
Some typographical errors have been corrected; a list follows the text. (In certain versions of this etext, in certain browsers,
clicking on this symbol Contents. (etext transcriber's note) |

Popular Science Library
EDITOR-IN-CHIEF
GARRETT P. SERVISS
AUTHORS
WILLIAM J. MILLER · HIPPOLYTE GRUENER · A. RUSSELL BOND
D. W. HERING LOOMIS HAVEMEYER · ERNEST G. MARTIN
ARTHUR SELWYN-BROWN · ROBERT CHENAULT GIVLER
ERNEST INGERSOLL · WILFRED MASON BARTON
WILLIAM B. SCOTT · ERNEST J. STREUBEL
NORMAN TAYLOR · DAVID TODD
CHARLES FITZHUGH TALMAN
ROBIN BEACH
ARRANGED IN SIXTEEN VOLUMES
WITH A HISTORY OF SCIENCE, GLOSSARIES
AND A GENERAL INDEX
ILLUSTRATED
![]()
![]()
VOLUME NINE
P. F. COLLIER & SON COMPANY
NEW YORK
Copyright 1922
By P. F. Collier & Son Company
MANUFACTURED IN U. S. A.
The Science of the Body
BY
ERNEST G. MARTIN
Professor of Physiology, Stanford University
![]()
![]()
P. F. COLLIER & SON COMPANY
NEW YORK
WHEN Alexander Pope wrote “The proper study of mankind is man,” he was thinking rather of man as a social being than as the possessor of an amazingly complex and interesting body. It is nevertheless true that to one who finds enjoyment in the study of intricate mechanisms or to one for whom that amazing sequence of events which we call life has appeal there is no more fascinating study than the study of the living body. That part of the study of the body which concerns itself primarily with activity and only secondarily with form and structure, makes up the science of Physiology. The way the body works is the central theme.
The practical value of Physiology to the general reader lies in the fact that it forms the basis of all sound rules of hygiene. Life is made up of bodily activities which may be carried on correctly or incorrectly. Carried on correctly they mean health, carried on incorrectly, unhealth. The world is flooded with health-preserving or health-restoring systems, urged upon the public, for the most part, by promoters in search of gain. Such of these as have merit are based on definite physiological principles, and anyone who has a common-sense working knowledge of his own body can order his life in accordance with them, at little or no expense. Moreover, a sound appreciation of Physiology drives home the truth that when the body is really out of order its restoration can be safely intrusted only to the properly trained physician: the man or woman who through years of painstaking study has won insight into the intricacies of the human mechanism and whose honest appreciation of the difficulties of his profession, and courageous sincerity in grappling with them, justify to the full the confidence in which he is held by his community.
Ernest G. Martin.
| CHAPTER | PAGE | |
| I. | The Signs of Life | 9 |
| II. | The Maintaining of Life | 18 |
| III. | The Sources of Food | 24 |
| IV. | The Uses of Food | 31 |
| V. | Body Cells | 41 |
| VI. | The Supporting Framework | 53 |
| VII. | Motion | 73 |
| VIII. | Sensation—Internal and Contact Senses | 86 |
| IX. | Sensation—Distance Senses | 96 |
| X. | The Nervous System and Simple Nervous Actions | 122 |
| XI. | The Brain and Complex Nervous Actions | 139 |
| XII. | Some Special Nervous Actions; Smooth Muscle and Gland Control | 155 |
| XIII. | The Body Fluids | 173 |
| XIV. | The Conveyer System of the Body | 191 |
| XV. | The Service of Supply of Food | 226 |
| XVI. | The Service of Supply of Oxygen | 253 |
| XVII. | The Service of Removal of Waste | 271 |
| XVIII. | More About the Use of Food by the Body | 277 |
| XIX. | Keeping Warm in Winter and Cool in Summer | 305 |
| XX. | The Perpetuation of the Race | 324 |
| XXI. | Child and Man | 345 |
Muscular Coordination—Weissmuller’s Horizontal Plunge into a Swimming Pool | Frontispiece |
| FACING PAGE | |
Factory’s Cooperation in Aiding Physical Fitness, Underweight Employes Are Given a Milk Lunch | 32 |
Weight Test in a Child’s Physical Examination to Determine How Nearly the Child Approaches Correct Standards of Physical Development | 33 |
French Method of Measuring Vertical Conformation, Considered Valuable in Advising and Training Athletes | 64 |
Curve of the Spine Measured and Drawn for Study of an Athlete’s Possibilities | 65 |
Muscular Development of an Athlete—The Discus Thrower of Myron | 80 |
A Modern “Victory”—Miss Sabie at Practice | 81 |
Hospital Designed to Supply the Maximum of Light and Air—Chief Requisites for Health—to Each Room | 96 |
Architect’s Plan Made with the Need for Light and Air in Mind | 97 |
Soft, Restful Colors of a Hospital Room | 112 |
Magnet Put to the Service of Removing Iron Filings from the Eye | 113 |
Test for Blood Pressure | 192 |
Indican Test in Urinalysis | 193 |
Microscopic Study of the Blood | 208 |
| 209 | |
X-Raying the Teeth and Jaw—Health Often Depends on Their Good Condition | 224 |
Throat Examination—Important in Determining Physical Condition | 225 |
Lunch Hour in French School Where Exercise, Out-of-Door Life, Sunshine and Fresh Air are Considered Essentials | 288 |
Diet Kitchen on a U. S. Hospital Ship | 289 |
Caring for the Ears—Important for Adults and Babies Alike | 336 |
Baby’s Footprint—a Means of Positive Identification | 336 |
Babies, Physically and Mentally Active | 337 |
PHYSIOLOGY is the study of living things, so the first thing to be asked when we begin to think about physiology is how we are to know whether anything is alive or not. It is usually pretty easy to tell whether a dog or cat is alive or dead, although sometimes when a dog is stretched out on the road we have to look closely to tell whether he has already met his end or is merely courting it by sleeping in the public highway. There are in the world hosts of animals with which we are not familiar, and to tell whether these are alive or dead is often a puzzle. More than one picnicker has been thoroughly surprised by seeing what looked like a bit of dead twig begin to walk away, and recognized the walking stick. On the whole we will agree that the sign of life which we find most reliable is motion of some sort on the part of the living animal. If the stretched-out dog makes breathing movements, we pronounce him alive; if not, we decide that he is dead. It is because the walking stick moves off when disturbed that we know it is not a twig. But while motion is the thing we look for in living animals we know perfectly well that it would be foolish to assert that anything that moves is alive. When the wind is blowing the air may be full of dead leaves and butterflies, all moving, but only part, the butterflies, alive. Unless the motion is produced by the animal itself it is not good as a sign of life. So widespread among animals is the making of movements, either on their own account, or when disturbed, that we shall not often find ourselves mistaken if we decide that an object which remains quiet indefinitely is not a living animal. Now how about the other side of the question? Is anything that moves on its own account or when disturbed to be judged alive?
Suppose that the inhabitants of Mars have finally succeeded in perfecting a flying boat that can be hermetically sealed and shot across space from that planet to our earth. Suppose further, that the first exploring party has set forth on a voyage of discovery, and has reached a point high in our sky from which objects on the earth’s surface begin to be distinguishable. Of course the huge landmarks, cities, lakes, and rivers, have been in view for a long while, and now the explorers are on the lookout for signs of living things. They are watching, just as we would be, for moving objects. The first moving thing that they see will probably be a train, and we can imagine their speculations as to whether they are actually looking or not at an inhabitant of the earth. As their craft sinks toward the surface the make-up of the train becomes perceptible as also the fact that it runs on rails, showing that it is a mechanical contrivance and not a living being. As smaller objects come into view black, shiny specks are seen moving about. These show every appearance of life; they start and stop; pass each other without interference; in fact conduct themselves about as animals usually do. If their apparent great power has the effect of discouraging the exploring party, so that they give up further investigation and fly away to Mars, the inhabitants of that planet will always suppose the earth to be populated by automobiles. We know that automobiles are not alive, yet, as this little allegory shows, they behave enough like living beings to deceive distant observers. There must be some sign of life which will apply to an animal and not to an automobile; what is it? Evidently what the Martian explorers missed was the fact that the automobile does not really start or stop itself, or guide itself past obstructions. If it had been alive, it would have done these things of itself. It is not so much the power of motion, then, that proves that the thing is alive as the power of making motions that are under the control of the animal itself.
The sight of an automobile which is not alive behaving as though it were because it is under control of a driver who is alive may lead us to ask whether the animal that we know to be living is actually alive in all its parts, or is a dead mechanism of some sort which has somewhere within it a living controller, corresponding to the living driver of the car. The animals with which we are most familiar are ourselves; how is it with our own bodies? Are they alive in all their parts, or is the brain the only part of us which is living? When a patient goes under ether on the operating table, or even when he is sound asleep, the signs of life are not conspicuously present; the heart goes on beating, to be sure, but so does the engine of an automobile go on running when the driver is away, provided he has not shut it off. A favorite belief among the Hindus is that when they go into a trance the body actually becomes lifeless while the living spirit soars among the heights. How are we to decide whether the Hindus are right or not? Evidently we shall have to look deeper than we have thus far, and learn something of what is actually going on in the different parts of our bodies when we are asleep and when we are awake.
Nearly everyone learns in school the main facts about the construction of the body; that there is a bony skeleton which supports the softer parts; that motions are made by muscles; that sense organs inform us as to what is going on in the world around; that the brain is the seat of the mind; that heart, lungs, stomach, kidneys, and other organs contribute in various ways to our well-being. Not so many go into detail as to the make-up of these organs, or into the way in which they do their work. This is not a simple matter, for several reasons. The first is that the construction units are so tiny that they cannot be seen by the unaided eye, but must be studied under the high magnification of a first-class microscope. It is much harder to make out the manner of the working of tiny pieces of machinery than of those that are of convenient size. When the parts are as small as those that make up our bodies, the task of finding out how they operate is so difficult that even now, after years of study, there are many details about which we know very little.
The construction units have been named cells. In some tiny animals the whole body consists of but one cell; all higher animals, including ourselves, have millions of cells making up the body. Undoubtedly some cells are alive; our question is as to whether all of them are, or whether there are some that are alive and some that are not. There are parts of our bodies, and of the bodies of nearly all other kinds of animals, as well, that are certainly not alive. Examples are the hair, the nails, the enamel of the teeth, and the hard parts of the bones. Actual living stuff is very soft and liquid. It is too fragile to hold its own structure except in the very tiniest animals; those that are larger need some additional supporting framework. In a body the size of a man’s the supporting framework amounts to a very considerable percentage of the entire weight (25 per cent). Not only is there the large bony skeleton, but between and among the individual cells is a framework made up of fine fibers and sheets which hold the cells in place. This latter framework is called connective tissue; we run across it in the gristly parts of meat. It makes up the stringy mass that clings to the cutter of the meat grinder when beef is being ground for Hamburg steak. We shall consider later how all this supporting material is made and put in place. Just now we are interested in the cells, and in determining whether all of them are alive or not.
There are many different kinds of cells in the body; some are muscle cells, others nerve cells, still others gland cells, and so on. Careful study shows, however, that at bottom all cells are alike. All are composed of one kind of substance to which has been given the name of protoplasm, meaning first or primary flesh. It is because some, at least, of this protoplasm is alive that our bodies are alive, and our physical life consists of nothing more than the combined life of all the living protoplasm which our bodies contain. Is there any way by which to tell whether any particular mass of protoplasm is alive or not? In other words, what are the signs of life of protoplasm as contrasted with the signs of life of whole animals?
We shall scarcely expect it to be as simple a matter to tell whether the tiny mass of protoplasm that we call a cell is alive or not as to decide whether a dog is dead or alive. For one thing, our most useful test of life, namely motion, cannot always be applied to single cells. We have in our bodies a great many cells, those in the brain, that we know are alive if any part of us is, but aside from the exceedingly gradual shifts in position that take place during growth the brain cells never make any motions at all, so far as anyone has ever been able to find out. Of course in the body of any ordinary animal most of the cells are hidden from view beneath the skin, but there are enough small transparent animals whose internal parts can be watched through the microscope to let us say with certainty that some of the cells which we know to be alive do not move. Tests of life that can be applied to all kinds of cells will necessarily be difficult to use, and we shall have to take the word of experts as to whether they have found particular cells alive or not, but the principle on which the tests are based is simple enough so that we can examine it. To do this, it will be necessary to turn our attention for a little while to some of the very tiniest of all living animals, those whose whole bodies consist of but one cell.
When these tiny one-celled animals are watched through the microscope as they swim about it can be seen that in one important feature they behave just as we do ourselves; that is in their care not to neglect mealtime. To be sure, mealtime comes for them whenever they happen to hit against any tinier particle than themselves, which they can take in and digest. But for them, as for us, the taking of food from time to time is a necessity of life. Only a small part of the food thus taken in is added permanently to the bulk of the animal. In other words, the growth does not go on as fast as does the taking of food. Of course in ourselves, after we have reached full size, there is little or no increase in permanent bulk even though we do keep right on eating. Evidently in the tiny one-celled animal, and in us as well, food is constantly being used for something besides growth. It can be proved that this food is used for precisely the same purpose that gasoline is used in the automobile, namely to run the machine. In a very real sense every living thing is a machine, and will no more run without a supply of power than will any other machine. From the engineering standpoint animals can be classified along with automobiles and locomotives as “prime movers,” namely, as machines which develop their power within themselves. There are not many kinds of power which can be developed by prime movers. By far the commonest is that seen in locomotives and automobiles, namely the burning of some kind of fuel. We have always known that the locomotive operates by the burning of coal or oil in the fire box. A moment’s thought will show us, if we have not realized it before, that the explosion of the air-gas mixture in the automobile cylinders is also a burning. Every steam-driven power plant depends on burning fuel for its power. Evidently fuel materials contain abundant power, if it can be extracted, and burning is a good method for doing the extracting. The word “burning” is the common name for a chemical process known technically as “oxidation,” meaning the union of oxygen with the fuel. The air is one-fifth oxygen, so there is plenty available, and fuel will usually oxidize readily after it is properly started.
Not only do animals correspond with other machines in using fuel as their source of power; they correspond also in that the power is extracted through the process of oxidation. To be sure, the oxidation in animals is not accompanied by flame and smoke as it usually is in power plants, nor do any parts of the animal get as hot as does the furnace where fuel is ordinarily burned; but in spite of these differences the fundamental fact is the same, namely that the extraction of power is by means of oxidation. What this shows is that great heat, flame, and smoke are not necessary in oxidation, but only in the kinds of oxidation with which we are most familiar.
As soon as we have described one more feature of animal power development, we shall be ready to apply what has been said to the topic in hand, namely the signs of life in single cells. The point that remains to be made is that in living cells power development has to go on all the time whether the cell is active or not. This means that fuel is constantly being burned, and oxygen is constantly being taken in to do the burning. There has been, and still is, a great deal of debate as to how much the oxidation can be reduced in living cells without destroying life. It is evident that it can be cut down to a very low level indeed, for seeds remain alive for years without using up, or even noticeably depleting, the store of fuel material which they contain. Most botanists of the present time doubt the truth of the tale that grains of wheat have sprouted after being taken from the wrappings of mummies, where they had lain for thousands of years. Careful efforts have been made to preserve wheat under as favorable conditions as existed in the mummy wrappings, but in every case the power to sprout was lost within a comparatively few years. So far as experiment enables us to judge, the complete cessation of power development in cells, either of plants or animals, means their death.
Here we have our sign of life that is applicable to all kinds of cells wherever they are located, whether making up the whole of a microscopic animal or deeply imbedded in the body of a large animal which consists of millions of them. If power development is going on, the cell is alive; if no power is being developed, the cell is not alive. When this test is applied it is found that all the protoplasmic cell masses which are present in the body of a plant or animal are alive, and since such masses are everywhere throughout the body, life is present in all parts of it, and not confined to the brain or to any other single region. We might admit that the Hindus are correct in assuming that the spirit can sometimes soar away and leave the lifeless body behind, but we cannot accept the possibility that it can return and establish life within it again. When life is resumed after a trance, that fact is proof positive that life continued throughout the trance itself.
EQUAL in importance to being alive is the power to go on living; therefore, having described the signs of life, our next task is to consider how that life is maintained. When the primary fact of life was given as continuous power development, the foundation was laid for this topic, for life cannot fail to go on if continuous power development is maintained.
Power development in living animals as in locomotives depends on fuel and oxygen; evidently continuous supplies of these must be provided if life is to go on. The living animal differs from the locomotive in this: that while some one attends to supplying the locomotive with fuel, most living animals, except the very young, have to attend to providing themselves. There are exceptions to this rule. The tapeworms that inhabit the intestines of animals, and sometimes of men, live in a stream of food; they are put to no trouble to obtain it. The same is true of many kinds of parasites. Except for these, however, it holds true that animals must attend to their own wants. We shall now begin to see the utility of the most conspicuous sign of life spoken of in the first chapter, namely, motion, for food must be gotten where it is; only tapeworms and similar animals swim in it. All the rest, including ourselves, must go to where the food is. Even animals like oysters, that are anchored to the rocks, have to use motion in getting food. In their case the motion consists in setting up a current in the sea water into and through their bodies, from which current they sift out the tiny food particles which abound in the ocean.
If an animal happens to live in the ocean, where every drop furnishes its particle or two of foodstuff, and especially if the animal is small, or sluggish, like the oyster, almost any kind of motion will serve to bring the animal all the food it needs. The simplest of the one-celled animals, that must be watched through the microscope to see how they behave, blunder about aimlessly, and in the course of their blundering bump up against food particles often enough to keep themselves fed. If an animal happens to live where food is scarcer, or if it is big and active, and so must have large quantities of food, aimless blundering about will never get it enough to keep it alive. It must have some means of finding out where the food is. Since we ourselves come under the head of animals whose food needs are so large that we must locate food supplies, and not depend on happening onto them, we can identify in ourselves the means which are used for doing this. We all know that our sense organs, the eyes, ears, nose, and finger tips are what we depend on for telling us where food is to be found. The same is true of all animals that are able to hunt for food; they have some sort of sense organs to help in guiding them to where the food is.
The story of the machinery for finding food is not yet quite complete, for the muscles which actually make the movements by which the animal gets about are in one part of the body, while the sense organs which are to furnish the information by which the movements are guided are in quite a different part, and in animals as large as ourselves, some distance away. From our eyes to our leg muscles is quite a space, and it is evident that this space must be bridged somehow if our legs are to move in obedience to information which our eyes bring in. In ourselves and in almost all other animals this space is bridged by means of special machinery for the purpose. We are familiar with it under the name of the nervous system. We may not have been in the habit of thinking of the nervous system in just this way, but at bottom this is exactly what the nervous system does for us, namely, guides our muscles according to the information brought in by our sense organs. There is more to nervous activity than just this, but this is the starting point and groundwork for all the rest, as we shall try to show presently.
Continuous food supplies are the main necessity for continued life, but there are some other things that have to be looked out for in addition. The favorite food for large numbers of animals, and, indeed, in many cases the only food, consists of the bodies of other animals. All the flesh-eating sorts prey on other animals for their food. This places the other animals on the defensive, so that a large part of their activity consists in escaping the attacks of the beasts that wish to eat them. For most kinds of animals the greater part of their waking life is taken up with movements which serve either to get them food or to prevent them from becoming food for others. If we add to these the movements that are necessary to preserve the animals against other kinds of danger than the danger of being eaten, and those connected with the propagation and care of the young, we shall have about covered the list of what we may call the serious activities of animals, and of men as well. Many kinds are active besides in play. This is particularly true of young animals, although grown-ups, both among animals and men, find play both agreeable and beneficial when not overindulged.
Protective motions need to be even more accurately made than those whose purpose is the getting of food, for if the food is missed at one effort another trial may be more successful, but if an attempt to escape fails there will probably be no more chances to try. The sense organs and the nervous system are just as deeply concerned, therefore, in avoiding harm as in finding food materials, and it is as important for them to do their work well in the one case as in the other. When we think of the activities of animals, for whatever purpose they are carried on, we must think of them as made up of the combined actions of the muscles, the nerves, and the sense organs, and not of any of them working by themselves.
These parts of us that are so closely concerned in the maintaining of our life by getting us food and keeping us safe from harm make up, also, the only parts of us which really share in what we may call conscious living. When we come right down to it we could spare our other organs—heart, lungs, stomach, and the rest—and never miss them so far as adding anything to our happiness is concerned. In fact, the less these organs intrude themselves into our attention the better off we are; only when we are ailing do we begin to think about them. Of course, they are absolutely necessary to us, and we should die instantly if one of the more important of them were to stop working, but the part they play is not one which enters actively into our consciousness, as do the muscles, nerves, and sense organs.
Naturally, we will ask what all these other organs are for if they do not share in our conscious life. Why can we not get along with just those that we use for getting food, for avoiding harm, for play, and for the other activities which they carry on? The answer to this question is found in the fact emphasized above that continuous power development is necessary to continued life. By themselves the muscles, nerves, and sense organs cannot carry on power development; they require the aid of a great many other organs to do this. Just how these other organs work will be described later; at present it will be enough to recall that every muscle, every nerve, and every sense organ is actually made up of a great many of the tiny construction units—the cells about which we were talking a few pages back—and that every one of these cells must be developing power all the time if it is to go on living. In order to be able to do this they must, every one, be able to oxidize fuel continuously, and this means that they must receive constant supplies, both of the fuel itself and of the oxygen with which it combines. Some system of delivering these materials must be in operation, and in case the materials have to be prepared for use beforehand this must be provided for also. The heart, the lungs, the stomach, and the various other organs that are useful but not conspicuous, are concerned in these necessary jobs. In an automobile factory we have a similar situation; the men that stand at the machines actually make the parts that go into the finished automobiles, but unless other men are hard at work preparing the castings, and bringing them to where the machine operators can get them, not many automobiles will be turned out. So in the body, unless the various organs are carrying on their work of preparing and delivering materials to the muscles, the nerves, and the sense organs, these latter cannot perform their tasks of getting the food for the whole body and of securing the body against harm, nor can they carry on the pleasant, but not absolutely necessary, activities of play and recreation.
WE have talked about the necessity of power development in all living things, and have seen that power development depends on the oxidation of fuel. Of course, our fuel is the food that we eat. No substance is suitable for fuel unless it contains power which can be gotten out by oxidation, and unless, in addition, it is suited to the particular kind of oxidation that goes on in the body, and can be handled by the body. Wood is excellent fuel for some purposes, but as a food for man it has no value, even when ground fine and mixed with flour as was done in some European countries during the Great War, because wood cannot be handled by the body in the way in which a usable fuel must be. Although wood is not good food, closely related materials are, and in fact make up the bulk of it. All fuel food is either vegetable or animal. All animal food traces back finally to the vegetable world, and it is an interesting fact that we do not usually care to eat flesh that is more than one remove from the vegetable kingdom. Animals that are flesh eaters are not considered fit for food, except in the case of fish and birds, and the flesh that these eat is not commonly thought of as being such, since it consists mostly of the flesh of insects, frogs, and fish themselves.
The real sources of food, then, are in the vegetable world. Of the countless thousands of kinds of plants that exist a few dozen have proven to be of enough use for human food, or for food for the animals on which human beings feed, to justify us in taking the trouble to raise them on our farms and in our gardens. There must be something about these particular plants to make us prefer them. If we look into the reason for the preference we shall understand something of the qualities which make plants good for food in the first place. At the beginning of the chapter were set down the things which make a substance fit for food. These are: the ability to yield power by oxidation, and a composition suitable to be used by the body. The ability to yield power involves the possession of a store of it. Power, or energy, which means the same thing in our present use of the words, is never present anywhere except as the result of an earlier exhibition of power. It is not made out of nothing. The sun is a reservoir of energy on which the earth draws, and we do not know with any certainty from whence the sun got its power. The heated center of the earth itself is a reservoir of power on which we may draw at some time in the future, when cheaper sources are used up. Except for energy from these sources and for trifling amounts that may be brought in by meteorites, there is none on the earth’s surface that has not always been here. On the other hand, the earth is constantly losing energy into space. The amount that reaches us from the sun balances our losses into space, so that the total energy present holds fairly steady. The energy that comes to us from the sun is chiefly in the two forms of heat and light. In actual horsepower the heat far outweighs the light, but in importance to mankind one stands about on a par with the other, for while without the sun’s heat the earth would become so cold that we would all die, without its light there would be no food and we would all starve. This is another way of saying that the energy that plants store up, and that we get when we eat them, comes originally from the light of the sun. Plants, like animals, are made up of cells. Those with which we are familiar consist of a great many cells, of a good many different kinds. Some are in the roots, others in the stems, still others in the leaves; the blossoms, fruits, and seeds are made up, likewise, of cells. The cells near the surface of the leaves, and, in many kinds of plants, near the surface of the stems as well, contain a green substance known as chlorophyll. This substance enables the cells in which it is present, although we do not know just how, to manufacture sugar, utilizing the energy of the sunlight for the purpose.
Sugar is composed of three very common chemical elements, carbon, hydrogen, and oxygen. As we all know, hydrogen and oxygen in combination of two atoms of hydrogen to one of oxygen form water; the most familiar of chemical symbols is that expressing this combination, namely H{2}O. Carbon, which we know in an almost pure state in anthracite coal, and in even purer form in diamonds, forms a combination with oxygen known as carbon dioxide. This is a gas; it makes up a small fraction of the air. The amount in the air is increased whenever coal or any other carbon-containing material is burned, since carbon dioxide is the product of the oxidation of carbon. Except in the arid regions of the earth there is always some water in the soil a greater or less distance below the surface of the ground. Water and carbon dioxide between them contain all the elements of which sugar is composed. The chlorophyll-containing cells are the factory; the sunlight is the power; and the carbon dioxide and water are the raw materials. Sugar is the finished product, and wherever sunlight is falling on green plants, whether directly or through a layer of cloud, its manufacture is going on. Sugar will oxidize readily, and in so doing will yield abundant power. The energy which it contains was derived by transformation from the energy of the sunlight. With the exception of a few kinds of bacteria every living thing on the earth depends for its food, and so for its energy, either directly or indirectly on the sugar which green plants manufacture. Since sugar dissolves in water it cannot easily be held in storage, so by a simple chemical process the plant changes it to starch, and it is in this latter form that we get it, except in the case of a few plants, like sugar cane.
The green parts of plants are the only places where sugar is made. We eat a certain amount of green food in lettuce and asparagus and similar vegetables, but for the most part the sugar or starch we eat comes from parts of plants that are not green. There is evidently a transportation from the point of manufacture to points of storage. The means of transport is in the sap; since starch is not soluble in water, it must be changed back into sugar. This is done, and then, by the movement of sap the dissolved sugar is carried to the points of storage, roots in such vegetables as beets, underground stems in potatoes, above ground stems in sugar cane, fruits or seeds in orchard and grain crops. In such of these as are sweet, the sugar itself is held in storage; in most kinds it is changed back into starch. Where the storage is in the form of starch the vegetable ordinarily keeps better than when sugar is the substance on deposit.
A few kinds of plants—olives, peanuts, and cocoanuts, for example—convert the sugar into oil and store their surplus material in that form. The chemical elements in oils and fats are the same as in starch and sugar, although the proportions are not the same. Weight for weight oil has more than twice the energy value of sugar; in making a given amount of peanut oil the peanut vine used up more than twice the amount of starch or sugar; but since energy value is what counts rather than bulk the plant is just as well off, and perhaps better on account of the smaller bulk occupied by the stored material. One of the very interesting examples of oil storage is found in the very tiny plants, called diatoms, which abound in the water of the ocean. Each tiny diatom stores within itself an even more tiny drop of oil. Although by themselves single oil drops would make no impression, if enough of them could be brought into one place a respectable accumulation of oil would result. This is precisely what the geologists tell us has happened in past ages; the bodies of diatoms have accumulated through thousands of years, and finally the oil accumulations have been covered over with sediment of one kind or another. When we tap through the sediment we strike into the “oil sand,” which contains this residue of the diatoms, and an oil well results.
Since we depend for our food, and so for our life, on the sugar-making activities of green plants, it will be worth our while to think for a moment of the slowness with which the process goes on. The slice of bread which we may eat in a dozen bites represents the result of a season’s growth of several wheat plants, every one of which was absorbing the sun’s energy and laying up starch grains during every daylight minute throughout the growing season. From the standpoint of the plant which does the storing the material which serves us as food is the excess over the plant’s own daily needs. In most cases it would be utilized at the beginning of the next season’s growth before the plant had put out a leaf system, if the course of events were not disturbed to satisfy the needs of man.
In addition to starch, sugar, and fat there is another kind of food material manufactured by plants, known as protein. This substance is much more complex chemically than any of the others; it contains, in addition to the three chemical elements—carbon, oxygen, and hydrogen—that are found in them, the element nitrogen, and usually some phosphorus and sulphur. These materials are dissolved in the soil water in the form of simple chemical substances, and are taken up by the plant along with the water which enters the roots and flows as sap up to the leaves. The same cells of the plant that make sugar have the power to make protein, using as raw materials some of the sugar along with the substances brought in with the soil water. The energy for the manufacture of protein comes from the oxidation of some of the sugar or starch in the leaf. The finished protein has about the same energy value, weight for weight, as has the starch from which it was mainly derived.
When an animal eats a plant or part of one, he is eating for the sake of the sugar or sugar products which the plant has made. There is one sugar product that is useful as food for many animals, but not for man, except possibly to a very slight extent. That is the woody substance, cellulose, which is formed in plants mainly as a support to the delicate living protoplasm. Cotton fiber is nearly pure cellulose. Cellulose is very similar to starch chemically, and is an excellent fuel wherever it can be burned. The human digestive tract is unable to handle it in a manner to make it usable, although grazing animals do so quite efficiently. A good many plants make products that are either disagreeable in flavor or actually poisonous. Of course, in such cases the plants become useless as food unless a treatment can be devised that will remove the objectionable material or convert it into something harmless. The few dozen kinds of plants that we raise for food are those that are free from harmful substances and that yield large quantities of stored food materials, or in some cases that taste especially good, even though they may not have much food value. Tomatoes, lettuce, and the like, come in this latter class. The world has been pretty well ransacked for food grains, fruits, and herbs, but probably there are others yet to be found besides those we now have.
WE have had a good deal to say thus far about power development in living animals, and have talked about food in connection with its use as fuel for the purpose. While we are on the topic it may be as well to say something about other uses to which food is put in animals besides that of serving as fuel, and also something about what is done with the power that is developed by the burning of such food as is used for fuel. To begin with, it is evident that one use that is made of food is to build the body itself. The new-born infant usually weighs somewhere between 5 and 12 pounds. From birth until the body gets its growth there is an almost continuous gain in weight until a total which may range anywhere between 90 and 250 pounds is reached. Of course, every bit of this additional material came into the body in the form of food. The whole mass of the body divides itself, as has been said before, into living protoplasm and nonliving substance. We do not know accurately what proportion of the whole weight is made up by protoplasm; it has been estimated at about 60 per cent, but any estimate can be only very rough because about half of the nonliving substance consists of fat deposits which vary greatly in different people.
In any case, that part of the food which goes to make gain in weight is passed over to the living
cells. If we accept the rough estimate given above, about 60 per cent is then used for the actual manufacture of new protoplasm; the remainder is worked over by cells specially devoted for the purpose and put into place to serve as supporting structure, or to be held in reserve as fat. Living protoplasm is chemically a very complex mixture. In consistency it resembles a rather thin, transparent jelly; the thickness of the jelly depends on how much water it contains and this varies greatly in different kinds of protoplasm. The watery part of the protoplasm has dissolved in it several substances; among them may be mentioned ordinary table salt; also salts of potash and lime. Only tiny amounts of these are present, but it is a curious fact that without these tiny amounts of salts protoplasm cannot live. The chief solid substance in protoplasm is protein; this

material, which is one of the most complex substances known to chemistry, has certain peculiarities which seem to fit it specially to serve as the chemical basis of life. Evidently of all the foodstuffs protein is the most important for the manufacture of new protoplasm, in other words for growth. In the case of a tiny one-celled animal, whose body is made up of protoplasm, not much else would be needed, but any animal that has a bony skeleton has to build this up to keep pace with the growth of the soft parts of the body. For this purpose mineral substances are needed, chiefly lime salts.
In addition to these foods which are actually used for making new body substance it has recently been discovered that proper growth in the higher animals, including man, depends on the presence in the diet of certain dietary accessories, whose use is not at all understood, although there is no doubt of their importance. These materials, to which has been given the rather cumbersome name of “growth-promoting vitamines,” are found dissolved in certain food fats. Apparently they are insoluble in water and soluble in oil. Most animal fats appear to contain them in small amounts, while most vegetable fats do not. Milk and eggs, which are growth foods in an especial sense, are richer in these accessories than any other articles of the diet. The discovery of these facts has emphasized the importance of including animal fats in the diet of growing children, milk and eggs particularly. Since milk is also rich in the lime salts which are necessary for bone formation it forms the best single foodstuff for children that there is. When very young children have to be fed on cow’s milk, which differs somewhat in proportion from mother’s milk, it is often found necessary to feed the top milk diluted with water, instead of the whole milk. When this is done, lime water is usually used in part for diluting the milk, instead of all ordinary water. In this way the proportion of lime is brought up enough to insure that the child will get plenty of it.
In addition to the use of protein as a growth food it has another use which no other kind of foodstuff can share. This is also because protein is the foundation material of living protoplasm. We do not know a great deal about what goes on in living protoplasm to make up what we call the life processes, but we do know that these processes are of a chemical nature, and that in connection with them there is a steady wastage of protein. The protein that thus goes to waste is broken down into simpler chemical compounds which are expelled from the cells. Why this occurs we do not know, but since it does it is evident that unless the wastage is made good the time will presently come when so much protein will have been lost from the protoplasm that it can no longer exist as such and must die. As a matter of fact, one might go on a diet excessively rich in starchy foods and fats and still starve to death if there were no protein present. This use of protein is called cell maintenance to distinguish it from the other special use of protein in cell growth. Evidently, whatever may be missing from the diet, protein must not be left out. Fortunately most of our common foods contain protein. It is especially abundant in lean meat, in dried beans and peas, and in grain. Potatoes and most garden vegetables are deficient in protein, as are almost all common fruits. Bread and meat are our chief stand-bys as furnishers of protein.
Just as there are vitamines that are important for growth, so are there vitamines that are necessary for cell maintenance. Many years ago Dr. Sylvester Graham made himself prominent by arguing that the outer coats of wheat grains contain something that is needed in the diet, which is removed in the process of manufacturing white flour. He accordingly invented a form of flour, familiar to us all under his name, which includes some of the bran from the outer layers of the wheat. This idea, which originated with Dr. Graham, has since been substantiated, although not precisely as Graham had it. We know that there are necessary accessories to the diet, but we know, also, that they are much more widespread than Graham thought. They occur in so many kinds of foodstuffs that anyone who eats a mixed diet usually gets enough of them for his needs. The ill effects of their lack are most evident when the diet is restricted to a few kinds of food which happen not to contain them. A striking example of bodily injury directly due to the absence of these vitamines from the food is seen among Orientals whose diet is apt to be made up of rice plus small amounts of other substances. Of recent years the natives of Japan and China and the Philippines have suffered much from a disease of the nerves known as beriberi. Investigation has shown that this disease is due to the absence from the diet of needed vitamines, and dates from the time when rice-milling machinery was introduced. The old hand methods of milling rice were so imperfect that much of the hull was left clinging to the grains, but machinery polishes the rice clean of every trace of hull. The hulls of rice contain the accessory that is wanting from the polished grains. Wherever it has been possible to bring about the use of unpolished (brown) rice instead of the usual polished kind, beriberi has disappeared. Or the same result can be secured by adding small amounts of beans to the diet. It is probable, also, that the hulls of most grains, including wheat, contain some of the same, or a similar accessory, so to that extent Dr. Graham was right in emphasizing the importance of adding hulls to the flour. Quite recently it has been shown that raw foods are richer in these accessories than cooked, and that ordinary compressed yeast contains more of them than any other easily obtainable material. Many people are being benefited by taking part or all of a yeast cake daily in a glass of milk.
For growth, or the making of new protoplasm, and for maintenance, or the repair of protoplasmic wastage, then, we must eat protein-containing foods, also foods containing various kinds of salts, and foods containing the necessary vitamines. All these are to provide required materials; the actual substances built into the protoplasm. There remains the requirement of power, for both growth and maintenance represent chemical activity on the part of the cell, and this activity depends on power just as does any other activity. In saying this we are merely saying over again in different words what was set down at the very beginning of the book as the chief sign of life, namely, the necessity on the part of living cells of continuous power development. The use of food as a source of energy or power has been talked about already, but it is necessary to say something about the different sorts of power development that may go on in cells, and since we shall have to talk about this a good deal, right here is a good place to bring in for the first time a word that has come to be used whenever the matter of the chemical activities of living cells is being mentioned. The word is metabolism; when we speak of cell metabolism we mean the chemical processes that are going on in the cells. Hereafter, instead of saying power development, the word metabolism will be used as meaning practically the same thing.
First of all, in describing the various kinds of metabolism that cells may show, we have the metabolism of rest. By this we mean the power development that is going on when the cell is doing nothing more than keeping alive; neither growing nor showing any special activity. This is evidently the minimum amount that any cell can show, so it is often referred to as the basic metabolism. We know of at least two things that may change the amount of basic metabolism; the first of these is a change in temperature; when a cell is cold, its basic metabolism is less than when it is warm. There is a very simple chemical reason for this, namely, that chemical processes as a rule go on more slowly the lower the temperature. Since all metabolism consists of chemical processes, this rule applies not only to basic metabolism, but to all other kinds as well, and, as we shall see, explains why the lower animals show such marked differences in behavior in cold and warm weather. The second thing that influences the amount of basic metabolism is the percentage of water in the protoplasm of the cell. Highly organized animals, like ourselves, are destroyed if the cells lose more than a small fraction of their water, but there are many of the lower animals that can be dried until their bodies contain only a very little water and still live. This applies to microscopic forms that live in puddles and similar places; when the puddle dries up the animal dries up too, until all that is left of it is a tiny particle of highly concentrated protoplasm. But this tiny particle preserves all the original cells, or at least enough of them to make a fresh start, and a very sluggish metabolism goes on in each cell. Of course, the advantage of this is that the stored food materials will not be used up as rapidly as they would if metabolism went on at the usual rate, and so there is a better chance that the animal may survive until more water falls or drains into the puddle, or until the particle of dust which the animal has become may be blown by the wind where it will fall into another one. Whenever either of these things happens the protoplasm takes up water again and the former rate of metabolism is resumed. It is only by means of this reduction in rate of metabolism that many kinds of animals are able to persist, for in large parts of the globe there is a period of each year when conditions become so unfavorable that the usual rate of metabolism could not possibly be maintained.
Next in order to basic metabolism comes the metabolism of growth, by which we mean the energy necessary for the making of new protoplasm. Not a great deal is known about growth metabolism; in fact, about the only reason for believing that it requires any energy at all is that the metabolism of young animals, whenever it has been studied, has been found to be greater in proportion than that of animals that are fully grown. It is hard to account for this, unless the growth process itself, namely, the making of new protoplasm, requires energy. When we think of the extreme complexity of living protoplasm, we can easily believe that its formation involves the expenditure of energy, perhaps in considerable amounts.
The last kind of power development to be considered is the metabolism of special activity. Most kinds of cells, particularly in highly organized animals, have some special kind of work to do. For example, the muscle cells have the task of making the motions; the gland cells of manufacturing the secretions, and so on. These we speak of as the particular functions of the cells, and the metabolism by which they are performed as functional metabolism. In some of the lower animals one can scarcely tell where basic metabolism leaves off and functional begins. There is a small shrimp, about a half inch long, that is found quite commonly in small ponds. This little animal has several pairs of legs by which he swims about, and the strokes of these legs go on continuously, day and night, with almost no interruptions, at the rate of a hundred or more a minute, for days or even weeks. It looks as though this, and other animals, that are continuously on the move, were organized without any sharp line between basic and functional metabolism; their protoplasm liberates energy by the oxidation of food, and various things happen as the result; among them are the maintenance of the protoplasm and the making of motions. In the higher animals the distinction between basic and functional metabolism is sharp, and, necessarily so, for the well-being of any of the higher animals requires that he shall have pretty complete control over the activities of his protoplasm, and this he could not have if the functional metabolism were blended in with the basic. In other words, it is as important for bodily well-being that the cells be able to become inactive as that they be capable of activity.
A GOOD deal has been said thus far about living cells without anything at all having been said to tell what they look like, or how they are made up, beyond the statement that they consist of living protoplasm, which is of a jellylike consistency. To look at living cells through a microscope would almost surely be a disappointment at first, for protoplasm is so transparent that not much of its form can be seen on direct inspection. Fortunately for our knowledge of how cells are made up, protoplasm that has been properly killed and preserved takes stain very well, and different chemical substances in the protoplasm stain differently. Thus features that could not be made out at all in the living cells become clearly visible after killing and staining. The first thing that attracts the attention when cells thus prepared are studied is that every cell has somewhere within it, and usually near its middle, a spot which is more deeply stained than any other part of the cell. This indicates the presence of a substance or substances that take stain more readily than the mass of the protoplasm. This peculiarity led to the naming of the deeply staining portion of the protoplasm chromatin, referring to the ease of staining. The part of the cell which contains chromatin is called the nucleus. In many kinds of cells the nucleus can be made out by an expert observer without resorting to stains, although the details of structure cannot be seen in that way.
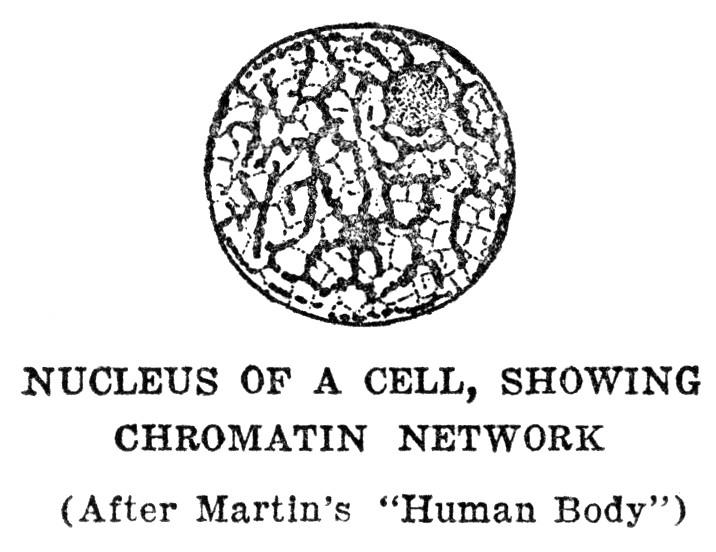
We now know that the nucleus, or rather the chromatin that it contains, plays a remarkable and interesting rôle in the life of the cell. To this we shall return presently. The remainder of the protoplasm, outside of the nucleus, shows the greatest possible variety of form, according to the kind of cell at which we happen to be looking. In some of the simpler types this part of the protoplasm seems to be merely a nearly uniform mass, perhaps with tiny particles scattered through it. In other types the protoplasm is drawn out into long slender threads, and these threads may have many branches; or the protoplasm may be distorted into a thin shell inclosing a mass of fat; or it may be subdivided into dense and thin portions with sharp lines of division between them. These various forms are related to the special functions which the cells have, and we shall learn more about them as we take up the different functions in order. On the whole, study of cell structure shows clearly that the protoplasm outside the nucleus carries on the greater part of the metabolism or power development, and is correspondingly important as the seat of the special functions shown by the cell. If it is a muscle cell, this is the part that does the moving; if a gland cell, this is the part that secretes. Nevertheless, the nucleus is a vital part of the cell. It has been definitely proven that a cell from which the nucleus is lost cannot survive more than a brief time. To gain some idea of the actual part played by the nucleus, we shall have to return to it in some detail.
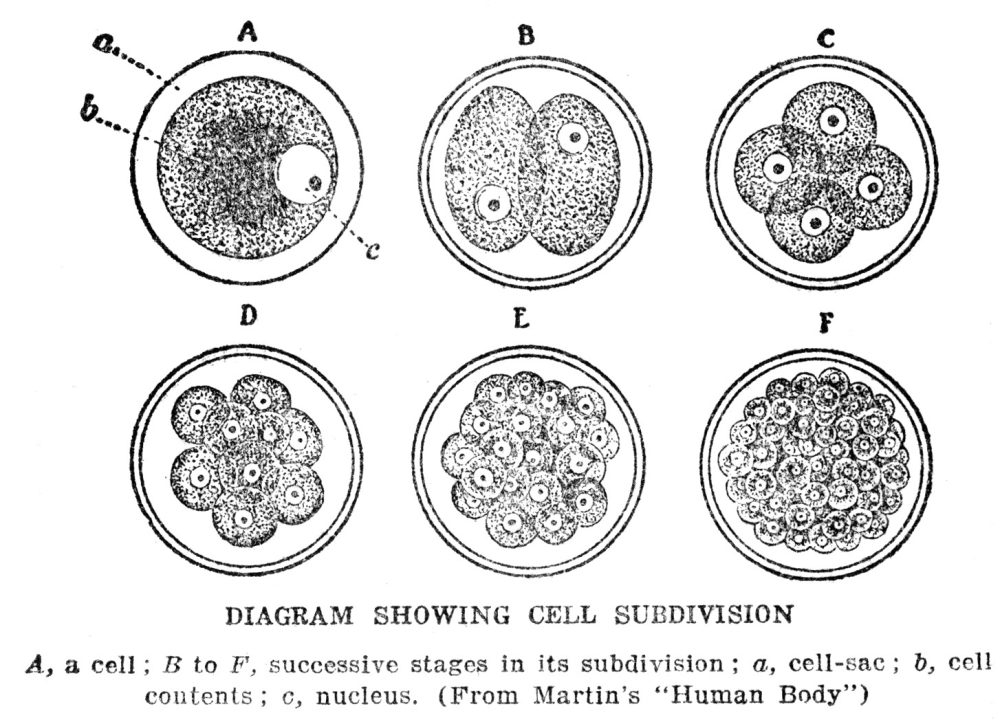
Before undertaking a further description of the nucleus itself, we shall be helped to an understanding of its function if we trace briefly the history of the cells which make up our body. At the beginning, as we probably all know, we start life as a single cell. This cell, after a series of events which will be described in a later chapter, begins the process known as development. Development consists of a series of subdivisions of cell material. At first the single cell divides into two; each of these then divides, giving four. At the next stage eight are formed, then sixteen and so on, until finally the millions of cells that make up the body are produced, all derived from the original single cell. We know that in the adult body there are very many different kinds of cells. Since they are all derived from a single cell, these differences must have put in their appearance during the course of the various cell divisions. In fact, this happens all along; at definite points in the process the two cells that come from the subdivision of some particular one will not be alike. The special kinds of cells that are thus produced become the starting points for whole masses of similar cells in the fully developed body. In human beings, and probably in most other kinds of animals, the very first subdivision does not result in any difference between the cells. The proof of this is that sometimes, in fact fairly often, the two cells become separated. When this happens twinning results, and the twins are exactly alike, being known as “identical twins.” Not only are they alike in all other respects, but they are always of the same sex, a fact that has escaped the attention of some writers of fiction, who have made twins, identical in all other features, brother and sister, instead of both boys or both girls. Twins that are not identical come from different original cells that happened to start developing together. Such twins need have no more resemblance than any members of the same family, and may or may not be of the same sex.
In every cell division the first step consists in a division of the chromatin of the nucleus, which is followed by a division of the rest of the protoplasm. The process by which the chromatin is subdivided is so curious as to be worth a brief description. The
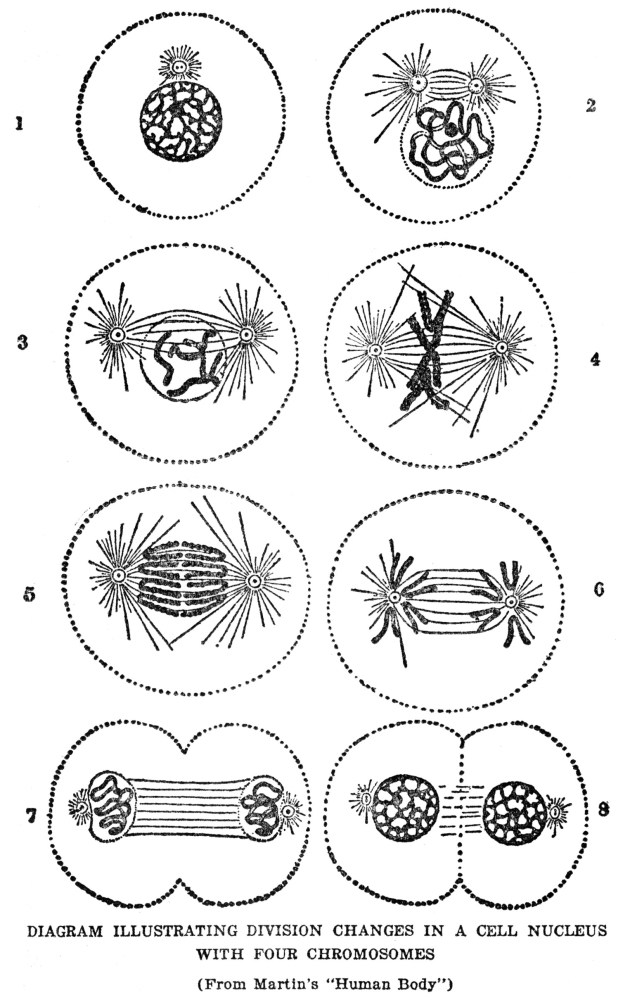
chromatin material is not a simple lump in the nucleus. It looks rather like a tiny string of beads thrown down carelessly, so as to become all mixed together. Each bead is a single bit of chromatin, and these bits are strung on a tiny thread. In an ordinary cell the beads are so mixed together that no order can be distinguished among them, but if a cell that is about to begin dividing is looked at it is found that the string has straightened itself out, and also that it has broken into pieces. The individual pieces are called chromosomes and their number is always the same for any one kind of animal or plant. There is a parasitic worm whose cells have only four chromosomes, and the number ranges from this up to as many as forty-eight in human beings. It may be that other species have even more, but they become so hard to count when there are as many as forty-eight that the number cannot be stated with certainty. So far as can be judged, the number of chromosomes has little to do with the complexity of the animal or plant, for some complex forms have few chromosomes, and some simple forms many.
At the same time as the chromatin is breaking up into chromosomes two tiny spots put in their appearance in the protoplasm of the cell on opposite sides of the nucleus, and tiny threads extend from one spot to the other through the nucleus. There are as many threads as there are chromosomes, the whole group making up a spindle-shaped figure. The chromosomes now become arranged at the middle of the spindle, and apparently each chromosome becomes fastened to a thread. Next each chromosome splits lengthwise through the middle and by what looks like a shortening of the threads the split halves are pulled apart and drawn to opposite tips of the spindle. The purpose of this elaborate scheme seems to be to insure an exactly equal division of the chromosomes between the cells, and the necessity of such an equal division will become clear when we learn something of what the chromatin is for. Meanwhile the description of cell division can be finished by saying that after the halves of the chromosomes are pulled apart the whole mass of protoplasm divides through the middle. As we stated above, sometimes the cells thus produced are alike and sometimes they are different, according to whether they are destined to become parts of similar or of different structures. In either case the chromatin material that goes into the two cells is exactly alike, so that if the cells themselves become different there must have developed a difference in the protoplasm at the two ends of the cell from which they came. Our bodies are made up of millions of cells, of a great many different kinds, but however different they may be the chromatin of each exactly duplicates that of every other one, or did when the cells were first formed; there is reason to believe that the chromatin may become changed during the lifetime of the cells, at least in some cases.
We may be interested in inquiring how long this process of cell division keeps up. Many children do not get through growing until they are twenty years old or more. Does cell division keep on during all this time? More than that; are there any cases of cell division that continue after full growth is reached? The answer to both these questions can be given in a brief paragraph. There are some tissues, particularly the outer layer of the skin, the connective tissues, the blood-corpuscle-forming tissues, and the reproductive tissues, in which cell division continues during all or most of life. The others finish at birth or shortly thereafter. We are born with the precise number of muscle cells with which we shall die, unless accident deprives us of some meanwhile; and if this happens no new ones will be formed to replace those that are lost. The same is true of gland cells. The last cell divisions among nerve cells are believed to occur within a few months after birth. As most of us have observed in our own cases, bodily injuries, if at all severe, are followed by the formation of scars. This means that connective tissue has grown in to fill the place of the cells destroyed by the injury, which cannot be replaced by cells of their own sort, since they have lost the power of cell division.
We have tried, in the above paragraphs, to get some idea of what living cells are like, and how they are derived, but have not attempted any detailed picture of particular kinds of cells. That will have to wait till we reach the story of the different kinds of bodily activity, when the cells that carry on each kind will have to be described more exactly. Something has also been told of the chromosomes, but the full account of them and their meaning is to be taken up in a later chapter, devoted to the matter of heredity and reproduction. In what remains of the present chapter we wish to talk about the conditions in which cells live so that we shall easily picture how they carry on their metabolism.
As an introduction to this topic a word may be said about the wide differences of complexity that are found in animals. They range from the simplest imaginable, a single cell with its nucleus and with protoplasm that appears almost uniform throughout, to a highly organized body like that of man, composed of millions of cells of many different kinds. Between these extremes almost every possible form is seen. The one-celled animals themselves show a wide range of complexity, and as soon as animals begin to be formed of numbers of cells grouped together the possibilities of complexity increase in proportion. One important difference between one-celled and many-celled animals needs to be emphasized; that is the matter of size. There are definite limits to the size that a single cell may attain; these limits are just over the boundary of naked eye vision. If animals are to attain larger sizes, they must necessarily be composed of many cells. The life of a single-celled animal presents no special problem, since it has only to take in through its outer layer from the surrounding water the various food materials and the oxygen which its metabolism requires, and to discharge into the same water any chemical products that may result from that same metabolism, and the question of whether it will live or die depends only on whether the water in which it happens to be contains sufficient materials and is otherwise suitable as a place to live. A many-celled animal, whose cells are arranged in not more than two layers, is in practically the same situation, for every cell has a frontage on the water and so can carry on interchanges of material directly; but the moment complexity reaches a stage where any cells are buried beneath other cells some special arrangement must be provided so that the buried cells can obtain the needed substances for their metabolism. The arrangement consists, in general, of furnishing what may be called an internal water frontage for the buried cells. In other words, complex animals have spaces all through their bodies, and these spaces are filled with fluid. There are no living tissues so dense that the cells of which they are composed are completely cut off from contact with body fluid. In thinking of our own bodies we should realize that this same arrangement applies; every one of our millions of living cells has contact with the fluid with which all the spaces of our bodies are filled, and it is from this fluid that the cells obtain the materials for their metabolism, and into this same fluid they discharge whatever substances their metabolism may produce.
The total amount of body fluid is not large, for the spaces among the cells are in most cases extremely tiny; it follows that with all the millions of cells absorbing food materials and oxygen from this fluid and discharging waste materials into it the time will soon come when no more food or oxygen will be left to be absorbed and there will be no more capacity for holding waste substances. If this state of affairs were actually to happen, metabolism would come to an end and death would be the result; evidently there must be some means of keeping the body fluids constantly renewed in respect to the things which the cells need for their metabolism, and constantly drained of the waste substances which the cells pour out. The way in which this renewal is accomplished is simple; part of the body fluid is separated off from the rest in a system of pipes, known to us as the blood vessels, and this part is kept in motion; at intervals along the system are stations at which the moving fluid can exchange substances with the fluid which actually comes in
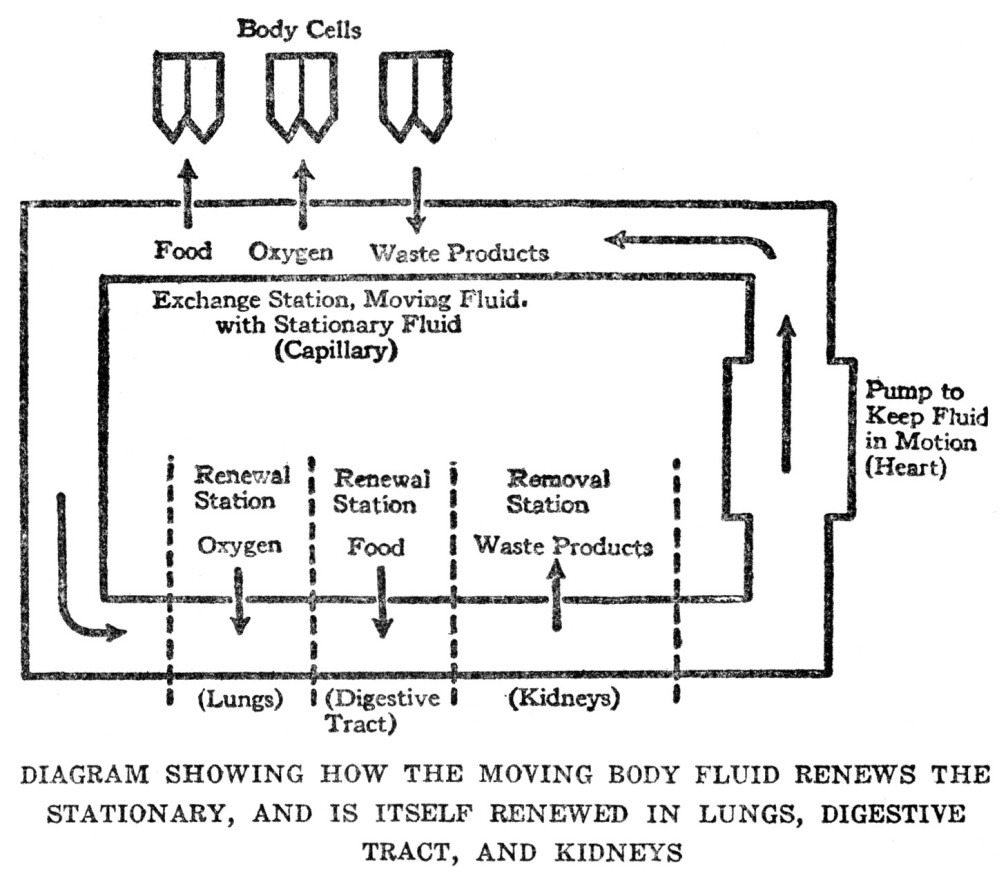
contact with the cells; thus the stationary fluid can obtain from the moving fluid the materials which the cells, in turn, are constantly withdrawing from it, and can pass on to the moving fluid the products with which the cells are continuously charging it. All that is necessary to complete the successful operation of the system is to have additional stations at which the moving fluid can obtain supplies of food materials and of oxygen, and stations where it can get rid of the wastes which it accumulates from the stationary fluid, and there must be a pump by which the moving fluid is kept in motion. We are familiar with the moving fluid under the name of blood; the system of pipes in which it moves are the blood vessels; the pump which keeps it in motion is the heart; the various supply stations include the digestive organs, the lungs, and the kidneys. In later chapters the operation of all these stations will be described in detail. The present outline has been given to show in a general way how the problem of metabolism is handled in highly organized bodies in which the individual cells have no direct access to food or oxygen supplies.
