In this transcription a black dotted underline indicates a hyperlink to a page or illustration; hyperlinks are also marked by aqua highlighting when the mouse pointer hovers over them. Page numbers are shown in the right margin.
Archaic spellings such as ‘sedementary’ and ‘millepedes’ have not been altered, and the following spelling inconsistencies have also been left as in the original text: omnivorous/omnivorus, eleagnus/elæagnus, silverfish/silver-fish, woodlice/wood-lice, blowfly/blow-fly.
The following typographic errors have been corrected:
kindom —> kingdom
aphis lion —> aphis-lion
Jose —> José
ocurring —> occurring
necesary —> necessary
Crytolæmus —> Cryptolæmus
Crape —> Grape
pupuli —> populi
CAWTHRON INSTITUTE MONOGRAPHS
| Page | |
Introduction | |
| Chapter i.: | General Review of the Animal Kingdom7 |
| Chapter ii.: | Soil Organisms and Soil Fertility12 |
| Chapter iii.: | Structure of Insects17 |
| Chapter iv.: | Life Histories of Insects22 |
| Chapter v.: | Sucking Insects29 |
| Chapter vi.: | Sucking Insects (Concluded)42 |
| Chapter vii.: | Leaf-feeding Insects50 |
| Chapter viii.: | Boring and Underground Insects61 |
| Chapter ix.: | Miscellaneous Pests67 |
| Chapter x.: | Principles of Pest Control74 |
Index |
This work deals with the insects and other animals having a detrimental or beneficial influence upon horticulture in New Zealand. Its purpose is to supply such general information as will enable the common animal inhabitants of the garden to be identified and controlled, to act as a popular guide for the use of practical gardeners and schools, and at the same time serve as a source from which the examination requirements set out in the syllabus of the New Zealand Institute of Horticulture may be met.
As this work is for the benefit of the gardening public, and an endeavour to diffuse some knowledge of certain natural problems, the language of the scientist—which, unfortunately, tends to guard what is known of these problems from the general reader—has been avoided as much as possible; at times, however, this ideal cannot be adhered to, but in such cases the reader should find no difficulty, and should be prepared to become familiar, with the few terms used. To know the scientific names of animals without being acquainted with the animals themselves is a habit to be avoided, and is just about as instructive as memorising the names of people in a town or telephone directory. But animals must be named; though their popular names are used in the following pages and as such names are very often misleading, the scientific names are given in brackets in order to avoid confusion.
In such a work as this, illustrations are of great value, and these are given wherever possible. One drawback to illustrations is that the relative proportions of animals may be lost; for example, a microscopic organism might require magnification by some 4,000 times its natural size and so become equal to that of some of the most conspicuous insects. Even with the best illustrations, however, it is essential that the reader becomes familiar with the animals themselves. This should present no difficulty to the reader, since he will find in his garden all of the animals with which he is concerned—mostly insects and their near relatives. Further, of very great assistance to him, he will find the several excellent public museums throughout the country, as well as the specialists at such research institutions as the Cawthron Institute at Nelson.
To keep a work for the general reader in a readable form, the desire of the author to cite the sources from which he derives his information must be suppressed. If this were not done, the text would rapidly become littered with endless references, much to the weariness and confusion of the reader. Therefore, it should be remembered that a work of this kind is a compilation from the publications of many scientists, to which is added what little original information the writer himself might possess.
Opportunity must be taken here to express one’s appreciation of the assistance given by Mr. W. C. Davies and Mr. L. J. Dumbleton in the preparation of the photographs and drawings, respectively.
At the outset it is advisable, by reviewing the animal kingdom as a whole, to secure in perspective the relationships of the animals with which the horticulturist has to deal.
To most people the animal kingdom is comprised chiefly of those animals commonly met with in everyday life or in general reading—the game and domestic animals and the fishes, all of which are similar in that they possess a backbone or vertebral column, and are consequently known as the vertebrates. Popularly, however, they are generally classed as the “lower” and “higher” animals; there is certainly some accuracy in such a haphazard classification, since, though all the vertebrates are, strictly speaking, the “higher” animals, some are “lower” (e.g., fish, frog, and bird) than others (e.g., kangaroo, dog, and man, the highest of all).
But when it comes to the true “lower” animals, that vast assemblage of less conspicuous creatures, the jelly-fish and corals, worms of all kinds, sea-urchins, crayfish, wood-lice, spiders and insects, shell-fish and snails, all characterised by the absence of a vertebral column and known as the invertebrates, they are not collectively visualised in a general sense as are the vertebrates. As a rule, these invertebrates are known individually as independent units, except, perhaps, in the case of worms, insects, spiders, wood-lice, etc., which are very often collectively and haphazardly referred to as “insects,” a term, in this sense, as ill-defined as it is unlimited.
That the average person should be more conversant with the vertebrates than the invertebrates is, to a great extent, the natural outcome of association and training; a possible influence is to be found at the outset of one’s career in the many illustrated nursery books depicting game and domestic animals, but seldom, if ever, any of the invertebrates; and this impression tends to be further fostered in later life by visits to the zoo, where we meet in person most of the nursery book animals, and perhaps some of the lower forms, such as insects; but the latter, in most cases, are there by chance, not design, and against the will of the authorities.
In recent years, however, more public attention has been given to the lower animals owing to the detrimental influence of many upon agricultural development as well as upon public health. That such animals are capable of ranking as fundamental factors hindering human progress, may be realised when it is considered that, of the invertebrates, insects alone comprise nearly four-fifths of the whole animal kingdom! This has been graphically illustrated as follows by F. E. Lutz, of the American Museum of Natural History:—Extend the arms and fingers at right angles to the body, and let the distance from the tip of the middle finger of one hand to that of the other represent the number of different kinds of living animals; then the last joint of the middle finger of the right hand will be proportionate to the number of mammals (kangaroos, hoofed animals, rabbits, man, etc.), the second joint to the reptiles and their relations, the first joint to the birds, and the distance between the knuckles and the wrist to the fishes. “In other words, you can hold the so-called zoological gardens and their aquarium annexes in one hand.” Finally, the distance between the wrist of the right arm and the tip of the middle finger of the left will proportionately represent all the known species of invertebrates, and of this section of the extended arms all except between a wrist and an elbow will be insects.
The zoologist classifies the animals under twelve main divisions, of which eleven contain the invertebrates and one the vertebrates; these divisions are arranged in a series, the first containing the simplest or lower animals, and the last the most complex or highest. A glance at this classification will serve to give some idea of the relative position in the animal kingdom of the animals which will be dealt with in the following pages. The very lowest forms, belonging to the first division, are micro-organisms known as the Protozoa; they inhabit water and soil, and live upon their own kind or upon minute plants, including bacteria, or are parasitic upon the higher animals, some of these parasites causing such diseases as malaria. The Protozoa are single units of living matter (protoplasm), and may be referred to as the one-celled animals; they are mostly microscopic, and lead an independent life, or are associated in colonies, but are capable, as a rule, of carrying on independently all the functions of life, though there are no organs such as those of digestion, respiration, and circulation, as we know them in the higher animals. It is amongst such simple forms that the distinction between the lowest animals and plants ceases to be clear. As will be discussed later, there is evidence that certain Protozoa have an important influence on soil fertility.
The remaining eleven divisions contain all other animals, ranging in size from mere specks to the mass of the elephant; the bodies of these are built up of a complex aggregate of countless cells of protoplasm arranged in groups to form the organs of digestion, circulation, respiration, reproduction, etc., each having its definite function in the animals’ lives. The following are some typical or well-known examples of each of these divisions, the technical names, with the exception of the Protozoa, not being given:—
The Protozoa (reference should be made here to Fig. 1) are followed by (2) sponges; (3) jelly-fish, sea-anemones, corals; (4) flat worms (tape-worms, etc.); (5) round worms (thread-worms, eel-worms); (6) sea-mats, lamp-shells; (7) wheel-animalcules; (8) star-fish, sea-urchins; (9) segmented worms (earthworms); (10) crayfish, woodlice, centipedes, millepedes, spiders, mites, insects; (11) shell-fish, slugs, snails; (12) fish, frogs, lizards, birds, hedgehogs, rabbits, man.
So far we have reviewed the animal kingdom from one aspect only—that of classification, based on the resemblances and differences of the individuals. It is now necessary to look at the subject from the viewpoint of the horticulturist—that is, the relationships of the animals to their surroundings, or environment, and to the welfare of man. Of the two great life-groups—animals and plants—the plants are of fundamental importance; without them no animal could exist, since, of all living things, it is the green plants alone that are able to convert the inorganic chemical constituents in soil, air and water into living matter or protoplasm; and all animals, either directly or indirectly, are dependent upon plants for their food supply. Plants, therefore, may be looked upon as the primary producers of life, and animals as the consumers. It is in this respect that the horticulturist becomes interested, in that certain of these consumers destroy too many of the plants grown by him for other purposes; fortunately, not all of the consumers are destructive; many are of very great use to the horticulturist and mankind in general.
The last point is well illustrated by the following classification of the animal kingdom based upon the part it plays in human welfare; this is a modification of the scheme adopted by the British Museum of Natural History:—
Group I.—Wild or domesticated animals used by man as beasts of burden, source of food, or in the manufacture of various products—e.g., sponges, crayfish, bees, silk-worms, shell-fish, and various vertebrates, as fish, birds and mammals.
Group II.—Animals detrimental to man’s welfare, attacking man himself; animals and plants of value to him, or the products derived therefrom—e.g., Protozoa, parasitic worms, mites, insects, and such vertebrates as certain birds and mammals.
Group III.—Animals aiding man’s welfare, as scavengers, or by pollinating flowers, or by attacking and checking such animals as are included in Group II.—e.g., Protozoa, parasitic worms, earthworms, parasitic insects, spiders, and such vertebrates as certain birds and mammals.
An analysis of the above classification shows that animals both aid and hinder the progress of man, hence the use of the terms “beneficial” and “destructive.” In nature, however, these terms are not altogether applicable in the same sense, since the balance maintained between animals and plants under natural conditions is an extremely fluctuating one, though sufficient for natural purposes; with man, however, the case is different. In order to compete in the world’s markets, and to supply the growing demands of increasing population, a much higher and dependable standard of productivity is required than is found in nature. Consequently, whilst utilising, and increasing the efficiency of the so-called natural enemies as auxiliaries in his fight against destructive animals, man has found it necessary to develop an effective system of artificial control, involving chemicals, resistant plants, cultivation, crop rotation, etc., for the purpose of maintaining a more stringent balance to meet his requirements.
The animal population of European New Zealand is very different from that of pre-European times, a position brought about naturally enough by the changes resulting from agricultural development as practised in the Old World, and the consequent creation of an environment foreign to the country.
Though the official date of the settlement of New Zealand by Europeans is 1840, the influences, inaugurating that upheaval of the natural conditions which was later to have such a marked effect on the economic development of the country, had commenced many years earlier.
When the first Europeans set foot in New Zealand, they must have been impressed by their unique surroundings, totally different from anything to be met with in the Old World. They found the country dominated by a forest quite unlike the forests of any other land, and inhabited by an animal population presenting many unusual features. This terrestrial population was characterised by an abundance of insects and spiders, and a paucity of vertebrates excepting the birds; the vertebrates consisted of a species or two of frogs, a few species of lizards, some 200 species of birds, and two species of bats, the last being the only terrestrial mammals. In fact, the insects, spiders and birds were the dominant animals, a feature common to other parts of the world, but the scanty vertebrate population, other than birds, was a characteristic of primeval New Zealand.
New Zealand being a country fitted for agriculture, settlement by Europeans naturally resulted in extensive and rapid changes, since the settlers brought with them the knowledge, implements, animals and plants of the civilised world; and to make way for settlement, it was necessary to remove the forests and drain the swamps, and to replace them with cultivated crops and pastures. These activities have been so thorough, that, within a period of some 90 years practically the whole of the original North Island forests, and the greater part of those of the South Island, have been cleared.
An outstanding feature of these changes is that many of the pests associated with the agricultural animals and plants have been brought to New Zealand with the animals and plants they infest, and these exotic pests comprise by far the greater proportion of the destructive animal population, there being but few native species forming the balance. For example, 71 per cent. of the destructive insects are exotic, and 29 per cent. native, while all the parasitic worms of economic importance, all the destructive birds (e.g., sparrows) and mammals (e.g., deer, wild pigs, and goats) are introduced.
The exotic factors that have set up this new environment may be summarised as follows:—
(1) Clearing of the native vegetation.
(2) Introduced plants: e.g., grasses, forage crops, trees, etc.
(3) Introduced game animals: e.g., deer, pigs, rabbits, birds, etc.
(4) Introduced destructive animals, infesting animals and plants of economic value: e.g., parasitic worms, insects, etc.
(5) Animals imported to control pests, but which have become destructive themselves: e.g., weasels, birds.
In the first chapter the plants were referred to as the primary producers of life, and the animals as the consumers; the former not only furnish nourishment for their own growth, but also for the support of the animal world as a whole. Living plants (in reference to green plants) utilise the sun’s energy in the manufacture of their complex food materials from comparatively simple chemical compounds. These latter compounds are carbon dioxide, derived from the air through the agency of leaves, and a weak solution of various chemical compounds in water, derived by means of the roots from the soil, and carried up through the plant to the leaves, where the elaboration into the complex compounds to be utilised by the plants as food takes place.
These comparatively simple compounds from which the plants elaborate their nourishment are the raw food materials, and that they must always be available for plant growth, is evident when one considers the vast areas of vegetation that cover, with the exception of desert regions, the surface of the earth. Under moist climatic conditions it has been calculated that some 500 tons of carbon dioxide and 1,000,000 tons of water, having the raw food materials in solution, are used annually by one square mile of dense forest. For their development, therefore, plants require:—
(1) Sunlight as the source of energy for the carrying on of their life functions;
(2) Air for the supply of carbon dioxide, oxygen, and, indirectly, nitrogen;
(3) An ample supply of water required for the living tissues and as a vehicle for the transport from the soil of
(4) The raw food materials, in the form of various chemical compounds.
With the exception of the carbon dioxide derived from the air, all the raw food materials—water, nitrates, phosphates, sulphates, potassium, calcium, magnesium, iron, etc.—are present in the soil, though only a part of them is in a form suitable for imbibition by plants. In the formation of these food materials, which render the soil fertile, physical forces and the activities of living organisms play a leading part. Our immediate concern is with the influence of these organisms upon soil fertility, but it is advisable to give some consideration to the soil itself, since it is the environment in which the organisms live, and with which their existence is intimately associated; in this respect attention will be confined to the type of soil usually cultivated by the horticulturist, and to the uppermost layers—that is, approximately, within one foot of the surface.
Soil is the product of disintegrated and weathered rocks with which are mixed the residues of organic matter. Apart from the particles of disintegrated rocks, which form the matrix, soil contains chemical compounds of two kinds: those of a purely mineral nature derived from the inorganic components of the original rocks, and those of an organic origin derived either from the ancient remains of organisms, which, in the case of sedementary deposits, became incorporated in the rocks at the time of their origin, or from the remains of present-day plants and animals decomposed by soil organisms. In addition, there is the humus, which has a fundamental physical influence, and for the production of which soil organisms are responsible.
In the initial stages of soil formation during the disintegration and decomposition of rocks, the first type of soil to be formed is suitable for the growth of only certain plants; it is of a purely mineral nature, containing raw food materials derived mainly from the rocks and not from organic matter, unless from such organic residues as were incorporated in the rocks during their formation in ancient times. Such soil cannot sustain the higher types of green plants, nor is it populated by soil organisms; it furnishes suitable pabulum, however, for the nourishment and growth of the more lowly types of vegetation, which are able to convert to their benefit the limited supply of food materials available. The complex organic compounds that such primitive plants elaborate from these food materials of purely mineral origin, and incorporate in their tissues, are, after death, returned to the soil, which becomes correspondingly enriched, and a favourable environment for the establishment of organisms; the latter reduce these plant residues to humus, and during this process of decomposition produce food materials of an organic origin suitable for the nutrition of the sequential plant covering. So the process proceeds until a soil is formed of sufficient extent and quality for the support of a more extensive and increasingly complex vegetation; thus, in the cycle of life and decay, stores of organic compounds are elaborated by plants and returned to the soil, which they enrich, and where they are decomposed by organisms, and so maintain the supplies of food materials suitable for the maintenance of vegetation.
These phenomena of plant establishment and succession, correlated with soil formation, were clearly demonstrated by the re-establishment of vegetation after the soil and plant life had been destroyed by the historic eruption in 1883 of Krakatoa, a volcanic island in the Straits of Sunda, between Java and Sumatra. The first plants to be established on the volcanic deposits were species of terrestrial algæ, which gradually spread and built up soil suitable for the development of soil organisms and for the growth of seeds brought to the island by birds and ocean currents. So rapid were the changes brought about by these influences, that within a period of twenty years after the eruption the barren ground was reclothed by a dense and varied plant covering.
Organisms that form part of the organic complex of the soil range from the more conspicuous species, such as slugs and snails, insects, spiders, wood lice, millepedes, earthworms and eelworms, to such microscopic forms as protozoa, fungi, algæ and bacteria, the last three being members of the plant kingdom. These organisms may be grouped as follows:—
(1) Temporary inhabitants that enter the soil for shelter, or to feed as scavengers on decaying organic matter, or both—e.g., slugs, snails, wood lice, certain insects and some eelworms.
(2) Permanent inhabitants that are dependent on the soil for their development and supplies of food, either throughout or during most of their lives—e.g., certain insects and spiders, millepedes, earthworms, eelworms, protozoa, fungi, algæ and bacteria.
The organisms in the first group play a comparatively minor part in soil development, and influence its fertility to an almost negligible extent, the temporary scavengers, perhaps, being of more importance since they aid in the reduction of vegetable residues. The forms in the second group, however, are invaluable as soil-making agents and in the production of plant food materials, the least important among them being the insects, spiders and millepedes. Many are merely scavengers, but some insects, such as grass-grubs and the caterpillars of certain moths, and millepedes, feed upon living plants and so add organic matter to the soil in their excreta, which also contains quantities of soil swallowed with the food, this latter mechanical action aiding in the pulverising and opening up of the soil; certain eelworms, too, that attack living plants play a somewhat similar part, in that they are primary causative agents in the decay of healthy tissues. Other forms of insects, together with spiders and some eelworms, are predaceous upon their fellows, the remains of the latter being added to the soil residual complex. Apart from the activities of all these organisms, however, it is the earthworms, protozoa, fungi, algæ and bacteria that have the most fundamental influence upon soil fertility.
Earthworms may be correctly called the great soil builders; they burrow through it, allowing the free passage of air and water; they swallow large quantities, which they eject on the surface in the form of “worm-casts,” the soil materials being well mixed in the process; they pull underground leaves and other parts of plants from the surface and so increase the supply of organic matter for the action of the micro-organisms that bring about decomposition. Further, by depositing their “casts” on the surface, earthworms soon cover the accumulations of dead vegetable matter, as has been illustrated by Darwin in his classic work on these animals. Without the aid of earthworms—e.g., in sour soils in which they do not abound—the plant residues accumulate on the surface, to form a partially decomposed, peaty mass, which only a limited number of plants can tolerate.
The protozoa, fungi, algæ and bacteria are all microscopic organisms, and are the agents responsible for the decomposition of the organic residues in the soil; they do not act as independent units, the processes of one group being dependent upon and intimately related with those of the others. During the activities of these organisms various organic and mineral substances are decomposed or transformed into materials, such as humus and the inorganic compounds of nitrogen, phosphorus, potassium, etc., necessary or helpful for the growth of plants.
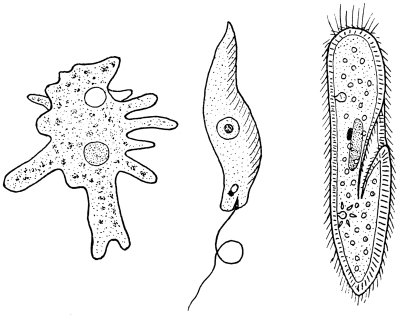
The protozoa (see Chapter I.) are the lowest and simplest forms of animal life, being mere specks of living matter. Three different groups of soil protozoa occur (Fig. 2). Some, like the amœba, progress by streaming movements, extruding temporary extensions of their substance in the form of finger or thread-like processes; the bodies of such protozoa may be naked, or enclosed in a shell-like covering secreted by the organism itself, or protected by an accumulation of particles of foreign matter. Some have a body of more definite shape and progress by means of the whip-like action of one or two thread-like processes, or flagella, arising from one end of the body. Such forms are the most numerous in the soil. Others, also of definite shape, control their movements by means of short, hair-like processes, or cilia, either distributed over the body or restricted to definite regions.
