OUR PRESENT KNOWLEDGE OF THE DESCENT OF MAN
BY
ERNST HAECKEL
(JENA)
WITH NOTES AND BIOGRAPHICAL SKETCHES
BY
HANS GADOW, F.R.S.
(CAMBRIDGE)
LONDON
ADAM AND CHARLES BLACK
1898
| page | |
| THE LAST LINK | |
| INTRODUCTORY | 1 |
| COMPARATIVE ANATOMY | 8 |
| PALÆONTOLOGY | 20 |
| OTHER EVIDENCE | 42 |
| STAGES RECAPITULATED | 47 |
| BIOGRAPHICAL SKETCHES: | |
| LAMARCK, SAINT-HILAIRE, CUVIER, BAER, | |
| MUELLER, VIRCHOW, COPE, KOELLIKER, GEGENBAUR, | |
| HAECKEL | 80 |
| THEORY OF CELLS | 115 |
| FACTORS OF EVOLUTION | 117 |
| GEOLOGICAL TIME AND EVOLUTION | 135 |
The address I delivered on August 26 at the Fourth International Congress of Zoology at Cambridge, 'On our Present Knowledge of the Descent of Man,' has, I find, from the high significance of the theme and the general importance of the questions connected with it, excited much interest, and has led to requests for its publication. Hence this volume, edited by my friend Dr. H. Gadow, my pupil in earlier days, who has not only revised the text, but has also enriched it by many valuable additions and notes.
ERNST HAECKEL.
Jena, December, 1898.
At the end of the nineteenth century, the age of 'natural science,' the department of knowledge that has made most progress is zoology. From zoology has arisen the study of transformism, which now dominates the whole of biology. Lamarck[1] laid its foundation in 1809, and forty years ago Charles Darwin obtained for it a recognition which is now universal. It is not my task to repeat the well-known principles of Darwinism. I am not concerned to explain the scientific value of the whole theory of descent. The whole of our biological study is pervaded by it. No general problem in zoology and[Pg 2] botany, in anatomy and physiology, can be discussed without the question arising, How has this problem originated? What are the real causes of its development?
This question was almost unknown seventy years ago, when Charles Darwin, the great reformer of biology, began his academical career at Cambridge as a student of theology. In the same year, 1828, Carl Ernst von Baer[2] published in Germany his classical work on the embryology of animals, the first successful attempt to elucidate by 'observation and reflection' the mysterious origin of the animal body from the egg, and to explain in every respect the 'history of the growing individuality.' Darwin at that time had no knowledge of this great advance, and he could not divine that forty years later embryology would be one of the strongest supports of his own life's work—of that very theory of transformism which, founded by Lamarck in the year of Darwin's birth,[Pg 3] was accepted with enthusiasm by Charles's grandfather Erasmus. There is no doubt that of all the celebrated naturalists of the nineteenth century Darwin achieved the greatest success, and we should be justified in designating the last forty years as the Age of Darwin.
In searching for the causes of this unexampled success, we must clearly separate three sets of considerations: first, the comprehensive reform of Lamarck's transformism, and its firm establishment by the many arguments drawn from modern biology; secondly, the construction of the new theory of selection, as established by Darwin, and independently by Alfred Wallace (a theory called Darwinism in the proper sense); thirdly, the deduction of anthropogeny, that most important conclusion of the theory of descent, the value of which far surpasses all the other truths in evolution.
It is the third point of Darwin's theory that I shall discuss here; and I shall discuss[Pg 4] it chiefly with the intention of examining critically the evidence and the different conclusions which at present represent our scientific knowledge of the descent of man and of the different stages of his animal pedigree.
It is now generally admitted that this problem is the most important of all biological questions. Huxley was right when in 1863 he called it the question of questions for mankind. The problem which underlies all others, and is more deeply interesting than any other, is as to the place which man occupies in nature and his relations to the universe of things. 'Whence our race has come; what are the limits of our power over nature, and of nature's power over us; to what goal are we tending—these are the problems which present themselves anew and with undiminished interest to every man born into the world.' This impressive view was explained by Huxley thirty-five years ago in his three celebrated essays on 'Evidence[Pg 5] as to Man's Place in Nature.' The first is entitled 'On the Natural History of the Man-like Apes'; the second, 'On the Relations of Man to the Lower Animals'; the third, 'On some Fossil Remains of Man.' Darwin himself felt the burden of these problems as much as Huxley; but in his chief work, 'On the Origin of Species,' in 1859, he had purposely only just touched them, suggesting that the theory of descent would shed light upon the origin of man and his history. Twelve years later, in his celebrated work on 'The Descent of Man, and Selection in Relation to Sex,' Darwin discussed fully and ingeniously all the different sides of this 'question of questions' from the morphological, historical, physiological, and psychological points of view. As early as 1866 I myself had applied in the Generelle Morphologie der Organismen the theory of transformism to anthropology, and had shown that the fundamental law of biogeny claims the same value for man as for all the other animals. The intimate causal[Pg 6] connection between ontogeny and phylogeny, between the development of the individual and the history of its ancestors, enables us to gain a safe and certain knowledge of our ancestral series. I had at that time distinguished in this series ten chief degrees of vertebrate organization. I attributed the highest importance to the logical connection of anthropogeny with transformism. If the latter be true, the truth of the former is absolute. 'Our theory that man is descended from lower vertebrates, and immediately from apes or primates, is a case of special deduction which follows with absolute certainty from the general induction of the theory of descent.' The full proof and detailed explanation of this view was afterwards given in my 'History of Natural Creation,' and especially in my 'Anthropogeny.'[3] Lastly, it has received an ample scientific and critical foundation in the third part of my 'Systematic Phylogeny.'[3]
During the forty years which have elapsed since Darwin's first publication of his theories an enormous literature, discussing the general problems of transformism as well as its special application to man, has been published. In spite of the wide divergence of the different views, all agree in one main point: the natural development of man cannot be separated from general transformism. There are only two possibilities. Either all the various species of animals and plants have been created independently by supernatural forces (and in this case the creation of man also is a miracle); or the species have been produced in a natural way by transmutation, by adaptation and progressive heredity (and in this case man also is descended from other vertebrates, and immediately from a series of primates). We are absolutely convinced that only the latter theory is fully scientific. To prove its truth, we have to examine critically the strength of the different arguments claimed for it.
First, we have to consider the relative place which comparative anatomy concedes to man in the 'natural system' of animals, for the true value of our 'natural classification' is based upon its meaning as a pedigree. All the minor and major groups of the system—the classes, legions, orders, families, genera, and species—are only different branches of the same pedigree. For man himself, his place in the pedigree has been fixed since Lamarck,[5] in 1801, defined the group of vertebrates. The most perfect[6] of these are[Pg 9] the Mammalia; and at the head of this class stands the order of Primates, in which Linnæus, in 1735, united four 'genera'—Homo, Simia, Lemur, and Vespertilio. If we exclude the last-named, the Chiroptera of modern zoology, there remain three natural groups of Primates—the Lemures, the Simiæ, and the Anthropi or Hominidæ. This is the classification[Pg 10] of the majority of zoologists; but if we compare man with the two chief groups of monkeys—the Eastern monkeys (or Catarrhinæ) and the Western or American monkeys (Platyrrhinæ)—there can be no doubt that the former group is much more closely related to man than is the latter. In the natural order of the Catarrhinæ we find united a long series of lower and higher forms. The lowest, the Cynopitheci, appear still closely related to the Platyrrhinæ and to the Lemures; while, on the other hand, the tailless apes (Anthropomorphæ) approach man through their higher organization. Hence one of our best authorities on the Primates, Robert Hartmann,[7] proposed to subdivide the whole order of the Simiæ into three groups:
(1) Primarii, man together with the other Anthropomorphæ, or tailless apes; (2) Simiæ, all the other monkeys; (3) Prosimiæ, or Lemurs. This arrangement has received[Pg 11] strong support from the interesting discovery by Selenka that the peculiar placentation of the human embryo is the same as in the great apes, and different from that of all the other monkeys. Our choice between these different classifications of Primates is best determined by the important thesis of Huxley, in which, in 1863, he carried out a most careful and critical comparison of all the anatomical gradations within this order. In my opinion, this ingenious thesis—which I have called the Huxleyan Law, or the 'Pithecometra-thesis of Huxley'—is of the utmost value. It runs as follows: 'Thus, whatever system of organs be studied, the comparison of their modifications in the ape-series leads to one and the same result—that the structural differences which separate man from the gorilla and the chimpanzee are not so great as those which separate the gorilla from the lower apes.' If we accept the Huxleyan law without prejudice, and apply it to the natural classification of the Primates, we[Pg 12] must concede that man's place is within the order of the Simiæ. On examining this relation with care, and judging with logical persistence, we may even go a step further. Instead of the wider conception of 'Simiæ,' we must use the restricted term of Catarrhinæ, and our Pithecometra-thesis has then to be formulated as follows: The comparative anatomy of all organs of the group of Catarrhine Simiæ leads to the result that the morphological differences between man and the great apes are not so great as are those between the man-like apes and the lowest Catarrhinæ. In fact, it is very difficult to show why man should not be classed with the large apes in the same zoological family. We all know a man from an ape; but it is quite another thing to find differences which are absolute and not of degree only. Speaking generally, we may say that man alone combines the four following features: (1) Erect walk; (2) extremities differentiated accordingly; (3) articulate speech; (4) higher[Pg 13] reasoning power. Speech and reason are obviously relative distinctions only—the direct result of more brains and more brain-power, the so-called mental faculties. The erect walk is not an absolutely distinguishing characteristic: the large apes likewise walk on their feet only, supporting their bodies by touching the ground with the backs of their hands—in fact, with their knuckles—and this is a mode of progression very different from that of the tailed monkeys, which walk upon the palms of their hands. There are, however, two obvious differences in the development of the muscles. In man alone the gastrocnemius and the soleus muscle are thick enough to form the calf of the leg, and the glutæus maximus is enlarged into the buttocks. A fourth glutæal muscle occurs occasionally in man, while it is constantly present in apes as the so-called musculus scansorius. Concerning the muscles of the whole body, we cannot do better than quote Testut's summary: 'The mass of recorded[Pg 14] observations upon the muscular anomalies in man is so great, and the agreement of many of these with the condition normal in apes is so marked, that the gap which usually separates the muscular system of man from that of the apes appears to be completely bridged over.'
There are, for example, the muscles of the
ear. In most people the majority, or even
all of them, are no longer movable at will,
while in the apes they are still in use. The
important point, however, is that these
muscles are still present in man, although
often in a reduced condition. They are the
following: (1) Musculus auricularis anterior
or attrahens auris, which is frequently much
reduced and no longer reaches the ear at all,
being then absolutely useless; (2) Musculus
auricularis superior or attollens auris, more
constant than the former; (3) Musculus
auricularis posterior or retrahens auris, likewise
often functional. Occasionally smaller
slips differentiated from these three muscles
are present, and as so-called intrinsic muscles[Pg 15]
[Pg 16]
are restricted to the ear itself; their function
is, or was, that of curling up or opening the
external ear.
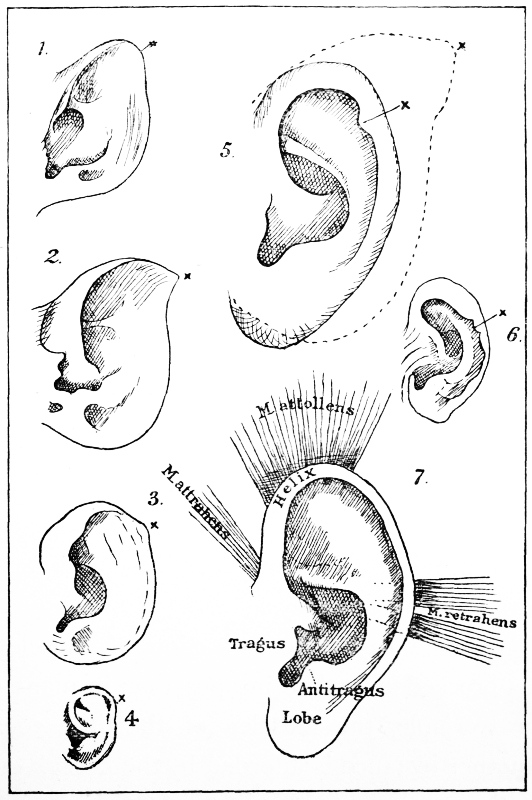
Outlines of the Left Ear of—
1. Lemur macaco; 2. Macacus rhesus, the Rhesus monkey; 3. Cercopithecus, a macaque; 4. human embryo of six months; 5. man, with Darwin's point well retained: the dotted outline is that of the ear of a baboon; 6. orang-utan (after G. Schwalbe):[8] x the original tip of the ear; 7. human ear with the principal muscles.
In connection with the ear, I may touch upon another interesting and most suggestive little feature which is present in many individuals—namely, 'Darwin's point.' This is the last remnant of the original tip of the ear, before the outer, upper, and hinder rim became doubled up or folded in. It is a feature quite useless, and absolutely impossible of interpretation, excepting as the vestige of such previous ancestral conditions as are normal in the monkeys.
In some cases the reduction of muscles has proceeded further in apes than in man—for example, the muscles of the little toe. Another instance is afforded by the coccyx or vestige of the tail; this is still furnished with muscles which are now in man, as well as in the apes, quite useless, and vary considerably with every sign of degeneration, most so in the orang-utan.
Darwin has mentioned the frequent action of the 'snarling muscle,' by which, in sneering, our upper canine teeth are exposed, like those of a dog prepared to fight.
Monkeys and apes possess vocal sacs, especially large in the orang-utan; survivals of them, although no longer used, persist in man in the shape of a pair of small diverticula, the pouches of Morgagni, between the true and the false vocal cords.
'In the native Australians, the dental formula appears least removed from the hypothetical original type, for in it are still found complete rows of splendid teeth, with powerfully-developed canines and molars, the latter being either uniform, or even increasing in size, as we proceed backwards, in such a way that the wisdom tooth is the largest of the series. This is decidedly a pithecoid characteristic which is always found in apes. The upper incisors of the Malay, apart from their prognathous disposition, have occasionally a distinctly pithecoid form, their anterior[Pg 18] surface being convex, and their lingual surface slightly concave. The ancestors of Europeans seem to have had the same form of teeth, for the oldest existing fragments of skulls from the Mammoth age (e.g., the jaws from La Naulette, in Belgium) reveal tooth-forms which must be classed with those of the lowest races of to-day.'[9]
Now we are able to apply this fundamental Pithecometra-thesis directly to the classification of the Primates and to the phylogeny of man, which is intimately connected with it, because in this order, as in all the other groups of animals, the natural system is the clear expression of true phylogenetic affinity. Four results follow from our thesis: (1) The Primates, as the highest legion or order of mammals, form one natural, monophyletic group. All the Lemures, Simiæ, and Homines descend from one common ancestral[Pg 19] form, from a hypothetical 'Archiprimas.' (2) The Lemures are the older and lower of the natural groups of the Primates; they stand between the oldest Placentalia (Prochoriata) and the true Simiæ. (3) All the Catarrhinæ, or Eastern Simiæ, form one natural monophyletic group. Their hypothetical common ancestor, the Archipithecus, may have descended directly or indirectly from a branch of the Lemures. (4) Man is descended directly from one series of extinct Catarrhine ancestors. The more recent ancestors of this series were tailless anthropoids (similar to the Anthropopithecus), with five sacral vertebræ. The more remote ancestors were tailed Cercopitheci, with three or four sacral vertebræ.
These four theses possess, in my opinion, absolute certainty. They are independent of all future anatomical, embryological, and palæontological discoveries which may possibly throw more light upon the details of our phyletic anthropogenesis.
The next question is, how the facts of palæontology agree with these most important results of comparative anatomy and ontogeny. The fossils are the true historical 'medals of creation,' the palpable evidence of the historical succession of all those innumerable organic forms which have peopled the globe for many millions of years. Here the question arises, If the known fossil specimens of Mammalia, and particularly of Primates, give proof of these Pithecometra-theses, do they confirm directly the descent of man from ape-like creatures? The answer to this question is, in my opinion, affirmative.
It is true that the gaps in the palæontological evidence, here as elsewhere, are[Pg 21] many and keenly felt. In the order of the Primates they are greater than in many other orders, chiefly because of the arboreal life of our ancestors. The explanation is very simple. It is really due to a long chain of favourable coincidences if the skeleton of a vertebrate, covered as it was with flesh and skin, and containing still more perishable viscera, is petrified at all. The body may be devoured by other creatures, and its bones scattered about; or it rots away and crumbles to pieces. Many animals hide in thick undergrowth when death approaches them; and, leading an almost entirely arboreal life, the Primates are especially likely to disappear without being fossilized. It is only when the body is quickly covered with sand, or is embedded in suitable lime or silica containing mud, that the process of petrifaction can come to pass. Even then it is only by great good luck that we come across such a fossil. Very few countries have been searched systematically, and the areas that have been[Pg 22] searched amount to little in comparison with the whole surface of the land, even if we leave out of account the fact that more than two-thirds of the globe are covered by water.
