
Transcriber Note
All corrections listed in the "Errata" have been made in the text. Linking text for the plates has been added to the "Contents".
THE ORNITHOSAURIA:
AN ELEMENTARY STUDY
OF
THE BONES OF PTERODACTYLES.
PRINTED BY G. J. CLAY, M.A.
AT THE UNIVERSITY PRESS.
THE ORNITHOSAURIA:
AN ELEMENTARY STUDY
OF
THE BONES OF PTERODACTYLES.
MADE FROM FOSSIL REMAINS FOUND IN THE
CAMBRIDGE UPPER GREENSAND,
AND
ARRANGED IN THE WOODWARDIAN MUSEUM OF THE
UNIVERSITY OF CAMBRIDGE
BY
HARRY GOVIER SEELEY,
OF ST. JOHN'S COLLEGE, CAMBRIDGE.
WITH TWELVE PLATES.
"And when the appointed end comes, they lie not dishonoured in
forgetfulness,"—Xenoph. Memor. Book 2, c. 1, § 83.
DEIGHTON, BELL, AND CO.
LONDON: BELL AND DALDY.
1870.
[All Rights reserved.]
The expense of printing this volume has been defrayed out of the Funds of the Syndics of the University Press; and Professor Sedgwick hereby expresses his grateful thanks to them for this great favour.
PREFACE.
This memoir is a portion of the Catalogue of the Woodwardian Museum which has been made at Professor Sedgwick's request and at his cost. When the Professor laid upon me his commands to prepare a Catalogue of the Museum, it was planned in three distinct works. First, a series of indexes to the specimens in the great divisions into which the Museum is arranged; secondly, a series of memoirs upon the orders and classes of animals concerning which new knowledge is given by fossils in the Museum; and, thirdly, memoirs descriptive of those species contained in the arranged collections which are at present unknown in scientific writings.
For the convenience of students the Catalogue is made in parts. The Syndics of the University Press printed last autumn as an example of the "Indexes to the Museum," an Index to the Pterodactyles, Birds, and Reptiles from the Secondary Strata. And this memoir is an example of the second kind of Catalogue, which explains the structures of the Pterodactyles of the Cambridge Greensand. In its progress « viii » questions have arisen which necessitated an examination both of the method, of research in comparative anatomy and of its results in classification. And in so far as the views here advanced differ from those commonly taught, the discrepancy is due to the writer's imperfect faith in the results of the inductive method of research, as commonly used by modern writers on Palæontology. It has not been consistent with the plan of this little work to do more than scatter through it a few hints upon method, a subject which will more fitly be discussed with a part of the Catalogue which forms a synopsis of the osteology of the fossil animals usually named Reptiles. The views here urged have however but little of novelty. The name Ornithosauria was proposed by the distinguished naturalist Prince Charles Bonaparte in 1838. The group as an order was recognized by Von Meyer in 1830. The affinities of the brain appear to have been detected by Oken, and the bird-like character of the respiratory system was expounded by Von Meyer. And most of whatever this memoir contains has been already thought or discovered by the German philosophers, who have had the Pterodactyles as fossils of their fatherland, though my own conclusions were arrived at separately and from different materials.
The oldest Ornithosaurians are from the Muschelkalk of Germany. In England the oldest are from the Lias,—several species of Dimorphodon—a genus in some respects nearly resembling the Pterosaurians of the Cambridge Upper Greensand. In the Oolite of Stonesfield are several species of Rhamphorhynchus or a similar genus. The great Pælolithic period from the Oxford Clay to the Kimeridge Clay, has yielded in its several divisions small Pterodactyles of « ix » new species. And the Psammolithic period from the Portland Sand to the Lower Greensand has afforded many excellent remains both of true Pterosaurians in the Purbeck, Wealden, and Potton Sands, and of animals which indicate a new order of Ornithosauria having affinities with Von Meyer's thick footed saurians, the Dinosauria. In the Cretaceous series, Galt, Upper Greensand, and Chalk all have representatives of the Pterosauria; but no English stratum has hitherto yielded so many as the Cambridge Upper Greensand. From this formation the collection accumulated during Prof. Sedgwick's long professoriate is unequalled; though, excepting a few fine bones from the Chalk and the Purbeck Limestone, the Woodwardian Museum is as yet deficient in Ornithosaurians from the other Secondary Rocks. Until descriptions of these animals shall have been published a classification of the Ornithosauria must necessarily be provisional. And it cannot be expected that descriptions of the structure of Cretaceous Pterosaurians here given will hold good for all the Ornithosaurian sub-class.
Finally, I have gratefully to express my thanks to the many friends, English and German, who have aided me with specimens and with their writings; to the chiefs and officers of the English museums, especially Prof. Owen, Prof Humphry, Prof Newton, Prof Phillips, Prof Flower, and Prof. Huxley; to the officers of the University Library, especially Mr Bradshaw, and Mr Crotch, for aid in consulting books; but chiefly to Prof Sedgwick, who while employing me as his paid Assistant to aid him in his Museum work, has generously encouraged me to carry on for several years, without restraint and as part of my daily labour, an investigation of which this treatise is the first fruit. Prof. Sedgwick « x » has placed at my disposal an ample number of copies for distribution among those who take an interest in the Museum; and especially among those who have contributed to the Ornithosaurian collections, and aided me in my work.
January 3, 1870.
CONTENTS.
| Page | |
| Introduction | 1 |
| Materials | 1 |
| History | 3 |
| Organization | 7 |
| Cuvier | 7 |
| Sömmerring | 10 |
| Oken | 10 |
| Wagler | 11 |
| Goldfuss | 11 |
| Wagner | 14 |
| Quenstedt | 17 |
| Burmeister | 17 |
| Von Meyer | 17 |
| Another view of the Ornithosauria | 24 |
Osteological collection illustrative of modifications of Ornithosauria in the Cambridge Greensand, pp. 28-94.
| Sternum | 28 |
| Coracoid | 32 |
| Scapula | 35 |
| Humerus | 38 |
| Radius and Ulna | 42 |
| Proximal carpal | 48 |
| Distal carpal | 50 |
| Lateral carpal | 51 |
| Metacarpal bone of wing-finger | 53 |
| First phalange of wing-finger | 56 |
| Second phalange of wing-finger | 57 |
| Claw phalange | 59 |
| Os innominatum | 59 |
| Femur « xii » | 62 |
| Tibia | 62 |
| Tarso-metatarsus | 63 |
| Atlas and axis | 64 |
| Cervical vertebræ | 65 |
| Dorsal vertebræ | 69 |
| Sacrum | 73 |
| Caudal vertebræ | 75 |
| Bones of the head | 77 |
| Basi-oocipital bone | 78 |
| Back of the cranium | 80 |
| Back of another cranium | 84 |
| Ethmo-sphenoid | 85 |
| Mould of the brain-cavity | 87 |
| ?Vomer | 88 |
| Quadrate bone | 89 |
| ?Pterygoid end of palatine bone | 91 |
| Premaxillary bones | 91 |
| Lower jaw | 91 |
| Teeth | 92 |
| Conclusion | 94 |
| A summing up | 94 |
| Restoration | 103 |
| Speculations on habits and aspect | 104 |
| Notes on German specimens | 106 |
| Classification | 108 |
| Synopsis of species | 112 |
| Appendix | 129 |
| Index | 133 |
| Plates, and explanation of Plates. | Plates |
| PAGE | LINE | ||
| 4, | 2, | from bottom, for procælian read procœlian. | |
| 7, | 13, | for Ossements read Ossemens. | |
| 8, | last line, paragraph (2), for outermost read innermost. | ||
| 10, | 21, | for Sömmering read Sömmerring. | |
| 11, | 5, | ||
| 11, | 13, | ||
| 14, | note, for Beyerischen read Bayerischen. | ||
| 15, | 5, | for ?zygapophyses read spinous-processes. | |
| 17, | 6, | from bottom, for Herman read Hermann. | |
| 37, | 5, | from bottom, after "spine as" insert "are" | |
| 92, | line above 'the Dentary Bone,' for Pterodactyle read Pterodactyles. | ||
| 97, | 11, | for Günter read Günther. | |
| 99, | 4, | for Ichthyopteria read Ichthyopterygia. | |
| 101, | 11, | from bottom, for procælous read procœlous. | |
| 102, | 15, | for procælous read procœlous. | |
| 111, | 8, | for Sömmering read Sömmerring. |
For epipubic bone read prepubic bone, pp. 61, 102, 109, 110, 111, and pl. 8.
INTRODUCTION
TO THE
OSTEOLOGY OF THE ORNITHOSAURIA FROM THE CAMBRIDGE UPPER GREENSAND.
The Cambridge Upper Greensand has yielded to collectors bones which illustrate nearly every part of the skeleton of the animals that are commonly named Pterodactyles. Large collections have been acquired for the Woodwardian Museum. A series of more than 500 bones have been arranged to exemplify the osteology and organization of the Ornithosauria in the area when the Cambridge Greensand was deposited. And this memoir is written to explain briefly some of the structures of the soft and hard parts of those animals which are exhibited or demonstrated by these relics. Another collection of nearly 400 bones has been arranged, which displays in association, as they were found entombed in the old Greensand sea-bed, the remains of the skeletons of thirty-three animals of the Pterodactyle kind. The whole of the remains from this formation hitherto gathered cannot be computed to have pertained to fewer than 150 individuals, which indicate a new sub-class of animals, two new genera and at least twenty-five new species.
The bones were mostly of a paper or card-like thinness, and were originally hollow like the thin bones of birds. In the jaws of other animals, and in the sea, they were easily fractured, so that proximal ends and distal ends and shafts and split bones abound, « 2 » while perfect bones are almost unknown. Even those bones like the carpals, which almost retain their entirety, invariably show indications of having been rolled on the sea-shore among the nodules of phosphate of lime with which they now occur, in their angular margins being rounded, and in the removal of slender processes. The rock in which these fossils are found is a thin bed of chalky marl which is heavily charged with dark-green grains of Glauconite, and is quarried largely, and entirely dug away to be deprived of the dark-brown nodules of phosphate of lime with which it is stored. In digging and in the subsequent washing, the workmen, stimulated by an ample reward, pick out the fossils as they are discovered. They are separated easily from the matrix of investing marl, so that every aspect of each bone is seen, except for the occasionally adherent oysters and the masses of phosphate of lime, with which material the bones are also filled. Hence these remains afford facilities for the study of the joints such as no other specimens have presented; and from their large size and comparatively great numbers, render easy the labour of the student who seeks to contrast them with the bones of other animals.
The osteological collection has been formed without regard to species or genera, and arranged to exhibit the structure and organization of the tribe of animals. So far as possible each bone, as humerus, femur, &c., has its variations of structures and form contrasted on a single tablet. The series comprises the following bones:
Fore-part of sternum.
Coracoid (perfect).
Scapula (nearly perfect).
Humerus (perfect).
?Radius (proximal end).
Radius (distal end).
?Ulna (proximal end).
Ulna (distal end).
Proximal carpal.
Distal carpal.
Lateral carpal.
Wing-metacarpal (proximal and distal ends).
First phalange (proximal and distal ends).
Second phalange (proximal end).
Metacarpal or metatarsal (distal end).« 3 »
Claw phalange.
Os innominatum (parts of ilium, ischium, and pubis).
Femur (perfect).
?Tibia (proximal end).
Atlas and axis.
Cervical vertebræ.
Dorsal vertebræ.
Sacrum and sacral vertebræ.
Caudal vertebræ.
Lower jaw (dentary and articular ends).
Premaxillary bones, &c.
Teeth.
Quadrate bone (distal end with quadrato-jugal).
Ethmoid with basi-sphenoid.
Occipital and parietal segments of skulls.
Basi-occipital and basi-temporal.
Cast of brain-cavity.
They are exhibited in Compartments a, b, c of the Table-case of Cabinet J. The letter F in a circle is placed against figured specimens.
History.
The Cambridge Pterodactyles first attain prominence in scientific literature in the year 1859. Professor Owen had figured (plate 32, fig. 6-8) fragments of bones in the Palæontographical Society's Monograph for 1851; the distal end of a large ulna (fig. 6); the shaft of a phalange of the wing-finger, probably the first (fig. 7); and the upper portion of the shaft of a small humerus showing part of the radial crest (fig. 8). Inadvertently the last specimen was referred to the Lower Greensand. But although fragments of humerus of Pterodactyle and vertebræ of Pterodactyloid animals have in the last few years been gathered from the Potton Sands, those deposits were believed to be barren of fossils when Prof. Owen wrote; and all the Pterodactyles yet made known from near Cambridge were collected from the Cambridge Upper Greensand.
Among the earliest successful collectors were Mr James Carter, « 4 » the Rev. H. G. Day, St John's Coll.; Prof. G. D. Liveing, St John's Coll.; the Rev. T. G. Bonney, St John's Coll.; and Mr Lucas Barrett, Trin. Coll.; and the Rev. Prof. Sedgwick, Trin. Coll., on behalf of the Woodwardian Museum. Mr Day and Mr Bonney both presented every specimen from their cabinets which could enrich the University collection. And in the last ten years the Woodwardian Museum has acquired, through the skillful collecting of the Messrs Farren, the present materials. The associated sets of bones were formed by William and Robert Farren, who, obtaining the specimens from day to day as they were discovered, were enabled to put together such parts of the skeleton as remained together on the sea-bottom. These collections will hereafter be used for the elucidation of species. They are the only materials which can give the proportions of the Cambridge Ornithosaurians, and the contrast of aspect which distinguished the living animals from those from other rocks.
The other collections of these fossils are those of Mr William Reed and Mr J. F. Walker at York, the Museum of Practical Geology, and the British Museum.
The Woodwardian specimens as collected were placed in the hands of Prof. Owen, and were first made known in the Professor's lectures on reptiles and birds delivered at the Museum of Practical Geology in 1858. In that year Prof. Owen communicated to the British Association for the Advancement of Science, and printed in their Report, the matter of the memoir which was published with plates by the Palæontographical Society in 1859. In this latter year Prof. Owen communicated to the Royal Society an account of the vertebral column of Pterodactyles. In 1859 Prof. Owen also produced a classification of recent and fossil reptiles at the meeting of the British Association, in which the order Pterosauria appears with new characters—such as the pneumatic structure of most of the bones—drawn from Cambridge specimens. In 1860 Prof. Owen produced another memoir on Pterodactyles, which was published by the Palæontographical Society. A brief account of the tribe appeared about the same time in the Professor's Palæontology.
In these writings are descriptions of the various parts of the vertebral column. Their procœlian centra are described, and the pneumatic foramina are noticed and supposed to have communicated « 5 » with air-cells. They are compared with birds, and distinguished from birds; but although the order is classed with reptiles no contrast with reptiles is made. Other bones described are a basi-occipital, and a doubtful bone, then thought to be a frontal, but which is more like the neural region of the sacrum.
The sternum is compared with the sternum of the birds Apteryx and Aptenodytes, is stated to be formed, in the main, on the Ornithic type, and to possess distinct synovial articular cavities for the coracoids such as only occur in birds. The inter-coracoid process of the sternum is compared with that of Bats, Birds, and Crocodiles.
The mechanism of the framework of the wings is said to be much more bird-like than bat-like, the anchylosed scapula and coracoid being remarkably similar to those of a bird of flight. The coracoid is shorter and straighter in birds than in Pterodactyles, but no comparisons are made with reptiles.
The humerus is known only by the proximal end. It is said to conform at its proximal end more with the Crocodilian than with the Avian type, but to have the radial crest much more developed than in either Crocodile or Bird. The bone is, however, chiefly compared with birds, and is figured between corresponding bones of a Vulture and a Crocodile. The pneumatic texture is said to be as well marked as in any bird of flight.
Of the carpus it is said, the Pterodactyle, in the complete separation of the metacarpus from the antibrachium by two successive carpals answering to the two rows, adheres more closely to the reptilian type than to that of birds. But the row which was regarded as proximal is the distal row, while the supposed distal row is proximal.
The claw-phalange and distal end of the wing-metacarpal, the mandible, teeth, and jaw are the other bones described, but their comparative osteology is not discussed. In the Professor's account of a fragment of a jaw it is said, "The evidence of the large and obviously pneumatic vacuities now filled with matrix, and the demonstrable thin layer of compact bone forming their outer wall, permit no reasonable doubt as to the Pterosaurian nature of this fossil. All other parts of the flying reptile being in proportion, it must have appeared with outstretched pinions like the soaring Roc of Arabian romance, but with the demoniacal features of the « 6 » leathern wings with crooked claws, and of the gaping mouth with threatening teeth, superinduced."
When the specimens on which Prof. Owen had founded the foregoing views of the osteology and classification of these animals were at length returned to the Woodwardian Museum, it became a duty of the present writer to arrange and name them. And in a Memoir on Pterodactyles which was communicated to the Cambridge Philosophical Society and read March 7 and May 2 and 16, 1864, a position was claimed for them, distinct from reptiles, as a separate sub-class of Sauropsida, nearly related to birds.
In September of the same year a communication was made to the British Association "On the Pterodactyle as evidence of a new sub-class of vertebrata (Sauromia)," with enlarged drawings of the skull and some of the other bones, in which the conclusions arrived at were that, excepting the teeth, there is little in such parts of the head as are preserved to distinguish the Cambridge Pterodactyles from birds; and that the remainder of the skeleton gives a general support to the inference from the skull.
Papers were communicated to the Cambridge Philosophical Society on February 17, 1868, on indications of Mammalian affinities in Pterodactyles in the pelvis and femur, and February 22, 1869, on the bird-like characters of the brain and metatarsus in the Pterodactyls from the Cambridge Greensand. The other references to Cambridge specimens are in a paper "On the literature of English Pterodactyles" in the Annals and Magazine of Natural History for Feb. 1865, and in "An epitome of the evidence that Pterodactyles are not reptiles, but a new sub-class of vertebrate animals allied to birds," in the same magazine for May, 1866.
In the meantime Prof. Owen's views have somewhat changed. In the first volume of the Comparative Anatomy and Physiology of the Vertebrata (1866), the Pterosauria are classed as the highest group of reptiles, and take rank above the Dinosauria. In the second volume of that work (1866), occurs the following passage:
"Derivatively the class of birds is most closely connected with the Pterosaurian order of cold-blooded air-breathers. In equivalency it is comparable rather with such a group than with the Reptilia in totality, or with the Mammalia."
Organization.
Nearly every writer on Pterodactyles, who has expressed any opinion at all, has formed an estimate of his own of their organization. They have been assigned to almost all possible positions in the vertebrate province, by great anatomists who all had before them very similar materials. An account of these views is given by von Meyer in his monograph of the Pterodactyles of the Lithographic Slate. It will not be necessary to discuss these conclusions here, for the materials from the Lithographic Slate and those from the Cambridge Greensand are so different that no light would be thrown on the organization of the animals by an exposition of any fallacious inferences from German specimens. In England they are classed with Reptilia, chiefly through the influence of the discourse upon them given by Baron Cuvier in his Ossemens Fossiles[A]. It therefore may conduce to a clear view of the subject to quote in Cuvier's words the passages in that memoir which have been supposed to establish their position among reptiles. He says,—"Ayant encore porté mon attention sur le petit os cylindrique marqué g (i.e. os quadratum) qui va du crâne à l'articulation des mâchoires, je me crus muni de tout ce qui étoit nécessaire pour classer ostéologiquement notre animal parmi les reptiles." The exact relations of the quadrate bone are not seen in either Cuvier's or Goldfuss' or von Meyer's figures of this Pterodactyle, the P. longirostris; but in von Meyer's figures of P. crassirostris, P. longicollum, and P. Kochi it appears to be a free bone articulated to the squamosal and petrosal region of the skull and with the lower jaw. This is not the case with either Chelonians or Crocodiles, which have the quadrate bone firmly packed in the skull; nor is it paralleled even among those lizards and serpents which have the bone as free; while, on the contrary, it is characteristic of the whole class of birds. The form of the bone is not more Lacertian than Avian, while its direct attachment to the bone of the brain-case finds no parallel among lizards, but is exactly paralleled in all birds.
[A] Tome V. Part a, pp. 358, 383. Edition, 1814.
Cuvier then goes on to say, "Ce n'étoit pas non plus un oiseau, quoiqu'il eût été rapporté aux oiseaux palmipèdes par un grand naturaliste[B]." Which position he supports as follows:—
[B] Blumenbach.
(1) "Un oiseau auroit des côtes plus larges, et munies chacune d'une apophyse récurrente[C]; son metatarse n'auroit formé qu'un seul os, et n'auroit pas été composé d'autanut d'os qu'il a de doigts." These, though they may not be characters which are those of birds, are certainly not eminently reptilian. The elongated form of the tarsals in birds is peculiar, but quite functional, as may be seen among the Penguins, where, when the so-called tarso-metatarsal bone is no longer erect, it becomes much shorter, and is nearly separated into three distinct bones. The cretaceous Pterodactyles appear to have this bone exactly like that of birds.
[C] This shown in other specimens since figured, and in the specimen from Stonesfield in the Oxford Museum.
(2) "Son aile n'auroit eu que trois divisions après l'avantbras, et non pas cinq comme celle-ce." This is a difference, but a difference of detail only, and not a reptilian character. The creatures have wings; and no reptile known, from recent or fossil specimens, has wings. The general plan of the wing, though very unlike, approximates to that of a bird. Most birds have two phalanges in the long finger, though some have three. One Pterodactyle is described as having only two phalanges in the wing-finger, while most of the German specimens appear to have four phalanges. In birds the longest finger appears to be the middle one, while in Pterodactyles it is the innermost one.
(3) "Son bassin auroit eu une toute autre étendue et sa queue osseuse un toute autre forme; elle seroit élargie, et non pas grêle et conique." The pelvis of Pterodactyle is not reptilian, and no living reptile has a pelvis like it. It is not unlike the pelvis of a Monotreme, but the ilium is more Avian. It resembles the pelvis of Dicynodon. And the discovery of a long-tailed bird-like the Archæopteryx shows that the tail is like that of old birds, even if it also presents some analogy in form to that of certain reptiles and mammals.
(4) "Il n'y auroit pas eu de dents au bec; les dents des harles ne tiennent qu'à l'enveloppe cornée, et non à la charpente osseuse." This is not a reptilian character. Among reptiles some tribes have teeth, others want them; and among mammals some animals are without teeth, though they are so characteristic of the class. It is an anomaly that birds should all be toothless. And so, without citing the supposed teeth of Archæopteryx, it may « 9 » be affirmed that it would be no more remarkable for some birds to have teeth than it is for some mammals and reptiles to be without them.
(5) "Les vertèbres du cou auroient été plus nombreuses. Aucun oiseau n'en a moins de neuf; les palmipèdes, en particulier, en ont depuis douze jusqu'à vingt-trois, et l'on n'en voit ici que six ou tout au plus sept." This is a variation of detail such as, had it occurred among birds, would hardly have been deemed evidence of their affinities. When the variation of the neck-vertebræ ranges from 23 to 9, the further reduction of the number to 7 becomes insignificant, and does not show that the animal was a reptile.
(6) "Au contraire, les vertèbres du dos l'auroient été beau-coup moins. Il semble qu'il y en ait plus de vingt, et les oiseaux en ont de sept à dix, ou tout au plus onze." This modification is obviously the result of smaller development of the pelvic bones from front to back, and hence of the small number of vertebræ in the sacrum. It does not support the reference of Pterodactyles to the class of reptiles.
Speaking of the teeth, it is said, "Elles sont toutes simples, coniques, et à peu près semblables entre elles comme dans les crocodiles, les monitors, et d'autres lézards." The teeth of Pterodactyles are (in the skull) for the most part in the premaxillary bones, in which it is so characteristic for the teeth of animals to, be merely conical and simple. Therefore it would have been difficult to imagine the teeth to have been anything but what they are, whatever the affinities of the Pterodactyle might be.
It is remarked, "La longueur du cou est proportionée à celle de la téte. On y voit cinq vertèbres grandes et prismatiques comme celles des oiseaux à long cou, et une plus petite se montre à chaque extrémité." This adds nothing to the evidence for its reptilian character.
"Ce qui est le plus fait pour étonner, c'est que cette longue téte et ce long cou soient portés sur un si petit corps; les oiseaux seuls offrent de semblable proportions, et sans doute c'est, avec la longueur du grand doigt, ce qui avoit determiné quelques naturalistes à rapporter notre animal à cette classe." Nor is this evidence that the animal was a reptile. And in many minor matters Cuvier is careful to show how their modifications resemble « 10 » those of birds; and when this is not so, birds are the only animals from which he finds them varying. And the few suggestions which are thrown out respecting their affinities with lizards are upon points which are also common to birds.
Thus what Cuvier did was to distinguish these animals from birds, and incidentally to show that their organization was a modification of that of the Avian class. And the legitimate inference would have been that their systematic place was near the birds, and not that they were reptiles.
But in Germany Cuvier's views on Pterodactyles have by no means been submissively received; and great anatomists, since he wrote, have propounded and defended views as various as those of the anatomists who preceded him, and with no less confidence in the results of their science. In the brief space at my command it would be impossible to do justice to the works of this array of philosophers, and therefore I present in a somewhat condensed version the epitome of their conclusions given by Hermann von Meyer in his Reptilien aus dem Lithographischen Schiefer der Jura. They form a commentary on the casts of Solenhofen Pterodactyles contained in the Woodwardian Museum.
Sömmerring
regarded the Pterodactyle as an unknown kind of bat, and thought that Cuvier was misled by Collini's imperfect description. He believed that he found in them different kinds of teeth as in mammals; and regarded them as differing from bats chiefly in having larger eye-holes, a longer neck, four fingers and four toes, a longer metatarsus, and in having but one elongated finger; and found the closest analogue of the fingers in Pteropus marginatus of Bengal. And although inclined to place the Pterodactyle between Pteropus and Galeopithecus, he suspects from the bird-like characters of the head and feet that its true place is intermediate between mammals and birds.
Oken[D].
[D] Isis, 1818, p. 551.
Oken reasoned carefully so far as his materials went. He dwells much on the analogy of the wing to that of a bat, and seems to suspect that the marsupial bones would hereafter be « 11 » found; and, excepting the head, finds that the other parts of the skeleton have their corresponding bones among mammals.
Afterwards, when he saw the specimens at Munich, he was so much struck at finding the quadrate bone of Lacertian form, though Sömmerring could not detect it even with a microscope, that he is shaken in his mammalian faith, and inclines to consider the animal a reptile.
Wagler[E].
[E] System der Amphibien, 1830, p. 75.
Wagler was impressed with the resemblance of the jaws and the rounded back part of the skull to those of Dolphins, and so far as the head went conceives it to have had nothing in common with Lizards. He recognizes mammalian characters in the pelvis and sternum, and fails, like Sömmerring, to detect a quadrate bone, and finds the sum of the characters like those of other extinct animals, such as Ichthyosaurus and Plesiosaurus, suggesting for it a position between mammals and birds. He supposed it unable to fly, that it never left the water, but swam about on the surface like a swan, and sought its food on the sea-bottom. He imagined the long arms to have been used after the fashion of turtles and penguins to row the body along; while to the claws he attributes the function of holding the females in the generative process.
Goldfuss[F].
[F] Nova Acta Acad. Leopold., 1831, Vol. XV. Pt. I. p. 103.
sees in Pterodactyle an indication of the course that nature took in changing the reptilian organization to that of birds and mammals. The less important organs, those of motion, assimilate partly to those of the bird and partly to those of the bats, but always preserve the fundament reptile type and reptile number of bones. The skull, fluctuating in character between the monitor and crocodile, hides its reptile nature under the outer form of the bird, but retains the teeth. To change the skull into a bird's skull it would only be necessary that a few separate elements should be blended together, and that a few peculiar bones should be removed. The length of the neck, varying only in a few species, is a deviation from the reptile type, and indicates an approximation to the structure of 'the bird; but the number of « 12 » the vertebræ remains constant notwithstanding the increased length. The fundamental plan of the crocodile may be recognised in all the important parts of the vertebræ. The body of the reptile, to be enabled to fly, would need a larger breast and a stronger structure of the fore-limbs. The shoulder-blade of the reptile, with its extremities forming the glenoid cavity, is necessarily smaller and prolonged backward, and altered to resemble that of a bird. The scapula only formed the back part of the glenoid cavity, but it is thick and strong, suggesting an affinity with the bats.
The breast-bone, in the form of a shield, is changing into that of a bird; as are the ribs, which are attached in a peculiar way to the vertebral column. It is really the strong sternum of the Chameleon, with moveable dorsal vertebræ. The whole chest is supported by the peculiar continuation of the wings of the pubic bones (Schambein). The ischiac and pubic bones resemble those of the Chameleon, but the ilium runs a little down, like that of a bird, and is only slightly connected with two sacral vertebræ, as in reptiles, prolonging itself a little upward and forward, as in mammals. The wings of the pubic bones exist in the Turtle and Monitor, but of small extent; they are also represented in the mammals by the upward development of the pubic bones in those families, genera, and species, in which nature has indicated by variety of shape, or peculiarities of development, or by affinities with reptiles, quite a new type and capacity for variation within certain limits, which is especially the case with certain Rodents and Opossums, and Monotremes. It would not be astonishing to find in Pterodactyles the marsupial bones. And indeed the Pterodactylus crassirostris has a small tongue-shaped bone, probably belonging to the pelvis. The less important part of the skeleton, the tail, is formed precisely as in mammals, and is identical with that of the bats. Both the thigh and shin are mammalian, and only the foot retains the same number of parts as in reptiles.
This animal was enabled by means of the pelvic bones and the long hind-legs to sit like the squirrels.
We should regard this position as natural but for the long wing-finger hanging far down the sides. If it were to creep along it would have the same difficulties as a bat, and the length and weight of the head, as well as the proportional weakness of the « 13 » bind limb, make it improbable that they progressed by leaping. These animals made use of their claws only to hang on to rocks and trees and to climb up steep cliffs. They could fly with their wings, and keep themselves aloft in order to catch insects or sea animals. The wide throat and the weak and high supports of the jaw-bone make it probable that they only used their teeth to capture their prey and not to mince it. By means of their long neck, which they usually bore curved backward in order to keep their balance, they could stretch out their head to their prey and change their centre of gravity, and so fly in different positions. The fundamental type of the Crocodile and Monitor leads us to suspect that they had a skin covered with scales. The approximation to the shape of the Bird makes it probable that they were feathered. And the whole outline, similar to that of the Bat, leads to the supposition that they were covered with hair, like the Monotremes. Goldfuss thinks he has got a clear insight into the covering of the body and the whole condition of the wing in examining the Pt. crassirostris. And the soft state of the stone near the bones he attributes to the presence of the soft parts of the animal; and supposes that on the original folds of the wing-membrane are to be seen tufts and bunches of curved hair directed downward and sideway[G]. And on the principal slab he finds evidence that the Pterodactyle had a mane on the neck like a horse. The tufts on the counter slab have some similarity with the feathers of the ostrich. Some very tender impressions on both plates still more resemble feathers. He recognizes the outline and faint diverging rays of a bird's feather, but never sees a strong quill. The microscope, instead of making the image clearer, makes it, on the contrary, vanish, because then the rough parts become prominent. Also on the slab which contains the Pterodactylus medius[H], are seen numerous lines and fibres diverging like a bird's feathers. And on the upper part of the belly is the appearance of a scanty texture of hairs and feathers. The visible marks of two cylinders of the thickness of a quill, made of thin substance and filled with limestone, he would regard as quills if there were clearer marks of their feathers to be seen. As a note upon this von Meyer says, after examining the slabs, that the particles considered by Goldfuss « 14 » to be hairs and feathers rest upon appearances not only to be seen in the vicinity of Pterodactyles, but which occur upon many other kinds of petrifactions that have nothing in common with the Pterodactyle; and that the roughnesses of the slab have nothing to do with the folds of the wing or the muscles.
[G] This is represented in Pl. 7, 8 of his memoir, loc. cit.
[H] Pl. 6, Nova Afta Acad. Leopold, Vol. XV. Pt. 1.
