
The cover image was created by the transcriber and is placed in the public domain.
GUIDE TO THE STUDY OF FISHES
BY
DAVID STARR JORDAN
President of Leland Stanford Junior University
With Colored Frontispieces and 427 Illustrations
IN TWO VOLUMES Vol I.
"I am the wiser in respect to all knowledge and the better qualified for all fortunes for knowing that there is a minnow in that brook."—Thoreau

NEW YORK
HENRY HOLT AND COMPANY
1905
Copyright, 1905
BY
HENRY HOLT AND COMPANY
Published March, 1905
To
Theodore Gill,
Ichthyologist, Philosopher, Critic, Master in Taxonomy,
this volume is dedicated.
This work treats of the fish from all the varied points of view of the different branches of the study of Ichthyology. In general all traits of the fish are discussed, those which the fish shares with other animals most briefly, those which relate to the evolution of the group and the divergence of its various classes and orders most fully. The extinct forms are restored to their place in the series and discussed along with those still extant.
In general, the writer has drawn on his own experience as an ichthyologist, and with this on all the literature of the science. Special obligations are recognized in the text. To Dr. Charles H. Gilbert, he is indebted for a critical reading of most of his proof-sheets; to Dr. Bashford Dean, for criticism of the proof-sheets of the chapters on the lower fishes; to Dr. William Emerson Ritter, for assistance in the chapters on Protochordata; to Dr. George Clinton Price, for revision of the chapters on lancelets and lampreys, and to Mr. George Clark, Secretary of Stanford University, for assistance of various kinds, notably in the preparation of the index. To Dr. Theodore Gill, he has been for many years constantly indebted for illuminating suggestions, and to Dr. Barton Warren Evermann, for a variety of favors. To Dr. Richard Rathbun, the writer owes the privilege of using illustrations from the "Fishes of North and Middle America" by Jordan and Evermann. The remaining plates were drawn for this work by Mary H. Wellman, Kako Morita, and Sekko Shimada. Many of the plates are original. Those copied from other authors are so indicated in the text.
No bibliography has been included in this work. A list of writers so complete as to have value to the student would make[Pg viii] a volume of itself. The principal works and their authors are discussed in the chapter on the History of Ichthyology, and with this for the present the reader must be contented.
The writer has hoped to make a book valuable to technical students, interesting to anglers and nature lovers, and instructive to all who open its pages.
David Starr Jordan.
Palo Alto, Santa Clara County, Cal.,
October, 1904.
| CHAPTER I. | |
|---|---|
| THE LIFE OF THE FISH (Lepomis megalotis). | |
| PAGE | |
| What is a Fish?—The Long-eared Sunfish.—Form of the Fish.—Face of the Fish.—How the Fish Breathes.—Teeth of the Fish.—How the Fish Sees.—Color of the Fish.—The Lateral Line.—The Fins of the Fish.—The Skeleton of the Fish.—The Fish in Action.—The Air-bladder.—The Brain of the Fish.—The Fish's Nest. | 3 |
| CHAPTER II. | |
| THE EXTERIOR OF THE FISH. | |
| Form of Body.—Measurement of the Fish.—The Scales or Exoskeleton.—Ctenoid and Cycloid Scales.—Placoid Scales.—Bony and Prickly Scales.—Lateral Line.—Function of the Lateral Line.—The Fins of Fishes.—Muscles. | 16 |
| CHAPTER III. | |
| THE DISSECTION OF THE FISH. | |
| The Blue-green Sunfish.—The Viscera.—Organs of Nutrition.—The Alimentary Canal.—The Spiral Valve.—Length of the Intestine. | 26 |
| CHAPTER IV. | |
| THE SKELETON OF THE FISH. | |
| Specialization of the Skeleton.—Homologies of Bones of Fishes.—Parts of the Skeleton.—Names of Bones of Fishes.—Bones of the Cranium.—Bones of the Jaws.—The Suspensorium of the Mandible.—Membrane Bones of Head.—Branchial Bones.—The Gill-arches.—The Pharyngeals.—The Vertebral Column.—The Interneurals and Interhæmals.—The Pectoral Limb.—The Shoulder-girdle.—The Posterior Limb.—Degeneration.—The Skeleton in Primitive Fishes.—The Skeleton of Sharks.—The Archipterygium.[Pg xii] | 34 |
| CHAPTER V. | |
| MORPHOLOGY OF THE FINS OF FISHES. | |
| Origin of the Fins of Fishes.—Origin of the Paired Fins.—Development of the Paired Fins in the Embryo.—Evidences of Palæontology.—Current Theories as to Origin of Paired Fin.—Balfour's Theory of the Lateral Fold.—Objections.—Objections to Gegenbaur's Theory.—Kerr's Theory of Modified External Gills.—Uncertain Conclusions.—Forms of the Tail in Fishes.—Homologies of the Pectoral Limb.—The Girdle in Fishes other than Dipnoans. | 62 |
| CHAPTER VI. | |
| THE ORGANS OF RESPIRATION. | |
| How Fishes Breathe.—The Gill Structures.—The Air-bladder.—Origin of the Air-bladder.—The Origin of Lungs.—The Heart of the Fish.—The Flow of Blood. | 91 |
| CHAPTER VII. | |
| THE NERVOUS SYSTEM. | |
| The Nervous System.—The Brain of the Fish.—The Pineal Organ.—The Brain of Primitive Fishes.—The Spinal Cord.—The Nerves. | 109 |
| CHAPTER VIII. | |
| THE ORGANS OF SENSE. | |
| The Organs of Smell.—The Organs of Sight.—The Organs of Hearing.—Voices of Fishes.—The Sense of Taste.—The Sense of Touch. | 115 |
| CHAPTER IX. | |
| THE ORGANS OF REPRODUCTION. | |
| The Germ-cells.—The Eggs of Fishes.—Protection of the Eggs.—Sexual Modification. | 124 |
| CHAPTER X. | |
| THE EMBRYOLOGY AND GROWTH OF FISHES. | |
| Post-embryonic Development.—General Laws of Development.—The Significance of Facts of Development.—The Development of the Bony Fishes.—The Larval Development of Fishes.—Peculiar Larval Forms.—The Development of Flounders.—Hybridism.—The Age of Fishes.—Tenacity of[Pg xiii] Life.—Effect of Temperature on Fishes.—Transportation of Fishes.—Reproduction of Lost Parts.—Monstrosities among Fishes. | 131 |
| CHAPTER XI. | |
| INSTINCTS, HABITS, AND ADAPTATIONS. | |
| The Habits of Fishes.—Irritability of Animals.—Nerve-cells and Fibers.—The Brain or Sensorium.—Reflex Action.—Instinct.—Classification of Instincts.—Variability of Instincts.—Adaptations to Environment.—Flight of Fishes.—Quiescent Fishes.—Migratory Fishes.—Anadromous Fishes.—Pugnacity of Fishes.—Fear and Anger in Fishes.—Calling the Fishes.—Sounds of Fishes.—Lurking Fishes.—The Unsymmetrical Eyes of the Flounder.—Carrying Eggs in the Mouth. | 152 |
| CHAPTER XII. | |
| ADAPTATIONS OF FISHES. | |
| Spines of the Catfishes.—Venomous Spines.—The Lancet of the Surgeon-fish.—Spines of the Sting-ray.—Protection through Poisonous Flesh of Fishes.—Electric Fishes.—Photophores or Luminous Organs.—Photophores in the Iniomous Fishes.—Photophores of Porichthys.—Globefishes.—Remoras.—Sucking-disks of Clingfishes.—Lampreys and Hogfishes.—The Swordfishes.—The Paddle-fishes.—The Sawfishes.—Peculiarities of Jaws and Teeth.—The Angler-fishes.—Relation of Number of Vertebræ to Temperature, and the Struggle for Existence.—Number of Vertebræ: Soft-rayed Fishes; Spiny-rayed Fishes; Fresh-water Fishes; Pelagic Fishes.—Variations in Fin-rays.—Relation of Numbers to Conditions of Life.—Degeneration of Structures.—Conditions of Evolution among Fishes. | 179 |
| CHAPTER XIII. | |
| COLORS OF FISHES. | |
| Pigmentation.—Protective Coloration.—Protective Markings.—Sexual Coloration.—Nuptial Coloration.—Coral-reef Fishes.—Recognition Marks.—Intensity of Coloration.—Fading of Pigments in Spirits.—Variation in Pattern. | 226 |
| CHAPTER XIV. | |
| GEOGRAPHICAL DISTRIBUTION OF FISHES. | |
| Zoogeography.—General Laws of Distribution.—Species Absent through Barriers.—Species Absent through Failure to Maintain Foothold.—Species Changed through Natural Selection.—Extinction of Species.—Barriers[Pg xiv] Checking Movements of Marine Species.—Temperature the Central Fact in Distribution.—Agency of Ocean Currents.—Centers of Distribution.—Distribution of Marine Fishes.—Pelagic Fishes.—Bassalian Fishes.—Littoral Fishes.—Distribution of Littoral Fishes by Coast Lines.—Minor Faunal Areas.—Equatorial Fishes most Specialized.—Realms of Distribution of Fresh-water Fishes.—Northern Zone.—Equatorial Zone.—Southern Zone.—Origin of the New Zealand Fauna. | 237 |
| CHAPTER XV. | |
| ISTHMUS BARRIERS SEPARATING FISH FAUNAS. | |
| The Isthmus of Suez.—The Fish Fauna of Japan.—Fresh-water Faunas of Japan.—Faunal Areas of Marine Fishes of Japan.—Resemblance of Japanese and Mediterranean Fish Faunas.—Significance of Resemblances.—Differences between Japanese and Mediterranean Fish Faunas.—Source of Faunal Resemblances.—Effects of Direction of Shore Lines.—Numbers of Genera in Different Faunas.—Significance of Rare Forms.—Distribution of Shore-fishes.—Extension of Indian Fauna.—The Isthmus of Suez as a Barrier to Distribution.—Geological Evidences of Submergence of Isthmus of Suez.—The Cape of Good Hope as a Barrier to Fishes.—Relations of Japan to the Mediterranean Explained by Present Conditions.—The Isthmus of Panama as a Barrier to Distribution.—Unlikeness of Species on the Shores of the Isthmus of Panama.—Views of Dr. Günther on the Isthmus of Panama.—Catalogue of Fishes of Panama.—Conclusions of Evermann & Jenkins.—Conclusions of Dr. Hill.—Final Hypothesis as to Panama. | 255 |
| CHAPTER XVI. | |
| DISPERSION OF FRESH-WATER FISHES. | |
| The Dispersion of Fishes.—The Problem of Oatka Creek.—Generalizations as to Dispersion.—Questions Raised by Agassiz.—Conclusions of Cope.—Questions Raised by Cope.—Views of Günther.—Fresh-water Fishes of North America.—Characters of Species.—Meaning of Species.—Special Creation Impossible.—Origin of American Species of Fishes. | 282 |
| CHAPTER XVII. | |
| DISPERSION OF FRESH-WATER FISHES. (Continued.) | |
| Barriers to Dispersion of Fresh-water Fishes: Local Barriers.—Favorable Waters Have Most Species.—Watersheds.—How Fishes Cross Watersheds.—The Suletind.—The Cassiquiare.—Two-Ocean Pass.—Mountain Chains.—Upland Fishes.—Lowland Fishes.—Cuban Fishes.—Swampy Watersheds.—The Great Basin of Utah.—Arctic Species in Lakes.—Causes of Dispersion still in Operation.[Pg xv] | 297 |
| CHAPTER XVIII. | |
| FISHES AS FOOD FOR MAN. | |
| The Flesh of Fishes.—Relative Rank of Food-fishes.—Abundance of Food-fishes.—Variety of Tropical Fishes.—Economic Fisheries.—Angling. | 320 |
| CHAPTER XIX. | |
| DISEASES OF FISHES. | |
| Contagious Diseases: Crustacean Parasites.—Myxosporidia or Parasitic Protozoa.—Parasitic Worms: Trematodes, Cestodes.—The Worm of the Yellowstone.—The Heart Lake Tape-worm.—Thorn-head Worms.—Nematodes.—Parasitic Fungi.—Earthquakes.—Mortality of Filefish. | 340 |
| CHAPTER XX. | |
| THE MYTHOLOGY OF FISHES. | |
| The Mermaid.—The Monkfish.—The Bishop-fish.—The Sea-serpent. | 359 |
| CHAPTER XXI. | |
| THE CLASSIFICATION OF FISHES. | |
| Taxonomy.—Defects in Taxonomy.—Analogy and Homology.—Coues on Classification.—Species as Twigs of a Genealogical Tree.—Nomenclature.—The Conception of Genus and Species.—The Trunkfishes.—Trinomial Nomenclature.—Meaning of Species.—Generalization and Specialization.—High and Low Forms.—The Problem of the Highest Fishes. | 367 |
| CHAPTER XXII. | |
| THE HISTORY OF ICHTHYOLOGY. | |
| Aristotle.—Rondelet.—Marcgraf.—Osbeck.—Artedi.—Linnæus.— Forskål.—Risso.—Bloch.—Lacépède.—Cuvier.—Valenciennes.— Agassiz.—Bonaparte.—Günther.—Boulenger.—Le Sueur.—Müller.— Gill.—Cope.—Lütken.—Steindachner.—Vaillant.—Bleeker.— Schlegel.—Poey.—Day.—Baird.—Garman.—Gilbert.—Evermann.— Eigenmann.—Zittel.—Traquair.—Woodward.—Dean.—Eastman.—Hay.— Gegenbaur.—Balfour.—Parker.—Dollo. | 387 |
| CHAPTER XXIII.[Pg xvi] | |
| THE COLLECTION OF FISHES. | |
| How to Secure Fishes.—How to Preserve Fishes.—Value of Formalin.—Records of Fishes.—Eternal Vigilance. | 429 |
| CHAPTER XXIV. | |
| THE EVOLUTION OF FISHES. | |
| The Geological Distribution of Fishes.—The Earliest Sharks.—Devonian Fishes.—Carboniferous Fishes.—Mesozoic Fishes.—Tertiary Fishes.—Factors of Extinction.—Fossilization of a Fish.—The Earliest Fishes.—The Cyclostomes.—The Ostracophores.—The Arthrodires.—The Sharks.—Origin of the Shark.—The Chimæras.—The Dipnoans.—The Crossopterygians.—The Actinopteri.—The Bony Fishes. | 435 |
| CHAPTER XXV. | |
| THE PROTOCHORDATA. | |
| The Chordate Animals.—The Protochordates.—Other Terms Used in Classification.—The Enteropneusta.—Classification of Enteropneusta.—Family Harrimaniidæ.—Balanoglossidæ.—Low Organization of Harrimaniidæ. | 460 |
| CHAPTER XXVI. | |
| THE TUNICATES, OR ASCIDIANS. | |
| Structure of Tunicates.—Development of Tunicates.—Reproduction of Tunicates.—Habits of Tunicates.—Larvacea.—Ascidiacea.—Thaliacea.—Origin of Tunicates.—Degeneration of Tunicates. | 467 |
| CHAPTER XXVII. | |
| THE LEPTOCARDII, OR LANCELETS. | |
| The Lancelet.—Habits of Lancelets.—Species of Lancelets.—Origin of Lancelets. | 482 |
| CHAPTER XXVIII.[Pg xvii] | |
| THE CYCLOSTOMES, OR LAMPREYS. | |
| The Lampreys.—Structure of the Lamprey.—Supposed Extinct Cyclostomes.—Conodontes.—Orders of Cyclostomes.—The Hyperotreta, or Hagfishes.—The Hyperoartia, or Lampreys.—Food of Lampreys.—Metamorphosis of Lampreys.—Mischief Done by Lampreys.—Migration or "Running" of Lampreys.—Requisite Conditions for Spawning with Lampreys.—The Spawning Process with Lampreys.—What Becomes of Lampreys after Spawning? | 486 |
| CHAPTER XXIX. | |
| THE CLASS ELASMOBRANCHII, OR SHARK-LIKE FISHES. | |
| The Sharks.—Characters of Elasmobranchs.—Classification of Elasmobranchs.—Subclasses of Elasmobranchs.—The Selachii.—Hasse's Classification of Elasmobranchs.—Other Classifications of Elasmobranchs.—Primitive Sharks.—Order Pleuropterygii.—Order Acanthodii.—Dean on Acanthodii.—Order Ichthyotomi. | 506 |
| CHAPTER XXX. | |
| THE TRUE SHARKS. | |
| Order Notidani.—Family Hexanchidæ.—Family Chlamydoselachidæ.—Order Asterospondyli.—Suborder Cestraciontes.—Family Heterodontidæ.—Edestus and its Allies.—Onchus.—Family Cochliodontidæ.—Suborder Galei.—Family Scyliorhinidæ.—The Lamnoid, or Mackerel-sharks.—Family Mitsukurinidæ, the Goblin-sharks.—Family Alopiidæ, or Thresher-sharks.—Family Pseudotriakidæ.—Family Lamnidæ.—Man-eating Sharks.—Family Cetorhinidæ, or Basking Sharks.—Family Rhineodontidæ.—The Carcharioid Sharks, or Requins.—Family Sphyrnidæ, or Hammer-head Sharks.—The Order of Tectospondyli.—Suborder Cyclospondyli.—Family Squalidæ.—Family Dalatiidæ.—Family Echinorhinidæ.—Suborder Rhinæ.—Family Pristiophoridæ, or Saw-sharks.—Suborder Batoidei, or Rays.—Pristididæ, or Sawfishes.—Rhinobatidæ, or Guitar-fishes.—Rajidæ, or Skates.—Narcobatidæ, or Torpedoes.—Petalodontidæ.—Dasyatidæ, or Sting-rays.—Myliobatidæ.—Family Psammodontidæ.—Family Mobulidæ. 523 | |
| CHAPTER XXXI. | |
| THE HOLOCEPHALI, OR CHIMÆRAS. | |
| The Chimæras.—Relationship of Chimæras.—Family Chimæridæ.—Rhinochimæridæ.—Extinct Chimæroids.—Ichthyodorulites. | 561 |
| CHAPTER XXXII. | |
| THE CLASS OSTRACOPHORI. | |
| Ostracophores.—Nature of Ostracophores.—Orders of Ostracophores.—Order Heterostraci.—Order Osteostraci.—Order Antiarcha.—Order Anaspida. | 568 |
| CHAPTER XXXIII. | |
| ARTHRODIRES. | |
| The Arthrodires.—Occurrence of Arthrodires.—Arthrognathi.—Anarthrodira.—Stegothalami.— Arthrodira.—Temnothoraci.—Arthrothoraci.—Relations of[Pg xviii] Arthrodires.—Suborder Cycliæ.—Palæospondylus.—Gill on Palæospondylus.—Views as to the Relationships of Palæospondylus: Huxley, Traquair, 1890. Traquair, 1893. Traquair, 1897. Smith Woodward, 1892. Dawson, 1893. Gill, 1896. Dean, 1896. Dean, 1898. Parker & Haswell, 1897. Gegenbaur, 1898.—Relationships of Palæospondylus | 581 |
| CHAPTER XXXIV. | |
| THE CROSSOPTERYGII. | |
| Class Teleostomi.—Subclass Crossopterygii.—Order of Amphibians.—The Fins of Crossopterygians.—Orders of Crossopterygians.—Haplistia.—Rhipidistia.—Megalichthyidæ.—Order Actinistia.—Order Cladistia.—The Polypteridæ | 598 |
| CHAPTER XXXV. | |
| SUBCLASS DIPNEUSTI, OR LUNGFISHES. | |
| The Lungfishes.—Classification of Dipnoans.—Order Ctenodipterini.—Order Sirenoidei.—Family Ceratodontidæ.—Development of Neoceratodus.—Lepidosirenidæ.—Kerr on the Habits of Lepidosiren | 609 |
[1] For most of this list of errata I am indebted to the kindly interest of Dr. B. W. Evermann.
| PAGE | |
|---|---|
| Lepomis megalotis, Long-eared Sunfish | 2 |
| Lepomis megalotis, Long-eared Sunfish | 4 |
| Eupomotis gibbosus, Common Sunfish | 7 |
| Ozorthe dictyogramma, a Japanese Blenny | 9 |
| Eupomotis gibbosus, Common Sunfish | 13 |
| Monocentris japonicas, Pine-cone Fish | 16 |
| Diodon hystrix, Porcupine-fish | 17 |
| Nemichthys avocetta, Thread-eel | 17 |
| Hippocampus hudsonius, Sea-horse | 17 |
| Peprilus paru, Harvest-fish | 18 |
| Lophius litulon, Anko or Fishing-frog | 18 |
| Epinephelus adscensionis, Rock-hind or Cabra Mora | 20 |
| Scales of Acanthoessus bronni | 21 |
| Cycloid Scale | 22 |
| Porichthys porosissimus, Singing-fish | 23 |
| Apomotis cyanellus, Blue-green Sunfish | 27 |
| Chiasmodon niger, Black Swallower | 29 |
| Jaws of a Parrot-fish, Sparisoma aurofrenatum | 30 |
| Archosargus probatocephalus, Sheepshead | 31 |
| Campostoma anomalum, Stone-roller | 33 |
| Roccus lineatus, Striped Bass | 35 |
| Roccus lineatus. Lateral View of Cranium | 36 |
| Roccus lineatus. Superior View of Cranium | 37 |
| Roccus lineatus. Inferior View of Cranium | 38 |
| Roccus lineatus. Posterior View of Cranium | 40 |
| Roccus lineatus. Face-bones, Shoulder and Pelvic Girdles, and Hyoid Arch | 42 |
| Lower Jaw of Amia calva, showing Gular Plate | 43 |
| Roccus lineatus. Branchial Arches | 46 |
| Pharyngeal Bone and Teeth of European Chub, Leuciscus cephalus | 47 |
| Upper Pharyngeals of Parrot-fish, Scarus strongylocephalus | 47 |
| Lower Pharyngeal Teeth of Parrot-fish, Scarus strongylocephalus | 47 |
| Pharyngeals of Italian Parrot-fish, Sparisoma cretense | 48 |
| Roccus lineatus, Vertebral Column and Appendages | 48 |
| Basal Bone of Dorsal Fin, Holoptychius leptopterus | 49 |
| Inner View of Shoulder-girdle of Buffalo-fish, Ictiobus bubalus[Pg xx] | 51 |
| Pterophryne tumida, Sargassum-fish. | 52 |
| Shoulder-girdle of Sebastolobus alascanus. | 52 |
| Cranium of Sebastolobus alascanus. | 53 |
| Lower Jaw and Palate of Sebastolobus alascanus. | 54 |
| Maxillary and Premaxillary of Sebastolobus alascanus. | 55 |
| Part of Skeleton of Selene vomer. | 55 |
| Hyostylic Skull of Chiloscyllium indicum, a Scyliorhinoid Shark. | 56 |
| Skull of Heptranchias indicus, a Notidanoid Shark. | 56 |
| Basal Bones of Pectoral Fin of Monkfish, Squatina. | 56 |
| Pectoral Fin of Heterodontus philippi. | 57 |
| Pectoral Fin of Heptranchias indicus. | 57 |
| Shoulder-girdle of a Flounder, Paralichthys californicus. | 58 |
| Shoulder-girdle of a Toadfish, Batrachoides pacifici. | 59 |
| Shoulder-girdle of a Garfish, Tylosurus fodiator. | 59 |
| Shoulder-girdle of a Hake, Merluccius productus. | 60 |
| Cladoselache fyleri, Restored. | 65 |
| Fold-like Pectoral and Ventral Fins of Cladoselache fyleri. | 65 |
| Pectoral Fin of a Shark, Chiloscyllium. | 66 |
| Skull and Shoulder-girdle of Neoceratodus forsteri, showing archipterygium. | 68 |
| Acanthoessus wardi. | 69 |
| Shoulder-girdle of Acanthoessus. | 69 |
| Pectoral Fin of Pleuracanthus. | 69 |
| Shoulder-girdle of Polypterus bichir. | 70 |
| Arm of a Frog. | 71 |
| Pleuracanthus decheni. | 74 |
| Embryos of Heterodontus japonicas, a Cestraciont Shark. | 75 |
| Polypterus congicus, a Crossopterygian Fish with External Gills. | 78 |
| Heterocercal Tail of Sturgeon, Acipenser sturio. | 80 |
| Heterocercal Tail of Bowfin, Amia calva. | 82 |
| Heterocercal Tail of Garpike, Lepisosteus osseus. | 82 |
| Coryphænoides carapinus, showing Leptocercal Tail. | 83 |
| Heterocercal Tail of Young Trout, Salmo fario. | 83 |
| Isocercal Tail of Hake, Merluccius productus. | 84 |
| Homocercal Tail of a Flounder, Paralichthys californicus. | 84 |
| Gephyrocercal Tail of Mola mola. | 85 |
| Shoulder-girdle of Amia calva. | 86 |
| Shoulder-girdle of a Sea-catfish, Selenaspis dowi. | 86 |
| Clavicles of a Sea-catfish, Selenaspis dowi. | 87 |
| Shoulder-girdle of a Batfish, Ogcocephalus radiatus. | 88 |
| Shoulder-girdle of a Threadfin, Polydactylus approximans. | 89 |
| Gill-basket of Lamprey. | 92 |
| Weberian Apparatus and Air-bladder of Carp. | 93 |
| Brain of a Shark, Squatina squatina. | 110 |
| Brain of Chimæra monstrosa. | 110 |
| Brain of Polypterus annectens.[Pg xxi] | 110 |
| Brain of a Perch, Perca flavescens. | 111 |
| Petromyzon marinus unicolor. Head of Lake Lamprey, showing Pineal Body. | 111 |
| Chologaster cornutus, Dismal-swamp Fish. | 116 |
| Typhlichthys subterraneus, Blind Cavefish. | 116 |
| Anableps dovii, Four-eyed Fish. | 117 |
| Ipnops murrayi. | 118 |
| Boleophthalmus chinensis, Pond-skipper. | 118 |
| Lampetra wilderi, Brook Lamprey. | 120 |
| Branchiostoma lanceolatum, European Lancelet. | 120 |
| Pseudupeneus maculatus, Goatfish. | 122 |
| Xiphophorus helleri, Sword-tail Minnow. | 124 |
| Cymatogaster aggregatus, White Surf-fish, Viviparous, with Young. | 125 |
| Goodea luitpoldi, a Viviparous Fish. | 126 |
| Egg of Callorhynchus antarcticus, the Bottle-nosed Chimæra. | 127 |
| Egg of the Hagfish, Myxine limosa. | 127 |
| Egg of Port Jackson Shark, Heterodontus philippi. | 128 |
| Development of Sea-bass, Centropristes striatus. | 135 |
| Centropristes striatus, Sea-bass. | 137 |
| Xiphias gladius, Young Sword-fish. | 139 |
| Xiphias gladius, Sword-fish. | 139 |
| Larva of the Sail-fish, Istiophorus, Very Young. | 140 |
| Larva of Brook Lamprey, Lampetra wilderi, before Transformation. | 140 |
| Anguilla chrisypa, Common Eel. | 140 |
| Larva of Common Eel, Anguilla chrisypa, called Leptocephalus grassii. | 141 |
| Larva of Sturgeon, Acipenser sturio. | 141 |
| Larva of Chætodon sedentarius. | 142 |
| Chætodon capistratus, Butterfly-fish. | 142 |
| Mola mola, Very Early Larval Stage of Headfish, called Centaurus boöps. | 143 |
| Mola mola, Early Larval Stage called Molacanthus nummularis. | 144 |
| Mola mola, Advanced Larval Stage. | 144 |
| Mola mola, Headfish, Adult. | 146 |
| Albula vulpes, Transformation of Ladyfish from Larva to Young. | 147 |
| Development of the Horsehead-fish, Selene vomer. | 148 |
| Salanx hyalocranius, Ice-fish. | 149 |
| Dallia pectoralis, Alaska Blackfish. | 149 |
| Ophiocephalus barca, Snake-headed China-fish. | 150 |
| Carassius auratus, Monstrous Goldfish. | 151 |
| Jaws of Nemichthys avocetta. | 156 |
| Cypsilurus californicus, Flying-fish. | 157 |
| Ammocrypta clara, Sand-darter. | 158 |
| Fierasfer acus, Pearl-fish, issuing from a Holothurian. | 159 |
| Gobiomorus gronovii, Portuguese Man-of-war Fish. | 160 |
| Tide Pools of Misaki. | 161 |
| Ptychocheilus oregonensis, Squaw-fish. | 162 |
| [Pg xxii] Ptychocheilus grandis, Squaw-fish, Stranded as the Water Falls. | 164 |
| Larval Stages of Platophrys podas, a Flounder of the Mediterranean, showing Migration of Eye. | 174 |
| Platophrys lunatus, the Wide-eyed Flounder. | 175 |
| Young Flounder Just Hatched, with Symmetrical Eyes. | 175 |
| Pseudopleuronectes americanus, Larval Flounder. | 176 |
| Pseudopleuronectes americanus, Larval Flounder (more advanced stage). | 176 |
| Face View of Recently-hatched Flounder. | 177 |
| Schilbeodes furiosus, Mad-Tom. | 179 |
| Emmydrichthys vulcanus, Black Nohu or Poison-fish. | 180 |
| Teuthis bahianus, Brown Tang. | 181 |
| Stephanolepis hispidus, Common Filefish. | 182 |
| Tetraodon meleagris. | 183 |
| Balistes carolinensis, the Trigger-fish. | 184 |
| Narcine brasiliensis, Numbfish. | 185 |
| Torpedo electricus, Electric Catfish. | 186 |
| Astroscopus guttatus, Star-gazer. | 187 |
| Æthoprora lucida, Headlight-fish. | 188 |
| Corynolophus reinhardti, showing Luminous Bulb. | 188 |
| Etmopterus lucifer. | 189 |
| Argyropelecus olfersi. | 190 |
| Luminous Organs and Lateral Line of Midshipman, Porichthys notatus. | 192 |
| Cross-section of Ventral Phosphorescent Organ of Midshipman, Porichthys notatus. | 193 |
| Section of Deeper Portion of Phosphorescent Organ, Porichthys notatus. | 194 |
| Leptecheneis naucrates, Sucking-fish or Pegador. | 197 |
| Caularchus mæandricus, Clingfish. | 198 |
| Polistotrema stouti, Hagfish. | 199 |
| Pristis zysron, Indian Sawfish. | 200 |
| Pristiophorus japonicus, Saw-shark. | 201 |
| Skeleton of Pike, Esox lucius. | 203 |
| Skeleton of Red Rockfish, Sebastodes miniatus. | 214 |
| Skeleton of a Spiny-rayed Fish of the Tropics, Holacanthus ciliaris. | 214 |
| Skeleton of the Cowfish, Lactophrys tricornis. | 215 |
| Crystallias matsushimæ, Liparid. | 218 |
| Sebastichthys maliger, Yellow-backed Rockfish. | 218 |
| Myoxocephalus scorpius, European Sculpin. | 219 |
| Hemitripterus americanus, Sea-raven. | 220 |
| Cyclopterus lumpus, Lumpfish. | 220 |
| Psychrolutes paradoxus, Sleek Sculpin. | 221 |
| Pallasina barbata, Agonoid-fish. | 221 |
| Amblyopsis spelæus, Blindfish of the Mammoth Cave. | 221 |
| Lucifuga subterranea, Blind Brotula. | 222 |
| Hypsypops rubicunda, Garibaldi. | 227 |
| Synanceia verrucosa, Gofu or Poison-fish. | 229 |
| [Pg xxiii] Alticus saliens, Lizard-skipper. | 230 |
| Etheostoma camurum, Blue-breasted Darter. | 231 |
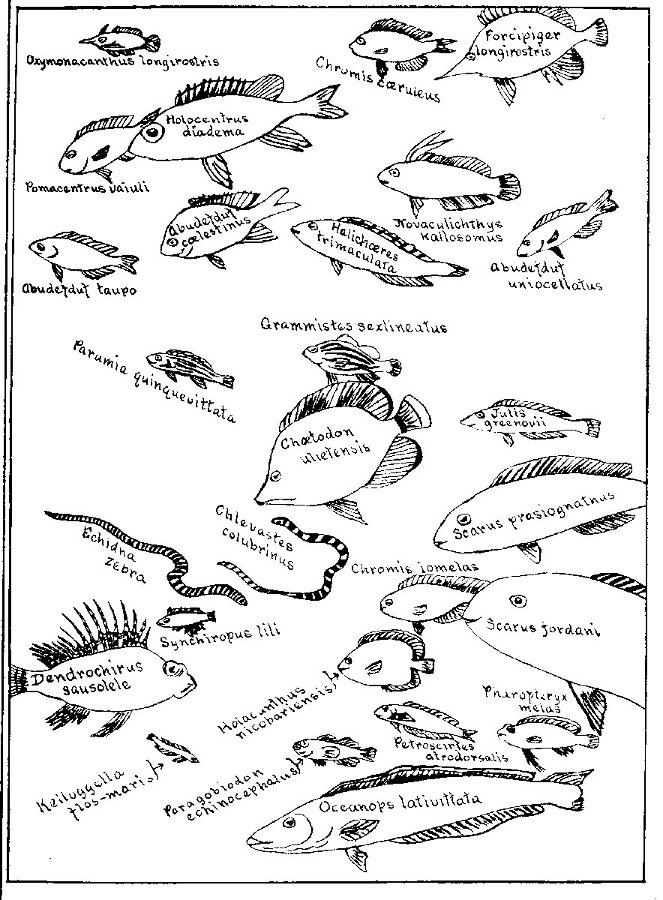
| Liuranus semicinctus and Chlevastes colubrinus, Snake-eels. | 233 |
| Coral Reef at Apia. | 234 |
| Rudarius ercodes, Japanese Filefish. | 241 |
| Tetraodon setosus, Globefish. | 244 |
| Dasyatis sabina, Sting-ray. | 246 |
| Diplesion blennioides, Green-sided Darter. | 247 |
| Hippocampus mohnikei, Japanese Sea-horse. | 250 |
| Archoplites interruptus, Sacramento Perch. | 258 |
| Map of the Continents, Eocene Time. | 270 |
| Caulophryne jordani, Deep-sea Fish of Gulf Stream. | 276 |
| Exerpes asper, Fish of Rock-pools, Mexico. | 276 |
| Xenocys jessiæ. | 279 |
| Ictalurus punctatus, Channel Catfish. | 280 |
| Drawing the Net on the Beach of Hilo, Hawaii. | 281 |
| Semotilus atromaculatus, Horned Dace. | 285 |
| Leuciscus lineatus, Chub of the Great Basin. | 287 |
| Melletes papilio, Butterfly Sculpin. | 288 |
| Scartichthys enosimæ, a Fish of the Rock-pools of the Sacred Island of Enoshima, Japan. | 294 |
| Halichœres bivittatus, the Slippery Dick. | 297 |
| Peristedion miniatum. | 299 |
| Outlet of Lake Bonneville. | 303 |
| Hypocritichthys analis, Silver Surf-fish. | 309 |
| Erimyzon sucetta, Creekfish or Chub-sucker. | 315 |
| Thaleichthys pretiosus, Eulachon or Ulchen. | 320 |
| Plecoglossus altivelis, the Japanese Ayu. | 321 |
| Coregonus clupeiformis, the Whitefish. | 321 |
| Mullus auratus, the Golden Surmullet. | 322 |
| Scomberomorus maculatus, the Spanish Mackerel. | 322 |
| Lampris luna, the Opah or Moonfish. | 323 |
| Pomatomus saltatrix, the Bluefish. | 324 |
| Centropomus undecimalis, the Robalo. | 324 |
| Chætodipterus faber, the Spadefish. | 325 |
| Micropterus dolomieu, the Small-mouthed Black Bass. | 325 |
| Salvelinus fontinalis, the Speckled Trout. | 326 |
| Salmo irideus, the Rainbow Trout. | 326 |
| Salvelinus oquassa, the Rangeley Trout. | 326 |
| Salmo gairdneri, the Steelhead Trout. | 327 |
| Salmo henshawi, the Tahoe Trout. | 327 |
| Salvelinus malma, the Dolly Varden Trout. | 327 |
| Thymallus signifer, the Alaska Grayling. | 328 |
| Esox lucius, the Pike. | 328 |
| Pleurogrammus monopterygius, the Atka-fish. | 328 |
| [Pg xxiv] Chirostoma humboldtianum, the Pescado blanco. | 329 |
| Pseudupeneus maculatus, the Red Goatfish. | 329 |
| Pseudoscarus guacamaia, Great Parrot-fish. | 330 |
| Mugil cephalus, Striped Mullet. | 330 |
| Lutianus analis, Mutton-snapper. | 331 |
| Clupea harengus, Herring. | 331 |
| Gadus callarias, Codfish. | 331 |
| Scomber scombrus, Mackerel. | 332 |
| Hippoglossus hippoglossus, Halibut. | 332 |
| Fishing for Ayu with Cormorants. | 333 |
| Fishing for Ayu. Emptying Pouch of Cormorant. | 335 |
| Fishing for Tai, Tokyo Bay. | 338 |
| Brevoortia tyrannus, Menhaden. | 340 |
| Exonautes unicolor, Australian Flying-fish. | 341 |
| Rhinichthys atronasus, Black-nosed Dace. | 342 |
| Notropis hudsonius, White Shiner. | 343 |
| Ameiurus catus, White Catfish. | 344 |
| Catostomus ardens, Sucker. | 348 |
| Oncorhynchus tschawytscha, Quinnat Salmon. | 354 |
| Oncorhynchus tschawytscha, Young Male. | 355 |
| Ameiurus nebulosus, Catfishes. | 358 |
| "Le Monstre Marin en Habit de Moine". | 360 |
| "Le Monstre Marin en Habit d'Évêque". | 361 |
| Regalecus russelli, Oarfish. | 362 |
| Regalecus glesne, Glesnæs Oarfish. | 363 |
| Nemichthys avocetta, Thread-eel. | 365 |
| Lactophrys tricornis, Horned Trunkfish. | 373 |
| Ostracion cornutum, Horned Trunkfish. | 376 |
| Lactophrys bicaudalis, Spotted Trunkfish. | 377 |
| Lactophrys bicaudalis, Spotted Trunkfish (Face). | 377 |
| Lactophrys triqueter, Spineless Trunkfish. | 378 |
| Lactophrys trigonus, Hornless Trunkfish. | 378 |
| Lactophrys trigonus, Hornless Trunkfish (Face). | 379 |
| Bernard Germain de Lacépède. | 399 |
| Georges Dagobert Cuvier. | 399 |
| Louis Agassiz. | 399 |
| Johannes Müller. | 399 |
| Albert Günther. | 403 |
| Franz Steindachner. | 403 |
| George Albert Boulenger. | 403 |
| Robert Collett. | 403 |
| Spencer Fullerton Baird. | 407 |
| Edward Drinker Cope. | 407 |
| Theodore Nicholas Gill. | 407 |
| George Brown Goode. | 407 |
| [Pg xxv] Johann Reinhardt. | 409 |
| Edward Waller Claypole. | 409 |
| Carlos Berg. | 409 |
| Edgar R. Waite. | 409 |
| Felipe Poey y Aloy. | 413 |
| Léon Vaillant. | 413 |
| Louis Dollo. | 413 |
| Decio Vinciguerra. | 413 |
| Bashford Dean. | 417 |
| Kakichi Mitsukuri. | 417 |
| Carl H. Eigenmann. | 417 |
| Franz Hilgendorf. | 417 |
| David Starr Jordan. | 421 |
| Herbert Edson Copeland. | 421 |
| Charles Henry Gilbert. | 421 |
| Barton Warren Evermann. | 421 |
| Ramsay Heatley Traquair. | 425 |
| Arthur Smith Woodward. | 425 |
| Karl A. Zittel. | 425 |
| Charles R. Eastman. | 425 |
| Fragment of Sandstone from Ordovician Deposits. | 435 |
| Fossil Fish Remains from Ordovician Rocks. | 436 |
| Dipterus valenciennesi. | 437 |
| Hoplopteryx lewesiensis. | 438 |
| Paratrachichthys prosthemius, Berycoid fish. | 439 |
| Cypsilurus heterurus, Flying-fish. | 440 |
| Lutianidæ, Schoolmaster Snapper. | 440 |
| Pleuronichthys decurrens, Decurrent Flounder. | 441 |
| Cephalaspis lyelli, Ostracophore. | 444 |
| Dinichthys intermedius, Arthrodire. | 445 |
| Lamna cornubica, Mackerel-shark or Salmon-shark. | 447 |
| Raja stellulata, Star-spined Ray. | 448 |
| Harriotta raleighiana, Deep-sea Chimæra. | 449 |
| Dipterus valenciennesi, Extinct Dipnoan. | 449 |
| Holoptychius giganteus, Extinct Crossopterygian. | 451 |
| Platysomus gibbosus, Ancient Ganoid fish. | 452 |
| Lepisosteus platystomus, Short-nosed Gar. | 452 |
| Palæoniscum macropomum, Primitive Ganoid fish. | 453 |
| Diplomystus humilis, Fossil Herring. | 453 |
| Holcolepis lewesiensis. | 454 |
| Elops saurus, Ten-pounder. | 454 |
| Apogon semilineatus, Cardinal-fish. | 455 |
| Pomolobus æstivalis, Summer Herring. | 455 |
| Bassozetus catena. | 456 |
| Trachicephalus uranoscopus. | 456 |
| [Pg xxvi] Chlarias breviceps, African Catfish. | 457 |
| Notropis whipplii, Silverfin. | 457 |
| Gymnothorax moringa. | 458 |
| Seriola lalandi, Amber-fish. | 458 |
| Geological Distribution of the Families of Elasmobranchs. | 459 |
| "Tornaria" Larva of Glossobalanus minutus. | 463 |
| Glossobalanus minutus. | 464 |
| Harrimania maculosa. | 465 |
| Development of Larval Tunicate to Fixed Condition. | 471 |
| Anatomy of Tunicate. | 472 |
| Ascidia adhærens. | 474 |
| Styela yacutatensis. | 475 |
| Styela greeleyi. | 476 |
| Cynthia superba. | 476 |
| Botryllus magnus, Compound Ascidian. | 477 |
| Botryllus magnus. | 478 |
| Botryllus magnus, a Single Zooid. | 479 |
| Aplidiopsis jordani, a Compound Ascidian. | 479 |
| Oikopleura, Adult Tunicate of Group Larvacea. | 480 |
| Branchiostoma californiense, California Lancelet. | 484 |
| Gill-basket of Lamprey. | 485 |
| Polygnathus dubium. | 488 |
| Polistotrema stouti, Hagfish. | 489 |
| Petromyzon marinus, Lamprey. | 491 |
| Petromyzon marinus unicolor, Mouth Lake Lamprey. | 492 |
| Lampetra wilderi, Sea Larvæ Brook Lamprey. | 492 |
| Lampetra wilderi, Mouth Brook Lamprey. | 492 |
| Lampetra camtschatica, Kamchatka Lamprey. | 495 |
| Entosphenus tridentatus, Oregon Lamprey. | 496 |
| Lampetra wilderi, Brook Lamprey. | 505 |
| Fin-spine of Onchus tenuistriatus. | 509 |
| Section of Vertebræ of Sharks, showing Calcification. | 510 |
| Cladoselache fyleri. | 514 |
| Cladoselache fyleri, Ventral View. | 515 |
| Teeth of Cladoselache fyleri. | 515 |
| Acanthoessus wardi. | 515 |
| Diplacanthus crassissimus. | 517 |
| Climatius scutiger. | 518 |
| Pleuracanthus decheni. | 519 |
| Pleuracanthus decheni, Restored. | 520 |
| Head-bones and Teeth of Pleuracanthus decheni. | 520 |
| Teeth of Didymodus bohemicus. | 520 |
| Shoulder-girdle and Pectoral Fins of Cladodus neilsoni. | 521 |
| Teeth of Cladodus striatus. | 522 |
| Hexanchus griseus, Griset or Cow-shark. | 523 |
| [Pg xxvii] Teeth of Heptranchias indicus. | 524 |
| Chlamydoselachus anguineus, Frill-shark. | 525 |
| Heterodontus francisci, Bullhead-shark. | 526 |
| Lower Jaw of Heterodontus philippi. | 526 |
| Teeth of Cestraciont Sharks. | 527 |
| Egg of Port Jackson Shark, Heterodontus philippi. | 527 |
| Tooth of Hybodus delabechei. | 528 |
| Fin-spine of Hybodus basanus. | 528 |
| Fin-spine of Hybodus reticulatus. | 528 |
| Fin-spine of Hybodus canaliculatus. | 529 |
| Teeth of Cestraciont Sharks. | 529 |
| Edestus vorax, Supposed to be a Whorl of Teeth. | 529 |
| Helicoprion bessonowi, Teeth of. | 530 |
| Lower Jaw of Cochliodus contortus. | 531 |
| Mitsukurina owstoni, Goblin-shark. | 535 |
| Scapanorhynchus lewisi, Under Side of Snout. | 536 |
| Tooth of Lamna cuspidata. | 537 |
| Isuropsis dekayi, Mackerel-shark. | 537 |
| Tooth of Isurus hastalis. | 538 |
| Carcharodon mega odon. | 539 |
| Cetorhinus maximus, Basking-shark. | 540 |
| Galeus zyopterus, Soup-fin Shark. | 541 |
| Carcharias lamia, Cub-shark. | 542 |
| Teeth of Corax pristodontus. | 543 |
| Sphyrna zygæna, Hammer-head Shark. | 544 |
| Squalus acanthias, Dogfish. | 545 |
| Etmopterus lucifer. | 546 |
| Brain of Monkfish, Squatina squatina. | 547 |
| Pristiophorus japonicus, Saw-shark. | 548 |
| Pristis pectinatus, Sawfish. | 550 |
| Rhinobatus lentiginosus, Guitar-fish. | 551 |
| Raja erinacea, Common Skate. | 552 |
| Narcine brasiliensis, Numbfish. | 553 |
| Teeth of Janassa linguæformis. | 554 |
| Polyrhizodus radicans. | 555 |
| Dasyatis sabina, Sting-ray. | 556 |
| Aëtobatis narinari, Eagle-ray. | 558 |
| Manta birostris, Devil-ray or Sea-devil. | 559 |
| Skeleton of Chimæra monstrosa. | 564 |
| Chimæra colliei, Elephant-fish. | 565 |
| Odontotodus schrencki, Ventral Side. | 570 |
| Odontotodus schrencki, Dorsal Side. | 570 |
| Head of Odontotodus schrencki, from the Side. | 571 |
| Limulus polyphemus, Horseshoe Crab. | 572 |
| Lanarkia spinosa. | 574 |
| [Pg xxviii] Drepanaspis gmundenensis. | 575 |
| Pteraspis rostrata. | 575 |
| Cephalaspis lyelli, Restored. | 576 |
| Cephalaspis dawsoni. | 577 |
| Pterichthyodes testudinarius. | 578 |
| Pterichthyodes testudinarius, Side View. | 579 |
| Birkenia elegans. | 579 |
| Lasianius problematicus. | 580 |
| Coccosteus cuspidatus, Restored. | 582 |
| Jaws of Dinichthys hertzeri. | 583 |
| Dinichthys intermedius, an Arthrodire. | 584 |
| Palæospondylus gunni. | 591 |
| Shoulder-girdle of Polypterus bichir. | 600 |
| Arm of a Frog. | 601 |
| Polypterus congicus, a Crossopterygian Fish. | 602 |
| Basal Bone of Dorsal Fin, Holoptychius leptopterus. | 603 |
| Gyroptychius microlepidotus. | 604 |
| Cœlacanthus elegans, showing Air-bladder. | 604 |
| Undina gulo. | 605 |
| Lower Jaw of Polypterus bichir, from Below. | 606 |
| Polypterus congicus. | 607 |
| Polypterus delhezi. | 607 |
| Erpetoichthys calabaricus. | 608 |
| Shoulder-girdle of Neoceratodus forsteri. | 609 |
| Phaneropleuron andersoni. | 613 |
| Teeth of Ceratodus runcinatus. | 614 |
| Neoceratodus forsteri. | 614 |
| Archipterygium of Neoceratodus forsteri. | 614 |
| Upper jaw of Neoceratodus forsteri. | 615 |
| Lower Jaw of Neoceratodus forsteri. | 616 |
| Adult Male of Lepidosiren paradoxa. | 619 |
| Lepidosiren paradoxa. Embryo Three Days before Hatching; Larva Thirteen Days after Hatching. | 620 |
| Larva of Lepidosiren paradoxa Forty Days after Hatching. | 621 |
| Larva of Lepidosiren paradoxa Thirty Days after Hatching. | 621 |
| Larva of Lepidosiren paradoxa Three Months after Hatching. | 621 |
| Protopterus dolloi. | 622 |

Fig. 1.—Long-eared Sunfish, Lepomis megalotis (Rafinesque). (From life by R. W. Shufeldt.)—Page 2.
A POPULAR ACCOUNT OF THE LIFE OF THE LONG-EARED SUNFISH, LEPOMIS MEGALOTIS

What is a Fish?—A fish is a back-boned animal which lives in the water and cannot ever live very long anywhere else. Its ancestors have always dwelt in water, and most likely its descendents will forever follow their example. So, as the water is a region very different from the fields or the woods, a fish in form and structure must be quite unlike all the beasts and birds that walk or creep or fly above ground, breathing air and being fitted to live in it. There are a great many kinds of animals called fishes, but in this all of them agree: all have some sort of a back-bone, all of them breathe their life long by means of gills, and none have fingers or toes with which to creep about on land.
The Long-eared Sunfish.—If we would understand a fish, we must first go and catch one. This is not very hard to do, for there are plenty of them in the little rushing brook or among the lilies of the pond. Let us take a small hook, put on it an angleworm or a grasshopper,—no need to seek an elaborate artificial fly,—and we will go out to the old "swimming-hole" or the deep eddy at the root of the old stump where the stream has gnawed away the bank in changing its course. Here we will find fishes, and one of them will take the bait very soon. In one part of the country the first fish that bites will be different from the first one taken in some other. But as we are fishing in the United States, we will locate our brook in the centre of population of our country. This will be to the northwest of Cincin[Pg 4]nati, among the low wooded hills from which clear brooks flow over gravelly bottoms toward the Ohio River. Here we will catch sunfishes of certain species, or maybe rock bass or catfish: any of these will do for our purpose. But one of our sunfishes is especially beautiful—mottled blue and golden and scarlet, with a long, black, ear-like appendage backward from his gill-covers—and this one we will keep and hold for our first lesson in fishes. It is a small fish, not longer than your hand most likely, but it can take the bait as savagely as the best, swimming away with it with such force that you might think from the vigor of its pull that you have a pickerel or a bass. But when it comes out of the water you see a little, flapping, unhappy, living plate of brown and blue and orange, with fins wide-spread and eyes red with rage.

Fig. 2.—Long-eared Sunfish, Lepomis megalotis (Rafinesque). (From Clear Creek, Bloomington, Indiana.) Family Centrarchidæ.
Form of the Fish.—And now we have put the fish into a bucket of water, where it lies close to the bottom. Then we take it home and place it in an aquarium, and for the first time we have a chance to see what it is like. We see that its body is almost elliptical in outline, but with flat sides and shaped on the lower parts very much like a boat. This form we see is such as to enable it to part the water as it swims. We notice that its progress comes through the sculling motion of its broad, flat tail.
Face of a Fish.—When we look at the sunfish from the front we see that it has a sort of face, not unlike that of higher animals. The big eyes, one on each side, stand out without eyelids, but the fish can move them at will, so that once in a while he seems to wink. There isn't much of a nose between the eyes, but the mouth is very evident, and the fish opens and shuts it as it breathes. We soon see that it breathes water, taking it in through the mouth and letting it flow over the gills, and then out through the opening behind the gill-covers.
How the Fish Breathes.—If we take another fish—for we shall not kill this one—we shall see that in its throat, behind the mouth-cavity, there are four rib-like bones on each side, above the beginning of the gullet. These are the gill-arches, and on each one of them there is a pair of rows of red fringes called the gills. Into each of these fringes runs a blood-vessel. As the water passes over it the oxygen it contains is absorbed through the skin of the gill-fringe into the blood, which thus becomes purified. In the same manner the impurities of the blood pass out into the water, and go out through the gill-openings behind. The fish needs to breathe just as we do, though the apparatus of breathing is not the same. Just as the air becomes loaded with impurities when many people breathe it, so does the water in our jar or aquarium become foul if it is breathed over and over again by fishes. When a fish finds the water bad he comes to the surface to gulp air, but his gills are not well fitted to use undissolved air as a substitute for that contained in water. The rush of a stream through the air purifies the water, and so again does the growth of water plants, for these in the sunshine absorb and break up carbonic acid gas, and throw out oxygen into the water.
Teeth of the Fish.—On the inner side of the gill-arch we find some little projections which serve as strainers to the water. These are called gill-rakers. In our sunfish they are short and thick, seeming not to amount to much but in a herring they are very long and numerous.
Behind the gills, at the opening of the gullet, are some roundish bones armed with short, thick teeth. These are called pharyngeals. They form a sort of jaws in the throat, and they are useful in helping the little fish to crack shells. If we look at the mouth of our live fish, we shall find that when it breathes or bites it moves[Pg 6] the lower jaw very much as a dog does. But it can move the upper jaw, too, a little, and that by pushing it out in a queer fashion, as though it were thrust out of a sheath and then drawn in. If we look at our dead fish, we shall see that the upper jaw divides in the middle and has two bones on each side. On one bone are rows of little teeth, while the other bone that lies behind it has no teeth at all. The lower jaw has little teeth like those of the upper jaw, and there is a patch of teeth on the roof of the mouth also. In some sunfishes there are three little patches, the vomer in the middle and the palatines on either side.
The tongue of the fish is flat and gristly. It cannot move it, scarce even taste its food with it, nor can it use it for making a noise. The unruly member of a fish is not its tongue, but its tail.
How the Fish Sees.—To come back to the fish's eye again. We say that it has no eyelids, and so, if it ever goes to sleep, it must keep its eyes wide open. The iris is brown or red. The pupil is round, and if we could cut open the eye we should see that the crystalline lens is almost a perfect sphere, much more convex than the lens in land animals. We shall learn that this is necessary for the fish to see under water. It takes a very convex lens or even one perfectly round to form images from rays of light passing through the water, because the lens is but little more dense than the water itself. This makes the fish near-sighted. He cannot see clearly anything out of water or at a distance. Thus he has learned that when, in water or out, he sees anything moving quickly it is probably something dangerous, and the thing for him to do is to swim away and hide as swiftly as possible.
In front of the eye are the nostrils, on each side a pair of openings. But they lead not into tubes, but into a little cup lined with delicate pink tissues and the branching nerves of smell. The organ of smell in nearly all fishes is a closed sac, and the fish does not use the nostrils at all in breathing. But they can indicate the presence of anything in the water which is good to eat, and eating is about the only thing a fish cares for.
Color of the Fish.—Behind the eye there are several bones on the side of the head which are more or less distinct from the skull itself. These are called membrane bones because they are formed of membrane which has become bony by the deposition[Pg 7] in it of salts of lime. One of these is called the opercle, or gill-cover, and before it, forming a right angle, is the preopercle, or false gill-cover. On our sunfish we see that the opercle ends behind in a long and narrow flap, which looks like an ear. This is black in color, with an edging of scarlet as though a drop of blood had spread along its margin. When the fish is in the water its back is dark greenish-looking, like the weeds and the sticks in the bottom, so that we cannot see it very plainly. This is the way the fish looks to the fishhawks or herons in the air above it who may come to the stream to look for fish. Those fishes which from above look most like the bottom can most readily hide and save themselves. The under side of the sunfish is paler, and most fishes have the belly white. Fishes with white bellies swim high in the water, and the fishes who would catch them lie below. To the fish in the water all outside the water looks white, and so the white-bellied fishes are hard for other fishes to see, just as it is hard for us to see a white rabbit bounding over the snow.

Fig. 3.—Common sunfish, Eupomotis gibbosus (Linnæus). Natural size. (From life by R. W. Shufeldt.)
But to be known of his own kind is good for the sunfish, and we may imagine that the black ear-flap with its scarlet edge helps his mate and friends to find him out, where they swim on his own level near the bottom. Such marks are called recognition-marks, and a great many fishes have them, but we have no certain knowledge as to their actual purpose.
We are sure that the ear-flap is not an ear, however. No fishes have any external ear, all their hearing apparatus being buried in the skull. They cannot hear very much: possibly a great jar or splash in the water may reach them, but whenever they hear any noise they swim off to a hiding-place, for any disturbance whatever in the water must arouse a fish's anxiety. The color of the live sunfish is very brilliant. Its body is covered with scales, hard and firm, making a close coat of mail, overlapping one another like shingles on a roof. Over these is a thin skin in which are set little globules of bright-colored matter, green, brown, and black, with dashes of scarlet, blue, and white as well. These give the fish its varied colors. Some coloring matter is under the scales also, and this especially makes the back darker than the lower parts. The bright colors of the sunfish change with its surroundings or with its feelings. When it lies in wait under a dark log its colors are very dark. When it rests above the white sands it is very pale. When it is guarding its nest from some meddling perch its red shades flash out as it stands with fins spread, as though a water knight with lance at rest, looking its fiercest at the intruder.
When the sunfish is taken out of the water its colors seem to fade. In the aquarium it is generally paler, but it will sometimes brighten up when another of its own species is placed beside it. A cause of this may lie in the nervous control of the muscles at the base of the scales. When the scales lie very flat the color has one appearance. When they rise a little the shade of color seems to change. If you let fall some ink-drops between two panes of glass, then spread them apart or press them together, you will see changes in the color and size of the spots. Of this nature is the apparent change in the colors of fishes under different conditions. Where the fish feels at its best the colors are the richest. There are some fishes, too, in which the male grows very brilliant in the breeding season through the deposition of red, white, black,[Pg 9] or blue pigments, or coloring matter, on its scales or on its head or fins, this pigment being absorbed when the mating season is over. This is not true of the sunfish, who remains just about the same at all seasons. The male and female are colored alike and are not to be distinguished without dissection. If we examine the scales, we shall find that these are marked with fine lines and concentric striæ, and part of the apparent color is due to the effect of the fine lines on the light. This gives the bluish lustre or sheen which we can see in certain lights, although we shall find no real blue pigment under it. The inner edge of each scale is usually scalloped or crinkled, and the outer margin of most of them has little prickly points which make the fish seem rough when we pass our hand along his sides.
Fig. 4.—Ozorthe dictyogramma (Herzenstein). A Japanese blenny, from Hakodate: showing increased number of lateral lines, a trait characteristic of many fishes of the north Pacific.
The Lateral Line.—Along the side of the fish is a line of peculiar scales which runs from the head to the tail. This is called the lateral line. If we examine it carefully, we shall see that each scale has a tube from which exudes a watery or mucous fluid. Behind these tubes are nerves, and although not much is known of the function of the tubes, we can be sure that in some degree the lateral line is a sense-organ, perhaps aiding the fish to feel sound-waves or other disturbances in the water.
The Fins of the Fish.—The fish moves itself and directs its course in the water by means of its fins. These are made up of stiff or flexible rods growing out from the body and joined together by membrane. There are two kinds of these rays or rods in the fins. One sort is without joints or branches, tapering to a sharp point. The rays thus fashioned are called spines, and they are in the sunfish stiff and sharp-pointed. The others,[Pg 10] known as soft rays, are made up of many little joints, and most of them branch and spread out brush-like at their tips. In the fin on the back the first ten of the rays are spines, the rest are soft rays. In the fin under the tail there are three spines, and in each fin at the breast there is one spine with five soft rays. In the other fins all the rays are soft.
The fin on the back is called the dorsal fin, the fin at the end of the tail is the caudal fin, the fin just in front of this on the lower side is the anal fin. The fins, one on each side, just behind the gill-openings are called the pectoral fins. These correspond to the arms of man, the wings of birds, or the fore legs of a turtle or lizard. Below these, corresponding to the hind legs, is the pair of fins known as the ventral fins. If we examine the bones behind the gill-openings to which the pectoral fins are attached, we shall find that they correspond after a fashion to the shoulder-girdle of higher animals. But the shoulder-bone in the sunfish is joined to the back part of the skull, so that the fish has not any neck at all. In animals with necks the bones at the shoulder are placed at some distance behind the skull.
If we examine the legs of a fish, the ventral fins, we shall find that, as in man, these are fastened to a bone inside called the pelvis. But the pelvis in the sunfish is small and it is placed far forward, so that it is joined to the tip of the "collar-bone" of the shoulder-girdle and pelvis attached together. The caudal fin gives most of the motion of a fish. The other fins are mostly used in maintaining equilibrium and direction. The pectoral fins are almost constantly in motion, and they may sometimes help in breathing by starting currents outside which draw water over the gills.
The Skeleton of the Fish.—The skeleton of the fish, like that of man, is made up of the skull, the back-bone, the limbs, and their appendages. But in the fish the bones are relatively smaller, more numerous, and not so firm. The front end of the vertebral column is modified as a skull to contain the little brain which serves for all a fish's activities. To the skull are attached the jaws, the membrane bones, and the shoulder-girdle. The back-bone itself in the sunfish is made of about twenty-four pieces, or vertebræ. Each of these has a rounded central part, concave in front and behind. Above this is a[Pg 11] channel through which the great spinal cord passes, and above and below are a certain number of processes or projecting points. To some of these, through the medium of another set of sharp bones, the fins of the back are attached. Along the sides of the body are the slender ribs.
The Fish in Action.—The fish is, like any other animal, a machine to convert food into power. It devours other animals or plants, assimilates their substance, takes it over into itself, and through its movements uses up this substance again. The food of the sunfish is made up of worms, insects, and little fishes. To seize these it uses its mouth and teeth. To digest them it needs its alimentary canal, made of the stomach with its glands and intestines. If we cut the fish open, we shall find the stomach with its pyloric cæca, near it the large liver with its gall-bladder, and on the other side the smaller spleen. After the food is dissolved in the stomach and intestines the nutritious part is taken up by the walls of the alimentary canal, whence it passes into the blood.
The blood is made pure in the gills, as we have already seen. To send it to the gills the fish has need of a little pumping-engine, and this we shall find at work in the fish as in all higher animals. This engine of stout muscle surrounding a cavity is called the heart. In most fishes it is close behind the gills. It contains one auricle and one ventricle only, not two of each as in man. The auricle receives the impure blood from all parts of the body. It passes it on to the ventricle, which, being thick-walled, is dark red in color. This passes the blood by convulsive action, or heart-beating, on to the gills. From these the blood is collected in arteries, and without again returning to the heart it flows all through the body. The blood in the fish flows sluggishly. The combustion of waste material goes on slowly, and so the blood is not made hot as it is in the higher beasts and birds. Fishes have relatively little blood; what there is is rather pale and cold and has no swift current.
If we look about in the inside of a fish, we shall find close along the lower side of the back-bone, covering the great artery, the dark red kidneys. These strain out from the blood a certain class of impurities, poisons made from nerve or muscle waste which cannot be burned away by the oxygen of respiration.
The Air-bladder.—In the front part of the sunfish, just above the stomach, is a closed sac, filled with air. This is called the air-bladder, or swim-bladder. It helps the fish to maintain its place in the water. In bottom fishes it is almost always small, while fishes that rise and fall in the current generally have a large swim-bladder. The gas inside it is secreted from the blood, for the sunfish has no way of getting any air into it from the outside.
But the primal purpose of the air-bladder was not to serve as a float. In very old-fashioned fishes it has a tube connecting it with the throat, and instead of being an empty sac it is a true lung made up of many lobes and parts and lined with little blood-vessels. Such fishes as the garpike and the bowfin have lung-like air-bladders and gulp air from the surface of the water.
In the very little sunfish, when he is just hatched, the air-bladder has an air-duct, which, however, is soon lost, leaving only a closed sac. From all this we know that the air-bladder is the remains of what was once a lung, or additional arrangement for breathing. As the gills furnish oxygen enough, the lung of the common fish has fallen into disuse and thrifty Nature has used the parts and the space for another and a very different purpose. This will serve to help us to understand the swim-bladder and the way the fish came to acquire it as a substitute for a lung.
The Brain of the Fish.—The movements of the fish, like those of every other complex animal, are directed by a central nervous system, of which the principal part is in the head and is known as the brain. From the eye of the fish a large nerve goes to the brain to report what is in sight. Other nerves go from the nostrils, the ears, the skin, and every part which has any sort of capacity for feeling. These nerves carry their messages inward, and when they reach the brain they may be transformed into movement. The brain sends back messages to the muscles, directing them to contract. Their contraction moves the fins, and the fish is shoved along through the water. To scare the fish or to attract it to its food or to its mate is about the whole range of the effect that sight or touch has on the animal. These sensations changed into movement constitute what is called reflex action, performance without thinking of[Pg 13] what is being done. With a boy, many familiar actions may be equally reflex. The boy can also do many other things "of his own accord," that is, by conscious effort. He can choose among a great many possible actions. But a fish cannot. If he is scared, he must swim away, and he has no way to stop himself. If he is hungry, and most fishes are so all the time, he will spring at the bait. If he is thirsty, he will gasp, and there is nothing else for him to do. In other words, the activities of a fish are nearly all reflex, most of them being suggested and immediately directed by the influence of external things. Because its actions are all reflex the brain is very small, very primitive, and very simple, nothing more being needed for automatic movement. Small as the fish's skull-cavity is, the brain does not half fill it.

Fig. 5.—Common Sunfish, Eupomotis gibbosus (Linnæus). Natural size. (From life by R. W. Shufeldt.)—Page 13.
The vacant space about the little brain is filled with a fatty fluid mass looking like white of egg, intended for its protection. Taking the dead sunfish (for the live one we shall look after carefully, giving him every day fresh water and a fresh worm or snail or bit of beef), if we cut off the upper part of the skull we shall see the separate parts of the brain, most of them lying in pairs, side by side, in the bottom of the brain-cavity. The largest pair is near the middle of the length of the brain, two nerve-masses (or ganglia), each one round and hollow. If we turn these over, we shall see that the nerves of the eye run into them. We know then that these nerve-masses receive the impressions of sight, and so they are called optic lobes. In front of the optic lobes are two smaller and more oblong nerve-masses. These constitute the cerebrum. This is the thinking part of the brain, and in man and in the higher animals it makes up the greater part of it, overlapping and hiding the other ganglia. But the fish has not much need for thinking and its fore-brain or cerebrum is very small. In front of these are two small, slim projections, one going to each nostril. These are the olfactory lobes which receive the sensation of smell. Behind the optic lobes is a single small lobe, not divided into two. This is the cerebellum and it has charge of certain powers of motion. Under the cerebellum is the medulla, below which the spinal cord begins. The rest of the spinal cord is threaded through the different vertebræ back to the tail, and at each joint it sends[Pg 15] out nerves of motion and receives nerves of sense. Everything that is done by the fish, inside or outside, receives the attention of the little branches of the great nerve-cord.
The Fish's Nest.—The sunfish in the spawning time will build some sort of a nest of stones on the bottom of the eddy, and then, when the eggs are laid, the male with flashing eye and fins all spread will defend the place with a good deal of spirit. All this we call instinct. He fights as well the first time as the last. The pressure of the eggs suggests nest-building to the female. The presence of the eggs tells the male to defend them. But the facts of the nest-building and nest protection are not very well understood, and any boy who can watch them and describe them truly will be able to add something to science.

Form of Body.—With a glance at the fish as a living organism and some knowledge of those structures which are to be readily seen without dissection, we are prepared to examine its anatomy in detail, and to note some of the variations which may be seen in different parts of the great group.
In general fishes are boat-shaped, adapted for swift progress through the water. They are longer than broad or deep and the greatest width is in front of the middle, leaving the compressed paddle-like tail as the chief organ of locomotion.
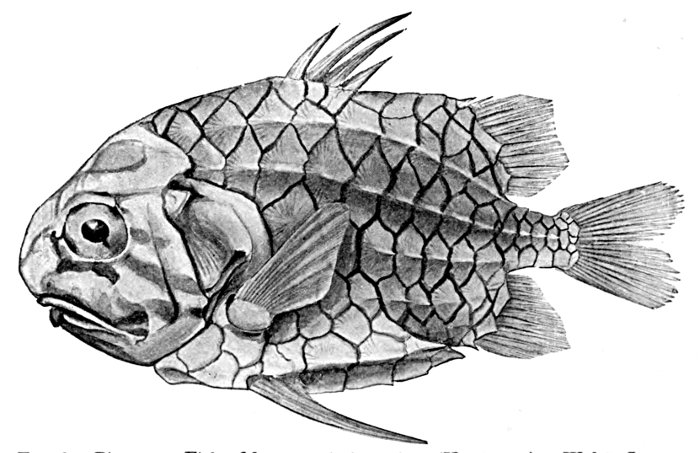
Fig. 6.—Pine-cone Fish, Monocentris japonicus (Houttuyn). Waka, Japan.
But to all these statements there are numerous exceptions. Some fishes depend for protection, not on swiftness, but on the thorny skin or a bony coat of mail. Some of these are almost globular in form, and their outline bears no resemblance to that of a boat. The trunkfish (Ostracion) in a hard bony box has no need of rapid progress.
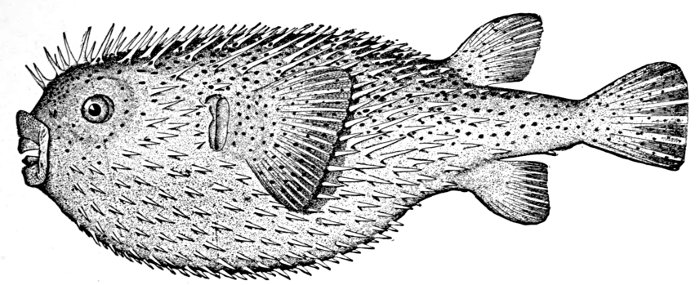
Fig. 7.—Porcupine-fish, Diodon hystrix (Linnæus). Tortugas Islands.

Fig. 8.—Thread-eel, Nemichthys avocetta Jordan and Gilbert. Vancouver Island.
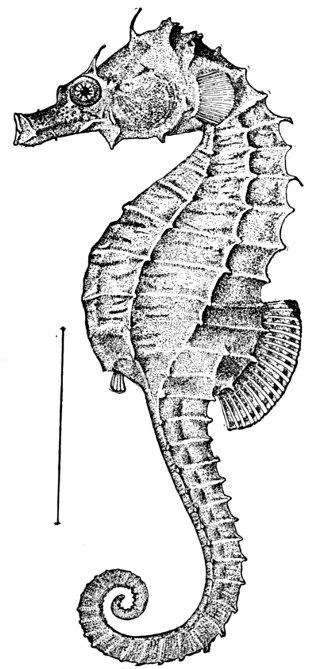
Fig. 9.—Sea-horse, Hippocampus hudsonius Dekay. Virginia.
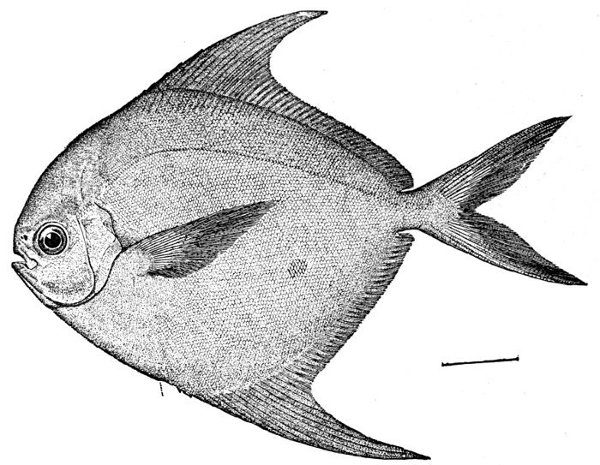
Fig. 10.—Harvest-fish, Peprilus paru (Linnæus). Virginia.
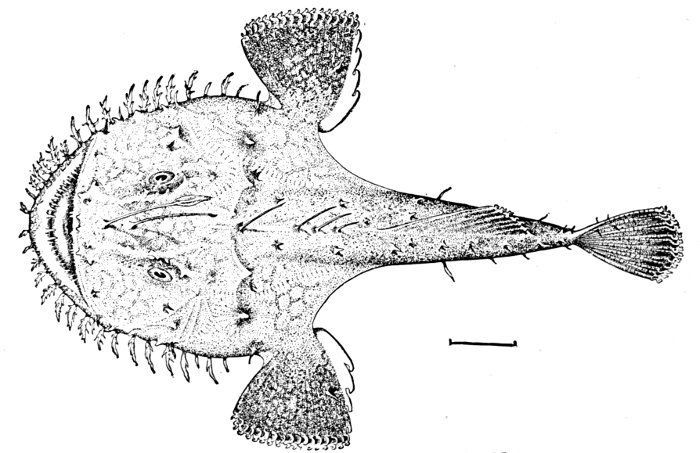
Fig. 11.—Anko or Fishing-frog, Lophius litulon (Jordan). Matsushima Bay, Japan. (The short line in all cases shows the degree of reduction; it represents an inch of the fish's length.)
The pine-cone fish (Monocentris japonicus) adds strong fin-spines to its bony box, and the porcupine fish (Diodon hystrix) is covered with long prickles which keep away all enemies.
Among swift fishes, there are some in which the body is much deeper than long, as in Antigonia. Certain sluggish fishes seem to be all head and tail, looking as though the body by some accident had been omitted. These, like the headfish (Mola mola) are protected by a leathery skin. Other fishes, as the eels, are extremely long and slender, and some carry this elongation to great extremes. Usually the head is in a line with the axis of the body, but in some cases, as the sea-horse (Hippocampus), the head is placed at right angles to the axis, and the body itself is curved and cannot be straightened without injury. The type of the swiftest fish is seen among the mackerels and tunnies, where every outline is such that a racing yacht might copy it.
The body or head of the fish is said to be compressed when it is flattened sidewise, depressed when it is flattened vertically. Thus the Peprilus (Fig. 10) is said to be compressed, while the fishing-frog (Lophius) (Fig. 11) has a depressed body and head. Other terms as truncate (cut off short), attenuate (long-drawn out), robust, cuboid, filiform, and the like may be needed in descriptions.
Measurement of the Fish.—As most fishes grow as long as they live, the actual length of a specimen has not much value for purposes of description. The essential point is not actual length, but relative length. The usual standard of measurement is the length from the tip of the snout to the base of the caudal fin. With this length the greatest depth of the body, the greatest length of the head, and the length of individual parts may be compared. Thus in the Rock Hind (Epinephelus adscensionis), fig. 12, the head is contained 2-3/5 times in the length, while the greatest depth is contained three times.
Thus, again, the length of the muzzle, the diameter of the eye, and other dimensions may be compared with the length of the head. In the Rock Hind, fig. 12, the eye is 5 in head, the snout is 4-2/5 in head, and the maxillary 2-3/5. Young fishes have the eye larger, the body slenderer, and the head larger in proportion than old fishes of the same kind. The mouth grows larger[Pg 20] with age, and is sometimes larger also in the male sex. The development of the fins often varies a good deal in some fishes with age, old fishes and male fishes having higher fins when such differences exist. These variations are soon understood by the student of fishes and cause little doubt or confusion in the study of fishes.
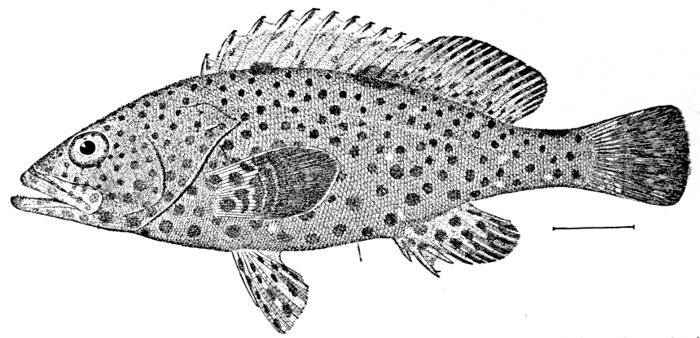
Fig. 12.—Rock Hind or Cabra Mora of the West Indies, Epinephelus adscensionis (Osbeck). Family Serranidæ.
The Scales, or Exoskeleton.—The surface of the fish may be naked as in the catfish, or it may be covered with scales, prickles, shagreen, or bony plates. The hard covering of the skin, when present, is known as the exoskeleton, or outer skeleton. In the fish, the exoskeleton, whatever form it may assume, may be held to consist of modified scales, and this is usually obviously the case. The skin of the fish may be thick or thin, bony, horny, leathery, or papery, or it may have almost any intermediate character. When protected by scales the skin is usually thin and tender; when unprotected it may be ossified, as in the sea-horse; horny, as in the headfish; leathery, as in the catfish; or it may, as in the sea-snails, form a loose scarf readily detachable from the muscles below.
The scales themselves may be broadly classified as ctenoid, cycloid, placoid, ganoid, or prickly.
Ctenoid and Cycloid Scales.—Normally formed scales are rounded in outline, marked by fine concentric rings, and crossed on the inner side by a few strong radiating ridges and folds.[Pg 21] They usually cover the body more or less evenly and are imbricated like shingles on a roof, the free edge being turned backward. Such normal scales are of two types, ctenoid or cycloid. Ctenoid scales have a comb-edge of fine prickles or cilia; cycloid scales have the edges smooth. These two types are not very different, and the one readily passes into the other, both being sometimes seen on different parts of the same fish. In general, however, the more primitive representatives of the typical fishes, those with abdominal ventrals and without spines in the fins, have cycloid or smooth scales. Examples are the salmon, herring, minnow, and carp. Some of the more specialized spiny-rayed fishes, as the parrot-fishes, have, however, scales equally smooth, although somewhat different in structure. Sometimes, as in the eel, the cycloid scales may be reduced to mere rudiments buried in the skin.
Ctenoid scales are beset on the free edge by little prickles or points, sometimes rising to the rank of spines, at other times soft and scarcely noticeable, when they are known as ciliate or eyelash-like. Such scales are possessed in general by the more specialized types of bony fishes, as the perch and bass, those with thoracic ventrals and spines in the fins.

Fig. 13.—Scales of Acanthoessus bronni (Agassiz). (After Dean.)
Placoid Scales.—Placoid scales are ossified papillæ, minute, enamelled, and close-set, forming a fine shagreen. These are characteristic of the sharks; and in the most primitive sharks the teeth are evidently modifications of these primitive structures. Some other fishes have scales which appear shagreen-like to sight and feeling, but only the sharks have the peculiar structure to which Agassiz gave the name of placoid. The rough prickles of the filefishes and some sculpins are not placoid, but are reduced or modified ctenoid scales, scales narrowed and reduced to prickles.
Bony and Prickly Scales.—Bony and prickly scales are found in great variety, and scarcely admit of description or classification. In general, prickly points on the skin are modifications of ctenoid scales. Ganoid scales are thickened and covered with bony enamel, much like that seen in teeth, otherwise[Pg 22] essentially like cycloid scales. These are found in the garpike and in many genera of extinct Ganoid and Crossopterygian fishes. In the line of descent the placoid scale preceded the ganoid, which in turn was followed by the cycloid and lastly by the ctenoid scale. Bony scales in other types of fishes may have nothing structurally in common with ganoid scales or plates, however great may be the superficial resemblance.
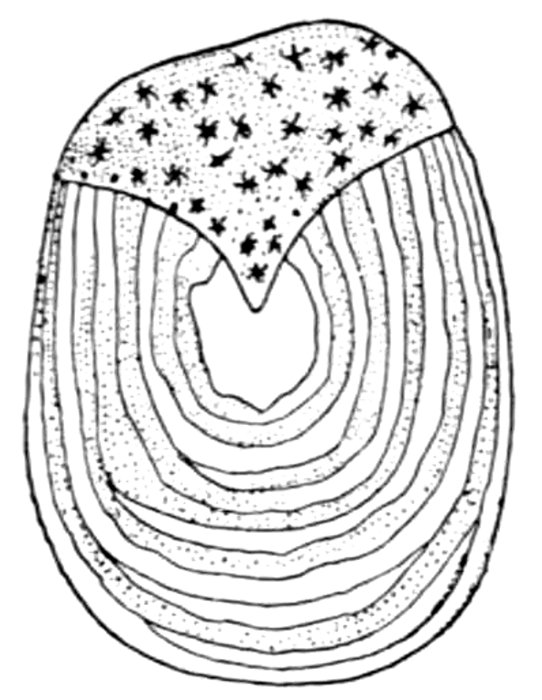
Fig. 14.—Cycloid Scale.
The distribution of scales on the body may vary exceedingly. In some fishes the scales are arranged in very regular series; in others they are variously scattered over the body. Some are scaly everywhere on head, body, and fins. Others may have only a few lines or patches. The scales may be everywhere alike, or they may in one part or another be greatly modified. Sometimes they are transformed into feelers or tactile organs. The number of scales is always one of the most valuable of the characters by which to distinguish species.
Lateral Line.—The lateral line in most fishes consists of a series of modified scales, each one provided with a mucous tube extending along the side of the body from the head to the caudal fin. The canal which pierces each scale is simple at its base, but its free edge is often branched or ramified. In most spiny-rayed fishes it runs parallel with the outline of the back. In most soft-rayed fishes it follows rather the outline of the belly. It is subject to many variations. In some large groups (Gobiidæ, Pæciliidæ) its surface structures are entirely wanting. In scaleless fishes the mucous tube lies in the skin itself. In some groups the lateral line has a peculiar position, as in the flying-fishes, where it forms a raised ridge bounding the belly. In many cases the lateral line has branches of one sort or another. It is often double or triple, and in some cases the whole back and sides of the fish are covered with lateral lines and their ramifications. Sometimes peculiar sense-organs and occasionally eye-like luminous spots are developed in connection with the lateral line, enabling the fish to see in the black depths of the sea. These will be noticed in another chapter.
The Lateral Line as a Mucous Channel.—The more primitive[Pg 23] condition of the lateral line is seen in the sharks and chimæras, in which fishes it appears as a series of channels in or under the skin. These channels are filled with mucus, which exudes through occasional open pores. In many fishes the bones of the skull are cavernous, that is, provided with cavities filled with mucus. Analogous to these cavities are the mucous channels which in primitive fishes constitute the lateral line.
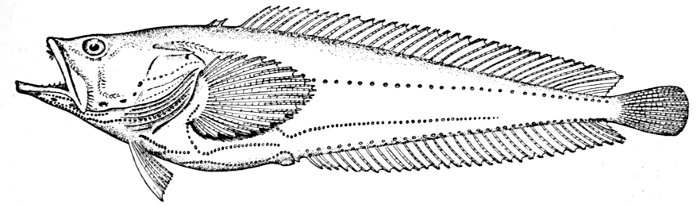
Fig. 15.—Singing Fish (with many lateral lines), Porichthys porosissimus (Cuv. and Val.). Gulf of Mexico.
Function of the Lateral Line.—The general function of the lateral line with its tubes and pores is still little understood. As the structures of the lateral line are well provided with nerves, it has been thought to be an organ of sense of some sort not yet understood. Its close relation to the ear is beyond question, the ear-sac being an outgrowth from it.
"The original significance of the lateral line," according to Dr. Dean,[2] "as yet remains undetermined. It appears intimately if not genetically related to the sense-organs of the head and gill region of the ancestral fish. In response to special aquatic needs, it may thence have extended farther and farther backward along the median line of the trunk, and in its later differentiation acquired its metameral characters." In view of its peculiar nerve-supply, "the precise function of this entire system of organs becomes especially difficult to determine. Feeling, in its broadest sense, has safely been admitted as its possible use. Its close genetic relationship to the hearing organ suggests the kindred function of determining waves of vibration. These are transmitted in so favorable a way in the aquatic medium that from the side of theory a system of [Pg 24]hypersensitive end-organs may well have been established. The sensory tracts along the sides of the body are certainly well situated to determine the direction of the approach of friend, enemy, or prey."
The Fins of Fishes.—The organs of locomotion in the fishes are knows as fins. These are composed of bony or cartilaginous rods or rays connected by membranes. The fins are divided into two groups, paired fins and vertical fins. The pectoral fins, one on either side, correspond to the anterior limbs of the higher vertebrates. The ventral fins below or behind them represent the hinder limbs. Either or both pairs may be absent, but the ventrals are much more frequently abortive than the pectorals. The insertion of the ventral fins may be abdominal, as in the sharks and the more generalized of the bony fishes, thoracic under the breast (the pelvis attached to the shoulder-girdle) or jugular, under the throat. When the ventral fins are abdominal, the pectoral fins are usually placed very low. The paired fins are not in general used for progression in the water, but serve rather to enable the fish to keep its equilibrium. With the rays, however, the wing-like pectoral fins form the chief organ of locomotion.
The fin on the median line of the back is called the dorsal, that on the tail the caudal, and that on the lower median line the anal fin. The dorsal is often divided into two fins or even three. The anal is sometimes divided, and either dorsal or anal fin may have behind it detached single rays called finlets.
The rays composing the fin may be either simple or branched. The branched rays are always articulated, that is, crossed by numerous fine joints which render them flexible. Simple rays are also sometimes articulate. Rays thus jointed are known as soft rays, while those rays which are neither jointed nor branched are called spines. A spine is usually stiff and sharp-pointed, but it may be neither, and some spines are very slender and flexible, the lack of branches or joints being the feature which distinguishes spine from soft ray.
The anterior rays of the dorsal and anal fins are spinous in most fishes with thoracic ventrals. The dorsal fin has usually about ten spines, the anal three, but as to this there is much variation in different groups. When the dorsal is di[Pg 25]vided all the rays of the first dorsal and usually the first ray of the second are spines. The caudal fin has never true spines, though at the base of its lobes are often rudimentary rays which resemble spines. Most spineless fishes have such rudiments in front of their vertical fins. The pectoral, as a rule, is without spines, although in the catfishes and some others a single large spine may be developed. The ventrals when abdominal are usually without spines. When thoracic each usually, but not always, consists of one spine and five soft rays. When jugular the number of soft rays may be reduced, this being a phase of degeneration of the fin. In writing descriptions of fishes the number of spines may be indicated by Roman numerals, those of the soft rays by Arabic. Thus D. XII-I, 17 means that the dorsal is divided, that the anterior portion consists of twelve spines, the posterior of one spine and seventeen soft rays. In some fishes, as the catfish or the salmon, there is a small fin on the back behind the dorsal fin. This is known as the adipose fin, being formed of fatty substance covered by skin. In a few catfishes, this adipose fin develops a spine or soft rays.
Muscles.—The movements of the fins are accomplished by the muscles. These organs lie along the sides of the body, forming the flesh of the fish. They are little specialized, and not clearly differentiated as in the higher vertebrates.
With the higher fishes there are several distinct systems of muscles controlling the jaws, the gills, the eye, the different fins, and the body itself. The largest of all is the great lateral muscle, composed of flake-like segments (myocommas) which correspond in general with the number of the vertebræ. In general the muscles of the fish are white in color. In some groups, especially of the mackerel family, they are deep red, charged with animal oils. In the salmon they are orange-red, a color also due to the presence of certain oils.
In a few fishes muscular structures are modified into electric organs. These will be discussed in a later chapter.
[2] Fishes Recent and Fossil, p. 52.

The Blue-green Sunfish.—The organs found in the abdominal cavity of the fish may be readily traced in a rapid dissection. Any of the bony fishes may be chosen, but for our purposes the sunfish will serve as well as any. The names and location of the principal organs are shown in the accompanying figure, from Kellogg's Zoology. It represents the blue-green sunfish, Apomotis cyanellus, from the Kansas River, but in these regards all the species of sunfishes are alike. We may first glance at the different organs as shown in the sequence of dissection, leaving a detailed account of each to the subsequent pages.
The Viscera.—Opening the body cavity of the fish, as shown in the plate, we see below the back-bone a membranous sac closed and filled with air. This is the air-bladder, a rudiment of that structure which in higher vertebrates is developed as a lung. The alimentary canal passes through the abdominal cavity extending from the mouth through the pharynx and ending at the anus or vent. The stomach has the form of a blind sac, and at its termination are a number of tubular sacs, the pyloric cæca, which secrete a digestive fluid. Beyond the pylorus extends the intestine with one or two loops to the anus. Connected with the intestine anteriorly is the large red mass of the liver, with its gall-bladder, which serves as a reservoir for bile, the fluid the liver secretes. Farther back is another red glandular mass, the spleen.
In front of the liver and separated from it by a membrane is the heart. This is of four parts. The posterior part is a thin-walled reservoir, the sinus venosus, into which blood enters through the jugular vein from the head and through the cardinal vein from the kidney. From the sinus venosus it passes forward into a large thin-walled chamber, the auricle.
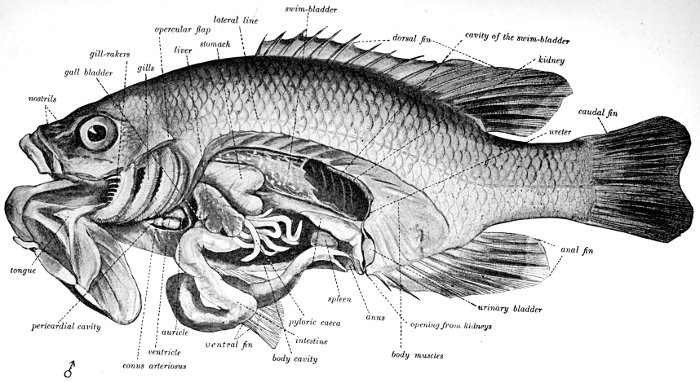
Fig. 16.—Dissection of the Blue-green Sunfish, Apomotis cyanellus Rafinesque. (After Kellogg.)—27.
[Pg 28] Next it flows into the thick-walled ventricle, whence by the rhythmical contraction of its walls it is forced into an arterial bulb which lies at the base of the ventral aorta, which carries it on to the gills. After passing through the fine gill-filaments, it is returned to the dorsal aorta, a large blood-vessel which extends along the lower surface of the back-bone, giving out branches from time to time.
The kidneys in fishes constitute an irregular mass under the back-bone posteriorly. They discharge their secretions through the ureter to a small urinary bladder, and thence into the urogenital sinus, a small opening behind the anus. Into the same sinus are discharged the reproductive cells in both sexes.
In the female sunfish the ovaries consist of two granular masses of yellowish tissue lying just below and behind the swim-bladder. In the spring they fill much of the body cavity and the many little eggs can be plainly seen. When mature they are discharged through the oviduct to the urogenital sinus. In some fishes there is no special oviduct and the eggs pass into the abdominal cavity before exclusion.
In the male the reproductive organs have the same position as the ovaries in the female. They are, however, much smaller in size and paler in color, while the minute spermatozoa appear milky rather than granular on casual examination. A vas deferens leads from each of these organs into the urogenital sinus.
The lancelets, lampreys, and hagfishes possess no genital ducts. In the former the germ cells are shed into the atrial cavity, and from there find their way to the exterior either through the mouth or the atrial pore; in the latter they are shed directly into the body cavity, from which they escape through the abdominal pores. In the sharks and skates the Wolffian duct in the male, in addition to its function as an excretory duct, serves also as a passage for the sperm, the testes having a direct connection with the kidneys. In these forms there is a pair of Müllerian ducts which serve as oviducts in the females; they extend the length of the body cavity, and at their anterior end have an opening which receives the eggs which have escaped from the ovary into the body cavity. In some bony fishes as the eels and female salmon the germ cells are shed into the body cavity and escape through genital pores, which, however, may[Pg 29] not be homologous with abdominal pores. In most other bony fishes the testes and ovaries are continued directly into ducts which open to the outside.
Organs of Nutrition.—The organs thus shown in dissection we may now examine in detail.
Fig. 17.—Black Swallower, Chiasmodon niger Johnson, containing a fish larger than itself. Le Have Bank.
The mouth of the fish is the organ or series of structures first concerned in nutrition. The teeth are outgrowths from the skin, primarily as modified papillæ, aiding the mouth in its various functions of seizing, holding, cutting, or crushing the various kinds of food material. Some fishes feed exclusively on plants, some on plants and animals alike, some exclusively on animals, some on the mud in which minute plants and animals occur. The majority of fishes feed on other fishes, and without much regard to species or condition. With the carnivorous fishes, to feed represents the chief activity of the organism. In proportion to the voracity of the fish is usually the size of the mouth, the sharpness of the teeth, and the length of the lower jaw.
The most usual type of teeth among fishes is that of villiform bands. Villiform teeth are short, slender, even, close-set, making a rough velvety surface. When the teeth are larger and more widely separated, they are called cardiform, like the teeth of a wool-card. Granular teeth are small, blunt, and sand-like. Canine teeth are those projecting above the level of the others, usually sharp, curved, and in some species barbed. Sometimes[Pg 30] the canines are in front. In some families the last tooth in either jaw may be a "posterior canine," serving to hold small animals in place while the anterior teeth crush them. Canine teeth are often depressible, having a hinge at base.
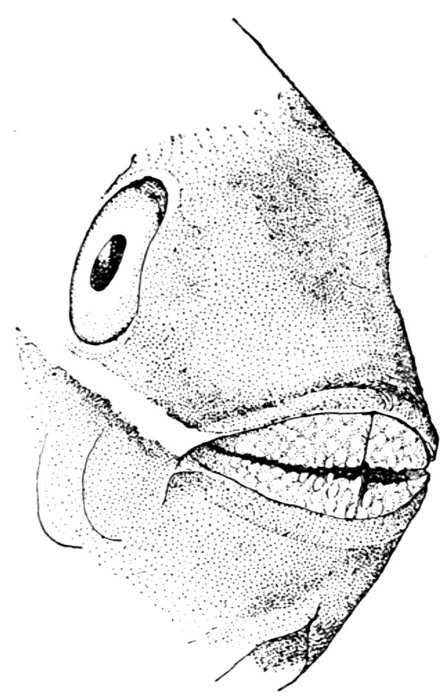
Fig. 18.—Jaws of a Parrot-fish, Sparisoma aurofrenatum (Val.). Cuba.
Teeth very slender and brush-like are called setiform. Teeth with blunt tips are molar. These are usually enlarged and fitted for crushing shells. Flat teeth set in mosaic, as in many rays and in the pharyngeals of parrot-fishes, are said to be paved or tessellated. Knife-like teeth, occasionally with serrated edges, are found in many sharks. Many fishes have incisor-like teeth, some flattened and truncate like human teeth, as in the sheepshead, sometimes with serrated edges. Often these teeth are movable, implanted only in the skin of the lips. In other cases they are set fast in the jaw. Most species with movable teeth or teeth with serrated edges are herbivorous, while strong incisors may indicate the choice of snails and crabs as food. Two or more of these different types may be found in the same fish. The knife-like teeth of the sharks are progressively shed, new ones being constantly formed on the inner margins of the jaw, so that the teeth are marching to be lost over the edge of the jaw as soon as each has fulfilled its function. In general the more distinctly a species is a fish-eater, the sharper are the teeth. Usually fishes show little discrimination in their choice of food; often they devour the young of their own species as readily as any other. The digestive process is rapid, and most fishes rapidly increase in size in the process of development. When food ceases to be abundant the fishes grow more slowly. For this reason the same species will grow to a larger size in large streams than in small ones, in lakes than in brooks. In most cases there is no absolute limit to growth, the species growing as long as it lives. But while some species endure many years, others are certainly very short[Pg 31]lived, and some may be even annual, dying after spawning, perhaps at the end of the first season.
Teeth are wholly absent in several groups of fishes. They are, however, usually present on the premaxillary, dentary, and pharyngeal bones. In the higher forms, the vomer, palatines, and gill-rakers are rarely without teeth, and in many cases the pterygoids, sphenoids, and the bones of the tongue are similarly armed.
No salivary glands or palatine velum are developed in fishes. The tongue is always bony or gristly and immovable. Sometimes taste-buds are developed on it, and sometimes these are found on the barbels outside the mouth.

Fig. 19.—Sheepshead (with incisor teeth), Archosargus probatocephalus (Walbaum). Beaufort, N. C.
The Alimentary Canal.—The mouth-cavity opens through the pharynx between the upper and lower pharyngeal bones into the œsophagus, whence the food passes into the stomach. The intestinal tract is in general divided into four portions—œsophagus, stomach, small and large intestines. But these divisions of the intestines are not always recognizable, and in the very lowest forms, as in the lancelet, the stomach is a simple straight tube without subdivision.
In the lampreys there is a distinction only of the œsophagus with many longitudinal folds and the intestine with but[Pg 32] one. In the bony fishes the stomach is an enlarged area, either siphon-shaped, with an opening at either end, or else forming a blind sac with the openings for entrance (cardiac) and exit (pyloric) close together at the anterior end. In the various kinds of mullets (Mugil) and in the hickory shad (Dorosoma), fishes which feed on minute vegetation mixed with mud, the stomach becomes enlarged to a muscular gizzard, like that of a fowl. Attached near the pylorus and pouring their secretions into the duodenum or small intestine are the pyloric cæca. These are tubular sacs secreting a pale fluid and often almost as long as the stomach or as wide as the intestine. These may be very numerous as in the salmon, in which case they are likely to become coalescent at base, or they be few or altogether wanting.
Besides these appendages which are wanting in the higher vertebrates, a pancreas is also found in the sharks and many other fishes. This is a glandular mass behind the stomach, its duct leading into the duodenum and often coalescent with the bile duct from the liver. The liver in the lancelet is a long diverticulum of the intestine. In the true fishes it becomes a large gland of irregular form, and usually but not always provided with a gall-bladder as in the higher vertebrates. Its secretions usually pass through a ductus cholodechus to the duodenum.
The spleen, a dark-red lymphatic gland, is found attached to the stomach in all fish-like vertebrates except the lancelet.
The lining membrane of the abdominal cavity is known as the peritoneum, and the membrane sustaining the intestines from the dorsal side, as in the higher vertebrates, is called the mesentery. In many species the peritoneum is jet black, while in related forms it may be pale in color. It is more likely to be black in fishes from deep water and in fishes which feed on plants.
The Spiral Valve.—In the sharks or skates the rectum or large intestine is peculiarly modified, being provided with a spiral valve, with sometimes as many as forty gyrations. A spiral valve is also present in the more ancient types of the true fishes as dipnoans, crossopterygians, and ganoids. This valve greatly increases the surface of the intestine, doing away with the necessity for length. In the bowfin (Amia) and the garpike (Lepi[Pg 33]sosteus) the valve is reduced to a rudiment of three or four convolutions near the end of the intestine. In the sharks and skates the intestine opens into a cloaca, which contains also the urogenital openings. In all fishes the latter lie behind the orifice of the intestine. In the bony fishes and the ganoids there is no cloaca.

Fig. 20.—Stone-roller, Campostoma anomalum (Rafinesque). Family Cyprinidæ. Showing nuptial tubercles and intestines coiled about the air-bladder.
Length of the Intestine.—In all fishes, as in the higher vertebrates, the length of the alimentary canal is coordinated with the food of the fish. In those which feed upon plants the intestine is very long and much convoluted, while in those which feed on other fishes it is always relatively short. In the stone-roller, a fresh-water minnow (Campostoma) found in the Mississippi Valley, the excessively long intestines filled with vegetable matter are wound spool-fashion about the large air-bladder. In all other fishes the air-bladder lies on the dorsal side of the intestinal canal.

Specialization of the Skeleton.—In the lowest form of fish-like vertebrates (Branchiostoma), the skeleton consists merely of a cartilaginous rod or notochord extending through the body just below the spinal cord. In the lampreys, sharks, dipnoans, crossopterygians, and sturgeons the skeleton is still cartilaginous, but grows progressively more complex in their forms and relations. Among the typical fishes the skeleton becomes ossified and reaches a very high degree of complexity. Very great variations in the forms and relations of the different parts of the skeleton are found among the bony fishes, or teleostei. The high degree of specialization of these parts gives to the study of the bones great importance in the systematic arrangement of these fishes. In fact the true affinities of forms is better shown by the bones than by any other system of organs. In a general way the skeleton of the fish is homologous with that of man. The head in the one corresponds to the head in the other, the back-bone to the back-bone, and the paired fins, pectoral and ventral, to the arms and legs.
Homologies of Bones of Fishes.—But this homology does not extend to the details of structure. The bones of the arm of the specialized fish are not by any means identical with the humerus, coracoid, clavicle, radius, ulna, and carpus of the higher vertebrates. The vertebrate arm is not derived from the pectoral fin, but both from a cartilaginous shoulder-girdle with undifferentiated pectoral elements bearing fin-rays, in its details unlike an arm and unlike the pectoral fin of the specialized fish.
The assumption that each element in the shoulder-girdle and the pectoral fin of the fish must correspond in detail to the arm of man has led to great confusion in naming the different[Pg 35] bones. Among the many bones of the fish's shoulder-girdle and pectoral fin, three or four different ones have successively borne the names of scapula, clavicle, coracoid, humerus, radius, and ulna. None of these terms, unless it be clavicle, ought by rights apply to the fish, for no bone of the fish is a true homologue of any of these as seen in man. The land vertebrates and the fishes have doubtless sprung from a common stock, but this stock, related to the crossopterygians of the present day, was unspecialized in the details of its skeleton, and from it the fishes and the higher vertebrates have developed the widely diverging lines.

Fig. 21.—Striped Bass, Roccus lineatus (Bloch). Potomac River.
Parts of the Skeleton.—The skeleton may be divided into the head, the vertebral column, and the limbs. The very lowest of the fish-like forms (Branchiostoma) has no differentiated head or skull, but in all the other forms the anterior part of the vertebral column is modified to form a cranium for the protection of the brain. In the lampreys there are no jaws or other appendages to the cranium.
In the sharks, dipnoans, crossopterygians, ganoids, and teleosts or bony fishes, jaws are developed as well as a variety of other bones around the mouth and throat. The jaw-bearing forms are sometimes known by the general name of gnathostomes. In the sharks and their relatives (rays, chimæras, etc.) all the skeleton is composed of cartilage. In the more specialized bony fishes, besides these bones we find also series of membrane bones, more or less external to the skull and composed of[Pg 36] ossified dermal tissues. Membrane bones are not found in the sharks and lampreys, but are developed in an elaborate coat of mail in some extinct forms.
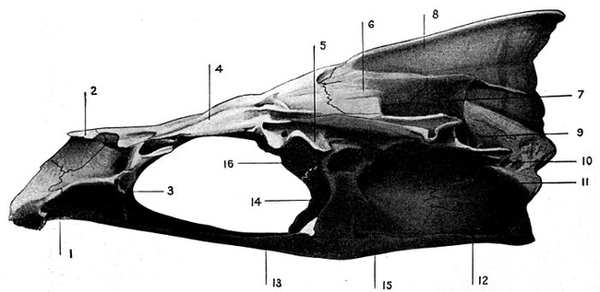
Fig. 22.—Roccus lineatus. Lateral view of cranium.
1. Vomer. 3. Prefrontal. 5. Sphenotic. 7. Epiotic. 9. Pterotic. 11. Exoccipital. 13. Parasphenoid. 15. Prootic. 2. Ethmoid. 4. Frontal. 6. Parietal. 8. Supraoccipital. 10. Opisthotic. 12. Basioccipital. 14. Basisphenoid.
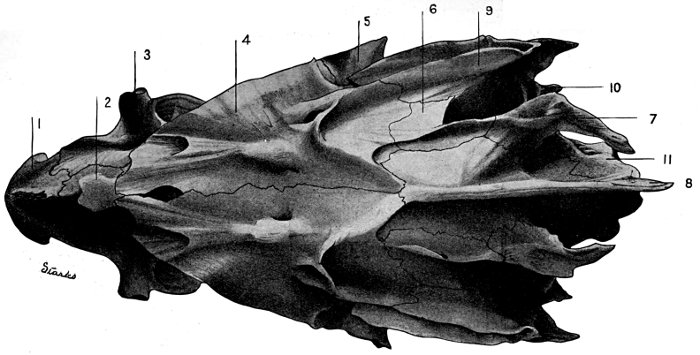
Fig. 23.—Roccus lineatus. Superior view of cranium.
1. Vomer. 3. Prefrontal. 5. Sphenotic. 7. Epiotic. 9. Pterotic. 11. Exoccipital. 2. Ethmoid. 4. Frontal. 6. Parietal. 8. Supraoccipital. 10. Opisthotic.
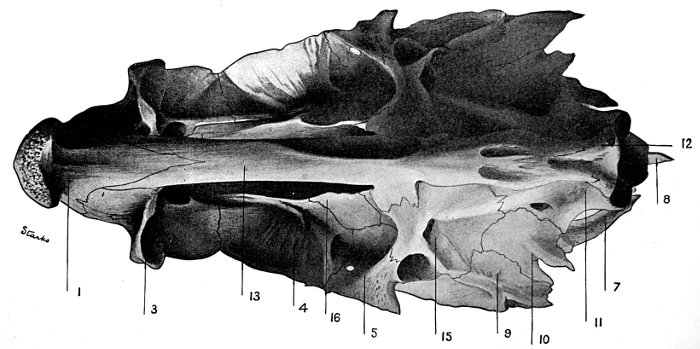
Fig. 24.—Roccus lineatus. Inferior view of cranium.
1. Vomer. 4. Frontal. 7. Epiotic. 9. Pterotic. 11. Exoccipital. 13. Parasphenoid. 16. Alisphenoid. 3. Prefrontal. 5. Sphenotic. 8. Supraoccipital. 10. Opisthotic. 12. Basioccipital. 15. Prootic.
Names of Bones of Fishes.—In the study of the names of the bones of fishes it will be more convenient to begin with a highly specialized form in which each of the various structures is present and in its normal position.
To this end we present a series of figures of a typical form, choosing, after Starks, the striped bass (Roccus lineatus) of the Atlantic coast of the United States. For this set of plates, drawn from nature by Mrs. Chloe Lesley Starks, we are indebted to the courtesy of Mr. Edwin Chapin Starks. The figures of the striped bass illustrate a noteworthy paper on "The Synonymy of the Fish Skeleton," published by the Washington Academy of Sciences in 1901.
Bones of the Cranium.—The vomer (1) is the anterior part of the roof of the mouth, armed with small teeth in the striped bass and in many other fishes, but often toothless. The ethmoid (2) lies behind the vomer on the upper surface of the skull, and the prefrontal (3) projects on either side and behind the ethmoid, the nostrils usually lying over or near it and near the nasal bone (51). Between the eyes above are the two frontal (4) bones joined by a suture. On the side behind the posterior angle of the frontal is the sphenotic (5) above the posterior part of the eye. Behind each frontal is the parietal (6). Behind the parietal and more or less turned inward over the ear-cavity is the epiotic (7). Between the parietals, and in most fishes rising into a thin crest, is the supraoccipital (8), which bounds the cranium above and behind, its posterior margin being usually a vertical knife-like edge. The pterotic (9) forms a sort of wing or free margin behind the epiotic and over the ear-cavity. The opisthotic (10) is a small, hard, irregular bone behind the pterotic. The exoccipital (11) forms a concave joint or condyle on each side of the basioccipital (12), by which the vertebral column is joined to the skull. The parasphenoid (13) forms a narrow ridge of the roof of the mouth, connecting the vomer with the basioccipital. In some fishes of primitive structure (Salmo, Beryx) there is another bone, called orbitosphenoid, on the middle line above and between the eyes. The basisphe[Pg 40]noid (14) is a little bone above the myotome or tube in which runs the rectus muscle of the eye. It descends toward the parasphenoid and is attached to the prootic. The prootic (15) is an irregular bone below the ear region and lying in advance of the opisthotic. The alisphenoid (16) is a small bone in the roof of the mouth before the prootic. These sixteen bones (with a loose bone of specialized form, the otolith, within the ear-cavity) constitute the cranium. All are well developed in the striped bass and in most fishes. In some specialized forms they are much distorted, coossified, or otherwise altered, and their relations to each other may be more or less changed. In the lower forms they are not always fully differentiated, but in nearly all cases their homologies can be readily traced. In the sharks and lampreys the skull constitutes a continuous cartilaginous box without sutures. In the dipnoans and other forms having a bony casque the superficial bones outside the cranium may not correspond to the cartilaginous elements of the soft skull itself.
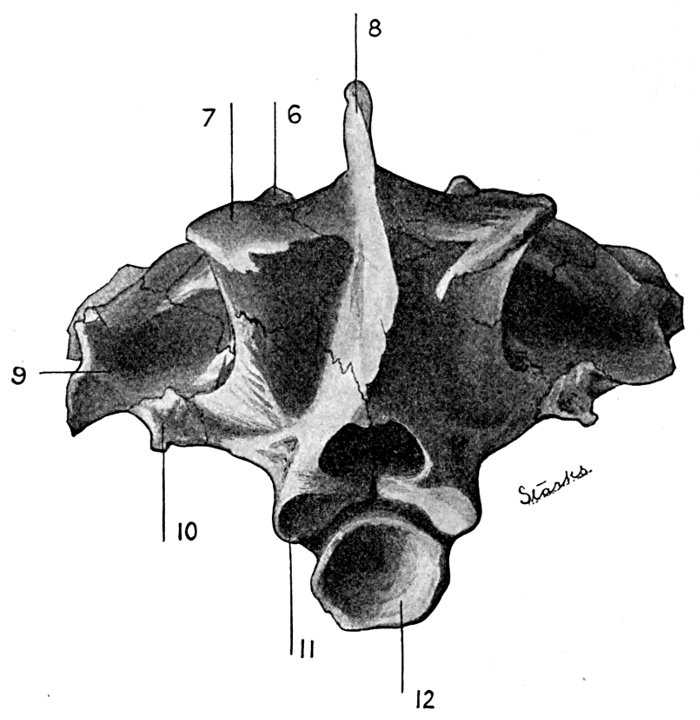
Fig. 25—Roccus lineatus. Posterior view of cranium.
Bones of the Jaws.—The bones of the jaws are attached to the cranium by membranes only, not by sutures, except in a few peculiarly specialized forms.
The Upper Jaw.—The premaxillary (32) lies on either side and forms the front of the upper jaw. Its upper posterior tip or premaxillary spine projects backward almost at right angles with the rest of the bone into a groove on the ethmoid. There is often a fold in the skin by which this bone may be thrust out or protracted, as though drawn out of a sheath. When the spines of the premaxillary are very long the upper jaw may be thrust out for a considerable distance. The premaxillary is also often known as intermaxillary.
Lying behind the premaxillary, its anterior end attached within the angle of the premaxillary, is the maxillary (31), or supramaxillary, a flattened bone with expanded posterior tip. In the striped bass this bone is without teeth, but in many less specialized forms, as the salmon, it is provided with teeth and joined to the premaxillary in a different fashion. In any case its position readily distinguishes it. In some cases the maxillary is divided by one or more sutures, setting off from it one or more extra maxillary (supplemental maxillary) bones. This suture is absent in the striped bass, but distinct in the black bass, and more than one suture is found in the shad and herring. The roof of the mouth above is formed by a number of bones, which, as they often possess teeth, may be considered with the jaws. These are the palatine bones (21), one on either side flanking the vomer, the pterygoid (20), behind it and articulating with it, the mesopterygoid (22), on the roof of the mouth toward the median line, and the metapterygoid (23), lying behind this. Although often armed with teeth, these bones are to be considered of the general nature of the membrane bones. In some degraded types of fishes (eels, morays, congers) the premaxillary is indistinguishable, being united with the vomer and palatines.
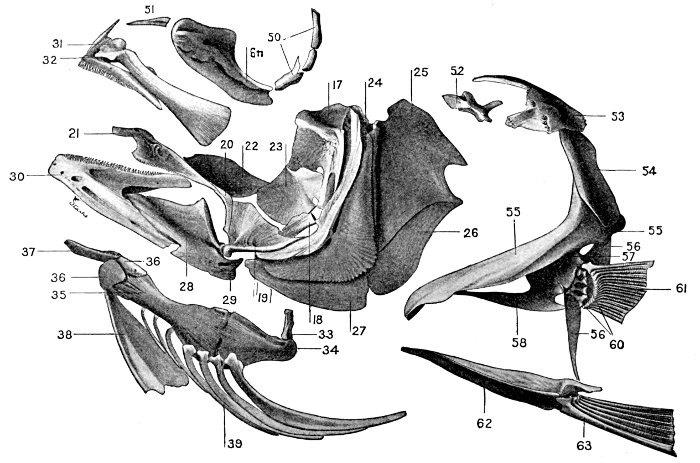
Fig. 26.—Roccus lineatus. Face-bones, shoulder and pelvic girdles, and hyoid arch.
The upper jaw of the shark is formed from the anterior portion of the palatine bones, which are not separate from the quadrate, the whole forming the palato-quadrate apparatus. In the himæra and the dipnoans this apparatus is solidly united with the cranium. In these fishes the true upper jaw, formed of maxillary and premaxillary, is wanting.
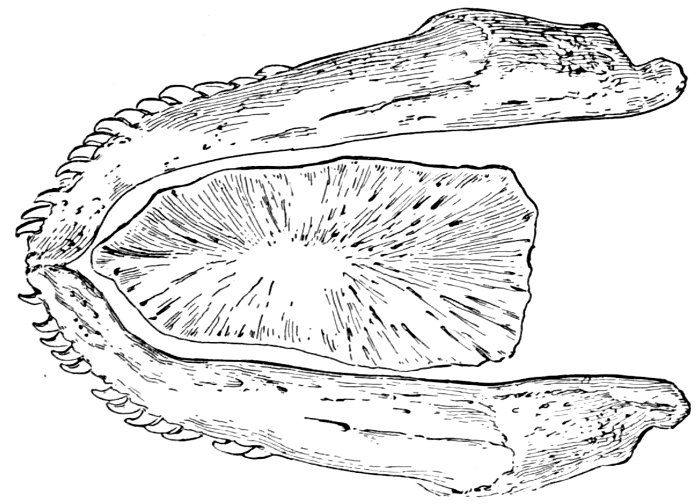
Fig. 27.—Lower jaw of Amia calva (Linnæus), showing the gular plate.
The Lower Jaw.—The lower jaw or mandible is also complex, consisting of two divisions or rami, right and left, joined in front by a suture. The anterior part of each ramus is formed by the dentary bone (30), which carries the teeth. Behind this is the articular bone (28), which is connected by a joint to the quadrate bone (19). At the lower angle of the articular bone is the small angular bone (29). In many cases another small bone, which is called splenial, may be found attached to the inner surface of the articular bone. This little bone has been called coronoid, but it is doubtless not homologous with the coronoid bone of reptiles. In a few fishes, Amia, Elopidæ, and certain fossil dipnoans, there is a bony gular plate, a membrane bone across the throat behind the chin on the lower jaw.
The Suspensorium of the Mandible.—The lower jaw is attached to the cranium by a chain of suspensory bones, which vary a good deal with different groups of fishes. The articular is jointed with the flat quadrate bone (19), which lies behind the pterygoid. A slender bone passes upward (18) under the preopercle and the metapterygoid, forming a connection above[Pg 44] with a large flattish bone, the hyomandibular (17), which in turn joins the cranium. The slender bone which thus keys together the upper and lower elements, hyomandibular and quadrate, forming the suspensorium of the lower jaw, is known as symplectic (18). The hyomandibular is thought to be homologous with the stapes, or stirrup-bone, of the ear in higher animals. In this case the symplectic may be homologous with its small orbicular bone, and the malleus is a transformation of the articular. The incus, or anvil-bone, may be formed from part of Meckel's cartilage. All these homologies are however extremely hypothetical. The core of the lower jaw is formed of a cartilage called Meckel's cartilage, outside which the membrane bones, dentary, etc., are developed. This cartilage forms the lower jaw in sharks, true jaw-bones not being developed in these fishes. In lampreys and lancelets there is no lower jaw.
Membrane Bones of Face.—The membrane bones lie on the surface of the head, when they are usually covered by thin skin and have only a superficial connection with the cranium. Such bones, formed of ossified membrane, are not found in the earlier or less specialized fishes, the lancelets and lampreys, nor in the sharks, rays, and chimæras. They are chiefly characteristic of the bony fishes, although in some of these they have undergone degradation.
The preorbital (49) lies before and below the eye, its edge more or less parallel with that of the maxillary. It may be broad or narrow. When broad it usually forms a sheath into which the maxillary slips. The nasal (51) lies before the preorbital, a small bone usually lying along the spine of the premaxillary. Behind and below the eye is a series of about three flat bones, the suborbitals (50), small in the striped bass, but sometimes considerably modified. In the great group of loricate fishes (sculpins, etc.), the third suborbital sends a bony process called the suborbital stay backward across the cheek toward the preopercle. The suborbital stay is present in the rosefish. In some cases, as in the gurnard, this stay covers the whole cheek with a bony coat of mail. In some fishes, but not in the striped bass, a small supraorbital bone exists over the eye, forming a sort of cap on an angle of the frontal bone.
The largest uppermost flat bone of the gill-covers is known[Pg 45] as the opercle (25). Below it, joined by a suture, is the subopercle (26). Before it is the prominent ridge of the preopercle (24), which curves forward below and forms a more or less distinct angle, often armed with serrations or spines. In some cases this armature is very highly developed. The interopercle (27) lies below the preopercle and parallel with the lower limb.
Branchial Bones.—The bones of the branchial apparatus or gills are very numerous and complex, as well as subject to important variations. In many fishes some of these bones are coossified, and in other cases some are wanting. The tongue may be considered as belonging to this series, as the bones of the gills are attached to its axis below.
In the striped bass, as in most fishes, the tongue, gristly and immovable, is formed anteriorly by a bone called the glossohyal (37). Behind this are the basihyals (36), and still farther back, on the side, is the ceratohyal (35). To the basihyals is attached a bone extending downward and free behind the urohyal (38). Behind the ceratohyal and continuous with it is the epihyal (34), to which behind is attached the narrow interhyal (33). On the under surface of the ceratohyal and the epihyal are attached the branchiostegals (39). These are slender rays supporting a membrane beneath the gills, seven in number on each side in the striped bass, but much more numerous in some groups of fishes. The gill membranes connecting the branchiostegals are in the striped bass entirely separate from each other. In other fishes they may be broadly joined across the fleshy interspace between the gill-openings, known as the isthmus, or again they may be grown fast to the isthmus itself, so that the gill-openings of the two sides are widely separated.
The Gill-arches.—The gills are attached to four bony arches with a fifth of the same nature, but totally modified by the presence of teeth, and very rarely having on it any of the gill-fringes. The fifth arch thus modified to serve in mastication instead of respiration is known collectively as the lower pharyngeals (46). Opposite these are the upper pharyngeals (45).
The gill-arches are suspended to the cranium from above by the suspensory pharyngeal (44). Each arch contains three parts—the epibranchial (43), above, the ceratobranchial (42), forming the middle part, and the hypobranchial (41), the lower part[Pg 46] articulating with the series of basibranchials (40) which lie behind the epihyal of the tongue. On the three bones forming the first gill-arch are attached numerous appendages called gill-rakers (47). These gill-rakers vary very greatly in number and form. In the striped bass they are few and spear-shaped. In the shad they are very many and almost as fine as hairs. In some fishes they form an effective strainer in separating the food, or perhaps in keeping extraneous matter from the gills. In some fishes they are short and lumpy, in others wanting altogether.
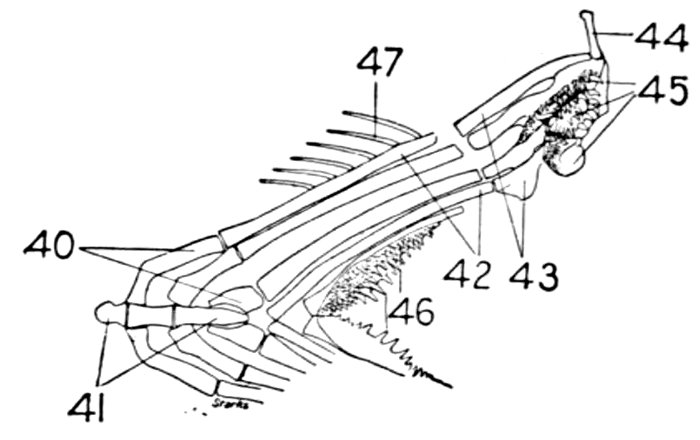
Fig. 28.—Roccus lineatus. Branchial arches. (After Starks.)
The Pharyngeals.—The hindmost gill-arch, as above stated, is modified to form a sort of jaw. The tooth-bearing bones above, 2 to 4 pairs, are known as upper pharyngeals (45), those below, single pair, as lower pharyngeals (46). Of these the lower pharyngeals are most highly specialized and the most useful in classification. These are usually formed much as in the striped bass. Occasionally they are much enlarged, with large teeth for grinding. In many families the lower pharyngeals are grown together in one large bone. In the suckers (Catostomidæ) the lower pharyngeal preserves its resemblance to a gill-arch. In the carp family (Cyprinidæ) retaining this resemblance, it possesses highly specialized teeth.
Vertebral Column.—The vertebral column is composed of a series of vertebræ, 24 in number in the striped bass and in many of the higher fishes, but varying in different groups from 16 to 18 to upwards of 400, the higher numbers being evidence of unspecialized or more usually degenerate structure.
Each vertebra consists of a double concave body or centrum (66). Above it are two small projections often turned backward, zygapophyses (71), and two larger ones, neurapophyses (67), which join above to form the neural spine (68) and thus form the neural canal, through which passes the spinal cord from end to end of the body.
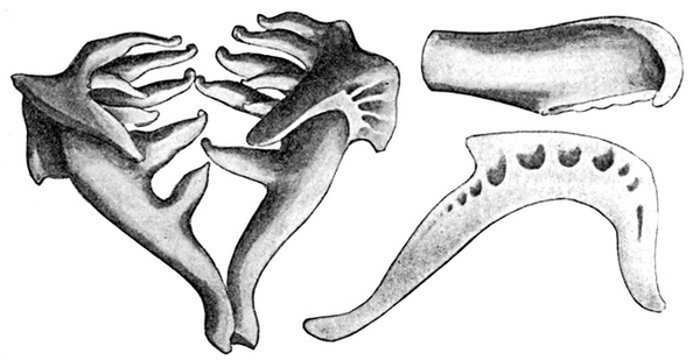
Fig. 29.—Pharyngeal bone and teeth of European Chub, Leuciscus cephalus (Linnæus). (After Seelye.)
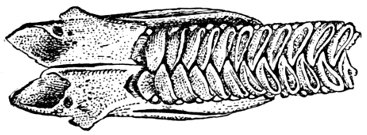
Fig. 30.—Upper pharyngeals of a Parrot-fish, Scarus strongylocephalus.
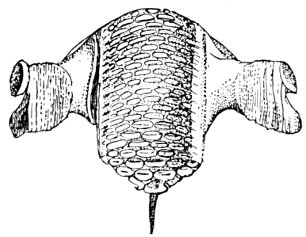
Fig. 31.—Lower pharyngeals of a Parrot-fish, Scarus strongylocephalus (Bleeker).
Below in the vertebræ of the posterior half of the body the hæmapophyses (69) unite to form the hæmal spine (70), and through the hæmal canal thus formed passes a great artery. The vertebræ having hæmal as well as neural spines are known as caudal vertebræ, and occupy the posterior part of the body, usually that behind the attachment of the anal fin (78).
The anterior vertebræ known as abdominal vertebræ, bounding the body-cavity, possess neural spines similar to those of[Pg 48] the caudal vertebræ. In place, however, of the hæmapophyses are projections known as parapophyses (72), which do not meet below, but extend outward, forming the upper part of the wall of the abdominal cavity.

Fig. 32.—Pharyngeals of Italian Parrot-fish, Sparisoma cretense (L.). a, upper; b, lower.
To the parapophyses, or near them, the ribs (73) are rather loosely attached and each rib may have one or more accessory branches (74) called epipleurals.
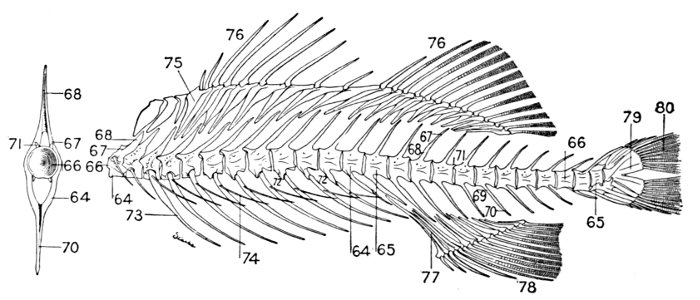
Fig. 33.—Roccus lineatus. Vertebral column and appendages, with a typical vertebra. (After Starks.)
In the striped bass the dorsal vertebræ are essentially similar in form, but in some fishes, as the carp and the catfish, 4 or 5 anterior vertebræ are greatly modified, coossified, and so arranged as to connect the air-bladder with the organ of hearing. Fishes with vertebræ thus altered are called plectospondylous.
In the garpike the vertebræ are convex anteriorly, concave[Pg 49] behind, being joined by ball-and-socket joints (opisthocœlian). In most other fishes they are double concave (amplicœlian). In sharks the vertebræ are imperfectly ossified, a number of terms, asterospondylous, cyclospondylous, tectospondylous, being applied to the different stages of ossification, these terms referring to the different modes of arrangement of the calcareous material within the vertebra.
The Interneurals and Interhæmals.—The vertical fins are connected with the skeletons by bones placed loosely in the flesh and not joined by ligament or suture. Below the dorsal fin (76) lies a series of these bones, dagger-shaped, with the point downward. These are called interneurals (75) and to these the spines and soft rays of the fin are articulated.
In like fashion the spines and rays of the anal fin (18) are jointed at base to bones called interhæmals (77). In certain cases the second interhæmal is much enlarged, made hollow and quill-shaped, and in its concave upper end the tip of the air-bladder is received. This structure is seen in the plume-fishes (Calamus). These two groups of bones, interneural and interhæmal, are sometimes collectively called inter-spinals. The flattened basal bone of the caudal fin (80) is known as hypural (79).
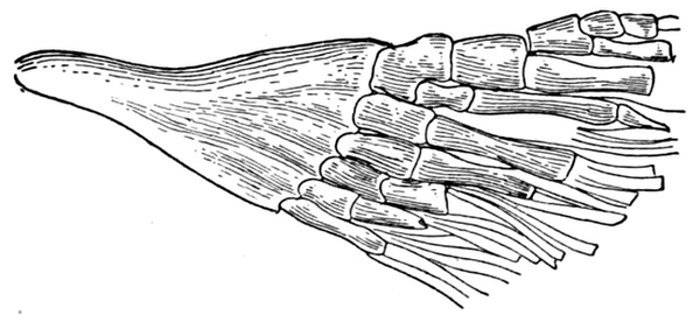
Fig. 34.—Basal bone of dorsal fin, Holoptychius leptopterus (Agassiz). (After Woodward.)
The tail of the striped bass, ending in a broad plate which supports the caudal, is said to be homocercal. In more primitive forms the tail is turned upward more or less, the fin being largely thrown to its lower side. Such a tail as in the sturgeon is said to be heterocercal. In the isocercal tail of the codfish and its relatives the vertebræ are progressively smaller behind and the hypural plate is obsolete or nearly so, the vertebræ remaining in the line of the axis of the body and dividing the caudal fin equally. The simplest form of tail, called diphycercal,[Pg 50] is extended horizontally, tapering backward, the fin equally divided above and below, without hypural plate. In any form of the tail, it may through degeneration be attenuate or whip-like, a form called leptocercal.
The Pectoral Limb.—The four limbs of the fish are represented by the paired fins. The anterior limb is represented by the pectoral fin and its basal elements with the shoulder-girdle, which in the bony fishes reaches a higher degree of complexity than in any other vertebrates. It is in connection with the shoulder-girdle that the greatest confusion in names has occurred. This is due to an attempt to homologize its parts with the shoulder-girdle (scapula, coracoid, and clavicle) of higher vertebrates. But it is not evident that a bony fish possesses a real scapula, coracoid, or even clavicle. The parts of its shoulder-girdle are derived by one line of descent from the undifferentiated elements of the cartilaginous shoulder-girdle of ancestral crossopterygian or dipnoan forms. From a similar ancestry by another line of differentiation has come the amphibian and reptilian shoulder-girdle and its derivative, the girdle of birds and mammals.
The Shoulder-girdle.—In the higher fishes the uppermost bone of the shoulder-girdle is called the post-temporal (suprascapula) (53). In the striped bass and in most fishes this bone is jointed to the temporal region of the cranium. Sometimes, as in the trigger-fishes, it is grown fast to the skull, but it usually rests lightly with the three points of its upper end. In sharks and skates the shoulder-girdle, which is formed of a continuous cartilage, does not touch the skull. In the eels and their allies, it has, by degradation, lost its connection and the post-temporal rests in the flesh behind the cranium.
The post-temporal sometimes projects behind through the skin and may bear spines or serrations. In front of the post-temporal and a little to the outside of it is the small supratemporal (52) also usually connecting the shoulder-girdle with the skull. Below the post-temporal, extending downward and backward, is the flattish supraclavicle (posterotemporal) (54). To this is joined the long clavicle (proscapula) (55), which runs forward and downward in the bony fishes, meeting its fellow on the opposite side in a manner suggesting the wishbone of a[Pg 51] fowl. Behind the base of the clavicle, the sword-shaped post-clavicle (56) extends downward through the muscles behind the base of the pectoral fin. In some fishes, as the stickleback and the trumpet-fish, a pair of flattish or elongate bones called interclavicles (infraclavicles) lie between and behind the lower part of the clavicle. These are not found in most fishes and are wanting in the striped bass. They are probably in all cases merely extensions of the hypocoracoid.
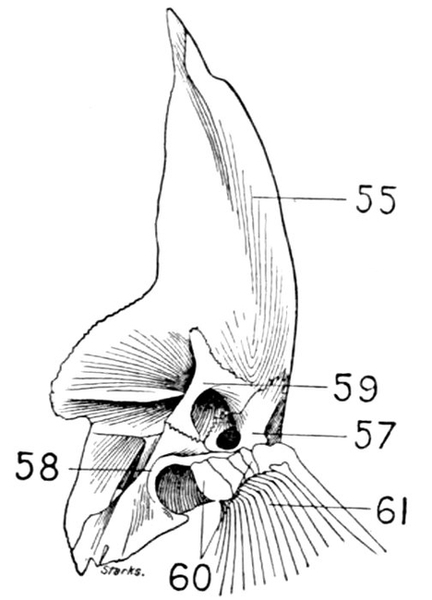
Fig. 35.—Inner view of shoulder-girdle of the Buffalo-fish, Ictiobus bubalus Rafinesque, showing the mesocoracoid (59). (After Starks.)
Two flat bones side by side lie at the base of the pectoral fin, their anterior edges against the upper part of the clavicle. These are the hypercoracoid (57), above, and hypocoracoid (58), below. These have been variously called scapula, coracoid, humerus, radius, and ulna, but being found in the higher fishes only and not in the higher vertebrates, they should receive names not used for other structures. The hypercoracoid is usually pierced by a round foramen or fenestra, but in some fishes (cods, weavers) the fenestra is between the two bones. Attached to the hypercoracoid in the striped bass are four little bones shaped like an hour-glass. These are the actinosts (60) (carpals or pterygials), which support the rays of the pectoral fin (61). In most bony fishes these are placed much as in the striped bass, but in certain specialized or aberrant forms their form and position are greatly altered.
In the anglers (Pediculati) the "carpals" are much elongated, forming a kind of arm, by which the fish can execute a motion not unlike walking.
In the Alaska blackfish (Dallia pectoralis) the two coracoids are represented by a thin, cartilaginous plate, imperfectly divided, and there are no actinosts. In almost all bony fishes, however, these bones are well differentiated and distinct. In most of the soft-rayed fishes an additional V-shaped bone or arch exists on the inner surface of the shoulder-girdle near the insertion of the hypercoracoid. This is known as the meso[Pg 52]coracoid (59). It is not found in the striped bass, but is found in the carp, catfish, salmon, and all their allies.