These deplorable deficiencies of empirical palæontology are balanced on the other side by a growing number of positive facts, which possess an inestimable value in human phylogeny. The most interesting and most important of these is the celebrated fossil Pithecanthropus erectus, discovered in Java in 1894 by Dr. Eugène Dubois.[10] Three years ago this now famous ape-like man provoked an animated discussion at the third International Zoological Congress at Leyden. I may therefore be allowed to say a few words as to its scientific significance. Unfortunately, the fossil remains of this creature are very scanty: the skull-cap, a femur, and two teeth. It is obviously impossible to form[Pg 23] from these scanty remains a complete and satisfactory reconstruction of this remarkable Pliocene Primate.
The more important points are the following: The remains in question rested upon a conglomerate which lies upon a bed of marine marl and sand of Pliocene age. Together with the bones of Pithecanthropus were found those of Stegodon, Leptobos, Rhinoceros, Sus, Felis, Hyæna, Hippopotamus, Tapir, Elephas, and a gigantic Pangolin. It is remarkable that the first two of these genera are now extinct, and that neither hippopotamus nor hyæna exists any longer in the Oriental region. If we may judge from these fossil remains, the bones of Pithecanthropus are not younger than the oldest Pleistocene, and probably belong to the upper Pliocene. The teeth are like those of man. The femur, also, is very human, but shows some resemblances to that of the gibbons. Its size, however, indicates an animal which stood when erect not less than 5 feet 6 inches high. The[Pg 24] skull-cap also is very human, but with very prominent eyebrow ridges, like those of the famous Neanderthal cranium. It is certainly not that of an idiot. It had an estimated cranial capacity of about 1,000 cubic centimetres—that is to say, much more than that of the largest ape, which possesses not more than 600 c.c. The crania of female Australians and Veddahs measure not more than 1,100, some even less than 1,000 c.c.; but, as these Veddah women stand only about 4 feet 9 inches high, the computed cranial capacity of the much taller Pithecanthropus is comparatively very low indeed.[11]
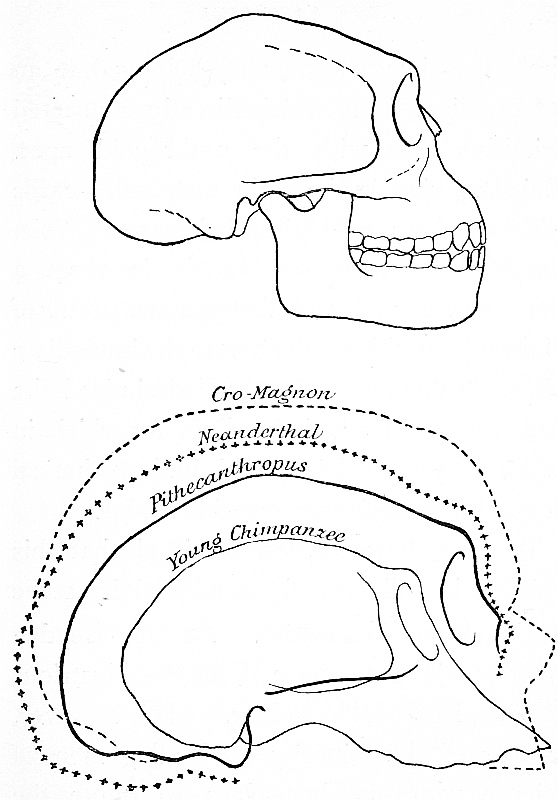
The upper figure represents the outlines of the skull of Pithecanthropus, as restored by Manouvier.[12] The lower figure shows the comparative size and shape of Pithecanthropus, the Neanderthal skull, a specimen of the Cro-Magnon race of neolithic France, and a Young Chimpanzee before the full development of the supraorbital crests.
The final result of the long discussion at
Leyden was that, of twelve experts present,
three held that the fossil remains belonged
to a low race of man; three declared them
to be those of a man-like ape of great size;[Pg 25]
[Pg 26]
the rest maintained that they belonged to an
intermediate form, which directly connected
primitive man with the anthropoid apes.
This last view is the right one, and accords
with the laws of logical inference. Pithecanthropus
erectus of Dubois is truly a
Pliocene remainder of that famous group of
highest Catarrhines which were the immediate
pithecoid ancestors of man. He is, indeed, the
long-searched-for 'missing link,' for which, in
1866, I myself had proposed the hypothetical
genus Pithecanthropus, species Alalus.
It must, however, be admitted that this opinion is still strongly combated by some distinguished authorities. At the Leyden Congress it was attacked by the illustrious pathologist Rudolf Virchow.[13] He, however, is one of the minority of leading men of science who set themselves to refute the theory of Evolution in every possible way. For thirty years he has defended the thesis: 'It is quite certain that man is not a[Pg 27] descendant of apes.' He declares any intermediate form to be unimaginable save in a dream.
Virchow went to the Leyden Congress with the set purpose of disproving that the bones found by Dubois belonged to a creature which linked together apes and man. First, he maintained that the skull was that of an ape, while the thigh belonged to man. This insinuation was at once refuted by the expert palæontologists, who declared that without the slightest doubt the bones belonged to one and the same individual. Next, Virchow explained that certain exostoses or growths observable on the thigh proved its human nature, since only under careful treatment the patient could have healed the original injury. Thereupon Professor Marsh, the celebrated palæontologist, exhibited a number of thigh-bones of wild monkeys which showed similar exostoses and had healed without hospital treatment. As a last argument the Berlin pathologist declared that[Pg 28] the deep constriction behind the upper margin of the orbits proved that the skull was that of an ape, as such never occurred in man. It so happened that a few weeks later Professor Nehring of Berlin demonstrated exactly the same formation on a human prehistoric skull received by him from Santos, in Brazil.
Virchow was, in fact, just as unlucky in Leyden in his fight with our pliocene ancestor as he had been unfortunate in his opinion on the famous skulls of Neanderthal, Spy, La Naulette, etc., every one of which he explained as a pathological abnormality. It would be a very curious coincidence indeed if all these and other fossil human remains were those of idiots or otherwise abnormal individuals, provided they are old and low enough in their organization to be of phylogenetic value to the unbiassed zoologist.
As the sworn adversary of Evolution, transformism, and Darwinism in particular, but a believer in the constancy of species, the[Pg 29] great and renowned pathologist has been driven to the incredible contention that all variations of organic forms are pathological.
Four years ago, as honorary president of the Anthropological Congress at Vienna, he attacked Darwinism in the severest manner, and declared that 'man may be as well descended from the elephant or from the sheep as from the ape.' Such attacks on the theory of transformism indicate a failure to understand the principles of the theory of Evolution and to appreciate the significance of palæontology, comparative anatomy, and ontogeny.
The thousands of other objections which have been made during the last forty years (chiefly by outsiders) may be passed over in silence. They do not require serious refutation. In spite of, or perhaps because of, these attacks, the theory of Evolution stands established more firmly than ever.
It is easy for the outsider to exult over the difficulties which our problem implies—diffi[Pg 30]culties which we who have given our lives to the study understand likewise, and try our best not only to bridge over, but also to point out. Anyhow, we do not conceal them; while those who reject the explanation offered by Evolution make the most of the gaps, and pass silently over the far more numerous points favourable to our theory.
How fruitful during the last thirty years the astonishing progress in our palæontological knowledge has been for our Pithecometra-thesis is best shown by a short glance at the growth of our knowledge of fossil Primates. Cuvier,[14] the founder of palæontology, continued up to the time of his death, in 1832, to assert that fossil remains of monkeys and lemurs did not exist. The only skull of a fossil lemuroid which he described (namely, Adapis) he declared to be that of an ungulate. Not until 1836 were the first fragments of extinct monkeys found in India; it was two years later, near Athens,[Pg 31] that the skeleton of Mesopithecus penthelicus was discovered. Other remains of lemurs were found in 1862. But during the last twenty years the number of fossil Primates has been augmented by the remarkable discoveries of Gaudry, Filhol, Milne Edwards, Seeley, Schlosser, and others in Europe; of Marsh, Cope, Osborn, Leidy, Ameghino, in South America; and Forsyth Major in Madagascar.[15] These tertiary remains, chiefly[Pg 32] of Eocene and Miocene date, fill many gaps between existing genera of Primates, and afford us quite a clear insight into the phyletic development of this order during the millions of years of the Cænozoic age.
The most important difference between the two groups of existing monkeys is indicated by their dentition. Adult man possesses, like all the other Catarrhine Simiæ, thirty-two teeth, whilst the American monkeys (the Platyrrhinæ) have thirty-six teeth—namely, one pair of premolars more in the upper and lower jaws. Comparative odontology leads us to the phylogenetic conclusion that this number has been produced by reduction from a still older form with forty-four teeth. This typical dental formula (three incisors, one canine, four premolars, and three molars, in each half-jaw) is common to all those most important older mammals which in the beginning of the Eocene period constituted the four large groups of Lemuravida, Condylarthra, Esthonychida, and Ictopsida. These are the[Pg 33] four ancestral groups of the four main orders of Placentalia—namely, of the Primates, Ungulata, Rodentia, and Carnassia. They seem to be so closely related by their primitive organization that they may be united in one common super-order, Prochoriata.
With a considerable degree of probability, we are led to formulate the further hypothesis that all the orders of Placentalia—from the lowest Prochoriata upwards to man—have descended from some unknown common ancestor living in the Cretaceous period, and that this oldest placental form originated from some Jurassic group of marsupials.
Among these numerous fossil Lemures which have been discovered within the last twenty years, there exist, indeed, all the connecting forms of the older series of Primates, all the 'missing links' sought for by comparative odontology.
The oldest Lemures of the tertiary age are the Eocene Pachylemures, or Hyopso[Pg 34]dina. They possess the complete dentition of the Prochoriata—namely, forty-four teeth (3.1.4.3/3.1.4.3). Then follow the Eocene Palæolemures, or Adapida, with forty teeth, they having lost one pair of incisors in each jaw. To these are attached the younger Autolemures, or Stenopida, with thirty-six teeth, they thus possessing already the same dentition as the Platyrrhinæ. The characteristic dentition of the Catarrhinæ is derived from this formula by the loss of another premolar.
These relations are so clear and so closely connected with a gradual transformation of the whole skull, and with the progressive differentiation of the Primate-form, that we are justified in saying that the pedigree of the Primates, from the oldest Eocene Lemures upwards to man, is now so well known, its principal features so firmly fixed within the Tertiary age, that there is no missing link whatever.
Quite different, and much more incom[Pg 35]plete, is the palæontological evidence, if we go further back into the Secondary or Mesozoic age, and look there for the older ancestors of the mammalian series. There we meet everywhere with wide gaps, and the scarce fragments of fossil Mesozoic mammals (excessively rare in the Cretaceous formation) are too poor to permit definite conclusions as to their systematic position. Indeed, comparative anatomy and ontogeny lead us to the hypothesis that the oldest Cretaceous Mammalia—the Prochoriata—are descended from Jurassic marsupials, and these again from Monotremes. We may also suppose with high probability that among the unknown Cretaceous Prochoriata there have been Lemuravida and forms intermediate between these and the Jurassic Amphitheriidæ, and that these marsupials in their turn are descendants of Pantotheria or similar monotreme-like creatures of the Triassic age. Any certain evidence for these hypotheses is at present still wanting. One important fact,[Pg 36] however, is established—namely, that these interesting and oldest Mammalia—the Pantotheria of Marsh, the Triassic Dromatheriidæ, and the Jurassic Triconodontidæ of Osborn—were small insectivorous mammals with a very primitive organization. Probably they were Monotremes, and may be derived directly from Permian Sauromammalia, an ill-defined mixture of Mammalia and Reptilia.
This generalized characteristic supports our view that the whole class of Mammalia is monophyletic, and that all its members, from the oldest Monotremes upwards to man, have descended from one common ancestor living in the older Triassic, or perhaps in the Permian, age. To acquire full conviction of this important conception, we have only to think of the hair and the glands of our human skin, of our diaphragm, the heart and the blood corpuscles without a nucleus, our skull with its squamoso-mandibular articulation. All these singular and striking modifications of the vertebrate organization are[Pg 37] common to mammals, and distinguish them clearly from the other Craniota. This characteristic combination and correlation proves that they have been developed only once in the history of the vertebrate stem, and that they have been transferred by heredity from one common ancestor to all the members of the class of Mammalia.
The next step, as we trace our human phylogeny to its origin, leads us further back into the lower Vertebrata, into that obscure Palæozoic age the immeasurable length of which (much greater than that of the Mesozoic) may, according to one of the newest geological calculations, have comprised about one thousand millions of years.[16]
The first important fact we have to face here is the complete absence of mammalian remains. Instead of these we find in the later Palæozoic period, the Permian, air-breathing reptiles as the earliest representatives of Amniota. They belong to the[Pg 38] most primitive order of that class, the Tocosauria; and besides them there were the Theromorpha, which approach the Mammalia in a remarkable manner. These reptiles in turn were preceded, in the Carboniferous period, by true Amphibia, most of them belonging to the armour-clad Stegocephali. These interesting Progonamphibia were the oldest Tetrapoda, the first vertebrates which had adapted themselves to the terrestrial mode of life; in them the swimming fin of fishes and Dipneusta was transformed into the pentadactyle extremities characteristic of quadrupeds.
To appreciate the high importance of this metamorphosis, we need only compare the skeleton of our own human limbs with that of the living Amphibia. We find in the latter the same characteristic composition as in man: the same shoulder and pelvic girdle; the same single bone, the humerus or the femur, followed by the same pair of bones in the forearm and leg; then the same[Pg 39] skeletal elements composing the wrist and the ankle regions; and, lastly, the same five fingers and toes.
The arrangement of these bones, peculiar and often complicated, but everywhere essentially the same in all the Tetrapoda, is a striking evidence that man is a descendant from the oldest pentadactyle Amphibia of the Carboniferous period. In man the pentadactyle type has been better preserved by constant heredity than in many other Mammalia, notably the Ungulata.
The oldest Carboniferous Amphibia, the armour-clad Stegocephali, and especially the remarkable Branchiosauri discovered by Credner, are now regarded by all competent zoologists as the indubitable common ancestral group of all Tetrapoda, comprising both Amphibia and Amniota. But whence this most remote group of Tetrapoda? That difficult question is answered by the marvellous progress of modern palæontology, and the answer is in complete harmony with[Pg 40] the older results arrived at by comparative anatomy and ontogeny. Thirty-four years ago Carl Gegenbaur,[17] the great living master of comparative anatomy, had demonstrated in a series of works how the skeletal parts of the various classes of Vertebrata, especially the skull and the limbs, still represent a continuous scale of phyletic gradations. Apart from the Cyclostomes, there are the fishes, and among them the Elasmobranchi (sharks and rays), which have best preserved the original structure in all its essential parts of organization. Closely connected with the Elasmobranchi are the Crossopterygii, and with these the Dipneusta or Dipnoi. Among the latter the highest importance attaches to the ancient Australian Ceratodus. Its organization and development is now, at last, becoming well known. This transitional group of Dipnoi, 'fishes with lungs' but without pentadactyle limbs, is the morphological bridge which joins the[Pg 41] Ganoids and the oldest Amphibia. With this chain of successive groups of Vertebrata, constructed anatomically, the palæontological facts agree most satisfactorily. Selachians and Ganoids existed in the Silurian times, Dipnoi in the Devonian, Amphibia in the Carboniferous, Reptilia in the Permian, Mammalia in the Trias. These are historical facts of first rank. They connote in the most convincing manner that remarkable ascending scale in the series of vertebrates for our knowledge of which we are indebted to the works of Cuvier and Blainville, Meckel, Johannes Mueller and Gegenbaur, Owen and Huxley. The historical succession of the classes and orders of the Vertebrata in the course of untold millions of years is definitely fixed by the concordance of those leading works, and this invaluable acquisition is much more important for the foundation of our human pedigree than would be a complete series of all possible skeletons of Primates.
Greater and more frequent difficulties arise if we penetrate further into the most remote part of the human phylogeny, and attempt to derive the vertebrate stem from an older stem of invertebrate ancestors. None of those had a skeleton which could be petrified; and the same remark applies to the lowest classes of Vertebrata—to the Cyclostomes and the Acrania. Palæontology, therefore, can tell us nothing about them; and we are limited to the other two great documents of phylogeny—the results of comparative anatomy and ontogeny. The value of their evidence is, however, so great that every competent zoologist can perceive the most important features of the most remote portion of our phylogeny.