Wagner[I]
[I] Abhandl. Bayerischen Acad. 1852, Vol. VI.
is so convinced that the Pterodactyles are Amphibians approximating to the Saurians, that he does not think it necessary to go into any controversy in the matter; but he acknowledges that their forms sometimes present peculiarities of bird and mammal. The head especially shows a blending of the bird and reptile types. Its outline, particularly when seen from above, is that of a long-beaked water-bird. And the long interval between the nose-holes and the tip of the jaw, and the peculiar fact of a hole between the nose and eye-holes, and the want of the continuation of the coronoid of the lower jaw, rather resemble a water-bird than a Saurian. But the presence and the form of the teeth show it to be a Saurian; and not only the teeth, but the configuration of the whole back part of the skull, reproduces the type of the Monitor. The sclerotic circle is a peculiar mark of birds and saurians. Very peculiar, however, is the extremely short back part of the skull; and the articulation of the lower jaw, stretched far forward and united just under the middle of the eye-hole. The more or less long neck, which may assume the form of an S, deviates very much from the short stiff neck of reptiles, and is quite bird-like, the neck-vertebræ of which those of the Pterodactyle closely resemble in shape; while their constant number of seven reminds us of mammals and crocodiles. The neck has the same flexibility as in a bird. The short and weak trunk-vertebræ are in such disproportion to the length and strength of the neck-vertebræ as is never met with even in the birds and mammals which have the longest necks. The trunk-vertebræ are completely separated from each other, and may be divided into dorsal, lumbar, and sacral vertebræ. The transverse processes of the back-vertebræ are notched out like those of the crocodile. The tail is short in most species, and this is a deviation from the type of the Saurians, and an approximation to birds and to many mammals. But « 15 » there are some kinds with very long tails, as is the case with mammals and usually with Saurians. But the vertebræ of these long-tailed Pterodactyles deviate very much from those of Saurians. And while the Saurian vertebræ are provided with long transverse processes and upper and lower spinous-processes (Dorn-Fortsätzen), they seem in the Pterodactyle to be almost devoid of processes and resemble those of mammals, on the tails of which these processes soon disappear. In a certain point of view we could say of the vertebral column of the Pterodactyle, that it has borrowed the neck from the bird, the trunk from the reptile, and the tail from the mammal. The ribs are connected to the transverse processes as in crocodiles, except with the atlas and axis. Quite in the type of the Saurians are the abdominal ribs, which are wanting to all birds and mammals, but often occur in the Lacertian order. The structure of the shoulder and breast-bone separate the Pterodactyle from the mammal, these parts being formed after the type of the Birds and Saurians, the characters of which are blended together. The small and elongated shoulder-blade, like the coracoid bone, belongs to the type of the bird rather than to that of the Saurians, of which, in reference to the last-named bone, only crocodiles have a similar one. The breast-bone, by its large expansion, points to the crocodiles, but at the same time, by the want of the keel, points to the ostrich-like birds, save that it is proportionally larger and wider than in these. The Pterodactyle, in common with the crocodile, wants the patella. The pelvis is formed on the type of the Saurians, although the ilium, by length and form, points somewhat to the mammals. The length and delicate form of the long bones of the limb, as well as the larger development of the fore-arm than of the upper-arm, and larger development of the lower thigh than of the upper thigh, and the thinness and elegance and shortness of the ?fibula (Wadenbein) have the characters of birds. The length of the middle hand [metacarpals] resembles that of birds, but its form in Pterodactyle is conformable to that of mammals. The first three fingers have the form and condition of the phalanges of lizards. The phalanges form the series 2, 3, and 4. The fourth, or air-finger, on the contrary, is of a peculiar type, of which no analogue is found in other animals, unless a somewhat similar arrangement be accredited to the bats. It is of enormous length, composed of four « 16 » parts and without a claw. The hind-leg is, in proportion to the fore-leg, weak, and in general does not take the bird-form, but that of a Saurian. It has five toes, with unusual arrangement of the phalanges into the series 1, 5, 4, 3, 2. One toe has no nail, and the others have claws weaker than those of the hand. It can hardly be supposed that the animal lived in the water. All Saurians that live either in the water or on land are short-legged; it is the same with the swimming birds. But the Pterodactyle has its hind-legs as long as a land or air-bird; and as in these, the shin especially exceeds the length of the thigh. At the same time the toes, when they are in their natural position, were so close together that we may suppose the animal not to have been web-footed. The great development of the hand, by means of the long middle hand and especially of the enormous length of the air-finger, makes it probable that it was the chief organ of flight, as in birds and bats; also deviating in a peculiar manner from both these types, the long air-finger served to expand the wing-membrane, which extended from the upper part of the finger to the trunk, and which in all probability did not touch the hind-legs. This we infer from the circumstance that the animal, in a position with the organs of flight folded up, was not supported like the bat on its four feet, but stood upright on its hind-legs like a bird. Such a position presumes the same freedom in moving the hind extremities as with birds; only in such a position could the animal walk on without being hindered by its flying organs when they were folded up like those of a bird. Only in such an upright position could the animal keep upright its unusually long head with the long and strong neck and be kept in balance, the neck being able to take a sigmoid curve like that of a bird.
Wagner concludes: "By these means we have recognised in the Pterodactyle a Saurian, but of a habitude which greatly removes him from all others of his kind, and approximates him to birds. Excepting in ability to fly, he has nothing in common with the birds. The opinion 'that the animal is half crocodile half monitor disguised as a bird, but intending to be a bird,' is therefore not only a paradox but also false. With more truth, but less phantasy, we could say that the Pterodactyle was a Saurian in transition to the Birds."
Quenstedt[J].
[J] Ueber Pterodactylus Suevicus. Tubingen 1855.
In the long thigh, with the long neck, Quenstedt sees evidence that the animal was able to walk upright, being probably still more upright than birds, since the great disproportion between the neck on the one hand, and the thigh on the other, could not have allowed a more appropriate position. At the same time he makes a question, Did it go on four feet? But a little later, in his book, Sonst und Jetzt, 1856, he gives a sketch of the animal resting on its four legs; and remarks, "The position upon four feet is however hypothetical, but is probable. It had its wings folded back. The slightly curved and thin bones of the middle hand probably served to support the flying-membrane, and had therefore the same function as the spur-bone in the bats." Finally, he says in his book, der Jura, p. 813, "Perhaps this animal walked from time to time on four legs, being then supported by the fore-end of the metatarsal bone."
Burmeister[K].
[K] Beleuchtung uniger Pterodactylus arten. 1855.
entirely rejects Quenstedt's opinions with regard to their upright position. He makes the following remarks: 'The animal walked on the free fore-toes and bore the wings like a bat, though with the body not in an upright position like a bird, but four-footed. The hind-foot is much too small for such an upright position, and the fore-foot much too strongly developed. I therefore believe that the Pterodactyle could much better have walked four-footed than a bat, because it possessed so much better developed fore-feet.' In the length of the tibia Burmeister sees no reason for the upright position, but, as he says, only a means for the wide expansion of the flying-membrane;—and an endeavour in walking on four feet to bring the leg into the necessary harmony with the arm, which is so much elongated with the flat-hand.
Hermann von Meyer[L].
[L] Fauna der Vorwelt. Reptilien aus dem Lithographischen schiefer. Frankfurt am Main. 1859. pp. 15-23.
The skull of the Pterodactyle can only be compared with those « 18 » of birds and lizards. The form is essentially Avian, and the sutures are indistinct or obliterated as in birds, while in reptiles they are persistent The temporal bone enters into the formation of the reservoir for the brain, which is eminently characteristic of birds and quite different from anything found in lizards. The snout resembles a bird in being chiefly formed by the intermaxillary bone, which bounds the front of the anterior nares; and, as in birds, the bone extends backward between the eye-cavities to the frontal bone. The corresponding intermaxillary ridge of the Monitor is of less extent.
The frontal-bone forms the highest part of the skull, and is similar to that of birds. The principal frontal is double, and forms the upper and hind part of the cavity for the eye, and covered the greater part of the large brain, composed of two hemispheres, in which Oken long ago saw a similarity to the higher animals. The arched form of the back part of the skull is bird-like. The double parietal adjoins the principal frontal, and is conditioned like the parietal in birds. The supra-occipital is single as in birds, expanded, and forms the part of the skull which extends furthest back. From the form of the back part of the skull it may be concluded that the foramen magnum was situated as in birds, and that the head and neck were moved as in birds, and not as in reptiles and mammals.
The temporal bone rests upon the parietal and frontal, and forms much of the temporal foss. Its anterior border does not appear to enter into the margin of the orbital cavity as in birds, but seems to be replaced by the post-frontal, which resembles that of the Chameleon. Its hindmost branch, which can hardly be supposed to be the jugal, forms the outer boundary of the temporal foss by uniting with a process which is probably part of the mastoid. A similar closing of the cavity for the temporal muscles is also to be found in birds. The jugal and maxillary do not follow the bird type. The jugal consists of a single bone which forms the greater part of the anterior and inferior boundary of the cavity of the eye, which is surrounded with bones, as in Dragons and Iguana. In those birds in which the cavity of the eye is surrounded with bones the jugal does not enter into it. As in lizards, at its upper end the jugal is commonly connected with the lachrymal, which bone is like that of a bird. « 19 » A bone, which appears to be the pre-frontal, enters into the back of the nasal aperture.
The nostril is double and often of large size.
The perforation in the skull between the orbit and nares is bird-like.
The quadrate bone is not quadratic as in birds, but cylindrical and shaft-like, as in the Chameleon. The articulation of the quadrate with the lower jaw is placed further forward than in birds and reptiles. The lower jaw, but for the teeth, has great similarity with that of a bird. Among reptiles its nearest resemblance is with Chameleons and Turtles. The hyoid is more bird-like than reptile-like.
Ribs and vertebræ.
It is uncertain whether the Pterodactyle had lumbar vertebræ. If they are wanting, therein the animals resemble birds, of which we are reminded in the short and stiff back and moveable neck. Pterodactyles possess a smaller number of neck-vertebræ and a larger number of back-vertebræ than birds. The long neck-vertebræ are paralleled by those of water-birds, by the Giraffe, the Camel, Protosaurus and Tanystrophæus. There are 7 cervical vertebræ, the 1st very short, 2nd not longer, but rather shorter than those which follow. There are in Pterodactyles from 12 to 16 dorsal vertebræ, while birds have never more than 11. It is not certain whether all Pterodactyles have an os sacrum; most have it, and therein resemble Mammals, Birds, and some fossil Saurians. In Pterodactylus dubius and P. grandipelvis and P. Kochi there are 5 or 6 vertebræ in the sacrum. In birds the sacral vertebræ vary from 5 to 22; in bats the number is from 5 to 6.
The short tails of Pterodactyles are more like those of mammals than birds; they include from 10 to 15 tail-vertebræ. In birds there are from 6 to 10 tail-vertebræ. Rhamphorhynchus has 38-40 tail-vertebræ, secured between thread-bones like those in the tail of rats.
The dorsal ribs are reptile-like. In herbivorous mammals and birds they are broader. A few species have the first pair of ribs large. The abdominal ribs belong neither to birds nor mammals, but are reptilian. In Rhamphorhynchus Gemmingi there are 6 pairs of sternal ribs.
The sternum
is bird-like, somewhat resembling lizards. It consists of a simple flat bone, but without the keel of a bird's sternum. It is relatively smaller than in birds, is broader than long, and therefore comparable with Struthious birds. They were not able flyers, since the part to which the muscles for flight should be affixed is wanting. And for the same reason they could not have been wandering animals. But Moles possess a keel on the breast-bone, which therefore is no evidence of flight. And in swimming-birds which do not fly the keel is much developed; and in swimming-birds the sternum is also long, so that neither length nor keel prove flight. So far as the evidence from the sternum goes, they were neither water-birds, nor diggers, but denizens of the air. In Rhamphorhynchus Gemmingi, besides the usual breast-bone, there is a plate with breast-ribs uniting the sternum with the dorsal ribs; they are cartilaginous, or horny, as in birds.
The scapula and coracoid
present the closest resemblance with those of a bird, and only deviate in the coracoid not being inserted in the breast-bone in the manner of birds[M]. It at first seemed that Rhamphorhynchus differed from Pterodactyle in having the scapula and coracoid anchylosed. In R. Gemmingi the bones are either separated or only slightly united.
[M] See however Pl. 1 and 2 of this memoir.
Oken and Goldfuss thought that the scapula consists of an upper and under part, as in lizards. Von Meyer sees nothing of the kind.
The humerus
presents no striking similarity with birds, and differs from bats.
The carpus
is more reptile-like. It consists of two rows of small bones. In birds there is one row made up of two bones.
The pteroid bone.
Von Meyer regards it as having supported the wing-membrane in flight. There has been a good deal of difference of opinion « 21 » about it, some thinking it, with Quenstedt, an ossified tendon; others, like Wagner and Burmeister, regarding it as an essential part of the Pterodactyle skeleton. Von Meyer regards its extent as indicating the extent of the wing-membrane. See p. 42.
Metacarpus.
In length the metacarpus resembles that of the Ruminants, in which however it consists of but one bone; while in Pterodactyles there is a separate bone for each of the four fingers; they are closely united together without being blended. In some Pterodactyles the metacarpals of the short fingers are as fine as hairs, so that it is impossible that they should have articular facets on the carpus. In Ornithopterus the metacarpus has some resemblance with that of the bird, but the articulation with the phalanges of the finger for flight is stiff. In Pterodactylus and Rhamphorhynchus there is a free articulation.
Burmeister remarks that the chief articulation of the wing in bats is with the carpus, while in Pterodactyle the articulation is with the end of the metacarpus.
The hand.
Von Meyer finds four fingers. It was formerly supposed that the order of the phalanges was 2, 3, 4, 4, but in the fly-finger this is not the case, Ornithopterus having but two. The number of joints in the other fingers is quite as irregular.
In Pt. longicollum the thumb consists of but one joint.
The Ilium
is more mammalian and avian than reptilian.
Pubis.
The pubis appears to have been excluded from the glenoid cavity, as in Crocodiles. It is more mammal-like than bird-like, and is to be compared with the marsupial bones.
The femur.
In certain Pterodactyles the proximal condyle of the femur resembles birds; but in other Pterodactyles the bone is more mammal-like in its straightness, and development of the upper condyle, and in the presence of a trochanter.
The tibia and fibula
may be compared, from their great length, with birds and flying vertebrate animals.
The fibula is style-shaped, like that of a bird, the lower part being wanting; while in bats the upper part is wanting.
The tarsus,
of two rows, is best compared with that of reptiles. The number of constituent bones has not been definitely determined.
The metatarsus
shows a certain return from the bird type to that of reptiles.
Foot.
Von Meyer never finds more than four toes, and sometimes a stump of a fifth. As a whole, the foot is Saurian-like. It differs from lizards in the number of toes, and approximates to Crocodiles. In Pterodactylus longirostris the formula of the toes is 2, 3, 4, 5, with a stump of two joints;—like lizards, if we abstract the outer toe; and like birds with four toes; but they are liable to variations.
In Pterodactylus scolopaciceps and P. Kochi the formula is 2, 3, 3, 4 joints. In Winkler's specimen of P. Kochi there is also a stump of three joints.
In Pterodactylus micronyx the formula is 2, 3, 3, 3, and a stump of two joints. In P. longicollum the number appears to be different from all the foregoing.
The stump was attached to the side of the outer toe. Wagner, in P. Kochi, supposed it to be on the inner side, and so gave a reverse arrangement to the toes. The stump may be compared with that of some Chelonians, in which it is not furnished with a claw.
There is a difference from birds in the claws being much less developed. It has a true reptile foot. In bats the toes are of equal length. Von Meyer thinks the hind-legs did not enable it to walk on the land.
In some Pterodactyles the flying-membrane is faintly seen. The presence of feathers might be inferred from there being but « 23 » one finger for flighty as in birds; but the function of feathers is subserved by the long and stiff finger. If it had been covered with scales, as was supposed by Cuvier, some traces of them would be found. The skin was probably naked, and had no connection with the hind-legs as it has in bats; in this respect resembling birds.
The condition of the several parts of the skeleton completely proves that the Pterodactyle was a reptile. Its head, neck, shoulder, and back, resemble a bird; while there are, on the other hand, some striking resemblances with the reptile in the pelvis, tail, and articular parts of the limbs. Sometimes the characters of the two classes run side by side, as in the skull, the fore-limbs, and especially in the hind-limbs, where the shin of a Bird is connected with the foot of a Saurian. The parts in which it corresponds with birds show that Pterodactyles also were flying animals. That we should be entitled to conclude, from the hollow state of the bones, that they belonged to flying animals, is sufficiently proved by Blumenbach, Buckland, Mantell, Owen having mistaken them for bones of birds.
The most absolute proof that it was a flying animal is the pneumatic character of its bones. This condition was discerned by me in some Pterodactyle bones from the Lias of Franken (Jahrb. für Mineral, 1837, p. 316), and was afterwards established by Owen in the Pterodactyles from the Chalk of England. This structure was previously only known in birds. And the supposition readily follows that in the respiratory process there was some similarity between the Pterodactyle and the Birds. They have the proportions of upper-arm and fore-arm which characterize birds of great flight, the humerus short and the fore-arm long; hence it may be presumed that Pterodactyles could fly well. From the absence and presence of the bony sclerotic ring in the eye, it may be supposed that the Pterodactyles were active in the day-time, while Rhamphorhynchus was nocturnal.
After this statement von Meyer gives a discursive summary, in which his views of the classification of reptiles in general and of Pterodactyles in particular are epitomized. And then goes on to combat the views of people who have departed from his classification and attempted to set up classifications of their own; and cites a number of authors who, labouring at the vertebrata, « 24 » have endeavoured to find a resting-place in their systems for the Pterodactyle. But the chief thing we learn of von Meyer's own views is, that in 1830 he published a classification of extinct Saurians, dividing them into those with limbs like the larger and heavier land-mammals, those with fin-like limbs, and those with a flying-finger. Which divisions have been widely adopted, though authors have sometimes given them other names than those by which they were first made known.
Von Meyer has freely stated the facts about the Pterodactyle, and draws the conclusion that the animal was a reptile; but how such a conclusion was obtained from such facts is a matter on which his pages are silent. One seems to hear the chirrup of the bird in almost every paragraph. The head is in the main a bird's head; the pectoral girdle and the sternal ribs are those of a bird; and very few are the structures in which some reminder of the bird is not present; and in their bones he discovered the pneumatic characteristic and inferred, for the animals bird-like lungs. How, then, comes it that the Pterodactyle is a reptile? We can only suppose the answer to be, Because if the head and pectoral girdle and other bones had been reptilian it would have been a bird.
* * * * *
In the views here epitomized it is difficult always to make out the logical foundations of the conclusions arrived at. Sometimes they have no foundations, and sometimes they represent the different aspects in which a truth presents itself to minds differently constituted or differently conversant with the structures of living animals. In now stating my own views I shall avail myself of the example of some previous writers, and attempt to investigate the Pterodactyle as though they had not written. And then, having placed before him all the theories that are known, the reader will be able to choose the theory that pleases him best, if indeed he needs one.
Much of the discrepancy of opinion that exists is probably due to the use of the inductive method of thought for the discovery of fundamental principles in classification. In palæontology, where the types are more generalized than are living forms, it must always be difficult to reason from the known to the unknown. The known is always more or less incomparable with the unknown; and there can be no reason for inferring that the specialities of « 25 » structure which now accompany specialities in organization would justify us in inferring for the animal, in which the structures formerly were united, the combined organizations of the living animals in which they are now found. On any hypothesis of evolution it would be allowed that the special modifications of a group were attained subsequently to the common plan of the larger group to which it belongs, and are entirely to be attributed to the function which the necessities or organization of the animal caused its structures to subserve. Inductive thought may sometimes discover function from structure, but never makes more than an approximate guess when it endeavours to determine fundamental organization from osseous structures which are not fundamental. And before a naturalist can say, since an animal has for instance a tail like a mammal that in so far it must be affiliated to the mammalia, he must have determined why the mammalian tail has its peculiar characters, and whether it is compatible with any other common plan of organization. And perhaps it might with equal reason be considered reptilian.
Therefore I prefer at firsts instead of reasoning from the details of structure, to adopt the à priori method, and ask, not what the Pterodactyle is like in its several bones, but what common plan it had whereon its hard structures were necessarily moulded. For I imagine, if it can be determined what the nervous and respiratory and circulatory structures of the Pterodactyle were, it becomes a secondary matter to know whether the phalanges are like a lizard's, or the pelvis like that of a mammal. If the animal is asserted to be a mammal, a reptile, or a bird, we ought to be able to adduce evidence that it had the soft parts which are deemed distinctive of the selected class. This no one has done or attempted to do.
Hereafter it will be necessary to describe the Pterodactyle's brain.
There is no organ more distinctive between hot-blooded animals on the one hand, and cold-blooded animals on the other, than the brain. In the cold-blooded groups, or those in which respiration is feeble and circulation imperfect, that is to say, in existing fishes, amphibians, and reptiles, the parts of the brain are arranged one behind another, so that when looked upon from above, a portion called the optic lobes intervenes between the anterior masses called the cerebrum and the posterior mass called « 26 » the cerebellum. In the hot-blooded groups, or those with an enormous extent of lung-surface for oxidation of the blood and a four-celled heart for its rapid circulation, that is to say, in birds and mammals, the front part of the brain called the cerebrum is immensely developed in proportion to the other parts, and abuts against the cerebellum and more or less completely covers the optic lobes, which in birds are squeezed out to the sides. The Pterodactyle brain is of this latter kind. And it being taken as a postulate that this kind of brain is the product of the organization which produces hot blood, it follows that the Pterodactyle was a hot-blooded animal.
Again, the Pterodactyle has perforations for pneumatic cells in many of the bones.
There is no structure in the animal kingdom more distinctive of a Class of animals than air-cells perforating the limb-bones. They are connected with a peculiar kind of lung and heart—those of the bird; for in this Class the bronchial tubes open on the outer surface of the lungs into air-cells, which are prolonged through the body into the bones. They follow the blood-vessels, and are most developed in the part of the body most used. In some lizards, as the Chameleon, the sack-like lung at its distal termination is as simple as the air-cells of a bird; but those air-cells are not comparable with the bird's air-cells, since they are not prolongations of the bronchial tubes through the walls of the lungs. And it cannot be inferred that a reptile with wings would develop air-cells like those of a bird: in the first place, because those mammals which have wings do not develop air-cells; and, in the second place, because there is nothing in existing nature to lead any one to think that reptiles might have wings. The mammalian lung is better comparable to that of a bird than is the Chameleon lung, and therefore the air-cell structure might with better reason have been anticipated to occur in the Chiroptera than in a Lizard-ally, if it were dependent on the development of wings. Moreover, among Struthious birds the legs have more of the air-cell prolongations than the wings. Therefore, being a peculiar Avian structure which only exists in association with the Avian heart and lung, it follows that because the Pterodactyle had the pneumatic foramina it also had the structures of which they are the evidence, viz. lung and heart formed on the bird plan.
Thus Pterodactyles have a nervous system of the bird type. That kind of brain only exists in association with a four-celled heart and hot blood.
They have a respiratory organization which is only met with among birds.
With that respiratory apparatus is always associated a four-celled heart and hot blood, which it would necessarily produce.
And with that respiratory organization is always associated a brain of the type that the Pterodactyle is found to possess.
Therefore it is firmly indicated that the general plan of the most vital and important of the soft structures was similar to that of living birds.
This proposition will be incidentally proved in the following memoir, in which it will be seen that with such a common plan, is associated a diversity of details sufficient to demonstrate that these animals are not birds, but constitute a new group of vertebrata of equal value with the birds—the sub-class, Ornithosauria.
ILLUSTRATIVE OF THE MODIFICATIONS OF THE
ORNITHOSAURIA (OR PTERODACTYLES) IN THE
CAMBRIDGE UPPER GREENSAND.
| Case. | Comp. | Tablet. |
| J | a | 1 |
The Sternum is the key to the bony apparatus supporting the anterior limbs. In the Pterodactyles from the Cambridge Greensand it has been well figured and described by Professor Owen, who enunciated its resemblance to the sternum of birds. The sternum in Pterodactyles from the Lithographic Slate, shows its proportional size to the body. The examples found in the Cambridge Greensand have as yet shown no evidence of a composite character like that attributed to Rhamphorhynchus Gemmingi.
The sternum consists of an expanded symmetrical shield having its lateral halves, which are inclined to each other at a large angle (about 150°), contracted superiorly, behind and immediately below the synovial cavities for the coracoids. The vertical angular ridge in which the lateral portions of the sternum unite becomes elevated as it is followed anteriorly, into a strong keel. This keel or interpectoral process is highest in front of the articulations for the coracoids; but the degree of elevation varies with the species. It is prolonged upward and in front of the coracoids for some distance, becoming very massive, and the prolonged mass which is flattened from side to side, reaches « 29 » laterally to the outer margins of the coracoid articulations, and on the visceral side a little between and over them. The anterior crest of the keel shows the attachment of powerful muscles.
Professor Owen has observed that only in birds are distinct synovial cavities provided for the coracoids, and that no reptile has a sternum showing characters like those seen in the Pterodactyle. These coracoid cavities are placed as in birds, close together, behind the manubrium, which forms the hindermost part of the keel. They are convex transversely, concave from front to back as in birds, and look upward at an angle of 35°, their main direction being outward and a little backward. Professor Owen recognises the function of the shield-shaped sternum in relation to the mechanism of respiration on the one hand, and on the other hand, for the attachment of pectoral muscles of great bulk and strength.
As is well known, the muscles of the breast in most birds consist chiefly of the 1st, 2nd, and 3rd pectoral muscles, and the coraco-brachialis.
The peculiar form of the bird's sternum appears to be due to the vertical development of the second pectoral muscle, since when the 1st and 3rd muscles are dissected off, the appearance presented nearly resembles that of the sternum in Pterodactyles. There can however be no doubt but that the third pectoral muscle, which in most birds is but feebly developed, attained a far greater bulk in the Pterodactyle, because there is evidence of its powerful insertion in the distal anterior face of the coracoid, as well as of the great lateral extension of the sternal shield to which such a muscle must—by the analogy of birds—have been attached. The peculiar lateral emargination of the sternum appears to be due to the anterior sternal termination of this muscle, caused by the outward direction of the coracoid bone.
Since the coracoids were developed outward and backward so much more than in birds, it would happen, from the apparent different direction of the second pectoral muscle, that the first pectoral muscle which in birds skirts the furculum, must have passed over the coracoid, probably pulling on its inside in opposition to the third pectoral. Either a subdivision of this muscle or a distinct muscle in the same place, in function corresponding to the subclavius muscle, appears to have been powerfully attached from « 30 » the anterior prolongation of the keel of the sternum to the front face of the coracoid. It is improbable that the second pectoral muscle was undeveloped, but merely directed differently to what it is in birds, since, as will be seen, there is a process at the proximal end of the coracoid homologous with that which forms the pulley round which this muscle in birds works.
Professor Owen concludes his remarks by observing that the Pterosaurian breast-bone is in the main formed on the ornithic type. The muscles also appear to be similar to those of birds.
All the specimens are much mutilated, but all show the distinctive post-coracoid lateral emarginations, but as these are not seen in German Pterodactyles they are to be regarded as characters of a peculiar sub-order and not as characteristics of the sub-class.
The example figured in this memoir and by Professor Owen is 25/8 inches in antero-posterior measurement, probably about one third its entire length.
A small example in the collection of Mr Reed of York extends 11/4 inch in the same measurement, and by the analogy of P. suevicus was more than twice that length when perfect. It is remarkable in that the coracoid facets look much less outward and much more backward than in the larger species.
The mammalian sternum is usually in many consecutive pieces like the vertebral column. The types in which it attains any size as an expanded shield are Cetaceans and the Manatee, but in these groups it has no keel and is not connected with the other bones of the pectoral girdle. The proximal portion of the sternum of the Mole is elongated and bird-like, with the shield narrower than in the typical gallinaceous birds, and with the keel similarly developed. It is connected with the humerus by small sub-quadrate bones named clavicles placed at the sides of the proximal end. The sternum in Bats usually consists of a proximal and a distal part. It is narrow except at the proximal-termination where it widens like the letter T or Y; and to the sides of the lateral prolongations are attached the long, slender, curved bones named clavicles, and a pair of ribs. This sternum develops a bird-like keel. Both Mole and Bat are regarded as differing from Pterodactyles in the bone giving attachment to the clavicles instead of to the coracoids. The proximal part of the sternum in both the living animals, gives attachment to but one pair of sternal « 31 » ribs. The Pterodactyle sternum otherwise differs from the Bats in having the articulations for the coracoids close together, of a peculiar concavo-convex character, with a massive portion or keel prolonged forward in front of the coracoid articulations. The Bat cannot be said to resemble the Pterodactyle closely. The sternum of the Mole differs from that of the Pterodactyle in having a less developed shield, and in having a more developed keel which is not prolonged in front of the coracoid articulations. These examples demonstrate that resemblance in conformation is functional, and no proof of affinity.
Pterodactyles make some approach in the proportions of their sternum to Struthious birds. But the Struthionidæ have the bone thick, do not develop a keel, nor, have they an inter-coracoid process while the coracoid articulations are singularly long and narrow instead of being ovate. With other birds the Pterodactyle sternum agrees in giving attachment to the coracoid bones by synovial articulations, in the bone being shield-shaped, and supporting a more or less developed keel. The keel is chiefly developed at the proximal end, as in the Albatross, which has the bone broad; and it is prolonged in front of the coracoids exactly as in Mergus merganser, which sternum if a little broader in the shield and thicker in the keel would very nearly reproduce the sternum of the Pterodactyle, even to the "post-coracoid lateral emargination" of Cambridge specimens. Among reptiles the only form which suggests comparison is the Chameleon, in which however the sternum consists of an anterior and a posterior part as in the Bats, the back part narrow, and the front part a long lozenge shape, with a keel made by inclination of the sides of the bone to each other as in the Dodo, but the keel such as it is, is at the back part of the bone, and there is no prolongation in front of the coracoids as in Pterodactyle. The coracoids are broad, and are applied to the two anterior sides of the lozenge. The Crocodile has a narrow flat sternum which is prolonged anteriorly between the coracoids.
The resemblance is greater with mammals than with reptiles. From birds the Pterodactyle sternum makes no essential difference, and in the Merganser finds a close ally.
| Case. | Comp. | Tablet. | Specimen. |
| J | a | 2 | 1—23 |
Commonly the coracoid in the Cambridge Pterodactyles is anchylosed to the scapula: occasionally the bones are separate, though the separation has hitherto only been observed in the largest species. In 1851 Professor Owen, when figuring the anchylosed ends of the scapula and coracoid in Pterodactylus giganteus (Bowerbank), observed that in no part of the skeleton does the Pterodactyle more nearly resemble a bird than in the scapular arch; a view again urged emphatically in 1859 when similar fragments were described from the Cambridge Greensand. Since then perfect examples of the coracoid have occurred, which show the characters given in the following description.
The bone is long, with sub-parallel sides, sub-triangnlar in section, with the proximal end expanded exteriorly and posteriorly, resembling in form the coracoid of a bird. The front surface looks forward and outward; it is flattened, is a little convex transversely, and a little convex in length; it is rugose with muscular attachments, which terminate in a tubercle on the uppermost fourth of the front, usually near to the inner side. The middle third of the slightly concave inside margin of the front aspect, is sharply angular; the parts above and below it have the angularity rounded off. The outside margin, a little more concave than the inside margin, is sharply angular in its distal third, in which the front gradually widens to near the sternal articulation, when it contracts—the whole sternal termination of the bone being directed a little inward towards the manubrium of the sternum. The inside, which faces the opposite coracoid, is convex transversely in the lower half or two-thirds; its distal termination is carried inward. The expanded proximal end of the inside is flattened, or channelled, by the developement inwardly, at the proximal end of the ridge formed with the front side, of a long strong process homologous with that on the inner side of the coracoid in birds. The channel so formed rounds on to the proximal surface of the bone, and extends backward to the limit of the scapula; over it the second pectoral muscle may be presumed to have worked[N]. The third side of the « 33 » bone is much more concave in length than either of the others; it looks backward, outward, and downward, the proximal end being turned outward and downward more than the distal end; it is a little concave transversely at the expanded proximal end. Near the distal end there are sometimes visible a few faint marks of the insertion of muscular fibres, but they are much less distinct than those made by the coraco-brachialis muscle in the corresponding region of the coracoid in birds. Throughout its length it rounds into the inner side, and the upper third rounds convexly into the front. On the most posterior part of this aspect of the proximal end is a groove terminating in a long pneumatic foramen, partly in the coracoid, partly in the scapula.
[N] The homologous process is more developed in Pterodactylus giganteus. See f. 7. pl. XXXI. Owen, Cret. Rept.
The muscular attachments on the front aspect of the coracoid appear to be two; one large and long inserted into the inner half of the middle third of the bone, terminating at the proximal end in a tubercle. No specimen shows the distal end of the insertion. This may indicate a subdivision of the first pectoral muscle. The other insertion, if it be distinct, is long and much narrower and at the distal end of the bone. This, according to the analogy of birds, should be the third pectoral muscle; if the insertion should be but part of that to which it is distally adjacent, then the third pectoral muscle must have had an enormous developement unparalleled in birds.
The distal end of the bone terminates in a synovial articulation concave transversely, convex from front to back, in form transversely ovate: the narrow side of the articulation, like the thin edge of the coracoid, being exterior. The articulation is about three fourths of the transverse diameter of the distal end; it is at right angles with the long axis of the bone, and looks downward and a little backward.
The proximal end, massively enlarged outward and backward, presents on the proximal surface three well defined regions. The largest of these is an irregular flattened surface half ovate in form, inclined to the axis of the bone at about 45°, looking backward, and upward also, when the bone is held vertically; the mesial hindermost half of the radius of this area is occupied by a pneumatic cavity: to this surface is applied the scapula. The next largest surface is rectangular and oblong, looking upward, outward, and a little forward. The transverse aspect which looks outward « 34 » being nearly half as long again as the antero-posterior aspect which looks forward; in the latter direction the area is slightly concave, in the former direction it is slightly convex; its posterior boundary is parallel with the front of the bone: this area forms the anterior moiety of the glenoid cavity, to which the proximal end of the humerus is applied.
The remaining surface of the proximal end is sub-quadrate, adjoins the two other surfaces as well as the front and the inside of the shaft, it is conically concave.
The entire bone when applied to the sternum looked outward, backward and upward.
Professor Owen remarked (1859) that the "coracoid is shorter and straighter in birds than in Pterodactyles, but is commonly broader, and with a longer and stronger anterior process."
The points in which the Pterodactyle coracoid resembles that of birds (e. g. Gallinaceæ) are the long slender triangular shaft; the concavo-convex articulation to the sternum; the convexity of the distal end in front, and its concavity behind; the posterior aspect of its scapular surface, and the pneumatic foramen.
The points in which it is distinct from birds are that the bone is not produced proximally beyond the glenoid cavity for the humerus, which, instead of being lateral as in birds, and looking outward, in Pterodactyles forms the proximal-termination of the bone. The sternal articulation is proportionally much shorter transversely in Pterodactyles, terminating in a convex margin which rounds up into the thin outer margin, as in the immature coracoid of the common Cock. It is bow-shaped in front instead of being straight, and is commonly longer than in birds. The usual ossified connection with the scapula is not entirely unparalleled in birds, the whole pectoral girdle being sometimes anchylosed into a bony mass as in the frigate bird.
In the monotremata, the only mammals in which the coracoids are separate bones, they rather recall those of Ichthyosaurus than those of any other animals, and have no connection with the sternum. The bone which represents it functionally in placental mammals is the clavicle.
In no reptile is there any structure resembling the Ornithosaurian coracoid. The nearest approximation is made by the Crocodile, in which as in the Chameleon the pectoral girdle is « 35 » formed as in pterodactyles and struthious birds by scapula, coracoid and sternum. But in the Crocodile the coracoid is compressed, and expanded from side to side both proximally and distally. Distally it has no synovial articulation with the sternum; and proximally a wide process of the bone extends beyond the articulation for the humerus as in birds, only the scapula unites with the prolonged part, and the glenoid cavity looks forward and inward.