Fig. 36.—Sargassum-fish, Pterophryne tumida (Osbeck). One of the Anglers. Family Antennariidæ.
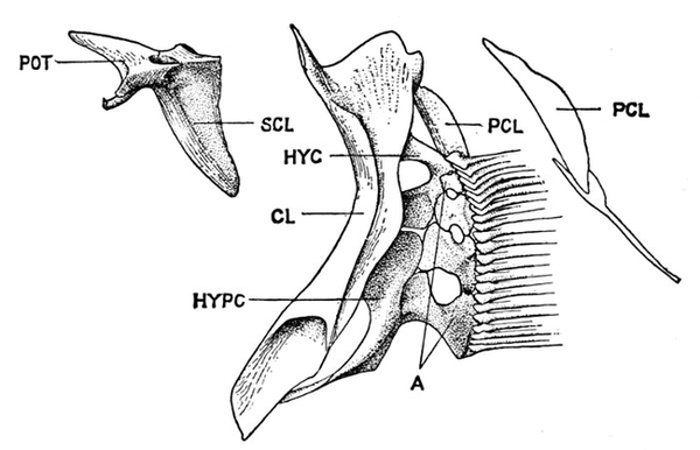
Fig. 37.—Shoulder-girdle of Sebastolobus alascanus Gilbert. (After Starks.)
The Posterior Limbs.—The posterior limb or ventral fin (63) is articulated to a single bone on either side, the pelvic girdle (62).
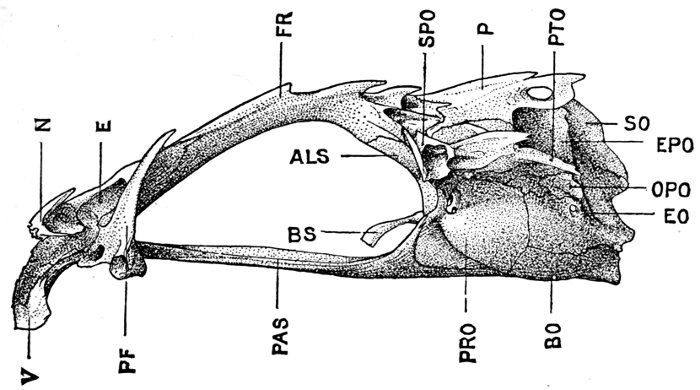
Fig. 38.—Cranium of Sebastolobus alascanus Gilbert. (After Starks.)
In the shark the pelvic girdle is rather largely developed, but in the more specialized fishes it loses its importance. In the less specialized of the bony fishes the pelvis is attached at a distance from the head among the muscles of the side, and free from the shoulder-girdle and other parts of the skeleton. The ventral fins are then said to be abdominal. When very close to the clavicle, but not connected with it, as in the mullet, the fin is still said to be abdominal or subabdominal. In the striped bass the pelvis is joined by ligament between the clavicles, near their tip. The ventral fins thus connected, as seen in most spiny-rayed fishes, are said to be thoracic. In certain forms the pelvis is thrown still farther forward and attached at the throat or even to the chin. When the ventral fins are thus inserted before the shoulder-girdle, they are said to be jugular.[Pg 54] Most of the fishes with spines in the fins have thoracic ventrals. In the fishes with jugular ventrals these fins have begun a process of degeneration by which the spines or soft rays or both are lost or atrophied.
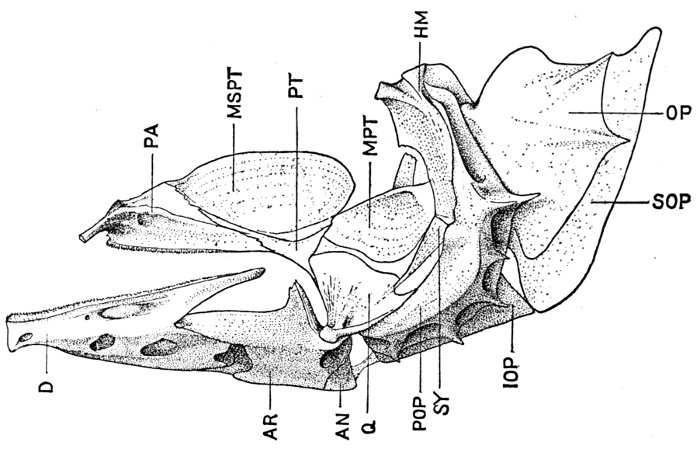
Fig. 39.—Lower jaw and palate of Sebastolobus alascanus. (After Starks.)
Degeneration.—By degeneration or degradation in biology is meant merely a reduction to a lower degree of complexity or specialization in structure. If in the process of development of the individual some particular organ loses its complexity it is said to be degenerate. If in the geological history of a type the same change takes place the same term is used. Degeneration in this sense is, like specialization, a phase of adaptation. It does not imply disease, feebleness, or mutilation, or any tendency toward extinction. It is also necessary to distinguish clearly phases of primitive simplicity from the apparent simplicity resulting from degeneration.
The Skeleton in Primitive Fishes.—To learn the names of bones we can deal most satisfactorily with the higher fishes, those in[Pg 55] which the bony framework has attained completion. But to understand the origin and relation of parts we must begin with the lowest types, tracing the different stages in the development of each part of the system.
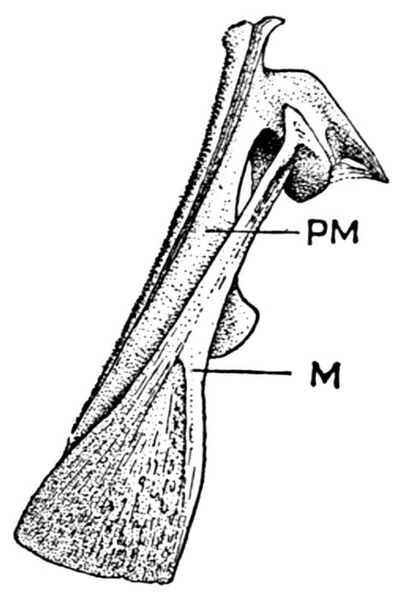
Fig. 40.—Maxillary and premaxillary of Sebastolobus alascanus. M, maxillary; PM, premaxillary.
In the lancelets (Leptocardii), the vertebral column consists simply of a gelatinous notochord extending from one end of the fish to the other, and pointed at both ends, no skull being developed. The notochord never shows traces of segmentation, although cartilaginous rods above it are thought to forecast apophyses. In these forms there is no trace of jaws, limbs, or ribs.
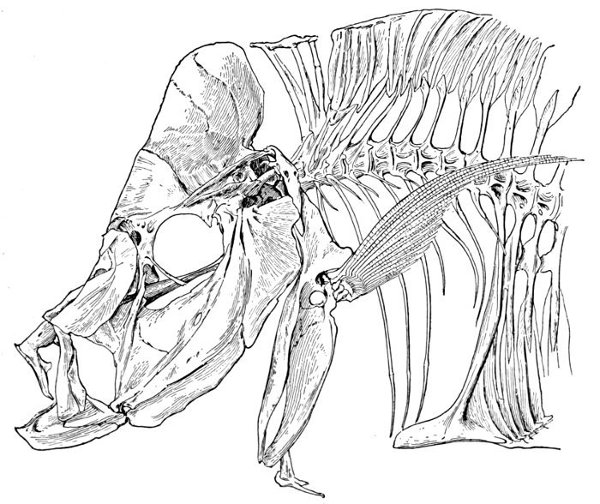
Fig. 41.—Part of skeleton of Selene vomer (Linnæus).
In the embryo of the bony fish a similar notochord precedes the segmentation and ossification of the vertebral column. In most of the extinct types of fishes a notochord more or less[Pg 56] modified persisted through life, the vertebræ being strung upon it spool fashion in various stages of development. In the Cyclostomi (lampreys and hagfishes) the limbs and lower jaw are still wanting, but a distinct skull is developed. The notochord is still present, but its anterior pointed end is wedged into the base of a cranial capsule, partly membranous, partly cartilaginous. There is no trace of segmentation in the notochord itself in these or any other fishes, but neutral arches are foreshadowed in a series of cartilages on each side of the spinal chord. The top of the head is protected by broad plates. There are ring-like cartilages supporting the mouth and other cartilages in connection with the tongue and gill structures.
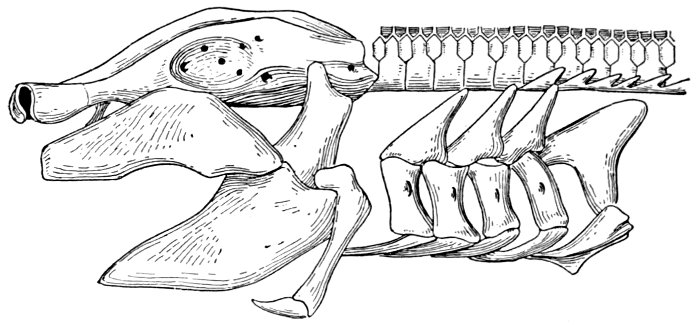
Fig. 42.—Hyostylic skull of Chiloscyllium indicum, a Scyliorhinoid Shark. (After Parker and Haswell.)
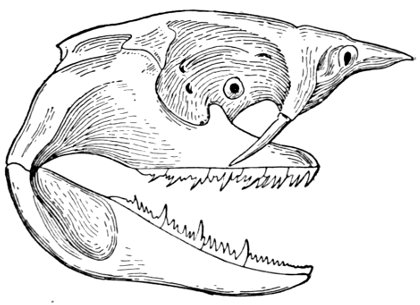
Fig. 43.—Skull of Heptranchias indicus (Gmelin), a notidanoid shark. (After Parker and Haswell.)
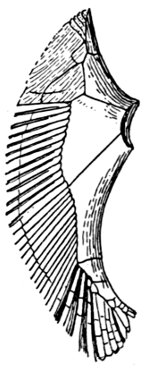
Fig. 44.—Basal bones of pectoral fin of Monkfish, Squatina. (After Zittel.)
The Skeleton of Sharks.—In the Elasmobranchs (sharks, rays, chimæras) the tissues surrounding the notochord are segmented and in most forms distinct vertebræ are developed. Each of these has a conical cavity before and behind, with a central canal through which the notochord is continued. The form and degree of ossification of these vertebræ differ materially in the different groups. The skull in all these fishes is cartilaginous, forming a continuous undivided box containing the brain and lodging the organs of sense. To the skull in the shark is attached a suspensorium of one or two pieces supporting the mandible and the hyoid structures. In the chimæra the mandible is articulated directly with the skull, the hyomandibular and quadrate elements being fused with the cranium. The skull in such case is said to be autostylic, that is, with self-attached mandible. In the shark it is said to be hyostylic, the hyomandibular intervening. The upper jaw in the shark consists not of maxillary and premaxillary but of palatine elements, and the two halves of the lower jaw are representatives of Meckel's cartilage, which is the cartilaginous centre of the dentary bone in the bony fishes. These jaw-bones in the higher fishes are in the nature of membrane bones, and in the sharks and their relatives all such bones are undeveloped. The hyoid structures are in the shark relatively simple, as are also the gill-arches, which vary in number. The vertical fins are supported by interneural and interhæmal cartilages, to which the soft fin-rays are attached without articulation.
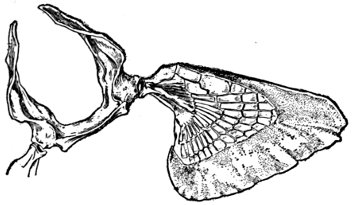
Fig. 45.—Pectoral fin of Heterodontus philippi. (From nature.)
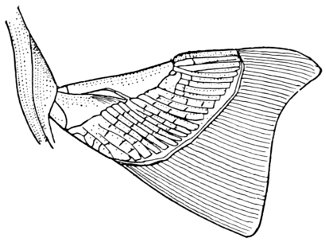
Fig. 46.—Pectoral fin of Heptranchias indicus (Gmelin). (After Dean.)
The shoulder-girdle is made of a single cartilage, touching[Pg 58] the back-bone at a distance behind the head. To this cartilage three smaller ones are attached, forming the base of the pectoral fin. These are called mesopterygium, propterygium, and metapterygium, the first named being in the middle and more distinctly basal. These three segments are subject to much variation. Sometimes one of them is wanting; sometimes two are grown together. Behind these the fin-rays are attached. In most of the skates the shoulder-girdle is more closely connected with the anterior vertebræ, which are more or less fused together.
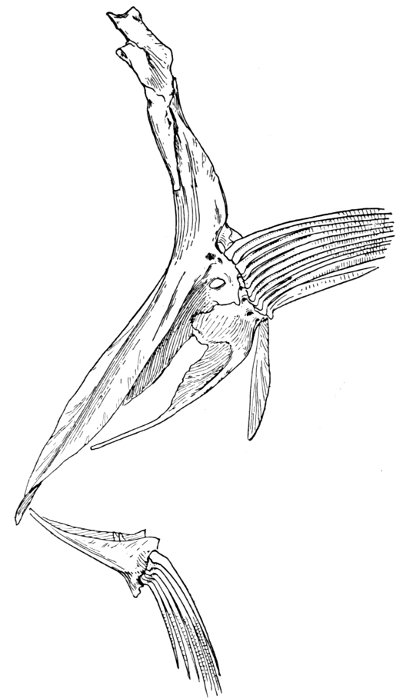
Fig. 47.—Shoulder-girdle of a Flounder, Paralichthys californicus (Ayres).
The pelvis, remote from the head, is formed, in the shark, of a single or paired cartilage with smaller elements at the base of the fin-rays. In the males a cartilaginous generative organ, known as the clasper, is attached to the pelvis and the ventral fins. In the Elasmobranchs the tail vertebræ are progressively smaller backward. If a caudal fin is present, the last vertebræ are directed upward (heterocercal) and the greater part of the fin is below the axis. In other forms (sting-rays) the tail degenerates into a whip-like organ (leptocercal), often without fins. In certain primitive sharks (Ichthyotomi), as well as in the Dipnoi and Crossopterygii, the tail is diphycercal, the vertebræ growing progressively smaller backward and not bent upward toward the tip.
In the chimæras (Holocephali) the notochord persists and is surrounded by a series of calcified rings. The palate with the suspensorium is coalesced with the skull, and the teeth are grown together into bony plates.
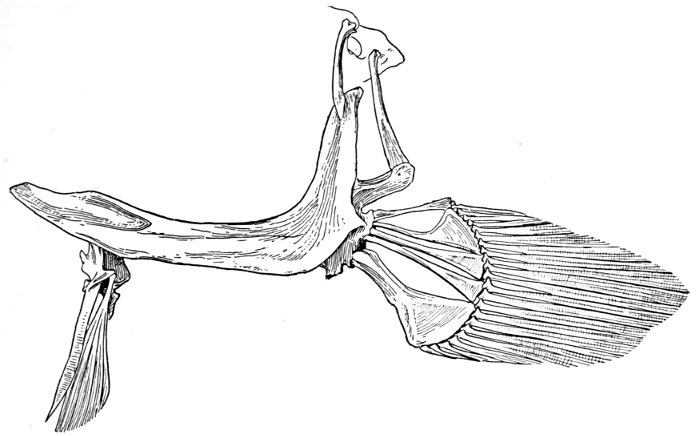
Fig. 48.—Shoulder-girdle of a Toadfish, Batrachoides pacifici (Günther).
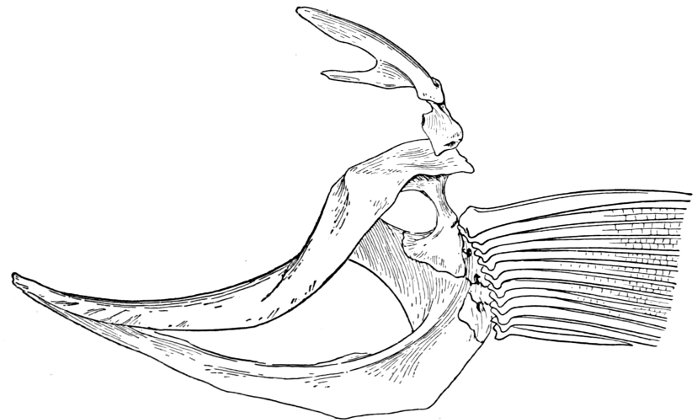
Fig. 49.—Shoulder-girdle of a Garfish, Tylosurus fodiator (Jordan and Gilbert).
The Archipterygium.—The Dipnoans, Crossopterygians, and Ganoids represent various phases of transition from the ancient cartilaginous types to the modern bony fishes.
In the Ichthyotomous sharks, Dipnoans, and Crossopterygians the segments of the pectoral limb are arranged axially, or one beyond another. This type of fin has been called archipterygium by Gegenbaur, on the theory that it represents the condition shown on the first appearance of the pectoral fin. This theory is now seriously questioned, but it will be convenient to retain the name for the pectoral fin with segmented axis fringed on one or both sides by soft rays.
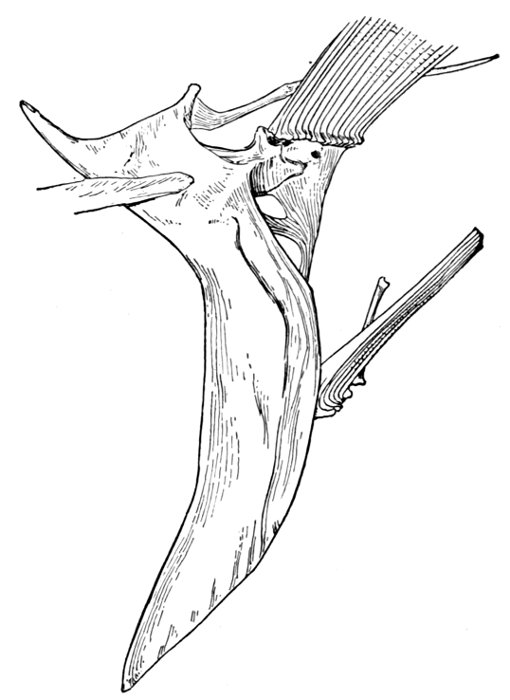
Fig. 50.—Shoulder-girdle of a Hake, Merluccius productus (Ayres).
The archipterygium of the Dipnoan genus Neoceratodus is thus described by Dr. Günther ("Guide to the Study of Fishes," p. 73): "The pectoral limb is covered with small scales along the middle from the root to the extremity, and is surrounded by a rayed fringe similar to the rays of the vertical fins. A muscle split into numerous fascicles extends all the length of the fin, which is flexible in every part and in every direction. The cartilaginous framework supporting it is joined to the scapular arch by a broad basal cartilage, generally single, sometimes[Pg 61] showing traces of a triple division. Along the middle of the fin runs a jointed axis gradually becoming smaller and thinner towards the extremity. Each joint bears on each side a three-, two-, or one-jointed branch."
In the genus Lepidosiren, also a Dipnoan, the pectoral limb has the same axial structure, but is without fin-rays, although in the breeding season the posterior limb or ventral fin in the male is covered with a brush of fine filaments. This structure, according to Prof. J. G. Kerr,[3] is probably without definite function, but belongs to the "category of modifications so often associated with the breeding season (cf. the newts' crest) commonly called ornamental, but which are perhaps more plausibly looked upon as expressions of the intense vital activity of the organisms correlated with its period of reproductive activity." Professor Kerr, however, thinks it not unlikely that this brush of filaments with its rich blood-supply may serve in the function of respiration, a suggestion first made by Professor Lankester.
[3] Philos. Trans., Lond., 1900.

Origin of the Fins of Fishes.—One of the most interesting problems in vertebrate morphology, and one of the most important from its wide-reaching relations, is that of the derivation of the fins of fishes. This resolves itself at once into two problems, the origin of the median fins, which appear in the lancelets, at the very bottom of the fish-like series, and the origin of the paired fins or limbs, which are much more complex, and which first appear with the primitive sharks.
In this study the problem is to ascertain not what theoretically should happen, but what, as a matter of fact, has happened in the early history of the fish-like groups. That these structures, with the others in the fish body, have sprung from simple origins, growing more complex with the demands of varied conditions, and then at times again simple, through degeneration, there can be no doubt. It is also certain that each structure must have had some element of usefulness in all its stages. In such studies we have, as Hæckel has expressed it, "three ancestral documents, paleontology, morphology, and ontogeny"—the actual history as shown by fossil remains, the sidelight derived from comparison of structures, and the evidence of the hereditary influences shown in the development of the individual. As to the first of these ancestral documents, the evidence of paleontology is conclusive where it is complete. But in very few cases are we sure of any series of details. The records of geology are like a book with half its leaves torn out, the other half confused, displaced, and blotted. Still each record actually existing represents genuine history, and in paleontology we must in time find our final court of appeal in all matters of biological origins.
The evidence of comparative anatomy is most completely secured, but it is often indecisive as to relative age and primi[Pg 63]tiveness of origin among structures. As to ontogeny, it is, of course, true that through heredity "the life-history of the individual is an epitome of the life-history of the race." "Ontogeny repeats phylogeny," and phylogeny, or line of descent of organisms and structures, is what we are seeking. But here the repetition is never perfect, never nearly so perfect in fact as Hæckel and his followers expected to find it. The demands of natural selection may lead to the lengthening, shortening, or distortion of phases of growth, just as they may modify adult conditions. The interpolation of non-ancestral stages is recognized in several groups. The conditions of the individual development may, therefore, furnish evidence in favor of certain theories of origins, but they cannot alone furnish the absolute proof.
In the process of development the median or vertical fins are doubtless older than the paired fins or limbs, whatever be the origin of the latter. They arise in a dermal keel which is developed in a web fitting and accentuating the undulatory motion of the body. In the embryo of the fish the continuous vertical fin from the head along the back and around the tail precedes any trace of the paired fins.
In this elementary fin-fold slender supports, the rudiments of fin-rays, tend to appear at intervals. These are called by Ryder ray-hairs or actinotrichia. They are the prototype of fin-rays in the embryo fish, and doubtless similarly preceded the latter in geological time. In the development of fishes the caudal fin becomes more and more the seat of propulsion. The fin-rays are strengthened, their basal supports are more and more specialized, and the fin-fold ultimately divides into distinct fins, the longest rays developed where most needed.
That the vertical fins, dorsal, anal, and caudal, have their origin in a median fold of the skin admits of no question. In the lowest forms which bear fins these structures are dermal folds, being supported by very feeble rays. Doubtless at first the vertical fins formed a continuous fold, extending around the tail, this fold ultimately broken, by atrophy of parts not needed, into distinct dorsal, anal, and caudal fins. In the lower fishes, as in the earlier sharks, there is an approach to this condition of primitive continuity, and in the embryos[Pg 64] of almost all fishes the same condition occurs. Dr. John A. Ryder points out the fact that there are certain unexplained exceptions to this rule. The sea-horse, pipefish, and other highly modified forms do not show this unbroken fold, and it is wanting in the embryo of the top-minnow, Gambusia affinis. Nevertheless the existence of a continuous vertical fold in the embryo is the rule, almost universal. The codfish with three dorsals, the Spanish mackerel with dorsal and anal finlets, the herring with one dorsal, the stickleback with a highly modified one, all show this character, and we may well regard it as a certain trait of the primitive fish. This fold springs from the ectoblast or external series of cells in the embryo. The fin-rays and bony supports of the fins spring from the mesoblast or middle series of cells, being thrust upward from the skeleton as supports for the fin-fold.
Origin of the Paired Fins.—The question of the origin of the paired fins is much more difficult and is still far from settled, although many, perhaps the majority of recent writers favor the theory that these fins are parts of a once continuous lateral fold of skin, corresponding to the vertical fold which forms the dorsal, anal, and caudal. In this view the lateral fold, at first continuous, became soon atrophied in the middle, while at either end it is highly specialized, at first into an organ of direction, then into fan-shaped and later paddle-shaped organs of locomotion. According to another view, the paired fins originated from gill structures, originally both close behind the head, the ventral fin migrating backward with the progress of evolution of the species.
Evidence of Paleontology.—If we had representations of all the early forms of fishes arranged in proper sequence, we could decide once for all, by evidence of paleontology, which form of fin appears first and what is the order of appearance. As to this, it is plain that we do not know the most primitive form of fin. Sharks of unknown character must have existed long before the earliest remains accessible to us. Hence the evidence of paleontology seems conflicting and uncertain. On the whole it lends most support to the fin-fold theory. In the later Devonian, a shark, Cladoselache fyleri, is found in which the paired fins are lappet-shaped, so formed and placed as to suggest[Pg 65] their origin from a continuous fold of skin. In this species the dorsal fins show much the same form. Other early sharks, constituting the order of Acanthodei, have fins somewhat similar, but each preceded by a stiff spine, which may be formed from coalescent rays.

Fig. 51.—Cladoselache fyleri (Newberry), restored. Upper Devonian of Ohio. (After Dean.)
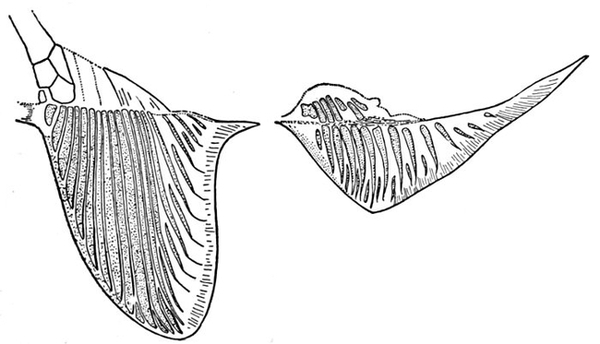
Fig. 52.—Fold-like pectoral and ventral fins of Cladoselache fyleri. (After Dean.)
Long after these appears another type of sharks represented by Pleuracanthus and Cladodus, in which the pectoral fin is a jointed organ fringed with rays arranged serially in one or two rows. This form of fin has no resemblance to a fold of skin, but accords better with Gegenbaur's theory that the pectoral limb was at first a modified gill-arch. In the Coal Measures are found also teeth of sharks (Orodontidæ) which bear a strong resemblance to still existing forms of the family of Heterodontidæ, which originates in the Permian. The existing Heterodontidæ have the usual specialized form of shark-fin, with three of the basal segments especially enlarged and placed side[Pg 66] by side, the type seen in modern sharks. Whatever the primitive form of shark-fin, it may well be doubted whether any one of these three (Cladoselache, Pleuracanthus, or Heterodontus) actually represents it. The beginning is therefore unknown, though there is some evidence that Cladoselache is actually more nearly primitive than any of the others. As we shall see, the evidence of comparative anatomy may be consistent with either of the two chief theories, while that of ontogeny or embryology is apparently inconclusive, and that of paleontology is apparently most easily reconciled with the theory of the fin-fold.
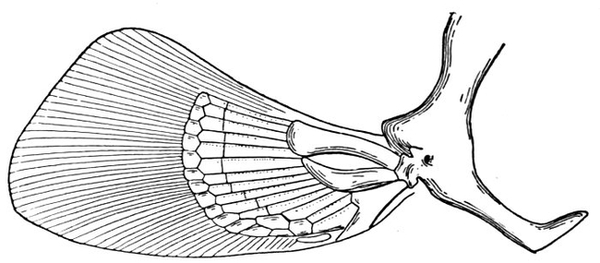
Fig. 53.—Pectoral fin of shark, Chiloscyllium. (After Parker and Haswell.)
Development of the Paired Fins in the Embryo.—According to Dr. John A. Ryder ("Embryography of Osseous Fishes," 1882) "the paired fins in Teleostei arise locally, as short longitudinal folds, with perhaps a few exceptions. The pectorals of Lepisosteus originate in the same way. Of the paired fins, the pectoral or anterior pair seems to be the first to be developed, the ventral or pelvic pair often not making its appearance until after the absorption of the yolk-sac has been completed, in other cases before that event, as in Salmo and in Gambusia. The pectoral fin undergoes less alteration of position during its evolution than the posterior pair."
In the codfish (Gadus callarias) the pectoral fin-fold "appears as a slight longitudinal elevation of the skin on either side of the body of the embryo a little way behind the auditory vesicles, and shortly after the tail of the embryo begins to bud out. At the very first it appears to be merely a dermal fold, and in some forms a layer of cells extends out underneath it from the sides of the body, but does not ascend into it. It[Pg 67] begins to develop as a very low fold, hardly noticeable, and, as growth proceeds, its base does not expand antero-posteriorly, but tends rather to become narrowed, so that it has a pedunculated form. With the progress of this process the margin of the fin-fold also becomes thinner at its distal border, and at the basal part mesodermal cells make their appearance more noticeably within the inner contour-line. The free border of the fin-fold grows out laterally and longitudinally, expanding the portion outside of the inner contour-line of the fin into a fan-shape. This distal thinner portion is at first without any evidence of rays; further than that there is a manifest tendency to a radial disposition of the histological elements of the fin."
The next point of interest is found in the change of position of the pectoral fin by a rotation on its base. This is associated with changes in the development of the fish itself. The ventral fin is also, in most fishes, a short horizontal fold and just above the preanal part of the median vertical fold which becomes anal, caudal, and dorsal. But in the top-minnow (Gambusia), of the order Haplomi, the ventral first appears as "a little papilla and not as a fold, where the body-walls join the hinder upper portion of the yolk-sac, a very little way in front of the vent." "These two modes of origin," observes Dr. Ryder, "are therefore in striking contrast and well calculated to impress us with the protean character of the means at the disposal of Nature to achieve one and the same end."
Current Theories as to Origin of Paired Fins.—There are three chief theories as to the morphology and origin of the paired fins. The earliest is that of Dr. Karl Gegenbaur, supported by various workers among his students and colleagues. In his view the pectoral and ventral fins are derived from modifications of primitive gill-arches. According to this theory, the skeletal arrangements of the vertebrate limb are derived from modifications of one primitive form, a structure made up of successive joints, with a series of fin-rays on one or both sides of it. To this structure Gegenbaur gives the name of archipterygium. It is found in the shark, Pleuracanthus, in Cladodus, and in all the Dipnoan and Crossopterygian fishes, its primitive form being still retained in the Australian genus of Dipnoans, Neoceratodus. This biserial archipterygium with its limb-girdle is[Pg 68] derived from a series of gill-rays attached to a branchial arch. The backward position of the ventral fin is due to a succession of migrations in the individual and in the species.
As to this theory, Mr. J. Graham Kerr observes:
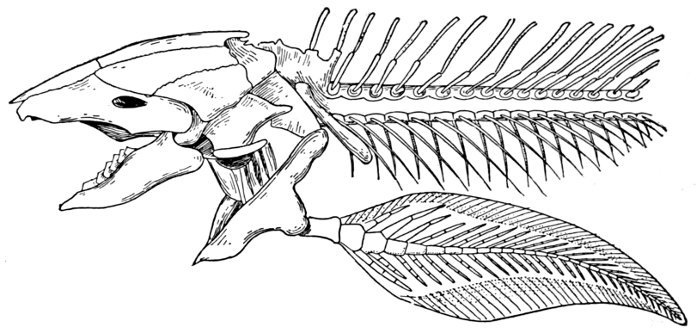
Fig. 54.—Skull and shoulder-girdle of Neoceratodus forsteri (Günther), showing the archipterygium.
"The Gegenbaur theory of the morphology of vertebrate limbs thus consists of two very distinct portions. The first, that the archipterygium is the ground-form from which all other forms of presently existing fin skeletons are derived, concerns us only indirectly, as we are dealing here only with the origin of the limbs, i.e., their origin from other structures that were not limbs.
"It is the second part of the view that we have to do with, that deriving the archipterygium, the skeleton of the primitive paired fin, from a series of gill-rays and involving the idea that the limb itself is derived from the septum between two gill-clefts.
"This view is based on the skeletal structures within the fin. It rests upon (1) the assumption that the archipterygium is the primitive type of fin, and (2) the fact that amongst the Selachians is found a tendency for one branchial ray to become larger than the others, and, when this has happened, for the base of attachment of neighboring rays to show a tendency to migrate from the branchial arch on to the base of the larger or, as we may call it, primary ray; a condition coming about which, were the process to continue rather farther than it is known to do in actual fact, would obviously result in a struc[Pg 69]ture practically identical with the archipterygium. Gegenbaur suggests that the archipterygium actually has arisen in this way in phylogeny."

Fig. 55.—Acanthoessus wardi (Egerton). Carboniferous. Family Acanthoessidæ. (After Woodward.)
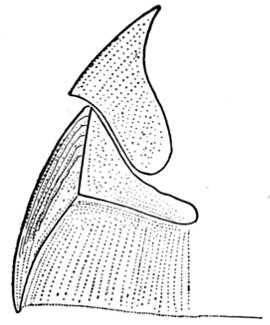
Fig. 56.—Shoulder-girdle of Acanthoessus. (After Dean.)
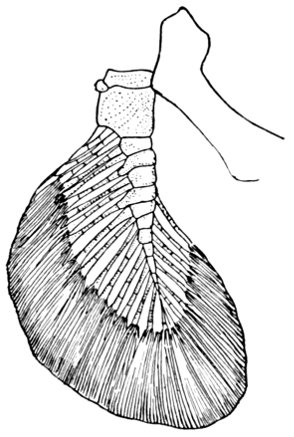
Fig. 57.—Pectoral fin of Pleuracanthus. (After Dean.)
The fin-fold theory of Balfour, adopted by Dohrn, Weidersheim, Thacher, Mivart, Ryder, Dean, Boulenger, and others, and now generally accepted by most morphologists as plausible, is this: that "The paired limbs are persisting and exaggerated portions of a fin-fold once continuous, which stretched along each side of the body and to which they bear an exactly similar phylogenetic relation as do the separate dorsal and anal fins to the once continuous median fin-fold."
"This view, in its modern form, was based by Balfour on his observation that in the embryos of certain Elasmobranchs[Pg 70] the rudiments of the pectoral and pelvic fins are at a very early period connected together by a longitudinal ridge of thickened epiblast—of which indeed they are but exaggerations. In Balfour's own words referring to these observations: 'If the account just given of the development of the limb is an accurate record of what really takes place, it is not possible to deny that some light is thrown by it upon the first origin of the vertebrate limbs. The facts can only bear one interpretation, viz., that the limbs are the remnants of continuous lateral fins.'
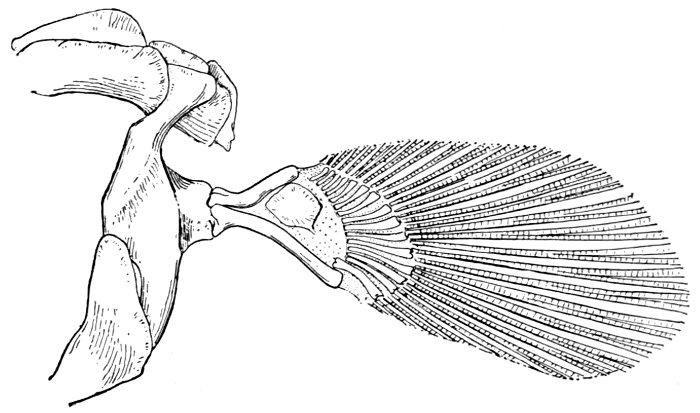
Fig. 58.—Shoulder-girdle of Polypterus bichir. Specimen from the White Nile.
"A similar view to that of Balfour was enunciated almost synchronously by Thacher and a little later by Mivart—in each case based on anatomical investigation of Selachians—mainly relating to the remarkable similarity of the skeletal arrangements in the paired and unpaired fins."
A third theory is suggested by Mr. J. Graham Kerr (Cambridge Philos. Trans., 1899), who has recently given a summary of the theories on this subject. Mr. Kerr agrees with Gegenbaur as to the primitive nature of the archipterygium, but believes that it is derived, not from the gill-septum, but from an external gill. Such a gill is well developed in the young of all the living sharks, Dipnoans and Crossopterygians, and in the latter types of fishes it has a form analogous to that of the archipterygium, although without bony or cartilaginous axis.
We may now take up the evidence in regard to each of the different theories, using in part the language of Kerr, the para[Pg 71]graphs in quotation-marks being taken from his paper. We may first consider Balfour's theory of the lateral fold.
Balfour's Theory of the Lateral Fold.—"The evidence in regard to this view may be classed under three heads, as ontogenetic, comparative anatomical, and paleontological. The ultimate fact on which it was founded was Balfour's discovery that in certain Elasmobranch embryos, but especially in Torpedo (Narcobatus), the fin rudiments were, at an early stage, connected by a ridge of epiblast. I am not able to make out what were the other forms in which Balfour found this ridge, but subsequent research, in particular by Mollier, a supporter of the lateral-fold view, is to the effect that it does not occur in such ordinary sharks as Pristiurus and Mustelus, while it is to be gathered from Balfour himself that it does not occur in Scyllium (Scyliorhinus).
"It appears to me that the knowledge we have now that the longitudinal ridge is confined to the rays and absent in the less highly specialized sharks greatly diminishes its security as a basis on which to rest a theory. In the rays, in correlation with their peculiar mode of life, the paired fins have undergone (in secondary development) enormous extension along the sides of the body, and their continuity in the embryo may well be a mere foreshadowing of this.
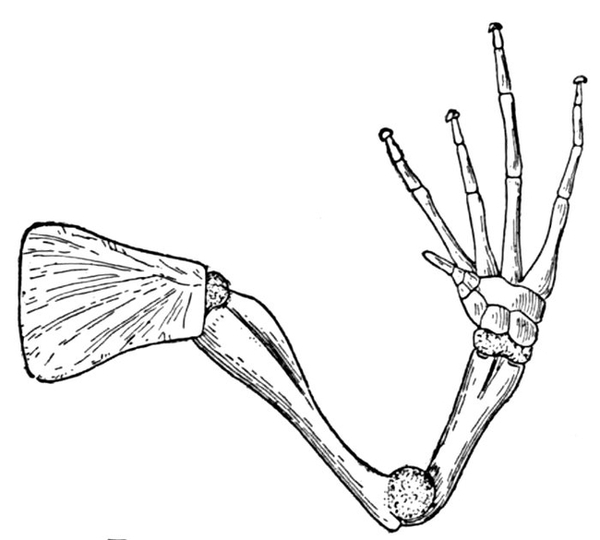
Fig. 59.—Arm of a frog.
"An apparently powerful support from the side of embryology came in Dohrn and Rabl's discoveries that in Pristiurus all the interpterygial myotomes produce muscle-buds. This, however, was explained away by the Gegenbaur school as being merely evidence of the backward migration of the hind limb—successive myotomes being taken up and left behind again as the limb moved farther back. As either explanation seems an adequate one, I do not think we can lay stress upon this body of facts as supporting either one view or the other. The[Pg 72] facts of the development of the skeleton cannot be said to support the fold view; according to it we should expect to find a series of metameric supporting rays produced which later on become fused at their bases. Instead of this we find a longitudinal bar of cartilage developing quite continuously, the rays forming as projections from its outer side.
"The most important evidence for the fold view from the side of comparative anatomy is afforded by (1) the fact that the limb derives its nerve supply from a large number of spinal nerves, and (2) the extraordinary resemblance met with between the skeletal arrangements of paired and unpaired fins. The believers in the branchial arch hypothesis have disposed of the first of these in the same way as they did the occurrence of interpterygial myotomes, by looking on the nerves received from regions of the spinal cord anterior to the attachment of the limb as forming a kind of trail marking the backward migration of the limb.
"The similarity in the skeleton is indeed most striking, though its weight as evidence has been recently greatly diminished by the knowledge that the apparently metameric segmentation of the skeletal and muscular tissues of the paired fins is quite secondary and does not at all agree with the metamery of the trunk. What resemblance there is may well be of a homoplastic character when we take into account the similarity in function of the median and unpaired fins, especially in such forms as Raja, where the anatomical resemblances are especially striking. There is a surprising dearth of paleontological evidence in favor of this view."
The objection to the first view is its precarious foundation. Such lateral folds are found only in certain rays, in which they may be developed as a secondary modification in connection with the peculiar form of these fishes. Professor Kerr observes that this theory must be looked upon and judged: "Just as any other view at the present time regarding the nature of the vertebrate limb, rather as a speculation, brilliant and suggestive though it be, than as a logically constructed theory of the now known facts. It is, I think, on this account allowable to apply to it a test of a character which is admittedly very apt to mislead, that of 'common sense.'
"If there is any soundness in zoological speculation at all, I think it must be admitted that the more primitive vertebrates were creatures possessing a notochordal axial skeleton near the dorsal side, with the main nervous axis above it, the main viscera below it, and the great mass of muscle lying in myotomes along its sides. Now such a creature is well adapted to movements of the character of lateral flexure, and not at all for movements in the sagittal plane—which would be not only difficult to achieve, but would tend to alternately compress and extend its spinal cord and its viscera. Such a creature would swim through the water as does a Cyclostome, or a Lepidosiren, or any other elongated vertebrate without special swimming organs. Swimming like this, specialization for more and more rapid movement would mean flattening of the tail region and is extension into an at first not separately mobile median tail-fold. It is extremely difficult to my mind to suppose that a new purely swimming arrangement should have arisen involving up-and-down movement, and which, at its first beginnings, while useless as a swimming organ itself, must greatly detract from the efficiency of that which already existed."
Objections to Gegenbaur's Theory.—We now return to the Gegenbaur view—that the limb is a modified gill-septum.
"Resting on Gegenbaur's discovery already mentioned, that the gill-rays in certain cases assume an arrangement showing great similarity to that of the skeletal elements of the archipterygium, it has, so far as I am aware, up to the present time received no direct support whatever of a nature comparable with that found for the rival view in the fact that, in certain forms at all events, the limbs actually do arise in the individual in the way that the theory holds they did in phylogeny. No one has produced either a form in which a gill-septum becomes the limb during ontogeny, or the fossil remains of any form which shows an intermediate condition.
"The portion of Gegenbaur's view which asserts that the biserial archipterygial fin is of an extremely primitive character is supported by a large body of anatomical facts, and is rendered further probable by the great frequency with which fins apparently of this character occur amongst the oldest known fishes. On the lateral-fold view we should have to[Pg 74] regard these as independently evolved, which would imply that fins of this type are of a very perfect character, and in that case we may be indeed surprised at their so complete disappearance in the more highly developed forms, which followed later on."
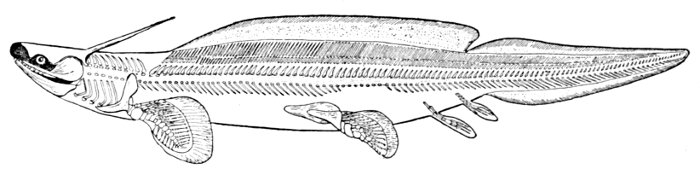
Fig. 60.—Pleuracanthus decheni (Goldfuss). (After Dean.)
As to Gegenbaur's theory it is urged that no form is known in which a gill-septum develops into a limb during the growth of the individual. The main thesis, according to Professor Kerr, "that the archipterygium was derived from gill-rays, is supported only by evidence of an indirect character. Gegenbaur in his very first suggestion of his theory pointed out, as a great difficulty in the way of its acceptance, the position of the limbs, especially of the pelvic limbs, in a position far removed from that of the branchial arches. This difficulty has been entirely removed by the brilliant work of Gegenbaur's followers, who have shown from the facts of comparative anatomy and embryology that the limbs, and the hind limbs especially, actually have undergone, and in ontogeny do undergo, an extensive backward migration. In some cases Braus has been able to find traces of this migration as far forward as a point just behind the branchial arches. Now, when we consider the numbers, the enthusiasm, and the ability of Gegenbaur's disciples, we cannot help being struck by the fact that the only evidence in favor of this derivation of the limbs has been that which tends to show that a migration of the limbs backwards has taken place from a region somewhere near the last branchial arch, and that they have failed utterly to discover any intermediate steps between gill-rays and archipterygial fin. And if for a moment we apply the test of common sense we cannot but be impressed by the improbability of the evolution of a gill-septum, which in all the lower forms of fishes is fixed[Pg 75] firmly in the body-wall, and beneath its surface, into an organ of locomotion.

Fig. 61.—Embryos of Heterodontus japonicus Maclay and Macleay, a Cestraciont shark, showing the backward migration of the gill-arches and the forward movement of the pectoral fin. a, b, c, representing different stages of growth. (After Dean.)
"May I express the hope that what I have said is sufficient to show in what a state of uncertainty our views are regarding the morphological nature of the paired fins, and upon what an exceedingly slender basis rest both of the two views which at present hold the field?"
As to the backward migration of the ventral fins, Dr. Bashford Dean has recently brought forward evidence from the embryo of a very ancient type of shark (Heterodontus japonicus) that this does not actually occur in that species. On the other[Pg 76] hand, we have a forward migration of the pectoral fin, which gradually takes its place in advance of the hindmost gill-arches. The accompanying cut is from Dean's paper, "Biometric Evidence in the Problem of the Paired Limbs of the Vertebrates" (American Naturalist for November, 1902). Dean concludes that in Heterodontus "there is no evidence that there has ever been a migration of the fins in the Gegenbaurian sense." "The gill region, at least in its outer part, shows no affinity during proportional growth with the neighboring region of the pectoral fin. In fact from an early stage onward, they are evidently growing in opposite directions."
Kerr's Theory of Modified External Gills.—"It is because I feel that in the present state of our knowledge neither of the two views I have mentioned has a claim to any higher rank than that of extremely suggestive speculations that I venture to say a few words for the third view, which is avowedly a mere speculation.
"Before proceeding with it I should say that I assume the serial homology of fore and hind limbs to be beyond dispute. The great and deep-seated resemblances between them are such as to my mind seem not to be adequately explicable except on this assumption.
"In the Urodela (salamanders) the external gills are well-known structures—serially arranged projections from the body-wall near the upper ends of certain of the branchial arches. When one considers the ontogenetic development of these organs, from knob-like outgrowth from the outer face of the branchial arch, covered with ectoderm and possessing a mesoblastic core, and which frequently if not always appear before the branchial clefts are open, one cannot but conclude that they are morphologically projections of the outer skin and that they have nothing whatever to do with the gill-pouches of the gut-wall. Amongst the Urodela one such gill projects from each of the first three branchial arches. In Lepidosiren there is one on each of the branchial arches I-IV. In Polypterus and Calamoichthys (Erpetoichthys) there is one on the hyoid arch. Finally, in many Urodelan larvæ we have present at the same time as the external gills a pair of curious structures called balancers. At an early stage of my work on Lepidosiren,[Pg 77] while looking over other vertebrate embryos and larvæ for purposes of comparison, my attention was arrested by these structures, and further examinations, by section or otherwise, convinced me that there were serial homologues of the external gills, situated on the mandibular arch. On then looking up the literature, I found that I was by no means first in this view. Rusconi had long ago noticed the resemblance, and in more recent times both Orr and Maurer had been led to the same conclusion as I had been. Three different observers having been independently led to exactly the same conclusions, we may, I think, fairly enough regard the view I have mentioned of the morphological nature of the balancers as probably a correct one.
"Here, then, we have a series of homologous structures projecting from each of the series of visceral arches. They crop up on the Crossopterygii, the Dipnoi, and the Urodela, i.e., in three of the most archaic of the groups of Gnathostomata. But we may put it in another way. The groups in which they do not occur are those whose young possess a very large yolk-sac (or which are admittedly derived from such forms). Now wherever we have a large yolk-sac we have developed on its surface a rich network of blood-vessels for purposes of nutrition. But such a network must necessarily act as an extraordinarily efficient organ of respiration, and did we not know the facts we might venture to prophesy that in forms possessing it any other small skin-organ of respiration would tend to disappear.
"No doubt these external gills are absent also in a few of the admittedly primitive forms such as, e.g., (Neo-) Ceratodus. But I would ask that in this connection one should bear in mind one of the marked characteristics of external gills—their great regenerative power. This involves their being extremely liable to injury and consequently a source of danger to their possessor. Their absence, therefore, in certain cases may well have been due to natural selection. On the other hand, the presence in so many lowly forms of these organs, the general close similarity in structure that runs through them in different forms, and the exact correspondence in their position and relations to the body can, it seems to me, only be adequately explained by looking on them as being homologous structures[Pg 78] inherited from a common ancestor and consequently of great antiquity in the vertebrate stem."
As to the third theory, Professor Kerr suggests tentatively that the external gill may be the structure modified to form the paired limbs. Of the homology of fore and hind limbs and consequently of their like origin there can be no doubt.
The general gill-structures have, according to Kerr, "the primary function of respiration. They are also, however, provided with an elaborate muscular apparatus comprising elevators, depressors, and adductors, and larvæ possessing them may be seen every now and then to give them a sharp backward twitch. They are thus potentially motor organs. In such a Urodele as Amblystoma their homologues on the mandibular arch are used as supporting structures against a solid substratum exactly as are the limbs of the young Lepidosiren.

Fig. 62.—Polypterus congicus, a Crossopterygian fish from the Congo River. Young, with external gills. (After Boulenger.)
"I have, therefore, to suggest that the more ancient Gnathostomata possessed a series of potentially motor, potentially supporting structures projecting from their visceral arches; it was inherently extremely probable that these should be made use of when actual supporting, and motor appendages had to be developed in connection with clambering about a solid substratum. If this had been so, we should look upon the limb as a modified external gill; the limb-girdle, with Gegenbaur, as a modified branchial arch.
"This theory of the vertebrate paired limb seems to me, I confess, to be a more plausible one on the face of it than either of the two which at present hold the field. If untrue, it is so dangerously plausible as to surely deserve more consideration than it appears to have had. One of the main differences between it and the other two hypotheses is that, instead of deriving[Pg 79] the swimming-fin from the walking and supporting limb, it goes the other way about. That this is the safer line to take seems to me to be shown by the consideration that a very small and rudimentary limb could only be of use if provided with a fixed point d'appui. Also on this view, the pentadactyle limb and the swimming-fin would probably be evolved independently from a simple form of limb. This would evade the great difficulties which have beset those who have endeavored to establish the homologies of the elements of the pentadactyle limb with those of any type of fully formed fin."
Uncertain Conclusions.—In conclusion we may say that the evidence of embryology in this matter is inadequate, though possibly favoring on the whole the fin-fold theory; that of morphology is inconclusive, and probably the final answer may be given by paleontology. If the records of the rocks were complete, they would be decisive. At present we have to decide which is the more primitive of two forms of pectoral fin actually known among fossils. That of Cladoselache is a low, horizontal fold of skin, with feeble rays, called by Cope ptychopterygium. That of Pleuracanthus is a jointed paddle-shaped appendage with a fringe of rays on either side. In the theory of Gegenbaur and Kerr Pleuracanthus must be, so far as the limbs are concerned, the form nearest the primitive limb-bearing vertebrate. In Balfour's theory Cladoselache is nearest the primitive type from which the other and with it the archipterygium of later forms may be derived.
Boulenger and others question even this, believing that the archipterygium in Pleuracanthus and other primitive sharks and that in Neoceratodus and its Dipnoan and Crossopterygian allies and ancestors have been derived independently, not the latter from the former. In this view there is no real homology between the archipterygium in the sharks possessing it and that in the Dipnoans and Crossopterygians. In the one theory the type of Pleuracanthus would be ancestral to the other sharks on the one hand, and to Crossopterygians and all higher vertebrates on the other. With the theory of the origin of the pectoral from a lateral fold, Pleuracanthus would be merely a curious specialized offshoot from the primitive sharks, without descendants and without special significance in phylogeny.
As elements bearing on this decision we may note that the tapering unspecialized diphycercal tail of Pleuracanthus seems very primitive in comparison with the short heterocercal tail of Cladoselache. This evidence, perhaps deceptive, is balanced by the presence on the head of Pleuracanthus of a highly specialized serrated spine, evidence of a far from primitive structure. Certainly neither the one genus nor the other actually represents the primitive shark. But as Cladoselache appears in geological time, long before Pleuracanthus, Cladodus, or any other shark with a jointed, archipterygial fin, the burden of proof, according to Dean, rests with the followers of Gegenbaur. If the remains found in the Ordovician at Cañon City referred to Crossopterygians are correctly interpreted, we must regard the shark ancestry as lost in pre-Silurian darkness, for in sharks of some sort the Crossopterygians apparently must find their remote ancestry.
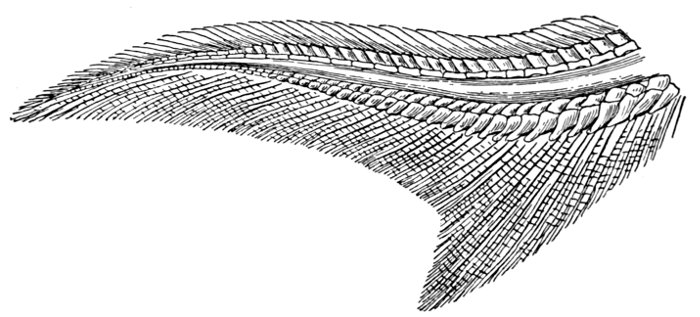
Fig. 63.—Heterocercal tail of Sturgeon, Acipenser sturio (Linnæus). (After Zittel.)
Forms of the Tail in Fishes.—In the process of development the median or vertical fins are, as above stated, older than the paired fins or limbs, whatever be the origin of the latter. They arise in a dermal keel, its membranes fitting and accentuating the undulatory motion of the body.
In this elementary fin-fold slender supports (actinotrichia), the rudiments of fin-rays, appear at intervals. In those fins of most service in the movement of the fish, the fin-rays are strengthened, and their basal supports specialized.
Dean calls attention to the fact that in fishes which swim,[Pg 81] when adult, by an undulatory motion, the paired fins tend to disappear, as in the eel and in all eel-like fishes, as blennies and eel-pouts.
The form of the tail at the base of the caudal fin varies in the different groups. In most primitive types, as in most embryonic fishes, the vertebræ grow smaller to the last (diphycercal). In others, also primitive, the end of the tail is directed upward, and the most of the caudal fin is below it. Such a tail is seen in most sharks, in the sturgeon, garpike, bowfin, and in the Ganoid fishes. It is known as heterocercal, and finally in ordinary fishes the tail becomes homocercal or fan-shaped, although usually some trace of the heterocercal condition is traceable, gradually growing less with the process of development.
Since Professor Agassiz first recognized, in 1833, the distinction between the heterocercal and homocercal tail, this matter has been the subject of elaborate investigation and a number of additional terms have been proposed, some of which are in common use.
A detailed discussion of these is found in a paper by Dr. John A. Ryder "On the Origin of Heterocercy" in the Report of the U. S. Fish Commissioner for 1884. In this paper a dynamic or mechanical theory of the causes of change of form is set forth, parts of this having a hypothetical and somewhat uncertain basis.
Dr. Ryder proposes the name archicercal to denote the cylindroidal worm-like caudal end of the larva of fishes and amphibians before they acquire median fin-folds. The term lophocercal is proposed by Ryder for the form of caudal fin which consists of a rayless fold of skin continuous with the skin of the tail, the inner surfaces of this fold being more or less nearly in contact. To the same type of tail Dr. Jeffries Wyman in 1864 gave the name protocercal. This name was used for the tail of the larval ray when it acquires median fin-folds. The term implies, what cannot be far from true, that this form of tail is the first in the stages of evolution of the caudal fin.
To the same type of tail Mr. Alexander Agassiz gave, in 1877, the name of leptocardial, on the supposition that it represented the adult condition of the lancelet. In this creature,[Pg 82] however, rudimentary basal rays are present, a condition differing from that of the early embryos.
The diphycercal tail, as usually understood, is one in which the end of the vertebral column bears "not only hypural but also epural intermediary pieces which support rays." The term is used for the primitive type of tail in which the vertebræ, lying horizontally, grow progressively smaller, as in Neoceratodus, Protopterus, and other Dipnoans and Crossopterygians. The term was first applied by McCoy to the tails of the Dipnoan genera Diplopterus and Gyroptychius, and for tails of this type it should be reserved.
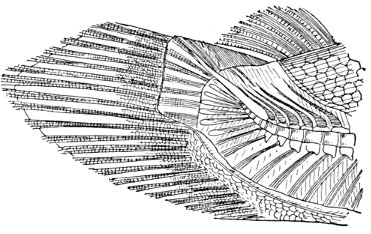
Fig. 64.—Heterocercal tail of Bowfin, Amia calva (Linnæus). (After Zittel.)
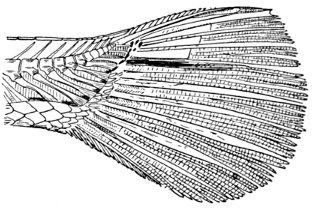
Fig. 65.—Heterocercal tail of Garpike, Lepisosteus osseus (Linnæus).
The heterocercal tail is one in which the hindmost vertebræ are bent upwards. The term is generally applied to those fishes only in which this bending is considerable and is externally evident, as in the sharks and Ganoids. The character disappears by degrees, changing sometimes to diphycercal or leptocercal by a process of degeneration, or in ordinary fishes becoming homocercal. Dr. Ryder uses the term heterocercal for all cases in which any up-bending of the axis takes place, even though it involves the modification of but a single vertebra. With this definition, the tail of salmon, herring, and even of most bony fishes would be considered heterocercal, and most or all of these pass through a heterocercal stage in the course of development. The term is, however, usually restricted to those forms in which the curving of the axis is evident without dissection.
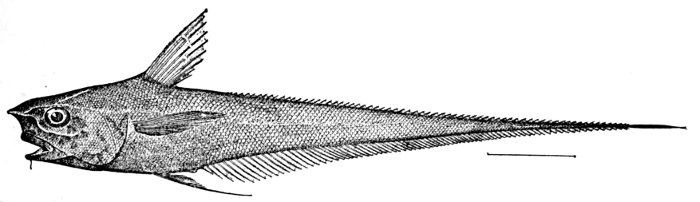
Fig. 66.—Coryphænoides carapinus (Goode and Bean), showing leptocercal tail. Gulf Stream.
The homocercal tail is the fan-shaped or symmetrical tail common among the Teleosts, or bony fishes. In its process of development the individual tail is first archicercal, then lophocercal, then diphycercal, then heterocercal, and lastly homocercal. A similar order is indicated by the sequence of fossil fishes in the rocks, although some forms of diphycercal tail may be produced by degeneration of the heterocercal tail, as suggested by Dr. Dollo and Dr. Boulenger, who divide diphycercal tails into primitive and secondary.
The peculiar tapering tail of the cod, the vertebræ growing progressively smaller behind, is termed isocercal by Professor Cope. This form differs little from diphycercal, except in its supposed derivation from the homocercal type. A similar form is seen in eels.
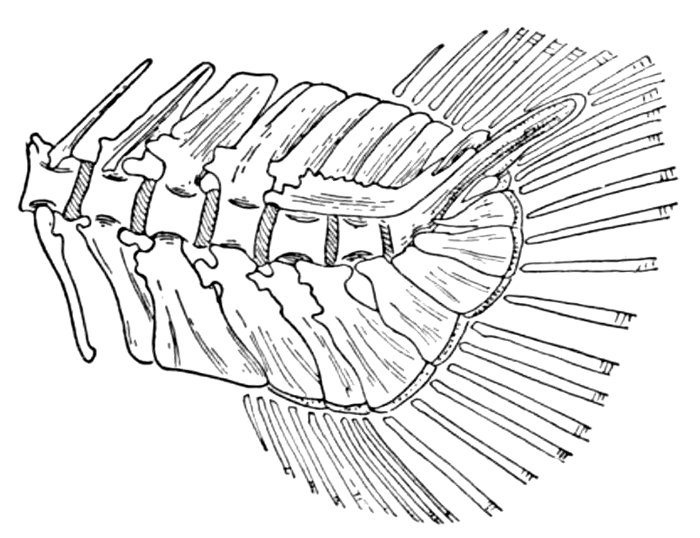
Fig. 67.—Heterocercal tail of Young Trout, Salmo fario (Linnæus). (After Parker and Haswell.)
The term leptocercal has been suggested by Gaudry, 1883, for those tails in which the vertebral column ends in a point. We may, perhaps, use it for all such as are attenuate, ending in a long point or whip, as in the Macrouridæ, or grenadiers, the sting-rays, and in various degenerate members of almost every large group.
The term gephyrocercal is devised by Ryder for fishes in which the end of the vertebral axis is aborted in the adult, leaving the caudal elements to be inserted on the end of this axis, thus bridging over the interval between the vertical fins,[Pg 84] as the name (γεφύρος, bridge; κέρκος, tail) is intended to indicate. Such a tail has been recognized in four genera only, Mola, Ranzania, Fierasfer, and Echiodon, the head-fishes and the pearl-fishes.
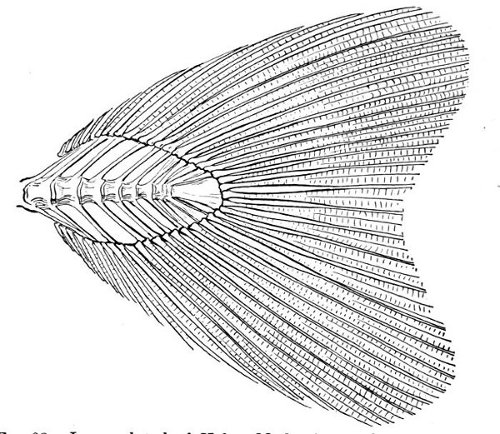
Fig. 68.—Isocercal tail of Hake, Merluccius productus (Ayres).
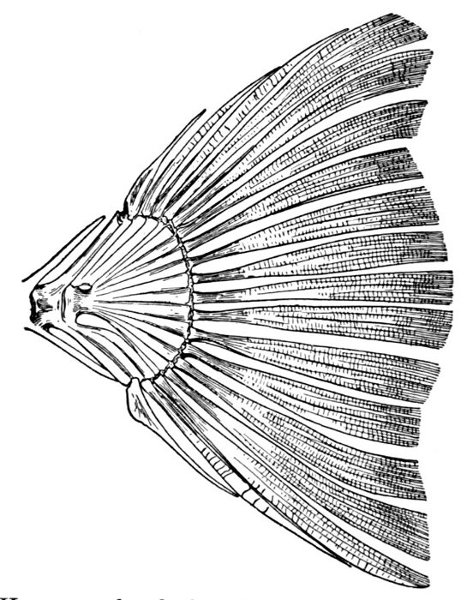
Fig. 69.—Homocercal tail of a Flounder, Paralichthys californicus.
The part of the body of the fish which lies behind the vent is known as the urosome. The urostyle is the name given to a modified bony structure, originally the end of the notochord, turned upward in most fishes. The term opisthure is suggested by Ryder for the exserted tip of the vertebral column, which in some larvæ (Lepisosteus) and in some adult fishes (Fistularia, Chimæra) projects beyond the caudal fin. The urosome, or posterior part of the body, must be regarded as a product of evolution and specialization, its function being largely that of locomotion. In the theoretically primitive fish there is no urosome, the alimentary canal, as in the worm, beginning at one end of the body and terminating at the other.
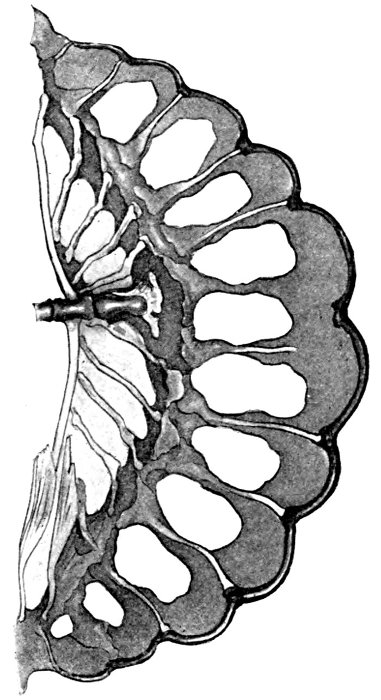
Fig. 70.—Gephyrocercal tail of Mola mola (Linnæus). (After Ryder.)
Homologies of the Pectoral Limb.—Dr. Gill has made an elaborate attempt to work out the homologies of the bones of the pectoral limb.[4] From his thesis we take the following:
"The following are assumed as premises that will be granted by all zootomists:
"1. Homologies of parts are best determinable, ceteris paribus, in the most nearly related forms.
"2. Identification should proceed from a central or determinate point outwards.
"The applications of these principles are embodied in the following conclusions:
"1. The forms that are best comparable and that are most nearly related to each other are the Dipnoi, an order of fishes at present represented by Lepidosiren, Protopterus, and Ceratodus, [Pg 86]and the Batrachians as represented by the Ganocephala, Salamanders, and Salamander-like animals.
"2. The articulation of the anterior member with the shoulder-girdle forms the most obvious and determinable point for comparison in the representatives of the respective classes.
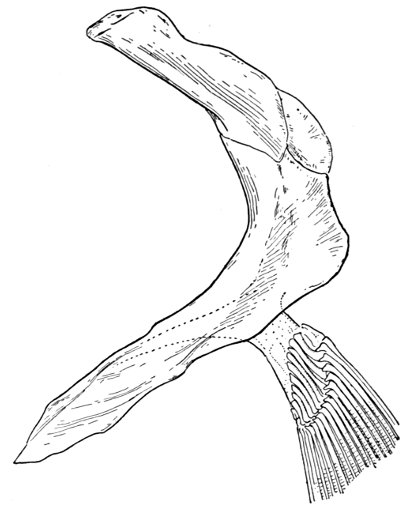
Fig. 71.—Shoulder-girdle of Amia calva (Linnæus).
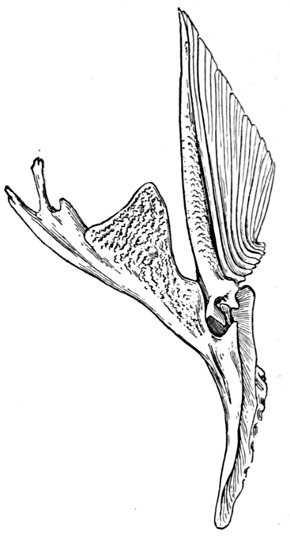
Fig. 72.—Shoulder-girdle of a Sea Catfish, Selenaspis dowi.
The Girdle in Dipnoans.—"The proximal element of the anterior limb in the Dipnoi has almost by common consent been regarded as homologous with the humerus of the higher vertebrates.
"The humerus of Urodele Batrachians, as well as the extinct Ganocephala and Labyrinthodontia, is articulated chiefly with the coracoid. Therefore the element of the shoulder-girdle with which the humerus of the Dipnoi is articulated must also be regarded as the coracoid (subject to the proviso hereinafter stated), unless some specific evidence can be shown to the contrary. No such evidence has been produced.
"The scapula in the Urodele and other Batrachians is entirely or almost wholly excluded from the glenoid foramen, and above[Pg 87] the coracoid. Therefore the corresponding element in Dipnoi must be the scapula.
"The other elements must be determined by their relation to the preceding, or to those parts from or in connection with which they originate. All those elements in immediate connection with the pectoral fin and the scapula must be homologous as a whole with the coraco-scapular plate of the Batrachians; that is, it is infinitely more probable that they represent, as a whole or as dismemberments therefrom, the coraco-scapular element than that they independently originated. But the homogeneity of that coraco-scapular element forbids the identification of the several elements of the fish's shoulder-girdle with regions of the Batrachian's coraco-scapular plate.
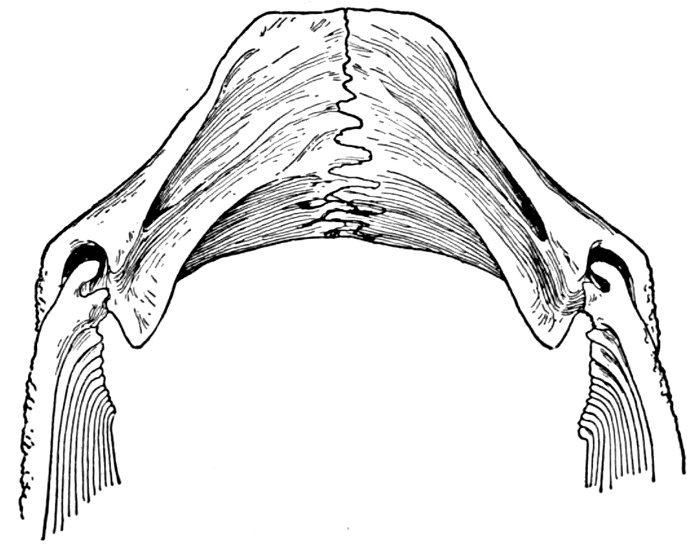
Fig. 73.—Clavicles of a Sea Catfish, Selenaspis dowi (Gill).
"And it is equally impossible to identify the fish's elements with those of the higher reptiles or other vertebrates which have developed from the Batrachians. The elements in the shoulder-girdles of the distantly separated classes may be (to use the terms introduced by Dr. Lankester) homoplastic, but they are not homogenetic. Therefore they must be named accordingly. The element of the Dipnoan's shoulder-girdle, continuous downward from the scapula, and to which the coracoid is closely applied, may be named ectocoracoid.
"Neither the scapula in Batrachians nor the cartilaginous extension thereof, designated suprascapula, is dissevered from the coracoid. Therefore there is an a priori improbability[Pg 88] against the homology with the scapula of any part having a distant and merely ligamentous connection with the humerus-bearing element. Consequently, as an element better representing the scapula exists, the element named scapula (by Owen, Günther, etc.) cannot be the homologue of the scapula of Batrachians. On the other hand, its more intimate relations with the skull and the mode of development indicate that it is rather an element originating and developed in more intimate connection with the skull. It may therefore be considered, with Parker, as a post-temporal.
"The shoulder-girdle in the Dipnoi is connected by an azygous differentiated cartilage, swollen backwards. It is more probable that this is the homologue of the sternum of Batrachians, and that in the latter that element has been still more differentiated and specialized than that it should have originated de novo from an independently developed nucleus."
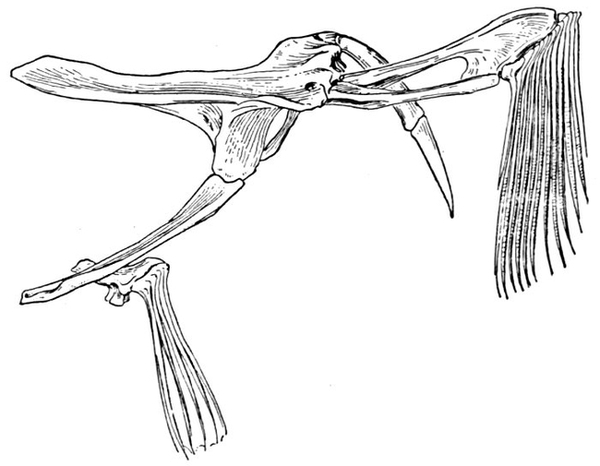
Fig. 74.—Shoulder-girdle of a Batfish, Ogcocephalus radiatus (Mitchill).
The Girdle in Fishes Other than Dipnoans.—"Proceeding from the basis now obtained, a comparative examination of other types of fishes successively removed by their affinities from the Lepidosirenids may be instituted.
"With the humerus of the Dipnoans, the element of the Polypterids (single at the base, but immediately divaricating and with its limbs bordering an intervening cartilage which supports the pectoral and its basilar ossicles) must be homologous. But it is evident that the external elements of the[Pg 89] so-called carpus of the teleosteoid Ganoids are homologous with that element in Polypterids. Therefore those elements cannot be carpal, but must represent the humerus.
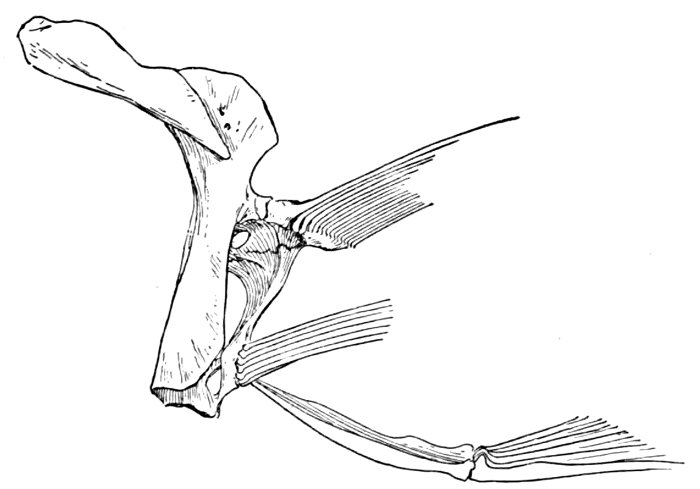
Fig. 75.—Shoulder-girdle of a Threadfin, Polydactylus approximans (Lay and Bennett).
"The element with which the homologue of the humerus, in Polypterids, is articulated must be homologous with the analogous element in Dipnoans, and therefore with the coracoid. The coracoid of Polypterids is also evidently homologous with the corresponding element in the other Ganoids, and the latter consequently must be also coracoid. It is equally evident, after a detailed comparison, that the single coracoid element of the Ganoids represents the three elements developed in the generalized Teleosts (Cyprinids, etc.) in connection with the basis of the pectoral fin, and, such being the case, the nomenclature should correspond. Therefore the upper element may be named hypercoracoid; the lower, hypocoracoid; and the transverse or median, mesocoracoid.
"The two elements of the arch named by Parker, in Lepidosiren, 'supraclavicle' (scapula) and 'clavicle' (ectocoracoid) seem to be comparable together, and as a whole, with the single element carrying the humerus and pectoral fin in the Crossopterygians (Polypterus and Calamoichthys) and other fishes, and therefore not identical respectively with the 'supraclavicle' and 'clavicle' (except in part) recognized by him in other fishes. As this compound bone, composed of the scapula and ectocoracoid fused together, has received no name which is not ambig[Pg 90]uous or deceptive in its homologous allusions, it may be designated as proscapula.
"The post-temporal of the Dipnoans is evidently represented by the analogous element in the Ganoids generally, as well as in the typical fishes. The succeeding elements (outside those already alluded to) appear from their relations to be developed from or in connection with the post-temporal, and not from the true scapular apparatus; they may therefore be named post-temporal, posterotemporal, and teleo-temporal. It will be thus seen that the determinations here adopted depend mainly (1) on the interpretation of the homologies of the elements with which the pectoral limbs are articulated, and (2) on the application of the term 'coracoid.' The name 'coracoid,' originally applied to the process so called in the human scapula and subsequently extended to the independent element homologous with it in birds and other vertebrates, has been more especially retained (e.g., by Parker in mammals, etc.) for the region including the glenoid cavity. On the assumption that this may be preferred by some zootomists, the preceding terms have been applied. But if the name should be restricted to the proximal element, nearest the glenoid cavity, in which ossification commences, the name paraglenal given by Dugès to the cartilaginous glenoid region can be adopted, and the coracoid would then be represented (in part) rather by the element so named by Owen. That eminent anatomist, however, reached his conclusion (only in part the same as that here adopted) by an entirely different course of reasoning, and by a process, as it may be called, of elimination; that is, recognizing first the so-called 'radius' and 'ulna,' the 'humerus,' the 'scapula,' and the 'coracoid' were successively identified from their relations to the elements thus determined and because they were numerically similar to the homonymous parts among higher vertebrates."
[4] Catalogue of the Families of Fishes, 1872.

How Fishes Breathe.—The fish breathes the air which is dissolved in water. It cannot use the oxygen which is a component part of water, nor can it, as a rule, make use of atmospheric air. The amount of oxygen required for the low vegetative processes of the fish is comparatively small. According to Dr. Günther, a man consumes 50,000 times as much oxygen as a tench. But some fishes demand more oxygen than others. Some, like the catfish or the loach, will survive long out of water, while others die almost instantly if removed from their element or if the water is allowed to become foul. In most cases the temperature of the blood of the fish is but little above that of the water in which they live, but in the mackerel and other muscular fishes the temperature of the body may be somewhat higher.
Some fishes which live in mud, especially in places which become dry in summer, have special contrivances by which they can make use of atmospheric air. In a few primitive fishes (Dipnoans, Crossopterygians, Ganoids) the air-bladder retains its original function of a lung. In other cases some peculiar structure exists in connection with the gills. Such a contrivance for holding water above the gills is seen in the climbing perch of India (Anabas scandens) and other members of the group called Labyrinthici.
In respiration, in fishes generally, the water is swallowed through the mouth and allowed to pass out through the gill-openings, thus bathing the gills. In a few of the lower types a breathing-pore takes the place of the gill-openings.
The gills, or branchiæ, are primarily folds of the skin lining the branchial cavity. In most fishes they form fleshy fringes or laminæ throughout which the capillaries are distributed. In the embryos of sharks, skates, chimæras, lung-fishes,[Pg 92] and Crossopterygians external gills are developed, but in the more specialized forms these do not appear outside the gill-cavity. In some of the sharks, and especially the rays, a spiracle or open foramen remains behind the eye. Through this spiracle, leading from the outside into the cavity of the mouth, water is drawn downwards to pass outward over the gills. The presence of this breathing-hole permits these animals to lie on the bottom without danger of inhaling sand.
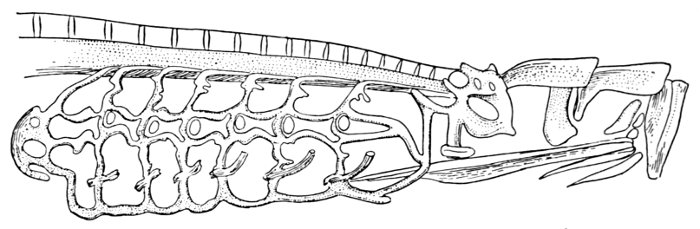
Fig. 76.—Gill-basket of Lamprey. (After Dean.)
The Gill-structures.—The three main types of gills among fishes are the following: (a) the purse-shaped gills found in the hagfishes and lampreys, known as a class as Marsipobranchs, or purse-gills. These have a number (5 to 12) of sac-like depressions on the side of the body, lined with gill-fringes and capillaries, the whole supported by an elaborate branchial basket formed of cartilage. (b) The plate-gills, found among the sharks, rays, and chimæras, thence called Elasmobranchs, or plate-gills. In these the gill-structures are flat laminæ, attached by one side to the gill-arches. (c) The fringe-gills found in ordinary fishes, in which the gill-filaments containing the capillaries are attached in two rows to the outer edge of each gill-arch. The so-called tuft-gills (Lophobranchs) of the sea-horse and pipefish are like these in structure, but the filaments are long, while the arches are very short. In most of the higher fishes a small accessory gill (pseudobranchia) is developed in the skin of the inner side of the opercle.
The Air-bladder.—The air-bladder, or swim-bladder, must be classed among the organs of respiration, although in the higher fishes its functions in this regard are rudimentary, and in some cases it has taken collateral functions (as a hydrostatic[Pg 93] organ of equilibrium, or perhaps as an organ of hearing) which have no relation to its original purpose.
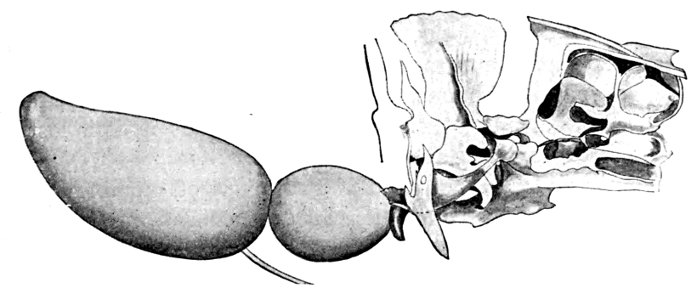
Fig. 77.—Weberian apparatus and air-bladder of Carp. (From Günther, after Weber.)
The air-bladder is an internal sac possessed by many fishes, but not by all. It lies in the dorsal part of the abdominal cavity above the intestines and below the kidneys. In some cases it is closely adherent to the surrounding tissues. In others it is almost entirely free, lying almost loose in the cavity of the body. In some cases it is enclosed in a bony capsule. In the allies of the carp and catfish, which form the majority of fresh-water fishes, its anterior end is connected through a chain of modified vertebræ to the ear. Sometimes its posterior end fits into an enlarged and hollow interhæmal bone. Sometimes, again, a mass of muscle lies in front of it or is otherwise attached to it. Sometimes it is divided into two or three parts by crosswise constrictions. Sometimes it is constricted longitudinally, and at other times it has attached to it a complication of supplemental tubes of the same character as the air-bladder itself. In still other cases it is divided by many internal partitions into a cellular body, similar to the lung of the higher vertebrates, though the cells are coarser and less intricate. This condition is evidently more primitive than that of the empty sac.
The homology of the air-bladder with the lung is evident. This is often expressed in the phrase that the lung is a developed air-bladder. This is by no means true. To say that the air-bladder is a modified and degenerate lung is much nearer the[Pg 94] truth, although we should express the fact more exactly to say that both air-bladder and lung are developed from a primitive cellular breathing-sac, originally a diverticulum from the ventral walls of the œsophagus.
The air-bladder varies in size as much as in form. In some fishes it extends from the head to the tail, while in others it is so minute as to be scarcely traceable. It often varies greatly in closely related species. The common mackerel (Scomber scombrus) has no air-bladder, while in the closely related colias or chub mackerel (Scomber japonicus) the organ is very evident. In other families, as the rockfishes (Scorpænidæ), genera with and those without the air-bladder are scarcely distinguishable externally. In general, fishes which lie on the bottom, those which inhabit great depths, and those which swim freely in the open sea, as sharks and mackerel, lack the air-bladder. In the sharks, rays, and chimæras there is no trace of an air-bladder. In the mackerel and other bony fishes without it, it is lost in the process of development.
The air-bladder is composed of two layers of membrane, the outer one shining, silvery in color, with muscular fibres, the inner well supplied by blood-vessels. The gas within the air-bladder must be in most cases secreted from the blood-vessels. In river fishes it is said to be nearly pure nitrogen. In marine fishes it is mostly oxygen, with from 6 to 10 per cent of carbonic-acid gas, while in the deep-sea fishes oxygen is greatly in excess. In Lopholatilus, a deep-sea fish, Professor R. W. Tower finds 66 to 69 per cent of oxygen. In Trigla lyra Biot records 87 per cent. In Dentex dentex, a shore fish of Europe, 40 per cent of oxygen was found in the air-bladder. Fifty per cent is recorded from the European porgy, Pagrus pagrus. In a fish dying from suffocation the amount of carbonic-acid gas (CO2) is greatly increased, amounting, according to recent researches of Professor Tower on the weak-fish, Cynoscion regalis, to 24 to 29 per cent. This shows conclusively that the air-bladder is to some degree a reservoir of oxygen secreted from the blood, to which channel it may return through a kind of respiration.
The other functions of the air-bladder have been subject to much question and are still far from understood. The following summary of the various views in this regard we copy[Pg 95] from Professor Tower's paper on "The Gas in the Swim-bladder of Fishes":
"The function of the swim-bladder of fishes has attracted the attention of scientists for many centuries. The rôle that this structure plays in the life of the animal has been interpreted in almost as many ways as there have been investigators, and even now there is apparently much doubt as to the true functions of the swim-bladder. Consequently any additional data concerning this organ are of immediate scientific value.
"Aristotle, writing about the noises made by fishes, states that 'some produce it by rubbing the gill-arches ...; others by means of the air-bladder. Each of these fishes contains air, by rubbing and moving of which the noise is produced.' The bladder is thus considered a sound-producing organ, and it is probable that he arrived at this result by his own investigations.
"Borelli (De Motu Animalium, 1680) attributed to the air-bladder a hydrostatic function which enabled the fish to rise and fall in the water by simply distending or compressing the air-bladder. This hypothesis, which gives to the fish a volitional control over the air-bladder—it being able to compress or distend the bladder at pleasure—has prevailed, to a greater or less degree, from the time of Borelli to the present. To my knowledge, however, there are no investigations which warrant such a theory, while, on the other hand, there are many facts, as shown by Moreau's experiment, which distinctly contradict this belief. Delaroche (Annales du Mus. d'Hist. Nat., tome XIV, 1807-1809) decidedly opposed the ideas of Borelli, and yet advanced an hypothesis similar to it in many respects. Like Borelli, he said that the fish could compress or dilate the bladder by means of certain muscles, but this was to enable the fish to keep the same specific gravity as the surrounding medium, and thus be able to remain at any desired depth (and not to rise or sink). This was also disproved later by Moreau. Delaroche proved that there existed a constant exchange between the air in the air-bladder and the air in the blood, although he did not consider the swim-bladder an organ of respiration.
"Biot (1807), Provençal and Humboldt (1809), and others made chemical analyses of the gas in the swim-bladder, and[Pg 96] found 1 to 5 per cent of CO2, 1 to 87 per cent of O2, and the remainder nitrogen. The most remarkable fact discovered about this mixture was that it frequently consisted almost entirely of oxygen, the per cent of oxygen increasing with the depth of the water inhabited by the fish. The reasons for this phenomenon have never been satisfactorily explained.
"In 1820 Weber described a series of paired ossicles which he erroneously called stapes, malleus, and incus, and which connected the air-bladder in certain fishes with a part of the ear—the atrium sinus imparis. Weber considered the swim-bladder to be an organ by which sounds striking the body from the outside are intensified, and these sounds are then transmitted to the ear by means of the ossicles. The entire apparatus would thus function as an organ of hearing. Weber's views remained practically uncontested for half a century, but recently much has been written both for and against this theory. Whatever the virtues of the case may be, there is certainly an inviting field for further physiological investigations regarding this subject, and more especially on the phenomena of hearing in fishes.
"Twenty years later Johannes Müller described, in certain Siluroid fishes, a mechanism, the so-called 'elastic-spring' apparatus, attached to the anterior portion of the air-bladder, which served to aid the fish in rising and sinking in the water according as the muscles of this apparatus were relaxed or contracted to a greater or lesser degree. This interpretation of the function of the 'elastic-spring' mechanism was shown by Sörensen to be untenable. Müller also stated that in some fish, at least, there was an exchange of gas between blood and air-bladder—the latter having a respiratory function—and regarded the gas in the air-bladder as the result of active secretion. In Malapterurus (Torpedo electricus) he stated that it is a sound-producing organ.
"Hasse, in 1873, published the results of his investigations on the functions of the ossicles of Weber, stating that their action was that of a manometer, acquainting the animal with the degree of pressure that is exerted by the gases in the air-bladder against its walls. This pressure necessarily varies with the different depths of water which the fish occupies. Hasse[Pg 97] did not agree with Weber that the ear is affected by the movements of these ossicles.
"One year later Dufosse described in some fishes an air-bladder provided with extrinsic muscles by whose vibration sound was produced, the sound being intensified by the air-bladder, which acted as a resonator. He also believed that certain species produced a noise by forcing the gas from the air-bladder through a pneumatic duct.
"At about the same time Moreau published his classical work on the functions of the air-bladder. He proved by ingenious experiments that many of the prevailing ideas about the action of the air-bladder were erroneous, and that this organ serves to equilibrate the body of the fish with the water at any level. This is not accomplished quickly, but only after sufficient time for the air in the bladder to become adjusted to the increase or decrease in external pressure that has taken place. The fish, therefore, makes no use of any muscles in regulating the volume of its air-bladder. The animal can accommodate itself only gradually to considerable changes in depth of water, but can live equally comfortably at different depths, provided that the change has been gradual enough. Moreau's experiments also convinced him that the gas is actually secreted into the air-bladder, and that there is a constant exchange of gas between it and the blood. In these investigations he has also noticed that section of the sympathetic-nerve fibres supplying the walls of the air-bladder hastens the secreting of the gas into the empty bladder. Since then Bohr has shown that section of the vagus nerve causes the secretion to cease. Moreau noticed in one fish (Trigla) having an air-bladder supplied with muscles that the latter served to make the air-bladder produce sound.
"Again, in 1885, the Weberian mechanism was brought to our attention with a new function attributed to it by Sagemehl who stated that this mechanism exists not for any auditory purposes, nor to tell the fish at what level of the water it is swimming, but to indicate to the fish the variations in the atmospheric pressure. Sörensen tersely contrasts the views of Hasse and Sagemehl by saying that 'Hasse considers the air-bladder with the Weberian mechanism as a manometer; Sagemehl regards it as a barometer.' The theory of Sagemehl has, naturally[Pg 98] enough, met with little favor. Sörensen (1895) held that there is but little evidence for attributing to the air-bladder the function of a lung. It is to be remembered, however, that, according to Sörensen's criterion no matter what exchange of gases takes place between blood and air-bladder, it cannot be considered an organ of respiration, 'unless its air is renewed by mechanical respiration.'
"Sörensen also refutes, from anatomical and experimental grounds, the many objections to Weber's theory of the function of the ossicles. He would thus attribute to the air-bladder the function of hearing; indeed in certain species the only reason for the survival of the air-bladder is that 'the organ is still of acoustic importance; that it acts as a resonator.' This idea, Sörensen states, is borne out by the anatomical structure found in Misgurnus and Chlarias, which resembles the celebrated 'Colladon resonator.' This author attributes to the air-bladder with its 'elastic spring' and various muscular mechanisms the production of sound as its chief function."
Origin of the Air-bladder.—In the more primitive forms, and probably in the embryos of all species, the air-bladder is joined to the œsophagus by an air-duct. This duct is lost entirely in the adult of all or nearly all of the thoracic and jugular fishes, and in some of the abdominal forms. The lancelets, lampreys, sharks, rays, and chimæras have no air-bladder, but in the most primitive forms of true fishes (Dipnoans and Crossopterygians), having the air-bladder cellular or lung-like, the duct is well developed, freely admitting the external air which the fish may rise to the surface to swallow. In most fishes the duct opens into the œsophagus from the dorsal side, but in the more primitive forms it enters from the ventral side, like the windpipe of the higher vertebrates. In some of the Dipnoans the air-bladder divides into two parts, in further resemblance to the true lungs.
The Origin of the Lungs.—The following account of the function of the air-bladder and of its development and decline is condensed from an article by Mr. Charles Morris:[5]
"If now we seek to discover the original purpose of this [Pg 99]organ, there is abundant reason to believe that it had nothing to do with swimming. Certainly the great family of the sharks, which have no bladder, are at no disadvantage in changing their depth or position in the water. Yet if the bladder is necessary to any fish as an aid in swimming, why not to all? And if this were its primary purpose, how shall we explain its remarkable variability? No animal organ with a function of essential importance presents such extraordinary modifications in related species and genera. In the heart, brain, and other organs there is one shape, position, and condition of greatest efficiency, and throughout the lower forms we find a steady advance towards this condition. Great variation, on the other hand, usually indicates that the organ is of little functional importance, or that it has lost its original function. Such we conceive to be the case with the air-bladder. The fact of its absence from some and its presence in other fishes of closely related species goes far to prove that it is a degenerating organ; and the same is shown by the fact that it is useless in some species for the purpose to which it is applied in others. That it had, at some time in the past, a function of essential importance there can be no question. That it exists at all is proof of this. But its modern variations strongly indicate that it has lost this function and is on the road towards extinction. Larval conditions show that it had originally a pneumatic duct as one of its essential parts, but this has in most cases disappeared. The bladder itself has in many cases partly or wholly disappeared. Where preserved, it seems to be through its utility for some secondary purpose, such as an aid in swimming or in hearing. That its evolution began very long ago there can be no question; and the indications are that it began long ago to degenerate, through the loss of its primitive function.
"What was this primitive function? In attempting to answer this question we must first consider the air-bladder in relation to the fish tribe as a whole. No shark or ray possesses the air-bladder. In some few sharks, indeed, there is a diverticulum of the pharynx which may be a rudimentary approach to the air-bladder; but this is very questionable. The conditions of its occurrence in the main body of modern fishes, the Teleostean, we have already considered. But in the most ancient living orders[Pg 100] of fishes it exists in an interesting condition. In every modern Dipnoan, Crossopterygian, and Ganoid the air-bladder has an effective pneumatic duct. This in the Ganoids opens into the dorsal side of the œsophagus, but in the Dipnoans and Crossopterygians, like the windpipe of lung-breathers, it opens into the ventral side. In the Dipnoans, also survivors from the remote past, the duct not only opens ventrally into the œsophagus, but the air-bladder does duty as a lung. Externally it differs in no particular from an air-bladder; but internally it presents a cellular structure which nearly approaches that of the lung of the batrachians. There are three existing representatives of the Dipnoans. One of these, the Australian lung-fish (Neoceratodus) has a single bladder, which, however, is provided with breathing-pouches having a symmetrical lateral arrangement. It has no pulmonary artery, but receives branches from the arteria cœliaca. In the other two forms, Lepidosiren and Protopterus, the kindred 'mudfishes' of the Amazon basin and tropical Africa, the bladder or lung is divided into two lateral chambers, as in the land animals, and is provided with a separate pulmonary artery.
"The opinion seems to have been tacitly entertained by physiologists that this employment of the air-bladder by the Dipnoans as a lung is a secondary adaptation, a side issue from its original purpose. It is more likely that this is the original purpose, and that its degeneration is due to the disappearance of the necessity of such a function. As regards the gravitative employment of the bladder, the Teleostean fishes, to which this function is confined, are of comparatively modern origin; while the Dipnoans are surviving representatives of a very ancient order of fishes, which flourished in the Devonian age of geology, and in all probability breathed air then as now; and the Crossopterygians and Ganoids, which approach them in this particular, are similarly ancient in origin, and were the ancestors of the Teleosteans. The natural presumption, therefore, is that the duty which it subserved in the most ancient fishes was its primitive function.
"The facts of embryology lend strong support to this hypothesis. For the air-bladder is found to arise in a manner very similar to the development of the lung. They each begin as an[Pg 101] outgrowth from the fore part of the alimentary tract, the only difference being that the air-bladder usually rises dorsally and the lung ventrally. The fact already cited, that the pneumatic duct is always present in the larval form in fishes that possess a bladder, is equally significant. All the facts go to show that the introduction of external air into the body was a former function of the air-bladder, and that the atrophy of the duct in many cases, and the disappearance of the bladder in others, are results of the loss of this function.
"Such an elaborate arrangement for the introduction of air into the body could have, if we may judge from analogy, but one purpose, that of breathing, to which purpose the muscular and other apparatus for compressing and dilating the bladder, now seemingly adapted to gravitative uses, may have been originally applied. The same may be said of the great development of blood-capillaries in the inner tunic of the bladder. These may now be used only for the secretion of gas into its interior, but were perhaps originally employed in the respiratory secretion of oxygen. In fact all the circumstances mentioned—the similarity in larval development between the bladder and lung, the larval existence of the pneumatic duct, the arrangements for compressing and dilating the bladder, and the capillary vessels on its inner tunic—point to the breathing of air as its original purpose.
"It is probable that the Ganoid, as well as the Dipnoan, air-bladder is to some extent still used in breathing. The Dipnoans have both lungs and gills, and probably breathe with the latter in ordinary cases, but use their lungs when the inland waters in which they live become thick and muddy, or are charged with gases from decomposing organic matter. The Ganoid fishes to some extent breathe the air. In Polypterus the air-bladder resembles the Dipnoan lung in having lateral divisions and a ventral connection with the œsophagus, while in Lepisosteus (the American garpike) it is cellular and lung-like. This fish keeps near the surface, and may be seen to emit air-bubbles, probably taking in a fresh supply of air. The American bowfin, or mudfish (Amia), has a bladder of the same lung-like character, and has been seen to come to the surface, open its jaws widely, and apparently swallow a large quantity of air. He[Pg 102] considers that both Lepisosteus and Amia inhale and exhale air at somewhat regular intervals, resembling in this the salamanders and tadpoles, 'which, as the gills shrink and the lungs increase, come more frequently to the surface for air.'
"As the facts stand there is no evident line of demarcation between the gas-containing bladders of many of the Teleosteans, the air-containing bladders of the others and the Ganoids, and the lung of the Dipnoans, and the indications are in favor of their having originally had the same function, and of this being the breathing of air.
"If now we ask what were the conditions of life under which this organ was developed, and what the later conditions which rendered it of no utility as a lung, some definite answer may be given. The question takes us back to the Devonian and Silurian geological periods, during which the original development of the bladder probably took place. In this era the seas were thronged with fishes of several classes, the Elasmobranchs among others, followed by the Dipnoi and Crossopterygians. The sharks were without, the Dipnoans and Crossopterygians doubtless with, an air-bladder—a difference in organization which was most likely due to some marked difference in their life-habits. The Elasmobranchs were the monarchs of the seas, against whose incursions the others put on a thick protective armor, and probably sought the shallow shore waters, while their foes held chief possession of the deeper waters without.
"We seem, then, to perceive the lung-bearing fishes, driven by their foes into bays and estuaries, and the waters of shallow coasts, ascending streams and dwelling in inland waters. Here two influences probably acted on them. The waters they dwelt in were often thick with sediment, and were doubtless in many instances poorly aerated, rendering gill-breathing difficult. And the land presented conditions likely to serve as a strong inducement to fishes to venture on shore. Its plant-life was abundant, while its only animal inhabitants seem to have been insects, worms, and snails. There can be little doubt that the active fish forms of that period, having no enemies to fear on the land, and much to gain, made active efforts to obtain a share of this vegetable and animal food. Even to-day, when they have numerous foes to fear, many fishes seek food on the shore, and[Pg 103] some even climb trees for this purpose. Under the conditions of the period mentioned there was a powerful inducement for them to assume this habit.
"Such conditions must have strongly tended to induce fishes to breathe the air, and have acted to develop an organ for this purpose. In addition to the influences of foul or muddy water and of visits to land may be named that of the drying-out of pools, by which fishes are sometimes left in the moist mud till the recurrence of rains, or are even buried in the dried mud during the rainless season. This is the case with the modern Dipnoi, which use their lungs under such circumstances. In certain other fresh-water fishes, of the family Ophiocephalidæ, air is breathed while the mud continues soft enough for the fish to come to the surface, but during the dry period the animal remains in a torpid state. These fishes have no lungs, but breathe the air into a simple cavity in the pharynx, whose opening is partly closed by a fold of the mucous membrane. Other Labyrinthici, of similar habits, possess a more developed breathing organ. This is a cavity formed by the walls of the pharynx, in which are thin laminæ, or plates, which undoubtedly perform an oxygenating function. The most interesting member of this family is Anabas scandens, the climbing perch. In this fish, which not only leaves the water, but is said to climb trees, the air-breathing organ is greatly developed. The labyrinthici, moreover, have usually large air-bladders. As regards the occasional breathing of air by fishes, even in species which do not leave the water, it is quite common, particularly among fresh-water species. Cuvier remarks that air is perhaps necessary to every kind of fish; and that, particularly when the atmosphere is warm, most of our lacustrine species sport on the surface for no other purpose.
"It is not difficult to draw a hypothetical plan of the development of the air-bladder as a breathing organ. In the two families of fishes just mentioned, whose air-bladders indicate that they once possessed the air-breathing function and have lost it, we perceive the process of formation of an air-breathing organ beginning over again under stress of similar circumstances. The larval development of the air-bladder points significantly in the same direction. In fact we have strong reason to believe that[Pg 104] air-breathing in fishes was originally performed, as it probably often is now, by the unchanged walls of the œsophagus. Then these walls expanded inwardly, forming a simple cavity, partly closed by a fold of membrane, like that of the Ophiocephalidæ. A step further reduced this membranous fold to a narrow opening, leading to an inner pouch. As the air-breathing function developed, the opening became a tube, and the pouch a simple lung, with compressing muscles and capillary vessels. By a continuation of the process the smooth-walled pouch became sacculated, its surface being increased by folding into breathing cells. Finally, a longitudinal constriction divided it into two lateral pouches, such as we find in the lung of the Dipnoans. This brings us to the verge of the lung of the amphibians, which is but a step in advance, and from that the line of progress is unbroken to the more intricate lung of the higher land animals.
"The dorsal position of the bladder and its duct would be a difficulty in this inquiry, but for the fact that the duct is occasionally ventral. This dorsal position may have arisen from the upward pressure of air in the swimming fish, which would tend to lift the original pouch. But in the case of fishes which made frequent visits to the shore new influences must have come into play. The effect of gravity tended to draw the organ and its duct downward, as we find in the Crossopterygians and in all the Dipnoans, and its increased use in breathing required a more extended surface. Through this requirement came the pouched and cellular lung of the Dipnoans. Of every stage of the process here outlined examples exist, and there is great reason to believe that the development of the lung followed the path above pointed out.
"When the carboniferous era opened there may have been many lung- and gill-breathing fishes which spent much of their time on land, and some of which, by a gradual improvement of their organs of locomotion, changed into batrachians. But with the appearance of the latter, and of their successors, the reptiles, the relations of the fish to the land radically changed. The fin, or the simple locomotor organ, of the Dipnoans could not compete with the leg and foot as organs of land locomotion, and the fish tribe ceased to be lords of the land, where, instead of feeble prey, they now found powerful foes, and were driven back to[Pg 105] their native habitat, the water. Nor did the change end here. In time the waters were invaded by the reptiles, numerous swimming forms appearing, which it is likely were abundant in the shallower shore-line of the ocean, while they sent many representatives far out to sea. These were actively carnivorous, making the fish their prey, the great mass of whom were doubtless driven into the deeper waters, beyond the reach of their air-breathing foes.
"In this change of conditions we seem to perceive an adequate cause for the loss of air-breathing habits in those fishes in which the lung development had not far progressed. It may indeed have been a leading influence in the development of the Teleostean or bony fishes, as it doubtless was in the loss of its primitive function by, and the subsequent changes of, the air-bladder.
"Such of the Crossopterygians and Dipnoans as survived in their old condition had to contend with adverse circumstances. Most of them in time vanished, while their descendants which still exist have lost in great measure their air-breathing powers, and the Dipnoans, in which the development of the lung had gone too far for reversal, have degenerated into eel-like, mud-haunting creatures, in which the organs of locomotion have become converted into the feeble paddle-like limbs of Neoceratodus and the filamentary appendages of the other species.
"As regards the presence of a large quantity of oxygen in the bladders of deep-swimming marine fishes, it not unlikely has a respiratory purpose, the bladder being, as suggested by Semper, used as a reservoir for oxygen, to serve the fish when sleeping, or when, from any cause, not actively breathing. The excess of oxygen is not due to any like excess in the gaseous contents of sea-water, for the percentage of oxygen decreases from the surface downward, while that of nitrogen remains nearly unchanged. In all cases, indeed, the bladder may preserve a share of its old function, and act as an aid in respiration. Speaking of this, Cuvier says: 'With regard to the presumed assistance which the swim-bladder affords in respiration, it is a fact that when a fish is deprived of that organ, the production of carbonic acid by the branchiæ is very trifling,' thus strongly indicating that the bladder still plays a part in the oxygenation of the blood.
"Under the hypothesis here presented the process of evolution involved may be thus summed up. Air-breathing in fishes was originally performed by the unchanged walls of the œsophagus perhaps at specially vascular localities. Then the wall folded inward, and a pouch was finally formed, opening to the air. The pouch next became constricted off, with a duct of connection. Then the pouch became an air-bladder with respiratory function, and finally developed into a simple lung. These air-breathing fishes haunted the shores, their fins becoming converted into limbs suitable for land locomotion, and in time developed into the lung- and gill-breathing batrachia, and these in their turn into the lung-breathing reptilia, the locomotor organs gradually increasing in efficiency. Of these pre-batrachia we have existing representatives in the mud-haunting Dipnoi, with their feeble limbs. In the great majority of the Ganoid fishes the bladder served but a minor purpose as a breathing organ, the gills doing the bulk of the work. In the Teleostean descendants of the Ganoids the respiratory function of the bladder in great measure or wholly ceased, in the majority of cases the duct closing up or disappearing, leaving the pouch as a closed internal sac, far removed from its place of origin. In this condition it served as an aid in swimming, perhaps as a survival of one of its ancient uses. It gained also in certain cases some connection with the organ of hearing. But these were makeshift and unimportant functions, as we may gather from the fact that many fishes found no need for them, the bladder, in these cases, decreasing in size until too small to be of use in swimming, and in other cases completely disappearing after having travelled far from its point of origin. In some other cases, above cited, the process seems to have begun again, in modern times, in an eversion of the wall of the œsophagus for respiratory purposes. The whole process, if I have correctly conceived it, certainly forms a remarkable organic cycle of development and degeneration, which perhaps has no counterpart of similarly striking character in the whole range of organic life."
The Heart of the Fish.—The heart of the fish is simple in structure, small in size, and usually placed far forward, just behind the branchial cavity, and separated from the abdominal[Pg 107] cavity by a sort of "diaphragm" formed of thickened peritoneum. In certain eels the heart is remote from the head.
The heart consists of four parts, the sinus venosus, into which the veins enter, the auricle or atrium, the ventricle, and the arterial bulb at the base of the great artery which carries the blood to the gills. Of these parts the ventricle is deepest in color and with thickest walls. The arterial bulb varies greatly in structure, being in the sharks, rays, Ganoids, and Dipnoans muscular and provided with a large number of internal valves, and contracting rhythmically like the ventricle. In the higher fishes these structures are lost, the walls of the arterial bulb are not contractile, and the interior is without valves, except the pair that separate it from the ventricle.
In the lancelet there is no proper heart, the function of the heart being taken by a contractile blood-vessel situated on the ventral side of the alimentary canal. In the Dipnoans, which are allied to the ancestors of the higher vertebrates, there is the beginning of a division of the ventricle, and sometimes of the auricle, into parts by a median septum. In the higher vertebrates this septum becomes more and more specialized, separating auricle and ventricle into right and left cavities. The blood in the fish is not returned to the heart after purification, but is sent directly over the body.
The Flow of Blood.—The blood in fishes is thin and pale red (colorless in the lancelet) and with elliptical blood-corpuscles. It enters the sinus venosus from the head through the jugular vein, from the kidney and body walls through the cardinal vein, and from the liver through the hepatic veins. Hence it passes to the auricle and ventricle, and from the ventricle through the arterial bulb, or conus arteriosus to the ventral aorta. Thence it flows to the gills, where it is purified. After passing through the capillaries of the gill-filaments it is collected in paired arteries from each pair of gills. These vessels unite to form the dorsal aorta, which extends the length of the body just below the back-bone. From the dorsal aorta the subclavian arteries branch off toward the pectoral fins. From a point farther back arise the mesenteric arteries carrying blood to the stomach, intestine, liver, and spleen. In the tail the caudal vein carries blood to the kidneys. These secrete impurities arising from[Pg 108] waste of tissues, after which the blood again passes to the heart through the cardinal vein. From the intestine the blood, charged with nutritive materials in solution, is carried by the portal vein to the liver. Here it again passes by the hepatic sinus to the sinus venosus and the heart.
The details of the circulatory system vary a good deal in the different groups, and a comparative study of the direction of veins and arteries is instructive and interesting.
The movement of the blood in fishes is relatively slow, and its temperature is raised but little above that of the surrounding water.
[5] The Origin of Lungs: A Chapter in Evolution. American Naturalist, December, 1892.

The nerves of the Fish.—The nervous system in the fish, as in the higher vertebrates, consists of brain and spinal cord with sensory, or afferent, and motor, or efferent, nerves. As in other vertebrates, the nerve substance is divided into gray matter and white matter, or nerve-cells and nerve-fibres. In the fish, however, the whole nervous system is relatively small, and the gray matter less developed than in the higher forms. According to Günther the brain in the pike (Esox) forms but 1/1305 part of the weight of the body; in the burbot (Lota) about 1/720 part.
The cranium in fishes is relatively small, but the brain does not nearly fill its cavity, the space between the dura mater, which lines the skull-cavity, and the arachnoid membrane, which envelops the brain, being filled with a soft fluid containing a quantity of fat.
The Brain of the Fish.—It is most convenient to examine the fish-brain, first in its higher stages of development, as seen in the sunfish, striped bass, or perch. As seen from above the brain of a typical fish seems to consist of five lobes, four of them in pairs, the fifth posterior to these and placed on the median line. The posterior lobe is the cerebellum, or metencephalon, and it rests on the medulla oblongata, the posterior portion of the brain, which is directly continuous with the spinal cord.
In front of the cerebellum lies the largest pair of lobes, each of them hollow, the optic nerves being attached to the lower surface. These are known as the optic lobes, or mesencephalon. In front of these lie the two lobes of the cerebrum, also called the hemispheres, or prosencephalon. These lobes are usually smaller than the optic lobes and solid. In some fishes they are crossed by a furrow, but are never corrugated as in the brain[Pg 110] of the higher animals. In front of the cerebrum lie the two small olfactory lobes, which receive the large olfactory nerve from the nostrils. From its lower surface is suspended the hypophysis or pituitary gland.
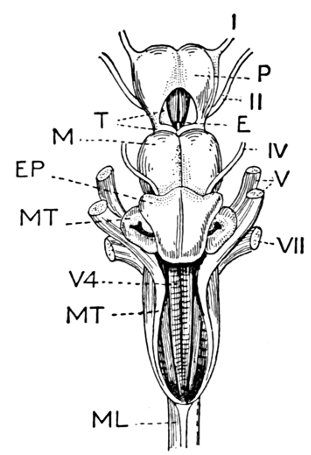
Fig. 78.—Brain of a Shark (Squatina squatina L.). (After Dean.)
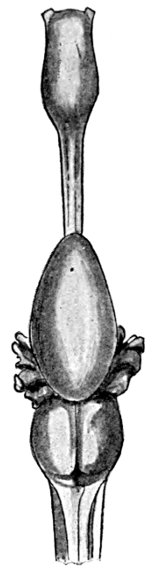
Fig. 79.—Brain of Chimæra monstrosa. (After Wilder per Dean.)
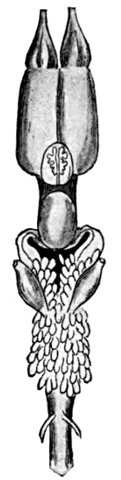
Fig. 80.—Brain of Protopterus annectens. (After Burckhardt per Dean.)
In most of the bony fishes the structure of the brain does not differ materially from that seen in the perch. In the sturgeon, however, the parts are more widely separated. In the Dipnoans the cerebral hemispheres are united, while the optic lobe and cerebellum are very small. In the sharks and rays the large cerebral hemispheres are usually coalescent into one, and the olfactory nerves dilate into large ganglia below the nostrils. The optic lobes are smaller than the hemispheres and also coalescent. The cerebellum is very large, and the surface of the[Pg 111] medulla oblongata is more or less modified or specialized. The brain of the shark is relatively more highly developed than that of the bony fishes, although in most other regards the latter are more distinctly specialized.
The Pineal Organ.—Besides the structures noted in other fishes the epiphysis, or pineal organ, is largely developed in sharks, and traces of it are found in most or all of the higher vertebrates. In some of the lizards this epiphysis is largely developed, bearing at its tip a rudimentary eye. This leaves no doubt that in these forms it has an optic function. For this reason the structure wherever found has been regarded as a rudimentary eye, and the "pineal eye" has been called the "unpaired median eye of chordate" animals.
Fig. 81.—Brain of a Perch, Perca flavescens. (After Dean.)
Fig. 82.—Petromyzon marinus unicolor (Dekay). Head of Lake Lamprey, showing pineal body. (After Gage.)
It has been supposed that this eye, once possessed by all vertebrate forms, has been gradually lost with the better de[Pg 112]velopment of the paired eyes, being best preserved in reptiles as "an outcome of the life-habit which concealed the animal in sand or mud, and allowed the forehead surface alone to protrude, the median eye thus preserving its ancestral value in enabling the animal to look directly upward and backward." This theory receives no support from the structures seen in the fishes.
In none of the fishes is the epiphysis more than a nervous enlargement, and neither in fishes nor in amphibia is there the slightest suggestion of its connection with vision. It seems probable, as suggested by Hertwig and maintained by Dean that the original function of the pineal body was a nervous one and that its connection with or development into a median eye in lizards was a modification of a secondary character. On consideration of the evidence, Dr. Dean concludes that "the pineal structures of the true fishes do not tend to confirm the theory that the epiphysis of the ancestral vertebrates was connected with a median unpaired eye. It would appear, on the other hand, that both in their recent and fossil forms the epiphysis was connected in its median opening with the innervation of the sensory canals of the head. This view seems essentially confirmed by ontogeny. The fact that three successive pairs of epiphyseal outgrowths have been noted in the roof of the thalamencephalon[6] appears distinctly adverse to the theory of a median eye."[7]
The Brain of Primitive Fishes.—The brain of the hagfish differs widely from that of the higher fishes, and the homologies of the different parts are still uncertain. The different ganglia are all solid and are placed in pairs. It is thought that the cerebellum is wanting in these fishes, or represented by a narrow commissure (corpus restiforme) across the front of the medulla. In the lamprey the brain is more like that of the ordinary fish.
In the lancelet there is no trace of brain, the band-like spinal cord tapering toward either end.
The Spinal Cord.—The spinal cord extends from the brain to the tail, passing through the neural arches of the different vertebræ when these are developed. In the higher fishes it is cylindrical [Pg 113]and inelastic. In a few fishes (headfish, trunkfish) in which the posterior part of the body is shortened or degenerate, the spinal cord is much shortened, and replaced behind by a structure called cauda equina. In the headfish it has shrunk into "a short and conical appendage to the brain." In the Cyclostomes and chimæra the spinal cord is elastic and more or less flattened or band-like, at least posteriorly.
The Nerves.—The nerves of the fish correspond in general in place and function with those of the higher animals. They are, however, fewer in number, both large nerve-trunks and smaller nerves being less developed than in higher forms.
The olfactory nerves, or first pair, extend through the ethmoid bone to the nasal cavity, which is typically a blind sac with two roundish openings, but is subject to many variations. The optic nerves, or second pair, extend from the eye to the base of the optic lobes. In Cyclostomes these nerves run from each eye to the lobe of its own side. In the bony fishes, or Teleostei, each runs from the eye to the lobe of the opposite side. In the sharks, rays, chimæras, and Ganoids the two optic nerves are joined in a chiasma as in the higher vertebrates.
Other nerves arising in the brain are the third pair, or nervus oculorum motorius, and the fourth pair, nervus trochlearis, both of which supply the muscles of the eye. The fifth pair, nervus trigeminus, and the seventh pair, nervus facialis, arise from the medulla oblongata and are very close together. Their various branches, sensory and motor, ramify among the muscles and sensory areas of the head. The sixth pair, nervus abducens, passes also to muscles of the eye, and in sharks to the nictitating membrane or third eyelid.
The eighth pair, nervus acousticus, leads to the ear. The ninth pair, glosso-pharyngeal, passes to the tongue and pharynx, and forms a ganglion connected with the sympathetic system. The tenth pair, nervus vagus, or pneumogastric nerve, arises from strong roots in the corpus restiforme and the lower part of the medulla oblongata. Its nerves, motor and sensory, reach the muscles of the gill-cavity, heart, stomach, and air-bladder, as well as the muscular system and the skin. In fishes covered with bony plates the skin may be nearly or quite without sen[Pg 114]sory nerves. The eleventh pair, nervus accessorius, and twelfth pair, nervus hypoglossus, are wanting in fishes.
The spinal nerves are subject to some special modifications, but in the main correspond to similar structures in higher vertebrates. The anterior root of each nerve is without ganglionic enlargement and contains only motor elements. The posterior or dorsal root is sensory only and widens into a ganglionic swelling near the base.
A sympathetic system corresponding to that in the higher vertebrates is found in all the Teleostei, or bony fishes, and in the body of sharks and rays in which it is not extended to the head.
[6] The thalamencephalon or the interbrain is a name given to the region of the optic thalami, between the bases of the optic lobes and cerebrum.
[7] Fishes Recent and Fossil, p. 55.

The Organs of Smell.—The sense-organs of the fish correspond in general to those of the higher vertebrates. The sense of taste is, however, feeble or wanting, and that of hearing is muffled and without power of acute discrimination, if indeed it exists at all. According to Dr. Kingsley (Vert. Zool., p. 75), "recent experiments tend to show that in fishes the ears are without auditory functions and are solely organs of equilibration."
The sense of smell resides in the nostrils, which have no relation to the work of breathing. No fish breathes through its nostrils, and only in a few of the lowest forms (hagfishes) does the nostril pierce through the roof of the mouth. In the bony fishes the nostril is a single cavity, on either side, lined with delicate or fringed membrane, well provided with blood-vessels, and with nerves from the olfactory lobe. In most cases each nasal cavity has two external openings. These may be simple, or the rim of the nostril may be elevated, forming a papilla or even a long barbel. Either nostril may have a papilla or barbel, or the two may unite in one structure with two openings or with sieve-like openings, or in some degenerate types (Tropidichthys) with no obvious openings at all, the olfactory nerves spreading over the skin of a small papilla. The openings may be round, slit-like, pore-like, or may have various other forms. In certain families of bony fishes (Pomacentridæ, Cichlidæ, Hexagrammidæ), there is but one opening to each nostril. In the sharks, rays, and chimæras there is also but one opening on either side and the nostril is large and highly specialized, with valvular flaps controlled by muscles which are said to enable them "to scent actively as well as to smell passively."
In the lancelet there is a single median organ supposed to[Pg 116] be a nostril, a small depression at the front of the head, covered by ciliated membrane. In the hagfish the single median nostril pierces the roof of the mouth, and is strengthened by cartilaginous rings, like those of the windpipe. In the lamprey the single median nostril leads to a blind sac. In the Barramunda (Neoceratodus) there are both external and internal nares, the former being situated just within the upper lip. In all other fishes there is a nasal sac on either side of the head. This has usually, but not always, two openings.
There is little doubt that the sense of smell in fishes is relatively acute, and that the odor of their prey attracts them to it. It is known that flesh, blood, or a decaying carcass will attract sharks, and other predatory fish are drawn in a similar manner. At the same time the strength of this function is yet to be tested by experiments.
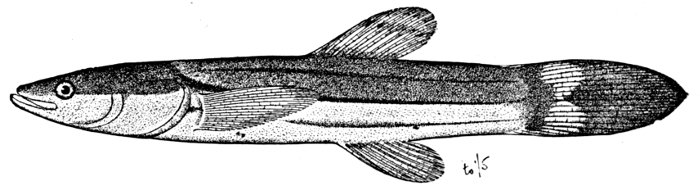
Fig. 83.—Dismal Swamp Fish, Chologaster cornutus Agassiz. Supposed ancestor of Typhlichthys. Virginia.
Fig. 84.—Blind Cavefish, Typhlichthys subterraneus Girard. Mammoth Cave, Kentucky.
The Organs of Sight.—The eyes of fishes differ from those of the higher vertebrates mainly in the spherical form of the crystalline lens. This extreme convexity is necessary because the lens itself is not very much denser than the fluid in which the fishes live. The eyes vary very much in size and somewhat in form and position. They are larger in fishes living at a moderate depth than in shore fishes or river fishes. At great depths,[Pg 117] as a mile or more, where all light is lost, they may become aborted or rudimentary, and may be covered by the skin. Often species with very large eyes, making the most of a little light or of light from their own luminous spots, will inhabit the same depths with fishes having very small eyes or eyes apparently useless for seeing, retained as vestigial structures through heredity. Fishes which live in caves become also blind, the structures showing every possible phase of degradation. The details of this gradual loss of eyes, whether through reversed selection or hypothetically through inheritance of atrophy produced by disuse, have been given in a number of memoirs on the blind fishes of the Mississippi Valley by Dr. Carl H. Eigenmann.
In some fishes the eye is raised on a short, fleshy stalk and can be moved about at the will of the fish. It is said that the vision of the pond-skipper, Periophthalmus, when hunting insects on the mud flats of Japan or India is "quite equal to that of a frog." It is known also that trout possess keen eyesight, and that they show a marked preference for one sort or another of real or artificial fly. Nevertheless the vision of fishes in general is probably not very precise. They apparently notice motion rather than outline, changes rather than objects, while the extreme curvature of the crystalline lens would seem to render them all near-sighted.

Fig. 85.—Four-eyed Fish, Anableps dovii Gill. Tehuantepec, Mexico.
In the eyes of the fishes there is no lachrymal gland. True eyelids no fishes possess; the integuments of the head pass over the eye, becoming transparent as they cross the orbit. In some fishes part of this integument is thickened, covering the eye fully although still transparent. This forms the adipose eyelid characteristic of the mullet, mackerel, and ladyfish. Many of the sharks possess a distinct nictitating membrane or special eyelid, moved by a set of muscles. The iris in most fishes surrounds a[Pg 118] round pupil without much power of contraction. It is frequently brightly colored, red, orange, black, blue, or green. In fishes, like rays or flounders, which lie on the bottom, a dark lobe covers the upper part of the pupil—a curtain to shut out light from above. The cornea is little convex, leaving small space for aqueous humor. In two genera of fishes, Anableps, Dialommus, the cornea is divided by a horizontal partition into two parts. This arrangement permits these fishes, which swim at the surface of the water, to see both in and out of the medium. Anableps, the four-eyed fish, is a fresh-water fish of tropical America, which swims at the surface like a top-minnow, feeding on insects. Dialommus is a marine blenny from the Panama region, apparently of similar habit.
Fig. 86.—Ipnops murrayi Günther.
In one genus of deep-sea fishes, Ipnops, the eyes are spread out to cover the whole upper surface of the head, being modified as luminous areas. Whether these fishes can see at all is not known.
Fig. 87.—Pond-skipper, Boleophthalmus chinensis (Osbeck). Bay of Tokyo, Japan; from nature. K. Morita. (Eye-stalks shrunken in preservation.)
The position of the optic nerves is described in a previous chapter.
In ordinary fishes there is one eye on each side of the head, but in the flounders, by a distortion of the cranium, both ap[Pg 119]pear on the same side. This side is turned uppermost as the fish swims in the water or when it lies on the bottom. This distortion is a matter of development. The very young flounder swims with its broad axis vertical in the water, and it has one eye on either side. As soon as it rests on the bottom it begins to lean to one side. The lower eye changes its axis and by degrees travels across the face of the fish, part of the bony interorbital moving with it across to the other side. In some soles it is said to pass through the substance of the head, reappearing on the other side. In all species which the writer has examined the cranium is twisted, the eye moving with the bones; and the frontal bone is divided, a new orbit being formed by this division. In most northern flounders the eyes are on the right side in the adult, in tropical forms more frequently on the left, these distinctions corresponding with others in the structure of the fish.
In the lowest of the fish-like forms, the lancelet, the eye is simply a minute pigment-spot situated in the anterior wall of the ventricle at the anterior end of the central nervous system. In the hagfishes, which stand next highest in the series, the eye, still incomplete, is very small and hidden by the skin and muscles. This condition is very different from that of the blind fishes of the higher groups, in which the eye is lost through atrophy, because in life in caves or under rocks the function of seeing is no longer necessary.
The Organs of Hearing.—The ear of the typical fish consists of the labyrinth only, including the vestibule and usually three semicircular canals, these dilating into sacs which contain one or more large, loose bones, the ear-stones or otoliths. In the lampreys there are two semicircular canals, in the hagfish but one. There is no external ear, no tympanum, and no Eustachian tube. The ear-sac on each side is lodged in the skull or at the base of the cranial cavity. It is externally surrounded by bone or cartilage, but sometimes it lies near a fontanelle or opening in the skull above. In some fishes it is brought into very close connection with the anterior end of the air-bladder. The latter organ it is thought may form part of the apparatus for hearing. The arrangement for this purpose is especially elaborate in the carp and the catfish families. In these fishes and their relatives[Pg 120] (called Ostariophysi) the two vestibules are joined in a median sac (sinus impar) in the substance of the basioccipital. This communicates with two cavities in the atlas, which again are supported by two small bones, these resting on a larger one in connection with the front of the air-bladder. The system of bones is analogous to that found in the higher vertebrates, but it connects with the air-bladder, not with an external tympanum. The bones are not homologous with those of the ear of higher animals, being processes of the anterior vertebræ. The tympanic chain of higher vertebrates has been thought homologous with the suspensory of the mandible.

Fig. 88.—Brook Lamprey, Lampetra wilderi Jordan and Evermann. (After Gage.) Cayuga Lake.
The otoliths, commonly two in each labyrinth, are usually large, firm, calcareous bodies, with enamelled surface and peculiar grooves and markings. Each species has its own form of otolith, but they vary much in different groups of fishes.
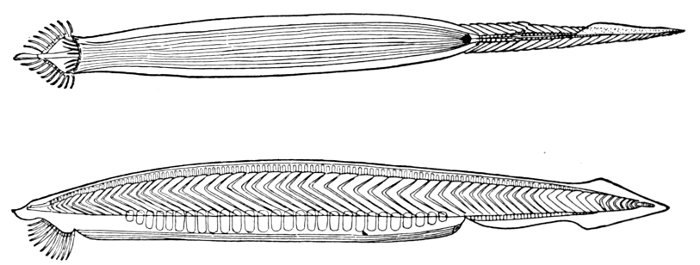
Fig. 89.—European Lancelet, Branchiostoma lanceolatum (Pallas). (After Parker and Haswell.)
In the Elasmobranchs (sharks and rays) and in the Dipnoans the ear-sac is enclosed in the cartilaginous substance of the skull. There is a small canal extending to the surface of the skull, ending sometimes in a minute foramen. The otoliths in these fishes are soft and chalk-like.
The lancelet shows no trace of an ear. In the cyclostomes, hagfishes, and lampreys it forms a capsule of relatively simple structure conspicuous in the prepared skeleton.
The sense of hearing in fishes cannot be very acute, and is at the most confined to the perception of disturbances in the water. Most movements of the fish are governed by sight rather than by sound. It is in fact extremely doubtful whether fishes really hear at all, in a way comparable to the auditory sense in higher vertebrates. Recent experiments of Professor G. H. Parker on the killifish tend to show a moderate degree of auditory sense which grades into the sense of touch, the tubes of the lateral line assisting in both hearing and touch. While the killifish responds to a bass-viol string, there may be some fishes wholly deaf.
Voices of Fishes.—Some fishes make distinct noises variously described as quivering, grunting, grating, or singing. The name grunt is applied to species of Hæmulon and related genera, and fairly describes the sound these fishes make. The Spanish name ronco or roncador (grunter or snorer) is applied to several fishes, both sciænoid and hæmuloid. The noise made by these fishes may be produced by forcing air from part to part of the complex air-bladder, or it may be due to grating one on another of the large pharyngeals. The grating sounds arise, no doubt, from the pharyngeals, while the quivering or singing sounds arise in the air-bladder. The midshipman, Porichthys notatus, is often called singing fish, from a peculiar sound it emits. These sounds have not yet been carefully investigated.
The Sense of Taste.—It is not certain that fishes possess a sense of taste, and it is attributed to them only through their homology with the higher animals. The tongue is without delicate membranes or power of motion. In some fishes certain parts of the palate or pharyngeal region are well supplied with nerves, but no direct evidence exists that these have a function of discrimination among foods. Fishes swallow their food very rapidly, often whole, and mastication, when it takes place, is a crushing or cutting process, not one likely to be affected by the taste of the food.
The Sense of Touch.—The sense of touch is better developed among fishes. Most of them flee from contact with actively[Pg 122] moving objects. Many fishes use sensitive structures as a means of exploring the bottom or of feeling their way to their food. The barbel or fleshy filament wherever developed is an organ of touch. In some fishes, barbels are outgrowths from the nostrils. In the catfish the principal barbel grows from the rudimentary maxillary bone. In the horned dace and gudgeon the little barbel is attached to the maxillary. In other fishes barbels grow from the skin of the chin or snout. In the goatfish and surmullet the two chin barbels are highly specialized. In Polymixia the chin barbels are modified branchiostegals. In the codfish the single beard is little developed. In the gurnards and related forms the lower rays of the pectoral are separate and barbel-like. Detached rays of this sort are found in the thread-fins (Polynemidæ), the gurnards (Triglidæ), and in various other fishes. Barbels or fleshy flaps are often developed over the eyes and sometimes on the scales or the fins.
Fig. 90.—Goatfish, Pseudupeneus maculatus (Bloch). Woods Hole.
The structure of the lateral line and its probable relation as a sense-organ is discussed on page 23. It is probable that it is associated with sense of touch, and hearing as well, the internal ear being originally "a modified part of the lateral-line system," as shown by Parker,[8] who calls the skin the lateral line and the ear "three generations of sense-organs."
The sense of pain is very feeble among fishes. A trout has been known to bite at its own eye placed on a hook, and similar insensibility has been noted in the pike and other fishes. "The Greenland shark, when feeding on the carcass of a whale, allows itself to be repeatedly stabbed in the head without abandoning its prey." (Günther.)
[8] See Parker, on the sense of hearing in fishes, American Naturalist for March, 1903.

The Germ-cells.—In most fishes the germ-cells are produced in large sacs, ovaries or testes, arranged symmetrically one on either side of the posterior part of the abdominal cavity. The sexes are generally but not always similar externally, and may be distinguished on dissection by the difference between the sperm-cells and the ova. The ovary with its eggs is more yellow in color and the contained cells appear granular. The testes are whitish or pinkish, their secretion milk-like, and to the naked eye not granular.
Fig. 91.—Sword-tail Minnow, male, Xiphophorus helleri Heckel. The anal fin modified as an intromittent organ. Vera Cruz.
In a very few cases both organs have been found in the same fish, as in Serranus, which is sometimes truly hermaphrodite. All fishes, however, seem to be normally diœcious, the two sexes in different individuals. Usually there are no external genital organs, but in some species a papilla or tube is developed at the end of the urogenital sinus. This may exist in the breeding season only, as in the fresh-water lampreys, or it may persist through life as in some gobies. In the Elasmobranchs, cartilaginous claspers, attached to the ventral fins in the male, serve as a conduit for the sperm-cells.
The Eggs of Fishes.—The great majority of fishes are oviparous, the eggs being fertilized after deposition. The eggs are laid in gravel or sand or other places suitable for the species, and the milt containing the sperm-cells of the male is discharged over or among them in the water. A very small quantity of the sperm-fluid may impregnate a large number of eggs. But one sperm-cell can enter a particular egg. In a number of families the species are ovoviviparous, the eggs being hatched in the ovary or in a dilated part of the oviduct, the latter resembling a real uterus. In some sharks there is a structure analogous to the placenta of higher animals, but not of the same structure or origin. In the case of viviparous fishes actual copulation takes place and there is usually a modification of some organ to effect transfer of the sperm-cells. This is the purpose of the sword-shaped anal fin in many top-minnows (Pæciliidæ), the fin itself being placed in advance of its usual position. In the surf-fishes (Embiotocidæ) the structure of part of the anal fin is modified, although it is not used as an intromittent organ. In the Elasmobranchs, as already stated, large organs of cartilage (claspers) are developed from the ventral fins.
Fig. 92.—White Surf-fish, viviparous, with young, Cymatogaster aggregatus Gibbons. San Francisco.
In some viviparous fishes, as in the rockfishes (Sebastodes) and rosefishes (Sebastes), the young are very minute at birth.

Fig. 93.—Goodea luitpoldi (Steindachner). A viviparous fish from Lake Patzcuaro, Mexico. Family Pæciliidæ. (After Meek.)
[Pg 127] In others, as the surf-fishes (Embiotocidæ), they are relatively large and few in number. In the viviparous sharks, which constitute the majority of the species of living sharks, the young are large at birth and prepared to take care of themselves.
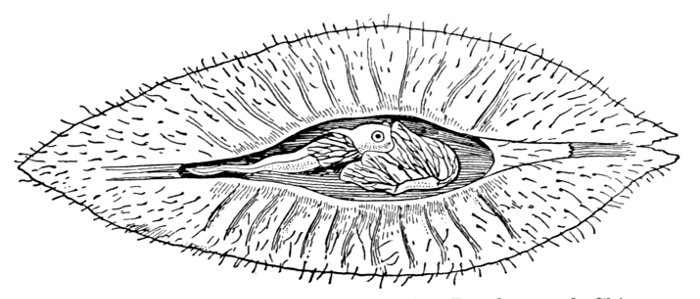
Fig. 94.—Egg of Callorhynchus antarcticus, the Bottle-nosed Chimæra. (After Parker and Haswell.)
The eggs of fishes vary very much in size and form. In those sharks and rays which lay eggs the ova are deposited in a horny egg-case, in color and texture suggesting the kelp in which they are laid. The eggs of the bullhead sharks (Heterodontus) are spirally twisted, those of the cat-sharks (Scyliorhinidæ) are quadrate with long filaments at the angles. Those of rays are wheelbarrow-shaped with four "handles." One egg-case of a ray may sometimes contain several eggs and develop several young. The eggs of lancelets are small, but those of the hagfishes are large, ovate, with fibres at each side, each with a triple hook at tip. The chimæra has also large egg-cases, oblong in form.

Fig. 95.—Egg of the Hagfish, Myxine limosa Girard, showing threads for attachment. (After Dean.)
In the higher fishes the eggs are spherical, large or small according to the species, and varying in the firmness of their[Pg 128] outer walls. All contain food-yolk from which the embryo in its earlier stages is fed. The eggs of the eel (Anguilla) are microscopic. According to Günther 25,000 eggs have been counted in the herring, 155,000 in the lumpfish, 3,500,000 in the halibut, 635,200 in the sturgeon, and 9,344,000 in the cod. Smaller numbers are found in fishes with large ova. The red salmon has about 3500 eggs, the king salmon about 5200. Where an oviduct is present the eggs are often poured out in glutinous masses, as in the bass. When, as in the salmon, there is no oviduct, the eggs lie separate and do not cohere together. It is only with the latter class of fishes, those in which the eggs remain distinct, that artificial impregnation and hatching is practicable. In this regard the value of the salmon and trout is predominant. In some fishes, especially those of elongate form, as the needle-fish (Tylosurus), the ovary of but one side is developed.
Fig. 96.—Egg of Port Jackson Shark, Heterodontus philippi (Lacépède). (After Parker and Haswell.)
Protection of the Young.—In most fishes the parents take no care of their eggs or young. In some catfishes (Platystacus) the eggs adhere to the under surface of the female. In a kind of pipefish (Solenostomus), a large pouch for retention of the eggs is formed on the belly of the female. In the sea-horses and pipefishes a pouch is formed in the skin, usually underneath the tail of the male. Into this the eggs are thrust, and here the young fishes hatch out, remaining until large enough to take care of themselves. In certain sea catfishes (Galeichthys, Conorhynchos) the male carries the eggs in his mouth, thus protecting them from the attacks of other fishes. In numerous cases the male constructs a rough nest, which he defends against all intruders, against the female as well as against outside enemies. The nest-building habit is especially developed in the stickle[Pg 129]backs (Gasterosteidæ), a group in which the male fish, though a pygmy in size, is very fierce in disposition.
In a minnow of Europe (Rhodeus amarus) the female is said to deposit her eggs within the shells of river mussels.
Sexual Modification.—In the relatively few cases in which the sexes are unlike the male is usually the brighter in color and with more highly developed fins. Blue, red, black, and silvery-white pigment are especially characteristic of the male, the olivaceous and mottled coloration of the female. Sometimes the male has a larger mouth, or better developed crests, barbels, or other appendages. In some species the pattern of coloration in the two sexes is essentially different.
In various species the male develops peculiar structures not found in the female, and often without any visible purpose. In the chimæra a peculiar cartilaginous hook armed with a brush of enamelled teeth at the tip is developed on the forehead in the male only. In the skates or true rays (Raja) the pectoral fin has near its edge two rows of stout incurved spines. These the female lacks. In the breeding season, among certain fishes, the male sometimes becomes much brighter by the accumulation of bright red or blue pigment accompanied by black or white pigment cells. This is especially true in the minnows (Notropis), the darters (Etheostoma), and other fresh-water species which spawn in the brooks of northern regions in the spring. In the minnows and suckers horny excrescences are also developed on head, body, or fins, to be lost after the deposition of the spawn.
In the salmon, especially those of the Pacific, the adult male becomes greatly distorted in the spawning season, the jaws and teeth being greatly elongated and hooked or twisted so that the fish cannot shut its mouth. The Atlantic salmon and the trout show also some elongation of the jaws, but not to the same extent.
In those fishes which pair the relation seems not to be permanent, nor is there anything to be called personal affection among them so far as the writer has noticed.
There is no evidence that the bright colors or nuptial adornments of the males are enhanced by sexual selection. In most species the males deposit the sperm-cells in spawning-grounds[Pg 130] without much reference to the preference of the females. In general the brightest colors are not found among viviparous fishes. None of the groups in which the males are showily colored, while the females are plain, belong to this class. The brightest colors are found on the individuals most mature or having greatest vitality.

