HANS DRIESCH, Ph.D.
HEIDELBERG
Transcriber’s notes:
In this transcription, page numbers are shown in the right margin, and page footnotes (renumbered in consecutive order) are grouped together at the end of the book. Hyperlinks to footnotes and page references are indicated by black dotted underlines plus aqua highlighting when the mouse pointer hovers over them. The footnotes are themselves hyperlinked back to the originating marker to facilitate easy return to the text. A red dashed underline as shown here indicates the presence of a transcriber’s comment; scrolling the mouse pointer over such text will reveal the comment.
The rare spelling typos noted in the original text have been corrected silently (e.g. invividual-->individual, hyberbola-->hyperbola) but inconsistent use of the ligature æ/ae (e.g. palæontology/palaeontology), inconsistent use of alternative spellings (e.g. learned/learnt), and occasional inconsistencies of hyphenation have been left as in the original. Minor punctuation typos have been corrected silently (e.g. index entries with missing commas). The abbreviation viz. appears in both roman and italic font.
Formatting of entries in the Table of Contents does not accurately match that of the corresponding headings in the text, particularly the heading Pt.I-B-3 which contains an extraneous α.
In Figure 12 caption, multiple ditto marks have been replaced by the relevant text for greater clarity.
THE SCIENCE AND PHILOSOPHY
OF THE ORGANISM
| AGENTS | |
| America | The Macmillan Company 64 & 66 Fifth Avenue, New York |
| Australasia | The Oxford University Press, Melbourne |
| Canada | The Macmillan Company of Canada, Ltd. 27 Richmond Street West, Toronto |
| India | Macmillan & Company, Ltd. Macmillan Building, Bombay 309 Bow Bazaar Street, Calcutta |
THE GIFFORD LECTURES DELIVERED BEFORE
THE UNIVERSITY OF ABERDEEN
IN THE YEAR 1907
BY
HANS DRIESCH, Ph.D.
HEIDELBERG
![]()
LONDON
ADAM AND CHARLES BLACK
1908
All rights reserved
This work is not a text-book of theoretical biology; it is a systematic presentment of those biological topics which bear upon the true philosophy of nature. The book is written in a decidedly subjective manner, and it seems to me that this is just what “Gifford Lectures” ought to be. They ought never to lose, or even try to lose, their decidedly personal character.
My appointment as Gifford Lecturer, the news of which reached me in February 1906, came just at the right moment in the progress of my theoretical studies. I had always tried to improve my previous books by adding notes or altering the arrangement; I also had left a good deal of things unpublished, and thus I often hoped that I might have occasion to arrange for a new, improved, and enlarged edition of those books. This work then is the realisation of my hopes; it is, in its way, a definitive statement of all that I have to say about the Organic.
The first volume of this work, containing the lectures for 1907—though the division into “lectures” has not been preserved—consists of Parts I. and II. of Section A, “The Chief Results of Analytical Biology.” It gives in Part I. a shortened, revised, and, as I hope, improved account of what was published in my Analytische Theorie der organischen Entwickelung (1894), Die Localisation morphogenetischer Vorgänge; ein Beweis Vitalistischen Geschehens (1899), and Die organischen Regulationen (1901), though for the professed biologist the two last-named books are by no means superseded by the new work. Part II. has never been published in any systematic form before, though there are many remarks on Systematics, Darwinism, etc., in my previous papers.
The second volume—to be published in the autumn, after the delivery of the 1908 lectures—will begin with the third and concluding part of the scientific section, which is a very carefully revised and rearranged second edition of my book, Die “Seele” als elementarer Naturfactor (1903). The greater part of this volume, however, will be devoted to the “Philosophy of the Organism,” i.e. Section B, which, in my opinion, includes the most important parts of the work.
Some apology is needed for my presuming to write in English. I was led to do so by the conviction, mistaken perhaps, that the process of translation would rob the lectures of that individual and personal character which, as I said before, seems to me so much to be desired. I wished nothing to come between me and my audience. I accordingly wrote my manuscript in English, and then submitted it to linguistic revision by such skilled aid as I was able to procure at Heidelberg. My reviser tells me that if the result of his labours leaves much to be desired, it is not to be wondered at, but that, being neither a biologist nor a philosopher, he has done his best to make me presentable to the English reader. If he has failed in his troublesome task, I know that it is not for want of care and attention, and I desire here to record my sense of indebtedness to him. He wishes to remain anonymous, but I am permitted to say that, though resident in a foreign university, he is of Scottish name and English birth.
My gratitude to my friends at Aberdeen, in particular to Professor and Mrs. J. A. Thomson, for their hospitality and great kindness towards me cannot be expressed here; they all know that they succeeded in making me feel quite at home with them.
I am very much obliged to my publishers, Messrs. A. and C. Black, for their readiness to fulfil all my wishes with respect to publication.
The lectures contained in this book were written in English by a German and delivered at a Scottish university. Almost all of the ideas discussed in it were first conceived during the author’s long residence in Southern Italy. Thus this book may be witness to the truth which, I hope, will be universally recognised in the near future—that all culture, moral and intellectual and aesthetic, is not limited by the bounds of nationality.
HANS DRIESCH.
Heidelberg, 2nd January 1908.
THE PROGRAMME
| PAGE | |
| On Lord Gifford’s Conception of “Science” | 1 |
| Natural Sciences and “Natural Theology” | 3 |
| Our Philosophical Basis | 5 |
| On Certain Characteristics of Biology as a Science | 9 |
| The Three Different Types of Knowledge about Nature | 13 |
| General Plan of these Lectures | 15 |
| General Character of the Organic Form | 19 |
SECTION A.—THE CHIEF RESULTS OF ANALYTICAL BIOLOGY
PART I.—THE INDIVIDUAL ORGANISM WITH REGARD TO
FORM AND METABOLISM
| A. ELEMENTARY MORPHOGENESIS— | |
| Evolutio and Epigenesis in the old Sense | 25 |
| The Cell | 27 |
| The Egg: its Maturation and Fertilisation | 31 |
| The First Developmental Processes of Echinus | 33 |
| Comparative Embryology | 44 |
| The First Steps of Analytical Morphogenesis | 45 |
| The Limits of Pure Description in Science | 50 |
| B. EXPERIMENTAL AND THEORETICAL MORPHOGENESIS— | |
| 1. The Foundations of the Physiology of Development. “Evolutio” and “Epigenesis” | 52 |
| The Theory of Weismann | 52 |
| Experimental Morphology | 56 |
| The Work of Wilhelm Roux | 58 |
| The Experiments on the Egg of the Sea-urchin | 59 |
| On the Intimate Structure of the Protoplasm of the Germ | 65 |
| On some Specificities of Organisation in Certain Germs | 70 |
| General Results of the First Period of “Entwickelungsmechanik” | 71 |
| Some New Results concerning Restitutions | 74 |
| 2. Analytical Theory of Morphogenesis | 76 |
| α. THE DISTRIBUTION OF MORPHOGENIC POTENCIES | 76 |
| Prospective Value and Prospective Potency | 76 |
| The Potencies of the Blastomeres | 79 |
| The Potencies of Elementary Organs in General | 80 |
| Explicit and Implicit Potencies: Primary and Secondary Potencies | 83 |
| The Morphogenetic Function of Maturation in the Light of Recent Discoveries | 85 |
| The Intimate Structure of Protoplasm: Further Remarks | 88 |
| The Neutrality of the Concept of “Potency” | 89 |
| β. THE “MEANS” OF MORPHOGENESIS | 89 |
| β′. The Internal Elementary Means of Morphogenesis | 90 |
| Some Remarks on the Importance of Surface Tension in Morphogenesis | 91 |
| On Growth | 93 |
| On Cell-division | 94 |
| β″. The External Means of Morphogenesis | 95 |
| The Discoveries of Herbst | 96 |
| γ. THE FORMATIVE CAUSES OR STIMULI | 99 |
| The Definition of Cause | 99 |
| Some Instances of Formative and Directive Stimuli | 102 |
| δ. THE MORPHOGENETIC HARMONIES | 107 |
| ε. ON RESTITUTIONS | 110 |
| A few Remarks on Secondary Potencies and on Secondary Morphogenetic Regulations in General | 110 |
| The Stimuli of Restitutions | 113 |
| 3. The Problem of Morphogenetic Localisation: The Theory of the Harmonious-Equipotential system—First Proof of the Autonomy of Life | 118 |
| The General Problem | 118 |
| The Morphogenetic “System” | 119 |
| The “Harmonious-equipotential System” | 122 |
| Instances of “Harmonious-equipotential Systems” | 126 |
| The Problem of the Factor E | 132 |
| No Explanation offered by “Means” or “Formative Stimuli” | 132 |
| No Explanation offered by a Chemical Theory of Morphogenesis | 134 |
| No Machine Possible Inside the Harmonious Systems | 138 |
| The Autonomy of Morphogenesis proved | 142 |
| “Entelechy” | 143 |
| Some General Remarks on Vitalism | 145 |
| The Logic of our First Proof of Vitalism | 146 |
| 4. On Certain other Features of Morphogenesis Advocating its Autonomy | 150 |
| Harmonious-equipotential Systems formed by Wandering Cells | 151 |
| On Certain Combined Types of Morphogenetic Systems | 153 |
| The “Morphaesthesia” of Noll | 157 |
| Restitutions of the Second Order | 158 |
| On the “Equifinality” of Restitutions | 159 |
| Remarks on “Retro-Differentiation” | 163 |
| C. ADAPTATION— | |
| Introductory Remarks on Regulations in General | 165 |
| 1. Morphological Adaptation | 168 |
| The Limits of the Concept of Adaptation | 168 |
| Adaptations to Functional Changes from Without | 172 |
| True Functional Adaptation | 176 |
| Theoretical Conclusions | 179 |
| 2. Physiological Adaptation | 184 |
| Specific Adaptedness not “Adaptation” | 186 |
| Primary and Secondary Adaptations in Physiology | 188 |
| On Certain Pre-requisites of Adaptations in General | 189 |
| On Certain Groups of Primary Physiological Adaptations | 190 |
| General Remarks on Irritability | 190 |
| The Regulation of Heat Production | 193 |
| Primary Regulations in the Transport of Materials and Certain Phenomena of Osmotic Pressure | 194 |
| Chromatic Regulations in Algae | 197 |
| Metabolic Regulations | 198 |
| Immunity the only Type of a Secondary Physiological Adaptation | 204 |
| No General Positive Result from this Chapter | 209 |
| A few Remarks on the Limits of Regulability | 212 |
| D. INHERITANCE. SECOND PROOF OF THE AUTONOMY OF LIFE— | |
| The Material Continuity in Inheritance | 214 |
| On Certain Theories which Seek to Compare Inheritance to Memory | 216 |
| The Complex-Equipotential System and its Rôle in Inheritance | 219 |
| The Second Proof of Life-Autonomy. Entelechy at the Bottom of Inheritance | 224 |
| The Significance of the Material Continuity in Inheritance | 227 |
| The Experimental Facts about Inheritance | 228 |
| The Rôle of the Nucleus in Inheritance | 233 |
| Variation and Mutation | 237 |
| Conclusions from the First Main Part of these Lectures | 240 |
PART II.—SYSTEMATICS AND HISTORY
| A. THE PRINCIPLES OF SYSTEMATICS— | |
| Rational Systematics | 243 |
| Biological Systematics | 246 |
| B. THE THEORY OF DESCENT— | |
| 1. Generalities | 250 |
| The Covert Presumption of all Theories of Descent | 253 |
| The Small Value of Pure Phylogeny | 255 |
| History and Systematics | 257 |
| 2. The Principles of Darwinism | 260 |
| Natural Selection | 261 |
| Fluctuating Variation the Alleged Cause of Organic Diversity | 264 |
| Darwinism Fails all along the Line | 269 |
| 3. The Principles of Lamarckism | 271 |
| Adaptation as the Starting-Point | 272 |
| The Active Storing of Contingent Variations as a Hypothetic Principle | 273 |
| Criticism of the “Inheritance of Acquired Characters” assumed by Lamarckism | 275 |
| Other Principles Wanted | 281 |
| Criticism of the Hypothesis of Storing and Handing Down Contingent Variations | 282 |
| 4. The Real Results and the Unsolved Problems of Transformism | 290 |
| 5. The Logical Value of the Organic Form according to the different Transformistic Theories | 293 |
| The Organic Form and Entelechy | 294 |
| C. THE LOGIC OF HISTORY | 297 |
| 1. The Possible Aspects of History | 299 |
| 2. Phylogenetic Possibilities | 304 |
| 3. The History of Mankind | 306 |
| Cumulations in Human History | 308 |
| Human History not an “Evolution” | 311 |
| The Problem of the “Single” as such | 315 |
| Conclusions about Systematics and History in General | 322 |
On Lord Gifford’s Conception of “Science”
This is the first time that a biologist has occupied this place; the first time that a biologist is to try to carry out the intentions of the noble and high-minded man to whom this lectureship owes its foundation.
On such an occasion it seems to be not undesirable to inquire what Lord Gifford’s own opinions about natural science may have been, what place in the whole scheme of human knowledge he may have attributed to those branches of it which have become almost the centre of men’s intellectual interest.
And, indeed, on studying Lord Gifford’s bequest with the object of finding in it some reference to the natural sciences, one easily notes that he has assigned to them a very high place compared with the other sciences, at least in one respect: with regard to their methods.
There is a highly interesting passage in his will which leaves no doubt about our question. After having formally declared the foundation of this lectureship “for Promoting, Advancing, Teaching and Diffusing the study of Natural Theology in the widest sense of that term,” and after having arranged about the special features of the lectures, he continues: “I wish the lecturers to treat their subject as a strictly natural science, the greatest of all possible sciences, indeed, in one sense, the only science, that of Infinite Being. . . . I wish it considered just as astronomy or chemistry is.”
Of course, it is not possible to understand these words of Lord Gifford’s will in a quite literal sense. If, provisionally, we call “natural theology” the ultimate conclusions which may be drawn from a study of nature in connection with all other results of human sciences, there cannot be any doubt that these conclusions will be of a rather different character from the results obtained in, say, the special field of scientific chemistry. But, nevertheless, there are, I think, two points of contact between the wider and the narrower field of knowledge, and both of them relate to method. Lord Gifford’s own phrase, “Infinite Being,” shows us one of these meeting-points. In opposition to history of any form, natural sciences aim at discovering such truths as are independent of special time and of special space, such truths as are “ideas” in the sense of Plato; and such eternal results, indeed, always stand in close relation to the ultimate results of human knowledge in general. But besides that there is still another feature which may be common both to “natural theology” and to the special natural sciences, and which is most fully developed in the latter: freedom from prepossessions. This, at least, is an ideal of all natural sciences; I may say it is the ideal of them. That it was this feature which Lord Gifford had in view in his comparison becomes clear when we read in his will that the lectures on natural theology are to be delivered “without reference to or reliance upon any supposed special exceptional or so-called miraculous revelation.”
So we might say that both in their logical and their moral methods, natural sciences are to be the prototype of “Natural Theology” in Lord Gifford’s sense.
Natural Sciences and “Natural Theology”
But now let us study in a more systematic manner the possible relations of the natural sciences to natural theology as a science.
How is it possible for a natural scientist to contribute to the science of the highest and ultimate subject of human knowledge?
Almost all natural sciences have a sort of naïveté in their own spheres; they all stand on the ground of what has been called a naïve realism, as long as they are, so to say, at home. That in no way prejudices their own progress, but it seems to stand in the way of establishing contact with any higher form of human knowledge than themselves. One may be a first-rate organic chemist even when looking upon the atoms as small billiard balls, and one may make brilliant discoveries about the behaviour of animals even when regarding them in the most anthropomorphic manner—granted that one is a good observer; but it can hardly be admitted that our chemist would do much to advance the theory of matter, or our biologist to solve the problem of the relations between body and mind.
It is only by the aid of philosophy, or I would rather say by keeping in constant touch with it, that natural sciences are able to acquire any significance for what might be called the science of nature in the most simple form. Unhappily the term “natural philosophy” is restricted in English to theoretical physics. This is not without a high degree of justification, for theoretical physics has indeed lost its naïveté and become a philosophy of nature; but it nevertheless is very unfortunate that this use of the term “natural philosophy” is established in this country, as we now have no proper general term descriptive of a natural science that is in permanent relation to philosophy, a natural science which does not use a single concept without justifying it epistemologically, i.e. what in German, for instance, would simply be called “Naturphilosophie.”
Let us call it philosophy of nature; then we may say that only by becoming a true philosophy of nature are natural sciences of all sorts able to contribute to the highest questions which man’s spirit of inquiry can suggest.
These highest questions themselves are the outcome of the combination of the highest results of all branches of philosophy, just as our philosophy of nature originated in the discussion of the results of all the separate natural sciences. Are those highest questions not only to be asked, are they to be also solved? To be solved in a way which does not exceed the limits of philosophy as the domain of actual understanding?
The beginning of a long series of studies is not the right place to decide this important question; and so, for the present certainly, “natural theology” must remain a problem. In other words: it must remain an open question at the beginning of our studies, whether after all there can be any final general answer, free from contradictions, applicable to the totality of questions asked by all the branches of philosophy.
But let us not be disturbed by this problematic entrance to our studies. Let us follow biology on its own path; let us study its transition from a “naïve” science to a real branch of the philosophy of nature. In this way we perhaps shall be able to understand what its part may be in solving what can be solved.
That is to be our subject.
Our Philosophical Basis
We call nature what is given to us in space.
Of course we are not obliged in these lectures to discuss the psychological and epistemological problems of space with its three dimensions, nor are we obliged to develop a general theory of reality and its different aspects. A few epistemological points will be considered later at proper times, and always in connection with results of theoretical biology.
At present it must suffice to say that our general philosophical point of view will be idealistic, in the critical meaning of the word. The universe, and within the universe nature, in the sense just defined, is my phenomenon. That is what I know. I know nothing more, either positively or negatively; that is to say, I do not know that the world is only my phenomenon, but, on the other hand, I know nothing about its “absolute reality.” And more, I am not even able to describe in intelligible words what “absolute reality” might mean. I am fully entitled to state: the universe is as truly as I am—though in a somewhat different sense of “being”—and I am as truly as the universe is; but I am not entitled to state anything beyond these two corresponding phrases. You know that, in the history of European philosophy at least, Bishop Berkeley was the first clearly to outline the field of idealism.
But my phenomenon—the world, especially nature—consists of elements of two different kinds: some of them are merely passive, some of them contain a peculiar sort of activity in themselves. The first are generally called sensations, but perhaps would be better called elements or presentations; the others are forms of construction, and, indeed, there is an active element embraced in them in this sense, that they allow, by their free combination, the discovery of principles which are not to be denied, which must be affirmed, whenever their meaning is understood. You know that I am speaking here of what are generally called categories and synthetic judgments a priori, and that it was Kant who, on the foundations laid by Locke, Hume, and Leibnitz, first gave the outlines of what may be called the real system of critical philosophy. Indeed, our method will be to a great extent Kantian, though with certain exceptions; it is to be strictly idealistic, and will not in the Kantian way operate with things in themselves; and it regards the so-called “synthetic judgment a priori” and the problem of the relation between categorical principles and experience in a somewhat different manner. We think it best to define the much disputed concept “a priori” as “independent of the amount of experience”; that is to say, all categories and categorical principles are brought to my consciousness by that fundamental event which is called experience, and therefore are not independent of it, but they are not inferences from experience, as are so-called empirical laws. We almost might say that we only have to be reminded of those principles by experience, and, indeed, we should not, I think, go very far wrong in saying that the Socratic doctrine, that all knowledge is recollection, holds good as far as categories and categorical principles are in question.
But enough at present about our general philosophy.
As to the philosophy of nature, there can be no doubt that, on the basis of principles like those we have shortly sketched, its ultimate aim must be to co-ordinate everything in nature with terms and principles of the categorical style. The philosophy of nature thus becomes a system; a system of which the general type is afforded by the innate constructive power of the Ego. In this sense the Kantian dictum remains true, that the Ego prescribes its own laws to nature, though, of course, “nature,” that is, what is given in space, must be such as to permit that sort of “prescription.”
One often hears that all sciences, including the science of sciences, philosophy, have to find out what is true. What, then, may be called “true” by an idealistic philosopher, for whom the old realistic formula of the conformity between knowledge and the object cannot have any meaning? Besides its ordinary application to simple facts or to simple judgments, where the word truth only means absence of illusion or no false statement, truth can be claimed for a philosophical doctrine or for a system of such doctrines only in the sense that there are no contradictions amongst the parts of the doctrine or of the system themselves, and that there are no features in them which impel our categorical Ego to further analysis.
Those of you who attended Professor Ward’s lectures on “Naturalism and Agnosticism,” or who have read his excellent book on that subject, will know what the aims of a theory of matter are. You will also be aware that, at present, there does not exist any theory of matter which can claim to be “true”; there are contradictions in every theory of matter, and, moreover, there are always some points where we are obliged to ask for further information and receive no answer. Experience here has not yet aroused all the categorical functions which are needed in order to form one unity out of what seem to be incompatibilities at the present day. Why is that? Maybe because experience is not yet complete in this field, but maybe also because the whole subject is so complicated that it takes much time to attach categorical functions to what is experienced.
But it is not our object here to deal either with epistemology proper or with ontology: a full analysis of biological facts is our problem. Why, then, all these introductions? why all these philosophical sketches in fields of knowledge which have quite another relation to philosophy than biology has? Biology, I hear some one say, is simply and solely an empirical science; in some sense it is nothing but applied physics and chemistry, perhaps applied mechanics. There are no fundamental principles in biology which could bring it in any close contact with philosophy. Even the one and only principle which might seem to be an innate principle of our experience about life, the principle of evolution, is only a combination of more simple factors of the physical and chemical type.
It will be my essential endeavour to convince you, in the course of these lectures, that such an aspect of the science of biology is wrong; that biology is an elemental natural science in the true sense of the word.
But if biology is an elemental science, then, and only then, it stands in close relations to epistemology and ontology—in the same relations to them, indeed, as every natural science does which deals with true elements of nature, and which is willing to abandon naïve realism and contribute its share to the whole of human knowledge.
And, therefore, a philosophical sketch is not out of place at the beginning of lectures on the Philosophy of the Organism. We may be forced, we, indeed, shall be forced, to remain for some time on the ground of realistic empiricism, for biology has to deal with very complicated experiences; but there will be a moment in our progress when we shall enter the realm of the elemental ontological concepts, and in that very moment our study of life will have become a part of real philosophy. It was not without good reasons, therefore, that I shortly sketched, as a sort of introduction to my lectures, the general point of view which we shall take with regard to philosophical questions, and to questions of the philosophy of nature in particular.
On Certain Characteristics of Biology as a Science
Biology is the science of life. Practically, all of you know what a living being is, and therefore it is not necessary to formulate a definition of life, which, at the beginning of our studies, would be either provisional and incomplete, or else dogmatic. In some respects, indeed, a definition should rather be the end of a science than its opening.
We shall study the phenomena of living organisms analytically, by the aid of experiment; our principal object will be to find out laws in these phenomena; such laws will then be further analysed, and precisely at that point we shall leave the realm of natural science proper.
Our science is the highest of all natural sciences, for it embraces as its final object the actions of man, at least in so far as actions also are phenomena observable on living bodies.
But biology is also the most difficult of all natural sciences, not only from the complexity of the phenomena, which it studies, but in particular for another reason which is seldom properly emphasised, and therefore will well repay us for a few words devoted to it.
Except so far as the “elements” of chemistry come into account, the experimenter in the inorganic fields of nature is not hampered by the specificity of composite objects: he makes all the combinations he wants. He is always able to have at his disposal red rays of a desired wave length when and where he wants, or to have, at a given time and place, the precise amount of any organic compound which he wishes to examine. And he forces electricity and electromagnetism to obey his will, at least with regard to space, time, and intensity of their appearance.
The biologist is not able to “make” life, as the physicist has made red rays or electromagnetism, or as the chemist has made a certain compound of carbon. The biologist is almost always in that strange plight in which the physicist would be if he always had to go to volcanoes in order to study the conductivity of heat, or if he had to wait for thunderstorms in order to study electricity. The biologist is dependent on the specificity of living objects as they occur in nature.
A few instances may show you what great inconveniences may hence arise to impede practical biological research. We later on shall have to deal with experiments on very young embryos: parts of the germ will have to be destroyed in order to study what will happen with the rest. Now almost all germs are surrounded by a membrane; this membrane has to be detached before any operation is possible. But what are we to do if it is not possible to remove the membrane without killing the embryo? Or what if, as for instance in many marine animals, the membrane may be removed but the germs are killed by contact with sea-water? In both cases no experiments at all will be possible on a sort of germ which otherwise, for some special circumstances of its organisation, might have given results of importance. These results become impossible for only a practical, for a very secondary reason; but enough: they are impossible, and they might have thrown light on problems which now must remain problems. Quite the same thing may occur in experiments on physiology proper or functional physiology: one kind of animals survives the operation, the other kind does not, and therefore, for merely extrinsic reasons, the investigations have to be restricted to the first, though the second might have given more important results. And thus the biological experimenter always finds himself in a sort of dependence on his subjects, which can hardly be called pleasant. To a great extent the comparatively slow advance of biological sciences is due to this very fact: the unalterable specific nature of biological material.
But there is still another feature of biology dependent on the same fact. If a science is tied down to specific objects in every path it takes, it first, of course, has to know all about those objects, and that requires nothing else but plain description. We now understand why pure description, in the most simple sense of the word, takes up such an enormous part of every text-book of biological science. It is not only morphology, the science of form, that is most actively concerned with description; physiology also, in its present state, is pure description of what the functions of the different parts of the body of animals and plants actually are, at least for about nine-tenths of its range. It seems to me important to press this point very emphatically, since we often hear that physiology is from the very beginning a much higher sort of knowledge than morphology, inasmuch as it is rational. That is not at all true of the beginning of physiology: what the functions of the liver or of the root are has simply to be described just as the organisation of the brain or of the leaf, and it makes no difference logically that one species of description has to use the experimental method, while the other has not. The experiment which only discovers what happens here or what happens there, possesses no kind of logical superiority over pure description at all.
But there will be another occasion in our lectures to deal more fully with the logic of experiment and with the differences of descriptive knowledge and real rational science.
The three Different Types of Knowledge about Nature
Natural sciences cannot originate before the given phenomena of nature have been investigated in at least a superficial and provisional manner, by and for the practical needs of man. But as soon as true science begins in any limited field, dealing, let us say, with animals or with minerals, or with the properties of bodies, it at once finds itself confronted by two very different kinds of problems, both of them—like all “problems”—created in the last resort by the logical organisation of the human mind, or, to speak still more correctly, of the Ego.
In any branch of knowledge which practical necessities have separated from others, and which science now tries to study methodically, there occur general sequences in phenomena, general orders of events. This uniformity is revealed only gradually, but as soon as it has shown itself, even in the least degree, the investigator seizes upon it. He now devotes himself chiefly, or even exclusively, to the generalities in the sequences of all changes. He is convinced that there must be a sort of most general and at the same time of most universal connection about all occurrences. This most universal connection has to be found out; at least it will be the ideal that always will accompany the inquiring mind during its researches. The “law of nature” is the ideal I am speaking about, an ideal which is nothing less than one of the postulates of the possibility of science at all.
Using for our purposes a word which has been already introduced into terminology by the philosopher Windelband, though in a somewhat different sense, we shall call that part of every branch of natural sciences which regards the establishment of a law of nature as its ideal, “nomothetic,” i.e. “law-giving.”
But while every natural science has its nomothetic side, it also has another half of a very different kind. This second half of every natural science does not care for the same general, the same universal, which is shown to us in every event in a different and specified kind: it is diversity, it is specification, that constitutes the subject of its interest. Its aim is to find a sufficient reason for the types of diversities, for the types of specifications. So in chemistry there has been found a systematic order in the long series of the compounds and of the elements; crystallography also has its different systems of crystals, and so on.
We have already employed the word by which we shall designate this second half of every natural science: it is the “systematic” side of science.
Nomothetic work on the one side and systematics on the other do, in fact, appear in every natural science, and besides them there are no other main parts. But “science” as a whole stands apart from another aspect of reality which is called “history.” History deals with particulars, with particular events at such and such a place, whilst science always abstracts from the particular, even in its systematic half.1
General Plan of these Lectures
Turning now to a sort of short outline of what is to be discussed in the whole of our future lectures, this summer and next, it seems clear, without further analysis, that biology as a science has its nomothetic and its systematic part also; respiration and assimilation, for instance, have proved to be types of natural laws among living phenomena, and that there is a “system” of animals and plants is too commonly known to require further explanation here. Therefore we might study first biological laws, and after that biological systematics, and in the third place perhaps biological history. But that would hardly correspond to the philosophical aims of our lectures: our chief object is not biology as a regular science, as treated in text-books and in ordinary university lectures; our chief object is the Philosophy of the Organism, as aided and supported by scientific biology. Therefore a general acquaintance with biology must be assumed in these lectures, and the biological materials must be arranged according to their bearing on further, that is on philosophical, analysis.
That will be done, not, of course, to the extent of my regarding every one of my audience as a competent biologist; on the contrary, I shall explain most fully all points of biology proper, and even of the most simple and descriptive kind of biology, which serve as bases for philosophical analysis. But I shall do so only if they indeed do serve as such bases. All our biology will be not for its own sake, but for the sake of philosophy.
Whilst regarding the whole of the biological material with such aims, it seems to me best to arrange the properly scientific material which is to be the basis of my discussions, not along the lines which biology as an independent science would select,2 but to start from the three different kinds of fundamental phenomena which living bodies offer to investigation, and to attach all systematics exclusively to one of them. For there will not be very much for philosophy to learn from biological systematics at present.
Life is unknown to us except in association with bodies: we only know living bodies and call them organisms. It is the final object of all biology to tell us what it ultimately means to say that a body is “living,” and in what sorts of relation body and life stand one to the other.
But at present it is enough to understand the terms “body” and “living” in the ordinary and popular sense.
Regarding living bodies in this unpretentious manner, and recollecting what the principal characters are of all bodies we know as living ones, we easily find that there are three features which are never wanting wherever life in bodies occurs. All living bodies are specific as to form—they “have” a specific form, as we are accustomed to say. All living bodies also exhibit metabolism; that is to say, they stand in a relation of interchange of materials with the surrounding medium, they take in and give out materials, but their form can remain unchanged during these processes. And, in the last place, we can say that all living bodies move; though this faculty is more commonly known among animals only, even elementary science teaches the student that it also belongs to plants.
Therefore we may ask for “laws of nature” in biology about form, about metabolism, and about movements. In fact, it is according to this scheme that we shall arrange the materials of the biological part of our lectures, though, as we cannot regard the three divisions as equally important in their bearing on our ultimate purposes, we shall not treat them quite on equal terms. It will appear that, at least in the present state of science, the problems of organic form and of organic movement have come into much closer relation to philosophical analysis than have most of the empirical data on metabolism.
It is form particularly which can be said to occupy the very centre of biological interest; at least it furnishes the foundation of all biology. Therefore we shall begin our scientific studies with a full and thorough analysis of form. The science of living forms, later on, will afford us a key to study metabolism proper with the greatest advantage for our philosophical aims, and therefore the physiology of what is usually called the vegetative functions will be to us a sort of appendix to our chapters on form; only the theory of a problematic “living substance” and of assimilation in the most general meaning of the word will be reserved for the philosophical part; for very good reasons, as I hope to show. But our chapters on the living forms will have yet another appendix besides the survey of the physiology of metabolism. Biological systematics almost wholly rests on form, on “morphology”; and what hitherto has been done on the metabolical side of their problems, consists of a few fragments, which are far from being an equivalent to the morphological system; though, of course it must be granted that, logically, systematics, in our general meaning of the word, as the sum of problems about the typically different and the specific, may be studied on the basis of each one of the principal characteristics of living bodies, not only on that of their forms. Therefore, systematics is to be the second appendix to the chief part of our studies in morphology, and systematics, in its turn, will later on lead us to a short sketch of the historical side of biology, to the theory of evolution in its different forms, and to the logic of history in general.
So far will our programme be carried out during this summer. Next year the theory of movements will conclude our merely scientific analysis, and the remaining part of the course next summer will be devoted to the philosophy of living nature. I hope that nobody will be able to accuse our philosophy of resting on unsound foundations. But those of you, on the other hand, who would be apt to regard our scientific chapters as a little too long compared with their philosophical results, may be asked to consider that a small clock-tower of a village church is generally less pretentious but more durable than the campanile of San Marco has been.
Indeed, these lectures will afford more “facts” to my hearers, than Gifford Lectures probably have done, as a rule. But how could that be otherwise on the part of a naturalist? Scientific facts are the material that the philosophy of nature has to work with, but these facts, unfortunately, are not as commonly known as historical facts, for instance, generally are; and they must be known, in order that a philosophy of the organism may be of any value at all, that it may be more than a mere entertainment.
Goethe once said, that even in so-called facts there is more “theory” than is usually granted; he apparently was thinking of what might be called the ultimate or the typical facts in science. It is with such typical or ultimate facts, of course, that we must become acquainted if our future philosophy is to be of profit to us.
Certainly, there would be nothing to prevent us from arranging our materials in a manner exactly the reverse of that which we shall adopt; we could begin with a general principle about the organic, and could try to deduce all its special features from that principle, and such a way perhaps would seem to be the more fascinating method of argument. But though logical it would not be psychological, and therefore would be rather unnatural. And thus our most general principle about the organic will not come on the scene before the eighteenth of these twenty lectures, although it is not a mere inference or deduction from the former lectures: it will be a culmination of the whole, and we shall appreciate its value the better the more we know what that whole really is.
General Character of the Organic Form
Our programme of this year, it was said, is to be devoted wholly to organic forms, though one of its appendixes, dealing with some characteristics of the physiology of metabolism, will lead us on to a few other phenomena. What then are the essentials of a living form, as commonly understood even without a special study of biology?
Living bodies are not simple geometrical forms, not, like crystals, merely a typical arrangement of surfaces in space, to be reduced theoretically, perhaps, to an arrangement of molecules. Living bodies are typically combined forms; that is to say, they consist of simpler parts of different characters, which have a special arrangement with regard to one another; these parts have a typical form of their own and may again be combinations of more simple different parts. But besides that, living bodies have not always the same typically combined form during the whole of their life: they become more complicated as they grow older; they all begin from one starting point, which has little form at all, viz., the egg. So the living form may be called a “genetic form,” or a form considered as a process, and therefore morphogenesis is the proper and adequate term for the science which deals with the laws of organic forms in general; or, if you prefer not to use the same word both for a science and for the subjects of that science, the physiology of morphogenesis.
Now there are different branches of the physiology of morphogenesis or physiology of form. We may study, and indeed we at first shall study, what are the laws of the morphogenetic processes leading from the egg to the adult: that may be properly called physiology of development. But living forms are not only able to originate in one unchangeable way: they may restore themselves, if disturbed, and thus we get the physiology of restoration or restitution as a second branch of the science of morphogenesis. We shall draw very important data, some of the foundations indeed of our philosophical discussions, from the study of such restitutions. Besides that, it is to them that our survey of the problems of the physiology of metabolism is to be appended.
Living forms not only originate from the egg and are able to restore themselves, they also may give origin to other forms, guaranteeing in this way the continuity of life. The physiology of heredity therefore appears as the counterpart to those branches of the physiology of form which deal with individual form and its restitutions. And our discussion on heredity may be followed by our second appendix to this chief section on form, an appendix regarding the outlines of systematics, evolution and history.
Theoretical considerations on biology generally start, or at least, used to start, from the evolution theory, discussing all other problems of the physiology of form by the way only, as things of secondary importance. You see from our programme, that we shall go just the opposite way: evolution will come last of all, and will be treated shortly; but the morphogenesis of the individual will be treated very fully, and very carefully indeed.
Why then this deviation from what is the common practice? Because we do not know very much about evolution at all, because in this field we are just at the very beginning of what deserves the name of exact knowledge. But concerning individual morphogenesis we really know, even at present, if not very much, at least something, and that we know in a fairly exact form, aided by the results of experiments.
And it will not be without its reward, if we restrict our aims in such a manner, if we prefer to deal more fully with a series of problems, which may seem at the first glance to be of less interest than others. After a few lectures we shall find already that we may decide one very important question about life merely by an analysis of individual form production, and without any regard to problematic and doubtful parts of biology: that we may decide the question, whether “life” is only a combination of chemical and physical events, or whether it has its elemental laws, laws of its own.
But to prepare the road that is to lead to such results we first have to restrict our aims once more, and therefore the next lecture of this course, which eventually is to touch almost every concept of philosophy proper, will begin with the pure description of the individual development of the common sea-urchin.
Evolutio and Epigenesis in the old Sense
The organism is a specific body, built up by a typical combination of specific and different parts. It is implied in the words of this definition, that the organism is different, not only from crystals, as was mentioned in the last lecture, but also from all combinations of crystals, such as those called dendrites and others, which consist of a typical arrangement of identical units, the nature of their combination depending on the forces of every single one of their parts. For this reason dendrites, in spite of the typical features in their combination, must be called aggregates; but the organism is not an aggregate even from the most superficial point of view.
We have said before, what must have been familiar to you already, that the organism is not always the same in its individual life, that it has its development, leading from simpler to more complicated forms of combination of parts; there is a “production of visible manifoldness” carried out during development, to describe the chief character of that process in the words of Wilhelm Roux. We leave it an open question in our present merely descriptive analysis, whether there was already a “manifoldness,” in an invisible state, before development, or whether the phrase “production of manifoldness” is to be understood in an absolute sense.
It has not always been granted in the history of biology, and of embryology especially, that production of visible manifoldness is the chief feature of what is called an organism’s embryology or ontogeny: the eighteenth century is full of determined scientific battles over the question. One school, with Albert von Haller and Bonnet as its leading men, maintained the view that there was no production of different parts at all in development, this process being a mere “evolutio,” that is, a growth of parts already existing from the beginning, yes, from the very beginning of life; whilst the other school, with C. F. Wolff and Blumenbach at its head, supported the opposite doctrine of so-called “epigenesis,” which has been proved to be the right one.
To some extent these differences of opinion were only the outcome of the rather imperfect state of the optical instruments of that period. But there were also deeper reasons beyond mere difficulties of description; there were theoretical convictions underlying them. It is impossible, said the one party, that there is any real production of new parts; there must be such a production, said the other.
We ourselves shall have to deal with these questions of the theory of organic development; but at present our object is narrower, and merely descriptive. It certainly is of great importance to understand most clearly that there actually is a “production of visible manifoldness” during ontogenesis in the descriptive sense; the knowledge of the fact of this process must be the very foundation of all studies on the theory of development in any case, and therefore we shall devote this whole lecture to studies in merely descriptive embryology.
But descriptive embryology, even if it is to serve merely as an instance of the universality of the fact of epigenesis, can only be studied successfully with reference to a concrete case. We select the development of the common sea-urchin (Echinus microtuberculatus) as such a case, and we are the more entitled to select this organism rather than another, because most of the analytical experimental work, carried out in the interests of a real theory of development, has been done on the germs of this animal. Therefore, to know at least the outlines of the individual embryology of the Echinus may indeed be called the conditio sine qua non for a real understanding of what is to follow.
The Cell3
You are aware that all organisms consist of organs and that each of their organs has a different function: the brain, the liver, the eyes, the hands are types of organs in animals, as are the leaves and the pistils in plants.
You are also aware that, except in the lowest organisms, the so-called Protista, all organs are built up of cells. That is a simple fact of observation, and I therefore cannot agree with the common habit of giving to this plain fact the title of cell-“theory.” There is nothing theoretical in it; and, on the other hand, all attempts to conceive the organism as a mere aggregate of cells have proved to be wrong. It is the whole that uses the cells, as we shall see later on, or that may not use them: thus there is nothing like a “cell-theory,” even in a deeper meaning of the word.
The cell may have the most different forms: take a cell of the skin, of a muscle, of a gland, of the wood in plants as typical examples. But in every case two parts may be distinguished in a cell: an outside part, the protoplasm, and an inside part, the nucleus, to leave out of special account several others, which, by the way, may only be protoplasmatic modifications.
Protoplasm is a mere name for what is not the nucleus; in any case it is not a homogeneous chemical compound; it consists of many such compounds and has a sort of architecture; all organic functions are based upon its metabolism. The nucleus has a very typical structure, which stands in a close relation to its behaviour during the most characteristic morphological period of the cell: during its division. Let us devote a few words to a consideration of this division and the part the nucleus plays in it; it will directly bear on future theoretical considerations about development.
There is a certain substance in every nucleus of a cell which stains most markedly, whenever cells are treated with pigments: the name of “chromatin” has been given to it. The chromatin always gives the reaction of an acid, while protoplasm is basic; besides that it seems to be a centre of oxidation. Now, when a division of a cell is to occur, the chromatin, which had been diffusely distributed before, in the form of small grains, arranges itself into a long and very much twisted thread. This thread breaks, as it were by sections, into almost equal parts, typical in number for each species, and each of these parts is split at full length. A certain number of pairs of small threads, the so-called “chromosomes,” are the ultimate result of this process, which intentionally has been described a little schematically, the breaking and the splitting in fact going on simultaneously or occasionally even in reverse order. While what we have described is performing in the nucleus, there have happened some typical modifications in protoplasm, and then, by an interaction of protoplasmatic and nuclear factors, the first step in the actual division of the cell begins. Of each pair of the small threads of chromatin one constituent is moved to one side of the cell, one to the other; two daughter-nuclei are formed in this way; the protoplasm itself at the same time forms a circular furrow between them; the furrow gets deeper and deeper; at last it cuts the cell in two, and the division of the cell is accomplished.
Not only is the growth of the already typically formed organism carried out by a series of cell-divisions, but also development proper in our sense, as a “production of visible manifoldness,” is realised to a great extent by the aid of such divisions, which therefore may indeed be said to be of very fundamental importance (Fig. 1).
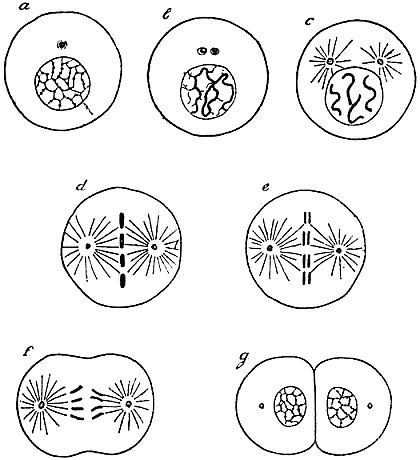
Fig. 1.—Diagram of Cell-Division (after Boveri).
| a. | Resting cell; the chromatin distributed in the form of small granules inside the nucleus. Outside the nucleus is the “centrosome,” not mentioned in the text. |
| b. | Beginning of division; the chromatin arranged in the form of a long thread. Centrosome divided in two. |
| c. | The thread of chromatin cut into four parts, the “chromosomes.” |
| d. | The four parts of the chromatin arranged symmetrically between the centrosomes and the star-like “spheres.” |
| e. | Each of the chromosomes split at full length. |
| f. | Beginning of division of protoplasm; the two parts of each chromosome separated. |
| g. | End of cell-division. |
Each cell-division which promotes growth is followed by the enlargement of the two daughter-cells which result from it; these two daughter-elements attain the exact size of the mother-cell before division, and as soon as this size is reached a new division begins: so the growth of the whole is in the main the result of the growth of the elements. Cell-divisions during real organ-formation may behave differently, as will be described at a proper occasion.
The Egg: its Maturation and Fertilisation
We know that all the organs of an animal or plant consist of cells, and we know what acts a cell can perform. Now there is one very important organ in all living beings, which is devoted to reproduction. This organ, the so-called ovary in animals, is also built up of cells, and its single cells are called the eggs; the eggs originated by cell-division, and cell-division is to lead from them to the new adult.
But, with a very few exceptions, the egg in the ovary is not able to accomplish its functions, unless certain typical events have occurred, some of which are of a merely preparatory kind, whilst the others are the actual stimulus for development.
The preparatory ones are generally known under the name of “maturation.” The egg must be “mature,” in order that it may begin development, or even that it may be stimulated to it. Maturation consists of a rather complicated series of phenomena: later on we shall have occasion to mention, at least shortly, what happens in the protoplasm during its course; as to the nuclear changes during maturation it may be enough for our purposes to say, that there occur certain processes among the chromosomes, which lead to an extension of half of them in the form of two very small cells, the “directive cells” or “directive or polar bodies,” as they have been somewhat cautiously called.
