: 113), Merritt (W.W.D.) and Maryhill (M.V.Z.).
Remarks.—A series of shrews from Moses Lake, Grant County, differs from monticola in larger size and darker color. They agree rather closely with Sorex r. amoenus from California and Nevada. Because they are isolated from that subspecies it seems best to consider them a microgeographic race referable to monticola.
Description.—The dusky shrew is similar in form of body to the cinereous shrew. The length of head and body is about 2-1/4 inches. The tail is about 2-1/2 inches. The upper parts are rusty or reddish brown. The underparts are brownish gray. The dusky shrew differs from the wandering shrew in possessing a slightly longer body and longer tail, but in eastern Washington the two species are almost indistinguishable.
Dusky shrews range from northern Alaska to southern New Mexico, and from the Rocky Mountains to the Pacific Ocean. Thirteen subspecies are recognized by Jackson (1928: 115), of which two occur in Washington.
Records of the dusky shrew are not available from the arid subdivision of the Transition or the Upper Sonoran life-zones. It occurs sparingly in the humid subdivision of the Transition and is common in the Canadian and Hudsonian life-zones.
The habitat of the dusky shrew is varied. Near Seattle, King County, several specimens were trapped in marshes where Sorex vagrans was abundant. Near Stevens Pass, King County, two were taken in a marsh; two others were trapped in a dry, coniferous forest; one was taken in a small bed of heather on a barren mountain top; and another was found dead in a pan of pancake batter in camp. Two specimens were trapped along a small stream at Dewey Lake, Mt. Rainier, Yakima County. Four specimens were caught in traps set in a talus slope on a dry hillside at the North Fork of the Quinault River, Jefferson County. Three others were trapped in dense, rain-forest thickets along the ocean at La Push, Jefferson County. These records indicate that the dusky shrew has a wider environmental range than other Washington shrews. Despite this wide range of tolerance the dusky shrew is common only locally, except in the Hudsonian Life-zone.
Little is known of the habits of dusky shrews but they seem to be as diurnal as they are nocturnal. At Wolf Bar, North Fork of the Quinault River, Jefferson County, a dusky shrew was seen on the packed-earth floor of an old trapper's cabin. A hat was carefully dropped over the live animal but, when the hat was lifted, the shrew was found dead. In an old cabin at Stevens Pass, King County, a dusky shrew was found dead one morning in a pan of flapjack batter prepared the evening before. As this pan was on a table about three feet from the floor, the shrew must have climbed to the table by way of the rough cabin wall, but how the animal managed to scale the side of the pan is a mystery.
Slipp (1942: 211) discovered the nest of a dusky shrew between Round Pass and Lake George, 4200 feet elevation, in Mt. Rainier National Park, on July 25, 1937. The nest was in a rotten fir log 20 inches in diameter. The nest, a ball of dry grass the size of a man's fist, had no central cavity or passages, the occupants "merely pushed through wherever they wished." Seven young shrews were found in and near the nest. Though the eyes of the young were still closed, they were able to creep about and squeal.
A specimen obtained 5 miles west-southwest of Guler, Skamania County, contained 4 embryos on July 10, 1939.
Fig. 30. Distribution of the dusky shrew in Washington. A. Sorex obscurus obscurus. B. Sorex obscurus setosus.
Sorex vagrans similis Merriam, N. Amer. Fauna, 5: 34, July 30, 1891 (not of Hensel, 1855).
Type.—Obtained on Timber Creek, 8,200 ft., Lemhi Mountains, Lemhi County, Idaho, by V. Bailey and B. H. Dutcher on August 26, 1890; type in United States National Museum.
Racial characters.—Tail relatively short, color pale.
Measurements.—Thirteen males and 10 females from Indian-point Lake, British Columbia, average, respectively: total length 105, 107; length of tail 45, 46; hind foot 13.3, 13.3.
Distribution.—Northeastern Washington west, according to Jackson (1928: 122), to Pasayten River, Stehekin and Wenatchee. Because specimens from Tye, King County, and Mt. Stuart, Easton and Lake Keechelus are clearly referable to S. o. setosus, the specimens recorded by Jackson (1928: 122), from Easton and Signal Peak, are mapped in the range of setosus.
Sorex setosus Elliot, Field Columb. Mus. Publ. 32. zoöl, ser., 1:274. March, 1899.
Sorex obscurus setosus Jackson, Proc. Biol. Soc. Washington. 31:127. November 29, 1918.
Sorex obscurus bairdi Jackson, N. Amer. Fauna, 51:140. July 24, 1928.
Type.—Obtained at Happy Lake, Clallam County, Washington, by D. G. Elliot on August 18, 1898; type in Field Museum of Natural History.
Racial characters.—Tail long, color dark.
Measurements.—Twelve males and 17 females from southwestern Washington average, respectively: total length 118, 119; length of tail 53, 53; hind foot 13.8, 13.7; weight 6.6, 5.2 grams.
Distribution.—Western Washington, east through the Cascades to Barron (Jackson, 1928: 137), Cascade River (Jackson 1928: 137) and Satus Pass (W. W. D.).
Fig. 31. Distribution of the mountain water shrew. Sorex palustris navigator, in Washington.
Neosorex navigator Baird, Rept. Pacific R. R. Survey, 8 (pt. I): 11, 1857.
Sorex (Neosorex) palustris navigator Merriam, N. Amer. Fauna, 10:92, December 31, 1895.
Sorex palustris navigator Stephens, California Mammals, p. 254, June, 1906.
Type.—Obtained at head of Yakima River, Kittitas County, Washington, by J. G. Cooper on August 31, 1853; type in United States National Museum.
Measurements.—Two males and 4 females from Washington average, respectively: total length 150, 150; length of tail 70, 74; hind foot 18.5, 20; weight?, 11.0 grams.
Distribution.—Mountainous areas of entire state, including the Olympic Mountains, from Elwah (Jackson, 1928: 188) south to Quinault River (Jackson, 1928: 189); the Cascades from Tomyhoi Lake (W. W. D.) south to 15 mi. N. Carson (Jackson, 1928: 189); northeastern Washington from Shovel Creek (W. W. D.) south to Gifford (Jackson, 1928: 189); the Blue Mountains at Godman Springs (M. V. Z.) and Hompeg Falls (M. V. Z.).
Description.—In general form of body the mountain water shrew resembles the cinereous shrew but is perhaps more stocky. It is a large shrew, nearly as large as a house mouse. The head and body measure about 3 inches; the tail also is about 3 inches long. The fur is exceedingly soft. The upper parts are blackish in color, lightly frosted with paler hairs. The underparts, from throat to vent, are whitish tinged with gray or brown. The stiff, curved fringe of bristles on the outer part of the hind foot serves as an aid in swimming.
Mountain water shrews range over much of Canada and in mountainous areas of the western United States extend south to Arizona. The subspecies found in Washington ranges over all of the western United States.
The mountain water shrew is primarily a mammal of the Hudsonian and Canadian life-zones. It sometimes descends to the Transition Life-zone along clear, cold streams where conditions are similar to those in the Canadian Life-zone.
The favored habitats of the mountain water shrew are the clear, cold streams of the alpine cirques and gushing streams on the mountain sides. In the pools and waterfalls, and among the rocks and mosses that border them, the larvae of aquatic insects, upon which the mountain water shrew feeds, are abundant. Svihla (1934: 45) observed that the fur of a swimming water shrew gathered air bubbles and "it had difficulty in forcing its way down to the bottom [of an aquarium]. On reaching the bottom it literally stood on its long flexible nose which was thrust into the sand and debris, searching for food, its feet kicking rapidly in order to maintain this position. A change in direction was brought about by a twist of the body. To come to the surface again it merely stopped kicking and immediately rose like a cork." The buoyancy of the water shrew allows it to float in the water, like a duck. Jackson (1928: 9) observed a water shrew run across the surface of a small pool. Near Stevens Pass a water shrew dashed from under a stone and ran, did not swim, across the surface of a small, deep pool to escape in a burrow on the other side. According to Jackson, an air bubble held in each foot supports the shrew on the surface of the water. A mountain water shrew observed at Shovel Creek, Ferry County, was as agile on land as any other species of shrew. In summary, water shrews are able to swim, dive, float like a duck, and walk on the surface of the water as well as walk on land.
The food of the mountain water shrew includes snails, leeches, and the larvae of aquatic insects. Mice caught in traps are sometimes eaten by water shrews. Svihla (loc. cit.) found a captive water shrew unable to capture pollywogs and minnows kept in the same aquarium.
Description.—The Bendire water shrew is similar in form of body to the cinereous shrew but possesses a more stocky body which, with the head, measures about 3-1/2 inches long; the tail is about 2-3/4 inches long. The Bendire water shrew closely resembles also the mountain water shrew but has a longer body and shorter tail. The upper parts are blackish in color, not lightly frosted with gray hairs. The hind feet lack the fringe of stiff, curved bristles characteristic of the mountain water shrew. The underparts are black in the race S. b. bendirii. In the race albiventer the throat is blackish but the abdomen is pale gray tinged with brownish.
Fig. 32. Distribution of the Bendire water shrew and the pigmy shrew in Washington. A. Sorex bendirii bendirii. B. Sorex bendirii albiventer. C. Microsorex hoyi washingtoni.
Bendire water shrews are restricted to the Pacific Coast of North America from southern British Columbia to northern California. [Pg 144] Generally they are found at elevations lower than are mountain water shrews. They are typically mammals of the humid division of the Transition Life-zone but often occur in the Canadian Life-zone. They occupy marshes, swamps, damp ravines, and the banks of slow-moving streams. Little is known of their habits, except what has been deduced from the circumstances of their capture. They seem less aquatic than the mountain water shrew. Near Jackson Guard Station on the Hoh River, Jefferson County, one was taken by setting traps on dense beds of water cress that floated in a slow-moving stream. The animal must have swum or walked on the surface of the mat of vegetation. Near Paradise Lake, King County, several were caught in a deep, dark, red cedar swamp. One was caught in a marsh nearby. Nothing is known of the food habits of the Bendire water shrew.
Atophyrax bendirii Merriam, Trans. Linn. Soc. New York, 2:217, August 28, 1884.
Atophyrax Bendirei True, Proc. U. S. Nat. Mus., 7 (1884):606, 1885.
Sorex bendirii Dobson, Monog. Insectivora, part 3, fasc. 1, pl. 23, 1890.
Neosorex bendirii bendirii Miller, U. S. Nat. Mus. Bull., 79:22, December 31, 1912.
Type.—Obtained approximately 1 mile from Williamson River, 18 miles southeast of Fort Klamath, Klamath County, Oregon, by C. C. Bendire on August 1, 1882; type in United States National Museum.
Racial characters.—Underparts everywhere sooty black.
Measurements.—Ten males and 10 females from southwestern Washington average, respectively: total length 163.9, 161.0; length of tail 71.0, 72.6; hind foot 20.5, 20.3; weight 16.8, 14.5 grams.
Distribution.—The southern Cascades and the lowlands of western Washington, exclusive of the Olympic Peninsula. Marginal localities on the west include Mt. Vernon (Jackson, 1928: 196), Bothell (W.S.M.), Renton (M.V.Z.), Puyallup (W.W.D.), Steilacoom (Jackson, 1928: 196) and Oakville (Jackson, 1928: 196).
Sorex (Atophyrax) bendirii albiventer Merriam, N. Amer. Fauna, 10:97, December 31, 1895.
Neosorex bendirii albiventer Miller, U. S. Nat. Mus. Bull., 79:22, December 31, 1912.
Sorex bendirii albiventer Jackson, N. Amer. Fauna, 51:198, July 24, 1928.
Type.—Obtained at Lake Cushman, Mason County, Washington, by C. P. Streator on July 7, 1894; type in United States National Museum.
Racial characters.—Area on abdomen whitish.
Measurements.—A male from near the type locality measures: total length 167; length of tail 69; hind foot 22. One from Potlatch, Mason County, measures 167; 69; 22.
Distribution.—The Olympic Peninsula of Washington. Marginal localities on the south are: Potlatch (M.V.Z.) and Lake Quinault (Jackson, 1928: 199).
Microsorex hoyi washingtoni Jackson, Proc. Biol. Soc. Washington, 38:125, November 13, 1925.
Type.—Obtained at Loon Lake, Stevens County, Washington, by V. Bailey on September 26, 1897; type in United States National Museum.
Measurements.—Of type: total length 89; length of tail 27; hind foot 9.
Distribution.—In Washington, known only from the type specimen which was "found dead in a trail in dry pine woods" (Jackson, 1928: 4).
Description.—The pigmy shrew is similar in form of body to the cinereous shrew but smaller. The head and body are about 1-3/4 inches in length; the tail is about 1 inch long. The upper parts are reddish brown and the underparts are gray.
These tiny mammals range widely across central Canada and northern United States from the Atlantic nearly to the Pacific, and north to central Alaska. A single species is known, one race of which occurs in Washington. The subspecies is known from but two specimens: the type and an individual from Montana (Koford, 1938: 372.)
Description.—The genus Myotis may be separated from all other bats that occur in Washington by the presence of 38 teeth (dental formula i. 2-2/3-3, c. 1-1/1-1, p. 3-3/3-3, m. 3-3/3-3 = 38). Their small size separates them from all other genera save Pipistrellus, from which Myotis may be distinguished by the straight, rather than hooked, anterior border of the tragus. Species of Myotis found in Washington vary considerably in size, but all are less than 100 mm. in total length. The upper parts are various shades of brown in color. The ears, when laid forward, always extend to the nostrils or beyond.
This genus is one of the most widely ranging groups of Recent mammals. It occurs on all continents, including Australia and many of the larger islands. Of the 19 American species recognized by Miller and Allen (1928), eight occur in the state of Washington. They are low-flying forms and as a rule appear relatively late in the evening. Their flight is rapid and erratic. They often hunt over the surfaces of streams, pools, and lakes. Some kinds hunt in the shade of forest trees and these are especially difficult to collect. Others hunt the brushy canyons and coulees of the desert areas of eastern Washington.
Grinnell (1918: 241-242) points out that, although bats are not subject to isolation by topographic barriers, as most wingless mammals are, they may be restricted by ecologic barriers in the same way as are other small mammals. This is particularly true of Myotis in Washington. Of the eight species in the state, five are represented by one subspecies west of the Cascade Mountains and another, paler subspecies in the more arid country east of the Cascades.
The mobility of bats makes it difficult to determine their origin and migrational history. Five of the Myotis found in Washington seem to belong to the Pacific Coastal Fauna, and to have been isolated south of the last continental glacier. Complete isolation is unlikely as these species occur in the Cascade Mountains as well as in the Pacific Coastal Faunal Area, and three occur also in the Blue Mountains of southeastern Washington. The differentiation of the Coastal type of Myotis may have come about through habitat selection, of the type discussed by Miller (1942: 25). One western [Pg 147] Washington Myotis (M. keenii) seems to belong to a northern fauna, and to have extended its range south to Washington. All seven species of the desert-living Myotis found in eastern Washington have subspecies which seem to have been derived from the Great Basin Faunal Area.
Myotis lucifugus is represented by two geographic races in Washington. The species ranges across Canada and the United States, from the Atlantic to the Pacific and from the northern limit of tree growth to southern Mexico.
Fig. 33. Distribution of the big myotis in Washington. A. Myotis lucifugus alascensis. B. Myotis lucifugus carissima.
It usually proves rather difficult to separate Myotis lucifugus, on the basis of external features, from other species with which it may occur. Its large foot (9-10 mm.), short ear (when laid forward not extending past nose) and the absence of a keel on the calcar separate it from all species except Myotis yumanensis. From the latter species, lucifugus may be distinguished by the gradually [Pg 148] rather than abruptly rising forehead, as seen in cleaned skulls, and by more shiny, metallic color of fur.
Little is known of the habits of this bat in Washington. It usually appears after dusk, and most specimens are shot over ponds or lakes, where the reflection of light from the sky on the water allows the hunter enough light to sight a gun. A few specimens were collected in deep forests. Its flight and feeding habits are not known to differ from those of other species with which it was associated, except at the south end of Lake Chelan, Chelan County, where two individuals were shot as they hovered near the tops of pine trees and seemed to be picking insects from the branches. I have never found this bat in its daytime retreat.
A specimen taken at Sportman's Lake, San Juan County, held one embryo on June 26, 1938.
Myotis (Leuconoë) carissima Thomas, Ann. and Mag. Nat. Hist., 13 (ser. 7): 383, May, 1904.
Myotis lucifugus carissima Cary, N. Amer. Fauna, 42:43, October 3, 1917.
Type.—Obtained at Yellowstone Lake, Yellowstone National Park, Wyoming by J. Darling in September, 1903; type in British Museum.
Racial characters.—Color pale, rather "brassy" in tone; distal border of interfemoral membrane paler than proximal part.
Measurements.—Four specimens from eastern Washington average: total length 77; length of tail 33; hind foot 11; ear 13; height of tragus 7.3.
Distribution.—East of the eastern base of the Cascade Mountains, save for the Blue Mountains of southeastern Washington. Western records are Stehekin (Miller and Allen, 1928: 52) and Vantage (W. W. D.)
Myotis lucifugus alascensis Miller, N. Amer. Fauna, 13:63, October 16, 1897.
Vespertilio gryphus lucifugus Allen, U. S. Nat. Mus. Bull., 43:78, March 14, 1894 (part specimens from Washington).
Type.—Obtained at Sitka, Alaska, by C. P. Streator on August 5, 1895; type in United States National Museum.
Racial characters.—Color dark, almost bronze; wing and tail membranes uniformly dark in color.
Measurements.—Five specimens from San Juan County, Washington, average: total length 80.9; length of tail 32.1; hind foot 12; ear 12; height of tragus 7; weight 5.4 grams.
Distribution.—From the eastern base of the Cascade Mountains west of the Pacific, and the Blue Mountains of southeastern Washington. Marginal occurrences listed by Miller and Allen (1928:49) are Chilliwack River, Lake Wenatchee, and Lyle.
Description.—The present species closely resembles Myotis lucifugus and specimens in worn pelage can not be distinguished from that species unless the cleaned skulls are examined. In fresh pelage, yumanensis is duller than lucifugus.
Myotis yumanensis ranges from southern British Columbia to central Mexico west of the Mississippi River. Four races are recognized by Miller and Allen (1928: 62).
Fig. 34. Distribution of the Yuma myotis in Washington. A. Myotis yumanensis saturatus. B. Myotis yumanensis sociabilis.
The habits of Myotis yumanensis and Myotis lucifugus appear to be the same. In Washington the two species are commonly found together. In western Washington, Myotis yumanensis seems to be more common than Myotis lucifugus.
In the San Juan Islands a yumanensis was found hiding in the attic of an old cabin on Blakeley Island. A specimen of long-eared bat was taken at the same place. Another Yuma myotis was caught behind a door of a mill on Blakeley Island (Dalquest, 1940: 4).
This species shares with Myotis californicus the habit of apparently drinking salt water.
A specimen obtained at Sportsmans Lake, San Juan County, held one embryo on June 27, 1938. One from Peavine Pass, Blakeley Island, San Juan County, held one embryo on June 22, 1939.
Myotis yumanensis sociabilis H. W. Grinnell, Univ. California Publ. Zoöl., 12:318, December 4, 1914.
Type.—Obtained at old Fort Tejon, Kern County, California, by J. Grinnell on July 23, 1904; type in Museum of Vertebrate Zoölogy.
Racial character.—Color pale.
Measurements.—Two males and 2 females from Selah, Yakima County, average: total length 78; length of tail 36; hind foot 10; ear 14; height of tragus 7.
Distribution.—Eastern Washington generally. Marginal records on the west are: Stehekin (Miller and Allen, 1928: 69), and Selah (W.W.D.).
Fig. 35. Distribution of the fringe-tailed myotis and the Keen myotis in Washington. A. Myotis thysanodes thysanodes. B. Myotis keenii keenii.
Myotis yumanensis saturatus Miller, N. Amer. Fauna, 13:68, October 16, 1897.
Type.—Obtained at Hamilton, Skagit County, Washington, by T. S. Palmer on September 13, 1889; type in United States National Museum.
Racial character.—Color dark.
Measurements.—Eighteen adults of both sexes from San Juan County, Washington, average: Total length 78.2; length of tail 34.4; hind foot 10.1; ear 15; height of tragus 7.4; weight 5.9 grams.
Distribution.—From the eastern base of the Cascade Mountains to the Pacific. This is the commonest Myotis found in western Washington. Marginal localities are: Hamilton (Miller and Allen, 1928: 71), and Goldendale (Miller and Allen, 1928: 71).
Vespertilio subulatus keenii Merriam, Amer. Nat., 29:860, September, 1895.
Myotis subulatus keenii Miller, N. Amer. Fauna, 13:77, October 16, 1897.
Type.—Obtained at Masset, Graham Island, Queen Charlotte Islands, British Columbia, by J. H. Keen in 1894; type in United States National Museum.
Measurements.—Miller and Allen (1928: 109) list the measurements of a male from Sol Duc Hot Springs, Clallam County, and a specimen of unknown sex from Lake Cushman, Jefferson County, as, respectively: total length 89, 87; length of tail 34, 36; hind foot 8.4, 7.4; ear?, 14.6.
Distribution.—Only the Olympic Peninsula, where it has been recorded by Miller and Allen (1928: 104) from Sol Duc Hot Springs and Lake Cushman.
Description.—Myotis keenii is similar, in general, to Myotis lucifugus and Myotis yumanensis, but the ears are longer and when laid forward reach about 4 mm. past the nose rather than ending at the nostrils. The foot is of medium size (about 8 mm.) and no keel is present on the calcar.
The distribution of this species is given by Miller and Allen (1928: 101) as "northern North America from the limits of tree growth south in the east to South Carolina and Arkansas, and in the west to northwestern Washington."
I have not observed this bat in Washington and know nothing of its habits. Its distribution is most unusual. Its range seems to lie only in the glaciated area of western British Columbia and northern Washington.
Description.—The distinguishing feature of Myotis evotis is its long ears, which, when laid forward, reach 5 mm. in front of the nose. Myotis thysanodes and Myotis keenii, other species in which the ears are rather long, have the ears ending less than 5 mm. anterior to the nose when laid forward. The foot of Myotis evotis is of moderate size (8 to 9 mm.).
This species ranges over the western United States, from British Columbia to central Mexico. Two subspecies of this interesting bat are recognized, both of which occur in Washington.
Though I have hunted for this species of bat in Washington on numerous occasions, I have taken no specimens. In the summer of 1939, Myotis identified as this species because of their large ears, were seen flying at midnight in the light of searchlights over Lake Washington Canal at Seattle. Mary Greer gave us a specimen which was struck by her auto near Baker Lake, Whatcom County. According to Miss Greer, the specimen was seen "hovering in the road, like a large moth." The time was about midnight. This evidence indicates that the species does its hunting late at night, when ordinary methods of hunting bats are useless, and may account for the scarcity of specimens from the state. Nevertheless, collectors from the California Museum of Vertebrate Zoölogy took specimens in the Blue Mountains where the bats flew slowly, in rather straight courses, 20 to 25 feet from the ground.
Fig. 36. Distribution of the long-eared myotis in Washington. A. Myotis evotis evotis. B. Myotis evotis pacificus.
This species has not, so far as is known, been taken in Washington in its daytime retreat. Daniel Bonell saved two specimens from under slabs of loose bark on old, dead snags near Tillamook, Oregon. Davis (1939: 214) reported them as hiding in the daytime in a cave in [Pg 153] Craters of the Moon National Monument, Idaho. Whitlow and Hall (1933: 241) report specimens found in an old cabin near Pocatello, Idaho, two of them containing one embryo each.
Vespertilio evotis Allen, Smithsonian Misc. Coll., 7 (no. 165):48, June, 1864.
Myotis evotis Miller, N. Amer. Fauna, 13:77, October 16, 1897.
Type.—Description based on a series of specimens, one of which came from Monterey, California. This locality was designated the type locality by Miller (1897: 78).
Racial character.—Color pale.
Measurements.—Two males and 2 specimens of unknown sex from the Blue Mountains, Columbia County, average: total length 87; length of tail 40; hind foot 7.5; ear 20; height of tragus 11; weight 5.4 grams.
Distribution.—The Blue Mountains area, of southeastern Washington; recorded from South Touchet (Miller and Allen, 1928: 116) and Godman Springs (W. S. M.).
Remarks.—Miller and Allen (1928: 116) record the dark race of Myotis evotis from the Blue Mountains. Specimens examined by me are much paler than pacificus and most of them are indistinguishable from specimens of evotis from California.
Vespertilio evotis Allen, Smithsonian Misc. Coll., 7 (no. 165):48, June, 1864 (part specimens from Puget Sound).
Myotis evotis pacificus Dalquest, Proc. Biol. Soc. Washington, 56:2, February 25, 1943.
Type.—Obtained from 3-1/2 miles east and 5 miles north of Yacolt, Clark County, Washington, by John Chattin on August 3, 1940; type in Museum of Vertebrate Zoölogy.
Racial character.—Color dark.
Measurements.—Five specimens from the type locality average: Total length 85; length of tail 41; hind foot 7.4; ear 19.4; height of tragus 10; weight 5.5 grams.
Distribution.—Western Washington from the Cascade Mountains westward. Marginal occurrences are: Baker Lake (W. W. D.) and Easton (Miller and Allen, 1928: 116).
Myotis thysanodes Miller, N. Amer. Fauna, 13:80, October 16, 1897.
Type.—Obtained at Old Fort Tejon, Kern County, California, by T. S. Palmer on July 5, 1891; type in United States National Museum.
Measurements.—Two males and 3 females from Vernon, British Columbia, average, respectively: total length 90.5, 82; length of tail 41.5, 37; hind foot 10.5, 10; ear 18.5, 16; height of tragus 14, 13.
Distribution.—In Washington known only from the southeastern border of the state, namely from Dayton (W. S. M.) and Anatone (Miller and Allen, 1928: 127).
Description.—Myotis thysanodes resembles Myotis evotis, but differs in larger size, smaller ear (reaching less than 5 mm. past nose when laid forward), and in possessing a well developed fringe of hairs along the border of the caudal membrane.
This species of bat ranges over western North America from southern British Columbia to southern Mexico. Two geographic races are currently recognized, only one of which occurs in the United States. This bat is not recorded by Davis (1939) as occurring in Idaho and has been found only once in Oregon. The only published account of the habits of thysanodes seems to be that of Palmer (in Miller, 1897: 84, also Grinnell, 1918) who found adults and young of various sizes in company with Myotis yumanensis in the attic of an old adobe building near Old Fort Tejon, California, in July, 1891. The specimens obtained in Washington and British Columbia came from dry areas of pine forest.
Description.—This species, in Washington, may be easily recognized by its relatively large size and the presence of a distinct keel on the side of the calcar, posterior to the foot.
Four subspecies of Myotis volans are recognized by Miller and Allen (1928: 136). These range over western North America from southern Alaska to southern Mexico. Two subspecies occur in Washington.
The record stations in Washington for the pale, southern race are all in arid places and the dark, coastal race is a forest animal. Most of the specimens taken by me (all of the dark race) were in clearings or along roads through timber near the crests of hills. They appeared relatively late in the evening, after the big-brown and the silver-haired bats had been in the air for some time. Often they were taken in company with Myotis lucifugus and Myotis yumanensis. They were appreciably larger than those species and their flight was slower and less erratic. They usually flew in relatively straight lines or large circles at from ten to forty feet from the ground. At Lake Kapowsin, Pierce County, they were attracted by swishing a long pole in the air. At Renton, King County, one was shot as it hunted insects at a city street light several hours after dark.
Vespertilio longicrus True, Science, 8:588, 1886.
Vespertilio nitidus longicrus H. Allen, U. S. Nat. Mus. Bull., 43:103, March 14, 1894.
Myotis lucifugus longicrus Miller, N. Amer. Fauna, 13:64, October 16, 1897.
Myotis longicrus Lyon and Osgood, U. S. Nat. Mus. Bull., 62:271, January 28, 1909.
Type.—Obtained in the "vicinity of Puget Sound, Washington" by D. S. Jordan, and catalogued in the U. S. National Museum on December 16, 1886.
Racial character.—Color dark.
Measurements.—A female from 6 miles northeast of Kelso, Cowlitz County, measures: total length 95; length of tail 39; hind foot 8; ear 13; height of tragus 8.
Distribution.—From the eastern base of the Cascade Mountains to the Pacific. Marginal records (from Miller and Allen, 1928: 142) are Oroville, Entiat, and Carson.
Fig. 37. Distribution of the hairy-winged myotis in Washington. A. Myotis volans longicrus. B. Myotis volans interior.
Myotis longicrus interior Miller, Proc. Biol. Soc. Washington, 27:211, October 31, 1914.
Type.—Obtained 5 miles south of Twining, Taos County, New Mexico, by Vernon Bailey on July 23, 1904; type in United States National Museum.
Racial characters.—Smaller and paler than Myotis v. longicrus.
Measurements.—Four males and a female from the Blue Mountains, Columbia County, average: total length 93; length of tail 41.5; hind foot?; ear 12; height of tragus 6.3.
Distribution.—Known only from the Blue Mountains area of the southeastern part of the state, from Walla Walla (E. S. B.) east to Anatone (Miller and Allen, 1928: 144).
Remarks.—Of 5 specimens available from the Blue Mountains, 4 are like interior and 1 is like longicrus.
Fig. 38. Distribution of the California myotis in Washington. A. Myotis californicus californicus. B. Myotis californicus caurinus.
Description.—Myotis californicus may be separated from all Myotis that occur in Washington, except Myotis subulatus, by its small foot (about 6 mm.). It is the only small-footed bat found in western Washington. In eastern Washington, where Myotis subulatus occurs, the cleaned skulls of the two species must be compared before certain identification of some specimens is possible. The skull of M. californicus possesses a higher cranium and more abruptly rising forehead than that of M. subulatus.
Four geographic races of this bat recognized by Miller and Allen (1928: 149) range from southern Alaska southward over western [Pg 157] North America to southern Mexico. Two subspecies occur in Washington.
Myotis californicus, in western Washington, often occurs in company with Myotis yumanensis, lucifugus, and volans. In flight it cannot be distinguished from M. yumanensis or M. lucifugus. Most of our specimens were collected over water, for these bats usually fly rather late and can be shot most easily where their reflection on the water assists the hunter in aiming. They are usually not common, one or two being taken at a single locality. This species, like Myotis yumanensis, seems to drink salt water. On May 9, 1936, a living specimen was caught under a loose piece of bark on a dead tree.
Vespertilio nitidus H. Allen, Proc. Acad. Nat. Sci. Philadelphia, p. 247, 1862 (part of the specimens were from Fort Steilacoom, Pierce Co., Washington).
Myotis californicus caurinus Miller, N. Amer. Fauna, 13:72, October 16, 1897.
Type.—Obtained at Masset, Graham Island, Queen Charlotte Islands, British Columbia, by J. H. Keen in 1895; type in United States National Museum.
Racial character.—Color reddish-brown.
Fig. 39. Distribution of the small-footed myotis, Myotis subulatus melanorhinus, in Washington.
Measurements.—Eight specimens, including both sexes, from the San Juan Islands, San Juan and Skagit counties, average: total length 77.8; length of tail 36.7; hind foot 6.7; ear?; height of tragus 7.6; weight 5.2 grams.
Distribution.—Western Washington east through the northern Cascades to Chelan, Blue Creek and Colville (Miller and Allen, 1928: 156) in northeastern Washington, and, farther south, east to Mount Rainier (Miller and Allen, 1928: 156) and Carson (Miller and Allen, 1928: 156).
Vespertilio californicus Audubon and Bachman, Jour. Acad. Nat. Sci., Philadelphia, 8 (ser. 1, ser. 1, pt. 2):285, 1842.
Myotis californicus Miller, N. Amer. Fauna, 13:69, October 16, 1897.
Type.—None designated. Type locality fixed at Monterey, Monterey County, California, by Miller and Allen (1928: 153).
Racial character.—Color pale.
Measurements.—A female from Crooked River, Crook County, Oregon, measures: Total length 80; length of tail 40; hind foot 7; ear 13; height of tragus 5; weight 3.2 grams.
Distribution.—Recorded only from the eastern part of the state. Westernmost records, according to Miller and Allen (1928: 155) are: Orondo, Goldendale and Lyle.
Vespertilio melanorhinus Merriam, N. Amer. Fauna, 3:46, September 11, 1890.
Type.—Obtained on San Francisco Mountain, 8250 ft. elevation, Coconino County, Arizona, by C. H. Merriam and V. Bailey on August 4, 1889; type in United States National Museum.
Measurements.—Three males and 2 females from eastern Washington average: total length 77; length of tail 37; hind foot 7; ear 15; height of tragus 8. One weighed 5.4 grams.
Distribution.—Desert areas of eastern Washington, including the Columbian Plateau and the lower Columbia River Valley. Marginal records are: 5 mi. S Grand Coulee Dam (W. W. D.) in the north, Wenatchee (W. W. D.) in the northwest, Lyle (W. W. D.) in the southwest, and Bly (Miller and Allen, 1928: 171) in the southeast.
Description.—Myotis subulatus is closely similar to Myotis californicus, but is more orange in color and has the skin on the face more nearly black. Specimens cannot be identified with certainty until the cleaned skulls are examined. The more flattened cranium and less abruptly rising forehead separate Myotis subulatus from M. californicus.
This species is confined to the United States and northern Mexico. Two races are recognized by Miller and Allen (1928), of which one occurs in Washington. It is the commonest bat in the desert of [Pg 159] eastern Washington. It lives far from trees on plains and in sandy or rocky areas, emerging rather early in the evening. It is not difficult to shoot. Its flight is erratic. The animal usually hunts in large, irregular circles at 10 to 25 feet from the ground. It has not been found in its daytime retreat but may hide in crevices in rocky outcrops. Near Vantage, Grant County, individuals were shot as they hung up in a concrete underpass to digest food. The stomachs of specimens taken were so crammed with the remains of insects that their abdomens were greatly distended. The underpass seemed to be only a resting place, not inhabited by day. Others were taken when they came to rest in the loft of a barn at Selah, Yakima County. They usually did not arrive at the barn until an hour after sunset and were still present there at midnight.
V[espertilio]. noctivagans Le Conte, McMurtrie's Cuvier, Anim. Kingd., 1:431, 1831.
Vesperugo noctivagans True, Proc. U. S. Nat. Mus., 7:602, 1885.
Lasionycteris noctivagans Allen, U. S. Nat. Mus. Bull., 43:105, March 14, 1894.
Type.—None designated; described from a specimen obtained in the "eastern United States."
Measurements.—Five males from San Juan County, Washington, average: total length 96.2; length of tail 46.4; hind foot 8.5; ear 16; height of tragus 7.2; weight 8.2 grams.
Distribution.—Forested areas of the entire state. This species is migratory and first appears about the middle of May. September 15th is the latest recorded occurrence, when one was seen at Seattle, King County. Marginal records are: Sportsmans Lake (W. W. D.), in the northwest; Carson (Taylor and Shaw, 1929: 9) in the southwest; Pass Creek Pass (W. W. D.) in the northeast; and Bly (Taylor and Shaw, 1929: 9) in the southeast.
Remarks.—The chocolate-brown color phase of the silver-haired bat is most common east of the Cascade Mountains, while almost all specimens from western Washington represent the black phase.
This medium-sized bat is the darkest-colored species living in the state, ranging from chocolate-brown to nearly black. Numerous white-tipped hairs give the upper parts a frosted appearance. The upper surface of the interfemoral membrane is well furred, a character shared only with Lasiurus. The dental formula (i. 2-2/3-3, c. 1-1/1-1, p. 2-2/3-3, m. 3-3/3-3 = 36) is the same as that of Corynorhinus. The small ears and short tragus immediately distinguish the silver-haired bat from the long-eared bat.
The genus Lasionycteris contains but a single species, of which [Pg 160] no geographic races have been described. It ranges across North America from coast to coast and from central Canada southward, in forested areas, nearly to Mexico. This species is known to be migratory, and southern records probably do not represent breeding individuals.
In Washington these bats have been taken in the Transition, Canadian, and Hudsonian life-zones. They were found near clearings in forests of open pine woods in the arid section of eastern Washington, near mountain hemlock thickets in the high mountains, and in the dense rain-forests of the Pacific Coastal area.
Fig. 40. Distribution of the silver-haired bat. Lasionycteris noctivagans, in Washington.
The size and flight of the silver-haired bats are distinctive, and after some experience it is possible to identify them in the air. It is an early flier, usually appearing just after the swallows roost. They fly at a considerable height, rarely coming within forty feet of the ground. The wings are moved with a "fluttery" motion, and their flight is interrupted by frequent short glides. They fly more rapidly than the big brown-bats, and twist and dart sideways more frequently. Compared with big brown-bats, silver-haired bats are relatively gregarious, and six to a dozen individuals were seen in the [Pg 161] same area. They generally hunt in sweeping circles, from fifty to one hundred yards in diameter. In the daytime the silver-haired bats hide beneath slabs of loose bark on dead trees. Near Cottage Lake, King County, two individuals were found beneath the bark on an old, lightning-blasted stub. The dark color of the bats blended with the charred surface of the stub.
This species feeds mainly on forest insects and for this reason is probably of considerable value to man. The stomachs of specimens were usually crammed with the remains of small, soft-bodied insects.
Description.—Diagnostic characters of the long-eared bat are: medium size (total length about 4 inches); dull, grayish-brown color; exceptionally long ears (over 1 inch from notch); thin, tissue-like membranes; and paired "lumps" on the rostrum. The dental formula is: i. 2-2/3-3, c. 1-1/1-1, p. 2-2/3-3, m. 3-3/3-3 = 36.
Fig. 41. Long-eared bat (Corynorhinus rafinesquii intermedius), female with young; Boulder Cave, Kittitas County, Washington, July 20, 1928. (Fish and Wildlife Service photo by T. H. Scheffer, No. B-33332.)
Long-eared bats range from southern British Columbia to southern Mexico. Three species are listed by Miller (1924: 82), one of which (rafinesquii) is divisible into five geographic races.
This bat is colonial and is not uncommonly found in caves, mine shafts, and darkened attics of old buildings. It is of scattered distribution throughout the state. In Washington its distribution in winter is unknown. A specimen from Friday Harbor, San Juan County, taken in March, 1936, indicates that it hibernates in the [Pg 162] state. Whitlow and Hall (1933: 245) give a detailed account of individuals found hibernating in winter near Pocatello, Idaho.
Fig. 42. Entrance to Boulder Cave, inhabited by long-eared bats (Corynorhinus); Kittitas County, Washington, May 26, 1938. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 56.)
John K. Townsend (1839: 325) mentions that the "great-eared bat" at the forts of the Columbia River district (Fort Vancouver) were protected by the "gentlemen of the Hudson's Bay Company for their services in destroying the dermestes which abound in their fur establishments." Townsend mentions also that the long-eared bats seldom left the "storehouses attached to the forts," even at night. My own observations are similar, in that at Boulder Cave, Kittitas County, on July 7, 1936, when not less than 100 long-eared bats were present in the cave, I watched the entrances to the cave until an hour after dark but no bats were seen to emerge. All of the specimens available from Washington were caught in their daytime hiding place. The number of long-eared bats at Boulder Cave has decreased in recent years. On July 12, 1930, bats were so abundant that 90 were captured with a single sweep of a butterfly net (T. H. Scheffer, 1930: 11). On July 7, 1936, it was estimated that there were slightly more than 100 in the cave. On June 11, 1937, the number was less, probably about 75.
Little is known of the food habits of the long-eared bat. The stomach of a specimen from Blakeley Island, San Juan County, was [Pg 163] crammed with the remains of insects, including the wing scales of Lepidoptera and the wings of small Diptera.
Scheffer noted that the long-eared bats at Boulder Cave were nearly ready to give birth to young on July 12, 1930, and were carrying naked young a week later. On July 7, 1936, at the same locality, females contained nearly full-term embryos.
Fig. 43. Distribution of the long-eared bat in Washington. A. Corynorhinus rafinesquii townsendii. B. Corynorhinus rafinesquii intermedius.
Plecotus townsendii Cooper, Ann. Lyc. Nat. Hist. New York, 4:73, November, 1837.
Corynorhinus macrotis townsendii Miller, N. Amer. Fauna, 13:53, October 16, 1897.
Corynorhinus megalotis townsendii G. M. Allen, Bull. Mus. Comp. Zoöl., 60:344, April, 1916.
Corynorhinus rafinesquii townsendii Miller, U. S. Nat. Mus. Bull., 128:82, April 29, 1924.
Type.—Probably obtained at Fort Vancouver, Clark County, Washington; type not now in existence.
Racial character.—Dark color.
Measurements.—A male from Blakeley Island, San Juan County, measured: total length 83; length of tail 43; hind foot 8.5; ear 37.4; tragus 15.1; weight 10 grams.
Distribution.—Western Washington, from Blakeley Island (W. W. D.) in the north, south to Seattle (W. W. D.) and Fort Vancouver.
Corynorhinus rafinesquii intermedius H. W. Grinnell, Univ. California Publ. Zoöl., 12:320, December 4, 1914.
Corynorhinus rafinesquii townsendii Dalquest, Jour. Mamm., 19:213, May 14, 1938.
Type.—Obtained at Auburn, Placer County, California, by J. C. Hawver on July 31, 1909; type in Museum of Vertebrate Zoölogy.
Racial characters.—Paler and duller than townsendii.
Measurements.—One male and 6 females from 15 miles east of Tonasket, Okanogan County, average: total length 96, length of tail 47; hind foot 11; ear 35.5; tragus 14.
Distribution.—Scattered localities in the arid subdivision of the Transition Life-zone of eastern Washington, from 15 mi. E Tonasket (W. W. D.) on the north to Boulder Cave (W. W. D.) on the west and Spokane (W. S. C.) on the east.
Remarks.—Of specimens in the University of Kansas, Museum of Natural History, those from Selah (not plotted on distribution map), Yakima County, are paler than those from Boulder Cave, Yakima County, but both series are paler than specimens from the coast of Oregon.
Fig. 44. Distribution of the western pipistrelle, Pipistrellus hesperus hesperus, in Washington.
Scotophilus hesperus H. Allen, Smithsonian Misc. Coll., 7 (no. 165):43, June, 1864.
Vesperugo hesperus True, Proc. U. S. Nat. Mus., 7:602, 1885.
Pipistrellus hesperus Miller, N. Amer. Fauna, 13:88, October 16, 1897.
Type.—Obtained at Old Fort Yuma, Imperial County, California, by G. H. Thomas; catalogued in U. S. National Museum on October 31, 1861.
Measurements.—A specimen from Maryhill, Klickitat County, measures: total length 68; length of tail 27; hind foot 7; ear 10; height of tragus 3; weight 4.2 grams. Two males from Vantage, Grant County, average: 68.5; 27.5; 6; 11; 4.
Distribution.—Known only from along the Snake and Columbia rivers of south-central Washington; recorded from Vantage (W.W.D.), south to Maryhill (M.V.Z.), and east to Almota (Taylor and Shaw, 1929: 9).
Description.—This is the smallest bat found in Washington, its body being approximately 1-3/4 inches long and the tail 1-1/4. It may be separated from Myotis by the bent tragus and by the possession of 34 rather than 38 teeth. The dental formula is: i. 2-2/3-3, c. 1-1/1-1, p. 2-2/2-2, m. 3-3/3-3 = 34.
The genus Pipestrellus is cosmopolitan in distribution. The few records for the single subspecies found in Washington indicate that it is a casual, though probably regular, summer visitant from the south.
Two western pipistrelles were shot at Vantage, Grant County, on July 23, 1937. They flew in slow circles about 50 feet from the ground. No breeding records are known from the state.
Eptesicus fuscus bernardinus Rhoads, Proc. Acad. Nat. Sci. Philadelphia, 1901:619, February 6, 1902.
Eptesicus fuscus pallidus Engels, Amer. Midland Nat., 17:656, May, 1936 (part specimens from Washington).
Type.—Obtained near San Bernardino, San Bernardino County, California, by R. B. Herron on May 26, 1893; type in Academy of Natural Sciences of Philadelphia.
Measurements.—Three males and 5 females from Washington average: total length 117; length of tail 48; hind foot 11.5; ear 18.3; height of tragus 9.
Distribution.—Forested areas of the entire state of Washington. Marginal localities are San Juan Island (W.W.D.) in the northwest, Carson (Taylor and Shaw, 1929: 9), in the southwest, Newport (W.W.D.) in the northeast, and Grand Ronde River (Taylor and Shaw, 1929: 9) in the southeast.
Remarks.—Specimens from both eastern and western Washington vary greatly in color, and series of specimens from eastern Washington average little, if any, paler than series from western Washington. Washington specimens most closely resemble specimens from California of the race bernardinus and average darker than pallidus.
Description. The big brown-bat may be distinguished from other bats by its large size (about 4-1/2 inches), rich, brown color, and small ears (reaching only to nostril when laid forward). It possesses 32 teeth, the dental formula being: i. 2-2/3-3, c. 1-1/1-1, p. 1-1/2-2, m. 3-3/3-3 = 32.
Bats of the genus Eptesicus are cosmopolitan in distribution. A single species occurs in North America, of which Engels (1936) recognizes four races in western United States.
Fig. 45. Distribution of the big brown-bat, Eptesicus fuscus bernardinus, in Washington.
The big brown-bat appears early in the evening, often before the swallows have retired. This, and its large size, makes it relatively easy to study. It is principally a forest bat but also is found in towns and cities. It is common in Seattle and hunts around street lights and about the trees in the city parks. In its more natural habitat it flies over trees and clearings. Big brown-bats were repeatedly timed, with a car's speedometer, at 17 miles an hour as they flew down a road lined with tall trees. This is a greater speed [Pg 167] than that at which they usually hunt. When the speed of the car was increased the bats dodged sideways, around the car. At lesser speeds they pulled ahead and escaped.
Big brown-bats are less gregarious, when hunting, than some bats. One or two may be found in a small area, the limits of which are definitely fixed. We noted this repeatedly near Cottage Lake, King County, where the big brown-bats hunted along roads through second-growth conifers. The bats patrolled back and forth along a section of a road about a quarter of a mile in length. When a bat reached the end of its personal territory, it would wheel and return. On six successive trips a bat turned, to retrace its course, at points less than 50 feet distant from the point of the first turn. When the bat approached the turning point on the seventh trip, a bat from the adjoining strip of road approached the area. The two animals fluttered about each other with shrill squeaks, audible 50 feet away. The fluttering and squeaking continued for nearly a minute, after which both bats resumed their hunting. Darkness concluded the observations.
On several occasions we stood on a road patrolled by a big brown-bat. The bat immediately detected the watcher and fluttered about his head and face. Big brown-bats ignore other species of bats hunting on their territory. Indeed, such comparatively slow flyers could scarcely drive the more speedy bats away.
Two Eptesicus, kept in captivity for several weeks, exhibited very different personalities. One was shy and sullen. It cowered in the back of its cage and attempted to bite when handled. It would eat only small quantities of finely scraped liver tendered to it on a spoon. If a bit of liver adhered to its face it would shake violently and refuse to eat more. When it had eaten its fill, it would seize the spoon in its teeth and spill the remaining food. The second specimen was tame and greedy. It ate insects, liver, earthworms, and even pieces of flesh. After two days in captivity it learned to come to the door of its cage at a person's approach and open its mouth, in anticipation of food. If it dropped bits of liver on which it was chewing it would descend from the top of its cage to retrieve them, squeaking indignantly while doing so. It also descended to the floor of the cage to lap water from a dish. It habitually hung head downward from the roof of its cage, but reversed its position when evacuating urine or feces. Engler (1943: 96) discovered that big brown-bats will kill and eat smaller bats, at least in captivity.
Vespertilio cinereus (misspelled linereus) Beauvois, Catal. Raisonné Mus. Peale, Philadelphia, 1796:18 (p. 15 of English edition by Peale and Beauvois).
Lasiurus cinereus H. Allen, Smithsonian Misc. Coll., 7 (no. 165): 21, 1864.
Atalapha cinerea True, Proc. U. S. Nat. Mus., 7:602, 1885.
Type.—None designated. Type locality Pennsylvania, probably near Philadelphia.
Measurements.—Ten specimens from California average: Total length 130.5; length of tail 54.8; hind foot 10.7; ear 16; height of tragus 9.5; weight (of 3) 20.4 grams.
Distribution.—Recorded from Seattle (W.S.M.) to the north, Westport (W.S.M.) to the west and Pullman (W.S.M.) to the east.
Fig. 46. Record stations for the hoary bat, Lasiurus cinerea, in Washington.
Remarks.—The hoary bat is the largest and most distinctively marked kind of bat in the state. Adults are usually more than 5 inches in total length. The fur is exceptionally long and soft. The [Pg 169] wing-membranes are thick and leathery. The posterior half of the wing-membrane is black; the anterior half is pale. The interfemoral membrane is furred. Dorsally the color of the fur is mottled white and seal-brown, giving a silvery-gray effect. The ears are short and thick; the feet short and wide. The dental formula is: i. 1-1/3-3, c. 1-1/1-1, p. 2-2/2-2, m. 3-3/3-3 = 32.
The genus Lasiurus is found in North America, South America, and the Hawaiian Islands. Lasiurus cinereus ranges from British Columbia to southern South America. Osgood (1943: 53) records two subspecies from Chile.
Information on the natural history of the hoary bat is meager. It was observed in the Kettle River Mountains, but none was obtained. It flew erratically and rapidly and did not appear until darkness had set in. At least two were seen above a road through a forest of western larch and ponderosa pine. Probably it breeds in the mountains of northeastern Washington and in the northern Cascades.
The hoary bat is migratory, leaving the state in August and September, rarely lingering until early October. It winters along the coast of central and southern California (Dalquest, 1943: 23).
Antrozous pallidus cantwelli Bailey, N. Amer. Fauna, 55:391, August 29, 1936.
Type.—Obtained at Rogersburg, Asotin County, Washington, by G. G. Cantwell on May 28, 1918; type in United States National Museum.
Measurements.—A male from the south bank of the Columbia River, opposite Fallbridge, Klickitat County, measures: total length 113; length of tail 40; hind foot 14; ear 36; height of tragus 13; weight 19.8 grams.
Distribution.—Recorded only from localities near the Columbia and Snake rivers in eastern Washington; from Wenatchee (W.W.D.) south and east to Bly and Rogersburg (Taylor and Shaw, 1929: 9).
Description.—Antrozous pallidus is more likely to be confused with Corynorhinus than with any other kind of bat and can be distinguished from Corynorhinus by its larger size (4-1/2 to 5 inches), lighter color, thick and leathery membranes, and shorter, wider ears. Antrozous lacks the prominent "lumps" on the rostrum, characteristic of the long-eared bat. The dental formula is: i. 1-1/2-2, c. 1-1/1-1, p. 1-1/2-2, m. 3-3/3-3 = 28.
The genus Antrozous is confined to western United States and northwestern Mexico. Two species are recognized by Miller (1924: 84). One of these (pallidus) includes three geographic races. A single race is known to occur in Washington.
Fig. 47. Distribution of the pallid bat, Antrozous pallidus cantwelli, in Washington.
The pallid bat chooses a crevice in a cliff, cave, or building as its daytime hiding place. Like Corynorhinus, it is colonial. The basalt cliffs of eastern Washington offer such ideal hiding places that pallid bats are seldom seen and consequently little is known of their habits. In the southwestern United States, pallid bats are more abundant and better known. H. W. Grinnell (1918: 355) notes that the floor of a roost of pallid bats was strewn each morning with the heads, wings, and legs of insects. Most numerous were the remains of the Jerusalem cricket (Stenopelmatus), a flightless insect that the bats must have secured on the ground. Engler (1943: 96) found that in captivity, pallid bats would kill and eat smaller bats and lizards confined with them.
Bailey (1936: 392) believes that the young of this bat are born in late June and early July. One or two young constitute a litter.
Description.—The black bear is the largest carnivore found in Washington if the grizzly is extinct there. The exact size of the adult male black bear is somewhat in question. Few actual weights are on record of Washington bears. Grinnell, Dixon and Linsdale (1937: 101) paid special attention to the maximum weight of black bears in California and concluded that few ever exceeded 500 pounds. The total length of the largest black bear among specimens from the entire Pacific Coastal area, especially from California and Alaska, they give as 5 feet, 10 inches (op. cit., p. 102). It may safely be assumed that few individuals ever reach a length of six feet. Females are smaller; recorded weights of adults rarely exceed 350 pounds. The largest males have hind feet from 10 to 10-1/2 inches in length.
Fig. 48. Female black bear (Ursus americanus altifrontalis), and two cubs, near Big Four Inn, Mt. Baker National Forest, Washington, July, 1939. (Forest Service photo by R. L. Fromme.)
The black bear, including its closely related species, the Mexican bear, Ursus machetes, ranges over Alaska, Canada, the United States and northern Mexico. The distinction between the black and grizzly bears has been listed under the account of Ursus chelan.
The fur of black bears in the Cascade Mountains in the fall is [Pg 172] long, sleek and glossy but rather stiff. In the spring and early summer the fur of animals at Mount Rainier often appeared coarse, wooly and patchy or rubbed on older animals. Some smaller bears possessed sleek, well-groomed pelts. Bears from the lowland areas are said to possess poor pelts because of constant abrasion in the forests. In the fall, when the salmon are spawning, the bears are said by trappers to roll in decayed fish until they "smell so bad you can't come near them" and their fur is matted and "crawling with lice."
It is now known that the cinnamon bear, sometimes called brown bear, is merely a color phase of the black bear. The brown phase varies in color from a rich, dark reddish brown to a pale cinnamon brown. Cowan (1938: 204) has recorded the geographic variation in the brown and black coats of the bears of British Columbia and, in part, of Washington. On the Olympic Peninsula the brown phase is rare. Old residents have told me of seeing but a few brown bears in their life. I have seen only black bears in the lowlands of western Washington. Cowan (loc. cit.) lists 1,197 black and 79 brown bears from Fort Nisqually, Pierce County, between 1834 and 1852. In the Cascade Mountains the brown phase is not uncommon, perhaps one out of five bears seen being brown. In northeastern Washington the brown and black phases are about equal in number, and some trappers state that the browns are more numerous. Cowan lists 3,813 black and 2,871 brown bears from Fort Colville between 1826 and 1856.
The black bear occurs in a variety of habitats in Washington. It seems to be absent only from the treeless areas of eastern Washington, and is most abundant in the Cascade Mountains and Olympic Mountains where food is abundant and men are few. It is not uncommon throughout the timbered lowlands of northeastern Washington and western Washington, however, and shows a surprising ability to exist unnoticed near the larger cities. In the dense, junglelike forests of southwestern Washington it is numerous. Jackson (1944: 1) estimates that 13,679 black bears live in Washington, more than in any other state.
The habits of the black bear have changed greatly where it has come into contact with man. At Mount Rainier National Park black bears now ignore people, save to beg for food. In the Cascades the bears are wild but so seldom see men that in many places they live much as they did hundreds of years ago. In the lowlands of western Washington they are in daily contact with evidence of [Pg 173] man or his guns. As a result they are silent and shy, rarely being seen.
Fig. 49. Black bear (Ursus americanus altifrontalis), in "hibernation," Mt. Baker region, Washington, about 1936. (John E. Candle photo, courtesy "Field and Stream.")
The black bear in the mountains is active sometimes by day and sometimes by night. Probably it is principally nocturnal but active by day only when the food obtained at night is insufficient to meet the needs of the animal. In the lowlands it is almost completely nocturnal. In the mountains it enters upon its winter sleep with the first snows or, if the snows are late, when the huckleberries are gone. Ordinarily black bears are in "hibernation" by the middle of November. In the lowlands of western Washington they are active until after the salmon spawning season, and probably do not "hibernate" before the middle of December and may not do so at all. In the mountains they emerge from the winter sleep in May; in the lowlands they emerge in February or March.
The black bear is omnivorous in the truest sense of the word. Because the animal is of large size it requires much nourishment. This is obtained by eating large quantities of material with low food value. Material eaten passes rapidly through the digestive tract and often only easily digested parts seem to be utilized. Feces commonly contain complete and undigested berries and seeds or almost undigested pieces of apples or other fruit.
The principal food of the black bear in Washington is berries. Many species are eaten, but the huckleberry (Vaccinium sp.) is favored. Other food items are: Gaultheria shallon (pulpy fruit); Oregon grape, Berberis nervosa (flowers and fruit); salmonberry, Rubus parviflorus (leaves and fruit); leaves of several plants, including Rubus macropetalus and other thorny kinds; grass, succulent plants, and roots. Insects are eaten and most feces show remains of a few. In June, 1938, near Cle Elum, Kittitas County, a species of sword-tailed cricket fully two inches in length was abundant, and droppings of a bear there were composed entirely of the remains of these crickets. Fish, especially spawning salmon, are important food. When salmon are spawning the bears in the vicinity feed on nothing else. Warm-blooded vertebrates probably are eaten but none of the many bear droppings examined by me contained remains of birds or mammals. Bennett, English and Watts (1943: 30) found few mammals to be eaten by bears studied by them in Pennsylvania. Locally bears kill pigs and sheep, smash bee-hives and raid fruit trees.
The large size and sometimes awkward appearance of black bears combine to give a comic note to some of their natural actions. A bear in full flight, lumbering along a trail with head swinging from side to side and hind feet stretching past the forelegs at each stride, is more apt than not to amuse the observer. Near Stevens Pass, a large black bear was observed lying on its back in a mud wallow. This was an oval opening in the ground in a meadow of heather and huckleberry near a small stream. The wallow was approximately 5 feet long, 3 feet deep and 4 feet wide. The bear was immersed in muddy water with only its head and feet visible. It seemed well content and comfortable, shifting its bulk occasionally and waving its paws. When a stray breeze brought to the bear the scent of my companion and myself the animal's contentment vanished and it hastily made a frantic effort to depart. It had wedged itself deeply in the hole, and as it twisted and turned in attempting to sit up, waves of water gushed from the wallow. Eventually escaping [Pg 175] from its wallow, the bear half ran and half rolled to the cover of alders 100 feet away. This particular wallow had been in use by bears for some time, as was attested by old droppings at its edge and by its well-worn condition.
Fig. 50. Distribution of the black bear in Washington. A. Ursus americanus altifrontalis. B. Ursus americanus cinnamomum.
The fact that Washington has the largest population of black bears of the states in the union is due, largely, I feel, to abundant natural cover, food, such as salmon and huckleberries, and an intelligent game code. In California and some other states the black bear has been considered a fur bearer, to be trapped with steel traps. This is permitted in spite of the fact that their pelts bring but a few dollars, usually less than ten. Their sale scarcely repays the trapper for his labor in preparing the hide. Townsend (1887: 182) remarked on the ease with which bears could be trapped in California although in the same area they were so shy that they could seldom be shot. As a result of trapping, the number of bears in some states has been dangerously reduced. In Washington the bear is a game animal, to be hunted with a rifle for a few weeks in the fall when the pelt and flesh are at their best. As a result bears are numerous and can be hunted with fair chances of success.
Ursus altifrontalis Elliot, Field Columb. Mus. Publ. 80, zoöl. ser. 3:234, June, 1903.
Euarctos altifrontalis Miller, U. S. Nat. Mus. Bull., 128:90, April 29, 1924.
Ursus americanus americanus Taylor and Shaw, Mammals and Birds of Mount Rainier National Park, U. S. Nat. Park Service, Washington, p. 37, 1927.
Euarctos americanus altifrontalis Bailey, N. Amer. Fauna, 55:321, August 29, 1936.
Ursus americanus altifrontalis Hall, Univ. California Publ. Zoöl., 30:232, March 2, 1928.
Type.—Obtained at Lake Crescent, Olympic Peninsula, Clallam County, Washington, by D. G. Elliot in 1898; type in Field Museum of Natural History.
Racial characters.—Color dark, almost always black instead of brown; skull wide, high and heavy; molar teeth wide and heavy.
Distribution.—From the eastern base of the Cascade Mountains westward to the Pacific. Marginal occurrences are Chelan (W.W.D.) and Signal Peak (Taylor and Shaw, 1929: 10).
Ursus americanus var. cinnamomum Audubon and Bachman, Quadrupeds of North America, 3:125, 1854.
Euarctos cinnamomum Miller, U. S. Nat. Mus. Bull., 128:91, April 29, 1924.
Euarctos americanus cinnamomum Bailey, N. Amer. Fauna, 55:319, August 29, 1936.
Ursus americanus cinnamomum Hall, Univ. California Publ. Zoöl., 30:232, March 2, 1928.
Type.—Obtained near the mouth of Jim Ford Creek, Lower Clearwater River, western Idaho (Bailey, 1936: 319) by Lewis and Clark on May 31, 1806.
Racial characters.—Resembling altifrontalis but skull and molar teeth narrower; color even in black phase paler and browner; brown and black phases of approximately equal incidence.
Distribution.—Northeastern Washington and the Blue Mountains of southeastern Washington, occurring westward as far as Republic (Taylor and Shaw, 1929: 10).
Ursus chelan Merriam, Proc. Biol. Soc. Washington, 29:136, September 6, 1916.
Type.—Obtained in Township 30 N. Range 16 E. Willamette Meridian, Wenatchee National Forest, Chelan County, Washington, by D. S. Rice on September 1, 1913; type in United States National Museum.
Measurements.—The skull of the type, an old male, measures: basal length 314; occipito-nasal length 323; palatal length 170; zygomatic breadth 225; interorbital breadth 86.
Distribution.—Probably once from the Stevens Pass area north to British Columbia. Possibly still remaining in the mountains between Mount Baker and Lake Chelan.
Remarks.—Grizzly bears occurred in the lowlands of Oregon (Bailey, 1936: 324) and California (Grinnell, Dixon and Linsdale, 1937: 67) but there is no record of their ever having occurred in the lowlands of western Washington.
Bears of the genus Ursus range over Europe, Asia and North America. No less than 75 kinds of grizzly and big brown bears are [Pg 177] recognized for North America by Merriam (1918). They are closely related to the brown bears of Asia. The 13 kinds of black bears of North America are now thought to belong to but one species, namely americanus, and have no close relatives in Eurasia.
Fig. 51. Probable past distribution of grizzly bears in Washington. A. Ursus chelan. B. Ursus canadensis. C. Ursus idahoensis.
The grizzly is the largest carnivorous mammal in North America. From the black bear the grizzly may be distinguished in life by the prominent muscular hump on the shoulders. Another feature is the long, slim, gently curved claws of the forefoot as compared with the shorter, stouter and sharply curved or hooked foreclaws of the black bear. The grizzly is much larger than the black bear. Cranially, the grizzly differs in possessing a larger skull with lower frontal area, higher, more extensive sagittal crest, more posteriorly jutting occipital condyles and much longer tooth row. The color of both species is variable. Some grizzly bears from Alaska and British Columbia are dark brown, almost as black as a black bear. Others are pale cinnamon with the longer guard hairs pale gray. Individuals of this frosted appearance popularly are known as "silver tips." In Washington, the black bear varies in color from glossy black, through various shades of brown to a pale cinnamon.