Here the first place belongs to the invaluable results which modern comparative ontogeny has gained by the aid of the biogenetic law or the theory of recapitulation. The foundation-stones of vertebrate embryology had been laid by the works of Von[Pg 43] Baer, Bischoff,[18] Remak, and Koelliker;[19] but the clearest light was thrown upon it by the famous discoveries of Kowalevsky[20] in 1866. He proved the identity of the first developmental stages of Amphioxus and the Ascidians, and thereby confirmed the divination of Goodsir, who had already announced the close affinity of Vertebrates and Tunicates. The acknowledgment of this affinity has proved of increasing importance, and has abolished the erroneous hypothesis that the Vertebrata may have arisen from Annelids or from other Articulata. Meanwhile, from 1860 to 1872, I myself had been studying the development of the Spongiæ, Medusæ, Siphonophora, and other Cœlenterata. Their comparison led me to the statements embodied in the[Pg 44] 'Gastræatheorie,' the first abstract of which was published in 1872 in my monograph of the Calcispongiæ.
These ideas were carried on and expanded during the subsequent ten years by the help of many excellent embryologists—first of all by E. Ray Lankester and Francis Balfour. The most fruitful result of these widely extended researches was the conclusion that the first stages of embryonic development are essentially the same in all the different Metazoa, and that we may derive from these facts certain views on the common descent of all from one ancestral form. The unicellular egg[21] repeats the stage of our Protozoan ancestors; the Blastula is equivalent to an ancestral cœnobium of Magosphæra or Volvox; the Gastrula is the hereditary repetition of the Gastræa, the common ancestor of all the Metazoa.
Man agrees in all these respects with the other vertebrates, and must have descended with them from the same common root.
Particularly obscure is that part of our phylogeny which extends from the Gastræa to Amphioxus. The morphological importance of this last small creature had been perceived by Johannes Mueller, who in 1842 gave the first accurate description of it. It would not, of course, be correct to proclaim the modern Amphioxus the common ancestor of all the vertebrates; but he must be regarded as closely related to them, and as the only survivor of the whole class of Acrania. If the Amphioxidæ had through some unfortunate accident become extinct, we should not have been able to gain anything like a positive glimpse at our most remote vertebrate ancestor. On the one hand, Amphioxus is closely connected with the early larva of the Cyclostomes, which are the oldest Craniota, and the pre-Silurian ancestors of the fishes. On the other hand, the ontogeny of Amphioxus is in harmony with that of the Ascidians, and if this agreement is not merely coincidental, but due to relationship, we are justified in reconstructing for both Ascidians[Pg 46] and Amphioxus one common ancestral group of chordate animals, the hypothetical Prochordonia. The modern Copelata give us a remote idea of their structure. The curious Balanoglossus, the only living form of Enteropneusta, seems to connect these Prochordonia with the Nemertina and other Vermalia, which we unite in one large class—Frontonia.
No doubt these pre-Cambrian Vermalia, and the common root of all Metazoa, the Gastræades, were connected during the Laurentian period by a long chain of intermediate forms, and probably among these were some older forms of Rotatoria and Turbellaria; but at present it is not possible to fill this wide gap with hypotheses that are satisfactory, and we have to admit that here indeed are many missing links in the older history of the Invertebrata. Still, every zoologist who is convinced of the truth of transformism, and is accustomed to phylogenetic speculations, knows very well that their results are most unequal, often incomplete.
Let us now recapitulate the ancestral chain of man, as it is set forth in the accompanying diagram (p. 55), which represents our present knowledge of our descent. For simplicity's sake the many side-issues or branches which lead to groups not in the main line of our descent have been left out, or have been indicated merely. Many of the stages are of course hypothetical, arrived at by the study of comparative anatomy and ontogeny; but an example for each of them has been taken from those living or fossil creatures which seem to be their nearest representatives.
1. The most remote ancestors of all living organisms were living beings of the simplest imaginable kind, organisms without organs,[Pg 48] like the still existing Monera. Each consisted of a simple granule of protoplasm, a structureless mass of albuminous matter or plasson, like the recent Chromaceæ and Bacteriæ. The morphological value of these beings is not yet that of a cell, but that of a cytode, or cell without a nucleus. Cytoplasm and nucleus were still undifferentiated.
I assume that the first Monera owe their existence to spontaneous creation out of so-called anorganic combinations, consisting of carbon, hydrogen, oxygen, and nitrogen. An explanation of this hypothesis I have given in my 'Generelle Morphologie.'
The Monera probably arose early in the Laurentian period. The oldest are the Phytomonera, with vegetable metabolism. They possessed the power (characteristic of plants) of forming albumin by synthesis from carbon, water, and ammonia. From some of these plasma-forming Monera arose the plasmophagous Zoomonera with animal metabolism, living directly upon the produce of[Pg 49] their plasmodomous or plasma-forming sisters. This is the first instance of the great principle of division of labour.
2. The second stage is that of the simple and single cell, a bit of protoplasm with a nucleus. Such unicellular organisms are still very common. The Amœbæ are their simplest representatives. The morphological value of such beings is the same as that of the egg of any animal. The naked egg cells of the sponges creep about in an amœboid fashion, scarcely distinguishable from Amœba. The same remark applies to the egg-cell of man himself in its early stages before it is enclosed in a membrane. The first unicellular organisms arose from Monera through differentiation of the inner nucleus from the outer protoplasm.
3. Repeated division of the unicellular organism produces the Synamœbium, or community of Amœbæ, provided the divisional products, or new generations of the original cell, do not scatter, but remain[Pg 50] together. The existence of such a Cœnobium, a number of equal and only loosely-connected cells, as a separate stage in the ancestral history of animals, is made highly probable by the fact that the eggs of all animals undergo after fertilization such a process of repeated self-division, or 'cleavage,' until the single egg cell is transformed into a heap of cells closely packed together, not unlike a mulberry (morula)—hence morula stage in ontogeny.
4. The morula of most animals further changes into a Blastula, a hollow ball filled with fluid, the wall being formed by a single layer of cells, the blastoderm or germinal layer. This modification is brought about by the action of the cells—they conveying nourishing fluid into the interior of the whole cell colony and thereby being themselves forced towards the surface. The Blastula of most Invertebrata, and even that of Amphioxus, is possessed of fine ciliæ, or hair-like processes, the vibrating motion of which causes the whole organism to rotate and advance in[Pg 51] the water. Living representatives of such Blastæads, namely, globular gelatinous colonies of cells enclosing a cavity, are Volvox and Magosphæra.
5. The Blastula of most animals assumes a new larval form called Gastrula, in which the essential characteristics are that a portion of the blastoderm by invagination converts the Blastula into a cup with double walls, enclosing a new cavity, the primitive gut. This invagination or bulging-in obliterates the original inner cavity of the Blastula. The outer layer of the Gastrula is the ectoderm, the inner the endoderm; both pass into each other at the blastoporus, or opening of the gut cavity. The Gastrula is a stage in the embryonic development of the various great groups of animals, and some such primitive form as ancestral to all Metazoa is thus indicated. This hypothetical Gastræa is still very essentially represented by the lower Cœlenterates—e.g., Olynthus, Hydra.
6. The sixth stage—that of the Platodes,[Pg 52] or flat-worms—is very hypothetical. They are bilateral gastræads, with a flattened oblong body, furnished with ciliæ, with a primitive nervous system, simple sensory and reproductive organs, but still without appendages, body cavity, vent, and blood-vessels. The nearest living representatives of such creatures are the acœlous Turbellarians—e.g., Convoluta, a free-swimming, ciliated creature.
7. The next higher stage is represented by such low animals as the Gastrotricha—e.g., Chætonotus among the Rotatoria, which differ from the rhabdocœlous Turbellarians chiefly by the formation of a vent and the beginnings of a cœlom, or cavity, between gut and body wall. The addition of a primitive vascular system and a pair of nephridia, or excretory organs, is first met with in the Nemertines.
8. These, together with the Enteropneusta (Balanoglossus), are comprised under the name of Frontonia, or Rhynchelminthes, and form the highest group of the Vermalia.
The Enteropneusta especially fix our attention, because they alone, although essentially 'worms,' exhibit certain characteristics which make it possible to bridge over the gulf which still separates the Invertebrata from the vertebrate phylum. The anterior portion of the gut is transformed into a breathing apparatus—hence Gegenbaur's term of Enteropneusta, or Gut-breathers. Moreover, Balanoglossus and Cephalodiscus possess another modification of the gut—namely, a peculiar diverticulum, which, in the present state of our knowledge, may be looked upon as the forerunner of the chorda dorsalis.
9. Stage of Prochordonia, as indicated by the larval form, called Chordula, which is common to the Tunicata and all the Vertebrata. These two groups possess three most important features: (a) A chorda dorsalis, a stiff rod lying in the long axis of the body, dorsally from the gut and below the central nervous system. This latter, for the first time in the animal kingdom, appears in[Pg 54] the shape of a spinal cord. (b) The use of the anterior portion of the gut for respiratory purposes. (c) The larval development of the Tunicata is essentially the same as that of the Vertebrata in its early stages. Only the free-swimming Copelata or Appendicularia among the Tunicates retain most of these features. The others, which become sessile—namely, the Ascidiæ, or sea-squirts—degenerate and specialize away from the main line.
ANCESTRAL TREE OF THE VERTEBRATA
Abridged from 'Systemat. Phylogenie,' § 15.
Names underlined refer to hypothetical groups.
| Aves | Mammalia | |
| | | Reptilia | | |
| '——— | ———| | | |
| |——— | ———' | |
| | | ||
| Proreptilia | ||
| Pisces | | | Amphibia |
| | | |——— | ———' |
| | | | | |
| | | Stegocephali | Dipnoi |
| | | |——— | ———' |
| '——— | ———| | |
| Proselachii | Cyclostomata | |
| | | | | |
| |——— | ———' | |
| | | ||
| Tunicata | Archicrania | Acrania |
| | | |——— | ———' |
| | | Prospondylia | |
| | | | | |
| '——— | ———| | |
| Prochordonia |
10. Stage of the Acrania, represented by Amphioxus. The early development of this little marine creature agrees closely with that of the Tunicates; but one important feature is added to its organization—namely, metamerism, segmentally arranged mesoderm. Amphioxus still possesses neither skull nor vertebræ, neither ribs nor jaws, and no limbs. But it is a member of the Vertebrata if we define these as follows: Bilateral symmetrical animals with segmentally arranged mesoderm, with a chorda dorsalis between the [Pg 55] [Pg 56] tubular nervous system and the gut, and with respiratory organs which arise from the anterior portion of the gut. We do not assume that Amphioxus stands in the direct ancestral line; it is probably much specialized, partly degenerated, and represents a side-branch; but it is, nevertheless, the only creature, hitherto known, which satisfactorily connects the Vertebrata with their invertebrate ancestors. Many other efforts have been made to solve the mystery of the origin of the Vertebrata—all less satisfactory than the present suggestion, or even absolutely futile. This remark applies especially to the attempts to derive them from either Articulata or Echinoderms. The other great and highly developed phylum, the Mollusca, is quite out of the question. We have to go back to a level at which all these principal phyla meet, and there we find the Vermalia, the lower of which alone permit connection in an upward direction with the higher phyla.
11. Stage of Cyclostomata. This now[Pg 57] small group of Lampreys and Hagfishes represents the lowest Craniota; and although much specialized as a side-branch of the main-stem from which the other Craniota have sprung, they give us an idea of what the direct ancestors of the latter must have been like:—still without visceral arches, without jaws and without paired limbs; with a persistent pronephros; the ear with one semicircular canal only; mouth suctorial; cranium very primitive; and the metamerism of the vertebral column indicated only by little blocks of cartilage in the perichordal sheath. Such creatures must have existed at least as early as the Lower Silurian epoch; but until 1890 fossil Cyclostomes were unknown. Their life in the mud, or as endoparasites of fishes, coupled with their soft structure, makes them very unfit for preservation. This gives all the greater importance to Traquair's discovery, in 1890, of many little creatures, called by him Palæospondylus gunni, in the Old Red Sandstone of[Pg 58] Caithness, which seem to be very closely allied to Cyclostomata.
12. The Elasmobranchi (sharks and skates), with their immediate forerunners, the Acanthodi of the Devonian and Carboniferous age, are the first typical fishes. That they existed as far back as the Silurian age is proved by many enamelled spines of the dermal armour, chiefly from the dorsal fins. This higher stage is characterized by the possession of typical jaws, by visceral or gill-bearing arches, and by two pairs of limbs. None of the Elasmobranchs, fossil or recent, stands in the direct ancestral line; but they are the lowest Gnathostomata, jaw-and-limb-possessing creatures, known.
13. Closely connected with the Elasmobranchs in a wider sense are the Crossopterygii, which begin in the Devonian age as a large group, but have left only two survivals, the African Polypterus and Calamoichthys. They are possessed of dermal bones and other ossifications, and are characterized by[Pg 59] their lobate paired fins, which have a thick axis beset with biserial fin rays. Their gill-clefts are covered by an operculum, and they have a well-developed air-bladder. Whilst they are in many respects more highly developed than the Elasmobranchs, and are intimately connected with the typical Ganoids and other bony fishes (all of which form a great, manifold side-branch of the general vertebrate stem), they stand in many other respects (notably, the structure of the paired fins, the vertebral column, and the air-bladder) nearer the main-stem of our own ancestral line.
14. This is shown by their intimate relation to the Dipnoi, which are still represented by the Australian, African, and South American mud-fishes: Ceratodus, Protopterus, and Lepidosiren. The genus Ceratodus existed in the Upper Trias, whence various other unmistakably dipnoous forms lead down through the Carboniferous (e.g., Ctenodus) to the Devonian strata—e.g., Dipterus. They[Pg 60] are characterized as follows: The paired fins still retain the archipterygial form (namely, one axis with biserial rays); the heart is already trilocular, and receives blood which is mixed arterial and venous, owing to the gills being retained, while the air-bladder has been modified into a lung. In fact, the generalized Dipnoi form the actual link between fishes and Amphibia.
15. Amphibia. The earliest amphibian fossils occur in the Carboniferous strata. They alone—the Stegocephali or Phractamphibia—stand in the ancestral line, while the Lissamphibia, to which all the recent forms belong, are side-branches. The Stegocephali are the earliest Tetrapoda, the archipterygial paired fins having been transformed into the pentadactyle fore and hind limbs, which are so characteristic of all the higher Vertebrata. The cranium is roofed over by dermal bones, of which, besides others, supra-occipitals, supra-orbitals, and supra-temporals are always present. The lowest members[Pg 61] (Branchiosauri) still retained gills besides the lungs, while others (Microsauri) have lost the gills. Be it remembered that all the recent Amphibia still undergo the same metamorphosis during their ontogenetic development.
In the very important Temnospondyli, a subgroup of the Stegocephali—e.g., Trimerorhachis of the Lower Red Sandstone or Lower Permian—the component cartilaginous or bony units which compose the vertebræ still remained in a separate, unfused state, showing at the same time an arrangement whence has arisen that which is typical of the Amniota. The same applies to the limbs and their girdles. In fact, the Stegocephali, taken as a whole, lead imperceptibly to the Proreptilia.
16. Proreptilia are represented by the Permian genera Eryops and Cricotus. Until quite recently these and many other fossils from the Carboniferous strata were looked upon as Amphibia, while many undoubted[Pg 62] fossil Amphibia were mistaken for reptiles, as indicated by the frequent termination '-saurus' in their names.
The nearest living representative of these extinct Proreptilia is the New Zealand reptile Hatteria, or Sphenodon, close relations of which are known from the Upper Trias; while others—e.g., Palæohatteria—have been discovered in the Permian. Anyhow, Sphenodon is the reptile which stands nearest to the main stem of our ancestry.
The most important characteristics of the Reptilia, which mark a higher stage or level, are (1) The entire suppression of the gills—although during the embryonic development the gill-clefts still appear in all reptiles, birds, and mammals; (2) The development of an amnion and an allantois, both for the embryonic life only, but so characteristic that all these animals are comprised under the name of Amniota; (3) The articulation of the skull with the first neck vertebræ by well-developed condyles, either single (really triple) or[Pg 63] double (such a condylar arrangement begins with the Amphibia, but only the two lateral condyles are developed, while the middle portion, belonging to the basi-occipital element, remains rudimentary[22]); (4) The formation of centra, or bodies of the vertebræ, mainly by a ventral pair of the original quadruple constituents, or arcualia.
17. Between the Proreptilia and the Mammalia, which latter occur in the Upper Triassic epoch, we have necessarily to intercalate a group of very low reptiles, which are still so generalized that their descendants could branch off either into the Reptilia proper or into the Mammalia. The changes concerned chiefly the brain and the heart; of the skele[Pg 64]ton, the skull and the pelvis; and, of the tegumentary structures, the formation of a hairy covering. Many such creatures existed in the Triassic epoch—namely, the Theromorpha—some of which indeed possess so many characteristics which otherwise occur in the Mammalia only, that these creatures have been termed Sauro-Mammalia. However, it has to be emphasized that none of the Theromorpha hitherto discovered fulfils all the requirements which would entitle them to this important linking position. They only give us an approximate idea of what this link was like.