Segmentation of the Egg.—The egg of the fish develops only after fertilization (amphimixis). This process is the union of its nuclear substance with that of the sperm-cell from the male, each cell carrying its equal share in the function of heredity. When this process takes place the egg is ready to begin its segmentation. The eggs of all fishes are single cells containing more or less food-yolk. The presence of this food-yolk affects the manner of segmentation in general, those eggs having the least amount of food-yolk developing most typically. The simplest of all fish like vertebrates, the lancelet (Branchiostoma) has very small eggs, and in their early development it passes through stages that are typical for all many-celled animals. The first stage in development is the simple splitting of the egg into two halves. These two daughter cells next divide so that there are four cells; each of these divides, and this division is repeated until a great number of cells is produced. The phenomenon of repeated division of the germ-cell is called cleavage, and this cleavage is the first stage of development in the case of all many-celled animals. Instead of forming a solid mass the cells arrange themselves in such a way as to form a hollow ball, the wall being a layer one cell thick. The included cavity is called the segmentation cavity, and the whole structure is known as a blastula. This stage also is common to all the many-celled animals. The next stage is the conversion of the blastula into a double-walled cup, known as a gastrula by the pushing in of one side. All the cells of the blastula are very small, but those on one side are somewhat larger than those of the other, and here the wall first flattens and then bends in until finally the larger cells come into contact with the smaller and the segmentation cavity is entirely obliterated. There is now[Pg 132] an inner layer of cells and an outer layer, the inner layer being known as the endoblast and the outer as the ectoblast. The cavity of the cup thus formed is the archenteron and gives rise primarily to the alimentary canal. This third well-marked stage is called the gastrula stage; and it is thought to occur either typically or in some modified form in the development of all metazoa, or many-celled animals. In the lampreys, the Ganoids, and the Dipnoans the eggs contain a much greater quantity of yolk than those of the lancelet, but the segmentation resembles that of the lancelet in that it is complete; that is, the whole mass of the egg divides into cells. There is a great difference, however, in the size of the cells, those at the upper pole being much smaller than those at the lower. In Petromyzon and the Dipnoans blastula and gastrula stages result, which, though differing in some particulars from the corresponding stages of the lancelet, may yet readily be compared with them. In the hagfishes, sharks, rays, chimæras, and most bony fishes there is a large quantity of yolk, and the protoplasm, instead of being distributed evenly throughout the egg, is for the most part accumulated upon one side, the nucleus being within this mass of protoplasm. When the food substance or yolk is consumed and the little fish is able to shift for itself, it leaves the egg-envelopes and is said to be hatched. The figures on page 135 show some of the stages by which cells are multiplied and ultimately grouped together to form the little fish.
Post-embryonic Development.—In all the fishes the development of the embryo goes on within the egg long after the gastrula stage is passed, and until the embryo becomes a complex body, composed of many differing tissues and organs. Almost all the development may take place within the egg, so that when the young animal hatches there is necessary little more than a rapid growth and increase of size to make it a fully developed mature animal. This is the case with most fishes: a little fish just hatched has most of the tissues and organs of a full-grown fish, and is simply a small fish. But in the case of some fishes the young hatches from the egg before it has reached such an advanced state of development, and the young looks very different from its parent. It must yet undergo considerable change before it reaches the structural condition of a fully developed[Pg 133] and fully grown fish. Thus the development of most fishes is almost wholly embryonic development—that is, development within the egg or in the body of the mother—while the development of some of them is to a considerable degree post-embryonic or larval development. There is no important difference between embryonic and post-embryonic development. The development is continuous from egg-cell to mature animal and, whether inside or outside of an egg, it goes on with a degree of regularity. While certain fishes are subject to a sort of metamorphosis, the nature of this change is in no way to be compared with the change in insects which undergo a complete metamorphosis. In the insects all the organs of the body are broken down and rebuilt in the process of change. In all fishes a structure once formed maintains a more nearly continuous integrity although often considerably altered in form.
General Laws of Development.—The general law of development may be briefly stated as follows: All many-celled animals begin life as a single cell, the fertilized egg-cell; each animal goes through a certain orderly series of developmental changes which, accompanied by growth, leads the animal to change from single-cell to many-celled, complex form characteristic of the species to which the animal belongs; this development is from simple to complex structural condition; the development is the same for all individuals of one species. While all animals begin development similarly, the course of development in the different groups soon diverges, the divergence being of the nature of a branching, like that shown in the growth of a tree. In the free tips of the smallest branches we have represented the various species of animals in their fully developed condition, all standing clearly apart from each other. But in tracing back the development of any kind of animal we soon come to a point where it very much resembles or becomes apparently identical with some other kind of animal, and going farther back we find it resembling other animals in their young condition, and so on until we come to that first stage of development, that trunk stage where all animals are structurally alike. Any animal at any stage in its existence differs absolutely from any other kind of animal, in this respect: it can develop into only its own kind. There is something inherent in each develop[Pg 134]ing animal that gives it an identity of its own. Although in its young stages it may be indistinguishable from some other species of animal in its young stages, it is sure to come out, when fully developed, an individual of the same kind as its parents were or are. The young fish and the young salamander may be alike to all appearance, but one embryo is sure to develop into a fish, and the other into a salamander. This certainty of an embryo to become an individual of a certain kind is called the law of heredity. Viewed in the light of development, there must be as great a difference between one egg and another as between one animal and another, for the greater difference is included in the less.
The Significance of Facts of Development.—The significance of the process of development in any species is yet far from completely understood. It is believed that many of the various stages in the development of an animal correspond to or repeat the structural condition of the animal's ancestors. Naturalists believe that all animals having a notochord at any stage in their existence are related to each other through being descended from a common ancestor, the first or oldest chordate or back-boned animal. In fact it is because all these chordate animals—the lancelets, lampreys, fishes, batrachians, the reptiles, the birds, and the mammals—have descended from a common ancestor that they all develop a notochord, and those most highly organized replace this by a complete back-bone. It is believed that the descendants of the first back-boned animal have, in the course of many generations, branched off little by little from the original type until there came to exist very real and obvious differences among the back-boned animals—differences which among the living back-boned animals are familiar to all of us. The course of development of an individual animal is believed to be a very rapid and evidently much condensed and changed recapitulation of the history which the species or kind of animal to which the developing individual belongs has passed through in the course of its descent through a long series of gradually changing ancestors. If this is true, then we can readily understand why the fish and the salamander and the tortoise and bird and rabbit are all alike in their earlier stages of development, and gradually[Pg 135] come to differ more and more as they pass through later and later developmental stages.
Development of the Bony Fishes.[9] The mode of development of bony fishes differs in many and apparently important regards from that of their nearest kindred, the Ganoids. In their eggs a large amount of yolk is present, and its relations to the embryo have become widely specialized. As a rule, the egg of a Teleost is small, perfectly spherical, and enclosed in delicate but greatly distended membranes. The germ disc is especially small, appearing on the surface as an almost transparent fleck. Among the fishes whose eggs float at the surface during development, as of many pelagic Teleosts, e.g., the sea-bass, Centropristes striatus, the yolk is lighter in specific gravity than the germ; it is of fluid-like consistency, almost transparent. In the yolk at the upper pole of the egg an oil globule usually occurs; this serves to lighten the relative weight of the entire egg, and from its position must aid in keeping this pole of the egg uppermost.
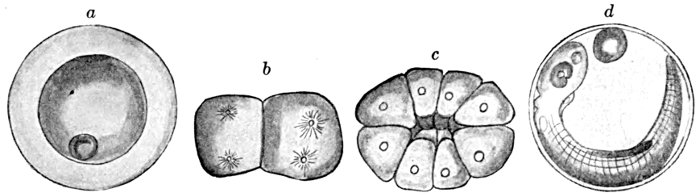
Fig. 97.—Development of Sea-bass, Centropristes striatus (Linnæus). a, egg prior to germination; b, germ-disk after first cleavage; c, germ-disk after third cleavage; d, embryo just before hatching. (After H. V. Wilson.)
In the early segmentation of the germ the first cleavage plane is established, and the nuclear divisions have taken place for the second; in the latter the third cleavage has been completed. As in other fishes these cleavages are vertical, the third parallel to the first. A segmentation cavity occurs as a central space between the blastomeres, as it does in the sturgeon and garpike.
In stages of late segmentation the segmentation cavity is [Pg 136]greatly flattened, but extends to the marginal cells of the germ-disk; its roof consists of two tiers of blastomeres, its floor of a thin film of the unsegmented substance of the germ; the marginal blastomeres are continuous with both roof and floor of the cavity, and are produced into a thin film which passes downward, around the sides of the yolk. Later the segmentation cavity is still further flattened; its roof is now a dome-shaped mass of blastomeres; the marginal cells have multiplied, and their nuclei are seen in the layer of the germ, below the plane of the segmentation cavity. These are seen in the surface view of the marginal cells of this stage; they are separated by cell boundaries only at the sides; below they are continuous in the superficial down-reaching layer of the germ. The marginal cells shortly lose all traces of having been separate; their nuclei, by continued division, spread into the layer of germ flooring the segmentation cavity, and into the delicate film of germ which now surrounds the entire yolk. Thus is formed the periblast of the Teleost development, which from this point onward is to separate the embryo from the yolk; it is clearly the specialized inner part of the germ, which, becoming fluid-like, loses its cell-walls, although retaining and multiplying its nuclei. Later the periblast comes into intimate relations with the growing embryo; it lies directly against it, and appears to receive cell increments from it at various regions; on the other hand, the nuclei of the periblast, from their intimate relations with the yolk, are supposed to subserve some function in its assimilation.
Aside from the question of periblast, the growth of the
blastoderm appears not unlike that of the sturgeon. From
the blastula stage to that of the early gastrula, the changes
have been but slight; the blastoderm has greatly flattened out
as its margins grow downward, leaving the segmentation cavity
apparent. The rim of the blastoderm has become thickened
as the 'germ-ring'; and immediately in front of the dorsal lip
of the blastopore its thickening marks the appearance of the
embryo. The germ-ring continues to grow downward, and
shows more prominently the outline of the embryo; this now
terminates at the head region; while on either side of this point
[Pg 137]spreads out tail-ward on either side the indefinite layer of outgrowing
mesoderm. In the next stage the closure of the blastopore
is rapidly becoming completed; in front of it stretches
the widened and elongated form of the embryo. The yolk-plug
is next replaced by periblast, the dorsal lip by the tail-mass, or
more accurately the dorsal section of the germ-rim; the cœlenteron
under the dorsal lip has here disappeared, on account of
the close approximation of the embryo to the periblast; its last
remnant, the Kupffer's vesicle, is shortly to disappear. The
germ-layers become confluent, but, unlike the sturgeon, the
flattening of the dorsal germ-ring does not permit the formation
of a neurenteric canal.

Fig. 98. Sea-bass, Centropristes striatus, natural size. (From life, by R. W. Shufeldt.)—Page 137.
The process of the development of the germ-layers in Teleosts appears as an abbreviated one, although in many of its details it is but imperfectly known. In the development of the medullary groove, as an example, the following peculiarities exist: the medullary region is but an insunken mass of cells without a trace of the groove-like surface indentation. It is only later, when becoming separate from the ectoderm, that it acquires its rounded character; its cellular elements then group themselves symmetrically with reference to a sagittal plane, where later, by their dissociation, the canal of the spinal cord is formed. The growth of the entoderm is another instance of specialized development. In an early stage the entoderm exists in the axial region, its thickness tapering away abruptly on either side; its lower surface is closely apposed to the periblast; its dorsal thickening will shortly become separate as the notochord. In a following stage of development the entoderm is seen to arch upward in the median line as a preliminary stage in the formation of the cavity of the gut. Later, by the approximation of the entoderm-cells in the median ventral line, the condition is reached where the completed gut-cavity exists.
The formation of the mesoderm in Teleosts is not definitely understood. It is usually said to arise as a process of 'delamination,' i.e., detaching itself in a mass from the entoderm. Its origin is, however, looked upon generally as of a specialized and secondary character.
The mode of formation of the gill-slit of the Teleost does not differ from that in other groups; an evagination of the entoderm coming in contact with an invaginated tract of[Pg 139] ectoderm fuses, and at this point an opening is later established.
The late embryo of the Teleost, though of rounded form, is the more deeply implanted in the yolk-sac than that of the sturgeon; it is transparent, allowing notochord, primitive segments, heart, and sense-organs to be readily distinguished; at about this stage both anus and mouth are making their appearance.
Fig. 99.—Young Sword-fish, Xiphias gladius (Linnæus). (After Lütken.)
The Larval Development of Fishes.[10]—"When the young fish has freed itself from its egg-membranes it gives but little suggestion of its adult form. It enters upon a larval existence, which continues until maturity. The period of change of form varies widely in the different groups of fishes, from a few weeks' to longer than a year's duration; and the extent of the changes that the larva undergoes are often surprisingly broad, investing every organ and tissue of the body, the immature fish passing through a series of form stages which differ one from the other in a way strongly contrasting with the mode of growth of amniotes; since the chick, reptile, or mammal emerges from its embryonic membranes in nearly its adult form.

Fig. 100.—Sword-fish, Xiphias gladius (Linnæus). (After Day.)
The fish may, in general, be said to begin its existence as [Pg 140]a larva as soon as it emerges from its egg-membranes. In some instances, however, it is difficult to decide at what point the larval stage is actually initiated: thus in sharks the excessive amount of yolk material which has been provided for the growth of the larva renders unnecessary the emerging from the egg at an early stage; and the larval period is accordingly to be traced back to stages that are still enclosed in the egg-membranes. In all cases the larval life may be said to begin when the following conditions have been fulfilled: the outward form of the larva must be well defined, separating it from the mass of yolk, its motions must be active, it must possess a continuous vertical fin-fold passing dorsally from the head region to the body terminal, and thence ventrally as far as the yolk region; and the following structures, characteristic in outward appearance, must also be established: the sense-organs—eye, ear, and nose—mouth and anus, and one or more gill-clefts.
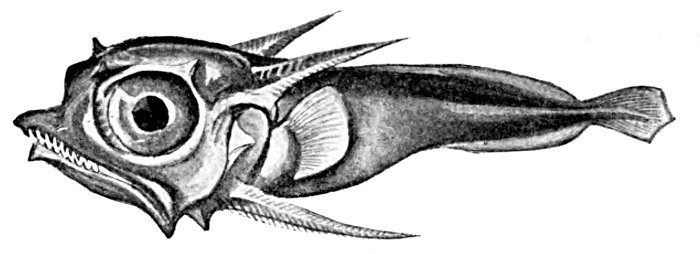
Fig. 101.—Larva of the Sail-fish, Istiophorus, very young. (After Lütken.)

Fig. 102.—Larva of Brook Lamprey, Lampetra wilderi, before transformation, being as large as the adult, toothless, and more distinctly segmented.

Fig. 103.—Common Eel. Anguilla chrisypa Rafinesque. Family Anguillidæ.
Among the different groups of fishes the larval changes are brought about in widely different ways. These larval peculiarities appear at first of far-reaching significance, but may ultimately be attributed, the writer believes, to changed environmental conditions, wherein one process may be lengthened, another shortened. So, too, the changes from one stage to another may occur with surprising abruptness. As a rule, it may be said the larval stage is of longest duration in the Cyclostomes, and thence diminished in length in sharks, lung-fishes, Ganoids, and Teleosts; in the last-named group a very much curtailed (i.e., precocious) larval life may often occur.
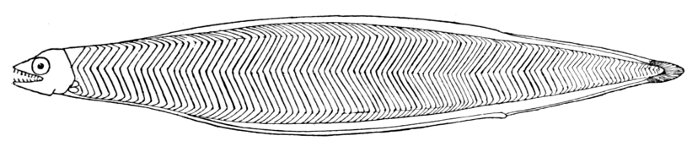
Fig. 104.—Larva of Common Eel, Anguilla chrisypa (Rafinesque), called Leptocephalus grassii. (After Eigenmann.)
The metamorphoses of the newly hatched Teleost must finally be reviewed; they are certainly the most varied and striking of all larval fishes, and, singularly enough, appear to be crowded into the briefest space of time; the young fish, hatched often as early as on the fourth day, is then of the most immature character; it is transparent, delicate, easily injured, inactive; within a month, however, it may have assumed almost every detail of its mature form. A form hatching three millimeters in length may acquire the adult form before it becomes much longer than a centimeter.
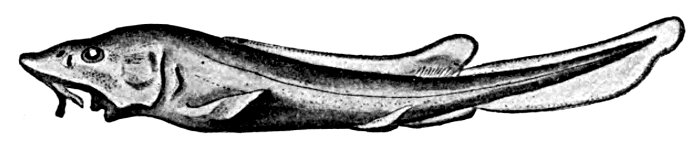
Fig. 105.—Larva of Sturgeon, Acipenser sturio (Linnæus). (After Kupffer, per Dean.)

Fig. 106.—Larva (called Tholichthys) of Chætodon sedentarius (Poey). Cuba. (After Lütken,)

Fig. 107.—Butterfly-fish, Chætodon capistratus Linnæus. Jamaica.
Peculiar Larval Forms.—The young fish usually differs from the adult mainly in size and proportions. The head is larger in the young, the fins are lower, the appendages less developed, and the body more slender in the young than in the adult. But to most of these distinctions there are numerous exceptions, and in some fish there is a change so marked as to be fairly called a metamorphosis. In such cases the young fish in its first condition is properly called a larva. The larva of the lamprey (Petromyzon) is nearly blind and toothless, with slender head, and was long supposed to belong to a different genus (Ammocœtes) from the adult. The larva of sharks and rays, and also of Dipnoans and Crossopterygians, are provided with bushy external gills, which disappear in the process of development. In most soft-rayed fishes the embryonic fringe which precedes the[Pg 143] development of the vertical fins persists for a considerable time. In many young fishes, especially the Chætodontidæ and their allies (butterfly-fishes), the young fish has the head armed with broad plates formed by the backward extension of certain membrane-bones. In other forms the bones of the head are in the young provided with long spines or with serrations, which vanish totally with age. Such a change is noticeable in the swordfish. In this species the production of the bones of the snout and upper jaw into a long bony sword, or weapon of offense, takes place only with age. The young fish have jaws more normally formed, and armed with ordinary teeth. In the headfish (Mola mola) large changes take place in the course of growth, and the young have been taken for a different type of fishes. Among certain soft-rayed fishes and eels the young is often developed in a peculiar way, being very soft, translucent, or band-like, and formed of large or loosely aggregated cells. These peculiar organisms, long known as leptocephali, have been shown to be the normal young of fishes when mature very different. In the ladyfish (Albula) Dr. Gilbert has shown, by a full series of specimens, that in their further growth these pellucid fishes shrink in size, acquiring greater compactness of body, until finally reaching about half their maximum length as larvæ. After this, acquiring essentially the form of the adult fish, they begin a process of regular growth. This leptocephalous condition is thought by Günther to be due to arrest of growth in abnormal individuals, but this is not the case in Albula, and it is probably fully normal in the conger and other eels. In the surf-fishes the larvæ have their vertical fins greatly elevated, much higher than in the adult, while the body is much more closely compressed. In the deal-fish (Trachypterus) the form of the body and fins changes greatly with age, the body becoming more elongate and the fins lower. The differences between different stages of the same fish seem greater than the differences between distinct species. In fact with this and with other forms which change with age, almost the only test of species is found in the count of the fin-rays. So far as known the numbers of these structures do not change. In the moonfishes (Carangidæ) the changes with age are often very considerable. We copy Lütken's figure of the changes in the genus Selene (fig. 113). Similar changes take place in Alectis, Vomer, and other genera.

Fig. 108.—Mola mola (Linnæus). Very early larval stage of the Headfish, called Centaurus boöps. (After Richardson.)

Fig. 109.—Mola mola (Linnæus). Early larval stage, called Molacanthus nummularis. (After Ryder.)

Fig. 110.—Mola mola (Linnæus). Advanced larval stage. (After Ryder.)
The Development of Flounders.—In the great group of flounders and soles (Heterosomata) the body is greatly compressed and the species swim on one side or lie flat on the bottom, with one side uppermost. This upper side is colored like the bottom, sand-color, gray, or brown, while the lower side is mostly white. Both eyes are brought around to the upper side by a twisting of the cranium and a modification or division of the frontal bones. When the young flounder is hatched it is translucent and symmetrical, swimming vertically in the water, with one eye on either side of the head. After a little the young fish rests the ventral edge on the bottom. It then leans to one side, and as its position gradually becomes horizontal the eye on the lower side moves across with its frontal and other bones to the other side. In most species it passes directly under the first interneurals of the dorsal fin. These changes are best observed in the genus Platophrys.
Hybridism.—Hybridism is very rare among fishes in a state of nature. Two or three peculiar forms among the snappers (Lutianus) in Cuba seem fairly attributable to hybridism, the single specimen of each showing a remarkable mixture of characters belonging to two other common species. Hybrids may be readily made in artificial impregnation among those fishes with which this process is practicable. Hybrids of the different salmon or trout usually share nearly equally the traits of the parent species.
The Age of Fishes.—The age of fishes is seldom measured by a definite period of years. Most of them grow as long as they live, and apparently live until they fall victims to some stronger species. It is reputed that carp and pike have lived for a century, but the evidence needs verification. Some fishes, as the salmon of the Pacific (Oncorhynchus), have a definite period[Pg 146] of growth (usually four years) before spawning. After this act all the individuals die so far as known. In Japan and China the Ice-fish (Salanx), a very long, slender, transparent fish allied to the trout, may possibly be annual in habit, all the individuals perhaps dying in the fall to be reproduced from eggs in the spring. But this alleged habit needs verification.
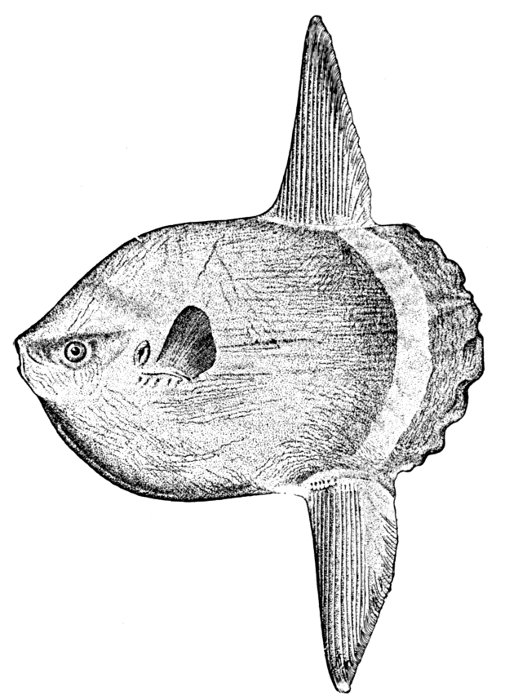
Fig. 111.—Headfish (adult), Mola mola (Linnæus). Virginia.
Tenacity of Life.—Fishes differ greatly in tenacity of life. In general, fishes of the deep seas die at once if brought near the surface. This is due to the reduction of external pressure. The internal pressure forces the stomach out through the mouth and may burst the air-bladder and the large blood-vessels. Marine fishes usually die very soon after being drawn out from the sea.
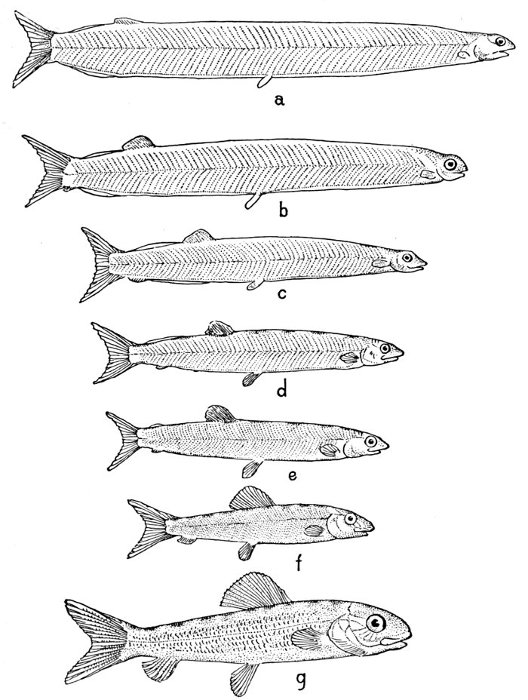
Fig. 112.—Albula vulpes (Linnæus). Transformation of the Ladyfish, from the translucent, loosely compacted larva to the smaller, firm-bodied young. Gulf of California. (After Gilbert.)
Fig. 113.—Development of the Horsehead-fish, Selene vomer (Linnæus). Family Carangidæ. (After Lütken.)
[Pg 149] Some fresh-water fishes are very fragile, dying soon in the air, often with injured air-bladder or blood-vessels. They will die even sooner in foul water. Other fishes are extremely tenacious of life. The mud-minnow (Umbra) is sometimes ploughed up in the half-dried mud of Wisconsin prairies. The related Alaskan blackfish (Dallia) has been fed frozen to dogs, escaping alive from their stomachs after being thawed out. Many of the catfishes (Siluridæ) will live after lying half-dried in the dust for hours. The Dipnoan, Lepidosiren, lives in a ball of half-dried mud during the arid season, and certain fishes, mostly Asiatic, belonging to the group Labyrinthici, with accessory breathing organ can long maintain themselves out of water. Among these is the China-fish (Ophiocephalus), often kept alive in the Chinese settlements in California and Hawaii. Some fishes can readily endure prolonged hunger, while others succumb as readily as a bird or a mammal.

Fig. 114.—Ice-fish, Salanx hyalocranius Abbott. Family Salangidæ. Tientsin, China.
Fig. 115.—Alaska Blackfish, Dallia pectoralis (Bean). St. Michaels, Alaska.
The Effects of Temperature on Fish.—The limits of distribution of many fishes are marked by changes in temperature. Few marine fishes can endure any sudden or great change in this regard, although fresh-water fishes adapt themselves to the seasons. I have seen the cutlass-fish (Trichiurus) benumbed with cold off the coast of Florida while the temperature was still above the frost-line. Those fishes which are tenacious of life and little sensitive to changes in climate and food are most successfully acclimatized or domesticated. The Chinese carp[Pg 150] (Cyprinus carpio) and the Japanese goldfish (Carassius auratus) have been naturalized in almost all temperate and tropical river basins. Within the limits of clear, cold waters most of the salmon and trout are readily transplanted. But some similar fishes (as the grayling) are very sensitive to the least change in conditions. Most of the catfish (Siluridæ) will thrive in almost any fresh waters except those which are very cold.

Fig. 116.—Snake-headed China-fish, Ophiocephalus barca. India. (After Day.)
Transportation of Fishes.—The eggs of species of salmon, placed in ice to retard their development, have been successfully transplanted to great distances. The quinnat-salmon has been thus transferred from California to Australia. It has been found possible to stock rivers and lakes with desirable species, or to restock those in which the fish-supply has been partly destroyed, through the means of artificially impregnated eggs.
The method still followed is said to be the discovery of J. L. Jacobi of Westphalia (about 1760). This process permits the saving of nearly all the eggs produced by the individuals taken. In a condition of nature very many of these eggs would be left unfertilized, or be destroyed by other animals. Fishes are readily kept in captivity in properly constructed aquaria. Unless injured in capture or transportation, there are few species outside the deep seas which cannot adapt themselves to life in a well-constructed aquarium.
Reproduction of Lost Parts.—Fishes have little power to reproduce lost parts. Only the tips of fleshy structures are thus restored after injury. Sometimes a fish in which the tail has been bitten off will survive the injury. The wound will heal, leaving the animal with a truncate body, fin-rays sometimes arising from the scars.

Fig. 117.—Monstrous Goldfish (bred in Japan), Carassius auratus (Linnæus). (After Günther.)
Monstrosities among Fishes.—Monstrosities are rare among fishes in a state of nature. Two-headed young are frequently seen at salmon-hatcheries, and other abnormally divided or united young are not infrequent. Among domesticated species monstrosities are not infrequent, and sometimes, as in the goldfish, these have been perpetuated to become distinct breeds or races. Goldfishes with telescopic eyes and fantastic fins, and with the green coloration changed to orange, are reared in Japan, and are often seen in other countries. The carp has also been largely modified, the changes taking place chiefly in the scales. Some are naked (leather-carp), others (mirror-carp) have a few large scales arranged in series.
[9] This account of the normal development of the Teleost fishes is condensed from Dr. Dean's "Fishes Living and Fossil," in which work the details of growth in the Teleost are contrasted with those of other types of fishes.
[10] This paragraph is condensed from Dean's "Fishes Living and Fossil."

The Habits of Fishes.—The habits of fishes can hardly be summarized in any simple mode of classification. In the usual course of fish-life the egg is laid in the early spring, in water shallower than that in which the parents spend their lives. In most cases it is hatched as the water grows warmer. The eggs of the members of the salmon and cod families are, however, mostly hatched in cooling waters. The young fish gathers with others of its species in little schools, feeds on smaller fishes of other species or of its own, grows and changes until maturity, deposits its eggs, and the cycle of life begins again, while the old fish ultimately dies or is devoured.
Irritability of Animals.—All animals, of whatever degree of organization, show in life the quality of irritability or response to external stimulus. Contact with external things produces some effect on each of them, and this effect is something more than the mere mechanical effect on the matter of which the animal is composed. In the one-celled animals the functions of response to external stimulus are not localized. They are the property of any part of the protoplasm of the body. In the higher or many-celled animals each of these functions is specialized and localized. A certain set of cells is set apart for each function, and each organ or series of cells is released from all functions save its own.
Nerve-cells and Fibres.—In the development of the individual animal certain cells from the primitive external layer or ectoblast of the embryo are set apart to preside over the relations of the creature to its environment. These cells are highly specialized, and while some of them are highly sensitive, others are adapted for carrying or transmitting the stimuli received by the sensitive cells, and still others have the function of receiv[Pg 153]ing sense-impressions and of translating them into impulses of motion. The nerve-cells are receivers of impressions. These are gathered together in nerve-masses or ganglia, the largest of these being known as the brain, the ganglia in general being known as nerve-centres. The nerves are of two classes. The one class, called sensory nerves, extends from the skin or other organ of sensation to the nerve-centre. The nerves of the other class, motor nerves, carry impulses to motion.
The Brain, or Sensorium.—The brain or other nerve-centre sits in darkness, surrounded by a bony protecting box. To this main nerve-centre, or sensorium, come the nerves from all parts of the body that have sensation, the external skin as well as the special organs of sight, hearing, taste, and smell. With these come nerves bearing sensations of pain, temperature, muscular effort—all kinds of sensation which the brain can receive. These nerves are the sole sources of knowledge to any animal organism. Whatever idea its brain may contain must be built up through these nerve-impressions. The aggregate of these impressions constitute the world as the organism knows it. All sensation is related to action. If an organism is not to act, it cannot feel, and the intensity of its feeling is related to its power to act.
Reflex Action.—These impressions brought to the brain by the sensory nerves represent in some degree the facts in the animal's environment. They teach something as to its food or its safety. The power of locomotion is characteristic of animals. If they move, their actions must depend on the indications carried to the nerve-centre from the outside; if they feed on living organisms, they must seek their food; if, as in many cases, other living organisms prey on them, they must bestir themselves to escape. The impulse of hunger on the one hand and of fear on the other are elemental. The sensorium receives an impression that food exists in a certain direction. At once an impulse to motion is sent out from it to the muscles necessary to move the body in that direction. In the higher animals these movements are more rapid and more exact. This is because organs of sense, muscles, nerve-fibres, and the nerve-cells are all alike highly specialized. In the fish the sensation is slow, the muscular response sluggish, but the method remains the same. This is simple reflex action, an impulse from the[Pg 154] environment carried to the brain and then unconsciously reflected back as motion. The impulse of fear is of the same nature. Reflex action is in general unconscious, but with animals, as with man, it shades by degrees into conscious action, and into volition or action "done on purpose."
Instinct.—Different animals show differences in method or degree of response to external influences. Fishes will pursue their prey, flee from a threatening motion, or disgorge sand or gravel swallowed with their food. Such peculiarities of different forms of life constitute the basis of instinct.
Instinct is automatic obedience to the demands of conditions external to the nervous system. As these conditions vary with each kind of animal, so must the demands vary, and from this arises the great variety actually seen in the instincts of different animals. As the demands of life become complex, so do the instincts. The greater the stress of environment, the more perfect the automatism, for impulses to safe action are necessarily adequate to the duty they have to perform. If the instinct were inadequate, the species would have become extinct. The fact that its individuals persist shows that they are provided with the instincts necessary to that end. Instinct differs from other allied forms of response to external condition in being hereditary, continuous from generation to generation. This sufficiently distinguishes it from reason, but the line between instinct and reason and other forms of reflex action cannot be sharply drawn.
It is not necessary to consider here the question of the origin of instincts. Some writers regard them as "inherited habits," while others, with apparent justice, doubt if mere habits or voluntary actions repeated till they become a "second nature" ever leave a trace upon heredity. Such investigators regard instinct as the natural survival of those methods of automatic response which were most useful to the life of the animal, the individual having less effective methods of reflex action perishing, leaving no posterity.
Classification of Instincts.—The instincts of fishes may be roughly classified as to their relation to the individual into egoistic and altruistic instincts.
Egoistic instincts are those which concern chiefly the individual animal itself. To this class belong the instincts of feed[Pg 155]ing, those of self-defense and of strife, the instincts of play, the climatic instincts, and environmental instincts, those which direct the animal's mode of life.
Altruistic instincts are those which relate to parenthood and those which are concerned with the mass of individuals of the same species. The latter may be called the social instincts. In the former class, the instincts of parenthood, may be included the instinct of courtship, reproduction, home-making, nest-building, and care for the young. Most of these are feebly developed among fishes.
The instincts of feeding are primitively simple, growing complex through complex conditions. The fish seizes its prey by direct motion, but the conditions of life modify this simple action to a very great degree.
The instinct of self-defense is even more varied in its manifestations. It may show itself either in the impulse to make war on an intruder or in the desire to flee from its enemies. Among carnivorous forms fierceness of demeanor serves at once in attack and in defense.
Herbivorous fishes, as a rule, make little direct resistance to their enemies, depending rather on swiftness of movement, or in some cases on simple insignificance. To the latter cause the abundance of minnows, anchovies, and other small or feeble fishes may be attributed, for all are the prey of carnivorous fishes, which they far exceed in number.
The instincts of courtship relate chiefly to the male, the female being more or less passive. Among many fishes the male makes himself conspicuous in the breeding season, spreading his fins, intensifying his pigmented colors through muscular tension, all this supposedly to attract the attention of the female. That this purpose is actually accomplished by such display is not, however, easily proved. In the little brooks in spring, male minnows can be found with warts on the nose or head, with crimson pigment on the fins, or blue pigment on the back, or jet-black pigment all over the head, or with varied combination of all these. Their instinct is to display all these to the best advantage, even though the conspicuous hues lead to their own destruction.
The movements of many migratory animals are mainly con[Pg 156]trolled by the impulse to reproduce. Some pelagic fishes, especially flying fishes and fishes allied to the mackerel, swim long distances to a region favorable for a deposition of spawn. Some species are known only in the waters they make their breeding homes, the individuals being scattered through the wide seas at other times. Many fresh-water fishes, as trout, suckers, etc., forsake the large streams in the spring, ascending the small brooks where they can rear their young in greater safety. Still others, known as anadromous fishes, feed and mature in the sea, but ascend the rivers as the impulse of reproduction grows strong. An account of these is given in a subsequent paragraph.
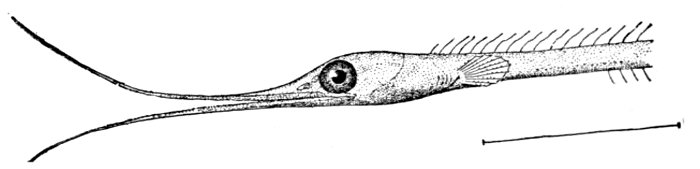
Fig. 118.—Jaws of Nemichthys avocetta. Jordan and Gilbert.
Variability of Instincts.—When we study instincts of animals with care and in detail, we find that their regularity is much less than has been supposed. There is as much variation in regard to instinct among individuals as there is with regard to other characters of the species. Some power of choice is found in almost every operation of instinct. Even the most machine-like instinct shows some degree of adaptability to new conditions. On the other hand, in no animal does reason show entire freedom from automatism or reflex action. "The fundamental identity of instinct with intelligence," says Dr. Charles O. Whitman, "is shown in their dependence upon the same structural mechanism (the brain and nerves) and in their responsive adaptability."
Adaptation to Environment.—In general food-securing structures are connected with the mouth, or, as in the anglers, are hung as lures above it; spines of offense and defense, electric organs, poison-glands, and the like are used in self-protection; the bright nuptial colors and adornments of the breeding season are doubtfully classed as useful in rivalry; the egg-sacs, nests, and other structures or habits may serve to defend the young, while skinny flaps, sand or weed-like markings, and[Pg 157] many other features of mimicry serve as concessions to the environment.
Each kind of fishes has its own ways of life, fitted to the conditions of environment. Some species lie on the bottom, flat, as a flounder, or prone on their lower fins, as a darter or a stone-roller. Some swim freely in the depths, others at the surface of the depths. Some leap out of the water from time to time, as the mullet (Mugil) or the tarpon (Tarpon atlanticus).

Fig. 119.—Catalina Flying Fish, Cypsilurus californicus (Cooper). Santa Barbara.
Flight of Fishes.—Some fishes called the flying-fishes sail through the air with a grasshopper-like motion that closely imitates true flight. The long pectoral fins, wing-like in form, cannot, however, be flapped by the fish, the muscles serving only to expand or fold them. These fishes live in the open sea or open channel, swimming in large schools. The small species fly for a few feet only, the large ones for more than an eighth of a mile. These may rise five to twenty feet above the water.
The flight of one of the largest flying fishes (Cypsilurus californicus) has been carefully studied by Dr. Charles H. Gilbert and the writer. The movements of the fish in the water are extremely rapid. The sole motive power is the action under the water of the strong tail. No force can be acquired while the fish is in the air. On rising from the water the movements[Pg 158] of the tail are continued until the whole body is out of the water. When the tail is in motion the pectorals seem in a state of rapid vibration. This is not produced by muscular action on the fins themselves. It is the body of the fish which vibrates, the pectorals projecting farthest having the greatest amplitude of movement. While the tail is in the water the ventral fins are folded. When the action of the tail ceases the pectorals and ventrals are spread out wide and held at rest. They are not used as true wings, but are held out firmly, acting as parachutes, enabling the body to skim through the air. When the fish begins to fall the tail touches the water. As soon as it is in the water it begins its motion, and the body with the pectorals again begins to vibrate. The fish may, by skimming the water, regain motion once or twice, but it finally falls into the water with a splash. While in the air it suggests a large dragon-fly. The motion is very swift, at first in a straight line, but is later deflected in a curve, the direction bearing little or no relation to that of the wind. When a vessel passes through a school of these fishes, they spring up before it, moving in all directions, as grasshoppers in a meadow.
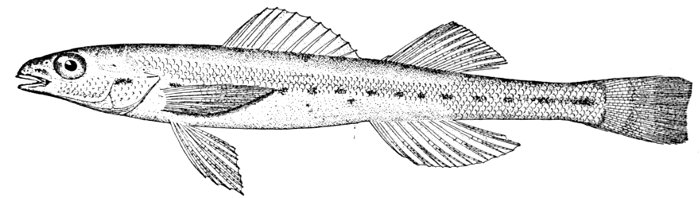
Fig. 120.—Sand-darter, Ammocrypta clara (Jordan and Meek). Des Moines River.
Quiescent Fishes.—Some fishes, as the lancelet, lie buried in the sand all their lives. Others, as the sand-darter (Ammocrypta pellucida) and the hinalea (Julis gaimard), bury themselves in the sand at intervals or to escape from their enemies. Some live in the cavities of tunicates or sponges or holothurians or corals or oysters, often passing their whole lives inside the cavity of one animal. Many others hide themselves in the interstices of kelp or seaweeds. Some eels coil themselves in the crevices of rocks or coral masses, striking at their prey like snakes. Some sea-horses cling by their tails to gulfweed or sea-wrack. Many[Pg 159] little fishes (Gobiomorus, Carangus, Psenes) cluster under the stinging tentacles of the Portuguese man-of-war or under ordinary jellyfishes. In the tide-pools, whether rock, coral, or mud, in all regions multitudes of little fishes abound. As these localities are neglected by most collectors, they have proved of late years a most prolific source of new species. The tide-pools of Cuba, Key West, Cape Flattery, Sitka, Unalaska, Monterey, San Diego, Mazatlan, Hilo, Kailua and Waiahæ in Hawaii, Apia and Pago-Pago in Samoa, the present writer has found peculiarly rich in rock-loving forms. Even richer are the pools of the promontories of Japan, Hakodate Head, Misaki, Awa, Izu, Waka, and Kagoshima, where a whole new fish fauna unknown to collectors in markets and sandy bays has been brought to light. Some of these rockfishes are left buried in the rock weeds as the tide flows, lying quietly until it returns. Others cling to the rocks by ventral suckers, while still others depend for their safety on their powers of leaping or on their quickness of their movements in the water. Those of the latter class are often brilliantly colored, but the others mimic closely the algæ or the rocks. Some fishes live in the sea only, some prefer brackish-water. Some are found only in the rivers, and a few pass more or less indiscriminately from one kind of water to another.
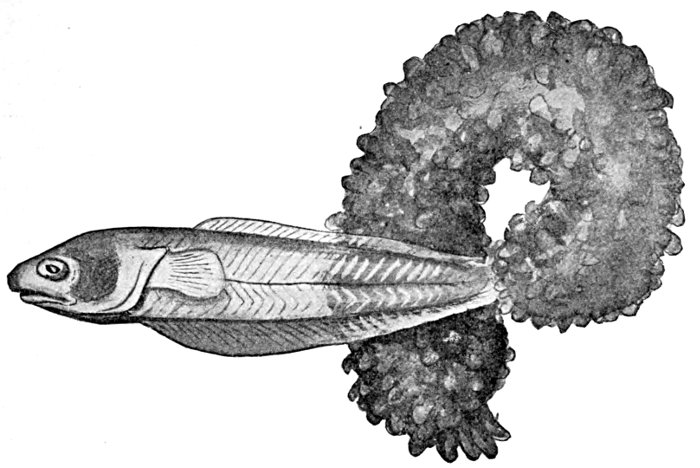
Fig. 121.—Pearl-fish, Fierasfer acus (Linnæus), issuing from a Holothurian. Coast of Italy. (After Emery.)
Fig. 122.—Portuguese Man-of-war Fish, Gobiomorus gronovii. Family Stromateidæ.
Migratory Fishes.—The movements of migratory fishes are mainly controlled by the impulse of reproduction. Some pelagic fishes, especially those of the mackerel and flying-fish families, swim long distances to a region favorable for the deposition of spawn. Others pursue for equal distances the schools of menhaden or other fishes which serve as their prey. Some species are known mainly in the waters they make their breeding homes, as in Cuba, Southern California, Hawaii, or Japan, the individuals being scattered at other times through the wide seas.
Anadromous Fishes.—Many fresh-water fishes, as trout and suckers, forsake the large streams in the spring, ascending the small brooks where their young can be reared in greater safety. Still others, known as anadromous fishes, feed and mature in the sea, but ascend the rivers as the impulse of reproduction grows strong. Among such fishes are the salmon, shad, alewife, sturgeon, and striped bass in American waters. The most remarkable case of the anadromous instinct is found in the king salmon or quinnat (Oncorhynchus tschawytscha) of the Pacific Coast. This great fish spawns in November, at the age of four years and an average weight of twenty-two pounds. In the Columbia River it begins running with the spring freshets in March and April. It spends the whole summer, without feeding, in the ascent of the river. By autumn the individuals have reached the mountain streams of Idaho, greatly changed in appearance,[Pg 161] discolored, worn, and distorted. The male is humpbacked, with sunken scales, and greatly enlarged, hooked, bent, or twisted jaws, with enlarged dog-like teeth. On reaching the spawning beds, which may be a thousand miles from the sea in the Columbia, over two thousand in the Yukon, the female deposits her eggs in the gravel of some shallow brook. The male covers them and scrapes the gravel over them. The female salmon does as much as the male in covering the eggs. Then both male and female drift tail foremost helplessly down the stream; none, so far as certainly known, ever survive the reproductive act. The same habits are found in the five other species of salmon in the Pacific, but in most cases the individuals do not start so early nor run so far. The blue-back salmon or redfish, however, does not fall far short in these regards. The salmon of the Atlantic has a similar habit, but the distance traveled is everywhere much less, and most of the hook-jawed males drop down to the sea and survive to repeat the acts of reproduction.

Fig. 123.—Tide-pools of Misaki. The Misaki Biological Station, from the north side.
Catadromous fishes, as the true eel (Anguilla), reverse this order, feeding in the rivers and brackish estuaries, apparently finding their usual spawning-ground in the sea.

Fig. 124.—Squaw-fish, Ptychocheilus oregonensis (Richardson). Columbia River.
Pugnacity of Fishes.—Some fishes are very pugnacious, always ready for a quarrel with their own kind. The sticklebacks show this disposition, especially the males. In Hawaii the natives take advantage of this trait to catch the Uu (Myripristis murdjan), a bright crimson-colored fish found in those waters. The species lives in crevices in lava rocks. Catching a live one, the fishermen suspend it by a string in front of the rocks. It remains there with spread fins and flashing scales, and the others come out to fight it, when all are drawn to the surface by a[Pg 163] concealed net. Another decoy is substituted and the trick is repeated until the showy and quarrelsome fishes are all secured.
In Siam the fighting-fish (Betta pugnax) is widely noted. The following account of this fish is given by Cantor:[11]
"When the fish is in a state of quiet, its dull colors present nothing remarkable; but if two be brought together, or if one sees its own image in a looking-glass, the little creature becomes suddenly excited, the raised fins and the whole body shine with metallic colors of dazzling beauty, while the projected gill membrane, waving like a black frill round the throat, adds something of grotesqueness to the general appearance. In this state it makes repeated darts at its real or reflected antagonist. But both, when taken out of each other's sight, instantly become quiet. The fishes were kept in glasses of water, fed with larvæ of mosquitoes, and had thus lived for many months. The Siamese are as infatuated with the combats of these fish as the Malays are with their cock-fights, and stake on the issue considerable sums, and sometimes their own persons and families. The license to exhibit fish-fights is farmed, and brings a considerable annual revenue to the king of Siam. The species abounds in the rivulets at the foot of the hills of Penang. The inhabitants name it 'Pla-kat,' or the 'fighting-fish'; but the kind kept especially for fighting is an artificial variety cultivated for the purpose."
A related species is the equally famous tree-climber of India (Anabas scandens). In 1797 Lieutenant Daldorf describes his capture of an Anabas, five feet above the water, on the bark of a palm-tree. In the effort to do this, the fish held on to the bark by its preopercular spines, bent its tail, inserted its anal spines, then pushing forward, repeated the operation.
Fear and Anger in Fishes.—From an interesting paper by Surgeon Francis Day[12] on Fear and Anger in Fishes we may make the following extracts, slightly condensed and with a few slight corrections in nomenclature. The paper is written in amplification of another by Rev. S. J. Whitmee, describing the behavior of aquarium fishes in Samoa.

Fig. 125.—Squaw-fish, Ptychocheilus grandis Agassiz. Running up a stream to spawn, the high water, after a rain, falling, leaves the fishes stranded. Kelsey Creek, Clear Lake, California, April 29, 1899. (Photograph by O. E. Meddaugh.)—Page 164.
The means of expression in animals adverted to by Mr. Darwin (excluding those of the ears, which would be out of place in fishes) are: sounds, vocally or otherwise produced; the erection of dermal appendages under the influence of anger or terror, which last would be analogous to the erection of scales and fin-rays among fishes. Regarding special expressions, as those of joy, pain, astonishment, etc., we could hardly expect such so well marked in fishes as in some of the higher animals, in which the play of the features often affords us an insight into their internal emotions. Eyes[13] destitute of movable eyelids, cheeks covered with scales, or the head enveloped in dermal plates, can scarcely mantle into a smile or expand into a broad grin. We possess, however, one very distinct expression in fishes which is absent or but slightly developed in most of the higher animals, namely, change of color. All are aware that when a fish sickens, its brilliant colors fade, but less so how its color may be augmented by anger, and a loss of it be occasioned by depression, the result of being vanquished by a foe. Some forms also emit sounds when actuated by terror, and perhaps in times of anger; but of this last I possess no decided proofs.
Similar to the expression of anger in Betta is that of the three-spined stickleback (Gasterosteus aculeatus).[14] After a fight between two examples, according to Couch, "a strange alteration takes place almost immediately in the defeated party: his gallant bearing forsakes him; his gay colors fade away; he becomes again speckled and ugly; and he hides his disgrace amongst his peaceable companions who occupy together that part of the tub which their tyrants have not taken possession of; he is, moreover, for some time the constant object of his conqueror's persecution."
Fear is shown by fish in many ways. There is not an angler unacquainted with the natural timidity of fishes, nor a keeper in [Pg 166]charge of a salmon-pass, who does not know how easy it is for poachers to deter the salmon from venturing along the path raised expressly for his use.
Among the coral reefs of the Andaman Islands I found the little Chromis lepisurus abundant. As soon as the water was splashed they appeared to retire for safety to the branching coral, where no large fish could follow them; so frightened did they become that on an Andamanese diving from the side of the boat, they at once sought shelter in the coral, in which they remained until it was removed from the sea. In Burma I observed, in 1869, that when weirs are not allowed to stretch across the rivers (which would impede navigation), the open side as far as the bank is studded with reeds; these, as the water passes over them, cause vibration, and occasion a curious sound alarming the fishes, which, crossing to the weired side of the river, become captured.
Hooker, alluding to gulls, terns, wild geese, and pelicans in the Ganges Valley, observes: "These birds congregate by the sides of pools and beat the water with violence, so as to scare the fish, which then become an easy prey—a fact which was, I believe, first indicated by Pallas during his residence on the banks of the Caspian Sea."[15] Fishes, under the influence of terror, dash about with their fins expanded, and often run into places which must destroy them. Thus droves and droves of sardines in the east, impelled by the terror of pursuing sharks, bonitos, and other voracious fishes, frequently throw themselves on the shores in enormous quantities. Friar Odoric, who visited Ceylon about 1320, says: "There are fishes in those seas which come swimming towards the said country in such abundance, that for a great distance into the sea nothing can be seen but the backs of fishes, which, casting themselves on the shore, do suffer men for the space of three days to come, and to take as many of them as they please, and then they return again into the sea."[16]
Pennant tells us that the river bullhead (Cottus gobio) "deposits its spawn in a hole it forms in the gravel, and quits it with great reluctance." General Hardwicke tells how the [Pg 167]gouramy (Osphromenus gouramy), in the Mauritius, forms a nest amongst the herbage growing in the shallow water in the sides of tanks. Here the parent continues to watch the place with the greatest vigilance, driving away any interloping fish. The amphibious walking-fish of Mysore (Ophiocephalus striatus) appears to make a nest very similar to that of the gouramy, and over it the male keeps guard; but should he be killed or captured, the vacant post is filled by his partner. (Colonel Puckle.) When very young the fishes keep with and are defended by their parents, but so soon as they are sufficiently strong to capture prey for themselves they are driven away to seek their own subsistence. (See Fishes of India, p. 362.) But it is not only these monogamous amphibious fishes which show an affection for their eggs and also for their fry, but even the little Etroplus maculatus has been observed to be equally fond of its ova. "The eggs are not very numerous and are deposited in the mud at the bottom of the stream, and, when hatched, both parents guard the young for many days, vigorously attacking any large fish that passes near them."[17]
Although the proceedings of the members of the marine and estuary genus of sea-cat (Tachysurus) and its allies show not quite so distinctly signs of affection, still it must be a well-developed instinct which induces the male to carry about the eggs in its mouth until hatched, and to remove them in this manner when danger is imminent. I have taken the ova just ready for the young to come forth out of the mouth and fauces of the parent (male) fish; and in every animal dissected there was no trace of food in the intestinal tract.
Calling the Fishes.—At many temples in India fishes are called to receive food by means of ringing bells or musical sounds. Carew, in Cornwall, is said to have called the gray mullet together by making a noise like chopping with a cleaver. Lacépède relates that some fishes, which had been kept in the basins out of the Tuileries for more than a century, would come when called by their names, and that in many parts of Germany trout, carp, and tench are summoned to their food by the sound of a bell. These instances are mostly due to the [Pg 168]fishes having learned by experience that on the hearing certain sounds they may expect food. But Lacépède mentions that some were able to distinguish their individual names; and the same occurs in India. Lieutenant Connolly[18] remarked upon seeing numerous fishes coming to the ghaut at Sidhnath to be fed when called; and on "expressing our admiration of the size of the fish, 'Wait,' said a bystander, 'until you have seen Raghu.' The Brahmin called out his name in a peculiar tone of voice; but he would not hear. I threw in handful after handful of ottah (flour) with the same success, and was just leaving the ghaut, despairing and doubting, when a loud plunge startled me. I thought somebody had jumped off the bastion of the ghaut into the river, but was soon undeceived by the general shout of 'Raghu, raghu,' and by the fishes, large and small, darting away in every direction. Raghu made two or three plunges, but was so quick in his motions that I was unable to guess at his species." [It may be said in relation to these stories quoted by Dr. Day, that they probably belong to the mythology of fishes. It is very doubtful if fishes are able to make any such discrimination among sounds in the air.]
Sounds of Fishes.—Pallegoix states that in Siam the dog's-tongue (Cynoglossus) is a kind of sole; it attaches itself to the bottom of boats, and makes a sonorous noise, which is more musical when several are stuck to the same boat and act in concert (vol. i. p. 193). These noises can scarcely be due to anger or fear. Sir J. Bowring (vol. ii. p. 276) also remarks upon having heard this fish, "which sticks to the bottoms of the boats, and produces a sound something like that of a jew's-harp struck slowly, though sometimes it increases in loudness, so as to resemble the full tones and sound of an organ. My men have pointed me out a fish about four inches long as the author of the music."
Some years since, at Madras, I (Dr. Day) obtained several specimens of a fresh-water Siluroid fish (Macrones vittatus) which is termed the "fiddler" in Mysore. I touched one which was on the wet ground, at which it appeared to become very irate, erecting its dorsal fin, making a noise resembling the buzzing of [Pg 169]a bee. Having put some small carp into an aquarium containing one of these fishes, it rushed at a small example, seized it by the middle of its back, and shook it like a dog killing a rat; at this time its barbels were stiffened out laterally like a cat's whiskers.
Many fish when captured make noises, perhaps due to terror. Thus the Carangus hippos, Tetraodon, and others grunt like a hog. Darwin (Nat. Journ., vol. vii) remarks on a catfish found in the Rio Paraná, and called the armado, which is remarkable for a harsh grating noise when caught by hook and line; this noise can be distinctly heard when the fish is beneath the water.
The cuckoo-gurnard (Trigla pini) and the maigre (Pseudosciæna aquila) utter sounds when taken out of the water; and herrings, when the net has been drawn over them, have been observed to do the same: "this effect has been attributed to an escape of air from the air-bladder; but no air-bladder exists in the Cottus, which makes a similar noise."
The lesser weaver (Trachinus) buries itself in the loose soil at the bottom of the water, leaving only its head exposed, and awaits its prey. If touched, it strikes upwards or sideways; and Pennant says it directs its blows with as much judgment as a fighting-cock. (Yarrell, vol. i. p. 26.) Fishermen assert that wounds from its anterior dorsal spines are more venomous than those caused by the spines on its gill-covers.
As regards fighting, I should suppose that, unless some portion of the body is peculiarly adapted for this purpose, as the rostrum of the swordfish, or the spine on the side of the tail in the lancet-fishes, we must look chiefly to the armature or covering of the jaws for weapons of offense.
Lurking Fishes.—Mr. Whitmee supposes that most carnivorous fish capture their prey by outswimming them; but to this there are numerous exceptions; the angler or fishing-frog (Lophis piscatorius), "while crouching close to the ground, by the action of its ventral and pectoral fins stirs up the sand and mud; hidden by the obscurity thus produced, it elevates its anterior dorsal spines, moves them in various directions by way of attraction as a bait, and the small fishes, approaching either to examine or to seize them, immediately become the[Pg 170] prey of the fisher." (Yarrell.) In India we find a fresh-water Siluroid (Chaca lophioides) which "conceals itself among the mud, from which, by its lurid appearance and a number of loose filamentous substances on its skin, it is scarcely distinguishable; and with an immense open mouth it is ready to seize any small prey that is passing along." (Ham. Buchanan.) In March, 1868, I obtained a fine example of Ichthyscopus lebeck (Fishes of India, p. 261), which I placed in water having a bed of mud; into this it rapidly worked itself, first depressing one side and then another, until only the top of its head and mouth remained above the mud, whilst a constant current was kept up through its gills. It made a noise, half snapping and half croaking, when removed from its native element.
In the Royal Westminster Aquarium, says Dr. Day, is a live example of the electric eel (Electrophorus electricus) which has in its electric organs the means of showing when it is affected by anger or terror. Some consider this curious property is for protection against alligators: it is certainly used against fishes for the purpose of obtaining food; but when we remember how, when the Indians drive in horses and mules to the waters infested by the eels, they immediately attack them, we must admit that such cannot be for the purpose of preying upon them, but is due to anger or terror at being disturbed. (Day.)
Carrying Eggs in the Mouth.—Many catfishes (Siluridæ) carry their eggs in the mouth until hatched. The first and most complete account of this habit of catfishes is that by Dr. Jeffries Wyman, which he communicated to the Boston Society of Natural History at its meeting on September 15, 1857. In 1859, in a paper entitled "On Some Unusual Modes of Gestation," Dr. Wyman published a full account of his observations as follows, here quoted from a paper on Surinam fishes by Evermann and Goldsborough:
"Among the Siluroid fishes of Guiana there are several species which, at certain seasons of the year, have their mouths and branchial cavities filled either with eggs or young, and, as is believed, for the purpose of incubation. My attention was first called to this singular habit by the late Dr. Francis W.[Pg 171] Cragin, formerly United States consul at Paramaribo, Surinam. In a letter dated August, 1854, he says:
"'The eggs you will receive are from another fish. The different fishermen have repeatedly assured me that these eggs in their nearly mature state are carried in the mouths of the parent till the young are relieved by the bursting of the sac. Do you either know or believe this to be so, and, if possible, where are the eggs conceived and how do they get into the mouth?'
"In the month of April, 1857, on visiting the market of Paramaribo, I found that this statement, which at first seemed to be very improbable, was correct as to the existence of eggs in the mouths of several species of fish. In a tray of fish which a negro woman offered for sale, I found the mouths of several filled with either eggs or young, and subsequently an abundance of opportunities occurred for repeating the observation. The kinds most commonly known to the colonists, especially to the negroes, are jara-bakka, njinge-njinge, kœpra, makrede, and one or two others, all belonging either to the genus Bagrus or one nearly allied to it. The first two are quite common in the market, and I have seen many specimens of them; for the last two I have the authority of negro fishermen, but have never seen them myself. The eggs in my collection are of three different sizes, indicating so many species, one of the three having been brought to me without the fish from which they were taken.
"The eggs become quite large before they leave the ovaries, and are arranged in three zones corresponding to three successive broods, and probably to be discharged in three successive years; the mature eggs of a jara-bakka 18 inches long measure three-fourths of an inch in diameter; those of the second zone, one-fourth; and those of the third are very minute, about one-sixteenth of an inch.
"A careful examination of eight specimens of njinge-njinge about 9 inches long gave the following results:
"The eggs in all instances were carried in the mouths of the males. This protection, or gestation of the eggs by the males, corresponds with what has been long noticed with regard to other fishes, as, for example, Syngnathus, where the mar[Pg 172]supial pouch for the eggs or young is found in the males only, and Gasterosteus, where the male constructs the nest and protects the eggs during incubation from the voracity of the females.
"In some individuals the eggs had been recently laid, in others they were hatched and the fœtus had grown at the expense of some other food than that derived from the yolk, as this last was not proportionally diminished in size, and the fœtus weighed more than the undeveloped egg. The number of eggs contained in the mouth was between twenty and thirty. The mouth and branchial cavity were very much distended, rounding out and distorting the whole hyoid and branchiostegal region. Some of the eggs even partially protruded from the mouth. The ova were not bruised or torn as if they had been bitten or forcibly held by the teeth. In many instances the fœtuses were still alive, though the parent had been dead for many hours.
"No young or eggs were found in the stomach, although the mouth was crammed to its fullest capacity.
"The above observations apply to njinge-njinge. With regard to jarra-bakka, I had but few opportunities for dissection, but in several instances the same conditions of the eggs were noticed as stated above; and in one instance, besides some nearly mature fœtuses contained in the mouth, two or three were squeezed apparently from the stomach, but not bearing any marks of violence or of the action of the gastric fluid. It is probable that these found their way into that last cavity after death, in consequence of the relaxation of the sphincter which separates the cavities of the mouth and the stomach. These facts lead to the conclusion that this is a mouth gestation, as the eggs are found there in all stages of development, and even for some time after they are hatched.
"The question will be very naturally asked, how under such circumstances these fishes are able to secure and swallow their food. I have made no observations bearing upon such a question. Unless the food consists of very minute particles it would seem necessary that during the time of feeding the eggs should be disgorged. If this supposition be correct, it would give a very probable explanation of the only fact which might be considered at variance with the conclusion stated above, viz., that[Pg 173] we have in these fishes a mouth gestation. In the mass of eggs with which the mouth is filled I have occasionally found the eggs, rarely more than one or two, of another species. The only way in which their presence may be accounted for, it seems to me, is by the supposition that while feeding the eggs are disgorged, and as these fishes are gregarious in their habits, when the ova are recovered the stray eggs of another species may be introduced into the mouth among those which naturally belong there."
One of the earliest accounts of this curious habit which we have seen is that by Dr. Günther, referring to specimens of Tachysurus fissus from Cayenne received from Prof. R. Owen:
"These specimens having had the cavity of the mouth and of the gills extended in an extraordinary manner, I was induced to examine the cause of it, when, to my great surprise, I found them filled with about twenty eggs, rather larger than an ordinary pea, perfectly uninjured, and with the embryos in a forward state of development. The specimens are males, from 6 to 7 inches long, and in each the stomach was almost empty.
"Although the eggs might have been put into the mouth of the fish by their captor, this does not appear probable. On the other hand, it is a well-known fact that the American Siluroids take care of their progeny in various ways; and I have no doubt that in this species and in its allies the males carry the eggs in their mouths, depositing them in places of safety and removing them when they fear the approach of danger or disturbance."
The Unsymmetrical Eyes of Flounders.—In the two great families of flounders and soles the head is unsymmetrically formed, the cranium being twisted and both eyes placed on the same side. The body is strongly compressed, and the side possessing the eyes is uppermost in all the actions of the fish. This upper side, whether right or left, is colored, while the eyeless side is white or very nearly so.
It is well known that in the very young flounder the body rests upright in the water. After a little there is a tendency to turn to one side and the lower eye begins its migration to the other side, the interorbital bones or part of them moving before[Pg 174] it. In most flounders the eye seems to move over the surface of the head, before the dorsal fin, or across the axil of its first ray. In the tropical genus Platophrys the movement of the eye is most easily followed, as the species reach a larger size than do most flounders before the change takes place. The larva, while symmetrical, is in all cases transparent.
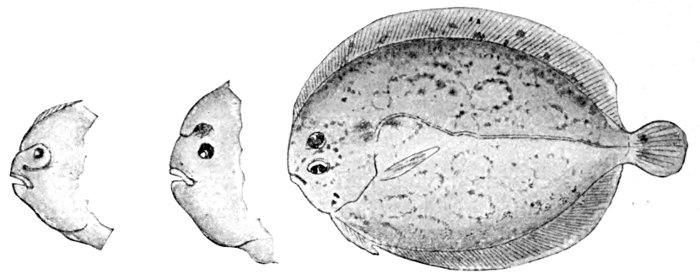
Fig. 126.
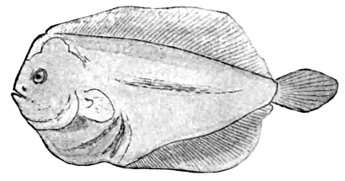
Fig. 127.
Figs. 126, 127.—Larval stages of Platophrys podas, a flounder of the Mediterranean, showing the migration of the eye. (After Emery.)
In a recent study of the migration of the eye in the winter flounder (Pseudopleuronectes americanus) Mr. Stephen R. Williams reaches the following conclusions:
1. The young of Limanda ferruginea (the rusty dab) are probably in the larval stage at the same time as those of Pseudopleuronectes americanus (the winter flounder).
2. The recently hatched fish are symmetrical, except for the relative positions of the two optic nerves.
3. The first observed occurrence in preparation for metamorphosis in P. americanus is the rapid resorption of the part of the supraorbital cartilage bar which lies in the path of the eye.
4. Correlated with this is an increase in distance between[Pg 175] the eyes and the brain, caused by the growth of the facial cartilages.
5. The migrating eye moves through an arc of about 120 degrees.
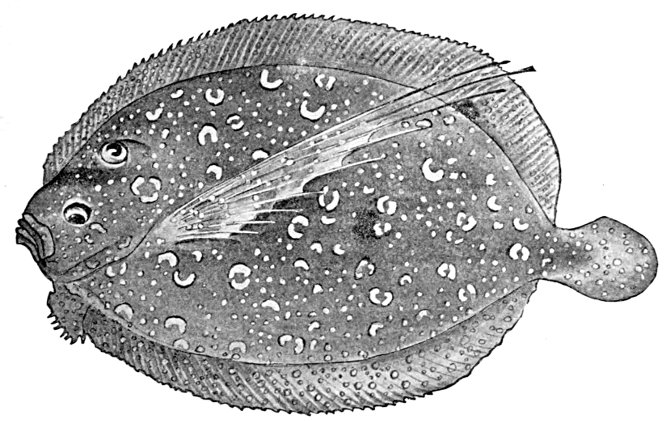
Fig. 128.—Platophrys lunatus (Linnæus), the Wide-eyed Flounder. Family Pleuronectidæ. Cuba. (From nature by Mrs. H. C. Nash.)
6. The greater part of this rotation (three-fourths of it in P. americanus) is a rapid process, taking not more than three days.
7. The anterior ethmoidal region is not so strongly influenced by the twisting as the ocular region.
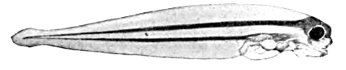
Fig. 129.—Young Flounder, just hatched, with symmetrical eyes. (After S. R. Williams.)
8. The location of the olfactory nerves (in the adult) shows that the morphological midline follows the interorbital septum.
9. The cartilage mass lying in the front part of the orbit of the adult eye is a separate anterior structure in the larva.
10. With unimportant differences, the process of metamorphosis in the sinistral fish is parallel to that in the dextral fish.
11. The original location of the eye is indicated in the adult by the direction first taken, as they leave the brain, by those cranial nerves having to do with the transposed eye.
12. The only well-marked asymmetry in the adult brain is due to the much larger size of the olfactory nerve and lobe of the ocular side.
13. There is a perfect chiasma.
14. The optic nerve of the migrating eye is always anterior to that of the other eye.

Fig. 130.—Larval Flounder, Pseudopleuronectes americanus. (After S. R. Williams.)

Fig. 131.—Larval Flounder, Pseudopleuronectes americanus. (After S. R. Williams.)
"The why of the peculiar metamorphosis of the Pleuronectidæ is an unsolved problem. The presence or absence of a swim-bladder can have nothing to do with the change of habit of the young flatfish, for P. americanus must lose its air-bladder before metamorphosis begins, since sections showed no evidence of it, whereas in Lophopsetta maculata, 'the windowpane flounder,' the air-sac can often be seen by the naked eye up to the time when the fish assumes the adult coloration, and long after it has assumed the adult form.
"Cunningham has suggested that the weight of the fish acting upon the lower eye after the turning would press it toward the upper side out of the way. But in all probability the planktonic larva rests on the sea-bottom little if at all before metamorphosing. Those taken by Mr. Williams into the laboratory showed in resting no preference for either side until the eye was near the midline.
"The fact that the change in all fishes is repeated during the development of each individual fish has been used to support the proposition that the flatfishes as a family are a comparatively recent product. They are, on the other hand, comparatively ancient. According to Zittel flatfishes of species referable to genera living at present, Rhombus (Bothus) and Solea, are found in the Eocene deposits. These two genera are notable in that Bothus is one of the least and Solea the most unsymmetrical of the Pleuronectidæ.
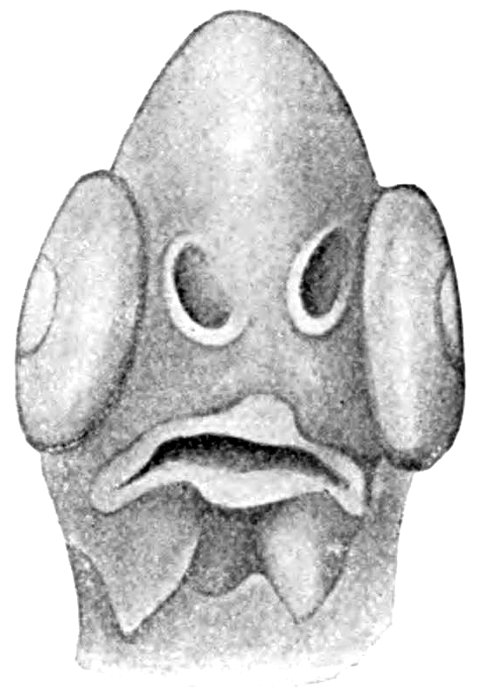
Fig. 132.—Face view of recently hatched Flounder. (After S. R. Williams.)
"The degree of asymmetry can be correlated with the habit of the animal. Those fishes, such as the sole and shore-dwelling flounders, which keep to the bottom are the most twisted representatives of the family, while the more freely swimming forms, like the sand-dab, summer flounder, and halibut, are more nearly symmetrical. Asymmetry must be of more advantage to those fishes which grub in the mud for their food than to those which capture other fishes; of the latter those which move with the greatest freedom are the most symmetrical.
"This deviation from the bilateral condition must have come about either as a 'sport' or by gradual modification of the adults. If by the latter method—the change proving to be advantageous—selection favored its appearing earlier and earlier in ontogeny, until it occurred in the stages of planktonic life. Metamorphosis at a stage earlier than this would be a distinct disadvantage, because of the lack of the customary planktonic food at the sea-bottom. At present some forms of selection are probably continually at work fixing the limit of the period of metamorphosis by the removal of those individuals which attempt the transformation at unsuitable epochs; for instance, at the time of hatching. That there are such individuals is shown by Fullarton, who figures a fish just hatched 'anticipating the twisting and subsequent unequal development exhibited by the head of Pleuronectids.' Those larvæ which remain pelagic until better able to compete at the sea-bottom[Pg 178] become the adults which fix the time of metamorphosis on their progeny." (S. R. Williams.)
So far as known to the writer, the metamorphosis of flounders always occurs while the individual is still translucent and swimming at the surface of the sea before sinking to the bottom.
[11] Cantor, Catal. Malayan Fishes, 1850, p. 87. Bowring, Siam, p. 155, gives a similar account of the battles of these fishes.
[12] Francis Day, on Fear and Anger in Fishes, Proc. Zool. Society, London, Feb. 19, 1878, pp. 214-221.
[13] Couch (Illustrations, etc., p. 305) says: "The faculty of giving forth brilliant light from the eyes is said to have been observed by fishermen in the blue shark, as in a cat."
[14] Couch, "British Fishes," 1865, vol. iv. p. 172.
[15] Himalayan Journals, vol. i. p. 80.
[16] Hakluyt, vol. ii. p. 37.
[17] Jerdon, "Madras Journal of Literature and Science," 1849, p. 143.
[18] "Observations on the Past and Present Condition of Onjein," Journal of the Asiatic Society of Bengal, vi, p. 820.
Fig. 133.—Mad-tom, Schilbeodes furiosus Jordan and Meek. Showing the poisoned pectoral spine. Family Siluridæ. Neuse River.

Spines of the Catfishes.—The catfishes or horned pouts (Siluridæ) have a strong spine in the pectoral fin, one or both edges of this being jagged or serrated. This spine fits into a peculiar joint and by means of a slight downward or forward twist can be set immovably. It can then be broken more easily than it can be depressed. A slight turn in the opposite direction releases the joint, a fact known to the fish and readily learned by the boy. The sharp spine inflicts a jagged wound. Pelicans which have swallowed the catfish have been known to die of the wounds inflicted by the fish's spine. When the catfish was first introduced into the Sacramento, according to Mr. Will S. Green, it caused the death of many of the native "Sacramento perch" (Archoplites interruptus). This perch (or rather bass) fed on the young catfish, and the latter erecting their pectoral spines in turn caused the death of the perch by tearing the walls of its stomach. In like manner the sharp dorsal and ventral spines of the sticklebacks have been known to cause the death of fishes who swallow them, and even of ducks. In Puget Sound the stickleback is often known as salmon-killer.
Certain small catfishes known as stone-cats and mad-toms (Noturus, Schilbeodes), found in the rivers of the Southern and Middle Western States, are provided with special organs of offense. At the base of the pectoral spine, which is sometimes very jagged, is a structure supposed by Professor Cope to be a poison gland the nature of which has not yet been fully ascertained. The wounds made by these spines are exceedingly painful like those made by the sting of a wasp. They are, however, apparently not dangerous.
Fig. 134.—Black Nohu, or Poison-fish, Emmydrichthys vulcanus Jordan. A species with stinging spines, showing resemblance to lumps of lava among which it lives. Family Scorpænidæ. From Tahiti.
Venomous Spines.—Many species of scorpion-fishes (Scorpæna, Synanceia, Pelor, Pterois, etc.), found in warm seas, as well as the European weavers (Trachinus), secrete poison from under the skin of each dorsal spine. The wounds made by these spines are very exasperating, but are not often dangerous. In some cases the glands producing these poisons form an oblong bag excreting a milky juice, and placed on the base of the spine.
In Thalassophryne, a genus of toad-fishes of tropical America, is found the most perfect system of poison organs known among fishes. The spinous armature of the opercle and the two spines of the first dorsal fin constitute the weapons. The details are known from the dissections of Dr. Günther. According to his[19] observations, the opercle in Thalassophryne "is very narrow, [Pg 181]vertically styliform and very mobile. It is armed behind with a spine eight lines long and of the same form as the hollow venom-fang of a snake, being perforated at its base and at its extremity. A sac covering the base of the spine discharges its contents through the apertures and the canal in the interior of the spine. The structure of the dorsal spines is similar. There are no secretory glands imbedded in the membranes of the sacs and the fluid must be secreted by their mucous membrane. The sacs are without an external muscular layer and situated immediately below the thick, loose skin which envelops the spines at their extremity. The ejection of the poison into a living animal, therefore, can only be effected as in Synanceia, by the pressure to which the sac is subjected the moment the spine enters another body."
Fig. 135.—Brown Tang, Teuthis bahianus (Ranzani). Tortugas, Florida.
The Lancet of the Surgeon-fish.—Some fishes defend themselves by lashing their enemies with their tails. In the tangs, or surgeon-fishes (Teuthis), the tail is provided with a formidable weapon, a knife-like spine, with the sharp edge directed forward. This spine when not in use slips forward into a sheath. The fish, when alive, cannot be handled without danger of a severe cut.
In the related genera, this lancet is very much more blunt and immovable, degenerating at last into the rough spines of Balistapus or the hair-like prickles of Monacanthus.
Spines of the Sting-ray.—In all the large group of sting-rays the tail is provided with one or more large, stiff, barbed spines, which are used with great force by the animal, and are capable of piercing the leathery skin of the sting-ray itself. There is no evidence that these spines bear any specific poison, but the ragged wounds they make are always dangerous and often end in gangrene. It is possible that the mucus on the surface of the spine acts as a poison on the lacerated tissues, rendering the wound something very different from a simple cut.
Fig. 136.—Common Filefish, Stephanolepis hispidus (Linnæus). Virginia.
Protection Through Poisonous Flesh of Fishes.—In certain groups of fishes a strange form of self-protection is acquired by the presence in the body of poisonous alkaloids, by means of which the enemies of the species are destroyed in the death of the individual devoured.
Such alkaloids are present in the globefishes (Tetraodontidæ), the filefishes (Monacanthus), and in some related forms, while members of other groups (Batrachoididæ) are under suspicion in this regard. The alkaloids produce a disease known as ciguatera, characterized by paralysis and gastric derangements. Severe cases of ciguatera with men, as well as with lower animals, may end fatally in a short time.
The flesh of the filefishes (Stephanolepis tomentosus), which[Pg 183] the writer has tested, is very meager and bitter, having a decidedly offensive taste. It is suspected, probably justly, of being poisonous. In the globefishes the flesh is always more or less poisonous, that of Tetraodon hispidus, called muki-muki, or death-fish, in Hawaii, is reputed as excessively so. The poisonous fishes have been lately studied in detail by Dr. Jacques Pellegrin, of the Museum d'Histoire Naturelle at Paris. He shows that any species of fish may be poisonous under certain circumstances, that under certain conditions certain species are poisonous, and that certain kinds are poisonous more or less at all times. The following account is condensed from Dr. Pellegrin's observations.
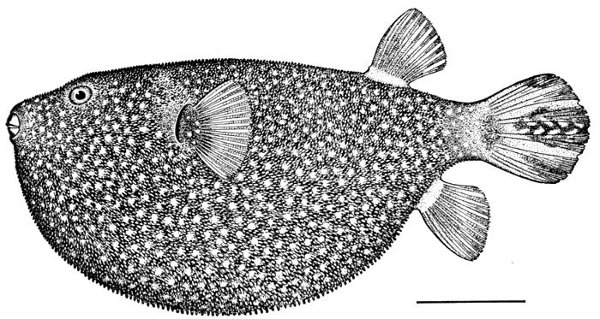
Fig. 137.—Tetraodon meleagris (Lacépède). Riu Kiu Islands.
The flesh of fishes soon undergoes decomposition in hot climates. The consumption of decayed fish may produce serious disorders, usually with symptoms of diarrhœa or eruption of the skin. There is in this case no specific poison, but the formation of leucomaines through the influence of bacteria. This may take place with other kinds of flesh, and is known as botulism, or allantiasis. For this disease, as produced by the flesh of fishes, Dr. Pellegrin suggests the name of ichthyosism It is especially severe in certain very oily fishes, as the tunny, the anchovy, or the salmon. The flesh of these and other fishes occasionally produces similar disorders through mere indigestion. In this case the flesh undergoes decay in the stomach.
In certain groups (wrasse-fishes, parrot-fishes, etc.) in the tropics, individual fishes are sometimes rendered poisonous by feeding on poisonous mussels, holothurians, or possibly polyps, species which at certain times, and especially in their spawning season, develops alkaloids which themselves may cause ciguatera. In this case it is usually the very old or large fishes which are liable to be infected. In some markets numerous species are excluded as suspicious for this reason. Such a list is in use in the fish-market of Havana, where the sale of certain species, elsewhere healthful, or at the most suspected, was rigidly prohibited under the Spanish régime. A list of these suspicious fishes has been given by Prof. Poey.
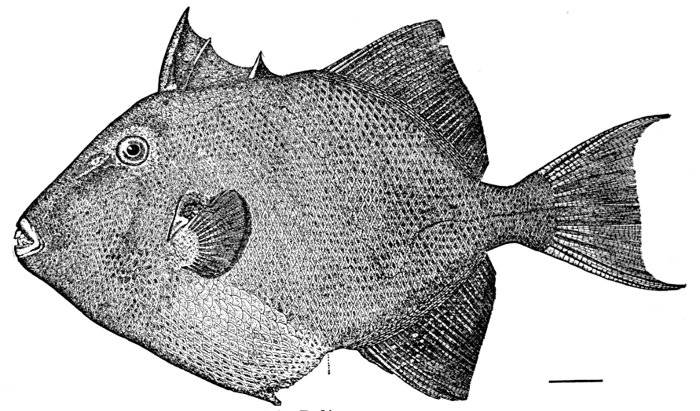
Fig. 138.—The Trigger-fish, Balistes carolinensis Gmelin. New York.
In many of the eels the serum of the blood is poisonous, but its venom is destroyed by the gastric juice, so that the flesh may be eaten with impunity, unless decay has set in. To eat too much of the tropical morays is to invite gastric troubles, but no true ciguatera. The true ciguatera is produced by a specific poisonous alkaloid. This is most developed in the globefishes or puffers (Tetraodon, Spheroides, Tropidichthys, etc.). It is present in the filefishes (Monacanthus, Alutera, etc.), probably in some toad-fishes (Batrachoides, etc.), and similar compounds are found in the flesh of sharks and especially in sharks' livers.
These alkaloids are most developed in the ovaries and testes, and in the spawning season. They are also found in the liver and sometimes elsewhere in the body. In many species otherwise innocuous, purgative alkaloids are developed in or about the eggs. Serious illness has been caused by eating the roe of the pike and the barbel. The poison is less virulent in the species which ascend the rivers. It is also much less developed in cooler waters. For this reason ciguatera is almost confined to the tropics. In Havana, Manila, and other tropical ports it is of frequent occurrence, while northward it is practically unknown as a disease requiring a special name or treatment. On the coast of Alaska, about Prince William Sound and Cook Inlet, a fatal disease resembling ciguatera has been occasionally produced by the eating of clams.
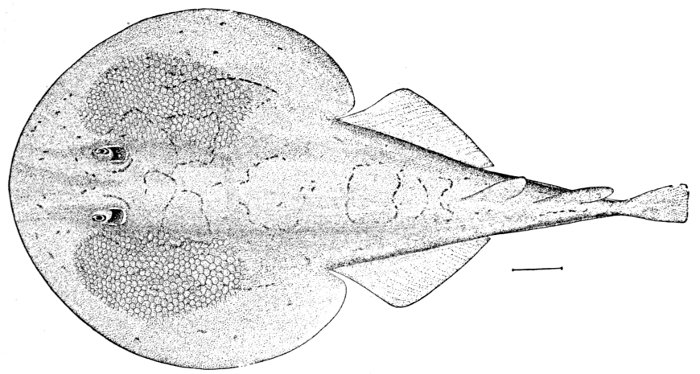
Fig. 139.—Numbfish, Narcine brasiliensis Henle, showing electric cells. Pensacola, Florida.
The purpose of the alkaloids producing ciguatera is considered by Dr. Pellegrin as protective, saving the species by the poisoning of its enemies. The sickness caused by the specific poison must be separated from that produced by ptomaines and leucomaines in decaying flesh or in the oil diffused through it. Poisonous bacteria may be destroyed by cooking, but the alkaloids which cause ciguatera are unaltered by heat.
It is claimed in tropical regions that the germs of the bubonic plague may be carried through the mediation of fishes which feed on sewage. It is suggested by Dr. Charles B. Ash[Pg 186]mead that leprosy may be so carried. It is further suggested that the custom of eating the flesh of fishes raw almost universal in Japan, Hawaii, and other regions may be responsible for the spread of certain contagious diseases, in which the fish acts as an intermediate host, much as certain mosquitoes spread the germ of malaria and yellow fever.
Electric Fishes.—Several species of fishes possess the power to inflict electric shocks not unlike those of the Leyden jar. This is useful in stunning their prey and especially in confounding their enemies. In most cases these electric organs are evidently developed from muscular substance. Their action, which is largely voluntary, is in its nature like muscular action. The power is soon exhausted and must be restored by rest and food. The effects of artificial stimulation and of poisons are parallel with the effect of similar agents on muscles.

Fig. 140.—Electric Catfish, Torpedo electricus (Gmelin). Congo River. (Alter Boulenger.)
In the electric rays or torpedos (Narcobatidæ) the electric organs are large honeycomb-like structures, "vertical hexagonal prisms," upwards of 400 of them, at the base of the pectoral fins. Each prism is filled "with a clear trembling jelly-like substance." These fishes give a shock which is communicable through a metallic conductor, as an iron spear or the handle of a knife. It produces a peculiar and disagreeable sensation not at all dangerous. It is said that this living battery shows all the known qualities of magnetism, rendering the needle magnetic, decomposing chemical compounds, etc. In the Nile is an electric catfish (Torpedo electricus) having similar powers. Its electric organ extends over the whole body, being thickest below. It consists of rhomboidal cells of a firm gelatinous substance.
The electric eel (Electrophorus electricus), the most powerful[Pg 187] of electric fishes, is not an eel, but allied rather to the sucker or carp. It is, however, eel-like in form and lives in rivers of Brazil and Guiana. The electric organs are in two pairs, one on the back of the tail, the other on the anal fin. These are made up of an enormous number of minute cells. In the electric eel, as in the other electric fishes, the nerves supplying these organs are much larger than those passing from the spinal cord for any other purpose. In all these cases closely related species show a no trace of the electric powers.

Fig. 141.—Star-gazer (Astroscopus guttatus) settling in the sand. (From life by R. W. Shufeldt.)
Dr. Gilbert has described the electric powers of species of star-gazer (Astroscopus y-græcum and A. zephyreus), the electric cells lying under the naked skin of the top of the head. Electric power is ascribed to a species of cusk (Urophycis regius), but this perhaps needs verification.
Photophores or Luminous Organs.—Many fishes, chiefly of the deep seas, develop organs for producing light. These are known as luminous organs, phosphorescent organs, or photophores. These are independently developed in four entirely unrelated groups of fishes. This difference in origin is accompanied by corresponding difference in structure. The best-known type is found in the Iniomi, including the lantern-fishes and their many relatives. These may have luminous spots, differ[Pg 188]entiated areas round or oblong which shine star-like in the dark. These are usually symmetrically placed on the sides of the body. They may have also luminous glands or diffuse areas which are luminous, but which do not show the specialized structure of the phosphorescent spots. These glands of similar nature to the spots are mostly on the head or tail. In one genus, Æthoprora, the luminous snout is compared to the headlight of an engine.
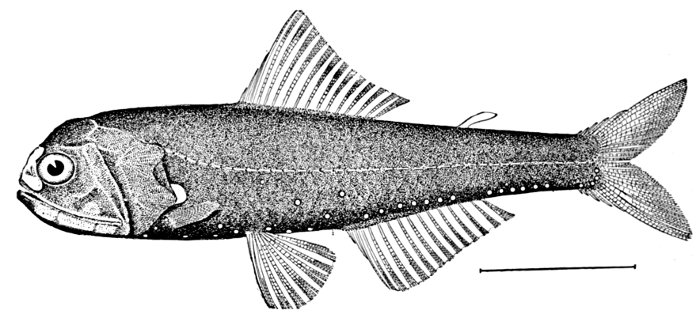
Fig. 142.—Headlight Fish, Æthoprora lucida Goode and Bean. Gulf Stream.

Fig. 143.—Corynolophus reinhardti (Lütken), showing luminous bulb (modified after Lütken). Family Ceratiidæ. Deep sea off Greenland.
Entirely different are the photophores in the midshipman or singing-fish (Porichthys), a genus of toad-fishes or Batrachoididæ. This species lives near the shore and the luminous spots are outgrowths from pores of the lateral line.
In one of the anglers (Corynolophus reinhardti) the complex bait is said to be luminous, and luminous areas are said to occur on the belly of a very small shark of the deep seas of Japan (Etmopterus lucifer). This phenomenon is now the subject of study by one of the numerous pupils of Dr. Mitsukuri. The structures in Corynolophus are practically unknown.
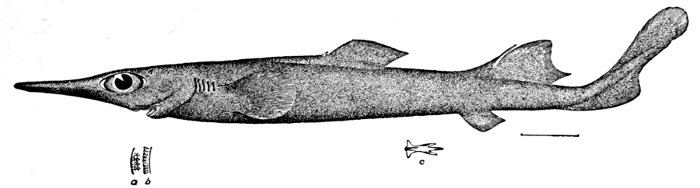
Fig. 144.—Etmopterus lucifer Jordan and Snyder. Misaki, Japan.
Photophores in Iniomous Fishes.—In the Iniomi the luminous organs have been the subject of an elaborate paper by Dr. R. von Lendenfeld (Deep-sea Fishes of the Challenger. Appendix B). These he divides into ocellar organs of regular form or luminous spots, and irregular glandular organs or luminous areas. The ocellar spots may be on the scales of the lateral line or on other definite areas. They may be raised above the surface or sunk below it. They may be simple, with or without black pigment, or they may have within them a reflecting surface. They are best shown in the Myctophidæ and Stomiatidæ, but are found in numerous other families in nearly all soft-rayed fishes of the deep sea.
The glandular areas may be placed on the lower jaw, on the barbels, under the gill cover, on the suborbital or preorbital, on the tail, or they may be irregularly scattered. Those about the eye have usually the reflecting membrane.
In all these structures, according to Dr. von Lendenfeld, the whole or part of the organ is glandular. The glandular part is at the base and the other structures are added distally. The primitive organ was a gland which produced luminous slime.[Pg 190] To this in the process of specialization greater complexity has been added.
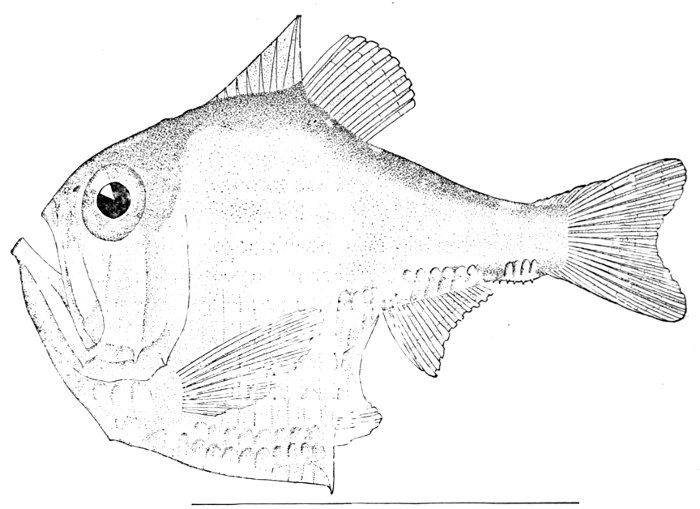
Fig. 145.—Argyropelecus olfersi Cuvier. Gulf Stream.
The luminous organs of some fishes resemble the supposed original structure of the primitive photophore, though of course these cannot actually represent it. The simplest type of photophore now found is in Astronesthes, in the form of irregular glandular luminous patches on the surface of the skin. There is no homology between the luminous organs of any insect and those of any fish.
Photophores of Porichthys.—Entirely distinct in their origin are the luminous spots in the midshipman (Porichthys notatus), a shore fish of California. These have been described in detail by Dr. Charles Wilson Greene (late of Stanford University, now of the University of Missouri) in the Journal of Morphology, xv., p. 667. These are found on various parts of the body in connection with the mucous pores of the lateral lines and about the mucous pores of the head. The skin in Porichthys is naked, and the photophores arise from a modification of its epidermis. Each is spherical, shining white, and consists of four parts—the[Pg 191] lens, the gland, the reflector, and the pigment. As to its function Prof. Greene observes:
"I have kept specimens of Porichthys in aquaria at the Hopkins Seaside Laboratory, and have made numerous observations on them with an effort to secure ocular proof of the phosphorescence of the living active fish. The fish was observed in the dark when quiet and when violently excited, but, with a single exception, only negative results were obtained. Once a phosphorescent glow of scarcely perceptible intensity was observed when the fish was pressed against the side of the aquarium. Then, this is a shore fish and quite common, and one might suppose that so striking a phenomenon as it would present if these organs were phosphorescent in a small degree would be observed by ichthyologists in the field, or by fishermen, but diligent inquiry reveals no such evidence.
"Notwithstanding the fact that Porichthys has been observed to voluntarily exhibit only the trace of phosphorescence mentioned above, still the organs which it possesses in such numbers are beyond doubt true phosphorescent organs, as the following observations will demonstrate. A live fish put into an aquarium of sea-water made alkaline with ammonia water exhibited a most brilliant glow along the location of the well-developed organs. Not only did the lines of organs shine forth, but the individual organs themselves were distinguishable. The glow appeared after about five minutes, remained prominent for a few minutes, and then for twenty minutes gradually became weaker until it was scarcely perceptible. Rubbing the hand over the organs was followed always by a distinct increase in the phosphorescence. Pieces of the fish containing the organs taken five and six hours after the death of the animal became luminous upon treatment with ammonia water.
"Electrical stimulation of the live fish was also tried with good success. The interrupted current from an induction coil was used, one electrode being fixed on the head over the brain or on the exposed spinal cord near the brain, and the other moved around on different parts of the body. No results followed relatively weak stimulation of the fish, although such currents produced violent contractions of the muscular system[Pg 192] of the body. But when a current strong enough to be quite painful to the hands while handling the electrodes was used then stimulation of the fish called forth a brilliant glow of light apparently from every well-developed photophore. All the lines on the ventral and lateral surfaces of the body glowed with a beautiful light, and continued to do so while the stimulation lasted. The single well-developed organ just back of and below the eye was especially prominent. No luminosity was observed in the region of the dorsal organs previously described as rudimentary in structure. I was also able to produce the same effect by galvanic stimulation, rapidly making and breaking the current by hand.
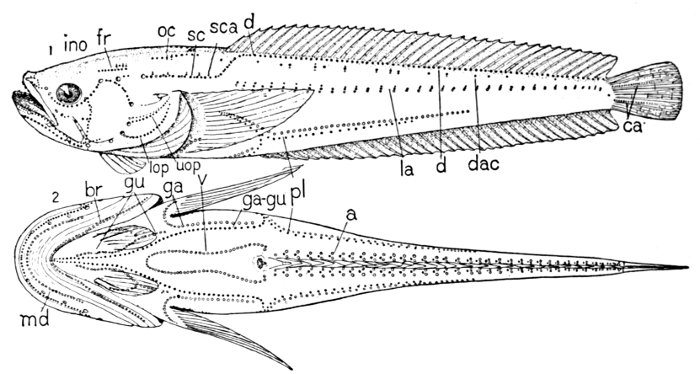
Fig. 146.—Luminous organs and lateral line of Midshipman, Porichthys notatus Girard. Family Batrachoididæ. Monterey, California. (After Greene.)
"The light produced in Porichthys was, as near as could be determined by direct observation, a white light. When produced by electric stimulation it did not suddenly reach its maximal intensity, but came in quite gradually and disappeared in the same way when the stimulation ceased. The light was not a strong one, only strong enough to enable one to quite easily distinguish the apparatus used in the experiment.
"An important fact brought out by the above experiment is that an electrical stimulation strong enough to most violently stimulate the nervous system, as shown by the violent contractions of the muscular system, may still be too weak to produce phosphorescence. This fact gives a physiological con[Pg 193]firmation of the morphological result stated above that no specific nerves are distributed to the phosphorescent organs.
"I can explain the action of the electrical current in these experiments only on the supposition that it produces its effect by direct action on the gland.

Fig. 147.—Cross-section of a ventral phosphorescent organ of the Midshipman, Porichthys notatus Girard. l, lens; gl, gland; r, reflector; bl, blood; p, pigment. (After Greene.)
"The experiments just related were all tried on specimens of the fish taken from under the rocks where they were guarding the young brood. Two specimens, however, taken by hooks from the deeper water of Monterey Bay, could not be made to show phosphorescence either by electrical stimulation or by treatment with ammonia. These specimens did net have the high development of the system of mucous cells of the skin exhibited by the nesting fish. My observations were, how[Pg 194]ever, not numerous enough to more than suggest the possibility of a seasonal high development of the phosphorescent organs.
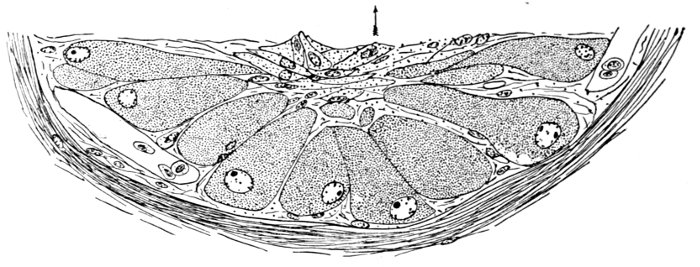
Fig. 148.—Section of the deeper portion of phosphorescent organ of Porichthys notatus, highly magnified. (After Greene.)
"Two of the most important parts of the organ have to do with the physical manipulation of light—the reflector and the lens, respectively. The property of the reflector needs no discussion other than to call attention to its enormous development. The lens cells are composed of a highly refractive substance, and the part as a whole gives every evidence of light refraction and condensation. The form of the lens gives a theoretical condensation of light at a very short focus. That such is in reality the case, I have proved conclusively by examination of fresh material. If the fresh fish be exposed to direct sunlight, there is a reflected spot of intense light from each phosphorescent organ. This spot is constant in position with reference to the sun in whatever position the fish be turned and is lost if the lens be dissected away and only the reflector left. With needles and a simple microscope it is comparatively easy to free the lens from the surrounding tissue and to examine it directly. When thus freed and examined in normal saline, I have found by rough estimates that it condenses sunlight to a bright point a distance back of the lens of from one-fourth to one-half its diameter. I regret that I have been unable to make precise physical developments.
"The literature on the histological structure of known phosphorescent organs of fishes is rather meager and unsatisfactory. Von Lendenfeld describes twelve classes of phosphorescent organs from deep-sea fishes collected by the Challenger expe[Pg 195]dition. All of these, however, are greater or less modifications of one type. This type includes, according to von Lendenfeld's views, three essential parts, i.e., a gland, phosphorescent cells, and a local ganglion. These parts may have added a reflector, a pigment layer, or both; and all these may be simple or compounded in various ways, giving rise to the twelve classes. Blood-vessels and nerves are distributed to the glandular portion. Of the twelve classes direct ocular proof is given for one, i.e., ocellar organs of Myctophum which were observed by Willemoes-Suhm at night to shine 'like a star in the net.' Von Lendenfeld says that the gland produces a secretion, and he supposes the light or phosphorescence to be produced either by the 'burning or consuming' of this secretion by the phosphorescent cells, or else by some substance produced by the phosphorescent cells. Furthermore, he says that the phosphorescent cells act at the 'will of the fish' and are excited to action by the local ganglion.
"Some of these statements and conclusions seem insufficiently grounded, as, for example, the supposed action of the phosphorescent cells, and especially the control of the ganglion over them. In the first place, the relation between the ganglion and the central nervous system in the forms described by von Lendenfeld is very obscure, and the structure described as a ganglion, to judge from the figures and the text descriptions, may be wrongly identified. At least it is scarcely safe to ascribe ganglionic function to a group of adult cells so poorly preserved that only nuclei are to be distinguished. In the second place, no structural character is shown to belong to the 'phosphorescent cells' by which they may take part in the process ascribed to them.[20]
"The action of the organs described by him may be explained on other grounds, and entirely independent of the so-called 'ganglion cells' and of the 'phosphorescent cells.'
"Phosphorescence as applied to the production of light by a living animal is, according to our present ideas, a chemical action, an oxidation process. The necessary conditions for producing it are two—an oxidizable substance that is luminous on oxidation, i.e., a photogenic substance on the one hand, and the presence of free oxygen on the other. Every phosphorescent organ must have a mechanism for producing these two conditions; all other factors are only secondary and accessory. If the gland of a firefly can produce a substance that is oxidizable and luminous on oxidation, as shown as far back as 1828 by Faraday and confirmed and extended recently by Watasé, it is conceivable, indeed probable, that phosphorescence in Myctophum and other deep-sea forms is produced in the same direct way, that is, by direct oxidation of the secretion of the gland found in each of at least ten of the twelve groups of organs described by von Lendenfeld. Free oxygen may be supplied directly from the blood in the capillaries distributed to the gland which he describes. The possibility of the regulation of the supply of blood carrying oxygen is analogous to what takes place in the firefly and is wholly adequate to account for any 'flashes of light' 'at the will of the fish.'
"In the phosphorescent organs of Porichthys the only part the function of which cannot be explained on physical grounds is the group of cells called the gland. If the large granular cells of this portion of the structure produce a secretion, as seems probable from the character of the cells and their behavior toward reagents, and this substance be oxidizable and luminous in the presence of free oxygen, i.e., photogenic, then we have the conditions necessary for a light-producing organ. The numerous capillaries distributed to the gland will supply free oxygen sufficient to meet the needs of the case. Light produced in the gland is ultimately all projected to the exterior, either directly from the luminous points in the gland or reflected outward by the reflector, the lens condensing all the rays into a definite pencil or slightly diverging cone. This explanation of the light-producing process rests on the assumption of a secretion product with certain specific characters. But comparing the organ with structures known to produce such a substance, i.e., the glands of the firefly or the photospheres of Eu[Pg 197]phausia, it seems to me the assumption is not less certain than the assumption that twelve structures resembling each other in certain particulars have a common function to that proved for one only of the twelve.
"I am inclined to the belief that whatever regulation of the action of the phosphorescent organ occurs is controlled by the regulation of the supply of free oxygen by the blood-stream flowing through the organ; but, however this may be, the essential fact remains that the organs in Porichthys are true phosphorescent organs." (Greene.)
Other species of Porichthys with similar photophores occur in Texas, Guiana, Panama, and Chile. The name midshipman alludes to these shining spots, compared to buttons.
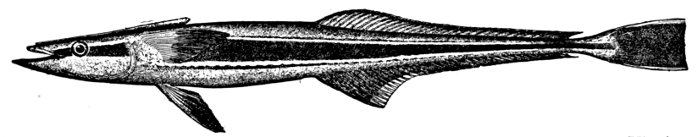
Fig. 149.—Sucking-fish, or Pegador, Leptecheneis naucrates (Linnæus). Virginia.
Globefishes.—The globefishes (Tetraodon, etc.) and the porcupine-fishes have the surface defended by spines. These fishes have an additional safeguard through the instinct to swallow air. When one of these fishes is seriously disturbed it rises to the surface, gulps air into a capacious sac, and then floats belly upward on the surface. It is thus protected from other fishes, although easily taken by man. The same habit appears in some of the frog-fishes (Antennarius) and in the Swell sharks (Cephaloscyllium).
The writer once hauled out a netful of globefishes (Tetraodon hispidus) from a Hawaiian lagoon. As they lay on the bank a dog came up and sniffed at them. As his nose touched them they swelled themselves up with air, becoming visibly two or three times as large as before. It is not often that the lower animals show surprise at natural phenomena, but the attitude of the dog left no question as to his feeling.
Remoras.—The different species of Remora, or shark-suckers, fasten themselves to the surface of sharks or other fishes and are carried about by them often to great distances. These[Pg 198] fishes attach themselves by a large sucking-disk on the top of the head, which is a modified spinous dorsal fin. They do not harm the shark, except possibly to retard its motion. If the shark is caught and drawn out of the water, these fishes often instantly let go and plunge into the sea, swimming away with great celerity.
Sucking-disks of Clingfishes.—Other fishes have sucking-disks differently made, by which they cling to rocks. In the gobies the united ventrals have some adhesive power. The blind goby (Typhlogobius californiensis) is said to adhere to rocks in dark holes by the ventral fins. In most gobies the adhesive power is slight. In the sea-snails (Liparididæ) and lumpfishes (Cyclopteridæ) the united ventral fins are modified into an elaborate circular sucking-disk. In the clingfishes (Gobiesocidæ) the sucking-disk lies between the ventral fins and is made in part of modified folds of the naked skin. Some fishes creep over the bottom, exploring it with their sensitive barbels, as the gurnard, surmullet, and goatfish. The suckers (Catostomus) test the bottom with their thick, sensitive lips, either puckered or papillose, feeding by suction.
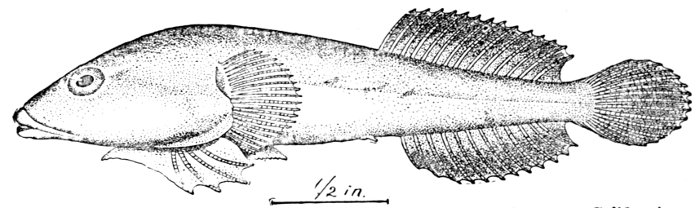
Fig. 150.—Clingfish, Caularchus mæandricus (Girard). Monterey, California.
Lampreys and Hagfishes.—The lampreys suck the blood of other fishes to which they fasten themselves by their disk-like mouth armed with rasping teeth.
The hagfishes (Myxine, Eptatretus) alone among fishes are truly parasitic. These fishes, worm-like in form, have round mouths, armed with strong hooked teeth. They fasten themselves at the throats of large fishes, work their way into the muscle without tearing the skin, and finally once inside devour all the muscles of the fish, leaving the skin unbroken and the viscera undisturbed. These fishes become living hulks before[Pg 199] they die. If lifted out of the water, the slimy hagfish at once slips out and swims quickly away. In gill-nets in Monterey Bay great mischief is done by hagfish (Polistotrema stouti). It is a curious fact that large numbers of hagfish eggs are taken from the stomachs of the male hagfish, which seems to be almost the only enemy of his own species, keeping the numbers in check.

Fig. 151.—Hagfish, Polistotrema stouti (Lockington).
The Swordfishes.—In the swordfish and its relatives, the sailfish and the spearfish, the bones of the anterior part of the head are grown together, making an efficient organ of attack. The sword of the swordfish, the most powerful of these fishes, has been known to pierce the long planks of boats, and it is supposed that the animal sometimes attacks the whale. But stories of this sort lack verification.
The Paddle-fishes.—In the paddle-fishes (Polyodon spatula and Psephurus gladius) the snout is spread out forming a broad paddle or spatula. This the animal uses to stir up the mud on the bottoms of rivers, the small organisms contained in mud constituting food. Similar paddle-like projections are developed in certain deep-water Chimæras (Harriottia, Rhinochimæra), and in the deep-sea shark, Mitsukurina.

Fig. 152.—Indian Sawfish, Pristis zysron Latham. River mouths of Hindustan. (After Day.)
The Sawfishes.—A certain genus of rays (Pristis, the sawfish) and a genus of sharks (Pristiophorus, the saw-shark), possess a similar spatula-shaped snout. But in these fishes the snout is provided on either side with enamelled teeth set in sockets and standing at right angles with the snout. The animal swims through schools of sardines and anchovies, strikes right and left with this saw, destroying the small fishes, who thus become an easy prey. These fishes live in estuaries and river mouths, Pristis in tropical America and Guinea, Pristiophorus in Japan and Australia. In the mythology of science, the sawfish attacks the whale, but in fact the two animals never come within miles of each other, and the sawfish is an object of danger only to the tender fishes, the small fry of the sea.

Fig. 153.—Saw-shark, Pristiophorus japonicus Günther. Specimen from Nagasaki.
Peculiarities of Jaws and Teeth.—The jaws of fishes are subject to a great variety of modifications. In some the bones are joined by distensible ligaments and the fish can swallow other fishes larger than itself. In other cases the jaws are excessively small and toothless, at the end of a long tube, so ineffective in appearance that it is a marvel that the fish can swallow anything at all.
In the thread-eels (Nemichthys) the jaws are so recurved that they cannot possibly meet, and in their great length seem worse than useless.
In some species the knife-like canines of the lower jaw pierce through the substance of the upper.
In four different and wholly unrelated groups of fishes the teeth are grown fast together, forming a horny beak like that of the parrot. These are the Chimæras, the globefishes (Tetraodon), and their relatives, the parrot-fishes (Scarus, etc.), and the stone-wall perch (Oplegnathus). The structure of the beak varies considerably in these four cases, in accord with the difference in the origin of its structures. In the globefishes the[Pg 202] jaw-bones are fused together, and in the Chimæras they are solidly joined to the cranium itself.
The Angler-fishes.—In the large group of angler-fishes the first spine of the dorsal fin is modified into a sort of bait to attract smaller fishes into the capacious mouth below. This structure is typical in the fishing-frog (Lophius), where the fleshy tip of this spine hangs over the great mouth, the huge fish lying on the bottom apparently inanimate as a stone. In other related fishes this spine has different forms, being often reduced to a vestige, of little value as a lure, but retained in accordance with the law of heredity. In a deep-sea angler the bait is enlarged, provided with fleshy streamers and a luminous body which serves to attract small fishes in the depths.
The forms and uses of this spine in this group constitute a very suggestive chapter in the study of specialization and ultimate degradation, when the special function is not needed or becomes ineffective.
Similar phases of excessive development and final degradation may be found in almost every group in which abnormal stress has been laid on a particular organ. Thus the ventral fins, made into a large sucking-disk in Liparis, are lost altogether in Paraliparis. The very large poisoned spines of Pterois become very short in Aploactis, the high dorsal spines of Citula are lost in Alectis, and sometimes a very large organ dwindles to a very small one within the limits of the same genus. An example of this is seen in the poisoned pectoral spines of Schilbeodes.
Relation of Number of Vertebræ to Temperature and the Struggle for Existence.—One of the most remarkable modifications of the skeleton of fishes is the progressive increase of the number of vertebræ as the forms become less specialized, and that this particular form of specialization is greatest at the equator.[21]
It has been known for some years that in several groups of [Pg 203]fishes (wrasse-fishes, flounders, and "rock-cod," for example) those species which inhabit northern waters have more vertebræ than those living in the tropics. Certain arctic flounders, for example, have sixty vertebræ; tropical flounders have, on the average, thirty. The significance of this fact is the problem at issue. In science it is assumed that all facts have significance, else they would not exist. It becomes necessary, then, to find out first just what the facts are in this regard.
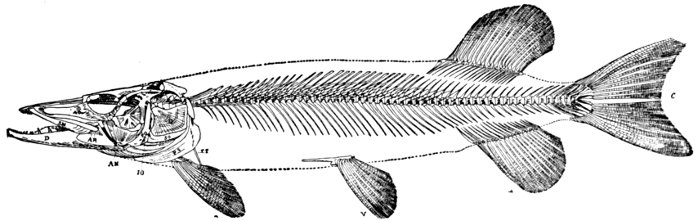
Fig. 154.—Skeleton of Pike, Esox lucius Linnæus, a river fish with many vertebræ.
Going through the various groups of non-migratory marine fishes we find that such relations are common. In almost every group the number of vertebræ grows smaller as we approach the equator, and grows larger again as we pass into southern latitudes. Taking an average netful of fishes of different kinds at different places along the coast, the variation would be evident. At Point Barrow or Cape Farewell or North Cape a seineful of fishes would perhaps average eighty vertebræ each, the body lengthened to make room for them; at Sitka or St. Johns or Bergen, perhaps sixty vertebræ; at San Francisco or New York or St. Malo, thirty-five; at Mazatlan or Pensacola or Naples, twenty-eight; and at Panama or Havana or Sierra Leone, twenty-five. Under the equator the usual number of vertebræ in shore fishes is twenty-four. Outside tropical and semi-tropical waters this number is the exception. North of Cape Cod it is virtually unknown.
Number of Vertebræ.—The numbers of vertebræ in different groups may be summarized as follows:
Lancelets.—Among the lancelets the numbers of segments range from 50 to 80, there being no vertebræ.
Lampreys.—In this group the number of segments ranges from 100 to 150.
Elasmobranchs.—Among sharks and skates the usual number of segments is from 100 to 150 and upwards. In the extinct species as far as known the numbers are not materially different. The Carboniferous genus, Pleuracanthus, has about 115 vertebræ. The Chimæras have similar numbers; Chimæra monstrosa has about 100 in the body and more than as many more in the filamentous tail.
Cycliæ.—Palæospondylus has about 85 vertebræ.
Arthrodires.—There are about 100 vertebræ in Coccosteus.
Dipnoans.—In Protopterus there are upwards of 100 vertebræ, the last much reduced in size. Figures of Neoceratodus show about 80.
Crossopterygians.—Polypterus has 67 vertebræ; Erpetichthys, 110; Undina, about 85.
Ganoids.—In this group the numbers are also large—95 in Amia, about 55 in the short-bodied Microdon. The Sturgeons all have more than 100 vertebræ.
Soft-rayed Fishes.—Among the Teleostei, or bony fishes, those which first appear in geological history are the Isospondyli, the allies of the salmon and herring. These have all numerous vertebræ, small in size, and none of them in any notable degree modified or specialized. They abound in the depths of the ocean, but there are comparatively few of them in the tropics. The Salmonidæ which inhabit the rivers and lakes of the northern zones have from 60 to 65 vertebræ. The Myctophidæ, Stomiatidæ, and other deep-sea forms have from 40 upwards in the few species in which the number has been counted. The group of Clupeidæ is nearer the primitive stock of Isospondyli than the salmon are. This group is essentially northern in its distribution, but a considerable number of its members are found within the tropics. The common herring (Clupea harangus) ranges farther into the arctic regions than any other. Its vertebræ are 56 in number. In the shad (Alosa sapidissima), a northern species which ascends the rivers, the same number is recorded. The sprat (Clupea sprattus) and sardine (Sardinia pilchardus), ranging farther south, have from 48 to 50, while in certain small herrings (Sardinella) which are strictly confined to tropical shores the number is but 40. Allied to the herring are the anchovies, mostly tropical. The northern[Pg 205]most species, the common anchovy of Europe (Engraulis enchrasicolus), has 46 vertebræ. A tropical species (Anchovia browni) has 41.
There are, however, a few soft-rayed fishes confined to the tropical seas in which the numbers of vertebræ are still large, an exception to the general rule. Among these are Albula vulpes, the bonefish, with 70 vertebræ, Elops saurus, the ten-pounder, with 72, the tarpon (Tarpon atlanticus), with about 50, and the milkfish, Chanos chanos, with 72.
In a fossil Eocene herring from the Green River shales (Diplomystus) I count 40 vertebræ; in a bass-like fish (Mioplosus) from the same locality 24—these being the usual numbers in the present tropical members of these groups.
The great family of Siluridæ, or catfishes, is represented in all the fresh waters of temperate and tropical America, as well as in the warmer parts of the Old World. One division of the family, containing numerous species, abounds on the sandy shores of the tropical seas. The others are all fresh-water fishes. So far as the vertebræ in the Siluridæ have been examined, no conclusions can be drawn. The vertebræ in the marine species range from 35 to 50; in the North American forms, from 37 to 45; and in the South American fresh-water species, where there is almost every imaginable variation in form and structure, the numbers range from 28 to 50 or more. The Cyprinidæ (carp and minnows), confined to the fresh waters of the northern hemisphere, and their analogues, the Characinidæ of the rivers of South America and Africa, have also numerous vertebræ, 36 to 50 in most cases.
In general we may say of the soft-rayed fishes that very few of them are inhabitants of tropical shores. Of these few, some which are closely related to northern forms have fewer vertebræ than their cold-water analogues. In the northern species, the fresh-water species, and the species found in the deep sea the number of vertebræ is always large, but the same is true of some of the tropical species also.
The Flounders.—In the flounders, the halibut and its relatives, arctic genera (Hippoglossus and Atheresthes), have from 49 to 50 vertebræ. The northern genera (Hippoglossoides, Lyopsetta, and Eopsetta) have from 43 to 45; the members of[Pg 206] a large semi-tropical genus (Paralichthys) of wide range have from 35 to 41; while the tropical forms have from 35 to 37.
In the group of turbots and whiffs none of the species really belong to the northern fauna, and the range in numbers is from 35 to 43. The highest number, 43, is found in a deep-water species (Monolene), and the next, 40, in species (Lepidorhombus, Orthopsetta) which extend their range well toward the north. Among the plaices, which are all northern, the numbers range from 35 to 65, the higher numbers, 52, 58, 65, being found in species (Glyptocephalus) which inhabit considerable depths in the arctic seas. The lowest numbers (35) belong to shore species (Pleuronichthys) which range well toward the south.
Spiny-rayed Fishes.—Among the spiny-rayed fishes the facts are more striking. Of these, numerous families are chiefly or wholly confined to the tropics, and in the great majority of all the species the number of vertebræ is constantly 24,—10 in the body and 14 in the tail (10+14). This is true of all or nearly all the Berycidæ, Serranidæ, Sparidæ, Sciænidæ, Chætodontidæ, Hæmulidæ, Gerridæ, Gobiidæ, Acanthuridæ, Mugilidæ, Sphyrænidæ, Mullidæ, Pomacentridæ, etc.
In some families in which the process of reduction has gone on to an extreme degree, as in certain Plectognath fishes, there has been a still further reduction, the lowest number, 14, existing in the short inflexible body of the trunkfish (Ostracion), in which the vertebral joints are movable only in the base of the tail. In all these forms the process of reduction of vertebræ has been accompanied by specialization in other respects. The range of distribution of these fishes is chiefly though not quite wholly confined to the tropics.
Thus Balistes, the trigger-fish, has 17 vertebræ; Monacanthus and Alutera, foolfishes, about 20; the trunkfish, Ostracion, 14; the puffers, Tetraodon and Spheroides, 18; Canthigaster, 17; and the headfish, Mola, 17. Among the Pediculates, Malthe and Antennarius have 17 to 19 vertebræ, while in their near relatives, the anglers, Lophiidæ, the number varies with the latitude. Thus, in the northern angler, Lophius piscatorius, which is never found south of Cape Hatteras, there are 30 vertebræ. In a similar species, inhabiting the north of Japan (Lophius litulon), there are 27. In another Japanese species, ranging[Pg 207] farther south, Lophiomus setigerus, the vertebræ are but 19. Yet in external appearance these two fishes are almost identical. It is, however, a notable fact that some of the deep-water Pediculates, or angling fishes, have the body very short and the number of vertebræ correspondingly reduced. Dibranchus atlanticus, from a depth of 3600 fathoms, or more than 4 miles, has but 18 vertebræ, and others of its relatives in deep waters show also small numbers. These soft-bodied fishes are simply animated mouths, with a feeble osseous structure, and they are perhaps recent offshoots from some stock which has extended its range from muddy bottom or from floating seaweed to the depths of the sea.
A very few spiny-rayed families are wholly confined to the northern seas. One of the most notable of these is the family of viviparous surf-fishes (Embiotocidæ), of which numerous species abound on the coasts of California and Japan, but which enter neither the waters of the frigid nor of the torrid zone. The surf-fishes have from 32 to 42 vertebræ, numbers which are never found among tropical fishes of similar appearance or relationship.
The facts of variation with latitude were first noticed among the Labridæ. In the northern genera (Labrus, Tautoga, etc.) there are 38 to 41 vertebræ; in the semi-tropical genera (Crenilabrus, Bodianus, etc.), 30 to 33; in the tropical genera (Halichœres, Xyrichthys, Thalassoma, etc.), usually 24.
Equally striking are the facts in the great group of Pareioplitæ, or mailed-cheek fishes, composed of numerous families, diverging from each other in various respects, but agreeing in certain peculiarities of the skeleton.
Among these fishes the family most nearly related to ordinary fishes is that of the Scorpænidæ (scorpion-fishes, etc.).
This is a large family containing many species, fishes of local habits, swarming about the rocks at moderate depths in all zones. The species of the tropical genera have all 24 vertebræ. Those genera chiefly found in cooler waters, as in California, Japan, Chile, and the Cape of Good Hope, have in all their species 27 vertebræ, while in the arctic genera there are 31.
Allied to the Scorpænidæ, but confined to the tropical or semi-tropical seas, are the Platycephalidæ, with 27 vertebræ, and[Pg 208] the Cephalacanthidæ (flying gurnards), with but 22. In the deeper waters of the tropics are the Peristediidæ, with 33 vertebræ, and extending farther north, belonging as much to the temperate as to the torrid zone, is the large family of the Triglidæ (gurnards) in which the vertebræ range from 25 to 38.
The family of Agonidæ (sea-poachers), with 36 to 40 vertebræ, is still more decidedly northern in its distribution. Wholly confined to northern waters is the great family of the Cottidæ (sculpins), in which the vertebræ ascend from 30 to 50. Entirely polar and often in deep waters are the Liparididæ (sea-snails), an offshoot from the Cottidæ, with soft, limp bodies, and the vertebræ 35 to 65. In these northern forms there are no scales, the spines in the fins have practically disappeared, and only the anatomy shows that they belong to the group of spiny-rayed fishes. In the Cyclopteridæ (lumpfishes), likewise largely arctic, the body becomes short and thick, the back-bone inflexible, and the vertebræ are again reduced to 28. In most cases, as the number of vertebræ increases, the body becomes proportionally elongate. As a result of this, the fishes of arctic waters are, for the most part, long and slender, and not a few of them approach the form of eels. In the tropics, however, while elongate fishes are common enough, most of them (always excepting the eels) have the normal number of vertebræ, the greater length being due to the elongation of their individual vertebræ and not to their increase in number. Thus the very slender goby, Gobionellus oceanicus, has the same number (25) of vertebræ as its thick-set relative Gobius soporator or the chubby Lophogobius cyprinoides. In the great group of blenny-like fishes the facts are equally striking. The arctic species are very slender in form as compared with the tropical blennies, and this fact, caused by a great increase in the number of their vertebræ, has led to the separation of the group into several families. The tropical forms composing the family of Blenniidæ have from 28 to 49 vertebræ, while in the arctic genera the numbers range from 75 to 100.
Of the true Blennidæ, which are all tropical or semi-tropical, Blennius has 28 to 35 vertebræ; Salarias, 35 to 38; Lepisoma, 34; Clinus, 49; Cristiceps, 40. A fresh-water species of Cristiceps found in Australia has 46. Blennioid fishes in the arctic seas are Anarrhichas, with 76 vertebræ; Anarrhichthys, with[Pg 209] 100 or more; Lumpenus, 79; Pholis, 85; Lycodes, 112; Gymnelis, 93. Lycodes and Gymnelis have lost all the dorsal spines.
In the cod family (Gadidæ) the number of vertebræ is usually about 50. The number is 51 in the codfish (Gadus callarias), 58 in the Siberian cod (Eleginus navaga), 54 in the haddock (Melanogrammus æglifinus), 54 in the whiting (Merlangus merlangus), 54 in the coalfish (Pollachius virens), 52 in the Alaskan coalfish (Theragra chalcogramma), 51 in the hake (Merluccius merluccius). In the burbot (Lota lota), the only fresh-water codfish, 59; in the deep-water ling (Molva molva), 64; in the rocklings (Gaidropsarus), 47 to 49. Those few species found in the Mediterranean and the Gulf of Mexico have fewer fin-rays and probably fewer vertebræ than the others, but none of the family enter warm water, the southern species living at greater depths.
In the deep-sea allies of the codfishes, the grenadiers or rat-tails (Macrouridæ), the numbers range from 65 to 80.
Fresh-water Fishes.—Of the families confined strictly to the fresh waters the great majority are among the soft-rayed or physostomous fishes, the allies of the salmon, pike, carp, and catfish. In all of these the vertebræ are numerous. A few fresh-water families have their affinities entirely with the more specialized forms of the tropical seas. Of these the Centrarchidæ (comprising the American fresh-water sunfish and black bass) have on the average about 30 vertebræ, the pirate perch 29, and the Percidæ, perch and darters, etc., 35 to 45, while the Serranidæ or sea-bass, the nearest marine relatives of all these, have constantly 24. The marine family of damsel-fishes (Pomacentridæ) have 26 vertebræ, while 30 to 40 vertebræ usually exist in their fresh-water analogues (or possibly descendants), the Cichlidæ, of the rivers of South America and Africa. The sticklebacks (Gasterosteidæ), a family of spiny fishes, confined to the rivers and seas of the north, have from 31 to 41 vertebræ.
Pelagic Fishes.—Among the free-swimming or migratory pelagic fishes, the number of vertebræ is usually greater than among their relatives of local habits. This fact is most evident among the scombriform fishes, the allies of the mackerel and tunny. All of these belong properly to the warm seas, and the reduction of the vertebræ in certain forms has no evident rela[Pg 210]tion to the temperature, though it seems to be related in some degree to the habits of the species. Perhaps the retention of many segments is connected with that strength and swiftness in the water for which the mackerels are preeminent.
The variations in the number of vertebræ in this group led Dr. Günther to divide it into two families, the Carangidæ and Scombridæ.
The Carangidæ or Pampanos are tropical shore fishes, local or migratory to a slight degree. All these have from 24 to 26 vertebræ. In their pelagic relatives, the dolphins (Coryphæna), there are from 30 to 33; in the opah (Lampris), 45; in Brama, 42; while the great mackerel family (Scombridæ), all of whose members are more or less pelagic, have from 31 to 50.
The mackerel (Scomber scombrus) has 31 vertebræ; the chub mackerel (Scomber japonicus), 31; the tunny (Thunnus thynnus), 39; the long-finned albacore (Germo alalonga), 40; the bonito (Sarda sarda), 50; the Spanish mackerel (Scomberomorus maculatus), 45.
Other mackerel-like fishes are the cutlass-fishes (Trichiuridæ), which approach the eels in form and in the reduction of the fins. In these the vertebræ are correspondingly numerous, the numbers ranging from 100 to 160. Aphanopus has 101 vertebræ; Lepidopus, 112; Trichurus, 159.
In apparent contradiction to this rule, however, the pelagic family of swordfishes (Xiphias), remotely allied to the mackerels, and with even greater powers of swimming, has the vertebræ in normal number, the common swordfish having but 24.
The Eels.—The eels constitute a peculiar group of soft-rayed ancestry, in which everything else has been subordinated to muscularity and flexibility of body. The fins, girdles, gill-arches, scales, and membrane bones are all imperfectly developed or wanting. The eel is perhaps as far from the primitive stock as the most highly "ichthyized" fishes, but its progress has been of another character. The eel would be regarded in the ordinary sense as a degenerate type, for its bony structure is greatly simplified as compared with its ancestral forms, but in its eel-like qualities it is, however, greatly specialized. All the eels have vertebræ in great numbers. As the great majority of the species are tropical, and as the vertebræ in very few of[Pg 211] the deep-sea forms have been counted, no conclusions can be drawn as to the relation of their vertebræ to the temperature.
It is evident that the two families most decidedly tropical in their distribution, the morays (Murænidæ) and the snake-eels (Ophichthyidæ), have diverged farthest from the primitive stock. They are most "degenerate," as shown by the reduction of their skeleton. At the same time they are also most decidedly "eel-like," and in some respects, as in coloration, dentition, muscular development, most highly specialized. It is evident that the presence of numerous vertebral joints is essential to the suppleness of body which is the eel's chief source of power.
So far as known the numbers of vertebræ in eels range from 115 to 160, some of the deep-sea eels (Nemichthys, Nettastoma, Gordiichthys) having much higher numbers, in accord with their slender or whip-like forms.
Among the morays, Muræna helena has 140; Gymnothorax meleagris, 120; G. undulatus, 130; G. moringa, 145; G. concolor, 136; Echidna catenata, 116; E. nebulosa, 142; E. zebra, 135. In other families the true eel, Anguilla anguilla, has 115; the conger-eel, Leptocephalus conger, 156; and Murænesox cinereus, 154.
Variations in Fin-rays.—In some families the number of rays in the dorsal and anal fins is dependent on the number of vertebræ. It is therefore subject to the same fluctuations. This relation is not strictly proportionate, for often a variable number of rays with their interspinal processes will be interposed between a pair of vertebræ. The myotomes or muscular bands on the sides are usually coincident with the number of vertebræ. As, however, these and other characters are dependent on differences in vertebral segmentation, they bear the same relations to temperature or latitude that the vertebræ themselves sustain.
Thus in the Scorpænidæ, Sebastes, and Sebastolobus arctic genera have the dorsal rays xv, 13, the vertebræ 12+19. The tropical genus Scorpæna has the dorsal rays xii, 10, the vertebræ 10+14, while the genus Sebastodes of temperate waters has the intermediate numbers of dorsal rays xii, 12, and vertebræ 12+15.
Relation of Numbers to Conditions of Life.—Fresh-water fishes have in general more vertebræ than marine fishes of shallow waters. Pelagic fishes and deep-sea fishes have more than those which live along the shores, and more than localized or non-migratory forms. To each of these generalizations there are occasional partial exceptions, but not such as to invalidate the rule.
The presence of large numbers of vertebræ is noteworthy among those fishes which swim for long distances, as, for example, many of the mackerel family. Among such there is often found a high grade of muscular power, or even of activity, associated with a large number of vertebræ, these vertebræ being individually small and little differentiated. For long-continued muscular action of a uniform kind there would be perhaps an advantage in the low development of the vertebral column. For muscular alertness, moving short distances with great speed, the action of a fish constantly on its guard against enemies or watching for its prey, the advantage would be on the side of a few vertebræ. There is often a correlation between the free-swimming habit and slenderness and suppleness of the body, which again is often dependent on an increase in numbers of the vertebral segments. These correlations appear as a disturbing element in the problem rather than as furnishing a clew to its solution. In some groups of fresh-water fishes there is a reduction in number of vertebræ, not associated with any degree of specialization of the individual bone, but correlated with simple reduction in size of body. This is apparently a phenomenon of degeneration, a survival of dwarfs, where conditions are unfavorable in full growth.
All these effects should be referable to the same group of causes. They may, in fact, be combined in one statement. All other fishes now extant, as well as all fishes existing prior to Cretaceous times, have a larger number of vertebræ than the marine shore fishes of the tropics of the present period. There is good reason to believe that in most groups of spiny-rayed fishes, those with the smaller number of segments are at once the most highly organized and the most primitive. This is true among the blennies, the sculpins, the flounders, the perches, and probably the labroid fishes as well. The present writer once[Pg 213] held the contrary view, that the forms with the higher numbers were primitive, but the evidence both from comparative anatomy and from palæontology seems to indicate that among spiny-rayed fishes the forms most ancient, most generalized, and most synthetic are those with about 24 vertebræ. The soft-rayed fishes without exception show larger numbers, and these are still more primitive. This apparent contradiction is perhaps explained by Dr. Boulenger's suggestion that the prevalence of the same number, 24, in the vertebræ of various families of spiny-rayed fishes is due to common descent, probably from Cretaceous berycoids having this number. In this theory, perches, sparoids, carangoids, chætodonts, labroids, parrot-fishes, gobies, flounders, and sculpins must be regarded as having a common origin from which all have diverged since Jurassic times. This view is not at all unlikely and is not inconsistent with the facts of palæontology. If this be the case, the members of these and related families which have larger numbers of vertebræ must have diverged from the primitive stock. The change has been one of degeneration, the individual vertebræ being reduced in size and complexity, with a vegetative increase in their number. At the same time, the body having the greater number of segments is the more flexible though the segments themselves are less specialized.
The primitive forms live chiefly along tropical shores, while forms with increased numbers of vertebræ are found in all other localities. This fact must be considered in any hypothesis as to the causes producing such changes. If the development of large numbers be a phase of degeneration the causes of such degeneration must be sought in the colder seas, in the rivers, and in the oceanic abysses. What have these waters in common that the coral reefs, the lava crags, and tide-pools of the tropics have not?
It is certain that the possession of fewer vertebræ indicates the higher rank, the greater specialization of parts, even though the many vertebræ be a feature less primitive. The evolution of fishes is rarely a movement of progress toward complexity. The time movement in some groups is accompanied by degradation and loss of parts, by vegetative repetition of structures, and often by a movement from the fish-form toward the [Pg 214]eel-form. Water life is less exacting than land life, having less variation of conditions. It is, therefore, less effective in pushing forward the differentiation of parts. When vertebræ are few in number each one is relatively larger, its structure is more complicated, its appendages larger and more useful, and the fins with which it is connected are better developed. In other words, the tropical fish is more intensely and compactly a fish, with a better fish equipment, and in all ways better fitted for the business of a fish, especially for that of a fish that stays at home.
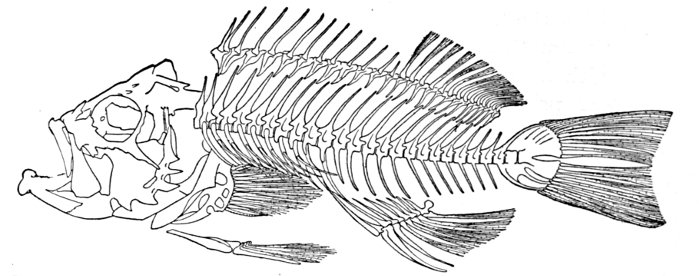
Fig. 155.—Skeleton of Red Rockfish, Sebastodes miniatus Jordan and Gilbert. California.
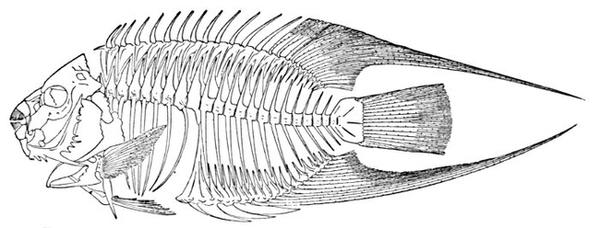
Fig. 156.—Skeleton of a spiny-rayed fish of the tropics, Holacanthus ciliaris (Linnæus).
In the center of competition no species can afford to be handicapped by a weak back-bone and redundant vertebræ. Those who are thus weighted cannot hold their own. They must change or perish.
The conditions most favorable to fish life are among the rocks[Pg 215] and reefs of the tropical seas. About the coral reefs is the center of fish competition. A coral archipelago is the Paris of fishes. In such regions is found the greatest variety of surroundings, and therefore the greatest number of possible adjustments. The struggle is between fish and fish, not between fishes and hard conditions of life. No form is excluded from the competition. Cold, darkness, and foul water do not shut out competitors, nor does any evil influence sap the strength. The heat of the tropics does not make the sea-water hot. It is never sultry or laden with malaria.
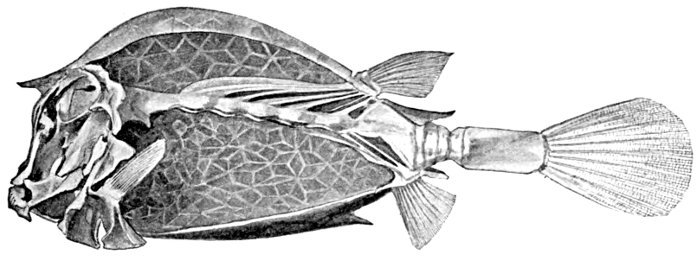
Fig. 157.—Skeleton of the Cowfish, Lactophrys tricornis (Linnæus).
From conditions otherwise favorable in arctic regions the majority of competitors are excluded by their inability to bear the cold. River life is life in isolation. To aquatic animals river life has the same limitations that island life has to the animals of the land. The oceanic islands are far behind the continents in the process of evolution in so far as evolution implies specialization of parts. In a like manner the rivers are ages behind the seas, so far as progress is concerned, though through lack of competition the animals in isolation may be farthest from the original stock.
Therefore the influences which serve as a whole to intensify fish life, to keep it up to its highest effectiveness, and which tend to rid the fish of every character or structure it cannot "use in its business," are most effective along the shores of the tropics. One phase of this is the retention of low numbers of vertebræ, or, more accurately, the increase of stress on each individual bone.
Conversely, as the causes of these changes are still in opera[Pg 216]tion, we should find that in cold waters, deep waters, dark waters, fresh waters, and inclosed waters the strain would be less, the relapses to less complex organization more frequent, the numbers of vertebræ would be larger, while the individual vertebræ would become smaller, less complete, and less perfectly ossified.
This in a general way is precisely what we do find in examining the skeletons of a large variety of fishes.
The cause of the increased numbers of vertebræ in cold waters or extratropical waters is as yet unknown. Several guesses have been made, but these can scarcely rise to the level of theories. To ascribe it to natural selection, as the present writer has done, is to do little more than to restate the problem.
As a possible tentative hypothesis we may say that the retention of the higher primitive traits in the tropics is due to continuous selection, the testing of individuals by the greater variety of external conditions. The degeneration of extratropical fishes may be due to isolation and cessation or reversal of selection. Thus fresh waters, the arctic waters, the oceanic abysses are the "back woods" of fish life, localities favorable to the retention of primitive simplicity, equally favorable to subsequent degeneration. Practically all deep-sea fishes are degenerate descendants of shore fishes of various groups. Monotony and isolation permit or encourage degeneration of type. Where the struggle for existence is most intense the higher structures will be retained or developed. Among such facts as these derived from natural selection the cause of the relation of temperature to number of vertebræ must be sought. How the Cretaceous berycoids first acquired their few vertebræ and the high degree of individual specialization of these structures we may not know. The character came with the thoracic ventrals with reduced number of rays, the ctenoid scales, the toothless maxillary, and other characters which have long persisted in their subsequent descendants.
An exception to the general rule in regard to the number of vertebræ is found in the case of the eel. Eels inhabit nearly all seas, and everywhere they have many vertebræ. The eels of the tropics are at once more specialized and more degraded. They are better eels than those of northern regions, but, as the[Pg 217] eel is a degraded type, they have gone farther in the loss of structures in which this degradation consists.
It is not well to push this analogy too far, but perhaps we can find in the comparison of the tropics and the cities some suggestion as to the development of the eel.
In the city there is always a class which follows in no degree the general line of development. Its members are specialized in a wholly different way. By this means they take to themselves a field which others have neglected, making up in low cunning what they lack in humanity or intelligence.
Thus, among fishes, we have in the regions of closest competition this degenerate and non-fish-like type, lurking in holes among the rocks, or creeping in the sand; thieves and scavengers among fishes. The eels thus fill a place otherwise left unfilled. In their way they are perfectly adapted to the lives they lead. A multiplicity of vertebral joints is useless to the tropical fish, but to the eel strength and suppleness are everything. No armature of fin or scale or bone is so desirable as its power of escaping through the smallest opening. With the elongation of the body and its increase in flexibility there is a tendency toward the loss of the paired fins, the ventrals going first, and afterwards the pectorals. This tendency may be seen in many groups. Among recent fishes, the blennies, the eel-pouts, and the sea-snails furnish illustrative examples.
Degeneration of Structures.—In the lancelet, which is a primitively simple organism, the various structures of the body are formed of simple tissues and in a very simple fashion. It is probable from the structure of each of these that it has never been very much more complex. As the individual develops in the process of growth each organ goes as it were straight to its final form and structure without metamorphosis or especial alterations by the way. When this type of development occurs, the organism belongs to a type which is primitively simple. But there are other forms which in their adult state appear feeble or simple, in which are found elements of organs of high complexity. Thus in the sea-snail (Liparis), small, weak, with feeble fins and flabby skin, we find the essential anatomy of the sculpin or the rosefish. The organs of the latter are there, but each one is reduced or degenerate, the bones as soft as membranes, the spines[Pg 218] obsolete or buried in the skin. Such a type is said to be degenerate. It is very different from one primitively simple, and it is likely in its earlier stages of development to be more complex than when it is fully grown.
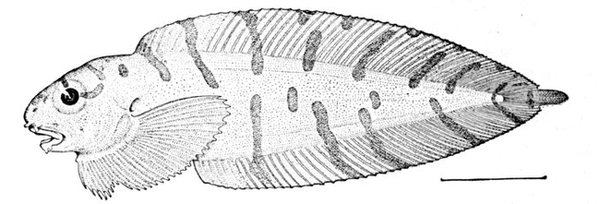
Fig. 158.—Liparid, Crystallias matsushimæ (Jordan and Snyder). Family Liparididæ. Matsushima Bay, Japan.
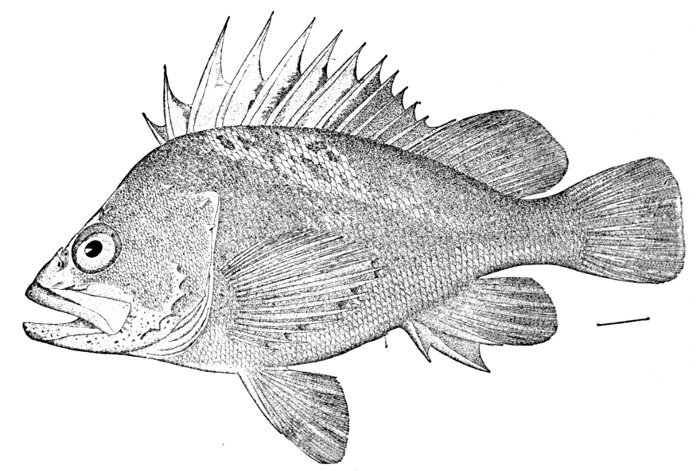
Fig. 159.—Yellow-backed Rockfish, Sebastichthys maliger Jordan and Gilbert. Sitka, Alaska.
In the evolution of groups of fishes it is a common feature that some one organ will be the center of a special stress, in view of some temporary importance of its function. By the process of natural selection it will become highly developed and highly specialized. Some later changes in conditions will ren[Pg 219]der this specialization useless or even harmful for at least a part of the species possessing it. The structure then undergoes degeneration, and in many cases it is brought to a lower estate than before the original changes. An example of this may be taken from the loricate or mailed-cheek fishes. One of the primitive members of this group is the rockfish known as priestfish (Sebastodes mystinus). In this fish the head is weakly armed, covered with ordinary scales. A slight suggestion of cranial ridges and a slight prolongation of the third suborbital constitute the chief suggestions of its close affinity with the mailed-cheek fishes. In other rockfishes the cranial ridges grow higher and sharper. The third suborbital extends itself farther and wider. It becomes itself spinous in still others. Finally it covers the whole cheek in a coat of mail. The head above becomes rough and horny and at last the whole body also is enclosed in a bony box. But while this specialization reaches an extraordinary degree in forms like Agonus and Peristedion, it begins to abate with Cottus, and thence through Cottunculus, Psychrolutes, Liparis, and the like, and the mailed cheek finds its final degradation in Parliparis. In this type no spines are present anywhere, no hard bone, no trace of scales, of first dorsal, or of ventral fins, and in the soft, limp structure covered with a fragile, scarf-like skin we find little suggestion of affinity with the strong rockfish or the rough-mailed Agonus. Yet a study of the skeleton shows that all these loricate forms constitute a continuous divergent series. The forms figured constitute only a few of the stages of specialization and degradation which the members of this group represent.
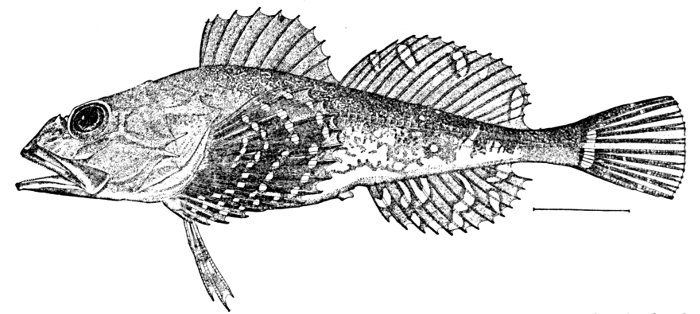
Fig. 160.—European Sculpin, Myoxocephalus scorpius (Linnæus). Cumberland Gulf, Arctic America
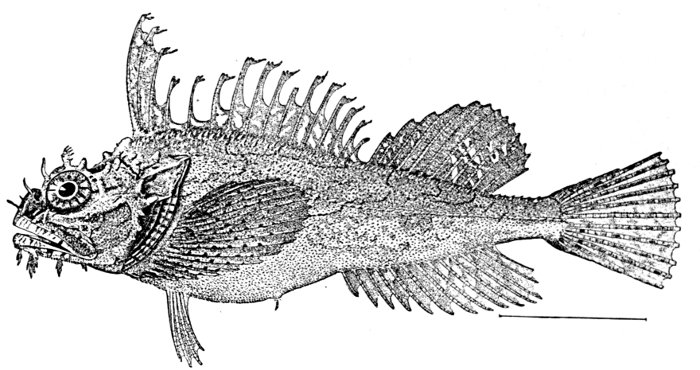
Fig. 161.—Sea-raven, Hemitripterus americanus (Gmelin). Halifax, Nova Scotia.
Some of the features of the habits and development of certain fresh-water fishes are mentioned in the following chapter.
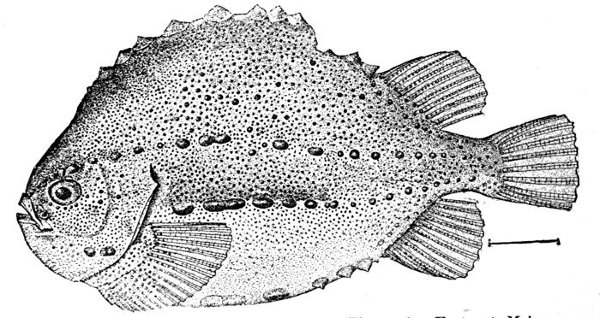
Fig. 162.—Lumpfish, Cyclopterus lumpus (Linnæus). Eastport, Maine.
The degeneration of the eye of the blind fishes of the caves of the Mississippi Valley, Amblyopsis, Typhlichthys, and Troglichthys, have been very fully studied by Dr. Carl H. Eigenmann.
According to his observations
"The history of the eye of Amblyopsis spelæus may be divided into four periods:
Fig. 163.—Sleek Sculpin, Psychrolutes paradoxus (Günther). Puget Sound.
"(a) The first extends from the appearance of the eye till the embryo is 4-5 mm. long. This period is characterized by a normal palingenic development, except that the cell division is retarded and there is very little growth.
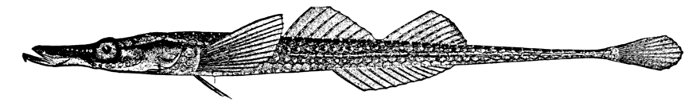
Fig. 164.—Agonoid-fish, Pallasina barbata (Steindachner). Port Mulgrave, Alaska.
"(b) The second period extends till the fish is 10 mm. long. It is characterized by the direct development of the eye from the normal embryonic stage reached in the first period to the highest stage reached by the Amblyopsis eye.