The grizzly bear is extinct over most of Washington. A few may remain in remote parts of the northern Cascades, and are regularly listed on the game census reports of the Mount Baker National Forest. Nevertheless, I have found no one who has seen an undoubted grizzly bear in Washington. The type specimen was obtained well up in the Cascade Mountains where the animals, like those in British Columbia, feed on roots, berries, marmots, pikas and other vegetable and animal life.
Description.—The raccoon is a relatively large carnivore with a stout, heavy body. Large adults measure three feet in length and weigh up to 20 pounds. The legs are of moderate length. The broad head tapers sharply to a pointed muzzle. The ears are erect, broad, low and rounded; the tail is long, about one-half the length of the head and body, bushy and round; the hind feet are large, flat and naked-soled but the forefeet are smaller with long, slim, handlike toes. The color of the body is a grizzled gray heavily washed with black on the back and sides. A jet-black "mask" across the eyes, sharply outlined by white muzzle and forehead, is the most distinctive feature of the head. The tail is alternately marked with six or seven black and five or six yellowish gray bands. The long, loose guard hairs give the body a shaggy appearance. The fur of the wrists is short, smooth, coarse, and directed downward.
The raccoon is not an aquatic mammal, yet it is closely associated with water, whether it be streams, rivers, lakes or the ocean. The favored habitat of the raccoon is the shore line. It is a wader and, if it can be considered as specialized for any occupation, it is wading in mud and shallow water. Its long toes, naked feet, and short-haired wrists are admirable adapted for walking on muddy bottoms or in shallow water. The coon is also at home on land. It moves swiftly and silently and when chased by dogs can cover miles in a few hours. It is an agile climber and lives in dens well up in trees.
The raccoon is almost completely nocturnal. Individuals are occasionally seen in the morning or evening, especially, when the tide is low along the ocean beach or Puget Sound. Near Fall City, King County, a small raccoon was seen eating a crayfish at 3:30 p.m. on a warm, sunny June day. In the eastern United States raccoons sleep during much of the winter, and probably they do the same in eastern Washington. In western Washington they are active most of the winter. Some trappers stated that the animals "hole up" in spells of unusually cold weather. Along the Tolt River, 10 miles southeast of Duvall, King County, their tracks were seen daily in January, 1936, although the temperature fell well below freezing each night.
Washington is near the northern limit of the range of the raccoon. The animal is reasonably common in western Washington, and [Pg 180] ranges well up into the Cascade Mountains. The highest altitudinal record available is Longmire, Mount Rainier National Park (Taylor and Shaw, 1927). Raccoons are not uncommon in southeastern Washington and in the Yakima Valley. They follow the Columbia River northward, as shown by tracks at Wenatchee on two occasions. The river valleys that flow into the Columbia in northeastern Washington seem admirably adapted for raccoons but the animal is rare there. Trappers who have lived and trapped in northeastern Washington for many years tell of seeing tracks at intervals of years along the Okanogan, San Poil, Colville and Kettle rivers. Several stated that tracks had been seen more often in recent years. Northeastern Washington seems to be the peripheral range of the species, occupied at rare intervals by animals wandering north from the Columbia River.
The food habits of raccoons are almost as varied as those of black bears. Animal matter forms their diet over most of the year; along stream courses crayfish, fish, thin-shelled fresh-water mussels, frogs (Hyla and Rana), and aquatic insects are eaten. Along beaches fish brought in by the tide, crustaceans, and mussels (Mytilus edulis) are taken; small sharks appear not to be eaten. A recently dead dogfish that lay on the beach at Whidby Island, Island County, was ringed by the tracks of a raccoon but had not been eaten. The shore crabs (Hemigrapsus nudus and Hemigrapsus oregonensis) are a favored food and regular items of diet. The edible crab (Cancer productus) is also eaten and in the San Juan Islands the porcelain crab (Petrolisthes eriomerus) was commonly eaten. Small mammals and birds are eaten regularly by this race of raccoon in California (Grinnell, Dixon and Linsdale, 1937: 157), and the eggs and young of wild birds are sought in the spring (op. cit.: 158). The ability of the raccoon as a climber makes it a particular menace to nesting birds. Berries, including domestic blackberries and salmonberries, are eaten in quantity when available. Apples are dearly loved and the trees and orchards of abandoned ranches are regularly visited. The fondness of the raccoon for green corn is well known. Insects are present in small quantities in most droppings and in the late summer some feces were composed entirely of the remains of grasshoppers. Chickens, ducks, young turkeys and eggs are stolen and individuals become extremely adept at raiding hen-houses.
At one time, about 1920, the raccoon became scarce in western Washington as a result of heavy trapping and high price of the pelts. A closed season was strictly enforced until it again became common. [Pg 181] The price received by the trapper for raw furs of raccoon has since been rather low, from two to ten dollars. With fur prices in this range, the number of raccoons probably will not be reduced to a dangerously low level by trapping, but instead may be expected to furnish a regular winter income to the trappers who do trap for it.
Raccoons are known to breed in their first year of life (Pope, 1944: 91).
Procyon psora Gray, Ann. and Mag. Nat. Hist., 10: 261. December, 1842.
Procyon psora pacifica Merriam, N. Amer. Fauna, 16: 107, October 28, 1899 (type from Lake Keechelus, Kittitas County, Washington).
Procyon proteus Brass, Aus dem Reiche der Pelze, p. 564, 1911.
Procyon lotor psora Grinnell, Dixon, and Linsdale, Fur-bearing Mamm. California, Univ. California Press, Berkeley, p. 137, July 22, 1937.
Type locality.—Sacramento, Sacramento County, California.
Racial characters.—Medium size; dark color; smoothly rounded skull.
Measurements.—A female from Forks, Clallam County, measures: total length 905; length of tail 355; hind foot 125; ear 50; weight 14-1/2 lbs. A female and 6 males, young animals of the year, taken between November 15 and December 15, average: 772; 284; 114; weight (of 3) 8-2/3 lbs.
Fig. 52. Distribution of the raccoon in Washington. A. Procyon lotor psora. B. Procyon lotor excelsus.
Distribution.—From the western slope of the Cascade Mountains westward. Marginal occurrences are Lake Keechelus (type locality) and Mount Rainier (Taylor and Shaw, 1927: 45).
Remarks.—The range of variation in color and cranial characters of coastal raccoons is large. I am unable to find any character or average difference to separate the raccoons of western Washington from those of northern California.
Type.—Obtained on upper Owyhee River, near the mouth of the North Fork in southeastern Oregon by J. W. Fisk on April 15, 1920; type in United States National Museum.
Racial characters.—Similar to psora but larger; body paler and grayer; skull larger, heavier and more angular.
Distribution.—Southeastern Washington, the valleys of the Snake and Yakima rivers and the valley of the Columbia River north to northeastern Washington. Marginal records, from reports of trappers, include the Okanogan River, Okanogan County, Wenatchee, Chelan County, and Selah, Yakima County.
Remarks.—The assignment of the raccoons of northeastern Washington to excelsus is tentative for I have seen no specimens.
Description.—The marten is slightly smaller and slimmer than a house cat, and at first glance resembles a large squirrel. The legs are longer, the body stouter and the fur more fluffy than those of a mink or weasel. Adult males weigh from two to two and a half pounds, and females from one and a half to two pounds. Males are slightly more than two feet in total length and females about 18 inches, the tail comprising one-third or more of the total length. The head is broad and narrows rapidly to a sharp muzzle. The ears are large, erect and prominent. The feet are large with stout toes and long, sharply curved pinkish-white claws. The body and head are rich golden brown, the tail, wrists, feet and muzzle being darker.
The western marten ranges from British Columbia southward through Idaho and Washington to California. A closely related species, americana, is found in Alaska, the eastern United States and Canada. Martens and fishers may be distinguished from weasels and minks by the presence of 18 rather than 16 teeth in the upper jaw and 20 instead of 18 teeth in the lower jaw.
The western marten is arboreal. Its principal habitat in Washington is the Canadian Life-zone forests of the Olympic, Cascade and Blue mountains and the various ranges in the northeastern part of the state. At one time it ranged near sea level along the densely forested coastal belt and may still do so in the more rugged parts of the Willapa Hills.
The marten is both diurnal and nocturnal. In Mount Rainier National Park the species has become quite tame and may be seen in the daytime. Many of the small mammals upon which it feeds are diurnal, but others are nocturnal. The marten is active throughout the year. Trappers report that during a storm the marten "holes up in rock slides" where it lives on conies and chipmunks until the storm passes. It spends a large part of its time in trees, and travels through them for long distances. It climbs more skillfully than the tree squirrels upon which it feeds. On the ground or on snow the marten travels in bounds, a yard at a leap, and its characteristic bounding gait forms tracks that are distinctive and easily followed.
Mammals of the weasel family mostly are not gregarious but the marten is exceptional in that in the winter it travels in bands of 6 [Pg 184] to 10 animals. Individuals composing these bands are inclined to wander but nevertheless the whole band travels in a definite general direction at a good rate of speed. Travel-ways or "runs" may be used by more than one band, and a run may extend for many miles, perhaps for as many as 50. A band of martens may take two weeks to complete the circuit, but usually returns to the starting point in less than a week. Most runs are about "half-way up the mountain," or midway between the crest of the hill or timber-line and the floor of the stream valley below. In summer the marten ranges higher; it lives in the trees just below timber-line and in the talus slides near timber-line. When the snows are unusually late the martens may keep to these higher areas until November.
The food of the marten consists principally of small mammals and probably birds; the staple food in winter is the Douglas squirrel. In summer they feed on pikas, mantled ground squirrels and chipmunks. Mice, also, are eaten. The deer mouse, Peromyscus maniculatus, is usually abundant about old cabins and is successfully used as bait by trappers. Wood rats and flying squirrels are also eaten, the latter being especially important in certain areas. The tracks of martens that had been following snowshoe rabbits were seen on several occasions but the martens had turned off before a kill was made. In every instance the rabbit tracks indicated that the animals were hopping leisurely and browsing; apparently the tracks were made some time before the marten began to follow them.
Martens are inquisitive, and to judge from their tracks in snow they investigate almost every object that they pass; a fallen mound of snow, branch, bit of moss, log or isolated tree is apt to be visited. Failing to find a meal in or around one of these objects, the marten visits the next object that catches its eye. Seemingly the animal always is giving concentrated attention to some definite object although the attention can shift in an instant when a more interesting object comes in to view. As a result the trail of a marten in the snow is an intricate affair composed of numerous straight lines and sharp turns.
The trapping of martens is specialized work, engaged in by professional trappers that follow trap lines many miles in length. Trappers commonly have a base cabin and one or two shelters situated a day's march apart, The trap-line is set in a circle, requiring one to three days' travel by the trapper to complete the circuit.
The standard "set" for marten in the Cascades consists of an [Pg 185] opening 8 to 10 inches wide, 4 inches high and 6 inches deep chopped into the side of a dead stub. The hole is cut as high as the trapper can reach. A trap is set in the opening and bait is placed in the back of the hole. A sapling 1 to 3 inches in diameter inclined at an angle of 45° from the ground to the hole completes the set. As the snow deepens, new holes are chopped higher up on the stub. Such sets seen in the summer may consist of six or seven holes spaced a foot apart. The chips from the holes are left lying on the snow and are said to attract the animals. Bait commonly consists of flying squirrel, red squirrel or deer mouse.
Fig. 53. Distribution of the western marten in Washington. A. Martes caurina caurina. B. Martes caurina origenes.
The value of the marten's pelt fluctuates from year to year. In recent years average pelts have sold at from ten to twenty dollars each. The number of individuals taken by a trapper varies with the trapper's skill and energy and the location of the trap line. The largest catch made by one trapper in a single winter, of which I know, was 300 animals, taken near Mt. Adams.
Mustela caurina Merriam, N. Amer. Fauna, 4:27, October 8, 1890.
Martes caurina caurina Miller, U. S. Nat. Mus. Bull., 79:93, December 31, 1912.
Type.—Obtained near Grays Harbor, Grays Harbor County, Washington, by L. C. Toey on February 4, 1886; type in United States National Museum.
Racial characters.—Color dark; throat patch bright orange or brownish.
Distribution.—From the Cascade Mountains westward. Marginal localities are (from Taylor and Shaw, 1929: 10) head of Cascade River, Riverside, Chelan, Easton and Trout Lake. The record from Chelan seems doubtful, and the specimen may have come from somewhere around Lake Chelan, rather than from the town of that name.
Remarks.—Skulls of martens from many localities in western North America were studied in an effort to determine the relationship of the eastern pine marten (Martes americana) and the western marten. East of the Rocky Mountains the ranges of the two species approach closely, but each retains distinctive characters, notably the shape of the auditory bullae. The two species have not been taken together and the possibility of intergradation exists. On the basis of the evidence at hand, the two should be regarded as full species until positive proof of intergradation is established.
In the absence of sufficient material, the marten of the Cascades is referred to Martes caurina caurina. Fur graders distinguish between a dark "coast marten" and a paler "Cascade marten."
Mustela caurina origenes Rhoads, Proc. Acad. at Sci. Philadelphia, 1902:458, September 30, 1902.
Martes caurina origenes Miller, U. S. Nat. Mus. Bull., 79:93, December 31, 1912.
Type.—Obtained at Marvine Lodge, Garfield County, Colorado, by E. T. Seton on September 16, 1901; type in Academy of Natural Sciences of Philadelphia.
Racial characters.—Paler than caurina with grayer head and yellow or white rather than deep orange or brown throat patch.
Distribution.—Mountainous areas of northeastern Washington and the Blue Mountains of southeastern Washington. Trappers have reported this marten from the mountains near Republic, Ferry County.
[Mustela] pennanti Erxleben, Syst. Regni. Anim., 1:470, 1777.
Martes pennanti pennanti Miller, U. S. Nat. Mus. Bull., 79:94, December 31, 1912.
Mustela canadensis pacifica Rhoads, Trans. Amer. Philos. Soc., n.s., 19:435, September, 1898 (type from Lake Keechelus, Kittitas County, Washington).
Martes pennanti pacifica Miller, U. S. Nat. Mus. Bull., 79:94, December 31, 1912.
Martes pennanti Grinnell, Dixon and Linsdale, Fur-bearing Mamm. California, Univ. California Press, Berkeley, p. 211, July 22, 1937.
Type locality.—Eastern Canada.
Distribution.—Originally forested areas from the eastern base of the Cascades westward and possibly the Blue Mountains of southeastern Washington and the mountains of northeastern Washington; now probably confined to the Cascade and Olympic mountains.
Fig. 54. Fisher (Martes pennanti). 2-year-old male in captivity; New Westminster, B. C., March 7, 1939. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 598.)
Remarks.—The fisher is the size of a large cat. In general proportions it resembles the marten. Adult males measure about 3-1/2 feet in length; the tail comprises 16 inches of this. Adult females are slightly less than 3 feet in length of which the tail makes up approximately 15 inches. Males weigh up to 10 lbs. and females about 5-1/2 pounds (Grinnell, Dixon and Linsdale, 1937: 213). The fisher has a slim body, bushy tail, short legs, large feet, and wide, low and triangular head. The ears are low, wide, rounded and erect. The fur is an ashy, brownish gray in color with an overwash of [Pg 188] blackish caused by long, dark hairs. The head is slightly paler than the body. The feet, rump and tail are darkest. The claws are strong and sharply curved.
The fisher is found in wooded parts of North America, extending southward in the Cascade-Sierra Nevada Chain to central California. Its near relatives are the martens. It is active all year. Like the marten, it is active by day and probably also by night. In spite of absolute protection for several years, the fisher is rare in Washington, and seems never to have been common. In consequence relatively little is known of its habits, and the little that is known has been reported by fur-trappers.
Fig. 55. Distribution of the fisher, Martes pennanti, in Washington.
The fisher seems not to live in bands as does the marten. Most of the actual records of fishers trapped are at higher altitudes but are misleading because most trappers agree that the fisher occupied a lower zone, altitudinally, than does the marten. There are old records of its occurrence near sea level (Scheffer, 1938: 9). The animals are usually taken in marten sets or in traps set especially for fishers by trappers who find their tracks on their marten trap lines. [Pg 189] Since marten trappers are almost the only persons who travel in the mountains in winter, and since they operate mostly above the areas where fishers live, relatively few fishers are reported.
Fishers are said to feed on chipmunks, squirrels, mice, birds and other small, warm-blooded animals, and to climb trees and catch squirrels in their natural habitat. Also, fishers are said to catch and kill martens. Their tracks in the snow resemble the marten's in that the hind feet land in the same places as the forefeet; both animals bound rather than walk.
The pelt of the fisher commands a high price. The smaller sized, females, are the more valuable. The price paid for pelts fluctuates widely and has ranged from as low as twelve dollars to as high as one hundred dollars in recent years.
In Washington more fishers live in the Olympic Peninsula and the northern Cascade Mountains than elsewhere. A few may occur in northeastern Washington, the Blue Mountains and the Willapa Hills.
The name pacifica was regarded by Grinnell, Dixon, and Linsdale (1937: 217) as a synonym of pennanti.
Description.—Though it is similar in general characters and proportions to the long-tailed weasel, the ermine is much smaller and has a relatively shorter tail. It is darker and less reddish or yellowish in summer. Adult males measure about 10 inches in length, of which 3-1/2 inches is the length of the tail. Females measure about 8 inches and have tails 2 inches long. The upper parts are chocolate brown; the underparts are white or pale yellow. Along the coast of Washington, the pale color of the underparts is more restricted in the ermine than in the long-tailed weasel. The dark brown tail has a black tip. Ermines east of the summit of the Cascades become pure white in winter, save for the black tail tip. West of the summit of the Cascades the winter pelt is similar to the summer pelt but is slightly paler with denser underfur.
Ermines in America range from the Arctic southward, in mountainous areas, to the southern end of the Sierra Nevada in California, and in the Rocky Mountains to northern New Mexico. In Washington they are found over the entire state except the arid parts of eastern Washington, where only the long-tailed weasel occurs. So far as my observations go, the ermine is principally nocturnal in Washington; I have seen only one abroad in the daytime. It dashed from a roadside thicket near Glacier, Whatcom County, and was crushed beneath the wheels of a car. Taylor and Shaw (1927: 53) note several instances of diurnal activity of the ermines at Mount Rainier National Park.
The ermine seems to feed principally upon mice. Its small size adapts it to entering burrows that larger weasels cannot enter. It probably eats chipmunks, birds, and other small, warm-blooded animals. Ermines climb readily and are often taken in traps set in trees for martens. Near Skykomish, King County, William Hoffman took two ermines in traps set in burrows of mountain beavers. The mountain beavers were needed for use as bait on his trap line. The traps were reset and later caught mountain beavers. Seemingly the ermines were traveling through the burrows, perhaps to catch the mice which utilize the burrows as highways, rather than to prey upon mountain beavers. It is difficult to see how the tiny ermine could kill an adult mountain beaver, which outweighs it many times. Should the incisors of the mountain beaver close even once upon an ermine it would most certainly be seriously wounded or killed. Judging by the forest habitat occupied by the ermine in [Pg 191] Washington, one might expect its principal food to consist of deer mice (Peromyscus maniculatus), red-backed mice (Clethrionomys) and meadow mice (Microtus).
In observing the activities and habits of mammals in their natural habitat, I have often relied on tracking in fresh snow. Strangely enough, tracks of ermines were seldom found, and the few that were seen came from beneath a log, bush or wind-fall and disappeared beneath similar cover, rarely extending 20 feet on the surface of the snow. The larger, long-tailed weasels often traveled for miles on the surface of the snow. Possibly the ermines were following the burrows of mice through the snow, or perhaps they kept beneath the surface from fear of owls. This suggests a reason why ermines are so seldom seen abroad. They may be following burrows and runways of mice and seldom come to the surface of the ground.
On November 18, 1936, we saw three ermines and two long-tailed weasels taken by a trapper on Deception Creek near Stevens Pass, King County. All were in the white winter coat. In December, 1938, we obtained two ermines at Skykomish, King County, 18 miles west of Stevens Pass. These were in the brown winter coat. In that area the break between the brown and white winter coat seems to come just west of the main Cascade Summit, or at the same point that the break occurs between the brown and white winter coats of the long-tailed weasel.
Ermine skins have little value and usually bring from 10 to 35 cents. They are saved incidentally by trappers, for it takes but a few moments to skin and prepare them. They possess the strong, musk odor so typical of the long-tailed weasel.
Mustela erminea invicta Hall, Jour. Mamm., 26:75, February 27, 1945.
Type.—Obtained at Benewah, Benewah County, Idaho, by W. T. Shaw on October 24, 1926; type in Museum of Vertebrate Zoölogy.
Racial characters.—Largest of the Washington ermines; winter coat entirely white; upper lips white; black of tail more than half length of tail vertebrae.
Measurements.—Ten males and 5 females from central Idaho average, respectively (Hall, 1945): total length 291, 255; length of tail 86, 71; hind foot 40, 32. 3.
Distribution.—Northeastern Washington and the northeastern Cascades, west to Chilliwack River and Hannegan Pass (Hall, 1945: 78).
Mustela erminea fallenda Hall, Jour. Mamm., 26:79, February 27, 1945.
Type.—Obtained at Huntingdon, British Columbia, by C. H. Young on May 21, 1927; type in National Museum of Canada.
Racial characters.-Size large; winter coat usually brown; pale color of underparts much restricted; color of upper parts dark; color of lips variable; tail with more than distal half black.
Measurements.—Seven males and 2 females average, respectively (Hall, 1945: 79): total length 278, 232; length of tail 77, 60; hind foot 36.5, 27.
Distribution.—The extreme north coast of Washington, from the Canadian boundary south through Whatcom County (Hall, 1945: 80-81).
Remarks.—The range of this ermine is similar to that of the yellow-pine chipmunk, Eutamias amoenus felix.
Fig. 56. Distribution of the ermine in Washington. A. Mustela erminea invicta. B. Mustela erminea murica. C. Mustela erminea gulosa. D. Mustela erminea fallenda. E. Mustela erminea streatori. F. Mustela erminea olympica.
Mustela erminea olympica Hall, Jour. Mamm., 26:81, February 27, 1945.
Type.—Obtained near head of Sol Duc River, Clallam County, Washington, by V. Bailey, on April 28, 1897; type in United States National Museum.
Racial characters.—Similar to streatori but smaller, especially females.
Measurements.—Twelve males and 6 females average, respectively: total length 243, 196; length of tail 65, 52; hind foot 31, 23.4 (Hall, 1945: 81).
Distribution.—The Olympic Peninsula, extending southeastward to Olympia.
Putorius streatori Merriam, N. Amer. Fauna, 11:13, June 30. 1896.
Mustela streatori streatori Miller, U. S. Nat. Mus. Bull., 79:96, December 31, 1912.
Mustela cicognanii streatori Taylor and Shaw, Occ. Pap. Chas. R. Conner Mus., no. 2:11, December, 1929.
Mustela erminea streatori Hall, Jour. Mamm., 26:76, February 23, 1945.
Type.—Obtained at Mount Vernon, Skagit County, Washington, by D. R. Lucky on February 29, 1896; type in United States National Museum.
Racial characters.—Similar to fallenda but pale color of underparts less restricted; tail with less than distal half black.
Measurements.—Two males and 2 females from extreme southwestern Washington average, respectively: total length 245, 210; length of tail 72, 54; hind foot 31.5, 25.5; ear 17, 14; weight 72.3, 46.
Distribution.—The lowlands of western Washington; north to Skagit County and Whidby Island (Taylor and Shaw, 1929: 11), southward and westward to Ilwaco (M.V.Z.) and eastward to Carson (U.S.N.M.).
Mustela erminea gulosa Hall, Journ. Mamm., 26:84, February 27, 1945.
Type.—Obtained at Trout Lake, Klickatat County, Washington, by P. Schmid on February 3, 1897; type in United States National Museum.
Racial characters.—Similar to invicta but smaller; winter coat usually white; tail less than one-half black; upper lips white.
Measurements.—Five males from Mount Rainier, and 4 females from the Cascade Mountains, average, respectively (Hall, 1945: 84): total length 253, 208; length of tail 76, 54; hind foot 30.2, 24.3.
Distribution.—The Cascade Mountains. Known from Skykomish (W.W.D.) and southward to Mount Adams (Trout Lake, Hall, 1945: 85).
Putorius (Arctogale) muricus Bangs, Proc. New England Zoöl. Club, 1:71, July 31, 1899.
Mustela muricus Miller, U. S. Nat. Mus. Bull., 79:96, December 31, 1912.
Mustela cicognanii muricus Bailey, N. Amer. Fauna, 55:292, August 29, 1936.
Mustela erminea murica Hall, Jour. Mamm., 26:77, February 27, 1945.
Type.—Obtained at Echo, El Dorado County, California, by W. W. Price and E. M. Nutting on July 15, 1897; type in Museum of Comparative Zoölogy.
Racial characters.—Similar to invicta but smaller and paler.
Measurements.—A young female from Stay-a-while Spring, Columbia County, measured: total length 201; length of tail 46; hind foot 24; ear 14; weight 45.8. A female from Butte Creek, Columbia County, measured: 185; 50; 26. A male from Baker Creek, White Pine County, Nevada, measured: 220; 56; 26; 14.5; weight 57.7.
Distribution.—The Blue Mountains of southeastern Washington.
Remarks.—Two specimens available from the Blue Mountains are darker than typical murica.
Description.—Male long-tailed weasels measure about 16 inches in total length of which 6 inches is the length of the tail. Females are smaller, measuring about 14 inches in length with a tail 5 inches long. The body is long and exceedingly thin. The legs are short and stout with rather large feet and strong, curved claws. The tail is well-furred, not tapered, but lacks the bushy appearance of the tail of the fisher. The head is low, wide at the base and abruptly tapering to the muzzle. The ears are erect, low and rounded. The fur is short but dense and rather soft. In summer the color of the head, back, sides and outside of the legs is brown. The throat, chest, underside of body and insides of legs are yellow or orange. In winter they may be entirely white, save for the black tip of the tail. The tail is slightly darker than the back in summer and possesses a long black tip.
Long-tailed weasels do not hibernate even in the coldest parts of Washington. They are active both by day and by night, apparently doing their hunting at the time of day or night when they can most easily obtain food. In the Cascade Mountains where diurnal mammals such as squirrels, chipmunks and pikas are common, weasels are not uncommonly seen by day. In the lowlands of western Washington, where they feed on more nocturnal mammals, they are seldom seen.
The long-tailed weasel is relatively unspecialized in habits. They climb readily and skillfully. They are active on the surface of the ground and follow the burrow systems of fossorial animals such as gophers and mountain beavers.
Weasels seen in the wild rarely exhibit fear of man but rather are curious and apt to watch his actions. Weasels are also often hit by cars and the number so killed seems to me to be out of proportion to their actual numbers.
In moving on the surface of the ground the weasel arches its back and contracts the body until the four feet are rather close together. When the long neck and small head are held upright the animal presents a surprisingly giraffelike appearance. When climbing, the long, slim body has a snakelike appearance. A weasel travels swiftly and erratically in a series of bounds and seems always to know where the next hole is situated.
The weasel has been accused of killing birds and doubtless does [Pg 195] so when opportunity presents itself. However, in Washington I have no actual evidence of its killing birds other than domestic fowls. At Republic, Ferry County, a companion and I saw a weasel enter the burrow of a ground squirrel (Citellus columbianus). The following day we returned to the area. The weasel was not seen but a ground squirrel dashed into the hole at our approach. Seemingly the ground squirrel had eluded the weasel. At Conconully, Okanogan County, we set a number of gopher traps in an alfalfa field. The following morning an adult male long-tailed weasel was found in a trap but not a gopher was taken. Near Moses Lake, Grant County, an adult male weasel was caught in a gopher trap, but no gophers were taken. At Shelton, Mason County, 50 gopher traps were set. At daylight the following morning the first trap visited was found to be pulled into the burrow. When a gentle tug was given the wire fastening the trap, a decisive jerk at the other end showed that the catch was alive. If trapped gophers that pull the traps back into their burrows are pulled out by main strength, their skins are often torn and damaged. Therefore an attempt was made to reach into the burrow and pull back the sod. An adult female long-tailed weasel promptly fastened its teeth into my forefinger and clung on, bulldog fashion, to be lifted into the air with the attached trap swinging. When the left hand was used to force the animal to release its grip, it fastened onto the left thumb. With right thumb and forefinger I forced it to release its grip, but was unable to elude its teeth which again fastened to my right forefinger. Only by laying it on the ground and crushing its chest with my foot could I free myself from the vicious little beast. No gophers were taken in traps set less than 150 feet from where the weasel was trapped. In the three instances mentioned above, weasels had seemingly killed all the gophers in their immediate vicinity. As regards the gophers near Moses Lake, none was found a year later in the area where the weasel was taken and only old, abandoned burrows were seen. T. H. Scheffer (1932: 54) records other instances of the capture of weasels in gopher burrows.
At the northern limits of the city of Seattle, steel traps were set for mountain beavers in a rather dense colony of these mammals. Well-used burrows indicated that approximately 10 individuals were present. The following morning an exceptionally large male weasel was found in a trap, but all others were empty. The traps were left out for two additional nights but no mountain beavers were taken. A month later the colony seemed abandoned and no evidence [Pg 196] of recent digging was noted. It could only be concluded that the weasel had killed the animals comprising the colony. A large mountain beaver weighs three or four pounds, which is 6 or 8 times as much as a weasel. Edson (1933: 76) recounts trapping 7 weasels in burrows of mountain beavers near Bellingham, Whatcom County.
Near Forks, Clallam County, a weasel was seen pursuing a young snowshoe rabbit (Lepus americanus washingtonii) along the edge of a concrete highway. As our car approached and passed the animals, they separated, the weasel retreating to the cover of horsetail (Equisetum) beside the road. The car was stopped 50 feet ahead. As we emerged the weasel dashed from cover to intercept the rabbit in the center of the road. The weasel knocked the rabbit to its side and, placing its feet on the rabbit's shoulders, bit fiercely at its neck. It then dashed back to the cover of the horsetails. The rabbit stood up, made two hops and died. It was approximately two weeks old.
Fig. 57. Distribution of the long-tailed weasel in Washington. A. Mustela frenata washingtoni. B. Mustela frenata altifrontalis. C. Mustela frenata nevadensis. D. Mustela frenata effera.
The pelts of weasels bring the trapper from twenty-five cents to a dollar and a half. Only skins in the white winter coat command [Pg 197] the higher price. They are usually taken in traps set for other animals.
In the western part of the state, long-tailed weasels do not turn white in the winter; the back is slightly less reddish than in summer and the underparts are pale yellow or white or may be both yellow and white. From the summit of the Cascades eastward weasels become white. The break in winter color seems to occur slightly west of the main summit of the Cascades. Specimens taken by marten trappers at Tye and Scenic, near Stevens Pass, were white on November 15, 1936, but specimens from Skykomish and Baring, 18 miles to the west, were brown.
The long-tailed weasel possesses a distinctive odor. It results from a glandular secretion and, although it has no great carrying power or lasting quality, it does affect some persons strongly. It is a heavy, rather sickening scent.
The gestation period of Mustela f. nevadensis has been recorded as more than 131 days (Hall, 1938B: 250). The gestation period of the long-tailed weasel of eastern United States has been recorded as more than 70 days. Three to five young are born in a litter, usually four.
Mustela washingtoni Taylor and Shaw, in part, Occ. Pap. Chas. R. Conner Mus., no. 2:11, December, 1929.
Mustela frenata nevadensis Hall, Carnegie Inst. Washington, Publ. 473:91, November 20, 1936.
Type.—Obtained 3 miles east of Baker, White Pine County, Nevada, by E. R. Hall and W. C. Russell on May 30, 1929; type in Museum of Vertebrate Zoölogy.
Racial characters.—Size medium; colors pale; back near Brussels Brown in summer, chin white and underparts yellow, rarely orange; color in winter white with black tip on tail.
Measurements.—A male from Neppel, Grant County, measures: total length 412; length of tail 151; hind foot 43; ear 19. Two males from Yakima, Yakima County, average: 379; 135; 42.5; 21; weight 176.5. A female from Ellensburg and one from 4 miles east of Ellensburg, Kittitas County, average: 284; 98; 33; 17.
Distribution.—From the high Cascades eastward, save for the Blue Mountains area. Marginal occurrences are Barron (Hall, 1936: 93) and Easton (W. W. D.).
Remarks.—Specimens from the northern Cascades are intergrades between nevadensis and the form to the west, altifrontalis. Specimens from extreme northeastern Washington might be referred to oribasa Bangs as readily as to nevadensis.
Mustela frenata effera Hall, Carnegie Inst. Washington, Publ. 473:93, November 20, 1936.
Type.—Obtained at Ironside, Malheur County, Oregon, by H. E. Anthony on September 8, 1912; type in American Museum of Natural History.
Racial characters.—Similar to nevadensis in color but smaller, with smaller, lighter skull. Color in winter white with black tip on tail.
Measurements.—A female from Prescott, Walla Walla County, measures: total length 310; length of tail 105; hind foot 34.
Distribution.—Southeastern Washington, south of the Snake River. Specimens from Walla Walla and Prescott have been examined.
Putorius washingtoni Merriam, N. Amer. Fauna, 11:18, June 30, 1896.
Mustela washingtoni Miller, U. S. Nat. Mus. Bull., 79:98, December 31, 1912.
Mustela frenata washingtoni Hall, Carnegie Inst. Washington, Publ. 473:106, November 20, 1936.
Type.—Obtained at Trout Lake, Skamania County, Washington, by D. N. Kaegi on December 15, 1895; type in United States National Museum.
Racial characters.—Color rich, dark; hind feet free of color of underparts.
Measurements.—A male from Spray Park, Pierce County, measures: total length 423; length of tail 164; hind foot 52.
Distribution.—Higher Cascades from Mt. Rainer (M. V. Z.) south to Mt. Adams (Taylor and Shaw, 1929: 11).
Mustela frenata altifrontalis Hall, Carnegie Inst. Washington, Publ. 473:94, November 20, 1936.
Type.—Obtained at Tillamook, Tillamook County, Oregon, by A. Walker on July 10, 1928; type in Museum of Vertebrate Zoölogy.
Racial characters.—Similar to nevadensis but darker above in summer and richer orange beneath; winter color of upper parts dark brown, underparts pale yellow or white.
Measurements.—Four males and 2 females from western Washington average, respectively: total length 411.5, 267; length of tail 145, 137.5; hind foot 50, 43; ear 26, 21.5; weight 227, 136.7 grams.
Distribution.—From the Cascade Mountains to the Pacific. Marginal occurrences are Rockport (Hall, 1936: 95) and Tye (W. W. D.).
Putorius vison energumenos Bangs, Proc. Boston Soc. Nat. Hist., 27:5, March, 1896.
Mustela vison energumenos Miller, U. S. Nat. Mus. Bull., 79:101, December 31, 1912.
Type.—Obtained at Sumas, British Columbia, by A. C. Brooks on September 23, 1895; type in Museum of Comparative Zoölogy.
Measurements.—A male from Seattle, King County, measures: total length 540; length of tail 180; hind foot 66; weight 1 pound, 15 ounces.
Distribution.—Throughout the state save for the Columbian Plateau; recorded from Neah Bay (Taylor and Shaw, 1929: 11) in the northwest, Ilwaco (M.V.Z.) in the southwest, Metaline (W.W.D.) in the northeast, and the Blue Mountains (Dice, 1919: 12) in the southeast.
Fig. 58. Distribution of the mink, Mustela vison energumenos, in Washington.
Description.—Because of the value and lasting popularity of its fur, the mink is known to all. Few persons, however, recognize the animal in the wild. The mink possesses the long, slim body and short, rather stout legs of a weasel but has a more bushy tail. The mink is larger than the weasel. Large males weigh up to 3 pounds; females 1-1/2 pounds. Males measure about 2 feet in length, of [Pg 200] which the tail comprises eight inches. Females measure about 20 inches and have tails 7 inches long. The color is rich, dark reddish or chocolate brown. The underparts are slightly paler than the back. There are usually small white markings on the chin, chest or other part of the ventral surface. These markings sometimes take the form of narrow white lines.
The mink ranges from the Atlantic to the Pacific and from Alaska south to Florida, New Mexico and central California. It is active throughout the year and is principally, though not exclusively, nocturnal. McMurry (1940: 47) records three mink seen at 3:30 p. m. on August 8, 1939, at Packwood Lake, Lewis County, one of which was carrying a garter snake two feet in length. This mustelid is semiaquatic, living along rivers, streams, lakes and salt water and spends most of its time along the shore, on muddy or sandy beaches. In this respect it resembles the raccoon. Whereas the raccoon spends a part of its time on the land, the mink spends a corresponding amount of its time in the water. It is an excellent swimmer, able to overtake and capture fish in the water.
In the San Juan Islands minks have forsaken the shoreline and roam over the uplands, feeding on the abundant, feral domestic rabbits. I found evidence of their presence far inland, miles from water, in grassy and bushy wastes; along the beaches their tracks were rarely seen. Along Puget Sound, minks spend part of their time on the beaches, feeding on dead fish and other marine animal life. These animals, however, seem to live along the rivers and streams flowing into the sound. Along the ocean coast, some minks seem to live exclusively in the marine shoreline habitat.
The mink ascends some distance into the Cascade and Olympic mountains along the larger watercourses. There are reliable reports of minks from Heart Lake and Lake Dorothy, King County, high in the Cascades. These animals were seen in the summer but they may live in higher parts of the Cascades, at least about some of the larger lakes throughout the year. As far as is known there are no minks on the Columbian Plateau.
The food probably varies with locality. Along the ocean beaches they eat dead sea birds, stranded fish, crustaceans, and mollusks. Along Puget Sound their tracks are commonly seen following the caked, decaying seaweed and debris that collects at the high tide mark. Svihla and Svihla (1931: 22) captured a mink that was feeding on the beach of the Olympic Peninsula. This individual, in captivity, was able to open and feed on clams. Fur trappers report that minks feed on mountain beavers, and that the flesh of the [Pg 201] mountain beaver is the best bait for attracting minks. Along streams and lakes, minks are thought to feed on mice, birds, fish, crayfish and thin-shelled, fresh-water mussels. The muskrat forms an important item of diet near the larger lakes and streams. Muskrats trapped near Seattle were often attacked by minks and either eaten or so slashed and torn that their pelts were worth but a fraction of what they would bring in an undamaged condition.
The tracks of a mink noted in freshly fallen snow about a garbage dump on the shore of Lake Washington, Seattle, indicated that the animal had been hunting house rats, which were numerous there. Minks are not uncommon in the marshes along Lake Washington near the University of Washington campus, Seattle, where I discovered evidence of their preying on ducks and coots. The ducks included mallards and green-winged teal, species most apt to alight in small pools in the rushes where a mink might find cover. I found the fresh tracks of one mink about a half-eaten golden tench 8 inches long. The tench is a fish of the sucker tribe, introduced into the lake. Tracks of minks are not uncommonly seen along rivers and streams under conditions that indicate they were hunting crayfish.
The raw pelt of the mink sells usually at from 10 to 20 dollars. The fur is in constant demand and fluctuates in price less than most furs. The average trapper takes a few minks each year along with his catch of muskrats, raccoons, and skunks but some trappers have taken as many as 100 minks in a winter in areas where the animal is especially numerous.
Gulo luteus Elliot, Field Columb. Mus., Publ. 87, zoöl. ser. 3:260, December, 1903.
Gulo luscus luteus Grinnell, Dixon, and Linsdale, Fur-bearing Mamm. California, Univ. California Press, Berkeley, p. 251, July 22, 1937.
Type.—Obtained on Mt. Whitney, Tulare County, California, by E. Heller; type in Field Museum of Natural History.
Measurements.—The dried skin of a wolverine taken 3 miles south of Riverside, Okanogan County, was 4 feet in length. The animal weighed 40 pounds (Scheffer, 1941: 37).
Distribution.—Timber-line region of the Cascades Mountains. From Robinson Creek (Scheffer, 1938: 8) south to Mount Rainier (Taylor and Shaw, 1929: 12).
Remarks.—Specimens from Washington are intermediate in character between luteus and the race inhabiting British Columbia, although available skulls are closer to luteus.
Fig. 59. Wolverine (Gulo luscus), mounted specimen, male, trapped by Billy Robinson about 1902 on Billy Robinson Creek, Okanogan County, Washington. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 76.)
Description.—Large males measure more than three feet in length, of which the tail makes up 10 inches; they weigh as much as 50 pounds (Grinnell, Dixon and Linsdale, 1937: 253). The body is wide and stocky; the tail is short; the legs, especially the forelegs, are short, thick and powerful; the feet are large and are armed with long, curved claws; the head is wide; the jaws are powerful with heavy teeth. The pelage is long and rather shaggy. Face, legs, [Pg 203] back and terminal portion of the tail are dark, blackish brown but the forehead is grayish, contrasting with the dark color of face and top of the head. Two broad, yellowish-brown bands begin on the side of the shoulders and pass back, low on the sides, to the hips where they join across the back and base of the tail.
The wolverines, nominally of several species, form a compact group that ranges over the boreal regions of the Old and New world. In North America they range southward from the Arctic to California and Colorado.
Fig. 60. Wolverine (Gulo luscus), male, trapped by Reuben R. Lamb, March 22, 1941, three miles south of Riverside, Washington; reported to weigh 40 pounds; dried pelt 3 × 4 feet. (Reuben R. Lamb photo.)
Fig. 61. Distribution of the wolverine, Gulo luscus luteus, in Washington.
The wolverine is adapted to boreal conditions and is most abundant in the arctic. In the Cascades it occurs only at or near timber-line. Except the wolf and the possibly extinct grizzly bear, the wolverine is the rarest carnivore in Washington. Probably it has been rare since the retreat of the continental glaciers, for suitable habitat for wolverines is not abundant and each animal requires a large area over which to range. An unusual record, possibly of a wandering animal, is given by Scheffer (1941: 37). This is of an adult male taken in the Okanogan Valley 3 miles south of Riverside, Okanogan County, on March 22, 1941.
Description.—The otter has the long body of many mustelids, but is specialized for aquatic life. Males are slightly larger than females. Large adults measure more than 3-1/2 feet in length, of which the tail comprises 18 inches. The largest adults may weigh as much as 25 pounds. The body is elongate but rather plump. The forelimbs are small and laterally placed. The hind legs are the heavier, and are situated posteriorly. The feet are large and webbed for swimming. The tail is long, thick at the base and gradually tapering to a narrow tip. The head is small and rounded. The ears are tiny and the eyes are of moderate size. The upper lips are large, somewhat spaniel-like, and support a mustache of stiff vibrissae. The fur is short, dense and soft. The tail is sleek with short, posteriorly-directed fur, and is not bushy. The fur is rich, dark chocolate brown in color, slightly paler on the animal's underparts than on its upper parts.
Otters range over Eurasia, North America and South America. Those occurring in North America north of Mexico are thought to belong to a single species, canadensis. Several species from tropical America may eventually be shown to also be races of canadensis.
The aquatic habits of the otter allow it to exist in several life-zones, in both marine and fresh-water habitats. The principal environment is the streams, rivers and lakes of the Transition Life-zone, but along the larger rivers, otters range up into the Canadian Life-zone and along the Snake and Columbia rivers they enter the Upper Sonoran Life-zone. They are active throughout the year. As we have observed them, they are principally nocturnal.
Otters are extremely powerful swimmers. Tracks along the North Fork of the Tolt River, King County, showed where otters had entered water that flowed over a stony bottom at the velocity of rapids. Their occurrence in the Tolt, Skykomish and Snoqualmie rivers where these are swift mountain streams indicates exceptional swimming ability. After emerging from the water, an otter often follows along the shore, sometimes for miles.
In the San Juan Islands the otters have taken to a marine existence. In the summer of 1938, abundant opportunity offered to observe the otters at Thatcher Bay, Blakely Island. Here, in the evening, bats were shot as they flew over the calm water of the bay. Specimens killed were retrieved by rowing out to them with a dory. For the first two nights bats were collected and retrieved without [Pg 206] incident. On the third night, several of the bats vanished between the time they were killed and the time that the boat was launched from the rocky beach. The presence of a shark or other large fish was at first suspected, but observation showed a group of three or possibly four otters to be the thieves. On succeeding nights the animals became bolder. A dead bat became the object of a race between otters and collector. They completely ignored shouts, dodged stones hurled at them, and stole almost all the bats shot. Only a sense of humor and desire to study the animals saved them from a load of fine shot. Bat collecting was given up in disgust.
These otters were remarkably seallike in many actions. In swimming their heads, shoulders and part of their back were exposed. In resting in the water, only the round head remained above the surface. They were never seen to float prone on the surface or rest on their backs, as does the sea otter. They were noisy swimmers, splashing with paws or heads as they dived. Their eyesight was remarkable. Stones the size of a walnut were hurled at them from distances as short as 25 feet. All were dodged with little effort. In this connection the experience of two trappers who caught an otter in the Samamish River near Woodinville, King County, is interesting. The trap holding the otter had been attached by a wire 6 feet long to a pole on the bank. The wire was detached from the pole; while one trapper held the wire, the other tried to strike the otter with an oar as the boat drifted over deep water. The otter was free to swim and dive for the length of the wire and trap chain. It detected, and evaded by dodging or diving, every blow directed at it and not until the animal tired, a half hour later, could it be killed. The trappers then noted that the splashing of oars and otter had nearly filled the boat with water. The trappers were exhausted.
The otters of Blakely Island were not unique in occupying a marine habitat. At Strawberry Bay, Cypress Island, on July 5, 1938, fishermen brought in a "strange animal" caught by them at Black Rock, a tiny bare and isolated islet 5 miles to the west. The heavy box holding the animal was opened, disclosing a very frightened young otter. Questioned, the fishermen stated that four young and one adult had been seen in the surf. They had thought the animals a species of seal, and were somewhat surprised at being successful in their attempts to catch one. The otter was too young to fare for itself and was kept in camp in the hope that it might be reared and released. It ate a few fresh herring and candlefish and drank some condensed milk. It refused whole salmon and sea bass but [Pg 207] ate some skinned and boned strips of these fish. It died a week after it was captured. This young otter made a ticking sound, almost a chirp. A variety of crying and whimpering sounds were also made, and when petted it grunted in satisfaction. If surprised or when first picked up it gave a deep harsh growl, unexpectedly vicious in sound, for such a small animal.
The otters at Blakely Island fed on a great variety of food. The only food they were actually seen to eat was candlefish, a slim silvery fish 6 inches in length. On several occasions an otter was seen swimming with the head of a candlefish held fast in its mouth and the fish's body extending out in a silvery bow. Many feces of these otters were composed entirely of the feathers of grebes and scoters. These birds were probably not killed by the otters but died a natural death or were shot by men. At that camp scarcely a day passed in which at least one person was not seen firing a .22 caliber rifle from a cabin cruiser into the flocks of scoters at the mouth of the bay. Dead sea birds were common on the shore and doubtless furnished food for otters as they did for crows and eagles. The majority of the otter feces examined at Blakely Island were composed of the remains of invertebrates. The small mussel (Mytilus edulis) was most abundant. Crabs formed an important part of their diet. Strangely enough the shore crab (Hemigrapsus nudus) and the porcelain crab (Petrolisthes eriomerus) were seldom eaten though they were abundant beneath the rocks upon which the feces were found, and formed the principal food item of raccoons. These species of invertebrates live beneath rocks and the raccoon probably reaches beneath the rocks to catch the crabs with its handlike forefeet, as the otter is unable to do. The edible crab (Cancer productus) and kelp crab (Telmessus cheiragonus) were most often eaten by otters. Also eaten were snails (Margarites, Littorina), bitter oysters (Pododesmus macroschisma), unidentified pelecypods, barnacles (Balanus), one chiton (Mopalia muscosa), and once a starfish. The invertebrate remains, save for the snails, were crushed and broken.
Fish of many species were abundant in these waters. Salmon were often seen leaping near the playing otters. Yet no scales or other remains of fish were detected in the feces. Candlefish possess tiny scales that could probably be seen by microscopic examination only. As for the bird remains, the quantity of feathers consumed is notable. The otters apparently do not pluck birds, as do minks. Indeed, of the material eaten, the food value by volume seems extraordinarily low. A great quantity must be eaten to nourish an animal as large and as active as an otter. The number of fecal droppings seen each morning indicates that this is the case.
Fig. 62. River otter (Lutra canadensis pacifica), trapped in Columbia National Forest, Washington, February, 1937. (Adolph Roth photo.)
The favored food of the otter in fresh water streams and lakes is the crayfish. The greater part, perhaps 80 per cent, of otter feces seen in the lowlands of western Washington, were composed of crayfish remains. Some fish are also eaten and locally otters may do damage among the steelhead trout. Even worse, from the fisherman's standpoint, they frighten and scatter the schools of steelhead trout. Frogs are eaten, as well as the thin-shelled mussels. The feathers of coots were twice seen in otter feces near Cottage Lake, King County.
The fur of the otter is dense and beautiful but the skin is rather heavy. Changes in styles cause great fluctuation in the value of the otter's pelt.
Fig. 63. Distribution of the river otter in Washington. Unshaded Lutra canadensis pacifica. Shaded Lutra canadensis vancouverensis.
Lutra hudsonica pacifica Rhoads, Trans. Amer. Philos. Soc., n. s., 19: 429, September, 1898.
Lutra canadensis pacifica Allen, Bull. Amer. Mus. Nat. Hist., 10: 460, November 10, 1898.
Type.—Obtained at Lake Keechelus, Kittitas County, Washington (altitude 3,000, rather than 8,000 feet as stated in original citation); type in Academy of Natural Sciences of Philadelphia.
Measurements.—An adult male from the Lower Calawah River, near Forks, Clallam County, measured (skinned carcass): total length 1205; length of tail 490; weight (entire) 23 pounds. A male from Puget Island, Wahkiakum County, measured (skinned carcass): total length 1250; length of tail 490. A female from Satsop, Grays Harbor County, measured (skinned carcass): total length 1205; length of tail 440. A female from Calawah River near Forks, Clallam County, measured: total length 1062; length of tail 430; hind foot 120; ear 23; weight 14-3/4 pounds.
Distribution.—Watercourses throughout the state, scarcer east of the Cascades; recorded at Forks (V. B. S.) in the northwest, Puget Island (V. B. S.) in the southwest, Colville (Taylor and Shaw, 1929: 12) in the northeast, and Touchet (Taylor and Shaw, 1929: 12) in the southeast.
Lutra vancouverensis Goldman, Proc. Biol. Soc. Washington, 48: 186, November 15, 1935.
Type.—Obtained at Quatsino, Vancouver Island, British Columbia, by H. O. Berg in 1905; type in United States National Museum.
Racial characters.—No adults, referred to this race from Washington, have been examined. Said to have broader skull than pacifica.
Measurements.—The only available specimen, a young female from Black Rock, San Juan County, measured: total length 792; length of tail 247; hind foot 103; ear 22.
Distribution.—The San Juan Islands of northern Puget Sound and adjacent marine waters.
Remarks.—The otters of the San Juan Islands are referred to this race on geographical grounds. Numerous islands connect Vancouver Island with the San Juan Islands, and intervening channels are not too wide for otters to swim. Adult material may show that the otters of the San Juan Islands are referable to pacifica or to an undescribed race rather than to vancouverensis.
Latax lutris nereis Merriam, Proc. Biol. Soc. Washington, 17: 159, October 6, 1904.
Enhydra lutris nereis Grinnell, Univ. California Publ. Zoöl., 21: 316, January 27, 1923.
Type.—Obtained on San Miguel Island, Santa Barbara Islands, by G. M. McGuire in 1904; type in United States National Museum.
Distribution.—Formerly occurred along the ocean coast of Washington. Now extinct there.
Description.—Adult sea otters reach a length of six feet. The body is stout, long and round; the tail is short, flattened and about a foot in length. The legs are short and thick. The hind feet are webbed for swimming. The head is rounded with small eyes and ears, and a seallike mustache of stiff bristles on the muzzle. The upper lip is thick and heavy, drooping over the sides of the mouth in a spaniel-like fashion. The fur is long, soft and extremely dense. The color of the body is various shades of rich, dark brown. Scattered white hairs often give the coat a frosted appearance. The heads of the adults are pale grey, occasionally almost white.
The sea otter once ranged from the Gulf of California north along the west coast of North America to Alaska, the Aleutian Islands, Siberia and Japan. It is now found only in a small area along the coast of California and in remote islands of the north Pacific. Once abundant along the coast of Washington, it is now completely extinct there.
The habits of sea otters living on the California Coast in the vicinity of Carmel have been described by Fisher (1939: 21-36). Available information on the sea otter in Washington has been given by Scheffer (1940B: 369-388). The search for sea-otter skins influenced the exploration and settlement of the northwest. Captain James Cook obtained skins of sea otter at Nootka Sound, Vancouver Island, in 1778. The mouth of the Columbia River was discovered by Captain Robert Gray on May 11, 1792, while trading for sea otter skins.
According to Scheffer, the bulk of the sea-otter population on the Washington coast was concentrated between the mouth of the Columbia River and Point Grenville at the mouth of the Quinault River. The otters were found several miles from land in extensive kelp beds. The principal food of the sea otter in Washington seems to have been the short-spined purple sea urchin (Strongylocentrotus purpuratus). The animals were social, living in herds of up to 400 individuals. They never came out on land, living, sleeping, and bringing forth their young on the kelp beds of the open ocean. The single young was born at any month of the year.
Description.—The civet cat is slightly less than 18 inches in total length, of which the tail constitutes 6 inches. Large, fat animals may weigh more than a pound. The body is long and moderately stout, heavier than that of the weasel but slimmer than that of the striped skunk. The tail is long with long, plumelike fur. The legs are slender and of moderate length. The feet are small with long claws. The head is small and triangular with low, erect ears and large, bright eyes. The fur is short but soft and silky. The ground color of the civet cat is black. Wavy lines and spots of white or salmon intercept the black in a complex pattern. Variations in color patterns include width of stripes, waviness of stripes, and breaking up of the stripes into spots.
Spilogale is restricted to America; it ranges from southern British Columbia south into Central America. Several species are now recognized, but additional material probably will show intergradation between some of them and reduce the number.
The name "spotted skunk" is seldom used by trappers or other persons familiar with the animal. Nor is it appropriate, for the distinctive color pattern is composed of short stripes. Neither is the term "civet cat" appropriate, for the true civet is found in Asia. The name civet cat is well established, however, and will doubtless remain in general use.
The civet cat is principally a lowland animal, but has been recorded from the Nisqually entrance of Mt. Rainier National Park (Brockman, 1939: 70). Its rather generalized habits allow it to exist in areas that will not support larger carnivores or species with specialized food habits. It dwells in areas of thickets, brushy tangles or deep woods. In southeastern Washington it lives in rocky places as well as in river-side thickets of willow and cottonwood. Where conditions are suitable it ranges up into the Canadian Life-zone but is most abundant in the Transition Life-zone.
The civet cats, in contrast to the striped skunks, are exclusively nocturnal. They are active the year around in western Washington but move about less in spells of unusually cold weather. We have taken them, however, in freezing weather.
In spite of their abundance and wide range, civet cats are known to few persons other than trappers, perhaps because of their exclusively nocturnal habits. They do occasionally make people [Pg 213] aware of themselves by taking up residence about old buildings. They are rather noisy at times. At the Tolt River, 10 miles southeast of Duvall, King County, we were asleep in an old building when a stamping noise from the next room awakened us. Investigation revealed a civet cat indulging in a series of short, stiff-legged hops. The forefeet were held slightly ahead and six or eight hops made. The animal would then relax, turn, and prance off in a new direction. The amount of noise made by its feet striking the board floor was surprising. The civet cat seemed to enjoy the noise it made, and the prancing may have been a method of playing. Trapped civet cats have been seen to make a similar but shorter series of hops at the trapper's approach, which might have been a warning. In the case described, however, the noise of the animal's actions was heard before the animal was seen and presumably when it was unaware of human presence. The civet cat also makes a drumming noise similar to the drumming of a wood rat. This was heard twice from wild animals that did not suspect an observer's presence, and once from a captive animal. While an animal was drumming I never had the opportunity to watch the actions closely enough to describe them. Apparently the drumming is done with the forefeet.
The civet cat is said to be a good climber and to do some of its hunting in trees. One trapped civet cat climbed to the limit of a trap chain and wire in a low bush near Cottage Lake, King County, but, in Washington, I have no other evidence of this animal climbing. Near Cottage Lake, we took a civet cat in a trap set beneath 2 inches of water at the side of a stream. The animal had either been swimming or wading.
Trapped civet cats rarely dig holes near the traps, as striped skunks often do. One caught near Ocean Park, Pacific County, did dig a hole beneath a log and forced earth and leaves over the entrance. I walked within a few feet of the animal in searching for the trap. Not until the trap chain was seen did I find the civet cat. None was taken in traps set in mountain beaver burrows, although I have caught both striped skunks and weasels in such burrows.
Some trappers state that the musk of the civet cat smells different from that of the striped skunk. I think the odor is slightly more acrid and that it does not carry so far as the skunk musk. The civet cat is far more active and nervous than the striped skunk. Whereas the striped skunk almost never throws its scent when trapped, the civet cat almost invariably does so, apparently when the trap closes [Pg 214] about its legs. Striped skunks in traps move slowly and steadily but civet cats jump, roll and squirm erratically.
Fig. 64. Distribution of the civet cat in Washington. A. Spilogale gracilis latifrons. B. Spilogale gracilis saxatilis.
In the winter of 1934 a large male civet cat was taken near Cottage Lake, King County. It had been killed and partially eaten in the trap. The ground about the set was torn up, indicating that the civet cat had put up a fight. The area for many feet about smelled of skunk musk. The body of the civet cat was used for bait and the trap reset. The following morning a large male striped skunk was in the trap. Its skin bore superficial cuts in several places about its neck and rump, and it was doubtless the animal that had killed the civet cat. Under ordinary circumstances a striped skunk could scarcely catch the far swifter and more agile civet cat. One civet cat in a trap was killed and partly eaten by a horned owl. Save for dislodging a few feathers, the civet cat seemed to have done little damage to the owl. The thick cover inhabited by the civet cat ordinarily protects it from owls. Dead civet cats run over by cars on the highway are rarely seen.
The stomach of a specimen from Ocean Park, Pacific County, contained the remains of three red-backed mice (Clethrionomys [Pg 215] californicus). The stomachs of most specimens trapped were empty. Mice, birds and insects probably constitute the bulk of the food.
Notes on the early life of Spilogale interrupta, a species related to the one occurring in Washington, have been published by Crabb (1944: 213-221).
The fur of the civet cat is of little value; in recent years trappers have received from fifteen cents to a dollar for large pelts. Because of its habit of throwing scent when in the traps, most trappers discard the animals without skinning them.
Spilogale saxatilis Merriam, N. Amer. Fauna, 4:13, October 8, 1890.
Spilogale gracilis saxatilis Howell, N. Amer. Fauna, 26:23, November 24, 1906.
Type.—Obtained at Provo, Utah County, Utah, by V. Bailey on November 13, 1890; type in United States National Museum.
Racial characters.—Size small; white areas extensive, especially on tail but white triangle on head small; white areas less often tinged with salmon or orange than latifrons.
Measurements.—Howell (1906: 32) gives the measurements of a male from Harney, Oregon, and the average of 3 females from Oregon as, respectively: total length 455, 360; length of tail 155, 129; hind foot 50, 40.
Distribution.—Southeastern Washington, north to Kamiak Butte (Taylor and Shaw, 1929: 12).
Spilogale phenax latifrons Merriam, N. Amer. Fauna, 4:15, October 8, 1890.
Spilogale olympica Elliot, Field Columb. Mus., Publ. 32, zoöl. ser. 1:270, March, 1899 (type from Lake Sutherland, Clallam County, Washington).
Spilogale phenax olympica Howell, N. Amer. Fauna, 26:33, November 24, 1906.
Spilogale gracilis latifrons Grinnell, Dixon and Linsdale, Fur-bearing Mamm. California, Univ. California Press, Berkeley, p. 301, July 22, 1937.
Type.—Obtained at Roseburg, Douglas County, Oregon, by T. S. Palmer on July 13, 1889; type in United States National Museum.
Racial characters.—Larger than saxatilis with less extensive white markings, especially on tail, but white markings on head larger; white usually tinged with salmon or pale orange.
Measurements.—Means for four males, from Pacific County, are total length 411; length of tail 136; hind foot 47.5; ear 27. One weighed 784 grams.
Distribution.—Western Washington from the western edge of the Cascades westward. Marginal records, given by Taylor and Shaw (1929: 12) are Hamilton, on the north; Lake Keechelus, on the east; and Carson, on the south.
Remarks.—Comparison of adult specimens of civet cats from western Washington and western Oregon shows no reliable character for recognition of two races. No difference in tail length exists. Narrowness of rostrum was the only diagnostic character found by Howell (1906: 34) to separate latifrons and olympica. We have carefully compared civet cats from western Oregon, southwestern Washington and the area about Seattle. Some local variation exists but overlapping is great and specimens from Washington do not have rostra that average narrower than specimens from Oregon.
Description.—The striped skunk is a heavy-bodied animal about the size of a house cat. The legs are of moderate length and stout and the hind feet are large. The claws of the forefeet are long, strong and curved. The head is small and pointed, with small eyes and ears. The tail is long, nearly equal in length to the head and body. The fur is long, soft and shiny, and is jet black with sharply contrasting white markings. These consist of a narrow stripe on the forehead, a broad band on the neck that diverges into two stripes on the back. The two lateral stripes fuse on the rump. The tail has long black hairs some of which are white at the base.
Striped skunks range over North America from central Canada southward to southern Mexico. Two species are recognized by Hall (1936: 64), namely mephitis and macroura. The latter species is found in Mexico and parts of the southwestern United States.
Skunks are principally nocturnal but are sometimes active in the morning and evening, especially on cloudy days. They prefer relatively open country such as logged-over land, old fields and river-side and streamside thickets. Their dens usually consist of old Aplodontia burrows or burrows which they, themselves, dig under stumps or log jams, the floors of old buildings or among rocks. They feed on a variety of animals and wild fruit. Along Puget Sound they wander over the beaches when the tide is out, eating stranded fish, crustaceans and other marine animals. The purple shore crab (Hemigrapusus nudus) forms a staple article of diet. Along streams and rivers they wander along low, muddy banks and sand bars searching for fish, crayfish, insects and insect larvae. The larger water beetles (Dytiscidae) are often eaten; feces are at times composed entirely of their shells. For the most part, however, skunks have no regular food habits but eat such insects, small mammals, birds or refuse as are available.
The skunk is famous for the musk which it uses as a defensive weapon. This highly volatile liquid is ejected from two small, nipplelike ducts situated in the edge of the anus. The consistency, color, and distance to which the musk can be discharged varies with the amount thrown. The first discharge or two is usually a fine, pale yellow spray, which can be accurately directed to a distance of 25 feet. The third discharge consists of small drops of heavy, bright-yellow liquid that travels an arching curve, 5 feet high, reaching [Pg 217] a maximum distance of about 10 feet. Later discharges consist of heavy yellow mucus and can be thrown only a few feet. As many as seven or eight discharges are possible. Skunk musk is acrid and pungent in order. In quantity or at close range it is choking. In small quantities it is not unpleasant. It is extremely lasting, sometimes being noticeable for months on clothes or buildings. It is soluble in gasoline and clothes may be de-scented by several washings in that fluid.