The ripe or mature egg is capable of being fertilised.
Before turning to this important fact, which, by the way, will bring us to our specially chosen type, the Echinus, a few words may be devoted to the phenomenon of “parthenogenesis,” that is to say, the possibility of development without fertilisation, since owing to the brilliant discoveries of the American physiologist, Jacques Loeb, this topic forms one of the centres of biological interest at present. It has long been known that the eggs of certain bees, lice, crayfishes, and other animals and also plants, are capable of development without fertilisation at all. Now Richard Hertwig and T. H. Morgan already had shown, that at least nuclear division may occur in the eggs of other forms—in the egg of the sea-urchin for instance—when these eggs are exposed to some chemical injuries. But Loeb4 succeeded in obtaining a full development by treating the eggs of echinoderms with chloride of magnesium; thus artificial parthenogenesis had been discovered. Later researches have shown that artificial parthenogenesis may occur in all classes of the animal kingdom and may be provoked by all sorts of chemical or physical means. We do not know at present in what the proper stimulus consists that must be supposed here to take the place of fertilisation; it seems, of course, highly probable that it is always the same in the last resort.5
But enough about processes, which at present are of a highly scientific, but hardly of any philosophic interest.
By fertilisation proper we understand the joining of the male element, the spermatozoon or the spermia, with the female element, the egg. Like the egg, the spermatozoon is but a cell, though the two differ very much from one another in the relation between their protoplasm and nucleus: in all eggs it is the protoplasm which is comparatively very large, if held together with somatic cells, in the spermatozoon it is the nucleus. A large amount of reserve material, destined for the growth of the future being, is the chief cause of the size of the egg-protoplasm. The egg is quite or almost devoid of the faculty of movement, while on the contrary, movement is the most typical feature of the spermia. Its whole organisation is adapted to movement in the most characteristic manner: indeed, most spermatozoa resemble a swimming infusorium, of the type of Flagellata, a so-called head and a moving tail are their two chief constituents; the head is formed almost entirely of nuclear substance.
It seems that in most cases the spermatozoa swim around at random and that their union with the eggs is assured only by their enormous number; only in a few cases in plants have there been discovered special stimuli of a chemical nature, which attract the spermia to the egg.
But we cannot enter here more fully into the physiology of fertilisation, and shall only remark that its real significance is by no means clear.6
The First Development Process of Echinus
Turning now definitively to the special kind of organism, chosen of our type, the common sea-urchin, we properly begin with a few words about the absolute size of its eggs and spermatozoa. All of you are familiar with the eggs of birds and possibly of frogs; these are abnormally large eggs, on account of the very high amount of reserve material they contain. The almost spherical egg of our Echinus only measures about a tenth of a millimetre in diameter; and the head of the spermatozoon has a volume which is only the four-hundred-thousandth part of the volume of the egg! The egg is about on the extreme limit of what can be seen without optical instruments; it is visible as a small white point. But the number of eggs produced by a single female is enormous and may amount to hundreds of thousands; this is one of the properties which render the eggs of Echinus so very suitable for experimental research; you can obtain them whenever and in any quantity you like; and, moreover, they happen to be very clear and transparent, even in later stages, and to bear all kinds of operations well.
The spermia enters the egg, and it does so in the open water—another of the experimental advantages of our type. Only one spermia enters the egg in normal cases, and only its head goes in, the tail is left outside. The moment that the head has penetrated the protoplasm of the egg a thin membrane is formed by the latter. This membrane is very soft at first, becoming much stronger later on; it is very important for all experimental work, that by shaking the egg in the first minutes of its existence the membrane can easily be destroyed without any damage to the egg itself.
And now occurs the chief phenomenon of fertilisation: the nucleus of the spermatozoon unites with the nucleus of the egg. When speaking of maturation, we mentioned that half of the chromatin was thrown out of the egg by that process: now this half is brought in again, but comes from another individual.
It is from this phenomenon of nuclear union as the main character of fertilisation that almost all theories of heredity assume their right to regard the nuclei of the sexual cells as the true “seat” of inheritance. Later on we shall have occasion to discuss this hypothesis from the point of view of logic and fact.
After the complete union of what are called the male and the female “pronuclei,” the egg begins its development; and this development, in its first steps, is simply pure cell-division. We know already the chief points of this process, and need only add to what has been described, that in the whole first series of the cell-divisions of the egg, or, to use the technical term, in the whole process of the “cleavage” or “segmentation” of it, there is never any growth of the daughter-elements after each division, such as we know to occur after all cell-divisions of later embryological stages. So it happens, that during cleavage the embryonic cells become smaller and smaller, until a certain limit is reached; the sum of the volumes of all the cleavage cells together is equal to the volume of the egg.
But our future studies will require a more thorough knowledge of the cleavage of our Echinus; the experimental data we shall have to describe later on could hardly be properly understood without such knowledge. The first division plane, or, as we shall say, the first cleavage plane, divides the eggs into equal parts; the second lies at right angles to the first and again divides equally: we now have a ring of four cells. The third cleavage plane stands at right angles to the first two; it may be called an equatorial plane, if we compare the egg with a globe; it also divides equally, and so we now find two rings, each consisting of four cells, and one above the other. But now the cell-divisions cease to be equal, at least in one part of the egg: the next division, which leads from the eight- to the sixteen-cell stage of cleavage, forms four rings, of four cells each, out of the two rings of the eight-cell stage. Only in one half of the germ, in which we shall call the upper one, or which we might call, in comparison with a globe, the northern hemisphere, are cells of equal size to be found; in the lower half of the egg four very small cells have been formed at one “pole” of the whole germ. We call these cells the “micromeres,” that is, the “small parts,” on the analogy of the term “blastomeres,” that is, parts of the germ, which is applied to all the cleavage cells in general. The place occupied by the micromeres is of great importance to the germ as a whole: the first formation of real organs will start from this point later on. It is sufficient thus fully to have studied the cleavage of our Echinus up to this stage: the later cleavage stages may be mentioned more shortly. All the following divisions are into equal parts; there are no other micromeres formed, though, of course, the cells derived from the micromeres of the sixteen-cell stage always remain smaller than the rest. All the divisions are tangential; radial cleavages never occur, and therefore the process of cleavage ends at last in the formation of one layer of cells, which forms the surface of a sphere; it is especially by the rounding-up of each blastomere, after its individual appearance, that this real surface layer of cells is formed, but, of course, the condition, that no radial divisions occur, is the most important one in its formation. When 808 blastomeres have come into existence the process of cleavage is finished; a sphere with a wall of cells and an empty interior is the result. That only 808 cells are formed, and not, as might be expected, 1024, is due to the fact that the micromeres divide less often than the other elements; but speaking roughly, of course, we may say that there are ten steps of cleavage-divisions in our form; 1024 being equal to 210.
We have learned that the first process of development, the cleavage, is carried out by simple cell-division. A few cases are known, in which cell-division during cleavage is accompanied by a specific migration of parts of the protoplasm in the interior of the blastomeres, especially in the first two or first four; but in almost all instances cleavage is as simple a process of mere division as it is in our sea-urchin. Now the second step in development, at least in our form, is a typical histological performance: it gives a new histological feature to all of the blastomeres: they acquire small cilia on their outer side and with these cilia the young germ is able to swim about after it has left its membrane. The germ may be called a “blastula” at this stage, as it was first called by Haeckel, whose useful denominations of the first embryonic stages may conveniently be applied, even if one does not agree with most, or perhaps almost all, of his speculations (Fig. 2).
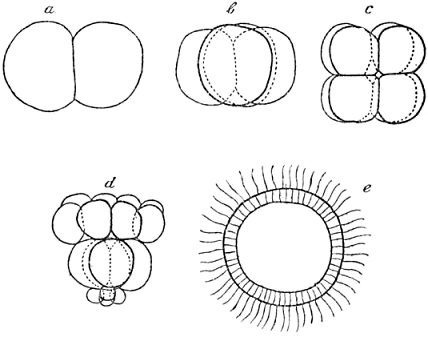
Fig. 2.—Early Development of Echinus, the Common Sea-urchin.
| a. | Two cells. |
| b. | Four cells. |
| c. | Eight cells, arranged in two rings of four, above one another. |
| d. | Sixteen cells, four “micromeres” formed at the “vegetative” pole. |
| e. | Optical section of the “blastula,” a hollow sphere consisting of about one thousand cells, each of them with a small cilium. |
It is important to notice that the formation of the “blastula” from the last cleavage stage is certainly a process of organisation, and may also be called a differentiation with regard to that stage. But there is in the blastula no trace of one part of the germ becoming different with respect to others of its parts. If development were to go on in this direction alone, high organisatory complications might occur: but there would always be only one sort of cells, arranged in a sphere; there would be only one kind of what is called “tissue.”
But in fact development very soon loads to true differences of the parts of the germ with respect to one another, and the next step of the process will enable us to apply different denominations to the different parts of the embryo.
At one pole of the swimming blastula, exactly at the point where the descendants of the micromeres are situated, about fifty cells lose contact with their neighbours and leave the surface of the globe, being driven into the interior space of it. Not very much is known about the exact manner in which these changes of cellular arrangement are carried out, whether the cells are passively pressed by their neighbours, or whether, perhaps, in a more active manner, they change their surface conditions; therefore, as in most ontogenetic processes, the description had best be made cautiously in fairly neutral or figurative words.
The cells which in the above manner have entered the interior of the blastula are to be the foundation of important parts of the future organism; they are to form its connective tissue, many of its muscles, and the skeleton. “Mesenchyme,” i.e. “what has been infused into the other parts,” is the technical name usually applied to these cells. We now have to learn their definite arrangement. At first they lie as a sort of heap inside the cell wall of the blastula, inside the “blastoderm,” i.e. skin of the germ. But soon they move from one another, to form a ring round the pole at which they entered, and on this ring a process takes place which has a very important bearing upon the whole type of the organisation of the germ. You will have noticed that hitherto the germ with regard to its symmetry has been a monaxial or radial formation; the cleavage stages and the blastula with its mesenchyme were forms with two different poles, lying at the ends of one single line, and round this line everything was arranged concentrically. But now what is called “bilateral symmetry” is established; the mesenchyme ring assumes a structure which can be symmetrically divided only by one plane, but divided in such a way, that one-half of it is the mirror image of the other. A figure shows best what has occurred, and you will notice (Fig. 3) two masses of cells in this figure, which have the forms of spherical triangles: it is in the midst of these triangles that the skeleton of the larva originates. The germ had an upper and a lower side before: it now has got an upper and lower, front and back, right and left half; it now has acquired that symmetry of organisation which our own body has; at least it has got it as far as its mesenchyme is concerned.
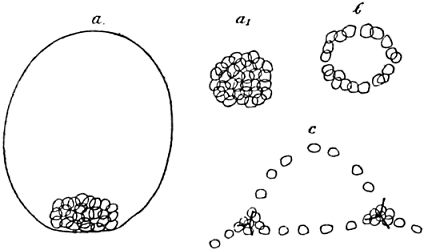
Fig. 3.—Formation of Mesenchyme in Echinus.
| a. | Outlines of blastula, side-view; mesenchyme forms a heap of cells at the “vegetative” pole. |
| a1. | Heap of mesenchyme-cells from above. |
| b. | Mesenchyme-cells arranged in a ring round the vegetative pole. |
| c. | Mesenchyme-cells arranged in a bilateral-symmetrical figure; primordia of skeleton in the midst of two spherical triangles. |
We leave the mesenchyme for a while and study another kind of organogenesis. At the very same pole of the germ where the mesenchyme cells originated there is a long and narrow tube of cell growing in, and this tube, getting longer and longer, after a few hours of growth touches the opposite pole of the larva. The growth of this cellular tube marks the beginning of the formation of the intestine, with all that is to be derived from it. The larva now is no longer a blastula, but receives the name of “gastrula” in Haeckel’s terminology; it is built up of the three “germ-layers” in this stage. The remaining part of the blastoderm is called “ectoderm,” or outer layer; the newly-formed tube, “endoderm,” or inner layer; while the third layer is the “mesenchyme” already known to us.
The endoderm itself is a radial structure at first, as was the whole germ in a former stage, but soon its free end bends and moves against one of the sides of the ectoderm, against that side of it where the two triangles of the mesenchyme are to be found also. Thus the endoderm has acquired bilateral symmetry just as the mesenchyme before, and as in this stage the ectoderm also assumes a bilateral symmetry in its form, corresponding with the symmetrical relations in the endoderm and the mesenchyme, we now may call the whole of our larva a bilateral-symmetrical organisation.
It cannot be our task to follow all the points of organogenesis of Echinus in detail. It must suffice to state briefly that ere long a second portion of the mesenchyme is formed in the larva, starting from the free end of its intestine tube; that the formation of the so-called “coelum” occurs by a sort of splitting off from this same original organ; and that the intestine itself is divided into three parts of different size and aspect by two circular sections.
But we must not, I think, dismiss the formation of the skeleton so quickly. I told you already that the skeleton has its first origin in the midst of the two triangular cell-masses of the mesenchyme; but what are the steps before it attains its typical and complicated structure? At the beginning a very small tetrahedron, consisting of carbonate of calcium, is formed in each of the triangles; the four edges of the tetrahedron are produced into thin rods, and by means of a different organogenesis along each of these rods the typical formation of the skeleton proceeds. But the manner in which it is carried out is very strange and peculiar. About thirty of the mesenchyme cells are occupied in the formation of skeleton substance on each side of the larva. They wander through the interior space of the gastrula—which at this stage is not filled with sea water but with a sort of gelatinous material—and wander in such a manner that they always come to the right places, where a part of the skeleton is to be formed; they form it by a process of secretion, quite unknown in detail; one of them forms one part, one the other, but what they form altogether, is one whole.
When the formation of the skeleton is accomplished, the typical larva of our Echinus is built up; it is called the “pluteus” (Fig. 4). Though it is far from being the perfect adult animal, it has an independent life of its own; it feeds and moves about and does not go through any important changes of form for weeks. But after a certain period of this species of independent life as a “larva,” the changes of form it undergoes again are most fundamental: it must be transformed into the adult sea-urchin, as all of you know. There are hundreds and hundreds of single operations of organogenesis to be accomplished before that end is reached; and perhaps the strangest of all these operations is a certain sort of growth, by which the symmetry of the animal, at least in certain of its parts—not in all of them—is changed again from bilateral to radial, just the opposite of what happened in the very early stages.
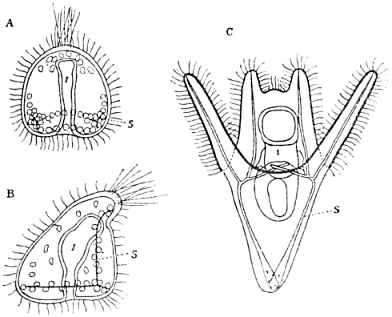
Fig. 4.—Larval Development of Echinus.
| A. | The gastrula. |
| B. | Later stage, bilateral-symmetrical. Intestine begins to divide into three parts. |
| C. | Pluteus larva. S = Skeleton. I = Intestine. |
But we cannot follow the embryology of our Echinus further here; and indeed we are the less obliged to do so, since in all our experimental work we shall have to deal with it only as far as to the pluteus larva. It is impossible under ordinary conditions to rear the germs up to the adult stages in captivity.
You now, I hope, will have a general idea at least of the processes of which the individual development of an animal consists. Of course the specific features leading from the egg to the adult are different in each specific case, and, in order to make this point as clear as possible, I shall now add to our description a few words about what may be called a comparative descriptive embryology.
Comparative Embryology
Even the cleavage may present rather different aspects. There may be a compact blastula, not one surrounded by only one layer of cells as in Echinus; or bilaterality may be established as early as the cleavage stage—as in many worms and in ascidians—and not so late as in Echinus. The formation of the germ layers may go on in a different order and under very different conditions: a rather close relative of our Echinus, for instance, the starfish, forms first the endoderm and afterwards the mesenchyme. In many cases there is no tube of cells forming the “endoderm,” but a flat layer of cells is the first foundation of all the intestinal organs: so it is in all birds and in the cuttlefish. And, as all of you know, of course, there are very many animal forms which have no proper “larval” stage: there is one in the frog, the well-known “tadpole,” but the birds and mammals have no larvae; that is to say, there is no special stage in the ontogeny of these forms which leads an independent life for a certain time, as if it were a species by itself, but all the ontogenetical stages are properly “embryonic”—the germ is always an “embryo” until it becomes the perfect young organism. And you also know that not all skeletons consist of carbonate of calcium, but, that there are skeletons of silicates, as in Radiolaria, and of horny substance, as in many sponges. And, indeed, if we were to glance at the development of plants also, the differences would seem to us probably so great that all the similarities would seem to disappear.
But there are similarities, nevertheless, in all development, and we shall now proceed to examine what they are. As a matter of fact, it was especially for their sake that we studied the ontogeny of a special form in such detail; one always sees generalities better if one knows the specific features of at least one case. What then are the features of most general and far-reaching importance, which may be abstracted from the individual history of our sea-urchin, checked always by the teachings of other ontogenies, including those of plants?
The First Steps of Analytical Morphogenesis
If we look back upon the long fight of the schools of embryologists in the eighteenth century about the question whether individual development was to be regarded as a real production of visible manifoldness or as a simple growth of visibly pre-existing manifoldness, whether it was “epigenesis” or “evolutio,” there can be no doubt, if we rely on all the investigations of the last hundred and fifty years, that, taken in the descriptive sense, the theory of epigenesis is right. Descriptively speaking there is a production of visible manifoldness in the course of embryology: that is our first and main result. Any one possessed of an average microscope may any day convince himself personally that it is true.
In fact, true epigenesis, in the descriptive sense of the term, does exist. One thing is formed “after” the other; there is not a mere “unfolding” of what existed already, though in a smaller form; there is no “evolutio” in the old meaning of the word.
The word “evolution” in English usually serves to denote the theory of descent, that is of a real relationship of all organisms. Of course we are not thinking here of this modern and specifically English meaning of the Latin word evolutio. In its ancient sense it means to a certain degree just the opposite; it says that there is no formation of anything new, no transformation, but simply growth, and this is promoted not for the race but for the individual. Keeping well in mind these historical differences in the meaning of the word “evolutio,” no mistakes, it seems to me, can occur from its use. We now shall try to obtain a few more particular results from our descriptive study of morphogenesis, which are nevertheless of a general bearing, being real characteristics of organic individual development, and which, though not calculated of themselves to further the problem, will in any case serve to prepare for a more profound study of it.
The totality of the line of morphogenetic facts can easily be resolved into a great number of distinct processes. We propose to call these “elementary morphogenetic processes”; the turning in of the endoderm and its division into three typical parts are examples of them. If we give the name “elementary organs” to the distinct parts of every stage of ontogeny which are uniform in themselves and are each the result of one elementary process in our sense, we are entitled to say that each embryological stage consists of a certain number of elementary organs. The mesenchyme ring, the coelum, the middle-intestine, are instances of such organs. It is important to notice well that the word elementary is always understood here with regard to visible morphogenesis proper and does not apply to what may be called elementary in the physiological sense. An elementary process in our sense is a very distinct act of form-building, and an elementary organ is the result of every one of such acts.
The elementary organs are typical with regard to their position and with regard to their histological properties. In many cases they are of a very clearly different histological type, as for instance, the cells of the three so-called germ-layers; and in other cases, though apparently almost identical histologically, they can be proved to be different by their different power of resisting injuries or by other means. But there are not as many different types of histological structure as there are typically placed organs: on the contrary there are many elementary organs of the same type in different typical parts of the organism, as all of you know to be the case with nerves and muscles. It will not be without importance for our future theory of development, carefully to notice this fact, that specialisation in the position of embryonic parts is more strict than in their histology.
But elementary organs are not only typical in position and histology, they are typical also with regard to their form and their relative size. It agrees with what has been said about histology being independent of typical position, that there may be a number of organs in an embryonic stage, all in their most typical positions, which though all possessing the same histology, may have different forms or different sizes or both: the single bones of the skeleton of vertebrates or of adult echinoderms are the very best instances of this most important feature of organogenesis. If we look back from elementary organs to elementary processes, the specialisation of the size of those organs may also be said to be the consequence of a typical duration of the elementary morphogenetic process leading to them.7
I hardly need to say, that the histology, form, and size of elementary organs are equally an expression of their present or future physiological function. At least they prepare for this function by a specific sort of metabolism which sets in very early.
The whole sequence of individual morphogenesis has been divided by some embryologists into two different periods; there is a first period, during which the foundations of the organisation of the “type” are laid down, and a second period, during which the histo-physiological specifications are modelled out (von Baer, Götte, Roux). Such a discrimination is certainly justified, if not taken too strictly; but its practical application would encounter certain difficulties in many larval forms, and also, of course, in all plants.
Our mention of plants leads us to the last of our analytical results. If an animal germ proceeds in its development from a stage d to the stage g, passing through e and f, we may say that the whole of d has become the whole of f, but we cannot say that there is a certain part of f which is d, we cannot say that f is d + a. But in plants we can: the stage f is indeed equal to a + b + c + d + e + a in vegetable organisms; all earlier stages are actually visible as parts of the last one. The great embryologist, Carl Ernst von Baer, most clearly appreciated these analytical differences between animal and vegetable morphogenesis. They become a little less marked if we remember that plants, in a certain respect, are not simple individuals but colonies, and that among the corals, hydroids, bryozoa, and ascidia, we find analogies to plants in the animal kingdom; but nevertheless the differences we have stated are not extinguished by such reasoning. It seems almost wholly due to the occurrence of so many foldings and bendings and migrations of cells and complexes of cells in animal morphogenesis, that an earlier stage of their development seems lost in the later one; those processes are almost entirely wanting in plants, even if we study their very first ontogenetic stages. If we say that almost all production of surfaces goes on outside in plants, inside in animals, we shall have adequately described the difference. And this feature again leads to the further diversity between animals and plants which is best expressed by calling the former “closed,” the latter “open” forms: animals reach a point where they are finished, plants never are finished, at least in most cases.
I hope you will allow that I have tried to draw from descriptive and comparative embryology as many general analytical results as are possibly to be obtained. It is not my fault if there are not any more, nor is it my fault if the results reached are not of the most satisfactory character. You may say that these results perhaps enable you to see a little more clearly and markedly than before a few of the characters of development, but that you have not really learnt anything new. Your disappointment—my own disappointment—in our analysis is due to the use of pure description and comparison as scientific methods.
The Limits of Pure Description in Science
We have analysed our descriptions as far as we could, and now we must confess that what we have found cannot be the last thing knowable about individual morphogenesis. There must be something deeper to be discovered: we only have been on the surface of the phenomena, we now want to get to the very bottom of them. Why then occurs all that folding, and bending, and histogenesis, and all the other processes we have described? There must be something that drives them out, so to say.
There is a very famous dictum in the Treatise on Mechanics by the late Gustav Kirchhoff, that it is the task of mechanics to describe completely and in the most simple manner all the motions that occur in nature. These words, which may appear problematic even in mechanics, have had a really pernicious influence on biology. People were extremely pleased with them. “‘Describing’—that is just what we always have done,” they said; “now we see that we have done just what was right; a famous physicist has told us so.” They did not see that Kirchhoff had added the words “completely and in the most simple manner”; and moreover, they did not consider that Kirchhoff never regarded it as the ultimate aim of physics to describe thunderstorms or volcanic eruptions or denudations; yet it only is with such “descriptions” that biological descriptions of given bodies and processes are to be compared!
Physicists always have used both experiment and hypothetical construction—Kirchhoff himself did so in the most gifted manner. With these aids they have gone through the whole of the phenomena, and what they found to be ultimate and truly elemental, that alone may they be said to have “described”; but they have “explained” by the aid of elementalities what proved to be not elemental in itself.8
It is the method of the physicists—not their results—that morphogenesis has to apply in order to make progress; and this method we shall begin to apply in our next lectures. Physiology proper has never been so short-sighted and self-satisfied as not to learn from other sciences, from which indeed there was very much to be learned; but morphology has: the bare describing and comparing of descriptions has been its only aim for about forty years or more, and lines of descent of a very problematic character were its only general results. It was not seen that science had to begin, not with problematic events of the past, but with what actually happens before our eyes.
But before saying any more about the exact rational and experimental method in morphology, which indeed may be regarded as a new method, since its prevalence in the eighteenth century had been really forgotten, we first shall have to analyse shortly some general attempts to understand morphogenesis by means of hypothetic construction exclusively. Such attempts have become very important as points of issue for really exact research, and, moreover, they deserve attention, because they prove that their authors at least had not quite forgotten that there were still other problems to be solved in morphology than only phylogenetical ones.
THE THEORY OF WEISMANN
Of all the purely hypothetic theories on morphogenesis that of August Weismann9 can claim to have had the greatest influence, and to be at the same time the most logical and the most elaborated. The “germ-plasma” theory of the German author is generally considered as being a theory of heredity, and that is true inasmuch as problems of inheritance proper have been the starting-point of all his hypothetic speculations, and also form in some respect the most valuable part of them. But, rightly understood, Weismann’s theory consists of two independent parts, which relate to morphogenesis and to heredity separately, and it is only the first which we shall have to take into consideration at present; what is generally known as the doctrine of the “continuity of the germ-plasm” will be discussed in a later chapter.
Weismann assumes that a very complicated organised structure, below the limits of visibility even with the highest optical powers, is the foundation of all morphogenetic processes, in such a way that, whilst part of this structure is handed over from generation to generation as the basis of heredity, another part of it is disintegrated during the individual development, and directs development by being disintegrated. The expression, “part” of the structure, first calls for some explanation. Weismann supposes several examples, several copies, as it were, of his structure to be present in the germ cells, and it is to these copies that the word “part” has been applied by us: at least one copy has to be disintegrated during ontogeny.
The morphogenetic structure is assumed to be present in the nucleus of the germ cells, and Weismann supposes the disintegration of his hypothetic structure to be accomplished by nuclear division. By the cleavage of the egg, the most fundamental parts of it are separated one from the other. The word “fundamental” must be understood as applying not to proper elements or complexes of elements of the organisation, but to the chief relations of symmetry; the first cleavage, for instance, may separate the right and the left part of the structure, the second one its upper and lower parts, and after the third or equatorial cleavage all the principal eighths of our minute organisation are divided off: for the minute organisation, it must now be added, had been supposed to be built up differently in the three directions of space, just as the adult organism is. Weismann concedes it to be absolutely unknown in what manner the proper relation between the parts of the disintegrated fundamental morphogenetic structure and the real processes of morphogenesis is realised; enough that there may be imagined such a relation.
At the end of organogenesis the structure is assumed to have been broken up into its elements, and these elements, which may be chemical compounds, determine the fate of the single cells of the adult organism.
Here let us pause for a moment. There cannot be any doubt that Weismann’s theory resembles to a very high degree the old “evolutio” doctrines of the eighteenth century, except that it is a little less crude. The chick itself is not supposed to be present in the hen’s egg before development, and ontogeny is not regarded as a mere growth of that chick in miniature, but what really is supposed to be present in the egg is nevertheless a something that in all its parts corresponds to all the parts of the chick, only under a somewhat different aspect, while all the relations of the parts of the one correspond to the relations of the parts of the other. Indeed, only on such an hypothesis of a fairly fixed and rigid relation between the parts of the morphogenetic structure could it be possible for the disintegration of the structure to go on, not by parts of organisation, but by parts of symmetry; which, indeed, is a very strange, but not an illogical, feature of Weismann’s doctrine.
Weismann is absolutely convinced that there must be a theory of “evolutio,” in the old sense of the word, to account for the ontogenetic facts; that “epigenesis” has its place only in descriptive embryology, where, indeed, as we know, manifoldness in the visible sense is produced, but that epigenesis can never form the foundation of a real morphogenetic theory: theoretically one pre-existing manifoldness is transformed into the other. An epigenetic theory would lead right beyond natural science, Weismann thinks, as in fact, all such theories, if fully worked out, have carried their authors to vitalistic views. But vitalism is regarded by him as dethroned for ever.
Under these circumstances we have a good right, it seems to me, to speak of a dogmatic basis of Weismann’s theory of development.
But to complete the outlines of the theory itself: Weismann was well aware that there were some grave difficulties attaching to his statements: all the facts of so-called adventitious morphogenesis in plants, of regeneration in animals, proved that the morphogenetic organisation could not be fully disintegrated during ontogeny. But these difficulties were not absolute: they could be overcome: indeed, Weismann assumes, that in certain specific cases—and he regarded all cases of restoration of a destroyed organisation as due to specific properties of the subjects, originated by roundabout variations and natural selection—that in specific cases, specific arrangements of minute parts were formed during the process of disintegration, and were surrendered to specific cells during development, from which regeneration or adventitious budding could originate if required. “Plasma of reserve” was the name bestowed on these hypothetic arrangements.
Almost independently another German author, Wilhelm Roux,10 has advocated a theoretical view of morphogenesis which very closely resembles the hypothesis of Weismann. According to Roux a minute ultimate structure is present in the nucleus of the germ and directs development by being divided into its parts during the series of nuclear divisions.
But in spite of this similarity of the outset, we enter an altogether different field of biological investigation on mentioning Roux’s name: we are leaving hypothetic construction, at least in its absoluteness, and are entering the realms of scientific experiment in morphology.
EXPERIMENTAL MORPHOLOGY
I have told you already in the last lecture that, while in the eighteenth century individual morphogenesis had formed the centre of biological interest and been studied experimentally in a thoroughly adequate manner, that interest gradually diminished, until at last the physiology of form as an exact separate science was almost wholly forgotten. At least that was the state of affairs as regards zoological biology; botanists, it must be granted, have never lost the historical continuity to such a degree; botany has never ceased to be regarded as one science and never was broken up into parts as zoology was. Zoological physiology and zoological morphology indeed were for many years in a relationship to one another not very much closer than the relation between philology and chemistry.
There were always a few men, of course, who strove against the current. The late Wilhelm His,11 for instance, described the embryology of the chick in an original manner, in order to find out the mechanical relations of embryonic parts, by which passive deformation, as an integrating part of morphogenesis, might be induced. He also most clearly stated the ultimate aim of embryology to be the mathematical derivation of the adult form from the distribution of growth in the germ. To Alexander Goette12 we owe another set of analytical considerations about ontogeny. Newport, as early as 1850, and in later years Pflüger and Rauber, carried out experiments on the eggs of the frog, which may truly be called anticipatory of what was to follow. But it was Wilhelm Roux,13 now professor of anatomy at Halle, who entered the field with a thoroughly elaborated programme, who knew not only how to state the problem analytically, but also how to attack it, fully convinced of the importance of what he did. “Entwickelungsmechanik,”—mechanics of development—he called the “new branch of anatomical science” of which he tried to lay the foundations.
I cannot let this occasion pass without emphasising in the most decided manner how highly in my opinion Roux’s services to the systematic exploration of morphogenesis must be esteemed. I feel the more obliged to do so, because later on I shall have to contradict not only many of his positive statements but also most of his theoretical views. He himself has lately given up much of what he most strongly advocated only ten years ago. But Roux’s place in the history of biological science can never be altered, let science take what path it will.
It is not the place here to develop the logic of experiment; least of all is it necessary in the country of John Stuart Mill. All of you know that experiment, by its method of isolating the single constituents of complicated phenomena, is the principal aid in the discovery of so-called causal relations. Let us try then to see what causal relations Wilhelm Roux established with the aid of morphogenetic experiment.
THE WORK OF WILHELM ROUX
We know already that an hypothesis about the foundation of individual development was his starting-point. Like Weismann he supposed that there exists a very complicated structure in the germ, and that nuclear division leads to the disintegration of that structure. He next tried to bring forward what might be called a number of indicia supporting his view.
A close relation had been found to exist in many cases between the direction of the first cleavage furrows of the germ and the direction of the chief planes of symmetry in the adult: the first cleavage, for instance, very often corresponds to the median plane, or stands at right angles to it. And in other instances, such as have been worked out into the doctrine of so-called “cell-lineages,” typical cleavage cells were found to correspond to typical organs. Was not that a strong support for a theory which regarded cellular division as the principal means of differentiation? It is true, the close relations between cleavage and symmetry did not exist in every case, but then there had always happened some specific experimental disturbances, e.g. influences of an abnormal direction of gravity on account of a turning over of the egg, and it was easy to reconcile such cases with the generally accepted theory on the assumption of what was called “anachronism” of cleavage.
But Roux was not satisfied with mere indicia, he wanted a proof, and with this intention he carried out an experiment which has become very celebrated.14 With a hot needle he killed one of the first two blastomeres of the frog’s egg after the full accomplishment of its first cleavage, and then watched the development of the surviving cell. A typical half-embryo was seen to emerge—an organism indeed, which was as much a half as if a fully formed embryo of a certain stage had been cut in two by a razor. It was especially in the anterior part of the embryo that its “halfness” could most clearly be demonstrated.
That seemed to be a proof of Weismann’s and Roux’s theory of development, a proof of the hypothesis that there is a very complicated structure which promotes ontogeny by its disintegration, carried out during the cell divisions of embryology by the aid of the process of nuclear division, the so-called “karyokinesis.”
To the dispassionate observer it will appear, I suppose, that the conclusions drawn by Roux from his experiment go a little beyond their legitimate length. Certainly some sort of “evolutio” is proved by rearing half the frog from half the egg. But is anything proved, is there anything discovered at all about the nucleus? It was only on account of the common opinion about the part it played in morphogenesis that the nucleus had been taken into consideration.
Things soon became still more ambiguous.
THE EXPERIMENTS ON THE EGG OF THE SEA-URCHIN
Roux’s results were published for the first time in 1888; three years later I tried to repeat his fundamental experiment on another subject and by a somewhat different method. It was known from the cytological researches of the brothers Hertwig and Boveri that the eggs of the common sea-urchin (Echinus microtuberculatus) are able to stand well all sorts of rough treatment, and that, in particular, when broken into pieces by shaking, their fragments will survive and continue to segment. I took advantage of these facts for my purposes. I shook the germs rather violently during their two-cell stage, and in several instances I succeeded in killing one of the blastomeres, while the other one was not damaged, or in separating the two blastomeres from one another.15
Let us now follow the development of the isolated surviving cell. It went through cleavage just as it would have done in contact with its sister-cell, and there occurred cleavage stages which were just half of the normal ones. The stage, for instance, which corresponded to the normal sixteen-cell stage, and which, of course, in my subjects was built up of eight elements only, showed two micromeres, two macromeres and four cells of medium size, exactly as if a normal sixteen-cell stage had been cut in two; and the form of the whole was that of a hemisphere. So far there was no divergence from Roux’s results.
The development of our Echinus proceeds rather rapidly, the cleavage being accomplished in about fifteen hours. I now noticed on the evening of the first day of the experiment, when the half-germ was composed of about two hundred elements, that the margin of the hemispherical germ bent together a little, as if it were about to form a whole sphere of smaller size, and, indeed, the next morning a whole diminutive blastula was swimming about. I was so much convinced that I should get Roux’s morphogenetical result in all its features that, even in spite of this whole blastula, I now expected that the next morning would reveal to me the half-organisation of my subject once more; the intestine, I supposed, might come out quite on one side of it, as a half-tube, and the mesenchyme ring might be a half one also.
But things turned out as they were bound to do and not as I had expected; there was a typically whole gastrula on my dish the next morning, differing only by its small size from a normal one; and this small but whole gastrula was followed by a whole and typical small pluteus-larva (Fig. 5).
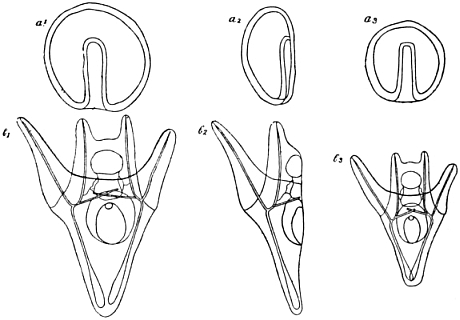
Fig. 5.—Illustration of Experiments on Echinus.
| a1 and b1. | Normal gastrula and normal pluteus. |
| a2 and b2. | “Half”-gastrula and “half”-pluteus, that ought to result from one of the first two blastomeres, when isolated, according to the theory of “evolutio.” |
| a3 and b3. | The small but whole gastrula and pluteus that actually do result. |
That was just the opposite of Roux’s result: one of the first two blastomeres had undergone a half-cleavage as in his case, but then it had become a whole organism by a simple process of rearrangement of its material, without anything that resembled regeneration, in the sense of a completion by budding from a wound.
If one blastomere of the two-cell stage was thus capable of performing the morphogenetical process in its totality, it became, of course, impossible to allow that nuclear division had separated any sort of “germ-plasm” into two different halves, and not even the protoplasm of the egg could be said to have been divided by the first cleavage furrow into unequal parts, as the postulate of the strict theory of so-called “evolutio” had been. This was a very important result, sufficient alone to overthrow at once the theory of ontogenetical “evolutio,” the “Mosaiktheorie” as it had been called—not by Roux himself, but according to his views—in its exclusiveness.
After first widening the circle of my observations by showing that one of the first four blastomeres is capable of performing a whole organogenesis, and that three of the first four blastomeres together result in an absolutely perfect organism, I went on to follow up separately one of the two fundamental problems which had been suggested by my first experiment: was there anything more to find out about the importance or unimportance of the single nuclear divisions in morphogenesis?16
By raising the temperature of the medium or by diluting the sea-water to a certain degree it proved at first to be possible to alter in a rather fundamental way the type of the cleavage-stages without any damage to the resulting organism. There may be no micromeres at the sixteen-cell stage, or they may appear as early as in the stage of eight cells; no matter, the larva is bound to be typical. So it certainly is not necessary for all the cleavages to occur just in their normal order.
But of greater importance for our purposes was what followed. I succeeded in pressing the eggs of Echinus between two glass plates, rather tightly, but without killing them; the eggs became deformed to comparatively flat plates of a large diameter. Now in these eggs all nuclear division occurred at right angles to the direction of pressure, that is to say, in the direction of the plates, as long as the pressure lasted; but the divisions began to occur at right angles to their former direction, as soon as the pressure ceased. By letting the pressure be at work for different times I therefore, of course, had it quite in my power to obtain cleavage types just as I wanted to get them. If, for instance, I kept the eggs under pressure until the eight-cell stage was complete, I got a plate of eight cells one beside the other, instead of two rings, of four cells each, one above the other, as in the normal case; but the next cell division occurred at right angles to the former ones, and a sixteen-cell stage, of two plates of eight cells each, one above the other, was the result. If the pressure continued until the sixteen-cell stage was reached, sixteen cells lay together in one plate, and two plates of sixteen cells each, one above the other, were the result of the next cleavage.
We are not, however, studying these things for cytological, but for morphogenetical purposes, and for these the cleavage phenomenon itself is less important than the organogenetic result of it: all our subjects resulted in absolutely normal organisms. Now, it is clear, that the spatial relations of the different nuclear divisions to each other are anything but normal, in the eggs subjected to the pressure experiments; that, so to say, every nucleus has got quite different neighbours if compared with the “normal” case. If that makes no difference, then there cannot exist any close relation between the single nuclear divisions and organogenesis at all, and the conclusion we have drawn more provisionally from the whole development of isolated blastomeres has been extended and proved in the most perfect manner. There ought to result a morphogenetic chaos according to the theory of real “evolutio” carried out by nuclear division, if the positions of the single nuclei were fundamentally changed with regard to one another (Fig. 6). But now there resulted not chaos, but the normal organisation: therefore it was disproved in the strictest way that nuclear divisions have any bearing on the origin of organisation; at least as far as the divisions during cleavage come into account.
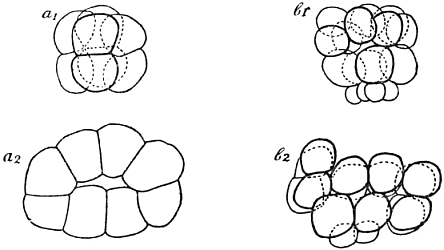
Fig. 6.—Pressure-experiments on Echinus.
| a1 and b1. | Two normal cleavage stages, consisting of eight and sixteen cells. |
| a2 and b2. | Corresponding stages modified by exerting pressure until the eight-cell stage was finished. See text. |
On the egg of the frog (O. Hertwig), and on the egg of annelids (E. B. Wilson), my pressure experiments have been carried out with the same result.17
ON THE INTIMATE STRUCTURE OF THE PROTOPLASM OF THE GERM
Nuclear division, as we have seen, cannot be the basis of organogenesis, and all we know about the whole development of isolated blastomeres seems to show that there exists nothing responsible for differentiation in the protoplasm either.
But would that be possible? It cannot appear possible on a more profound consideration of the nature of morphogenesis, it seems to me: as the untypical agents of the medium cannot be responsible in any way for the origin of a form combination which is most typical and specific, there must be somewhere in the egg itself a certain factor which is responsible at least for the general orientation and symmetry of it. Considerations of this kind led me, as early as 1893,18 to urge the hypothesis that there existed, that there must exist, a sort of intimate structure in the egg, including polarity and bilaterality as the chief features of its symmetry, a structure which belongs to every smallest element of the egg, and which might be imagined by analogy under the form of elementary magnets.19 This hypothetic structure could have its seat in the protoplasm only. In the egg of echinoderms it would be capable of such a quick rearrangement after being disturbed, that it could not be observed but only inferred logically; there might, however, be cases in which its real discovery would be possible. Indeed Roux’s frog-experiment seems to be a case where it is found to be at work: at least it seems very probable to assume that Roux obtained half of a frog’s embryo because the protoplasm of the isolated blastomere had preserved the “halfness” of its intimate structure, and had not been able to form a small whole out of it.
Of course it was my principal object to verify this hypothesis, and such verification became possible in a set of experiments which my friend T. H. Morgan and myself carried out together,20 in 1895, on the eggs of ctenophores, a sort of pelagic animals, somewhat resembling the jelly-fish, but of a rather different inner organisation. The zoologist Chun had found even before Roux’s analytical studies, that isolated blastomeres of the ctenophore egg behave like parts of the whole and result in a half-organisation like the frog’s germ does. Chun had not laid much stress on his discovery, which now, of course, from the new points of view, became a very important one. We first repeated Chun’s experiment and obtained his results, with the sole exception that there was a tendency of the endoderm of the half-larva of Beroë to become more than “half.” But that was not what we chiefly wanted to study. We succeeded in cutting away a certain mass of the protoplasm of the ctenophore egg just before it began to cleave, without damaging its nuclear material in any way: in all cases, where the cut was performed at the side, there resulted a certain type of larvae from our experiments which showed exactly the same sort of defects as were present in larvae developed from one of the first two blastomeres alone.
The hypothesis of the morphogenetic importance of protoplasm had thus been proved. In our experiments there was all of the nuclear material, but there were defects on one side of the protoplasm of the egg; and the defects in the adult were found to correspond to these defects in the protoplasm.
And now O. Schultze and Morgan succeeded in performing some experiments which directly proved the hypothesis of the part played by protoplasm in the subject employed by Roux, viz., the frog’s egg. The first of these investigators managed to rear two whole frog embryos of small size, if he slightly pressed the two-cell stage of that form between two plates of glass and turned it over; and Morgan,21 after having killed one of the first two blastomeres, as was done in the original experiment of Roux, was able to bring the surviving one to a half or to a whole development according as it was undisturbed or turned. There cannot be any doubt that in both of these cases, it is the possibility of a rearrangement of protoplasm, offered by the turning over, which allows the isolated blastomere to develop as a whole. The regulation of the frog’s egg, with regard to its becoming whole, may be called facultative, whilst the same regulation of the egg of Echinus is obligatory. It is not without interest to note that the first two blastomeres of the common newt, i.e. of a form which belongs to the other class of Amphibia, after a separation of any kind, always develop as wholes, their faculty of regulation being obligatory, like that of Echinus.