The coracoid is essentially avian in its affinities, though with peculiar characters of its own. In the German genera it closely resembles specimens from the Cambridge Greensand.
23 specimens are exhibited. Nos. 4, 10, 12, are the middle parts of shafts of left coracoids. Nos. 3-12, 22, are the middle parts of shafts of right coracoids. Nos. 2, 5, 14, are proximal ends of left coracoids. Nos. 1, 6, 8, 9, 23, are proximal ends of right coracoids. Nos. 15, 16, 17, 18, 19, 20, 21, are distal ends of left coracoids. No. 13 is a nearly perfect left coracoid, and No. 7 is the glenoid cavity for the humerus formed by a right coracoid with the anchylosed scapula.
| Case. | Comp. | Tablet. | Specimen. |
| J | a | 3 | 1—17 |
| 4 | 1— 6 | ||
| 5 | 1— 4 |
Professor Owen described the scapula of Pterodactylus giganteus in 1851, and added further particulars regarding the Species from the Cambridge Greensand, in 1859; but, as with the coracoid, only the humeral end has hitherto been figured. The only example sufficiently perfect to give the length and proportions of the bone is preserved in the collection of Mr Reed, of York. This left scapula is a stout strong bone, short in proportion to its strength, of flattened ovate form in section, expanding at the humeral end into an irregular sub-rhomboid mass. It is smaller in the middle, contracting both from side to side and from back to front till the back to front measurement is 7/16 of an inch, and the side to side measurement is 11/16 of an inch, and it expands a little at the free end, which terminates in a smooth heart-shaped surface, convex in the long diameter, which measures 7/8 of an inch, and flat in the short « 36 » one, which measures nearly 5/8 of an inch; it is at right angles with the inside of the bone. The sharp superior lateral outline is concave, but less so than the inferior lateral outline; into that inferior aspect of the bone the sides are more fully rounded. The flattened inner surface applied to the ribs is concave in the length of the bone, which measures 31/2 inches; the posterior half of which is convex transversely, the anterior humeral half is concave transversely so as to be cup-shaped, and measures in extreme width 111/16 inch; the outline of the transversely convex outer side in length is nearly straight, but the exterior part and glenoid cavity of the proximal end is broken away, and there only remains a small median proximal surface broken at both ends, a little concave in length, measuring 5/8 of an inch, and convex in breadth measuring 1/4 of an inch.
As there is no specimen in the Woodwardian Museum showing clearly the connection of the proximal with the distal end, the specimens are arranged on separate tablets.
Humeral End of Scapula.
The humeral end of the scapula exhibits in the different species much diversity of form, spreading laterally from the shaft, and terminating in an elongated articular surface truncating the bone nearly at right angles. On its inferior border it throws out a large convex tuberosity, separated from the humeral articular surface by a deep emargination. From the tuberosity usually arises a crescentic row of muscular insertions, which is continued inward and forward over the most compressed part of the scapula towards the middle of the humeral articulation. From the superior margin, interior to the coracoid, arises a prominent ridge, the spine of the scapula, which is directed diagonally backward and downward, terminating in the middle of the outer surface, where it is bordered on the anterior aspect by a long narrow muscular attachment. Between this spine and the elevated margin of the glenoid cavity the bone is much compressed and concave.
On the inside surface of the bone there appear to be small muscular attachments in front of and behind the great tuberosity. The area between the spine and the inner surface is sometimes flattened, sometimes gently convex.
With well-marked distinctive characters in the inferior tuberosity, « 37 » the pre-tuberous emargination and the thick rounded form of the bone, the Pterodactyle scapula is intermediate in character between that of a mole, a bird, and the crocodile; wanting the sabre shape of the bird's scapula, it also wants the wide expanded form of the scapula of the Crocodile, but resembles the latter in the direction and degree of developement of the spine. This modification is probably due to the outward direction and clavicular function of the coracoid, as well as to the raptorial habit of the organism.
In no living Reptile is there a scapula to be compared with that of the Pterodactyle, for besides the free end being expanded, in the crocodile, it is also thin and squamous and the bone makes a continuous curve with the coracoid as in struthious birds, and not a sharp angle as in Pterodactyles. The "spine" in crocodiles is on the anterior border of the bone and directed upward and backward, while in Pterodactyles it is on the posterior border and directed upward and forward. In the Chameleon the scapula is more elongated and narrow, narrower in proportion to its length than in Pterodactyle, but becomes rapidly wide at its union with the coracoid. It is curved in length so as to fit on to convex ribs. A scapula presenting some resemblance to Pterodactyle is found in certain Liassic Ichthyosaurs.
Among mammals a straight elongated narrow scapula is rare. The mole however has a scapula of this kind somewhat cylindrical in its proximal half and not much expanded at the free end, on which there is a small spine. The anterior emargination above the glenoid cavity in Pterodactyle is entirely mammalian, as is the anterior tuberosity above the emargination, for it entirely corresponds with what in ruminants, pachyderms and many mammals would be named the coracoid process. If that process is accurately determined it is difficult to say what this is.
In birds there is often a prolonged process on the inner side of the coracoid, which however extends interior to other parts of the scapula, and to this the furculum is attached. Such traces of a spine as are to be detected in the swan conform to the Pterodactyle.
No bird has the scapula cylindrical, even struthious birds only making an approximation to such a condition; and no birds have the scapula so straight. The bone is more avian and mammalian « 38 » than reptilian; and more avian than mammalian but with strong distinctive characters of its own.
17 specimens of the humeral ends of scapulæ are exhibited. Nos. 1, 4, 6, 7, 8, 9, 11, 13, 14, 15, 17 are left scapulæ. Nos. 2, 3, 5, 10, 12, 16 are right scapulæ.
The tablet of the distal ends of scapulæ comprises 6 specimens.
| Case. | Comp. | Tablet. | Specimen. |
| J | a | 6 | 1—46 |
| 7 | 1— 3 | ||
| 8 | 1 |
There are among the fossils of the Cambridge Greensand at least two well-marked types of Pterodactyle humerus, readily recognised by the forms of the proximal and of the distal ends, and by the positions of the pneumatic foramina. In the group having the ulnar ridge developed the pneumatic foramen is on the posterior aspect of the bone[O] under the ulnar ridge, as in birds; but in some of the small Pterodactyles the foramen is on the anterior surface, and on its radial side. This latter kind of humerus has the distal end more or less divided into three convex surfaces, while the radial crest is enormously developed and terminates in a smooth oblong flattened surface nearly as large as the proximal articular surface, and looking anteriorly. The distal articular surfaces are not as in birds parallel to that of the proximal end, though they agree with those of birds in being at right angles to the radial crest; this ridge in Pterodactyles being directed much further outward and backward than in birds.
[O] Professor Owen states (p. 16, 3d Supt.) that the foramen is palmar. Fig. 15. T. III. 2d Supt. shows it to be anconal.
The largest forms of Pterodactyle all have the distal articular surface flatter, and the proximal articulation less bent back so as to look more upwards. No specimen of this kind of humerus has occurred with the radial crest preserved; but it is apparently carried farther down the shaft and not so far forward as in the other group. This latter kind of bone is shown by Prof. Owen in T. III. figs. 1, 2, 3rd Sup. Cret. Reptiles; the former kind has been illustrated in figure 5 of the same plate.
Some of the most gigantic Pterodactyles appear to have had « 39 » the limb-bones as solid as those of crocodiles, and unpermeated by air; and there is no evidence that the high Avian characteristics of most of these Greensand fossils also pertained to all the previously known types from the lower secondary rocks.
The osteological series comprises 46 specimens. No. 30 is a nearly perfect right humerus. Nos. 1, 2, 5, 6, 7, 8, 9, 11, 18, 22, 23, 25, 39 are examples of the proximal ends of left humeri. Nos. 3, 4, 10, 12, 13, 14, 15, 16, 17, 19, 24, 26, 27, 28, 38, 40, 41 are examples of the proximal ends of right humeri. Nos. 20, 21, 32, 33, 34, 35, 37, 42, 44, 45, are examples of the distal ends of left humeri. Nos. 29, 31, 36, 43 and 46 are distal ends of right humeri.
No. 30 shows the entire length of the humerus to be 21/2 inches. It has a nearly circular shaft with a diameter of a little more than a quarter of an inch, being more slender than the corresponding bone of Pt. suevicus, which has the same length. The proximal articular surface is crescentic, the anterior concavity corresponding with the concave anterior aspect of the proximal end, while the convex border corresponds to the convex posterior side of the bone, which it overhangs: it is worn, but appears to measure half an inch from the radial to the ulnar side. The ulnar ridge (which is worn) has not extended more than a quarter of an inch beyond the articular surface. The thin bird-like radial crest, arising rather more distally than the ulnar ridge, is flat on its posterior surface, and extends anteriorly for a distance nearly half as far again as the length of the proximal articular surface of the humerus. On the proximal third of the posterior face are two contiguous long narrow oblique muscular insertions. The proximal ends Nos. 22, 23, 24, 25 are examples of this kind of bone, having the pneumatic foramen radially situated on the anterior aspect near to the articular surface, as may be seen in No. 24. No. 25 shows the termination of the radial crest in an oblique oblong smooth surface, slightly convex in length and breadth, directed distally towards the ulnar side.
No. 6, 7, 13, 27, are examples of another kind of proximal end, where the pneumatic foramen is an oval hole on the ulnar side of the posterior surface. The radial crest arises more distally, and the ulnar ridge more proximally, than in the small species, like No. 30.
Nos. 4, 11, 14, 16 are examples of other species with the foramen placed as in the last group, only less near to the proximal end, while it enters obliquely, being directed distally from the broad concave area proximal to it. The largest proximal ends known, such as No. 2, which though very imperfect measures 23/4 inches over what remains of the articular surface, appear to conform to this latter type.
Distally the humerus No. 30 enlarges, widening rapidly on the radial side, which is bordered near the distal end by a sharp ridge showing a muscular attachment, while the ulnar side is rounded and rather inflated. The articular surface looks downward and in the direction of the radial process. There is a mesial concavity on the radial side which is bordered on the right and on the left by a prominent rounded condyle, and behind by a condyloid convexity. On that side which in conformity with the nomenclature applied to birds' bones, has here been named the ulnar side, the ulnar and mesial condyles are impressed with a flattened slightly concave sub-rhomboid area, which looks downward, backward, and towards the ulnar side. These characters are not well seen in No. 30, but may be effectively studied in their specific variations in Nos. 36, 37, 42, 43, 44, 45, and 46.
Nos. 20, 21, 29, 31, 32, 33, 34, 35, are examples of the distal ends of humeri of a different type. They are mostly larger than the preceding group, and correspond in characters with the large proximal ends, but appear to be separable into two groups, namely those with a pneumatic foramen on the anterior radial side near to the articular surface, and those where no pneumatic foramen is seen. Unlike the previously considered type, the ulnar side is sometimes more inflated than the radial side.
The mesial condyle in this group appears in every case to be an epiphysis, which is wanting. The radial condyle becomes a large flattened slightly convex surface looking downwards, which in some of the species, as Nos. 21 and 32 (in other respects remarkable species), shows an approach to a trochlear character on its anterior side. In Nos. 33, 34 and 35 the mesial anterior concavity becomes flattened and abuts at an angle against the flattened radial condyle. No. 20 shows the rhomboid impression on the ulnar side to be more concave and more ovate. The ulnar condyle remains a smaller but prominent tubercle directed distally. Nos. 21, 22 and 34 show a « 41 » ridge developed on the ulnar side of the shaft like that on the radial side in the other group, while the radial ridge is not so near to the articular surface. The largest and smallest distal ends of humeri known, both show the characters here enumerated. The great distal end of a left humerus, figured by Prof. Owen, Pl. IV. f. 1, 2, 3 of the 1st Supplement to the Cretaceous Pterosauria, is of this kind, and though imperfect measures more than three inches over what remains of the articular surface. In the small humerus, No. 30, the width over the distal articular surface is 5/8ths of an inch. If it is assumed that the large bone was no more than 5 times the length of the small one, the entire length of the humerus would have been about twelve inches. The smallest humerus, No. 29, measures over the shaft rather more than one eighth of an inch.
The Ornithosaurian humerus has but little in common with that of any mammal. Most mammals have the proximal head of the bone hemispherical, and a pit at the distal end for the olecranon process of the ulna, while there is usually little indication of a radial crest, and the proximal and distal ends are in the same plane. In the Bat however the bone is twisted a little so that the slight radial crest looks in the same direction as the distal end, here also there is no pit for the olecranon; but the bone is sigmoid and proportionally much longer than in Pterodactyles. In the horse, hippopotamus, &c., the radial process becomes more developed but never resembles that of a Pterodactyle.
Among reptiles, the bone may be compared with lizards and crocodiles. In crocodiles the proximal and distal ends are nearly in the same plane, the distal end has two condyles, the head is convex from side to side, and the radial crest is moderately developed and never extends so far outward or so far proximally as in Pterodactyle. In the Chameleon the bone is more twisted than in Crocodile, and as in Pterodactyle the distal end is compressed on the radial side to a sharp margin. In Iguana, Scink, and Monitor both proximal and distal ends are much expanded, and the radial process makes no approximation to that of a Pterodactyle.
The bird humerus does not approximate more closely in form to that of the Pterodactyle than does the Chameleon humerus, though it has the cardinal distinction of pneumatic foramina, and these sometimes corresponding in position in the two groups.
The bird humerus is commonly longer, though in the parrots the proportions and straightness are not unlike Pterodactyle. In some respects a nearer resemblance is seen in the raptorial bird Gypogeranus serpentarius, in which the radial process is rather more developed than in the Crocodile, and extends further proximally though still much smaller than in Pterodactyle; here too the superior surface is concave from side to side, and the distal articulation is not unlike that of some Pterodactyles. But no Pterodactyle has the head of the humerus convex from the radial to the ulnar sides, and the bird is distinctive in having the ulnar crest developed on the inferior side of the head: a faint approximation to a similar development is seen in Crocodile, but there is no trace of such a process in Pterodactyle. The distal end is more Bird-like than Lacertian in form, but is twisted to a greater angle with the proximal end than in birds.
Altogether the bone is distinctive. The points in which it is unlike birds and reptiles are those in which Birds and Lizards resemble each other; it would not be easy to say that in form it resembles one group more than the other. But it is linked with birds by the pneumatic foramina.
| Case. | Comp. | Tablet. | Specimen. |
| J | a | 9 | 5— 6 |
| 10 | 1—10 | ||
| 11 | 1— 7 | ||
| 12 | 1— 4 | ||
| 13 | 5— 6 |
Of neither of these bones has a perfect specimen been found. While fragments of humeri are met with frequently, fragments of these bones are rare. In accordance with the analogy with birds the Ulna might be presumed to be the larger bone of the two. But from a study of German specimens the larger bone is found to be the Radius, which according to the mammalian plan is placed in front of the ulna. As a whole, the fore-arm of Ornithosaurians is only to be compared with the insectivorous mammal Chrysochloris Capensis, in which there are also three bones in the fore-arm,—the third bone like the Pteroid bone in Ornithosaurians, extending about half-way from the carpus to the humerus, and holding, relatively, a similar position and development to the fibula in bats.
The pteroid bone articulated with a separate carpal, and was placed on the side of the arm, adjacent to the radius, which at the distal end extended in German specimens more inward than the ulna. In Chrysochloris the third bone appears to be behind the other bones, and adjacent to the ulna[P].
[P] See D'Alton and Pander Chiropteren und Insectivoren, Bonn, 1831, pl. 5, Chrysochloris.
Among neither birds nor reptiles is any comparable modification of the fore-arm to be found. Then by examining the proximal surface of the proximal carpal, the characters of the distal end of the Radius are readily discovered. The proximal carpal shows on the same surface another articular facets with which however only one fragmentary distal end of a bone corresponds. That accordingly is identified as the ulna. Besides these, three other articular ends of bones occur, one of which fits on to the distal end of the femur. The remaining two are both large bones, with epiphyses which formed portions of the articular surfaces, and are usually wanting. One of these bones corresponds in form with the ulna of a bird, and would fit the facet on the ulnar side of the distal end of the Pterodactyle humerus. The other bone is massive with a sub-quadrate articular end, and might well be the proximal end of the radius. Some specimens are among the largest fragments of Pterodactyle bone known. The only other bone that either of these could be is the distal end of the tibia, a bone not yet known, but probably not unlike that of a bird.
I. Distal End of Ulna.
Four specimens which show articular ends such as the ulna should have, are mounted together. They are compressed bones with the section of the fracture elongately oval; and the shaft widens from the fracture to the articulation without increasing in thickness. The outer surface is gently convex, becoming concave mesially near the articulation; the inner surface has the same characters, only the concavity at the extreme distal end reaches from side to side of the bone. The two short sides both look outward as well as laterally; one of them flattened so as to thicken the bone, is concave in vertical outline owing to the extreme distal end turning suddenly outward; the other side a little convex, compresses the bone and inflects its inner margin. The longest « 44 » specimen measures 15/8 inch; 5/8 inch wide at the fracture, and 11/8 inch wide at the distal end. The greatest thickness at the distal end is half an inch, the thickness of the fractured shaft is 5/8 of an inch.
The articular surface appears to have an elongated sub-reniform shape, the part at the compressed side of the bone being narrower than the broad ovate part on the thick side of the bone, to the lateral limit of which it extends, while the narrow part does not extend laterally nearly so far as the inflected border, which appears to give attachment to powerful muscles. There is also a strong muscular attachment at the corresponding diagonal corner of the bone where the outer surface on its right meets the side of the bone in an elevated ridge.
In its long diameter the articulation is a little convex; transversely it is very convex in the ovate part, but more flattened in its narrower continuation. Where widest it measures about 4/10ths of an inch.
Nos. 5 and 6 on another tablet appear to be distal ends of ulna of another kind of Pterodactyle. They are less compressed, more quadrate in section, and have the sides more nearly parallel The flattened side similarly has a concave border, but instead of having its distal termination developed laterally, has it thickened behind. The opposite side of the bone which in the other specimens was compressed is here thick and well rounded, and not at all inflected. There is an absence of the concavity noticed on the outer surface of the bone in the compressed specimens. The articular surface is much flatter, and a little concave in length instead of being convex; as in the other examples it looks downward. The largest fragment. No. 5, measures 13/8 inch long; it is 6/8 inch wide at the fracture, and 4/8 inch thick. The sub-quadrate distal end is more than an inch long, more than 4/8ths inch thick on the thick side, and nearly 4/8ths inch thick on the compressed side.
II. Distal End of Radius.
The best preserved of the 10 specimens here exhibited is 3 inches long, No. 2. The shaft is oval, flattened on one side; measuring at an inch from the fractured end 7/10ths of an inch in the least diameter, and one inch in the wide diameter. It widens « 45 » distally at first slowly, then rapidly, till at the articular end its greatest width is two inches. But while expanding laterally it contracts from side to side, the more convex side of the two at about an inch from the articular end, beginning to approximate to the flatter side till the articular end has a short diameter of less than half an inch.
On the left-hand corner of the convex inner side of the bone is an elevated flattened disc for muscular attachments, fully half an inch in diameter, there is a slight muscular attachment interior to this, nearer the middle of the bone. The left-hand corner of the flattened outer side of the distal end of the radius is marked by a vertical ridge bordering a similarly elevated oval muscular attachment. Parallel to this nearer the middle of the side is a much stronger and acutely elevated ridge.
The articulation is made up of three distinct parts, all in a straight line. The portion of bone adjacent to the large muscular disc is compressed and rounded on the distal end; then first there is a rather deep circular cup 3/8ths of an inch wide, nearer to the more convex than to the flatter side of the bone; adjacent to this cup is a convex ball of about the same size; while the remainder of the articulation is concave in length, convex from side to side, and looks downward and a little towards the inner convex side of the bone. The specimens are arranged so as to display these characters.—The example described is of nearly the same size as that figured for the humerus in fig. 1, T. XXIV. of the Cretaceous Reptilia. The less well preserved bone in that figure exhibits the Ulna in its true position behind the Radius.
III. Proximal End of Ulna.
This bone has much the proportions of the Ulna in birds, the smaller specimens nearly resembling the ulna of the Heron. The specimen (No. 1) with the shaft best preserved is 21/4 inches long, cylindrical at the fracture, where it measures in diameter 3/16ths of an inch. It gradually enlarges proximally widening to about 7/10ths of an inch; near the proximal end it is a little curved, the side which is concave in length being a little flattened, while there is a lateral elevation on the opposite side, apparently corresponding to the quill-ridge on the convex side of the bird-ulna. There is a separate ossification for the olecranon, which is an irregular sub-oblong « 46 » bone forming the outer part of the articulation; it is only preserved in No. 1. Nos. 4, 5 and 6 show the concave transverse groove from which it has come away.
The articular surface looks upward and forward, in which aspect it has a trapezoidal form. Sometimes, as in No. 2, the great sigmoid area is divided into two parts by a vertical ridge, the more elevated part of the articulation on the radial side of the bone being concave, while the outer part, as in the heron, besides being concave, has its border on the concave side of the bone produced and rounded. There is a small triangular elevation on the radial aspect of the proximal end like that on the corresponding part of the ulna of the heron. On this aspect the bone is flattened, on the opposite and outward aspect it is compressed and produced as in the bird. No. 2 measures 11/8 inch over the articular end. The series includes 6 specimens.
IV. Proximal End of Radius.
This bone terminated in an epiphysis which formed part of the articular surface, and has disappeared from all the 7 specimens mounted. So much of the articulation as remains does not oppose the idea of its having been attached to the humerus, while the large size of the example No. 7, which could not have measured less than 21/2 inches from side to side over the articulation, is more in accordance with what is at present known of the dimensions attained by the distal end of the humerus than with the size that would be expected in the distal end of the tibia, which is the only other unknown bone to which these specimens could be referred.
The longest specimen, No. 3, is 3 inches long; broadly ovate at the fracture, measuring in the long diameter 1 inch, and in the short diameter more than 3/4ths of an inch. Nearer the articular end the bone becomes in section sub-quadrate or rather sub-rhomboid. No. 1 shows these terminal characters extremely well. On the posterior aspect of the specimen the surface is divided into two flattened slightly convex parts by a median vertical well-rounded angular bend. In front the side is similarly divided into two parts, both of them a little concave proximally, by a sharp median vertical ridge, which does not reach to the articulation by a varying distance, never so long as the bone is wide. The ridge « 47 » terminates in, and is pierced by, a vertical groove apparently for a nutritive vessel. Where the anterior and posterior aspects of the bone converge laterally the sides are well rounded.
Only a small part of the articular surface is preserved, looking upward and a little forward; it terminates the wider of the halves of the bone laterally and in front. The remainder of the articular surface, from which the epiphysis has come away, may be divided principally in the majority of specimens into a posterior flattened median rhomboid space and an oblong cup-shaped anterior space divided from it by an elevated ridge. The extreme lateral termination appears to have been a ball-shaped convexity.
The great length of the fore-arm relatively to the humerus, characteristic of German Ornithosaurians, from the fragmentary condition of Cambridge specimens is not seen.
Although the fore-arm resembles Chrysochloris in plan the resemblance is not close in the details of form. In many Mammals it is characteristic for the radius to be the principal bone of the fore-arm, and among Ruminants in which this is especially the case the radius is altogether in front and the ulna behind as is the position with Birds and Crocodiles. And among mammals with claws, as in the Lion, Bear, &c., and in the Chameleon, it is characteristic, for the radius also to be on the inside of the limb at the distal end, as in Ornithosaurians. In form, ridges, and muscular attachments (see pl. 3) the distal end of the radius approximates closely to the Bear and the Lion, and may also be compared with the Bats and Birds, though with Birds it is a small bone. From the epiphysis of the proximal end apparently being wanting it would be difficult to compare closely. But though not like any particular mammal, it might have pertained to a mammal since it has the large perforation for the nutritive vessel near to the proximal end as in the Camel and many of the mammalia.
The ulna of the Pterodactyles is at the proximal end altogether distinguished from mammals by the slight development of the olecranon, nor can the distal end, especially in its relation to the carpus, be paralleled.
Among birds and reptiles the ulna is the large bone, and here a general resemblance in form to the ulna of Pterodactyles is seen at the proximal end. It is not compressed from side to side as in the Crocodile, Iguana, Monitor, &c., but from back to fronts in this « 48 » rather resembling Birds than the Chameleon. It however at the distal end is more crocodilian.
The fore-arm in plan is mammalian. The Pteroid bone is mammalian, the Radius is mammalian and avian; the Ulna is avian, and crocodilian in form, but mammalian in proportion. The pneumatic foramen of the ulna is peculiarly avian.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 1 | 1—13 |
| 2 | 1—18 | ||
| 3 | 1— 4 | ||
| 4 | 1— 8 |
The pterodactyle wrist is made up of three bones, arranged as a proximal carpal, a distal carpal, and a lateral carpal. Two of them are figured by Professor Owen, who regarded the distal carpal of this description as the scapho-cuneiform; while A very imperfect example of the proximal carpal is named the unciform: neither of these determinations, the reverse of those which follow, were given as more than probable guesses.
I. Proximal Carpal.
No. 10 shows the proximal surface well; portions of it are seen in Nos. 4, 5, 6, 8, 9, 11, and 12. The distal surface is best exhibited in No. 1; portions of it are shown in Nos. 2, 3, 5, 7, 8. No. 13 is an impression taken from the proximal surface of a distal carpal to show its correspondence with distal surface of the proximal carpal. The bone is proximally of an irregular oblong form, being five sided, and much broader towards the inner end than towards the outer end. The two ends are sub-parallel, and rather obliquely connected on one side by a nearly straight border more than twice as long as the shorter end. The other limits of the sub-parallel ends are connected by two concave borders meeting in a well rounded convexity, which is near to the broader inner end.
The proximal surface of the bone is flattened, and may be divided into a sub-rhomboid space, adjacent to the shorter of the sub-parallel ends, which is moderately concave in the long axis of the bone and slightly convex transversely, and an oblong space adjacent to the longer of the two ends. This is separated from the sub-rhomboid « 49 » space, toward the straight side of the bone, by an elevated ridge sub-parallel with the ends. It is directed towards the convexity on the opposite side, in which the long and short concave parts meet, but after half crossing the bone it becomes forked in a U shape, and less elevated; the smooth unarticular included space shows an oval pneumatic foramen, which varies in size with the different species. The region between this Y-shaped ridge and the longer of the two ends, is sub-reniform, slightly concave in its long diameter, and deeply concave in the short diameter, exactly corresponding in form with the articular surface already described as the distal end of the ulna. Also parallel with the long end of the bone are marks of an articular surface exactly corresponding with those described as the distal end of the radius; that is, at the convex angle of the angulated side is placed a hemispherical boss,' interior to which is a hemispherical concavity, and extending toward the straight side is the oblique smooth border of the sub-rhomboid area described. There still remains a space to be accounted for. This consists of a sub-quadrate area forming the corner of the bone made by the concave side and the shorter outer end; it is made up of an inner concave part separated from the radial articulation by a ridge, and an outer convex part constituting the shorter end of the bone.
This carpal is moderately compressed from the proximal to the distal side, except towards the shorter end of the bone, being there prolonged distally into a wedge-shaped process, showing at its termination marks of a powerful muscular attachment.
The outer lateral surface is of variable antero-posterior extent.
The distal articular surface is placed entirely toward the narrow end of the bone, leaving at the proximal end a large smooth rhomboid unarticular area, of which every side is a little concave: it connects obliquely the proximal with the distal articular surfaces. The distal articular area is divided by a diagonal ridge into a long oblong area of which the inner and outer sides are sub-parallel and the ends rounded: it is slightly concave in length as well as transversely, and is slightly twisted like the flukes of a screw. Adjacent to this region laterally is the other and sub-triangular part of the articulation. The broad end of the triangle is toward the broad end of the bone; it is concave in length and flattened transversely. The two parts of the articulation « 50 » are inclined to each other at a large angle, both looking downward and outward, but on opposite sides of the bone.
II. Distal Carpal.
The tablets of this bone comprise 22 specimens. Nos. 2, 3, 4, 5, 6, 8, 9, 10, 15, 16, 19 and 22 are so mounted as to exhibit the proximal surface. Nos. 7, 11, 12, 13, 14, 15, 16, 20 and 21 show the distal surface of the bone. No. 17 is a cast from the distal surface of a proximal carpal for comparison with the proximal surface of the distal carpal. No. 16 is a cast from the proximal end of the wing-metacarpal for comparison with the distal surface of the distal carpal. No. 20 is a distal carpal of unusual type, 19 is a cast from its proximal surface, and 21 is a cast from the distal surface of the same specimen.
The proximal aspect of this bone is rather narrower than the distal aspect; each is sub-triangular in outline, the sides being convexly curved. In the long axis from the apex on the inside to the short outer[Q] side the bone is convex proximally with an oblique transverse depression; in the short axis, that is, between the two longer sides, the middle of the bone is hollow, but the oblique transverse depression makes both sides of the hollow convex,—so that excepting the smooth unarticular triangular area adjacent to the apex, the sub-quadrate articular surface is shaped somewhat like two cones put side by side in such manner that the apex of each touches the base of the other: the apex of that cone which should touch the short side or base of the triangle formed by the bone, is truncated by a depression which exhibits an oval pneumatic foramen. Towards the apex, on the same side as the pneumatic foramen, the margin of the bone is rounded for a small terminal oval articulation which looks outward and upward.
[Q] Outer and inner are here used in accordance with the usual interpretation, and the better to compare with birds.
The lateral aspects of the bone are at right angles to the proximal and distal surfaces. They are smooth, a little concave in antero-posterior extent, and convex in the opposite direction. That one on to which the marginal articular surface impinges is except for that surface sub-quadrate in outline; the opposite side has a slightly crescentic form, the flattened outline being distal. They show several small foramina.
The distal aspect of the bone is comparatively flat. The distal surface consists of a smooth unarticular part adjacent to the apex, rather smaller than the corresponding area on the proximal aspect of the bone. Between this part and the sub-crescentic articular surface, which occupies the remainder of the distal area, is a large circular pit, furthest removed from the side of the bone which forms the sub-apical marginal articulation. The pit on the apical side shows several small foramina; on the outer side of the bone the roughened articular surface extends down the pit side. The articulation is flattened from side to side of the bone. Its outer margin is slightly prominent, and the margin of the pit is slightly convex and prominent, so that the intervening articular surface in the direction between these limits is concave. It is commonly divided into two parts by a median band limiting a depressed half, which is in a slightly different plane from the other half of the articulation. Where the depressed part terminates towards the marginal articulation, which does not extend so far distally, there is between the two a small step-like roughened articular portion.
The large crescentic articulation described gave attachment to the wing-metacarpal bone; if there was a second metacarpal terminating in a claw, it must have been attached to the small articulation last referred to. In No. 20 the pit is extremely small, the impressed part of the articulation is small and deeply sunk, while the apicular articulation is widened and shortened so as to make the outline of the bone quadrate.
III. Lateral Carpal.
The tablet exhibits eight examples of a bone which at its distal end is attached to the marginal apicular articulation of the distal carpal, thence extending proximally, and terminating in an articular facet for the third bone of the fore-arm, so as to overlap laterally both of the other carpals. The bone is compressed, is three times as wide as thick, and in outline sub-quadrate with a distal talon. On the inner side it is flat, and on the outer side above the talon it is concave vertically and convex transversely in such way that the side of the bone to which the distal articulation is adjacent is thicker than the other side, and sometimes bent at a sharp angle. The talon on the inner aspect of the bone is « 52 » flat and continuous with the quadrate side, but on the outer aspect it is separated from the side by an elevated transverse thickening, distally to which the bone is compressed, and rounded into the adjacent parts. The talon extends over more than half of the distal end of the bone, and constitutes with the remainder of the distal end, the distal articulation, which is flat from front to back, and concave from side to base. The proximal articulation is cupped and extends over the whole proximal surface; it is at right angles with the sides of the bone. Both the inner and outer sides exhibit small pneumatic foramina. No. 8 differs from the other specimens in its sub-triangular lateral outline, and general less complex modifications.
The Carpus of the Cambridge Ornithosaurians at first sight is not easily compared with that of Birds; Birds having but one bone between the radius and the metacarpus. But that one bone in the Ostrich, for instance, is not unlike in form to the proximal carpal of Pterodactyle; while the proximal end of the metacarpus presents so close an analogy with the distal carpal of the Pterodactyle, that even were it not easily demonstrated that the bone in Birds commonly called the metacarpus is a carpo-metacarpus[R], it would be strong evidence for such a determination. In Birds there is a small lateral bone between the ulna and carpo-metacarpus which is evidently homologous with the lateral carpal of our Pterodactyles, and so, since this lateral carpal of the Ostrich is the pisiform bone, it results that the lateral carpal of Pterodactyle is the pisiform bone also. From this follows a conclusion of the first importance in the interpretation of the hand. The fine hair-like metacarpals of the Pterodactyle are on the side towards the pisiform bone, while the great wing-metacarpal is on the side towards the index finger.
[R] They separate without difficulty in the Chicken.
In Birds the rudimentary thumb (or second finger, according to Owen) has no connection with the carpus. In the Penguin, Aptenodytes Patagonica, it has disappeared altogether, and there then remain two fingers of which the outer one (seen from the front as we have placed our animal) is the larger, and has the greatest number of phalanges, precisely as in Ornithosaurians. Moreover the wing-metacarpal, in the Penguin especially, is seen to unite with the carpus directly under the radius, as is the case with the « 53 » Cambridge Ornithosaurians. Hence it follows that in Pterodactyles the thumb is not developed, and that the wing-finger is not the little finger, but the index finger, precisely as in Birds. If Goldfuss gave a reverse arrangement it was because the hand in his specimen, as is proved by the claws, was upside down. In the immature state the distal carpal of Pterodactyle appears to have been composite.