The striped skunk is an even-tempered animal. Its defensive fluid is discharged only when it is cornered or attacked. A trapped skunk rarely releases musk. A man, by speaking softly and moving slowly, can come within a distance of six feet of a trapped skunk. Experienced trappers utilize this fact to approach and shoot trapped skunks through the head or neck and so produce odorless furs.
The skunk is potentially a source of considerable income to trappers in Washington. The value of their furs varies with the demand but large prime skins usually bring from $1 to $4. In eastern Washington, where trapping for coyotes and other terrestrial mammals is carried on, the skunks taken incidentally are an important source of revenue. In western Washington they are often abundant but are seldom trapped. The most sought pelts in western Washington are the mink, muskrat and raccoon; all semiaquatic species. Skunks are rarely taken in traps set for these mammals and few trappers bother to set overland trap lines for skunks.
The young of the striped skunk usually number four to six. They are born in late May or early June in western Washington; possibly later in eastern Washington.
Mephitis americana var. hudsonica Richardson, Fauna Boreali-Americana, 1:55, 1829.
Mephitis hudsonica Bangs, Proc. Boston Soc. Nat. Hist., 26:536, July 31, 1895.
Chincha hudsonica Howell, N. Amer. Fauna, 20:24, August 31, 1901.
Mephitis mephitis hudsonica Hall, Carnegie Inst. Washington, Publ. 473:65, November 20, 1936.
Type.—Obtained on the "plains of the Saskatchewan, Canada."
Racial characters.—Size moderate; stripes diverging anteriorly on back of neck; tail long with white stripe reaching well out, nearly to tip; zygomatic arches nearly parallel.
Measurements.—Howell (1901: 24) gives the average of 3 males from Saskatchewan, Montana and Wyoming, and of 3 females from Montana and Idaho, as, respectively: total length 726, 602; length of tail 268, 250; hind foot 82, 71.
Distribution.—Northeastern Washington and the eastern edge of the northern Cascades, south probably to the Wenatchee Mountains. Recorded west to Oroville (W. W. D.) and Timentwa (W. W. D.) and south to Spokane (Taylor and Shaw, 1929: 12).
Chincha occidentalis major Howell, N. Amer. Fauna, 20:37, August 31, 1901.
Mephitis mephitis major Hall, Univ. California Publ. Zoöl., 37:2, April 10, 1931.
Fig. 65. Distribution of the striped skunk in Washington. A. Mephitis mephitis hudsonica. B. Mephitis mephitis major. C. Mephitis mephitis notata. D. Mephitis mephitis spissigrada.
Type.—Obtained at Fort Klamath, Klamath County, Oregon, by B. L. Cunningham on January 5, 1898; type in United States National Museum.
Racial characters.—Slightly larger than hudsonica with white stripes diverging on shoulders rather than neck.
Measurements.—A young male from 1 mile north of Burbank, Walla Walla County, measures: total length 474; length of tail 205; hind foot 68; ear 30; weight 815 grams.
Distribution.—Southeastern Washington, south of the Snake River and east of the Columbia River, occurring west to Burbank (M. V. Z.).
Chincha occidentalis notata Howell, N. Amer. Fauna, 20:36, August 31, 1901.
Mephitis mephitis notata Hall, Carnegie Inst. Washington, Publ. 473:67, November 20, 1936.
Type.—Obtained at Trout Lake, south base of Mt. Adams, Klickitat County, Washington, by P. Schmid on March 22, 1897; type in United States National Museum.
Racial characters.—Similar to hudsonica but larger; stripes narrower, diverging anteriorly on neck or back of head; tail shorter, sometimes without long, white hairs.
Measurements.—Howell (1901:37) gives the average of 3 adult males from Trout Lake, Klickitat County, as: total length 633; length of tail 249; hind foot 76.
Distribution.—The Columbia River Valley of the southern Cascades from the Wind River east to the Snake River and the Yakima Valley area (trappers' reports).
Mephitis spissigrada Bangs, Proc. Biol. Soc. Washington, 12:31, March 24, 1898.
Mephitis foetulenta Elliot, Field Columb. Mus., Publ. 32, zoöl. ser., 1:269, March, 1899 (type from Laguna, near Port Angeles, Clallam County, Washington).
Chincha occidentalis spissigrada Howell, N. Amer. Fauna, 20:35, August 31, 1901.
Mephitis mephitis spissigrada Hall, Carnegie Inst. Publ. 473:67, November 20, 1936.
Type.—Obtained at Sumas [prairie], British Columbia, by A. Brooks on September 30, 1895; type in Museum of Comparative Zoölogy.
Racial characters.—Similar to hudsonica but larger; stripes broader, usually diverging on shoulders; hairs of stripes often cream color or yellowish near base; end of tail usually white.
Measurements.—An adult male from 2-1/2 miles southeast of Chinook, Pacific County, measures: total length 578; length of tail 260; hind foot 30. A female from the same place and one from Renton, King County, average: 575; 233; 72; 27.
Distribution.—Lowlands of western Washington. This form rarely goes far into the mountains except along the valleys of the larger rivers. Marginal records are Skykomish (W. W. D.) and Washougal River (W. W. D.).
Ursus taxus Schreber, Saugethiere, 3:520, 1778.
Taxidea taxus Rhoads, Amer. Nat., 28:524, June, 1894.
Taxidea taxus taxus Hall, Carnegie Inst. Washington, Publ. 473:78, November 20, 1936.
Type.—None. Type locality probably southwest of Hudson Bay (Hall, 1936: 78).
Measurements.—Two males from northern Nevada measure, respectively: total length 780, 762; length of tail 113, 150; hind foot 136, 120; ear 55, 54; weight 15, 11 pounds.
Distribution.—Of general distribution over open country east of the Cascades. Marginal records are Wauconda (Taylor and Shaw, 1929: 12), Chelan (Taylor and Shaw, 1929: 13), Goldendale (W. W. D.) and "Divide above Trout Lake" (trapper's photograph).
Fig. 66. Badger (Taxidea taxus taxus), tame animal in Seattle, Washington. January 29, 1938. Captured at Lakeside, Chelan County, and photographed at approximate age of ten months; subadult male. (Eloise Kuntz photo).
Description.—The badger is the size of a small dog, measuring up to 32 inches in total length and weighing up to 20 pounds. The body [Pg 221] is heavy, powerful and remarkably flat and compressed. The tail and legs are short. The forelegs are thick and strong, armed with long heavy claws for digging. The ears are wide and low. The color of the upper parts is a grizzled yellowish brown, not unlike the color of the yellow-bellied marmot. The underparts are buffy, often with a white area on the abdomen. The legs, feet, top of head, ears, and small areas on the cheeks are blackish. Triangular areas about the eyes are buffy. A white stripe extends from the nose pad backwards, between the eyes, to the shoulders and serves as the best recognition mark.
Fig. 67. Distribution of the badger, Taxidea taxus taxus, in Washington.
Badgers are found over central and western United States, Canada and northern Mexico. They are commonest on the plains and desert, principally because the burrowing mammals upon which they feed are most abundant there. The badger is a powerful and rapid digger, being able to overtake and capture mice, ground squirrels, and even pocket gophers. Perry (1939: 49-53) in her interesting accounts of the habits of a pet badger obtained at Lakeside, Chelan County, found the animal powerful enough to dig through a concrete floor! Evidence of badger's activities are usually [Pg 222] seen at any ground squirrel colony in eastern Washington. This evidence consists of large holes in the ground. Rarely a horizontal tunnel begins at the depth of two to four feet and extends for an unknown distance. Earth removed in excavating is heaped beside one or both of the narrower sides of the surface opening.
In examining badger workings in ground squirrel colonies I have been impressed by the fact that most of the holes ended not more than four or five feet from the entrances—perhaps at the places where the ground squirrel nests were located, although it may be that the digging of the badger so terrified the squirrels that they dashed out in an attempt to escape past the badger, before he reached the nests. Kangaroo rats and pocket mice often attempt to escape by dashing past a person when he is excavating their burrows.
The power of the short, thick and slightly bowed foreleg of the badger is tremendous. The claws are stout and fully an inch long. The animal is able to break up and remove at a scoop, the baked, claylike loess of the Columbian Plateau. Clods of this same material are so firm that only by twisting and exerting considerable power was I able to break them. The soil a foot beneath the sunbaked crust is softer and more easily worked.
It is difficult to estimate the size of badger populations. In the arid land of eastern Washington their diggings may exist almost unchanged for many years. Interviews with professional trappers serve to indicate their range and numbers as follows: southeastern edge of the Cascades and Yakima Valley, not common, rarely straying up into mountains—average trapper's catch, only one to three a year; Okanogan Valley and northeastern edge of Cascades, not common—average trapper's catch is six to ten a year, occasional trappers catching as many as 35; Columbia, Kettle River, and other valleys in northeastern Washington, uncommon, a few records only; Columbian Plateau, fairly common—average professional trapper's yearly catch includes 10 to 20 badgers; southeastern Washington, now rare because of overtrapping, formerly common.
Trappers state that the badgers taken in northeastern Washington are usually classified as "hair badger" by fur buyers and bring only a dollar or two. The badgers of the eastern Cascades are "fur" badgers but do not bring top prices. The badgers of the Columbian Plateau bring the best prices. Seemingly some geographic variation exists among badgers in Washington. Those from the more humid [Pg 223] areas of northeastern Washington and the eastern Cascades are darker and bring poorer prices than the paler "silver" badgers of the more open desert areas.
The principal food of the badger in Washington seems to be ground squirrels, Citellus washingtoni, townsendii, columbianus, or saturatus, depending on locality. Pocket mice, gophers, and other mammals are also eaten, as are grasshoppers, sword-tailed crickets, other insects, and birds.
Young of badgers number 3 to 5 and are born in late April, May, or early June.
Vulpes cascadensis Merriam, Proc. Washington Acad. Sci., 2:665, December 28, 1900.
Vulpes fulvus cascadensis Bailey, N. Amer. Fauna, 55:281, August 29, 1936.
Type.—Obtained at Trout Lake, Klickitat County, Washington, by P. Schmid on March 3, 1898; type in United States National Museum.
Racial characters.—Skull heavy, narrow; color yellowish.
Measurements.—A male from Crater Lake, Klamath County, Oregon, measures: total length 1113; length of tail 441; hind foot 180; ear 112; weight 9 pounds.
Distribution.—From Trout Lake northward, through the higher Cascades, to Loomis (Taylor and Shaw, 1929: 13).
Description.—The red fox of the Cascades is large and measures about 4 feet in total length, of which the tail comprises 15 inches. The body is slender and doglike; the legs long and slim; the tail thick and bushy, and the ears are large and erect. In the red phase the red fox of the Cascades is distinctly more yellowish than the red fox of the eastern United States; the head is especially yellow. The body has more red on the shoulder area than posteriorly, and is darkest on the rump. The tail is rather pale with a dusky, not black, area distally and a white tip. The ears are dusky. The lower legs and feet are black. The throat, chest and underparts are white. The "cross" phase, according to Cowan (1938: 202-206), is rather common in the Cascades. In cross foxes the color is darker, brown rather than yellowish, and the area from the nape of the neck back between the shoulders, including a bar across the shoulders, is deep blackish or grayish brown. In a pelt that is stretched out a cross is formed by the dark areas. The black and silver phases of the red fox are also said by Cowan to be relatively common in the Cascades, constituting 48 per cent of the population. Of 3,163 foxes traded at Fort Colville, in northeastern Washington, only 19 per cent were silver or cross. One fox, trapped in Okanogan County, is said by its captor to have been black above and straw colored beneath, with no white on the body.
Red foxes range from Alaska and northern Canada well southward into the United States. Related forms occur in Eurasia.
The red fox of Washington is an alpine animal, ranging at or slightly below timber-line. Here food is abundant in summer and fall but must be scarce in winter. In winter its habitat is difficult for man to reach and few persons penetrate the dangerous terrain where the fox lives. A few professional trappers regularly catch foxes in the Cascades but know little of their habits.
The feces of red foxes examined by Taylor and Shaw on Mt. Rainier contained remains of insects and berries (Taylor and Shaw, 1927: 43).
The red fox is rare in Washington; it lives in inaccessible territory and its fur is not especially valuable. The animal is of relatively little economic importance.
The brood den of a red fox found by Livezey and Evendan (1943: 500) near Corvallis, Oregon, was two-thirds of the way up a 300-foot hill in a strip of oaks (Quercus garryana). Well-packed trails led to an entrance concealed in poison oak (Rhus diversiloba). Remains of a turkey, 5 ground squirrels, and a jack rabbit were found near the den. The entrance was 8 inches wide and 15 inches high. The tunnel tapered to 5 inches in diameter and was 47 feet long. Seven pups, 4 males and 3 females, were found in the den.
Fig. 68. Distribution of the red fox in Washington. A. Vulpes fulva cascadensis. B. Vulpes fulva macroura. (See p. 450.)
Description.—The coyote is a large carnivore, about the size and general proportions of a small collie dog but with bushier tail, and more slender body. Adults measure about four feet in length, of which the tail comprises a fourth. Adult males weigh about 30 pounds. The color is somewhat variable, yellowish, buffy, or grayish. The muzzle and backs of the ears are more reddish, and the legs brown. The tail is yellowish gray with a dusky tip. The throat, chest and underparts are white.
Fig. 69. Coyote (Canis latrans lestes), in trap, 5 miles southeast of McKenna, Washington, April 10, 1924. (Fish and Wildlife Service photo by G. R. Bach, No. 26901.)
The coyote ranges from Alaska southward, over western North America, to Central America.
At the present time the coyote ranges over almost all of Washington, except for the highest parts of the mountains and the dense forest areas of western Washington. According to available [Pg 227] information it was not found about Puget Sound or the Olympic Peninsula until relatively recent times. Some old trappers are of the opinion that coyotes did not come into western Washington until the timber wolves became rare there. The scarcity of the timber wolves seems to coincide with settlement, clearing and lumbering. Probably coyotes did not become common in western Washington until lumbering provided extensive clearings and open areas more suitable to them than forest. Certainly coyotes and wolves existed together in eastern Washington.
It is difficult to determine whether coyotes were completely absent from western Washington in the early days or simply scarce. The glacial prairies of the Puget Sound area provide suitable habitat for coyotes and coyotes are abundant there at the present time. If coyotes were present at all in western Washington in the early days it is reasonable to suppose that they would have been common on the prairies. Yet I was told by an old trapper who had lived near Scotts Prairie, Mason County, for many years, that he had never seen or heard coyotes there until about 1910. Although a coyote might be mistaken for a small wolf, the call, as this trapper pointed out, is distinctive.
The principal habitat requirement of the coyote seems to be extensive areas of open country. This it finds in the desert area of the Columbian Plateau, the open forests of northeastern Washington and the eastern Cascades, and in the extensive timbered and burned-over lands in western Washington. In summer coyotes range well up into the Hudsonian Life-zone of the Cascade and Olympic mountains. Their principal range is lower, in the Transition and Upper Sonoran life-zones.
The coyotes are both nocturnal and diurnal. In the vicinity of human habitations they are most active at night. In the heat of the day they take refuge in brushy areas or small gullies. Many actions of the coyote are doglike, and were it not for the large, round, bushy tail, a coyote might easily be mistaken for a dog. The tail is carried in a drooped position with the tip bowed slightly backwards. When badly frightened and running at full speed the tail is stretched out straight behind. The ordinary gait is a purposeful trot with the head held erect, the ears pricked up, and the legs moving smoothly and effortlessly. Near Moses Lake, Grant County, I watched a coyote trot along the side of one of a series of small sand dunes. At the report of the small shotgun fired at it, the coyote's dignified trot changed to terrified bounds, its feet dug into the dune, throwing showers of sand into the air, as it crossed [Pg 228] several successive dunes in full flight before turning to take advantage of the shelter of a draw between two dunes. A coyote chased by an automobile attained a speed of 43 miles an hour for a short distance (Zimmerman, 1943: 400).
I have not watched a coyote stalk game, but as described to me it creeps up to within a few yards of its prey and catches it with a sudden dash. In hunting a jack rabbit, the coyote is said to pursue it, taking advantage of short cuts, until close enough to seize it.
Better known than the coyote itself is its howl—several doglike barks, each successive one of a series shorter and higher pitched, the last one ending in a long, drawn-out howl. In the winter of 1936 the coyotes near Cottage Lake, King County, were especially vociferous. They ordinarily began to howl about 9 p.m., but could be induced to howl earlier by imitating their call. Their howling was a signal for all nearby ranchers' dogs to howl in reply. In the desert areas of eastern Washington I heard coyotes most often just at dusk or at dawn.
The coyote is principally carnivorous, feeding on any mammals and birds easy to kill. It willingly eats carrion, even when much decayed. Large insects such as grasshoppers and crickets are eaten when they are abundant and easily caught. Fruit and berries are eaten when available.
Sperry (1941) reported on the analysis of the contents of 8,339 coyote stomachs from various places in the United States, 1,186 of the stomachs being from Washington. The following information is from his report on all of the 8,339 stomachs: rabbits formed one-third of the food; Microtus was found in 7 per cent of the stomachs; Peromyscus in 6 per cent; Neotoma in 4 per cent. Reithrodontomys were found in 53 of the 8,339 stomachs and in insignificant numbers. Onychomys occurred in 11 of the 8,339 stomachs; Clethrionomys in 8. Ondatra occurred in 8 of the 8,339. Two muskrats were in stomachs obtained from Washington. Citellus (exclusive of beecheyi, lateralis, and saturatus) were found in 4 per cent and were locally important; Citellus beecheyi occurred in 84 stomachs, including 1 from Washington; Citellus lateralis and saturatus occurred in 50 stomachs, including 5 from Washington. Marmota were found in 1 per cent of the stomachs. They were included in stomachs from Washington but the number was not reported. Tamias (= Eutamias) were found in 43 stomachs from western United States and were present in 1 per cent of the 1,186 stomachs from Washington. Sciurus and Tamiasciurus occurred in [Pg 229] 33 stomachs. Included were 3 Tamiasciurus douglasii, a Tamiasciurus hudsonicus, and a Sciurus griseus from Washington. Glaucomys occurred in 6 Washington-taken coyotes, of a total of 11 from the entire United States. Perognathus occurred in 3 per cent of the stomachs, and 274 individuals were represented. They were found in 10 per cent of the 1,186 Washington stomachs. Thomomys occurred in 4 per cent of the stomachs examined and in 7 per cent of the stomachs from Washington. Erethizon appeared in 2 per cent of the stomachs (135 records), including some from Washington. Aplodonta occurred in only 11 stomachs, all taken in Washington. Locally, it is concluded, mountain beavers are important coyote food. House mice occurred in but five stomachs, including 2 from Washington. Zapus did not occur in coyote stomachs from Washington. Domestic sheep and goats formed 7 per cent of the food of Washington coyotes. Calves occurred in 3 stomachs of coyotes from Washington. Pigs occurred in 8. Deer formed 3 per cent, by volume, of coyote food in Washington. A part of the stock and deer reported was doubtless carrion.
One shrew was found in the stomach of a coyote from Washington, and two stomachs contained moles. A house cat was eaten by one Washington coyote, and another coyote had eaten a raccoon. Birds occurred in 13 per cent of the stomachs examined but formed only a small part by volume. Poultry formed one-fourth of this volume. Reptiles formed 0.08 per cent of the food eaten by coyotes and occurred in 3 per cent of the stomachs. A coyote from Washington had eaten a garter snake. No frogs were found in coyote stomachs. A coyote from Washington had eaten a salamander. Another had eaten a fish. Insects formed 1.08 per cent of the total food eaten by coyotes. Fruit formed 3.63 per cent. Carrion constituted 25.2 per cent of the total food eaten.
It is extremely difficult to draw conclusions regarding the economic value of any species. Much depends on the outlook of the individual, his occupation, the locality where the animal occurs and local conditions there. Furthermore it is impossible to understand and fairly weigh all the factors involved.
Coyotes destroy game and stock. They also eat carrion and destroy sick animals, thus preventing the spread of disease. They eat jack rabbits which are pests, and snowshoes and cottontails which are game. They eat mice, which are a pest in agricultural areas, a benefit on rangelands where they destroy weed seeds, and of neutral importance elsewhere. To analyze the economic value [Pg 230] of the coyote, the economic importance of all animals on which it feeds must be considered and the "good" and "bad" balanced—a well-nigh impossible task.
In any event, the coyote has been judged and found guilty. Coyotes are controlled by poison, trapping, and bounty. In spite of control measures the coyote is holding its own in numbers or increasing.
Stimulated in part by the bounty, professional trappers take many coyotes each year in Washington. The pelts fluctuate in value from year to year but a large, prime skin usually brings from 5 to 10 dollars.
Fig. 70. Distribution of the coyote in Washington. A. Canis latrans lestes. B. Canis latrans incolatus.
Canis lestes Merriam, Proc. Biol. Soc. Washington, 11:25, March 15, 1897.
Type.—Obtained in the Toyabe Mountains near Cloverdale, Nye County, Nevada, by V. Bailey on November 21, 1890; type in United States National Museum.
Racial characters.—Frontal region of skull only slightly concave.
Measurements.—A male from 20 miles south of Ephrata, Grant County, on the Columbian Plateau, measures: total length 1185; length of tail 365; hind foot 198; ear 125; weight 26-1/2 lbs. A female from 10 miles northeast of Goldendale, Klickitat County, in the southern Cascades, measured: 1105; 280; 197; weight 19 lbs. A female from Cashmere, Chelan County, in the northern Cascades, measured: 1209; 410; 210; 131. A female from the middle fork of the Nooksack River, Whatcom County, in western Washington, measured: total length 1185; length of tail 358.
Distribution.—Suitable areas of the entire state, except for northeastern Washington.
Remarks.—Pending a revision of the coyotes, those of western Washington are referred to lestes.
Canis latrans incolatus Hall, Univ. California Publ. Zoöl., 40:369, November 5, 1934.
Type.—Obtained at Isaacs Lake, 3,000 ft. elevation, Bowron Lakes Region, British Columbia, by T. T. and E. B. McCabe on October 23, 1928; type in Museum of Vertebrate Zoölogy.
Racial characters.—Similar to lestes but frontal region more concave.
Measurements.—A male from 20 miles east of Tonasket, Okanogan County, measures: total length 1033; length of tail 291; hind foot 163; ear 110.
Distribution.—Northeastern Washington, west to Okanogan and Conconully (W. W. D.).
Remarks.—Several series of coyote skulls from northeastern Washington are more variable than series of lestes from California and incolatus from British Columbia. The "dish-faced" character of incolatus is more strongly represented in some coyotes from northeastern Washington than in topotypes of incolatus but others are more like lestes. In average measurements they are nearer incolatus.
Canis lupus var. fusca Richardson, Mammalia, Zoölogy, Captain Beechey's voyage of the Blossom, p. 5, 1839.
Lupus gigas Townsend, Jour. Acad. Sci. Philadelphia, 2:75, November, 1850 (type from near Vancouver, Clark County, Washington).
Canis gigas Miller, Smithsonian Misc. Coll., 59 (no. 15):4, June 8, 1912.
Canis lycaon gigas Bailey, N. Amer. Fauna, 55:272, August 29, 1936.
Type locality.—Banks of the Columbia below The Dalles in Oregon or Washington.
Racial characters.—A large, relatively dark colored wolf with wide skull and heavy dentition.
Distribution.—Probably occurred from the eastern base of the Cascades westward to the Pacific. Now extinct over most of its range.
Fig. 71. Probable past distribution of the wolf in Washington. A. Canis lupus fuscus. B. Canis lupus columbianus. C. Canis lupus irremotus.
Description.—The wolf is a large carnivore of the general proportions of a large collie dog. Wolves closely resemble coyotes but are larger, with stouter body, larger feet, thicker muzzle and more massive, powerful skull and teeth. The fur of wolves is long and [Pg 233] rather stiff. In general color of the body they resemble coyotes, but the underparts are less white and the legs and feet are more contrastingly reddish.
Wolves of the species Canis lupus range over the northern parts of both the Old and New World. In America they are found from the Arctic south into Mexico.
Wolves occurred in western, northeastern and southeastern Washington. They seem not to have occurred on the Columbian Plateau. Wolves are generally associated with the larger, hooved mammals upon which they feed. In Washington these probably once included the elk, deer, mountain sheep and mountain goat. Hooved animals, except mountain sheep and deer, may not have occurred on the Columbian Plateau in historic times, and even the deer and sheep probably were scarce. There was probably little food for wolves on the Plateau.
The early settlers found wolves to be common and a serious pest. By 1900 they had nearly disappeared. In the settled parts of western Washington they were doubtless exterminated at an early date but it is difficult to account for their disappearance on the Olympic Peninsula. To the best of my knowledge, two wolves killed on the north fork of the Quinault River in 1920, or a few years earlier, were the last ones from that peninsula. Previously they were common and I doubt that man killed them all; perhaps some introduced disease, such as rabies, brought about their extinction.
In the Cascade area they probably still exist in small numbers and in remote places. There are said to be some near Mount Adams. They have been reported from Mount Rainier as recently as 1920. Trappers state that there are a few in the northern Cascades, between Lake Chelan and Mount Baker. There are no recent records for southeastern Washington. The last wolves killed on the Columbian Plateau were two taken at Wahluke, Grant County, on September 17, 1917. Only rumors—no authentic reports—of wolves are available from northeastern Washington in recent years. The reappearance of the caribou there may attract wolves from British Columbia.
Any report of wolves, even from experienced trappers, is open to some question. An unusually large coyote often is mistaken for a wolf. A large, wild and wolflike dog is even more likely to be mistaken for a wolf.
The wolf no longer is an important element in the mammalian fauna of Washington and will probably never be so again. The species, like the grizzly bear, is nearly extinct in the state.
Canis familiaris Linnaeus, Syst. Nat., ed. 12, 1:56, 1766.
Type locality.—Sweden.
Dogs were present with aboriginal man in Washington, previous to the arrival of the white man. For greater detail see Bailey (1936), Suckley and Gibbs (1860) and Allen (1920).
Fig. 72. Cougar or mountain lion (Felis concolor missoulensis), skin mounted as a rug; shot on Mill Creek, Pend Oreille County, Washington, February 13, 1935, by Ralph Johnson. Skin now measures: snout to tip of tail 252 cm., span between tips of forepaws 157 cm., tail 89 cm.; male. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 66.)
Description.—The cougar is a large cat and has the general proportions of the house cat. Large cougars measure more than seven feet in length of which the tail comprises two feet. Large males [Pg 235] weigh more than 150 pounds. Females are smaller and lighter than males. In color the head, back, tail and outside of the legs are reddish brown. The throat, underparts and inside of the legs are white. The tip of the tail, or area near the tip, is black or blackish.
Cougars, often called mountain lions, pumas and panthers, range from Canada to southern South America.
The cougar feeds extensively upon deer and its habits and habitat are accordingly specialized. It is active in both winter and summer and zonally ranges from the Transition through the Canadian to the Hudsonian life-zones.
Fig. 73. Cougar or mountain lion (Felis concolor oregonensis), adults and young taken by Dewey Schmid in White Salmon Valley, Washington, about December 1, 1937, and January 1, 1938. Two bobcats show at extreme right. (Dewey Schmid photo.)
The number of deer killed by the average cougar is unknown. Some trappers believe that a cougar kills a deer at least every other day while others feel that only one a week is taken. Other food is eaten when available. Because cougars kill deer, they are incessantly warred upon by hunters and sportsmen. Bounties, often generous, have long been paid in Washington. Although many cougars are killed each year they are still numerous in many areas. So many remote areas in the mountains are not easily accessible to man and dogs that the cougar, as a species, probably will persist for many years.
A cougar is one of the most secretive animals in the wilds. Rarely are individuals seen by man. They are difficult to trap, principally because they are such wide-ranging animals and partly because they prefer to feed on fresh deer meat and hence are not [Pg 236] apt to be attracted to trap bait. For these same reasons they are difficult to poison. By using poisoned hamburger, a government agent did poison one near Leavenworth, Chelan County. The principal method of killing cougar is to hunt them with packs of especially trained hounds which pursue the cougar until it takes refuge in a tree or other supposed place of safety, where it is shot.
A common gait is a swift, smooth trot in which the body is kept low and the tail droops with the terminal part bent backwards. A wild cougar seen near Leavenworth, Chelan County, traveled with effortless speed until fired upon. At the sound of the shot it made two great bounds and disappeared into the brush fringing a canyon.
Fig. 74. Distribution of the cougar in Washington. A. Felis concolor oregonensis. B. Felis concolor missoulensis.
Although the cougar is a large and powerful carnivore it almost never makes unprovoked attacks upon man. An exception was the cougar that partially devoured a thirteen-year old boy near Lake Chelan, Chelan County (Finley, 1925: 197-199).
The hunting range of an individual cougar is many miles in extent. These ranges are traveled periodically and any particular area may be visited regularly every few days. The cougars may [Pg 237] travel many miles each night in search of deer. Their ranges must overlap to a certain extent for as many as 12 have been taken from a single drainage area. Over most of the year they are solitary but breeding females may attract several males and hunters occasionally capture a female and several males at one locality. Breeding occurs in almost any month of the year. The young are cared for by the female and follow her for perhaps a year. Young individuals have been taken at the same time and in the same locality as an adult female and several adult males that supposedly were breeding. There are from one to three young in a litter. For the first months of their life cougar kittens are spotted. In this they differ from the young of the bobcat which are plain or slightly mottled and do not become spotted until later in life. Cougar kittens differ further from bobcat kittens in possessing a long tail.
The pelage of the cougar is short and of no value as fur although the skins often are sold at a good price as trophies when prepared as rugs.
Felix [sic] oregonensis Rafinesque, Atlantic Journal, 1:62, 1832.
Felis hippolestes olympus Merriam, Proc. Biol. Soc. Washington, 11:220, July 15, 1897 (type from Lake Cushman, Mason County, Washington).
Felis oregonensis Stone, Science, n. s., 9:35, January 6, 1899.
Felis oregonensis oregonensis Miller, U. S. Nat. Mus. Bull., 128:158, April 29, 1924.
Type locality.—"Oregon Mountains, or east or west of them."
Racial characters.—Light, rounded skull, dark color, extensive black on tip of tail and short, rather coarse fur.
Distribution.—From the eastern base of the Cascade Mountains westward to the Pacific. Marginal records are (trappers' specimens): Ruby, Leavenworth, and Goldendale.
Felis concolor missoulensis Goldman, Jour. Mamm., 24:229, June 8, 1943.
Type.—Obtained 10 miles southwest of Missoula, Missoula County, Montana, by R. and C. Thompson, on December 30, 1936; type in United States National Museum.
Racial characters.—A large cougar with heavy, wide skull, pale color and rather long, soft fur.
Distribution.—Northeastern Washington west at least to Republic (W.W.D.) and the Blue Mountains of southeastern Washington.
Remarks.—Goldman (1943: 229) states "Between the Rocky Mountains and the Cascade Range missoulensis intergrades with oregonensis." My own comparison of specimens reveals differences of considerable magnitude between the cougars of western and northeastern Washington. I suppose that intergradation takes place in a limited area in, and west of, the Okanogan River Valley in Washington.
Fig. 75. Canadian lynx, Lynx canadensis, male, taken February 16, 1939, on Baldy Mountain, northwest Idaho, by Lloyd Robinson of Sandpoint. (Ross A. Hall photo.)
Lynx canadensis Kerr, Anim. Kingd., 1: systematic catalogue inserted between pages 32 and 33 (description, p. 157), 1792.
Lynx borealis canadensis True, Proc. U. S. Nat. Mus., 7:611, 1885.
Lynx canadensis canadensis Miller, U. S. Nat. Mus. Bull., 128:160, April 29, 1924.
Type locality.—Eastern Canada.
Measurements.—A female from Buttermills Creek, Twisp River, Okanogan County, measured: total length 900; length of tail 95; hind foot 205.
Distribution.—Higher parts of the Cascade Mountains, Blue Mountains and mountains of northeastern Washington, recorded from Oroville on the north (W. W. D.) to Mount Adams (Taylor and Shaw, 1929: 13) on the south.
Fig. 76. Canadian lynx (Lynx canadensis), catch of nine, with two coyotes, taken by Lester Fairbrother in hills west of Oroville, Washington, March, 1938. (Lester Fairbrother photo.)
Description.—The Canadian lynx weighs about 20 pounds and is catlike in general proportions but differs in possessing longer, stouter legs, much larger feet, a short tail, tufted ears and long, very soft fur. The pelage of the upper parts is soft gray with a slightly yellowish tone; the ears and tail are black; and the underparts are gray with indistinct black spots. The pelage is shorter and more reddish in summer. Bailey (1936: 271) records a maximum weight of 28 pounds.
Fig. 77. Distribution of the Canadian lynx, Lynx canadensis, in Washington.
The lynx ranges over the forested parts of North America from the Arctic south into the northern United States. It has a restricted range in Washington, occurring in the same areas as does the red fox. Although the lynx is an important fur bearer in Canada and Alaska, it is unimportant in Washington because only a few are trapped each winter. Most of the natural range is in the remote and wilder parts of the mountains. Here, each of several trappers regularly takes a dozen or more each year. Mr. Lester Fairbrother of Oroville, Okanogan County, regularly traps lynxes in the northern Cascades. They are taken in wooded areas where snowshoe rabbits, their principal food in winter, are abundant. In the more accessible parts of the animal's range, such as the Blue Mountains and the mountains of northeastern Washington, lynxes are rare. As much as sixty dollars each is offered for large skins.
Description.—The average male bobcat weighs approximately 20 pounds. The female is about one-fourth lighter. A bobcat has longer, stouter legs and larger feet than a house cat and a short tail. The ears are short, with pointed tufts of hair.
Fig. 78. Bobcat (Lynx rufus fasciatus), two-year-old male captured as a kit near Lyman, Washington, in the spring of 1937, by Earl Scott; photographed March 9, 1939. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 588.)
Bobcats range from southern Canada south to central Mexico. Whereas the Canadian lynx occupies the boreal region, the bobcat occupies the temperate region. It is thought to be principally nocturnal but is occasionally active by day. One that I watched near Lake Samamish, King County, when it was unaware of my presence, [Pg 242] was decidedly uncatlike as it strolled with a smooth but stiff-legged gait on a forest trail, with head held up, short tail erect and wagging back and forth with each step. The general impression was of a large, extremely long-legged animal. There was nothing stealthy in its movements. Another individual seen in the same locality on a later date saw me. As it bounded away the body was kept low and the legs were bent with the forelegs appearing almost bowlegged.
Like many other carnivores, each bobcat has a home range which varies with the available food supply. The range may include deep forest, dense thickets and open grasslands, but country with considerable edge-environment seems to be preferred to dense cover, and rocky areas to smooth soil. Perhaps the abundance of mice and wood rats attracts bobcats to the rocks, but the cover afforded is also a factor.
The bobcat is almost universally regarded as a predator and the state of Washington now offers a bounty on it. The animals doubtless do kill a certain amount of game, but kill also other animals which man regards as pests because they interfere with reforestation and growing of food crops. One of the best natural checks on these pests is the bobcat which, in certain areas, does more good than harm. Where doing damage to game or livestock bobcats may be eliminated by trapping or hunting with dogs.
Trappers report that bobcats have a poor sense of smell but very keen sight. They are easily trapped. The fur is soft and handsome but does not wear well. Fur buyers designate the large, pale bobcats of eastern Washington as lynx cats and reserve the name bobcat for the more reddish race of western Washington. Immature and unprime pelts from eastern Washington also are classed as "bobcat."
The difference between the pale bobcat of eastern Washington and the dark race of western Washington is greater than that which separates many subspecies. Dewey Smith of Guler, Klickitat County, showed me skins of bobcats taken on his trap-line along the White Salmon River, which drains southward to the Columbia, and over into the watershed of the Lewis River. Bobcats from the White Salmon River were pale and gray, and those from along the Lewis River were more reddish. The difference was striking. A very few intermediate skins indicated that interbreeding occurs. The geographic variation between the two races is reminiscent of that in the snowshoe rabbits of the western Cascades.
Lynx fasciatus Rafinesque, Amer. Monthly Mag., 2:46, November, 1817.
Lynx fasciatus fasciatus Miller, U. S. Nat. Mus. Bull., 128:160, April 29, 1924.
Lynx rufus fasciatus Bailey, N. Amer. Fauna, 55:269, August 29, 1936.
Type locality.—Near mouth of the Columbia River on "Netul" River (Lewis and Clark River) near Astoria (Bailey, 1936: 269).
Racial characters.—Size moderate; fur short; color distinctly reddish.
Measurements.—A male from Forks, Clallam County, measured: total length 890; length of tail 190; hind foot 167; ear 80; weight 24-1/2 pounds.
Distribution.—From the Cascade Mountains westward, including the valleys of rivers draining westward in the Cascades. Marginal occurrences are: Skykomish (W. W. D.), Mt. Rainier (Taylor and Shaw, 1927: 60) and headwaters of Lewis River (W. W. D.).
Fig. 79. Distribution of the bobcat in Washington. A. Lynx rufus fasciatus. B. Lynx rufus pallescens.
Lynx fasciatus pallescens Merriam, N. Amer. Fauna, 16:104, October 28, 1899.
Lynx rufus uinta Bailey, N. Amer. Fauna, 55:267, August 29, 1936.
Lynx rufus pallescens Bailey, N. Amer. Fauna, 55:268, August 29, 1936.
Type.—Obtained at Trout Lake, Klickitat County, Washington, by D. Kaegi on January 10, 1895; type in United States National Museum.
Racial characters.—Size large; skull heavy with extensive crests and ridges; fur long and soft; color pale, more grayish and less reddish than in fasciatus.
Distribution.—From the Cascade Mountains eastward, including the valleys of rivers in the Cascades which drain southward and eastward. Marginal records of occurrences are Oroville (W. W. D.), Lake Keechelus (Taylor and Shaw, 1929: 15) and Trout Lake (W. W. D.).
Otaria californiana Lesson, Dict. Class Nat. Hist., 13:420, 1828.
Zalophus californianus Allen, Monogr. N. Amer. Pinnipeds, U. S. Geol. and Geogr. Surv. Terr., Misc. Publ., 12:276, 1880.
Type locality.—California.
Distribution.—Rare or casual along the coast of Washington.
Description.—Adult males are 7 to 8 feet in length and females about 6 feet. Bulls weigh as much as 1,000 pounds, and females up to 600 pounds. Both fore and hind limbs are modified for swimming; they are flippers directed posteriorly. The body is cylindrical and streamlined, the neck thick and the head small. Adult males possess a high sagittal crest resulting in a high forehead. Eyes and ears are small. The reddish brown pelage is short and coarse. Females are darker than the males.
The California sea lion occurs along the Pacific Coast from Mexico to northern California and has been recorded from Oregon (Bailey, 1936: 332), Washington and British Columbia (Greenwood, Newcombe, and Fraser, 1918: 1-39). It is of rare or casual occurrence along the coast of Washington.
Phoca jubata Schreber, Säugthiere, 3:300, 1776.
Eumetopias Stelleri True, Proc. U. S. Nat. Mus., 7 (1884):607, 1885.
Eumetopias jubata Allen, Bull. Amer. Mus. Nat. Hist., 16:113, March 15, 1902.
Type locality.—North Pacific Ocean.
Distribution.—Ocean coast, breeding on small, rocky islets such as the Quillayute Needles.
Description.—The Steller sea lion resembles the California sea lion but is larger. Old bulls are from 10 to 12 feet in length and weigh from 1,200 to 1,500 pounds, perhaps even a ton. Females are 8 to 9 feet in length and may weigh up to 1000 pounds. The female Steller sea lion is nearly as large as the male of the California sea lion. The bull Steller sea lion lacks the high forehead characteristic of the California sea lion, and the body is heavier and stouter, especially in the neck and chest. Bulls are distinctly bicolored, the head, neck and chest being cinnamon and the rest of the body darker. Females are a uniform dark brown color. The Steller sea lion makes a loud, deep roaring sound.
Steller sea lions occur along the coast of Asia south to Japan and the coast of North America from the Bering Strait south to central California. The range thus meets and overlaps that of the California sea lion.
Steller sea lions are splendid swimmers and spend much of their time in the surf. They often lie in the water a few yards from [Pg 245] where the waves pound some jagged cliff, retaining their dangerous position without discernible effort. They are curious and will raise their head and neck vertically out of the water to observe a passing boat. Much time is spent beneath the surface, presumably hunting fish. They sometimes leap up out of the water in an arching dive or may simply lower their heads beneath the surface and appear several minutes later at another place. They may come to the surface with a porpoiselike roll, breathe, and again dive. Many of their actions seem inspired by playfulness—a means of working off excess energy.
Sea lions haul out on a number of rocky islands along the coast. They are slow and clumsy on land. Their rear flippers can be directed forward and so give them some assistance in moving about. When startled they are able, with much struggling, to move with fair speed to the water. They can dive from considerable heights.
The studies of Greenwood, Newcombe and Fraser (1918: 1-39) show that the Steller sea lions eat a great variety of marine fish and other sea life. Included in their diet are squid, starfish, crabs, clams, mussels, salmon, herring, flounder, rock cod, sea bass and dogfish. They seem not to be selective in their food habits but eat the food most easily available at the time. Unfortunately this is occasionally salmon in nets or traps and for this reason fishermen usually kill sea lions on sight. The number of sea lions along the coast of Washington has been greatly reduced by shooting and dynamiting the animals on their hauling-out grounds. There is no doubt that sea lions do occasionally eat salmon, especially in traps or nets. They also become entangled in the nets, and damage them. On the other hand, investigations of their food habits have shown that they do not eat any great number of salmon or other important food fishes. They are a relatively harmless and exceedingly interesting animal and might well be preserved in numbers exceeding their present population.
Until relatively recent times the sea lion was an important source of food to the Indians living along the ocean coast. The bones of sea lions are often the most numerous vertebrate remains in shell mounds.
The Steller sea lion now breeds only in a few places along the coast of Washington. They are polygamous and each of the stronger bulls has eight or ten cows in his harem. Fierce battles between bulls are said to take place in the breeding season, late in May. A single pup is born after a gestation period of about one year. Pups are darker in color than adults.
Siren cynocephalus Walbaum, Petri Artedi Sueci Genera Pisc., p. 560, 1792.
Callorhinus ursinus True, Proc. U. S. Nat. Mus., 7 (1884):607, 1885.
Callorhinus alascanus Jordan and Clark, Fur Seals and Fur Seal Islands of the North Pacific, pt. 3, p. 2, November, 1899.
Callotaria ursina cynocephala Stejneger, George Wilhelm Steller, Harvard Univ. Press, p. 285, 1936.
Type locality.—Pribilof Islands, Alaska.
Distribution.—Breeds on the Pribilof Islands, Alaska. Abundant off the coast of Washington on migration.
Description.—The difference in the size of male and female fur seals is great. Males are about 8 feet in length when fully grown but the females are only about 4 feet long. Males weigh up to 700 pounds but females only about 100 pounds. In general appearance the fur seal resembles the sea lion but the fur is longer, denser and softer. Males are very dark brown in color. Females and young are grayish brown. The fur seal, like the sea lion, can reverse its hind flippers and use them to a certain extent in walking.
The Alaskan fur seal breeds on the Pribilof Islands, Alaska. The seals emerge from the water to certain favored hauling-out places in May and June. They leave the Islands in November to begin their long migration. The bulls spend the winter off the coast of Alaska south of the Aleutians but the females and young travel south to the ocean off California. In the spring they start north, arriving off the coast of Washington in February, March and April. Young, rarely adults, are washed up on the ocean beaches of Washington (Scheffer, 1939: 43).
The United States Government, by international treaty, manages the fur seal herds on a sustained yield basis and pelagic sealing by independent hunters is prohibited. Indians are allowed to take the fur seals on migration but are subject to certain restrictions. Modern boats, outboard motors and guns are prohibited. Indians living on the coast of Washington hunt the fur seals from dugout canoes. The weapons are double-headed harpoons with long cedar shafts. One harpoon head is at the tip of the shaft and the other is on a short fork that projects downward and forward at a 30° angle from the main shaft. The harpoon is thrown with the aid of hand grips at the end of the shaft. The seal is recovered by rawhide lines connected to the harpoon heads. The hunters leave shore at dawn and travel ten to twenty miles from land, at which distance the seal herds are usually encountered. The hunting is dangerous work and is carried on only by skilled and brave men. Relatively calm [Pg 247] weather is required. The flesh of the seals is eaten and the skins sold.
Schultz and Rafn (1936: 13-15) examined the stomachs of 41 fur seals taken within 30 miles of La Push, Washington, in March, April and May, 1930. Food found included squids, shrimps, herring and lampreys.
Halicyon richardii Gray, Proc. Zoöl. Soc. London, p. 28, 1864.
Phoca richardii Allen, Bull. Amer. Mus. Nat. Hist., 16:491, December 12, 1902.
Phoca richardii richardii Miller, U. S. Nat. Mus. Bull., 128:164, April 29, 1924.
Phoca vitulina richardii Doutt, Ann. Carnegie Mus., 29:117, May 12, 1942.
Type.—Described from a specimen obtained at the Fraser River, British Columbia, probably on March 23, 1861, by C. B. Wood and another obtained by Wood at Queen Charlotte Sound, British Columbia, in 1862 (see Scheffer and Slipp, 1944: 374); type in British Museum of Natural History.
Distribution.—Puget Sound, the Strait of Juan De Fuca and the coast of Washington, extending at times up the Columbia River to The Dalles. Has been recorded in Lake Washington, Seattle (Bonham, 1942: 76).
Description.—Adults are about 5 feet in length and weigh up to 250 pounds; males are approximately a quarter larger than females. The hair seals differ from the sea lions and fur seals in a number of respects. The body is widest in the midsection rather than in the chest. The neck is short and slim. The eyes are large and there are no external ears. The hind flippers are not reversible but are permanently directed posteriorly. The pelage is short, stiff, and directed posteriorly. The ground color is silver gray or yellow; usually it is yellow, blotched and marbled with black, dusky or gray.
Hair seals range over the northern Atlantic and Pacific oceans, occurring southward along the Pacific Coast of North America to central Mexico. Six races are recognized by Doutt (1942: 115).
The hair seal is the common seal in Puget Sound, the Strait of Juan De Fuca and on the ocean coast. Scheffer and Slipp (1944: 373) estimate that 5,000 live along the coast of Washington. Hair seals are social to some degree but are often seen singly. In Puget Sound, where there are relatively few hauling-out areas, they rarely leave the water but on the ocean coast they emerge to bask on the reefs. On Destruction Island, Jefferson County, V. B. Scheffer and I watched a herd of about 20 animals that lay on a reef exposed by low tide. According to the lighthouse keeper the reef was occupied daily by these seals. Scheffer and Slipp (loc. cit., p. 388) report herds of up to 200 hair seals. In Puget Sound 10 to [Pg 248] 20 seals seem to be the usual number in a herd, but occasionally there are as many as 50.
Hair seals often exhibit curiosity concerning small boats; the seals lie in the water with only their round heads above the surface, staring at the boat for many minutes. Constant shooting has made them shy and any quick movement will cause them to dive; several dived instantly when I pointed either a stick or gun at them.
Hair seals are less spectacular than sea lions. When basking on rocks the seals are silent. When an observer is yet a long distance away the seals raise their small heads and remain on the alert. When they take to the water, they travel with a jerking motion. In the water one rarely sees more than their heads. When they dive they usually sink below the surface, never making the spectacular arching dive so typical of the sea lions.
At Useless Bay, Whidby Island, Island County, a herd of five or six hair seals was studied in July, 1936. These animals played and slept in the shallow water beneath a high, forested bluff and could be watched from the bluff above. The seals seemed to gather here in the early afternoon. Much of their time was spent sleeping on the surface with the body in a bowed position and drifting freely. Occasional waving of the rear flippers kept them from drifting away. They played a great deal, splashing, diving and swimming in circles or spirals. Occasionally one would dive down to swim slowly along just above the bottom. At times one would put on a sudden burst of speed, apparently in pursuit of some fish, rarely continued for more than 30 or 40 feet after which the seal usually rose to the surface to resume play. Never did I see one with a fish in its mouth and these pursuits, if pursuits they were, seemed to have been prompted more by a spirit of play than by hunger.
A young seal was kept as a pet for several weeks at Friday Harbor, San Juan County. It was tame and affectionate but decidedly temperamental. It demanded constant attention and whimpered, cried or moaned if left alone. It was more active by night than by day and made a characteristic mooing cry, which continued at intervals throughout the night. This habit led to its eventual disposal. The temper of this young seal was shown by its actions when it encountered strands of kelp while swimming. If the kelp strands held back its progress it would turn back, seize the kelp in its teeth and bite viciously. Never did it attempt to bite persons.
The hair seal was generally thought to feed almost exclusively on salmon, but the work of Scheffer (1928: 10-16) showed this view to be incorrect. Of 14 hair seal stomachs examined, all of which [Pg 249] were full, only two contained salmon. In a later study (Scheffer and Sperry, 1931: 214-226), only two of 100 stomachs examined contained salmon. Other food items were tomcod, flounder, sculpin, herring, shiner, hake, skate, blenny, unidentified fish, squid, octopus, shrimp, crab and starfish. In spite of Scheffer's work, hair seals are killed at every opportunity by fisherman and boatman. Bounties were paid on their scalps for many years, and more than 1,000 bounties were paid for each of several years previous to 1930. Their small, round heads bobbing on the waves offer a poor target and many seals, after having been fired at, become extremely shy. They seem to be holding their own in numbers at the present time.
Scheffer and Slipp (1944: 401) found that the young were born in late May along the ocean coast and in June and July in Puget Sound. The young seal mentioned as having been kept captive at Friday Harbor was obtained from an Indian on July 26, 1938, and was said to be two weeks old at the time. The Indian said that he had watched the birth of the young and then killed the mother for bounty. On July 28 the young seal weighed approximately 20 pounds and was in good health. The seal could swim well. It was said to have been born "on the rocks" at Long Island, San Juan County. The seal drank milk from a baby's bottle but refused fresh scallops, clams and fish of several species. When put into a large, screened box sunk in the water it at once investigated the other animals in the box. It showed no fear of a large bull cod weighing 50 pounds, or of a 20-pound skate and several sharks 5 feet long but seemed to be frightened by a large octopus weighing about 30 pounds. In swimming, the front flippers were held flat against the body and the actual swimming was accomplished by the vertically-held rear flippers and the rapid swinging of the hips. Its eyes were very dark brown, almost black, but soft and appealing. The bases of the vibrissae were thick and soft. The belly was silvery white and unspotted. The sides and back were iron gray spotted with dark, bluish gray, the whole overlaid with a silvery tint. The claws were long, round, and sharply pointed.
When sleeping, the seal usually lay on its side, occasionally upon its back or belly. The front flippers were held tight to its sides but the back flippers were held straight back with the digits bent inward at right angles and laid so that the right digits were against the left. In moving on land the front flippers were folded into fists and used to push the animal forward while the body was moved by snakelike motions of the hips. It breathed in short gasps.
The chipmunks of Asia and western North America have usually been separated under the generic name Eutamias from those of the genus Tamias of eastern America. Ellerman (1940: 428) placed both in the same genus and Bryant (1945: 257-390) reached the same conclusions after intensive study of American sciurids. Bryant's treatment is followed here. The sciurid genera as they occur in Washington, are listed by Bryant as follows: Tamias, Marmota, Citellus, Sciurus, Tamiasciurus, Glaucomys. This order, rather than that of Miller (1924) is used here. Four species of Tamias are listed for Washington: minimus represented by two subspecies; amoenus, by six; ruficaudus by one; and townsendii, by two.
Chipmunks from Washington vary in size from less than 8 inches in total length to more than 10 inches in total length. Some race of chipmunk occurs in almost every part of Washington. Their striped color pattern serves as a universal recognition mark. The somewhat similarly striped mantled ground squirrel is often mistakenly called chipmunk. The mantled ground squirrel is larger than any chipmunk, has but two dark stripes as compared with five dark stripes of chipmunks, and has a plain, reddish head unlike the distinctly striped head of Tamias.
Like most members of the squirrel family, chipmunks are active by day and are therefore better known to man than are most of the other kinds of small mammals, most of which are nocturnal. The attractive color and sprightly actions of chipmunks make them a delightful feature of the outdoors. They feed on fruit, seeds, and fungus and eagerly eat food that can be begged or stolen from man. They have been known to kill mice and they have been accused of destroying nests and eggs of birds. They often eat insects and occasionally eat the flesh of mice or other chipmunks held in collector's traps.
Chipmunks climb trees and bushes readily but only townsendii can be called arboreal, and even it prefers to climb on stumps and dead trunks rather than in living trees. All species are fond of climbing about rocky outcrops and talus slides.
The call of the chipmunks is a birdlike cheep. In minimus it is shrill and uttered rapidly, but it is low-pitched and is uttered by townsendii with longer intervals between the notes. The call of amoenus is of an intermediate nature.
Young of chipmunks vary in number from four to six. Nests are constructed of dry grass and are placed under rocks, logs, and in burrows in the ground. There are four pairs of mammae, one pectoral, two abdominal, and one inguinal.
Description.—The least chipmunk is the smallest chipmunk found in Washington. The head and body of adults measure about 3-1/2 inches; the tail about 3-1/2 inches. Its fur is short and sleek. The dorsal stripe is black; the upper pale stripe is buffy gray; the lower dark stripe is rich brown; the lower stripe is white. The sides are pale buff and the head, rump and thighs are buffy gray. The tail is brownish above, yellowish beneath.
Tamias minimus has a wide range, being found from the Cascade-Sierra Nevada Chain to the Great Lakes and from northern Canada to central Arizona and New Mexico. Two races occur in Washington, both in the sagebrush desert area.
Least chipmunks are only locally common in Washington. I have found them in areas where the soil was firmly packed and sagebrush the dominant vegetation. All were far from water. Two miles west of Vantage, Kittitas County, several were found near an old sheep corral, where one took shelter in a pile of boards. In my experience, least chipmunks are wary and difficult to collect. Many times as I crept silently through the sagebrush chipmunk after chipmunk scampered with tail aloft into a hole at the base of same sage bush, each far out of gunshot, voicing alarmed chirps. The extreme caution of least chipmunks, as compared with other species, is doubtless a necessary adaptation to living in an exposed situation. The open sagebrush desert is a favored hunting place of hawks and eagles; also coyotes, wildcats, and badgers sometimes abound there. All these probably find the least chipmunk a suitable food item and only the most cautious chipmunk survives to reproduce. The least chipmunk has been timed at a speed of 10 miles per hour (Cottam and Williams, 1943: 262).
The food of the least chipmunk in eastern Washington is almost entirely seeds of the annuals that flourish briefly in the spring. Insects probably are eaten and one specimen had the remains of two scorpions in its stomach.
Eutamias minimus pictus Howell, N. Amer. Fauna, 52:39, November 30, 1929.
Eutamias minimus scrutator Hall and Hatfield, Univ. California Publ. Zoöl., 40:321, February 12, 1934.
Tamias minimus scrutator Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 432, June 8, 1940.
Fig. 80. Distribution of the least chipmunk in Washington. A. Tamias minimus scrutator. B. Tamias minimus grisescens.
Type.—Obtained near Blanco Mountain, 10500 ft. elevation, Mono County, California, by J. Grinnell on July 28, 1917; type in Museum of Vertebrate Zoölogy.
Racial characters.—Buffy color; wide dark stripes and narrow pale stripes.
Measurements.—A male and a female from Sunnyside, Yakima County, measure respectively; total length 186, 186; length of tail 81, 82; hind foot 31, 31; ear 9, 10.
Distribution.—The sagebrush areas west of the Columbia River. Present in scattered and widely separated areas, ranging, according to Howell (1929: 41), north to Ellensburg and south to Wiley City.
This species is a member of the Great Basin Fauna that entered the state from Oregon. The population in Washington is now isolated north and west of the Columbia River but seems not to differ from least chipmunks from Oregon and Nevada.
Eutamias minimus grisescens Howell, Jour. Mamm., 6:52, February 9, 1925.
Tamias minimus grisescens Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 431, June 8, 1940.
Type.—Obtained at Farmer, Douglas County, Washington, by J. A. Loring, on July 31, 1897; type in United States National Museum.
Racial characters.—Similar to scrutator but smaller; more grayish, less buffy in color, dark stripes narrower and pale stripes wider.
Measurements.—Seven topotypes and near topotypes average: Total length 177; length of tail 78.7; hind foot 26.8; ear 10.6.
Distribution.—Known only from the Columbian Plateau. This race is rare and though we hunted for it in localities where specimens have been collected, including the type locality, it was found but twice. Ranchers living in the area know the chipmunk but see individuals only occasionally. Marginal occurrences are Douglas (Howell, 1929: 41), Vantage (V. B. S.) and Pasco (Howell, 1929: 41).
Remarks.—The geographic range of this race is separated from that of scrutator by the Columbia River and many miles of country uninhabited by chipmunks of this species.
Description.—The yellow-pine chipmunk resembles the least chipmunk but is larger. The underside of the tail is more ochraceous, less yellowish. The color of the sides varies considerably in the various races. The stripes are narrow and sharply delineated, the dorsal one being black. Next lower is a grayish stripe, followed by one of brownish black. The lower stripe is white. The underparts, in most races, are white but in luteiventris are buffy.
The geographic range of the species is west of the Great Plains from central British Columbia to central California. There are twelve races, six of them occurring in Washington.
The yellow-pine chipmunk is a small animal, being but little larger than the least chipmunk, and much smaller than townsendii. Externally it may be separated from the Townsend chipmunk by its small size, sleek, appressed pelage and brighter color. Separation from minimus is more difficult but, in the Washington races of minimus, the colored fur of the underside of the tail is pale yellowish while in amoenus it is more ochraceous or buffy. So far as is known, the two species do not occur together in Washington.
The yellow-pine chipmunks live in open woods, brushy areas, clearings, and rocky outcrops. Suitable conditions are abundant in mountainous areas and the distribution of mountain ranges affects the distribution of these chipmunks. Where yellow pine forests descend to relatively low altitudes, the chipmunks enter the lowlands. [Pg 254] Such conditions are present in some places along the eastern base of the Cascades, the inter-mountain river valleys of northeastern Washington, and along the central-eastern border of the state.
Yellow-pine chipmunks are sprightly and active. They seem always to be moving restlessly about, running, investigating for food, and watching for enemies. They are far tamer than the least chipmunks, usually allowing the observer to approach within twenty feet or closer. Some, after coaxing, will take food from a person's hand.
Near Stevens Pass, King County, numbers of Tamias townsendii cooperi and Tamias amoenus ludibundus, apparently on good terms, were feeding together on blue huckleberries which grew in abundance on an extensive snowslide area. The yellow-pine chipmunks had been drawn from surrounding open areas by the berries, while the Townsend chipmunks had been attracted from the forest by the same food. The nervous movements of amoenus contrasted strongly with the more sedate behavior of townsendii. At the observer's close approach the yellow-pine chipmunks went scampering off through the brush and tangles of logs and branches, to emerge again and watch from a hundred feet away. The same individuals, when repeatedly followed, always remained in sight. Most of the Townsend chipmunks, when frightened, ran into the nearest dense cover and vanished, not to appear again. A goodly number, perhaps ten per cent, climbed high up in fir trees. None of the yellow-pine chipmunks took refuge in trees.
The yellow-pine chipmunk is usually found at considerable altitude and consequently there is deep snow and bitter cold in winter where it lives. Hibernation is probably complete. Seemingly these chipmunks depend on stored food rather than on accumulated fat to tide them over the winter, for animals collected in autumn are no fatter than those taken in the spring. Svihla (1936B: 290) found that Tamias a. canicaudus, hibernating in captivity at Pullman, Whitman County, awoke at intervals to eat stored food. The time of retirement of yellow-pine chipmunks for the winter seems to coincide with the coming of winter weather. In mid-November of one year, when no snow had yet fallen in the Cascades, yellow-pine chipmunks were common near Stevens Pass, although their actions were noticeably slow. In another autumn, when the winter snows came early, I looked in vain for chipmunks in October where they had been common earlier. George C. Cantwell noted a yellow-pine chipmunk at Republic, Ferry County, on November 9, 1903, after the ground was "well frozen," but apparently free of snow (Howell, [Pg 255] 1929: 7). Like other species that hibernate, they, at times, seem to become active in winter; J. B. Flett reported seeing a yellow-pine chipmunk at Longmire, Mt. Rainier, on February 14, 1920, and again on March 31 (Howell, loc. cit., p. 7). At Deer Park, Clallam County, at timber-line, several Tamias townsendii cooperi were active in early April, 1938, but only one Tamias amoenus was seen. In the previous June they were abundant there and townsendii was scarce.
I have watched these chipmunks eat the berries of the red huckleberry (Vaccinium parvifolium), salmonberry (Rubus spectabilis), thimbleberry (Rubus parviflorus), devil's club (Fatsia horrida), and mountain ash (Sorbus cascadensis and S. occidentalis). Of these, the blue huckleberry (Vaccinum occidentale and V. membranaceum) are probably the most important to the chipmunks. At Sherman Creek Pass, in the Kettle River Mountains, Ferry County, I watched a pika (Ochotona) busily harvesting wild raspberry plants (Rubus leucodermis), and laying them in a pile under a rock. A yellow-pine chipmunk waited under the rock and ate the ripe berries from each branch as it was laid away.
A brood nest of the yellow-pine chipmunk was discovered by Shaw (1944: 274) at Hurricane Ridge, Clallam County. The entrance was a hole 1-1/2 inches in diameter constructed among the grass and alpine flowers of a meadow at 6,450 feet elevation. The burrow itself was 2 inches in diameter, and had a turning-around pocket 9 inches from the entrance. The nest was situated just beneath the sod, 4 feet from the entrance. The nest chamber was 7 inches high by 7-1/2 inches in diameter and in the shape of a "round-bottomed flask." It was filled snugly with nest material composed of a grasslike sedge (Carex spectabilis) mixed with feathers of the blue grouse. Earth excavated from the nest cavity had been forced upwards through the sod in the manner of a mole in forming mounds. The single burrow was unbranched. Seven young of about 16 to 18 days of age were found in the nest.
Eutamias caurinus Merriam, Proc. Acad. Nat. Sci. Philadelphia, p. 352, October 4, 1898.
Eutamias amoenus caurinus Howell, Jour. Mamm., 3:184, August 4, 1922.
Tamias amoenus caurinus Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 432, June 8, 1940.
Type.—Obtained at timber-line, head of Sol Duc River, Olympic Mountains, Clallam County, Washington, by C. H. Merriam and Vernon Bailey on August 27, 1897; type in United States National Museum.
Racial characters.—Small size and pale color.
Measurements.—Seven males and 3 females from Deer Park, Clallam County, average: total length 207.4; length of tail 93.2; hind foot 32.5; ear 16.