The protozoa are widely distributed, being most abundant in the richer types of soil, especially during the spring and autumn. A great amount of research has been undertaken at Rothamsted, England, and elsewhere, on the part played by protozoa in soil fertility; the evidence thus secured points to the probability that some of these organisms may be detrimental in that they devour certain kinds of bacteria responsible for the production of nitrates and other substances of nutritive value to plants. The extent of this may be realised from the fact that in a definite weight of soil (about 1-28th of an ounce) the micro-population was calculated to include not only about 1,550,000 protozoa, of which 430,000 were amœbæ (Fig. 2), but also some 6,000,000,000 bacteria. Observations showed that a single bacteria-destroying amœba required about 400 organisms for its nourishment, so that the amœbæ, to say nothing of the other protozoa, present in the weight of soil above-mentioned, would be capable of destroying about 172,000,000 of the bacterial population. Since the partial sterilisation of soil by steam results in an increase of fertility, it is thought, on account of the sterilisation destroying the protozoa, being more susceptible, and not the bacteria, that protozoa inhibit the activities of the bacteria to such an extent as to reduce the fertility of the soil; but this is a subject as yet open to argument. Apart from the bacteria-destroying protozoa, there are other forms that are thought to have something to do with the decomposition of organic substances.
The fungi, algæ and bacteria are amongst the lowest forms of plant life, and hold somewhat the same position in the plant kingdom as the protozoa do among animals; they are, especially the fungi and bacteria, of primary importance in the maintenance of soil fertility. The role of algæ lies mainly in increasing the organic content of the soil, and they are invaluable in developing favourable conditions for the establishment of vegetation on purely mineral soils. The fungi and bacteria are responsible for setting up the intricate reactions involved in the decomposition of organic matter, the bacteria being concerned in practically all of the chemical processes going on in the soil. Both fungi and bacteria are of two kinds: those that bring about decomposition, and those that live in a reciprocal relationship with plants upon the roots of the latter. Such relationship, which benefits both organisms and plants, is called symbiosis, the fungi being known as mycorrhiza, while the bacteria form nodules on the roots of such plants as the legumes.
Although insects present a great variety of forms, they nevertheless agree in general features; thus by studying the structure of some generalised species, which will give a broad idea of the main characteristics, one is enabled to recognise different structural modifications assumed by various species. For this purpose a weta, grasshopper, or cockroach may be taken as a type.
Just as in the case of the crayfish, so the body of an insect is completely covered and protected by a continuous “shell,” very solid in some insects, more or less pliable in others, but even in the most delicate forms tending to become rigid and brittle after death. This shell acts as a skeleton and as a very effective armour-plating, protecting and supporting the soft body within. Unlike the shell of the crayfish, which is mainly calcareous, that of insects consists of a horny substance called chitin, secreted by the underlying skin, and constitutes what is known as a cuticle. It is due to this horny cuticle or shell that the form and colour of most insects are preserved after death, though the enclosed body tissues decay unless preserved in some suitable medium.
The cuticle, though forming a complete covering, does not enclose the body in an inflexible shell; flexibility is allowed by the cuticle being formed of a segmented series of strongly-chitinised sections alternating with skin-like, feebly-chitinised, and very elastic sections; this arrangement gives freedom of movement to the enclosed body, as is readily seen in the movements of a caterpillar.
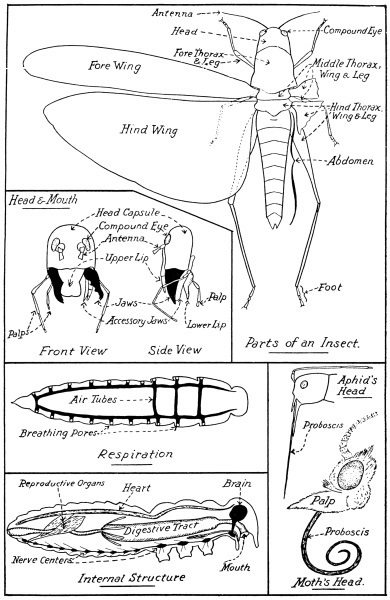
There are three distinctly separated divisions of the insect body—head, thorax, and abdomen—each consisting of a varying number of segments (Fig. 3). The head segments are so closely fused as to be practically untraceable, the cuticle forming a rigid capsule; the thorax, to which the head is attached, carries the wings (when present) and the legs, and consists of three segments; posterior to the thorax is the abdomen, comprised of several segments, which show the typical segmentation of insects better than any other part of the body.
The head capsule is more or less freely movable on the thorax, and bears certain sensory organs, together with the mouth appendages. The sensory organs are the eyes and the feelers, or antennæ. On each side is a compound eye of varying size, according to the insect; each eye consists of a variable number (from a comparative few to several thousand) of microscopic, hexagonal lenses, each of which records a separate image. Between the compound eyes, on top of the head, are three simple eyes in some insects, but in others one or all of these may be absent. Between the compound eyes on the front aspect of the head is a pair of feelers, or antennæ; they consist of a variable number of joints, are freely movable and highly sensory, thread-like or hair-like, short, or longer even than the whole body, and may be bare or clothed to a varying degree with hair or bristles. On the antennæ are the organs of touch, smell, and sometimes hearing.
When the head of a weta, grasshopper, or cockroach is removed from the body and boiled for a few minutes in a 10 per cent. solution of caustic potash, and then washed in water in order to remove the muscles and other tissues, a large opening will be seen on the posterior surface where the head was attached to the thorax; also, if the mouth appendages are pulled apart, they will be seen to surround another opening on the lower aspect of the head capsule, marking the position of the mouth. The digestive canal passes from the mouth through the posterior opening into the thorax.
The mouth appendages are as follows (Fig. 3):—Suspended from the fore aspect of the mouth opening is a more or less conspicuous movable flap, which forms the upper lip, while from the posterior aspect of the same opening is another suspended appendage forming the lower lip; this latter appendage is really a complicated one, and bears a pair of short, jointed appendages—the palps—which are sensory organs, while on its inner surface—i.e., within the mouth—is a swollen area or tongue, an organ very greatly modified in certain insects. Between the upper and lower lips, and suspended from both sides of the mouth opening, is a pair of true jaws immediately behind the upper lip, followed by a pair of accessory jaws immediately before the lower lip; these jaws do not move up and down, but have a side-wise action, closing and opening like scissor blades. While the true jaws are each of one piece, the accessory jaws consist of several parts, and each bears in addition a jointed palp, as in the case of the lower lip. The upper and lower lips serve to hold the food in the mouth, the true jaws nibble or tear off portions of the food and masticate it (if the term can be used), while the accessory jaws, aided by the lower lip, manipulate the food during the process of feeding.
The comparatively simple arrangement of mouth parts found in the weta, grasshopper, and cockroach, as described above, is characteristic of all insects that gnaw or chew their food—e.g., earwigs, beetles and their larvæ or grubs, the caterpillars of moths, and so on. There is, however, a vast number of insects that has developed more or less complex variations of this generalised pattern, according to the manner of feeding.
The mouth parts of the worker honey-bee, for example, have the jaws adapted for eating pollen and moulding wax for the comb; the accessory jaws, however, are lengthened, though their palps are reduced to mere vestiges in contrast with the elongated palps of the lower lip; the most remarkable modification is that of the greatly elongated tongue, with its spoon-like tip adapted for reaching nectar of flowers having deep-seated nectaries. For the same purpose, the mouth parts are modified in a moth (Fig. 3) to form a long proboscis, which lies curled up in a spiral beneath the head when not in use; in this case the proboscis is the modified accessory jaws, the remaining mouth parts, with the exception of the well-developed palps of the lower lip, being greatly reduced. In a blood-sucking insect, such as the female mosquito, all the mouth parts are well developed, but are very delicate and greatly lengthened and suited for piercing the skin. The greatest modification is found in the blow-fly proboscis, which is a soft, sucking tube, with no outward resemblance to the generalised plan, except for the palps of the accessory jaws. The mouth parts of insects (e.g., aphids) which feed on the nutrient sap of plants, just in the same way as mosquitoes do on blood, are modified for puncturing the tissues of plants; in such insects the upper lip is short, and both pairs of palps are atrophied, but the jaws and accessory jaws are greatly lengthened in the form of bristle-like stylets, which lie in a groove along the equally lengthened lower lip (Fig. 3). The manner in which insects feed is of great importance in controlling them with insecticides, and the two types to bear in mind are those that chew their food and those that suck the sap of plants, reached by puncturing the tissues.
As already stated, the thorax consists of the three segments immediately behind the head, and carries the organs of locomotion; its three segments are distinct, and may be referred to, respectively, as the fore, middle, and hind thorax. The cuticle of each thoracic segment consists of a number of chitinised plates connected by membranous areas; these plates are arranged in three series—the back or dorsal; the lower or ventral, forming the sternum; and the lateral, or side-pieces, connecting the dorsal and ventral ones.
At the lower surface of each thoracic segment is attached a pair of legs, the members of each pair being separated by the sternum of the segment to which they belong. The presence of three pairs of legs is a character by which insects can be distinguished from all other animals; indeed, on account of this feature, insects are sometimes called the hexapods, or six-legged animals. Each leg is covered by a continuation of the body cuticle, and is five-jointed; the first two joints at the attachment to the body are small; the next two are long, and form the greater part of the limb; while the fifth, or foot, consists of a varying number of small joints, the terminal one bearing a pair of claws.
In the typical winged insects there are two pairs of wings: one pair attached to the middle thorax, and the other to the hind thorax; owing to the development of muscles controlling flight, the middle and hind thorax of winged insects are usually better developed than the fore thorax; this is especially noticeable in the thorax of two-winged flies (daddy-long-legs and blow-flies), where the hind wings are reduced to vestiges, the power of flight being thus confined to the middle thorax, which forms by far the greater portion of the whole thorax.
Each wing, arising from the junction of the dorsal and lateral thoracic plates, is a bag-like extension of the cuticle, flattened leaf-like, so as to form a double flexible membrane. The wing membrane is supported by several ribs or veins, which may be very numerous (grasshopper) or few (aphid), while the fore edge, where it cuts the air in flight, is bordered by a stouter vein, ensuring rigidity. The fore and hind wings of some insects work independently, but in agreement of movement, while in others the fore and hind wings of each side are coupled along their adjoining margins, giving greater rigidity during flight.
The abdomen of insects consists of a varying number of visible segments; each segment is covered by an upper and lower chitinous plate connected by membrane, there being no side plates as are found in the thorax. There are no organs of locomotion (except in a very few cases), the only appendages being those connected with reproduction; the latter are well developed in the female weta, where the egg-laying apparatus, or ovipositor, projects blade-like from the apex of the abdomen. In very many insects, however, the external reproductive organs are not readily seen without special study.
All insects, from the largest to the most minute, contain internally a well-formed heart and a digestive, reproductive, respiratory, and nervous system (Fig. 3), while the spaces surrounding these organs are, for the most part, packed with a complex system of muscles. The heart is a delicate tube lying along the middle of the back or dorsal surface of the body, immediately under the skin, and extends almost from one end of the insect to the other; in an almost similar position, close to the lower or ventral surface of the body, the nervous system is situated, and consists of a chain of nerve centres, or ganglia, connected by a double nerve cord, the most anterior of these ganglia being in the head and forming the brain, the following three lying in the thorax, one to each segment, while the remainder are confined to the abdomen, one ganglion to each segment, as in the thorax. In many insects the number of nerve centres is reduced, owing to the fusion of two or more. The reproductive organs are located in the abdomen.
The digestive system consists of a tube (Fig. 3), with its appendages, opening at the mouth and at the posterior end of the body; this alimentary canal may be straight and simple, or convoluted and complex, according to the insect and the nature of its food. Respiration in insects is carried on by means of a system of air tubes (Fig. 3), which branch and re-branch to form an intricate system of delicate tubular airways, carrying the atmosphere to all tissues of the body; the main air tubes open at the surface by a series of breathing pores normally arranged along each side of the body, except on the head; these pores are best seen on a caterpillar or on the abdomen of adult insects.
No doubt owing to the endless assortment of sizes, from mere specks to giants of a few inches, a widespread idea has arisen, particularly in regard to such insects as have a general resemblance to one another, that the smaller individuals are the younger stages of the larger. Though gradation in size may be a sign of successive ages in certain insects, the presence of functioning wings denotes that growth has ceased; in the case of wingless insects, the characters of maturity may be less conspicuous. Although there may be at times a fairly wide range in size among fully-grown individuals of the one species, such variation is not due to age, but to certain factors influencing the insect during growth, such as the abundance or scarcity of food supply, and favourable or unfavourable climatic conditions. On the other hand, the sex to which an individual belongs is often responsible for difference in size, males very frequently being smaller than females. Size, therefore, is by no means a sign of age, and the smaller winged insects must not be regarded as the young of the larger ones, no matter how close is the resemblance.
Insects, with the exception of certain species giving birth to living young, are reproduced from eggs laid by the females; with few exceptions, the latter take no further interest in the eggs beyond placing them in surroundings offering the most favourable conditions for their well-being, and a sufficient food supply for the forthcoming young; each egg is protected by a delicate shell, through which the young insect makes its way on hatching.
On emerging from the egg, the young insect commences to feed and grow in size, until very soon a stage is reached when the cuticle or shell becomes too small for the enclosed insect; a fluid then collects between the cuticle and the underlying skin, and a new and more roomy cuticle is secreted by the latter; on this process being completed, the old chitinous covering splits, and the insect withdraws itself. This moulting takes place several times, until the body is fully grown, when the cuticle formed at the last moult is retained by the now adult insect for the rest of its life.
The different stages through which an insect passes from egg to adult constitute its life history, or life cycle, and the relation of the latter to the seasons, its seasonal history. According to the species, a full twelve months or even more may be necessary for the complete life cycle, or the cycle may be repeated several times within the year; when the cycle occupies twelve months, the insect is single-brooded; but two, three, or four-brooded, etc., when the cycle is repeated two, three, or four times, respectively, in the year. Climatic and food-supply conditions have a distinct influence on the number of broods, the one species in many cases being single-brooded in colder, and two or three-brooded in warmer climates. During the winter, when the temperature is low enough, insects are more or less dormant in some stage of their life cycle; such a state is the period of hibernation.
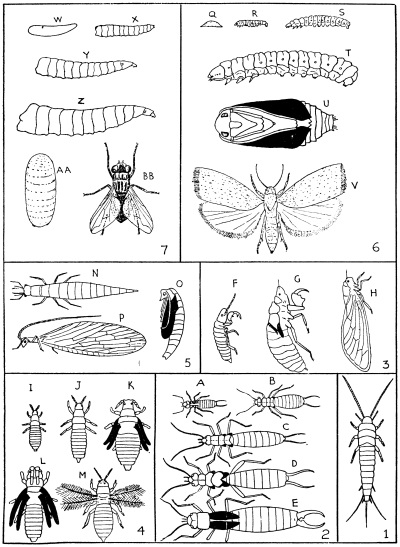
FIGURE 4.
1, Silverfish. 2, Earwig; a, young larva; b–d, later stages; e, adult. 3, Cicada; f, young larva; g, resting stage prior to emergence of adult; h, adult. 4, Thrips; i and j, larvae; k, first stage pupa; l, second stage pupa; m, adult. 5, Aphis-lion; n, larva; o, pupa; p, adult. 6, Moth; q, egg; r–t, larvae; u, pupa; v, adult. 7, House-fly; w, egg; x–z, larvae; aa, puparium; bb, adult. NOTE: Developing wings shown in black.
All insects do not follow the same method of development from egg to adult, and the adaptations of structure and habit are many and varied as well as simple and complex. Species having a complex development, during which they pass through stages, each differing in form from its predecessor, undergo what is known as a metamorphosis; contrasted with such insects are those developing in a simple manner without pronounced differences in the form of successive stages, the young resembling the adult in most features except size and maturity—these insects are without a metamorphosis. Intermediate between these two extremes are other insects with a partial metamorphosis.
A consideration of the life cycle of some common insects will serve to illustrate the principles of development discussed above. Firstly, will be taken examples of complex development or complete metamorphosis; secondly, examples of simple development or absence of metamorphosis, followed by a review of species having a partial metamorphosis, thus linking the first two types.
A convenient type of insect undergoing a complete metamorphosis is any common moth (Fig. 4); one of the most suitable, most easily obtained in all stages and commonest in any part of the country from spring to autumn, is the magpie moth (Nyctemera annulata) and its caterpillar, the “woolly bear.” The moth, unlike most of its kind, is a day-flying species, and is very conspicuous owing to its black colour relieved by white wing spots, and orange-yellow bands on the abdomen; the equally conspicuous caterpillar, feeding on groundsel, ragwort and cineraria, is black, with a very hairy body marked with narrow brick-red lines.
The eggs are laid in clusters by the female moth on the under side of the leaves of the caterpillars’ food-plant; at first the eggs are of a pale green colour, but assume a darker yellowish tint within a few hours, and finally a leaden colour some time later. These colour changes are due to the developing embryo, and just before the young insect (the caterpillar in this case) hatches, its outline as it lies curled within the egg is easily seen through the transparent egg-shell; near the top of the egg is a black spot marking the position of the caterpillar’s head, while the numerous delicate black lines below the egg surface are the black hairs with which the caterpillar is clothed. According to temperature and humidity, the incubation period—that is, the period between egg-laying and the hatching of the young caterpillar—varies from eight days to three weeks. The process of hatching occupies about two hours, the young insect using its jaws to eat an exit hole through the egg. The caterpillar stage—indeed, the first stage of all insects—is known as the larva.
At first the larva of the magpie moth, measuring about one-sixteenth of an inch long, is pale yellow in colour, except for the black head and hairs clothing the body; very soon, however, the body becomes characteristically black, and develops the reddish lines. During growth the larva feeds continuously day and night, undergoing from five to ten moults before becoming fully grown. During a moult the cuticle of the head is cast separately from that of the body.
The body of the larva is worm-like, not only in general form, but also in its segmented appearance; it is, however, a very different animal from a worm. The larva has a distinct head, a pair of eyes, and short antennæ, and a set of mouth parts, similar to those of the weta or grasshopper, well adapted for devouring foliage; the first three segments behind the head correspond to the thorax of the moth, and each bears a pair of short feet; the remaining segments are those of the abdomen, and have no true feet, but six pairs of sucker-like appendages called pro-legs. The number of pro-legs varies from four to six pairs, according to the species of moth, and are found only on the larva.
The time occupied by larval development of the magpie moth varies from forty to eighty days in summer and autumn; but if winter intervenes, causing the larvæ to hibernate before completing their development, the larval period may be as long as two hundred and forty-eight days; normally this insect hibernates in the larval state, completing its development during the following spring. Throughout winter the larvæ hibernate singly or in colonies under loose bark, in leaf axils, or any suitable crevice.
The fully-grown larva measures about one and a-half inches long. Prior to the final moult it ceases to feed, and wanders in search of a suitable place in which to undergo the next transformation, usually among stones, rubbish, or under loose bark, etc. There it spins a white silken cocoon, among the strands of which are entangled the long black body hairs; herein the larva undergoes the final moult, the cast cuticle being easily seen at one end inside the cocoon.
The insect, however, has now assumed a form quite different from that of the larva; this form is the chrysalis or pupa, and as such is incapable of locomotion and feeding. The pupa measures about three-quarters of an inch long, is yellowish at first, but soon becomes black with yellow markings, while the form of the future moth (head, antennæ, thorax, legs, wings and abdomen) can be traced on the pupal cuticle. After from about two to five weeks, the pupa opens by a cross-shaped slit on the back just behind the head, and the moth draws itself out. At first the moth is comparatively helpless after having been confined within the limited space of the pupal cuticle; soon, however, the body hardens, the wings smooth out, and the insect is ready for flight.
Metamorphosis is carried to a much higher state of perfection in the case of such insects as blowflies and houseflies (Fig. 4). The larva, or maggot, is without any external sign of head and legs, though these, together with the wings of the future fly, develop from rudiments within the body of the maggot. At the final moult the larval cuticle is not discarded, as in the case of the moth, but hardens to form a case—the puparium—within which the pupa lies.
The life-cycle of the magpie moth is illustrative of the principles of metamorphosis characterising the development of a great many insects, such as all moths and butterflies, beetles, flies, bees and wasps, etc.; but, although the general characters of the larva, pupa, and adult moth are common, with but slight variation, to corresponding stages of moths and butterflies as a whole, these stages in other insects, though readily recognised, have their own characteristics.
Outstanding features in a life-cycle involving metamorphosis are that growth takes place only in the larval state, and that the insect parades through life in different guises—egg, larva, pupa, and adult—each with its own peculiarity of habit and form, although the adult and pupa resemble one another much more than do the adult and larva; but no matter how dissimilar the larva, pupa, and adult may outwardly seem, structures common to them all may be traced throughout. Make, for example, a comparative study of the larva, pupa, and moth of the magpie moth; the head, thorax, and abdomen can be seen in each stage, while counterparts of the larval antennæ, eyes, mouth-parts and feet persist in the moth, though more or less profoundly modified during pupal transformation. Although there are no external signs of wings in the larva, these appendages are developing, nevertheless, in concealed “pockets” within the larval thorax, and, at the time of pupal formation, become extruded and lie ensheathed with the legs and antennæ in the pupal cuticle along the sides of the pupal body. Apart from these changes, the larval mouth parts undergo a most profound metamorphosis; apparently, though there is no similarity between the long “tongue” or proboscis of the moth and the jaws and accessory jaws of the caterpillar, the proboscis, adapted for sipping the nectar of flowers, is nothing but the accessory jaws of the leaf-chewing larva greatly elongated; with the exception of the palps of the accessory jaws, the other larval mouth parts are either absent in the moth or reduced to vestiges.