SINCE protoplasm is so very soft and fragile it must be supported in all animals and plants except the very tiniest. The nature of the supporting framework has a great deal to do with both the form and the working of the body, so it is desirable that we become familiar with it before trying to go further in the examination of the living protoplasm itself.
A large heavy body like that of man requires an arrangement for support that shall meet several conditions. In the first place there must be strength and stiffness, combined with flexibility, so that the body as a whole shall be firm, yet not rigid. The weight, also, must be kept as small as possible. Then every single cell, and every grouping of cells that we call an organ, must be supported in its place securely but without hindering the free performance of its function. Not only must the protoplasm be held in place, but on account of its fragility it has also to be protected against injury; the vital parts require more careful protection than those that are less immediately essential to life. Finally, bodily motions of all sorts depend on the framework to give purchase to the muscles, which are the actual organs of motion, and so to make their movements effective. For support, for protection, and for motion, then, the framework is important.
The material that does the real supporting is not, of course, alive, for living protoplasm lacks the necessary qualities needed here. It is manufactured and put in place, however, by living cells. They do this by withdrawing the special materials needed from the body fluid which surrounds them; in large part what they get from the fluid is not the finished substance but material from which the living cells make the finished substance. It is then passed outside their bodies and deposited in the surrounding space. Of course this is a gradual process. Bit by bit the structure, bone, cartilage, or connective tissue, as the case may be, is built up by the combined activities of many cells.
Of the three kinds of supporting material mentioned above, bone is the most familiar. No description of its appearance is necessary, for everyone has seen it as it appears in meat animals and in poultry, and it looks precisely the same in man. There are several things about bone, however, that are worth describing. One is the arrangement by which the very hard, compact material is deposited in large masses without cutting off the cells which are doing the depositing from their contact with the body fluid, and so destroying them and bringing their work to an end. The way this is managed can be made out by examination of the figure, showing the structure of bone. At the beginning the bone cells are lying near one of the tiny blood vessels known as capillaries, which are the exchange stations for material between blood and the stationary part of the body fluid. Thus these cells are favorably located for obtaining materials from which bone can be constructed. As they proceed with the formation of bone they always leave tiny passages open between themselves and the blood capillary. Finally the capillary may become completely surrounded by bone, but all along it will be left the passages through which fluid can make its way from the blood to where the cells are imprisoned within the bony walls of their own construction. The metabolism of bone cells is not on a very active scale; the amount of bone substance that a single bone cell has to produce in a day is only a fraction of the amount of saliva, for instance, that a single cell of the salivary gland turns out in the same time; so the bone cell can manage even though its supply of material has to come to it through a few very tiny passages in the bone.
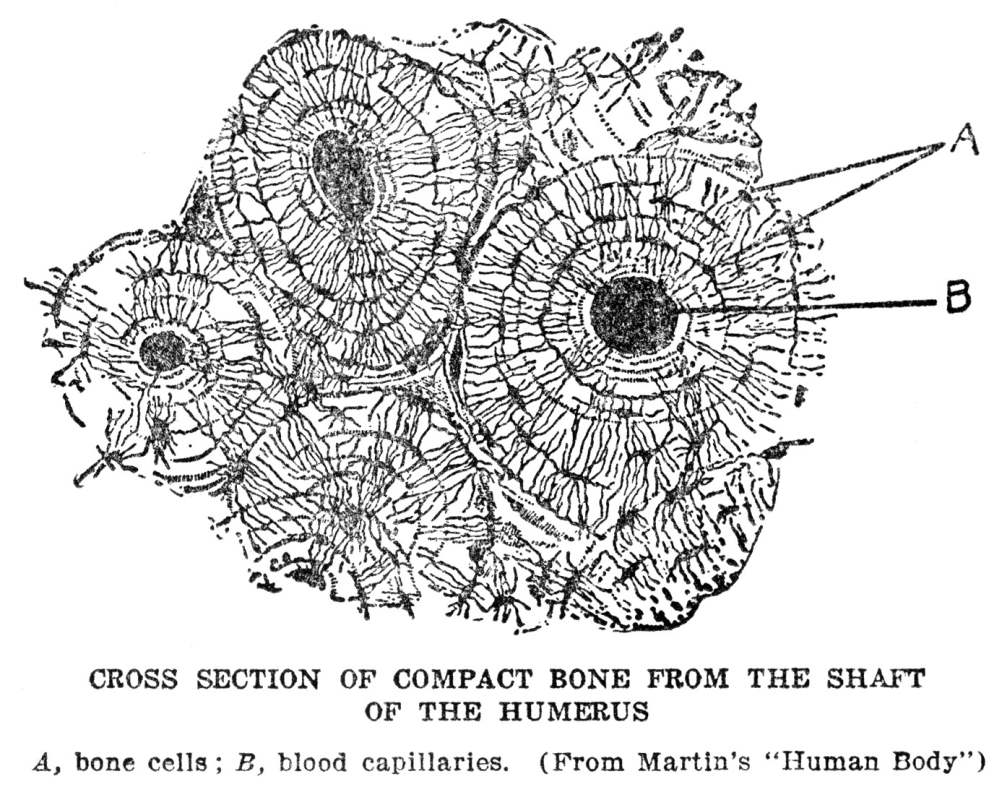
Another interesting feature of bone is the ease with which it can be remodeled. We are apt to think of bone as permanent, after it has once been formed, but as a matter of fact bone is about as subject to change as any of the softer tissues. This is because there are in and around the bones, in addition to the bone-forming cells, a great many cells of different appearance which may be named bone-destroying cells. These latter have the ability to dissolve out the hard material which the bone-forming cells have deposited. Good examples of their work are seen in the hollows of the long bones. We know, of course, that the bones in a child’s leg are so much smaller than those in the leg of an adult that they could almost be fitted into the hollows of the latter. Evidently the bone substance has been moved bodily outward during the course of growth. As the bone-forming cells add material to the outer surface of the bone, the bone-destroying cells dissolve it away from the inner surface. The same thing happens all over the body. A child’s face grows by an increase in size of the bones. Again the inner surfaces are dissolved away. Apparently one condition which makes the bone-destroying cells active is constant pressure. A good example of this is seen in what is known as a gumboil. If a tooth becomes ulcerated, gas and pus are formed at its root, and cannot escape since this is completely surrounded by bone. They accordingly press upon the surrounding bone, and also upon the sensitive tissues, resulting in extreme pain. The pressure upon the bone starts the bone-destroying cells into great activity and in the course of a few days they will dissolve a hole right through the bone, allowing the gas and pus to escape to the outside, and relieving the pain.
Of recent years school authorities have had much to say about the importance of adjusting school seats and desks so that they shall be at the proper height for the particular children that are to occupy them. This is because if the feet hang clear of the ground for hours at a time, as they will if the seat is too high, or if the body must be screwed around to enable the child to work at his desk, as happens when the desk is too low, there is real danger that some of the bones may become misshapen. Most of the stoop shoulders and many of the crooked backs that we see are the result of the habitual taking of wrong postures. Children, and adults as well, should form habits of standing and sitting so straight that none of the bones are put under a pressure that may tend to distort them.
After the teeth are lost the bony sockets in which they lie are dissolved away, making the jaws much shallower than formerly, a fact that accounts for the shortening of the distance between chin and nose in aged people. An important result of this dissolving away of bone by the bone-destroying cells is that the bones are kept as light as possible, without undue sacrifice of strength.
A second kind of supporting material is cartilage. This is both softer and more flexible than bone. It is found in places where flexibility is more important than great strength, as in the ears, the parts of the nose just below the bridge, the Adam’s apple and wind pipe. The chief difference in make-up between bone and cartilage is that while in bone about three-fourths of the nonliving substance consists of lime salts, in cartilage there is almost none of this material, organic substances making up the entire mass. There are no living cells in the body that are more poorly located with respect to obtaining supplies from the body fluids than the cartilage cells, for as these deposit the cartilage around themselves they leave no definite passages through which fluid may pass; the material incloses the cells completely. Although cartilage looks as though it were altogether nonporous, there must be some degree of sponginess present, since the cells do succeed in getting the materials on which their life depends. Cartilage seems to be a more primitive kind of supporting substance than bone. This is shown by the fact that it makes up the entire skeleton in the lowest fishes, and also by the fact that in the higher animals, including man, the bony skeleton starts, in large part, as cartilage. In the parts in which this happens a mass of cartilage is deposited in the place which is later to be occupied by bone. Then at certain points the cartilage begins to be dissolved away by cartilage-destroying cells, which are precisely like bone-destroying cells, and the bone-forming cells come in and build up the real bone as fast as the cartilage is removed. This process of replacing cartilage by bone is practically completed at birth, except in the long bones of legs and arms. These bones, which will about double in length during the growth of the body to adult size, as well as the other bones, which grow to some extent, retain plates of cartilage near each end during all the growth period, and the increase in length is obtained by a continuous formation of new cartilage, which is continuously replaced by bone.
The third kind of supporting material is connective tissue. This is composed of tiny threads or fibers, some of which are inelastic, others are very elastic. The inelastic fibers are found in places where a flexible, but unyielding support is required; the elastic fibers are located where elasticity is particularly important. Either kind of fiber may be grouped into sheets, or into loose networks, or into stout cords. A good example of inelastic connective tissue in sheet form is in the mesentery which holds the organs of the abdominal cavity in place. Just under the skin, anchoring it loosely to the underlying muscles, is connective tissue in network form. The tendons by which most of the muscles are attached to the bones upon which they pull are made up of inelastic connective tissue in the form of cords. The best example of elastic connective tissue is in the large arteries, which are just as elastic as best quality rubber tubing. Another good example is the large and strong elastic cord which passes along the back of the neck in cattle and sheep, and helps to support the weight of the head. Connective tissue fibers are deposited by living connective tissue cells. Since connective tissue is of open and relatively loose construction, there is no problem presented in supplying the cells with material. The meshes among the fibers are filled with fluid, and this fluid has ready connection, in turn, with the blood. Use is made of the abundance of body fluid in the connective tissue spaces whenever a subcutaneous injection is given, for what is done is to inject the desired material into the fluid which fills the spaces in the connective tissue just beneath the skin, trusting that it will pass from there to the blood, which it does rather gradually, and so is distributed about the whole body.
While we are on the topic of the supporting framework, something must be said about the grouping of the bones into what we know as the skeleton. Of course it is evident that the effectiveness of the bony part of the framework depends almost altogether on the way in which the individual bones are grouped together. If the whole skeleton were
composed of one great bone, or of different bones anchored solidly together, the body would be perfectly rigid; since motion is necessary to life, flexible connections between some of the bones are absolutely essential. Our movements are actually made by means of muscles, but nearly all of them become effective through the motions of bones to which the muscles are fastened. The bones are often very irregular in shape; careful study shows that the irregularities are due either to provision for the contact of one bone with another in the joints, a contact that must allow in most cases for motion of one on the other, or to provision of places to which muscles can be fastened in such a way as to make their pull effective. It is, of course, out of the question for us to examine the skeleton bone by bone. Figures are given of a number of typical bones: all that we can do in addition is to mention some of the interesting features of the skeleton.
The skeleton of the head is called the skull; its chief features are the brain case, the eye sockets, and the parts about the nose and mouth. The brain case is made up of eight bones firmly joined together by saw-tooth margins to make up a roughly spherical box which holds the brain, and protects this delicate and vitally important organ from all injury except the most severe. There are a number of small openings out from the brain case through which nerves pass, and one large opening below and at the back through which the spinal cord merges into the brain. The bones which make up the sides of the brain case are much thickened just behind the ears. A hollow extends from each ear into the bone, and within this hollow, securely protected from harm, is the actual organ of hearing. There are extensions of the hollow backward which are not occupied by any organs, and which communicate with the cavity of the ear. These sometimes become infected from the ear, causing the condition known as mastoiditis. Not only is this condition excruciatingly painful, but on account of the thin layer of bone which separates it from the brain itself it is highly dangerous. For this reason any ear trouble should be carefully watched lest it develop into mastoid trouble.
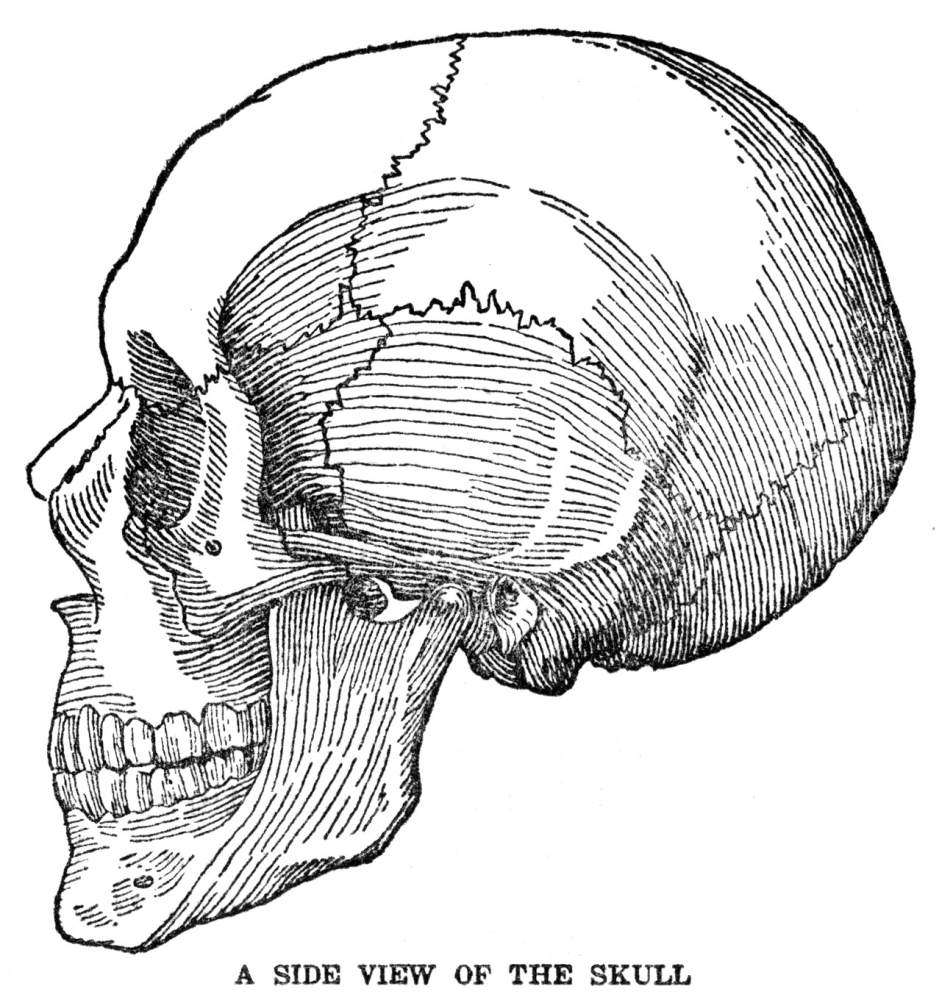
Of the bones that make up the eye sockets not much need be said, except that they have a great deal to do with determining the shape of the upper part of the face and so the appearance. There are bones within the nostrils that are very irregular in outline. Their effect is to increase greatly the surface over which the air that is breathed must pass, enabling it to become both warm and moist before entering the lungs. The jaw bones serve as receptacles for the teeth; the lower jaw, which is the only movable bone of the head, except for the tiny bones within the ears, has also the duty of operating as a mill in reducing the food to suitable form for swallowing. To aid in this function the lower jaw is hinged to the rest of the skull in such a way that it not only opens and closes but can slide forward and back or from side to side. All these motions are used in chewing. There are twenty-two bones altogether in the skull, not counting the three tiny ones in each ear which will be described later.
The body consists of trunk and limbs, and each part has its skeleton. The skeleton of the trunk consists of the spinal column, the rib cage, the shoulder girdle, and the hip girdle. The skeletons
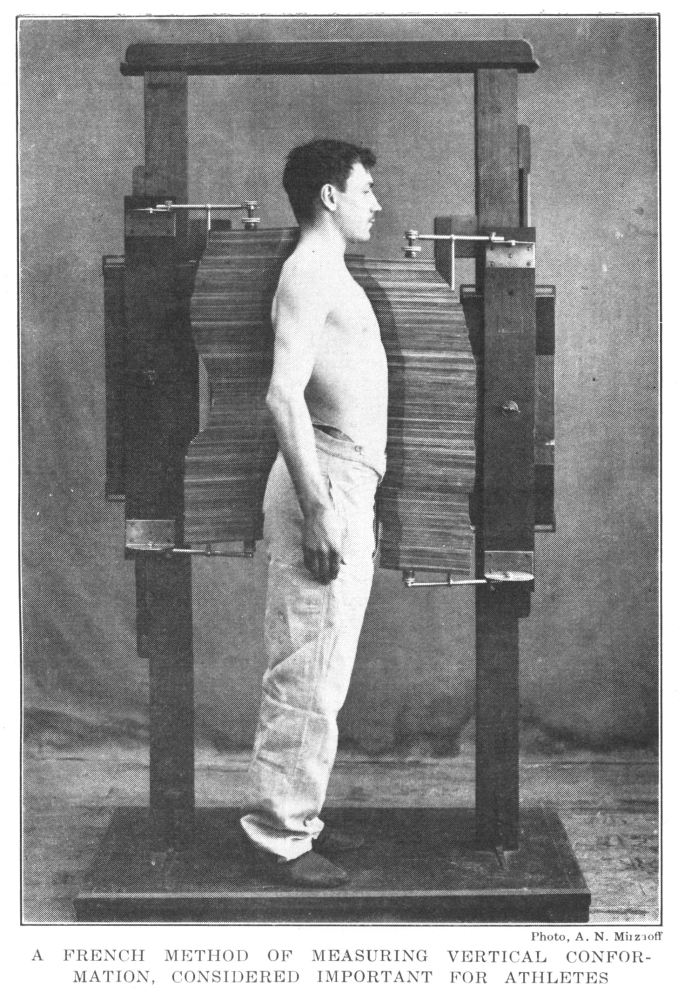
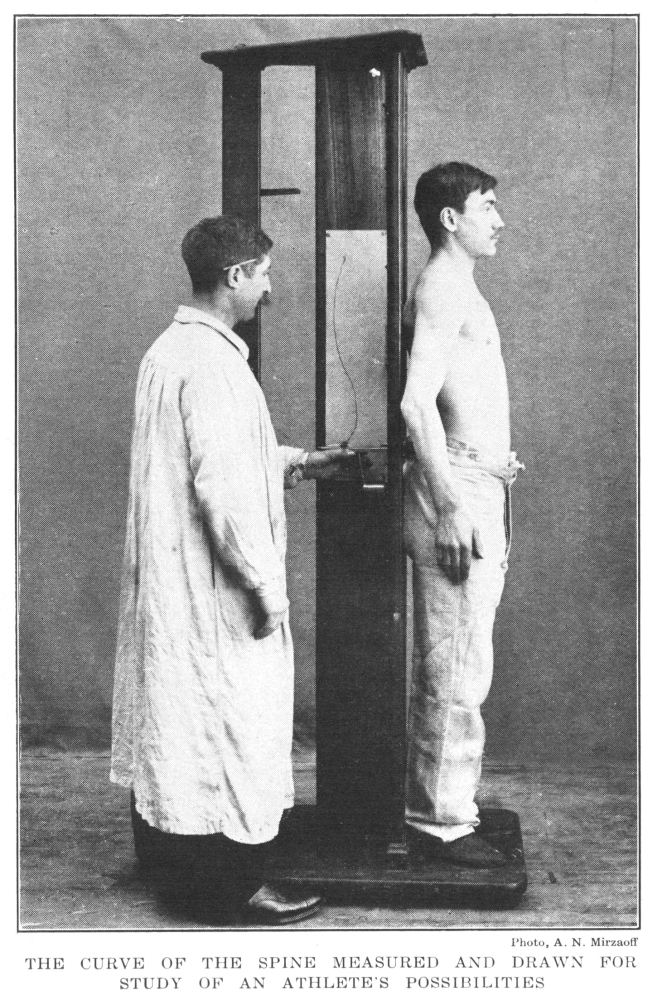
of the limbs are all according to a single plan to be described in a moment. The spinal column is a remarkable example of strength combined with flexibility and elasticity. It is made up of thirty-three bones or vertebræ; each of these has a disk-shaped part known as the body, and these disks are placed in line as shown in the figure. Between each disk and its neighbor is an elastic pad composed of a mixture of cartilage and elastic connective tissue. There is a small amount of give in each pad and this, taken along the whole length of the spinal column, is enough to give it the great flexibility which it enjoys. During the day the weight of the body packs these pads down hard, so that it is said that a man may be as much as an inch shorter at night than in the morning. Behind the disk each vertebra has an arch of bone, and beyond and beside this arch most of them have projections. All the arches together make up a bony canal which contains and protects the spinal cord. The projections serve for the attachment of the ribs and the back muscles by which the bending motions of the body are made.
The rib cage includes the breastbone and twelve pairs of ribs. It serves two purposes: to protect the heart and lungs from injury; and to take part in the movements of breathing. The latter function involves some degree of motion of the rib cage. All the ribs are attached behind to the vertebral column in a fashion that permits of a little motion up and down. All except the last two are fastened in front, seven pairs to the breastbone, three pairs each to the rib above it. In breathing the breastbone and ribs are moved up and down by muscles attached to them.
The shoulder girdle is made up of the collar bones and shoulder blades. Each collar bone is fastened at its inner end to the upper edge of the breastbone; this is the only direct contact the shoulder girdle has with any other part of the skeleton of the trunk; at the point where collar bone and shoulder blade meet, there is a shallow cup into which the head of the skeleton of the arm fits. The arrangement is favorable to great freedom of movement of the arm. Not only is the shoulder joint very flexible owing to the shallowness of the cup into which the arm bone fits, but the shoulder blade itself is capable of a considerable range of movement. This is because it is imbedded in and held in place by muscles. If one watches a person with bare shoulders while he raises his arms, it will be seen that the shoulder blades do not move much while the arms are being lifted to the horizontal position, but as that point is passed they begin to swing outward rapidly, so that when the arms are high above the head the shoulder blades are in a quite different position from that which they have when the arms are down.
The hip girdle consists of five bones of the vertebral column welded firmly together to make up what is called the sacrum, and two other large bones known as the innominate bones, each of which, in turn, is made up of three bones tightly fused together. The innominate bones are firmly joined to the sacrum at the back and they meet in front, also in a firm joint. The hip girdle or pelvis is rigid, suiting it to bear the strains that come upon it on account of its position at the junction of the legs with the trunk. At the outer side of each innominate bone is a cup, much deeper than the corresponding cup of the shoulder girdle, and into this fits the head of the skeleton of the leg. The arrangement is a typical ball and socket, and has been much copied in machinery where a flexible joint is required. In a good many people the union of the innominate bones to the sacrum is not so firm but that it yields somewhat when strains are put on it. Ordinary strains in these cases produce severe backache. Heavy strains may cause an excessively painful as well as disabling dislocation. In either case medical attention is needed.
Each arm can be subdivided into upper arm, forearm, wrist, and hand. The skeleton of the upper arm is a single long bone. The forearm has two bones, one of which is hinged at the elbow to the bone of the upper arm in a way to limit the movement to the single back and forth swing of which the elbow is capable. The other bone of the forearm can be rolled over the one which is fast at the elbow; this is what happens whenever the hand is changed from the palm up to the palm down position. There are eight bones in the wrist; these are irregular in shape, and are so grouped as to permit of a very wide range of movement. The bones of the hand and fingers make up five rows numbering four bones each for the fingers and three for the thumb. The joints are all practically simple hinges except for the one where the thumb joins the wrist, which is a much more flexible joint; flexible enough, in fact, to allow the thumb to be brought opposite any of the fingers. No animal except man enjoys this degree of flexibility in the thumb, so no animal equals man in the nicety of the grasp, particularly of small tools. When we recall how constantly we take advantage of this property of our hands we can realize how greatly our superiority over the lower animals has been aided by this rather slight structural difference between our hands and theirs.
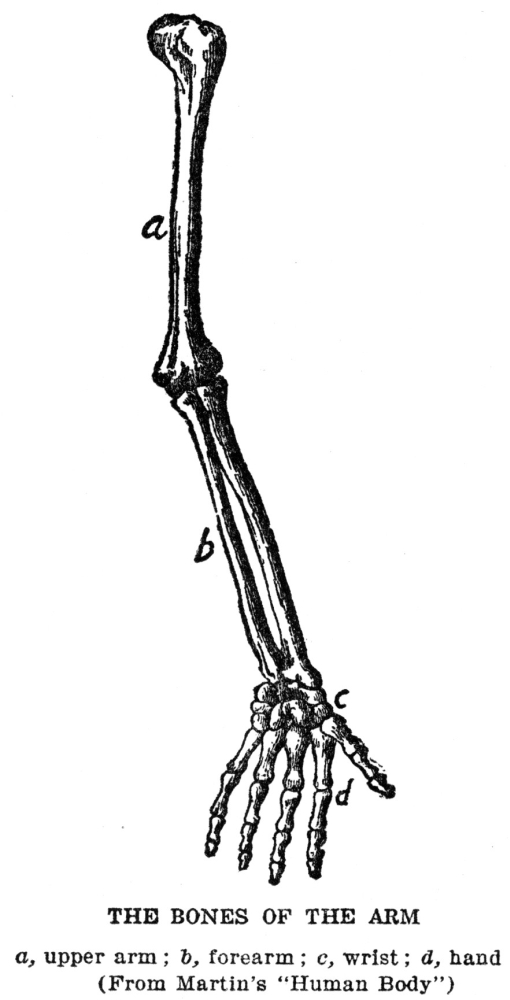
The leg subdivides along the same lines as the arm into upper leg, or thigh, lower leg, or shin, and foot. The order of bones is, on the whole, the same; one in the thigh; two in the lower leg. Instead of a flexible wrist the corresponding bones of the foot are grouped into a less flexible, but much stronger, heel and upper instep. Two of the bones of this group are fused together into one, reducing the total number from eight to seven. The bones of the lower instep and toes correspond in number and arrangement to those of the hand and fingers, but the great toe does not have superior flexibility as does the thumb. There is one bone in the leg, the knee cap or patella, that does not correspond to any bone in the arm, although it does correspond to a part of a bone, namely, the projection, at the elbow, of the long bone of the forearm. A feature of the skeleton of the foot that is worth a word is the arching of the instep. This undoubtedly adds greatly to the ease of walking. The natural position for the foot is, of course, with both the heel and the ball of the foot on the ground. For some reason it has become the universal custom among civilized people to raise the heel off the ground by adding a heel to the shoe. This does not seem to make much difference as long as the heel is not too high. In fact soldiers wearing properly fitted heel shoes can march as far and fast as can be expected. Excessively high heels throw the weight too much on the ball of the foot, thus doing away with the benefits that come from the arching of the instep. The effect on the gait is very apparent in any one who walks in high-heeled shoes. The foot itself does not appear to be greatly harmed by the wearing of high heels provided the shoes are otherwise well fitting. Whether the heels are high or low, the fit of the shoe is of utmost importance to the preservation of the feet. Crowding the feet into shoes that are too small in any direction is a fruitful means of bringing on foot trouble. Wearing shoes that are loose enough to allow the foot to turn over inside the shoe is nearly as bad. If the shoes are properly fitted in the beginning and then the heels are kept squared up, so that the feet will always stand straight on the ground, there will usually be little trouble with fallen arches or other foot disturbances.
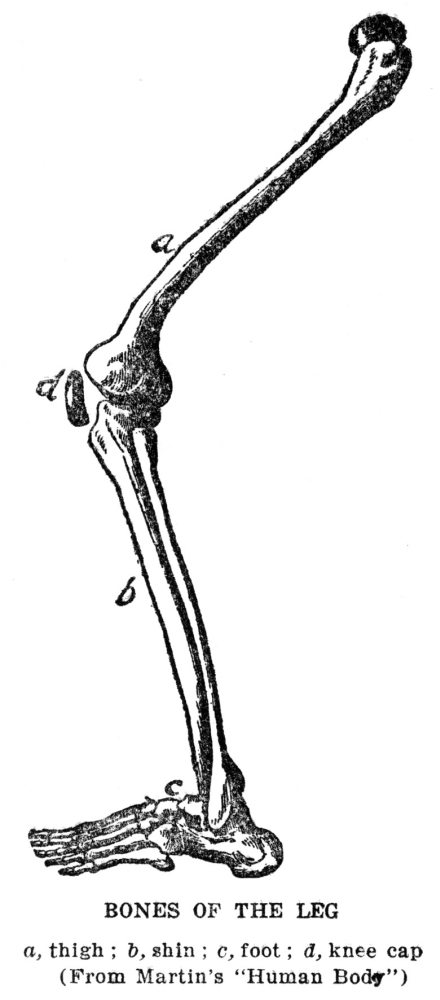
The bones are fastened together at the movable joints by stout sheets or bands of connective tissue known as ligaments. These hold them in place very securely and as additional support the muscles which surround every joint help to prevent the bones from slipping out of place. At nearly all the joints of the body the combined action of ligaments and muscles is sufficient to guarantee the joint against dislocation; the shoulder joint, and to a less extent the hip joint, is more likely to suffer this accident. The reason is that in obtaining flexibility of movement security of attachment is somewhat lessened. If the ligaments at the shoulder were tight enough to prevent the joint from ever becoming dislocated they would bind it to a serious degree. Most of the ligaments are of inelastic connective tissue, but those that fasten the separate vertebræ of the spinal column together are elastic, allowing of the bending in every direction which makes our backs as flexible as they are. The only movable joints which are bound by other means than ligaments are the connections of the ribs with the breastbone. These are of cartilage, but the movement here is so slight that the cartilage yields enough to permit it.
This completes our account of the bony skeleton. We shall finish the description of the supporting framework by a word about what may be called the connective tissue skeleton. The bony skeleton serves to support the body as a whole and to permit the muscles to do their work; the individual organs and the cells which make them up are held in place by sheets and bands of connective tissue. These are coarse and strong when their purpose is to support a large and heavy organ like the stomach; they become finer and finer as the parts to be supported become smaller, and when the individual cells are reached the connective tissue which surrounds them is almost inconceivably delicate. So completely does connective tissue permeate the whole body that it has been said that if everything else could be dissolved away, leaving only this tissue in place, there would still remain a model of the body, complete to the last detail.
OUR account of the body has now reached the point where we can take up in detail the special activities of the different kinds of cells. The first to be considered is motion, both because it is the familiar sign of life, as pointed out in the first chapter, and because it has so much to do with everything that enters into life. There are probably no animals that live out their entire lives without making any active motions, although some parasites, like the tapeworm, are stationary most of the time. There are a number of different ways in which movements are brought about. The very simplest animals, which consist of nothing but a bit of protoplasm, move by causing the protoplasm to flow bodily in one direction or another, a projection of part of the protoplasm being balanced by withdrawal of an equal part on the opposite side, and the whole mass progresses in the direction of the first projection. Next beyond this simplest means comes motion by tiny threads of protoplasm which project beyond the surface of the cell and whip back and forth. The stroke of these threads or cilia, as they are called, is stronger in one direction than in the other, so the effect of their beating is to propel the cell of which they are part in one direction through the water; or if they are on a surface which is stationary they set up a current in the water itself. This latter is the means by which oysters and similar animals which are anchored to the rocks get their food supplies. In some one-celled animals there are only one or two large cilia at one end; these beat back and forth, propelling the animal much as a fish swims.
The commonest, as well as the most effective, means of making motions is by cells specially developed for that purpose. These are called muscle cells, and every highly organized animal depends on them for most if not all of the motions which take place in its body. In muscle cells the functional metabolism takes the form of forcible changes in shape of the cells by which bodily motions are brought about. A muscle cell might be described as a mechanical device for transforming the chemical energy of burning fuel into the energy of motion. We have something comparable in the automobile cylinder, where the energy obtained from the explosive burning of the air-gas mixture drives the piston and so propels the car. Of course the two devices are not even remotely alike in the actual way in which they operate; their resemblance is purely the general one of converting one type of energy (chemical) into another type (motion).
There are three kinds of muscle cells in our bodies. The simplest are those that are found in the wall of the stomach and intestines and other internal organs that are capable of movements; the next kind is found only in the heart; the third, and most complex, makes up the bulk of our muscular tissue; it is the muscle that moves the bones. The first kind, because it shows no particular markings when examined through the microscope, is usually called smooth muscle; the second kind is known as heart muscle; the third kind, because it moves the skeleton, is named skeletal muscle. We shall devote most of our attention to this third kind of muscle, because it is a much more efficient machine than the others, and also because it has to do with our general bodily movements instead of with motions of internal organs.
A single skeletal muscle cell is an exceedingly slender fiber, much smaller than the finest thread; it may also be very short, not more than a twenty-fifth of an inch long, or it may be as much as an inch long. A muscle is made up of many of these fibers grouped side by side in bundles, and also, if the muscle is long, placed end to end. The fibers are held in place and fastened together by connective tissue. Lean meat consists of thousands of these muscle cells with their connective tissue fastenings. In coarse meat there is relatively more connective tissue and less actual muscle tissue than in the finer grades. In every muscle the connective tissue is loose enough to allow body fluid to penetrate among the muscle cells. Blood vessels are also distributed through the mass of the muscles between and among the cells; thus their nutrition is provided for.
Although not all muscle cells are exactly equal in power, on the whole the force that muscle can show is the force of one cell multiplied by the number of cells that can join in the pull. A strong muscle must have many cells side by side; in other words it must be thick. Also, the distance through which muscles can make movements depends on their length, so a muscle that has to pull for a considerable way must be long, and since single muscle cells are short there will have to be a good many cells end to end to make the whole muscle long enough for its task. The actual make-up and arrangement of muscles in the body depends in part, therefore, on the thickness and length needed for the particular work to be done, and in part on the architecture of the part of the body where the muscle is located. For example, the strongest muscle in the body is that by which one rises on the toes. This muscle operates by pulling upward at the back of the heel. If it were located right at the ankle, where it would have to be if attached directly to the place where its force is exerted, the resulting clumsiness can easily be imagined. By shifting it up to the middle of the lower leg room is found for the large mass of muscle needed for the work. The connection with the heel is made by means of a long and very strong tendon, known as the tendon of Achilles, because that was the part Achilles’ mother failed to immerse when she was dipping the infant in the river Styx to make him invulnerable. Other equally good examples are the muscles for operating the fingers. If placed in the hands, the latter would be too bulky and clumsy for any kind of efficient use. By placing them up in the forearms out of the way, and connecting them with the fingers by long tendons, delicacy is secured for the hands.
The muscles of the arms and legs are arranged in groups about the joints, and these groups always include opposing sets. Thus if the joint is a simple hinge, as at the elbow, where the only motion possible is back and forth, there will be one muscle to bend the joint and another opposing muscle to straighten it out again. The first is known as a flexor; the second as an extensor. In the arm the biceps, on the upper surface, is the flexor and the triceps, on the under side, the extensor. Joints that permit of motion in several directions have correspondingly more opposing sets of muscles acting upon them. The same scheme applies to the trunk, but since in the trunk instead of a few very movable joints we have the whole row of slightly movable vertebræ, the grouping of muscles is more complicated. Not all the skeletal muscles work about joints. The tongue, the muscles of the lips and about the eyes, those along the front of the abdomen, and some others are attached to bones only at one end or not at all, and do their work by pulling upon one another.
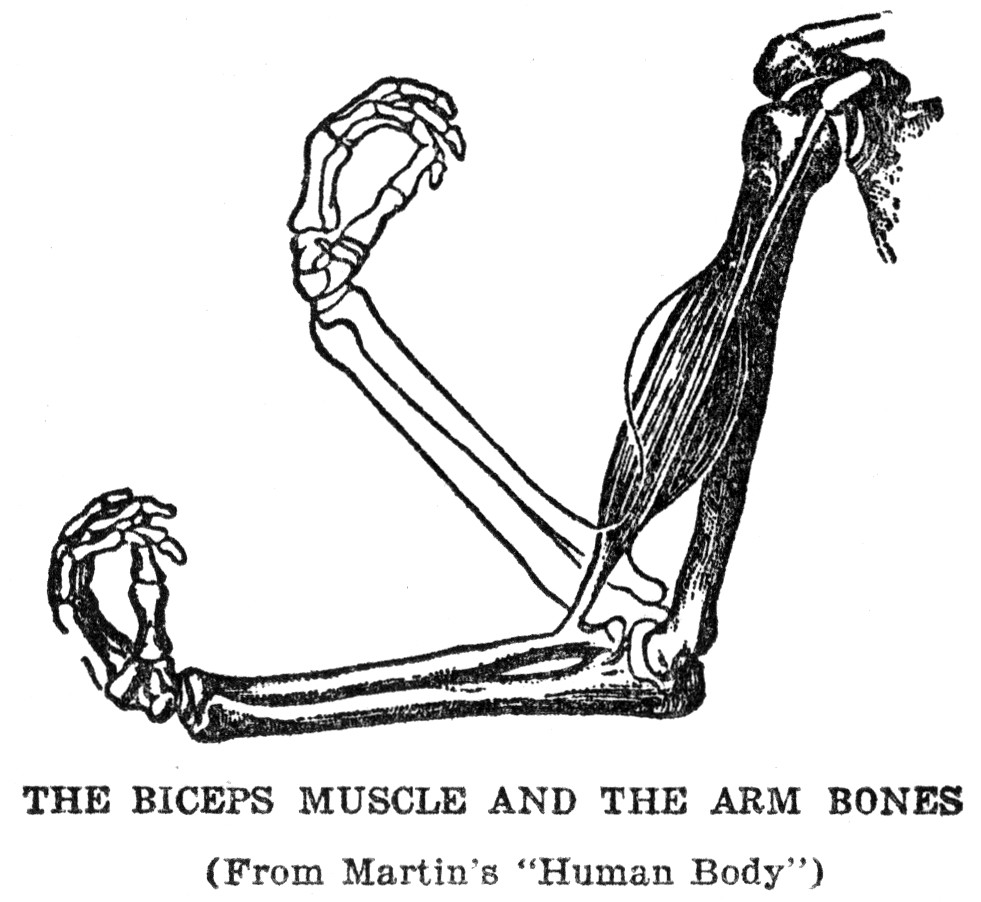
In earlier paragraphs we have seen that the movements made by muscles represent their functional metabolism, and also that the actions of whole muscles are merely the sum of the actions of the individual cells. Our present task is to see how muscles act; in other words to examine their functional metabolism. One feature that must be in mind from the very beginning is that the functional metabolism of muscle cells is under control; they do not go off at random, but only when started. This is more or less true of the functional metabolism of all the cells in highly organized animals. The agency that starts them off is named a stimulus. To picture how stimuli act we shall have to think for a moment of the state of affairs in cells at rest. As we have tried to make clear, cells at rest are not stagnating; a more or less active basic metabolism goes on within them all the time. This metabolism is of such a sort that it does not disturb the balance existing within the cell. The various chemical processes go on, using up material and producing wastes, but without arousing the additional chemical processes of functional metabolism. Meanwhile the substances that are required for this latter are present in the cell, so that when the disturbance that we call a stimulus comes along there is an increase in the total amount of metabolism, the extra chemical processes being those which perform the special function of the cell. In the case of muscle cells the stimulus ordinarily reaches them by way of the nervous system, although electric shocks, sharp blows, some irritating chemicals, and perhaps one or two other kinds of disturbance can act as stimuli. The effect of the stimulus is to start certain chemical processes; these in turn bring about the forcible shortening which is the thing that happens in active muscle. In skeletal muscle the shortening may be very rapid; the muscle can contract and relax again more quickly than the eye can follow. This is true at the temperature of our bodies. In cold-blooded animals, like fish or frogs, muscles become sluggish when they are cold. We see here one of the advantages we enjoy in having bodies that stay at the same temperature the year around; if our bodies cooled off in cold weather as do those of frogs, we should have to do as they do, become inactive whenever the weather becomes cold. As each muscle cell shortens it pulls upon the connective tissue that surrounds it; this communicates with the connective tissue of other cells, and all the connective tissue within the mass of the muscle fastens to the very stout sheets or cords of the same at the ends which are called tendons, by which the muscles are attached to the bones. Thus, although the pull of any single cell is so feeble as to be scarcely measurable, when hundreds or thousands of them pull all at once the effect may be very powerful.
We are familiar with the very wide range of effort that our muscles can show. They may contract with utmost delicacy, as when we hold a humming bird’s egg in our fingers, or they may pull with a force, in our largest muscles, of several hundred pounds. Of course this possibility of variation is of great advantage in our use of our muscles. It depends upon the very large number of individual fibers of which even our smallest muscles are made up. Whenever any single fiber contracts, it pulls to its full extent; if only a few become active, the pull of the whole muscle will be slight; as more come into action, more force will be exerted; the muscle will show its utmost power when all the fibers are contracting at once. We are conscious of greater mental effort when we make a powerful muscular contraction. This can be explained as due to the greater nervous discharge required to excite all the muscle fibers at once.
One feature of muscular action calls for an additional word. This is the temporary loss of power, resulting from too long-continued use, which is called fatigue. We know that a well-constructed machine can operate day in and day out without having to stop to rest; why cannot our muscles do the same? Evidently the necessity of resting cuts down the possibilities of life more than any other one thing; our real life is only two-thirds as long as it counts up in years because we have to spend one-third of the time in sleep. Of course muscular fatigue is not the only kind; there is nervous fatigue, as well, about which something will be said later. The activity of our muscles is based on functional metabolism; it follows, therefore, that fatigue is also due to metabolism. We can think of two ways in which metabolism might cause fatigue; the first of these is by using up the materials which furnish energy; clearly no cells can go on working after they have exhausted their supplies of fuel. The second results from the fact that metabolism produces waste products. It is a familiar fact of chemistry that when the substances formed in chemical processes are not removed they interfere with the processes themselves. In active muscles very rapid metabolism is going on and large quantities of waste substance are being formed; these have to be discharged from the cells into the surrounding fluid, and removed from there in turn by the blood. We can easily imagine that this might not take place as fast as necessary to keep the cells from becoming more or less clogged; in fact this clogging is exactly what happens, so that muscles begin to show fatigue some time before their supplies of fuel material are used up.
One familiar fact of muscular fatigue is that soreness, which indicates that fatigue has really


been present in large amount, occurs much more often when we use our muscles in ways to which we are not accustomed than when they are exercised according to habit. It is the experience of every one who does manual labor that when he gets a new job, one that calls for different use of the muscles than he has been in the habit of, his muscles are very sore until he is “broken in.” After that, although he does as much or even more work than at first, he no longer becomes sore. This is explained as being due to two things. First, whenever we make an unaccustomed movement we overstimulate our muscles; that is, we call more fibers into action than are necessary to do the job; as the motion becomes familiar we cut down the action to that which just meets the demand. Thus there is a great deal more metabolism than necessary when unfamiliar motions are being made. Then, secondly, there is a spot in every muscle cell, just at the point where the nerve makes its connection with the muscle, that is more easily fatigued than any other part of the muscle cell. This spot, by becoming fatigued first, tends to cause metabolism to stop in time to prevent the rest of the cell from being seriously fatigued. Only when we are so much interested in what we are doing that we pay no attention to the fatigue of this safety spot, or when necessity keeps us at work after we would quit if we had our own way, do we push the metabolism so far that muscular soreness results. Other types of fatigue, including feelings of exhaustion, are due to effects on the nervous system, and will be described when we have that system before us.
Before we leave the subject of the skeletal muscles it will be interesting to say a word about the different kinds of motions that they bring about. We have already seen that they work by pulling at the joints, and we have no intention of enlarging on that topic. What we want to do here is to group the bodily motions into a few classes, regardless of what joints are actually moved. First, and most important, comes locomotion; by that we mean any motions that move the body from one place to another. Under that head we have walking, running, swimming, jumping; in birds, flying. Next in order comes grasping; this includes all motions by which we take hold of anything. We can realize the importance of this group of movements when we think that our fore limbs are specifically grasping organs, while in the great majority of animals they are organs of locomotion along with the hind limbs. Originally grasping had to do, undoubtedly, with the taking of food and not much else. In civilized man we have in addition the use of all kinds of tools from the coarsest to the finest. In most four-legged animals the chief organ of grasping is the mouth. We still use our mouths to some extent as grasping organs, and could probably learn to make even more use of them in that direction if forced to it. Chewing and swallowing make up a group of movements concerned primarily with the handling of food after it has been grasped. Not much need be said about them. Of small extent but great importance are the motions connected with sense perception; these include chiefly the motions of the body, neck, and eyes in vision; we are constantly turning to look at something; in such animals as horses movements of the ears help greatly in hearing; and both man and animals make sniffing motions to increase the keenness of smelling. There is a group of motions devoted to voice production (including breathing). In man the vocal cords, tongue, and muscles of the cheeks are the chief muscles that have to do with the voice, not including the muscles of breathing, which, of course, are essential. The interesting things about the vocal cords are the excessive fineness of their operation, enabling expert singers to produce tones that vary by only a few vibrations a second, and the amazing exactitude of the control that the nerves have over them, so that good singers can set them at the tension needed for producing a particular tone with absolute certainty. The tongue is not a single muscle, but a mass of several muscles working one upon the other. It plays a part both in voice production and in the chewing of food. As an organ of voice production it helps by changing the shape of the mouth cavity. Speech depends very largely on this, since not the tension of the vocal cords but the shape of the mouth and throat determines the making of letters and syllables.
In addition to these familiar uses of the muscles there is a use which is just as important but about which we are apt to think less. This is their use in connection with posture, the taking and holding of particular bodily positions. Posture is unlike other muscular activities in several things. In the first place there is a steady, but rather feeble, tension which can be held without marked fatigue for long periods; all other forms of muscular contraction become severely fatiguing rather quickly if held steadily. In the second place the nervous control of posture seems to be different in some respects from our ordinary control of our muscles. Finally there is some doubt as to whether the contractions of the muscles themselves are the same. Measurements of the functional metabolism of posture show that it is much less than would be expected if the muscular action were of the ordinary type. This, of course, explains why posture is less fatiguing than other forms of activity.
The other two kinds of muscle, heart muscle and smooth muscle, must have a word of description. Heart muscle contracts quickly and powerfully, as does skeletal. It differs from skeletal in not depending on nervous stimulation to make it contract; the heart can be cut clean out of the body and will go on beating for a short time; in cold-blooded animals, like frogs or turtles, for a long time. This could not be true if the heart muscle had to be aroused to activity by nerves. Besides being automatic, heart muscle shows the peculiarity that whenever it contracts all the fibers join. We do not have a varying strength of pull shown by heart muscle as we do in skeletal. As we shall see, it would be a serious disadvantage rather than an advantage if heart muscle were to be like skeletal in this respect.
Smooth muscle has the duty of operating the internal organs. For this function no great strength is required; the motions do not have to be powerful. Nor is rapid motion important. Smooth muscle does not have to be so highly developed, then, as is skeletal. It is sluggish and rather feeble in its actions. There are, however, two points of superiority about smooth muscle, which fit in well with its special task. The first of these is its freedom from fatigue. There are in the body numerous smooth muscle masses that are in contraction practically all the time. This would be impossible if fatigue were to develop. These masses make up what are called the sphincters, rings of muscle surrounding openings like that from the esophagus to the stomach or from the stomach to the small intestine. It is the duty of these sphincters to hold the openings closed all the time except occasionally when they open for just an instant to let material through. The second point about smooth muscle which fits it for its work is that it is capable of stretching out greatly or contracting sharply without much difference in the force with which it is pulling. For example, at the beginning of a meal the walls of the stomach are drawn up, so that the food that is swallowed enters a small space. With the progress of the meal the stomach enlarges, so that at the end it has a much greater bulk than at the beginning. But the actual pressure of the stomach upon its contents is about the same as at the beginning. If the stomach were an ordinary elastic bag this could not happen; the walls would have to stretch as the stomach filled, and the stretching would mean greater pressure. Since the stomach walls are of smooth muscle they adjust themselves to the progress of the meal. It is important to note that there is a limit to this possibility of adjustment. If one is so greedy as to keep on stuffing after the stomach has reached its full size, stretching does occur, and if this is repeated it may lead to a diseased condition known as “dilated stomach,” which will cause much digestive trouble.
WE have talked a good deal about muscles and the different sorts of activities they can perform. We have also mentioned the fact that the skeletal muscles are under accurate nervous control. Our next task is to investigate the control of this nervous control; in other words to find out just what it is that causes the nerves to stimulate the muscles so that they shall perform as skillfully and usefully as they do. In Chapter II we saw that our bodily movements are adjusted to our needs through the sense organs. These bring information of the situation and we act accordingly. We may group the kinds of information which the sense organs furnish under three heads; first, what is going on inside our bodies; second, what is happening at the surface of the body, and third, what is happening at a distance from us. The senses which bring the first kind of information are called the internal senses; the second group are the contact senses; and the third are the distance senses. We need to remember that the primary purpose of the senses is to guide our muscles, and that our muscles are to find food for us, to keep us from bodily harm, and to assist in the perpetuation of life by propagating and caring for the young. By keeping these facts in mind we shall have no difficulty in understanding the way in which the various senses do their work.
Pain, hunger, and thirst are the internal senses with which we are most familiar. Pain is evidently a protective sense. It is never aroused unless something is amiss; for that reason pain should never be neglected. Of course, in the majority of cases the pain is due to some simple disturbance which can be located, and if no permanent harm is to follow, or if no relief is possible, the heroic bearing of the pain is meritorious; but thousands of women, thinking mistakenly that to complain of suffering is a sign of weakness, or hoping to spare loved ones distress, bear in secret or make light of pains that are the signs of insidious disease, curable if taken in hand early enough, but sure to cause acutest suffering and untimely death if allowed to go on unchecked. Unfortunately our most dangerous internal enemies, the organisms of infectious disease, do not give warning of their attack by causing pain until the disease itself is so far advanced that there is no escaping it. In this respect pain falls short of being efficient as a means of warning us against impending injury.
Hunger and thirst are the stimuli which drive us to the taking of food and water. It is interesting to think that of all the living things that roam the earth only men have discovered the connection between the taking of food and the avoidance of starvation; all other animals are impelled to nourish themselves wholly through the operation of these senses. There are two distinct phases to hunger. The first is appetite, and this by itself seems not to be a sense in the strict meaning of the word, but rather a memory of agreeable experiences associated with the taking of food. In man appetite is often sufficient by itself to lead to eating, as is proved by the frequency with which food is eaten between meals when there cannot possibly be any genuine hunger, but probably in animals it acts to arouse genuine hunger, rather than to cause eating by itself. Genuine hunger is a sense as definite as any other. It is aroused by spasmodic contractions of the stomach. These contractions cannot occur except when the wall of the stomach is in a certain state of tension. Various things can influence the coming on of this degree of tension in the stomach, and so the possibility of hunger. Appetite itself probably does this very effectively. Habit seems also to have something to do with it. Hunger is usually felt just as mealtime draws near, and it is often much keener at noon or night than before breakfast, although the stomach has been longer empty at breakfast than at any other meal. A curious fact about hunger is that it may disappear completely after a few days of complete starvation. Contrary to the popular idea that hunger becomes more and more acute as starvation continues, the testimony of practically all persons who have starved for more than a few days is that all sensations of hunger, as well as all strong longings for food, subside and do not return. This is especially true if the body is kept quiet and if the mind is diverted, so that recollection of meals particularly enjoyed shall not come up.
Thirst is due to actual drying of the throat. When the cells lining that region become deficient in moisture the sense is aroused. The drying may occur from without or from within. When it occurs from without, as in sleeping with the mouth open, relief can usually be obtained by merely swallowing saliva copiously. The same treatment helps for the moment when the lack of moisture is due to deficiency in the amount in the body, but in this latter case no permanent relief can be had except by the taking of water. When the amount in the body falls below the proper level no comfort can be had until the loss has been made good. An interesting thing about thirst is that it is the only sense which is said never to be lost or seriously impaired by disease.
In addition to these familiar internal senses we have some that are less well known. They are for the purpose of what may be described as the routine guidance of the muscles. The act of walking, as we well know, is made up of a series of muscular movements which are both accurately timed and accurately graded. We obtain startling realization of this when we come to the bottom step on our way down stairs without noticing that we have arrived there. This timing and grading are done for us by our bodies without our having to attend to it. The amount of labor that is saved is shown by walking upon railroad ties. These are irregularly spaced, and on that account it is necessary for us to pay attention to every step. There is no comparison between the fatigue of this kind of walking and ordinary progress along a smooth path. The senses that keep track of the position of the body and of individual muscles are known respectively as the equilibrium sense and the muscle-and-joint sense. The equilibrium sense has as its organ a part of the internal ear. Deeply imbedded in the bone is a series of chambers and canals lined with a delicate membrane and filled with liquid. The canals, which are three in number in each ear, are semicircular in shape, and accordingly have been named the semicircular canals. One of them is horizontal; the other two are vertical, and the two vertical canals lie at right angles to one another. This arrangement makes it inevitable that any movement of the head, in any direction whatsoever, will register differently on the canal system than any other movement, which is exactly what is required to make the apparatus efficient as an organ by which motions of the body are kept track of and guided. Along with the semicircular canals is a structure known as the vestibule which registers the position of the head, and so indirectly of the body, when no movements are being made. We are not ordinarily conscious of the working of these senses; they carry on their guidance of muscular movement without our attention. We can, however, pay attention to what they show if we wish. For example, one who is swimming under water is never in doubt as to whether his head is turned up or down, even though his eyes may be shut. His knowledge of position in such a place is obtained from his equilibrium organ, even though he may not be aware of the fact. Sometimes the organ becomes diseased. The results, so far as the victim is concerned, are highly distressing. He usually has to stay in bed because he cannot balance himself well enough to get about.
The organs for muscle-and-joint sense consist of tiny spindles distributed around the joints and embedded within the mass of the muscles. They are arranged so as to be affected by every motion of a joint or every contraction of a muscle. They register not only the fact of motion but also the extent. There is a disease, commonly known as locomotor ataxia, in which the muscle-and-joint sense is impaired or lost, particularly in the legs. The result is that walking becomes difficult and unsteady, and usually impossible when the eyes are shut or the room is dark. This is because the victim learns to make his sight serve instead of his muscle-and-joint sense for guiding his muscular movements, and when this also is withdrawn all knowledge of where his legs are or what they are doing fails, and the only course is to fall down or lie down as quickly as possible.
We have some additional bodily sensations, such as nausea, repletion, fatigue, ill feeling or malaise, which guide our conduct more or less, and are not very different in consciousness from hunger or thirst. So far as is known there are no sense organs by which these sensations are aroused. They are not strictly senses, therefore. We do not know enough about how they originate to say anything more about them.
The contact senses are touch, warmth, cold, and taste. Pain that comes as the result of bodily injury might also be classified as a contact sense, since its cause is something that comes in direct contact with the body from outside, but it differs from internal pain only in its source and not at all in the sensations it arouses, so there is no need of describing it over again. The sense of touch is the fundamental sense; the very lowest animals, even those that have no specially developed sense organs, and few organs of any kind, react to the contact of objects with their bodies just as the highest animals react to the sense of touch. When no other information is available, that of simple contact guides the animal in its securing of food and its avoidance of harm. In accordance with this primitive character of the touch sense, the psychologists tell us that we interpret the information from our more highly developed sense organs, sight particularly, in terms of the feel of objects. When we look at anything our judgment of it actually consists in an idea of how it would feel if we were to take hold of it. Our touch organs consist of tiny spots scattered all over the surface of the body. They are much closer together on some parts than on others. The total number is estimated at a half million or more. A good way to test their sensitiveness is by pressing down on different parts of the skin with fine hairs. When this is done it is found that the most sensitive regions—the tip of the tongue, for instance—are fifty or sixty times as sensitive as the dullest regions, like the small of the back. To obtain sensations of touch it is necessary that there be unaffected points alongside those that are affected. If all are acted on alike, there will be no more sensation than if none is acted upon. This can be shown by dipping the hand into quicksilver. The very heavy liquid presses on all the touch points hard enough to affect them, but since it presses on all alike nothing at all can be felt except along the line where the hand enters the quicksilver where the pressure is strongly marked. It is this feature of the touch sense that makes the wearing of clothing bearable. If we had to feel the contact of the clothes constantly we should presently find them so trying that we could no longer endure them. We do feel rough places and are often seriously annoyed by them, so we can judge what would be the effect if the whole surface were felt as plainly.
Closely related to touch is the sensation of tickling or itching. Curious facts about this sensation are the violence of the feeling that may be aroused by very delicate irritation, drawing a thread along the corner of the nose, for example; the persistence of the feeling beyond the actual irritation; and the effectiveness of scratching as a means of alleviating the condition. Almost nothing is known in explanation of any of these peculiarities.
In addition to organs of touch the skin contains two kinds of organs for perceiving differences of temperature. The first of these detects warmth; the second cold. It is by means of these senses that we judge whether the place where we are is of a suitable temperature in which to remain; whether we should be quiet or active; whether special provisions, like changes in the clothing, are necessary. In the case of both senses the temperature of the skin is the comparison point. We judge that an object is warm or cold according as its temperature is above or below that of the skin which touches it. The ears are usually a few degrees cooler than the hands; thus it is possible for one and the same object to feel cold to the hands and warm to the ears. The two kinds of temperature organs are side by side in the skin, although there are many more “cold” spots than “warmth” spots. Very warm objects affect both kinds, and then we get the sensation that we call “hot,” as distinguished from merely warm. The cold spots are a little nearer the surface of the skin than are the warmth spots; for this reason a hot bath may feel cold at the very instant of stepping into it, although the sensation changes to hot almost at once. We need to remember that our sensations of warmth or cold depend altogether on the state of the skin, and tell us nothing at all about whether our bodies as a whole are warm or cold. Because the blood is always warm a flushed skin always feels warm, and to produce flushing by means of alcohol has long been used as a means of making the body feel warm and comfortable. This may be a serious mistake in cold weather, for to drive the blood to the surface then may mean that the body as a whole will cool off to the point of actual injury. It is better to feel cold and conserve the body’s heat than to feel warm and waste it.