18. Stage of the Promammalia, or Prototheria. The only surviving members are the famous duck-bill, Ornithorhynchus, and the spiny ant-eaters, Echidna and Proechidna, of the Australian region. These few genera, however, differ so much from one another in various important respects that they cannot but be remnants of an originally much larger group. Indeed, many fossils from the Upper[Pg 65] Triassic and from the Jurassic strata have without much doubt to be referred to the Prototheria. The Prototheria are typical mammals, because they possess the following characteristics: The heart is completely quadrilocular; the blood is warm, and its red corpuscles have, owing to the loss of their nucleus, been modified from biconvex into biconcave discs; they have a hairy coat and sweat glands, and two occipital condyles; the ilio-sacral connection is preacetabular; the ankle-joint is cruro-tarsal; the quadrate bone of the Reptilia has ceased to carry the under jaw, which now articulates directly with the squamosal portion of the skull. Their low position is shown by the retention of the following reptilian features: Complete coracoid bones and a T-shaped interclavicle; a cloaca, or common chamber for the passage of the fæces, the genital and the urinary products; they are still oviparous; the embryo develops without a chorion, and is therefore not nourished through a placenta. Even the[Pg 66] milk glands, which are absolutely peculiar to the Mammalia, are still in a very primitive stage, and do not yet produce milk proper; and there is only a temporary shallow marsupium.
19. Stage of Metatheria, or Marsupialia, are direct descendants of Prototheria; but they show higher development by the reduction of the coracoid bones and the interclavicle. The original cloaca is divided into a rectal chamber and a uro-genital sinus, completely separated, at least in the males; they are viviparous; the young are received into a permanent marsupium, in the walls of which are formed typical milk glands and nipples, but the embryo is still devoid of a placenta, although some recent marsupials show indications of such an organ. The corpus callosum in the brain is still very weak.
Most of the marsupials are extinct. They occur from the Upper Trias onwards, and had in the Jurassic epoch attained a wide distribution both in Europe and in America.[Pg 67] Since the Tertiary epoch they have been restricted to America and to the Australian region, and are now represented by about 150 species.
20. Stage of Prochoriata, or early Placentalia: a further development of the Metatheria by the development of a placenta, loss of the marsupium and the marsupial bones, complete division by the perineum of the anal and uro-genital chambers, stronger development of the corpus callosum, or chief commissure of the two hemispheres of the brain.
Placentalia must have come into existence during the Cretaceous epoch. Up to that time all the Mammalia seem to have belonged to either Prototheria or to Metatheria; but in the early Eocene we can distinguish the main groups of Placentalia—namely, (1) Trogontia, now represented by the rodents; (2) Edentata, or sloths, armadilloes, etc.; (3) Carnassia, or Insectivora and Carnivora; (4) Chiroptera, or bats;[Pg 68] (5) Cetomorpha, or whales and dugongs; (6) Ungulata; (7) Primates. Of these groups, the first and second, third and fourth, fifth and sixth, can perhaps, to judge from palæontological evidence, be combined into three greater groups, as indicated by the fossil Esthonychida, Ictopsida, and Condylarthra, in addition to the ancestral Primates, or Lemuravida, as the fourth large branch of the ancestral-tree where this has reached the placental level. Among none of the first three branches can we look for the ancestors of the Primates. The Lemuravida, therefore, represent a branch equivalent to the three other branches.
21. Stage of Lemures, or Prosimiæ, comprising the older members of the Primates, consequently approaching most nearly to the Lemuravida. The limbs are modified into pentadactyle hands and feet of the arboreal type, and are protected by nails. The dentition is of the frugivorous or omnivorous type, with an originally complete series of teeth,
with milk teeth and with permanent. The orbit is surrounded by a complete bony ring, posteriorly by a fronto-jugal arch, but still widely communicating with the temporal fossa. The placenta is diffuse and non-deciduous.
ANCESTRAL TREE OF THE MAMMALIA.
'Systematische Phylogenie,' § 386.
| Perissodactyla | Homo | Carnivora | ||||
| Artiodactyla | | | (Litopterna) | | | | | Pinnipedia | |
| | | | | | | Anthropoidae | |—— | ——' | |
| | | | —— | —— ' | | | | | ||
| '—— | ——| | Catarhinæ | Carnassia | |||
| '—— | —— , | | | | | Chiroptera | ||
| (Amblypoda) | Proboscidea | | | Platyrhinæ | Insectivora | | | | |
| '—— | ——| | | | | | |—— | ——' | | |
| ' —— | —— | | Simiæ | | | | | ||
| Cetacea | Sirenia | | | | | |—— | ———— | ——' |
| |—— | ——' | | | Lemures | | | Rodentia | |
| Cetomorpha | | | | | | | | | ||
| | | Hyracoidea | | | | | Ictopsales | (Tillodontia) | |
| | | | | | | Lemuravidæ | | | | | |
| '—— | ——'—— | —— | | | | | | Trogontia | Edentata |
| Condylarthrales | | | | | |—— | ——' | ||
| | | | | | | Esthonychales | |||
| ' —— | ——|—— | ——'—— | ——' | |||
| | | ||||||
| Eutheria s. Placentalia | ||||||
| | | ||||||
| Marsupialia diprotodontia | | | Marsupialia polyprotodontia | ||||
| '—— | ———— | ——|—— | ———— | ——' | ||
| Metatheria | ||||||
| | | Monotremata | (Allotheria) | ||||
| Prototheria | | | | | ||||
| |—— | ——'—— | ———— | ——' | |||
| Hypotheria s. Promammalia | ||||||
Names in brackets indicate extinct groups.
Names underlined indicate hypothetical groups or combinations.
22. Stage of Simiæ. Orbit completely separated from the temporal fossa by an inward extension of the frontal and malar bones meeting the alisphenoid. Placenta consolidated into a disc, and with a maternal deciduous portion. Mammæ pectoral only. The dental formula is 2.1.3.3. All the fingers and toes are protected by flat nails. The tail is long. The American prehensile-tailed monkeys are a lower side-branch.
23. Stage of Catarrhinæ Cercopithecidæ. The dental formula is 2.1.2.3, owing to the loss of one pair of premolars in each jaw. The frontal and alisphenoid bones are in contact, separating the parietal from the malar bone; this feature is correlated with the enlarged brain. The internarial septum[Pg 71] is narrow, and the nostrils look forwards and downwards instead of sidewards—hence the term 'Catarrhinæ.' The external auditory meatus is long and bony. The tail is long, with the exception of Macacus inuus. The body is covered with a thick coat of furry hair. Catarrhine monkeys have existed, we know with certainty, since the Miocene.
24. Stage of Catarrhinæ Anthropoidæ, or Apes. Now represented by the large apes—namely, the Hylobates or gibbon of South-Eastern Asia, Simia satyrus, the orang-utan of Sumatra and Borneo, Troglodytes gorilla, T. niger and T. calvus, the gorilla and the chimpanzees from Western Equatorial Africa. Of fossils are to be mentioned Pliopithecus and Dryopithecus from European Miocene, and Troglodytes sivalensis from the Pliocene of the Punjaub. The tail is reduced to a few caudal vertebræ, which are transformed into a coccyx, not visible externally; but in the embryos of apes and man the tail is still a conspicuous feature. The walk is semi[Pg 72]erect; in adaptation to the prevailing arboreal life, the arms are longer than the legs. The hair of the body is considerably more scanty than in the tailed monkeys. Troglodytes calvus, a species or variety of chimpanzee, is bald-headed. None of the recent genera of apes can lay claim to a place in the ancestry of mankind.
25. Stage of Pithecanthropi. Hitherto the only known representative is Pithecanthropus erectus, from the Upper Pliocene of Java. In adaptation to a more erect gait, the legs have become stronger and the hind-hand has been turned into a flat-soled walking 'foot.' The brain is considerably enlarged. Presumably it is still devoid of so-called articulate speech; this is indicated by the fact that children have to learn the language of their parents, and by the circumstance that comparative philology declares it impossible to reduce the chief human languages to anything like one common origin.
26. Man. Known with certainty to have[Pg 73] existed as an implement-using creature in the last Glacial epoch. His probable origin cannot, therefore, have been later than the beginning of the Plistocene. The place of origin was probably somewhere in Southern Asia.
Whilst we have to admit that there are great defects in the older (invertebrate) portion of our pedigree, we have all the more reason to be satisfied with the positive results of our investigation of the more recent (vertebrate) part of it. All modern researches have confirmed the views of Lamarck, Darwin, and Huxley, and they allow of no doubt that the nearest vertebrate ancestors of mankind were a series of Tertiary Primates.
Particularly valuable are the admirable attempts of the two zoologists, Paul and Fritz Sarasin,[23] to throw light upon the human phylogeny by painstaking comparison of all the skeletal parts of man with those of the[Pg 74] anthropoid apes. They have shown that among the lower races of man the primitive Veddahs of Ceylon approach the apes most nearly, and that among the latter the chimpanzee stands nearest to man.
The direct descent of man from some extinct ape-like form is now beyond doubt, and admits of being traced much more clearly than the origin of many another mammalian order. The pedigrees of the Elephants, the Sirenia, the Cetacea, and, above all, of the Edentata, for example, are much more obscure and difficult to explain. In many parts of their organization—for example, in the number and structure of his five digits and toes—man and monkeys have remained much more primitive than most of the Ungulata.
The immense significance of this positive knowledge of the origin of man from some Primate does not require to be enforced. Its bearing upon the highest questions of philosophy cannot be exaggerated. Among modern philosophers no one has perceived this more[Pg 75] deeply than Herbert Spencer.[24] He is one of those older thinkers who before Darwin were convinced that the theory of development is the only way to solve the 'enigma of the world.' Spencer is also the champion of those evolutionists who lay the greatest weight upon progressive heredity, or the much combated heredity of acquired characters. From the first he has severely attacked and criticised the theories of Weismann, who denies this most important factor of phylogeny, and would explain the whole of transformism by the 'all-sufficiency of selection.' In England the theories of Weismann were received with enthusiastic acclamation, much more so than on the Continent, and they were called 'Neo-Darwinism,' in opposition to the older conception of Evolution, or 'Neo-Lamarckism.' Neither of those expressions is correct. Darwin himself was convinced of the fundamental importance of progressive heredity[Pg 76] quite as much as his great predecessor Lamarck; as were also Huxley and Spencer.
Three times I had the good fortune to visit Darwin at Down, and on each occasion we discussed this fundamental question in complete harmony. I agree with Spencer in the conviction that progressive heredity is an indispensable factor in every true monistic theory of Evolution, and that it is one of its most important elements. If one denies with Weismann the heredity of acquired characters, then it becomes necessary to have recourse to purely mystical qualities of germ-plasm. I am of the opinion of Spencer, that in that case it would be better to accept a mysterious creation of all the various species as described in the Mosaic account.
If we look at the results of modern anthropogeny from the highest point of view, and compare all its empirical arguments, we are justified in affirming that the descent of man from an extinct Tertiary series of Primates is not a vague hypothesis, but an historical fact.
Of course, this fact cannot be proved exactly. We cannot explain all the innumerable physical and chemical processes, all the physiological mutations, which have led during untold millions of years from the simplest Monera and from the unicellular Protista upwards to the chimpanzee and to man. But the same consideration applies to all historical facts. We all believe that Aristotle, Cæsar, and King Alfred did live; but it is impossible to give a proof within the meaning of modern exact science. We believe firmly in the former existence of these and other great heroes of thought, because we know well the works they have left behind them, and we see their effects in the history of human culture. These indirect arguments do not furnish stronger evidence than those of our history as vertebrates. We know of many Jurassic mammals only a single bone, the under jaw. We all believe that these mammals possessed also an upper jaw, a skull, and other bones. But[Pg 78] the so-called 'exact school,' which regards the transformation of species as a hypothesis not proven, must suppose that the mandibula was the only bone in the body of these curious animals.
Looking forward to the twentieth century, I am convinced that it will universally accept our theory of descent, and that future science will regard it as the greatest advance made in our time. I have no doubt that the influence of the study of anthropogeny upon all other branches of science will be fruitful and auspicious. The work done in the present century by Lamarck and Darwin will in all future times be considered one of the greatest conquests made by thinking man.
EVOLUTIONARY STAGES OF THE PRINCIPAL GROUPS OF VERTEBRATA.[25]
| STAGES OF THE | CLASSES. | STAGES OF THE HEART. |
| PAIRED LIMBS. | ||
| {1. Acrania. | I. Leptocardia. | |
| I. Adactylia | { | Cold-blooded; heart |
| s. Impinnata. | { | with one chamber; |
| Without jaws | { | without lungs. |
| and limbs. | { | |
| {2. Cyclostomata. | }II. Ichthyocardia. | |
| } Cold-blooded; heart | ||
| } two-chambered, with | ||
| } one atrium and one | ||
| } ventricle; heart | ||
| } containing venous | ||
| II. Polydactylia | {3. Pisces. | } blood only; without lungs |
| s. Pinnata. | { | |
| With two | { | |
| pairs of fins. | { 4. Dipnoi. | }III. Amphicardia. |
| }Cold-blooded; heart | ||
| } with three complete | ||
| } chambers, namely, with | ||
| } two atria and one | ||
| { 5. Amphibia. | } ventricle, or (Reptilia) | |
| { | } two ventricles with still | |
| { | } incomplete septum; heart | |
| { | } containing mixed venous | |
| { | } and arterialized | |
| III. Pentadactylia | { 6. Reptilia. | } blood; with lungs. |
| s. Tetrapoda. | { | |
| With two | { | {IV. Thermocardia. |
| pairs of | { | { Warm-blooded; heart |
| pentadactyle | { | { with four complete |
| limbs (unless | {7. Aves. | { chambers, namely, two |
| they have | { | { auricles and two |
| been lost by | { | { ventricles; right half |
| reduction). | { | { of the heart with venous, |
| { | { left half with arterialized, | |
| {8. Mammalia. | { blood; with lungs. |
Jean Baptiste de Monet, Chevalier de Lamarck, was born on August 1, 1744, in Picardy, where his father owned land. Originally educated for the Church, he soon enlisted, and distinguished himself in active service. Owing to an accident affecting his health, the young Lieutenant gave up the military career, and, without means, studied medicine and natural sciences at Paris. In 1778 appeared his 'Flore française.' In 1793 he was appointed to a Chair of Zoology at the newly-formed Musée d'Histoire Naturelle. He had the misfortune to become gradually blind, and the last years of his life were spent amid straitened circumstances. He died in 1829.
In 1794 Lamarck divided the whole animal kingdom into vertebrate and invertebrate animals, and founded successively the groups of Crustacea, Arachnida, Annelida, and Radiata. Between 1816 and 1822 he published his celebrated 'Histoire naturelle des Animaux sans Vertèbres.'
His most famous work is the 'Philosophie zoologique,' 1809.
Assuming the spontaneous origin of life, he propounded the doctrine that all animals and plants have arisen from low forms through incessant modifications and changes. In this respect he was in absolute opposition to Cuvier, who upheld the immutability of species, and did his best by absolute silence to suppress the spread of the new doctrine.
Lamarck has explained his views of transformism chiefly in the seventh chapter of the first volume of his 'Philosophie zoologique.'
Organisms strive to accommodate or adapt themselves to new circumstances, or to satisfy new requirements—e.g., climate, mode of[Pg 82] procuring food, escape from enemies. The continued function of parts of an organism changes the old and produces new organs. The acquirements are inherited by the offspring, and thus are produced the more complicated from simpler organisms. Continued disuse brings about degeneration and ultimate loss of an organ.
Lamarck consequently sees in the adaptability, or power of adaptation, which he assumes for all living matter the ultimate cause of variation; and, as he was certainly the first to point out that acquired characters are inherited by the progeny, he has given a working explanation of Evolution.
But his doctrine did not spread—partly because he was misunderstood. His theory, that a new want, by making itself felt, exacts from the animal new exertions, perhaps from parts hitherto not used, until the want is satisfied—this way of putting it sounds too teleological to explain the yearned-for change in a mechanical or natural way. Moreover,[Pg 83] many of his examples lacked the exact basis of experiment and observation necessary for their acceptance. Witness that of the neck of the giraffe,—a never-failing source of ridicule to men who cannot see the deeper purpose underlying the well-meant attempt at an explanation, which failed from want of complete knowledge of the intricate circumstances.
However, the theory of transformism was, so to speak, in the air; and various authors have written on the subject, filling the gap between Lamarck and Darwin, especially Goethe, Treviranus, Leopold von Buch, and Herbert Spencer. But it is Darwin's immortal merit to have opened our eyes by his theory of natural selection, which is, at least, the first attempt to explain some of the causes and incidents of organic Evolution in a natural mechanical way. Moreover, he was the first clearly to express the fundamental principles of the theory of descent, to elaborate what had been at best a general sketch of an ill-defined problem, and to enter[Pg 84] into detail, supported by a host of painstaking observations, the making of which had taken him half a lifetime. Darwin, without going further than cursorily into the causes of variation, argued as follows: We know that variations do occur in every kind of living creatures. Some of these variations lead to something, while others do not. An enormously greater number of animals and plants are born than reach maturity and can in their turn continue the race. What is the regulating factor? His answer is, The struggle for existence—in other words, the weeding out of the less fit, or rather of the owners of those variations which are not so well adapted to their surroundings.