In the case of insects that develop without a metamorphosis, the life-cycle is one of comparative simplicity. An example of such an insect is the so-called “silverfish” (Lepisma saccharina), common in dwellings, especially in damp places, dark and dusty corners, flour and sugar bins, while not uncommonly it causes some considerable damage by devouring the paste and glaze from wallpapers and the binding and leaves of books.
The silverfish (Fig. 4), wingless throughout life, measures about one-quarter of an inch long when full grown; it is silver-white in colour, due to a clothing of glistening scales that rub off as a silky powder when the insect is handled. It glides rapidly about, especially after dark, and is one of the most primitive insects, there being minute leg-like processes attached in pairs to the under side of the abdomen; the normal thoracic legs are well developed. The body is wedge-shaped, tapering to the posterior end, from which three tail-like appendages project, while anteriorly a pair of long, delicate antennæ arises from the head.
All stages of the silverfish, from the minute, freshly-hatched individuals to fully-grown ones, may be found in the one place, the smaller ones being immature developing stages. In the case of another species allied to the common silverfish, the female lays from six to ten eggs at one time in sheltered crevices, and the young hatch forty-five to sixty days later, when the temperature ranges from 65 degrees to 68 degrees Fahrenheit.
Unlike the moth larva, that of the silverfish throughout its growth resembles the adult both in habit and form, the only marked differences being that of size and the absence of the abdominal leg-like appendages. During growth several moults take place, and at the final one the adult appears with all its characteristics. Some species take two years to reach maturity. In this type of insect there is, therefore, no pupal or resting stage, and the larval habits and food are the same as those of the adult insect, while there is but little difference in structure throughout all the stages.
There are many winged insects (e.g., cockroaches, crickets and earwigs) that show a slight advance toward a metamorphosis. Though their larvæ differ from the adults principally in the absence of wings, there are stages between the younger larvæ and the adults in which the wing rudiments appear. These rudiments first appear after one of the moults as small bud-like structures on each side of the thorax (earwig, Fig. 4), becoming larger after each succeeding moult, when the developing wings may be seen enclosed in a sheath of the cuticle; at the final moult the wings, no longer enclosed in their coverings, straighten out and become functional. A very pronounced difference is here noted between the wing development of such insects and that of a moth, in that the wing rudiments of the former develop externally and those of the latter internally.
A decided advance toward a metamorphosis is exhibited by insects known as thrips (Fig. 4). Though readily overlooked on account of their minute size (one-twenty-fourth of an inch and less), they are nevertheless conspicuous on green foliage and white flowers owing to their blackish or yellowish colour. Thrips, when magnified, are easily recognised by their peculiar wings; each is feather-like, being formed of a narrow rib-like membrane clothed along the margins with long and delicate stiff hairs. Thrips’ eggs are laid upon the plant surface or within the tissues, according to the species, and are very minute (about one-twenty-third of an inch long). The larvæ puncture the plant tissues and feed upon the juices just as do the parents, which they resemble in general form, except that there are no wings and the antennæ are very short and the eyes small. There are two or three larval moults, after which the insect is more like the adult, though still resembling the larva. It now differs from the latter, however, in the antennæ being considerably shortened, and in the appearance of a pair of finger-like processes on each side of the body attached to the thorax and lying along the sides of the abdomen; these processes are the sheaths enclosing the wing rudiments of the future adult. The insect again moults, changing to a form resembling the preceding stage in many respects, but differing in the wing sheaths being much longer, and in having the antennæ, enclosed in sheaths of cuticle, turned back over the head. Although during these two stages the insect is capable of moving about, it is nevertheless sluggish and does not feed; from this second semi-quiescent stage the adult emerges. In the thrip’s cycle, therefore, although the habits of the larva and adult are similar, the presence of the two intermediate semi-quiescent stages, during which feeding ceases, shows a decided advance toward a true metamorphosis and represents a pupal stage.
In the case of those insects not involved by a metamorphosis, as discussed above, the structure and habit of both adult and the immature stages differ but little, the development of wings being the principal change, except in the case of the thrips, where there is a definite tendency toward a pupa. However, passing on to a consideration of the common cicada (wrongly called a locust), a change in both structure and habit occurs during the life-cycle, the immature stages being adapted to a subterranean life, while the winged adult frequents the foliage of trees; all stages agree, however, in puncturing plant tissues with their proboscis and sucking up the nutrient juices from the roots by the larva and from the stems and leaves by the adult.
The female cicada (Fig. 4) lays its eggs in colonies beneath the young bark of trees and shrubs; the larvæ, on hatching, drop to the ground, into which they burrow; the antennæ and soft body are comparatively long, while the fore legs are greatly modified for grasping plant roots and as digging tools. After a number of moults, the body shortens, the antennæ come to resemble those of the adult, and the rudiments of the wings appear. Growth and the activities of the developing insect continue until finally the larva constructs an earthen underground chamber, in which it lies torpid until ready to undergo the final moult; in this inactive state, though still resembling the later larval stages, the insect corresponds to the pupa of the moth. For the final moult the pupa leaves the ground, crawls up some support (a tree trunk or post), where the winged adult emerges, leaving the empty pupal husk attached to the support. Besides the change in habit and the possession of functional wings, the adult cicada differs in many structural features from the immature stages. Outstanding differences are the normal fore legs, the development of a “voice-box” in the male, and an ovipositor in the female.
An insect that shows some linkage between those having a true metamorphosis and those having a partial metamorphosis is the aphis-lion (Micromus tasmaniæ), though undergoing a true metamorphosis itself. The larvæ are predaceous and feed upon aphids (Fig. 4). Its larva, pupa, and adult are distinct forms, as in the moth, but the larva is not of the specialised caterpillar or grub type, rather resembling in general appearance the silverfish, or the type of young larva peculiar to such insects as the earwig or thrips before the wing rudiments develop. Furthermore, the pupa, though one in the strict sense, is capable of great freedom of movement, its head, mouth-parts, antennæ, legs and wings, ensheathed by the cuticle, being freely movable, and not rigidly attached to the body.
A review of the early larval stages of the earwig, thrips and cicada, prior to wing development, and of the aphis-lion larva, shows a conformity to a generalised type exemplified by the primitive silverfish. On the other hand, the moth caterpillar exhibits another larval type more highly specialised, though still retaining a modified semblance to the silverfish type, while specialisation is carried to the highest degree in the blowfly maggot, where all outward sign of the primitive larval type is lost. Regarding the pupæ, there are three types; the most simple is the free pupa, like that of the aphis-lion, and some moths, beetles, etc., where the appendages are freely movable. The most complex is the pupa of the blowfly, enclosed in its puparium, while intermediate between these two extremes are many moth pupæ that have the appendages firmly attached to the body, but nevertheless visible.
The term “sucking insect” is applied to all insects that have the mouth parts modified as delicate stylets, by means of which the plant tissues are punctured and the nutrient sap sucked up. Not only may such insects weaken the infested plants, but they also cause the destruction of chlorophyll, interfere with the normal functioning of the stomata, and have a toxic effect upon the tissues; further, many serious plant diseases are carried and spread by sucking insects, whilst the punctures made when feeding may allow the entry of disease spores.
Among sap-sucking insects are scale insects, mealy-bugs, aphids, leaf-hoppers, white-flies, thrips, etc. Infestation by most of these insects (especially in the case of scale insects, mealy-bugs, and aphids) is very often detected by the sticky nature and blackened appearance of the plants; this is due to the fact that the insects excrete a sweet, sticky substance known as “honey-dew,” which collects on the foliage and branches, whilst upon it grows a black, sooty mould.
Scale insects and mealy-bugs, collectively known as coccids, are of very great economic importance on account, not only of their widespread depredations upon plants, few being free from infestation, but also of the commercial value of some species—e.g., in the production of lac, cochineal, Chinese wax, etc.; it is with the injurious forms that the New Zealand horticulturist is concerned. The term “scale insects” is derived from the appearance of many of the species that are protected by a scale-like covering, which forms a conspicuous scaly incrustation when a plant is heavily infested.
Of the several kinds of insects injurious to vegetation, the coccids as a family are undoubtedly of major importance, because they infest not one group, or allied group, of plants, as do so many other injurious insects, but an extensive range of widely different plants. Some coccids are much more injurious than others, the San José Scale, for example, having a very virulent toxic influence, while the Greedy Scale may cause but little damage, even when the plant is completely encrusted by it; further, some plants may be more susceptible to injury than others by the same species of coccid.
Coccids, as a whole, are highly specialised insects, and among themselves exhibit a great variety of forms. Throughout the group the sexes differ to a marked degree. The adult males, which vary but little in all the coccids, are usually minute, and, with few exceptions, two-winged (Fig. 5); none has mouth parts, these appendages having become atrophied during metamorphosis, which is complete, while many have one or more hair-like tail appendages. On the other hand, females are never winged; some are comparatively large; all have well-developed mouth parts throughout life, and undergo incomplete metamorphosis, while in many forms the legs and antennæ are lost before maturity.
In all cases coccids secrete a protective covering, which assumes different forms; this fact, together with the chief methods of female development, is utilised for the purpose of this work to arrange the coccids under three main types as follows:—
1. Less Specialised Forms.—Examples are the mealy-bugs and cottony-cushion scale, which belong to the more generalised or least specialised representatives. The protective body covering is in the form of a powdery or mealy secretion; the legs and antennæ are retained throughout life, and the insect remains freely mobile.
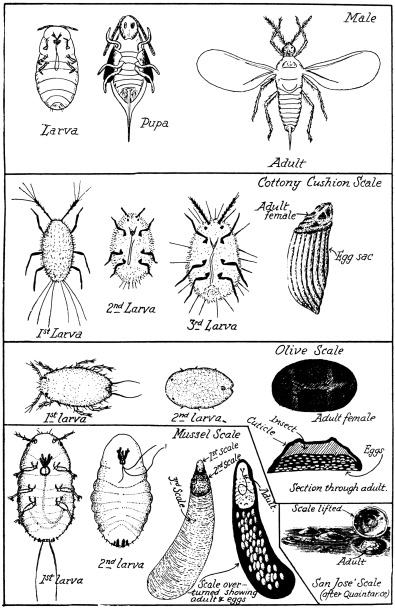
A typical-form life-cycle may be studied in that of the cottony-cushion scale (Figs. 5 and 6a). During development the female insect passes through three larval stages; each of these stages is, on the whole, similar, except for size and minor structural changes, and the white powdery secretion that covers the reddish body of the adult.
2. Intermediate Forms.—An example is the olive scale (Fig. 5). In such forms there is a tendency to specialisation, owing to more or less sedentary habits in later life, and protection is afforded by a thickening and toughening of the cuticle on the upper surface of the body. Unlike the cottony-cushion scale, the female olive scale passes through two larval stages; the minute first stage larva is active and very flat; it soon settles upon a leaf and commences to feed, when it becomes much flatter and a little larger; the second stage differs from the first in size and in the development of a dorsal longitudinal ridge, which eventually forms the cross-bar of the two transverse ridges that are characteristic of the third or adult stage, when the insect swells and assumes the shape of the mature form. After settling in the first larval stage, the insect becomes very sluggish, and does not move, except to migrate, as most do, from the leaves to the twigs, there to take up a permanent position. The legs and antennæ are retained throughout life, but in the adult are functionless, being folded against the body; in some species of intermediate forms the appendages become atrophied during development. In the olive scale, and related forms, the toughened cuticle not only serves as a protection to the insect, but also as a receptacle for the eggs (Fig. 5); as these are laid and increase in numbers, the body of the parent diminishes and is crowded against the dome-shaped cuticle.
3. Specialised Forms.—The apple mussel-scale (Figs. 5 and 7, Nos. 2 and 6) is a representative of this group, the members of which are markedly specialised, the legs and antennæ of the adult female becoming completely atrophied during development, and the shape of the body profoundly altered; protection is afforded by a scale-like covering not attached to the body. In the mussel-scale development there are two larval stages: the first, like all coccids, has the legs and antennæ well developed and is active.
On settling to feed, this first larva commences to produce a covering of white threads that mat together to form the first scale; the second stage larva presents profound changes in the absence of legs and antennæ, while the body has become pear-shaped, the head, thorax and abdomen seeming as one; a second more waxy scale is now formed. After a second moult, the adult appears, and resembles the second stage larva in form; the adult constructs a third scale, very much larger than the earlier ones, to which it remains attached by its anterior end.
Though many of the specialised coccids form elongate scales, as in the case of the mussel-scale, numerous others construct circular scales, as does the San José (Fig. 5); in the latter, the second and third scales are constructed round the first, so that the first and second appear as pimple-like structures in the centre, or slightly to one side of the completed covering. As with the olive scale, the covering of the specialised forms serves as a receptacle for the eggs (Fig. 5).
Some of the more important coccids occurring in New Zealand will now be discussed.
Cottony Cushion Scale (Icerya purchasi).—This insect (Fig. 6a) is a native of Australia, but has now become established in many other countries, including New Zealand. For a time it was a serious pest of citrus, until the introduction and establishment of its natural enemy, the ladybird beetle (Novius cardinalis).
The adult female is more or less oval, and covered with a yellowish powder, partly concealing the reddish-brown ground colour and dark spots along the sides of the body; the legs are black. A characteristic feature is the white corrugated egg-sac attached to the end of the body (Fig. 5). As the eggs are laid, this sac increases in size, until it may measure fully 2 ½ times the length of the parent, which becomes tilted up. The eggs are orange-yellow, and as many as 800 may be produced by a single female. The eggs hatch in about a fortnight during summer, and the period of development to the adult ranges from three to five months. The larvæ most frequently congregate along the mid-ribs of leaves, and as development advances they usually migrate to the twigs and branches. There are two generations each year. A considerable variety of plants is attacked by this insect, chief among which are citrus, acacia, gorse, wattle, and Douglas fir.
Control is effected by the agency of the ladybird, but epidemics sometimes occur with which the beetle cannot immediately cope; in such a case fumigation in the glass-house, or spraying with red oil in the open, should be resorted to.
Mealy Bugs.—Mealy bugs are characterised in the female by the nature of the waxy protective secretion which forms a powdery meal-like covering over the body, but is developed as a fringe of leg-like processes at the side (Fig. 6b); these processes at the posterior end of the insect may be prolonged as longer or shorter tail-like appendages in some species, or they may be no longer than those fringing the body margins in others. Immediately after each moult the larvæ are devoid of mealy covering and lateral processes, which are secreted anew each time the cuticle is shed. In a mealy bug colony are numerous small, narrow cocoons, in each of which a developing male insect lies.
Most mealy bugs produce eggs, which are laid in a spacious, cottony sac secreted at the posterior end of the female; the female insects, egg sacs, and male cocoons together form characteristic woolly masses on infested plants.
The injury caused by mealy bugs may be considerable, not only through the drainage of plant sap, but also owing to the production of honey-dew and its consequent sooty mould. All parts of plants are subject to mealy bug attack, and the insects are frequently attended by ants.

FIG. 6.
(a) Cottony cushion scale. (b) Mealy bug. (c) The black olive scale. (d) Gum tree scale: On right, females on twig; upper left, male scales; lower left, the ladybird beetle; centre, scales destroyed by beetle. (e) Hemispherical scale. (f) Fruit lecanium scale.
Photographs by W. C. Davies, Cawthron Institute.
Mealy bugs are controlled to a great extent by natural enemies, among which are the Tasmanian lace wing (Micromus tasmaniæ) and the Cryptolæmus ladybird (Cryptolæmus montrouzieri), but the influence of these is insufficient for commercial purposes. Attempts are now being made at the Cawthron Institute, Nelson, to establish other parasites recently imported from California.
Control under glass is effective by means of fumigation, but in the open is a more difficult matter, though red oil and lime-sulphur give some satisfactory results, together with the practice of removing rough bark on trees where the insects hibernate. In New Zealand are several species of mealy bugs, of which the following are of interest to the horticulturist:—
Long-tailed Mealy Bug (Pseudococcus adonidum).—This species is readily recognised by the long tail-like appendages of the female. It is widely distributed and commonly met with under glass, where it infests almost any plant; in the warmer and moister districts of the Dominion it occurs out of doors. Its list of host plants is a lengthy one, and includes grape vine, passion vine, wistaria, fig, oleander, Phormium, cineraria, begonia, apple, plum, palms, ferns, etc. Considerable injury may be caused by the insect when it occurs in dense masses on the under side of foliage and upon young, succulent growth.
No eggs are produced by this insect, the young being born alive; the production of young lasts for a period of from two to three weeks at the rate of about twelve each day; the time taken to reach maturity varies considerably, according to climatic conditions, the range being from one to three months. There are comparatively few generations each year out of doors, but under glass there may be several.
Citrophilus Mealy Bug (Pseudococcus gahani).—In New Zealand this species is met with on grape vines and begonia in glass-houses, where it becomes epidemic if left uncontrolled; out of doors it infests apple and potato, and no doubt other plants are attacked. It is characterised by the mealy covering being coarse and distributed unevenly over the body, while the marginal fringe is short, the processes being comparatively thick, particularly the tail-like ones, which are much shorter than the body, though conspicuous.
Egg-laying covers a period of about two weeks, from 394 to 679 eggs being deposited by each female; development to the adult is completed in about six weeks, though this will vary according to the conditions. In California four generations in the year have been noted.
Apple Mealy Bugs (Pseudococcus maritimus and P. comstocki).—Both these species occur upon apple, pear and potato in New Zealand, the former species originating in America, and the latter in Japan; the injury to the host itself is not severe, but the presence of these insects on the fruit is responsible for apples and pears being rejected for export.
Both species are very similar in appearance, and are of the short-tailed type; they differ from the citrophilus mealy bug in having the mealy covering evenly distributed over the body, while the marginal fringe is delicate and thread-like. The eggs hatch in from one to three weeks, and the larvæ migrate freely, the insects reaching maturity one or two months later, according to climatic conditions. In the open the winter is passed in the egg stage, but under glass or in mild climates activity among the different stages occurs throughout the year.
Apart from apple and pear, these insects have been recorded from many plants: Baker’s mealy bug (maritimus) on lemon, orange, walnut, willow, elder, ivy, iris; and Comstock’s mealy bug on citrus, elder, euonymus, gooseberry, grape, horse chestnut, hydrangea, mulberry, peach, persimmon, plum, poplar, wistaria.
The Gum Scale (Eriococcus coriaceus).—This is one of the most spectacularly destructive scale insects now established in the Dominion. It is a native of Australia, and its normal hosts are the several species of eucalyptus, though it is sometimes found on apricot and willow. A characteristic feature of infected eucalyptus is their blackened appearance, due to sooty mould growing on the copious honey-dew secreted by the scale.
On an infested twig or branch, the insects may be so closely packed as to conceal the bark (Fig. 6, d); each female lies in a pear-shaped sac of felted secretion, reddish-brown, tawny, or sometimes white in colour, measuring about three-twenty-fifths of an inch long, and having a circular aperture at one end. The enclosed insect is somewhat flattened, oval, and blood-red in colour; when crushed, it leaves a reddish and sticky smear. The developing males are to be found forming white patches of innumerable individuals on the tree trunks under the loose bark (Fig. 6, d).
The female is viviparous; during spring, mid-summer and autumn immense numbers of young are produced, which escape through the opening at one end of the female sac, and are carried long distances by the wind. These young insects first settle on the eucalypt leaves, whence they migrate, the females to take up their final position on the twigs and smaller branches, and the males to continue their development on the trunk of the tree.
The gum tree scale occurs throughout the districts east of the Southern Alps and in the vicinity of Nelson, in the South Island, and over the southern half of the North Island; it is, however, spreading rapidly northward.
This pest is held in control by means of the black-ladybird beetle (Rhizobius ventralis)—Fig. 6, d—which was imported for the purpose from Australia; birds such as the tui, wax-eye, fantail, blackbird and thrush congregate on infested trees and eat the insect.
Olive Scale (Saissetia oleæ).—This insect has a world-wide distribution, and is one of the most important pests of citrus in New Zealand, although it occurs on a wide range of plants; in all cases it infests the fruit, bark, and the under side of leaves. The host plants include citrus, apple, pear, apricot, plum, almond, fig, grape-vine, wistaria, pepper tree, oleander, holly, laurel, palms, camellia, rose.
The injury caused by the insect is not so much on account of its weakening influence upon the infested plants as of the fact that it copiously secretes honey-dew, so that black mould develops to a marked degree, necessitating the washing of herbaceous plants and fruit.
The adult female (Fig. 6, c) is hemispherical, and measures about one-fifth of an inch in diameter, a characteristic distinguishing feature being the three ridges forming the letter H on its upper surface (Fig. 5). According to age, the colour varies from brownish or greyish to jet black, the insect being conspicuous against the lighter background of bark or leaf; the small, immature individuals are light brown or yellowish, and almost flat.
In New Zealand the winter is passed in both egg and larval stages, though a few adults may be found at that time; on turning over what appears to be an adult, it will usually be found that the female has died and her place taken by numerous eggs (Fig. 5). The average number of eggs produced has been estimated at from 1,500 to 2,000 per female; at first the eggs are white, but prior to hatching they turn a deep orange-red. Development is slow, the adult state being reached about three months after time of hatching; egg laying commences about five weeks after maturity, and continues for a period of about six weeks. There is only one generation each year, and all stages may be met with on the one plant; the greatest activity occurs during the summer months. An important natural enemy of this scale is the steel-blue ladybird beetle (Orcus chalybæus), introduced from Australia.