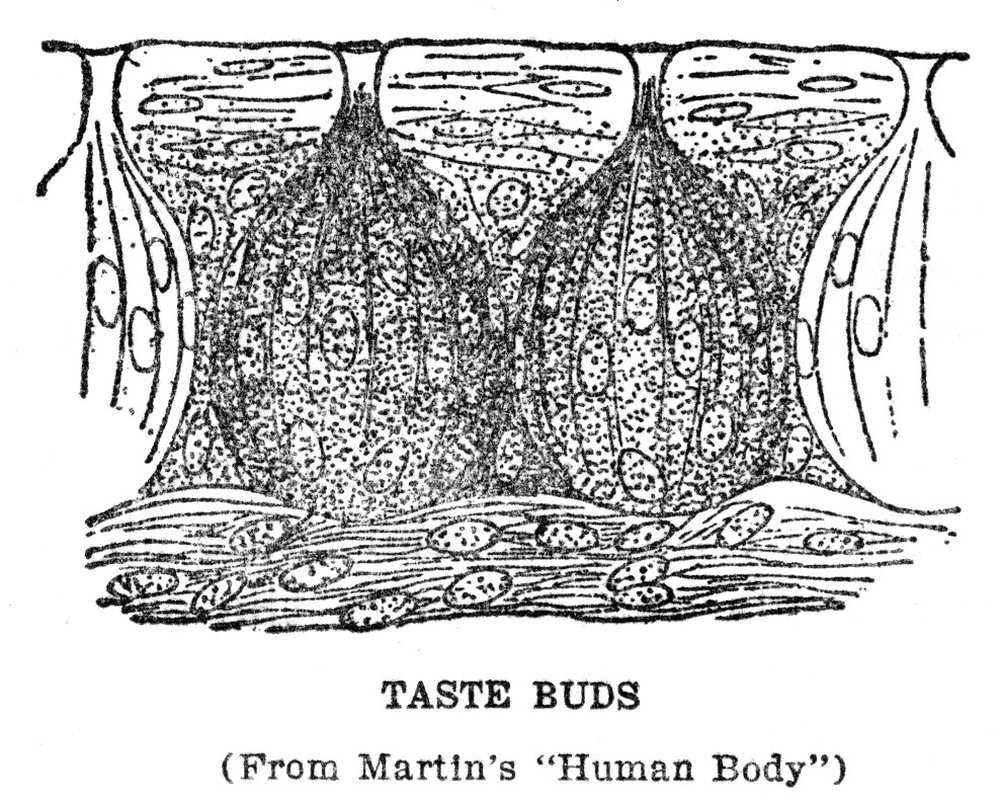
The last of the contact senses is that of taste. This is found only on the tongue. Scattered about on that organ are many tiny sense organs known as taste buds. These are usually in little hollows, so they cannot be affected unless liquids which can enter the hollows are on the surface of the tongue. If the tongue is wiped dry and then dry sugar is sprinkled on it, no sweet taste will develop so long as the dryness continues. The purpose of taste is evidently to give final information about the food after it has passed the inspection of the other senses and has been inserted into the mouth, but before it is swallowed. In the higher animals there has been a subdivision of this sense into two. The other is the sense of smell. In large part smell is a distance sense, and will be treated when we are talking about the distance senses. Smell has monopolized most of the properties of food-judging, so there is left for taste proper only four kinds of perception. These are sweet, sour, salty and bitter. We have, apparently, four kinds of taste buds, one for each of these kinds of taste. All the other sensations that we call taste are flavors, and are really smells. Of the four tastes sour and bitter would probably be called warning and sweet and salty recommending. Only by practice do we come to care for bitter foods, and children usually object just as strongly to those that are sour. Tropical savages, for whom salt is a rarity, esteem it much more highly than sugar, which they can usually get in abundance.
In concluding this chapter we need to remember that the contact senses make up the court of last resort; by the time anything comes close enough to the body to act upon any of them it is so close that the effect in guiding the muscles must be immediate; there is no time for deliberation; whatever the muscles are going to do in response to information thus obtained must be done at once. Later we shall see how this affects our whole bodily make-up.
THE three senses that give us information of what is happening beyond the surface of our bodies are smell, hearing, and sight. Since smell is closely related to taste, which was talked about in the last chapter, we shall take it up first. Smell is like taste in that it is aroused by chemical substances, but to be smelled these must be in gaseous form, not dissolved in water, as for taste. The organ for smell is in the upper part of the nasal chamber. There are really two of them, one in each nostril. They are made up simply of little patches of mucous membrane, as the membrane that lines the nose is called, in which are many of the particular kind of cells that are affected by odors. An interesting thing about these patches is that they are not in the part of the nostrils through which the main current of air sweeps in breathing, but in a little pocket off this main channel. If air containing an odorous substance is breathed in or out, a little of it works its way into the side pocket and is smelled. If we wish to get more of the odor we do it by sniffing, which is changing the shape of the nostrils to throw the air current more directly against the smell organs.
These organs are amazingly sensitive. It is hard to appreciate the minuteness of the amounts of material that can be smelled. Especially is this true of those animals that have a really keen sense of

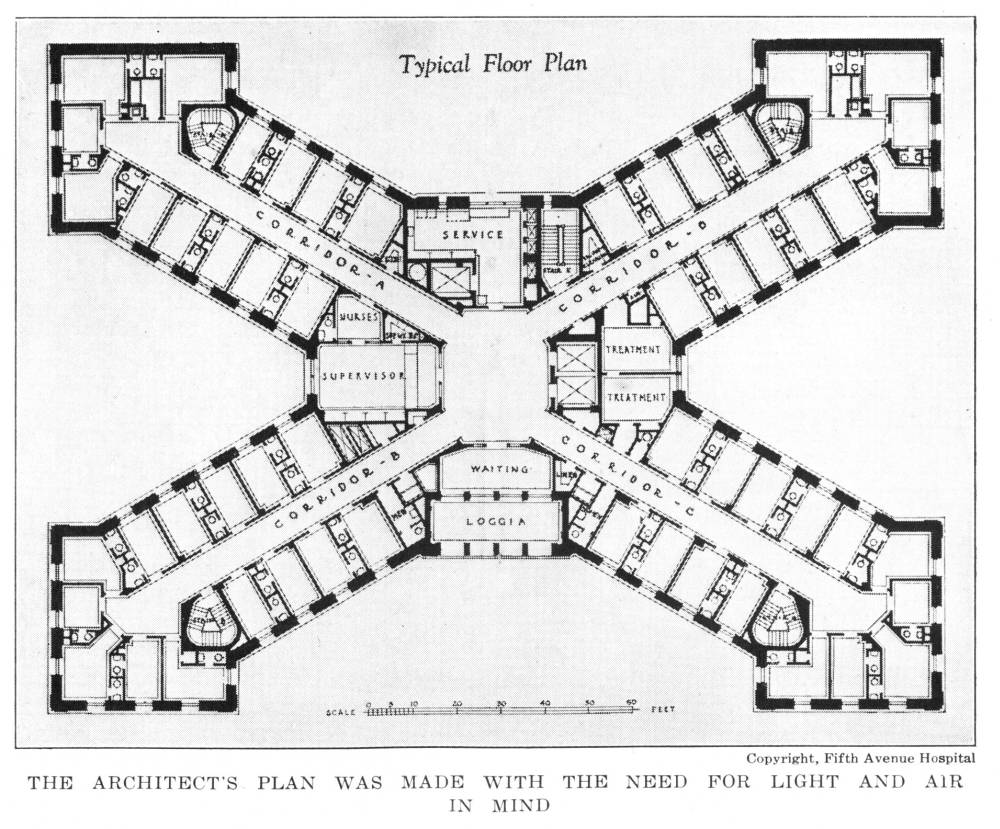
smell. The amount of substance that rubs off from a rabbit’s feet onto the ground at each step cannot be much to begin with; yet this continues for hours to give off gas into the air, and a dog coming along at any time meanwhile will get enough of the gas into his nostrils to smell it. Fortunately for our comfort the sense of smell fatigues very rapidly. An odor that is excessively disagreeable at first presently no longer troubles us. If it were not for this, it would be almost, if not quite, impossible to obtain laborers in those industries where the odor is necessarily bad. There is, however, a source of danger in this quick subsidence of smell perception. About our only method of judging offhand as to the ventilation of a room is by the smell, and this fails as a guide when we have been in the room for a time. Persons coming in from outside are often struck by the bad state of the air in rooms whose occupants are not conscious that there is anything amiss. Because of this the ventilation of schoolrooms usually is not, and never should be, left merely to the judgment of the teacher, but definite rules are laid down as to opening of ventilators or windows.
When the gas that is smelled is part of an inward air current we recognize it as coming from the outside and call it an odor; when it is part of an outward current we call it a flavor. On account of the rapid fatigue of the sense of smell we are unconscious of the smell of our own breath, but can get fresh smells from within, and these come, practically always, from materials that have just been taken into the mouth. In comparison with taste flavor furnishes great variety of perception. As persons become connoisseurs in food their enjoyment depends more and more on flavor and less and less on taste. The sensation from spices is a combination of flavor with irritation of the tongue that is partly pain and partly touch. The sense of smell has evidently a two-fold use; it makes us aware that there is food in the vicinity, or sometimes that disagreeable things are near at hand; and it shares with taste the duty of enabling us to judge of the food as it is being eaten. Agreeable tastes, flavors, and odors add much to the enjoyment of life. Within reasonable limits it is well to cultivate this kind of enjoyment, for while there is no doubt that it can be overdone, as in the excessive lengths to which the decadent Romans went to gratify their taste and smell, neither is there any doubt that bodily health in general, and the bodily function of digestion in particular, benefit definitely from the kind of enjoyment that savory food and delightful odors bring. It is the duty of those charged with the responsibility of preparing and serving food to take pains that full advantage is taken of the possibilities present in what food is to be prepared. This does not mean expensive food; what it does call for is skillful handling of all food, whether cheap or costly.
In introducing the subject of hearing we shall have to say a few words about that which is heard, namely sound. Any object that has any degree of elasticity at all is apt, if struck or rubbed or otherwise set suddenly in motion, to start vibrating back and forth; the vibrations will nearly always be regular, and will occur at a rate that is the same for that particular object whenever it vibrates. The rate depends on the size, the character, and the degree of stretch of the object. Air is to all intents and purposes perfectly elastic; it is set vibrating by any object that is vibrating in it, but since it has no particular size nor degree of stretch it takes the vibration rate of the object that started it going in the first place. The vibrations once started in air spread in all directions, just as waves spread from a stone thrown into a pond, and when these air waves strike upon another object that is free to vibrate they will set it going at the same rate. The human hearing apparatus is a device which is set in vibration by air waves, and the result is called sound. The ear is limited in its ability to respond to vibrations; they must be neither too fast nor too slow; if slower than 16 a second, most people will fail to hear them, and the same is true if they are more rapid than about 40,000 a second. Between these limits vibrations that strike upon the ear are heard as sounds.
Differences in vibration rate between one sound and another can be recognized by the ear; the difference is a matter of pitch. By the pitch of a tone we mean the vibration rate which it has. More rapid vibrations give tones of higher pitch; a slow rate means a low pitch. Middle C on the piano has a rate of either 256 or 261 a second according to the system used by the tuner. The human voice has an extreme range starting with the lowest note that the bass voice can compass with a rate of about 80 vibrations a second, to the highest note that famous sopranos can attain at about 1,400 a second. There is a record of a singer who could achieve a tone with a rate of 2,100 a second, but this has not been duplicated so far as is known. Of course no single voice can cover more than a fraction of this range. Most men produce all their tones at rates of between 90 and 500 a second, and women between 200 and 800 a second. Not every different vibration rate is heard as a tone of different pitch; the ear is not sensitive enough for that. The interval between one note and the next includes several vibrations, more the higher one goes in the scale. A perfectly true tone has exactly the rate called for; a departure of one or two vibrations a second may not be noticed, but if the error is greater the singer is sharping or flatting his tone, according as he is above or below the true rate. A note that has just double the rate of another one is said to be its octave. For convenience the interval of the octave has been split up into twelve tones, and all our music is constructed on that basis.
It is evident that the ear must be a very complicated organ; not only must it perceive differences in pitch, as just indicated, but differences in loudness must also register differently. More than that, the ear has to be able to deal with sounds made up of a great many tones coming into it all at once. When we listen to an orchestra or band, the waves that strike our ears represent the commotion set up in the air by all the instruments together. It is a remarkable fact that in this case, instead of getting a meaningless jumble, we actually get a blend of tones from which, if we are sufficiently musical, we can pick out the individual elements.
The ear consists, in the first place, of a vibrator that will respond accurately to any vibration rate or combination of vibration rates within its range, and secondly of a sensitive apparatus that is acted upon by the vibrator. The vibrator must respond freely to feeble impulses, and, what is of prime importance, to any vibration rate as readily as to any other. Almost all elastic bodies have a preferred vibration rate; that is, they will respond better to some rates than to others. About the only exception to this rule
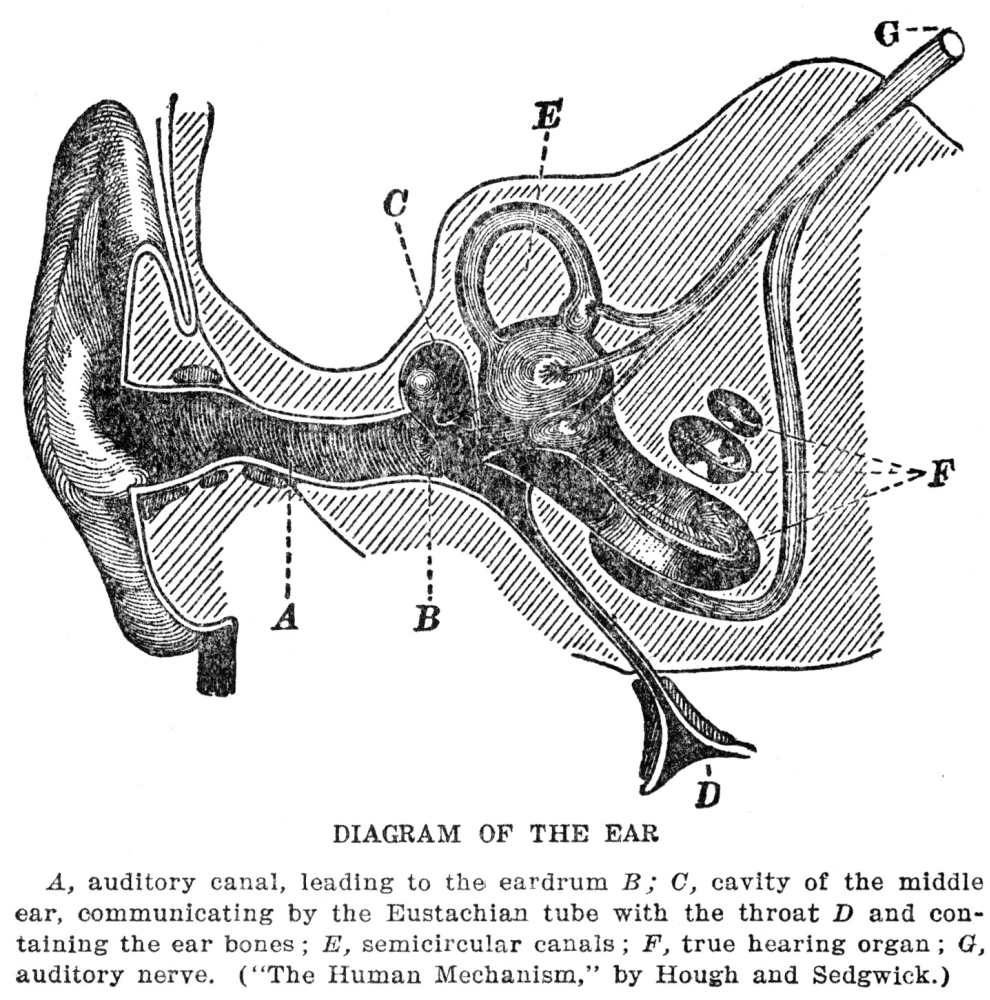
is in the case of membranes that are not tightly stretched. A stretched membrane, like a drumhead, has its own vibration rate, but one that is not on the stretch is able to vibrate at almost any rate. This fact is taken advantage of in the telephone and the phonograph, both of which depend on being able to vibrate at various rates almost equally well. In the ear, also, an unstretched membrane is the vibrator. We are familiar with it as the eardrum. It is located at the bottom of the ear canal, but cannot be seen by looking therein, because of a slight curve at about the middle. When the ear specialist wants to examine the eardrum he thrusts a small metal tube into the canal. This straightens it out enough to bring the drum into view.
The eardrum does not act directly upon the sensitive hearing apparatus, but its vibrations are transmitted across a space known as the middle ear. The necessity for this space is found in the fact that atmospheric pressure is not constant, but changes frequently from day to day, besides falling off as one ascends higher above sea level. The free action of the eardrum depends on its not being stretched; if there were no means of readjustment it might be properly set for one air pressure, but greater pressures would bulge it inward, putting it on the stretch, and so cutting down its ability to respond to a wide range of tones. The middle ear, which is the space behind the drum, connects with the outside air by a tube leading from it to the back of the throat, which latter communicates freely with the air through the nose, as well as through the mouth whenever it is open. The tube is known as the Eustachian tube. Its walls are ordinarily collapsed, so it is not an open passage, but every time one swallows the tube is pulled open, thus allowing differences in air pressure on the two sides of the eardrum to equalize. Whenever one ascends a high hill quickly, as by train or automobile, or even in going to the top of a high building by elevator, the difference in air pressure behind and in front of the eardrum can be felt. The sensation is disagreeable, and there is definite impairment of hearing. Repeated swallowing gives relief.
The vibrations of the eardrum are transmitted across the space of the middle ear by a chain of three tiny bones; these are very irregular in shape, and are attached to one another in such a way that every movement of the drum is followed exactly as to time and direction, but with reduced size and increased power. The hearing apparatus, which is part of the internal ear, but not the same part as makes up the equilibrium organ, contains liquid which is set moving by the last of the chain of bones, and this liquid acts upon the actual sensitive cells which make up the sense organ proper. These latter are arranged in an exceedingly complicated fashion. There are various theories as to the precise manner in which the vibrations of the liquid in the inner ear arouse the sensitive cells. It is thought that different cells respond to tones of different pitch, but exactly how this is accomplished is not known.
Deafness may result from failure of the sensitive inner ear to respond, or from poor transmission of vibrations across the middle ear by the chain of bones, or by interference with the freedom of action of the eardrum, or by preventing the air waves from striking upon the drum. Injury to the inner ear is rare, because of its secure position within the bone. A common form of deafness is the result of hardening of the connections between the ear bones, so that the chain no longer follows well the vibrations of the drum. This usually begins to come on during the twenties or thirties, and causes almost complete deafness by the age of fifty. In most if not all cases it is hereditary. Interference with the action of the eardrum may be due to the partial destruction of the drum itself. Scarlet fever and measles are particularly likely to leave the drum in a delicate condition, and any strain upon it then may rupture it beyond repair. Continuous closing of the Eustachian tube, by preventing equalization of air pressure on the two sides of the drum, causes partial deafness. Inflammation accompanying a cold may cause this, or the growth of adenoids in the back of the throat. Adenoids will be described later; here they are mentioned only because they may press upon and close the Eustachian tubes. There is danger from the closure of the tubes by inflammation, because the inflammation may creep up to the middle ear and cause serious trouble, both there and in the mastoid region adjoining. Earache in children should be carefully watched, since it usually means that the middle ear has been invaded by the same inflammatory condition that is present in the throat in colds, and may do serious damage to the delicate structures there. Often in children, and sometimes in adults, the hearing is impaired by the accumulation of wax in the ear canal. This wax is a sticky secretion that serves to catch particles entering the ear canal and to prevent them from striking against the drum. Unless the ear canals are washed out frequently with hot water the wax accumulates and hardens into a plug which closes the ear canal, shutting off faint sounds. The wax dissolves in hot water, but not in cold, so its accumulation is to be prevented by taking pains that hot water actually gets to the bottom of the ear canal once in a while. Digging the wax out with hard instruments should be done only by an expert with greatest caution, lest the drum be injured.
The last of the senses to be described is sight; this is the one of which we make the most use ordinarily, and curiously enough is the only one that we can turn on and off. Loud noises or penetrating smells must be endured, but by shutting our eyes tightly we can escape sight whenever we want to. Altogether there are three kinds of information which the sense of sight brings to us; the first is the knowledge simply of light and darkness; the second is the knowledge of the shape and size of objects; the third is the knowledge of color. Nearly all animals seem to have some power of distinguishing between light and shade. Even the one-celled kinds, that have no eyes or anything corresponding to eyes, behave in a way that proves them to have this power. A lot of them can be put in a dish of water that is well lighted on one side and in shade on the other and in a few minutes all of them will be found to have traveled to one side or the other according as they happen to be a light-seeking kind or a dark-seeking species. As we go higher up the animal scale we find that parts of the body show this same power of distinguishing light from darkness. In the case of the common angleworm, or earthworm as it is more properly called, the front end has the power but the rear end has not. In very highly organized animals only the special organs known as eyes possess the power; all the rest of the body has lost it. The next feature of sight, the ability to perceive the shape and size of objects, requires a special apparatus, the eye, so animals that lack eyes cannot perceive objects, although they may be able to tell light from darkness.
In order to see an object it is necessary that a pattern or image of it be thrown on a sensitive surface; this surface registers the details of the pattern, and so the object is seen. What we have to do here is to find out how these patterns or images are formed in our eyes. In the first place we must realize that every visible object has rays of light going off from every part of its surface in every direction. So-called self-luminous objects, like lamps or the sun, produce the light within themselves; all others merely reflect light that falls on them from some source. Whenever light falls on any object, unless it is a perfect mirror, part is absorbed by the object and the rest is reflected; that which is reflected presently strikes against another object and is again in part absorbed and in part reflected; as this process is repeated over and over again the light becomes so broken up that rays of it are traveling in every direction from every point on every object. Any spot so protected that no rays can strike it evidently cannot reflect any out again, and such a spot will be absolutely dark.
For the formation of an image of any object all that is necessary is that some of the rays of light from every point on the object be caused to fall in exactly corresponding positions on the image. The simplest possible means of doing this takes advantage of the fact that rays of light travel in straight lines. If we inclose an incandescent bulb in a tight box with a round hole in one side of it, every spot on the incandescent filament will be giving off light in every direction, but all the light will be cut off by the box except that which happens to have the direction which takes it out through the hole. From every incandescent spot, then, there will be a beam of light in the form of a cone escaping from the box through the hole. The tip of the cone will be the incandescent spot; the slope of its sides will depend upon the size of the hole. If a screen is placed in front of the hole, all these cones of light will strike on it, and it will be illuminated in a pattern which is made up of all the cones from all the incandescent spots which make up the filament, but these will overlap so much that one cannot be told from another. Now if the hole in
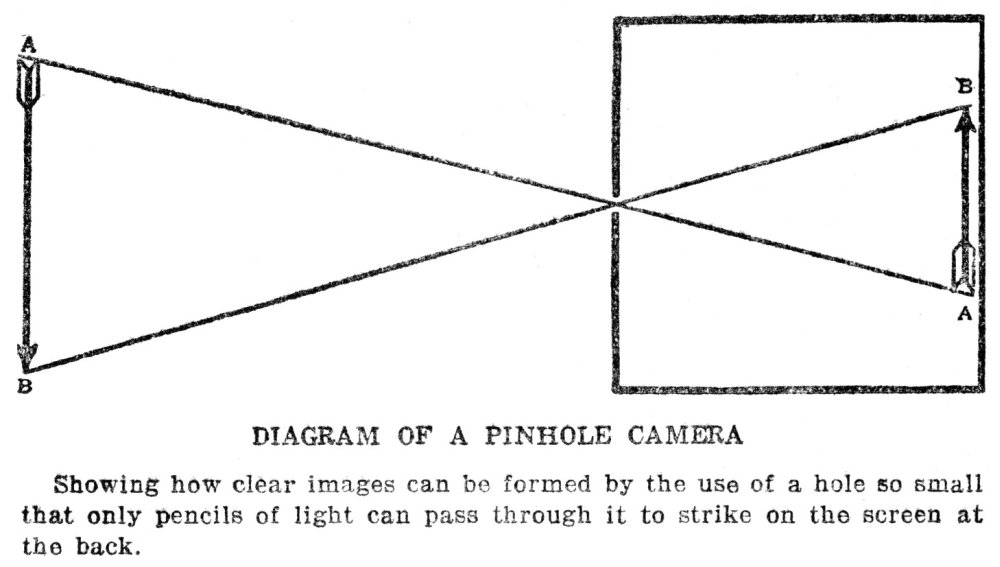
the box is made small enough, a pinhole, in fact, and if the screen is placed close to the hole, the cones of light from the different incandescent spots become so narrow that when they strike the screen they overlap scarcely at all, and what we get is a tiny spot of light on the screen corresponding to every incandescent spot on the filament and straight in line with it through the hole. Here we have exactly what we have been talking about, namely a pattern or image of an object. The image will be upside down, because those rays from the top of the bulb that strike the tiny hole will be below on the outside, and those from the bottom will be above. The same thing can be worked exactly in reverse; we can place a box with a pinhole in it in front of any object and get an image inside the box on the back; by placing a photographic plate or film there an excellent picture can be taken. There is just one reason why this scheme is not used in all cameras; that is that unless the object is very brightly illuminated indeed the amount of light that passes through the pinhole is not enough to affect the plate or film except on long exposure. Perfect pictures can be taken with a pinhole camera wherever long exposures are possible, or wherever the object shines brightly enough. This difficulty is gotten around in the ordinary camera by gathering up all the light in a wide cone from each spot on the object and condensing it again on the plate or film. The image is formed just as before, but now each spot on the image includes, not only the beam of light that comes in a straight line from the corresponding spot on the object, but in addition that in a wide cone surrounding the straight beam. It is naturally brighter the wider the cone; which explains why in poor light we open the diaphragm of the camera wider than when the light is good; a brightly illuminated object will pass enough light through a narrow opening, but a dimmer object must have as wide an opening as possible in order that enough may get through.
The method of condensing the light in a spreading cone so that it shall come back to a point again is by means of a lens; not only is this true of cameras, but also of the eye; in fact everything that has been said thus far about cameras applies perfectly to the eye. There is one thing about the way in which light is brought to a point by a lens that makes the formation of images by this method troublesome in comparison with their formation in pinhole cameras. That is that the cone of light which strikes the lens is condensed as an opposite cone on the other side, and since the formation of an image requires that every point of the object shall be reproduced as a point in the image, there is only one place where the image can come, which is where the tips of the cones of light are. This place is spoken of as the focus. Unless the screen or film is exactly there the image will be made up of overlapping circles of light instead of points, and so will be blurred. In a pinhole camera the cones are so small that they cannot overlap, so there is no one place where the image is better than elsewhere; in other words, there is no necessity of focusing. The chief reason that focusing gives trouble is that the farther away from the lens the object is the closer to the lens will be its image; hence if the field of view consists of several objects at different distances they will not all focus at the same level; the distant objects will focus near the lens; the near objects farther back. In practice this trouble is met by using a thick lens, which has a very short focus to begin with, so that a considerable range of distances will be covered without serious blurring, and for finer work by adjusting the distance between the lens and the back of the camera, where the film or plate is, so that there shall be a good focus of the particular object that is desired.
In the eye the clear part that projects between the lids, and is called the cornea, is the important lens. Just behind is the arrangement that corresponds to the diaphragm of the camera; this is the colored part with a round hole in the center; it is called the iris and the round hole is the pupil. Behind the iris, and resting right against it, is the secondary lens of the eye, known as the crystalline lens. The eye as a whole is a globe just under an inch in diameter; at the back of it, straight behind the lenses and pupil, is the sensitive surface upon which images are formed. This is the retina; it extends pretty well around to the sides, but the part we use most in seeing is the small portion straight in line with the pupil. The cornea by itself is a lens whose focus is
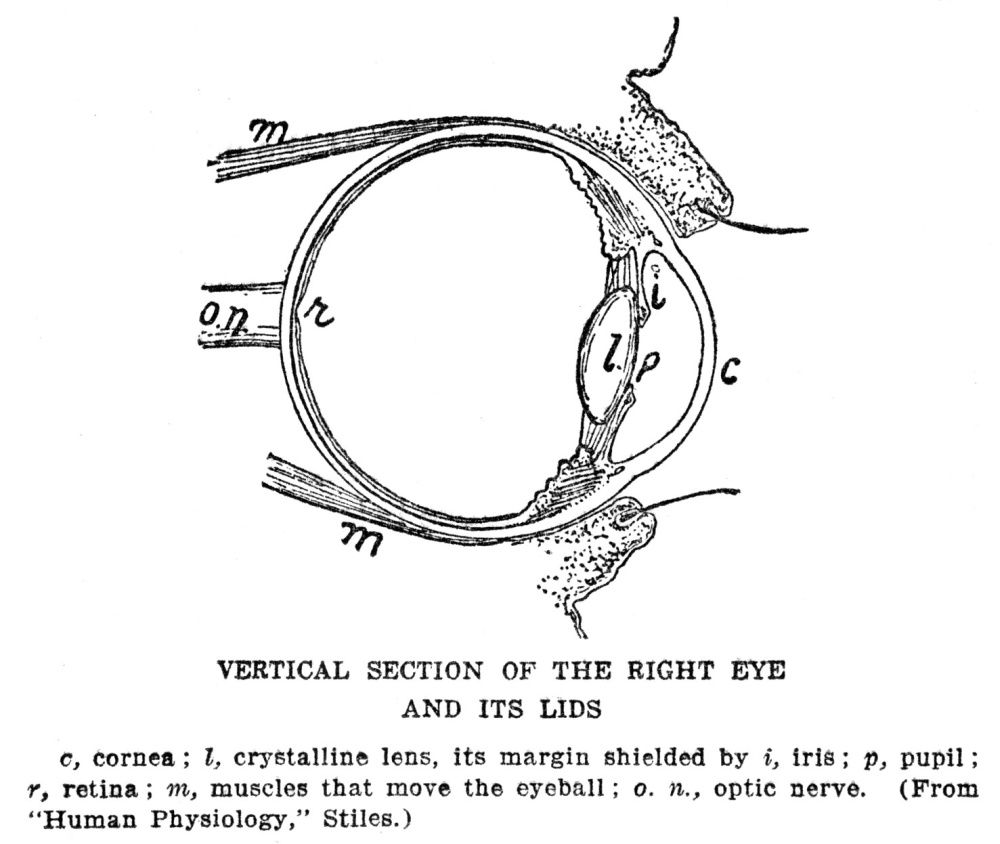
longer than the length of the eyeball, and the crystalline lens by itself also has a long focus, but the two in combination give a focus that just corresponds with the length of the eyeball, so that the images of all objects at a distance of eighteen feet or more fall sharply on the retina. Since near objects focus farther away from the lens than far objects, the effect of this is to make objects nearer than eighteen feet out of focus. For them a longer eyeball would be needed, and it would have to become longer and longer the nearer the object was brought to the eye. We all know that when we look at near objects we make an adjustment in the eyes. This is known as accommodation; for a long time it was supposed that accommodation was actually secured by lengthening or shortening the eyeball to bring the focus right, but we now know that the eyeball does not change in shape when we accommodate. The same result is secured by another means, namely, by letting the crystalline lens bulge out and become thicker. It was stated incidentally a few pages back that thick lenses have shorter focuses than thin. So when the crystalline lens thickens it shortens the focus of the eye, and throws the image forward. This will locate the image of near objects on the retina, instead of behind it, as in the unaccommodated eye. The crystalline lens is not stiff like glass, but rather like a thick jelly; it is in a transparent bag, or capsule, which is fastened to the inside of the eyeball all around, and the pull on the capsule stretches it out pretty flat, making the lens thin. There are tiny muscles inside the eyeball, known as the ciliary muscles. These are so fastened that when they contract they pull the eyeball forward and inward, loosening the tension on the capsule of the lens. This then bulges out, taking up all the slack. When we learn, as babies, to accommodate for near objects we find out just how much the capsule must be loosened to give the proper adjustment for any distance. As people get along in years the crystalline lens often loses its elasticity so that it does not bulge when the capsule is loosened. After this happens vision for near objects is no longer clear. Relief is obtained by wearing reading glasses. These are merely glass lenses worn in front of the eyes selected so that their focus fits in with that of the lenses of the eye itself to bring the image of objects at reading distance sharply on the retina. It is usually necessary to replace these glasses from time to time as the crystalline lens becomes stiffer and stiffer and ordinary accommodation fails more and more.
There is a disease known as cataract in which the crystalline lenses become cloudy and finally completely opaque. Of course this means blindness, since the passage of light to the retina is interfered with. Relief is obtained by the simple expedient of removing the opaque lenses bodily. This is possible merely because the crystalline lens is not the chief lens of the eye. To be sure the cornea by itself will not focus on the retina, but a glass lens can be placed in front of it which will add itself to the cornea and the combined lenses will. There is no possibility of accommodation in a case like this, so the patient has to be furnished with bifocal lenses; the main part gives clear distance vision; the lower section gives clear vision at the reading distance. The patient has to get along with blurred vision in the regions between.
Not all eyes are exactly the right size so that distant objects shall focus sharply on the retina. In fact a large proportion of them are either too long or too short. It is clear that in an eyeball that is too long the image of distant objects will fall in front of the retina, but near objects that happen to be at just the right distance will focus exactly on the retina. The distance at which this happens depends, of course, on how much too long the eyeball is. Persons that have unduly long eyeballs are, therefore, nearsighted. The condition is called myopia. The correction for it consists in the use of lenses in front of the eyes that instead of shortening the focus shall lengthen it. Concave lenses, namely, those that are thick at the edge and thin in the middle, will do this, and these are the kind that are worn by near-sighted people.
When the eyeball is too short the image of distant objects falls behind the retina, and of course that of near objects tends to fall farther back yet. Since by
accommodation the focus can be thrown forward, most persons with short eyeballs can see distant objects clearly by accommodating for them, and near objects that are not too near by extreme accommodation. For this reason hyperopia, as this condition is called, is usually not discovered until the person begins to feel the strain of the constant accommodation that is necessary whenever the eyes are open. Eyestrain usually shows itself in headaches; in fact, so large a proportion of headaches come from this cause that anyone who suffers from them at all frequently should have his eyes examined by a competent oculist. Relief for hyperopia is by means of glasses that shorten the focus and thus bring the image of distant objects forward to where the retina is in the short eyeball.
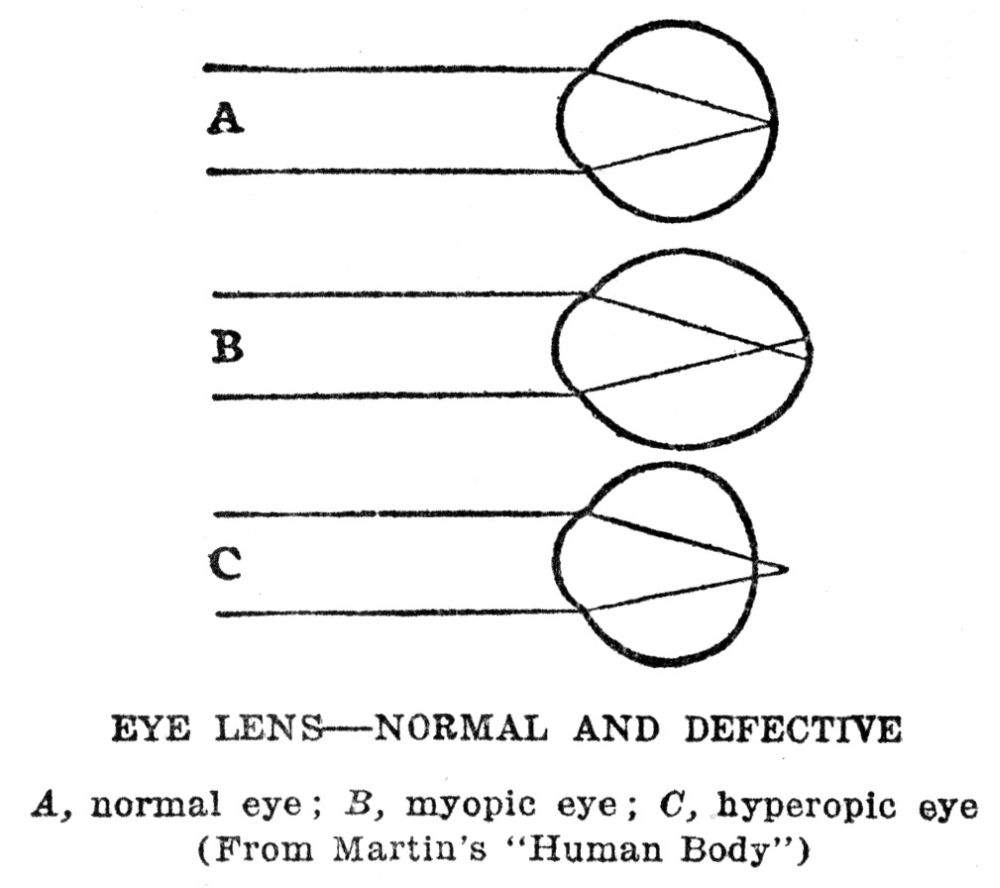
There is one other defect of vision that is so common as to call for a word; this is astigmatism. It is the condition in which the cornea is not curved equally in all directions; the vertical curvature may be greater or less than the horizontal. The effect is that points on objects do not focus sharply as points in the image, but as little elliptical spots. If one adjusts the accommodation so that the top and bottom edges of objects are sharp the sides will be blurred and vice versa. Usually the blurring is not great enough to be noticeable, but only enough to make the person unconsciously dissatisfied with the accommodation. He, therefore, constantly tries to improve it by changing the tension of his ciliary muscles, and so brings on eyestrain. The correction for this condition is glasses that are not equally curved in all directions, but so selected that their less curvatures shall fit in with the greater curvatures of the cornea. In fact, glasses that are curved only in one direction are usually used, this curvature being just enough to bring up the total curvature in that direction to equal the other curvatures of the cornea. It ought not be necessary to give warning that only competent persons should be allowed to examine and prescribe for the eyes. Great skill is needed to determine accurately just how far from correct the eyeball is, and unless this is known there is no means except guesswork by which to decide on a prescription.
The third feature of vision is the perception of color. Color is to light what pitch is to sound; that is, it depends on the vibration rate of the light waves. Light, as already explained, is one of the forms in which energy reaches the earth from the sun. Heat is another form. Both are portions of a great energy stream to which we give the name of radiant energy. This, as it comes from the sun, is made up of a mixture of vibrations having almost every imaginable rate, except that the slowest are many times faster than the highest pitched sound. At a certain rate, and one that for these vibrations ranks as slow, the energy is what we know as heat, and over a considerable range it continues to be called heat; more rapid vibrations, and they are so rapid that they have to be expressed in trillions per second, cause the effect on our retinas that we call light. The slowest that we can see give the sensation of red; the most rapid the sensation of violet; the other colors of the rainbow, which in order after red are orange, yellow, green and blue, are vibration rates between those that give red and those that give violet. It will be noticed that only six colors are given for the rainbow instead of seven. This is because there is not enough real difference between blue and indigo to justify making them separate colors. The distinction was made at the time when it was supposed that there was something specially wonderful about the number seven, which made it necessary that every important feature in nature should show that number. We now know of no reason why seven should have virtue over any other number. When all the vibration rates are mixed together, as they are in the sunlight, the sensation is white. There are other mixtures of colors that give white also, but they are not exactly equivalent to the white of sunlight, as is proven when one tries to match colors under artificial light. A great deal of labor has been devoted to the attempt to get an artificial light that shall be practically equivalent to sunlight, and only lately have good results been obtained. When no light enters the eye the sensation is of black, and it is worth while to note that so far as our sensations are concerned black is as much a color as white or any other, notwithstanding the fact that no light falls on the retina when the black sensation is being felt. Contrary to ordinary belief, blind persons who are blind because their eyes have been destroyed do not see black all the time; they simply have no sensations at all from the eyes. On the other hand, persons blind because of cataract do see black, because in them the retinas are still present, but no light falls on them.
The perception of color is very complicated, and not at all well understood. Persons who do not have the same color perception as most of us are called color-blind, and by learning some things about color-blindness we shall best get an idea of color perception itself. About four men in one hundred have defective color sense; the proportion in women is only about a tenth as great. By far the commonest type is one in which neither red nor green is seen correctly, but both are seen as neutral tints, and in many cases look so much alike that the person cannot tell one from the other. The practical importance of knowing whether or not this defect is present is seen when we think that red and green lights are used more than any other colors in signaling, so that railroad men and others who work by signals must have normal color sense. It has been discovered that even people that have normal color vision are color-blind in the margins of the retina. This can easily be demonstrated by bringing a red or green disk slowly around in front of the eye of a person who, meanwhile, keeps looking straight ahead. He will see the disk out of the corner of his eye some time before he can tell what color it is. In fact, if a red or green disk is used, he usually will not be certain as to the color until it is almost straight in front. Blue or yellow disks can be told with certainty much farther out, but even these colors are not perceived clear to the edge of the field of vision. These experiments show that the retina becomes more and more highly developed as an organ for perceiving color as we get closer and closer to the center; at the extreme edges there is no color sense at all, but only the primitive ability to tell light from darkness; closer in blue and yellow are distinguished and not red and green; only in the central part are all colors clearly seen.
In addition to the kinds of information which have been described thus far, that the distance senses bring in, there is another kind, fully as important as any in our actual use of our senses; that is information as to the “direction from us” of the object or objects which are arousing the sense. We can get this through all the distance senses, but much more perfectly in the case of sight than in the others. We locate the direction of objects that we smell by turning the head this way and that, sniffing meanwhile, and noting the position in which the odor is caught most clearly. Animals with a keen sense of smell, like dogs, can locate directions very accurately by this means. In the case of hearing the method is to turn the head until the sound is equally loud in both ears. We would expect that a person who was hard of hearing in one ear would never be able to locate sounds by this method; but, as a matter of fact, such persons unconsciously allow for the difference in hearing in the two ears, and so can judge the direction of sounds about as well as any of us. Animals, like horses or rabbits, that have very movable outer ears, undoubtedly can locate sounds much more accurately than we can. Our outer ears are of almost no use in hearing; persons who have had the misfortune to lose them hear practically as well as anyone.
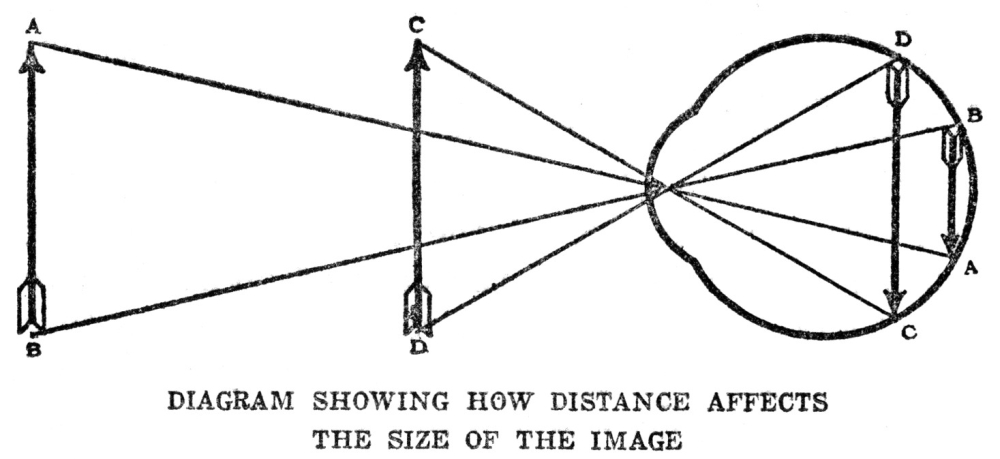
We locate directions with the sense of sight with perfect accuracy, because unless the image of the object we are looking at falls on the center of the retina it is not seen clearly. The only way to make the image fall just there is to look directly at the object. The muscle sense in the eye muscles is extremely delicate, so that if the eyes are rolled at all in looking at anything we know it and can judge, also, how much they are turned from the straight position. In this way we are able to tell exactly the direction from us of any object we can see.
We can judge the distance of a near object very accurately by noting the degree to which the two eyes have to be turned in in order to see it clearly with both. We are quite unconscious of this means of making the judgment; all we know is that we can tell. It is easy to prove that it depends on the two eyes by closing one and trying to make movements that depend on accurate knowledge of distance. A good example is threading a needle sideways. With both eyes open this can be done fairly easily, but with one shut it cannot be done at all, except by chance. Objects so far away that the eyes are not turned in perceptibly in looking at them are judged as to distance wholly on the basis of their size. It is clear that the actual size of any image on the retina will depend in part on how large the object is, and in part on how far away it is. If an object that we know to be large casts a small image on the retina, we conclude that it is far away. It follows that unless there are some familiar objects in view, judgments of distance are not at all trustworthy. A good illustration is in looking up a bare hillside, and trying to estimate the distance to the top. If, while this estimation is being made, a man or horse suddenly comes into view at the top, the man or horse will nearly always appear unexpectedly large, showing that the top of the hill is not actually as far away as it was judged to be.
The possession of two eyes instead of one is an advantage to us in another way, in addition to helping in the estimation of near distances. This is in making objects appear solid, or in other words, in helping the estimation of depth. When we look about us we have no difficulty in realizing that some objects are near and others far, and that the objects themselves have some parts that are nearer to us than other parts. A great many things assist us in this realization. First and foremost comes that which is known in art as perspective, namely the tendency of distant objects or distant parts of objects to appear smaller than those that are near. This can best be illustrated in the case of parallel lines extending away from the eye, as when one stands on a straight railroad track and looks along it. Although we know perfectly well that the rails are the same distance apart all along, if we were to believe our eyesight implicitly we should think that they came gradually together. It is on account of this matter of perspective that drawings of solid objects must show supposedly parallel sides nearer together at the far end than at the near. Besides perspective there are the shadows to be taken into account. Only solid objects cast shadows, so if we see a shadow apparently cast by any object we naturally conclude that it is solid. Both perspective and shadows can be and are used by artists in making drawings and paintings look real. In fact, they have almost no other means of doing this. As we all know, even the cleverest paintings do not give an impression of depth equal to that which comes from actually looking at solid objects. This is because of help we get from the two eyes in the latter case. The reason for the difference is that when we look at a solid object with both eyes the view we get with one eye is not exactly the same as with the other; we see a little farther around on the left side with the left eye, and on the right side with the right eye. The combined view with the two eyes gives us an impression of solidity that cannot possibly be had when the view with the two eyes is exactly the same, as when we look at a picture. The only way in which the impression of depth can really be gotten in a picture is by using the familiar method of the stereoscope, where two pictures are taken simultaneously by two cameras, placed a little farther apart than the two eyes; and then the two are looked at together through a special pair of prisms.
By means of the three distance senses, smell, hearing, and sight, we are informed pretty completely as to what is around us. All three give an idea as to the direction from us of objects; although sight does this better than either of the others. Sight, also, lets us know accurately as to the distance away of objects, provided they are fairly near. Smell and hearing, as well as sight, may give us some idea as to the distance of far objects, but only when we are dealing with familiar sensations. A very faint smell, or faint sound, means a distant object provided we know enough about the source to know that if the object were near the sensations would be keener. Our judgment of distant objects by sight is better than this, but not by any means perfect. When we contrast the distance senses with the contact senses we see at once that the great advantage coming from the possession of distance senses is in the time that is permitted for action. In the chapter on contact senses we emphasized the fact that the response to them must be immediate; there is no time to pick and choose. When we can learn of the presence of objects before they reach us, and something of their direction and distance as well, we can usually take time to select the most fitting course of action. We do not have to jump into a mud puddle to escape an automobile if we see it soon enough. We shall learn in a later chapter how this opportunity of choice is wrapped up with our development into highly intelligent animals.
IN the second chapter we saw that to make our muscles act in accordance with the information brought in by the sense organs some means of communication between them is necessary; we saw, also, that this means consists of the nervous system. Now that we have learned something about both muscles and sense organs we are ready to look into the way in which communication between them is carried on. First of all, we must realize that living protoplasm does this. The nerve cells are alive and have their basic metabolism just as do all other living cells. They also have their functional metabolism; but in them, instead of taking the form of forcible motion, as in muscles, or of the manufacture of special materials, as in gland cells, it takes the form of the transmission of a disturbance from one part of the cell to another. An interesting and important fact about this transmission of disturbances is that the actual amount of functional metabolism required by it is very small. Only by the most careful measurements has it been shown that nerve cells that are functioning have a greater metabolism than those that are at rest. For a long time it was thought that a nerve cell acted very much like a telephone or a telegraph wire, transmitting some kind of a disturbance which was set up in it, but not having any active part itself in the process. We now know that the special activity of nerve cells is a form of functional metabolism, just as is the special activity of muscle cells or gland cells.
The nerve cells have to make communication between sense organs and muscles, and, as we have already seen, these are often quite a distance apart. It is necessary, therefore, that the nerve cells be long enough to reach over these distances. As a matter of fact, it is not necessary for single cells to have this great length, because it is possible for them to be arranged end to end, making a path of living protoplasm consisting not of one cell but of a chain of them. If we look at a nerve cell under the microscope we see that it is made up of a little central mass of protoplasm to which has been given the name of “cell body.” From this cell body extends a tiny thread of living protoplasm. This thread is called the axon. It is so very slender that it cannot be seen except under a powerful microscope, and yet in our own bodies and the bodies of all large animals many of these are three feet or more in length without a break. This tiny protoplasmic thread, the axon, was formed originally by growing out from the cell body. As it grew it became surrounded by a sheath, which probably gives it strength and decreases the danger of its being broken. Another thing which helps to keep the axons from being injured is that they are always in bundles. Instead of one of these very slender axons lying all by itself, it will be bound up with several hundred others; the arrangement is similar to that in a telephone cable, where a great many single wires are bound together in the large and very strong cable. The living protoplasm of the nerve cell has a gray color, so that wherever this shows we have what is commonly called gray matter. We saw a moment ago that every axon is inclosed in a sheath. Some of these sheaths are transparent, so that the gray color can be seen underneath, but most of them have a layer of white material, which makes them look white instead of gray. The bundles of axons corresponding to the telephone cables make up what we call the nerves. Nearly all nerves are white in color because of the white material in the sheaths.
The sense organs, as we have seen in Chapters VIII and IX, are some of them inside the body, others spread over the surface of the skin, and the rest in the special sense organs, like the eyes or the ears. A very complex organ, like the eye or the ear, has thousands of axons leading from it. In the case of the eye these are grouped into a large nerve leading away from it at the back, which is called the optic nerve. There is a similar large nerve leading from the ear. When any sense organ is acted upon, as when light falls in the eye or sound on the ear, it starts a disturbance in some or all of the axons leading away from it.
As we have said over and over, the purpose of the nervous system is to arouse the muscles to activity, and to guide them in that activity. We must ask next, then, how the nerves are distributed to the muscles. If we dissect the body of any animal or bird, we can find nerves passing to various parts. For example, a large nerve goes down each leg; this nerve subdivides here and there. Since we know that the nerve consists of a great many axons bundled together, we will realize that this subdivision is not a real branching, but simply a passing of some of the axons away from the main trunk along the smaller stem. Some of these smaller stems can be traced to endings in the skin; these contain the axons connecting with sense organs. Others lead directly into muscles. Some of these axons may also connect with sense organs, since, as we have already seen, every muscle has embedded in it the organs of muscle sense, but in addition any nerve that leads to a muscle contains a great many axons which pass directly to the muscle fibers. These are the axons by which the muscles are aroused to activity. It is a general rule of the nervous system that no nerve cell extends without a break from any sense organ to any muscle fiber. The axon which communicates with the sense organ belongs to one nerve cell; the axon which connects with the muscle fiber belongs to a different nerve cell. The first is called a sensory nerve cell, the second a motor nerve cell. Some idea of the appearance of these cells can be gotten from the figures on page 126.
It will be seen that the cell body of the sensory cell appears to be off on a little side branch. As a matter of fact, the branch is double, so that when a nervous disturbance passing along from the sense organ comes to the beginning of this branch it can pass up to the cell body and then out from the cell body along the second part of the branch, and so along the other part of the axon. This part of the axon is seen in the figure to have several branches; these are really branches of protoplasm and not separate axons coming off, as in the case of the nerve trunk. The use of these branches we shall see in a moment. Also at the tips of each branch there is a tiny feathering. We shall explain this presently. Let us look first at the figure of the motor nerve cell. This has a cell body and long axon, and, besides these, has a great many short protoplasmic branches sticking out in all directions from the cell body. Since a nervous disturbance to get from a sense organ to a muscle has to pass over a sensory nerve cell, and also over a motor nerve cell, evidently there will have to be some point at which it leaves the sensory cell and gets into the motor. This is accomplished by having the tiny feathering at the tip of the sensory cell interwoven with the fine processes projecting from the body of the motor cell. This arrangement we may call a nerve junction. In the whole body there are, of course, millions of these nerve junctions.
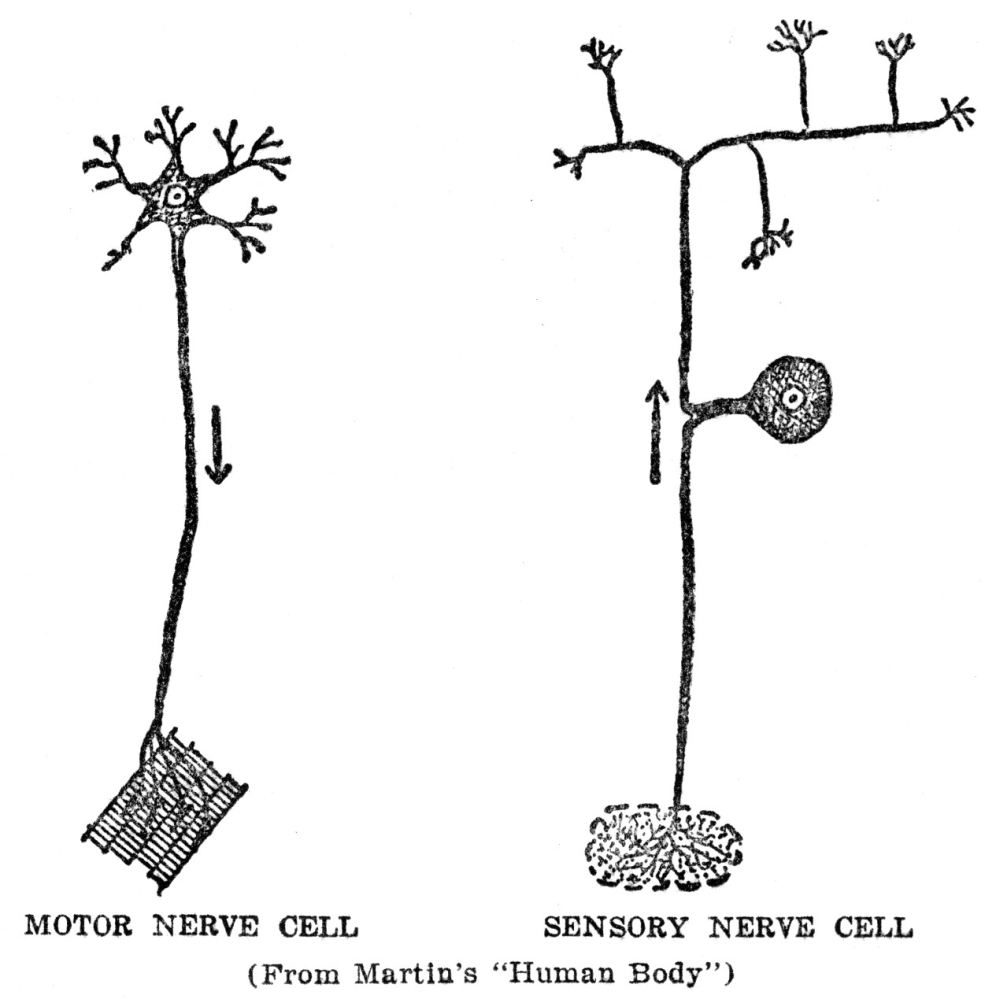
We have just described the simplest arrangement of a nerve path from a sense organ to a muscle; it consists of the sensory nerve cell, a nerve junction, and a motor nerve cell. This arrangement will answer where the sense organ and the muscle are in the same part of the body, but it may happen that the sense organ is in one part of the body and the muscle is in a distant part, as, for example, the eye and the muscles of the hand. To make these distant connections there must be additional nerve cells, and these we find in the body in the form of a kind of nerve cell that serves as a connecting link between sensory cells and motor cells. It may be called simply a connecting nerve cell. In appearance it is like the motor nerve cell, except that it has many branches which do not terminate in muscle fibers, but in fine feathering like that at the tips of the sensory nerve cell. When these connecting cells are present in the chain, the arrangement is as follows: from the sense organ the sensory nerve cell will pass just as previously described, the feathering at the tip will form a nerve junction with a connecting cell instead of with a motor cell. This connecting cell also has feathering at the tips of its branches, and these featherings will form a nerve junction with another nerve cell. This may be a motor cell, in which case the pathway is completed, or it may be another connecting cell, which in turn may lead into motor cells or connecting cells.
The nervous system is made up, then, of chains of nerve cells. Now why is this arrangement present? In very many cases sense organs and muscles are side by side or within a short distance of each other; why does not a single nerve cell reach directly from the sense organ to the muscle? The
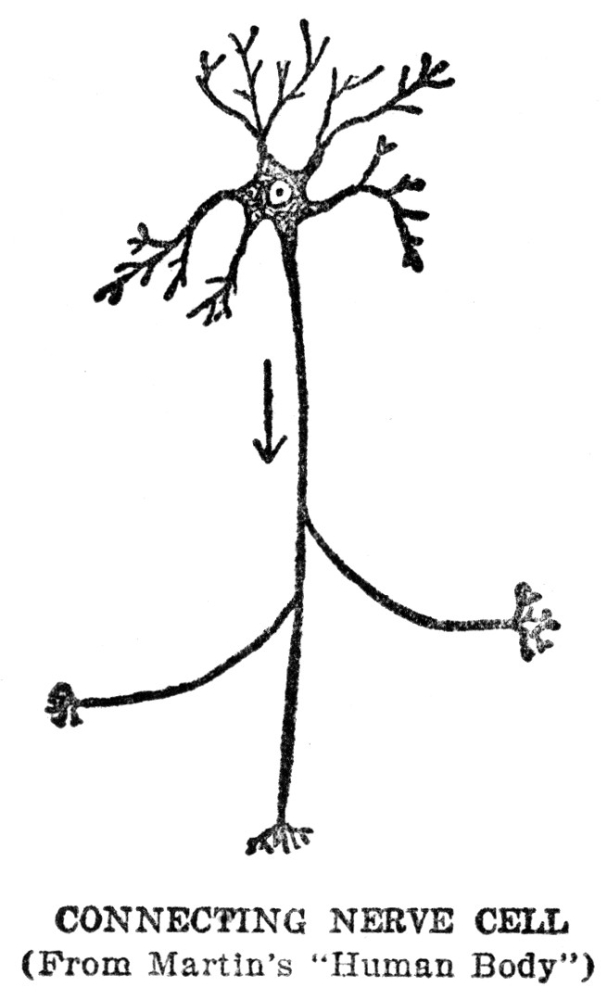
answer is simple, if we think for a moment of how the body works. The information that comes in by way of particular sense organs cannot always be used to arouse particular muscles to activity. Things that we see will sometimes cause us to move one hand, sometimes another hand, sometimes the legs, sometimes muscles of the head, and so on. This means that the eye is able to make connection with a very large number of different muscles. The same thing is true of the other sense organs. The body could not possibly work as it does, if certain sense organs connected with certain muscles and no others. As a matter of fact, it is not an exaggeration to say that the proper working of the body requires any sense organ to be able to make connection with any muscle. It might be possible to do this by giving every sense organ as many sensory nerve cells as there are muscles, but this would be as bunglesome as to attempt to provide every business house in a large city with a separate telephone wire to every other business house.