Notwithstanding the opinions of eminent German philosophers to the contrary no reptile has a carpus comparable to that of the Pterodactyle. If some of them have two rows of bones and a pisiform bone, so have mammals, and the mammalian arrangement is not more like the Ornithosauria than is that in Reptiles.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 5 | 1 3 |
The illustrations of this bone comprise 31 specimens. Nos. 1 to 15 are examples of the proximal end, and Nos. 16 to 31 show the distal trochlear end of the bone. No. 1, which is nearly entire, gives the form and proportions of the wing-metacarpal in one species, but a knowledge of its variableness in German forms would guard against an assumption that all the other Greensand species were to be restored on the plan of this example. It is 35/8 inches long, to which three-eighths of an inch may be added for the distal articulation, making the length up to 4 inches. The proximal end is not well preserved, but in its wide measurement is 5/8ths of an inch; the distal end in the same measurement is about 3/8ths of an inch. A large example from the Chalk, in the Museum of C. Moore Esq. of Bath, shows the bone more attenuated distally. No. 1 is compressed so as to be oblong in section at the proximal end, and ovate in the middle of the shaft, which is slightly smaller than the distal end. One of the lateral outlines is straight; the other is concave. The bone is straight. In No. 30 the shaft where thickest measures less than 1/4 of an inch, becoming nearly circular in section. The shaft of No. 31 measures nearly an inch in width at its distal end, rather more than half an inch in thickness. No. 10 is 11/2 inch wide at the proximal end and 7/8ths of an inch thick. No. 9 would not have measured less when perfect than 21/2 inches over the proximal end, so that if it had the « 54 » proportions of No. 1 it would have measured when entire not less than 16 inches in length.
The Proximal End.
The proximal end has never been figured. Prof. Owen's figure pl. IV. fig, 4-5, First Supt. Cret. Rep. is probably part of a jaw, and not the wing-metacarpal. The articular surface is oblong with one corner rounded off so that the adjacent long and short sides become confluent on the exterior surface of the bone.
In the middle of the flat inside margin and extending proximally
is a semi-cylindrical process, which is prolonged a short
distance down the side of the bone as an elevated ridge. On the
flattened articular end this process is bordered by a semicircular
furrow which extends half-way across the bone, outside of which is
a slightly convex semicircular band which extends to the outer margin
of the bone, except towards the short side opposite to that one
which rounds into the outer side, where there appears to be a
narrow unarticular area. On the inside of the bone where the two
ends of the semicircular proximal furrow terminate are two deep
grooves which extend a short distance distally; they are both
limited by inward extensions of the short sides of the bone, that
crest being most developed in height and length which is toward
the flattened short side. The outline which these modifications
give to the inner side of the proximal surface is intermediate
in form between the letters S and ![]() .
.
The Distal End.
The distal end has been figured by Prof. Owen in the British Fossil Mammals, p. 545; in Dixon's Geology of Sussex, Pl. XXIX. fig. 12; Cret. Reptiles, Pl. XXXII. figs. 4 and 5, First Supt. Pl. IV, fig. 9-11, and other places, and fully described. It closely resembles the distal end of a bird's tibia; and consists of a pulley-shaped end set obliquely on to the compressed shaft, which just above the junction is reniform in section, owing to the development of a median rounded ridge on the same inner side of the bone which bears the median ridge at the proximal end, while on the opposite side there is a corresponding median depression which does not extend far proximally. In this depression is « 55 » an oval pneumatic foramen; on the right of the median ridge of the other side, but placed more distally, is another pneumatic foramen. The median ridge has sometimes a slight furrow on each side. It terminates proximally in strong muscular insertions, which extend round the right side of the bone; and distally, becoming more elevated and rounded, it curves obliquely to the rights and forming one of the sides of the pulley, passes round the base as three quarters of a spiral, the termination extending laterally beyond the shaft. On this side of the bone, distal to the median depression, arises another ridge strong and well rounded, which is directed to the right, similarly passes round the base as a spiral, and forms the other side of the pulley. It is not so prominent as the border previously described. While the spirals approximate at their origin, they become widely separated at the base, making the articulation wider than the shaft. In No. 31 the three inches of the shaft which remain show both pairs of its sides sub-parallel; the widest measures nearly an inch; the base of the articulation is less than a quarter of an inch wider.
Limited to the base, between the two outer ridges of the pulley, is a short median ridge slightly developed; so as to flatten the middle of the concavity between the ridges, and divide it into two grooves. The degree to which the middle ridge is developed varies in different species. In No. 30, the smallest pterodactyle, remarkable for a long wing-metacarpal bone, it is not to be detected. The exterior sides of the trochlear articulation are broad, flattened, and a little concave.
There is some variation in the way in which the shaft is set on the trochlear end. It being often in the middle, but not unfrequently inclined more to one side than to the other.
The metacarpus finds no close parallel among living animals. The thread-like metacarpal bones suggest the condition of the hind-foot in the Kangaroo. The predominant metacarpal suggests the ruminants. But the nearest approximation is found among birds where the bone for the middle finger is large and the bone for the third finger is slender. This may be observed (among other examples) in the Penguin and the Swan. But here the parallel ends. The proximal end in Birds, we have already seen to be hidden by the anchylosed distal row of the carpus, and the distal end though often convex from side to side never presents the trochlear joint of « 56 » the Pterodactyle. Consequently so far as regards the form of the articular ends the resemblance is closer with Reptiles and clawed Mammals than with Birds. In Birds the small metacarpal is usually of similar length with the large one as is the case with Pterodactyles.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 6 | 1—10 |
No perfect specimen of the first phalange has been found in the Cambridge Greensand. Ten bones are mounted to illustrate it, all of them less perfect than others in the series of associated bones. No. 1 shows the heel of the proximal end; Nos. 9 and 10 are portions of the proximal articulation from which the epiphysis which forms the articular heel-part seen in No. 1 has come away. Nos. 2 to 8 are the distal articular ends of first phalanges. It is improbable that any of them belong to the second phalange, since they agree in form, and show muscular attachments which correspond.
Prof. Owen has figured the shaft of a fine example of this bone in Dixon's Geology of Sussex, Pl. XXXIX. fig. 11. A good proximal end is shown in Pl. XXXII. fig. 2, of Prof. Owen's monograph of the Cretaceous Reptilia, but the figure appears to have been previously given in Pl. XXIV. fig. 2 of the same monograph. By far the grandest specimens are drawn in Pl. XXX. Prof. Owen names these wing bones. In the "Literature of English Pterodactyles" the loss of the proximal epiphysis from the specimen represented in Prof Owen's fig. 1 and 2 led me to interpret the bone as an ulna. Figs. 1 to 4 represent the proximal ends and greater portions of the shafts of first phalanges. The lower bone in fig. 5 is neither radius nor ulna, as stated in the text of the Cretaceous Reptilia, but the shaft and distal end of a first phalange; the upper bone being the second phalange.
The Proximal End.
The straight shaft throughout its length is triangular in section. One side of the bone is gently convex; this may be named for convenience the outside. The two parts which make up the other « 57 » side are inclined, and have the angle in which they meet rounded; one part looks upward and inward, the other downward and inward. Towards the proximal end the bone widens and thickens, and the moiety of the inner side which is away from the heel becomes cleft, and has the sides of the depression rounded to form a large pneumatic foramen. The articular surface looks upward and a little outward on the side of the pneumatic foramen. It consists of two semicircular concave grooves, separated by an intervening low convexity. The outer of these grooves extends from the margin of the extreme proximal point of the heel to the widest point of the bone; the other groove more deeply concave, is a third shorter, extending from inside the pneumatic foramen to the heel. Here both grooves converge, terminating in a point, exterior to which a little distally is a hemispherical mammilate eminence. On the distal side of the eminence there is a depression so as to make the angle behind the heel almost hemispherically rounded. This articulation fits on to the distal articulation of the wing-metacarpal.
When the proximal epiphysis forming the heel comes away, it leaves a large sub-circular pit with a depressed narrow border.
Distal End.
On nearing the distal end, the angle of the inner side of the shaft becomes more depressed; and the articulation becomes an elongated oval, slightly convex transversely and convex in length so as to extend distally in a curve in such way that the articulation looks downward and outward from an aspect of the bone exactly opposite to the aspect from which the proximal articulation looks upward and inward. Hence the two articular surfaces are sub-parallel; but the distal one at its distal termination is bent inward, so as to make the adjacent lateral outline of the bone concave on the inside at its termination. The articulation does not cover the most proximal part of the distal surface.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 7 | 1—14 |
On this tablet are mounted 14 specimens. Nos. 1 to 9 are examples of the proximal end of the second phalange. If there « 58 » were more than two phalanges, of which there is no osteological evidence, it is possible that proximal ends of succeeding phalanges may be included with these. They all however resemble each, other so closely as to lend no support to such a supposition. Nos. ?10 to ?14 have been mounted with the proximal ends because they appear to be portions of the middle of the shaft of the second phalange; they indicate a rapid distal attenuation, favouring the idea of there being but two phalanges.
The proximal end of the shaft has the outer side flattened, rarely concave, commonly slightly convex; the inner side being much more inflated, and not dissimilar in form to the inner side of the first phalange. Proximally the bone widens and one lateral outline extends outward in a curve, on the inner side of which, under the proximal articulation, is placed the pneumatic foramen. The elongated oval articular surface is concave from side to side and concave in length; it does not extend in length so far as the straight side outline, exterior to it being a crescentic flattened or convex area. The distal end attenuates more rapidly in some specimens than others, and appears in Nos. 11, 12, and 14 eventually to become cylindrical; but none of the specimens show its distal termination.
The phalanges of the wing-finger attain a grand development in length which is not paralleled in Birds, nor surpassed in Bats. In the Ostrich there are three phalanges in the wing-finger, while in Ornithopterus there are two joints, and in other German Pterodactyles four joints. The terminal joint in the Ostrich is a claw, but in Pterodactyle the terminal joint appears to be unarmed as in ordinary birds. The form of the bones in being compressed from side to side is more bird-like than bat-like. But the claws in their compression from side to side are more like the bat than the bird.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 7 | 1—14 |
Sub-cylindrical bones, apparently elongated, and a little compressed obliquely, terminating distally in a slightly expanded trochlear articulation. Some of them show on one side marks of an osseous adhesion: this has led to their being regarded as « 59 » claw metacarpals rather than as the distal ends of tibiæ. But on the supposition of their being claw metacarpals, they are as compared with the same bones in Pt. Suevicus, out of all proportion large, since wing-metacarpals from the Cambridge Greensand would not as a rule have a diameter more than twice that of these bones. The trochlear articulation is smaller in proportion to the shaft than in the wing-metacarpal, and usually shows a pit at the side and grooves above for ligaments; the mesial pulley groove is shallow and broad. Seven specimens are mounted in illustration, of which No. 3 may be regarded as doubtful. It is possible that they may be metatarsals.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 8 | 1—3 |
These three sub-triangular bones, which supported the claws, are much compressed from side to side, and consequently deep. The superior outline is convex from front to back and rounded from side to side. The inferior outline is concave from front to back, sometimes narrower, sometimes broader than the upper part of the bone, while the inferior aspect is always more flattened than the superior aspect. On each side on the lower half of the bone is a deep groove. The articular end is divided into an upper articular part, which extends as far down as the lateral groove and a lower non-articular part with ligament markings. The articulation is concave from above downward, and is divided into two lateral parts by a mesial vertical ridge. The articular end is about half as deep as the bone is long.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 10 | 1—9 |
Nine specimens are mounted in illustration of the pelvic girdle: Nos. 1, 3, 4, 5 and 6 show the acetabular or femoral aspects. The right os innominatum is exemplified by Nos. 1, 4 and 5; the left by Nos. 3 and 6. No. 2 shows the sacral aspect of a left ischium, and its attachments with the pubis and ilium. No. 8 is the sacral aspect of a left os pubis. No. 9 is the femoral aspect of a right OS pubis. None of the specimens are sufficiently complete « 60 » to give the form of any of the bones. The only known example of an entire or nearly entire pelvis at all comparable in form, is seen in the original specimen of Dimorphodon macronyx figured by Buckland, Trans. Geol. Soc. Ser. 2. vol. III. p. 217. In nearly all the fossils from the Cambridge Greensand the bones of the pelvis are anchylosed together.
The ossa innominata have been determined as right and left on the supposition that the pelvis of the Dimorphodon is in situ, and from the general correspondence of the form of the constituent elements with elements of the pelvis in the lower mammals, reptiles, and birds.
Each os innominatum shows a hemispherical acetabulum which is slightly elongated in antero-posterior extent In the Dimorphodon the bone which is superior to the cup, that is to say, which extends dorsally along the sacral vertebræ is prolonged anteriorly as a strong narrow straight style, the base of which is seen in the parts marked Ilium in Nos. 1 and 6. A more perfect example may be studied in a pelvis from the Cambridge Greensand preserved in the collection of the Geological survey. Posterior to the acetabulum a similar but stronger bony style extends for more than the length of the acetabulum, curving slightly downward at its posterior part. The dorsal outline of this portion of the bone is slightly concave. The posterior horn like the anterior horn forms part of the ilium which constitutes the upper half of the acetabular cup. The os innominatum contracts in antero-posterior extent below the acetabulum, and immediately widens again in a thin concave bony expansion. The anterior or pubic outline is comparatively straight, and at right angles with the ilium; the posterior or ischiac outline is deeply cupped where the ischium unites with the ilium, and becoming straight extends backward at a considerable angle. The ischium contributes less to the pelvic cup than either the ilium or pubis; it is flat in front and convex on the visceral side, rounding into the narrow flattened posterior side. The pubis is separated from the ischium by a suture extending vertically through the obturator foramen. The obturator foramen [seen in No. 9] is small and oval, less than half the diameter of the acetabulum, situated below its ventral border. It passes obliquely downward and a little forward, and its opening makes the exterior aspect of the pubis concave; the visceral aspect of the pubis is convex from side to « 61 » side like the ischium. The sacral aspect of so much of the os innominatum as is seen, is concave from the dorsal to the ventral margins, and is cupped behind and below the acetabulum, the surface being rough. Among reptiles the ilium is chiefly behind the acetabulum, in mammals it is chiefly in front. In the over-lapping group, Aves, it extends both ways. Among the Amphibia the ilium is chiefly anterior to the acetabulum. In Crocodiles it has a slight extension both ways, in Dinosaurs the extensions are more marked and the whole arrangement approximates to birds. But among animals which have been affiliated with reptiles the Dicynodonts are the only order in which there is a pelvis so mammalian and massive. If the ilium of the Monotreme genus Echidna had a posterior extension, the pelvis would be altogether comparable with the pelvis of this Pterodactyle, and would differ chiefly in the larger obturator foramen, the perforated acetabulum and the unanchylosed condition of the pelvic elements. The pelvis of Apteryx does not make any near approximation.
Moreover specimens Nos. 3 and 4 show on the anterior pubic border, about the base of the acetabulum, a slight pit or roughness to which something has been attached, and in the original specimen of Dimorphodon associated with the pelvis are two triangular bones which recall something of the form of the prepubic bones of Echidna. Most German Pterodactyles show on the OS pubis an enormous prepubic bone. In Iguana the pubis forms at its anterior border, a sharp angular process. In Chelydra the process is long and narrower, and arises from the middle of the border. In Echidna this prepubic process has become a distinct prepubic bone and is more elongated. Unlike the marsupial bones it is attached to the pubis by a wide base. The anterior pubic roughness of Cambridge specimens, and the loose bones of the Dimorphodon, &c. indicate the existence of structures in the Ornithosauria homologous with the prepubic bones of the Ornithodelphia.
So far as it is comparable with living animals, the ilium is altogether avian, differing in being narrower; and the pubis and ischium are mammalian.
The upper anterior corner is the most elevated part of the acetabular border, as in the great Auk and some birds of vertical position of body, and many mammals.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 11 | 1—16 |
Twenty-six specimens are mounted to illustrate the Femur. 10 are proximal ends; 16 distal ends. But in the series illustrative of species is an entire specimen of a right Femur 4 inches long. Fragments Nos. 3 and 12 show proximal and distal ends twice as large, but most of the examples are about the size of the entire femur.
It is a straight sub-cylindrical bone, flattened in front, a little compressed from front to back distally, and (in one type) compressed proximally from side to side behind. The distal articulation has a broad shallow channel passing down from the front and imperfectly separating two condyloid parts, which extend a little backward and are divided behind. The outer condyle extends a little outward, and so gives the outer side of the bone at the distal end an oblique compressed aspect like that which prevails among birds and many mammals. Proximally the shaft contracts suddenly and is produced upward, forward, and inward as a rounded neck, as long as in the femur of any mammalian carnivore, which expands rapidly at the end to form the hemispherical ball, which articulates with the pelvic acetabulum.
No. 1 shows a well-marked pit for the ligamentum teres at the back part of the ball. At the proximal end of the shaft below the neck is a large pit for the obturator muscle, and at the outer front angle a great trochanter. Proximally the bone can only be compared with the mammalian Carnivora, Quadrumana and Man; distally it is avian and mammalian.
In one genus exemplified by specimens 5-10 the obturator pit is not developed.
Sometimes the shaft is curved a little convexly, outward and forward.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 12 | 1—11 |
Eleven specimens are mounted to illustrate the tibia, of which 1 to 9 are regarded as proximal ends; and 10, 11 with less confidence are regarded as distal ends from which the distal epiphysis « 63 » has come away. It is to be anticipated that the distal end of the tibia in Pterodactyle will when found approximate to the distal end of the tibia in the bird.
The bone is at the proximal end straight and sub-cylindrical, slowly enlarging proximally; convex behind, except for an elevated boss some little way below the proximal articulation for the attachment of powerful muscles. In front the shaft is a little flattened proximally, with a mesial groove dividing two prominences which are apparently homologous with the ridges below the patella in birds. The proximal articular surface truncates the shaft at right angles except at what is regarded as the outer front aspect, where it rises into a small patelloid prominence.
It shows the impressions of two condyles, which correspond in form with the distal end of the Femur.
Nos. 3 and 6 are regarded as left tibia; Nos. 4, 5, 7, 8, 9 as examples of right tibia.
No specimen likely to be a fibula has been found. In Dimorphodon and in German Genera the fibula is Avian in form. The Crocodile offers some approximation to the Pterodactyle shape in the proximal end of the Tibia, but the Pterodactyle has Avian characters in addition. Its straightness and length, ridges on the front and patelloid prominence, are Avian.
| Case. | Comp. | Tablet. | Specimen. |
| J | b | 13 | 1 |
This fragment, which may be the distal end of the bone corresponding to that called in birds the tarsus or tarso-metatarsus, is badly preserved. Yet so close is its resemblance in form, structure, and apposition of the constituent bones to what obtains among birds, that it may probably be identified as the tarsus; while the peculiar characteristics of Pterodactyle bones which it shows, demonstrate that it is not from a bird, but from an Ornithosaurian skeleton.
The bones are of paper thinness, and consist of a strong bone behind which distally appears on the inner side to be compressed and thrown backward and flattened at the side, exactly like the inner toe in Natatorial birds. On the front of this strong support, confluent with it, and confluent together, so that « 64 » the places of union are only seen at the distal end and in transverse section, are three bones, together as wide as the bone on which they rest. It does not appear possible that the distal articulations could have supported more than three digits.
This bone, if correctly determined, offers points of affinity with birds as pronounced and as important as any thing shown by the extremities, for among reptiles a welding of the (tarsal or) tarso-metatarsal bones is unknown, and here it is as absolute as in any bird, and takes a characteristic bird shape. In the Rodent Jerboa the metatarsus has much the same form as in a bird.
No phalanges have been recognised.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 1 | 1—15 |
Fifteen specimens are mounted to exemplify the structures of the Pterodactyle atlas and axis. Nos. 1, 11, and 2 have already been figured, and described by Prof Owen, the latter as a section of a cervical vertebra.
The atlas centrum, a saucer-shaped disk of bone, commonly united more or less intimately with the centrum of the axis, but sometimes free. It presents in front a hemispherical cup for the basi-occipital, and is flattened or slightly convex behind. Its neural arch is seen in Nos. 2, 10, and 12; but the only specimen with the arch entire is in the museum of James Carter, Esq. The neurapophyses vary in form and size, but always are small obliquely flattened lamellar bones, which extend upward and backward to meet the neural arch of the axis, just above the neural canal, where a thin and small cross piece connects them together.
The distinctive aspect of these bones is given by the neural arch of the axis, which is very much elevated, and is formed by two flattened sides, which meet in a vertical ridge above the neural canal, and look forward, outward and upward; extending laterally more and more beyond the side of the centrum, but not reaching so far back as the posterior articulation of the centrum. Each side of the neural arch at its middle part behind is produced into a thick obliquely flattened process, the under portion of which « 65 » shows the small posterior zygapophysial facets, which look downward, outward and backward. The lateral outline of the part of the neural arch above this process is concave; as is the lateral outline between it and the centrum. Behind, the neural arch is concave, and looks a little backward. The neural canal is stirrup-shaped in front, but is higher and sub-ovate behind. The neural arch of this, as of all the other vertebræ, except a few dorsals, is inseparably united to the centrum, without a trace of the line of union. In the middle of the side of the vertebra, and at what may be presumed to be the union between the neural arch and the centrum, in a concavity, is the pneumatic foramen. It is round or oval, and varies in form and size though not in position. In No. 8 it exhibits the subdivided reticular structure characteristic of the pneumatic foramina of birds. In No. 10, which has a short centrum, the pneumatic cavities are reduced to a few small perforations, no larger than would be made with fine needles.
The centrum is shorter than in cervical vertebræ, commonly convex (No. 8) on the visceral surface; often with a slight longitudinal hypapophysial ridge (Nos. 1; 7; 12) rarely flattened (No. 10). Towards the hinder part the centrum widens, and becomes concave on the visceral surface, sending off as do the other cervicals, below the transversely elongated posterior articulation, a pair of short strong apophyses.
The posterior articulation can only by a modification of the idea be said to conform to the cup-and-ball plan, for though convex from above downward and convex from side to side, the elongated transverse measurement would be three times the depth. On the under side an impressed transverse line divides the articulation from the concave part of the centrum below.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 2 | 1—43 |
Forty-three specimens are mounted to exemplify the variations in size and characters of cervical vertebræ. These for the most part are specific characters; and between the axis and the first dorsal vertebra the variations in an individual were slight. [Those nearest to the back, as in birds, are widest in front, and have the highest neural arches.] The associated series show commonly « 66 » four cervical vertebræ behind the axis, and in two cases apparently five; never more. So that as seven appears to be the number of true cervical vertebræ in most if not all of the German Pterodactyles, it may be presumed that the Cretaceous Ornithosaurians also had this character in common, with Mammals, and probably as persistent. In Iguana there are 6, in Monitor 7, and in Crocodile 8.
The centrum is united to the neural arch as in birds, without a trace of suture; sometimes the neural arch is no wider than the centrum, sometimes it extends over the centrum on each side. Those forms with a narrow neural arch have the neural spine high, and its sides look forward as well as outward. The pneumatic foramen is oblique. An example is figured by Prof. Owen, in the memoir on Pterodactylus simus, pl. 2, fig. 4. The forms with a wide neural arch have the neural spine rising from the middle of the dorsal surface, erect and equally compressed from side to side. The pneumatic foramen is horizontal. An example is figured in Prof. Owen's memoir on Pt. simus, pl. 2, fig. 1. These two forms of cervical vertebræ may be regarded as typifying two genera.
In both forms many characters occur in common, and as the specimens illustrative of special modifications will be described hereafter, the following description has been made to embrace the chief characteristics of these vertebræ in Cretaceous Ornithosaurians.
The inferior aspect of the centrum is oblong (being narrower than long), or quadrate; when quadrate the additional lateral expansions are external to the pneumatic foramina, and are formed by the neural arch and zygapophyses. The centrum proper is a little wider in front than behind, and the side outlines are concave. The base of the centrum is flattened, or more or less hollow, or more or less tumid and regularly convex; in front there is often a mesial ridge, which never reaches the posterior articulation, and forms a prominent tubercle at the base of the anterior articulation. At the posterior end the outline of the centrum is concave, and mesially the bone has a hollow corresponding to the tubercle in front of the adjacent vertebra; and the part of the centrum on each side is prolonged slightly into a strong rounded or flattened tubercle below the side borders « 67 » of the posterior articulation; these posterior processes, in vertebræ in situ, fitted, on each side of the mesial anterior process of the vertebra behind, on to concavities more or less marked. Analagous processes are developed in the cervical vertebræ of many birds.
All the Cretaceous Pterodactyles have the articular surfaces of the centrum transversely oblong, as have some birds. The posterior articulation is convex from side to side, and convex from above downward, and appears to extend a little further on this neural than on the hæmal surface; in outline it is commonly an elongated oval, but sometimes attends on the upper surface of the inferior lateral tubercles. The anterior articulation is transversely elongated, concave in both directions, and sub-triangular in outline; that is to say, the superior outline is more or less convex, and from its limits to the mesial tubercle at the base, the inferior outlines are more or less concave.
The neural canal is sub-circular or ovate in outline, and quite as large as the neural canal in vertebræ of Dinornis of similar size.
The neural arch like the centrum has commonly a depressed appearance. It always has a neural spinous process which is directed upward. In the depressed type the neural surface of the vertebra is in outline usually sub-quadrate, but concave at each side, and concave in front and behind; the four corners are the processes which support the zygapophysial facets, the surface is divided into two lateral parts by the strong neural spine. These lateral parts are from front to back flat, or slightly concave, or slightly convex; and from the neural spine outward they are always concave. The neural spine is commonly sharp in front and flattened behind. The neural arch is placed well forward, so that while a third of the neural canal remains uncovered by it behind, rarely a sixth would be uncovered in front.
The anterior and posterior zygapophyses are commonly connected by a more or less rounded ridge, undefined above, but well defined below, since under its posterior part at about the middle of the side of the centrum is placed the pneumatic foramen.
The anterior zygapophysial processes are separated from the anterior articular surface of the centrum by a more or less oblique « 68 » channel. Towards the base of this channel in many vertebræ may be seen a small and short flattened antero-posterior perforation corresponding in position with the usually large perforation for the vertebral artery. If the passages are to be regarded as having subserved such a function, it will not be without interest to remark the small relative size of the cerebellum in these animals; since the vertebral artery conveys the blood to that region of the brain.
The anterior zygapophyses are strong processes directed forward and outward, compressed a little from side to side; they are placed at the outer sides of the anterior articular face of the centrum, and extend in front of it.
The zygapophysial facet is commonly oval and looks upward and inward and forward.
The posterior zygapophyses are short and massive, but otherwise correspond closely with the anterior zygapophyses, only with all the parts reversed, and except that necessarily they are relatively to the neural canal a little higher.
A sharp and well defined angular ridge, commencing at the back of the zygapophysis, is directed inward, and forward, and upward along the posterior margin of the neural arch to the top of the neural spine. The posterior aspect of the neural arch is concave from side to side, and makes a right angle with the superior lateral aspect.
The part of the centrum exposed behind the neural arch is convex above from side to side.
The pneumatic foramen between the centrum and the neural arch varies greatly in size; it is oval and longitudinal.
The largest specimens have the centrum 21/2 inches long; in the smallest the centrum measures 5/8ths of an inch in length.
In the second type of cervical vertebra the side of the centrum makes a right angle with the base, and is separated from it by a sharp angle as in struthious birds. The side of the centrum is concave, with a few small pneumatic perforations; and the side of the centrum, which is high posteriorly, rounds over the oblique ridge connecting the zygapophyses, into the oblique lateral face of the neural arch. The anterior zygapophyses are very large and the posterior zygapophyses small and placed as high as the top of the neural canal.
Every region of the vertebral column displays pneumatic foramina, situated as in the vertebræ of birds.
The large proportional size of the neck-vertebræ is common to some birds, and is conspicuous in many mammals, like the Llama. In most mammals where the vertebræ have a cup-and-ball articulation, the ball is in front, as it is in the dorsal vertebræ of the penguin, so that those vertebræ are not comparable with Pterodactyles, although on the under side of the centrum they similarly give off a mesial process below the cup, and a lateral process below the ball on each side. The neural spine in Pterodactyle is commonly more developed than is the case with long-necked birds or mammals. Reptiles such as Crocodiles and Lizards have the neural spines of the neck-vertebræ well developed. Birds differ from Pterodactyles in the peculiar articulation of their vertebræ. In both the centrum is often depressed, in both it is concave from side to side in front, and convex from side to side behind, but in birds it is also convex from above downward in front, and concave from above downward behind, while the reverse arrangement obtains in Pterodactyles. A similar condition to that of the bird is seen in the neck-vertebræ of the Kangaroo, of Man, and several mammals, only the vertical curves are less marked. Vertebræ concave in front, and convex behind, and devoid of cervical ribs, are met with among the Lizards, but neither Monitor nor Iguana offer any parallel to the form of the cervical vertebræ of Pterodactyle, which is best matched among Marsupials and Birds. Birds commonly have more vertebræ in the neck than have Pterodactyles, which in that respect resemble mammals and some Lizards.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 3 | 1—20 |
Twenty specimens are mounted to exemplify pectoral and dorsal vertebræ. Like the cervical vertebræ, they include two types of form, one with the centrum flat, figured in pl. 2. fig. 20-22 of the memoir on Pterodactylus Sedgwicki, and regarded by Prof Owen as anterior dorsal; and the other form with a convex centrum, figured 24-25 of the same plate of the same memoir, regarded by Prof. Owen as posterior dorsal. Following the analogy of birds such determination is as well « 70 » supported as the similar reference of the two types of cervical vertebræ to anterior and posterior parts of the neck, but fuller materials compel a reference of the two types of dorsal vertebræ to two different genera.
Nos. 1, 3, 6, 7, 8, 9, 10, 14, 15, 19 belong to the flat type. Nos. 2, 4, 5, 11, 12, 16, 17, 18 exemplify the convex type.
Dorsal vertebræ are rare fossils; and in the associated sets of bones never more than four dorsal vertebræ are found, rarely more than one. No specimen of the type with a convex centrum occurs in the associated sets.
The dorsal vertebræ with convex centra have all lost their neural arches except No. 2. The form of the centrum is half a cylinder, as long, or longer than wide, but sometimes depressed, and wider behind than in front. The exterior surface is smooth, convex from side to side, and slightly concave from front to back. The neural surface is mesially excavated. Both anterior and posterior articular surfaces are semicircular or sub-ovate, being wide from side to side.
The anterior articulation is cupped, concave from the neural to the hæmal surface, and concave from side to side. The posterior articulation is convex from the neural to the hæmal surface, in which direction, it usually shows striations, and from side to side has a gentle convexity, sometimes so slight as to be nearly flat.
The neural canal is large, ovate, and as high as is the centrum.
The neural arch is strong, compressed from back to front, and placed as usual on the anterior part of the centrum. In outline it is sub-rhomboid with the sides concave. There is a strong process on each side above the neural canal for a rib, and apparently a neural spine, but all are broken. The transverse processes for the ribs are directed outward, and a little forward, flattened in front and behind, the surfaces being sub-parallel, so that in front the neural arch is concave from side to side. Behind, the neural spine is directed between the transverse processes so as to over-hang the exposed part of the superior surface of the centrum. At the points where the neural spine bends back from the transverse processes are the posterior zygapophyses, high above the neural canal, and parted from each other by an interspace as wide as the « 71 » canal is high. They look downward, outward, and backward. The lateral surface below the transverse process is narrow, flattened, bends at a right angle with the posterior surface, rounds into the anterior surface, is a continuous curve with the side of the centrum, and is concave from below upward. The superior surfaces of the neural arch have the sides sub-parallel, they are each concave from side to side; and these surfaces are excavated for pneumatic foramina.
Dorsal vertebræ of the type with the centrum flattened closely resemble cervical vertebræ with the centrum flattened, differing chiefly in the less length of the centrum. Occasionally (as in No. 3) the neural arch comes away from the body of the vertebra.
The centrum is very depressed, sub-quadrate, and wider than long; the base is flat, or slightly concave, with occasionally a slight longitudinal mesial ridge; the lateral outlines are concave, so that the bone is pinched in from side to side. The neural surface of the centrum is flat and parallel with the base, and, as usual, wider behind than in front, but the centrum is not there so high. The surfaces for the neural arch are flat, and extend nearly to the base of the centrum in front, so that they look upward, outward and a little forward.
The articular ends are remarkable for their depressed oblong character, still preserving the anterior concavity with a small mesial process below, as in cervical vertebræ, and similar but smaller processes at the inferior outer angles of the posterior sub-semicylindrical convexity. The middle third of the anterior cup is made by the trapezoidal anterior end of the centrum; sometimes the sutures between it and the neural arch are well marked.
The neural arch is large, commonly with a sub-circular neural canal. The neural spine is high, compressed so as to have the lateral surfaces sub-parallel and rounding into each other superiorly; and it has a less antero-posterior extent than the centrum. At its base behind it widens rapidly, and forms massive quadrate processes, extending outward and backward, which on the outside each have a flattened ovate zygapophysial facet, which also looks downward. Above the facet and separated from it by a groove is a tubercle. Between the zygapophyses behind the bone is concave from side to side; the facets are placed above the neural canal.
The posterior zygapophyses are placed considerably higher than the anterior zygapophyses, and the part of the neural arch between is rather constricted from front to back. The neural arch steadily widens in front down to the base of the anterior zygapophysial processes in such way that the more or less flattened lateral surface looks outward and is gently concave from above downward. A ridge commencing at the tubercle over the posterior zygapophysial facet descends in a curve forward and downward, to form the posterior border of the anterior zygapophysial process. This is separated by a groove from the anterior articular surface, and anterior part of the base of the centrum, and has the aspect of a compressed part of the neural arch, extending obliquely downward, and forward, over and beyond the articular surface of the centrum. The anterior zygapophysial facets are oblong, narrow from side to side, and long from front to back; they are directed forward and a little outward, and are flattened, make nearly a right angle behind with the front of the neural arch, and look upward and inward. They are sometimes placed as high as the top of the neural canal, but are commonly lower. Around the neural canal the bone is conically impressed.
Minute pneumatic foramina are in the usual position, between the centrum and the neural arch; and sometimes others behind the anterior zygapophysial process.
The largest specimen known has the centrum nearly an inch and a half long.