Whole or partial development may thus be dependent on the power of regulation contained in the intimate polar-bilateral structure of the protoplasm. Where this is so, the regulation and the differences in development are both connected with the chief relations of symmetry. The development becomes a half or a quarter of the normal because there is only one-half or one-quarter of a certain structure present, one-half or one-quarter with regard to the very wholeness of this structure; the development is whole, in spite of disturbances, if the intimate structure became whole first. We may describe the “wholeness,” “halfness,” or “quarterness” of our hypothetic structure in a mathematical way, by using three axes, at right angles to one another, as the base of orientation. To each of these, x, y, and z, a certain specific state with regard to the symmetrical relations corresponds; thence it follows that, if there are wanting all those parts of the intimate structure which are determined, say, by a negative value of y, by minus y, then there is wanting half of the intimate structure; and this halfness of the intimate structure is followed by the halfness of organogenesis, the dependence of the latter on the intimate structure being established. But if regulation has restored, on a smaller scale, the whole of the arrangement according to all values of x, y and z, development also can take place completely (Fig. 7).
Fig. 7.—Diagram illustrating the intimate Regulation of Protoplasm from “Half” to “Whole.”
The large circle represents the original structure of the egg. In all cases where cleavage-cells of the two-cell stage are isolated this original structure is only present as “half” in the beginning, say only on the right (+y) side. Development then becomes “half,” if the intimate structure remains half; but it becomes “whole” (on a smaller scale) if a new whole-structure (small circle!) is formed by regulatory processes.
I am quite aware that such a discussion is rather empty and purely formal, nevertheless it is by no means without value, for it shows most clearly the differences between what we have called the intimate structure of germs, responsible only for the general symmetry of themselves and of their isolated parts, and another sort of possible structure of the egg-protoplasm which we now shall have to consider, and which, at the first glance, seems to form a serious difficulty to our statements, as far at least as they claim to be of general importance. The study of this other sort of germinal structure at the same time will lead us a step farther in our historical sketch of the first years of “Entwickelungsmechanik” and will bring this sketch to its end.
ON SOME SPECIFICITIES OF ORGANISATION IN CERTAIN GERMS
It was known already about 1890, from the careful study of what has been called “cell-lineage,” that in the eggs of several families of the animal kingdom the origin of certain organs may be traced back to individual cells of cleavage, having a typical histological character of their own. In America especially such researches have been carried out with the utmost minuteness, E. B. Wilson’s study of the cell-lineage of the Annelid Nereis being the first of them. If it were true that nuclear division is of no determining influence upon the ontogenetic fate of the blastomeres, only peculiarities of the different parts of the protoplasm could account for such relations of special cleavage cells to special organs. I advocated this view as early as in 1894, and it was proved two years later by Crampton, a pupil of Wilson’s, in some very fine experiments performed on the germ of a certain mollusc.22 The egg of this form contains a special sort of protoplasm near its vegetative pole, and this part of it is separated at each of the first two segmentations by a sort of pseudo-cleavage, leading to stages of three and five separated masses instead of two and four, the supernumerary mass being the so-called “yolk-sac” and possessing no nuclear elements (Fig. 8). Crampton removed this yolk-sac at the two-cell stage, and he found that the cleavage of the germs thus operated upon was normal except with regard to the size and histological appearance of one cell, and that the larvae originating from these germs were complete in every respect except in their mesenchyme, which was wanting. A special part of the protoplasm of the egg had thus been brought into relation with quite a special part of organisation, and that special part of the protoplasm contained no nucleus.
Fig. 8.—The Mollusc Dentalium (after E. B. Wilson).
| a. | The egg, consisting of three different kinds of protoplasmatic material. |
| b. | First cleavage-stage. There are two cells and one “pseudo-cell,” the yolk-sac, which contains no nucleus. This was removed in Crampton’s experiment. |
GENERAL RESULTS OF THE FIRST PERIOD OF “ENTWICKELUNGSMECHANIK”
This experiment of Crampton’s, afterwards confirmed by Wilson himself, may be said to have closed the first period of the new science of physiology of form, a period devoted almost exclusively to the problem whether the theory of nuclear division or, in a wider sense, whether the theory of a strict “evolutio” as the basis of organogenesis was true or not.
It was shown, as we have seen, that the theory of the “qualitatively unequal nuclear division” (“qualitativ-ungleiche Kernteilung” in German) certainly was not true, and that there also was no strict “evolutio” in protoplasm. Hence Weismann’s theory was clearly disproved. There certainly is a good deal of real “epigenesis” in ontogeny, a good deal of “production of manifoldness,” not only with regard to visibility but in a more profound meaning. But some sort of pre-formation had also been proved to exist, and this pre-formation, or, if you like, this restricted evolution, was found to be of two different kinds. First an intimate organisation of the protoplasm, spoken of as its polarity and bilaterality, was discovered, and this had to be postulated for every kind of germs, even when it was overshadowed by immediate obligatory regulation after disturbances. Besides that there were cases in which a real specificity of special parts of the germ existed, a relation of these special parts to special organs: but this sort of specification also was shown to belong to the protoplasm.
It follows from all we have mentioned about the organisation of protoplasm and its bearing on morphogenesis, that the eggs of different animals may behave rather differently, in this respect, and that the eggs indeed may be classified according to the degree of their organisation. Though we must leave a detailed discussion of these topics to morphology proper, we yet shall try shortly to summarise what has been ascertained about them in the different classes of the animal kingdom. A full regulation of the intimate structure of isolated blastomeres to a new whole, has been proved to exist in the highest degree in the eggs of all echinoderms, medusae, nemertines, Amphioxus, fishes, and in one class of the Amphibia (the Urodela); it is facultative only among the other class of Amphibia, the Anura, and seems to be only partly developed or to be wanting altogether among ctenophora, ascidia, annelids, and mollusca. Peculiarities in the organisation of specific parts of protoplasm have been proved to occur in more cases than at first had been assumed; they exist even in the echinoderm egg, as experiments of the last few years have shown; even here a sort of specification exists at the vegetative pole of the egg, though it is liable to a certain kind of regulation; the same is true in medusae, nemertines, etc.; but among molluscs, ascidians, and annelids no regulation about the specific organisation of the germ in cleavage has been found in any case.
The differences in the degree of regulability of the intimate germinal structure may easily be reduced to simple differences in the physical consistency of their protoplasm.23 But all differences in specific organisation must remain as they are for the present; it will be one of the aims of the future theory of development to trace these differences also to a common source.
That such an endeavour will probably be not without success, is clear, I should think, from the mere fact that differences with regard to germinal specific pre-formation do not agree in any way with the systematic position of the animals exhibiting them; for, strange as it would be if there were two utterly different kinds of morphogenesis, it would be still more strange if there were differences in morphogenesis which were totally unconnected with systematic relationship: the ctenophores behaving differently from the medusae, and Amphioxus differently from ascidians.
SOME NEW RESULTS CONCERNING RESTITUTIONS
We now might close this chapter, which has chiefly dealt with the disproof of a certain sort of ontogenetic theories, and therefore has been almost negative in its character, did it not seem desirable to add at least a few words about the later discoveries relating to morphogenetic restorations of the adult. We have learnt that Weismann created his concept of “reserve plasma” to account for what little he knew about “restitutions”: that is, about the restoration of lost parts: he only knew regeneration proper in animals and the formation of adventitious buds in plants. It is common to both of these phenomena that they take their origin from typically localised points of the body in every case; each time they occur a certain well-defined part of the body is charged with the restoration of the lost parts. To explain such cases Weismann’s hypothesis was quite adequate, at least in a logical sense. But at present, as we shall discuss more fully in another chapter, we know of some very widespread forms of restitution, in which what is to be done for a replacement of the lost is not entrusted to one typical part of the body in every case, but in which the whole of the morphogenetic action to be performed is transferred in its single parts to the single parts of the body which is accomplishing restoration: each of its parts has to take an individual share in the process of restoration, effecting what is properly called a certain kind of “re-differentiation” (“Umdifferenzierung”), and this share varies according to the relative position of the part in each case. Later on these statements will appear in more correct form than at present, and then it will become clear that we are fully entitled to emphasise at the end of our criticism of Weismann’s theory, that his hypothesis relating to restorations can be no more true than his theory of development proper was found to be.
And now we shall pass on to our positive work.
We shall try to sketch the outlines of what might properly be called an analytical theory of morphogenesis; that is, to explain the sum of our knowledge about organic form-production, gained by experiment and by logical analysis, in the form of a real system, in which each part will be, or at least will try to be, in its proper place and in relation with every other part. Our analytical work will give us ample opportunity of mentioning many important topics of so-called general physiology also, irrespective of morphogenesis as such. But morphogenesis is always to be the centre and starting-point of our analysis. As I myself approach the subject as a zoologist, animal morphogenesis, as before, will be the principal subject of what is to follow.
Prospective Value and Prospective Potency
Wilhelm Roux did not fail to see that the questions of the locality and the time of all morphogenetic differentiations had to be solved first, before any problem of causality proper could be attacked. From this point of view he carried out his fundamental experiments.
It is only in terminology that we differ from his views, if we prefer to call our introductory chapter an analysis of the distribution of morphogenetic potencies. The result will be of course rather different from what Roux expected it would be.
Let us begin by laying down two fundamental concepts. Suppose we have here a definite embryo in a definite state of development, say a blastula, or a gastrula, or some sort of larva, then we are entitled to study any special element of any special elementary organ of this germ with respect to what is actually to develop out of this very element in the future actual course of this development, whether it be undisturbed or disturbed in any way; it is, so to say, the actual, the real fate of our element, that we take in account. I have proposed to call this real fate of each embryonic part in this very definite line of morphogenesis its prospective value (“prospective Bedeutung” in German). The fundamental question of the first chapter of our analytical theory of development may now be stated as follows: Is the prospective value of each part of any state of the morphogenetic line constant, i.e. is it unchangeable, can it be nothing but one; or is it variable, may it change according to different circumstances?
We first introduce a second concept: the term prospective potency (“prospective Potenz” in German) of each embryonic element. The term “prospective morphogenetic potency” is to signify the possible fate of each of those elements. With the aid of our two artificial concepts we are now able to formulate our introductory question thus: Is the prospective potency of each embryonic part fully given by its prospective value in a certain definite case; is it, so to say, identical with it, or does the prospective potency contain more than the prospective value of an element in a certain case reveals?
We know already from our historical sketch that the latter is true: that the actual fate of a part need not be identical with its possible fate, at least in many cases; that the potency of the first four blastomeres of the egg of the sea-urchin, for instance, has a far wider range than is shown by what each of them actually performs in even this ontogeny. There are more morphogenetic possibilities contained in each embryonic part than are actually realised in a special morphogenetic case.
As the most important special morphogenetic case is, of course, the so-called “normal” one, we can also express our formula in terms of special reference to it: there are more morphogenetic possibilities in each part than the observation of the normal development can reveal. Thus we have at once justified the application of analytical experiment to morphogenesis, and have stated its most important results.
As the introductory experiments about “Entwickelungsmechanik” have shown already that the prospective potency of embryonic parts, at least in certain cases, can exceed their prospective value—that, at least in certain cases, it can be different from it—the concept of prospective potency at the very beginning of our studies puts itself in the centre of analytical interest, leaving to the concept of prospective value the second place only. For that each embryonic part actually has a certain prospective value, a specified actual fate in every single case of ontogeny, is clear from itself and does not affirm more than the reality of morphogenetic cases in general; but that the prospective value of the elements may change, that there is a morphogenetic power in them, which contains more than actuality; in other words, that the term “prospective potency” has not only a logical but a factual interest: all these points amount to a statement not only of the most fundamental introductory results but also of the actual problems of the physiology of form.
If at each point of the germ something else can be formed than actually is formed, why then does there happen in each case just what happens and nothing else? In these words indeed we may state the chief problem of our science, at least after the fundamental relation of the superiority of prospective potency to prospective value has been generally shown.
We consequently may shortly formulate our first problem as the question of the distribution of the prospective morphogenetic potencies in the germ. Now this general question involves a number of particular ones. Up to what stage, if at all, is there an absolutely equal distribution of the potencies over all the elements of the germ? When such an equal distribution has ceased to exist at a certain stage, what are then the relations between the parts of different potency? How, on the other hand, does a newly arisen, more specialised sort of potency behave with regard to the original general potency, and what about the distribution of the more restricted potency?
I know very well that all such questions will seem to you a little formal, and, so to say, academical at the outset. We shall not fail to attach to them very concrete meanings.
The Potencies of the Blastomeres
At first we turn back to our experiments on the egg of the sea-urchin as a type of the germ in the very earliest stages. We know already that each of the first two, or each of the first four, or three of the first four blastomeres together may produce a whole organism. We may add that the swimming blastula, consisting of about one thousand cells, when cut in two quite at random, in a plane coincident with, or at least passing near, its polar axis, may form two fully developed organisms out of its halves.25 We may formulate this result in the words: the prospective potency of the single cells of a blastula of Echinus is the same for all of them; their prospective value is as far as possible from being constant.
But we may say even a little more: what actually will happen in each of the blastula cells in any special case of development experimentally determined depends on the position of that cell in the whole, if the “whole” is put into relation with any fixed system of co-ordinates; or more shortly, “the prospective value of any blastula cell is a function of its position in the whole.”
I know from former experience that this statement wants a few words of explanation. The word “function” is employed here in the most general, mathematical sense, simply to express that the prospective value, the actual fate of a cell, will change, whenever its position in the whole is different.26 The “whole” may be related to any three axes drawn through the normal undisturbed egg, on the hypothesis that there exists a primary polarity and bilaterality of the germ; the axes which determine this sort of symmetry may, of course, conveniently be taken as co-ordinates; but that is not necessary.
The Potencies of Elementary Organs in General
Before dealing with other very young germs, I think it advisable to describe first an experiment which is carried out at a later stage of our well-known form. This experiment will easily lead to a few new concepts, which we shall want later on, and will serve, on the other hand, as a basis of explanation for some results, obtained from the youngest germs of some other animal species, which otherwise would seem to be rather irreconcilable with what our Echinus teaches us.
You know, from the second lecture, what a gastrula of our sea-urchin is. If you bisect this gastrula, when it is completely formed, or still better, if you bisect the gastrula of the starfish, either along the axis or at right angles to it, you get complete little organisms developed from the parts: the ectoderm is formed in the typical manner in the parts, and so is the endoderm; everything is proportionate and only smaller than in the normal case. So we have at once the important results, that, as in the blastula, so in the ectoderm and in the endoderm of our Echinus or of the starfish, the prospective potencies are the same for every single element: both in the ectoderm and in the endoderm the prospective value of each cell is a “function of its position” (Fig. 9).
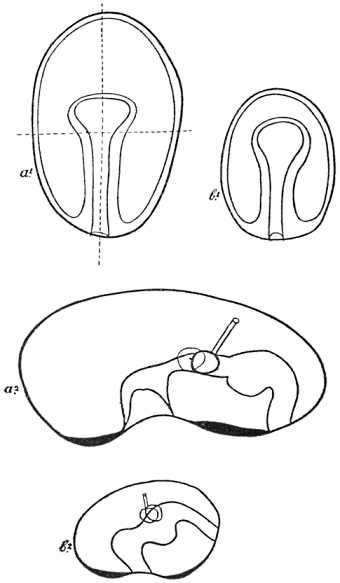
Fig. 9.—The Starfish, Asterias.
| a1. | Normal gastrula; may be bisected along the main axis or at right angles to it (see dotted lines). |
| a2. | Normal larva, “Bipinnaria.” |
| b1. | Small but whole gastrula that results by a process of regulation from the parts of a bisected gastrula. |
| b2. | Small but whole “Bipinnaria,” developed out of b1. |
But a further experiment has been made on our gastrula. If at the moment when the material of the future intestine is most distinctly marked in the blastoderm, but not yet grown into a tube, if at this moment the upper half of the larva is separated from the lower by an equatorial section, you will get a complete larva only from that part which bears the “Anlage” of the endoderm, while the other half will proceed in morphogenesis very well but will form only ectodermal organs. By another sort of experiment, which we cannot fully explain here, it has been shown that the endoderm if isolated is also only able to form such organs as are normally derived from it.
And so we may summarise both our last results by
saying: though ectoderm and endoderm have their potencies
equally distributed amongst their respective cells, they possess
different potencies compared one with the other. And the
same relation is found to hold for all cases of what we call
elementary organs: they are “equipotential,” as we may say,
in themselves, but of different potencies compared with each
other.
Explicit and Implicit Potencies: Primary and Secondary Potencies
We shall first give to our concept of “prospective potency” a few words of further analytical explanation with the help of our newly obtained knowledge.
It is clear from what we have stated that the prospective potencies of the ectoderm and of the endoderm, and we may add, of every elementary organ in relation to every other, differ between themselves and also in comparison with the blastoderm, from which they have originated. But the diversity of the endoderm with respect to the ectoderm is not of the same kind as its diversity in respect to the blastoderm. The potency of the endoderm and that of the ectoderm are both specialised in their typical manner, but compared with the potency of the blastoderm they may be said not only to be specialised but also to be restricted: the potency of the blastoderm embraces the whole, that of the so-called germ-layer embraces only part of the whole; and this species of restriction becomes clearer and clearer the further ontogeny advances: at the end of it in the “ultimate elementary organs” there is no prospective potency whatever.
A few new terms will serve to state a little more accurately what happens. Of course, with regard to all morphogenesis which goes on immediately from the blastoderm, the potency of the blastoderm is restricted as much as are the potencies of the germ layers. We shall call this sort of immediate potency explicit, and then we see at once that, with regard to their explicit potencies, there are only differences among the prospective potencies of the elementary organs; but with respect to the implicit potency of any of these organs, that is with respect to their potency as embracing the faculties of all their derivations, there are also not only differences but true morphogenetic restrictions lying at the very foundations of all embryology.
But now those of you who are familiar with morphogenetic facts will object to me, that what we have stated about all sorts of restrictions in ontogeny is not true, and you will censure me for having overlooked regeneration, adventitious budding, and so on. To some extent the criticism would be right, but I am not going to recant; I shall only introduce another new concept. We are dealing only with primary potencies in our present considerations, i.e. with potencies which lie at the root of true embryology, not with those serving to regulate disturbances of the organisation. It is true, we have in some way disturbed the development of our sea-urchin’s egg in order to study it; more than that, it would have been impossible to study it at all without some sort of disturbance, without some sort of operation. But, nevertheless, no potencies of what may properly be called the secondary or restitutive type have been aroused by our operations; nothing happened except on the usual lines of organogenesis. It is true, some sort of regulation occurred, but that is included among the factors of ontogeny proper.
We shall afterwards study more fully and from a more general point of view this very important feature of “primary regulation” in its contrast to “secondary regulation” phenomena. At present it must be enough to say that in speaking of the restriction of the implicit potencies in form-building we refer only to potencies of the primary type, which contain within themselves some properties of a (primary) regulative character.
The Morphogenetic Function of Maturation in the Light of Recent Discoveries
Turning again to more concrete matters, we shall first try, with the knowledge acquired of the potencies of the blastoderm and the so-called germ layers of Echinus, to understand certain rather complicated results which the experimental morphogenetic study of other animal forms has taught us. We know from our historical sketch that there are some very important aberrations from the type, to which the Echinus germ belongs,27 i.e. the type with an equal distribution of the potencies over all the blastomeres. We know not only that in cases where a regulation of the intimate structure of the protoplasm fails to occur a partial development of isolated cells will take place, but that there may even be a typical disposition of typical cells for the formation of typical organs only, without any regulability.
Let us first consider the last case, of which the egg of mollusca is a good type: here there is no equal distribution of potencies whatever, the cleavage-cells of this germ are a sort of real “mosaic” with regard to their morphogenetic potentialities. Is this difference between the germ of the echinoderms and the molluscs to remain where it is, and not to be elucidated any further? Then there would be rather important differences among the germs of different animals, at least with regard to the degree of the specification of their cleavage cells, or if we ascribe differences among the blastomeres to the organisation of the fertilised egg ready for cleavage, there would be differences in the morphogenetic organisation of the egg-protoplasm: some eggs would be more typically specialised at the very beginning of morphogenesis than others.
In the first years of the study of “Entwickelungsmechanik” I pointed out that it must never be forgotten that the egg itself is the result of organogenesis. If, therefore, there are real mosaic-like specifications in some eggs at the beginning of cleavage, or during it, there may perhaps have been an earlier stage in the individual history of the egg which did not show such specifications of the morphogenetic structure. Two American authors share the merit of having proved this hypothesis. Conklin showed, several years ago, that certain intracellular migrations and rearrangements of material do happen in the first stages of ovogenesis in certain cases, but it is to E. B. Wilson28 that science owes a proper and definitive elucidation of the whole subject. Wilson’s researches, pursued not only by descriptive methods,29 but also by means of analytical experiment, led him to the highly important discovery that the eggs of several forms (nemertines, molluscs), which after maturation show the mosaic type of specification in their protoplasm to a more or less high degree, fail to show any kind of specification in the distribution of their potencies before maturation has occurred. In the mollusc egg a certain degree of specification is shown already before maturation, but nothing to be compared with what happens afterwards; in the egg of nemertines there is no specification at all in the unripe egg.
Maturation thus becomes a part of ontogeny itself; it is not with fertilisation that morphogenesis begins, there is a sort of ontogeny anterior to fertilisation.
These words constitute a summary of Wilson’s researches. Taken together with the general results obtained about the potencies of the blastula and the gastrula of Echinus, they reduce what appeared to be differences of degree or even of kind in the specification of the egg-protoplasm to mere differences in the time of the beginning of real morphogenesis. What occurs in some eggs, as in those of Echinus, at the time of the definite formation of the germ layers, leading to a specification and restriction of their prospective potencies, may happen very much earlier in other eggs. But there exists in every sort of egg an earliest stage, in which all parts of its protoplasm are equal as to their prospectivity, and in which there are no potential diversities or restrictions of any kind.
So much for differences in the real material organisation of the germ and their bearing on inequipotentialities of the cleavage cells.
The Intimate Structure of Protoplasm: Further Remarks
Where a typical half- or quarter-development from isolated blastomeres happens to occur, we know already that the impossibility of a regulation of the intimate polar-bilateral structure may account for it. As this impossibility of regulation probably rests on rather simple physical conditions30 it may properly be stated that equal distribution of potencies is not wanting but is only overshadowed here. In this respect there exists a logical difference of fundamental importance between those cases of so-called “partial” or better, “fragmental” development of isolated blastomeres in which a certain embryonic organ is wanting on account of its specific morphogenetic material being absent, and those cases in which the “fragmental” embryo lacks complete “halves” or “quarters” with regard to general symmetry on account of the symmetry of its intimate structure being irregularly disturbed. This logical difference has not always received the attention which it undoubtedly deserves. Our hypothetical intimate structure in itself is, of course, also a result of factors concerned in ovogenesis. Only in one case do we actually know anything about its origin: Roux has shown that in the frog it is the accidental path of the fertilising spermatozoon in the egg which, together with the polar axis, normally determines the plane of bilateral symmetry; but this symmetry may be overcome and replaced by another, if gravity is forced to act in an abnormal manner upon the protoplasm; the latter showing parts of different specific gravity in the eggs of all Amphibia.
The Neutrality of the Concept of “Potency”
Now we may close our rather long chapter on the distribution of potencies in the germ; it has been made long, because it will prove to be very important for further analytical discussion; and its importance, in great measure, is due to its freedom from prepossessions. Indeed, the concept of prospective potency does not prejudice anything; we have said, it is true, that limitations of potencies may be due to the presence of specific parts of organisation in some cases; that, at least, they may be connected therewith; but we have not determined at all what a prospective potency really is, what the term really is to signify. It may seem that such a state of things gives an air of emptiness to our discussions, that it leaves uncertain what is the most important. But, I think, our way of argument, which tries to reach the problems of greatest importance by degrees, though it may be slow, could hardly be called wrong and misleading.
We now proceed to an analysis of what may properly be called the means of morphogenesis, the word “means” being preferable to the more usual one “conditions” in this connection, as the latter would not cover the whole field. It is in quite an unpretentious and merely descriptive sense that the expression “means” should be understood at present; what is usually called “conditions” is part of the morphogenetic means in our sense.
We know that all morphogenesis, typical or atypical, primary or secondary, goes on by one morphogenetic elementary process following the other. Now the very foundation of these elementary processes themselves lies in the elementary functions of the organism as far as they result in the formation of stable visible products. Therefore the elementary functions of the organism may properly be called the internal “means” of morphogenesis.
Secretion and migration are among such functions; the former happening by the aid of chemical change or by physical separation, the latter by the aid of changes in surface tension. But hardly anything more concrete has been made out about these or similar points at present.
We therefore make no claim to offer a complete system of the internal elementary means of morphogenesis. We shall only select from the whole a few topics of remarkable morphogenetic interest, and say a few words about each.
But, first of all, let us observe that the elementary means of morphogenesis are far from being morphogenesis themselves. The word “means” itself implies as much. It would be possible to understand each of these single acts in morphogenesis as well as anything, and yet to be as far from understanding the whole as ever. All means of morphogenesis are only to be considered as the most general frame of events within which morphogenesis occurs.
Some Remarks on the Importance of Surface Tension in Morphogenesis.—There are a few purely physical phenomena which have a special importance in organic morphology, all of them connected with capillarity or surface tension. Soap-lather is a very familiar thing to all of you: you know that the soap-solution is arranged here in very thin planes separated by spaces containing air: it was first proved by Berthold31 that the arrangement of cells in organic tissues follows the same type as does the arrangement of the single bubbles of a soap-lather, and Bütschli32 added to this the discovery that the minute structure of the protoplasm itself is that of a foam also. Of course it is not one fluid and one gas which make up the constituents of the structure in the organisms, as is the case in the well-known inorganic foams, but two fluids, which do not mix with one another. One general law holds for all arrangements of this kind: the so-called law of least surfaces, expressed by the words that the sum of all surfaces existing is a minimum; and it again is a consequence of this law, if discussed mathematically, that four lines will always meet in one point and three planes in one line. This feature, together with a certain law about the relation of the angles meeting in one line to the size of the bubbles, is realised most clearly in many structures of organic tissues, and makes it highly probable, at least in some cases, that capillarity is at work here. In other cases, as for instance in many plants, a kind of outside pressure, the so-called tissue tension, may account for the arrangement in surfaces minimae areae. Cleavage stages are perhaps the very best type in which our physical law is expressed: and here it may be said to have quite a simple application whenever all of the blastomeres are of the same physical kind, whilst some complications appear in germs with a specialised organisation and, therefore, with differences in the protoplasm of their single blastomeres. In such instances we may say that the physical law holds as far as the conditions of the system permit, these conditions ordinarily consisting in a sort of non-homogeneity of the surfaces.
It seems, from the researches of Dreyer,33 that the formation of organic skeletons may also be governed by the physically conditioned arrangement of protoplasmatic or cellular elements, and some phenomena of migration and rearrangement among cleavage cells, as described by Roux, probably also belong here.
But let us never forget that the laws of surface tension only give us the most general type of an arrangement of elements in all these cases, nothing else. A physical law never accounts for the Specific! Capillarity gives us not the least clue to it. As the organic substance, at least in many cases, is a fluid, it must of course follow the general laws of hydrostatics and hydrodynamics, but life itself is as little touched by its fluid-like or foam-like properties as it is by the fact that living bodies have a certain weight and mass.
All indeed that has been described may be said to belong, in the broadest meaning of the word, to what is called by Roux “correlation of masses,” though this author originally intended to express by this term only some sorts of passive pressure and deformation amongst embryonic parts as discovered especially by His.
We must be cautious in admitting that any organic feature has been explained, even in the most general way, by the action of physical forces. What at first seems to be the result of mechanical pressure may afterwards be found to be an active process of growth, and what at first seems to be a full effect of capillarity among homogeneous elements may afterwards be shown to depend on specialised metabolic conditions of the surfaces as its principal cause.34
There are other physical phenomena too, which assist morphogenesis; osmotic pressure for instance, which is also well known to operate in many purely physiological processes. But all these processes are only means of the organism, and can never do more than furnish the general type of events. They do not constitute life; they are used by life; let it remain an open question, for the present, how the phenomenon of “life” is to be regarded in general.35
On Growth.—Among the internal morphogenetical means which are of a so-called physiological character, that is, which nobody claims to understand physically at present, there is in the first place growth, which must be regarded as a very essential one.
Analytically we must carefully discriminate between the increase in the size of the cavities of an organism by a passive extension of their surfaces and the proper growth of the individual cells, which again may be due either to mere extension or to real assimilation. Osmotic pressure, of course, plays an important part both in the growth of the body-cavities and in simple cellular extension. We repeat the caution against believing too much to be explained by this phenomenon: it is the organism which by the secretion of osmotic substances in the cavities or the protoplasm of the cells prepares the ground for growth even of this osmotic sort. The real cellular growth which proceeds on the basis of assimilation cannot, of course, be accounted for by osmotic events, not even in its most general type.
Ontogenetical growth generally sets in, both in animals and in plants, after the chief lines of organisation are laid out; it is only the formation of the definite histological structures which usually runs parallel to it.
On Cell-division.—We have already said a good deal about the importance of cell-division in ontogeny: it accompanies very many of the processes of organisation in all living beings. But even then, there are the Protozoa, in the morphogenesis of which it does not occur at all, and there have also become known many cases of morphogenesis in higher animals, mostly of the type of regulation, in which cellular division is almost or wholly wanting. Therefore, cellular division cannot be the true reason of differentiation, but is only a process, which though necessary in some cases, cannot be essential to it. It must be conceded, I believe, that the same conclusion can be drawn from all our experiments on very young stages of the germ.
The investigations of the last few years have made it quite clear that even in organisms with a high power of morphogenetic regulation it is always the form of the whole, but not the individual cell, which is subjected to the regulation processes. Starting from certain results obtained by T. H. Morgan, I was able to show that in all the small but whole larvae, reared from isolated blastomeres, the size of the cells remains normal, only their number being reduced; and Boveri has shown most clearly that it is always the size of the nucleus—more correctly, the mass of the chromatin—which determines how large a cell of a certain histological kind is to be. In this view, the cell appears even more as a sort of material used by the organism as supplied, just as workmen can build the most different buildings with stones of a given size.
We now know what internal means of morphogenesis are, and so we may glance at some of the most important “outer means” or “conditions” of organisation.
Like the adult, the germ also requires a certain amount of heat, oxygen, and, when it grows up in the sea, salinity in the medium. For the germ, as for the adult, there exists not only a minimum but also a maximum limit of all the necessary factors of the medium; the same factor which at a certain intensity promotes development, disturbs it from a certain other intensity upwards.
Within the limits of this minimum and this maximum of every outside agent there generally is an increase in the rate of development corresponding to the increase of intensity of the agent. The acceleration of development by heat has been shown to follow the law of the acceleration of chemical processes by a rise of temperature; that seems to prove that certain chemical processes go on during the course of morphogenesis.
Almost all that has been investigated of the part played by the external conditions of development has little bearing on specific morphogenesis proper, and therefore may be left out of account here: we must, however, lay great stress on the general fact that there is a very close dependence of morphogenesis on the outside factors, lest we should be accused afterwards of having overlooked it.
Of course all “external” means or conditions of morphogenesis can actually relate to morphogenetic processes only by becoming in some way “internal,” but we unfortunately have no knowledge whatever how this happens. We at present are only able to ascertain what must necessarily be accomplished in the medium, in order that normal morphogenesis may go on, and we can only suppose that there exist certain specific internal general states, indispensable for organogenesis but inaccessible to present modes of investigation.36
The Discoveries of Herbst.—There are but few points in the doctrine of the external means or conditions of organogenesis which have a more special bearing on the specification of proper form, and which therefore require to be described here a little more fully. All these researches, which have been carried out almost exclusively by Herbst,37 relate to the effect of the chemical components of sea-water upon the development of the sea-urchin. If we select the most important of Herbst’s results, we must in the first place say a few words on the part taken by lime or calcium, not only in establishing specific features of form, but in rendering individual morphogenesis possible at all. Herbst has found that in sea-water which is deprived of calcium the cleavage cells and many tissue cells also completely lose contact with each other: cleavage goes on quite well, but after each single division the elements are separated; at the end of the process you find the 808 cells of the germ together at the bottom of the dish, all swimming about like infusoria. There seems to be some influence of the calcium salts upon the physical state of the surfaces of the blastomeres.
It is not without interest to note that this discovery has an important bearing on the technical side of all experiments dealing with the isolation of blastomeres. Since the separation of the single cleavage elements ceases as soon as the germs are brought back from the mixture without lime into normal sea-water, it of course is possible to separate them up to any stage which it is desired to study, and to keep them together afterwards. Thus, if for instance you want to study the development of isolated cells of the eight-cell stage, you will leave the egg in the artificial mixture containing no calcium until the third cleavage, which leads from the four- to the eight-cell stage, is finished. The single eight cells brought back to normal sea-water at this point will give you the eight embryos you want. All researches upon the development of isolated blastomeres since the time of Herbst’s discovery have been carried out by this method, and it would have been quite impossible by the old method of shaking to pursue the study into such minute detail as actually has been done. It may be added that calcium, besides its cell-uniting action, is also of primary importance in the formation of the skeleton.
Among all the other very numerous studies of Herbst we need only mention that potassium is necessary for the typical growth of the intestine, just as this element has been found necessary for normal growth in plants, and that there must be the ion SO4, or in other terms, sulphur salts present in the water, in order that the germs may acquire their pigments and their bilateral symmetry. This is indeed a very important result, though it cannot be said to be properly understood. It is a fact that in water without sulphates the larvae of Echinus retain the radial symmetry they have had in the very earliest stages, and may even preserve that symmetry on being brought back to normal sea-water if they have spent about twenty-four hours in the artificial mixture.
We may now leave the subject of Herbst’s attempts to discover the morphogenetic function of the single constituents of normal sea-water, and may devote a few words to the other branch of his investigations, those dealing with the morphogenetic effects of substances which are not present in the water of the sea, but have been added to it artificially. Here, among many other achievements, Herbst has made the most important discovery that all salts of lithium effect radical changes in development.38 I cannot describe fully here how the so-called “lithium larva” originates; let me only mention that its endoderm is formed outside instead of inside, that it is far too large, that there is a spherical mass between the ectodermal and the endodermal part of the germ, that a radial symmetry is established in place of the normal bilateralism, that no skeleton exists, and that the mesenchyme cells are placed in a quite abnormal position. All these features, though abnormal, are typical of the development in lithium. The larvae present no really pathological appearance at all, and, therefore, it may indeed be said that lithium salts are able to change fundamentally the whole course of morphogenesis. It detracts nothing from the importance of these discoveries that, at present, they stand quite isolated: only with lithium salts has Herbst obtained such strange results, and only upon the eggs of echinids, not even upon those of asterids, do lithium salts act in this way.
The Definition of Cause
We cannot begin the study of the “causes” of the differentiation of form without a few words of explanation about the terminology which we shall apply. Causality is the most disputed of all categories; many modern scientists, particularly in physics, try to avoid the concept of cause altogether, and to replace it by mere functional dependence in the mathematical meaning of the term. They claim to express completely by an equation all that is discoverable about any sort of phenomena constantly connected.
I cannot convince myself that such a very restricted view is the right one: it is very cautious, no doubt, but it is incomplete, for we have the concept of the acting “cause” in our Ego and are forced to search for applications of it in Nature. On the other hand, it does not at all escape me that there are many difficulties, or rather ambiguities, in applying it.
We may call the “cause” of any event, the sum total of all the constellations of facts which must be completed in order that the event may occur; it is in this meaning, for instance, that the first principle of energetics applies the term in the words causa aequat effectum. But, by using the word only in this very general sense, we deprive ourselves of many conveniences in the further and more particular study of Nature. Would it be better to say that the “cause” of any event is the very last change which, after all the constellations necessary for its start are accomplished, must still take place in order that the event may actually occur? Let us see what would follow from such a use of the word causality. We here have an animal germ in a certain stage, say a larva of Echinus, which is just about to form the intestine; all the internal conditions are fulfilled, and there is also a certain temperature, a certain salinity, and so on, but there is no oxygen in the water: the intestine; of course, will not grow in such a state of things, but it soon will when oxygen is allowed to enter the dish. Is, therefore, oxygen the cause of the formation of the intestine of echinus? Nobody, I think, would care to say so. By such reasoning, indeed, the temperature, or sodium, might be called the “cause” of any special process of morphogenesis. It, therefore, seems to be of little use to give the name of cause to that factor of any necessary constellation of events which accidentally happens to be the last that is realised. But what is to be done then?
Might we not say that the cause of any morphogenetic process is that typical property, or quality, or change, on which its specific character depends, on which depends for example, the fact that now it is the intestine which appears, while at another time it is the lens of the eye? We might very well, but we already have our term for this sort of cause, which is nothing else than our prospective potency applied to that elementary organ from which the new process takes its origin. The prospective potency indeed is the truly immanent cause of every specification affecting single organogenetic processes. But we want something more than this.
We may find what we want by considering that each single elementary process or development not only has its specification, but also has its specific and typical place in the whole—its locality. Therefore we shall call the “cause” of a single morphogenetic process, that occurrence on which depends its localisation, whether its specific character also partly depends on this “cause” or not.39
This definition of “cause” in morphology may be artificial; in any case it is clear. And at the same time the concepts of the prospective potency and of the “means” of organogenesis now acquire a clear and definite meaning: potency is the real basis of the specific character of every act in morphogenesis, and “means,” including conditions, are the sum of all external and internal general circumstances which must be present in order that morphogenetic processes may go on, without being responsible for their specificity or localisation.
It is implied in these definitions of cause and potency, that the former almost always will be of that general type which usually is called a stimulus or “Auslösung,” to use the untranslatable German word. There is no quantitative correspondence between our “cause” and the morphogenetic effect.
Some Instances of Formative and Directive Stimuli
Again it is to Herbst that we owe not only a very thorough logical analysis of what he calls “formative and directive stimuli”40 but also some important discoveries on this subject. We cannot do more here than barely mention some of the most characteristic facts.
Amongst plants it has long been known that the direction of light or of gravity may determine where roots or branches or other morphogenetic formations are to arise; in hydroids also we know that these factors of the medium may be at work41 as morphogenetic causes, though most of the typical architecture of hydroid colonies certainly is due to internal causes, as is also much of the organisation in plants.
Light and gravity are external formative causes; beside that they are merely “localisers.” But there also are some external formative stimuli, on which depends not only the place of the effect, but also part of its specification. The galls of plants are the most typical organogenetic results of such stimuli. The potencies of the plant and the specific kind of the stimulus equally contribute to their specification; for several kinds of galls may originate on one sort of leaves.
Scarcely any exterior formative stimuli are responsible for animal organisation; and one would hardly be wrong in saying that this morphogenetic independence in animals is due to their comparatively far-reaching functional independence of those external agents which have any sort of direction. But many organogenetic relations are known to exist between the single parts of animal germs, each of these parts being in some respect external to every other; and, indeed, it might have been expected already a priori, that such formative relations between the parts of an animal embryo must exist, after all we have learned about the chief lines of early embryology. If differentiation does not go on after the scheme of Weismann, that is, if it is not carried out by true “evolutio” from within, how could it be effected except from without? Indeed, every embryonic part may in some respect be a possible cause for morphogenetic events, which are to occur on every other part: it is here that the very roots of epigenesis are to be found.
Heliotropism and geotropism are among the well-known physiological functions of plants: the roots are seen to bend away from the light and towards the ground; the branches behave just in the opposite way. It now has been supposed by Herbst that such “directive stimuli” may also be at work among the growing or wandering parts of the embryo, that their growth or their migration may be determined by the typical character of other parts, and that real morphogenetic characters can be the result of some such relation; a sort of “chemotropism” or “chemotaxis” may be at work here. Herbst himself has discussed theoretically several cases of organogenesis in which the action of directive stimuli is very probable. What has become actually known by experiment is not very much at present: the mesenchyme cells of Echinus are directed in their migration by specified places in the ectoderm, the pigment cells of the yolk-sac of the fish fundulus are attracted by its blood vessels, and nerves may be forced to turn into little tubes containing brain substance; but of course only the first two instances have any bearing on typical morphogenesis.
The first case of an “internal formative stimulus” in the proper sense, that is, of one embryonic part causing another to appear, was discovered by Herbst himself. The arms of the so-called pluteus of the sea-urchin are in formative dependence on the skeleton—no skeleton, no arms; so many skeleton primordia,42 in abnormal cases, so many arms; abnormal position of the skeleton, abnormal position of the arms: these three experimental observations form the proof of this morphogenetic relation.
It may be simple mechanical contact, or it may be some chemical influence that really constitutes the “stimulus” in this case; certainly, there exists a close and very specific relation of the localisation of one part of the embryo to another. Things are much the same in another case, which, after having been hypothetically stated by Herbst on the basis of pathological data, was proved experimentally by Spemann. The lens of the eye of certain Amphibia is formed of their skin in response to a formative stimulus proceeding from the so-called primary optic vesicle. If this vesicle fails to touch the skin, no lens appears; and, on the other hand, the lens may appear in quite abnormal parts of the skin if they come into contact with the optic vesicle after transplantation.
But formative dependence of parts may also be of different types.
We owe to Herbst the important discovery that the eyes of crayfishes, after being cut off, will be regenerated in the proper way, if the optic ganglion is present, but that an antenna will arise in their place if this ganglion has also been removed. There must in this case be some unknown influence of the formative kind on which depends, if not regeneration itself, at least its special character.
In other cases there seems to be an influence of the central nervous system on the regenerative power in general. Amphibia, for instance, are said to regenerate neither their legs (Wolff), nor their tail (Godlewski), if the nervous communications have been disturbed. But in other animals there is no such influence; and in yet others, as for instance, in Planarians, it must seem doubtful at present whether the morphogenetic influence of the nervous system upon processes of restoration is more than indirect; the movements of the animal, which become very much reduced by the extirpation of the ganglia, being one of the main conditions of a good regeneration.
Of course, all we have said about the importance of special materials in the ripe germ, as bearing on specifically localised organisations, might be discussed again in our present chapter, and our intimate polar-bilateral structure of germs may also be regarded as embracing formative stimuli, at any rate as far as the actual poles of this structure are concerned. This again would bring us to the problem of so-called “polarity” in general, and to the “inversion” of polarity, that is to a phenomenon well known in plants and in many hydroids and worms, viz., that morphogenetic processes, especially of the type of restitutions, occur differently, according as their point of origin represents, so to speak, the positive or the negative, the terminal or the basal end of an axis, but that under certain conditions the reverse may also be the case. But a fuller discussion of these important facts would lead us deeper and deeper into the science of morphogenesis proper, without being of much use for our future considerations.
And so we may close this section43 on formative stimuli or “causes” of morphogenesis by shortly adding, more on account of its factual than of its logical interest, that the phenomenon of the determination of sex,44 according to the latest researches, seems to depend on cytological events occurring in the very earliest embryonic stages, say even before ontogeny, and not on formative stimuli proper45: it seems, indeed, as if the sexual products themselves would account for the sex of the individual produced by them, particularly if there were differences in their chromatin.46
Let us now turn again to considerations of a more abstract kind: we have become acquainted with some morphogenetic interactions among the parts of a developing embryo; and, indeed, we can be sure that there exist far more of such interactions than we know at present.
But it is far from being true that the development of each embryonic part depends on the existence or development of every other one.
On the contrary, it is a very important and fundamental feature of organogenesis that it occurs in separate lines, that is to say, in lines of processes which may start from a common root, but which are absolutely independent of one another in their manner of differentiation. Roux has coined the term “self-differentiation” to denote this phenomenon, and we admit that this term may be conveniently used for the purpose, if only it can be kept in mind that its sense is always relative, and that it is also negative. Suppose a part, A, shows the phenomenon of self-differentiation: this means that the further development of A is not dependent on certain other parts, B, C, and D; it does not mean at all that A has not been formatively dependent on some other parts, E or F at the time of its first appearance, nor does it imply that there might not be many formative actions among the constituents of A itself.
We indeed are entitled to say that the ectoderm of Echinus shows “self-differentiation” with regard to the endoderm; it acquires its mouth, for instance, as has been shown by experiment, even in cases where no intestine is present at all (Fig. 10); but ectoderm and endoderm both are formatively dependent on the intimate and the material organisation of the blastoderm. It further seems from the most recent experiments that the nerves and the muscles of the vertebrates are independent of each other in their differentiation, but that their fate is probably determined by formative processes in the very earliest stages of ontogeny.