The arrangement of the nervous system is very much like that of a city telephone system. The sensory nerve cells all lead into a part of the nervous system to which is given the name of the Central Nervous System, just as the telephone wires all lead into a “central exchange.” From this central nervous system or “exchange” all the motor nerve cells extend to the muscles. There is one important difference between the arrangement of the nervous system and that of the telephone exchange; namely, that the nervous system is a “one-way” system. As we all know, a telephone instrument can be used either for sending or receiving and the same wires conduct the messages in both directions. This is not true of the nervous system. The messages from the sense organs pass into the center by way of the sensory nerves and out from the center by way of the motor nerves. The central nervous system operates as an “exchange,” connections can be made from any sensory cell to any of the motor cells.
Since this ability of the central nervous system to make connections here and there within itself is about the most important of all our nervous activities, as we shall see shortly, we must try to form an idea of how it is done. If we look again at the figures of the sensory and connecting nerve cells, we shall note that both kinds are branched; the tip of every branch makes a nerve junction. This means that every sensory cell, for example, has as many outlets as it has branches. If every one of these outlets were to communicate directly with a motor nerve cell, the number of connections that
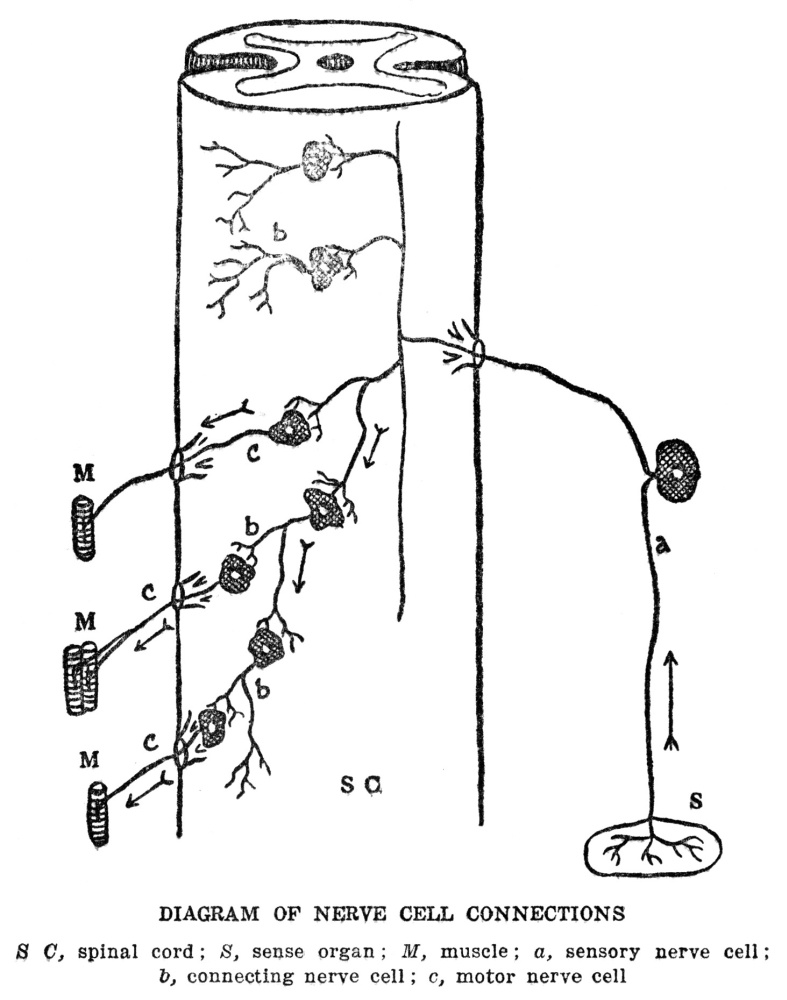
this sensory cell could make would depend on the number of its branches; but, as a matter of fact, most of the branches from the sensory cells make nerve junctions with connecting cells, and not with motor cells directly. These connecting cells in turn are branched and many of these branches lead again to connecting cells, so we see that the number of connections that a single sensory cell can make quickly becomes very large. The arrangement is shown in the accompanying diagram. Not only does one sensory cell have in this way the possibility of nerve connection with all the muscles, but the reverse is also true; namely, that every muscle has the possibility of being connected with every sense organ. This means that there must be a number of nerve junctions connecting with the cell body of each of the motor nerve cells. If we look back at the diagram of the motor nerve cell, we shall see that it has a great many tiny branches leading off it. These are numerous enough to enable the feathery tips of a great many sensory or connecting nerve cells to interweave with them, and so enable any motor cell to be acted upon from a great many different directions.
The central nervous system, which is the place where all these much-branched pathways are and where all the nerve junctions are located, is made up of two chief parts, the brain and the spinal cord. The brain is inside the skull and the spinal cord is an extension down the back. It lies in the tunnel made up of the arches of the bones of the vertebral column as described in Chapter VI. The cables which contain the axons of both sensory and motor nerve cells extend from the brain or from the spinal cord out to the different parts of the body where the sense organs and the muscles are located. They start as large nerve trunks which divide and subdivide as they get farther and farther away from the central nervous system. The large nerve trunks are arranged in pairs—those that spring from the brain are called cranial nerves; those that spring from the spinal cord are called spinal nerves. The cranial nerves, with the exception of one, lead only to points in the head or neck, and so are short; the spinal nerves on the other hand reach to the various parts of the trunk or down the arms and legs, so that some of these may be three feet long or more. An idea of the arrangement of the nervous system is given in the accompanying figure.
A good way to realize the actual working of the nervous system is to take a particular action and follow it through. Suppose a barefooted boy steps on a sharp thorn. The thorn arouses some of the sense organs in the sole of the foot. These in turn start a disturbance in the sensory nerve cells which pass up the leg to the lower end of the spinal cord in the small of the back. Within the spinal cord the sensory cells branch and the disturbance set up by the prick of the thorn spreads all over these branches to their tips. Some of the nerve junctions thus affected lead to muscles which will cause the foot to be jerked up; others communicate with connecting nerve cells which extend all the way up the spinal cord and into the brain and make the boy aware of the fact that he has stepped on the thorn; still others may make connection with muscles which would cause him to sit down and look at the bottom of his foot; still others may lead to the vocal muscles and the tear glands, causing the boy to cry. Of course these are not the only possible nerve connections; every muscle in the body might theoretically be aroused to action as the result of the stepping on the thorn. As a matter of fact, there is a condition in which this will happen. The drug strychnine has such an effect upon the nervous system that the stimulation of any sense organ actually does arouse all
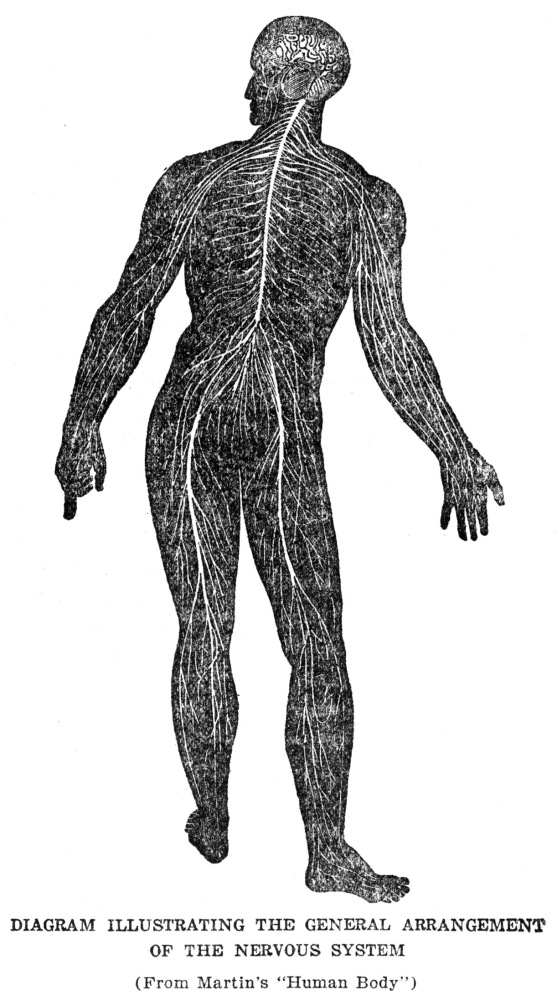
the muscles in the body, giving what is called a convulsion. There are some other poisons which may act similarly. Convulsions are not at all uncommon in young children. The way in which a convulsion is produced is by the spreading of the disturbance all over from a single sense organ. What we have, then, in the nervous system is an arrangement whereby under special conditions a nervous disturbance can pass from any sense organ to any or all of the muscles, but under ordinary conditions the disturbance spreads to a particular muscle or group of muscles only. It is evident that the nervous system would be of no use at all, if this latter arrangement did not exist. In order for our muscles to serve us, they must act in obedience to information brought in by the sense organs, and this can happen only when certain groups of muscles work in accordance with the information brought in by certain sense organs or groups of sense organs. We explain the behavior of the central nervous system by saying that there are preferred pathways through it, or, to put it in a slightly different way, when a nervous disturbance spreads over any nerve cell it extends equally over all parts of it, but does not pass with equal ease over all the nerve junctions. Some of the nerve junctions allow the disturbance to pass more readily than others, and it is this difference in the ease of passing the nerve junctions that determines which pathway the disturbance shall follow. We know no other means by which this picking out of particular paths from the huge number of possible paths could be accomplished.
What we are describing now is the simple foundation on which all our nervous activities rest. For that reason we are not taking up at this point the working of the brain, but only the direct connections between sense organs and muscles. In some very low animals the whole nervous system is made up of such simple connections.
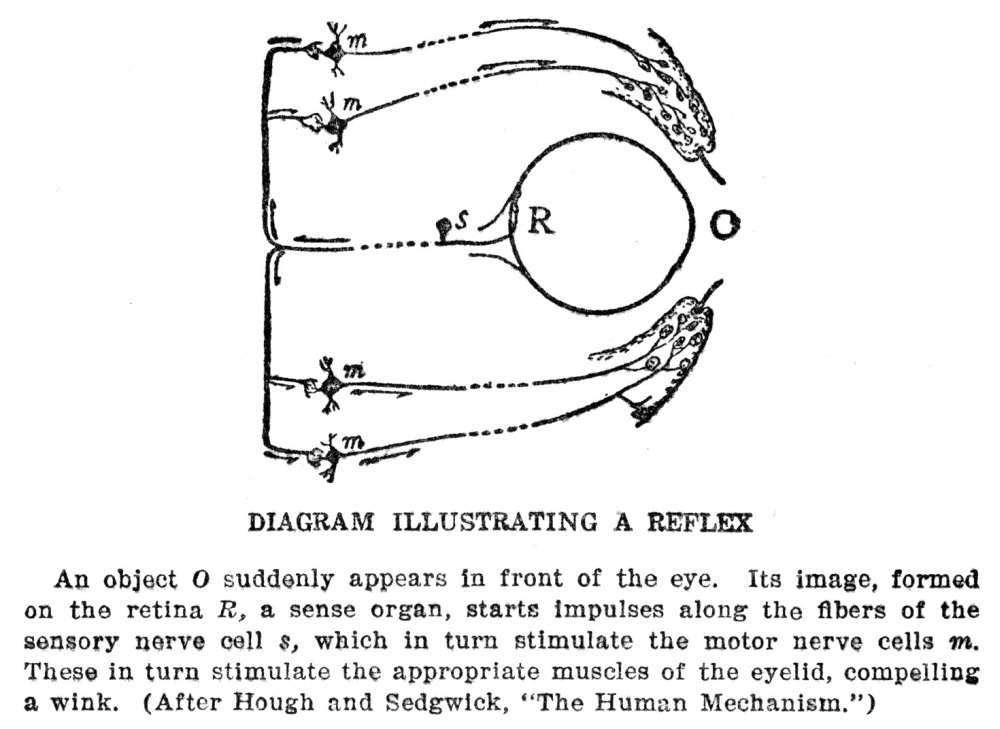
An object O suddenly appears in front of the eye. Its image, formed on the retina R, a sense organ, starts impulses along the fibers of the sensory nerve cell s, which in turn stimulate the motor nerve cells m. These in turn stimulate the appropriate muscles of the eyelid, compelling a wink. (After Hough and Sedgwick, “The Human Mechanism.”)
A nervous activity which consists of the passage of a disturbance from a sense organ to a muscle is called a reflex. We have many examples in ourselves; if we inadvertently touch something hot, the hand is jerked away. Tickling the soles of the feet in one who is asleep will cause them to be drawn up; irritation in the throat causes us to cough, or in the nose to sneeze; the flashing of a bright light into the eye compels us to wink; all these are examples of the direct passage of nervous disturbances from sense organs to muscles. In every case the path is from the sense organ over the sensory nerve cell to some point in the central nervous system, then either by a direct nerve junction to a motor nerve cell or over one or more connecting nerve cells to a motor nerve cell and so to the appropriate muscles. These reflex actions follow the arousing of the sense organ with no more delay than is required for the passage of the disturbance over the nervous pathway. There is a delay of a small fraction of a second at every nerve junction, which makes some reflexes slower in their action than others. For example, there is a reflex known as the knee jerk; this is an outward kick which results from a sharp blow on the front of the leg just below the knee. The kick follows so closely after the blow that there cannot be more than one or at the most two nerve junctions in the pathway. The reflex of winking, on the other hand, takes several times longer, although the eye is much nearer the central nervous system than is the place on the leg which is struck in arousing the knee jerk. Since the actual length of nerve to be passed by the disturbance is much shorter in the winking than in the knee jerk, while the time for the reflex is a good deal longer, we conclude that the nerve pathway which is used in arousing winking contains a great many more nerve junctions, and therefore includes a great many more connecting cells than does the path for the knee jerk. In this particular case the reason why the pathway of winking contains so many connecting cells is that it is a brain pathway, while that for the knee jerk includes only the lower end of the spinal cord, where the arrangement of nerve cells is very much simpler.
It is important for us to get clearly in mind the working of the reflexes in order to be able to understand the more complex nervous actions which will be described in the next chapter. We need to remember that the ordinary way of starting nervous disturbances is from the sense organs. With a few exceptions, to be described later, whenever a nervous action occurs anywhere in our bodies it can be traced back, although often very indirectly, to the sense organ from which the disturbance originally came. The part played in this by the brain and by what we call our mental processes will be described in the next chapter.
Before going on to that topic we have a word to say about nervous fatigue. We mentioned the fact in Chapter VII that much of our actual feeling of fatigue is nervous rather than muscular. Not as much is known about nervous fatigue as about the fatigue that comes on when the muscles are overworked. One thing that seems pretty evident is that the place where the fatigue actually is located is in the very delicate nerve junctions. These junctions offer some resistance to the passing of nervous disturbances over them, and if they are compelled to submit too often to this passing they appear to offer still more resistance; in other words, to become fatigued. We must remember that the nerve junctions are exceedingly delicate things, consisting, as they do, of the interweaving of the almost inconceivably tiny featherings at the tips of sensory or connecting nerve cells with the equally tiny featherlike branches from the cell bodies of connecting or motor nerve cells. It is likely, also, that on account of their delicacy they are easily affected by the waste products that may be circulating about in the blood stream from the active muscles. In either case, the way to recover from nervous fatigue is simply by resting. It is not hard for the delicate nerve junctions to throw off fatigue if given a chance. The way to give them this chance is not to use them. As we shall see in the next chapter, mental processes are made up of nervous disturbances passing here and there in the brain. If we allow ourselves to be occupied too continuously with the same lines of thought, we are evidently sending nervous disturbances over the same nerve junctions over and over again. In order to give those nerve junctions a chance to rest, what we have to do is to think about something entirely different. The word that best expresses what we have in mind is “diversion.” In the strict sense diversion means a turning aside from what we have been doing to something different, and that is the best way to allow the brain to rest. The man who takes his business home with him, and dwells on it during the hours that are supposed to be set aside for rest, may be able to achieve more for the moment than if he were really to rest, although even that is doubtful; but in the long run there is no doubt that continuous efficiency depends on allowing the fatigued nerve junctions ample opportunity to recover, which means that the thoughts must be directed into entirely different channels.
WE have just seen that the underlying arrangement of the nervous system is one which makes communication possible between any sense organ in the body and any muscle; also that in the working of the nervous system there are certain paths from given sense organs to particular muscles which we may call the selected paths, and over which nervous disturbances easily pass. These paths work out in such a way that the muscles that are thrown into activity as the result of the arousing of any sense organ are those most likely to serve the needs of the animal. There are a number of respects in which a nervous system, having no more than we have thus far described, would fall short of meeting the requirements of any animal except perhaps the very simplest. The thing that is missing that is of most importance, perhaps, is the ability to perform actions in response to information received at some past time; in other words, the ability to profit by experience. As the nervous system has been described up to this point, only immediate effects upon the sense organs are transmitted to the muscles to throw them into activity. As a matter of fact, we know that we ourselves, as well as many of the higher animals, are able to profit by past experience. That is to say, we can make our muscles move in accordance with information received at some time in the past. There must be in the nervous system, then, some arrangement for doing this, and our present task is to describe it. In the chapter on the distance sense organs considerable emphasis was placed on the fact that the advantage of having distance sense organs is to allow time for a choice among the various responses that the animal might make to the information received by the sense organs. It is this opportunity for choice that lies at the foundation of the higher nervous activities that we are now beginning to describe. As soon as the element of choice enters, the nervous disturbance cannot pass from the sense organ to the muscle in the least possible time. In order for there to be a choice there must be a delay while the selection of the muscle to be aroused to activity is being made. We have, then, a new feature in connection with the operation of distance sense organs in that the nervous disturbance can come to a stop at some point in the course of its progress from sense organs to muscles. Every animal that has distance sense organs shows this feature of possible delay in the movement of the nervous disturbance over the nervous pathway, and in every one the particular part of the nervous system in which this occurs is known as the brain. In fact no other nerve cells except some of the nerve cells of the brain have this ability of stopping the nervous disturbance and holding it for a time before sending it on.
One thing about the information that comes in through the distance sense organs is that much of it does not require an immediate activity. For example, a hungry fox, seeing a fowl roosting in an accessible place, might pounce upon it at once, but if he happened at the moment to be either fully satisfied or to be carrying a fowl which he had captured a moment ago, there would be no occasion for him to seize this one; rather would it be desirable for him to set it aside in his mind to be captured in the future. In the case used in the illustration the nervous action goes on in the beginning just as we have been picturing it; that is, the sense organ is aroused and this in turn starts a disturbance over the nerve path; when it comes to the brain, however, it does not go on from there to cause immediate muscular activity, but instead is held in the brain and can cause activity at some future time, as for example if the fox, after taking the fowl which he is carrying home to his young, comes back in search of the one that he saw roosting. We are perfectly familiar with this ability to stop nervous disturbances; we know it under the name of memory, although we may not have thought of memory in exactly this sense. If we consider what memory really is, we shall see that it is just what we have been describing, namely the stopping of nervous disturbances that come in from the sense organs and holding them, so that they may in turn set up at some future time a nervous disturbance that shall produce activity. If we try to picture what actually goes on in the nerve cells where this memory is located, we can say only that when the nervous disturbance strikes upon these cells, it does not pass on through at once leaving them very much as they were before, but stops in them and brings about in them some kind of permanent change. An interesting thing about memory is its persistence. We may act upon memories that we have held for years. More than one person moving away from a particular town early in life, and going back to it in later years has remembered things he saw in childhood well enough to find his way to them again.
Since memory is registered in the brain cells as a permanent or fairly permanent change, it can become the source of nervous disturbance over and over again. This in fact is one of its greatest advantages, because when we have once learned a thing, we can make use of it a great many times, and do not have to have the sense organs freshly aroused every time it is desirable to use this particular bit of information.
In order that the brain cells may receive and store up memories, it is evidently necessary that nerve paths from the sense organs should lead into the brain; so, if we go back to the sensory nerve cells, we shall find that in nearly every one of them, if not in every one, one of the branches makes a nerve junction with a connecting cell, which either extends directly into the brain, or links with other connecting cells which do. This is true not only of the distance sense organs, but also of the contact sense organs and to some extent of those inside the body. The path by which the brain is reached from the sense organs that are located in the body is over the sensory nerve cells to the spinal cord, and along the spinal cord by way of connecting nerve cells to the brain. All the distance sense organs are located in the head, so their nerve paths lead into the brain directly over cranial nerves; for the eye, the optic nerve; for the ear, the auditory nerve; and for the organ of smell, the olfactory nerve. One very interesting fact about the connection of the sense organs with the brain is that the nerve paths in every case cross from the left-hand
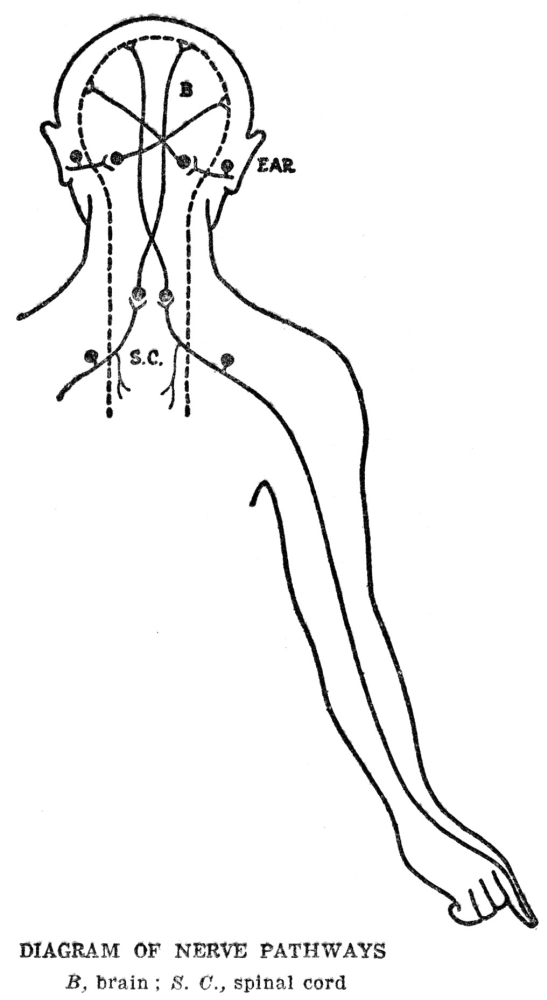
side to the right-hand side, and vice versa, so that all the sense organs in the left half of the body have their connection with the right half of the brain, and those in the right half of the body with the left half of the brain. We know of no reason why this should be so. It is merely an interesting fact. A diagram showing the path from a sense organ in the body and from one of the head senses to the brain is given in the accompanying figure.
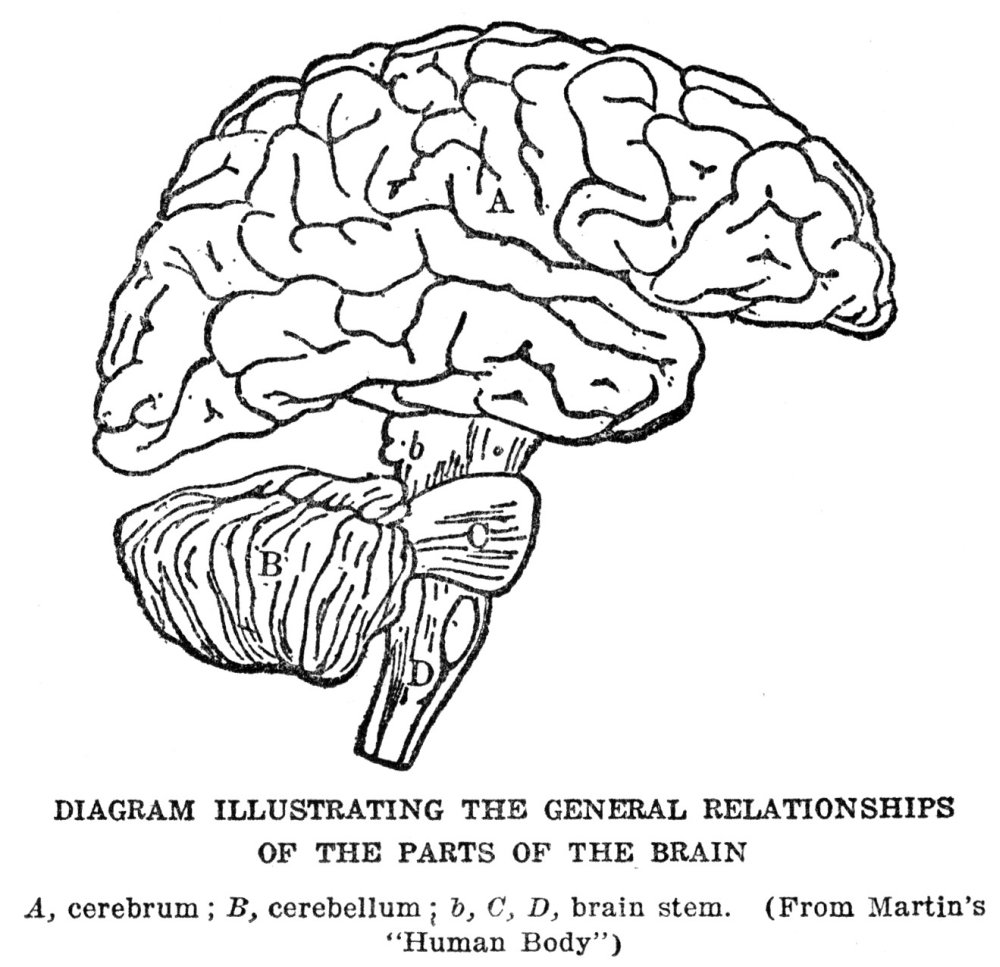
The brain in man and in all the higher animals is a very complicated organ made up of a number of different parts. To simplify the description as much as possible we shall omit all account of smaller subdivisions and speak only of the most important parts. These are three in number, named the cerebrum, the cerebellum, and the brain stem. Their location with reference to one another is shown herewith.
As the figure shows, the cerebrum is the main part of the brain. It in fact is the part we ordinarily think of, when we have the brain in mind. The brain stem is really the upper extension of the spinal cord within the head. It is a very important part of the brain, because all the nerve pathways in from the sense organs, and from the brain out to the muscles, pass through it. Besides that it has some activities of its own, which will be described a little later. The cerebellum is a subdivision of the brain which plays a very important part, but is not concerned in those complicated nervous actions which make up our mental processes. All these are conducted wholly in the cerebrum. In fact, this section of the brain is the only part which has the ability to stop the progress of nervous disturbances; the property of memory, which was described a moment ago, is found only in the cerebrum. Since the cerebrum is the seat of memory and of our mental life in general, it is both the most interesting and the most important part of the whole body, and a very large amount of study has been given to it. Many years ago attempts were made to show that the brain is subdivided into a number of parts, each of which has control over certain mental characteristics. Nothing very important came of these attempts, although they gave rise to the false science of phrenology, which has been widely exploited by fakers. Notwithstanding the failure of these early attempts at locating particular mental activities in particular parts of the brain, recent studies have shown that there is something of the sort, although it is not at all what the phrenology charts would indicate. It has been shown, however, that the various sense organs do make connections with particular parts of the cerebrum. Comparing the pathways over which the nervous disturbances pass to railroad tracks these places in the brain are often spoken of as the “arrival platforms” of the various senses. Thus the arrival platform for the sense of
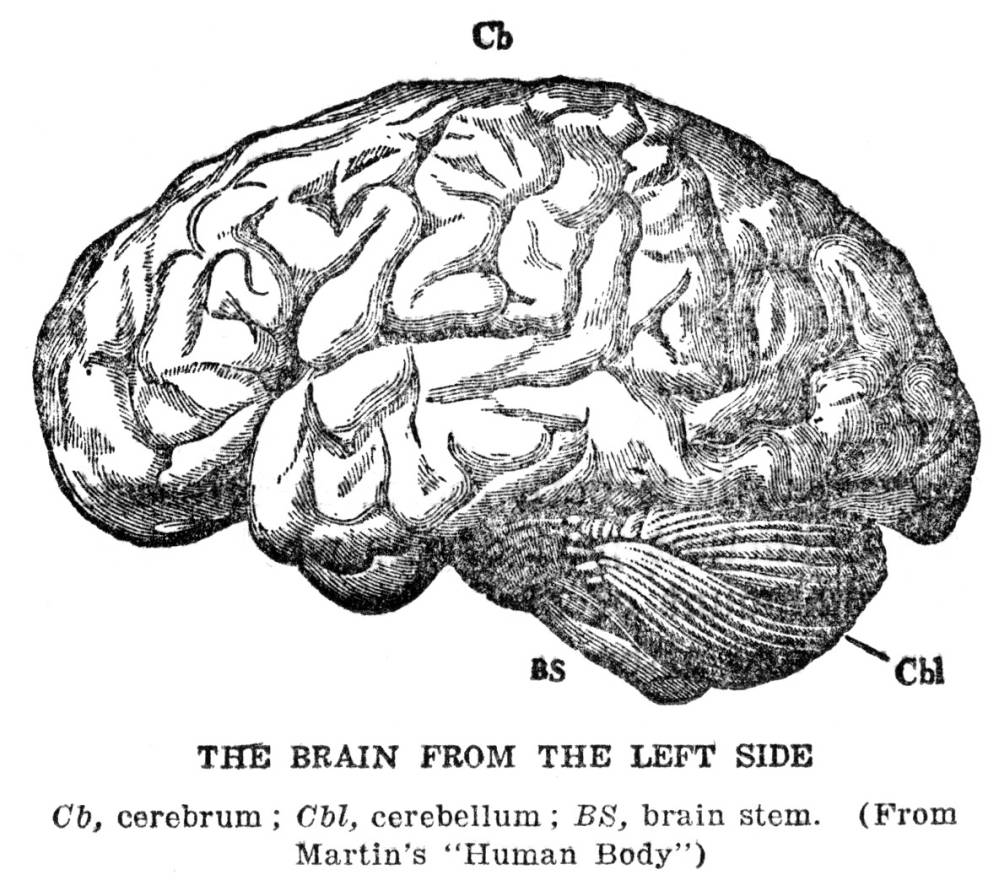
sight is at the back of the cerebrum down at its lower margin; the arrival platform for the senses of touch, pain, and the like are just about at the top of the brain. The arrival platform for the sense of hearing is down at the side. The location of these arrival platforms has been worked out by studying the effects of brain diseases. It has been found, for example, that persons who are blind, although there is nothing the matter with their eyes, are so because of a disease of the lower back part of the cerebrum, and persons who are deaf, even though their ears are perfect, are so because of diseases in the parts of the cerebrum at the sides. The only way to explain these findings is by supposing that the arrival platforms of these senses are located in the parts that are found diseased. The cells which stop the nervous disturbances coming in from the sense organs and so serve as the seat of memories of sensations are in the arrival platforms, so that when we recall how something looked, for example, the nerve cells which are active in this recollection are those at the lower back part of the cerebrum. All these arrival platforms taken together occupy only a very small fraction of the whole of the cerebrum, so that evidently it has more to do than simply to register these memories.
One additional thing we shall look for in the cerebrum is a pathway from the brain to the muscles whereby the memories that are stored in the brain can make themselves effective in arousing the muscles to activity. There is such a pathway, and it is one of the best marked of all the nerve pathways of the body. It is made up of nerve cells which start just about at the top of the cerebrum, a little in front of the middle and pass down into the brain stem; these cross over from the side in which they started to the opposite side; and go on down the spinal cord to make connection by nerve junctions directly with the motor nerve cells. Thus from the brain to the muscles the nerve pathway is made up of just two nerve cells. Since this pathway crosses over from one side to the other in the brain stem, the whole right half of the body, both sensory and motor, connects with the left half of the brain, and vice versa. The part of the cerebrum in which this nerve pathway starts is called the motor area of the brain.
We have now seen how nervous disturbances from the various sense organs can come into the brain and be registered there as memories. We have also seen that there is a nerve pathway by which nervous disturbances starting in the motor area can pass out to the muscles, and arouse them to activity. We have left to see how the connections are made between the incoming and the outgoing disturbances, or in
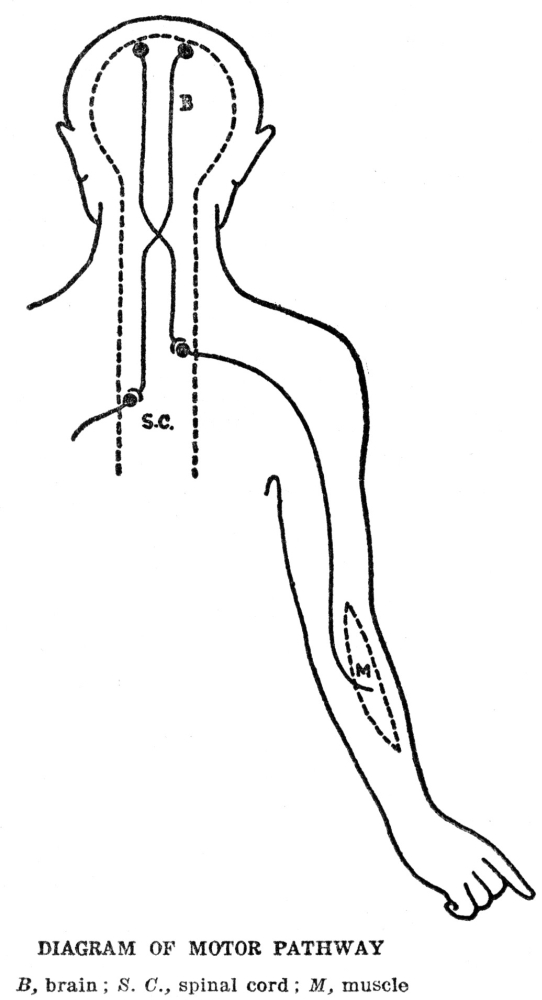
other words to see how the memories that are registered in the brain act upon the outgoing pathways. We shall expect to find connecting nerve cells reaching across from the various arrival platforms to the motor area, and such connecting cells exist, but they are not simple and direct connections for a reason which we shall now try to make clear. We all know from our own experience that our memories are never the pure registering of a single sense. What we mean is that the sight of something by itself or the sound of something by itself never remains as a separate memory, but is always worked in with some other memories from some of the other sense organs. This putting together of the simple sensory memories began in earliest childhood, long before we were old enough to think about our mental processes, and see how they are carried on, so that unless our attention has been called to it, we have probably failed to realize how complicated our simplest memories are. A good example of this is in the experience of a baby with its mother’s voice. So far as the baby is concerned, the voice is an influence affecting the organ of hearing, arousing nervous disturbances which pass to the arrival platform for hearing in the brain and are registered there as memories, but it does not take the infant long to learn that some other memories that have come to it by way of other sense organs belong with this particular memory and always go with it. For example there is the sight of the mother, or the feel of the mother’s face and hands, all these are influences affecting different sense organs and registering in the child’s brain in different arrival platforms. Yet within his brain they become fused into a single composite memory of the mother, and after this fusion has once occurred, the arousing of any part of the memory brings up the whole of it, so that the child may hear the mother’s voice from the next room, but the memory that will be aroused as the result will be not simply of the voice but of the mother as a whole. This is an illustration of how our pure sense memories are fused into complex memories. After one of these memory complexes is once started, we add to it any time any sense organ is acted upon by anything that has relationship to the complex. For example, the child’s idea of the mother at first is a very simple one made up of a few sensory impressions, but as time goes on and more and more sensory impressions of the mother are received, the child’s idea of her becomes more and more complex. This process of memory fusion is called association or sometimes association of ideas. Strictly it is an association of memories and this is the method by which all our mental activities are carried on. The thought process consists of putting together various memories in various ways and so building up associations of different kinds. Of course adults of wide experience can form associations which are made of literally hundreds or perhaps thousands of separate memories of sense impressions. We receive sense impressions at the rate of hundreds every day, and very many of these, perhaps all of them, are registered as memories and are fitted into their proper associations.
The nervous machinery for carrying on different processes of association consists of nerve cells of the brain. These are all of the kind known as connecting cells. It will be remembered that the connecting cells may have many nerve junctions leading into them and they in turn are much branched so that many nerve junctions can lead out from them. The connecting cells of the brain are richer in these respects than those in any other part of the body, and the cells of the human brain than the brain of any of the lower animals. We suppose that the associations are formed by the passage of nervous disturbances from the arrival platforms where the sense memories are registered over various connecting nerve cells to a common meeting point in some cell or in some group of cells, where the associated memories are all brought together into a single memory complex. It is supposed that the large areas of the brain which are not taken up either by the arrival platforms or by the motor areas are the regions in which these associations take place.
There is one particular group of associations that are so interesting as to call for special mention. These are the associations concerned with language. We are so in the habit of using language that we are likely never to have thought of it in its real meaning as a part of our mental activity. We know that we have two kinds of language, spoken and written, and that spoken language consists of certain sounds, and written language of certain visual symbols. Two of the distance senses then are concerned, sight for written language, hearing for spoken, and language itself consists simply of sense impressions coming in through one or the other of these sensory channels. The important feature of language is that mankind has selected arbitrarily certain sounds or certain written symbols to stand for particular things. When a child is learning to understand what is said, what he is really doing is fitting a particular set of arbitrary sense impressions into their proper places in his associations. We can illustrate this by the same example that was used a moment ago. The child becomes thoroughly familiar with its mother, so far as sense impressions are concerned that come directly from her. In course of time it adds to the associations thus formed an additional one made up of the sound of the word “mother.” Of course there is no particular reason why this sound should have that meaning rather than any other. The proof is that different languages have different sounds which stand for the same thing. After this sound has once been selected and learned, it becomes as much a part of the idea of mother as any other of the associations concerned, and thereafter, whenever that particular sound strikes upon the child’s ear, the association of mother is aroused. Precisely the same sort of thing is true of written language. Arbitrarily selected symbols act through the sense of sight to arouse nervous disturbances, which are built into particular associations, and here again it makes no particular difference what the symbol is; one will do as well as another, provided a number of people have agreed to use that symbol to stand for the same thing. One very useful feature of language is that we can make a single word stand for a complex group of sensory impressions or even associations. To illustrate, the word physiology, whether spoken or written, is in itself simply a sound or a visible symbol, but it stands for a highly complex group of associations in the human mind. In this respect language is a kind of shorthand.
It is hard to overestimate the importance of language to the human race. This is because of the enormous extension it gives to the ability to profit by experience. The lower animals are able to profit by experience, but the only experience that they can get is that which comes to them individually or to another animal which they may observe. Human beings, on the other hand, may profit by the experience of their contemporaries through learning of them by word of mouth, or may profit by the experience of present or past generations as recorded in writing. It is not too much to say that the progress of civilization depends on our ability to profit by the experience of past generations as made known to us in writing. We are further advanced than our remote ancestors, not because we are actually superior to them in mental power, but because we have a much larger background of experience than they had, and this background depends altogether on our ability to use language.
The complex nervous activities that have been described up to this point are purely a matter of the association of memories, or what we speak of as intellectual processes. Besides these our mental life includes what we call the feelings or emotions, and these make up so large a part of our mental life, that we cannot leave the consideration of the brain without saying a word about them. For convenience we may subdivide the feelings into the two classes of agreeable and disagreeable. If we try to get at the meaning of these, we can perceive that the agreeable feelings are fundamentally associated with one of two things: either a condition in which bodily well-being is assured, or conditions associated with the perpetuation of the race. An example of this latter is the pleasure that parents find in the care of children. The disagreeable feelings are fundamentally concerned with the immediate preservation of the body from injury. Thus fear and anger are two of our most disagreeable emotions, and both of these are aroused ordinarily only when danger threatens or when bodily well-being is otherwise interfered with. One other disagreeable emotion, worry or anxiety, is interesting in that it applies to ill-being that is expected in the future, and while it is well to be on guard against trouble, this emotion is really for the most part futile, because the expected ill-being commonly fails to materialize. Not only do the feelings share with the intellectual processes in making up our mental life as a whole, but they also show themselves in certain bodily changes, which are so important that we shall return to them at greater length in the next chapter.
The feelings as well as the intellectual processes are made up of nervous disturbances passing over the brain cells. We do not know whether those which are concerned with the emotions are in different parts of the brain from those that are concerned with the simple associations or not. There are some differences between the two kinds of nervous activity; particularly is this true of the vividness of the memories concerned. Every intellectual process, presumably, results in the formation of some associations that were not previously present, and the memory of the new associations should be and usually is reasonably sharp. Emotions on the other hand, although they may be remembered, do not register themselves in such a way that a new emotion is aroused every time the old one is remembered. It is probably true that for a while after any strong emotion the recollection of it will tend to bring up the emotional state again, but in course of time there is a dulling, so that although we may remember clearly the occasion of the emotion, the feelings are no longer actively aroused.
IN the last two chapters we have talked about the arrangement of the nervous system and its working as seen in simple reflex actions, and the special activities of the brain which make up the intellectual processes and the emotions. Before we can leave the subject of nervous activity, there are some special kinds of reflexes to be spoken of, and there are also some additional things to say about the working of the emotions. When we speak of special reflex actions we have in mind some of the most useful of our own reflexes. They are not different from the simple reflexes that are described in Chapter X, except that they involve a larger number of sense organs and many muscles, and are correspondingly more complicated. Furthermore, on account of this complication a special part of the nervous system is devoted to carrying them on. Back in Chapter VIII, under the heading of motion, the different types of bodily movement were listed. Chief of these was the act of locomotion; that is, the means by which we get from one place to another, an act which is, of course, of prime importance in maintaining life, since it is concerned both with the securing of food and the escaping of harm. Locomotion is performed in a number of different ways; in ourselves, walking, running, leaping, swimming, riding a bicycle are all forms of locomotion. Any of these acts, as stated in Chapter VIII, requires the accurate timing and accurate grading of the contractions of a great many different muscles. As we saw in that same chapter there are special sense organs which furnish the information upon which this accurate control of the muscles of locomotion is based. We have now to consider the part played by the nervous system in carrying it on. Whenever one starts to walk or to perform any of the other acts of locomotion, nervous disturbances will be set up in great quantity from the organs of muscle and joint sense in the parts of the body that are active, and also from the organs of equilibrium in the ear. It is necessary that the whole volume of nervous disturbance that is pouring into the central nervous system from these sources be translated into a series of streams of disturbances passing out over the motor nerves to cause the muscles to work just as they should in performing the act. All this is reflex in the strict sense. That is to say, the muscular action follows the arousing of the sense organs with no more delay than is necessary for the passage of the nervous disturbance. The work of transforming the incoming stream into a suitable outgoing stream is performed by a part of the brain about which little has been said thus far, namely the cerebellum. This is connected with other parts of the central nervous system in such a way that pathways from organs of equilibrium and from the organs of muscle and joint sense lead into it. From it in turn come a great many connecting nerve cells whose axons pass along the spinal cord and form nerve junctions with the motor nerve cells; and so nervous disturbances coming from the cerebellum can arouse the muscles to activity. We all know that walking or any other locomotor act, after it has once been learned, is carried on successfully without our performing any intellectual work in connection with it. What we do have to do is to start the act, and this is done through a discharge from the cerebrum; either some sense organ is aroused and sends a disturbance into the cerebrum, which arouses in us an association which indicates the necessity for going somewhere, or else a pure memory not stimulated from outside has the same effect. In either case the nervous disturbance passes over from the cell where the association resides to the motor area, and from there down to start the act of walking; but as soon as it is started, the reflex takes hold and carries it on without any further thought on our part except as some additional associations enter to cause us to change or perhaps to stop the action. It is possible that the complex muscular acts which we group under the head of grasping are operated in similar fashion. For example, the act of writing is essentially a modified grasping movement. When we are writing we think not about the muscular movements but about the sense of what is being written, and it may be that the actual control of the muscles in this case is something like that which we know exists in locomotion, although this is by no means certain. It is of course a very great advantage to us to have these complicated activities, which we are performing so constantly, carried on as reflexes without conscious effort on our part. If we had to think about the movement of every muscle, whenever we started to walk, our minds would be so taken up with the act itself that there would be no freedom for anything else; but since the reflex machinery takes care of these muscular movements for us, our minds are left free to deal with other matters at the same time as the muscular actions are going on.
Not only is the simple act of walking carried on for us without mental effort on our part, but to a very large extent the guidance through the sense of sight becomes reflex also. Thus, as we go along the street, we step down from or up on to the curb at the crossings without noticing particularly what we are doing. The approach of the curb is registered through the eyes, and the sensory disturbances thus set up are carried into the brain and intermingled with those from the organs of muscle sense and equilibrium, so that the motions are adjusted to carry us safely down or up. This reflex guidance through the eye becomes so perfect that one who habitually walks over a certain course, as a man in going from his home to his place of business, ordinarily goes the entire distance with scarcely any conscious attention to what he is doing; in fact, persons who move from one locality to another frequently, when they start to walk home, unless they pay attention, find themselves going to the old home instead of the new. The reflex machinery which guided them to their old home works so well that only by conscious interference with it are they able to direct themselves to the new one. Whenever an act of locomotion is learned, it is carried on at first by nervous disturbances from the cerebrum, just as are muscular actions of all sorts; but, as time goes on, there seems to be a transference of the control from the cerebrum to the cerebellum, and as this happens the act is more accurately performed. We have an amusing illustration of this in persons who are learning to ride a bicycle. So long as they are making movements of the handle bars and at the same time pedaling by pure acts of the will, they proceed with the very greatest difficulty; after these actions become habitual, they are carried on easily and skillfully. The great effort of athletic trainers is to develop what is called “form.” What this really means is that they try to teach the athlete the best way of making the desired motions, and then drill him in that particular way, so that it becomes completely reflex with him. The more fully this is done, the better will the athlete perform.
All that has been said about the nervous system thus far has applied to its duty of controlling the activities of skeletal muscles; namely, the muscles by which the body carries on its principal motions. These, as already stated in Chapter VII, are not the only kind of muscles in the body. There are also the smooth muscles and heart muscle. There is also another kind of functional metabolism besides the metabolism of muscular motion over which the nerves have control. This is the metabolism of secretion, or the manufacture of special chemical substances by gland cells. In later sections of the book we shall hear more about what the glands do, but here we have to say something about the way they are controlled through the nervous system. We speak of them along with the smooth muscles, because it happens that the same nervous arrangements are found in both cases; the smooth muscles and the glands have a precisely similar kind of nervous control. In appearance the nerves which lead from the central nervous system to the smooth muscles and the glands do not differ much from other nerves. They are composed of nerve cells, just as are all the other nerve pathways. It happens to be true that the sheaths which inclose the axons of these particular nerves usually have little or no white material in them, so that nerve trunks which are made up of these fibers look gray instead of white, as do the other nerves. Just why there should be this difference, we do not know. Of more importance than this difference in appearance is the fact that the connection from the central nervous system to the smooth muscle cell and the gland cell in no case is made up of a single nerve cell, but always of a chain of two. In the case of nerve paths to skeletal muscles the cell body of the motor nerve cell lies either in the spinal cord or the brain and its axon extends all the way from there directly to the muscle fibers. In the case of smooth muscles and glands a connecting nerve cell extends out from the central nervous system some distance and makes its nerve junction with the motor nerve cell part way between the central nervous system and the muscle or gland. These connecting cells are much branched, as are all connecting cells; also the motor cells which connect with smooth muscles or glands are very much branched; the result is that a single nerve cell starting from the spinal cord finally connects with a large number of smooth muscle cells or gland cells. We do not have, then, in these organs the fine subdivision of control that there is in the skeletal muscles, for a single skeletal motor cell in the spinal cord connects with one or, at the most, a very few muscle fibers. If we consider the uses to which smooth muscles are put, we shall see that finely subdivided control is not important, as it is in the case of the skeletal muscles, where the movements have to be graded very accurately. Smooth muscles are present in the stomach and intestines and other abdominal organs which make movements. They are found also in the walls of the small blood vessels where their contractions or relaxations can change the caliber of the vessels, and so affect the amount of blood passing through them. A good example of this action is seen in the color of the face; when one is pale, it is because the small blood vessels just under the skin of the face are much contracted, and very little blood can pass through; when flushed, on the other hand, the blood vessels are relaxed and there is a large amount of blood passing. These contractions and relaxations of the blood vessels are performed by movements of the smooth muscles. Another interesting place where there are smooth muscles is at the roots of the hairs. Everybody knows that the hair is supposed to stand on end when one is frightened. Most of us have thought of this as a picturesque figure of speech, but as a matter of fact it actually does happen. A human being has such a small equipment of hair that its rising on end does not make much of a showing, but in an angry dog or frightened cat we see it in all its glory. If one looks at the forearm when it is so chilly that “goose-fleshing” occurs, he sees his hair then standing clearly on end. In fact, goose-fleshing is caused by contractions of these smooth muscles just under the skin. Every hair has a few smooth muscle cells at its base and they are attached in such a way that, when they contract, the hair is pulled from the slanting to the erect position. The surface of the skin acts as the fulcrum on which the motion is made. Besides these places where smooth muscle is found there are some in the eye. The muscles of accommodation and also those by which the size of the pupil is changed are smooth muscles. There are some also in the small bronchial passages. In persons who suffer from asthma these smooth muscles become contracted, and the difficulty in breathing experienced is the result of the partial closing of the air passages caused by this contraction. Glands which are under the control of the same kind of nerves as control the smooth muscles are found in many places in the body; in the head we have the tear glands and the salivary glands; in the trunk there are the gastric glands, the pancreatic, and various others about which we shall hear later. With the single exception of the muscles of accommodation there is no case either among the smooth muscles or the glands where there is any occasion for a finely subdivided control. When the muscles of the stomach or intestines become active, a great many fibers work together. When the hair stands on end, a great many hairs become affected at once. A flushing or paling of the skin is also a matter of a large surface. The same is true of gland secretion. In an active gland there is no picking out of a few cells to do the secretion, but a large part of the gland, if not the whole of it, becomes active, whenever any of it is. The advantage of the construction of the nerve paths from the spinal cord to the smooth muscles or to the glands is that a single nervous disturbance from the central nervous system can affect many muscle or gland cells all at once. In the case of the skeletal muscles, on the other hand, the only way a great many fibers can be made active at once, is by sending out nervous disturbances through a great many nerve cells. There is undoubted economy in having the control of the smooth muscles and glands carried on by means of comparatively few discharges from the central nervous system.
A peculiarity of the structures that we are talking about now is that we have very little voluntary control over them. This contrasts sharply with the completeness of our control over the activities of our skeletal muscles. With very few exceptions we can start, stop, or regulate skeletal muscle activities at pleasure, but the activities of our smooth muscles and of our glands are not under our control in the same way. We cannot cause ourselves to turn pale by an act of the will, nor can we set the stomach to working when we wish. This latter inability is a real misfortune, for there are many cases of acute indigestion that are due to stagnation of the stomach, and if we could set the stomach into operation at will, these would be avoided. Since we have not this voluntary control, it is of the utmost importance for us to learn how the control is really carried on, so that the motions necessary to health can be induced in a normal manner.
To a very large extent the smooth muscles and glands are subject to reflex control; that is, nervous disturbances coming in from the sense organs pass over by way of connecting nerve cells to the nerve pathways leading out to the smooth muscles or to the glands. Examples of this kind of control are seen in the paling of the skin, when one is chilly, or its flushing, when one is warm; in the copious secretion of saliva into the mouth that one can get by chewing a rubber band; or in the secretion of tears that comes when the front of the eye is hit.
A second kind of control of the smooth muscles and glands is their control through the emotions. We have already given illustrations of this in the paling of the skin when one is frightened, or the erection of the hairs in the tail of an angry cat. For a long time it has been known that most of our emotions, if not all of them, are accompanied by some sort of bodily change; laughter and crying are familiar illustrations. To a certain extent, as in these instances, skeletal muscles share in the bodily effects of emotions, but their most important examples are found in the smooth muscles and glands. Every serious display of emotion is accompanied by a discharge of nervous disturbances into some of the smooth muscles or some of the glands. This fact has long been known, but its meaning has become clear only very recently. Before attempting to explain this, an additional word will be necessary about the nervous control of smooth muscles. These muscles differ from skeletal muscles in one important particular, which is this: when a skeletal muscle is aroused to contraction, it will relax as soon as the stream of nervous disturbances stops. To keep it in contraction a continuous stream must come into it through the nervous system. This is not true of smooth muscle. When a smooth muscle has been caused to contract, it is very likely to stay contracted without further action on the part of the nervous system. In fact, in very many cases another kind of nervous disturbance must be sent into it in order to make it relax. This means that the smooth muscles, or at least most of them, have two sets of nerves coming to them; one set which causes them to contract, the other set which causes them to relax. Both these sets of nerves are of the type described above as supplying the smooth muscles and glands. A good illustration of the double action is found in the stomach. There is one set of nerves whose activity causes the walls of the stomach to perform the churning motions which go on ordinarily during digestion. There is another set of nerves whose effect upon the stomach is to cause all motions in it to stop. This same double arrangement of nerves is found in many of our most important smooth muscle organs. We said a moment ago that every emotional display is accompanied by an outflow of nervous disturbances to the smooth muscle or gland structures. We now see that what happens in the body is determined by which of the possible pathways is followed by the nervous disturbance. One turns pale, when frightened, because in the emotion of fright the nervous discharge is over the nerves that cause the blood vessels of the face to contract. The mouth waters at the sight of delicious food, because the pleasurable emotions aroused thereby cause the nerves to the salivary glands to become active. As was said a moment ago, for a long time there seemed to be no rhyme or reason about these emotional reactions. Why, for example, should worry cause the muscles of the stomach to stop completely, thus bringing on thousands of attacks of acute indigestion? We now have a clue to the meaning of these facts, which is about as follows: The emotions are to be looked upon as part of the machinery by which bodily well-being is promoted. Emotions of pleasure signify that matters are progressing favorably, and the nervous discharges accompanying these emotions are those in accord with well-being. For example, the activities of the alimentary tract in digestion are of a kind that prosper when the body as a whole is well off. Disagreeable emotions, on the other hand, are associated with the necessity for self-preservation. Thus anger or fear arise in the presence of an emergency calling either for combat or flight. The nervous discharges that accompany these emotions are found on analysis to bring about bodily changes whose effect is to prepare the body successfully for one or the other. This is best shown by actually naming over some of the bodily changes that occur. We may begin with the top of the head and work down. The hair stands on end; this is of no importance in human beings, but in many animals contributes materially to the ferocity of the appearance. The pupils are dilated, aiding vision; the face is pale, and the mouth is dry; not because these in themselves are of importance but because they represent a diversion of energy from unimportant to important parts of the body. The blood that ordinarily flows through the face is shunted out of it into the brain and muscles, where it will really help in the emergency. The activity of the salivary glands is stopped to free the energy that they would consume for use in the brain or muscles. The heart beat is quickened; this again, by making the circulation more efficient, helps the animal to fight effectively. The activity of the stomach is suspended for the same reason as is that of the salivary glands. What we have is a marshaling of the bodily forces into the organs which are of immediate importance in meeting the emergency; namely, the nervous system and the muscles. The other parts of the body, which are necessary in the long run, are allowed to stop functioning for the moment. This behavior of the body in an emergency illustrates better than anything else could the point that our muscles, nerves, and sense organs make up the really important parts of us and the rest of the body has simply the duty of keeping these in good order. In time of emergency all the energies are devoted to the task of keeping these parts of us at highest efficiency, however bad may be the effect on the other less necessary organs; so it is that one who worries himself into an attack of indigestion is really preparing for an emergency by sacrificing the less essential for the more essential activities. The practical difficulty in this is that the worry almost always fails to be of use, either because the expected emergency does not arise at all or else arises so far in the future that the immediate bodily changes caused by the worry do no good. Evidently worry is an emotion to be combated. The old adage about not crossing the bridge until it is reached has really a great deal more to do with the preservation of health than most of us realize.
We have described the control of smooth muscles and glands through reflexes and in connection with the emotions. We have still to mention briefly a third way in which nervous disturbances leading to these organs may be originated. In the last chapter the brain was described as consisting of three chief parts: the cerebrum, which is the seat of memory and association, and so of all thought processes; the cerebellum, which governs the reflex of locomotion; and the brain stem about which nothing was said except that it is a region through which all nervous disturbances have to pass on their way into or out from the brain. The brain stem has a special function of its own in connection with the control of what are often called the “vital processes.” Since most of these vital processes are concerned with smooth muscle or gland action they are of interest to us here. In the brain stem are located groups of nerve cells to which are given the name of “centers.” From these extend nervous pathways to various bodily structures. For example, there are two centers having to do with the activity of the heart, and both of these have nerve pathways leading to that organ. Over one of them pass the discharges which cause the heart to beat more rapidly, over the other the discharges which slow it down. There are also centers for controlling the caliber of the various small blood vessels and so the amount of blood that flows through the various organs of the body. Still another center has control of the secretion of sweat. These are a few of the centers which are present in the brain stem. It is evident that these centers must be played upon directly from the sense organs in the reflex control of the smooth muscles and glands and also from the cerebrum in their emotional control. The third way in which these centers can be affected is by a direct action of the blood which circulates through them. Of course, every center is made up of nerve cells which are alive and which share with all other living cells the necessity of being sufficiently nourished. This means that there must be small blood vessels here and there among the nerve cells, so that interchanges of material can take place between the blood and the fluid immediately surrounding the cells. Among the materials which may come from the blood are some special chemical substances which have the ability to arouse these cells to activity. For example, the very rapid beating of the heart following vigorous muscular exercise is due, in part at least, to the presence of chemical substances that are poured out into the blood from the laboring muscles and which have the effect of arousing the center whose action speeds up the heart. In at least one case, that of the center controlling the sweat glands, the nerve cells are aroused by an increase in temperature; on a very hot day or when one exercises briskly the temperature of the blood begins to go up; this arouses the center which controls the secretion of sweat; nervous disturbances are poured out to the sweat glands and they become active. The effect of their activity is to remove heat from the body, and so help to lower its temperature. Here we have an example of an automatic regulating device in which nerve cells are controlled through the blood. Except for this one instance of the effect of blood temperature the actions of this class are thought to be wholly by means of chemical substances, and for that reason we speak commonly of the third method of control of the smooth muscles and glands as chemical control. We have then altogether three methods of control; through reflexes, through the emotions, and through chemical substances in the blood.