For 'adapted' we had better read 'adaptable,' because a variation which does not answer, which cannot be made use of, or, still more notably, is a hindrance or disadvantage, does not become an adapted feature. There is often a confusion between adaptation as an accomplished fact, a feature,[Pg 85] or resultant condition, and adaptation as the mode of fitting the organism to, or making the best of, the prevailing surroundings or circumstances.
Étienne Geoffroy Saint-Hilaire was born in 1772 at Étampes, Seine-et-Oise. He was originally brought up for the Church; but when already ordained he attended lectures on natural science and medicine in Paris. He managed to get the place of assistant in the Musée d'Histoire Naturelle; he became Professor of Zoology in 1793, and took the opportunity of encouraging young Cuvier. Later he became Professor of Zoology of the Faculté des Sciences, and in 1818 he published his remarkable 'Philosophie anatomique.' He died in 1844.
He had conceived the 'unity of organic composition,' meaning that there is only one plan of construction,—the same principle, but varied in its accessory parts. In 1830, when Geoffroy proceeded to apply to the Inverte[Pg 86]brata his views as to the uniformity of animal composition, he found a vigorous opponent in Cuvier. Geoffroy, like Goethe, held that there is in Nature a law of compensation, or balancing of growth, so that if one organ take on an excess of development, it is at the expense of another part; and he maintained that, since Nature takes no sudden leaps, even organs which are superfluous in any given species, if they have played an important part in other species of the same family, are retained as rudiments, which testify to the permanence of the general plan of creation. It was his conviction that, owing to the conditions of life, the same forms had not been perpetuated since the origin of all things, although it was not his belief that existing species were becoming modified. Cuvier, on the other hand, maintained the absolute invariability of species, which, he declared, had been created with regard to the circumstances in which they were placed, each organ con[Pg 87]trived with a view to the function it had to fulfil,—thus putting the effect for the cause ('Encyclopædia Britannica,' 9th edition, vol. xxi., p. 171).
George Cuvier in the department of Doubs, which at that time belonged to Württemberg. He was educated at Stuttgart, and studied political economy. While acting as private tutor to a French family in France he followed his favourite pursuit, the study of natural sciences. Geoffroy Saint-Hilaire heard of him, and appointed him assistant in the department of comparative anatomy in the Musée d'Histoire Naturelle. In 1799 he was elected Professor of Natural History at the Collège de France, and soon after he became Perpetual Secretary of the Institut National. In 1831, a year before his death, Louis Philippe raised him to the rank of a peer of France.
Cuvier was the first to indicate the true[Pg 88] principle upon which the natural classification of animals should be based—namely, their structure. It is the study of the anatomy of the creatures and their comparison which affords the only sound basis of a classification. The work which had the greatest influence upon the scientific public is his 'Règne animal distribué d'après son Organisation,' 1817. The system which he propounded in this book gradually came to have almost world-wide fame, and, in spite of its many obvious deficiencies, still lingers in some of our most recent text-books.
A standard work is his 'Leçons d'Anatomie comparée,' and, in truth, he is the founder of that kind of comparative anatomy which was brought to such a high state by his pupil, the late Sir Richard Owen. Cuvier discovered the law of 'correlation of growth,' and was the first to apply this law to the reconstruction of animals from fragments: see his monumental work entitled 'Recherches sur les Ossemens fossiles,' 1812.
Cuvier, however, as a strict matter-of-fact man, was incapable of appreciating the speculative conclusions which were drawn by his contemporaries Saint-Hilaire and Lamarck. On the contrary, he firmly stuck to the doctrine of the immutability of species; and, in order to account for the existence of animals whose kind exists no longer, he invented the famous doctrine of successive cataclysms.
Karl Ernst von Baer was born in 1792 in Esthonia, studied at Dorpat and then at Würzburg, where Döllinger introduced him to comparative anatomy. For a few years he was a Privat-docent at Berlin; then he went to Königsberg as Professor of Zoology and Embryology. In 1834 he became an Academician at St. Petersburg, where for many years he was occupied with the most varied studies, chiefly geographical and ethnological. The last years of his long, active life he spent in contemplative retire[Pg 90]ment on his paternal estate, and he died at Dorpat in 1876.
While still at Würzburg he induced his friend Pander, a young man of means, to study the development of the chick; and Pander was the first to start the theory of the germinal layers from which all the organs arise. Baer, however, continued these researches in Königsberg, and after nine years' labour produced his epoch-making work, 'Ueber Entwicklungsgeschichte der Thiere: Beobachtung und Reflexion,' Königsberg, 1828. Nine years later he completed the second volume. He established upon a firm basis the theory of the germinal layers, and by further 'reflexions' arrived at the elucidation of some of the most fundamental laws of biology. For example, in the first volume he made the following prophetic statement: 'Perhaps all animals are alike, and nothing but hollow globes at their earliest developmental beginning. The farther back we trace their development, the more resemblance we find[Pg 91] in the most different creatures. And this leads to the question whether at the beginning of their development all animals are essentially alike, and referable to one common ancestral form. Considering that the "germ" (which at a certain stage appears in the shape of a hollow globe or bag) is the undeveloped animal itself, we are not without reason for assuming that the common fundamental form is that of a simple vesicle, from which every animal is evolved, not only theoretically, but historically.'
This statement is all the more wonderful when we consider that the cells, the all-composing individual units, were not discovered until ten years later.
In 1829 Baer discovered the human egg, and later the chorda dorsalis. In an address delivered in 1834, entitled 'The Most Universal Law of Nature in all Development,' he explained that only from a most superficial point of view can the various species be looked upon as permanent and immutable types;[Pg 92] that, on the contrary, they can be nothing but passing stages, or series of stages, of development, which have been evolved by transformation out of common ancestral forms.
Johannes Mueller, born at Coblenz in 1801, established himself as Privat-docent at Bonn, where in 1830 he became Professor of Physiology. In 1833 he accepted the Chair of Anatomy and Physiology at Berlin, where he died in 1858.
He was one of the most distinguished physiologists and comparative anatomists. By summarizing the labours and discoveries already made in the field of physiology, by reducing them to order, and abstracting the general principles, he became the founder of modern physiology. But he was scarcely less distinguished by his researches in comparative anatomy. His 'Vergleichende Anatomie der Myxinoiden,' in Abhandlungen der Berliner Akademie, 1835-45, and 'Ueber die Grenzen der Ganoiden' (ibid., 1846), are standard works of lasting value.
Mueller exercised a stimulative influence as a teacher. Many well-known men—such as Helmholtz, Gegenbaur, Bruecke the physiologist, Guenther the zoologist, Virchow the pathologist, Koelliker and Haeckel—have been his pupils.
Rudolph Virchow Schievelbein, a small town in Eastern Pomerania. He studied medicine in Berlin as a pupil of Johannes Mueller, and went in 1849 to Würzburg, where, under the influence of Koelliker, and Leydig the pathologist, he laid the foundation of an entirely new branch of medical science—that of 'cellular pathology.' Since 1856 he has filled the principal Chair of Pathology at Berlin. In 1892 he received the Copley medal of the Royal Society.
'His contributions to the study of morbid anatomy have thrown light upon the diseases of every part of the body; but the broad and philosophical view he has taken of the[Pg 94] processes of pathology has done more than his most brilliant observations to make the science of disease.
'In pathology, strictly so called, his two great achievements—the detection of the cellular activity which lies at the bottom of all morbid as well as normal physiological processes, and the classification of the important group of new growths on a natural histological basis—have each of them not only made an epoch in medicine, but have also been the occasion of fresh extension of science by other labourers' (Proc. Royal Soc., 1892).
Virchow has not confined himself to medicine. He takes the keenest interest in anthropology and ethnology, on which subjects he has contributed many papers. Together with his colleagues Helmholtz the physicist, and Du Bois Reymond the physiologist, he has taken a leading place in the spreading of natural science; but, unfortunately, he did not take to the doctrine of[Pg 95] Evolution, and for the last thirty years has been its declared antagonist, rarely missing an opportunity of denouncing everything but descriptive anatomy and zoology as the unsound speculations of dreamers. This has on more than one occasion brought him into sharp conflict with Haeckel. His activity is astonishing, especially if it be remembered that Virchow has for many years been one of the most conspicuous leaders of the Progressists and Radicals in the German Parliament and Berlin town-council.
Edward Drinker Cope was born at Philadelphia, Pa. After studying at several Continental Universities, especially at Heidelberg, he became first Professor of Natural Science at Haverford College, and later Professor of Geology and Mineralogy. He died at an early age in 1897. As a member of various geological expeditions and other surveys, he explored chiefly Kansas, Wyoming, and Colorado; and he published many[Pg 96] most suggestive papers on the fossil vertebrate fauna of North America, and on classification especially of Amphibia and Reptiles.
Among works of a more general philosophical scope may be mentioned 'The Origin of the Fittest,' 1887, and his latest work, 'The Primary Factors of Organic Evolution,' 1896.
Albert von Koelliker became Professor of Anatomy at Würzburg. His earlier studies and discoveries contributed considerably to the systematic development of the cell theory. In 1844 he observed the division and further multiplication of the original egg cell. Next year he showed the continuity between nerve cells and nerve fibres in the Vertebrata; later, that the non-striped or smooth muscular tissue is composed of cellular elements. He demonstrated that the Gregarinæ are unicellular creatures. In 1852 he went with his younger friend Gegenbaur to Messina, where he studied especially[Pg 97] the development of the Cephalopoda (cuttlefishes and allies); and he produced a magnificent work on Alcyonaria, Medusæ, and other allied forms. He elucidated the development of the vertebral column, especially with reference to the notochord.
In 1848 he founded, together with Th. von Siebold, the famous Zeitschrift für wissenschaftliche Zoologie.
A standard work on mammalian embryology is his 'Entwicklungsgeschichte des Menschen und der höheren Thiere,' a text-book of which the second edition appeared in 1879.
At the anniversary meeting of 1897 he received the Copley medal, the highest honour which the Royal Society can bestow.
Carl Gegenbaur was born on August 21, 1826, in Bavaria. He studied medicine and kindred subjects in Würzburg, and as a pupil of Johannes Mueller in Berlin.
In 1852 he went with Koelliker to Messina[Pg 98] to study the structure and development of the marine fauna. Important papers on Siphonophora, Echinoderms, Pteropoda, and, later, Hydrozoa and Mollusca, were the result. Soon after his return he was offered the chair of Anatomy at Jena, and at this retired spot he produced his most important works, devoting himself more and more to the study of the Vertebrata. Since 1875 he has held the Chair of Anatomy at Heidelberg.
In 1859 he published his 'Principles of Comparative Anatomy'; but in 1870 he remodelled it completely, the theory of descent being the guiding principle. These 'Grundzüge' were followed by a somewhat more condensed 'Grundriss,' the second edition of which was published in 1878, and has been translated into French and English. In the meantime he had broken new ground by the development and treatment of certain problems concerning the composition and origin of the limbs, the shoulder-girdle and the skull, researches which are embodied[Pg 99] in his 'Untersuchungen zur vergleichenden Anatomie der Wirbelthiere,' 1864-65-72.
In 1883 he brought out a text-book on human anatomy. This also marked a new epoch, because for the first time, not only the nomenclature, but also the general treatment of human anatomy, was put upon a firm comparative anatomical basis. The success of this work is indicated by the fact that it reached the sixth edition in 1897.
Lastly, in 1898, appeared the first volume of what may be called his crowning work, 'Vergleichende Anatomie der Wirbelthiere.'
Gegenbaur is universally recognised, not only as the greatest living comparative anatomist, but also as the founder of the modern side of this science, by having based it on the theory of descent.
In 1896 he received from the Royal Society the Copley medal 'for his pre-eminence in the science of comparative anatomy or animal morphology.'
His marvellously powerful influence as a[Pg 100] teacher and investigator has made Heidelberg a centre whence many pupils have spread his teaching, and above all his method of research.
Ernst Heinrich Haeckel was born on February 16, 1834, at Potsdam. He carried out his academical studies alternately at Berlin and Würzburg, attracted by such men as Johannes Mueller, Koelliker, and Virchow. For years he was undecided what his career should be, whether that of botanist, collector, or geographical traveller. Certainly that of medicine attracted him least, although in deference to his father's wishes he qualified and settled down for a year's practice in Berlin. As he himself has told us, he might perhaps have proved rather successful as a physician, to judge from the fact that he did not lose a single patient. But 'I had only three patients all told, and the reason of this is perhaps that I had given on my plate the hours of consultation as from 5 to 6 a.m.'
During the year 1859 he travelled as medical man and artist in Sicily. In 1861 he was induced by Gegenbaur, whose acquaintance he had made in Würzburg, to establish himself as a Privat-docent for comparative anatomy in Jena. And there he has remained ever since, filling the Chair of Zoology, and having declined several much more tempting offers from the Universities of Würzburg, Vienna, Strassburg, and Bonn.
Within one year, 1865, he wrote the two volumes of his 'Generelle Morphologie der Organismen,' as he himself relates, in order to master his sorrow over the loss of his first wife. But he broke down, and went to the Canaries to recruit health and strength. The 'Morphologie,' which has long been out of print,[26] made scarcely any impression. It[Pg 102] was ignored, probably because he had placed the old-fashioned study of zoology and morphology upon a thoroughly Darwinistic basis.
On the advice of his friend Gegenbaur, he gave a more popularly written abstract of his 'Generelle Morphologie'—in fact, the substance of a series of his lectures—in the shape of his 'Natürliche Schöpfungsgeschichte.' This 'History of Natural Creation,' which in 1898 has reached the ninth edition (first edition translated into English in 1873), had the desired effect. So also had his 'Anthropogenie oder Entwicklungsgeschichte des Menschen,' the fourth edition of which appeared in 1891.
It was a lucky coincidence that Haeckel had just finished his preliminary academical studies, was entirely at leisure, and undetermined to which branch of natural science he should devote his genius, when Darwin's great work was given to the world. Haeckel embraced the new doctrine fervently, and, as Huxley was doing in England, he spread[Pg 103] it and fought for it with ever-increasing vigour in Germany.
With marvellous vigour and quickness of perception he applied the principles of Evolution or the theory of descent to the whole organic world, and not only opened entirely new vistas for the study of morphology, but also worked them out and fixed them. He was the first to draw up pedigrees of the various larger groups of animals and plants, filling the gaps by fossils or with hypothetical forms (the necessary existence of which he arrived at by logical deductions); and thus he reconstructed the first universal pedigree, a gigantic ancestral tree, from the simple unicellular Amœba to Man. Of course these pedigrees were entirely provisional, as he himself has over and over again avowed; but they are, nevertheless, the ideal which all systematists and morphologists working upon the basis of Evolution have since been seeking to establish.
Naturally he was vigorously attacked, not[Pg 104] only by anti-Darwinians, or rather anti-Evolutionists, but also by many of those who, having accepted the principle of transformism, ought to have known better. Perhaps they thought they did know better. Imperfections or mistakes in details of the grand attempt,—and these, naturally, were many,—were singled out as samples of the whole, which was ridiculed as the romance of a dreamer.
In the end, however, this hostility, narrow-minded and unfair in many respects, has done good to the cause. There has arisen an ever-increasing school of workers in favour of the new doctrine. Owing to renewed research, criticism, corrections in all directions, we now know considerably more about natural classification (and this is pedigree) than when Haeckel first opened out the whole problem.
Owing to his fearless mode of exposition, regardless of the indignant wrath which the new doctrine aroused in certain ecclesiastical[Pg 105] quarters, Haeckel bore the brunt of almost endless attacks, and had to write polemical essays. The result has been that friend and foe alike are now working on the lines which he has laid down; most of the ideas which he was the first to conceive, and to formulate by inventing a scientific terminology for them, have become important branches, or even disciplines, of the science.
Most morphologists of the younger generations now take these terms for granted, without remembering the name of their founder. It is, therefore, perhaps not quite superfluous to mention some of them:
Phylum, or stem, the sum total of all those organisms which have probably descended from one common lower form. He distinguished eight such phyla—Protozoa, Cœlenterata, Helminthes or Vermes, Tunicata, Mollusca, Articulata, and Vertebrata. The phyla are more or less analogous to 'super-classes,' large branches or 'circles,' or principal groups of other zoologists.
Phylogeny, the history of the development of the various phyla, classes, orders, families,and species.
Ontogeny, the history or study of the development of the individual, generally called embryology. In reality the scope of embryology is the ontogenetic study of the various species, and this branch of developmental study alone can be checked by direct, 'exact' observation, for the simple reason that the individuals alone are entities, while the species, genera, families, etc., are abstract ideas.
The ontogenesis of any given living organism is a short, condensed recapitulation of its ancestral history or of its phylogenesis. This is Haeckel's 'fundamental biogenetic law.'
A complete proof of the phylogeny of any creature would be given by the preservation of an unbroken series of all its fossil ancestors. Such a series will in most cases, for obvious reasons, always remain a desideratum. In a few cases, however, the desideratum is nearly[Pg 107] met: for example, the ancestral line of the one-toed digitigrade horse from a four-or five-toed plantigrade and still very generalized Ungulate is approaching completion.