The dorsal vertebræ in Cambridge specimens would appear to make a nearer approximation in number to birds than to Mammals or Lizards or Crocodiles, though Chelonians have few vertebræ in the back. Among Reptiles the form of the vertebra makes some approach to that of the Monitor, Chameleon and Scink. In most Mammals the dorsal vertebræ have the centrum convex, though in the lumbar region its visceral surface often becomes flattened. But though very unlike there is a nearer resemblance to the lower dorsal vertebræ of a Struthious bird.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 4 | 1—7 |
Seven specimens are mounted to exemplify the ordinary structures of the Ornithosaurian sacrum.
Nos. 1 and 2 have the centrum convex, exactly as in the dorsal vertebræ of the convex type. Nos. 3-7 have the centrum flattened, following in general features the plan of the dorsal vertebræ with flat centra.
No. 1 is a vertebra from a sacrum, where perfect anchylosis had not been induced; it has the neural arch well preserved, and shows the sharp suture which united it to the preceding vertebra.
No. 2 shows two entire vertebræ and part of a third, which have lost the neural arches but have the centra perfectly anchylosed together. The middle vertebra measures 5/8 of an inch in length, and at the suture from side to side measures more. The surface is smooth, regularly convex from side to side, and gently concave from back to front. The last vertebra shows the articular vertebral surface; it is convex in both directions, and oblique, so that a large part looks upward. The anterior of the three vertebræ is pinched in at the lower part of the sides of the centrum. No. 1 shows that the neural surface of the centrum is deeply excavated, making the neural canal an elongated upright oval. Above the centrum, which forms only the middle third of the articular surface, the neural arch expands on each side into a wedge-shaped transverse process, the lower surfaces are flattened, and continuous with the centrum, while the upper surfaces are flat and horizontal as in birds and Dinosaurs, and form the platform from which arises the massive neural spine.
In front the transverse wedge is flattened and compressed, so as to look forward and outward, and in the middle shows a large ovate pneumatic foramen. Behind, the wedge is compressed so as to look backward and downward.
The neural spine is massive and forms rather more than half the height of the vertebra. It is flattened with a ridge rising near its base in front and ascending in a concave curve obliquely backward and upward. The anterior parts approximate a little in front, while the small parts posterior to the ridge approximate « 74 » a little behind. The sides of the neural spine approximate superiorly, and appear to round into each other.
There is a notch on each side in front at the base of the neural spine, and another above the central articulation. The neural spines appear to have been united by suture. It may be instructive to compare the neural spine just described with the specimens J. c. 10.
Of the second type or genus No. 4 to 7 all show the anterior cup for the last lumbar vertebra. No. 3, 5 and 6 all show two entire vertebræ and part of a third preserved, but no specimen shows the posterior termination of the sacrum. No. 7 has the articular face of the centrum very broad, and greatly depressed. In No. 6 it is ovate and has the neural arch preserved; above a semicircular neural canal it sends out on each side a short horn-like zygapophysial process. No. 4 is remarkable for the small size of the circular neural canal, the centrum when entire measuring an inch from side to side, while the neural canal only measures 5/16 of an inch. No. 5 is figured by Prof. Owen. No. 4-6 appear to have given off transverse processes from the sides of the centra. No. 7 appears to widen into transverse processes at the point of suture between the centra.
In No. 3 the base of the sacrum is flattened, and its sides pinched in, and concave in outline from back to front. In this hollow are small pneumatic foramina, and between the hollows the vertebræ widen in the line of the suture so as to send out strong short transverse processes or tubercles. Above the hollows are given out the strong horizontal quadrate pyramidal transverse processes. All their sides are flattened or a little concave, and the under side displays a large ovate pneumatic foramen. Each of the four angles of the transverse process gives off a ridge. The lower ones descend obliquely to the anterior and posterior intersutural tubercles. The upper two ascend obliquely, in front and behind, and form rounded ridges on the neural spine. The neural spine is flattened, moderately compressed from side to side, and cupped a little over each transverse process. In front the neural spine is flattened transversely and perpendicular; the transverse processes are also flattened and a little in advance of the neural spine.
The sacrum in its general aspect is Mammalian. In the Bird « 75 » the vertebræ are much more numerous and do not retain their individuality so well. In Reptiles properly so called, the sacrum never includes more than two or three vertebræ, and those commonly remain unanchylosed. But in almost any placental Mammal in which several vertebræ are anchylosed together, a sacrum similar to that of the Pterodactyle is met with. No mammalian sacrum, however, is furnished with pneumatic foramina.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 5 | 1—13 |
Thirteen specimens are mounted to exemplify the osteology of caudal vertebræ. No. 7 has been figured by Prof. Owen in the memoir on Pterodactylus simus, pl. 2 fig. 13-16. The centrum of the largest specimens measures one inch and a quarter long, and the vertebra is half an inch wide from side to side in the middle. The smallest specimen No. 13 has the centrum 3/4 of an inch long. The vertebræ vary in proportions, some being much more slender than others. They present a close approximation in form to the first type of cervical vertebræ, differing chiefly in being more elongated.
They are elongated bones constricted in the middle, so that the outlines of the sides seen from above or below are gently concave; the outline of the anterior end is sub-rhomboid, the outline of the posterior end is sub-pentagonal, as would be a transverse section of the vertebra. The long outlines of the base of the centrum and of the top of the neural arch are sub-parallel.
The two sides of the upper surface of the neural arch are smooth, flattened, a little concave from back to front; they are inclined to each other pent-house wise at about a right angle, and are separated throughout their length by a narrow slightly elevated neural spine. Behind, the neural arch is truncated transversely so as to expose the posterior neural surface of the centrum, which is convex from side to side. The outermost lateral angles of the neural arch are the posterior zygapophysial processes, short and strong above the centrum, with a tubercle on the upper surface, and showing the sub-circular zygapophysial facets behind; they look backward and downward, and « 76 » are separated by a groove from the region of the centrum. Under the sharp ridge which connects these zygapophyses behind, the neural arch is excavated, and the cup shows the termination of three canals. The largest one is the upright oval of the neural canal in the middle, on each of its sides separated by a narrow bony wall is another perforation, very variable in size and shape, sometimes b& large as the neural canal, but usually small and circular. The anterior end of the neural arch is cut into, so that as seen from above, the straight sharp anterior margins diverge mesially from each other at a right angle, and so expose to view a small anterior part of the neural surface of the centrum. These lines are prolonged forward and outward, to form the upper margin of the anterior zygapophyses, which are compressed and prolonged over and beyond the sides of the anterior articulation, from which they are separated by a slight groove; the anterior and posterior zygapophyses are connected by a rounded ridge. The anterior end of the neural arch is excavated, but less so than the posterior end; in the middle is the oval perforation of the neural canal, and at the sides other perforations corresponding to those behind are placed a little in advance of the neural canal. The anterior and posterior articular surfaces differ in no respect from those of cervical vertebræ.
The inferior surface of the centrum is separated from the sides by two ridges parallel to the lateral concave outlines of the neural arch; they extend from sides of the front, more or less well marked, to the tubercular processes at the base of the sides of the centrum behind. The dice-box shaped area of the centrum so inscribed is usually concave from front to back, and concave from side to side behind, and convex from side to side in the middle; this convexity is only broken in front by the development of the slight mesial hypapophysial ridge.
The sides are narrow, flattened, look downward and outward, are a little concave from front to back, round into the centrum and into the neural arch, and show at about the middle a small pneumatic foramen, which is variable in size, but largest in No. 8, and sometimes a mere puncture.
The caudal vertebræ differ in many ways from other animals. « 77 » They have neither transverse processes, neural spines, hypapophyses or hæmapophyses. In the persistence of the neural arch down the tail they resemble reptiles and birds rather than mammals, in which nothing but the centrum persists to the end of the tail. The vertebræ are furnished with vertebral arteries which run through the neural arch parallel to the neural canal, in exactly the same position as do the vertebral arteries in the neck-vertebræ of the Llama.
The skull of Dimorphodon differs in form and in many important details of structure from that of Rhamphorhynchus; and both of these types of skull are strikingly unlike that of the short-tailed animals named Pterodactyle. Hence, as it will be shown that the Cretaceous fossils of this class belong to very distinct new genera, there is no reason for assigning to them by anticipation any class of cranial structures. The cranium of this type of animal has never been critically described, and for all that is yet known to the contrary Pterodactyles may differ between themselves as much as birds or mammals. Their affinities have been unknown. Therefore, before describing bones it may be desirable to state the grounds on which the several specimens are referred to the Ornithosauria. The fossils on which this section of the memoir is founded are, the basi-occipital and basi-temporal bones, the anterior portion of a cranium, the back parts of four crania, facial bones, and the quadrate and quadrato-jugal.
The crania are all no larger than that of the Heron; though from the Greensand are bones and jaws indicating Pterodactyles both smaller and larger. The skulls are mostly remarkable for wanting both basi-occipital and basi-temporal bones. And the specimen of basi-temporal and basi-occipital corresponds posteriorly with the Pterodactyle atlas, anteriorly with these crania; it is hence concluded to have belonged to a similar animal. Being relatively twice as large, it indicates that in these animals the basi-occipital condyle was proportionally larger than in known birds; and that animals of a cognate kind had skulls probably twice the size of these. The anterior basal part of the hinder « 78 » sphenoid terminates in a remarkable triangular surface, with two perforations, which are separated by a median ridge. Almost entirely corresponding with this is the basal surface of the anterior part of a cranium, fractured in front of the pituitary fossa. Therefore, and as it indicates a similar capacity of brain, it is regarded as belonging to the same kind of animal as the others ; but being five times the size, it must, if the proportions of the Heron were preserved, have been part of a head a yard long.
Now, as there is no other animal with the same texture of bone, or exhibiting with high organization the same diversity of size, these cranial fragments are referred to the jaws and bones of Pterodactyle. So marked are their structures that many quarry-men refer vertebrate fossils to their several orders with almost as much accuracy as would a practised anatomist.
| Case. | Comp. | Tablet. |
| J | c | 7 |
Basi-occipital, Owen, Sup. Cret. Rep. p. 6, T. 1, figs. 11, 12, 13.
This bone was not found associated with any set of fossils that would induce us to refer it to one species more than to another. Its Ornithosaurian character was probable; and Prof. Owen described it in his last memoir on the Greensand Pterodactyles.
But though indubitably basi-occipital, it is so anomalous in some respects that the Professor regarded the under as the upper surface; since then the investing phosphate of lime has been removed, and the bone is now described in what appears to be its natural position.
Viewed from above the fossil divides into two parts; the occipital condyle, and an anterior, wide, transversely oblong extension terminating at each side in a strong short horn. The posterior half of the condyle shows large cancelli as though so much of it had been covered by the articular cartilage. The sides of the condyle converge, so that posteriorly it is only two-thirds of the width it has at the foramen magnum, which would appear to indicate a comparatively slight lateral motion of the head. The condyle is hemispherical posteriorly and superiorly; there is a depression between it and the great foramen of the skull; inferiorly it is flat.
It is 7/16 of an inch long; posteriorly 9/16 wide, nearly 6/16 of an inch high anteriorly. It terminates in front superiorly in an elevated transverse ridge.
On removing the matrix, the anterior surface of this occipital bone was found to be concave; yet as nothing but cancellous structure is seen it may be but imperfectly ossified or more probably, imperfectly preserved. And the bottom of this cup expands forward in a thin sheet of bone a quarter of an inch long and half an inch wide, which on the under side is continuous with the base of the condyle.
On each side of this floor and partly extending in front of it, and below it, is an irregular piece of bone, half an inch long, resembling anterior zygapophyses of cervical vertebræ.
Though in most vertebrates the basi-occipital enters into the basal floor of the skull, the median bones are either so placed that they rest one upon another from before backwards or abut against one another nearly perpendicular, so that the basi-sphenoid comes commonly to underlap and partly hide the basi-occipital. Nowhere among Amphibia or Reptilia do I know of the reverse position occurring. In some fishes there is an approach to it. Thus a slight anterior bony expansion of the basi-occipital in the Cod fits partly into a horizontal slit in the basi-sphenoid[A]. In the Carp the basi-occipital has a spathulate basal expansion like that of Pterodactyle, but it is underlapped by the basi-sphenoid[S]. In some mammals the under side of the basi-occipital extends further forward than does the neural side, as for example in the Sheep and Goat; while in a few others, as in the Walrus, the reverse positions obtain.
[S] Parasphenoid of Prof. Huxley.
But it is among Birds that the structure described in Pterodactyle is evident and characteristic. For although the bony plate under the sphenoid,—Mr Parker's basi-temporals,—is mostly anchylosed to the bones about it, and less with the occipital than with others, its position and relations are quite the same as those of the expanded flap of this Pterodactyle basi-occipital. Therefore it is identified with the basi-temporal bones.
| Case. | Comp. | Tablet. | |
| J | c | 8 | 1 |
This fossil is an inch high, rather wider, and half an inch long. It well shows the bones at the back of the skull, the basi-cranial bones, and the bones posterior to the frontals, which roof in the Cranium. There are in it striking resemblances to the back of the skull of some Natatores, as the Grannet and Cormorant, and of some Grallatores as the Heron, and Gallinaceous birds as the Cock.
The base of the skull. The bones here indicated are the basi-occipital, basi-temporal, and basi-sphenoid. The former two have come away as from an articular joint, and are wanting. The basi-occipital does not enter into the floor of the cranial cavity, and only rims the foramen magnum. But its basi-temporal expansion rests beneath the posterior part of the basi-sphenoid forming the base of the skull; its long convex anterior end fits into the concave groove at the back of the anterior part of the sphenoid. The squamous basi-temporal bone appears in this species to have been as long as the foramen magnum is wide, and to have been relatively thicker than in the other form already described.
The basi-sphenoid is a thin expanded bone forming the floor for the cerebellum, and terminating anteriorly in a triangular mass, while the slightly convex part behind, covered with the basi-temporals, is nearly square. It enters into the foramen magnum, forming its lower part; and is confluent with the ex-occipitals behind, with the periotic, alisphenoid and perhaps with the squamosal at the side; and as in birds all these sutures are obliterated. This is probably the only instance in the Animal Kingdom in which the basi-sphenoid takes so important and singular a share in the functions of the basi-occipital bone. The anterior part of the basi-sphenoid projects below the posterior part, is nearly flat on the basal surface, and forms an equilateral triangle with the apex in front and base behind. In the middle of the triangular bone is a slight longitudinal ridge, and behind the middle of each outer side a rather large foramen which appears to be the inferior opening for the carotid artery. The triangular part is hollow and as long as the quadrate portion. The lateral « 81 » parts of this anterior bone are nearly flat. They converge upwards and are rounded in front to form the boundary of the pituitary fossa, and do not appear to have terminated in a spine. Above are the alisphenoids.
The upper part of the skull is divided into two segments by a strong straight transverse ridge, which leaves the occipital bones behind, and the parietal &c. in front.
The occipital bones anchylosed together are about two-thirds the width of the foramen magnum, and of the parietal bones, with which latter the supra-occipital makes an angle of 45°. The surface is irregular, and especially is marked by a deep concavity just above each ex-occipital. The supra-occipital projects slightly over the plane of the foramen magnum, to which the strong ridge bounding the segment in front is parallel. The great foramen is nearly round, being slightly compressed at the upper part of the sides: it measures 3/8 of an inch high and is nearly as wide.
The occipital bones make with those at the base of the skull an angle of about 145° or 150°. In outline they are a transverse diamond shape. The mastoid portion is not to be distinguished from the other bones, but appear to terminate the sides of the strong occipital crest, which by posterior compression of the squamosals and parietals, becomes very strong, and makes the backward boundary of the temporal foss. This crest is in the same plane with the anterior border of the basi-temporals.
The parietals meet above in a slight ridge. They are two rectangular bones twice as wide as long, forming a semicircular roof for the brain, which looks outward and a little backward. Anteriorly these bones unite with the frontals in a slightly flexuous transverse line; and inferiorly they are connected with the periotic, the squamosal, and perhaps with the anterior point of the alisphenoid: they do not descend to the plane of the articulations of the free quadrate bones. The surface is smooth, and on the upper part flat, but concave below from side to side.
Below these parietals are the squamosals and alisphenoids, but the suture between them is not seen. They are in form a trapezium where the short side is anterior, and the lower third is folded inward so as to be confluent with the anterior part of the sphenoid. The fold forms a ridge, which I suppose « 82 » may run obliquely over the alisphenoid. The unfolded squamosal part is a flat and smooth oblong, with parallel sides, the bones are in parallel planes and nearly perpendicular to the base of the skull. Where the alisphenoid joins the sphenoid, there is a considerable concavity, above which is a small circular impression. These strips approximate inferiorly, so that the width of the skull there is rather more than half what it is at their outer margins. They shut off the pituitary body in front of them, and appear to form part of the wall for the orbit of the eye.—The slightly convex, lateral, squamosal parts above the fold continue the circular transverse outline of which the parietals are the upper half. They extend anterior to the parietals, and on the inside give attachment to the frontals. Like the parietals, they make a sharp bend outward at their hinder border, and form the lateral terminations of the occipital ridge, which is the widest part of this fossil.
The only portion of the specimen now to be described is the large region at each side looking downward, which extends from the occipital ridge to the sphenoid. It is an irregular pentangular hollow with many cavities, the hinder of which are for the ear. Two cavities above these, under the widest part of the skull, appear to be a double articulation for the quadrate bone. The outer transverse one with the squamosal is separated by a deep groove from the inner and more vertical one, which may therefore be regarded as with the petrosal bone. These excavations form the posterior half of the pentagon. The anterior half is a smooth rhombus not separable from the basi-sphenoid.
Such is the external appearance of the occipital and parietal segments of the skull of a Cambridge Pterodactyle. Each segment forms a large ring of thin bone, inclosing part of a brain-cavity as large as that of a bird and shaped like that of a bird; and which moreover is made up of the same bones as the cranium of a bird; and these are in almost exactly the same proportions as those of the Common Cock.
My own investigations do not substantiate Wagner's discovery, that the back part of the skull resembles that of the Monitor. Iguana would have offered a slightly nearer comparison, but they both differ from Cambridge specimens of Pterodactyles in characters like these.
In the lizard,
The cranial bones do not enclose the brain.
There is no division of the back of the skull into an occipital segment and a parietal segment by a girdling crest.
The squamosal bone does not enter into the cranial wall.
The quadrate bone does not articulate with the wall of the brain-case.
While the peculiar backward development of wings of the parietal in a diverging V form, give the Lizard skull an aspect of its own.
So that it must be asserted that the differences of these Pterodactyles from Lizards are so wide as to preclude comparison.
With the Crocodile, in which the cranial bones are massive, and the quadrate bone firmly packed in the skull, comparison would be no less difficult.
The Delphinidæ, in both the form of the jaws and of the back of the head, give some support to Wagler's fancy, in putting the Pterodactyle into his curious creation, the Gryphi[T]. But in the porpoises the parietal bones form as narrow a band as they do in the Duck; and are quite unlike the bones here described. In the Dolphin the two condyles almost unite into one semicircular condyle (in young specimens), owing to the enormous development of the ex-occipitals, which almost if not entirely exclude the basi-occipital from the foramen magnum. The dolphin moreover has no quadrate bone. But notwithstanding the absence of a division into occipital and parietal segments, the form and arrangement of the bones in the skull of the porpoises approximate more to the Cambridge Pterodactyles than is the case with Lizards.
[T] The Gryphi are a class of animals intermediate between Birds and Mammals according to Wagler, and including Pterodactyles, Ichthyosaurus, Plesiosaurs, Ornithorhynchus, and Myrmecophaga.
But with Birds the correspondence is so close that it would be difficult to discover differences. That one of the condition of the occipital bone seems to be the most important; another is, that from the relatively smaller size of the cerebellum the parietal bones appear to cover a larger part of the cerebrum; and a third is the strong triangular condition of the sphenoid in front of the sella tursica. With these exceptions there is nothing to distinguish the fossil described from the cranium of a bird.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 8 | 2 |
Another cranium has occurred which must be referred to a different genus. Its preservation is less perfect, but it similarly exhibits the occipital and parietal segments of the skull. All the bones are blended together without a trace of a suture.
The occipital region is flat. Its outline is not defined owing to the extent to which the sharp crest, in which it terminated outwardly, has been broken away. The occipital condyle is broken off. The foramen magnum is of an ovate form—flattened at the base. The ex-occipitals at its sides are impressed as though from contact with the neurapophyses of the atlas. Mesially, over the foramen magnum is a vertical elevated crest (now rubbed away), which may have given attachment to a bone like that post-superoccipital crest described by Quenstedt in the Pterodactylus suevicus. The occipital region makes a great angle with the flat basi-temporal region, as in birds.
The parietal region is convex from below upward, the lateral parts converging towards the crown, which however presents a broken and worn surface. From side to side the squamosal and parietal bones are concave, owing to the extended occipital crest behind, and the rapid widening of the skull in front caused by the large size of the brain.
In front is seen a section of the brain-cavity. It is very like in form to the two halves of a pear put together side by side with the stalk downward. I have removed some of the phosphate of lime from the brain-cavity, and although it has not been excavated to the cerebellum, the great depth of the brain is well seen, and the convex character of the cerebral lobes, between which a crest of bone descends mesially as in the ethmo-sphenoid mass next described. At each of the lower outer angles of the brain, extending into the cancellous brain-walls to the outermost film, is an ovoid convexity, covered with a thin film of bone. They entirely correspond with the optic lobes, being in exactly the same position as in birds, only relatively rather small. Underneath the optic lobe on the outside is a small concavity, apparently the articulation for the quadrate « 85 » bone. The basi-sphenoid mass below the brain is of considerable height, the upper half flat and smooth, the lower half fractured and cancellous.
In the main this skull is like the other one, differing chiefly in the depth of the sphenoid, in the mesial ridge between the cerebral lobes, in showing the optic lobes, and in having anchylosed basi-temporal bones. There would hence appear to have been considerable variations in the skulls of Pterodactyles even in the Cambridge Greensand.
| Case. | Comp. | Tablet. |
| J | c | 9 |
The symmetrical bone which I have so named is a wedge-like mass tapering in front, keeled above; flattened below, and cupped behind on each side. It belonged to a very much larger animal than the last fossil, and probably to a very different genus.
The inferior surface is triangular, an inch and an eighth wide behind, at the base, and an inch and a quarter long; but it is broken at both ends. In its longitudinal median line is a strong keel stopping short in front, dying away behind, and forming with the compressed margins a considerable hollow on each side, at the back part of which is a large oval foramen. This surface, though five times the size, corresponds in form, ridges, and foramina with the anterior part of the sphenoid described in the article on the back of the cranium.
The posterior surface is at right angles to the inferior one, but its lower third shows only fractured phosphate of lime filling perhaps the anterior part of the pituitary fossa. Its upper part also is broken. But on each side is a large concavity measuring in the fractured fossil an inch and a quarter high, three quarters of an inch wide, and half an inch deep from the unbroken median ridge where the cups become confluent at their base. The whole specimen is two and a quarter inches high. From the determination of the under side it follows that these smooth hollows, over each of which an impressed mesial line descends obliquely outward, are a part of the anterior boundary of the brain.
From the middle of the outer convex border of the oval remains of these cups for the cerebral hemispheres, a strong blunt ridge descends obliquely down the sides of the bone to « 86 » terminate the compressed anterior end of the bone just in front of the hypapophysial ridge of the sphenoid. Above this ridge the bone is much compressed anteriorly, forming a strong straight mesial keel above, which rapidly approximates to the base; the height of the bone in front being one inch and a half, which is also its extreme length.
The region below the oblique ridge is a concavity, but it is a little compressed from side to side behind, and has the same anterior compression, so that the elongated oval of the fracture at the anterior end of the bone is only three-eighths of an inch wide.
The superior ridge will probably have supported the frontals, and the anterior end would terminate in the orbito-sphenoid.
The lateral ridges appear to correspond with what Prof. Huxley has described in the Ostrich as the ridge indicative of a supra-presphenoid ossification pointed out by Kölliker. The groove which is here noticed on the cerebral surface may indicate the same division. If so, the upper and anterior part of the mass would be the ethmoid.
This mass offers a considerable resemblance to the frontal portion of the skull of a dolphin (e. g. Delphinus delphis) from which the maxillary, premaxillary, palatine and nasal bones have been removed. But in the Porpoise the mesial ridge dividing the cerebral hemispheres is not prolonged so far forward as in the Pterodactyle; the cranial bones are often as smooth on the inside. Notwithstanding Wagner's assurance that the Pterodactyle skull is very like a Monitor's, he would have looked in vain for an ossification in Monitor, Iguana, or other Lizards, comparable with this mass. And although the brain is closed in front by bones in Serpents, it is by the frontal bones, which form a covering for nearly the whole of the conical cerebrum. Nor in the Crocodile is there any ossified mass in front of the brain, although the brain approximates nearer to Birds than is the case with other living Reptiles. Among Birds such a structure as that of the Pterodactyle is characteristic, but no bird has it so massive and mammal-like, though an approximation is made in some thick-skulled birds like Ciconia marabou. And in birds it usually is prolonged much further forward than appears to have been the case with Pterodactyle, where from the rapid tapering of the mass in front it appears to have ended in a vertical ridge « 87 » like that in Parrots and Birds with a moveable beak. In Birds there is usually a median ridge dividing the cerebral hemispheres, but there is also often a small olfactory lobe prolonged in front of the cerebrum, to which nothing analogous is indicated in these fossils.
[U] For the opportunity of making this description, I am indebted to the kindness of John Francis Walker, Esq., M.A., F.G.S., F.C.S., &c., who some time since forwarded to me the whole of his rarer Ornithosaurian remains for description in the Geological Magazine,
The original specimen is in the collection of J. F. Walker, Esq., of Sidney Sussex College. When found it only displayed the front of the cerebral hemispheres, and Mr. Walker generously gave me permission to remove the investing cancellous bone and phosphate of lime, and thus exhibit the form of the cerebrum and its relations to the cerebellum. The lower part of the brain is not preserved. But adherent to the sides of the fossil are still left parts of the temporal bones, and part of the bone at the back of the orbit which closes in the brain. The form of the cerebellum is not quite perfect behind, but it must have been unusually small.
The cerebral lobes taken together are much wider from side to side than from back to front, and have a transverse elliptical outline, except for the mesial notch behind for the cerebellum. The lobes are a little flattened above, and divided from each other by a deep mesial groove, which makes each lobe convex from side to side. They are well rounded at the front and at the sides, and are a little compressed towards each other below in the region of the orbits. Behind they become covered superiorly as in birds with a greatly thickened part of the squamosal and parietal bones. The surface of the cerebrum is smooth. There is no indication of a pineal gland. The cerebellum is small, like a pea between two filberts. It is sub-hemisphercal, is placed close against the cerebrum, and appears to give off narrow lateral parts, like those seen in many birds, only that they abut against the back of the cerebral lobes as in the Hare and some Mammals. « 88 » In no reptile is there a brain in which the cerebrum embraces the front of the cerebellum, or in which it attains to such an enormous size. Fœtal Mammals (e. g. the horse and the sheep), even when they have attained to a considerable bulk, and many adult mammals, still have the optic lobes dividing the cerebrum from the cerebellum as in Reptiles.
The only Mammal which shows any near approximation to this brain is the Ornithorhynchus, in which the cerebellum is very small, but the cerebrum is not so well rounded in front. The form approximates to the brain in Man. But with Birds the resemblance is so close—with the owl and the goose—that there is no character to distinguish the brain of the fossil animal from those of the recent ones. A section of the cerebrum in this specimen entirely corresponds with a section of the brain-cavity in the second skull described, as does the backward extension of the cerebrum with the extent of the cerebral cavity, and the narrow cerebellum with the narrow channel parallel to the walls of the foramen magnum, as in Gallus domesticus and Birds. The front of the brain corresponds with the cast of the front of the cerebral lobes taken from the Ethmo-sphenoid mass. Thus the specimens agree among themselves, and prove the Pterodactyle to have had a brain indistinguishable from that of a Bird. And when it is remembered how distinctive this kind of brain is, and that it approximates rather towards the higher Mammals than towards Reptiles, the fact attains unusual importance in determining the Pterodactyle's place in nature.
?NEURAL ARCH OF SACRAL VERTEBRA, ?VOMER.
Pl. 12.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 10 | 1—3 |
In 1859 Prof. Owen described with doubt as the Frontal of Pterodactyle, a symmetrical bone. A smaller but more perfect specimen has since been obtained for the Woodwardian Museum; and a fragment of intermediate size is in the rich collection of the Rev. T. G. Bonney. From the descriptions already given it is impossible for it to be the frontal. There « 89 » is no proof that it is a skull bone. If of Pterodactyle the compressed lateral spaces could only be part of the nasal passages, or the impressions of a palatine or pterygoid articulation. And as the external surface of every specimen is keeled, and as the palatal surface of the upper jaw of every known Greensand Pterodactyle is keeled, and as the concavities slightly converge to the keel, it might be a bone from the under side of the head,—the vomer.
The smallest specimen is a compressed sub-semicircular bone 11/4 inch long, 9/16 inch high, and a 1/4 inch thick. The under surfaces converge to form a strong keel, which is flattened off behind. Above this, the posterior third of the bone is compressed obliquely to half the thickness, as though a bone had over-lapped this area on each side. If the oval spaces are nares, that bone might have been the pterygoid or palatine. Three-fifths of the remainder of the bone are taken up by the smooth oval depressions, which might be the inner walls of the nares; and above this is a margin of bone widening into the triangular compressed part in front, which, if the fossil is rightly determined, must have fitted into the posterior end of the maxillary or anterior end of the palatine bones.
A specimen collected by the Rev. T. G. Bonney is preserved on the sacral side of a left os innominatum with the keel downward. It appears to show a sutural surface from which an anterior part has come away. And if this specimen is compared with the neural arch of the sacral vertebra J.c.4.1, it will be found to correspond entirely. It is not impossible that c.10.1, 2 may be vomerine, and c.10.3 sacral, but there are no distinctive characters between the specimens to warrant such a determination.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 11 | 1—4 |
In the Woodwardian Museum are two distal ends of the quadrate bone and two other fragments showing the quadrato-jugal with it.
Quadrate. The smallest specimen is 1/2 an inch over the articular surface for the lower jaw and a quarter of an inch thick. It is concave from side to side in front where it shows a large pneumatic foramen near the basal end; it is bent from the articulation a little backward. It is convex behind; and between the foramen and the articulation sends inward and forward a great wing like that of the quadratum in birds. The specimens are broken short off and do not show any articulation above, where the bone contracts.
The distal articulation is double, like two long cones placed together; that in front having the base outward, while the hinder one has the base on the inner side. The largest specimen, which is much broken, shows the articulation half an inch thick.
Quadrato-jugal. This is a thin flat squamous bone, apparently of a transverse diamond shape, which is anchylosed to the anterior lateral margin of the quadrate, at right angles to the articulation. The lower margin is straight, as is the upper anterior margin, which appears to have received the malar bone above.
The upper posterior side is broken, but shows a large foramen near to the side of the quadrate. The base of the diamond is at the articulation, and at its apex is a small fragment of smooth surface, either part of a foramen, or the orbit of the eye.
In this specimen the articulation, which is broken, is about 3/4 of an inch wide, 3/8 of an inch thick. The remaining piece of the quadrate is an inch long. The quadrato-jugal is an inch and 3/16 high, and between its broken ends 13/4 inch long. It is thick and strong where joining the quadrate, but the rest of the bone is about an 1/8th of an inch thick.
The quadrate bone is Avian in possessing a pneumatic foramen, and Avian in the form of so much of the distal end as is preserved, and in the articulation for the lower jaw. The process which it sends inward on the inside is probably for the pterygoid bone, after the manner of Birds. Before anchylosis with the quadrato-jugal bone set in, as may be seen in J.c.11.4, the union was made by a hemispherical knob on the outside of the quadrate, as in Gallus domesticus. The squamose quadrato-jugal is a distinctive character.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 14 | 1—2 |
This determination is conjectural. Its form is such as would make it probable that it is part of the head. A more perfect specimen is seen in J.c.1.2.7.
The best specimen is a compressed sub-quadrate fragment of bone terminating at one end in a long reniform articular surface, and at the other end in a fracture where the bone is rapidly thickening. A side, regarded as the outer one, is flattened, being slightly concave in length, and slightly convex from side to side. The form of the inner side of the bone is determined by the inward curve of the thick part of the articular surface, which sends a rounded ridge obliquely on to the side, so that while it is concave from side to side at the articulation, at the fracture it is convex from side to side. All the specimens are large, the articulation being not less than an inch long.
appear to be developed as in birds. An account of their structure will be found in the notes on the species, page 112.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 12 | 1—6 |
Prof. Owen has described in a 'Palæontographical' monograph the proximal end of a mandible in which the sutures are obliterated. But there is one specimen of a young right ramus showing the inner and under part of the mandible to be the surangular bone which unites with the angular or outer bone by a longitudinal and vertical suture traversing on the inner side the great upper groove; and on the surangular the greater part of the articular bone rests. The articulation is strong and double, consisting of a deep transverse hollow, bounded by a strong over-locking ridge in front and a slight ridge behind; and this area is divided into two tapering furrows by a strong oblique and « 92 » rounded crest, which passes from behind inward and forward. Just behind the articulation is a ?pneumatic aperture, and then the upper surface tapers to the under surface, forming a heel, of which one specimen measuring an inch deep on the inside of the articulation has 3/4 of an inch still left and is more than 1/4 inch thick at the fracture. In a specimen belonging to the Rev. T. G. Bonney the outside of the jaw is 11/16 of an inch deep, and under the articulation 5/16 of an inch deep. The articular area is 3/4 of an inch wide and 6/16 of an inch long.
Seven specimens indicate four species.
The proximal end of the lower jaw is entirely Avian. The pneumatic aperture, as in birds, is placed behind the articulation, which is shaped as in many birds. Commonly in Ornithosaurians the bones are anchylosed and all trace of sutures obliterated, as in most birds. In the Goose, however, the six elements of each side are sometimes as readily separated as in reptiles. And in some Pterodactyles the bones separate.