Fig. 165.—Blindfish of the Mammoth Cave, Amblyopsis spelæus (De Kay). Mammoth Cave, Kentucky.
"(c) The third, from 10 mm. to about 80 or 100 mm. It is characterized by a number of changes which are positive as con[Pg 222]trasted with degenerative. There are also distinct degenerative processes taking place during this period.
"(d) The fourth, 80-100 mm. to death. It is characterized by degenerative processes only.
"The eye of Amblyopsis appears at the same stage of growth as in normal fishes developing normal eyes. The eye grows but little after its appearance.
"All the developmental processes are retarded and some of them give out prematurely. The most important, if the last, is the cell division and the accompanying growth that provide material for the eye.
"The lens appears at the normal time and in the normal way, but its cells never divide and never lose their embryonic character.
"The lens is first to show degenerative steps and disappears entirely before the fish is 10 mm. long.

Fig. 166.—Blind Brotula, Lucifuga subterranea (Poey), showing viviparous habit. Joignan Cave, Pinar del Rio, Cuba. Photographed by Dr. Eigenmann.
"The optic nerve appears shortly before the fish reaches 5 mm. It does not increase in size with the growth of the fish and disappears in old age.
"The scleral cartilages appear when the fish is 10 mm. long; they grow very slowly, possibly till old age.
"There is no constant ratio between the extent and degree of ontogenic and phylogenic degeneration.
"The eye is approaching the vanishing point through the route indicated by the eye of Troglichthys rosæ.
"There being no causes operative or inhibitive, either within the fish or in the environment, that are not also operative or[Pg 223] inhibitive in Chologaster agassizii, which lives in caves and develops well-formed eyes, it is evident that the causes controlling the development are hereditarily established in the egg by an accumulation of such degenerative changes as are still notable in the later history of the eye of the adult.
"The foundations of the eye are normally laid, but the superstructure, instead of continuing the plan with additional material, completes it out of the material provided for the foundations. The development of the foundation of the eye is phylogenic; the stages beyond the foundations are direct."
Conditions of Evolution among Fishes.—Dr. Bashford Dean ("Fishes, Living and Fossil") has the following observations on the processes of adaptation among fishes:
"The evolution of groups of fishes must accordingly have taken place during only the longest periods of time. Their aquatic life has evidently been unfavorable to deep-seated structural changes, or at least has not permitted these to be perpetuated. Recent fishes have diverged in but minor regards from their ancestors of the Coal Measures. Within the same duration of time, on the other hand, terrestrial vertebrates have not only arisen, but have been widely differentiated. Among land-living forms the amphibians, reptiles, birds, and mammals have been evolved, and have given rise to more than sixty orders.
"The evolution of fishes has been confined to a noteworthy degree within rigid and unshifting bounds; their living medium, with its mechanical effects upon fish-like forms and structures, has for ages been almost constant in its conditions; its changes of temperature and density and currents have rarely been more than of local importance, and have influenced but little the survival of genera and species widely distributed; its changes, moreover, in the normal supply of food organisms cannot be looked upon as noteworthy. Aquatic life has built few of the direct barriers to survival, within which the terrestrial forms appear to have been evolved by the keenest competition.
"It is not, accordingly, remarkable that in their descent fishes are known to have retained their tribal features, and to have varied from each other only in details of structure. Their[Pg 224] evolution is to be traced in diverging characters that prove rarely more than of family value; one form, as an example, may have become adapted for an active and predatory life, evolving stronger organs of progression, stouter armoring, and more trenchant teeth; another, closely akin in general structures, may have acquired more sluggish habits, largely or greatly diminished size, and degenerate characters in its dermal investiture, teeth and organs of sense or progression. The flowering out of a series of fish families seems to have characterized every geological age, leaving its clearest imprint on the forms which were then most abundant. The variety that to-day maintains among the families of bony fishes is thus known to be paralleled among the carboniferous sharks, the Mesozoic Chimæroids, and the Palæozoic lung-fishes and Teleostomes. Their environment has retained their general characters, while modelling them anew into forms armored or scaleless, predatory or defenseless, great, small, heavy, stout, sluggish, light, slender, blunt, tapering, depressed.
"When members of any group of fishes became extinct, those appear to have been the first to perish which were the possessors of the greatest number of widely modified or specialized structures. Those, for example, whose teeth were adapted for a particular kind of food, or whose motions were hampered by ponderous size or weighty armoring, were the first to perish in the struggle for existence; on the other hand, the forms that most nearly retained the ancestral or tribal characters—that is, those whose structures were in every way least extreme—were naturally the best fitted to survive. Thus generalized fishes should be considered those of medium size, medium defenses, medium powers of progression, omnivorous feeding habits, and wide distribution, and these might be regarded as having provided the staples of survival in every branch of descent.
"Aquatic living has not demanded wide divergence from the ancestral stem, and the divergent forms which may culminate in a profusion of families, genera, and species do not appear to be again productive of more generalized groups. In all lines of descent specialized forms do not appear to regain by regression or degeneration the potential characters of their ancestral condition. A generalized form is like potter's clay, plastic in the[Pg 225] hands of nature, readily to be converted into a needed kind of cup or vase; but when thus specialized may never resume unaltered its ancestral condition: the clay survives; the cup perishes." (Dean.)
[19] Günther, Introd. to the Study of Fishes, p. 192.
[20] The cells which von Lendenfeld designates 'phosphorescent cells' have as their peculiar characteristic a large, oval, highly refracting body imbedded in the protoplasm of the larger end of the clavate cells. These cells have nothing in common with the structure of the cells of the firefly known to be phosphorescent in nature. In fact the true phosphorescent cells are more probably the 'gland-cells' found in ten of the twelve classes of organs which he describes.
[21] See a more technical paper on this subject entitled "Relations of Temperature to Vertebræ among Fishes," published in the Proceedings of the United States National Museum for 1891, pp. 107-120. Still fuller details are given in a paper contained in the Wilder Quarter-Century Book, 1893. The substance is also included in Chapter VIII of foot-notes to Evolution: D. Appleton & Co.

Pigmentation.—The colors of fishes are in general produced by oil sacs or pigment cells beneath the epidermis or in some cases beneath the scales. Certain metallic shades, silvery blue or iridescent, are produced, not by actual pigment, but, as among insects, by the deflection of light from the polished skin or the striated surfaces of the scales. Certain fine striations give an iridescent appearance through the interference of light.
The pigmentary colors may be divided into two general classes, ground coloration and ornamentation or markings. Of these the ground color is most subject to individual or local variation, although usually within narrow limits, while the markings are more subject to change with age or sex. On the other hand, they are more distinctive of the species itself.
Protective Coloration.—The ground coloration most usual among fishes is protective in its nature. In a majority of fishes the back is olivaceous or gray, either plain or mottled, and the belly white. To birds looking down into the water, the back is colored like the water itself or like the bottom below it. To fishes in search of prey from below, the belly is colored like the surface of the water or the atmosphere above it. In any case the darker colored upper surface casts its shadow over the paler lower parts.
In shallow waters or in rivers the bottom is not uniformly colored. The fish, especially if it be one which swims close to the bottom, is better protected if the olivaceous surface is marked by darker cross streaks and blotches. These give the fish a color resemblance to the weeds about it or to the sand and stones on which it lies. As a rule, no fish which lies on the bottom is ever quite uniformly colored.
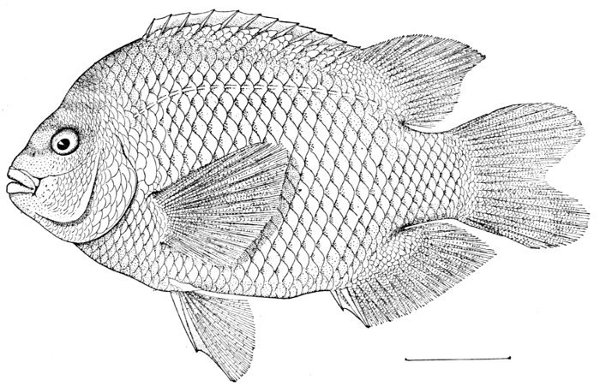
Fig. 167.—Garibaldi (scarlet in color), Hypsypops rubicunda (Girard). La Jolla, San Diego, California.
In the open seas, where the water seems very blue, blue colors, and especially metallic shades, take the place of olivaceous gray or green. As we descend into deep water, especially in the warm seas, red pigment takes the place of olive. At a moderate depth a large percentage of the fishes are of various shades of red. Several of the large groupers of the West Indies are represented by two color forms, a shore form in which the prevailing shade is olive-green, and a deeper-water form which is crimson. In several cases an intermediate-color form also exists which is lemon-yellow. On the coast of California is a band-shaped blenny (Apodichthys flavidus) which appears in three colors, according to its surroundings, blood-red, grass-green, and olive-yellow. The red coloration is also essentially protective, for the region inhabited by such forms is the zone of the rose-red algæ. In the arctic waters, and in lakes where rose-red algæ are not found, the red-ground coloration is almost unknown, although red may appear in markings or in nuptial colors. It is possible that the red, both of fishes and algæ, in deeper water is related to the effect of water on the waves of light, but whether this should make fishes red or violet has never been clearly under[Pg 228]stood. It is true also that where the red in fishes ceases violet-black begins.
In the greater depths, from 500 to 4000 fathoms, the ground color in most fishes becomes deep black or violet-black, sometimes with silvery luster reflected from the scales, but more usually dull and lusterless. This shade may be also protective. In these depths the sun's rays scarcely penetrate, and the fish and the water are of the same apparent shade, for black coloration is here the mere absence of light.
In general, the markings of various sorts grow less distinct with the increase of depth. Bright-red fishes of the depths are usually uniform red. The violet-black fishes of the oceanic abysses show no markings whatever (luminous glands excepted), and in deep waters there are no nuptial or sexual differences in color.
Ground colors other than olive-green, gray, brown, or silvery rarely appear among fresh-water fishes. Marine fishes in the tropics sometimes show as ground color bright blue, grass-green, crimson, orange-yellow, or black; but these showy colors are almost confined to fishes of the coral reefs, where they are often associated with elaborate systems of markings.
Protective Markings.—The markings of fishes are of almost every conceivable character. They may be roughly grouped as protective coloration, sexual coloration, nuptial coloration, recognition colors, and ornamentation, if we may use the latter term for brilliant hues which serve no obvious purpose to the fish itself.
Examples of protective markings may be seen everywhere. The flounder which lies on the sand has its upper surface covered with sand-like blotches, and these again will vary according to the kind of sand it imitates. It may be true sand or crushed coral or the detritus of lava, in any case perfectly imitated.
Equally closely will the markings on a fish correspond with rock surroundings. With granite rocks we find an elaborate series of granitic markings, with coral rocks another series of shades, and if red corals be present, red shades of like appearance are found on the fish. Still another kind of mark indicates rock pools lined with the red calcareous algæ called corallina. Black species are found in lava masses, grass-green ones[Pg 229] among the fronds of ulva, and olive-green among Sargassum or fucus, the markings and often the form corresponding to the nature of the algæ in which the species makes its home.
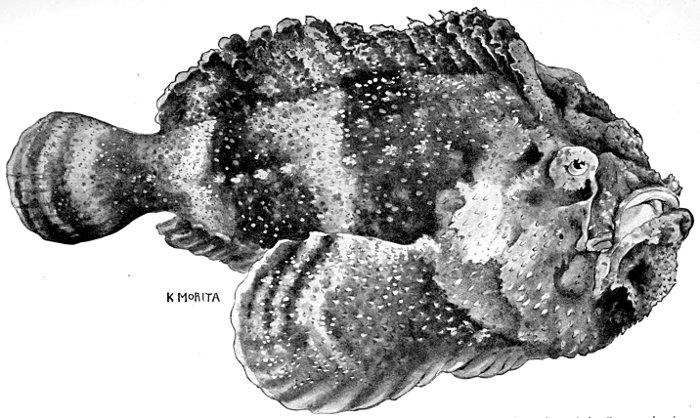
Fig. 168.—Gofu, or Poison Fish, Synanceia verrucosa (Linnæus). Family Scorpænidæ. Specimen from Apia, Samoa, showing resemblance to coral masses, in the clefts of which it lives.
Sexual Coloration.—In many groups of fishes the sexes are differently colored. In some cases bright-red, blue, or black markings characterize the male, the female having similar marks, but less distinct, and the bright colors replaced by olive, brown, or gray. In a few cases, however, the female has marks of a totally different nature, and scarcely less bright than those of the male.

Fig. 169.—Lizard-skipper, Alticus saliens (Forster). A blenny which lies out of water on lava-rocks, leaping from one to another with great agility. From nature; specimen from Point Distress, Tutuila Island, Samoa. (About one-half size.)
Nuptial Coloration.—Nuptial colors are those which appear on the male in the breeding season only, the pigment afterwards vanishing, leaving the sexes essentially alike. Such colors are found on most of the minnows and dace (Cyprinidæ) of the rivers and to a less degree in some other fresh-water fishes, as the darters (Etheostominæ) and the trout. In the[Pg 231] minnows of many species the male in spring has the skin charged with bright pigment, red, black, or bright silvery, for the most part, the black most often on the head, the red on the head and body, and the silvery on the tips of the fins. At the same time other markings are intensified, and in many species the head and sometimes the body and fins are covered with warty excrescences. These shades are most distinct on the most vigorous males, and disappear with the warty excrescences after the fertilization of the eggs.
Fig. 170.—Blue-breasted Darter, Etheostoma camurum (Cope), the most brilliantly colored of American river-fishes. Cumberland Gap, Tennessee.
Nuptial colors do not often appear among marine fishes, and in but few families are the sexes distinguishable by differences in coloration.
Recognition-marks.—Under the head of "recognition-marks" may be grouped a great variety of special markings, which may be conceived to aid the representatives of a given species to recognize each other. That they actually serve this purpose is a matter of theory, but the theory is plausible, and these markings have much in common with the white tail feathers, scarlet crests, colored wing patches, and other markings regarded as recognition-marks among birds.
Among these are ocelli, black- or blue-ringed with white or yellow, on various parts of the body; black spots on the dorsal fin; black spots below or behind the eye; black, red, blue, or yellow spots variously placed; cross-bars of red or black or green, with or without pale edges; a blood-red fin or a fin of shining blue among pale ones; a white edge to the tail; a yellow, blue, or red streamer to the dorsal fin, a black tip to the pectoral[Pg 232] or ventral; a hidden spot of emerald in the mouth or in the axil; an almost endless variety of sharply defined markings, not directly protective, which serve as recognition-marks, if not to the fish itself, certainly to the naturalist who studies it.
These marks shade off into an equally great variety for which we can devise no better name than "ornamentation." Some fishes are simply covered with brilliant spots or bars or reticulations, their nature and variety baffling description, while no useful purpose seems to be served by them, unless we stretch still more widely the convenient theory of recognition-marks.
In many cases the markings change with age, certain bands, stripes, or ocelli being characteristic of the young and gradually disappearing. In such cases the same marks will be found permanent in some related species of less differentiated coloration. In such cases it is safe to regard them as ancestral.
In case of markings on the fins and of elaborate ornamentation in general, it is best defined in the oldest and most vigorous individuals, becoming intensified by degrees. The most brilliantly colored fishes are found about the coral reefs. Here may be found species of which the ground color is the most intense blue, others are crimson, grass-green, lemon-yellow, jet-black, and each with a great variety of contrasted markings. The frontispiece of this volume shows a series of such fishes drawn from nature from specimens taken in pools of the great coral reef of Apia in Samoa. These colors are not protective. The coral masses are mostly plain gray, and the fishes which lie on the bottom are plain gray also. Nothing could be more brilliant or varied than the hues of the free-swimming fishes. What their cause or purpose may be, it is impossible to say. It is certain that their intense activity and the ease with which they can seek shelter in the coral masses enable them to defy their enemies. Nature seems to riot in bright colors where her creatures are not destroyed by their presence.
Intensity of Coloration.—In general, coloration is most intense and varied in certain families of the tropical shores, and especially about coral reefs. But in brilliancy of individual markings some fresh-water fishes are scarcely less notable, especially the darters (Etheostominæ) and sunfishes (Centrarchidæ) of the streams of eastern North America. The bright[Pg 233] hues of these fresh-water fishes are, however, more or less concealed in the water by the olivaceous markings and dark blotches of the upper parts.

Fig. 171.—Snake-eels, Liuranus semicinctus (Lay and Bennett), and Chlevastes colubrinus (Boddaert), from Riu Kiu Islands, Japan.

Fig. 172.—Coral Reef at Apia.
Coral-reef Fishes.—The brilliantly colored fishes of the tropical reefs seem, as already stated, to have no need of protective coloration. They save themselves from their enemies in most cases by excessive alertness and activity (Chætodon, Pomacentrus), or else by busying themselves in coral sand (Julis gaimard), a habit more frequent than has been suspected. Every large mass of branching coral is full of lurking fishes, some of them often most brilliantly colored.
Fading of Pigments in Spirits.—In the preservation of specimens most red and blue pigments fade to whitish, and it requires considerable care to interpret the traces which may be left of red bands or blue markings. Yet some blue pigments are absolutely permanent, and occasionally blood-red pigments persist through all conditions. Black pigment seldom changes in spirits, and olivaceous markings simply fade a little without material alteration. It is an important part of the work of the systematic ichthyologist to learn to interpret the traces of the faded pigment left on specimens he may have occasion to examine. In such cases it is more important to trace the markings than to restore the ground color, as the ground color is at once more variable with individuals and more constant in large groups.
Variation in Pattern.—Occasionally, however, a species is found in which, other characters being constant, both ground color and markings are subject to a remarkable range of variation. In such cases the actual unity of the species is open to serious question. The most remarkable case of such variation known is found in a West Indian fish, the vaca, which bears the incongruous name of Hypoplectrus unicolor. In the typical vaca the body is orange with black marks and blue lines, the fins checkered with orange and blue. In a second form the body is violet, barred with black, the head with blue spots and bands. In another form the blue on the head is wanting. In still another the body is yellow and black, with blue on the head only. In others the fins are plain orange, without checks, and the body yellow, with or without blue stripes and spots, and[Pg 236] sometimes with spots of black or violet. In still others the body may be pink or brown, or violet-black, the fins all yellow, part black or all black. Finally, there are forms deep indigo-blue in color everywhere, with cross bands of indigo-black, and these again may have bars of deeper blue on the head or may lack these altogether. I find, no difference among these fishes except in color, and no way of accounting for the differences in this regard.
Certain species of puffer (Tetraodon setosus, of Panama, and Tetraodon nigropunctatus, of Polynesia) show similar remarkable variations, being dark gray with white spots, but varying to indigo-blue, lemon-yellow, or sometimes having coarse blotches of either. Lemon-yellow varieties of several species are known, and these may be due to a failure of pigment, a sort of semi-albinism. True albinos, individuals wholly without pigment, are rare among fishes. In some cases the markings, commonly black, will be replaced by a deep crimson which does not fade in alcohol. This change happens most frequently among the Scorpænidæ. An example of this is shown in the frontispiece of Volume II of this work. The Japanese okose or poison-fish (Inimicus) is black and gray about lava-rocks. In deeper water among red algæ it is bright crimson, the color not fading in spirits, the markings remaining the same. In still deeper water it is lemon-yellow.

Zoogeography.—Under the head of distribution we consider the facts of the actual location of species of organisms on the surface of the earth and the laws by which their location is governed. This constitutes the subject-matter of the science of zoogeography. In physical geography we may prepare maps of the earth or of any part of it, these bringing to prominence the physical features of its surface. Such maps show here a sea, there a plateau, here a mountain chain, there a desert, a prairie, a peninsula, or an island. In political geography the maps show their physical features of the earth as related to the people who inhabit them and the states or powers which receive or claim their allegiance. In zoogeography the realms of the earth are considered in relation to the species or tribes of animals which inhabit them. Thus series of maps could be drawn representing those parts of North America in which catfishes or trout or sunfishes are found in the streams. In like manner the distribution of any particular fish as the muskallonge or the yellow perch could be shown on the map. The details of such a map are very instructive, and their consideration at once raises a series of questions as to the cause behind each fact. In science it must be supposed that no fact is arbitrary or meaningless. In the case of fishes the details of the method of diffusion of species afford matters of deep interest. These are considered in a subsequent chapter.
The dispersion of animals may be described as a matter of space and time, the movement being continuous but modified by barriers and other conditions of environment. The tendency of recent studies in zoogeography has been to consider[Pg 238] the facts of present distribution as the result of conditions in the past, thus correlating our present knowledge with the past relations of land and water as shown through paleontology. Dr. A. E. Ortmann well observes that "Any division of the earth's surface into zoogeographical regions which starts exclusively from the present distribution of animals without considering its origin must always be unsatisfactory." We must therefore consider the coast-lines and barriers of Tertiary and earlier times as well as those of to-day to understand the present distribution of fishes.
General Laws of Distribution.—The general laws governing the distribution of all animals are reducible to three very simple propositions.
Each species of animal is found in every part of the earth having conditions suitable for its maintenance, unless
(a) Its individuals have been unable to reach this region through barriers of some sort; or,
(b) Having reached it, the species is unable to maintain itself, through lack of capacity for adaptation, through severity of competition with other forms, or through destructive conditions of environment; or else,
(c) Having entered and maintained itself, it has become so altered in the process of adaptation as to become a species distinct from the original type.
Species Absent through Barriers.—The absence from the Japanese fauna of most European or American species comes under the first head. The pike has never reached the Japanese lakes, though the shade of the-lotus leaf in the many clear ponds would suit its habits exactly. The grunt[22] and porgies[23] of our West Indian waters have failed to cross the ocean and therefore have no descendants in Europe or Asia.
Species Absent through Failure to Maintain Foothold.—Of species under (b), those who have crossed the seas and not found lodgement, we have, in the nature of things, no record. Of the existence of multitudes of estrays we have abundant evidence. In the Gulf Stream off Cape Cod are every year taken many young fishes belonging to species at home in the Bahamas and which find no permanent place in the New England fauna. In [Pg 239]like fashion, young fishes from the tropics drift northward in the Kuro Shiwo to the coasts of Japan, but never finding a permanent breeding-place and never joining the ranks of the Japanese fishes. But to this there have been, and will be, occasional exceptions. Now and then one among thousands finds permanent lodgement, and by such means a species from another region will be added to the fauna. The rest disappear and leave no trace. A knowledge of these currents and their influence is eventual to any detailed study of the dispersion of fishes.
The occurrence of the young of many shore fishes of the Hawaiian Islands as drifting plankton at a considerable distance from the shores has been lately discovered by Dr. Gilbert. Each island is, in a sense, a "sphere of influence," affecting the fauna of neighboring regions.
Species Changed through Natural Selection.—In the third class, that of species changed in the process of adaptation, most insular forms belong. As a matter of fact, at some time or another almost every species must be in this category, for isolation is a source of the most potent elements in the initiation and intensification of the minor differences which separate related species. It is not the preservation of the most useful features, but of those which actually existed in the ancestral individuals, which distinguish such species. Natural selection must include not only the process of the survival of the fittest, but also the results of the survival of the existing. This means the preservation through heredity of the traits not of the species alone, but those of the actual individuals set apart to be the first in the line of descent in a new environment. In hosts of cases the persistence of characters rests not on any special usefulness or fitness, but on the fact that individuals possessing these characters have, at one time or another, invaded a certain area and populated it. The principle of utility explains survivals among competing structures. It rarely accounts for qualities associated with geographical distribution.
Extinction of Species.—The extinction of species may be noted here in connection with their extension of range. Prof. Herbert Osborn has recognized five different types of elimination.
1. That extinction which comes from modification or progressive evolution, a relegation to the past as the result of a transmutation into more advanced forms. 2. Extinction from changes of physical environment which outrun the powers of adaptation. 3. The extinction which results from competition. 4. The extinction from extreme specialization and limitation to special conditions the loss of which means extinction. 5. Extinction as a result of exhaustion. As an illustration of No. 1, we may take almost any species which has a cognate species on the further side of some barrier or in the tertiary seas. Thus the trout of the Twin Lakes in Colorado has acquired its present characters in the place of those brought into the lake by its actual ancestors. No. 2 is illustrated by the disappearance of East Indian types (Zanclus, Platax, Toxotes, etc.) in Italy at the end of the Eocene, perhaps for climatic reasons. Extinction through competition is shown in the gradual disappearance of the Sacramento perch (Archoplitis interruptus) after the invasion of the river by catfish and carp. From extreme specialization certain forms have doubtless disappeared, but no certain case of this kind has been pointed out among fishes, unless this be the cause of the disappearance of the Devonian mailed Ostracophores and Arthrodires. It is not likely that any group of fishes has perished through exhaustion of the stock of vigor.
Barriers Checking Movement of Marine Fishes.—The limits of the distribution of individual species or genera must be found in some sort of barrier, past or present. The chief barriers which limit marine fishes are the presence of land, the presence of great oceans, the differences of temperature arising from differences in latitude, the nature of the sea bottom, and the direction of oceanic currents. That which is a barrier to one species may be an agent in distribution to another. The common shore fishes would perish in deep waters almost as surely as on land, while the open Pacific is a broad highway to the albacore or the swordfish.
Again, that which is a barrier to rapid distribution may become an agent in the slow extension of the range of a species. The great continent of Asia is undoubtedly one of the greatest of barriers to the wide movement of species of fish, yet its long shore-line enables species to creep, as it were, from bay to bay,[Pg 241] or from rock to rock, till, in many cases, the same species is found in the Red Sea and in the tide-pools or sand-reaches of Japan. In the North Pacific, the presence of a range of half-submerged volcanoes, known as the Aleutian and the Kurile Islands, has greatly aided the slow movement of the fishes of the tide-pools and the kelp. To a school of mackerel or of flying-fishes these rough islands with their narrow channels might form an insuperable barrier.
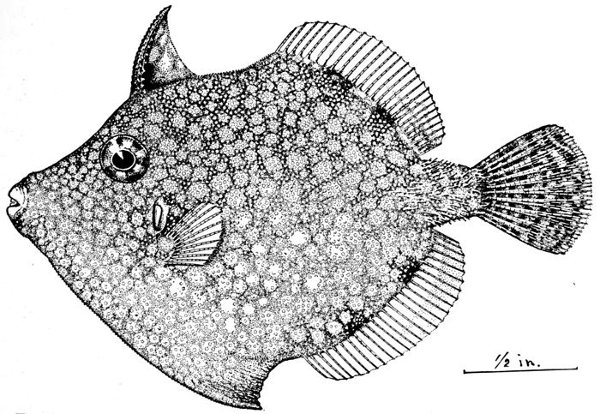
Fig. 173.—Japanese filefish, Rudarius ercodes Jordan and Snyder. Wakanoura, Japan. Family Monacanthidæ.
Temperature the Central Fact in Distribution.—It has long been recognized that the matter of temperature is the central fact in all problems of geographical distribution. Few species in any group freely cross the frost-line, and except as borne by oceanic currents, not many extend their range far into waters colder than those in which the species is distinctively at home. Knowing the average temperature of the water in a given region we know in general the types of fishes which must inhabit it. It is the similarity in temperature and physical conditions which chiefly explains the resemblance of the Japanese fauna to that of the Mediterranean or the Antilles. This fact alone[Pg 242] must explain the resemblance of the Arctic and Antarctic faunæ, there being in no case a barrier in the sea that may not some time be crossed. Like forms lodge in like places.
Agency of Ocean Currents.—We may consider again for a moment the movements of the great currents in the Pacific as agencies in the distribution of species.
A great current sets to the eastward, crossing the ocean just south of the equator. It extends past Samoa and passes on nearly to the coast of Mexico, touching the Galapagos Islands, Clipperton Island, and especially the Revillagigedos. This may account for the number of Polynesian species found on these islands, about which they are freely mixed with immigrants from the mainland of Mexico.
From the Revillagigedos[24] the current moves northward and westward, passing the Hawaiian Islands and thence onward to the Ladrones. The absence in Hawaii of most of the characteristic fishes of Polynesia and Micronesia may be in part due to the long detour made by these currents, as the conditions of life in these groups of islands are not very different. Northeast of Hawaii is a great spiral current, moving with the hands of the watch, forming what is called Fleurieu's Whirlpool. This does not reach the coast of California. This fact may help to account for the almost complete distinction in the shore fishes of Hawaii and California.[25]
No other group of islands in the tropics has a fish fauna so isolated as that of Hawaii. The genera are largely the ordinary tropical types. The species are largely peculiar to these islands.
The westward current from Hawaii reaches Luzon and Formosa. It is deflected to the northward and, joining a northward current from Celebes, it forms the Kuro Shiwo or Black Stream of Japan, which strews its tropical species in the rock pools along the Japanese promontories as far as Tokio. Then, turning into the open sea, it passes northward to the Aleutian Islands, across to Sitka. Thence it moves southward as a cold [Pg 243]current, bearing Ochotsk-Alaskan types southward as far as the Santa Barbara Islands, to which region it is accompanied by species of Aleutian origin. A cold return current seems to extend southward in Japan, along the east shore perhaps as far as Matsushima. A similar current in the sea to the west of Japan extends still further to the southward, to Noto, or beyond.
It is, of course, not necessary that the movements of a species in an oceanic current should coincide with the direction of the current. Young fishes, or fresh-water fishes, would be borne along with the water. Those that dwell within floating bodies of seaweed would go whither the waters carry the drifting mass. But free-swimming fishes, as the mackerel or flying-fishes, might as readily choose the reverse direction. To a free-swimming fish the temperature of the water would be the only consideration. It is thus evident that a current which to certain forms would prove a barrier to distribution, to others would be a mere convenience in movement.
In comparing the Japanese fauna with that of Australia, we find some trace of both these conditions. Certain forms are perhaps excluded by cross-currents, while certain others seem to have been influenced only by the warmth of the water. A few Australian types on the coast of Chile seem to have been carried over by the cross-currents of the South Atlantic.
It is fair to say that the part taken by oceanic currents in the distribution of shore fishes is far from completely demonstrated. The evidence that they assist in such distribution is, in brief, as follows:
1. The young of shore fishes often swim at the surface.
2. The young of very many tropical fishes drift northward in the Gulf Stream and the Japanese Kuro Shiwo.
3. The faunal isolation of Hawaii may be correlated with the direction of the oceanic currents.
Centers of Distribution.—We may assume, in regard to any species, that it has had its origin in or near that region in which it is most abundant and characteristic. Such an assumption must involve a very large percentage of error or of doubt, but in considering the mass of species, it may represent essential truth. In the same fashion we may regard a genus as being autochthonous or first developed in the region where it shows[Pg 244] the greatest range or variety of species. Those regions where the greatest number of genera are thus autochthonous may be regarded as centers of distribution. So far as the marine fishes are concerned, the most important of these supposed centers are found in the Pacific Ocean. First of these in importance is the East-Indian Archipelago, with the neighboring shores of India. Next would come the Arctic Pacific and its bounding islands, from Japan to British Columbia. Third in importance in this regard is Australia. Important centers are found in temperate Japan, in California, the Panama region, and in New Zealand, Chili, and Patagonia. The fauna of Polynesia is almost entirely derived from the Indies; and the shore fauna of the Red Sea, the Bay of Bengal, and Madagascar, so far as genera are concerned, seems to be not really separable from the Indian fauna generally.
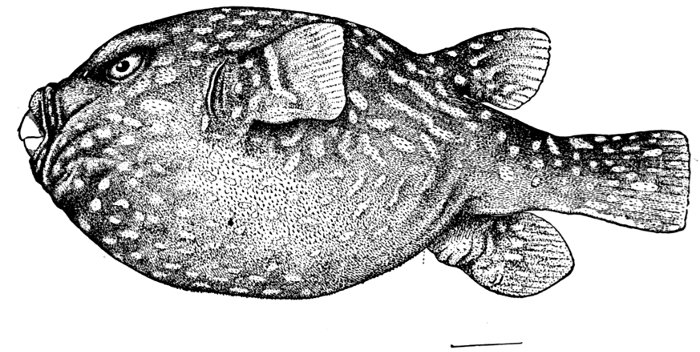
Fig. 174.—Globefish, Tetraodon setosus Rosa Smith. Clarion Island, Mexico.
I know of but six genera which may be regarded as autochthonous in the Red Sea, and nearly all of these are of doubtful value or of uncertain relation. The many peculiar genera described by Dr. Alcock, from the dredgings of the Investigator in the Bay of Bengal, belong to the bathybial or deep-water series, and will all, doubtless, prove to be forms of wide distribution.
In the Atlantic, the chief center of distribution is the West Indies; the second is the Mediterranean. On the shores to the northward or southward of these regions occasional genera have[Pg 245] found their origin. This is true especially of the New England region, the North Sea, the Gulf of Guinea, and the coast of Argentina. The fish fauna of the North Atlantic is derived mainly from the North Pacific, the differences lying mainly in the relative paucity of the North Atlantic. But in certain groups common to the two regions the migration must have been in the opposite direction, exceptions that prove the rule.
Distribution of Marine Fishes.—The distribution of marine fishes must be indicated in a different way from that of the fresh-water forms. The barriers which limit their range furnish also their means of dispersion. In some cases proximity overbalances the influence of temperature; with most forms questions of temperature are all-important.
Pelagic Fishes.—Before consideration of the coast-lines we may glance at the differences in vertical distribution. Many species, especially those in groups allied to the mackerel family, are pelagic—that is, inhabiting the open sea and ranging widely within limits of temperature. In this series some species are practically cosmopolitan. In other cases the genera are so. Each school or group of individuals has its breeding place, and from the isolation of breeding districts new species may be conceived to arise. The pelagic types have reached a species of equilibrium in distribution. Each type may be found where suitable conditions exist, and the distribution of species throws little light on questions of distribution of shore fishes. Yet among these species are all degrees of localization. The pelagic fishes shade into the shore fishes on the one hand and into the deep-sea fishes on the other.
Bassalian Fishes.—The vast group of bassalian or deep-sea fishes includes those forms which live below the line of adequate light. These too are localized in their distribution, and to a much greater extent than was formerly supposed. Yet as they dwell below the influence of the sun's rays, zones and surface temperatures are nearly alike to them, and the same forms may be found in the Arctic or under the equator. Their differences in distribution are largely vertical, some living at greater depths than others, and they shade off by degrees from bathybial into semi-bathybial, and finally into ordinary pelagic and ordinary shore types. Apparently all of the bassalian fishes[Pg 246] are derived from littoral types, the changes in structure being due to degeneration of the osseous and muscular systems and of structures not needed in deep-sea life.
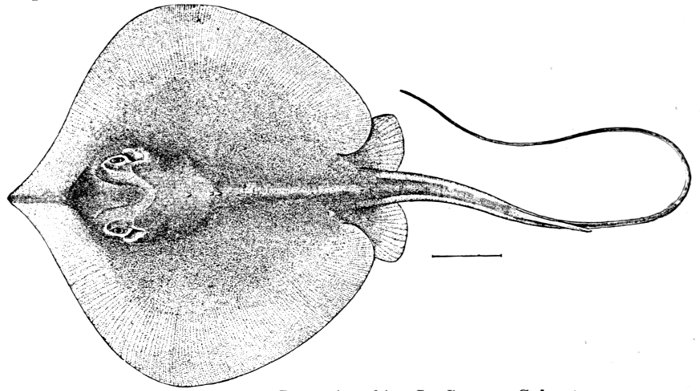
Fig. 175.—Sting-ray, Dasyatis sabina Le Sueur. Galveston.
The fishes of the great depths are soft in substance, some of them blind, some of them with very large eyes, all black in color, and very many are provided with luminous spots or areas. A large body of species of fishes are semi-bathybial, inhabiting depths of 20 to 100 fathoms, showing many of the characters of shore fishes, but far more widely distributed. Many of the remarkable cases of wide distribution of type belong to this class. In moderate depths red colors are very common, corresponding to the zone of red algæ, and the colors in both cases are perhaps determined from the fact that the red rays of light are the least refrangible.
A certain number of species are both marine and fresh water, inhabiting estuaries and brackish waters, while some more strictly marine ascend the rivers to spawn. In none of these cases can any hard and fast line be drawn, and some groups which are shore fishes in one region will be represented by semi-bathybial or fluviatile forms in another.[26]
Littoral Fishes.—The shore fishes are in general the most highly specialized in their respective groups, because exposed to the greatest variety of selecting conditions and of competition. Their distribution in space is more definite than that of the pelagic and bassalian types, and they may be more definitely assigned to geographical areas.
Distribution of Littoral Fishes by Coast-lines.—Their distribution is best indicated, not by realms or areas, but as forming four parallel series corresponding to the four great north and south continental outlines. Each of these series may be represented as beginning at the north in the Arctic fauna, practically identical in each of the four series, actually identical in the two Pacific series. Passing southward, forms are arranged according to temperature. One by one in each series, the Arctic types disappear; subarctic, temperate, and semi-tropical types take their places, giving way in turn to south-temperate and Antarctic forms. The distribution of these is modified by barriers and by currents, yet though genera and species may be different, each isotherm is represented in each series by certain general types of fishes.
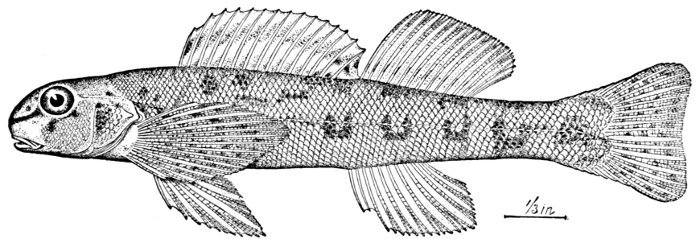
Fig. 176.—Green-sided Darter, Diplesion blennioides Rafinesque. Clinch River. Family Percidæ.
Passing southward the two American series, the East Atlantic and the East Pacific, pass on gradually through temperate to Antarctic types. These are analogous to those of the Arctic, and in a few cases they are generally identical. The West Pacific (East Asian) series is not a continuous line on account of the presence of Australia, the East Indies, and Polynesia. The irregularities of these regions make a number of subseries, which break up the simplicity expressed in the idea of four[Pg 248] parallel series. Yet the fauna of Polynesia is strictly East Indian, modified by the omission or alteration of species, and that of Australia is Indian at the north, and changes to the southward much as that of Africa does. In its marine fishes, it does not constitute a distinct "realm." The East Atlantic (Europe-African) series follows the same general lines of change as that of the West Atlantic. It extends, however, only to the South Temperate Zone, developing no Antarctic elements. The relative shortness of Africa explains in large degree, as already shown, the similarity between the tropical elements in the two Old-World series, as the similarity in tropical elements in the two American series must be due to a former depression of the connecting Isthmus. The practical unity of the Arctic marine fauna needs no explanation in view of the present shore lines of the Arctic Ocean.
Minor Faunal Areas.—The minor faunal areas of shore fishes may be grouped as follows:
East Atlantic.
West Atlantic.
East Pacific.
West Pacific.
Equatorial Fishes Most Specialized.—In general, the different types are most highly specialized in equatorial waters. The processes of specific change, through natural selection or[Pg 249] other causes, if other causes exist, take place most rapidly there and produce most far-reaching modification. As elsewhere stated, the coral reefs of the tropics are the centers of fish-life, the cities in fish economy. The fresh waters, the arctic waters, the deep sea and the open sea represent forms of ichthyic backwoods, regions where change goes on more slowly, and in them we find survivals of archaic or generalized types. For this reason the study in detail of the distribution of marine fishes of equatorial regions is in the highest degree instructive.
Realms of Distribution of Fresh-water Fishes.—If we consider the fresh-water fishes alone we may divide the land areas of the earth into districts and zones not differing fundamentally with those marked out for mammals and birds. The river basin, bounded by its shores and the sea at its mouth, shows many resemblances, from the point of view of a fish, to an island considered as the home of an animal. It is evident that with fishes the differences in latitude outweigh those of continental areas, and a primary division into Old World and New World would not be tenable.
The chief areas of distribution of fresh-water fishes we may indicate as follows, following essentially the grouping proposed by Dr. Günther:[27]
Northern Zone.—With Dr. Günther we may recognize first the Northern Zone, characterized familiarly by the presence of sturgeon, salmon, trout, whitefish, pike, lamprey, stickleback, and other species of which the genera and often the species are identical in Europe, Siberia, Canada, Alaska, and most of the United States, Japan, and China. This is subject to cross-division into two great districts, the first Europe-Asiatic, the second North American. These two agree very closely to the northward, but diverge widely to the southward, developing a variety of specialized genera and species, and both of them passing finally by degrees into the Equatorial Zone.
Still another line of division is made by the Ural Mountains in the Old World and by the Rocky Mountains in the New. In both cases the Eastern region is vastly richer in genera and species, as well as in autochthonous forms, than the Western. The reason for this lies in the vastly greater extent of the river [Pg 250]basins of China and the Eastern United States, as compared with those of Europe or the Californian region.
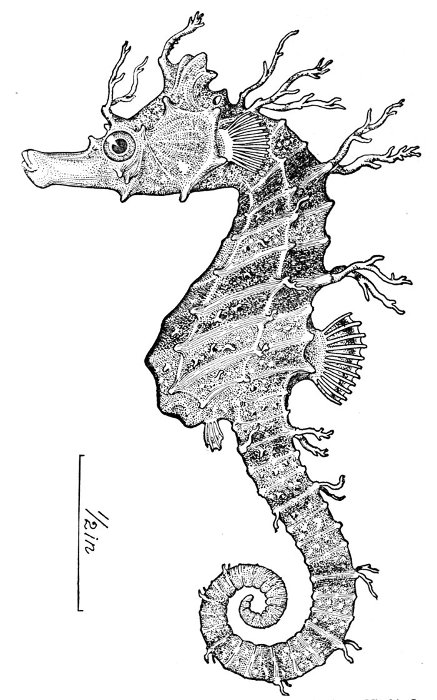
Fig. 177.—Japanese Sea-horse, Hippocampus mohnikei Bleeker. Misaki, Japan.
Minor divisions are those which separate the Great Lake region from the streams tributary to the Gulf of Mexico; and in Asia, those which separate China from tributaries of the Caspian, the Black, and the Mediterranean.
Equatorial Zone.—The Equatorial Zone is roughly indicated by the tropics of Cancer and Capricorn. Its essential feature is that of the temperature, and the peculiarities of its divisions are caused by barriers of sea or mountains.
Dr. Günther finds the best line of separation into two divisions to lie in the presence or absence of the great group of dace or minnows,[28] to which nearly half of the species of fresh-water fishes the world over belong. The entire group, now spread everywhere except in the Arctic, South America, Australia, and the islands of the Pacific, seems to have had its origin in India, from which region its genera have radiated in every direction.
The Cyprinoid division of the Equatorial Zone forms two districts, the Indian and the African. The Acyprinoid division includes South America, south of Mexico, and all the islands of the tropical Pacific lying to the east of Wallace's line. This line, separating Borneo from Celebes and Bali from Lompoe, marks in the Pacific the western limit of Cyprinoid fishes, as well as that of monkeys and other important groups of land animals. This line, recognized as very important in the distribution of land animals, coincides in general with the ocean current between Celebes and Papua, which is one of the sources of the Kuro Shiwo.
In Australia, Hawaii, and Polynesia generally, the fresh-water
fishes are derived from marine types by modification of
one sort or another. In no case, so far as I know, in any island
to the eastward of Borneo, is found any species derived from
fresh-water families of either the Eastern or the Western Continent.
Of course, minor subdivisions in these districts are formed
by the contour lines of river basins. The fishes of the Nile differ
from those of the Niger or the Congo, or of the streams of Madagascar
[Pg 252]or Cape Colony, but in all these regions the essential
character of the fish fauna remains the same.
Southern Zone.—The third great region, the Southern Zone, is scantily supplied with fresh-water fishes, and the few it possesses are chiefly derived from modifications of the marine fauna or from the Equatorial Zone to the north. Three districts are recognized—Tasmania, New Zealand, and Patagonia.
Origin of the New Zealand Fauna.—The fact that certain peculiar groups are common to these three regions has attracted the notice of naturalists. In a critical study of the fish fauna of New Zealand,[29] Dr. Gill discusses the origin of the four genera and seven species of fresh-water fishes found in these islands, the principal of these genera (Galaxias) being represented by nearly related species in South Australia, in Patagonia,[30] the Falkland Islands, and in South Africa.
According to Dr. Gill, we can account for this anomaly of distribution only by supposing, on the one hand, that their ancestors were carried for long distances in some unnatural manner, as (a) having been carried across entombed in ice, or (b) being swept by ocean currents, surviving their long stay in salt water, or else that they were derived (c) from some widely distributed marine type now extinct, its descendants restricted to fresh water.
On the other hand, Dr. Gill suggests that as "community of type must be the expression of community of origin," the presence of fishes of long-established fresh-water types must imply continuity or at least contiguity of land. The objections raised by geologists to the supposed land connection of New Zealand and Tasmania do not appear to Dr. Gill insuperable. It is well known, he says, "that the highest mountain chains are of comparatively recent geological age. It remains, then, to consider which is the more probable, (1) that the types now common in distant regions were distributed in some unnatural manner by the means referred to, or (2) that they are descendants of forms once wide-ranging over lands now submerged." After considering questions as to change of type in other groups, Dr. Gill is inclined to postulate, from the occurrence of species of the [Pg 253]trout-like genus Galaxias, in New Zealand, South Australia, and South America, that "there existed some terrestrial passage-way between the several regions at a time as late as the close of the Mesozoic period. The evidence of such a connection afforded by congeneric fishes is fortified by analogous representatives among insects, mollusca, and even amphibians. The separation of the several areas must have occurred little later than the late Tertiary, inasmuch as the salt-water fishes of corresponding isotherms found along the coast of the now widely separated lands are to such a large extent specifically different. In general, change seems to have taken place more rapidly among marine animals than fresh-water representatives of the same class."
In this case, when one guess is set against another, it seems to me that the hypothesis first suggested, rather than the other, lies in the line of least logical resistance. I think it better to adopt provisionally some theory not involving the existence of a South Pacific Antarctic Continent, to account for the distribution of Galaxias. For this view I may give five reasons:
1. There are many other cases of the sort equally remarkable and equally hard to explain. Among these is the presence of species of paddle-fish and shovel-nosed sturgeon,[31] types characteristic of the Mississippi Valley, in Central Asia. The presence of one and only one of the five or six American species of pike[32] in Europe; of one of the three species of mud-minnow in Austria,[33] the others being American. Still another curious case of distribution is that of the large pike-like trout of the genus Hucho, one species (Hucho hucho) inhabiting the Danube, the other (Hucho blackistoni) the rivers of northern Japan. Many such cases occur in different parts of the globe and at present admit of no plausible explanation.
2. The supposed continental extension should show permanent traces in greater similarity in the present fauna, both of rivers and of sea. The other fresh-water genera of the regions in question are different, and the marine fishes are more [Pg 254]different than they could be if we imagine an ancient shore connection. If New Zealand and Patagonia were once united other genera than Galaxias would be left to show it.
3. We know nothing of the power of Galaxias to survive submergence in salt water, if carried in a marine current. As already noticed, I found young and old in abundance of the commonest of Japanese fresh-water fishes in the open sea, at a distance from any river. Thus far, this species, the hakone[34] dace, has not been recorded outside of Japan, but it might well be swept to Korea or China. Two fresh-water fishes of Japanese origin now inhabit the island of Tsushima in the Straits of Korea.
4. The fresh-water fishes of Polynesia show a remarkably wide distribution and are doubtless carried alive in currents. One river-goby[35] ranges from Tahiti to the Riu Kiu Islands. Another species,[36] originally perhaps from Brazil through Mexico, shows an equally broad distribution.
5. We know that Galaxias with its relatives must have been derived from a marine type. It has no affinity with any of the fresh-water families of either continent, unless it be with the Salmonidæ. The original type of this group was marine, and most of the larger species still live in the sea, ascending streams only to spawn.
When the investigations of geologists show reason for believing in radical changes in the forms of continents, we may accept their conclusions. That geological evidence exists which seems to favor the existence of a former continent, Antarctica, is claimed on high authority. If this becomes well established we may well explain the distribution of Galaxias with reference to it. But we cannot, on the other hand, regard the anomalous distribution of Galaxias alone constituting proof of shore connection. There can be no doubt that almost every case of anomalies in the distribution of fishes admits of a possible explanation through "the slow action of existing causes."
Real causes are always simple when they are once known. All anomalies in distribution cease to be such when the facts necessary to understand them are at our disposal.
[22] Hæmulon.
[23] Calamus.
[24] Clarion Island and Socorro Island.
[25] A few Mexican shore fishes, Chætodon humeralis, Galeichthys dasycephalus, Hypsoblennius parvipinnis, have been wrongly accredited to Hawaii by some misplacement of labels.
[26] The dragonets (Callionymus) are shore fishes of the shallowest waters in Europe and Asia, but inhabit considerable depths in tropical America. The sea-robins (Prionotus) are shore fishes in Massachusetts, semi-bathybial fishes at Panama. Often Arctic shore fishes become semi-bathybial in the Temperate Zone, living in water of a given temperature. A long period of cold weather will sometimes bring such to the surface.
[27] "Introduction to the Study of Fishes."
[28] Cyprinidæ.
[29] "A Comparison of Antipodal Faunæ," 1887.
[30] Galaxias, Neochanna, Prototroctes, and Retropinna.
[31] The shovel-nosed sturgeon (Scaphirynchus and Kessleria) and the paddle-fish (Polyodon and Psephurus).
[32] Esox lucius.
[33] Umrba, the mud-minnow.
[34] Leuciscus hakuensis.
[35] Eleotris fusca.
[36] Awaous genivittatus.

The Isthmus of Suez.—In the study of the effect of the Isthmus of Suez on the distribution of fishes we may first consider the alleged resemblance between the fauna of the Mediterranean and that of Japan. Dr. Günther claims that the actual identity of genera and species in these two regions is such as to necessitate the hypothesis that they have been in recent times joined by a continuous shore-line. This shore-line, according to Prof. A. Ortmann and others, was not across the Isthmus of Suez, but farther to the northward, probably across Siberia.
The Fish Fauna of Japan.—For a better understanding of the problem we may give a brief analysis of the fish fauna of Japan.
The group of islands which constitute the empire of Japan is remarkable for the richness of its animal life. Its variety in climatic and other conditions, its nearness to the great continent of Asia and to the chief center of marine life, the East Indian Islands, its relation to the warm Black Current or Kuro Shiwo from the south and to the cold currents from the north, all tend to give variety and richness to the fauna of its seas. Especially is this true in the group of fishes. In spite of the political isolation of the Japanese Empire, this fact has been long recognized and the characteristic types of Japanese fishes have been well known to naturalists.
At present about 900 species of fishes are known from the four great islands which constitute Japan proper—Hondo, Hokkaido, Kiusiu, and Shikoku. About 200 others are known from the volcanic islands to the north and south. Of these 1100 species, about fifty belong to the fresh waters. These are all closely allied to forms found on the mainland of Asia, from which re[Pg 256]gion all of them were probably derived. In general the same genera appear in China and with a larger range of species.
Fresh-water Faunas of Japan.—Two faunal areas of fresh waters may be fairly distinguished, although broadly overlapping. The northern region includes the island of Hokkaido and the middle and northern part of the great island of Hondo. In a rough way, its southern boundary may be defined by Fuji Yama, and the Bay of Matsushima. It is characterized by the presence of salmon, trout, and sculpins, and northward by sturgeon and brook lampreys. The southern area loses by degrees the trout and other northern fishes, while in its clear waters abound various minnows, gobies, and the famous ayu, or Japanese dwarf salmon, one of the most delicate of food fishes. Sculpins and lampreys give place to minnows, loaches, and chubs. Two genera, a sculpin[37] and a perch,[38] besides certain minnows and catfishes, are confined to this region and seem to have originated in it, but, like the other species, from Chinese stock.
Origin of Japanese Fresh-water Fishes.—The question of the origin of the Japanese river fauna seems very simple. All the types are Asiatic. While most of the Japanese species are distinct, their ancestors must have been estrays from the mainland. To what extent river fishes may be carried from place to place by currents of salt water has never been ascertained. One of the most widely distributed of Japanese river fishes is the large hakone dace or chub.[39] This has been repeatedly taken by us in the sea at a distance from any stream. It would evidently survive a long journey in salt water. An allied species[40] is found in the midway island of Tsushima, between Korea and Japan.
Faunal Areas of Marine Fishes in Japan.—The distribution of the marine fishes of Japan is mainly controlled by the temperature of the waters and the motion of the ocean currents. Five faunal areas may be more or less clearly recognized, and these may receive names indicating their scope—Kurile, Hokkaido, Nippon, Kiusiu, Kuro Shiwo, and Riu Kiu. The first or Kurile district is frankly subarctic, containing species characteristic of the Ochotsk Sea on the one hand, and of Alaska on [Pg 257]the other. The second or Hokkaido[41] district includes this northern island and that part of the shore of the main island of Hondo[42] which lies to the north of Matsushima and Noto. Here the cold northern currents favor the development of a northern fauna. The herring and the salmon occupy here the same economic relation as in Norway, Scotland, Newfoundland, and British Columbia. Sculpins, blennies, rockfish, and flounders abound of the rocky shores and are seen in all the markets.
South of Matsushima Bay and through the Island Sea as far as Kobe, the Nippon fauna is distinctly one of the temperate zone. Most of the types characteristically Japanese belong here, abounding in the sandy bays and about the rocky islands.
About the islands of Kiusiu and Shikoku, the semi-tropical elements increase in number and the Kiusiu fauna is less characteristically Japanese, having much in common with the neighboring shores of China, while some of the species range northward from India and Java. But these faunal districts have no sharp barriers. Northern fishes[43] unquestionably of Alaskan origin range as far south as Nagasaki, while certain semi-tropical[44] types extend their range northward to Hakodate and Volcano Bay. The Inland Sea, which in a sense bounds the southern fauna, serves at the same time as a means of its extension. While each species has a fairly definite northern or southern limit, the boundaries of a faunal district as a whole must be stated in the most general terms.
The well-known boundary called Blackiston's Line, which passes through the Straits of Tsugaru, between the two great islands of Hondo and Hokkaido, marks the northern boundary of monkeys, pheasants, and most tropical and semi-tropical birds and mammals of Japan. But as to the fishes, either marine or fresh water, this line has no significance. The northern fresh-water species probably readily cross it; the southern rarely reach it.
We may define as a fourth faunal area that of the Kuro
Shiwo district itself, which is distinctly tropical and contrasts strongly with that of the inshore bays behind it. This warm "Black Current," analogous to our Gulf Stream, has its origin in part from a return current from the east which passes westward through Hawaii, in part from a current which passes between Celebes and New Guinea. It moves northward by way of Luzon and Formosa, touching the east shores of the Japanese islands Kiusiu and Shikoku, to the main island of Hondo, flooding the bays of Kagoshima and Kochi, of Waka, Suruga, and Sagami. The projecting headlands reach out into it and the fauna of their rock-pools is distinctly tropical as far to the northward as Tokio.

Fig. 178.—Sacramento Perch, Archoplites interruptus Girard. Family Centrarchidæ. Sacramento River.
These promontories of Hondo, Waka, Ise, Izu, Misaki, and Awa have essentially the same types of fishes as are found on the reefs of tropical Polynesia. The warmth of the off-shore currents gives the fauna of Misaki its astonishing richness, and the wealth of life is by no means confined to the fishes. Corals, crustaceans, worms, and mollusks show the same generous profusion of species.
A fifth faunal area, closely related to that of the Black Current, is formed by the volcanic and coral reefs of the Riu Kiu Archipelago. This fauna, so far as known, is essentially East Indian, the genera and most of the species being entirely identical with those of the islands about Java and Celebes.
Resemblance of the Japanese and Mediterranean Fish Faunas.—It has been noted by Dr. Günther that the fish fauna of Japan bears a marked resemblance to that of the Mediterranean. This likeness is shown in the actual identity of genera and species, and in their relation to each other. This resemblance he proposes to explain by the hypothesis that at some recent period the two regions, Japan and the Mediterranean, have been united by a continuous shore-line. The far-reaching character of this hypothesis demands a careful examination of the data on which it rests.
The resemblance of the two faunal areas, so far as fishes are concerned, may be stated as follows: There are certain genera[45] of shore fishes, tropical or semi-tropical, common to the Mediterranean and Japan, and wanting to California, Panama, and the West Indies, and in most cases to Polynesia also. Besides these, certain others found in deeper water (100 to 200 fathoms) are common to the two areas,[46] and have been rarely taken elsewhere.
Significance of Resemblance.—The significance of these facts can be shown only by a fuller analysis of the fauna in question, and those of other tropical and semi-tropical waters. If the resemblances are merely casual, or if the resemblances are shown by other regions, the hypothesis of shore continuity would be unnecessary or untenable. It is tenable if the resemblances are so great as to be accounted for in no other way.
Of the genera regarded as common, only two[47] or three are represented in the two regions by identical species, and these have a very wide distribution in the warm seas. Of the others, nearly all range to India, to the Cape of Good Hope, to Australia, or to Brazil. They may have ranged farther in the past; they may even range farther at present. Not one is confined to the two districts in question. As equally great resemblances exist between Japan and Australia or Japan and the West Indies, the case is not self-evident without fuller comparison. I shall [Pg 260]therefore undertake a somewhat fuller analysis of the evidence bearing on this and similar problems with a view to the conclusions which may be legitimately drawn from the facts of fish distribution.
Differences between Japanese and Mediterranean Fish Faunas.—We may first, after admitting the alleged resemblances and others, note that differences are equally marked. In each region are a certain number of genera which we may consider as autochthonous. These genera are represented by many species or by many individuals in the region of their supposed origin, but are more scantily developed elsewhere. Such genera in Mediterranean waters are Crenilabrus, Labrus, Spicara, Pagellus, Mullus, Boops, Spondyliosoma, Oblata. None of these occurs in Japan, nor have they any near relatives there. Japanese autochthonous types, as Pseudoblennius, Vellitor, Duymæria, Anoplus, Histiopterus, Monocentrus, Oplegnathus, Plecoglossus, range southward to the Indies or to Australia, but all of them are totally unknown to the Mediterranean. The multifarious genera of Gobies of Japan show very little resemblance to the Mediterranean fishes of this family, while blennies, labroids, scaroids, and scorpænoids are equally diverse in their forms and alliances. To the same extent that likeness in faunas is produced by continuity of means of dispersion is it true that unlikeness is due to breaks in continuity. Such a break in continuity of coast-line, in the present case, is the Isthmus of Suez, and the unlikeness in the faunas is about what we might conceive that such a barrier should produce.
Sources of Faunal Resemblances.—There are two main sources of faunal resemblances: first, the absence of any barriers permitting the actual mingling of the species; second, the likeness of temperature and shore configuration on either side of an imperfect barrier. Absolute barriers do not exist and apparently never have existed in the sea. If the fish faunas of different regions have mingled in recent times, the fact would be shown by the presence of the same species in each region. If the union were of a remote date, the species would be changed, but the genera might remain identical.
In case of close physical resemblances in different regions, as in the East Indies and West Indies, like conditions would favor[Pg 261] the final lodgement of like types, but the resemblance would be general, the genera and species being unlike. Without doubt part of the resemblance between Japan and the Mediterranean is due to similarity of temperature and shores. Is that which remains sufficient to demand the hypothesis of a former shore-line connection?
Effects of Direction of Shore-line.—We may first note that a continuous shore-line produces a mingling of fish faunas only when not interrupted by barriers due to climate. A north and south coast-line, like that of the East Pacific, however unbroken, permits great faunal differences. It is crossed by the different zones of temperature. An east and west shore-line lies in the same temperature. In all cases of the kind which now exist on the earth (the Mediterranean, the Gulf of Mexico, the Caribbean Sea, the shores of India), even species will extend their range as far as the shore-line goes. The obvious reason is because such a shore-line rarely offers any important barrier to distribution, checking dispersion of species. We may, therefore, consider the age and nature of the Isthmus of Suez and the character of the faunas it separates.
Numbers of Genera in Different Faunas.—For our purposes the genera must be rigidly defined, a separate name being used in case of each definable difference in structure. The wide-ranging genera of the earlier systematists were practically cosmopolitan, and their geographical distribution teaches us little. On the other hand, when we come to the study of geological distribution, the broad definition of the genus is the only one usually available. The fossil specimens are always defective. Minor characters may be lost past even the possibility of a guess, and only along broad lines can we achieve the classification of the individual fossil.
Using the modern definition of genus, we find in Japan 483 genera of marine fishes; in the Red Sea, 225; in the Mediterranean, 231. In New Zealand 150 are recorded; in Hawaii, 171; 357 from the West Indies, 187 from the Pacific coast of tropical America, 300 from India, 450 from the East-Indian islands, and 227 from Australia.
Of the 483 genera ascribed to Japan, 156 are common to the Mediterranean also, 188 to the West Indies and Japan, 169 to[Pg 262] the Pacific coast of the United States and Mexico. With Hawaii Japan shares 90 genera, with New Zealand 62; 204 are common to Japan and India, 148 to Japan and the Red Sea, most of these being found in India also. Two hundred genera are common to Japan and Australia.
From this it is evident that Japan and the Mediterranean have much in common, but apparently not more than Japan shares with other tropical regions. Japan naturally shows most likeness to India, and next to this to the Red Sea. Proportionately less is the resemblance to Australia, and the likeness to the Mediterranean seems much the same as that to the West Indies or to the Pacific coast of America.
But, to make these comparisons just and effective, we should consider not the fish fauna as a whole; we should limit our discussion solely to the forms of equatorial origin. From the fauna of Japan we may eliminate all the genera of Alaskan-Aleutian origin, as these could not be found in the other regions under comparison. We should eliminate all pelagic and all deep-sea forms, for the laws which govern the distribution of these are very different from those controlling the shore fishes, and most of the genera have reached a kind of equilibrium over the world.
Significance of Rare Forms.—We may note also, as a source of confusion in our investigation, that numerous forms found in Japan and elsewhere are very rarely taken, and their real distribution is unknown. Some of these will be found to have, in some unexpected quarter, their real center of dispersion. In fact, since these pages were written, I have taken in Hawaii representatives of three[48] genera which I had enumerated as belonging chiefly to Japan and the West Indies. Numerous other genera common to the two regions have since been obtained by Dr. Gilbert. Such species may inhabit oceanic plateaus, and find many halting places in their circuit of the tropical oceans. We have already discovered that Madeira, St. Helena, Ascension, and other volcanic islands constitute such halting places. We shall find many more such, when the deeper shore regions are explored, the region between market-fishing and the deep-sea dredgings of the Challenger and [Pg 263]the Albatross. In some cases, no doubt, these forms are verging on extinction and a former wide distribution has given place to isolated colonies.
The following table shows the contents, so far as genera are concerned, of those equatorial areas in which trustworthy catalogues of species are accessible. It includes only those fishes of stationary habit living in less than 200 fathoms. It goes without saying that considerable latitude must be given to these figures, to allow for errors, omissions, uncertainties, and differences of opinion.
Distribution of Shore Fishes.—
| A. Japan and the Mediterranean. | |
|---|---|
| Genera[49] chiefly confined to these regions | 2 |
| Genera of wide distribution | 77 |
| Total of common genera | 79 |
| Total in both regions | 399 |
| Genera above included, found in all equatorial regions | 55 |
| Genera[50] found in most equatorial regions | 11 |
| Genera more or less restricted | 13 |
| 79 | |
| B. Japan and the Red Sea. | |
| Genera[51] chiefly confined to these two regions | 2 |
| Genera of wide distribution | 109 |
| Total genera common | 111 |
| Total in both regions | 424 |
| C. Japan and Hawaii. | |
| Genera chiefly confined to these regions | 3 |
| Genera of wide distribution | 79 |
| Total genera common | 82 |
| Total in both regions | 396 |
| D. Japan and Australia. | |
| Genera chiefly confined to these regions | 13 |
| Genera of wide distribution (chiefly East Indian) | 122 |
| Total genera common | 135 |
| Total in both regions | 533 |
| E. Japan and Panama. | |
| Genera chiefly confined to these regions | 2 |
| Genera of wide distribution | 89 |
| Total genera common | 91 |
| Total in both regions | 499 |
| F. Japan and the West Indies. | |
| Genera chiefly confined to these regions | 5 |
| Genera of wide distribution | 108 |
| Total genera common | 113 |
| Total in both regions | 520 |
| G. The Mediterranean and the Red Sea. | |
| Genera confined to the Suez region | 0 |
| Genera of wide distribution (chiefly Indian) | 40 |
| Total genera common | 40 |
| Total in both regions | 295 |
| H. West Indies and the Mediterranean. | |
| Genera chiefly confined to the equatorial Atlantic | 11 |
| Genera of wide distribution | 59 |
| Total | 70 |
| Total in both regions | 373 |
| I. West Indies and Panama. | |
| Genera chiefly confined to equatorial America | 68 |
| Genera of wide distribution | 101 |
| Total genera common | 169 |
| Total in equatorial America | 376 |
| J. Hawaii and Panama. | |
| Genera chiefly confined to the regions in question | 3 |
| Genera of wide distribution | 74 |
| Total genera common | 77 |
| Total in both regions | 323 |
| K. Hawaii and the East Indies. | |
| Genera chiefly confined to Hawaii | 4 |
| Genera of wide distribution in the equatorial Pacific | 123 |
| Genera confined to Hawaii and the West Indies | 1 |
| Summary. | |
| Genera (shore fishes only) in the Mediterranean Sea. | 144 |
| Genera in the Red Sea | 191 |
| Genera in India | 280 |
| Genera in Japan (exclusive of northern forms) | 334 |
| Genera in Australia | 344 |
| Genera in New Zealand | 108 |
| Genera in Hawaii | 144 |
| Genera about Panama | 256 |
| Genera in West Indies | 299 |
Extension of Indian Fauna.—From the above tables it is evident that the warm-water fauna of Japan, as well as that of Hawaii, is derived from the great body of the fauna of the East Indies and Hindostan; that the fauna of the Red Sea is derived in the same way; that the fauna of the Mediterranean bears no especial resemblance to that of Japan, rather than to other[Pg 266] elements of the East Asiatic fauna in similar conditions of temperature, and no greater than is borne by either to the West Indies; that the faunas of the sides of the Isthmus of Suez have relatively little in common, while those of the two sides of the Isthmus of Panama show large identity of genera, although few species are common to the two sides. Of the 255 genera recorded from the Panama region, 179, or over 70 per cent., are also in the West Indies, while 68, or more than 30 per cent. of the number, are limited to the two regions in question.
The Isthmus of Suez as a Barrier to Distribution.—With the aid of the above table we may examine further the relation of the fauna of Japan to that of the Mediterranean. If a continuity of shore-line once existed, it would involve the obliteration of the Isthmus. With free connection across this isthmus the fauna of the Red Sea must have been once practically the same as that of the Mediterranean. The present differences must be due to later immigrations to one or the other region, or to the extinction of species in one locality or the other, through some kind of unfitness. In neither region is there evidence of extensive immigration from the outside. The present conditions of water and temperature differ a little, but not enough to explain the difference in faunæ. The Red Sea is frankly tropical and its fauna is essentially Indian, much the same, so far as genera are concerned, as that of southern Japan. The Mediterranean is at most not more than semi-tropical and its fishes are characteristically European. Its tropical forms belong rather to Guinea than to the East Indies. With the Red Sea the Mediterranean has very little in common, not so much, for example, as has Hawaii. Forty genera of shore fishes (and only fifty of all fishes) are identical in the two regions, the Mediterranean and the Red Sea. Of those, every one is a genus of wide distribution, found in nearly all warm seas. Of shore fishes, only one genus in seven is common to the two regions. Apparently, therefore, we cannot assume a passage across the Isthmus of Suez within the lifetime of the present genera. Not one of the types alleged to be peculiar to Japan and the Mediterranean is thus far known in the Red Sea. Not one of the characteristically abundant[Pg 267] Mediterranean types[52] crosses the Isthmus of Suez, and the distinctive Red Sea and Indian types[53] are equally wanting in the Mediterranean. The only genera which could have crossed the Isthmus are certain shallow-water or brackish-water forms, sting-rays, torpedoes, sardines, eels, and mullets, widely diffused through the East Indies and found also in the Mediterranean. The former channel, if one ever existed, had, therefore, much the same value in distribution of species as the present Suez Canal.
Geological Evidence of Submergence of the Isthmus of Suez.—Yet, from geological data, there is strong evidence that the Isthmus of Suez was submerged in relatively recent times. The recognized geological maps of the Isthmus show that a broad area of post-Pliocene or Pliocene deposits constitutes the Isthmus and separates the nummulitic hills of Suez from their fellows about thirty miles to the eastward. The northern part of the Isthmus is alluvium from the Nile, and its western part is covered with drifting sands. The Red Sea once extended farther north than now and the Mediterranean farther to the southeast. Assuming the maps to be correct, the Isthmus must have been open water in the late Pliocene or post-Pliocene times.
Admitting this as a fact, the difference in the fish fauna would seem to show that the waters over the submerged area were so shallow that the rock-loving forms did not and could not cross it. Moreover, the region was very likely overspread with silt-bearing fresh waters from the Nile. To such fishes as Chætodon, Holocentrus, Thalassoma of the Red Sea, or to Crenilabrus, Boops, and Zeus of the Mediterranean, such waters would form a barrier as effective as the sand-dunes of to-day.
Conclusions as to the Isthmus of Suez.—We are led, therefore, to these conclusions:
1. There is no evidence derivable from the fishes of the recent submergence of the Isthmus of Suez.
2. If the Isthmus was submerged in Pliocene or post-Pliocene times, the resultant channel was shallow and muddy, so [Pg 268]that ordinary marine fishes or fishes of rock bottoms or of deep waters did not cross it.
3. It formed an open water to brackish-water fishes only.
4. The types common to Japan and the Mediterranean did not enter either region from the other by way of the Red Sea.
5. As most of these are found also in India or Australia or both, their dispersion was probably around the south coast of Africa or by the Cape of Good Hope.
6. In view of the fact that numerous East Indian genera, as Zanclus, Enoplosus, Toxotes, Ephippus, Platax, Teuthis, Acanthurus (Monoceros), Myripristis occur in the Eocene rocks of Tuscany, Syria, and Switzerland, we may well suppose that an open waterway across Africa then existed. Perhaps these forms were destroyed in European waters by a wave of glacial cold, perhaps after the Miocene. As our knowledge of the Miocene fish faunæ of Europe is still imperfect, we cannot locate accurately the period of their disappearance. About half the species found in the Eocene of Italy belong to existing genera, and these genera are almost all now represented in the Indian fauna, and those named above with others are confined to it.
The study of fishes alone furnishes no adequate basis for mapping the continental masses of Tertiary times. The known facts in regard to their distribution agree fairly with the provisional maps lately published by Dr. Ortmann (Bull. Philos. Soc., XLI). In the Eocene map (Fig. 179) the Mediterranean extends to the northward of Arabia, across to the mouth of the Ganges. This extension would account for the tropical, Eocene, and Miocene fish fauna of Southern Europe.
The Cape of Good Hope as a Barrier to Fishes.—The fishes of the Cape of Good Hope are not well enough known for close comparison with those of other regions. Enough is known of the Cape fauna to show its general relation to those of India and Australia. The Cape of Good Hope lies in the South Temperate Zone. It offers no absolutely impassable barrier to the tropical fishes from either side. It bears a closer relation to either the Red Sea or the Mediterranean than they bear to each other. It is, therefore, reasonable to conclude that the transfer of tropical shore fishes of the Old World between the Atlantic and Pacific, in recent times, has taken place mainly[Pg 269] around the southern point of Africa. To pelagic and deep-sea fishes the Cape of Good Hope has offered no barrier whatever. To ordinary fishes it is an obstacle, but not an impassable one. This the fauna itself shows. It has, however, not been passed by many tropical species, and by these only as the result of thousands of years of struggle and point-to-point migration.
Relations of Japan to Mediterranean Explainable by Present Conditions.—We may conclude that the resemblance of the Mediterranean fish fauna to that of Japan or India is no more than might be expected, even had the present contour of the continents been permanent for the period of duration of the present genera and species. An open channel in recent times would have produced much greater resemblances than actually exist.
The Isthmus of Panama as a Barrier to Distribution.—Conditions in some regards parallel with those of the Isthmus of Suez exist in but one other region—the Isthmus of Panama. Here the first observers were very strongly impressed by the resemblance of forms. Nearly half the genera found on the two sides of this isthmus are common to both sides. Taking those of the Pacific shore for first consideration, we find that three-fourths of the genera of the Panama fauna occur in the West Indies as well.
This identity is many times greater than that existing at the Isthmus of Suez. Moreover, while the Cape of Good Hope offers no impassable barrier to distribution, the same is not true of the southern part of South America. The subarctic climate of Cape Horn has doubtless formed a complete check to the movements of tropical fishes for a vast period of geologic time.
Unlikeness of Species on the Shores of the Isthmus of Panama.—But, curiously enough, this marked resemblance is confined chiefly to the genera and does not extend to the species on the two shores.
Of 1400 species of fishes recorded from tropical America north of the Equator, only about 70 are common to the two coasts. The number of shore fishes common is still less. In this 70 are included a certain number of cosmopolitan types which might have reached either shore from the Old World.
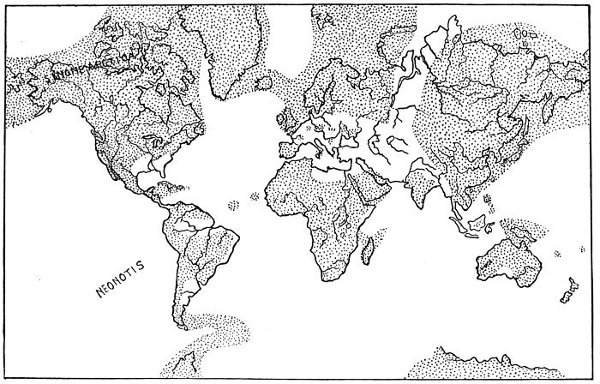
Fig. 179.—Map of the Continents, Eocene time. (After Ortmann.)
A few others invade brackish or fresh waters and may possibly have found their way, in one way or another, across the Isthmus of Nicaragua. Of fishes strictly marine, strictly littoral, and not known from Asia or Polynesia, scarcely any species are left as common to the two sides. This seems to show that no waterway has existed across the Isthmus within the lifetime, whatever that may be, of the existing species. The close resemblance of genera shows apparently with almost equal certainty that such a waterway has existed, and within the period of existence of the groups called genera. How long a species of fish may endure unchanged no one knows, but we know that in this regard great differences must exist in different groups. Assuming that different species crossed the Isthmus of Panama in Miocene times, we should not be surprised to find that a few remain to all appearances unchanged; that a much larger number have become "representative" species, closely related forms retaining relations to the environment to those of the parent form, and, finally, that a few species have been radically altered.
This is exactly what has taken place at the Isthmus of Panama with the marine shore fishes. Curiously enough, the movement of genera seems to have been chiefly from the Atlantic to the Pacific. Certain characteristic genera[54] of the Panama region have not passed over to the Pacific. On the other hand, most of the common genera[55] show a much larger number of species on the Atlantic side. This may be held to show their Atlantic origin.
Of the relatively small number of genera which Panama has received from Polynesia[56] few have crossed the Isthmus to appear in the West Indian fauna.
Views of Earlier Writers on the Fishes of the Isthmus of Panama.—The elements of the problem at Panama may be better understood by a glance at the results of previous investigations.
In 1869 Dr. Günther, after enumerating the species examined by him from Panama, reaches the conclusion that nearly one-third of the marine fishes on the two shores of tropical America will be found to be identical. He enumerates 193 such species as found on the two coasts; 59 of these, or 31 per cent. of the total, being actually identical. From this he infers that there must have been, at a comparatively recent date, a depression of the Isthmus and intermingling of the two faunas.[57]
Catalogue of Fishes of Panama.—In an enumeration of the fishes of the Pacific coast in 1885,[58] the present writer showed that Dr. Günther's conclusions were based on inadequate data.
In my list 407 species were recorded from the Pacific coast of tropical America—twice the number enumerated by Dr. Günther. Of these 71 species, or 17½ per cent., were found also in the Atlantic. About 800 species are known from the Caribbean and adjacent shores, so that out of the total number of 1,136 species but 71, or 6 per cent. of the whole, are common to the two coasts. This number does not greatly exceed that of the species common to the West Indies and the Mediterranean, or even the West Indies and Japan. It is to be noted also that the number 71 is not very definitely ascertained, as there must be considerable difference of opinion as to the boundaries of species, and the actual identity in several cases is open to doubt.
This discrepancy arises from the comparatively limited representation of the two faunas at the disposal of Dr. Günther. He enumerates 193 marine or brackish-water species as found on the two coasts, 59 of which are regarded by him as specifically identical, this being 31 per cent. of the whole. But in 30 of these 59 cases I regard the assumption of complete identity as erroneous, so that taking the number 193 as given I would reduce the percentage to 15. But these 193 species form but a fragment of the total fauna, and any conclusion based on such narrow data is certain to be misleading.
Of the 71 identical species admitted in our list, several (e.g., Mola, Thunnus) are pelagic fishes common to most warm seas.
Still others (e.g., Trachurus, Carangus, Diodon sp.) are cosmopolitan in the tropical waters. Most of the others (e.g., Gobius, Gerres, Centropomus, Galeichthys sp., etc.) often ascend the rivers of the tropics, and we may account for their diffusion, perhaps, as we account for the dispersion of fresh-water fishes on the Isthmus, on the supposition that they may have crossed from marsh to marsh at some time in the rainy season.
In very few cases are representatives of any species from opposite sides of the Isthmus exactly alike in all respects. These differences in some cases seem worthy of specific value, giving us "representative species" on the two sides. In other cases the distinctions are very trivial, but in most cases they are appreciable, especially in fresh specimens.
Further, I expressed the belief that "fuller investigation will not increase the proportion of common species. If it does not, the two faunas show no greater resemblance than the similarity of physical conditions on the two sides would lead us to expect." This similarity causes the same types of fishes to persist on either side of the Isthmus while through isolation or otherwise these have become different as species.
This conclusion must hold so far as species are concerned, but the resemblance of the genera on the sides has a significance of its own.
In 1880[59] Dr Günther expressed his views in still stronger language, claiming a still larger proportion of the fishes of tropical America to be identical on the two sides of the continent. He concluded that "with scarcely any exceptions the genera are identical, and of the species found on the Pacific side, nearly one-half have proved to be the same as those of the Atlantic. The explanation of this fact has been found in the existence of communications between the two oceans by channels and straits which must have been open till within a recent period. The isthmus of Central America was then partially submerged, and appeared as a chain of islands similar to that of the Antilles; but as the reef-building corals flourished chiefly north and east of these islands and were absent south and west of them, reef fishes were excluded from the Pacific shores when the communications were destroyed by the upheaval of land."
Conclusions of Evermann and Jenkins.—This remark led to a further discussion of the subject on the part of Dr. B. W. Evermann and Dr. O. P. Jenkins. From their paper on the fishes of Guaymas[60] I make the following quotations:
"The explorations since 1885 have resulted (1) in an addition of about 100 species to one or other of the two faunas; (2) in showing that at least two species that were regarded as identical on the two shores[61] are probably distinct; and (3) in the addition of but two species to those common to both coasts.[62]
"All this reduces still further the percentage of common species.
"Of the 110 species obtained by us, 24, or less than 21 per cent., appear to be common to both coasts. Of these 24 species, at least 16, from their wide distribution, would need no hypothesis of a former waterway through the Isthmus to account for their presence on both sides. They are species fully able to arrive at the Pacific shores of the Americas from the warm seas west. It thus appears that not more than eight species, less than 8 per cent. of our collection, all of which are marine species, require any such hypothesis to account for their occurrence on both coasts of America. This gives us, then, 1,307 species that should properly be taken into account when considering this question, not more than 72 of which, or 5.5 per cent., seem to be identical on the two coasts. This is very different from the figures given by Dr. Günther in his 'Study of Fishes.'
"Now, if from these 72 species, admitted to be common to both coasts, we subtract the 16 species of wide distribution—so wide as to keep them from being a factor in this problem—we have left but 56 species common to the two coasts that bear very closely upon the waterway hypothesis. This is less than 4.3 per cent. of the whole number.
"But the evidence obtained from a study of other marine life of that region points to the same conclusion.
"In 1881, Dr. Paul Fischer discussed the same question in his 'Manual de Conchyliologie,' pp. 168, 169, in a section on the Molluscan Fauna of the Panamic Province, and reached the same general conclusions. He says: 'Les naturalistes Américians se sont beaucoup preéoccupés des espèces de Panama qui paraissent identiques avec celles des Antilles, ou qui sont représentatives. P. Carpenter estime qu'il en existe 35. Dans la plupart des cas, l'identite absolue n'a pu être constantée et on a trouvé quelques caractères distinctifs, ce qui n'a rien d'ètonnant, puisque dans l'hypothèse d'une origine commune, les deux races pacifique et atlantique sont séparée depuis la periode Miocène. Voici un liste de ces espèces représentatives ou identiques.' Here follows a list of 20 species. 'Mais ces formes semblables,' he says, 'constituent un infime minorité (3 per cent.).'
"These facts have a very important bearing upon certain geological questions, particularly upon the one concerning the cold of the Glacial period.
"In Dr. G. Frederick Wright's recent book, 'The Ice Age in North America,' eight different theories as to the cause of the cold are discussed. The particular theory which seems to him quite reasonable is that one which attributes the cold as due to a change of different parts of the country, and a depression of the Isthmus of Panama is one of the important changes he considers. He says: 'Should a portion of the Gulf Stream be driven through a depression across the Isthmus of Panama into the Pacific, and an equal portion be diverted from the Atlantic coast of the United States by an elevation of the sea-bottom between Florida and Cuba, the consequences would necessarily be incalculably great, so that the mere existence of such a possible cause for great changes in the distribution of moisture over the northern hemisphere is sufficient to make one hesitate before committing himself unreservedly to any other theory; at any rate, to one which has not for itself independent and adequate proof.'
"In the appendix to the same volume Mr. Warren Upham, in discussing the probable causes of glaciation, says: 'The quaternary uplifts of the Andes and Rocky Mountains and of the West Indies make it nearly certain that the Isthmus of Panama[Pg 276] has been similarly elevated during the recent epoch.... It may be true, therefore, that the submergence of this isthmus was one of the causes of the Glacial period, the continuation of the equatorial oceanic currents westward into the Pacific having greatly diminished or wholly diverted the Gulf Stream, which carries warmth from the tropics to the northern Atlantic and northwestern Europe.'
Fig. 180.—Caulophryne jordani Goode and Bean, a deep-sea fish of the Gulf Stream. Family Ceratiidæ.
Fig. 181.—Exerpes asper Jenkins and Evermann, a fish of the rock-pools, Guaymas, Mexico. Family Blenniidæ.
"Any very recent means by which the fishes could have passed readily from one side to the other would have resulted in making the fish faunas of the two shores practically identical; but the time that has elapsed since such a waterway could have[Pg 277] existed has been long enough to allow the fishes of the two sides to become practically distinct. That the mollusks of the two shores are almost wholly distinct, as shown by Dr. Fischer, is even stronger evidence of the remoteness of the time when the means of communication between the two oceans could have existed, for 'species' among the mollusks are probably more persistent than among fishes.
"Our present knowledge, therefore, of the fishes of tropical America justifies us in regarding the fish faunas of the two coasts as being essentially distinct, and believing that there has not been, at any comparatively recent time, any waterway through the Isthmus of Panama."
It is thus shown, I think, conclusively, that the Isthmus of Panama could not have been depressed for any great length of time in a recent geological period.
Conclusions of Dr. Hill.—These writers have not, however, considered the question of generic identity. To this we may find a clue in the geological investigations of Dr. Robert T. Hill.
In a study of "The Geological History of the Isthmus of Panama and Portions of Costa Rica," Dr. Hill uses the following language:
"By elimination we have concluded that the only period of time since the Mesozoic within which communication between the seas could have taken place is the Tertiary period, and this must be restricted to the Eocene and Oligocene epochs of that period. The paleontologic evidence upon which such an opening can be surmised at this period is the occurrence of a few California Eocene types in the Atlantic sides of the tropical American barrier, within the ranges of latitude between Galveston (Texas) and Colon, which are similar to others found in California. There are no known structural data upon which to locate the site of this passage, but we must bear in mind, however, that this structure has not been completely explored.
"Even though it was granted that the coincidence of the occurrence of a few identical forms on both sides of the tropical American region, out of the thousands which are not common, indicates a connection between the two seas, there is still an absence of any reason for placing this connection at the Isthmus of Panama, and we could just as well maintain that the[Pg 278] locus thereof might have been at some other point in the Central American region.
"The reported fossil and living species common to both oceans are littoral forms, which indicate that if a passage existed it must have been of a shallow and ephemeral character.
"There is no evidence from either a geologic or a biologic standpoint for believing that the oceans have ever communicated across the Isthmian regions since Tertiary time. In other words, there is no evidence for these later passages which have been established upon hypothetical data, especially those of Pleistocene time.
"The numerous assertions, so frequently found in literature, that the two oceans have been frequently and recently connected across the Isthmus, and that the low passes indicative of this connection still exist, may be dismissed at once and forever and relegated to the domain of the apocryphal. A few species common to the waters of both oceans in a predominantly Caribbean fauna of the age of the Claiborne epoch of the Eocene Tertiary is the only paleontologic evidence in any time upon which such a connection may be hypothesized.
"There has been a tendency in literature to underestimate the true altitude of the isthmian passes, which, while probably not intentional, has given encouragement to those who think that this Pleistocene passage may have existed. Maack has erroneously given the pass at 186 feet. Dr. J. W. Gregory states 'that the summit of the Isthmus at one locality is 154 feet and in another 287 feet in height.' The lowest isthmian pass, which is not a summit, but a drainage col, is 287-295 feet above the ocean.
"If we could lower the isthmian region 300 feet at present, the waters of the two oceans would certainly commingle through the narrow Culebra Pass. But the Culebra Pass is clearly the headwater col of two streams, the Obispo flowing into the Chagres, and the Rio Grande flowing into the Pacific, and has been cut by fluviatile action, and not by marine erosion, out of a land mass which has existed since Miocene time. Those who attempt to establish Pleistocene interoceanic channels through this pass on account of its present low altitude must not omit from their calculations the restoration of former rock[Pg 279] masses which have been removed by the general levelling of the surface by erosion."

Fig. 182.—Xenocys jessiæ Jordan and Bollman. Galapagos Islands. Family Lutianidæ.
In conclusion, Dr. Hill asserts that "there is considerable evidence that a land barrier in the tropical region separated the two oceans as far back in geologic history as Jurassic time, and that that barrier continued throughout the Cretaceous period. The geological structure of the Isthmus and Central American regions, so far as investigated, when considered aside from the paleontology, presents no evidence by which the former existence of a free communication of oceanic waters across the present tropical land barriers can be established. The paleontologic evidence indicates the ephemeral existence of a passage at the close of the Eocene period. All lines of inquiry—geologic, paleontologic, and biologic—give evidence that no connection has existed between the two oceans since the close of the Oligocene. This structural geology is decidedly opposed to any hypothesis by which the waters of the two oceans could have been connected across the regions in Miocene, Pliocene, Pleistocene, or recent times."
Final Hypothesis as to Panama.—If we assume the correctness of Dr. Hill's conclusions, they may accord in a remarkable degree with the actual facts of the distribution of the fishes about the Isthmus. To account for the remarkable identity of genera and divergence of species I may suggest the following hypothesis:
During the lifetime of most of the present species, the Isthmus has not been depressed. It was depressed in or before Miocene time, during the lifetime of most of the present genera. We learn from other sources that few of the extant species of fishes are older than the Pliocene. Relatively few genera go back to the Eocene, and most of the modern families appear to begin in the Eocene or later Cretaceous. In general the Miocene may be taken as the date of the origin of modern genera. The channel formed across the Isthmus was relatively shallow, excluding forms inhabiting rocky bottoms at considerable depths. It was wide enough to permit the infiltration from the Caribbean Sea of numerous species, especially of shore fishes of sandy bays, tide pools, and brackish estuaries. The currents set chiefly to the westward, favoring the transfer of Atlantic rather than Pacific types.

Fig. 183.—Channel Catfish, Ictalurus punctatus (Rafinesque). Illinois River. Family Siluridæ.
Since the date of the closing of this channel the species left on the two sides have been altered in varying degrees by the processes of natural selection and isolation. The cases of actual specific identity are few, and the date of the establishment as species, of the existing forms, is subsequent to the date of the last depression of the Isthmus.
We may be certain that none of the common genera ever found their way around Cape Horn. Most of them disappear to the southward, along the coasts of Brazil and Peru.
While local oscillations, involving changes in coast-lines, have doubtless frequently taken place and are still going on, the past and present distribution of fishes does not alone give adequate data for their investigation.
Further, it goes without saying that we have no knowledge of the period of time necessary to work specific changes in a body of species isolated in an alien sea. Nor have we any data as to the effect on a given fish fauna of the infiltration of many species and genera belonging to another. All such forces and results must be matters of inference.
The present writer does not wish to deny that great changes have taken place in the outlines of continents in relatively recent times. He would, however, insist that the theory of such changes must be confirmed by geological evidence, and evidence from groups other than fishes, and that likeness in separated fish faunas may not be conclusive.

Fig. 184.—Drawing the net on the beach of Hilo, Hawaii. Photograph by Henry W. Henshaw.
[37] Rheopresbe.
[38] Bryttosus.
[39] Leuciscus hakuensis Günther.
[40] Leuciscus jouyi.
[41] Formerly, but no longer, called Yeso in Japan.
[42] Called Nippon on foreign maps, but not so in Japan, where Nippon means the whole empire.
[43] Pleuronichthys cornutus, Hexogrammos otakii, etc.
[44] As Halichœres, Tetrapturus, Callionymus, Ariscopus, etc.
[45] Of these, the principal ones are Oxystomus, Myrus, Pagrus, Sparus, Macrorhamphosus, Cepola, Callionymus, Zeus, Uranoscopus, Lepidotrigla, Chelidonichthys.
[46] Among these are Beryx, Helicolenus, Lotella, Nettastoma, Centrolophus, Hoplostethus, Aulopus, Chlorophthalmus, Lophotes.
[47] Beryx, Hoplostethus.
[48] Antigonia, Etelis, Emmelichthys.
[49] Lepadogaster, Myrus; Lophotes, thus far recorded from Japan, the Mediterranean, and the Cape of Good Hope, is bassalian and of unknown range. Beryx, Trachichthys, Hoplostethus, etc., are virtually cosmopolitan as well as semi-bassalian.
[50] In this group we must place Cepola, Callionymus, Pagrus, Sparus, Beryx, Zeus, all of which have a very wide range in Indian waters.
[51] Cryptocentrus, Asterropteryx. The range of neither of these genera of small shore fishes is yet well known.
[52] As Crenilabrus, Labrus, Symphodus, Pagellus, Spondyliosoma, Sparisoma.
[53] As Chætodon, Lethrinus, Monotaxis, Glyphisodon, etc.
[54] Hoplopagrus, Xenichthys, Xenistius, Xenocys, Microdesmus, Cerdale, Cratinus, Azevia, Microlepidotus, Orthostœchus, Isaciella, etc.
[55] Hæmulon, Anisotremus, Gerres, Centropomus, Galeichthys, Hypoplectrus, Mycteroperca, Ulæma, Stellifer, Micropogon, Bodianus, Microspathodon.
[56] Among these are perhaps Teuthis (Acanthurus), Ilisha, Salarias, Myripristis, Thalassoma. Some such which have not crossed the Isthmus are Cirrhitus, Sectator, Sebastopsis, and Lophiomus.
[57] "Fishes of Central America," 1869, 397.
[58] Proc. U. S. Nat. Mus., 1885, 393.
[59] Introduction to the "Study of Fishes," 1880, p. 280.
[60] Proc. U. S. Nat. Mus., 1891, pp. 124-126.
[61] Citharichthys spilopterus and C. gilberti.
[62] Hæmulon steindachneri and Gymnothorax castaneus of the west coast probably being identical with H. schranki and Gymnothorax funebris of the east coast.

Dispersion of Fishes.—The methods of dispersion of fishes may be considered apart from the broader topic of distribution or the final results of such dispersion. In this discussion we are mainly concerned with the fresh-water fishes, as the methods of distribution of marine fishes through marine currents and by continuity of shore and water ways are all relatively simple.
The Problem of Oatka Creek.—When I was a boy and went fishing in the brooks of western New York, I noticed that the different streams did not always have the same kinds of fishes in them. Two streams in particular in Wyoming County, not far from my father's farm, engaged in this respect my special attention. Their sources are not far apart, and they flow in opposite directions, on opposite sides of a low ridge—an old glacial moraine, something more than a mile across. The Oatka Creek flows northward from this ridge, while the East Coy runs toward the southeast on the other side of it, both flowing ultimately into the same river, the Genesee.
It does not require a very careful observer to see that in these two streams the fishes are not quite the same. The streams themselves are similar enough. In each the waters are clear and fed by springs. Each flows over gravel and clay, through alluvial meadows, in many windings, and with elms and alders "in all its elbows." In both streams we were sure of finding trout,[64] and in one of them the trout are still abundant. In both we used to catch the brook chub,[65] or, as we [Pg 283]called it, the "horned dace"; and in both were large schools of shiners[66] and of suckers.[67] But in every deep hole, and especially in the millponds along the East Coy Creek, the horned pout[68] swarmed on the mucky bottoms. In every eddy, or in the deep hole worn out at the root of the elm-trees, could be seen the sunfish,[69] strutting in green and scarlet, with spread fins keeping intruders away from its nest. But in the Oatka Creek were found neither horned pout nor sunfish, nor have I ever heard that either has been taken there. Then besides these nobler fishes, worthy of a place on every schoolboy's string, we knew by sight, if not by name, numerous smaller fishes, darters[70] and minnows,[71] which crept about in the gravel on the bottom of the East Coy, but which we never recognized in the Oatka.
There must be a reason for differences like these, in the streams themselves or in the nature of the fishes. The sunfish and the horned pout are home-loving fishes to a greater extent than the others which I have mentioned; still, where no obstacles prevent, they are sure to move about. There must be, then, in the Oatka some sort of barrier, or strainer, which keeping these species back permits others more adventurous to pass; and a wider knowledge of the geography of the region showed that such is the case. Farther down in its course, the Oatka falls over a ledge of rock, forming a considerable waterfall at Rock Glen. Still lower down its waters disappear in the ground, sinking into some limestone cavern or gravel-bed, from which they reappear, after some six miles, in the large springs at Caledonia. Either of these barriers might well discourage a quiet-loving fish; while the trout and its active associates have some time passed them, else we should not find them in the upper waters in which they alone form the fish fauna. This problem is a simple one; a boy could work it out, and the obvious solution seems to be satisfactory.
Generalizations as to Dispersion.—Since those days I have been a fisherman in many waters,—not an angler exactly, but one who fishes for fish, and to whose net nothing large or small ever comes amiss; and wherever I go I find cases like this.
We do not know all the fishes of America yet, nor all those well that we know by sight; still this knowledge will come with time and patience, and to procure it is a comparatively easy task. It is also easy to ascertain the more common inhabitants of any given stream. It is difficult, however, to obtain negative results which are really results. You cannot often say that a species does not live in a certain stream. You can only affirm that you have not yet found it there, and you can rarely fish in any stream so long that you can find nothing that you have not taken before. Still more difficult is it to gather the results of scattered observations into general statements regarding the distribution of fishes. The facts may be so few as to be misleading, or so numerous as to be confusing, and the few writers who have taken up this subject in detail have found both these difficulties to be serious. Whatever general propositions we may maintain must be stated with the modifying clause of "other things being equal"; and other things are never quite equal. The saying that "Nature abhors a generalization" is especially applicable to all discussions of the relations of species to environment.
Still less satisfactory is our attempt to investigate the causes on which our partial generalizations depend,—to attempt to break to pieces the "other things being equal" which baffle us in our search for general laws. The same problems, of course, come up on each of the other continents and in all groups of animals or plants; but most that I shall say will be confined to the question of the dispersion of fishes in the fresh waters of North America. The broader questions of the boundaries of faunæ and of faunal areas I shall bring up only incidentally.
Questions Raised by Agassiz.—Some of the problems to be solved were first noticed by Prof. Agassiz in 1850, in his work on Lake Superior. Later (1854), in a paper on the fishes of the Tennessee River,[72] he makes the following statement:
"The study of these features [of distribution] is of the greatest importance, inasmuch as it may eventually lead to a better understanding of the intentions implied in this seemingly arbitrary disposition of animal life....
"There is still another very interesting problem respecting the geographical distribution of our fresh-water animals which may be solved by the further investigation of the fishes of the Tennessee River. The water-course, taking the Powell, Clinch, and Holston Rivers as its head waters, arises from the mountains of Virginia in latitude 37°; it then flows S.W. to latitude 34° 25', when it turns W. and N.W., and finally empties into the Ohio, under the same latitude as its source in 37°.

Fig. 185.—Horned Dace, Semotilus atromaculatus (Mitchill). Aux Plaines River, Ills. Family Cyprinidæ.
"The question now is this: Are the fishes of this water system the same throughout its extent? In which case we should infer that water communication is the chief condition of geographical distribution of our fresh-water fishes. Or do they differ in different stations along its course? And if so, are the differences mainly controlled by the elevation of the river above the level of the sea, or determined by climatic differences corresponding to differences of latitude? We should assume that the first alternative was true if the fishes of the upper course of the river differed from those of the middle and lower courses in the same manner as in the Danube, from its source to Pesth, where this stream flows nearly for its whole length under the same parallel. We would, on the contrary, suppose the second alternative to be well founded if marked differences were observed between the fish of such tracts of the river as do not[Pg 286] materially differ in their evolution above the sea, but flow under different latitudes. Now, a few collections from different stations along this river, like that sent me by Dr. Newman from the vicinity of Huntsville, would settle at once this question, not for the Tennessee River alone, but for most rivers flowing under similar circumstances upon the surface of the globe. Nothing, however, short of such collections, compared closely with one another, will furnish a reliable answer.... Whoever will accomplish this survey will have made a highly valuable contribution to our knowledge."
Conclusions of Cope.—Certain conclusions were also suggested by Prof. Cope in his excellent memoir on the fishes of the Alleghany region[73] in 1868. From this paper I make the following quotations:
"The distribution of fresh-water fishes is of special importance to the questions of the origin and existence of species in connection with the physical conditions of the waters and of the land. This is, of course, owing to the restricted nature of their habitat and the impossibility of their making extended migrations. With the submergence of land beneath the sea, fresh-water fish are destroyed in proportion to the extent of the invasion of salt water, while terrestrial vertebrates can retreat before it. Hence every inland fish fauna dates from the last total submergence of the country.
"Prior to the elevation of a given mountain chain, the courses of the rivers may generally have been entirely different from their later ones. Subsequent to this period, they can only have undergone partial modifications. As subsequent submergences can rarely have extended to the highlands where such streams originate, the fishes of such rivers can only have been destroyed so far as they were unable to reach those elevated regions, and preserve themselves from destruction from salt water by sheltering themselves in mountain streams. On the other hand, a period of greater elevation of the land, and of consequent greater cold, would congeal the waters and cover their courses with glaciers. The fishes would be driven to the neighborhood of the coast, though no doubt in more [Pg 287]southern latitudes a sufficient extent of uncongealed fresh waters would flow by a short course into the ocean, to preserve from destruction many forms of fresh-water fishes. Thus, through many vicissitudes, the fauna of a given system of rivers has had opportunity of uninterrupted descent, from the time of the elevation of the mountain range, in which it has its sources....
"As regards the distinction of species in the disconnected basins of different rivers, which have been separated from an early geologic period, if species occur which are common to any two or more of them, the supporter of the theory of distinct creations must suppose that such species have been twice created, once for each hydrographic basin, or that waters flowing into the one basin have been transferred to another. The developmentalist, on the other hand, will accept the last proposition, or else suppose that time has seen an identical process and similar result of modification in these distinct regions.

Fig. 186.—Chub of the Great Basin, Leuciscus lineatus (Girard). Heart Lake, Yellowstone Park. Family Cyprinidæ.
"Facts of distribution in the eastern district of North America are these. Several species of fresh-water fishes occur at the same time in many Atlantic basins from the Merrimac or from the Hudson to the James, and throughout the Mississippi Valley, and in the tributaries of the Great Lakes. On the other hand, the species of each river may be regarded as pertaining to four classes, whose distribution has direct reference to the character of the water and the food it offers: first, those of the tide-waters, of the river channels, bayous, and sluggish waters near them, or in the flat lands near the coast; second, those of the river channels of its upper course, where the currents are[Pg 288] more distinct; third, those of the creeks of the hill country; fourth, those of the elevated mountain streams which are subject to falls and rapids."
In the same paper Prof. Cope reaches two important general conclusions, thus stated by him:
"I. That species not generally distributed exist in waters on different sides of the great watershed.
"II. That the distribution of the species is not governed by the outlet of the rivers, streams having similar discharges (Holston and Kanawha, Roanoke and Susquehanna) having less in common than others having different outlets (Kanawha, or Susquehanna and James).
Fig. 187.—Butterfly-sculpin, Melletes papilio Bean, a fish of the rock-pools. St. Paul, Pribilof Islands.
"In view of the first proposition, and the question of the origin of species, the possibility of an original or subsequent mingling of the fresh waters suggests itself as more probable than that of distinct origin in the different basins."
Questions Raised by Cope.—Two questions in this connection are raised by Prof. Cope. The first question is this: "Has any destruction of the river faunæ taken place since the first elevation of the Alleghanies, when the same species were thrown into waters flowing in opposite directions?" Of such destruction by submergence or otherwise, Prof. Cope finds no evidence. The second question is, "Has any means of communication[Pg 289] existed, at any time, but especially since the last submergence, by which the transfer of species might occur?" Some evidence of such transfer exists in the wide distribution of certain species, especially those which seek the highest streamlets in the mountains; but except to call attention to the cavernous character of the Subcarboniferous and Devonian limestones, Prof. Cope has made little attempt to account for it.
Prof. Cope finally concludes with this important generalization:
"It would appear, from the previous considerations, that the distribution of fresh-water fishes is governed by laws similar to those controlling terrestrial vertebrates and other animals, in spite of the seemingly confined nature of their habitat."
Views of Günther.—Dr. Günther[74] has well summarized some of the known facts in regard to the manner of dispersion of fishes:
"The ways in which the dispersal of fresh-water fishes has been affected were various. They are probably all still in operation, but most work so slowly and imperceptibly as to escape direct observation; perhaps they will be more conspicuous after science and scientific inquiry shall have reached a somewhat greater age. From the great number of fresh-water forms which we see at this present day acclimatized in, gradually acclimatizing themselves in, or periodically or sporadically migrating into, the sea, we must conclude that under certain circumstances salt water may cease to be a barrier at some period of the existence of fresh-water species, and that many of them have passed from one river through salt water into another. Secondly, the headwaters of some of the grandest rivers, the mouths of which are at opposite ends of the continents which they drain, are sometimes distant from each other a few miles only. The intervening space may have been easily bridged over for the passage of fishes by a slight geological change affecting the level of the watershed or even by temporary floods; and a communication of this kind, if existing for a limited period only, would afford the ready means of an exchange of a number of species previously peculiar to one or the other of these river or lake systems. Some fishes provided with gill-openings [Pg 290]so narrow that the water moistening the gills cannot readily evaporate, and endowed, besides, with an extraordinary degree of vitality, like many Siluroids (Chlarias, Callichthys), eels, etc., are enabled to wander for some distance over land, and may thus reach a water-course leading them thousands of miles from their original home. Finally, fishes or their ova may be accidentally carried by water-spouts, by aquatic birds or insects, to considerable distances."
Fresh-water Fishes of North America.—We now recognize about six hundred species[75] of fishes as found in the fresh waters [Pg 291]of North America, north of the Tropic of Cancer, these representing thirty-four of the natural families. As to their habits, we can divide these species rather roughly into the four categories proposed by Prof. Cope, or, as we may call them,
(1) Lowland fishes; as the bowfin,[76] pirate-perch,[77] large-mouthed black bass,[78] sunfishes, and some catfishes.
(2) Channel-fishes; as the channel catfish,[79] the mooneye,[80] garpike,[81] buffalo-fishes,[82] and drum.[83]
(3) Upland fishes; as many of the darters, shiners, and suckers, and the small-mouthed black bass.[84]
(4) Mountain-fishes; as the brook trout and many of the darters and minnows.
To these we may add the more or less distinct classes of (5) lake fishes, inhabiting only waters which are deep, clear, and cold, as the various species of whitefish[85] and the Great Lake trout;[86] (6) anadromous fishes, or those which run up from the sea to spawn in fresh waters, as the salmon,[87] sturgeon,[88] shad,[89] and striped bass;[90] (7) catadromous fishes, like the eel,[91] which pass down to spawn in the sea; and (8) brackish-water fishes, which thrive best in the debatable waters of the river-mouths, as most of the sticklebacks and the killifishes.
As regards the range of species, we have every possible gradation from those which seem to be confined to a single river, and are rare even in their restricted habitat, to those which are [Pg 292]in a measure cosmopolitan,[92] ranging everywhere in suitable waters.
Characters of Species.—Still, again, we have all degrees of constancy and inconstancy in what we regard as the characters of a species. Those found only in a single river-basin are usually uniform enough; but the species having a wide range usually vary much in different localities. Such variations have at different times been taken to be the indications of as many different species. Continued explorations bring to light, from year to year, new species; but the number of new forms now discovered each year is usually less than the number of recognized species which are yearly proved to be untenable. Four complete lists of the fresh-water fishes of the United States (north of the Mexican boundary) have been published by the present writer. That of Jordan and Copeland,[93] published in 1876, enumerates 670 species. That of Jordan[94] in 1878 contains 665 species, and that of Jordan and Gilbert[95] in 1883, 587 species. That of Jordan and Evermann[96] in 1898 contains 585 species, although upwards of 130 new species were detected in the twenty-two years which elapsed between the first and the last list. Additional specimens from intervening localities are often found to form connecting links among the nominal species, and thus several supposed species become in time merged in one. Thus the common channel catfish[97] of our rivers has been described as a new species not less than twenty-five times, on account of differences real or imaginary, but comparatively trifling in value.
Where species can readily migrate, their uniformity is preserved; but whenever a form becomes localized its representatives assume some characters not shared by the species as a whole. When we can trace, as we often can, the disappearance by degrees of these characters, such forms no longer represent to us distinct species. In cases where the connecting forms are extinct, or at least not represented in collections, each form which is apparently different must be regarded as a distinct species.
The variations in any type become, in general, more marked as we approach the tropics. The genera are represented, on the whole, by more species there, and it would appear that the processes of specific change go on more rapidly under the easier conditions of life in the Torrid Zone.
We recognize now in North America twenty-five distinct species of fresh-water catfishes,[98] although nearly a hundred (93) nominal species of these fishes have been from time to time described. But these twenty-five species are among themselves very closely related, and all of them are subject to a variety of minor changes. It requires no strong effort of the imagination to see in them all the modified descendants of some one species of catfish, not unlike our common "bullhead,"[99] an immigrant probably from Asia, and which has now adjusted itself to its surroundings in each of our myriad of catfish-breeding streams.
Meaning of Species.—The word "species," then, is simply a term of convenience, including such members of a group similar to each other as are tangibly different from others, and are not known to be connected with these by intermediate forms. Such connecting links we may suppose to have existed in all cases. We are only sure that they do not now exist in our collections, so far as these have been carefully studied.
When two or more species of any genus now inhabit the same waters, they are usually species whose differentiation is of long standing,—species, therefore, which can be readily distinguished from one another. When, on the other hand, we have "representative species,"—closely related forms, neither of which is found within the geographical range of the other,—we [Pg 294]can with some confidence look for intermediate forms where the territory occupied by the one bounds that inhabited by the other. In very many such cases the intermediate forms have been found; and such forms are considered as subspecies of one species, the one being regarded as the parent stock, the other as an offshoot due to the influences of different environment. Then, besides these "species" and "subspecies," groups more or less readily recognizable, there are varieties and variations of every grade, often too ill-defined to receive any sort of name, but still not without significance to the student of the origin of species. Comparing a dozen fresh specimens of almost any kind of fish from any body of water with an equal number from somewhere else, one will rarely fail to find some sort of differences,—in size, in form, in color. These differences are obviously the reflex of differences in the environment, and the collector of fishes seldom fails to recognize them as such; often it is not difficult to refer the effect to the conditions. Thus fishes from grassy bottoms are darker than those taken from over sand, and those from a bottom of muck are darker still, the shade of color being, in some way not well understood, dependent on the color of the surroundings. Fishes in large bodies of water reach a larger size than the same species in smaller streams or ponds. Fishes from foul or sediment-laden waters are paler in color and slenderer in form than those from waters which are clear and pure. Again, it is often true that specimens from northern waters are less slender in body than those from farther south; and so on. Other things being equal, the more remote the localities from each other, the greater are these differences.
Fig. 188.—Scartichthys enosimæ Jordan and Snyder, a fish of the rock-pools of the sacred island of Enoshima, Japan. Family Blenniidæ.
In our fresh-water fishes each species on an average has been described as new from three to four times, on account of minor variations, real or supposed. In Europe, where the fishes have been studied longer and by more different men, upwards of six or eight nominal species have been described for each one that is now considered distinct.
Special Creation Impossible.—It is evident, from these and other facts, that the idea of a separate creation for each species of fishes in each river-basin, as entertained by Agassiz, is wholly incompatible with our present knowledge of the specific distinctions or of the geographical distribution of fishes. This is an unbroken gradation in the variations from the least to the greatest,—from the peculiarities of the individual, through local varieties, geographical subspecies, species, sub-genera, genera, families, super-families, and so on, until all fish-like vertebrates are included in a single bond of union.
Origin of American Species of Fishes.—It is, however, evident that not all American types of fishes had their origin in America, or even first assumed in America their present forms. Some of these are perhaps immigrants from northern Asia, where they still have their nearest relatives. Still others are evidently modified importations from the sea; and of these some are very recent immigrants, land-locked species which have changed very little from the parent stock.
The problems of analogous variation or parallelism without homology are very often met with among fishes. In shallow, swift brooks in all lands there are found small fishes which hug the bottom—large-finned, swift of movement, with speckled coloration, and with the air-bladder reduced in size. In the eastern United States these fishes are darters, dwarf perches; in northern India they are catfishes; in Japan, gobies or loaches; in Canada, sculpins; in South America, characins. Members of various groups may be modified to meet the same conditions of life. Being modified to look alike, the thought of mutual affinity is naturally suggested, but in such cases the likeness is chiefly external. The internal organs show little trace of such modifications. The inside of an animal tells what it really is, the outside where it has been. In other words, it is the external characters which are most readily affected by the environ[Pg 296]ment. Throughout all groups of animals and plants, there are large branches similarly affected by peculiarities of conditions.
This is the basis of the law of "Adaptive Radiation." Prof. H. F. Osborn thus states this law:
"It is a well-known principle of zoological evolution that an isolated region, if large and sufficiently varied in its topography, soil, climate, and vegetation, will give rise to a diversified fauna according to the law of adaptive radiation from primitive and central types. Branches will spring off in all directions to take advantage of every possible opportunity of securing food. The modifications which animals undergo in this adaptive radiation are largely of mechanical nature; they are limited in number and kind by hereditary stirp or germinal influences, and thus result in the independent evolution of similar types in widely separated regions under the law of parallelism or homoplasy."
[63] This chapter and the next are in substance reprinted from an essay published by the present writer in a volume called Science Sketches. A. C. McClurg & Co., Chicago.
[64] Salvelinus fontinalis Mitchill.
[65] Semotilus atromaculatus Mitchill.
[66] Notropis cornutus Rafinesque.
[67] Catostomus commersoni (Lacépède).
[68] Ameiurus melas Rafinesque.
[69] Eupomotis gibbosus Linnæus.
[70] Etheostoma flabellare Rafinesque.
[71] Rhinichthys atronasus Mitchill.
[72] On Fishes from Tennessee River, Alabama. American Journal of Science and Arts, xvii., 2d series, 1854, p. 26.
[73] On the Distribution of Fresh-water Fishes in the Alleghany Region of Southwestern Virginia. Journ. Acad. Nat. Sci., Phila., 1868, pp. 207-247.
[74] Introduction to the Study of Fishes, 1880, p. 211.
[75] The table below shows approximately the composition of the fresh-water fish fauna of Europe, as compared with that of North America north of the Tropic of Cancer.
| Families. | Europe. | N. America. | |
|---|---|---|---|
| Lamprey | Petromyzonidæ | 3 species. | 8 species. |
| Paddle-fish | Polyodontidæ | — " | 1 " |
| Sturgeon | Acipenseridæ | 10 " | 6 " |
| Garpike | Lepisosteidæ | — " | 3 " |
| Bowfin | Amiidæ | — " | 1 " |
| Mooneye | Hiodontidæ | — " | 3 " |
| Herring | Clupeidæ | 2 " | 5 " |
| Gizzard-shad | Dorosomidæ | — " | 1 " |
| Salmon | Salmonidæ | 12 " | 28 " |
| Characin | Characinidæ | — " | 1 " |
| Carp | Cyprinidæ | 61 " | 230 " |
| Loach | Cobiridæ | 3 " | — " |
| Sucker | Catostomidæ | — " | 51 " |
| Catfish | Siluridæ | 1 " | 25 " |
| Trout-perch | Percopsidæ | — " | 2 " |
| Blindfish | Amblyopsidæ | — " | 6 " |
| Killifish | Cyprinodontidæ | 3 " | 52 " |
| Mud-minnow | Umbridæ | 1 " | 2 " |
| Pike | Esocidæ | 1 " | 5 " |
| Alaska blackfish | Dalliidæ | — " | 1 " |
| Eel | Anguillidæ | 2 " | 1 " |
| Stickleback | Gasterosteidæ | 3 " | 7 " |
| Silverside | Atherinidæ | 2 " | 2 " |
| Pirate perch | Aphredoderidæ | — " | 1 " |
| Elassoma | Elassomidæ | — " | 2 " |
| Sunfish | Centrarchidæ | — " | 37 " |
| Perch | Percidæ | 11 " | 72 " |
| Bass | Serranidæ | 1 " | 4 " |
| Drum | Sciænidæ | — " | 1 " |
| Surf-fish | Embiotocidæ | — " | 1 " |
| Cichlid | Cichlidæ | — " | 2 " |
| Goby | Gobiidæ | 2 " | 6 " |
| Sculpin | Cottidæ | 2 " | 21 " |
| Blenny | Blenniidæ | 3 " | — " |
| Cod | Gadidæ | 1 " | 1 " |
| Flounder | Pleuronectidæ | 1 " | — " |
| Sole | Soleidæ | 1 " | 1 " |
Total: Europe, 21 families; 126 species. North America, 34 families; 590 species. A few new species have been added since this enumeration was made.
According to Dr. Günther (Guide to the Study of Fishes, p. 243), the total number of species now known from the temperate regions of Asia and Europe is about 360. The fauna of India, south of the Himalayas, is much more extensive, numbering 625 species. This latter fauna bears little resemblance to that of North America, being wholly tropical in its character.
[76] Amia calva Linnæus.
[77] Aphredoderus sayanus Gilliams.
[78] Micropterus salmoides Lacépède.
[79] Ictalurus punctatus Rafinesque.
[80] Hiodon tergisus Le Sueur.
[81] Lepisosteus osseus Linnæus.
[82] Ictiobus bubalus, cyprinella, etc.
[83] Aplodinotus grunniens Rafinesque.
[84] Micropterus dolomieu Lacépède.
[85] Coregonus clupeiformis, Argyrosomus artedi, etc.
[86] Cristivomer namaycush Walbaum.
[87] Salmo salar Linnæus.
[88] Acipenser sturio and other species.
[89] Alosa sapidissima Wilson.
[90] Roccus lineatus Bloch.
[91] Anguilla chrysypa Raf.
[92] Thus the chub-sucker (Erimyzon sucetta) in some of its varieties ranges everywhere from Maine to Dakota, Florida, and Texas; while a number of other species are scarcely less widely distributed.
[93] Check List of the Fishes of the Fresh Waters of North America, by David S. Jordan and Herbert E. Copeland. Bulletin of the Buffalo Society of Natural History, 1876, pp. 133-164.
[94] A Catalogue of the Fishes of the Fresh Waters of North America. Bulletin of the United States Geological Survey, 1878, pp. 407-442.
[95] A Catalogue of the Fishes Known to Inhabit the Waters of North America North of the Tropic of Cancer. Annual Report of the Commissioner of Fish and Fisheries for 1884 and 1885.
[96] Check List of the Fishes of North and Middle America. Report of the U. S. Commissioner of Fisheries for 1895.
[97] Ictalurus punctatus Rafinesque.
[98] Siluridæ.
[99] Ameiurus nebulosus.