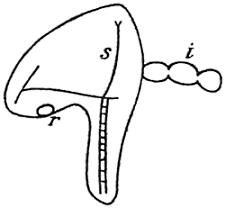
Fig. 10.—Pluteus-larva of Sphaerechinus.
The Intestine (i) is developed outside instead of inside (by means of raising the temperature); but the mouth (r) is formed in its normal place. S = Skeleton.
The phenomenon of self-differentiation, properly understood, now may help to the discovery of one most general character of all development. If the phenomenon of self-differentiation really occurs in ontogeny in its most different aspects, and if, on the other hand, in spite of this relative morphogenetic independence of embryonic parts, the resulting organism is one whole in organisation and in function, some sort of harmony of constellation, as it may properly be styled, must be said to be one of the most fundamental characters of all production of individual form. In establishing this harmony we do nothing more than describe exactly what happens: the harmony is shown by the fact that there is a whole organism at the end, in spite of the relative independence of the single events leading to it.
But still another sort of harmony is revealed in morphogenesis, by an analysis of the general conditions of the formative actions themselves. In order that these actions may go on properly the possibility must be guaranteed that the formative causes may always find something upon which to act, and that those parts which contain the potencies for the next ontogenetic stage may properly receive the stimuli awaking these potencies: otherwise there would be no typical production of form at all. This, the second species of harmonious relations to be described in ontogeny, may be called causal harmony; the term simply expresses the unfailing relative condition of formative causes and cause-recipients.
Finally, in functional harmony we have an expression descriptive of the unity of organic function, and so we may state, as the latest result of our analytical theory of development up to this point, that individual morphogenesis is marked by a threefold harmony among its parts.
At this stage we leave for a while our analytical studies of ontogeny proper. We must not forget that typical ontogenesis is not the only form in which morphogenesis can occur: the organic form is able to restore disturbances of its organisation, and it certainly is to be regarded as one of the chief problems of analytical morphogenesis to discover the specific and real stimulus which calls forth the restoring processes. For simply to say that the disturbance is the cause of the restoration would be to evade the problem instead of attacking it. But there are still some other problems peculiar to the doctrine of restitutions.
A few Remarks on Secondary Potencies and on Secondary Morphogenetic Regulations in General
We have only briefly mentioned in a previous chapter that there exist many kinds of potencies of what we call the secondary or truly restitutive type, and that their distribution may be most various and quite independent of all the potencies for the primary processes of ontogeny proper. Let us first add a few words about the concept of “secondary restitution” and about the distribution of secondary potencies in general.
Primary ontogenetic processes founded upon primary potencies may imply regulation, or more correctly, restitution in many cases: so it is, when fragments of the blastula form the whole organism, or when the mesenchyme cells of Echinus reach their normal final position by an attraction on the part of specific localities of the ectoderm in spite of a very abnormal original position enforced upon them by experiment. In these cases we speak of primary regulations or restitutions; disturbances are neutralised by the very nature of the process in question. We speak of secondary restitution whenever a disturbance of organisation is rectified by processes foreign to the realm of normality; and these abnormal lines of events are revealed to us in the first place by the activity of potencies which remain latent in ontogeny proper.
We know already that a certain kind of secondary restitution has been discovered lately, very contradictory to the theoretical views of Weismann; the process of restoration being carried out not by any definite part of the disturbed organisation, but by all the single elements of it. The problem of the distribution of secondary potencies in these cases of so-called “re-differentiation” is to form our special study in the next chapter. In all other cases restoration processes start from specific localities; if they occur on the site of the wound which caused the disturbance, we speak of regeneration; if they occur at some distance from the wound, we call them adventitious processes. Besides these three types of processes of restitution there may be mentioned a fourth one, consisting in what is generally called compensatory hypertrophy; the most simple case of such a compensatory process is when one of a pair of organs, say a kidney, becomes larger after the other has been removed.48 Finally, at least in plants, a change of the directive irritability, of so-called “geotropism” for instance, in certain parts may serve to restore other more important parts.
In two of these general types of restitution, in regeneration proper and in the production of adventitious organs, the potencies which underlie these processes may be said to be “complex.” It is a complicated series of events, a proper morphogenesis in itself, for which the potency has to account, if, for instance, a worm newly forms its head by regeneration, or if a plant restores a whole branch in the form of an adventitious bud.
Such generalisations as are possible about the distribution of complex potencies are reserved for a special part of our future discussion.
Secondary restitution is always, like ontogeny, a process of morphogenesis, and therefore all the questions about single formative stimuli, and about internal and external conditions or means, occur again. But of course we cannot enter into these problems a second time, and may only say that, especially in regeneration proper, the specific type of the regenerative formation of any part may differ very much from the ontogenetic type of its origin: the end of both is the same, but the way can be even fundamentally different in every respect.
The Stimuli of Restitutions49
But now we turn to the important question: what is the precise stimulus50 that calls forth processes of restitution; or, in other words, what must have happened in order that restitution may occur?
That the operation in itself, by its removing of mechanical obstacles, cannot be the true stimulus of any restitutions, is simply shown by all those restitutions that do not happen at the place of the wound. If we took a narrower point of view, and if we only considered regeneration proper from the wound itself, we might probably at first be inclined to advocate the doctrine that the removing of some obstacles might in fact be the stimulus to the process of restoration; but, even then, why is it that just what is wanted grows out? Why is there not only growth, but specific growth, growth followed by specification? The removing of an obstacle could hardly account for that. But, of course, taking account of all the adventitious restitutions—that is, all restorations not beginning at the wound itself—the theory that the removing of obstacles is the stimulus to restoration becomes, as we have said, quite impossible.51
But where then is the stimulus to be found? There is another rather simple theory of the “Auslösung” of restitutions,52 which starts from the phenomena of compensatory hypertrophy and some occurrences among plants. The removal of some parts of the organism, it is said, will bring its other parts into better conditions of nutrition, and therefore these parts, particularly if they are of the same kind, will become larger. Granted for the moment that such a view may hold in cases when one of a pair of glands becomes larger after the other has been removed, or when pruning of almost all the leaves of a tree leads to the rest becoming larger, it certainly must fail to explain the fact that in other cases true new formations may arise in order to restore a damaged part, or that the latter may be regenerated in its proper way. For merely quantitative differences in the mixture of the blood or of the nourishing sap in plants can never be a sufficient reason for the highly typical and qualitative structure of newly-formed restitutions. And even in the most simple cases of a mere increase in the size of some parts, that is, in the simplest cases of so-called compensatory hypertrophy,53 it is at least doubtful, if not very improbable, that the compensation is accomplished in such a purely passive way, because we know that in other cases it is usually the growth of the young parts that actively attracts the nourishment: there is first differentiation and growth, and afterwards there is a change in the direction of the nourishing fluids.
The process of true regeneration, beginning at the locality of the wound itself, has been shown by Morgan, even as regards its rate, to occur quite irrespectively of the animal being fed or not.54 There could hardly be a better demonstration of the fundamental fact that food assists restitution, but does not “cause” it in any way.
But in spite of all we have said, there seems to be some truth in regarding the nutritive juices of animals and plants as somehow connected with the stimulus of restitutions: only in this very cautious form, however, may we make the hypothesis. It has been shown for both animals and plants, that morphogenesis of the restitutive type may be called forth even if the parts, now to be “regenerated” have not been actually removed; e.g. in the so-called super-regeneration of legs and tails in Amphibia, of the head in Planarians, of the root-tip in plants and in some other cases. Here it has always been a disturbance of the normal connection of some parts with the rest of the organism which proved to be the reason of the new formation. This shows that something to do with the communication among parts is at least connected with restitution, and this communication may go on either by the unknown action of specific tissues or by the aid of the blood or sap.55 But in what this change or break of specific communication consists, is absolutely unknown. One might suppose that each part of the organisation constantly adds some sort of ferment to the body fluids outside or inside the cells, that the removing of any part will change the composition of these fluids in this particular respect, and that this change acts as a sort of communication to summon the restituting parts of the whole to do their duty.56
But I see quite well that such a theory is very little satisfactory; for what has to be done in restitution in each case is not a simple homogeneous act, for which one special material might account, but is a very complicated work in itself. It was the defect of the theory of “organ-forming substances” as advocated by Sachs, that it overlooked this point.
So all we know about the proper stimuli of restitutions is far from resting on any valid grounds at all; let us not forget that we are here on the uncertain ground of what may be called the newest and most up-to-date branch of the physiology of form. No doubt, there will be something discovered some day, and the idea of the “whole” in organisation will probably play some part in it. But in what manner that will happen we are quite unable to predict.
This is the first time that, hypothetically at least, the idea of the whole has entered into our discussion. The same idea may be said to have entered it already in a more implicit form in the statement of the threefold harmony in ontogeny.
Let us now see whether we can find the same problem of the “whole” elsewhere, and perhaps in more explicit and less hypothetical form. Let us see whether our analytical theory of development is in fact as complete as it seemed to be, whether there are no gaps left in it which will have to be filled up.
We have come to the central point of the first part of these lectures; we shall try in this chapter to decide a question which is to give life its place in Nature, and biology its place in the system of sciences. One of the foundation stones is to be laid upon which our future philosophy of the organism will rest.
The General Problem
Our analytical theory of morphogenesis has been founded upon three elementary concepts: the prospective potency, the means, and the formative stimulus. Its principal object has been to show that all morphogenesis may be resolved into the three phenomena expressed by those concepts; in other terms, that morphogenesis may be proved to consist simply and solely of what is expressed by them. Have we indeed succeeded in attaining this object? Has nothing been left out? Is it really possible to explain every morphogenetic event, at least in the most general way, by the aid of the terms potency, means, and stimulus?
All of these questions are apt to lead us to further considerations. Perhaps these considerations will give us a very clear and simple result by convincing us that it is indeed possible to analyse morphogenesis in our schematic way.
But if the answer were a negative one? What would that suggest?
The full analysis of morphogenesis into a series of single formative occurrences, brought about by the use of given means and on the basis of given potencies, might assure us, perhaps, that, though not yet, still at some future time, a further sort of analysis will be possible: the analysis into the elemental facts studied by the sciences of inorganic nature. The organism might prove to be a machine, not only in its functions but also in its very origin.
But what are we to say if even the preliminary analysis, which possibly might lead to such an ultimate result, fails?
Let us then set to work. Let us try to consider most carefully the topic in which our concept of the formative cause or stimulus may be said to be centred, the localisation of all morphogenetic effects. Is it always possible in fact to account for the typical localisation of every morphogenetic effect by the discovery of a single specific formative stimulus? You will answer me, that such an analysis certainly is not possible at present. But I ask you again, are there any criteria that it is possible, at least in principle; or are there any criteria which will render such an aim of science impossible for all future time?
The Morphogenetic “System”
We know from our experimental work that many, if not all, of the elementary organs in ontogeny show one and the same prospective potency distributed equally over their elements. If we now borrow a very convenient term from mechanics, and call any part of the organism which is considered as a unit from any morphogenetic point of view, a morphogenetic “system,” we may sum up what we have learnt by saying that both the blastoderm of the echinoderms, at least around its polar axis, and also the germ-layers of these animals, are “systems” possessing an equal potentiality in all of their elements, or, in short, that they are equipotential systems.
But such a term would not altogether indicate the real character of these systems.
Later on we shall analyse more carefully than before the distribution of potencies which are the foundation both of regeneration proper and of adventitious growth, and then we shall see that, in higher plants for instance, there is a certain “system” which may be called the organ proper of restitutions, and which also in each of its elements possesses the same restoring potency; I refer to the well-known cambium. This cambium, therefore, also deserves the name of an “equipotential system.” But we know already that its potencies are of the complex type, that they consist in the faculty of producing the whole, of such a complicated organisation as a branch or a root, that the term “equipotential system” is here only to signify that such a complicated unit may arise out of each of the cells of the cambium.
The potencies we have been studying in the blastula or gastrula of echinoderms are not of the complex type: our systems are equipotential to the extent that each of their elements may play every single part in the totality of what will occur in the whole system; it is to this single part that the term “function of the position” relates. We therefore might call our systems equipotential systems with single potencies; or, more shortly, singular-equipotential systems.
But even this terminology would fail to touch precisely the very centre of facts: it is not only the simplicity or singularity of their potencies which characterises the rôle of our systems in morphogenesis,57 but far more important with respect to the production of form are two other leading results of the experimental researches. The proper act to be performed by every element in each actual case is in fact a single one, but the potency of any element as such consists in the possibility of many, nay of indefinitely many, single acts: that then might justify us in speaking of our systems as “indefinite equipotential,” were it not that another reason makes another title seem still more preferable. There are indeed indefinite singular potencies at work in all of our systems during ontogeny: but the sum of what happens to arise in every case out of the sum of the single acts performed by all of the single equipotential cells is not merely a sum but a unit; that is to say, there exists a sort of harmony in every case among the real products of our systems. The term harmonious-equipotential system therefore seems to be the right one to denote them.
We now shall try first to analyse to its very extremes the meaning of the statement that a morphogenetic system is harmonious-equipotential.
The “Harmonious-Equipotential System”
We have an ectoderm of the gastrula of a starfish here before us; we know that we may cut off any part of it in any direction, and that nevertheless the differentiation of the ectoderm may go on perfectly well and result in a typical little embryo, which is only smaller in its size than it would normally be. It is by studying the formation of the highly complicated ciliary band, that these phenomena can be most clearly understood.
Now let us imagine our ectoderm to be a cylinder instead of being approximately a sphere, and let us imagine the surface of this cylinder unrolled. It will give us a plane of two definite dimensions, a and b. And now we have all the means necessary for the analytical study of the differentiation of an harmonious-equipotential system.
Our plane of the dimensions a and b is the basis of the normal, undisturbed development; taking the sides of the plane as fixed localities for orientation, we can say that the actual fate, the “prospective value” of every element of the plane stands in a fixed and definite correlation to the length of two lines, drawn at right angles to the bordering lines of the plane; or, to speak analytically, there is a definite actual fate corresponding to each possible value of x and of y. Now, we have been able to state by our experimental work, that the prospective value of the elements of our embryonic organ is not identical with their “prospective potency,” or their possible fate, this potency being very much richer in content than is shown by a single case of ontogeny. What will be the analytical expression of such a relation?
Let us put the question in the following way: on what factors does the fate of any element of our system depend in all possible cases of development obtainable by means of operations? We may express our results in the form of an equation:—
p.v. (X) = f( . . . )
i.e. “the prospective value of the element X is a function of . . .”—of what?
We know that we may take off any part of the whole, as to quantity, and that a proportionate embryo will result, unless the part removed is of a very large size. This means that the prospective value of any element certainly depends on, certainly is a function of, the absolute size of the actually existing part of our system in the particular case. Let s be the absolute size of the system in any actual experimental case of morphogenesis: then we may write p.v. (X) = f(s . . . ). But we shall have to add still some other letter to this s.
The operation of section was without restriction either as to the amount of the material removed from the germ, or as to the direction of the cut. Of course, in almost every actual case there will be both a definite size of the actual system and a definite direction of the cut going hand-in-hand. But in order to study independently the importance of the variable direction alone, let us imagine that we have isolated at one time that part of our system which is bounded by the lines a1 b1, and at another time an equal amount of it which has the lines a2 b2 as its boundaries. Now since in both cases a typical small organism may result on development, we see that, in spite of their equal size the prospective value of every element of the two pieces cut out of the germ may vary even in relation to the direction of the cut itself. Our element, X, may belong to both of these pieces of the same size: its actual fate nevertheless will be different. Analytically, it may be said to change in correspondence to the actual position of the actual boundary lines of the piece itself with regard to the fundamental lines of orientation, a and b; let this actual position be expressed by the letter l, l marking the distance of one58 of the actual boundary lines of our piece from a or b: then we are entitled to improve our formula by writing p.v. (X) = f(s, l . . . ) (Fig. 11).
Fig. 11.—Diagram to show the Characteristics of an “Harmonious-equipotential System.”
The element X forms part of the systems a b or a1 b1 or a2 b2; its prospective value is different in each case.
But the formula is not yet complete: s and l are what the mathematicians call variables: they may have any actual value and there will always be a definite value of p.v., i.e. of the actual fate which is being considered; to every value of s and l, which as we know are independent of each other, there corresponds a definite value of the actual prospectivity. Now, of course, there is also a certain factor at work in every actual case of experimental or normal development, which is not a variable, but which is the same in all cases. This factor is a something embraced in the prospective potency of our system, though not properly identical with it.
The prospective potency of our system, that is to say of each of its elements, is the sum total of what can be done by all; but the fact that a typically proportionate development occurs in every possible case, proves that this sum comes into account, not merely as a sum, but as a sort of order: we may call this order the “relation of localities in the absolutely normal case.” If we keep in mind that the term “prospective potency” is always to contain this order, or, as we may also call it, this “relative proportionality,” which, indeed, was the reason for calling our systems “harmonious,” then we may apply it without further explanation in order to signify the non-variable factor on which the prospective value of any element of our systems depends, and, if we denote the prospective potency, embracing order, by the letter E, we are now able to complete our formula by saying p.v. (X) = f(s, l, E).
So far the merely analytical study of the differentiation of harmonious-equipotential systems.59
Instances of “Harmonious-Equipotential Systems”
We must try at first to learn a few more positive facts about our systems, in order that we may know how important is the part which they play in the whole animal kingdom, and in order that our rather abstract analysis may become a little more familiar to us. We know already that many of the elementary morphogenetic organs have been really proved to be harmonious-equipotential systems, and that the same probably is true of many others; we also know that the immature egg of almost all animals belongs to this type, even if a fixed determination of its parts may be established just after maturation. Moreover, we said, when speaking about some new discoveries on form-restitution, that there are many cases in which the processes of restitution do not proceed from single localities, the seat of complex potencies in the organism, but in which each single part of the truncated organism left by the operation has to perform one single act of restoration, the full restitution being the result of the totality of all. These cases must now be submitted to a full analysis.
All of you have seen common sea-anemones or sea-roses, and many of you will also be familiar with the so-called hydroid polyps. Tubularia is one genus of them: it looks like a sea-anemone in miniature placed on the top of a stem like a flower. It was known already to Allman that Tubularia is able to restore its flower-like head when that is lost, but this process was taken to be an ordinary regeneration, until an American zoologist, Miss Bickford, succeeded in showing that there was no regeneration process at all, in the proper sense of the word, no budding of the missing part from the wound, but that the new tubularian head was restored by the combined work of many parts of the stem. Further analysis then taught us that Tubularia indeed is to be regarded as the perfect type of an harmonious-equipotential system: you may cut the stem at whatever level you like: a certain length of the stem will always restore the new head by the co-operation of its parts. As the point of section is of course absolutely at our choice, it is clear, without any further discussion, that the prospective value of each part of the restoring stem is a “function of its position,” that it varies with its distance from the end of the stem; and so at once we discover one of the chief characteristics of our systems. But also the second point which enters into our formula can be demonstrated in Tubularia: the dependence of the fate of every element on the actual size of the system. You would not be able to demonstrate this on very long stems, but if you cut out of a Tubularia stem pieces which are less than ten millimetres in length, you will find the absolute size of the head restored to be in close relation to the length of the stem piece, and this dependence, of course, includes the second sort of dependence expressed in our formula.
The figures will serve to show you a little more concretely what has been described. The head of Tubularia consists of a sort of broad base with a thin proboscis upon it, both bearing a large number of tentacles; these tentacles are the first things to be seen as primordia (“Anlagen”) in the process of restitution. You notice two rings of longitudinal lines inside the stem; the lines will become walls and then will separate from the stem until they are only connected with it at their basal ends; the new tentacles are ready as soon as that has happened, and a process of growth at the end will serve to drive the new head out of the so-called perisarc or horny skeleton, which surrounds the stem. By comparing the two figures, 12 e, and g, you easily find out that the absolute lengths of the two tentacle rings are very different, and that both are in proportion60 to the actual size of the stem (Fig. 12).
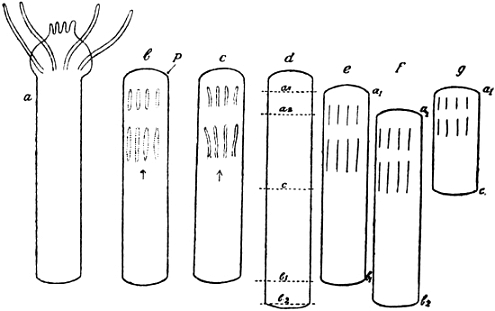
Fig. 12.—Tubularia.
| a. | Diagram of the “Hydranth,” with its short and long tentacles. |
| b. | Restitution of a new hydranth inside the perisarc (p). |
| c. | The same—later stage; the tentacles are complete; the whole hydranth will be driven out of the perisarc by a process of growth that occurs at the locality marked ↑. |
| d. | A stem of Tubularia cut either at a1b1 or at a2b2 or at a1c. |
| e. | Position of tentacles in the piece cut at a1b1. |
| f. | Position of tentacles in the piece cut at a2b2 which is equal in length to a1b1. |
| g. | Position of tentacles in the piece cut at a1c, which is half as long as a1b1. |
So we find our formula p.v. (X) = f(s, l, E) very well illustrated in Tubularia. The formula indeed may help us to predict, in any case, where a certain part of the polyp’s organisation is to originate, at least if we know all that is included under our letter E, i.e. the normal proportion of our form. Of course such prediction would not have much practical importance in all our cases of morphogenesis, but nevertheless I should like to state here that it is possible; for many scientific authors of recent times have urged the opinion that prediction of, and domination over, what will happen, can be the only true aims of sciences at all. I myself judge these aims to be of second or third-rate importance only, but, if they may be reached by what our purely theoretical study teaches, so much the better.
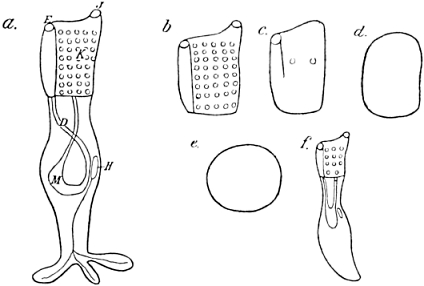
Fig. 13.—Clavellina.
| a. | Diagram of the normal animal: E and J = openings; K = branchial apparatus; D = intestine; M = stomach; H = heart. |
| b. | The isolated branchial apparatus. |
| c-e. | Different stages of reduction of the branchial apparatus. |
| f. | The new whole little ascidian. |
Another very typical case of a morphogenetic system of the harmonious type is supplied by the phenomena of restoration in the ascidian Clavellina. I cannot fully describe the organisation of this form (Fig. 13a), and it must suffice to say that it is very complicated, consisting of two very different chief parts, the branchial apparatus and the so-called intestinal sac; if these two parts of the body of Clavellina are separated one from the other, each may regenerate the other in the typical way, by budding processes from the wound. But, as to the branchial apparatus, there may happen something very different: it may lose almost all of its organisation and become a small white sphere, consisting only of epithelia corresponding to the germ-layers, and of mesenchyme between them, and then, after a certain period of rest, a new organisation will appear. Now this new organisation is not that of a branchial apparatus but represents a very small but complete ascidian (Fig. 13). Such a fact certainly seems to be very important, not to say very surprising; but still another phenomena may be demonstrated on the animal which seems to be even more important. You first isolate the branchial apparatus from the other part of the body, and then you cut it in two, in whatever direction you please. Provided they survive and do not die, as indeed many of them do, the pieces obtained by this operation will each lose their organisation, as did the whole branchial apparatus, and then will each acquire another one, and this new organisation is also that of a complete little Clavellina. So we see that not only is the branchial apparatus of our animal capable of being transformed into a whole animal by the co-operative work of all its parts, but even each part of it may be transformed into a small whole, and it is quite at our disposal how large this part shall be, and what sort of a fragment of the original branchial apparatus it shall represent.
We could hardly imagine a better instance of an harmonious-equipotential system.
I cannot give you a description of all the other types of our systems subservient to restitution, and I can only mention here that the common hydra and the flatworm Planaria are very fine examples of them. But to one special case of harmonious equipotentiality you must allow me to direct your further attention.
It has been known for many years that the Protozoa are also capable of a restoration of their form and organisation after disturbances, if at least they contain a certain amount of their nuclear substance. This process of restoration used to be regarded as belonging to the common type of regeneration proper, until T. H. Morgan succeeded in showing that in the genus Stentor it follows just the very lines which we know already from our study of embryonic organs or from Tubularia; that an harmonious-equipotential system is at the basis of what goes on. Now, you know that all Protozoa are but one highly organised cell: we have therefore here an instance where the so-called “elements” of our harmonious-morphogenetic system are not cells, but something inside of cells; and this feature must appear to be of very great moment, for it first shows, as we have already pointed out on another occasion, that morphogenesis is not dependent on cell-division, and it states at the same time that our concept of the harmonious-equipotential system may cover a very great area—that, in fact, it is a scheme of a very wide extent.
The Problem of the Factor E
We turn back again to considerations of a more abstract form. We left our analysis of the differentiation of the harmonious-equipotential systems, and particularly of the phenomena of localisation during this differentiation, at the point where we had succeeded in obtaining an equation as the expression of all those factors on which the prospective value, the actual fate, of any element of our systems depends, p.v. (X) = f(s, l, E) was the short expression of all the relations involved; s and l, the absolute size of the system and the relative position of the element with respect to some fixed points, were independent variables; E was a constant, namely, the prospective potency, with special regard to the proportions embraced by it.
We shall now study the significance of the factor E.
What does this E mean? Is it a short expression merely for an actual sum of elemental agents having a common resultant? And, if so, of what kind are these agents? Or what may E mean, if it can be shown not to be a short sign for a mere sum?
No Explanation Offered by “Means” or “Formative Stimuli”
For practical purposes it seems better if we modify the statement of our question. Let us put it thus: E is one of the factors responsible, among variables, for the localisation of organic differentiation; what then do we actually know about the causal factors which play a localising part in organogenesis? We, of course, have to look back to our well-studied “formative stimuli.” These stimuli, be they “external” or “internal,” come from without with respect to the elementary organ in which any sort of differentiation, and therefore of localisation, occurs: but in our harmonious systems no localising stimulus comes from without, as was the case, for instance, in the formation of the lens of the eye in response to the optical vesicle touching the skin. We know absolutely that it is so, not to speak of the self-evident fact that the general “means” of organogenesis have no localising value at all.61
So we see there is nothing to be done, either with the means or with the formative stimuli; both are entirely unable to account for those kinds of localisation during differentiation which appear in our harmonious systems.
But is there no possibility of explaining the phenomena of organogenetic localisation by any other sort of interaction of parts? Two such possibilities may at the first glance seem to exist.
No Explanation Offered by a Chemical Theory of Morphogenesis
Though never set forth, in the form of a properly worked-out theory, the view has sometimes been advocated by biologists, that a chemical compound of a very high degree of complication might be the very basis of both development and inheritance, and that such a chemical compound by its disintegration might direct morphogenesis.
Let us first examine if such a view may hold for the most general features of organic morphogenesis. It seems to me that from the very beginning there exists one very serious objection to every chemical theory of form-building, in the mere fact of the possibility of the restoration of form starting from atypical localities. The mere fact, indeed, that there is such a thing as the regeneration of a leg of a newt—to say nothing about restitution of the harmonious type—simply contradicts,62 it seems to me, the hypothesis, that chemical disintegration of one compound may govern the course of morphogenetic events: for whence comes the re-existence of the hypothetical compound, newly to be disintegrated, after disintegration has been completed once already? And we even know that regeneration may go on several times running from the same locality!
But, if we intentionally disregard this difficulty, in spite of its fundamental character, how could the hypothesis of chemical disintegration give the reason for the differentiation of our harmonious-equipotential systems, with special regard to the localisation of it; how could it account, in other words, for the appearance of typically localised specifications in an organ for which no external localising causes can be predicated?
Let us remember that a few original intimate differences exist in our harmonious systems: the main directions of the intimate protoplasmic structure including polarity and bilaterality. There are therefore three times two specified poles in each of these systems, at least in bilateral organisms, but no other differences are present in them. A few very simple cases of harmonious differentiation might indeed be understood on the theory of a disintegrating chemical compound in connection with these few differences. Imagine that the original compound, of the quantity a, is disintegrated to the amount of a1; from a1 are formed the two more simple compounds, b and c, both of them in definite quantities; then we have the three chemical individuals, a-a1, b and c, as the constituents of our harmonious system; and it now might be assumed, without any serious difficulty, though with the introduction of some new hypotheses, that the two poles of one of the fundamental axes of symmetry attract b and c respectively, a-a1 remaining unattracted between them. We thus should have the three elementary constituents of the system separated into three parts, and as they all three are of a definite quantity, their separation would mean that the system had been divided into three parts, a-a1, b and c, also with regard to its proper form. It is clear, that by taking away any part of the original system, by means of operations, there would be taken away a certain amount of the original compound; say that a/n is left; then, of course, the three constituents after the partial disintegration would be a-a1/n, b/n and c/n, and so it follows that the proportionality of localisation would really be preserved in any case.
But these considerations, evident as they seem to be in the most simple case, fail to satisfy in a really general sense: for two different reasons. First, they could never account for the fact that the differentiated organism by no means consists of so many different compounds as it shows single parts of its differentiation, but that, on the contrary, it only consists, as we know, of a certain rather limited number of true different morphogenetic elements, these elements occurring again and again—as for instance, nervous or muscular elements—but typical each time in locality, quantity, and form. And in the second place, the very form of elementary organs, their form as such, does not at all go hand-in-hand with chemical differences; this feature alone would absolutely overthrow any sort of a chemical morphogenetic theory to account for the problem of localisation. Take the typically arranged ring of the mesenchyme cells in our Echinus-gastrula, with its two spherical triangles, so typically localised; look at any sort of skeleton, in Radiolaria, or in starfishes, or in vertebrates: here you have form, real form, but form consisting of only one material. Not only is the arrangement of the elements of form typical here, e.g. the arrangement of the single parts of the skeleton of the hand or foot, but also the special form of each element is typical, e.g. the form of each single bone of the foot; and, on a purely chemical theory of morphogenesis the sufficient reason for the production of typical form in such a sense would be wanting. For atoms or molecules by themselves can only account for form which is arranged, so to speak, according to spatial geometry—as in fact they do in crystallography; but they can never account for form such as the skeleton of the nose, or hand, or foot. You will answer me perhaps, that there may be non-chemical agents in the germ,63 responsible for typical form-localisation, but by such reasoning you would be departing from a purely chemical theory. Our next paragraph will be devoted to this side of the question.
That is the principal reason for rejecting all sorts of chemical morphogenetic theories put forward to explain the problem of localisation; it is more explicit, and therefore, I suppose, still more convincing than the more general consideration that the very fact of restitutions in itself must contradict the hypothesis that a disintegration of compounds might be the directive agency in morphogenesis. To sum up: Specificity of organic form does not go hand-in-hand with specificity of chemical composition, and therefore cannot depend on it; and besides that, specific organic form is such that it can never be explained by atomic or molecular arrangement in the chemical sense; for, to state it in a short but expressive manner, the “form” of an atom or molecule can never be that of a lion or a monkey. To assume that would be to go beyond the limits of chemistry in chemistry itself.
No Machine Possible Inside the Harmonious Systems
And now we turn to the last possibility which is left to us in our endeavour to “understand” the localisation of the differentiation in our harmonious-equipotential systems by the means of physics and chemistry. Outside causes have failed to account for it, chemical disintegration of a compound has failed too. But could there not exist some sort of complicated interactions amongst the parts of the harmonious system themselves? Could there not exist some kind of a real machine in the system, which, if once set going, would result in the differentiations that are to take place? Then we might say that the “prospective potency” of the system is in fact that machine; we should know what the letter E of our equation stood for: viz., a resultant action of many complicated elemental interactions, and nothing more.
Weismann, we know already, had assumed that a sort of machine was the prime mover of morphogenesis. We have seen that his theory cannot be true; the results of experiments most strongly contradict it. But, of course, the experiments only showed us that such a machine as he had imagined to exist could not be there, that development could not be governed by the disintegration of a given complicated structure into its simplest parts. But might not some other machine be imaginable?
We shall understand the word “machine” in a most general sense. A machine is a typical configuration of physical and of chemical constituents, by the acting of which a typical effect is attained. We, in fact, lay much stress upon embracing in our definition of a machine the existence of chemical constituents also; we therefore understand by the word “machine” a configuration of a much higher degree of complication than for instance a steam-engine is. Of course a machine, whose acting is to be typical with regard to the three dimensions in space, has to be typically constructed with regard to these three dimensions itself; a machine that was an arrangement of elements in a strict plane could never have typical effects at right angles to that plane. This is a point which must well be kept in mind in all hypothetical considerations about machines that claim to explain morphogenesis.
It must be granted that a machine, as we understand the word, might very well be the motive force of organogenesis in general, if only normal, that is to say, if only undisturbed development existed, and if a taking away of parts of our systems led to fragmental development.
But we know that, at least in our harmonious-equipotential systems, quite another process occurs after parts have been taken away: the development that occurs is not fragmental but whole, only on a smaller scale.
And we know, further, that this truly whole development sets in irrespective of the amount and direction of the separation. Let us first consider the second of these points. There may be a whole development out of each portion of the system—above certain limits—which is, say, of the volume V. Good! Then there ought to exist a machine, like that which exists in the whole undisturbed system, in this portion V also, only of smaller dimensions; but it also ought to exist in the portion V1 which is equal to V in amount, and also in V2, in V3, V4 and so on. Indeed, there do exist almost indefinitely many Vn all of which can perform the whole morphogenesis, and all of which therefore ought to possess the machine. But these different portions Vn are only partly different from each other in spatial relation. Many parts of V2 are also parts of V1 and of V3 and of V4 and so on; that is to say, the different volumes Vn overlap each other successively and in such a manner that each following one exceeds the preceding one in the line by a very small amount only. But what then about our machines? Every volume which may perform morphogenesis completely must possess the machine in its totality. As now every element of one volume may play any possible elemental rôle in every other, it follows that each part of the whole harmonious system possesses any possible elemental part of the machine equally well, all parts of the system at the same time being constituents of different machines.
A very strange sort of machine indeed, which is the same in all its parts (Fig. 14)!
Fig. 14.—An “Harmonious-equipotential System” of whatever kind.
According to the “machine-theory” of life this system ought to possess a certain unknown very complicated machine in its completeness:
(a) in its total length,
and (b) in each of the equal volumes v, v1, v2, v3 and so on,
and (c) in each of the unequal volumes w, x, y, and so on,
and (d) in every imaginable volume, no matter of what size.
Therefore the “machine-theory” of life is absurd.
But we have forgotten, I see, that in our operation the absolute amount of substance taken away from the system was also left to our choice. From this feature it follows that not only all the different Vn, all of the same size, must possess the hypothetic machine in its completeness, but that all amounts of the values Vn-n, n being variable, must possess the totality of the machine also: and all values Vn-n, with their variable n, may again overlap each other.
Here we are led to real absurdities!
But what is the conclusion of our rather wild considerations?
It seems to me that there is only one conclusion possible. If we are going to explain what happens in our harmonious-equipotential systems by the aid of causality based upon the constellation of single physical or chemical factors and events, there must be some such thing as a machine. Now the assumption of the existence of a machine proves to be absolutely absurd in the light of the experimental facts. Therefore there can be neither any sort of a machine nor any sort of causality based upon constellation underlying the differentiation of harmonious-equipotential systems.
For a machine, typical with regard to the three chief dimensions of space, cannot remain itself if you remove parts of it or if you rearrange64 its parts at will.
Here we see that our long and careful study of morphogenesis has been worth while: it has afforded us a result of the very first importance.
The Autonomy of Morphogenesis Proved
No kind of causality based upon the constellations of single physical and chemical acts can account for organic individual development; this development is not to be explained by any hypothesis about configuration of physical and chemical agents. Therefore there must be something else which is to be regarded as the sufficient reason of individual form-production. We now have got the answer to our question, what our constant E consists in. It is not the resulting action of a constellation. It is not only a short expression for a more complicated state of affairs, it expresses a true element of nature. Life, at least morphogenesis, is not a specialised arrangement of inorganic events; biology, therefore, is not applied physics and chemistry: life is something apart, and biology is an independent science.
All our results at present, indeed, are negative in their form; our evidence was throughout what is called per exclusionem, or indirect or apagogic. There were excluded from a certain number of possibilities all except one; a disjunctive proposition was stated in the form: E is either this, or that, or the other, and it was shown that it could not be any of all these except one, therefore it was proved to be that one. Indeed, I do not see how natural science could argue otherwise; no science dealing with inorganic phenomena does; something new and elemental must always be introduced whenever what is known of other elemental facts is proved to be unable to explain the facts in a new field of investigation.
We shall not hesitate to call by its proper name what we believe we have proved about morphogenetic phenomena. What we have proved to be true has always been called vitalism, and so it may be called in our days again. But if you think a new and less ambitious term to be better for it, let us style it the doctrine of the autonomy of life, as proved at least in the field of morphogenesis. I know very well that the word “autonomy” usually means the faculty of giving laws to oneself, and that in this sense it is applied with regard to a community of men; but in our phrase autonomy is to signify the being subjected to laws peculiar to the phenomena in question. This meaning is etymologically defensible, and besides that I perhaps may remind you of a certain chapter of Professor Ward’s Gifford Lectures, in which he holds the view that, psychologically and epistemologically, there is more than a mere verbal relation between the civil and the natural “law.”
Vitalism then, or the autonomy of life, has been proved by us indirectly, and cannot be proved otherwise so long as we follow the lines of ordinary scientific reasoning. There can indeed be a sort of direct proof of vitalism, but now is not the time to develop this proof, for it is not of the purely scientific character, not so naïve as our present arguments are, if you choose to say so. An important part of our lectures next summer will be devoted to this direct proof.
“Entelechy”
But shall we not give a name to our vitalistic or autonomous factor E, concerned in morphogenesis? Indeed we will, and it was not without design that we chose the letter E to represent it provisionally. The great father of systematic philosophy, Aristotle, as many of you will know, is also to be regarded as the founder of theoretical biology. Moreover, he is the first vitalist in history, for his theoretical biology is throughout vitalism; and a very conscious vitalism indeed, for it grew up in permanent opposition to the dogmatic mechanism maintained by the school of Democritus.
Let us then borrow our terminology from Aristotle, and let that factor in life phenomena which we have shown to be a factor of true autonomy be called Entelechy, though without identifying our doctrine with what Aristotle meant by the word έντελέχεια. We shall use this word only as a sign of our admiration for his great genius; his word is to be a mould which we have filled and shall fill with new contents. The etymology of the word ἐντελέχεια allows us such liberties, for indeed we have shown that there is at work a something in life phenomena “which bears the end in itself,” ὃ ἔχει ἐν ἑαυτᾣ τὸ τέλος.
Our concept of entelechy marks the end of our analysis of individual morphogenesis. Morphogenesis, we have learned, is “epigenesis” not only in the descriptive but also in the theoretical sense: manifoldness in space is produced where no manifoldness was, real “evolutio” is limited to rather insignificant topics. But was there nothing “manifold” previous to morphogenesis? Nothing certainly of an extensive character, but there was something else: there was entelechy, and thus we may provisionally call entelechy an “intensive manifoldness.” That then is our result: not evolutio, but epigenesis—“epigenesis vitalistica.”
Some General Remarks on Vitalism
We now shall leave entelechy where it stands: next summer we shall turn back to it and shall make its full logical and ontological analysis our chief study. At present we are satisfied with having proved its existence in nature, with having laid some of the foundations of a doctrine to be based upon it. I hope that these foundations will evince themselves strong: that is all-important.65 It indeed has been the fault of all vitalism in the past that it rested on weak foundations. Therefore the discussion of the basis underlying our doctrine of the autonomy of life is to occupy us still a considerable time. We shall devote to it two more of this year’s lectures and three of the next; we shall examine all sorts of phenomena of life in order to find out if there are any further proofs of vitalism, independent perhaps, of what we way call our first proof, which is based upon the analysis of the differentiation of harmonious-equipotential systems. We shall find some more independent proofs; and besides that we shall find many kinds of phenomena upon which future times perhaps may erect more of such independent proofs.
For we shall be chary of bestowing the name “proof” except on what is a proof indeed, of course according to our critical conviction. Vitalistic views in biology have arisen in rather numerous forms during the last fifteen years, especially in Germany—though in very strong contrast to the so-called official German biology—but I can only admit that one of all the arguments of “neo-vitalism” has proved its statements. I refer to the theory of “morphaesthesia” as developed by Noll, which we shall study briefly in the next lecture. I cannot concede that Reinke or Schneider or Pauly have really proved what they believe, and I cannot even allow to the most original thinker in this field, Gustav Wolff, that he has given a real demonstration of his views. He states that the existence of so-called “primary purposefulness,” that is, the existence of adaptive processes, which cannot be imagined to have arisen on Darwinian principles, is able to prove vitalism; but I say that it only proves teleology, which is a broader concept than vitalism.
The possibility of a machine at the root of the phenomena in question always has to be excluded in order that vitalism may be proved, and I cannot grant that the necessity of such an exclusion has been actually shown by any of my fellow-combatants against so-called mechanism, except Noll.66
The Logic of our First Proof of Vitalism
Let us devote the end of our present lecture to an account of the logical means by which it has been possible to develop what we hope will be regarded as a true proof of life autonomy.
Firstly, we have looked upon the phenomena of morphogenesis without any prepossessions; we may say that we have fully surrendered ourselves to them; we have not attacked them with any sort of dogmatism except the inherent dogmatism of all reasoning. But this dogmatism, if it may be called so, does not postulate that the results of the inorganic doctrines must hold for the organic world, but only that both the inorganic and the organic must be subject to certain most general principles.
By studying life as a given phenomenon, by fully devoting ourselves to our problem, we not only have analysed into its last elements what was given to us as our subject, but we also, more actively, have created new combinations out of those elements: and it was from the discussion of these positive constructions that our argument for vitalism was derived.
We have analysed morphogenesis into elementary processes, means, potency, formative stimulus, just as the physicist analyses mechanics into time, velocity, mass, and force; we have then rearranged our elements into “systems”—the equipotential systems, the harmonious-equipotential system in particular, just as the physicist composes his elements into the concepts of momentum or of kinetic energy or of work. And finally, we have discussed our compositions and have obtained our result, just as the physicist gets his ultimate results by discussing work and kinetic energy and momentum.
Of course the comparison is by no means intended to show that mechanics and biology are sciences of the same kind. In my opinion, they are not so at all; but nevertheless there do exist similarities of a logical kind between them.
And it is not the formal, logical character alone which allows us to compare biology with other natural sciences: there is still something more, there is one kind of assumption or postulate, or whatever you may choose to call it, without which all science whatever would be altogether impossible. I refer to the concept of universality. All concepts about nature which are gained by positive construction out of elements resulting from analysis, claim to be of universal validity; without that claim there could indeed be no science.
Of course this is no place for a lecture on methodology, and it therefore must suffice to make one remark with special regard to our purpose, which we should like to emphasise. Our concept of the harmonious-equipotential system—say rather, our concept of the prospective potency itself—presumes the understanding that indeed all blastomeres and all stems of Tubularia, including those upon which we have not carried out our experiments, will behave like those we have experimented with; and those concepts also presume that a certain germ of Echinus, A, the blastomeres of which were not separated, would have given two whole larvae, if separation had taken place, while another germ, B, which actually gave us two larvae after separation, would only have given one without it. Without this presumption the concept of “potency” is meaningless, and, indeed, every assumption of a “faculty” or a “possibility” would be meaningless in the whole area of science.
But this presumption can never be proved; it can only be postulated. It therefore is only with this postulate that our first proof of vitalism holds; but this restriction applies to every law of nature.
I cannot force you to agree with this postulate: but if you decline you are practically saying that there exists a sort of pre-established harmony between the scientific object and the scientist, the scientist always getting into his hands such objects only as have been predestinated from the very beginning to develop two larvae instead of one, and so on.
Of course, if that is so, no proof of natural laws is possible at all; but nature under such views would seem to be really dæmonic.
And so, I hope, you will grant me the postulate of the universality of scientific concepts—the only “hypothesis” which we need for our argument.
Our next studies on the physiology of form will be devoted in the first place to some additional remarks about our harmonious-equipotential systems themselves, and about some other kinds of morphogenetic “systems” which show a certain sort of relationship with them. For it is of the greatest importance that we should become as familiar as possible with all those facts in the physiology of form upon the analysis of which are to be based almost all of the future theories that we shall have to develop in biology proper and philosophical. Our discussions, so far as they relate to questions of actual fact, will contain only one other topic of the same importance.