Since we have spoken of chemical control it will be appropriate to close this chapter with just a further word about it. In addition to the regulation of smooth muscle and gland activity there are a number of other bodily processes which are affected more or less through chemical agents brought to them by the blood. Many of these processes are of a kind that cannot well be controlled through the nervous system. For example, growth is something which goes on very slowly and yet which distinctly requires some kind of control. It has not perhaps occurred to us that it is more than chance that our ears are substantially the same size, our arms about the same length, our legs the same length, our feet about the same size, and so on. Yet if we pause a moment we realize that as the ears grow there would seem to be no particular reason why they should both stop when they have reached the same size, unless there is some definite regulation. This regulation is present in the form of chemical substances. Of late years a very large amount of study has been given to the materials which have the power of acting as chemical regulators, and a good deal has been learned about them, although there is every reason to believe that a great deal more remains to be found out. It has been necessary to invent a name for substances which act as chemical regulators, because, without such a name, talking about them becomes too cumbersome. The name that has been adopted is hormone, from a Greek word meaning to arouse or to stimulate, referring to the ability these substances have of acting upon living cells. Thus far we have spoken of hormones only in connection with their ability to excite those nerve cells by which smooth muscles and glands are controlled. As a matter of fact, their action is much wider than that. They have the power of affecting the metabolism of very many body cells. Some of them appear to have the special function of regulating the rate of metabolism. These we shall talk about in detail, when we return to the study of metabolism in a later chapter. One of the hormones is interesting in connection with the relation of emotions to smooth muscle and gland activity. This is a secretion manufactured by a pair of small glands located in the abdomen near the kidneys. They are known as the adrenal bodies. These glands secrete a substance to which has been given the name of adrenalin, which has been shown to be a very efficient chemical excitant to a number of bodily processes. We are particularly interested here in a property it has of arousing smooth muscles and glands in precisely the same way as they are aroused in connection with the disagreeable emotions, such as fright or anger. For example the injection of a little of this adrenalin into the veins of a person or animal will cause his heart beat to be quickened, the blood flow into the brain and the muscles to be increased at the expense of the flow through the skin and abdominal organs, and the other effects to occur that were described above as accompanying these emotions. These facts about the effects of adrenalin had been known for some time before it was realized that they fit in with the reaction by which the body prepares itself to operate efficiently in time of stress. Now we have learned to look upon adrenalin as the “emergency hormone,” meaning that it is a chemical substance which does the same things to the body that are done through the emotions in time of emergency. We look upon this as a reenforcement of the nervous action, indicating the efficiency with which the fitting of the body for time of stress works out. The adrenal bodies are themselves glands and are acted upon like other glands through the part of the nervous system that we have just been describing. In time of emergency a stream of nervous discharges pours into them; they secrete an abundant supply of adrenalin which passes out into the blood and is carried by the blood to all parts of the body, exerting its special functions wherever it comes. There is no doubt that the ability of the body to care for itself in time of need is helped by the outpouring of adrenalin. In one respect, however, this arrangement does not work out perfectly. This is because chemical substances that are poured out into the blood cannot be gotten rid of instantly, and are likely to linger for a shorter or longer time after the actual emergency subsides, so the bodily effects persist for a time after the occasion for them has disappeared. We are all familiar with the fact, for example, that the heart goes on thumping for a long time after a sudden shock. It may not have occurred to us to wonder what kept it going at that rate after the immediate disturbance was passed, but we now see that it must do so until the adrenalin which was poured out in connection with the shock has been gotten rid of, which is a more or less gradual process.
WE have learned to think of the cells which make up the body as dependent on the fluid which surrounds them for the various materials they require, and as a place into which they discharge the products of their metabolism. We have seen furthermore that the fluids which bathe the cells directly must be constantly renewed. The renewal is accomplished by interchanges between this fluid and the blood, which constantly flows through the tiny blood vessels that are everywhere present in the body. In its course, in turn, it passes through the blood vessels of the organs in which it is to be itself renewed; the digestive organs for food supplies, the lungs for oxygen, the kidneys for the discharge of waste material. We must now look further into the nature and action of the various body fluids. Of course the foundation of all of them is water. In this water must be dissolved everything that is used by any of the cells for food or anything that any of these cells produces. Under this latter head we have the waste products of ordinary metabolism, or in the case of some cells special products of functional metabolism. The presence of all these various materials would be bound to make the body fluid an extremely complex mixture. In addition to these various materials there are certain substances present besides water to make up what we may call the structure of the body fluids as distinguished from the materials which they are carrying from one place to another. These structural materials include a number of salts of which ordinary table salt (sodium chloride) is the most abundant as well as the most familiar. In addition there are salts of lime and potash and magnesium; all these latter in very small proportions. Just why the body fluids should contain these salts is not very clear. We know that if they were not present the cells of our bodies could not live, yet it is true that there are a great many kinds of living cells that get along perfectly well in fresh water, which may have no salts dissolved in it at all. On the other hand there are more kinds living in the ocean and exposed to the rather strong concentration of salts which make up ocean water, and there are even some kinds of animals that live in strong brine, so that evidently living protoplasm can adjust itself to surroundings in which the strength of the salt in solution is widely different. It is true that very few kinds of animals can endure being changed suddenly from ocean water to fresh water or the reverse. One of the best ways to clean the bottom of a ship that has become foul through long sailing about the sea is to transfer it into a fresh water lake or stream, where the accumulation of living animals and plants will be killed and will drop off. Most kinds of ocean animals die rather promptly if changed to fresh water, or fresh water animals if put into the ocean. There are a few kinds of fish, like the salmon and shad, which live in the ocean but lay their eggs in rivers, and these are able to endure the change from the one kind of water to the other without being destroyed. They, to be sure, make the transition gradually, swimming up from the ocean into water that is less and less salty, until they finally reach the fresh water stream itself. There are other kinds of living things which can endure a much more abrupt change from salt to fresh water and back again. In various parts of the world are large rivers emptying into the ocean and so situated relative to towns and cities that steamers make regular round trips from the ocean up to the fresh water of the river and back again, and it will be found that on the bottoms of these steamers are various plants and water animals which endure the frequent shift from salt water to fresh and back again without harm.
The percentage of salt in the body fluids of all the higher animals including ourselves is only about one-fifth that in the ocean. Furthermore the fluids of most kinds of land animals have about the same percentage of salts dissolved in them. Naturally there has been much speculation as to why there should be this percentage of salt in preference to any other. One ingenious theory supposes that back in the beginning of things, when the earth cooled down below the boiling point of water, so that it was possible for water to collect on the earth, the water in all the oceans was fresh. This would have to be true, since water must have fallen in the form of rain; but in course of time some of the salts in the earth’s crust would be dissolved, making the water salty, and as time went on the ocean would become saltier and saltier. This is still happening, for every river that discharges into the ocean carries with it materials that it has dissolved from the underlying soil during its passage from its source, and such material, when it once enters the ocean, must stay there until the ocean water becomes saturated with that particular substance. According to this theory there was a time, then, when the ocean water was just about as salty as our body fluids at the present time, and it supposes also that that was the time when the ancestors of present land animals crawled out of the ocean and took up their abode on land. Of course there is no way to prove that this is so, but it does account for the particular percentage of salts in our bodies as well as any other explanation we know anything about.
In addition to these various salts our body fluids contain in solution moderate amounts of very complex chemical substances belonging to the class of proteins. A fact about proteins which has not yet been emphasized is that they make liquids in which they are dissolved sticky or gelatinous. An excellent example of this is ordinary raw white of egg, which is a solution of protein in water, and which shows the gelatinous character very strikingly. Because of the protein that is in solution in the body fluids, they have also this gelatinous character, although to a much less extent than in the white of egg, because the amount of protein in solution is so much less. As we shall show later, this sticky quality of the fluid is of a good deal of importance in its actual use in the body. It may be that the protein in the body fluids serves other purposes as well. One interesting fact that needs to be emphasized is that it is not used for fuel or building material for the cells. The protein that comes to them as part of their food supply is entirely distinct from that about which we are now talking, which is part of the permanent structure of the body fluids. We shall speak of the protein that serves to nourish the cells in a moment, when we are talking about the relations of the fluids to the transportation of food material.
In addition to the salts and proteins we have also dissolved a great many very complex materials which may be looked upon as permanent or relatively permanent constituents of the fluid, but about which we know practically nothing chemically. We are sure that they are present, because of certain effects which they produce, but the substances themselves have never been made out by chemical analysis. These are materials which are concerned with the resistance of the body to infectious disease, and it will be necessary to say just a word about infection to make clear the part played by them.
What we call an infection is the invasion of the body from the outside by minute living organisms, either plant or animal, and the establishing of them within the body, so that they grow and multiply. They carry on their metabolism just as do all other living cells and produce various chemical products as a result. There are many organisms living within our bodies whose metabolic products apparently do us no harm, and so we serve as hosts for these unbidden guests year in and year out without even knowing of their existence. The products from other kinds of organisms are poisonous to us, and when some of these organisms multiply within us we discover it, because we are poisoned and become ill. Only organisms whose products of metabolism are poisonous are counted ordinarily as causing infection. In the strict sense we might be said to be infected by the harmless organisms of which so many thrive within us, but in the usual use of the word we speak of one as having an infection only when his body has been invaded by organisms that produce poison. In the case of most kinds of injurious organisms their multiplication within the body unchecked would lead finally to its destruction. It is necessary, therefore, that the body have some means either of checking the development of the organisms or of neutralizing the poisons which they produce. The body has this power, and the machinery for it consists very largely of substances or structures in the body fluids. The detailed story of these is too complex to be told here. We shall content ourselves by saying that when an infection becomes established, as of scarlet fever, for example, the poisons that are produced are poured out by the organisms into the body fluids of the part where they happen to be located, and are taken up from there by the blood and distributed all over the body. The fever, headache, and other disagreeable symptoms are due to these poisons. The interesting thing about it is that the poisons themselves act toward the body as chemical regulators or hormones, exciting some or perhaps all of the cells of the body to a special kind of functional metabolism, which results in the manufacture of materials which neutralize the poisons. Thus, one of the very important properties of living protoplasm is to respond to the poisons from the metabolism of other cells by producing neutralizing material. Whether one dies from an infection or recovers from it depends on whether the cells are able to produce enough neutralizing material to prevent themselves from being killed or whether the poison is so abundant or so malignant that the cells are destroyed in spite of their activity in pouring out the neutralizing material.
Still another interesting thing about this whole matter is that every kind of infecting organism has its own kind of poison, which differs from that of the other kinds, and so the chemical effect of the poison upon the cells is not the same for one infection as for another. The functional metabolism of the cells in turn is adjusted to the kind of poison, so that the material they pour out is suitable to neutralize the particular poison which aroused them to activity in the first place, and in most cases no other. If one gets well from any infection, there is a surplus of the neutralizing material left in his body fluids, and, as long as it remains, he is secure from another infection of the same kind. This condition is defined as immunity. Since the neutralizing materials are different for different infections, immunity against one is in most cases of no avail against another. One may be immune against scarlet fever, but be just as likely to catch pneumonia as a person who has never suffered from any infection at all. It follows that an individual who has had and recovered from a great many infections has a correspondingly large assortment of neutralizing materials in his body fluids. Some of these appear to persist throughout life, others disappear fairly soon.
The next group of permanent constituents to be described consists of some materials which seem to have nothing at all to do as long as everything is going well: the body fluids bathing the cells, or, in the case of the blood, circulating about from part to part through the blood vessels. These come into play only when, as the result of injury, the fluids begin to escape; namely, in the case of bleeding, or, as it is technically called, hemorrhage. It is clear that if any injury is started sufficient to allow the fluid to escape, there would be no more reason why it should stop running out, unless prevented, than there is why water should stop running out of an open faucet of itself. Since, as we know, we do not bleed to death every time we get a slight cut, but after a long or shorter time the bleeding stops of itself, there must be some automatic arrangement by which the opening is plugged. All of us are familiar with the way in which this is done. We know that in the course of a few minutes after the bleeding begins, the blood tends to set into a firm jelly, which is called the clot. This clotting is the result of a chemical transformation which goes on in the blood as the result of its escape from the blood vessels and its exposure to the outside. The details of the chemical processes are too complicated to be described here; all we need to remember is that the blood within the body contains certain special materials which are soluble and therefore float in solution along with the other soluble materials. When any blood vessel is injured, so that the blood begins to escape, a series of chemical changes is started automatically by which this soluble material is changed and becomes insoluble, so that instead of remaining dissolved in the blood it is precipitated out. In this form it is called fibrin. Fibrin is a very sticky, stringy mass which forms a spongy network, spreads itself over the injury, and clings firmly to the edges, thus plugging the opening, unless the rush of blood is so strong that it keeps washing the fibrin away as fast as it is formed. Bleeding from small wounds will presently stop of itself, but if the hemorrhage is too great for this, it is necessary that the blood flow be slowed down or stopped artificially. This is done by locating and pressing upon the large blood vessel through which the blood is escaping. In order to do this successfully, one has to know something about the course of the blood vessels, and this will be described in a later chapter.
There are a few people whose blood lacks some of the necessary chemical substances to enable it to clot; such persons are known technically as “bleeders.” Even a slight injury in one of them will cause serious, or even fatal, hemorrhage, unless the escape of blood is stopped artificially, since it will not stop of itself. An interesting fact about this condition is that it runs in families; in other words it is hereditary.
In addition to all these constituents of the body fluids which are dissolved in them, there are in that part of the fluids confined to the blood vessels, which we call the blood, three kinds of structures floating; these we have next to describe briefly. The first of these are the red corpuscles. They give the blood its red color, although if looked at singly they appear yellowish rather than red. Red corpuscles are almost inconceivably tiny. They are red flexible disks, a little bit thinner in the middle than at the edges, about one three-thousandth of an inch in diameter. Some idea of the enormous numbers in the body can be gathered, when we say that a drop of blood the size of the head of a pin would contain four or five million of them. The red corpuscles are made up of a sort of framework of protein within which is inclosed a red coloring matter or pigment, known as hemoglobin. It is this pigment that gives the blood its color, and in some respects it is one of the most important of the nonliving substances in
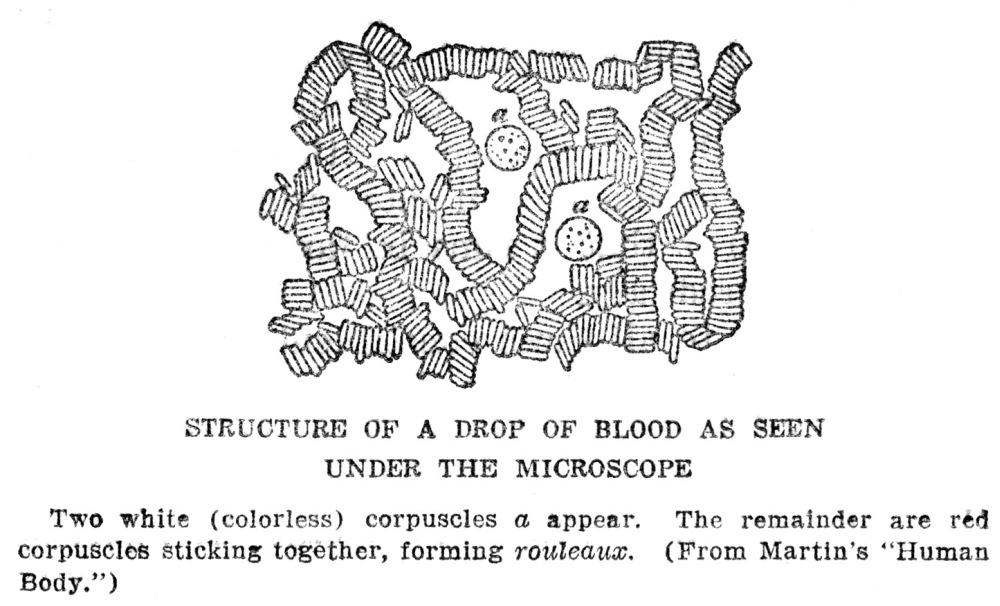
Two white (colorless) corpuscles a appear. The remainder are red corpuscles sticking together, forming rouleaux. (From Martin’s “Human Body.”)
the body. This is because it is the means by which the cells obtain sufficient supplies of oxygen. As we have already seen, every cell is constantly drawing from the body fluids about it the oxygen which is required for carrying on its metabolism. The fluids in turn get oxygen from the blood. It is necessary, therefore, for the blood to convey abundant supplies. Oxygen will dissolve in water, as is proven by the fact that fish and other aquatic animals are able to get enough oxygen from the water in which they live to serve their needs; but it is not sufficiently soluble to supply the needs of an active body like that of man. It is necessary, therefore, to have a special additional means of conveying oxygen besides its simply dissolving in the blood. This additional means is furnished by the hemoglobin, which is an “oxygen-carrying” pigment. What this means is that the hemoglobin has the property of taking up oxygen chemically, whenever it is exposed to a region where there is oxygen in abundance, and of giving it up again whenever it passes through a region where oxygen is scarce. It is thus that oxygen is conveyed from the lungs to the active tissues of the body. We shall have more to say about this in the chapter devoted especially to the matter of the oxygen supply.
We said a moment ago that the red corpuscles consist of a protein framework inclosing hemoglobin. They are not living. They must, therefore, have been made by living cells and poured out into the blood stream. We might suppose that this was done once for all and that the same red corpuscles are floating in our blood now that started floating there when the blood was first formed; but, as a matter of fact, this is not the case. There is a continuous breaking down of red corpuscles which must be made good by a continuous manufacture of new ones. Most of the larger bones in our bodies have a sort of spongy framework by which the ends, where the joints are, are made stronger. Within the space of these frameworks is a kind of marrow, known as red marrow, because it has such a very abundant blood supply. It is in this red marrow that the manufacture of red corpuscles goes on. There are throughout the red marrow living cells which are constantly dividing and subdividing, forming more and more so-called daughter cells. Within these daughter cells hemoglobin is presently deposited; a little later they lose the nucleus and probably the remainder of the living protoplasm as well, leaving just the framework of nonliving protein with its contained store of hemoglobin. This is the finished red corpuscle, and it breaks loose from the red marrow and floats out into the blood stream. The rate of manufacture of red corpuscles is very rapid; undoubtedly millions of them are formed daily in the various red marrow regions of the body. The total number of red corpuscles does not increase correspondingly, because they are broken down at the same rate as they are formed. It is now believed that in the spleen, which is a large organ of the abdomen whose function has always been obscure, those red corpuscles which are destined for destruction are picked out of the blood and broken down. We commonly suppose that this fate overtakes corpuscles that are worn out and are no longer efficient oxygen carriers, but we do not know, as a matter of fact, that the corpuscles do lose their effectiveness in course of time, nor have we any idea how the spleen could select out of the millions in the blood those particular ones which are no longer useful.
What the spleen does to the corpuscle is to break it up so that the protein and the hemoglobin in it are set free in the blood stream. We do not know what becomes of the protein; probably it is taken up and utilized. We do know that the hemoglobin is decomposed in the liver. One constituent of hemoglobin, in fact the constituent which gives it its ability to carry oxygen, is the element iron. Iron is not particularly abundant in living things, and we find that the body is thrifty with regard to it. When the liver decomposes the hemoglobin, the iron is saved in some way which enables the blood to carry it back to the red marrow, where it can be used over again. There are also some portions of the hemoglobin which are valuable as food material; the remainder, which is of no further use, is discharged from the body as a part of the bile.
Besides the red corpuscles the blood contains what are known as the white or, better, the colorless corpuscles. These, instead of being dead structures, as are the red corpuscles, are actual living cells, which float along in the blood stream or have the power of clinging to the walls of the blood vessels and crawling along them; in places where the blood vessels are very thin, they work their way right through, and so get out into the spaces among the tissues. There are not nearly so many colorless as red corpuscles, in fact, the latter are eight or nine hundred times as numerous as the former. A great deal of study has been devoted to colorless corpuscles to find out just what they do. We are by no means certain that we understand fully all their activities, but we do know that one very important thing that they do is to absorb into their own bodies tiny foreign particles that may be present in the blood stream or in the spaces among the tissues. There are not many kinds of foreign particles that can get into these places. In fact, about the only sort that can are the tiny plant or animal cells which are responsible for disease. When these foreign organisms invade the body, the colorless corpuscles may engulf them into their own bodies and destroy them. In this way a great deal of infection is prevented. Of course it may happen that the invading organisms are so numerous that the colorless corpuscles can not get rid of all of them, or the corpuscles themselves may show a diminished activity. It is an interesting fact that both the number of colorless corpuscles in the body and the vigor with which each colorless corpuscle attacks foreign organisms vary from time to time, so that we are much more secure against infection at some times than at others. In general we may say that when the body is in a condition of vigorous health the colorless corpuscles will be efficient. When we are run down, on the other hand, these cells share with all other body cells this state of low vitality. This explains why people who allow themselves to become run down are more likely to fall victims to infection than those who are in vigorous health, and emphasizes, of course, the importance of a habit of life that shall keep the body vigorously healthy at all times. It should be understood that the colorless corpuscles do not show the same effectiveness against all kinds of foreign organisms; some kinds of disease germs are able to bring about infection in the body quite regardless of the activity of the colorless corpuscles. From other kinds, on the other hand, they give the body very complete protection.
The most familiar example of the action of the colorless corpuscles is in the formation of what we all know as pus. There are a few kinds of organisms that, instead of getting into the body and becoming scattered through its fluids, establish themselves at certain points and by growth and multiplication accumulate at those places in large numbers. Examples are pimples and boils. In these cases the pus-forming organisms have located just under the skin and are multiplying there at a great rate. They produce poison which is absorbed from the place where they are and distributed through the body. This poison appears to have some sort of chemical attraction for the colorless corpuscles; at any rate the corpuscles gather from all around to the place where these organisms are located and engulf as many of them as they are able, but in so doing they themselves are destroyed, and pus, as we know it, is simply a mass made up of the dead bodies of the colorless corpuscles along with the organisms which they have destroyed and which in turn have destroyed them. In the ordinary pimple or boil the colorless corpuscles ultimately win the combat and the infection is completely overcome. It happens occasionally, however, that a pus-forming infection becomes established in some place where it manages to maintain itself in spite of the attacks of the colorless corpuscles. This happens very frequently in the tonsils, so that persons who have infected tonsils may have pus formation going on month in and month out. This may also take place at the roots of the teeth. In fact it is now commonly believed that whenever the nerve to the tooth dies, infection is certain sooner or later to become established at the tip of the root. These places where pus formation is going on continually are known as “pus pockets.” For a long time very little attention was paid to them. Persons occasionally suffered acute distress from gumboils or had attacks of sore throat owing to the infected condition of the tonsils, but beyond these no particular attention was paid to the pus-forming organisms, unless, as occasionally happened, an especially virulent type became established which brought on the condition commonly known as blood poisoning. Within the last few years the discovery has been made that there is a steady discharge of poison from every pus pocket into the body fluids and so to the blood stream. The amount is ordinarily so small that the body as a whole is not seriously affected, but now and then, either because of a larger production of poison or because of a lower resistance on the part of the body, serious ill effects are produced. Among them may be mentioned acute (inflammatory) rheumatism. This is not only an extremely painful condition, but it is very likely to leave serious after-effects, as, for example, injured heart valves, to give trouble for the rest of life. The discovery of these facts has given us great respect for pus pockets, so we no longer treat them carelessly. Infected tonsils are looked after, not simply because they bring about sore throat now and then, but even more because of the poison which they are likely to send through the body. It is probably not too much to say that the practice of dentistry has been revolutionized since the significance of pus pockets has been discovered. Formerly the killing of nerves to relieve aching teeth or to insure them against future aching was a common practice. The modern dentist, on the other hand, kills nerves to teeth with the very greatest reluctance and only as a last resort, because he knows full well that in so doing he is opening the way for the establishment of pus pockets with the train of ills that is likely to follow. At the present time methods of curing pus pockets at the base of the teeth are not very adequate, so that the extraction of the tooth has to be resorted to, but there is every reason to believe that the dental profession will shortly find methods by which pus pockets can be controlled without having to sacrifice the teeth.
The third kind of structures which are present in the blood stream are much smaller than the red corpuscles, but are nowhere near so numerous. They are called the platelets and are disk-shaped bodies composed chiefly of protein material, and probably, although not certainly, living cells. Their presence in the blood remained unsuspected up to about the end of the last century, not so much because of their very small size, as because they go to pieces very quickly after the blood is shed. By the time a drop of blood could be gotten under the microscope they would be all gone. They were discovered only as the result of the adoption of a method of treating the blood which preserved them long enough so that they would still be present when the blood was looked at under the microscope. They seem to have something to do with the changes that take place in the blood by which it is caused to clot. Whether this is their only function, we do not know.
We have now described the substances which are present in the blood and in the other tissue fluids as a fairly permanent part of their make-up. In addition to these there are present all the materials that are in transit to the cells or from them. These include all kinds of foodstuffs on their way from the digestive organs to the cells. As we shall see in detail later, the digestive organs work over the food that we eat before passing it on to the blood, so that the actual food materials that are being transported by the blood are the digested products of the food rather than the food itself. For example, there are to be found in the blood, in addition to the regular blood proteins which were described a moment ago, the digestion products of the food proteins on their way to serve the needs of the various cells of the body. There will be found also the digestion products of the other classes of foodstuffs. The body fluids contain also the waste products of cell metabolism on the way to be discharged from the body and the special products, such as the hormones, which are manufactured by certain cells and carried through the blood stream to act upon other cells. Of course we realize that not all of the materials that are manufactured by cells are poured out into the blood stream. Such materials as saliva, gastric juice and the like are passed directly from the cells in which they are manufactured into tubes by which they are conveyed to the digestive canal, where they carry on their special work of digesting the food.
Among the things which are present in the body fluids should be mentioned the two gases oxygen and carbon dioxide. We should expect oxygen to be present in the body fluids, because it is necessary for the metabolism of cells and can get to them only by being carried in the blood stream. We have seen in the red corpuscles the special method by which an abundance of oxygen is transported. Carbon dioxide is the gaseous product of the oxidation of carbon and is found in large amounts wherever there is burning, since carbon is the chief constituent of all fuel and whenever carbon is burned, carbon dioxide is formed. Since the fuel materials that are burned in the living cells consist largely of carbon, carbon dioxide is produced in them as well. They have to get rid of it, and the only way they can do so is by passing it out into the fluids that surround them, which in turn pass it on to the blood. The way in which the body handles these two gases makes up a special subdivision of the subject of physiology and will be so treated in the chapter on respiration.
IN the last chapter we talked about the body fluids and saw that they can be subdivided into the tissue fluids, which surround the cells, and the blood, which is inclosed in a system of pipes and which carries the materials to and from the tissue fluids. We now have to take up this matter of transporting the material in more detail. The first step will be to see how materials that are in the blood get from it to the tissue fluids, and how materials that are in the tissue fluids get from them into the blood. Unless these interchanges can take place freely, there is no way in which the blood can serve as a conveyer system. In order to see how the interchanges are carried on, we shall have to look first at some features of the construction of the system of pipes through which the blood flows. As we all know, the large blood vessels are either arteries or veins, the arteries being blood vessels which are carrying blood away from the heart, and the veins vessels that are carrying blood toward the heart. If we start with an artery and trace it through the body, we find that it is continually giving off branches which in turn give off smaller branches, until finally the subdivisions are so small that we cannot trace them any further with the naked eye. In the days before the microscope was discovered, there was a great deal of question as to how these finest branches ended. At first no one suspected that there was connection between the fine subdivisions of the arteries and the fine subdivisions of the veins through which the blood could pass.
About the beginning of the seventeenth century William Harvey became convinced that there must be fine vessels leading across from the smallest arteries to the smallest veins and that the blood must pass through these. He came to this conclusion without ever having seen these small vessels, since at that time there was no microscope by which such tiny structures could be seen. Before Harvey’s time it was supposed that the blood ebbed and flowed in the arteries and in the veins. He showed that the blood flows in one direction constantly, leaving the heart by way of the arteries and coming back into it by the veins. Harvey was, therefore, the discoverer of the circulation, one of the most important discoveries in physiology. After the microscope was perfected the tiny tubes connecting the finest arteries with the finest veins were made out. They were found to be very small in diameter and to have very thin and delicate walls. They were also found to be extremely numerous. The finest subdivisions of the arteries that can be seen with the naked eye are scattered very thickly through all the tissues of the body which have a blood supply, and they go on subdividing microscopically, so that the finest vessels are scattered more and more thickly through the mass of living substance. The very finest of all, which are the tubes connecting the smallest arteries with the smallest veins, are called capillaries, from a Latin word meaning a hair, to indicate their very small size. They are so close together in most parts of the body that it would be difficult to thrust a
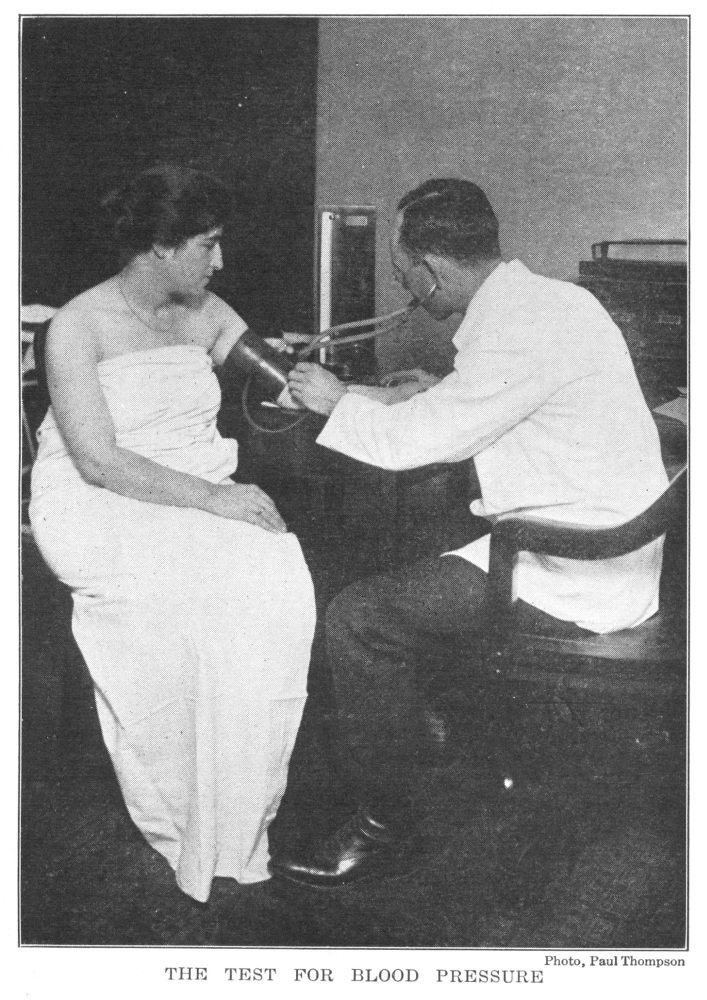

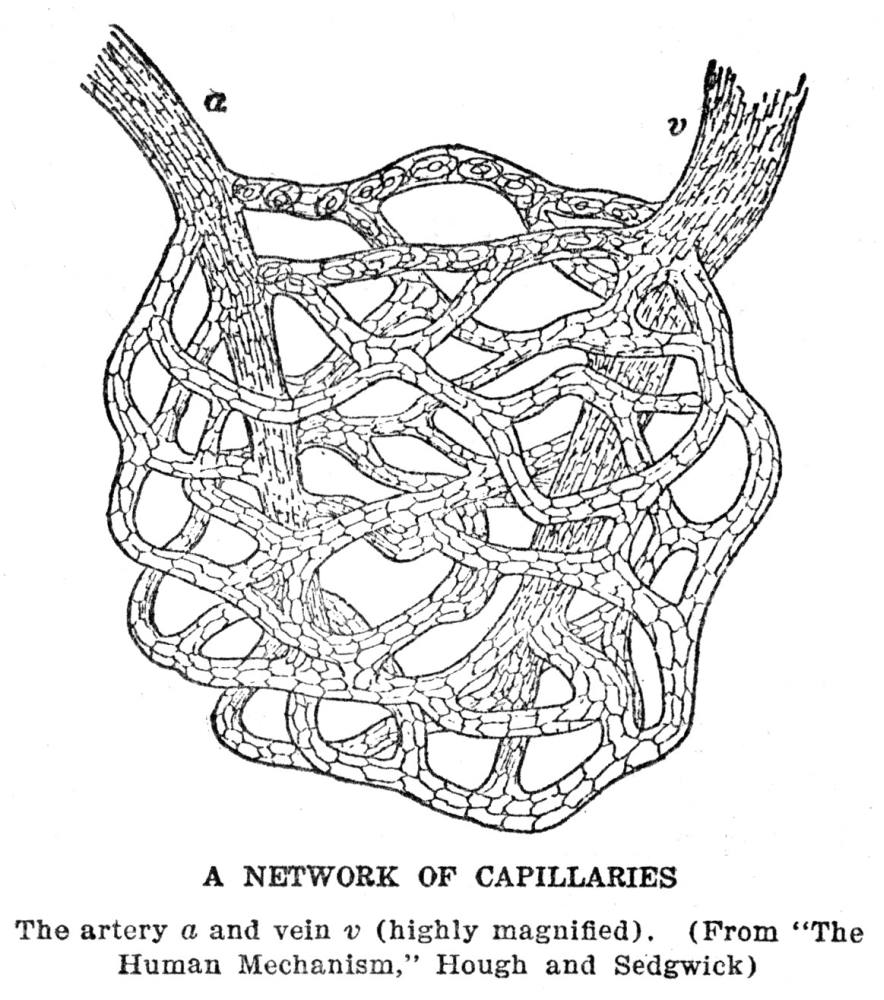
pin in anywhere for any distance without striking against one or more of them. The capillaries are spread so thickly that there are not many places in the body where living cells are more than a very small fraction of an inch from one of them. The cells do not, however, lie right against the capillaries, but are separated from them by tiny spaces filled with tissue fluid. In order for material to get from the blood to any living cell, then, it must pass through the wall of the capillary into the fluid which fills the tissue space and from that in turn to the cell itself. The wall of the capillaries is so delicate that if the blood flowing through any capillary contains more of any substance than is present in the tissue fluid surrounding that capillary, some of it will pass out through the wall and into the tissue fluid, just about as freely as though there were no wall there at all. The arrangement can be illustrated by a familiar example; if a drop of ink is allowed to fall into a glass of water, it will color only a small part of the water at first, but quickly spreads out until each part of the water is as deeply colored as any other part. If the glass of water were to be divided in half by a very delicate membrane, and the ink dropped in on one side, it would spread out in the same way, passing through the membrane in so doing, until again all the water in the glass was equally colored. Of course, the quickness with which the ink could pass through the membrane would depend on how delicate the membrane was. We could imagine membranes which would not let any ink at all through, and every degree from that up to membranes so delicate as to offer no obstruction at all to the passage of the ink. The walls of the capillaries rank as membranes of such delicacy as apparently to offer almost no obstruction to the passage of materials through them. They hold back the red corpuscles and the platelets fairly well, so that they do not pass out of the blood and into the tissue spaces, unless the capillaries are actually injured. The colorless corpuscles are able to make their way through the capillary walls and so also do nearly all the substances that are dissolved in the blood. It is an interesting fact that the blood proteins do not pass freely through the capillary walls, although the digestion products of food proteins, do. It will be remembered that in the chapter on Body Fluids the importance of the sticky quality of the blood proteins was spoken of. It is now believed that it is because of this gelatinous nature that the blood proteins are not able to pass out through the capillary walls, and this is supposed to be important in the proper working of the circulation. In fact there is a condition of greatly lowered vitality to which the name “shock” is applied, in which the blood proteins escape through the capillary walls to so great an extent as to interfere with the proper working of the body. It has been found possible to prevent this in a very large measure by the simple expedient of injecting some substance like mucilage directly into the blood stream. We are to think of the capillary walls, then, as allowing materials to pass freely through them in either direction, from the blood into the tissue spaces or from the tissue spaces into the blood, with the exception of the red corpuscles, platelets, and the blood proteins, and as thus keeping the tissue fluids supplied with whatever materials the blood contains or taking from the tissue fluids the waste products of cell metabolism, which the cells are pouring out. With this arrangement clearly in mind, all that remains for the understanding of the conveyer system is to see how the blood is kept in motion and distributed among the various organs of the body, and then to consider where the blood in turn gets its supplies of materials which it can pass on to the tissue fluids, or how it gets rid of the substances which the tissue fluids have passed on to it from the cells.
At the beginning of the chapter we said something about the arteries and veins and their branching into smaller and smaller subdivisions with the final connecting link between the smallest arteries and the smallest veins in the form of capillaries. We are now to consider in detail the movement of the blood through these tubes, and to do that it will be necessary to speak again of this arrangement. In describing the circulation we usually begin with the heart. The heart itself will be taken up presently. First let us trace the blood vessels from the heart through the body and back to it again. The large main artery leading from the heart is known as the aorta. This springs from the upper side of the heart, bends over in an arch, and passes down through the chest into the abdomen.
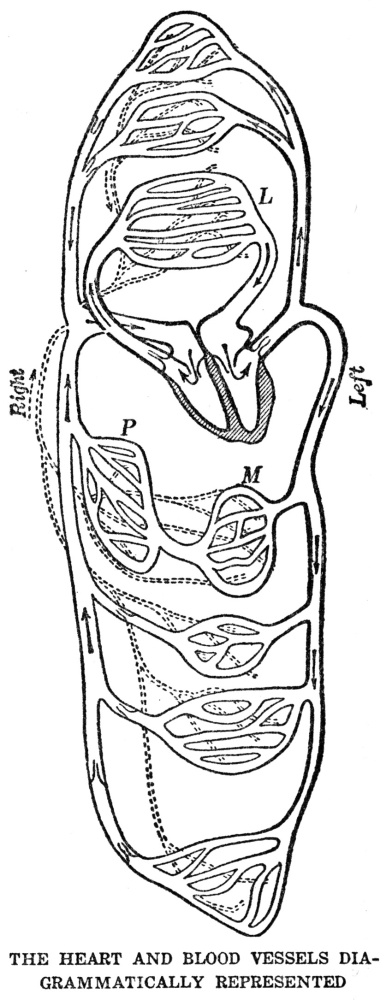
Branches are given off from the aorta all along its length. The very first of these come off before the aorta gets away from the heart, and are the arteries by which the tissues of the heart itself are supplied with blood. A little farther on are large arteries, one for the left arm with a large branch running up the left side of the neck, another for the right arm with a large branch running up the right side of the neck. Each of these in turn gives off branches all along to provide for the tissues of the arms, neck, and head. It is worth while noting that the arteries running up the neck to the head are large in proportion to the size of the head itself; this is because the brain, as the most important organ in the body, requires and receives a disproportionately large blood supply. Besides the brain the head contains numerous muscles and also the salivary and tear glands, all of which carry on active metabolism and therefore require abundant blood supply. The main branches of the aorta to the head and arms are given off from the arch; as the aorta passes down through the chest it gives off small branches to the muscles of the chest wall, and then passes into the abdomen. Here are located two of the three important arrangements for renewing the blood; namely, the digestive organs and the organs of excretion (kidneys). Large branches from the aorta pass to the digestive organs and others to the kidneys; smaller branches lead to the muscles of the abdominal wall, and also to the various secreting glands that are located in the abdomen. At the lower end of the abdomen the aorta divides, giving one large branch for each leg. As we have already seen, if we follow any of these subdivisions through its finer and finer branchings, we shall finally be able, with the aid of a microscope, to trace it to capillaries, where the interchanges between blood and tissue fluid occur, and beyond the capillaries to tiny veins which unite with other tiny veins from other capillaries into larger veins. These again continue to come together into main veins corresponding in every part of the body with the main arteries. All these veins finally unite into two, one for the lower part of the body, called the inferior vena cava, and one for the upper part of the body, called the superior vena cava. These two come together just at the entrance to the heart. One special feature of the blood supply to the digestive organs may as well be mentioned here; it is that the blood which flows through the capillaries of the stomach and intestines is all reassembled into a vein known as the portal vein, which instead of passing directly into the inferior vena cava goes first to the liver, where the vein breaks up into another set of capillaries, the liver capillaries, beyond which is another vein which leads into the inferior vena cava. The result of this arrangement is that all blood passing into the capillaries of the stomach or intestines is obliged to pass again through the capillaries of the liver before going on into the main stream of the circulation. This is an important feature of the renewal of the food supplies of the blood.
We have now traced the blood from the heart through the body back to the heart again, and have seen how in its course some of it will pass through such active tissues as muscles or brain or glands, so that the interchanges can go on by which the fluids in these active tissues can take up needed materials and give off wastes. Also a part of it flows through the digestive organs, where food materials can be taken up, and another part flows into the kidneys, where wastes can be gotten rid of. This leaves us to consider only the passage of the blood through the lungs, where the supply of oxygen is to be taken up and the gaseous waste product, carbon dioxide, is to be disposed of.
The most urgent requirement of the body is the requirement for oxygen. There is under ordinary circumstances at all times some surplus of food materials stored in the cells, so that even though the renewal of their surrounding fluids from the blood should stop, they could keep on going for a time on the material that is stored within them; but there is no such storage of oxygen. The cells in the body lead almost a hand-to-mouth existence so far as their oxygen supply is concerned. They are constantly withdrawing oxygen from the tissue fluids surrounding them, and these fluids are just as constantly withdrawing it from the blood; therefore any failure of the blood to be properly supplied with oxygen results very promptly in a condition of oxygen hunger in the cells. This means prompt cutting off of metabolism, since metabolism is a matter of oxidizing fuel, and oxidation cannot go on unless the oxygen is provided. This urgent need for oxygen is met in the body by having the arrangement for supplying it to the blood much more perfect than the other renewal arrangements. We saw a moment ago that only part of the whole blood stream passes through the digestive organs at any given time and only part of the stream passes through the kidneys. The whole stream, on the other hand, passes through the capillaries of the lungs. This is brought about by having an arrangement whereby the combined venæ cavæ after entering the heart communicate with an outlet in the form of an artery leading to the lungs. This artery, which is called the pulmonary artery, breaks up into capillaries in the lungs, which reunite into the pulmonary vein which comes back to the heart again. It is from the pulmonary vein that connection is made with the aorta, starting the blood on its course through the body again. We see then that the blood passes through the heart twice in each complete round, once as it comes in from the body at large on its way to the lungs, and again as it comes in from the lungs on its way to the body at large.
We have spoken of the heart thus far as a single organ; it is actually two hearts side by side, and these would work just as well if they were at a distance instead of being built into one organ. There has probably been more misunderstanding of the heart by people in general than of any of the other parts of the body. This is because from the earliest times the heart has been looked upon as the seat of the affections, and so powers and properties have been attributed to it to which it is not at all entitled. As we tried to make perfectly clear in a former chapter all intelligence and all feelings are located in the brain; the heart cannot possibly take any more active part in these than can the stomach, liver, kidneys or any of the other parts of the body which are concerned with the maintaining of the tissues in good working order. Probably no one really knows how it came about originally that the heart was endowed with these peculiar gifts. It is true that in time of strong emotion there are changes in the activity of the heart which we can perceive. These occur because the heart is under the same kind of nervous control as are the smooth muscles and glands, and shares with them in the disturbances which accompany emotion; but the real seat of these emotions is, of course, the brain. As a matter of fact the heart is nothing but a muscular pump whose sole function is to keep the blood in motion. From what has already been said it is clear that the heart cannot relax its activity for more than an instant without disastrous results; the pressing need of the tissues for oxygen requires that the blood be kept moving. If there were any other way in which the needs of the cells could be supplied except through the movement of the blood, the heart could be dispensed with perfectly well. We have emphasized this about the heart because it is much easier to understand its working if one thinks of it simply as a pumping organ than if one is attributing to it mystic functions connected with our higher emotions.
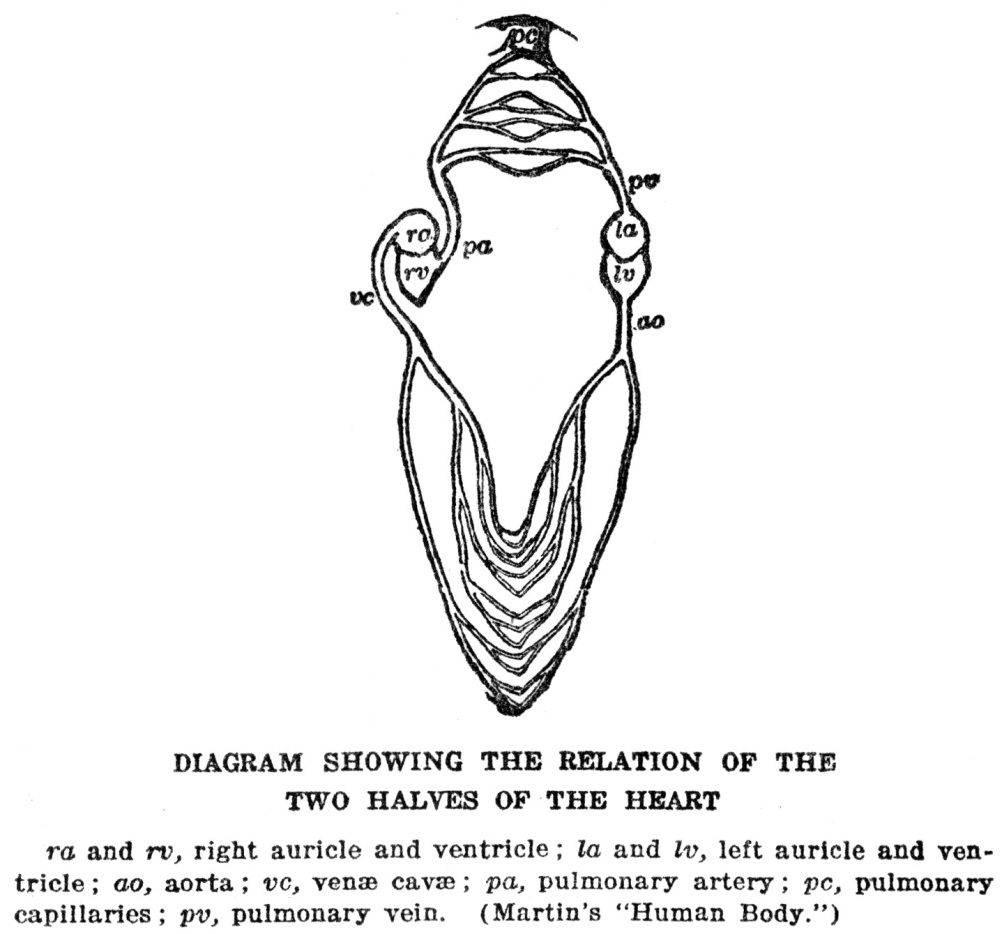
We showed a moment ago that the heart is really a double pump. The relation of the two halves is shown in the diagram. One of the two pumps, that on the right side of the heart, receives the blood from the body at large and pumps it out into the pulmonary artery and through the capillaries of the lungs; the pump on the left side of the heart receives the blood from the lungs through the pulmonary vein and pumps it out into the aorta and so through all the other capillaries of the body. Since the circuit of the body is much more extensive than the circuit of the lungs, the work of pumping is correspondingly greater, and we find the left part of the heart a much more powerful pump than the right. The heart operates as a reciprocating pump, by which we mean that it alternately fills and empties. In this respect it is like ordinary pumps except those of the rotary variety. Any reciprocating pump must have a chamber which will fill and which can then be emptied forcibly. In order that it shall not empty itself back through the pipe from which it filled there must be a valve in the intake pipe which shall close as the pump is being emptied. If, as is the case in the heart, it is emptying itself into a system which permits backflow, there must be another valve in the outlet pipe to prevent the fluid that has been expelled from running back in. Each of the heart pumps consists, then, of a chamber, which alternately fills and empties itself, and two valves, one on the intake and one on the outflow side. In ordinary pumps the forcible emptying is performed by a piston which moves through the pump chamber expelling the liquid ahead of it and then has to draw back, making room for the chamber to fill again. In the heart the forcible emptying is accomplished by muscular action. The wall of the heart consists of a great many muscle fibers so arranged that when they contract they pull the walls of the heart together, making the cavity smaller, or even obliterating it completely. The contraction of these fibers makes up what we are familiar with as the beat of the heart. The frequency with which they contract varies a good deal in different individuals. The average is about seventy-two a minute; but it may be as slow as forty-eight or fifty, or may run up to one hundred and forty or one hundred and fifty a minute. Whatever the rate, in every case there is an alternation of contraction and relaxation; during the relaxation the cavity is filling with blood through the intake valve, the outflow valve being closed, so that no blood that has once been pumped out can rush back in again. By the contraction of the muscles the heart is emptied, the outflow valve being open, and the intake valve being closed to prevent an escape of blood backward into the veins through which it flowed in. The part of the heart that carries on this active pumping work is known as the ventricle; that on the right side, which receives the blood from the body and pumps it to the lungs, is the right ventricle; and that on the left side, which receives the blood from the lungs and pumps it to the body, is the left ventricle.
In addition to the ventricles, which are the active pumps, each side of the heart has an additional chamber known as the auricle, whose purpose is to serve as a reservoir into which blood can flow during the time that the ventricles are emptying themselves. If it were not for the auricles, the movement of blood into the heart would have to stop with every beat, because while the ventricles are contracting the intake valves are closed and there would be no place to which blood could flow, but since each side has its auricle, the flow of blood goes on during the beat of the ventricles, the auricles filling up. The intake valve, in order to operate properly, should be located between the auricle and ventricle, and this is where it is. The vein opens directly into the auricle without any valve between; the auricle opens into the ventricle with the intake valve at the point of junction. The intake valves are given rather formidable names; they are sometimes spoken of as the auriculo-ventricular valves; that on the right side of the heart between the right auricle and the right ventricle is composed of three flaps of membrane, and has therefore been named the tricuspid valve. The intake valve on the left side of the heart, which is composed of but two flaps, is known as the mitral valve. As soon as the beat of the ventricles is over and the ventricular muscle relaxes, the blood which has accumulated in the auricles presses the intake valve open and blood begins to flow through it directly into the ventricle. Both the intake and outflow valves are composed of stout but thin sheets of membrane, so that very little pressure is required to operate them. The weight of the blood that is accumulated in the auricles during the beat of the ventricle is more than sufficient to force the valve open and allow the blood to flow on through into the ventricle. In a heart that is beating seventy-two times a minute, there cannot be much time occupied either in filling or emptying. As a matter of fact both these intervals are measured in tenths of seconds. If we take a heart that is beating at the average rate of seventy-two times a minute the whole of a single beat amounts to eight-tenths of a second. The beat of the ventricle takes about three-tenths of a second or three-eighths of the whole time; the period of relaxation of the ventricle, during which it is filling with blood through the open intake valve, is about five-tenths of a second or five-eighths of the whole time. The movement of blood is rapid enough so that this five-tenths of a second allows the ventricle to fill; in fact much less time than this is required, for in a heart that is beating at twice the average rate, the ventricle still fills with blood between beats.
A word remains to be said about the beat of the auricle. During most of the period when the ventricle is relaxed the auricle is also quiet and blood is pouring directly through it from the veins into the ventricle; but just an instant before the ventricular beat begins, one-tenth of a second to be exact, the auricle contracts, emptying what blood it contains into the nearly filled ventricle; thus, when the ventricle beats, which it does immediately, closing the intake valve at the same time, the auricle is empty and so is able to accommodate the inflow of blood from the vein during the three-tenths of a second that the intake valve is shut. Both sides of the heart work exactly together, the two auricles beating simultaneously, and the two ventricles. Of course it is necessary that this be so, for if they did not keep pace exactly, one with the other, there would be a piling up of the blood either in the lungs or in the veins leading from the body to the heart, and the efficiency of the circulation would be seriously impaired.
We can get a good deal of information about the way our hearts are behaving simply by holding the hand against the chest directly over the heart or by pressing the ear against the chest of some one else and listening to the heart’s action. The physician makes use of a stethoscope, which is merely an apparatus for conducting clearly the sounds which the heart makes, so that it is not necessary to apply the ear to the chest. When one listens thus to the heart he finds that with every beat there are two distinct sounds: the first is a rather dull sound which comes just at the beginning of the beat of the ventricle, the second is a sharp sound occurring just at the end of the ventricular beat. As we saw in Chapter IX, sound is always the result of vibrations, and a great deal of study has been devoted to an attempt to find out where the vibrations come from that cause the heart sounds. It is now generally believed that the first sound is partly the result of vibrations set up in the contracting heart muscle and partly due to vibrations from the sharp closing of the intake valves. The second sound is known to be wholly due to the sudden closing of the outflow valves. The sounds are chiefly of importance in that they enable the physician to determine whether the valves are holding tight or whether there is a leakage of blood through them. In case the intake valve leaks, there will be a backward jet of blood from the ventricle into the auricle with every beat of the heart. This will cause a sort of hissing or murmur which can be heard with the stethoscope in connection with the first sound. If the outflow valve is the one that leaks, blood will squirt back into the ventricle from the aorta, while the ventricle is relaxing. The murmur in this case will come just after the second sound. The skillful physician by comparing the loudness of the murmur when the stethoscope is pressed at different points on the chest and back can determine whether the leaky valves are on the right side or the left side. Thus an accurate diagnosis of imperfect valves can be obtained. Of course the heart will not work well if its valves are not tight any more than will an ordinary pump, so that persons suffering from this trouble cannot have as good a circulation as those whose valves have nothing the matter with them. It is true that in most cases of imperfect valve action there is a compensation in the form of an increase in the size and strength of the heart muscle, so that the circulation is maintained in spite of poor valve action by harder work on the part of the heart. It is evident that in a case of this kind exceptional strains on the heart are more dangerous than if the heart is normal to begin with, so that persons with faulty valve action must avoid physical strains, such as sharp running after street cars or trains, which would be borne with impunity by ordinary individuals. Since faulty valves are a frequent result of acute rheumatism, which in turn comes from pus pockets, and since no way is known to cure a defective valve, once the trouble has developed, it is evident that prevention is of the utmost importance. Physical efficiency is very seriously hampered by poor heart action.
One feature of the heart action with which we are all perfectly familiar is that both the rate and the vigor of the heartbeat vary greatly from time to time. When one is lying quietly, the heartbeat is at its lowest point. It becomes more rapid as one sits up, still more rapid upon standing, increasing still more with the taking of any form of muscular exercise, and in vigorous muscular exercise attains its greatest rapidity and force. The rate in this latter case may be fully double that of the quiet standing position, and, as the vigorous thumping tells us, the force is also very much increased. As we saw in Chapter VII the heart muscle works automatically, contracting and relaxing without being stimulated through the nervous system. The variations in rate and force, however, are the result of nervous action. The heart muscle, as we have already seen, is under the same sort of nervous control as the smooth muscles and glands. It has passing to it two sets of nerves, one to slow it down, the other to speed it up. Both these sets of nerves arise from centers in the brain stem, and both these centers appear to be discharging continuously. So it works out that the heart muscle is under the constant influence of two opposing sets of nerves, and its actual rate and vigor depend upon the balance between them. This has the effect of making the heart extremely responsive to nervous influences. The slightest relaxation on the part of the nerves whose function is to cause slowing will lead to a prompt increase of rate, since the nerves that tend to cause increase are active all the time. Various things may bring about changes in the nervous balance governing the heart; chief of these are muscular activity and emotional disturbance. Practically all the changes in the heart action that we observe from moment to moment can be explained as being due to one or the other of these causes. There are, however, two additional points to be noted briefly; the first is that after muscular exercise the heart slows down very gradually, not returning to its ordinary resting rate for a half hour to an hour after the exercise is over, depending on how long the exercise was kept up. The explanation of this long-continued rapid beat is found in the great outpouring of waste products as the result of the exercise. We have already learned that the functional metabolism of muscular work involves the oxidation of a large amount of energy-yielding material and therefore brings about the production of large amounts of oxidation products. Their presence in the blood serves as a stimulus to

the nerve center in the brain stem, which acts to quicken the heart, and this keeps the rapid beat going until these products are gradually gotten rid of from the body. Another somewhat similar case is the prolonged rapid heartbeat following a violent emotion. The explanation of this we saw a couple of chapters ago in the outpouring of adrenalin that accompanies emotion. One property of adrenalin, as already noted, is to quicken the heart; so, as long as any adrenalin remains in the blood stream, the heartbeat will be faster than normal.
In the above paragraphs we have tried to make clear that the blood is kept in motion through the body by the work of the heart, and that the heart’s activity varies in accordance with the needs of the body; in muscular exercise there is a great increase in metabolism, which means a greatly increased demand both for food supplies and for oxygen. To meet this increased demand it is necessary that the blood circulate more abundantly, and in the automatic speeding up of the heart through the nervous system we have the means by which this is done. In the case of strong emotion, as already emphasized, the bodily reactions are such as put the body into the best possible condition for meeting the emergency. Evidently a quickened heartbeat, by insuring abundant supplies of oxygen and of foodstuffs, contributes to this end. The slowing of the heartbeat, when one lies down, is evidently helpful in enabling the heart itself to recover from any strains that may have been put upon it. The heart is a muscle, and like any other muscle carries on its functional metabolism, which means that it is oxidizing fuel materials and producing waste products. Since it is absolutely necessary that the heart go on beating regularly year in and year out for perhaps eighty or a hundred years, any relief from activity that it can get by slowing down during sleep is evidently an advantage. It has been calculated that the heart muscle really enjoys an “eight-hour day,” by which is meant that on the average the functional metabolism of contraction is going on in heart muscle only about one-third of the time, eight hours out of each twenty-four. During the active waking time the metabolism takes a larger percentage than that, but during sleep enough less to even up.
The heart empties itself into the large arteries; the left heart into the aorta, the right heart into the pulmonary artery. Both these arteries, as well as their subdivisions, are highly elastic. The very best quality of rubber tubing is not superior to our arteries as samples of elastic tubes. The blood, as we have already seen, is quite sticky, and the capillaries through which it must pass in its course around the body are microscopically tiny. The heart pumps the blood out of itself at the rate of four or five quarts a minute or more, according to whether it is working moderately or at high speed. To force this amount of the sticky blood through the tiny capillaries evidently requires very considerable force. As a matter of fact, the force is sufficient so that if it were applied to working a fountain it would force a jet to the height of nearly eight feet. Evidently pumping blood into elastic arteries with this force and against the resistance offered by the tiny capillaries causes the arteries themselves to be not only filled but overfilled, so that their walls are greatly stretched. This fact, that our arteries are elastic and are kept on the stretch by the pressure of the blood within them, is of the very greatest importance to the proper flow of blood and this in turn is so important to our well-being that some of our most serious chronic diseases are traceable to the loss of elasticity on the part of the arteries.