The dentary bone consists of a single piece, as in birds and chelonians; and differs from both in being provided with teeth. It is described under the species O. machærorhynchus, page 113.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 17 | 1—39 |
The first three teeth are usually larger than those which are placed behind them, in this respect rather resembling some fossil reptiles than Dolphins, and presenting a character like that seen in the Dimorphodon. They are placed in oblique oval sockets. They have a single fang like Cetaceans, Edentates, Reptiles, and like the premaxillary teeth of Mammals. Cambridge specimens of jaws are not sufficiently perfect to show whether the teeth are limited to the premaxillary bone; but this appears to be the case in Pterodactylus crassirostris (Goldf.), and probably in Ornithocheirus compressirostris (Owen), [Palæontographical Society, 1851, Pl. 27], and is so regarded by Professor Owen in his later writings. Yet the significance of this fact seems to have been forgotten, and Cuvier's dictum about their teeth still has influence. He says, « 93 » "The teeth, by which the examination of an animal ought always to be commenced, here present nothing equivocal. They are all simple, conical, and nearly alike, as in the crocodiles, monitors, and other lizards." But, on the one hand, the Dolphins demonstrate that a mammal might have similar teeth even in the maxillary bone; and, on the other hand, since teeth in the premaxillary bone always are single-fanged, and commonly have a simple sub-conical crown, there is absolutely no evidence in the teeth of the affinities of the animal, which, so far as this portion of its economy went, might as well have been a fish or a mammal as anything else. In the succession there is nothing very distinctive. In the Crocodile one tooth comes up under another, as is commonly the case with mammals; and in mammals the fangs of the old teeth are often partially absorbed so that the teeth drop out into the mouth. In the Pterodactyle the new teeth came up on the inner side, as in the Ichthyosauria—a tribe of animals as singular in their affinities as the Ornithosauria. Occasionally specimens show a small furrow on the inner side of the fang, indicating absorption, but there is nothing to show how many times the teeth were renewed: in the Dolphins there is but one set, and in Crocodiles the teeth are replaced many times. In form and size the teeth are very variable. They are directed obliquely forward, and are curved backward and inward. They taper in an elongate cone, compressed from side to side, flattened on the outside, moderately convex on the inside; rarely the sides meet in a ridge after the plan of Pliosaurus, Megalosaurus, Dakosaurus, &c.; more frequently the lateral margins round into each other. Usually the enamel is quite smooth, sometimes, as in No. 1, it is finely striated and wrinkled. Some teeth are nearly circular and some quite straight. The ovate fang contracts below, conically, and is closed, leaving a long hollow pulp-cavity in its interior. Nos. 9, 10 show the marks of the successional teeth on their inner sides. No. 11 appears to have had the crown slightly worn at the tip during the animal's lifetime. In transverse section of the crown the tooth structure resembles Ichthyosaurus, Cetaceans, and Bats. The dentine is filled with calciferous tubes which radiate as in Ichthyosaurus, and towards the centre of the tooth are seen in transverse section to present many angles, almost like radiated corpuscles. They are separated by interspaces of their own width, and run towards the « 94 » circumference, sometimes straight and sometimes wavy, parallel to each other. They send off branches usually at right angles which anastomose with the adjoining tubes. The dentine is in concentric layers, and shows layers of sub-circular cells as in the teeth of Mammals. The enamel is a thin transparent layer with fewer and finer tubes than the dentine.
The story of the structure of the Ornithosaurians of the Cambridge Greensand has now been told, and it only remains to gather up the threads of their affinities and determine the Pterodactyle's place in nature. But before doing so, so various in importance are the characters enumerated, that I would first offer a few remarks on the classificational value of characters among the Reptilia, with which Pterodactyles have been most commonly grouped.
The naturalist who only examines organisms now living on the earth, symbolizes to himself, by the term Reptile, a definite sum of characters, with definite subdivisions and subordinate grouping, to which the extinct types of life extricated from the rocks cannot entirely be adapted. When the fragmentary, and often isolated or ill-associated, bones of fossilized animals are contrasted with corresponding bones in the skeletons of Serpents, Crocodiles, Lizards and Turtles, not infrequently it is found that the characters attributed to different Ordinal groups are interlaced in a single individual with a type of organization peculiar to itself, and important as are the modifications of existing orders. These characters occasionally are grouped with others which in living animals had been deemed characteristic of Fishes, Amphibia, Birds, and Mammals.
The Reptilia of the Palæontologist is therefore a vast and provisional group, ever acquiring new characters, to which no diagnosis can be applied. And although certain empirical characters have served to allocate the specimens in their several orders, in general with sufficient accuracy, yet from the imperfect preservation of some of the remains, or the imperfect extent to which their structures are known, and the want of recognised canons by which to measure their relative values, it has not been possible to « 95 » discuss the relations of the several orders to each other or with the larger groups on which some of them impinge.
Classifications represent more or less faithfully the gradational increase in the sum of the characters of an organism, as well as the increase in importance that those character severally attain. Thus gathering, so far as may be, from the chaos of individuals, a common plan of structures on which the genus, order, or class is moulded from a less specialized group of organs. The fundamental structures of a vertebrate animal, so far as their persistent importance can be measured, are, those connected with
| I. | Perpetuating the race. |
| II. | Construction of the brain. |
| III. | Circulation and oxidation of the blood. |
| IV. | Locomotion, i.e. skeleton, muscles, &c. |
And these characteristics are for the most part so interlinked, that it becomes difficult to assign to one order of animals a relative superiority over another order; since when a single set of organs is prominently developed in one group it often happens that another set of organs has a like pre-eminence in an allied group. Thus among reptiles it might be considered that
And since these structures in their functions severally modify and determine the use of other structures, the meaning that terms like Crocodilian and Chelonian really have is that they represent the aspect of Reptilian organization when seen through the specialization of respiration, or circulation of the blood. The soft parts thus determining the nutrition and function of the muscles and skeleton, anatomists in examining the bones of extinct animals are accustomed to reverse the order of their inferences, and infer from modifications of the skeleton what had been the characters of the soft and more vital structures.
On the presumed accuracy of this method of research rest many results of Comparative Anatomy. But since the shapes of bones are determined by the muscles as well as by inheritance, it is always to be remembered that a similar form of bone may obtain in different orders or classes of animals, as the result of a similar function in a special region of the body. Such resemblances are « 96 » familiar to anatomists. Hence much caution is required from the Palæontologist to distinguish between the characteristics of a group, and the extent to which they may be modified by function. This distinction is the first principle of classification. But it is always difficult to estimate the importance of characters in fragments of bones or parts of skeletons, and the difficulty is increased by the fact that if what appears to be but a functional modification should pervade all the species, it becomes a characteristic of the group, and its power of modifying the other organs in a peculiar way has to be considered.
Thus for all practical purposes birds may be said to be characterized by wings, which almost acquire the dignity of class characters from their influence on the respiratory function. But in some birds it has been thought that no bone of the fore-limb was ever developed[V]; and the difference between such a phenomenon and the wing of a Swift, for example, is one almost of infinity, as compared with any other aspect that the anterior limb might have assumed. Therefore, since a bird may part with its fore-limbs and yet remain a bird, I infer that it might apply its fore-limbs to the ground, become a quadruped, and be a bird still. And if in this process the other structures remained unchanged, no one would regard the modification as more than an ordinal one. But should the vertebræ change also, or the pelvis, or the covering of the integument, or the jaws become toothed, then, although the heart and lungs and brain of the imaginary animal retained their class characters, the functional differences being more than those of an order would constitute it a sub-class.
[V] According to Prof. Owen, in Dinornis.
In the same way it is conceivable that serpents may have existed with well-developed limbs, and if they retained their other characters the limbed forms would constitute a sub-order of serpents; but if to these characters they added a closed palate united to the cranium, they would constitute a new order of reptiles. A chelonian might be entirely deprived of its bony covering, and it would still be a chelonian, differing only as a separate family. So that structures which to the eye appear fundamental may be lost without affecting an animal's systematic position, just as animals while resembling each other in form may possess dissimilar organization.
Even with the living or typical Reptilia, naturalists are divided as to the number of ordinal groups into which they naturally fall. It is however generally agreed that the Amphibia or Dipnoa of Fitzinger, have no near affinity with the true reptiles. Milne-Edwards, Van der Hoeven and Agassiz make the remainder into three orders, as did Cuvier:
Stannius, Gray, Owen and Huxley, on the other hand, by dividing the Saurians make four orders, to which Dr Günther by his description of Sphenodon has given evidence of a fifth:
De Blainville in a remarkable classification (1816), made three orders, Chelonians, Emydosaurians [crocodiles], and Saurophidians; the latter group being subdivided into Saurians and Ophidians.
In his "Handbuch der Anatomie der Wirbelthiere" Stannius unites the Crocodilia and Chelonia into a group called Monimostylica; while of the Sauria and Ophidia he makes another group called Streptostylica. Similar groups were made by Dr Gray, and named Cataphracta and Squamata. They are identical with the "cuirassed" and "scaly" reptiles of Dumeril and Bibron.
The Astylica (Sphenodon) have no penis.
The Streptostylica have a double penis, lungs simplified at the distal end into a mere air-bladder, brains with a moderately elongated cerebrum, the palate mesially open, scales, leathery shell to the egg cut through by a tooth on the premaxillary bone.
The Monimostylica have a single penis, lungs well subdivided, ventricle of heart partly [turtles] or entirely divided [crocodiles], brains having the cerebrum broad or high, a closed palate, scutes, a calcareous shell to the egg.
Thus the chief differences between Turtles and Crocodiles on the one hand, and Lizards and Serpents on the other hand, are not so much in the fundamental vital structures, though these « 98 » undergo changes even in the families, as in the different ways in which the muscles and skeleton are modified. The typical lizards diverge widely from the crocodiles, and in those osteological features which admit of comparison they make at least as near an approach to the Chelonians. But leaving the limbs and pectoral and pelvic girdles out of consideration, lizards find their natural place side by side with the serpents.
Attempts have been made by Palæontologists to incorporate the new ordinal groups which they have been compelled to create for some fossils, along with the true Reptilia; but such a proceeding destroys the value of the term Reptile as a measure of a known organization. In the absence of knowledge of the brains of Dinosaurs, Ichthyosaurs, and Dicynodonts, their union with the Reptilia can only have a stagnating effect on Palæontology, for there is no proof that they are Reptiles in the same sense as are Crocodiles or Chameleons: while their bones being used as standards of Reptilian structure in comparisons, they adjudicate the place in nature of other animals by an authority which has never been established.
Before any inference can be drawn from the forms of bones in extinct animals, their relations to vital structures and to way of life must be known in animals which still live. This may give some clue both to their functional significance and to the extent to which they are inherited and not directly attributable to function. But an idea of the morphological value of the bones of living animals is only gained by comparing them with the remains of their extinct allies, tracing the now imitative structure back to its origin in a function which has ceased to be displayed.
Professor Owen in his "Comparative Anatomy of the Vertebrates" (1866) admits nine orders of Reptiles, five of which are extinct, some of the extinct orders being supposed to rank lower, while others are higher than the living types. They are arranged in this way,
| * | Pterosauria, |
| * | Dinosauria, |
| Crocodilia, | |
| Ophidia, | |
| Lacertilia, | |
| Chelonia | |
| * | Anomodontia, |
| * | Sauropterygia, |
| * | Ichthyopterygia.+ |
| * | Extinct. |
| + | Prof. Owen, Comp. Anat. Vol. I. p. 7-9, defines his
sub-classes. At p. 15, in the details of the orders, he puts
Ichthyosaurus in the 5th sub-class Monopnoa. But at p. 50,
treating of the vertebral column of Ichthyosaurus, it is written of
as an extinct order of Dipnoal reptiles. The Dipnoa then
would include
Ichthyosauria,
But Ichthyosaurus obviously belongs to Haeckel's group Monocondylia.Batrachia, Labyrinthodontia, Ganocephala. |
In what characters the Ichthyosaurs are lower than living reptiles I have been unable to discover. The palate may be better compared with a struthious bird than with a reptile; and the pectoral girdle may be better compared with the Ornithodelphia than with a reptile, while all the trunk-vertebræ have ribs such as are associated in living animals with a four-celled heart. But if it is a lower animal type than living reptilia, the student will ask, how much lower? does it descend to the Dipnoa, and prove to be the missing link between the Amphibia and Reptilia? and wherein is the evidence? Or does it not with Dicynodonts and Dinosaurs rather form an outlying class uniting the reptiles with the mammals.
In the same way, when Pterosauria and Dinosauria are placed above living reptiles, we are compelled to ask how much are they above, or what are the characters which bind them to the Reptilia at all? No satisfactory evidence has ever been adduced to show that the Dinosauria are Reptiles. And of the claim of the Pterodactyles to such a position, the facts detailed and now summarised will be the best evidence.
The highest structure shown in these remains is the brain-case. The cavity for the brain is in every respect like that in the skull of a bird. It resembles brains of a high type in having the cerebral lobes convex in front; since, in the lower mammals, there is a resemblance to reptiles in the conical form of the cerebrum; while the brains even of some of the placental mammals are not well distinguished from those of reptiles. Although the brain of the Ornithorhynchus is entirely mammalian, « 100 » it is more like the brain of a reptile than is the brain of the Pterodactyle. No evidence of affinities could be adduced which would outweigh this. Taken by itself it would lead us to anticipate for the Pterodactyle those vital structures which birds have in common.
Next in importance to the brain are the pneumatic perforations in the bones. They are seen in the lower jaw, the quadrate bone, in the whole of the vertebral column, in all the bones of the fore-limb, excepting one or two fragments, in the scapula and coracoid, in the os innominatum, in the femur and in the tibia. In such of the bones as can be compared, the pneumatic perforation is usually situated in Birds as it is in Pterodactyles. In Birds the bones are filled with air through these perforations, and as a principle the greater the motion of the animal, the greater is the number of bones filled with air. This air is received from the air-sacs which receive it from the lungs, and return it through the lungs again. There is thus in birds a sort of supplemental lung-system, which circulates air through the body. Nothing of the kind exists in any other class of animals. The respiratory system in birds is more perfect and complex than in the other vertebrata, and, as a result, the temperature of the blood on the whole is hotter.
In Pterodactyles the reticulate character of the perforations proves that they were pneumatic, and supplied the bones with air. The fact that the bones were supplied with air, necessitates an elaborate system of air-sacs to furnish the supply. And the existence of these air-sacs speaks incontestably to bronchial tubes opening on the surface of the lungs to supply them, and to the existence of lungs essentially like those of birds. The outward and backward direction of the coracoid bones may indicate that the lungs were larger than in a bird.
The circulation of air through the bird's body has relation to rapid motion through the atmosphere, which necessarily produces more rapid respiration than would comparative quiescence. The same inference must be applied to the Pterodactyles. But rapid respiration only means more rapid oxidation of the blood, and conversion of the purple cruorine into scarlet cruorine,—that is, the conversion of venous blood into arterial blood. And if venous blood is converted into arterial blood by a lung-apparatus « 101 » like that of a bird, and with a rapidity like that in a bird, there must be a circulation of the blood as rapid as in birds. Such a circulation is only maintained by a heart with two auricles and two ventricles. Therefore Pterodactyles had the heart like that of birds and mammals.
Now, since the temperature of the blood is chiefly dependent on respiration and circulation, and Pterodactyles had respiratory and circulatory organs which in living animals produce hot blood, it results that they were hot-blooded animals.
Thus the heart and lungs are exactly such as would have been inferred from the brain, and, like it, they are avian. And so important are these vital structures all taken together, that the inference from them upon an animal's affinities would overbear all other evidence that could be adduced except reproduction; for they demonstrate the plan on which an animal was built, and are the motor power which enabled it to use its skeleton in a way that stamped upon it a peculiar form.
In the head such structures as are preserved conform with slight variations to the avian plan. Other Ornithosaurians show in the parts which are not preserved in Cambridge specimens some characters which are not avian; they are in part as much mammalian as reptilian, and in a few points entirely reptilian. But it might be misleading to take German specimens into consideration in forming an estimate of the Pterodactyles of the Cambridge Greensand, which were probably a different ordinal group, and may have had material differences in structure.
The vertebral column as a whole is distinctive.
The neck and sacrum are mammalian, and the tail reptilian. The procœlous vertebræ are characteristic of reptiles, but in some animals, as Chelonians, they vary in different regions of the body; and among amphibians the character is inconstant in genera nearly allied.
The hind-limb is in part mammalian and in part avian; if there be any reptilian characters in the foot, they are not less mammalian.
The os innominatum is avian and mammalian.
The pectoral girdle is avian.
The fore-limb is avian and mammalian.
The wing-finger is distinctive, though formed on the avian plan.
Thus, if with an avian basis some parts of the skeleton present points of agreement with reptiles, in other points there are resemblances with mammals not less characteristic. These phænomena do not show that in so far the animal is a mammal or a reptile, but only that mammals, ornithosaurians, and reptiles have had a common origin, and that while they have been differentiated so as to form separate classes they have severally retained characters which formerly were united in one class. It is a skeleton intermediate between reptiles and mammals, and well distinguished by mammalian, reptilian, and peculiar characters, from birds. It therefore forms a parallel group with birds, displaying the ornithic organization in a differently modified skeleton. Yet it differs more from existing birds than they differ among themselves, for the discrepancies are in points of structure in which all existing birds agree: they are in having teeth, in the procœlous centrum, in the separate condition of the carpal and metacarpal (and of the tarsal and metatarsal) bones; in having more than two bones in the fore-arm, in the sacrum formed of few vertebræ, in the expanded pubic (and prepubic) bones, in a long neck to the femur, and in the modification of the wing by the great development of the phalanges of one finger.
I therefore regard the Pterodactyles as forming a group of equal value with birds, for which group the name Ornithosauria is here used. It cannot form a separate class, because they have a fundamental organization in common; and it cannot form an order of birds, because its differences from birds are greater than those of an order. It is a group which itself probably includes several orders, and must constitute a sub-class, which finds its place in nature side by side with birds and between mammals and reptiles, thus:—

Restoration.
Of the form and size[W] of the animals from the Cambridge Greensand, an idea will best be given by a few measurements.
[W] There are Ornithosaurians hereafter to be described compared with which the largest at present known will seem diminutive. A vertebra of one such, from the Wealden, is contained in the British Museum (numbered 28632). The centrum alone is between 9 and 10 inches long and 8 inches deep. It is named Streptospondylus, but constitutes a new group of Ornithosaurians. Nothing so gigantic exists in the Woodwardian Museum. Another vertebra of the same or an allied genus has been figured by Prof. Owen as the tympanic bone of ?Iguonodon (Fossil Reptilia of the Wealden, Part 2, pl. 10).
In the species Ornithocheirus nasutus (Seeley), J.c.2.11.1:
The premaxillary extends for 6 inches without reaching the nares.
The lower jaw is 3/4 of an inch deep at the articulation.
The four cervical vertebræ are each 11/2 inch long.
The sternum measures 11/2 inch over the facets for the coracoids.
The humerus is 21/16 inches over the proximal end, the radial crest not being preserved.
The coracoid is 11/4 inch over the proximal end.
The scapula is about 31/2 inches long.
The proximal carpal (imperfect) is 15/8 inch wide.
The distal carpal is 11/2 inch wide.
The lateral carpal is 11/4 inch long.
The wing-metacarpal is 11/4 inch wide at the proximal end, and 7/8 inch wide at the distal end.
The proximal end of the first phalange is about 15/8 inch wide.
The proximal end of the second phalange is less than an inch wide.
The claw-phalange (imperfect) is about 11/4 inch long.
The femur is 4 inches long.
Putting the animal together, the bones give this size :
| Head | 1 ft. | 3 in. | long. |
| Neck | 9 | ” | |
| (Back and sacrum) | ? | 8 | ” |
| (Tail) | ? | 10 | ” |
With the hypothetical parts, this would give a length of about 3 ft. 6 in. from the tip of the snout to the tip of the tail. Then
| Humerus | 8 in. | long. | |
| (Fore-arm) | ?1 ft. | 0 | ” |
| Carpus | 2 | ” | |
| Metacarpus | 10 | ” |
Which, if the fore-limbs were kept together as in ordinary quadrupeds, would give a height to the body of about 2 ft. 6 in., but as the limbs probably spread in walking as among the bats, the hind-limb would give a better idea of the height of the animal.
| (Flesh, sacrum, os innominatum). | 2 | in. |
| Femur | 4 | ” |
| (Tibia) | 6 | ” |
| (Metatarsus, &c.) | 1 | ” |
Which would give a height of about 13 inches; and, standing in the position of a bird, the height to the crown of the head would be about 2 feet. The majority of the Ornithosaurians of the Cambridge Greensand are of this size.
The spread of the wings, if there were 4 phalanges, would be
| Body | 10 | in. wide. | |
| Two arms | 5 ft. | 2 | ” |
| Two wing fingers | 7 | 0 | ” |
Giving a total expanse of about 13 feet. But, from the indications of the wing-finger, I should incline to think an expanse of 10 feet a truer estimate. The largest species attained to twice this size, and the smallest was a fourth as large. Another memoir will present descriptions and restorations of the Greensand species.
Habits.
The varying organization of different Ornithosaurians probably depends on the different habits of the tribes. That they could all fly is probable from the enormous radial crest to the humerus and the great development of the wing-bones, to which a wing-membrane was stretched, comparable to that of a Bat in texture, but more comparable to a Bird in its extent. The groups with long hind-legs probably had the membrane limited to the bones of the arm, while in the species with small hind-legs it may have attained even as great a development as in Bats, though there is no reason « 105 » for suspecting that it extended to the tail. A Pterodactyle cannot be supposed to have hung itself up by the hind-legs as does a Bat, because the hind-claws appear invariably to be directed forward. A Bat walks upon four legs with considerable elegance and speed; the wing is folded in, close to the side, so as to be scarcely noticed; and the outer claw is free to climb with. There can be little doubt but that Pterodactyles walked in a similar way. The thickened mammilate knob at the proximal end of the first phalange is well calculated for contact with the ground. And if it were supposed that the large wing-metacarpal bone were only used to support the wing, and the small metacarpals only used to support the claws by which the creature has sometimes been pictured suspending itself, it would be difficult to believe that the forces of pressure and tension in flying so exactly corresponded to the forces manifested in suspension as to cause the large and the small metacarpals invariably to attain the same length. A correspondence of this kind may be presumed to indicate a correspondence in function; and since the animal did not fly by means of its claws, the inference is that it walked by means of the metacarpal bones. In no other way could the bones have been used equally. The avian ilium would suggest a probability that they also at times stood erect like birds, from which position they could with more ease expand their wings; nor is such an idea opposed by the resemblance of some bones of the hind-limb to what obtains in birds, and of the neck of the femur to what is seen in mammals of great power in the hind-legs.
That they lived exclusively upon land and in air is improbable, considering the circumstances under which their remains are found. It is likely that they haunted the sea-shores, and, while sometimes rowing themselves over the water with their powerful wings, used the wing-membrane as does the Bat to enclose their prey and bring it to the mouth. But the superior development of the pneumatic foramina suggest that their activity was greater than in ordinary sea-birds.
The large Cambridge Pterodactyles probably pursued a more substantial prey than dragon-flies. Their teeth are well suited for fish, but probably fowl and small mammal, and even fruits, made a variety in their food. As the lord of the cliff, it may be presumed to have taken toll of all animals that could be conquered « 106 » with tooth and nail. From its brain it might be regarded as an intelligent animal. The jaws present indications of having been sheathed with a horny covering, and some of the species show a rugose anterior termination of the snout suggestive of fleshy lips like those of the Bat, and which may have been similarly used to stretch and clean the wing-membrane.
The high temperature, coupled with the sub-aerial life, are opposed to the idea of the animal having been naked. The undisturbed condition of the skeleton and some points of structure are opposed to the idea of their having had large feathers. The absence of such remains does not favour the hypothesis of their having been covered with scales, though in the legs of birds a scaly covering is met with. I should anticipate for them a filamentous downy feather, or hair, like a Bat's. The Bat combs its hair with its claws, and the Ornithosaurians may have used their claws in a similar way.
They cannot be supposed to have been gregarious, from the large number of species relatively to specimens. The reproduction may have been much the same as in birds; and the young were probably reared with affectionate care[X].
[X] Mr Carruthers has shown me crushed Turtle-like eggs from the Stonesfield slate, which in the external pitting of the egg-shell are not so different from some birds as to preclude a suspicion that they might possibly be Ornithosaurian.
The following notes indicate structures in perfect specimens from the Lithographic slate which supplement the fragmentary remains from the Cambridge Greensand[Y].
[Y] The German animals form different family groups. And it cannot be inferred that the structures seen in them pertained to Cambridge specimens.
In the head, Cambridge specimens show no trace of the parts which are between the brain-cavity and the fore-part of the jaw. The form and condition of the orbits, nares, and of the space between them, vary in German specimens. Some Birds and certain Ruminants, such as deer, the giraffe, &c., have an interspace between the orbits and nares corresponding to that in some Pterodactyles, but no such perforation is found in living reptiles. « 107 » In mammals it appears to be surrounded by the frontal, nasal, lachrymal, and often by the maxillary bone. In birds the bones appear to be the lachrymal, nasal, maxillary and premaxillary, as is the case with Pterodactyles, except that the nasal bones would seem sometimes to be excluded. The chief peculiarity of the Pterodactyle skull in this region is made by the malar bone (and, according to some authors, the maxillary also) sending up a process to meet the lachrymal. This is not seen in birds, but is characteristic of many mammals and reptiles.
The premaxillary bone is single, as in birds and Iguana; but it appears to attain as great a development as in birds, and to occupy the portion of the jaw which among reptiles and mammals is made by the maxillary bone. Owing to the great development of the premaxillary bones, the exterior nares are placed far back toward the middle of the skull as in birds, and not near the tip of the snout as in living reptiles and most mammals.
The orbits in Pterodactyles are surrounded with bone, as is commonly the case with mammals and reptiles. Among birds a complete orbit is seen among the parrots, in which it is completed below by a prolongation of the outer posterior corner of the frontal, which would correspond to the post-frontal bone, and by the lachrymal bone. Thus the malar bone, which in most mammals and reptiles forms an important part of the lower margin of the orbit, is in birds entirely excluded. In Pterodactyles the malar bone is placed between the lachrymal and the post-frontal process of the frontal bone.
The quadrate bone in German Pterodactyles, instead of being vertical as in birds, stretches obliquely forward below the malar bone, so that the articulation for the lower jaw is brought forward to be under the middle of the orbit. In Pterodactylus Kochi and in other species there appears to be a process, or small separate triradiate bone, comparable to a diminished lacertian post-frontal, and homologous with the post-frontal process of the parrots. Its upper branch meets the frontal. In some genera the front appears to meet the malar. The lower branch goes to the front of the quadrate bone, and the backward branch goes to the squamosal immediately above the articulation for the quadrate bone. Thus it is a post-frontal bone resembling that of the Iguana, but modified and adapted to a cranium like that of a bird. Its form and « 108 » size in the different genera are very variable. No similar development is seen among mammals, where the post-frontals have probably ceased to exist. It is a carious point of resemblance, but from the other resemblances to Iguana being so few it is robbed of much of its force as a mark of affinity, and becomes of interest chiefly as an evidence of independent persistence of structures.
The pterygoid and palatine bones approximate to those of bird and lizard in Pterodactylus crassirostris. And the bones in Pterodactylus suevicus, which Quenstedt names vomera, should rather have been named palatines. There is a bone in Goldfuss' specimen, between the malar and palatine, which he identifies with the transverse bone, but it is not seen in any other specimen.
The ribs sometimes appear to articulate by single heads, but in P. crassirostris they are apparently articulated as in the Crocodile. Some species show abdominal ribs like those of some reptiles; but the segments of the mammalian sternum and abdominal ribs are to be regarded as homologous structures. The vertebræ offer considerable variety in size and shape, but the greatest variation in number is seen in the tail, which is sometimes stiff and long, and sometimes short. The pelvic bones show a large amount of variation in different genera, often appearing to be crocodilian, sometimes lacertian, sometimes mammalian. In the aim the humerus is variable in the length of the radial crest, and the metacarpus also varies in length.
When the external similarity of the skeletons of birds is borne in mind, it is impossible, without disregard of classification altogether, to place animals differing so widely as do the different Ornithosaurians in the few genera in which they are at present packed.
The orders of Ornithosaurians may be established hereafter. Under the name Pterosauria, Prof. Owen founded one order which has for its type the Pterodactylus longirostris.
Von Meyer proposed to separate this order into two groups, one with two phalanges in the wing-finger, of which Ornithopterus is the only example, forming his Diathri; while the other group, « 109 » Tetrathri, or those "with four fingers, comprised all other Pterosaurians. The Tetrathri he again subdivided, following out, as he states, the suggestion of Munster and Goldfuss, into Dentirostres or such Pterodactyles as have the jaws furnished with teeth to their anterior termination; and the Subulirostres, or such as want teeth at the extremities of the jaws. To the former group he left the name Pterodactylus, and to the latter was given the name Rhamphorhynchus. Von Meyer says that he might easily have made a few more species, as will be evident to those who inspect his plates, but he "believes that the students of living animals go too far in their tendency to subdivide:" a fancy that, if indulged in by Palæontologists, would have the effect of restoring the old Linnæan groups; and a complaint which, although often heard, has usually come from those who do not readily discern and appraise classificational characters. In Palæontology genera are sometimes co-extensive with orders, while species often mean genera. It may be wearisome to the collector to be lured on to follow the devious ways of a science, but Palæontology, the source whence the mysteries of existing nature must unravel their meaning, is the handmaid of all nature's truths which have been buried in evolving the existing creation; and a duty devolves upon Palæontologists to make the past an inseparable part of the present, by applying to the two the same scientific method.
A year previous to the formation of Owen's Pterosauria, Bonaparte named the Order Ornithosaurii, and divided it into a family—Pterodactylæ, and a sub-family Pterodactylinæ.
Fitzinger (Systema Reptilium, 1843) also used the same ordinal name, and recognized three genera—
Pachyrhamphus, of which the type is Pterodactylus crassirostris (Gold.).
Pterodactylus, with the type P. longirostris (Cuv.).
And Ornithocephalus, with the type O. brevirostris (Sömm.).
These and other attempts at classification all endeavour to subdivide Ornithosaurians by the head or by the tail. Other characters for primary divisions may be obtained from the pelvis.
In the majority of German Pterodactyles the ilium extends for a long distance in front of the os pubis, and only for a very short distance behind the large ischium; and the small pubis from its anterior margin gives attachment to a large prepubic bone, which « 110 » resembles in form the os pubis of the Crocodile[Z], and is unlike that of the Monotreme. These appear to include the long-legged animals with short tails, at present called Pterodactyles, and form a well-marked family or order.
[Z] Prof. Haughton, from a study of the bones and muscles, came to the conclusion that the pubic bones of Crocodiles are the marsupial bones.
Another kind of pelvis is that in which the ilium extends a short way in front of the acetabulum, in which the pelvic bones inclose a much larger space. These include the Cambridge Ornithosaurians, the Rhamphorhynchus, and the Dimorphodon, and form another well-marked family.
These long-tailed Pterodactyles subdivide into three sub-families—Rhamphorhynchæ, Dimorphodontæ, and Ornithocheiræ. The four families may then be defined thus:
Pterodactylæ. Tail short. Hind-legs long. Ilium narrow, extending far anterior to the acetabulum; ischium extending behind the acetabulum. Epipubic bones ficiform. Head with the middle holes large, often confluent with the exterior nares. Jaws toothed to the anterior extremity.
Rhamphorhynchæ. Tail long and stiff. Hind-legs short. Pubis and ischium small, oblique to ilium, which extends less far anteriorly than in Pterodactylæ. Epipubic bones narrow and bent; they unite mesially and form a three-sided bow in front of the pelvis. Head with the middle holes and nares both small. Jaws never toothed to the anterior extremity.
Dimorphodontæ. Tail long and stiff. Hind-legs long. Pubis and ischium forming an expanded sheet of bone at right angles with the narrow ilium, which extends as far behind as in front [prepubic bones triangular (?) attached by the apex of the triangle]. Head with the nares and middle holes large. Quadrate bone large. Jaws with large teeth at the extremities, and small teeth behind. No sacrum.
Ornithocheiræ. Tail long and flexible. Hind-legs short. Pelvis as in Dimorphodontæ. [Epipubic bones with a small attachment, form unknown.] Head with the quadrate bone small. Sacrum of not fewer than three vertebræ.
In the Pterodactylæ the genera are—
Pterodactylus (Cuvier), in which the exterior nares are at the sides of the face, very large, and only partially, if at all, separated by bone from the small middle hole of the head. The head is elongated. The neck is long. Among others, it includes the species P. longirostris, P. Kochi, P. scolopaciceps, P. longicollum.
Ornithocephalus (Sömmerring), in which the anterior nares are entirely separated from the middle holes of the head, both being small, and the latter exceedingly small. The head is short The neck is short. The large ischium appears to be excluded from the acetabulum, and the ilium appears to extend less far forward than in Pterodactylus[AA].
[AA] So far as can be judged from figures, it appears to have but three bones in the wing-finger: what Cuvier regarded as a terminal and fourth joint, the bone n, Pl. XXIII. fig. 7, Oss. Foss., appearing to me to be the fibula of the tibia marked e. s in the same figure would be the terminal phalange, and r the first phalange, as may be proved by measuring them with those of the other hand, so that a phalange is missing from between them. Both the terminal phalanges appear to be hooked at the termination. Goldfuss figures the phalanges so as to make the bone which appears to be fibula in Sömmerring and Cuvier look like a fourth phalange.
Pachyrhamphus (Fitzinger). The nares are entirely separated from the middle holes of the head; both are large. The head is thick and massive. The prepubic bones meet mesially. No evidence of the number of phalanges in the wing-finger. The quadrate bone is massive, but has small attachment to the skull. Two sacral vertebræ. Wing-metacarpal very short. The type is P. crassirostris (Goldfuss).
Cycnorhamphus (Seeley). Nares very small, looking upward from a swan-like beak. The middle hole of the skull very large and elongated and lateral. Neck long. Wing-metacarpal long. Four joints in the wing-finger. Ilium widening in front. Epipubic bones meeting mesially. The type is Pterodactylus suevicus (Quenstedt).
In the Rhamphorhynchæ at present there appears to be but one genus known:
Rhamphorhynchus (von Meyer). The nares and middle holes are both small, ovate, of nearly equal size, and close together at the side of the head in front of the orbit.