But though it is designed to complete and to deepen our analysis, the present considerations may yet be said to mark a point of rest in the whole of our discussions: we have followed one single line of argumentation from the beginning until now; this line or this stream of thought, as you might call it, is now to break into different branches for a while, as if it had entered from a rocky defile into a plain. It seems to me that such a short rest will be not unconducive to a right understanding of all we have made out; and such a full and real conceiving again, such a realising of our problems of morphogenesis and their solutions, will be the best preparation for the philosophical part of these lectures.
HARMONIOUS-EQUIPOTENTIAL SYSTEMS FORMED BY WANDERING CELLS
All of the harmonious-equipotential systems which we have studied so far were the bases of histological differentiation; that is to say, the processes of their differentiation consisted in specifically localised elements of theirs becoming different in situ. Now we know at least one type of systems which also may be called harmonious-equipotential, but the differentiation of which does not simply relate to elements at a fixed place. An additional phenomenon enters here into the sphere of the others. The elements not only become different where they are, but a specific changing of locality, a specific kind of wandering, goes hand-in-hand with differences relating to the prospective value to be attained. I am speaking of the formation of the larval skeleton of our well-known Echinus. We know that the mesenchyme cells, which have left the blastoderm and are arranged in a sort of ring of bilateral structure, are the starting-point of this skeleton: it indeed originates in a sort of secretive process on the part of the cells; the cells are moving about and are secreting carbonate of lime during their wandering. The experiments now have shown, as we know, that a whole, though smaller, skeleton may also be formed, if only a half or a quarter of the mesenchyme cells are present, as happens to be the case in all experiments with isolated blastomeres of the two or four-cell stage of cleavage. It is clear that in these cases the performance of each single cell must be different from what it is in the normal case, and that the same sort of differences in the morphogenetic performances appears again, if the two- and the four-cell stage are compared with each other. And there are still some other phenomena showing the possibility of different performances being carried out by the individual cells. Peter has shown that the number of mesenchyme cells may vary enormously under certain conditions; but, in spite of that, the skeleton always will be complete. It may be said that this line of research is only of a relative value to our own questions, as, of course, variability relates to different individuals: but it seems to me that it adds a very good supplementary instance to what the experiment on the individual itself has established.
We should only be repeating ourselves if we were to analyse again what happens here as the expression of the harmonious-equipotentiality itself. But indeed there occurs something new in this instance: the single mesenchyme cell not only has to perform in each case that single act of specific secretion which the case requires, but it also has to wander to the right place in order to perform it; there must be some order, not only about the acts of secretion after wandering, but also in the migrations themselves. If undisturbed ontogeny alone were possible, and if therefore a theory like that of Weismann were in place, we might say perhaps that each mesenchyme-cell is specified not only as to its performance in secretion, but also with regard to its chemotactical irritability, the latter being typically localised, so that its effect becomes typical, thanks to the typical arrangement of all the cells with respect to each other. But that is certainly not the case. Now, you may ask yourselves if you could imagine any sort of a machine, which consists of many parts, but not even of an absolutely fixed number, all of which are equal in their faculties, but all of which in each single case, in spite of their potential equality, not only produce together a certain typical totality, but also arrange themselves typically in order to produce this totality. We are indeed familiar with certain occurrences in nature where such curious facts are observed, but I doubt if you would speak of “machines” in these cases. The mesenchyme-cells, in fact, behave just as a number of workmen would do who are to construct, say, a bridge. All of them can do every single act, all of them also can assume every single position: the result always is to be a perfect bridge; and it is to be a perfect bridge even if some of the workmen become sick or are killed by an accident. The “prospective values” of the single workman change in such a case.
I well know that it is only an analogy which I am offering to you. The mesenchyme-cells have not “learned,” have no “experience.” All that is to occupy us next summer. But in spite of it, there is truth in the analogy; and perhaps you will prefer it to the merely abstract consideration.
ON CERTAIN COMBINED TYPES OF MORPHOGENETIC SYSTEMS
For the sake of completeness it may be remarked, only by the way, that the type of the proper harmonious-equipotential system may go hand in hand with another type of “systems” which play a part in morphogenesis; a type which we have shortly mentioned already and which will be studied fully a few chapters later. We know that there are equipotential systems with complex potencies: that is to say, systems which may produce a whole organism equally well from any one of their elements; we know the cambium of Phanerogams to be such a system. Now it is easily understood that the germ of our Echinus, say in the stage of two or four or eight cleavage cells, is not only an harmonious-equipotential system, but a complex-equipotential system too. Not only may there arise a whole organism out of 2/4 or 3/4; or 3/8, 4/8, 5/8, 6/8, 7/8 of its elements, in which cases the harmonious rôle of the single element with regard to its single performance in a totality is variable, but there may also arise four whole single larvae out of the four cells of the four-cell stage, or eight single whole larvae out of the eight-cell stage.67 In these cases, of course, each of the four or eight elements has performed not a part of the totality, changing with its “position,” but the totality itself. With respect to these possible performances the “systems” present in the four or eight-cell stages of cleavage must be called complex-equipotential ones.
We propose to give the name of mixed-equipotential systems to all those equipotential systems which, at the same time, may be regarded as belonging to the harmonious or to the complex type. It is not only among cleavage-stages that they are to be found; you may also find them very clearly exhibited in our ascidian Clavellina for instance. We know already that the branchial apparatus of this form is typically harmonious-equipotential, but it is complex-equipotential too, for it also may regenerate what is wanting in the proper way, by a budding from the wound; and the same is true of many other cases, the flatworm Planaria for instance.
Another type of systems, which might be said to be of a higher degree, is exhibited in some very strange phenomena of regeneration. It was first shown most clearly by some experiments of Godlewski’s that a whole tail may be regenerated from a wound inflicted on the body of a newt, even if this wound involves section of only a portion of the body-diameter. Section of the whole of the body-diameter of course would cause the formation of the whole tail also; but it was found that even an incomplete cross-section of the body is capable of performing the whole on a smaller scale. The series of possible cross-sections which are all capable of regeneration would have to be called a system of the complex type in this case; but, now we learn that every single cross-section is of the harmonious type, we must speak of complex-harmonious systems. What we have described is not the only instance of our new type of morphogenetic systems. Some other instances had been discovered a few years earlier, though nobody had pointed out their true significance. In the flatworm Planaria a partial cross-section is also capable of forming a whole structure, say a head, and all cases of so-called “super-regeneration” after the infliction of a complicated wound probably belong here also.
You may say that our two additions to the theory of systems are merely formal, and indeed I am prepared to concede that we shall not learn anything altogether new from their discussion: their analysis would lead either to what was our “first proof” of the autonomy of life-phenomena or to what will be our “second” one. But the mere descriptions of the facts discovered here will interest you, I think, and will fill your minds with more vivid pictures of the various aspects of form-autonomy.
While dealing with our harmonious-equipotential systems as the starting-points of processes of restitution, e.g. in Tubularia, Clavellina, the flatworms, and other instances, we always have regarded cross-sections of the body as constituting the elements of equipotentiality. Now cross-sections, of course, are by no means simple in themselves, but are made up of very different tissues, which are derivates of all three of the original germ layers—ectoderm, mesoderm, and endoderm. Owing to this composite character of the cross-sections, taken as elements of harmonious systems, a special phenomenon of morphogenesis is presented to us, which teaches somewhat more than the mere concept of harmonious-equipotentiality can express. If composite elements concerned in morphogenesis result in one whole organisation in spite of the development of the single tissues of these elements going on independently, then there must be a sort of correspondence or reciprocity of the harmonious development among these tissue constituents themselves; otherwise a proportionate form could not be the final result. We may conveniently speak of a reciprocity of harmony as existing between the single tissues or germ layers which constitute many harmonious-equipotential systems, and there can be little doubt that we have here an important feature with regard to general morphogenesis.68
A few other groups of morphogenetic facts may find their proper place here, though they are not properly to be regarded as additions to the theory of harmonious systems but as forming a sort of appendix to it.
THE “MORPHAESTHESIA” OF NOLL69
We may briefly mention that group of botanical phenomena, by which the botanist Noll has been led to the concept of what he calls “morphaesthesia,” or the “feeling” for form; a concept, the full discussion of which would lead to almost the same conclusions as our analysis of the harmonious systems has done. In the Siphoneae, a well-known order of marine algae with a very complicated organisation as to their exterior form, the protoplasm which contains the nuclei is in a constant state of circulation round the whole body, the latter not being divided by proper cell-walls. On account of this constant movement it is certainly impossible to refer morphogenetic localisation to definite performances of the nuclei. Nor can any sort of structure in the outer protoplasmic layer, which is fixed, be responsible for it, for there is no such structure there: hence there must be a sort of feeling on the part of the plant for its relative body localities, and on account of this feeling morphogenesis occurs. This “feeling” is styled “morphaesthesia” by Noll, and to it he tries to refer all sorts of different botanical form-phenomena,70 for instance what is called “autotropism,” that is, the fact that branches of plants always try to reassume their proper angle with regard to their orientation on the main axis, if this orientation has been disturbed. It may be an open question if this particular application of the theory is right: certainly there seems to be much truth in the establishment of the concept of morphaesthesia, and we only have to object to its psychological name. But that may be done in a more general form on a later occasion.
RESTITUTIONS OF THE SECOND ORDER
In the hydroid polyp Tubularia, already familiar to us as being a most typical representative of the harmonious-equipotential systems, a very interesting phenomenon has been discovered71, almost unparalleled at present but nevertheless of a general importance, a phenomenon that we may call a restitution of a restitution, or a restitution of the second order. You know that the first appearance of the new head of Tubularia, after an operation, consists in the formation of two rings of red lines, inside the stem, these rings being the primordia of the new tentacles. I removed the terminal ring by a second operation soon after it had arisen, disturbing in this way the process of restitution itself: and then the process of restitution itself became regulated. The organism indeed changed its course of morphogenesis, which was serving the purposes of a restitution, in order to attain its purpose in spite of the new disturbance which had occurred. For instance, it sometimes formed two rings out of the one that was left to it, or it behaved in a different way. As this difference of morphogenetic procedure is a problem by itself, to be discussed farther on, we shall postpone a fuller description of this case of a restitution of the second degree.
At present I do not see any way of proving independently the autonomy of life by a discussion of these phenomena; their analysis, I think, would again lead us to our problem of localisation and to nothing else; at least in such an exact form of reasoning as we demand.
ON THE “EQUIFINALITY” OF RESTITUTIONS72
I have told you already that Tubularia in the phenomena of the regulation of restitutions offers us a second problem of a great general importance, the problem of the Equifinality of Restitutions. There indeed may occur restitutions, starting from one and the same initial state and leading to one and the same end, but using very different means, following very different ways in the different individuals of one and the same species, taken from the same locality, or even colony.
Imagine that you have a piece of paper before you and wish to sketch a landscape. After drawing for some time you notice that you have miscalculated the scale with regard to the size of the paper, and that it will not be possible to bring upon the paper the whole of the landscape you want. What then can you do? You either may finish what you have begun to draw, and may afterwards carefully join a new piece of paper to the original one and use that for the rest of the drawing; or you may rub out all you have drawn and begin drawing to a new scale; or lastly, instead of continuing as you began, or erasing altogether, you may compromise as best you can by drawing here, and erasing there, and so you may complete the sketch by changing a little, according to your fancy, the proportions as they exist in nature.
This is precisely analogous to the behaviour of our Tubularia. Tubularia also may behave in three different ways, if, as I described to you, the terminal one of its two newly arisen rings of tentacle primordia is removed again. It may complete what is left, say the basal tentacle ring, then put forth from the horny skeleton (the “perisarc”) the new head as far as it is ready, and finally complete this head by a regular process of budding regeneration. But it also may behave differently. It may “erase” by a process of retro-differentiation all that has been left of what had already been formed, and then may form de novo the totality of the primordia of a new head. Or, lastly, it may remove a part of the middle of the one ring of tentacle rudiments which was left, and may use this one ring for the formation of two, which, of course, will not be quite in the normal relations of place with regard to each other and to the whole, but will be regulated afterwards by processes of growth. Thus, indeed, there is a sort of equifinality of restitution: one starting-point, one end, but three different means and ways.
It would, of course, contradict the principle of univocality, as we shall see more fully later on, to assume that there actually are different ways of regulation whilst all the conditions and stimuli are the same. We are obliged to assume, on the contrary, that this is not the case, that there are certain differences in the constellation, say of the general conditions of age or of metabolism, which are responsible for any given individual choosing one process of restitution instead of another; but even then the phenomenon of equifinality remains very striking.
It has long been known that restitution in general does not always follow the same lines of morphogenesis as are taken by ontogeny, and it was this feature that once led Roux to point out that the adult forms of organisms seem to be more constant than their modes of origin. But, comparing ontogeny with restitution in general, we see that only the ends are the same, not the points of starting; the latter are normal or non-typical in ontogeny, atypical in restitution. In the new discoveries of an equifinality of restitutions we have the same starting-point, which is decidedly non-typical but atypical, i.e. dependent on our arbitrary choice, leading by different ways always to the same end.
There may be many who will regard the fact of equifinality as a proof of vitalism. I should not like to argue in this easy way; I indeed prefer to include part of the phenomena of equifinality in our first proof of autonomy, and part in the second one, which is to follow.
Another important phenomenon of the equifinality of regulation was discovered by Morgan. A species of the flatworm Planaria was found to restore its totality out of small pieces either by regeneration proper, if the pieces were fed, or by a sort of rearrangement of material, on the basis of its harmonious-equipotentiality, if they were kept fasting. It is important to note that here we see one of the conditions determining the choice of the way to restoration, as we also do in the well-known equifinal restitutions of the root in plants, where the behaviour of the organism depends on the distance of the operation-wound from the tip.73 In Tubularia the actual stage of restitution that has been already reached by the stem when the second operation takes place, may account for the specification of its future organogenesis, but this is not at all clearly ascertained at present.
Clavellina also shows equifinality in its restitution, as has already been shortly mentioned. The isolated branchial apparatus may restitute itself by retro-differentiation to an indifferent stage followed by renovation; or it may regenerate the intestine-sac in the proper way. Nothing is known here about the conditions, except perhaps that young individuals seem more apt to follow the first of these two ways, older ones the second; but there are exceptions to this rule.
The discussion of other instances of equifinality, though important in themselves, would not disclose anything fundamentally new, and so we may close the subject with the remark that nothing can show better than the fact of the equifinality of restitutions how absolutely inadequate all our scientific conceptions are when confronted with the actual phenomena of life itself. By analysis we have found differences of potencies, according as they are simple or complex; by analysis we have found differences of “systems,” differences of means, and indeed we were glad to be able to formulate these differences as strictly as possible: but now we see how, in defiance of our discriminations, one and the same species of animals behaves now like one sort of our “systems,” and now like the other; how it uses now one sort of “potencies,” now another.
But even if it is granted that, in the presence of such phenomena of life, our endeavour seems to be like a child’s play on the shores of the ocean, I do not see any other way for us to go, so long, at least, as our goal is human science—that is, a study of facts as demanded by our mental organisation.
REMARKS ON “RETRO-DIFFERENTIATION”
We shall finish this part of our studies by mentioning a little more explicitly one fundamental fact which has already entered incidentally into our considerations, viz. retro- or back-differentiation.74 We know that it occurs in Clavellina and in Tubularia; we may add that it also happens in Hydra, and that in the flatworm Planaria the pharynx, if it is too large for a piece that is cut out, may be differentiated back and be replaced by a new pharynx, which is smaller.
It is not death and sloughing of parts that occurs in these cases,75 but a real process of active morphogenesis; not, however, a process consisting in the production of visible manifoldness, but the opposite. Loeb was the first to lay much stress upon this topic, and indeed, there may appear a very strange problem in its wake: the problem, whether all morphogenesis might be capable perhaps of going backwards under certain conditions.
It is important to note that in most76 cases retro-differentiation occurs in the service of restitution: it goes on wherever restitution requires it. This fact alone would show that not very much could be explained here by the discovery of modern chemistry, important as it is, that one and the same “ferment” or “enzyme” may affect both the composition and the decomposition of the same compound. We could regard what is called “catalysis” solely as an agent in the service of entelechy. But this point also will become clearer in another part of the work.
We have finished our long account of individual morphogenesis proper. If we look back upon the way we have traversed, and upon those topics in particular which have yielded us the most important general results, the material for the higher analysis which is to follow, it must strike us, I think, that all these results relate to regulations. In fact, it is “secondary” form-regulations, according to our terminology, that we have been studying under the names of equifinality, back-differentiation, restitution of the second order, and so on, and our harmonious-equipotential systems have figured most largely in processes of secondary form-regulations also. But even where that has not been the case, as in the analysis of the potencies of the germ in development proper, form-regulations of the other type have been our subject, regulations of the primary or immanent kind, the connection of normal morphogenetic events being regulatory in itself. It was not the phenomenon of organic regulation as such that afforded us the possibility of establishing our proof of the autonomy of morphogenesis: that possibility was afforded us by the analysis of the distribution of potencies; but upon this distribution regulation is based, and thus we may be said to have studied some types of regulation more or less indirectly when analysing potencies.
It therefore seems to me that we shall have hopes of a successful issue to our inquiries, if we now, on passing to what is called the physiology of the vegetative functions, proceed to focus our attention on the concept of regulation as such. And that is what we shall do: on our way through the whole field of physiology, we shall always stop at any occurrence that has any sort of regulatory aspect, and shall always ask ourselves what this feature has to teach us.
But let us first try to give a proper definition of our concept. We shall understand by “regulation” any occurrence or group of occurrences on a living organism which takes place after any disturbance of its organisation or normal functional state, and which leads to a reappearance of this organisation or this state, or at least to a certain approach thereto. Organisation is disturbed by any actual removal of parts; the functional state may be altered by any change among the parts of the organism on the one hand, by any change of the conditions of the medium on the other; for physiological functioning is in permanent interaction with the medium. It is a consequence of what we have said that any removal of parts also changes the functional state of the organism, but nevertheless organisation is more than a mere sum of reactions in functional life. All regulations of disturbances of organisation may be called restitutions, while to regulations of functional disturbances we shall apply the name adaptations. It is with adaptations that we have to deal in the following.
Let us begin our studies of adaptations in a field which may justly be called a connecting link between morphogenesis and physiology proper, not yet wholly separated from the science of the organic form, morphology.
Morphological adaptation is a well-established fact, and I need only mention the striking differences between the land and water form of amphibious plants, or the differences between the same species of plants in the Alps and in the plains, or the very different aspect of the arms of an athlete and of an ascetic, to recall to your memory what is meant by this term.
Morphological adaptation is no part of individual morphogenesis proper, but occurs at the end of it; at least it never occurs previous to the full individual life of an organism, previous to its true functional life; for it relates to the functions of the complete organism.
THE LIMITS OF THE CONCEPT OF ADAPTATION
It is especially, though by no means exclusively, among plants that morphological adaptation assumes its most marked forms; and this topic, indeed, may very easily be understood if we remember that plant-life is in the very closest permanent dependence on the medium, and that this medium is liable to many changes and variations of all kinds. In order to elucidate our problem, it therefore seems convenient to restrict our considerations for a while to the study of plants. There exist very many external formative stimuli in the morphogenesis of vegetation: would it then be possible to regard every effect of such an external formative stimulus as a real morphological adaptation? No; for that would not meet the point. The general harmony of form is indeed concerned if gravity forces roots to shoot forth below at a spot where they can enter the ground, or if light induces branches and leaves to originate at places where they can obtain it for assimilation; but gravity and light themselves are mere formative stimuli—of the localising type—in these instances, for they relate only to the individual production of form, not to the functioning of already existing form. We therefore are warned not to confuse the effects of formative stimuli from without with real adaptive effects until we have fully analysed the particular case.
We have drawn a sharp line between causes and means of morphogenesis, applying the term “means” to those conditions of the morphogenetic process which relate neither to the specificity nor to the localisation of its constituents, though they are necessary for the accomplishment of the process in the most thorough manner. Would it be possible to connect our new concept of an adaptation with our well-established concept of a means of morphogenesis in such a way that we might speak of a morphological “adaptation” whenever any specific feature about morphogenesis proves to be immediately dependent for its success on some specific means, though it does not owe its localisation to that means as its “cause”? It seems to me that such a view would also fall wide of the mark. It is well known, for instance, that the flowers of many plants never fully develop in the dark; light is necessary for their morphogenesis. Is, therefore, their growth in the presence of light to be called a morphological “adaptation” to light? Certainly not: they simply cannot originate without light, because they require it for some reason. It is precisely here that our conception of light as a “means” of morphogenesis is most fully justified. There are many77 such cases; and there are still others of an apparently different type, but proving the same. All pathological forms produced in plants by animal parasites or by parasitic fungi could hardly be called adaptations, but must be attributed to some abnormality of means or of stimuli. It may be that the organism reacts as well as possible in these cases, and that if it reacted otherwise it would die—we know absolutely nothing about this question. But even then there would only be some sort of regulation in the process of pathological morphogenesis, but the process itself could hardly be called adaptive.
So far we have only learned what is not to be regarded as morphological adaptation. No response to external formative stimuli is in itself an example of adaptation, nor are processes dependent for their existence on any kind of condition or means to be called, simply because they are dependent on them, adaptations to those agents. What then, after all, is a morphological adaptation?
Let us remember what the word adaptation is really to mean in our discussions: a state of functioning is adapted—a state of functioning must therefore have been disturbed; but as functioning itself, at least in plants, certainly stands in close relations to the medium, it follows that all adaptations are in the last resort connected with those factors of the medium which affect functioning. In being correctives to the disturbances of functioning they become correctives to the disturbing factors themselves.
But again, the question seems to arise whether these factors of the medium, when they provoke an adaptation by some change that is followed by functional disturbance, do so in the capacity of “causes” or of “means,” and so it might seem that we have not gained very much so far by our analysis. The reproach, however, would not be quite justified, it seems to me: we indeed have gained a new sort of analytical concept, in the realm of causal concepts in general, by clearly stating the point that adaptations are related directly to functionality, and only indirectly, through functionality, to external changes. By the aid of this logical formulation we now are entitled to apply the term “cause,” in our restricted sense of the word, to every change of the medium which is followed by any sort of adaptation in regard to itself. Our definition stated that a “cause” is any one of the sum of necessary factors from without that accounts either for the localisation or for the specification of the effect, and the definition holds very well in this case. Indeed, the specification of the effect is determined by the outside factor in every case of an adaptation to it, by the mere fact of its being a specific adaptation to this specific factor.
We must not forget that in this chapter we are not studying real individual morphogenesis as the realisation of what has been inherited, but that at present we regard morphogenesis proper as an accomplished fact. Morphogenesis proper has laid the general lines of organisation; and now adaptation during the functional life, so to speak, imposes a second kind of organisation upon the first. It is for that reason that the meaning of the word “cause” is now becoming a little different from what it was before.
In order to study a little more in detail what has been discovered about morphological adaptation in animals and plants, let us separate our materials into two groups, one of them embracing adaptations with regard to functional changes from without, the other adaptations to those functional changes which come from the very nature of functioning. Almost all of our previous general considerations have applied to the former group, with which we shall now proceed to deal.
ADAPTATIONS TO FUNCTIONAL CHANGES FROM WITHOUT78
The differences between plants grown in very dry air, very moist air, and water, respectively, are most distinctly seen in all the tissues that assist in what is called transpiration, that is, the exchange of water-vapour between the plant and the medium, but especially in the epidermis and the conductive fibres, both of which are much stronger in plants grown in the dry. Indeed, it seems from experiments that transpiration is the most essential factor to which “adaptation” occurs in amphibious plants, though the changes of the mechanical conditions according to the medium also seem to have some sort of structural effect. If plants stand very deeply in water, the conditions of illumination, so important for assimilation in plants, may have been altered, and therefore much of the structural change can be attributed also to them. It is unimportant in our general question what is due to one of these factors and what to the other. That there is a real sort of adaptation cannot be doubtful; and the same is true, as experimental observations of the last few years have shown, with regard to the structural differences between so-called sun-leaves and shade-leaves of plants grown in the air: it has been actually shown here that the functional life of the former goes on better in the sun, of the latter better in the shade.
It is very important to emphasise this point, as the adaptive character of all sorts of structural differences in plants dependent on light and on moisture has lately been denied, on the supposition that there is only a stopping of organogenesis in the case of the more simple, a continuance in the case of the more complicated modification, but nothing else. Indeed, all morphological adaptation has been conceived as only consisting in differences dependent upon the absence or the presence of necessary means or causes of development, and as offering no problem of its own. We have gained the right position from which to oppose this argument, it seems to me, in our formula that all adaptations do relate not directly to the agents of the medium, but to changes of functional states induced by those agents; that adaptations only are “adaptations” by being correctives to the functional state.
There simply is an “adaptation” of structure in such a sense in all the cases we have mentioned. We can say neither more nor less. Granted that one of the outside factors which comes into account is merely a necessary “means”: then why is the histological consequence of the presence of the means an actual adaptation to it as far as its relation to functioning is concerned—why is the consequence of its absence also an adaptation to this absence in its relation to functioning? Why, to complete the series, is the degree of the consequence of its presence an adaptation to the degree of its presence?
All these relationships, which are so many facts, have been absolutely overlooked by those who have been pleased to deny morphological adaptation to functional changes from without.
To do full justice to them we may speak of “primary” regulative adaptations in all the cases mentioned above, applying the word “primary,” just as was done with regard to restitutions, to the fact that there is some sort of regulation in the normal connection of processes. We reserve the title of “secondary adaptations” for cases such as those described, for instance, by Vöchting,79 where not merely one and the same tissue originates adaptively with regard to the degree of its normal functioning, but where a profound disturbance of all functioning connections, due to the removal of portions of the organisation, is followed by histological changes at absolutely abnormal localities; that is, where a real change of the kind of functioning is the consequence of the adaptation. It, of course, will be found very difficult to discriminate such phenomena from real restitutions, though logically there exists a very sharp line between them.
A few more concrete instances may now close this account of adaptation to functional changes coming from without. Though almost all the adaptive characters in the aquatic forms of amphibious plants represent a less complicated state of organisation than the corresponding structures in their terrestrial forms, and therefore have wrongly been regarded as simply due to a stopping of morphogenesis for want of necessary means, yet there are a few of them that are positive complications in comparison with the land-forms: the so-called aërenchyme, especially well developed in the water-form of Jussiaea is such an instance. This tissue stands in the direct service of respiration, which is more difficult to be accomplished under water than ordinarily, and represents a true adaptation to the altered function.
Among animals there is only one well-studied instance of our first type of adaptive morphological characters. Salamandra atra, the black salamander, a species which only inhabits regions at least two thousand feet above sea-level, does not bring forth its young until metamorphosis has taken place. The larvae, however, may be removed from the mother’s body at an earlier stage and forced to complete their development in water. Under these circumstances, as was shown in an excellent memoir by Kammerer,80 they will change the whole histological type of their gills and skin in order to meet the new functional conditions. The change of the conditions of functioning is very severe here, for whereas the gills had served for nutrition and respiration in the uterus—by a process of endosmosis—they now serve for respiration only, and, of course, are surrounded by quite an abnormal chemical medium.
TRUE FUNCTIONAL ADAPTATION81
But all other cases of morphological adaptation among animals, and several in the vegetable kingdom too, belong to our second group of these phenomena, which in our analytical discussion we have called adaptations to functional changes that result from the very nature of functioning, and which we shall now call by their ordinary name, “functional adaptation.”
It was Roux who first saw the importance of this kind of organic regulation and thought it well to give it a distinguishing name. By functioning the organisation of organic tissues becomes better adapted for functioning. These words describe better than any others what happens. It is well known that the muscles get stronger and stronger the more they are used, and that the same holds for glands, for connective tissue, etc. But in these cases only quantitative changes come into account. We meet with functional adaptations of a much more complicated and important kind, when for instance, as shown by Babák,82 the intestine of tadpoles changes enormously in length and thickness according as they receive animal or vegetable food, being nearly twice as long in the second case. Besides this the so-called mechanical adaptations are of the greatest interest.
It has long been known, especially from the discoveries of Schwendener, Julius Wolff, and Roux, that all tissues whose function it is to resist mechanical pressure or mechanical tension possess a minute histological structure specially suitable to their requirements. This is most markedly exhibited in the stem of plants, in the tail of the dolphin, in the arrangements of the lime lamellae in all bones of vertebrates. All these structures, indeed, are such as an engineer would have made them who knew the sort of mechanical conditions they would be called upon to encounter. Of course all these sorts of mechanically adapted structures are far from being “mechanically explained,” as the verbal expression might perhaps be taken to indicate, and as indeed has sometimes been the opinion of uncritical authors. The structures exist for mechanics, not by it. And, on the other hand, all these structures, which we have called mechanically “adapted” ones, are far from being mechanical “adaptations,” in our meaning of the word, simply because they are “adapted.” Many of them indeed exist previous to any functioning, they are for the most part truly inherited, if for once we may make use of that ambiguous word.
But, the merely descriptive facts of mechanical adaptedness having been ascertained, there have now been discovered real mechanical processes of adaptations also. They occur among the statical tissues of plants, though not in that very high degree which sometimes has been assumed to exist; they also occur in a very high perfection in the connective tissue, in the muscles and in the bone tissue of vertebrates. Here indeed it has proved possible to change the specific structure of the tissue by changing the mechanical conditions which were to be withstood, and it is in cases of healing of broken bones that these phenomena have acquired a very great importance, both theoretically and practically: the new joints also, which may arise by force of circumstances, correspond mechanically to their newly created mechanical function.
So far a short review of the facts of “functionelle Anpassung.” They seem to prove that there does exist a morphological adaptation to functional changes which result from the very nature of functioning. In fact, the actual state of all functioning tissue, the intensity of its state of existence, if you care to say so, may be said to be due to the functioning itself: the so-called atrophy by inactivity being only one extreme of a very long line of correspondences.83
We now, of course, have to ask ourselves if any more intimate analysis of these facts is possible, and indeed we easily discover that here also, as in the first of our groups of morphological adaptations, there are always single definite agents of the medium, which might be called “causes” or “means” of the adaptive effects, the word “medium” being taken as embracing everything that is external to the reacting cells. But of course also here the demonstration of single formative agents does not detract in the least from the adaptive character of the reaction itself. So we may say, perhaps, that localised pressure is the formative stimulus for the secretion of skeleton substance at a particular point of the bone tissue, or of the fibres of the connective tissue; the merely quantitative adaptations of muscles might even allow of a still more simple explanation.84 But adaptations remain adaptations in spite of that; even if they only deserve the name of “primary” regulations.
THEORETICAL CONCLUSIONS
We have stated in the analytical introduction to this chapter and elsewhere, that functional changes, which lead to morphological adaptations of both of our groups, may arise not only from changes of factors in the medium, but also from a removal of parts. As such removal is generally followed by restitution also, it is clear that restitutions and adaptations very often may go hand in hand, as is most strikingly shown in a fine series of experiments carried out by Vöchting, which we have already alluded to. Here again I should like to lay the greatest stress upon the fact that, in spite of such actual connections, restitutions and adaptations always have been separated from another theoretically, and that the forms are never to be resolved into sums of the latter. Such a view has been advocated by some recent authors, especially by Klebs, Holmes, and Child:85 it is refuted I think by the simple fact that the first phase of every process of restitution, be it regeneration proper or be it a sort of harmonious differentiation, goes on without functioning at all, and only for future functioning.86
And there has been advocated still another view in order to amplify the sphere of adaptation: all individual morphogenesis, not only restitution, is adaptation, it has been said. In its strictest form such an opinion of course would simply be nonsense: even specific adaptive structures, such as those of bones, we have seen to originate in ontogeny previous to all specific functions, though for the help of them, to say nothing of the processes of the mere outlining of organisation during cleavage and gastrulation. But they are “inherited” adaptations, it has been answered to such objections. To this remark we shall reply in another chapter. It is enough to state at present that there is a certain kind of, so to speak, architectonic morphogenesis, both typical and restitutive, previous to specific functioning altogether.
If now we try to resume the most general results from the whole field of morphological adaptations, with the special purpose of obtaining new material for our further philosophical analysis, we have reluctantly to confess that, at present at least, it does not seem possible to gather any new real proof of life-autonomy, of “vitalism,” from these facts, though of course also no proof against it.
We have stated that there is in every case of both our types of adaptive events a correspondence between the degree of the factor to which adaptation occurs, and the degree of the adaptive effect. We here may speak of an answering between cause and effect with regard to adaptation, and so perhaps it may seem as if the concept of an “answering reaction” (“Antwortsreaktion”), which was introduced into science by Goltz87 and which is to play a great part in our discussions of next summer, may come into account: but in our present cases “answering” only exists between a simple cause and a simple effect and relates almost only to quantity and locality. There is therefore lacking the most important feature, which, as will be seen, would have made the new concept of value.
We only, I believe, can state the fact that there are relations between morphogenetic causes and effects which are adaptations, that functional disturbances or changes are followed by single histogenetic reactions from the organism, which are compensations of its disturbed or changed functional state. We are speaking of facts here, of very strange ones indeed. But I feel unable to formulate a real proof against all sorts of mechanism out of these facts: there might be a machine, to which all is due in a pre-established way. Of course we should hardly regard such a machine as very probable, after we have seen that it cannot exist in other fields of morphogenesis. But we are searching for a new and independent proof; and that is indeed not to be found here.88
At present it must be taken as one of the fundamental facts of the organogenetic harmony, that the cells of functioning tissues do possess the faculty of reacting to factors which have changed the state of functioning, in a way which normalises this state histologically. And it is a fact also that even cells, which are not yet functioning but are in the so-called embryonic or indifferent condition contributing to the physiological completion of the tissue, react to factors embracing new functional conditions of the whole in a manner which leads to an adaptation of that whole to those conditions.
This is a very important point in almost all morphological adaptation, whether corresponding to functional changes from without or resulting from the very nature of functioning. In fact, such cells as have already finished their histogenesis are, as a rule, only capable of changing their size adaptively, but are not able to divide into daughter-cells or to change their histological qualities fundamentally; in technical terms, they can only assist “hypertrophy” but not “hyperplasia.” Any adaptive change of a tissue therefore, that implies an increase in the number of cellular elements or a real process of histogenesis, has to start from “indifferent” cells, that is to say, cells that are not yet functioning in the form that is typical of the tissue in question; and, strange to say, these “embryonic” cells—i.e. the “cambium” in higher plants and many kinds of cells in animals—can do what the functional state requires. It is to be hoped that future investigations will lay a greater stress upon this very important feature of all adaptation.
It is but a step from morphological adaptations to adaptations in physiology proper. The only difference between regulations of the first type and those which occur in mere functioning is, that the resulting products of the regulation are of definite shape and therefore distinctly visible in the first case, while they are not distinctly visible as formed materials but are merely marked by changes in chemical or physical composition in the latter.
Metabolism, it must never be forgotten, is the general scheme within which all the processes of life in a given living organism go on; but metabolism means nothing else, at least if we use the word in its descriptive and unpretentious meaning, than change in the physical or chemical characteristics of the single constituents of that organism. In saying this, we affirm nothing about the physical or chemical nature of the actual processes leading to those physical or chemical characteristics, and by no means are these “processes” a priori regarded as being physical or chemical themselves: indeed, we have learned that in one large field, in the differentiation of our harmonious systems they certainly are not. Now, if the metabolism does not end in any change of visible form, then true physiological processes, or more particularly physiological regulations, are going on before us. But we are dealing with morphogenetic events or regulations, if the result of metabolism is marked by any change in the constituents of form. This however may depend on rather secondary differences as to the nature of regulation itself, and any kind of metabolism may really be of the regulatory type, whether we actually see its result as a constituent of form, e.g. owing to the production of some insoluble compound, or whether we do not.
I do not mean to say that these are the only differences between mere physiological activities or regulations and organogenesis proper, as an originating of typical form-combination; but if we regard, as we do in this chapter, the given organisation of a living being as a substratum of its functional life, morphological and physiological adaptations are indeed of almost the same logical order.
We had best therefore begin our discussions with a recapitulation of our problem. We are studying adaptations in functioning—that means we want to know how the organism behaves with regard to any change which may take place in its functional state. We apply the term regulation, or in particular adaptation, to any kind of reaction on the part of the organism which re-establishes the normal state of functioning, and we now want to learn to what degree such adaptations exist in the field of physiology.
SPECIFIC ADAPTEDNESS NOT “ADAPTATION”
It is important to keep well in mind our strictly formulated theme, as by doing so we shall be able to exclude at once from our materials a large group of phenomena which occasionally have been called regulations by physiological authors, but which, in fact, are not of the adaptation type and therefore cannot be said to afford those problems which possibly might have been expected. Typical peculiarities in functional life cannot be called “regulations” for this very reason. If, for instance, the organism selects specific amounts of specific kinds of organic food or of salts out of the combinations of salts or organic food normally offered to it in the medium, as indeed is most typically shown for instance by the roots of plants, there cannot be said to occur a “regulation” or “adaptation” with regard to the permeability of the cell, nor is it strictly a case of “regulation,” if so-called selective qualities are discovered in the processes of secretion, say of the epithelium of the kidney.
All these facts are typical and specific peculiarities in functioning which are duly to be expected, where a very typical and specific organisation of the most elaborated kind exists. Indeed, after studying such an organisation we must not be astonished that functions in organisms follow lines which certainly they would not have taken without it. Take the fact which is quoted very often, that the migration of compounds or of ions in the organisms can happen quite contrary to all the laws of osmosis, from the less concentrated to the more concentrated side of a so-called “membrane.” There is no simple “membrane” in the organism, but a complicated organisation of an almost unknown character takes its place, and nothing, indeed, is against the assumption that this organisation may include factors which actually drive ions or compounds to the side of higher concentration, which indeed drive them by “doing work,” if we like to speak in terms of energy; and these factors included in organisation may very well be of a true physical or chemical nature.90
I lay great stress upon these statements, as I should like to be as careful as possible in the admission of anything like a “proof” of vitalism. It was want of scientific criticism and rigid logic that discredited the old vitalism; we must render our work as difficult as possible to ourselves, we must hold the so-called “machine theory” of life as long as possible, we must hold it until we are really forced to give it up.
In a more general form we now can sum up our discussion by saying: There never are adaptations in physiology, requiring any special analysis, where there are only complications or even apparent deviations from the purely physico-chemical type of events which are, so to say, statical, i.e. fixed in quantity or quality, however peculiar or typically complicated they may be; all such peculiarities indeed, may properly be called “adapted,” that is to say, very well fitted to perform a specific part in the service of normal general functioning, and they are “adapted” to their part by virtue of a certain “adaptedness” of the organisation; but they are not “adaptations” in any sense of the word.
PRIMARY AND SECONDARY ADAPTATIONS IN PHYSIOLOGY
We approach the subject of true adaptations, that is, of adapting processes, as soon as any kind of variation in functioning occurs which corresponds to a variation of any factor of the medium in the widest sense. But even here our work is by no means done by simply showing such a correspondence of outer and inner variations. We know very well already, from our former studies, that now we are faced by a further problem, that we are faced by the question whether we have to deal with simple primary kinds of adaptations or with the far more important secondary ones.
As the discrimination between primary and secondary regulations proves indeed to be of first-rate importance, you will allow me, I hope, to summarise our chief analytical statements regarding them in a most general form. We call primary regulatory any kind of morphogenetic or functional performance, which, by its very intimate nature, always serves to keep the whole of organisation or of functions in its normal state. We call secondary regulations all features in the whole of morphogenesis or of functioning which serve to re-establish the normal state after disturbances along lines which are outside the realm of so-called normality. This analytical discrimination will help us very much to a proper understanding of physiology. But before we turn to apply our definitions to actual facts, another preliminary problem has to be solved.
ON CERTAIN PRE-REQUISITES OF ADAPTATIONS IN GENERAL
We are thinking of the general and important question, what types of adaptations may be expected in the field of physiology and whether there may be certain classes of regulatory events which possibly might be expected to occur in the organism on a priori grounds, but which, nevertheless, are to be regarded as impossible after a more intimate analysis of its nature, even at the very beginning? Or, in other words, to what kinds of changes of the medium will an organism be found able or unable to adapt itself?
We know that the state of functioning must be altered in order to call forth any sort of adaptation at all. Now, there can be no doubt that a priori it would seem to be very useful for the organism, if it never would let enter into its blood, lymph, etc., be it through the skin or through the intestine, any chemical compound that would prove to be a poison afterwards. In fact, a man, judging on the principle of the general usefulness of all the phenomena of the living, might suppose that there would exist a sort of adaptation against all poisons to the extent that they would never be allowed to enter the real interior of the body. We know that such reasoning would be incorrect. But we also can understand, I suppose, that an a priori analysis of a more careful kind would have reasoned differently. How could the functional state of the organism be changed, and how, therefore, could adaptation be called forth by any factor of the medium which had not yet entered the organism, but was only about to enter it? Not at all therefore is such a regulation to be expected as we have sketched; if there is to be any adaptation to poisons, it only can occur after the poison has really acted in some way, and in this case we shall indeed find regulations.
You may perhaps regard this discussion as a little too academical and hair-splitting, but here again it was for the sake of ensuring a perfectly sound foundation of our chief principles that I undertook it. Very often, indeed, the question has been raised by the defenders of a mechanistic theory of life, Why then did the organisms not reject all poisons from the very beginning? We now may reply to that only—how could they do so? How could they “know” what is a poison and what is not, unless they had experienced it?—if we are allowed for a moment to use very anthropomorphistic language.
We repeat, therefore, that the functional conditions of the organism must have been actually changed in order that an adaptation may occur. Nothing is more essential to a clear understanding of our problems than to keep fully in mind the exact sense of this definition.
ON CERTAIN GROUPS OF PRIMARY PHYSIOLOGICAL ADAPTATIONS
General Remarks on Irritability.—Turning now to more special groups of problems concerning physiological adaptations, let us begin with the primary class of them, and let us first say a few words on a subject which occasionally has been regarded as the basis of physiological regulation in general. I refer to a most important fact in the general physiology of irritability. Irritability of any kind is known to be re-established, after it has been disturbed by the process of reacting to the stimulus, and in certain cases, in which two different—or rather two opposite—kinds of reactions are possible on the same substratum, which increase with regard to one process whilst decreasing at the same time with regard to the other. The irritability of the muscle or of the leaves of Mimosa is a very good instance of the first case, whilst the second more complicated one cannot be illustrated better than by what all experience has taught us about the irritability of the retina. The retina is more irritable by green rays and less by red ones the more it has been stimulated by the latter, and more sensitive to light in general the more it has been exposed to darkness; and something very similar is true, for instance, as regards phototactic irritability in plants, all these phenomena being in relation to the so-called law of Weber.91
It seems to me that there would be little difficulty in harmonising the phenomenon of the inversion of irritability with the so-called principle of the “action of masses” and with the laws of certain “reversible” processes well known in chemistry. As to the simple fact of the re-establishment of irritability after stimulation has occurred, or, in certain other cases, the fact that in spite of permanent stimulation irritability seems to exist permanently also, physical analogies or even explanations might very well be found.92
If now we ask whether there is anything like an adaptation appearing in the general characteristics of irritation and irritability, it seems to me that we may answer the question in an affirmative manner, as far as primary regulation comes into account. We, certainly, have not studied any abnormal regulatory lines of general functioning, we only have studied general functioning itself; but, indeed, there was a certain sort of regulation in functioning. Of course, by showing that one of the most general features of all functioning is primary-regulatory in itself, we do not deny the possibility of many specific functions in which real secondary regulations actually do exist. Nothing indeed is asserted about the specific character of functioning in its different types, by proving that one of the general features of all functioning may comparatively easily be understood. It seems to me that this important logical point has not always received the attention it deserved.
The Regulation of Heat Production.93—Having finished our introductory remarks we now turn to the proper study of special physiological functioning with regard to its adaptive side, and begin with the most simple cases.