Hemispherical Scale (Saissetia hemispherica).—This world-wide species is commonly met with in New Zealand, and, though not a serious pest, has a wide range of host plants, both in the open and under glass; some of the commoner hosts are citrus, fig, oleander, palms, japonica, camellia, asparagus, and orchids.
Both leaves and stems are infested by the insect, which resembles the olive scale (Fig. 6, e); from the latter it may be distinguished by its light brown colour and smooth surface, there being no ridges; the longest diameter of the adult female is one-seventh of an inch. Between 500 and 1,000 eggs are laid by each female, and the life-cycle is completed in about six months; the young insects settle along the main leaf-veins.
Turtle Scale (Coccus hesperidum).—This widely-distributed insect, though common in hot-houses and out of doors in the warmer parts of the Dominion, is not especially injurious, except for the copious honey-dew secreted and the consequent sooty mould; it occurs on holly, ivy, camellia, citrus, laurel, myrtle, oleander, and japonica.
The insect infests leaves and stems, and is especially abundant on succulent growth. The adult female is rather reddish-brown in colour, dome-shaped, but with the margins flattened on the host plant; on each side the margin is notched by a shallow depression, and there is a deeper one at one end; over the surface is a reticulation of ridges, resembling the pattern on the back of a turtle; fully-developed individuals measure from one-sixth to one-eighth inch in diameter. This species is viviparous, and development to the adult occupies about nine weeks; there may be three or four generations each year.
Fruit Lecanium Scale (Eulecanium corni).—This European insect is common throughout the Dominion, where occasionally it becomes epidemic and causes some temporary damage; with it are associated honey-dew and sooty mould. Among the plants infested are apricot, peach, nectarine, plum, pear, grape-vine, wistaria, raspberry, mulberry, blackberry, gooseberry, black currant, ferns.
Leaves and bark are infested, and a narrow twig may be partly encircled by the margins of the scale. The adult female (Fig. 6f) is oval and dome-shaped, some individuals measuring one-sixth of an inch in length; the surface is smooth, except toward the margins, parallel to which are some wrinkles. The general colour is dark brown, but just prior to egg-laying there are numerous transverse and longitudinal markings of a lighter colour over the surface. The winter is passed in the egg stage or as partly-grown young.
Another, but larger, species, closely resembling the preceding, and found on grape-vines, wistaria, eleagnus, etc., is Eulecanium berberidis. It is reddish-brown in colour, and measures up to one-third of an inch in length.
Golden Oak Scale (Asterolecanium variolosum).—This insect is very common upon English oak trees in parts of New Zealand. In many cases so badly are the trees infested, that they become sickly in appearance, and at times the greater part, or even the whole, of the tree is killed through the agency of the pest.
The individual scale (Fig. 7, 1) is more or less circular, and about one-sixteenth of an inch in diameter; it is of a greenish-yellow colour, with a narrow paler circumference, though some, with the exception of the rim, are partly or wholly brownish. Each scale forms and lies in a depression of the bark. The insect is viviparous. A minute parasite, Habrolepis dalmanni (note the exit holes made by the parasite during emergence from some of the scales shown in the photograph) has recently been established as a means of control and is proving effective.
Camellia Scale (Pulvinaria camelicola).—This European scale sometimes heavily infests camellias and euonymus in New Zealand, but is not a very serious pest, though more so in glass-houses than out of doors. After the female has produced her eggs, she drops off the plant, so that, though the latter shows evidence of injury, there may be no sign of the insect.
The adult female is oval and about one-third of an inch at its longest length; in shape it resembles a rather flattened turtle scale, but when laying eggs the body shrivels and numerous transverse wrinkles develop, although the margins of the scale remain smooth. There is at least one generation each year, and in warmer parts probably a second, which may reach maturity before winter or not till the following spring. The eggs are laid in an elongate, white, cottony sac secreted at one end of the female; this sac is sometimes as much as four to five times the length of the insect. The eggs continue to hatch over a period of from four to six weeks, and the larvæ rapidly spread; the latter settle along the leaf mid-rib, margin, or lower surface.
Apple Mussel Scale (Lepidosaphes ulmi).—The apple mussel scale is now established throughout the temperate regions of the world. It is commonly met with on apple, but has a long list of host plants, among which are pear, hawthorn, willow, poplar, gooseberry, and currant.
The insect (Fig. 7, Nos. 2 and 6) forms incrustations on bark and fruit, and is commonly met with at the stalk end of the apple; the individual scale is chocolate-brown in colour, is shaped like the shell of the salt water mussel—hence the name “mussel scale”—and when full grown measures one-eighth of an inch long.
A single female is capable of laying up to 700 eggs, in which stage the winter is passed. The eggs hatch in the spring, and the young insects swarm over the host plant in search of a suitable place to settle. A continuous warm spell of weather in the spring will allow all the eggs to hatch almost at one time, but alternating cold spells will retard development, so that emergences take place over a longer period. After emerging from the egg until maturity, when egg-laying again takes place, a period of three months elapses; the insect is a slow breeder, and produces only one brood a year in colder climates, but is two-brooded in warm districts, such as Auckland.
A small hymenopterous parasite (Aphelinus mytilaspidis), less than one-twenty-fifth of an inch long, attacks this scale, but does not serve as an efficient control; individual scales that have been killed by the parasite show a small hole through which the adult parasite has emerged. The most effective control is secured by treating infested trees with red oil or lime-sulphur during winter.
Cabbage Tree Scales (Leucaspis cordylinidis and Leucaspis stricta).—Cabbage trees and also New Zealand flax often have the leaves encrusted by the white masses of these two native scales. The adult female of one species (L. cordylinidis) measures one-eighth of an inch long, is very narrow and straight as a rule, and white in colour, except for the yellow anterior end (Fig. 7, 4). The other species (L. stricta) resembles the former, except that the adult is one-eleventh of an inch long, and has the anterior half blackish. In the case of ornamental cabbage trees and flax, control can be effected by removing all dead and scale-infested leaves, thus allowing access to sunlight.
San José Scale (Aspidiotus perniciosus).—Of all scale insects of major importance, the San José (Fig. 7, 5) is outstanding, in that it is one of the insects most destructive to deciduous trees and shrubs, a considerable number of which are liable to attack. It is of Chinese origin, and first came into prominence when it became established at San José, in California, hence its name. Owing to its small size, it is easily overlooked, except when epidemic, and is readily transported upon plants from one country to another.
The list of plants attacked is a long one, but the following may be mentioned:—Acacia, hawthorn, quince, privet, poplar, almond, apricot, cherry, plum, peach, pear, apple, gooseberry, currant, roses, willow, ash, elm.
The female San José scale is circular in outline, having a diameter of about one-twenty-fifth of an inch; in profile it has the form of a flat cone with a crater-like depression at the apex, in the centre of which lies a minute pimple-like prominence; the immature scales are smaller and whitish in colour, while the male scale is elongate-oval in outline, with the crater-like depression toward one end. The individual scales are greyish and are readily overlooked, but when well established upon a tree they form an incrustation giving a characteristic dull silver-grey appearance to the tree; bark, fruit and leaves are infested. A characteristic feature of San José scale infection is the discolouration of the plant tissues immediately surrounding each insect, which turn a distinct red or purple, giving at once an indication that this scale is present.
The winter is passed by the insect in almost a mature state; on the advent of spring, development to maturity continues, when, after mating, the females give birth to living young over a period of several weeks. The young reach maturity and commence to reproduce five to six weeks from birth, there being several generations in the course of a season. The average number of young produced by each female has been found to be about 400.
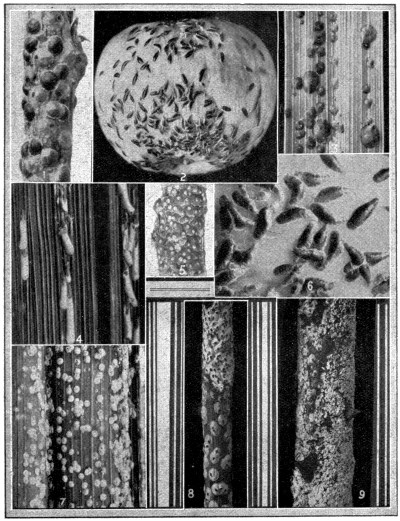
FIG. 7.
(1) Golden Oak Scale; (2) Apple Mussel Scale; (3) Black Scale; (4) Cabbage Tree Scale; (5) San José Scale; (6) Apple Mussel Scale; (7) Oleander Scale; (8) and (9) Rose Scale.
Photographs by W. C. Davies, Cawthron Institute.
Natural enemies in New Zealand are two species of hymenopterous parasites, Aphelinus fuscipennis and A. mytilaspidis, the latter also attacking the apple mussel scale. Ladybird beetles also feed upon the insect.
Control requires close attention, and can be effected by the application of lime-sulphur in the dormant season, when it is essential to apply a strong wash to kill off as many scales as possible before reproduction commences in the spring. At bud movement further applications are necessary to destroy the young insects.
Red Orange Scale (Chrysomphalus aurantii).—The red orange scale is distributed throughout the world, and is especially abundant in tropical and sub-tropical regions, the most southern limit being New Zealand. As a major pest it is peculiar to citrus, but infests to a minor extent other plants—e.g., plum, apple, pear, quince, grape, fig, euonymus and rose. So far it has been found only on citrus in New Zealand, it being well established in the Auckland province, and also in the South Island on Banks Peninsula. It is very often found on imported oranges and lemons.
This scale is a circular one, with a central pimple-like prominence, as in the case of the San José, but is flatter, about half as large again, and is of a characteristic reddish colour. The damage done to citrus trees by this insect is of a serious nature, as the entire tree or part of it may be killed in severe infestations. A characteristic feature of this species is that no honey-dew is secreted, and hence there is a total absence of sooty mould on infested trees.
Like the San José scale, the red scale is viviparous, and over-winters as partially mature adults, completing development in early spring, when the young insects make their appearance. An average of about 55 young is produced by each female, and development to maturity takes from two or two and a-half months; about one month later young are produced, and their production continues over a period of one or two months; climatic conditions, however, have a direct influence on development.
An important natural enemy is the steel blue ladybird (Orcus chalybæus), imported from Australia; but the most efficient control is cyanide fumigation, or spraying with red oil or lime-sulphur.
The Black Scale (Chrysomphalus rossi).—Foliage of palms, oleander and citrus is often infested by this reddish-black to black circular scale (Fig. 7, 3); it is almost flat, with a central whitish spot, and measures up to one-tenth of an inch in diameter; when many individuals are crowded together, their outline becomes irregular. This species is not especially injurious, though common.
Oleander Scale (Aspidiotus hederæ).—This cosmopolitan insect occurs on orchids, oleander, ivy, camellia, palms, citrus, coprosma, and karaka, infesting stems, leaves and fruit. In the case of citrus, this insect delays colouring of the fruit, which becomes blotched with yellow or green. The insect may be so numerous, that it completely covers the whole plant, giving to the latter a white appearance; this is due to the preponderance of white male scales, the female being slightly yellow, with a purplish tint.
The female scale is almost circular (Fig. 7, 7), having a diameter of from one-twenty-fifth of an inch to two-twenty-fifths of an inch, and is rather flat; the male is more oval and of the same size, and in both cases there is a central orange-yellow spot. The eggs are comparatively large, and hatch soon after being deposited. The insect reaches maturity in from four to six weeks.
Greedy Scale (Aspidiotus rapax).—This European insect is now widespread, and in New Zealand is common on apple, pear, quince, and wattle; it has a wide range of hosts. The adult female scale is convex and of a general grey colour, though sometimes yellowish. The winter is passed in all stages of development.
Rose Scale (Aulacaspis rosæ).—This is a very common insect, forming white incrustations on the bark of roses, briar, raspberry, loganberry, blackberry, and sometimes pear. The adult female (Fig. 7, 8), which is from one-twelfth of an inch to one-eighth of an inch in diameter, is rather thin and flat, circular or oval in outline, but irregular when crowded; the general colour is white or slightly yellowish. The male (Fig. 7, 9) differs, in being elongated and narrow. This insect can withstand severe winters, and is to be controlled by the use of red oil.
The small, soft-bodied plant-lice, or aphides, usually found forming dense colonies on all sorts of plants, are pests well known to every gardener; they attack plants by inserting into the tissues their delicate piercing mouth-parts, and drain the nutrient sap (Fig. 8, 1g). All parts of a plant may be infested, and the insects, owing to their ability to reproduce abundantly and rapidly, may destroy the plant, or at least injure it by stunting its growth, curling the leaves, or deforming the flowers and fruit. In many cases aphides copiously secrete honey-dew, upon which sooty mould grows, rendering the plant unsightly; on this honey-dew ants feed, and are frequently seen associated with aphides. Apart from their direct injurious effects, aphides are of outstanding importance, in that they transmit some of the most serious plant diseases. Of all the species occurring in New Zealand, only one species is supposed to be a native.
Most aphides live exposed upon the host plant (e.g., Rose Aphis), but some (e.g., Woolly Aphis) secrete a protective covering, while others cause a malformation of the plant tissues which form a partial protection as a semi-gall (e.g., Elm-leaf Aphis), or a complete protection as a true gall (e.g., Leaf-petiole Gall-aphis of Poplar).
Aphides present certain variations in structure, and, generally speaking, the one species presents four or five types (Fig. 8, 1): the asexual (parthenogenetic) wingless and winged females that give birth to living young (viviparous) in the absence of males, and the sexual forms, both males and females, the latter producing eggs (oviparous).
The best character by which the New Zealand aphides are to be recognised is to be found in the pair of longer or shorter horn-like processes, or “cornicles,” projecting from the upper surface of the abdomen; in some species, however, the “cornicles” are reduced and inconspicuous (e.g., Woolly Aphis), or altogether absent (e.g., Grape Phylloxera). The “cornicles” are frequently called “honey-tubes,” since for many years it was thought that they secreted the honey-dew; it has been shown, however, that the honey-dew is secreted from the rectum, and that the function of the “cornicles” is to secrete a waxy protective substance, which may take the form of a powder or woolly threads. The wings, when present, are membranous, the front pair being much larger than the hind ones, and when not in use usually close roof-like over the body.
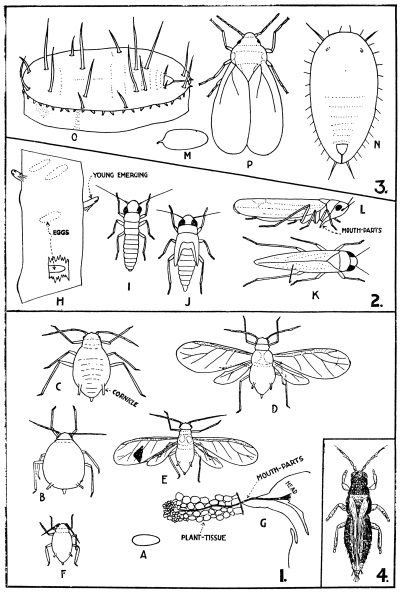
FIG. 8.
(1) Life History of an Aphis: A, egg; B, C, and F, wingless females; D, winged female; E, male; G, section of head and plant tissue to show method of attack. (2) Life History of a Leaf-Hopper: H, eggs under bark of twig; I, first stage hopper; J, later stage hopper with developing wings; K, adult from above; L, adult from side. (3) Life History of a White Fly: M, egg; N, first stage larva; O, pupal stage under scale covering; P, adult. (4) An adult Thrips.
In their life-histories and habits aphides present many variations, sometimes of considerable complexity, but fundamentally the processes are as follows:—Eggs are laid on the host plant during the autumn, and give rise to wingless females in the spring; these females (being asexual or parthenogenetic, since they reproduce without being fertilised) are viviparous, producing living forms similar to themselves. Some of these forms remain wingless, while others may develop wings, upon which a wider dispersal of the species depends, but in both cases such females are asexual and viviparous. Several such generations may develop until the autumn, when males and females appear, the latter being oviparous, producing the over-wintering eggs when fertilised by the males. Very often, however, the life-cycle is considerably complicated by the winged forms flying to other host plants and establishing there colonies differing in many respects from the parent stock; from these secondary hosts there is a return migration to the original species of plant. Again, the migrations may be restricted to different parts of the same plant, from the leaves or branches to the roots, for example. Most aphides are readily controlled by means of insecticides, such as nicotine-sulphate, or kerosene-emulsion. They are also very often held in check by natural enemies, such as aphis-lions, hover-flies, ladybirds, and numerous forms of hymenoptera. The following species are some of the commoner aphides met with in New Zealand:—
Black Peach—aphis (Aphis persicæ-niger).—From early spring, even before the foliage develops, this aphis may be found heavily infesting the young, succulent shoots of peach; it also occurs on cherry, plum and nectarine. The adult insects are black and the immature stages pale reddish-brown, dull brown, or lemon-yellow. During the winter the insect lies underground about the roots of the host plant, and thence migrates to the young growth in spring. At first only wingless forms are seen, but as the season advances the winged migratory aphides develop; at that time the foliage is so severely attacked that it becomes crumpled and functionless (Fig. 9, 1), while the developing fruit is distorted and rendered useless. The heat of the late summer destroys the aphides still on the foliage, but sufficient numbers descend underground for protection, where they live over winter.
Green Peach—aphis (Rhophalosiphum persicæ).—This aphid occurs on a wide range of plants, including the peach, and, as a rule, is most abundant during summer and autumn; as the name implies, the general colour is green, though some individuals are reddish or brownish-yellow; the wingless forms have black-tipped “cornicles,” and on the abdomen of the winged insects are dark markings.
Black Cherry—aphis or fly (Myzus cerasi).—This aphid has now a world-wide distribution. In New Zealand it has been found on cherry and plum, though in other countries its hosts include peaches, red and black currants, and cruciferous plants, such as common mustard, shepherd’s purse, etc. This species exudes copious honey-dew, upon which sooty mould develops, thus rendering fruit unfit for use. The principal injury, however, is due to the destruction of shoots and leaves, the latter frequently curling up when the insect clusters in dense colonies upon the infested plant. The complete life-cycle has not been followed under New Zealand conditions, but the shiny black eggs occur on the bark and buds of cherry trees during the winter. In spring the eggs hatch, and the insects, rapidly reproducing, attack the young shoots and leaves. Observers in other countries have noted that there is a summer migration of winged females to cruciferous plants, where colonies are established, and whence there is a return migration during the autumn to the original host. The wingless females are black, with part of the legs yellow, while the young individuals are pale in colour; the winged females have a green abdomen, from which arise the black “honey-tubes.” Since all the over-wintering eggs have hatched by the time the buds open, the insect can be then controlled by applications of nicotine-sulphate.
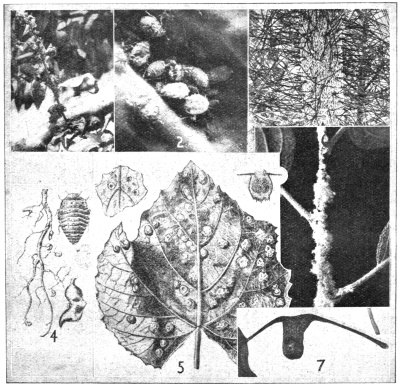
FIG. 9.
(1) Peach leaves attacked by Black Peach aphis. (2) Colony of Cabbage aphis on leaf. (3) Stem of insignis pine attacked by Chermes. (4) Grape Phylloxera and galls on vine roots. (5) Grape Phylloxera galls on vine leaf. (6) Woolly aphis on apple twig. (7) Galls of Poplar aphis. (Figs. 1, 2 and 6 by W. C. Davies; Fig. 4. after U.S. Dept. Agric.; Fig. 5. after N.Z. Dept. Agric.)
Cabbage Aphis (Brevicoryne brassicæ).—The cabbage aphis, or cabbage green fly, is widely distributed throughout the world, and has become a serious pest in New Zealand, causing considerable damage to cruciferous crops; it infests rape, turnip, cabbage, Brussels sprouts, cauliflower, as well as related weeds, such as wild mustard, shepherd’s purse and watercress. Most damage is done during dry seasons, when the plants succumb more readily to attack; if the insects are numerous, they cause the leaves to curl, and give a greyish appearance to infested plants, which may become flaccid and sticky from the copious honey-dew of the insect. The wingless forms are bluish in colour and coated with a greyish powder, but the winged females have the head and thorax black and the abdomen greenish (Fig. 9, 2). In New Zealand all stages may be found throughout the year on winter crucifers or on weeds, though reproduction is retarded during the winter; in the spring the winged females fly to young crops. In very cold climates eggs are laid in the autumn, and these survive the winter. The cabbage aphis is attacked by a number of parasites, and usually the brownish empty shells of a large number that have been destroyed by a small parasite are to be found at any time; other important enemies are the hover-flies, the eleven-spotted ladybird beetle, and the Tasmanian aphis-lion. The insect can be controlled by spraying with nicotine-sulphate to which soap has been added.