We must remember that once every second, or oftener, the heart is shooting a jet of blood into the large artery which is already stretched with blood and which can empty itself only through the tiny capillaries at the tips of its finest subdivisions. On account of the inertia of the blood stream, room is made for this additional jet of blood by stretching the arteries near the heart more than they were stretched before. The result is that there is an inequality in the amount to which the arteries are stretched, those near the heart being stretched more than those farther along. As quickly as possible this inequality of stretch will be equalized by a spreading of the additional tension out over all the arteries in the form of a wave. This wave makes up what we know as the pulse. It can be felt in any artery that is near enough to the surface so that the finger tips can press upon it. The radial artery at the wrist is the one commonly used by physicians for feeling it. There is a large artery in the neck in which the pulse can also be felt readily, and if one takes the pulse of another person in the neck with one hand and in the wrist with the other he can easily satisfy himself that the pulse in the neck always comes an instant earlier than that in the wrist. This is simply because the pulse spreads from the heart as a wave, and the distance to the neck is not so great as that to the wrist. By the time the finest subdivisions have been reached, the tension is equalized throughout the arterial system, and there is no more pulse. The advantage of this is that the blood flows through the capillaries in a steady stream and not in a series of jerks. This, in fact, is the chief, but not the only, benefit we derive from having elastic arteries. Since the heart operates as an intermittent pump, it is evident that unless the arteries were elastic and so could take up the shock, the blood would have to pass through the capillaries in a series of jerks, exactly corresponding with the beats of the heart. There is abundant proof, which we shall return to in a moment, that to have the blood move through the capillaries in this jerky fashion would be disastrous. Before taking that up, however, we wish to show that by having elastic arteries the actual work of the heart is less than it would be if the arteries were stiff. The reason is really very simple. As was stated a few pages back, the heart is actually emptying itself only during three-eighths of every beat. If the arteries were stiff tubes, and therefore not able to take up any of the blood within themselves, exactly as much would have to pass out through the capillaries during this three-eighths of the beat as was pumped in by the heart. In other words, if the heart were pumping five quarts a minute, five quarts would have to pass through the capillaries in three-eighths of the minute instead of having the whole minute in which to do it. Since the arteries are actually elastic, they are able, by stretching a little more, to make room for part of the blood and so spread the time of its passage through the capillaries out over the whole time instead of confining it just to the period when the ventricle is contracting. Evidently it would take more work to pump five quarts of blood through the capillaries in three-eighths of a minute than in a whole minute.
We measure the work of the heart by what we call blood pressure, about which we are hearing so much these days, so that it will be well to explain as clearly as possible just what is meant by it. The blood pressure really means the pressure of the blood within the large arteries. It could be measured with any ordinary pressure gauge, if it were not for the fact that we cannot very well cut into our bodies to apply gauges to the arteries. For this reason it has been necessary to invent means of finding out what the blood pressure is from the outside. The way it is done is to put a band around the arm, press this band down upon the arm until it squeezes the large arm artery shut, and then, by means of a suitable gauge, find out how much pressure was required. Of course, it is necessary to be able to tell when the artery has actually been squeezed shut, so that the determination of blood pressure in human beings is the work of an expert. Furthermore, blood pressure, as should be clear from what has already been said, varies with every heartbeat. It is at its maximum the instant the heart finishes emptying itself into the artery, and falls off steadily, reaching a minimum just before the next beat comes. The more elastic the arteries, the less difference there will be between the maximum and the minimum blood pressure. The reason for this will be clear when we think that if the arteries were entirely rigid there would have to be a very high pressure during the time the heart was actually beating to force the blood out through the capillaries, but that between beats the pressure would fall off to zero. The more elastic the arteries are, the more nearly do they exert a steady pressure on the blood within them, and so the less will be the difference between the maximum and the minimum pressure. When the physician determines blood pressure, he really determines both the maximum and the minimum pressure, in order that he may be able to judge whether or not the arteries are as elastic as they should be. High blood pressure just by itself might not mean much more than that the heart was beating more rapidly than it should, but a high maximum pressure and a low minimum pressure means nonelastic arteries. This in turn means that the blood is forced through the capillaries in jets rather than in a steady stream, and we may judge of the importance of having a steady flow through the capillaries when we recall the well-known medical proverb that “a man is as old as his arteries.” It is an actual fact that the chronological age of an individual need not have much to do with his physical age. If his arteries continue elastic over a long period of years, he will be physiologically youthful, while if his arteries become rigid he will be physiologically aged, no matter how few his actual years upon earth may have been. Unfortunately we do not know as much as we would like to about the causes of loss of elasticity in the arteries. It does appear, however, that self-indulgence of various kinds is apt to lead to loss of elasticity. For example, even the moderate use of alcohol is now generally recognized by the medical profession as a cause of impairment of elasticity in the arteries. It is probable that intemperance in the use of various foods and drugs leads also to this same condition.
We have just seen that the heart is obliged to maintain high blood pressure in order to force the blood through the tiny capillaries. It will be clear that the actual amount of pressure will depend in part upon how much blood is forced through in a minute and in part upon the extent to which the capillaries offer resistance. It is a familiar law of friction that the smaller the tube the greater will be the resistance it will offer to the passage of liquid through it, so that if the capillaries change in size their resistance to the flow of blood through them is bound to vary. The walls of the capillaries are very sparsely provided with muscle fibers, but the very finest subdivisions of the arteries, which are really no larger in diameter than the capillaries, have much more smooth muscle in their walls. These muscles, as we have already seen, can by their contraction or relaxation make the tiny vessels smaller or larger. We have examples of this in the flushing and pallor of the skin. What we wish to do now is to show how the flow of blood through different parts of the body is controlled by changes in the caliber of these tiny tubes. The muscles in the vessel walls have the double nervous control commonly found in smooth muscles, and both sets of nerves trace back to centers in the brain stem. One of these centers causes the muscles to contract and the vessels to become smaller; the effect of this is, of course, to increase the resistance to the passage of blood through them. The nervous center which brings this about is called vasomotor, or, more properly, the vasoconstrictor center. Vasoconstriction means literally causing contraction of the vessels, which is exactly what this center does. Not all the blood vessels in the body are acted upon through the vasoconstrictor center. Those of the skin and of the abdominal organs are under its control, but those of the skeletal muscles are not. The result of activity of this center, then, is to make it more difficult for blood to flow through the skin and through the abdominal organs, but the ease of flow of blood through the muscles is not affected. Of course, it will follow automatically that the blood stream will be diverted in a large part from the former regions into the latter. The skin and abdomen together make up so large a part of the whole body that marked constriction of the blood vessels in these two regions is bound to cause a considerable increase in blood pressure.
A fairly high blood pressure is necessary for bodily well-being, because only thereby is the brain assured of sufficient nourishment. To see why this is so, we have only to remember that the brain is unfavorably situated for receiving ample supplies of blood. It is at the top of the body, so that the influence of gravity has to be overcome in forcing blood up to it. Also it is completely inclosed in the bony skull, which it in turn fills so completely that there is almost no room for the accommodation of extra blood in it. In all other parts of the body a rise in blood pressure stretches the arteries and so leads to there being actually more blood within them, but there is no room for the arteries in the brain to stretch, so that the total quantity of blood in the brain cannot vary greatly from time to time. The only way in which an increased blood supply can be obtained is by causing the blood to flow more rapidly. This is precisely what happens every time the blood pressure in the body rises. Whenever the vasoconstrictor center sends nervous discharges into the blood vessels of the skin and abdominal organs, causing them to contract, there is a diversion of blood from them directly into the skeletal muscles and also a rise in blood pressure due to the increase in resistance to the circulation, which causes blood to flow more rapidly through the brain. The net result, then, is improvement in the blood supply to the skeletal muscles and to the brain. So important is the blood supply to the brain that the vasoconstrictor center discharges actively throughout the waking part of the day. If for any cause the activity of this center diminishes, there will be an increased circulation in the skin and in the abdominal organs, the blood pressure will fall, the circulation through the brain will decrease, and along with it there will be a decrease in brain function. After this passes a certain point unconsciousness will result. This is what happens when one faints. For some reason or other the vasoconstrictor center becomes inactive, and the series of events just described is set in motion. Fainting ordinarily cures itself automatically, because when consciousness is lost, the individual falls over; this brings his head down on the level with the rest of his body, makes it easier for the blood to flow through it, and so in a moment or two consciousness will be regained. It is a mistake to try to scramble to one’s feet immediately, because until the vasoconstrictor center recovers its ordinary activity, raising of the head above the level of the rest of the body is bound to result in its failure to receive sufficient blood, and so faintness will come on again. It has long been a practice to dash cold water in the face of a fainting person. The physiological value of this is in the sudden stimulation of the sensory nerves in the face by which is set up a stream of nervous discharges which will play upon the vasomotor center, and arouse it again to activity. Almost any vigorous sensory stimulation may have the same effect.
During sleep the vasoconstrictor center is usually not very active; the cause of sleep is not completely understood, but one of the most satisfactory theories regarding it is that during the period of waking the vasoconstrictor center becomes gradually more and more fatigued, and so requires more and more stimulation to keep it active. This stimulation may come in part through the ordinary channels of the sense organs and in part from the higher brain centers, as when one keeps awake by an effort of the will. Upon going to bed sensory stimulation is cut off to a very large extent, also the will to remain awake is no longer present. Under these circumstances the fatigued vasoconstrictor center is under a minimum of stimulation and tends, therefore, to lessen its activity. The result is that the blood pressure falls and presently the circulation through the brain drops below the level of consciousness and the individual is asleep. During the period of sleep the fatigued center recuperates, so that it becomes more susceptible to sensory stimulations and, in course of time, is aroused by such stimuli as accompany the returning day to sufficient activity to restore the brain to consciousness. Of course it will be perceived that there are many things about sleep which are not satisfactorily explained on this theory; in fact no one at the present time pretends that we can account for it completely on this basis of changes of circulation through the brain. It is believed, however, that they have a good deal to do with it and it is certainly true that in healthy individuals the course of sleep follows very closely the activity of the vasoconstrictor center. Undue wakefulness in persons not suffering from disease can nearly always be explained on the basis of excessive activity on the part of this center. The activity may be the result of chemical stimulation, as when persons are kept awake by drinking coffee or strong tea in the evening, or by the persistence of adrenalin in the blood following a period of great excitement. Mental activity, itself, tends to keep the vasoconstrictor center whipped up, so that one who allows his mind to work actively during the time when he should be asleep is very apt to find sleep refusing to come when it is desired. The center is often stimulated from the digestive tract; gastric irritation, even though not acute enough to be recognized as indigestion, may cause wakefulness by arousing nervous disturbances which play upon the vasoconstrictor center. Flatulence, namely the presence of large volumes of gas in the digestive tract, frequently acts as a mechanical source of irritation by which wakefulness is induced. Evidently the factors favoring healthy sleep are the inducing of fatigue, preferably by physical exercise, the avoidance of chemical irritation or of excitement in the hours just before going to bed, and finally the adoption of dietary habits which shall insure good digestion. If an individual in whom all these precautions are combined still continues chronically wakeful, the trouble is sufficiently deep-seated to call for competent medical attention.
Besides the vasoconstrictor center about which we have just been talking there is in the brain stem a center which relaxes the tension of the muscles in the walls of the blood vessels. This is known as the vasodilator center, and it acts in opposition to the vasoconstrictor. In most parts of the body it does not appear to have any very great importance for the simple reason that the blood is under such high pressure that any relaxation of effort on the part of the vasoconstrictor center leads at once to a forcing open of the blood vessels. There are a few regions, however, in which the action of the vasodilator center is of real importance; one of these is in the skeletal muscles. These, as should be perfectly clear by this time, are the seats of our most active functional metabolism. When the muscles are active, great amounts of oxygen and food are being withdrawn from the blood and large amounts of waste material, including carbon dioxide, poured out into it. Only a very rapid circulation will take care of this situation. At the same time the skeletal muscles are the most compact of our living tissues. The cells are crowded together in making up the very strong muscular machine by which our movements are performed. As the muscles contract, they squeeze hard on the blood vessels passing through them. In view of this situation it is very important that the blood vessels be opened as widely as possible during muscular activity, and so we find that the vasodilator center acts to improve the circulation through the muscles, while they are active. In time of special emergency, as we saw a moment ago, the vasoconstrictor center is at the same time engaged in cutting down the blood supply to the skin and to the abdominal organs, thus insuring to the muscles the maximum possible nourishment through the blood stream.
Besides the skeletal muscles there is active functional metabolism in the various secreting glands; a feature of their activity is that it must be very great at certain times, but falls to little or nothing at others; thus during the actual taking and digesting of food the various glands which secrete digestive juices are exceedingly active, but in the intervals they may be doing little or no work. They require a very copious blood supply when they are functioning, but need very little between times. The blood vessels flowing through all these glands are under the influence of the vasodilator center, so that they are opened as widely as possible while the glands are active. They, therefore, receive much more blood in proportion to their size than they would if it were not for the action of this center. Between times their blood supply falls off to that which suffices for inactive tissues generally. In time of emergency the action of the vasoconstrictor center upon the vessels through these glands is such as to cut off the blood supply to them almost completely. This is well illustrated in the dry mouth of the frightened man, showing that the salivary glands have suspended activity, a suspension due largely, if not wholly, to the cutting down of the blood supply through them.
In the section just completed we have tried to give some idea of the way in which the circulating blood provides the various tissues with the materials they require and is adjusted automatically to meet variations in demand from different tissues. Just one more point needs to be noted in completing the account of the conveyer system. The tissue fluids which serve as the connecting links between the circulating blood and the individual cells are necessarily full of waste products, because these come directly from the cells to the tissue fluids and afterward pass on from them to the blood. The result is that the cells are constantly bathed in a solution of their own waste products. There is only one way in which relief from this condition can be obtained and that is by moving the used tissue fluid away bodily and letting it be replaced by fresh fluid. As a matter of fact, this happens; there is an oozing of fluid through the walls of the capillaries from the blood into the tissue spaces; this of course pushes the fluid already in those spaces on ahead of it. If there were no place to which the fluid could go, the result would be a swelling, as the tissue became more and more filled with fluid. This is avoided ordinarily by a drainage system whereby the tissue fluids are carried off as fast as more fluid comes in from the blood, but when liquid is poured out faster than it will drain off, as from a bruise or wrench, we do get a swelling.
The drainage system consists of very delicate vessels known as the lymphatics, which come together into larger and larger vessels, just as the veins do, and finally empty into the vena cava just at the point where it enters the heart. There is no back pressure of blood here, so that the movement of fluid through the lymphatics is not hindered. There is no pump for forcing the lymph to move along; the very gradual motion, which is all that is necessary to keep the fluid from accumulating in the tissue spaces, is brought about by pressure upon the lymphatics resulting from the bodily movements. The lymphatic vessels are like the veins in having valves here and there along them. Whenever by any bodily movement either a lymphatic vessel or a vein is squeezed, the valves insure that the liquid shall be passed along in the direction toward the heart and never in the reverse direction. In the case of the veins this action is not absolutely necessary, since the heart itself is able to maintain the circulation, although it does help, particularly in bringing back the blood from the extremities. In the lymphatics this is the only way by which movement of fluid is brought about. The result is that when the body is perfectly quiet, there is very little movement of fluid through the lymphatics; muscular activity, on the other hand, leads to rather active movement through these vessels. The fact is well illustrated in ourselves. One who sits for a long time humped over a desk finds himself feeling very dull; to obtain relief he stirs about, stretches, and yawns. The dullness was merely the result of the stagnation of fluid in his tissues, causing the cells to be more or less poisoned by their own waste products. By making active movements, these stagnating fluids were forced along to be replaced by fresh liquid direct from the blood and the beneficial effect is felt immediately. Massage properly applied has very much the same effect, although it is doubtful whether as good results can ever be obtained thus as by actual vigorous exercise.
At various places along the lymphatics are little spongelike lumps of tissue known as lymph nodes; the particular spongy substance of which they are composed is called adenoid tissue. This adenoid tissue acts as a filter for the fluid passing through it. Any foreign particles, living or nonliving, that get into the stream are caught in the lymph nodes and held there more or less permanently. Most of the nonliving particles that get into our tissue fluids are from the dust that we inhale, which works its way through the mucous membrane of the respiratory passages and so into the body fluids. The lymphatics that drain the lungs carry along these dust particles and they lodge in lymph nodes at the base of the neck. Persons who have lived in dusty regions or have pursued a dusty livelihood, such as coal heaving, will have by the end of their lives lymph nodes which are literally black with dirt.
The tonsils are lymph nodes at the base of the tongue. Unfortunately they are so near the surface of the throat that they frequently become infected from the throat itself, and so become the seats of pus pockets, as already noted. Closely related to the tonsils are the masses of adenoid tissue at the back of the throat which frequently grow to an undue size in children, and are then known as adenoids. The harm done by adenoids is chiefly mechanical; they may block the Eustachian tubes, and so cause deafness, as already mentioned in Chapter IX, or they may interfere with the free movement of the air through the nasal passages. Children in whom this condition exists are mouth breathers. Adenoids, like tonsils, are subject to infection, and so may give trouble by becoming the sites of poison formation. Adenoids represent always an overgrowth and for that reason may be removed without any possibility of hampering the proper working of the body. Experience has shown that children whose development appears to be hindered by the presence of adenoids are almost invariably benefited by having them removed. The tonsils are normal parts of the bodily structure and as such undoubtedly have a regular work to do, but here again experience has shown that harm from persistent pus pockets is so much greater than harm from loss of function following their removal as to justify taking them out, whenever pus pockets develop in them. There are enough lymph nodes in the region about the throat, so that if tonsils or adenoids are removed any foreign matters that get into the body fluids will still be filtered out.
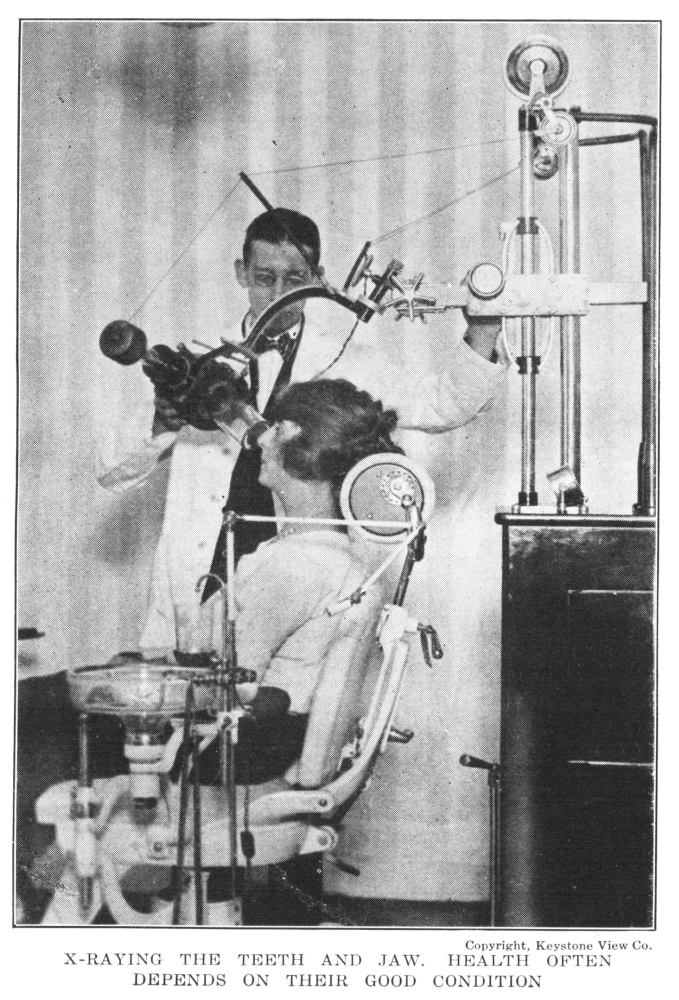
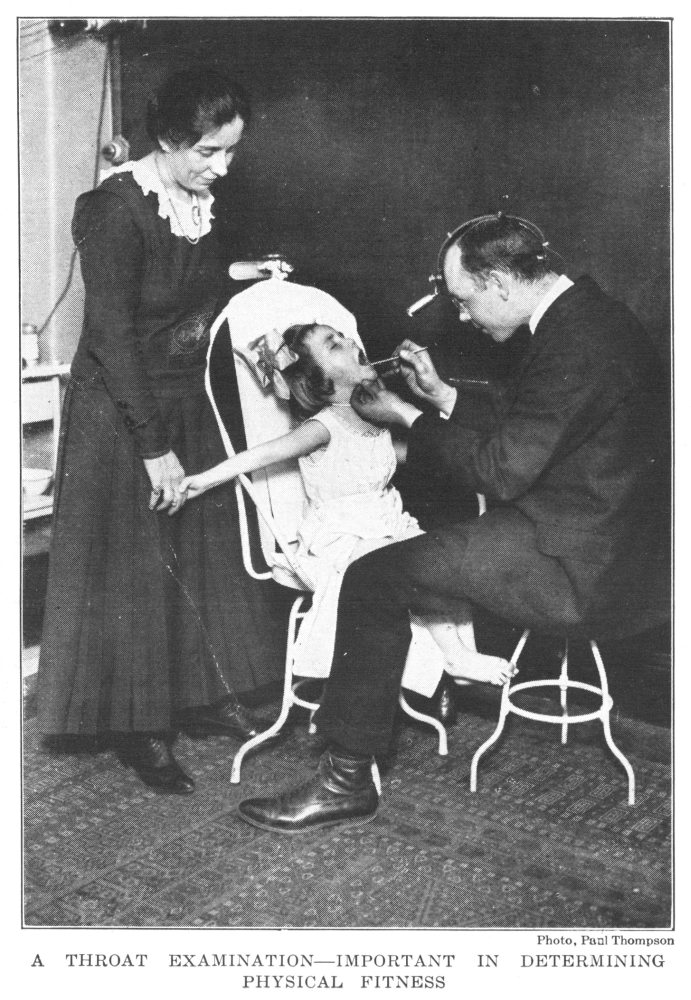
One more function of the lymph nodes must be mentioned; this is their property in preventing the spread of cancer cells. We now know that so-called secondary cancers are the result of the spread of cancer cells from the original seat of the cancer to other parts of the body, and that this spread is much hindered by the ability of the lymph nodes to catch the cancer cells and hold them. Unfortunately sooner or later some of the cells will escape beyond the lymph nodes and so spread the malignant growth throughout the body, but until this happens the cancer is confined to the region where it started, and it is during this period that complete cure by surgical means is possible. It is because of the imminent danger of the escape of cancer cells beyond the restraining lymph nodes that relief by surgery should be sought at the very earliest possible moment. Delay, whether due to carelessness or any other cause, is as certainly fatal in the case of cancer as in any other disease for which a cure is known.
IN order for the blood that circulates through the body to pass on to the body cells the materials which they need, it is evident that the blood itself must have some source from which to obtain the materials. Our present task is to examine this source. In Chapter IV we talked about food and its uses; here we are concerned with the way in which the foodstuffs are taken into the body and prepared for use. We know that the material which we take in is part solid and part liquid, giving rise to the familiar distinction between food and drink. For our present purposes this distinction has no importance and will not be made. What we do have to note is that of the complex mixture of materials which makes up any ordinary meal, some of the substances are ready to be taken up at once by the blood and to be distributed around the body; others have to go through a preliminary course of preparation. All of the dietary accessories, so far as we know, pass from the digestive organs into the blood without change. All or nearly all of the energy-yielding foods, on the other hand, must have a preliminary preparation to which we give the name of digestion; the operation of digestion being to break down the complex foodstuffs that are eaten into simpler materials of which the body can make use. Digestion is made up of a series of chemical changes by which the large molecules of the original foodstuffs are broken into smaller and simpler molecules. This breaking down of large molecules into smaller is a very common kind of chemical process. A feature of it is that under ordinary circumstances the breaking down of the large molecule goes on very slowly, but if the right conditions are provided the breaking down proceeds rapidly. The particular condition which is necessary is the presence in the solution where the large molecules are breaking down of something which will hurry up the process. There are a good many kinds of substances which have this ability to hasten molecular decomposition; those that do it in carrying on digestion are given the name of enzymes. We do not know just how the enzymes act; we merely know that when they are put along with the substance which is to be decomposed, it goes to pieces very much more rapidly than if none of them was present. The enzymes themselves are not used up in the process, so if time enough is allowed, a very small amount of enzyme can bring about the decomposition of a very large amount of material. In our bodies the whole digestive process consists of a succession of decompositions of complex materials into simpler ones under the speeding-up influence of enzymes. We have a number of different kinds of foodstuffs to be decomposed and a corresponding number of enzymes.
The process begins in the mouth; here the food is chewed and moistened with saliva before being swallowed; both the chewing and the moistening are important to insure good digestion later on. The enzymes have no particular ability to penetrate a mass of food material; what they do is to attack it from the outside and work in as it decomposes. Since enzyme action is thus a surface action, it is evident that the larger the surface the more efficient will be the action. Chewing is nothing in the world but a mechanical breaking up of the food to get the largest possible surface. We have sufficient proof of its importance in the digestive disturbances that arise as the direct consequence of improper chewing. Of recent years it has been realized that undernutrition, particularly in children, is often a result of the failure to chew the food properly; it has been found, furthermore, that bad teeth or improperly shaped mouths are very frequently responsible. For this reason in most of our large cities dental clinics are being established for the purpose of inspecting and, if necessary, caring for the mouths of school children. The result of this work is to improve the general average of health among children simply by increasing the extent to which the food is chewed. This fact, together with that described in Chapter XIII as to pus pockets, should impress upon both parents and teachers the fundamental importance of proper care of the teeth. This includes not only the prevention of decay by daily thorough cleaning and the securing at intervals of not more than six months of dental inspection with treatment where necessary, but also in the case of children with deformed jaws the special treatment necessary to bring the teeth into position for effective chewing. Too much stress can scarcely be laid on the importance of these precautions.
The moistening of the food with saliva is likewise important to good digestion. As a matter of fact, without it even the act of swallowing would be impossible. One can easily prove that insufficient moistening prevents swallowing by eating a rather large quantity of a very dry food, like crackers. A common habit of people is to moisten the food with water which they drink instead of waiting for it to be properly moistened by saliva. Contrary to popular belief there is absolutely no objection to drinking water at mealtime. In fact, the presence of the additional liquid in the alimentary tract is probably beneficial rather than the reverse; there is, however, a very serious objection to using water for washing down half-chewed food. The best possible way to judge whether the food is sufficiently chewed is by observing whether it has been sufficiently moistened so that it will be swallowed easily. If so, the chances are that the chewing has been sufficient. Between mouthfuls as much water may be taken as one desires, although if ice water is drunk, it should be held in the mouth until the worst chill is taken off before being swallowed, so that it will not chill the stomach. Since the comfort of drinking ice water is in the cooling of the mouth and throat and not at all in the cooling of the stomach, this increases rather than diminishes the enjoyment one gets from a glass of it on a hot day. So much satisfaction can be obtained by proper drinking habits, that it is a pity to allow the health to be injured by improper habits to such an extent as to necessitate, as frequently happens, the complete abandonment of water drinking with meals.
The saliva is primarily for the purpose of moistening the food, but besides this it is also a definite digestive juice, because it contains one of the digestive enzymes. This enzyme, to which is given the name ptyalin, acts upon starch, changing it to sugar, but not to the particular sugar of which the body makes use. Perhaps we should digress for a moment to say that there are chemically a number of sugars. These are in many respects alike, although they also vary a good deal among themselves. For example, some are much sweeter than others; glucose, which is a sugar that can be made from cornstalks and other plant products, is not nearly so sweet as cane or beet sugar, although one is fully equal to the other so far as nutritive value is concerned. Of all the various kinds of sugar that exist, only those in the glucose class can actually be used by the body, so that all other sugars have to be changed by digestion into a glucoselike sugar before passing on into the blood. The enzyme of saliva converts starch not into glucose but into a more complex sugar to which is given the name of maltose. It, therefore, starts digestion, but does not carry it through to completion. Starch digestion begins during the course of chewing and mixing the food with saliva; it goes on while the food is being swallowed, and for a short time after it enters the stomach, being stopped there sooner or later by means which will be described in a moment. No other real digestion occurs in the mouth, although the chewing and moistening are of very great importance in preparing the food for the digestion that is to come further along.
The food that is swallowed passes down the esophagus and enters the stomach. Before speaking of this it will be necessary to recall what was said in Chapter VII about the behavior of the stomach walls. Between meals these are in a relaxed and flabby state, with the opposite walls lying more or less in contact. There is usually a little liquid and some swallowed air in the stomach, so that it is not actually empty, even when we speak of it as being so. Just before mealtime the walls of the stomach draw up so that instead of a flabby bag we have a fairly tense tubular organ. It is at this time that the contractions of the stomach wall, which we recognize as hunger, begin to come on. There is a sphincter muscle between the esophagus and stomach which closes the opening tightly and so prevents the pressure within the stomach from forcing gas or liquid back up into the esophagus. This sphincter opens automatically only in connection with the act of swallowing. Every time we make a swallowing movement a sort of wave passes down the esophagus, and when this wave arrives at the stomach the sphincter relaxes, allowing whatever was moving down the esophagus to enter. If one watches a horse or cow in the act of swallowing, the rather deliberate progress of this wave down the neck can be followed. What happens as we eat a meal is that every mouthful, as it is swallowed, is passed through the sphincter into the stomach and room is made for it by a gradual relaxation of the stomach wall, so that, as we saw in Chapter VII, the pressure of the stomach against its contents stays fairly steady, in spite of the fact that more and more material is being taken into the stomach from the esophagus. One result of this behavior of the stomach is that the first food that is swallowed is nearest to the stomach walls, and that which is swallowed afterward is nearer the middle, being inclosed on all sides by the previously swallowed food. This is fairly important because the gastric juice is secreted by glands in the wall of the stomach and so will get in contact first with the food that is swallowed first, and only afterward will reach the food that was swallowed later. Occasionally the sphincter between the stomach and the esophagus relaxes unexpectedly; this is said to happen more often in smokers than in nonsmokers. The result is that some of the sharply acid stomach contents are forced up into the esophagus and vigorous swallowing is necessary to crowd them back down into the stomach. The burning sensation which accompanies this is known as heartburn, although, as we have just seen, it is really entirely a matter of the esophagus and has nothing whatever to do with the heart.
Although the stomach carries on a certain part of the work of digestion, its primary purpose, as we shall see presently, is to serve as a storage place into which a considerable amount of food can be placed in a few moments and so enable us to do our eating at three or four definite meals instead of little by little throughout the day. When we rise from table after any meal, we have in our stomachs roughly one-third of our total food supply for the day, the exact proportion depending, of course, on our individual habits as regards our distribution of food-taking among the three meals. During the two to four hours following the meal this accumulated material in the stomach will be passed along little by little to the small intestine, which is the real digestive organ of the body. In the course of this time there will be some additional digestion within the stomach, but not enough to prepare any foodstuffs for actual use by the tissues. The outlet from the stomach to the small intestine is guarded by
another sphincter similar to that which closes the opening between the esophagus and stomach. This sphincter is ordinarily closed tightly, relaxing occasionally to let a little food through into the small intestine. It is interesting to note, however, that the sphincter does not hold against some materials; for example, water that is drunk between meals passes rapidly from the stomach on into the small intestine. It is said that raw oysters, raw white of egg, and other materials of a similar consistency also pass the sphincter immediately. Most other kinds of food material cannot pass unless the sphincter actually relaxes.
Shortly after food begins to enter the stomach churning movements are set up. These consist of regular contractions of the smooth muscle beginning at about the middle of the stomach in the form of a ring and so causing a deep depression to form right around the stomach, which then travels along toward the sphincter at the outlet. This, of course, crowds the food up toward that end; but if the sphincter does not relax, the food instead of escaping simply squirts back through the space left in the middle of the ringlike construction and so is actively churned. A regular procession of these waves travels over this part of the stomach during the whole time that gastric digestion is going on. The outer half of the stomach, the side toward the esophagus, does not join in this churning motion. The walls here remain quiet, merely pressing upon the food to crowd it up into the part where the churning is going on as fast as room is made for it there. At intervals during this process the sphincter relaxes and a small mass of food is crowded through into the small intestine; then the sphincter closes again, preventing more from passing until the former mass has had time to be acted upon by the digestive juices in the intestine. The way in which this sphincter is controlled is one of the very interesting facts of physiology, but before describing it we shall have to say something about actual gastric digestion.
The juice which the glands in the stomach wall secrete is called gastric juice. This contains three important constituents, first of which is hydrochloric acid. From the standpoint of chemistry it is a very interesting thing that the gastric glands should he able to manufacture a mineral acid like hydrochloric acid. That they do so, however, is proven by the facts of digestion in everybody. In addition to hydrochloric acid gastric juice contains two enzymes; the first is pepsin, which begins the digestion of protein foods, although it does not carry it through to completion. The other constituent is the enzyme rennin, which clots milk; this property appears to be useful in that clotted milk will not pass the sphincter into the small intestine as quickly as it would if it were liquid, and so the digestive processes can go on in milk after clotting just as it goes on in any other of our foods. Since milk is the chief, and in many cases the only, food of the young it is of course very important that its digestion should be very efficiently carried on. As the food is churned by the stomach muscles, it becomes mixed with gastric juice. Any milk that is present will be clotted and any protein will begin to be digested. The food will also be mixed with hydrochloric acid. It happens that the salivary enzyme, ptyalin, will not act in the presence of an acid, so that as soon as any of the food in the stomach comes in contact with hydrochloric acid of the gastric juice, the digestion of starch by ptyalin stops. Since gastric juice is mixed only with that part of the food which is being churned, the other portion, that in the part of the stomach toward the esophagus, goes right on undergoing salivary digestion. So, contrary to the old idea that salivary digestion is unimportant because there is not time enough during the chewing and swallowing of the food for it to go on effectively, we now know that the latter parts of the meal particularly may be very thoroughly acted upon by ptyalin before the action is stopped through the contact of this part of the food with the acid of the gastric juice.
The sphincter between the stomach and small intestine is operated by hydrochloric acid, making up a remarkably ingenious arrangement for securing the emptying of the stomach as fast as the food is ready to be passed on into the small intestine. In order that gastric digestion may have full opportunity, it is necessary that the food should be thoroughly mixed with the gastric juice. As soon as this has happened, however, it is proper that the food should be passed along to be acted on by the rest of the digestive juices. Until that part of the food which is next to the outlet sphincter is thoroughly mixed with gastric juice there will be no surplus of hydrochloric acid by which to operate the sphincter, but as soon as the mixing is complete there will be a surplus. We have here a simple case of stimulation by a chemical substance; the mere contact of the acid with the part of the sphincter which fronts on the stomach is sufficient to stimulate the smooth muscles in it to relax, and, of course, as soon as relaxation occurs, the pressure of the churning movements upon the food will crowd that part of it which is nearest to the sphincter through into the small intestine.
Now we encounter the second part of the action which is just as interesting as the first part. The small intestine is a rather narrow tube; its cavity is not much more than three-quarters of an inch or so in diameter. If the outlet sphincter from the stomach were to stay relaxed after it once let go, the rapid pressing of the food from the stomach would fill the small intestine quickly to a distance of several feet. It happens, however, that the next of the digestive juices to act upon the food are the pancreatic juice and bile, which are secreted respectively by the pancreas and liver, and are poured into the small intestine by ducts from these organs just a few inches beyond the outlet sphincter from the stomach. In order for these juices to mix well with the food it is important that only a little food come into the small intestine at a time. Otherwise a large part of it would pass the opening of these ducts so quickly that there would be no chance for it to become mixed with the juice from them. What actually happens is that the sphincter relaxes under the stimulus of surplus hydrochloric acid and then as soon as a little food has passed through closes again. The closing as well as the opening is operated by hydrochloric acid, but there is this difference: the opening is the result of the stimulation of acid upon the stomach side of the sphincter; when the same acid comes in contact with the intestinal side of the sphincter, its presence there causes the sphincter to contract. Of course, this means that whenever any food passes out from the stomach into the small intestine the sphincter is stimulated by acid simultaneously from both sides, but under that condition the stimulus which causes the sphincter to contract is more potent than that which causes it to relax, so that whenever there is acid on both sides the sphincter will be shut. The only way in which it can be opened to allow more food to pass out is to get rid of the acid on the intestinal side. This is fairly quickly accomplished, because both the pancreatic juice and bile are strongly alkaline liquids, so that as fast as they mix with the food they tend to neutralize the acid; the result is, of course, that by the time these digestive juices have impregnated the food mass thoroughly and so have accomplished their purpose, the stimulus which keeps the sphincter closed has disappeared and there is left only the stimulus to open it, due to the presence of acid in the stomach. As soon as this condition is reached, the sphincter will open again, another mass of food will pass through bringing its acid with it, which, of course, closes the sphincter promptly, and so the whole story goes on again. This action has been described in detail because it is one of the very best examples we have of the remarkably ingenious arrangements by which the complex bodily functions are carried on automatically.
From what we have just said it is evident that the proper emptying of the stomach and, therefore, the proper carrying on of the whole process of digestion, depends upon the formation and outpouring of ample supplies of acid-containing gastric juice. A failure of gastric secretion is bound to be followed by a failure of the stomach to empty itself. Instead of going on to the small intestine where digestion is completed and the digestion products passed on to the blood stream to be used by the body, the food mass lies inert in the stomach. After a few hours, if not immediately, the acute symptoms of indigestion are certain to develop. We are describing not a theoretical possibility but an actual happening in the life of thousands of people, sometimes almost daily. Indigestion is one of the great causes of misery and of impaired efficiency throughout the civilized world. It is said that Napoleon lost the Battle of Waterloo on account of an acute indigestion which befell him on the morning of the crucial day.
There are several causes of indigestion. Among them should be mentioned food poisoning; that is, the inadvertent taking of some material which instead of being a food proves to be actively poisonous. A second type about which we shall have something to say presently is the result of poisoning by substances which are products of the putrefaction of the intestinal contents. A third type, and the one in which we are particularly interested at this moment, is the one which results from the failure of gastric juice to be properly secreted. A fourth occasional cause of indigestion, which should be mentioned in passing, is that which results from eyestrain, and is avoided by properly fitted glasses. There is comparatively little food poisoning; most persons who are susceptible to poisoning from particular kinds of foods find it out promptly and learn to avoid the article which poisons them. A good deal of indigestion is the direct result of intestinal sluggishness; this usually comes on gradually enough to give warning of its approach. Sharp attacks for which there seems to be no justification are nearly always the result of the failure of gastric secretion. Because of the practical importance of this topic a very large amount of study has been devoted to it, and its story makes one of the very fascinating chapters of physiology.
Our space permits us to touch on it only very lightly. It begins with the time back in the middle of the eighteenth century when a couple of Italian investigators fed sponges inside perforated metal capsules to birds and animals and after recovering and squeezing the sponges demonstrated that the juice which had been soaked up would dissolve meat. The scene then shifts from Italy in the middle of the eighteenth century to our own country at the close of the first quarter of the nineteenth, when a French-Canadian lad, by the name of Alexis St. Martin, was brought in to the frontier army post on Mackinac Island with a gaping gunshot wound in his side. The victim of this accident recovered, but meanwhile a permanent opening into his stomach was left. The army doctor on duty at that post, William Beaumont by name, perceiving the unique opportunity for the study of digestion that was in his hands, employed the young man in his family and for many years studied the digestive process as it went on in his stomach. Next we turn to Russia, where an eminent physiologist, who was still living at last reports, demonstrated by making artificially an opening into the stomach of animals, similar to the one which was made by accident in the case of St. Martin, that the secretion of gastric juice is under the control of the nervous system and, furthermore, that it is under the control of that part of the nervous system which we have already learned regulates the functioning of smooth muscles and glands. Our present knowledge of the nervous system would lead us to expect nothing else, but at the time when these discoveries were made much less was known than now about how the glands, as well as the smooth muscles, are controlled. This Russian investigator showed that it is not necessary for food to enter the stomach in order for gastric juice to be secreted. The enjoyment that accompanies the taking of food into the mouth and so getting its taste and flavor is sufficient to arouse nervous disturbances which excite the gastric glands to activity. On the other hand, unfavorable emotions such as anger, fright, or worry, prevent the discharge of these nervous disturbances and so the glands remain inactive. Hand in hand with this, as we saw in Chapter XII, there will be a complete absence of muscular activity in the stomach; not only does the food fail to be churned, but gastric juice fails to be poured out. No digestion goes on in the stomach nor is the food passed on into the small intestine to be digested there. We can expect nothing else than that indigestion will come on. It appears that in human beings, with their high nervous organization, simple weariness may suffice to hinder the outpouring of the nervous disturbances upon which gastric secretion depends, so that one may not be worried at mealtime, but may be simply overtired and still fail of digesting the food. Fortunately we have means of avoiding this; it has been shown that meat extracts have a definite chemical effect upon the gastric glands, arousing them directly to activity. A hot, well-flavored soup at the beginning of a meal is a great aid to digestion, both because it tends to arouse us to active enjoyment of the meal and also because of the chemical influence it exerts directly upon our gastric glands. Cheerful conversation at table is an aid to digestion, because it contributes to our enjoyment of our food. Heated or angry discussion is to be avoided because of the danger that unfavorable emotional disturbances may be aroused, hampering or even putting a stop to the digestive activities.
We have here a powerful argument in favor of good cookery; some may be inclined to think that food is food and that one kind is about as nourishing as another. Not only theoretically, but in practice, food that is attractive both in appearance and in flavor benefits the body more than unattractive and badly cooked food. Of course, we must recognize that no statement of this sort has absolutely general application. The old proverb which says that “hunger is the best sauce” continues true. Hard-working manual laborers and actively playing children enjoy and digest food that is indifferently attractive. Brain workers and people of moderate health are better for an attractively prepared dietary.
We have now traced the food into the small intestine and will remember that up to this point the actual act of digestion has not gone very far. The enzyme of saliva has decomposed part of the starch, or perhaps all of it, into a complex sugar; the enzyme of gastric juice has split some of the very complex proteins into simpler ones. Of the final products of digestion none has yet been formed; their formation is the work of the small intestine. Into the upper end of it, as we have already seen, pancreatic juice and bile are poured from the pancreas and liver respectively. The pancreatic juice contains three enzymes; one of them is identical with that of saliva and completes the decomposition of any starch that the saliva has failed to act upon; the second is an enzyme which acts upon proteins, differing from pepsin of gastric juice in that it carries the splitting of proteins to completion; the third is the enzyme which acts upon fats. Fats are the most troublesome of all the foodstuffs from the digestive standpoint, because they are insoluble in water. Not only do they themselves offer difficulty, but they smear over the other food masses and make it hard for the digestive juices to get at them. Although the enzyme for digesting fat is secreted by the pancreas, its successful working depends on the presence of bile from the liver. If there is an obstruction of the bile passages, so that bile cannot be poured out into the small intestine, the digestion of fats stops and, along with it to a considerable extent, all the other digestive processes of the intestine. There is a familiar common condition known as jaundice, in which the bile passages become stopped, usually as the result of inflammation. When one is suffering from this condition, it is very important that not much fat be included in the diet; otherwise there is likely to be trouble with indigestion.
In addition to the pancreatic juice and bile there is a secretion known as intestinal juice which is poured out by small glands scattered along the wall of the small intestine. These secrete enzymes which convert the various kinds of sugar into the particular sugar, glucose, which is the one the body can use. The glands also secrete an enzyme which can carry on to completion protein digestion. Thus there is ample insurance that all the foodstuffs shall be made ready for the use of the body.
The next thing in order is for the digested foods to pass from the digestive cavity into the blood. This process is known as absorption. The first part of the alimentary tract from which absorption might occur is the stomach, but, as we have just seen, the process of digestion is not carried far enough in the stomach to fit the foods for going on into the blood stream. Absorption from the stomach is, therefore, undesirable rather than desirable. This is interesting because many condiments, particularly the hot spices and mustard, irritate the stomach lining in such a way as to promote absorption through it. Formerly this was looked upon as an argument in favor of the use of these condiments, but we now realize that it is rather an argument against them. The same applies to alcohol. Alcohol irritates the lining of the stomach and produces absorption through it. Unfortunately for those who desire a physiological argument in favor of the use of alcohol at meals this is a disadvantage rather than an advantage. The great region of absorption is the small intestine, where the digestive process is completed and where, therefore, absorption can properly go on. As to the method of absorption not a great deal needs to be said; there is a very delicate membrane lining the intestine, and just beneath this are the very numerous capillaries through which the blood is flowing; also in the spaces between the capillaries is the tissue fluid which drains into the lymphatic system. The products of protein digestion and glucose pass through the lining membrane of the intestine and through the capillary walls directly into the blood stream and are carried along in it. The digested fats, on the other hand, after passing through the intestinal lining enter the tissue fluids instead of the blood. Incidentally it should be noted that they are rebuilt into tiny fat droplets during the process of absorption, so that what we find in the tissue fluids of the intestine are true fats and not digestion products of fat. These tiny fat droplets pass away from the intestine by way of the lymphatic system. This, as we saw in the last chapter, drains finally through large lymphatic vessels into the great veins just at the entrance of the heart, and so the fat finally reaches the blood stream, but not so directly as do glucose and the digestion products of protein.
There is a reason for this difference in the handling of fats and the other foodstuffs which will be clear when we recall the point made in the last chapter that all the blood which flows through the walls of the intestine is collected in the portal vein and must pass again through the capillaries of the liver before entering the circulation at large. What happens is that all the digestion products, except the fats, pass through the capillaries of the liver, but the fat gets into the blood stream by another way and does not go through the liver en route; for some reason or other it seems to be to the advantage of the body that the fat should not be allowed to circulate through the liver capillaries. It is, as we shall see in a later chapter, a distinct advantage to have the glucose pass through the liver, and there is probably also a good reason for having the digestion products of protein take that course.
The digesting food is moved along the small intestine by contractions of its muscular wall, which travel along slowly in the form of a wave, very much as the wave of swallowing passes down the neck of a cow or horse. These waves do not occur regularly but only now and then; what happens is that several inches or two or three feet of the small intestine will be filled with a food mass which stays for a while at that place, digestion going on in it, and the digested food being absorbed through the wall. Both the digestion and the absorption are aided by a kind of churning motion made up of a series of contractions spaced about an inch apart along the part of the intestine where the food mass lies. These contractions form and disappear quite regularly at the rate of several a minute. They have the effect of squeezing the food mass rhythmically, but do not move it away from the place where it is lying. After this churning has been going on for a while it subsides, leaving that part of the intestine entirely quiet; then a contraction wave sets in at the end of the food mass toward the stomach and pushes the mass bodily along the intestine to a new section where the wave dies out and the churning motion begins again. This process is repeated with every food mass as it comes through from the stomach into the intestine until, finally, usually three or four hours after the first food has commenced to come into the intestine from the stomach, the whole mass has been propelled the length of the small intestine and what is left has been passed on through into the large intestine. The small intestine is about twenty-five feet long in man; flesh-eating animals of about the same size have intestines only about one-half as long; grazing animals the same size, on the other hand, have intestines two or three times longer than those of man; this intermediate length of the human intestine is ordinarily looked upon as indicating that man is adapted to a mixed diet, being neither strictly a flesh-eating nor strictly a vegetable-eating animal.
During the slow passage of the food mass through the small intestine the processes of digestion and absorption are completed, so that very little material enters the large intestine except the indigestible parts of the original food. These are made up mostly of cellulose and similar indigestible vegetable materials, but they include also indigestible fragments of gristle and other meat remnants. It is also worthy of note that the amount of water does not decrease much during the passage of material through the small intestine; in other words the material that enters the large intestine is about as liquid as that which passes from the stomach in to the small intestine. This does not mean that there is no absorption of water from the small intestine; in fact, water that we drink is probably absorbed quite rapidly; what it does mean is that enough water is secreted into the intestine from the blood in the lower parts to keep the whole mass liquid; this is of course important as an aid both to digestion and absorption.
The large intestine, as its name implies, is a tube of much larger diameter than the small; it begins low down on the right-hand side, passes up to about the level of the stomach, then across the body and down on the left-hand side, making thus a sort of inverted “U” in the abdomen; the space within the “U” is filled up by the loops and coils of the small intestine. The first part of the large intestine is devoted to the absorption of water; the result of this absorption is to leave the contents of the large intestine in a semidry condition; absorption of water goes on continuously, so that the longer the material remains in the large bowel, the drier will it become. In the ordinary course of events the material, as it is dried out, is passed along through the large intestine and at intervals, which should not exceed twenty-four hours, the accumulated material is discharged from the body.
Various facts about the functioning of the intestine which have seemed to many people mysterious are easily explained on the basis of what we have next to talk about, which is the presence both in the small and in the large intestine of flourishing colonies of bacteria. It has perhaps not occurred to most of us that we are in the position of involuntary hosts to enormous numbers of these microscopic plants, but that is exactly what we are. There is no possibility of preventing this, since the combination of warmth, moisture, and abundant food, which is afforded within the intestine, is the most favorable possible situation for many kinds of bacteria, and no effort on our part would enable us to be free from them. Fortunately, in the ordinary course of events, we are not affected one way or the other by their presence. In the small intestine their chief diet consists of the sugars which are produced as the result of starch digestion or are eaten directly. In connection with nourishing themselves from these sugars, they bring about what is known as alcoholic fermentation; that is, some of the sugar is converted into alcohol with a by-product of carbon dioxide. Exactly the same thing happens when yeast develops in a mass of dough; in the latter case the bubbles of carbon dioxide are what we are after, since they are what makes the bread light, the alcohol being driven off by the heat of cooking. In the alcoholic fermentation that goes on in our small intestines the carbon dioxide is a waste product and is passed on out of the body, but the alcohol is absorbed and used as fuel. Little has been said thus far about the physiological effects of alcohol. At this point all we care to say is that alcohol can be oxidized by the body tissues with the liberation of energy and so is a perfectly good fuel food. In the minute amounts in which it is absorbed from the small intestine it is utilized completely and there is not the slightest reason to suppose that it has any ill effects whatever. The objections to the use of alcohol, which have led to its abolition as a beverage in this country, depend on certain definite pernicious drug effects which it shows when consumed in any but the smallest amounts, and which destroy the usefulness of what would otherwise be a valuable article of human diet. Fermentation within the small intestine ordinarily goes on entirely unperceived by us. Certain foods, or the invasion of the intestinal tract by certain species of bacteria, may change the fermentation in such a way that irritating substances are produced which cause the movements of the small intestine to be very greatly increased; its contents are swept on through in very much less than the usual time and we have the condition known as diarrhea.
Within the large intestine bacterial action goes on fully as vigorously as in the small, and because of the smaller relative amounts of digestible food and of water a much larger proportion of the intestinal contents consists of the bodies of the bacteria and of the products of their metabolism. Since the sugars have been completely absorbed out by the time the food reaches the large intestine and the remaining materials upon which the bacteria can feed are more largely of the protein class, the bacterial action changes from fermentation to that which we commonly describe as putrefaction. This type of action, instead of giving rise to alcohol and carbon dioxide, produces a number of highly offensive compounds; many of these can pass through the lining membrane of the large intestine into the blood and so can circulate around the body. Of late years we have realized that these products of intestinal putrefaction are highly poisonous, especially if present in the blood stream in any considerable amount. The term “auto-intoxication,” which we run across frequently in health literature, means strictly the poisoning of the body by the products of intestinal putrefaction. Of course, the condition is much aggravated if putrefaction is allowed to go on for too long a time. The ill feelings which result from constipation and which in the life of very many people constitute a really serious impairment of health are the direct result of poisoning by these putrefaction products. The obvious remedy is the avoidance of intestinal sluggishness; unfortunately this is easier in theory than it often is in practice. Probably two conditions of modern life are chiefly responsible: the first is our tendency to make the diet more and more highly concentrated; that is, to leave out of it more and more the indigestible parts. The result is that not enough indigestible material enters the large intestine to make a sufficient bulk upon which the intestinal muscle can work in moving the mass forward. While bulk is accumulating, both absorption of water and putrefaction are going on, until by the time a sufficient mass is present, it is so dry that the muscles are not able to move it along and it has produced undue quantities of poisons. Obviously the way to treat this condition is by eating more indigestible material. For this purpose there is nothing better than apples in their season. The old proverb “An apple a day keeps the doctor away” has very sound common sense back of it. Apples are not ideal for this purpose for all people, since they frequently cause a distressing evolution of gas or even headache. Nearly everybody, though, by experimenting, can find a time of day at which an apple can be eaten without any digestive trouble whatever, and frequently with considerable benefit. Apples are valuable simply for their bulk of indigestible substance. The process of cooking converts most of this into digestible material, so that for this particular purpose they must be eaten raw. There are various other fruits such as figs, prunes, and raisins which function similarly and in addition have a direct stimulating effect upon the intestinal movements and so favor the discharge of material. Some people can eat popcorn to good advantage, although others suffer from gas distress if they do so.
The second condition of modern life which favors intestinal sluggishness is the sedentary habit which so many of us, both men and women, have. A vigorous outdoor existence is practically never complicated by auto-intoxication. Anyone who can maintain habits of active exercise will usually find himself troubled little by this condition. One further point should be made, and this can scarcely be over-emphasized, since it probably has as much to do as any other single factor in the avoidance of auto-intoxication; this is the development of regular habits in the matter of evacuation of the bowel. It is a general fact of the operation of smooth muscle that it readily develops certain habits. We have already seen an example of this in the behavior of the stomach in connection with mealtime. As we have noted, the stomach lies flabby between meals and enters into a state of tension just about the time that we are in the habit of eating. This adjustment is made no matter what particular habits individuals may have. Those who habitually eat only two meals a day will have this tightening of the stomach twice a day; others whose habit it is to eat five times a day will have a similar tightening five times in the twenty-four hours. Similarly the large intestine can establish a regular habit with regard to evacuation. This is best done in childhood, so that parents by insisting upon regularity in their children in this respect can usually assure them of a lifetime in which there will be little trouble from auto-intoxication. On the other hand, parents who are neglectful of their children’s welfare in this respect are laying up for them a lifetime of trouble and very much discomfort.
In concluding this part of our discussion we should note that artificial stimulation to intestinal activity should be regarded as a measure of last resort and under no circumstances as a habitual means of inducing evacuation. Persons who allow themselves to become dependent upon laxatives are laying up for themselves trouble, since these invariably become less and less effective, making it necessary to increase the dose and finally establishing a condition in which only the vigorous efforts of a physician will restore the body to normal. A safe general rule is that anyone who finds himself becoming dependent upon laxatives should immediately put himself under competent medical care for the purpose of restoring his system to normal functioning in which he will not be dependent upon drugs.
EVERY cell must have oxygen for its metabolism. This it must get from the tissue fluid upon which it fronts, and tissue fluid in turn must get it from the circulating blood. The blood in turn has to get it from somewhere and the place from which the blood gets it is called a lung or a gill, according as the animal breathes air directly or gets its oxygen out of the water. The purpose of the present chapter is to trace oxygen from the air through the blood to the tissue fluids and so to the living cells. We saw in the chapter on “Blood” that we have a special substance, the hemoglobin, which helps in the transportation of oxygen. We shall have occasion here to see how it does this. It is well to bear in mind that for practical purposes we include the methods by which carbon dioxide is gotten rid of along with the study of the supply of oxygen, so that although this chapter carries the heading “The Service of Supply of Oxygen,” we shall also study in it how the carbon dioxide is carried away. This is a convenient way of dealing with the subject, because the two gases are handled in very much the same manner; it is also made necessary by the fact that the control of the apparatus by which these gases are handled is so interwoven that the transportation of one could not well be studied without giving attention also to the transportation of the other.
The problem of the oxygen supply and of the removal of the carbon dioxide is in theory very simple; the air contains a large percentage of oxygen and a very small percentage (three parts in ten thousand) of carbon dioxide. If the blood is exposed to air with nothing between but a very delicate membrane, oxygen will diffuse from the air into the blood until the blood has taken up all that it is capable of holding. As the blood circulates around the body and comes to the tissue spaces where there is little or no oxygen, because the living cells are constantly taking it up and using it, the oxygen which previously diffused into the blood will diffuse out into the tissue spaces. The only special arrangements that have to be provided are a sufficiently great surface of exposure to the air, so that no matter how rapidly the blood may be flowing it shall be able to take up all the oxygen it can hold, and an arrangement for making sure that the blood can hold and carry as much oxygen as the tissues need. The first of these requirements is met by the special construction of the lung or gill; the second by having present in the blood stream a chemical substance (hemoglobin) which automatically takes up large quantities of oxygen and so insures that sufficient shall be transported.
In principle the structure of a gill corresponds with that of a lung; since we are particularly interested in the working of our own bodies we shall content ourselves with describing only a lung. We saw in the chapter on the Circulation that the pulmonary artery which leads away from the right side of the heart breaks up into a system of capillaries. These capillaries are thousands in number and they are spread out over the whole lung surface. The lung itself consists of a hollow bag with very thin and very elastic walls connecting with the throat by means of the windpipe. In reality the bag is double, for the windpipe splits at its lower end into two tubes, known as the chief bronchial tubes, and these subdivide repeatedly until their fine terminals end in the elastic lung sacs themselves. We spoke of the lung as a bag; in reality it is a system of thousands upon thousands of separate tiny bags. The structure is comparable to that of a bunch of grapes, the stem representing the chief bronchial tube, the smaller stems the subdivisions and the individual grapes the lung sacs proper. The advantage of this arrangement is, of course, in the very large surface which it gives; every one of the individual lung sacs has its wall filled with capillaries and there are so many of the tiny individual sacs that the total surface over which the blood is spread is measured in hundreds of square feet. We cannot imagine any other arrangement by which so large a surface of exposure could be packed away into a cavity the size of the human chest.
Of course, we see immediately one serious defect of this arrangement of the lung surface; every one of the individual sacs is full of air and so the blood vessels which line its wall have exposure to air, but between the individual lung sac and the outside atmosphere is, first, the very tiny bronchial tube with which the sac connects, then the somewhat larger one into which that opens, then a still larger one, and so on until we come by way of the chief bronchial tube and windpipe up to the throat, and so through the mouth or nose to the outside. It is quite evident that this system of passages does not permit of a very free movement of air. We must realize also that the blood which flows through the walls of the lung sacs must constantly take up oxygen from the air within the sacs, if it is to meet the needs of the body tissues. Simple diffusion through the narrow bronchial tubes could not possibly bring oxygen into the lung sacs fast enough to supply the requirements of the blood flowing through their walls. The situation is met by active lung ventilation; that is, by forcibly changing the air in the lungs at frequent intervals. The way in which this is done is, as we all know, by breathing. Breathing is nothing but a bellows movement of the chest by which air is alternately expelled and allowed to enter. It does not require a very active lung ventilation to keep the air in the lung sacs sufficiently supplied with oxygen under conditions of bodily quiet. Our ordinary breathing movements are gentle, less than a quart of air is breathed in and out again in every breath, and we breathe only fifteen or sixteen times a minute. Of course, when there is vigorous functional metabolism, as in brisk muscular exercise, the oxidation processes in the tissues go on at a very much more rapid rate, and correspondingly larger amounts of oxygen must be carried by the blood to meet the demand. Under these circumstances there is an improvement in lung ventilation, the movements of the chest are greater and also happen more times in a minute.