The so-called “regulation of heat” in warm-blooded vertebrates is an instance of a special function which can be said to be regulatory in itself. There exists a normal blood heat for each species, which is maintained no matter whether the temperature of the medium rise or fall. It might seem at first as if in this case there were a little more of an adaptive regulation than only its well-known primary type; no reversion, one might say, of the direction of one and the same process occurs in the regulation of heat production, but one kind of process is called into action if it is necessary to raise the temperature, and another whenever it is necessary to lower it. Even in the dilatation and constriction of capillary vessels there are different nerves serving for each operation respectively, and far more important are the increasing of transpiration for cooling, the increasing of combustion for heating—two radically different processes. But, nevertheless, there is a certain unity in these processes, in so far as a specific locality of the brain has been proved to be the “centre” of them all; it is to this centre of course that the analysis of heat production considered as a kind of regulation or adaptation must be directed. Such an ultimate analysis, it seems to me, would have to classify heat regulation under the primary type of adaptations in physiology without any restriction. The centre acts in one sense or in the other, if stimulated by any temperature beyond a very limited range, and it is in the action of the centre that the “regulation” of heat consists.94
Primary Regulations in the Transport of Materials and Certain Phenomena of Osmotic Pressure.—Very similar phenomena of regulation are present in many processes concerned in the whole of metabolism. Let us consider for a moment the migration of materials in plants. Whenever any compound is used at a certain place, a permanent afflux of this compound to that place sets in from all possible directions. No doubt this is a “regulation,” but it is also the function itself, and besides that, a very simple function based almost entirely on well-known laws of physical chemistry. And in other cases, as in the ascent of water to the highest tops of our trees, which purely physical forces are said to be insufficient to explain, we can appeal to the unknown organisation of many cells, and there is nothing to prevent our attributing to these cells certain functions which are, if you like to say so, regulatory in themselves. Among other facts of so-called regulations there is the stopping of metabolic processes by an accumulation of their products: as, for instance, the transformation of starch into sugar is stopped, if the sugar is not carried away. Of course that is a regulation, but it again is an intrinsic one, and it is one of the characteristics of reversible chemical processes to be stopped in that way. I know very well that in this particular case a certain complication is added by the fact that it is a so-called ferment, the diastase, which promotes the transformation of starch into cane-sugar, and that this ferment is actively produced by the organism: but even its production would not prove that any real kind of secondary regulation exists here, if nothing more were known about such an active production than this single case.
In a special series of experiments almost all carried out in Wilhelm Pfeffer’s botanical laboratory at Leipzig, an attempt has been made to discover in what manner the cells of plants are able to withstand very high abnormalities of the osmotic pressure of the medium—that is to say, very great changes in the amount of its salinity. That many, particularly the lower plants, are able to stand such changes had been ascertained already by the careful examinations of Eschenhagen; but recent years have given us a more profound insight into what happens. Von Mayenburg95 has found that sundry of the species of Aspergillus, the common mould, are able to live in very highly concentrated solutions of several salts (KNO3 and Na2SO4). They were found to regulate their osmotic pressure not by taking in the salts themselves, but by raising the osmotic pressure of their own cell sap, producing a certain amount of osmotically active substances, probably carbohydrates. If in this case it were possible to assume that the osmotic pressure of the medium were the real stimulus for the production of the osmotic substances in the cell, stimulus and production both corresponding in their degree, we should be entitled to speak of a primary though physiological96 regulation only; and it seems to me that despite the discoveries of Nathansohn that certain algae and cells of higher plants are able to change the permeability of their surfaces in a way which regulates the distribution of single salts or ions in the sap of their cells without any regard to pure osmotic equilibrium, such a simple explanation might be possible.97
There are many regulation phenomena connected with osmotic pressure and permeability in animal physiology also, though at present they are not worked out as fully as possible. The works of Frédéricq, J. Loeb, Overton and Sumner98 would have to be taken into account by any one who wished to enter more deeply into these problems. We can only mention here that permeability to water itself also plays its part, and that, according to Overton’s experiments, it is a kind of solubility of the media in the very substance of the cell surface on which all permeability and its regulation depend.
Chromatic Regulations in Algae.—The phenomena of osmotic pressure and its regulation may be said to be the preliminaries of metabolism proper, conditions necessary for it to take place. Now there is another branch of such preliminaries to metabolism, in which the most interesting regulation phenomena have been lately discovered. It is well known that what is called assimilation in plants, that is, the formation of organic compounds out of carbon dioxide (CO2) and water, occurs only in the light by means of certain pigments. This pigment is in all higher plants and in many others the green chlorophyll, but it may be different in certain species of algae, and can generally be said99 to be of the colour complementary to the colour of those rays which especially are to be absorbed and to be used for assimilation. But here we have “adaptedness,” not adaptation. It was in some species of primitive algae, the Oscillariae, that Gaidukow100 found a very interesting instance of an active regulation in the formation of pigments. These algae always assume a colour which corresponds to the accidental colour of the rays of the medium and is complementary to it; they become green in red light, yellow in blue light, and so on—that is, they always actively take that sort of colouring which is the most suitable to the actual case.101 There indeed occurs a sort of complementary photography in these algae; but, though adaptive, it could hardly be said to exceed the limits of “primary phenomena.”
Metabolic Regulations.—And now we enter the field of regulations in metabolism itself. There are two kinds of outside factors of fundamental importance for all metabolic processes: food is one, and oxygen is the other. And metabolism as a whole is of two different aspects also: it both serves for assimilation proper—that is, building up—and it supplies the energy for driving the functional machine. It is clear that food alone—together of course with the assimilating means of the organism, can account for the first type of metabolism, while both food and oxygen, or some sort of substitute for the latter, as in certain bacteria, supply functional energy. Of course we are not entitled to say that the importance of so-called oxidation or respiration is exhausted by its energetic rôle: it certainly is not, for if it were, the organism would only be stopped in its functions if deprived of oxygen but would not die. It seems that certain substances always arise in the metabolism, in the processes of decomposition, which have to be burnt up in order not to become poisonous. But we shall return to the phenomena of organic oxidation in another chapter of the book, and shall deal with them from a more general point of view.102
Let us now try to take a short survey of all the regulations discovered relating to the substitution of one kind of food for another. We have said that food serves in the first place as building material, in the second place as fuel. It only deserves brief mention that, as all recent investigations have shown, fats, carbohydrates, and albumen are equally well able to serve as fuel.103
It is in the state of fasting, i.e. in the case of a real absence of all nourishing materials, that the organism has proved to be capable of regulations of the most marked nature, with regard to the combustion of its own materials. Respiration, we know, must go on if death is to be avoided, and now indeed it has been found that this process attacks the different tissues of the organism subjected to fasting in such an order that, after the combustion of the reserves, the most unimportant tissues with regard to life in general are destroyed first, the most important ones last. Thus in vertebrates the nerve cells and the heart are preserved as long as possible; in infusoria it is the nucleus; in flatworms, as the very careful studies of E. Schultz104 have lately shown, it is the nerve cells and the sexual cells which longest resist destruction, whilst almost all the rest of the organisation of these animals may disappear. I should not say that we can do very much with these facts at present in our theoretical discussion, but they are certainly witness of very astonishing adaptive powers.105
We now turn to study the cases of a compensation of nourishments serving for the real building up of the organism. Albumen, we know, is absolutely indispensable for animals, even for adults, though nothing is known about the purpose it serves in the latter; its place can be taken of course by those less complicated compounds which result from its first decomposition, effected by pepsin and trypsin, but nothing else will do. The salts of sea-water, according to Herbst’s experiments, may only vary to a very small degree if the development of marine animals is to go on well; potassium may be replaced by caesium or rubidium, and that is all. Much the same is true of the salts necessary to plants. It will not surprise us very much to hear that algae can also be successfully fed with the potassium salts of organic compounds, and higher plants with acid amides or glucoses instead of carbonic acid, as those products are normal steps in their assimilation; and it may also be fairly easily understood that nitrogen can be offered in organic form instead of as a nitrate.
It was in the group of fungi that really important adaptations with regard to the proper form-producing alimentation were first discovered, and these are of a very complicated kind indeed. Fungi are known to be satisfied with one single organic compound instead of the group of three—fat, carbohydrate and albumen—necessary for animals. Now Pfeffer showed that the most different and indeed very abnormal compounds were able to bring his subjects to a perfect growth and morphogenesis; and, moreover, he found that, if several kinds of such food were offered together, they were consumed quite indifferently as to their chemical constitution, but only with regard to their nutritive value: that sort of food which had produced a better growth than another when both were offered separately was found to save the latter from consumption whenever both were offered together.
Here we are faced by one of the most typical cases of regulations in metabolic physiology: the organism is able to decompose compounds of the most different constitution, which have never been offered to it before; but nevertheless, it must remain an open question whether real “secondary” regulation has occurred, as nothing is known in detail about the single steps of metabolism in these fungi. There might be some ferments equally able to destroy different classes of compounds,106 and that the most nutritive compound is used up first may be a question of physico-chemical equilibrium.
That is almost all107 that is actually known of adaptation with regard to the use of an abnormal food supply. Though important, it cannot be said to be very much. But could we expect very numerous regulations here at all after what we laid down in a former paragraph about the possibilities of adaptive regulation in general? The functional state must have been altered in order that such regulations may occur. Now there is no doubt that this state may be really altered only if an abnormal food has first been taken in altogether by the cell-protoplasm of the body-surfaces, but never if it has only entered the cavity of the intestine, which, strictly speaking, is a part of the exterior medium. Fungi indeed not only take in the abnormal food, but also know what to do with it, but all animals are obliged to treat first with their chemical secretions what happens to be present in their intestine, in order that it may be taken up by their living cells, and one hardly can wonder that these secretions are only formed in correspondence to a limited number of outside stimuli. In fact, as soon as we look upon what adaptive or regulatory work happens in metabolism inside the body interior, we meet, even in animals, regulations of a far more developed type.
Discoveries of the last few years have taught us that almost all metabolic processes in the organism, including oxidation, are carried out by the aid of special materials, the so-called enzymes or ferments. These are known to exist in the most different forms even in the inorganic world. They are simply chemical compounds, of specific types, that bring about chemical reactions between two other chemical materials, which in their absence would either not go on at all or would go on very slowly. We cannot enter here into the much disputed chemical theory of what is called “catalysis”: we can only say that there is no objection to our regarding almost all metabolic processes inside the organism as due to the intervention of ferments or catalytic materials, and that the only difference between inorganic and organic ferments is the very complicated character of the latter and the very high degree of their specification.
Such a statement, of course, does not say that all metabolism has proved to be of a chemical nature: the action of the ferment when produced is chemical, but we do not know at all how the ferment is produced; we only know that a high degree of active regulation is shown in this production. In fact, it has been proved in some cases, and probably will be proved in a great many more in the near future, that all metabolic ferments, whether they promote oxidation or assimilation proper or chemical decomposition, are produced in a regulatory manner with regard to the specific compound to be dissociated or to be built up. In this way the whole field of metabolism is really covered by “regulations.” Are they real “secondary” ones? Of course the regulatory correspondence applies to the process of secretion in the first place, not to the actual formation of the ferment inside the cell. The correspondence as to secretion, no doubt, is of the primary type; is there any secondary regulation with regard to the real production of the ferment? I am sorry that I cannot answer this question affirmatively. Nothing is known at present, even here, that really proves the existence of adaptation of the secondary type: there might be a sort of statical “harmony” at the base of it all, established before all functioning for functioning.108
The only facts of secondary metabolic regulations which are known at present have been found in combination with phenomena of restitution after real disturbances of organisation, where, indeed, numbers and numbers of regulatory changes of metabolism, both in animals and plants, have also been recorded. But there is not one case of a secondary regulation really known to affect pure metabolism alone.109 This is a new indicium of the primacy of form in the organism.
IMMUNITY THE ONLY TYPE OF A SECONDARY PHYSIOLOGICAL ADAPTATION
There is only one class of physiological processes in which the type of the real secondary regulation occurs. The discoveries of the last twenty years have proved beyond all doubt, and future discoveries will probably prove even more conclusively, that the so-called immunity against diseases is but one case out of numerous biological phenomena in which there is an adaptive correspondence between abnormal chemical stimuli and active chemical reactions on the part of the organism and in its interior, exceeding by far everything that was formerly supposed to be possible in organic regulation.
The adaptive faculty of the organism against inorganic poisonous substances110 is but small comparatively, and is almost always due not to a real process of active regulation but to the action of substances pre-existing in the organism—that is, to a sort of adaptiveness but not adaptation. Metallic poisons, for instance, may be transformed into harmless compounds by being combined with albumen or sulphuric acid and thus becoming insoluble, or free acids may be neutralised, and so on; but all these processes go on to a certain extent only, and, as was mentioned already, are almost always the result of reactions with pre-existing materials. Only in a few cases is there any sort of true adaptation to metallic substances, such as sublimate and, in a very small degree, arsenic, comparable in some respects with the adaptation to abnormally high temperatures. The organism which has been accustomed to receive at first very small amounts, say, of sublimate, and then receives greater and greater amounts of this substance by degrees, will at the end of this treatment be able to stand a quantity of the poison that would have been instantly fatal if administered at the first dose.111 But the explanation of this adaptation is not known in any case; there seems to be some similarity between it and the so-called histogenetic immunity against organic poisons.
It is in the fight against animal and vegetable poisons, such as those produced by bacteria, by some plants and by poisonous snakes, that the true adaptation of the organism reaches its most astonishing degree. The production of so-called “anti-bodies” in the body fluids is not the only means applied against noxious chemical substances of this kind: the existence of so-called histogenetic immunity is beyond all doubt, and Metschnikoff112 certainly was also right in stating that the cells of the organism themselves repel the attack of living bacteria. Cells of the connective tissue and the white blood cells, being attracted by them as well as by many other foreign bodies, take them in and kill them. This process, called “phagocytosis” is of special frequency among lower animals, but it also contributes to what is called inflammation in higher ones.113 And there are still other kinds of defence against parasites, as for instance the horny or calcareous membranes, employed to isolate trichinae and some kinds of bacteria. But all this is of almost secondary importance as compared with the adaptive faculties of the warm-blooded vertebrates, which produce anti-poisonous substances in their lymph and blood.
It is impossible to say here114 more than a few words about the phenomena and the theory of immunity proper, which have attained the dimensions of a separate science. Let me only mark those general points which are of the greatest theoretical interest. Discoveries of the most recent years have shown not only that against the “toxins” of bacteria, snakes, and some plants, the organism is able actively to produce so-called “anti-toxins”—that is, soluble substances which react with the toxins and destroy their poisonous character—whenever required, but that against any foreign body of the albumen group a specific reaction may occur, resulting in the coagulation of that body. But the destruction of the noxious substance or foreign albumen actually present is not all that is accomplished by the organism. “Acquired immunity” proper, that is, security against the noxious material for a more or less extensive period of the future, depends on something more. Not only is there produced as much of the so-called “anti-body” as is necessary to combine with the noxious, or at least foreign substances which are present, but more is produced than is necessary in the actual case. On this over-production depends all active immunity, whether natural or, as in some kinds of vaccination, artificial; and so-called “passive” immunity, obtained by the transfusion of the serum of an actively immune organism into another also depends upon this feature.115
This phenomenon in particular—the production of more of the antitoxin or the “precipitin” than is actually necessary—seems to render almost impossible any merely chemical theory of these facts. The reaction between toxin and antitoxin, albumen and precipitin is indeed chemical; it may in fact be carried out in a test-tube; but whether the production of the anti-body itself is also chemical or not could hardly be ascertained without a careful and unbiassed analysis. There can be no doubt that the well-known theory of Ehrlich,116 the so-called theory of side-chains (“Seitenkettentheorie”) has given a great impulse to the progress of science; but even this theory, irrespective of its admissibility in general, is not a real chemical one: the concept of a regeneration of its so-called haptophore groups is a strictly biological concept.117
And, indeed, here if anywhere we have the biological phenomenon of adaptation in its clearest form. There are very abnormal changes of the functional state of the organism, and the organism is able to compensate these changes in their minutest detail in almost any case. The problem of the specification of the reactions leading to immunity seems to me, as far as I can judge as an outsider, to stand at present in the very forefront of the science. There cannot be the slightest doubt that especially against all sorts of foreign albumens the reaction is as strictly specific as possible; but there are some typical cases of specificity in the production of antitoxins also. It is, of course, the fact of specific correspondence between stimulus and reaction, that gives to immunity its central position among all adaptations, no matter whether the old hypothesis of the production of specific anti-bodies proves tenable, or whether, as has been urged more recently by some authors, the anti-body is always the same but reacts differently according to the medium. In the latter case it would be the medium that is regulated in some way by the organism in order to attain a specific adaptedness.
NO GENERAL POSITIVE RESULT FROM THIS CHAPTER
But now let us look back to the sum of all the physiological reactions studied, and let us see if we have gained a new proof of the autonomy of life from our long chapter.
We freely admit we have not gained any really new proof, but we may claim, I think, to have gained many indicia for the statement that the organism is not of the type of a machine, in which every single regulation is to be regarded as properly prepared and outlined.
It is precisely in the field of immunity that such a machine-like preparation of the adaptive effects seems almost impossible to be imagined. How indeed could there be a machine, the chemical constituents of which were such as to correspond adaptively to almost every requirement?—to say nothing of the fact that the production of more of the protecting substance than is actually necessary could hardly be said to be “chemical.”
In fact, we are well entitled to say that we have reached here the very heart of life and of biology. If nevertheless we do not call the sum of our facts a real proof of vitalism, it is only because we feel unable to formulate the analysis of what happens in such a manner as to make a machine as the basis of all reactions absolutely unimaginable and unthinkable. There might be a true machine in the organism producing immunity with all its adaptations. We cannot disprove such a doctrine by demonstrating that it would lead to a real absurdity, as we did in our analysis of differentiation of form; there is only a very high degree of improbability in our present case. But an indirect proof must reduce to absurdity all the possibilities except one, in order to be a proof.
Mechanistic explanations in all branches of functional physiology proper, so much in vogue twenty years ago, can indeed be said to have failed all along the line: the only advantage they have brought to science is the clearer statement of problems to which we are now accustomed. But we are not fully entitled to say118 that there never will be any mechanistic explanation of physiological functions in the future. It may seem as improbable as anything can be; but we wish to know not what is improbable but what is not possible.
Now of course you might answer me that after we have indeed shown that the production of form, as occurring on the basis of harmonious-equipotential systems, is a fact that proves vitalism, the acts taking place on the basis of that form after its production would have been proved to be vitalistic also, or at least to be in some connection with vitalistic phenomena. Certainly they would, and I myself personally should not hesitate to say so. But that is not the question. We have to ask: Is any new proof, independent of every other, to be obtained from the facts of physiological adaptation in themselves? And there is really none. Mere regulatory correspondence between stimuli and reactions, even if it be of the adaptive type and occur in almost indefinite forms, never really disproves a machine as its basis so long as the stimuli and reactions are simple and uniform. Next summer, however, we shall see that vitalism may be proved by such a correspondence if the two corresponding factors are not simple and not uniform.
We most clearly see at this point what it really was in our analysis of differentiation that allowed us to extract a real proof of vitalism from it. Not the mere fact of regulability, but certain specific relations of space, of locality, lay at the very foundation of our proof. These relations, indeed, and only these relations, made it possible to reduce ad absurdum any possible existence of a machine as the actual basis of what we had studied. In our next chapter again it will be space-relations, though analysed in a different manner, that will enable us to add a second real proof of vitalism to our first one.
With this chapter we conclude the study of organic regulation in all its forms, as far as morphogenesis and metabolism are in question.
But our analysis of these regulations would be incomplete and indeed would be open to objections, if we did not devote at least a few words to two merely negative topics, which will be taken more fully into consideration later on.
A FEW REMARKS ON THE LIMITS OF REGULABILITY
There has never been found any sort of “experience” in regulations about morphogenesis or in adaptations of the proper physiological type. Nothing goes on “better” the second time than it did the first time;119 everything is either complete, whenever it occurs, or it does not occur at all.
That is the first of our important negative statements about regulations; the second relates to the phrase just used, “or it does not occur at all.” There are indeed limits of regulability; adaptations are not possible to every sort of change of the physiological state: sickness and death could not exist if they were; nor is restitution possible in all cases where it might be useful. It is a well-known fact, that man is only able to heal wounds but is altogether destitute of the faculty of regeneration proper. But even lower animals may be without this faculty, as are the ctenophores and the nematodes for instance, and there is no sort of correspondence between the faculty of restitution and the place in the animal kingdom. It is not altogether impossible that there may be found, some day, certain conditions under which every organism is capable of restoring any missing part; but at present we know absolutely nothing about such conditions.120
But no amount of negative instances can disprove an existing positive—which is what we have been studying. Our analysis based upon the existence of regulations is as little disparaged by cases where no regulability exists as optical studies are by the fact that they cannot be undertaken in absolute darkness.
All organisms are endowed with the faculty of re-creating their own initial form of existence.
In words similar to these Alexander Goette, it seems to me, has given the shortest and the best expression of the fact of inheritance. Indeed, if the initial form in all its essentials is re-created, it follows from the principle of univocality, that, ceteris paribus, it will behave again as it did when last it existed.
By the fact of inheritance life becomes a rhythmic phenomenon, that is to say, a phenomenon, or better, a chain of phenomena, whose single links reappear at constant intervals, if the outer conditions are not changed.
The Material Continuity in Inheritance
It was first stated by Gustav Jaeger and afterwards worked out into a regular theory by Weismann, that there is a continuity of material underlying inheritance. Taken in its literal meaning this statement is obviously self-evident, though none the less important on that account. For as all life is manifested on bodies, that is on matter, and as the development of all offspring starts from parts of the parent bodies, that is from the matter or material of the parents, it follows that in some sense there is a sort of continuity of material as long as there is life—at least in the forms we know of. The theory of the continuity of “germ-plasm” therefore would be true, even if germ-cells were produced by any and every part of the organism. That, as we know, is not actually the case: germ-cells, at least in the higher animals and in plants, are produced at certain specific localities of the organism only, and it is with regard to this fact that the so-called theory of the “continuity of germ-plasm” acquires its narrower and proper sense. There are distinct and specific lines of cell-lineage in ontogenesis, so the theory states, along which the continuity of germ-protoplasm is kept up, which, in other words, lead from one egg to the other, whilst almost all other lines of cell-lineage end in “somatic” cells, which are doomed to death. What has been stated here is a fact in many cases of descriptive embryology, though it can hardly be said to be more than that. We know already, from our analytical and experimental study of morphogenesis, that Weismann himself had to add a number of subsidiary hypotheses to his original theory to account for the mere facts of regeneration proper and the so-called vegetative reproduction in plants and in some animals, and we have learned that newly discovered facts necessitate still more appendixes to the original theory. In spite of that, I regard it as very important that the fact of the continuity of some material as one of the foundations of inheritance has clearly been stated, even if the specialised form of the theory, as advocated by Weismann in the doctrine of the “germ-lineages” (“Keimbahnen”) should prove unable to stand against the facts.
The important problem now presents itself: What is the material, the matter, which is handed down from generation to generation as the basis of inheritance? Weismann, as we know, regarded it as a very complicated structure, part of which by its disintegration became the foundation of individual embryology. We have disproved, on the authority of many facts, the latter part of this assumption; but of course the first part of it may turn out to be true in spite of this. We have no means at present to enable us to say a priori anything positive or negative about the important question of the nature of that matter, the continuity of which in inheritance is in some sense a self-evident fact, and we therefore shall postpone the answer until a later point of our analytical discussion.
On Certain Theories which Seek to Compare Inheritance to Memory
It will be advisable first to study some other theoretical views which have been put forward with regard to inheritance. The physiologist Hering, as early as 1876, compared all heredity to the well-known fact of memory, assuming, so to say, a sort of remembrance of all that has happened to the species in the continuity of its generations; and several German authors, especially Semon, have lately made this hypothesis the basis of more detailed speculation.
It is not clear, either from Hering’s paper121 or from Semon’s book,122 what is really to be understood here by the word “memory,” and, of course, there might be understood by it very different things, according to the author’s psychological point of view. If he is a “parallelist” with regard to so-called psychical phenomena, he would use the word memory only as a sort of collective term to signify a resultant effect of many single mechanical events, as far as the material world of his parallel system comes into account, with which of course the problem of inheritance alone deals; but if he maintains the theory of so-called psycho-physical interaction, the psychical would be to him a primary factor in nature, and so also would memory. As we have said, it is by no means clear in what sense the word “memory” is used by our authors, and therefore the most important point about the matter in question must remain in dubio.
But another topic is even more clear in the theory of inheritance, as stated in Hering’s and Semon’s writings. The hypothetical fact that so-called “acquired characters” are inherited is undoubtedly the chief assumption of that theory. Indeed, it would be difficult to understand the advantage of the ambiguous word memory, had it not to call attention to the hypothetic fact that the organism possesses the faculty of “remembering” what once has happened to it or what it once has “done,” so to speak, and profiting by this remembering in the next generation. The zoologist Pauly indeed has stated this view of the matter in very distinct and clear terms.
As we soon shall have another occasion to deal with the much-discussed problem of the “inheritance of acquired characters,” we at present need only say a few words about the “memory-theory” as a supposed “explanation” of heredity. Undoubtedly this theory postulates, either avowedly or by half-unconscious implication, that all the single processes in individual morphogenesis are the outcome either of adaptations of the morphological type, which happened to be necessary in some former generation, or of so-called contingent “variations,” of some sort or other, which also happened once in the ancestral line. Such a postulate, of course, is identical with what is generally called the theory of descent in any of its different forms. This theory is to occupy us in the next lectures; at present we only analyse the “memory-theory” as a theory of heredity in itself. In any case, to regard memory as the leading point in inheritance, at least if it is to signify what is called memory in any system of psychology, would be to postulate that either adaptation or contingent “variation” has been the origin of every morphogenetic process. Indeed, the American physiologist Jennings did not hesitate to defend such a view most strongly, and many others seem to be inclined to do the same.
But such an assumption most certainly cannot be true.
It cannot be true, because there are many phenomena in morphogenesis, notably all the phenomena akin to restitution of form, which occur in absolute perfection even the very first time they happen. These processes, for the simple reason of their primary perfection, cannot be due either to “learning” from a single adaptation, or to accidental variation. We shall afterwards employ a similar kind of argument to refute certain theories of evolution. It therefore may be of a certain logical interest to notice that at present, combating the memory-theory of inheritance, and hereafter, combating certain theories of descent, we select not “adaptation” or “variation” as the central points to be refuted, but the assumed contingency of both of them.
The word “memory,” therefore, may be applied to the phenomena of inheritance only in a very figurative meaning, if at all. We do not wholly deny the possibility of an inheritance of acquired characters, as will be seen later on, and to such a fact there might perhaps be applied such a term as “memory” in its real sense, but we simply know that there is something in inheritance which has no similarity whatever to what is called “memory” in any species of psychology. A primary perfection of processes occurring quite abnormally proves that there is a “knowing” of something—if we may say so—but does not prove at all that there is a “remembering.”
The Complex-Equipotential System and its Rôle in Inheritance123
But we thus far have reached only negative results. Is the question necessarily to remain at this point, which could hardly be said to be very satisfying; or could we perhaps get better, that is, positive results about inheritance by a change of our analytic methods? Let us try to analyse the facts that occur in inheritance instead of beginning with hypotheses which claim to be complete explanations. Perhaps we shall gain, if but small, yet certainly fixed results by an analysis which goes from the facts to the theory and not from the theory to the facts.
Let the discussions that are to follow be placed upon a basis as broad as possible.
Our studies of morphogenetic restitution have shown us that besides the harmonious-equipotential systems another and widely different type of morphogenetic “systems” (i.e. unities consisting of elements equal in morphogenetic faculty) may also be the basis of restitution processes. Whilst in the harmonious system the morphogenetic acts performed by every single element in any actual case are single acts, the totality of all the single acts together forming the harmonious whole, in the other type of systems now to be examined, complex acts, that is, acts which consist of a manifoldness in space and in time, can be performed by each single element, and actually are performed by one or the other of them. We therefore have given the title of “complex-equipotential systems” to the systems in question, as all our denominations are based on the concept of the prospective morphogenetic potency, that is of the possible fate of the elements.
The cambium of the Phanerogams may be regarded as the very type of a complex-equipotential system, promoting restitution of form. It runs through the whole stem of our trees, in the form of a hollow tube, placed between the inner and the outer cell-layers of the stem, and either branch or root may originate from any single one of its cells, just as circumstances require. We might call the cambium a system of the “complex” type of course, even if every one of its constituents were able to form only a root or only a branch by way of restitution. But in fact one and the same element can form both of these complex-structures; it depends only on its relative position in the actual part of the stem isolated for the purposes of experiment, what will be accomplished in every case. Here we have a state of affairs, which we shall encounter again when studying regeneration in animals: every element of the system may be said to contain potencies for the “ideal whole,” though this ideal whole will never be realised in its proper wholeness.124
But there is no need to recur to the “ideal whole” in many other cases of adventitious restitution in plants. On isolated leaves of the well-known begonia, a whole plant, containing all the essential parts, may arise from any single cell125 of the epidermis, at least along the veins, and in some liverworts it has been shown by Vöchting, that almost every cell of the whole is able to reproduce the plant, as is also the case in many algae.
In the animal kingdom it is chiefly and almost solely the phenomena of regeneration proper which offer typical instances of our systems, since adventitious restitution, though occurring for instance in the restitution of the lens of vertebrates from the iris, and though connected also with the events in regeneration proper,126 is of but secondary importance in animal restitution, at least, if compared with restitution in plants. If we study the regeneration of a leg in the common newt, we find that it may take place from every section, the point of amputation being quite at our choice. Without regarding here the exact order of the regeneration phenomena, which is almost unknown at present, we in any case can say without any doubt that the line of consecutive possible cross-sections forms a complex-morphogenetic system, as every one of them is able to give rise to a complex organ, viz. the foot and part of the leg. It is an open question whether this complex system is to be called “equipotential” or not. It indeed seems to be inequipotential at the first glance, for each single section has to form a different organogenetic totality, namely, always that specific totality which had been cut off; but if we assume hypothetically that the real “Anlage” which is produced immediately by the cells of the wounded surface is the very same for all of them, and that it is the actual state of organisation which determines to what result this Anlage is to lead,127 we may say that the series of consecutive cross-sections of a newt’s leg does form a morphogenetic system of the complex-equipotential type, promoting secondary regulations of form.
Now all these difficulties vanish, if we consider the regeneration of animals, such for instance as many worms of the annelid class or our familiar ascidian Clavellina, in which regeneration in both directions is possible. The wound at the posterior end of the one half which results from the operation forms a posterior body half, the wound at the anterior end of the other half forms an anterior one. Again, it is the ideal whole which we meet here: each section of the body indeed may be said to contain the potencies for the production of the totality, though actually this totality is always realised by the addition of two partial organisations. The title of complex-equipotential systems thus seems to be fully justified as applied to the systems which are the basis of regeneration: each section of the regenerating body may in fact produce the same complex whole, or may, if we prefer to say so, at least prepare the ground for that complex Anlage, out of which the complex totality is actually to arise, in the same manner.
It often occurs in science, that in rather strange and abnormal conditions something becomes apparent which might have been found everywhere, which is lying before our eyes quite obviously. Are we not in just such a condition at present? In order to study the complex-equipotential systems, we turn to the phenomena of regeneration and of restitution in general; we occasionally even introduce hypotheses to render our materials more convenient for our purposes; and all the time there is one sort of complex-equipotential system in the body of every living being, which only needs to be mentioned in order to be understood as such, and which indeed requires no kind of preliminary discussion. The system of the propagation cells, in other words the sexual organ, is the clearest type of a complex-equipotential system which exists. Take the ovary of our sea-urchin for instance, and there you have a morphogenetic system every element of which is equally capable of performing the same complex morphogenetic course—the production of the whole individual.
Further on we shall deal exclusively with this variety of our systems, and in doing so we shall be brought back to our problem of heredity. But it had its uses to place our concept of the complex-equipotential system upon such a broad basis: we at once gave a large range of validity to all that is to follow—which, indeed, does not apply to inheritance alone, though its significance in a theory of heredity may be called its most important consequence.
The Second Proof of Life-Autonomy. Entelechy at the Bottom of Inheritance
After we had established the concept of the harmonious-equipotential system in a former chapter, we went on to study the phenomena of the differentiation of it, and in particular the problem of the localisation of all differentiations. Our new concept of the complex-equipotential system is to lead us to an analysis of a different kind: we shall pay special attention to the origin, to the genesis of our complex systems that show equipotentiality.
If we review the process of ontogenesis, we are able to trace back every complex system to a very small group of cells, and this small group of cells again to one single cell. So in plants the cambium may be shown to have originated in a sort of tissue-rudiment, established at a very early period, and the ovary may be demonstrated to be the outcome of a group of but a few cells, constituting the first visible “Anlage” of the reproductive organs. At the end then, or from another point of view at the beginning, a single cellular element represents the very primordial egg-cell.
The whole cambium, there can be no doubt, must be regarded as the result of a consecutive number of cell-divisions of the one cell from which it originates. So must it be with the ovary. The primordial egg-cell has undergone a long line of consecutive divisions; the single eggs are the last result of them.
We now proceed to some considerations which have a certain logical similarity to those which inaugurated our analysis of the differentiation of the harmonious-equipotential systems, though the facts in question are very different.
Viewed by itself without any kind of prepossessions, as it might be by any one who faces a new problem with the single postulate of introducing new natural entities—to use the scholastic phrase—as little as possible, the development of the single egg might be regarded as proceeding on the foundation of a very complicated sort of machine, exhibiting a different kind of construction in the three chief dimensions of space, as does also the organism which is to be its result.
But could such a theory—irrespective of all the experimental facts which contradict it—could such a theory stand before the one fact, that there occurs a genesis of that complex-equipotential system, of which our one single egg forms a part? Can you imagine a very complicated machine, differing in the three dimensions of space, to be divided hundreds and hundreds of times and in spite of that to remain always the same whole? You may reply that during the period of cell-divisions there is still no machine, that the machine is established only after all the divisions are complete. Good; but what then constructs this machine in the definitive cells of our systems, say in the eggs? Another sort of machine perhaps? That could hardly be said to be of much use. Or that entelechy of which we have spoken? Then you would recur to our first proof of vitalism and would burden entelechy with a specific performance, that is with the construction of the hypothetic machine which you are postulating in every single egg. But of course you would break the bounds of physics and chemistry even then.
It seems to me that it is more simple, and so to say more natural, not to recur to our first proof of life-autonomy in order to keep to the “machine theory” in this new branch of inquiry, but to consider facts as they offer themselves to analysis.
But then indeed we are entitled to draw an independent second proof of the autonomy of life from our analysis of the genesis of systems of the complex-equipotential type. We say it is a mere absurdity to assume that a complicated machine, typically different in the three dimensions of space, could be divided many many times, and in spite of that always be the whole: therefore there cannot exist any sort of machine as the starting-point and basis of development.
Let us again apply the name entelechy to that which lies at the very beginning of all individual morphogenesis.
Entelechy thus proves to be also that which may be said to lie at the very root of inheritance,128 or at least of the outcome of inheritance; the individual formation of the next generation is shown not to be performed by a machine but by a natural agent per se.
The Significance of the Material Continuity in Inheritance
But what about the material continuity appearing in inheritance, which we have said to be almost self-evident, as life is only known to exist on material bodies? Is there not, in fact, a serious contradiction in admitting at the same time entelechy on the one side and a sort of material condition on the other as the basis of all that leads to and from inheritance? Next summer the relation between matter and our autonomous agent of life will be studied more fully; at present it must be enough to state in a more simple and realistic way, what we hold this relation to be. There is no contradiction at all in stating that material continuity is the basis of inheritance on the one side, and entelechy on the other. It would be very inconvenient for us if there were any: for the material continuity is a mere fact and our entelechy we hope we have proved to exist also; if now there were any sort of contradiction in assuming the existence of both of them, of course it would be fatal to our proof.
Let us try to comprehend what is meant by the statement that entelechy and something material are at work in inheritance at the same time. Entelechy has ruled the individual morphogenesis of the generation which is regarded as being the starting-point for inheritance, and will rule also the morphogenesis of the generation which is to follow; entelechy determines the egg to be what it is, and the morphogenesis starting from this egg to be what it is also. Entelechy, at present, is not much more for us than a mere word, to signify the autonomous, the irreducible of all that happens in morphogenesis with respect to order, in the one generation and in the next. But may not the material continuity which exists in inheritance account perhaps for the material elements which are to be ordered? In such a way, indeed, I hope we shall be able to reconcile entelechy and the material basis of heredity. May it not be that there exist some “means” for morphogenesis, which are handed down from generation to generation, always controlled by entelechy, and which constitute the real significance of the continuity of matter during inheritance?
The Experimental Facts about Inheritance
Discoveries of the last few years do seem to show that such means of a material character, though not the foundation of that order of processes which is inherited, are nevertheless among the most necessary conditions for the accomplishment of inheritance in general. It is scarcely necessary to remind you that for very many years all concrete research on heredity proper—that is, the actual comparison of the various specific characters in the generations of the grandfather, the father, and the child—was due to Galton. You may also be aware that in spite of Galton’s inestimable services it was not till 1900 that one of the active principles concerned in inheritance was found independently by de Vries, Correns, and Tschermak, and that this principle happened to be one that had been discovered already, stated with the utmost clearness and precision by the Augustinian monk, Gregor Mendel,129 as early as 1865, though it had been completely forgotten ever since.
The so-called “rule of Mendel” is based upon experiments with hybrids, that is, with the offspring of parents belonging to different species, or, at least, varieties, but it relates not to the characters of the generation resulting immediately from hybridisation, the “first” generation of hybrids, as we shall call it, but to the characters of that generation which is the result of crossing the hybrids with each other, provided that this leads to any offspring at all. There are many cases indeed, both amongst animals and plants, where the offspring of the hybrids, or in other terms the “second” generation, is found to consist of individuals of three different types—the mixed130 type of the hybrids themselves, and the two pure types of the grandparents. Whenever the individuals of the “second” generation are separated into these three different types, hybrids are said to “split.” It is the fact of this splitting on the one hand, and on the other hand a certain statement about the numbers of individuals in the three different types of the “second” generation, that gives its real importance to Mendel’s rule.
Before discussing what may follow from Mendel’s discovery for the theory of heredity, we must lay stress on the fact that there are many exceptions to his rule. In quite a number of cases the hybrids are of one or more types, which remain constant: there is no splitting at all in the second generation. But that does not affect the rule of Mendel in those cases where it is true. Where there is a “splitting” in the second generation, there also are the numerical proportions stated by Mendel; there never are other relations among the numbers of individuals of the mixed and of the two pure types than those given by his rule. I regard it as very important that this real meaning of Mendel’s principle should be most clearly understood.
From the fact of the splitting of hybrids in the second generation most important consequences may be drawn for the theory of inheritance; the split individuals, if crossed with each other, always give an offspring which remains pure; there is no further splitting and no other change whatever. The germ-cells produced by the split individuals of the second generation may therefore be said to be “pure,” as pure as were those of the grandparents. But that is as much as to say that the pureness of the germ-cells has been preserved in spite of their passing through the “impure” generation of the hybrids, and from this fact it follows again that the union of characters in the hybrids must have been such as to permit pure separation: in fact, the germ-cells produced by Mendelian hybrids may hypothetically be regarded as being pure themselves.131
We have not yet considered one feature of all experiments in hybridisation, which indeed seems to be the most important of all for the theory of inheritance, if taken together with the fact of the pureness of the germs. The rule of Mendel always relates to one single character of the species or varieties concerned in hybridisation, and if it deals with more than one character, it regards every one of them separately; indeed, the rule holds for every one of them irrespective of the others. We cannot study here how this most important fact of the independence of the single characters of a species with regard to inheritance leads to the production of new races, by an abnormal mixture of those characters. We only take advantage of the fact theoretically, and in doing so, I believe, we can hardly escape the conclusion that the independence of the single characters in inheritance, taken together with the pureness of the germ-cells in the most simple form of hybrids, proves that there occurs in inheritance a sort of handing over of single and separate morphogenetic agents which relate to the single morphogenetic characters of the adult. We may use Bateson’s word “allelomorphs” for these agents, or units, as they may be called, thereby giving expression to the fact that the single and separate units, which are handed over in inheritance, correspond to each other in nearly related species without being the same.
And so we have at least an inkling of what the material continuity of inheritance is to mean, though, of course, our “single and separate morphogenetic agents,” or “units” or “allelomorphs” are in themselves not much more than unknown somethings described by a word; but even then they are “somethings.”
Besides the researches relating to the rule of Mendel and its exceptions, founded, that is, upon a study of the “second” generation of hybrids, there is another important line of research lately inaugurated by Herbst, which investigates the first generation in hybridisation. The hybrids themselves are studied with the special purpose of finding out whether the type of the single hybrid may change according to the conditions of its development, both outer and inner. The discoveries thus made may lead some day to a better understanding of the intimate nature of the “units” concerned in heredity, and perhaps to some knowledge of the arranging and ruling factor in morphogenesis also.
Starting from the discovery of Vernon, that the hybrids of sea-urchins are of different types according to the season, Herbst132 was able to show that differences among the hybrids with regard to their being more of the paternal or more of the maternal type, are in part certainly due to differences in temperature. But there proved to be still another factor at work, and Herbst has succeeded in discovering this factor by changing the internal conditions of morphogenesis. Whenever he forced the eggs of Sphaerechinus to enter into the first133 phase of artificial parthenogenesis and then fertilised them with the sperm of Echinus, he was able to approximate the offspring almost completely to the maternal type, whilst under ordinary conditions the hybrids in question follow the paternal far more than the maternal organisation.
What is shown, in the first place, by these discoveries is the importance of an arranging and ruling factor in spite of all units. The organism is always one whole whether the paternal properties prevail or the more complicated maternal ones; in other words, all so-called properties that consist in the spatial relations of parts have nothing to do with “units” or “allelomorphs,” which indeed cannot be more than necessary means or materials, requiring to be ordered. As to the character of the morphogenetic single and separate units themselves Herbst is inclined to regard them as specific chemical substances which unite correspondingly during nuclear conjugation, forming a sort of loose chemical compound. It would depend on the constitution of this compound whether germ-cells of hybrids could become pure or not.
The Rôle of the Nucleus in Inheritance
At the end of our studies on heredity we hardly can avoid saying a few words about the problem of the localisation of the morphogenetic units in the germ-cells themselves. Is it in the protoplasm or in the nucleus that they are placed? You all know that this question was for a long time regarded as more important than any other, and perhaps you have already blamed me for not raising it until now. But in my opinion results gained by the purely analytical method and carefully established, are always superior to those which are of a merely descriptive nature and doubtful besides. The famous problem of the part played by the nucleus in inheritance is both descriptive and doubtful: it is only, so to say, of factual, not of analytical importance, and quite insoluble at present.
As for our second proof of vitalism, stating that no kind of machine inside the germ-cells can possibly be the foundation of their morphogenesis, it is clear that the protoplasm and the nucleus may both come into account here on equal terms. If you prefer to say so, it is to the nucleus and to its division in particular that the second proof of autonomy relates, while the first, though not over-looking the presence of nuclei,134 deals “especially” with the protoplasmic nature of its “systems.”
What then can we say, on the basis of actual facts, about the part taken by the protoplasm and by the nucleus in inheritance, now that we have learnt from our analytical discussion that both of them cannot be any kind of morphogenetic machine, but can only be means of morphogenesis? Let us state our question in the following way: whereabouts in the germ-cells are those “means” of morphogenesis localised, the existence of which we infer from the material continuity in the course of generations in general and from the facts discovered about hybridisation in particular?
The first of the facts generally said to support the view that the nucleus of the germ-cells exerts a specified influence upon the processes of development and inheritance, relates to the proportion between protoplasm and nuclear material in the egg and in the spermiae. This proportion is very different in the two sexual products, as we know, there being an enormous preponderance of the protoplasm in the egg, of the nucleus in the spermatozoon. This seems to indicate that the proportion between protoplasm and nucleus is fairly indifferent for inheritance, as all the facts go to show that inheritance from the father is as common as inheritance from the mother. It is in the nucleus, and in the nucleus alone, that any similarity of organisation exists between the two sexual products, so very different in all other respects: therefore the nucleus should be the organ of inheritance. The phenomena of nuclear division, of karyokinesis, which are quite equal in both sexual cells, are certainly well fitted to support this hypothesis.