Phylogenetic study has to rely upon other help. This is afforded by comparative anatomy and by the study of ontogeny. If the latter were a faithful, unbroken recapitulation of all the stages through which the ancestors have passed, the whole matter would be very simple; but we know for certain that in the individual development many stages are left out (or, rather, are hurried through, and are so condensed by short-cuts being taken that we cannot observe them), while other features which have been introduced obscure, and occasionally modify beyond recognition, the original course.
Again, the sequence of the appearance of the various organs is frequently upset (heterochronism). Some organs are accelerated in their development, while others,[Pg 108] which we know to be phylogenetically older, are retarded in making their reappearance in the embryo.
These disturbing or distorting newly introduced features or factors show themselves chiefly in connection with the embryonic conditions of growth—for example, yolk-sac, placenta, amnion. They all come within the category of cænogenesis: they are cænogenetic, while the true, undisturbed recapitulation is palingenetic.
Lastly, some features, so-called rudimentary or vestigial organs, instead of disappearing, are most tenacious in their recurrence, while others of originally fundamental importance scarcely leave recognisable traces, and are, so to speak, only hinted at during the embryonic growth of the creature we happen to study. Hence arises the philosophical study of 'Dysteleology.'
Among other terms invented by Haeckel, and now in general use, are Metamere, Metamerism, Cœlom, Gonochorism, Gastrula,[Pg 109] Metazoa, Gnathostomata, Acrania, Craniota, and Amniota.
Hitherto we have dealt with his general work only, a résumé of which he gave for many years in a course of thirty lectures before an audience composed of 'all sorts and conditions of men.' Students of biology and of medicine side by side with theologians, incipient and ordained, jurists, political economists, and philosophers, crowded his lecture-room during the 'seventies to hear the master explaining the 'natural history of creation' or the mysteries of anthropogenesis. Another course of eighty lectures during the winter semester was, and still is, devoted to a systematic treatment of zoology, while practical classes are reserved for the more select.
His winning personality and fascinating eloquence, combined with a clear and concise delivery, have gained the enthusiastic admiration of many a student who went to the quiet University town in order to learn with his own ears and eyes.
List of Separate Publications by Professor Haeckel.
'Biologische Studien. I.: Studien ueber die Moneren und andere Protisten.' Leipzig, 1870 (out of print). He was the first to make observations on the natural history of the Monera, living bits of protoplasm, devoid even of a nucleus—e.g., Protogenes primordialis, Protomyxa aurantiaca.
'Monographie der Radiolarien.' Berlin, 1862-88. With 171 plates.
'Entwicklungsgeschichte der Siphonophoren.' Utrecht, 1869.
'Plankton-Studien. Vergleichende Untersuchungen ueber die Bedeutung und Zusammensetzung der pelagischen Fauna und Flora.' Jena, 1880.
'Metagenesis und Hypogenesis von Aurelia aurita.' Jena, 1881.
'Monographie der Geryoniden oder Ruesselquallen.' Leipzig, 1865.
'Generelle Morphologie der Organismen.' 2 vols. Berlin, 1866.
'Anthropogenie oder Entwicklungsgeschichte des Menschen,' 1874; 4th edition, 1891.
'Natuerliche Schoepfungs-Geschichte.' 2 vols. Berlin, 1st edition, 1868; 9th edition, 1898. This work has been translated into most European languages (the first edition in English, under the title 'Natural History of Creation' in 1873; the eighth in 1892).
'Monographie der Kalkschwaemme.' 3 vols. Berlin, 1872 (out of print). With the subtitle, 'An Attempt to solve analytically the Problem of the Origin of Species.' In this work, illustrated by sixty plates, he showed that the Calcispongia are individually so yielding, so adaptive to external influences, that it is practically impossible to break up the whole group into anything like satisfactory species or genera. According to predilection, we can distinguish either 1 genus with only 3 species, or 3, 21, 43 genera, with 21, 111, 181, or 289 species respectively.
In this work, in 1872, Haeckel established[Pg 112] the homology of the two primary layers, ecto- and endoderm, throughout the Metazoa. The attempt to do the same for the four secondary layers, as made in the second part of his 'Gastræa-theory,' failed. It caused an enormous amount of research, hitherto without a satisfactory solution of the problem.
'Studien zur Gastræa-Theorie.' Jena, 1874. The transformation of the single primitive egg-cell by cleavage into a globular mass of cells (Morula)—which latter, becoming hollow (and then known as the Blastula), turns ultimately by invagination or by delamination into the Gastrula—is a series of processes which applies to all Metazoa. The Gastrula is, therefore, the ancestral form of the Metazoa; and the Gastræa-theory, founded by Haeckel, throws light, on the one hand, upon the mystery of the phyletic connection of the various animal groups, while, on the other hand, it connects the Metazoa, or multicellular organisms, with the lowest Protozoa. We come to this conclusion because[Pg 113] the Gastrula arises from and passes through stages which exist as independent, permanent organisms among the Protozoa.
Needless to say this Gastræa-theory has been violently attacked in detail, with the result that various modifications of the Gastrula, until then undreamed of, have become known.
'Monographie der Medusen.' Jena, 1879-81. With 72 coloured plates.
'Reports on the Scientific Results of the Voyage of H.M.S. Challenger.' With 230 plates:
1. Deep-sea Medusæ. 1881.
2. Radiolaria. 1887.
3. Siphonophoræ. 1888.
4. Deep-sea Keratosa. 1889.
A short holiday spent on the coasts of the Red Sea produced the volume 'Arabische Korallen' (Berlin, 1876); and a longer trip to Ceylon has been described in 'Indische Reisebriefe,' of which the third edition[Pg 114] appeared in 1893. The English translation (1883) is entitled 'A Visit to Ceylon.'
'Monism as connecting Religion and Science: the Confession of Faith of a Man of Science.' 1894.
Haeckels latest work is the 'Systematische Phylogenie' (Berlin, 1896), three volumes dealing with Protistæ and Plants, Invertebrata and Vertebrata. They contain the author's views on the natural system of the organic world, both living and extinct. Notable in the work are the many reconstructions of ancestral forms which, provided Evolution is true, must have existed—hypothetical until they, or something like them, are found in a fossil state. Everybody who works systematically, and upon the basis of Evolution, does, sometimes unconsciously, reconstruct such links, although he may perhaps not see the necessity, or have the courage to fix his vision, by assigning to it all those attributes or characters which are indicated by deductions from comparative anatomy, palæontology, and embryology.
The vegetable cell was discovered by Schleiden, Professor of Botany at Jena, in 1838. Next year Schwann found the animal cell.
In 1844 Koelliker discovered that the egg cell, by division and multiplication, becomes an aggregation—a heap of new cells.
In 1849 Huxley found the two primary layers (observed long before by Pander and Baer in the chick) also in certain Invertebrata, the Medusæ; and he called these layers 'ectoderm' and 'endoderm' respectively.
In 1851 Remak, in his 'Untersuchungen über die Entwicklung der Thiere,' showed the egg to be a simple cell, and that from it, by repeated division or multiplication, arise[Pg 116] the germinal layers, and that by differentiation of the cells of these layers are formed all the tissues of the body.
Kowalevsky, of St. Petersburg, found the two primary germinal layers also in Worms, Echinoderms, Articulata, and other animals.
Haeckel, in 1872, found the same in the Sponges. He stated that these two germinal layers occur in all animals, except in the Protozoa; and that they are homologous, or equivalent, in all the groups of animals, from the Sponges up to Man. In 1873, in his 'Gastræa-theorie,' he explained the phylogenetic significance, and tried to show the homology, of the four secondary germinal layers.
An organism, as living matter, does not stand in opposition to, or outside of, the rest of the world. It is part of the world. It receives matter from its surroundings, and gives some back; therefore it is influenced by its surroundings. It is acted upon, and it reacts upon the latter, and if these change (and they are nowhere and never strictly the same) the organism also varies. It adapts itself, and if it does not, or, rather, cannot, do so, it dies, because it is unfit to live in the world, or, rather, in those particular surroundings and conditions in which it happens to be. That organism which yields most easily, accommodates itself most quickly, has the best chance of existence—survival of the fittest.[Pg 118] 'Fitness' in this case does not mean fitness to live, but rather a particular condition which happens to fit into the new circumstances.
Adaptation and variation are simultaneous: they are fundamentally the same. If there were no adaptability and no variability, those simplest of organisms which we suppose to have sprung into existence in the pre-Cambrian period would long ago have ceased to exist.
It is the physiological momentum which models the organism, and, by causing its adaptations, has produced its organs by change of function. Gegenbaur illustrates this most important fundamental truth by an excellent example. Suppose that, in an absolutely simple organism, all the parts of its exterior are under the same functional conditions, so that each part of the surface can take in food, and that this is digested, assimilated, in the interior. There is, in this condition, not yet any definite organ. If this organism sinks to the bottom and[Pg 119] becomes sessile, this part is excluded from taking in nourishing matter, while the opposite surface alone remains, or becomes more, fit for this function. Thus, a simple variation and adaptation has been produced, and if the same organism continues in this position, its bottom cells will estrange themselves from their original function, while those on the top will convey the food into the interior, where a cavity will be formed, ultimately with a permanent opening, the primitive gut and mouth, both very different from the 'foot.'
Thus, by adaptation and variation the organism acquires new functions, organs, features, and it gives up and eventually loses others. Its offspring is like it. Like produces like. This is the principle of heredity. Adaptation, when going on generation after generation on the same lines in the same direction, becomes continuous, and has an intensifying, cumulative effect. By always weeding out from a flock of pigeons those birds[Pg 120] which possess more dark feathers than the rest, we ultimately produce an entirely white race. We hurry on what Nature does slowly.
The inheritance of acquired characters becomes very obvious in the following example: The Monera are the lowest living organisms known; they consist of a mass of protoplasm, and are still devoid of even a nucleus. They multiply simply by division; each half is like the other, and like the parent (which by this process has ceased to exist), except that each is smaller and has to grow. A certain Moneron, Protomyxa aurantiaca, is orange-coloured, and its offspring is from the beginning of the same colour, and this colour has been acquired by that kind of Monera-like protoplasm which thereby has become the species called Aurantiaca. We have no reason for assuming that there existed from the beginning of life not only colourless, but also red, orange, and other kinds of protoplasm. In these simplest of organisms the whole process of heredity[Pg 121] seems very obvious; but in the higher ones, in those which propagate by eggs, the problem is infinitely more complicated. It is true that the egg is, strictly, nothing but a small part of the parental organism, and we know from everyday experience that this single egg-cell has in it all the attributes and characteristics of the parent; but these attributes and characteristics make their appearance successively, just as the egg cell of a chick has neither wings nor feathers, not even a backbone, but develops these organs because its parents have them.
The theory that acquired characters are hereditary has often been vigorously attacked; but the champions of the negative position have not given us anything satisfactory instead. They question, also, the principle of adaptation as a factor in Evolution, and substitute 'variation,' coupled with 'natural selection.'
They point to Darwin's argument: (1) It is a fact that animals and plants produce a[Pg 122] much greater number of young than in their turn grow up to propagate the race; (2) no two of the frequently many individuals of the same breed are exactly alike, although the differences may be hidden to our perception (this is quite true, because no two entities can live in absolutely the same place and conditions); (3) through heredity the offspring takes over the faculties and features of the parents; (4) what decides which of the many individuals (each one possessing some aberration or variation) are to live and to propagate the race?—obviously those individual variations which happen to make the lucky possessors most fit for the struggle for life.
So far, well; but the 'Neo-Darwinians' imagine that 'adaptation' is not the cause, but the result, the effect, of the formation of species. According to them, the species are neither adapted by, nor do they adapt themselves to, their surroundings. Adaptation is to them an accomplished fact, a condition which a[Pg 123] species happens to be in because its particular variation is the one which, to the exclusion of others, suits or fits into its surroundings. Such a view simply takes variation for granted, and stipulates it as a something a priori, without raising the further necessary question, why there should be any variations at all. Why, indeed, unless they are caused by external influences? Haeckel elucidated this by the conception of adaptation as explained in the foregoing pages.
These and kindred speculations have produced some rather curious discussions, which not infrequently end in conundrums. If we speak of a case of adaptation as a condition, a fact, we easily run the risk of getting into confusion about cause and effect. For example: Is the stag swift because he has long and slender legs, or are his legs long because he is swift? In reality, swiftness and length of legs are cause and effect in one. His legs have been so modified as to make him swift, because he has put them continuously to whatever was[Pg 124] his full speed, which in his thick-footed ancestors was probably a very slow one. The above question reads, therefore, more sensibly as follows: Has the stag become swift because his legs have become long and slender, or have his legs become long and slender because he has attained swiftness? Now, we see that both halves of the double question are practically the same and instantly suggest the answer.
A fundamental difference between artificial machines and living organisms is that the former are worn out by use, while the latter not only repair the loss caused by use, but are also stimulated to further increase. On the other hand, organs which are not put into function, or are not used, degenerate. The various cells of the organ react upon external stimuli by increased activity. Why this should be so is another question—perhaps because those which do not would soon be not fit to survive. Each cell has a function; the more specialized the more intense it is. Every[Pg 125] external stimulus, every contact with the outer surroundings, is an insult, necessarily of detrimental effect, as it disturbs the equilibrium of the cell body. It must, therefore, be of advantage to the cells' well-being to return as soon as possible to the status quo ante, and this can only be done by increased activity.
In the present state of our knowledge, we can approach only the simplest cases of acquisition of characteristics. Mostly they are so complicated, subject to so many unthought-of conditions, that we do not know from which end to approach the problem. Frequently the supposed use of certain obvious features is the merest guesswork. This applies especially to features to which we are not accustomed (although wrongly so) to assign a function—for example, coloration. A green tree-frog will with predilection rest on green leaves. The advantages of concealment are obvious, and in this case he 'adapts himself' to the surroundings by[Pg 126] making for green localities: if he did not he would be eaten up sooner than his more circumspect comrades. But this making for, and sitting in, the green has not necessarily made him of that colour. Extreme advocates of one view would argue as follows: Once upon a time there were among the offspring of ancestral tree-frogs some which, among other colours, exhibited green, not much, perhaps not even perceptible to our eyes. The occurrence of this colour, according to them, was spontaneous, a freak—as if in reality there were anything spontaneous in the sense of being causeless. The descendants of these more greenish creatures, provided they did not pair with frogs of the ordinary set, became still greener (by accumulative inheritance), and so on, until the green was pronounced sufficient to be of advantage when competition could set in.
With this view there is always the difficulty of understanding how the initial very small changes can be useful, unless we have[Pg 127] to deal with extremely simple organisms. Is it likely in the case of our frogs that an almost imperceptible variation in colour makes them more fit to live? We have to assume that 'luck' or chance kept them for generations out of harm's reach, until the accumulation of green, hitherto quite ineffective, neither harmful nor useful, became strong enough to be effective. Such cases undoubtedly happen.
But we can also argue out this problem in a somewhat different way, which goes nearer to the root of the whole process. The original slight, imperceptible change in pigmentation is not a spontaneous freak; it was caused by the direct influence of the surroundings in which the particular frogs happened to live, be this factor light or temperature or food. Thus it stands to reason that the offspring, living under similar conditions, will be acted upon in the same way. That factor which has added green to the parents will add green to the children,[Pg 128] until by accumulative inheritance a more decidedly green race is produced.
The offspring of green plants do not become green when grown in the dark; the young plants inherit not the green, but the capacity of becoming green when acted upon by sunlight. This as an instance of direct influence of the surroundings on a substance (chlorophyll), which has not yet performed a function. But the kittens of a pair of black cats produce black hair before they are born, and we have no reason to doubt that the black pigment in their tegumentary structures is ultimately referable to the action of the sunlight. In many instances creatures living for generations in darkness become white, pigmentless, and they regain it when exposed to light. For example, the white, colourless Proteus from the caves of Adelsberg becomes clouded grey, and ultimately jet black, when kept in a tank whence light is not strictly excluded.
Blindness is a very general characteristic[Pg 129] of creatures which dwell in darkness. There are all stages between total blindness and weak eyes. Now, do these blind creatures live in darkness because they are blind, or have they become first weak-eyed and then blind because of the continuous disuse of their eyes? The former explanation has actually been suggested! Individuals not smitten, but spontaneously, as a freak, born with sore eyes, have crept into the darkness for relief and have produced a blind race! To carry such a notion to the bitter end leads to absurdities. Anyhow, it is not understandable where the benefit of losing the eyesight arises. It can be explained only by continued disuse: witness Spalax typhlus, the blind mole, and, above all, the Endoparasites.