Distribution.—Higher parts of the Olympic Mountains, from Deer Park (W. W. D.) south to head of Dosewallips River (Howell, 1929:77).
Tamias quadrivittatus felix Rhoads, Amer. Nat., 29:941, October, 1895.
Eutamias quadrivittatus felix Miller and Rehn, Proc. Boston Soc. Nat. Hist., 30:44, December 27, 1901.
Eutamias amoenus felix Howell, Jour. Mamm., 3:184, August 4, 1922.
Tamias amoenus felix Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 432, June 8, 1940.
Fig. 81. Distribution of the yellow-pine chipmunk in Washington. A. Tamias amoenus luteiventris. B. Tamias amoenus canicaudus. C. Tamias amoenus affinis. D. Tamias amoenus ludibundus. E. Tamias amoenus felix. F. Tamias amoenus caurinus.
Type.—Obtained at Church Mountain, British Columbia, near the United States boundary by Allan Brooks on August 13, 1895; type in Philadelphia Academy of Natural Sciences.
Racial characters.—Large size and rich, dark color.
Measurements.—Ten topotypes average: total length 224.7; length of tail 98.8; hind foot 34.1; ear 14.4.
Distribution.—The extreme northwestern Cascades, north and west of Mt. Baker.
Remarks.—This richly-colored coastal race barely enters Washington. It is abundant near Goldrun Pass and Tomyhoi Lake, Whatcom County, just south of the international boundary.
Eutamias ludibundus Hollister, Smithsonian Misc. Coll., 56 (no. 26):1, December 5, 1911.
Eutamias amoenus ludibundus Howell, Jour. Mamm., 3:184, August 4, 1922.
Tamias amoenus ludibundus Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 432, June 8, 1940.
Type.—Obtained at Yellowhead Lake, 3,700 ft., British Columbia, by N. Hollister on August 29, 1911; type in United States National Museum.
Racial characters.—Moderate size, brownish rump, ochraceous sides.
Measurements.—Five males and 4 females from the higher parts of the Cascades average, respectively: total length 210, 217; length of tail 89, 90; hind foot 33, 33; ear 17.4, 16.7; weight 50, 59.7 grams.
Distribution.—The higher Cascade Mountains. Marginal records are: Barron (Howell, 1929:75), Lyman Lake (Howell, 1929:75), Cascade Tunnel (W. W. D.), Mt. Stuart (W. W. D.), Lake Kachees (W. W. D.), Boulder Cave (W. W. D.), and Mt. St. Helens (Howell, 1929:75).
Remarks.—At the higher altitudes this race seems to be the equivalent of affinis. The latter race lives in relatively arid yellow pine forests and ludibundus occupies more moist and varied habitats higher in the mountains.
Tamias quadrivittatus affinis Allen, Amer. Mus. Nat. Hist. Bull., 3:103, June, 1890.
Eutamias quadrivittatus affinis Miller and Rehn, Proc. Boston Soc. Nat. Hist., 30:44, December 27, 1901.
Eutamias amoenus affinis Howell, Jour. Mamm., 3:184, August 4, 1922.
Tamias amoenus affinis Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 432, June 8, 1940.
Type.—Obtained at Ashcroft, British Columbia, by C. P. Streator on July 3, 1889; type in United States National Museum.
Racial characters.—Small size, grayish color including rump, and white underparts.
Measurements.—Fourteen males and 7 females from Washington average: total length 201.5; length of tail 86.5; hind foot 31.7; ear 17.2.
Distribution.—The eastern slope of the Cascade Mountains. Marginal records on the west are: Bald Mountain (Howell, 1929:73), Mazama (Howell, 1929:73), Hart Lake (Howell, 1929:73), Lake Wenatchee (W. W. D.), 10 mi. S. Dryden (W. W. D.), Blewett Pass (W. W. D.), 10 mi. N. W. Ellensburg (W. W. D.), Wenas Creek (W. W. D.), Mt. Adams (Howell, 1929:73), and Lyle (Howell, 1929:73). Marginal occurrences on the east are: Mt. Chopaka (Howell, 1929:73), 20 mi. E. Tonasket (W. W. D.) and Omak Lake (Howell, 1929:73).
Eutamias canicaudus Merriam, Proc. Biol. Soc. Washington, 16:77, May 29, 1903.
Eutamias amoenus canicaudus Howell, Jour. Mamm., 3:184, August 4, 1922.
Tamias amoenus canicaudus Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 432, June 8, 1940.
Type.—Obtained at Spokane, Spokane County, Washington, by C. P. Streator, on April 11, 1891; type in United States National Museum.
Racial characters.—Large size, pale color, grayish tail, white or buffy underparts.
Measurements.—Thirteen topotypes average: total length 227.2; length of tail 104.4; hind foot 33.7; ear 14.
Distribution.—The pine-covered lowlands along the central-eastern border of the state, ranging, according to Howell (1929: 71), from Spokane County south to Pullman.
Tamias quadrivittatus luteiventris Allen, Amer. Mus. Nat. Hist. Bull., 3:101, June, 1890.
Eutamias quadrivittatus luteiventris Miller and Rehn, Proc. Boston Soc. Nat. Hist., 30:44, December 27, 1901.
Eutamias amoenus luteiventris, Howell, Jour. Mamm., 3:179, August 4, 1922.
Tamias amoenus luteiventris Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 432, June 8, 1940.
Type.—Obtained at Chief Mountain Lake (Waterton Lake), Alberta (3-1/2 mi. N. United States boundary) by Elliott Coues on August 24, 1874; type in United States National Museum.
Racial characters.—Small size, rich color, buffy underparts.
Measurements.—Twelve males and 12 females from the Blue Mountains, Columbia County, average respectively: total length 212, 219; length of tail 96.7, 101; hind foot 31.7, 32.5; ear 17.3, 18; weight 46.5, 52.8 grams.
Distribution.—The Blue Mountains of southeastern Washington, and the Pend Oreille Mountains of northeastern Washington, west to Eureka, in the Kettle River Mountains, Ferry County (Howell, 1929: 69), and south to Newport (W. W. D.).
Eutamias ruficaudus simulans Howell, Jour. Mamm., 3:179, August 4, 1922.
Tamias ruficaudus simulans Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., p. 434, June 8, 1940.
Type.—Obtained at Coeur d'Alene, Kootenai County, Idaho, by C. P. Streator on June 1, 1891.
Measurements.—Six males and 3 females from northeastern Washington average: total length 234; length of tail 109; hind foot 31.6; ear 18.
Distribution.—Northeastern Washington, reported from Pend Oreille, Stevens and Ferry counties by Howell (1929: 98).
Description.—The red-tailed chipmunk closely resembles Tamias amoenus. From T. a. luteiventris and T. a. canicaudus it differs principally in larger size, wider brain case and especially in its pure white underparts. From T. a. affinis it differs in richer coloration, especially the brownish rather than gray rump. The differences separating it from amoenus are slight, and only adult specimens can be identified in the field.
According to Howell (1929: 81) this species occurs in northern Idaho, western Montana, northeastern Washington, southeastern British Columbia and extreme southwestern Alberta. Two subspecies are recognized, only one of which occurs in Washington.
A number of large, white-bellied chipmunks have been taken in northeastern Washington that answer well to the description of ruficaudus. Also, there are a number of specimens that I cannot definitely identify as either amoenus or ruficaudus. Some buff-bellied chipmunks from Idaho are as large, and possess brain cases as wide, as specimens from Washington unhesitatingly called ruficaudus, while some white-bellied individuals match amoenus luteiventris in all other characters. When all the specimens available from Pend Oreille, Stevens and Ferry counties are separated into amoenus and ruficaudus and the skulls are examined, it is noticeable that all the ruficaudus are old, fully adult animals and that most of the amoenus are younger, showing less wear on the teeth. Perhaps the buffy underparts are lost with increasing age.
This leads to the suspicion that ruficaudus, as applied to chipmunks in Washington, is a synonym of amoenus. Until considerable material is collected in northeastern Washington, showing individual and age variation, it seems best to retain the name ruficaudus.
Most of the specimens referable to ruficaudus were taken in talus slides high in the Pend Oreille Mountains. A smaller series was collected at a lower altitude in open pine forests near Pend Oreille Lakes, Stevens County. I could detect no difference between amoenus and ruficaudus in habitat or habits.
Description.—The Townsend chipmunk is the largest of the chipmunks that occur in Washington. Head and body measure about 5-1/2 inches; the tail about 4-1/2 inches. The fur of the Townsend chipmunk is more lax and less sleek than that of other species. The upper parts are duller and darker ochraceous. Stripes are wide and not sharply delineated. The dark stripes are deep chestnut or blackish. The upper pale stripe is pale buffy gray; the lower is whitish. The tail is blackish frosted with white above and rich ochraceous below. Underparts are dull white.
The Townsend chipmunk ranges from the Fraser River in southern British Columbia through western Washington and Oregon, to central California. Like other members of the Pacific Coastal Fauna which extend southward to California, its geographic range [Pg 260] extends farther inland to the south and geographic variation is greater; 3 races are listed by Johnson (1943: 114) in California.
The Townsend chipmunk is the largest and darkest chipmunk in Washington. Over much of its range it is the only chipmunk found although in some mountainous areas both townsendii and amoenus occur together. The larger size and richer coloration, especially the rich tawny color of the underside of the tail, separate Townsend chipmunks from amoenus.
Fig. 82. Townsend chipmunk (Tamias townsendii cooperi), captured on Goat Creek, 3,000 feet, western Cascade Mountains near Chinook Pass, Washington, September 16, 1940, by Earl J. Larrison; photographed February 1, 1941. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 1139.)
Townsend chipmunks are closely associated with the coniferous forest where they live in clearings and tangles of underbrush such as on steep hillsides, fire slashings, snowslide areas or mountain glades. Though capable of swift movement, they are less nervous and active than other chipmunks. Rarely are they as abundant, even locally, as other species, and the home range of an individual seems to be larger than that of an amoenus or a minimus. Rarely are more than two seen in a locality. Their more sedentary habits make them less conspicuous than other species. In the fall, when berries are dried out and seeds are ripe, they are most often seen. The Townsend chipmunk seems to be the most arboreal species found in Washington and individuals are not uncommonly seen in trees.
The brushy cover inhabited by Townsend chipmunks protects them from most hawks while their diurnal habits prevent owls from feeding on them. Predatory mammals probably constitute their [Pg 261] greatest menace. A weasel (Mustela frenata) was seen to follow a Townsend chipmunk into a pile of timbers near Cottage Lake, King County, and another was seen carrying a dead Townsend chipmunk at Stevens Pass, King County. Tracks around an area of blood and fur showed where a mink (Mustela vison) had killed a Townsend chipmunk near Cottage Lake, King County.
Fig. 83. Distribution of the Townsend chipmunk in Washington. A. Tamias townsendii townsendii. B. Tamias townsendii cooperi.
Shaw (1944: 278) discovered the brood nest of a Townsend chipmunk on Hurricane Ridge, Clallam County, 4,500 feet elevation. The burrow was in a cool, damp area among surface runs of moles (Scapanus), and led to an underground nest among the roots of a tree. The nest was formed of the gray, moss-like lichen (Usnea), lined within with sedge leaves (Carex spectabilis) and covered outside with leaves of the same sedge. A turning-about chamber was constructed near the entrance. The burrow was single and not branched. Three young, only two or three days old, were in the nest.
Tamias townsendii Bachman, Jour. Acad. Nat. Sci. Philadelphia, 8 (pt. 1):68, 1839.
Tamias quadrivittatus townsendii Allen, Proc. Boston Soc. Nat. Hist., 16:290, 1874.
Tamias asiaticus var. townsendii Allen, Monog. N. Amer. Rodentia, Rept. U. S. Geol. Surv. Terr., 11:794, 1877.
Eutamias townsendii Merriam, Proc. Biol. Soc. Washington, 11:195, July 1, 1897.
Type.—Lectotype obtained near the lower mouth of the Willamette River, Multnomah County, Oregon, by J. K. Townsend in 1834; in Philadelphia Academy of Natural Sciences.
Racial Characters.—Color of sides rich tawny; dark stripes black or dark brown, and pale stripes cinnamon; underside of tail tawny.
Measurements.—Fifteen males and 10 females from western Washington average, respectively: total length 254.7, 258.6; length of tail 116, 122; hind foot 36.7, 37.1; ear 20.5, 20.4; weight 72, 81.2 grams.
Distribution.—The humid coastal belt of western Washington, from the western base of the Cascade Mountains to the Pacific, exclusive of the Olympic Mountains. When A. H. Howell revised the chipmunks in 1929, he employed a concept of a subspecies different from the writer's own. The locality records listed by Howell (1929: 109-112) for Tamias townsendii townsendii and T. t. cooperi are not in agreement with Howell's own distribution map (op. cit.: 107). When the localities listed by Howell are plotted on a map of Washington, the ranges of the two races overlap in some critical areas. Not all of the material examined by Howell was seen by the writer, and, consequently, the ranges shown in Fig. 83 are plotted, in part, on geographic grounds.
Marginal localities on the east, so plotted, for T. t. townsendii, are: Hamilton (U. S. N. M.), 5 mi. E. Monroe (W. W. D.), Redmond (W. W. D.), Roy (U. S. N. M.), and Vancouver (U. S. N. M.).
Tamias cooperi Baird, Proc. Acad. Nat. Sci. Philadelphia, 7:334, 1855.
Tamias townsendii var. cooperi Baird, Mamm. N. Amer., p. 737, 1857.
Eutamias cooperi Lyon, Smithsonian Misc. Coll., 50:89, June 27, 1907.
Eutamias townsendii cooperi Howell, Jour. Mamm., 3:184, August 4, 1922.
Type.—Cotypes obtained at Klickitat Pass, 4,500 ft., Skamania County, Washington, by J. S. Cooper in July, 1853; in United States National Museum.
Racial Characters.—Similar to T. t. townsendii but paler with pale stripes whitish rather than cinnamon.
Measurements.—Ten males and 10 females from the Cascades average, respectively: total length 246.4, 246.8; length of tail 111.7, 107.6; hind foot 35, 35.4; ear 20, 20; weight 77, 89.9 grams.
Distribution.—The higher and eastern Cascade Mountains and the Olympic Mountains. Marginal localities along the Cascades probably include: Swamp Creek (U. S. N. M.), Index (W. S. C.), North Bend (U. S. N. M.), Mt. St. Helens (U. S. N. M.), and Yacolt (M. V. Z.).
Marmota monax petrensis Howell, N. Amer. Fauna, 37:33, April 7, 1915.
Type.—Obtained at Revelstoke, British Columbia, by W. Spreadborough on May 12, 1890; type in United States National Museum.
Measurements.—Howell (1915: 33) gives the measurements of an adult male (the type) as: total length 540; length of tail 127; hind foot 76. An adult female from Barkerville, British Columbia, measured: total length 505; length of tail 125; and hind foot 68.
Distribution.—The Pend Oreille Mountains. Pend Oreille County. The woodchuck was seen and positively identified in northeastern Washington but no specimens were collected.
Description.—The woodchuck is the smallest member of the genus Marmota that occurs in Washington. Adults are about 22 inches in length, of which the tail comprises 5 inches. The body is stout and plump. The legs are short. The ears are low and rounded. The eyes are large but not prominent. The fur is rather stiff but dense. The upper parts are cinnamon, frosted with white-tipped guard hairs. The underparts are tawny. There is no white bar across the nose.
Woodchucks occur from Alaska to Idaho and eastward to the Atlantic, extending southward in the eastern United States. Miller (1924: 173-175) lists seven subspecies, one of which enters the extreme northeastern corner of Washington.
The habits of the eastern woodchuck (Marmota monax rufescens) have been studied by Hamilton (1934: 85-178), but the northern races are less well known. Cowan (1939: 77-79) gives observations on the habits, nests and burrows of Marmota monax canadensis.
Arctomys flaviventer avarus Bangs, Proc. New England Zoöl. Club, 1:68, July 31, 1899.
[Marmota flaviventer] avarus Trouessart, Catal. Mamm., viv. foss., suppl., p. 344, 1904.
Marmota flaviventris avara Miller, U. S. Nat. Mus. Bull., 128:175, April 29, 1924.
Type.—Obtained at Okanogan, British Columbia, by A. C. Brooks on July 17, 1897; type in Museum of Comparative Zoölogy.
Measurements.—A male from 5 miles north of Entiat, Chelan County, measured: total length 610; length of tail 182; hind foot 70; ear 28.
Distribution.—From the eastern edge of the Cascade Mountains eastward, except, apparently, extreme northeastern Washington. Marginal records are:
Okanogan (W. W. D.) in the north, Wenas (W. W. D.) in the west, Pasco (M. V. Z.) in the south, and 14 mi. S. W. Pullman (Howell, 1915: 42) in the east.
Description.—The yellow-bellied marmot is similar to the woodchuck but is slightly larger. It is decidedly paler in color, less reddish, but possesses white-tipped hairs on the dorsal surface, as does the woodchuck. The yellow-bellied marmot also differs from the woodchuck in having a distinct white bar on the nose. Its pelage is coarse and rather thin.
The yellow-bellied marmot is typically an animal of the basalt talus of eastern Washington but occurs in mountainous areas in northeastern Washington. These animals are usually found near streams, ponds, lakes, or rivers. They wander considerably, however, and are often found far from water. Their wandering habits probably account for their presence near temporary streams and ponds on the Columbian Plateau. When these temporary sources of water dry up in July or early August, the marmots go into hibernation. Edson (1935: 68) records a marmot from Bellingham, Whatcom County, far west of the usual range of the species.
The "ground hog" is often hunted for sport and sometimes for food. Near centers of human population the yellow-bellied marmots are extremely shy. Along the highways of the Columbia River on any Sunday in June, it is not unusual to see a dozen cars in an hour, moving slowly past a talus slide while eager hunters scan the rocks for marmots. In the late afternoon, when the marmots leave the protection of the talus slides to drink at the river, they fall easy prey to rifles with telescope sights. Near cities in eastern Washington yellow-bellied marmots have become partially nocturnal.
Couch (1930: 2-6) attempted to excavate several dens of yellow-bellied marmots, but decided to leave the task "to some future road-building crew." Embryos found by Couch numbered three to six. Couch thought the young were born about March 15 in the Snake River area and about April 15 in the upper Okanogan area. The young appear above ground approximately 30 days after birth.
The yellow-bellied marmots enter aestivation from late June to early August, depending on the locality and local conditions. They are active longer in northeastern Washington. Couch records a yellow-bellied marmot seen in Okanogan County on October 10, but regards this as exceptional. The marmots near Wenatchee, Chelan County, emerge from their burrows in early March (March 5 to 10, from reports of residents in 1937, 1938). Couch (1930: 5) gives February 20 to March 15 as the date of appearance.
A principal requirement for marmots is the presence of rocks. At Cle Elum, Kittitas County, I took a marmot from an alfalfa field where a farmer had placed all the surface stones in a loose pile. Fifteen miles east of Tonasket, Okanogan County, marmots were living in the stones piled by road builders to support the ends of a small bridge. A hundred feet away another marmot was living under an abandoned building. A high, convenient rock near their burrow serves the yellow-bellied marmots as a look-out post. These look-out posts seem, in many cases, to have been used by many generations of marmots, for their feces sometimes fill nearby crevices to a depth of several feet and cover the look-out rock itself. The glacial boulders on the plateau between the Okanogan River and Omak Lake, Okanogan County, furnish the best examples of look-out posts. These numerous isolated boulders, ten to fifty feet in diameter and ten to thirty feet high, each seem to furnish shelter to one or more marmots. Well-worn trails lead from the boulders to burrows and feeding areas.
The food of the yellow-bellied marmot includes grasses and succulent plants found in their habitat. Fondness for alfalfa makes them a serious pest in some areas, for their large size enables them to make considerable inroads on a field. Natural enemies probably include most larger predaceous mammals. Hawks and eagles probably kill their young. Near Tonasket, Okanogan County, Robert Dalquest surprised a coyote as it ran across a small wash. A shot caused the coyote to drop a half-grown marmot which it had been carrying.
Marmota caligata cascadensis Howell, Proc. Biol. Soc. Washington, 27:17, February 2, 1914.
Type.—Obtained on Mt. Rainier, 6,000 ft., Pierce County, Washington, by A. K. Fisher on August 11, 1897; type in United States National Museum.
Measurements.—A young adult male from Stevens Pass, King County, measures: total length 773; length of tail 221; hind foot 93; ear 27.
Distribution.—The Cascade Mountains, ranging from Mt. Baker (W. W. D.) and Mt. Chopaka (Taylor and Shaw, 1929: 15) south to Mt. Adams (Taylor and Shaw, 1929: 15).
Description.—The hoary marmot is the largest of the American marmots; adults are 28 inches or more in length, of which the tail makes up about 8 inches. The pelage is dense and rather woolly. The upper parts are not frosted with white-tipped hairs, but are gray washed with blackish. The head is blackish with white facial [Pg 266] markings, and the shoulders, legs and underparts are gray. The tail is dark reddish brown.
Hoary marmots range from Alaska south to Washington and Idaho. Howell (1915: 57-67) recognizes seven races of this species.
Fig. 84A. Distribution of marmots in Washington. A. Marmota monax petrensis. B. Marmota flaviventris avara. C. Marmota caligata cascadensis. D. Marmota olympus.
This mammal of the higher altitudes rarely goes below the Hudsonian Life-zone. It is most common in the talus slides at the lower edge of the Arctic-Alpine Life-zone. Like other marmots, it prefers to live amid loose boulders. The steep talus or "scab rock" slides in the glacial cirques provide an ideal habitat. The crevices and caves beneath the rocks offer concealment for young and adults. A large boulder with a flat top is usually selected as a look-out. Well-worn trails lead from the talus slides to nearby grassy slopes. When surprised in the open, the hoary marmot exhibits a peculiar bounding run, reminiscent of that of tree squirrels. The short tail "follows through" in the leaps. In the rock slides the hoary marmot is surprisingly agile.
The clear, shrill whistle of the hoary marmot is familiar to all who penetrate its haunts. The whistle is remarkably similar to the whistle of a person. Locally the hoary marmot is known as "whistler" or "whistle pig." Individuals emerge from hibernation early in June; most adults retire again by the middle of September. On September 14, 1937, a young of the year was shot and few were seen where they had been common in June. All were shy. Only one adult was seen.
Large hawks and eagles commonly hunt over the rock slides inhabited by marmots, and probably kill very young individuals. Only the larger predators such as bear, cougar, wolf, coyote, lynx, and bobcat would be expected to kill an adult hoary marmot.
Arctomys olympus Merriam, Proc. Acad. Nat. Sci. Philadelphia, p. 352, October 4, 1898.
[Marmota] olympus Trouessart, Catal. Mamm., viv. foss. suppl., p. 344, 1904.
Type.—Obtained at timber-line, head of Sol Duc River, Clallam County, Washington, by C. H. Merriam, on August 27, 1897; type in United States National Museum.
Measurements.—Two small but adult females from Deer Park, Clallam County, measure respectively: total length 758, 691; length of tail 163, 161; hind foot 106, 97; ear 31, 29.
Distribution.—This species is confined to the Olympic Mountains.
Description.—The Olympic marmot closely resembles the hoary marmot, and differs in being slightly larger, and reddish or rusty brown where the hoary marmot is gray. It lacks the blackish overwash of the hoary marmot. The nose is whitish.
Although the Olympic marmot belongs to the hoary marmot group it is a distinct species, most closely related to the marmot of the mountains of Vancouver Island. Its habits differ but little from those of the hoary marmot. It lives in burrows in talus slides and boulder piles near timber line. A few burrows are dug beneath logs. Well worn trails lead from burrows to feeding grounds on nearby grassy slopes and heather meadows. High rocks or logs serve as lookouts. The alarm whistle is similar to that of the hoary marmot.
Spermophilus townsendii Bachman, Jour. Acad. Nat. Sci. Philadelphia, 8:61, 1839.
Spermophilus mollis yakimensis Merriam, Proc. Biol. Soc. Washington, 12:70, March 24, 1898.
[Citellus mollis] yakimensis Trouessart, Cat. Mamm., Sup., p. 339, 1904.
Citellus townsendii townsendii Howell, N. Amer. Fauna, 56:60, May 18, 1938.
Fig. 84B. Distribution of the Townsend and Washington ground squirrels in Washington. A. Citellus townsendii townsendii. B. Citellus washingtoni.
Type.—Obtained on the western bank of the Columbia River "about 300 miles above its mouth" (probably near the mouth of the Walla Walla or Touchet river; more exactly, opposite Wallula, in Benton County, Washington) by J. K. Townsend in July, 1836; type in Academy of Natural Sciences of Philadelphia.
Measurements.—Ten adults from Mabton and North Yakima, Yakima County, average: total length 212.2; length of tail 45.7; hind foot 33.9.
Distribution.—The Upper Sonoran Life-zone area of the Yakima Valley from Ellensburg (Howell, 1938: 63) south to the Columbia at Kennewick (W. W. D.).
Remarks.—This race is probably derived from C. t. mollis of Oregon, from which it is but slightly differentiated. It was known for many years as Citellus mollis yakimensis. For use of the name townsendii see Howell (1938: 62).
The head and body are about 6-1/2 inches long and the tail is about 2 inches in length. The head is large with low, rounded ears and large eyes. The neck is distinctly constricted and the body is plump. Adults are "pot-bellied." The legs are short and the feet small. The tail is short, round at the base but bushy throughout the rest of its length. The pelage is short and rather harsh. In color the upper parts are yellowish gray flecked with tiny, pale dots. The underparts are buffy. The face, thighs and tail are reddish buff.
Ground squirrels occur in parts of Europe, Asia, and much of western North America. Howell (1938: 36-37) recognizes eight subgenera and thirty-one species in North America. Three subgenera occur in Washington, namely: Citellus, the spotted ground squirrels, represented by Citellus townsendii, Citellus washingtoni and Citellus columbianus; Otospermophilus, the long-tailed ground squirrels, represented by Citellus beecheyi; and Callospermophilus, the striped ground squirrels, represented by Citellus lateralis and Citellus saturatus.
All ground squirrels are diurnal and consequently are familiar to man. Most species bear local names and the striped or mantled ground squirrels are often mistaken for chipmunks. Some species are destructive to crops, especially grain. In addition they harbor sylvatic plague. Economically the genus Citellus is of great importance in Washington.
All Washington ground squirrels live in burrows which they construct themselves. The life histories of the two striped species are less well known than those of the more economically important species. All species hibernate. The striped species disappear in October and reappear the following March. This can be called true hibernation. The long-tailed ground squirrels probably hibernate at high altitudes but aestivate in more arid localities. In Washington the Douglas ground squirrel occupies an area that is relatively temperate and humid. They may hibernate or aestivate, depending on local conditions, or they may remain active all year. The spotted ground squirrels disappear in midsummer and sleep (aestivate) until the following spring, for in their arid habitat the [Pg 270] disappearance of green food in late summer and fall makes living conditions almost as unfavorable as in winter.
Townsend ground squirrels range from central Washington south to southern Nevada and from the Cascade Mountains east to eastern Idaho and central Utah. Five races are recognized, only one of which occurs in Washington, where it is confined to the Upper Sonoran Life-zone. It occupies the sagebrush area, being most common where the sage is in scattered clumps separated by grassy areas. Occasionally these squirrels occupy extensive grasslands where sage is scarce and in the Yakima area may enter pastures and fields. They live in colonies, often with more than ten separate burrows to the acre.
Burrows are dug in dusty ground, either near sage bushes or in openings among them. Well-worn trails lead from feeding places to openings of burrows. Mouths of burrows are usually situated on flat ground and are surrounded by a rim of excavated earth four to six inches high. If located on a slope, the excavated earth collects as a mound on the lower side, and serves as a look-out post for the squirrels. Burrows excavated by Scheffer are described by Howell (1938: 5). One reached a depth of 5-1/4 feet at a point where it branched 11 feet from the entrance. One branch led to the nest chamber three feet to the right of the main tunnel. The other reached a depth of six feet at a point 14 feet from the entrance, then turned upwards at a 70 degree angle and reached the surface through a partly obstructed entrance, 12 feet from the original entrance. The nest chamber was 6-1/2 inches in diameter and filled with a perfectly dry nest of fine grass, partly broken and shredded.
Food consists of soft green vegetation and seeds. Foods listed by Howell (1938: 5) include: Sphaeralida munroana, Plantago purshii, Bromus tectorum, Agropyron pauciflorum, Oryzopsis hymenoides, Norta altissima, Artemisiae spinescens, sunflower, alfalfa, wheat, barley, potato, beets, carrots, lettuce, and insects (grasshoppers, cicadas). In 1917, these squirrels were reported to have practically destroyed a ten-acre field of beets at White Swan, Yakima County.
Aestivation of older individuals begins in late May and the last young disappear in early June. The squirrels appear again in late January, before the snow has disappeared (Scheffer, 1941: 272).
The voice of the Townsend ground squirrel is a faint, high pitched "pe-eee-ep," of remarkable carrying power for so faint a sound, and extremely difficult to trace to its source. Scheffer (in Howell, [Pg 271] op. cit., p. 6) mentions also a chirping sound and a chatter of alarm which I have not heard.
Five to seven embryos were found in specimens collected near Yakima. Scheffer (1941: 270) found the number of fetuses in 52 pregnant females from the Kennewick area to vary from 4 to 16 with an average of 8.6. Francis (1922: 5) reported tularemia in this species. They probably harbor also sylvatic plague.
Citellus washingtoni washingtoni Howell, N. Amer. Fauna, 56:69, May 18, 1938.
Citellus washingtoni loringi Howell, N. Amer. Fauna, 56:71, May 18, 1938 (type from Douglas, Douglas County, Washington).
Type.—Obtained at Touchet, Walla Walla County, Washington, by C. P. Streator on May 18, 1891; type in United States National Museum.
Distribution.—The Columbian Plateau and southward into Oregon, ranging from Farmer (Howell, 1938: 71) and Moses Coulee (W. W. D.) south to Wallula (M. V. Z.).
Measurements.—Fifteen specimens from the vicinity of the type locality average: total length 229.2; length of tail 50; hind foot 35.3. A male from 4 miles west of Pasco, Franklin County, weighed 201 grams.
Remarks.—Citellus w. loringi allegedly differs from typical washingtoni in smaller size. The specimens collected by us from within the range of loringi are as large as those from farther south. C. w. loringi is regarded as a synonym of C. w. washingtoni.
Description.—The Washington ground squirrel closely resembles the Townsend ground squirrel in size and appearance. It differs principally in color. The upper parts are brownish gray marked with distinct white spots, and the underparts are buffy. The face, thighs and tail are dull reddish.
This species is closely related to the Townsend ground squirrel but its habitat is more varied. It is most common in areas of low sage bushes surrounded by grasslands and on extensive grasslands, but occurs also in sandy places, wheat fields, and rocky hillsides. The animals live in colonies, in some places 50 or more individuals to the acre. They occur also as scattered individuals or small colonies in smaller areas of suitable habitat. For example, along the highway from Farmer, Douglas County, to Waterville, a distance of 15 miles, a band of natural grass, 50 feet or less in width, lies between the road and the extensive wheat fields beyond. This strip of natural cover possessed about one Citellus to each 200 feet along most of its length. The squirrels were often seen sitting on rolls of wire drift fence, used to keep snow drifts from the road in [Pg 272] winter but in summer rolled into bundles 3 feet in diameter and left standing at intervals of one each 100 feet. The squirrels had constructed their burrow entrances beneath these rolls and used the rolls as look-out posts. At my approach they would dodge into the rolls of wire and, unless scared, remained in the wire roll. By approaching carefully and overturning the rolls I was able to trap and capture a good series of living specimens.
Burrows, nests, habits, and food of this species seem identical to those of townsendii. Aestivation dates seem to be the same (Scheffer, 1941: 270-279). The hibernation of this species has been discussed in detail by Svhila (1939: 6-10). Food species listed by Scheffer at Wallula (Howell, 1938: 8) are identical to those of townsendii at Kennewick, across the Columbia River (see account of townsendii).
Scheffer (1941: 270-279) examined 26 pregnant females of this species and found the fetuses to vary from 5 to 11 in number, with an average of 8.
Description.—The Columbian ground squirrel has the general body proportions, large head, large eyes, low, rounded ears, plump body and short tail of the Townsend ground squirrel, but is much larger. The head and body of an adult measure about 10 inches and the tail about 4 inches. The upper parts are grayish buff mottled with round white dots. The underparts and forefeet are pale ochraceous. The back of the head and neck are gray. The face, thighs and tail are reddish.
The Columbian ground squirrel occupies inland mountainous areas from central British Columbia to central Oregon and Idaho and from eastern Washington and Oregon to western Montana and southwestern Alberta. It is closely related to the arctic ground squirrels of the Citellus parryii group, which are in turn related to the Siberian forms, buxtoni and stejnegeri.
The habitat of the Columbian ground squirrel is varied but is usually more humid than that of the Townsend and Washington ground squirrels. The most arid of the lands occupied by them in Washington are the grasslands and wheat fields of the Poulouse country and the eastern border of the state. In northeastern Washington they live in meadows and grasslands in the valleys, in openings in the coniferous forest at higher altitudes and in parks and alpine meadows almost to timber-line on the mountains. Some individuals [Pg 273] were in brushy places or even in forests, far from grassy clearings. Near Republic, Ferry County, a Columbian ground squirrel was killed as it ran along a log in dense larch woods. Its burrows were found in a tangle of brush and fallen logs. No clearings, meadow or grassland existed nearby. Narrow bands of cleared land beside roads and railroad tracks are extensively used by these squirrels. Where small meadows or pastures occur, this species lives in dense colonies. If grasslands are extensive it lives in loose colonies, often with considerable distances between individual burrow systems. In the mountains the squirrels are scattered or live in small groups.
Burrows of the Columbian ground squirrel are often constructed in the open. Mouths of burrows, especially if the animals are living in colonies, usually are marked by a large pile of excavated earth. If, however, there is some large object on the surface of the ground, such as a stone, stump, or log, the entrance to the burrow is located beside this. Such objects are often undermined and made to settle, eventually becoming buried. The squirrels live under houses, and the yards of abandoned ranch buildings are often infested with them.
The burrows of the Columbian ground squirrel have been described by Shaw (1919, 1924B, 1925, A, B, E, 1926) and Bailey (1918: 47). Two types of burrows are constructed, summer burrows and hibernation dens. Summer burrows are used year after year, perhaps by succeeding generations of squirrels. They vary considerably but usually are 3 or 4 inches in diameter and possess several entrances. The depth to which the burrow descends, as well as the number of forks and subsidiary burrows, depends to some extent on the whims of the individual squirrel and the length of time the burrow has been occupied. Usually the burrow reaches a depth of four feet and two or more branches are present. A turning-about chamber is present, not far from the entrance. Here a startled squirrel can turn about and watch the entrance and, if the alarm was false, the squirrel resumes its activities above ground. Nests are above the level of the main burrow system and thus are protected from flooding by rain or melting snow. Stored food and excrement are deposited in separate compartments. Escape entrances, concealed in grass and weeds, for use in case the burrow is invaded, are dug from beneath the surface of the ground and thus are unmarked by any telltale mound of earth.
Separate dens are used for aestivation. While the squirrel is in aestivation the entrances to the summer burrow are solidly plugged with tamped earth. Aestivation dens are short and may or may not communicate with the summer burrow. Shaw (1925B: 58) measured 50 such dens and found the aestivation cell to average 2 feet 6 inches beneath the surface with extreme depths of 6 inches and 4 feet 11 inches. Ordinarily a drainage burrow is dug beneath the level of the nest.
Fig. 85. Distribution of the Columbian and Beechey ground squirrels in Washington. A. Citellus columbianus columbianus. B. Citellus columbianus ruficaudus. C. Citellus beecheyi douglasii.
The time and length of aestivation is variable. Old males enter aestivation before the females and young, and emerge earlier in the spring. In the lowlands, as about Pullman, Whitman County, the squirrels begin to disappear in mid July. In the mountains of northeastern Washington they are active until late August. They emerge again in late February or early March. Shaw (1925B) mentions that food is occasionally stored in aestivation dens of male squirrels. This food is not eaten in the winter but in the spring, when the squirrels awaken.
The gestation period was determined by Shaw (1925C: 108) as 24 days. Two to five, rarely seven, young are born. The average litter is about 3.5. Mating occurs in late March and the young are born about the middle of April.
Most green vegetation occurring in its habitat is food for the Columbian ground squirrel. Bulbs, seeds, fruit, berries, grain, clover, alfalfa, and garden truck are eagerly eaten. The squirrels are especially fond of wheat and great damage results from their depredations. In northeastern Washington some wheat fields are almost entirely destroyed. Shaw (1925G) showed that one squirrel destroyed an average of more than 50 pounds of wheat in a season. When populations range as high as ten squirrels to the acre, little wheat is left.
In Montana the Columbian ground squirrel is a known reservoir of Rocky Mountain spotted fever (Birdseye, 1912: 1-46) while sylvatic plague has been reported in this species in Oregon (Meyer, 1936: 965). Control of the Columbian ground squirrel by traps, poison, and shooting is only locally successful.
Arctomys columbianus Ord, Guthrie's Geography, 2d American Edition, 2:292 (description, p. 303), 1815.
Spermophilus columbianus Merriam, N. Amer. Fauna, 5:39, July 30, 1891.
Citellus columbianus Allen, Bull. Amer. Mus. Nat. Hist., 19:536, October 10, 1903.
Type.—None designated. Description based on Lewis and Clark's account of animals taken by them between the forks of the Clearwater and Kooskooskie rivers, Idaho.
Racial characters.—Smaller size and paler color than in ruficaudus.
Measurements.—Five females from northeastern Washington average: total length 346; length of tail 77; hind foot 48.6; ear 20.
Distribution.—Northeastern Washington, from 15 mi. E. Tonasket (W. W. D.) east to Pass Creek Pass (U. S. N. M.) and south to Pullman (W. S. C.) along the eastern edge of the state.
Citellus columbianus ruficaudus Howell, Proc. Biol. Soc. Washington, 41:212, December 18, 1928.
Type.—Obtained at Wallowa Lake, Wallowa County, Oregon, by G. G. Cantwell on April 13, 1919; type in United States National Museum.
Racial characters.—Similar to Citellus columbianus columbianus but tail redder, especially above, and red color everywhere deeper.
Distribution.—Restricted to the Blue Mountains of extreme southeastern Washington.
Remarks.—This is at best a slightly differentiated race and upon further study, animals assigned to it may prove unworthy of recognition as a distinct subspecies.
Arctomys? (Spermophilus?) douglasii Richardson, Fauna Boreali-Americana, 1:172, 1829.
Spermophilus douglasii F. Cuvier, Sup. a l'hist. natur. Buffon, 1:333, 1831.
[Spermophilus grammurus] var. douglasii Allen, Proc. Boston Soc. Nat. Hist., 16:293, 1874.
Citellus v[ariegatus]. douglasii Elliot, Field Columb. Mus. Publ. 76, zoöl. ser. 3:183, May, 1903.
Citellus beecheyi douglasi Grinnell, Proc. California Acad. Sci., 3 (ser. 4):345, August 28, 1913.
Otospermophilus grammurus douglasii Miller, U. S. Nat. Mus. Bull., 128:18, April 29, 1924.
Citellus beecheyi douglasii Howell, N. Amer. Fauna, 56:150, May 18, 1938.
Type.—None. Description based on a hunter's skin from "Banks of the Columbia," probably near The Dalles, Wasco County, Oregon.
Measurements.—A male and 6 females from Klickitat County, average, respectively: total length 469, 480; length of tail 181, 198; hind foot 58, 62; ear 26, 26.
Distribution.—The Columbia River Valley from Goldendale (W. W. D.) west to the White Salmon River (W. W. D.).
Remarks.—Ground squirrels have been common on the Oregon side of the Columbia River for a long time. The specimen upon which the description was based was presumably obtained there in the early 1800's. Yet the species did not become established in Washington until 100 years later, in about 1915.
The head is large with eyes and ears of moderate size. The body is rather stout; thicker than that of a tree squirrel but slimmer than that of the Columbian ground squirrel. Head and body are about 11 inches in length and the tail is about 7 inches long. The upper parts are dark brownish or blackish gray heavily marked with white spots. A triangular area at the shoulders is clear black outlined in whitish. The head and neck are grayish brown. The underparts are buffy. The tail is gray above and buffy beneath.
This species belongs to the subgenus Otospermophilus. Externally the species beecheyi differs from Citellus washingtoni, townsendii, and columbianus, all of which belong to the subgenus Citellus, in possessing more slender limbs, a longer, thinner body, a rather bushy tail that is nearly two-thirds, rather than less than half, the length of the body, and larger, more prominent ears. The Beechey ground squirrel lacks also the hazel color of nose and thigh that is characteristic of the subgenus Citellus in Washington.
Citellus beecheyi ranges from the southern edge of the state of Washington southward through western Oregon and California into northern Lower California. It is a western coastal species and [Pg 277] reaches eastward only as far as Nevada. A related species, Citellus variegatus, occurs farther inland, from Utah, Colorado, and Texas, southward to central Mexico. Eight subspecies of Citellus beecheyi are recognized by Howell (1938), all but three of which are restricted to California.
At present the Beechey ground squirrel occupies a limited area of Washington, which it has invaded in recent years (Scheffer and Dalquest, 1939: 44). However, it is extending its range and may be expected eventually to occupy a considerable part of the state.
The habitat of this ground squirrel is varied. It occupies a more humid terrain than do most members of the genus. At the present time it is most common in grassy fields and rocky outcrops along the Columbia River. It is common also in the open oak groves on the hillsides back from the river valley and some individuals were seen near Guler, well up on the slopes of Mount Adams. As observed in Washington, its favored habitat is about rock outcrops and talus slides near extensive fields or grasslands. In California and Oregon, however, it occurs among other places, in scattered fields and meadows of the coniferous forests in the humid subdivision of the Transition Life-zone. The range of tolerance of the species is such that it might extend its range into much of western Washington, including the Puget Sound area. It may be expected to move northward through the eastern Cascades.
Because of its limited range and the short time of its establishment, the Beechey ground squirrel has been studied little in Washington. According to Howell (1938: 28), this race is less prolific and abundant than other members of the species. Of the Beechey ground squirrel, certainly the subspecies C. b. douglasii, is less common, locally and over wide areas, in its range in California and Oregon, than are the subspecies C. b. beecheyi and C. b. fisheri in California. In Washington, on the other hand, douglasii is locally abundant, perhaps more so than elsewhere.
The habits of this race in California have been studied by Grinnell and Dixon (1919: 595-807). Burrows have been excavated near Eugene, Oregon, and the findings published (Edge, 1934: 189-193). Burrows were dug on sloping or well-drained ground. Mounds of earth at the entrances were usually inconspicuous but well-worn trails led to the burrows. The burrows entered the ground at an angle of 35° for about two feet, then flattened out horizontally for six feet or more. The burrows were from 1 to 4 feet deep, usually about 2 feet. Ordinary burrows had a single entrance but some [Pg 278] older burrows had two or more. Burrows branched frequently. Nest chambers were of the shape of a flattened sphere 10 inches in diameter. Frequently more than one nest was found in a burrow, but only one was in use at a time. Some nests were infested with fleas. Nests frequently contained food. Most of the burrows observed by me had their entrances beside rocks or oak roots. In alfalfa fields near Bingen and Lyle, Klickitat County, the burrows were in open fields and the entrances were marked by large mounds of earth. In actual diameter the burrows seemed smaller than those of Citellus columbianus.
Aestivation and hibernation of the ground squirrels in the lower Columbia River Valley seem unnecessary, for the climate is temperate and humid. It is not established that douglasii hibernates in other parts of its range where the climate is mild. Specimens shot at White Salmon in early March were lean and gave no evidence of having hibernated.
The Beechey ground squirrel is an excellent climber; I have seen individuals in oak trees on a number of occasions. They like to sit on fence posts and when alarmed descend head foremost, with speed and agility. In running on the ground their movements are more graceful than those of Citellus columbianus but are unlike the flowing, bounding run of tree squirrels.
Near White Salmon and Lyle, these ground squirrels were eating burr clover (Medicago), grasses (Bromus), and alfalfa. In some places they do considerable damage to alfalfa fields. They probably eat acorns and are said to eat some insects.
In California the young are born in May and number five to seven.
No disease has been recorded for this subspecies but a related form in California (Citellus b. beecheyi) is known to harbor plague (Kellogg, 1935: 857) and tularemia (McCoy, 1911: 53-71).
Description.—The head and body measure about 6 inches and the tail about 4 inches. The head is large and rounded with eyes and ears of moderate size. The body is stouter than that of a chipmunk. The tail is long and well furred. Upper parts are buffy gray with one pale stripe bordered by two black stripes extending from shoulders to rump, and the underparts are buffy. The head and shoulders are tawny. The tail is buffy beneath and darker above.
Externally golden-mantled ground squirrels somewhat resemble chipmunks but actually are no more closely related to chipmunks [Pg 279] than are other ground squirrels. They are larger than chipmunks and their stripes differ in that there is but one pale stripe on each side bordered with black stripes rather than two pale stripes, and in that the stripes end at the shoulders rather than continuing on to the face.
These ground squirrels are distributed in mountainous areas of western North America from central British Columbia south to central Arizona and New Mexico. A closely related species (Citellus madrensis) occupies a limited area in northern Mexico and another lives in the Cascades of Washington. There are two subspecies of lateralis in Washington, each with a major part of its range outside of Washington.
This species inhabits mountain clearings, parks and talus slides, where it is most common about rocks, stumps, and logs. Its fondness for talus slides and outcrops has earned it the common name of "rock squirrel" in some places.
Entrances to burrows are usually beside a rock, stump, or root, or are concealed under talus slides. A burrow excavated by Hatt (1927) in Colorado was three inches in diameter at the entrance and uniformly two inches in diameter for the rest of its length. Most of the burrow was 8 inches beneath the surface. The tunnel branched twice and contained one pocket, probably a turning or passing chamber, in addition to the nest. "A runway surrounded the nest on three sides, from which there were four passages leading in. The nest cavity was 4 inches deep, the nest not filling the space available, but occurring more as a mat in the bottom of a cup."
The habits of this ground squirrel in Washington are little known. Since it occupies areas that are cold and under deep snow in winter, it probably hibernates. According to Howell (1938: 32) these animals become fat in the fall and must retire for the winter about the middle of September.
The food habits of these ground squirrels in Washington are little known. In the Pend Oreille Mountains, Pend Oreille County, I found them eating the fruit of the western raspberry (Rubus leucodermis). Howell lists, as food of this species, the seeds of yellow pine and douglas fir, serviceberries, other berries and seeds, grain, mushrooms, and several kinds of insects.
Callospermophilus lateralis tescorum Hollister, Smithsonian Misc. Coll., 56 (no. 26):2, December 5, 1911.
Citellus lateralis tescorum Elliot, Check-list Mamm. N. Amer., Supp., p. 29, 1917.
Type.—Obtained at the head of Moose Pass Branch, Smoky River, Alberta (near Moose Pass, British Columbia), 7,000 ft., by N. Hollister on August 2, 1911; type in United States National Museum.
Racial characters.—Large size, gray color.
Measurements.—A female from Pass Creek Pass, Pend Oreille County, measures: total length 257; length of tail 90; hind foot 40; ear 14.
Distribution.—Known only from the Pend Oreille Mountains, Pend Oreille County, at Pass Creek Pass (W. W. D.).
Fig. 86. Distribution of golden-mantled ground squirrels in Washington. A. Citellus lateralis tescorum. B. Citellus lateralis connectens. C. Citellus saturatus.
Callospermophilus chrysodeirus connectens Howell, Jour. Mamm., 12:161, May 14, 1931.
Citellus lateralis connectens Howell, N. Amer. Fauna, 56:205, May 18, 1938.
Type.—Obtained at Homestead, Oregon, by H. H. Sheldon on June 1, 1916; type in United States National Museum.
Racial Characters.—Similar to C. l. tescorum but browner, more buffy and less grayish, mantle brighter, size slightly smaller.
Measurements.—Seven males from northeastern Oregon average (Howell, 1938: 206): total length 266; length of tail 92; hind foot 41.7; ear 14.8 (dry). An adult female from Godman Springs, Columbia County, measures: total length 257; length of tail 87; hind foot 37; ear 14.
Distribution.—The Blue Mountains of extreme southeastern Washington.
Remarks.—This race differs markedly from the subspecies that occupies northeastern Washington, but differs but little from Citellus l. chrysodeirus of the Cascades of Oregon.
Tamias lateralis saturatus Rhoads, Proc. Acad. Nat. Sci. Philadelphia, 1895:43, April 9, 1895.
[Spermophilus lateralis] saturatus Elliot, Field Columb. Mus. Publ. 45, zoöl. ser., 2:83, 1901.
Citellus lateralis saturatus Elliot, Field Columb. Mus. Publ. 105, zoöl. ser., 6:106, 1905.
Callospermophilus lateralis saturatus Miller, U. S. Nat. Mus. Bull., 79:316, December 31, 1912.
Citellus saturatus Howell, N. Amer. Fauna, 56:212, May 18, 1936.
Type.—Obtained at Lake Keechelus, 3000 ft., Kittitas County, Washington, by A. Rupert, in September, 1893; type in Academy of Natural Sciences of Philadelphia.
Measurements.—Ten males from the Cascade Mountains average: total length 305; length of tail 110.9; hind foot 46.5; ear 17 (dry). The weights of 3 males and 5 females average, respectively: 281 grams; 259.4 grams.
Distribution.—The higher and eastern Cascade Mountains from Barron (Howell, 1938: 213) and Bauerman Ridge (Howell, 1938: 213) south to Cleveland (Howell, 1938: 213) and Goldendale (W. W. D.).
Remarks.—The golden-mantled ground squirrel of the Cascades is similar to Citellus lateralis but is larger and duller-colored. Head and body are about 8 inches in length and the tail is about 4-1/2 inches long. The upper parts are brownish gray with one pale and two dark stripes on each side. Head and shoulders are tawny. The underparts are dull buffy gray.
Citellus saturatus occupies the Cascade Mountains of Washington and southern British Columbia. It inhabits talus slides and clearings. The rock embankment of the Great Northern Railroad between the Cascade Tunnel and Leavenworth, Chelan County, is a favored habitat and the population along the railroad probably averages one squirrel each hundred yards. To the east it extends well into the Transition Life-zone, being abundant in clearings in the open forest of yellow pine. In places its range nearly reaches the Upper Sonoran Life-zone. A few miles east of Leavenworth, Chelan County, we found this squirrel in chapparal and brush, principally composed of Ceanothus creneatus. Here the animals were unusually abundant and formed a colony almost as dense as colonies of Citellus washingtoni.
Entrances of burrows are usually placed beside rocks, stumps or logs. In the area near Leavenworth, mentioned above, entrances [Pg 282] to burrows were in the open or among roots of bushes. The entrances to many burrows are doubtless concealed under talus slides. No complete records of excavations of burrows are available. One burrow, uncovered by road construction operations three miles east of Scenic, Chelan County, was dug in the earth-filled cleft of a great granite boulder. The cleft was 5 feet wide at the surface of the ground but narrowed until at a depth of six feet the stones were in contact. The cleft was at least 10 yards long. The burrow descended at an angle of 45 degrees, to a depth of three feet. Here the construction work had passed the cleft, but digging into the almost vertical wall uncovered the nest at the end of a horizontal tunnel two feet farther on. It was a matted cup of dry grass with two fresh, green fern fronds lying loose in the cup. Two divergent burrows emerged on opposite sides of the nest cavity, but a fall of the loose, gravelly soil prevented further observation. The body of the occupant, crushed and mangled by the steam-shovel scoop, was that of a small male.
Fig. 87. Golden-mantled ground squirrel (Citellus saturatus), captured when young at Tye, Washington, by Earl J. Larrison, June 20, 1940; photographed February 1, 1941. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 1139.)
In spite of their vivacious appearance, these squirrels are rather sedentary. When undisturbed they move leisurely over rocks and stumps, pausing often, and occasionally sitting on their hindquarters to gaze about for minutes at a time. They are good climbers and [Pg 283] often ascend smooth-barked trees to heights of 20 feet or more. At the approach of danger they descend and enter the nearest burrow. In August these ground squirrels become exceedingly fat and by late September only a few may be seen, basking in the midday sun. These take alarm at the slightest excuse. All have usually gone into hibernation by October 15.
The food includes the berries of salal, huckleberry, mountain ash, and seeds of lupine. Near Liberty, Kittitas County, a squirrel killed by a car had its cheek pouches stuffed with garden peas. The source of its loot was doubtless the garden of a farmhouse 100 feet away. Numerous individuals are run over by cars, and on many occasions I have found other individuals eating the sun-dried flesh of their mates, parents, or young. These cannibals are often run over while so engaged, and it is not unusual to find two or three dead on a section of pavement 20 feet in length.
Economically this species is of little importance. Its principal enemies probably include hawks, weasels, martens, bobcats and coyotes.
Sciurus griseus Ord, Jour. de phys., 87:152, 1818.
Sciurus griseus griseus Miller, U. S. Nat. Mus. Bull., 128:222, April 29, 1924.
Type.—None. Described from a squirrel seen by Lewis and Clark. Type locality, The Dalles, Wasco County, Oregon.
Measurements.—A female from midway between Satus Pass and Goldendale, Klickitat County, measured: total length 560; length of tail 264; hind foot 76; weight 897 grams.
Distribution.—Southwestern Washington north to the glacial prairies near Tacoma (W. W. D.); the Columbia River Valley of the southern Cascades; the eastern edge of the Cascades north to Lake Chelan (Taylor and Shaw, 1929: 18).
Fig. 88. Distribution of the western gray squirrel, Sciurus griseus, in Washington.
Description.—This is the largest tree squirrel found in Washington. The head and body measure about 12 inches, and the tail about 11 inches. The body is long and slender. The fur is long and soft, that on the tail being exceptionally long. The upper parts are silvery gray and the underparts white.
The western gray squirrel is restricted to the region of the Pacific Coast and ranges from central Washington south to northern Lower California. Three subspecies are recognized, only one of which occurs in Washington. Like the red squirrels, the western gray squirrel is arboreal. Its favored habitat is the oak woods rather than coniferous forest. Its range in Washington is largely regulated by the distribution of oaks, especially the garry oak (Quercus garryana). Altitudinally it ranges from near sea level at Puget Sound to above 2,500 feet elevation in the eastern Cascade Mountains. It seems to be restricted to the Transition Life-zone.
The habits of the western gray squirrel are modified by its arboreal existence. Homes consist of holes in hollow trees or outside nests of sticks and twigs. It is extremely active in trees and travels from tree to tree on branches that seem scarcely strong enough to bear the weight of so large an animal. The oak woods in Washington are usually rather open and the trees more scattered than is the case in coniferous forests. Consequently gray squirrels must more often descend to the ground than Douglas and red squirrels. On the ground, western gray squirrels travel in smooth leaps each of two feet or more in length. The long tail is held out behind and "follows through" the animal's leaps in a remarkable way. The tip of the tail may be descending from the previous leap while the animal's fore parts are already at the height of a new jump. This results in a "flowing" movement that is extremely graceful. The squirrel may stop momentarily to dig in the ground or search for an acorn. At such times the tail is immediately switched up, over the back.
The western gray squirrel is able to drop unharmed for considerable distances. Near Fort Lewis, Pierce County, a gray squirrel was cornered in a tall fir tree and an agile friend volunteered to climb the tree. The squirrel ascended to the very topmost branches. When only a few feet separated them the squirrel leaped far out into the air. Its legs were stretched out stiffly, the tail was extended and the body slightly arched. It struck the ground with an audible thud and bounced fully 18 inches. At the height of its bounce, the squirrel's legs began moving rapidly, and it struck the ground the second time at a full run.
The principal food is acorns, although the seeds of the Douglas fir and probably other conifers are eaten. Four embryos were found by Victor B. Scheffer in a specimen from Klickitat County on March 20, 1939.
Sciurus carolinensis hypophaeus Merriam, Science, 7:351, April 16, 1886.
Type.—Obtained at Elk River, Sherburne County, Minnesota.
Description.—Size large, slightly smaller than the western gray squirrel; color of upper parts less silvery, more reddish, especially on the dorsal area and top of the tail.
Remarks.—The eastern gray squirrel was introduced at Woodland Park, Seattle, in 1925. The original stock (7 pairs) came from Minneapolis, Minnesota. It has spread through the nearby woods, around Green Lake, Cowan Park, and to the woods on the University of Washington Campus. Occasional individuals are found outside the city limits, but the species seems not to spread away from the city.
[Sciurus] niger Linnaeus, Syst. Nat., 1 (10th ed.):64, 1758.
Description.—Similar in size and appearance to the eastern gray squirrel but upper parts more reddish and underparts reddish orange rather than white.
Remarks.—Occasional fox squirrels are encountered near Seattle where they have been introduced from the southeastern United States. The source and date of the introduction are unknown.
Description.—The head and body of the red squirrel measure about 7-3/4 inches, the tail about 5 inches. It may be recognized by its trim body, bushy tail and white underparts. The upper parts are reddish gray, reddest on the dorsal area. The red color of the center of the back extends to the tail. A black line separates the dark upper parts from the white underparts.
Red squirrels range over North America from the northern limit of tree growth south through the United States to Tennessee and North Carolina. A related species, douglasii, is found along the Pacific coast from British Columbia to California. The genus Tamiasciurus differs from Sciurus of Washington in lacking a penis bone or baculum. There are other fundamental differences in anatomy (see Mossman, Lowlah and Bradley, 1932: 89-155).
The habitat of the red squirrels is the coniferous forests from which they rarely stray. Zonally they range through the Transition and Canadian life-zones into the Hudsonian Life-zone. Red squirrels are arboreal and most of their habits are modified by arboreal existence. They are swift and agile climbers, able to travel from tree to tree on slender twigs or by leaping as much as ten feet to span the distance from one branch to another. They ascend and descend trees head first. They hang by their hind feet, high in the air, to clip the cones of conifers. If cornered in a tree they leap far out and, by extending the legs and tail stiffly, fall to the ground unharmed by leaps of fifty feet or more.
Fig. 89. Douglas squirrel (Tamiasciurus douglasii douglasii): feeding station with remnants of Douglas fir cone, Longmire, Washington, elevation 2,700 feet, June 25, 1937. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 268.)
The homes of red squirrels are woodpecker holes or other holes in hollow trees. More rarely they build outside nests of twigs and branches, about two feet in diameter, or add onto old nests of crows or jays. Most nests and holes are some distance from the ground but some holes are between roots on the ground itself.
The food consists principally of the seeds of coniferous trees, especially Douglas fir and various species of pines. Cones are clipped when green or just before ripening and are either allowed to fall to the ground to be retrieved later or are at once carried to a favored feeding place to be consumed. Cones are held between the forepaws while the squirrel sits on its hind feet, tail curved up over its back, and rapidly clips the cone apart to get the seeds [Pg 288] it contains. The cone is rotated between the paws and a steady stream of husks drops to the ground. Soon only the core is left and this too joins the husks on the ground. Favored feeding stations are used continually, perhaps by generation after generation of squirrels, and debris from thousands of cones accumulates in great piles.
Cones are stored. In the vicinity of a favored feeding place, at times virtually every hiding place is filled with green cones. Cones may be jammed into cracks or crevices in logs or stumps without effort at concealment or may be carefully covered with leaves or dry needles. Many are placed in craterlike pits dug in the ground. Most of these pits probably are later covered over but many are left open to the weather. Hollows in trees are probably also used for storage, as are holes dug into the piles of accumulated cone debris beneath feeding stations. Other food eaten by squirrels includes hazelnuts, berries, maple seeds, and mushrooms. A variety of fruits and seeds are doubtless eaten when opportunity offers.
Red squirrels do not hibernate. In the lowlands they are active all winter long but are noticeably shy and quiet. In the mountains they disappear after the snow falls but tracks may be seen in the snow about their dens and occasional individuals are seen. Specimens collected at this time are not fat, as would be the case if hibernation had been interrupted. Seemingly they stay close to their homes and feed on stored food.
The call of the red squirrel is harsher, more metallic, than that of the Douglas squirrel. The common call is a rolling "bur-r-r," starting loud but fading out entirely in a half minute. A lower-pitched "pauf" is uttered at intervals when the squirrel is going about its business. The danger cry is a loud "pee-ee," not unlike the call of a red-tailed hawk but less shrill. A low pitched "chauf-chauf-chauf," repeated at intervals of about two seconds, is occasionally given in the fall of the year.
Sciurus hudsonicus streatori Allen, Bull. Amer. Mus. Nat. Hist., 10:267, July 22, 1898.
T[amiasciurus]. h[udsonicus]. streatori Howell, Proc. Biol. Soc. Washington, 49:135, August 22, 1936.
Type.—Obtained at Ducks, British Columbia; type in American Museum of Natural History.
Racial characters.—Similar to richardsoni but smaller and darker, redder above.
Measurements.—Seven males from north-central Washington average: total length 330; length of tail 129; hind foot 51; ear 23.
Distribution.—Northeastern Washington. Marginal occurrences are (from Taylor and Shaw, 1929: 17): Beaver Creek, Ruby Creek, and head of Lake Chelan.
Remarks.—This race reaches Washington from the interior of British Columbia. Intergradation between hudsonicus streatori and h. richardsoni takes place over much of northeastern Washington.
Fig. 90. Distribution of the red squirrel and Douglas squirrel in Washington. A. Tamiasciurus hudsonicus richardsoni. B. Tamiasciurus hudsonicus streatori. C. Tamiasciurus douglasii douglasii.
Sciurus richardsoni Bachman, Proc. Zoöl. Soc. London, p. 100, 1838.
Sciurus hudsonius Richardsoni True, Proc. U. S. Nat. Mus., 7:595, 1884.
Sciurus hudsonius Richardsoni Allen, Bull. Amer. Mus. Nat. Hist., 10:265, July 22, 1898.
Sciurus hudsonicus richardsoni Miller, U. S. Nat. Mus. Bull., 128:211, April 29, 1924.
Type.—Obtained at the head of Big Lost River, Custer County, Idaho.
Racial characters.—Color paler, especially on back.
Measurements.—Four males and 4 females average, respectively: total length 339, 339; length of tail 133, 131; hind foot 54, 51; ear 24, 24; weight 256, 266.
Distribution.—The Blue Mountains of southeastern Washington.
Sciurus douglasii Bachman, Proc. Zoöl. Soc. London, p. 99, 1838.
Sciurus hudsonius Douglasii True, Proc. U. S. Nat. Mus., 7:595, 1884.
Sciurus douglasii douglasii Miller, U. S. Nat. Mus. Bull., 128:212, April 29, 1924.
Sciurus douglasii cascadensis Taylor and Shaw, Occ. Papers Chas. R. Conner Mus., no. 2:18, December, 1929.
Tamiasciurus douglasii Holdenried, Jour. Mamm., 21:406, November 14, 1940.
Type.—Obtained near the mouth of the Columbia River by David Douglas.