There seems indeed to be some truth in this reasoning, but nevertheless it must remain hypothetical; and it must never be forgotten that there may be very probably some sort of morphogenetic importance in protoplasm also. Rauber and afterwards Boveri135 have tried to prove experimentally that it is on the nuclear chromatic substance only that inheritance depends, but the first of these authors failed to get any results at all, and the latter obtained only ambiguous ones. Godlewski, on the contrary, has fertilised purely protoplasmic egg-fragments of the sea-urchin with the sperm of quite another group of Echinoderms, and obtained in spite of that a few stages of development of the pure maternal type. This experiment seems to place the morphogenetic importance of protoplasm beyond all doubt.
I should prefer not to make any definite statement about our problem at present. Our actual knowledge of the organisation and metabolism of both nucleus and protoplasm is so extremely small and may relate to such very insignificant topics, that any definite decision is impossible. I myself believe that the nucleus plays an important part in heredity, perhaps even a greater one than protoplasm, but this is only my belief.136
The discovery of Gruber and others, that Protozoa are only capable of restitution if they contain at least a fragment of the nucleus, has also been used occasionally as a proof of the morphogenetic importance of the nucleus. But might not this absence of restitution where nuclear material is lacking be understood equally well on the hypothesis of Loeb and R. S. Lillie that the nucleus is a centre of oxidation in the cell? Remove the heart from a vertebrate and the animal will not digest any more; but in spite of that the heart is not the organ of digestion.
And so we lay stress once more upon this point: that the experimental results of hybridisation and the analytical results obtained by the discussion of the complex-equipotential systems are of greater value to the theory of heredity than all speculation about the importance or unimportance of special constituents of the cell, of whose organisation, chemistry, and physics, scarcely anything is known at present.137
Variation and Mutation
Heredity, it has been said, may be understood as resting upon the fact that each organism forms its own initial stage again, and that this initial stage always encounters conditions of the same kind.
If this statement were quite correct, all the individuals of a given species would be absolutely alike everywhere and for ever. But they are not alike; and that they are not alike everywhere and for ever is not merely the only real foundation of the so-called theory of descent we possess, but also forces us to change a little our definition of heredity, which now proves to have been only a sort of approximation to the truth, convenient for analytical discussion.
In the first place, the conditions which surround the initial stages of morphogenesis are not quite equal in every respect: and indeed the offspring of a given pair of parents, or better, to exclude all complications resulting from sexual reproduction, or amphimixis, as Weismann called it—the offspring of one given parthenogenetic female are not all equal among themselves. The individuals of each generation are well known to vary, and it is especially in this country that the so-called individual or fluctuating variation has been most carefully studied by statistical methods, Galton and Weldon being the well-known pioneers in this field.138 In fact, if we are allowed to assume that this sort of variation is the outcome of a variation of conditions—in the most general meaning of the word—we only follow the opinion which has almost universally been adopted by the biologists139 that are working at this branch of the subject. Variation proper is now generally allowed to be the consequence of variations in nutrition; the contingencies of the latter result in contingencies of the former, and the law of contingencies is the same for both, being the most general law of probability. Of course under such an aspect fluctuating variation could hardly be called an exception, but rather an addition to inheritance.
But there are other restrictions of our definition of heredity. The initial stage which is formed again by an organism is not always quite identical in itself with the initial stage of its own parent: Bateson and de Vries were the first to study in a systematic way these real exceptions140 to true inheritance. As you know, de Vries has given them the name of “mutations.” What is actually known on this subject is not much at present, but nevertheless is of great theoretical value, being the only real foundation of all theories of descent, as we shall see in the next lectures. “Mutations” are known to exist at present only among some domesticated animals and plants. Nothing of a more general character can be said about their law or meaning.141
In finishing our chapter on inheritance, we at the same time have finished the first main part of our lectures; that part of them which has been devoted exclusively to the study of the morphogenesis of the individual, including the functioning of the adult individual form. We now turn to our second part, which is to deal with the problems of the diversities of individual forms, with morphological systematics. The end of our chapter on inheritance has already led us to the threshold of this branch of biological science.
The chief result of the first main part of our lectures has been to prove that an autonomy of life phenomena exists at least in some departments of individual morphogenesis, and probably in all of them; the real starting-point of all morphogenesis cannot be regarded as a machine, nor can the real process of differentiation, in all cases where it is based upon systems of the harmonious equipotential type. There cannot be any sort of machine in the cell from which the individual originates, because this cell, including both its protoplasm and its nucleus, has undergone a long series of divisions, all resulting in equal products, and because a machine cannot be divided and in spite of that remain what it was. There cannot be, on the other hand, any sort of machine as the real foundation of the whole of an harmonious system, including many cells and many nuclei, because the development of this system goes on normally, even if its parts are rearranged or partly removed, and because a machine would never remain what it had been in such cases.
If our analytical discussions have thus led us to establish a typical kind of vitalism, it follows that we can by no means agree with Wilhelm Roux in his denomination of the analytical science of the individual form and form-production as “Entwickelungsmechanik,” “developmental mechanics,” a title, which, of course, might easily be transformed into that of “morphogenetic mechanics,” to embrace not only normal development, but restitution and adaptation too. We feel unable to speak of “mechanics” where just the contrary of mechanics, in the proper meaning of the word, has been proved to exist.
Names of course are of comparatively small importance, but they should never be allowed to be directly misleading, as indeed the term “Entwickelungsmechanik” has already proved to be. Let us rather say, therefore, that we have finished with this lecture that part of our studies in biology which has had to deal with morphogenetic physiology or physiological morphogenesis.
Once more we repeat, at this resting-point in our discussions, that both of our proofs of life-autonomy have been based upon a careful analysis of certain facts about the distribution of morphogenetic potencies in two classes of morphogenetic systems, and upon nothing else. To recall only one point, we have not said that regeneration, merely because it is a kind of restitution of the disturbed whole, compels us to admit that biological events happen in a specific and elemental manner, but, indeed, regeneration does prove vitalism, because it is founded upon the existence of certain complex-equipotential systems, the analysis of the genesis of which leads to the understanding of life-autonomy. This distinction, in fact, is of the greatest logical importance.
Rational Systematics
All systematics which deserves the predicate “rational” is founded upon a concept or upon a proposition, by the aid of which a totality of specific diversities may be understood. That is to say: every system claiming to be rational gives us a clue by which we are able to apprehend either that there cannot exist more than a certain number of diversities of a certain nature, or that there can be an indefinite number of them which follow a certain law with regard to the character of their differences.
Solid geometry, which states that only five regular bodies are possible, and points out the geometrical nature of these bodies, is a model of what a rational system should be. The theory of conic sections is another. Take the general equation of the second degree with two unknowns, and study all the possible forms it can assume by a variation of its constants, and you will understand that only four different types of conic sections are possible—the circle, the ellipse, the hyperbola, and the parabola.
In physics and chemistry no perfect rational systems have been established hitherto, but there are many systems approaching the ideal type in different departments of these sciences. The chemical type of the monohydric saturated alcohols, for instance, is given by the formula CnH2n+1OH, and in this formula we not only have an expression of the law of composition which all possible alcohols are to follow,—but, since we know empirically the law of quantitative relation between n and various physical properties, we also possess in our formula a general statement with respect to the totality of the properties of any primary alcohol that may be discovered or prepared in the future. But chemistry has still higher aims with regard to its systematics: all of you know that the so-called “periodic law of the elements” was the first step towards a principle that may some day give account of the relation of all the physical and chemical properties of any so-called element with its most important constant, the atomic weight, and it seems to be reserved for the present time to form a real fundamental system of the “elements” on the basis of the periodic law by the aid of the theory of electrons. Such a fundamental system of the elements would teach us that there can only be so many elements and no more, and only of such a kind. In crystallography a similar end has been reached already by means of certain hypothetic assumptions, and systematics has here accounted for the limited number and fixed character of the possible forms of crystalline symmetry.
It is not difficult to understand the general logical type of all rational systems, and logic indeed can discover it without appealing to concrete sciences or to geometry. Rational systematics is always possible whenever there exists any fundamental concept or proposition which carries with it a principle of division; or to express it somewhat differently, which would lead to contradictions, if division were to be tried in any but one particular manner. The so-called “genus,” as will easily be perceived, then embraces all its “species” in such a manner that all peculiarities of the species are represented already in properties of the genus, only in a more general form, in a form which is still unspecified. The genus is both richer in content and richer in extent than are the species, though it must be added that its richness in content is, as it were, only latent: but it may come into actuality by itself and without any help from without.
We are dealing here with some of the most remarkable properties of the so-called synthetic judgments a priori in the sense of Kant, and, indeed, it seems that rational systematics will only be possible where some concept of the categorical class or some proposition based upon such concept lies at the root of the matter or at least is connected with it in some way. In fact, all rational systems with regard to the relations of symmetry in natural bodies deal ultimately with space; or better, all systems in such fields are able to become rational only if they happen to turn into questions of spatial symmetry.
All other genera and species, whether of natural bodies or of facts, can be related only on the basis of empirical abstraction, i.e. can never attain rationality: here, indeed, the genus is richer in extent and poorer in content than are the species. The genus is transformed into the species, not by any inherent development of latent properties, but by a mere process of addition of characteristic points. It is impossible to deduce the number or law or specifications of the species from the genus. Mere “classification,” if we may reserve the honorable name of systematics for the rational type, is possible here, a mere statement in the form of a catalogue, useful for orientation but for nothing more. We may classify all varieties of hats or of tables in the same way.
Biological Systematics
At this point we return from our logical excursion to our proper subject of biology; for I am sorry to say biological systematics is at present of our second type of systematics throughout: it is classification pure and simple. We have a catalogue in our hands, but nothing more.
Such a statement of fact conveys not a particle of censure, casts not the least reflection on the gifted men who created the classification of animals or plants. It is absolutely necessary to have such a catalogue, and indeed the catalogue of the organisms can be said to have been improved enormously during the advance of empirical and descriptive biological science. Any classification improves as it becomes more “natural,” as the different possible schemes of arrangement, the different reasons of division, agree better and better in their results; and, in fact, there has been a great advance of organic classification in this direction. The “natural” system has reached such perfection, that what is related from one point of view seems nearly related also from almost all points of view which are applicable, at least from those which touch the most important characteristics. There has been a real weighing of all the possible reasons of division, and that has led to a result which seems to be to some extent final.
But, nevertheless, we do not understand the raison d’être of the system of organisms; we are not at all able to say that there must be these classes or orders or families and no others, and that they must be such as they are.
Shall we ever be able to understand that? Or will organic systematics always remain empirical classification? We cannot answer this question. If we could, indeed, we should have what we desire! As simple relations of space are certainly not the central point of any problematic rational organic systematics even of the future, the question arises, whether there could be found any principle of another type in the realm of synthetic a priori judgments which could allow an inherent sort of evolution of latent diversities, as do all judgments about spatial symmetry. At the end of the second course of these lectures, which is to be delivered next summer, we shall be able to say a few more words about this important point.
The concept of what is called “a type,” due almost wholly to Cuvier and Goethe, is the most important of all that classification has given to us. Hardly second in importance is the discovery of the “correlation of parts,” as a sort of connection which has the character of necessity without being immediately based upon causality. Rádl seems to be the only modern author who has laid some stress on this topic. The harmony which we have discovered in development is also part of this correlation. When, later on, we come to discuss analytically our well established entelechy as the ultimate basis of individual organisation, we shall be able to gain more satisfactory ideas with respect to the meaning of the non-causal but necessary connection, embraced in the concepts of type and of correlation of parts.
The type is a sort of irreducible arrangement of different parts; the correlation deals with the degree and the quality of what may be called the actual make of the parts, in relation to one another: all ruminants, for instance, are cloven-footed, the so-called dental formulae are characteristic of whole groups of mammals. Of course all such statements are empirical and have their limits: but it is important that they are possible.142
It has been the chief result of comparative embryology to show that the type as such is more clearly expressed in developmental stages than it is in the adults, and that therefore the embryological stages of different groups may be very much more similar to each other than are the adults: that is the truth contained in the so-called “biogenetisches Grundgesetz.” But the specific differences of the species are not wanting in any case of ontogeny, in spite of such similarities in different groups during development.
We have applied the name “systematics” or, if rationality is excluded, “classification” to all that part of a science which deals with diversities instead of generalities: in such a wide meaning systematics, of course, is not to be confused with that which is commonly called so in biology, and which describes only the exterior differences of form. Our systematics is one of the two chief parts of biology; what are called comparative anatomy and comparative embryology are its methods. For it must be well understood that these branches of research are only methods and are not sciences by themselves.
It is most generally conceded at the present time that the actually existing state of all organisms whatsoever is the result of their history. What does that mean? What are the foundations upon which the assumption rests? What is the relation of systematics to history? In raising such questions and considerations we are treading the ground sacred to the theory of descent.
I well know that you prefer the name “theory of evolution” for what I am speaking of: but it may be misleading in various respects. We already know that quite a determinate meaning has been given to the word “evolutio” as applied to individual morphogenesis, “evolutio” being here opposed to “epigenesis.” Now there would be nothing against the use of the word evolution in a wider sense—indeed it is often applied nowadays to denote the fact that a something is actually “evolved” in embryology—if only our entelechy had taken the place of the machine of the mechanists. But that is the very point: there must be a real “evolving” of a something, in order that the word evolution may be justified verbally: and that is not the case in so-called phylogeny. At least we know nothing of an evolutionary character in the problematic pedigree of the organisms, as we shall see more fully hereafter. The term “theory of descent” is therefore less open to objection than is the usual English term. The word transformism, as used by the French, would also be a very good title.
The theory of descent is the hypothetic statement that the organisms are really allied by blood among each other, in spite of their diversities.143 The question about their so-called monophyletic or polyphyletic origin is of secondary importance compared with the statement of relationship in general.
There are two different groups of facts which have suggested the idea of transformism: none of these facts can be said to be conclusive, but there certainly is a great amount of probability in the whole if taken together.
The first group of evidences which lead to the hypothesis of the real relationship of organisms consists of facts relating to the geographical distribution of animals and plants and to palæontology. As to geography, it seems to me that the results of the floral and faunal study of groups of islands are to be mentioned in the first place. If, indeed, on each of the different islands, A B C and D, forming a group, the species of a certain genus of animals or plants are different in a certain respect, and show differences also compared with the species living on the neighbouring continent, of which there is geological evidence that the islands once formed a part, whilst there is no change in the species on the continent itself for very wide areas, then, no doubt, the hypothesis that all these differing species once had a common origin, the hypothesis that there is a certain community among them all, will serve to elucidate in some way what would seem to be very abstruse without it. And the same is true of the facts of palaeontology. In the geological strata, forming a continuous series, you find a set of animals, always typical and specific for every single stratigraphical horizon, but forming a series just as do those horizons. Would not the whole aspect of these facts lose very much of its peculiarity if you were to introduce the hypothesis that the animals changed with the strata? The continuity of life, at least, would be guaranteed by such an assumption.
The geographical and geological evidences in favour of the theory of descent are facts taken from sciences which are not biology proper; they are not facts of the living but only facts about the living. That is not quite without logical importance, for it shows that not biology alone has led to the transformism hypothesis. Were it otherwise, transformism might be said to be a mere hypothesis ad hoc; but now this proves to be not the case, though we are far from pretending that transformism might be regarded as resting upon a real causa vera.
But let us study the second group of facts which support the theory of descent. It is a group of evidences supplied by biology itself that we meet here, there being indeed some features in biology which can be said to gain some light, some sort of elucidation, if the theory of descent is accepted. Of course, these facts can only be such as relate to specific diversities, and indeed are facts of systematics; in other words, there exists something in the very nature of the system of organisms that renders transformism probable. The system of animals and plants is based upon a principle which might be called the principle of similarities and diversities by gradation; its categories are not uniform but different in degree and importance, and there are different kinds of such differences. No doubt, some light would be shed upon this character of the system, if we were allowed to assume that the relation between similarities and diversities, which is gradual, corresponded to a blood-relationship, which is gradual also.
THE COVERT PRESUMPTION OF ALL THEORIES OF DESCENT
We have used very neutral and somewhat figurative words, in order to show what might be called the logical value of the theory of descent, in order to signify its value with respect to so-called “explanation.” We have spoken of the “light” or the “elucidation” which it brings, of the “peculiarity of aspect” which is destroyed by it. We have used this terminology intentionally, for it is very important to understand that a specific though hidden addition is made almost unconsciously to the mere statement of the hypothesis of descent as such, whenever this hypothesis is advocated in order to bring light or elucidation into any field of systematic facts. And this additional hypothesis indeed must be made from the very beginning, quite irrespective of the more detailed problems of the law of transformism, in order that any sort of so-called explanation by means of the theory of descent may be possible at all. Whenever the theory that, in spite of their diversities, the organisms are related by blood, is to be really useful for explanation, it must necessarily be assumed in every case that the steps of change, which have led the specific form A to become the specific form B, have been such as only to change in part that original form A. That is to say: the similarities between A and B must never have become overshadowed by their diversities.
Only on this assumption, which indeed is a newly formed additional subsidiary hypothesis, joined to the original hypothesis of descent in general—a hypothesis regarding the very nature of transformism—only on this almost hidden assumption is it possible to speak of any sort of “explanation” which might be offered by the theory of transformism to the facts of geography, geology, and biological systematics. Later on we shall study more deeply the logical nature of this “explanation”; at present it must be enough to understand this term in its quasi-popular meaning.
What is explained by the hypothesis of descent—including the additional hypothesis, that there always is a prevalence of the similarities during transformism—is the fact that in palaeontology, in the groups of island and continent faunae and florae taken as a whole, as well as in the single categories of the system, the similarities exceed the diversities. The similarities now are “explained”; that is to say, they are understood as resting on but one principle: the similarities are understood as being due to inheritance;144 and now we have but one problem instead of an indefinite number. For this reason Wigand granted that the theory of descent affords what he calls a numerical reduction of problems.
Understanding then what is explained by the theory of descent with its necessary appendix, we also understand at once what is not elucidated by it: the diversities of the organism remain as unintelligible as they always were, even if we know that inheritance is responsible for what is similar or equal. Now there can be no doubt that the diversities are the more important point in systematics; if there were only similarities there would be no problem of systematics, for there would be no system. Let us be glad that there are similarities in the diversities, and that these similarities have been explained in some way; but let us never forget what is still awaiting its explanation. Unfortunately it has been forgotten far too often.
THE SMALL VALUE OF PURE PHYLOGENY
And so we are led to the negative side of the theory of transformism, after having discussed its positive half. The theory of descent as such, without a real knowledge of the factors which are concerned in transformism, or of the law of transformism, in other terms, leaves the problem of systematics practically where it was, and adds really nothing to its solution. That may seem very deplorable, but it is true.
Imagine so-called historical geology, without any knowledge of the physical and chemical factors which are concerned in it: what would you have except a series of facts absolutely unintelligible to you? Or suppose that some one stated the cosmogenetic theory of Kant and Laplace without there being any science of mechanics: what would the theory mean to you? Or suppose that the whole history of mankind was revealed to you, but that you had absolutely no knowledge of psychology: what would you have but facts and facts and facts again, with not a morsel of real explanation?
But such is the condition in which so-called phylogeny stands. If it is based only on the pure theory of transformism, there is nothing explained at all. It was for this reason that the philosopher Liebmann complained of phylogeny that it furnishes nothing but a “gallery of ancestors.” And this gallery of ancestors set up in phylogeny is not even certain; on the contrary, it is absolutely uncertain, and very far from being a fact. For there is no sound and rational principle underlying phylogeny; there is mere fantastic speculation. How could it be otherwise where all is based upon suppositions which themselves have no leading principle at present? I should not like to be misunderstood in my polemics against phylogeny. I fully grant you that it may be possible in a few cases to find out the phylogenetic history of smaller groups with some probability, if there is some palaeontological evidence in support of pure comparative anatomy; and I also do not hesitate to allow that such a statement would be of a certain value with regard to a future discovery of the “laws” of descent, especially if taken together with the few facts known about mutations. But it is quite another thing with phylogeny on the larger scale. Far more eloquent than any amount of polemics is the fact that vertebrates, for instance, have already been “proved” to be descended from, firstly, the amphioxus; secondly, the annelids; thirdly, the Sagitta type of worms; fourthly, from spiders; fifthly, from Limulus, a group of crayfishes; and sixthly, from echinoderm larvae. That is the extent of my acquaintance with the literature, with which I do not pretend to be specially familiar. Emil du Bois-Reymond said once that phylogeny of this sort is of about as much scientific value as are the pedigrees of the heroes of Homer, and I think we may fully endorse his opinion on this point.
HISTORY AND SYSTEMATICS
A few words should be devoted to the relations between history and systematics in biology. Is there no contradiction between historical development and a true and rational system which, we conceded, might exist some day in biological sciences, even though it does not at present? By no means. A totality of diversities is regarded from quite different points of view if taken as the material of a system, and if considered as realised in time. We have said that chemistry has come very near to proper rational systematics, at least in some of its special fields; but the compounds it deals with at the same time may be said to have originated historically also, though not, of course, by a process of propagation. It is evident at once that the geological conditions of very early times prohibited the existence of certain chemical compounds, both organic and inorganic, which are known at present. None the less these compounds occupy their proper place in the system. And there may be many substances theoretically known to chemical systematics which have never yet been produced, on account of the impossibility of arranging for their proper conditions of appearance, and nevertheless they must be said to “exist.” “Existence,” as understood in systematics, is independent of special space and of special time, as is the existence of the laws of nature: we may speak of a Platonic kind of existence here. Of course it does not contradict this sort of ideal existence if reality proper is added to it.
Thus the problem of systematics remains, no matter whether the theory of descent be right or wrong. There always remains the question about the totality of diversities in life: whether it may be understood by a general principle, and of what kind that principle would be. As, in fact, it is most probably by history, by descent, that organic systematics is brought about, it of course most probably will happen some day that the analysis of the causal factors concerned in the history will serve to discover the principle of systematics also.
Let us now glance at the different kinds of hypotheses which have been established in order to explain how the descent of the organisms might have been possible. We have seen that the theory of transformism alone is not worth very much as a whole, unless at least a hypothetical picture can be formed of the nature of the transforming factors: it is by some such reasoning that almost every author who has defended the theory of descent in its universality tries to account for the manner in which organisms have acquired their present diversities.
There is no need in our times and particularly in this country, to explain in a full manner the theory known under the name of Darwinism. All of you know this theory, at least in its outlines, and so we may enter at once upon its analytic discussion. A few words only I beg you to allow me as to the name of “Darwinism” itself. Strange to say, Darwinism, and the opinion of Charles Darwin about the descent of organisms, are two different things. Darwin, the very type of a man devoted to science alone and not to personal interests,—Darwin was anything but dogmatic, and yet Darwinism is dogmatism in one of its purest forms. Darwin, for instance, gave the greatest latitude to the nature of the variations which form the battleground of the struggle for existence and natural selection; and he made great allowances for other causal combinations also, which may come into account besides the indirect factors of transformism. He was Lamarckian to a very far-reaching extent. And he had no definite opinion about the origin and the most intimate nature of life in general. These may seem to be defects but really are advantages of his theory. He left open the question which he could not answer, and, in fact, he may be said to be a good illustration of what Lessing says, that it is not the possession of truth but the searching after it, that gives happiness to man. It was but an outcome of this mental condition that Darwin’s polemics never left the path of true scientific discussions, that he never in all his life abused any one who found reason to combat his hypotheses, and that he never turned a logical problem into a question of morality.
How different is this from what many of Darwin’s followers have made out of his doctrines, especially in Germany; how far is “Darwinism” removed from Darwin’s own teaching and character!
It is to Darwinism of the dogmatic kind, however, that our next discussions are to relate, for, thanks to its dogmatism, it has the advantage of allowing the very sharp formulation of a few causal factors, which a priori might be thought to be concerned in organic transformism, though we are bound to say that a really searching analysis of these factors ought to have led to their rejection from the very beginning.
The logical structure of dogmatic Darwinism reveals two different parts, which have nothing at all to do with one another.
NATURAL SELECTION
We shall first study that part of it which is known under the title of natural selection, irrespective of the nature of the causes of primary differences, or, in other words, the nature of variability. This part may be said to belong to Darwin’s personal teachings and not only to “Darwinism.” The offspring of a certain number of adults show differences compared with each other; there are more individuals in the offspring than can grow up under the given conditions, therefore there will be a struggle for existence amongst them, which only the fittest will survive; these survivors may be said to have been “selected” by natural means.
It must be certain from the very beginning of analysis that natural selection, as defined here, can only eliminate what cannot survive, what cannot stand the environment in the broadest sense, but that natural selection never is able to create diversities. It always acts negatively only, never positively. And therefore it can “explain”—if you will allow me to make use of this ambiguous word—it can “explain” only why certain types of organic specifications, imaginable a priori, do not actually exist, but it never explains at all the existence of the specifications of animal and vegetable forms that are actually found. In speaking of an “explanation” of the origin of the living specific forms by natural selection one therefore confuses the sufficient reason for the non-existence of what there is not, with the sufficient reason for the existence of what there is. To say that a man has explained some organic character by natural selection is, in the words of Nägeli, the same as if some one who is asked the question, “Why is this tree covered with these leaves,” were to answer “Because the gardener did not cut them away.” Of course that would explain why there are no more leaves than those actually there, but it never would account for the existence and nature of the existing leaves as such. Or do we understand in the least why there are white bears in the Polar Regions if we are told that bears of other colours could not survive?
In denying any real explanatory value to the concept of natural selection I am far from denying the action of natural selection. On the contrary, natural selection, to some degree, is self-evident; at least as far as it simply states that what is incompatible with permanent existence cannot exist permanently, it being granted that the originating of organic individuals is not in itself a guarantee of permanency. Chemical compounds, indeed, which decompose very rapidly under the conditions existing at the time when they originated may also be said to have been eliminated by “natural selection.” It is another question, of course, whether in fact all eliminations among organic diversities are exclusively due to the action of natural selection in the proper Darwinian sense. It has been pointed out already by several critics of Darwinism and most clearly by Gustav Wolff, that there are many cases in which an advantage with regard to situation will greatly outweigh any advantage in organisation or physiology. In a railway accident, for instance, the passengers that survive are not those who have the strongest bones, but those who occupied the best seats; and the eliminating effect of epidemics is determined at least as much by localities, e.g. special houses or special streets, as by the degree of immunity. But, certainly, natural selection is a causa vera in many other cases.
We now may sum up our discussion of the first half of Darwinism. Natural selection is a negative, an eliminating factor in transformism; its action is self-evident to a very large degree, for it simply states that things do not exist if their continuance under the given conditions is impossible. To consider natural selection as a positive factor in descent would be to confound the sufficient reason for the non-existence of what is not, with the sufficient reason of what is.
Natural selection has a certain important logical bearing on systematics, as a science of the future, which has scarcely ever been alluded to. Systematics of course has to deal with the totality of the possible, not only of the actual diversities; it therefore must remember that more forms may be possible than are actual, the word “possible” having reference in this connection to originating, not to surviving. Moreover, systematics is concerned not only with what has been eliminated by selection, but also with all that might have originated from the eliminated types. By such reasoning natural selection gains a very important aspect—but a logical aspect only.
FLUCTUATING VARIATION THE ALLEGED CAUSE OF ORGANIC DIVERSITY
The second doctrine of dogmatic Darwinism states that all the given diversities among the organisms that natural selection has to work upon are offered to natural selection by so-called fluctuating variation; that is, by variation as studied by means of statistics. This sort of variation, indeed, is maintained to be indefinite in direction and amount, at least by the most conservative Darwinians; it has occasionally been called a real differential; in any case it is looked upon as being throughout contingent with regard to some unity or totality; which, of course, is not to mean that it has not had a sufficient reason for occurring.
It could hardly be said to be beyond the realm of possibility that such differences among organic species as only relate to degree or quantity and perhaps to numerical conditions also, might have been “selected” out of given contingent variations, if but one postulate could be regarded as fulfilled. This postulate may appropriately be stated as the fixation of new averages of variation by inheritance. Let the average value of a variation, with regard to a given property of a given species be n and let the value n + m—m being variable—which is represented in fewer individuals of course than is n, be such as to offer advantages in the struggle for existence; then the individuals marked by n + m will have the greater chance of surviving. Our postulate now states that, in order that a permanent increase of the average value of the variation in question may be reached, n + m in any of its variable forms must be able to become the average value of the second generation, as n was the average value of the first. Out of the second generation again it would be the few individuals marked by n + m + o, which would be selected; n + m + o would be the new average; afterwards n + m + o + p would be selected, would become the new average, and so on. A black variety for instance might be selected by such a series of processes out of a grey-coloured one without difficulty.
But our postulate is not beyond all doubt: certain experiments, at least, which have been carried out about the summation of variations of the true fluctuating type by any kind of selection seem to show that there may be a real progress for a few generations, but that this progress is always followed by a reversion. Of course our experience is by no means complete on this subject, and, indeed, it may be shown in the future that positive transforming effects of fluctuating variability, in connection with selective principles, are possible in the case of new quantitative differences (in the widest sense), but we are not entitled to say so at present.
And this is the only condition on which we can give credit to the second doctrine of dogmatic Darwinism. Its second principle, indeed, proves to be absolutely inadequate to explain the origin of any other kind of specific properties whatever.
I cannot enter here into the whole subject of Darwinian criticism.145 Our aims are of a positive character, they desiderate construction and only use destruction where it is not to be avoided. So I shall only mention that dogmatic Darwinism has been found to be unable to explain every kind of mutual adaptations, e.g. those existing between plants and insects; that it can never account for the origin of those properties that are indifferent to the life of their bearer, being mere features of organisation as an arrangement of parts; that it fails in the face of all portions of organisation which are composed of many different parts—like the eye—and nevertheless are functional units in any passive or active way; and that, last not least, it has been found to be quite inadequate to explain the first origin of all newly formed constituents of organisation even if they are not indifferent: for how could any rudiment of an organ, which is not functioning at all, not only be useful to its bearer, but be useful in such a degree as to decide about life or death?
It is only for one special feature that I should like to show, by a more full analysis, that dogmatic Darwinism does not satisfy the requirements of the case. The special strength of Darwinism is said to lie in its explaining everything that is useful in and for organisms; the competitive factor it introduces does indeed seem to secure at least a relative sort of adaptedness between the organism and its needs. But in spite of that, we shall now see that Darwinism fails absolutely to explain those most intimate organic phenomena which may be said to be the most useful of all.
Darwinism in its dogmatic form is not able to explain the origin of any sort of organic restitution; it is altogether impossible to account for the restitutive power of organisms by the simple means of fluctuating variation and natural selection in the struggle for existence. Here we have the logical experimentum crucis of Darwinism.
Let us try to study in the Darwinian style the origin of the regenerative faculty, as shown in the restitution of the leg of a newt. All individuals of a given species of the newt, say Triton taeniatus, are endowed with this faculty; all of them therefore must have originated from ancestors which acquired it at some time or other. But this necessary supposition implies that all of these ancestors must have lost their legs in some way, and not only one, but all four of them, as they could not have acquired the restitutive faculty otherwise. We are thus met at the very beginning of our argument by what must be called a real absurdity, which is hardly lessened by the assumption that regeneration was acquired not by all four legs together, but by one after the other. But it is absolutely inevitable to assume that all the ancestors of our Triton must have lost one leg, or more correctly, that only those of them survived which had lost one! Otherwise not all newts at the present day could possess the faculty of regeneration! But a second absurdity follows the first one; out of the ancestors of our newt, which survived the others by reason of having lost one of their legs, there were selected only those which showed at least a very small amount of healing of their wound. It must be granted that such a step in the process of selection, taken by itself, would not at all seem to be impossible; since healing of wounds protects the animals against infection. But the process continues. In every succeeding stage of it there must have survived only those individuals which formed just a little more of granulative tissue than did the rest: though neither they themselves nor the rest could use the leg, which indeed was not present! That is the second absurdity we meet in our attempt at a Darwinian explanation of the faculty of regeneration; but I believe the first one alone was sufficient.
If we were to study the “selection” of the faculty of one of the isolated blastomeres of the egg of the sea-urchin to form a whole larva only of smaller size, the absurdities would increase. At the very beginning we should encounter the absurdity, that of all the individuals there survived only those which were not whole but half; for all sea-urchins are capable of the ontogenetical restitution in question, all of their ancestors therefore must have acquired it, and they could do that only if they became halved at first by some accident during early embryology. But we shall not insist any further on this instance, for it would not be fair to turn into ridicule a theory which bears the name of a man who is not at all responsible for its dogmatic form. Indeed, we are speaking against Darwinism of the most dogmatic form only, not against Darwin himself. He never analysed the phenomena of regeneration or of embryonic restitution—they lay in a field very unfamiliar to him and to his time. I venture to say that if he had taken them into consideration, he would have agreed with us in stating that his theory was not at all able to cover them; for he was prepared to make great concessions, to Lamarckism for instance, in other branches of biology, and he did not pretend, to know what life itself is.
Darwin was not a decided materialist, though materialism has made great capital out of his doctrines, especially in Germany. His book, as is well known, is entitled “The Origin of Species,” that is of organic diversities, and he himself possibly might have regarded all restitution as belonging to the original properties of life, anterior to the originating of diversities. Personally he might possibly be called even a vitalist. Thus dogmatic “Darwinism” in fact is driven into all the absurdities mentioned above, whilst the “doctrine of Darwin” can only be said to be wrong on account of its failing to explain mutual adaptation, the origin of new organs, and some other features in organic diversities; the original properties of life were left unexplained by it intentionally.
DARWINISM FAILS ALL ALONG THE LINE
The result of our discussion then must be this: selection has proved to be a negative factor only, and fluctuating variation as the only way in which new properties of the organisms might have arisen has proved to fail in the most marked manner, except perhaps for a few merely quantitative instances. Such a result betokens the complete collapse of dogmatic Darwinism as a general theory of descent: the most typical features of all organisms remain as unexplained as ever.
What then shall we put in the place of pure Darwinism? Let us first try a method of explanation which was also adopted occasionally by Darwin himself: let us study that form of transformation theories which is commonly known under the title of Lamarckism.
As the word “Darwinism” does not signify the proper theoretical system of Charles Darwin, so Lamarckism as commonly understood nowadays is a good deal removed from the original views of Jean Baptiste Lamarck. Lamarckism is generally regarded as reducing all organic diversities to differences in the needs of individual life, but Lamarck himself, as must be emphasised from the very beginning, did not at all maintain the opinion that the great characteristics of the types were only due to such accidental factors. He supposed a sort of law of organisation to be at the root of systematics, as developed in history, and the needs of life were only responsible, according to him, for splitting the given types of organisation into their ultimate branches. Thus Lamarck, to a great extent at any rate, belongs to a group of authors that we shall have to study afterwards: authors who regard an unknown law of phylogenetic development as the real basis of transformism. Modern so-called Neo-Lamarckism, on the other hand, has indeed conceded the principle of needs to be the sole principle of transformism. Let us then study Lamarckism in its dogmatic modern form.
ADAPTATION AS THE STARTING-POINT
All facts of morphological adaptations—facts which we have analysed already from a different point of view, as being among the most typical phenomena of organic regulation—form the starting-point of this theory, and it must be granted that they form a very solid foundation, for they are facts. The theory only has to enlarge hypothetically the realm of these facts, or rather the realm of the law that governs them. Indeed, it is assumed by Lamarckism that the organism is endowed with the faculty of responding to any change of the environment which may change its function by a morphologically expressed alteration of its functional state and form, which is adapted to the state of conditions imposed from without. Of course, as stated in this most general form, the assumption is not true, but it is true within certain limits, as we know; and there seems to be no reason why we should not believe that there are many more cases of adaptation than we actually know at present, or that, in former phylogenetic times, the organisms were more capable of active adaptation than they are now. So to a certain extent, at least, Lamarckism can be said to rest upon a causa vera.
It is important to notice that this causa vera would imply vitalistic causality when taken in the wide meaning which Lamarckism allows to it: indeed, the power of active adaptation to indefinite changes would imply a sort of causal connection that is nowhere known except in the organism. Lamarck himself is not very clear about this point, he seems to be afraid of certain types of uncritical vitalism in vogue in his days; but modern writers have most clearly seen what the logical assumptions of pure Lamarckism are. Next to Cope, August Pauly146 may be said to be the most conscious representative of a sort of so-called psychological vitalism, which indeed Lamarckism as a general and all-embracing theory must have as its basis.
THE ACTIVE STORING OF CONTINGENT VARIATIONS AS A HYPOTHETIC PRINCIPLE
This point will come out more fully, if now we turn to study a certain group of principles, upon which dogmatic Lamarckism rests: I say principles and not facts, for there are no facts but only hypothetic assumptions in this group of statements. We do know a little about adaptations, at least to a certain extent, and it was only about the sphere of the validity of a law, which was known to be at work in certain cases, that hypothetical additions were made. In the second group of the foundations of Lamarckism we know absolutely nothing; accidental variations of form are supposed to occur, and the organism is said to possess the faculty of keeping and storing these variations and of handing them down to the next generation, if they happen to satisfy any of its needs.
But these needs are not of the actual type, brought forth by a change of the functional state of the individual, as in the case of adaptations: they are of a somewhat mysterious nature. A glance at the theory of the origin of the movements which are called acts of volition in the human child may serve to elucidate what is meant.
Acts of volition are said thus to originate in random movements of the new-born infant: certain of these accidental motions which happen to relieve some pain or to afford some pleasure are “remembered,” and are used another time quite consciously to bring forth what is liked or to remove what is disliked. So much for the present on a very difficult subject, which will occupy us next year at much greater length. It is clear that at least three fundamental phenomena are concerned in this theory of the origin of acts of volition: the liking and disliking, the keeping in mind, and the volition itself. The real act of volition, indeed, is always based upon a connection of all these factors, these factors now being connected in such a way that even their kind of connection may be said to be a fourth fundamental principle. In order that the particular effect may be obtained which is wanted because it is liked, the possible ways leading to it, which appeared among the random movements in the very beginning, are now regarded as “means” and may now be said to be “used.” But that is as much as to say that the “means” are judged with respect to their usefulness for the actual purpose, and therefore judgment is the fourth foundation of the act of volition.
In fact, Pauly does not hesitate to attribute judgment, along with the other psychological elements, to the organisms whilst undergoing their transformation. There has been formed, for instance, by accidental variation some pigment which by its chemical nature brings the organism into a closer connection with the light of the medium; the individual likes that, keeps the pigment for itself and produces it again in the next generation; and indeed it will safeguard any sort of improvement which chance may effect in this primitive “eye.” Such a view is said to hold well with respect to the origin of every new organ. And this psychological argument is also said to afford the real explanation of adaptation proper. Adaptation also is regarded not as a truly primary faculty of the organism, but as a retention or provoking of metabolic states which occurred by accident originally and were then found to be useful; now they are reproduced either in every single case of individual morphogenesis, without regard to actual requirements, or else only in response to such: in the first case they are “inherited,” in the second they only occur as regulations. Thus the process of judgment, together with all the other elemental factors of psychical life concerned in it, has been made to account for adaptation proper. The whole theory has accordingly become very uniform and simple.
CRITICISM OF THE “INHERITANCE OF ACQUIRED CHARACTERS” ASSUMED BY LAMARCKISM
In addressing ourselves to the criticism of Neo-Lamarckism we shall neglect as far as possible all the different psychological principles concerned in it—which in any case would need rather a great amount of epistemological sifting—and shall keep to those hypothetic facts which are supposed to be such as may be actually observed in nature.
All of you know that the so-called inheritance of acquired characters lies at the root of Lamarckism; and from this hypothesis our critical analysis is to start, disregarding a larger or smaller number of psychological principles that are brought into the field.
The name of “acquired characters” may a priori be given to three different types of phenomena: firstly, variations including mutations; secondly, disease or injuries; and thirdly, the results of the actual process of adaptation of every kind.
In the first of these groups, the true problem of the inheritance of “acquired” characters appears only with certain restrictions. All variations and mutations are indeed “acquired” by one generation so far as the earlier generation did not possess them, but mutations, at least, cannot be said to be acquired by the actual adult personality: they are innate in it from its very beginning, and therefore may better be called congenital.147 Congenital properties of the mutation type are, in fact, known to be inherited: their inheritance does not present any problem of its own, but is included in the changes of the hereditary condition to which they are due altogether.148 All properties of the variation type, on the other hand, having been studied statistically, are known to be inherited, to a certain small extent, as we have seen already whilst studying Darwinism, though they are possibly always liable to reversion. Modern science, as we know,149 regards them as due to changes of nutrition, in the most general meaning of the word. Under such a view variations might indeed be said to belong to the acquired group of organic specifications; their inheritance, as will be seen later on, would hardly be quite a pure instance of what we are searching for. In no case can true variations claim to be of great importance in problems of transformism.
But what is known about the inheritance of those properties which beyond any doubt may be said to have originated in the adult individual as such, and of which lesions and adaptations proper, as shown for instance among amphibious plants, are instances of the two most typical groups?150 Weismann did good service by putting an end to the scientific credulity which prevailed with regard to this subject. Weismann was led by his theory of the germ plasm to deny the inheritance of acquired characters of the typical kinds. He could not imagine how the effect of any agent upon the adult, be it of the merely passive or of the adaptive kind, could have such an influence upon the germ as to force it to produce the same effect in spite of the absence of that agent. In fact, that is what the inheritance of acquired characters would render necessary, and a very strange phenomenon it would be, no doubt. But, of course, taken alone, it could never be a decisive argument against such inheritance. I fully agree, that science is obliged to explain new facts by what is known already, as long as it is possible; but if it is no longer possible, the theory of course has to be changed, and not the facts. On this principle one would not neglect the fact of an inheritance of acquired properties, but on the contrary one perhaps might use it as a new evidence of vitalism.
But are there any facts?
At this point we come to speak about the second group of Weismann’s reasonings. He not only saw the difficulty of understanding inheritance of acquired characters on the principles of the science of his time, but he also criticised the supposed facts; and scarcely any of them stood the test of his criticism. Indeed, it must fairly be granted that not one case is known which really proves the inheritance of acquired characters, and that injuries certainly are never found to be inherited. In spite of that, I do not believe that we are entitled to deny the possibility of the inheritance of a certain group of acquired characters in an absolute and dogmatic manner, for there are a few facts which seem at least to tend in the direction of such an inheritance, and which seem to show that it might be discovered perhaps one day, if the experimental conditions were changed.
I am not referring here to the few cases in which bacteria were made colourless or non-virulent by outside factors, or in which certain fungi were forced to permanent agamic reproduction by abnormal external conditions and were shown to retain their “acquired properties” after the external conditions had been restored. In these cases only reproduction by simple division occurred, and that does not imply the true problem of inheritance. Nor am I referring to the few cases of non-adaptive “modifications” found by Standfuss and Fischer, in which butterflies that had assumed an abnormal kind of pigmentation under the influence of abnormal temperature acting upon the pupa, were seen to form this same kind of pigmentation in the next generation under normal conditions of temperature. These cases, though important in themselves, are capable perhaps of a rather simple explanation, as in fact has been suggested. Some necessary means both of inheritance and of morphogenesis, the former being present in the propagation cells, may be said to have been changed or destroyed by heat, and therefore, what seems to be inherited after the change of the body only, would actually be the effect of a direct influence of the temperature upon the germ itself.151 Let me be clearly understood: I do not say that it is so, but it may be so. What seems to me to be more important than everything and to have a direct bearing on the real discovery of the inheritance of acquired characters in the future, is this. In some instances plants which had been forced from without to undergo certain typical morphological adaptations, or at least changes through many generations, though they did not keep the acquired characters permanently in spite of the conditions being changed to another type, were yet found to lose the acquired adaptations not suddenly but only in the course of three or more generations. A certain fern, Adiantum, is known to assume a very typical modification of form and structure, if grown on serpentine; now Sadebeck,152 while cultivating this serpentine modification of Adiantum on ordinary ground, found that the first generation grown in the ordinary conditions loses only a little of its typical serpentine character, and that the next generation loses a little more, so that it is not before the fifth generation that all the characters of the serpentine modification have disappeared. There are a few more cases of a similar type relating to plants grown in the plains or on the mountains. There also it was found to take time, or rather to take the course of several generations, until what was required by the new conditions was reached. Of course these cases are very very few compared with those in which a sudden change of the adaptive character, corresponding to the actual conditions, sets in; but it is enough that they do exist.