Let us now take an example to explain the influence of a tangible external stimulus. Repeated pressure produces callosities. Although they are not exactly beneficial in the shape of corns on our toes, they are[Pg 130] so on our hands. At any rate, the morphologist can trace the development of the footpads, nails, hoofs, and horns, step by step from small beginnings. The cells of the Malpighian stratum, of the inner, active portion of our epidermis, are excited to extra activity, and by continually producing more horn cells than peel off the surface of the skin in the normal process of wear and tear cause the formation of the pad. It need scarcely be mentioned that hypertrophic growths are not necessarily useful; they are often harmful, and in that case pathological.
Lastly, a few words about the very difficult question of teleology. In trying to explain Evolution in a mechanical—sometimes called monistic, but in reality natural—way, we exclude anything like a set purpose, a goal, or ideal, a final condition which the organism strives to attain. Unknown, however, to many morphologists, especially embryologists, their writings are full of this teleological notion. Indeed, there are many cases[Pg 131] in which an organism becomes changed, and quickly, too, in a way which cannot but be called reasonable. It starts modifications, be they outgrowths, alterations in shape or colour, or the making good of injuries received, which by 'short-cuts' produce the only advantageous result that can reasonably satisfy the new requirement or altered circumstances.
Trees growing in precarious positions, after part of the supporting rock has slipped away, throw out new roots, and rearrange some of the old ones in the only way which could save the tree. In animals which have lost part of a limb the wound closes up, and what is left is turned into a serviceable stump—for example, in water-tortoises (creatures in which reproduction of lost limbs does not happen). In frogs and newts the lost part is reproduced, not correctly, but in a good semblance. Tortoises which have had their shell smashed can throw off an astonishingly large portion and renew the bone as well as the over[Pg 132]lapping scutes; but this mending is not neatly done. It serves the requirement, but it is patchwork; the new shell is such as no tortoise ever possessed before.
Mammals transported into colder countries, or subjected to continued exposure, grow a thicker coat; and the same kind of tree which in a sheltered valley is tall, large-leaved, and soft-wooded, assumes a very different aspect, although perhaps growing into a healthy specimen, when planted on a wind-exposed hill.
There is no room, or, rather, no time, to apply to these cases the principle of many variations or the long-continued accumulation of infinitely small changes. The thing is to be done quickly, or not at all. Nor can we explain the mending of a wound, which implies an activity of countless cells, simply as a case of, or similar to, the reproduction of a lost part; against such an explanation militates the almost absolute unlikelihood of that precise injury having[Pg 133] happened before to any of the creature's ancestors.
Still, I think we are brought near the solution of the mystery by such considerations. We see no difficulty in the regeneration of a few cells, or in the making good of the disturbance suffered by one of the most simple organisms; but we become suspicious when we see that countless cells, not of one kind, but of the most varied tissues and parts of the body, make common cause in remedying a defect in a serviceable way.
We must assume that since the beginning of life organisms have been subjected to countless insults. We can scarcely speak of a wound in an Amæba; but these insults have always been made good, and whenever this was not the case, that particular organism came to an end. As these organisms developed into more complicated ones, the possible insults became more serious, more complicated; and the organisms took adaptive measures so as to be superior to them. This[Pg 134] action, I have no hesitation in declaring, became by heredity a habit. The whole creature became so thoroughly 'imbued' (for want of a better word) with the finding of ways and means for meeting sudden, serious conditions, that it now acts directly, and produces by a short-cut, with the least amount of time and with the smallest possible waste of material, that which meets the occasion, thereby saving the life of the individual and that of the race. This we cannot but call reasonable and to the purpose, although it is all carried out by causæ efficientes without there being any causæ finales.
One million years is a stretch of time beyond our conception. We can arrive at a more or less adequate understanding of what a million individuals or concrete things means. Several Continental nations can put more than a million men into the field. We can gaze at a building which contains as many bricks; and we know that our own body is composed of millions of millions of cells. No such help applies to time, because that itself is an entirely relative, abstract conception. We can imagine what one hundred years are like—a span of time seemingly short to the hale and hearty octogenarian, enormous to the child, totally inapplicable to certain[Pg 136] animals whose whole life is crowded into one single day.
Astronomers have long ceased to reckon distances by miles or any other understandable unit. They express the distances between us and the stars and nebulæ by 'years of light.' Try to imagine a unit of length equal to that which is passed through by light (186,000 miles per second) in one year. Not so very long ago the enormous distances resulting from astronomical calculations were looked upon as the most serious objection to the correctness of the astronomers' views as to the distances which separate our globe from the nearest fixed stars. We have not yet accustomed ourselves to reckoning time by some similar broadly-conceived standard—say æons of so many thousand years each.
Unfortunately, we possess no data whatever for calculating the age of the successive geological strata. Thanks to Lyell, the theory of violent universal cataclysms has[Pg 137] been done away with. It is more probable that the same agencies have acted which are now changing the aspect of the globe; and these changes are slow, as far as we know them—at least, as far as the formation of sedimentary strata is concerned, and these alone we have to deal with. Various calculations have been made, based upon the denudation of the mountains, the filling up of the valleys by the débris, the formation of deltas, etc. The results give enormous stretches of time, but all of them unsatisfactory, because the methods are so very local in their application.
The least objectionable attempt is that which, based upon astronomical calculations, tried to fix the height of the last Glacial epoch[27] at about 200,000 years ago, and asserted that since its beginning in the Pliocene epoch as many as 270,000 years have elapsed. The duration of the whole[Pg 138] Tertiary period has by the same authorities been fixed approximately at 3,000,000 to 4,000,000 years. Beyond this we cannot venture without the wildest speculation; but we know to a certain extent the thickness of the various sedimentary strata, which amount in all to from 100,000 to 175,000 feet—on the average perhaps 130,000 feet, or about twenty miles.
Unless we prefer giving up all attempt at calculation as absolutely hopeless, and thus resign the whole problem, we must at least try to arrive at some results, and then see if these cannot reasonably be made use of.
Neither geologist nor physicist, and no zoologist, would accept the suggestion that these 130,000 feet of stratified rocks have been deposited within only as many years, although the average rate of deposit would in that case be not more than 1 foot per year. On the other hand, an indignant protest is raised against the assumption of 1,000,000,000 years.
Lord Kelvin[28] has come to the conclusion (from data which various other authorities regard as very unsatisfactory) that not much more than 100,000,000 years can have elapsed since the molten globe acquired a consolidated crust. Further time must have passed before the surface had become stable and cool enough to allow the temperature of the collecting oceans to fall below boiling-point, and it is obvious that life cannot possibly have begun until after this had happened.
Wallace, in his 'Island Life,' by making use of Professor A. Geikie's results as to the rate of denudation of matter by rivers from the area of their basins, and estimating the average rate of deposition, concludes that 'the time required to produce this thickness of rock [Professor Haughton's maximum of 177,000 feet] at the present rate of denudation and deposition is only 28,000,000 years.'[Pg 140] Our lower assumption of 130,000 feet thickness would give only 20,000,000 years—a rate of 1 foot in 154 years.
Again, if we prefer round numbers to start with, we have only to assume that the age of the whole Tertiary period, with its 3,000 feet thickness, is 3,000,000 years (i.e., 1,000 feet in 1,000,000 years, or 1 foot in 1,000 years, surely an excessively slow rate); then 130,000,000 years would bring us to the bottom of the Laurentian or pre-Cambrian deposits. Of course, it is a pure assumption that the same rate of destruction and sedimentation applies to the whole of the strata; but we know nothing to the contrary, especially if we consider the average periods, the quick periods of extra activity, taken with the slow periods or those of standstill.
Dana estimated the length of the whole Tertiary period at one-fifteenth of the Mesozoic and Palæozoic combined. If we take the duration of the Tertiary period, as before, as 3,000,000 to 4,000,000 years, the total[Pg 141] will amount to from 45,000,000 to 60,000,000 years.
Lastly, Walcott[29] has estimated the duration of the Palæozoic, Mesozoic, and Cænozoic or Tertiary epochs at about 17,000,000, 7,000,000 and 3,000,000 years respectively, giving 27,700,000 years from the beginning of the Cambrian; and Williams[30] has calculated the relative duration of the smaller epochs. See the table on p. 149.
The results of all these calculations fall surprisingly well within the limits of Lord Kelvin's allowance. Of course they are based upon assumptions, but none of them is inherently unreasonable; and it was my purpose to draw attention to the surprising coincidence in the closeness of these results, perhaps too good to be true. Such calculations are considered close enough if they range within a few multiples of each other.
Zoologists have fallen into the habit of requiring enormous lengths of time for the evolution of the animal kingdom. We know that Evolution is at best a slow process, and the conception of the changes necessary to evolve man from monkey-like creatures, these from the lowest imaginary mammals, these from some reptilian stock, thence descending to Dipnoan fish-like creatures, and so on back into Invertebrata, down to the simple Monera—this conception is indeed gigantic. Innumerable, almost endless, slow changes require seemingly unlimited time, and as time is endless, why not draw upon it ad libitum?
Huxley pointed out that it took nearly the whole of the Tertiary epoch to produce the horse out of the four-toed Eohippos, and that, if we apply this rate to the rest of its pedigree, enormous times would be required. This is, however, a very misleading statement, which necessitates considerable reduction, in conformity with our increased palæontological knowledge. Animals of the[Pg 143] genus Equus—namely, Ungulata, with one toe, and with a certain tooth pattern—from the Upper Miocene of India are now known. Moreover, it is not simply a question of the gradual loss of the side-toes. The change from the fox-sized little Eohippos and Hyracotherium, so far as skull, teeth, vertebral column, and limbs are concerned (about the soft parts we know next to nothing), is a very great one indeed.
Elephants and mammoths seem to have developed very rapidly. None are known from Eocene strata; but towards the end of the Miocene they had spread over Asia, Europe, and North America, and that in great numbers. The Eocene Amblypoda are still so different that we hesitate to connect them ancestrally with the elephants.
The Pinnipedia (seals and walruses) are strongly modified fissiped Carnivora, and have existed since at least the Upper Miocene; the transformation must have been accomplished within the Miocene period.
We cannot shut our eyes to the fact that various groups have from the time of their first appearance burst out into an exuberant growth of modifications in form, size, and numbers, into all possible—and one might almost say impossible—shapes; and they have done this within comparatively short periods, after which they have died out not less rapidly. It seems almost as if these go-ahead creatures had, by accepting every possible modification and carrying the same to the extreme, too quickly exhausted their plasticity—which, after all, must have limits—thereby becoming unable to meet successfully the requirements of further changes in their surroundings. The slowly developing groups, keeping within main lines of Evolution, and not being tempted into aberrant side-issues, had, after all, a much better chance of onward evolution.
A good example of the former are the Dinosaurs. We do not know their ancestors; but we have here to deal only with their[Pg 145] range of transformation. The oldest known forms occur in the Upper Trias; they attain their most stupendous development in the Upper Jurassic and in the Wealden; and they have died out with the Cretaceous epoch. But already some of their earliest forms had assumed bipedal gait, and the Oolitic Compsognathus had developed almost bird-like hind-limbs.
On the other hand, there are many instances of extremely slow development—facts which raise the difficult question of 'persistent types.' Are these due to a state of perfection which cannot be improved upon? Or are they due to a kind of morphological consolidation (not necessarily specialization) which can no longer yield easily, so that therefore through changes in their surroundings they may come to an end sooner than more plastic groups?
Struthio, the ostrich; Orycteropus, the Cape ant-eater; Tapirus, and many others, existed in the Miocene age practically as[Pg 146] they are now; but pre-Pliocene dolphins, cats, monkeys, stags, all belong to closely-allied and well-defined 'genera,' but different from the living forms.
Alligators and crocodiles are known from the Upper Chalk; Tomistoma since the Miocene; Gavialis since the Pliocene.
The oldest surviving reptile is Sphenodon, the Hatteria of New Zealand, a fair representative of what generalized reptiles of the later Triassic period seem to have been like; and to the same period belongs Ceratodus, the Australian mud-fish, hitherto the oldest known surviving genus of a very ancient and low type so far as Vertebrata are concerned.
Now let us see if the above estimates of geological time are so utterly inapplicable to animal evolution. On purpose we take one of the lowest estimates, about 28,000,000 years, and apportion them equally to the various strata or epochs.
The original owner of the famous Trinil skull, a Pithecanthropus erectus, lived,[Pg 147] according to some, in the Late Pliocene, according to others in the Early Plistocene, period—that is to say, somewhere about the beginning of our last Glacial epoch, some 270,000 years ago. Assuming that he and his like reached puberty at sixteen to twenty years of age, about 17,000 generations would lie between him and ourselves, or, to put it more forcibly, between him and the lowest living human races—say the Ceylonese Veddahs. Only 250 generations, at twenty years, carry us back to 3000 B.C. (i.e., beyond the ken of history); and if it be objected that the differences between the oldest inhabitants of Egypt, the Naquada, and the present Fellahin are very slight, we are welcome to multiply these differences sixty or seventy fold, in order to arrive at the Pithecanthropus level. But these Naquada had no metal implements, and there cannot be the slightest doubt that the development of the human race went on by leaps and bounds after certain discoveries had been made—to[Pg 148] wit, the use of implements and that of fire. That creature which first took up a stone or a branch and wielded it thereby got such an enormous advantage over his fellow-creatures that his mental and bodily development went on apace. The same applies to the improvement of speech. We assume the single, monophyletic origin of mankind at one place, in one district; and the differences between some of the races of man are great enough to constitute what we might call species. Compare the Venus of Milo, that noble expression of the ancient Greeks' notion of female beauty, with the 'products of art' of the Veddahs or the dwarfs of Central Africa, or think of the beau-idéal which a Michael Angelo could possibly have evolved if he had never seen any but such people.
| I. | II. | III. | IV. | V. | VII. | VII. |
| Recent | } | Adam and Eve | 250 | |||
| Plistocene | } 5 | } | } | Man, contemp- | 3,500 | |
| } | } | } 270,000 | orary with Reindeer | |||
| } | } | in France | ||||
| Pliocene - | } | } 3,000,000 | Pithecanthropus | 16 | 17.000 | |
| } | } | } 600,000 | erectus | |||
| Miocene - | }10 | } | Anthropoid | 10 | 60,000 | |
| } | } | } 2,100,000 | Apes | |||
| Eocene - | } | } | Lemurs | 5 | 420,000 | |
| Cretaceous - | 10 | } | 3,600,000 | |||
| Jurassic - | 5 | } | 1,800,000 | |||
| Rhætic - | } | } | } | Prototheria, or | 3 | 1,800,000 |
| } | } | } | first Mammalia | |||
| } | } 7,200,000 | } | ||||
| Keuper - | } 5 | } | }1,800,000 | |||
| Muschelkalk - | } | } | } | |||
| New Red | } | } | } | Theomorpha | 4 | 425,000 |
| Sandstone | ||||||
| Magnesian | } | } | } | |||
| Limestone | } | } | } | |||
| Lower Red | } | } | } | Proreptilia | 4 | 250,000 |
| Sandstone | }15 | } | }4,000,000 | |||
| Coal-measures | } | } | } | Eotetrapoda | 4 | 500,000 |
| Mountain | } | } | } | |||
| Limestone | } | }17,500,000 | } | |||
| Devonian - | 15 | } | 4,000,000 | Dipnoi and | 5 | 1,000,000 |
| } | Crossopterygii | |||||
| Silurian - | 10 | } | 2,700,000 | First fishlike | 3 | 900,000 |
| } | } | creatures | ||||
| Ordovician - | 10 | } | 2,700,000 | |||
| Cambrian - | 15 | } | 4,000,000 | Sum total of | ||
| Laurentian - | generations | ———— | ||||
| Archean or | (about) | 5,375,000 | ||||
| Metamorphic |
Explanation of the Table on p. 149.
Column I. contains the names of the successive sedimentary strata.
" II. contains the percentage of the duration of the various epochs, according to Williams, the time from the Cambrian until recent times being taken as 100.
" III. gives the estimated duration in years of the Palæozoic, Mesozoic, and Cænozoic periods, according to Walcott.
" IV. gives in years the duration of the various smaller epochs, as computed from Walcott and Williams' statements.
" V. Representatives of stages of the ancestral line of man. The names stand in the level of the stratum in which they have made their first appearance.
" VI. contains the number of years which, in the present calculation, have been assumed necessary for the animal to reach puberty.
" VII. contains the number of generations which can have elapsed from stage to stage. For example, 60,000 generations separate the earliest known anthropoid apes from Pithecanthropus.
Let us follow the descent of man further back. The next stage, reckoning backwards, is that from Pithecanthropus to bonâ-fide anthropoid apes. They are represented in the Miocene by various genera—e.g., Pliopithecus and Dryopithecus. According to Croll and Wallace, 850,000 years ago carry us[Pg 151] into the Miocene epoch. Assuming that these apes lived about 600,000 years before Pithecanthropus, namely, in the later half of the Miocene, and taking puberty at ten years of age, a high estimate, we get not less than 60,000 generations.
2. From Apes back to lowest Lemurs in the lowest Eocene. The date of Eocene being fixed at 3,000,000, we have about 2,100,000 years for this stage; assuming as much as five years for puberty, this results in 420,000 generations.