Pine Tree Chermes (Chermes pini).—This is a widely-distributed species, occurring upon both Austrian and insignis pine in New Zealand. The insect lives in colonies upon the cones, twigs and branches, as well as around the bases of the needles; each aphis exudes a woolly covering, which forms conspicuous white masses when the trees are heavily infested (Fig. 9, 3). Young trees seem to be the more subject to infestation, from which they may recover as they grow, but some damage is caused by the insect by a weakening of the trees, especially where grown in unsuitable localities. It is frequently noticed that individual trees in a plantation are heavily infested, while adjacent trees of the same species are not. The wingless form of the insect, covered by its mat of white threads, is brownish in colour and ornamented with numerous dark spots; there are no “honey-tubes” on the abdomen. The life-cycle of this insect becomes complicated, when it develops on two types of conifers; in the latter case the primary host is a species of spruce upon which the insect forms galls, and the secondary host may be larch, Douglas fir or pine, upon which gall formation is unusual. So far as is known, only the pine-infesting form of the aphis occurs in New Zealand.
Grape Phylloxera (Phylloxera vastatrix).—This destructive aphis, sometimes called the grape louse, is a native of North America, where it normally infests grape vines. It was accidentally introduced into the grape-growing districts of France, where it became very destructive. It later made its appearance in New Zealand. The insect infests both the leaves and roots of grape vines, the root-feeding stages being the most destructive, in consequence of which vines are now grown on resistant root stocks. The leaf-infesting stages of the insect cause pocket-like galls to form, which open on the upper surface of the leaf by a narrow aperture concealed under a tuft of delicate hairs (Fig. 9, 5). In each gall the aphid matures and deposits several hundreds of eggs, from which wingless females hatch; these wander to other leaves, and each insect forms a new gall for itself. Several generations develop thus, but later many of the offspring migrate underground and join the root-infesting colonies. The irritation set up by the latter causes yellow flabby nodules to develop on the roots (Fig. 9, 4). These nodules, or galls, later decay. The root-feeding aphides are wingless, and reproduce by means of eggs for several generations. Although they may go on developing thus for many years, it usually happens that, toward autumn, some of the insects transform to winged females, which fly to other vines or are carried thence by the wind. There each female feeds on the lower leaf surface, and deposits two kinds of eggs, some larger and some smaller; from the larger develop wingless females, and from the smaller wingless males, which are unable to feed. After fertilisation, each of these females deposits a single egg upon the older bark of the vine; such eggs do not hatch until the spring, when they give rise to the wingless females that start the galls on the leaves. Control depends on the use of phylloxera-resistant stocks, since it is from the root colonies of the aphis that the foliage is re-infested in the spring. An important feature is to prevent the scion from sending down roots where the union of the scion and root stock is close to the soil; if such scion roots form, they should be cut away and the soil removed from the union.
Rose Aphis (Macrosiphum rosæ).—The rose aphis is perhaps one of the best-known insects of the garden, mainly owing to its prevalence upon the young growth of all kinds of roses; it sometimes occurs on apple, tomato and rhododendrons. In a colony some of the insects are pink, and others bright green, though in the winged forms the head, antennæ, thorax, a row of spots on each side of the abdomen, and the “honey-tubes” are black; in both winged and wingless forms the eyes are red. In the case of severe infestations, plant growth is retarded and the leaves and flowers become distorted. Control can be effected by applications of nicotine-sulphate, kerosene, or soap solution.
Apple Woolly Aphis (Eriosoma lanigerum).—Although frequently called “American Blight,” the apple woolly aphis is probably a native of Europe. It occurs throughout New Zealand, and was a very serious pest until controlled by the Aphelinus parasite. The presence of this insect is made apparent by the characteristic white woolly patches (Fig. 9, 6) which appear upon the apple trees, due to the woolly material secreted by the aphid. Another feature is that the part of the tree attacked, even after the insects have disappeared, is disfigured by gnarled swellings, due to abnormal thickening of the inner bark. This species also infests apple tree roots, which become similarly malformed. However, root infestation has been overcome by using root stocks, such as Northern Spy, that are immune. The individuals comprising a colony of woolly aphis are variously coloured, yellow, green and red predominating; a considerable amount of honey-dew is secreted. This species has been found to migrate to the foliage of the elm and mountain ash, but in New Zealand the elm-infesting form has not been found to occur. The insect becomes active in spring, and rapidly increases until the autumn. Under favourable climatic conditions, winged females develop and produce males and females, the latter laying eggs. The woolly aphis is preyed upon by the nine-spotted ladybird, but, as this beetle is itself the victim of another insect, its utility is greatly minimised. The most important check to the aphis is the Aphelinus parasite (Aphelinus mali), the influence of which has been spectacular under New Zealand conditions.
Plum Aphis (Rhophalosiphum nymphææ).—This insect is sometimes very common during spring upon the shoots and leaves of plum in New Zealand; in other countries it has been found to migrate to and infest the flowers and leaves of water lilies. The insects assume various shades of green, the winged females having the head, thorax, and legs blackish; the “honey-tubes” vary in colour, and may be reddish, blackish or yellowish.
Poplar Gall Aphis (Pemphigus pupuli-transversus).—Upon the leaf stems of poplar trees in many parts of New Zealand sac-like growths (Fig. 9, 7), measuring anything from half an inch to one inch in length, may be found. These are the galls formed by the North American poplar gall aphis. In each gall are colonies of the aphis surrounded by a mass of flocculent secretion. The walls of the gall are thick and tough, with the outer surface wrinkled, while at the end, toward one side, is a slit-like, or sometimes circular, opening surrounded by a thickened rim, presenting much the same appearance as the mouth of a sack gathered together and tied. For the most part, these insects are wingless females only, but during the summer, and particularly toward the end of autumn, winged females develop and migrate to cruciferous plants, such as cabbage, rape, mustard and turnips, or weeds allied to these cultivated forms, upon the roots of which they establish colonies surrounded by a woolly secretion. In spring a return migration to the poplar takes place, and galls are again established.
Leaf-hoppers form a group of small, narrow-bodied, sap-sucking insects; as the name implies, they infest the foliage of a variety of plants, and when disturbed have the habit of suddenly leaping or hopping to safety; the species present in New Zealand are usually of a greenish or yellowish colour. The adult insect is winged (Fig. 8, K, L), and the female lays her eggs in the plant tissues (H); from these eggs the young wingless hoppers (I) hatch and attack the plant; as they grow, wings develop (J), but until then the insect depends for locomotion upon its hopping powers.
The most outstanding species in New Zealand is the apple leaf-hopper (Typhlocyba australis). This insect causes considerable damage to apple trees unless controlled, which can be effected by spraying with nicotine-sulphate against the young insects in the spring.
White-flies, or mealy-wings, are minute sap-sucking insects, having the body and wings covered with mealy wax. The female (Fig. 8, P) lays her eggs, frequently in circular batches, upon foliage, and the young insects (N) are active, but settle down and commence feeding soon after hatching. Later the insects change to another form (O), without legs and antennæ, and so resemble scale insects to a certain extent; from the latter, however, they may be distinguished by the waxy covering, bearing spine-like processes, and by being surrounded by a distinct marginal area. An important species in New Zealand is the greenhouse white-fly (Trialeurodes vaporariorum), against which fumigation with calcium cyanide is the best as a check.
The foliage of many plants is sometimes infested by very minute black insects, known as thrips. A species commonly met with is that found upon ripe peaches. Thrips are readily identified by the structure of the wings (Fig. 8, 4), which are but narrow strips fringed with long, rigid hairs. These insects, by puncturing the plant tissues and sucking up the nutrient sap, very often are responsible for infecting healthy plants with disease, such as mosaic.
According to the species of thrips, the female lays her eggs either in the plant tissues or upon the surface. The young insects are wingless, but attack the plant in the same manner as does the adult; as development proceeds, the insect transforms to a pupa, from which the adult ultimately emerges. A characteristic symptom of thrips infestation is a silvering of the foliage, while the leaves are further rendered unsightly by the minute specks of hardened excreta ejected by the insects. Many thrips pass their whole development upon the host plants, while others pass part of their lives underground. One of the commonest species met with under glass and out of doors is the greenhouse thrips (Heliothrips hæmorrhoidalis). Thrips are readily controlled by means of nicotine-sulphate.
Leaf-feeding insects have their mouth-parts developed for the biting off and mastication of their food; such insects are, in general, earwigs, crickets and grasshoppers, the caterpillars of moths and butterflies, beetles and their grubs, and the grubs of saw-flies. Such insects vary, not only in their period of activity, some feeding at night, others during the day, but also in the manner under which they set about it. Many feed exposed upon the surface of the plant, while others require protection, such as is afforded by the webbing together of leaves. Some feed upon the leaf epidermis only; some eat holes in the leaf-surface, or gnaw irregular notches from the leaf-edge; while the grosser feeders completely devour the whole.
In many parts of New Zealand the European earwig (Forficula auricularia) causes considerable damage in gardens, while in Central Otago it sometimes ruins the stone fruits. During the winter this insect lies underground, where the female will be found with her cluster of eggs. In the spring these eggs hatch, and the small whitish young earwigs (Fig. 4, 2) emerge from the ground to feed largely upon the pollen and pistils of flowers. At that time the insects and the injury they do are not very noticeable, but as the earwigs grow in size they become conspicuous and extend their depredations to the foliage of plants and to fruit. Earwigs are nocturnal in their habits, and during the day take shelter among fallen leaves, under stones, sacking, or boards, etc., lying on the ground, and may even burrow into the soil itself.
In the control of the earwig, a great deal can be done by what may be called clean gardening—that is, the removal of all places likely to shelter the insect above ground during the day. Another important means is systematic trapping, one of the simplest methods being to place crumpled newspapers on the ground at nightfall, in which many of the insects will seek shelter, the papers being collected and burned next day. But the best method is the use of the following poison bait:—With 12lb. of bran mix 6 quarts of water, to which has been added 5oz. of glycerine and 6oz. of sodium fluoride; to this mash add 4lb. of treacle, taking care to thoroughly mix the whole.
This bait is spread at nightfall in places frequented by earwigs, and should be repeated regularly, especially after wet weather. It is obvious, if satisfactory results are to be secured, that there should be a co-operative campaign organised among the residents of an earwig-infected district.
Fortunately, neither crickets nor grasshoppers (Fig. 10, 1 and 2) are a serious menace to the New Zealand horticulturist, though at times, especially in the warmer parts of the country, crickets may do some extensive damage. The control of these pests is a difficult matter, since they are mobile insects, and breed in places outside the boundaries of the horticulturist’s activities. Some benefits can be secured, however, by thorough cultivation, which breaks up the egg-masses which are placed in the ground. In the case of serious outbreaks, the use of a poisoned bait would have to be resorted to, and the following is recommended from the several recipes in use:—With 25lb. of bran mix 3 or 4 gallons of water in order to make a thin mash; to this, add 2 quarts of molasses and 1lb. of Paris green, thoroughly mixing the whole. If crickets alone are to be dealt with, then use a little more of the Paris green. This mash is spread on the ground invaded by the insects.
Of the leaf-feeding insects, the caterpillars of moths are the most commonly met with, there being a considerable number of destructive species. Caterpillars (Fig. 10, 3) can be readily distinguished by their structure from the grubs of other insects. They resemble short earthworms in shape, and in having the body divided into several segments, of which there are usually thirteen; but here the resemblance to worms stops. There is a distinct head—the first segment—provided with jaws, and on the under side of each of the next three segments, or thorax, is a pair of short feet. The remaining segments comprise the abdomen, and possess sucker-like feet, varying in number according to the kind of caterpillar; in some forms there may be as many as five pairs of such feet, in some three pairs, and in others two, but in all the pair on the terminal segment persists. Many caterpillars are more or less hairy, and others comparatively nude. The following are amongst the most injurious kinds:—
Leaf Rollers.—It is a common sight to see small greenish caterpillars sheltering between two or more leaves of plants that have been tied together by the silken threads spun by the caterpillars; protected thus, the insects feed more or less in security. These caterpillars belong to several species of the tortricid moths, which are themselves comparatively small and drab in colour. Of these species, the most abundant one, comprising over 84 per cent. of the leaf-roller population, is the Australian apple-leaf roller (Tortrix postvittana); the caterpillars of this insect by no means confine their attacks to the apple, but feed equally well upon pear, orange, grape, rose, insignis pine, oak, pelargoniums, etc. Apart from attacking the foliage, the caterpillars frequently tie a leaf to the surface of apple and stone fruits, and feed upon the skin of the latter, causing a blemish.
The apple-leaf roller passes the winter in the caterpillar stage between two leaves. In the spring these caterpillars transform to pupæ, which give rise to moths from the end of August to about the end of October; there are at least two broods of caterpillars during the year, but the limits of these broods are not clearly defined. The caterpillars are attacked by several species of parasites.
Leaf-rollers are easily controlled by the arsenical sprays used against codlin moth, but these sprays must be continued into the late summer after their need against codlin moth is past.
Diamond-backed Moth (Plutella maculipennis).—The caterpillars of this moth (Fig. 10, 4) are commonly found attacking the leaves of cabbages, rape and other cruciferous crops and weeds. These caterpillars are small and greenish, and, if disturbed, suddenly drop suspended by a silken thread attached to the plant. The damage they do is very often extensive, considerable areas of the foliage being devoured. When fully developed, each caterpillar spins a silken cocoon on the under side of the leaf, and there transforms to the pupæ, from which a moth eventually emerges. The insect is small, narrow, and has a light-coloured, diamond-shaped marking along the back. The moth is nocturnal, and shelters amongst the denser foliage during the day; it emerges at night, and lays its eggs upon the leaves. The life-cycle from eggs to adult occupies some 36 days, more or less, according to the season, and there may be six or seven generations during the year.
In control, an important point to note is that the diamond-backed moth breeds upon cruciferous weeds—watercress, shepherd’s purse, and hedge-mustard—as well as on the old plants of a crop left in the ground; it is from such places that infestation of future crops arises, and the clearing up of such breeding places should be given close attention. Under garden conditions, control can be secured by spraying the plants with arsenate of lead (to which a spreader must be added in the case of cabbage), which should be done especially when the plants are young.
Kowhai Moth (Mecyna maorialis).—The caterpillar of this native moth sometimes becomes epidemic, when it does considerable damage to kowhai, broom, lupins, and sometimes clover. The caterpillar, which measures about an inch when mature, is of a greenish colour, having rows of black tubercles with white centres along the sides, and a double row of white spots along the back; from the black tubercles black bristle-like hairs arise. The caterpillar spins a silken cocoon, in which it pupates. The moth is comparatively small, the fore wings being yellowish-brown with darker markings, and the hind wings orange-yellow with a blackish border. There are at least two broods of caterpillars annually: the first in the spring, and the second during autumn. Arsenate of lead will give effective control on garden legumes.
Cut-worms.—This term is applied to the caterpillars of a number of night-flying noctuid moths; these caterpillars are smooth-bodied and rather worm-like, in some cases measuring from one and a-half to two inches in length when full grown. They feed at night, and their method of attack is characteristic in that they nip off young plants close to the ground (Fig. 10, 5), so that the latter fall over, when they are devoured by the caterpillars; this habit has given rise to the name “cut-worms.” During the day the cut-worms are to be found curled up in the ground close to the plants they have been attacking. The moths of these caterpillars are rather stout-bodied, and measure about three-quarters of an inch long. One of the commonest species is the cosmopolitan greasy-cut-worm (Agrotis ypsilon).
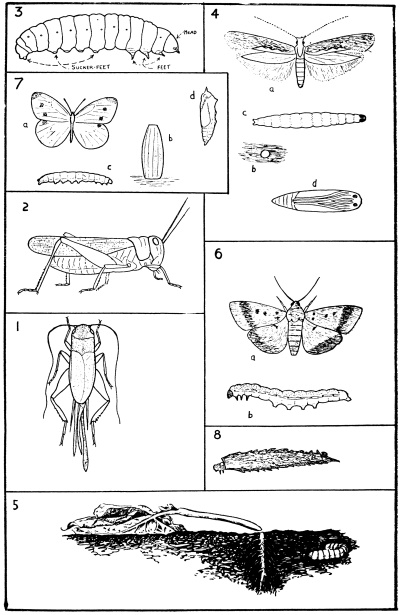
FIG. 10.
(1) Cricket. (2) Grasshopper. (3) Caterpillar. (4) Diamond-backed Moth—a, adult moth; b, egg; c, larva; d, pupa. (5) Cut-worm lying by damaged plant. (6) Tomato-worm Caterpillar—a, adult; b, larva. (7) Cabbage White Butterfly—a, adult; b, egg; c, larva; d, pupa. (8) Larva case of Bag-moth.
Though cut-worms are active throughout the growing period of plants, most damage is done to young and tender plants at the time of establishment, and this is particularly noticeable in the spring. When plants are grown isolated in rows, and the area is not too large, complete protection from cut-worms can be secured by enclosing each plant in a tin collar pushed into the ground and projecting a few inches from the surface; these collars are removed when the plant is well established. In localities where cut-worms are very troublesome it is advisable to reduce their numbers by means of a poison bait made as follows:—50lb. of bran and 1lb. of Paris green are thoroughly mixed in a dry state; when this is done, and just before being used, the bran is moistened with water, sweetened with molasses, until the bait reaches a crumbly, but not saturated, condition. This bait may be broadcast over the infected area or laid around each plant as a barrier. This bait must be applied every few days until the plants have reached a stage when they are able to withstand cut-worm attack.
A great deal can be done to check cut-worms by removing dense growths of weeds and rough herbage growing in unused parts of the garden; in such places the insects breed, and are a source of infestation. Another point to consider is that thorough cultivation will destroy many pupæ that are lying underground, and which would otherwise give rise to another generation of moths.
“Army-worms.”—These caterpillars are similar in their appearance and general habits to the cut-worms, but differ in their method of attack. When present in numbers, they move through a crop—especially cereals—eating as they go, and leaving nothing but devastation in their wake, much as does an invading army on the march. They are not of so much interest to the horticulturist as to the farmer.
Tomato-worm (Heliothis armigera).—This caterpillar (Fig. 10, 6) is one of the most conspicuous caterpillars met with in the garden. Its habit of boring into and eating the contents of tomatoes gives it the name of “tomato-worm.” It is a cosmopolitan insect, and is especially destructive to flower buds and fruit, a wide range of plants being attacked. The caterpillars vary in colour, some being greenish and others brownish, with reddish, yellowish or white markings. The moth, which belongs to the noctuid group, is on the wing both day and night, mostly during the earlier part of the year; it is a stoutly-built insect, measuring somewhat over half-an-inch long; its colour is a brownish-orange, with oblique darker bands on the wings. As the insect passes the winter and spring as a pupa in the ground, thorough cultivation will help to destroy a considerable number. The use of arsenate of lead sprays, however, is the most effective control for the caterpillars.
Hawk or Sphinx Moth (Sphinx convolvuli).—This conspicuous insect and its caterpillars are most abundant in the Auckland province, though found as far south as Christchurch. The caterpillars feed on convolvulus, but do considerable damage to the foliage of the kumara and sometimes tobacco. The caterpillar is the largest met with in the garden; it is stout in form, and measures up to 3 ½ inches when fully grown. It is to be recognised at once on account of the dark red, horn-like process arising from the end of the body. The caterpillar may be of two colours—the one green, with diagonal yellow bars on the sides; the other, brownish-yellow, with dark lines on the back and sides. From about February to November the insect lies in the ground as a pupa. The latter can be recognised by a curved process arising from the head and lying along the body. The moth flies rapidly during the last and earlier months of the year; it is a large, conspicuous insect, about 1 ½ inches long, with greyish-brown mottled wings, while the abdomen is conspicuously barred with white, red and brown. Arsenate of lead against the young caterpillars during November to February would act as an efficient control.
Cabbage White Butterfly (Pieris rapæ).—This butterfly (Fig. 10, 7) is a recent importation, having been first noted at Napier in 1930. Since then it has spread with marvellous rapidity throughout the North Island, and has appeared in the South Island, in the vicinity of Timaru.
The caterpillars of this insect are particularly severe in their attacks upon the foliage of cabbages and cauliflowers, though they also feed upon many other related plants, such as lettuce and radish, besides cruciferous weeds. The caterpillars of the white butterfly are not to be confused with those of the diamond-backed moth, already described. The full-grown white butterfly caterpillar is a conspicuous insect, and measures up to an inch and a-quarter in length; it is easily distinguished by its leaf-green colour and velvet-like appearance, while down the centre of the back is a narrow orange stripe, and on each side a brownish line formed of little spots. The chrysalis measures about three-quarters of an inch long, having a pointed process from the head, and a keel-like ridge on its back, while the colour varies according to the surroundings with which the chrysalis blends; it is not protected by a cocoon of silk, and may be found upon the food plant or any other support near by.
The butterfly itself is a very conspicuous insect, measuring about two inches across the expanded wings. The female is of a yellowish-white colour, with darker to blackish markings at the fore-angles of the front wings, while there are two similar spots on the surface of the same wings, and one on the hind pair. The male is whitish, with a dull greyish-green or bluish hue, marked much as the female, except that there is only a single spot on the surface of each wing.
The eggs (Fig. 10, 7b) are bottle-shaped, and stand erect upon the leaf surface, where they are laid singly, and not in batches; they are visible to the naked eye. The eggs hatch within a week after being laid. There are several generations each year.
The cabbage butterfly can be controlled by the use of lead arsenate. It has been found effective when planting out to first dip the seedlings in lead arsenate at the rate of 1lb. in 50 gallons of water, to which 1lb. of laundry soap is added as a spreader. During the growth of the crop the same strength of arsenate and soap can be applied as a spray.