The Process of Natural Selection.—We can say, in general, that in all waters not absolutely uninhabitable there are fishes. The processes of natural selection have given to each kind of river or lake species of fishes adapted to the conditions of life which obtain there. There is no condition of water, of bottom, of depth, of speed of current, but finds some species with characters adjusted to it. These adjustments are, for the most part, of long standing; and the fauna of any single stream has as a rule been produced by immigration from other regions or from other streams. Each species has an ascertainable range of distribution, and within this range we may be reasonably certain to find it in any suitable waters.
Fig. 189.—Slippery-dick or Doncella, Halichœres bivittatus Bloch, a fish of the coral reefs, Key West. Family Labridæ.
But every species has beyond question some sort of limit to its distribution, some sort of barrier which it has never passed in all the years of its existence. That this is true becomes evident when we compare the fish fauna of widely separated rivers. Thus the Sacramento, Connecticut, Rio Grande, and[Pg 298] St. John's Rivers have not a single species in common; and with one or two exceptions, not a species is common to any two of them. None of these[100] has any species peculiar to itself, and each shares a large part of its fish fauna with the water-basin next to it. It is probably true that the faunas of no two distinct hydrographic basins are wholly identical, while on the other hand there are very few species confined to a single one. The supposed cases of this character, some twenty in number, occur chiefly in the streams of the South Atlantic States and of Arizona. All of these need, however, the confirmation of further exploration. It is certain that in no case has an entire river fauna[101] originated independently from the divergence into separate species of the descendants of a single type.
The existence of boundaries to the range of species implies, therefore, the existence of barriers to their diffusion. We may now consider these barriers and in the same connection the degree to which they may be overcome.
Local Barriers.—Least important to these are the barriers which may exist within the limits of any single basin, and which tend to prevent a free diffusion through its waters of species inhabiting any portion of it. In streams flowing southward, or across different parallels of latitude, the difference in climate becomes a matter of importance. The distribution of species is governed very largely by the temperature of the water. Each species has its range in this respect,—the free-swimming fishes, notably the trout, being most affected by it; the mud-loving or bottom fishes, like the catfishes, least. The latter can reach the cool bottoms in hot weather, or the warm bottoms in cold weather, thus keeping their own temperature more even than that of the surface of the water. Although water communication is perfectly free for most of the length of the Mississippi, there is a material difference between the faunæ of the stream in Minnesota and in Louisiana. This difference is caused chiefly by the difference in temperature occupying the difference in latitude. That a similar difference in longitude, with free [Pg 299]water communication, has no appreciable importance, is shown by the almost absolute identity of the fish faunæ of Lake Winnebago and Lake Champlain. While many large fishes range freely up and down the Mississippi, a majority of the species do not do so, and the fauna of the upper Mississippi has more in common with that of the tributaries of Lake Michigan than it has with that of the Red River or the Arkansas. The influence of climate is again shown in the paucity of the fauna of the cold waters of Lake Superior, as compared with that of Lake Michigan. The majority of our species cannot endure the cold. In general, therefore, cold or Northern waters contain fewer species than Southern waters do, though the number of individuals of any one kind may be greater. This is shown in all waters, fresh or salt. The fisheries of the Northern seas are more extensive than those of the tropics. There are more fishes there, but are far less varied in kind. The writer once caught seventy-five species of fishes in a single haul of the seine at Key West, while on Cape Cod he obtained with the same net but forty-five species in the course of a week's work. Thus it comes that the angler, contented with many fishes of few kinds, goes to Northern streams to fish, while the naturalist goes to the South.
Fig. 190.—Peristedion miniatum Goode and Bean, a deep-red colored fish of the depths of the Gulf Stream.
But in most streams the difference in latitude is insignificant, and the chief differences in temperature come from differences in elevation, or from the distance of the waters from the colder source. Often the lowland waters are so different in character as to produce a marked change in the quality of their fauna. These lowland waters may form a barrier to the free movements[Pg 300] of upland fishes; but that this barrier is not impassable is shown by the identity of the fishes in the streams[102] of the uplands of middle Tennessee with those of the Holston and French Broad. Again, streams of the Ozark Mountains, similar in character to the rivers of East Tennessee, have an essentially similar fish fauna, although between the Ozarks and the Cumberland range lies an area of lowland bayous, into which such fishes are never known to penetrate. We can, however, imagine that these upland fishes may be sometimes swept down from one side or the other into the Mississippi, from which they might ascend on the other side. But such transfers certainly do not often happen. This is apparent from the fact that the two faunas[103] are not quite identical, and in some cases the same species are represented by perceptibly different varieties on one side and the other. The time of the commingling of these faunæ is perhaps now past, and it may have occurred only when the climate of the intervening regions was colder than at present.
The effect of waterfalls and cascades as a barrier to the diffusion of most species is self-evident; but the importance of such obstacles is less, in the course of time, than might be expected. In one way or another very many species have passed these barriers. The falls of the Cumberland limit the range of most of the larger fishes of the river, but the streams above it have their quota of darters and minnows. It is evident that the past history of the stream must enter as a factor into this discussion, but this past history it is not always possible to trace. Dams or artificial waterfalls now check the free movement of many species, especially those of migratory habits; while conversely, numerous other species have extended their range through the agency of canals.[104]
Every year fishes are swept down the rivers by the winter's floods; and in the spring, as the spawning season approaches, almost every species is found working its way up the stream. In some cases, notably the Quinnat salmon[105] and the blue-back salmon,[106] the length of these migrations is surprisingly great. To some species rapids and shallows have proved a sufficient barrier, and other kinds have been kept back by unfavorable conditions of various sorts. Streams whose waters are always charged with silt or sediment, as the Missouri, Arkansas, or Brazos, do not invite fishes; and even the occasional floods of red mud such as disfigure otherwise clear streams, like the Red River or the Colorado (of Texas), are unfavorable. Extremely unfavorable also is the condition which obtains in many rivers of the Southwest, as, for example, the Red River, the Sabine, and the Trinity, which are full from bank to bank in winter and spring, and which dwindle to mere rivulets in the autumn droughts.
Favorable Waters have Most Species.—In general, those streams which have conditions most favorable to fish life will be found to contain the greatest number of species. Such streams invite immigration; and in them the struggle for existence is individual against individual, species against species, and not a mere struggle with hard conditions of life. Some of the conditions most favorable to the existence in any stream of a large number of species of fishes are the following, the most important of which is the one mentioned first: Connection with a large hydrographic basin; a warm climate; clear water; a moderate current; a bottom of gravel (preferably covered by a growth of weeds); little fluctuation during the year in the volume of the stream or in the character of the water.
Limestone streams usually yield more species than streams flowing over sandstone, and either more than the streams of regions having metamorphic rocks. Sandy bottoms usually are not favorable to fishes. In general, glacial drift makes a suitable river bottom, but the higher temperature usual in regions beyond the limits of the drift gives to certain Southern streams conditions still more favorable. These conditions are all well [Pg 302]realized in the Washita River in Arkansas, and in various tributaries of the Tennessee, Cumberland, and Ohio; and in these, among American streams, the greatest number of species has been recorded.
The isolation and the low temperature of the rivers of New England have given to them a very scanty fish fauna as compared with the rivers of the South and West. This fact has been noticed by Professor Agassiz, who has called New England a "zoological island."[107]
In spite of the fact that barriers of every sort are sometimes crossed by fresh-water fishes, we must still regard the matter of freedom of water communication as the essential one in determining the range of most species. The larger the river basin, the greater the variety of conditions likely to be offered in it, and the greater the number of its species. In case of the divergence of new forms by the processes called "natural selection," the greater the number of such forms which may have spread through its waters; the more extended any river basin, the greater are the chances that any given species may sometimes find its way into it; hence the greater the number of species that actually occur in it, and, freedom of movement being assumed, the greater the number of species to be found in any one of its affluents.
Of the six hundred species of fishes found in the rivers of the United States, about two hundred have been recorded from the basin of the Mississippi. From fifty to one hundred of these species can be found in any one of the tributary streams of the size, say, of the Housatonic River or the Charles. In the Connecticut River there are but about eighteen species permanently resident; and the number found in the streams of Texas is not much larger, the best known of these, the Rio Colorado, having yielded but twenty-four species.
The waters of the Great Basin are not rich in fishes, the

Fig. 191.—Ancient Outlet of Lake Bonneville, Great Salt Lake, in Idaho. (Photograph by Prof. J. M. Aldrich.)
[Pg 304] species now found being evidently an overflow from the Snake River when in late glacial times it drained Lake Bonneville. This postglacial lake once filled the present basin of the Great Salt Lake and Utah Lake, its outlet flowing northwest from Ogden into Snake River. The same fishes are now found in the upper Snake River and the basins of Utah Lake and of Sevier Lake. In the same fashion Lake Lahontan once occupied the basin of Nevada, the Humboldt and Carson sinks, with Pyramid Lake. Its drainage fell also into the Snake River, and its former limits are shown in the present range of species. These have almost nothing in common with the group of species inhabiting the former drainage of Lake Bonneville. Another postglacial body of water, Lake Idaho, once united the lakes of Southeastern Oregon. The fauna of Lake Idaho, and of the lakes Malheur, Warner, Goose, etc., which have replaced it, is also isolated and distinctive. The number of species now known from this region of these ancient lobes is about 125. This list is composed almost entirely of a few genera of suckers,[108] minnows,[109] and trout.[110] None of the catfishes, perch, darters, or sunfishes, moon-eyes, pike, killifishes, and none of the ordinary Eastern types of minnows[111] have passed the barrier of the Rocky Mountains.
West of the Sierra Nevada the fauna is still more scanty, only about seventy species being enumerated. This fauna, except for certain immigrants[112] from the sea, is of the same general character as that of the Great Basin, though most of the species are different. This latter fact would indicate a considerable change, or "evolution," since the contents of the two faunæ were last mingled. There is a considerable difference between the fauna of the Columbia and that of the Sacramento. The species which these two basins have in common are chiefly those which at times pass out into the sea. The rivers of Alaska contain but few species, barely a dozen in all, most of these being found also in Siberia and Kamchatka. In the scantiness [Pg 305]of its faunal list, the Yukon agrees with the Mackenzie River, and with Arctic rivers generally.
There can be no doubt that the general tendency is for each species to extend its range more and more widely until all localities suitable for its growth are included. The various agencies of dispersal which have existed in the past are still in operation. There is apparently no limit to their action. It is probable that new "colonies" of one species or another may be planted each year in waters not heretofore inhabited by such species. But such colonies become permanent only where the conditions are so favorable that the species can hold its own in the struggle for food and subsistence. That the various modifications in the habitat of certain species have been caused by human agencies is of course too well known to need discussion here.
Watersheds.—We may next consider the question of watersheds, or barriers which separate one river basin from another.
Of such barriers in the United States, the most important and most effective is unquestionably that of the main chain of the Rocky Mountains. This is due in part to its great height, still more to its great breadth, and most of all, perhaps, to the fact that it is nowhere broken by the passage of a river. But two species—the red-throated or Rocky Mountain trout[113] and the Rocky Mountain whitefish[114]—are found on both sides of it, at least within the limits of the United States; while many genera, and even several families, find in it either an eastern or a western limit to their range. In a few instances representative species, probably modifications or separated branches of the same stock, occur on opposite sides of the range, but there are not many cases of correspondence even thus close. The two faunas are practically distinct. Even the widely distributed red-spotted or "dolly varden" trout[115] of the Columbia River and its affluents does not cross to the east side of the mountains, nor does the Montana grayling[116] ever make its way to the West. In Northern Mexico, however, numerous Eastern river fishes have crossed the main chain of the Sierra Madre.
How Fishes Cross Watersheds.—It is easy to account for this separation of the faunæ; but how shall we explain the almost universal diffusion of the whitefish and the trout in suitable waters on both sides of the dividing ridge? We may notice that these two are the species which ascend highest in the mountains, the whitefish inhabiting the mountain pools and lakes, the trout ascending all brooks and rapids in search of their fountainheads. In many cases the ultimate dividing ridge is not very broad, and we may imagine that at some time spawn or even young fishes may have been carried across by birds or other animals, or by man, or more likely by the dash of some summer whirlwind. Once carried across in favorable circumstances, the species might survive and spread.
The following is an example of how such transfer of species may be accomplished, which shows that we need not be left to draw on the imagination to invent possible means of transit.
The Suletind.—There are few watersheds in the world better defined than the mountain range which forms the "back-bone" of Norway. I lately climbed a peak in this range, the Suletind. From its summit I could look down into the valleys of the Lära and the Bägna, flowing in opposite directions to opposite sides of the peninsula. To the north of the Suletind is a large double lake called the Sletningenvand. The maps show this lake to be one of the chief sources of the westward-flowing river Lära. This lake is in August swollen by the melting of the snows, and at the time of my visit it was visibly the source of both these rivers. From its southeastern side flowed a large brook into the valley of the Bägna, and from its southwestern corner, equally distinctly, came the waters which fed the Lära. This lake, like similar mountain ponds in all northern countries, abounds in trout; and these trout certainly have for part of the year an uninterrupted line of water communication from the Sognefjord on the west of Norway to the Christianiafjord on the southeast,—from the North Sea to the Baltic. Part of the year the lake has probably but a single outlet through the Lära. A higher temperature would entirely cut off the flow into the Bägna, and a still higher one might dry up the lake altogether. This Sletningenvand, with its two[Pg 307] outlets on the summit of a sharp watershed, may serve to show us how other lakes, permanent or temporary, may elsewhere have acted as agencies for the transfer of fishes. We can also see how it might be that certain mountain fishes should be so transferred while the fishes of the upland waters may be left behind. In some such way as this we may imagine that various species of fishes have attained their present wide range in the Rocky Mountain region; and in similar manner perhaps the Eastern brook trout[117] and some other mountain species[118] may have been carried across the Alleghanies.
The Cassiquiare.—Professor John C. Branner calls my attention to a marshy upland which separates the valley of the La Plata from that of the Amazon, and which permits the free movement of fishes from the Paraguay River to the Tapajos. It is well known that through the Cassiquiare River the Rio Negro, another branch of the Amazon, is joined to the Orinoco River. It is thus evident that almost all the waters of eastern South America form a single basin, so far as the fishes are concerned.
As to the method of transfer of the trout from the Columbia to the Missouri, we are not now left in doubt.
Two-Ocean Pass.—To this day, as the present writer and later Evermann and Jenkins[119] have shown, the Yellowstone and Snake Rivers are connected by two streams crossing the main divide of the Rocky Mountains from the Yellowstone to the Snake across Two-Ocean Pass.
Prof. Evermann has described the locality as follows:
"Two-Ocean Pass is a high mountain meadow, about 8,200 feet above the sea and situated just south of the Yellowstone National Park, in longitude 110° 10' W., latitude 44° 3' N. It is surrounded on all sides by rather high mountains except where the narrow valleys of Atlantic and Pacific creeks open [Pg 308]out from it. Running back among the mountains to the northward are two small canyons down which come two small streams. On the opposite is another canyon down which comes another small stream. The extreme length of the meadow from east to west is about a mile, while the width from north to south is not much less. The larger of the streams coming in from the north is Pacific Creek, which, after winding along the western side of the meadow, turns abruptly westward, leaving the meadow through a narrow gorge. Receiving numerous small affluents, Pacific Creek soon becomes a good-sized stream, which finally unites with Buffalo Creek a few miles above where the latter stream flows into Snake River.
"Atlantic Creek was found to have two forks entering the pass. At the north end of the meadow is a small wooded canyon down which flows the North Fork. This stream hugs the border of the flat very closely. The South Fork comes down the canyon on the south side, skirting the brow of the hill a little less closely than does the North Fork. The two, coming together near the middle of the eastern border of the meadow, form Atlantic Creek, which after a course of a few miles flows into the Upper Yellowstone. But the remarkable phenomena exhibited here remain to be described.
"Each fork of Atlantic Creek, just after entering the meadow, divides as if to flow around an island, but the stream toward the meadow, instead of returning to the portion from which it had parted, continues its westerly course across the meadow. Just before reaching the western border the two streams unite and then pour their combined waters into Pacific Creek; thus are Atlantic and Pacific creeks united and a continuous waterway from the Columbia via Two-Ocean Pass to the Gulf of Mexico is established.
"Pacific Creek is a stream of good size long before it enters the pass, and its course through the meadow is in a definite channel, but not so with Atlantic Creek. The west bank of each fork is low and the stream is liable to break through anywhere and thus send part of its water across to Pacific Creek. It is probably true that one or two branches always connect the two creeks under ordinary conditions, and that following heavy rains or when the snows are melting, a much greater[Pg 309] portion of the water of Atlantic Creek crosses the meadow to the other side.
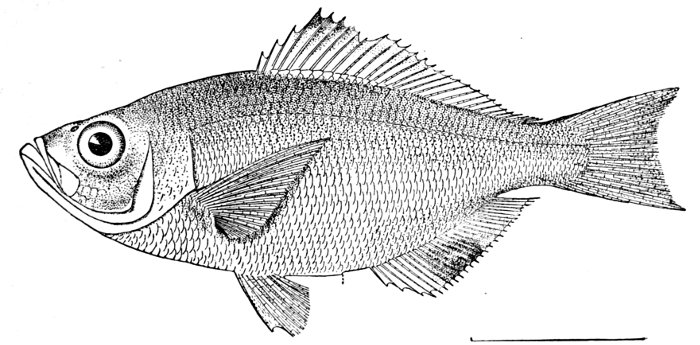
Fig. 192.—Silver Surf-fish (viviparous), Hypocritichthys analis (Agassiz). Monterey.
"Besides the channels already mentioned, there are several more or less distinct ones that were dry at the time of our visit. As already stated, the pass is a nearly level meadow covered with a heavy growth of grass and many small willows one to three feet high. While it is somewhat marshy in places it has nothing of the nature of a lake about it. Of course, during wet weather the small springs at the borders of the meadow would be stronger, but the important facts are that there is no lake or even marsh there and that neither Atlantic nor Pacific Creek has its rise in the meadow. Atlantic Creek, in fact, comes into the pass as two good-sized streams from opposite directions and leaves it by at least four channels, thus making an island of a considerable portion of the meadow. And it is certain that there is, under ordinary circumstances, a continuous waterway through Two-Ocean Pass of such a character as to permit fishes to pass easily and readily from Snake River over to the Yellowstone, or in the opposite direction. Indeed, it is quite possible, barring certain falls in the Snake River, for a fish so inclined, to start at the mouth of the Columbia, travel up that great river to its principal tributary, the Snake, thence on through the long, tortuous course of that stream, and, under the shadows of the Grand Teton, enter the cold waters of Pacific Creek, by which it could journey on up to[Pg 310] the very crest of the great continental divide,—to Two-Ocean Pass; through this pass it may have a choice of two routes to Atlantic Creek, in which the down-stream journey is begun. Soon it reaches the Yellowstone, down which it continues to Yellowstone Lake, then through the lower Yellowstone out into the turbid waters of the Missouri; for many hundred miles it may continue down this mighty river before reaching the Father of Waters, which will finally carry it to the Gulf of Mexico—a wonderful journey of nearly 6,000 miles, by far the longest possible fresh-water journey in the world.
"We found trout in Pacific Creek at every point where we examined it. In Two-Ocean Pass we found trout in each of the streams and in such positions as would have permitted them to pass easily from one side of the divide to the other. We also found trout in Atlantic Creek below the pass, and in the upper Yellowstone they were abundant. Thus it is certain that there is no obstruction, even in dry weather, to prevent the passage of trout from the Snake River to Yellowstone Lake; it is quite evident that trout do pass over in this way; and it is almost certain that Yellowstone Lake was stocked with trout from the west via Two-Ocean Pass."—Evermann.
Mountain Chains.—The Sierra Nevada constitutes also a very important barrier to the diffusion of species. This is, however, broken by the passage of the Columbia River, and many species thus find their way across it. That the waters to the west of it are not unfavorable for the growth of Eastern fishes is shown by the fact of the rapid spread of the common Eastern catfish,[120] or horned pout, when transported from the Schuylkill to the Sacramento. The catfish is now one of the important food fishes of the San Francisco markets, and with the Chinaman its patron, it has gone from California to Hawaii. The Chinese catfish, described by Bleeker as Ameiurus cantonensis, was doubtless carried home by some Chinaman returning from San Francisco. In like fashion the small-mouthed black bass is now frequent in California streams, as is also the blue-green sunfish, Apomotis cyanellus, introduced as food for the bass.
The mountain mass of Mount Shasta is, as already stated, a considerable barrier to the range of fishes, though a number of species find their way around it through the sea. The lower and irregular ridges of the Coast Range are of small importance in this regard, as the streams of their east slope reach the sea on the west through San Francisco Bay. Yet the San Joaquin contains a few species not yet recorded from the smaller rivers of southwestern California.
The main chain of the Alleghanies forms a barrier of importance separating the rich fish fauna of the Tennessee and Ohio basins from the scantier faunæ of the Atlantic streams. Yet this barrier is crossed by many more species than is the case with either the Rocky Mountains or the Sierra Nevada. It is lower, narrower, and much more broken,—as in New York, in Pennsylvania, and in Georgia there are several streams which pass through it or around it. The much greater age of the Alleghany chain, as compared with the Rocky Mountains, seems not to be an element of any importance in this connection. Of the fish which cross this chain, the most prominent is the brook trout,[121] which is found in all suitable waters from Hudson's Bay to the head of the Chattahoochee.
Upland Fishes.—A few other species are locally found in the head waters of certain streams on opposite sides of the range. An example of this is the little red "fallfish,"[122] found only in the mountain tributaries of the Savannah and the Tennessee. We may suppose the same agencies to have assisted these species that we have imagined in the case of the Rocky Mountain trout, and such agencies were doubtless more operative in the times immediately following the glacial epoch than they are now. Prof. Cope calls attention also to the numerous caverns existing in these mountains as a sufficient medium for the transfer of many species. I doubt whether the main chains of the Blue Ridge or the Great Smoky can be crossed in that way, though such channels are not rare in the subcarboniferous limestones of the Cumberland range. In the brooks at the head waters of the Roanoke River about Alleghany Springs in Virginia, fishes of the Tennessee Basin are found, instead of those characteristic [Pg 312]of the lower Roanoke. In this case it is likely that we have to consider the results of local erosion. Probably the divide has been so shifted that some small stream with its fishes has been cut off from the Holston and transferred to the Roanoke.
The passage of species from stream to stream along the Atlantic slope deserves a moment's notice. It is under present conditions impossible for any mountain or upland fish, as the trout or the miller's thumb,[123] to cross from the Potomac River to the James, or from the Neuse to the Santee, by descending to the lower courses of the rivers, and thence passing along either through the swamps or by way of the sea. The lower courses of these streams, warm and muddy, are uninhabitable by such fishes. Such transfers are, however, possible farther north. From the rivers of Canada and from many rivers of New England the trout does descend to the sea and into the sea, and farther north the whitefish does this also. Thus these fishes readily pass from one river basin to another. As this is the case now everywhere in the North, it may have been the case farther south in the time of the glacial cold. We may, I think, imagine a condition of things in which the snow-fields of the Alleghany chain might have played some part in aiding the diffusion of cold-loving fishes. A permanent snow-field on the Blue Ridge in western North Carolina might render almost any stream in the Carolinas suitable for trout, from its source to its mouth. An increased volume of colder water might carry the trout of the head streams of the Catawba and the Savannah as far down as the sea. We can even imagine that the trout reached these streams in the first place through such agencies, though of this there is no positive evidence. For the presence of trout in the upper Chattahoochee we must account in some other way.
It is noteworthy that the upland fishes are nearly the same in all these streams until we reach the southern limit of possible glacial influence. South of western North Carolina the faunæ of the different river basins appear to be more distinct from one another. Certain ripple-loving types are represented by closely related but unquestionably different species in each [Pg 313]river basin, and it would appear that a thorough mingling of the upland species in these rivers has never taken place.
The best examples of this are the following: In the Santee basin are found Notropis pyrrhomelas, Notropis niveus, and Notropis chloristius; in the Altamaha, Notropis xænurus and Notropis callisemus; in the Chattahoochee, Notropis hypselopterus and Notropis eurystomus; in the Alabama, Notropis cœruleus, Notropis trichroistius, and Notropis callistius. In the Alabama, Escambia, Pearl, and numerous other rivers is found Notropis cercostigma. This species descends to the sea in the cool streams of the pine woods. Its range is wider than that of the others, and in the rivers of Texas it reappears in the form of a scarcely distinct variety, Notropis venustus. In the Tennessee and Cumberland, and in the rivers of the Ozark range, is Notropis galacturus; and in the upper Arkansas Notropis camurus,—all distinct species of the same general type. Northward, in all the streams from the Potomac to the Oswego, and westward to the Des Moines and the Arkansas, occurs a single species of this type, Notropis whipplei, varying eastward into Notropis analostanus. But this species is not known from any of the streams inhabited by any of the other species mentioned, although very likely it is the parent stock of them all.
Lowland Fishes.—With the lowland species of the Southern rivers it is different. Few of these are confined within narrow limits. The streams of the whole South Atlantic and Gulf Coast flow into shallow bays, mostly bounded by sand-spits or sand-bars which the rivers themselves have brought down. In these bays the waters are often neither fresh nor salt; or, rather, they are alternately fresh and salt, the former condition being that of the winter and spring. Many species descend into these bays, thus finding every facility for transfer from river to river. There is a continuous inland passage in fresh or brackish waters, traversable by such fishes, from Chesapeake Bay nearly to Cape Fear; and similar conditions exist on the coasts of Louisiana, Texas, and much of Florida. In Perdido Bay I have found fresh-water minnows[124] and silversides[125] living together with marine gobies[126] and salt-water eels.[127] Fresh-water alligator [Pg 314]gars[128] and marine sharks compete for the garbage thrown over from the Pensacola wharves. In Lake Pontchartrain the fauna is a remarkable mixture of fresh-water fishes from the Mississippi and marine fishes from the Gulf. Channel-cats, sharks, sea-crabs, sunfishes, and mullets can all be found there together. It is therefore to be expected that the lowland fauna of all the rivers of the Gulf States would closely resemble that of the lower Mississippi; and this, in fact, is the case.
The streams of southern Florida and those of southwestern Texas offer some peculiarities connected with their warmer climate. The Florida streams contain a few peculiar fishes;[129] while the rivers of Texas, with the same general fauna as those farther north, have also a few distinctly tropical types,[130] immigrants from the lowlands of Mexico.
Cuban Fishes.—The fresh waters of Cuba are inhabited by fishes unlike those found in the United States. Some of these are evidently indigenous, derived in the waters they now inhabit directly from marine forms. Two of these are eyeless species,[131] inhabiting streams in the caverns. They have no relatives in the fresh waters of any other region, the blind fishes[132] of our caves being of a wholly different type. Some of the Cuban fishes are common to the fresh waters of the other West Indies. Of Northern types, only one, the alligator gar,[133] is found in Cuba, and this is evidently a filibuster immigrant from the coasts of Florida.
Swampy Watersheds.—The low and irregular watershed which separates the tributaries of Lake Michigan and Lake Erie from those of the Ohio is of little importance in determining the range of species. Many of the distinctively Northern fishes are found in the headwaters of the Wabash and the Scioto. The considerable difference in the general fauna of the Ohio Valley as compared with that of the streams of Michigan is due to the higher temperature of the former region, rather than [Pg 315]to any existing barriers between the river and the Great Lakes. In northern Indiana the watershed is often swampy, and in many places large ponds exist in the early spring.
At times of heavy rains many species will move through considerable distances by means of temporary ponds and brooks. Fishes that have thus emigrated often reach places ordinarily inaccessible, and people finding them in such localities often imagine that they have "rained down." Once, near Indianapolis, after a heavy shower, I found in a furrow in a corn-field a small pike,[134] some half a mile from the creek in which he should belong. The fish was swimming along in a temporary brook, apparently wholly unconscious that he was not in his native stream. Migratory fishes, which ascend small streams to spawn, are especially likely to be transferred in this way. By some such means any of the watersheds in Ohio, Indiana, or Illinois may be passed.

Fig. 193.—Creekfish or Chub-sucker, Erimyzon sucetta (Lacépède). Nipisink Lake, Illinois. Family Catostomidæ.
It is certain that the limits of Lake Erie and Lake Michigan were once more extended than now. It is reasonably probable that some of the territory now drained by the Wabash and the Illinois was once covered by the waters of Lake Michigan. The cisco[135] of Lake Tippecanoe, Lake Geneva, and the lakes of the Oconomowoc chain is evidently a modified descendant of the so-called lake herring.[136] Its origin most likely [Pg 316]dates from the time when these small deep lakes of Indiana and Wisconsin were connected with Lake Michigan. The changes in habits which the cisco has undergone are considerable. The changes in external characters are but trifling. The presence of the cisco in these lakes and its periodical disappearance—that is, retreat into deep water when not in the breeding season—have given rise to much nonsensical discussion as to whether any or all of these lakes are still joined to Lake Michigan by subterranean channels. Several of the larger fishes, properly characteristic of the Great Lake region,[137] are occasionally taken in the Ohio River, where they are usually recognized as rare stragglers. The difference in physical conditions is probably the sole cause of their scarcity in the Ohio basin.
The Great Basin of Utah.—The similarity of the fishes in the different streams and lakes of the Great Basin is doubtless to be attributed to the general mingling of their waters which took place during and after the Glacial Epoch. Since that period the climate in that region has grown hotter and drier, until the overflow of the various lakes into the Columbia basin through the Snake River has long since ceased. These lakes have become isolated from each other, and many of them have become salt or alkaline and therefore uninhabitable. In some of these lakes certain species may now have become extinct which still remain in others. In some cases, perhaps, the differences in surroundings may have caused divergence into distinct species of what was once one parent stock. The suckers in Lake Tahoe[138] and those in Utah Lake are certainly now different from each other and from those in the Columbia. The trout[139] in the same waters can be regarded as more or less tangible species, while the whitefishes[140] show no differences at all. The differences in the present faunas of Lake Tahoe and Utah Lake must be chiefly due to influences which have acted since the Glacial Epoch, when the whole Utah Basin was part of the drainage of the Columbia.
Arctic Species in Lakes.—Connected perhaps with changes [Pg 317]due to glacial influences is the presence in the deep waters of the Great Lakes of certain marine types,[141] as shown by the explorations of Professor Sidney I. Smith and others. One of these is a genus of fishes,[142] of which the nearest allies now inhabit the Arctic Seas. In his review of the fish fauna of Finland,[143] Professor A. J. Malmgren finds a number of Arctic species in the waters of Finland which are not found either in the North Sea or in the southern portions of the Baltic. These fishes are said to "agree with their 'forefathers' in the Glacial Ocean in every point, but remain comparatively smaller, leaner, almost starved." Professor Lovén[144] also has shown that numerous small animals of marine origin are found in the deep lakes of Sweden and Finland as well as in the Gulf of Bothnia. These anomalies of distribution are explained by Lovén and Malmgren on the supposition of the former continuity of the Baltic through the Gulf of Bothnia with the Glacial Ocean. During the second half of the Glacial Period, according to Lovén, "the greater part of Finland and of the middle of Sweden was submerged, and the Baltic was a great gulf of the Glacial Ocean, and not connected with the German Ocean. By the gradual elevation of the Scandinavian Continent, the Baltic became disconnected from the Glacial Ocean and the Great Lakes separated from the Baltic. In consequence of the gradual change of the salt water into fresh, the marine fauna became gradually extinct, with the exception of the glacial forms mentioned above."
It is possible that the presence of marine types in our Great Lakes is to be regarded as due to some depression of the land which would connect their waters with those of the Gulf of St. Lawrence. On this point, however, our data are still incomplete.
To certain species of upland or mountain fishes the depression of the Mississippi basin itself forms a barrier which cannot be passed. The black-spotted trout,[145] very closely related species [Pg 318]of which abound in all waters of northern Asia, Europe, and western North America, has nowhere crossed the basin of the Mississippi, although one of its species finds no difficulty in passing Bering Strait. The trout and whitefish of the Rocky Mountain region are all species different from those of the Great Lakes or the streams of the Alleghany system. To the grayling, the trout, the whitefish, the pike, and to arctic and subarctic species generally, Bering Strait has evidently proved no serious obstacle to diffusion; and it is not unlikely that much of the close resemblance of the fresh-water faunæ of northern Europe, Asia, and North America is due to this fact. To attempt to decide from which side the first migration came in regard to each group of fishes might be interesting; but without a wider range of facts than is now in our possession, most such attempts, based on guesswork, would have little value. The interlocking of the fish faunas of Asia and North America presents, however, a number of interesting problems, for migrations in both directions have doubtless taken place.
Causes of Dispersion Still in Operation.—One might go on indefinitely with the discussion of special cases, each more or less interesting or suggestive in itself, but the general conclusion is in all cases the same. The present distribution of fishes is the result of the long-continued action of forces still in operation. The species have entered our waters in many invasions from the Old World or from the sea. Each species has been subjected to the various influences implied in the term "natural selection," and under varying conditions its representatives have undergone many different modifications. Each of the six hundred fresh-water species we now know in the United States may be conceived as making every year inroads on territory occupied by other species. If these colonies are able to hold their own in the struggle for possession, they will multiply in the new conditions, and the range of the species becomes widened. If the surroundings are different, new species or varieties may be formed with time; and these new forms may again invade the territory of the parent species. Again, colony after colony of species [Pg 319]after species may be destroyed by other species or by uncongenial surroundings.
The ultimate result of centuries on centuries of the restlessness of individuals is seen in the facts of geographical distribution. Only in the most general way can the history of any species be traced; but could we know it all, it would be as long and as eventful a story as the history of the colonization and settlement of North America by immigrants from Europe. But by the fishes each river in America has been a hundred times discovered, its colonization a hundred times attempted. In these efforts there is no co-operation. Every individual is for himself, every struggle a struggle of life and death; for each fish is a cannibal, and to each species each member of every other species is an alien and a savage.
[100] Except possibly the Sacramento.
[101] Unless the fauna of certain cave streams in the United States and Cuba be regarded as forming an exception.
[102] For example, Elk River, Duck River, etc.
[103] There are three species of darters (Cottogaster copelandi Jordan, Hadropterus evides Jordan and Copeland, Hadropterus scierus Swain) which are now known only from the Ozark region or beyond and from the uplands of Indiana, not yet having been found at any point between Indiana and Missouri. These constitute perhaps isolated colonies, now separated from the parent stock in Arkansas by the prairie districts of Illinois, a region at present uninhabitable for these fishes. But the non-occurrence of these species over the intervening areas needs confirmation, as do most similar cases of anomalous distribution.
[104] Thus, Dorosoma cepedianum Le Sueur and Pomolobus chrysochloris Rafinesque have found their way into Lake Michigan through canals.
[105] Oncorhynchus tschawytscha Walbaum.
[106] Oncorhynchus nerka Walbaum.
[107] "In this isolated region of North America, in this zoological island of New England, as we may call it, we find neither Lepidosteus, nor Amia, nor Polyodon, nor Amblodon (Aplodinotus), nor Grystes (Micropterus), nor Centrarchus, nor Pomoxis, nor Ambloplites, nor Calliurus (Chænobryttus), nor Carpiodes, nor Hyodon, nor indeed any of the characteristic forms of North American fishes so common everywhere else, with the exception of two Pomotis (Lepomis), one Boleosoma, and a few Catostomus."—Agassiz, Amer. Journ. Sci. Arts, 1854.
[108] Catostomus, Pantosteus, Chasmistes.
[109] Gila, Ptychocheilus, etc.
[110] Salmo clarkii and its varieties.
[111] Genera Notropis, Chrosomus, etc.
[112] As the fresh-water surf-fish (Hysterocarpus traski) and the species of salmon.
[113] Salmo clarki Richardson.
[114] Coregonus williamsoni Girard.
[115] Salvelinus malma (Walbaum).
[116] Thymallus tricolor Cope.
[117] Salvelinus fontinalis Mitchill.
[118] Notropis rubricroceus Cope, Rhinichthys atronasus Mitchill, etc.
[119] Evermann, A Reconnoissance of the Streams and Lakes of Western Montana and Northwestern Wyoming, in Bull. U. S. Fish. Comm., XI, 1891, 24-28, pls. I and II; Jordan, The Story of a Strange Land, in Pop. Sci. Monthly, Feb., 1892, 447-458; Evermann, Two-Ocean Pass, in Proc. Ind. Ac. Sci., 1892, 29-34, pl. I; Evermann, Two-Ocean Pass, in Pop. Sci. Monthly, June, 1895, with plate.
[120] Ameiurus nebulosus Le Sueur: Ameiurus catus Linnæus.
[121] Salvelinus fontinalis.
[122] Notropis rubricroceus Cope.
[123] Cottus ictalops Rafinesque.
[124] Notropis cercostigma, Notropis xænocephalus.
[125] Labidesthes sicculus.
[126] Gobiosoma molestum.
[127] Myrophis punctatus.
[128] Lepisosteus tristœchus.
[129] Jordanella, Rivulus, Heterandria, etc.
[130] Heros, Tetragonopterus.
[131] Lucifuga and Stygicola, fishes allied to the cusk, and belonging to the family of Brotulidæ.
[132] Amblyopsis, Typhlichthys.
[133] Lepisosteus tristœchus.
[134] Esox vermiculatus Le Sueur.
[135] Argyrosomus sisco Jordan.
[136] Argyrosomus artedi Le Sueur.
[137] As Lota maculosa; Percopsis guttata; Esox masquinongy.
[138] Catostomus tahoensis, in Lake Tahoe; Catostomus macrocheilus and discobolus, in the Columbia; Catostomus fecundus; Catostomus ardens; Chasmistes liorus and Pantosteus generosus, in Utah Lake.
[139] Salmo henshawi and virginalis.
[140] Coregonus williamsoni.
[141] Species of Mysis and other genera of Crustaceans, similar to species described by Sars and others, in lakes of Sweden and Finland.
[142] Triglopsis thompsoni Girard, a near ally of the marine species Oncocottus quadricornis L.
[143] Kritisk Öfversigt of Finlands Fisk-Fauna, Helsingfors, 1863.
[144] See Günther, Zoological Record for 1864, p. 137.
[145] Salmo fario L., in Europe; Salmo labrax Pallas, etc., in Asia; Salmo gairdneri Richardson, in streams of the Pacific Coast; Salmo perryi, in Japan; Salmo clarki Richardson, throughout the Rocky Mountain range to the Mexican boundary and the headwaters of the Kansas, Platte, and Missouri.

The Flesh of Fishes.—Among all races of men, fishes are freely eaten as food, either raw, as preferred by the Japanese and Hawaiians, or else as cooked, salted, dried, or otherwise preserved.
The flesh of most fishes is white, flaky, readily digestible, and with an agreeable flavor. Some, as the salmon, are charged with oil, which aids to give an orange hue known as salmon color. Others have colorless oil which may be of various consistencies. Some have dark-red flesh, which usually contains a heavy oil which becomes acrid when stale. Some fishes, as the sharks, have tough, coarse flesh. Some have flesh which is watery and coarse. Some are watery and tasteless, some dry and tasteless. Some, otherwise excellent, have the muscular area, which constitutes the chief edible part of the fish, filled with small bones.
Relative Rank of Food-fishes.—The writer has tested most of the noted food-fishes of the Northern Hemisphere. When properly cooked (for he is no judge of raw fish) he would place first in the ranks as a food-fish the eulachon, or candle-fish (Thaleichthys pacificus).

Fig. 194.—Eulachon, or Ulchen. Thaleichthys pretiosus Girard. Columbia River. Family Argentinidæ.
This little smelt, about a foot long, ascends the Columbia River, Frazer River, and streams of southern Alaska in the spring in great numbers for the purpose of spawning. Its flesh is white, very delicate, charged with a white and very agreeable oil, readily digested, and with a sort of fragrance peculiar to the species.

Fig. 195.—Ayu, or Japanese Samlet, Plecoglossus altivelis Schlegel. Tanagawa, Tokyo, Japan.
Next to this he is inclined to place the ayu (Plecoglossus altivelis), a sort of dwarf salmon which runs in similar fashion in the rivers of Japan and Formosa. The ayu is about as large as the eulachon and has similar flesh, but with little oil and no fragrance.

Fig. 196.—Whitefish, Coregonus clupeiformis Mitchill. Ecorse, Mich.
Very near the first among sea-fishes must come the pampano[Pg 322] (Trachinotus carolinus) of the Gulf of Mexico, with firm, white, finely flavored flesh.
The red surmullet of Europe (Mullus barbatus) has been long famed for its delicate flesh, and may perhaps be placed next. Two related species in Polynesia, the munu and the kumu (Pseudupeneus bifasciatus and Pseudupeneus porphyreus), are scarcely inferior to it.
Fig. 197.—Golden Surmullet, Mullus auratus Jordan & Gilbert. Woods Hole, Mass.

Fig. 198.—Spanish Mackerel, Scomberomorus maculatus Mitchill. Family Scombridæ. Key West.
Side by side with these belongs the whitefish of the Great Lakes (Coregonus clupeiformis). Its flesh, delicate, slightly gelatinous, moderately oily, is extremely agreeable. Sir John Richardson records the fact that one can eat the flesh of this fish longer than any other without the feeling of cloying. The salmon cannot be placed in the front ranks because, however excellent, the stomach soon becomes tired of it. The Spanish mackerel (Scomberomorus maculatus), with flesh at once rich and delicate, the great opah (Lampris luna), still richer and still[Pg 323] more delicate, the bluefish (Pomatomus saltatrix) similar but a little coarser, the ulua (Carangus sem), the finest large food-fish of the South Seas, the dainty California poppy-fish, miscalled "Pampano" (Palometa simillima), and the kingfish firm and well-flavored (Scomberomorus cavalla), represent the best of the fishes allied to the mackerel.

Fig. 199.—Opah, or Moonfish, Lampris luna (Gmelin). Specimen in Honolulu market weighing 317½ lbs. (Photograph by E. L. Berndt.)—Page 323.

Fig. 200.—Bluefish, Pomatomus saltatrix (L.). New York.

Fig. 201.—Robalo, Centropomus undecimalis (Bloch). Florida.
The shad (Alosa sapidissima), with its sweet, tender, finely oily flesh, stands also near the front among food-fishes, but it sins above all others in the matter of small bones. The weak-fish (Cynoscion nobilis) and numerous relatives rank first among those with tender, white, savorous flesh. Among the bass and perch-like fishes, common consent places near the first the striped bass (Roccus lineatus), the bass of Europe (Dicentrarchus labrax), the susuki of Japan (Lateolabrax japonicus), the red tai of Japan (Pagrus major and P. cardinalis), the sheep's-head (Archosargus probatocephalus), the mutton-fish or Pargo Criollo of Cuba (Lutianus analis), the European porgy (Pagrus pagrus),[Pg 325] the robalo (Centropomus undecimalis), the uku (Aprion virescens) of Hawaii, the spadefish (Chætodipterus faber), and the black bass (Micropterus dolomieu).

Fig. 202.—Spadefish, Chætodipterus faber (L.). Virginia.

Fig. 203.—Small-mouthed Black Bass, Micropterus dolomieu (Lacépède). Potomac River.

Fig. 204.—Speckled Trout (male), Salvelinus fontinalis (Mitchill). New York.

Fig. 205.—Rainbow Trout, Salmo irideus Gibbons. Sacramento River, California.

Fig. 206.—Rangeley Trout, Salvelinus oquassa (Girard). Lake Oquassa, Maine.
The various kinds of trout have been made famous the world over. All are attractive in form and color; all are gamey; all have the most charming of scenic surroundings, and, finally, all are excellent as food, not in the first rank perhaps, but well above the second. Notable among these are the European charr (Salvelinus alpinus), the American speckled trout or charr (Salvelinus fontinalis), the Dolly Varden or malma (Salvelinus malma), and the oquassa trout (Salvelinus oquassa). Scarcely[Pg 327] less attractive are the true trout, the brown trout, or forelle (Salmo fario), in Europe, the rainbow-trout (Salmo irideus), the steelhead (Salmo gairdneri), the cut-throat trout (Salmo clarkii), and the Tahoe trout (Salmo henshawi), in America, and the yamabe (Salmo perryi) of Japan. Not least of all these is the flower of fishes, the grayling (Thymallus), of different species in different parts of the world.

Fig. 207.—Steelhead Trout, Salmo gairdneri Richardson. Columbia River.

Fig. 208.—Tahoe Trout, Salmo henshawi Gill & Jordan. Lake Tahoe, California.

Fig. 209.—The Dolly Varden Trout, Salvelinus malma (Walbaum). Lake Pend d'Oreille, Idaho. (After Evermann.)

Fig. 210.—Alaska Grayling, Thymallus signifer Richardson. Nulato, Alaska.

Fig. 211.—Pike, Esox lucius L. Ecorse, Mich.
Fig. 212.—Atka-fish, Pleurogrammus monopterygius (Pallas). Atka Island.
Other most excellent food-fishes are the eel (Anguilla species), the pike (Esox lucius), the muskallonge (Esox Roccus), the sole of Europe (Solea solea), the sardine (Sardinella pilchardus), the atka-fish (Pleurogrammus monopterygius) of Bering Sea, the pescado blanco of Lake Chapala (Chirostoma estor and other species), the Hawaiian mullet (Mugil cephalus), the channel catfish (Ictalurus punctatus), the turbot (Scophthalmus maximus), the barracuda (Sphyræna), and the young of various sardines and herring, known as whitebait. Of large fishes, probably the[Pg 329] swordfish (Xiphias gladius), the halibut (Hippoglossus hippoglossus), and the king-salmon, or quinnat (Oncorhynchus tschawytscha), may be placed first. Those people who feed on raw fish prefer in general the large parrot-fishes (as Pseudoscarus jordani in Hawaii), or else the young of mullet and similar species.
Fig. 213.—Pescado blanco, Chirostoma humboldtianum (Val.). Lake Chalco, City of Mexico.

Fig. 214.—Red Goatfish, or Salmonete, Pseudupeneus maculatus Bloch. Family Mullidæ (Surmullets).
Abundance of Food-fishes.—In general, the economical value of any species depends not on its toothsomeness, but on its abundance and the ease with which it may be caught and preserved. It is said that more individuals of the herring (Clupea harengus in the Atlantic, Clupea pallasi in the Pacific) exist than of any other species. The herring is a good food-fish and whenever it runs it is freely sought. According to Björnsön, wherever the school of herring touches the coast of Norway, there a village springs up, and this is true in Scotland, Newfoundland, and[Pg 330] from Killisnoo in Alaska to Otaru in Japan, and to Strielok in Siberia. Goode estimates the herring product of the North Atlantic at 1,500,000,000 pounds annually. In 1881 Professor Huxley used these words:

Fig. 215.—Great Parrot-fish, or Guacamaia, Pseudoscarus guacamaia Bloch & Schneider. Florida.

Fig. 216.—Striped Mullet, Mugil cephalus (L.). Woods Hole, Mass.
"It is said that 2,500,000,000 or thereabout of herrings are every year taken out of the North Sea and the Atlantic. Suppose we assume the number to be 3,000,000,000 so as to be quite safe. It is a large number undoubtedly, but what does it come to? Not more than that of the herrings which may be contained in one shoal, if it covers half a dozen square miles, and shoals of much larger size are on record. It is safe to say that scattered through the North Sea and the Atlantic, at one and the same time, there must be scores of shoals, any one of[Pg 331] which would go a long way toward supplying the whole of man's consumption of herrings."
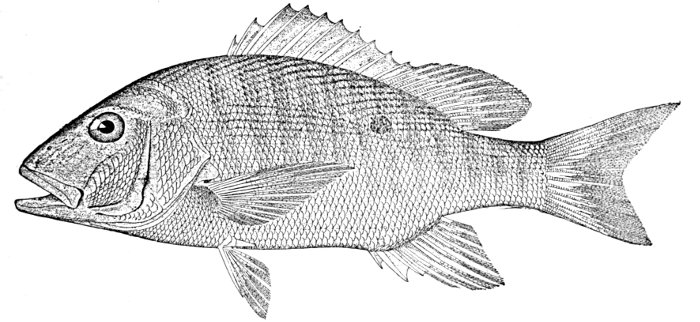
Fig. 217.—Mutton-snapper, or Pargo criollo, Lutianus analis (Cuv. & Val.). Key West.

Fig. 218.—Herring, Clupea harengus L. New York.
Fig. 219.—Codfish, Gadus callarias L. Eastport, Maine.
The codfish (Gadus callarias in the Atlantic; Gadus macrocephalus in the Pacific) likewise swarms in all the northern seas, takes the hook readily, and is better food when salted and dried than it is when fresh.
Next in economic importance probably stands the mackerel of the Atlantic (Scomber scombrus), a rich, oily fish which bears salting better than most.

Fig. 220.—Mackerel, Scomber scombrus L. New York.
Not less important is the great king-salmon, or quinnat (Oncorhyanchus tschawytscha), and the still more valuable blue-back salmon, or redfish (Oncorhynchus nerka).

Fig. 221.—Halibut, Hippoglossus hippoglossus (Linnæus). St. Paul Island, Bering Sea. (Photograph by U. S. Fur Seal Commission.)
The salmon of the Atlantic (Salmo salar), the various species of sturgeon (Acipenser), the sardines (Sardinella), the halibut (Hippoglossus), are also food-fishes of great importance.
Variety of Tropical Fishes.—In the tropics no one species is represented by enormous numbers of individuals as is the case in colder regions. On the other hand, the number of species regarded as food-fishes is much greater in any given port. In Havana, about 350 different species are sold as food in the markets, and an equal number are found in Honolulu. Upward of 600 different species appear in the markets of Japan. In England, on the contrary, about 50 species make up the list of fishes commonly used as food. Yet the number of individual fishes is probably not greater about Japan or Hawaii than in a similar stretch of British coast.
Economic Fisheries.—Volumes have been written on the economic value of the different species of fishes, and it is not the purpose of the present work to summarize their contents.

Fig. 222.—Fishing for Ayu with Cormorants in the Tanagawa, near Tokyo. (After Photograph by J. O. Snyder by Sekko Shimada.)
Equally voluminous is the literature on the subject of catching fishes. It ranges in quality from the quaint wisdom of the "Compleat Angler" and the delicate wit of "Little Rivers" to elaborate discussions of the most economic and effective forms and methods, of the beam-trawl, the purse-seine, and the cod[Pg 334]fish hook. In general, fishes are caught in four ways—by baited hooks, by spears, by traps, and by nets. Special local methods, such as the use of the tamed cormorant[146] in the catching of the ayu, by the Japanese fishermen at Gifu, may be set aside for the moment, and all general methods of fishing come under one of these four classes. Of these methods, the hook, the spear, the seine, the beam-trawl, the gill-net, the purse-net, the sweep-net, the trap and the weir are the most important. The use of the hook is again extremely varied. In the deep sea long, sunken lines, are sometimes used for codfish, each baited with many hooks. For pelagic fish, a baited hook is drawn swiftly over the surface, with a "spoon" attached which looks like a living fish. In the rivers a line is attached to a pole, and when fish are caught for pleasure or for the joy of being in the woods, recreation rises to the dignity of angling. Angling may be accomplished with a hook baited with an earthworm, a grasshopper, a living fish, or the larva of some insect. The angler of to-day, however, prefers the artificial fly, as being more workmanlike and also more effective than bait-fishing. The man who fishes, not for the good company of the woods and brooks, but to get as many fish as possible to eat or sell, is not an angler but a pot-fisher. The man who kills all the trout he can, to boast of his skill or fortune, is technically known as a trout-hog. Ethically, it is better to lie about your great catches of fine fishes than to make them. For most anglers, also, it is more easy.
Fisheries.—With the multiplicity of apparatus for fishing, there is the greatest variety in the boats which may be used. The fishing-fleet of any port of the world is a most interesting [Pg 335]object, as are also the fishermen with their quaint garb, plain speech, and their strange songs and calls with the hauling in of the net.

Fig. 223.—Fishing for Ayu in the Tanagawa, Japan. Emptying the pouch of the cormorant. (Photograph by J. O. Snyder.)
For much information on the fishing apparatus in use in America the reader is referred to the Reports of the Fisheries in the Tenth Census, in 1880, under the editorship of Dr. George Brown Goode. In these reports Goode, Stearns, Earle, Gilbert, Bean, and the present writer have treated very fully of all economic relations of the American fishes. In an admirable work entitled "American Fishes," Dr. Goode, with the fine literary[Pg 336] touch of which he was master, has fully discoursed of the game- and food-fishes of America with especial reference to the habits and methods of capture of each. To these sources, to Jordan and Evermann's "Food and Game Fishes of North America," and to many other works of similar purport in other lands, the reader is referred for an account of the economic and the human side of fish and fisheries.
Angling.—It is no part of the purpose of this work to describe the methods or materials of angling, still less to sing its praises as a means of physical or moral regeneration. We may perhaps find room for a first and a last word on the subject; the one the classic from the pen of the angler of the brooks of Staffordshire, and the other the fresh expression of a Stanford student setting out for streams such as Walton never knew, the Purissima, the Stanislaus, or perchance his home streams, the Provo or the Bear.
"And let me tell you, this kind of fishing with a dead rod, and laying night-hooks, are like putting money to use; for they both work for the owners when they do nothing but sleep, or eat, or rejoice, as you know we have done this last hour, and sat as quietly and as free from cares under this sycamore as Virgil's Tityrus and his Melibœus did under their broad beech-tree. No life, my honest scholar,—no life so happy and so pleasant as the life of a well-governed angler; for when the lawyer is swallowed up with business and the statesman is preventing or contriving plots, then we sit on the cowslip-banks, hear the birds sing, and possess ourselves in as much quietness as these silent silver streams which we now see glide so quietly by us. Indeed, my good scholar, we may say of angling, as Dr. Boteler said of strawberries, 'Doubtless God could have made a better berry, but doubtless God never did'; and so, if I might be judge, 'God never made a more calm, quiet, innocent recreation than angling.'
"I'll tell you, scholar, when I sat last on this primrose-bank, and looked down these meadows, I thought of them as Charles the Emperor did of Florence, 'That they were too pleasant to be looked on but only on holidays.'
"Gentle Izaak! He has been dead these many years, but his disciples are still faithful. When the cares of business lie[Pg 337] heavy and the sound of wheels jarring on cobbled streets grows painful, one's fingers itch for the rod; one would away to the quiet brook among the pines, where one has fished so often. Every man who has ever got the love of the stream in his blood feels often this longing.
"It comes to me each year with the first breath of spring. There is something in the sweetness of the air, the growing things, the 'robin in the greening grass' that voices it. Duties that have before held in their performance something of pleasure become irksome, and practical thoughts of the day's work are replaced by dreamy pictures of a tent by the side of a mountain stream—close enough to hear the water's singing in the night. Two light bamboo rods rest against the tent-pole, and a little column of smoke rising straight up through the branches marks the supper fire. Jack is preparing the evening meal, and, as I dream, there comes to me the odor of crisply browned trout and sputtering bacon—was ever odor more delicious? I dare say that had the good Charles Lamb smelled it as I have, his 'Dissertation on Roast Pig' would never have been written. But then Charles Lamb never went a-fishing as we do here in the west—we who have the mountains and the fresh air so boundlessly.
"And neither did Izaak Walton for that matter. He who is sponsor for all that is gentle in angling missed much that is best in the sport by living too early. He did not experience the exquisite pleasure of wading down mountain streams in supposedly water-proof boots and feeling the water trickling in coolingly; nor did he know the joy of casting a gaudy fly far ahead with a four-ounce rod, letting it drift, insect-like, over that black hole by the tree stump, and then feeling the seaweed line slip through his fingers to the whirr of the reel. And, at the end of the day, supper over, he did not squat around a big camp-fire and light his pipe, the silent darkness of the mountains gathering round, and a basketful of willow-packed trout hung in the clump of pines by the tent. Izaak's idea of fishing did not comprehend such joy. With a can of worms and a crude hook, he passed the day by quiet streams, threading the worms on his hook and thinking kindly of all things. The day's meditations over, he went back to the village, and, may[Pg 338]hap, joined a few kindred souls over a tankard of ale at the sign of the Red Lobster. But he missed the mountains, the water rushing past his tent, the bacon and trout, the camp-fire—the physical exaltation of it all. His kind of fishing was angling purely, while modern Waltons, as a rule, eschew the worm.

Fig. 224.—Fishing for Tai, Tokyo Bay. (Photograph by J. O. Snyder.)
"To my mind, there is no real sport in any kind of fishing except fly-fishing. This sitting on the bank of a muddy stream with your bait sunk, waiting for a bite, may be conducive to gentleness and patience of spirit, but it has not the joy of action in which a healthy man revels. How much more sport is it to clamber over fallen logs that stretch far out a-stream, to wade slipping over boulders and let your fly drop caressingly on ripples and swirling eddies and still holes! It is worth all the work to see the gleam of a silver side as a half-pounder rises, and, with a flop, takes the fly excitedly to the bottom. And then the nervous thrill as, with a deft turn of the wrist, you hook him securely—whoever has felt that thrill cannot forget it. It will come back to him in his law office when he should be thinking of other things; and with it will come a longing for that dear remembered stream and the old days. That is the hold trout-fishing takes on a man.
"It is spring now and I feel the old longing myself, as I always do when life comes into the air and the smell of new growth is sweet. I got my rod out to-day, put it together, and have been looking over my flies. If I cannot use them, I can at least muse over days of the past and dream of those to come." (Waldemar Young.)
[146] The cormorant is tamed for this purpose. A harness is placed about its wings and a ring about the lower part of its neck. Two or three birds may be driven by a boy in a shallow stream, a small net behind him to drive the fish down the river. In a large river like that of Gifu, where the cormorants are most used, the fishermen hold the birds from the boats and fish after dark by torchlight. The bird takes a great interest in the work, darts at the fishes with great eagerness, and fills its throat and gular pouch as far down as the ring. Then the boy takes him out of the water, holds him by the leg and shakes the fishes out into a basket. When the fishing is over the ayu are preserved, the ring is taken off from the bird's neck, and the zako or minnows are thrown to him for his share. These he devours greedily.

Contagious Diseases.—As compared with other animals the fishes of the sea are subject to but few specific diseases. Those in fresh waters, being more isolated, are more frequently attacked by contagious maladies. Often these diseases are very destructive. In an "epidemic" in Lake Mendota, near Madison, Wis., Professor Stephen A. Forbes reports a death of 300 tons of fishes in the lake. I have seen similar conditions among the land-locked alewife in Cayuga and Seneca Lakes, the dead fishes being piled on the beaches so as to fill the air with the stench of their decay.

Fig. 225.—Menhaden, Brevoortia tyrannus (Latrobe). Woods Hole, Mass.
Crustacean Parasites.—The external parasites of fishes are of little injury. These are mainly lernæans and other crustaceans (fish-lice) in the sea, and in the rivers different species of leeches. These may suck the blood of the fish, or in the case of certain crustaceans which lie under the tongue, steal the food as it passes along, as is done by Cymothoa prægustator, the "bug" of the mouth of the menhaden (Brevoortia tyrannus).

Fig. 226.—Australian Flying-fish, Exonautes unicolor (Valenciennes). Specimen from Tasman Sea, having parasitic lernæan crustaceans, to which parasitic barnacles are attached. (After Kellogg.)
[Pg 342] The relation of this crustacean to its host suggested to Latrobe, its discoverer, the relation of the "foretaster" in Roman times to the tyrant whom he served. A similar commensation exists in the mouth of a mullet (Mugil hospes) at Panama. The writer has received, through the courtesy of Mr. A. P. Lundin, a specimen of a flying-fish (Exonautes unicolor) taken off Sydney, Australia. To this are attached three large copepod crustaceans of the genus Penella, the largest over two inches long, and to the copepods in turn are attached a number of barnacles (Conchoderma virgatum) so joined to the copepods as to suggest strange flowers, like orchids, growing out of the fish.

Fig. 227.—Black-nosed Dace, Rhinichthys atronasus (Mitchill). East Coy Creek, W. N. Y. Showing black spots of parasitic organisms. (From life by Mary Jordan Edwards.)
Myxosporidia, or Parasitic Protozoa.—Internal parasites are very numerous and varied. Some of them are bacteria, giving rise to infectious diseases, especially in ponds and lakes. Others are myxosporidia, or parasitic protozoans, which form warty appendages, which burst, discharging the germs and leaving ulcers in their place. In the report of the U. S. Fish Commissioner for 1892, Dr. R. R. Gurley has brought together our knowledge of the protozoans of the subclass Myxosporidia, to which these epidemics are chiefly due. These creatures belong to the class of Sporozoa, and are regarded as animals, their nearest relatives being the parasitic Gregarinida, from which they differ in having the germinal portion of the spore consisting of a single protoplasmic mass instead of falciform protoplasmic rods as in the worm-like Gregarines. The Myxosporidia are parasitic on fishes, both fresh-water and marine, especially beneath the epidermis of the gills and fins and in the gall-bladder and urinary bladder. In color these protozoa are[Pg 343] always cream-white. In size and form they vary greatly. The cyst in which they lie is filled with creamy substance made up of spores and granule matter.
Dr. Gurley enumerates as hosts of these parasites about sixty species of fishes, marine and fresh-water, besides frogs, crustaceans, sea-worms, and even the crocodile. In the sharks and rays the parasites occur mainly in the gall-ducts, in the minnows within the gill cavity and epidermis, and in the higher fishes mainly but not exclusively in the same regions. Forty-seven species are regarded by Gurley as well defined. The diseases produced by them are very obscurely known. These parasites on American fishes have been extensively studied by Charles Wardall Stiles, Edwin Linton, Henry B. Ward, and others.
According to Dr. Linton the parasitism which results from infection with protozoan parasites will, of all kinds, be found to be the most important. Epidemics among European fish have been repeatedly traced to this source. The fatality which attends infection with psorosperms appears to be due to a secondary cause, however, namely, to bacilli which develop within the psorosperms (Myxobolus) tumors and give rise to ulceration. The discharge of these ulcers then disseminates the disease.

Fig. 228.—White Shiner, Notropis hudsonius (Clinton), with cysts of parasitic psorosperms. (After Gurley.)
"Brief mention of the remedies there proposed may appropriately be repeated here. Megnin sees no other method than to collect all the dead and sick fishes and to destroy them by fire. Ludwig thinks that the waters should be kept pure, and that the pollutions of the rivers by communities or industrial establishments should be interdicted. Further he says:
"That most dangerous contamination of the water by the Myxosporidia from the ulcers cannot of course be stopped entirely, but it is evident that it will be less if all fishermen are impressed with the importance of destroying all diseased and dead fish instead of throwing them back into the water. Such destruction must be so effected as to prevent the re-entry of the germs into the water.
"Railliet says that it is expedient to collect the diseased fish and to bury them at a certain depth and at a great distance from the water-course. He further states that this was done on the Meuse with success, so that at the end of some years the disease appeared to have left no trace."

Fig. 229.—White Catfish, Ameiurus catus (Linnæus), from Potomac River, infested by parasitic protozoa, Ichthyophthirus multifilis Fouquet. (After C. W. Stiles.)
Parasitic Worms: Trematodes.—Parasitic worms in great variety exist in the intestinal canal or in the liver or muscular substance of fishes.
Trematode worms are most common in fresh-water fishes. These usually are sources of little injury, especially when found in the intestines, but they may do considerable mischief when encysted within the body cavity or in the heart or liver. Dr. Linton describes 31 species of these worms from 25 different species of American fishes. In 20 species of fishes from the Great Lakes, 102 specimens, Dr. H. B. Ward found 95 specimens infected with parasites, securing 4000 trematodes, 2000 acanchocephala, 200 cestodes, and 200 nematodes. In the bowfin (Amia calva), trematodes existed in enormous numbers.
Cestodes.—Cestode worms exist largely in marine fishes, the adults, according to Dr. Linton, being especially common in the spiral valve of the shark. It is said that one species of[Pg 345] human tape-worm (Bothriocephalus tænia) has been got from eating the flesh of the European tench (Tinca tinca).
The Worm of the Yellowstone.—The most remarkable case of parasitism of worms of this type is that given by the trout of Yellowstone Lake (Salmo clarki). This is thus described by Dr. Linton:
"One of the most interesting cases of parasitism in which direct injury results to the host, which has come to my attention, is that afforded by the trout of Yellowstone Lake (Salmo clarki). It was noticed by successive parties who visited the lake in connection with government surveys that the trout with which the lake abounded were, to a large extent, infested with a parasitic worm, which is most commonly in the abdominal cavity, in cysts, but which in time escapes from the cyst and tunnels into the flesh of its host. Fish, when thus much afflicted, are found to be lacking in vitality, weak, and often positively emaciated.
"It was my good fortune, in the summer of 1890, to visit this interesting region for the purpose of investigating the parasitism of the trout of Yellowstone Lake. The results of this special investigation were published in the Bulletin of the U. S. Fish Commission for 1889, vol. ix., pp. 337-358, under the title 'A Contribution to the Life-history of Dibothrium cordiceps, a Parasite Infesting the Trout of Yellowstone Lake.'
"I found the same parasite in the trout of Heart Lake, just across the great continental divide from Yellowstone Lake, but did not find any that had tunneled into the flesh of its host, while a considerable proportion of the trout taken in Yellowstone Lake had these worms in the flesh. Some of these worms were as much as 30 centimeters in length when first removed; others which had lain in water a few hours after removal before they were measured were much longer, as much as 54 centimeters. They are rather slender and of nearly uniform size throughout, 2.5 to 3 millimeters being an average breadth of the largest. I found the adult stage in the intestine of the large white pelican (Pelecanus erythrorhynchus), which is abundant on the lake and was found breeding on some small islands near the southern end of the lake.
"In the paper alluded to above I attempted to account for[Pg 346] two things concerning this parasitism among the trout of Yellowstone Lake: First, the abundance of parasitized trout in the lake; second, the migration of the parasite into the muscular tissue of its host. The argument cannot be well summarized in as short space as the requirements of this paper demand. It is sufficient to say that what appear to me to be satisfactory explanations are supplied by the peculiar conditions of distribution of fish in the lakes of this national park. Until three or four years ago, when the U. S. Fish Commission stocked some of the lakes and streams of the park, the conditions with relation to fish life in the three principal lakes were as follows: Shoshone Lake, no fish of any kind; Heart Lake, at least three species, Salmo clarki, Leuciscus lineatus, and Catostomus ardens; Yellowstone Lake, one species, Salmo clarki. Shoshone and Yellowstone Lakes are separated from the river systems which drain them by falls too high for fish to scale. Heart Lake has no such barrier. The trout of Yellowstone Lake are confined to the lake and to eighteen miles of river above the falls. Whatever source of parasitism exists in the lake, therefore, must continue to affect the fish all their lives. They cannot be going and coming from the lake as the trout of Heart Lake may freely do. If their food should contain eggs of parasites, or if the waters in which they swim should contain eggs or embryos of parasites, they would be continually exposed to infection, with no chance for a vacation trip for recuperation. To quote from my report:
"'It follows, therefore, from the peculiar conditions surrounding the trout of Yellowstone Lake, that if there is a cause of parasitism present in successive years the trout are more liable to become infested than they would be in waters where they had a more varied range. Trout would become infested earlier and in greater relative numbers, and the life of the parasites themselves—that is, their residence as encysted worms—must be of longer duration than would be the rule where the natural conditions are less exceptional.... There are probably not less than one thousand pelicans on the lake the greater part of the time throughout the summer, of which at any time not less than 50 per cent. are infested with the adult form of the parasite, and, since they spend the greater part of[Pg 347] their time on or over the water, disseminate millions of tape-worm eggs each in the waters of the lake. It is known that eggs of other dibothria hatch out in the water, where they swim about for some time, looking much like ciliated infusoria. Donnadieu found in his experiments on the adult dibothria of ducks that the eggs hatched out readily in warm water and very slowly in cold. If warm water, at least water that is warmer than the prevailing temperature of the lake, is needed for the proper development of these ova, the conditions are supplied in such places as the shore system of geysers and hot springs on the west arm of the lake, where for a distance of nearly three miles the shore is skirted by a hot spring and geyser formation, with numerous streams of hot water emptying into the lake, and large springs of hot water opening in the floor of the lake near shore.
"'Trout abound in the vicinity of these warm springs, presumably on account of the abundance of food there. They do not love the warm water, but usually avoid it. Several persons with whom I talked on the subject while in the park assert that diseased fish—that is to say, those which are thin and affected with flesh worms—are more commonly found near the warm water; that they take the bait readily but are logy. I frequently saw pelicans swimming near the shore in the vicinity of the warm springs on the west arm of the lake. It would appear that the badly infested or diseased fish, being less active and gamy than the healthy fish, would be more easily taken by their natural enemies, who would learn to look for them in places where they most abound. But any circumstances which cause the pelican and the trout to occupy the same neighborhood will multiply the chances of the parasites developing in both the intermediate and final host. The causes that make for the abundance of the trout parasite conspire to increase the number of adults. The two hosts react on each other and the parasite profits by the reaction. About the only enemies the trout had before tourists, ambitious to catch big strings of trout and photograph them with a kodak, began to frequent this region, were the fish-eating birds, and chief among these in numbers and voracity was the pelican. It is no wonder, therefore, that the trout should have become seriously parasitized.[Pg 348] It may be inferred from the foregoing statements that the reason why the parasite of the trout of Yellowstone Lake migrates into the muscular tissue of its host must be found in the fact that the life of the parasite within the fish is much more prolonged than is the case where the conditions of life are less exceptional.
"The case just cited is probably the most signal one of direct injury to the host from the presence of parasites that I have seen. I shall enumerate more briefly a few additional cases out of a great number that I have encountered in my special investigations on the entozoa of fishes for the U. S. Fish Commission."
Many worms of this type abound in codfishes, bluefishes, striped bass, and other marine fishes, rendering them lean and unfit for food.
The Heart Lake Tape-worm.—Another very interesting case of parasitism is that of the large tape-worm (Ligula catostomi) infecting the suckers, Catostomus ardens, in the warm waters of Witch Creek, near Heart Lake, in the Yellowstone Park. Of this Dr. Linton gives the following account:
Fig. 230.—Sucker, Catostomus ardens (Jordan & Gilbert), from Heart Lake, Yellowstone Park, infested by a flatworm, Ligula catostomi Linton, itself probably a larva of Dibothrium. (After Linton.)
"In the autumn of 1889 Dr. David Starr Jordan found an interesting case of parasitism in some young suckers (Catostomus ardens) which he had collected in Witch Creek, a small stream which flows into Heart Lake, in the Yellowstone National Park. Specimens of these parasites were sent to me for identification. They proved to be a species of ligula, probably identical with the European Ligula simplicissima Rud., which is[Pg 349] found in the abdominal cavity of the tench. On account of its larval condition in which it possesses few distinctive characters, I described it under the name Ligula catostomi. These parasites grow to a very large size when compared with the fish which harbors them, often filling the abdominal cavity to such a degree as to give the fish a deceptively plump appearance. The largest specimen in Dr. Jordan's collection measured, in alcohol, 28.5 centimeters in length, 8 millimeters in breadth at the anterior end, 11 millimeters at a distance of 7 millimeters from the anterior end, and 1.5 millimeters near the posterior end. The thickness throughout was about 2 millimeters. The weight of one fish was 9.1 grams, that of its three parasites 2.5 grams, or 27½ per cent. the weight of the host. If a man weighing 180 pounds were afflicted with tape-worms to a similar degree, he would be carrying about with him 50 pounds of parasitic impedimenta.
"In the summer of 1890 I collected specimens from the same locality. A specimen obtained from a fish 19 centimeters in length measured while living 39.5 centimeters in length and 15 millimeters in breadth at the anterior end. Another fish 15 centimeters in length harbored four parasites, 12, 13, 13, and 20 centimeters long, respectively, or 58 centimeters aggregate. Another fish 10 centimeters long was infested with a single parasite which was 39 centimeters in length.
"These parasites were found invariably free in the body cavity. Dr. Jordan's collections were made in October and mine in July of the following year. Donnadieu has found that this parasite most frequently attains its maximum development at the end of two years. It is probable, therefore, that Dr. Jordan and I collected from the same generation. Since these parasites, in this stage of their existence, develop, not by levying a toll on the food of their host, after the manner of intestinal parasites, but directly by the absorption of the serous fluid of their host, it is quite evident that they work a positive and direct injury. Since, however, they lie quietly in the body cavity of the fish and possess no hard parts to cause irritation, they work their mischief simply by the passive abstraction of the nutritive juices of their host, and by crowding the viscera into confined spaces and unnatural positions. The worms, in[Pg 350] almost every case, had attained such a size that they far exceeded in bulk the entire viscera of their host.
"From the fact that the examples obtained were of comparatively the same age, it may be justly inferred that the period of infection to which the fish are subjected must be a short one. I did not discover the final host, but it is almost certain to be one or more of the fish-eating species of birds which visit that region, and presumably one of which, in its migrations, pays but a brief visit to this particular locality. This parasite was found only in the young suckers which inhabit a warm tributary of Witch Creek. They were not found in the large suckers of the lake. These young Catostomi were found in a single school, associated with the young of the chub (Leuciscus lineatus), in a stream whose temperature was 95° F. near where it joined a cold mountain brook whose temperature was 46° F. We seined several hundred of these young suckers and chubs, ranging in length from 6 to 19 centimeters. The larger suckers were nearly all infested with these parasites, the smaller ones not so much, and the smallest scarcely at all. Or, to give concrete examples: Of 30 fish ranging in length from 14 to 19 centimeters, only one or two were without parasites; of 45 specimens averaging about 10 centimeters in length, 15 were infested and 30 were not; of 65 specimens averaging about 9 centimeters in length, 10 were infested and 55 were not; of 62 specimens less than 9 centimeters in length, 2 were infested and 60 were not. None of the chubs were infested with this parasite.
"The conditions under which these fish were found are worthy of passing notice. The stream which they occupied flowed with rather sluggish current into a swift mountain stream, which it met almost at right angles. The school of young chubs and suckers showed no inclination to enter the cold water, even to escape the seine, but would dart around the edge of the seine, in the narrow space between it and the bank, in preference, apparently, to taking to the colder water. When not disturbed by the seine they would swim up near to the line which marked the division between the cold and the warm water, and seemed to be gazing with open mouth and eyes at the trout which occasionally darted past in the cold stream. The trout appeared to avoid the warm water, while[Pg 351] the chubs and suckers appeared to avoid the cold water. It may be that what the latter really avoided was the special preserve of the trout, since large chubs and suckers are found in abundance in the lake, which is quite cold, a temperature of 40° F. having been taken by us at a depth of 124 feet.
"Since the eggs of this parasite, after the analogy of closely related forms, in all probability are discharged into the water from the final host and hatch out readily in warm water, where they may live for a longer or shorter time as free-swimming planula-like forms, it will be observed that the sluggish current and high temperature of the water in which these parasitized fish occur give rise to conditions which are highly favorable to infection.
"It may be of passing interest to state here what I have recorded elsewhere, that ligulæ, probably specifically identical with L. catostomi, form an article of food in Italy, where they are sold in the markets under the name maccaroni piatti; also in southern France, where they are less euphemistically but more truthfully called the ver blanc. So far as my information goes, this diet of worms is strictly European.
"It is not necessary to prove cases of direct injury resulting from the presence of parasites in order to make out a case against them. In the sharp competition which nature forces on fishes in the ordinary struggle for existence, any factor which imparts an increment either of strength or of weakness may be a very potent one, and in a long term of years may determine the relative abundance or rarity of the individuals of a species. In most cases the interrelations between parasite and host have become so adjusted that the evil wrought by the parasite on its host is small. Parasitic forms, like free forms, are simply developing along the lines of their being, but unlike most free forms they do not contribute a fair share to the food of other creatures."
Thorn-head Worms.—The thorn-head worms called Acanthocephala are found occasionally in large numbers in different kinds of fishes. They penetrate the coats of the intestines, producing much irritation and finally waxy degeneration of the tissues.
According to Linton, there is probably no practical way of[Pg 352] counteracting the bad influences of worms of this order, since their larval state is passed, in some cases certainly, and in most cases probably, in small crustacea, which constitute a constant and necessary source of food for the fish. The same remark which was made in another connection with regard to the disposal of the viscera of fish applies here. In no case should the viscera of fish be thrown back into the water. In this order the sexes are distinct, and the females become at last veritable sacs for the shelter and nourishment of enormous numbers of embryos. The importance, therefore, of arresting the development of as many embryos as possible is at once apparent.
Nematodes.—The round worms or nematodes are very especially abundant in marine fishes, and particularly in the young. The study of these forms has a large importance to man. Dr. Linton pertinently observes:
"Where there is exhaustive knowledge of the thing itself the application of that knowledge toward getting good out of it or averting evil that may come from it first becomes possible. For example, a knowledge of the life-history of Trichina spiralis and its pathological effects on its host has taught people a simple way of securing immunity from its often deadly effects. A knowledge of the life-histories of the various species of tæniæ which infest man and the domestic animals, frequently to their serious hurt, has made it possible to diminish their numbers, and may, in time, lead to their practical extinction.
"So with the parasites of fishes. Whenever for any reason or reasons parasitism of any sort becomes so prevalent with any species as to amount to a disease, the remedy will be suggested, and in some cases may be practically applied. If, for example, it were thought desirable to counteract the influences which are at work to cause the parasitism of the trout of Yellowstone Lake, it could be very largely accomplished by breaking up the breeding-places of the pelican on the islands of the lake. With regard to parasitism among the marine food-fishes, the remedy while plainly suggested by the circumstances, might be difficult of application. Yet something could be done even there, if it were thought necessary to lessen the amount of parasitism. If such precautions as the destruction of the parasites which abound in the viscera of fish before throwing them[Pg 353] back into the water, and if no opportunity be lost of killing those sharks which feed on the food-fishes, two sources of the prevalence of parasites would be affected and the sum total of parasitism diminished. These remarks are made not so much because such precautions are needed as to suggest possible applications of knowledge which is already available."
Parasitic Fungi.—Fishes are often subject to wounds. If not too serious these will heal in time, with or without scars. Some lost portions may be restored, but not those including bone fin-rays or scales. In the fresh waters, wounds are usually attacked by species of fungus, notably Saprolegnia ferox, Saprolegnia mixta, and others, which makes a whitish fringe over a sore and usually causes death. This fungus is especially destructive in aquaria. This fungus is not primarily parasitic, but it fixes itself in the slime of a fish or in an injured place, and once established the animal is at its mercy. Spent salmon are very often attacked by this fungus. In America the spent salmon always dies, but in Scotland, where such is not the case, much study has been given to this plant and the means by which it may be exterminated. Dr. G. P. Clinton gives a useful account of the development of Saprolegnia, from which we take the following:
"The minute structure and life-history of such fungous forms have been so thoroughly made out by eminent specialists that no investigation along this line was made, save to observe those phenomena which might be easily seen with ordinary microscopic manipulations. The fungus consists of branched, hyaline filaments, without septa, except as these are found cutting off the reproductive parts of the threads. It is made up of a root-like or rhizoid part that penetrates the fish and a vegetative and reproductive part that radiates from the host. The former consists of branched tapering threads which pierce the tissues for a short distance, but are easily pulled out. The function of this part is to obtain nourishment for the growth of the external parts. Prostrate threads are found running through the natural slime covering the fish, and from these are produced the erect radiating hyphæ so plainly seen when in the water. The development of these threads appears to be very rapid when viewed under the microscope, although the[Pg 354] growth made under favorable conditions in two days is only about a third of an inch. From actual measurements of filaments of the fungus placed in water and watched under the microscope, it was found that certain threads made a growth of about 3000 microns in an hour. Two others, watched for twenty minutes, gave in that time a growth of 90 and 47 microns respectively; and yet another filament, observed during two periods of five minutes each, made a growth of 28 microns each time. In ordinary cultures the rate of growth depends upon the condition of the medium, host, etc."

Fig. 231.—Quinnat Salmon, Oncorhynchus tschawytscha (Walbaum). Monterey Bay. (Photograph by C. Rutter.)
Professor H. A. Surface thus speaks of the attacks of Saprolegnia on the lamprey:
"The attack that attends the end of more lampreys than does any other is that of the fungus (Saprolegnia sp.). This looks like a gray slime and eats into the exterior parts of the animal, finally causing death. It covers the skin, the fins, the eyes, the gill-pouches, and all parts, like leprosy. It starts where the lamprey has been scratched or injured or where its mate has held it, and develops very rapidly when the water is warm. It is found late in the season on all lampreys that have spawned out, and it is almost sure to prove fatal, as we have repeatedly seen with attacked fishes or lampreys kept in tanks or aquaria. With choice aquarium fishes a remedy, or at least a palliative, is to be found in immersion in salt water for a few minutes or in bathing the affected parts with listerine. Since these creatures complete the spawning process before the fungoid attack proves serious to the individual, it can be seen that it affects no injury to the race, as the fertilized eggs are left to[Pg 355] come to maturity. Also, as it is nature's plan that the adult lampreys die after spawning once, we are convinced that death would ensue without the attack of the fungus; and in fact this is to be regarded as a resultant of those causes that produce death rather than the immediate cause of it. Its only natural remedy is to be found in the depths of the lake (450 feet) where there is a uniform or constant temperature of about 39° Fahr., and where the light of the noon-day sun penetrates with an intensity only about equal to starlight on land on a clear but moonless night.

Fig. 232.—Young Male Quinnat Salmon, Oncorhynchus tschawytscha, dying after spawning. Sacramento River. (Photograph by Cloudsley Rutter.)
"As light and heat are essential to the development of the fungus, which is a plant growth and properly called a water mold, and as their intensity is so greatly diminished in the depth of the lake, it is probable that if creatures thus attacked should reach this depth they might here find relief if their physical condition were otherwise strong enough to recuperate. However, we have recently observed a distinct tendency on the part of fungus-covered fishes to keep in the shallower, and consequently warmer, parts of the water, and this of course results in the more rapid growth of the sarcophytic plant, and the death of the fishes is thus hastened.
"All kinds of fishes and fish-eggs are subject to the attacks of such fungus, especially after having been even slightly scratched or injured. As a consequence, the lamprey attacks on fishes cause wounds that often become the seat of a slowly spreading but fatal fungus. We have seen many nests of the bullhead, or horned pout (Ameiurus nebulosus), with all the eggs thus destroyed, and we have found scores of fishes of various kinds thus killed or dying. It is well known that in many rivers this is the apparent cause of great mortality among adult salmon. Yet we really doubt if it ever attacks uninjured fishes that are in good strong physical condition which have not at least had the slime rubbed from them when captured. It is contagious, not only being conveyed from one infested fish to another, but from dead flies to fishes." (For a further discussion of this subject see an interesting and valuable Manual of Fish Culture, by the U. S. Fish Commission, 1897.)
Earthquakes.—Occasionally an earthquake has been known to kill sea-fishes in large numbers. The Albatross obtained[Pg 357] specimens of Sternoptyx diaphana in the Japanese Kuro Shiwo, killed by the earthquakes of 1896, which destroyed fishing villages of the coast of Rikuchu in northern Japan.
Mortality of Tilefish.—Some years ago in the Gulf Stream off Newfoundland an immense mortality of the filefish (Lopholatilus chamæleonticeps) was reported by fishermen. This handsome and large fish, inhabiting deep waters, died by thousands. For this mortality, which almost exterminated the species, no adequate cause has been found.
As to the destruction of fresh-water fishes by larger enemies, we may quote from Professor H. A. Surface. He says there is no doubt that these three species, the lake lamprey (Petromyzon marinus unicolor), the garpike (Lepidosteus osseus), and the mud-puppy (Necturus maculosus), named "in order of destructiveness, are the three most serious enemies of fishes in the interior of this State [New York], each of which surely destroys more fishes annually than are caught by all the fishermen combined. The next important enemies of fishes in order of destructiveness, according to our observations and belief, are spawn-eating fishes, water-snakes, carnivorous or predaceous aquatic insects (especially larvæ), and piscivorous fishes and birds." The lamprey attaches itself to larger fishes, rasping away their flesh and sucking their blood, as shown in the accompanying plate.

Fig. 233.—Catfishes, Ameiurus nebulosus Le Sueur, destroyed by lampreys (Petromyzon marinus unicolor De Kay). Cayuga Lake, N. Y. (Modified from photograph by Prof. H. A. Surface.)

The Mermaid.—A word may be said of the fishes which have no existence in fact and yet appear in popular literature or in superstition.
The mermaid, half woman and half fish, has been one of the most tenacious among these, and the manufacture of their dried bodies from the head, shoulders, and ribs of a monkey sealed to the body of a fish has long been a profitable industry in the Orient. The sea-lion, the dugong, and other marine mammals have been mistaken for mermaids, for their faces seen at a distance and their movements at rest are not inhuman, and their limbs and movements in the water are fish-like.
In China, small mermaids are very often made and sold to the curious. The head and torso of a monkey are fastened ingeniously to the body and tail of a fish. It is said that Linnæus was once forced to leave a town in Holland for questioning the genuineness of one of these mermaids, the property of some high official. These monsters are still manufactured for the "curio-trade."
The Monkfish.—Many strange fishes were described in the Middle Ages, the interest usually centering in some supposed relation of their appearance with the affairs of men. Some of these find their way into Rondelet's excellent book, "Histoire Entière des Poissons," in 1558. Two of these with the accompanying plate of one we here reproduce. Other myths less interesting grew out of careless, misprinted, or confused accounts on the part of naturalists and travelers.
"In our times in Norway a sea-monster has been taken after a great storm, to which all that saw it at once gave the name of[Pg 360] monk; for it had a man's face, rude and ungracious, the head shorn and smooth. On the shoulders, like the cloak of a monk, were two long fins instead of arms, and the end of the body was finished by a long tail. The picture I present was given me by the very illustrious lady, Margaret de Valois, Queen of Navarre, who received it from a gentleman who gave a similar one to the emperor, Charles V., then in Spain. This gentleman said that he had seen the monster as the portrait shows it in Norway, thrown by the waves and tempests on the beach at a place called Dieze, near the town called Denelopoch. I have seen a similar picture at Rome not differing in mien. Among the sea-beasts, Pliny mentions a sea-mare and a Triton as among the creatures not imaginary. Pausanias also mentions a Triton."

Fig. 234.—"Le monstre marin an habit de Moine." (After Rondelet.)
Rondelet further says:
The Bishop-fish.—"I have seen a portrait of another sea-monster at Rome, whither it had been sent with letters that affirmed for certain that in 1531 one had seen this monster in a bishop's garb, as here portrayed, in Poland. Carried to the king of that country, it made certain signs that it had a great desire to return to the sea. Being taken thither it threw itself instantly into the water."

Fig. 235.—"Le monstre marin en habit d'Évêque." (After Rondelet.)
The Sea-serpent.—A myth of especial persistency is that of the sea-serpent. Most of the stories of this creature are seaman's yarns, sometimes based on a fragment of wreck, a long strip of kelp, the power of suggestion or the incitement of alcohol. But certain of these tales relate to real fishes. The sea-serpent with an uprearing red mane like that of a horse is the oarfish (Regalecus), a long, slender, fragile fish compressed like a ribbon and reaching a length of 25 feet. We here present a photograph of an oarfish (Regalecus russelli) stranded on the California coast at Newport in Orange County, California. A figure of a European species (Regalecus glesne) is also given showing the fish in its uninjured condition. Another reputed sea-serpent is the frilled shark (Chlamydoselachus angineus), which has been occasionally noticed by seamen. The struggles of the great killer (Orca orca) with the whales it attacks and destroys has also given rise to stories of the whale struggling in the embrace of some huge sea-monster. This description is correct, but the mammal is a monster itself, a relative of the whale and not a reptile.

Fig. 236.—Oarfish, Regalecus russelli, on the beach at Newport, Orange Co., Cal. (Photograph by C. P. Remsberg.)

Fig. 237.—Glesnæs Oarfish, Regalecus glesne Ascanius. Newcastle, England. (After Day.)
It is often hard to account for some of the stories of the sea-serpent. A gentleman of unquestioned intelligence and sincerity lately described to the writer a sea-serpent he had seen at short range, 100 feet long, swimming at the surface, and with a head as large as a barrel. I do not know what he saw, but I do know that memory sometimes plays strange freaks.
Little venomous snakes with flattened tails (Platyurus, Pelamis) are found in the salt bays in many tropical regions of the Pacific (Gulf of California, Panama, East Indies, Japan), but these are not the conventional sea-serpents.
Certain slender fishes, as the thread-eel (Nemichthys) and the wolf-eel (Anarrhichthys), have been brought to naturalists as young sea-serpents, but these of course are genuine fishes.
Whatever the nature of the sea-serpent may be, this much is certain, that while many may be seen, none will ever be caught. The great swimming reptiles of the sea vanished at the end of Mesozoic time, and as living creatures will never be known of man.
As a record of the Mythology of Science, we may add the following remarks of Rafinesque on the imaginary garpike (Litholepis adamantinus), of which a specimen was painted for him by the wonderful brush of Audubon:
"This fish may be reckoned the wonder of the Ohio. It is only found as far up as the falls, and probably lives also in the Mississippi. I have seen it, but only at a distance, and have been shown some of its singular scales. Wonderful stories are related concerning this fish, but I have principally relied upon the description and picture given me by Mr. Audubon. Its length is from 4 to 10 feet. One was caught which weighed 400 pounds. It lies sometimes asleep or motionless on the surface of the water, and may be mistaken for a log or snag. It is impossible to take it in any other way than with the seine or a very strong hook; the prongs of the gig cannot pierce the scales, which are as hard as flint, and even proof against lead balls! Its flesh is not good to eat. It is a voracious fish. Its vulgar names are diamond-fish (owing to its scales being cut like diamonds), devil-fish, jackfish, garjack, etc. The snout is large, convex above, very obtuse, the eyes small and black; nostrils small, round before the eyes; mouth beneath the eyes,[Pg 365] transversal with large angular teeth. Pectoral and abdominal fins trapezoidal. Dorsal and anal fins equal, longitudinal, with many rays. The whole body covered with large stone scales, lying in oblique rows; they are conical, pentagonal pentædral, with equal sides, from half an inch to one inch in diameter, brown at first but becoming the color of turtle-shell when dry. They strike fire with steel and are ball-proof!"

Fig. 238.—Thread-eel, Nemichthys avocetta Jordan & Gilbert. Puget Sound.

Taxonomy.—Classification, as Dr. Elliott Coues has well said,[147] is a natural function of "the mind which always strives to make orderly disposition of its knowledge and so to discover the reciprocal relations and interdependencies of the things it knows. Classification presupposes that there do exist such relations, according to which we may arrange objects in the manner which facilitates their comprehension, by bringing together what is like and separating what is unlike, and that such relations are the result of fixed inevitable law. It is therefore taxonomy (τάξις, away; νόμος, law) or the rational, lawful disposition of observed facts."
A perfect taxonomy is one which would perfectly express all the facts in the evolution and development of the various forms. It would recognize all the evidence from the three ancestral documents, palæontology, morphology, and ontogeny. It would consider structure and form independently of adaptive or physiological or environmental modifications. It would regard as most important those characters which had existed longest unchanged in the history of the species or type. It would regard as of first rank those characters which appear first in the history of the embryo. It would regard as of minor importance those which had arisen recently in response to natural selection or the forced alteration through pressure of environment, while fundamental alterations as they appear one after another in geologic time would make the basal characters of corresponding groups in taxonomy. In a perfect taxonomy or natural system of classification animals would not be divided into groups nor ranged in linear series. We should imagine [Pg 368]series variously and divergently branched, with each group at its earlier or lower end passing insensibly into the main or primitive stock. A very little alteration now and then in some structure is epoch-making, and paves the way through specialization to a new class or order. But each class or order through its lowest types is interlocked with some earlier and otherwise diverging group.
Defects in Taxonomy.—A sound system of taxonomy of fishes should be an exact record of the history of their evolution. But in the limitations of book-making, this transcript must be made on a flat page, in linear series, while for centuries and perhaps forever whole chapters must be left vacant and others dotted everywhere with marks of doubt. For science demands that positive assertion should not go where certainty cannot follow. A perfect taxonomy of fishes would be only possible through the study, by some Artedi, Müller, Cuvier, Agassiz, Traquair, Gill, or Woodward, of all the structures of all the fishes which have ever lived. There are many fishes living in the sea which are not yet known to any naturalist, many others are known from one or two specimens, but not yet accessible to students in other continents. Many are known externally from specimens in bottles or drawings in books, but have not been studied thoroughly by any one, and the vast multitude of species have perished in Palæozoic, Mesozoic, and Tertiary seas without leaving a tooth or bone or fin behind them. With all this goes human fallibility, the marring of our records, such as they are, by carelessness, prejudice, dependence, and error. Chief among these defects are the constant mistaking of analogy for homology, and the inability of men to trust their own eyes as against the opinion of the greater men who have had to form their opinions before all evidence was in. Because of these defects, the current system of classification is always changing with each accession of knowledge.
The result is, again to quote from Dr. Coues, "that the natural classification, like the elixir of life or the philosopher's stone, is a goal far distant."
Analogy and Homology.—Analogy, says Dr. Coues, "is the apparent resemblance between things really unlike—as the wing of a bird and the wing of a butterfly, as the lungs of a bird and[Pg 369] the gills of a fish. Homology is the real resemblance, or true relation between things, however different they may appear to be—as the wing of a bird and the foreleg of a horse, the lungs of a bird and the swim-bladder of a fish. The former commonly rests upon mere functional, i.e. physiological, modifications; the latter is grounded upon structural, i.e., morphological, identity or unity. Analogy is the correlative of physiology, homology of morphology; but the two may be coincident, as when structures identical in morphology are used for the same purposes, and are therefore physiologically identical. Physiological diversity of structure is incessant, and continually interferes with morphological identity of structure, to obscure or obliterate the indications of affinity the latter would otherwise express clearly.... We must be on our guard against those physiological appearances which are proverbially deceptive!"
"It is possible and conceivable that every animal should have been constructed upon a plan of its own, having no resemblance whatever to the plan of any other animal. For any reason we can discover to the contrary, that combination of natural forces which we term life might have resulted from, or been manifested by, a series of infinitely diverse structures; nor would anything in the nature of the case lead us to suspect a community of organization between animals so different in habit and in appearance as a porpoise and a gazelle, an eagle and a crocodile, or a butterfly and a lobster. Had animals been thus independently organized, each working out its life by a mechanism peculiar to itself, such a classification as that now under contemplation would be obviously impossible; a morphological or structural classification plainly implying morphological or structural resemblances in the things classified.
"As a matter of fact, however, no such mutual independence of animal forms exists in nature. On the contrary, the members of the animal kingdom, from the highest to the lowest, are marvelously connected. Every animal has something in common with all its fellows—much with many of them, more with a few, and usually so much with several that it differs but little from them.
"Now, a morphological classification is a statement of these[Pg 370] gradations of likeness which are observable in animal structures, and its objects and uses are manifold. In the first place, it strives to throw our knowledge of the facts which underlie, and are the cause of, the similarities discerned into the fewest possible general propositions, subordinated to one another, according to their greater or less degree of generality; and in this way it answers the purpose of a memoria technica, without which the mind would be incompetent to grasp and retain the multifarious details of anatomical science."
Coues on Classification.—It is obvious that fishes like other animals may be classified in numberless ways, and as a matter of fact by numberless men they have been classified in all sorts of fashions. "Systems," again quoting from Dr. Coues, "have been based on this and that set of characters and erected from this or that preconception in the mind of the systematist.... The mental point of view was that every species of bird (or of fish) was a separate creature, and as much of a fixture in nature's museum as any specimen in a naturalist's cabinet. Crops of classifications have been sown in the fruitful soil of such blind error, but no lasting harvest has been reaped.... The genius of modern taxonomy seems to be so certainly right, to be tending so surely even if slowly in the direction of the desired consummation, that all differences of opinion we hope will soon be settled, and defect of knowledge, not perversity of mind, is the only obstacle in the way of success. The taxonomic goal is not now to find the way in which birds (or other animals) may be most conveniently arranged, but to discover their pedigree, and so construct their family tree. Such a genealogical table, or phylum (φῦλον, tribe, race, stock), as it is called, is rightly considered the only taxonomy worthy the name—the only true or natural classification. In attempting this end, we proceed upon the belief that, as explained above, all birds, like all other animals and plants, are related to each other genetically, as offspring are to parents, and that to discover their generic relations is to bring out their true affinities—in other words, to reconstruct the actual taxonomy of nature. In this view there can be but one 'natural' classification, to the perfecting of which all increase in our knowledge of the structure of birds infallibly and inevitably tends. The classification now in use[Pg 371] or coming into use is the result of our best endeavors to accomplish this purpose, and represents what approach we have made to this end. It is one of the great corollaries of that theorem of evolution which most naturalists are satisfied has been demonstrated. It is necessarily a morphological classification; that is, one based solely upon considerations of structure or form (μορφή, form, morphe), and for the following reasons: Every offspring tends to take on precisely the form or structure of its parents, as its natural physical heritage; and the principle involved, or the law of heredity, would, if nothing interfered, keep the descendants perfectly true to the physical characters of their progenitors; they would 'breed true' and be exactly alike. But counter influences are incessantly operative, in consequence of constantly varying external conditions of environment; the plasticity of organization of all creatures rendering them more or less susceptible of modifications by such means, they become unlike their ancestors in various ways and to different degrees. On a large scale is thus accomplished, by natural selection and other natural agencies, just what man does in a small way in producing and maintaining different breeds of domestic animals. Obviously, amidst such ceaselessly shifting scenes, degrees of likeness or unlikeness of physical structure indicate with the greatest exactitude the nearness or remoteness of organisms in kinship. Morphological characters derived from the examination of structure are therefore the surest guides we can have to the blood relationships we desire to establish; and such relationships are the 'natural affinities' which all classification aims to discover and formulate."
Species as Twigs of a Genealogical Tree.—In another essay Dr. Coues has compared species of animals to "the twigs of a tree separated from the parent stem. We name and arrange them arbitrarily in default of a means of reconstructing the whole tree according to nature's ramifications." If one had a tree, all in fragments, pieces of twig and stem, some of them lost, some destroyed, and some not yet separated from the mass not yet picked over, and wished to place each part where he could find it, he would be forced to adopt some system of natural classification. In such a scheme he would lay those parts together which grew from the same branch. If he were com[Pg 372]pelled to arrange all the fragments in a linear series, he would place together those of one branch, and when these were finished he would begin with another. If all this were a matter of great importance and extending over years or over many lifetimes, with many errors to be made and corrected, a set of names would be adopted—for the main trunk, for the chief branches, the lesser branches, and on down to the twigs and buds.
A task of this sort on a world-wide scale is the problem of systematic zoology. There is reason to believe that all animals and plants sprang from a single stock. There is reasonable certainty that all vertebrate animals are derived from a single origin. These vertebrate animals stand related to each other, like the twigs of a gigantic tree of which the lowermost branches are the aquatic forms to which we give the name of fishes. The fishes are here regarded as composed of six classes or larger lines of descent. Each of these, again, is composed of minor divisions called orders. The different species or ultimate kinds of animals are grouped in genera. A genus is an assemblage of closely related species grouped around a central species as type. The type of a genus is, in common usage, that species with which the name of the genus was first associated. The name of the genus as a noun, often with that of the species which is an adjective in signification if not in form, constitutes the scientific name of the species. Thus Petromyzon is the genus of the common large lamprey, marinus is its species, and the scientific name of the species is Petromyzon marinus. Petromyzon means stone-sucker; marinus, of the sea, thus distinguishing it from a species called fluviatilis, of the river. In like fashion all animals and plants are named in scientific record or taxonomy. Technical names are necessary because vernacular names fail. Half a million kinds of animals are known, while not half a thousand vernacular names exist in any language. And these are always loosely used, half a dozen of them often for the same species, one name often for a dozen species.
In the same way, whenever we undertake an exact description, we must use names especially devised for that purpose. We cannot use the same names for the bones of the head of a fish and those of the head of a man, for a fish has a different series of bones, and this series is different with different fishes.
Nomenclature.—A family in zoology is an assemblage of related genera. The name of a family, for convenience, always ends in the patronymic idæ, and it is always derived from the leading genus, that is, the one best known or earliest studied. Thus all lampreys constitute the family Petromyzonidæ. An order may contain one or more families. An order is a division of a larger group; a family an assemblage of related smaller groups. Intermediate groups are often recognized by the prefixes sub or super. A subgenus is a division of a genus. A subspecies is a geographic race or variation within a species; a super-family a group of allied families. Binomial nomenclature, or the use of the name of genus and species as a scientific name, was introduced into science as a systematic method by Linnæus. In the tenth edition of his Systema Naturæ, published in 1758, this method was first consistently applied to animals. By common consent the scientific naming of animals begins with this year, and no account is taken of names given earlier, as these are, except by accident, never binomial. Those authors who wrote before the adoption of the rule of binomials and those who neglected it are alike "ruled out of court." The idea of genus and species was well understood before Linnæus, but the specific name used was not one word but a descriptive phrase, and this phrase was changed at the whim of the different authors.
Fig. 239.—Horned Trunkfish, Cowfish, or Cuckold, Lactophrys tricornis (Linnæus). Charleston, S. C.
Nomenclature of Trunkfishes.—Examples of such names are those of the West Indian trunkfish, or cuckold (Ostracion tricorne, Linnæus). Lister refers to a specimen in 1686 as "Piscis triangularis capiti cornutu cui e media cauda cutanea[Pg 374] aculeus longus erigitus." This Artedi alters in 1738 to Ostracion triangulatus aculeis duobus in capite et unico longiore superne ad caudam. This is more accurately descriptive and it recognizes the existence of a generic type, Ostracion, or trunkfish, to cover all similar fishes. French writers transformed this into various phrases beginning "Coffre triangulaire à trois cornes," or some similar descriptive epithet, and in English or German it was likely to wander still farther from the original. But Linnæus condenses it all in the word tricornis, which, although not fully descriptive, is still a name which all future observers can use and recognize.
It is true that common consent fixes the date of the beginning of nomenclature at 1758. But to this there are many exceptions. Some writers date genera from the first recognition of a collective idea under a single name. Others follow even species back through the occasional accidental binomials. Most British writers have chosen the final and completed edition of the Systema Naturæ, the last work of Linnæus, in 1766, in preference to the earlier volume. But all things considered, justice and convenience alike seem best served by the use of the edition of 1758.
Synonymy and Priority.—Synonymy is the record of the names applied at different times to the same group or species. With characteristic pungency Dr. Coues defines synonymy as "a burden and a disgrace to science." It has been found that the only way to prevent utter confusion is to use for each genus or species the first name applied to it and no other. The first name, once properly given, is sacred because it is the right name. All other later names whatever their appropriateness are wrong names. In science, of necessity, a name is a name without any necessary signification. For this reason and for the further avoidance of confusion, it remains as it was originally spelled by the author, obvious misprints aside, regardless of all possible errors in classical form or meaning. The names in use are properly written in Latin or in Latinized Greek, the Greek forms being usually preferred as generic names, the Latin adjectives for names of species. Many species are named in honor of individuals, these names being usually given the termination of the Latin genitive, as Sebas[Pg 375]todes gillii, Liparis agassizi. In recent custom all specific names are written with the small initial; all generic names with the capital.
One class of exceptions must be made to the law of priority. No generic name can be used twice among animals, and no specific name twice in the same genus. Thus the name Diabasis has to be set aside in favor of the next name Hæmulon, because Diabasis was earlier used for a genus of beetles. The specific name Pristipoma humile is abandoned, because there was already a humile in the genus Pristipoma.
The Conception of Genus.—In the system of Linnæus, a genus corresponds roughly to the modern conception of a family. Most of the primitive genera contained a great variety of forms, as well as usually some species belonging to other groups disassociated from their real relationships.
As greater numbers of species have become known the earlier genera have undergone subdivision until in the modern systems almost any structural character not subject to intergradation and capable of exact definition is held to distinguish a genus. As the views of these characters are undergoing constant change, and as different writers look upon them from different points of view, or with different ideas of convenience, we have constant changes in the boundaries of genera. This brings constant changes in the scientific names, although the same specific name should be used whatever the generic name to which it may be attached. We may illustrate these changes and the burden of synonymy as well by a concrete example.
The Trunkfishes.—The horned trunkfish, or cuckold, of the West Indies was first recorded by Lister in 1686, in the descriptive phrase above quoted. Artedi, in 1738, recognized that it belonged with other trunkfishes in a group he called Ostracion. This, to be strictly classic, he should have written Ostracium, but he preferred a partly Greek form to the Latin one. In the Nagg's Head Inn in London, Artedi saw a trunkfish he thought different, having two spines under the tail, while Lister's figure seemed to show one spine above. This Nagg's Head specimen Artedi called "Ostracion triangulatus duobus aculeis in fronte et totidem in imo ventre subcaudalesque binis."
Next came Linnæus, 1758, who named Lister's figure and[Pg 376] the species it represented, Ostracion tricornis, which should in strictness have been Ostracion tricorne, as ὀστρακίον, a little box, is a neuter diminutive. The Nagg's Head fish he named Ostracion quadricornis. The right name now is Ostracion tricornis, because the name tricornis stands first on the page in Linnæus' work, but Ostracion quadricornis has been more often used by subsequent authors because it is more truthful as a descriptive phrase. In 1798, Lacépède changed the name of Lister's fish to Ostracion listeri, a needless alteration which could only make confusion.

Fig. 240.—Horned Trunkfish, Ostracion cornutum Linnæus. East Indies. (After Bleeker.)
In 1818, Dr. Samuel Latham Mitchill, receiving a specimen from below New Orleans, thought it different from tricornis and quadricornis and called it Ostracion sexcornutus; Dr. Holard, of Paris, in 1857, named a specimen Ostracion maculatus, and at about the same time Bleeker named two others from Africa which seem to be the same thing, Ostracion guineensis and Ostracion gronovii. Lastly, Poey calls a specimen from Cuba Acanthostracion polygonius, thinking it different from all the rest, which it may be, although my own judgment is otherwise. This brings up the question of the generic name. Among trunkfishes there are four-angled and three-angled kinds, and of each form there are species with and without horns and spines. The original Ostracion of Linnæus we may interpret as being Ostracion cubicus of the coasts of Asia, a species similar to the Ostracion rhinorhynchus. This species, cubicus, we call the type species of the genus, as the Nagg's Head specimen of Artedi was the type specimen of the species quadricornus, and the one that was used for Lister's figure the type specimen of tricornis.
Ostracion cubicus is a four-angled species, and when the[Pg 377] trunkfishes were regarded as a family (Ostraciidæ), the three-angled ones were set off as a separate genus. For this two names were offered, both by Swainson in 1839. For trigonus, a species without horns before the eyes, he gave the name Lactophrys, and for triqueter, a species without spines anywhere, the name of Rhinesomus. Most recent American authors have placed the three-cornered species which are mostly American in one genus, which must therefore be called Lactophrys. Of this name Rhinesomus is a synonym, and our species should stand as Lactophrys tricornis. The fact that Lactophrys as a word (from Latin lætus, smooth; Greek ὀφρύς, eyebrow; or else from lactoria, a milk cow, and ὀφρύς) is either meaningless or incorrectly written makes no difference with the necessity for its use.
Fig. 241.—Spotted Trunkfish, Lactophrys bicaudalis (Linnæus). Cozumel Island, Yucatan.

Fig. 242.—Spotted Trunkfish (face view), Lactophrys bicaudalis (Linnæus).
In 1862, Bleeker undertook to divide these fishes differently. Placing all the hornless species, whether three-angled or four-angled, in Ostracion, he proposed the name Acanthostracion for the species with horns, tricornis being the type. But Acanthostracion has not been usually adopted except as the name of a section under Lactophrys. The three-angled American species are usually set apart from the four-angled species of Asia, and our cuckold is called Lactophrys tricornis. But it may be with perfect correctness called Ostracion tricorne, in the spirit called conservative. Or with the "radical" systematists we may[Pg 378] accept the finer definition and again correctly call it Acanthostracion tricorne. But to call it quadricornis or listeri or maculatus with any generic name whatever would be to violate the law of priority.
Fig. 243.—Spineless Trunkfish, Lactophrys triqueter (Linnæus). Tortugas.
Trinomial Nomenclature.—By trinomial nomenclature we mean the use of a second subordinate specific name to designate a geographic subspecies, variety, or other intergrading race. Thus Salmo clarki virginalis indicates the variety of Clark's trout, or the cut-throat trout, found in the lakes and streams of the Great Basin of Utah, as distinguished from the genuine Salmo clarkii of the Columbia. Trinomials are not much used among fishes, as we are not yet able to give many of the local forms correct and adequate definition such as is awarded to similar variations among birds and mammals. Usually varieties in ichthyology count as species or as nothing.
Fig. 244.—Hornless Trunkfish, Lactophrys trigonus (Linnæus). Tortugas, Florida.

Fig. 245.—Hornless Trunkfish (face-view), Lactophrys trigonus (Linnæus). Charleston, S. C.
Meaning of Species.—Quoting once more from the admirable essay of Dr. Coues on the taxonomy of birds: "The student cannot be too well assured that no such things as species, in the old sense of the word, exist in nature any more than have genera or families an actual existence. Indeed they cannot be, if there is any truth in the principles discussed in our earlier paragraphs. Species are simply ulterior modifications, which once were, if they be not still, inseparably linked together; and their nominal recognition is a pure convention, like that of a genus. More practically hinges upon the way we regard them than turns upon our establishment of higher groups, simply because upon the way we decide in this case depends the scientific labeling of specimens. If we are speaking of a robin, we do not ordinarily concern ourselves with the family or order it belongs to, but we do require a technical name for constant use. That name is compounded of its genus, species, and variety. No infallible rule can be laid down for determining what shall be held to be a species, what a conspecies, subspecies, or variety. It is a matter of tact and experience, like the appreciation of the value of any other group in zoology. There is, however, a convention upon the subject, which the present workers in ornithology in this country find available; at any rate we have no better rule to go by. We treat as "specific" any form, however little different from the next, that we do not know or believe to intergrade with that next one, between which and the next one no intermediate equivocal specimens are forthcoming, and none, consequently, are supposed to exist. This is to imply that differentiation is accomplished, the links are lost and the characters actually become "specific." We treat as "varietal" of each other any forms, however different in their extreme manifestation, which we know to intergrade, having the intermediate specimens before us, or which we believe with any good reason do intergrade. If the links still exist, the differentiation[Pg 380] is still incomplete, and the characters are not specific, but only varietal, in the literal sense of these terms."
Generalization and Specialization.—A few terms in common use may receive a moment's discussion. A type or group is said to be specialized when it has a relatively large number of peculiarities or when some one peculiarity is carried to an extreme. A sculpin is a specialized fish having many unusual phases of development, as is also a swordfish, which has a highly peculiar structure in the snout. A generalized type is one with fewer peculiarities, as the herring in comparison with the sculpin. In the process of evolution generalized types usually give place to specialized ones. Generalized types are therefore as a rule archaic types. The terms high and low are also relative, a high type being one with varied structure and functions. Low types may be primitively generalized, as the lancelet in comparison with all other fishes, or the herring in comparison with the perch, or they may be due to degradation, a loss of structures which have been elaborately specialized in their ancestry. The sea-snail (Liparis), an ally of the sculpin, with scales lost and fins deteriorated is an example of a low type which is specialized as well as degraded.
High and Low Forms.—In the earlier history of ichthyology much confusion resulted from the misconception of the terms "high" and "low." Because sharks appeared earlier than bony fishes, it was assumed that they should be lower than any of their subsequent descendants. That the brain and muscular system in sharks was more highly developed than in most bony fishes seemed also certain. Therefore it was thought that the teleost series could not have had a common origin with the series of sharks. It is now understood that evolution means chiefly adaptation. The teleost is adapted to its mode of life, and to that end it is specialized in fin and skeleton rather than in brain and nerves. All degeneration is associated with specialization. The degeneration of the blindfish is a specialization for better adaptation to life in the darkness of caves; the degeneration of the deep-sea fish meets the demands of the depths, the degeneration of the globefish means the sinking of one line of functions in the extension of some other.
Referring to his own work on the fossil fishes in the early[Pg 381] forties, Professor Agassiz once said to the writer: "At that time I was on the verge of anticipating the views of Darwin, but it seemed to me that the facts were contrary to the theories of evolution. We had the highest fishes first." This statement leads us to consider what is meant by high and low. Undoubtedly the sharks are higher than the bony fishes in the sense of being nearer to the higher vertebrates. In brain, muscle, teeth, and reproductive structures they are also more highly developed. In all skeletal and cranial characters the sharks stand distinctly lower. But the essential fact, so far as evolution is concerned, is not that the sharks are high or low. They are, in almost all respects, distinctly generalized and primitive. The bony fishes are specialized in various ways through adaptation to the various modes of life they lead. Much of this specialization involves corresponding degeneration of organs whose functions have ceased to be important. As a broad proposition it is not true that "we had our highest fishes first," for in a complete definition of high and low, the specialized perch or bass stands higher. But whether true or not, it does not touch the question of evolution which is throughout a process of adaptation to conditions of life.
Referring to the position of Agassiz and his early friend and disciple, Hugh Miller, Dr. Traquair (1900) uses these words in an address at Bradford, England:
"It cannot but be acknowledged that the paleontology of fishes is not less emphatic in the support of descent than that of any other division of the animal kingdom. But in former days the evidence of fossil ichthyology was by some read otherwise.
"It is now a little over forty years since Hugh Miller died: he who was one of the first collectors of the fossil fishes of the Scottish old red sandstone, and who knew these in some respects better than any other man of his time, not excepting Agassiz himself. Yet his life was spent in a fierce denunciation of the doctrine of evolution, then only in its Lamarckian form, as Darwin had not yet electrified the world with his 'Origin of Species.' Many a time I wonder greatly what Hugh Miller would have thought had he lived a few years longer, so as to have been able to see the remarkable revolution which was wrought by the publication of that book.
"The main argument on which Miller rested was the 'high' state of organization of the ancient fishes of the Paleozoic formations, and this was apparently combined with a confident assumption of the completeness of the geological record. As to the first idea, we know of course that evolution means the passage from the more general to the more special, and that as the general result an onward advance has taken place; yet 'specialization' does not always or necessarily mean 'highness' of organization in the sense in which the term is usually employed. As to the idea of the perfection of the geological record, that of course is absurd.
"We do not and cannot know the oldest fishes, as they would not have had hard parts for preservation, but we may hope to come to know many more old ones, and older ones still than we do at present. My experience on the subject of fossil ichthyology is that it is not likely to become exhausted in our day.
"We are introduced at a period far back in geological history to certain groups of fishes, some of which certainly are high in organization as animals, but yet of generalized type, being fishes and yet having the potentiality of higher forms. But because their ancestors are unknown to us, that it is no evidence that they did not exist, and cannot overthrow the morphological testimony in favor of evolution with which the record actually does furnish us. We may therefore feel very sure that fishes or 'fish-like vertebrates' lived long ages before the oldest forms with which we are acquainted came into existence.
"The modern type of bony fishes, though not so 'high' in many anatomical points as that of the Selachii, Crossopterygii, Dipnoi, Acipenseroidei, and Lepidosteoidei of the Palæozoic and Mesozoic eras, is more specialized in the direction of the fish proper, and, as already indicated, specialization and 'highness' in the ordinary sense of the word are not necessarily coincident. But ideas about these things have undergone a wonderful change since those pre-Darwinian days, and though we shall never be able fully to unravel the problems concerning the descent of animals, we see many things a great deal more clearly now than we did then."
Dr. Gill observes: "Perhaps there are no words in science that have been productive of more mischief and more retarded the progress of biological taxonomy than those words pregnant with confusion, High and Low, and it were to be wished that they might be erased from scientific terminology. They deceive the person to whom they are addressed. They insensibly mislead the one who uses them. Psychological prejudices and fancies are so inextricably associated with these words that the use of them is provocative of such ideas. The words, generalized and specialized, having become almost limited to the expression of the ideas which the scientific biologist wishes to unfold by the others, can with great gain be employed in their stead." ("Families of Fishes," 1872.)
The Problem of the Highest Fishes.—As to which fishes should be ranked highest and which lowest, Dr. Gill gives ("Families of Fishes," 1872) the following useful discussion: "While among the mammals there is almost universal concurrence as to the forms entitled to the first as well as the last places, naturalists differ much as to the 'highest' of the ichthyoid vertebrates, but are all of one accord respecting the form to be designated as the 'lowest.' With that admitted lowest form as a starting-point, inquiry may be made respecting the forms which are successively most nearly related.
"No dissent has ever been expressed from the proposition that the Leptocardians (Branchiostoma) are the lowest of the vertebrates; while they have doubtless deviated much from the representatives of the immediate line of descent of the higher vertebrates, and are probably specialized considerably, in some respects, in comparison with those vertebrates from which they (in common with the higher forms) have descended, they undoubtedly have diverged far less, and furnish a better hint as to the protovertebrates than any other form.
"Equally undisputed it is that most nearly related to the Leptocardians are the Marsipobranchiates (Lampreys, etc.), and the tendency has been rather to overlook the fundamental differences between the two, and to approximate them too closely, than the reverse.
"But here unanimity ends, and much difference of opinion has prevailed with respect to the succession in the system of the[Pg 384] several subclasses (by whatever name called) of true fishes: (1) Some (e.g., Cuvier, J. Müller, Owen, Lütken, Cope) arranging next to the lowest the Elasmobranchiates, and, as successive forms, the Ganoids and Teleosteans; (2) while others (e.g., Agassiz, Dana, Duméril, Günther) adopt the sequence Leptocardians, Marsipobranchiates, Teleosteans, Ganoids, and Elasmobranchiates. The source of this difference of opinion is evident and results partly from metaphysical or psychological considerations, and partly from those based (in the case of the Ganoids) on real similarities and affinities.
"The evidence in favor of the title of the Elasmobranchiates to the 'highest' rank is based upon (1) the superior development of the brain; (2) the development of the egg, and the ovulation; (3) the possession of a placenta; and (4) the complexity of the organs of generation.
"(1) It has not been definitely stated wherein the superior development of the brain consists, and as it is not evident to the author, the vague claim can only be met by this simple statement; it may be added, however, that the brains comparable in essentials and most similar as a whole to those of the Marsipobranchiates are those of the sharks. In answer to the statement that the sharks exhibit superior intelligence, and thus confirm the indications of cerebral structure, it may be replied that the impression is a subjective one, and the author has not been thus influenced by his own observations of their habits. Psychological manifestations, at any rate, furnish too vague criteria to be available in exact taxonomy.
"(2) If the development of the eggs, their small number, and their investment in cases are arguments in favor of the high rank of the Elasmobranchiates, they are also for the Marsipobranchiates, and thus prove too much or too little for the advocates of the views discussed. The variation in number of progeny among true fishes (e.g., Cyprinodonts, Embiotocids) also demonstrates the unreliability of those modifications per se.
"(3) The so-called placenta of some Elasmobranchiates may be analogous to that of mammals, but that it is not homologous (i.e., homogenetic) is demonstrable from the fact that all the forms intervening between them and the specialized pla[Pg 385]cental mammals are devoid of a placenta, and by the variation (presence or want) among the Elasmobranchiates themselves.
"(4) The organs of generation in the Elasmobranchiates are certainly more complex than in most other fishes, but as the complexity results from specialization of parts sui generis and different from those of the higher (quadruped) vertebrates, it is not evident what bearing the argument has. If it is claimed simply on the ground of specialization, irrespective of homological agreement with admitted higher forms, then are we equally entitled to claim any specialization of parts as evidence of high rank, or at least we have not been told within what limits we should be confined. The Cetaceans, for example, are excessively specialized mammals, and, on similar grounds, would rank above the other mammals and man; the aye-aye exhibits in its dentition excessive specialization and deviation from the primitive type (as exhibited in its own milk teeth) of the Primates, and should thus also rank above man. It is true that in other respects the higher primates (even including man) may be more specialized, but the specialization is not as obvious as in the cases referred to, and it is not evident how we are to balance irrelative specializations against each other, or even how we shall subordinate such cases. We are thus compelled by the reductio ad absurdum to the confession that irrelative specialization of single organs is untrustworthy, and are fain to return to that better method of testing affinities by the equation of agreement in whole and after the elimination of special teleological modifications.
"The question then recurs, What forms are the most nearly allied to the Marsipobranchiates, and what show the closest approach in characteristic features? And in response thereto the evidence is not undecisive. Wide as is the gap between Marsipobranchiates and fishes, and comparatively limited as is the range of the latter among themselves, the Elasmobranchiates are very appreciably more like, and share more characters in common with them, than any other; so much is this the case that some eminent naturalists (e.g., Pallas, Geoffroy, St. Hilaire, Latreille, Agassiz, formerly Lütken) have combined the two forms in a peculiar group, contradistinguished from the other fishes. The most earnest and extended argument in English,[Pg 386] in favor of this combination has been published by Professor Agassiz in his 'Lake Superior,' but that eminent naturalist subsequently arrived at the opposite conclusions already indicated.
"The evidences of the closer affinity of the Elasmobranchiates (than of any other fishes) with the Marsipobranchiates are furnished by (1) the cartilaginous condition of the skeleton; (2) the post-cephalic position of the branchiæ; (3) the development of the branchiæ and their restriction to special chambers; (4) the larger number of the branchiæ; (5) the imperfect development of the skull; (6) the mode of attachment of the teeth; (7) the slight degree of specialization of the rays of the fins; and (8) the rudimentary condition of the shoulder-girdle."
[147] Key to North American Birds.

Science consists of human experience, tested and placed in order. The science of ichthyology represents our knowledge of fishes, derived from varied experiences of man, tested by methods or instruments of precision and arranged in orderly sequence. This science, in common with every other, is the work of many persons, each in his own field, and each contributing a series of facts, a series of tests of the alleged facts of others, or some improvement in the method of arrangement. As in other branches of science, this work has been done by sincere, devoted men, impelled by a love for this kind of labor, and having in view, as "the only reward they asked, a grateful remembrance of their work." And in token of this reward it is well sometimes, in grateful spirit, to go over the names of those who made even its present stage of completeness possible.
We may begin the history of ichthyology with that of so many others of the sciences, with the work of Aristotle (383-322 B.C.). This wonderful observer recorded many facts concerning the structure and habits of the fishes of Greece, and in almost every case his actual observation bears the closest modern test. These observations were hardly "set in order." The number of species he knew was small, about 118 in all, and it did not occur to him that they needed classification. His ideas of species were those of the fishermen, and the local vernacular supplied him with the only names needed in his records.
As Dr. Günther wisely observes, "It is less surprising that Aristotle should have found so many truths as that none of his followers should have added to them." For nearly 1800 years the scholars of the times copied the words of Aristotle, confus[Pg 388]ing them by the addition of fabulous stories and foolish superstitions, never going back to nature herself, "who leads us to absolute truth whenever we wander." A few observations were made by Caius Plinius, Claudius Ælianus, Athenæus and others. Theophrastus (370-270 B.C.) wrote on the fishes which may live out of water. About 400 A.D., Decius Magnus Ausonius wrote a pleasing little poem on the Moselle, setting forth the merits of its various fishes. It was not, however, until the middle of the seventeenth century that any advance was made in the knowledge of fishes. At that time the development of scholarship among the nations of Europe was such that a few wise men were able to grasp the idea of species.
In 1553, Pierre Bélon (1518-64) published his octavo volume of 448 pages, entitled "De Aquatilibus," in which numerous (110) species of fishes of the Mediterranean were described, with tolerable figures, and with these is a creditable attempt at classification. At about this time Ulysses Aldrovandi, of Bologna, founded the first museum of natural history and wrote on the fishes it contained. In 1554-58, Ippolito Salviani (1513-72), a physician at Rome, published a work entitled "Aquatilium Animalium Historia," with good figures of most of the species, together with much general information as to the value and habits of animals of the sea.
More important than these, but almost simultaneous with them, is the great work of Guillaume Rondelet (1507-57), "De Piscibus Marinus" (1554-55), at first written in Latin, later translated into French and enlarged under other titles. In this work, 244 different species, chiefly from the Mediterranean, are fairly described, and the various fables previously current are subjected to severe scrutiny. Recognizable woodcuts represent the different species. Classification, Rondelet had none, except as simple categories for purposes of convenience. More than usual care is given to the vernacular names, French and Greek. He closes his book with these words:
"Or s'il en i a qui prennent les choses tant à la rigueur, qui ne veulent rien apparouver qui ne soit du tout parfait, je les prie de bien bon cueur de traiter telle, ou quelque autre histoire parfaitement, sans qu'il i ait chose quelconque à redire et la receverons é haut louerons bien vouluntiers. Cependant[Pg 389] je scai bien, et me console . . . avec grand travail . . . qu'on pourra trouver plusieurs bones choses e dignes de louange ou proufit é contentement des homes studieux é à l'honneur é grandissime admiration des tres excellens é perfaits œuvres de Dieu."
And with the many "bones choses" of the work of Rondelet, men were too long satisfied, and it was not until the impulse of commerce had brought them face to face with new series of animals not found in the Mediterranean that the work of investigating fishes was again resumed. About 1640, Prince Moritz (Maurice) of Nassau (1604-79) visited Brazil, taking with him two physicians, Georg Marcgraf (1610-44) and Wilhelm Piso. In the great work "Historia Naturalis Brasiliæ," published at Leyden (1648), Marcgraf described about one hundred species, all new to science, under Portuguese names and with a good deal of spirit and accuracy. This work was printed by Piso after Marcgraf's death, and his colored drawings—long afterward used by Bloch—are in the "History of Brazil" reduced to small and crude woodcuts. This is the first study of a local fish fauna outside the Mediterranean region and it reflects great credit on Marcgraf and on the illustrious prince whose assistant he was.
There were no other similar attempts of importance in ichthyology for a hundred years, when Per Osbeck, an enthusiastic student of Linnæus, published (1757) the records of his cruise to China, under the name of "Iter Chinensis." At about the same time another of Linnæus' students, Fredrik Hasselquist, published, in his "Iter Palestinum" the account of his discoveries of fishes in Palestine and Egypt. More pretentious than these and of much value as an early record is Mark Catesby's (1679-1749) "Natural History of Carolina and the Bahamas," published in 1749, with large colored plates which are fairly correct except in those cases in which the drawing was made from memory.
At about the same time, Hans Sloane (1660-1752) published his large volume on the "Fishes of Jamaica," Patrick Browne (1720-90) wrote on the fishes of the same region, while Father Charles Plumier (1646-1704) made paintings of the fishes of Martinique, long after used by Bloch and Lacépède. Dr. Alex[Pg 390]ander Garden (1730-91), of Charleston, S. C., collected fishes for Linnæus, as did also Dr. Pehr Kalm in his travels in the northern parts of the American colonies.
With the revival of interest in general anatomy several naturalists took up the structure of fishes. Among these Günther mentions Borelli, Malpighi, Swammerdam, and Duverney. Other anatomists of later dates were Albrecht von Heller (1708-77), Peter Camper (1722-89), Felix Vicq d'Azyr (1748-94), and Alexander Monro (1783).
The basis of classification was first fairly recognized by John Ray (1628-1705) and Francis Willughby (1635-72), who, with other and varied scientific labors, undertook, in the "Historia Piscium," published in Oxford in 1686, to bring order out of the confusion left by their predecessors. This work, edited by Ray after Willughby's death, is ostensibly the work of Willughby with additions by Ray. In this work 420 species were recorded, 180 of which were actually examined by the authors, and the arrangement chosen by them pointed the way to a final system of nomenclature.
Direct efforts in this direction, with a fairly clear recognition of genera as well as species, were made by Lorenz Theodor Gronow, called Gronovius, a German naturalist of much acumen, and by Jacob Theodor Klein (1685-1757), whose work, "Historic Naturalis Piscium," published about 1745, is of less importance, not being much of an advance over the catalogue of Rondelet.
Far greater than any of these investigators, and earlier than either Klein or Gronow, was he who has been justly called the Father of Ichthyology, Petrus (Peter) Artedi (1705-35). Artedi was born in Sweden. He was a fellow student of Linnæus at Upsala, and he devoted his short life wholly to the study of fishes. He went to Holland to examine the collection of East and West Indian fishes of a rich Dutch merchant in Amsterdam named Albert Seba, and there at the age of twenty-nine he was, by accident, drowned in one of the Dutch canals. "His manuscripts were fortunately rescued by an Englishman, Cliffort," and they were edited and published by Linnæus in a series of five parts or volumes.
Artedi divided the class of fishes into orders, and these orders[Pg 391] again into genera, the genera into species. The name of each species consisted of that of the genus with a descriptive phrase attached. This cumbersome system, called polynomial, used by Artedi, Gronow, Klein, and others, was a great advance on the shifting vernacular, of which it now took the place. But the polynomial method as a system was of short duration. Linnæus soon substituted for it the convenient, in fact inevitable binomial system which has now endured for 150 years, and which with certain modifications must form the permanent substructure of the nomenclature in systematic zoology and botany.
The genera of Artedi are in almost all cases natural groups, corresponding essentially equivalent to the families of to-day. Families in ichthyology were first clearly recognized and defined by Cuvier.
The following is a list of Artedi's genera and their arrangement:
ORDER MALACOPTERYGII.
ORDER ACANTHOPTERYGII.
ORDER BRANCHIOSTEGI.
ORDER CHONDROPTERYGII.
In all 47 genera and 230 species of fishes were known from the whole world in 1738.
The cetaceans, or whales, constitute a fifth order, Plagiuri, in Artedi's scheme.
As examples of the nomenclature of species I may quote:
"Zeus ventre aculeato, cauda in extremo circinata." This polynomial expression was shortened by Linnæus to Zeus faber. The species was called by Rondelet "Faber sive Gallus Marinus" and by other authors "Piscis Jovii." "Jovii" suggested Zeus to Artedi, and Rondelet's name faber became the specific name.
"Anarhichas Lupus marinus nostras." This became with Linnæus "Anarhichas lupus."
"Clupea, maxilla inferiore longiore, maculis nigris carens: Harengus vel Chalcis Auctorum, Herring vel Hering Anglis, Germanis Belgis." This became Clupea harengus in the convenient binomial system of Linnæus.
The great naturalist of the eighteenth century, Carl von Linné, known academically as Carolus Linnæus, was the early associate and close friend of Artedi, and from Artedi he obtained practically all his knowledge of fishes. Linnæus, professor in the University of Upsala and for a time its rector, primarily a botanist, was a man of wonderful erudition, and his great strength lay in his skill in the orderly arrangement of things. In his lifetime, his greatest work, the "Systema Naturæ," passed through twelve editions. In the tenth edition, in 1758, the binomial system of nomenclature was first consistently applied to all animals. For this reason most naturalists use the date of its publication as the beginning of zoological nomenclature, although the English naturalists have generally preferred the more complete twelfth edition, published in 1766. This difference in the recognized starting-point has been often a source of confusion, as in several cases the names of species were needlessly changed by Linnæus and given differently in the twelfth edition. In taxonomy it is not nearly so important that a name be pertinent or even well chosen as that it be stable. In changing his own established names, the father of classification set a bad example to his successors, one which they did not fail to follow.
In Linnæus' system (tenth and twelfth editions) all of[Pg 393] Artedi's genera were retained save Lepturus, which name was changed to Trichiurus. The following new genera were added: Chimæra, Tetraodon, Diodon, Centriscus, Pegasus, Callionymus, Uranoscopus, Cepola, Mullus, Teuthis, Loricaria, Fistularia, Atherina, Mormyrus, Polynemus, Amia, Elops. The classification was finally much altered: the Chondropterygia and Branchiostegi (with Syngnathus) being called Amphibia Nantes, and divided into two groups—Spiraculis compositis and Spiraculis solitariis. The other fishes were more naturally distributed according to the position of the ventral fins into Pisces Apodes, Jugulares, Thoracici, and Abdominales. The Apodes of Linnæus do not form a homogeneous group, as members of various distinct groups have lost their ventral fins in the process of evolution. But the Jugulares, the Thoracici, and the Abdominales must be kept as valid categories in any natural system.
Linnæus' contributions to zoology consisted mainly of the introduction of his most ingenious and helpful system of bookkeeping. By it naturalists of all lands were able to speak of the same species by the same name in whatever tongue. Unfortunately, ignorance, carelessness, and perversity brought about a condition of confusion. For a long period many species were confounded under one name. This source of confusion began with Linnæus himself. On the other hand, even with Linnæus, the same species often appeared under several different names; in this matter it was not the system of naming which was at fault. It was the lack of accurate knowledge, and sometimes the lack of just and conscientious dealing with the work of other men. No system of naming can go beyond the knowledge on which it rests. Ignorance of fact produces confusion in naming. The earlier naturalists had no conception of the laws of geographical distribution. The "Indies," East or West, were alike to them, and "America" or "India" or "Africa" was a sufficiently exact record of the origin of any specimen.
Moreover, no thought of the geological past of groups and species had yet arisen, and without the conception of common origin, the facts of homology had no significance. All classification was simply a matter of arbitrary pigeon-holing the records of forms, rather than an expression of actual blood rela[Pg 394]tionship. To this confusion much was added through love of novelty. Different authors changed names to suit their personal tastes regardless of rights of priority. Amia was altered to Amiatus by Rafinesque in 1815 because it was too short a name. Hiodon was changed to Amphiodon because it sounded too much like Diodon, Batrachoides to Batrictius because βατράχος means a frog, not a fish, and other changes even more wanton were introduced, to be condemned and discarded by the more methodical workers of a later period. With all its abuses, however, the binomial nomenclature made possible systematic zoology and botany, and with the "Systema Naturæ" arose a new era in the science of living organisms.
In common with most naturalists of his day, the spirit of Linnæus was essentially a devout one. Admiration for the wonderful works of God was breathed on almost every page. "O Jehovah! quam ampla sunt opera Tua" is on the title-page of the "Systema Naturæ," and the inscription over the door of his home at Hammarby was to Linnæus the wisdom of his life. This inscription read: "Innocue vivito: Numen adest" (Live blameless: God is here).
The followers of Linnæus are divided into two classes, explorers and compilers. To the first class belonged his own students and others who ransacked all lands for species to be added to the lists of the "Systema Naturæ." Those men, mostly Scandinavian and Dutch, worked with wonderful zeal, enduring every hardship and making great contributions to knowledge, which they published in more or less satisfactory forms. To these men we owe the beginnings of the science of geographical distribution. Among the most notable of these are Pehr Osbeck and Fredrik Hasselquist, already noted; Otto Fabricius (1744-1822), author of an excellent "Fauna of Greenland"; Carl Peter Thunberg (1743-), successor of Linnæus as rector of the University of Upsala, who collected fishes about Nagasaki, intrusting most of the descriptive work to the less skillful hands of his students, Jonas Nicolas Ahl and Martin Houttuyn; Martin Th. Brünnich, who collected at Marseilles the materials for his "Pisces Massiliensis"; Petrus Forskål (1736-63), whose work on the fishes of the Red Sea ("Descriptio Animalium," etc.), published posthumously in 1775, is one of the most accu[Pg 395]rate of faunal lists, and one which shows a fine feeling for taxonomic distinctions scarcely traceable in any previous author. Georg Wilhelm Steller (1709-45), naturalist of Bering's expedition, gathered amid incredible hardships the first knowledge of the fishes of Alaska and Siberia, his notes being printed after his tragic death, by Pallas and Krascheninnikov. Petrus Simon Pallas (1741-1811) gives the account of his travels in the North Pacific in his most valuable volumes, "Zoographia Russo-Asiatica"; Johann Georg Gmelin (1709-55) with Samuel Theophilus Gmelin (1745-84), and Johann Anton Güldenstädt (1745-91), like Steller, crossed Siberia, recording its animals. Johann David Schöpf (1752-1800), a Hessian surgeon stationed at Long Island in the Revolutionary War, gave an excellent account of the fishes about New York.
Still other naturalists accompanied navigators around the globe, collecting specimens and information as opportunity offered. John Reinhold Forster (1729-98), with his son, John George Adam Forster (1754-94), and Daniel Solander (1736-81), a student of Linnæus, and Sir Joseph Banks (1743-1820), sailed with Captain James Cook. Philibert Commerson (1727-73) accompanied the explorer, Louis Antoine de Bougainville, and furnished nearly all the original material used by Lacépède. Other noted travelers of the early days were Pierre Sonnerat and Mungo Park.
Still other naturalists, scarcely less useful, gave detailed accounts of the fauna of their own native regions. Ablest of these was Anatole Risso, an apothecary of Nice, who published in 1810 the "Ichthyologie de Nice," an excellent work, afterward (1826) expanded by him into a "Histoire Naturelle de l'Europe Méridionalé."
Contemporary with Risso was a man of very different character, Constantine Samuel Rafinesque (1784-1842), who wrote at Palermo in 1810 his "Caratteri di Alcuni Nuovi Generi" and his "Ittiologia Siciliana." Later he went to America, where he was for a time professor in the Transylvania University at Lexington, Ky. Brilliant, erudite, irresponsible, fantastic, he wrote of the fishes of Sicily and later ("Ichthyologia Ohiensis," 1820) of the fishes of the Ohio River, with wide knowledge, keen taxonomic insight, and a hopeless disregard of the[Pg 396] elementary principles of accuracy. Always eager for novelties, restless and credulous, his writings have been among the most difficult to interpret of any in ichthyology.
Earlier than Risso and Rafinesque, Thomas Pennant (1726-58) wrote of the British fishes; Otto Fredrik Müller of the fishes of Denmark; J. E. Gunner, Bishop of Thröndhjem, of fishes of Norway; Francis Valentijn (1660-1730), Jan Nieuhof (1600-1671), Renard, and Castour of the fishes of the Dutch East Indies; Duhamel du Monceau of the fisheries of France; Francesco Cette of the fishes of Sicily; José Cornide of the fishes of Spain; Ignacio Molina of the fishes of Chile; and Meidinger of those of Austria. Some of these writers lived before Linnæus. Others knew little of the Linnæan system, and their records are generally in the vernacular. Most important of this class is the work of Antonio Parra, "Descripcion de Diferentes Piezas de Historia Natural de la Isla de Cuba," published in Havana in 1787. In 1803, Patrick Russell gave a valuable account, non-binomial, of "Two Hundred Fishes Collected at Vizagapatam and on the Coast of Coromandel."
Papers on the fishes of Bering Sea and Japan by Wilhelm Theophilus Tilesius (1775-1835), are published in the transactions of the early societies of Russia. The collections of the traveler Krusenstern were recorded by Tilesius. Stephen Krascheninnikov (1786) wrote a history of Russia in Asia.
Other notable names among the early writers are those of Pierre Marie Auguste Broussonet, of Montpelier, whose work (1780), too soon cut short, showed marked promise; Fr. Faber, who wrote of the fishes of Iceland; E. Blyth, who studied the fishes of the Andamans; A. G. Desmarest, who made excellent studies of the fishes of Cuba; J. T. Kölreuter and Everard Home in the East Indies; Geoffrey Saint-Hilaire, who recorded the fishes of Egypt at the command of Napoleon. Others equally notable were B. A. Euphrasen, Iwan Lepechin (1750-1802), John Latham, W. E. Leach, George Montagu, C. Quensel, Jean-Antoine Scopoli, Peter Ascanius, Francois Etienne de la Roche (1789-1812), Hans Ström, M. Vahl and Zuieuw.
The compilers who followed Linnæus belonged to a wholly different class. These were men of extensive learning, methodi[Pg 397]cal ways, sometimes brilliant, occasionally of deep insight, but more often, on the whole, dull, plodding, and mechanical.
Earliest of those is Antoine Gouan, whose "Historia Piscium" was published in Paris in 1770. In this work, which is of fair quality, only genera were included, and the three new ones which he introduces into the "System" (Lepadogaster, Lepidopus, and Trachypterus) are still retained with his definition of them.
Johann Friedrich Gmelin (1748-1804), a relative of the explorers of Siberia, published in 1788 a thirteenth edition of the "Systema Naturæ" of Linnæus, adding to it the discoveries of Forskål, Forster, and others who had written since Linnæus' time. This work was useful as bringing the compilation of Linnæus to a later date, but it is not well done, the compiler having little knowledge of the animals described and little penetration in matters of taxonomy. Very similar in character, although more lucid in expression, is the French compilation of the same date (1788), "Tableau Encyclopédique et Méthodique des Trois Règnes de la Nature," by the Abbé J. P. Bonnaterre. Another volume of the "Encyclopédie Méthodique," of still less merit, was published as a dictionary in Paris in 1787 by Réné Just Haüy. Another dictionary in 1817 even poorer was the work of Hippolyte Cloquet.
In 1792, Johann Julius Walbaum (1721-1800), a German compiler of a little higher rank, gathered together the records of all known species, using the work of Artedi as a basis and giving binominal names in place of the vernacular terms used by Schöpf, Steller, Pennant, and Krascheninnikov.
Far more pretentious and more generally useful, as well as containing a large amount of original material, is the "Ichthyologia" of Mark Eliezer Bloch, published in Berlin in various parts from 1782 to 1785. It was originally in German and divided into two portions—"Oeconomische Naturgeschichte der Fische Deutschlands" and "Naturgeschichte der auslandischen Fische." Bloch was a Jewish physician, born at Anspach in 1723, and at the age of fifty-six began to devote himself to ichthyology. In his great work is contained every species which he had himself seen, every one which he could purchase[Pg 398] from collections, and every one of which he could find drawings made by others.
That part which relates to the fishes of Germany is admirably done. In the treatment of East Indian and American fishes there is much guesswork and many errors of description and of fact, for which the author was not directly responsible. To learn to interpret the personal equation in the systematic work of other men is one of the most delicate of taxonomic arts.
After the publication of these great folio volumes of plates, Dr. Bloch began a systematic catalogue to include all known species. This was published after his death by his collaborator, the philologist, Dr. Johann Gottlob Schneider. This work, "M. E. Blochii Systema Ichthyologia," contains 1519 species of fishes, and is the most creditable compilation subsequent to the death of Linnæus.
Even more important than the work of Bloch is that of the Comte de La Cépède, who became with the progress of the French Revolution, "Citoyen Lacépède," his original full name being Bernard Germain Etienne de la Ville-sur-Illon, Comte de La Cépède. His great work, "Histoire Naturelle des Poissons," was published originally in five volumes, in Paris, from 1798 to 1803. It was brought out under great difficulties, his materials being scattered, his country in a constant tumult. For original material he depended largely on the collections and sagacious notes of the traveler Commerson. Dr. Gill sums up the strength and weakness of Lacépède's work in these terms:
"A work by an able man and eloquent writer even prone to aid rhetoric by the aid of the imagination in absence of desirable facts, but which because of undue confidence in others, default of comparison of material from want thereof and otherwise, and carelessness generally is entirely unreliable."
The work of Lacépède had a great influence upon subsequent investigators, especially in France. A considerable number of the numerous new genera of Rafinesque were founded on divisions made in the analytical keys of Lacépède.