The act of breathing is carried on by ordinary skeletal muscles. This is the only act connected with bodily maintenance of which this is true. Our other “vital” organs are operated by means of smooth muscles. On account of this difference we have a certain degree of control over our movements of breathing. As we all know, we can hold the breath for a short time without difficulty, or can breathe more quickly or more deeply any time we choose. In this respect breathing differs strikingly from either the heart action or the movements of the digestive organs, over which we have no direct control at all. Our control of the muscles of breathing is, however, rather limited; we cannot hold the breath indefinitely. This means that the nervous mechanism which causes the muscles to contract will work in spite of the efforts of our will to prevent it. The actual machinery is very much like that which has already been talked about in connection with other “vital processes.” We have a “center” in the brain stem from which the nervous discharges come. This center is located immediately adjoining the vasoconstrictor center about which we learned in Chapter XIV. Because of the location of these two important centers in a single very small space, the spot where they are was named by a French physiologist more than a hundred years ago “the vital knot”; the point of this is that death can be induced more quickly and with less actual tissue destruction here than anywhere else in the body.
The center which controls breathing has been named the “respiratory center.” It discharges automatically about fifteen or sixteen times a minute, causing the muscles of breathing to contract and so the bellows action of the chest to be carried on. Like the other centers in the brain stem this one can be acted upon by nervous disturbances passing into the brain stem from the sense organs. Perhaps the best example of this is the modification of breathing that comes as the result of a dash of cold water on the skin. Most of us have noticed that we give a sort of gasp upon stepping suddenly under a cold shower or plunging into cold water. It may not have occurred to us that this gasp is entirely involuntary, but we can easily prove that it is by trying to breathe with perfect regularity at the moment of stepping under a cold shower. We shall easily convince ourselves that this modification of the breathing is something over which we have no control. It is, as a matter of fact, an excellent example of a simple reflex. Coughing and sneezing are other reflexes in which sensory irritation of some sort acts upon the respiratory center modifying its discharges. In addition to these reflex changes in breathing we have also the familiar effects of muscular exercise. We know that after even moderate exercise the breathing is quickened somewhat, and after vigorous exercise it becomes very rapid and deep, and that after very severe exertion, particularly in an untrained person, the puffing and blowing is not only pronounced but even distressful. We shall see presently how muscular exercise brings these changes about, but before doing so it will be necessary for us to take up the movement of the gases between the lungs and the tissues, by way of the blood; oxygen from lungs to tissues, carbon dioxide from tissues to lungs.
By lung ventilation the air in the tiny individual lung sacs is kept supplied with oxygen and also measurably free from carbon dioxide. From this air there is a continuous diffusion of oxygen into the blood. The first oxygen that diffuses in may dissolve in the blood liquid just as oxygen will dissolve in any water to which it is exposed, but as this goes on the hemoglobin of the red corpuscles begins to take up oxygen, forming a chemical compound to which is given the name of oxyhemoglobin. If there is enough of the gas present, every molecule of hemoglobin will take up oxygen to its full capacity. The amount that will dissolve directly in blood is so slight that to all intents and purposes the ability of the blood to carry oxygen depends on the extent to which hemoglobin can combine with it. It is important to emphasize this, because it means that there is a definite limit to the amount of oxygen that the blood can carry, a limit which is reached as soon as the hemoglobin is saturated. Hemoglobin has so great a power of combining with oxygen that the moderate lung ventilation which ordinary quiet breathing gives suffices usually to saturate it. Now and then we encounter statements which give the impression that there is a virtue in deep breathing in improving the amount of oxygen which becomes available for our tissues. As a matter of fact, this is not the case; ordinary quiet breathing when the body is at rest saturates the blood with oxygen, which means that it is carrying its full cargo, and evidently no amount of deep breathing can make it do more than that. We should not be understood as intimating that deep breathing is not a valuable exercise; the point which we wish to emphasize is that its value does not lie in affording an increased supply of oxygen.
Oxyhemoglobin is of a bright scarlet color; hemoglobin itself, not combined with oxygen, is a very dark purplish color; partial combinations are brighter and brighter as they contain more oxygen, so that an expert can judge of the degree to which any specimen of hemoglobin is combined with oxygen by noting its color in comparison with fully saturated hemoglobin. The combination of hemoglobin with oxygen takes place as the blood is passing through the capillaries of the lungs; therefore the blood which leaves the lungs has the bright scarlet color characteristic of oxyhemoglobin. This blood is called arterial blood, the reason is that it is the kind that is found in the arteries of the body in general. It happens that it makes its first appearance in the pulmonary vein, by which it is conveyed from the lungs to the left side of the heart, so that the expression arterial blood does not mean anything in particular except to describe blood in which the hemoglobin is saturated with oxygen. This blood is pumped out by the left side of the heart to all the parts of the body; in its passage through the capillaries it is in a region where there is active utilization of oxygen by the living cells. These are steadily taking up oxygen from the tissue fluids about them, so the blood in the capillaries, which is carrying an abundant supply of oxygen, is brought in contact with tissue fluids containing little or none, with only the delicate wall of the capillaries between. Under these circumstances rapid diffusion of oxygen from the blood into the tissue fluids takes place and accompanying this there is a breakdown of oxyhemoglobin, so that not only most of the oxygen which was dissolved in the blood passes out, but also a considerable part of that which was formerly in combination with hemoglobin. Under ordinary circumstances only from one-fourth to one-third of the oxyhemoglobin decomposes during the rapid passage of the blood through the capillaries; thus the blood that goes on into the veins will still be carrying two-thirds or more of the total oxygen cargo. The color of venous blood will be darker than that of arterial blood, because it contains a good deal less oxyhemoglobin, but it is nowhere near so dark as is blood in which all the oxyhemoglobin has been decomposed. This is well recognized in melodramatic fiction where the wounds of persons who have met death by strangulation are described as oozing black blood. It is an actual fact that blood from which all the oxygen has been withdrawn is so much darker than ordinary venous blood that it gives the impression of being black, although it is really a dark purple.
If we should be moved to inquire why so small a fraction of its whole store of oxygen is given up by the blood to the tissues ordinarily, we shall find the answer in remembering that the demand of the body for oxygen is extremely variable; every increase in functional metabolism means an increase in the amount of fuel that is oxidized and therefore an increase in the amount of oxygen that is required. Actual measurements have shown that in very vigorous muscular exercise the oxygen consumption may be approximately ten times as great as in complete rest. In order that this very greatly increased metabolism may be carried on it must be possible for the blood to deliver approximately ten times as much oxygen to the active tissues as it delivers to them when quiet. There are just two ways in which this can be done; one is by a more complete decomposition of the oxyhemoglobin by which all its oxygen is set free; the other is by a more rapid movement of the blood. It is by a combination of these two that the oxygen requirements of the body in times of vigorous metabolism are taken care of. As has already been said, the heart rate is just about doubled in vigorous exercise. There has also been shown to be some increase in the amount of blood that it pumps with every beat. The result is that more than twice as much blood leaves the heart in a minute under these circumstances as in time of rest. The oxyhemoglobin is also completely decomposed when the tissues are active, and these two facts together are sufficient to account for the great increase in the oxygen supply.
Hand in hand with the increased consumption of oxygen, there is of course an increased production of carbon dioxide and of water, since the oxidation of fuel substances produces these waste products. The cells are always pouring both out into the tissue fluids, but to a very much greater extent when they are actively functioning. We need make no effort to keep track of the water, since it merely adds itself to the water already present, and we shall consider later how the water supplies of the body are handled. The carbon dioxide, however, must be gotten rid of, and the mechanism for getting rid of it must work efficiently, otherwise metabolism itself will be hampered, since it is a familiar law of chemical action that if the products of an action are allowed to accumulate they interfere with its further progress. The method of getting rid of carbon dioxide is by simple diffusion from the cells into the tissue fluids and from the tissue fluids into the blood. Carbon dioxide is many times as soluble as oxygen, so that a great deal more of it can be handled by merely dissolving. This is not sufficient, however, to take care of all the carbon dioxide; the remainder must go into chemical combination with some substances that are in the blood. There is no single conspicuous material for carrying carbon dioxide like the hemoglobin which transports the oxygen. There are, however, a number of compounds in the blood which are able to combine with carbon dioxide, among them the blood proteins of which much was made in Chapter XIII. The carbon dioxide distributes itself among these various substances and so is transported. It should be noted that the blood does not become saturated with carbon dioxide as it does with oxygen. Arterial blood ordinarily carries practically all the oxygen it is able to take up; venous blood on the other hand probably never comes anywhere near being as fully charged with carbon dioxide as it is able to be.
During the passage of the blood through the capillaries of the lungs an outward diffusion of carbon dioxide into the air in the lung sacs is going on simultaneously with the inward movement of oxygen from this air into the blood. The diffusion is never so complete as to deprive the blood of all its carbon dioxide; there is in fact only a little less of it in arterial blood than in venous, although the diffusion is sufficiently rapid so that as much carbon dioxide as is produced in the whole body in a minute is passed out into the air of the lung sacs in the same time. The effect of this outward diffusion is naturally to increase the amount of carbon dioxide in the air of the lung sacs, and if this increase is allowed to go on unhampered, there will presently be so much carbon dioxide there as to stop further outward diffusion, and so to put an end to the escape of carbon dioxide from the blood. This is avoided by lung ventilation. Every time a breath is drawn some air that is almost free from carbon dioxide enters the lung spaces to replace the carbon dioxide-laden air that was expelled at the previous exhalation.
The description of gas transportation that we have just given opens the way for an account of the control of breathing. From what has just been said it should be clear that the amount of carbon dioxide in the blood corresponds closely with the amount that is in the air of the lung sacs. As the percentage of carbon dioxide in this air goes up, outward diffusion becomes less free, and so the amount of carbon dioxide in the blood will have to increase. The tissues are all the time producing and pouring out carbon dioxide, and so there will be a steady increase in the amount of carbon dioxide in the blood. This applies to the arterial blood as well as to the venous, since, as we saw a moment ago, there is nearly as much carbon dioxide in the former as in the latter. This is the fact which is utilized in the body for operating the breathing machinery. The respiratory center in the brain stem is susceptible to carbon dioxide; the more of this gas there is in the blood, the more tendency there will be for the center to discharge. There is a certain level of carbon dioxide below which it is entirely inactive, but when this level is passed nervous discharges begin and become more and more powerful as the amount of carbon dioxide in the blood goes up. Now we can see what makes us breathe. Let us imagine that there is not very much carbon dioxide in our blood, but that the tissues are constantly producing it and giving it off. Since we are supposing the amount is not enough to excite the respiratory center, there will be no movements of breathing. There will be a steady increase of the amount of carbon dioxide in the blood and at the same time a corresponding increase of the amount of carbon dioxide in the air sacs of the lungs; presently there will be enough in the blood to arouse the respiratory center to discharge. This will cause a breath to be drawn; the effect of this will be to sweep out much of the accumulated carbon dioxide from the lung sacs; this in turn enables more rapid diffusion of carbon dioxide from the blood to occur, and so the amount of it in the blood may fall below the level at which the respiratory center is made active. In a moment, of course, the continued outpouring of carbon dioxide from the tissues will raise the level again to the point of exciting the respiratory center, and so we will have a rhythmically recurring discharge of that center causing a rhythmic drawing of breath.
According to the account just given the activity of the respiratory center is determined exclusively by the carbon dioxide in the blood; it could be so regulated, but, as a matter of fact, in all higher animals, including man, the carbon dioxide control of the respiratory center is interwoven with a complicated nervous control whose effect is to make us breathe more often in a minute, but to make the individual breaths shallower than they would be if the control of breathing were exclusively by means of carbon dioxide. The net result in lung ventilation is exactly the same, but the rapid shallow breaths are advantageous in that they avoid large fluctuations in the amount of carbon dioxide in the blood, while they do serve fully to provide sufficient oxygen.
The rate and vigor of breathing are ordinarily adjusted automatically to the amount of carbon dioxide in the blood stream, but, as we know, we can, of our own will, breathe quite differently. Let us see what will happen if while we are sitting quietly we begin to breathe deeply and rapidly, overventilating the lungs. So far as oxygen is concerned, this will make no difference at all, since, as we have already seen, the ordinary automatic breathing keeps the blood charged with all the oxygen it can hold. What overventilation does is to sweep out the carbon dioxide from the lung sacs more rapidly than usual and this permits of a correspondingly more rapid outward diffusion of carbon dioxide from the blood. The result will be that carbon dioxide will leave the blood faster than it is being poured into it from the tissues, and so the total amount of the gas in the body will be cut down. The first effect of this we would expect to be the removal of the automatic stimulation of the respiratory center, so that, after a period of excessive breathing, one would not at once resume breathing spontaneously. This, as a matter of fact, is the case; anyone can easily prove on himself, by breathing deeply and rapidly for a minute or two, that the automatic control of breathing is temporarily suspended immediately after. It follows naturally that one can hold the breath a good deal longer if the lungs are overventilated for a short time just before the attempt is made. This also can be easily proved. Prolonged overventilation of the lungs has, likewise, a number of other effects, all of which are due to cutting down the total amount of carbon dioxide in the body. The most conspicuous is a feeling of dizziness or light-headedness that comes on. If pushed to excess, there is a very definite feeling as though one were about to soar away into space, and this is followed by unconsciousness. Certain religious cults in India have interpreted this sensation resulting from deep breathing as an actual severance of soul from body, and maintain that during the time of unconsciousness the spirit really floats freely in space. Without venturing any statement as to the relation between the soul and the body during either consciousness or unconsciousness, we would point out that these bodily sensations are definitely due to the very simple fact that there is less carbon dioxide in the blood than is normal on account of the overventilation of the lungs, and just as soon as the metabolism that goes on all of the time in the tissues pours out enough carbon dioxide to bring the amount up to normal, consciousness will return and the ordinary condition of affairs will be resumed.
Although this finishes what we have to say about the movements of gases into and out of the body, the general subject cannot be completed without a word concerning the conditions that should be maintained in the air immediately surrounding us. This makes up the topic of ventilation. We all know that some air is much more fit to breathe than other; until very recently, however, our ideas as to the conditions which make air fit or unfit to breathe have been hazy or entirely erroneous. Fortunately, of late years, the subject of ventilation has been actively investigated and we now have a satisfactory knowledge of its laws.
There are, of course, two things that must be true of any air that is to be breathed; these are that it must contain enough oxygen and must not contain too much carbon dioxide. So far as the oxygen supply is concerned we may state that only with the greatest difficulty are conditions reached in which there is not enough oxygen in the air. As we all know, the air becomes thinner the higher we go above the surface of the earth; both mountain climbers and aviators have attained heights at which the amount of oxygen in the air is only about one-third that of ordinary air and have been able to obtain enough oxygen for their bodily needs even under those extreme conditions. It is quite evident that a room could scarcely be so poorly ventilated as to bring the oxygen supply down below this figure, so that no attention need be paid to the oxygen supply in working out practical methods of ventilation. Air which contains carbon dioxide to the extent of four per cent could not be breathed because the carbon dioxide being produced in the body would not diffuse out fast enough into an atmosphere containing that amount of carbon dioxide to keep the body alive. This again is a percentage of carbon dioxide that is practically never reached. Probably the most famous case in history of death from poor ventilation is the “Black Hole of Calcutta,” a dungeon room about twenty feet square with only two small windows, in which one hundred and fifty British soldiers were imprisoned over one night; all but twenty-three of these died, but it is doubtful whether their death was actually due either to deficiency of oxygen or to excess of carbon dioxide. This is because there were enough other factors which would make the air unbreathable to bring on death before either of these could come into play. The modern science of ventilation concerns itself with these other factors; chief among them is the factor of moisture. As we shall see in the next chapter our bodies are constantly giving off from the lungs and by evaporation from the sweat glands water vapor into the air. This causes the humidity to go up rapidly in rooms where people are congregated, and particularly so where there are many people present. Also everyone gives off a great deal of heat. We now know that the feeling of closeness which we ascribe to a poorly ventilated room is due to the combination of warmth and moisture. We also know that the discomfort which comes from being in such rooms is due to the same causes. Actual vitiation of the air is much less disagreeable than is the accumulation of heat and moisture. In theory, of course, the best ventilation is secured by keeping rooms flooded with outdoor air. In practice, however, this does not always work out; for example, in many cities the air is so laden with dust and smoke as to be bad for everybody and even dangerous for sick people. Before such air is breathed the smoke and dirt should be gotten out of it. This is done sometimes by forcing it through fine mesh cloth bags, or the most modern scheme is by passing it through a thin screen of water and so washing the dirt and smoke out of it. The second practical difficulty with flooding rooms with outside air is the expense in cold weather of warming the large volumes that would be required. For this reason it has been found feasible in churches and public halls that are occupied only occasionally to use the same air over and over by keeping down the temperature and moisture. Of course, this cuts down very greatly the expense of heating.
There is one other source of harmful effect from bad air besides the high humidity and undue warmth; this is the presence in it of ammonia and other poisonous compounds that are given off from the bodies of people. It used to be believed that organic poisons were exhaled from the lungs with every breath, but we now know that the amount of these, if any are present, is too small to be important in comparison with the very much larger amounts that come off from the evaporating sweat, from decaying teeth, and from the digestive tract; there is no doubt that in any assemblage of people the air will be vitiated by organic poisons from these sources. The more cleanly the individuals are, the less will be the contamination. It is generally believed, although perhaps not absolutely proven, that the bad health found in sweatshops and crowded slums generally is due largely to chronic poisoning from the constant breathing of effluvia from the unwashed bodies and clothing of the inhabitants. The obvious remedy is insistence upon personal cleanliness, although this does not lessen the desirability of breathing as pure air as can be gotten. The point to be emphasized is that where personal cleanliness prevails, the closeness of rooms is chiefly due to excessive moisture ordinarily accompanied by too high a temperature. Ventilation measures should be carried out with this in mind.
THE metabolism that goes on in all our body cells results in the formation of waste products. The chief of these, carbon dioxide, is taken care of by means which were described in the last chapter. Besides this gaseous substance there are produced a number of compounds which are soluble in water and which are poured out from the cells into the tissue spaces surrounding them and which pass thence into the blood stream. The special places in the body wherein these substances leave the blood stream make up the apparatus for the removal of waste. This apparatus must be able to take out of the blood stream all the waste products of cell metabolism except carbon dioxide, also any materials which may have been absorbed into the blood stream from the digestive tract, but not used by the tissues, and finally all the accumulations of water. The discharge of water makes up a topic by itself which will be considered after a word has been said about the apparatus by which the removal of wastes is carried on. Sharing in this function we have first the kidneys, second the liver, and third the sweat glands.
The kidneys are the most important of the organs for removing waste. They are a pair of bean-shaped bodies lying in the small of the back. Each

one is made up of thousands of tiny tubes; each of these tubes starts as a little sac in which is a knot of blood vessels which is really a tiny filter, filtering out from the blood water and the inorganic salts that are dissolved in it.
All the other materials that are in the blood are held back by the filter, so that nothing escapes from it in these places except water and salts. Further along each of the tiny tubes is a section which is surrounded by a network of fine blood vessels where two things take place; the first of these is the taking out from the blood of the various kinds of waste products; the second is the absorbing back into the blood of part of the water which filtered out through the knot at the tip of the tube. Beyond the point where these things happen the tube does nothing except to convey the finished kidney secretion, or urine, down to the outlet where opens a very large tube, the ureter, by which the kidney secretion is carried down to the bladder. The formation of urine is made up then of three stages; first the filtration of water and salts out of the blood, second the escape from the blood into the kidney tube of the various waste products of cell activity, and third, the reabsorption into the blood of part of the water. It is estimated that about four times as much water filters out from the thousands of tiny filters at the tips of the kidney tubes as comes out at their ends to be carried down to the bladder. We do not understand exactly why this should happen; one result of it is to make the urine contain about four times as much salt as the blood; the water that filters out at the beginning of the kidney tube carries with it just the amount of salt that is dissolved; after the water is reabsorbed the salt is left behind in the kidney tube, so that all the salt that filters out stays in the urine, but only about a quarter of the water. The discharge of waste products is easier to understand than the way in which the water is handled; all organic substances which happen to be in the blood and which do not belong there pass out from the blood during its passage through the fine network surrounding the kidney tubes. Of course we do not know just how this is carried on; there must be some special features about the protoplasm in this part of the kidney to cause these substances to pass out from the blood here and nowhere else. Not only waste products of cellular activity, but also all or nearly all the organic accessories of the diet and all drugs taken as medicine leave the blood in this region.
An interesting thing about the removal of water from the blood by the kidneys is the relation it shows to the activity of the sweat glands. The total amount of water gotten rid of in any day is the amount taken in in drinking during the same time. The amount we drink is controlled chiefly by the sense of thirst, assuming that we are in a place where water can be had. The amount of water that passes out in the form of sweat is not under our control; in a later chapter we shall see how the formation of sweat is regulated. There is also a constant and fairly steady loss of water from the lungs, since every breath that we exhale carries with it all the water vapor which it can hold. What the kidneys have to do is to get rid of the water that is not passed out either from the sweat glands or the lungs. The amount of urine that is formed in a day is, therefore, extremely variable; on days that are warm and when a great deal of exercise is taken there will be so very much sweat formed that in spite of copious drinking comparatively little water will be discharged from the kidneys. On the other hand on cold days in which not much exercise is taken there will be a large amount of urine formed. It is interesting to note that some substances like asparagus, and to a less extent coffee, cause a large increase in the amount of urine. We do not know certainly whether these act by increasing filtration from the tips of the kidney tubes or by diminishing the reabsorption in the part farther along, but so far as our information goes either or both may occur.
The kidney is not the only organ by which wastes are removed; the liver has a part in this as well. The action of the liver is not, however, simply to assist the kidney; there are certain special substances which are taken care of by the liver and which are never gotten rid of through the kidneys. The first of these are the broken-down blood pigments. We have already seen that after the red corpuscles become worn out they are decomposed in the spleen and the hemoglobin carried to the liver where the iron and other usable parts are saved and the useless remainder is passed on into the bile to be conveyed out of the body. Bile is green in vegetable-eating animals and golden brown in flesh-eating; the difference depends on the so-called bile pigments which are the waste remains of hemoglobin. The difference in color is the result of a slight chemical difference in the pigments which in turn depends on the diet, so that in man the bile is either green or golden brown according to whether the diet is vegetarian or chiefly of flesh. The liver also takes out from the blood a waste substance which has the formidable name of cholesterin. The interesting thing about this substance is that unless the bile is altogether normal chemically it will not be dissolved and so will be thrown down in the form of little grains or lumps. So long as these grains are small they can pass down the bile duct to the small intestine and be carried out of the body, but after they become bunched together into lumps it is difficult or even impossible for them to be carried down the narrow bile duct. These lumps make up what are commonly called gallstones. The acute pain known as gallstone colic is due to the stretching of the bile duct by the passage of one or more of these stones down it. At the present time no way is known to prevent the formation of gallstones in persons in whom it starts, nor of softening them after they have once been formed; the only relief is through surgery, which, fortunately, is entirely adequate, so that thousands of persons are living in great comfort to-day who would be dead or living in acute suffering if the surgeon had not been able to remove the accumulation of gallstones from the passages in their livers.
The sweat glands, as we have already seen, make up part of the service of the removal of waste in that they help to carry away water from the body. To a very slight extent, but much slighter than is commonly believed, they get rid also of waste products of cell activity; for example ammonia, which is one such product, can frequently be detected in the sweat by its odor; practically, however, the importance of sweat glands is not in their discharge of waste organic substances, but only in their discharge of water. In this they are playing a very important part in the regulation of body temperature, so that the further account of the sweat glands as well as of the skin of which they form a part is left for the chapter in which the regulation of temperature is described.
WE have now brought our description of the various things that happen in our bodies up to the point where we may begin to make some kind of summary of them; particularly in respect to what goes on within our individual cells. We have seen how some of our cells are muscle cells by which we make motions; others are the cells of sense organs by which we get the necessary information for guiding our activities; still others are in the nervous system and carry on the adjustments by which we act in accordance with the indications of our surroundings and also of our past experience; finally we have the cells which manufacture materials, as in the various digestive glands or in the glands which make hormones. We have also looked into the kind of materials which these cells require, where they come from, and how they are prepared in the digestive tract, and we have seen how these prepared materials and oxygen are conveyed to the different parts of the body where the cells can get them and finally how the waste products which all these cells give off are gotten rid of. We are now to turn to the process of metabolism itself as it takes place within the cells. In Chapters III and IV we looked into the use of food for power development and for repairing the wastage of protoplasm, as well as for the making of new protoplasm in growth. We have seen also that the body contains much nonliving material, as in the bones and teeth, which must come from the food and which must be put in place as the result of metabolism on the part of living cells.
The first thing which we wish to take up here is the use of protein in the repair of protoplasmic wastage and in growth. We saw in Chapter III that protein is manufactured originally by the living cells of green plants. We have also seen that protein is the only material that can repair protoplasmic wastage or that can make new protoplasm. We have omitted to say thus far that the most important place from which we can obtain protein is from living protoplasm. It is true that most seeds store up within themselves nonliving protein to be used by the young sprout as it forms, and seeds make up a large part of our diet; but except in grain and other seed foods we obtain our supplies of protein by eating protoplasm. This protein is to be used by us for repairing our protoplasmic wastage or, in parts of us that are growing, for making new protoplasm. We have already seen that the protein which we eat must go through a process of digestion before we can use it for these purposes, and our present task is to explain just why this is necessary and to show how the protein is actually used in our bodies.
A thing about protein which fits it specially to be the chief material of living protoplasm is that it is very much the same sort of substance wherever we find it and yet can differ enough to account for the differences that exist among animals and plants. In spite of the fact that protoplasm analyzes about the same, no matter where it comes from, we are bound to believe that the difference between a dog and an oak tree is at bottom a chemical difference; they are unlike because the protoplasm of one is not the same substance chemically as the protoplasm of the other, and the difference is a difference in the proteins. We can come even nearer home than that and say that the differences between the races of mankind are probably chemical differences between their proteins. Human protein is undoubtedly different from the protein of beef or pork or mutton. What we have then is a substance which can be at the same time similar and different; also since we can make human protein out of beef protein or any other kind which we happen to eat it must be fairly easy to change one into the other.
Protein is about the most complex substance that we know anything about; it is made up of a number of organic acids combined chemically. These organic acids all contain nitrogen, which puts them into a class to which is given the name of amino acids. To the chemist the name amino acid shows a certain kind of chemical formation; to us it need mean no more than an organic acid which contains nitrogen. The proteins which are in our bodies are as complex as any that exist and some of them are made up of as many as eighteen different amino acids. The same eighteen acids are present in the proteins of all the higher animals. When we eat lean beef or pork we get exactly the same eighteen amino acids that are in our own proteins, but not put together in precisely the same way. What we have to do with these proteins is to break them up into the amino acids of which they are composed and then put these together again in the combination which makes up human protein. The breaking up of the proteins is carried on in the digestive organs; we have said a good deal about it in Chapter XV; what were called in that chapter digestion products of proteins we now see are amino acids. These are taken up into the blood stream, carried around the body to the tissue fluids and by the living cells taken up to be built into human protein. Exactly the same thing happens to any proteins that we eat.
One of the great differences between animal protein and plant protein is that the percentages of the different amino acids are very different. Some of the amino acids that make up a large proportion of animal protein are very scantily represented in plant protein. To make human protein we must not only have all the amino acid ingredients, but we must also have enough of every one. On a purely vegetarian diet to get enough of those amino acids which are scantily present in plants we have to eat a large surplus of those amino acids which are specially abundant. In this respect plants as providers of amino acids are less economical than animals, because animal proteins have more nearly the same proportions of the different amino acids as do our own human proteins. Of course, as we will see at once, in theory cannibalism is the most economical way of getting protein; if we were to eat human protein we would have exactly the correct proportion of the different amino acids and so could get along with a minimum amount. This is not to be interpreted as an argument for the practice of cannibalism among human beings, although we may as well face the fact that there is no physiological or dietary reason for avoiding the practice. In some of the lower animals, particularly in rats, cannibalism is a regular part of the life habit. Rats do not have the instinct of storing up food supplies as do squirrels and some other kinds of animals. When food is abundant they multiply very rapidly and then when food becomes scarce the stronger feed upon the weaker. It is largely for this reason that the unsanitary and extremely expensive rat nuisance is so hard to abate.
The amino acids that are circulating in the blood stream after every meal are primarily to be used for repairing protoplasmic wastage; also they serve for the manufacture of new protoplasm, provided growth is going on. In theory an adult who is through with all his growth except in the skin and one or two other minor tissues should be able to get along with just the amount of protein which will make good his protoplasmic wastage. Since protein is an expensive food and likely to be hard to get in times of scarcity, the question of how much protein should be eaten is of great practical importance. There are several ways of studying the problem; one is by the actual study of diets to find out how much protein people do habitually eat; another is by finding out how much the daily protoplasmic wastage amounts to. If no more protein is being eaten than is necessary for the protoplasmic wastage, these two figures should be about the same. The way of finding out how much protoplasmic wastage there is is to go on a diet which contains abundant starch and fat for energy supplies, but no protein or amino acids. When one is on that kind of diet he knows that he will not have to burn up any of his own tissues to supply him with the energy for his metabolism; whatever breakdown of protoplasm occurs on such a diet is the natural wastage of the body and not the result of using tissues for fuel, as is the case in complete starvation. It is fairly easy to keep track of the decomposition of protein in the body, because protein contains nitrogen and the nitrogen is given off almost wholly in the waste products that are passed out from the kidneys. By collecting the urine and analyzing it for nitrogen the amount of protoplasmic breakdown in the body can be determined, provided no nitrogen-containing compounds were taken in with the food. Otherwise, of course, one could not be sure that nitrogen appearing in the urine had actually come from the wastage of protoplasm. The fact is that when an average-sized human being goes on a diet which contains no protein, but is ample in other respects, he loses daily from his body about an ounce of protein. This is proven by the occurrence in the urine of an amount of nitrogen which stands for that much protein. The person may be gaining or losing weight meanwhile; if his consumption of fats, starches, and sugars is excessive, he may deposit some fat, in which case he might gain weight in spite of the loss of some of his actual living protoplasm. Usually though, in experiments of this kind, there is a steady loss of weight made up of the ounce of protein and of three or four ounces of water. We have to remember that living protoplasm is three-fourths or more water, so that whenever any protoplasm wastes away, some water will be lost as well as protein. It is a very interesting fact that this protoplasmic wastage goes on steadily at the rate of about an ounce a day whether the body is active or inactive; this means that the wastage is a matter of the basic metabolism and not of the functional metabolism. The former goes on all the time day and night, in sleep and in waking, and in connection with it the living protoplasm shows this small amount of wastage. Functional metabolism does not, at least under ordinary conditions, increase the amount. Speaking of the body as though it were a machine, we would say that it rusts out just as fast as it wears out. This is one of the features in which the living machine differs from most mechanical devices of human manufacture.
Although the loss of protein due to wastage is only about an ounce a day, nobody can get along on a diet which contains no more than that amount. Between three and four ounces of protein is the average daily consumption of adults in this country. We should not forget that our diet consists of meat, bread, vegetables, fruits, etc., which are mixtures of proteins with the other food materials and with a large percentage of water, so that in order to get three or four ounces of protein we have to eat four or five times that weight of ordinary foodstuffs. There has been much debate as to whether it is necessary or even desirable for adults to eat three or four times as much protein as the body requires for making up its wastage. The decision will have to rest in part on what use the body makes of the surplus. Since from time immemorial human beings have habitually eaten every day this large surplus, it is evident that they have been wasting enormous amounts of good food or else that some use is made of it even though it does not serve its purpose of repairing the body waste. The surplus materials are present in the body in the form of amino acids, since what the cells do in repairing their wastage is to take up from the whole quantity of amino acids in the tissue fluids as much as they require for making good their loss. The mixture of amino acids that is left over will make perfectly good fuel provided the nitrogen that is in it is gotten rid of, and this is what happens in the body. All the amino acids in excess of the amount needed for restoring the tissues are decomposed in such a way that the nitrogen is abstracted in the form of ammonia and the substance that is left, which is a starchlike compound, joins with the other starch products and fats to be burned in the body as a source of energy. We do not know certainly which tissues have the power of decomposing the surplus amino acids. At the present time it is believed that all or nearly all of them can do it, so that as they take up from the tissue fluids the particular amino acids which they need for making good their own wastage they take up also the surplus which they decompose, utilizing the starchlike part for fuel and turning the ammonia back into the body fluid as a waste product.
Ammonia is a very poisonous substance and it quickly poisons the body if allowed to accumulate in the tissue fluids. This is prevented by the action of the liver in changing the poisonous ammonia into a harmless substance known as urea. This urea is carried by the blood stream from the liver to the kidneys where it is passed out to become the chief organic substance in urine. The more protein one eats the more surplus amino acids will there be, and so the more urea will be formed and passed out of the body. Flesh-eating animals and men (Eskimos for an example of the latter) eat a very large surplus of proteins, the fuel for their metabolism being furnished almost altogether either from the usable remains of the decomposed amino acids or the fats that were in the flesh they ate. Some people have been inclined to believe the production of so much ammonia and its subsequent conversion into correspondingly large amounts of urea to be injurious. As a matter of fact, there is no particular reason for thinking this to be the case; it is part of the duty of the liver to change all the ammonia that comes to it to urea and of the kidneys to pass out all the urea that comes to it; so long as these organs are healthy they are able to fulfill these duties effectively, so this does not seem to be a good reason for cutting down the percentage of meat in the diet. It is generally believed that meat has special effects upon the nervous system, such as to incite to cruelty and bloodthirstiness. There is no real scientific proof as to whether this is true or not. The scientific fact is that man is fitted for a mixed diet, neither exclusively of flesh nor exclusively vegetarian. He has lived for thousands of years on that kind of diet and can apparently go on for thousands of years more. We need to remember that the various dietary fads which come into great prominence from time to time are rarely based on a well-established scientific foundation nor have any of them any long experience back of them. On the other hand, the common mixed diet which all of us eat in accordance with custom and the dictates of our appetites has the sanction of thousands of years of successful maintenance of the human race. It is quite true that one can get along on almost any kind of a diet provided it contains enough protein to make good the daily body wastage and enough fuel material to provide for the demands of metabolism. Anyone who is disposed to adopt for himself a dietary fad will rarely suffer seriously from it; on the other hand, those who prefer to eat as our fore-fathers have eaten need not feel conscience-stricken because there is agitation against the commonly accepted diet.
While on the topic of diet, a word should be said about cooking. In a previous chapter the advantage of good cookery as an aid to digestion was emphasized. We would add here merely the comment that in defending the ancestral diet we do not intend to imply that their cookery was always what it should have been. Over most of America there has prevailed from pioneer days a habit of frying food in preference to other means of cooking it. Our hardy pioneer ancestors throve on fried meats; an outdoor life of muscular toil makes almost any kind of cookery both acceptable and digestible. As labor-saving machines tend more and more to diminish the amount of muscular labor that most of us do, we find it harder and harder to maintain good digestion on fried foods. The objection to frying is simple; fats are the hardest of all foods to digest and fried foods are smeared all over with fat. It is only logical to expect fried foods to be harder to digest than other kinds; it is undeniable that the flavor of many fried foods is so agreeable that we would be unwilling to omit them from the diet altogether. What is realty objectionable is the practice of smearing all the food with fat in the process of cooking it.
We have finished what we have to say about the use of food for the repair of bodily wastage. While we are on the topic, a few words about the use of food in growth will come in well, since the growth process is closely related to the process of tissue repair. The chief difference between them is that the process of growth comes to an end in all but a few of our body tissues as soon as we become adults. The tissues which go on growing are the layers of the skin just under the surface, the reproductive tissues, and the blood-corpuscle-forming tissues. Connective tissue has the power to grow at any time during life, although it does not actually keep on growing as does the skin. Whenever an injury is suffered which actually destroys muscular tissue or the deep layers of the skin, there is no growth of new tissue to take the place of either. Repair is made by a growth of connective tissue to fill up the space. The result is the formation of a scar. If the edges of the injury can be brought together skillfully enough, the outer layers of the skin which do have the power of growth may bridge across the space so that no scar will result.
A second thing to be mentioned about growth is a discovery which has attracted a great deal of attention of late years; we will realize of course that the chief thing in the making of new protoplasm is the building together of protein out of amino acids. It is evident that for the manufacture of new protein all the eighteen amino acids must be present in sufficient proportion to give enough of each. There are some plant proteins which lack one or two of the amino acids that are present in human proteins and when a growing animal goes on a diet in which these are the only proteins present it at once stops growing. Most of the experiments proving this have been done on white rats, and it has been found possible to keep a rat for more than a year at the size it was when only a few weeks old simply by feeding it proteins in which one or two amino acids were lacking. The fact that the animal lived during this time proves that his protoplasmic wastage was made good and therefore that there are proteins which can replace the body wastage but cannot manufacture new protein. There is only one conclusion to be drawn from this, namely that the daily wastage which the protein suffers does not include all of it; some amino acids when once built in are there for good; others, on the other hand, are constantly being lost in the process of wastage, and these are the ones which must be replaced. There is a common protein, gelatin, which is used a great deal for food, but which by itself will not serve either for repair or growth because it not only lacks some of the amino acids that are in living protoplasm, but also some that are lost in the process of wastage. Gelatin is useful as a food, therefore, only in combination with other proteins that contain the necessary amino acids; or after the nitrogen has been taken out, it becomes a good fuel.
One interesting question that was settled by the experiments described above was whether the ability to grow is exclusively a matter of youth; as we know, under ordinary conditions only young animals grow. When they reach a certain age, they become mature and thereafter no increase in size takes place. In the case of these white rats which were kept for more than a year at the size of partially grown rats it was found that as soon as their diet was changed to one in which all the necessary amino acids were present they would begin at once to grow and grow to full size, when they would stop growing just as they would have when young. This proves that growth is not a matter of age, but is a matter of achieving a certain size, and is controlled by factors which we do not at present understand. An animal that has not been able to attain


full growth because it has been denied the amino acids necessary for making new protein retains the power of growth, so that even though it may be long past the ordinary age of maturity it can go on growing as soon as the necessary materials are provided. The dependence of growth on certain dietary accessories was spoken of in Chapter IV and need not be repeated here.
The final use of food is as a source of energy for carrying on metabolism. A good deal was said about that in an earlier chapter, but there are a few additional points to be brought out here. The energy for metabolism can come from any of the foodstuffs; these are present in the blood stream in the form of sugar or fat or amino acids. A moment ago we saw that in the case of the amino acids the nitrogen is removed before they are ready for use as fuel. After this has happened the part that remains is so similar to sugar that it can be thought of as the same material and will be so considered by us. We have then to follow only the two food substances, sugar and fat, through their use by the body cells. Fat will be dealt with first since we have less to say about it. It passes from the digestive tract into the blood stream in the form known as an emulsion; all this means is that it is broken up into very tiny particles which are kept from running together by some sort of a film. In the case of the fats in the blood it is likely that this film is composed of ordinary soap. In this state the fat circulates in the blood stream until it is taken out by the tissues and burned. In the process of this burning some very poisonous substances are likely to be produced, but this happens only when large amounts of fat are being burned by themselves. If sugar is present and is being burned at the same time, there is no danger. Ordinarily in the body sugar is always present, but in a certain disease, diabetes, about which more will be said later, the body is not able to burn sugar as well as it ordinarily does, and under these circumstances poisoning from the products of the fat burning is apt to happen. This makes up, in fact, the serious danger in diabetes. As we all know, fat is chiefly important as the form in which food is stored in the body against a time of future need. We shall return to the way in which the body does this after we have spoken of the use of sugar as fuel.
Sugar is the great fuel substance of the body. About two-thirds or our food ordinarily consists of starchy materials which are digested into sugar. When we add to this the sugarlike remains of all the protein food which is not actually used for the repair of tissues we see that this substance makes up the great bulk of the material which is carried by the blood to the tissues. This material is handled in the body in an interesting way which depends on the curious fact that although sugar is the most important fuel for living cells they cannot endure its presence in them or in the fluids surrounding them except in very small amounts. Sugar, as we all know, is very soluble in water, and it would be perfectly possible for the blood to dissolve all the sugar that enters it from the digestive tract and simply carry it in solution until the tissues withdrew it for their needs; but this would mean that immediately after every meal the percentage of sugar in the blood would mount up to a high point from which it would gradually sink as the sugar was taken out, to mount again after the next meal when absorption began again. This does not actually happen because it is prevented by the liver, which has as the most important of its many functions that of storing the sugar that is taken up by the blood from the digestive tract and dealing it out little by little as the cells of the body need it. Back in the chapter on digestion we saw that all the blood that passes through the intestinal tract is gathered up by the portal vein and passes through the capillaries of the liver. It is during this passage through the liver that the sugar is taken out of the blood and stored in the form of a less easily dissolved material known as animal starch or glycogen. The liver cells have the ability to convert sugar into glycogen and they do this whenever the amount of sugar in the blood passing through them is greater than the very small amount which is suitable for the body cells. The blood that leaves the liver carries in it only this small percentage. The liver cells have the ability to change glycogen back into sugar, and this they do whenever the blood that enters them is deficient in it, so that the blood leaving the liver tends always to have the same amount of sugar in it. Whenever it enters with more, there is a conversion of sugar into glycogen; whenever it enters with less, there is a conversion of glycogen back into sugar.
So important is the protection of the body cells against having too much sugar in the fluids surrounding them that the kidney acts to prevent undue accumulation; this it does by withdrawing from the blood and passing out into the urine any sugar that may be in the blood in excess of the small amount which the tissues are able to endure. Thus we see that if the liver did not have its function of converting the sugar into glycogen we would have to change our eating habits completely, taking only a little food at a time instead of eating it in three meals, since otherwise most of our food would be wasted by being passed out from the kidneys as fast as it was poured into the blood from the digestive organs. There is a limit to the ability of the liver to change sugar into glycogen. If the amount in the portal vein at any one time goes above a certain figure, not all will be saved: a part will escape into the blood stream, and since this will raise the percentage, the kidney comes into action and passes it out. In order for this to happen, there must be a very large amount of digested sugar in the small intestine leading to rapid absorption. Since some starchy foods are easier to digest than others some diets are more likely to result in the appearance of sugar in the urine than others. Some of our common foods, notably honey and corn sirup, consist largely of the kind of sugar the body uses. These require no digestion at all, but are ready for absorption as soon as they enter the small intestine. Naturally, if they are eaten in any quantity, they are likely to flood the liver with sugar beyond the amount which it can change to glycogen. Common table sugar and the sugar of milk have to be digested before they are absorbed and so are less likely to flood the liver. It is true, however, that either of them if taken in very large amounts may do this. The digestion of starch goes on much more slowly and so is absorbed more gradually and it is doubtful whether the liver is ever flooded on a starch diet. Since the presence of sugar in the urine is a common indication of diabetes, it is necessary to know that other conditions may bring it about. Obviously, as in the case of an examination for life insurance, a perfectly healthy person might be rejected on account of the presence of sugar in his urine, if it were not that the examining physician knows of this other possibility and is on his guard against it.
The liver ordinarily turns glycogen back into sugar at just the rate necessary to keep the amount in the blood constant. This means that when functional metabolism is going on glycogen is being turned into sugar more rapidly than when the body is quiet. One of the very interesting discoveries of recent years is that in times of strong excitement leading to the outpouring of adrenalin into the blood the rate of change of glycogen into sugar is much increased, so that instead of the usual small amount there is present in the blood a large concentration of sugar. This is evidently advantageous in insuring ample fuel supplies to the muscles at the time of an emergency. It is wasteful in that a large part of the surplus sugar is passed out by the kidneys. The fact is illustrated that in marshaling the bodily functions for meeting an emergency economy is lost sight of.
In addition to these healthy conditions in which sugar may appear in the urine there is the disease diabetes, in which the presence of sugar in the urine is a conspicuous symptom. In diabetes there is a serious disturbance of the whole fuel-supplying mechanism; the liver does not carry on its function of changing sugar to glycogen and glycogen back to sugar as perfectly as it should, and what is of much more importance the muscles which are the chief users of sugar as fuel cannot use sugar in anything like their usual manner. In fact in severe cases they appear to be almost wholly unable to use sugar as fuel. Since the protein from which the nitrogen has been removed classes itself with sugar in this regard, the muscles are thrown back upon fat as their only source of fuel, and this confronts the body with the danger already mentioned that in the burning of fat when little or no sugar is being burned along with it very poisonous products may be formed. Medical investigators have devoted a vast amount of labor to the attempts to find a diet that can be successfully eaten by diabetics. Starches and, almost equally, proteins are not serviceable because they simply flood the tissues with sugar, making an environment which is not good for the cells, and keeping the kidneys busy getting rid of the surplus. Fats are dangerous for the reason just stated. Quite recently real progress has been made by means of the discovery that when the body is living on its own tissue there will be no accumulation of sugar in the body fluids nor outpouring of it from the kidneys. One who is being starved is living on his own tissues and so by simply starving a diabetic his symptoms can be relieved. This is not in itself a very promising expedient, since evidently without food one cannot go on living very long. The point of the treatment is that after starvation has proceeded until the body is actually living on its own tissue it is possible to begin feeding proteins cautiously until little by little a protein diet can be established in which there is little or no indication of surplus sugar. In other words it seems that when the body is compelled to live upon its own tissue it uses proteins efficiently and will then go on using them efficiently when they are supplied to it in the diet.
In an earlier section of the chapter we talked about the amount of protein that the body requires; now we have to take up the matter of the amounts of energy-yielding foods. It is evident that the amounts of these depend upon the amount of metabolism; the total metabolism is made up of the basic metabolism which is steady, shifting little day in and day out, plus the functional metabolism which depends upon how actively the body works. The main functional metabolism is that of the muscles and it is that which varies from day to day. In the case of children there is the additional metabolism of growth for which energy is required and for which food must be eaten. In order to talk intelligently about the use of foods for metabolism we must have a word by which to express a definite amount of it. The word that we use for this is “calory.” Primarily this word stands for a certain amount of energy in the form of heat; since one kind of energy can be transformed into another kind without changing the actual energy value we can use this word as a measure of all kinds of energy and this has become the custom. The calory as a unit of energy means really the amount of energy in the form of heat required to raise the temperature of 1,000 grams of water by 1 degree centigrade. We can translate it into more familiar terms by stating that it equals almost exactly the amount of energy required to raise a weight of 300 pounds to a height of 10 feet, or 30 pounds to a height of 100 feet, or any other combination of weight in pounds multiplied by distance in feet which figures up to 3,000.
The total metabolism, as we said a moment ago, varies greatly day by day because the extent to which we use our muscles is so different. The basic metabolism is very steady and the functional metabolism of the vital processes like breathing, the heartbeat, etc., is also pretty steady, so that the metabolism for one who makes no use of his muscles is fairly uniform. Of course it differs in large people as compared with small, although curiously enough the difference is not proportional to the weight but to the body surface. Just why this is so we do not know. The average figure for the total metabolism of a resting man is about 1,900 calories a day; this includes the motions that are necessary for eating and swallowing food, since life cannot go on indefinitely without making those motions, but supposes that all other activities are done away with. This figure, as we said a moment ago, holds pretty steady day in and day out. To find the total metabolism on any day we have simply to add to it the figure for the energy expenditure from the use of the muscles, which will vary with the amount of work that is done. In figuring this we have to reckon with the fact that the muscles are like other machines in working at what is called in engineering a low efficiency; by that we mean that the amount of energy that can be gotten out in the form of useful work is less than the energy that is actually consumed; there is a waste of energy which takes the form of heat. Our muscles are under ordinary circumstances about twenty per cent efficient, which means that for every calory-worth of actual muscular work we do we use up five calories-worth of fuel; the energy of four wasted calories takes the form of heat and we all know from experience that the amount of heat thus produced inside our bodies warms us up very quickly, when we are using our muscles actively.
The actual muscular work done in a day by individuals varies from almost nothing in the case of invalids confined to bed through about forty calories for a person of decidedly sedentary habits and about 120 calories for the average clerical or professional man up to from 300 to 400 in the case of manual laborers. These figures are for the actual muscular work done; to obtain the energy expenditure we have to multiply each of them by five on account of the inefficiency of the muscles. If we do this and then add to each product the constant figure of 1,900 for the metabolism of rest we obtain for the total metabolism of a decidedly sedentary person an average of about 2,100 calories; for an average clerical or professional man 2,500 calories, for manual laborers from 3,200 to 4,000 calories. Of course, individuals may exceed even these latter figures. It is believed that athletes in extreme competitions such as for example a six-day bicycle race may liberate energy at the rate of 10,000 calories a day, although they probably cannot keep this up long enough actually to do that amount in a single day.
In order to satisfy the requirements of metabolism the food that is eaten must yield corresponding amounts of energy. If it does not do so enough of the tissues will be consumed to make up the deficiency. Of these the first to be drawn upon will be the stored glycogen in the liver and secondly the body fat. Only when the deficiency is great enough so that all these are used up, does the protoplasm itself begin to be drawn upon as a source of fuel. This happens in cases of prolonged starvation and it is interesting to note that in this case the tissues drawn upon are the muscles. When one wastes away as the result of starvation, the only tissues that suffer serious loss at first are the muscles; the rest of the body is fed at their expense. In this process no muscle cells are actually destroyed—apparently each can sacrifice a little of its protoplasm without being itself injured; the material thus obtained is converted into amino acids and from most of these the nitrogen is removed, leaving a fuel material which can be burned in the cells all over the body to keep them going. It is only after extreme starvation, when the muscles can no longer yield of their substance without being themselves destroyed, that the other tissues begin to show serious wastage. This explains why the brain of a starving man remains clear almost up to the end.
If a surplus of food is eaten over the energy requirements, the liver will store it in the form of glycogen so far as it is able; but if this will not suffice, the excess will be changed into fat and stored in the body in what are called adipose tissues. These are located in various regions, one of the most important being directly under the skin. It is the loading of this with fat that causes the bodily enlargement of fat people. We do not know exactly how the fat is made; we do know that it is not ordinarily food fat that has been simply transported to these tissues and deposited there. There is abundant proof that the body can manufacture fat even if there is none in the diet. Grazing cows that get no fat of any kind produce milk with its regular percentage of fat in the form of cream and do this day in and day out, showing that the fat that the body makes does not have to come from fat in the food. Since body fat represents ordinarily a storage of fuel against a future need, we ordinarily think of it as made whenever the temporary storage in the form of glycogen becomes inadequate to take care of the surplus of food over the amount consumed in carrying on the metabolism. Since the vast majority of people are neither gaining nor losing weight, the amount of food that they take in each day must balance the average metabolism. This is interesting because the amount of food that is eaten is regulated chiefly by the sense of hunger, or by the hunger and appetite together, and it is remarkable that these should cause us to eat so accurately just the amount of food our metabolism requires. In order that we may get some idea of how much energy is furnished in our common foods a table is given below. The figures are for the numbers of calories in a pound of the food material as purchased in the market. In most vegetables and meats there is a loss of about ten per cent in preparing them for the table, or, in the case of meat, in the bones. The figures were prepared by officials of the United States Government in arranging dietaries for the Army.
| CALORIES PER POUND AS PURCHASED | |
| Apples, fresh | 219 |
| Bacon | 2,979 |
| Bananas | 298 |
| Beans, dried | 1,603 |
| Beef, fresh | 1,009 |
| Bread | 1,300 |
| Butter | 3,478 |
| Cabbage | 124 |
| Celery | 70 |
| Cheese, American | 1,948 |
| Chocolate | 2,858 |
| Eggs | 614 |
| Fish, salmon, canned | 679 |
| Fish, fresh | 368 |
| Flour | 1,651 |
| Lard and substitutes | 4,218 |
| Milk | 302 |
| Pork, salt | 2,948 |
| Potatoes, white | 311 |
| Rice | 1,631 |
| Sugar | 1,860 |
| Tomatoes | 106 |
Very few of our foodstuffs are exclusively of one kind of material; that is they are not exclusively protein or exclusively fat or exclusively of sugar or starch compounds. Most vegetables are mixtures of starch with protein, fruits are mixtures of starch and sugar with a little protein, both fruits and vegetables contain so much water that their actual fuel value per pound amounts to little. Meats are mixtures of proteins with fats. Milk is a mixture of proteins, sugar, and fat. Table sugar and butter are as near pure unmixed foodstuffs as any of the things we commonly eat. In an earlier part of the chapter we saw that about two-thirds of our ordinary diet is of starch or starchlike materials. This figure is given, not in weight of material, but in the energy value. What it means is that about two-thirds of the energy for our metabolism comes from the starch and sugar that we eat; the other one-third is divided between fat and protein in the proportion of about two to one. The energy we get from fat being about double that which we get from protein—since a given weight of fat has about twice the energy value of the same weight of protein—we actually eat about the same amount of protein as of fat. The combined weight of the starch and sugar is between four and five times that of the protein. These, of course, are average figures representing not what we eat at any one meal, but the way in which the foodstuffs are found to be divided in our diet taken as a whole. Where meals have to be planned on a large scale, as in armies or in institutions where the persons to eat the food have not much choice in selecting them, it is necessary that the diets be arranged both to give the proper amounts of material and also to furnish them in about the correct proportions. The dietary experts who have charge of these matters do this with tables similar to the one given above. In domestic feeding arrangements, although usually the choice of foods is determined by the state of the markets, the preferences of the various members of the family, and the abilities of the cook, it is astonishing how closely the result will correspond in the long run to the figures here given. Most of us, without any effort on our part to do so, eat a diet which is made up week in and week out in just about the proportions here given. The experience of ourselves and of all our ancestors indicates that these are the correct proportions for the human race.
In talking about metabolism thus far we have spoken as though the metabolism of the body at rest were about the same day in and day out regardless of conditions. This is true in the main for healthy persons; there is one condition which may bring about a change in the resting metabolism of health which must be mentioned, and one or two coming under the category of disease about which also something must be said. The change in resting metabolism that comes about in health is one that is seen when the diet contains an especially large percentage of protein. For some reason which we do not understand the digestion, absorption, and utilization of protein stimulate the resting metabolism so that during the time that this protein is being used the metabolism is higher than at other times. The curious thing about it is that the increase of metabolism is not just enough to take care of the protein itself, but goes so far to cause sugars or even fats to be burned at a more rapid rate than usual. This fact is taken advantage of in treatment for reducing flesh; where one lays on flesh it is evident that more food has been taken than was required for metabolism so that the surplus has been stored in the form of fat. The only way to reduce the weight is to compel the body to burn up that stored fat. In theory the simplest way to do this is simply to starve. If starvation is combined with very vigorous exercise, the reduction of weight is bound to be rapid, since metabolism cannot be carried on without fuel, and if not enough fuel is supplied in the form of food, the body will have to furnish it and the stored fat is the place from which it will be taken. Unfortunately this is much simpler in theory than it is in practice. A good deal of discomfort and sometimes even disturbance of health results from too drastic efforts to reduce the weight by starvation. It is perfectly feasible to do it by adopting and sticking to a practice of never eating quite enough. This is a perfectly successful method, but requires great strength of will to carry it out. Probably the easiest way to reduce weight is to combine self-denial with a diet which consists chiefly of protein. Thus the stimulating effect upon metabolism is obtained and the result will be a gradual burning away of the body fat. The selection of a diet to fit any particular individual can best be made under competent medical advice, since personal peculiarities have to be taken into account in selecting among the various foods those best adapted for the purpose.
The variations in resting metabolism that fall under the head of disease are, first, the increase of metabolism in fever, about which we shall speak in detail in the next chapter, and, secondly, variations in metabolism that result from variations in the activity of the thyroid gland. The thyroid gland is an organ at the front of the neck; when it is enlarged, as it is in some people, we have the condition known as goiter. This gland is now known to manufacture and pour out into the blood a hormone which is a regulator of metabolism. When it is produced in normal amounts, the metabolism goes on at the rate that we find in healthy people. If the gland is inactive and does not secrete enough of the hormone, there is a reduction in the metabolism. Since this implies a lowering of the vigor of the life processes, we might expect it to have important effects. The most marked of these are in the nervous system. Persons whose thyroid glands are relatively inactive are mentally sluggish; the less active the gland is, the more marked is this sluggishness, and in case of practically complete absence of the hormone the condition amounts to idiocy. Occasionally a child is born without an active thyroid gland; this child is doomed for life, unless artificial aid can be procured, not only to complete idiocy but to all the other results of lowered metabolism; these show themselves in dwarfishness and a misshapen body. One of the conspicuous and beneficent discoveries of medicine in comparatively recent years has been that extracts of the thyroid glands of meat animals, when eaten by persons whose own thyroids are not sufficiently active, supply the lack, and so they may be restored to the normal condition. There are many people alive to-day who are in all respects normal, but who, if they were to discontinue taking thyroid extract, would relapse rapidly into a condition of idiocy.
The thyroid gland may sometimes become overactive as well as underactive; when the former happens we have an increase in the resting metabolism and a group of symptoms that indicate, so far as the nervous system is concerned, a condition of overexcitability. Unfortunately this does not mean exceptional mental power but rather mental instability. The victims of this condition are exceptionally quick nervously, but they are quick to take offense and quick to be disturbed by all sorts of conditions. If the overactivity of the gland becomes too pronounced, mental instability or even marked insanity results. Another fact of the heightened metabolism is that large amounts of food must be eaten to carry it on. Sufferers from overactivity of the thyroid gland eat voraciously, but, in spite of doing so, are thin or even emaciated. They have rapid heartbeat, high blood pressure, and other symptoms indicative of too great activity of the gland. The successful treatment of this condition depends on the removal by surgery of enough of the thyroid gland to reduce the outpouring of the hormone to the normal amount. This feat is now accomplished successfully by our most skillful surgeons, and the result has been the restoring to health and happiness of large numbers of people whose lives were rendered miserable through no fault of their own, but because their thyroid glands had become unduly active. We do not know how the thyroid gland itself is controlled; there is evidently something which causes it in the vast majority of us to produce its hormone at the rate which keeps the metabolism steady at what we look upon as the level of health. Deficient thyroid is, at least in some cases, hereditary. Excessive thyroid activity seems to be rather the secondary result of some preceding disturbance of the nervous system, but as to that we cannot say with confidence.
A GOOD deal of what has been said thus far in the book applies to nearly all kinds of animals about as well as it does to man. We have now to take up a feature found only in two great groups of the animal kingdom—birds, and the four-legged animals, to which are given the name of mammals. This peculiarity is commonly called warm-bloodedness. What we really mean when we say that an animal is warm-blooded is that the temperature of its body runs about the same summer and winter, day in and day out. An animal that we call cold-blooded, on the other hand, cools down when in a cold place, but is warm when in a warm place. It therefore has a very variable temperature as compared with the almost constant temperature of warm-blooded animals. The maintaining of the warm-blooded condition, which means really the maintaining of a constant temperature, involves only one thing, namely that the amount of heat lost from the body shall exactly balance the amount of heat produced in it. If more heat is produced than is lost, the temperature must rise; if more heat is lost than is produced, the temperature must fall. These are simple facts of physics which apply as well to the body of an animal as to any other source of heat.