In the Dimorphodontæ the only genus is
Dimorphodon (Owen). It has the nares enormously large. The middle holes are also large.
In the Ornithocheiræ the genus is
Ornithocheirus (Seeley), in which teeth are prolonged anterior to the muzzle, and the palate has a longitudinal ridge.
With the osteological illustrations of the Ornithosauria are arranged some premaxillary bones, which show varieties of form of the snout. These variations of shape serve easily to indicate different species. And the following memoranda from those specimens and other specimens in the drawers form a synopsis of the species of the Cambridge genera, which may hereafter be fully elucidated from the copious materials in the series of associated remains.
I.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 13 | 2 |
The fragment is 27/8ths inches long, with the elliptical teeth opposite to each other, 6 on a side on the palate, and one pair in front. The first three teeth are large; behind these the teeth are about half the size. The palate is gently convex, with a faint median ridge, and measures from side to side over the fourth and subsequent sockets 13/16ths of an inch. The height of the jaw at the fourth socket 11/4 inch. The sides converge to an acute rounded rostral keel. The jaws appear to have been long. The anterior termination is vascular.
The rostral keel figured by Owen Pl. I, fig. 1 d, in the 1st Supt. Cret. Reptiles, is not square as represented there, but rounded; the sides converge more acutely, and at the ridge the keel is not half so wide as the figure makes it. The enormous size of the third tooth-socket is partly due to the cracked bone having absorbed more phosphate of lime than it could hold, and extended the cracks to fissures. The type specimen shows that there was another pair of sockets in front of, but quite close to, those which appear to terminate the lower jaw.
II.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 15 | 1—3 |
A portion of a premaxillary bone fractured at both ends, and two inches long, corresponds with Dr Bowerbank's fossil figured Pl. XXVII. fig. 1, 3, 4, in the Palæontographical volume for 1851. The palate is just as wide; the median ridge, the same; the teeth the same in shape and as far apart. The jaw is of the same depth, but does not deepen so rapidly behind. The only other difference is that the sockets of the teeth are less prominent on the sides, and appear to look more directly down.
The ridge in which the converging sides meet is well rounded in a dentary bone which may have pertained to this species. In the space of two inches and a quarter are 5 teeth, the posterior four extending over two inches, the other pair being in front. The palatal surface is 3/4 of an inch broad behind the third tooth, and rather more than 5/8 of an inch broad behind the fourth tooth. The length of the 4th or of the 5th sockets is two-thirds that of the second or third. In front of the 5th tooth, the jaw is an inch deep, and it tapers in a curve to the anterior end. The teeth behind the third have interspaces greater than the length of the sockets; that between the 4th and 5th being 3/8 of an inch, while the socket only measures a quarter of an inch long. Behind the 2nd socket commences the palatal groove, broad in fronts but narrowing behind; and its sides instead of diverging as in the type, are concave so as to form a channel like a straightened Siliquaria shell. The halves of the palate bevel off so as to make a right angle with each other, and greater angles with the flat sides.
III.
| Case. | Comp. | Tablet. | Specimen. |
| J | c6 | 35 | 1 |
Dentary bone. Broken at both ends, and wanting all its teeth, this interesting fossil shows the suture where its whole length rests on the angular bone which almost reached to the termination of the beak, quite unlike what is seen in any German Pterodactyle.
It is a narrow mandible, less than three quarters of an inch wide, with the alveolar margins parallel. The palatal surface 11/2 inch long, is divided into 3 equal strips; the middle one being a deep glossal groove, slightly narrowing in front, and deepening behind, made by two inclined flat surfaces. The lateral strips are horizontal behind, and in front slope a little outward. The tooth-sockets are oval, directed outward, and as long as the interspaces, though these seem to get longer behind. In an inch and a quarter there are four teeth. Below the teeth, the sides of the jaw are compressed: though nearly parallel at the hinder fracture, the flattened surfaces approximate in front till they meet in a sharp keel, which appears to make an acute angle of about 45° with the palate; and below, where the jaw is an inch deep extends for half an inch in front of the suture with the angular bone: this suture is straight and irregularly concave, and in an inch and a quarter approximates to within 5/8ths of an inch of the palate.
IV.
| Case. | Comp. | Tablet. | Specimen. |
| J | c2 | 12 | 1 |
Middle part of a premaxillary bone fractured behind and in front, slightly distorted by compression; it is 21/8th inches long, and nearly resembles O. compressirostris (Owen). The palate is about 1/2 an inch wide in front, and 5/8ths of an inch wide behind; it is compressed mesially into a strong angular keel, between which and the teeth there is a shallow groove on each side. The groove dies away behind, and the converging parts of the keel occupy the whole space between the teeth. The teeth-sockets are small, elliptical, not opposite to each other, and placed along a distinct flattened tooth area, which looks downward and outward and separates the palate from the side of the jaw. The first pair of sockets preserved are almost 3/16ths of an inch long and 1/16th of an inch wide. The interspace between that tooth and the next tooth behind is 7/16 of an inch. Separated by similar interspaces, behind these on one side are two sockets, and on the other side one socket. The sides are flattened in front, and convex behind, (making the section of the jaw lanceolate); they are compressed and round into a narrow rostral keel. The height from the « 115 » palatal ridge to the rostral keel in front is 11/16ths of an inch; behind it is fractured, but the height was probably 14/16ths of an inch.
The palatal keel, distance of the teeth, and proportions of the jaw, distinguish it from O. compressirostris (Owen).
V.
| Case. | Comp. | Tablet. |
| J | c | 20 |
The small piece of premaxillary on which this species is founded indicates a small animal, and nearly resembles the jaw of O. microdon.
It is scarcely an inch long; nearly 9/16ths inch high behind, and nearly 7/16ths of an inch high in front, so that it tapers very rapidly, and could scarcely have been an inch longer in front.
The nose is well rounded, but the sides are a little concave, and become well pinched in in the middle, behind, showing the near approach as I think to the nostril.
The palate half an inch broad, is divided into two concave channels by the strong and sharp median ridge, which projects below the alveolar margins. The dental margins are not rounded as in C. microdon, but flattened, making more than a right angle with both the outer side-wall and palate. The interspaces between the teeth are rough, looking as though they had supported minute teeth. The alveolar margin is a tenth of an inch wide; along it are the perfectly circular sockets, a sixteenth of an inch in diameter. There are 3 sockets between 5/8 of an inch, so that they are separated by 3 times their diameter. The palate is obliquely impressed with blood-vessels running forward to the teeth from the median ridge.
The points in which this jaw differs from that of O. microdon are that in this species the teeth are circular instead of being oval; that the interspaces here are as long as in that species, though this jaw is only two-thirds the width; that instead of having a sharp keel on the upper surface, this has a well rounded roof. That though the jaw is scarcely higher than it is wide, it shows strong furrows running up to the nares, while in O. microdon, though the proportions are the « 116 » same, the sides are perfectly flat without trace of pinching in, while the line of the nasal opening is indicated by a faint furrow running all along the jaw. And lastly it differs in size, which, where the sutures are lost, may be important in discriminating forms.
VI.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 29 | 1—2 |
Premaxillary bone. The fossil is nearly 13/4ths inch long, and at the proximal end, where it is less than 3/4ths of an inch high, has flat sides, which converge to form a keel which is depressed anteriorly and rounded so that where fractured in front the bone is 7/16ths of an inch deep. The palatal surface contains two wide concave channels, between which descends a sharp median ridge, which behind becomes more prominent than the alveolar border.
The palate is 5/8ths of an inch wide. The alveolar margins are compressed and rounded. The small tooth-sockets are oval, and four are contained in 11/8th inch; they look downward.
There is a small tip of a jaw associated with this fossil, which is so like that it might be part of the bone broken off before fossilization. It corresponds in every way except that the teeth are closer. In this terminal lanceolate fragment there are in 5/8ths of an inch four teeth. The snout is terminated by two, which are close together.
VII.
Ornithocheirus Huxleyi (Seeley).
The only specimen of this species yet known is the greater part of a dentary bone contained in the Museum of the Geological Survey. An inch and 1/4 long and 3/4ths of an inch wide, it is less than half an inch deep: the sides slowly converge towards the front, and it appears to have had an obtusely lanceolate beak. The under surface is convex, too inflated for trace of a keel, and tapers to the end of the beak, which, with the left alveolar margin is abraded. The palatal surface is smooth at its front end, but two diverging ridges soon arise and form the boundary of a posteriorly deepening mesial channel, which is a quarter of an inch « 117 » wide at the fracture. These ridges too, which are parallel with the compressed and rounded alveolar margins, convert the lateral spaces into shallow channels. The right side shows the sockets of 3 small oval teeth separated by interspaces wider than teeth. A tooth and two interspaces measure 7/16ths of an inch.
The only cretaceous Pterodactyle which this at all resembles is O. microdon, but the palate is wider than in that species; the sides converge towards each other more rapidly, as though it belonged to a species with a shorter snout.
I am indebted to Prof. Huxley for the opportunity of making a notice of this species.
VIII.
| Case. | Comp. | Series. | Specimen. |
| J | c2 | 13 | 1 |
This well-marked species is a portion of a premaxillary bone 11/4 inch long, fractured behind and in front. The palate is half an inch wide; its two halves are inclined to each other at a considerable angle, and where they meet form a more prominent keel. The tooth-sockets look more outward than downward, are nearly circular, separated by interspaces as long as the sockets; three sockets and two interspaces measure one inch. The jaw is about 5/8ths of an inch high in front, and about 1/16th of an inch higher behind. The sides are flat and converge like the sides of a wedge to a sharp rostral keel.
IX.
Ornithocheirus xyphorhynchus (Seeley).
I have seen but one example of this form. It has lost much of the outer layer of bone, and shows on the sides impressions like tooth-marks from an eater of Pterodactyles. A groove which has some appearance of being due to fracture traverses each side, but the specimen is symmetrical, and has its characters in no way changed by the accident.
It is a portion of a lower jaw of a long-beaked Pterodactyle of the O. Sedgwicki type, with parallel sides, and the rounded basal ridge nearly parallel with the palate.
The fragment is two inches long, showing four large and « 118 » obliquely set sockets in If inch. The tooth-sockets are on the outer two-thirds of the palate, and looked forward, upward, and outward The interspaces each measure 5/16ths of an inch.
Each half of the palatal surface which is 5/16 of an inch wide, inclines to the other half at a right angle, being parted by a narrow groove; the diameter of the jaw is half an inch.
The depth of the jaw is 5/8ths of an inch in front, and 3/4ths of an inch behind. The sides are flat and approximate below to a sharp keel. This species is one of many in the collection of W. Reed, Esq. of York, kindly placed in my hands for the elucidation of those in the Woodwardian Museum.
X.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 14 | 1, 2 |
The fragment is 11/2 inch long, with two large elliptical tooth-sockets on each side of the flattened palate, and one pair in front. The third socket is separated from the fourth by a considerable interspace. Between the third sockets arises the median palatal ridge, and from the inner margin of each socket a lateral ridge appears to be continued. Behind the third socket the jaw measures 11/16ths of an inch from side to side, and 10/16ths of an inch high. The sides converge and round convexly into each other. The jaws appear to have been long; It is only known by upper jaws. The type specimen shows the socket of another tooth in front of the last one figured by Prof. Owen. It is directed outward at a greater angle, and separated from the hinder one by a wall not 1/16th of an inch thick, and the teeth of this pair must have been parted from each other by a film equally thin. There is no truncation of the snout as in O. Woodwardi.
Another specimen shows some variations. This fragment of a premaxillary bone is fractured through the third pair of tooth-sockets in front and through the seventh pair behind. It is about 21/8th inches long; the palate is 11/16ths of an inch wide behind the great tooth, and maintains the same width. The jaw is 11/16ths of an inch high behind, and 10/16ths high in front. The sides are gently convex, and imperceptibly unite to form the well-rounded depressed mesial ridge of the beak. From the front of the third « 119 » to the back of the fifth socket measures 13/8ths inch. The sockets are ovate, rather smaller, and closer together than in the type of O. Fittoni; margins elevated. The variations from types are so many, and often so considerable, as to suggest the idea that the fossil groups called species may in the living animals have often been genera.
In all the specimens the end of the palate is a little reflected upward.
XI.
| Case. | Comp. | Series. | Specimen. |
| J | c1 | 9 | 1 |
A fragment of premaxillary bone two inches long, fractured behind the socket for the seventh tooth. It most nearly resembles O. Sedgwicki and O. Cuvieri. Behind the second tooth the palate is 1/2 an inch wide; behind the sixth socket it is 5/8ths of an inch wide; the distance between these points is nearly 11/2 inch. The palate is flattened, with a sharp slight mesial keel and a wide concave channel on each side which dies away in front. The first pair of teeth are in front of the snout, rather small, and look forward. In this specimen the large third tooth is not developed on the left side. The second and third sockets are large and close together; the succeeding teeth are parted from each other by interspaces equal to their own diameter. They are gibbously elliptical. The sides of the jaw are gently convex from above downward; they round into each other to form a narrow rostral keel. Behind the second socket the jaw is 1/2 an inch high; behind the sixth it is nearly 7/8ths of an inch high.
The grooved and relatively wider palate, and the relatively smaller teeth, abundantly distinguish this species from O. Sedgwicki (Owen).
The smaller, more circular teeth, placed closer together, distinguish it from O. Cuvieri (Bowerbank).
XII.
| Case. | Comp. | Tablet. |
| J | c | 22 |
This fragment of premaxillary bone is 11/2 inch long. The palate is 1/2 an inch wide behind, and the jaw is rather more than 1/2 an inch high; behind the second tooth it is nearly 5/8ths of an « 120 » inch high. The sides converge superiorly to form a well-rounded keel. The palate is flattened, with a slightly elevated blunt median keel. There appears to be a pair of small teeth in front of the snout as usual, and six on the palate, with an indication of another at the posterior fracture. The teeth are of moderate size and almost circular. In the form of the bone it is readily distinguished from all the species enumerated.
XIII.
| Case. | Comp. | Series. | Specimen. |
| J | c6 | 32 | 1 |
An ill-preserved fragment fractured in front and behind, yet indicating a distinct species. The palate is flat, with the faintest median ridge, and the sides are flat and round into a narrow rostral keel, which in front approximates rapidly towards the palate. The first pair of sockets are missing; what appears to be the second pair are about 1/8th of an inch long, separated from the pair behind by an interspace of 1/4th of an inch. These are ovate and less than 1/4th of an inch long, and separated from the next pair by an interspace of not less than 1/4th of an inch. The height of the jaw over the first pair of sockets preserved is 9/16ths of an inch; over the second pair it is 14/16ths of an inch; the space between these points is 9/16ths of an inch. Behind the second pair of teeth the palate is nearly 5/8ths of an inch wide.
The only species which it resembles is O. brachyrhinus, but differs from that in the flatter, narrower palate, which makes a greater angle with the rostral keel, and in the smaller teeth, which are separated by wider interspaces.
XIV.
| Case. | Comp. | Series. | Specimen. |
| J | c2 | 11 | 1 |
A fragment of a premaxillary bone 6 inches long. It somewhat resembles O. Cuvieri in the aspect of the palate, but the jaw is more elongated, and expands from side to side at the anterior end. The teeth are opposite to each other in front, but become irregular after the sixth. The palate measures behind the second pair of sockets 3/4ths of an inch, behind the third pair it is a sixteenth of an inch wider, behind the ninth pair half an inch, and in the « 121 » last two inches it begins to widen again. A sharp keel arises behind the second pair of sockets and becomes more prominent to behind the tenth pair, when the channel which accompanies it on each side seems to disappear. The first pair of teeth, which look forward, is smaller than the second and third pairs; they are closer together than those which follow. The third sockets are 7/8ths of an inch from the tip of the snout. Then follow three smaller, more circular teeth, which are separated from each other by interspaces as long as the sockets. The back of the sixth sockets are 21/4 inches from the tip of the snout. Then follow two larger more elliptical sockets; after which the sockets become smaller and are separated by longer distances, that between the 10th and 11th pairs is nearly 3/4ths of an inch.
The height of the jaw behind the second pair of sockets is 5/8ths of an inch, behind the sixth sockets 15/16ths, behind the tenth sockets 11/4 inch. In front, the nose has the aspect of being compressed from above downward, and behind it is compressed from side to side. The sides are flattened and round into a narrow rostral ridge which is depressed at the anterior end.
XV.
| Case. | Comp. | Tablet. |
| J | c | 21 |
This species is founded on the anterior end of a premaxillary bone; in form not unlike O. Fittoni. It is 5/8ths of an inch wide; the lateral margins approximate very slowly, and in front it appears to be truncated. It is an inch and a quarter long, and in that space were on each side six large round teeth, almost as close together as they could be, five on the palate and a pair in front. The terminal two are no wider apart than the rest, and point more forward. A moderate, sharp, median ridge descends in the flattened palate, making its lateral halves a little concave. The front termination of the palate is slightly reflected upward. The jaw, which is 1/2 an inch deep behind, tapers to its termination more rapidly than does O. Fittoni. The flat sides similarly converge, and form a well-rounded ridge, which does not get blunter in front. From their close approximation, it results that the tooth-sockets are entirely above the palatal surface, so that they are better seen from the side of the jaw than from the palate.
It is a clearly marked species, as well distinguished from O. Fittoni by the closeness of its teeth, as O. Sedgwicki is from O. Cuvieri.
XVI.
| Case. | Comp. | Series. | Tablet. |
| J | c5 | 28 | 1 |
This is a species which can only be confounded with O. polyodon. It is a fragment of premaxillary bone 13/4 inch long, fractured through the seventh socket. It differs from O. polyodon in having larger teeth, which are wider apart, look more downward, have a narrower palatal interspace between each pair, and a rostral keel, which is more compressed from side to side behind and from above downward in front, and makes a greater angle with the palate.
The sockets are more uniform in size and closer together than usual, the second and third pairs being but slightly larger than the others; all are broadly elliptical. The palatal keel becomes sharp and prominent behind the fourth sockets. Behind the second pair of sockets the height of the jaw is nearly 7/16ths of an inch, behind the fourth sockets the height is 10/16ths of an inch; the distance between these points is about 10/16ths of an inch.
XVII.
| Case. | Comp. | Series. | Tablet. |
| J | c1 | 2 | 2 |
This is a fragment of a ?premaxillary bone, fractured behind through the socket for the fourth tooth. It approximates to O. colorhinus, but differs chiefly in the nose not extending in front of the first pair of teeth; in there not being any lunate area above the first pair of teeth; in there being but one tooth in front, which is relatively large; in the socket for the fourth tooth being quite close to that for the third tooth, and in the palatal sockets looking much more outward. The nose also appears to be better rounded.
The fragment is 17/8 inch long. The second and third sockets, with their interspace, measure 11/8 inch. On the opposite side the first socket is intermediate in position between the first and second.
Though not likely, it is just possible that this might be the premaxillary bone of O. eurygnathus.
XVIII.
| Case. | Comp. | Tablet. |
| J | c | 24 |
This fragment of a premaxillaiy bone is fractured behind the sockets for the third pair of teeth. It is 11/8 inch long, and shows one pair of small teeth in front and two pairs of large ovate teeth on the palate. The first pair are divided from each other and from the second pair by films of bone; and the second pair are separated from the third by rather more than half the length, of the third socket. Behind the third pair of sockets the palate is 5/8ths of an inch wide; it is flattened, and has a blunt moderately elevated mesial ridge. Behind the second pair of sockets the jaw is 5/8ths of an inch high; behind the third pair of sockets it is 3/4ths of an inch high; the distance between the places of measurement is 1/2 an inch. The sides are flat and converge to a rounded nose. The jaw is rounded from side to side in front, and the outline of the top of the nose rounds over the blunt termination of the snout above the teeth on to the palate.
In the shortness of the nose it somewhat resembles the ?P. giganteus (Bowerbank), but the jaw attenuates less rapidly, is truncated, and has larger teeth.
XIX.
| Case. | Comp. | Tablet. |
| J | c | 25 |
This species nearly resembles O. brachyrhinus, from which it differs in larger size, with a relatively wider palate, which is without a keel, and in a larger front pair of teeth. It approximates towards O, colorhinus, but is smaller, and wants the rugose lunate area over the front pair of teeth characteristic of that species. There are many varieties or species nearly related to this type, but from their imperfect preservation and the small part of the head which they represent, it is not possible to give descriptions of them.
XX.
| Case. | Comp. | Series. | Tablet. |
| J | c3 | 16 | 1 |
A fragment of a ?dentary bone, fractured behind through the socket for the third tooth. The sockets are nearly circular. It « 124 » measures about an inch long,, and behind the socket for the second tooth 13/4 inch high. The sides of the jaw are gently concave from above downward, having a pinched aspect and approximating; they round into a narrow rostral ridge, which widens towards the tip of the snout and is truncated by a small sub-circular [or sub-pentagonal] rugose area at right angles with the part of the palate behind the first pair of sockets. The first pair of sockets are nearly as large as the second, and from the steep incline of the jaw look more than usually upward; they are 7/16ths of an inch long, are separated from each other by an interspace of 6/16 ths, and from the second sockets by an interspace of more than 1/8th of an inch, while the second socket is separated from the third by about 1/4th of an inch. The palatal space between the second pair is about 3/4ths of an inch.
XXI.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 17 | 1, 2 |
Fragments of premaxillary bones. The largest portion is 21/2 inches long, and is fractured behind the socket for the fourth tooth, and the upper part of the nose is also broken away. The palate is flattened, with the median part slightly convex. The sides of the jaw converge upward, but not rapidly; in front they round into each other, but there is a slight mesial depression. The front pair of teeth are large, separated from each other and from the second pair by films of bone. Above the first pair of sockets, so as to look downward and forward, is an impressed lunate area 9/16ths of an inch wide and 5/16ths of an inch high, to which a soft lip may have been attached. This area is in the same plane with the first pair of teeth and at right angles with the upper outline of the nose. The sockets of the first pair of teeth are a little smaller than the second pair; they are both about half an inch in diameter and nearly circular. An interspace of 3/16ths of an inch separates the second socket from the third. The tooth is elliptical, the socket being narrower and longer than that of the second. The palatal interspace between the third pair is more than 3/4ths of an inch. The interspace between the third and fourth sockets is about 3/8ths of an inch. The diameter of the nearly circular fourth socket is 1/4th of an inch.
The overhanging lunate lip space, with the size of the teeth and width of the palate, abundantly distinguish this species.
XXII.
| Case. | Comp. | Tablet. | Specimen. |
| J | c | 18 | 1—4 |
I regard the fragment on which this species was founded as being the terminal end, and not a section of a jaw; partly from the rounding of the lateral surfaces to the front, and chiefly from the snapped off teeth in the middle of the truncated anterior end, for they are smaller than the pair behind them, and look forward at a greater angle, so that the converging sockets of both pairs meet behind. These characters are well shown in Mr Dinkel's excellent figure, Pl. II. fig. 3a. Second Sup. Palæont. The palate is destroyed, and gives no clue to the bone being either lower or upper.
Another specimen, rather smaller, shows the rostrum well rounded; the front is truncated at right angles to it: there is the same rounding of its lower part into the sides, and the stumps of the front pair of teeth are visible though they are again worn level with the rugose front of the snout.
But the finest fragment of this species is a rostral end, (perhaps of the upper jaw) three inches long, two inches deep, and with the palate as wide. It indicates 5 teeth on a side: the front pair small, 2nd and 3rd much larger, and two pairs behind, which are smaller. The palate is flat, and attains its greatest width at the third tooth, behind which it contracts noticeably. The third tooth is more than half an inch in diameter, the fourth is 5/16ths of an inch long. The spaces between teeth seem equal to the long diameter of the sockets, which are oval and straight. The sides round into the front of the muzzle more gradually in this specimen than in the others. An impressed line runs along the median ridge of the upper surface. Just as the jaw gets narrower behind, so the well-rounded upper surface becomes more acute behind.
Behind the third socket the palate measures 17/8 inch from side to side, and the jaw is there nearly 2 inches high.
This is the most massive Pterodactyle jaw known. In the recent state it may have indicated a creature sufficiently distinguished « 126 » from those to which the smaller fossils belonged, but now the divergence of characters is so slight as to be for zoological purposes of no value.
It is related to O. Fittoni; the chief points of difference being the truncated muzzle, the compression behind the third tooth, the much sharper (?) dorsal ridge, and the large size of the head.
XXIII.
| Case. | Comp. | Series. | Tablet. |
| J | c3 | 14 | 1 |
A fragment of premaxillary bone, well distinguished from every other specimen, except one in the collection of Mr Reed of York, which is here named O. Reedi. It is a large head, with larger teeth than any known species. The jaw is truncated in front, with a rugose vertical area in front reaching 13/4 inch high from the palate, on which the usual front pair of teeth are not seen. At the angle of this front area with the palate is a large elliptical tooth 9/16ths of an inch wide, and behind it, with an interspace of 3/16ths of an inch, is a socket measuring 10/16ths of an inch in length; the next interspace is about 1/8th of an inch, and the next nearly circular socket is 5/16ths long; then another interspace of 1/8th of an inch, and another and a smaller tooth. The palate appears to have been channelled. The sides of the jaw are flat, or slightly concave, and where fractured above, are 3 inches high. Above the rugose vertical area of the snout, is an area, concave from back to front, reaching up to the rostral keel; it is flat from side to side behind, and convex from side to side in front. So much as is preserved measures 13/4 inch in length, and appears to be relatively narrower than in O. Reedi.
XXIV.
Ornithocheirus Reedi (Seeley).
The anterior part of an upper jaw has flattened slightly concave sides, which converge above so as to form boundaries of (1) a flat triangular area which looks anteriorly, and of (2) an oblong area, traversed by a mesial groove, which looks upward and forward and is concave from back to front. In the lower half of the truncated triangular anterior termination are the « 127 » remains of the stumps of the two anterior teeth; they are oval in outline, 9/16ths of an inch high, and 7/16ths of an inch wide; they are parted by an interspace nearly 1/4 of an inch wide, which becomes concave vertically as it rounds on to the palatal surface. All the front triangular surface above the teeth is rough: its entire height is about 11/4 inch, and is nearly as wide across the base. The side rounds a little into the concave median upper surface, and into the triangular front; so much as is preserved measures 21/2 inches high, and 13/8 inch long. The palatal surface, which is very small and badly preserved, is 13/4 inch wide behind, but gives no indication of further widening. On its outer border are seen two large circular teeth 5/8ths of an inch in diameter; they are separated by a median palatal interspace of 7/8ths of an inch. Where it is fractured behind, the specimen shows the sockets of another pair of teeth behind these, with an interspace of 1/4 of an inch in the antero-posterior direction. The palate is convex.
The superior oblong area is concave in length as well as transversely. It makes a great angle with the triangular front of which it is the upward continuation; so much as is preserved extends 11/2 inch in length; it is about 1/2 an inch wide.
I am indebted to W. Reed, Esq. of York, for the opportunity of making a notice of this species, which closely resembles O. capito.
The species which follow were separated in the "Index to the Ornithosauria," &c. as a different genus. That proposal might still be sustained, for these massive truncated jaws are unlike the spear-shaped jaws of many of the species. And to the minds of some readers the forms already described will arrange themselves in groups which not improbably indicate genera. But a re-examination of the type Pterodactylus simus (Owen) has convinced me that it is a lower jaw, and therefore it affords no evidence of the presence or absence of the peculiar front premaxillary teeth which characterize nearly all the Cretaceous species.
XXV.
| Case. | Comp. | Specimen. | |
| J | c | c | 16 |
The palate is 23/4 inches long, and at the second pair of teeth about 7/8ths of an inch wide. It is fractured at the end through the fifth socket, and at the side along the palatal groove. The first pair of teeth is smaller and closer together than the others. The palatal interspace between the second pair is 3/8ths of an inch; « 128 » between the third pair, which are large teeth, it is 1/2 an inch. The sockets are sub-circular, and are not separated from each other by wider interspaces than their own length. In front is a long triangular rugose area, convex from above downward, a distance of 11/2 inch; and concave from side to side, a width above of rather more than 1/2 an inch. Below this the flattened sides converge to a blunt keel; where, fractured, the jaw is 21/2 inches deep. There are several fragments of species allied to the last; one has the triangular area in front very small, only half as high as in the type and very narrow, for the sides are gently rounded into it. It is marked by short longitudinal furrows, impressed vessels I think, while in O. simus the surface is irregularly rough. The first pair of teeth are much larger than in O. simus; they are longer, more conical, and circular, and separated by as wide a space as the second pair. There is not much to found a species on, but as it appears to be quite distinct from O. simus, it is named O. Carteri. Another fragment, with the area very long, is marked O. platyrhinus. But a sufficiency of species has been indicated to make known the Ornithosaurian fauna of the Cambridge Greensand. And the detailed description of critical types and of the other parts of the skeletons is beyond the general osteology of the tribe, and will rather belong to a memoir in which this flock of Pterodactyles will be restored to their living forms.
A fragment of the lower jaw of a large Ornithocheirus has been obtained from an outlier of the Upper Greensand at Rocken End in the Isle of Wight. It appears to indicate a distinct species. It is 21/2 inches long, and shows three large teeth still preserved in their sockets. The extreme width outside the third pair of sockets is nearly 2 inches. The sides, which are slightly concave from above downward, converge so as to give the broken end a triangular outline. In front is a small sub-triangular area, deeply scored with vascular markings; below this the outline slopes obliquely backward, and the two sides there round convexly into each other. The first socket is 7/16ths of an inch long, the tooth coarsely striated, and like the others elliptical; the interspace between the first and the second teeth is 5/16ths of an inch. The second tooth, probably immature, is an inch in length, smooth, and like the third traversed in front and behind by a slight lateral ridge; at the base it measures 5/16ths of an inch from front to back. The third tooth is rather less than 5/8ths of an inch from front to back. The interspace between the first and second sockets, which the teeth do not entirely fill, is more than 1/4 of an inch. The posterior margin of each socket is elevated into a sort of collar.
Enumeration of some of the principal writings on the Ornithosauria (selected chiefly from Von Meyer's Reptilien aus dem Lithographischen Schiefer), with references to the shelves in the Cambridge University Library, where the books may be consulted.
| Agassiz (Louis).—Memoires Soc. Nat. Neuchâtel, Vol. 1, p. 19, paragraph notice in a memoir, "Résumé des travaux de la section d'histoire naturelle, et de celle des sciences medicales pendant l'année, 1833" | B. 3. 66. |
| A briefer notice in a paper, "A Period in the History of our Planet," in Edinburgh New Phil. Journal, 1843, Vol. 35, p. 9, quoted by Von Meyer | XXVIII. 36. 65. |
| de Blainville (D.).—Osteographie; Palæotherium, p. 9 (Vol. 2), quoted by v. Meyer | AF. 5. 9. |
| Bonaparte (C. L.).—Nuovi Annali delle Scienze Naturali Bologna[1], Vol. 1, 1838, p. 391; Vol. 4, 1840, 24 Sept. p. 91. | |
| Blumenbach.—Manuel d'Histoire naturelle, éd. 1803, Vol. 2, p. 408 | B. 12. 20. |
| (Vergleichende Anatomie, 1805, p. 75), § 44, Translation, 1807 | Tt. 18. 51. |
| Handbuch der Naturgeschichte, 1825, p. 620 | Yy. 39. 6. |
| Burmeister.—Gesellsch. zu Halle, Vol. 3, Part 2, 1855; Viertel-jahrsbericht, 28 April, p. 11 | XXVI. 50. 2. |
| Collini.—Acta Acad. Theod. Palat. 1784, Vol. 5, p. 58, pl. 1. | 17. 5. 34. |
| Cuvier.—Ossemens fossiles, Vol. 5, Pt. 2, p. 359, ed. 1824, pl. 23. | VII. 1. 36. |
| Annales du Museum, 1809, Vol. 13, p. 424 « 130 » | B. 42. 13. |
| Règne Animal, ed. 1850, Vol. Rept. p. 62. | XVIII. 15. 15. |
| Dumeril et Bibron.—Erpétologie générale, Vol. 4, p. 549. | B. 37. 33. |
| Fischer.—Bibliotheca Palæontologica, Moscow, 1834, p. 163. | LR. 15. 58. |
| Fitzinger.—Systema Reptilium[1], 1843, p. 35. | |
| Fraas.—Württemb. naturw. Jahreshefte, XI. 1855, p. 102. | XIII. 24. 25. |
| Giebel.—Jahresbericht des naturwiss. Vereins zu Halle[1], 1849-50. Fauna der Vorwelt, 1847 (Vögel und Amphib. p. 89). | B. 46. 17. |
| Allgemeine Palæontologie[1], 1852, p. 231. | |
| Goldfuss.—Nova Acta Leopold., XV. Part 1, p. 63, pl. 7-10. | 23. 4. 63. |
| Van der Hoeven.—Verslagen en Mededeelingen van het K, Nederl. Institut over den Jare, 1846, p. 430. | 23. 6. 136. |
| Merk.—Bald. Medic. Journ. Stück. 1787, Vol. 4, p. 74. | XVIII. 23. 10. |
| H. von Meyer.—Reptilien aus dem Lithograph. Schiefer, 1859 (Fauna der Vorwelt) | KK. 1. 55. |
| Nova Acta Leopold., XV. Part 2, 1831, p. 198, pl. 60 | 23. 4. 64. |
| Palæologica, 1832, pp. 115, 228 | X. 20. 39. |
| Jahrb. für Mineral. 1837, p. 316 | XIII. 14. 32. |
| 1838, pp. 415, 667 | XIII. 14. 33. |
| 1843, p. 583 | XIII. 14. 38. |
| Palæontographica, Vol. 1, p. 1846 | B. 40. 53. |
| Jahrb. für Mineral. 1854, p. 51 | XIII. 14. 50. |
| 1855, p. 328 | XIII. 14. 61. |
| 1856, p. 826 | XIII. 14. 52. |
| 1857, p. 535 | XIII. 14. 53. |
| 1858, p. 62 | XIII. 14. 62. |
| Von Munster.—Jahrb. für Mineral. 1832, p. 412 | XIII. 14. 27. |
| Nova Acad. Leopold., XV. Part 1, p. 49, pl. 6 | 23. 4. 63. |
| Jahrb. für Mineral. 1836, p. 580 | XIII. 14. 31. |
| Beiträge zur Petrefaktenkunde, i.; p. 83, 1839 | XIII. 11. 49. |
| Jahrb. für Mineral. 1839, p. 677 | XIII. 14. 34. |
| 1842, p. 35 | XIII. 14. 37. |
| Oken.—Isis, 1818, p. 246, pl. 4 « 131 » | XXII. 5. 2. |
| 1819, p. 1788 | XXII. 5. 3. |
| A. Oppel.—Württemb. naturw. Jahreshefte, XII. 1856, p. 326. | XIII. 24. 25. |
| Württemb. naturw. Jahreshefte, XIV. 1858, p. 55 | XIII. 24. 26. |
| Quenstedt.—Jahrb. für Mineral. 1854, p. 570 | XIII. 14. 50. |
| Ueber Pterodactylus Suevicus[1], 4to. 1855. | |
| Sonst und Jetzt[1], 1856, p. 130. | |
| Württemb. naturw. Jahreshefte, XIII. 1857, p. 41; XIV. 1858, p. 299 | XIII. 24. 26. |
| Der Jura, 1858, p. 812 | B. 44. 48. |
| Ritgen.—Nova Acta Leopold., XIII. Part 1, 1826, p. 329, pl. 16. | 23. 4. 68. |
| Th. von Sömmerring.—Denkschriften Akad. München, | |
| 1812, Vol. IV. p. 89, pl. 5-7[2] | 23. 3. 28. |
| 1820, Vol. VI. pp. 89, 102, pl. | 23. 3. 31. |
| Spix.—Denkschriften Akad. München, VI. 1820, p. 59 | 23. 3. 31. |
| Theodori.—Notiz für Nat. u. Heilk. 1830, No. 623, p. 101. | 24. 2. 28. |
| Bericht des naturforschenden Vereins in Bamberg, 1852, p. 17. | |
| Wagler.—System der Amphibien, 1830, p. 61, figs. 1, 2. | |
| Wagner (A.).—Abhandl. Bayerischen Akad., | |
| II. 1837, p. 163, pl. | 23. 3. 41. |
| VI. Part 1, 1851, p. 129, pl. 5, 6; Part 3, 1852, p. 690, pl. 19 | 23. 3. 45. |
| VIII. Part 2, 1858, p. 439, pl. 15-17 | 23. 3. 61. |
[1] May be consulted on application to the Librarian.