Measurements.—Fifteen males and 10 females from the lowlands of western Washington average, respectively: total length 317, 320; length of tail 121, 126.7; hind foot 50, 49.4; ear 23.2, 22.6; weight 204, 202 grams.
Distribution.—From the eastern base of the Cascade Mountains westward to the Pacific. Marginal occurrences are: Nooksack River (Taylor and Shaw, 1929: 18), Lake Wenatchee (W. W. D.) and Wenatchee (Taylor and Shaw, 1929: 18).
Remarks.—Squirrels from the Puget Sound area are slightly less intensely colored than specimens from the ocean coast and the Olympic Mountains. Those from the higher and eastern Cascade Mountains are paler still. When, however, squirrels from the Cascades of Washington are compared with individuals of the race inhabiting the Cascades of Oregon, it is apparent that the Washington squirrels are nearest to douglasii. The pale tips of the hairs in the tail of the Oregon race are strikingly white while in Washington specimens they are orange or yellow.
The Douglas squirrel resembles the red squirrel in size and proportions but differs in being dusky olive (less reddish) on the upper parts and orange instead of white on the underparts.
Douglas squirrels range from southern British Columbia southward to Lower California. They are confined to the Pacific Coastal region. Although closely related to Tamiasciurus hudsonicus, no intergradation with that species has been found.
The rich coloration of the Douglas squirrel matches the background of the humid forests it inhabits. It occurs at sea level along the ocean coast, the shores of Puget Sound and on some islands in Puget Sound. It ranges through the Transition and Canadian life-zones well into the Hudsonian.
It makes its home in holes in trees or in nests constructed of conifer twigs, needles and bark. Old nests of birds may be modified and taken over, but most outside nests seem to have been made entirely by the squirrel. Nests are usually placed near the top of denser fir and cedar trees. Nests are hemispherical in shape, 12 [Pg 291] inches or more in diameter, and open on top. The central cup, 4 inches in diameter, is lined with strips of inner bark of red cedar coiled but unshredded. Nests in holes are usually some distance from the ground, but the entrances to some are at ground level.
The habits of the Douglas squirrels vary with the season. In spring they are shy. They rarely call and are skillful in keeping branches or tree trunks between themselves and persons. In June and July they become bolder, calling more frequently and moving about more. In September and October they become exceptionally bold. They call almost incessantly from late morning until early afternoon, and spend considerable time on the ground.
The call notes of the Douglas squirrel are similar to those of the red squirrel but are softer, more slurred. The common call is the long, trilling "burr," loud at first and becoming gradually softer. On hot afternoons in late summer a barking "pauf" note is given. This sound is repeated several times at intervals of a minute or so. It has remarkable carrying power. The danger signal is a loud, explosive "pe-ee." Another common call is a short barking or chirping "bauf" with a musical, questioning sound. This is repeated at intervals of a few seconds when the presence of danger is suspected. At times it, rather than the loud danger signal, is given also when a person is sighted. A low chirping note is often given while the squirrel is busily at work. This is made while the squirrel is moving about. Other calls described are made from a standing or sitting position and are accompanied by a jerk or a flick of the tail.
The Douglas squirrel eats seeds of the Douglas fir, seeds of maple, hazelnuts, dogwood berries and mushrooms. The berries of forest shrubs, such as the red huckleberry, salal and Oregon grape, may also be eaten.
Two broods of young may be born in a year. The first is born in early June. Embryos found from June 11 to 28 varied in number from 6 to 8. A nursing female was taken as early as June 10, 1938, and one was taken as late as October 10, 1938.
Douglas squirrels commonly have a few fleas and 2 to 5 ticks at the bases of the ears. No lice, cases of mange, or serious infestations of parasites have been seen.
Fig. 91. Northern flying squirrel (Glaucomys sabrinus), probably from near Seattle, in Woodland Park Zoo, Seattle, Washington, January 28, 1941. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 1137.)
Description.—The northern flying squirrel is slightly smaller than the red squirrel (Tamiasciurus). The head and body measure about 7 inches and the tail about 5 inches. The most distinctive feature is the loose fold of skin that stretches from the wrist of the foreleg to the ankle of the hind leg. The fur is extremely soft and plushlike. It is not separated into guard hair and underfur, and because all hairs are of approximately the same length, the fur appears extremely sleek. The fur of the tail is of the same texture as that on the body but is dorso-ventrally compressed. The eyes are large and [Pg 293] dark. The color of the upper parts varies from reddish brown in some races to brownish gray in others. The underparts are buffy gray.
Flying squirrels range over the forested parts of North America from Guatemala to Alaska and northern Canada and from the Atlantic to the Pacific. A closely related genus (Pteromys) occurs in Europe. Two species are found in North America. The small volans ranges in eastern United States, Mexico, and Guatemala, while the larger sabrinus occurs in parts of the western United States, most of Canada, and central Alaska. Near the Great Lakes the two species occur in the same area, probably in different habitats.
Howell (1918: 16) recognized 18 subspecies of sabrinus, five of which were recorded in the state of Washington. One of these (olympicus) proves to be a synonym of an earlier-named subspecies but another form (bangsi), not recorded for Washington by Howell, has been found in the Blue Mountains in the southeastern corner of the state (Taylor and Shaw, 1929: 18).
Flying squirrels are strictly arboreal and occur only in woods and forests. They prefer areas where trees grow close together and to considerable heights. Older woods with dead stubs and hollow trees are preferred. They occasionally invade attics of cabins and other habitations of man. The old Forestry Building on the University of Washington Campus, later the home of the Washington State Museum, was inhabited by flying squirrels for many years. Several specimens preserved in the Museum prove the poor judgment of the animal in choosing a natural history museum as a home.
Relatively little is known of the detailed habits of Glaucomys sabrinus although the life history of the eastern Glaucomys volans has been described. Flying squirrels are active all winter, even in the high mountains. They are often caught in traps set for fur bearers and, where trapping is an important means of livelihood, they are serious pests. Flying squirrels do not actually fly but only glide through the air. The loose fold of fur between their limbs is stretched by extending the legs. With it the flying squirrel is able to sail 50 yards or more. The flat tail serves as a rudder and allows the squirrel to change direction while in flight. Glides end with an upward swoop, allowing the squirrel to alight, head upwards, on a tree trunk slightly lower than their starting point.
Unlike other members of the squirrel family, flying squirrels are completely nocturnal. They are seen in the daytime only when frightened from their retreats. Homes consist principally of old [Pg 294] woodpecker holes or other cavities in trees. They are said to build outside nests, similar to those of tree squirrels, but I have found none of these in Washington. The animals can be frightened from their holes by pounding on the bases of trees in which their nests are situated. Certain holes seem to be preferred nesting places. Near Cottage Lake, King County, two flying squirrels were taken from a woodpecker hole in succeeding years. Cowan (1936B: 58) discovered remains of 14 nests of flying squirrels in a single hollow tree near Alta Lake, British Columbia. According to Cowan, hollow trees are used in winter but the young are born in outside nests of shredded bark and lichens. The young usually number 3 and are born in May and June.
Fig. 92. Distribution of flying squirrel in Washington. A. Glaucomys sabrinus oregonensis. B. Glaucomys sabrinus fuliginosus. C. Glaucomys sabrinus columbiensis. D. Glaucomys sabrinus latipes. E. Glaucomys sabrinus bangsi.
Only once have I heard the voice of a flying squirrel. Near Dewey Lake, Yakima County, the squirrels were unusually abundant in the black hemlock and Douglas fir forests of the mountain sides. Here a birdlike twittering note caught my attention and occasional dark, sailing shapes were glimpsed against the clear sky. Eight traps set [Pg 295] in the afternoon and visited at 11 p. m. held two flying squirrels. In the morning another was in a trap. Seemingly the squirrels are active most of the night.
The food consists principally of conifer seeds and probably other nuts, seeds, and fruit. They eat the meat bait of traps set for fur bearers and probably eat the eggs of birds.
Pteromys oregonensis Bachman, Jour. Acad. Nat. Sci. Philadelphia, 8:101, 1839.
Sciuropterus alpinus oregonensis Rhoads, Proc. Acad. Nat. Sci. Philadelphia, p. 324, June, 1897.
Sciuropterus alpinus olympicus Elliot, Field Columb. Mus. Publ. 30, zoöl. ser., 1:225, February 1, 1899 (type from Happy Lake, Clallam County, Washington).
Glaucomys sabrinus oregonensis Howell, N. Amer. Fauna, 44:44, June 13, 1918.
Glaucomys sabrinus olympicus Howell, N. Amer. Fauna, 44:49, June 13, 1918.
Type.—Obtained "in pine [= conifer] woods of the Columbia near the sea" by J. K. Townsend in 1839. Probably near St. Helen, Columbia County, Oregon (Rhoads, 1897:324).
Racial characters.—Small size, rich color.
Measurements.—A male from Cottage Lake, King County, measured: total length 287; length of tail 125; hind foot 38. A female from 5 miles southeast of Sequim, Clallam County, measured: 303; 133; 41; ear 27. A male from Quilcene, Jefferson County, measured: 311; 140; 41.
Distribution.—Western Washington west of the Cascade Mountains. The locality records for flying squirrels in the northern Cascade Mountains, given by Taylor and Shaw (1929: 18), when plotted on a distribution map, show overlapping of ranges in this area. The ranges of the three races involved have been drawn on the accompanying map (fig. 92) on the basis of geographic probability and are subject to revision because the specimens from this area have not been examined.
Marginal records that may apply to G. s. oregonensis are, from Taylor and Shaw (1929: 18): Nooksack River, Rockport, North Bend and Skamania.
Remarks.—Of all the races occurring in Washington, oregonensis is the most distinct. Glaucomys s. olympicus Elliot must be regarded as a synonym of oregonensis.
Sciuropterus alpinus bangsi Rhoads, Proc. Acad. Nat. Sci. Philadelphia, p. 321, June, 1897.
Glaucomys sabrinus bangsi Howell, N. Amer. Fauna, 44:38, June 13, 1918.
Type.—Obtained in Idaho County, Idaho, by Harbison and Bargamin on March 8, 1897; type in Philadelphia Academy of Natural Sciences.
Racial characters.—Similar to oregonensis but larger and paler throughout.
Measurements.—A female from Wildcat Spring, Columbia County, measured: total length 333; length of tail 147; hind foot 45; ear 29; weight 151 grams.
Distribution.—Found only in the Blue Mountains of extreme southeastern Washington.
Glaucomys sabrinus columbiensis Howell, Proc. Biol. Soc. Washington, 28:111, May 27, 1915.
Type.—Obtained at Okanogan, British Columbia, by Allan Brooks on May 9, 1898; type in United States National Museum.
Racial characters.—Larger than oregonensis and paler. Similar to bangsi but paler, especially beneath, and less reddish above.
Measurements.—Howell (1918: 46) gives the average of two subadult topotypes as: total length 313; length of tail 143; hind foot 42.
Distribution.—Northeastern Washington, probably extending, from records of Taylor and Shaw (1929: 18), west to Mazama and Stehekin and east to Molson.
Glaucomys sabrinus latipes Howell, Proc. Biol. Soc. Washington, 28:112, May 27, 1915.
Type.—Obtained at Glacier, British Columbia, by J. A. Loring, on August 13, 1894; type in United States National Museum.
Racial characters.—Similar to bangsi and columbiensis but with larger feet and grayer color.
Measurements.—Howell (1918: 49) gives the average of 10 specimens as: total length 342; length of tail 153; hind foot 41.5.
Distribution.—The Pend Oreille Mountains of extreme northeastern Washington, north (from records of Taylor and Shaw, 1929: 18-19, as revised) to Sullivan Lake and south to Loon Lake.
Sciuropterus alpinus fuliginosus Rhoads, Proc. Acad. Nat. Sci. Philadelphia, p. 321, June, 1897.
Glaucomys sabrinus fuliginosus Howell, N. Amer. Fauna, 44:47, June 13, 1918.
Type.—Obtained at Martin Station, Kittitas County, Washington, by Allan Rupert in March, 1893; type in Philadelphia Academy of Natural Sciences.
Racial characters.—Similar to columbiensis but underparts darker and tail paler. Larger and paler than oregonensis.
Measurements.—Three females from the Cascades average: total length 327; length of tail 145; hind foot 40.7; ear 25.
Distribution.—The Cascade Mountains, according to Taylor and Shaw (1929: 18), as revised here, east to Entiat River and south to Carson.
Description.—Great Basin pocket mice are slightly larger than house mice. The ears are tiny and round; the tail is relatively long, slightly longer than the head and body; the forefeet are small but the hind feet are large and powerful, more than one-fourth as long as the head and body. They have relatively large, fur-lined, external cheek pouches. The color of the upper parts varies with the subspecies from blackish olive-gray to buffy olive-gray; a line of clear buff or buffy-olive extends along the lower part of side, separating the darker color of the upper parts from the white underparts; the tail is short-haired, blackish above, olive beneath.
Pocket mice of the genus Perognathus range from Mexico northward to British Columbia. Only one species, parvus, occurs in Washington where it is represented by three subspecies. Pocket mice are most common in the Upper Sonoran Life-zone in sandy areas dotted with desert shrubs. They are found occasionally in dry, grassy places in the Arid Transition Life-zone. They may be locally abundant in rocky areas and are often trapped high on talus slides, many yards from the nearest soil. Gray (1943: 191-193) estimates their numbers on the sagebrush areas of the Yakima Valley at 32 per acre. They are completely nocturnal. Their ordinary movements are rather slow and specimens studied by the aid of a searchlight usually crept quietly into the protection of the nearest desert shrub. The trail of a pocket mouse in soft sand may be recognized by the distinctive mark left by the dragging tail.
Burrows of pocket mice usually are at the bases of shrubs where tough roots furnish protection. They are kept closed during the day by means of a plug of fresh earth or sand. Openings may often be recognized by a fan-shaped pile of fresh sand before the hole. Burrows excavated were usually less than four feet in length and branched from two to four times. No nests were found in the burrows but one contained a few fresh stalks of desert annuals. The air in the burrows seemed warm and humid.
The pouches of pocket mice from Washington often contain the fresh, green tips of desert plants, grass seeds, seeds of plants other than grasses, and plant leaves. Because of their occasional great abundance, pocket mice may be a menace to agriculture. Fortunately much of their habitat is unsuited to farming.
In Washington pocket mice breed in March and April. From 4 to 8 embryos were found in pregnant females.
Cricetodipus parvus Peale, U. S. Explor. Exped., 8 (mamm. and ornith.):53, 1848.
Perognathus parvus Cassin, U. S. Explor. Exped., 8 (mamm. and ornith.):48, 1858.
Perognathus parvus parvus Miller, Bull. U. S. Nat. Mus., 128:278, April 29, 1924.
Fig. 93. Distribution of the Great Basin pocket mouse in Washington. A. Perognathus parvus parvus. B. Perognathus parvus columbianus. C. Perognathus parvus lordi.
Type.—Probably obtained in the neighborhood of The Dalles, Wasco County, Oregon.
Racial characters.—Size small; upper parts brownish-buff washed with blackish or, in gray phase, ashy gray washed with blackish; sides buffy yellow; a buffy spot often present on throat; tail blackish above, olive below; facial markings usually brownish-buff but in older animals indistinct and washed with blackish.
Measurements.—Thirty-one males and 19 females from Washington average, respectively: total length 169, 164; length of tail 90, 86; hind foot 22.6, 21.8; ear 5, 5.
Distribution.—Southeastern Washington south of the Snake River, the area on the north side of the Columbia River in Klickitat County, and the Yakima Valley area as far north as the Vantage, Kittitas County (W. W. D.). Other marginal records are: Kennewick (W. W. D.), Atilla (W. W. D.) and Walla Walla (E. S. B.).
Remarks.—Dichromatism seems to be rather common in Perognathus p. parvus—so common that Osgood (1900: 35) recognized both a "red" and a "gray" phase. Anderson (1932: 102) found no dichromatism in P. p. lordi in British Columbia. A single specimen from the Grand Coulee at Dry Falls, Grant County, does show dichromatism. It is even more red than the reddest topotypes of parvus examined.
Seemingly the recessive gene for red is still present in lordi, or has mutated anew. When present, the color is deeper and brighter than in the parent population, in keeping with the heavier pigmentation of the race lordi.
Abromys lordi Gray, Proc. Zoöl. Soc. London, p. 202, 1868.
Perognathus lordi Merriam, N. Amer. Fauna, 1:28, October 25, 1889.
Perognathus lordi lordi Miller, U. S. Nat. Mus. Bull., 128:279, April 29, 1924.
Perognathus parvus lordi Davis, Recent Mamm. of Idaho, p. 266, Caxton Printers, Caldwell, Idaho, April 5, 1939.
Type.—Obtained in southern British Columbia (probably near Lake Osoyoos) by J. K. Lord, probably in 1860.
Racial characters.—Size large; color of upper parts buffy-olive, washed with blackish; sides buff-olive; facial markings absent or, if present, indistinct and pale olive; tail blackish above, olive below.
Measurements.—Twenty-nine males and 10 females average, respectively: total length 175, 171; length of tail 93, 89; hind foot 23.4, 22.9; ear 5.3, 5.1.
Distribution.—Okanogan Valley and the Columbian Plateau, except for the southwestern part. Marginal localities are: Vantage, Grant County (W. W. D.), 10 mi. S. Moses Lake (W. W. D.), Washtucna (M. V. Z.) and Pullman (M. V. Z.).
Remarks.—There are constant differences between Perognathus parvus parvus and Perognathus parvus lordi. The latter is larger, darker, and differs slightly in average cranial measurements. The Snake River, the Columbia River, and the Wenatchee Mountains separate the geographic ranges of the two subspecies and prevent intergradation between them. Nevertheless their close similarity and probable common origin indicate that both belong to one species, namely parvus.
Perognathus columbianus Merriam, Proc. Acad. Nat. Sci. Philadelphia, p. 263, September 27, 1894.
Perognathus lordi columbianus, Osgood, N. Amer. Fauna, 18:40, September 20, 1900.
Type.—Obtained at Pasco, Franklin County, Washington, by Clark P. Streator on May 9, 1891; type in United States National Museum.
Racial characters.—Size and color as in lordi but skull considerably wider in mastoid region.
Measurements.—Twenty-one male and 9 female topotypes average, respectively: total length 173, 168; length of tail 91, 89; hind foot 23.9, 22.8; ear 5, 5.
Distribution.—The vicinity of the type locality and the part of the Columbian Plateau north of the Snake and east of the Columbia rivers.
Remarks.—This race is separated from parvus by river barriers and the two do not intergrade. It differs significantly from lordi only in the wider mastoid region. No barrier separates the range of lordi from that of parvus, and the two races intergrade over a wide area (north to Moses Lake, east to Washtucna). The race columbianus must have originated in situ from lordi. The habitat of columbianus seems not to differ from that of parvus or lordi.
Perodipus ordi columbianus Merriam, Proc. Biol. Soc. Washington, 9:115, June 21, 1894.
Dipodomys ordii columbianus Grinnell, Jour. Mamm., 2:96, May 2, 1921.
Type.—Obtained at Umatilla, Umatilla County, Oregon, by C. P. Streator on October 18, 1890; type in United States National Museum.
Measurements.—Fourteen males and 9 females from Walla Walla County average, respectively: total length 261.5, 248.4; length of tail 137.2, 139.1; hind foot 40.6, 40.8; ear 13.1, 13.0; weight 52.1, 49.0 grams.
Distribution.—Western Walla Walla County from the Oregon boundary north to the Snake River and on Blalock Island in the Columbia River, Benton County.
Fig. 94. Distribution of the Ord kangaroo rat, Dipodomys ordii columbianus, in Washington.
Description.—The kangaroo rat with a body about the size of that of a chipmunk has an exceptionally large head and large, black eyes. The forelegs and forefeet are tiny but the hind feet and legs are large and powerful. The hind foot is almost a third as long as the head and body. The tail is long, longer than the head and body. Kangaroo rats possess fur-lined, external cheek pouches, like those of [Pg 301] the pocket mice. The upper parts are soft buff in color. The underparts and a stripe on each flank are white. The tail is dusky above and below, with white sides; it is tufted at the tip.
Kangaroo rats are typical of the desert regions of the southwestern United States, where numerous species and subspecies are found. A single subspecies of the wide-ranging species ordii occurs into southeastern Washington, where it is restricted to sandy areas in the Upper Sonoran Life-zone. In the soft, drifted sand along the Columbia River where sagebrush and other desert shrubs are low and widely spaced kangaroo rats are abundant.
These rats are strictly nocturnal. When individuals are dug from their burrows in the daytime they usually hop about in a dazed manner and appear to be blinded by sunlight. Near Wallula, Walla Walla County, these rats were caught at night with a butterfly net as they stood "paralyzed" in the beam of a powerful searchlight. Such night hunting was unsuccessful on cloudy or windy nights when kangaroo rats seem not to move about.
As might be guessed from their powerful hind legs, kangaroo rats travel in bounds. Near Wallula, where we watched them in their natural habitat, they traveled, when unfrightened, in slow hops, each hop followed by a pause. As they struck the surface of the ground an audible thud could be heard for a distance of several feet. After each jump they paused for a second or so, perhaps to allow a pursuing enemy to over-run them.
Near Wallula the burrows of kangaroo rats were dug in large mounds of wind-blown sand. The burrows entered these natural mounds horizontally and branched two or three times. Their average length was about five feet. No nests or food stores were discovered although several kangaroo rats were caught as they burst from entrances at sides of the mounds. All entrances to burrows were plugged with soft sand. The air in the burrows seemed warm and humid.
Food found in the cheek pouches of kangaroo rats from Washington included the seeds of desert annuals, short sections of sprouts of an unidentified plant, grass seeds, and the leaves of the hop-sage.
A female taken March 22, 1939, contained 3 embryos.
Description.—The pocket gopher is a fossorial animal, being but slightly less adapted to an underground existence than the moles. The body is stout, the legs short and the head broad. The tail is short, sparsely haired, cylindrical and blunt-ended. The fur is soft and dense. The eyes are small and the ears tiny and naked. The incisor teeth are external, being separated from the mouth cavity by a furry strip of skin. Like the pocket mouse and kangaroo rat, the pocket gopher possesses external, fur-lined cheek pouches. The openings of these begin just below and posterior to the nostrils, sweep out and down in a semicircle, and end at the chin posterior to the base of the lower incisors. They extend laterally to the shoulders and easily accommodate a fifty-cent piece.
Fig. 95. Northern pocket gopher (Thomomys talpoides yelmensis), from two miles southwest of Tenino, Washington, January 28, 1941. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 1133.)
The family Geomyidae is composed of eight genera, so similar in appearance that the name "pocket gopher" is applied to all of them. The family is confined to North and Central America. Three genera occur in the United States but only one, Thomomys, occurs in Washington. Thomomys is restricted to western North America [Pg 303] where it ranges from central Canada south to the southern edge of the table land of Mexico.
Several hundred kinds of Thomomys have been described and as systematic work with this genus has been continued, more and more kinds, originally thought to be species, are found to intergrade and to be only subspecies. All of the 17 kinds of pocket gophers occurring in Washington belong to a single species.
The pocket gopher is principally nocturnal or crepuscular but sometimes it is active at midday, especially if the day be dark and cloudy. Pocket-gopher activity is indicated by fresh mounds of earth on the surface of the ground. Rarely, an observer may see movement of plants as the gopher molests the roots of the plants, or even see the head and shoulders of an animal that partly emerges from an open burrow. The ordinary gopher mound consists of less than a cubic foot of earth. The earth is forced up from a single opening and usually is pushed out in one direction. In consequence it forms in a fan-shaped pile about the opening, and the last load forms a circular plug above and to one side of the burrow opening. When so much earth has been forced out of one opening that expulsion of additional loads of earth is overly difficult, the burrow is extended slightly to one side, or even extended into the newly formed mound, and another fan formed. Usually not more than three coalesced fans form a mound, but where the soil is exceedingly soft and fluffy, hundreds of fans may form a composite mound and the one mound may include a cubic yard of earth. Large composite mounds probably are formed gradually over a period of weeks or even months.
The earth in a fresh gopher mound is usually "scratched," and gives the appearance of having been sieved. Pebbles weighing more than 100 grams are included in material ejected from burrows. The entrances to the burrows of gophers are usually solidly plugged with earth. The plug may be from a few inches to more than a foot in length. At times a burrow entrance may appear to be open, but in such cases investigation will usually reveal it to be plugged some distance back—sometimes several feet.
In contrast to the gopher mounds described above, the mounds of moles are not fan-shaped but volcano-shaped. The earth from a mole's burrow is forced straight upwards, whence it falls to either side. Later loads are pushed up from beneath, raising the entire mound, with the last material ejected at the center and bottom. The earth of a fresh mole-mound is not of fine texture but instead [Pg 304] is "clotted" and, if damp, gives the mound a fractured appearance. When mounds are older, perhaps changed by rain and sun, their identity as of mole-origin or pocket gopher-origin is more difficult to establish. In such cases, if no fresh mounds can be found, the observer must rely on the spacing of the mounds. Mole-mounds are spaced along a burrow, about as far apart as a man can step. Gopher mounds are irregularly spaced, and the course of the burrow cannot be traced merely by observing the arrangement of the mounds as can that of the mole.
In addition to mounds, gopher burrows have plugged openings where a gopher has come to the surface, probably to cut plants. Such entrances are marked by a plug of earth several inches long. Mounds and feeding entrances of the gopher burrow are usually not constructed in the main burrow system itself, but at the ends of lateral burrows of varying length. If one traces the burrow back from the mound, a junction with the main, better-constructed burrow is found. The junction is usually T-shaped, with the lateral burrow at right angles to the main burrow. More rarely the junction has a Y-shape.
Fig. 96. Giant mounds raised by pocket gophers on Mima Prairie, Thurston County, Washington, July 13, 1941. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 1209.)
The burrow system of the pocket gopher may be divided into three main parts. These are the laterals, just discussed, the main burrow, and the deep nesting burrow. The main burrow is a sinuous [Pg 305] tube or tunnel at a relatively uniform depth, that marks the extent of the gopher's home territory. This tunnel may branch, or even intersect. As it is extended in one direction, the earth excavated by the gopher may in part be thrust into an unused portion of the burrow.
The deep, nesting burrows may be used only in the breeding season. They are connected with the main tunnel system but descend to a greater depth. Usually they descend into the harder, consolidated layers of soil below the zone where plant roots penetrate. Here chambers are constructed in which nests and food are stored. Usually a vertical shaft is dug in the burrow ahead of the nest to lead rain water away from the nest.
In areas where gophers live in a thin layer of soil underlain by a more or less impenetrable layer of rock, clay or gravel, it has been suggested that they form unique structures known as Mima Mounds. The formation of these mounds has been discussed in detail elsewhere (Dalquest and Scheffer, 1942: 68-84). At least in the breeding season the gophers work mainly and make their nest where the soil is deepest. In the vicinity of this nest, considerable mounding and cultivation of soil ensues. This stimulates plant growth in the area. Much observation indicates that cultivation of earth by gophers stimulates plant growth to a greater extent than the depredations of the gopher deplete the ensuing growth. Thus the gopher, by cultivation of the soil in the area about its burrow, stimulates the growth of vegetation and so increases his own food supply. Consequently there is but little incentive for the gopher to leave the vicinity of the nest. The gopher does, however, construct lateral tunnels into surrounding areas. Earth from these lateral tunnels is, in part, thrown to the surface in mounds and in part transported back to fill the abandoned burrows near the nest. The earth from the burrows about the nest was earlier ejected on the surface. Slowly, then, earth is transported from surrounding areas to burrows in the vicinity of the original nest. Each succeeding generation finds in the vicinity of the original nest, better food and deeper soil, while areas surrounding the nest possess thinner soil and less vegetation. Over a period of thousands of generations of gophers, large mounds, known as Mima Mounds, are formed. Since the removal of earth from the surrounding areas and its accumulation in the Mima Mound are chance affairs, the contours of the mound are smooth and flowing while the contours of the intermound areas are smoothly convex.
The pocket gophers in Washington are economically important. In truck and flower gardens they are a pest, especially if the crop be bulb plants. In grain fields they are a pest because their mounds cover considerable grain and are apt to clog or dull the knife of the mower. In fields of young alfalfa they are apt to crop back the plant more rapidly than it can grow. Once the alfalfa plant is well established, however, the cultivation resulting from activities of gophers, some persons believe, stimulates the plant to such an extent that it grows larger and healthier in spite of the gopher's feeding on it. In the White Salmon Valley, Klickitat County, I examined numerous alfalfa fields. The most luxuriant growth was invariably in fields where gophers were common. In these fields, the largest plants were those in the immediate vicinity of gopher activity. The commensal relation between the gopher and alfalfa was understood by many farmers, who forbade us to take gophers for specimens from their fields. Several told us that they always trapped the gophers from the fields of young alfalfa and from hay meadows but encouraged their presence in fields of older alfalfa.
Fig. 97. Food cache of northern pocket gopher (Thomomys talpoides tacomensis), from chamber four inches below surface of ground, Tacoma, Washington, December 1, 1940. Contents 575 grams (about 2 liters) of roots, principally quackgrass, Agropyron repens. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 1119.)
Another economic factor is the gopher's removal of certain weeds from grazing land. A number of introduced weeds form dense rosettes that prevent the growth of grass from several square inches of ground and themselves lie so close to the ground as to be unavailable as food to grazing stock. These weeds seem to be favored food of gophers which cut not only the rosettes but the roots of the weeds as well.
In irrigated parts of eastern Washington the gopher is a serious pest. It burrows in the banks of the main ditches, causing cave-ins and occasional breaks with resultant water loss. The mounds of gophers fill in the smaller channels and divert the streams. Constant attention is necessary to keep the ditches free of gopher mounds.
In uncultivated land the gopher is a distinct asset unless the land is near enough to cultivated land to serve as a reservoir of pests. In the mountains and on the desert the gopher cultivates and keeps the soil soft and stimulates the growth of water-retaining vegetation, thus preventing rapid run-off and erosion and keeping the flow of springs and streams constant. Boulders, logs, and other obstructions are undermined and, as a result of gopher activity, in time sink under the surface of the ground. Thus a larger surface is available for plant growth. Lastly the gopher furnishes an important food source for certain fur-bearing mammals and eases the predator pressure on game species.
The subspecies of pocket gopher occupying the Puget Sound area are highly restricted in their habitat preference; they occur only on the grassy prairies of the glacial outwash aprons. They do not occur in woods, brush or even small openings on the borders of the prairies. The alpine forms occupy the mountain meadows and are slightly less selective in their habits. It should be noted that in western Washington the forests are far more open at higher altitudes than at low elevations. The races inhabiting the desert are found in open areas, often in sandy places. They occur more rarely in areas where the soil is baked and claylike, and then usually in the vicinity of springs or watercourses. The race T. t. fuscus has a wide range of tolerance as regards environmental factors; it occurs near Wenatchee in essentially desert conditions, in alpine meadows of northeastern Washington, and in many habitats at intermediate elevations. It occurs also in brushy areas and is often abundant in open pine forests.
Since the gopher usually has a narrow range of tolerance as regards [Pg 308] its environmental adaptations, this has resulted in considerably more isolation than is the case with other mammals, and has probably contributed to the formation of the many subspecies. Within the range of almost every race, microgeographic races, or local populations with distinctive characters, are found. Many subspecies of Thomomys are probably the result of chance fixation of genetic characters already present in a more genetically variable ancestral population, and the loss of other genetic factors. Such races might be considered degenerative (see Dalquest and Scheffer, 1944: 24).
Fig. 98. Distribution of the northern pocket gopher in Washington. A. Thomomys talpoides devexus. B. Thomomys talpoides columbianus. C. Thomomys talpoides aequalidens. D. Thomomys talpoides wallowa. E. Thomomys talpoides fuscus. F. Thomomys talpoides yakimensis. G. Thomomys talpoides shawii. H. Thomomys talpoides immunis. I. Thomomys talpoides limosus. J. Thomomys talpoides douglasii. K. Thomomys talpoides pugetensis. L. Thomomys talpoides tacomensis. M. Thomomys talpoides glacialis. N. Thomomys talpoides tumuli. O. Thomomys talpoides yelmensis. P. Thomomys talpoides couchii. Q. Thomomys talpoides melanops.
The history of the pocket gophers of Washington has been traced previously (Dalquest and Scheffer, 1942, 1944). It may be briefly summarized as follows: At the close of Vashon-Wisconsin times, [Pg 309] gophers were found in the southern Cascades, on the Simcoe Bridge, the Columbian Plateau, and in southeastern Washington. Following the retreat of the ice, the gophers in the Mount Rainier area spread westward on the outwash of the Nisqually and perhaps other glaciers to the Vashon Outwash about southern Puget Sound and thence to the Olympic Mountains. In the southern Cascades, gophers spread westward on glacial terraces of the Columbia River to the vicinity of Vancouver, Clark County. The establishment and growth of the forests split up the original populations, and continued spread of forest has exterminated many units. All the races in the lowlands of western Washington face extermination as the prairies are reclaimed by forest.
Pocket gophers also invaded northeastern Washington from Idaho and spread westward to the Cascades, thence southward until the native gophers were encountered north of Mount Rainier and in the Yakima Valley. The invading gophers nearly surrounded the Columbian Plateau.
T. H. Scheffer (1938B: 220-224) found the gestation period of the pocket gopher to be approximately 28 days. No second brood is raised in Washington. Near Kennewick, Yakima County, the young are born from February to April. The average number of embryos found in 76 female gophers was 6.3. Near Olympia, Thurston County, the young are born from March to June. The average number of embryos from 312 females was 5.0.
Thomomys talpoides ericaeus Goldman, Jour. Mamm., 20:243, May 15, 1939 (type from Badger Mountains, Douglas County, Washington).
Type.—Obtained 1 mile west-southwest of Neppel (now Moses Lake), Grant County, Washington, by W. W. Dalquest on May 30, 1938; type in Museum of Vertebrate Zoölogy.
Racial characters.—Size medium; ears tiny; color of upper parts pale brownish gray; underparts white; postauricular spots dark.
Measurements.—Two male and 4 female topotypes average, respectively: total length 184, 184; length of tail 54, 55; hind foot 25, 25; weight 89, 71 grams.
Distribution.—The Columbian Plateau. Marginal occurrences of the races of pocket gophers occurring in Washington are not listed here because they have recently been placed on record (Dalquest and Scheffer, 1944: 308-333, 423-450).
Remarks.—This is the smallest and palest race of pocket gopher found in Washington.
Thomomys fuscus columbianus Bailey, Proc. Biol. Soc. Washington, 27:117, July 10, 1914.
Thomomys columbianus Bailey, N. Amer. Fauna, 39:106, November 15, 1915.
Thomomys talpoides columbianus Goldman, Jour. Mamm., 20:234, May 15, 1939.
Type.—Obtained at Touchet, Walla Walla County, Washington, by C. P. Streator on September 10, 1890; type in United States National Museum.
Racial characters.—Larger and brighter in color than devexus. Near ochraceous orange in color.
Measurements.—Five male and three female topotypes average, respectively: total length 213, 209; length of tail 60, 58; hind foot 29, 28.
Distribution.—Walla Walla County, between the Snake River and the Oregon boundary and from the Columbia River east to the Columbia County line.
Thomomys talpoides aequalidens Dalquest, Murrelet, 23:3, May 14, 1942.
Type.—Obtained at Abel Place, 2,200 ft., 6 miles south-southeast of Dayton, Columbia County, Washington, by S. H. Lyman, on April 6, 1934; type in Museum of Vertebrate Zoölogy.
Racial characters.—Large size, very dark color.
Measurements.—The average of four male topotypes and the measurements of one female topotype are, respectively: total length 202, 201; length of tail 57, 59; hind foot 26, 27.
Distribution.—Southeastern Washington east of the range of columbianus and north of the higher parts of the Blue Mountains.
Thomomys talpoides wallowa Goldman, Jour. Mamm., 20:234, May 15, 1939.
Type.—Obtained at Catherine Creek, 7 miles east of Telocaset, 3,500 ft., Union County, Oregon, by R. T. Orr on June 29, 1932; type in Museum of Vertebrate Zoölogy.
Racial characters.—Similar to devexus but much darker. Similar (in Washington) to aequalidens but much smaller.
Measurements.—Four males and 4 females from Mountain Top and Stay-a-while Spring, Columbia County, average, respectively: total length 191, 180; length of tail 56, 52; hind foot 26, 25.
Distribution.—The higher parts of the Blue Mountains.
Remarks.—Washington specimens referred to this race are intermediate between wallowa and aequalidens but are colored like aequalidens.
Thomomys clusius fuscus Merriam, N. Amer. Fauna, 5:69, July 30, 1891.
Thomomys myops Merriam, Proc. Biol. Soc. Washington, 14:112, July 19, 1901 (type from Conconnully, Okanogan County, Washington).
Thomomys fuscus fuscus Bailey, N. Amer. Fauna, 39:126, November 15, 1915.
Type.—Obtained at the head of Big Lost River, Custer County, Idaho, by B. H. Dutcher on September 23, 1890; type in United States National Museum.
Racial characters.—Similar to devexus but upper parts rich tawny in color.
Measurements.—Three males and 2 females from Newport, Pend Oreille County, average, respectively: total length 189, 186; length of tail 54, 57; hind foot 27, 26.
Distribution.—Along the eastern border of the state to the north of the Snake River, northeastern Washington, and the northeastern Cascades.
Thomomys talpoides badius Goldman, Jour. Mamm., 20:242, May 15, 1939 (type from Wenatchee, Chelan County, Washington).
Type.—Obtained at Selah, Yakima County, Washington, by P. Burgner, on November 27, 1938; type in Museum of Vertebrate Zoölogy.
Racial characters.—Similar to fuscus but more orange, less red.
Measurements.—Four male and 3 female topotypes average, respectively: total length 209, 191; length of tail 60, 56; hind foot 27, 26.
Distribution.—The eastern edge of the Cascades from the Wenatchee Mountains south to the Simcoe Anticline.
Thomomys douglasii shawi Taylor, Proc. Biol. Soc. Washington, 34:121, June 30, 1921.
Type.—Obtained at Owyhigh Lakes, Mount Rainier, Pierce County, Washington, by G. G. Cantwell, on August 9, 1919; type in United States National Museum.
Racial characters.—A large, tan-colored pocket gopher, similar to aequalidens but paler.
Measurements.—Two male and 7 female topotypes average, respectively: total length 227, 213; length of tail 72, 64; hind foot 32, 30.
Distribution.—The higher Cascade Mountains from Mount Rainier southward. Southern limits of range unknown.
Type.—Obtained 5 miles south of Trout Lake, Klickitat County, Washington, by W. W. Dalquest on July 27, 1937; type in Museum of Vertebrate Zoölogy.
Racial characters.—Similar to shawi but paler and grayer.
Measurements.—Two males and 2 females from Morrison Springs Ranger Station, Skamania County, average, respectively: total length 211, 212; length of tail 64, 58; hind foot 28, 29.
Distribution.—The Cascade Mountains from the vicinity of Mount Adams north. The zone of intergradation between shawi and immunis is in the rugged, inaccessible mountains between Mount Rainier and Mount Adams.
Thomomys limosus Merriam, Proc. Biol. Soc. Washington, 14:116, July 19, 1901.
Thomomys talpoides limosus Goldman, Jour. Mamm., 20:235, May 15, 1939.
Type.—Obtained at White Salmon, Klickitat County, Washington, by J. A. Loring on June 26, 1897; type in United States National Museum.
Racial characters.—Similar to immunis but darker with smaller, shorter skull.
Measurements.—One male and 13 female topotypes average, respectively: total length 210, 198; length of tail 55, 56; hind foot 30, 28.
Distribution.—The lower Columbia River Valley, from the type locality east to the Columbian Plateau.
Geomys douglasii Richardson, Fauna Boreali-American, 1:200, 1829.
Geomys fuliginosus Schinz, Syn. Mamm., 2:136, 1846 (type from "Habitat ad fluvium Columbia").
Thomomys douglasii Allen, Bull. Amer. Mus. Nat. Hist., 5:66, April 28, 1893.
Thomomys douglasi douglasi Bailey, N. Amer. Fauna, 39:116, November 15, 1915.
Thomomys talpoides douglasii Goldman, Jour. Mamm., 20:234, May 15, 1939.
Type.—Obtained at Fort Vancouver (now the city of Vancouver), Clark County, Washington, by David Douglas, probably in 1825. Probably not now in existence.
Racial characters.—A medium-sized, yellowish gopher with tiny, pointed ears and very small postauricular patches.
Measurements.—Two male and 10 female topotypes average, respectively: total length 227, 213; length of tail 68, 63; hind foot 30, 30; and weight 148, 117 grams.
Distribution.—Known only from Clark County.
Thomomys talpoides glacialis Dalquest and Scheffer, Proc. Biol. Soc. Washington, 55:97, August 13, 1942.
Type.—Obtained 2 miles south of Roy, Pierce County, Washington, by W. W. Dalquest, on December 19, 1941; type in United States National Museum.
Racial characters.—A dark, yellowish-brown gopher with orange-tinged underparts.
Measurements.—Twenty male and 17 female topotypes average, respectively: total length 225, 220; length of tail 72, 71; hind foot 30, 30; and weight 128, 116 grams.
Distribution.—Known only from Roy Prairie, Pierce County.
Thomomys douglasii tacomensis Taylor, Proc. Biol. Soc. Washington, 32:169, September 30, 1919.
Thomomys talpoides tacomensis Goldman, Jour. Mamm., 20:235, May 15, 1939.
Type.—Obtained 6 miles south of Tacoma, Pierce County, Washington, by G. G. Cantwell on December 24, 1918; type in United States National Museum.
Racial characters.—A large, dark form; rich hazel in color with large, black postauricular patches and ochraceous underparts.
Measurements.—Thirteen male and 15 female topotypes average, respectively: total length 224, 196; length of tail 71, 57; hind foot 31, 29; and weight 127, 104 grams.
Distribution.—Restricted to the area about Steilacoom and Tacoma, Pierce County, Washington.
Thomomys talpoides pugetensis Dalquest and Scheffer, Proc. Biol. Soc. Washington, 55:96, August 13, 1942.
Type.—Obtained 4 miles south of Olympia, Thurston County, Washington, by W. W. Dalquest, on December 31, 1941; type in United States National Museum.
Racial characters.—Similar to glacialis but underparts not tinged with orange and conspicuous dusky areas present on sides of neck.
Measurements.—Fourteen male and 19 female topotypes average, respectively: total length 223, 205; length of tail 62, 59; hind foot 30, 29; and weight 123, 96 grams.
Distribution.—Known only from the type locality.
Thomomys talpoides tumuli Dalquest and Scheffer, Proc. Biol. Soc. Washington, 55:96, August 13, 1942.
Type.—Obtained on Rocky Prairie, 7 miles north of Tenino, Thurston County, Washington, by W. W. Dalquest on January 2, 1942; type in United States National Museum.
Racial characters.—Similar to pugetensis but grayer, less yellow.
Measurements.—Eleven male and 14 female topotypes average, respectively: total length 225, 216; length of tail 60, 64; hind foot 31, 30; and weight 140, 118 grams.
Distribution.—Known only from the type locality.
Thomomys douglasi yelmensis Merriam, Proc. Biol. Soc. Washington, 13:21, January 31, 1899.
Thomomys douglasii yelmensis Taylor, Proc. Biol. Soc. Washington, 32:169, September 30, 1919.
Thomomys talpoides yelmensis Goldman, Jour. Mamm., 20:235, May 15, 1939.
Type.—Obtained at Tenino, Thurston County, Washington, by C. P. Streator on October 24, 1891; type in United States National Museum.
Racial characters.—A rather small, yellowish race with whitish underparts.
Measurements.—Twenty-one male and 21 female topotypes average, respectively; total length 213, 202; length of tail 64, 61; hind foot 29, 28; and weight 121, 101 grams.
Distribution.—Known only from Grand Mound Prairie (type locality), Vail Prairie, near Vail, and Rochester Prairie, near Rochester, all in Thurston County.
Thomomys talpoides couchi Goldman, Jour. Mamm., 20:243, May 15, 1939.
Type.—Obtained on Scotts Prairie, 4 miles north of Shelton, Mason County, Washington, by L. K. Couch, on June 27, 1922; type in United States National Museum.
Racial characters.—A small race; rich hazel in color.
Measurements.—Thirteen male and 9 female topotypes average, respectively: total length 196, 191; length of tail 55, 53; hind foot 27, 27; and weight 87, 79 grams.
Distribution.—Known only from the type locality and from Lost Lake Prairie, near Satsop, Mason County.
Thomomys melanops Merriam, Proc. Biol. Soc. Washington, 13:21, January 31, 1899.
Thomomys douglasi melanops Bailey, N. Amer. Fauna, 39:119, November 15, 1915.
Thomomys talpoides melanops Goldman, Jour. Mamm., 20:235, May 15, 1939.
Type.—Obtained at head of Sol Duc River, Clallam County, Washington, by V. Bailey on August 28, 1897; type in United States National Museum.
Racial characters.—Most like shawi but smaller, with larger postauricular patches and dusky areas on side of head.
Measurements.—A male topotype measures: total length 211; length of tail 67; hind foot 28.
Distribution.—Higher Olympic Mountains.
Description.—The beaver is the largest rodent occurring in Washington. Large individuals weight approximately 50 pounds. They are heavily-built, robust animals with large heads and short necks. The large, flat, naked tail immediately separates them from all other mammals occurring in the state. The forelegs are short and the forefeet handlike. The hind legs are long, thick and powerful. The hind feet are large and webbed for swimming. The ears are small and the eyes, although of moderate size, are not prominent. The incisors are large and prominent with a distinct yellow or orange color. The close, dense underfur is overlaid with long, lax, rather stiff overfur or guardhairs.
Fig. 99. Beaver (Castor canadensis). Silvana, Washington, August 15, 1921. (Fish and Wildlife Service by Leo K. Couch, No. B-21912.)
Beavers are found in Europe, Asia and North America. In America they range from Alaska to Mexico and from the Atlantic to the Pacific. Many American subspecies have been described, all thought to belong to a single species.
Beavers live in streams and lakes of Washington under diverse climatic conditions. They are perhaps most abundant in the western Washington lowlands where there are numerous watercourses. In [Pg 316] the Cascade Mountains they are found well up into the Canadian Life-zone, where the streams are swift and clear with stony bottoms. The highest altitudinal record known to us is Reflection Lake, Mt. Rainier, 4,861 feet (Brockman, 1939: 71). Farther east, in the timbered regions of the eastern Cascades and in northeastern Washington, beavers live in deeper, more sluggish streams. Abundant beaver signs were noted at the junction of the Snake and Columbia rivers where the animals were occupying one of the hottest, most desertlike portions of the state. They are found in Moses Lake, in the center of the arid Columbian Plateau. Beavers once occurred in the San Juan Islands and have recently been reintroduced there.
Fig. 100. Beaver (Castor canadensis): lodge and beaver pond, Elbe, Washington, August 24, 1926. (T. H. Scheffer photo.)
The part the beaver played in man's exploration of the state of Washington is a story in itself. The establishment of Fort Vancouver and Fort Spokane and other settlements was principally due to the trade in beaver pelts. These forts served as headquarters for Douglas, Suckley, Townsend, Nuttall and other early naturalists who contributed so greatly to the knowledge of the mammalian fauna of the western United States.
Fig. 101. Cottonwood pole carved by beaver, Lake Wenatchee, Washington, May 13, 1938. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 33.)
Trapping for fur drastically reduced the number of beavers in Washington. After trapping was prohibited they were slow in recovering and, until about 1930, they were uncommon. After that date they seemed to increase rapidly, becoming abundant about 1940. The present system of removing beavers only from areas where they are doing damage, and then only under strict supervision from the State Department of Game, has held their numbers at a high level.
The food of the beaver varies greatly with locality. Along the Columbia River, in eastern Washington, cottonwood (Populus hastata) and willow (Salix sp.) seem to constitute favorite foods. Few other trees grow in this arid region. On Puget Island, near the mouth of the Columbia, willow, alder (Alnus oregonus) and probably other shrubs are eaten. In the many streams that run from the western Cascade Mountains to Puget Sound, various species of willow seem to be the principal food eaten. Some alder, cascara, Douglas fir, red huckleberry and salmonberry are eaten. In some streams on the eastern slope of the Cascades, coniferous trees, including Douglas fir, yellow pine (Pinus ponderosa), and red cedar (Thuja plicata) are used.
Beaver dams are abundant in certain sections of Washington, notably the area between the Cascade Mountains and Puget Sound. Dams several hundred feet in length were seen in the valley of Cherry Creek, 7 miles northeast of Duvall, King County. However, in most parts of the state few dams are built. No dams are necessary in the larger streams and lakes while the smaller mountain streams possess a gradient too steep for successful damming. Spring floods and high water following rains would destroy such dams.
True beaver houses, or lodges, are seldom seen in Washington. In the larger streams, rivers and lakes the beavers are "bank beavers," living in burrows with underwater entrances. In the mountain streams, log jams furnish protection. Great piles of logs and debris accumulate at bends of streams in periods of high water. These jams commonly contain large, well-anchored key logs and persist for years. Back-cutting by the stream often forms deep pools before the jams while the sloping bank opposite may support willows and alders. Thus beavers may find a home, food, and a protecting pool of deep water at a log jam. In periods of low water, and when meandering of the stream causes it to leave the log jam, canals are constructed by the beavers to connect the log jam with the stream.
Where dams are built a few lodges may occur. For the most part, however, the beavers dig holes in the bank or in small islands that [Pg 319] result from their dam-building activities. Burrows are occasionally easy to locate because of the mud and sticks laid on the ground above. At best these sticks form a layer only a few inches thick and are too loosely packed to furnish any additional protection. They probably represent a concession to the beavers' lodge-building instincts.
The beaver probably breeds in January. A specimen from Cle Elum, taken on March 1, contained 4 embryos and one from the Teanaway River near Cle Elum held 2 embryos on February 28. One from Swauk Creek, Kittitas County, on the eastern slope of the Cascades, had 3 embryos on February 12. A young beaver was born to a captive animal on May 18. Its mother was taken at Soos Creek in southeastern King County. This baby weighed 884 grams.
The majority of the beavers trapped are small animals, measuring from 725 to 900 mm. (total length) in February, and weighing from 10 to 20 pounds. These are kits, probably one and two years old. It is rather difficult to divide these animals into two age groups but probably the first year young (7 to 11 months old) weigh from 10 to 15 pounds and measure between 725 and 800 mm. in total length. Two-year olds (19 to 23 months) measure from 800 to 925 mm. and weigh from 15 to 25 pounds. They measure approximately 1050 mm. the third year, when they become adult, and weigh in the neighborhood of 45 pounds. Individuals occasionally weigh more than 50 pounds; these are probably old adults.
Damage by beavers is limited. Their dams occasionally flood roads and rarely fields. In some parts of eastern Washington, where fruit raising is an important industry, an individual beaver may cause considerable damage by cutting valuable trees. Ornamental trees may be damaged near the larger cities. Beavers living in Lake Washington in almost the center of the city of Seattle, caused some damage to ornamental trees on the University of Washington Golf Course. Canals and dams in small streams and ditches in areas where stock raising is practiced, flood stock food and sometimes form mudholes or bogs where stock might be trapped. Where individual beavers are doing damage they are trapped and killed, or are live-trapped and transported to more favorable habitats. In places where any beaver may be potentially dangerous, as in the fruit-growing areas of eastern Washington or stock-raising areas in western Washington, their numbers should be kept down by continued trapping.
Fig. 102. Forest road flooded by beavers which built a dam on the upstream (left) side of a bridge; Tronson Creek, Blewett Pass, Wenatchee National Forest, May 13, 1938. (Fish and Wildlife Service by Victor B. Scheffer, No. 30.)
Probably the greatest value of beavers lies in the impounding of water by their dams. This is especially true in the arid mountains of eastern Washington and in the logged-off or burned-over parts of western Washington. These ponds prevent erosion, raise the water table and so stimulate the growth of vegetation, insure greater regularity of stream flow, provide suitable water for trout and muskrats as well as some breeding grounds for waterfowl. On burned-over or logged-off land, the greatest, and often the only, growth of new conifers is in the vicinity of beaver ponds. In the vicinity of beaver dams, rich growths of willows and other deciduous vegetation provides food and cover for deer and fur-bearing mammals.
Fig. 103. Distribution of the beaver in Washington. A. Castor canadensis leucodonta. B. Castor canadensis idoneus.
Trapping of beaver for their fur at present is not a private enterprise but controlled trapping by the state adds to public funds.
Castor canadensis leucodonta Gray, Ann. and Mag. Nat. Hist., 4 (ser. 4):293, October, 1869.
Castor canadensis pacificus Rhoads, Trans. Amer. Philos. Soc., 19 (n. s.):422, September, 1898 (type from Lake Keechelus, Kittitas County, Washington).
Type.—Obtained by Dr. Robert Brown, probably on Vancouver Island, British Columbia.
Racial characters.—Large size; reddish color; elongate skull.
Distribution.—Found over all but the southwestern corner of the state. The most westward locality from which specimens have been examined is Lake Crescent (U. S. N. M.).
Remarks.—The good series of beaver skulls available from several areas of Washington has led to careful comparison of Washington material with specimens from Vancouver Island. Skulls from extreme southwestern Washington are small and decidedly short, being referable to the race idoneus. No satisfactory way of distinguishing between the others could be found and all are consequently referred to the earliest named form, leucodonta.
Castor canadensis idoneus Jewett and Hall, Jour. Mamm., 21:87, February 15, 1940.
Type.—Obtained at Foley Creek, tributary to Nehalem River, Tillamook County, Oregon, by C. Leach on December 15, 1914; type in California Museum of Vertebrate Zoölogy.
Description.—Similar to leucodonta but smaller, darker, with shorter and wider skull.
Distribution.—Lowlands near the mouth of the Columbia River. Specimens from Puget Island (U. S. N. M.) have been examined.
Onychomys leucogaster fuscogriseus Anthony, Bull. Amer. Mus. Nat. Hist., 32: 11, March 7, 1913.
Type.—Obtained at Ironside, Malheur County, Oregon, by H. E. Anthony, in 1912; type in American Museum of Natural History.
Measurements.—Nine males and 5 females from Franklin, Benton, and Walla Walla counties, Washington, average, respectively: total length 132.3, 133.0; length of tail 35.8, 35.6; hind foot 19.2, 19.6; ear 17.6, 17.6; weight 23.3, 24.1 grams.
Distribution.—The Columbian Plateau, southeastern Washington, and the Yakima Valley, ranging north to Douglas (Taylor and Shaw, 1929: 21), west to Yakima (M. V. Z.), east to Asotin (Taylor and Shaw, 1929: 21) and south to Wallula (Taylor and Shaw, 1929: 21).
Fig. 104. Distribution of the northern grasshopper mouse, Onychomys leucogaster fuscogriseus, in Washington.
Description.—A mature grasshopper mouse is slightly larger than a house mouse; the head and body measure about 4 inches and the tail about 1-1/4 inches; it may be recognized by the rather plump [Pg 324] body, naked ears, short, thick and tapering tail and soft, dense fur. The upper parts of adults are buffy. Younger animals are blue-gray above. Underparts and tail are white.
Grasshopper mice of two species range over much of northern Mexico and the western United States. The species occurring in Washington, leucogaster, is found from southern Manitoba to northern Mexico, and from the Pacific states east to North Dakota. This species is characteristic of the Upper Sonoran Life-zone, and usually is associated with sagebrush and greasewood. It prefers open areas of sand and avoids heavy cover, and seems to be strictly nocturnal. It eats vegetation, seeds, insects, or the flesh of other mice. According to Bailey (1936: 178), grasshopper mice pursue, catch and kill other mice. They hunt by scent and follow tracks like a wolf. They make a sound, or "call," like a miniature wolf howl. They are said to be rather docile and make friends with humans quickly.
On some occasions grasshopper mice are locally abundant, but for the most part they are rather uncommon over their range. Usually one is taken in about 200 traps set in suitable habitats. They do not hibernate; one was caught at Moses Lake, Grant County, on a freezing winter night.
The grasshopper mouse is almost always associated with the pocket mouse (Perognathus parvus), deer-mouse (Peromyscus maniculatus), and often the harvest mouse (Reithrodontomys megalotis). The same snakes, owls, and carnivorous mammals that prey on these mice doubtless eat also the grasshopper mouse.
R. D. Svihla (1936: 172) determined the gestation period of 3 lactating grasshopper mice captured at Lind, Adams County, as 33, 39 and 47 days. A non-lactating female gave birth to a litter in 32 days. Young varied from 2 to 5 in number and were pink and hairless at birth, with eyes and ears closed.
Reithrodon megalotis Baird, Mamm. N. Amer., p. 451, 1857.
Reithrodontomys megalotis nigrescens Howell, N. Amer. Fauna, 36:32, June 5, 1914 (type from Payette, Idaho).
Reithrodontomys megalotis megalotis Bailey, N. Amer. Fauna, 55:189, August 29, 1936.
Type.—Obtained between Janos, Chihuahua, and San Luis Springs, New Mexico, by C. B. R. Kennerly, in 1855; type in United States National Museum.
Measurements.—Five males and 6 females, showing moderate wear on the third upper molars, from Walla Walla County, average, respectively: total length 141, 137.5; length of tail 71.8, 68.1; hind foot 17.2, 16.7; weight (males only) 10.5 grams. A female, 136 mm. long, from Grant County, weighed 9.5 grams. Over the range of megalotis in the United States, males average consistently larger than females in length of the hind foot and consistently less in length of head and body.
Distribution.—The Columbian Plateau, the Upper Sonoran Life-zone area west of the Columbia River and south of the Wenatchee Mountains in Kittitas, Yakima, and Benton counties, the north side of the Columbia River in Klickitat County, and Okanogan County east of the Okanogan River. Marginal localities are: Timentwa (W. W. D.) on the north, Maryhill (M. V. Z.) on the west, Wallula (M. V. Z.) on the south, and Colfax (Taylor and Shaw, 1929: 21) on the east.
Fig. 105. Distribution of the western harvest mouse, Reithrodontomys megalotis megalotis, in Washington.
Remarks.—Harvest mice from the states of Washington, Oregon, Idaho, Utah, Nevada, California, Arizona, and New Mexico were examined to ascertain the validity of the name Reithrodontomys megalotis nigrescens Howell (type from Payette County, Idaho). No cranial characters or measurements could be found to separate nigrescens from megalotis. Specimens from Washington, Oregon, and west-central Idaho, within the range ascribed to nigrescens, averaged slightly darker in color with a more distinct, blackish dorsal line than specimens from New Mexico. This difference is slight, and specimens from any one locality vary greatly in color. Some Washington specimens are as pale as the palest material from New Mexico, and some of the darkest nigrescens can be matched by specimens of megalotis from Arizona and New Mexico. This slight color variant is not worthy of recognition as a distinct subspecies, and I agree with Howell (1939: 390) that nigrescens is a synonym of megalotis.
Description.—The western harvest mouse is about the size and shape of the common house mouse. The length of the tail is approximately equal to the length of the head and body. The tail is slim and lightly haired. The upper parts are brownish buff lightly washed with blackish. A faint but distinct dark dorsal stripe is usually present. The underparts are white. From Mus and Peromyscus, Reithrodontomys can be distinguished by the grooves on the anterior face of the upper incisors and the long brown hairs at the bases of the ears.
This genus reaches the extreme northern limit of its distribution in the state of Washington, where it ranges to the Canadian border. A single subspecies occurs in Washington. The harvest mouse is a resident of the Upper Sonoran Life-zone and it is most abundant in dense growths of grass, weeds, and other vegetation. Along the Yakima River, near Ellensburg, it was abundant in thick, river-side jungle. Near Yakima many specimens were taken in a marsh of cattails and tules. In the Grand Coulee they live in thick grass. Near Moses Lake several were found in their nests in a thick growth of tumbleweed. Harvest mice occur also, although never in great numbers, in relatively open areas. At the Potholes near Moses Lake, Grant County, a few were caught on sand dunes among scattered bushes of sage. They were taken on sagebrush flats in several localities. Gray (1943: 191) estimates their numbers in the sagebrush area of the Yakima Valley at 5 per acre.
Where harvest mice live in dense vegetation they make small runways and food piles similar to those of Microtus but in more open areas they leave no perceptible signs. Near Yakima the nest of a harvest mouse was found among the roots of an overturned apple tree. This nest was an irregular ball composed of fine grasses and shredded bark closed above, with a side entrance and a central cavity three inches in diameter. Near Moses Lake three nests of harvest mice were discovered on the ground under cover of a dense growth of tumbleweed, and six live harvest mice were captured in the same area. These nests were cuplike, open above, and were composed of dry grasses, bits of weeds, and newspaper. The smoothly rounded inner cups measured about three inches in diameter.
Embryos were found in harvest mice in Washington from March to April and numbered 3 to 6.
Description.—The deer mouse is slightly larger than the house mouse, the head and body measuring from 3-1/2 to 4 inches. The length of the tail varies considerably in the several races, ranging from approximately 2-1/2 inches in gambelii to 4 inches or more in some individuals of oreas. The eyes are large and protrude slightly. The ears are large, erect and naked. The tail is sparsely furred with short, posteriorly directed hairs. The color of the upper parts varies from yellowish buff in gambelii to reddish brown in oreas and dusky in austerus. Young deer mice are gray or bluish gray above. Underparts and feet are white. The tail is bicolored, being dusky above and white below.
Fig. 106. Deer mouse (Peromyscus maniculatus austerus), in captivity, Seattle, Washington, September 18, 1939. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 723.)
In Washington Peromyscus may be separated from other naked-eared mice by ungrooved teeth and slightly tapering, bicolor tail. Nearly every part of North America is inhabited by one or more kinds of Peromyscus. The single species, maniculatus, which occurs in Washington, ranges from the Atlantic to the Pacific and from the Arctic to tropical Mexico. Osgood (1909: 17) remarks: "... it is probable that a line, or several lines, could be drawn from Labrador to Alaska and thence to southern Mexico throughout which not a single square mile is not inhabited by some form of this species."
Deer mice are the most abundant mice in Washington. They occur at sea level on the ocean beaches and above timber-line, even breeding in the Arctic-alpine Life-zone. They occur on the islands in Puget Sound, the lowland marshes and deciduous thickets of western Washington, the great conifer forests, the alpine cirques and mountain parks, and the deserts of eastern Washington. Almost everywhere they are the commonest mammal encountered. They make their homes in city dwellings, under logs or in hollow trees in the forest, in the burrows of larger rodents and, if necessary, in burrows of their own construction.