3. From Lemures to Prototheria. The earliest known mammalian remains come from the Rhætic, or top formation of the Triassic epoch; allowing for the Rhætic only 100,000 years, we have to add the whole of the Jurassic and Cretaceous, in all about 5,500,000 years. Assuming three years for a generation, we get 1,800,000 generations.
4. From Prototheria to something like the Theromorpha at the bottom of the Triassic[Pg 152] strata. A duration of 1,700,000 years divided by four gives 425,000 generations.
5. From Theromorpha to Proreptilia, represented by Eryops and Cricotus from the Lower Permian of Texas. Allowing 1,000,000 years, each generation at four years, we obtain 250,000 generations.
6. From Proreptilia to Eotetrapoda, the first terrestrial Vertebrata, represented by something like the Stegocephali, the earliest of which are known from the Coal-measures. Assuming them to have come into existence at the bottom of the Coal-measures, for the duration of which we may guess 2,000,000 years, we get, with four years' allowance for puberty, 500,000 generations.
7. From Eotetrapoda to a not yet separated or differentiated group of Crossopterygian and Dipnoan fishes, both of which are known from Devonian strata. The duration of the latter has been computed at 4,000,000 years, which, with 1,000,000 for the Mountain Limestone formation, gives us[Pg 153] 5,000,000 for this stage. Assuming, for the sake of round numbers, as much as five years for a generation, we get 1,000,000 generations.
8. Earliest stage, down to the first fish-like creatures. Teeth and spines indicating the existence of fishes are known from the Upper Silurian. By carrying the earliest fishes down to the bottom of the Silurian, with 2,700,000 years' duration, and allowing three years for attaining puberty, the calculation results in 900,000 generations.
Further back we cannot go. We do not know of any Vertebrate remains from the Ordovician and Cambrian, which together represent 6,700,000 years, enough for at least half as many generations of Prochordate creatures. The pre-Cambrian or Laurentian epoch lies quite beyond the reach of calculation, nor have we any trustworthy fossil remains of living matter from these strata, to which, however, Haeckel and others refer the first beginnings of life.
All the above calculations are, of course, only approximate. What we do know is the existence of representatives of the stages, our proofs being the fossils; but when we refer the origin of the Eotetrapoda, for example, to the bottom and not somewhere to the middle of the Coal-measures, we are guessing merely. Alterations in the levels assumed for the various stage-representatives will, of course, alter the result of the number of generations; but the leading idea, as a whole, is not thereby upset. The fact remains that in the Upper Silurian we have fishes; from the Coal-measures onwards, fishes and Amphibia; since the Permian, fishes, Amphibia, and reptiles; since the end of the Trias these three classes and the Mammalia; and lastly, at least since the Plistocene, man himself. If Evolution is true at all, the transformation from early fish-like creatures to man has come about within these epochs. Being able to assign a time of duration to each of them, with an approximate total of[Pg 155] 21,000,000 years, we are also able to put the whole ancestral series to a test by expressing each great stage in generations. The result is very satisfactory. The whole enormous stretch from the lowest fish-like creatures to man has been resolved into more than 5,000,000 successive generations, and each of these means a little step forwards in onward Evolution.
Nothing is to be gained for the understanding of our problem of Evolution if we multiply this enormous number of generations by ten or any other multiple. We are not able to conceive changes so small as those which necessarily have existed between Pithecanthropus and man if the whole striking difference is analysed into 17,000 steps. Every one of these stages in the modifications of the muscles, the skeletal framework, increase of brain, shortening of the trunk, lengthening of the legs, improvement of the hands, loss of the hairy coat, etc., is truly microscopical, imperceptible, just as[Pg 156] the Evolutionist imagines the whole process to have been. Again, where is the difficulty implied by the change from an air-breathing, in many structural points half-amphibian, fish into a primitive land-crawling four-footed creature, if we are allowed to resolve the transformation into 1,000,000 stages? So far from there being any difficulty, rather does it appear questionable if so many infinitely small changes have been necessary to bring about this result.
One thousand years make apparently no difference in the evolution of animals, nor does one second change the aspect of the hands on the face of a clock, nor did Julius Cæsar's commission of scientific men appreciate the error of about eleven minutes in the length of the year beyond its real value; but now the Russians are, owing to this neglect, nearly two weeks behind the civilized nations.
THE END.
BILLING AND SONS, PRINTERS, GUILDFORD.
By PROFESSOR ERNST HAECKEL
MONISM;
OR,
The Confession of Faith of a Man of Science.
Translated from the German by J. D. F. GILCHRIST.
Crown 8vo., cloth. Price 1s. 6d. net.
'We may readily admit that Professor Haeckel has stated his case with the clearness and courage which we should expect of him, and that his lecture may be regarded as a fair and authoritative statement of the views now held by a large number of scientifically educated people.'—Times.
'The Monism, which is the substance of his faith, is thus defined by him: "Our conviction that there lives one spirit in all things, and that the whole cognizable world is constituted, and has been developed, in accordance with one common fundamental law." As the confession of a distinguished man of science, this little work deserves to be read.'—North British Daily Mail.
'This "Confession of Faith" was delivered by the great German scientist, its author, as an extemporaneous address at Altenburg rather more than two years ago. There are, no doubt, a large number of English readers who will welcome a translation, for this "connecting of religion and science" has long troubled many earnest students of modern science.'—Publisher's Circular.
'This is a little book of great daring, an example of the wild speculative flights of one of the very ablest and greatest of our contemporary men of science.'—Aberdeen Free Press.
'The address, whatever we may think of its conclusions, is, however, most interesting reading, and is admirably done into English by the translator.'—Literary World.
LONDON: ADAM & CHARLES BLACK, SOHO SQUARE.
Demy 8vo., price 7s. 6d. net.
SOURCES OF THE APOSTOLIC CANONS.
With a Treatise on the Origin of the Readership and other Lower Orders.
By Professor ADOLF HARNACK.
Translated by LEONARD A. WHEATLEY.
With an Introductory Essay on the Organization of the Early Church and the Evolution of the Reader.
By the Rev. John Owen, Author of 'Evenings with the Skeptics.'
'Dr. Adolf Harnack is at the present time undoubtedly the leading liberal authority in Germany on matters connected with early Christian history.'—The Times.
'Those who are interested in early Church history know how to prize anything from the pen of Prof. Harnack. They will not be disappointed with the present paper, in which, with his accustomed learning and acute criticism, he annotates and comments upon the fragments of primitive church law which partly form the basis of the Apostolic Canons.'—British Weekly.
'The wide circulation of this volume would be of the happiest augury for a more scientific and worthy conception of the organization of the primitive Church.'—Dr. Marcus Dods in The Bookman.
Crown 8vo., cloth, price 1s. 6d. net.
CHRISTIANITY AND HISTORY.
By ADOLF HARNACK.
Translated, with the Author's sanction, by THOMAS BAILEY SAUNDERS, with an Introductory Note.
'It is highly interesting and full of thought. The short introductory note with which Mr. Saunders prefaces it is valuable for its information and excellent in its tone.'—Athenæum.
'A singularly able exposition and defence of Christianity, as seen in the newer light, by one of the most learned and acute "evangelical" critics of Germany. The essay is a masterly one.'—Glasgow Herald.
' ... We hope the lecture will be widely read.'—Primitive Methodist Quarterly Review.
'The lecture itself is weighty in its every word, and should be read and re-read by those desiring to have in a nutshell the central positions of modern Christianity.'—Christian World.
LONDON: ADAM & CHARLES BLACK, SOHO SQUARE.
Third Edition. Crown 8vo., cloth, price 5s.
SKETCH OF THE HISTORY OF ISRAEL AND JUDAH.
By J. WELLHAUSEN,
PROFESSOR AT MARBURG.
'This work is now issued for the third time as an independent treatise. It admirably epitomizes the subject, and exhibits on almost every page evidences of Professor Wellhausen's profound study.'—Publishers' Circular.
'We would only say that those who differ from his critical views will yet do well to study them, and to read this history in which he applies them. Its separate publication, in a handy form and at a moderate price, makes it generally accessible.'—North British Daily Mail.
'The publication in a separate form of Professor Wellhausen's article in the "Encyclopædia Britannica" on "Israel" will be very warmly welcomed by many readers.'—Manchester Guardian.
'We are very glad to welcome an edition of Professor Wellhausen's "Sketch of the History of Israel and Judah" in a convenient and handy form. This is the first time it has appeared in a separate form. It is already known to students; it ought now to become popular. It is based on the learned author's studies in Hebrew literature and history, and, though not controversial in form, it differs totally from orthodox presentations of the subject.'—Westminster Review.
'A sketch which has created such widespread and profound interest as this could not be kept in the pages of a voluminous encyclopædia. Wellhausen's words necessarily have exceptional importance, even in the esteem of those who differ from him toto cœlo.'—Baptist Magazine.
'The profound scholarship of the author does not elevate his writing above the interest of the general reader, and a vivid idea of the involved Jewish history is obtainable from this volume.'—Christian Advocate.
LONDON: ADAM & CHARLES BLACK, SOHO SQUARE.
Demy 8vo., boards, price 3s. 6d. net.
A CLASSIFICATION OF VERTEBRATA,
RECENT AND EXTINCT.
With Diagnoses and Definitions, a Chapter on Geographical Distribution, and an Etymological Index.
By HANS GADOW, M.A., Ph.D., F.R.S.,
STRICKLAND CURATOR AND LECTURER ON ZOOLOGY TO THE UNIVERSITY, CAMBRIDGE.
'At the end of his work Dr. Gadow adds a useful chapter on the geographical distribution of the Vertebrata, with a table showing the approximate number of the known recent species. He also gives a fanciful though striking calculation to show how some groups are still in the ascendant, while others are distinctly declining. The little volume is indeed a welcome addition to the biological student's library, and it deserves the wide circulation which its author's eminence is likely to ensure for it.'—Natural Science.
'It is a book, it need hardly be said, for the student; it is simply a list of the principal sub-divisions of backboned animals, with just as much definition as is needed. It may be regarded as an exceedingly concentrated extract of a full text-book of the vertebrates.'—Daily Chronicle.
LONDON: ADAM & CHARLES BLACK, SOHO SQUARE.
Demy 8vo., cloth, price 21s.
IN NORTHERN SPAIN.
By Dr. HANS GADOW, M.A., Ph.D., F.R.S.
Containing Map and 89 Illustrations.
'Some years back "Wild Spain," one of the best books of its kind, made you desirous of knowing more of the country. And Hans Gadow has deepened this feeling in his excellent volume "In Northern Spain," and that to an enormous extent. Dwelling at inn or farm, or in their own tent, they saw the country as it has been seen but rarely, and they came to know the inhabitants as they can be known in no other fashion.'—Black and White.
'To persons visiting the provinces with which the author deals, this book will be invaluable, and will do more to point their attention to objects of interest than existing guide-books of Spain, most of which are out of date.'—The Field.
'About the best book of European travel that has appeared these many years.'—Literary World.
LONDON: ADAM & CHARLES BLACK, SOHO SQUARE.
[6] Perfect, in the sense of highest stage of evolution, may seem a petitio principii. Leaving aside the consideration that no living creature is absolutely perfect, in the sense that its organization cannot become more efficient or proficient, we have here to deal with relative perfection of the whole organization. A fish or a snake is in its way more specialized than a mammal; but specialization does not necessarily mean height of development: it generally means life in a comparatively narrow groove. The acts of giving birth and nourishing the young with the mother's milk is a much higher stage than the act of laying eggs and letting them run their chance. The development of a hairy coat goes along with heightened temperature of the blood, subsequent greater independence of the surrounding temperature, and increased steady activity of the brain and other nerve-centres. The brain of the Mammalia, in its minute structure, is much more complex. This rule applies to some of the principal sense organs, chiefly the nose and the ear. The skeleton, not so much as a whole as in the various bones and joints, is more neatly finished, and built up more in conformity with 'scientific principles,' than is the case even with birds, in spite of their marvellous specialization. The same is the case with the vascular system, notably the heart and the veins, and with the excretory organs. In all of these many imperfections, still to be found in the other classes, have been corrected in Mammalia. The Primates take an easy first by their hands, and among them the apes and man himself by their brains.
[7] 'Die menschenähnlichen Affen und ihre Organisation im Vergleich zur menschlichen.' 1883.
[8] G. Schwalbe, 'In wiefern ist die menschliche Ohrmuschel ein rudimentäres Organ?'—In what Respects is the Human Outer Ear a Rudimentary Organ? (Archiv f. Anatomie und Physiologie, 1889).
[9] Wiedersheim, 'Der Bau des Menschen als Zeugniss für seine Vergangenheit.' Freiburg, 1888. Translated: 'The Structure of Man an Index to his Past History.' London, 1895.
[10] Pithecanthropus erectus. 'Eine menschenähnliche Uebergangsform aus Java' ('A Human-like Transitional Form'). Batavia, 1894.
[11] On the day after the delivery of this address Dr. Dubois exhibited the cranium of Pithecanthropus, from which he had removed the stony matrix which filled the inside, in order to examine the impression left by the cerebral convolutions. He was able to show that they also are very human, and more highly developed than those of the recent apes.
[12] L. Manouvier: 'Deuxième étude sur le Pithecanthropus erectus comme précurseur présumé de l'homme.' (Bulletins de la Soc. d'Anthropologie de Paris, 1895.)
F. Ameghino: 'Contribucion al conocimiento de los mamíferos de la república Argentina.' In Actas de la Academia nacional de Sciencias en Cordoba, 1889.—Another article in Revista Argentina de Historia natural. Buenos Aires, 1891.
A. Gaudry: 'Animaux fossiles et géologie de l'Attique.' 1862.—'Le Dryopithèque.' Mém. Soc. géol. de France: 'Paléontologie.' 1890.
O. Marsh: 'Introduction and Succession of Vertebrate Life in America.' Address, Amer. Assoc. Adv. Sci., Nashville, 1887.
H. F. Osborn: 'The Rise of the Mammalia in North America.' Address, Amer. Assoc. Adv. Sci., Madison, 1893.
L. Ruetimeyer: 'Ueber die Herkunft unserer Thierwelt,' Basel, 1867.
C. S. Forsyth Major: 'Fossil Monkeys from Madagascar.' Geological Magazine, 1896.
M. Schlosser: 'Ueber die Beziehungen der ausgestorbenen Saeugethierfaunen und ihr Verhaeltniss zur Saeugethierfauna der Gegenwart.' Biolog. Centralblatt, 1888.
[18] Wilhelm Bischoff of Munich: works on the history of the development of the rabbit, dog, guinea-pig, roe-deer. 1840-1854.
[20] 'Ueber die Entwicklung der einfachen Ascidien,' Mém. Acad. St. Petersbourg, vii. ser., tome x. (1866). Other papers in 'Archiv f. Mikroskop. Anatomie,' vii. (1871); xiii. (1877).
[22] Similar conditions seem to have prevailed among the Proreptilia; but in those of their descendants which have specialized into Reptiles and Birds the basi-occipital element becomes more and more predominant in that formation which ultimately leads to the apparently single condyle. Hence it is misleading to divide the Tetrapoda into the two main groups of Amphi-and Mono-condylia, and therefrom to conclude that the two-condyled Mammalia are more closely related to the likewise amphicondylous Amphibia than to the so-called monocondylous Reptiles.
[23] 'Ergebnisse naturwissenschaftlicher Forschungen auf Ceylon,' vols. 4 and 5. (With an atlas of 84 plates; 1893.)
[24] 'Principles of Biology': 'The Factors of Organic Evolution'; 'The Inadequacy of Natural Selection.'
[25] Abridged from Haeckel's 'Systematische Phylogenie der Vertebraten,' § 14.
[26] That this great work is now comparatively rare, although still in the second-hand market, may perhaps be urged in excuse of the fact of so many attempts made by many authors, both professional and amateur, to find fault with or to explain the principles of adaptation, variation, heredity, cænogenesis, phylogeny, etc., in complete ignorance that all these and many more fundamental questions were fully discussed more than thirty years ago in the 'Generelle Morphologie.'
[27] James Croll: 'On Geological Time, and the Probable Date of the Glacial and Upper Miocene Period,' Philos. Magazine, xxxv., 1868, pp. 363-384; xxxvi., pp. 141-154; 362-386.
[28] William Thomson: 'On the Secular Cooling of the Earth,' Transact. R. S. Edinb., xxiii., 1864, pp. 157-169.
[29] 'Geological Time as indicated by the Sedimentary Rocks of North America.' Proc. Amer. Assoc. Adv. Sci., xlii., 1893, pp. 129-169.
[30] Henry Shaler Williams, 'Geological Biology.' New York, 1895.
Transcriber's Notes
Variations in spelling, punctuation and hyphenation have been retained
except in obvious cases of typographical errors.
Inconsistent hyphenation and spelling are as in the original.
The layout of the chart Ancestral Tree of The Mammalia has been changed
from the original to enhance clarity, the essential relationships have
been preserved.
The second reference to footnote 3, in the same paragraph as the first,
has been left blind as it is redundant.