Bernard Germain de Lacépède.
Georges Dagobert Cuvier.
Louis Agassiz.
Johannes Müller.
In 1803 and 1804, Dr. George Shaw published in London his "General Zoology," the fishes forming part of volumes IV and V. This is a poor compilation, the part concerning the fishes being mostly extracted from Bloch and Lacépède. Another weak compilation for the supposed use of students was the "Ichthyologie Analytique" of A. M. Constant Duméril. About 1815, Henri Ducrotay de Blainville wrote the "Faune Française" and contributed important studies to the taxonomy of sharks.
With Georges Léopold Chrétien Frédéric Dagobert Cuvier (1769-1832) and the "Règne Animal arrangé aprés son Organization" (1817; 1829-30) we have the beginning of a new era in ichthyology. This period is characterized by a recognition of the existence of a natural classification inevitable in proportion to the exactness of our knowledge, because based on the principles of morphology. The "Règne Animal" is, in the history of ichthyology, not less important than the "Systema Naturæ" itself, and from it dates practically our knowledge of families of fishes and the interrelations of the different groups. The great facts of homology were clearly understood by Cuvier. Their significance as indications of lines of descent were never grasped by him, and this notwithstanding the fact that Cuvier was almost the first to bring extinct forms into proper relations with those now living.
Dr. Günther well says that the investigation of anatomy of fishes was continued by Cuvier until he had succeeded in completing so perfect a framework of the system of the whole class that his immediate successors could content themselves with filling up those details for which their master had no leisure. Indefatigable in examining all the external and internal characters of the fishes of a rich collection, he ascertained the natural affinities of the infinite variety of fishes, and accurately defined the divisions, orders, families, and genera of the class as they appear in the two original editions of the "Règne Animal." His industry equaled his genius; he opened connections with almost every accessible part of the globe; not only French travelers and naturalists, but also Germans, Englishmen, Americans rivaled one another to assist him with collections; and for many years the Museum of the Jardin des Plantes was the Center where all ichthyological treasures were deposited. Thus Cuvier brought together a collection the like of which had never been seen before, and which, as it contains all the materials[Pg 401] on which his labors were based, must still be considered to be one of the most important in existence.
"Those little low rooms, five in number" (in the museum of the Jardin des Plantes), "they should be the Mecca of scientific devotees. Perhaps every great zoologist of the past hundred years has sat in them and discussed those problems of life which are always inviting solution and are never solved. The spirits of great naturalists still haunt these corridors and speak from the specimens their hands have set in order." (Theodore Lyman.)
Cuvier's studies of the different species of fishes are contained in the great "Histoire Naturelle des Poissons," the joint work of Cuvier and his pupil and successor, Achille Valenciennes (1794-1865). Of this work 22 volumes were published, from 1828 to 1849, containing 4514 nominal species, the greater portion being written after the death of Cuvier (1832). The work was finally left unfinished on account of a disagreement with the publisher. Dr. Gill tells me that at this time Valenciennes made an unsuccessful appeal to the Smithsonian Institution for assistance in the publication of the remaining chapters.
This is a most masterly work, indispensable to the student of fishes. Its descriptions are generally fairly correct, its plates accurate, and its judgments trustworthy. But with all this it is very unequal. Too often nominal species are based on variations due to age or sex or to the conditions of preservation of specimens. Many of the species are treated very lightly by Cuvier; many of the descriptions of Valenciennes are very mechanical, as though the author had grown weary of the endless process, "a failing commonly observed among zoologists when attention to descriptive details becomes to them a tedious task."
After the death of Valenciennes (1865) Dr. Auguste Duméril began another Natural History of the Fishes. Of this two volumes (1865-70) were published covering sharks, ganoids, and other fishes not treated by Cuvier and Valenciennes, his category beginning at the opposite end of the fish series. The death of Duméril left this catalogue also unfinished. Duméril's work is useful and carefully done, but his excessive trust in[Pg 402] slight differences has filled his book with nominal species. Thus among the living ganoid fishes he recognizes 135 species, the actual number being not far from 40.
We may anticipate the sequence of time by here referring to the remaining attempts at a record of all the fishes in the world, Dr. Albert C. L. G. Günther, a naturalist of German birth, but resident in London for many years, long the honored keeper of the British Museum, published in eight volumes the "Catalogue of the Fishes of the British Museum," from 1859 to 1870. In this monumental work, the one work most essential to all systematic study of fishes, 6843 species are described and 1682 doubtful species are mentioned. The book is a remarkable example of patient industry. Its great merits are at once apparent, and those of us engaged in the same line of study may pass by its faults with the leniency which we may hope that posterity may bestow on ours.
The publication of this work gave an immediate impetus to the study of fishes. The number of known species has been raised from 9000 to about 12,000 in the last thirty years, although meanwhile some hundreds of species even accepted by the conservatism of Günther have been erased from the system.
A new edition of this work has been long in contemplation, and in 1898 the first volume of it, covering the percoid fishes, was published by Dr. George Albert Boulenger. This volume is one of the most satisfactory in the history of ichthyology. It is based on ample material. Its accepted species have been subject to thorough criticism and in its classification every use has been made of the teachings of morphology and especially of osteology. Its classification is distinctly modern, and with the writings of the contemporary ichthyologists of Europe and America, it is fully representative of the scientific era ushered in by the researches of Darwin. The chief criticism which one may apply to this work concerns most of the publications of the British Museum. It is the frequent assumption that those species not found in the greatest museum of the world do not really exist at all. There are still many forms of life, very many, outside the series gathered in any or all collections.

Albert Günther.
Franz Steindachner.
George A. Boulenger.
Robert Collett.
We may now turn from the universal catalogues to the work on special groups, on local faunas, or on particular branches of the subject of ichthyology. These lines of study were made possible by the work of Cuvier and Valenciennes and especially by that of Dr. Günther.
Before taking up the students of faunal groups, we may, out of chronological order, consider the researches of three great taxonomists, who have greatly contributed to the modern system of the classification of fishes.
Louis Agassiz (born at Motiers in western Switzerland in 1807; died at Cambridge, Mass., in 1873) was a man of wonderful insight in zoological matters and possessed of a varied range of scientific information, scarcely excelled in any age—intellectually a lineal descendant of Aristotle. His first work on fishes was the large folio on the fishes collected by Jean Baptiste Spix (1781-1826) in Brazil, published at Munich in 1827. After his establishment in America in 1846, soon after which date, he became a professor in Harvard University, Agassiz published a number of illuminating papers on the fresh-water fishes of North America. He was the first to recognize the necessity of the modern idea of genera among fishes, and most of the groups designated by him as distinct genera are retained by later writers. He was also the first to investigate the structure of the singular viviparous surf-fishes of California, the names Embiotoca and Holconotus applied to these fishes being chosen by him.
His earlier work, "Recherches sur les Poissons des Eaux Douces," published in Europe, gave a great impetus to our knowledge of the anatomy and especially of the embryology of the fresh-water fishes. Most important of all his zoological publications was the "Recherches sur les Poissons Fossiles," published at Neufchatel from 1833 to 1843. This work laid the foundation of the systematic study of the extinct groups of fishes. The relations of sharks were first appreciated by Agassiz, and the first segregation of the ganoids was due to him. Although he included in this group many forms not truly related either to anything now called ganoids, nor even to the extinct mailed forms which preceded them, yet the definition of this order marked a distinct step in advance.
The great, genial, hopeful personality of Agassiz and his[Pg 405] remarkable skill as a teacher made him the "best friend that ever student had" and gave him a large following as a teacher. Among his pupils in ichthyology were Charles Girard (1822-1895), Frederick Ward Putnam, Alexander Agassiz, Samuel Garman, Samuel H. Scudder, and the present writer.
Johannes Müller (1808-1858), of Berlin, was one of the greatest of comparative anatomists. In his revision of Cuvier's "System of Classification" he corrected many errors in grouping, and laid foundations which later writers have not altered or removed. Especially important is his classical work, "Ueber den Bau and die Grenzen der Ganoiden." In this he showed some of the real fundamental characters of that group of archaic fishes, and took from it the most heterogeneous of the elements left in it by Agassiz. To Müller we also owe the first proper definition of the Leptocardii and the Cyclostomata, and, in association with Dr. J. Henle, Müller has given us one of the best general accounts of the sharks ("Systematische Beschriebungen der Plagiostomen"). To Müller we owe an accession of knowledge in regard to the duct of the air-bladder, and the groups called Physostomi, Physoclysti, Dipneusti (Dipnoi), Pharyngognathi, and Anacanthini were first defined by him.
In his work on Devonian fishes, the great British comparative anatomist, Thomas Henry Huxley, first distinguished the group of Crossopterygians, and separated it from the ganoids and dipnoans.
Theodore Nicholas Gill is the keenest interpreter of taxonomic facts yet known in the history of ichthyology. He is the author of a vast number of papers, the first bearing date of 1858, touching almost every group and almost every phase of relation among fishes. His numerous suggestions as to classification have been usually accepted in time by other authors, and no one has had a clearer perception than he of the necessity of orderly methods in nomenclature. Among the orders first defined by Gill are the Eventognathi, Nematognathi, Pediculati, Iniomi, Heteromi, Haplomi, Xenomi, and the group called Teleocephali, originally framed to include all the bony fishes except those which showed peculiar eccentricities or modifications. Dr. Gill's greatest excellence has been shown as a scientific critic. Incisive, candid, and friendly, there is[Pg 406] scarcely an investigator in biology, in America, who is not directly indebted to him for critical aid of the highest importance. The present writer cannot too strongly express his own obligations to this great teacher, his master in fish taxonomy. Dr. Gill's work is not centered in any single great treatise, but is diffused through a very large number of brief papers and catalogues, those from 1861 to 1865 mostly published by the Academy of Natural Sciences in Philadelphia, those of recent date by the United States National Museum. For many years Dr. Gill has been identified with the work of the Smithsonian Institution at Washington.
Closely associated with Dr. Gill was Dr. Edward Drinker Cope, of Philadelphia, a tireless worker in almost every field of zoology, and a large contributor to the broader fields of ichthyological taxonomy as well as to various branches of descriptive zoology. Cope was one of the first to insist on the close relation of the true ganoids with the teleost fishes, the nearest related group of which he defined as Isospondyli. At the same time he recognized the wide range of difference even among the forms which Johannes Müller had assembled under that name. In breadth of vision and keenness of insight, Cope ranked with the first of taxonomic writers. Always bold and original, he was not at all times accurate in details, and to the final result in classification his contribution has been less than that of Dr. Gill. Professor Cope also wrote largely on American fresh-water fishes, a large percentage of the Cyprinidæ and Percidæ of the eastern United States having been discovered by him, as well as much of the Rocky Mountain fauna. In later years his attention was absorbed by the fossil forms, and most of the species of Cretaceous rocks and the Eocene shales of Wyoming were made known through his ceaseless activity.

Spencer Fullerton Baird.
Edward Drinker Cope.
Theodore Nicholas Gill.
George Brown Goode.
The enumeration of other workers in the great field of ichthyology must assume something of the form of a catalogue. Part of the impulse received from the great works of Cuvier and Valenciennes and of Günther was spent in connection with voyages of travel. In 1824 Quoy and Gaimard published in Paris the great folio work on the fishes collected by the corvette l'Uranie and la Physicienne in Freycinet's voyages around the world, and in 1834 the same authors published the fishes collected in Duperrey's voyage of the Astrolabe. In 1826 Lesson published the fishes of Dumont D'Urville's voyage of the Coquille. These three great works lie at the foundation of our knowledge of the fishes of Polynesia. In 1839 Eydoux and Gervais published an account of the fishes of the voyage of La Favorite. In 1853, also in Paris, Hombron and Jacquinot gave an account of the fishes taken in Dumont D'Urville's expedition to the South Pole. In England, Sir John Richardson (1787-1865), a wise and careful naturalist, wrote of the fishes collected by the Sulphur (1845), the Erebus and Terror (1846), the Samarang, and the Herald. Lay and Bennett recorded the species taken by Beechey's voyage on the Blossom. A most useful work is the account of the species taken by Charles Darwin on the voyage of the Beagle, prepared by the conscientious hand of Rev. Leonard Jenyns. Still more important and far ranging is the voyage of the Challenger, including the first important work in the deep seas, one stately volume and parts of other volumes on fishes being the work of Dr. Günther. Other deep-sea work of equal importance has been accomplished in the Atlantic and the Pacific by the U. S. Fish Commission steamer Albatross. Its results in Central America, Alaska, Japan, Hawaii, as well as off both coasts of the United States, have been made known in different memoirs by Goode and Bean, Gilbert, Garman, Gill, Jordan, Cramer, Ryder, and others. The deep-sea fish collections of the Fish Hawk and the Blake have been studied by Goode and Bean and Garman.
The deep-sea work of other countries may be briefly noticed. The French vessels Travailleur and Talisman have made collections chiefly in the Mediterranean and along the coast of Africa, the results having been made known by Léon Valliant. The Hirondelle about the Azores and elsewhere has furnished material for Professor Robert Collett, of the University of Christiania. Dr. Decio Vinciguerra, of Rome, has reported on the collections of the Violante, a vessel belonging to the Prince of Monaco. Dr. A. Alcock, of Calcutta, has had charge of the most valuable deep-sea work of the Investigator in the Indian Seas. Edgar R. Waite and James Douglas Ogilby, of the Australian Museum at Sydney, have[Pg 409] described the collections of the Thetis, on the shores of the New South Wales.

Johann Reinhardt.
Edward Waller Claypole.
Carlos Berg.
Edgar R. Waite.
From Austria the voyage of the frigate Novara has yielded large material which has been described by Dr. Rudolph Kner. The cream of many voyages of many Danish merchant vessels has been gathered in the "Spolia Atlantica" and other truly classical papers of Christian Frederik Lütken, of the University of Copenhagen, one of the most accomplished naturalists of recent times.
F. H. von Kittlitz has written on the fishes seen by him in the northern Pacific, and earlier and more important we may mention the many ichthyological notes found in the records of travel in Mexico and South America by Alexander von Humboldt (1796-1859).
The local faunal work in various nations has been very extensive. In Great Britain we may note Parnell's "Natural History of the Fishes of the Firth of Forth," published in Edinburgh in 1838, William Yarrell's "History of British Fishes" (1859), the earlier histories of British fishes by Edward Donovan and by William Turton, and the works of J. Couch (1862) and Dr. Francis Day (1888), possessing similar titles. The work of Day, with its excellent plates, will long be the standard account of the relatively scant fish fauna of the British islands. H. G. Seeley has prepared (1886) also a useful synopsis of "The Fresh-water Fishes of Europe."
We may here notice without praise the pretentious work of William Swainson (1838-39). W. Thompson has written of the fishes of Ireland, and Rev. Richard T. Lowe and J. Y. Johnson have done most excellent work on the fishes of Madeira. F. McCoy, better known for work on fossil fishes, may be mentioned here.
The fish fauna of Scandinavia has been described more or less fully by S. Kröyer (1840), Robert Nilsson (1855), Fries and Ekström (1836), Robert Collett, Robert Lilljeborg, and F. A. Smitt, besides special papers by other writers, notably Reinhardt, L. Esmarck, Japetus Steenstrup, Lütken, and A. W. Malm. Reinhardt, Kröyer, Lütken, and A. J. Malmgren have written of the Arctic fishes of Greenland and Spitzbergen.
In Russia, Nordmann has described the fishes of the Black[Pg 411] Sea ("Ichthyologic Pontique," Paris, 1840) and Eichwald those of the Caspian. More recently, S. Herzenstein, Warpachowsky, K. Kessler, B. N. Dybowsky, and others have written of the rich fauna of Siberia, the Caucasus, and the scarcely known sea of Ochotsk. Stephan Basilevsky has written of the fishes of northern China. A. Kowalevsky has contributed very much to our knowledge of anatomy. Peter Schmidt has studied the fishes of the Japan Sea.
In Germany and Austria the chief local works have been those of Heckel and Kner on the fresh-water fishes of Austria (1858) and C. Th. von Siebold on the fresh-water fishes of Central Europe (1863). German ichthyologists have, however, often extended their view to foreign regions where their characteristic thoroughness and accuracy has made their work illuminating. The two memoirs of Eduard Rüppell on the fishes of the Red Sea and the neighboring parts of Africa, "Atlas zu der Reise im Nördlichen Afrika," 1828, and "Neue Wirbelthiere," 1837, rank with the very best of descriptive literature. Günther's illustrated "Fische der Südsee," published in Hamburg, may be regarded as German work. The excellent colored plates are mostly from the hand of Andrew Garrett. Other papers are those of Dr. Wilhelm Peters on Asiatic fishes, the most important being on the fishes of Mozambique. J. J. Heckel, Rudolph Kner, and Franz Steindachner, successively directors of the Museum at Vienna, have written largely on fishes. The papers of Steindachner cover almost every part of the earth and are absolutely essential to any systematic study of fishes. No naturalist of any land has surpassed Steindachner in industry or accuracy, and his work has the advantage of the best illustrations of fishes made by any artist, the noted Eduard Konopicky. In association with Dr. Döderlein, formerly of Tokyo, Dr. Steindachner has given an excellent account of the fishes of Japan. Other German writers are J. J. Kaup, who has worked in numerous fields, but as a whole with little skill, Dr. S. B. Klunzinger, who has given excellent accounts of the fishes of the Red Sea, and Dr. Franz Hilgendorf, of the University of Berlin, whose papers on the fishes of Japan and other regions have shown a high grade of taxonomic insight. A writer of earlier date is W. L. von Rapp,[Pg 412] who wrote on the "Fische den Bodensees." J. F. Brandt has written of the sturgeons of Russia, and Johann Marcusen, to whom we owe much of our knowledge, of the Mormyri of Africa.
In Italy, Charles Lucien Bonaparte, Prince of Canino, has published an elaborate "Fauna Italica" (1838) and in numerous minor papers has taken a large part in the development of ichthyology. Many of the accepted names of the large groups (as Elasmobranchii, Heterosomata, etc.) were first suggested by Bonaparte. The work of Rafinesque has been already noticed. O. G. Costa published (about 1850) a "Fauna of Naples." In recent times Camillo Ranzani, of Bologna, wrote on the fishes of Brazil and of the Mediterranean. Giovanni Canestrini, Decio Vinciguerra, Enrico Hillyer Giglioli, Luigi Döderlein, and others have contributed largely to our knowledge of Italian fishes, while Carlo F. Emery, F. de Filippi, Luigi Facciolá, and others have studied the larval growth of different species. Camillo Ranzani, G. G. Bianconi, Domenico Nardo, Cristoforo Bellotti, Alberto Perugia, and others have contributed to different fields of ichthyology.
Nicholas Apostolides and, still later, Horace A. Hoffman and the present writer, have written of the fishes of Greece.
In France, the fresh-water fishes are the subject of an important work by Emile Blanchard (1866), and Emile Moreau has given us a convenient account of the fish fauna of France. Léon Vaillant has written on various groups of fishes, his monograph of the American darters (Etheostominæ) being a masterpiece so far as the results of the study of relatively scanty material would permit. The "Mission Scientifique au Mexique," by Valliant and F. Bocourt, is one of the most valuable contributions to our knowledge of the fishes of that region. Dr. H. E. Sauvage, of Boulogne-sur-Mer, has also written largely on the fishes of Asia, Africa, and other regions. Among the most important of these are the "Poissons de Madagascar," and a monograph of the sticklebacks. Alexander Thominot and Jacques Pellegrin have also written, in the Museum of the Jardin des Plantes, on different groups of fishes. Earlier writers were Constant Duméril, Alphonse Guichenot, L. Brissot de Barneville, H. Hollard, an able anatomist, and Bibron, an associate of Auguste Duméril.

Felipe Poey y Aloy.
Léon Vaillant.
Louis Dollo.
Decio Vinciguerra.
In Spain and Portugal the chief work of local authors is that of J. V. B. Bocage and F. de Brito Capello on the fishes of Portugal. So far as the fishes of Spain are concerned, the most valuable memoir is Steindachner's account of his travels in Spain and Portugal. The principal studies of the Balkan region have also been made by Steindachner. José Gogorza y González, of the Museum of Madrid, has given a list of the fishes of the Philippines. A still more elaborate list, praiseworthy as a beginning, is the work of the Reverend Padre Casto de Elera, professor of Natural History in the Dominican College of Santo Tomas in Manila.
In Holland, the chief great works have been those of Schlegel and Pieter van Bleeker. Professor H. Schlegel, of the University of Leyden, described the fishes collected about Nagasaki by Ph. Fr. de Siebold and Bürger. His work on fishes forms a large folio illustrated by colored plates, a volume of the "Fauna Japonica," published in Leyden from 1843 to 1847. Schlegel's work in every field is characterized by scrupulous care and healthful conservatism, and the "Fauna Japonica" is a most useful monument to his rare powers of discrimination.
Pieter von Bleeker (1819-78), a surgeon in the Dutch East Indies, is the most voluminous writer in ichthyology. He began his work in Java without previous training and in a very rich field where almost everything was new. With many mistakes at first he rose to the front by sheer force of industry and patience, and his later work, while showing much of the "personal equation," is still thoroughly admirable. At his death he was engaged in the publication of a magnificent folio work, "Atlas Ichthyologique des Indes Orientales Neerlandaises," illustrated by colored plates. This work remains about two-thirds completed. The writings of Dr. Bleeker constitute the chief source of our knowledge of the fauna of the East Indies.
Dr. Van Lidth de Jeude, of the University of Leyden, is the author of a few descriptive papers on fishes.
To Belgium we may assign part at least of the work of the eminent Belgian naturalist, George Albert Boulenger, now long connected with the British Museum. His various valuable papers on the fishes of the Congo are published under the[Pg 415] auspices of the "Congo Free State." To Belgium also we may ascribe the work of Louis Dollo on the morphology of fishes and on the deep-sea fishes obtained by the "Expedition Antarctique Belge."
The fish fauna of Cuba has been the lifelong study of Dr. Felipe Poey y Aloy (1799-1891), a pupil of Cuvier, for a half century or more the honored professor of zoology in the University of Havana. Of his many useful papers, the most extensive are his "Memorias sobre la Historia Natural de la Isla de Cuba," followed by a "Repertorio" and an "Enumeratio" in which the fishes are elaborately catalogued. Poey devoted himself solely to the rich fish fauna of his native island, in which region he was justly recognized as a ripe scholar and a broad-minded gentleman. A favorite expression of his was "Comme naturaliste, je ne suis pas espagnol: je suis cosmopolite." Before Poey, Guichenot, of Paris, had written on the fishes collected in Cuba by Ramon de la Sagra (1810-60). His account was published in Sagra's "Historia de Cuba," and later Philip H. Gosse (1810-1888) wrote on the fishes of Jamaica. Much earlier, Robert Hermann Schomburgk (1804-65) wrote on the fishes of British Guiana. Other papers on the Caribbean fishes were contributed by Johannes Müller and F. H. Troschel, and by Richard Hill and J. Hancock.
Besides the work in South America of Marcgraf, Agassiz, Reinhardt, Lütken, Steindachner, Jenyns, Boulenger, and others already named, we may note the local studies of Dr. Carlos Berg in Argentina, Dr. R. A. Philippi, and Frederico T. Delfin in Chile, Miranda-Ribeiro in Brazil, with Garman, J. F. Abbott, and others in recent times. Carl H. Eigenmann and earlier Jordan and Eigenmann have studied the great collections made in Brazil by Agassiz. Steindachner has described the collections of Johann Natterer and Gilbert those made by Dr. John Casper Branner. The most recent examinations of the myriads of Brazilian river fishes have been made by Dr. Eigenmann. Earlier than any of these (1855), Francis de Castelnau (1800-65) described many Brazilian fishes and afterwards numerous fishes of Australia and southern Africa, Alphonse Guichenot, of Paris, contributed a chapter on fishes to Claude Gay's (1800-63) "History of Chile," and J. J. von[Pg 416] Tschudi, of St. Gallen, published an elaborate but uncritical "Fauna Peruana" with colored plates of Peruvian fishes.
In New Zealand, F. W. Hutton and J. Hector have published a valuable work on the fishes of New Zealand, to which Dr. Gill added useful critical notes in a study of "Antipodal Faunas." Later writers have given us a good knowledge of the fishes of Australia. Notable among them are Charles DeVis, William Macleay, H. de Miklouho-Maclay, James Douglas Ogilby, and Edgar R. Waite. Clarke has also written on "Fishes of New Zealand."
The most valuable work on the fishes of Hindustan is the elaborate treatise on the "Fishes of India" by Surgeon Francis Day. In this all the species are figured, the groups being arranged as in Günther's catalogue, a sequence which few non-British naturalists seem inclined to follow. Cantor's "Malayan Fishes" is a memoir of high merit, as is also McClelland's work on Indian fishes and the still earlier work of Francis Buchanan Hamilton on the fishes of the Ganges. We may here refer to Andrew Smith's papers on the fishes of the Cape of Good Hope and to R. I. Playfair and A. Günther's "Fishes of Zanzibar." T. C. Jerdon, John Edward Gray, E. Tyrwhitt Bennett, and others have also written on the fishes of India; J. C. Bennett has published several excellent papers on the fishes of Polynesia and the East Indies.
In Japan, following the scattering papers of Thunberg, Tilesius, and Houttuyn, and the monumental work of Schlegel, numerous species have been recorded by James Carson Brevoort, Günther, Gill, Eduard Nyström, Hilgendorf, and others. About 1884 Steindachner and Döderlein published the valuable "Fische Japans," based on the collections made about Tokyo by Dr. Döderlein. In 1881, Motokichi Namiye, then assistant curator in the Imperial University, published the first list of Japanese fishes by a native author. In 1900, Dr. Chiyomatsu Ishikawa, on the "Fishes of Lake Biwa," was the first Japanese author to venture to name a new species of fish (Pseudogobio zezera). This reticence was due not wholly to lack of self-confidence, but rather to the scattered condition of the literature of Japanese ichthyology. For this reason no Japanese author has ever felt that any given undetermined[Pg 417] species was really new. Other Japanese ichthyologists of promise are Dr. Kamakichi Kishinouye, in charge of the Imperial fisheries Bureau, Dr. Shinnosuke Matsubara, director of the Imperial Fisheries Institute, Keinosuke Otaki, S. Hatta, S. Nozawa, T. Kitahara, and Michitaro Sindo, and we may look for others among the pupils of Dr. Kakichi Mitsukuri, the distinguished professor of zoology in the Imperial University.

Bashford Dean.
Kakichi Mitsukuri.
Carl H. Eigenmann.
Franz Hilgendorf.
The most recent, as well as the most extensive, studies of the fishes of Japan were made in 1999 by the present writer and his associate, John Otterbein Snyder.
The scanty pre-Cuvieran work on the fishes of North America has been already noticed. Contemporary with the early work of Cuvier is the worthy attempt of Professor Samuel Latham Mitchill (1764-1831) to record in systematic fashion the fishes of New York. Soon after followed the admirable work of Charles Alexandre Le Sueur (1789-1840), artist and naturalist, who was the first to study the fishes of the Great Lakes and the basin of the Ohio. Le Sueur's engravings of fishes, in the early publications of the Academy of Natural Sciences in Philadelphia, are still among the most satisfactory representations of the species to which they refer. Constantine Samuel Rafinesque (1784-1842), the third of this remarkable but very dissimilar trio, published numerous papers descriptive of the species he had seen or heard of in his various botanical rambles. This culminated in his elaborate but untrustworthy "Ichthyologia Ohiensis." The fishes of Ohio received later a far more conscientious though less brilliant treatment at the hands of Dr. Jared Potter Kirtland (1793-1877), an eminent physician of Cleveland, Ohio. In 1842 the amiable and scholarly James Ellsworth Dekay (1799-1851) published his detailed report on the fishes of the "New York Fauna," and a little earlier (1836) in the "Fauna Boreali-Americana" Sir John Richardson (1787-1865) gave a most valuable and accurate account of the fishes of the Great Lakes and Canada. Almost simultaneously, Rev. Zadock Thompson (1796-1856) gave a catalogue of the fishes of Vermont, and David Humphreys Storer (1804-91) began his work on the fishes of Massachusetts, finally expanded into a "Synopsis of the Fishes of North America" (1846) and a "History of the Fishes of Massachu[Pg 419]setts" (1853-67). Dr. John Edwards Holbrook (1794-1871), of Charleston, published (1855-60) his invaluable record of the fishes of South Carolina, the promise of still more important work, which was prevented by the outbreak of the Civil War in the United States. The monograph on Lake Superior (1850) and other publications of Louis Agassiz (1807-73) have been already noticed. One of the first of Agassiz's students was Charles Girard (1822-95), who came with him from Switzerland, and, in association with Spencer Fullerton Baird (1823-87), described the fishes from the United States Pacific Railway Surveys (1858) and the United States and Mexican Boundary Surveys (1859). Professor Baird, primarily an ornithologist, became occupied with executive matters, leaving Girard to finish these studies of the fishes. A large part of the work on fishes published by the United States National Museum and the United States Fish Commission has been made possible through the direct help and inspiration of Professor Baird. Among those engaged in this work, James William Milner (1841-80), Marshall Macdonald (1836-95), and Hugh M. Smith may be noted.
Most eminent, however, among the students and assistants of Professor Baird was his successor, George Brown Goode (1851-96), one of the most accomplished of American naturalists, whose greatest work, "Oceanic Ichthyology," published in collaboration with his long associate, Dr. Tarleton Hoffman Bean, was barely finished at the time of his death. The work of Theodore Nicholas Gill and Edward Drinker Cope has been already noticed.
Other faunal writers of more or less prominence were William Dandridge Peck (1763-1822) in New Hampshire, George Suckley (1830-69) in Oregon, James William Milner (1841-80) in the Great Lake Region, Samuel Stehman Haldeman (1812-80) in Pennsylvania, William O. Ayres (1817-91) in Connecticut and California; Dr. John G. Cooper (died 1902), Dr. William P. Gibbons and Dr. William N. Lockington (died 1902) in California; Philo Romayne Hoy (1816-93) studied the fishes of Wisconsin, Charles Conrad Abbott those of New Jersey, Silas Stearns (1859-88) those of Florida, Stephen Alfred Forbes and Edward W. Nelson those of Illinois, Oliver Perry Hay,[Pg 420] later known for his work on fossil forms, those of Mississippi, Alfredo Dugés, of Guanajuato, those of Central Mexico.
Samuel Garman, at Harvard University, a student of Agassiz, is the author of numerous valuable papers, the most notable being on the sharks and on the deep-sea collections of the Albatross in the Galapagos region, the last illustrated by plates of most notable excellence. Other important monographs of Garman treat of the Cyprinodonts and the Discoboli.
The present writer began a "Systematic Catalogue of the Fishes of North America" in 1875 in association with his gifted friend, Herbert Edson Copeland (1849-76), whose sudden death, after a few promising beginnings, cut short the undertaking. Later, Charles Henry Gilbert (1860-), a student of Professor Copeland, took up the work and in 1883 a "Synopsis of the Fishes of North America" was completed by Jordan and Gilbert. Later, Dr. Gilbert has been engaged in studies of the fishes of Panama, Alaska, and other regions, and the second and enlarged edition of the "Synopsis" was completed in 1898, as the "Fishes of North and Middle America," in collaboration with another of the writer's students, Dr. Barton Warren Evermann. A monographic review of the Fishes of Puerto Rico was later (1900) completed by Dr. Evermann, together with numerous minor works. Other naturalists whom the writer may be proud to claim as students are Charles Leslie McKay (1854-83), drowned in Bristol Bay, Alaska, while engaged in explorations, and Charles Henry Bollman (1868-89), stricken with fever in the Okefinokee Swamps in Georgia. Still others are Dr. Carl B. Eigenmann, the indefatigable investigator of Brazilian fishes and of the blind fishes of the caves; Dr. Oliver Peebles Jenkins, the first thorough explorer of the fishes of Hawaii; Dr. Alembert Winthrop Brayton, explorer of the streams of the Great Smoky Mountains; Dr. Seth Eugene Meek, explorer of Mexico; John Otterbein Snyder, explorer of Mexico, Japan, and Hawaii; Edwin Chapin Starks, explorer of Puget Sound and Panama and investigator of fish osteology. Still other naturalists of the coming generation, students of the present writer and of his lifelong associate, Professor Gilbert, have contributed in various degrees to the present fabric of American ichthyology. Among them are[Pg 421] Mrs. Rosa Smith Eigenmann, Dr. Joseph Swain, Wilbur Wilson Thoburn (1859-99), Frank Cramer, Alvin Seale, Albert Jefferson Woolman, Philip H. Kirsch (1860-1902), Cloudsley Rutter (died 1903), Robert Edward Snodgrass, James Francis Abbott, Arthur White Greeley, Edmund Heller, Henry Weed Fowler, Keinosuke Otaki, Michitaro Sindo, and Richard Crittenden McGregor.

David Starr Jordan.
Herbert Edson Copeland.
Charles Henry Gilbert.
Barton Warren Evermann.
Other facts and conclusions of importance have been contributed by various persons with whom ichthyology has been an incident rather than a matter of central importance.
The Fossil Fishes.[148]—The study of fossil fishes was begun systematically during the first decades of the nineteenth century, for it was then realized that of fossils of back-boned animals, fishes were the only ones which could be determined from early Palæozoic to recent horizons, and that from the diversity of their forms they could serve as reliable indications of the age of rocks. At a later time, when the evolution of vertebrates began to be studied, fishes were examined with especial care with a view of determining the ancestral line of the Amphibians. The earliest work upon fossil fishes is, as one would naturally expect, of a purely systematic value. Anatomical observations were scanty and crude, but as the material for study increased, a more satisfactory knowledge was gained of the structures of the various major groups of fishes; and finally by a comparison of anatomical results important light came to be thrown upon more fundamental problems.
The study of fossil fishes can be divided for convenience into three periods: (I) That which terminated in the magnum opus of Louis Agassiz; (II) that of the systematists whose major works appeared between 1845 and the recent publication of the Catalogue of Fossil Fishes of the British Museum (from this period date many important anatomical observations); and (III) that of morphological work, roughly from 1870 to the present. During this period detailed consideration has been given to the phylogeny of special structures, to the probable lines of descent of the groups of fossil fishes, and to the relationships of terrestrial to aquatic vertebrates.
First Period.—The Work of Louis Agassiz.—The real beginning of our knowledge of fossil fishes dates from the publication of the classic volumes of Agassiz, "Recherches sur les Poissons Fossiles (Neuchâtel, 1833-44)." There had previously existed but a fragmentary and widely scattered literature; the time was ripe for a great work which should bring together a knowledge of this important vertebrate fauna and the museums throughout Europe had been steadily growing in their collections of fossils. Especially ripe, too, since the work of Cuvier (1769-1832) had been completed and the classic anatomical papers of J. Müller (1802-56) were appearing. And Agassiz (1807-73) was eminently the man for this mission. At the age of one and twenty he had already mapped out the work, and from this time he devoted sixteen active years to its accomplishment. One gets but a just idea of the personality of Agassiz when he recalls that the young investigator while in an almost penniless position contrived to travel over a large part of Europe, mingle with the best people of his day, devote almost his entire time to research, employ draughtsmen and lithographers, support his own printing-house, and in the end publish his "Poissons Fossiles" in a fashion which would have done credit to the wealthiest amateur. With tireless energy he collected voluminous notes and drawings numberless; he corresponded with collectors all over Europe and prevailed upon them to loan him tons of specimens; in the meanwhile he collated industriously the early but fragmental literature in such works as those of de Blainville, Münster, Murchison, Buckland, Egerton, Redfield, W. C. Williamson, and others. Hitherto less than 300 species of fossil fishes were known; at the end of Agassiz's work about 900 were described and many of them figured.
It is easy to see that such a work made a ready basis of future studies. Doubtless, too, much is owing to the personal energy of Agassiz that such keen interest was focused in the collection and study of fossil fishes during the middle of the nineteenth century. The actual value of Agassiz's work can hardly be overestimated; his figures and descriptions are usually clear and accurate. And it is remarkable, perhaps, that in view of the very wide field which he covered that his errors[Pg 424] are not more glaring and numerous. Upon the purely scientific side, however, one must confess that the "Poissons Fossiles" is of minor importance for the reason that as time has gone by it has been found to yield no generalizations of fundamental value. The classification of fishes advocated by Agassiz, based upon the nature of the scales, has been shown to be convenient rather than morphological. This indeed Agassiz himself appears to realize in a letter written to Humboldt, but on the other hand he regards his creation of the now discarded order of Ganoids, which was based upon integumental characters, as his most important contribution to the general study of ichthyology. And although there passed through his hands a series of forms more complete than has perhaps been seen by any later ichthyologist,[149] a series which demonstrates the steps in the evolution of the various families and even orders of fishes, he is nowhere led to such important philosophical conclusions as was, for example, his contemporary, Johannes Müller. And even to his last day, in spite of the light which palæontology must have given him, he denied strenuously the truth of the doctrine of evolution, a result the more remarkable since he has even given in graphic form the geological occurrence of the various groups of fishes in a way which suggests closely a modern phylogenetic table, and since at various times he has emphasized the dictum that the history of the individual is but the epitomized history of the race. The latter statement, which has been commonly attributed to Agassiz, is clearly of much earlier origin; it was definitely formulated by von Baer and Meckel, the former of whom even as early as 1834 pronounced himself a distinct evolutionist.

Ramsay Heatley Traquair.
Arthur Smith Woodward.
Karl A. Zittel.
Charles R. Eastman.
Second Period.—Systematic Study of Fossil Fishes.—On the ground planted by Agassiz, many important works sprang up within the next decades. In England a vigorous school of palæichthyologists was soon flourishing. Many papers of Egerton date from this time, and the important work of Owen on the structure of fossil teeth and the often-quoted papers of Huxley in the "British Fossil Remains." Among other workers may be mentioned James Powrie, author of a number of papers upon Scottish Devonian fossils; the enthusiastic Hugh Miller, stone-mason and geologist; Montague Brown, Thomas Atthey, J. Young, and W. J. Barkas, students upon Coal Measure fishes; E. Ray Lankester, some of whose early papers deal with pteraspids; E. T. Newton, author of important works on chimæroids. The extensive works of J. W. Davis deal with fishes of many groups and many horizons. Mr. Davis, like Sir Philip Gray Egerton, was an amateur whose devotion did much to advance the study of fossil fishes. The dean of British palæichthyology is at present Dr. R. H. Traquair, of the Edinburgh Museum of Science and Arts. During four decades he has devoted himself to his studies with rare energy and success, author of a host of shorter papers and numerous memoirs and reports. Finally, and belonging to a younger generation of palæontologists, is to be named Arthur Smith Woodward, curator of vertebrate palæontology of the British Museum. Dr. Woodward has already contributed many scores of papers to palæichthyology, besides publishing a four-volume Catalogue of the Fossil Fishes of the British Museum, a compendial work whose value can only be appreciated adequately by specialists.
In the United States the study of fossil fishes was taken up by J. H. and W. C. Redfield, father and son, prior to the work of Agassiz, and there has been since that time an active school of American workers. Agassiz himself, however, is not to be included in this list, since his interest in extinct fishes became almost entirely unproductive during his life in America. Foremost among these workers was John Strong Newberry (1822-92), of Columbia College, whose publications deal with fishes of many horizons and whose work upon this continent is not unlike that of Agassiz in Europe. He was the author of many state reports, separate contributions, and two monographs, one upon the palæozoic fishes of North America, the other upon the Triassic fishes. Among the earlier palæontologists were Orestes H. St. John, a pupil of Agassiz at Harvard, and A. H. Worthen (1813-88), director of the Geological Survey of Illinois; also W. Gibbes and Joseph Leidy. The late E. D. Cope (1840-97) devoted a considerable portion of his labors to the study of extinct fishes. E. W. Claypole, of Buchtel College, is next to be mentioned as having produced note[Pg 427]worthy contributions to our knowledge of sharks, palæaspids, and arthrodires, as has also A. A. Wright, of Oberlin College. Among other workers may be mentioned O. P. Hay, of the American Museum; C. R. Eastman, of Harvard, author of important memoirs upon arthrodires and other forms; Alban Stewart, a student of Dr. S. W. Williston at Kansas University, and Bashford Dean. Among Canadian palæontologists G. F. Matthew deserves mention for his work on Cyathaspis, Principal Dawson for interesting references to Mesozoic fishes, and J. F. Whiteaves for his studies upon the Devonian fishes of Scaumenac Bay.
Belgian palæontologists have also been active in their study of fishes. Here we may refer to the work of Louis Dollo, of Brussels, of Max Lohest, of P. J. van Beneden, of L. G. de Koninck, of T. C. Winckler, and of R. Storms, the last of whom has done interesting work on Tertiary fishes.
Foremost among Russian palæichthyologists is to be named C. H. Pander, long-time Academician in St. Petersburg, whose elaborate studies of extinct lung-fishes, ostracophores, and crossopterygians published between 1856 and 1860 will long stand as models of careful work. We should also refer to the work of H. Asmuss and H. Trautschold, E. Eichwald and of Victor Rohon, the last named having published many important papers upon ostracophores during his residence in St. Petersburg.
German palæichthyologists include Otto Jaekel, of Berlin; O. M. Reis of the Oberbergamt, in Munich; A. von Koenen, of Göttingen; A. Wagner, E. Koken, and K. von Zittel. Among Austro-Hungarians are Anton Fritsch, author of the Fauna der Gaskohleformations Boemens; Rudolf Kner, an active student of living fishes as well, as is also Franz Steindachner.
French palæichthyologists are represented by the veteran H. E. Sauvage, of Boulogne-sur-Mer, V. Thollière, M. Brongniart, and F. Priem. In Italy Francesco Bassani, of Naples, is the author of many important works dealing with Mesozoic and Tertiary forms; also was Baron Achille di Zigno. Robert Collett, of Bergen, and G. Lindström are worthy representatives of Scandinavia in kindred work.
Third Period.—Morphological Work on Fossil Fishes.—Among the writers who have dealt with the problems of the rela[Pg 428]tionships of the Ostracophores as well as Palæospondylus and the Arthrodires may be named Traquair, Huxley, Newberry, Smith Woodward, Rohon, Eastman, and Dean; most recently William Patten. Upon the phylogeny of the sharks Traquair, A. Fritsch, Hasse, Cope, Brongniart, Jaekel, Reis, Eastman, and Dean. On Chimæroid morphology mention may be made of the papers of A. S. Woodward, Reis, Jaekel, Eastman, C. D. Walcott, and Dean. As to Dipnoan relationships the paper of Louis Dollo is easily of the first value; of especial interest, too, is the work of Eastman as to the early derivation of the Dipnoan dentition. In this regard a paper of Rohon is noteworthy, as is also that of Richard Semon on the development of the dentition of recent Neoceratodus, since it contains a number of references to extinct types. Interest notes on Dipnoan fin characters have been given by Traquair. In the morphology of Ganoids, the work of Traquair and A. S. Woodward takes easily the foremost rank. Other important works are those of Huxley, Cope, A. Fritsch, and Oliver P. Hay.
Anatomists.—Still more difficult of enumeration is the long list of those who have studied the anatomy of fishes usually in connection with the comparative anatomy or development of other animals. Pre-eminent among these are Karl Ernst von Baer, Cuvier, Geoffroy St. Hilaire, Louis Agassiz, Johannes Müller, Carl Vogt, Carl Gegenbaur, William Kitchen Parker, Francis M. Balfour, Thomas Henry Huxley, Meckel, H. Rathke, Richard Owen, Kowalevsky, H. Stannius, Joseph Hyrtl, Gill, Boulenger, and Bashford Dean. Other names of high authority are those of Wilhelm His, Kölliker, Bakker, Rosenthal, Gottsche, Miklucho-Macleay, Weber, Hasse, Retzius, Owsjannikow, H. Müller, Stieda, Marcusen, J. A. Ryder, E. A. Andrews, T. H. Morgan, G. B. Grassi, R. Semon, Howard Ayers, R. R. Wright, J. P. McMurrich, C. O. Whitman, A. C. Eyclesheimer, E. Pallis, Jacob Reighard, and J. B. Johnston.
Besides all this, there has risen, especially in the United States, Great Britain, Norway, and Canada and Australia, a vast literature of commercial fisheries, fish culture, and angling, the chief workers in which fields we may not here enumerate even by name.
[148] For these paragraphs on the history of the study of fossil fishes the writer is indebted to the kind interest of Professor Bashford Dean.
[149] Dr. Arthur Smith Woodward excepted.

How to Secure Fishes.—In collecting fishes three things are vitally necessary—a keen eye, some skill in adapting means to ends, and some willingness to take pains in the preservation of material.
In coming into a new district the collector should try to preserve the first specimen of every species he sees. It may not come up again. He should watch carefully for specimens which look just a little different from their fellows, especially for those which are duller, less striking, or with lower fins. Many species have remained unnoticed through generations of collectors who have chosen the handsomest or most ornate specimens. In some groups with striking peculiarities, as the trunkfishes, practically all the species were known to Linnæus. No collector could pass them by. On the other hand, new gobies or blennies can be picked up almost every day in the lesser known parts of the world. For these overlooked forms—herrings, anchovies, sculpins, blennies, gobies, scorpion-fishes—the competent collector should be always on the watch. If any specimen looks different from the rest, take it at once and find out the reason why.
In most regions the chief dependence of the collector is on the markets and these should be watched most critically. By paying a little more for unusual, neglected, or useless fish, the supply of these will rise to the demand. The word passed along among the people of Onomichi in Japan, that "Ebisu the fish-god was in the village" and would pay more for okose (poison scorpion-fishes) and umiuma (sea-horses) than real fishes were worth soon brought (in 1900) all sorts of okose and umiuma into the market when they were formerly left neglected on the beach. Thus with a little ingenuity the markets in any country can be greatly extended.
The collector can, if he thinks best, use all kinds of fishing tackle for himself. In Japan he can use the "dabonawa" long lines, and secure the fishes which were otherwise dredged by the Challenger and Albatross. If dredges or trawls are at his hand he can hire them and use them for scientific purposes. He should neglect no kind of bottom, no conditions of fish life which he can reach.
Especially important is the fauna of the tide-pools, neglected by almost all collectors. As the tide goes down, especially on rocky capes which project into the sea, myriads of little fishes will remain in the rock-pools, the algæ, and the clefts of rock. In regions like California, where the rocks are buried with kelp, blennies will lie in the kelp as quiescent as the branches of the algæ themselves until the flow of water returns.
A sharp three-tined fork will help in spearing them. The water in pools can be poisoned on the coast of Mexico with the milky juice of the "hava" tree, a tree which yields strychnine. In default of this, pools can be poisoned by chloride of lime, sulphate of copper, or, if small enough, by formaline. Of all poisons the commercial chloride of lime seems to be most effective. By such means the contents of the pool can be secured and the next tide carries away the poison. The water in pools can be bailed out, or, better, emptied by a siphon made of small garden-hose or rubber tubing. On rocky shores, dynamite can be used to advantage if the collector or his assistant dare risk it and if the laws of the country do not prevent.
Most effective in rock-pool work is the help of the small boy. In all lands the collector will do well to take him into his pay and confidence. Of the hundred or more new species of rock-pool fishes lately secured by the writer in Japan, fully two-thirds were obtained by the Japanese boys. Equally effective is the "muchacho" on the coasts of Mexico.
Masses of coral, sponges, tunicates, and other porous or hollow organisms often contain small fishes and should be carefully examined. On the coral reefs the breaking up of large masses is often most remunerative.
The importance of securing the young of pelagic fishes by tow-nets and otherwise cannot be too strongly emphasized.
How to Preserve Fishes.—Fishes must be permanently preserved in alcohol. Dried skins are far from satisfactory, except as a choice of difficulties in the case of large species.
Dr. Günther thus describes the process of skinning fishes:
"Scaly fishes are skinned thus: With a strong pair of scissors an incision is made along the median line of the abdomen from the foremost part of the throat, passing on one side of the base of the ventral and anal fins to the root of the caudal fin, the cut, being continued upward to the back of the tail close to the base of the caudal. The skin of one side of the fish is then severed with the scalpel from the underlying muscles to the median line of the back; the bones which support the dorsal and caudal are cut through, so that these fins remain attached to the skin. The removal of the skin of the opposite side is easy. More difficult is the preparation of the head and scapulary region. The two halves of the scapular arch which have been severed from each other by the first incision are pressed toward the right and left, and the spine is severed behind the head, so that now only the head and shoulder bones remain attached to the skin. These parts have to be cleaned from the inside, all soft parts, the branchial and hyoid apparatus, and all smaller bones being cut away with the scissors or scraped off with the scalpel. In many fishes which are provided with a characteristic dental apparatus in the pharynx (Labroids, Cyprinoids), the pharyngeal bones ought to be preserved and tied with a thread to their specimen. The skin being now prepared so far, its entire inner surface as well as the inner side of the head are rubbed with arsenical soap; cotton-wool or some other soft material is inserted into any cavities or hollows, and finally a thin layer of the same material is placed between the two flaps of the skin. The specimen is then dried under a slight weight to keep it from shrinking.
"The scales of some fishes, as for instance of many kinds of herrings, are so delicate and deciduous that the mere handling causes them to rub off easily. Such fishes may be covered with thin-paper (tissue paper is the best) which is allowed to dry on them before skinning. There is no need for removing the paper before the specimen has reached its destination.
"Scaleless fishes, as siluroids and sturgeons, are skinned in[Pg 432] the same manner, but the skin can be rolled up over the head; such skins can also be preserved in spirits, in which case the traveler may save to himself the trouble of cleaning the head.
"Some sharks are known to attain to a length of thirty feet, and some rays to a width of twenty feet. The preservation of such gigantic specimens is much to be recommended, and although the difficulties of preserving fishes increase with their size, the operation is facilitated, because the skins of all sharks and rays can easily be preserved in salt and strong brine. Sharks are skinned much in the same way as ordinary fishes. In rays an incision is made not only from the snout to the end of the fleshy part of the tail, but also a second across the widest part of the body. When the skin is removed from the fish, it is placed into a cask with strong brine mixed with alum, the head occupying the upper part of the cask; this is necessary, because this part is most likely to show signs of decomposition, and therefore most requires supervision. When the preserving fluid has become decidedly weaker from the extracted blood and water, it is thrown away and replaced by fresh brine. After a week's or fortnight's soaking the skin is taken out of the cask to allow the fluid to drain off; its inner side is covered with a thin layer of salt, and after being rolled up (the head being inside) it is packed in a cask the bottom of which is covered with salt; all the interstices and the top are likewise filled with salt. The cask must be perfectly water-tight."
Value of Formalin.—In the field it is much better to use formalin (formaldehyde) in preference to alcohol. This is an antiseptic fluid dissolved in water, and it at once arrests decay, leaving the specimen as though preserved in water. If left too long in formalin fishes swell, the bones are softened, and the specimens become brittle or even worthless. But for ordinary purposes (except use as skeleton) no harm arises from two or three months' saturation in formalin. The commercial formalin can be mixed with about twenty parts of water. On the whole it is better to have the solution too weak rather than too strong. Too much formalin makes the specimens stiff, swollen, and intractable, besides too soon destroying the color.
Formalin has the advantage, in collecting, of cheapness and of ease in transportation, as a single small bottle will make[Pg 433] a large amount of the fluid. The specimens also require much less attention. An incision should be made in the (right) side of the abdomen to let in the fluid. The specimen can then be placed in formalin. When saturated, in the course of the day, it can be wrapped in a cloth, packed in an empty petroleum can, and at once shipped. The wide use of petroleum in all parts of the world is a great boon to the naturalist.
Before preservation, the fishes should be washed, to remove slime and dirt. They should have an incision to let the fluid into the body cavity and an injection with a syringe is a useful help to saturation, especially with large fishes. Even decaying fishes can be saved with formalin.
Records of Fishes.—The collector should mark localities most carefully with tin tags and note-book records if possible. He should, so far as possible, keep records of life colors, and water-color sketches are of great assistance in this matter. In spirits or formalin the life colors soon fade, although the pattern of marking is usually preserved or at least indicated. A mixture of formalin and alcohol is favorable to the preservation of markings.
In the museum all specimens should be removed at once from formalin to alcohol. No substitute for alcohol as a permanent preservative has been found. The spirits derived from wine, grain, or sugar is much preferable to the poisonous methyl or wood alcohol.
In placing specimens directly into alcohol, care should be taken not to crowd them too much. The fish yields water which dilutes the spirit. For the same reason, spirits too dilute are ineffective. On the other hand, delicate fishes put into very strong alcohol are likely to shrivel, a condition which may prevent an accurate study of their fins or other structures. It is usually necessary to change a fish from the first alcohol used as a bath into stronger alcohol in the course of a few days, the time depending on the closeness with which fishes are packed. In the tropics, fishes in alcohol often require attention within a few hours. In formalin there is much less difficulty with tropical fishes.
Fishes intended for skeletons should never be placed in formalin. A softening of the bones which prevents future[Pg 434] exact studies of the bones is sure to take place. Generally alcohol or other spirits (arrack, brandy, cognac, rum, sake "vino") can be tested with a match. If sufficiently concentrated to be ignited, they can be safely used for preservation of fishes. The best test is that of the hydrometer. Spirits for permanent use should show on the hydrometer 40 to 60 above proof. Decaying specimens show it by color and smell and the collector should be alive to their condition. One rotting fish may endanger many others. With alcohol it is necessary to take especial pains to ensure immediate saturation. Deep cuts should be made into the muscles of large fishes as well as into the body cavity. Sometimes a small distilling apparatus is useful to redistil impure or dilute alcohol. The use of formalin avoids this necessity.
Small fishes should not be packed with large ones; small bottles are very desirable for their preservation. All spinous or scaly fishes should be so wrapped in cotton muslin as to prevent all friction.
Eternal Vigilance.—The methods of treating individual groups of fishes and of handling them under different climatic and other conditions are matters to be learned by experience. Eternal vigilance is the price of a good collection, as it is said to be of some other good things. Mechanical collecting—picking up the thing got without effort and putting it in alcohol without further thought—rarely serves any useful end in science. The best collectors are usually the best naturalists. The collections made by the men who are to study them and who are competent to do so are the ones which most help the progress of ichthyology. The student of a group of fishes misses half the collection teaches if he has made no part of it himself.

The Geological Distribution of Fishes.—The oldest unquestioned remains of fishes have been very recently made known by Mr. Charles D. Walcott, from rocks of the Trenton period in the Ordovician or Lower Silurian. These are from Cañon City in Colorado. Among these is certainly a small Ostracophore (Asteraspis desideratus). With it are fragments (Dictyorhabdus) thought to be the back-bone of a Chimæra, but more likely, in Dean's view, the axis of a cephalopod, besides bony, wrinkled scales, referred with doubt to a supposed Crossopterygian genus called Eriptychius. This renders certain the existence of Ostracophores at this early period, but their association with Chimæras and Crossopterygians is questionable. Primitive sharks may have existed in Ordovician times, but thus far no trace of them has been found.
Fig. 246.—Fragment of Sandstone from Ordovician deposits, Cañon City, Colo., showing fragments of scales, etc., the earliest known traces of vertebrates. (From nature.)
The fish-remains next in age in America are from the Bloomfield sandstone in Pennsylvania of the Onondaga period in the[Pg 436] upper Silurian. The earliest in Europe are found in the Ludlow shales, both of these localities being in or near the horizon of the Niagara rocks, in the Upper Silurian Age.
It is, however, certain that these Lower Silurian remains do not represent the beginning of fish-life. Probably Ostracophores, and Arthrodires, with perhaps Crossopterygians and Dipnoans, existed at an earlier period, together perhaps with unarmed, limbless forms without jaws, of which no trace whatever has been left.
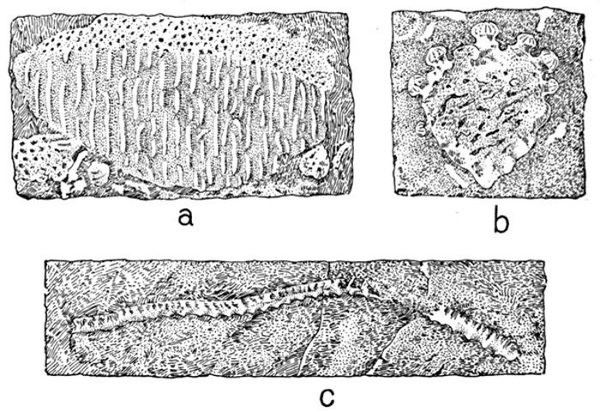
Fig. 247.—Fossil fish remains from Ordovician rocks, Cañon City, Colo. (After Walcott.) a. Scale of Eriptychius americanus Walcott. Family Holoptychiidæ? b. Dermal plate of Asteraspis desideratus Walcott. Family Asterolepidæ. c. Dictyorhabdus priscus Walcott, a fragment of uncertain nature, thought to be a chordal sheath of a Chimæra, but probably part of a Cephalopod (Dean). Chimæridæ?
The Earliest Sharks.—The first actual trace of sharks is found in the Upper Silurian in the form of fin-spines (Onchus), thought to belong to primitive sharks, perhaps Acanthodeans possibly to Ostracophores. With these are numerous bony shields of the mailed Ostracophores, and somewhat later those of the more highly specialized Arthrodires. Later appear the teeth of Cochliodontidæ, with Chimæras, a few Dipnoans, and Crossopterygians.
Devonian Fishes.—In the Devonian Age the Ostracophores increase in size and abundance, disappearing with the beginning[Pg 437] of the Carboniferous. The Arthrodires also increase greatly in variety and in size, reaching their culmination in the Devonian, but not disappearing entirely until well in the Carboniferous. These two groups (often united by geologists under the older name Placoderms) together with sharks and a few Chimæras made up almost exclusively the rich fish-fauna of Devonian times. The sharks were chiefly Acanthodean and Psammodont, as far as our records show. The supposed more primitive type of Cladoselache is not known to appear before the latter part of the Devonian Age, while Pleuracanthus and Cladodus, sometimes regarded as still more primitive, are as yet found only in the Carboniferous. It is clear that the records of early shark life are still incomplete, whatever view we may adopt as to the relative rank of the different forms. Chimæroids occur in the Devonian, and with them a considerable variety of Crossopterygians and Dipnoans. The true fishes appear also in the Devonian in the guise of the Ganoid ancestors and relatives of Palæoniscum, all with diamond-shaped enameled scales. In the Devonian, too, we find the minute creature Palæospondylus, our ignorance of which is concealed under the name Cycliæ.
Carboniferous Fishes.—In the Carboniferous Age the sharks increase in number and variety, the Ostracophores disappear, and the Arthrodires follow them soon after, the last being recorded from the Permian. Other forms of Dipnoans, Crossopterygians, and some Ganoids now appear giving the fauna a somewhat more modern aspect. The Acanthodei and the Ichthyotomi pass away with the Permian, the latest period of the Carboniferous Age.
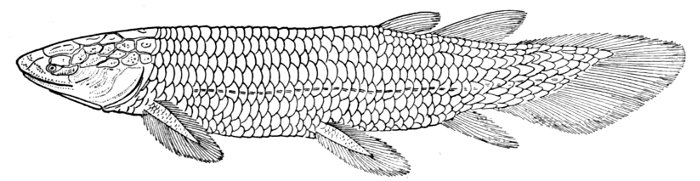
Fig. 248.—Dipterus valenciennesi Agassiz, a Dipnoan. (After Dean, from Woodward.)
Mesozoic Fishes.—In the Triassic period which follows the Permian, the earliest types of Ganoids give place to forms ap[Pg 438]proaching the garpike and sturgeon. The Crossopterygians rapidly decline. The Dipnoans are less varied and fewer in number; the primitive sharks, with the exception of certain Cestracionts, all disappear, only the family of Orodontidæ remaining. Here are found the first true bony fishes, doubtless derived from Ganoid stock, the allies and predecessors of the great group of herrings. Herring-like forms become more numerous in the Jurassic, and with them appear other forms which give the fish-fauna of this period something of a modern appearance. In the Jurassic the sharks become divided into several groups, Notidani, Scyllioid sharks, Lamnoid sharks, angel-fishes, skates, and finally Carcharioid sharks being now well differentiated. Chimæras are still numerous. The Acanthodei have passed away, as well as the mailed Ostrachopores and Arthrodires. The Dipnoans and Crossopterygians are few. The early Ganoids have given place to more modern types, still in great abundance and variety. This condition continues in the Cretaceous period. Here the rays and modern sharks increase in number, the Ganoids hold their own, and the other groups of soft-rayed fishes, as the smelts, the lantern-fishes, the pikes, the flying-fishes, the berycoids and the mackerels join the group of herring-like forms which represent the modern bony fishes. In the Cretaceous appear the first spiny-rayed fishes, derived probably from herring-like forms. These are allies or ancestors of the living genus Beryx.

Fig. 249.—Hoplopteryx lewesiensis (Mantell), restored. English Cretaceous. Family Berycidæ. (After Woodward.)
Dr. Woodward observes:
"As soon as fishes with a completely osseous endoskeleton began to predominate at the dawn of the Cretaceous period, specializations of an entirely new kind were rapidly acquired. Until this time the skull of the Actinopterygii had always been remarkably uniform in type. The otic region of the cranium often remained incompletely ossified and was never prominent or projecting beyond the roof bones; the supraoccipital bone was always small and covered with the superficial plates; the maxilla invariably formed the greater part of the upper jaw; the cheek-plates were large and usually thick; while none of the head or opercular bones were provided with spines or ridges. The pelvic fins always retained their primitive remote situation, and the fin-rays never became spines. During the Cretaceous period the majority of the bony fishes began to exhibit modifications in all these characters, and the changes occurred so rapidly that by the dawn of the Eocene period the diversity observable in the dominant fish-fauna was much greater than it had ever been before. At this remote period, indeed, nearly all the great groups of bony fishes, as represented in the existing world, were already differentiated, and their subsequent modifications have been quite of a minor character."

Fig. 250.—A living Berycoid fish, Paratrachichthys prosthemius Jordan & Fowler. Misaki, Japan. Family Berycidæ.

Fig. 251.—Flying-fish, Cypsilurus heterurus (Rafinesque). Family Exocætidæ Woods Hole, Mass.

Fig. 252.—The Schoolmaster Snapper, a Perch-like fish. Family Lutianidæ. Key West.
Tertiary Fishes.—With the Eocene or first period of the Tertiary great changes have taken place. The early families of bony fishes nearly all disappear. The herring, pike, smelt, salmon, flying-fish, and berycoids remain, and a multitude of other forms seem to spring into sudden existence. Among these are the globefishes, the trigger-fishes, the catfishes, the eels, the morays, the butterfly-fishes, the porgies, the perch, the bass, the pipefishes, the trumpet-fishes, the mackerels, and the John-dories, with the sculpins, the anglers, the flounders, the blennies, and the cods. That all these groups, generalized and specialized, arose at once is impossible, although all seem[Pg 441] to date from the Eocene times. Doubtless each of them had its origin at an earlier period, and the simultaneous appearance is related to the fact of the thorough study of the Eocene shales, which have in numerous localities (London, Monte Bolca, Licata, Mount Lebanon, Green River) been especially favorable for the preservation of these forms. Practically fossil fishes have been thoroughly studied as yet only in a very few parts of the earth. The rocks of Scotland, England, Germany, Italy, Switzerland, Syria, Ohio, and Wyoming have furnished the great bulk of all the fish remains in existence. In some regions perhaps collections will be made which will give us a more just conception of the origin of the different groups of bony fishes. We can now only say with certainty that the modern families were largely existent in the Eocene, that they sprang from ganoid stock found in the Triassic and Jurassic, that several of them were represented in the Cretaceous also, that the Berycoids were earliest of the spiny-rayed fishes, and forms allied to herring the earliest of the soft-rayed forms. Few modern families arose before the Cretaceous. Few of the modern genera go back to the Eocene, many of them arose in the Miocene, and few species have come down to us from rocks older than the end of the Pliocene. The general modern type of the fish-faunas being determined in the latter Eocene and the Miocene, the changes which bring us to recent times have largely concerned the abundance and variety of the individual species. From geological distribution we have arising the varied problems of geographical distribution and the still more complex conditions on which depend the extinction of species and of types.
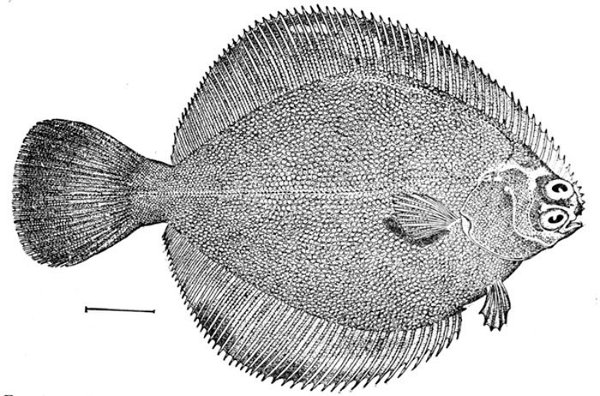
Fig. 253.—Decurrent Flounder, Pleuronichthys decurrens Jordan & Gilbert. San Francisco.
Factors of Extinction.—These factors of extinction have been recently formulated as follows by Professor Herbert Osborn. He considers the process of extinction as of five different types:
"(1) That extinction which comes from modification or progressive evolution, a relegation to the past as a result of the transmutation into more advanced forms. (2) Extinction from changes of physical environment which outrun the powers of adaptation. (3) The extinction which results from competition. (4) The extinction which results from extreme specialization and limitation to special conditions the loss of which means extinction. (5) Extinction as a result of exhaustion."
Fossilization of a Fish.—When a fish dies he leaves no friends. His body is at once attacked by hundreds of creatures ranging from the one-celled protozoa and bacteria to individuals of his own species. His flesh is devoured, his bones are scattered, the gelatinous substance in them decays, and the phosphate of lime is in time dissolved in the water. For this reason few fishes of the millions which die each year leave any trace for future preservation. At the most a few teeth, a fin-spine, or a bone buried in the clay might remain intact or in such condition as to be recognized.
But now and then it happens that a dead fish may fall in more fortunate conditions. On a sea bottom of fine clay the bones, or even the whole body, may be buried in such a way as to be sealed up and protected from total decomposition. The flesh will usually disappear and leave no mark or at the most a mere cast of its surface. But the hard parts, even the muscles may persist, and now and then they do persist, the salts of lime unchanged or else silicified or subjected to some other form of chemical substitution. Only the scales, the teeth, the bones, the spines, and the fin-rays can be preserved in the rocks of sea or[Pg 443] lake bottom. In a few localities, as near Green River in Wyoming, Monte Bolca, near Verona, and Mount Lebanon in Syria, the London clays, with certain quarries in Scotland and lithographic stones in Germany, many skeletons of fishes have been found pressed flat in layers of very fine rock, their structures traced as delicately as if actually drawn on the smooth stone. Fragments preserved in ruder fashion abound in the clays and even the sandstones of the earliest geologic ages. In most cases, however, fossil fishes are known from detached and scattered fragments, many of them, especially of the sharks, by the teeth alone. Fishes have occurred in all ages from the Silurian to the present time and probably the very first lived long before the Silurian.
The Earliest Fishes.—No one can say what the earliest fishes were like, nor do we know what was their real relation to the worm-like forms among which men have sought their presumable ancestors, nor to the Tunicates and other chordate forms, not fish-like, but still degenerate relatives of the primeval fish.
From analogy we may suppose that the first fishes which ever were bore some resemblance to the lancelet, for that is a fish-like creature with every structure reduced to the lowest terms. But as the lancelet has no hard parts, no bones, nor teeth, nor scales, nor fins, no traces of its kind are found among the fossils. If the primitive fish was like it in important respects, all record of this has probably vanished from the earth.
The Cyclostomes.—The next group of living fishes, the Cyclostomes, including the hagfishes and lampreys,—fishes with small skull and brain but without limbs or jaws,—stands at a great distance above the lancelet in complexity of structure, and equally far from the true fishes in its primitive simplicity. In fact the lamprey is farther from the true fish in structure than a perch is from an eagle. Yet for all that it may be an offshoot from the primitive line of fish descent. There is not much in the structure of the lamprey which may be preserved in the rocks. But the cartilaginous skull, the back-bone, fins, and teeth might leave their traces in soft clay or lithographic stone. But it is certain that they have not done so in any rocks yet explored, and it may be that the few existing lampreys owe their form and structure to a process of degradation from a more complex and more fish-like ancestry. The supposed[Pg 444] lamprey fossil of the Devonian of Scotland, Palæospondylus, has little in common with the true lampreys.
The Ostracophores.—Besides the lampreys the Devonian seas swarmed with mysterious creatures covered with an armor of plate, fish-like in some regards, but limbless, without true jaws and very different from the true fishes of to-day. These are called Ostracophori, and some have regarded them as mailed lampreys, but they are more likely to be a degenerate or eccentric offshoot from the sharks, as highly modified or specialized lampreys, a side offshoot which has left no descendants among recent forms. Recently Professor Patten has insisted that the resemblance of their head-plates to those of the horseshoe crab (Limulus) is indicative of real affinity.
Among these forms in mail-armor are some in which the jointed and movable angles of the head suggest the pectoral spines of some catfishes. But in spite of its resemblance to a fin, the spine in Pterichthyodes is an outgrowth of the ossified skin and has no more homology with the spines of fishes than the mailed plates have with the bones of a fish's cranium. In none of these fishes has any trace of an internal skeleton been found. It must have retained its primitive gelatinous character. There are, however, some traces of eyes, and the mucous channels of the lateral line indicate that these creatures possessed some other special senses.
Fig. 254.—An Ostracophore, Cephalaspis lyelli Agassiz, restored. Devonian. (After Agassiz, per Dean.)
Whatever the Ostracophores may be, they should not be included within the much-abused term Ganoidei, a word which was once used in the widest fashion for all sorts of mailed fishes, but little by little restricted to the hard-scaled relatives and ancestors of the garpike of to-day.
The Arthrodires.—Dimly seen in the vast darkness of Paleozoic time are the huge creatures known as Arthrodires. These are mailed and helmeted fishes, limbless so far as we know,[Pg 445] but with sharp, notched, turtle-like jaws quite different from those of the fish or those of any animal alive to-day. These creatures appear in Silurian rocks and are especially abundant in the fossil beds of Ohio, where Newberry, Claypole, Eastman, Dean and others have patiently studied the broken fragments of their armor. Most of them have a great casque on the head with a shield at the neck and a movable joint connecting the two. Among them was almost every variation in size and form.

Fig. 255.—An Arthrodire, Dinichthys intermedius Newberry, restored. Devonian, Ohio. (Family after Dean.)
These creatures have been often called ganoids, but with the true ganoids like the garpike they have seemingly nothing in common. They are also different from the Ostracophores. To regard them with Woodward as derived from ancestral Dipnoans is to give a possible guess as to their origin, and a very unsatisfactory guess at that. In any event these have all passed away in competition with the scaly fishes and sharks of later evolution, and it seems certain that they, like the mailed Ostracophores, have left no descendants.
The Sharks.—Next after the lampreys, but a long way after them in structure, come the sharks. With the sharks appear for the first time true limbs and the lower jaw. The upper jaw is, however, formed from the palate, and the shoulder-girdle is attached behind the skull. "Little is known," says Professor Dean, "of the primitive stem of the sharks, and even the lines of descent of the different members of the group can only be generally suggested. The development of recent forms has yielded few results of undoubted value to the phylogenist. It would appear as if paleontology alone could solve the puzzles of their descent."
Of the very earliest sharks in the Upper Silurian Age the remains are too scanty to prove much save that there were sharks in abundance and variety. Spines, teeth, fragments of shagreen, show that in some regards these forms were highly specialized. In the Carboniferous Age the sharks became highly varied and extensively specialized. Of the Paleozoic types, however, all but a single family seems to have died out, leaving Cestraciontes only in the Permian and Triassic. From these the modern sharks one and all may very likely have descended.
Origin of the Sharks.—Perhaps the sharks are developed from the still more primitive shark imagined as without limbs and with the teeth slowly formed from modification of the ordinary shagreen prickles. In determining the earliest among the several primitive types of shark actually known we are stopped by an undetermined question of theory. What is the origin of paired limbs? Are these formed, like the unpaired fins, from the breaking up of a continuous fold of skin, in accordance with the view of Balfour and others? Or is the primitive limb, as supposed by Gegenbaur, a modification of the bony gill-arch? Or again, as supposed by Kerr, is it a modification of the hard axis of an external gill?
If we adopt the views of Gegenbaur or Kerr, the earliest type of limb is the jointed archipterygium, a series of consecutive rounded cartilaginous elements with a fringe of rays along its length. Sharks possessing this form of limb (Ichthyotomi) appear in the Carboniferous rocks, but are not known earlier. It may be that from these the Dipnoans, on the one hand, may be descended and, on the other, the true sharks and the Chimæras; but there is no certainty that the jointed arm or archipterygium of the Dipnoans is derived from the similar pectoral fin of the Ichthyotomi.
On the other hand, if we regard the paired fins as parts of a lateral fold of skin, we find primitive sharks to bear out our conclusions. In Cladoselache of the Upper Devonian, the pectoral and the ventral fins are long and low, and arranged just as they might be if Balfour's theory were true. Acanthoessus, with a spine in each paired fin and no other rays, might be a specialization of this type or fin, and Climatius, with rows of spines in place of pectorals and ventrals, might be held to[Pg 447] bear out the same idea. In all these the tail is less primitive than in the Ichthyotomi. On the other hand, the vent in Cladoselache is thought by Dean to have been near the end of the tail. If this is the case, it should indicate a very primitive character. On the whole, though there is much to be said in favor of the primitive nature of the Ichthyotomi (Pleuracanthus) with the tapering tail and jointed pectoral fin of a dipnoan, and other traits of a shark, yet, on the whole, Cladoselache is probably nearer the origin of the shark-like forms.
The relatively primitive sharks called Notidani have the weakly ossified vertebræ joined together in pairs and there are six or seven gill-openings. This group has persisted to our day, the frilled shark (Chlamydoselachus) and the genera Hexanchus and Heptranchias still showing its archaic characters.
Here the sharks diverge into two groups, the one with the vertebræ better developed and its calcareous matter arranged star-fashion. This forms Hasse's group of Asterospondyli, the typical sharks. The earliest forms (Orodontidæ, Heterodontidæ) approach the Notidani, and so far as geological records go, precede all the other modern sharks. One such ancient type, Heterodontus, including the bullhead shark, and the Port Jackson shark, still persists. The others diverge to form the three chief groups of the cat-sharks (Scyliorhinus, etc.), the mackerel-sharks (Lamna, etc.), and the true sharks (Carcharhias, etc.).

Fig. 256.—Mackerel-shark or Salmon-shark, Lamna cornubica (Gmelin). Santa Barbara, Cal.
In the second group the vertebræ have their calcareous matter arranged in rings, one or more about the notochordal center. In all these the anal fin is absent, and in the process of speciali[Pg 448]zation the shark gradually gives place to the flattened body and broad fins of the ray. This group is called Tectospondyli. Those sharks of this group with one ring of calcareous matter in each vertebra constitute the most primitive extreme of a group representing continuous evolution.
From Cladoselache and Chlamydoselachus through the sharks to the rays we have an almost continuous series which reaches its highest development in the devil rays or mantas of the tropical seas, Manta and Mobula being the most specialized genera and among the very largest of the fishes. However different the rays and skates may appear in form and habit, they are structurally similar to the sharks and have sprung from the main shark stem.
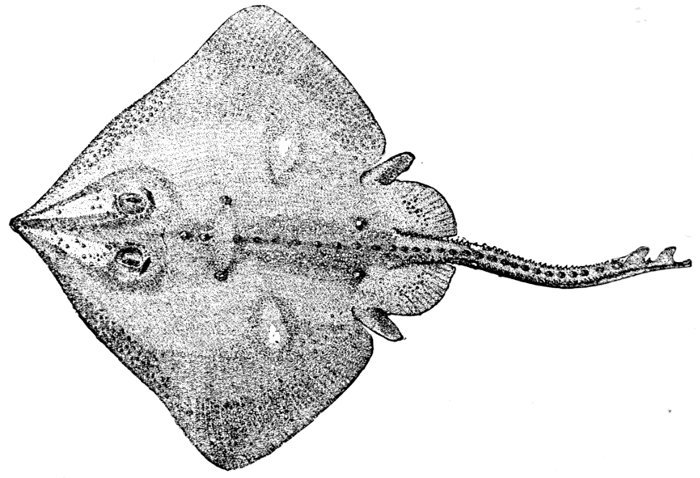
Fig. 257.—Star-spined Ray, Raja stellulata Jordan & Gilbert. Monterey, Cal.
The Chimæras.—The most ancient offshoot from the shark stem, perhaps dating from Silurian times and possibly separated at a period earlier than the date of any known shark, is the group of Holocephali or Chimæras, shark-like in essentials, but differing widely in details. Of these there are but few living forms and the fossil types are known only from dental plates and fin-spines. The living forms are found in the deeper seas the world over, one of the simplest in structure being the newly dis[Pg 449]covered Rhinochimæra of Japan. The fusion of the teeth into overlapping plates, the covering of the gills by a dermal flap, the complete union of the palato-quadrate apparatus or upper jaw with the skull and the development of a peculiar clasping spine on the forehead of the male are characteristic of the Chimæras. The group is one of the most ancient, but it ends with itself, none of the modern fishes being derived from Chimæras.
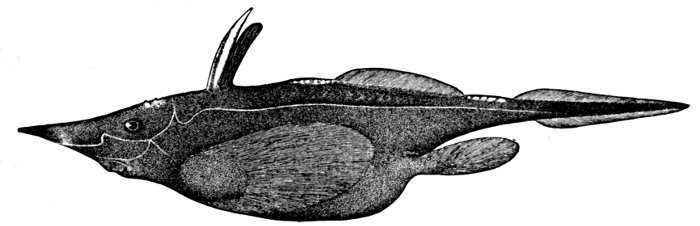
Fig. 258.—A Deep-sea Chimæra, Harriotta raleighiana Goode & Bean. Gulf Stream.

Fig. 259.—An extinct Dipnoan, Dipterus valenciennesi Agassiz. Devonian. (After Pander.)
The Dipnoans.—The most important offshoot of the primitive sharks is not the Chimæras, nor even the shark series itself, but the groups of Crossopterygians and Dipnoans, or lung-fishes, with the long chain of their descendants. With the Dipnoan appears the lung or air-bladder, at first an outgrowth from the ventral side of the œsophagus, as it still is in all higher animals, but later turning over, among fishes, and springing from the dorsal side. At first an arrangement for breathing air, a sort of accessory gill, it becomes the sole organs of respiration in the higher forms, while in the bony fishes its respiratory function is lost altogether. The air-bladder is a degenerate lung. In the Dipnoans the shoulder-girdle moves forward to the skull, and the pectoral limb, a jointed and fringed archipterygium, is its[Pg 450] characteristic appendage. The shark-like structure of the mouth remains.
The few living lung-fishes resemble the salamanders in many regards, and some writers have ranged the class as midway between the primitive sharks and the amphibians. These forms show their intermediate characters in the development of lungs and in the primitive character of the pectoral and ventral limbs. Those now extant give but little idea of the great variety of extinct Dipnoans. The living genera are three in number—Neoceratodus in Australian rivers, Lepidosiren in the Amazon, and Protopterus in the Nile. These are all mudfishes, some of them living through most of the dry season encased in a cocoon of dried mud. Of these forms Neoceratodus is certainly the nearest to the ancient forms, but its embryology, owing to the shortening of its growth stages due to its environment, has thrown little light on the question of its ancestry.
From some ally of the Dipnoans the ancestry of the amphibians, and through them that of the reptiles, birds, and mammals may be traced, although a good deal of evidence has been produced in favor of regarding the primitive crossopterygian or fringe fin as the point of divergence. It is not unlikely that the Crossopterygian gave rise to Amphibian and Dipnoan alike.
In the process of development we next reach the characteristic fish mouth in which the upper jaw is formed of maxillary and premaxillary elements distinct from the skull. The upper jaw of the shark is part of the palate, the palate being fused with the quadrate bone which supports the lower jaw. That of the Dipnoan is much the same. The development of a typical fish mouth is the next step in evolution, and with its appearance we note the decline of the air-bladder in size and function.
The Crossopterygians.—The fish-like mouth appears with the group of Crossopterygians, fishes which still retain the old-fashioned type of pectoral and ventral fin, the archipterygium. In the archaic tail, enameled scales, and cartilaginous skeleton the Crossopterygian shows its affinity with its Dipnoan ancestry. Thus these fishes unite in themselves traits of the shark, lung-fish, and Ganoid. The few living Crossopterygians, Polypterus and Erpetoichthys, are not very different from those which pre[Pg 451]vailed in Devonian times. The larvæ possess external gills with firm base and fringe-like rays, suggesting a resemblance to the pectoral fin itself, which develops from the shoulder-girdle just below it and would seem to give some force to Kerr's contention that the archipterygium is only a modified external gill. In Polypterus the archipterygium has become short and fan-shaped, its axis made of two diverging bones with flat cartilage between. From this type it is thought that the arm of the higher forms has been developed. The bony basis may be the humerus, from which diverge radius and ulna, the carpal bones being formed of the intervening cartilage.

Fig. 260.—An extinct Crossopterygian, Holoptychius giganteus Agassiz (1835). (After Agassiz, per Zittel.)
The Actinopteri.—From the Crossopterygians springs the main branch of the true fishes, known collectively as Actinopteri, or ray-fins, those with ordinary rays on the paired fins instead of the jointed archipterygium. The transitional series of primitive Actinopteri are usually known as Ganoids. The Ganoid differs from the Crossopterygian in having the basal elements of the paired fins small and concealed within the flesh. But other associated characters of the Crossopterygii and Dipnoans are preserved in most of the species. Among these are the mailed head and body, the heterocercal tail, the cellular air-bladder, the presence of valves in the arterial bulb, the presence of a spiral valve in the intestine and of a chiasma in the optic nerves. All these characters are found in the earlier types so far as is known, and all are more or less completely lost or altered in the teleosts or bony fishes. Among these early types is every variety of form, some of them being almost as long[Pg 452] as deep, others arrow-shaped, and every intermediate form being represented. An offshoot from this line is the bowfin (Amia calva), among the Ganoids the closest living ally of the bony fishes, showing distinct affinities with the great group to which the herring and salmon belong. Near relatives of the bowfin flourished in the Mesozoic, among them some with a forked tail, and some with a very long one. From Ganoids of this type the vast majority of recent fishes may be descended.
Fig. 261.—An ancient Ganoid fish, Platysomus gibbosus Blainville. Family Platysomidæ. (After Woodward.)

Fig. 262.—A living Ganoid fish, the Short-nosed Gar, Lepisosteus platystomus Rafinesque. Lake Erie.
Another branch of Ganoids, divergent from both garfish and bowfin and not recently from the same primitive stock, included the sturgeons (Acipenser, Scaphirhynchus, Kessleria) and the paddle-fishes (Polyodon and Psephurus). All these are regarded by Woodward as degenerate descendants of the earliest Ganoids, Palæoniscidæ, of Devonian and Carboniferous time.
Fig. 263.—A primitive Ganoid fish, Palæoniscum macropomum (Agassiz), restored. Permian. Family Potaconiscidæ. (After Traquair.)

Fig. 264.—A fossil Herring, Diplomystus humilis Leidy. (From a specimen obtained at Green River, Wyo.) The scutes along the back lost in the specimen. Family Clupeidæ.
The Bony Fishes.—All the remaining fishes have ossified instead of cartilaginous skeletons. The dipnoan and ganoid traits one by one are more or less completely lost. Through these the main line of fish development continues and the various groups are known collectively as bony fishes or teleosts.

Fig. 265.—A primitive Herring-like fish, Holcolepis lewesiensis Mantell, restored. Family Elopidæ. English Chalk. (After Woodward.)

Fig. 266.—Ten-pounder, Elops saurus L. An ally of the earliest bony fishes. Virginia.
The earliest of the true bony fishes or teleosts appear in Mesozoic times, the most primitive forms being soft-rayed fishes with the vertebræ all similar in form, allied more or less remotely to the herring of to-day, but connected in an almost unbroken series with the earliest ganoid forms. In these and other soft-rayed fishes the pelvis still retains its posterior insertion, the ventral fins being said to be abdominal. The next great stage in evolution brings the pelvis forward, attaching it to the shoulder-girdle so that the ventral fins are now thoracic as in the perch and bass. If brought to a point in front of the pectoral fins, a feature of specialized degradation, they become jugular as in the codfish. In the abdominal fishes the air-bladder still retains its rudimentary duct joining it to the œsophagus.
From the abdominal forms allied to the herring, the huge[Pg 455] array of modern fishes, typified by the perch, the bass, the mackerel, the wrasse, the globefish, the sculpin, the sea-horse, and the cod descended in many diverging lines. The earliest of the spine-rayed fishes with thoracic fins belong to the type of Berycidæ, a group characterized by rough scales, the retention of a primitive bone between the eyes, and the retention of the primitive larger number of ventral rays. These appear in the Cretaceous or chalk deposits, and show various attributes of transition from the abdominal to the thoracic type of ventrals.
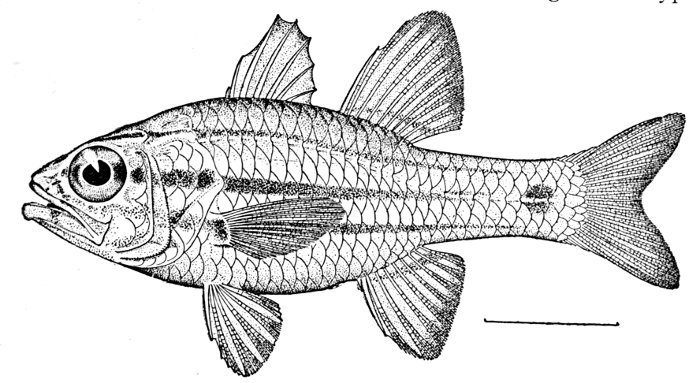
Fig. 267.—Cardinal-fish, a perch-like fish, Apogon semilineatus Schlegel. Misaki, Japan.
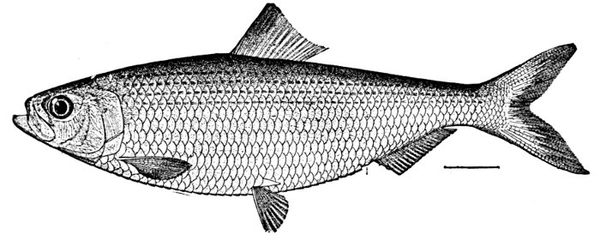
Fig. 268.—Summer Herring, Pomolobus æstivalis (Mitchill). Potomac River. Family Clupeidæ.
Another line of descent apparently distinct from that of the[Pg 456] herring and salmon extends through the characins to the loach, carps, catfishes, and electric eel. The fishes of this series have the anterior vertebræ coossified and modified in connection with the hearing organ, a structure not appearing elsewhere among fishes. This group includes the majority of fresh-water fishes. Still another great group, the eels, have lost the ventral fins and the bones of the head have suffered much degradation.
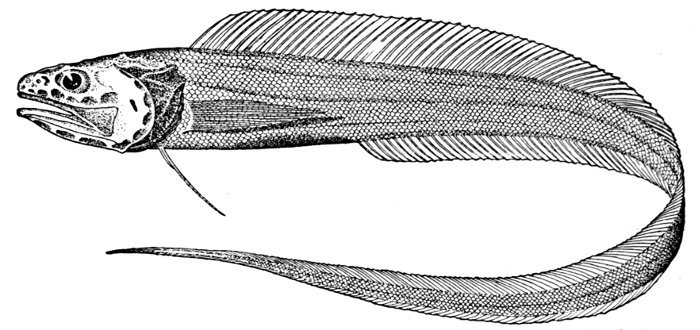
Fig. 269.—Fish with jugular ventral fins, Bassozetus catena Goode & Bean. Family Brotulidæ. Gulf Stream.
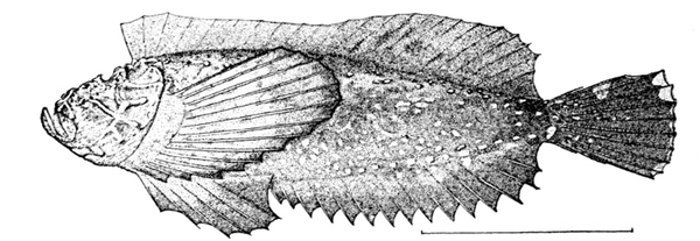
Fig. 270.—A specialized bony fish, Trachicephalus uranoscopus. Family Scorpænidæ. From Swatow, China.
The most highly developed fishes, all things considered, are doubtless the allies of the perch, bass, and sculpin. These fishes have lost the air-duct and on the whole they show the greatest development of the greatest number of structures. In these groups their traits one after another are carried to an extreme and these stages of extreme specialization give way one after another to phases of degeneration. The specialization of one[Pg 457] organ usually involves degeneration of some other. Extreme specialization of any organ tends to render it useless under other conditions and may be one step toward its final degradation.

Fig. 271.—An African Catfish, Chlarias breviceps Boulenger. Congo River. Family Chlariidæ. (After Boulenger.)

Fig. 272.—Silverfin, Notropis whipplii (Girard). White River, Indiana. Family Cyprinidæ.
We have thus seen, in hasty review, that the fish-like vertebrates spring from an unknown and possibly worm-like stock, that from this stock, before it became vertebrate, degenerate branches have fallen off, represented to-day by the Tunicates and Enteropneustans. We have seen that the primitive vertebrate was headless and limbless and without hard parts. The lancelet remains as a possible direct offshoot from it; the cyclostome with brain and skull is a possible derivative from archaic lancelets. The earliest fishes leaving traces in the rocks were mailed ostracophores. From an unknown but possibly lamprey-like stock sprang the sharks and chimæras. The sharks developed into rays in one right line and into the highest sharks along another, while by a side branch through lost stages the primitive sharks passed into Crossopterygians, into Dipnoans, or lung-fishes, and perhaps into Ostracophores. All these types[Pg 458] and others abound in the Devonian Age and the early records were lost in the Silurian. From the Crossopterygians or their ancestors or descendants by the specialization of the lung and limbs, the land animals, at first amphibians, after these reptiles, birds, and mammals, arose.
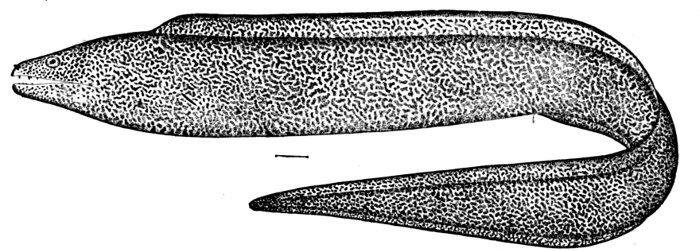
Fig. 273.—Moray, Gymnothorax moringa Bloch. Family Murænidæ Tortugas.

Fig. 274.—Amber-fish, Seriola lalandi (Cuv. & Val.). Family Carangidæ. Woods Hole.
In the sea, by a line still more direct, through the gradual emphasis of fish-like characters, we find developed the Crossopterygians with archaic limbs and after these the Ganoids with fish-like limbs but otherwise archaic; then the soft-rayed and finally the spiny-rayed bony fishes, herring, mackerel, perch, which culminate in specialized and often degraded types, as the anglers, globefishes, parrot-fishes, and flying gurnards; and from each of the ultimate lines of descent radiate infinite branches till the sea and rivers are filled, and almost every body of water has fishes fitted to its environment.
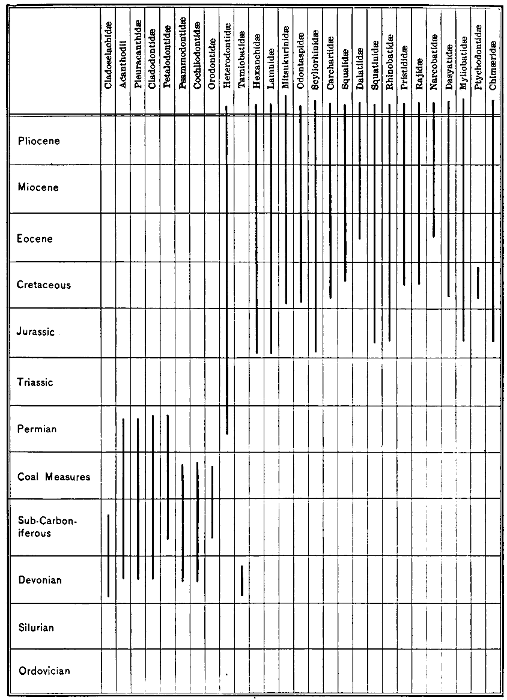
Geological Distribution of the Families of Elasmobranchs.
| Pliocene | Miocene | Eocene | Cretaceous | Jurassic | Triassic | Permian | Coal Measures | Sub-Carbonifero | Devonian | Silurian | Ordovician | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cladoselachidæ | █ | █ | ||||||||||
| Acanthodii | █ | █ | █ | █ | ||||||||
| Pleuracanthidæ | █ | █ | █ | █ | ||||||||
| Cladodontidæ | █ | █ | █ | █ | ||||||||
| Petalodontidæ | █ | █ | █ | |||||||||
| Psammodontidæ | █ | █ | █ | |||||||||
| Cochliodontidæ | █ | █ | █ | |||||||||
| Orodontidæ | █ | █ | ||||||||||
| Heterodontidæ | █ | █ | █ | █ | █ | █ | █ | |||||
| Tamiobatidæ | █ | |||||||||||
| Hexanchidæ | █ | █ | █ | █ | █ | |||||||
| Lamnidæ | █ | █ | █ | █ | █ | |||||||
| Mitsukurinidæ | █ | █ | █ | █ | ||||||||
| Odontaspidæ | █ | █ | █ | █ | ||||||||
| Scyliorhinidæ | █ | █ | █ | █ | █ | |||||||
| Carchariidæ | █ | █ | █ | █ | ||||||||
| Squalidæ | █ | █ | █ | █ | ||||||||
| Dalatiidæ | █ | █ | █ | |||||||||
| Squatinidæ | █ | █ | █ | █ | █ | |||||||
| Rhinobatidæ | █ | █ | █ | █ | █ | |||||||
| Pristididæ | █ | █ | █ | █ | ||||||||
| Rajidæ | █ | █ | █ | █ | ||||||||
| Narcobatidæ | █ | █ | █ | |||||||||
| Dasyatidæ | █ | █ | █ | █ | ||||||||
| Myliobatidæ | █ | █ | █ | █ | █ | |||||||
| Ptychodontidæ | █ | |||||||||||
| Chimæridæ | █ | █ | █ | █ | █ |

The Chordate Animals.—Referring to our metaphor of the tree with its twigs as used in the chapter on classification we find the fishes with the higher vertebrates as parts of a great branch from which the lower twigs have mostly perished. This great branch, phylum, or line of descent is known in zoology as Chordata, and the organisms associated with it or composing it are chordate animals.
The chordate animals are those which at some stage of life possess a notochord or primitive dorsal cartilage which divides the interior of the body into two cavities. The dorsal cavity contains the great nerve centers or spinal cord; the ventral cavity contains the heart and alimentary canal. In all other animals which possess a body cavity, there is no division by a notochord, and the ganglia of the nervous system if existing are placed on the ventral side or in a ring about the mouth.
The Protochordates.—Modern researches have shown that besides the ordinary back-boned animals certain other creatures easily to be mistaken for mollusks or worms and being chordate in structure must be regarded as offshoots from the vertebrate branch. These are degenerate allies, as is shown by the fact that their vertebrate traits are shown in their early or larval development and scarcely at all in their adult condition. As Dr. John Sterling Kingsley has well said: "Many of the species start in life with the promise of reaching a point high in the scale, but after a while they turn around and, as one might say, pursue a downward course, which results in an adult which displays but few resemblances to the other vertebrates." In the Tunicates or Ascidians (sea-squirts, sea-pears, and salpas), which constitute the class known as Tunicata or Urochordata,[Pg 461] there is no brain, the notochord is confined to the tail and is usually present only in the larval stage of the animal when it has the form of a tadpole. In later life the animal usually becomes quiescent, attached to some hard object, fixed or floating. It loses its form and has the appearance of a hollow, leathery sac, the body organs being developed in a tough tunic. There are numerous families of Tunicates and the species are found in nearly all seas. They suggest no resemblance to fishes and look like tough clams without shells. The internal cavity being usually filled with water it is squirted out through the two apertures when the animal is handled. The class Enteropneusta (Adelochorda, or Hemichordata), includes the rather rare worm-like forms related to Balanoglossus. Bateson has shown that these animals possess a notochord which is developed in the anterior part of the body. They have no fins and before the mouth is a long proboscis. Gill-slits are found in the larval tunicate. In Balanoglossus these persist through life as in the fishes.
The remaining chordate forms constitute the vertebrates proper, not worm-like nor mollusk-like, the notochord not disappearing with age, except as it gives way, by specialized segmentation to the complex structures of the vertebral column. These vertebrates, which are permanently aquatic, are known in a popular sense as fishes. The fish, in the broad sense, is a back-boned animal which retains the homologue of the back-bone throughout life, which does not develop jointed limbs, its locomotive members, if present, being developed as fins, and which breathes through life the air contained in water by means of gills. This definition excludes the Tunicates and Enteropneusta on the one hand and the Amphibia or Batrachia with the reptiles, birds, and mammals on the other. The Amphibia are much more closely related to certain fishes than the classes of fishes are to each other. Still for purposes of systematic study, the frogs and salamanders are left out of the domain of ichthyology, while the Tunicata and the Enteropneusta might well be included in it.
The known branchiferous or gill-bearing chordates living and extinct may be first divided into eight classes—the Enteropneusta, the Tunicata, the Leptocardii, or lancelets, the Cyclostomi,[Pg 462] or lampreys, the Elasmobranchii, or sharks, the Ostracophori the Arthrodira, and the Teleostomi, or true fishes. The first two groups, being very primitive and in no respect fish-like in appearance, are sometimes grouped together as Protochordata, the others with the higher Chordates constituting the Vertebrata.
Other Terms used in Classification.—The Leptocardii are sometimes called Acraniata (without skull), as distinguished from the higher groups, Craniota, in which the skull is developed. The Leptocardii, Cyclostomi, and Ostracophori are sometimes called Agnatha (without jaws) in contradistinction to the Gnathostomi (jaw mouths), which include the sharks and true fishes with the higher vertebrates. The sharks and Teleostomes are sometimes brought together as Pisces, or fishes, as distinguished from other groups not true fishes. To the sharks and true fishes the collective name of Lyrifera has been given, these fishes having the harp-shaped shoulder-girdle, its parts united below. The Ostracophores and Arthrodires agreeing in the bony coat of mail, and both groups now extinct and both of uncertain relationship, have been often united under the name of Placoderms, and these and many other fishes have been again erroneously confounded with the Ganoids. Again, the Teleostomi have been frequently divided into three classes—Crossopterygii, Dipneusti or Dipnoi, and Actinopterygii. The latter may be again divided into Ganoidei and Teleostei and all sorts of ranks have been assigned to each of these groups. For our purposes a division into eight classes is most convenient, and lowest among these we may place the Enteropneusta.
The Enteropneusta.—Most simple, most worm-like, and perhaps most primitive of all the Chordates is the group of worm-shaped forms, forming the class of Enteropneusta. The class of Enteropneusta, also called Adelochorda or Hemichordata, as here recognized, consists of a group of small marine animals allied to the genus Balanoglossus, or acorn-tongues (βάλανος, acorn; γλώσσα, tongue). These are worm-like creatures with fragile bodies buried in the sand or mud, or living under rocks of the seashore and in shallow waters, where they lie coiled in a spiral, with little or no motion. From the surface of the body[Pg 463] a mucous substance is secreted, holding together particles by which are formed tubes of sand. The animal has a peculiar odor like that of iodoform. At the front is a long muscular proboscis, very sensitive, capable of great extension and contraction, largely used in burrowing in the ground, and of a brilliant orange color in life. Behind this is a collar which overlaps the small neck and conceals the small mouth at the base of the proboscis. The gill-slits behind the collar are also more or less concealed by it.
The body, which is worm-like, extends often to the length of two or three feet. The gill-slits in the adult are arranged in regular pairs, there being upwards of fifty in number much like the gill-slits of the lancelet. As the animal grows older the slits become less conspicuous, their openings being reduced to small slit-like pores.
In the interior of the proboscis is a rod-like structure which arises as an outgrowth of the alimentary canal above the mouth. In development and structure this rod so resembles the notochord of the lancelet that it is regarded as a true notochord, though found in the anterior region only. From the presence of gill-slits and notochord and from the development and structure of the central nervous system Balanoglossus was recognized by William Bateson, who studied an American species, Dolichoglossus kowalevskii, at Hampton Roads in Virginia in 1885, and at Beaufort in North Carolina, as a member of the Chordate series. Unlike the Tunicates it represents a primitively simple, not a degenerate, type. It seems to possess real affinities with the worms, or possibly, as some have thought, with the sea-urchins.
Fig. 275.—"Tornaria" Larva of Glossobalanus minutus. (After Minot.)
A peculiar little creature, known as Tornaria, was once considered to be the larva of a starfish. It is minute and transparent, floating on the surface of the sea. It has no visible resemblance to the adult Balanoglossus, but it has been reared in aquaria and shown to pass into the latter or into the related genus Glossobalanus. No such metamorphosis was found by[Pg 464] Bateson in the more primitive genus Dolichoglossus, studied by him. This adult animal may be, indeed, a worm as it appears, but the presence of gill-slits, the existence of a rudimentary notochord, and the character of the central nervous system are distinctly fish-like and therefore vertebrate characters. With the Chordates, and not with the worms, this class, Enteropneusta (ἔντερον, intestine; πνεῖν, to breathe), must be placed if its characters have been rightly interpreted. It is possibly a descendant of the primitive creatures which marked the transition from the archaic worms, or possibly archaic Echinoderms, to the archaic Chordate type.
Fig. 276.—Glossobalanus minutus, one of the higher Enteropneustans. (After Minot.)
It is perhaps not absolutely certain that the notochord of Balanoglossus and its allies is a true homologue of the notochord of the lancelet. There may be doubt even of the homologies of the gill-slits themselves. But the balance of evidence seems to throw Balanoglossus on the fish side of the dividing line which separates the lower Chordates from the worms.
It may be noticed that Hubrecht regards the proboscis of various marine Nemertine worms as a real homologue of the notochord, and other writers have traced with more or less success other apparent or possible homologies between the Chordate and the Annelid series.
Classification of Enteropneusta.—Until recently the Enteropneusta have been usually placed in a single family or even in a single genus. The recent researches of Professor J. W. Spengel of Giessen and of Professor William Emerson Ritter of the University of California, have shown clearly that the group is much larger than had been generally supposed, with numerous species[Pg 465] in all the warm seas. In Spengel's recent paper, "Die Benennung der Enteropneusten-Gattungen," three families are recognized with nine genera and numerous species. At least seven species are now known from the Pacific Coast of North America.
Family Harrimaniidæ.—In Harrimania maculosa, lately described by Dr. Ritter from Alaska, the eggs are large, with much food yolk, and the process of development is probably, without Tornaria stage. A second species of Harrimania (H. kupferi) is now recognized from Norway and Greenland. This genus is the simplest in structure among all the Enteropneustans and may be regarded as the lowest of known Chordates, the most worm-like of back-boned animals.
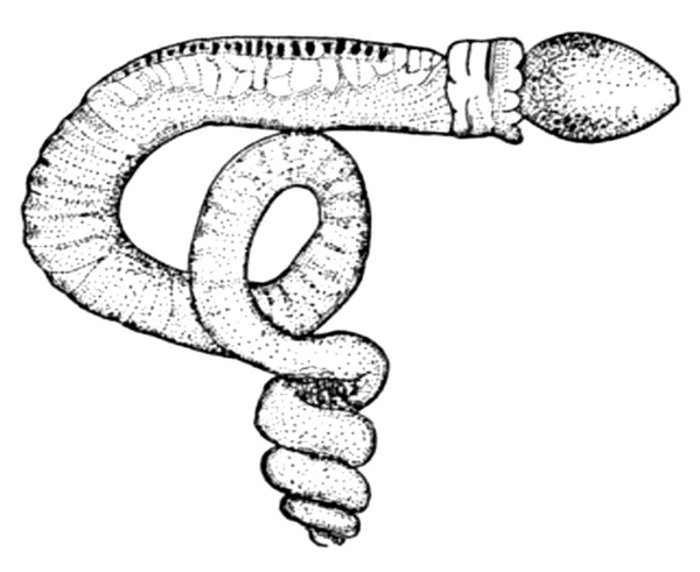
Fig. 277.—Harrimania maculosa (Ritter), the lowest of chordate animals. An Enteropneustan from Alaska. (After Ritter.)
In Dolichoglossus kowalevskii the species studied by Bateson on the Virginia coast, the same simplicity of development occurs. This genus, with a third, Stereobalanus (canadensis), constitutes in Spengel's system the family of Harrimaniidæ.
Balanoglossidæ.—The family Glandicepitidæ contains the genera Glandiceps, Spengelia, and Schizocardium. In the Balanoglossidæ (Ptychoderidæ of Spengel) the eggs are very small and numerous, with little food yolk. The species in this family pass through the Tornaria stage above described, a condition strikingly like that of the larval starfish. This fact has given rise to the suggestion that the Enteropneusta have a real affinity with the Echinoderms.
The Balanoglossidæ include the genera Glossobalanus, Balanoglossus, and Ptychodera, the latter the oldest known member of the group, its type, Ptychodera flava, having been described by Eschscholtz from the Pacific Coast in 1825, while Balanoglossus clavigerus was found by Della Chiaje in 1829.
Low Organization of Harrimaniidæ.—Apparently the Harrimaniidæ, with simpler structure, more extensive notochord, and direct development, should be placed at the bottom as the most primitive of the Enteropneustan series. Dr. Willey, however, regards its characters as due to degeneration, and considers the[Pg 466] more elaborate Balanoglossidæ as nearest the primitive type. The case in this view would have something in common with that of the Larvacea, which seems to be the primitive Tunicates, but which may have been produced by the degeneration of more complex forms.

Structure of Tunicates.—One of the most singular groups of animals is that known as Ascidians, or Tunicates. It is one of the most clearly marked yet most heterogeneous of all the classes of animals, and in no other are the phenomena of degeneration so clearly shown.
Among them is a great variety of form and habit. Some lie buried in sand; some fasten themselves to rocks; some are imbedded in great colonies in a gelatinous matrix produced from their own bodies, and some float freely in long chains in the open sea. All agree in changing very early in their development from a free-swimming or fish-like condition to one of quiescence, remaining at rest or drifting with the current.
Says Dr. John Sterling Kingsley: "Many of the species start in life with the promise of reaching a point high in the scale, but after a while they turn around and, as one might say, pursue a downward course which results in an adult which displays but few resemblances to the other vertebrates. Indeed, so different do they seem that the fact that they belong here was not suspected until about thirty-five years ago. Before that time, ever since the days of Cuvier, they were almost universally regarded as mollusks, and many facts were adduced to show that they belonged near the acephals (clams, oysters, etc.). In the later years when the facts of development began to be known, this association was looked on with suspicion, and by some they were placed for a short time among the worms. Any one who has watched the phases of their development cannot help believing that they belong here, the lowest of the vertebrate series."
The following account of the structure and development of the Tunicate is taken, with considerable modification and condensation, from Professor Kingsley's chapter on the group in the Riverside Natural History. For the changes suggested I am indebted to the kindness of Professor William Emerson Ritter:
The Tunicates derive their name from the fact that the whole body is invested with a tough envelope or "tunic." This tunic or test may be either gelatinous, cartilaginous, or leathery. In some forms it is perfectly transparent, in others it is translucent, allowing enough light to pass to show the colors of the viscera, while in still others it is opaque and variously colored. The tunic is everywhere only loosely attached to the body proper, except in the region of the two openings now to be mentioned. One of these openings occupies a more or less central position, while the other is usually at one side, or it may even be placed at the opposite end of the body. On placing one of the Ascidians in a glass dish and sprinkling a little carmine or indigo in the water, we can study some of the functions of the animal. As soon as the disturbance is over, the animals will open the two apertures referred to, when it will be seen that each is surrounded with blunt lobes, the number of which varies with the species. As soon as they are opened a stream of water will be seen to rush into the central opening, carrying with it the carmine, and a moment later a reddish cloud will be ejected from the other aperture. From this we learn that the water passes through the body. Why it does so is to be our next inquiry. On cutting the animal open we find that the water, after passing through the first-mentioned opening (which may be called the mouth) enters a spacious chamber, the walls of which are made up of fine meshes, the whole appearing like lattice-work. Taking out a bit of this network and examining it under the microscope, we find that the edges of the meshes are armed with strong cilia, which are in constant motion, forcing the water through the holes. Of course, the supply has to be made good, and hence more water flows in through the mouth. This large cavity is known as the branchial or pharyngeal chamber. It is, according to Professor Ritter, "as we know from the embryology of the animal, the greatly[Pg 469] enlarged anterior end of the digestive tract; and as the holes, or stigmata, as they are technically called, are perforations of the wall for the passage of water for purposes of respiration, they are both morphologically and physiologically comparable with the gill openings of fishes." There can be no doubt, therefore, that the pharyngeal sac of Ascidians is homologous with the pharynx of fishes.
Surrounding the mouth, or branchial orifice, just at its entrance into the branchial chamber is a circle of tentacles. These are simple in some genera, but elaborately branched in others.
In close connection with the cerebral ganglion, which is situated between the two siphons, there is a large gland with a short trumpet-shaped duct opening into the branchial sac a little distance behind the mouth. The orifice of the duct is just within a ring consisting of a ciliated groove that extends around the mouth outside the circle of branchial tentacles. On the opposite side of the mouth from the gland the ciliated groove joins another groove which is both ciliated and glandular, and which runs backward along the upper floor of the pharyngeal sac to its posterior extremity. This organ, called the endostyle, is concerned in the transportation of the animal's food through the pharyngeal sac to the opening of the œsophagus. Comparative embryology makes it almost certain that the subneural gland with its duct, described above, is homologous with the hypophesis cerebri of true vertebrates, and that the endostyle is homologous with the thyroid glands of vertebrates.
The water after passing through the branchial network is received into narrow passages and conducted to a larger cavity—the cloacal or atrial chamber. The general relations can he seen from our diagram, illustrating a vertical and horizontal section. From the atrial chamber the water flows out into the external world.
Now we can readily see how in the older works naturalists were misled as to the affinities of the Tunicates. They regarded the tunic as the equivalent of the mantle of the mollusks, while the incurrent and excurrent openings corresponded to the siphons. In one genus, Rhodosoma, the resemblance was even stronger, for there the tunic is in two parts, united[Pg 470] by a hinge line, and closed by an adductor muscle. How and why these views were totally erroneous will be seen when we come to consider the development of these animals.
At the bottom of the pharnygeal sac is the narrow œsophagus surrounded with cilia, which force a current down into the digestive tract. The branchial meshes serve as a strainer for the water, and the larger particles which it contains fall down until they are within reach of the current going down the œsophagus. After passing through the throat, they come to the stomach, where digestion takes place, and then the ejectamenta are carried out through the intestine and poured into the bottom of the atrial cavity.
The heart lies on the ventral side of the stomach and is surrounded by a well-developed pericardium. The most remarkable fact connected with the circulation is that the heart, after beating a short time, forcing the blood through the vessels, will suddenly stop for a moment and then resume its beats; but, strange to say, after the stoppage the direction of the circulation is reversed, the blood taking an exactly opposite course from that formerly pursued. This most exceptional condition was first seen in the transparent Salpa, but it may be witnessed in the young of most genera. We have already referred to the branchial chamber. The walls of this chamber, besides acting as a strainer, are also respiratory organs. The meshes of which they are composed are in reality tubes through which the blood circulates and thus is brought in contact with a constantly renewed supply of fresh water.
The central nervous system in the adults of all except the Larvacea is reduced to a single ganglion placed near the mouth thus indicating the dorsal side. In forms like Cynthia it holds the same relative position with regard to the mouth, but by the doubling of the body (to be explained further on) it is also brought near the atrial aperture, where it is shown in our first diagram.
Development of Tunicates.—The sexes are combined in the same individual, though usually the products ripen at different times. As a rule, the earlier stages of the embryo are passed inside the cloacal chamber, though in some the development occurs outside the body. As a type of the development we[Pg 471] will consider that of one of the solitary forms, leaving the many curious modifications to be noticed in connection with the species in which they occur. This will be best, since these forms show the relationship to the other vertebrates in the clearest manner.
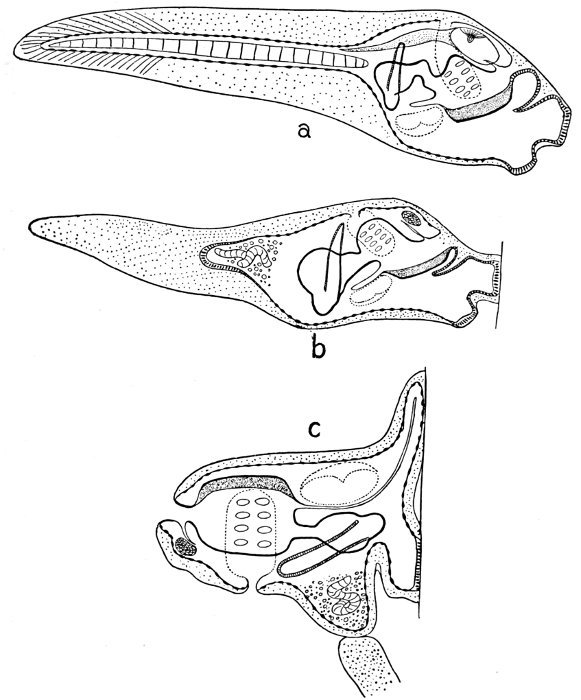
Fig. 278.—Development of the larval Tunicate to the fixed condition. (From Seeliger, per Parker & Haswell.) a, larva; b, intermediate stage; c, adult.
The egg undergoes a total segmentation and a regular gastrulation. Soon a tail appears, and under the microscope the young embryo, which now begins its free life, appears much[Pg 472] like the tadpole of the frog. It has a large oval body and a long tail which lashes about, forcing the animal forward with a wriggling motion. Nor is the resemblance superficial; it pervades every part of the structure, as may be seen from the adjacent diagram. The mouth is nearly terminal and communicates with a gill-chamber provided with gill-clefts. At the posterior end of the gill-chamber begins the alimentary tract, which pursues a convoluted course to the vent. In the tail, but not extending to any distance into the body, is an axial cylinder, the notochord, which here, as in all other vertebrates, arises from the hypoblast; and above it is the spinal cord (epiblastic in origin), which extends forward to the brain, above the gill-chamber. Besides, the animal is provided with organs of sight and hearing, which, however, are of peculiar construction and can hardly be homologized with the corresponding organs in vertebrates. So far the correspondence between the two types is very close, and if we knew nothing about the later stages, one would without doubt predict that the adult tunicate would reach a high point in the scale of vertebrates. These high expectations are never fulfilled; the animal, on the contrary, pursues a retrograde course, resulting in an adult whose relationship to the true vertebrates never would have been suspected had its embryology remained unknown.
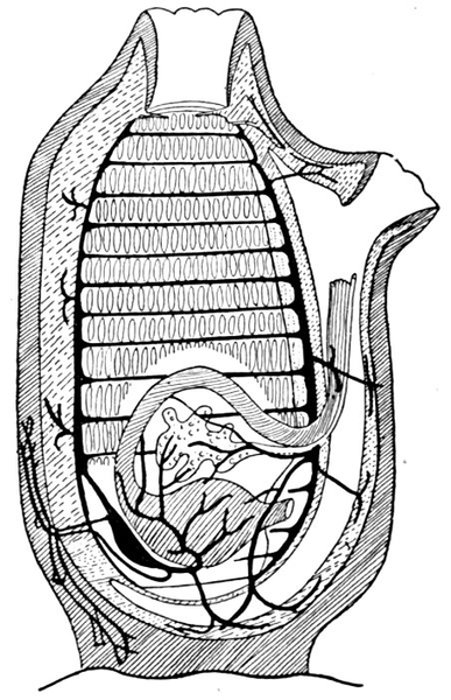
Fig. 279.—Anatomy of Tunicate. (After Herdman, per Parker & Haswell.)
After the stage described this retrograde movement begins. From various parts of the body lobes grow out, armed on their extremities with sucking-disks. These soon come in contact with some subaquatic object and adhere to it. Then the notochord breaks down, the spinal cord is absorbed, the tail follows suit, the intestine twists around, and the cloaca is formed, the result being much like the diagram near the head of this section. In forms[Pg 473] like Appendicularia, little degeneration takes place, so far as is known, the tail, with its notochord and neural chord, persisting through life.
Reproduction of Tunicates.—As to the reproduction of the Tunicates, Dr. Ritter writes: "In addition to the sexual method of reproduction, many tunicates reproduce asexually by budding. The capacity for bud reproduction appears to have been acquired by certain simple Ascidians in connection with, probably as a result of, their having given up the free-swimming life and become attached and consequently degenerate.
"Instructive as the embryonic development of the creatures is from the standpoint of evolution, the bud method of development is scarcely less so from the same point of view. The development of the adult zooid from the simple bud has been conclusively shown to be by a process in many respects fundamentally unlike that by which the individual is developed from the egg. We have then in these animals a case in which practically the same results are reached by developmental processes that are, according to prevailing conceptions of animal organizations, fundamentally different. This fact has hardly a parallel in the animal kingdom."
Habits of Tunicates.—The Tunicates are all marine, some floating or swimming freely, some attached to rocks or wharves, others buried in the sand. They feed on minute organisms, plants, or animals, occasional rare forms being found in their stomachs. Some of them possess a single median eye or eye-like structure which may not do more than recognize the presence of light. No fossil Tunicates are known, as they possess no hard parts, although certain Ostracoderms have been suspected, though on very uncertain grounds, to be mailed Tunicates, rather than mailed lampreys. It is not likely that this hypothesis has any sound foundation. The group is divided by Herdman and most other recent authorities into three orders, viz., the Larvacea, the Ascidiacea, and the Thaliacea.
Larvacea.—In the most primitive order the animals are minute and free-swimming, never passing beyond the tadpole stage. The notochord and the nervous chord persist through life, the latter with ganglionic segmentations at regular intervals. The species mostly float in the open sea, and some[Pg 474] of them form from their own secretions a transparent gelatinous envelope called a "house." This has two apertures and a long chamber "in which the tail has room to vibrate."
The order consists of a single small family, Appendiculariidæ. The lowest type is known as Kowalevskia, a minute creature without heart or intestine found floating in the Mediterranean. It is in many respects the simplest in structure among Chordate animals. Oikopleura (Fig. 288) is another genus of this group.
Ascidiacea.—In the Ascidiacea the adult is usually attached to some object, and the two apertures are placed near each other by the obliteration of the caudal area. The form has been compared to a "leathern bottle with two spouts."
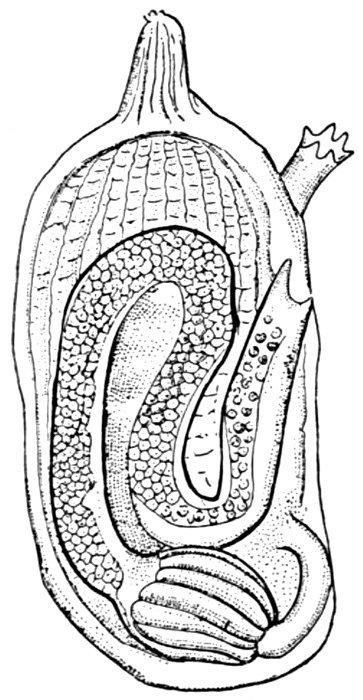
Fig. 280.—Ascidia adhærens Ritter. Glacier Bay, Alaska. (After Ritter.)
The suborder Ascidiæ simplices includes the solitary Ascidians or "sea-squirts," common on our shores, as well as the social forms in which an individual is surrounded by its buds. The common name arises from the fact that when touched they contract, squirting water from both apertures. The Ascidiidæ comprise the most familiar solitary forms, some of them the largest of the Tunicates and represented on most coasts. In the Molgulidæ and most Ascidiæ compositæ the young hatch out in the cloaca, from which "these tadpoles swim out as yellow atoms," while in a new genus, Euherdmania, described by Ritter, from the coast of California, the embryos are retained through their whole larval stage in the oviduct of the parent. They form, according to Kingsley, adhesive processes on the body, but those of Molgula cannot use them in becoming attached to rocks, since they are entirely inclosed in a peculiar envelope. This envelope is after a while very adhesive, and if the little tadpole happens to touch any part of himself to a stone or shell he is fastened for life. Thus "I have frequently seen them adhere by the tail, while the anterior part was making the most violent struggles to escape. Soon, however, they settle down contentedly, absorb the tail, and in a few weeks assume the adult structure."
In the family Cynthiidæ the brightly-colored red and yellow species of Cynthia are known as sea-peaches by the fishermen. The sea-pears, Boltenia, are fastened to long stalks. These have a leathery and wrinkled tunic, to which algæ and hydroids freely attach themselves. Into the gill-cavity of these forms small fishes, blennies, gobies, and pearl-fishes often retreat for protection.
Fig. 281.—Styela yacutatensis (Ritter), a simple Ascidian. Family Molgulidæ. Yakutat Bay, Alaska. (After Ritter.)
The social Ascidians constitute the Clavellinidæ. They are similar to the Ascidiidæ in form, but each individual sends out a bud which forms a stern bearing another individual at the end. By this means large colonies may be formed.
The suborder, Ascidiæ compositæ, contains the compound Ascidians or colonies enveloped in a common gelatinous "test." These colonies are usually attached to rock or seaweed, and the individuals are frequently regularly and symmetrically arranged. The bodies are sometimes complex in form.
Fig. 282.—Styela greeleyi Ritter. Family Molgulidæ. Lukanin, Pribilof Islands. (After Ritter.)
Fig. 283.—Cynthia superba Ritter. A Tunicate from Puget Sound. Family Cynthiidæ. (After Ritter.)
In the Botryllidæ and Polystyelidæ the individuals are not segmented and in the former family are arranged in star-shaped groups about a common cloaca, into which the atrial siphons of the different individuals open. The group springs by budding from the tadpole, or larva, which has attached itself to some object. These forms are often brightly colored. Botryllus gouldi is a species very common along our North Atlantic coast, forming gray star-shaped masses sometimes an inch across on eel-grass (Zostera) and on flat-leaved seaweeds. Goodsiria dura, a representative of the Polystyelidæ, is one of the most common Ascidians on the California coast southward, where the brick-red[Pg 477] masses incrusting on seaweeds of various kinds, and on other Ascidians, are frequently thrown ashore in great quantities during heavy storms.
Fig. 284.—Botryllus magnus Ritter. A compound Ascidian. Shumagin Islands, Alaska. (After Ritter.)
In Didemnidæ the body is more complex, of two parts, called the "thorax" and "abdomen." In Amarœcium, the "sea pork" of the fishermen, the body is in three parts and the individuals are very long. These sometimes form great masses a foot or more long, "colored like boiled salt pork, but more translucent." Other families of this type are the Distomidæ and the Polyclinidæ.
In the suborder Luciæ, including the family Pyrosomidæ, the colonies are thimble-shaped and hollow, the incurrent openings being on the outer surface of the thimble, the outgoing stream opening within. Pyrosoma is highly phosphorescent. In the tropical seas some colonies reach a length of two or three feet. It is said that a description of a colony was once written by a naturalist on a page illumined by the colony's own light. "Each of the individuals has a number of cells near the mouth the function of which is to produce the light."
Thaliacea.—In the order Thaliacea the Tunicates have the two orifices at opposite ends of the body. All are free-swimming and perfectly transparent. The principal family is that of Salpidæ. The gill-cavity in Salpa is much altered, the gills projecting into it dividing it into two chambers.
In these forms we have the phenomena of alternation of generations. A sexual female produces eggs, and from each hatches a tadpole larva which is without sex. This gives rise to buds, some at least of the individuals arising which in turn produce eggs.
Fig. 285.—Botryllus magnus Ritter. Part of colony. (After Ritter.)
In the family Salpidæ two kinds of individuals occur, the solitary salpa, or female, and the chain salpa, or bisexual males. The latter are united together in long bands, each individual forming a link in the chain held together by spurs extending from one to the next. From each solitary individual a long process or cord grows out, this dividing to form the chain. Each chain salpa produces male reproductive organs and each develops as well a single egg. The egg is developed within the body attached by a sort of placenta, while the spermatozoa are cast into the sea to fertilize other eggs. From each egg develops the solitary salpa and from her buds the chain of bisexual creatures. Dr. W. K. Brooks regards these as nursing males, the real source of the egg being perhaps the solitary female. Of this extraordinary arrangement the naturalist-poet Chamisso, who first described it, said: "A salpa mother is not like its daughter or its own mother, but resembles its sister, its granddaughter, and its grandmother." But it is misleading to apply such terms taken from the individualized human relationship to the singular communal system developed by these ultra-degenerate and strangely specialized Chordates.
Fig. 286.—Botryllus magnus Ritter, a single Zooid. Shumagin Islands, Alaska. (After Ritter.)
Fig. 287.—Aplidiopsis jordani Ritter, a compound Ascidian. Lukanin Beach, Pribilof Islands. (After Ritter.)
The Salpas abound in the warm seas, the chains often covering the water for miles. They are perfectly transparent, and the chains are often more than a foot in length. In Doliolum the body is barrel-shaped and the gills are less modified than in Salpa. The alternation of generations in this genus is still more complicated than in Salpa, for here we have not only a sexual and a non-sexual generation, the individuals of which differ from each other, but there is further a differentiation among the asexually produced individuals themselves; so that we have in all three instead of two sorts of animals in the complete life cycle. Besides the proliferating stolon situated on the ventral side, the bud-producing individual possesses a dorsal process larger than the stolon proper. The buds become completely severed from the true stolon at an early stage and actually crawl along the side of the parent up to the dorsal process, upon which they arrange themselves in three rows, two lateral and one median. The buds of the lateral rows become nutritive and respiratory zooids, while those of the[Pg 480] median row, ultimately at least, give rise in turn to the egg-producing individuals.
Origin of Tunicates.—There can be little doubt that the Tunicata form an offshoot from the primitive Chordate stock, and the structure of their larva in connection with that of the lancelet throws a large light on the nature of their common parents. "We may conclude," says Dr. Arthur Willey, "that the proximate ancestor of the Vertebrates was a free-swimming animal intermediate in organization between an Ascidian tadpole and Amphioxus, possessing the dorsal mouth, hypophysis, and restricted notochord of the former and the myotomes, cœlomic epithelium, and straight alimentary canal of the latter. The ultimate or primordial ancestor of the Vertebrates would, on the contrary, be a worm-like animal whose organization was approximately on a level with that of the bilateral ancestors of the Echinoderms."
Fig. 288.—Adult Tunicate of the group Larvacea, Oikopleura. Family Appendiculariidæ. (After Fol, per Parker & Haswell.)
Degeneration of Tunicates.—There is no question, furthermore, Professor Ritter observes, "that most of the group has undergone great degeneration in its evolutionary course. Just what the starting-point was, however, is a matter on which there is considerable difference of opinion among authorities. According to one view, particularly championed by Professor W. K. Brooks, Appendicularia is very near the ancestral form. The ancestor was consequently a small, marine, free-swimming creature. From this ancestor the Ascidiacea were evolved largely through the influence of the attached habit of life, and the tadpole stage in their development is a recapitulation of the ancestral form, just as the tadpole stage in the frog's life is a repetition of the fish ancestry of the frog.
"According to the most common view Appendicularia is not an ancestral form at all, but is the tadpole stage of the Ascidiacea that has failed to undergo metamorphosis and has become sexually mature in the larval condition, as the larva of certain Amphibians and insects are known to never pass into the adult state but reproduce their kind sexually in the larval condition. By this view the tadpole of such Ascidian as Ciona, for example, represents more closely the common ancestor of the group than does any other form we know. This view is especially defended by Professor K. Heider and Dr. Arthur Willey."