Nests are usually cup-shaped masses of soft material, 4 to 8 inches in diameter with central cavities 2 to 3 inches in diameter. Materials utilized include dry grass, shredded bark, rope, rags, newspaper, moss, cattail fluff, fur, wool, and feathers. One nest discovered in the attic of an old building near Coulee City, Grant County, consisted of the mummified body of a wood rat (Neotoma cinerea). The fur of the arched body of the rat formed one side of the nest, while the fur of the far side of the body had been plucked to form the remainder. In the extensive marshes along Lake Washington, King County, deer mice occupy the ball-shaped nests of tule wrens (Telamtodytes palustris). These are constructed entirely of fragments of cattail leaves and cattail fluff and are placed in the cattail rushes about four feet above the wet ground of the marsh. Other deer mouse nests were found in an unused typewriter, in the pocket of an old coat, in a window frame, under a piece of tar paper on the ground, in a cavity in the ground under a board, between a ceiling and a loose bit of roofing paper, under rocks and logs, in hollows in logs, and in an unused nest of a wood rat. Where plant cover is dense, nests are placed on the surface of the ground, as under thick growths of tumbleweed (Salsola) in eastern Washington.
Deer mice are definitely nocturnal. Of thousands of specimens trapped only a few were taken in the daytime. Two of these were taken in a rock slide at Pass Creek Pass, Pend Oreille County, on the same day, and some unusual conditions may have incited them to travel in daylight. Deer mice are active throughout the winter and their lacy footprints are seen on the snow in below-freezing weather. They are mainly terrestial but one race (oreas) is at least partly arboreal for it is often taken in traps set in trees for flying squirrels. One was caught in a trap set 50 feet from the ground. All are good climbers, and oreas is perhaps the best. One was watched as it climbed the rough cedar shake wall of a cabin, ran upside-down across three feet of rough ceiling, and descended [Pg 329] the opposite wall head first. They are equally agile in climbing rocks and cliffs.
The ordinary gait of the deer mouse is a steady run. When frightened it may leap a distance of one foot or more. When surprised in its nest it usually remains in the vicinity, hopping about in confusion, returning time after time to the exposed nest.
The food of deer mice consists principally of vegetable material such as seeds, nuts, berries, and the soft parts of fleshy plants. Mushrooms and lichens are eaten. Insect remains are present in small quantities in most stomachs examined. Flesh is also eaten and small mammals caught in traps are often eaten by deer mice. Cannibalism appears to be common only in captivity.
Fig. 107. Distribution of the deer mouse in Washington. A. Peromyscus maniculatus oreas. B. Peromyscus maniculatus austerus. C. Peromyscus maniculatus hollisteri. D. Peromyscus maniculatus gambelii. E. Peromyscus maniculatus artemisiae.
Deer mice are often heavily parasitized by tapeworms and roundworms. Virtually all of the deer mice taken on Jones Island in the San Juan Islands had livers so infected by the eggs of a nematode worm as to be swollen to several times their natural size, and had a yellow, crystalline appearance. The maggots of parasitic flies often occur in the anal and genital region of deer mice, effectively sterilizing some individuals.
Predatory snakes, birds, and mammals doubtless kill and eat deer mice. Strangely enough, although deer mice may be the commonest mammal in an area, the pellets of owls collected in the same area usually contain the remains of relatively few deer mice. The impact of owls seems to be greatest upon mice, such as Microtus, which follow definite runways.
The breeding season of the deer mouse extends over most of the spring and summer. Growth and development of the young of Peromyscus m. oreas have been discussed by A. Svihla (1936A). Embryos found varied in number from 2 to 8 with a mean of 4.5.
Peromyscus oreas Bangs, Proc. Biol. Soc. Washington, 12:84, March 24, 1898.
Peromyscus akeleyi Elliot, Field Columb. Mus. Publ. 30., zoöl. ser., 1:226. February 1, 1899 (type from Elwha River, Olympic Mountains, Washington).
Peromyscus maniculatus oreas Osgood, N. Amer. Fauna, 28:51, April 17, 1909.
Type.—Obtained on Church Mountain, 6,500 ft., Mt. Baker Range, near the U. S. boundary, British Columbia, by Allan Brooks on August 29, 1896; type in Museum of Comparative Zoölogy.
Racial characters.—Size large; tail more than 50 per cent of total length; color of upper parts reddish.
Measurements.—Twenty-five males and 15 females average: total length 202; length of tail 110; hind foot 22.6; ear 18.7.
Distribution.—From the Cascade Mountains west to the Pacific Ocean, save for the Puget Sound area and a narrow band extending southward nearly to the Columbia River. Marginal localities along the west side of Puget Sound are: Deer Park (W.W.D.), Lake Cushman (W.W.D.), and Kelso (M.V.Z.). Marginal localities on the west side of the Cascade Mountains are: Tomyhoi Lake (W.W.D.), Forks of Skykomish River (W.W.D.), North Bend (W.W.D.), Mt. St. Helens (V.B.S.) and Yacolt (M.V.Z.). Marginal localities on the east slope of the Cascade Mountains are: Grouse Creek (W.W.D.), Lake Wenatchee (W.W.D.), Easton (W.W.D.), Morrison Springs Ranger Station (W.W.D.) and Wind River C.C.C. Camp (W.W.D.).
Peromyscus maniculatus hollisteri Osgood, N. Amer. Fauna, 28:62, April 17, 1909.
Type.—Obtained at Friday Harbor, San Juan Island, San Juan County, Washington, by N. Hollister on October 23, 1903; type in United States National Museum.
Racial characters.—Similar to oreas but darker, less reddish, and with much shorter tail. Length of tail about 40 per cent of total length.
Measurements.—Populations of deer mice on the various islands vary slightly in average measurements. The average measurements of 10 males and 10 females from San Juan Island, 20 males and 15 females from Blakeley Island, and 20 males and 15 females from Cypress Island, are respectively: total length 189, 183, 179; length of tail 87, 82, 79; hind foot 22.8, 22, 19.5; ear 19.4, 18.7, 19.5; weight 33, 29, 32 grams.
Distribution.—The San Juan Islands, San Juan and Skagit counties, in northern Puget Sound.
Hesperomys austerus Baird, Proc. Acad. Nat. Sci. Philadelphia, 7:336, 1855.
Sitomys americanus austerus Allen, Bull. Amer. Mus. Nat. Hist., 5:192, August 18, 1893.
P[eromyscus]., austerus Bangs, Amer. Nat., 31:75, January 1, 1897.
Peromyscus maniculatus austerus Osgood, N. Amer. Fauna, 28:63, April 17, 1909.
Type.—Obtained at Old Fort Steilacoom, Pierce County, Washington, by J. G. Suckley, probably on January 20, 1854; type in United States National Museum.
Racial characters.—Similar to oreas but smaller with relatively shorter tail; tail narrower at base, less sharply bicolor; upper parts duller, browner, less reddish. In oreas the tail is usually more than 100 mm. In austerus the tail is usually less than 90 mm.
Measurements.—Twenty-seven males and 23 females from Seattle, King County, average: total length 180.5; length of tail 88.6; hind foot 21; ear 18. The average weight of 15 adults is 21 grams.
Distribution.—The Puget Sound area and south to the Columbia River in a narrow band through the lowlands. Marginal localities on the west side of Puget Sound are: Port Angeles (W.W.D.), Bremerton (W.W.D.), 4 miles north of Shelton (W.W.D.), Tenino (W.W.D.) and Mouth of Klama River (M.V.Z.). Marginal localities on the east side of Puget Sound are: Bellingham (W.W.D.), Arlington (W.W.D.), Everett (W.W.D.), 4 miles east of Monroe (W.W.D.), Fall City (W.W.D.), and Vancouver (W.W.D.).
Peromyscus oreas rubidus Osgood, Proc. Biol. Soc. Washington, 14:193, December 12, 1901.
Peromyscus maniculatus rubidus Osgood, N. Amer. Fauna, 28:65, April 17, 1909.
Type.—Obtained at Mendocino City, California, by J. A. Loring on November 17, 1897; type in United States National Museum.
Racial characters.—Closely similar to austerus but paler with longer skull. Specimens from Washington are almost purplish-brown in appearance.
Measurements.—Six males and 2 females from Puget Island, Wahkiakum County, average: total length 177; length of tail 89; hind foot 21.9; ear 18.7.
Distribution.—This is the deer mouse of the humid coastal belt of Oregon and California. It enters Washington only at Puget Island, Wahkiakum County, in the Columbia River.
Peromyscus gambelii Baird, Mamm. N. Amer., Pacific R. R. Reports, 8:464, 1857.
Sitomys americanus gambelii Allen, Bull. Amer. Mus. Nat. Hist., 5:190, August 18, 1893.
P[eromyscus]., t[exanus]. gambelii Mearns, Proc. U. S. Nat. Mus., 18:446, March 25, 1896.
Peromyscus maniculatus gambeli Osgood, N. Amer. Fauna, 28:67, April 17, 1909.
Type.—Obtained at Monterey, Monterey County, California, by W. P. Trowbridge; skin catalogued on January 4, 1853; type in United States National Museum.
Racial characters.—Size small; tail short; color pale, varying from buffy gray to rich buff; rarely dark reddish brown.
Measurements.—Twenty-five males and 15 females average: total length 160; length of tail 71; hind foot 19.8; ear 17.7.
Distribution.—The Columbian Plateau. Intergradation with oreas occurs between the Columbia River and the Cascade Mountains. Marginal records on the west are: Chelan (W.W.D.), Leavenworth (W.W.D.), Cle Elum (W.W.D.), Satus Pass (W.W.D.), Lyle (V.B.S.). Marginal localities on the north are: Chelan (W.W.D.), 5 miles north of Coulee (W.W.D.), Sprague (V.B.S.) and Steptoe Butte (M.V.Z.).
Sitomys americanus artemisiae Rhoads, Proc. Acad. Nat. Sci. Philadelphia, p. 260, October 23, 1894.
Peromyscus texanus artemisiae Miller and Rehn, Proc. Boston Soc. Nat. Hist., 30:84, December, 1901.
Peromyscus maniculatus artemisiae Osgood, N. Amer. Fauna, 28:58, April 17, 1909.
Type.—Obtained at Ashcroft, British Columbia, by S. N. Rhoads on July 5, 1892; type in Philadelphia Academy of Sciences.
Racial characters.—Similar to gambelii but darker, more reddish; often with longer tail.
Measurements.—Ten males and 10 females from northeastern Washington average: total length 167; length of tail 80; hind foot 20; ear 17.8.
Distribution.—Northeastern Washington and the Blue Mountains of southeastern Washington, extending west, in northeastern Washington, to Conconully (W.W.D.) and south to Duly Lake (W.W.D.).
Description.—The bushy-tailed wood rat is slightly larger than the common Norway or brown rat. The head and body measure about 12-1/2 inches and the tail about 3-1/2 inches. It resembles the deer mouse in general proportions. The ears are large and naked and the black eyes are large and protruding. The tail is bushy, squirrel-like. The feet are small and have furry soles. The fur is soft and silky. Adults are brownish gray above with white underparts and a gray tail. Young individuals have blue-gray upper-parts.
Wood rats range over most of North America. They exhibit great variation, especially in the southwestern United States. The bushy-tailed species occurs in the western United States and Canada. Wood rats are notorious for invading buildings in the mountains and in the desert. However, their natural habitat is broken rock or talus. This habitat preference accounts for their distribution in Washington, for talus is common except in the humid subdivision of the Transition Life-zone. Wood rats are probably most abundant in the talus slides of the Columbian Plateau, especially in the canyon of the Columbia River and in Moses and Grand coulees. They are common in all the mountainous areas in the state where high altitudes and steep slopes result in the accumulation of talus. Wood rats range from sea level to 10,000 feet elevation on Mount Rainier and from the Upper Sonoran to the Arctic-alpine life-zones. Great horned owls and probably all of the carnivores that share the range of the wood rat prey on it to some extent. Sperry (1941: 15) lists Neotoma in four per cent of 8,339 coyote stomachs gathered throughout the United States.
Wood rats in Washington are definitely nocturnal and are rarely seen in daylight. On January 10, 1939, however, near Colville, Stevens County, a companion and I were sitting on the porch of a deserted shack eating lunch. Suddenly a wood rat darted out of the open door into the full sunlight and tugged at a can, containing a few drops of tomato juice, which one of us was holding in his hand. When the rat looked up and saw the human, it hastily retreated. The wood rat has a habit of flattening its body tightly against the ground when observed and also of "drumming" with its feet when [Pg 334] excited. This habit is shared by the snowshoe rabbit and the spotted skunk. A captive specimen "drummed" by lifting its back feet, alternately, about one-half inch from the surface of a piece of wood and striking downward with surprising power. The agility of the wood rat was demonstrated near Wallula where we watched a specimen by the light of a flashlight as it climbed an almost vertical rock wall, taking advantage of small fractures for toe-holds.
Wood rats are far from noiseless in their nocturnal activities. Near Moses Coulee, Douglas County, wood rats were heard from a distance of 50 feet as they scampered back and forth through a concrete culvert under the highway.
Many species of wood rats build complicated stick houses in which they live. In Washington, elaborate stick houses are sometimes built but are usually not occupied. In the attic of an old building near Pend Oreille Lakes, Stevens County, two large and complicated houses were discovered. These were built of sticks, paper and other debris and measured more than three feet in diameter. Three small, cup-shaped nests resembling those of a bird, made of soft grasses, moss, and shredded paper were found on the attic floor ten feet or more from the stick houses. These nests gave every indication of being used, while the piles of sticks contained no nests and seemed never to have been occupied. Near Moses Lake, Grant County, a wood rat house built in the rafters of an old shed was constructed entirely of tumbleweeds (Salsola pestifer). This house was spherical in shape and measured more than five feet in diameter. It appeared to be unoccupied and a wood rat, probably its builder, was living under the floor of the shed. Most wood rat houses consist of a half-dozen sticks arranged in a crevice in a rock pile or a cave. Wood rats were placing fresh material on a house in the Wenatchee National Forest on January 17, 1939, when there was two feet of snow on the ground and the temperature was hovering around zero. The house measured about one cubic yard and consisted of Ceanothus leaves and steins, with some twigs of fir (Abies). It was built under a bunk in a broken-down cabin left by deer hunters.
The collecting habit of the wood rat has gained it the name "pack-rat." Taylor (1920A: 91) lists the following materials from a wood rat nest on Mt. Rainier: Nesting material: rags, leaves, paper, thumb of a glove, string, thongs, oakum; Food: apple core, onion peel, bacon rind, raisins, 10 bars of chocolate, figs, puff balls, bread crust, meat scraps, cantaloupe rind, potatoes, dried apricots, lemons, [Pg 335] mushrooms, beans, peanuts, banana, 15 lumps of sugar; Miscellaneous: dime, coffee can cover, paraffin from jelly glass, bones, 19 pieces of candles and several cakes of soap.
As a rule only one or two wood rats are trapped at a given locality, indicating that the species is not gregarious. The presence of wood rats in a building, cave, mine tunnel, or talus slide can be detected in several ways. The white, mineral-like incrustation formed by the urine of many generations of wood rats is usually conspicuous on rocks near their homes. A strong musky odor pervades the atmosphere in every habitat occupied by wood rats. Wood rat feces, consisting of hard, black cylinders one-half inch long and three-sixteenths of an inch in diameter, are invariably found scattered on stones or exposed areas of ground.
Fig. 108. Distribution of the bushy-tailed wood rat in Washington. A. Neotoma cinerea occidentalis. B. Neotoma cinerea alticola.
The food of the bushy-tailed wood rat is varied and includes insects and vegetation. A captive specimen that escaped in the University [Pg 336] of Washington Zoölogical Laboratories killed and ate a number of lizards.
Embryos found from April 18 to June 14 varied in number from 3 to 5.
Neotoma occidentalis Baird, Proc. Acad. Nat. Sci. Philadelphia, p. 335, 1855.
Neotoma cinerea occidentalis Merriam, N. Amer. Fauna, 5:58, July 30, 1891.
Type.—Obtained at Shoalwater Bay (Willapa Bay), Pacific County, Washington, by J. G. Cooper, in June, 1854; type in United States National Museum.
Racial characters.—Size large; color of upper parts cinnamon brown.
Measurements.—A male and a female from Glade Creek, 1/2 mile north of the Columbia River, Benton County, measure, respectively: total length 425, 395; length of tail 181, 176; hind foot 50, 44; ear 32, 31; weight 532.5, 330.0 grams.
Distribution.—Throughout the state except in the Blue Mountains.
Remarks.—Specimens from the Olympic Mountains and western Washington are slightly duller-colored than specimens from eastern Washington and young from western Washington are more bluish, less gray, than young individuals from the Columbian Plateau. The specimens from the Columbian Plateau show some resemblance to alticola. Specimens from northeastern Washington are more like typical occidentalis.
Neotoma cinerca alticola Hooper, Univ. California Publ. Zoöl., 42:409, May 17, 1940.
Type.—Obtained at Parker Creek, 5,500 ft., Warner Mountains, Modoc County, California, by N. B. Stern on June 22, 1916; type in Museum of Vertebrate Zoölogy.
Racial characters.—Similar to Neotoma c. occidentalis but slightly paler, with paler hind feet and narrower sphenopalatine vacuities.
Measurements.—A female from Stay-a-while Spring, Columbia County, measures: total length 387; length of tail 171; hind foot 43; ear 33; weight 330 grams.
Distribution.—Southeastern Washington, south of the Snake River.
Remarks.—The only adult specimen available from the Blue Mountains shows well the characters attributed to alticola.
Synaptomys (Mictomys) wrangeli Merriam, Proc. Biol. Soc. Washington, 10:63, March 19, 1896.
Synaptomys (Mictomys) truei Merriam, Proc. Biol. Soc. Washington, 10:62, March 19, 1896 (type from Skagit Valley, Skagit County, Washington).
Synaptomys borealis wrangeli Howell, N. Amer. Fauna, 50:26, June 30, 1927.
Type.—Obtained at Wrangel, Alexander Archipelago, Alaska, by C. P. Streator on September 6, 1895; type in United States National Museum.
Racial character.—Dark color.
Distribution.—The northwestern Cascades, from Mt. Baker northward.
Remarks.—The relationship of this race to artemisiae remains to be worked out.
Fig. 109. Distribution of the northern lemming mouse in Washington. A. Synaptomys borealis wrangeli. B. Possible range of Synaptomys borealis artemisiae.
Description.—Lemming mice may be recognized by their short, thick bodies, slightly larger than the bodies of the common house mouse (Mus); short tails, which are less than 20 per cent of their total length; small, inconspicuous ears; and grooved upper incisors.
Mice of the genus Synaptomys range over much of boreal North America. Two species and eleven races are recognized by Howell (1927B: 9). Synaptomys borealis ranges westward across Canada from Labrador to the Pacific Coast and from Alaska south to Washington. The lemming mice are terrestrial and inhabit runways similar to those of meadow mice (Microtus). They are alpine in distribution. Shaw (1930: 7-10) found them among typical annual herbs in a cold, boggy mountain valley. Other than this, nothing is known of the species in Washington. Indeed little is known of the life history of any member of the genus.
The lemming mouse possesses four pairs of mammae, two pectoral and two inguinal. It is the most primitive of the microtines. It seems to be of boreal origin but is not known from the Old World. It was evidently forced southward by the Pleistocene glaciers into Washington and is now retreating northward in the wake of the ice. The only specimens recorded from Washington are from the northern border of the state.
Description.—Heather voles are heavy-bodied, short-legged mice, closely similar in general appearance to other microtines that occur in Washington. Their dull, brownish upper parts, lacking a distinct reddish dorsal stripe, separate them from the red-backed mice (Clethrionomys) and the lack of grooves on their upper incisors separates them from Synaptomys. Their relatively short tail (30 per cent or less of their total length) separates them from most meadow mice (Microtus). There are no external characters which serve to separate them from all species of Microtus and the teeth must be examined certainly to identify Phenacomys. In Microtus the angles between the cusps of the inner and outer sides of the lower molars are of approximately equal depth, but in Phenacomys the angles on the inner side of the lower molars are at least twice as deep as those of the outer side of the jaw. In addition the molars of Phenacomys are rooted while those of Microtus are not.
The heather voles and their relatives are primitive microtines that range through boreal North America including the higher parts of the Rocky Mountains and the cool area along the Pacific Coast. Several species are included in the genus: an arctic form (ungava), a lowland Pacific Coastal species (albipes), two tree-inhabiting species (longicaudus and silvicola) and a mountain species (intermedius). Thus far only intermedius has been found in Washington [Pg 339] but further search may reveal that one or more of the arboreal species and possibly albipes are also present.
Heather voles are alpine animals, seldom occurring below the Hudsonian Life-zone. They are rare as compared with most microtines, and are seldom taken in traps, even by the experienced collector of small mammals. Evidence of their presence is most often found in the dense meadows of heather (Cassiope and Phyllodoce) high on the mountains. Here their feces, runways, and old winter nests are not uncommon, and heather voles may be far more abundant than trapping records indicate. They are not restricted to the vicinity of heather, however, for one of my specimens was taken in a marshy willow thicket near Stevens Pass in King County, one in a patch of skunk-cabbage in a grassy glacial cirque near Tomyhoi Lake in Whatcom County, and one at the edge of a snowbank on a steep hillside at Deer Park, Clallam County. In each of these places, however, there were heather meadows within a thousand yards. Shaw (1924A: 12-15) also found the heather mouse on "comparatively dry hill slopes" and reports that it "rather characteristically occurs in such log-tangled areas as former fire zones."
Fig. 110. Distribution of the heather vole in Washington. A. Phenacomys intermedius oramontis. B. Phenacomys intermedius intermedius.
The winter nests of Phenacomys intermedius are placed on the surface of the ground and in the season when occupied are deeply buried under the snow. They are spherical or oval in shape, six to eight inches in diameter and with a side entrance. One examined by Shaw was "formed of white heather twigs, the interstices being filled with leaves of huckleberry." It was "lined with finely shredded grass." All those examined by me were composed of grass and lichens.
The principal foods of the heather vole seem to be white heather (Cassiope mertensiana) and huckleberry (Vaccinium). Red heather (Phyllodoce empetriformis) is not eaten. Shaw also lists beargrass (Xerophyllum tenax) and lousewort (Pedicularis) as eaten.
A burrow excavated and figured by Shaw, was slightly more than a yard long and contained four young in a nest situated but a few inches from one of the three entrances. The nest was made of grass and moss. A female taken at Tye, King County, contained 4 embryos on May 28, 1939. One taken at Pass Creek Pass, Pend Oreille County, contained 3 small embryos on June 14, 1938.
Phenacomys intermedius Merriam, N. Amer. Fauna, 2:32, October 30, 1889.
Phenacomys intermedius intermedius Howell, N. Amer. Fauna, 48:15, October 12, 1926.
Type.—Obtained 20 miles north-northwest of Kamloops, 5,500 ft. elevation, British Columbia, by G. M. Dawson on October 2, 1888; type in United States National Museum.
Racial characters.—Size small; color of upper parts brownish gray; skull small.
Measurements.—A female from Pass Creek Pass, Pend Oreille County, measured: total length 127; length of tail 29; hind foot 17; weight 25 grams.
Distribution.—Known only from extreme northeastern Washington at Pass Creek Pass (W.W.D.) and the Blue Mountains.
Phenacomys oramontis Rhoads, Amer. Nat., 29:941, October, 1895.
Phenacomys olympicus Elliot, Field Columb. Mus. Publ. 30, zoöl. ser., 1:225, February 1, 1899 (type from Happy Lake, 5,000 ft., Clallam County, Washington).
Microtus (Lagurus) pumilus Elliot, Field Columb. Mus. Publ. 30, zoöl. ser., 1:226, February 1, 1899 (type from Happy Lake, 5,000 ft., Clallam County, Washington).
Phenacomys intermedius olympicus Howell, N. Amer. Fauna, 48:21, October 12, 1926.
Phenacomys intermedius oramontis Anderson, Canadian Field Nat., 56:59, June 8, 1942.
Type.—Obtained on Church Mountain, 6,000 ft., Mt. Baker Range, British Columbia, just north of international boundary, by A. C. Brooks on August 6, 1895; type in Philadelphia Academy of Natural Sciences.
Racial characters.—Similar to intermedius but darker, browner above and deeper gray beneath; skull large and heavy.
Measurements.—A female from Tye, King County, measures: total length 152; length of tail 40; hind foot 19; ear 12. A female from Tomyhoi Lake, Whatcom County, measures: 170; 45; 20; 15.
Distribution.—The Cascade and Olympic mountains, from Tomyhoi Lake (W. W. D.) on the north, to Mt. Adams (Taylor and Shaw, 1929:23) on the south.
Description.—Red-backed mice are heavy-bodied, short-tailed and short-legged mice, similar in general appearance to the meadow mice (Microtus). Unlike the meadow mice, they possess rooted molars, a primitive character. They do not have grooved incisors, like Synaptomys, or the great difference in the depth of the angles of the lower molars that characterizes both Synaptomys and Phenacomys. Externally Clethrionomys may be recognized by the broad red dorsal area from which they obtain their common name. Their sides are grayish or dusky and the undersides buffy white.
The red-backed mice, including more than one species, range over the boreal parts of the Old and New World. In America they are found in most of Alaska, Canada and the northern United States, and extend southward in the Rocky Mountains and along the Pacific Coast. The one species Clethrionomys gapperi ranges across southern Canada from the Atlantic to the Pacific, extending southward in forested areas to New Mexico. They usually live in clearings in the forest. In the Pend Oreille Mountains saturatus was common in damp, mossy talus slides, along with Microtus longicaudus and Phenacomys intermedius. The Clethrionomys outnumbered the two latter species combined by 25 to 1. In nearby forest, red-backed mice were scarce but no other microtines were found. In the Kettle River Mountains a week later, red-backed mice were rare. A single specimen was taken in a damp place in the forest; none was found in talus slides. Near Stevens Pass, King County, in the Cascades, cascadensis was taken in equal numbers in talus slides and under logs in the forest. Near Dewey Lake, Yakima County, in the Cascades, I took them only in an extensive grassy meadow. In the Blue Mountains I found idahoensis in the dense chaparral, far from forests.
Unlike Microtus, red-backed mice do not make runways, although they sometimes follow the runways of other mammals.
Taylor (1920B: 92) found red-backed mice breeding on Mount Rainier from early July to the middle of September. One female gave birth to four young in a nest in his duffle bag.
Fig. 111. Distribution of the Gapper and California red-backed mice in Washington. A. Clethrionomys gapperi saturatus. B. Clethrionomys gapperi idahoensis. C. Clethrionomys gapperi nivarius. D. Clethrionomys gapperi cascadensis. E. Clethrionomys californicus occidentalis.
Evotomys gapperi saturatus Rhoads, Proc. Acad. Nat. Sci. Philadelphia, p. 284, October 23, 1894.
[Clethrionomys gapperi] saturatus Whitlow and Hall, Univ. California Publ. Zoöl., 40:265, September 30, 1933.
Type.—Obtained at Nelson, British Columbia, by S. N. Rhoads on August 27, 1892; type in Philadelphia Academy of Sciences.
Racial characters.—Size small, about 140 mm. in total length; tail short, about one-third of total length; ears large, projecting above fur; color of head and sides gray tinged with yellowish; back with distinct red stripe; underparts whitish tinged with buff.
Measurements.—Five males and 10 females from Pass Creek Pass, Pend Oreille County, average: total length 147; length of tail 43; hind foot 18.3; ear 14.
Distribution.—Northeastern Washington from Sherman Creek Pass (W. W. D.) on the west to Pass Creek Pass on the east.
Evotomys idahoensis Merriam, N. Amer. Fauna, 5:66, July 30, 1891.
Clethrionomys gapperi idahoensis Whitlow and Hall, Univ. California Publ. Zoöl., 40:265, September 30, 1933.
Type.—Obtained at Sawtooth (Alturas) Lake, 7,200 ft., Blaine County, Idaho, by C. H. Merriam and V. Bailey, on October 4, 1890; type in United States National Museum.
Racial characters.—Similar to saturatus but brain case longer and narrower and dorsal area more reddish.
Measurements.—Seven males and 6 females from the Blue Mountains average, respectively: total length 138, 142; length of tail 41, 41; hind foot 18.7, 19.0; ear 13.6, 13.5; weight 20.5, 22.7 grams.
Distribution.—The Blue Mountains of southeastern Washington.
Evotomys gapperi saturatus Taylor and Shaw, Occ. Pap. Chas. R. Conner Mus., no. 2:23, December, 1929.
Clethrionomys gapperi cascadensis Booth, Murrelet, 26:27, August 10, 1945.
Type.—Obtained 2 miles south of Blewett Pass, 3,000 ft. elevation, Kittitas County, Washington, by G. G. Cantwell, on October 30, 1921; type in United States National Museum.
Racial characters.—Dark and dull with underparts dull buffy.
Measurements.—Ten adults from the Cascade Mountains average (Booth, 1945: 27): total length 162; length of tail 50; hind foot 19; ear 14.
Distribution.—The Cascade Mountains, according to Booth (loc. cit., p. 28), from Hannegan Pass south to Mount Adams. Marginal localities listed are: Swamp Creek, Glacier Peak, McKenna, and Mt. St. Helens. These, and other localities listed by Booth are not plotted on the accompanying distribution map (fig. 111).
Evotomys nivarius Bailey, Proc. Biol. Soc. Washington, 11:136, May 13, 1897.
Clethrionomys nivarius Svihla, Murrelet, 12:54, May, 1931.
Type.—Obtained on the northwest slope of Mt. Ellinor, 4,000 ft. elevation, Mason County, Washington, by C. P. Streator, on July 9, 1894; type in United States National Museum.
Racial characters.—Similar to saturatus but paler throughout.
Measurements.—Two males and a female from 1 mile northwest of Lake Cushman, Mason County, average: total length 140; length of tail 42; hind foot 18; ear 13.
Distribution.—The Olympic Mountains, from Sol Duc Hot Springs (W.S.M.) south and west at least to Staircase, on Lake Cushman (W.W.D.).
Evotomys occidentalis Merriam, N. Amer. Fauna, 4:25, October 8, 1890.
Evotomys pygmaeus Rhoads, Proc. Acad. Nat. Sci. Philadelphia, p. 284, October 23, 1894 (type from mouth of Nisqually River, Pierce County, Washington).
Clethrionomys occidentalis Hall, Murrelet, 13:79, September, 1932.
Type.—Obtained at Aberdeen, Grays Harbor County, Washington, by T. S. Palmer, on August 16, 1889; type in United States National Museum.
Measurements.—Ten males and 10 females average: total length 137; length of tail 45; hind foot 18; ear 12.3.
Distribution.—The lowlands of western Washington, east at least to Cottage Lake (W. W. D.).
Remarks.—Specimens from the type locality of occidentalis and other places along the ocean coast are larger and brighter in color than specimens from farther inland, but the difference appears to me to be insufficient to warrant subspecific separation of the two lots.
Clethrionomys californicus resembles Clethrionomys gapperi but is darker and duller in color with the red dorsal area more obscured and forming less of a stripe. C. californicus is found in the forested lowlands of western Washington, Oregon, and northern California, and in the Cascades of Oregon. In Washington, it is confined entirely to the forest where it is trapped under logs and on the layer of dead needles at the bases of conifers. Mice of this species were numerous in the forest along the Pacific Coast on the Long Beach Peninsula and at Aberdeen, Grays Harbor County. At Lost Lake Prairie, Mason County, at the southeastern base of the Olympic Mountains, they were rather scarce, but were the only mammals taken in the deep woods. Near Shelton, Mason County, at the eastern edge of the Olympic Mountains, a number of specimens were taken along with twice as many Peromyscus maniculatus. At Cottage Lake, King County, near Seattle, they were rare, comprising about two per cent of the mammals taken in two weeks' trapping.
Almost nothing was learned of the habits of these mice. They seem to be rigidly restricted to a habitat where few plants other than trees grow. The stomachs examined contained pasty masses of finely chewed white vegetation with occasional gray particles that might have been bits of lichens. The mice do not make runways like those of Microtus.
Description.—The upper parts are dark blackish brown and the underparts grayish or whitish. The tail comprises about a fourth of the total length and the foot does not exceed 21 mm. These two features separate it from most other species of meadow mice. From Microtus oregoni it may be separated by its larger size, blackish color and well-developed eyes. From Microtus montanus it differs in being larger, darker, and in having a closed posterior loop on the innerside of the second upper molar.
Fig. 112. Pennsylvania meadow mouse (Microtus pennsylvanicus kincaidi), from ten miles south of Moses Lake, Washington, April 28, 1940. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 937.)
Microtus pennsylvanicus is the most common meadow mouse of the eastern United States. It ranges westward through Canada and southward in the Rocky Mountains. A number of races occur in this extensive range. In northeastern Washington the eastern meadow mouse is locally common, being confined to marshes and damp meadows. Well-used runways are made through the dense vegetation and piles of feces and blades of grass are deposited therein. A number of broods are raised in a season, for young of many different ages are taken together in midsummer.
On the Columbian Plateau this mouse lives in marshy areas about lakes and potholes. Narrow trails are constructed through the [Pg 346] dense vegetation. It burrows in damp earth and some occupied burrows are half-full of seepage water. Cuttings found in runways show that the mice feed on a variety of plants, including sedge (Carex). In the Moses Lake area the breeding season begins in March. Embryos found March 24, 1940, varied from 4 to 7 in number. This species is cyclically abundant.
Fig. 113. Distribution of the Pennsylvania meadow mouse in Washington. A. Microtus pennsylvanicus funebris. B. Microtus pennsylvanicus kincaidi.
Microtus pennsylvanicus modestus Bailey, N. Amer. Fauna, 17:20, June 6, 1900.
Microtus drummondii Bailey, N. Amer. Fauna, 17:22, June 6, 1900.
Microtus pennsylvanicus funebris Dale, Jour. Mamm., 21:338, August 13, 1940.
Type.—Obtained at Coldstream. 1,450 ft., 3-1/2 miles southeast of Vernon. British Columbia, by T. P. Maslin. Jr., on August 2, 1937; type in Museum of Vertebrate Zoölogy.
Racial characters.—Size medium; color reddish brown; fur short, harsh; skull small and narrow.
Measurements.—Three males from Newport, Pend Oreille County, average: total length 165; length of tail 40; hind foot 20; ear 15.3.
Distribution.—Northeastern Washington, west to Conconully (Taylor and Shaw, 1929: 24) and east to Newport (W. W. D.).
Remarks.—Specimens from northeastern Washington are larger and darker, more reddish and less gray, than drummondii. They are larger, more reddish, and have relatively narrower skulls, than modestus from Colorado and Idaho. They closely resemble funebris from south-central British Columbia, and are best referred to that race. Rand (1943: 123) considers funebris a synonym of modestus but I feel is incorrect in doing so.
Microtus montanus kincaidi Dalquest, Proc. Biol. Soc. Washington, 54:145, September 30, 1941.
Type.—Obtained at The Potholes, 10 miles south of Moses Lake, Grant County, Washington, by W. W. Dalquest on March 24, 1940; type in Museum of Vertebrate Zoölogy.
Racial characters.—Large size; dark blackish color; long fur; wide, angular skull.
Measurements.—Six male and 12 female topotypes average, respectively: total length 176.6, 168.0; length of tail 45.1, 43.8; hind foot 20.8, 20.3; ear 14.0, 13.4.
Distribution.—The Columbian Plateau, in the Grand Coulee area.
Description.—Montane meadow mice have short tails, flaring zygomatic arches, and heavily ridged skulls. They are small, about 6-1/2 inches long, and buffy-gray in color. Their short tails, less than one-third the length of head and body, and gray color separate them from all other Washington meadow mice.
Montane meadow mice are widely distributed in the southwestern United States, where numerous races are found. They occupy damp meadows and marshes in the arid subdivision of the Transition Life-zone of eastern Washington. They seem to require deep, dense cover of grasses, reeds, or sedges, near water. They are usually associated with harvest mice (Reithrodontomys megalotis), wandering shrews (Sorex vagrans monticola), and muskrats (Ondatra zibethica). Hawks and owls are their principal enemies, but predatory mammals and snakes probably kill many.
Montane meadow mice evidently are both diurnal and nocturnal; specimens were taken in the daytime as well as at night, and captives were active periodically day and night. Captives were extremely quick in their movements. If disturbed they sat up on their haunches and chattered indignantly. It was almost impossible to handle them without being bitten.
The trails of montane meadow mice are narrower than those of other meadow mice, and appear to be used the year around. Burrows [Pg 348] are numerous where the mice are plentiful. A heap of damp earth marks the entrance to each burrow. One nest of M. m. canescens near Lake Chelan, Chelan County, was under a log on a small knoll in a marsh. The nest was round, about 8 inches in diameter, and was composed of cattails, grasses, and moss.
Fig. 114. Distribution of the montane and Townsend meadow mice in Washington. A. Microtus montanus nanus. B. Microtus montanus canescens. C. Microtus townsendii townsendii. D. Microtus townsendii pugeti.
Arvicola (Mynomes) nanus Merriam, N. Amer. Fauna, 5:63, July 30, 1891.
Microtus nanus Miller, Proc. Biol. Soc. Washington, 11:67, April 21, 1897.
Microtus nanus nanus Miller, Bull. U. S. Nat. Mus., 128:409, April 29, 1924.
Microtus montanus nanus Hall, Proc. Biol. Soc. Washington, 51:133, August 23, 1938.
Type.—Obtained in the Pahsimeroi Mountains, 9,300 ft., Custer County, Idaho, by C. H. Merriam and V. Bailey on September 16, 1890; type in United States National Museum.
Racial characters.—Size medium; color of upper parts brownish gray; sides paler; underparts grayish white.
Measurements.—A large male from Prescott, Walla Walla County, measures: total length 168; length of tail 48; hind foot 20. A female from Prescott, Walla Walla County, and a female from Pullman, Whitman County, average: 133; 31; 18.5.
Distribution.—Southeastern Washington, westward as far as 5 miles east of Wallula (M.V.Z.).
Microtus nanus canescens Bailey, Proc. Biol. Soc. Washington, 12:87, April 30, 1898.
Microtus montanus canescens Hall, Proc. Biol. Soc. Washington, 51:133, August 23, 1938.
Type.—Obtained at Conconully, Okanogan County, Washington, by J. A. Loring, on September 12, 1897; type in United States National Museum.
Racial characters.—Size small; color of upper parts pale brownish gray; sides yellowish gray. This race differs from Microtus m. nanus in generally paler, less brownish coloration. It does not differ from nanus in any distinctive cranial features.
Measurements.—Two male and 2 female topotypes average, respectively: total length 151, 143; length of tail 35, 33.5; hind foot, 19.5, 19.5. A series of 8 males and 6 females from Selah, Yakima County, average, respectively: total length 151.7, 150.5; length of tail 41.5, 40.1; hind foot 19.7, 18.8; ear 12.8, 13.0; weight 47.2, 36.8 grams.
Distribution.—The eastern foothills of the Cascade Mountains from the British Columbian boundary south, probably to the Columbia River. Recorded east to Benton City (Taylor and Shaw, 1929: 24).
Remarks.—A large series from Selah, Yakima County, is somewhat intermediate between nanus and canescens. These specimens from south of the Wenatchee Mountains are, however, more like canescens in color than they are like nanus.
Bailey (1900: 32) records a specimen of this race from North Yakima as Microtus canicaudus.
Description.—The Townsend meadow mouse is a large-bodied, long-furred mouse with a tail of moderate length. Head and body measure about 8 inches, the tail about 2-1/2 inches. The legs are short and the ears scarcely project through the fur on the head. In summer the color is dark reddish-brown. The winter color is dark brownish-black. Underparts are paler, more grayish brown, than are the upper parts. The tail is sparsely haired.
Townsend meadow mice occur west of the Cascade Mountains in California, Washington, Oregon, British Columbia, and on some British Columbian and Washington islands. The habitats of the two races of the Townsend meadow mice found in Washington are wholly in the humid subdivision of the Transition Life-zone but vary somewhat in nature. The race townsendii lives in marshes or damp meadows, under cover of deep, rank vegetation. These mice avoid forested areas or dry brush, but sometimes occur in dry grass when it is deep enough to conceal them from enemies. The race pugeti has been recorded from meadows, salt marshes, driftwood strewn on sea-beaches, areas of sparse, dry grass, and piles of rocks.
Townsend meadow mice are as diurnal as they are nocturnal; specimens have been trapped at almost all hours of the day and night. Their ordinary method of traveling is a slow run. When startled they make a dash for the nearest cover, into which they dive headlong. They make considerable noise while moving about and often may be heard from several feet away. Captives in the laboratory seemed rather dull and stupid as compared with other meadow mice.
Fig. 115. Runways of Townsend meadow mice (Microtus townsendii) worn to grooves in the damp soil at Seattle; May 18, 1938; dense cover of cattails has been burned off (W. W. Dalquest photo).
In the fall, winter, and early spring the Townsend meadow mice keep to their runways. These runways are used by successive generations of mice, and often are worn into ditches several inches deep. In the late spring and summer, when the grass and other [Pg 351] vegetation in their habitat is tall and rank, offering complete concealment, the runways are abandoned and cuttings of rejected food are found scattered over the surface of the ground.
The food of the Townsend meadow mice includes the succulent leaves and stems of many grasses and annuals. Near Seattle the staple summer food is the velvet grass (Holcus lanatus), although many other plants, including the horsetail (Equisetum arvense), are eaten. In winter the common cattail is eaten. Couch (1925: 200) found caches of the roots of mint (Mentha canadensis) stored by this species. As much as 14 quarts was found in a single cache.
The cup-shaped nests of Townsend meadow mice near Seattle were below ground. Embryos were found from May 4 to May 20 and varied in number from 5 to 8 with a mean of 7. In the San Juan Islands the nests of M. t. pugeti were under driftwood.
Arvicola townsendii Bachman, Jour. Acad. Nat. Sci. Philadelphia, 8 (pt. 1):60, 1839.
Arvicola occidentalis Peale, U. S. Expl. Exped., Mammalogy, p. 45, 1848 (type from Puget Sound).
M[icrotus]. townsendi Miller, N. Amer. Fauna, 12:66, July 23, 1896.
Type.—Obtained on the Columbia River (probably on or near Sauvie Island, Multnomah County, Oregon); type in Philadelphia Academy of Natural Sciences.
Racial characters.—Size large; skull narrow in interorbital region.
Measurements.—Eight males and 5 females from Clark and Pacific counties, southwestern Washington, average, respectively: total length 208.0, 208.4; length of tail 65.7, 66.0; hind foot 26.5, 25.4; ear 15.0, 14.6; weight 80.8, 76.7 grams. Thirteen males and 9 females from Seattle average, respectively: 211.8, 209.0; 71.2, 68.6; 25.7, 26.0; 15.8, 15.4.
Distribution.—The lowlands of western Washington from Bellingham (J.M.E.) south to Puget Island (V.B.S.).
Microtus townsendii pugeti Dalquest, Murrelet, 21:7, April 1, 1940.
Type.—Obtained at Neck Point, northwest corner of Shaw Island, San Juan County, Washington, by D. H. Johnson, on July 10, 1938; type in Museum of Vertebrate Zoölogy.
Racial characters.—Size small; skull wide in interorbital region, averaging about 4.0 mm. (3.8-4.2); basi-sphenoid truncate posteriorly; upper incisors strongly curved.
Measurements.—Two males and 6 females average: total length 182.6; length of tail 50.3; hind foot 23.2; ear 15.0.
Distribution.—Found only on the San Juan Islands, San Juan and Skagit counties.
Remarks.—The islands occupied by this race of mouse were heavily glaciated by the last continental glacier (Vashon). Mice of the species townsendii apparently migrated to the islands early in the Recent era, and under isolation developed the differences which now separate them from the mainland population.
Description.—The several races of this species vary from small to large in size. Their bodies are relatively longer and slimmer than those of the other meadow mice that occur in Washington. Their most distinctive feature is the long tail, only slightly shorter than the head and body. Their fur is rather coarse. The color varies from grayish brown to dull reddish brown with a brighter brown dorsal stripe. The tail is bicolor; black or dark brown above, yellowish below. The underparts are whitish gray.
Fig. 116. Distribution of the long-tailed meadow mouse in Washington. A. Microtus longicaudus macrurus. B. Microtus longicaudus halli.
Long-tailed meadow mice are widely distributed over the western United States, Canada, and Alaska. In Washington the long-tailed meadow mouse has been taken in many habitats. One specimen was taken along a small, temporary stream through sagebrush in the Upper Sonoran Life-zone. Others were found in marshes and near [Pg 353] water in the arid subdivision of the Transition Life-zone. In the humid subdivision of the Transition Life-zone they are not uncommon in damp areas along the ocean coast, but are rather rare in dry, grassy habitats. In the Canadian and Hudsonian life-zones they are fairly common in forest-free, grassy places, being most abundant near talus slides. Altitudinally they range from sea level to 6,000 feet.
Long-tailed meadow mice are rather rare. Several were taken in the daytime at Round Top Mountain, Pend Oreille County, and two others were taken in daytime two miles south of Tenino, Thurston County. However most of the specimens were caught at night.
Long-tailed meadow mice do not, at least ordinarily, make trails as do other meadow mice. Specimens are usually taken unexpectedly, and intensive trapping in the area where a specimen or two is taken rarely yields additional individuals.
Two specimens taken in the Cascade Mountains in September were pregnant; one contained two embryos and the other four.
Microtus mordax angustus Hall, Univ. California Publ. Zoöl., 37:13, April 10, 1931 (not of Thomas, 1908).
Microtus longicaudus angustus Goldman, Jour. Mamm., 19:491, November 14, 1938.
Microtus mordax halli Ellerman, Fam. and Genera of Living Rodents, British Mus. Nat. Hist., 2:603, March 21, 1941 (new name for Microtus mordax angustus Hall).
Type.—Obtained at Godman Springs, 5,700 ft., Blue Mountains, Columbia County, Washington, by S. H. Lyman, on September 1, 1927; type in Museum of Vertebrate Zoölogy.
Racial characters.—Size small; color of sides pale grayish brown; brown dorsal stripe conspicuous; tail relatively short.
Measurements.—Four males and 5 females from eastern Washington average, respectively: total length 164.8, 166.6; length of tail 55.8, 55.8; hind foot 22.0, 21.4.
Distribution.—From the Blue Mountains of southeastern Washington north, along the eastern edge of the state, to British Columbia, and thence west, north of the Columbia River, to the Cascades, and south along the eastern edge of the Cascades to the Wenatchee Mountains. Marginal occurrences are: Pasayten River (Taylor and Shaw, 1929: 24), Hart Lake (Taylor and Shaw, 1929: 24), Blewett Pass (W.W.D.), Boulder Cave (W.W.D.), and Satus Pass (W.W.D.).
Remarks.—Four specimens from Satus Pass, Klickitat County, are somewhat intermediate between this race and macrurus, and indicate that halli crossed the Columbia River when an alpine meadow land extended from the Simcoe Anticline to southeastern Washington.
One specimen from Selah, Yakima County, is colored somewhat like halli, and is smaller than macrurus. Possibly the halli type of meadow mouse spread northward to the Yakima Valley. This specimen was taken in the Upper Sonoran Life-zone. A specimen from the Arid Transition Zone at Naches, Yakima County (taken in almost Canadian Life-zone habitat), is like macrurus-halli intergrades from the higher Cascade Mountains.
Microtus macrurus Merriam, Proc. Acad. Nat. Sci. Philadelphia, p. 353, October 4, 1898.
Microtus mordax macrurus Dice, Murrelet, 13:49, May, 1932.
Microtus longicaudus macrurus Goldman, Jour. Mamm., 19:491, November 14, 1938.
Type.—Obtained at Lake Cushman, Mason County, Washington, by C. P. Streator on June 26, 1894; type in United States National Museum.
Racial characters.—Size large; tail relatively long, almost as long as head and body; color of upper parts dull brown; dorsal stripe obscure; underparts gray.
Measurements.—Two males and 4 females from the Olympic Mountains and the coast of Washington, average, respectively: total length 212.5, 220.5; length of tail 82.5, 86.8; hind foot 25.5, 24.7; ear 15.0, 13.7.
Distribution.—The Olympic Mountains, the coast region of western Washington, and the Cascade Mountains, save in the southeastern part. Marginal occurrences are: Sauk (Taylor and Shaw, 1929: 24), Tye (W.W.D.), Naches River (W.W.D.), and Wind River (W.W.D.).
Remarks.—Specimens from the Cascade Mountains are referred to macrurus but are intermediate between that race and halli.
Description.—This is the largest meadow mouse found in Washington. It is too large to be called a mouse and the term water rat, applied to it by Merriam, suits it well. It is the only Washington microtine measuring more than 8 inches in total length. The upper parts are dark, reddish brown in color; the underparts are grayish brown. The tail comprises about one-third of the total length.
This species of meadow mouse ranges from Canada south to Colorado in the Rocky Mountains and in the Cascades south into Oregon. The water rats are strictly alpine animals, occurring about streams, marshes and damp meadows. In the Cascade Mountains they are most common in the glacial cirques where tiny streams flow through grassy meadows to plunge over the lip of the cirque on a rocky course of cascades to the valley below. Here their broad trails occur along the stream banks, commonly entering the water where it is swift. These trails are well-worn roads, usually about four inches wide but often wider. The burrows of the water rat are about three inches in diameter and are constructed with no effort at concealment; large mounds of earth mark their entrances. Freshly dug [Pg 355] burrows are so abundant that it seems likely more are dug than are actually inhabited. Burrows are often dug beneath rocks.
The water rat is mainly nocturnal but not uncommonly is seen in the daytime. Twice I cornered a water rat away from its burrow and each time it escaped by swimming. The rats swam with great speed but with much splashing. One dived under the surface of a small pool and disappeared. Stones along the bank were pulled out until the rat was captured in a small chamber at the end of a burrow. The burrow entered the bank at the base of a large stone six inches beneath the surface. This burrow resembled a miniature muskrat burrow and apparently had been dug when the rat was under water.
Like Phenacomys, the water rat constructs sub-snow nests on the surface of the ground. These are recognizable by their large size and by piles of ovoid droppings a quarter of an inch in length. These nests are loosely built and fall apart soon after the snow melts.
The water rat is sometimes a pest to the mammal collector for they spring mouse traps set for other mammals without becoming caught. At times the greater part of an entire trap line was thus rendered ineffective by these mammals. Fully adult animals are uncommon in collections for a rat trap or steel trap is needed to take them and these items, when packed on back up mountains to water-rat habitat, are usually set for still larger animals. Racey (Racey and Cowan, 1935: H27) recounting his difficulty in securing specimens when no suitable traps were at hand, writes: "Killed one with my hands, shot another, and a third was stunned by a mouse trap."
Taylor and Shaw (1927: 76) list food eaten by the water rat on Mount Rainier as the avalanche lily, dogtooth violet, Ligusticum purpureum, Valeriana sitchensis, Polygonum bistortoides, Petasites frigida, Phyllodoce empetriformis, Potentilla flabellifolia, Aster sp., grass, wild clover, conifer seeds, two kinds of blueberry (Vacinnium) and Xerophyllum tenax. Racey and Cowan (1935) list foods eaten in the Cascades of southern British Columbia as Lupinus polyphyllus, Senecio balsamitae, Pedicularis bractiosa and Arnica alpinus.
A female from Dewey Lake, Yakima County, contained 4 embryos on September 1, 1940. One from Tye, King County, had 2 embryos on September 8, 1940.
Aulacomys arvicoloides Rhoads, Amer. Nat., 28:182, February, 1894.
Microtus richardsoni arvicoloides Bailey, N. Amer. Fauna, 17:62, June 6, 1900.
Type.—Obtained at Lake Keechelus, Kittitas County, Washington, by A. Rupert in September, 1893. Rhoads gives the altitude as 8,000 ft. This apparently is an error, for the elevation of the lake is 2,458 ft. and the summit of Snoqualmie Pass, to the west, is 3,100 ft. Probably 3,000 ft. was intended; type in Philadelphia Academy of Natural Sciences.
Racial characters.—Size large; color of upper parts dark reddish brown; underparts paler.
Fig. 117. Distribution of the water rat in Washington. A. Microtus richardsoni arvicoloides. B. Microtus richardsoni macropus.
Measurements.—A female from Tomyhoi Lake, Whatcom County, measured: total length 242; length of tail 68; hind foot 27; ear 17. A female from Tye, King County, measured: 257; 83; 26; 17.
Distribution.—The Cascade Mountains, from Tomyhoi Lake (W.W.D.) south to Potato Hill (Taylor and Shaw, 1929: 25).
Arvicola (Mynomes) macropus Merriam, N. Amer. Fauna, 5:60, July 30, 1891.
Microtus richardsoni macropus Bailey, N. Amer. Fauna, 17:61, June 6, 1900.
Microtus richardsonii macropus Taylor and Shaw, Occ. Pap. Chas. R. Conner Mus., no. 2:25, December, 1929.
Type.—Obtained in the Pashimeroi Mountains, 9,700 ft., Custer County, Idaho, by C. H. Merriam and V. Bailey in 1890; type in United States National Museum.
Racial characters.—Similar to arvicoloides but slightly smaller and redder.
Measurements.—A female from Stay-a-while Spring, Columbia County, measures: total length 228; length of tail 73; hind foot 25; ear 14.
Distribution.—The Blue Mountains of southeastern Washington.
Arvicola oregoni Bachman, Jour. Acad. Nat. Sci. Philadelphia, 8:60, 1839.
Microtus oregoni Miller, N. Amer. Fauna, 12:9, July 23, 1896.
Microtus morosus Elliot, Field Columb. Mus. Publ. 30, zoöl. ser., 1:227, February 1, 1899 (type from Boulder Lake, 5,000 ft., Clallam County, Washington).
Microtus oregoni oregoni Miller, U. S. Nat. Mus. Bull., 79:227, December 31, 1912.
Microtus oregoni cantwelli Taylor, Jour. Mamm., 1:180, August 24, 1920 (type from Glacier Basin, 5,935 ft., Mt. Rainier, Pierce County, Washington).
Fig. 118. Distribution of the creeping mouse, Microtus oregoni oregoni, in Washington.
Type.—Obtained at Astoria, Clatsop County, Oregon, by J. K. Townsend in 1836; type in Philadelphia Academy of Natural Sciences.
Measurements.—Ten males and 10 females from southwestern Washington, average, respectively: total length 133, 126; length of tail 35, 32; hind foot 16.9, 16.7; ear 9.8, 9.7; weight 19.3, 19.1 grams.
Distribution.—The Cascade Mountains, the Olympic Mountains, and the lowlands of western Washington.
Remarks.—A large series of topotypes of Microtus o. oregoni in the California Museum of Vertebrate Zoölogy, and the rather large series from Cowlitz County, Washington, show a wide range of variation in color, size, and cranial characters. Specimens from the Cascade and Olympic Mountains seem to average a bit paler than topotypes of oregoni, but are not worthy of recognition as distinct races. Too few topotypes of Microtus o. serpens from British Columbia are available to judge the status of that race with any certainty, but specimens from northwestern Washington are certainly oregoni.
Description.—The creeping mouse is a small species of general microtine form. The head and body measure about 4 inches and the tail about 1-1/2 inches. The ears are small and the eyes tiny, nearly buried in the fur. The fur is short and rather rough. It does not lie back smoothly, thereby giving a woolly appearance. The upper parts are reddish or grayish brown and the underparts are grayish white. The nondescript appearance suggests a young rather than adult mouse.
The creeping mouse (subgenus Chilotus) is restricted to the Pacific Coast and ranges from British Columbia to California. In Washington it occupies almost every conceivable "mouse" habitat in its range, including wet marshes, damp ravines, dry forest, damp, mossy forest, meadows, alpine meadows and fields of short grass. It is rare in all but the latter habitat. In fields of short or dry grass it is often abundant. In the Cascade Mountains it was in relatively dry places along streams or rock slides. Altitudinally it ranges from sea level to at least 6,000 feet, and from the Humid Transition well into the Hudsonian life-zones.
Creeping mice construct tiny tunnels among the grass roots and seldom venture out of them. In suitable habitat the surface of the ground beneath the grass is a maze of these tunnels, which cross, intersect, and divide in a complex network. An observer standing in a field occupied by creeping mice can scarcely conceive of the extent and perfection of the tiny tunnel system at his feet.
Creeping mice lived but a day or two in captivity. Save for the bits of grass blades left in their runways, little is known of their food or other life habits. Their nests are round balls of dry grasses placed in cavities under logs. None of the many examined possessed a lining of softer materials. Embryos found in pregnant females from April 10 to May 18 numbered from 2 to 4.
Arvicola pauperrima Cooper, Amer. Nat., 2:535, December, 1868.
Arvicola pauperrimus Merriam, N. Amer. Fauna, 5:64, July 30, 1891.
L[agurus]. pauperrimus Thomas, Ann. and Mag. Nat. Hist., ser. 8, 9:401, April, 1912.
Microtus pauperrimus Bailey, N. Amer. Fauna, 55:214, August 29, 1936.
Lemmiscus pauperrimus Davis, Recent Mamm. Idaho, Caxton Printers, p. 327, April 5, 1939.
Lemmiscus curtatus pauperrimus Goldman, Proc. Biol. Soc. Washington, 54:70, July 31, 1941.
Type.—Obtained on the "Plains of the Columbia" near the Snake River, southwestern Washington by J. G. Cooper on October 9, 1860. Probably from the Bunchgrass Hills near Wallula (Old Fort Walla Walla), Walla Walla County: type in United States National Museum.
Measurements.—Bailey (1900: 69) gives the average of 3 adults from the vicinity of Antelope, Oregon, as: total length 115; length of tail 20; hind foot 16.
Distribution.—Known in Washington only from the type and a specimen from Badger Mountains, 8 miles southwest of Waterville (Taylor and Shaw, 1929: 25).
Remarks.—This rare vole is the smallest microtine rodent in Washington. The head and body measure about 4 inches and the tail about 1 inch. The upper parts are grayish or yellowish brown and the underparts grayish. The upper incisors are not grooved and the inner and outer angles of the molars are about equal in length.
Mice of the genus Lagurus occur in Siberia and in prairie areas of the northwestern United States and Canada. The sagebrush vole inhabits the Upper Sonoran Life-zone. It prefers upland areas of low sagebrush with sparse grass. Poorly formed runways and small piles of feces indicate its presence. The type of Lagurus pauperrimus was obtained in southwestern Washington 80 years ago, and a single additional record has since been obtained. Nothing is known of its habits in Washington. The life history of related forms has been reported on by Hall (1928: 201-204) from Nevada and Moore (1943: 188-191) from Oregon.
Davis (1939: 326) raised the subgenus Lemmiscus Thomas (for American forms) to generic rank and Goldman (1941: 69) accepted this usage. Comparison of the Siberian and American species reveals but three impressive differences: the dorsal stripe of the Siberian mice, the greater development of prisms in the molars of the American species, and the presence of cement in the angles of the molars of the Siberian specimens and its lack in the American. These differences seem to be of no more than subgeneric [Pg 360] value. Goldman (1941: 69) showed that all American Lagurus belong to a single species.
Description.—The muskrat is a large aquatic rodent. Head and body measure about 14 inches; the tail about 10 inches. The body is plump and the head small. Eyes and ears are relatively small. The forefeet are small and handlike with furred wrists. The hind feet are large with webbed toes and naked wrists. The tail is narrow, constricted at the base and flattened vertically. It is scaled and possesses scattered, stiff hairs. The underfur is dense and soft. The guard hair is stiff and shiny. The upper parts are rich, dark brown. The underparts are gray washed with cinnamon.
Fig. 119. Muskrat (Ondatra zibethicus osoyoosensis), male, Lake Washington, Seattle, October 13, 1939; weight 906 grams on November 4, 1939. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 736.)
Muskrats are found in Canada and the United States. They are lowland animals, rarely ranging into the Canadian Life-zone. Their aquatic habitat makes them relatively independent of temperature, and consequently they occur in both the Transition and Upper Sonoran life-zones.
The muskrat is well adapted to aquatic life. The toes of the large hind feet are webbed at the base, the hind feet are turned slightly outward, a fringe of stiff bristles at the rear edge of each foot furnishes additional aid in swimming, and the laterally compressed tail makes it an efficient rudder. In the water the surface of the fur flattens down to entrap air in the dense underfur, keeping the body dry.
The typical habitat of the muskrat is slow-moving water or still [Pg 361] water, such as in lakes, ponds, marshes and sluggish rivers and streams. Muskrats occur commonly, though not in large numbers, in more swiftly moving streams. In Puget Sound and the San Juan Islands they occupy a marine habitat.
In the extensive marshes along Lake Washington, King County, muskrats are abundant. They occupy sluggish water, often water polluted by garbage and sewage. In these marshes, banks suitable for the construction of burrows are absent and houses are made of cattail stalks and leaves. The food of these marsh-living muskrats consists principally of cattail and other marsh vegetation.
In contrast to the marsh-occupying muskrats, muskrats along the open, marsh-free shores of the lake live in deep clear water where waves lap the shore. These muskrats live exclusively in burrows dug in the banks and feed upon fresh-water mussels.
In still greater contrast were muskrats living 20 miles away, near Cottage Lake, King County. Here we found them in small clear streams, 4 to 10 feet wide. Depth of the water varied from a few inches to three feet. The streams flowed through meadows, pastures and junglelike, deciduous woods. Muskrats were slightly more common along wooded stretches than in open areas. Some were trapped where the streams flowed at high velocity over shallow, gravel bottoms. The animals lived in burrows and fed upon fresh-water mussels and a variety of plants.
Near Richmond Beach, Snohomish County, muskrats took up residence in a small tidal pool along Puget Sound. The nearest fresh water stream large enough to support a muskrat was two miles away. Two muskrats were trapped here. Investigation of a tidal pool a mile to the north disclosed unmistakable muskrat signs. Traps set in the culvert connecting the pool with Puget Sound at high tide took several specimens. Study showed that the muskrats were not living in the pool but among the large boulders forming the breakwater for the Great Northern Railroad, along the sound itself. They were feeding on marine mussels (Mytilus). These mussels lived in the salt water of the sound, not in the tidal pool.
At Peavine Pass, Blakely Island, in the San Juan Islands, muskrats were living in the swift tidal current and deep, marine waters. Several were seen in late afternoon. All were swimming parallel to the shore about 50 feet out. Here also they fed on Mytilus, but their homes were not discovered. Certainly they were not living in the tidal pool at Flat Point, a half-mile away.
In the interior of Blakely Island a colony of muskrats was discovered [Pg 362] living in a marsh of about one acre. In the rainy season the ground of the marsh was covered with less than one inch of water. Residents said that in the dry season springs kept the ground moist. Muskrats were living in burrows whose entrances descended at a 45-degree angle and were filled with water. The ground about some occupied burrows was dry, the only water visible being in the burrow itself. A variety of marsh vegetation provided food.
Fig. 120. Distribution of the muskrat in Washington. A. Ondatra zibethicus osoyoosensis. B. Ondatra zibethicus occipitalis.
Burrows of muskrats always have entrances under water. Usually they enter a vertical bank 6 to 15 inches below water line and occasionally 3 feet below it. About half the burrows excavated near Lake Washington, King County, had a single entrance. About 40 per cent had double or triple entrances situated 2 to 3 feet apart and converging within a yard to a single burrow. About 10 per cent had double burrows more than 3 feet in length. Burrows were from 5 to 8 inches in diameter. Nest chambers were from 12 to 15 inches in diameter, spherical, and from 6 to 30 feet from the burrow entrance. The nests themselves were bulky, loose masses of cattail [Pg 363] leaves. Embryos found in late February and early March numbered 4 to 8.