[2] Good figure.
Chief English Writings on Ornithosaurians.
| J. S. Bowekbank.—Quart. Jour. Geol. Soc. 1846, p. 7. | VII. 3. 42. |
| Quart Jour. Geol. Soc. 1848, p. 2. | VII. 3. 44. |
| Proc. Zool. Soc. 1851, p. 14. | XVIII. 18. 3. |
| W. Buckland.—Geol. Trans. Ser. 2, Vol. III. p. 217. | XIII. 2. 8. |
| Geology and Mineralogy, Vol. I. p. 221, Vol. II. p. 31, pl. 21, 22. | Zz. 34. 10. |
| T. H. Huxley.—Quart. Jour. Geol. Soc. 1859, p. 658. | VII. 3. 35. |
| Introduction to Classification of Animals, 1869, p. 110. | B. 41. 70. |
| Proc. Zool. Soc. 1867, p. 417 | XVIII. 18. 19. |
| G. A. Mantell.—Geol. Trans. Ser. 2, Vol. V. p. 170. | XIII. 2. 10. |
| Quart. Jour. Geol. Soc. Vol. II. p. 104. | VII. 3. 42. |
| R. Owen.- Geol. Trans. Ser. 2, Vol. VI. 1840, p. 411. | XIII. 2. 12. |
| Brit. Assoc. Reports, 1841, p. 156. | II. 6. 10. |
| Quart. Jour. Geol. Soc. Vol. II. p. 96. | VII. 3. 42. |
| British Fossil Mammals and Birds, 1846, p. 545. | IX. 5. 15. |
| Odontography, Vol. I. p. 273. | IX. 10. 23. |
| Dixon's Geology of Sussex, 1850, p. 401. | VII. 1. 5. |
| Palæont. Soc. Monograph, Owen, 1851, p. 80. | XVIII. 14. 17. |
| Proc. Zool. Soc. 1851, p. 21. | XVIII. 18. 3. |
| British Assoc. Reports, 1858, p. 97, sec. | II. 6. 27. |
| Philosophical Trans. Royal Soc, 1859, Vol. 149, p. 161. | 15. 3. 61. |
| Palæontographical Soc. Monograph, 1859. | XVIII. 14. |
| 1860 | XVIII. 14. 9. |
| Palæontology, p. 244. | B. 46. 29. |
| Anat. Vertebrates, Vol. I. pp. 6, 18, 161, 175, 192, Vol. II. p. 13. | IX. 11. 22. |
| H. G. Seeley.—British Assoc. Reports, 1864, p. 69, sec. | II. 6. 33. |
| Annals of Natural History, 1865, Vol. XV. p. 148. | XIII. 30. 29. |
| 1866, Vol. XVII. p. 321. | XIII. 30. 31. |
| 1869, Vol. III. p. 465. | XIII. 30. 37. |
| Index to Aves Ornithosauria and Reptilia, p. 4, p. 89. | VII. 6. 71. |
Affinities, 24, 94
Albatross, 31
Alisphenoid, 81
Appendix, 129
Archæopteryx, 8
Aspect, 105
Astylica, 97
Atlas and axis, 64
Avian carpus, 52
Basi-occipital bone, 78
Basi-sphenoid, 80
Bat, 31, 105
Birds, 52
Blainville, 97
Body, 108
Bonaparte, 109
Brain, 25
Brain-cavity, 87
Buckland, 60
Burmeister, 17
Cambridge upper Greensand, 2
Camel, 47
Carp, 79
Carpus, 48
Carruthers (Mr), 106
Caudal vertebræ, 75
Cerebral lobes, 87
Cervical vertebræ, 65
Cetaceans, 30
Chameleon, 31, 34, 37, 41, 47, 72
Chelydra, 61
Chrysochloris, 42, 47
Ciconia marabou, 86
Circulation, 100
Classification, 108
Claw phalange, 59
Cod, 79
Coracoid, 32
Cranium, 80
Crocodile, 31, 35, 37, 41, 47, 63, 69, 83, 93, 95, 97
Cuvier, 7, 92
Cycnorhamphus, 111
Delphinidæ, 83
Dentary bone, 92
Dicynodonts, 61
Dimensions, 103
Dimorphodon, 112, 60
Dinornis, 67
Dinosaurs, 99, 61
Dipnoal reptiles, 99
Dorsal vertebræ, 69
Echidna, 61
?Eggs, 106
Epipubic bones (see prepubic bones)
Evidence that Pterodactyles were Reptiles, 5
Facial bones, 91
Families, 110
Femur, 62
« 134 »
Fibula, 63, 22
Food, 105
Fore-arm, 48
Gallus domesticus, 90, 82, 34
Genera, 111
German Pterodactyles, 106
Goldfuss, 11, 63
Goose, 92
Grouping of reptiles, 97
Gypogeranus serpentarius, 42
Habits, 104
Hand, 53
Hare, 87
Head 18, 77, 106
History, 3
Horse, 41
How the meaning of the word reptile is lost, 98
Humerus, 38
Huxley (Prof), 86, 116
Ichthyosaurus, 34, 37
Iguana, 41, 47, 69, 82
Ilium, 60
Ischium, 60
Jerboa, 64
Kangaroo, 55, 69
Ligamentum teres, 62
Lower jaw, 91
Llama, 77, 69
Malar bone, 107
Mammalian Affinities, 31, 34, 87, 41, 42, 61, 62, 69, 75, 79, 83, 86, 94, 105
Manubrium, 29
Marsupial bones, 61, 110
Materials, 1
Mergus merganser, 31
Metacarpus, 53
Metatarsus, 63
Meyer (H. von), 17, 109
Mole, 30, 37
Monimostylica, 97
Monitor, 41, 47, 69, 72, 86
Monotremata, 34
Mould of Brain-cavity, 87
Objections to Prof. Owen's grouping, 99
Occipital bones, 81
Oken, 10
Optic lobes, 84
Orbito-ethmo-sphenoid bone, 85
Orbits, 107
Organization, 7
Ornithocephalus, 111
Ornithocheirus, 112
Ornithorhynchus, 88
Ornithosauria, 27
Ossemens fossiles, 7
Ostrich, 52, 58, 86
Owen (Prof. R.), 3, 29, 32, 36, 48, 54, 56, 64, 66, 69, 75, 78, 88, 91, 92, 98, 108, 115
Pachyrhamphus, 111
Parrot, 87
Palæontology, 109
Parietal bones, 81
Parker (Mr W. K.), 79
Pectoral girdle, 28
Pelvis, 59
Penguin, 8, 69
Petrosal, 82
Phalange, 56
Plan of organisation, 25
Pneumatic cavities, 23, 26, 100
Porpoise, 86
Post frontal, 107
Premaxillary bones, 91, 107
Prepubic bones (prepubic), 61, 110, 111
« 135 »
Pterodactyle's place in nature, 102
Pterodactylus, 111
Pteroid bone, 48
Pterosauria, 99, 108
?Pterygoid end of palatine bone, 91
Quadrate bone, 89, 107
Quadrato-jugal, 90
Quenstedt, 17, 21
Radius, 42
Reptilia, 94
Respiration, 26, 100
Restoration, 103
Rhamphorhynchus, 111
Ribs, 108
Roc, 5
Sacrum, 73
Scapula, 35
Scink, 41, 72
Second phalange, 57
Sömmerring, 10
Species, 112
Squamosal bone, 81
Stannius, 97
Sternum, 28
Streptostylica, 97
Struthious birds, 31, 72
Tarso-metatarsus, 63
Teeth, 92
Tibia, 62
Ulna, 43
Vertebral column, 64
?Vomer, 88
Wagler, 11
Wagner, 14
Walker (Mr J. F.), 87
Walking, 105
Walrus, 79
Wing-finger, 66
THE END.
PRINTED BY C. J. CLAY, M.A.
AT THE UNIVERSITY PRESS.
PLATE I.[AB]
Sternum and Scapula.
Fig. 1. Fore-part of sternum showing the ovate synovial facet for the coracoid. J.a.1, p. 28.
2. Outside of the proximal end of a right scapula. Largest specimen. J.a.3, no. 2, p. 35.
3. Outside of greater portion of a left scapula. J.a.3, no. 13.
4. Inner side of a small right scapula. J.a.3, no. 12.
5. Outside of proximal end of a right scapula. J.a.3, no. 3.
6. Surface of J.a.3, no. 3. articulating with humerus.
7. Outside of distal end of a scapula. J.a.4, no. 1.
8. View of the distal termination of a scapula.
9. View of proximal end of left scapula looking from the distal toward the articular end. J.a.3, no. 17.
10. Proximal end of right scapula where united with coracoid, looking at the scapula from the articulation. J.c4.18.6. Compare fig. 6.
11. Inner surface of same specimen showing the pneumatic foramen at the union of scapula and coracoid.
12. Outer view of the same specimen.
[AB] For the Lithographic details of plates 1 to 3, the author is not answerable. Accidents happened to these plates in the printing, and they were replaced without his knowledge by good copies; which however have sometimes deprived the bones of their characters.
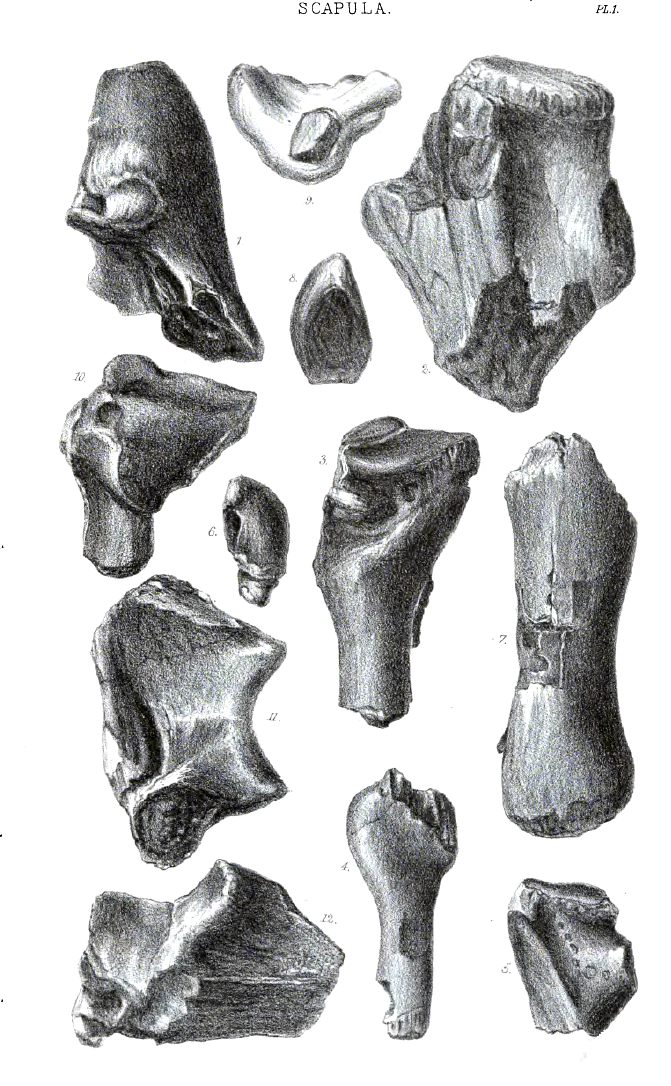 Click on image to view larger vesion
Click on image to view larger vesion
PLATE II.
Coracoid and Radius.
Fig. 1. Outer side view of left coracoid. J.c3.16.5, p. 32.
2. Back view of the same specimen showing the surface which unites with the scapula.
3. Outer side view of perfect right coracoid. J.c4.18. 5. Near the figure 3 is the pneumatic notch.
4. View of the proximal articular surface of a right coracoid. J.a.2, no. 23.
5. Inner view of distal end of left coracoid. J.a.2, no. 18.
6. The distal articulation of the same specimen.
7. Fragment of proximal end of radius 4/5 nat. size. J.a.11, no. 7, p. 46.
8. Proximal end of radius. J.a.11, no. 1.
9. Proximal articular surface of radius from the same specimen.
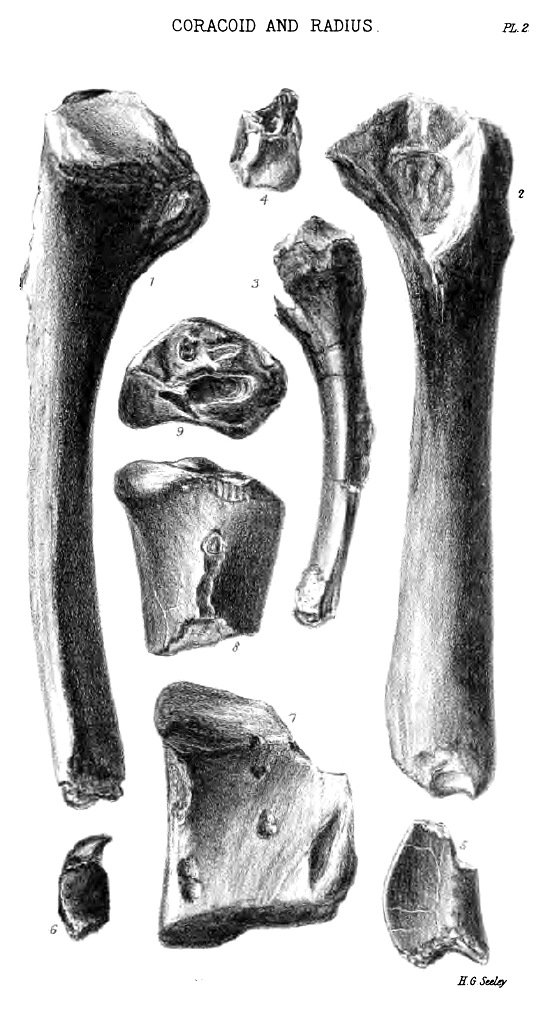 Click on image to view larger vesion
Click on image to view larger vesion
PLATE III.
Radius and Ulna.
Fig. 1. Inner view of distal end of right radius. J.a.10, no. 2, p. 44.
2. Outer view of distal end of right radius. J.a.10, no. 3.
3. Distal articulation of right radius. J.a.10, no. 6.
4. Inner view of proximal end of ulna with olecranon anchylosed, p. 45.
5. Side view of the same specimen. J.a.9, no. 1.
6. Proximal end of ulna from which the olecranon has come away. J.a.9, no. 5.
7. Proximal articular surface of same specimen.
8. Proximal articular surface of ulna. J.a.9, no. 4.
9. Proximal articular end of ulna from which the olecranon has come away.
10. Distal end of right ulna. J.a.13, no. 5, p. 43.
11. Distal articulation of the same specimen.
12. Distal end of left ulna. J.a.12, no. 3.
13. Distal articulation of the same specimen.
 Click on image to view larger vesion
Click on image to view larger vesion
PLATE IV.
Humerus.
Fig. 1. A nearly perfect right humerus, from Ashwell. J.a.6, no. 30, p. 38.
2. Same specimen seen from the proximal end, so as to display the distal end, twisted at right angles with the radial crest. The pneumatic foramen is on the anterior and radial side.
3. Proximal end of left humerus showing the radial crest perfect. J.a.6. 25.
4. Articular surface of same specimen showing the termination of the radial crest.
5. Posterior aspect of proximal end of right humerus. The pneumatic foramen is on the posterior and ulnar side.
6. Proximal articular surface of left humerus. J.a.6, no. 2.
7. Distal end of right humerus. J.a.6, no. 29.
8. Distal articulation of left humerus. J.a.6, no. 45.
9. Distal end of same specimen.
10. Distal end of left humerus. J.a.6.20.
11. Distal end of right humerus. J.a.6.46.
12. Distal end of left humerus. J.a.6.34.
13. Distal end of left humerus from a specimen lent by J. B. Lee, Esq.
14. Distal end of left humerus. J.a.6.35.
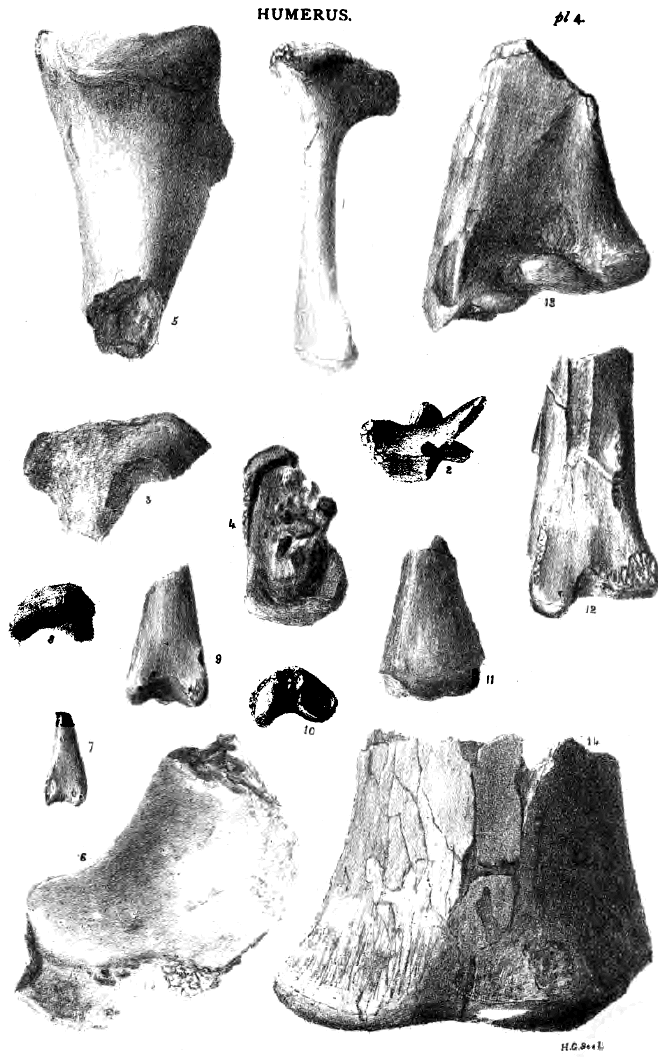 Click on image to view larger vesion
Click on image to view larger vesion
PLATE V.
Carpal Bones.
Fig. 1. Distal surface of right proximal carpal bone, p. 48.
2. Same specimen seen from outer end, showing the large unarticular surface, above is a part of the distal articulation. J.b.1, no. 1. (figured upside down).
3. Proximal articular surface of right proximal carpal bone. J.b.1, no. 7. The right upper part is for the radius, the left lower part for the ulna.
4. View of same specimen (upside down) from the ulnar side.
5. View of same specimen from the radial side.
6. Portion of distal articular surface of a right distal carpal bone. J.b.3, no. 23, 4/5 nat. size, p. 50.
7. Front radial side of right distal carpal. J.b.3.24.
8. Back ulnar side of the same specimen.
9. Proximal articular surface of the same distal carpal.
10. Distal articular surface of the same distal carpal.
11. View of the proximal articular surface of the same distal carpal, seen from the inside.
12. Perfect element of left distal carpal bone showing the distal carpal bone to be composite.
13. Distal surface of a right distal carpal of another genus. J.b.3, no. 20.
14. Lateral carpal or pisiform bone, seen from the inside, the distal articular talon partly broken. J.b.4, no. 2.
15. Lateral carpal seen from the outside. J.b.4.9.
16. Same bone showing the distal articulation, p. 51.
17. Lateral carpal bone of a different genus, seen from the inside.
 Click on image to view larger vesion
Click on image to view larger vesion
PLATE VI.
Wing Metacarpal Bone, &c.
Fig. 1. Fragment of the proximal end of a large wing-metacarpal bone. J.b.5, no. 9. It is figured upside down, a part of the surface articulating with the distal carpal bone being over the fig. 1, p. 53.
2. Aspect of the proximal articular sur&.ce of the wing-metacarpal bone. J.b.5, no. 3.
3. Exterior aspect of the same specimen.
4. Inner aspect of another proximal end. J.b.5, no. 4.
5. The greater part of a small wing-metacarpal bone. J.b.5, no. 1. Imperfect at the distal end.
6. Distal end of a wing-metacarpal bone. J.b.5, no. 31.
7. Front aspect of the same specimen.
8. Distal end of metatarsal bone or of a metacarpal bone of a small finger. J.b.8, no. 1.
9. Lateral aspect of a similar bone. J.b.8, no. 2.
10. Outline of the imperfect distal termination of a bone regarded as left metatarsus of an Ornithosaurian. J.b.13, p. 63.
11. Front aspect of the same specimen.
12. Articular aspect of proximal end of first phalange of the wing-finger, from which the terminal epiphysis has come away. J.b.6, no. 10.
13. Diagram outline of the same specimen, p. 56.
 Click on image to view larger vesion
Click on image to view larger vesion
PLATE VII.
Wing Finger.
Fig. 1. Exterior aspect of proximal end of first phalange of the wing-finger. J.c3.16.12, p. 56.
2. Inner aspect of proximal end of a small wing-metacarpal bone which has lost its proximal epiphysis; it shows the notch for the pneumatic foramen. J.c1.8.8.
3. Fragment of the proximal end of a large wing-metacarpal bone, showing near the fig. 3 part of the articular surface. J.c3.15. 10.
4. Distal end of 1 first phalange of the wing-finger. J.c6.31. 7, no. 1.
5. Distal articular surface of a first phalange.
6. Distal end of a first phalange. J.b.6, no. 4.
7. Proximal end of the second phalange of the wing-finger. J.c2.12.12, p. 57.
8. Proximal end of a small second phalange. J.b.7, no. 7.
9. Proximal end of a large second phalange. J.b.7, no. 4.
10. Side view of distal end of right femur. J.b.11, no. 11, p. 62.
 Click on image to view larger vesion
Click on image to view larger vesion
PLATE VIII.
Pelvis, Femur, Tibia, &c.
Fig. 1. Fragment of a large right os innominatum. The faint T-shaped lines in the acetabulum indicate the limits of the three component pelvic bones; fig. 1 is placed at the posterior border of the ischium. J.b.10, no. 1.
2. Imperfect right os innominatum, with the anterior and posterior wings of the ilium broken away. J.b.10, no. 4, p. 69.
3. Imperfect left os innominatum showing the small obturator foramen which divides the pubis from the ischium. On the anterior border of the pubis is seen a depression, which may have given attachment to the prepubic bone. J.b.10, no. 3.
4. Visceral aspect of an imperfect right ischium. J.c4.20.2.
5. Exterior side aspect of a right femur. J.c2.11. 20.
6. Front aspect of the same specimen, p. 62.
7. Posterior aspect of proximal end of right femur of a different genus, showing a pit for the obturator muscle. J.b.11, no. 1.
8. Front aspect of the same specimen.
9. Outline of the proximal articular end; the obturator pit is darkened.
10. Posterior aspect of distal end of right femur. J.b.11, no. 20.
11. Outline of the distal articular end of the same specimen.
12. Distal end of a large right femur. J.b.11, no. 12.
13. Proximal end of tibia (? front aspect). J.b.12, no. 8.
14. Another view of the same specimen, p. 62.
15. Outline of the articular aspect of the same tibia. The non-articular part is shaded.
16. Claw phalange. J.c1.2.5, p. 69.
17. Claw phalange. J.c.9, no. 4.
 Click on image to view larger vesion
Click on image to view larger vesion
PLATE IX.
Neck Vertebræ.
Fig. 1. Anterior aspect of an axis to which the atlas was not anchylosed. J.c3.15. 2, p. 64.
2. Anchylosed atlas and axis seen from the base of the vertebra. J.c.1, no. 8.
3. Anchylosed atlas and axis seen from above. J.c.1, no. 14.
4. Atlas, neural arch imperfect. J.c.1, no. 10.
5. Anchylosed atlas and axis seen from the side, the neural arch of the atlas is wanting. The light space in the centrum of the axis is the pneumatic foramen. J.c.1, no. 14.
6. Large cervical vertebra seen from below. J.c.2, no. 42, p. 65.
7. Small cervical vertebra seen from below. J.c.2, no. 43.
8. Cervical vertebra seen from behind. J.c.2, no. 5.
9. Cervical vertebra seen from above. J.c.2, no. 23.
10. Cervical vertebra seen from the left side. J.c6.27.1, no. 4.
11. Cervical vertebra of another genus seen from the left side. J.c.2, no. 13.
12. Base of the centrum of the last true cervical vertebra. J.c.2, no. 40.
13. Right side of cervical vertebra. J.c.2, no. 7.
 Click on image to view larger vesion
Click on image to view larger vesion
PLATE X.
Back and Tail Vertebræ.
Fig. 1. Centrum of a vertebra from the region between the neck and the back, called pectoral. J.c.3, no. 19, p. 69.
2. Dorsal vertebra seen from below. J.c2.12.3, no. 2.
3. The same specimen seen from behind.
4. Right side view of a dorsal vertebra showing the neural spine nearly perfect. J.c.3, no. 20.
5. The same specimen seen from behind.
6. Right side of dorsal vertebra showing anterior and posterior zygapophyses. The neural spine broken.
7. Front view of the same specimen. The centrum is seen to form but a small part of the anterior articular surface.
8. Bight side of a sacral vertebra J.c.4, no. 1, p. 73.
9. Front aspect of the same specimen. The neural arch forms part of the intervertebral articulation with the centrum.
10. Side view of the anterior part of a sacrum, presented by H. C. Raban Esq. J.c.4, no. 3.
11. The same specimen seen from below.
12. Inferior aspect of posterior part of sacrum of a different genus. J.c.4, no. 2.
13. Large caudal vertebra seen from above. J.c.5, no. 9.
14. The same specimen seen from beneath, p. 75.
15. Left side of the same specimen.
16. Anterior articulation of the same specimen.
17. Posterior aspect of the same specimen.
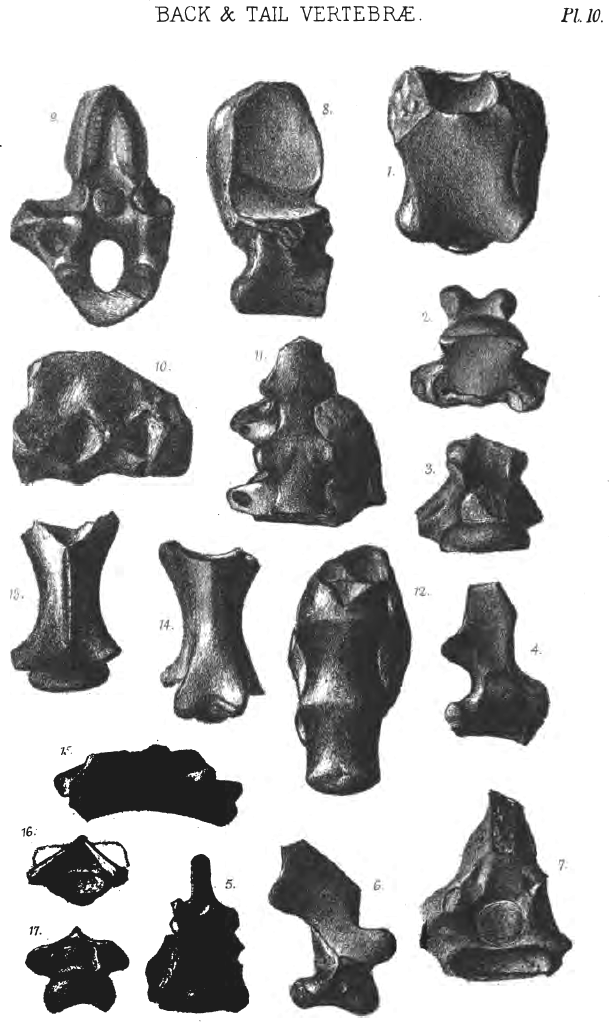 Click on image to view larger vesion
Click on image to view larger vesion
PLATE XI.
Cranium.
Fig. 1. Occipital aspect of the skull of a Pterosaurian. J.c.8, no. 2, p. 84.
2. Anterior aspect of the same skull, showing a transverse section of the brain cavity fractured through the parietal bones. At its base on each side are seen the optic lobes.
3. Anterior aspect of a Pterodactyle skull of a different genus. J.c.8, no. 1. The frontal bones have come away from the parietal at the suture, p. 80.
4. Superior aspect of the same specimen looking upon the parietal, supra-occipital, and ex-occipital bones.
5. Occipital aspect of the same specimen, showing the foramen magnum, the absence of the basi-occipital bone, and the basi-sphenoid mass.
6. Side view of the same specimen, showing below the girdling occipital crest the excavation for the quadrate bone's articulation with the skull, and the forward prolongation of the basi-sphenoid mass.
7. Palatal aspect of the basi-sphenoid bone. J.c.9. To be compared with the small triangular mass in fig. 5, p. 85.
8. Side view of the ethmo-sphenoid mass, J.c.9, showing the lateral boundary of the front of the cerebral hemispheres, p. 85.
9. Posterior aspect of the same specimen, showing parts of the cups which covered the anterior termination of the cerebral lobes.
10. Anterior view of the cerebral lobes in a natural mould of the brain, in the collection of J. F. Walker, Esq. It may be compared with figs. 2. and 9, p. 87.
11. Superior aspect of a natural mould of the brain, showing the outline of the cerebral lobes, and the cerebellum between them behind. Portions of bone in the temporal region are left attached, p. 87.
12. Side view of the same specimen; one cerebral lobe is seen behind the other. The anterior termination of this figure may be compared with the posterior outline of fig. 8.
13. Side view of basi-occipital bone, p. 78.
14. Palatal aspect of quadrate bone, showing the articulation for the lower jaw, and the thin quadrato-jugal attached to its outside, p. 89.
15. Exterior aspect of quadrato-jugal and quadrate bones. Above the articulation in German specimens is the outline of the orbit of the eye.
16. Anterior aspect of the distal end of a left quadrate bone.
17. Posterior aspect of the same specimen, showing the wing for the pterygoid articulation.
 Click on image to view larger vesion
Click on image to view larger vesion
PLATE XII.
Facial Bones and Lower Jaw.
Fig. 1. Side view of the dentary bone of Ornithocheirus machærorhynchus, showing its posterior attenuation towards the palate. J.c6.33.1, p. 113.
2. Superior aspect of the same specimen, showing the palatal groove and tooth sockets.
3. Articular end of left ramus of mandible, J.4, showing its posterior termination, p. 91.
4. Articular end of left ramus of mandible, J.c6.32. 2, fractured through the articulation.
5. Side view of anterior part of dentary bone of Ornithocheirus Cuvieri ? J.c.15, p. 113.
6. Side view of anterior part of premaxillary bone of Ornithocheirus microdon, fractured at both ends. J.c.29, p. 116.
7. Palatal aspect of the same specimen, showing the palatal ridge and tooth sockets.
8. Palatal aspect of anterior part of premaxillary bone of Ornithocheirus denticulatus. J.c5.28.1, p. 122.
9. Side view of the same specimen.
10. Tooth, showing absorption by the successional tooth, on the inner side of the fang. J.c.27, no. 10, p. 92.
11. Tooth. J.c1.1.4.
12. Fang of a large tooth. J.c.27, no. 34.
13. Undetermined [? pterygoid end of palatine bone]. J.c1.2.7, p. 91.
14. Other side of same specimen.
15. 1 Vomer, side view. J.c.10, no. 2, p. 88.
16. 1 Palatal view of the same specimen.
17. Pelvis with a bone attached like the middle part of J.c.10, no. 2. ?Neural arch of sacral vertebra.
 Click on image to view larger vesion
Click on image to view larger vesion
Minor typos were corrected and the Errata list changes were applied. Standardization of hyphenation was standardized to the most common form used. Headers for each genera's description was standardized to list the specimen information first. Cover artwork and plate images derived from materials made available on Google Books and The Internet Archive. All are placed in the Public Domain.