Magpie Moth (Nyctemera annulata).—One of the commonest and most conspicuous day-flying insects of the garden and field is the magpie moth. It is black in colour, relieved by an orange-banded abdomen and whitish spots on the wings, two on each of the front wings and one on each hind one. Its black, hairy caterpillars, commonly called “woolly bears,” have narrow brick-red lines along the body, and very often do some considerable damage to cinerarias; they also attack weeds, such as ragwort and groundsel.
The small globular eggs are laid in clusters on the leaves of the food plant. At first they are pale green, later becoming dark yellow, and just before the young caterpillars emerge from them they change to a leaden colour. When fully fed, the caterpillar seeks a sheltered place (beneath stones, under, bark, etc.), and there spins a loose cocoon, in which it transforms to the chrysalis; the latter becomes blackish or brownish in colour, with yellow markings. There are several generations during the year.
Cinerarias can be protected by spraying with lead arsenate, or, better, by removing the caterpillars by hand and destroying them.
Cabbage Tree Moth (Venusia verriculata).—The foliage of the cabbage tree is frequently holed on the surface and notched along the edges—this is the work of the cabbage tree moth caterpillars. The history of the insect is as follows:—The nocturnal moth measures about an inch and a-half across the expanded wings, which are characteristically coloured by alternating chocolate-brown and yellowish-white lines running from wing-tip to wing-tip across the body, so that the insect merges into the general pattern and colour of a dead leaf, upon which it usually rests. The eggs are green, and at first blend with the green leaf, on which they are often laid in batches; when on dead leaves they become conspicuous. Later the eggs change colour to brown, and finally red. The caterpillars congregate in the unopened foliage, and their injury becomes apparent as the leaves open. The larvæ transform to chrysalids in silken cocoons, loosely spun in any suitable crevice upon the trees. If it was necessary and practicable to protect ornamental cabbage trees from the attacks of this insect, it could be done by removing dead leaves from the crown and spraying with arsenate of lead to which laundry soap had been added.
Bag Moth (Œceticus omnivorus).—This is an insect that never fails to attract attention on account of its cigar-shaped bags (Fig. 10, 8), constructed by the larvæ, and are to be found attached to a variety of plants, upon the foliage of which the larvæ feed, though they are not serious pests. Each caterpillar spins its own tough silken bag, which it never leaves, and to the outside of which it frequently attaches fragments of leaves and twigs. Though the male is a normal moth, and flies about (it is practically black, and densely haired, with translucent smoky-black wings having an expanse of about an inch and a-quarter), the female develops in an abnormal manner, and assumes a grub-like form, never leaving the bag woven by its caterpillar.
If it should be found necessary, as sometimes happens, the only satisfactory way of controlling the bag-moth is to remove by hand and destroy.
Unlike the caterpillars of moths, there are very few beetles in New Zealand that are important leaf-feeders. Though few in numbers, however, the outstanding ones are very destructive. The beetles themselves, as well as their larvæ, according to the species, may attack foliage, but in other cases it is only the beetles that feed on foliage while their larvæ live underground on roots. The following species are outstanding:—
Cockchafers.—These are the adults of the grass grubs, and there are several species, all native to New Zealand. The commonest and most destructive one (Fig. 11, 1a) is the so-called brown beetle (Odontria zealandica), misnamed the “turnip fly,” which is on the wing for about six weeks each year, during November and early December as a rule. It swarms at dusk, creating a loud, droning sound, and is responsible for widespread damage by defoliating garden plants and field crops, as well as trees.

FIG. 11.
(1) a, Brown-chafer beetle; b, antenna of beetle, showing finger-like processes; c, larva or grass grub. (2) Bronze beetle. (3) a, Gum-tree weevil; b, egg capsule; c, larva. (4) Eucalyptus tortoise beetle. (5) a, Pear saw-fly; b, larva from the side; c, larva from above. (6) Pear midge.
This beetle is easily identified. It is rather plump-bodied, brownish, smooth, and measures about three-eighths of an inch long. Like all beetles, the front wings are hard, and form a cover over the body when closed; these hardened wings, or elytra, do not reach quite to the end of the abdomen, the tip of which remains uncovered. Another definite character is found in the antennæ, which terminate in finger-like processes (Fig. 11, 1b). There are several species of cockchafers, to which all these characteristics, except the colour, might be referred, but none is so abundant as the species under review. There is one, however, that is on the wing about the same time as, or a little earlier than, the brown beetle. This species is somewhat larger, about half an inch long; it is sparsely clothed with hair, and the elytra are marked by broad brown stripes, alternated with very narrow darker ones.
The brown beetle lays its spherical eggs in the ground, preferably amongst the roots of grass, strawberries, etc. The grubs (Fig. 11, 1c) are well known as grass grubs; they are whitish in colour, the swollen terminal segment of the abdomen being very often darker. These grubs are sometimes called “curl-grubs,” from their habit of lying doubled-up when at rest or feeding in the ground. Towards September each year the grubs of the brown beetle pupate prior to the beetles emerging in November. These grubs will be referred to later under the chapter dealing with subterranean insects.
In gardens and nurseries, the depredations of the beetles may be lessened by spraying with lead arsenate, or by the use of sulphur smudges. The use of smudges was developed very effectively as follows by Mr. D. J. Buchanan, forest ranger at the Tapanui State Forest nurseries. Sulphur is spread on strips of scrim, which are then rolled up and placed in containers, such as old paint pots. The latter are set about the nursery, and fired at evening, when they will burn throughout the night, the fumes acting as a deterrent to the beetles. When only a few plants are to be protected, such as bush roses, the beetles can be warded off by allowing a hose to play over the plants throughout the night.
Another common cockchafer which is on the wing most of the summer and autumn is the green manuka beetle (Pyronota festiva). This insect is capable of causing considerable damage as a defoliator. It is active both day and night. The general colour is bright green, with a dark stripe down the middle of the back, though some specimens are brown or coppery; the legs are orange-yellow, and the length of the insect is a little over a quarter of an inch.
Bronze Beetle (Eucolaspis brunneus).—This insect (Fig. 11, 2) is very often confused with the brown beetle, from which, however, it is easily distinguished. It is active during the day, and attacks the foliage and fruit of a great variety of plants, eating holes from leaves, so that the latter appear as if they had been subjected to a charge of shot, or devouring the epidermis from fruits and berries. This beetle is active during November to January; it measures about one-sixteenth of an inch long, is oval in outline, and varies in colour from yellowish, with darker markings, to greenish or bronzy-black; the antennæ are comparatively long, and do not terminate in any unusual manner, as do those of the cockchafers. A characteristic habit of the bronze beetle is to leap off the plant if disturbed; this habit has been responsible for the group to which this insect belongs being called “flea beetles.” The bronze beetle lays its eggs in the ground, where the larvæ feed, though they are not injurious in that stage. The beetles are to be controlled by the use of lead arsenate.
Gum Tree Weevil (Gonipterus scutellatus).—Both the adults and larvæ of this Australian weevil attack eucalyptus foliage, particularly bluegum, in most parts of New Zealand, the adult weevils eating from the leaf margin, as well as devouring tender shoots, while the larvæ cut elongated holes from the leaf surface.
The weevil (Fig. 11, 3a), which is of a tawny to brownish-black colour, and clothed with yellowish-white and golden hairs, measures from a quarter to one-third of an inch in length; it possesses a short snout on the head. The eggs are yellowish, and are packed in a hard, black capsule (Fig. 11, 3b), attached mainly to the surface of young leaves. The larvæ (Fig. 11, 3c) are legless, like those of all weevils, and yellowish at first, when they are studded with small black dots, and have a dark stripe along each side. Frequently these young larvæ are seen with a tail-like thread of blackish excrement projecting from the posterior end. The plump, fully-developed grub is yellowish-green, with a wrinkled skin, and is slug-like in general appearance. Pupation takes place in the ground. This insect over-winters in the adult stage, and large numbers of the weevils are very often to be found beneath loose bark on the tree trunks during the winter. Control depends upon the use of an egg parasite which has been established in certain localities of the Dominion. In the case of small ornamental trees, spraying with lead arsenate to which laundry soap has been added should be effective.
Eucalyptus Tortoise Beetle (Paropsis dilatata).—This is another Australian insect restricted so far to the East Coast districts of the South Island, where it attacks eucalyptus foliage in company with the weevil. The beetle (Fig. 11, 4) is tortoise-shaped, varies in colour from reddish-yellow to reddish-brown, with darker markings on the back, which is pitted by minute depressions, and has a length of from one-third to half an inch. Like the weevil, this beetle passes the winter beneath loose bark.
The eggs are conspicuous and cigar-shaped, being laid in clusters, lying more or less on their sides, upon the foliage. The larva is rather plump, and pointed posteriorly; it possesses legs, while at the tip of the body is a sucker-like false foot. The general colour is yellowish, varying to a rosy-pink, there being a darker stripe down the back, while along each side is a similar one above a row of black dots.
Pear and Cherry Slug, or Saw-fly (Caliroa limacina).—The slug-like larvæ of this insect are very abundant upon hawthorn foliage, and if not controlled do considerable damage to cherry, plum, pear, and peach. These larvæ (Fig. 11, 5b) are very often called leeches, and devour the epidermis, exposing the skeleton of the infested leaves; they are slimy, of a dark green, though orange-coloured immediately after a moult, and the head end is much enlarged, giving a clubbed shape to the body, along the under side of which are several false legs. Pupation takes place in the ground. The adult (Fig. 11, 5a) measures about a quarter of an inch long, is rather thickly set, black in colour, and possesses four transparent wings. The female deposits her eggs in the tissue of the foliage by means of a saw-like ovipositor—hence the name “saw-fly”—which is thrust through the lower epidermis of the leaf, when a pocket is formed to receive the egg; each egg pocket forms a little pimple on the upper surface. This insect is very easily controlled by spraying foliage infested by the larvæ with lead arsenate.
Another saw-fly closely related to the foregoing species is the willow saw-fly (Pontania proxima). This species has only recently appeared in New Zealand, and its larvæ live in galls, or swellings, on the foliage of willows.
Pear Midge (Perrisia pyri).—A serious pest of pear trees, which for some years retarded the culture of pears, especially in the Auckland district, is the pear midge. This is a minute, delicate, two-winged fly (Fig. 11, 6), measuring about one-twenty-fifth of an inch long; it has a blackish head and thorax, and an orange-red to brownish abdomen. The female alights upon young leaves just burst from the bud; and, while they are yet curled, lays her eggs between the folds. The larvæ, on hatching, live protected in the curled leaves, which they attack, and which never unfold. The result is that the infested leaves eventually turn black and brittle, and cease to function. The fully-developed larvæ drop to the ground, which they enter, and there pupate. The midges become abundant in early spring, when the first young pear foliage develops, and they keep on producing generation after generation until the autumn. The winter is passed in the larval stage underground beneath the trees.
A parasite has been established against the pest, and is doing good work. The insect can be reduced to a large extent by thorough winter cultivation, especially beneath the trees. The insect’s larvæ, being protected within the curled-up leaves, are not reached by ordinary sprays, but Dr. R. H. Makgill, of Henderson, secured some excellent results on young trees by the use of nicotine.
Olearia Gall Midge (Cecidomyia oleariæ).—In many parts of New Zealand where Olearia forsteri is grown as a hedge, it is very often disfigured by the formation of malformations, or galls. These are caused by a native midge known as the olearia gall midge. The midge itself resembles the pear midge in structure, but is larger, measuring from one-tenth to one-eighth of an inch long; it is conspicuous on account of its black thorax and blood-red abdomen. In early spring the midges appear and lay their conspicuous masses of bright red eggs upon the buds of the developing shoots. The larvæ, on hatching, set up an irritation in the rapidly-developing tissues, causing the latter to swell and become malformed into bunches of rosette-like galls. If the latter are cut open, a number of the yellowish larvæ will be found, each in its own compartment within the fleshy gall. There is only one brood of adults each year. Control can be effected to a great extent by cutting back and burning the badly-infested parts during winter, and by pruning the young growth carrying the eggs in the spring. Spraying with nicotine when the midges are active should also help to protect the plants.
CODLIN MOTH (Cydia pomonella).—The codlin moth caterpillar burrows in developing apples and pears, and such “wormy” fruit is known to everybody.
The moth itself is seldom seen, since it lies concealed until after nightfall, when it becomes active and lays its eggs. The insect (Fig. 12a) measures about three-quarters of an inch long, and is inconspicuously, though beautifully, coloured; the fore wings, which cover the body when closed, are light grey, crossed by fine bands of a darker hue, giving the appearance of watered-silk, while at the extremity of each wing is a large bronze spot; the hind wings, seen only when expanded, are of a light brown colour. The minute flat eggs are laid on the foliage of leaves, on the fruit, or even on young bark; they appear at first as glistening white specks, but, as development advances, a red ring develops, and finally a black spot just prior to the caterpillars hatching.
In some places the first larvæ developing in the spring enter the fruit by way of the calyx, but under New Zealand conditions it is more usual for entry to be made by boring through the skin of the apple. Having completed their development in the fruit, the caterpillars bore their way out and spin their cocoons beneath the loose bark of the tree trunks; in these cocoons pupation takes place, and from them the next generation of moths develops.
Although in New Zealand there is usually only one generation produced each year, three or even four develop in other countries. The winter is passed by the larvæ in their cocoons, and pupation takes place just prior to the period when the moths emerge in the spring. As the moths continue to emerge and lay their eggs for a period extending from November to February, it is essential that regular applications of arsenate of lead be made during that time in order to protect the fruit from the larvæ hatching from the eggs laid by the moths. In localities where the spring larvæ enter the calyx of the fruit, it is essential to apply the first spray just after the petals fall, so that the poison may lodge in the calyx before it closes. The removal of rough bark from the trunks of both apple and pear trees is a help in controlling the insect. Another method sometimes used is to band the tree trunks with strips of scrim; under these bands the larvæ collect, and the former can be later removed and destroyed with their tenants.
Currant Clear-wing Borer (Sesia tipuliformis).—This destructive moth has been carried to and established in New Zealand, as well as many other parts of the world. In currant gardens its larvæ cause the death of canes by eating out the pith. The moth (Fig. 12b) is a very conspicuous and beautiful insect; the wings, which expand to about three-quarters of an inch, are transparent and bordered with golden-purple, a bar of the same colour crossing the surface of the fore wings; the body (about half an inch long) is metallic-purple, the thorax having a yellow stripe on each side, while the abdomen, barred with golden bands, terminates in a fan-shaped tuft of purplish hairs.
The moths are active each year in the spring, when they lay their brownish, globular eggs singly on the bark of the currant canes. The resultant larvæ bore into the stem and destroy the pith, passing the winter in the damaged canes. In the following spring the larvæ become active once more and approach the surface, where pupation takes place shortly before the moths emerge.
There is only one generation each year, and control lies in the removal and burning of infested canes in late winter.
Tomato Stem Borer (Gnorimoschema plæsiosema).—Tomato growers are frequently faced with the problem of the destruction of tomato plants caused by the attacks of the larvæ of the tomato stem borer moth. This insect caused considerable damage for the first time in Auckland some fourteen years ago, though it was known in other parts of the country as well.
The moth itself (Fig. 12c) is a small one, measuring about a quarter of an inch with the wings closed. In this position the insect is wedge-shaped and conspicuous. Against the general greyish-brown colour is a dark brownish area on each side. The eggs are laid on the tomato leaves, in which the young caterpillars tunnel as they work toward the leaf petioles, down which they burrow into the main stems. In the damaged stems, pupation takes place. Under favourable conditions, this insect may pass through at least three generations during the season.
Control depends upon sanitation and the use of arsenate of lead sprays. All infested stems, together with plants after the crop has been removed, should be burned; as the insect is known to attack potato plants and tubers, care should be taken to destroy all potato tops after harvesting. Frequent applications of arsenate of lead are essential to protect the tomato plants, especially during the earlier part of the season.
When on this subject, mention should be made of the potato-tuber moth (Phthorimæa operculella), which is somewhat similar to the tomato-stem borer, both in appearance and habits. The larva of this insect is best known from its habit of boring through potato-tubers; these burrows become filled by a fungus after the larvæ have vacated them. The adult potato-tuber moth is a night-flyer, and lays its eggs upon the leaves of the plants; the larvæ burrow down the stems, and may even reach the tuber below ground. When seed is not properly buried, the moth will also lay its eggs in the “eyes,” and so directly infest the tuber; this danger applies also to potatoes in store or in bags.
In the control of the potato-tuber moth, the following points should be noted:—Select only sound seed and cover well when planted. On harvesting the crop, do not leave the bagged potatoes standing in the field overnight, as they are exposed to infestation; neither cover the open bags with the potato-tops, as is commonly done, since this will attract the moths. Destroy all tops immediately after harvesting. Dusting potatoes in store with slaked lime will tend to act as a protection against the moth.
FIGURE 12.
A—1, Codlin moth; 2, codlin larva in apple. B—1, Currant clear-wing moth; 2, clear-wing moth larva in stem. C—1, Tomato stem-borer moth; 2, larva of moth; 3, damaged tomato stem. D—1, A long-horn beetle; 2, larva of long-horn beetle. E—1, A leaf-mining fly; 2, leaf attacked by leaf-miner. F—1, Subterranean grass-caterpillar moth; 2, subterranean grass-caterpillar. G—1, A click beetle; 2, a wire-worm. H—1, Larger narcissus fly; 2, smaller narcissus fly. K—1, A subterranean spring-tail; 2, a leaf-eating spring-tail.
Round-headed Borers.—Apple, almond, and citrus trees, together with gooseberry and such ornamental and shelter trees as poplars, tree-lucerne, and goat-willow, are sometimes damaged by round-headed borers, which tunnel in the stems and branches. These borers (Fig. 12d) are white in colour, narrow-bodied, and cylindrical, the segments being usually well defined, and belong to a group of beetles known as long-horned beetles, a group of insects to which the common hu-hu beetle belongs. These beetles are narrow-bodied, and their antennæ are comparatively long and conspicuous.
To control these pests, the only thing to do is to cut out and burn the badly-infested parts. Where a borer is located (and this can be frequently done by the presence of the powdered wood ejected from the burrows), the culprit may be killed by injecting into the tunnel some carbon bisulphide and plugging up the openings with some clay or other similar substance.
Leaf-mining Flies.—Very often the leaves of cineraria and chrysanthemum are disfigured by the tortuous tunnellings of the maggots of minute flies (Fig. 12e). The adult insects are two-winged, and in structure resemble in many respects miniature houseflies. The eggs are laid in the leaf tissues, in which the whole development of the maggots and pupæ takes place. The white maggots are small, legless and headless. Spraying with black-leaf 40 would act as a deterrent to the flies, while infested leaves should be removed and destroyed before infestation becomes general.
Grass Grub (Odontria zealandica).—As explained in the preceding chapter, the grass grub is the larva of a native cockchafer beetle (Fig. 11, 1). This grub, by feeding upon roots, causes extensive damage to pastures and lawns, as well as to many garden plants, including strawberries. In the case of pasture and lawns, the presence of even a considerable number of grass grubs is not detrimental unless they occur concentrated in definite areas, when the damage is pronounced. With garden plants, however, which are isolated when compared with the dense root masses of grasses, the attacks of one or two grubs upon the roots of a single plant may cause serious injury.
Grass grub damage to grasses is not merely due to attack upon the roots. While feeding, the grubs swallow soil with the roots, rendering the former spongy, and so disturb the normal circulation of moisture about the grass roots. In the case of infested lawns, it is advantageous to roll infested areas in order to pack the soil pulverised by the grubs, and re-establish normal circulation of soil moisture. Another important feature in grub control is to stimulate root development by means of fertilisers. A recently-developed method of “grub-proofing” lawns is to broadcast over every thousand square feet of turf to be treated one bushel of screened sand or clean soil, in which 5lb. of lead arsenate powder have been intimately mixed. This is said to remain effective for a period of three years; but such fertilisers as nitrate of soda, superphosphate, sulphate of potash, and potassium chloride should not be used on “grub-proofed” turf, as they react with the lead arsenate, and reduce its effectiveness, though rotted manure or ammonia sulphate may be used.
The control of grass grubs damaging the roots of strawberry and other plants is a difficult matter, though some benefit is to be derived by making holes about four inches deep with a stick in the soil near to the infested plants and pouring in a little carbon bisulphide; the holes should be closed immediately. To protect strawberry beds, if they are not too extensive, the most satisfactory method is to cover the plants with scrim, stretched on frames, at dusk during November and early December, when the beetles are flying; this will prevent the insects from infesting the ground with their eggs. The use of sulphur smudges, already referred to, is of great importance in this respect.
Subterranean Grass Caterpillars.—These caterpillars are the larvæ of native moths (Fig. 12, f1) belonging to the genus Porina, and when they become epidemic they cause much more extensive damage to pasture and lawns than do the grass grubs. When full grown, the greyish-black caterpillars (Fig. 12, f2) reach a length of about three inches; they are soft-bodied and rather flaccid, and live in underground burrows of varying depth. After dark, these caterpillars come to the surface and devour the grass, eating it close to the ground, much soil being swallowed by the larvæ during the feeding. This soil is evacuated, and resembles earthworm castings, but is mixed with silk spun by the caterpillars; the emergence holes of the caterpillars, about the diameter of a lead pencil, are conspicuous on the surface denuded of its covering of grass. Pupation takes place underground, and when the moths emerge the pupæ first move to and project beyond the surface of the ground; these pupæ are large and easily recognised by the wing-cases, which are very short compared with the length of the body. The moths are on the wing during spring and summer, the rest of the year being spent in the larval stage. The moths are night-flyers, and are amongst the largest species in New Zealand, their wings having an expanse of from one to over two inches; they are heavy-bodied insects, and vary considerably in colour. One of the commonest, species is brownish-yellow, or sometimes a smoky-grey, with a white streak bordered with black on the fore wings; the hind wings may be pinkish.