The Lancelet.—The lancelet is a vertebrate reduced to its very lowest terms. The essential organs of vertebrate life are there, but each one in its simplest form unspecialized and with structure and function feebly differentiated. The skeleton consists of a cartilaginous notochord inclosed in a membranous sheath. There is no skull. No limbs, no conspicuous processes, and no vertebræ are present. The heart is simply a long contractile tube, hence the name Leptocardii (from λεπτός, slender; καρδία, heart). The blood is colorless. There is a hepatic portal circulation. There is no brain, the spinal cord tapering in front as behind. The water for respiration passes through very many gill-slits from the pharynx into the atrium, from which it is excluded through the atripore in front of the vent. A large chamber, called the atrium, extends almost the length of the body along the ventral and lateral regions. It communicates with the pharynx through the gill-slits and with the exterior through a small opening in front of the vent, the atripore. The atrium is not found in forms above the lancelets.
The reproductive organs consist of a series of pairs of segmentally arranged gonads. The excretory organs consist of a series of tubules in the region of the pharynx, connecting the body-cavity with the atrium. The mouth is a lengthwise slit without jaws, and on either side is a row of fringes. From this feature comes the name Cirrostomi, from cirrus, a fringe of hair, and στόμα, mouth. The body is lanceolate in form, sharp at either end. From this fact arises a third name, Amphioxus, from ἀμφί, both; ὀξύς, sharp. Dorsal and anal fins are developed as folds of the skin supported by very slender rays.[Pg 483] There are no other fins. The alimentary canal is straight, and is differentiated into pharynx and intestine; the liver is a blind sac arising from the anterior end of the intestine. A pigment spot in the wall of the spinal cord has been interpreted as an eye. Above the snout is a supposed olfactory pit which some have thought to be connected with the pineal structure. The muscular impressions along the sides are very distinct and it is chiefly by means of the variation in numbers of these that the species can be distinguished. Thus in the common lancelet of Europe, Branchiostoma lanceolatum, the muscular bands are 35+14+12=61. In the common species of the Eastern coasts of America, Branchiostoma caribæum, these are 35+14+9=58, while in the California lancelet, Branchiostoma californiense, these are 44+16+9=69.
Habits of Lancelets.—Lancelets are slender translucent worm-like creatures, varying from half an inch (Asymmetron lucayanum) to four inches (Branchiostoma californiense) in length. They live buried in sand in shallow waters along the coasts of warm seas. One species, Amphioxides pelagicus, has been taken at the depth of 1000 fathoms, but whether at the bottom or floating near the surface is not known. The species are very tenacious of life and will endure considerable mutilation. Some of them are found on almost every coast in semi-tropical and tropical regions.
Species of Lancelets.—The Mediterranean species ranges northward to the south of England. Others are found as far north as Chesapeake Bay, San Diego, and Misaki in Japan, where is found a species called Branchiostoma belcheri. The sands at the mouth of San Diego Bay are noted as producing the largest of the species of lancelets, Branchiostoma californiense. From the Bahamas comes the smallest, the type of a distinct genus, Asymmetron lucayanum, distinguished among other things by a projecting tail. Other supposed genera are Amphioxides (pelagicus), dredged in the deep sea off Hawaii and supposed to be pelagic, the mouth without cirri; Epigonichthys (cultellus), from the East Indies, and Heteropleuron (bassanum), from Bass Straits, Australia. These little animals are of great interest to anatomists as giving the clue to the primitive structure of vertebrates. While possibly these have diverged widely from[Pg 484] their actual common ancestry with the fishes, they must approach near to these in many ways. Their simplicity is largely primitive, not, as in the Tunicates, the result of subsequent degradation.

Fig. 289.—California Lancelet, Branchiostoma californiense Gill. (From San Diego.)
The lancelets, less than a dozen species in all, constitute a single family, Branchiostomidæ. The principal genus, Branchiostoma, is usually called Amphioxus by anatomists. But while the name Amphioxus, like lancelet, is convenient in vernacular use, it has no standing in systematic nomenclature. The name Branchiostoma was given to lancelets from Naples in 1834, by Costa, while that of Amphioxus, given to specimens from Cornwall, dates from Yarrell's work on the British fishes in 1836. The name Amphioxus may be pleasanter or shorter or more familiar or more correctly descriptive than Branchiostoma, but if so the fact cannot be considered in science as affecting the duty of priority.
The name Acraniata (without skull) is often used for the lower Chordates taken collectively, and it is sometimes applied to the lancelets alone. It refers to those chordate forms which have no skull nor brain, as distinguished from the Craniota, or forms with a distinct brain having a bony or cartilaginous capsule for its protection.
Origin of Lancelets.—It is doubtless true, as Dr. Willey suggests, that the Vertebrates became separated from their worm-like ancestry through "the concentration of the central nervous system along the dorsal side of the body and its conversion[Pg 485] into a hollow tube." Besides this trait two others are common to all of them, the presence of the gill-slits and that of the notochord. The gill-slits may have served primarily to relieve the stomach of water, as in the lowest forms they enter directly into the body-cavity. The primitive function of the notochord is still far from clear, but its ultimate use of its structures in affording protection and in furnishing a fulcrum for the muscles and limbs is of the greatest importance in the processes of life.
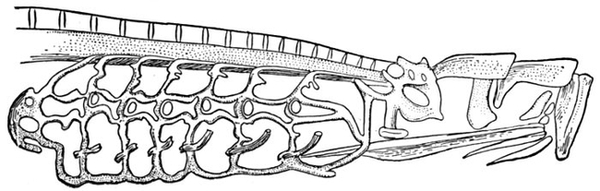
Fig. 289a.—Gill-basket of Lamprey.

The Lampreys.—Passing upward from the lancelets and setting aside the descending series of Tunicates, we have a long step indeed to the next class of fish-like vertebrates. During the period this great gap represents in time we have the development of brain, skull, heart, and other differentiated organs replacing the simple structures found in the lancelet.
The presence of brain without limbs and without coat-of-mail distinguishes the class of Cyclostomes, or lampreys (κυκλός, round; στόμα, mouth). This group is also known as Marsipobranchi (μαρσιπίον, pouch; βράγχος, gill); Dermopteri (δέρμα, skin; πτερόν, fin); and Myzontes (μυζάω, to suck). It includes the forms known as lampreys, slime-eels, and hagfishes.
Structure of the Lamprey.—Comparing a Cyclostome with a lancelet we may see many evidences of specialization in structure. The Cyclostome has a distinct head with a cranium formed of a continuous body of cartilage modified to contain a fish-like brain, a cartilaginous skeleton of which the cranium is evidently a differentiated part. The vertebræ are undeveloped, the notochord being surrounded by its membranes, without bony or cartilaginous segments. The gills have the form of fixed sacs, six to fourteen in number, on each side, arranged in a cartilaginous structure known as "branchial basket" (fig. 289a), the elements of which are not clearly homologous with the gill-arches of the true fishes. Fish-like eyes are developed on the sides of the head. There is a median nostril associated with a pituitary pouch, which pierces the skull floor. An ear-capsule is developed. The brain is composed of paired ganglia in general appearance resembling the brain of the true fish, but[Pg 487] the detailed homology of its different parts offers considerable uncertainty. The heart is modified to form two pulsating cavities, auricle and ventricle. The folds of the dorsal and anal fins are distinct, supported by slender rays.
The mouth is a roundish disk, with rasping teeth over its surface and with sharper and stronger teeth on the tongue. The intestine is straight and simple. The kidney is represented by a highly primitive pronephros and no trace exists of an air-bladder or lung. The skin is smooth and naked, sometimes secreting an excessive quantity of slime.
From the true fishes the Cyclostomes differ in the total absence of limbs and of shoulder and pelvic girdles, as well as of jaws. It has been thought by some writers that the limbs were ancestrally present and lost through degeneration, as in the eels. Dr. Ayers, following Huxley, finds evidence of the ancestral existence of a lower jaw. The majority of observers, however, regard the absence of limbs and jaws in Cyclostomes as a primitive character, although numerous other features of the modern hagfish and lamprey may have resulted from degeneration. There is no clear evidence that the class of Cyclostomes, as now known to us, has any great antiquity, and its members may be all degenerate offshoots from types of greater complexity of structure.
Supposed Extinct Cyclostomes.—No species belonging to the class of Cyclostomes has been found fossil. We may reason theoretically that the earliest fish-like forms were acraniate or lancelet-like, and that lamprey-like forms would naturally follow these, but this view cannot be substantiated from the fossils. Lancelets have no hard parts whatever, and could probably leave no trace in any sedimentary deposit. The lampreys stand between lancelets and sharks. Their teeth and fins at least might occasionally be preserved in the rocks, but no structures certainly known to be such have yet been recognized. It is however reasonably certain that the modern lamprey and hagfish are descendants, doubtless degraded and otherwise modified from species which filled the gap between the earliest chordate animals and the jaw-bearing sharks.
Conodontes.—Certain structures found as fossils have been from time to time regarded as Cyclostomes, but in all such[Pg 488] cases there is doubt as to the real nature of the fossil relic in question or as to the proper interpretation of its relationship.
Thus the Conodontes of the Cambrian, Silurian, and Devonian have been regarded as lingual teeth of extinct Cyclostomes. The Cycliæ of the Devonian have been considered as minute lampreys, although the vertebral segments are highly specialized, to a degree far beyond the condition seen in the lampreys of to-day. The Ostracophores have been regarded as monstrous lampreys in coat of mail, and the possibility of a lamprey origin even for Arthrodires has been suggested. The Cycliæ and Ostracophori were apparently without jaws or limbs, being in this regard like the Cyclostomes, but their ancestry and relationships are wholly problematical.
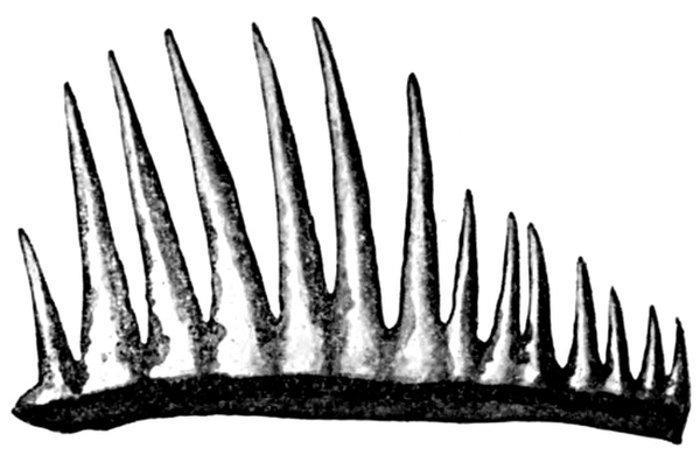
Fig. 290.—Polygnathus dubium Hinde. A Conodont from the New York Devonian. (After Hinde.)
The nature of the Conodontes is still uncertain. In form they resemble teeth, but their structure is different from that of the teeth of any fishes, agreeing with that of the teeth of annelid worms. Some have compared them to the armature of Trilobites. Some fifteen nominal genera are described by Pander in Russia, and by Hinde about Lake Erie and Lake Ontario. Some of these, as Drepaniodus, are simple, straight or curved grooved teeth or tooth-like structures; others, as Prioniodus, have numerous smaller teeth or denticles at the base of the larger one.
Orders of Cyclostomes.—The known Cyclostomes are naturally divided into two orders, the Hyperotreta, or hagfishes, and the Hyperoartia, or lampreys. These two orders are very distinct from each other. While the two groups agree in the general form of the body, they differ in almost every detail, and there is much pertinence in Lankester's suggestions that each should stand as a separate class. The ancestral forms of each, as well as the intervening types if such ever existed, are left unrecorded in the rocks.
The Hyperotreta, or Hagfishes.—The Hyperotreta (ὑπερῴα, pal[Pg 489]ate; τρετός, perforate), or hagfishes, have the nostril highly developed, a tube-like cylinder with cartilaginous rings penetrating the palate. In these the eyes are little developed and the species are parasitic on other fishes. In Polistotrema stouti, the hagfish of the coast of California, is parasitic on large fishes, rockfishes, or flounders. It usually fastens itself at the throat or isthmus of its host and sometimes at the eyes. Thence it works very rapidly to the inside of the body. It there devours all the muscular part of the fish without breaking the skin or the peritoneum, leaving the fish a living hulk of head, skin, and bones. It is especially destructive to fishes taken in gill-nets. The voracity of the Chilean species Polistotrema dombeyi is equally remarkable. Dr. Federico T. Delfin finds that in seven hours a hagfish of this species will devour eighteen times its own weight of fish-flesh. The intestinal canal is a simple tube, through which most of the food passes undigested. The eggs are large, each in a yellowish horny case, at one end of which are barbed threads by which they cling together and to kelp or other objects. In the California hagfish, Polistotrema stouti, great numbers of these eggs have been found in the stomachs of the males.

Fig. 291.—California Hagfish, Polistotrema stouti Lockington.
Similar habits are possessed by all the species in the two families, Myxinidæ and Eptatretidæ. In the Myxinidæ the gill-openings are apparently single on each side, the six gills being internal and leading by six separate ducts to each of the six branchial sacs. The skin is excessively slimy, the extensible tongue is armed with two cone-like series of strong teeth. About the mouth are eight barbels.
Of Myxine, numerous species are known—Myxine glutinosa, in the north of Europe; Myxine limosa, of the West Atlantic; Myxine australis, and several others about Cape Horn, and Myxine garmani in Japan. All live in deep waters and none have been fully studied. It has been claimed that the hagfish is male when young, many individuals gradually changing to female, but this conclusion lacks verification and is doubtless without foundation.
In the Eptatretidæ the gill-openings, six to fourteen in number, are externally separate, each with its own branchial sac as in the lampreys.
The species of the genus Eptatretus (Bdellostoma, Heptatrema, and Homea, all later names for the same group) are found only in the Pacific, in California, Chile, Patagonia, South Africa, and Japan. In general appearance and habits these agree with the species of Myxine. The species with ten to fourteen gill-openings (dombeyi: stouti) are sometimes set off as a distinct genus (Polistotrema), but in other regards the species differ little, and frequent individual variations occur. Eptatretus burgeri is found in Japan and Eptatretus forsteri in Australia.
The Hyperoartia, or Lampreys.—In the order Hyperoartia, or lampreys, the single nostril is a blind sac which does not penetrate the palate. The seven gill-openings lead each to a separate sac, the skin is not especially covered with mucus, the eyes are well developed in the adult, and the mouth is a round disk armed with rasp-like teeth, the comb-like teeth on the tongue being less developed than in the hagfishes. The intestine in the lampreys has a spiral valve. The eggs are small and are usually laid in brooks away from the sea, and in most cases the adult lamprey dies after spawning. According to Thoreau, "it is thought by fishermen that they never return, but waste away and die, clinging to rocks and stumps of trees for an indefinite period, a tragic feature in the scenery of the river-bottoms worthy to be remembered with Shakespeare's description of the sea-floor." This account is not far from the truth, as recent studies have shown.
The lampreys of the northern regions constitute the family of Petromyzonidæ. The larger species (Petromyzon, Entosphenus) live in the sea, ascending rivers to spawn, and often becoming[Pg 491] land-locked and reduced in size by living in rivers only. Such land-locked marine lampreys (Petromyzon marinus unicolor) breed in Cayuga Lake and other lakes in New York. The marine forms reach a length of three feet. Smaller lampreys of other genera six inches to eighteen inches in length remain all their lives in the rivers, ascending the little brooks in the spring, clinging to stones and clods of earth till their eggs are deposited. These are found throughout northern Europe, northern Asia, and the colder parts of North America, belonging to the genera Lampetra and Ichthyomyzon. Other and more aberrant genera from Chile and Australia are Geotria and Mordacia, the latter forming a distinct family, Mordaciidæ. In Geotria, a large and peculiar gular pouch is developed at the throat. In Macrophthalmia chilensis from Chile the eyes are large and conspicuous.
Food of Lampreys.—The lampreys feed on the blood and flesh of fishes. They attach themselves to the sides of the various species, rasp off the flesh with their teeth, sucking the blood till the fish weakens and dies. Preparations made by students of Professor Jacob Reighard in the University of Michigan show clearly that the lamprey stomach contains muscular tissue as well as the blood of fishes. The river species do a great deal of mischief, a fact which has been the subject of a valuable investigation by Professor H. A. Surface, who has also considered the methods available for their destruction. The flesh of the lamprey is wholesome, and the larger species, especially the great sea lamprey of the Atlantic, Petromyzon marinus, are valued as food. The small species, according to Prof. Gage, never feed on fishes.

Fig. 292.—Lamprey, Petromyzon marinus L. Woods Hole, Mass.
Metamorphosis of Lampreys.—All lampreys, so far as known, pass through a distinct metamorphosis. The young, known as the Ammocœtes form, are slender, eyeless, and with the mouth[Pg 492] narrow and toothless. From Professor Surface's paper on "The Removal of Lampreys from the Interior Waters of New York" we have the following extracts (slightly condensed):
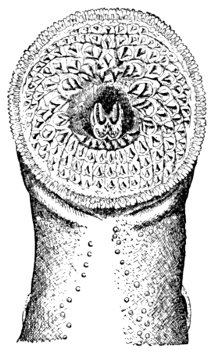
Fig. 293.—Petromyzon marinus unicolor (De Kay). Mouth of Lake Lamprey, Cayuga Lake. (After Gage.)
Fig. 294.—Lampetra wilderi Jordan & Evermann. Larval Brook Lamprey in its burrow in a glass filled with sand. (After Gage.)

Fig. 295.—Lampetra wilderi Jordan & Evermann. Mouth of Brook Lamprey. Cayuga Lake. (After Gage.)
"In the latter part of the fall the young lampreys, Petromyzon marinus unicolor, the variety land-locked in the lakes of Central New York, metamorphose and assume the form of the adult. They are now about six or eight inches long. The externally segmented condition of the body disappears. The eyes appear to grow out through the skin and become plainly visible and functional. The mouth is no longer filled with vertical membranous sheets to act as a sieve, but it contains nearly one hundred and fifty sharp and chitinous teeth, arranged in rows that are more or less concentric and at the same time presenting the appearance of circular radiation. These teeth are very strong, with sharp points, and in structure each has the appearance of a hollow cone of chitin placed over another cone or papilla. A little below the center of the mouth is the oral opening, which is circular and contains a flattened tongue which bears finer teeth of chitin set closely together and arranged in two interrupted (appearing as four) curved rows extending[Pg 493] up and down from the ventral toward the dorsal side of the mouth. Around the mouth is a circle of soft membrane finally surrounded by a margin of fimbriæ or small fringe. This completes the apparatus with which the lamprey attaches itself to its victims, takes its food, carries stones, builds and tears down its nest, seizes its mate, holds itself in position in a strong current, and climbs over falls."
Mischief Done by Lampreys.—"The most common economic feature in the entire life history of these animals is their feeding habits in this (spawning) stage, their food now consisting wholly of the blood (and flesh) of fishes. A lamprey is able to strike its suctorial mouth against a fish, and in an instant becomes so firmly attached that it is very rarely indeed that the efforts of the fish will avail to rid itself of its persecutor. When a lamprey attaches itself to a person's hand in the aquarium, it can only be freed by lifting it from the water. As a rule it will drop the instant it is exposed to the open air, although often it will remain attached for some time even in the open air, or may attach itself to an object while out of water.
"Nearly all lampreys that are attached to fish when they are caught in nets will escape through the meshes of the nets, but some are occasionally brought ashore and may hang on to their victim with bulldog pertinacity.
"The fishes that are mostly attacked are of the soft-rayed species, having cycloid scales, the spiny-rayed species with ctenoid scales being most nearly immune from their attacks. We think there may be three reasons for this: 1st, the fishes of the latter group are generally more alert and more active than those of the former, and may be able more readily to dart away from such enemies; 2d, their scales are thicker and stronger and appear to be more firmly imbedded in the skin, consequently it is more difficult for the lampreys to hold on and cut through the heavier coat-of-mail to obtain the blood of the victim; 3d, since the fishes of the second group are wholly carnivorous and in fact almost exclusively fish-eating when adult, in every body of water they are more rare than those of the first group, which are more nearly omnivorous. According to the laws and requirements of nature the fishes of the first group must be more abundant, as they become the food for those of the[Pg 494] second, and it is on account of their greater abundance that the lampreys' attacks on them are more observed.
"There is no doubt that the bullhead, or horned pout (Ameiurus nebulosus), is by far the greatest sufferer from lamprey attacks in Cayuga Lake. This may be due in part to the sluggish habits of the fish, which render it an easy victim, but it is more likely due to the fact that this fish has no scales and the lamprey has nothing to do but to pierce the thick skin and find its feast of blood ready for it. There is no doubt of the excellency of the bullhead as a food-fish and of its increasing favor with mankind. It is at present the most important food- and market-fish of the State (New York), being caught by bushels in the early part of June when preparing to spawn. As we have observed at times more than ninety per cent. of the catch attacked by lampreys, it can readily be seen how very serious are the attacks of this terrible parasite which is surely devastating our lakes and streams."
Migration or "Running" of Lampreys.—"After thus feeding to an unusual extent, their reproductive elements (gonads) become mature and their alimentary canals commence to atrophy. This duct finally becomes so occluded that from formerly being large enough to admit a lead-pencil of average size when forced through it, later not even liquids can pass through, and it becomes nearly a thread closely surrounded by the crowding reproductive organs. When these changes commence to ensue, the lampreys turn their heads against the current and set out on their long journeys to the sites that are favorable for spawning, which here may be from two to eight miles from the lake. In this migration they are true to their instincts and habits of laziness in being carried about, as they make use of any available object, such as a fish, boat, etc., that is going in their direction, fastening to it with their suctorial mouths and being borne along at their ease. During this season it is not infrequent that as the Cornell crews come in from practice and lift their shells from the water, they find lampreys clinging to the bottoms of the boats, sometimes as many as fifty at one time. They are likely to crowd up all streams flowing into the lake, inspecting the bed of the stream as they go. They do not stop until they reach favorable spawning sites, and if they[Pg 495] find unsurmountable obstacles in their way, such as vertical falls or dams, they turn around and go down-stream until they find another, up which they go. This is proved every spring by the number of adult lampreys which are seen temporarily in Pall Creek and Cascadilla Creek. In each of these streams, about a mile from its mouth, there is a vertical fall over thirty feet in height which the lampreys cannot surmount, and in fact they have never been seen attempting to do so. After clinging with their mouths to the stones at the foot of the falls for a few days, they work their way down-stream, carefully inspecting all the bottom for suitable spawning sites. They do not spawn in these streams because there are too many rocks and no sand, but finally enter the only stream (the Cayuga Lake inlet) in which they find suitable and accessible spawning sites.

Fig. 296.—Kamchatka Lamprey, Lampetra camtschatica (Tilesius). Kamchatka.
"The three-toothed lampreys (Entosphenus tridentatus) of the West Coast climb low falls or rapids by a series of leaps, holding with their mouths to rest, then jumping and striking again and holding, thus leap by leap gaining the entire distance.
"The lampreys here have never been known to show any tendency or ability to climb, probably because there are no rapids or mere low falls in the streams up which they would run. In fact, as the inlet is the only stream entering Cayuga Lake in this region which presents suitable spawning conditions and no obstructions, it can be seen at once that all the lampreys must spawn in this stream and its tributaries.

Fig. 297.—Oregon Lamprey, Entosphenus tridentatus, ascending a brook. (Modified from a photograph by Dr. H. M. Smith. Published by Prof. H. A. Surface.) Willamette River, Oregon.
"In 'running' they move almost entirely at night, and if they do not reach a suitable spawning site by daylight, they will cling to roots or stones during the day and complete their journey the next night. This has been proven by the positive observation of individuals. Of the specimens that run up early in the season, about four-fifths are males. Thus the males do not exactly precede the females, because we have found the latter sex represented in the stream as early in the season as the former, but in the earlier part of the season the number of the males certainly greatly predominates. This proportion of males gradually decreases, until in the middle of the spawning season the sexes are about equally represented, and toward the latter part of the season the females continue to come until they in turn show the greater numbers. Thus it appears very evident in general that the reproductive instinct impels the most of the males to seek the spawning ground before the most of the females do. However, it should be said that neither the males nor the females show all of the entirely sexually mature features when they first run up-stream in the beginning of the season, but later they are perfectly mature and 'ripe' in every regard when they first appear in the stream. When they migrate, they stop at the site that seems to suit their fancy, many stopping near the lake, others pushing on four or five miles farther up-stream. We have noted, however, that later in the season the lower courses become more crowded, showing that the late comers do not attempt to push up-stream as far as those that came earlier. Also it thus follows, from what was just said about late-running females, that in the latter part of the season the lower spawning beds are especially crowded with females. In fact, during the early part of the month of June we have found, not more than half a mile above the lowest spawning bed, as many as five females on a spawning nest with but one male; and in that immediate vicinity many nests indeed were found at that time with two or three females and but one male.
"Having arrived at a shoal which seems to present suitable conditions for a spawning nest, the individual or pair commences at once to move stones with its mouth from the centre to the margin of an area one or two feet in diameter. When many stones are thus placed, especially at the upper edge, and they are cleaned quite free of sediment and algæ, both by being moved and by being fanned with the tail, and when the proper condition of sand is found in the bottom of the basin thus formed,[Pg 498] it is ready to be used as a spawning bed or nest. A great many nests are commenced and deserted. This has been left as a mystery in publications on the subject, but we are well convinced that it is because the lampreys do not find the requisites or proper conditions of bottom (rocks, sand, etc., as given below) to supply all their needs and fulfill all conditions for ideal sites. This desertion of half-constructed nests is just what would be expected and anticipated in connection with the explanation of 'Requisite Conditions for Spawning,' given below, because some shallows contain more sand and fewer stones, and others contain many larger stones but no sand, while others contain pebbles lying over either rocks or sand. The lampreys remove some of the material, and if they do not find all the essentials for a spawning nest, the site is deserted and the creatures move on."
Requisite Conditions for Spawning with Lampreys.—"For a spawning site two conditions are immediately essential—proper conditions of water and suitable stream bed or bottom. Of course with these it is essential that no impassable barriers (dam or falls) exist between the lake and the spawning sites to prevent migration at the proper 'running' season. Lampreys will not spawn where there is no sand lying on the bottom between the rocks, as sand is essential in covering the eggs (see remarks on the 'Spawning Process'); neither will they spawn where the bottom is all sand and small gravel, as they cannot take hold of this material with their mouths to construct nests or to hold themselves in the current, and they would not find here pebbles and stones to carry over the nest while spawning, as described elsewhere. It can thus be seen that, as suggested above, the reason they do not spawn in Fall Creek and Cascadilla Creek, between the lake and the falls, is that the beds of these streams are very rocky, being covered only with large stones and no sand. There is no doubt that the lampreys find here suitable conditions of water, but they do not remain to spawn on account of the absence of the proper conditions of stream bed. Again, they do not spawn in the lower course of the inlet for a distance of nearly two miles from the lake, because near the lake the bed of the stream is composed of silt, while for some distance above this (up[Pg 499]-stream) there is nothing but sand. Farther up-stream are found pebbles and stones commingled with sand, which combination satisfies the demands of the lampreys for material in constructing nests and covering eggs. The accessibility of these sites, together with their suitable conditions, render the inlet the great and perhaps the only spawning stream of the lake; and, doubtless, all the mature lampreys come here to spawn, excepting a few which spawn in the lower part of Six-mile Creek, a tributary of the inlet.
"As the course of the stream where the beds abound is divided into pools, separated by stony ripples or shallows, the nests must be made at the ends of the pools. Of the spawning beds personally observed during several seasons, nine-tenths of the entire number were formed just above the shallows at the lower ends of the pools, while only a few were placed below them. An advantage in forming the nest above the shoals rather than below it is that in the former place the water runs more swiftly over the lower and middle parts of such a bed than at its upper margin, since the velocity decreases in either direction from the steeper part of the shallows; and any organic material or sediment that would wash over the upper edge of the nest is thus carried on rather than left as a deposit. When formed below the shallows, owing to the decreased velocity at the lower part of the nest compared with that at the upper, the sediment is likely to settle in the hollow of the nest, and, through the process of decay of the organic material, prove disastrous or unfavorable for the developing embryos.
"The necessity of sand in the spawning bed indicates the explanation of why we see so many shallows which have no spawning lampreys upon them, while there are others in the same vicinity that are crowded. There will be no nests formed if there is too little or too much sand, not enough or too many stones, or stones that are all too small or all too large. The stones must vary from the size of an egg to the size of a man's hand, and must be intermingled with sand without mud or rubbish.
"The lampreys choose to make their spawning nests just where the water flows so swiftly that it will carry the sand a short distance, but will not sweep it out of the nest. This[Pg 500] condition furnishes not only force to wash the sand over the eggs when laid, but also keeps the adult lampreys supplied with an abundance of fresh water containing the dissolved air needed for their very rapid respiration. Of course in such rapid water the eggs are likely to be carried away down-stream, but Nature provides against this by the fact that they are adhesive, and the mating lampreys stir up the sand with their tails, thus weighing down the freshly laid eggs and holding them in the nest. Hence the necessity of an abundance of sand at the spawning site."
The Spawning Process with Lampreys.—"There is much interest in the study of the spawning process, as it is for the maintenance of the race that the lampreys risk and end their lives; and as they are by far the lowest form of vertebrates found within the United States, a consideration of their actions and apparent evidences of instinct becomes of unusual attraction. Let us consider one of those numerous examples in which the male migrates before the female. When he comes to that portion of the stream where the conditions named above are favorable, he commences to form a nest by moving and clearing stones and making a basin with a sandy bottom about the size of a common wash-bowl. Several nests may be started and deserted before perfect conditions are found for the completion of one. The male may be joined by a female either before or after the nest is completed. There is at once harmony in the family; but if another male should attempt to intrude, either before or after the coming of the female, he is likely to be summarily dealt with and dismissed at once by the first tenant. As soon as the female arrives she too commences to move pebbles and stones with her mouth.
"Sometimes the nest is made large enough to contain several pairs, or often unequal numbers of males and females; or they may be constructed so closely together as to form one continuous ditch across the stream, just above the shallows. Many stones are left at the sides and especially at the upper margin of the nest, and to these both lampreys often cling for a few minutes as though to rest. While the female is thus quiet, the male seizes her with his mouth at the back of her head, clinging as to a fish. He presses his body as tightly as possible[Pg 501] against her side, and loops his tail over her near the vent and down against the opposite side of her body so tightly that the sand, accidentally coming between them, often wears the skin entirely off of either or both at the place of closest contact. In most observed instances the male pressed against the right side of the female, although there is no unvarying rule as to position. The pressure of the male thus aids to force the eggs from the body of the female, which flow very easily when ripe. The vents of the two lampreys are thus brought into close proximity, and the conspicuous genital papilla of the male serves to guide the milt directly to the issuing spawn. There appears to be no true intromission, although definite observation of this feature is quite difficult, and, in fact, impossible. During the time of actual pairing, which lasts but a few seconds, both members of the pair exhibit tremendous excitement, shaking their bodies in rapid vibrations and stirring up such a cloud of sand with their tails that their eggs are at once concealed and covered. As the eggs are adhesive and non-buoyant, the sand that is stirred up adheres to them immediately and covers most of them before the school of minnows in waiting just below the nest can dart through the water and regale themselves upon the eggs of these enemies of their race; but woe to the eggs that are not at once concealed. We would suggest that the function of the characteristic anal fin, which is possessed only by the female, and only at this time of year, may be to aid in this vastly important process of stirring up the sand as the eggs are expelled; and the explanation of the absence of such a fin from the ventral side of the tail of the male may be found in the fact that it could not be used for the same purpose at the instant when most needed, since the male is just then using his tail as a clasping organ to give him an essential position in pairing. As soon as they shake together they commence to move stones from one part of the nest to another, to bring more loose sand down over their eggs. They work at this from one to five minutes, then shake again, thus making the intervals between mating from one to five minutes, with a general average of about three and a half minutes.
"Although their work of moving stones does not appear to be systematic in reference to the placing of the pebbles, or[Pg 502] as viewed from the standpoint of man, it does not need to be so in order to perfectly fulfill all the purposes of the lampreys. As shown above in the remarks on the spawning habits of the brook lampreys, the important end which they thus accomplish is the loosening and shifting of the sand to cover their eggs; and the more the stones are moved, even in the apparently indiscriminate manner shown, the better is this purpose achieved. Yet, in general, they ultimately accomplish the feat of moving to the lower side of the nest all the stones they have placed or left at the upper margin. At the close of the spawning season when the nest is seen with no large pebbles at its upper margin, but quite a pile of stones below, it can be known that the former occupants completed their spawning process there; but if many small stones are left at the upper edge and at the sides, and a large pile is not formed at the lower edge, it can be known that the nest was forsaken or the lampreys removed before the spawning process was completed. The stones they move are often twice as heavy as themselves, and are sometimes even three or four times as heavy. Since they are not attempting to build a stone wall of heavy material, there is no occasion for their joining forces to remove stones of extraordinary size, and they rarely do so, although once during the past spring (1900) we saw two lake lampreys carrying the same large stone down-stream across their nest. Although this place was occupied by scores of brook lampreys, there were but three pairs of lake lampreys seen here. It is true that one of these creatures often moves the same stone several times, and many even attempt many times to move a stone that has already been found too heavy for it; but sooner or later the rock may become undermined so that the water will aid them, and they have no way of knowing what they can do under such circumstances until they try. Also, the repeated moving of one stone may subserve the same purpose for the lamprey in covering its eggs with sand as would the less frequent removal of many.
"When disturbed on the spawning nest, either of the pair will return to the same nest if its mate is to be found there; but if its mate is in another place, it will go to it, and if its mate is removed or killed, it is likely to go to any part of the[Pg 503] stream to another nest. When disturbed, they often start up-stream for a short distance, but soon dart down-stream with a velocity that is almost incredible. They can swim faster than the true fishes, and after they get a start are generally pretty sure to make good their escape, although we have seen them dart so wildly and frantically down-stream that they would shoot clear out on the bank and become an easy victim of the collector. This peculiar kind of circumstance is most likely to happen with those lampreys that are becoming blinded from long exposure to the bright light over the clear running water. If there is a solitary individual on a nest when disturbed, it may not return to that nest, but to any that has been started, or it may stay in the deep pool below the shallows until evening and then move some distance up-stream. When the nest is large and occupied by several individuals, those that are disturbed may return to any other such nest. We have never seen evidence of one female driving another female out of a spawning-nest; and from the great number of nests in which we have found the numbers of the females exceeding those of the males, we would be led to infer that the former live together in greater harmony than do the males.
"Under the subject of the number of eggs laid, we should have said that at one shake the female spawns from twenty to forty. We once caught in fine gauze twenty-eight eggs from a female at one spawning instant. In accordance with the frequency of spawning stated, and the number of eggs contained in the body of one female, the entire length of time given to the spawning process would be from two to four days. This agrees with the observed facts, although the lampreys spend much time in moving stones and thoroughly covering the nests with sand. Even after the work of spawning and moving stones is entirely completed, they remain clinging to rocks in various parts of the stream, until they are weakened by fungus and general debility, when they gradually drift down-stream.
"In forming nests there is a distinct tendency to utilize those sites that are concealed by overhanging bushes, branches, fallen tree-tops, or grass or weeds, probably not only for concealment, but also to avoid the bright sunlight, which sooner or later causes them to go blind, as it does many fishes when[Pg 504] they have to live in water without shade. Toward the end of the spawning season, it is very common to see blind lampreys clinging helplessly to any rocks on the bottom, quite unable to again find spawning-beds. However, at such times they are generally spent and merely awaiting the inevitable end.
"As with the brook lamprey, the time of spawning and duration of the nesting period depend upon the temperature of the water, as does also the duration of the period of hatching or development of the embryo. They first run up-stream when the water reaches a temperature of 45° or 48° Fahr., and commence spawning at about 50°. A temperature of 60° finds the spawning process in its height, and at 70° it is fairly completed. It is thus that the rapidity with which the water becomes heated generally determines the length of time the lampreys remain in the stream. This may continue later in the season for those that run later, but usually it is about a month or six weeks from the time the first of this species is seen on a spawning-nest until the last is gone."
What becomes of Lampreys after Spawning?—"There has been much conjecture as to the final end of the lampreys, some writers contending that they die after spawning, others that they return to deep water and recuperate, and yet others compromise these two widely divergent views by saying that some die and others do not. The fact is that the spawning process completely wears out the lampreys, and leaves them in a physical condition from which they could never recover. They become stone-blind; the alimentary canal suffers complete atrophy; their flesh becomes very green from the katabolic products, which find the natural outlet occluded; they lose their rich yellow color and plump, symmetrical appearance; their skin becomes torn, scratched, and worn off in many places, so that they are covered with sores, and they become covered with a parasitic or sarcophytic fungus, which forms a dense mat over almost their entire bodies, and they are so completely debilitated and worn out that recovery is entirely out of the question. What is more, the most careful microscopical examination of ovaries and testes has failed to reveal any evidence of new gonads or reproductive bodies. This is proof that reproduction could not again ensue without a practical rebuilding of[Pg 505] the animals, even though they should regain their vitality. A. Mueller, in 1865, showed that all the ova in the lamprey were of the same size, and that after spawning no small reproductive bodies remained to be developed later. This is strong evidence of death after once spawning.
"One author writes that an argument against the theory of their dying after spawning can be found in the fact that so few dead ones have been found by him. However, many can be found dead if the investigator only knows how and where to look for them. We should not anticipate finding them in water that is shallow enough for the bottom to be plainly seen, as there the current is strong enough to move them. It is in the deep, quiet, pools where sediment is depositing that the dead lampreys are dropped by the running water, and there they sink into the soft ooze.
"The absence of great numbers of dead lampreys from visible portions of the stream cannot be regarded as important evidence against the argument that they die soon after spawning once, as the bodies are very soon disintegrated in the water. In the weir that we maintained in 1898, a number of old, worn-out, and fungus-covered lampreys were caught drifting down-stream; some were dead, some alive, and others dying and already insensible, but none were seen going down that appeared to be in condition to possibly regain their strength."

Fig. 297a.—Brook Lamprey, Lampetra Wilderi. (After Gage.)

The Sharks.—The gap between the lancelets and the lampreys is a very wide one. Assuming the primitive nature of both groups, this gap must represent the period necessary for the evolution of brain, skull, and elaborate sense organs. The interspace between the lampreys and the nearest fish-like forms which follow them in an ascending scale is not less remarkable. Between the lamprey and the shark we have the development of paired fins with their basal attachments of shoulder-girdle and pelvis, the formation of a lower jaw, the relegation of the teeth to the borders of the mouth, the development of separate vertebræ along the line of the notochord, the development of the gill-arches, and of an external covering of enameled points or placoid scales.
These traits of progress separate the Elasmobranchs from all lower vertebrates. For those animals which possess them, the class name of Pisces or fishes has been adopted by numerous authors. If this term is to be retained for technical purposes, it should be applied to the aquatic vertebrates above the lampreys and lancelets. We may, however, regard fish as a popular term only, rather than to restrict the name to members of a class called Pisces. From the bony fishes, on the other hand, the sharks are distinguished by the much less specialization of the skeleton, both as regards form and substance, by the lack of membrane bones, of air-bladder, and of true scales, and by various peculiarities of the skeleton itself. The upper jaw, for example, is formed not of maxillary and premaxillary, but of elements which in the lower fishes would be regarded as belonging to the palatine and pterygoid series. The lower jaw is formed[Pg 507] not of several pieces, but of a cartilage called Meckel's cartilage, which in higher fishes precedes the development of a separate dentary bone. These structures are sometimes called primary jaws, as distinguished from secondary jaws or true jaws developed in addition to those bones in the Actinopteri or typical fishes. In the sharks the shoulder-girdle is attached, not to the skull, but to a vertebra at some distance behind it, leaving a distinct neck, such as is possessed or retained by the vertebrate higher than fishes. The shoulder-girdle itself is a continuous arch of cartilage, joining its fellow at the breast of the fish. Other peculiar traits will be mentioned later.
Characters of Elasmobranchs.—The essential character of the Elasmobranchs as a whole are these: The skeleton is cartilaginous, the skull without sutures, and the notochord more or less fully replaced or inclosed by vertebral segments. The jaws are peculiar in structure, as are also the teeth, which are usually highly specialized and found on the jaws only. There are no membrane bones; the shoulder-girdle is well developed, each half of one piece of cartilage, and the ventral fins, with the pelvic-girdle, are always present, always many-rayed, and abdominal in position. The skin is covered with placoid scales, or shagreen, or with bony bucklers, or else it is naked. It is never provided with imbricated scales. The tail is diphycercal, heterocercal, or else it degenerates into a whip-like organ, a form which has been called leptocercal. The gill-arches are 5, 6, or 7 in number, with often an accessory gill-slit or spiracle. The ventral fins in the males (except perhaps in certain primitive forms) are provided with elaborate cartilaginous appendages or claspers. The brain is elongate, its parts well separated, the optic nerves interlacing. The heart has a contractile arterial cone containing several rows of valves; the intestine has a spiral valve; the eggs are large, hatched within the body, or else deposited in a leathery case.
Classification of Elasmobranchs.—The group of sharks and their allies, rays, and Chimæras, is usually known collectively as Elasmobranchii (ἐλάσμος, blade or plate; βράγχος, gill). Other names applied to all or a part of this group are these: Selachii (σελαχός, a cartilage, the name also used by the Greeks for the gristle-fishes or sharks); Plagiostomi (πλαγιός, oblique; στόμα,[Pg 508] mouth); Chondropterygii (χόνδρος, cartilage; πτερύξ, fin); and Antacea (ἀντακαῖος, sturgeon). They represent the most primitive known type of jaw-bearing vertebrates, or Gnathostomi (γνάθος, jaw; στόμα, mouth), the Chordates without jaws being sometimes called collectively Agnatha (ἀ-γνάθος, without jaws). These higher types of fishes have been also called collectively Lyrifera, the form of the two shoulder-girdles taken together being compared to that of a lyre. Through shark-like forms all the higher vertebrates must probably trace their descent. Sharks' teeth and fin-spines are found in all rocks from the Upper Silurian deposits to the present time, and while the majority of the genera are now extinct, the class has had a vigorous representation in all the seas, later Palæozoic, Mesozoic, and Cenozoic, as well as in recent times.
Most of the Elasmobranchs are large, coarse-fleshed, active animals feeding on fishes, hunting down their prey through superior strength and activity. But to this there are many exceptions, and the highly specialized modern shark of the type of the mackerel-shark or man-eater is by no means a fair type of the whole great class, some of the earliest types being diminutive, feeble, and toothless.
Subclasses of Elasmobranchs.—With the very earliest recognizable remains it is clear that the Elasmobranchs are already divided into two great divisions, the sharks and the Chimæras. These groups we may call subclasses, the Selachii and the Holocephali, or Chismopnea.
The Selachii, or sharks and rays, have the skull hyostylic, that is, with the quadrate bone grown fast to the palate which forms the upper jaw, the hyomandibular, acting as suspensorium to the lower jaw, being articulated directly to it.
The palato-quadrate apparatus, the front of which forms the upper jaw in the shark, is not fused to the cranium, although it is sometimes articulated with it. There are as many external gill-slits as there are gill-arches (5, 6, or 7), and the gills are adnate to the flesh of their own arches, without free tips. The cerebral hemispheres are grown together. The teeth are separated and usually strongly specialized, being primitively modified from the prickles or other defences of the skin. There is no frontal holder or bony hook on the forehead of the male.
The subclass Holocephali, or Chimæras, differ from the sharks in all this series of characters, and its separation as a distinct group goes back to the Devonian or even farther, the earliest known sharks having little more in common with Chimæras than the modern forms have.
The Selachii.—There have been many efforts to divide the sharks and rays into natural orders. Most writers have contented themselves with placing the sharks in one order (Squali or Galei or Pleurotremi) having the gill-openings on the side, and the rays in another (Rajæ, Batoidei, Hypotrema) having the gill-openings underneath. Of far more importance than this superficial character of adaptation are the distinctions drawn from the skeleton. Dr. Gill has used the attachment of the palato-quadrate apparatus as the basis of a classification. The Opistharthri (Hexanchidæ) have this structure articulated with the postorbital part of the skull. In the Prosarthri (Heterodontidæ) it is articulated with the preorbital part of the skull, while in the other sharks (Anarthri) it is not articulated at all. But these characters do not appear to be always important. Chlamydoselachus, for example, differs in this regard from Heptranchias, which in other respects it closely resembles. Yet, in general, the groups thus characterized are undoubtedly natural ones.

Fig. 298.—Fin-spine of Onchus tenuistriatus Agassiz. (After Zittel.)
Hasse's Classification of Elasmobranchs.—In 1882, Professor Carl Hasse proposed to subdivide the sharks on the basis of the structure of the individual vertebræ. In the lowest group, a hypothetical order of Polyospondyli, possibly represented by the fossil spines called Onchus, an undivided notochord, perhaps swollen at regular intervals, is assumed to have represented the vertebral column. In the Diplospondyli (Hexanchidæ) the im[Pg 510]perfectly segmented vertebræ are joined in pairs, each pair having two neural arches. In the Asterospondyli or ordinary sharks each vertebra has its calcareous lamella radiating star-like from the central axis. In the Cyclospondyli (Squalidæ, etc.) the calcareous part forms a single ring about the axis, and in the Tectospondyli (Squatina, rays, etc.) it forms several rings. These groups again are natural and correspond fairly with those based on other characters. At the same time there is no far-reaching difference between Cyclospondyli and Tectospondyli, and the last-named section includes both sharks and rays.
Fig. 299.—Section of vertebræ of sharks, showing calcification. (After Hasse.) 1. Cyclospondyli (Squalus); 2. Tectospondyli (Squatina); 3. Asterospondyli (Carcharias).
Nothing is known of the Polyospondyli, and they may never have existed at all. The Diplospondyli do not differ very widely from the earlier Asterospondyli (Cestraciontes) which, as a matter of fact, have preceded the Diplospondyli in point of time, if we can trust our present knowledge of the geological record.
Other Classifications of Elasmobranchs.—Characters more fundamental may be drawn from the structure of the pectoral fin. In this regard four distinct types appear. In Acanthoessus this fin consists of a stout, stiff spine, with a rayless membrane attached behind it. In Cladoselache the fin is low, with a very long base, like a fold of skin (ptychopterygium), and composed of feeble rays. In Pleuracanthus it is a jointed axis of many segments, with a fringe of slender fin-rays, corresponding in structure to all appearance to the pectoral fin of Dipnoans and[Pg 511] Crossopterygians, the type called by Gegenbaur archipterygium on the hypothesis that it represents the primitive vertebrate limb.
In most sharks the fin has a fan-shape, with three of the basal segments larger than the others. Of these the mesopterygium is the central one, with the propterygium before it and the metapterygium behind. In the living sharks of the family of Heterodontidæ, this form of fin occurs and the teeth of the same general type constitute the earliest remains distinctly referable to sharks in the Devonian rocks.
Primitive Sharks.—Admitting that these four types of pectoral fin should constitute separate orders, we have next to consider which form is the most primitive and what is the line of descent. In this matter we have, in the phrase of Hæckel, only the "three ancestral documents, Palæontology, Morphology, and Ontogeny."
Unfortunately the evidence of these documents is incomplete and conflicting. So far as Palæontology is concerned, the fin of Cladoselache, with that of Acanthoessus, which may be derived from it, appears earliest, but the modern type of pectoral fin with the three basal segments is assumed to have accompanied the teeth of Psammodonts and Cochliodonts, while the fin of the Chimæra must have been developed in the Devonian. The jointed fin of Cladodus and Pleuracanthus may be a modification or degradation of the ordinary type of shark-fin.
Assuming, however, that the geological record is not perfect and that the fin of Cladoselache is not clearly shown to be primitive, we have next to consider the evidence drawn from morphology.
Those who with Balfour and others (see page 69) accept the theory that the paired fins are derived from a vertebral fold, will regard with Dean the fin of Cladoselache as coming nearest the theoretical primitive condition.
The pectoral fin in Acanthoessus Dean regards as a specialized derivative from a fin like that of Cladoselache, the fin-rays being gathered together at the front and joined together to form the thick spine characteristic of Acanthoessus. This view of the morphology of the fin of Acanthoessus is not accepted by[Pg 512] Woodward, and several different suggestions have been recorded.
If with Gegenbaur we regard the paired fins as derived from the septa between the gill-slits, or with Kerr regard them as modified external gills, the whole theoretical relation of the parts is changed. The archipterygium of Pleuracanthus would be the nearest approach to the primitive pectoral limb, and from this group and its allies all the other sharks are descended. This central jointed axis of Pleuracanthus is regarded by Traquair as the equivalent of the metapterygium in ordinary sharks. (See Figs. 44, 45, 46.)
According to Traquair: "The median stern [of the archipterygium], simplified, shortened up and losing all its radials on the postaxial side, except in sometimes a few near the tip, becomes the metapterygium, while the mesopterygium and propterygium are formed by the fusion into two pieces of the basal joints of a number of preaxial radials, which have reached and become attached to the shoulder-girdle in front of the metapterygium."
According to Dr. Traquair, the pectoral fin in Cladodus neilsoni, a shark from the Coal Measures of Scotland, is "apparently a veritable uniserial archipterygium midway between the truly biserial one of Pleuracanthus and the pectoral fin of ordinary sharks." Other authors look on these matters differently, and Dr. Traquair admits that an opposite view is almost equally probable. Cope and Dean would derive the tribasal pectoral of ordinary sharks directly from the ptychopterygium or fan-like fold of Cladoselache, while Fritsch and Woodward would look upon it as derived in turn from the Ceratodus-like fin of Pleuracanthus, itself derived from the ptychopterygium or remains of a lateral fin-fold.
If the Dipnoans are descended from the Crossopterygians, as Dollo has tried to show, the archipterygium of Pleuracanthus has had a different origin from the similar-appearing limb of the Dipnoans, Dipterus and Ceratodus.
In such case the archipterygium would not be the primitive pectoral limb, but a structure which may have been independently evolved in two different groups.
In the view of Gegenbaur, the Crossopterygians and Dipnoans with all the higher vertebrates and the bony fishes would[Pg 513] arise from the same primitive stock, ancestors, or allies of the Ichthyotomi, which group would also furnish the ancestors of the Chimæras. In support of this view, the primitive protocercal or diphycercal tail of Pleuracanthus may be brought in evidence as against the apparently more specialized heterocercal tail of Cladoselache. But this is not conclusive, as the diphycercal tail may arise separately in different groups through degeneration, as Dollo and Boulenger have shown.
The matter is one mainly of morphological interpretation, and no final answer can be given. On page 68 a summary of the various arguments may be found. Little light is given by embryology. The evidence of Palæontology, so far as it goes, certainly favors the view of Balfour. Omitting detached fin-spines and fragments of uncertain character, the earliest identifiable remains of sharks belong to the lower Devonian. These are allies of Acanthoessus. Cladoselache comes next in the Upper Devonian. Pleuracanthus appears with the teeth and spines supposed to belong to Cestraciont sharks, in the Carboniferous Age. The primitive-looking Notidani do not appear before the Triassic. For this reason the decision as to which is the most primitive type of shark must therefore rest unsettled for the present and perhaps for a long time to come.
The weight of authority at present seems to favor the view of Balfour, Wiedersheim, Boulenger, and Dean, that the pectoral limb has arisen from a lateral fold of skin. But weight of authority is not sufficient when evidence is confessedly lacking.
For our purpose, without taking sides in this controversy, we may follow Dean in allowing Cladoselache to stand as the most primitive of known sharks, thus arranging the Elasmobranchs and rays, recent and fossil, in six orders of unequal value—Pleuropterygii, Acanthodei, Ichthyotomi, Notidani, Asterospondyli, and Tectospondyli. Of these orders the first and second are closely related, as are also the fourth and fifth, the sixth being not far remote. The true sharks form the culmination of one series, the rays of another, while from the Ichthyotomi the Crossopterygians and their descendants may be descended. But this again is very hypothetical, or perhaps impossible; while, on[Pg 514] the other hand, the relation of the Chimæras to the sharks is still far from clearly understood.
Order Pleuropterygii.—The order of Pleuropterygii of Dean (πλεύρον, side; πτερύξ, fin), called by Parker and Haswell Cladoselachea, consists of sharks in which the pectoral and ventral fins have each a very wide horizontal base (ptychopterygium), without jointed axis and without spine. There are no spines in any of the fins. The dorsal fin is low, and there were probably two of them. The notochord is persistent, without intercalary cartilage, such as appear in the higher sharks. The caudal fin is short, broad, and strongly heterocercal. Apparently the ventral fin is without claspers. The gill-openings were probably covered by a dermal fold. The teeth are weak, being modified denticles from the asperities of the skin. The lateral line is represented by an open groove. The family of Cladoselachidæ consists of a single genus Cladoselache from the Cleveland shale or Middle Devonian of Ohio. Cladoselache fyleri is the best-known species, reaching a length of about two feet. Dean regards this as the most primitive of the sharks, and the position of the pectorals and ventrals certainly lend weight to Balfour's theory that they were originally derived from a lateral fold of skin. I am recently informed by Dr. Dean that he has considerable evidence that in Cladoselache the anus was subterminal. If this statement is verified, it would go far to establish the primitive character of Cladoselache.

Fig. 300.—Cladoselache fyleri (Newberry), restored. Upper Devonian of Ohio. (After Dean.)
Order Acanthodei.—Near the Pleuropterygii, although much more highly developed, we may note the strange group of Acanthodei (ἀκανθώδης, spinous). These armed fishes were once placed among the Crossopterygians, but there seems no doubt that Woodward is right in regarding them as a highly specialized aberrant offshoot of the primitive sharks. In this group the paired[Pg 515] fins consist each of a single stout spine, nearly or quite destitute of other rays. A similar spine is placed in front of the dorsal fin and one in front of the anal. According to Dean these spines are each produced by the growing together of all the fin-rays normally belonging to the fin, a view of their morphology not universally accepted.
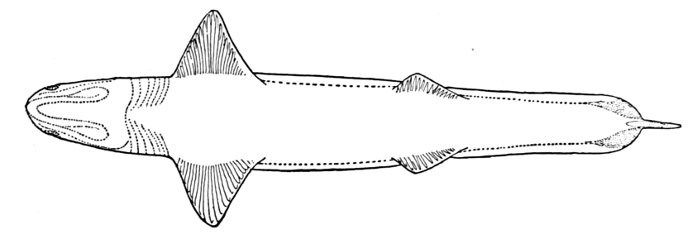
Fig. 301.—Cladoselache fyleri (Newberry), restored. Ventral view. (After Dean.)

Fig. 302.—Teeth of Cladoselache fyleri (Newberry). (After Dean.)

Fig. 303.—Acanthoessus wardi (Egerton). Carboniferous. Family Acanthoessidæ. (After Woodward.)
The dermal covering is highly specialized, the shagreen denticles being much enlarged and thickened, often set in little squares suggesting a checker-board. The skull is covered with small bony plates and membrane bones form a sort of ring about the eye. The teeth are few, large, and "degenerate[Pg 516] in their fibrous structure." Some of the species have certainly no teeth at all. The tail is always heterocercal, or bent upward at tip as in the Cladoselache, not diphycercal, tapering and horizontal as in the Ichthyotomi.
The lower Acanthodeans, according to Woodward, "are the only vertebrates in which there are any structures in the adult apart from the two pairs of fins which may be plausibly interpreted as remnants of once continuous lateral folds. In Climatius, one of the most primitive genera (see Fig. 305), there exists, according to Woodward, and as first noticed by Cope, between the pectoral and pelvic (or ventral) fins a close and regular series of paired spines, in every respect identical with those supporting the appendages that presumably correspond to the two pairs of fins in the higher genera. They may even have supported fin membranes, though specimens sufficiently well preserved to determine this point have not yet been discovered. However, it is evident that dermal calcifications attained a greater development in the Acanthodei than in any of the more typical Elasmobranchs, and we may look for much additional information on the subject when the great fishes to which the undetermined Ichthyodorulites pertained became known." (See Fig. 305.)
The Acanthodei constitute three families. In the Acanthoessidæ there is but one short dorsal fin opposite the anal, and clavicular bones are absent. The gill-openings being provided with "frills" or collar-like margins, perhaps resembled those of the living genus Chlamydoselachus, the frilled shark. The pectoral spine is very strong, and about the eye is a ring of four plates. The body is elongate, tapering, and compressed. Acanthoessus of Agassiz, the name later changed by its author to Acanthodes, is the principal genus, found in the Devonian and Carboniferous.
The species of Acanthoessus are all small fishes rarely more than a foot long, with very small teeth or none, and with the skin well armed with a coat-of-mail. Acanthoessus bronni is the one longest known. In the earliest species known, from the Devonian, the ventral fins are almost as large as the pectorals and nearly midway between pectorals and anal. In the later species the pectoral fins become gradually larger[Pg 517] and the ventrals move forward. In the Permian species the pectorals are enormous.
Traquairia pygmæa, from the Permian of Bohemia, is a diminutive sharklet three or four inches long with large scales, slender spines, and apparently no ventral fins.
In the genus Cheiracanthus the dorsal fin is placed before the anal. In Acanthodopsis the teeth are few, large, and triangular, and the fin-spines relatively large.
The Ischnacanthidæ have no clavicles, and two dorsal fins. Ischnacanthus gracilis of the Devonian has a few large conical teeth with small cusps between them.
The Diplacanthidæ, with two dorsal fins, possess bones interpreted as clavicles. The teeth are minute or absent. In Diplacanthus striatus and Diplacanthus longispinus of the Lower Devonian stout spines are attached to the shoulder-girdle between the pectoral spines below.

Fig. 304.—Diplacanthus crassissimus Duff. Devonian. Family Diplacanthidæ. (After Nicholson). (Restoration of jaws and gill-openings; after Traquair.)
In the very small sharks called Climatius the fin-spines are very strong, and a series of several free spines occurs, as above stated, on each side between the pectoral and ventral fins, a supposed trace of a former lateral fold. In Paraxus the first dorsal spine is enormously enlarged in size, the other spines remaining much as in Climatius.
Dean on Acanthodei.—In his latest treatise on these fishes, "The Devonian Lamprey," Dr. Dean unites the Pleuropterygii and Acanthodei in a single order under the former name, re[Pg 518]garding Acanthoessus as an ally and perhaps descendant of the primitive Cladoselache. Dr. Dean observes:
"In the foregoing classification it will be noted that the Acanthodia are regarded as included under the first order of sharks, Pleuropterygii. To this arrangement Smith Woodward has already objected that the spines of Acanthodians cannot be regarded as the homologues of the radial elements of the Cladoselachian fin (which by a process of concrescence have become fused in its interior margin), since he believes the structure to be entirely dermal in origin. His criticism, however, does not seem to me to be well grounded, for, although all will admit that Acanthodian spines have become incrusted, and deeply incrusted, with a purely dermal calcification, it does not follow that the interior of the spine has not had primitively a non-dermal core. That the concrescence of the radial supporting elements of the fin took place pari passu with the development of a strengthening dermal support of the fin margin was the view expressly formulated in my previous paper on this subject. It is of interest in this connection to recall that the earliest types of Acanthodian spines were the widest, and those which, in spite of their incasing dermal calcification, suggest most clearly the parallel elements representing the component radial supports. There should also be recalled the many features in which the Acanthodians have been shown to resemble Cladoselache."
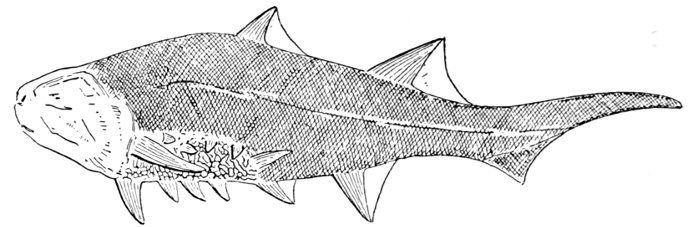
Fig. 305.—Climatius scutiger Egerton, restored. Family Diplacanthidæ. (After Powrie, per Zittel.)
From these primitive extinct types of shark we may proceed to those forms which have representatives among living fishes. From Cladoselache a fairly direct series extends through[Pg 519] the Notidani and Cestraciontes, culminating in the Lamnoid and Galeoid sharks.
Still another series, destitute of anal fin, probably arising near the Acanthodei, reaches its highest development in the side branch of the Batoidei or rays. The Holocephali and Dipneusti must also find their origin in some of these primitive types, certainly not in any form of more highly specialized sharks.
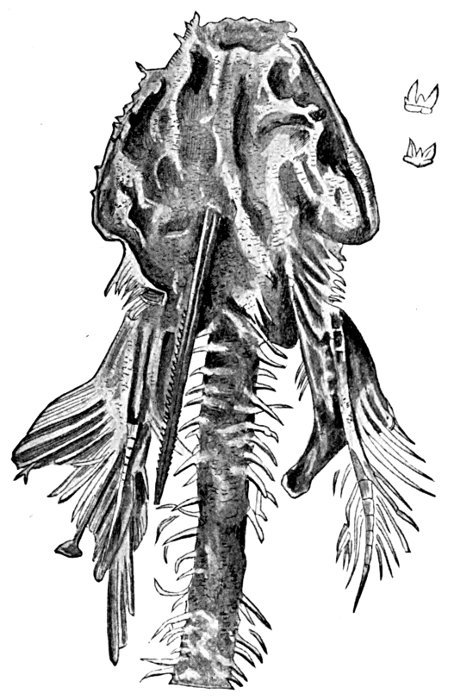
Fig. 306.—Pleuracanthus decheni Goldfuss. Family Pleuracanthidæ. (After Roemer, per Zittel.)
Woodward prefers to place the Tectospondyli next to the Ichthyotomi, leaving the specialized sharks to be treated later. There is, however, no linear system which can interpret natural affinities, and we follow custom in placing the dogfishes and rays at the end of the shark series.
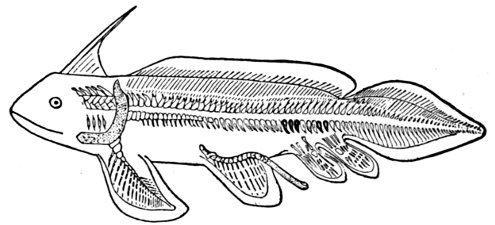
Fig. 307.—Pleuracanthus decheni, restored. (After Brongniart.) The anterior anal very hypothetical.
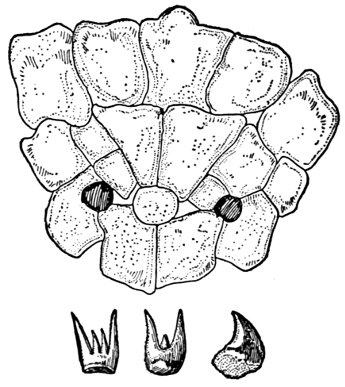
Fig. 308.—Head-bones and teeth of Pleuracanthus decheni Goldfuss. (After Davis, per Dean.)
Fig. 309.—Teeth of Didymodus bohemicus Quenstadt. Carboniferous. Family Pleuracanthidæ. (After Zittel)
Order Ichthyotomi.—In the order Ichthyotomi (ἰχθύς, fish; τομός, cutting; named by Cope from the supposed segmentation of the cranium; called by Parker and Haswell Pleuracanthea) the very large pectoral fins are developed each as an archipterygium. Each fin consists of a long segmented axis fringed on one or both sides with fin-rays. The notochord is very simple, scarcely or never constricted, the calcifications of its sheath "arrested at the most primitive or rhachitomous stage, except in the tail." This is the best defined of the orders of sharks, and should perhaps rank rather as a subclass, as the Holocephali. Two families of Ichthyotomi are recognized by Woodward, the Pleuracanthidæ and the Cladodontidæ. In the Pleuracanthidæ the dorsal fin is long and low, continuous from head to tail, and the pectoral rays are in two rows. There is a long barbed spine with two rows of serrations at the nape.[Pg 521] The body is slender, not depressed, and probably covered with smooth skin. The teeth have two or more blunt cusps, sometimes with a smaller one between and a blunt button behind. The interneural cartilages are more numerous than the neural spines. The genera are imperfectly known, the skeleton of Pleuracanthus decheni only being well preserved. This is the type of the genus called Xenacanthus which, according to Woodward, is identical with Pleuracanthus, a genus otherwise known from spines only. The denticles on the spine are straight or hooked backward, in Pleuracanthus (lævissimus), the spine being flattened. In Orthacanthus (cylindricus), the spine is cylindrical in section. The species called Dittodus and Didymodus are known from the teeth only. These resemble the teeth of Chlamydoselachus. It is not known that Dittodus possesses the nuchal spine, although detached spines like those of Pleuracanthus lie about in remains called Didymodus in the Permian rocks of Texas. In Dicranodus texensis the palato-quadrate articulates with the postorbital process of the cranium, as in the Hexanchidæ, and the hyomandibular is slender.
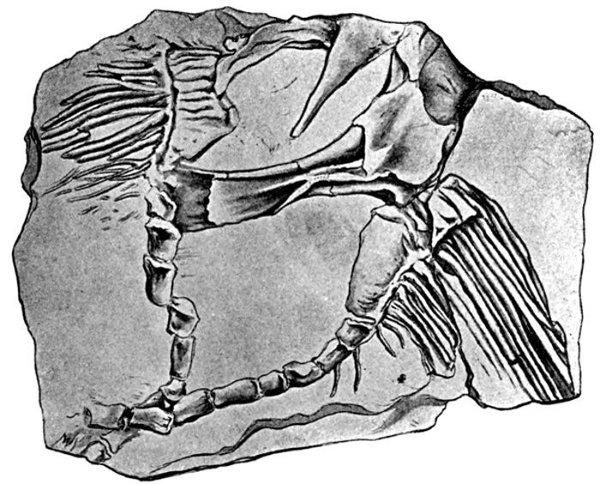
Fig. 310.—Shoulder-girdle and pectoral fins of Cladodus neilsoni Traquair.
A genus, Chondrenchelys, from the sub-Carboniferous of[Pg 522] Scotland, is supposed to belong to the Pleuracanthidæ, from the resemblance of the skeleton. It has no nuchal spine, and no trace of paired fins is preserved.
The Cladodontidæ differ in having the "pectoral fin developed in the form of a uniserial archipterygium intermediate between the truly biserial one of Pleuracanthus and the pectoral fin of modern sharks." The numerous species are known mainly from detached teeth, especially abundant in America, the earliest being in the Lower Carboniferous. One species, Cladodus nelsoni (Fig. 310), described by Traquair, from the sub-Carboniferous of Scotland shows fairly the structure of the pectoral fin.
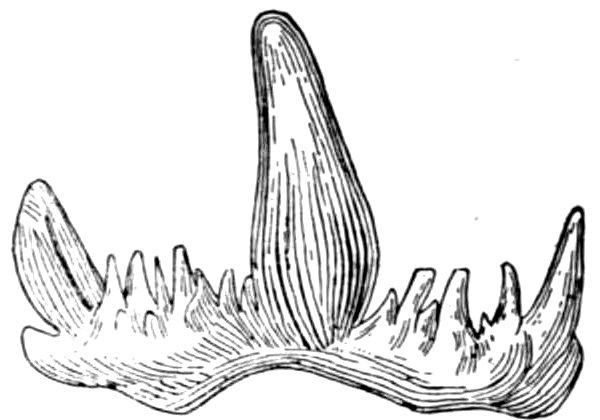
Fig. 311.—Teeth of Cladodus striatus Agassiz. (After Davis.) Carboniferous.
In Cladodus mirabilis the teeth are very robust, the crown consisting of a median principal cone and two or three large lateral cones on each side. The cones are fairly striate. In Lambdodus from Illinois there are no lateral cones. Other genera are Dicentrodus, Phœbodus, Carcharopsis, and Hybocladodus.

Order Notidani.—We may recognize as a distinct order, a primitive group of recent sharks, a group of forms finding its natural place somewhere between the Cladoselachidæ and Heterodontidæ, both of which groups long preceded it in geological time.
The name Notidani (Notidanus, νωτιδάνος, dry back, an old name of one of the genera) may be retained for this group, which corresponds to the Diplospondyli of Hasse, the Opistharthri of Gill, and the Protoselachii of Parker and Haswell. The Notidani are characterized by the primitive structure of the spinal column, which is without calcareous matter, the centra being imperfectly developed. There are six or seven branchial arches, and in the typical forms (not in Chlamydoselachus) the palato-quadrate or upper jaw articulates with the postorbital region of the skull. The teeth are of primitive character, of different forms in the same jaw, each with many cusps. The fins are without spines, the pectoral fin having the three basal cartilages (mesopterygium with propterygium and metapterygium) as usual among sharks.
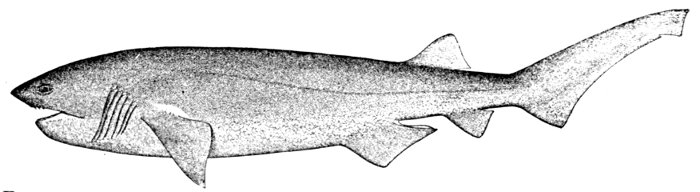
Fig. 312.—Griset or Cow-shark, Hexanchus griseus (Gmelin). Currituck Inlet, N. C.
The few living forms are of high interest. The extinct species are numerous, but not very different from the living species.
Family Hexanchidæ.—The majority of the living Notidanoid sharks belong to the family of Hexanchidæ. These sharks have six or seven gill-openings, one dorsal fin, and a relatively simple organization. The bodies are moderately elongate, not eel-shaped, and the palato-quadrate articulates with the postorbital part of the skull. The six or eight species are found sparsely in the warm seas. The two genera, Hexanchus, with six, and Heptranchias, with seven vertebræ, are found in the Mediterranean. The European species are Hexanchus griseus, the cow-shark, and Heptranchias cinereus. The former crosses to the West Indies. In California, Heptranchias maculatus and Hexanchus corinus are occasionally taken, while Heptranchias deani is the well known Aburazame or oil shark of Japan. Heptranchias indicus, a similar species, is found in India.
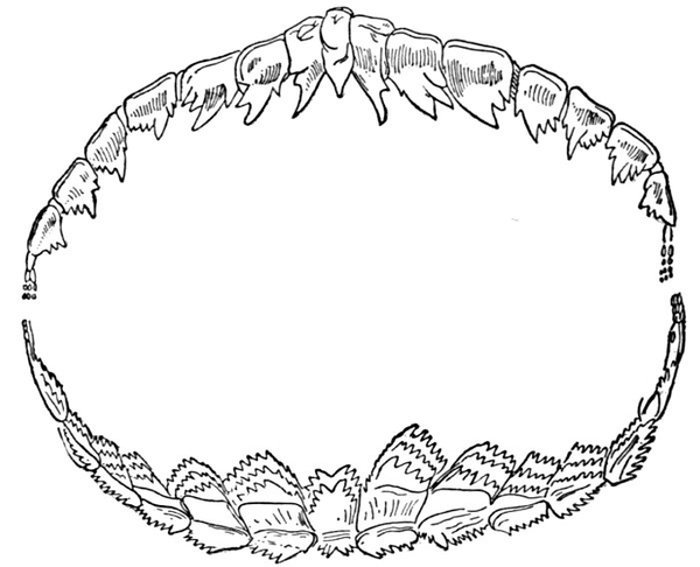
Fig. 313.—Teeth of Heptranchias indicus Gmelin.
Fossil Hexanchidæ exist in large numbers, all of them referred by Woodward to the genus Notidanus (which is a later name than Hexanchus and Heptranchias and intended to include both these genera), differing chiefly in the number of gill-openings, a character not ascertainable in the fossils. None of these, however, appear before Cretaceous time, a fact which may indicate that the simplicity of structure in Hexanchus and Heptranchias is a result of degeneration and not altogether a mark of primitive simplicity. The group is apparently much[Pg 525] younger than the Cestraciontes and little older than the Lamnoids, or the Squaloid groups. Heptranchias microdon is common in English Cretaceous rocks, and Heptranchias primigenius and other species are found in the Eocene.
Family Chlamydoselachidæ.—Very great interest is attached to the recent discovery by Samuel Garman of the frilled shark, Chlamydoselachus anguineus, the sole living representative of the Chlamydoselachidæ.
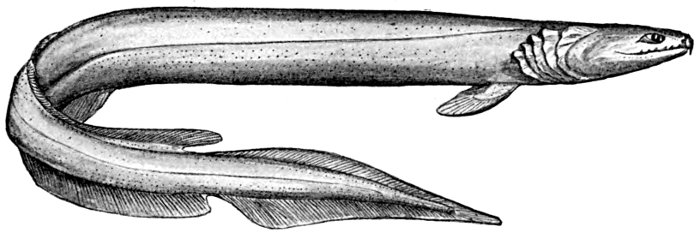
Fig. 314.—Frill-shark, Chlamydoselachus anguineus Garman. From Misaki, Japan. (After Günther.)
This shark was first found on the coast of Japan, where it is rather common in deep water. It has since been taken off Madeira and off the coast of Norway. It is a long, slender, eel-shaped shark with six gill-openings and the palato-quadrate not articulated to the cranium. The notochord is mainly persistent, in part replaced by feeble cyclospondylic vertebral centra. Each gill-opening is bordered by a broad frill of skin. There is but one dorsal fin. The teeth closely resemble those of Dittodus or Didymodus and other extinct Ichthyotomi. The teeth have broad, backwardly extended bases overlapping, the crown consisting of three slender curved cusps, separated by rudimentary denticles. Teeth of a fossil species, Chlamydoselachus lawleyi, are recorded by J. W. Davis from the Pliocene of Tuscany.
Order Asterospondyli.—The order of Asterospondyli comprises the typical sharks, those in which the individual vertebræ are well developed, the calcareous lamellæ arranged so as to radiate, star-fashion, from the central axis. All these sharks possess two dorsal fins and one anal fin, the pectoral fin is normally[Pg 526] developed, with the three basal cartilages; there are five gill-openings, and the tail is heterocercal.

Fig. 315.—Bullhead-shark, Heterodontus francisci (Girard). San Pedro, Cal.
Suborder Cestraciontes.—The most ancient types may be set off as a distinct suborder under the name of Cestraciontes or Prosarthri.
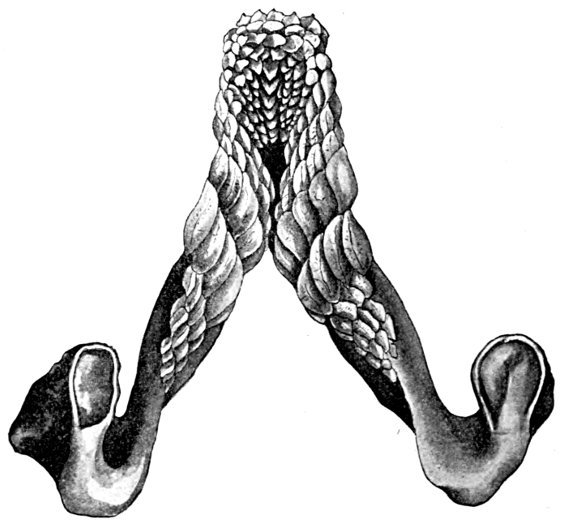
Fig. 316.—Lower jaw of Heterodontus philippi. From Australia. Family Heterodontidæ. (After Zittel.)
These forms find their nearest allies in the Notidani, which they resemble to some extent in dentition and in having the palato-quadrate articulated to the skull although fastened[Pg 527] farther forward than in the Notidani. Each of the two dorsal fins has a strong spine.
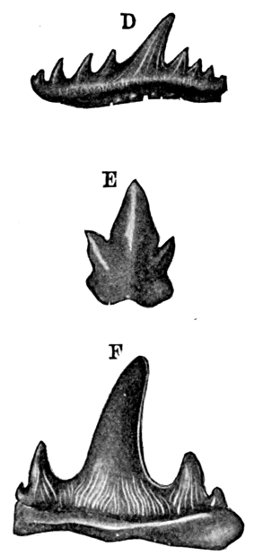
Fig. 317.—Teeth of Cestraciont Sharks. (After Woodward.) d, Synechodus dubrisianus Mackie; e, Heterodontus canaliculatus Egerton; f, Hybodus striatulus Agassiz. (After Woodward.)
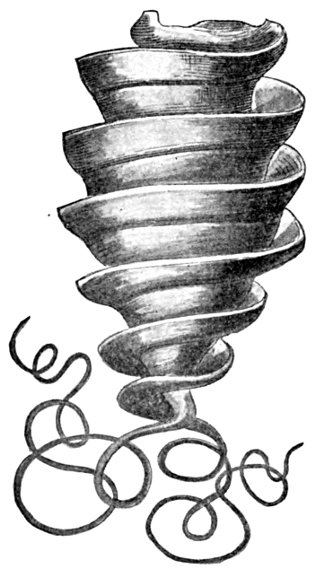
Fig. 318.—Egg of Port Jackson Shark, Heterodontus philippi (Lacépède). (After Parker & Haswell.)
Family Heterodontidæ.—Among recent species this group contains only the family of Heterodontidæ, the bullhead sharks, or Port Jackson sharks. In this family the head is high, with usually projecting eyebrows, the lateral teeth are pad-like, ridged or rounded, arranged in many rows, different from the pointed anterior teeth, the fins are large, the coloration is strongly marked, and the large egg-cases are spirally twisted. All have five gill-openings. The living species of Heterodontidæ are found only in the Pacific, the Port Jackson shark of Australia, Heterodontus philippi, being longest known. Other species are Heterodontus francisci, common in California, Heterodontus japonicus, in Japan, and Heterodontus zebra, in China. These small and harmless sharks at once attract attention by their peculiar forms. In the American species the jaws are less[Pg 528] contracted than in the Asiatic species, called Heterodontus. For this reason Dr. Gill has separated the former under the name of Gyropleurodus. The differences are, however, of slight value. The genus Heterodontus first appears in the Jurassic, where a number of species are known, one of the earliest being Heterodontus falcifer.
Three families of Cestraciontes are recognized by Hay. The most primitive of these is the group of Orodontidæ. Orodus, from the Lower Carboniferous, has the teeth with a central crown, its surface wrinkled. Of the Heterodontidæ, Hybodus, of the Carboniferous and Triassic, is one of the earliest and largest genera, characterized by elongate teeth of many cusps, different in different parts of the jaw, somewhat as in the Hexanchidæ, the median points being, however, always longest. The dorsal fins are provided with long spines serrated behind. The vertebræ with persistent notochord show qualities intermediate between those of Hexanchidæ and Heterodontidæ, and the same relation is shown by the teeth. In this genus two large hooked half-barbed dermal spines occur behind each orbit.
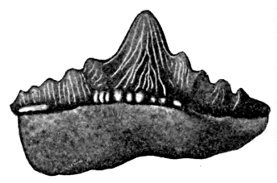
Fig. 319.—Tooth of Hybodus delabechei Charlesworth. (After Woodward.)

Fig. 320.—Fin-spine of Hybodus basanus Egerton. Cretaceous. Family Heterodontidæ. (After Nicholson.)

Fig. 321.—Fin-spine of Hybodus reticulatus Agassiz. (After Zittel.)
Palæospinax, with short stout spines and very large pectoral fins, formerly regarded as a dogfish, is placed near Heterodontus by Woodward. Acrodus, from the Triassic, shows considerable resemblance to Heterodontus. Its teeth are rounded and without cusps.
Most of these species belong to the Carboniferous, Triassic, and Jurassic, although some fragments ascribed to Cestraciont sharks occur in the Upper Silurian. Asteracanthus, known only from fin-spines in the Jura, probably belongs here.
It is a singular fact first noted by Dr. Hay, that with all the great variety of sharks, ten families in the Carboniferous age, representatives of but one family, Heterodontidæ, are found in the Triassic. This family may be the parent of all subsequent sharks and rays, six families of these appearing in the Jurassic and many more in the Cretaceous.
Edestus and its Allies.—Certain monstrous structures, hitherto thought to be fin-spines, are now shown by Dr. Eastman and others to be coalescent teeth of Cestraciont sharks.
Fig. 322.—Fin-spine of Hybodus canaliculatus Agassiz.
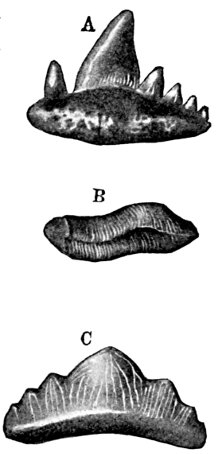
Fig. 323.—Teeth of Cestraciont Sharks. (After Woodward.) a, Hybodus lævis Woodward (after Woodward); b, Heterodontus rugosus Agassiz; c, Hybodus delabechei Charlesworth.
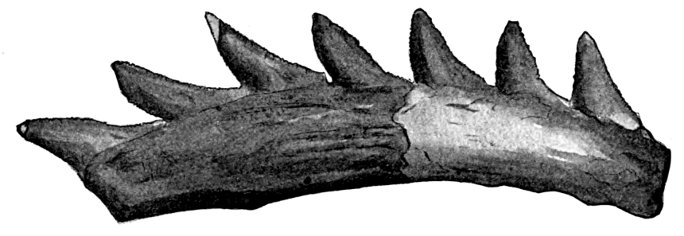
Fig. 324.—Edestus vorax Leidig, supposed to be a whorl of teeth. (After Newberry.)
These remarkable Ichthyodorulites are characteristic structures of sharks of unknown nature, but probably related to the Heterodontidæ. Of these the principal genera are Edestus, Helicoprion, and Campyloprion. Karpinsky regards these ornate serrated spiral structures as whorls of unshed teeth[Pg 530] cemented together and extending outside the mouth, "sharp, piercing teeth which were never shed but became fused in whorls as the animals grew."
Dr. Eastman has, however, shown that these supposed teeth of Edestus are much like those of the Cochliodontidæ, and the animals which bore them should doubtless find their place among the Cestraciont sharks, perhaps within the family of Heterodontidæ.
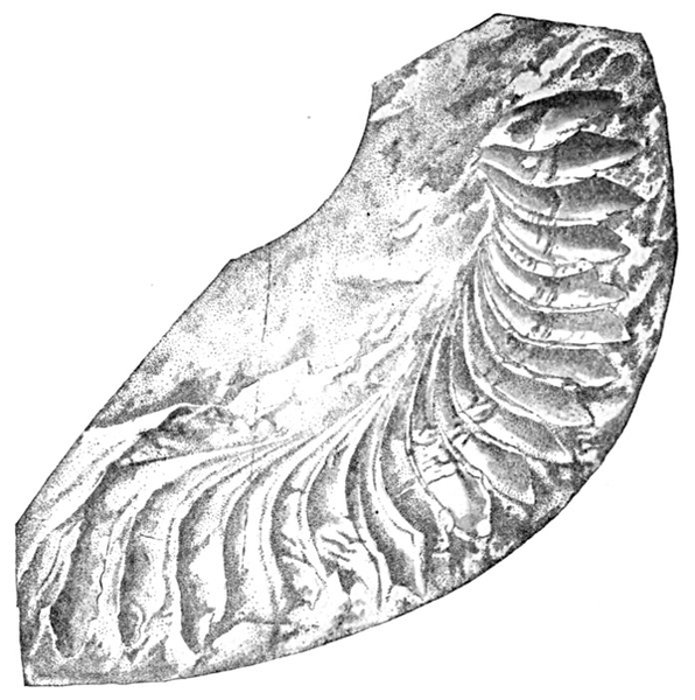
Fig. 325.—Helicoprion bessonowi Karpinsky. Teeth from the Permian of Krasnoufimsk, Russia. (After Karpinsky.)
Onchus.—The name Onchus was applied by Agassiz to small laterally compressed spines, their sides ornamented with smooth or faintly crenulated longitudinal ridges, and with no denticles behind. Very likely these belonged to extinct Cestraciont sharks. Onchus murchisoni and Onchus tenuistriatus occur in the Upper Silurian rocks of England, in the lowest strata in which sharks have been found.
To a hypothetical group of primitive sharks Dr. Hasse has given the name of Polyospondyli. In these supposed[Pg 531] ancestral sharks the vertebræ were without any ossification, a simple notochord, possibly swollen at intervals. The dorsal fin was single and long, a fold of skin with perhaps a single spine as an anterior support. The teeth must have been modified dermal papillæ, each probably with many cusps. Probably seven gill-openings were developed, and the tail was diphycercal, ending in a straight point. The finely striated fin-spines not curved upward at tip, called Onchus from the Upper Silurian of the Ludlow shales of England and elsewhere, are placed by Hasse near his Polyspondylous sharks. Such spines have been retained by the group of Chimæras, supposed to be derived from the ancestors of Onchus, as well as by the Heterodontidæ and Squalidæ.
Family Cochliodontidæ.—Another ancient family known from teeth alone is that of Cochliodontidæ. These teeth resemble those of the Heterodontidæ, but are more highly specialized. The form of the body is unknown, and the animals may have been rays rather than sharks. Eastman leaves them near the Petalodontidæ, which group of supposed rays shows a similar dentition. The teeth are convex in form, strongly arched, hollowed at base, and often marked by ridges or folds, being without sharp cusps. In each jaw is a strong posterior tooth with smaller teeth about. The elaborate specialization of these ancient teeth for crushing or grinding shells is very remarkable. The species are chiefly confined to rocks of the Carboniferous age. Among the principal genera are Helodus, Psephodus, Sandalodus, Venustodus, Xystrodus, Deltodus, Pœcilodus, and Cochliodus.
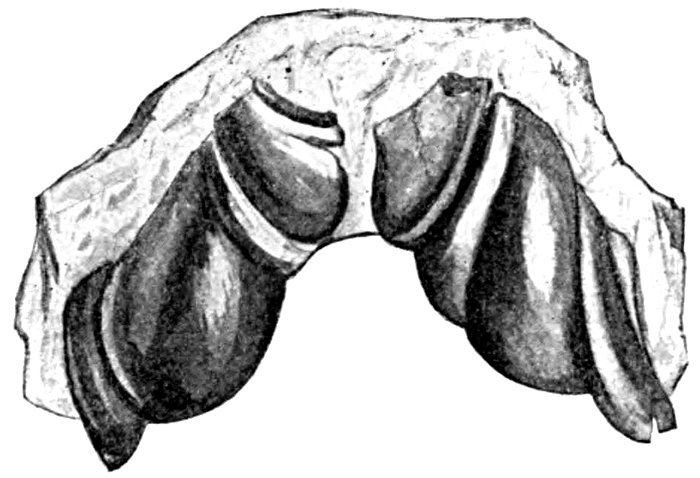
Fig. 326.—Lower jaw of Cochliodus contortus Agassiz. Carboniferous. (After Zittel.)
Concerning the teeth of various fossil sharks, Dr. Dean observes: "Their general character appears to have been primitive, but in structural details they were certainly specialized. Thus their dentition had become adapted to a shellfish diet, and they had evolved defensive spines at the fin margins, some[Pg 532]times at the sides of the head. In some cases the teeth remain as primitive shagreen cusps on the rim of the mouth, but become heavy and bluntish behind; in other forms the fusion of tooth clusters may present the widest range in their adaptations for crushing; and the curves and twistings of the tritoral surfaces may have resulted in the most specialized forms of dentition which are known to occur, not merely in sharks but among all vertebrates."
In this neighborhood belongs, perhaps, the family of Tamiobatidæ, known from the skull of a single specimen, called Tamiobatis vetustus, from the Devonian in eastern Kentucky. The head has the depressed form of a ray, but it is probably a shark and one of the very earliest known.
Suborder Galei.—The great body of recent sharks belong to the suborder Galei, or Euselachii, characterized by the asterospondylous vertebræ, each having a star-shaped nucleus, and by the fact that the palato-quadrate apparatus or upper jaw is not articulated with the skull. The sharks of this suborder are the most highly specialized of the group, the strongest and largest and, in general, the most active and voracious. They are of three types and naturally group themselves about the three central families Scyliorhinidæ, Lamnidæ, and Carchariidæ (Galeorhinidæ).
The Asterospondyli are less ancient than the preceding groups, but the modern families were well differentiated in Mesozoic times.
Among the Galei the dentition is less complex than with the ancient forms, although the individual teeth are more highly specialized. The teeth are usually adapted for biting, often with knife-like or serrated edges; only the outer teeth are in function; as they are gradually lost, the inner teeth are moved outward, gradually taking the place of these.
We may place first, as most primitive, the forms without nictitating membrane.
Family Scyliorhinidæ.—The most primitive of the modern families is doubtless that of the Scyliorhinidæ, or cat-sharks. This group includes sharks with the dorsal fins both behind the ventrals, the tail not keeled and not bent upward, the spiracles present, and the teeth small and close-set. The species[Pg 533] are small and mostly spotted, found in the warm seas. All of them lay their eggs in large cases, oblong, and with long filaments or strings at the corners. The cat-sharks, or roussettes, Scyliorhinus canicula and Catulus stellaris, abound in the Mediterranean. Their skin is used as shagreen or sandpaper in polishing furniture. The species of swell-sharks (Cephaloscylium) (C. uter, in California; C. ventriosus, in Chile; C. laticeps, in Australia; C. umbratile, in Japan) are short, wide-bodied sharks, which have the habit of filling the capacious stomach with air, then floating belly upward like a globefish. Other species are found in the depths of the sea. Scyliorhinus, Catulus, and numerous other genera are found fossil. The earliest is Palæoscyllium, in the Jurassic, not very different from Scyliorhinus, but the fins are described as more nearly like those of Ginglymostoma.
Close to the Scyliorhinidæ is the Asiatic family, Hemiscylliidæ, which differs in being ovoviviparous, the young, according to Mr. Edgar R. Waite, hatched within the body. The general appearance is that of the Scyliorhinidæ, the body being elongate. Chiloscyllium is a well-known genus with several species in the East Indies. Chiloscyllium modestum is the dogfish of the Australian fishermen. The Orectolobidæ are thick-set sharks, with large heads provided with fleshy fringes. Orectolobus barbatus (Crossorhinus of authors) abounds from Japan to Australia.
Another family, Ginglymostomidæ, differs mainly in the form of the tail, which is long and bent abruptly upward at its base. These large sharks, known as nurse-sharks, are found in the warm seas. Ginglymostoma cirrhatum is the common species with Orectolobus. Stegostoma tigrinum, of the Indian seas and north to Japan, one of several genera called tiger-sharks, is remarkable for its handsome spotted coloration. The extinct genus Pseudogaleus (voltai) is said to connect the Scyliorhinoid with the Carcharioid sharks.
The Lamnoid or Mackerel Sharks.—The most active and most ferocious of the sharks, as well as the largest and some of the most sluggish, belong to a group of families known collectively as Lamnoid, because of a general resemblance to the mackerel[Pg 534]-shark, or Lamna, as distinguished from the blue sharks and white sharks allied to Carcharias (Carcharhinus).
The Lamnoid sharks agree with the cat-sharks in the absence of nictitating membrane or third eyelid, but differ in the anterior insertion of the first dorsal fin, which is before the ventrals. Some of these sharks have the most highly specialized teeth to be found among fishes, most effective as knives or as scissors. Still others have the most highly specialized tails, either long and flail-like, or short, broad, and muscular, fitting the animal for swifter progression than is possible for any other sharks. The Lamnoid families are especially numerous as fossils, their teeth abounding in all suitable rock deposits from Mesozoic times till now. Among the Lamnoid sharks numerous families must be recognized.
The most primitive is perhaps that of the Odontaspididæ (called Carchariidæ by some recent authors), now chiefly extinct, with the tail unequal and not keeled, and the teeth slender and sharp, often with smaller cusps at their base. Odontaspis and its relatives of the same genus are numerous, from the Cretaceous onward, and three species are still extant, small sharks of a voracious habit, living on sandy shores. Odontaspis littoralis (also known as Carcharias littoralis) is the common sand-shark of our Atlantic coast. Odontaspis taurus is a similar form in the Mediterranean.
Family Mitsukurinidæ, the Goblin-sharks.—Closely allied to Odontaspis is the small family of Mitsukurinidæ, of which a single living species is known. The teeth are like those of Odontaspis, but the appearance is very different.
The goblin-shark, or Tenguzame, Mitsukurina owstoni, is a very large shark rarely taken in the Kuro Shiwo, or warm "Black Current" of Japan. It is characterized by the development of the snout into a long flat blade, extending far beyond the mouth, much as in Polyodon and in certain Chimæras. Several specimens are now known, all taken by Capt. Alan Owston of Yokohama in Sagami Bay, Japan. The original specimen, a young shark just born, was presented by him to Professor Kakichi Mitsukuri of the University of Tokyo. From this our figure was taken. The largest specimen now known is in the United States National Museum and is fourteen feet in[Pg 535] length. In the Upper Cretaceous is a very similar genus, Scapanorhynchus (lewisi, etc.), which Professor Woodward thinks may be even generically identical with Mitsukurina, though there is considerable difference in the form of the still longer rostral plate, and the species of Scapanorhynchus differ among themselves in this regard.
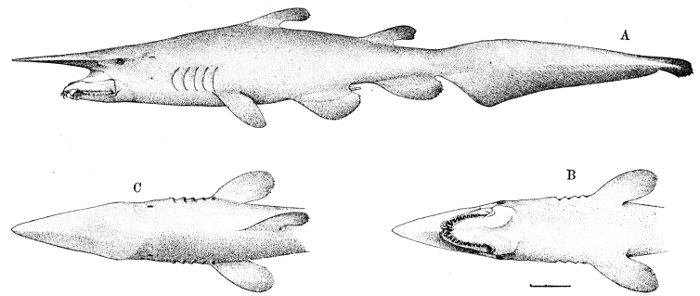
Fig. 327.—Goblin-shark (Tenguzame), Mitsukurina owstoni Jordan. From a young specimen in the Imperial University of Tokyo.
Mitsukurina, with Heterodontus, Heptranchias, and Chlamydoselache, is a very remarkable survival of a very ancient form. It is an interesting fact that the center of abundance of all these relics of ancient life is in the Black Current, or Gulf Stream, of Japan.
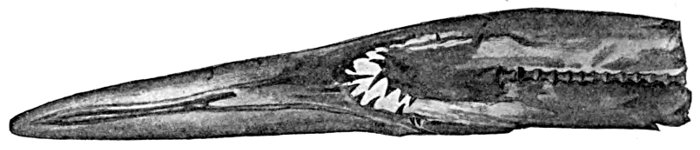
Fig. 328.—Scapanorhynchus lewisi Davis. Family Mitsukurinidæ. Under side of snout. (After Woodward.)
Family Alopiidæ, or Thresher Sharks.—The related family of Alopiidæ contains probably but one recent species, the great fox-shark, or thresher, found in all warm seas. In this species, Alopias vulpes, the tail is as long as the rest of the body and bent upward from the base. The snout is very short, and the teeth are small and close-set. The species reaches a length of about twenty-five feet. It is not especially ferocious, and the current stories of its attacks on whales probably arise from a mistake of the observers, who have taken the great killer, Orca, for a shark. The killer is a mammal, allied to the porpoise. It attacks the whale with great ferocity, clinging to its flesh by its strong teeth. The whale rolls over and over, throwing the killer into the air, and sailors report it as a thresher. As a matter of fact the thresher very rarely if ever attacks any animal except small fish. It is said to use its tail in rounding up and destroying schools of herring and sardines. Fossil teeth of thresher-sharks of some species are found from the Miocene.
Family Pseudotriakidæ.—The Pseudotriakidæ consist of two species. One of these is Pseudotriakis microdon, a large shark[Pg 537] with a long low tail, long and low dorsal fin, and small teeth. It has been only twice taken, off Portugal and off Long Island. The other, the mute shark, Pseudotriakis acrales, a large shark with the body as soft as a rag, is in the museum of Stanford University, having been taken by Mr. Owston off Misaki.
Family Lamnidæ.—To the family of Lamnidæ proper belong the swiftest, strongest, and most voracious of all sharks. The chief distinction lies in the lunate tail, which has a keel on either side at base, as in the mackerels. This form is especially favorable for swift swimming, and it has been independently developed in the mackerel-sharks, as in the mackerels, in the interest of speed in movement.
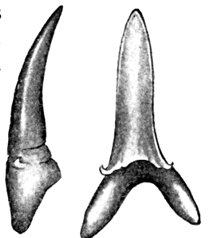
Fig. 329.—Tooth of Lamna cuspidata Agassiz. Oligocene. Family Lamnidæ. (After Nicholson.)
The porbeagle, Lamna cornubica, known as salmon-shark in Alaska, has long been noted for its murderous voracity. About Kadiak Island it destroys schools of salmon, and along the coasts of Japan, and especially of Europe and across to New England, it makes its evil presence felt among the fishermen. Numerous fossil species of Lamna occur, known by the long knife-like flexuous teeth, each having one or two small cusps at its base.

Fig. 330.—Mackerel-shark, Isuropsis dekayi Gill. Pensacola, Fla.
In the closely related genus, Isurus, the mackerel-sharks, this cusp is wanting, while in Isuropsis the dorsal fin is set farther back. In each of these genera the species reach a length of 20 to 25 feet. Each is strong, swift, and voracious.[Pg 538] Isurus oxyrhynchus occurs in the Mediterranean, Isuropsis dekayi, in the Gulf of Mexico, and Isuropsis glauca, from Hawaii and Japan westward to the Red Sea.
Man-eating Sharks.—Equally swift and vastly stronger than these mackerel-sharks is the man-eater, or great white shark, Carcharodon carcharias. This shark, found occasionally in all warm seas, reaches a length of over thirty feet and has been known to devour men. According to Linnæus, it is the animal which swallowed the prophet Jonah. "Jonam Prophetum," he observes, "ut veteris Herculem trinoctem, in hujus ventriculo tridui spateo bæsisse, verosimile est."

Fig. 331.—Tooth of Isurus hastalis (Agassiz). Miocene. Family Lamnidæ. (After Nicholson.)
It is beyond comparison the most voracious of fish-like animals. Near Soquel, California, the writer obtained a specimen in 1880, with a young sea-lion (Zalophus) in its stomach. It has been taken on the coasts of Europe, New England, Carolina, California, Hawaii, and Japan, its distribution evidently girdling the globe. The genus Carcharodon is known at once by its broad, evenly triangular, knife-like teeth, with finely serrated edges, and without notch or cusp of any kind. But one species is now living. Fossil teeth are found from the Eocene. One of these, Carcharodon megalodon (Fig. 332), from fish-guano deposits in South Carolina and elsewhere, has teeth nearly six inches long. The animal could not have been less than ninety feet in length. These huge sharks can be but recently extinct, as their teeth have been dredged from the sea-bottom by the Challenger in the mid-Pacific.
Fossil teeth of Lamna and Isurus as well as of Carcharodon are found in great abundance in Cretaceous and Tertiary rocks. Among the earlier species are forms which connect these genera very closely.
The fossil genus Otodus must belong to the Lamnidæ. Its massive teeth with entire edges and blunt cusps at base are common in Cretaceous and Tertiary deposits. The teeth are formed much as in Lamna, but are blunter, heavier, and much less effective as instruments of destruction. The extinct genus Corax is also placed here by Woodward.
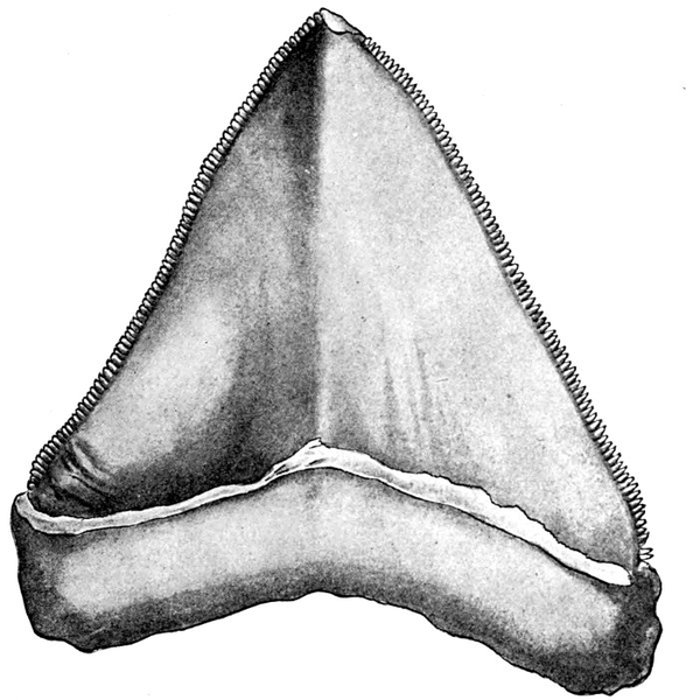
Fig. 332.—Carcharodon megalodon Charlesworth. Miocene. Family Lamnidæ. (After Zittel.)
Family Cetorhinidæ, or Basking Sharks.—The largest of all living sharks is the great basking shark (Cetorhinus maximus), constituting the family of Cetorhinidæ. This is the largest of all fishes, reaching a length of thirty-six feet and an enormous weight. It is a dull and sluggish animal of the northern seas, almost as inert as a sawlog, often floating slowly southward in pairs in the spring and caught occasionally by whalers for its liver. When caught, its huge flabby head spreads out wide on the ground, its weight in connection with the great size of the mouth-cavity rendering it shapeless. Although so clumsy and without spirit, it is said that a blow with its tail will crush[Pg 540] an ordinary whaleboat. The basking shark is known on all northern coasts, but has most frequently been taken in the North Sea, and about Monterey Bay in California. From this locality specimens have been sent to the chief museums of Europe. In its external characters the basking shark has much in common with the man-eater. Its body is, however, relatively clumsy forward; its fins are lower, and its gill-openings are much broader, almost meeting under the throat. The great difference lies in the teeth, which in Cetorhinus are very small and weak, about 200 in each row. The basking shark, also called elephant-shark and bone-shark, does not pursue its prey, but feeds on small creatures to be taken without effort. Fossil teeth of Cetorhinus have been found from the Cretaceous, as also fossil gill-rakers, structures which in this shark are so long as to suggest whalebone.
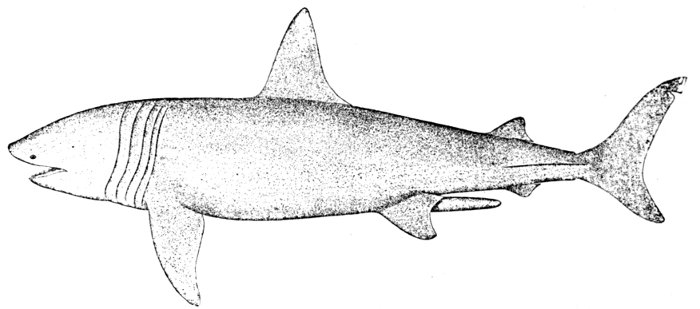
Fig. 333.—Basking Shark, Cetorhinus maximus (Gunner). France.
Family Rhineodontidæ.—The whale-sharks, Rhineodontidæ, are likewise sluggish monsters with feeble teeth and keeled tails. From Cetorhinus they differ mainly in having the last gill-opening above the pectorals. There is probably but one species, Rhineodon typicus, of the tropical Pacific, straying northward to Florida, Lower California, and Japan.
The Carcharioid Sharks, or Requins.—The largest family of recent sharks is that of Carchariidæ (often called Galeorhinidæ, or Galeidæ), a modern offshoot from the Lamnoid type, and especially characterized by the presence of a third eyelid, the nictitating membrane, which can be drawn across the eye from[Pg 541] below. The heterocercal tail has no keel; the end is bent upward; both dorsal fins are present, and the first is well in front of the ventral fins; the last gill-opening over the base of the pectoral, the head normally formed; these sharks are ovoviviparous, the young being hatched in a sort of uterus, with or without placental attachment.
Some of these sharks are small, blunt-toothed, and innocuous. Others reach a very large size and are surpassed in voracity only by the various Lamnidæ.
The genera Cynias and Mustelus, comprising the soft-mouthed or hound-sharks, have the teeth flat and paved, while well-developed spiracles are present. These small, harmless sharks abound on almost all coasts in warm regions, and are largely used as food by those who do not object to the harsh odor of shark's flesh. The best-known species is Cynias canis of the Atlantic. By a regular gradation of intermediate forms, through such genera as Rhinotriacis and Triakis with tricuspid teeth, we reach the large sharp-toothed members of this family. Galeus (or Galeorhinus) includes large sharks having spiracles, no pit at the root of the tail, and with large, coarsely serrated teeth. One species, the soup-fin shark (Galeus zyopterus), is found on the coast of California, where its fins are highly valued by the Chinese, selling at from one to two dollars for each set. The delicate fin-rays are the part used, these dissolving into a finely flavored gelatine. The liver of this and other species is used in making a coarse oil, like that taken from the dogfish. Other species of Galeus are found in other regions, Galeus galeus being known in England as tope, Galeus japonicus abounding in Japan.

Fig. 334.—Soup-fin Shark, Galeus zyopterus (Jordan & Gilbert). Monterey.
Galeocerdo differs mainly in having a pit at the root of the tail. Its species, large, voracious, and tiger-spotted, are found[Pg 542] in warm seas and known as tiger-sharks (Galeocerdo maculatus in the Atlantic, Galeocerdo tigrinus in the Pacific).
The species of Carcharias (Carcharhinus of Blainville) lack the spiracles. These species are very numerous, voracious, armed with sharp teeth, broad or narrow, and finely serrated on both edges. Some of these sharks reach a length of thirty feet. They are very destructive to other fishes, and often to fishery apparatus as well. They are sometimes sought as food, more often for the oil in their livers, but, as a rule, they are rarely caught except as a measure for getting rid of them. Of the many species the best known is the broad-headed Carcharias lamia, or cub-shark, of the Atlantic. This the writer has taken with a great hook and chain from the wharves at Key West. These great sharks swim about harbors in the tropics, acting as scavengers and occasionally seizing arm or leg of those who venture within their reach. One species (Carcharias nicaraguensis) is found in Lake Nicaragua, the only fresh-water shark known, although some run up the brackish mouth of the Ganges and into Lake Pontchartrain. Carcharias japonicus abounds in Japan.

Fig. 335.—Cub-shark, Carcharias lamia Rafinesque. Florida.
A closely related genus is Prionace, its species Prionace glauca, the great blue shark, being slender and swift, with the dorsal farther back than in Carcharias. Of the remaining genera the most important is Scoliodon, small sharks with oblique teeth which have no serrature. One of these, Scoliodon terræ-novæ, is the common sharp-nosed shark of our Carolina coast. Fossil teeth representing nearly all of these genera are common in Tertiary rocks.
Probably allied to the Carchariidæ is the genus Corax, containing large extinct sharks of the Cretaceous with broad[Pg 543]triangular serrate teeth, very massive in substance, and without denticles. As only the teeth are known, the actual relations of the several species of Corax are not certainly known, and they may belong to the Lamnidæ.

Fig. 336.—Teeth of Corax pristodontus.
Family Sphyrnidæ, or Hammer-head Sharks.—The Sphyrnidæ, or hammer-headed sharks, are exactly like the Carchariidæ except that the sides of the head are produced, so as to give it the shape of a hammer or of a kidney, the eye being on the produced outer edge. The species are few, but mostly widely distributed; rather large, voracious sharks with small sharp teeth.
The true hammer-head, Sphyrna zygæna, Fig. 337, is common from the Mediterranean to Cape Cod, California, Hawaii, and Japan. The singular form of its head is one of the most extraordinary modifications shown among fishes. The bonnet-head (Sphyrna tiburo) has the head kidney-shaped or crescent-shaped. It is a smaller fish, but much the same in distribution and habits. Intermediate forms occur, so that with all the actual differences we must place the Sphyrnidæ all in one genus. Fossil hammer-heads occur in the Miocene, but their teeth are scarcely different from those of Carcharias. Sphyrna prisca, described by Agassiz, is the primeval species.
The Order of Tectospondyli.—The sharks and rays having no anal fin and with the calcareous lamellæ arranged in one or more rings around a central axis constitute a natural group to which, following Woodward, we may apply the name of Tectospondyli. The Cyclospondyli (Squalidæ, etc.) with one ring only of calcareous lamellæ may be included in this order, as also the rays, which have tectospondylous vertebræ and differ from the sharks as a group only in having the gill-openings relegated to the lower side by the expansion of the pectoral fins. The group of rays and Hasse's order of Cyclospondyli we may consider each as a suborder of Tectospondyli. The origin[Pg 544] of this group is probably to be found in or near the Cestraciontes, as the strong dorsal spines of the Squalidæ resemble those of the Heterodontidæ.

Fig. 337.—Hammer-head Shark, Sphyrna zygæna L. Hindustan. (After Day.)
Suborder Cyclospondyli.—In this group the vertebræ have the calcareous lamellæ arranged in a single ring about the central axis. The anal fin, as in all the tectospondylous sharks and rays, is wanting. In all the asterospondylous sharks, as in the Ichthyotomi, Acanthodei, and Chimæras, this fin is present. It is present in almost all of the bony fishes. All the species have spiracles, and in all are two dorsal fins. None have the nictitating membrane, and in all the eggs are hatched internally. Within the group there is considerable variety of form and structure. As above stated, we have a perfect gradation among Tectospondyli from true sharks, with the gill-openings lateral, to rays, which have the gill-opening on the ventral side, the great expansion of the pectoral fins, a character of relatively recent acquisition, having crowded the gill-openings from their usual position.
Family Squalidæ.—The largest and most primitive family of Cyclospondyli is that of the Squalidæ, collectively known as dogfishes or skittle-dogs. In the Squalidæ each dorsal fin has a stout spine in front, the caudal is bent upward and not keeled, and the teeth are small and varied in form, usually not all alike in the same jaw.

Fig. 338.—Dogfish, Squalus acanthias L. Gloucester, Mass.
The genus Squalus includes the dogfishes, small, greedy sharks abundant in almost all cool seas and in some tropical waters. They are known by the stout spines in the dorsal fins and by their sharp, squarish cutting teeth. They are largely sought by fishermen for the oil in their livers, which is used to adulterate better oils. Sometimes 20,000 have been taken in one[Pg 546] haul of the net. They are very destructive to herrings and other food-fishes. Usually the fishermen cut out the liver, throwing the shark overboard to die or to be cast on the beach. In northern Europe and New England Squalus acanthias is abundant. Squalus sucklii replaces it in the waters about Puget Sound, and Squalus mitsukurii in Japan and Hawaii. Still others are found in Chile and Australia. The species of Squalus live near shore and have the gray color usual among sharks. Allied forms perhaps hardly different from Squalus are found in the Cretaceous rocks and have been described as Centrophoroides. Other genera related to Squalus live in greater depths, from 100 to 600 fathoms, and these are violet-black. Some of the deep-water forms are the smallest of all sharks, scarcely exceeding a foot in length. Etmopterus spinax lives in the Mediterranean, and teeth of a similar species occur in the Italian Pliocene rocks. Etmopterus lucifer,[150] a deep-water species of Japan, has a brilliant luminous glandular area along the sides of the belly. Other small species of deeper waters belong to the genera Centrophorus, Centroscymnus, and Deania. In some of these species the scales are highly specialized, pedunculate, or having the form of serrated leaves. Some species are Arctic, the others are most abundant about Misaki in Japan and the Madeira Islands, two regions especially rich in semi-bathybial types. Allied to the Squalidæ is the small family of Oxynotidæ with short bodies and strong dorsal spine. Oxynotus centrina is found in the Mediterranean, and its teeth occur in the Miocene.
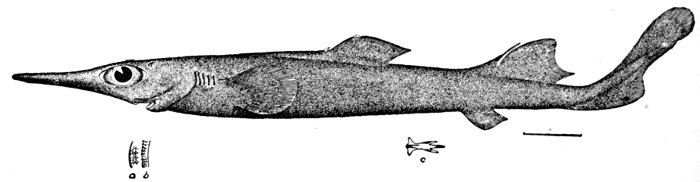
Fig. 339.—Etmopterus lucifer Jordan & Snyder. Misaki, Japan.
Family Dalatiidæ.—The Dalatiidæ, or scymnoid sharks, differ from the Squalidæ almost solely in the absence of dorsal spines. The smaller species belonging to Dalatias (Scymnorhinus, or Scymnus), Dalatias licha, etc., are very much like the dogfishes.
They are, however, nowhere very common. The teeth of Dalatias major exist in Miocene rocks. In the genus Somniosus the species are of very much greater size, Somniosus microcephalus attaining the length of about twenty-five feet. This species, known as the sleeper-shark or Greenland shark, lives in all cold seas and is an especial enemy of the whale, from which it bites large masses of flesh with a ferocity hardly to be expected from its clumsy appearance. From its habit of feeding on fish-offal, it is known in New England as "gurry-shark." Its small quadrate teeth are very much like those of the dogfish, their tips so turned aside as to form a cutting edge. The species is stout in form and sluggish in movement. It is taken for its liver in the north Atlantic on both coasts in Puget Sound and Bering Sea, and I have seen it in the markets of Tokyo. In Alaska it abounds about the salmon canneries feeding on the refuse.
Family Echinorhinidæ.—The bramble-sharks, Echinorhinidæ, differ in the posterior insertion of the very small dorsal fins, and in the presence of scattered round tubercles, like the thorns of a bramble instead of shagreen. The single species, Echinorhinus spinosus reaches a large size. It is rather scarce on the coasts of Europe, and was once taken on Cape Cod. The teeth of an extinct species, Echinorhinus richardi, are found in the Pliocene.
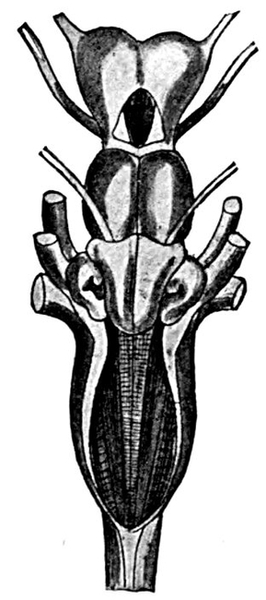
Fig. 340.—Brain of Monkfish, Squatina squatina L. (After Duméril.)
Suborder Rhinæ.—The suborder Rhinæ includes those sharks having the vertebræ tectospondylous, that is, with two or more series of calcified lamellæ, as on the rays. They are transitional forms, as near the rays as the sharks, although having the gill-openings rather lateral than inferior, the great pectoral fins being separated by a notch from the head.
The principal family is that of the angel-fishes, or monkfishes (Squatinidæ). In this group the body is depressed and flat like that of a ray. The greatly enlarged pectorals form a sort of shoulder in front alongside of the gill-openings, which has suggested the bend of the angel's wing.[Pg 548] The dorsals are small and far back, the tail is slender with small fins, all these being characters shared by the rays. But one genus is now extant, widely diffused in warm seas. The species if really distinct are all very close to the European Squatina squatina. This is a moderate-sized shark of sluggish habit feeding on crabs and shells, which it crushes with its small, pointed, nail-shaped teeth. Numerous fossil species of Squatina are found from the Triassic and Cretaceous, Squatina alifera being the best known.

Fig. 341.—Saw-shark, Pristiophorus japonicus Günther. Specimen from Nagasaki.
Family Pristiophoridæ, or Saw-sharks.—Another highly aberrant family is that of the sawsharks, Pristiophoridæ. These are small sharks, much like the Dalatiidæ in appearance, but with the snout produced into a long flat blade, on either side of which is a row of rather small sharp enameled teeth. These teeth are smaller and sharper than in the sawfish (Pristis), and the whole animal is much smaller than its analogue among the rays. This saw must be an effective weapon among the schools of herring and anchovies on which the sawsharks feed. The true teeth are small, sharp, and close-set. The few species of sawsharks are marine, inhabiting the shores of eastern Asia and Australia. Pristiophorus japonicus is found rather sparsely along the shores of Japan. The vertebræ in this group are also tectospondylous. Both the Squatina and Pristiophorus represent a perfect transition from the sharks and rays. We regard them as sharks only because the gill-openings are on the side, not[Pg 549] crowded downward to the under side of the body-disk. As fossil, Pristiophorus is known only from a few detached vertebræ found in Germany.
Suborder Batoidei, or Rays.—The suborder of Batoidei, Rajæ, or Hypotrema, including the skates and rays, is a direct modern offshoot from the ancestors of tectospondylous sharks, its characters all specialized in the direction of life on the bottom with a food of shells, crabs, and other creatures less active than fishes.
The single tangible distinctive character of the rays as a whole lies in the position of the gill-openings, which are directly below the disk and not on the side of the neck in all the sharks. This difference in position is produced by the anterior encroachment of the large pectoral fins, which are more or less attached to the side of the head. By this arrangement, which aids in giving the body the form of a flat disk, the gill-openings are limited and forced downward. In the Squatinidæ (angel-fishes) and the Pristiophoridæ (sawsharks) the gill-openings have an intermediate position, and these families might well be referred to the Batoidei, with which group they agree in the tectospondylous vertebræ.
Other characters of the rays, appearing progressively, are the widening of the disk, through the greater and greater development of the fins, the reduction of the tail, which in the more specialized forms becomes a long whip, the reduction, more and more posterior insertion, and the final loss of the dorsal fins, which are always without spine, the reduction of the teeth to a tessellated pavement, then finally to flat plates and the retention of the large spiracle. Through this spiracle the rays breathe while lying on the bottom, thus avoiding the danger of introducing sand into their gills, as would be done if they breathed through the mouth. In common with the cyclospondylous sharks, all the rays lack the anal fin. The rays rarely descend to great depths in the sea. The different members have varying relations, but the group most naturally divides into thick-tailed rays or skates (Sarcura) and whip-tailed rays or sting-rays (Masticura). The former are much nearer to the sharks and also appear earliest in geological times.
Pristididæ, or Sawfishes.—The sawfishes, Pristididæ, are long, shark-like rays of large size, having, like the sawsharks, the[Pg 550] snout prolonged into a very long and strong flat blade, with a series of strong enameled teeth implanted in sockets along either side of it. These teeth are much larger and much less sharp than in the sawsharks, but they are certainly homologous with these, and the two groups must have a common descent, distinct from that of the other rays. Doubtless when taxonomy is a more refined art they will constitute a small suborder together. This character of enameled teeth on the snout would seem of more importance than the position of the gill-openings or even the flattening and expansion of the body. The true teeth in the sawfishes are blunt and close-set, pavement-like as befitting a ray. (See Fig. 152.)
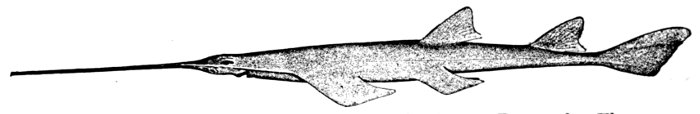
Fig. 342.—Sawfish, Pristis pectinatus Latham. Pensacola, Fla.
The sawfishes are found chiefly in river-mouths of tropical America and West Africa: Pristis pectinatus in the West Indies; Pristis zephyreus in western Mexico; and Pristis pectinatus in the Senegal. They reach a length of ten to twenty feet, and with their saws they make great havoc among the schools of mullets and sardines on which they feed. The stories of their attacks on the whale are without foundation. The writer has never found any of the species in the open sea. They live chiefly in the brackish water of estuaries and river-mouths.
Fossil teeth of sawfishes occur in abundance in the Eocene. Still older are vertebræ from the Upper Cretaceous at Maestricht. In Propristis schweinfurthi the tooth-sockets are not yet calcified. In Sclerorhynchus atavus, from the Upper Cretaceous, the teeth are complex in form, with a "crimped" or stellate base and a sharp, backward-directed enameled crown.
Rhinobatidæ, or Guitar-fishes.—The Rhinobatidæ (guitar-fishes) are long-bodied, shovel-nosed rays, with strong tails; they are ovoviviparous, hatching the eggs within the body. The body, like that of the shark or sawfish, is covered with nearly uniform shagreen. The numerous species abound in all warm seas; they are olive-gray in color and feed on small animals of the sea[Pg 551]bottoms. The length of the snout differs considerably in different species, but in all the body is relatively long and strong. Most of the species belong to Rhinobatus. The best-known American species are Rhinobatus lentiginosus of Florida and Rhinobatus productus of California. The names guitar-fish, fiddler-fish, etc., refer to the form of the body. Numerous fossil species, allied to the recent forms, occur from the Jurassic. Species much like Rhinobatus occur in the Cretaceous and Eocene. Tamiobatis vetustus, lately described by Dr. Eastman from a skull found in the Devonian of eastern Kentucky, the oldest ray-like fish yet known, is doubtless the type of a distinct family, Tamiobatidæ. It is more likely a shark however than a ray, although the skull has a flattened ray-like form.

Fig. 343.—Guitar-fish, Rhinobatus lentiginosus Garman. Charleston, S. C.
Closely related to the Rhinobatidæ are the Rhinidæ (Rhamphobatidæ), a small family of large rays shaped like the guitar-fishes and found on the coast of Asia. Rhina ancylostoma extends northward to Japan.
In the extinct family of Astrodermidæ, allied to the Rhinobatidæ, the tail has two smooth spines and the skin is covered with tubercles. In Belemnobatis sismondæ the tubercles are conical; in Astrodermus platypterus they are stellate.
Rajidæ, or Skates.—The Rajidæ, skates, or rays, inhabit the colder waters of the globe and are represented by a large number of living species. In this family the tail is stout, with two-rayed dorsal fins and sometimes a caudal fin. The skin is variously armed with spines, there being always in the male two series of specialized spinous hooks on the outer edge of the pectoral fin. There is no serrated spine or "sting," and in all the species the eggs are laid in leathery cases, which are[Pg 552] "wheelbarrow-shaped," with a projecting tube at each of the four angles. The size of this egg-case depends on the size of the species, ranging from three to about eight inches in length. In some species more than one egg is included in the same case.
Most of the species belong to the typical genus Raja, and these are especially numerous on the coasts of all northern regions, where they are largely used as food. The flesh, although rather coarse and not well flavored, can be improved by hot butter, and as "raie au beurre noir" is appreciated by the epicure. The rays of all have small rounded teeth, set in a close pavement.
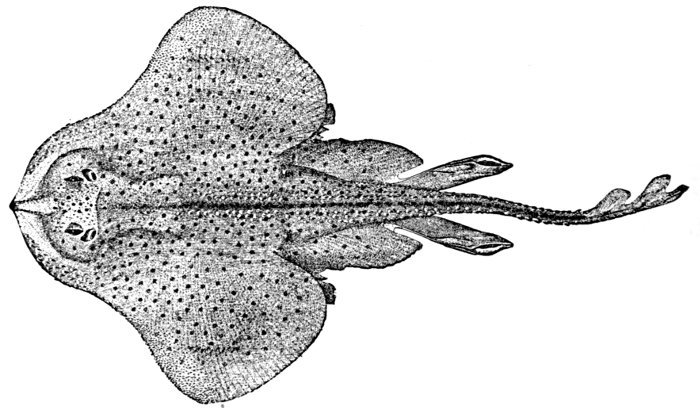
Fig. 344.—Common Skate, Raja erinacea Mitchill. Woods Hole, Mass.
Some of the species, known on our coasts as "barn-door skates," reach a length of four or five feet. Among these are Raja lævis and Raja ocellata on our Atlantic coast, Raja binoculata in California, and Raja tengu in Japan. The small tobacco-box skate, brown with black spots, abundant on the New England coast, is Raja erinacea. The corresponding species in California is Raja inornata, and in Japan Raja kenojei. Numerous other species, Raja batis, clavata, circularis, fullonica, etc., occur on the coasts of Europe. Some species are variegated in color, with eye-like spots or jet-black marblings. Still others, living in deep waters, are jet-black with the body very soft and[Pg 553] limp. For these Garman has proposed the generic name Malacorhinus, a name which may come into general use when the species are better known. In the deep seas rays are found even under the equator. In the south-temperate zone the species are mostly generically distinct, Psammobatis being a typical form, differing from Raja. Discobatus sinensis, common in China and Japan, is a shagreen-covered form, looking like a Rhinobatus. It is, however, a true ray, laying its eggs in egg-cases, and with the pectorals extending on the snout. Fossil Rajidæ, known by the teeth and bony tubercles, are found from the Cretaceous onward. They belong to Raja and to the extinct genera Dynatobatis, Oncobatis, and Acanthobatis. The genus Arthropterus (rileyi) from the Lias, known from a large pectoral fin, with distinct cylindrical-jointed rays, may have been one of the Rajidæ, or perhaps the type of a distinct family, Arthropteridæ.
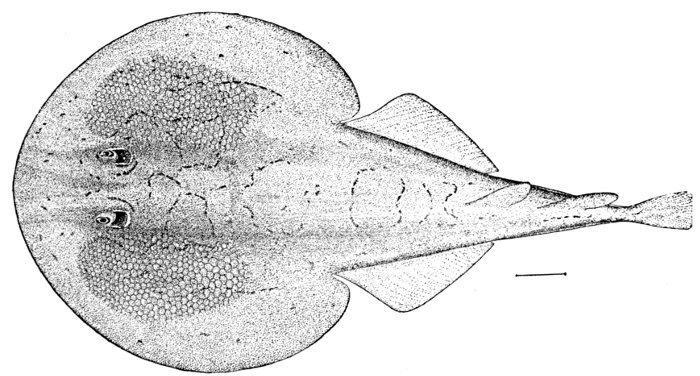
Fig. 345.—Numbfish, Narcine brasiliensis Henle, showing electric cells. Pensacola, Fla.
Narcobatidæ, or Torpedoes.—The torpedoes, or electric rays (Narcobatidæ), are characterized by the soft, perfectly smooth skin, by the stout tail with rayed fins, and by the ovoviviparous habit, the eggs being hatched internally. In all the species is developed an elaborate electric organ, muscular in its origin and composed of many hexagonal cells, each filled with soft fluid. These cells are arranged under the skin about the back[Pg 554] of the head and at the base of the pectoral fin, and are capable of benumbing an enemy by means of a severe electric shock. The exercise of this power soon exhausts the animal, and a certain amount of rest is essential to recovery.
The torpedoes, also known as crampfishes or numbfishes, are peculiarly soft to the touch and rather limp, the substance consisting largely of watery or fatty tissues. They are found in all warm seas. They are not often abundant, and as food they have not much value.
Perhaps the largest species is Tetronarce occidentalis, the crampfish of our Atlantic coast, black in color, and said sometimes to weigh 200 pounds. In California Tetronarce californica reaches a length of three feet and is very rarely taken, in warm sandy bays. Tetronarce nobiliana in Europe is much like these two American species. In the European species, Narcobatus torpedo, the spiracles are fringed and the animal is of smaller size. To Narcine belong the smaller numbfish, or "entemedor," of tropical America. These have the spiracles close behind the eyes, not at a distance as in Narcobatus and Tetronarce. Narcine brasiliensis is found throughout the West Indies, and Narcine entemedor in the Gulf of California. Astrape, a genus with but one dorsal fin, is common in southern Japan. Fossil Narcobatus and Astrape occur in the Eocene, one specimen of the former nearly five feet long. Vertebræ of Astrape occur in Prussia in the amber-beds.
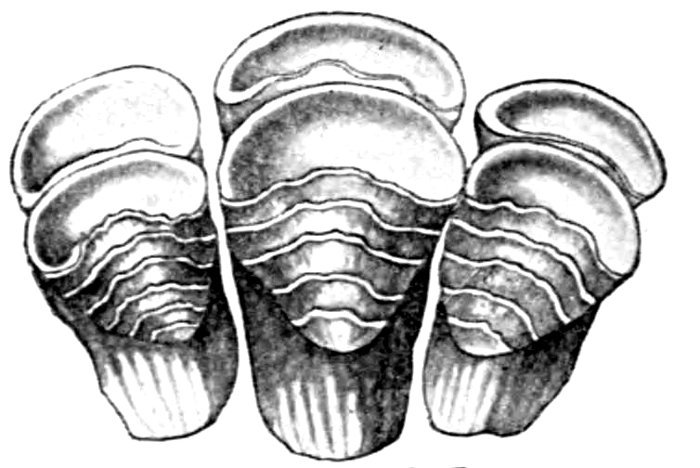
Fig. 346.—Teeth of Janassa linguæformis Atthey. Carboniferous. Family Petalodontidæ. (After Nicholson.)
Petalodontidæ.—Near the Squatinidæ, between the sharks and the rays, Woodward places the large extinct family of Petalodontidæ, with coarsely paved teeth each of which is elongate with a central ridge and one or more strong roots at base. The best-known genera are Janassa and Petalodus, widely distributed in Carboniferous time. Janassa is a broad flat shark, or, perhaps, a skate, covered with smooth shagreen. The large pectoral fins are grown to the head; the rather large ventral fins are separated from them. The tail is small,[Pg 555] and the fins, as in the rays, are without spines. The teeth bear some resemblance to those of Myliobatis. Janassa is found in the coal-measures of Europe and America, and other genera extend upward from the Subcarboniferous limestones, disappearing near the end of Carboniferous time. Petalodus is equally common, but known only from the teeth. Other widely distributed genera are Ctenoptychius and Polyrhizodus.
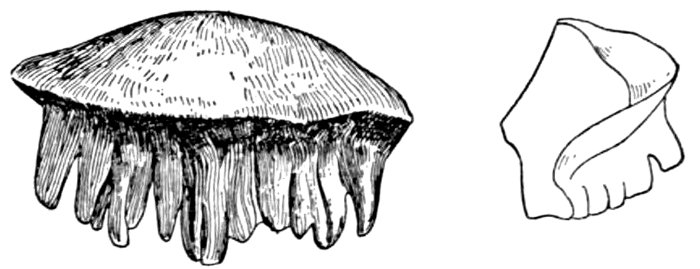
Fig. 347.—Polyrhizodus radicans Agassiz. Family Petalodontidæ. Carboniferous of Ireland. (After McCoy.)
These forms may be intermediate between the skates and the sting-rays. In dentition they resemble most the latter.
Similar to these is the extinct family of Pristodontidæ with one large tooth in each jaw, the one hollowed out to meet the other. It is supposed that but two teeth existed in life, but that is not certain. Nothing is known of the rest of the body in Pristodus, the only genus of the group.
Dasyatidæ, or Sting-rays.—In the section Masticura the tail is slender, mostly whip-like, without rayed dorsal or caudal fins, and it is usually armed with a very long spine with saw-teeth projecting backward. In the typical forms this is a very effective weapon, being wielded with great force and making a jagged wound which in man rarely heals without danger of blood-poisoning. There is no specific poison, but the slime and the loose cuticle of the spine serve to aggravate the irregular cut. I have seen one sting-ray thrust this spine through the body of another lying near it in a boat. Occasionally two or three of these spines are present. In the more specialized forms of sting-rays this spine loses its importance. It becomes very small and not functional, and is then occasionally or even generally absent in individuals.
The common sting-rays, those in which the caudal spine is most developed, belong to the family of Dasyatidæ. This group is characterized by the small skate-like teeth and by the non-extension of the pectoral rays on the head. The skin is smooth or more or less rough. These animals lie flat on the sandy bottoms in nearly all seas, feeding on crabs and shellfish. All hatch the eggs within the body. The genus Urolophus has a[Pg 556] rounded disk, and a stout, short tail with a caudal fin. It has a strong spine, and for its size is the most dangerous of the sting-rays. Urolophus halleri, the California species, was named for a young man who was stung by the species at the time of its first discovery at San Diego in 1863. Urolophus jamaicensis abounds in the West Indies, Urolophus mundus at Panama, and Urolophus fuscus in Japan. None of the species reach Europe. The true sting-ray (stingaree, or clam-cracker), Dasyatis, is more widely diffused and the species are very closely related. In these species the body is angular and the tail whip-like. Some of the species reach a length of ten or twelve feet. None have any economic value, and all are disliked by fishermen. Dasyatis pastinaca is common in Europe, Dasyatis centrura along our Atlantic coast, Dasyatis sabina ascends the rivers of Florida, and Dasyatis dipterura abounds in the bay of San Diego. Other species are found in tropical America, while still others (Dasyatis akajei, kuhlii, zugei, etc.) swarm in Japan and across India to Zanzibar.
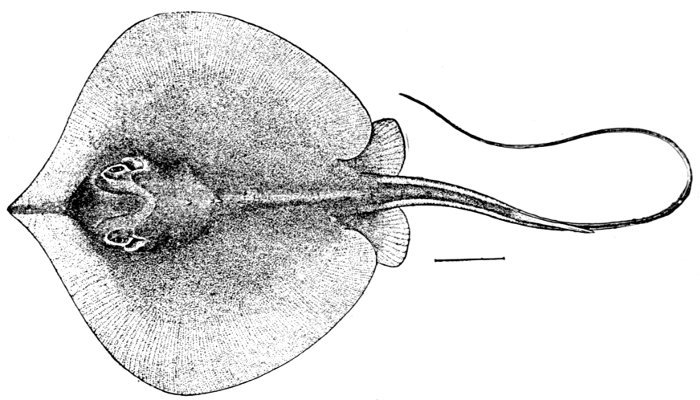
Fig. 348.—Sting-ray, Dasyatis sabina Le Sueur. Galveston.
Pteroplatea, the butterfly-ray, has the disk very much broader than long, and the trivial tail is very short, its little spine more often lost than present. Different species of this genus circle the globe: Pteroplatea maclura, on our Atlantic coast; Pteroplatea marmorata, in California; Pteroplatea japonica, in Japan;[Pg 557] and Pteroplatea altavela, in Europe. They are all very much alike, olive, with the brown upper surface pleasingly mottled and spotted.
Sting-rays of various types, Tæniura, Urolophus, etc., occur as fossils from the Eocene onward. A complete skeleton called Xiphotrygon acutidens, distinguished from Dasyatis by its sharp teeth, is described by Cope from the Eocene of Twin Creek in Wyoming. Vertebræ of Urolophus are found in German Eocene. Cyclobatis (oligodactylus), allied to Urolophus, with a few long pectoral rays greatly produced, extending over the tail and forming a rayed wreath-like projection over the snout, is known from the Lower Cretaceous.
Myliobatidæ.—The eagle-rays, Myliobatidæ, have the pectoral fins extended to the snout, where they form a sort of rayed pad. The teeth are very large, flat, and laid in mosaic. The whip-like tail is much like that in the Dasyatidæ, but the spine is usually smaller. The eagle-like appearance is suggested by the form of the skull. The eyes are on the side of the head with heavy eyebrows above them. The species are destructive to clams and oysters, crushing them with their strong flat teeth.
In Aëtobatus the teeth are very large, forming but one row. The species Aëtobatus narinari is showily colored, brown with yellow spots, the body very angular, with long whip-like tail. It is found from Brazil to Hawaii and is rather common.
In Myliobatis the teeth are in several series. The species are many, and found in all warm seas. Myliobatis aquila is the eagle-ray of Europe, Myliobatis californicus is the batfish of California, and Myliobatis tobijei takes its place in Japan.
In Rhinoptera the snout is notched and cross-notched in front so that it appears as if ending in four lobes at the tip. These "cow-nosed rays," or "whipparees," root up the soft bottoms of shallow bays in their search for clams, much as a drove of hogs would do it. The common American species is Rhinopterus bonasus. Rhinoptera steindachneri lives in the Gulf of California.
Teeth and spines of all these genera are common as fossils from the Eocene onwards, as well as many of the extinct genus, Ptychodus, with cyclospondylous vertebræ. Ptychodus mammilaris, rugosus, and decurrens are characteristic of the Creta[Pg 558]ceous of England. Myliobatis dixoni is common in the European Eocene, as is also Myliobatis toliapicus and Aëtobatis irregularis. Apocopodon seriacus is known from the Cretaceous of Brazil.
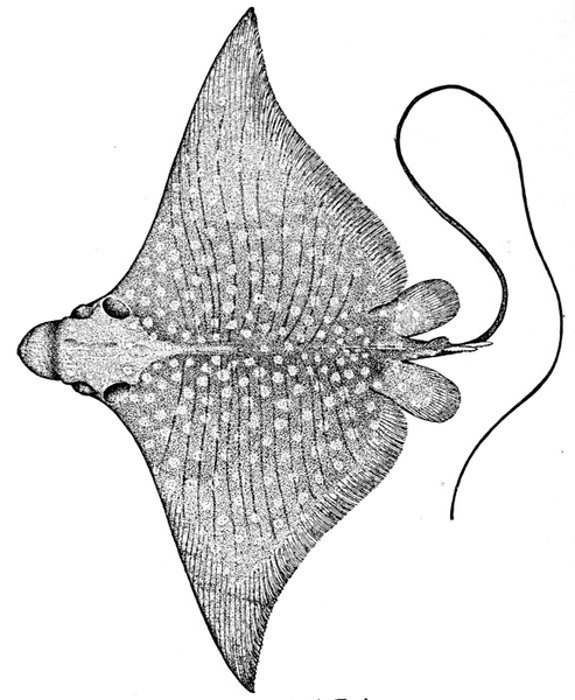
Fig. 349.—Eagle-ray, Aëtobatis narinari (Euphrasen). Cedar Keys, Fla.
Family Psammodontidæ.—The Psammodontidæ are known only from the teeth, large, flat, or rounded and finely dotted or roughened on the upper surface, as the name Psammodus (ψάμμος,[Pg 559] sand; ὀδούς, tooth) would indicate. The way in which the jaws lie indicates that these teeth belonged to rays rather than sharks. Numerous species have been described, mostly from the Subcarboniferous limestones. Archæobatis gigas, perhaps, as its name would indicate, the primeval skate, is from the Subcarboniferous limestone of Greencastle, Indiana. Teeth of numerous species of Psammodus and Copodus are found in many rocks of Carboniferous age. Psammodus rugosus common in Carboniferous rocks of Europe.
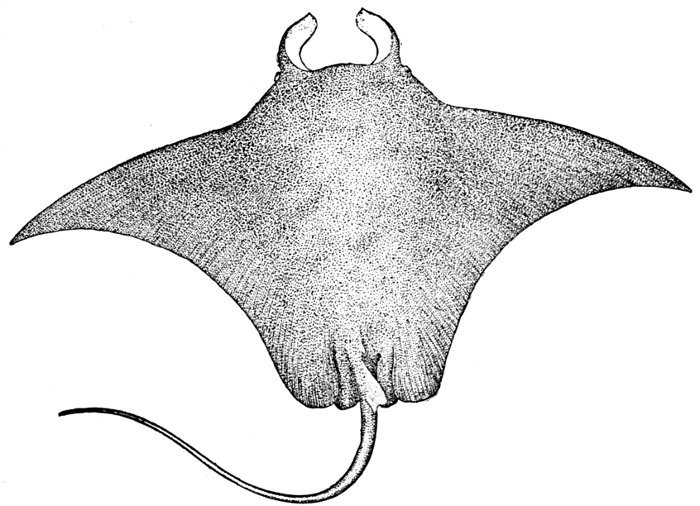
Fig. 350.—Devil-ray or Sea-devil, Manta birostris (Walbaum). Florida.
Family Mobulidæ.—The sea-devils, Mobulidæ, are the mightiest of all the rays, characterized by the development of the anterior lobe of the pectorals as a pair of cephalic fins. These stand up like horns or cars on the upper part of the head. The teeth are small and flat, tubercular, and the whip-like tail is with or without spine. The species are few, little known, and inordinately large, reaching a width of more than twenty feet and a weight, according to Risso, of 1250 pounds. When harpooned it is said that they will drag a large boat with great swiftness. The manta, or sea-devil, of tropical America is[Pg 560] Manta birostris. It is said to be much dreaded by the pearl-fishers, who fear that it will devour them "after enveloping them in its vast wings." It is not likely, however, that the manta devours anything larger than the pearl-oyster itself. Manta hamiltoni is a name given to a sea-devil of the Gulf of California. The European species Mobula edentula reaches a similarly enormous size, and Mobula hypostoma has been scantily described from Jamaica and Brazil. Mobula japonica occurs in Japan. A fœtus in my possession from a huge specimen taken at Misaki is nearly a foot across. In Mobula (Cephaloptera) there are teeth in both jaws, in Manta (Ceratoptera) in the lower jaw only. In Ceratobatis from Jamaica (C. robertsi) there are teeth in the upper jaw only. Otherwise the species of the three genera are much alike, and from their huge size are little known and rarely seen in collections. Of Mobulidæ no extinct species are known.
[150] Dr. Peter Schmidt has made a sketch of this little shark at night from a living example, using its own light.