Because it is abundant, widely spread and easy to trap, the muskrat is one of the most important fur bearers in the state. The fur is relatively stable in value. In recent years the average skin has brought the trapper slightly less than a dollar. Muskrat flesh is eaten and sold on the market in the eastern United States but has never been popular in Washington. The muskrat does little damage to agriculture, most complaints arising from its burrows which interfere with irrigation ditches.
Fiber osoyoosensis Lord, Proc. Zoöl. Soc. London, p. 97, 1863.
F[iber]. z[ibethicus]. osoyoosensis Hollister, Proc. Biol. Soc. Washington, 23:1, February 2, 1910.
Ondatra zibethica osoyoosensis Miller, U. S. Nat. Mus. Bull., 79:231, December 31, 1912.
Type.—Obtained at Osoyoose Lake, British Columbia, on British Columbia-Washington boundary at head of Okanogan River, by J. K. Lord, in 1861 or 1862.
Racial characters.—Color of the upper parts rich, dark brown.
Measurements.—Two males and a female from Seattle, King County, average and measure respectively: total length 565, 555; length of tail 262, 257; hind foot 80.5, 79; weight 2 pounds 13 ounces and 2 pounds 3 ounces.
Distribution.—Eastern Washington generally and all but the southern part of western Washington. Specimens from as far southwest as Tenino (W.W.D.) are typical of osoyoosensis.
Fiber occipitalis Elliot, Field Columb. Mus. Publ. 74, zoöl. ser., 3:162, April, 1903.
Ondatra zibethica occipitalis Miller, U. S. Nat. Mus. Bull., 79:231, December 31, 1912.
Type.—Obtained at Florence, Lane County, Oregon, by E. Heller, in 1901; type in Chicago Natural History Museum.
Racial characters.—Similar to osoyoosensis but redder; fur shorter and interpterygoid spaces of skull narrower.
Measurements.—A male from Chinook, Pacific County, measures: total length 580; length of tail 248; hind foot 78; ear 21; weight 863 grams.
Distribution.—The southwestern corner of the state, extending north to Aberdeen (V.B.S.) and east to Cathlamet (V.B.S.).
Description.—Size large, total length approximately 400 mm.; tail long, naked, comprising 50 per cent or more of total length; color sooty black or brown; in brown phase, whitish beneath.
The roof rat became established in Central America some 350 years ago, and entered the United States (English Colonies) late in the seventeenth century. Subsequent to the introduction of the Norway rat the roof rat decreased in numbers and is now found only in restricted areas.
[Mus] rattus Linnaeus, Syst. Nat., 1 (ed. 10):61, 1758.
Rattus rattus Hollister, Proc. Biol. Soc. Washington, 26:126, June 6, 1916.
Type.—From Uppsala, Sweden.
Remarks.—In Washington I have taken specimens of the roof rat in the San Juan Islands and in the coniferous forests on the west slope of the Cascades. In the latter area it seems to be widely spread and to live in the wild.
Mus alexandrinus Geoffroy, Catal. Mam. du Mus. Nat. d'Hist. Nat. Paris, p. 192, 1803.
R[attus]. rattus alexandrinus Hinton, Jour. Bombay Nat. Hist. Soc., 26:63, December 20, 1918.
Type.—From Alexandria, Egypt.
Remarks.—This subspecies seems to be rare in Washington. My specimens are all from small, isolated islands in the San Juan group. All were taken in the vicinity of human habitations. R. r. alexandrinus resembles R. r. rattus, differing only in brown color of upper parts.
[Mus] norvegicus Erxleben, Syst. Regni Anim., 1:381, 1777.
Rattus norvegicus Hollister, Proc. Biol. Soc. Washington, 29:126, June 6, 1916.
Type.—From Norway.
Description.—Larger, heavier-bodied and coarser-furred than the black rat or roof rat with shorter, heavier tail. The tail is less than 50 per cent of the total length. The color of the back is dull, reddish brown, the sides are paler and the underparts are dirty gray.
Remarks.—The Norway rat was absent from the Pacific Coast of the United States before 1851. It probably reached the coast slightly after that date. It is common about all large cities in Washington. In the western part of the state it lives along streams and marshes under feral conditions.
[Mus] musculus Linnaeus, Syst. Nat., 1 (ed. 10):62, 1758.
Description.—Size small; tail about 50 per cent of total length, naked; ears small, about 12 mm. in height; upper incisors not grooved; color of upper parts reddish or grayish brown; underparts brown or gray.
Remarks.—The house mouse became established in North America soon after its settlement by Europeans. It is now common throughout the state of Washington, principally near human habitations, but often lives in the wild.
Races of the house mouse have been dealt with by Schwartz and Schwartz (1943: 59-72), and by Nichols (1944: 82-89), but lack of adequate material prevents subspecific identification of house mice from Washington at this time.
Description.—The mountain beaver is a stout-bodied animal about 14 inches in length, with a tiny tail that is almost invisible externally. The head is large, wide and low with small eyes, small ears and long vibrissae. The legs are short and heavy, but the forefeet are small and handlike; the hind feet are large and powerful. The claws of both forefeet and hind feet are long and strong. The pelage is short, coarse and rough. The upper parts are dark reddish brown and the underparts are grayish brown. The feet are pink.
Fig. 121. Mountain beaver (Aplodontia rufa rufa), Seattle, Washington, March 19, 1940. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 919.)
Mountain beavers are confined to the Pacific Coast and range from southern British Columbia to central California. The genus contains a single species of which Taylor (1918) recognized nine races. The principal habitat of the mountain beaver is clearings at the edge of coniferous forests. The animals are most abundant near springs, streams and damp places, although they are not [Pg 367] aquatic. The tangled jungles of deciduous trees and shrubs that grow in the ravines and stream valleys of the Puget Sound area present optimum habitat. They occur also on hillsides, on logged-off land and along roadside clearings. In the mountains they occur in thickets and forests, always, in our experience, near streams.
The most conspicuous evidence of the presence of mountain beavers is their burrows. These are large tunnels, four to eight inches in diameter. To each set of tunnels there are numerous entrances, some partly concealed in brush or beside logs or stumps, and some are in the open. Those in the open are less used as entrances than as places for receiving the loose earth which the animals excavate. A pile may contain nearly a cubic yard of earth and stones. Many of the burrows are shallow, and cave-ins are common. Breaks in the roof of a burrow are not repaired, although debris is removed from the burrow itself. The burrows seem not to be constructed according to a system, but are extended to take in whatever brush, logs or other cover is available. They are commonly dug through damp or muddy soil. Small streams flow through some burrows. Such partly flooded runs seem to be favored by the animals.
The nest of a mountain beaver excavated by Scheffer (1929: 15), under the roots of a fallen tree, was oval in shape, twenty inches high and 13 inches wide. The nest was protected from flooding by a basal chamber, or basin, six inches beneath the nest. Two drainage tunnels lead away from this basal chamber. The nest was composed of the leaves and stems of bracken laced together with grass and fine twigs. Two other nests examined by Scheffer measured 17 by 18 inches and 19 by 17 inches. Both were about two feet beneath the surface.
Around Puget Sound the mountain beavers mate in early March. The young number two to three, rarely four, per litter and are born in early April.
Mountain beavers enter water readily but wade rather than swim. They are rather noisy, splashing in water and breaking twigs or rustling leaves on the ground. They climb bushes and saplings, clipping off branches for food as they ascend. According to Scheffer (1929: 15) they leave the stubs of branches attached to the trunk to facilitate their descent. Twice a mountain beaver was found several feet up in a sapling. In both saplings the animal had clipped the branches close to the trunk and was desperately reaching with its hind feet for missing branches. When the observer came near, one animal squealed, tumbled to the ground, and scurried frantically to its burrow.
Although principally nocturnal, mountain beavers are not infrequently active by day, especially in the fall. At this season they harvest food and spread it on logs to dry. The cured hay is removed to their burrows for nesting material and food. In winter mountain beavers are more restricted in habits and are rarely seen by day. Presumably they feed on stored food at this time but they forage somewhat. In winter they eat such evergreen shrubs as salal (Gaultheria shallon) and Oregon grape (Berberis nervosa). They eat also the bark of trees, especially that of the willow (Salix). Under cover of snow, in the mountains, they burrow to some extent and pack excavated earth in snow burrows. The melting of the snow in the spring reveals the earth core, six to eight inches in diameter and two to four feet long. Several such earth cores were forked, showing that part of the earth had been pushed into a branching burrow.
Fig. 122. Distribution of the mountain beaver in Washington. A. Aplodontia rufa rufa. B. Aplodontia rufa rainieri.
The mountain beaver holds its food in its forefeet, squirrel-like, when it eats. Its food consists of the leaves and bark of woody plants and entire herbs, including roots. The mountain beaver is the only mammal so far as known that eats the bracken fern. It feeds on the branches of coniferous trees, including Douglas fir, red cedar, and hemlock. Such thorny species as the blackberry, [Pg 369] blackcap and devil's club are eaten. The odiferous skunk cabbage and the stinging nettle are on its bill of fare. A list of its food would include most plants found in its habitat, and we know of no species that it refuses as food.
The mountain beaver is more of a nuisance than a pest. In most of its range there is but little farming although where crops are raised the mountain beaver may do some damage. It undermines roads and trails and defiles springs and streams. Control is simple for the animals readily enter steel traps set in their burrows.
Anisonyx? rufa Rafinesque, Amer. Monthly Mag., 2:45, November, 1817.
Haplodon rufus True, Proc. U. S. Nat. Mus., 7 (1884):596, 1885.
Aplodontia rufa Merriam, Ann. New York Acad. Sci., 3:316, May, 1886.
Aplodontia olympica Merriam, Proc. Biol. Soc. Washington, 13:20, January 31, 1899 (type from Lake Quiniault, Grays Harbor County, Washington).
Aplodontia rufa grisea Taylor, Univ. California Publ. Zoöl., 12:497, May 6, 1916 (type from Renton, King County, Washington).
Aplodontia rufa rufa Taylor, Univ. California Publ. Zoöl., 12:497, May 6, 1916.
Type.—None. Based on a description by Lewis and Clark. Taylor (1918: 455) regarded as typical specimens collected at "Marmot, Clackamas County, Oregon (western slope of Mount Hood, not far from the Columbia River)."
Racial characters.—Size small; skull small.
Measurements.—Eight males and 7 females, from the area about Puget Sound, average, respectively: total length 343, 338; length of tail 37.5, 33.5; hind foot 57.8, 56; ear 25, 24.5; weight 1342, 1300 grams.
Distribution.—Western Washington, between Puget Sound and the Cascade Mountains and southward. Marginal localities are: Bellingham (U.S.N.M.), Sauk (U.S.N.M.), Forks of Skykomish River (W.W.D.), North Bend (U.S.N.M.), and mouth of Klama River (M.V.Z.).
Remarks.—Individual variation in mountain beavers is considerable. Two weakly defined races are recognized in Washington.
Aplodontia major rainieri Merriam, Proc. Biol. Soc. Washington, 13:21, January 31, 1899.
[Haplodontia rufa] raineri Elliot, Field Columb. Mus. Publ. 45, zoöl. ser., 2:112, 1901.
[Aplodontia rufa] raineri Trouessart, Catal. Mamm., viv. foss., suppl. p. 348, 1904.
Aplodontia rufa columbiana Taylor, Univ. California Publ. Zoöl., 12:499, 1916 (type from Hope, British Columbia).
Type.—Obtained at Paradise Creek, 5, 200 ft., Mount Rainier, Pierce County, Washington, by V. Bailey on August 6, 1897; type in United States National Museum.
Racial characters.—Similar to rufa but larger with larger skull.
Measurements.—Three males and a female from Tye, King County, average and measure respectively: total length 352, 340; length of tail 35, 40; hind foot 60, 59; ear 24, 19.
Distribution.—The higher Cascade Mountains from the Columbia River northward to the Canadian boundary. Marginal localities are: Canyon Creek (U.S.N.M.), Cascade Pass (U.S.N.M.), Tye (W.W.D.), Mt. Rainier (U.S.N.M.), Mt. St. Helens (U.S.N.M.), and Yacolt (M.V.Z.).
Remarks.—Intergradation between rufa and raineri is seen in specimens from the area between Stevens Pass and Skykomish, King County.
Fig. 123. Big jumping mouse (Zapus princeps trinotatus) in hibernation. Puyallup, Washington, January 30, 1939. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 576.)
Description.—The big jumping mouse is a medium-sized mouse, slightly larger than the house mouse, with an exceptionally long, tapering tail. Head and body measure about 4 inches and the tail about 5 inches. The body is slender with a small head, small eyes and small, naked ears. The forefeet are small and handlike but the hind feet are large with long, powerful legs. The long, naked tail is smoothly tapering to a narrow point. The fur is short, posteriorly directed, stiff and bristlelike. The dorsal area is dusky, the sides are orange or yellow, and the underparts are creamy white. Each upper incisor tooth has a groove on its anterior face.
Jumping mice are boreal mammals occurring in wooded regions ranging from the Arctic region southward to North Carolina, New Mexico and California. Jumping mice are of rather general distribution in Washington, being only locally common. They are partial to damp, boggy areas but avoid true swamps. They occur in clearings in the forests in the Puget Sound area and in meadows in river bottoms and in jungles in ravines. They reach their greatest abundance in the boggy meadows and glacial cirques of the mountains and spread out from such areas to seemingly less favored habitat on dryer slopes and heather meadows. Unlike many species, they do not seem to inhabit talus slides. They are entirely absent from the eastern Washington desert.
Sometimes the jumping mouse walks on all four feet but the ordinary means of progression is by short hops on the hind feet alone. When startled they travel in great bounds, covering six feet or more at a jump. When jumping they make considerable noise, swishing or rustling through the grass and landing with an audible thud. The long tail serves as a balancing organ. A specimen whose tail had been lost was reported by Svihla and Svihla (1933: 133) to turn somersaults in the air and invariably to land on its back rather than its feet.
Jumping mice become very fat and hibernate in middle summer or early fall. In the lowlands they disappear by late July but in the mountains they remain active until the middle of September. They spend the winter in nests of grass several feet beneath the surface. A hibernating individual figured by Flahaut (1939: 17) was curled in a ball, head down with the tail wrapped completely around the greatest circumference of the ball.
Near Seattle the principal food of the jumping mouse was the velvet grass (Holchus lanatus), and the seeds of grasses and the broad-leaved dock. The fruit of the blackberry (Rubus macropetalus) is eaten and occasional individuals are seen with the chin stained a deep purple from the juice.
The new-born young of the jumping mouse were reported by Svihla and Svihla (1933: 132) to average 0.8 grams in weight. They are pink, hairless, lacking even the facial vibrissae, with eyes closed and ears folded.
Zapus trinotatus Rhoads, Proc. Acad. Nat. Sci. Philadelphia, 1894, p. 421, January 15, 1895.
Zapus imperator Elliot, Field Columb. Mus. Publ. 30, zoöl. ser., 1:228, February 1, 1899 (type from Sieg's Ranch, Elwha River, Olympic Mountains, Clallam County, Washington).
Type.—Obtained on Lulu Island, mouth of Fraser River, British Columbia, by S. N. Rhoads on May 31, 1892; type in Philadelphia Academy of Natural Sciences.
Racial characters.—Similar to oregonus but brighter; sides orange; underparts creamy white; buffy area often present on chest.
Measurements.—Twenty males and 15 females from western Washington average, respectively: total length 230, 233; length of tail 140, 140; hind foot 33, 32.8; ear 14.9, 16.3; weight 23.7, 25.7 grams.
Distribution.—Western Washington and the Cascade Mountains, east in the northern Cascades at least to Tomyhoi Lake (W.W.D.).
Remarks.—Actual intergrades between trinotatus and kootenayensis have not been examined but the differences separating the two forms are of the degree that usually distinguish subspecies. Since trinotatus occurs in the high Cascades as far east as Mount Baker, at least, and kootenayensis probably occurs in the northeastern Cascades, the two forms doubtless come together in the rugged, inaccessible area between these two localities. Further collecting will probably show a narrow zone of intergradation in extreme western Okanogan County.
Fig. 124. Distribution of the big jumping mouse in Washington. A. Zapus princeps oregonus. B. Zapus princeps idahoensis. C. Zapus princeps kootenayensis. D. Zapus princeps trinotatus.
Zapus princeps kootenayensis Anderson, Ann. Rept. Nat. Mus. Canada for 1931, p. 108, November 24. 1932.
Type.—Obtained on Green Mountain, 6,000 ft., 10 miles north of Rossland, West Kootenay District, British Columbia, by R. M. Anderson, on July 18, 1929; type in National Museum of Canada.
Racial characters.—Similar to oregonus but paler; yellow more faded.
Measurements.—Twenty females, including 15 topotypes, average (Anderson, 1932: 109): total length 245; length of tail 140; hind foot 30.5.
Distribution.—Northeastern Washington. Specimens from Sullivan Lake (E.S.B.) have been examined.
Zapus princeps idahoensis Davis, Jour. Mamm., 15:221, August 10, 1934.
Type.—Obtained 5 miles east of Warm Lake, 7,000 feet elevation, Valley County, Idaho, by W. B. Davis; type in Museum of Vertebrate Zoölogy.
Racial characters.—Similar to kootenayensis but brighter in color, more ochraceous. Similar to oregonus but paler, more yellowish.
Measurements.—Davis (1939:339) gives the measurements of six adult topotypes as: total length 240; length of tail 144; hind foot 31.
Distribution.—A single specimen in the Charles R. Conner Museum from Kamiak Butte, Whitman County, is referable to this race.
Zapus princeps oregonus Preble, N. Amer. Fauna. 15:24, August 8, 1899.
Type.—Obtained at Elgin, Union County, Oregon, by E. A. Preble, on May 29, 1896; type in United States National Museum.
Racial characters.—Small size, pale color.
Measurements.—Three males and 3 females from the Blue Mountains average, respectively: total length 233, 234; length of tail 138, 139; hind foot 31.8, 31.8; ear 16, 16; weight 29, 33 grams.
Distribution.—The Blue Mountains of southeastern Washington.
Description.—The porcupine is one of the largest rodents found in Washington, being exceeded in size only by the beaver. Its body is heavy and stocky, its legs short, its tail long and thick and its eyes small. It is best known for the modified hairs, or quills, of its tail and dorsal area. These vary in greatest diameter from one-sixteenth to three-sixteenths of an inch and from three-quarters of an inch to five inches in length. They are ivory-white with black tips. In addition to quills, the porcupine possesses wooly, black underfur and long, banded guard hair. The bands of the guard hairs are black and yellow, varying in width. Commonly they are of a single color, black, yellow or brown.
Porcupines range over virtually all wooded parts of North America north of Mexico, in and above the Transition Life-zone. The Canadian porcupines have been studied by Anderson and Rand (1943A) and intergradation between the eastern dorsatum and the western epixanthum has been shown.
The porcupines are commonly considered to be forest animals. However, they are rather rare in the denser coniferous forests. In the more open areas on the Cascade Mountains, especially on the eastern slopes, they are not uncommon. They are rather common in the coniferous forests of northeastern Washington and the Blue Mountains. They seem to be rather common also in desert areas at the southern edge of the Columbian Plateau.
The huge incisors of the porcupine are adapted to feeding on bark. They do feed on bark to a certain extent in Washington, but it is my observation that more herbs and bushes are eaten than bark. In areas where porcupines are common, trees are commonly girdled, usually close to the top. Trees girdled in this manner in the Kettle River Mountains included western larch, ponderosa pine, and grand fir. The tops of some trees were killed.
The ordinary walking gait of the porcupine is a slow deliberate walk in which he appears to waddle somewhat. They can increase their speed to a slow trot. They are slow, deliberate climbers, ascending and descending trees with head upward. They are able also to climb rocks and cliffs, sometimes being seen on the tops of large boulders.
Despite their large size, porcupines are not commonly seen. They are mainly nocturnal and, in the daytime, find concealment high in the branches of some conifer or a cave between the rocks in a talus slide.
While encamped near Sherman Creek Pass in the Kettle River Mountains my companion and I heard a crunching of gravel from the road fifty feet away. We listened intently, wondering what person would be abroad in the mountains at midnight. In the vicinity of our car, concealed from our view by trees, the noise stopped, to be followed a few minutes later by a rasping and clattering that could be heard far away. We raced to the car to discover a large porcupine crouched on the running board by a pile of "pick-up" antlers of the white-tailed deer left there by us. The "porkie" had been chewing on these, heedless of the noise made by the loose antlers clashing against the metal side of the car.
Fig. 125. Distribution of the porcupine, Erethizon epixanthum in Washington. Boundaries between ranges of subspecies are uncertain.
In Washington the single young is born late in May or early in June. There are two pairs of mammae, both pectoral, of which only the anterior are functional.
Erethizon epixanthus Brandt, Mem. Acad. Imp. Sci. St. Pétersbourg, ser. 6, 3 (Sci. Nat. vol. 1): 390, 1835.
Erethizon dorsatus epixanthus True, Proc. U. S. Nat. Mus., 7:600, 1885.
Erethizon epixanthum epixanthum Miller, U. S. Nat. Mus. Bull., 128:437, April 29, 1924.
Erethizon dorsatum epixanthum Anderson and Rand, Canadian Jour. Research, 21:293, September 24, 1943.
Type.—None. Type locality California.
Racial characters.—Size large, total length of adults approximately 30 inches; tail long (nearly one-third of total length), thick, heavy and spiny; body stout; legs short; claws long and curved; ears and eyes small; body spines short, thick and most abundant on posterior part of back, longer and more slender on sides and shoulders; guard hairs of shoulders and sides long, almost concealing spines; fur of underparts shorter; color variable, brown, black or yellow. In winter the fur is longer and woolly, concealing spines.
Distribution.—The Columbian Plateau and the Blue Mountains.
Remarks.—Anderson and Rand (1943A: 295) ascribe two races to Washington. With inadequate material myself to verify this ascription. I think it probable that the northern forest porcupine and the Great Basin animal are racially different. In consequence the available names, nigrescens and epixanthum, are here applied, pending a revision of the entire genus.
Erethizon epixanthus nigrescens Allen, Bull. Amer. Mus. Nat. Hist., 19:558, October 10, 1903.
Erethizon epixanthum nigrescens Miller, U. S. Nat. Mus. Bull., 128:437, April 29, 1924.
Erethizon dorsatum nigrescens Anderson and Rand, Canadian Jour. Research, 21:293, September 24, 1943.
Type.—Obtained on the Shesley River, British Columbia, by M. P. Anderson on August 23, 1902; type in American Museum of Natural History.
Measurements.—A female from Sherman Creek Pass, Ferry County, measured: total length 770; length of tail 250; hind foot 95; ear 37. A female from Tye, King County, measured: 930; 280; 125; weight 20 pounds.
Distribution.—Forested parts of the state, exclusive of the Blue Mountains.
Remarks.—Porcupines are extremely rare west of the Cascades but are occasionally reported from as far west as the Olympic Peninsula.
Mus coypus Molina, Sagg. Stor. Nat. Chili, p. 287, 1782.
Myocastor coypus Kerr, Anim. Kingd., p. 225, 1792.
Type locality.—Chile.
Description.—Size large, slightly smaller than a beaver; color rich, reddish brown; tail long, round; hind feet webbed for swimming.
Remarks.—The nutria, a native of South America, has been brought to the United States and raised commercially on "fur farms." The species has become established in the wild in several localities in western Washington and at the Colville Indian Reservation in northeastern Washington. For further details see Larrison (1943).
Description.—The pika is of guinea-pig size, with a short, chunky body about 200 mm. in length. The tail is represented externally merely by a tuft of white fur. The short, wide head has large, circular ears, large black eyes, and long whiskers. The legs are short and the soles of the feet are furred. The color of the Washington races varies from grayish-yellow to dark reddish brown. Like rabbits, all pikas have two pairs of upper incisors. The second pair, located just in back of the first, is small and delicate.
Ochotona is a wide-ranging genus with many more species in Asia and extreme eastern Europe than in North America. Three races of the species princeps occur in the state of Washington, where they are confined to the Cascade, Kettle River, and Pend Oreille Mountains. The ranges of all three races extend northward into British Columbia; one (brunnescens) occurs also in Oregon, and one (cuppes) in Idaho.
In Washington pikas live only in talus slides and rock piles, where they find refuge from most of the carnivores that prey on small mammals. Their distribution in the state seems to be regulated by the distribution of talus slides, and areas free of talus act as effective barriers to pikas. They are abundant throughout the Cascades but are absent from the Olympic Mountains although conditions there are well suited to them. Probably the lowlands of western Washington which, owing to moderate temperature and low relief, have little talus and exposed rock, serve as a barrier. The Columbian Plateau is also free of pikas. This may be due to the relative scarcity of talus as compared with mountainous areas, and the fact that much of the talus on the Plateau is composed of fragments of basalt too small to afford the shelter needed by pikas. The aridity of the Columbian Plateau may contribute to the absence of pikas, although this seems unlikely in view of the fact that they occur on arid lands in Nevada and elsewhere.
Altitudinally, pikas range from 300 feet, in Clark County, to 6,000 feet on Mt. Rainier, Pierce County, and on Round Top Mountain, Pend Oreille County. They occur from the arid subdivision of the Transition Life-zone, at Milk Creek, Kittitas County, to the upper edge of the Hudsonian Life-zone, at Glacier Basin, Mt. Rainier. Generally speaking, they are mammals of the mountains.
Common enemies of the pikas are the weasel (Mustela frenata), marten (Martes caurina), and hawks of several species. Pikas are active by day, especially in the early morning. Their call note is a short "eek!" which carries a long distance. This squeaking note is often heard throughout the night when rain threatens their drying hay.
Fig. 126. Distribution of the pika in Washington. A. Ochotona princeps brunnescens. B. Ochotona princeps fenisex. C. Ochotona princeps cuppes.
Vegetation used as food, either for immediate consumption or for winter use, includes almost all grasses, vines, shrubs, and trees available near the pika's home. The subalpine lupines are especially favored. Even such a thorny growth as the devil's club (Oplopanax horridum) is eaten. Heather (Phyllodoce, Cassiope) has not been found in any of the numerous hay piles examined, even when it is the commonest plant in the vicinity. Large bundles of plants are carried in the pika's mouth. The forefeet do not assist in transporting the load. If intended for immediate consumption, the plants are deposited on one of last year's hay piles and are eaten at leisure. The eating habits of the pika are rabbitlike. A large leaf is seized at the tip and drawn into the mouth with rapid chewing motions without assistance from the forefeet. Plants destined to become [Pg 379] hay are carefully spread out and exposed to the sun. In cloudy or rainy weather the exposed plants are gathered and stored under large rocks, to be reëxposed for curing when the weather improves. Large hay piles often include more than fifty pounds of perfectly cured grasses, annuals, bushes and evergreens.
No record of embryos is available for Washington pikas, but a male with enlarged testes was taken at Lake Keechelus, Kittitas County, on March 22, 1940. Half-grown young of fenisex were taken at Sawtooth Mountain, Skamania County, on July 13, 1939, and of brunnescens at Slate Creek, Whatcom County, on August 16, 1937. Nearly full-grown young of fenisex are reported taken at Bald Mountain, head of Ashnola River, Okanogan County, on September 16, 1920. Young cuppes of several sizes were taken at Sherman Creek Pass, Ferry County, on September 11, 1938. The breeding season possibly extends from March to August with a tendency to be earlier at lower elevations.
Ochotona cuppes Bangs, Proc. New England Zoöl. Club, 1:40, June 5, 1899.
Ochotona princeps cuppes A. H. Howell, N. Amer. Fauna, 47:27, August 21, 1924.
Type.—Obtained by Allan Brooks at the Monashee Divide, 4,000 feet, Gold Range, British Columbia, on August 2, 1897; type in Museum of Comparative Zoölogy.
Racial characters.—Smallest and palest of the Washington pikas; total length less than 8 inches; color of upper parts grayish-yellow, grayest on posterior third of back; underparts pale buff; skull small, but with relatively wide zygomatic and interorbital regions.
Measurements.—Eight males and 3 females from Round Top Mountain, Pend Oreille County, average, respectively: total length 183.5, 181.5; hind foot 30.7, 31.0; ear 22.4, 23.7; one adult male from the same locality weighed 141.6 grams.
Distribution.—This pika has been found at Round Top Mountain (W.W.D.) and Pass Creek Pass (W.W.D.) in northeastern Washington.
Lagomys minimus Lord, Proc. Zoöl. Soc. London, p. 98, 1863 (not of Schinz, 1821).
Ochotona minimus Bangs, Proc. New England Zoöl. Club, 1:39, June 5, 1899.
Ochotona fenisex Osgood, Proc. Biol. Soc. Washington, 26:80, March 22, 1913 (substitute for minimus Lord).
Ochotona princeps fenisex A. H. Howell, N. Amer. Fauna, 47:28, August 21, 1924.
Type.—Obtained by J. K. Lord at "Ptarmigan Hill," near head of Ashnola River, Cascade Range, British Columbia, in early fall of 1860 (?); type in British Museum.
Racial characters.—Size and color intermediate between brunnescens and cuppes; length about 8 inches; color of upper parts near Pinkish Cinnamon, becoming gray on posterior third of back; underparts washed with buff; skull of medium size and proportions.
Measurements.—Nine males and 5 females from Okanogan and Chelan counties average, respectively: total length 190.0, 197.4; hind foot 31.1, 32.8; ear 21.6, 22.0.
Distribution.—Specimens referable to this race occur from the British Columbian boundary south, through the eastern Cascade Mountains. Along the western border of its range, fenisex becomes larger and darker, merging into the race brunnescens. Marginal records are: Hidden Lakes (U.S.N.M.), Lyman Lake (U.S.N.M.), Mt. Stuart (W.W.D.), Easton (U.S.N.M.), Mt. Aix (U.S.N.M.), Steamboat Mt. (M.V.Z.).
Ochotona fenisex brunnescens A. H. Howell, Proc. Biol. Soc. Washington, 32:108, May 20, 1919.
Ochotona princeps brunnescens A. H. Howell, N. Amer. Fauna. 47:31, August 21, 1924.
Type.—Obtained by George G. Cantwell at Lake Keechelus, Kittitas County, Washington, on August 23, 1917; type in United States National Museum.
Racial characters.—Largest and darkest of the three races of Washington pikas; total length 8 inches or more; color of upper parts rich cinnamon, heavily washed with blackish; posterior part of back slightly paler; underparts buffy cinnamon; skull large and heavy with wide zygomatic arches but relatively narrow interorbital region and relatively narrow across maxillary tooth rows.
Measurements.—Eight males and 9 females from within three miles of Stevens Pass, King and Chelan counties, average, respectively: total length 201.8, 208.3; hind foot 33.3, 33.0; ear 22.3, 22.6. The average weights of 4 males and 3 females from 3 mi. S E Tumtum Mountain, Clark County, are 178.0 and 174.3 grams, respectively.
Distribution.—From the British Columbia boundary south to the Columbia River and from the western Cascades east to the area of intergradation with fenisex. Marginal occurrences are: Whatcom Pass (U.S.N.M.), Stevens Pass (W.W.D.), Keechelus (U.S.N.M.), Cowlitz Pass (U.S.N.M.), Tumtum Mountain (M.V.Z.).
Lepus townsendii Bachman, Jour. Acad. Nat. Sci. Philadelphia, 8 (pt. 1):90, pl. 2, 1839.
Lepus campestris townsendi Merriam, Proc. Biol. Soc. Washington, 17:132, July 14, 1904.
Lepus townsendii townsendii Hollister, Proc. Biol. Soc. Washington, 28:70, March 12, 1915.
Type.—Obtained by J. K. Townsend at old Fort Walla Walla (present town of Wallula), Walla Walla County, Washington; type in Philadelphia Academy of Natural Sciences.
Measurements.—A male from Miller's Island, Klickitat County, in the Columbia River, measured: total length 564; length of tail 117; hind foot 156; ear 110; weight 337.5 grams.
Distribution.—Previously found over the grasslands of eastern Washington. Now restricted and scarce except in the Okanogan Valley.
Remarks.—The white-tailed jack rabbit is the largest rabbit in the state, adults measuring 24 or more inches in length. The long legs and long ears accentuate the impression of large size. Its body is more bulky than that of its relative, the black-tailed jack rabbit. In summer the pelage of the upper parts is dark gray and in winter it is white over nearly the entire body.
The white-tailed jack rabbit occurs from southern Saskatchewan south to extreme northern New Mexico, and from eastern Washington east to Wisconsin. A single race occurs in Washington. In eastern Washington "whitetails" favor the hilly, bunchgrass territory of the arid subdivision of the Transition and Upper Sonoran life-zones. In winter they descend to the lower sagebrush valleys.
The principal enemies of the white-tailed jack rabbit are the eagle, coyote, and bobcat. Of 1,186 stomachs of coyotes from Washington, Sperry (1941: 11) found that 27 percent contained rabbits, including jack rabbits, snowshoe hares, and cottontails.
In the daytime, white-tailed jack rabbits hide in forms which consist of shallow holes dug at the bases of bushes or beside rocks. They feed in the morning, evening, and in the night along wide, well-defined trails through the bunchgrass. If startled from their forms they dash off in bounding, erratic leaps, skimming away until lost to sight. A whitetail has been timed at a speed of 34 miles per hour (Cottam and Williams, 1943: 262).
The early explorers and settlers found the white-tailed jack rabbits abundant in eastern Washington. With the invasion and spread of the black-tailed jack rabbit, and the reduction of native bunchgrass through overgrazing by livestock, the whitetail has become rare. In several years of field work on the Columbian Plateau, I saw none. Near Wallula, the type locality, residents had not seen whitetails for years, but thought there might be a few left "back in the hills." There are thought to be a few left near Ellensburg and Yakima.
Only in the Okanogan Valley are the whitetails holding their own; they are reasonably common there. In winter they come down from the hills on to the sagebrush flats along the Okanogan River in Okanogan County. In January it is not unusual to see as many as five in a day's drive. When, as will most certainly occur, the black-tailed jack rabbit enters the Okanogan Valley, the splendid whitetail may be expected to disappear from Washington.
Because this species has been so reduced in numbers, no distributional map has been included. Taylor and Shaw (1929: 28) give its range as: "north to Oroville, east to Pullman, south to Asotin, [Pg 382] Walla Walla, and Kennewick, and west to Lake Chelan (Manson), Yakima Valley, and Klickitat County." This range is similar to that of the Nuttall Cottontail (Fig. 129).
Description.—The appearance, size, and proportions of the snowshoe rabbit are similar to those of the Belgian hare. The body is about 16 inches in length, the ears are midway in size between those of the cottontail and the jack rabbit, and the feet are relatively long and the tail is short. In summer the color of the upper parts is reddish brown, varying with the subspecies. The winter pelage of Lepus a. washingtonii is a slightly paler brown than the summer coat. In the other three races in Washington the winter coat is entirely white, except for the dusky borders of the ears.
Snowshoe rabbits occur in Alaska, Canada, and the northern United States, from the Atlantic to the Pacific. They are absent in desert or prairie regions but range far southward in the United States in mountainous areas. They are found throughout Washington, except on the Columbian Plateau and in the Okanogan River Valley. None of the four races found in Washington is restricted exclusively to the state. Snowshoe rabbits live only in wooded areas. Their habitat varies from dense, impenetrable rain-forests along the ocean to the alpine parks, dotted with trees, of the Hudsonian Life-zone. They occur in humid and arid subdivisions of the Transition, Canadian, and Hudsonian life-zones. Altitudinally they range from sea level to 6,000 feet (Mt. Rainier).
Enemies of the snowshoe rabbit include the coyote, bobcat, lynx, long-tailed weasel, and great horned owl.
Snowshoe rabbits are largely nocturnal or crepuscular in habit. They are secretive and slip away quietly at the least threat of danger. Persons often live for years in localities where snowshoe rabbits are abundant without seeing a live individual. Those that are seen ordinarily have been startled from their forms at midday, or surprised while feeding on clover along a highway in the early morning. More commonly they are seen crossing a road in the lights of an automobile. Tracks, easily found after a fresh snowfall, give some indication of their numbers in any locality.
Little information is available on the fluctuations of numbers of snowshoe rabbits in Washington. Floyd Thornton, a trapper living at Forks, Clallam County, states that they were numerous in 1924, scarce in 1930-31, and fairly common in 1938-39. More rabbits [Pg 383] are seen shortly after the breeding season than at other times of the year. From April 8-10, 1941, I saw none on a highway extending about 100 miles along the west coast of the Olympic Peninsula, but on June 4-5 here counted 3 dead on the road and saw 3 running across it. One was about one-third grown and another two-thirds grown.
Fig. 127. Distribution of the snowshoe hare in Washington. A. Lepus americanus washingtonii. B. Lepus americanus cascadensis. C. Lepus americanus columbiensis. D. Lepus americanus pineus.
The snowshoe rabbits are classed as game animals in Washington but few people hunt them. Their winter food includes buds and needles of hemlock, Douglas fir, and probably other evergreens. Annuals, grasses, and shrubs, as well as Douglas fir needles, are eaten in the summer. Snowshoe rabbits do some damage by eating the bark of trees and the boughs of newly planted evergreens. Together with rodents they are responsible for serious damage to plantations of Douglas fir and hemlock on the Olympic Peninsula.
At least in summer, they are heavily parasitized by fleas and ticks and may in addition carry tularemia, or rabbit fever.
Scheffer (1933: 77-78) found that the young were born from May 5 to July 4 in the Puget Sound area and that there were from 2 to 5, usually 5 per litter.
Lepus washingtonii Baird, Proc. Acad. Nat. Sci. Philadelphia, 7:333, 1855.
[Lepus americanus] var. Washingtoni J. A. Allen, Proc. Boston Soc. Nat. Hist., 17:434, February 17, 1875.
Lepus americanus Washingtoni True, Proc. U. S. Nat. Mus., 7:601, 1885.
Type.—Obtained by G. Suckley at Steilacoom, Pierce County, Washington, on April 1, 1854; type in United States National Museum.
Racial characters.—Size small for a snowshoe rabbit; color of upper parts dark, in summer between Sayal Brown and Cinnamon, in winter slightly paler, near Pale Cinnamon Buff (capitalized color terms in the accounts of the lagomorphs are after Ridgway, Color Standards and Color Nomenclature, Washington, D. C., 1912); underparts white; soles of feet usually stained yellowish, brownish, or blackish.
Measurements.—Two males and 6 females from the Olympic Peninsula average, respectively: total length 407, 402.5; hind foot 114, 119. Weight of a 408 mm. male from the same locality 2-1/4 lbs.
Distribution.—This snowshoe rabbit occupies the humid subdivision of the Transition Life-zone of western Washington from the British Columbian boundary south to the Columbia River. Marginal occurrences are (from Dalquest, 1942: 175): Mt. Vernon, Paradise Lake, Lake Kapowsin, and White Salmon.
Lepus bairdi cascadensis Nelson, Proc. Biol. Soc. Washington, 20:87, December 11, 1907.
Lepus americanus cascadensis Racey and Cowan, Ann. Rept. Provincial Mus. British Columbia, p. H 18, 1935.
Type.—Obtained by W. C. Colt near Hope, British Columbia, on June 12, 1894; type in Museum of Comparative Zoölogy.
Racial characters.—Size medium for a snowshoe rabbit; color of upper parts in summer near Orange Cinnamon; head paler, sharply marked off from body; underparts white. Color in winter: entire body pure white except for dusky borders of ears and eyelids.
Measurements.—Three males and 5 females from Kittitas County, Washington, average, respectively: total length 405, 440; hind foot 124, 133.
Distribution.—The Cascade Mountains from the British Columbia boundary south to Mount Adams. Marginal occurrences are: Skykomish (Dalquest, 1942: 177), Vance (Dalquest, 1942: 177) and 7 mi. W Guler (W.W.D.).
Lepus americanus pineus Dalquest, Jour. Mamm., 23:178, May 14, 1942.
Type.—Obtained by P. G. Putnam at Cedar Mountain (now Moscow Mountain), Latah County, Idaho, on May 29, 1921; type in University of Michigan, Museum of Zoölogy.
Racial characters.—Size small for a snowshoe rabbit; in summer, upper parts Cinnamon Brown, with sides slightly paler and rump patch blackish; color of the head between Sayal Brown and Cinnamon; hips Light Olivaceous Buff; chest-band light Cinnamon; ears blackish, often edged with white. In winter the color of the entire body is white, save for the dusky edges of the ears and the blackish eyelids. The color of the underfur in winter is usually Pale Pinkish Cinnamon, rarely Light Vinaceous Cinnamon or Orange Cinnamon, with the basal portion slaty.
Measurements.—Two males and 5 females, from northeastern Washington, average, respectively: total length 419, 439; hind foot 138.5, 138. The average measurements of 3 males and 7 females from the Blue Mountains are: 407, 422; 128, 131.
Distribution.—The pine forests of the arid subdivision of the Transition Life-zone along the eastern border of Washington, in the Blue Mountains, and in the forested parts of northeastern Washington as far west as the Kettle River Range. Western records of occurrence are (Dalquest, 1942: 179): Deep Lake, Colville, Calispel Peak.
Lepus americanus columbiensis Rhoads, Proc. Acad. Nat. Sci. Philadelphia, p. 242, June, 1895.
Type.—Obtained by S. N. Rhoads at Vernon, British Columbia, on July 29, 1892; type in Philadelphia Academy of Natural Sciences.
Racial characters.—Size large; color in summer: upper parts near Cinnamon Brown, sides and head slightly paler; top of tail and small rump-patch blackish; chest-band pale Cinnamon Brown; hips light Ochraceous Buff; chin and belly white.
Measurements.—A male from Molson, Okanogan County, measures: total length 435; length of tail 150. A male and a female from Danville, Ferry County, measure, respectively: 460, 430; 150, 142.
Distribution.—Only a small part of the range of this rabbit lies within the state of Washington; the greater part is in British Columbia. In Washington it occupies the timbered areas north of the Columbia River, east of the Okanogan River, and west of the Kettle River Range. Records are (Dalquest, 1942: 182): Molson, Danville and Republic. In the 1942 paper, the latter locality was erroneously listed under pineus.
Lepus texianus deserticola Mearns, Proc. U. S. Nat. Mus., 18:564, June 24, 1896.
Lepus texianus wallawalla Merriam, Proc. Biol. Soc. Washington, 17:137, July 14, 1904 (type from Touchet, Walla Walla County, Washington).
Lepus californicus deserticola Nelson, N. Amer. Fauna, 29:137, August 31, 1909.
Type.—Obtained at western edge of Colorado desert, Imperial County, California; type in American Museum of Natural History.
Measurements.—Three males and 4 females from Union Gap, Yakima County, average, respectively: total length 538, 539; length of tail 72, 71; hind foot 128, 124; ear 120, 124. Two females from the same locality weighed 5 and 6-1/2 pounds, respectively.
Distribution.—The Columbian Plateau, southeastern Washington, and the Yakima Valley area. The northernmost locality record is Moses Coulee (W.W.D.).
Remarks.—The blacktail is the commonest jack rabbit of the West. It is larger than the domestic rabbit and its enormous ears, long, gangling legs, and bounding gait make it appear even larger than it really is. Its iron-gray color, black tail and black ear tips match its sagebrush habitat.
Fig. 128. Distribution of the black-tailed jack rabbit, Lepus californicus deserticola, in Washington.
Black-tailed jack rabbits reach the northern limit of their distribution on the Columbian Plateau of eastern Washington. They extend from Washington south to the Valley of Mexico and from the Pacific Coast east to Missouri (Nelson, 1909: 127). About 20 races are recognized, of which only one is native to Washington.
The blacktail is restricted to the Upper Sonoran Life-zone. Seldom is it found far from sagebrush (Artemisia) and rabbitbrush (Chrysothamus). It is well adapted to desert life, and is able to withstand the bitterly cold winters and hot, dry summers of eastern Washington. Blacktails are active in the evening, night, morning, [Pg 387] and cooler parts of the day. In the daytime they crouch in forms consisting of shallow depressions at the bases of shrubs.
Blacktails eat the twigs and leaves of sagebrush, rabbitbrush, other desert shrubs, and grasses. They are particularly fond of alfalfa and cultivated crops, of which they destroy great quantities. They are serious pests during periodic years of abundance. Some measure of their abundance may be gained by counting the bodies of jack rabbits killed by cars on well-traveled highways through sagebrush areas. In years of abundance these may number 50 to 100 per mile, while in years of scarcity these may number only one or two.
Black-tailed jack rabbits are susceptible to numerous parasites and diseases including tularemia. Diseased rabbits are especially noticeable during years of abundance. Jack rabbits are almost never used as food although many are shot for sport or to protect crops.
The black-tailed jack rabbit is a swift runner. Cottam and Williams (1943: 263) timed 6 individuals while running under varying conditions. Full speeds for 50 to 300 yards varied from 27 to 38 miles per hour. The maximum speed was attained by two individuals, each for 100 yards.
Embryos found in March numbered 4, 5 and 6.
Lepus nuttallii Bachman, Jour. Acad. Nat. Sci. Philadelphia, 7:345, 1837.
Lepus artemisia Bachman, Jour. Acad. Nat. Sci. Philadelphia, 8:94, 1839 (type from Wallula, Walla Walla County, Washington).
[Lepus sylvaticus] var. Nuttallii Allen, Proc. Boston Soc. Nat. Hist., 17:434, February 17, 1875.
Lepus sylvaticus Nuttalli True, Proc. U. S. Nat. Mus., 7:601, 1885.
Sylvilagus (Sylvilagus) nuttallii Lyon, Smithsonian Misc. Coll., 45 (no. 1456):336, June 15, 1904.
Sylvilagus nuttalli Nelson, N. A. Fauna, 29:201, August 31, 1909.
Type.—Obtained near mouth of Malheur River, Malheur County, Oregon. by T. Nuttall in August, 1834; type in Philadelphia Academy of Natural Sciences.
Measurements.—Two males and 4 females from Moses Lake, Grant County, average, respectively: total length 355, 348; length of tail 33, 35; hind foot 83, 83; ear 63.5, 60.
Distribution.—The Columbian Plateau, Okanogan Valley, Yakima Valley and Columbia Valley in southeastern Washington; in general, the sagebrush area of eastern Washington; north in the Okanogan River Valley to Oroville and in the Columbia Valley to Kettle Falls (W.W.D., records not all shown on map).
Remarks.—Nuttall cottontails are small, grayish-brown rabbits with relatively short, rounded ears and short legs. Their small size and small ears, which lack black tips, distinguish them from jack rabbits where the two occur together.
Fig. 129. Distribution of the Nuttall cottontail. Sylvilagus nuttallii nuttallii, in Washington.
The genus Sylvilagus is found in both North and South America. The species nuttallii, with three subspecies, is restricted to the western part of the United States. It ranges from southern Canada south to central New Mexico and from western South Dakota west to the Cascades. A single race occurs in Washington. Nuttall cottontails depend on cover for concealment from enemies. They frequent thick stands of tall sagebrush, riparian thickets, or rocky coulees. Seldom are they encountered in the open. In the sand-dune areas near Moses Lake cottontails were abundant in the dense, thorny thickets about potholes and in areas of tall sagebrush. They are especially common near the talus at the bases of the walls of Grand Coulee, and Moses Coulee where they do not hesitate to enter crevices in rock slides for protection. Indeed, cottontails are [Pg 389] abundant everywhere within their range in the state of Washington, where suitable cover and food are present. They seem to be confined to the Upper Sonoran Life-zone.
Cottontails are most active at night, as is attested by the number killed then by automobiles on highways. The greatest number are seen by observers in the morning and evening but it is not unusual to see an animal feeding at midday. When startled they dash for the nearest thicket or pile of rocks with their tiny white tails erect. They travel in relatively straight lines and do not dash from side to side in flight as do jack rabbits. They sometimes seek concealment by "freezing" motionlessly in plain sight. When feeding undisturbed they travel by slow hops.
The trails of cottontails are characteristic of thickets in sagebrush country. The trails are narrow, less than four inches wide, and often enter thickets of strong, thorny growths which can scarcely be penetrated by man. Near Okanogan Lake the trails of cottontails were found among greasewood bushes on hard-packed gravel. Trails are usually most abundant in thickets near water. Permanent trails are not made through low sagebrush or over sandy areas where the animals prefer to pick their way when traveling from one clump of cover to another.
Nuttall cottontails probably eat many desert grasses, annuals, and shrubs; observation indicates that sagebrush (Artemisiae tridentata) and rabbit brush (Chrysothamnus nauseosus) are particularly important as food.
The young of cottontails in Washington seem to vary from one to four per litter and are born between April and June.
Lepus sylvaticus floridanus Allen, Bull. Amer. Mus. Nat. Hist., 3:160, October 8, 1890.
Sylvilagus floridanus Lyon, Smithsonian Misc. Coll., 45 (no. 1456):322, June 15, 1904.
Description.—Slightly larger than the sagebrush cottontail; smaller than the snowshoe rabbit; ears small; color of upper parts pinkish cinnamon-brown; sides pale grayish-cinnamon; underparts white; nape of neck cinnamon; chest band paler cinnamon; tail brown above, white beneath; forefeet and sides of hind feet cinnamon.
Remarks.—The Florida cottontail is not native to the state of Washington but has been introduced at several localities. It is spreading rapidly at the present time. Points of introduction include [Pg 390] Pullman, Whitman County (1926-1927); Battleground, Clark County (1933); Auburn, King County (1927); and Whidby Island, Island County (1931). At least two subspecies (mearnsi, alacer) have been introduced and a third (similis) may have been introduced.
For further information on introduced cottontails see Dalquest (1941B: 408-411).
Lepus idahoensis Merriam. N. Amer. Fauna, 5:75, July 30, 1891.
Brachylagus idahoensis Lyon, Smithsonian Misc. Coll., 45 (no. 1456):323, June 15, 1904.
Sylvilagus idahoensis Grinnell, Dixon and Linsdale, Univ. California Publ. Zoöl., 35:553, October 10, 1930.
Type.—Obtained in the Pahsimeroi Valley, Custer County, Idaho, by V. Bailey and B. H. Dutcher on September 16, 1890; type in United States National Museum.
Distribution.—Known only from the central part of the Columbian Plateau.
Description.—The pigmy rabbit is a tiny species, differing from the cottontail in smaller size, paler, grayer color, shorter ears and smaller legs.
The pigmy rabbit is restricted to the Great Basin region. No subspecies has been described. It is rare and of local occurrence in Washington, having been recorded only from the central part of the Columbian Plateau. Orr (1940), who studied the species in California, found them only in stands of tall, dense sage (Artemisiae tridentata). It is a burrowing form, not straying far from its hole.
Description.—The elk, next to the moose, is our largest deer. The legs of the elk are slender. The tail is a short, pointed stub a few inches long. The neck is thick in proportion to the head. Both males and females possess the canine teeth familiar as "elk tooth charms." Only the males possess antlers. These are huge, slender beams that curve up, out and back with the basal tine or "dog killer" and four to six points on each antler. The antlers are deciduous and are shed annually. The body is grayish or tan in color. The head, neck, chest and legs are rich, dark brown, strongly contrasting with the paler body. The distinctive rump patch is pale tan or white.
In the past the elk was found over most of the forested areas of Washington. Lumbering, agriculture and settlement as well as excessive hunting removed it from parts of eastern Washington and all except the most inaccessible parts of the lowlands of western Washington. Only in the Cascade and Olympic Mountains and the coastal strip between the Columbia River and the Olympic Mountains, did the elk survive in appreciable numbers. Conservationists and a more enlightened game policy began to protect the elk at the turn of the century. It was already too late to save the species in eastern Washington, where it seems never to have been truly abundant and where relatively open country afforded little protection from the high-powered rifle. In the dense, rugged forests of western Washington a sizable number remained on the Olympic Peninsula and these, under protection, increased to their present numbers. The dense, tangled forests of the southwestern coastal area and the western Cascades lack conditions suitable to support truly large elk herds. These areas probably now have as large an elk population as can safely be supported and fed.
The elk of eastern Washington had disappeared or nearly disappeared by 1910. The race originally occurring there was the Rocky Mountain form; it has been reintroduced from Montana and Wyoming into northeastern Washington and the Blue Mountains area. These plantings have not been very successful. Introduced in the eastern Cascades, however, the Rocky Mountain elk thrived and increased on what was probably once the peripheral range of the coastal elk.
Fig. 130. Elk or wapiti (Cervus canadensis nelsoni), Banff, Alberta, October, 1939. (G. A. Thomas photo.)
The habits of the elk are best known from the herds in the Olympic Mountains and on the Rattlesnake Game Refuge in the eastern Cascade Mountains. Here the animals are numerous and relatively tame. Their habits seem to differ somewhat in the forests of the lowlands from those of the animals in the higher Olympics where the topography and climate are very different.
The elk is a social animal, gathering in herds over most of the year. The old males leave the herds in the spring but seem to stay in small bands while their antlers are growing. In the Olympic Mountains, herds of 100 or more animals have been seen. In the lowlands there are ordinarily from five to ten in a herd.
Fig. 131. Group of elk or wapiti (Cervus canadensis nelsoni), Banff, Alberta, October 10, 1939. (G. A. Thomas photo.)
The elk is a browsing animal, feeding on twigs and leaves of deciduous trees, shrubs and evergreens. In spring and summer it eats grasses and succulent annuals, but in winter twigs and needles of evergreens, perennial ferns, dry grass and even moss is utilized. To a certain extent the elk are migratory, ascending to the open meadows of the Hudsonian Life-zone in the early summer and returning to the dense forests of the Transition and lower Canadian Life-zones with the winter snows. The lowland elk make no such migrations, merely leaving the riverbottom jungles when the leaves [Pg 394] are off the deciduous plants growing there, and live in the coniferous timber.
In the forests the elk is capable of swift and almost silent movement. It is an eerie experience to trail a herd of elk through a dense forest in a winter rain, knowing that a number of the large animals are within a few feet, moving swiftly but silently away. When a herd is feeding and does not suspect the presence of an observer, the animals rustle branches, break twigs, snort and wheeze as they breathe.
The antlers of the males are shed in March. The bulls retire from the herd until the new horns are well grown, in late August or September. Breeding takes place in September or October and the young are born the following April or May.
Cervus roosevelti Merriam, Proc. Biol. Soc. Washington, 11:272, December 17, 1897.
Cervus canadensis occidentalis Taylor and Shaw, Occ. Papers Chas. R. Conner Mus., 2:29, December, 1929.
Type.—Obtained on Mount Elaine, Mason County, Washington, by H. and C. Emmet on October 4, 1897; type in United States National Museum.
Racial characters.—A large, dark elk with short, heavy antlers.
Measurements.—The type measured, in the flesh (Bailey, 1936:81): total length 2,490; tail 80; ear (dry) 208.
Distribution.—From the Cascade Mountains westward; possibly occurred formerly in the eastern Cascades, and perhaps still present there, in places, or mixed with introduced nelsoni.
Remarks.—For use of the name roosevelti rather than occidentalis see Bailey (1936:81).
Cervus canadensis canadensis Taylor and Shaw, Occ. Papers Chas. R. Conner Mus., 2:29, December, 1929.
Cervus canadensis nelsoni Bailey, Proc. Biol. Sec. Washington, 48:188, November 15, 1935.
Type.—Obtained at Yellowstone National Park, Wyoming, by J. Pitcher; died in captivity on September 21, 1904; type in United States National Museum.
Description.—Smaller and paler than roosevelti with longer, slimmer antlers.
Distribution.—Formerly occurred in northeastern Washington and the Blue Mountains of southeastern Washington. Since then exterminated and reintroduced to both areas from the Rocky Mountains. Also introduced and established in the eastern Cascades.
Description.—The white-tailed deer differs from the black-tailed and mule deer in several anatomical details. The tail is long, wide, and when the animal is frightened is carried upright with the long, white hair of the underside spread out. The antlers of the buck are not of a biramous system of branching but instead consist of a main beam which curves sharply out and forward, remaining low. All subsidiary tines, save the basal one, emerge from the dorsal side of the main beam. The basal tine is usually a spurlike point arising vertically from the main beam not far from the base of the antler. In exceptional specimens the basal tine is large and divided into several points. The body of the "whitetail" is more smoothly rounded, and more graceful, than that of the mule deer.
Fig. 132. White-tailed deer (Odocoileus virginianus leucurus), doe at least nine years old, kept as a pet by Mrs. Jack Hovis, Puget Island, Washington, December 16, 1939. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 750.)
White-tailed deer range from Canada south to Panama and from the Atlantic to the Pacific. Their range is much more extensive to the east and south than that of the "blacktail" and mule deer. [Pg 396] In western Washington the whitetail occupies a limited habitat near the mouth of the Columbia River. Its geographic range was probably somewhat greater in the past. Its habitat includes the low, damp, marshy islands and floodplain of the Columbia. Blacktail range in the wooded hills surrounding the whitetail's range but rarely enter it to compete with the whitetail (Scheffer, 1940A: 282).
In northeastern Washington the whitetail shows the same tendency to occupy low, marshy areas and valleys. The habitat preference of the whitetail in northeastern Washington is less exact than in western Washington, for the species ranges up from the valleys into the forests of larch and cottonwood of the Transition Life-zone. Its habitat includes denser forest and brushy areas. Rarely does it occur in the open type of forest occupied by the mule deer.
Fig. 133. White-tailed deer (Odocoileus virginianus ochrourus): fawn kept as a pet by L. E. Borud, Ione, Washington, June 13, 1938. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 69.)
In Washington the whitetail is not an important game animal. The race occurring in western Washington is characterized by small size and small antlers. Further, its total population is estimated at [Pg 397] but 400 to 900 individuals. The whitetail of northeastern Washington is larger and possesses larger antlers. Indeed, some antlers from Ferry County are the largest antlers of whitetail that I have ever seen. Nevertheless, the whitetail of northeastern Washington is smaller than the mule deer and far less common. The habitat is dense and difficult to traverse. The animal is shy and silent, fleeing soundlessly when approached. For these reasons most hunters in northeastern Washington prefer to hunt the mule deer.
Fig. 134. White-tailed deer (Odocoileus virginianus ochrourus): left antler found on ground at Park Rapids, Pend Oreille County, Washington, October 1, 1937. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 756.)
Odocoileus virginianus macrourus Taylor and Shaw, Occ. Papers Chas. R. Conner Mus., 2:30, December, 1929.
Odocoileus virginianus ochrourus Bailey, Proc. Biol. Soc. Washington, 45:43, April 2, 1932.
Fig. 135. White-tailed deer (Odocoileus virginianus leucurus), antlers of large buck killed in the fall of 1939 at Cathlamet, Washington, by Paul Lewis. (Fish and Wildlife Service photo by Victor B. Scheffer. No. 752.)
Type.—Obtained at Coolin, south end of Priest Lake, Bonner County, Idaho, by F. Lemmer on December 27, 1908; type in United States National Museum.
Racial characters.—A large, pale white tail with large, low antlers and distinctive cranial features.
Measurements.—The type, an adult male, measures (Bailey, 1936. p. 43): total length 1,752; tail 265; hind foot 483; ear (dry) 120.
Distribution.—Northeastern Washington. Probably once occurred in southeastern Washington.
Cervus leucurus Douglas, Zoöl. Jour., 4:330, 1829.
Odocoileus leucurus Thompson, Forest and Stream, 51:286, October 8, 1898.
Odocoileus virginianus leucurus Lydekker, Cat. Ung. Mamm. British Mus., 4:162, 1915.
Type.—Obtained on the North Umpqua River, Oregon, by David Douglas on October 17, 1826; type originally, and perhaps still, in British Museum.
Racial characters.—A small, handsome whitetail with slender, erect antlers and brownish or grayish color.
Measurements.—A young adult female from 1 mile south of Skamokawa, Wahkiakum County, measured: total length 1,545; length of tail 250; hind foot 458; ear 143; weight 88 pounds.
Distribution.—Previously occurred over much of southwestern Washington; now confined to the banks and islands of the Columbia River in Wahkiakum County.
Description.—The mule and black-tailed deer are among deer of medium to large size. Adult mule deer may weigh up to 400 pounds while some fully adult blacktail bucks may weigh as little as 100 pounds. The body is heavily muscled, the legs long and the tail only about 6 inches in length. The ears are long, from 6 to 8 inches from notch to tip. The head is long and the male has well-developed antlers. The first antlers are almost straight spikes. Those of the second year are bent slightly outward and forward and are equally or subequally forked with the anterior branch usually the stouter. In the third year the anteriolateral curvature is more pronounced and one or both of the forks again fork. In succeeding years forks are larger and more numerous but the essential biramous arrangement of forking is maintained. The "blacktail" and mule deer are strongly marked races. The blacktail differs from the mule deer in: smaller size; darker color, especially on face and tail; tail dark brown above with dark tip rather than whitish with black tip; tail haired beneath, not naked for half its length; antlers smaller and lighter; and skull and teeth smaller.
Mule deer and black-tailed deer range over western North America from southeastern Alaska southward into northern Mexico. They inhabit forested parts of the state of Washington. Blacktails occupy the San Juan Islands, the islands in Puget Sound, the Olympic Mountains, the lowlands of western Washington, and the Cascade Mountains. Mule deer occupy the Cascades, including their eastern slope, northeastern and southeastern Washington and parts of the Columbian Plateau. Over this large range there is considerable local geographic variation.
Jackson (1944: 1-56) estimated that 109,600 blacktail and 175,725 mule deer live in Washington. Thus Washington is second only to California in number of blacktail and ranks fifth in number of mule deer.
Individual variation over the range of the mule deer is considerable but no trends of variation are distinguishable. Mule deer from the Blue Mountains, northeastern Washington and the eastern Cascades are essentially similar.
There is geographic variation as well as great individual variation in the black-tailed deer of Washington. The deer of the San Juan Islands and the islands of Puget Sound are smaller and darker than those of the mainland and possess smaller, lighter antlers. The deer of Whidby Island are sometimes contemptuously referred to by residents as jackrabbit deer. Fully grown bucks on the Islands weigh in the neighborhood of 100 pounds, rarely exceeding 150 pounds, whereas bucks on the mainland commonly weigh more than 150 pounds dressed.
Fig. 136. Mule deer (Odocoileus hemionus hemionus), subadult male from Okanogan County, Washington, raised in captivity; photographed June 29, 1938, on Hurricane Ridge, Olympic National Park. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 99.)
The dividing line between the ranges of the black-tailed and mule deer in general is the summit of the Cascade Mountains. In summer the two races come together and doubtless mingle in the Cascades but as fall approaches they migrate. The mule deer moves eastward to the yellow-pine areas on the lower slopes while the [Pg 401] blacktails descend westward to the denser Douglas fir and hemlock forests on the western flanks of the Cascades. In the breeding season the two races are separated. However, as shown by Cowan (1936A: 219), in the Pemberton Valley, British Columbia, the two forms occur together during the breeding season and intergradation occurs there. Intergradation occurs also in the Lake Wenatchee area of Chelan County. Observation of hunters' specimens showed some undoubted intergrades among the more abundant, typical mule deer.