The most satisfactory method of controlling the insect is to roll infested lawns after dark, in order to crush the caterpillars whilst feeding on the surface. Flooding an infested lawn with water will bring most of the caterpillars to the surface, when they can be collected and destroyed. Spraying grass in spring and early summer with arsenate of lead will tend to poison the immature caterpillars. There are at least three species of insect parasites that attack these larvæ, and there is also a fungus which invades and destroys the whole body, taking the shape of the insect; such fungus-infested caterpillars are commonly called “vegetable caterpillars.”
Wireworms.—The roots of garden plants and germinating seeds are often damaged by hard, wiry beetle grubs, reddish-brown or whitish in colour, called “wireworms,” so named from their resemblance to short pieces of wire; they have three pairs of legs behind the head and a sucker-like appendage on the last body-segment (Fig. 12, g2). These grubs transform to narrow-bodied, brownish or blackish beetles, known as “click-beetles” (Fig. 12, g1) from their habit, when overturned, of righting themselves by a springing action, during which a distinct and sharp clicking sound is made; the spring apparatus consists of a spine, the tip of which fits into a notch on the under side of the thorax.
Practically nothing is known as yet in regard to the biology of the New Zealand click-beetles. They are extremely difficult to control, and the larval stage covers a period of two or more years.
Narcissus Flies.—There are two species of narcissus flies—the larger (Merodon equestris) and the smaller (Eumerus strigatus) both occur in New Zealand. The larvæ of these flies attack bulbs of various kinds, the hosts of the larger fly being narcissus, hyacinth, tulip, amaryllis, habranthus, vallota, galtonia, scylla, and leucojum; and of the smaller fly, narcissus, hyacinth, onion and shallot. These flies are two-winged insects, the hind wings being wanting as such, and belong to a group called the syrphid, or hover flies.
The larger narcissus fly (Fig. 12, h1) resembles somewhat a humble-bee (which, however, has four wings); its stout and very hairy body measures about half an inch long. There is considerable variation in colour, though black or brown predominates, with greyish or yellowish hairs, and bands of the same colour; the bands, however, are absent in some individuals.
During spring the insects fly about in the sun, and lay their eggs at the leaf bases of the host plants, or on the exposed neck of bulbs, or in the soil close by. The larvæ, which are legless, yellowish grubs, enter the bulb, and may completely destroy it. Infested bulbs may be detected by an unnatural softness near the neck when pressed between the fingers.
The smaller narcissus fly (Fig. 12, h2) is about half the length of the larger, of a shiny black colour, with metallic reflections, and is not clothed with hair. The eggs are laid in the ground, or at times upon the plant itself. Several larvæ of this fly may be found in the one bulb; the larvæ resemble those of the larger fly, but are smaller, and have three small processes at the end of the body. The smaller narcissus fly usually attacks the bulbs already damaged by some other agent, though it has been known to infect sound bulbs.
Control of both these flies depends upon the destruction of infested bulbs. Recent researches have shown that the flies themselves can be poisoned in large numbers by a spray made of 4oz. of sodium arsenate, 1lb. of crude glycerine, 2lb. of white sugar, and four gallons of water; this spray is applied during bright and warm weather.
Springtails.—These are very minute, soft-bodied insects, which are very active, and have a habit of springing with the agility of fleas. There are several species, but two are of interest to the horticulturist.
One of these (Fig. 12, k1) is white in colour, narrow-bodied, and lives underground, especially in damp places, where it damages germinating seeds, or the roots of seedlings; even older herbaceous garden plants are attacked. As a control, it is important to drain the soil in damp locations and to dig in calcium cyanide about two weeks before planting or sowing.
The second species is blackish and more or less spherical (Fig. 12, k2); at times it does considerable damage in the spring to the seed-leaves of young plants as soon as they appear above ground. Spraying small areas—e.g., of cucumbers, turnips, etc.—with black-leaf 40 would help to protect the plants; as the eggs are laid in the ground, and as these develop best under moist conditions, thorough cultivation prior to sowing the crop is an important controlling factor.
In this chapter will be grouped for convenience mites, woodlice, millepedes, slugs, snails, and eelworms.
Mites, together with spiders and ticks, belong to a group of animals distinct from the insects, from which, they differ in many respects; for example, they possess four, and not three, pairs of legs in the adult state, no head separated from the body as a movable, distinct region, while in many cases, especially in mites and ticks, the abdomen and thorax are continuous; in no case are wings developed.
Mites are of small size, some being microscopic, while others are just discernable by the unaided eye. All species have the mouth-parts developed for the purpose of feeding upon liquid food—e.g., blood (in the case of those species that attack animals), decaying vegetable matter, or the saps of plants. It is the last—that is, those parasitic upon plants—with which we are here concerned.
The life-history of mites presents some variability, and, though there are fundamentally four stages of development, additional stages have been developed by some species which tend to complicate the cycle. The principal stages in development are as follows (Fig. 13, 1–5):—In practically all cases eggs are deposited, but few species being viviparous. The larva, on hatching, possesses but six legs, and resembles an insect in this respect; the larva then becomes quiescent, and after moulting the eight-legged nymph appears. While in the nymphal state the mite may undergo one or more moults, giving rise to additional nymphal forms, that may complicate the life-history. From the final moult of the nymph the adult mite emerges.
Perhaps the best-known mite in New Zealand is the European red mite of apple trees (Paratetranychus pilosus), though it attacks a wide range of plants apart from deciduous fruit trees, which it favours; it has been found on grape vine, raspberry, rose, hawthorn, citrus, etc. This mite (Fig. 13, 6) occurs in Europe, Russia, British Isles, North America, Australia, and New Zealand, and it causes considerable injury to foliage, which assumes a brown appearance, owing to the tissues drying up where they have been punctured by the mouth-parts of the mite.
In the case of heavily-infested trees, the red eggs of this mite form conspicuous patches on the bark during winter; these winter eggs are laid from January onward till leaf-fall, and from them the young mites hatch in the spring, when the foliage is again attacked. The red mite develops rapidly, and reaches the adult stage in about two weeks; several generations are thus produced from spring to autumn, when the eggs are laid upon the foliage.
The eggs (Fig. 13, 7) are very small, globular, and ribbed on the surface; from the centre of each projects a hair-like stalk, somewhat bent at the tip. The colour is bright red, changing to a deep orange. The red mite lives freely upon the foliage, and does not produce a web, as do related species; the adult female is bright red to dark brownish-red, rather globular in shape, with comparatively stout legs and numerous spine-like hairs on the back. Although the eggs of the European red mite are exposed on bark and readily accessible to sprays during the winter, no effective winter wash for their control is yet known; the most satisfactory method for checking the pest is to spray the active stages of the mite with summer oil.
Another species of mite, having much the same habits and host plants as the European red mite, is the brown mite (Bryobia prætiosa). The eggs of this species are of a deep red, with a yellowish tinge in many cases, but differ from those of the European red mite in the absence of the polar-stalk and ribbed surface. The brown mites (Fig. 13, 8) are of a dull red or greenish colour, lack the spine-like hairs on the back, are decidedly flattened, and have the front pair of legs abnormally long.
The common red spider (Tetranychus telarius) is a species of mite frequently met with on a wide range of plants too numerous to mention here; in New Zealand it frequently injures violet, hop, currant, willow, and many weeds. This mite is to be found in all stages practically all the year round; during the spring it is mostly found on weeds and such cultivated plants as strawberry and violet. It is a web-spinning species, and the minute yellowish-red eggs are to be found scattered among a fine web attached to the lower surface of leaves as a rule. The adult mite (Fig. 13, 9) is very active; it is somewhat larger than the two foregoing species, and of a yellowish-green colour, with a pair of conspicuous dark spots on the back. Though this mite can be held in check by the application of lime-sulphur sprays, advantage should be taken of thorough cultivation during the dormant season, since the mite hibernates on weeds and among dead leaves and in the soil.
A mite very often met with by bulb growers is the bulb-mite (Rhizoglyphus hyacinthi), now found in most parts of the world. Although this mite may possibly be able to attack practically all tubers or bulbs, it is commonly found infesting narcissus, hyacinth, tulip, crocus, and Easter lily; it is especially abundant in bulbs with loose scales, and has been found to be capable of attacking healthy tissue. The life-history of this species is complicated at times by the development of additional stages; one of these—the hypopus—is of particular interest, as it shows more activity than the others, and attaches itself to the bodies of insects, and is so transported. The mite develops from egg to adult within a period of nine days under favourable conditions, or as long as six weeks at other times. All stages of the bulb-mite occur at the same time in infested bulbs, which become soft and rotten. The adult mites (Fig. 13, 10) are smooth, yellowish-white, tinged with pink, and have legs and mouth-parts reddish. Symptoms of their presence are to be found in stunted growth and yellowing leaves, failure of flower development, reddish spots on bulb scales, or a softening of the bulbs. All seriously-infested bulbs should be destroyed, and the ground where they were grown treated with calcium cyanide. For the treatment of bulbs, they should be immersed for ten minutes in a two per cent. solution of formalin heated to 122 deg. Fahr., or simply in water at a temperature of 131 deg. Fahr.

FIG. 13.
(1) Five stages in mite development: (1) Egg, (2) larva, (3) nymph, (4) older nymph, (5) adult mite. (6) European red mite and (7) egg of same. (8) Brown mite. (9) Common red spider. (10) Bulb mite. (11) Pear-leaf blister mite. (12) Common woodlouse. (13) Garden millepede. (14) Garden slug. (15) Garden snail. (16) Bulb eelworm. (17) and (18) Immature and mature beet eelworm. (19) and (20) Immature and mature root knot eelworm.
Another group of mites of importance to the horticulturist is that of the blister mites; they are so minute—measuring about a hundred-and-fiftieth of an inch long—as to be invisible to the unaided eye. Though so minute, however, their damage to foliage is characteristic and conspicuous, so that their presence is easily detected. The most important blister mite in New Zealand is the pear-leaf blister mite (Eriophyes pyri); it differs from the other mites described above in having a long and cylindrical body, with only two pairs of legs crowded at the head end, the elongate abdomen having the appearance of being composed of innumerable segments (Fig. 13, 11). This mite lives in colonies in blisters formed on the leaf, and sometimes on the leaf petioles. In the spring the yellowish-green blisters will give the upper surface of an infested leaf a spotted appearance, and as the season advances these blisters become reddish and finally brown; in the case of severe infestation, the blisters become so crowded as to merge into masses.
During the winter the mites lie in the shelter of the bud scales; as soon as the foliage begins to develop in the spring the over-wintering mites attack the leaves, each mite forming a blister, in which it produces a colony of young. The offspring then migrate from the parent blister and form blisters for themselves, and this goes on until autumn, when the last generation of mites migrates for the winter to the shelter of the bud scales.
Owing to the mites being protected within the leaf blisters, summer sprays are not effective as a means of control, which can be effected, however, by spraying with lime-sulphur in the autumn, when the mites are taking up their winter quarters, and again at bud movement in the spring.
Woodlice are so well known, that but little description is necessary here. However, the following features are of interest. They belong to the group of animals known as the Crustacea, which also includes the crabs; these animals breathe by means of gills, and are usually aquatic, but some forms, such as the woodlice, have become adapted to a life on land. In outline (Fig. 13, 12) the woodlice are more or less oval, with the upper surfaces somewhat arched, and the lower flat; the body is divided into several segments, which may enable the animals to curl up in the form of a pill. There is a distinct head, bearing a pair of antennæ and the mouth-parts, followed by seven large thoracic segments, to each of which a pair of legs is attached; finally, the remaining six segments are more or less crowded together, and constitute the abdomen.
Since woodlice are terrestrial gill-breathing animals, moisture is essential for them, and it is in moist places that they abound. They depend upon a mixed diet, being carnivorous, as well as herbivorous; though normally scavengers, their attacks upon seedlings and tender parts of plants bring them into the ranks of important garden pests.
Woodlice hibernate under any convenient shelter; in the spring, eggs are produced and carried by the female on the under side of the body until the young woodlice hatch. During growth the cuticle or shell is periodically cast, and a freshly-moulted woodlouse is white in colour.
The best method of control is garden sanitation, all rubbish likely to harbour the woodlice being removed. Since they are nocturnal, the woodlice can be trapped by means of moss laid on the ground; the moss in which the woodlice have taken shelter is collected during the day and burned, or immersed in hot water to kill the animals, when it can be used again. Some good results have been secured by means of sliced potatoes dipped in arsenate of lead or Paris green; the potatoes are placed within reach of the woodlice, which are attracted to and feed upon the poisoned bait. Horse manure should not be used in seed beds likely to be infested by woodlice.
Millepedes are short, worm-like animals, with a fringe of numerous short legs on each side (Fig. 13, 13), and have a characteristic habit of curling up when disturbed. Though scavengers for the most part, feeding upon decaying vegetation and on small organisms, they may do considerable damage to sprouting seeds, seedlings, and to tender plants; they are particularly abundant in damp and warm soil, where there is an abundance of rotting vegetable matter.
Having a keen sense of smell, millepedes are readily attracted to poisoned bait in the form of sliced potato spread with Paris green: another method is to place a piece of freshly-cut potato under an inverted flower pot, to which the millepedes will be attracted, when they can be collected and destroyed. A satisfactory control measure is to treat infested soil with black-leaf 40, using one part in one thousand parts of water.
Plants are very often greatly damaged by the depredations of slugs and snails; frequently young plants are devoured as soon as they appear above ground. These animals attack the plants after nightfall, and during the day seek cover. Though slugs will shelter in the soil, they, together with snails, will shelter in any convenient place, such as under old boards, sacking, bricks and stones upon the ground, or under large leaves or amongst rank herbage—indeed, in almost any place that affords cover and moisture. Slugs and snails are especially active during wet weather, and at such times, owing to the overcast conditions, they will continue their depredations in the daytime.
Though slugs are active throughout the year, and even during winter when the temperature is favourable, snails pass the winter, as well as hot, dry spells in summer, in a dormant state, often being found together in sheltered positions where the conditions are dry.
Both slugs and snails copiously secrete a slimy substance, that affords them protection against chemicals used for purposes of control. In the case of the slug (Fig. 13, 14), the shell is small and inconspicuous, but the large spiral shell of the snail (Fig. 13, 15) affords the animal adequate protection, into which it withdraws itself in times of danger. Both slugs and snails reproduce by means of eggs; these are white, spherical and opaque, and are deposited in the soil or under decaying vegetation.
One of the best means of control is to dust the plants with powdered tobacco. Another method is to treat infested plants with soot or lime, but this must be done at night, and the material used must come into actual contact with the pests. An effective poison bait, but one that requires to be carefully handled, owing to its poisonous nature, is a mash made of 6lb. of bran mixed with 1lb. of arsenate of lead and an equal weight of treacle; this is made into a stiff paste, water being added if necessary. Lumps of this mash are placed about the plants to be protected. As a barrier to prevent the inroads of slugs and snails, plants may be surrounded by a belt of calcium cyanide; this would have to be replaced each night, and the utmost care taken in handling, since the substance and the gas evolved from it are highly poisonous; out of doors, however, the gas, being diluted with air, would not be very injurious as long as one did not stand over the treated ground longer than was necessary for laying the cyanide.
Apart from the above methods, the key to the control of slugs and snails is “clean farming”—that is, the removal of all places, such as rubbish and rank vegetation, where the animals will find shelter; the compost heap is a favourite breeding place, and this should be turned over at intervals and dressed with lime.
Eelworms are minute, unsegmented worms, related to the parasitic thread-worms of animals, and are abundant in soil and water; it is usually the surface layers of the richer soils that are inhabited by them. Of the long list of species, only a few are destructive to vegetation, but these constitute one of the greatest problems of the horticulturist. It is thought that the injury caused to plants by eelworms is toxic rather than mechanical, and some plants apparently are capable of producing anti-toxins, which neutralise the toxins of the eelworms; such plants possess an immunity. There are three important species in New Zealand.
The so-called bulb-eelworm (Anguillulina dipsaci) attacks more than two hundred kinds of plants, but is of especial interest to the horticulturist on account of its attacks upon hyacinths, daffodil, narcissus, and gladiolus, causing deformity and rotting of the tissues (Fig. 13, 16). It has been found that this eelworm develops from egg to adult within a period of between three and four weeks; the eggs are capable of lying dormant in the soil for as long as seven years. Infested bulbs and corms should be treated by immersion for three hours in water heated to 110 deg. Fahr.
Potatoes are often damaged by the beet eelworm (Heterodera schachtii), which causes what is known as “potato sickness,” when the growth is retarded, and wilting takes place; the root-system shows an abnormal development of secondary or “hunger-roots.” The eggs are retained in the body of the female, which forms a protective sack or cyst (Fig. 13, 17 and 18), and in this state the eggs pass the winter in the ground, where they are known to remain dormant for a period of ten years; under favourable conditions in the spring, the larvæ emerge from the eggs and attack the rootlets of suitable host plants, entering them at the extreme tip. Satisfactory methods of control have not yet been developed under field conditions, but a four-year crop rotation following potatoes is suggested; seed potatoes from infested ground should not be used.
The roots of tomatoes are often found to be a mass of galls, due to attack by the root-knot eelworm (Heterodera radicicola), which also infests tobacco roots as well as other plants (Fig. 13, 19 and 20). All stages of this species are to be found in the root galls; the female lays her eggs in a gelatinous egg sack, which remains attached to the parent. The larvæ, on hatching, either remain within the parent gall or leave it and enter the soil, where they seek out and attack the roots of another plant. In tomato gardens steam sterilisation of the soil is the most effective means of control.
In dealing with the control of plant pests, the objective is to prevent attacks, or, when the attacks have established, to check them as much as possible. In the latter case the term “exterminate” is in too frequent use; it is not usually practicable to exterminate a pest, and the best that can be done is to check or control it.
In the control of animal pests, it should be borne in mind that the pests are usually associated with other factors inimical to plant life, such as unthrifty plants, due to injury or malnutrition, and fungous and bacterial diseases, any one of which might be either the primary or secondary cause of plant injury.
Though at times one method may serve as a means of control, generally it is a combination of methods that gives the most satisfactory results, rendering the conditions favourable for the plant and unfavourable for the pests and diseases. The principles underlying control are:—
(a) Garden management.
(b) Use of chemicals.
(c) Influence of natural enemies.
All parts of plants, both above and below ground, are subject to infestation by pests and diseases. Under garden conditions, cultivation is intensive, and plants are grown year after year on the same ground in surroundings much more sheltered and crowded than in the open field. Sound garden management is therefore an important control factor, and the following features are fundamental:—
Condition of Soil.—The vigour of plants is dependent on the soil, which therefore must be kept in the right state; it must be well tilled, and must contain the requisite nourishment and moisture available for plant use, and as far as possible be free of an abnormal population of root feeding pests, such as eelworms and the larvæ of many insects. Proper cultivation is therefore the important factor in bringing the soil into the state most favourable to plant life, as all inimical factors, including pests, are reduced. Wherever practicable, as in glass-houses, soil-inhabiting pests and diseases can be completely controlled by steam sterilisation.
Importation of Pests.—One of the readiest methods of infesting a garden is the importation of pests on plants, and every care should be taken to secure only pest-free stock. In this respect, also, must be mentioned the use of stable and barnyard manure, in which pests such as insect larvæ, woodlice and eelworms are introduced; artificial fertilisers are therefore safer.
Overcrowding.—The tendency to overcrowd, especially in household gardens, is to be avoided; a favourite habit is to plant something in every available space. Under such conditions pests and diseases will abound, and before attempting to spread over a large area, and so lessen the effect of their depredations, they concentrate in mass formation within the confines of the garden as long as the food supply lasts; further, plants tend to be less vigorous and more susceptible to infestation under crowded than under more open conditions.
Injury to Plants.—Care should be taken not to injure plants with garden tools during cultivation, and a clean cut should always be the object in pruning. Mechanical injury opens the way for infestation by diseases and some insects.
Garden Sanitation.—Clean gardening is an extremely important control factor. In most gardens there are rank growths of grass and weeds in out-of-the-way places, along boundaries, and bordering cultivated plots. Such growths, especially when the weeds are related to the garden plants, are always favourite breeding places for many pests that move on to cultivated plants immediately they appear above ground. If these growths are cut and burned in the winter, and the ground thoroughly dug, many a spring infestation will be suppressed by the control of hibernating pests; it is the control of spring infestations that will save a great deal of trouble throughout the summer and autumn.
The compost heap, where garden refuse is dumped until sufficiently rotted, may be a source of infestation; not only does it attract and breed many destructive underground pests, but it may be infested with the spores of diseases harboured by the plant refuse of which it is composed; it is thus a ready means of reinfesting the soil. Diseased and pest-infested refuse should be burned without delay, and only healthy refuse used for the compost heap if not dug into the ground, where it will rot.