The Chimæras.—Very early in geological times, certainly as early as the middle Silurian, the type of Chimæras diverged from that of the sharks. Hasse derives them directly from his hypothetical primitive Polyospondyli, by way of the Acanthodei and Ichthyotomi. In any event the point of divergence must be placed very early in the evolution of sharks, and this suggestion is as likely as any other. The chief character of Chimæras is found in the autostylic skull, which is quite different from the hyostylic skull of the sharks. In the sharks and in all higher fishes the mandible is joined to the skull by a suspensorium of bones or cartilages (quadrate, symplectic, and hyomandibular bones in the Teleost fishes). To this arrangement the name hyostylic is given. In the Chimæra there is no suspensorium, the mandible being directly attached to the cranium, of which the hyomandibular and quadrate elements form an integral part, this arrangement being called autostylic. The palato-quadrate apparatus, of which the upper jaw is the anterior part, is immovably fused with the cranium, instead of being articulated with it. This fact, gives the name to the subclass Holocephali (ὅλος, whole or solid; κεφαλή, head). Other characters are found in the incomplete character of the back-bone, which consists of a scarcely segmented notochord differing from the most primitive condition imagined only in being surrounded by calcareous rings, no lime entering into the composition of the notochord itself. The tail is diphycercal and usually prolonged in a filament (leptocercal). The shoulder-girdle, as in the sharks, is free from the skull. The pectoral fins are short and broad, without segmented axis or archiptery[Pg 562]gium and without recognizable analogue of the three large cartilages seen in the sharks, the propterygium, mesopterygium, and metapterygium. In the mouth, instead of teeth, are developed flat, bony plates called tritors or grinders, set endwise in the front of the jaws. The gills are fringe-like, free at the tips as in ordinary fishes, and there is a single external opening for them all as in true fishes, and they are covered with a flap of skin. These structures are, however, quite different from those of the true fishes and are doubtless independently developed. There is no spiracle. The skin is smooth or rough. In the living forms and most of the extinct species there is a strong spine in the dorsal fin. The ventral fin in the male has complex, usually trifid, claspers, and an analogous organ, the cephalic holder, is developed on the front of the head, in the adult male. This is a bony hook with a brush of glistening enameled teeth at the end. The eggs are large, and laid in oblong or elliptical egg-cases, provided with silky filaments. The eggs are fertilized after they are extruded. Mucous channels and lateral line are highly developed, being most complex about the head. The brain is essentially shark-like, the optic nerves form a chiasma, and the central hemispheres are large.
The teeth of the Chimæras are thus described by Woodward, vol. 2, pp. 36, 37:
"In all the known families of Chimæroids, the dentition consists of a few large plates of vascular dentine, of which certain areas ('tritors') are specially hardened by the deposition of calcareous salts within and around groups of medullary canals, which rise at right angles to the functional surface. In most cases there is a single pair of such plates in the lower jaw, meeting at the symphysis, while two pairs are arranged to oppose these above. As a whole, the dentition thus closely resembles that of the typical Dipnoi (as has often been pointed out); and the upper teeth may be provisionally named palatine and vomerine until further discoveries shall have revealed their precise homologies. The structures are sometimes described as 'jaws,' and regarded as dentaries, maxillæ, and premaxillæ, but the presence of a permanent pulp under each tooth is conclusive proof of their bearing no relation to the familiar membrane-bones thus named in higher fishes."
Relationship of Chimæras.—As to the origin of the Chimæras and their relation to the sharks, Dr. Dean has this recent ("The Devonian Lamprey") and interesting word:
"The Holocephali have always been a doubtful group, anatomy and palæontology contributing but imperfect evidence as to their position in the gnathostome phylum. Their embryology, however, is still undescribed, except in a brief note by T. J. Parker, and it is reasonably looked to to contribute evidence as to their line of descent. The problem of the relationships of the Chimæroids has long been of especial interest to me, and it has led me to obtain embryonic material of a Pacific species of one of these forms. It may be of interest in this connection to state that the embryology of this form gives the clearest evidence that the wide separation of the Selachii and Holocephali is not tenable. The entire plan of development in Chimæra colliei is clearly like that of a shark. The ovulation is closely like that of certain of the rays and sharks: the eggs are large, the segmentation is distinctly shark-like; the circular blastoderm overgrows the yolk in an elasmobranchian manner. The early embryos are shark-like; and the later ones have, as T. J. Parker has shown, external gills, and I note further that these arise, precisely as in shark-embryos, from the posterior margin of the gill-bar. A spiracle also is present. A further and most interesting developmental feature is the fact that the autostylism in Chimæra is purely of secondary nature and is at the most of ordinal value. It is found that in a larva of Chimæra measuring 45 mm. in length, the palato-quadrate cartilage is still separated from the skull by a wide fissure. This becomes gradually reduced by the confluence of the palato-quadrate cartilage with the skull, the fusion taking place at both the anterior and posterior ends of the mesal rim of the cartilage. The remains of the fissure are still well marked in the young Chimæra, four inches in length; and a rudiment of it is present in the adult skull as a passage-way for a nerve. Regarding the dentition: it may also be noted in the present connection that the growth of the dental plates in Chimæra suggests distinctly elasmobranchian conditions. Thus on the roof of the mouth the palatine plates are early represented by a series of small more or less conical[Pg 564] elements which resemble outwardly, at least, the 'anlagen' of the pavement teeth in cestraciont sharks."
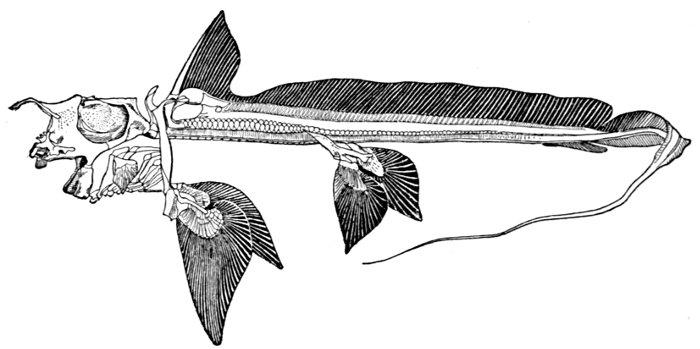
Fig. 351.—Skeleton of Chimæra monstrosa Linnæus. (After Dean.)
Family Chimæridæ.—The existing Chimæras are known also as spookfishes, ratfishes, and elephant-fishes. These are divided by Garman into three families, and in the principal family, the Chimæridæ, the snout is blunt, the skin without plates, and the dorsal fin is provided with a long spine. The flat tritors vary in the different genera. The single genus represented among living fishes is Chimæra, found in cold seas and in the oceanic depths. The best-known species, Chimæra colliei, the elephant-fish, or chimæra of California, abounds in shallow waters of ten to twenty fathoms from Sitka to San Diego. It is a harmless fish, useless except for the oil in its liver, and of special interest to anatomists as the only member of the family to be found when desired for dissection. This species was first found at Monterey by Mr. Collie, naturalist of Captain Beechey's ship, the Blossom. It is brown in color, with whitish spots, and reaches a length of 2½ feet. As a shallow-water form, with certain differences in the claspers and in the tail, Chimæra colliei is sometimes placed in a distinct genus, Hydrolagus. Other species inhabit much greater depths and have the tail produced into a long filament. Of these, Chimæra monstrosa, the sea-cat of the north Atlantic, has been longer known than any other Chimæra. Chimæra affinis has been dredged in the Gulf Stream and off Portugal. Chimæra phantasma and Chimæra mitsukurii are frequently taken in Japan,[Pg 565] and the huge jet-black Chimæra purpurascens in Hawaii and Japan. None of these species are valued as food, but all impress the spectator with their curious forms.
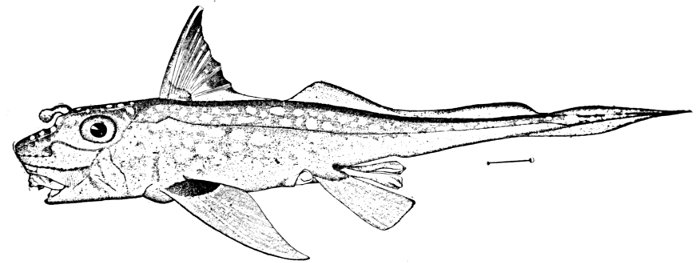
Fig. 352.—Elephant-fish, Chimæra colliei Lay & Bennett. Monterey.
The fossil Chimæridæ, although numerous from Triassic times and referred to several genera, are known chiefly by their teeth with occasional fin-spines, frontal holders, or impressions of parts of the skeleton. The earliest of chimæroid remains has been described by Dr. Charles D. Walcott[151] from Ordovician or Lower Silurian rocks at Cañon City, Colorado. Of the species called Dictyorhabdus priscus, only parts supposed to be the sheath of the notochord have been preserved. Dr. Dean thinks this more likely to be part of the axis of a cephalopod shell. The definitely known Chimæridæ are mainly confined to the rocks of the Mesozoic and subsequent eras. Ischyodus priscus (avitus) of the lower Jura resembles a modern chimæra. Granodus oweni is another extinct chimæra, and numerous fin-spines, teeth, and other fragments in the Cretaceous and Eocene of America and Europe are referred to Edaphodon. A species of Chimæra has been recorded from the Pliocene of Tuscany, and one of Callorhynchus from the greensand of New Zealand. Other American Cretaceous genera of chimæroids are Mylognathus, Bryactinus, Isotænia, Leptomylus, and Sphagepœa. Dental plates called Rhynchodus are found in the Devonian.
Rhinochimæridæ.—The most degenerate of existing chimæras belong to the family of Rhinochimæridæ, characterized by the long flat soft blade in which the snout terminates. This structure [Pg 566]resembles that seen in the deep-sea shark, Mitsukurina, and in Polyodon. In Rhinochimæra pacifica of Japan the teeth in each jaw form but a single plate. In Harriotta raleighana, of the Gulf Stream, they are more nearly as in Chimæra. Both are bathybial fishes, soft in texture, and found in great depths. The family of Callorhynchidæ, or Antarctic Chimæras, includes the bottle-nosed Chimæra (Callorhynchus callorhynchus) of the Patagonian region. In this species the snout is also produced, a portion being turned backward below in front of the mouth, forming a sensory pad well supplied with nerves.
Extinct Chimæroids.—According to Woodward, three other families are recognizable among the extinct forms.
The Ptyctodontidæ are known from the teeth only, a single pair of large, laterally compressed dental plates in each jaw, with a few hard tritoral areas. These occur in Silurian and Devonian rocks. Ptyctodus obliquus from the Devonian of Russia is the best-known species. Other genera are Rhynchodus and Palæomylus.
The Squalorajidæ have the head depressed and the snout produced in a flat rostrum, as in Harriotta. There is no dorsal spine, and the teeth are a few thin curved plates. The frontal holder of the male is well developed. The few species occur in the Lias. Squaloraja dolichognathos is known from numerous fragments from the Triassic in England and Scotland. Chalcodus permianus is found in German Permian.
The Myriacanthidæ have the body elongate, with dermal plates on the head and a long straight spine in the dorsal fin. The frontal holder is large. The species, few in number, are found in Mesozoic rocks. Myriacanthus paradoxus is the best-known species. Of another species, Chimæropsis paradoxa, a skeleton about three feet long has been found which shows a number of peculiar traits. The skin is covered with ribbed shagreen scales. The dorsal fin has a large spine with retrorse serrations behind. The tail is slim, and the pectoral and ventral fins are very large. Bony plates with conical spines protect the neck. The teeth are large and angular, of peculiar form.
Ichthyodorulites.—The term ichthyodorulite (ἰχθύς, fish; δόρυ, lance; λίθος, stone) is applied to detached fin-spines, dermal spines, and tubercles belonging to unrecognized species of[Pg 567] sharks and chimæras. Some of these are serrated, others entire, some straight, some curved, and some with elaborate armature or sculpture. Some doubtless belong to Cestraciontes, others to Pleuracanthidæ; some to Squalidæ, some to chimæras, and others, perhaps, to forms still altogether unknown.
[151] Bulletin Geol. Soc. America, 1892.

Ostracophores.—Among the earliest vertebrates actually recognized as fossils belongs the group known as Ostracophori (ὄστρακος, a box; φορέω, to bear). These are most extraordinary creatures, jawless, apparently limbless, and enveloped in most cases anteriorly in a coat of mail. In typical forms the head is very broad, bony, and horseshoe-shaped, attached to a slender body, often scaly, with small fins and ending in a heterocercal tail. What the mouth was like can only be guessed, but no trace of jaws has yet been found in connection with it. The most remarkable distinctive character is found in the absence of jaws and limbs in connection with the bony armature. The latter is, however, sometimes obsolete. The back-bone, as usual in primitive fishes, is developed as a persistent notochord imperfectly segmented. The entire absence of jaw structures, as well as the character of the armature, at once separates them widely from the mailed Arthrodires of a later period. But it is by no means certain that these structures were not represented by soft cartilage, of which no traces have been preserved in the specimens known.
Nature of the Ostracophores.—The Ostracophores are found in the Ordovician or Lower Silurian rocks, in the Upper Silurian, and in the Devonian. After the latter period they disappear. The species are very numerous and varied. Their real affinities have been much disputed. Zittel leaves them with the Ganoids, where Agassiz early placed them, but they show little homology in structure with the true Ganoids. Some have regarded them as aberrant Teleosts, possibly as freakish catfishes. Cope saw in them a huge mailed group of archaic Tunicates, while Patten has soberly and with considerable plausibility urged their affinity[153] to the group of spiders, especially to the horseshoe-crabs (Limulus) and their palæozoic ancestors, the Eurypteridæ and Merostomata.
The best guess as to the affinities of the Ostracophores is perhaps that given by Dr. Ramsey H. Traquair ("Fossil Fishes of the Silurian Rocks of the South of Scotland," 1899). Traquair regards them as highly aberrant sharks, or, more exactly, as being derived, like the Chimæras, from a primitive Elasmobranch [Pg 570]stock. In favor of this view is the character of their armature, the bony plates themselves to be regarded as formed by the fusion of shagreen grains or scales. According to Traquair: "Specialization from the most specialized form, Lanarkia, has been accompanied by (1) fusion of the spinelets (Lanarkia) or shagreen grains (Thelodus) into plates, scutes, and rhombic scales, supported by hard matter developed in a deeper layer of skin, and (2) alterations in the pectoral fin-flaps, which, becoming covered up by the postero-lateral plates in Drepanaspis, are finally no longer recognizable in the Pteraspidæ."
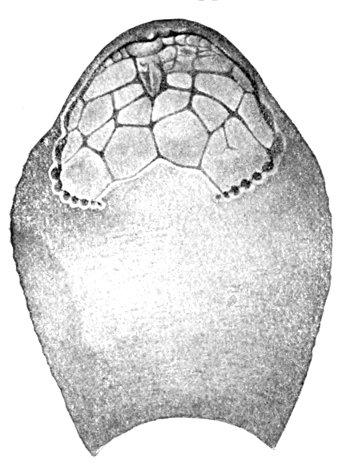
Fig. 353.—Odontotodus schrencki (Pander) (Tremataspis), ventral side. Island of Oesel. (After Patten.)
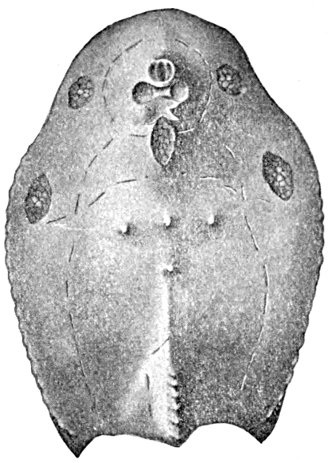
Fig. 354.—Odontotodus schrencki (Pander) (Tremataspis), dorsal side. Island of Oesel. (After Patten.)
Woodward leaves their exact relationship undefined, while others have regarded them as mailed lampreys, at any rate to be excluded from the Gnathostomi, or jaw-bearing series. The apparent absence of true jaws, true limbs, and limb-girdles certainly seems to separate them widely from true fishes, but these characters are negative only, perhaps due to degeneration, and at any rate they are not yet absolutely determined. Certainly they offer no positive proof of affinity with the modern Cyclostomes.
Dr. Traquair regards the Heterostraci or most primitive Ostracophores as most certainly derived from the Elasmobranchs. Other writers have attacked the integrity of the group of Ostracophores, questioning the mutual relationship of its component parts. Reiss, for example, regards the association of the Osteostraci with the Heterostraci as "unbegründet" and "unheilvoll," while Ray Lankester, as quoted by Traquair, affirms that "there is absolutely no reason for regarding Cephalaspis as allied to Pteraspis beyond that the two genera occur in the same rocks, and still less for concluding that either has any connection with Pterichthys." Elsewhere Lankester states that the Heterostraci are associated at present with the Osteostraci, "because they have, like Cephalaspis, a large head-shield, and because there is nothing else with which to associate them." Patten, on the other hand, seems inclined to deny the rank of Heterostraci and Osteostraci as even separate orders, regarding them as very closely related to each other as also to their supposed spider-like ancestors.

Fig. 355.—Head of Odontotodus schrencki Pander, from the side. (After Patten.)
But the consensus of opinion favors the belief that the four orders usually included under this head are distinct and at the same time are really related one to another. For our purposes, then, we may regard the Ostracophori as a distinct class of vertebrates. By placing it after the Elasmobranchs we may indicate its probable descent from a primitive shark-like stock.
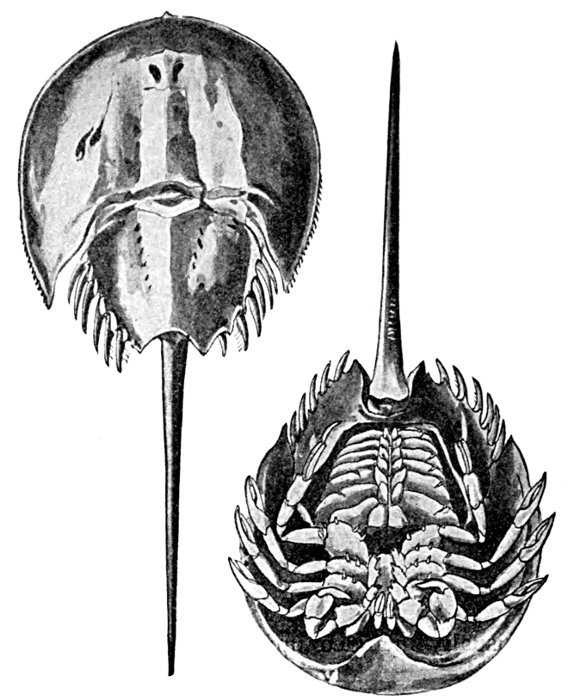
Fig. 356.—The Horseshoe Crab or King-crab, Limulus polyphemus Linnæus. Supposed by Professor Patten to be an ally of the Ostracophores; usually regarded as related to the Spiders.
On this subject Dr. Dean remarks: "The entire problem of the homology of the dermal plates and 'scales' in the Ostracophores and Arthrognaths is to the writer by no means as clear as previous writers have conceded. From the histological standpoint, admitting the craniote nature of the vasodentine and cancellous layers in the dermal plates, it nevertheless does not follow that they have been derived from the actual conditions of the dermal denticles of the ancestral Gnathostome, as were unquestionably the dermal plates of Teleostomes and Dipnoans. It seems equally if not more probable, on the other hand, that the dermal armoring of the distinct groups may have had an altogether different mode of origin, the product of a crude evolution which aimed to strengthen the skin by a general deposition of calcareous matter throughout its entire thickness. The tuberculation of plates thus acquired might have become an important step in the development of a more superficial type of armoring which is most preferably represented by the dermal denticles of Selachians. Nor, in passing, need the presence of a mucus-canal system in the early plated forms be of greater morphological importance than a foreshadowing of the conditions of Gnathostomes, for this system of organs[Pg 573] might serve as well as evidence, in a general way, of relationship with Marsipobranchs. Nor is this evidence the more conclusive when we reflect that no known type of Gnathostome, recent or fossil, possesses open sensory grooves in distinct dermal plates. The presence, furthermore, of a dorsal fin and a 'truly piscine heterocercal tail,' as noted by Traquair, is by no means as Gnathostome-like as these structures at first glimpse appear. For they lack not merely the characteristic radial supports of fishes, but even actinotrichia. Their mode of support, on the other hand, as Smith Woodward points out, is of a more generalized nature, bent scales, homologous with those of the adjacent body region, taking the place of the piscine external supports." The actual position in the system to be finally assigned to the Ostracophores is therefore still uncertain.
Orders of Ostracophores.—Four orders of Ostracophori are now usually recognized, known in the systems of Woodward and Traquair as Heterostraci, Osteostraci, Antiarcha, and Anaspida. The former is the most primitive and perhaps the most nearly allied to the sharks, the second is not very remote from it, the last two aberrant in very different directions. Hay places the Antiarcha with the Arthrodira under the superorder of Placodermi.
Order Heterostraci.—The Heterostraci (ἕτερος], different; ὀστράκος, box) have no bone-corpuscles in the coat of mail. This typically consists of a few pieces above, firmly united and traversed by dermal sense-organs or "lateral lines." The ventral shield is simple. Four families are recognized by Traquair as constituting the Heterostraci, these forming a continuous series from shark-like forms to the carapace-covered Pteraspis. In the most primitive family, the Thelodontidæ,[154] the head and trunk are covered with small scales or tubercles of dentine and not fused into large plates. The tail is slender and heterocercal, the caudal fin deeply forked. Until lately these tubercles were regarded as belonging to sharks, and they are still regarded by Traquair as evidence of the affinity of the Heterostraci with the Acanthodei. Dr. Traquair thinks that a flap or lappet-like projection behind the head may be [Pg 574]a pectoral fin. The three known genera are Thelodus, Lanarkia, and Ateleaspis. In Thelodus the scales consist of a base and a crown separated by a constriction or neck. Thelodus scoticus, Thelodus pagei, and Thelodus planus are found in the Silurian rocks of Scotland. Other species, as Thelodus tulensis of Russia, extend to the Upper Devonian.
In Lanarkia the large sharp scales have an expanded base like the mouth of a trumpet. Lanarkia horrida and L. spinulosa are found in the shire of Lanark in Scotland. In Ateleaspis (tesselatus) the skin is covered with small polygonal plates. The lateral flaps or possibly fins take the form of flat rhombic sculptured scales. In this genus the eyes seem to be on the top of the head.

Fig. 357.—Lanarkia spinosa Traquair. Upper Silurian. Family Thelodontidæ. (After Traquair.)
In the Psammosteidæ of the Devonian the head is covered with large plates which are not penetrated by the sense-organs. These plates are covered with minute, close-set tubercles, covered with brilliant ganoid enamel and with finely crimped edges. According to Dr. Traquair, these tubercles are shagreen granules which have coalesced and become united to plates formed in a deeper layer of the skin, as in Ateleaspis the minute scales have run together into polygonal plates. These creatures have been considered as "armored sharks," and Dr. Traquair regards them as really related to the acanthodean sharks. Nevertheless they are not really sharks at all, and they find their place with the Pteraspis and other longer known Heterostracans.
The family of Drepanaspidæ consists of a single recently known species, Drepanaspis gmundenensis, found in a pyritized condition[Pg 575] in purple roofing-slate in Gmünden, Germany. This fish, which reaches a length of about two feet, has a broad head, with eyes on its outer margin, with a slender body and heterocercal tail. The head has a broad median plate and smaller polygonal ones. The flaps, supposed to represent the pectoral fins, are here cased in immovable bone. No trace of internal skeleton is found by Traquair, who has given the restoration of this species, but the mouth has been outlined.
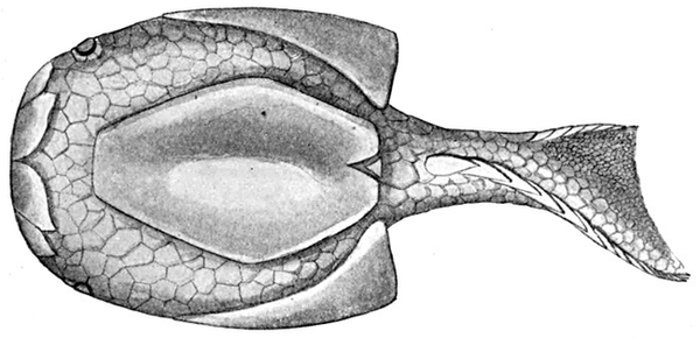
Fig. 358.—Drepanaspis gmundenensis Schlüter. Upper Silurian, Gmünden, Germany. (After Traquair.)
The best known of the Heterostracan families is that of Pteraspidæ. In this family the plates of the head are coalesced in a large carpace, the upper part originally formed of seven coalesced pieces. A stout dorsal spine fits into a notch of the carapace. The slender body is covered with small scales and ends in a heterocercal tail. The dermal sense-organs are well developed. Pteraspis rostrata occurs in the Lower Devonian. Other genera are Palæaspis and Cyrthaspis.
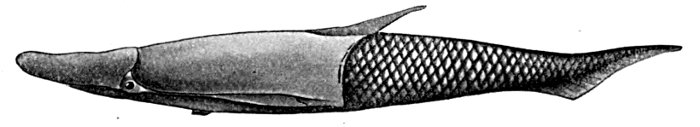
Fig. 359.—Pteraspis rostrata Agassiz. Devonian. Family Pteraspidæ. (After Nicholson.)
Order Osteostraci.—The Osteostraci (ὄστεον, bone; οστρακος, box) (called Aspidocephali by Rohon) have bone-corpuscles in the shields, and the shield of the back is in one piece without[Pg 576] lateral-line channels or sense-organs. Ventral shield single. The order includes three families. The Cephalaspidæ have the shields tuberculate, the one between the eyes fixed, and the anterior body-shields are not fused into a continuous plate. The best known of the numerous species is Cephalaspis lyelli from the Lower Devonian of England. Hemicyclaspis murchisoni occurs in the Upper Silurian of England, and the extraordinary Cephalaspis dawsoni in the Lower Devonian of Gaspé, Canada. Eukeraspis pustulifera has the head-shield very slender and armed with prickles. In the Thyestidæ the anterior body-scales are fused into a continuous plate. Thyestis and Didymaspis are genera of this type. The Odontotodontidæ (Tremataspidæ) have the shield truncate behind, its surface finely punctate, and the piece between the eyes not fixed. Odontotodus[155] schrenki is found in the Upper Silurian of the Island of Oesel in company with species of Thyestes. The Euphaneropidæ are represented in the Devonian of Quebec.
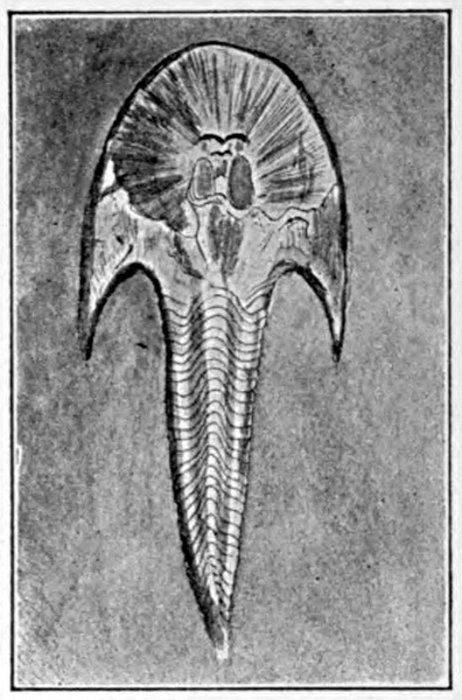
Fig. 360.—Cephalaspis lyelli Agassiz, restored. (After Agassiz.)
Order Antiarcha.—The Antiarcha (ἀντί, opposite; ἀρχός, anus) have also bone-corpuscles in the plates, which are also enameled. The sense-organs occupy open grooves, and the dorsal and ventral shields are of many pieces. The head is jointed on the trunk, and jointed to the head are paddle-like appendages, covered with bony plates and resembling limbs. There is no evidence that these erectile plates are real limbs. They seem to be rather jointed appendages of the head-plate, erectile on a hinge like a pectoral spine. There are traces of ear-cavities, gill-arches, and other fish-like structures, but nothing suggestive of mouth or limbs.
This group contains one family, the Asterolepidæ, with numerous species, mostly from Devonian rocks. The best known genus is Pterichthyodes,[156] in which the anterior median plate [Pg 577]of the back is overlapped by the posterior dorso-lateral. Pterichthyodes milleri from the Lower Devonian, named by Agassiz for Hugh Miller, is the best known species, although numerous others, mostly from Scottish quarries, are in the British Museum. Asterolepis maximus is a very large species from the same region, known from a single plate. Bothriolepis canadensis is from the Upper Devonian of Scaumenac Bay near Quebec, numerous specimens and fragments finely preserved having been found.
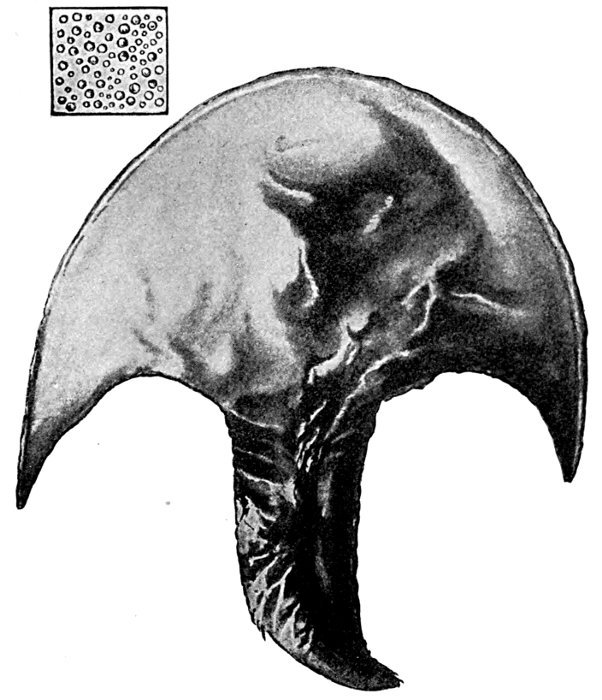
Fig. 361.—Cephalaspis dawsoni Lankester. Lower Devonian of Canada. Family Cephalaspidæ. (After Woodward.) In the square a portion of the tubercular surface is shown.
Microbrachium dicki with the pectoral appendages small occurs in the Devonian of Scotland.
The earliest remains of Ostracophori are found in Ordovician or Lower Silurian rocks of the Trenton horizon at Cañon[Pg 578] City, Colorado. These consist of enormous numbers of small fragments of bones mixed with sand. With these is a portion of the head carapace of a small Ostracophore which has been named by Dr. Walcott Asteraspis desiderata and referred provisionally to the family of Asterolepidæ, which belongs otherwise to the Lower Devonian.
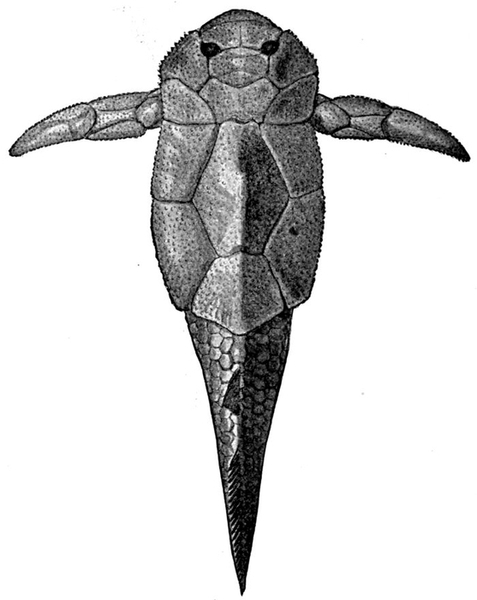
Fig. 362.—Pterichthyodes testudinarius (Agassiz), restored. Lower Devonian Family Asterolepidæ. (After Traquair and others.)
With these remains are found also scales possibly belonging to a Crossopterygian fish (Eriptychius). These remains make it evident that the beginning of the fish series lies far earlier than the rocks called Silurian, although fishes in numbers are not elsewhere known from rocks earlier than the Ludlow shales of the Upper Silurian, corresponding nearly to the Niagara period in America.
In the Ludlow shales we find the next appearance of the[Pg 579] Ostracophores, two families, Thelodontidæ and Birkeniidæ, being there represented.
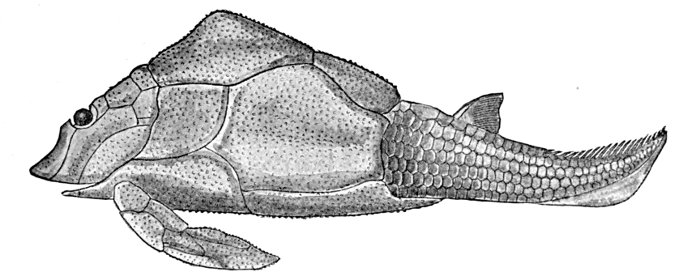
Fig. 363.—Pterichthyodes testudinarius Agassiz, side view. (After Zittel, etc.)
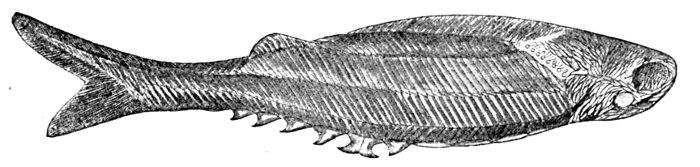
Fig. 364.—Birkenia elegans Traquair. Upper Silurian. (After Traquair.)
Order Anaspida.—Recently a fourth order, Anaspida (ἄ, without; ἀσπίς, shield), has been added to the Ostracophori through the researches of Dr. Traquair. This group occurs in the Upper Silurian in the south of Scotland. It includes the single family Birkeniidæ, characterized by the fusiform body, bluntly rounded head, bilobate, heterocercal tail, and a median row of hooked spinous scales along the ventral margin. No trace of jaws, teeth, limbs, or internal skeleton has been found. Unlike other Ostracophores, Birkenia has no cranial buckler with orbits on the top, nor have the scales and tubercles the microscopic structure found in other Ostracophores. In the genus Birkenia the head and body are completely covered by tubercular scutes. The gill-openings seem to be represented by a series of small perforations on the sides. A dorsal fin is present. Birkenia elegans is from the Ludlow and Downstonian rocks of southern Scotland. Lasianius problematicus from the same rocks is very similar, but is scaleless. It has a row of ventral plates like those of Birkenia, the only other hard parts it[Pg 580] possesses being a number of parallel rods behind the head, homologous with the lateral series of Birkenia. Lasianius is therefore a specialized and degenerate representation of Birkenia, differing somewhat as "the nearly naked Phanerosteon differs from other Palæoniscidæ whose bodies are covered with osseous scales."

Fig. 365.—Lasianius problematicus Traquair. Upper Silurian. (After Traquair.)
[152] This group was first called by Cope Ostracodermi—a name preoccupied for the group of bony trunkfishes, Ostracidæ. The still earlier name of Placodermi, chosen by McCoy (1848), was intended to include Arthrodires as well as Ostracophores. Rohon (1892) calls the group Protocephali, and to the two orders he assigns the names Aspidorhini and Aspidocephali. These groups correspond to Heterostraci and Osteostraci of Woodward. Another name of early date is that of Aspidoganoidei, given by Professor Gill in 1876, but not defined until 1896. These fishes are, however, not "Ganoids" and the name Ostracophori seems to receive general preference. The group Peltacephalata of Patten corresponds essentially to Ostracophori, as does also the order Hypostomata of Gadow.
[153] According to Professor Patten's view, the close resemblance of the shields of Pteraspis to those of contemporaneous Eurypterids indicates real affinity. But the Eurypterids are related to the spiders and to Limulus. The only reason for thinking that Pteraspis is a fish at all lies in its resemblance to Cephalaspis, which is in several ways fish-like, although its head shield is much like that of Limulus. All these resemblances in Patten's view indicate real affinity. Patten considers the Pteraspids as derived from primitive arachnid or spider-like forms having a bony carapace as Limulus has. From Pteraspis he derives the other Ostracophores, and from these the sharks and other vertebrates, all of which appear later in time than the earliest Ostracophores. This view of the origin of vertebrates is recently urged with much force by Professor Patten (Amer. Nat., 1904, 1827). Most naturalists regard such resemblances in specialized structures on the outside of an animal as parallelisms due to likeness in conditions of life. The external structure in forms of really different nature is often similarly modified. Thus certain catfishes, pipefishes, sea-moths, and agonoid fishes are all provided with bony plates not unlike those of ganoid fishes, although indicative of no real affinity with them. Commonly the ancestry of vertebrates is traced through enteropneustans to soft-bodied worms which have left no trace in the rocks.
In the same connection, Professor Patten suggests that the lateral fold from which many writers have supposed that the limbs or paired fins of vertebrates is evolved is itself a resultant of the fusion of the fringing appendages on the sides of the body. Such appendages are found in the primitive mailed arachnoids and in Limulus. They are shown very plainly in Patten's restoration of Cephalaspis. About thirty of them of a bony nature and jointed to the body occur on either side between the gill opening and the vent.
[154] Called Cœlolepidæ by Pander and Traquair, but Cœlolepis is a later synonym of Thelodus.
[155] This name, inappropriate or meaningless, is older than Tremataspis.
[156] The earlier name of Pterichthys has been already used for a genus of living fishes.

The Arthrodires.—Another large group of extinct fishes mailed and helmeted is included under the general name of Arthrodira[157] (ἄρθρος, joint; δεῖρα, neck), or Arthrognathi (ἄρθρος, γνάθος, jaw), the latter term recently framed by Dr. Dean with a somewhat broader application than the former.
These fishes differ from the Ostracophores, on the one hand, in the possession of jaws and in the nature of their armored covering. On the other hand, the nature of these jaws, the lack of differentiation of the skeleton, and the uncertain character of the limbs separate them still more widely from the true fishes. Their place in the system is still unknown, but their origin seems as likely to be traceable to Ostracophores as to any other group.
The head in all the species is covered with a great bony helmet. Behind this on the nape is another large shield, and [Pg 582]between the two is usually a huge joint which Dr. Dean compares to the hinge of a spring-beetle (Elater).
As to the presence of limbs, no trace of pectoral fin or anterior limb has been found. Dean denies the existence of any structures corresponding to either limb, but Woodward figures a supposititious posterior limb in Coccosteus, finding traces of basal bones which may belong to it.
These monstrous creatures have been considered by Woodward and others as mailed Dipnoans, but their singular jaws are quite unlike those of the Dipneusti, and very remote from any structures seen in the ordinary fish. The turtle-like mandibles seem to be formed of dermal elements, in which there lies little homology to the jaws of a fish and not much more with the jaws of Dipnoan or shark.
The relations with the Ostracophores are certainly remote, though nothing else seems to be any nearer. They have no affinity with the true Ganoids, to which vaguely limited group many writers have attached them. Nor is there any sure foundation to the view adopted by Woodward, that they are to be considered as armored offshoots of the Dipnoans.
According to Dean we might as well refer the Arthrodires to the sharks as to the Dipnoans. Dean further observes ("Fishes Living and Fossil"):
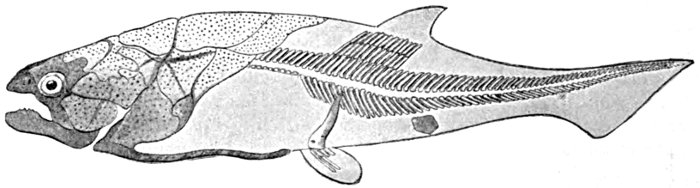
Fig. 366.—Coccosteus cuspidatus Agassiz, restored. Lower Devonian. (After Traquair, per Woodward.)
"The puzzling characters of the Arthrodirans do not seem to be lessened by a more definite knowledge of their different forms. The tendency, as already noted, seems to be at present to regard the group provisionally as a widely modified offshoot of the primitive Dipnoans, basing this view upon their general structural characters, dermal plates, dentition, autostylism. But only in the latter regard could they have differed more[Pg 583] widely from the primitive Elasmobranch or Teleostome, if it be admitted that in the matter of dermal structures they may be clearly separated from the Chimæroid. It certainly is difficult to believe that the articulation of the head of Arthrodirans could have been evolved after dermal bones had come to be formed, or that a Dipnoan could become so metamorphosed as to lose not only its body armoring, but its pectoral appendages as well. The size of the pectoral girdle is, of course, little proof that an anterior pair of fins must have existed, since this may well have been evolved in relation to the muscular supports of plastron, carapace, trunk, and head. The intermovement of the dental plates, seen especially in Dinichthys, is a further difficulty in accepting their direct descent from the Dipnoans."
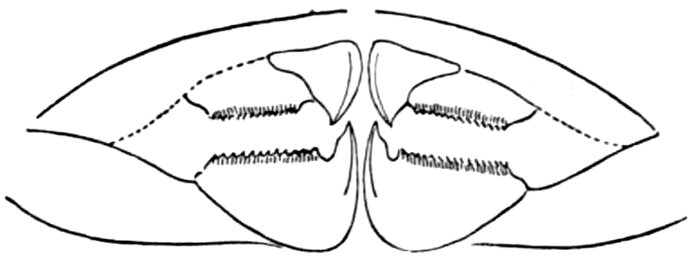
Fig. 367.—Jaws of Dinichthys hertzeri Newberry. Upper Devonian. Ohio. (After Newberry.)
Occurrence of Arthrodires.—These fishes occur in abundance from the Silurian times to the Mesozoic. In the Devonian their gigantic size and thick armor gave them the leading position among the hosts of the sea. Among the genera there occurred "series of forms most interesting as to their evolution." "It is found more and more evident," says Dr. Dean ("Fishes, Living and Fossil," pp. 135, 136) "that the Arthrodirans may have represented the dominant group in the Devonian period, as were the sharks in the Carboniferous, or as are the Teleosts in modern times. There were forms which, like Coccosteus, had eyes at the notches of the head-buckler; others, like Macropetalichthys, in which orbits were well centralized; some, like Dinichthys and Titanichthys, with the pineal foramen present; some with pectoral spines(?); some with elaborately sculptured dermal plates. Among their forms appear to have been those whose shape was apparently subcylindrical, adapted for swift swimming; others (Mylostoma) whose trunk was depressed to almost ray-like proportions. In size they varied from that of the perch to that of a basking shark. In dentition they presented the widest range in variation, from the formidable shear-like jaws of Dinichthys to the lip-like mandibles of Titanichthys, the tearing teeth of Trachosteus, the wonderfully forked tooth[Pg 584]-bearing jaw-tips of Diplognathus, to the Cestraciont type, Mylostoma. The latter form has hitherto been known only from its dentition, but now proves to be, as Newberry and Smith Woodward suggested, a typical Arthrodiran."
Classification of Arthrodira.—Our knowledge of the systematic relations of the Arthrodira is mostly of recent origin. Woodward refers most of the remains to the best known genus Coccosteus, and recognizes as families the Coccosteidæ, Mylostomidæ, Asterosteidæ, and Phyllolepidæ.

Fig. 368.—An Arthrodire, Dinichthys intermedius Newberry, restored. Devonian, Ohio. (After Dean.)
Dr. Bashford Dean in different papers has treated these fishes in great detail. In a recent paper on the "Relationships of the Arthrognathi"[158] he recognizes the group as a class coordinate with Cyclostomi and Elasmobranchii. This class, which he calls Arthrognathi, is first divided into two suborders, Anarthrodira, without joint at the neck, and Arthrodira, with such a joint. The former comprises one order, Stegothalami, and the latter two orders, Temnothoraci and Arthrothoraci. The following is Dr. Dean's definition of these orders and their component families:
Arthrognathi.—"Chordates whose anterior body region is encased in dermal elements, and divisible by a more or less definite partition into head and trunk. Dermal plates which surround the mouth function as jaws. No evidence of branchial arches. Column notochordal, showing no traces of centra; well-marked neural and hæmal elements. Paired limbs [absent or uncertain]. Dermal plates consisting typically of two layers, the superficial tuberculate, the inner bony with radiating lamellæ. [Pg 585]Orbits situated near or at the margin of the head-shield and separated from one another by fixed integumental plates. A pineal funnel present situated in a fixed plate. A mucous system whose canals radiate from the preoccipital region."
Anarthrodira.—"Arthrognaths in which the cranial and dorsal regions are separated by a fixed partition whose dorsal rim is overlapped and concealed by superficial plates. Of these a large median dorsal element is present which extends backward superficially from the region near the pineal funnel. Also a pair of elements which overlie the position of the external occipital joint. Suborbital plates apparently absent. Jaw elements undescribed."
Stegothalami (στέγος, roof; θάλαμος, chamber).—"Anarthrodires in which the cranio-dorsal septum is vertical and deep, its height equal apparently to that of the arch of the head-shield. By this deep partition the latter appears to inclose two chambers (whence the ordinal name). Orbits inclosed by pre- and postorbital plates. Mucous system lacks a postorbital canal."
One family, the Macropetalichthyidæ, thus defined:
"Stegothalami with large orbits and well-arched cranio-dorsal shield. Dorso-central shield long, wide, gomphoidal, extending backward to the hinder margin of the shield and bordered by all plates save the postorbitals and marginals. Pineal funnel small and obscure." Macropetalichthys sullivanti from Ohio Devonian rocks, and Macropetalichthys agassizi from the Devonian of Germany, are important species of this group.
The Asterosteidæ perhaps constitute a second family in this order. The single species Asterosteus stenocephalus is from the Devonian of Ohio.
Arthrodira.—"Arthrognaths in which the dorsal armoring is separated into dorsal and cranial elements, the latter attached to the former movably by means of a pair of peg-and-socket joints. The interval lying between cranial and dorsal armoring does not appear to have been protected by plates, and in the median line, instead of the cranio-central of the Anarthrodires, there are separate elements, median occipital, median dorsal, and perhaps others. Suborbital plates present. Jaws of three[Pg 586] pairs of elements. Ventral armoring of two pairs of lateral and two median elements."
Temnothoraci (τέμνω, to cut; θώραξ, thorax).—"Arthrodires whose cranial and dorsal shields are closely apposed, separated only by a transverse fissure-like interval (whence the ordinal name); interarticulation of cranial and dorsal shields little developed. Head-shield elliptical in outline as far as the line of the transverse division. The anterior rim of the shoulder-shield flattened at its sides, suggesting a rudiment of the vertical partition of the Anarthrodira. Suborbital plate is present, but takes no part, apparently, in the ventral boundary of the orbit, this being formed, as in the Anarthrodira, by the pre- and postorbital elements. Jaws, ventral armoring, and endoskeleton not definitely known."
One family, Chelonichthyidæ, thus defined:
"Temnothoraci with orbits relatively small in size and situated well forward in the head-shield. Occipital elements produced antero-posteriorly, the external occipital forming the posterior lateral angle of the head, no projection of the head occurring in the region of the marginal plate. Median occipital trapezoidal. Centrals take part in the median boundary of the orbits, and embrace the pineal plate. Median dorsal with poorly developed keel and terminal process."
Heterosteus asmussi (perhaps to be called Ichthyosauroides spinosus) is a gigantic species from the Lower Devonian of Livonia.
Allied to this species is Homostius milleri from Scotland, celebrated as the "Asterolepis of Stromness" in Hugh Miller's "Footsteps of the Creator." Another notable species is Homostius formosissimus from the Lower Devonian of Russia.
Arthrothoraci.—"Arthrodires whose dorsal shield articulates with the head-roof by a conspicuous and movable peg-and-socket joint, and leaves a definite interval (unprotected?) between the two armorings. Orbits marginal, bounded inferiorly not by the suborbital element. In the head-shield the postero-lateral angles formed by the marginal plate (Phlyctænaspis?), the occipital border concave. A dorsal fin is present, supported by endoskeletal elements." Five families, the most important being the Coccosteidæ, thus defined:
"Arthrothoraci with head-shield hexagonal in outline. Median occipital trapezoidal, margins underlapped conspicuously by the external occipitals. Prefrontals meet below pineal plates, thus occluding this element from contact with centrals. The median dorsal plate elongated, terminating in an acute heavy point; no definite ventral keel; its anterior border approaches the head-shield more closely than in related families. Cranio-dorsal joint relatively small. Postero-dorso-lateral large." (?A pair of spines occurs in the pectoral region.) The best-known species is Coccosteus cuspidatus (decipiens) of the Lower Red Sandstone or Devonian of Scotland.
The family of Dinichthyidæ consists of "Arthrothoraci with stout trenchant jaws, whose cutting surfaces have worn away marginal teeth. Plates heavy. Head-shield with conspicuous lateral indentation to form dorsal border of orbit. Preorbitals separated by rostral and pineal elements, the latter passing backward between the anterior ends of the centrals. Cranio-dorsal joint conspicuous. Median dorsal shovel-shaped, nearing a stout keel with a large neck and with heavy gouge-shaped terminal. Postero-dorso-lateral relatively small in size." Dinichthys hertzeri and numerous other species are described from the Devonian and Carboniferous rocks of Ohio.
The Titanichthyidæ are "Arthrothoraci with slender edentulous jaws bearing a longitudinal sulcus. Plates squamous. Head-shield wide, with indentations to form dorsal border of orbit. Cranio-dorsal joint complete, but of relatively small size. Median dorsal with lateral border indented with rudimentary keel and with flat and rounded terminal. Antero-dorso-lateral with an area of overlap on median border." Titanichthys agassizi is a gigantic mailed fish from the Lower Carboniferous of Cleveland, Ohio.
The Mylostomidæ are "Arthrothoraci with dental elements in the character of crushing plates. Cranial shield wide, rounded anteriorly, deeply indented in nuchal margin; orbital rim not apparent in dorsal aspect. Central separated from marginal." Mylostoma terrelli is based on jaws from Cleveland, Ohio.
The Selenosteidæ are "Arthrothoraci with jaws studded with cuspidate teeth; the mandibular rami rounding out anteriorly or presenting diverging tips, bearing teeth in the symphysis. Cranial[Pg 588] shield deeply concave on lateral margins, no orbital rim here apparent. Nuchal border deeply indented. (Centrals separate from marginals.) Cranio-dorsal hinges large in size. Dorsal armoring reduced antero-posteriorly, giving an almost zone-like appearance. Dorso-median crescent-shaped, with feeble keel and knob." Selenosteus glaber is described by Dean from the Cleveland shales.
Relations of Arthrodires.—To complete our account of the Arthrodira we may here summarize Dr. Dean's reasons for separating its members from true fishes on the one hand and from the Ostracophores on the other.
"First. The Arthrodira cannot be strictly included among the Pisces. According to the definition of the latter class its members are Craniotes possessing the following characters: a, dermal defenses which in their simplest terms can be reduced to the shagreen denticles of the Elasmobranch; b, a series of definite gill-arches whose foremost elements are metamorphosed into hyoid and mandibular apparatus; c, paired fins, or their equivalents. In the first of these regards I think it can be shown that the remarkable character of the dermal plates in the Arthrognaths approaches rather that of the Ostracophores than that of the Pisces. In certain of these forms, Trachosteus, for example, the tuberculated plates are made up of inner and outer elements, each with tubercles, which denote a distinctly different mode of origin from that of any known type of fish. The absence of remains of gill-arches in the Arthrognaths would be not a serious objection to including these forms among Pisces, especially in view of the fact that cartilaginous gill-arches are rarely preserved even in favorable fossils. But that their presence is more than doubtful is indicated by the peculiar character of the 'jaws' in these forms. For the character of these structures is such as to suggest that they are not homologous with the branchial arch jaws of the true fishes, but are rather parallel structures which owe their origin to distinctly exoskeletal elements, i.e., that they were derived from dermal plates surrounding the mouth, which became mobile, and whose edges became apposed as sectorial structures. I would in this connection call attention to the fact that the 'mandibles,' 'premaxillary,' and 'maxillary'[Pg 589] dental plates[159] were not fixed in the sense in which these elements are in the true Pisces. On the evidence of several types, Dinichthys, Titanichthys, Mylostoma, Trachosteus, Diplognathus, and other of the American forms, Macropetalichthys[160] excepted, there is the clearest proof that each element of the jaws had a considerable amount of independent movement. On account of the mobility of these elements the name Arthrognathi is suggested. Thus the mandibular rami could change the angle of inclination towards each other, as well as their plane with reference to the vertical axis. So, too, could the 'premaxillæ' be protracted like a pair of bent fingers, and it is more than probable that the 'maxillæ' had a considerable amount of independent movement. In connection with these characters it is also important to note that the blades of the 'mandible' show nowhere the faintest trace of an articular facet for attachment to the cranium. In short, the entire plan of the mandibular apparatus in these forms is strikingly unfish-like, although one will frankly confess that it is remarkable that these forms should have paralleled so strikingly the piscine conditions, to the extent of producing mandibular rami margined with teeth, and an arrangement of toothed elements on the 'upper jaw' which resembles superficially the premaxillary and maxillary structures of teleostomes, or the vomero-palatine structures of lung-fishes and chimæras.
"In the matter of paired fins there seems little evidence to conclude that either pectoral or pelvic fins were present. In spite of the researches upon these forms during the past half-century, no definite remains of pectoral fins have been described. The so-called pectoral spines described for Dinichthys by Newberry, whatever they may be, certainly are not, [Pg 590]as far as the present evidence goes, pterygial, nor are the similar structures in Brachydirus.[161] The sigmoid element, described as a 'pelvic girdle' by Smith Woodward, in Coccosteus, a structure which appears to occur in a small species of Dinichthys(?), may as reasonably be interpreted as a displaced element of the armor-plates of the trunk. In Coccosteus, as far as I am aware, it occurs in well-preserved condition in but a single specimen.
"In referring to the singular joint between the shoulder-plates and the hinder margin of the cranium Smith Woodward has called attention to one of the striking features of the group. It is one, however, which, as a functional structure, i.e., a joint, characterizes only a portion of its members; and in these the region in which vestiges of the joint are sought is overlaid and concealed by dermal plates. Such are the conditions in Macropetalichthys (with transitional characters in Trachosteus and in Mylostoma). For this form a special subclass (or order) may be created which we may term Anarthrodira.
"Second. The Arthrognathi cannot well be included in any other class. It would certainly be more convenient to retain the Arthrognaths among the Ostracophores, regarding them as a fourth subclass, were it not that they differ from them in so marked a way in the presence of well-marked vertebral arches, of supports for the unpaired fin, and in the possession of 'jaws.' In these regards—add to them the (probable if not certain) absence of the paired paddle-like 'spines'—they stand certainly further from the Antiarcha than these from the Osteostraci, or than the latter from the Heterostraci. It appears to me desirable, therefore, that the Arthrodira and the Anarthrodira be brought together as a separate class. Should subsequent researches demonstrate a closer affinity with the Ostracophores, the Arthrognathi can be regarded as of rank as a subclass, with the orders Anarthrodira and Arthrodira."[162]
In a recent paper Dr. Otto Jaekel unites Arthrodires and Ostracophores under the name Placodermi. He regards Pteraspis as a larval type, Asterolepis as one more specialized. In Coccosteus he claims to find a pelvic girdle as well as a more segmented skeleton. He regards all of these as true fishes, the Coccosteidæ as ancestral, related on the one hand to the Crossopterygians, and on the other to the Stegocephali and other ancestral Amphibians.
Suborder Cycliæ.—We may append to the Arthrodira as a possible suborder the group called Cycliæ by Dr. Gill, based on a single imperfectly known species. Few organisms discovered in recent times have excited as much interest as this minute fish-like creature, called Palæospondylus gunni, discovered in 1890 by Dr. R. H. Traquair in the flagstones of Caithness in Scotland. Many specimens have been obtained, none more than an inch and a half long. Its structure and systematic position have been discussed by Dr. R. H. Traquair, by Woodward, Gill, Gegenbaur, and recently by Dean, from whose valuable memoir on "The Devonian Lamprey" we make several quotations.
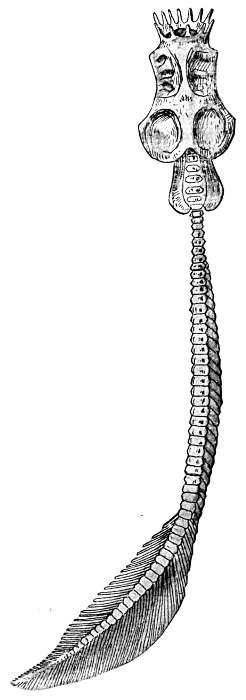
Fig. 369.—Palæospondylus gunni Traquair. Devonian. (After Traquair and Dean.)
Palæospondylus.—According to Dr. Traquair: "The Palæospondylus gunni is a very small organism, usually under one inch in length, though exceptionally large specimens occasionally measure one inch and a half.... It has a head and vertebral column, but no trace of jaws or limbs; and, strange to say, all the specimens are seen only from the ventral aspect, as is shown by the relation of the neural arches to the vertebral centra.
"The head is in most cases much eroded.... It is divided by a notch ... into two parts.... The anterior[Pg 592] part shows a groove the edges of which are elevated, while the surface on each side shows two depressions, like fenestræ, though perhaps they are not completely perforated, and also a groove partially divided off, posteriorly and externally, a small lobe. In front there is a ring-like opening ... surrounded by small pointed cirri, four ventrally, at least five dorsally, and two long lateral ones which seem to arise inside the margin of the ring instead of from its rim like the others. The posterior part of the cranium is flattened, but the median groove is still observable. Connected with the posterior or occipital aspect of the skull are two small narrow plates which lie closely alongside the first half-dozen vertebræ.
"The bodies of the vertebræ are hollow or ring-like, and those immediately in front are separated from each other by perceptible intervals; their surfaces are marked with a few little longitudinal grooves, of which one is median. They are provided with neural arches, which are at first short and quadrate, but towards the caudal extremity lengthen out into slender neural spines, which form the dorsal expansion of a caudal fin, while shorter hæmal ones are also developed on the ventral aspect."
Dr. Traquair concludes that "there seems to be no escape from the conclusion that the little creature must be classed as a Marsipobranch." "If Palæospondylus is not a Marsipobranch, it is quite impossible to refer it to any other existing group of vertebrates."
Gill on Palæospondylus.—In 1896 Dr. Gill proposed to regard Palæospondylus provisionally as the type of a distinct order of Cyclostomes to be called Cycliæ (κύκλος, circle), from the median ring on the head, whether nostril or mouth. Dr. Gill observes:
"Assuming the correctness of Dr. Traquair's description and figures, we certainly have a remarkable combination of characters. On the one hand, if the 'median opening or rim' is indeed nasal, the animal certainly cannot be referred to the class of Selachians or of Teleostomes. On the other hand, the cranium and the segmental vertebral column indicate a more advanced stage of development of the vertebrate line than that from the living Marsipobranchs must have originated. We[Pg 593] may, therefore, with propriety isolate it as the representative not only of a peculiar family (Palæospondylidæ), but of an order or even subclass (Cycliæ) of vertebrates which may provisionally (and only provisionally) be retained in the class of Marsipobranchs.
"The group may be defined as Monorrhines with a continuous (?) cranium, a median nasal (?) ring, and a segmented vertebral column.
"The differences between the Hyperoartia and Hyperotreta are very great, and Prof. Lankester did not go much too far when he elevated those groups to class rank. Among the numerous distinctive characters are the great differences in the auditory organs. Perhaps the organs of Palæospondylus might be worked out in some specimen and throw light on the subject of affinities. At present even the region of the auditory organs is not exactly known and we are now at a loss to orient the several parts of the cranium. In fact, the question of the relations of Palæospondylus is a very open one."
Views as to the Relationships of Palæospondylus.—Dr. Dean thus summarizes in a convenient and interesting fashion the views of different students of fossil fishes in regard to Palæospondylus:
Huxley.—A "baby Coccosteus."
Traquair, 1890.—"Certainly not a Placoderm, its resemblance to a supposed 'baby Coccosteus' being entirely deceptive. The appearance of the head does remind us in a strange way of the primitive skull of Myxine, a resemblance which is rendered still more suggestive by the apparent complete absence of the lower jaw, or of limbs or limb-girdles."
Traquair, 1893.—"It seems, indeed, impossible to refer the organism to any existing vertebrate class, unless it be the Marsipobranchs or Cyclostomata." Does not believe it a larval form, because the possible adult is unknown, and because of the highly differentiated vertebræ. Granting his interpretation of the parts of the fossil, "there seems no escape from the conclusion that the little creature must be classed as a Marsipobranch."
Traquair, 1897.—"The question of the affinities of Palæospondylus is left precisely where it was after I had written my last paper on the subject."
Smith Woodward, 1892.—"It seems to possess an unpaired nose, lip cartilages in place of functional jaws, and no paired limbs; thus agreeing precisely with the lampreys and hagfishes, of which the fossil representatives have long been sought. It is extremely probable, therefore, that Palæospondylus belongs to this interesting category."
Dawson, 1893.—Palæospondylus suggests "the smaller snake-like Batrachians of the Carboniferous and Permian; and I should not be surprised if it should come to be regarded as either a forerunner of the Batrachians or as a primitive tadpole."
Gill, 1896.—"The group to which Palæospondylus belongs may be defined as Monorrhines with a continuous (?) cranium, a median nasal (?) ring, and a segmented vertebral column." "The cranium and segmented vertebral column indicate a more advanced stage of development of the vertebrate line than that from which the living Marsipobranchs must have originated. We may, therefore, with propriety isolate it as the representative not only of a peculiar family (Palæospondylidæ), but of an order or even subclass (Cycliæ) of vertebrates which may provisionally (and only provisionally) be retained in the class of Marsipobranchs."
Dean, 1896.—"Place it with the Ostracoderms among the curiously specialized offshoots of the early Chordates, but this position would be at the best unsatisfactory."
Dean, 1898.—"Palæospondylus should not be given a place—even a provisional one—among the Marsipobranchs." To be accepted "as the representative of the new subclass (or class) Cycliæ constituted for it by Professor Gill."
Parker & Haswell, 1897.—"There is some reason to regard that Palæospondylus is referable to the Cyclostomes." "A distinctly higher type than recent forms."
Gegenbaur, 1898.—"Discovery of Palæospondylus one of the highest importance. If this organism stands in no way near the Cyclostomes, the tentacles lose their higher importance, since they also occur in other groups." "Through Palæospondylus came also the attempt (Pollard) to deduce the presence of the tentacular condition in the higher forms." (Mem.—In this Gegenbaur has not consulted the literature accurately. At[Pg 595] the time of founding his "Cirrhostomal Theory" Pollard was unaware of the discovery of Palæospondylus). "Ich muss sagen, das die positive Behauptung der einen wie der anderen Deutung mir sehr unsicher scheint, da auch an den übrigen Resten des Kopfskelets keine bestimmten Uebereinstimmungen mit anderen Organismen erweisbar sind. Es ist daher auch nicht zu vermuthen, dass sogar an Beziehung zu Froschlarven gedacht ward. Unter diesen Umständen möchte ich jene im Verhältniss zum Kopfe wie zum gesammten Körper bedeutende, von Cirren umstellte Eingangsöffnung als nicht einer Nase, sondern einem Munde oder beiden zugleich angehörig betrachten. Zu einem dem Cyclostomenriechorgan vergleichbaren Verhalten fehlen alle Bedingungen."
Relationships of Palæospondylus.—The arguments for and against the supposition that Palæospondylus is a Cyclostome may be here summed up after Professor Dean.
The vertebral column agrees with that of the lamprey in having the notochord in part persistent. On the other hand, the vertebræ have continuous centra, showing definite processes. Those of the different regions are differentiated. These conditions are quite unlike those seen in the lamprey.
The cranium is massive, over twice as large proportionally as that of the lamprey. In the latter type the cranium forms but a small portion of the bulk of the head; in Palæospondylus, on the other hand, the cranium bears every sign of having filled the contour of the head. Moreover, if the region adjacent to the structure is admitted to be that of the eye, and few, I believe, will doubt it, then the brain-cavity must, by many analogies, have been much larger than that of a Marsipobranch. Also the auditory capsules must have been of extraordinary size. In short, there is very little about the cranium to suggest the structures of Cyclostomes.
The "oral cirri" suggest somewhat the barbels of the nose and mouth of a hagfish. They, however, resemble even as much in arrangement and greater number the buccal cirri of Amphioxus. On the other hand, similar mouth-surrounding tentacles are evolved independently in many groups of fishes, siluroids, sharks, forms like Pogonias, Hemitripterus. A possi[Pg 596]bility further exists that the "cirri" may turn out to be remnants of cranial or facial structures of an entirely different nature. In fact the very uncertain preservation of these parts renders their evidence of little definite value. In but one specimen, as far as I am aware, is there any evidence of the presence of ventral cirri.
The jaw parts in Palæospondylus are unknown. It is possible that the ventral rim of the "nasal ring" may prove to be the remains of the Meckelian cartilage (the cartilaginous core of the lower jaw).
It is possible that certain very faint ray-like markings noted by Professor Dean may be the basalia of paired fins. In such case Palæospondylus can have no affinity with the lampreys. Dr. Dean asserts that the presence of these, in view of the wide dissimilarity in other and important structures, is sufficient to remove Palæospondylus from its provisional position among the Cyclostomes. The postoccipital plates may represent a pectoral arch. It is, however, much more likely, as Dr. Traquair has insisted, that the supposed rays are due to the reflection of light from striations on the stone, and that the creature had no pectoral limbs.
The caudal fin, with its dichotomous rays, is essentially like the tail of a lamprey. This condition is, however, found in other groups of fishes, as among sharks and lung-fishes. It is, moreover, doubtful whether the rays are really dichotomous.
It is possible that Palæospondylus may be, as Huxley suggests, a larval Arthrodire. It is not probable that this is the case, but, on the other hand, Palæospondylus seems to be an immature form. According to Dr. Dean, it is more likely to prove a larval Coccosteus, or the young of some other Arthrodire, than a lamprey. Against this view must be urged the fact that the tail of Palæospondylus is not heterocercal, a fact verified by Dr. Traquair on all of his many specimens. It is more like the tail of a lamprey than that of Coccosteus. It is, however, certain that it cannot be placed in the same class with the living Cyclostomes, and that it is far more highly specialized than any of them. In a still later paper (1904) Dr. Dean[Pg 597] shows that the fossil might as easily be considered a Chimæra as a lamprey, and repeats his conviction that it is a larval form of which the adult is still unrecognized.
We cannot go much farther than Dr. Dean's statement in 1896, that it belongs "among the curiously specialized offshoots of the early Chordates."
[157] "The name Arthrodira as given to Coccosteans, as distinguished from the Antiarcha, is not altogether a satisfactory one, since at least from the time of Pander the head of Pterichthys (Asterolepis) is known to be articulated with the armoring of the trunk in a way closely resembling that of Coccosteus. This term may, however, be retained as a convenient one for the order of Coccosteans, in which, together with other differentiating features, this structure is prominently evolved. A renewed examination of the subject has caused me to incline strongly to the belief, as above expressed, that Pterichthys and Coccosteans are not as widely separated in phylogeny as Smith Woodward, for example, has maintained. But, as far as present evidence goes, they appear to me certainly as distinct as fishes are from amphibia, or as reptiles are from birds or from mammals." (Dean.)
The name Placodermi used by McCoy in 1848 was applied to the Ostracophores as well as to the Arthrodires. Hay revives it as the name of a superorder to include the Antiarcha and the Arthrodira, the former being detached from the Ostracophores. This superorder is equivalent to the subclass Azygostei of Hay.
[158] Memoirs New York Academy of Sciences, 1901.
[159] It will be recalled that there is no ground for concluding that the "mandibular rami" possessed an endoskeletal core, and were comparable, therefore, to the somewhat mobile jaws of Elasmobranchs. On the other hand, there is the strongest evidence that they are entirely comparable to adjacent dermal plates. Histologically they are identical, and in certain cases their exposed surfaces bear the same tuberculation.
[160] The similarity of Macropetalichthys to Dinichthyids in the general matter of the dermal plates is so complete that I have had no hesitation in associating it with the Arthrognaths. (Cf. Eastman.) The circumstance that its "jaws" have not yet been found has to a large degree been due to the lack of energy on the part of local collectors. In the corniferous quarries near Delaware, Ohio, this fossil is stated to be relatively abundant.
[161] It is by no means impossible that there may ultimately be found pectoral elements to correspond in a general way with the paddle-like "spines" of the Antiarcha.
[162] The group Placodermi, created by McCoy (1848) as a "family" for the reception of Coccosteus and Pterichthys might then be justly elevated to rank as a class, superseding the Ostracophori of Cope (1891). The latter group might, however, be retained as a subclass, and include the Heterostraci and Osteostraci as ordinal divisions.

Class Teleostomi.—We may unite the remaining groups of fishes into a single class, for which the name Teleostomi (τέλεος, true; στόμα, mouth), proposed by Bonaparte in 1838, may be retained. The fishes of this class are characterized by the presence of a suspensorium to the mandible, by the existence of membrane-bones (opercles, suborbitals, etc.) on the head, by a single gill-opening leading to gill-arches bearing filamentous gills, and by the absence of claspers on the ventral fins. The skeleton is at least partly ossified in all the Teleostomi. More important as a primary character, distinguishing these fishes from the sharks, is the presence typically and primitively of the air-bladder. This is at first a lung, arising as a diverticulum from the ventral side of the œsophagus, but in later forms it becomes dorsal and is, by degrees, degraded into a swim-bladder, and in very many forms it is altogether lost with age.
This group comprises the vast majority of recent fishes, as well as a large percentage of those known only as fossils. In these the condition of the lung can be only guessed.
The Teleostomi are doubtless derived from sharks, their relationship being possibly nearest to the Ichthyotomi or to the primitive Chimæras. The Dipnoans among Teleostomi retain the shark-like condition of the upper jaw, made of palatal elements, which may be, as in the Chimæra, fused with the cranium. In the lower forms also the primitive diphycercal or protocercal form of tail is retained, as also the archipterygium or jointed axis of the paired fins, fringed with rays on one or both sides.
We may divide the Teleostomes, or true fishes, into three subclasses: the Crossopterygii, or fringe-fins; the Dipneusti, or lung-fishes; Actinopteri, or ray-fins, including the Ganoidei and the Teleostei, or bony fishes. Of these many recent writers are disposed to consider the Crossopterygii as most primitive, and to derive from it by separate lines each of the remaining subclasses, as well as the higher vertebrates. The Ganoidei and Teleostei (constituting the Actinopteri) are very closely related, the ancient group passing by almost imperceptible degrees into the modern group of bony fishes.
Subclass Crossopterygii.—The earliest Teleostomes known belong to the subclass or group called after Huxley, Crossopterygii (κρόσσος, fringe; πτερύξ, fin). A prominent character of the group lies in the retention of the jointed pectoral fin or archipterygium, its axis fringed by a series of soft rays. This character it shares with the Ichthyotomi among sharks, and with the Dipneusti. From the latter it differs in the hyostylic cranium, the lower jaw being suspended from the hyomandibular, and by the presence of distinct premaxillary and maxillary elements in the upper jaw. In these characters it agrees with the ordinary fishes. In the living Crossopterygians the air-bladder is lung-like, attached by a duct to the ventral side of the œsophagus. The lung-sac, though specialized in structure, is simple, not cellular as in the Dipnoans. The skeleton is more or less perfectly ossified. Outside the cartilaginous skull is a bony coat of mail. The skin is covered with firm scales or bony plates, the tail is diphycercal, straight, and ending in a point, the shoulder-girdle attached to the cranium is cartilaginous but overlaid with bony plates, and the branchiostegals are represented by a pair of gular plates.
In the single family represented among living fishes the heart has a muscular arterial bulb with many series of valves on its inner edge, and the large air-bladder is divided into two lobes, having the functions of a lung, though not cellular as in the lung-fishes.
The fossil types are very closely allied to the lung-fishes, and the two groups have no doubt a common origin in Silurian times. It is now usually considered that the Crossopterygian is more primitive than the lung-fish, though at the same time[Pg 600] more nearly related to the Ganoids, and through them to the ordinary fishes.
Origin of Amphibians.—From the primitive Crossopterygii the step to the ancestral Amphibia, which are likewise mailed and semi-aquatic, seems a very short one. It is true that most writers until recently have regarded certain Dipneustans as the Dipteridæ as representing the parents of the Amphibians. But the weight of recent authority, Gill, Pollard, Boulenger, Dollo, and others, seems to place the point of separation of the higher vertebrates with the Crossopterygians, and to regard the lobate pectoral member of Polypterus as a possible source of the five-fingered arm of the frog. This view is still, however, extremely hypothetical and there is still much to be said in favor of the theory of the origin of Amphibia from Dipnoans and in favor of the view that the Dipnoans are also ancestors of the Crossopterygians.
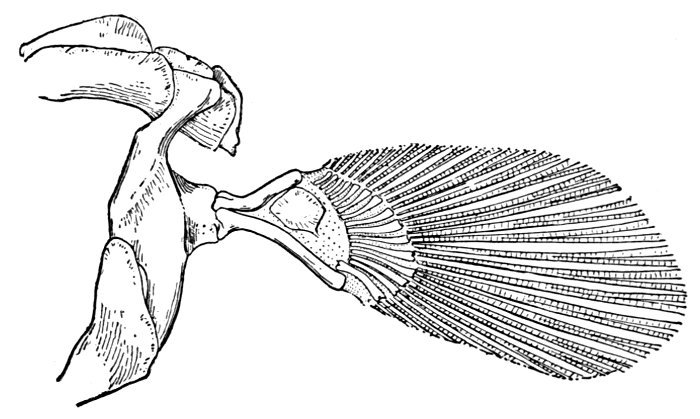
Fig. 370.—Shoulder-girdle of Polypterus bichir. Specimen from the White Nile.
In the true Amphibians the lungs are better developed than in the Crossopterygian or Dipnoan, although the lungs are finally lost in certain salamanders which breathe through epithelial cells. The gills lose, among the Amphibia, their primitive importance, although in Proteus anguineus of Austria and Necturus maculosus, the American "mud-puppy" or water-dog, these persist through life. The archipterygium, or primitive fin, gives place to the chiropterygium, or fingered arm. In[Pg 601] this the basal segment of the archipterygium gives place to the humerus, the diverging segments seen in the most specialized type of archipterygium (Polypterus) become perhaps radius and ulna, the intermediate quadrate mass of cartilage possibly becoming carpal bones, and from these spring the joints called metacarpals and phalanges. In the Amphibians and all higher forms the shoulder-girdle retains its primitive insertion at a distance from the head, and the posterior limbs remain abdominal.
The Amphibians are therefore primarily fishes with fingers and toes instead of the fringe-fins of their ancestors. Their relations are really with the fishes, as indicated by Huxley, who unites the amphibians and fishes in a primary group, Ichthyopsida, while reptiles and birds form the contrasting group of Sauropsida.
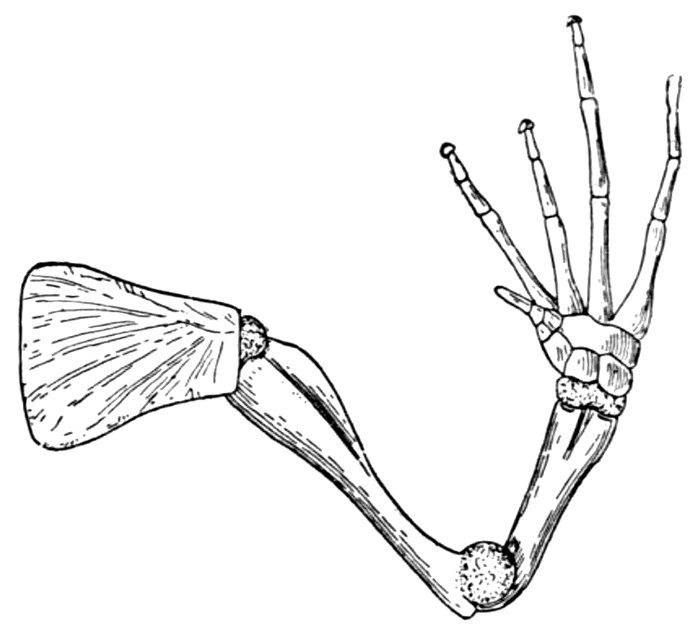
Fig. 371.—Arm of a frog.
The reptiles differ from the Amphibians through acceleration of development, passing through the gill-bearing stages within the egg. The birds bear feathers instead of scales, and the mammals nourish their young by means of glandular secretions. Through a reptile-amphibian ancestry the birds and mammals may trace back their descent from palæozoic Crossopterygians. In the very young embryo of all higher vertebrates traces of double-breathing persist in all species, in the form of rudimentary gill-slits.
The Fins of Crossopterygians.—Dollo and Boulenger regard the heterocercal tail as a primitive form, the diphycercal form being a result of degradation, connected with its less extensive use as an organ of propulsion. Most writers who adopt the theory of Gegenbaur that the archipterygium is the primitive form of the pectoral fin are likely, however, to consider the diphycercal tail found associated with it in the Ichthyotomi, Dipneusti, Crossopterygii as the more primitive form of the tail. From this form the heterocercal tail of the higher sharks and[Pg 602] Ganoids may be derived, this giving way in the process of development to the imperfectly homocercal tail of the salmon, the homocercal tail of the perch, and the isocercal tail of the codfish and its allies, the gephyrocercal and the leptocercal tail, tapering or whip-like, representing various stages of degeneration. Boulenger draws a distinction between the protocercal tail, the one primitively straight, and the diphycercal tail modified, like the homocercal tail, from an heterocercal ancestry.
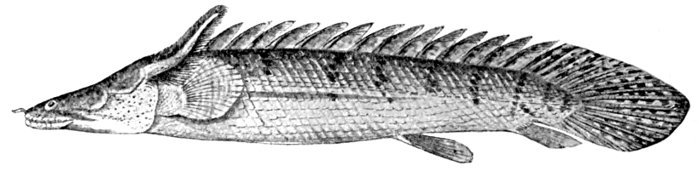
Fig. 372.—Polypterus congicus, a Crossopterygian fish from the Congo River. Young, with external gills. (After Boulenger.)
Orders of Crossopterygians.—Cope and Woodward divide the Crossopterygia into four orders or suborders, Haplistia, Rhipidistia, Actinistia, and Cladistia. To the latter belong the existing species, or the family of Polypteridæ, alone. Boulenger unites the three extinct orders into one, which he calls Osteolepida. In all three of these the pectorals are narrow with a single basal bone, and the nostrils, as in the Dipneustans, are below the snout. The differences are apparently such as to justify Cope's division into three orders.
Haplistia.—In the Haplistia the notochord is persistent, and the basal bones of dorsal and anal fins are in regular series, much fewer in number than the fin-rays. The single family Tarrassiidæ is represented by Tarrasius problematicus, found by Traquair in Scotland. This is regarded as the lowest of the Crossopterygians, a small fish of the Lower Carboniferous, the head mailed, the body with small bony scales.
Rhipidistia.—In the Rhipidistia the basal bones of the median fins ("axonosts and baseosts") are found in a single piece, not separate as in the Haplistia. Four families are recognized, Holoptychiidæ, Megalichthyidæ, Osteolepidæ, and Onychodontidæ, the first of these being considered as the nearest approach of the Crossopterygians to the Dipnoans.
The Holoptychiidæ have the pectoral fins acute, the scales cycloid, enameled, and the teeth very complex. Holoptychius nobilissimus is a very large fish from the Devonian. Glyptolepis leptopterus from the Lower Devonian is also a notable species. Dendrodus from the Devonian is known from detached teeth.
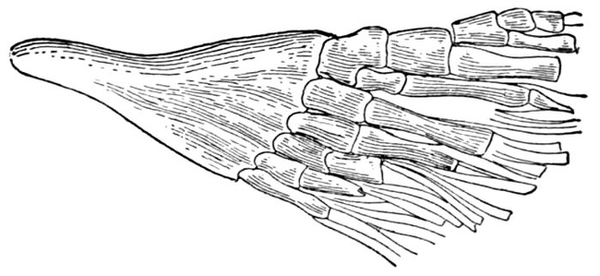
Fig. 373.—Basal bone of dorsal fin, Holoptychius leptopterus (Agassiz). (After Woodward.)
In the Ordovician rocks of Cañon City, Colorado, Dr. Walcott finds numerous bony scales with folded surfaces and stellate ornamentation, and which he refers with some doubt to a Crossopterygian fish of the family Holoptychiidæ. This fish he names Eriptychius americanus. If this identification proves correct, it will carry back the appearance of Crossopterygian fishes, the earliest of the Teleostome forms, to the beginning of the Silurian, these Cañon City shales being the oldest rocks in which remains of fishes are known to occur. In the same rocks are found plates of Ostracophores and other fragments still more doubtful. It is certain that our records in palæontology fall far short of disclosing the earliest sharks, as well as the earliest remains of Ostracophores, Arthrodires, or even Ganoids.
Megalichthyidæ.—The Megalichthyidæ (wrongly called "Rhizodontidæ") have the pectoral fins obtuse, the teeth relatively simple, and the scales cycloid, enameled. There are numerous species in the Carboniferous rocks, largely known from fragments or from teeth. Megalichthys, Strepsodus, Rhizodopsis, Gyroptychius, Tristichopterus, Eusthenopteron, Cricodus, and Sauripterus are the genera; Rhizodopsis sauroides from the coal-measures of England being the best-known species.
The Osteolepidæ differ from the Megalichthyidæ mainly in the presence of enameled rhomboid scales, as in Polypterus and[Pg 604] Lepisosteus. In Glyptopomus these scales are sculptured, in the others smooth. In Osteolepis, Thursius, Diplopterus, and Glyptopomus a pineal foramen is present on the top of the head. This is wanting in Parabatrachus (Megalichthys of authors). In Osteolepis, Thursius, and Parabatrachus the tail is heterocercal, while in Diplopterus and Glyptopomus it is diphycercal. Osteolepis macrolepidotus and numerous other species occur in the Lower Devonian. Diplopterus agassizii is common in the same horizon. Megalichthys hibberti is found in the coal-measures, and Glyptopomus minimus in the Upper Devonian. Palæosteus is another genus recently described.
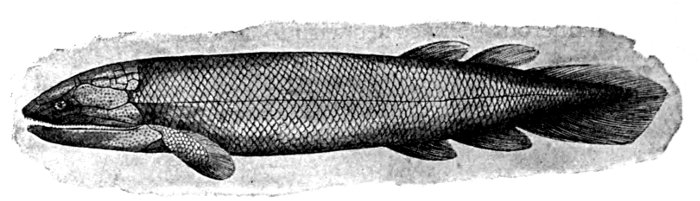
Fig. 374.—Gyroptychius microlepidotus Agassiz. Devonian. Family Megalichthyidæ. (After Pander.)
The Onychodontidæ are known from a few fragments of Onychodus sigmoides from the Lower Devonian of Ohio and Onychodus anglicus from England.

Fig. 375.—Cœlacanthus elegans Newberry. From the Ohio Carboniferous, showing air-bladder. (After Dean.)
Order Actinistia.—In the Actinistia there is a single fin-ray to each basal bone, the axonosts of each ray fused in a single piece. The notochord is persistent, causing the back-bone in fossils to appear hollow, the cartilaginous material leaving no trace in the rocks. The genera and species are numerous, ranging from the Subcarboniferous to the Upper Cretaceous, many of them belonging to Cœlacanthus, the chief genus of the[Pg 605] single family Cœlacanthidæ. In Cœlacanthus the fin-rays are without denticles. Cœlacanthus granulatus is found in the European Permian. Cœlacanthus elegans of the coal-measures is found in America also. In Undina the anterior fin-rays are marked with tubercles. Undina penicillata and Undina gulo from the Triassic are well-preserved species. In Macropoma (lewesiensis) the fin-rays are robust, long, and little articulated. Other genera are Heptanema, Coccoderma, Libys, Diplurus, and Graphiurus. Diplurus longicaudatus was found by Newberry in the Triassic of New Jersey and Connecticut.
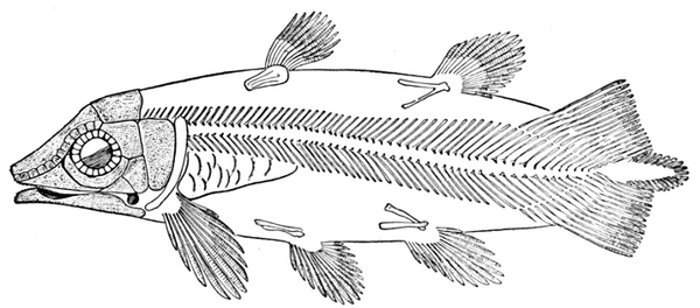
Fig. 376.—Undina gulo Egerton; Lias. Family Cœlacanthidæ. (After Woodward.)
Order Cladistia.—In the Cladistia the axis of the pectoral limb is fan-shaped, made of two diversified bones joined by cartilage. The notochord is restricted and replaced by ossified vertebræ. The axonosts of the dorsal and anal are in regular series, each bearing a fin-ray. The order contains the single family Polypteridæ. In this group the pectoral fin is formed differently from that of the other Crossopterygians, being broad, its base of two diverging bones with cartilage between. This structure, more specialized than in any other of the Crossopterygians or Dipneusti, has been regarded by Gill and others, as above stated, as the origin of the fingered hand (chiropterygium) of the frogs and higher vertebrates. The base of the diverging bones has been identified as the antecedent of the humerus, the bones themselves as radius and ulna, while the intervening non-ossified cartilage breaks up into carpal bones, from which metacarpals and digits ultimately diverge. This hypothesis is open to considerable doubt.[Pg 606] The nostrils, as in true fishes, are superior. The body in these fishes is covered with rhombic enameled scales, as in the garpike; the head is similarly mailed, but, in distinction from the garpike, the anterior rays of the dorsal are developed as isolated spines.
The young have a bushy external gill with a broad scaly base. The air-bladder is double, not cellular, with a large air-duct joining the ventral surface of the œsophagus. The intestine has a spiral valve.
The cranium, according to Boulenger ("Poissons du Bassin du Congo," p. 11), is remarkable for its generalized form, this character forming a trait of union between the Ganoids and the primitive Amphibia or Stegocephali. Without considering Polypterus, it is not possible to interpret the homologies of the cranium of the amphibians and the sharks.
The jaws are similar to those of the vertebrates higher than fishes. Tooth-bearing premaxillaries and dentaries are solidly joined at the front of the cranium, and united by a suture to the toothed maxillaries which form most of the edge of the mouth. Each half of the lower jaw consists of four elements, covering Meckel's cartilage, which is ossified at the symphysis. These are the articular, angular, dentary, and splenial (coronoid). Most of these bones are armed with teeth. The palato-suspensory consists of hyomandibular, quadrate, ectopterygoid, entopterygoid, metapterygoid, and palatine elements, the pterygoid elements bearing teeth. In Erpetoichthys only the opercle is distinct among the gill-covers. In Polypterus there is a subopercle also; the suborbital chain is represented by two small bones.
The gill-arches are four, but without lower pharyngeals. The teeth are conic and pointed, and in structure, according to Agassiz, they differ largely from those of bony fishes, approaching the teeth of reptiles.
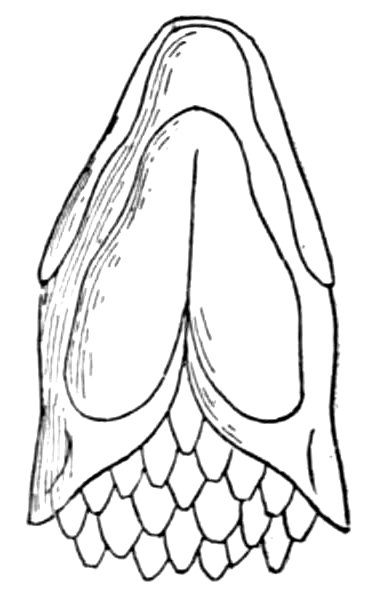
Fig. 377.—Lower jaw of Polypterus bichir, from below.
The external gill of the young, first discovered by Steindachner in 1869, consists of a fleshy axis bordered above and below by secondary branches, themselves fringed. In form and structure this resembles the external gills of amphibians.[Pg 607] It is inserted, not on the gill-arches, but on the hyoid arch. Its origin is from the external skin. It can therefore not be compared morphologically with the gills of other fishes, nor with the pseudobranchiæ, but rather with the external gills of larval sharks. The vertebræ are very numerous and biconcave as in ordinary fishes. Each of the peculiar dorsal spines is primitively a single spine, not a finlet of several pieces, as some have suggested. The enameled, rhomboid scales are in movable oblique whorls, each scale interlocked with its neighbors.

Fig. 378.—Polypterus congicus, a Crossopterygian fish from the Congo River. Young, with external gills. (After Boulenger.)

Fig. 379.—Polypterus delhezi Boulenger. Congo River.
The shoulder-girdle, suspended from the cranium by post-temporal and supraclavicle, is covered by bony plates. To the small hypercoracoid and hypocoracoid the pectoral fin is attached. Its basal bones may be compared to those of the sharks, mesopterygium, propterygium, and metapterygium, which may with less certainty be again called humerus, radius, and ulna. These are covered by flesh and by small imbricated scales. The air-bladder resembles the lungs of terrestrial vertebrates. It consists of two cylindrical sacs, that on the right the longer, then uniting in front to form a short tube, which enters the œsophagus from below with a slit-like glottis. Unlike the lung of the Dipneusti, this air-bladder is not cellular, and it receives only arterial blood. Its function is to assist the respiration by gills without replacing it.
The Polypteridæ.—All the Polypteridæ are natives of Africa. Two genera are known, no species having been found fossil. Of Polypterus, Boulenger, the latest authority, recognizes nine species: six in the Congo, Polypterus congicus, P. delhezi, P. ornatipinnis, P. weeksi, P. palmas, and P. retropinnis; one, P. lapradei, in the Niger; and two in the Nile, Polypterus bichir and P. endlicheri. Of these the only one known until very recently was Polypterus bichir of the Nile.
These fishes in many respects resemble the garpike in habits. They live close on the mud in the bottom of sluggish waters, moving the pectorals fan-fashion. If the water is foul, they rise to the surface to gulp air, a part of which escapes through the gill-openings, after which they descend like a flash. In the breeding season these fishes are very active, depositing their eggs in districts flooded in the spring. The eggs are very numerous, grass-green, and of the size of eggs of millet. The flesh is excellent as food.
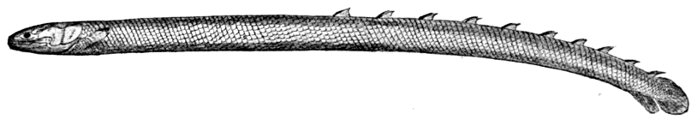
Fig. 380.—Erpetoichthys calabaricus Smith. Senegambia. (After Dean.)
The genus Erpetoichthys contains a single species, Erpetoichthys calabaricus,[163] found also in the Senegal and Congo. This species is very slender, almost eel-like, extremely agile, and, as usual in wriggling or undulating fishes, it has lost its ventral fin. It lives in shallow waters among interlaced roots of palms. When disturbed it swims like a snake.
[163] This genus was first called Erpetoichthys, but the name was afterwards changed by its author, J. A. Smith, to Calamoichthys, because there is an earlier genus Erpichthys among blennies, and a Herpetoichthys among eels. But these two names, both wrongly spelled for Herpetichthys, are sufficiently different, and the earlier name should be retained. "A name in science is a name without necessary meaning" and without necessarily correct spelling. Furthermore, if names are spelled differently, they are different, whatever their meaning. The efforts of ornithologists, notably those of Dr. Coues, to spell correctly improperly formed generic names have shown that to do so consistently would throw nomenclature into utter confusion. It is well that generic names of classic origin should be correctly formed. It is vastly more important that they should be stable. Stability is the sole function of the law of priority.

The Lungfishes.—The group of Dipneusti, or lung-fishes, is characterized by the presence of paired fins consisting of a jointed axis with or without rays. The skull is autostylic, the upper jaw being made as in the Chimæra of palatal elements joined to the quadrate and fused with the cranium, without premaxillary or maxillary. The dentary bones are little developed. The air-bladder is cellular, used as a lung in all the living species, its duct attached to the ventral side of the œsophagus. The heart has many valves in the muscular arterial bulb. The intestine has a spiral valve. The teeth are usually of large plates of dentine covered with enamel, and are present on the pterygo-palatine and splenial bones. The nostrils are concealed, when the mouth is closed, under a fold of the upper lip. The scales are cycloid, mostly not enameled.
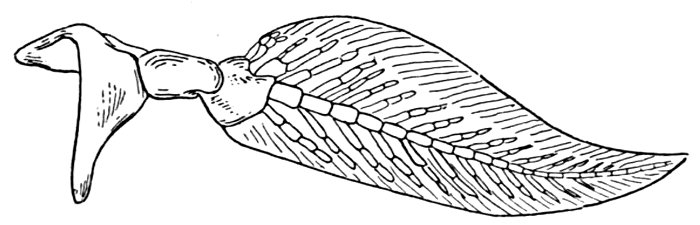
Fig. 381.—Shoulder-girdle of Neoceratodus forsteri Günther. (After Zittel.)
The lung-fishes, or Dipneusti (δίς, two; πνεῖν, to breathe), arise, with the Crossopterygians, from the vast darkness of [Pg 610]Palæozoic time, their origin with that or through that of the latter to be traced to the Ichthyotomi or other primitive sharks. These two groups are separated from all the more primitive fish-like vertebrates by the presence of lungs. In its origin the lung or air-bladder arises as a diverticulum from the alimentary canal, used by the earliest fishes as a breathing-sac, the respiratory functions lost in the progress of further divergence. Nothing of the nature of lung or air-bladder is found in lancelet, lamprey, or shark. In none of the remaining groups of fishes is it wholly wanting at all stages of development, although often lost in the adult. Among fishes it is most completely functional in the Dipneusti, and it passes through all stages of degeneration and atrophy in the more specialized bony fishes.
In the Dipneusti, or Dipnoans, as in the Crossopterygians and the higher vertebrates, the trachea, or air-duct, arises, as above stated, from the ventral side of the œsophagus. In the more specialized fishes, yet to be considered, it is transferred to the dorsal side, thus avoiding a turn in passing around the œsophagus itself. From the sharks these forms are further distinguished by the presence of membrane-bones about the head. From the Actinopteri (Ganoids and Teleosts) Dipnoans and Crossopterygians are again distinguished by the presence of the fringe-fin, or archipterygium, as the form of the paired limbs. From the Crossopterygians the Dipnoans are most readily distinguished by the absence of maxillary and premaxillary, the characteristic structures of the jaw of the true fish. The upper jaw in the Dipnoan is formed of palatal elements attached directly to the skull, and the lower jaw contains no true dentary bones. The skull in the Dipnoans, as in the Chimæra, is autostylic, the mandible articulating directly with the palatal apparatus, the front of which forms the upper jaw and of which the pterygoid, hyomandibular and quadrate elements form an immovable part. The shoulder-girdle, as in the shark, is a single cartilage, but it supports a pair of superficial membrane-bones.
In all the Dipnoans the trunk is covered with imbricated cycloid scales and no bony plates, although sometimes the scales are firm and enameled. The head has a roof of well[Pg 611]-developed bony plates made of ossified skin and not corresponding with the membrane-bones of higher fishes. The fish-like membrane-bones, opercles, branchiostegals, etc., are not yet differentiated. The teeth have the form of grinding-plates on the pterygoid areas of the palate, being distinctly shark-like in structure. The paired fins are developed as archipterygia, often without rays, and the pelvic arch consists of a single cartilage, the two sides symmetrical and connected in front. There is but one external gill-opening leading to the gill-arches, which, as in ordinary fishes, are fringe-like, attached at one end. In the young, as with the embryo shark, there is a bushy external gill, which looks not unlike the archipterygium pectoral fin itself, although its rays are of different texture. In early forms, as in the Ganoids, the scales were bony and enameled, but in some recent forms deep sunken in the skin. The claspers have disappeared, the nostrils, as in the frog, open into the pharynx, the heart is three-chambered, the arterial bulb with many valves, and the cellular structure of the skin and of other tissues is essentially as in the Amphibian.
The developed lung, fitted for breathing air, which seems the most important of all these characters, can, of course, be traced only in the recent forms, although its existence in all others can be safely predicated. Besides the development of the lung we may notice the gradual forward movement of the shoulder-girdle, which in most of the Teleostomous fishes is attached to the head. In bony fishes generally there is no distinct neck, as the post-temporal, the highest bone of the shoulder-girdle, is articulated directly with the skull. In some specialized forms (Balistes, Tetraodon) it is even immovably fused with it. In a few groups (Apodes, Opisthomi, Heteromi, etc.) this connection ancestrally possessed is lost through atrophy and the slipping backward of the shoulder-girdle leaves again a distinct neck. In the Amphibians and all higher vertebrates the shoulder-girdle is distinct from the skull, and the possession of a flexible neck is an important feature of their structure. In all these higher forms the posterior limbs remain abdominal, as in the sharks and the primitive and soft-rayed fishes generally. In these the pelvis or pelvic elements are attached toward the middle[Pg 612] of the body, giving a distinct back as well as neck. In the spiny-rayed fishes the "back" as well as the neck disappears, the pelvic elements being attached to the shoulder-girdle, and in a few extreme forms (as Ophidion) the pelvis is fastened at the chin.
Classification of Dipnoans.—By Woodward the Dipneusti are divided into two classes, the Sirenoidei and the Arthrodira. We follow Dean in regarding the latter as representative of a distinct class, leaving the Sirenoidei, with the Ctenodipterini, to constitute the subclass of Dipneusti. The Sirenoidei are divided by Gill into two orders, the Monopneumona, with one lung, and the Diplopneumona, with the lung divided. To the latter order the Lepidosirenidæ belong. To the former the Ceratodontidæ, and presumably the extinct families also belong, although nothing is known of their lung structures. Zittel and Hay adopt the names of Ctenodipterini and Sirenoidei for these orders, the former being further characterized by the very fine fin-rays, more numerous than their supports.
Order Ctenodipterini.—In this order the cranial roof-bones are small and numerous, and the rays of the median fins are very slender, much more numerous than their supports, which are inserted directly on the vertebral arches.
In the Uronemidæ the upper dentition comprises a cluster of small, blunt, conical denticles on the palatine bones; the lower dentition consists of similar denticles on the splenial bone. The vertical fins are continuous and the tail diphycercal. There is a jugular plate, as in Amia. The few species are found in the Carboniferous, Uronemus lobatus being the best-known species.
In Dipteridæ there is a pair of dental plates on the palatines, and an opposing pair on the splenials below. Jugular plates are present, and the tail is usually distinctly heterocercal.
In Phaneropleuron there is a distinct anal fin shorter than the very long dorsal; Phaneropleuron andersoni is known from Scotland, and Scaumenacia curta is found at Scaumenac Bay in the Upper Devonian of Canada.
In Dipterus there are no marginal teeth, and the tail is heterocercal, not diphycercal, as in the other Dipnoans generally. Numerous species of Dipterus occur in Devonian rocks.[Pg 613] In these the jugular plate is present, as in Uronemus. Dipterus valenciennesi is the best-known European species. Dipterus nelsoni and numerous other species are found in the Chemung and other groups of Devonian rocks in America.

Fig. 382.—Phaneropleuron andersoni Huxley; restored; Devonian. (After Dean.)
In the Ctenodontidæ the tail is diphycercal, and no jugular plates are present in the known specimens. In Ctenodus and Sagenodus there is no jugular plate and there are no marginal teeth. The numerous species of Ctenodus and Sagenodus belong chiefly to the Carboniferous age. Ctenodus wagneri is found in the Cleveland shale of the Ohio Devonian. Sagenodus occidentalis, one of the many American species, belongs to the coal-measures of Illinois.
As regards the succession of the Dipneusti, Dr. Dollo regards Dipterus as the most primitive, Scaumenacia, Uronemus, Ctenodus, Ceratodus, Protopterus, and Lepidosiren following in order. The last-named genus he thinks marks the terminus of the group, neither Ganoids nor Amphibians being derived from any Dipnoans.
Order Sirenoidei.—The living families of Dipneusti differ from these extinct types in having the cranial roof-bones reduced in number. There are no jugular plates and no marginal teeth in the jaws. The tail is diphycercal in all, ending in a long point, and the body is covered with cycloid scales. To these forms the name Sirenoidei was applied by Johannes Müller.
Family Ceratodontidæ.—The Ceratodontidæ have the teeth above and below developed as triangular plates, set obliquely each with several cusps on the outer margin. Nearly all the species, representing the genera Ceratodus, Gosfordia, and Conchopoma, are now extinct, the single genus Neoceratodus still existing in Australian rivers. Numerous fragments of Ceratodus are found in Mesozoic rocks in Europe, Colorado, and[Pg 614] India, Ceratodus latissimus, figured by Agassiz in 1838, being the best-known species.
The abundance of the fossil teeth of Ceratodus renders the discovery of a living representative of the same type a matter of great interest.
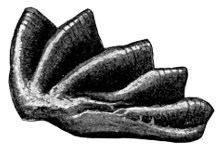
Fig. 383.—Teeth of Ceratodus runcinatus Plieninger. Carboniferous. (After Zittel.)

Fig. 384.—Neoceratodus forsteri (Günther). Australia. Family Ceratodontidæ. (After Dean.)
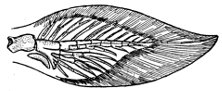
Fig. 385.—Archipterygium of Neoceratodus forsteri Günther.
In 1870 the Barramunda of the rivers of Queensland was described by Krefft, who recognized its relationship to Ceratodus and gave it the name of Ceratodus forsteri. Later, generic differences were noticed, and it was separated as a distinct group by Castelnau in 1876, under the name of Neoceratodus (later called Epiceratodus by Teller). Neoceratodus forsteri and a second species, Neoceratodus miolepis, have been since very fully discussed by Dr. Günther and Dr. Krefft.[Pg 615] They are known in Queensland as Barramunda. They inhabit the rivers known as Burnett, Dawson, and Mary, reaching a length of six feet, and being locally much valued as food. From the salmon-colored flesh, they are known to the settlers in Queensland as "salmon." According to Dr. Günther, "the Barramunda is said to be in the habit of going on land, or at least on mud-flats; and this assertion appears to be borne out by the fact that it is provided with a lung. However, it is much more probable that it rises now and then to the surface of the water in order to fill its lung with air, and then descends again until the air is so much deoxygenized as to render a renewal of it necessary. It is also said to make a grunting noise which may be heard at night for some distance. This noise is probably produced by the passage of the air through the œsophagus when it is expelled for the purpose of renewal. As the Barramunda has perfectly developed gills besides the lung, we can hardly doubt that, when it is in water of normal composition and sufficiently pure to yield the necessary supply of oxygen, these organs are sufficient for the purpose of breathing, and that the respiratory function rests with them alone. But when the fish is compelled to sojourn in thick muddy water charged with gases, which are the products of decomposing organic matter (and this must be the case very frequently during the droughts which annually exhaust the creeks of tropical Australia), it commences to breathe air with its lung in the way indicated above. If the medium in which it happens to be is perfectly unfit for breathing, the gills cease to have any function; if only in a less degree, the gills may still continue to assist in respiration. The Barramunda, in fact, can breathe by either gills or lung alone or by both simultaneously. It is not probable that it lives freely out of water, its limbs being much too flexible for supporting the heavy and unwieldy body and too feeble generally to be of much use in locomotion on land. However, it is quite possible that it is occasionally compelled to leave the water, although we cannot believe that it can exist without it in a lively condition for any length of time.
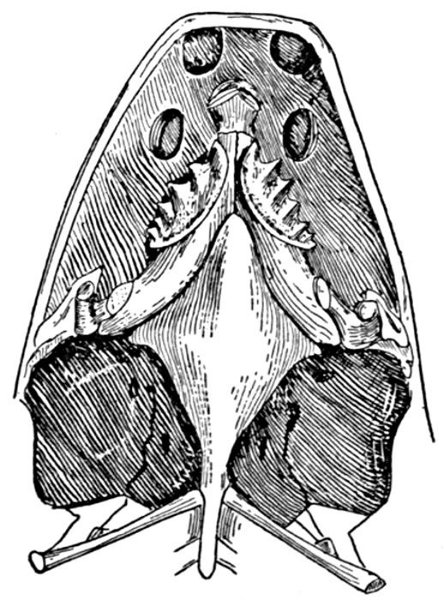
Fig. 386.—Upper jaw of Neoceratodus forsteri Günther. (After Zittel.)
"Of its propagation or development we know nothing except that it deposits a great number of eggs of the size of those of a newt, and enveloped in a gelatinous case. We may infer that the young are provided with external gills, as in Protopterus and Polypterus.
"The discovery of Ceratodus does not date farther back than the year 1870, and proved to be of the greatest interest, not only on account of the relation of this creature to the other living Dipneusti and Ganoidei, but also because it threw fresh light on those singular fossil teeth which are found in strata of Triassic and Jurassic formations in various parts of Europe, India, and America. These teeth, of which there is a great variety with regard to general shape and size, are sometimes two inches long, much longer than broad, depressed, with a flat or slightly undulated, always punctated, crown, with one margin convex, and with from three to seven prongs projecting on the opposite margin."
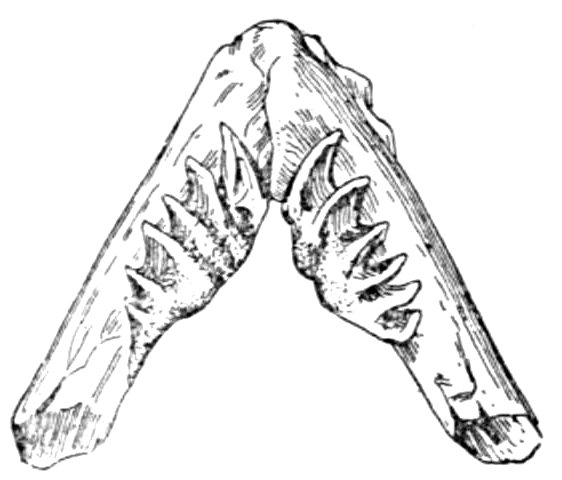
Fig. 387.—Lower jaw of Neoceratodus forsteri Günther. (After Günther.)
Development of Neoceratodus.—From Dean's "Fishes, Recent and Fossil," pp. 218-221, we condense the following account (after the observations of Dr. F. Semon) of the larval history of the Barramunda, Neoceratodus forsteri:
It offers characters of exceptional interest, uniting features of Ganoids with those of Cyclostomes and Amphibians.
The newly hatched Neoceratodus does not strikingly resemble the early larva of shark. No yolk-sac occurs, and the distribution of the yolk material in the ventral and especially the hinder ventral region is suggestive rather of lamprey or amphibian; it is, in fact, as though the quantum of yolk material had been so reduced that the body form had not been constricted off from it. The caudal tip in this stage appears, however, to resemble that of the shark, and, as far as can be inferred from surface views, a neurenteric canal persists. Like the[Pg 617] shark there then exists no unpaired fin; the gill-slits (five?) are well separated and there is an abrupt cephalic flexure. In this stage pronephros (primitive kidney) and primitive segments are well marked, and are outwardly similar to those structures in Ganoid; the mouth is on the point of forming its connection with the digestive cavity; the anus is the persistent blastophore; the heart, well established, takes a position, as in Cyclostomes, immediately in front of the yolk material.
In a later stage the unpaired fin has become perfectly established, the tail increasing in length; the gill-slits have now been almost entirely concealed by a surrounding dermal outgrowth, the embryonic operculum; a trace of the pectoral fin appears; the lateral line is seen proceeding down the side of the body; near the anal region the intestine[165] becomes narrower, and the beginnings of the spiral valve appear. In a larva of two weeks a number of developmental advances are noticed; the fish has become opaque; the primitive segments are no longer seen; the size of the yolk mass is reduced; the anal fin-fold appears; sensory canals are prominent in the head region; lateral line is completely established; the rectum becomes narrowed; and the cycloidal body-scales are already outlined. Gill-filaments may still be seen beyond the rim of the outgrowing operculum. In the ventral view of a somewhat later larva the following structures are to be noted: the pectoral fins, which have now suddenly budded out,[166] reminding one in their late appearance of the mode of origin of the anterior extremity of urodele; the greatly enlarged size of the opercular flap; external gills, still prominent; the internal nares, becoming constricted off into the mouth-cavity by the dermal fold of the anterior lip (as in some sharks); and finally (as in Protopterus and some batrachian larvæ) the one-sided position of the anus.
The larva of six weeks suggests the outline of the mature fish; head and sides show the various openings of the tubules of the insunken sensory canals; and the archipterygium of the pectoral fin is well defined. The oldest larva figured is ten weeks old; its operculum and pectoral fin show an increased size; the tubular mucous openings, becoming finely subdivided, are no longer noticeable; and although the basal supports of the remaining fins are coming to be established, there is as yet little more than a trace of the ventrals.
The early development of a lung-fish has thus far been described (Semon) only from the outward appearance of the embryo. The egg of Neoceratodus has its upper pole distinguished by its fine covering of pigment. From the first fine planes of cleavage it will be seen that the yolk material of the lower pole is not sufficient to prevent the egg's total segmentation. The first plane of cleavage is a vertical one, passing down the side of the egg as a shallow surface furrow, not appearing to entirely separate the substance of the blastomeres, although traversing completely the lower hemisphere. A second vertical furrow at right angles to the first is seen from the upper pole. The third cleavage is again a vertical one (as in all other fishes, but unlike Petromyzon), approximately meridional; its furrows appear less clearly marked than those of earlier cleavages, and seem somewhat irregular in occurrence. The fourth cleavage is horizontal above the plane of the equator. Judging from Semon's figure, at this stage the furrows of the lower pole seem to have become fainter, if not entirely lost. In a blastula showing complete segmentation the blastomeres of the upper hemisphere are the more finely subdivided. In the earlier stage the dorsal lip of the blastopore is crescent-like; in the later the blastopore acquires its oblong outline, through which the yolk material is apparent; its conditions may later be compared to those of a Ganoid.
The next change of the embryo is strikingly amphibian-like; the medullary folds rise above the egg's surface, and, arching over, fuse their edges in the median dorsal line. The medullary folds are seen closely apposed in the median line; hindward, however, they are still separate, and through this opening the blastopore may yet be seen. At this stage primitive segments[Pg 619] are shown; in the brain region the medullary folds are still slightly separated.
In an older embryo the fish-like form may be recognized. The medullary folds have completely fused in the median line, and the embryo is coming to acquire a ridge-like prominence; optic vesicles and primitive segments are apparent, and the blastopore appears to persist as the anus. The continued growth of the embryo above the yolk mass is apparent; the head end has, however, grown the more rapidly, showing gill-slits, auditory, optic, and nasal vesicles, at a time when the tail mass has hardly emerged from the surface. Pronephros has here appeared. It is not until the stage of the late embryo that the hinder trunk region and tail come to be prominent. The embryo's axis elongates and becomes straighter; the yolk mass is now much reduced, acquiring a more and more oblong form, lying in front of the tail in the region of the posterior gut. The head and even the region of the pronephros are clearly separate from the yolk-sac; the mouth is coming to be formed.
According to Eastman (Ed. Zittel), the skeleton of Neoceratodus is less developed and less ossified than that of its supposed Triassic ancestors. A similar rule holds with regard to the sturgeons and some Amphibians.

Fig. 388.—Adult male of Lepidosiren paradoxa Fitzinger. (After Kerr.)
Lepidosirenidæ.—The family Lepidosirenidæ, representing the suborder Diplopneumona, is represented by two genera of mudfishes found in streams of Africa and South America. Lepidosiren paradoxa was discovered by Natterer in 1837 in tributaries of the Amazon. It was long of great rarity in collections, but quite recently large numbers have been obtained, and Dr. J. Graham Kerr of the University of Cambridge has given a very useful account of its structure and development. From his memoir we condense the following record of its habits as seen in the swamps in a region known as Gran Chaco, which lies under the Tropic of Capricorn. These swamps[Pg 620] in the rainy season have a depth of from two to four feet, becoming entirely dry in the southern winter (June, July).
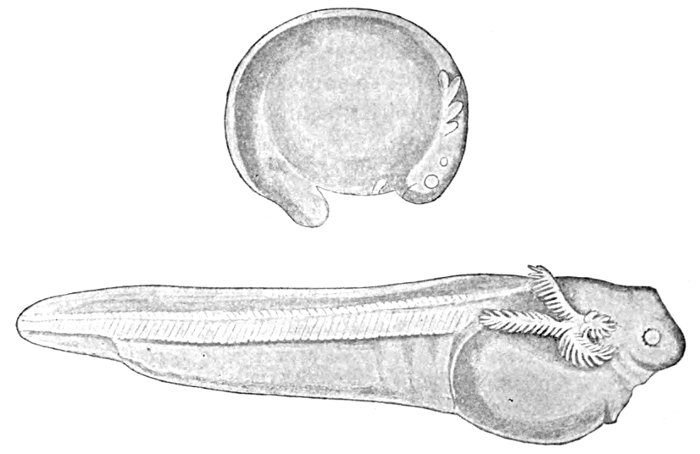
Fig. 389.—Embryo (3 days before hatching) and larva (13 days after hatching) of Lepidosiren paradoxa Fitzinger. (After Kerr.)
Kerr on the Habits of Lepidosiren.—The loalach, as the Lepidosiren is locally called, is normally sluggish, wriggling slowly about at the bottom of the swamp, using its hind limbs in irregular alternation as it clambers through the dense vegetation. More rapid movement is brought about by lateral strokes of the large and powerful posterior end of the body. It burrows with great facility, gliding through the mud, for which form of movement the shape of the head, with the upper lip overlapping the lower and the external nostril placed within the lower lip, is admirably adapted. It feeds on plants, algæ, and leaves of flower-plants. The gills are small and quite unable to supply its respiratory needs, and the animal must rise to the surface at intervals, like a frog. It breathes with its lungs as continuously and rhythmically as a mammal, the air being inhaled through the mouth. The animal makes no vocal sound, the older observation that it utters a cry like that of a cat being doubtless erroneous. Its strongest sense is that of smell. In darkness it grows paler in color, the black[Pg 621] chromatophores shrinking in absence of light and enlarging in the sunshine. In injured animals this reaction becomes much less, as they remain pale even in daylight.
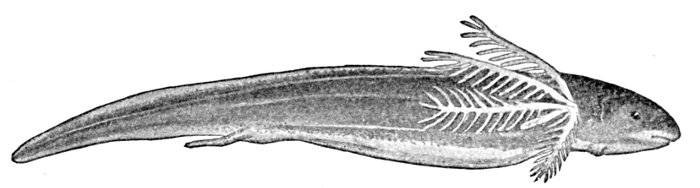
Fig. 390.—Larva of Lepidosiren paradoxa 30 days after hatching. (After Kerr.)

Fig. 391.—Larva of Lepidosiren paradoxa 40 days after hatching. (After Kerr.)
Fig. 392.—Larva of Lepidosiren paradoxa 3 months after hatching. (After Kerr.)
In the rainy season when food is abundant the Lepidosiren eats voraciously and stores great quantities of orange-colored fat in the tissues between the muscles. In the dry season it ceases to feed, or, as the Indians put it, it feeds on water. When the water disappears the Lepidosiren burrows down into the mud, closing its gill-openings, but breathing through the mouth. As the mud stiffens it retreats to the lower part of its burrow, where it lies with its tail folded over its face, the body surrounded by a mucous secretion. In its burrow there remains an opening which is closed by a lid of mud. At the end of the dry season this lid is pushed aside, and the animal comes out when the water is deep enough. When the waters rise the presence of Lepidosirens can be found only by a faint quivering movement of the grass in the bottom of the swamp. When taken the body is found to be as slippery as an eel and as muscular. The eggs are laid in underground burrows in the black[Pg 622] peat. Their galleries run horizontally and are usually two feet long by eight inches wide. After the eggs are laid the male remains curled up in the nest with them. In the spawning season an elaborate brush is developed in connection with the ventral fins.
Protopterus, a second genus, is found in the rivers of Africa, where three species, P. annectens, P. dolloi, and P. æthiopicus, are now known.
The genus has five gill-clefts, instead of four as in Lepidosiren. It retains its external gills rather longer than the latter, and its limbs are better developed. The habits of Protopterus are essentially like those of Lepidosiren, and the two types have developed along parallel lines doubtless from a common ancestry. No fossil Lepidosirenidæ are known.

Fig. 393.—Protopterus dolloi Boulenger. Congo River. Family Lepidosirenidæ. (After Boulenger.)
Just as the last page of this volume passes through the press, there has appeared a bold and striking memoir on the "Phylogeny of the Teleostomi," by Mr. C. Tate Regan of the British Museum of Natural History. In this paper Mr. Regan takes the view that the Chondrostean Ganoids (Palæoniscum, Chondrosteus, Polyodon, Psephurus, etc.) are the most primitive of the Teleostomous fishes; that the Crossopterygii, the Dipneusti, the Placodermi, and the Teleostei (as well as the higher vertebrates) are descended from these; that the Coccosteidæ (Arthrodires) are the most generalized of the Placoderms, the Osteostraci and most of the other forms called Ostracophores (Antiarcha, Anaspida) being allied to the Arthrodires, and to be included with them among the Placodermi; that the cephalic appendage of Pterichthyodes, etc., is really a pectoral fin; that the Heterostraci (Lanarkia, Pteraspis, etc.) are not Ostracophores or Placoderms at all, but mailed primitive sharks,[Pg 623] derived from the early sharks as the Chimæras are, and that the Holostean Ganoids (Lepisosteus, Amia, etc.) should be separated from the Chondrostei and referred to the Teleostei, of which they are the primitive representatives.
Mr. Regan especially calls attention to the very close similarity in structure of pectoral and ventral fins in the Chondrostean Ganoids, Psephurus and Polyodon, with that of the anal fin in the same fishes. From this he derives additional evidence in favor of the origin of paired fins from a lateral fold. In his view, the Chondrostei have sprung directly, through ancestors of the Lysopteri and Selachostomi, from pleuropterygian sharks (Cladoselache) of the Lower Silurian, and the true fishes on the one hand and the Crossopterygian-Dipneustan-Placoderm series on the other are descended from these. The absence of the lower jaw in fossil remains of Ostracophores may be due to its cartilaginous structure. "There is no justification for regarding the Crossopterygii as less specialized than the Chondrostei because they were the earlier dominant group."
These views are very suggestive and contain at least some elements of taxonomic advance, although few naturalists of to-day will regard the Chondrostean Ganoids as more primitive than the fishes called Crossopterygii and Placoderms.
These conclusions are summarized by Mr. Regan as follows:
(1) The Chondrostei are the most generalized Teleostomi.
(2) The Crossopterygii differ from them
(a) in the lobate pectoral fin;
(b) in the larger paired gular plates.
(3) The Placodermi (Coccosteidæ, Asterolepidæ, Cephalaspidæ) are a natural group, not related to the Heterostraci, which are Chondropterygii. They may probably be regarded as armored primitive Crossopterygii, this view being most in accordance with
(a) the arrangement of the cranial roof-bones in Coccosteus;
(b) the structure of the ventral fin in Coccosteus;
(c) the structure of the pectoral limb of the Asterolepidæ.
(4) The Dipneusti probably originated from more specialized Crossopterygii, e.g., from the neighborhood of the Holoptychiidæ.
(5) The Teleostei differ in so many respects from the Chondrostei that they should rank as an order, in which the Holostei are included.
[164] This group has been usually known as Dipnoi, a name chosen by Johannes Müller in 1845. But the latter term was first taken by Leuckart in 1821 as a name for Amphibians before any of the living Dipneusti were known. We therefore follow Boulenger in the use of the name Dipneusti, suggested by Hæckel in 1866. The name Dipnoan may, however, be retained as a vernacular equivalent of Dipneusti.
[165] The yolk appears to be contained in the digestive cavity, as in Ichthyophis and lamprey.
[166] The abbreviated mode of development of the fins is most interesting; from the earliest stage they assume outwardly the archipterygial form; the retarded development of the limbs seems curiously amphibian-like; the pectorals do not properly appear until about the third week, the ventrals not until after the tenth.
The Natural History of Plants
THEIR FORMS, GROWTH, REPRODUCTION AND DISTRIBUTION
FROM THE GERMAN OF
ANTON KERNER VON MARILAUN
Professor of Botany in the University of Vienna
By F. W. OLIVER
Quain Professor of Botany in University College, London
WITH THE ASSISTANCE OF
MARIAN BUSH AND MARY E. EWART
4to. New edition. 2 vols. The set—$11.00
A work for reference or continuous reading, at once popular and, in the modern sense, thoroughly scientific. The new edition is practically identical with the former four-volume edition except that the colored plates in the latter have been omitted. The wood-engravings, over two thousand in number, have been retained.
Prof. John M. Coulter, in The Dial: "Prof. Kerner has brought the most recent researches within reach of the intelligent reader, and in a style so charming that even the professional teacher may learn a lesson in the art of presentation.... It is such books as this that will bring botany fairly before the public as a subject of absorbing interest; that will illuminate the botanical lecture-room."
Prof. Chas. R. Barnes, in The Botanical Gazette: "This lucidity, and the excellent illustrations, not only will introduce the non-botanical reader to the science of botany, but should serve as a lesson to the professional botanist in the art of presentation."
The Nation: "He has succeeded in constructing a popular work on the phenomena of vegetation which is practically without any rival."
GUIDE TO THE STUDY OF INSECTS
AND A TREATISE ON THOSE
INJURIOUS AND BENEFICIAL TO CROPS
FOR THE USE OF
COLLEGES, FARM-SCHOOLS AND AGRICULTURISTS
By ALPHEUS S. PACKARD, M.D.
With 685 illustrations. Ninth edition. xii+715 pp., 8vo, $5.00 net
PLANT PHYSIOLOGY
By GEORGE J. PEIRCE
Professor in Leland Stanford University
vi+291 pages, 8vo $2.00
A modern and thoroughly scientific discussion of the general principles of plant physiology, intended for the student or general reader acquainted with the elements of botany.
Science: "The volume is full of original suggestions and differs quite markedly from the old-time works devoted to plant physiology."
William F. Ganong, Professor in Smith College: "I am much pleased with the clearness, proportion, and vigor with which it treats the subject. It seems to me an admirable exposition of the principles of plant physiology as they are understood at the present day, and it should have a wide use."

Henry Holt and Company
29 West 23d Street, New York
GEOLOGY
Vol I. "Geologic Processes and Their Results"
By Prof. THOMAS C. CHAMBERLIN
AND
Prof. ROLLIN D. SALISBURY
Heads of the Departments of Geology and Geography, University of Chicago; Members of the United States Geological Survey; Editors of the Journal of Geology
With numerous illustrations, including 24 colored maps and 3 tables. 654 pages, 8vo, $4.00 net
Vol. II. "Earth History." In preparation
Chas. D. Walcott, Director of U. S. Geological Survey: "I am impressed with the admirable plan of the work and with the thorough manner in which geological principles and processes and their results have been presented. The text is written in an entertaining style and is supplemented by admirable illustrations, so that the student cannot fail to obtain a clear idea of nature and the work of geological agencies, of the present status of the science, and of the spirit which actuates the working geologist."
T. A. Jaggar, Jr., Harvard University: "An excellent statement of modern American geology, with abundant new illustrative material based upon the most recent work of government and other surveys."
Henry S. Williams, Yale University: "It is the best treatise on this part of the subject which we have seen in America."
R. S. Woodward, Columbia University: "It is admirable for its science, admirable for its literary perfection, and admirable for its unequalled illustrations."
Israel C. Russell, University of Michigan. "I deem it an epoch-making book and one that will vastly extend the study of geology."
BUTTERFLIES
By S. H. SCUDDER
THEIR STRUCTURE, CHANGES, AND LIFE-HISTORIES
With Special Reference to American Forms. Being an Application of the "Doctrine of Descent" to the Study of Butterflies. With an Appendix of Practical Instruction
12mo $1.50 net
Brief Guide to the Commoner Butterflies of the Northern United States and Canada
Being an Introduction to the Knowledge of their Life-Histories
New edition. With 21 plates, containing in all 97 illustrations
12mo $1.50
THE LIFE OF A BUTTERFLY
A CHAPTER IN NATURAL HISTORY FOR THE GENERAL READER
16mo $1.00

Henry Holt and Company
29 West 23d Street New York
FERNS
A MANUAL FOR THE NORTHEASTERN STATES WITH ANALYTICAL KEYS BASED ON THE STALKS AND ON THE FRUCTIFICATION
With over two hundred illustrations from original drawings and photographs
By CAMPBELL E. WATERS
302 pages, square 8vo. Boxed, $3.00 net; by mail, $3.34
This book is thoroughly authoritative, and is written in popular style. It covers all the ferns in the region embraced either in Britton's or in Gray's Manuals.
"This book is likely to prove the leading popular work on ferns. No finer examples of fern photography have ever been produced. Dr. Waters brings to his work fifteen years of experience in field and herbarium study, and the book may be expected to prove of permanent scientific value, as well as to satisfy a want which existing treatises have but imperfectly filled."—Plant World.
"For all who study or wish to study our native ferns Dr. Waters has prepared a book which is sure to prove both helpful and inspiring. Especially charming and significant are the views showing typical habits and habitats."—The American Naturalist.
"There could hardly be a better book for those interested in the subject."—Boston Literary World.
OUR NATIVE FERNS
AND THEIR ALLIES
WITH SYNOPTICAL DESCRIPTION OF THE AMERICAN PTERIDOPHYTA NORTH OF MEXICO
By LUCIEN M. UNDERWOOD
Professor in Columbia University
Revised. xii+156 pages, 12mo $1.00
"The elementary part is clear and well calculated to introduce beginners to the study of the plants treated of. The excellent key makes the analysis of ferns comparatively easy. The writer cordially commends the book. It should be in the hands of all who are especially interested in the vascular cryptogams of the United States."—Bulletin of the Torrey Botanical Club, N. Y.

Henry Holt and Company
29 West 23d Street, New York
MUSHROOMS
By GEORGE FRANCIS ATKINSON
Professor of Botany in Cornell University, and Botanist of the Cornell University Experiment Station
Recipes for Cooking Mushrooms. By Mrs. SARAH TYSON RORER
Chemistry and Toxicology of Mushrooms. By J. F. CLARK
With 230 illustrations from photographs, including 15 colored plates
320 pages, 8vo. $3.00 net; by mail, $3.23
Educational Review:—"It would be difficult to conceive of a more attractive and useful book.... In addition to its general attractiveness and the beauty of its illustrations, it is written in a style well calculated to win the merest tyro."
Moulds, Mildews, and Mushrooms
By LUCIEN M. UNDERWOOD
Professor in Columbia University
iv+236 pages, 12mo $1.50
Bradley M. Davis, in the Botanical Gazette:—"Wonderfully free from the dry diagnoses of most systematic descriptions, and everywhere combined with interesting accounts of life-habits and activities.... A marvel in its compactness, with a wonderfully uniform tone throughout, condensed and yet very clear."
Flora of the Northern States and Canada
By Professor N. L. BRITTON
Director of the New York Botanical Garden
x+1080 pages, large 12mo $2.25
This manual is published in response to a demand for a handbook suitable for ordinary school use, which shall meet modern requirements and outline modern conceptions of the science. It is based on An Illustrated Flora prepared by Professor Britton in co-operation with Judge Addison Brown, in three volumes. The text has been revised and brought up to date, and much of novelty has been added, but all illustrations are omitted.
Conway MacMillan, Professor in the University of Minnesota, in Science:—"There is no work extant in the whole series of American botanical publications which deals with descriptions of the flowering plants that can for a moment be compared with it, either for a skillful and delightful presentation of the subject-matter or for modern, scientific, and accurate mastery of the thousandfold mass of detail of which such a work must consist."
V. M. Spalding, Professor in the University of Michigan:—"I regard the book as one that we cannot do without and one that will henceforth take its place as a necessary means of determination of the plant species within its range."

Henry Holt and Company
29 West 23d Street, New York
Transcriber's Notes:
Simple spelling, grammar, and typographical errors were corrected.
Punctuation normalized.
Anachronistic and non-standard spellings retained as printed.
The ERRATA on p. ix has been corrected in the text.
P. xxiii corrected "Salmo gairdneri, the Steelhead Trout. 326" to "Salmo irideus, the Rainbow Trout. 326" to agree with the actual illustration caption.
P. xxiii corrected "Salmo rivularis, the Steelhead Trout. 327" to "Salmo gairdneri, the Steelhead Trout. 327" to agree with the actual illustration caption.