All animals produce heat, because in all of them metabolism is going on, which means that in all of them energy is being liberated, and it is one of the fundamental laws of physics that whenever any energy is liberated all of it which does not take some other definite form is certain to appear as heat. The other possible forms that the energy liberation of the body may take are chemical energy in the manufacture of various materials and the energy of motion. Whenever either of these processes takes place a large amount of heat is produced in connection; this means, from the standpoint of mechanics, as we have already seen, that the body is an inefficient machine; if it were perfectly efficient, the energy that it liberates might all go into the form either of motion or of the manufacture of materials, with none left over to appear as heat; but since it is only 20 per cent efficient, four-fifths of all the energy that is displayed actually takes the form of heat. While this is a marked inefficiency from the mechanical standpoint, from the standpoint of bodily well-being it is by no means a bad thing, since it is this constant production of heat which makes possible in birds and mammals the maintenance of a constant and rather high temperature. That it is a great advantage to the body to be warm all of the time is clear when we compare the possibilities of our lives with those of cold-blooded animals, like insects or frogs. Whenever the weather cools down they necessarily become inactive since, as we saw in the third chapter, when protoplasm is cooled, its metabolism necessarily slows down, and when the cooling reaches a certain point the protoplasm becomes completely inactive. The temperature of 98½ degrees F., which our bodies maintain, is a temperature which is very well suited for an active metabolism; by keeping this temperature all the year around they are able to show this metabolism all the time instead of only in summer, as in the case of the cold-blooded animals.
As we said a moment ago, the maintaining of a constant temperature is altogether a matter of making the loss of heat balance the production of heat; the production of heat is altogether a matter of the metabolism; practically the whole of the resting metabolism takes the form of heat, since apparently the expenditure of energy in keeping the protoplasm alive is extremely wasteful; nearly all the energy that is actually liberated takes the form of heat, almost none of it being used in actual manufacture of material. This is, as has already been said, a very constant metabolism; the functional metabolism on the other hand is extremely variable depending on how hard we use our muscles; the total amount of heat produced from hour to hour is scarcely ever the same except when we are asleep; this means that the loss of heat, which has to balance the production of heat, must vary from hour to hour exactly parallel with the latter. In birds and in all mammals, except man, the adjustment of heat loss to heat production is almost wholly automatic; the animal or bird does very little to control it. We do see, however, a few examples of effort on the part of animals either to prevent heat from being lost too rapidly or to favor its more rapid loss. Thus, on a cold day, a cat or dog is apt to lie curled up or with its legs bunched under it; on a very warm day, on the other hand, it will lie stretched out as much as possible, and in the case of the dog, with the tongue hanging out full length. Birds sometimes are seen with feathers ruffled up apparently in an effort to keep warm. Both birds and mammals seek sheltered places in which to sleep, where they will be as little exposed as possible to cold winds or rains. In man the automatic adjustment of heat loss to heat production is very imperfect, in fact, it would not enable men to live naked except in the tropics. It is true that outside the arctic regions men might go naked and still make the heat production equal the heat loss during the waking hours by very vigorous exercise, but during the hours of sleep the loss of heat would be certain to be much more rapid than its production and so the body temperature would fall in severe weather enough to cause death. Man maintains himself outside the tropics, then, by using artificial aids for maintaining the balance between heat production and heat loss; these are of three sorts; first the use of clothing, second the use of shelter, and third the use of artificial heat. It is really a very curious fact when one stops to think of it that, although many animals enjoy artificial heat and gladly bask in it when opportunity offers, no animal has ever discovered the simple fact that throwing sticks on a fire will keep it going, and so no animals, except man, have ever made any real use of artificial heat. When we think that the progress of human civilization has been accomplished wholly in parts of the earth where clothing, shelter, and fire are necessary to human existence, we realize that these, instead of being mere incidents to our life, lie really at the very basis of advancement. Hairless man is evidently a tropical animal; if he had not devised means of maintaining himself outside the tropics, there is no reason to suppose that he would have behaved any differently than have the savages that inhabit those regions at the present time.
Clothing, shelter, and fire all operate to prevent us from losing heat too rapidly and so have their great value when the weather is cool; we also can and do bring about an increased heat production in cold weather partly by exercising more actively and partly by increasing the amount of protein in the diet, and so bringing into play the stimulating effect of that substance on metabolism. This latter is seen perhaps most strikingly in the experience of the Eskimos; when one of them comes in from a hunting trip in the depth of winter very much chilled, he scarcely stops to warm himself at all, but with the greatest haste gulps huge quantities of meat that are barely thawed out, not really cooked in our sense of the word; in fact, frequently the meat that is eaten is so cold that it makes the Eskimo chillier than before; as soon, however, as digestion and absorption have commenced, so that the stimulation of metabolism can begin, the rapid production of heat in the body warms it up to the point where complete comfort is obtained.
We have no other means of increasing heat production in our bodies except by muscular exercise or eating protein. It is interesting to note, however, that there is a form of involuntary muscular exercise which comes into play when we need to produce more heat in order to balance too rapid loss; this involuntary muscular action is the familiar shivering which one does when chilly. It is reflex in the strict sense, that is to say, the stimulation of cold on the skin arouses nervous disturbances which pass to the muscles in various parts of the body and set them into the violent movements which we call shivering. The functional metabolism of shivering is much greater than we might at first suppose, and the production of heat is correspondingly rapid. In fact, if one who feels inclined to shiver encourages it instead of attempting to keep from doing it, he will very quickly produce enough heat within his body so that he will no longer have a disposition to shiver.
We have seen above the various ways in which heat is produced in the body, and the artificial means we employ to prevent heat from being lost too rapidly. It is clear that all of these make up what we may call coarse adjustments. They tend on the whole to cause heat to be produced more rapidly when more is needed, or to prevent too rapid loss when conditions are of a sort to bring it about. Nothing that has yet been said accounts for the fine adjustment, that is, for the actual maintaining from moment to moment of a balance of the heat loss with the heat production. This fine adjustment is purely automatic and is carried on in us by two distinct means. The first of these is by an automatic regulation of the amount of blood flowing through the skin and so, within certain limits, of the temperature of the skin. It is evident that if the skin is warm heat will be given off from it more rapidly than if it is cool; a condition in which the balance is being upset by the failure of the body to lose heat rapidly enough can be corrected, at least in part, by causing the skin to become warmer and so more heat to be given off. The machinery for regulating the amount of blood flowing through the skin is the vasomotor system, about which a good deal has already been said. This system operates reflexly, stimulation of cold on the skin tends to diminish the flow of blood through it by contracting the blood vessels, while warmth on the skin has just the opposite effect, causing the blood vessels to become flushed and the blood flow to be more rapid. It is true that there is a reddening of exposed parts of the skin in extreme cold, as we all see frequently on our nose or ears, but this is a purely local effect and does not mean that the blood is flowing through the region rapidly enough to keep up its temperature and so favor the loss of heat. These changes in the amount of blood in the skin are very effective in the fine regulation of heat loss. The body can get rid of heat very much more effectively when the skin is flushed than when it is pale. There are, however, limits to the usefulness of this arrangement. Even when the skin is as pale as it can become it still is warm, and on a cold day it continues to lose heat more rapidly than is desirable. It is to prevent this that clothing is worn. Clothing operates for people as fur does for animals. It establishes a nonconducting layer between the skin and the outside. This nonconducting layer warms up to the temperature of the body and so hinders the escape of heat. The actual nonconducting material is the air which is caught in the meshes of the fabric, or in the case of fur-bearing animals among the different strands of the fur. The effectiveness of clothing for conserving heat depends altogether on the degree to which it imprisons air in its meshes. In this respect wool is the most effective of all fabrics, cotton next, silk and linen having very little effectiveness. This explains why wool is preferred for winter clothing, linen and silk for summer.
The temperature of the body as a whole is, as we have seen, about 98½ degrees F.; this figure represents the maximum temperature that the skin can reach, even when it is flushed to its utmost. In the warm weather of the summertime the temperature of the air frequently mounts to or even above that point and it is evident that under such circumstances the body cannot give heat directly from itself to the surrounding air, and no amount of flushing of the skin will enable it to do so. It is under these circumstances that the second method of getting rid of heat comes into play, namely, the evaporation of sweat. We have spoken of the sweat glands in a previous chapter, but have left their action and the description of the skin in which they lie for this point.
The skin serves a number of purposes; it is the great protective layer for all the parts beneath; in order that it may serve for this it must itself be reasonably free from injury. This is secured by having the outer layers of the skin composed of a horny dead material. Just beneath this dead layer is a layer of living cells tightly packed together and having exposure to the body fluids only at their under side. Cell multiplication goes on in these cells continuously and rapidly; as the cells divide, they grow, the result being that those that are underneath are constantly crowding those above them farther and farther out toward the surface; this cuts them off from their food supplies and so they die. This layer of dead cells becomes very closely packed and makes up what is commonly called the horny layer of the skin or the “cuticle.” The outer part of this cuticle is constantly rubbing off and, of course, as this happens any dirt that may be clinging is rubbed off as well. A large factor of cleanliness is the constant rubbing off from the surface of the body of its outermost layer. The extent to which this happens can be appreciated by anyone who has ever had a broken bone and has had to have an arm or leg put up in a splint or cast for several weeks. At the end of that time there is a great accumulation of dead skin waiting to be rubbed off.
Bathing is a very efficient aid to cleanliness; when baths are for this purpose, warm or hot water, with an abundance of soap, should be used; the choice of soap is an individual matter determined largely by the sensitiveness of the skin. Chemically there is no very great difference between one kind of soap and another. Some of them contain materials which are more irritating than those found in others and are to be avoided by persons whose skins are easily irritated. The common belief that a bath should not be taken immediately after eating rests on the feeling that the flushing of the skin that results from the contact with the warm water may divert blood unduly from the alimentary tract and so interfere with the process of digestion. This may very well be true if a very hot bath is taken; where the bath is only a degree or two above body temperature little trouble is to be feared from this cause. A cold bath shortly after eating sometimes gives rise to cramps either in the stomach or in the muscles. Not much is known about these cramps or about the causes which bring them about. In connection with its use as the protective surface of the body the skin has scattered over it numerous glands known as the sebaceous glands, which secrete an oily substance that is spread more or less completely over the skin. This oil probably helps to keep the skin soft and also to some degree to make the skin waterproof. One of the reasons why soap has to be used in bathing is to dissolve off this thin film of oil which is ordinarily over the surface of the body.
Another use of the skin is as a great sense organ; as we have already seen, the structures for touch, for temperature, and for pain are present in the deeper layers of the skin. These consist of tiny cell masses into which the tips of the sensory nerves pass. In the case of the nerves of pain there are believed to be no special end organs, but the nerves end nakedly among the cells of the skin.
The third use of the skin, and that with which we are particularly concerned here, is as the fine regulator of body temperature. We have already seen in part how the skin works in this connection, through being warmer when there is more blood flowing through it and cooler when there is less. Its other heat-regulating mechanism is made up of the sweat glands. These, as the name implies, are tiny glands in the skin; they consist of tubes opening on the surface and at their inner end coiled up into a sort of knot. In this knot are a great many fine blood vessels, so that each sweat gland has an abundant blood supply. So far as we can tell, the sweat glands act as filters, corresponding in that respect to the filtering tips of the kidney tubes. One interesting thing about them is that the filtering action is controlled by nerves. These nerves belong to the system which controls the smooth muscles and glands; when the nerves become active the sweat glands are also active, and there is rapid filtration of water and inorganic salts from the blood into the sweat glands and out to the outside of the body. Small amounts of organic materials come out also in the sweat, and these are responsible for its characteristic odor. They evidently differ in different people, since all dogs and some persons are able to recognize individuals by their odor, and body odor is largely the odor of sweat.
The sweat glands are carrying on their filtering activity most if not all of the time; we are not usually aware of it because the sweat comes out in exceedingly fine drops which evaporate as fast as they are formed; it is only when the sweat is poured out faster than it evaporates that we become aware of its presence. If the air is very full of moisture, sweat will stand on our bodies even though it is not being formed more abundantly than usual. This is because of its inability to evaporate into air already laden with moisture. Usually, however, when sweat appears on the surface of the body it is because the sweat glands have become more active. A couple of paragraphs ago we saw that the sweat glands are under the control of nerves; these arise like the other nerves that control smooth muscles and glands from a center in the brain stem; this particular one is called the sweat center. It may become active through various influences. The one most commonly arousing it is a rise in the temperature of the blood. Whenever we begin to exercise, heat is produced very rapidly, warming up the tissues in which the activity is going on and, since the blood is flowing through them rapidly, warming the blood up as well. This warm blood circulates all through the body, tending to raise all parts of it to the temperature of the active regions, or, as it passes through the skin, to cool by loss of heat from the surface. When this warm blood enters the brain stem it arouses the sweat center and an increased secretion of sweat results. The importance of this is that the evaporation of water, no matter at what temperature, requires a large amount of heat. This heat is abstracted from the nearest place, which in this case would be the body itself, so that the evaporation of sweat acts powerfully to cool off the body. In hot weather this is really our only effective means of getting rid of heat, for if the body is no warmer than its surroundings it cannot lose heat directly, but sweat can evaporate, taking heat with it, no matter how warm the surroundings may be. We said a moment ago that a high percentage of water in the air would hamper the evaporation of sweat. The practical workings of this fact are seen in connection with heat prostrations in the summertime. Careful scrutiny of the weather reports will show that these are much more numerous as well as more severe on days of high relative humidity than on days that are simply hot, but without much moisture in the air. As the sweat evaporates from the body, the resulting water vapor has a tendency to linger in the neighborhood and so to interfere with further evaporation. Under these circumstances it is desirable that this moisture-laden air be moved away and fresh air brought into its place. This is the benefit we obtain from fanning. The air that is brought upon us by a fan feels cooler than it really is; this we realize when we notice how comfortable the draft of a fan is even on the very hottest days. When one fans oneself in the old-fashioned way, the heat produced in working the fan is often almost enough to balance the loss through the improved evaporation of sweat. An electric fan, on the other hand, is a very powerful aid to comfort in hot weather. In fact it is hard to think of any mechanical device that has contributed more in recent years. An amusing illustration of the importance of moisture in the air in relation to bodily comfort is in the case of the man who thought he would obtain a cool-weather office in hot weather by installing a block of ice and causing his electric fan to play upon it. Very much to his surprise and quite contrary to his expectations the room quickly became unbearably sultry. The rapid evaporation of the ice filled the air so full of water vapor that evaporation of sweat could not take place and the persons in the room were much worse off than if they had contented themselves with merely keeping the warm and comparatively dry air of the room in rapid motion. Although the evaporation of sweat is a very effective means of getting rid of heat from the body it does not work perfectly in times of great muscular exertion in very hot weather. Careful studies of men in steel industries and other places where hard work is done in intense heat have shown that the production of heat during the course of the working day slightly out-strips its loss, so that by the end of the day it is the regular thing for the body temperature to be up two or three degrees, to 100 or 101 degrees Fahrenheit. Upon finishing the work the temperature quickly falls to normal, so that there is no reason at present for supposing this daily rise to be seriously harmful.
In fever the production of heat becomes greater than its loss for a time and so the temperature goes up. It must not be forgotten that presently the loss again balances the production; if it did not, the temperature would keep on rising until death resulted; usually the greater loss of heat from the body at the fever temperature is sufficient to restore the equilibrium, but it will be clear that the temperature cannot fall again until the heat loss exceeds the heat production. There has been a good deal of discussion as to whether fevers are caused by an increase of heat production or by an impairment of the machinery for getting rid of heat. There is no doubt that in fevers the resting metabolism is much greater than in health; it is not by any means always the case, however, that the total metabolism of an individual lying quietly in bed with a fever is greater than that of a healthy individual doing heavy manual labor. We must therefore look to an interference with the process of losing heat to account for the rise in temperature. The distribution of blood through the skin in fever does not differ from that at other times when the body is warm; it is the usual thing in fevers for the skin to be much flushed. There is, on the other hand, a definite impairment of the sweat-secreting mechanism. It is one of the most familiar facts about fever that the skin is not only hot but very dry. It is also well known that by inducing sweating the temperature can often be brought down. It is evident that the poisons which are responsible for the fever have at least two effects; to increase general resting metabolism to a high level and at the same time to interfere with the ordinary regulation of the sweat glands, so that, even though the temperature of the blood is several degrees above normal, the sweat glands are not stimulated to active production of sweat.
A fever is very uncomfortable to the person suffering from it, but is not in itself particularly dangerous, unless it mounts to a very high degree. The body wastes away rapidly, of course, because of the increased metabolism which is usually not balanced by any taking of food. Theoretically fever patients should eat as abundantly as laborers; practically this is impossible of accomplishment in most cases because of the depressed condition of mind and body of the patient. In some particularly wasting diseases, like typhoid, feeding has been carried on with some degree of success. The old practice with regard to fevers was to restore the patient to comfort as quickly as possible by getting rid of the fever, that is, by lowering his temperature to normal; at the present time the tendency in medicine is not so much to strive with the result of the poisoning as to seek to rid the body as quickly as possible of the poison and of its source. For that reason modern physicians are much less concerned with the degree of fever shown by the patient than with the extent to which the poisons in his body are being controlled or are becoming masters.
In all our discussion of the regulation of body temperature it is important to remember that our feelings of cold or of warmth depend altogether on the temperature of the outermost one-quarter inch of our body surface. No matter how cold or how warm we may feel, as soon as we get to that depth below the surface, we find the body temperature the same, namely 98½ degrees. We feel cold when the skin is cool; the skin is cool because not much blood is flowing through it; the stimulation of the surrounding cold on the skin has aroused reflex vasoconstriction and so diverted the blood out of the skin into other parts of the body. This means that the mechanism for preventing loss of heat is working as effectively as it can and, therefore, that the heat which the body produces is being conserved. Experience shows that in reality the feeling of cold is very largely a matter of the condition of the extremities. If the hands and feet are warm, one rarely finds the cold uncomfortable; on the other hand, cold extremities produce a very general feeling of discomfort, even though the rest of the body may be warm. The lesson to be drawn from this is to pay particular attention to keeping the extremities warm. Where workmen are standing on a floor it is much more important that the floor be warm than that the air of the room as a whole be so. Of course, it must not be cold enough to allow the fingers to be chilled and stiff where manual work is being done, but it has been found that a considerably lower factory temperature is endured when the floors are warm than when the floors are cold.
We feel cold in cold weather because the body is conserving heat; in warm weather, on the other hand, we feel warm because the skin is flushed and the body is losing heat as rapidly as it can. The practical bearing of this is that it might be disastrous to feel particularly warm in cold weather. Not, of course, if the feeling of warmth be due to a very rapid production of heat, as in vigorous exercise, but if it is due simply to a flushing of the skin when not much heat is being produced, the body is being deprived of what heat it has and a condition which cannot long be endured in cold weather will result. It happens that exactly this situation is met with in connection with the use of alcohol. One of the important effects of alcohol is to produce a flushing of the skin; this is a direct drug effect and has nothing whatever to do with the amount of heat that is being produced in the body. The result is that one who, by a few drinks, fortifies himself, when starting out for a long cold ride, feels very warm and comfortable as long as the alcoholic effect persists; but as soon as this passes off he is very much colder than he otherwise would be, since during all this time he has been losing heat rapidly instead of conserving it. It used formerly to be not at all uncommon in the cold parts of the country for persons to perish from exposure on long drives because they had attempted to keep themselves warm by taking alcohol, when if they had conserved their bodily heat instead of wasting it for the sake of comfort, they would probably have reached their destinations in safety.
While we are on the topic of temperature regulation and the part played by the skin generally in bodily activities a word should be said about bathing for other purposes than cleanliness. There is a good deal of discussion about whether the morning cold bath is really a health measure or is simply a fad; there can be very little doubt that persons who react well to the cold bath find that it contributes definitely to the enjoyment of life. It is particularly true that after a night of disturbed rest the morning cold bath makes one feel much more ready for the duties of the day, whether he actually is so or not. The value seems really to be in what is known as the reaction in which the first chill is followed by a warm glow. The bath should under no circumstances be prolonged beyond the time when a good reaction is obtained. Brisk rubbing with a harsh towel helps the oncoming of the reaction very much. For those who would like to take a cold bath but have found it too much of a hardship a hint may be helpful, namely, that if one enters the bath with the feet warm there is practically no discomfort connected with taking it. To step into a cold bath when the feet are already cold causes them to begin aching almost instantly and usually nullifies all the benefits the bath affords. One should either enter the bath immediately after rising, before there has been any time for the body to become chilly, or if one prefers to shave or carry on other activities first, it is a good plan to give the feet a momentary preliminary bath in hot water. When this condition is fulfilled, the cold bath will be both enjoyable and beneficial to the majority of people. Very hot baths have a distinct relaxing effect; they favor the onset of sleep and can be taken to best advantage just before retiring. There is nothing in the common idea that a hot bath opens the pores of the skin; the only pores the skin has are the openings of the sweat glands, and these do not vary in size. What a hot bath does do, is to cause a marked flushing of the skin as the result of which the body loses heat rapidly. It is necessary, therefore, after a hot bath to wrap oneself warmly so that excessive loss of heat shall not occur.
In finishing this topic, the subject of catching cold must be brought up, together with its relation to the regulation of body temperature. There is no doubt at the present time that common colds result from infection; in that sense they are acute diseases. It seems to be the fact, however, that the organisms which cause them are pretty constantly present in the air passages of our throats and lungs ready to invade the body whenever opportunity offers. Very recently it has been shown that rapid chilling of the skin is accompanied by contraction of the blood vessels in the linings of the throat and air passages. The effect of thus cutting down the blood supply to these linings seems to be to make them more ready of access by the organisms of colds. At any rate the fact that when the skin is suddenly chilled a cold frequently develops is best explained on this basis. Since colds are infectious, the organisms causing them may sometimes succeed in penetrating the linings of the air passages even when no change in the latter has occurred. It may therefore happen, and frequently does, that one develops a cold without being able to refer it to any time when there was a sudden chilling of the skin. Also it can easily happen that the chilling may not lead to a cold, since if the organisms do not happen to be abundant or if the general bodily resistance is high, the infection will not get a foothold, even though the linings of the air passages would permit the organisms to enter. One’s best precaution against colds would naturally be to avoid having the organisms which produce them present in the throat and lungs. Since this usually cannot be done, the remaining practical measures are to keep the general bodily health as good as possible and to avoid conditions which lead to a sudden rapid chilling, such as sitting in a cold draft when heated.
WE have tried to take up one by one the chief things that happen in the body, but thus far have emphasized their importance entirely in connection with bodily well-being; that is, we have seen how the body maintains itself against the competition of other living things and in a world full of hazards. Before leaving the subject entirely, a short account must be given of the way in which the race is maintained upon the earth as distinct from the maintenance of individuals. Early in the book we showed that every one of us starts life as a single cell which, by dividing and subdividing, along with continuous growth, finally develops into our large and complicated body. This cell is really the union of two cells; one furnished by each parent; in the body of the parent it formed part of what is known as the germinal tissue, this name being applied because the cell from which we start may be spoken of as the germ of life. As the cell from which the body is to come begins to develop, the cells formed from it quickly become different one from the other, so that very early it can be distinguished that certain cells are to form the future nervous system, others the future muscles, and so on. One of the earliest of these groups of cells to become distinguishable is that which is to become the germinal tissue in the newly formed individual. A great deal of importance has been attached of recent years to this fact that the germinal tissue is set aside almost from the beginning of development, and when we remember that the next generation will be derived from this germinal tissue and from no other tissue, we see that there is a very close relationship of the germinal tissue from generation to generation. The importance of this is in connection with heredity, namely, with the question of the resemblance of the child to the parent. We all know that sometimes children resemble their parents very closely, at other times there is almost no resemblance at all. The main fact of heredity is, as we have just seen, that the child comes from germinal tissue of the parent and not from the parent as a whole. We can think of an individual as a complex body carrying within itself and nourishing an independent group of cells which are to serve as the starting point for the next generation, but with which the individual himself has nothing to do except to provide nourishment. A little later we shall try to show just why we think of the germinal tissue as so little dependent on the rest of the body. What we want to do first is to point out how this notion must affect our views of heredity. If the germinal tissue is simply harbored and nourished by the body, but otherwise quite independent of it, it follows as a matter of course that things the parent does, provided that they do not disturb the nourishment of the germinal tissue, can have very little effect upon it. For example, anything that the parent may achieve during his lifetime cannot be passed on by heredity to his offspring. This is entirely contrary to the idea of heredity which is held by nearly everybody. Just a little over one hundred years ago a Frenchman by the name of Lamarck published writings in which he argued very strongly in favor of the inheritance of “acquired characters,” meaning by this that the achievements of parents could be and were transmitted to the offspring. It is evident that this would be a very great advantage; the progress of the race would be secure if the children could inherit directly the achievements of their parents. Of course, it might work the other way around and children might inherit unfavorable acquisitions of their parents as well as favorable. Since the time of Lamarck there has been not only hot discussion of his ideas but also a vast amount of experimentation to find out whether acquired characters actually can be inherited. One of the most famous experiments is that of a man who cut off the tails of mice at birth and did this generation after generation in the hope of being able to produce a race of tailless mice; after a great many generations he abandoned the attempt with the conclusion that the artificial removal of tails from parents would not cause the offspring to be born in the tailless condition. More recently similar experiments have been carried out on kinds of animals that have much shorter lives and in which the generations are correspondingly more frequent. In some cases thousands of generations have been studied without any indication that changes brought about during the lifetime of the parents can be transmitted to the offspring.
One of the first questions that is sure to be asked as soon as this inheritance of acquired characters is denied, is how changes can then be brought about; the answer to that question is found in the experience of animal and plant breeders. One fact of nature which should be emphasized, is that individuals are never exactly alike except perhaps in the case of so-called identical twins which have a special heredity spoken of in Chapter V. Even brothers and sisters always have pretty marked differences. These differences among individuals are some of them probably more or less accidental results of the way in which the germ develops into the complete body; others are the result of differences in the germ cells themselves. The distinction between these is that where the germ cells are different, the difference will be transmitted by heredity to future generations; if it is a modification that comes on in connection with development and is not due to a difference in the germ cell, it will not show itself in future generations. What the animal and plant breeders have to do is to watch for changes in their stock, and when the kinds that they are looking for appear, they breed them in the hope that the desired features will prove to be hereditary, so that a race can be established showing them. A very striking example of this kind of breeding is in the development of hornless cattle. Of late years it has been the practice to dehorn cattle regularly and this dehorning has never caused the offspring to be born without horns, but on the other hand it has happened occasionally that calves have been born which were naturally hornless and since this absence of horns was due to a difference in the germ tissue, the character was found to be hereditary and it has been possible to establish breeds of hornless cattle. Thus by selecting cases where a change occurred naturally in the germ tissue, a result has been obtained which could not possibly be gotten by any amount of work directly upon the parents.
What we have tried to make clear thus far is that heredity is absolutely a matter of the condition of the germinal tissue. What we have now to do is to see what the germinal tissue is like and how the process of heredity actually goes on. In Chapter V something was said about the nucleus; it will be remembered that the nucleus contains a substance known as chromatin, which, as seen under the microscope, looks like a tangled skein or network of fine threads, but is in reality a number of tiny structures known as chromosomes. We now know that these chromosomes are the actual controllers of heredity. Of course we do not know at all how they exert their control; what we do know is that when certain factors are present in the chromosomes of the germinal tissue of the parents, the offspring derived from that germinal tissue will have traits that would not be present if the chromosomes had been different.
Space does not permit us to tell at length how the facts that are now to be described were discovered. They date from the gardening experiments of an Austrian monk by the name of Mendel, who for many years grew ordinary garden peas and studied from season to season the varieties that appeared. The facts of heredity that he found to be true of peas, have since been shown to apply just as well to ourselves, and since this book deals with human beings we shall try to make the description apply directly to human heredity. The first thing to get clearly in mind is that every single thing about any one of us which can or does differ from the corresponding feature in another person, may be a hereditary trait, and, if it is, will have a factor controlling it somewhere among our chromosomes. Since the number of hereditary traits is legion, including not only the size and shape of all parts of our bodies inside and out, but our mental peculiarities and moral tendencies as well, there must be a huge number of controlling factors, or determiners, as they are often called. As a matter of fact, there are probably not quite so many determiners as traits, because a single determiner may govern more than one trait. But even so, the number of determiners is too large for comfort in trying to describe their working. The best way to go about it is by pretending that we are not so complicated as we really are. We shall do this by setting the number of different determiners human beings may possess at fifty-two, not because that is anywhere near their real number, but because it is the number of combined large and small letters in the alphabet, and we propose to use letters to stand for determiners, as an easy way to keep them separate in the description.
Every cell in our bodies has its nucleus with its equipment of chromosomes. An interesting fact already spoken of in Chapter V is that according to our best knowledge the chromosomes in any cell of any one of us are exactly like those in all the other cells of the same person. The chromosomes in the muscle cells are exactly like those in the nerve cells, and both correspond exactly with the chromosomes of the ordinary cells in the germinal tissue. In every one of these cells the chromosomes are arranged in pairs. It happens that in human beings the number of pairs is twenty-four, with one pair incomplete in some of us for a reason that will be explained presently; this is an awkwardly large number for our present purpose, since we have allotted only fifty-two determiners altogether, so we shall do some more pretending and set the number of pairs at nine. One more change will have to be made from the real state of affairs before we can go on with the description; this is to suppose that our various bodily features can show only one difference, instead of the many of which they are really capable. For example, we shall suppose that the hair can be either black or light, but never red; the nose can be Roman or Greek, but never Irish. By making this supposition, we can let the large letters stand for one set of hereditary determiners and the small for the same features but with exactly contrary traits. According to this arrangement, if we had two persons side by side one of whom had only large-letter determiners in his chromosomes and the other only small, they would have the same general human make-up, but in every possible detail one would be the exact opposite of the other.
It has been proven by complicated studies which we cannot take time to describe that the determiners are grouped in the different chromosomes in a definite plan. We saw a moment ago that the chromosomes are in pairs. This pairing is an essential part of the arrangement, for every hereditary bodily feature actually has two determiners governing it, which lie in corresponding positions in the two chromosomes of the pair. To illustrate how this works out, let us suppose that the chromosomes of the first pair contain determiners A, B, and C. Each of these three determiners will be present in both members of pair number one, and if this is true of any cell it will be true of all the cells, and in any other human being either they or the corresponding small-letter determiners, a, b, and c, will occupy pair one of the chromosomes. Furthermore, they will lie in a row within the chromosomes, always in the same order; thus if A and C are at the ends with B in the middle in one chromosome, every other chromosome that contains these three determiners will have them in the same order.
Thus far we have planned our diagrams as though only large letter determiners were concerned; but we saw a moment ago that there is a complete set also of small-letter determiners, which control a set of contrary hereditary traits, and we intimated that these will sometimes be found in the chromosomes in positions corresponding with those occupied by the equivalent large-letter determiners. It might happen, for example, that pair one of the chromosomes would contain large-letter determiners A, B, and C, while pair two would contain small-letter determiners a, b, and c. Evidently the person in whom this combination was present would differ from one all of whose chromosomes contained large-letter determiners, since part of his traits would be established by small-letter determiners.
We are now ready to go back to the germinal tissue and trace the process of heredity as it actually works out in the developing offspring. In the ordinary cells of the germinal tissue, as we have already seen, the chromosomes are exactly like those in the other cells of the body. Cell multiplication goes on actively in the germinal tissue of both parents; in the mother this leads to the production of germ cells which are called eggs, and in the father to the production of cells that are called sperm. During the process certain changes occur, so that neither eggs nor sperm are exactly like the original cells of the germinal tissue. If we look back at the description of cell division in Chapter V, we shall recall that the chromosomes split lengthwise and are pulled apart. Now that we have learned about determiners, we will realize that every determiner splits in half, because otherwise there would not be an equal distribution of determiners between the two cells. Much of the cell division in the germinal tissue is precisely like this, but at a certain stage in the production of both eggs and sperm there is one cell division in which the pairs of chromosomes are simply pulled apart, without there having been any previous lengthwise splitting. The result of this is to leave the resulting cells with only half as many chromosomes as the other cells of the body have. Some further changes take place in these cells before they become ripe eggs or ripe sperm, but there is no further disturbance of the chromosomes, so that eggs and sperm contain only one member of each pair, instead of both members, as do all other body cells.
The first step in development is the coming together of the egg cell with one sperm in the process that we call fertilization. The sperm penetrates the egg and its chromosomes line themselves up with the egg’s, restoring the pairing that is present regularly. Immediately afterward the cell divisions begin that make up development, and in all of these the usual lengthwise splitting of determiners takes place. Every cell in our body contains its pairs of chromosomes, one member of each pair tracing back directly to the egg cell while the other traces directly to the sperm. Thus half of our determiners came from the maternal germ tissue and half from the paternal.
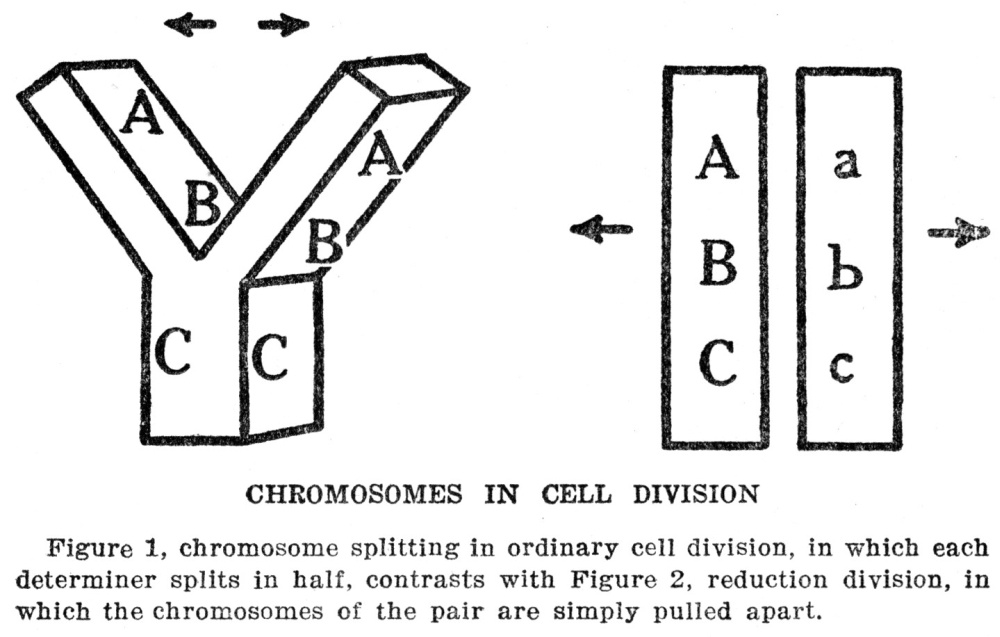
Figure 1, chromosome splitting in ordinary cell division, in which each determiner splits in half, contrasts with Figure 2, reduction division, in which the chromosomes of the pair are simply pulled apart.
We shall now begin to see how heredity works out. Suppose chromosome one in the egg has large-letter determiners A, B, and C, while the corresponding chromosome in the sperm has small-letter determiners a, b, and c. When these line up after fertilization, restoring pair one, we have A opposite a, B opposite b, and C opposite c. Since we have supposed the large and small letters to stand for contrary hereditary traits we introduce here a conflict and must ask at once how it is settled. One of the things the monk Mendel worked out in his studies of heredity in peas was this particular problem. He found that where there is conflicting heredity one of the determiners usually dominates over the other, and when this happens the trait in the offspring will be like that of the parent which contributed the dominant determiner. To illustrate: suppose A is dominant over a, then in the case in question the offspring will be like the mother in feature A. In some kinds of animals and plants conflicting characters blend in the offspring, producing an intermediate appearance. A good example of this latter case is in the common flower, the four-o’clock. In this plant white blossoms and red blossoms are due to determiners that occupy the same positions in the chromosomes; therefore, if white and red flowered plants are crossed, these conflicting determiners come together when the sperm and egg chromosomes pair. Since neither determiner dominates over the other, the color of the flowers in the offspring is neither white nor red, but pink.
Any individual whose chromosome pairs contain conflicting determiners is called a “hybrid”; there may be every degree of hybridism, from the simplest, in which all the chromosome pairs are alike except one, up to the most complete, in which all the pairs are unlike. It is easy to tell a hybrid by its appearance in the cases in which there is blending inheritance, but not so easy when one trait dominates over the other, for then the hybrid will look like the parent that furnished the dominant determiner. The sure way to detect hybrids is by the study of their offspring. Suppose we have two parents, both of whom are hybrids in respect to chromosome pair one. This pair in both will contain determiners A, B, and C lined up opposite a, b, and c. Since we have supposed the large letters to be dominant over the small, both will have the same appearance, dependent on the presence in their chromosomes of A, B, and C. Since in the formation of eggs and sperm the chromosome pairs are pulled apart, half the eggs produced by the mother will contain determiners A, B, and C, and the other half a, b, and c, and half the sperm of the father will similarly contain one set and half the other. Since it is an absolute matter of chance which sperm encounters which egg in fertilization we can safely conclude that in the long run all the chances will be realized equally. Calling, for convenience, the large-letter eggs E and the small-letter e, and, similarly, the large-letter sperm S and the small-letter s, this means that E can be fertilized either by S or s, and e also by either S or s. The possible combinations are ES, Es, eS, and es. In terms of actual determiners these combinations are ABCABC, ABCabc, abcABC, and abcabc.
The combinations just given represent the possible offspring from a pair of hybrid parents. If we look them over, we see at once that only half of them are hybrid, namely, combinations ABCabc and abcABC; the other half are pure breed, the chromosome pairs being exactly alike; but these pure breeds are of two kinds, one having only large-letter determiners, the other only small-letter. If it is a case where the large letters dominate over the small, the large letter pure breeds and the hybrids will look alike, but the small-letter pure breeds will look different, since in them the traits governed by the small-letter determiners have a chance to show themselves. A very good illustration of this is seen in eye color in human beings. Brown eyes are dominant over blue; in other words, the determiner that causes eyes to be brown dominates over that responsible for blueness in cases where both come together in hybrids. A person who has brown eyes may be either a pure breed in that respect or may be a hybrid; there is no way to tell the difference from the appearance; but if the brown-eyed person has offspring, and any of them turn up with blue eyes, it is proof positive that the parent is a hybrid so far as eye color is concerned. Moreover, the blue-eyed child is not a hybrid in this respect; his brown-eyed brethren may or may not be; in the long run two-thirds of them will prove so; the other third will be pure breed, having in their chromosome pairs only brown-eye determiners.
Where the hybrids differ from the pure breeds, as in the case of the four-o’clocks, given earlier, it is easy, of course, to tell which are pure and which are hybrid. When pink four-o’clocks are interbred, the chromosomes will combine just as described above for hybrids, since the plants that have pink flowers are hybrid. One-half the offspring will have pink flowers, showing that they are hybrid; one-fourth will have white flowers, proving that in them the white-flower determiners have separated out, and the other fourth will have red flowers, because in them the red-flower determiners are the only kind present. This and similar experiments have been tried hundreds of times, and whenever the numbers of offspring have been great enough to allow the chances to equalize, the proportion of different kinds of offspring has always agreed almost exactly with expectation. Of course in human beings the families are not large enough for this always to work out accurately, but even so the agreement is often striking.

In practical animal breeding the blended inheritance just described is not very useful, for even though a blended character might appear which is just what the breeder has been looking for, it will not occur in more than half the offspring and can not ever be depended on to show itself in any particular individual. This explains largely why pure-bred stock is always more desirable than hybrid, and why breeders strive so eagerly to obtain desired traits in pure-bred animals. In plants, blended characters are much more valuable, for two reasons; first, because the offspring are so numerous that even though half of them come out pure, and so lack the desired blend, there are enough left that have it to make the crop worth while; and second, because propagation by cuttings is possible in very many kinds of plants, which means that the same plant is kept going in hundreds of places, and for tens or even hundreds of years. A trait that is desirable can be perpetuated indefinitely by this means, even though it may be a blending of several hereditary traits, which would separate out in a few generations by ordinary means of propagation.
There are several more things in heredity that must be taken up while we are on the subject, so we shall have to return to the chromosomes for a while. We have seen that there are several determiners to each chromosome; for convenience, we assigned three apiece to our chromosomes, except the ninth, which has to get along with two; but in reality the number to each chromosome is often much greater. This grouping of the determiners, several to a chromosome, carries an interesting consequence with it, in that all the hereditary traits controlled by one chromosome have to go together in reproduction. In the example we have already used A, B, and C are together; therefore any individual that shows character A must show B and C as well. The most striking instances of this are certain traits that are bound up with sex, but we cannot describe these further until we have looked into the heredity of sex, which we shall do in a minute. First a word must be said about occasional exceptions that turn up to the rule that we have just stated. In the study of thousands of specimens now and then one has been found in which there has evidently been a swapping about of determiners. We can illustrate the situation by supposing chromosome one is found to contain determiners A, b, and C, instead of A, B, and C; one small-letter determiner has traded places with a large. Of course, the effect of this is to permit different combinations of hereditary traits than ordinarily occur, and at the present time students of heredity are actively engaged in following this up to see how it happens, and what advantage can be taken of it. This crossing over of determiners from one chromosome to another takes place only among such as are actually in contact at times within the nucleus as seen under the microscope, which confines it to the members of corresponding pairs.
In man, and in many of the lower animals, sex is a hereditary character. That means that there is a determiner for it which is grouped with other determiners in one of the chromosomes. In man the determiner is for femaleness; there is no special determiner for producing the male sex; it is produced whenever the female determiner is missing from one chromosome of the pair, and this is brought about by having the whole chromosome that should make up this pair absent. At the beginning of this chapter the fact was mentioned incidentally that a good many of us have only 47 chromosomes, instead of the 48 that are characteristic of human beings. The distribution is really almost exactly half and half, for all males have 47 and all females 48. This means that the cells of the germinal tissue of females have 24 complete pairs, while the corresponding cells in males have only 23 complete pairs and one chromosome over. This extra chromosome is the one that contains the determiner for femaleness; each of the chromosomes of pair 24 in females contains this determiner also. These are often spoken of as sex chromosomes.
Now when in the course of the production of egg cells within the mother’s germinal tissue the pairs of chromosomes are pulled apart, each separate cell, and so each egg, will contain the full number of chromosomes, 24, including the sex chromosome. But when the same thing happens in the course of the production of sperm only every other one will have the full number; the remaining half having only 23, and all of this half lacking the chromosome that contains the determiner for femaleness. There are, then, always equal numbers of two kinds of sperm, one with 24 and the other with only 23 chromosomes. If the egg is fertilized by a sperm containing 24, including the sex chromosome, the pairing of chromosomes is complete in the egg, and the offspring will be a female; if, on the other hand, the fertilizing sperm is one that contains only 23 chromosomes the pairing in the egg will be incomplete; the single sex chromosome of the egg will not be paired with a corresponding one from the sperm and the egg will develop into a male. Since it is a pure chance whether fertilization will be accomplished by a sperm of 24 or one of 23 chromosomes, we should expect the sexes to appear in exactly equal numbers, taking the world as a whole. As a matter of fact, whenever extensive birth data have been accumulated they have shown a very slight excess of male births over female. We are not able to explain this at the present time. It is possible that the 23 chromosome sperms are a little more vigorous for some reason than those that have 24, and so are able to fertilize slightly more than their share of eggs.
We spoke a moment ago of hereditary traits whose determiners are bound up in the sex chromosomes. All such behave interestingly in heredity for the simple reason that they can never be transmitted from father to son, but only from father to daughter. This is because, as we have just seen, the sex chromosome in the sperm always causes the egg which that sperm fertilizes to develop into a female. The single-sex chromosome which males possess invariably comes from the mother. An interesting example is the common type of color blindness known as Daltonism. Normal color vision is hereditary and the determiners which establish it are in the sex chromosomes. Occasionally a person is found in whom these determiners are defective. If this person is a male, he will be color blind, but if a female not, unless both sex chromosomes are defective in this regard, since normal color vision is dominant over color blindness; so if one sex chromosome is normal the vision will be also. The woman, in this case, will be a hybrid with respect to color vision; one of her sex chromosomes containing a normal determiner, the other a defective.
This works out in heredity as follows: If a color-blind man is married to a woman who has no color blindness in her heredity, none of his children will be color-blind because he cannot transmit the sex chromosomes which carry the determiners for color blindness to his sons, but only to his daughters; all the latter will be hybrid with respect to the character, since all of them come from fertilized eggs which received sex chromosomes from the sperm. If these daughters, in turn, marry men who are free from color blindness, some of their sons may be color-blind, but none of their daughters can be. The only way in which women can be color blind through inheritance is by descent from color-blind fathers and from mothers who are either themselves color-blind or are hybrid with respect to the trait. The result of this difference in the heredity of the two sexes is to make color blindness many times as frequent among men as among women. In round numbers four men out of every hundred have this type of color blindness, while only six or seven women in ten thousand show it.
We have left for discussion only one topic dealing with heredity, but this is the most baffling of all, since it deals with the problem of how the various kinds of determiners came into existence. It is evident that if given one parent with all large-letter determiners and the other with all small-letter, we might, in the course of many generations, get a great variety of combinations and so a great many different-appearing individuals. But unless we have various kinds of determiners to start with, there is no way in which this can be done. We do not pretend to know very much about how the innumerable determiners that are in existence came about, but we have one clue that is thought to point the way. In some animals, and many plants, descendants put in their appearance from time to time that are so different from their ancestors as not to be accountable according to ordinary laws of heredity. These have long been known, and the name of “sport” has been applied to them by breeders. Since the facts about determiners have been learned, it has been clear that these “sports” cannot have all their determiners like those present in their parents, and it has come to be believed that occasionally spontaneous changes take place within individual determiners. Since the determiners are undoubtedly complex chemical structures, we know of no reason why this might not happen. Probably it is much more common an occurrence in some kinds of plants and animals than in others. The name of “mutant” has been applied to the plant or animal in which this has happened, and the process is called “mutation.” It is important since it is the most likely way in which the innumerable kinds of determiners that are now in existence came into being.
We suppose that since life first put in its appearance on the earth there have been uncounted mutations, a vastly greater number than are now represented by determiners. Many of the mutants could not compete with their brothers and sisters of ordinary descent and so promptly died, but occasionally it might happen that a mutant would be as well fitted for life as its relatives, in which case it would establish itself, and in course of time become ancestor to a whole line like itself. If this happened often enough, and time were allowed for it to work out, all the kinds of plants and animals that are now in existence might have come by descent from a very few ancestors. The geological history of the earth shows that there has been plenty of time, even though valuable mutations did not occur oftener than once in a thousand years.
Our description of racial perpetuation should be finished by an account of the development of the fertilized egg. Snugly ensconced within the body of the mother, in an organ devoted solely to the purpose, the egg passes rapidly through the early stages, living at first on fats and proteins stored within itself. After these are exhausted it draws supplies from the body fluids of the mother. In the course of a few weeks it has developed its own conveyer system, with its own beating heart and its own stock of blood. There is never any actual mingling of the blood of the developing child with that of the mother; capillaries of the maternal circulation come into intimate contact with capillaries of the circulation of the child. Here interchange of all sorts of material goes on; food and oxygen pass from the mother’s blood to the child’s and waste materials from the child’s blood to the mother’s. During all this time the mother is eating, breathing, and excreting wastes for two. She cannot bring any nervous influences to bear on the developing child, since there is no connection between her nervous system and the child’s; she can, however, influence it chemically through the blood. Poisons that get into the blood of the mother can pass from it into the blood of the child. These may be the poisons of auto-intoxication, or drugs that the mother has taken. In either case they may do the child harm. We do not know very much about this, but it may be that a considerable percentage of children that are born with abnormalities that are not hereditary come by them through chemical influences received from the mother’s blood.
When the development of the child has gone far enough so that it can do its own breathing, feeding, and discharging of waste materials, it is expelled from the body of the mother in the process that we know as birth. This does not imply by any means that parental care and responsibility are at an end. Food, protection, and warmth must be provided. Education must be attended to, for the nervous system of the new-born infant is absolutely undeveloped. It has, through heredity, certain possibilities of achievement; their realization hinges upon the bringing to bear of worth-while influences. Upon the attainment of maturity the child will be expected to assume his place in society, and society has a right to the best that he is able to offer. In preparation for this it is the duty, both of the parents and of society itself, to provide throughout the formative years as nearly as possible the environment best suited to the development of those traits which make for usefulness. Environment cannot overcome the limitations of heredity, but environment can bring out the best that is in us.
EVERYONE is familiar with the beguilingly helpless picture the tiny baby presents. The disproportionately large head, with aimlessly rolling eyes and toothless mouth, the frail and delicate limbs, waving in the air or clutching spasmodically at anything within reach, the expressionless face, on which, for the first few days, only sensations of discomfort are registered, all mark a creature whose survival hinges absolutely on unremitting care; a far cry from the competent self-sufficiency of the average person of mature years. These surface marks of helplessness are by no means the most significant. Buried from view beneath the soft and velvety skin are characteristics of even greater meaning to those on whom falls the responsibility for the rearing of the new life.
At the time of birth the bony skeleton is very incomplete; there is a spot just above the forehead where the skull bones have not yet grown completely together, leaving a space where the brain is protected from injury only by the overlying skin. This spot can be detected easily in very young infants by its pulsations in time with the heartbeats. In most of the other bones the deposit of lime salts to which they owe their stiffness has gone only a little way, so that it would be impossible for the child to stand or walk even though it knew how. In this respect the contrast between the human infant and the new-born of such animals as horses or cattle is very striking, for the latter walk stumblingly from almost the moment of birth, and efficiently within a few hours thereafter.
Not only is the infant devoid of teeth, but in various other regards his digestive apparatus is undeveloped. Not only is he unable to chew solid food, but he could not digest most of it if it were served already chewed. During the early months of life the child is emphatically a one-diet being. His alimentary tract deals successfully with the mixture of proteins, fats, and sugars of which milk consists, provided the proportions are substantially those of mother’s milk and the quantity at a feeding is not too great. He can do this because the enzymes needed for digesting these particular substances are manufactured by his digestive glands from the very beginning, and because the muscles of his stomach and intestines can churn and propel the soft curd into which the milk is converted as soon as it enters the stomach. The fact that cow’s milk sets into a tough curd accounts for much of the difficulty some babies have in thriving upon cow’s milk.
There is no starch in milk, and neither the saliva nor the pancreatic juice of the infant contains the enzyme by which starch is digested. It is wholly useless to begin feeding starch-containing foods until this enzyme begins to be manufactured, which usually takes place when the child is about eight months old. Even then the introduction of starch into the diet should be gradual and cautious.
Modern science has discovered no better food for infants than mother’s milk, and no substitute more satisfactory in general than suitably modified cow’s or goat’s milk. Under modern conditions of life, particularly in cities where milk has to be transported over long distances, and where much time necessarily elapses before the milk can be placed in the hands of the consumer, there is serious danger of contamination of the milk with disease germs. In all enlightened communities this danger is fully recognized, and the entire milk supply, so far as possible, but that part of it destined for the feeding of children in particular, is safeguarded by all available means. A fairly reliable index to the degree of enlightenment of any community is the quality of the milk which its children receive. One recent discovery of considerable importance, and incidentally an interesting illustration of the way in which correct procedure may be hit upon in advance of the scientific knowledge which justifies it, is the finding that orange juice contains one of the dietary accessories, known as vitamines, essential to health, and present in raw milk, but destroyed when milk is heated. For years, physicians had recommended orange juice for babies. It is of vital importance when the milk must be pasteurized.
Both the heartbeat and the breathing in the young child are much more rapid than in the grown person. It is believed that this quickness of heart action and of breathing rate are related to the smaller size of the infant as compared with the adult and are of no very marked significance. At any rate it is true in general that the smaller the animal the more rapidly does its heart beat and the more quickly does it breathe. A very noticeable fact about young children is the susceptibility to all sorts of influences of the mechanism by which the breathing is controlled. Every passing interest reflects itself in heightened breathing. Violent emotion often leads to such extreme overbreathing as to drain the child’s blood of considerable of its store of carbon dioxide, whereupon the rapid breathing gives place to prolonged holding of the breath. Many a young mother has been seriously alarmed by the length of time her offspring in a tantrum is able to refrain from drawing its breath. Contrary to the appearance of things, which would indicate that the child is holding its breath out of spite, and in the hope of getting even with its parent, the cessation of breathing is largely or wholly automatic, indicating the way in which nervous and chemical influences have interacted to suspend the respiration.
The child is born with all its muscles in place, and all fully formed in that every muscle fiber the child will ever have has been produced previous to birth. In fact, as soon as the full equipment of muscle fibers has been laid down the body loses the power to form more, so that if, through injury, one is so unfortunate as to have some of his muscle fibers destroyed he will have to get along for the rest of his life with those that are left. The gaps in the muscle tissue that are produced by injuries are filled up by a kind of connective tissue known as scar tissue. The muscle fibers are all present, but smaller and weaker than they will be later.
The connections between muscles and nerves are also pretty well established at the time of birth, so that the body and limbs can be moved freely, even if not at all efficiently.
Not only are the motor nerves formed and in connection with the muscles at and even before birth, but the sensory nerves and most of the central nervous system are ready to begin functioning as well. Some reflex actions, including a few that require quite elaborate muscular and nervous coordination, are performed very shortly after birth. One famous example, that has been much cited as tending to reinforce the belief in man’s descent from tree-dwelling ancestors, is the curling of the fingers about a slender rod that is pressed against the palms. In most very tiny babies the grasp thus secured is strong enough so that the child can be raised and held in the air, supported wholly by its own grip. Sneezing, which is really a very complex act, requiring accurate cooperation on the part of many muscles, is done successfully by very young infants. The reflex of sucking, which is of paramount importance, in that without it the child would almost inevitably starve, is present practically from birth. Another important early reflex is that of crying. It is a curious thought that this reflex, by which bodily discomfort is made known, and through which relief may be summoned, is revealed at the very instant of birth, in connection with the drawing of the first breath, while the contrary reflex of laughter, by which bodily well-being is expressed, puts in an appearance only after some weeks or even months.
It is difficult to determine just how far the sense organs have arrived in their development at the time when the infant begins its independent existence. That touch and those senses related to bodily discomfort, of which pain is most important, are operative from the first is shown by the occurrence of the reflexes described above, which are brought into action by those particular senses. There is good reason to believe that the sense of hunger comes into play within two or three days at the latest. The sense of thirst does not have much chance to reveal itself early in life, for with a diet exclusively liquid, and feedings separated by only a few hours, there is no reason why the child should become thirsty. As the feedings become less frequent, and particularly as solid food begins to be added, there is real danger that the child may be insufficiently supplied with water.
Muscle sense and the equilibrium sense, if present at all, must be in a very imperfect condition at first. They seem to differ from the senses described thus far in that they depend on practice for their development. At any rate the bodily movements are largely aimless in the beginning, and it will be observed that the baby has the appearance of experimenting with its extremities, placing them repeatedly in particular positions and seeming to gain precision thereby. The eye movements, and especially those by which both eyes are focused on a single object, depend for their accuracy on the working of muscle sense. In the estimation of the parents a distinct mark of progress is registered the first time the baby follows a movement with its eyes. As soon as it does this accurately, and also brings both eyes to bear on any object, its muscle sense is known to be in efficient operation, so far, at least, as the eye muscles are concerned. Equilibrium sense first shows itself when real balancing motions of the body are made.
The senses of taste and smell may be operative to some degree in very young children, but it is doubtful whether they have either the breadth or the acuteness that will be shown later in life. Recognition of disagreeable tastes or smells seems to appear earlier than appreciation of agreeable. This is in line with the general fact that the self-protective reactions are developed very early.
Hearing and sight are probably in working order practically from birth. It is customary to test the sight of the new-born by passing a light directly in front of the eyes. If sight is present there will be some appreciable eye movement, suggesting that the eyes are attempting to follow the moving light. There is no reason to believe that there is any such thing as definite looking at objects thus early. So long as the eyes continue to roll aimlessly about, and before they begin to focus accurately, they are more likely concerned with distinctions between light and shadow, than with perceptions of form or size. In general we may say of the senses that those concerned immediately with bodily discomfort are about as fully developed at birth, or shortly thereafter, as they ever will be, while those that have to do with the general adjustments of the body to its environment reach full efficiency more gradually.
The higher parts of the brain, especially those concerned on the one hand with the mental life (the cerebrum), and on the other with the complicated reflex acts involved in locomotion (the cerebellum), are not ready to begin active functioning when the child is born. Indeed some parts of the cerebellum do not take on final form for from eight months to two years afterward. It is thought by some that the question of whether a child will learn to walk early or late depends, in part at least, on how soon his cerebellum reaches complete development.
Most parents are fully alive to the importance of abundant warm covering when their children are to be taken out into the cold, but there is much less appreciation of the harm that may be done by too much clothing in extremely hot weather. Of special importance is the avoidance of exposure to sharp drops in temperature. The adult adjusts himself more or less automatically to these, whereas the infant does so to only a limited extent.
Finally, the hold of the infant upon life, that quality that we know as ruggedness or vitality, is much slighter than it will be after a few years. Not only is the susceptibility to many kinds of infectious diseases very much greater, and the power of resisting them very much less, but the ill effects of poisons, whether taken in with the food or breathed in with the air, are more pronounced. Thus the vitiated air of slum dwellings, saturated with the effluvium from unwashed bodies and unclean clothing, while trying enough for the average grown person, is deadly for all but the toughest babies. Even in the ordinarily well-kept home, especially in the winter time when ventilation is apt to be neglected, the air within the house tends to become unsatisfactory from the standpoint of the infant’s welfare.
The wise parent, and wise he must be at this time, relaxes his care in just proportion as the child achieves ability to do things for himself. Since bodily development is more rapid than mental the close supervision of food, clothing, and physical occupation is necessary only during the early years, but the task of building up, through the slow processes of education, the sort of mind which will be able to do its proper share in dealing with the difficulties which confront the coming generations is one to which may well be devoted the best thought and effort not only of the parents, but of organized society as a whole.
| Typographical errors corrected by the etext transcriber: |
|---|
| that their less curvaturos=> that their less curvatures {pg 114} |
| as fast as it it formed.=> as fast as it is formed. {pg 180} |
| Russian invesigator showed=> Russian investigator showed {pg 240} |
| quantities of axygen=> quantities of oxygen {pg 254} |
| unless artificial air can be procured=> unless artificial aid can be procured {pg 303} |