Crop Rotation.—Growing the one type of crop on the same piece of ground for several seasons encourages the development of pests and diseases; but by a rotation of different kinds of plants the continuity of the conditions favourable for the pests and diseases is broken, and the latter do not have the chance of becoming thoroughly established.
Diseases Spread by Pests.—It should be borne in mind that the fewer the animal pests, the less chance there is for diseases to spread. It is now well known that many pests, though not necessarily epidemic themselves, are carriers from plant to plant of certain destructive fungous, bacterial and virus diseases.
Co-operation.—In a locality of many gardens a co-operative spirit is essential, since a single neglected garden in an otherwise well-managed locality will be responsible for discounting the labours of the neighbours.
Chemicals are essential in the control of pests and diseases, and are applied either in the form of sprays or dusts. The former method is the more usual in this country, but where the water supply is poor dusts tend to take the place of sprays. Chemicals used for horticultural purposes are of two distinct kinds—those for the control of animal pests and those for the control of diseases. The commercial horticulturist, however, finds it necessary to apply both in the one spray or dust for the dual purpose of controlling both pests and diseases. As the present work is concerned with the pests, and not diseases, only those types of chemicals for the control of the former will be referred to.
Sprays and dusts are of three kinds, and act upon pests accordingly: they are either stomach poisons, or act externally on the animal by actual contact and corrosion, or cause death by fumigation. The kind used is governed by the feeding habits of the pest; if the latter is possessed of jaws (woodlice, caterpillars, beetles, etc.), and feeds by chewing the plant tissues, then a stomach poison is applied and is swallowed with the food; if the food is the nutrient sap of plants, and so could not be poisoned, a spray acting by contact is used, as against such animals as aphids (green fly), scale insects, etc., in which the mouth-parts are not adapted for chewing, but for puncturing plant tissues to feed on the sap, much the same as a mosquito punctures one’s skin and sucks the blood. Fumigants can be used against both the chewing and sucking pests, the fumes passing into the breathing system.
Stomach Poisons.—The chief of these are arsenate of lead and Paris green, though the latter has practically gone out of use. Arsenate of lead is sold as a paste and as a powder, and is mixed with water to form a spray, 3lb. of paste, or 1½lb. of powder, to 100 gallons of water being the proportions used. For garden purposes, smaller quantities must be kept to this strength.
Contacts.—The chemicals used in contact control are red oil, kerosene and lime-sulphur, but all are also fumigants, lime-sulphur being also a stomach poison to a limited extent, though best known as a fungicide. Commercial red oils can be purchased ready for mixing with water without the necessity of emulsification, and the strength at which each brand should be used is given by the manufacturers. Though red oils have mostly replaced kerosene emulsion, many horticulturists still prefer the latter. It is prepared by dissolving 8oz. of soap in one gallon of hot water, and then adding two gallons of kerosene, stirring briskly until emulsification is complete. This is the stock emulsion, and must be diluted before use, the strengths being one part to six of water for use in the winter, and one part to fifteen of water for use in the growing season. Commercial brands of concentrated lime-sulphur are on the market, and the manufacturers’ directions for their dilution should be followed.
Fumigants.—The chief fumigants are black-leaf 40, carbon-bisulphide and calcium cyanide.
Black-leaf 40, in which nicotine sulphate is the effective principle, is the most useful fumigant on the market, and acts as a most effective control for sap-sucking, and even some chewing pests. The strength at which this fumigant is used is one part in 800 parts of water, and is applied as a spray.
Carbon-bisulphide is a liquid, the gas evolved from it being an effective fumigant. It is not used as a spray unless emulsified, its chief use in horticulture being for the fumigation of the soil, glass-houses, stored seeds and vegetables, and imported plants. It is very inflammable and extremely volatile, especially under higher temperatures, the heavy gas being highly explosive when mixed with air.
The amount of carbon-bisulphide to be used varies, according to circumstances. For soil fumigation a special type of “gun” is on the market for injecting the bisulphide into the soil, but for ordinary garden purposes it is sufficient to make holes in the ground with a stick, pour in the fumigant, and close up the holes. When holes are made about 18 in. apart, half an ounce of bisulphide to a hole is sufficient, the depth of the hole varying according to the depth of the pest to be controlled.
For the fumigation of seeds, bulbs, potatoes, etc., an airtight chamber is necessary. This is also of value in the control of pests of potted plants. The dimensions of a chamber will vary according to the demands made upon it. Carbon-bisulphide gas being heavy, the containers (shallow dishes) should be placed on a shelf near the top of the chamber during fumigation. The proportion of fumigant to the air space varies according to the plants and insects to be fumigated.
For lawn-infesting insects, carbon-bisulphide can also be used in an emulsion as a spray prepared as follows:—Fifty grams of powdered resin are gradually added to 135 cc. of a 7 per cent. solution of sodium hydroxide, previously warmed; 450 cc. of hot water is now added, and the whole agitated until the resin is completely dissolved, when 50 cc. of oleic acid is also added. To prepare the emulsion, three parts of this soap solution are thoroughly agitated with seven parts of carbon-bisulphide until emulsification is complete, which can be gauged by the creamy-white colour and viscosity. For use dilute in the proportions of 18 pints of the emulsion with 50 gallons of water, applying by means of a watering-can or spray-pump at the rate of one gallon to every square foot of lawn.
Calcium cyanide, on being exposed to the atmosphere, gives off hydrocyanic acid gas, the evolution of the gas being governed by temperature and humidity. Calcium cyanide has replaced the old method of generating the gas by the action of sulphuric acid on potassium cyanide, and is sold in the form of dusts or granules. In the use of this material very great care is necessary, since the gas is highly poisonous, and also scorching of the foliage of plants results if atmospheric conditions are not considered carefully. With ordinary care, however, calcium cyanide can be safely handled. It is extremely effective against all kinds of pests, and can be used to fumigate soil, glass-houses, or as a dust on plants in the open.
As stated in the first chapter, plants are to be looked upon as the primary producers of life (since all animals are directly or indirectly dependent upon them), and the animals as the consumers. Many of the latter are destructive to crops grown by man, and become pests, but others, fortunately, exist upon these pests, and are classed as beneficial animals; it is the purpose of this section to deal with the more important of these from a horticultural viewpoint. In New Zealand such beneficial animals are insects, birds, and the hedgehog.
There is a wide range of insects that live at the expense of their fellows, and without these plant production would be impossible, either by Nature or by man. These so-called beneficial insects or parasites are the greatest factor in maintaining within reasonable bounds the insects that destroy vegetation; they are of much greater value in this respect than birds. In recent times the utilising of beneficial insects as a means of pest control has developed as one of the most important branches of entomological research.
From a general viewpoint, the beneficial insects are to be found mainly among the groups, including wasps, beetles, flies (two-winged insects) and lace-wings. The following are some examples:—
Common examples of parasitic insects are the ichneumon wasps (Fig. 14a), chalcid wasps (Fig. 14b), and ensign wasps, the first being the most conspicuous, the others less so owing to the minute size of many of them. A characteristic feature of these forms is the stalk-like attachment of the abdomen to the thorax and the sting-like ovipositor of the female, which may be of short or moderate length, sometimes projecting as a tail-like appendage beyond the end of the abdomen. Parasites deposit their eggs either upon or within the body of their victims or hosts, which are eventually destroyed by the larvæ hatching from the parasites’ eggs. Destructive caterpillars and their pupæ, and also aphides, are attacked by these wasp-like parasites, which in many cases restrict their depredations to one or a limited number of host species, while others are more general in their selection. Another group, the predaceous wasps, should be mentioned here. These insects in the adult state are hunters, and capture and paralyse by stinging such insects as caterpillars and flies, as well as spiders, which are stored in nests or cells for the nourishment of the predators’ offspring.
Important natural enemies of aphides and young caterpillars are the hover-flies, which can be easily recognised by their manner of flight. They are two-winged insects (Fig. 14c), and when on the wing hang motionless, as if suspended by some unseen means, to suddenly dart off with marvellous rapidity, until they hang motionless as before. These flies lay their eggs upon the foliage of plants infested by aphids or caterpillars, and from these eggs legless and headless larvæ emerge (Fig. 14d), and commence to search for and feed upon their victims.
Another important group of two-winged flies is the tachinids. They are rather robust, usually very bristly (Fig. 14e); they vary in size from that of a large blue-bottle to comparatively minute forms. The tachinids lay their eggs either upon their hosts or on the food plants of the latter, where they can be swallowed; some tachinids give birth to living larvæ, which crawl about in search of their victims.
Among the beneficial beetles are the well-known ladybirds (Fig. 14f); they are mostly oval in outline, dome-shaped above and flat below, while many of them are spotted by yellow, red, or white in a characteristic manner, though others are of one uniform colour. The eggs are laid on plants infested by the aphides and scale insects upon which the beetles and their larvæ (Fig. 14g) feed. There are other kinds of beetles of importance as predators, such as the common tiger-beetle, but they are not especially selective in their types of victims.
A very valuable group of insects includes the lace-wings or aphis-lions. The adult insects (Fig. 14h) carry the seemingly over-large lace-veined wings roof-like over the small body; the larvæ are alligator-like (Fig. 14i), and possess a pair of caliper-shaped jaws, by means of which they capture their prey. The eggs are laid directly on plants or are attached at the end of long stalks.
It is generally recognised that birds are a very important aid in keeping destructive insects in check, though it is well-known that a great deal of damage can be done by these animals. Without a systematic study of the stomach contents of birds, it is not possible to decide when a species is beneficial or injurious, and in New Zealand no such study has been made; practically all the information we have is based on field observations, which are, unfortunately, influenced largely by the outlook of the observer, and are thus misleading. Though some species subsist for the most part on insects, most land-birds have a mixed diet of vegetable and animal food, but they specialise on an insect diet when rearing their young and when moulting.

FIGURE 14.
(a) An ichneumon (natural size 1¼ in); (b) a chalcid (natural size 1-25 in); (c) a hoverfly (natural size ⅛ in); (d) hoverfly larva (natural size ¼ in); (e) a tachinid fly (natural size ¼ in); (f) a ladybird beetle (natural size ⅕ in); (g) ladybird larva (natural size ¼ in); (h) lacewing (natural size ¼ in); (i) lacewing larva (natural size ⅕ in).
Based on the nature of their diet, birds fall into three principal groups: (1) those feeding almost solely upon seeds and fruits; (2) insectivorous birds feeding on insects and other animals; and (3) the omnivorous species feeding both on insects and vegetable matter. The seed-feeding birds are a potential menace to the agriculturist, though in New Zealand the native species are fundamental to the well-being of the native forests; the insectivorous birds are obviously beneficial, though they devour both destructive and useful insects; while the omnivorous birds may be either useful or harmful, according to the circumstances. It should be remembered that, no matter what the food of the adult bird may be, most species give their young a diet of insects or other animal matter. When it is realised that the weight of nestling birds increases from one-fifth to one-half each day, requiring at times more than half the weight of the nestling in food, one can better visualise the enormous quantities of insects daily destroyed for this purpose. Consider the common house sparrow, which is usually condemned: an analysis of the nestling diet has shown that it consisted of 40 per cent. grain and 60 per cent. insects and related forms, while that of the adult comprised 75 per cent. grain and 25 per cent. insects, etc.
To summarise the situation, it may be said that, on the whole, enormous numbers of insects are destroyed by birds each year, and, unless allowed to become abnormally abundant, the benefit derived from birds outweighs the damage they may cause.
The hedgehog was first introduced by the Canterbury Acclimatisation Society in 1870, and later by other societies and private individuals. The animal is now very abundant in many parts of the Dominion. Though condemned and destroyed by some people, who consider it a menace to eggs, chickens and even vegetables, the hedgehog is really a very useful animal, in that, being a night prowler itself, it destroys numerous nocturnal pests, such as slugs and snails, earwigs, grass caterpillars and cut-worms.
The hedgehog, on the approach of winter, constructs a nest in some suitable place, where it becomes torpid and hibernates. On the advent of spring, it becomes active once more, and during summer produces a litter of four young; a second litter is sometimes produced in the autumn.
| Page | Page | |
Iris35 | ||
Agrotis ypsilon52 | ||
Japonica36 | ||
Amaryllis66 | ||
American blight47 | Karaka40 | |
Amœbæ15 | Kerosene76 | |
Anguillulina dipsaci72 | Kowhai52 | |
Animal Kingdom, Divisions of8 | Kowhai moth52 | |
Aphelinus fuscipennis40 | Kumara54 | |
Aphelinus mali47 | ||
Aphides42 | Ladybird beetles78 | |
Larch46 | ||
Aphis persicæ-niger44 | Larger narcissus fly66 | |
Larva24 | ||
Apple leaf-hopper48 | ||
Apple leaf-roller51 | ||
Apple mealy-bug34 | Lead arsenate76 | |
Leaf-hoppers48 | ||
Apple red-mite57 | Leaf-mining flies64 | |
Leaf-rollers51 | ||
Army-worms54 | Leech59 | |
Arsenate of lead76 | ||
Asexual42 | Lepidosaphes ulmi37 | |
Ash38 | Lepisma saccharina26 | |
Asparagus36 | Lettuce55 | |
Aspidiotus hederæ40 | Leucaspis cordylinidis38 | |
Aspidiotus perniciosus38 | Leucaspis stricta38 | |
Asterolecanium variolosum37 | Leucojum66 | |
Austrian pine46 | Life-cycle of insects22 | |
Lime-sulphur76 | ||
Loganberry41 | ||
Bag-moth56 | Long-horned beetles64 | |
Baker’s mealy-bug35 | Long-tailed mealy-bug34 | |
Beet eelworm72 | Lupins52 | |
Beetles56 | ||
Begonia34 | Macrosiphum rosæ47 | |
Birds79 | Management of garden74 | |
Manure74 | ||
Blackbird35 | ||
Black cherry-aphis44 | Mealy-wings43 | |
Mecyna maorialis52 | ||
Black-leaf 4076 | Merodon equestris66 | |
Black peach-aphis44 | Metamorphosis24 | |
Black scale40 | ||
Blister-mites70 | Millepedes71 | |
Blue-gum59 | Mites67 | |
Borers, round-headed64 | Moulting22 | |
Breathing systems of insects21 | Mouth appendages of insects19 | |
Brevicoryne brassicæ45 | ||
Briar41 | ||
Bronze beetle58 | ||
Broods of insects22 | Myrtle36 | |
Broom52 | Myzus cerasi44 | |
Brown-beetle58 | ||
Brown-mite68 | ||
Brussels sprouts45 | Narcissus flies66 | |
Bryobia prætiosa68 | Natural enemies77 | |
Bulb-eelworm72 | ||
Bulb-mite68 | Nervous systems of insects21 | |
Bulb or Narcissus flies66 | Nicotine-sulphate76 | |
Northern Spy47 | ||
Novius cardinalis32 | ||
Cabbage aphis45 | ||
Cabbage green-fly45 | ||
Cabbage-tree56 | ||
Cabbage-tree moth56 | Oak-scale37 | |
Cabbage-tree scales38 | ||
Cabbage white butterfly55 | Oeceticus omnivorus56 | |
Calcium cyanide76 | ||
Caliora limacina59 | Oleander-scale40 | |
Olearia forsteri60 | ||
Camellia scale37 | Olearia gall-midge60 | |
Carbon-bisulphide76 | ||
Caterpillars51 | Onion66 | |
Caterpillars in lawns65 | ||
Orange-scale40 | ||
Cecidomyia oleariæ60 | ||
Cereals54 | ||
Chalcid wasps78 | Oviparous42 | |
Chemicals75 | Ovipositor21 | |
Chermes pini46 | ||
Cherry-aphis, black44 | Paratetranychus pilosus67 | |
Cherry slug59 | Paris green76 | |
Chitin17 | Paropsis dilatata59 | |
Chrysalis25 | Parthenogenetic42 | |
Chrysanthemum64 | Passion vine34 | |
Chrysomphalus aurantii40 | ||
Chrysomphalus rossi40 | ||
Cicada28 | Peach-aphis, black44 | |
Peach-aphis, green44 | ||
Citrophilus mealy-bug34 | ||
Pear and cherry saw-fly59 | ||
Click-beetles65 | Pear and cherry slug59 | |
Clover52 | Pear-leaf blister mite70 | |
Coccids29 | Pear-midge60 | |
Coccus hesperidum36 | Pelargonium51 | |
Cockchafers57 | Pemphigus populi-transversus48 | |
Pepper-tree35 | ||
Cocoon25 | Perrisia pyri60 | |
Codlin moth61 | Persimmon35 | |
Comstock’s mealy-bug35 | Phormium34 | |
Contact sprays and dusts76 | Phthorimæa operculella62 | |
Convolvulus54 | Phylloxera vastatrix46 | |
Coprosma40 | Pieris rapæ55 | |
Cornicles42 | Pine-tree chermes46 | |
Plant food12 | ||
Crayfish17 | Plant-lice42 | |
Crocus68 | Plum-aphis48 | |
Crop rotation75 | Plutella maculipennis52 | |
Cruciferous crops45 | Pontania proxima60 | |
Cryptolæmus ladybird34 | ||
Cryptolæmus montrouzieri34 | ||
Curl-grubs58 | Porina65 | |
Currant clear-wing borer61 | Potato sickness72 | |
Cuticle17 | Potato tuber-moth62 | |
Cut-worms52 | Predaceous wasps78 | |
Cydia pomonella61 | Privet38 | |
Daffodil72 | Pulvinaria camelicola37 | |
Development of insects22 | Pupa25 | |
Diamond-backed moth52 | Puparium25 | |
Digestive system of insects21 | Pyronota festiva58 | |
Dusts75 | ||
Earthworms15 | Radish55 | |
Easter lily68 | ||
Eggs of insects22 | Red currant44 | |
Elder35 | Red oil76 | |
Eleagnus37 | Red orange-scale40 | |
Red spider68 | ||
Rhizoglyphus hyacinthi68 | ||
Elytra58 | Rhododendron47 | |
Ensign wasps78 | Rhophalosiphum nymphææ48 | |
Eriococcus coriaceus35 | Rhophalosiphum persicæ44 | |
Eriophyes pyri70 | Root-knot eelworm of tomato73 | |
Eriosoma lanigerum47 | ||
Eucalyptus tortoise beetle59 | Rotation of crops75 | |
Eucolaspis brunneus58 | Round-headed borers64 | |
Eulecanium berberidis37 | ||
Eulecanium corni36 | Saissetia hemispherica36 | |
Eumerus strigatus66 | Saissetia oleæ35 | |
European earwig50 | Scale insects29 | |
European red-mite67 | Scylla66 | |
Seasonal history of insects22 | ||
Fantail35 | ||
Sesia tipuliformis61 | ||
Shallot66 | ||
Flax, New Zealand38 | ||
Flea-beetles58 | Silver-fish26 | |
Forficula auricularia50 | ||
Fruit lecanium scale36 | Smaller narcissus fly66 | |
Fumigants76 | ||
Soil fumigation76 | ||
Sphinx convolvuli54 | ||
Galls42 | Sphinx moth54 | |
Galtonia66 | ||
Gladiolus72 | Sprays75 | |
Gnorimoschema plæsiosema62 | Springtails66 | |
Goat-willow64 | Spruce46 | |
Golden oak-scale37 | Steam sterilisation74 | |
Gonipterus scutellatus59 | ||
Stomach poisons76 | ||
Gorse32 | ||
Grape louse46 | Subterranean grass caterpillars65 | |
Grape phylloxera46 | ||
Syrphids66 | ||
Grass caterpillars65 | ||
Tachinids78 | ||
Tetranychus telarius68 | ||
Greasy cut-worms52 | Thrush35 | |
Green-fly of cabbage45 | ||
Green-house thrips49 | ||
Green-house white-fly48 | Tomato root-knot eelworm73 | |
Green manuka beetle58 | Tomato stem-borer62 | |
Green peach-aphis44 | Tortoise beetle59 | |
Tortrix postvittana51 | ||
Grub-proofing lawns64 | Tree-lucerne64 | |
Gum-tree scale35 | Trialeurodes vaporariorum48 | |
Gum-tree weevil59 | Tui35 | |
Habranthus66 | ||
Habrolepis dalmanni37 | Turnip-fly57 | |
Hawk moth54 | Turtle-scale36 | |
Typhlocyba australis48 | ||
Hedgehog80 | ||
Heliothrips hæmorrhoidalis23 | Vallota66 | |
Hemispherical scale36 | Vegetable caterpillars65 | |
Heterodera radicola73 | Venusia verriculata56 | |
Heterodera schachtii72 | Vertebrates7 | |
Hibernation23 | Violet68 | |
Viviparous42 | ||
Honey-bee19 | ||
Walnut35 | ||
Honey-tubes42 | ||
Hop68 | Water lilies48 | |
Horse chestnut35 | ||
Wax-eye35 | ||
Weta17 | ||
Hydrangea35 | White butterfly55 | |
Hydrocyanic-acid gas77 | White-flies48 | |
Hypopus68 | ||
Willow saw-fly60 | ||
Icerya purchasi32 | Wings of insects20 | |
Ichneumon wasps78 | Wireworms65 | |
Importation of pests74 | ||
Insects, proportion of7 | ||
Invertebrates7 |