Both the mule deer and blacktail have a characteristic bounding gait, unlike the smooth run of the white-tailed deer. The tail is usually held down, rarely erect. When not frightened they move with dainty steps, making little noise.
Fig. 137. Black-tailed deer (Odocoileus hemionus columbianus), buck resting at midday, Van Trump Park, Mt. Rainier, Washington, August, 1931. (Fish and Wildlife Service photo by Victor B. Scheffer, No. 260.)
The black-tailed deer in certain areas is more nocturnal than diurnal. They feed principally in the early morning and evening. In the Olympic Mountains, on the North Fork of the Quinault River, in July, 1937, blacktails were watched feeding and playing until midnight. Also in December, 1939, deer, near the forks of the Skykomish River, were moving about, browsing as late as 11 p. m. while a light snow was falling. In the San Juan Islands, in the summer of 1939, deer were observed feeding at all hours of the day.
The blacktail lives in some of the most dense jungle areas of western Washington. On Whidby Island and some of the San Juan Islands the brush and vines grow so densely that a man can scarcely penetrate them. Were it not for the deer trails, formed by generations of deer, our small mammal investigations would have been far more difficult. The blacktail lives also in the dense forests where fir, hemlock and cedars grow to heights of more than 150 feet. Here lack of light allows only ferns and moss to form an understory vegetation. These forests often clothe the steep glacial hills and the trails of the deer on such hills show them to be adept climbers. Deer trails generally avoid fallen trees and other obstacles. When startled, however, a stump, fence or log is easily leaped by a blacktail. Often they are in small bands of 6 to 10 individuals, but almost as often are solitary, or in pairs.
The habitat of the mule deer is generally more open than that of the blacktail. In the summer, it may occupy rough and rugged country; in the higher Cascades individuals were found in the rocky and brushy country and in open glades and meadows. Farther east they were in the open yellow-pine forests where extensive grassy slopes, free of trees, existed. In some parts of northeastern Washington they lived in the larch and lodgepole pine forests almost dense enough to be "blacktail country." In the Okanogan Valley and on the northwestern corner of the Columbia Plateau mule deer lived in open prairie country where a few cottonwoods and willows were the only trees.
The mule deer seem more social than the blacktail. In the late summer they gather in bands of 10 to 20 or more. In winter, under pressure of hunger, they gather in herds and raid haystacks and pastures. Farmers in the Methow Valley, Okanogan County, report herds of 200 to 400 mule deer about a single haystack.
The mule deer and blacktail are the principal game mammals of the state of Washington. Thousands of hunters enter the woods each year in search of a buck. Eastern Washington is the favored hunting grounds for that is the home of the mule deer, the larger size of which makes it a more desirable trophy. Also the open country which it inhabits makes hunting more productive. Sums spent on equipment, gasoline and hunting licenses are tremendous but the feeling is that the return in recreational value and venison are worth the cost. A few casualties result each year among the army of deer hunters.
Cervus hemionus Rafinesque, Amer. Month. Mag., 1:436, October, 1817.
Cariacus macrotis True, Proc. U. S. Nat. Mus., 7:592, 1885.
Odocoileus hemionus Merriam, Proc. Biol. Soc. Washington, 12:100, April 30, 1898.
Odocoileus hemionus macrotis Bailey, National Geographic, 20:64, 1932.
Type locality.—Vicinity of Big Sioux River, South Dakota.
Racial characters.—Large size, pale color.
Measurements.—Two adult males from Stay-a-while Spring, Columbia County, measure respectively: total length 1,751, 1,559; length of tail 172, 205; hind foot 515, 485; ear 210, 211; length of metatarsal gland 150, 135.
Distribution.—Summit of Cascades eastward, in forested areas, exact limits not certainly known.
Cervus macrotis var. columbiana Richardson, Fauna Boreali-Americana, 1:257, 1829.
Cariacus columbianus True, Proc. U. S. Nat. Mus., 7 (1884):592, 1885.
Cervus columbianus Baird, U. S. Pacific R. R. Exp. and Surveys, p. 659, 1857.
Eucervus columbianus Gray, Ann. and Mag. Nat. Hist., 18 (ser. 3):338, 1866.
Odocoileus columbianus Merriam, Proc. Biol. Soc. Washington, 12:100, April 30, 1898.
Odocoileus columbianus columbianus Swarth, Univ. California Publ. Zoöl., 10:85, February 13, 1912.
Odocoileus hemionus columbianus Cowan, California Fish and Game, 22:215, July, 1936.
Type.—Obtained at Cape Disappointment, Pacific County, Washington, by Lewis and Clark on November 19, 1805 (Cowan, 1936A: 218).
Racial characters.—Small size, dark color.
Distribution.—Summit of Cascades westward, including islands in Puget Sound and San Juan Islands; exact limits of range uncertain.
Alces americanus shirasi Nelson, Proc. Biol. Soc. Washington, 27:72, April 25, 1914.
Type.—Obtained near the Snake River, Lincoln County, Wyoming, by J. Shire on December 11, 1913; type in United States National Museum.
Distribution.—Casual wanderer into northeastern Washington from Canada.
Remarks.—The moose is the largest North American deer. Its large size and huge, palmate antlers serve to separate it from any other member of the deer tribe.
Moose range from northern United States to central Canada and Alaska. The European elk and the American moose are subspecies of the same species. The moose is of only casual occurrence in Washington. The latest authentic record for Washington is a bull that wandered southward and westward from Canada until it was killed on an Indian reservation in Ferry County. Other moose are reported to have wandered into northeastern Washington from Canada in past years.
Dice (1919) was told that moose once occurred in the Blue Mountains of southeastern Washington but there is no confirmation of this report.
Rangifer montanus Seton-Thompson, Ottawa Naturalist, 13:129-30, August, 1899.
Rangifer arcticus montanus Jacobi, Erganzungsband, Zoöl. Anz., 96:92, November, 1931.
Type.—Obtained near Revelstoke, Selkirk Range, British Columbia.
Distribution.—Rare or casual along the Canadian boundary in northeastern Washington.
Remarks.—The caribou is a rather stout-bodied deer with large hoofs, short, rounded muzzle and long, erect, flattened antlers.
Caribou and their relatives, the reindeer, range over Arctic Europe, Asia, Greenland and America. In North America they range from the Atlantic to the Pacific and from the northern border of the United States northward into the Arctic. The caribou was, until recent years, a regular winter resident in small numbers in northeastern Washington near the Canadian Boundary. Their wintering grounds in Washington were said to have been destroyed by fire in 1915 and the species has appeared in the state only casually since then. Two were killed in 1940 by hunters who thought they were deer. Caribou are protected by law in Washington.
Bison bison oregonus Bailey, Proc. Biol. Soc. Washington, 45:48, April 2, 1932.
Type.—Skull and skeleton picked up at Malheur Lake, Oregon, by G. M. Benson in November, 1931; type in United States National Museum.
Distribution.—Perhaps casual in eastern Washington before coming of the white man.
Remarks.—Bison, or buffalo, occurred in southeastern Oregon but disappeared before white men reached the country (Bailey, 1936: 57). Gibbs (1860: 138) was told by an Indian hunter in 1853 that a lost bull had been killed in the Grand Coulee (state of Washington) 25 years before but that "this was an extraordinary occurrence, perhaps before unknown." In the days before horses reached the Indian tribes of eastern Washington and Oregon, wandering bison from herds in Oregon probably strayed into Washington, in somewhat the manner that the moose today stray in from Canada.
Description.—The mountain sheep is the size of a small deer. The horns of the males are massive spirals. Those of the females are smaller, curve upward and backward, rarely forming a semicircle. The horns are permanent structures, enlarged each year by growth at the base. Growth is irregular, probably as the result of seasonal reproductive activities. As a result the horns are "ringed" with concentric ridges. The color of the upper parts is dark, grayish brown. The face is paler, yellowish brown. The outer sides of the legs are dark brown. The rump, abdomen, and insides of legs are white.
Mountain sheep of the genus Ovis are abundantly represented in Asia. Two species occur in North America, Ovis dalli in Alaska, and Ovis canadensis in western North America. They range from Alaska south to northern Mexico.
In the past the mountain sheep inhabited most of the eastern Cascade Mountains, the Blue Mountains, Pend Oreille Mountains, and the cliffs of the Columbia River Valley in eastern Washington. They occurred on the eastern or Columbian Plateau side of the river and therefore probably occupied the cliffs of Moses Coulee and the Grand Coulee. Their habitat seems thus to have included rocky areas from the Upper Sonoran to the Hudsonian life-zones. At the present time they are extinct over most of their range. A small band still remains in the extreme northeastern Cascades near Mount Chopaka.
Little has been published concerning the habits of the mountain sheep in Washington. In caves along the Columbia River in Grant County, bones of sheep are found in association with stone arrowheads and other human artifacts. Presumably the sheep were killed and eaten by the Indians.
The history of the mountain sheep in North America is outlined by Cowan (1940: 506). The genus is thought to have crossed from Asia to America by the land bridge now under Bering Strait in the early Pleistocene and spread southward through the Rockies. The advance of the glacial ice forced them farther southward and the southern (canadensis) sheep were separated from their relatives farther north. The present differences between the Rocky Mountain and western sheep seem to have resulted from separation by glaciers during Wisconsin Time.
Ovis canadensis Shaw, Nat. Misc., vol. 15, text to pl. 610, about December, 1803.
Type.—Obtained in the mountains on Bow River; W. B. Davis (1939: 377) gives Dew River near Exshaw, Alberta.
Racial characters.—Large size; heavy, closely coiled horns.
Measurements.—Cowan (1940: 533) gives the measurements of a fully adult (6-year old) ram from Colorado as: total length 1,953; length of tail 127; hind foot 394; ear (dry) 63.
Distribution.—Now extinct in Washington. Cowan (op. cit.: 535) refers to this race as the sheep that formerly occurred in the Blue Mountains. This view seems logical but I feel that the sheep formerly occurring in the Pend Oreille Mountains of extreme northeastern Washington should, on the basis of geographic probability, also be referred to canadensis.
Ovis californianus Douglas, Zoöl. Jour., 4:332, 1829.
Ovis californica Richardson, Fauna Boreali-Americana, 1:272, 1829.
Ovis californiana Biddulph, Proc. Zoöl. Soc. London, p. 683, 1885.
Ovis canadensis californiana Lydekker, The Sheep and its Cousins, p. 288, 1912.
Type.—Obtained from near Mount Adams, Yakima County, Washington, by D. Douglas on August 27, 1826; type in British Museum.
Racial characters.—Similar to canadensis but smaller with more slender, spreading horns and horn tips less blunt.
Measurements.—Cowan (1940: 545) gives the measurements of a ram, five years old, from Owens Valley, California, as: total length, 1,582; length of tail, 110; hind foot, 240.
Distribution.—Formerly occupied the eastern Cascades, the valley of the Columbia and possibly the cliffs bordering Moses Coulee and The Grand Coulee. Now it is found only near Mt. Chopaka in the extreme northeastern Cascades.
Description.—The mountain goat is of deer-size, with a stout body and a pronounced hump on the shoulders. The legs are short but the hoofs are large with large dewclaws. The neck is short and thick. The head is large and goatlike in appearance. The tail is tiny. The horns are slim, round and curve up and slightly backwards. They are hollow and are permanent structures, added to each year. The body is snow white, consisting of long, soft wool, which is longer and coarser on forelegs, neck and chin than on the body. Males have a distinct beard.
Mountain goats range from Washington and Idaho north to Alaska. Their nearest relatives are the Old-World antelopes, especially the alpine species of Europe and Asia.
The mountain goat is an animal of the high mountains. Their habitat is the bare-rock cliffs and rock-strewn slopes of the Arctic-alpine and Hudsonian Life-zones. Where extensive, open rocky areas occur they descend to the Canadian Life-zone. Even in winter they keep to the high cliffs where steep slopes and strong winds keep the snow from the plants on which they feed.
Mountain goats are considered a game animal in Canada and Idaho. The species has been protected in Washington for many years. As a result they are common, although not present in numbers sufficient to withstand hunting. The high country which they occupy is unsuited to any of man's domestic animals and no reason is apparent at present why the mountain goat should not be protected and conserved for many years.
Fig. 138. Mountain goat (Oreamnos americanus americanus), old female, Lake Chelan, Washington, March 6, 1937. (Forest Service photo by Oliver T. Edwards, No. 348491.)
In spite of the protection accorded them, the mountain goat has not become tame. In driving over Stevens Pass, King County, one can, with the aid of glasses, usually pick out one or more mountain [Pg 408] goats on nearby mountains. Nevertheless, I have never seen one within 100 yards of a highway, nor heard of one being killed by cars on a highway.
The mountain goat does not, as a rule, allow close approach by man. At Goat Basin, Deception Creek, King County, circumstances are particularly favorable for goats and several are usually to be seen. While studying them on several occasions, I was never able to get truly close to them. On the few occasions when I came upon a band unexpectedly, they rapidly went over the mountain or up the nearest cliff.
Fig. 139. Group of mountain goats (Oreamnos americanus americanus), northern Cascade Mountains, Washington. (Forest Service photo, No. 348490.)
When unfrightened, these mountain goats spent much time standing, or lying with forefeet folded under them, looking into space. For such large animals they seemed to spend little time feeding. At intervals of about five minutes they plucked the ferns or other plants that grew in abundance on ledges or in tiny crevices in the cliffs. Some animals did browse for several minutes at a time. They seemed rather particular as to their food, plucking only one or two stems from a clump of vegetation.
The ordinary movements of the mountain goats are deliberate. They rarely move more than a few feet at a time. They climb with sure-footed ease but usually slowly. In climbing, the forelegs are spread and the knees are bent. The animal moves as if it were climbing steps. The greatest action is in the knee joints. If surprised in the open they run with a smoother gait, the legs moving from the shoulder and hip. If near a cliff they climb rapidly, jumping when necessary, and rarely stop while in sight. Evidence of the [Pg 409] presence of mountain goats is usually seen in the form of tufts and strands of wool. Their wool becomes caught in bushes and rough rocks and seemingly pulls out easily.
The range of the mountain goat in Washington includes the entire Cascade range from Mt. Adams and Mt. Saint Helens north to the Canadian boundary. They extend west to Mt. Baker, Mt. Higgins and Mt. Index and east to Lake Chelan and, in the Wenatchee Range, east of Mt. Stuart, perhaps almost to the Columbia River.
Dice (1919: 21) was told that mountain goats once occurred in the Blue Mountains of southeastern Washington. We know of no suitable goat country in the Blue Mountains of Washington and feel that the report was probably based on an erroneous identification.
There is a single record, supported by specimen, of a mountain goat from northeastern Washington. This individual probably wandered into Washington from northern Idaho, Montana, or adjacent British Columbia.
Ovis montanus Ord, Guthrie's Geol., 2d Amer. Ed., p. 292, 1815 (preoccupied).
R[upicapra]. americana Blainville, Bull. Sci. Soc. Philomath, Paris, p. 80, 1816.
Mazama dorsata Rafinesque, Amer. Month. Mag., 2:44, 1817 (new name for Ovis montanus Ord).
Mazama montana True, Proc. U. S. Nat. Mus., 7 (1884):592, 1885.
Oreamnos montanus Merriam, Science, n. s., 1:19, 1895.
Oreamnos montanus montanus Miller, U. S. Nat. Mus. Bull., 79:398, December 31, 1912.
Oreamnos americanus americanus Hollister, Proc. Biol. Soc. Washington, 25:186, December 24, 1912.
Type.—None. Descriptions all based on Ovis montanus Ord who, in turn, based his description on the account of skins seen by Lewis and Clark on the Columbia River of Washington or Oregon. As the mountain goat is not known to have ever occurred in Oregon in Recent times, the type locality is probably near Mt. Adams, Washington, the point where goats come nearest the Columbia.
Measurements.—The measurements of a large male killed "west of North Yakima" were reported by A. S. Harmer as: 8 feet 3 inches from tip of nose to tip of tail; horns 10 inches; weight 507 pounds (Outdoor Life, 1915: 459).
Distribution.—Throughout the higher Cascade Mountains.
Oreamnos montanus missoulae Allen, Bull. Amer. Mus. Nat. Hist., 20:20, February 10, 1904.
Oreamnos americanus missoulae Hollister, Proc. Biol. Soc. Washington, 25:186, December 24, 1912.
Type.—Obtained at Missoula, Missoula County, Montana; type in American Museum of Natural History.
Distribution.—A single record for northeastern Washington. Seemingly a rare wanderer from outside the state.
Because Victor B. Scheffer and John W. Slipp have in preparation a detailed account of Cetaceans properly ascribable to Washington, members of this order here are not treated in as much detail as are other native mammals.
Description.—Length about 40 feet; mandibles elongate, "beak-like"; one or two large teeth at tip of lower jaw; dorsal fin small and situated posteriorly; color black but abdomen occasionally grayish.
Remarks.—This rare whale is known from a few specimens recorded from Alaska south to California.
Description.—Similar to the previous species in proportions but smaller; length about 17 feet. Differs in possessing one large flat tusk in lower jaw.
Remarks.—Known only from a specimen from Alaska and another from the coast of Oregon.
Description.—A slender porpoise about 6 or 7 feet long; long, narrow beak with 80-120 conical teeth; color above black tinged with greenish; sides gray; belly and throat white.
Remarks.—Recorded from California and probably ranges into Washington in off-shore waters.
Description.—A small, beaked porpoise lacking a dorsal fin; color black with narrow white area from breast to tail; length about 4 feet.
Remarks.—A rare species described from waters 500 miles off the mouth of the Columbia River.
Description.—Length about 7 feet; beak not prominent; 44-46 conical teeth in each jaw; anterior edge of dorsal fin curved; color of upper parts greenish black; sides with one white or gray stripe posteriorly; underparts white.
Remarks.—This is the common porpoise in the Straits of Juan De Fuca from about the vicinity of Port Townsend to the ocean and in immediate off-shore waters. One specimen was obtained 100 miles off Grays Harbor. Rarely seen in Puget Sound.
Description.—A large porpoise, 20-30 feet in length; dorsal fin high and nearly straight; teeth large, conical, 12 above, 13 below; color black above with white patch on side anteriorly.
Remarks.—The killer is common in Puget Sound and off the ocean coast. Both the killer and the true blackfish are locally termed "blackfish."
Description.—A round-headed porpoise about 10 feet in length; dorsal fin high and narrow; 4 to 6 teeth in each jaw; color black, occasionally with white head or with body mottled with white and gray.
Remarks.—An off-shore species, probably not uncommon off the coast of Washington at times.
Description.—A large, stout-bodied, round-headed porpoise about 20 feet in length. Teeth large; 8-10 in lower jaw; 10-12 in upper jaw. Color black. Differs from the killer whale in possessing a low, long dorsal fin rather than a high, erect one.
Remarks.—The blackfish is a colonial species, often common in Puget Sound. It is frequently confused with the killer whale.
Description.—A small, stout-bodied porpoise with a rounded head. Length 5 to 6 feet. Teeth small, slightly flattened; about 25 above, 24 below; color blackish or brownish to liver color.
Remarks.—Small schools of harbor porpoises are common in Puget Sound, among the San Juan Islands, and in the Straits of Juan De Fuca. This is the commonest inshore porpoise but, because of its smaller size, is less well known to most persons than are the blackfish and killer whale.
Description.—A small porpoise, about 6 feet in length, with pointed head and moderately stout body. Teeth small, 23 above and 27 below. Color greenish black with flanks or posterior sides white.
Remarks.—A rare off-shore species.
Description.—A toothed whale about 60 feet long; head enormous, squarish; teeth conical, 50 or fewer in lower jaw.
Remarks.—Occurs off the coast of Washington and rarely enters the Straits of Juan De Fuca.
Description.—Similar to the sperm whale in proportions but less than 10 feet in length.
Remarks.—A rare species whose habits are little known. The available information concerning this species in the Atlantic has been summarized by Allen (1941).
Description.—A relatively slender whale, about 40 feet in length; dorsal fin absent; baleen short, yellow in color; color of body mottled gray or blackish.
Remarks.—The gray whale was once abundant and occurred off the coast of Washington in spring and summer migrations. Hunted until now rare.
Description.—A relatively slender whale, up to 82 feet in length; dorsal fin far posterior, prominent; baleen in two slabs, well developed, about two feet in length and gray in color; color of body blackish or brownish above, white below.
Remarks.—Recorded off-shore.
Description.—Similar to the finback whale but dorsal fin larger; baleen dark; body dark blue or brownish; belly with restricted white area.
Remarks.—Recorded off-shore.
Description.—A small whalebone whale, about 30 feet in length; body slender; head pointed; pectoral fin well developed and prominent; baleen short, white; color black above, white beneath.
Remarks.—A rare species that has been recorded from Washington.
Description.—Largest of the whales; length up to 100 feet; body slender; head flat above, rounded beneath; dorsal fin slender but prominent; baleen in two series, heavy and black; belly with numerous longitudinal ridges; color bluish black above, yellow beneath.
Remarks.—Probably uncommon off-shore.
Description.—A medium-sized, stout-bodied whale up to 54 feet in length; dorsal fin low, not prominent; head flat above, rounded beneath; prominent "hump" at back of head; belly with numerous longitudinal ridges; baleen small; color blackish or grayish above, paler below; body characteristically blotched with patches of whitish barnacles.
Remarks.—The humpback is the commonest whale off the coast of Washington, often coming into northern Puget Sound.
Description.—A large whale, 60-70 feet in length, with enormous, rounded head; dorsal fin absent; belly lacking longitudinal ridges; baleen blackish, 8 feet in length; color uniformly blackish.
Remarks.—A much hunted species now probably rare off shore.
Mammals Other Than Cetaceans Possibly Occurring in Washington
of Which Satisfactory Record Is Lacking
1. Ursus canadensis Merriam, grizzly bear.—This species has been recorded from Kootenay Lake, British Columbia, 30 miles northeast from the northeastern corner of the state of Washington and may have occurred in northeastern Washington.
2. Ursus idahoensis Merriam, grizzly bear.—Recorded by Merriam (1918: 54) from the Wallowa Mountains, Oregon. This form may have occurred in the Blue Mountains of southeastern Washington.
3. Vulpes fulva macroura Baird, red fox.—Reported from the Blue Mountains and northeastern Washington; no specimen recorded.
4. Canis lupus columbianus Goldman, wolf.—Possibly occurred in northeastern Washington in historic time, and perhaps is occasionally still found there.
5. Canis lupus irremotus Goldman, wolf.—Perhaps once occurred in southeastern Washington.
6. Mirounga angustirostris (Gill), sea elephant.—May occur as a casual wanderer off the coast of Washington. The home of this species is Lower California but a dead specimen was washed upon the shore of Prince of Wales Island, Alaska (Willett, 1943: 500).
7. Perognathus parvus laingi Anderson, Great Basin pocket mouse.—Probably present in the mountains east of Lake Osoyoos in Washington, but no specimens have yet been collected.
8. Synaptomys borealis artemisiae Anderson, northern lemming mouse.—Probably present in the Cascades of northern Okanogan County but no specimens have yet been obtained.
9. Antilocapra americana (Ord) subsp?, pronghorned antelope.—This species is restricted to North America and once ranged over much of the plains region of the western part of the continent. Taylor and Shaw (1929: 31) included the antelope in their list of Washington mammals with the statement "Now extirpated within State; the former range of the pronghorn included much of the plains country of eastern Washington." So far as known to me, there is no record by any of the early explorers of antelope killed or seen in what is now Washington. No bones of antelope have been discovered in caves in eastern Washington. Ogden (1909: 339) mentions an antelope killed in Oregon a day's journey south of The Dalles. This record is fairly close to Washington and indicates that antelope might have occurred at least in southeastern Washington in historic times.
1. The antelope (Antilocapra americana) has been introduced into the Yakima Valley by the state of Washington, Department of Game.
2. The elephant seal (Mirounga angustirostris) has been discovered off the coast of Washington (Seattle Times, April 15, 1947, p. 3).
3. The yellow-pine chipmunk of the Blue Mountains has been described as a new subspecies, Eutamias [Tamias] amoenus albiventris Booth, Murrelet, 28 (no. 1):7, 1947. Type locality Wickiup Spring, 23 miles west of Anatone, Asotin-Garfield County boundary.
| Allen, G. M. | ||
| 1920. | Dogs of the American aborigines. Harvard Col., Bull. Mus. Comp. Zoöl., 63:431-517, 12 pls. | |
| |
1941. |
Pigmy sperm whale in the Atlantic. Field Mus. Nat. Hist., zoöl.
ser., 27:17-36, 4 figs. in text. |
| Allen, J. A. | ||
| |
1893. |
On a collection of mammals from the San Pedro Martin region of
Lower California, with notes on other species, particularly of the
genus Sitomys. Bull. Amer. Mus. Nat. Hist., 5:181-202. |
| Anderson, R. M. | ||
| |
1932. |
Five new mammals from British Columbia. Ann. Rept. Canadian
Nat. Mus. for 1931, pp. 99-119, 1 pl. |
| Anderson, R. M., and Rand, A. L. | ||
| 1943A. | Variation in the porcupine (genus Erethizon) in Canada. Canadian Jour. Research, 21:292-309, 5 figs. in text. | |
| |
1943B. |
Status of the Richardson vole (Microtus richardsoni) in Canada.
Canadian Field-Nat., 57:106-107. |
| Bailey, V. | ||
| 1900. | Revision of the American voles of the genus Microtus. N. Amer. Fauna, 17:1-88, 5 pls., 17 figs. in text. | |
| 1918. | Wild animals of Glacier National Park: the mammals. U. S. Nat Park Serv. Bull., pp. 15-102, 21 pls., 18 figs. in text. | |
| |
1936. |
The mammals and life zones of Oregon. N. Amer. Fauna, 55:1-416,
51 pls., 102 figs. in text. |
| Baird, S. F. | ||
| |
1857. |
Mammals. Gen. Rept., Zoölogy of the Several Pacific R. R. Routes,
pp. xxv-xxxii, 1-737; xlviii + 757 pp., pls. xvii-lx, 35 figs. in text. |
| Bennett, L. J., English, P. F., and Watts, R. L. | ||
| |
1943. |
The food habits of the black bear in Pennsylvania. Jour. Mamm.,
24:25-31, 1 fig. in text. |
| Berry, E. W. | ||
| |
1931. |
A Miocene flora from Grand Coulee, Washington. In U. S. Geol.
Surv. Prof. Paper 170, pp. 31-42, pls. 11-13. |
| Birdseye, C. | ||
| |
1912. |
Some Common mammals of western Montana in relation to agriculture
and spotted fever. U. S. Dept. Agric., Farm. Bull. 484, pp.
1-46, 34 figs. in text. |
| Bonham, K. | ||
| |
1942. |
Records of harbor seals in lakes Washington and Union, Seattle.
Murrelet, 23:76. |
| Booth, E. S. | ||
| |
1945. |
A new red-backed mouse from Washington state. Murrelet,
26:27-28. |
| Bretz, J. H. | ||
| 1913. | Glaciation of the Puget Sound region. Washington Geol. Soc. Bull. 8, pp. 1-244, 24 pls., 27 figs. in text. | |
| [Pg 418]
|
1923. |
Glacial drainage on the Columbian Plateau. Bull. Geol. Soc. Amer.
vol. 34, pp. 573-608. 12 figs. in text. |
| Brockman, C. F. | ||
| |
1939. |
Items of interest from Mount Rainier. Murrelet, 20:70-71. |
| Broadbooks, H. E. | ||
| |
1939. |
Food habits of the vagrant shrew. Murrelet, 20:62-66. |
| Bryan, K. | ||
| |
1927. |
The "Palouse soil" problem. In U. S. Geol. Surv. Bull. 790, pp. 21-45,
pls. 4-7. |
| Bryant, M. D. | ||
| |
1945. |
Phylogeny of nearctic Sciuridae. Amer. Mid. Nat., vol. 33, pp. 257-390,
8 pls., 48 figs. in text. |
| Cottam, C., and Williams, C. S. | ||
| |
1943. |
Speed of some wild mammals. Jour. Mamm., 24:262.
|
| Couch, L. K. | ||
| 1925. | Storing habits of Microtus townsendii. Jour. Mamm., 6:200-201. | |
| 1927. | Migration of the Washington black-tailed jack rabbit. Jour. Mamm., 8:313-314. | |
| |
1930. |
Notes on the pallid yellow-bellied marmot. Murrelet, 11 (No. 2):2-6,
3 figs. |
| Cowan, I. M. | ||
| 1936A. | Distribution and variation in deer (genus Odocoileus) of the Pacific coastal region of North America. California Fish and Game, 22:155-246. figs. 51-63. | |
| 1936B. | Nesting habits of the flying squirrel Glaucomys sabrinus. Jour. Mamm., 17:58-60. | |
| 1937. | A new race of Peromyscus maniculatus from British Columbia. Proc. Biol. Soc. Washington, 50:215-216. | |
| 1938. | Geographic distribution of color phases of the red fox and black bear in the Pacific Northwest. Jour. Mamm., 19:202-206, 1 map. | |
| 1939. | The vertebrate fauna of the Peace River district of British Columbia. Occ. Papers British Columbia Prov. Mus., No. 1, pp. 1-102. | |
| |
1940. |
Distribution and variation in the native sheep of North America.
Amer. Mid. Nat., 24:505-580, 4 pls., 1 map. |
| Cowan, I. M., and Hatter, J. | ||
| |
1940. |
Two mammals new to the known fauna of British Columbia. Murrelet,
21:9. |
| Crabb, W. D. | ||
| 1944. | Growth, development, and seasonal weights of spotted skunks. Jour.
Mamm., vol. 25. pp. 213-221, 2 pls. |
|
| Culver, H. E. | ||
| 1936. | The geology of Washington: part 1, General features of Washington
geology (with map). Washington State Div. Geol., Bull. 32, pp. 1-70. |
|
| Dale, F. H. | ||
| 1940. | Geographic variation in the meadow mouse in British Columbia
and southeastern Alaska. Jour. Mamm., 21:332-340. |
|
| Dalquest, W. W. | ||
| [Pg 419] | 1938. | Bats in the state of Washington. Jour. Mamm., 19:211-213. |
| 1940. | Bats in the San Juan Islands, Washington. Murrelet, 21:4-5. | |
| 1941A. | Ecologic relationships of four small mammals in western Washington. Jour. Mamm., 22:170-173. | |
| 1941B. | Distribution of cottontail rabbits in Washington State. Jour. Wildlife Management, 5:408-411, 1 fig. | |
| 1942. | Geographic variation in northwestern snowshoe hares. Jour. Mamm., 23:166-183, 2 figs. in text. | |
| 1943. | Seasonal distribution of the hoary bat along the Pacific Coast. Murrelet, 24:20-24, 2 figs. | |
| 1944. | The moulting of the wandering shrew. Jour. Mamm., 25:146-148.
|
|
| Dalquest, W. W., and Burgner, R. L. | ||
| 1941. | The shrew-mole of western Washington. Murrelet, 22:12-14.
|
|
| Dalquest, W. W., and Orcutt, D. R. | ||
| 1942. | The biology of the least shrew-mole, Neurotrichus gibbsii minor.
Amer. Mid. Nat., 27:387-401, 4 figs. in text |
|
| Dalquest, W. W., and Scheffer, V. B. | ||
| 1942. | The origin of the mima mounds of western Washington. Jour. Geol., 50:68-84, 8 figs. in text. | |
| 1944. | Distribution and variation in pocket gophers, Thomomys talpoides,
in the state of Washington. Amer. Nat., part 1, 78:308-333, part 2,
78:423-550, 8 figs. in text. |
|
| Davis, W. B. | ||
| 1939. | The Recent Mammals of Idaho. The Caxton Printers, Caldwell,
Idaho, pp. 1-400. |
|
| Dice, L. R. | ||
| 1919. | The mammals of southeastern Washington. Jour. Mamm., 1:10-22, 2 pls. | |
| 1939. | Variation in the deer-mouse (Peromyscus maniculatus) of the Columbia Basin of southeastern Washington and adjacent Idaho and Oregon. Contrib. Lab. Vert. Genetics, Univ. Michigan, 12:1-22, 1 map. | |
| 1940. | Speciation in Peromyscus. Amer. Nat., 74:289-298.
|
|
| Dobzhansky, T. G. | ||
| 1937. | Genetics and the origin of species. Columbia Univ. Press, New York,
pp. xvi + 364. |
|
| Doutt, J. K. | ||
| 1942. | A review of the genus Phoca. Ann. Carnegie Mus., 29:61-125, 14 pls.,
11 figs. in text. |
|
| Edge, E. R. | ||
| 1934. | Burrows and burrowing habits of the Douglas ground squirrel. Jour.
Mamm., 15:189-193, 1 pl., 1 fig. in text. |
|
| Edson, J. M. | ||
| 1916. | Wild animals of Mount Baker. Mountaineer, 9:51-57. | |
| 1933. | A visitation of weasels. Murrelet, 14:76-77. | |
| 1935. | Yellow-bellied marmot out of bounds. Jour. Mamm., 16:68.
|
|
| [Pg 420] Ellerman, J. R. | ||
| 1940-1941. | The families and genera of living rodents. Jarrold and sons,
Ltd., Norwich, vol. 1 (1940), xxvi + 689, 189 figs. in text; vol. 2 (1941)
xii + 690, 49 figs, in text. |
|
| Elliot, D. G. | ||
| 1899. | Catalogue of mammals from the Olympic Mountains, Washington,
with descriptions of new species. Field Columb. Mus. Publ., 32,
zoöl. ser., 1:241-276, pls. 41-62, numerous figs. in text. |
|
| Engels, W. L. | ||
| 1936. | Distribution of races of the brown bat (Eptesicus) in western North
America. Amer. Mid. Nat., 17:653-660, 1 fig. in text. |
|
| Engler, C. H. | ||
| 1943. | Carnivorous activities of big brown and pallid bats. Jour. Mamm.,
24:96-97. |
|
| English, E. H. | ||
| 1930. | Mammals of Austin Pass, Mount Baker. Mazama, 12:34-43, illustrated.
|
|
| Finley, W. L. | ||
| 1919. | With the birds and animals of Rainier. Mazama, 5:319-326, pls. | |
| 1925. | Cougar kills a boy. Jour. Mamm., 6:197-199.
|
|
| Fisher, E. M. | ||
| 1939. | Habits of the southern sea otter. Jour. Mamm., 20:21-36.
|
|
| Flahaut, M. R. | ||
| 1939. | Unusual location of hibernating jumping mice. Murrelet, 20:17-18,
1 fig. |
|
| Flint, R. F. | ||
| 1935. | Glacial features of the southern Okanogan region. Bull. Geol. Soc. Amer., vol. 46, pp. 169-194, pls. 13-18, 2 figs. in text. | |
| 1937. | Pleistocene drift border in eastern Washington. Bull. Geol. Soc. Amer., vol. 48, pp. 203-232, 5 pls., 1 fig. in text. | |
| 1938. | Summary of late-Cenozoic geology of southeastern Washington.
Amer. Jour. Science, ser. 5, vol. 35, pp. 223-230. |
|
| Francis, E. | ||
| 1922. | The occurrence of tularaemia in nature as a disease of man. U. S.
Publ. Health Serv., Hyg. Lab. Bull. 130:1-8. |
|
| Furlong, E. L. | ||
| 1906. | The exploration of Samwel Cave. Amer. Jour. Sci., ser. 4, 22:235-247,
3 figs. in text. |
|
| Gibbs, G. | ||
| See Suckley, G., and Gibbs, G. 1860. |
||
| Gidley, J. W., and Gazin, C. L. | ||
| 1933. | New Mammalia in the Pleistocene fauna from Cumberland Cave. Jour. Mamm., 14:343-357. | |
| 1938. | The Pleistocene vertebrate fauna from Cumberland Cave, Maryland.
U. S. Nat. Mus. Bull. 171, pp. i-vi + 1-99, pls. 1-10, 50 figs. |
|
| [Pg 421] Goldman, E. A. | ||
| 1941. | Remarks on voles of the genus Lemmiscus, with one described as new. Proc. Biol. Soc. Washington, vol. 54, pp. 69-71. | |
| 1943. | Two new races of the puma. Jour. Mamm., 24:228-231.
|
|
| Gray, J. A. Jr. | ||
| 1943. | Rodent populations in the sagebrush desert of the Yakima Valley,
Washington. Jour. Mamm., 24:191-193. |
|
| Greenwood, W. H., Newcomb, F. C. and Fraser, C. M. | ||
| 1918. | Sea-lion question in British Columbia. Contr. Canadian Biol., Sessional
Paper No. 38a., pp. xv + 52, 36 photographs. |
|
| Grinnell, J. | ||
| 1923. | The burrowing rodents of California as agents in soil formation.
Jour. Mamm., 4:137-149, pls. 13-15. |
|
| Grinnell, J., and Dixon, J. | ||
| 1919. | Natural history of the ground squirrels of California. California
State Comm. Hort., Monthly Bull., 7:595-708, 5 col. pls., 30 figs. in
text. |
|
| Grinnell, J., Dixon, J., and Linsdale, J. M. | ||
| 1937. | Fur-bearing mammals of California. Univ. California Press, Berkeley,
2 vols., pp. xii + xiv + 777, 13 col. pls., 345 figs. in text. |
|
| Grinnell, H. W. | ||
| 1918. | A synopsis of the bats of California. Univ. California Publs. Zoöl.,
17:223-404, pls. 14-24, 24 figs. in text. |
|
| Hall, E. R. | ||
| 1928. | Notes on the life history of the sagebrush meadow mouse (Lagurus). Jour. Mamm., 9:201-204. | |
| 1936. | Mustelid mammals from the Pleistocene of North America with systematic notes on some Recent members of the genera Mustela, Taxidea and Mephitis. Carnegie Inst. Washington. Publ. 473, pp. 41-119, 5 pls., 6 figs. | |
| 1938A. | Variation among insular mammals of Georgia Strait, British Columbia. Amer. Nat., 72:453-463, 2 figs. | |
| 1938B. | Gestation period in the long-tailed weasel. Jour. Mamm., 19:249-250. | |
| 1944. | A new genus of American Pliocene badger, with remarks on the relationships of badgers of the northern hemisphere. Carnegie Inst. Washington, Publ. 551, pp. 9-23, 2 pls., 2 figs. in text. | |
| 1945. | Four new ermines from the Pacific Northwest. Jour. Mamm., 26:75-85,
1 fig. |
|
| Hamilton, W. J. | ||
| 1934. | The life history of the rufescent woodchuck, Marmota monax rufescens Howell. Carnegie Mus. Ann., 23:85-178, 6 pls., 9 figs. | |
| 1940. | The biology of the smoky shrew (Sorex fumeus fumeus Miller). Zoologica,
25:473-492, 4 pls. |
|
| Hanson, H. P. | ||
| 1939. | Pollen analysis of a bog near Spokane, Washington. Bull. Torrey Bot. Club., 66:215-220, 1 fig. in text. | |
| 1940. | Paleoecology of a montane peat deposit at Bonaparte Lake, Washington. Northwest Science, 14:60-69. | |
| [Pg 422] | 1941A. | Paleoecology of a peat deposit in west central Oregon. Amer. Jour. Botany, 28:206-212. 1 fig. in text. |
| 1941B. | Further studies of Post Pleistocene bogs in the Puget lowlands of Washington. Bull. Torrey Bot. Club., 68:133-148. 2 figs. in text. | |
| 1941C. | A pollen study of Post Pleistocene lake sediments in the Upper
Sonoran Life Zone of Washington. Amer. Jour. Sci., 239:503-522,
1 fig. |
|
| Hartman, C. | ||
| 1923. | Breeding habits, development, and birth of the opossum. Smithsonian
Rept. for 1921, pp. 347-363, 10 pls. |
|
| Hatt, R. T. | ||
| 1927. | Notes on the ground-squirrel, Callospermophilus. U. Michigan, Mus.
Zoöl., Occ. Papers 185, pp. 1-22. 1 pl., 2 figs. |
|
| Hay, O. P. | ||
| 1921. | Descriptions of species of Pleistocene Vertebrata, types or specimens
of most of which are preserved in the United States National Museum.
Proc. U. S. Nat. Mus., 59:599-642, pls. 116-124. |
|
| Hinton, M. A. C. | ||
| 1926. | Monograph of the voles and lemmings (Microtinae) living and extinct.
British Mus. Nat. Hist., London, pp. xvi + 1-488, 15 pls., 110
figs. in text. |
|
| Howell, A. B. | ||
| 1927A. | On the faunal position of the Pacific Coast of the United States. Ecology, 8:18-26. | |
| 1927B. | Revision of the American lemming mice (Genus Synaptomys).
N. Amer. Fauna, 50:1-38, 2 pls., 11 figs. |
|
| Howell, A. H. | ||
| 1901. | Revision of the skunks of the genus Chincha. N. Amer. Fauna, 20:1-62, 8 pls. | |
| 1906. | Revision of the skunks of the genus Spilogale. N. Amer. Fauna, 26:1-55, 10 pls. | |
| 1914. | Revision of the American harvest mice (Genus Reithrodontomys). N. Amer. Fauna, 36:1-97, 7 pls., 6 figs. | |
| 1915. | Revision of the American marmots. N. Amer. Fauna, 37:1-80, 15 pls., 3 figs. | |
| 1918. | Revision of the American flying squirrels. N. Amer. Fauna, 44:1-64, 7 pls., 4 figs. | |
| 1924. | Revision of the American pikas (Genus Ochotona). N. Amer. Fauna, 47:1-57, 6 pls., 4 figs. | |
| 1929. | Revision of the American chipmunks (Genera Tamias and Eutamias). N. Amer. Fauna, 52:1-157, 10 pls., 9 figs. | |
| 1938. | Revision of the North American ground squirrels with a classification of the North American Sciuridae. N. Amer. Fauna, 56:1-256, 32 pls., 10 figs. | |
| 1939. | |Review of "The Recent Mammals of Idaho" by William B. Davis|.
Jour. Mamm., 20:389-390. |
|
| Huxley, J. S. (edited by). | ||
| 1940. | The new systematics. Oxford Univ. Press, pp. viii + 583, 55 figs. in
text. |
|
| Jackson, H. H. T. | ||
| 1915. | A review of the American moles. N. Amer. Fauna, 38:1-100, 6 pls., 27 figs. | |
| 1928. | A taxonomic review of the American long-tailed shrews (Genera Sorex and Microsorex). N. Amer. Fauna, 51:i-vi + 1-238, 13 pls., 24 figs. | |
| [Pg 423] | 1944. | Big-game resources of the United States, 1937-1942. U. S. Dept.
Interior, Fish and Wildlife Research Rept. 8:1-56, 31 figs. in text. |
| Johnson, D. H. | ||
| 1943. | Systematic review of the chipmunks (genus Eutamias) of California.
Univ. California Publs. Zoöl., 48:63-148, 6 pls., 12 figs. |
|
| Jones, G. N. | ||
| 1936. | A botanical survey of the Olympic Peninsula, Washington. Univ. Washington Publs. Biol., 5:1-286, 9 pls. | |
| 1938. | The flowering plants and ferns of Mount Rainier. Univ. Washington
Publs. Biol., 7:1-192, 9 pls. |
|
| Kellogg, L. | ||
| 1912. | Pleistocene rodents of California. Univ. California Publs., Bull.
Dept. Geol., 7:151-168, 16 figs. in text. |
|
| Kellogg, W. H. | ||
| 1935. | Rodent plague in California. Jour. Amer. Med. Assoc. 105:856-859.
|
|
| Koford, C. B. | ||
| 1938. | Microsorex hoyi washingtoni in Montana. Jour. Mamm., 19:372.
|
|
| Larrison, E. J. | ||
| 1942. | Pocket gophers and ecological succession in the Wenas region of Washington. Murrelet, 23:34-41, 2 figs. | |
| 1943. | Feral coypus in the Pacific Northwest. Murrelet, 24:3-9, 1 fig.
|
|
| Livezey, R., and Evenden, F., Jr. | ||
| 1943. | Notes on the western red fox. Jour. Mamm., 24:500-501.
|
|
| Matthew, W. D. | ||
| 1902. | List of the Pleistocene Fauna from Hay Springs, Nebraska. Bull.
Amer. Mus. Nat. Hist., 16:317-322. |
|
| McCoy, G. W. | ||
| 1911. | A plague-like disease of rodents. U. S. Publ. Health and Marine-Hosp.
Serv., Pub. Health Bull., 43:53-71. |
|
| McMurry, F. B. | ||
| 1940. | Mink observations at Packwood Lake. Murrelet, 21:47.
|
|
| Merriam, C. H. | ||
| 1892. | The geographic distribution of life in North America. Ann. Rept. Smiths. Inst. for 1891, pp. 365-415. | |
| 1918. | Review of the grizzly and big brown bears of North America (genus
Ursus) with description of a new genus, Vetularctos. N. Amer. Fauna,
41:1-136, 16 pls. |
|
| Merriam, J. C. | ||
| 1911. | The fauna of Rancho La Brea, Pt. 1, Occurrence. Mem. Univ. California,
1:197-213, pls. 19-23. |
|
| Merriam, J. C., and Bulwalda, J. P. | ||
| 1917. | Age of strata referred to the Ellensburg Formation in the White
Bluffs of the Columbia River. Univ. California Publs. Bull. Dept.
Geol., 10:255-266, 1 pl. |
|
| Meyer, K. F. | ||
| 1936. | The sylvatic plague committee. Amer. Jour. Pub. Health, 26:961-969.
|
|
| [Pg 424] Miller, A. H. | ||
| 1942. | Habitat selection among higher vertebrates and its relation to intraspecific
variation. Amer. Nat., 76:25-35. |
|
| Miller, G. S., Jr. | ||
| 1897. | Revision of the North American bats of the family Vespertilionidae. N. Amer. Fauna, 13:1-140, 3 pls., 40 figs. | |
| 1924. | List of North American Recent mammals, 1923. U. S. Nat. Mus.
Bull. 128, pp. xvi + 673. |
|
| Miller, G. S., Jr., and Allen, G. M. | ||
| 1928. | American bats of the genera Myotis and Pizonyx. U. S. Nat. Mus.
Bull. 144, pp. 1-218, 1 pl., 1 fig., 13 maps. |
|
| Moore, A. W. | ||
| 1933. | Food habits of the Townsend and coast moles. Jour. Mamm., 14:36-40, 1 pl. | |
| 1940. | Wild animal damage to seed and seedlings on cut-over Douglas fir lands of Oregon and Washington. U. S. Dept. Agri., Tech. Bull. 706, pp. 1-28, 14 figs. in text. | |
| 1942. | Shrews as a check on Douglas fir regeneration. Jour. Mamm., 23:37-41, 1 pl. | |
| 1943. | Notes on the sage mouse in eastern Oregon. Jour. Mamm., 24:188-191.
|
|
| Mossman, H. W., Lowlah, J. W., and Bradley, J. A. | ||
| 1932. | The male reproductive tract of the Sciuridae. Amer. Jour. Anat.,
vol. 51, pp. 89-155, 7 pls., 16 figs. |
|
| Murie, O. J., and Murie, A. | ||
| 1931. | Travels of Peromyscus. Jour. Mamm., 12:200-209, 1 fig. | |
| 1932. | Further notes on travels of Peromyscus. Jour. Mamm., 13:78-79.
|
|
| Nelson, E. W. | ||
| 1909. | The rabbits of North America. N. Amer. Fauna, 29:1-314, 13 pls.,
19 figs. |
|
| Nichols, D. G. | ||
| 1944. | Further consideration of American house mice. Jour. Mamm., 25:82-84.
|
|
| Ogden, P. S. | ||
| 1909. | The Peter Skene Ogden Journals. Quart. Oregon Hist. Soc., 10:331-365.
|
|
| Orr, R. T. | ||
| 1940. | The rabbits of California. Occas. Papers, California Acad. Sci., 19:1-227,
10 pls., 30 figs. in text. |
|
| Osgood, W. H. | ||
| 1900. | Revision of the pocket mice of the genus Perognathus. N. Amer. Fauna, 18:1-72, 4 pls., 15 figs. | |
| 1909. | Revision of the mice of the American genus Peromyscus. N. Amer. Fauna, 28:1-285, 8 pls., 12 figs. | |
| 1943. | The mammals of Chile. Field Mus. Nat. Hist., Zoöl. Ser., vol. 30,
publ. 542, pp. 1-268, 33 figs., 10 maps. |
|
| Pardee, J. T., and Bryan, K. | ||
| 1926. | Geology of the Latah formation in relation to the lavas of the
Columbia Plateau near Spokane, Washington. In U. S. Geol. Surv.
Prof. Paper 140, pp. 1-16, 7 pls., 1 fig. in text. |
|
| [Pg 425] Paschall, S. E. | ||
| 1920. | Mountain beaver (Haplodontia or Aplodontia). Mountaineer, 13:40-43,
1 illustration. |
|
| Perry, M. L. | ||
| 1939. | Notes on a captive badger. Murrelet, 20:49-53, 1 fig.
|
|
| Piper, C. V. | ||
| 1906. | Flora of the state of Washington. Cont. U. S. Nat. Herb., 11:1-637,
22 pls., 1 map in pocket. |
|
| Pope, C. H. | ||
| 1944. | Attainment of sexual maturity in raccoons. Jour. Mamm., 25:91.
|
|
| Racey, K., and Cowan, I. M. | ||
| 1935. | Mammals of the Alta Lake region of southwestern British Columbia.
Ann. Rept. Prov. Mus. British Columbia 1935, pp. H15-H27, 5 pls.,
1 fig. in text. |
|
| Rand, A. L. | ||
| 1943. | Canadian forms of the meadow mouse (Microtus pennsylvanicus).
Canadian Field Nat., 57:115-123. |
|
| Rhoads. S. N. | ||
| 1897. | A revision of the west American flying squirrels. Proc. Acad. Nat.
Sci. Philadelphia 1897, pp. 314-327. |
|
| Russell, I. C. | ||
| 1893. | A geological reconnaisance in central Washington. Bull. U. S. Geol.
Surv. 108, pp. 1-108, 12 pls., 8 figs. in text. |
|
| Sampson, A. | ||
| 1906. | Wild animals of the Mt. Rainier National Park. Sierra Club Bull.,
6:32-38. |
|
| Scheffer, T. H. | ||
| 1922. | American moles as agriculture pests and as fur producers. U. S. Dept. Agri., Farm. Bull. 1247 (revised 1927), pp. 1-21, 18 figs. | |
| 1928. | Precarious status of the seal and sea-lion on our northwest coast. Jour. Mamm., 9:10-16. | |
| 1929. | Mountain beavers in the Pacific Northwest: their habits, economic status and control. U. S. Dept. Agric., Farm. Bull. 1598, pp. 1-18, 13 figs. | |
| 1930. | Bat matters. Murrelet, 11, (no. 2):11-13, 2 figs. | |
| 1931. | Habits and economic status of the pocket gophers. U. S. Dept. Agric., Tech. Bull. 224, pp. 1-26, 8 pls. | |
| 1932. | Weasels and snakes in gopher burrows. Murrelet, 13:54. | |
| 1933. | Breeding of the Washington varying hare. Murrelet, 14:77-78. | |
| 1938A. | Pocket mice of Oregon and Washington in relation to agriculture. U. S. Dept. Agric., Tech. Bull. 608, pp. 1-16, 6 pls., 1 fig. | |
| 1938B. | Breeding records of Pacific Coast pocket gophers. Jour. Mamm., 19:220-224. | |
| 1941. | Ground squirrel studies in the Four-rivers Country, Washington.
Jour. Mamm., 22:270-279, 2 pls. |
|
| Scheffer, T. H., and Sperry, C. C. | ||
| 1931. | Food habits of the Pacific harbor seal, Phoca richardii. Jour.
Mamm., 12:214-226. |
|
| [Pg 426] Scheffer, V. B. | ||
| 1938. | Notes on the wolverine and fisher in the state of Washington. Murrelet, 19:8-10, 2 figs. | |
| 1939. | Fur seal in Willapa Harbor. Murrelet, 20:43, 1 fig. | |
| 1940A. | A newly located herd of Pacific white-tailed deer. Jour. Mamm., 21:271-282, 1 pl. | |
| 1940B. | The sea otter on the Washington coast. Pacific Northwest Quart., October, 1940, pp. 369-388, 5 figs. | |
| 1941. | Wolverine captured in Okanogan County, Washington. Murrelet, 22:37, 1 fig. | |
| 1942. | A list of the marine mammals of the west coast of North America. Murrelet, 23:42-47. | |
| 1943. | The opossum settles in Washington State. Murrelet, 24:27-28.
|
|
| Scheffer, V. B., and Dalquest, W. W. | ||
| 1939. | Present distribution of the Douglas ground squirrel in Washington.
Murrelet, 20:44. |
|
| Scheffer, V. B., and Slipp, J. W. | ||
| 1944. | The harbor seal in Washington State. Amer. Mid. Nat., 32:373-416,
17 figs. |
|
| Schultz, L. P., and Rafn, A. M. | ||
| 1936. | Stomach contents of fur seals taken off the coast of Washington.
Jour. Mamm., 17:13-15. |
|
| Schwartz, E., and Schwartz, H. K. | ||
| 1943. | The wild and commensal stocks of the house mouse, Mus musculus
Linnaeus. Jour. Mamm., 24:59-72. |
|
| Scott, W. B. | ||
| 1937. | A history of land mammals in the western hemisphere. Macmillan
Co., New York, pp. xiv + 786, 420 figs. in text. |
|
| Shaw, W. T. | ||
| 1919. | The Columbian ground squirrel. (Citellus columbianus columbianus). California State Comm. Hort., Monthly Bull. 7, pp. 710-720, col. pl. vi. figs. 31-43. | |
| 1924A. | Alpine life of the heather vole (Phenacomys olympicus). Jour. Mamm., 5:12-15, pls. 2-4. | |
| 1924B. | The home life of the Columbian ground squirrel. Canadian Field Nat., 38:128-130, 4 figs. | |
| 1925A. | The seasonal differences of north and south slopes in controlling the activities of the Columbian ground squirrel. Ecology, 6:157-162, 2 figs. in text. | |
| 1925B. | Duration of the aestivation and hibernation of the Columbian ground squirrel (Citellus columbianus) and sex relations of the same. Ecology, 6:75-81, 2 figs. | |
| 1925C. | Breeding and development of the Columbian ground squirrel. Jour. Mamm., 6:106-113, pls. 11-14. | |
| 1925D. | The Columbian ground squirrel as a handler of earth. The Sci. Monthly, 20:483-490, 8 figs. in text. | |
| 1925E. | The food of ground squirrels. Amer. Nat., 59:250-264, 5 figs. in text. | |
| 1925F. | A life history problem and a means for its solution. Jour. Mamm., 6:157-162, pls. 15-17. | |
| 1925G. | Observations on the hibernation of ground squirrels. Jour. Agric. Research, 31:761-769, 7 figs. in text. | |
| 1925H. | The hibernation of the Columbian ground squirrel. Canadian Field Nat., 39:56-61, 79-82, 11 figs. | |
| [Pg 427] | 1925I. | The marmots of Hannegan Pass. Nat. Hist., 25:169-177, 6 unnumbered photographs. |
| 1926. | Age of the animal and slope of the ground surface, factors modifying the structure of hibernation dens of ground squirrels. Jour. Mamm., 7:91-96, 1 pl., 3 figs. | |
| 1930. | The lemming mouse in North America and its occurrence in the state of Washington. Murrelet, 11 (No. 2):7-10, 2 figs. | |
| 1944. | Brood nests and young of two western chipmunks in the Olympic
Mountains of Washington. Jour. Mamm., 25:274-284, 4 pls. 1 fig. |
|
| Sinclair, W. J. | ||
| 1903. | A preliminary account of the exploration of the Potter Creek Cave,
Shasta county, California. Science, 17:708-712. |
|
| Slipp, J. W. | ||
| 1942. | Nest and young of the Olympic dusky shrew. Jour. Mamm.,
23:211-212. |
|
| Smith, G. O. | ||
| 1903. | Ellensburg Folio (No. 86). U. S. Geol. Surv., Geol. Atlas of the U. S.
|
|
| Sperry, C. C. | ||
| 1941. | Food habits of the coyote. U. S. Dept. Interior, Wildlife Research
Bull. 4, pp. 1-70, 3 pls., 3 figs. in text. |
|
| Stager, K. E. | ||
| 1943. | Notes on the resting place of Pipistrellus hesperus. Jour. Mamm.
24:266-267. |
|
| St. John, H. | ||
| 1937. | Flora of southeastern Washington. Student Book Corp., Pullman,
Washington, pp. xxv + 531, front (map) illus. |
|
| St. John, H., and Jones, G. N. | ||
| 1928. | An annotated catalogue of the vascular plants of Benton County,
Washington. Northwest Science, 2:73-93, illustrated. |
|
| Stock, C. | ||
| 1918. | The Pleistocene fauna of Hawver Cave. Univ. California Publs. Bull. Dept. Geol., 10:461-515, 32 figs. in text. | |
| 1930. | Rancho La Brea: a record of Pleistocene life in California. Los
Angeles Mus. Publ. 1, pp. 1-82, 27 figs. in text. |
|
| Suckley, G., and Gibbs, G. | ||
| 1860. | Report upon the mammals collected on the survey. Repts. expls.
and surveys ... route for a railroad, from the Mississippi River
to the Pacific Ocean, ... 1853-1855, Pacific R. R. Rept., vol. 12,
book 2, pt. 3, zoöl. rept., No. 2, chap. 3, pp. 107-139, 6 pls. (for chaps.
1-3). |
|
| Sumner, F. B. | ||
| 1917A. | The role of isolation in the formation of a narrowly localized race of deer-mice (Peromyscus). Amer. Nat., 51:173-185. | |
| 1917B. | Several color "mutations" in mice of the genus Peromyscus. Genetics, 2:291-300, 1 fig. in text. | |
| 1932. | Genetic, distributional and evolutionary studies of the subspecies of
deer mice (Peromyscus). Bibliographia Genetica, 9:1-106, 24 figs. in
text. |
|
| [Pg 428] Svihla, A. | ||
| 1932. | A comparative life history study of the mice of the genus Peromyscus. Univ. Michigan Mus. Zoöl., Misc. Publs. 24, pp. 1-39. | |
| 1933. | Notes on the deer-mouse. Peromyscus maniculatus oreas (Bangs). Murrelet, 14:13-14. | |
| 1934. | The mountain water shrew. Murrelet, 15:44-45. | |
| 1936A. | Development and growth of Peromyscus maniculatas oreas. Jour. Mamm., 17:132-137, 2 figs. | |
| 1936B. | Notes on the hibernation of a western chipmunk. Jour. Mamm., 17:289-290. | |
| 1939. | Breeding habits of Townsend's ground squirrel. Murrelet, 20:6-10.
|
|
| Svihla, A., and Svihla, R. D. | ||
| 1931. | Mink feeding on clams. Murrelet, 12:22. | |
| 1933. | Notes on the jumping mouse Zapus trinotatus trinotatus Rhoads. Jour. Mamm., 14:131-134. | |
| 1940. | Annotated list of the mammals of Whitman County, Washington.
Murrelet, 21:53-58. |
|
| Svihla, R. D. | ||
| 1936. | Breeding and young of the grasshopper mouse (Onychomys leucogaster
fuscogriseus). Jour. Mamm., 17:172-173. |
|
| Tate, G. H. H. | ||
| 1942. | Review of the Vespertilionine bats, with special attention to genera
and species of the Archbold collections. Bull. Amer. Mus. Nat. Hist.,
80:221-297, 4 figs. in text. |
|
| Taylor, W. P. | ||
| 1918. | Revision of the rodent genus Aplodontia. Univ. California Publs. Zoöl., 17:435-504, pls. 25-29, 16 figs. in text. | |
| 1920A. | The wood rat as a collector. Jour. Mamm., 1:91-92. | |
| 1920B. | A novel nesting place of the red-backed mouse. Jour. Mamm., 1:92. | |
| 1921. | Some birds and mammals of Mount Rainier. The Mountaineer, 14:27-35, illustrated. | |
| 1922. | A distributional and ecological study of Mount Rainier, Washington.
Ecology, 3:214-236, 4 figs. in text. |
|
| Taylor, W. P., and Shaw, W. T. | ||
| 1927. | Mammals and birds of Mount Rainier National Park. U. S. Dept. Interior, Nat. Park Service, U. S. Govt. Printing Office, Washington, D. C., pp. 1-249. | |
| 1929. | Provisional list of the land mammals of the state of Washington.
Occ. Papers Charles R. Conner Mus. No. 2, pp. 1-32. |
|
| Timofeeff-Ressovsky, N. W. | ||
| 1932. | The genographical work with Epilachna chrysomelina, etc. Proc. 6th International Congress Genetics, 2:230. | |
| 1940. | Mutations and geographical variation, in The New Systematics, ed.
J. S. Huxley, Oxford Univ. Press, pp. 73-136, 38 figs. in text. |
|
| Townsend, C. H. | ||
| 1887. | Field-notes on the mammals, birds and reptiles of northern California.
Proc. U. S. Nat. Mus., 10:159-241. 1 pl., 4 unnumbered figs. in text. |
|
| [Pg 429] Townsend, J. K. | ||
| 1839. | Narrative of a journey across the Rocky Mountains, to the Columbia
River ... Reprint in Early Western Travels, 1748-1846, R. G.
Thwaits, ed., A. H. Clark Co., Cleveland, 21:107-369, 1 pl., 1905. |
|
| Weaver, C. E. | ||
| 1937. | Tertiary stratigraphy of western-Washington and northwestern Oregon.
Univ. Washington Publs. Geol., 4:1-266, 15 pls. |
|
| Whitlow, W. B., and Hall, E. R. | ||
| 1933. | Mammals of the Pocatello Region of southeastern Idaho. Univ.
California Publs. Zoöl., 40:235-275, 3 figs. |
|
| Wight, H. M. | ||
| 1928. | Food habits of the Townsend's mole, Scapanus townsendii (Bachman). Jour. Mamm., 9:19-33. | |
| Willett, G. | ||
| 1943. | Elephant seal in southeastern Alaska. Jour. Mamm., 24:500.
|
|
| Wilson, R. W. | ||
| 1933A. | Pleistocene mammalian fauna from the Carpinteria asphalt. Carnegie Inst. Washington, Publ., 440:59-76. | |
| 1933B. | A rodent fauna from the later Cenozoic beds of southwestern Idaho.
Carnegie Inst. Washington, Publ., 440:117-135, 2 pls., 8 figs. in text. |
|
| Wright, S. | ||
| 1932. | The roles of mutation, interbreeding, crossbreeding and selection in
evolution. Proc. 6th Internat. Congress Genetics, 1:356-366, figs. |
|
| Young, S. P., and Goldman, E. A. | ||
| 1944. | The wolves of North America. Amer. Wildlife Inst., Washington,
D. C., xx + 636 pp., 131 pls., 15 figs. in text. |
|
| Zimmerman, R. S. | ||
| 1943. | A coyote's speed and endurance. Jour. Mamm., 24:400.
|
|
| Transmitted August 14, 1947. | ||
Obvious typographical errors have been silently corrected, but other variations in spelling and punctuation remain unchanged.