Would it not be possible at least that adaptations which last for thousands of generations or more might in fact change the adaptive character into a congenital one? Then we not only should have inheritance of acquired characters, but should have a sort of explanation at the same time for the remarkable fact that certain histological structures of a very adapted kind are formed ontogenetically before any function exists, as is known to be the case with the structures in the bones of vertebrates, for instance. Experiments are going on at Paris, and perhaps in other places of scientific research also, which, it is hoped, will show that animals reared in absolute darkness for many generations will lose their perfectly formed eyes, and that animals from the dark with very rudimentary eyes will be endowed with properly functioning ones, after they have been reared in the light for generations. Such a result indeed would account for the many animals, of the most different groups, which live in dark caves and possess only rudiments of eyes: functional adaptation is no longer necessary, so-called atrophy by inactivity sets in, and the results “acquired” by it are inherited.153
But enough of possibilities. Let us be content at present to know at least a few real instances with regard to the slowness of the process of what might be said to be “re-adaptation” in some plants. This process shows us a way by which our problem may some day be solved; it allows us to introduce inheritance of acquired characters as a legitimate hypothesis at least, which not only will explain many of the diversities in systematics historically, but also can be called, though not a causa vera, yet certainly more than a mere fiction.
OTHER PRINCIPLES WANTED
We have only dealt with the probability of the inheritance of morphological or physiological154 adaptation. If that could really be considered as one of the factors concerned in the theory of descent, many, if not all of those congenital diversities among organic species which are of the type of a true structural correspondence to their future functional life, might be regarded as explained, that is, as reduced to one and the same principle. But nothing more than an explanation of this kind of diversities is effected by our principle, and very much more remains to be done, for organic diversities not only consist in specifications and differences as to histology, but are to a much more important degree, differences of organisation proper, that is, of the arrangement of parts, in the widest sense of the word.155
Would it be possible to interpret the origin of this sort of systematic diversities by a reasoning similar to that by which we have understood, at least hypothetically, congenital adaptedness?
Dogmatic Lamarckism, we know, uses two principles as its foundations; one of them, adaptation and its inheritance, we have studied with what may be called a partly positive result. The other is the supposed faculty of the organism to keep, to store, and to transfer those variations or mutations of a not properly adaptive sort which, though originating by chance, happen to satisfy some needs of the organism.
CRITICISM OF THE HYPOTHESIS OF STORING AND HANDING DOWN CONTINGENT VARIATIONS
Strange to say, this second hypothesis of dogmatic Lamarckism, invented with the express purpose of defeating Darwinism and taking the place of its fluctuating variability, which was found not to do justice to the facts—this second hypothesis of dogmatic Lamarckism is liable to just the same objections as dogmatic Darwinism itself.
As it is important to understand well the real logical nature of our objections to both of the great transformistic theories, we think it well to interrupt our argument for a moment, in order to consider a certain point which, though very important in itself, seems of only secondary importance to us in our present discussion. Dogmatic Darwinism—I do not say the doctrine of Charles Darwin—is materialistic at bottom, and indeed has been used by many to complete their materialistic view of the universe on its organic side. The word “materialism” must not necessarily be taken here in its metaphysical sense, though most materialists are dogmatic metaphysicians. It also can be understood as forming part of a phenomenological point of view. Materialism as a doctrine of science means simply this: that whether “nature” be reality or phenomenon, in any case there is but one ultimate principle at its base, a principle relating to the movements of particles of matter. It is this point of view which dogmatic Darwinism strengthens; on the theory of natural selection and fluctuating variations, due to accidental differences of nutrition, organisms are merely arrangements of particles of matter, nothing else; and moreover, their kinds of arrangement are understood, at least in principle. Lamarckism, on the other hand, is not materialistic, but most markedly vitalistic—psychistic even; it takes life for granted when it begins its explanations.
You may tell me that Darwin did the same, that he expressly states that his theory has nothing to do with the origin of life; that the title of his work is “The Origin of Species.” It would certainly be right to say so, at least with reference to Darwin personally; but in spite of that, it must be granted that Darwin’s doctrine contains a certain germ of materialism which has been fully developed by the Darwinian dogmatists, while Lamarckism is antimaterialistic by its very nature.
Now it is very important, I think, to notice that this difference between the two theories is unable to disguise one main point which is common to both: and it is to this point, and to this point only, that our chief objections against both these theories converge at present.
The contingency of the typical organic form is maintained by Darwinism as well as by Lamarckism: both theories, therefore, break down for almost the same reasons. The term “contingency” can signify very different relations, having but little in common; but it is sufficient for our present purpose to observe that there may be distinguished roughly two main classes of contingencies, which may provisionally be called the “contingency of being,” and the “contingency of occurring.” It is with the contingency of being that criticism of Darwinism and Lamarckism of the dogmatic type has to deal. Darwinism dealt with variations occurring at random; the organic form was the result of a fixation of only one kind of such variations, all others being extinguished by selection. In other terms, the specific organised form, as understood by Darwinism, was a unit only to the extent that all its properties related to one and the same body, but for the rest it was a mere aggregation or summation. It may be objected to this statement, that by being inherited in its specificity the Darwinian form proved to be a unit in a higher sense of the word, even in the opinion of dogmatic Darwinians; and this objection, perhaps, holds good as far as inheritance is concerned. But on the other hand, it must never be forgotten that the word “unit” had quite a vague and empty meaning even then, as indeed everything the organism is made up of is regarded as being in itself due to a contingent primary process, which has no relation to its fellow-processes. “Unit,” indeed, in spite of inheritance—which, by the way, is alleged also to be a merely materialistic event—means to Darwinians no more when applied to the organism than it does when applied to mountains or islands, where of course a sort of “unit” also exists in some sense, as far as one and the same body comes into account, but where every single character of this unit, in every single feature of form or of quality, is the result of factors or agents each of which is independent of every other.
To this sort of contingency of being, as maintained by Darwinians, criticism has objected, as we know, that it is quite an impossible basis of a theory of descent, since it would explain neither the first origin of an organ, nor any sort of harmony among parts or among whole individuals, nor any sort of restitution processes.
Now Lamarckism of the dogmatic kind, as will easily be seen, only differs from Darwinism in this respect, that what according to the latter happens to the organism passively by means of selection, is according to the former performed actively by the organism, by means of a “judgment”—by the retention and handing down of chance variations. The specificity of the form as a whole is contingent also according to Lamarckism. And, indeed, criticism must reject this contingency of being in exactly the same way as it rejected the contingency of form maintained by Darwinians.
As far as the inheritance of truly adaptive characters comes into account—that is, the inheritance of characters which are due to the active faculty of adaptation possessed by the organism, bearing a vitalistic aspect throughout—hardly anything could be said against Lamarckism, except that inheritance of acquired characters is still an hypothesis of small and doubtful value at present. But that specific organisation proper is due to contingent variations, which accidentally have been found to satisfy some needs of the individual and therefore have been maintained and handed down, this reasoning is quite an impossibility of exactly the same kind as the argument of Darwinism.
The process of restitution, perfect the very first time it occurs, if it occurs at all, is again the classical instance against this new sort of contingency, which is assumed to be the basis of transformism. Here we see with our eyes that the organism can do more than simply perpetuate variations that have occurred at random and bear in themselves no relation whatever to any sort of unit or totality. There exists a faculty of a certain higher degree in the organism, and this faculty cannot possibly have originated by the process which Lamarckians156 assume. But if their principle fails in one instance, it fails as a general theory altogether. And now, on the other hand, as we actually see the individual organism endowed with a morphogenetic power, inexplicable by Lamarckism, but far exceeding the organogenetic faculty assumed by that theory, would it not be most reasonable to conclude from such facts, that there exists a certain organising power at the root of the transformism of species also, a power which we do not understand, which we see only partially manifested in the work of restitutions, but which certainly is not even touched by any of the Lamarckian arguments? There does indeed exist what Gustav Wolff has called primary purposefulness (“primäre Zweckmässigkeit”), at least in restitutions, and this is equally unexplainable by Darwinism and by the dogmatism of the Lamarckians.
But before entering into this area of hypothesis, let us mention a few more objections to be made to the theory of the contingency of form as put forward by Lamarckians. In the first place, let us say a few words about the appropriateness of the term “contingency” as used in this connection. The forms are regarded as contingent by Lamarckians inasmuch as the variations which afterwards serve as “means” to the “needs” of the organism occur quite accidentally with regard to the whole organism. It might be said that these “needs” are not contingent but subject to an inherent destiny, but this plea is excluded by the Lamarckians themselves, when they say that the organism experiences no need until it has enjoyed the accidental fulfilment of the same. So the only thing in Lamarckian transformism which is not of a contingent character would be the psychological agent concerned in it, as being an agent endowed with the primary power of feeling needs after it has felt fulfilment, and of judging about what the means of future fulfilment are, in order to keep them whenever they offer. But these are characteristics of life itself, irrespective of all its specific forms, which alone are concerned in transformism. Now indeed, I think, we see as clearly as possible that Darwinism and Lamarckism, in spite of the great contrast of materialism and psychologism, shake hands on the common ground of the contingency of organic forms.
The whole anti-Darwinistic criticism therefore of Gustav Wolff for instance, may also be applied to Lamarckism with only a few changes of words. How could the origin of so complete an organ as the eye of vertebrates be due to contingent variations? How could that account for the harmony of the different kinds of cells in this very complicated organ with each other and with parts of the brain? And how is it to be understood, on the assumption of contingency, that there are two eyes of almost equal perfection, and that there are two feet, two ears? Islands and mountains do not show such symmetry in their structures.
We shall not repeat our deduction of the origin of restitutions, of regeneration for instance, on the dogmatic Lamarckian theory. As we have said already, it would lead to absurdities as great as in the case of dogmatic Darwinism, and indeed we already have mentioned that Lamarckians would hardly even attempt to explain these phenomena. It follows that dogmatic Lamarckism fails as a general theory about form.157
There is finally one group of facts often brought forward against Lamarckism by Darwinian authors158 which may be called the logical experimentum crucis of this doctrine, an experimentum destined to prove fatal. You know that among the polymorphic groups of bees, termites, and ants, there exists one type of individuals, or even several types, endowed with some very typical features of organisation, but at the same time absolutely excluded from reproduction: how could those morphological types have originated on the plan allowed by the Lamarckians? Of what use would “judgment” about means that are offered by chance and happen to satisfy needs, be to individuals which die without offspring? Here Lamarckism becomes a simple absurdity, just as Darwinism resulted in absurdities elsewhere.
We were speaking about dogmatic Darwinism then, and it is about dogmatic Lamarckism that we are reasoning at present; both theories must fall in their dogmatic form, though a small part of both can be said to stand criticism. But these two parts which survive criticism, one offered by Lamarck, the other by Darwin, are far from being a complete theory of transformism, even if taken together: they only cover a small area of the field concerned in the theory of descent. Almost everything is still to be done, and we may here formulate, briefly at least, what we expect to be accomplished by the science of the future.
What has been explained to a certain extent by the two great theories now current is only this. Systematic diversities consisting in mere differences as to intensity or number may perhaps owe their origin to ordinary variation. They may at least, if we are entitled to assume that heredity in some cases is able to hand on such variations without reversion, which, it must be again remarked, is by no means proved by the facts at present. Natural selection may share in this process by eliminating all those individuals that do not show the character which happens to be useful. That is the Darwinian part of an explanation of transformism which may be conceded as an hypothesis. On the other side, congenital histological adaptedness may be regarded hypothetically as due to an inheritance of adaptive characters which had been acquired by the organism’s activity, exerted during a great number of generations. That is the Lamarckian part in the theory of descent.
But nothing more is contributed to this theory either by the doctrine of Darwin or by that of Lamarck. So it follows that almost everything has still to be done; for no hypothesis at present accounts for the foundation of all systematics, viz., for the differences in organisation, in all that relates to the so-called types as such and the degree of complication in these types, both of which (types and degree of complication) are independent of histological adaptation and adaptedness.
What then do we know about any facts that might be said to bear on this problem? We have stated already at the end of our chapter devoted to the analysis of heredity that what we actually know about any deviation of inheritance proper, that is, about congenital differences between the parents and the offspring, relating to mere tectonics, is practically nothing: indeed, there are at our disposal only the few facts observed by de Vries or derived from the experience of horticulturalists and breeders. We may admit that these facts at least prove the possibility159 of a discontinuous variation, that is of “mutation,” following certain lines of tectonics and leading to constant results; but everything else, that is everything about a real theory of phylogeny, must be left to the taste of each author who writes on the theory of the Living. You may call that a very unscientific state of affairs, but no other is possible.
And, in fact, it has been admitted by almost all who have dealt with transformism without prepossessions that such is the state of affairs. Lamarck himself, as we have mentioned already, was not blind to the fact that a sort of organisatory law must be at the base of all transformism, and it is well known that hypothetical statements about an original law of phylogeny have been attempted by Nägeli, Kölliker, Wigand, Eimer, and many others. But a full discussion of all these “laws” would hardly help us much in our theoretical endeavour, as all of them, it must be confessed, do little more than state the mere fact that some unknown principle of organisation must have been at work in phylogeny, if we are to accept the theory of descent at all.
It is important to notice that even such a convinced Darwinian as Wallace, who is well known to have been an independent discoverer of the elimination principle, admitted an exception to this principle in at least one case—with regard to the origin of man. But one exception of course destroys the generality of a principle.
As we ourselves feel absolutely incapable of adding anything specific to the general statement that there must be an unknown principle of transformism, if the hypothesis of descent is justified at all, we may here close our discussion of the subject.
A few words only must be added about two topics: on the character of organic forms as regarded by the different transformistic theories, and on the relation of transformism in general to our concept of entelechy.
We have learnt that both Darwinism and Lamarckism, in their dogmatic shape, regard the specific forms of animals and plants as being contingent; in fact, it was to this contingency that criticism was mainly directed. We therefore are entitled to say that to Darwinism and Lamarckism organic forms are accidental in the very sense of the forma accidentalis of the old logicians. There are indefinite forms possible, according to these theories, and there is no law relating to these forms. Systematics, under such a view, must lose, of course, any really fundamental importance. “There is no rational system about organisms”: that is the ultimate statement of Darwinism and of Lamarckism on this doubtful question. Systematics is a mere catalogue, not at present only, but for ever, by the very nature of the organisms. It is not owing to the indefinite number of possible forms that both our theories came to deny the importance of systematics, but to the want of a law relating to this indefinite number: among chemical compounds indefinite possibilities also exist in some cases, but they obey the law of the general formula. It is very strange that Darwinians of all people are in the forefront of systematic research in all countries: do they not see that what they are trying to build up can only relate to accidental phenomena? Or have they some doubts about the foundations of their own theoretical views, in spite of the dogmatic air with which they defend them? Or is it the so-called historical interest which attracts them?
A new question seems to arise at this point: Have not we ourselves neglected history in favour of systematics and laws? Our next lecture, the last of this year, will give the answer to this question.
At present we continue our study of the possible aspects of systematics. It is not difficult to find out what meaning organic forms would assume under any phylogenetic theory opposed to the theories of contingency. It was their defence of contingency, that is, their lack of any law of forms, that caused these theories to be overthrown—reduced to absurdities even—and therefore, it follows that to assume any kind of transformistic law is at the same time to deny the accidental character of the forms of living beings.
There is no forma accidentalis. Does that mean that the forma essentialis is introduced by this mere statement? And what would that assert about the character of systematics?
THE ORGANIC FORM AND ENTELECHY
This problem is not as simple as it might seem to be at the first glance, and, in fact, it is insoluble at present. It is here that the relation of the hypothetic transformistic principle to our concept of entelechy is concerned.
We know that entelechy, though not material in itself, uses material means in each individual morphogenesis, handed down by the material continuity in inheritance. What then undergoes change in phylogeny, the means or the entelechy? And what would be the logical aspect of systematics in either case?
Of course there would be a law in systematics in any case; and therefore systematics in any case would be rational in principle. But if the transformistic factor were connected with the means of morphogenesis, one could hardly say that specific form as such was a primary essence. Entelechy would be that essence, but entelechy in its generality and always remaining the same in its most intimate character, as the specific diversities would only be due to a something, which is not form, but simply means to form. But the harmony revealed to us in every typical morphogenesis, be it normal or be it regulatory, seems to forbid us to connect transformism with the means of morphogenesis. And therefore we shall close this discussion about the most problematic phenomena of biology with the declaration, that we regard it as more congruent to the general aspect of life to correlate the unknown principle concerned in descent with entelechy itself, and not with its means. Systematics of organisms therefore would be in fact systematics of entelechies, and therefore organic forms would be formae essentiales, entelechy being the very essence of form in its specificity. Of course systematics would then be able to assume a truly rational character at some future date: there might one day be found a principle to account for the totality of possible160 forms, a principle based upon the analysis of entelechy.161 As we have allowed that Lamarckism hypothetically explains congenital adaptedness in histology, and that Darwinism explains a few differences in quantity, and as such properties, of course, would both be of a contingent character, it follows that our future rational system would be combined with certain accidental diversities. And so it might be said to be one of the principal tasks of systematic biological science in the future to discover the really rational system among a given totality of diversities which cannot appear rational at the first glance, one sort of differences, so to speak, being superimposed upon the other.
History, in the strictest sense of the word, is the enumeration of the things which have followed one another in order of time. History deals with the single, with regard both to time and space. Even if its facts are complex in themselves and proper to certain other kinds of human study, they are nevertheless regarded by history as single. Facts, we had better say, so far as they are regarded as single, are regarded historically, for what relates to specific time and space is called history.
Taken as a simple enumeration or registration, history, of course, cannot claim to be a “science” unless we are prepared to denude that word of all specific meaning. But that would hardly be useful. As a matter of fact, what has actually claimed to be history, has always been more than a mere enumeration, even in biology proper. So-called phylogeny implies, as we have shown, that every one of its actual forms contains some rational elements. Phylogeny always rests on the assumption that only some of the characters of the organisms were changed in transformism and that what remained unchanged may be explained by the fact of inheritance.
But this, remember, was the utmost we were able to say for phylogeny. It remains fantastic and for the most part unscientific in spite of this small degree of rationality, as to which it is generally not very clear itself. For nothing is known with regard to the positive factors of transformism, and we were only able to offer the discussion of a few possibilities in place of a real theory of the factors of descent.
In spite of that it will not be without a certain logical value to begin our analysis of history in general by the discussion of possibilities again. Biology proper would hardly allow us to do more: for the simple “fact” of history is not even a “fact” in this science, but an hypothesis, albeit one of some probability.
As discussions of mere possibilities should always rest on as broad a basis as possible, we shall begin our analysis by raising two general questions. To what kinds of realities may the concept of history reasonably be applied? And what different types of “history” would be possible a priori, if the word history is to signify more than a mere enumeration?
Of course, we could select one definite volume in space and call all the consecutive stages which it goes through, its history: it then would be part of its history that a cloud was formed in it, or that a bird passed through it on the wing. But it would hardly be found very suggestive to write the history of space-volumes. In fact, it is to bodies in space that all history actually relates, at least indirectly, for even the history of sciences is in some respect the history of men or of books. It may suffice for our analysis to understand here the word body in its popular sense.
Now in its relation to bodies history may have the three following aspects, as far as anything more than a simple enumeration comes into account. Firstly, it may relate to one and the same body, the term body again to be understood popularly. So it is when the individual history of the organism is traced from the egg to the adult, or when the history of a cloud or of an island or of a volcano is written. Secondly, the subject-matter of history may be formed by the single units of a consecutive series of bodies following each other periodically. To this variety of history the discoveries of Mendel and his followers would belong in the strictest sense, but so does our hypothetical phylogeny and a great part of the history of mankind. And lastly, there is a rather complicated kind of sequence of which the “history” has actually been written. History can refer to bodies which are in no direct relation with one another, but which are each the effect of another body that belongs to a consecutive series of body-units showing periodicity. This sounds rather complicated; but it is only the strict expression of what is perfectly familiar to you all. Our sentence indeed is simply part of the definition of a history of art or of literature for instance—or, say, of a phylogenetic history of the nests of birds. The single pictures are the subjects of the history of art, and nobody would deny that these pictures are the effects of their painters, and that the painters are individuals of mankind—that is, that they are bodies belonging to a consecutive series of body-units showing periodicity. Of course, it is only improperly that we speak of a history of pictures or of books or of nests. In fact, we are dealing with painters, and with men of letters or of science, and with certain birds, and therefore the third type of history may be reduced to the second. But it was not without value to pursue our logical discrimination as far as possible.
So far we have always spoken of history as being more than a mere enumeration, but we have not ascertained what this “more” signifies. It is not very difficult to do so: in fact, there are three different types of history, each of a different degree of importance with respect to the understanding of reality.
In the first place, history may start as a mere enumeration at the beginning, and at the end, in spite of all further endeavour, may remain that and nothing more. That may occur in the first as well as in the second group of our division of history with regard to its relation to bodies. Take a cloud and describe its history from the beginning to the end: there would never be much more than pure description. Or take one pair of dogs and describe them and their offspring for four generations or more: I doubt if you will get beyond mere descriptions in this case either. The only step beyond a mere enumeration which we can be said to have advanced in these instances, consists in the conviction, gained at the end of the analysis, that nothing more than such an enumeration is in any way possible.
Quite the opposite happens when “history” deals with the individual from the egg to the adult: here the whole series of historical facts is seen to form one whole. This case therefore we shall call not history, but evolution, an evolving of something; the word “evolution” being understood here in a much wider sense than on former occasions,162 and including, for instance, the embryological alternative “evolutio” or “epigenesis.”
And half-way between enumeration and evolution there now stands a type of history which is more than the one and less than the other: there is a kind of intelligible connection between the consecutive historical stages and yet the concept of a whole does not come in. The geological history of a mountain or of an island is a very clear instance of this class. It is easy to see here, how what has been always becomes the foundation of what will be in the next phase of the historical process. There is a sort of cumulation of consecutive phases, the later ones being impossible without the earlier. So we shall speak of the type of “historical cumulation” as standing between evolution and bare temporal sequence. By means of historical cumulations history may fairly claim to “explain” things. We “understand” a mountain or an island in all its actual characteristics, if we know its history. This “historical understanding” rests on the fact that what first appeared as an inconceivable complex has been resolved into a sequence of single events, each of which may claim to have been explained by actually existing sciences. The complex has been explained as being, though not a real “whole,” yet a sum of singularities, every element of which is familiar.
But you may tell me that my discussion of evolution and of cumulation, as the higher aspects of history, is by no means complete; nay, more—that it is altogether wrong. You would certainly not be mistaken in calling my analysis incomplete. We have called one type of history evolution, the other cumulation; but how have these higher types been reached? Has historical enumeration itself, which was supposed to stand at the beginning of all analysis, or has “history” itself in its strictest sense, as relating to the single as such, risen unaided into something more than “history”? By no means: history has grown beyond its bounds by the aid of something from without. It is unhistorical elements that have brought us from mere history to more than history. We have created the concept of evolution, not from our knowledge of the single line of events attendant on a single egg of a frog, but from our knowledge that there are billions or more of frogs’ eggs, all destined to the same “history,” which therefore is not history at all. We have created the concept of cumulation not from the historical study of a single mountain, but from our knowledge of physics and chemistry and so-called dynamical geology: by the aid of these sciences we “understood” historically, and thus our understanding came from another source than history itself.
Does history always gain its importance from what it is not? Must history always lose its “historical” aspect, in order to become of importance to human knowledge? And can it always become “science” by such a transformation? We afterwards shall resume this discussion on a larger scale, but at present we shall apply what we have learned to hypothetic phylogeny. What then are the possibilities of phylogeny, to what class of history would it belong if it were complete? Of course, we shall not be able to answer this question fully; for phylogeny is not complete, and scarcely anything is known about the factors which act in it. But in spite of that, so much, it seems to me, is gained by our analysis of the possible aspects of history and of the factors possibly concerned in transformism, that we are at least able to formulate the possibilities of a phylogeny of the future in their strict logical outlines.
Darwinism and Lamarckism, regarding organic forms as contingent, must at the same time regard organic history as a cumulation; they indeed might claim to furnish an historical explanation in the realm of biology—if only their statements were unimpeachable, which as we have seen, they are not.
But any transformistic theory, which locates the very principle of phylogeny in the organism itself, and to which therefore even organic forms would be not accidental but essential, might be forced to regard the descent of organisms as a true evolution. The singularities in phylogenetic history would thus become links in one whole: history proper would become more than history. But I only say that phylogeny might be evolution, and in fact I cannot admit more than this a priori, even on the basis of an internal transformistic principle, as has been assumed. Such a principle also might lead always from one typical state of organisation to the next: but ad infinitum.163 Then phylogeny, though containing what might in some sense be called “progress,” would not be “evolution”; it might even be called cumulation in such a case, in spite of the internal transforming principle, though, of course, cumulation from within would always mean something very different from cumulation from without.164
But we must leave this problem an open question, as long as our actual knowledge about transformism remains as poor as it is. We need only add, for the sake of logical interest, that phylogeny, as a true evolution, would necessarily be characterised by the possibility of being repeated.
We only assume hypothetically that phylogeny has happened, and we know scarcely anything about the factors concerned in it. Now, it certainly would be of great importance, if at least in a small and definite field of biology we were able to state a little more, if the mere fact of phylogeny, of “history,” were at least beyond any doubt within a certain range of our biological experience. And indeed there is one department of knowledge, where history, as we know, has happened, and where we also know at least some of the factors concerned in it.
I refer to the history of mankind; and I use the expression not at all in its anthropological or ethnographical sense, as you might expect from a biologist, but in its proper and common sense as the history of politics and of laws and of arts, of literature and of sciences: in a word, the history of civilisation. Here is the only field, where we know that there actually are historical facts: let us try to find out what these facts can teach us about their succession.
The theory of history in this narrower meaning of the word has been the subject of very numerous controversies in the last twenty years, especially in Germany, and these controversies have led very deeply into the whole philosophical view of the universe. We shall try to treat our subject as impartially as possible.
Hegel says, in the introduction to his Phänomenologie des Geistes: “Die Philosophie muss sich hüten erbaulich sein zu wollen” (“Philosophy must beware of trying to be edifying”). These words, indeed, ought to be inscribed on the lintel of the door that leads into historical methodology, for they have been sadly neglected by certain theoretical writers. Instead of analysing history in order to see what it would yield to philosophy, they have often made philosophy, especially moral philosophy, the starting-point of research, and history then has had to obey certain doctrines from the very beginning.
We shall try as far as we can not to become “erbaulich” in our discussions. We want to learn from history for the purposes of philosophy, and we want to learn from history as from a phenomenon in time and in space, just as we have learnt from all the other phenomena regarding life in nature. Every class of phenomena of course may be studied with respect to generalities as well as with respect to particulars. The particular, it is true, has not taught us much in our studies so far. Perhaps it may be successful in the domain of history proper.
If I take into consideration what the best authors of the last century have written about human history with respect to its general value, I cannot help feeling that none of them has succeeded in assigning to history a position where it would really prove to be of great importance for the aims of philosophical inquiry. Is that the fault of the authors or of human history? And what then would explain the general interest which almost every one takes, and which I myself take in history in spite of this unsatisfactory state of things?
CUMULATIONS IN HUMAN HISTORY
Let us begin our analytical studies of the value and the meaning of human history, by considering some opinions which deserve the foremost place in our discussion, not as being the first in time, but as being the first in simplicity. I refer to the views of men like Buckle, Taine, and Lamprecht, and especially Lamprecht, for he has tried the hardest to justify theoretically what he regards the only scientific aim of history to be. If we may make use of our logical scheme of the three possible aspects of history, it is clear from the beginning that the history of mankind, as understood by the three authors we have named, but most particularly by Lamprecht, is neither a mere enumeration nor a true evolution, but that it has to do with cumulations, in the clearest of their possible forms. The processes of civilisation among the different peoples are in fact to be compared logically with the origin of volcanoes or mountain-ranges in Japan, or in Italy, or in America, and show us a typical series of consecutive phases, as do these. There exists, for instance, in the sphere of any single civilisation an economic system, founded first on the exchange of natural products, and then on money. There are, or better, perhaps, there are said to be, characteristic phases succeeding one another in the arts, such as the “typical,” the “individualistic,” and the “subjective” phases. Any civilisation may be said to have its “middle ages,” and so on. All these are “laws” of course in the meaning of “rules” only, for they are far from being elemental, they are not “principles” in any sense. And there are other sorts of “rules” at work for exceptional cases: revolutions have their rules, and imperialism, for instance, has its rules also.
Now, as the consecutive phases of history have been shown to be true cumulations, it follows that the rules which are revealed by our analysis, are rules relating to the very origin of cumulations also. The real element upon which the cumulation-phases, and the cumulation-rules together rest, is the human individual as the bearer of its psychology. Nobody, it seems to me, has shown more clearly than Simmel that it is the human individual, qua individual, which is concerned in every kind of history.
History, viewed as a series of cumulations, may in fact claim to satisfy the intellect by “explaining” a good deal of historical facts. It explains by means of the elemental factor of individual psychology, which every one knows from himself, and by the simple concept that there is a cumulation, supported by language and by writing as its principal factors, which both of course rest on psychology again. Psychology, so we may say, is capable of leading to cumulation phenomena; the cumulations in history are such that we are able to understand them by our everyday psychology; and history, so far as it is of scientific value, consists exclusively of cumulations.
No doubt there is much truth in such a conception of history; but no doubt also, it puts history in the second rank as compared with psychology; just as geology stands in the second rank as compared with chemistry or physics. Geology and human history may lead to generalities in the form of rules, but these rules are known to be not elemental but only cumulative; and moreover, we know the elements concerned in them. The elements, therefore, are the real subjects for further studies in the realm of philosophy, but not the cumulations, not the rules, which are known to be due to accidental constellations. Of course, the “single” is the immediate subject of this sort of history, but the single as such is emphatically pronounced to be insignificant, and the cumulations and the cumulative rules, though “singles” in a higher sense of the word, are shown to be anything but elementalities.
Therefore, on a conception of human history such as that of Buckle, Taine, Lamprecht, and others, we, of course, ought to take an interest in history, because what is “explained” by historical research touches all of us most personally every day and every year. But our philosophy, our view of the world, would remain the same without history as it is with it. We only study history, and especially the history of our own civilisation, because it is a field of actuality which directly relates to ourselves, just as we study for practical purposes the railway time-tables of our own country, but not of Australia; just as we study the local time-table in particular.
If the mere rerum cognoscere causas is regarded as the criterium of science, history of Lamprecht’s type of course is a science, for its explanations rest upon the demonstration of the typical constellations and of the elemental factor or law from which together the next constellations are known necessarily to follow. But history of this kind is not a science in the sense of discovering den ruhenden Pol in der Erscheinungen Flucht.
HUMAN HISTORY NOT AN “EVOLUTION”
Quite another view of history has been maintained by Hegel, if his explanations about the Entwicklung des objectiven Geistes (“the development of the objective mind”) may be co-ordinated with our strictly logical categories of the possible aspects of history. But I believe we are entitled to say that it was a real evolution of mankind that Hegel was thinking of; an evolution regarding mankind as spiritual beings and having an end, at least ideally. One psychical state was considered by Hegel to generate the next, not as a mere cumulation of elemental stages, but in such a way that each of the states would represent an elementality and an irreducibility in itself; and he assumed that there was a continuous series of such stages of the mind through the course of generations. Is there any sufficient reason in historical facts for such an assumption?
The mind “evolves” itself from error to truth by what might be called a system of contradictions, according to Hegel, with respect to logic as well as to morality; the sum of such contradictions becoming smaller and less complicated with every single step of this evolution. No doubt there really occurs a process of logical and moral refining, so to say, in the individual, and no doubt also, the results of this process, as far as attained, can be handed down to the next generation by the spoken word or by books. But it is by no means clear, I think, that this process is of the type of a real evolution towards an end, so far as it relates to the actual series of generations as such. On the contrary, it seems to me that we have here simply what we meet everywhere in history—a sort of cumulation resting upon a psychological basis.
The dissatisfaction that exists at any actual stage of contradiction, both moral and logical, is one of the psychical factors concerned; the faculty of reasoning is the other. Now it is a consequence of the reasoning faculty that the dissatisfaction continually decreases, or at least changes in such a way that each partial result of the logical process brings with it the statement of new problems. The number of such problems may become less, as the logical process advances, and, indeed, there is an ideal state, both logical and moral, in which there are no more problems, but only results, though this ideal could hardly be regarded as attainable by the human mind. In the history of those sciences which are wholly or chiefly of the a priori type, this process of deliverance from contradictions is most advantageously to be seen. It is obvious in mechanics and thermodynamics, and the theory of matter is another very good instance. A certain result is reached; much seems to be gained, but suddenly another group of facts presents itself, which had been previously unknown or neglected. The first result has to be changed or enlarged; many problems of the second order arise; there are contradictions among them, which disappear after a certain alteration of what was the first fundamental result, and so on. And the same is true about morality, though the difficulties are much greater here, as a clear and well-marked standard of measurement of what is good and what is bad, is wanting, or at least, is not conceded unanimously. But even here there is a consensus on some matters: one would hardly go back to slavery again, for instance, and there are still other points in morality which are claimed as ideals at least by a great majority of moral thinkers.
But all this is not true “evolution,” and indeed, I doubt if such an evolution of mankind could be proved at present in the sense in which Hegel thought it possible. The process of logical and moral deliverance from contradictions might come to an end in one individual; at least that is a logical possibility, or it might come to an end in, say, six or ten generations. And there is, unfortunately for mankind, no guarantee that the result will not be lost again and have to be acquired a second time. All this proves that what Hegel regarded as an evolution of the race is only a cumulation. There is nothing evolutionary relating to the generations of mankind as such. At least, nothing is proved about such an evolution.165
You may call my view pessimistic, and indeed you may be right so far as the sum total of human beings as such is in question. But, be it pessimistic or not, we are here moving on scientific ground only, and have merely to study the probability or improbability of problematic facts, and with such a view in our mind, we are bound to say that a true logical and moral evolution of mankind is not at all supported by known facts. There is a process of logical and moral perfection, but this process is not one, is not “single” in its actuality; it is not connected with the one and single line of history, but only with a few generations each time it occurs, or even with one individual, at least ideally. And this process is not less a process of cumulation than any other sort of development or so-called “progress” in history is. Philosophers of the Middle Ages, in fact, sometimes regarded human history as one evolutionary unity, beginning with the Creation and ending with the Day of Judgment; but every one must agree, I think, that even under the dogmatic assumptions of orthodoxy history would by no means necessarily be an “evolution.” Even then the paths taken by different individuals or different branches of the human race on their way to redemption can be regarded as independent lines.
Thus Hegel’s conception of an evolution of mankind, it seems to me, fails to stand criticism. By emphasising that there are certain lines of development in history which bring with them a stimulus to perfection, and that these lines relate to all that is highest in culture, Hegel certainly rendered the most important service to the theory of history; but in spite of that he has revealed to us only a special and typical kind of cumulation process, and nothing like an evolution. We may say that the very essence of history lies in this sort of cumulation, in this “pseudo-evolution” as we might say; and if we like to become moral metaphysicians we might add, that it is for the sake of the possibility of this sort of cumulation that man lives his earthly life; the Hindoos say so, indeed, and so do many Christians. But even if we were to depart from our scientific basis in this way we should not get beyond the realm of cumulations.
All this, of course, is not to be understood to affirm that there never will be discovered any real evolutionary element in human history—in the so-called “subconscious” sphere perhaps—but at present we certainly are ignorant of such an element.
THE PROBLEM OF THE “SINGLE” AS SUCH
If history has failed to appear as a true evolution, and if, on the other hand, it reveals to us a great sum of different cumulations, some of very great importance, others of minor importance, what then remains of the importance of the single historical event in its very singleness? What importance can the description of this event have with regard to our scientific aims? We could hardly say at present that it appears to be of very much importance at all. The historical process as a whole has proved to be not a real elemental unit, as far as we know, and such elemental units as there are in it have proved to be of importance only for individual psychology but not as history. History has offered us only instances of what every psychologist knew already from his own experience, or at least might have known if he had conceived his task in the widest possible spirit.
But is no other way left by which true history might show its real importance in spite of all our former analysis? Can history be saved perhaps to philosophical science by any new sort of reasoning which we have not yet applied to it here.
As a matter of fact, such new reasoning has been tried, and Rickert,166 in particular, has laid much stress upon the point that natural sciences have to do with generalities, while historical sciences have to do with the single in its singleness only, and, in spite of that, are of the highest philosophical importance. He does not think very highly of so-called “historical laws,” which must be mere borrowings from psychology or biology, applied to history proper, and not touching its character as “history.” We agree with these statements to a considerable extent. But what then about “history proper,” what about “the single in its very singleness”?
Let us say at first a few words about this term “single” so very often applied by us. In the ultimate meaning of the word, of course, the series of actual sensations or “presentations” is the “single” which is given “historically” to each individual, and therefore to the writer of history also, and in fact, history as understood by Rickert is based to a great extent upon this primordial meaning of single “givenness.” The word “single,” in his opinion, relates to the actual and true specification of any event, or group of events, at a given time and at a given locality in space, these events possessing an identity of their own and never being repeated without change of identity. If the subject-matter of history is defined like this, then there are, indeed, “Grenzen der naturwissenschaftlichen Begriffsbildung” with regard to history, for natural sciences have nothing to do with the single in such an understanding of the word.
Rickert says somewhere that history as a real evolution, as one totality of a higher order, would cease to be proper history: and he is right. History, in fact, would soon lose the character of specific attachment to a given space and to a given time, and would lose its “non-repeatability,” in the logical sense at least, if it were one unit in reality: as soon as it was that, it would have become a logical generality, an element in nature, so to say, in spite of its factual singularity. But history is not obliged to become that, Rickert states; and we may add that history in fact cannot become that, because it simply proves not to be an evolution as far as we know at present.
But what importance does Rickert attach to his history specified and non-repeatably single?
History has a logic of its own, he says; the scheme of its logic is not the syllogism, but the relation to “values.” So far as the single historical facts can be related to values, they are of historical importance, and in such a way only does history in its proper sense become important in itself and through itself at the same time. Must history always lose its historical aspect to become of importance to human knowledge? That is the question we asked whilst considering the general logical types of the “evolution” and “cumulation” that arose out of the analysis of the historical facts of problematic phylogeny. It now might seem that this question may be answered, and that it may be answered by a clear and simple “No.” The history of mankind, according to Rickert, seems to be important in itself, and without borrowing from any other branch of study. But is his reasoning altogether cogent and convincing?
Has it really been able to attribute to history in the strictest sense such an importance for philosophy, for the theory of the universe, “für die Weltanschauung,” that history proper may in fact be allowed to take its place beside science proper?
The relation to values is not to include any kind of “Bewertung” of judgment, Rickert allows. In fact, history of any kind would hardly satisfy the reader, if moral judgment were its basis. Every reader, of course, has a moral judgment of his own, but, unfortunately, almost every reader’s judgment is different from his neighbour’s, and there is no uniformity of moral principles as there is of geometrical ones. We shall come back to this point. At present we only state the fact that indeed moral judgment can never be the foundation of history, and that Rickert was very right to say so: it is enough to put the names of Tolstoy and Nietzsche together to understand how devoid of even the smallest general validity would be a history resting upon moral principles.
But what then are the “values” of Rickert to which history has to relate, if moral values in their proper sense have to be excluded? It is here that his discussions begin to become obscure and unsatisfactory, and the reason is fairly intelligible. He is trying to prove the impossible; he wants to put history beside science in its real philosophical importance, in spite of the fact that all evidence to establish this is wanting.
These “values,” to which every historical act in its singularity has to be related in order to become an element of real history, are they after all nothing but those groups of the products of civilisation which in fact absorb the interest of men? Is it to groups of cultural phenomena, such as arts, science, the State, religion, war, economics, and so on, that “historical” facts have to be related? Yes, as far as I understand our author, it is simply to these or other even less important groups of cultural effects—cultural “cumulations,” to apply our term—that a single action of a man or a group of men must bear some relation in order to become important historically.
But what does that mean? Is the relation to such “values” to be regarded as really rendering history equal to the sciences of nature in philosophical importance?
In the first place, there is no more agreement about such “values” than there is in the field of morals. Imagine, for instance, a religious enthusiast or recluse writing history! I fancy there would be very little mention of warriors and politicians: war and politics would not be “values” in any sense to such a man. And we know that there are others to whom those products of civilised life rank amongst the first. Rickert well notes that there is one great objection to his doctrine—the character of universality167 is wanting to his history, or rather to the values forming its basis; for there cannot be, or at least there actually is not at present, a consensus omnium with regard to these “values.”
I am convinced that Rickert is right in his conception of real “history” as the knowledge of the single acts of mankind. But this conception proves just the contrary of what Rickert hoped to prove; for history in this sense is moulded by the actual products of culture, that is, by the effects which actually exist as groups of cultural processes, and it cannot be moulded by anything else; the historian correlates history with what interests him personally.
Here now we have met definitively the ambiguous word: history indeed is to end in “interest” and in being “interesting.” There is nothing like a real “value” in any sense underlying history; the word value therefore would better give place to the term “centre of interest”—a collection of stamps may be such a “centre.” History, then, as the knowledge of cultural singularities, is “interesting,” and its aspects change with the interests of the person who writes history: there is no commonly accepted foundation of history.168
And it follows that history as regarded by Rickert cannot serve as the preliminary to philosophy. It may be169 of use for personal edification or for practical life: granting that the “centres of interest” as referred to are of any real ethical or at least factual importance. But you may take away from history even the greatest personalities, and your view of the universe, your philosophy, would remain the same, except of course so far as these personalities themselves have contributed to philosophy in any way.
Now, on the other hand, it is worth noticing that, even if there were generally accepted “values,” history as the doctrine of singularities would be deprived of philosophical importance. Its single cases would then be merely instances of certain types of actions and occurrences which have been proved to be “valuable,” i.e. to be centres of interest, before-hand. Rickert has observed that the relation to any judgments about moral values would render history unhistorical, for the generalities to which it is related would be the main thing in such a case. But he did not notice, as far as I can see, that history, if related to any “values” whatever—if there were any generally conceded—would become “non-historical” just as well: for the generalities as expressed in the “values” would be the main thing in this case also. In fact, there is no escape from the dilemma:—either no general centres of interest, and therefore a mere subjective “history”; or general “values,” and therefore history a mere collection of instances.
The “limits of concepts in natural sciences” then are the same as the limits of intellectual concepts in general. For intellectual, i.e. logical, “values” are the only centres of interest that can lay claim to universality. There are indeed other groups of important concepts, the ethical ones, but they are outside intellectuality and may enter philosophy only as problems, not as solutions. Therefore, history in its true sense, even if related to the ethical group of concepts, has no bearing on philosophy. Philosophically it remains a sum of contingencies, in which certain laws of cumulation and certain series of cumulation may be discovered. But these series and these laws, if taken scientifically, only offer us instances of psychological elementalities. They also might be instances of primary ethical states and relations, if there were such relations of more than a mere subjective and personal validity, which at present at least seems not to be the case.
We have finished our analysis of the history of mankind as the only instance of an historical biological process that is actually known to exist and is not only assumed hypothetically.
What we have learnt from this analysis, though certainly important in itself, has not afforded us any new result for theoretical biology.
The history of mankind is proved to be of philosophical importance, at present, so far only as it offers instances to the science of psychology; besides that it may be of value and importance to many conditions of practical and emotional life.
There is only one science, and only one kind of logic too. “In one sense the only science”—that was the predicate attached to natural sciences by Lord Gifford, as you will remember from our first lecture. It is not without interest to note that at the end of our course of this year, we find occasion to realise on what a deep insight into logical and philosophical relations that sentence was grounded.
We now leave the theory of human history, which has been to us nothing more than a branch of biological phylogeny in general. We have dealt with it from quite a simple realistic point of view, not burdened by any epistemology. We have taken psychical states as realities, just as we have taken as realities all parts of the animal body; and it seems to me that we were entitled to do so, as it was only history about the actions of men we were dealing with, not their actions themselves. Next summer we shall begin with studying action as action, and then, in fact, a well-founded epistemology will be among our first requirements. And history also will come on the scene once more.
It is the main result of our last chapters, devoted to systematics, transformism, and human history in particular, that no conclusions really useful for further philosophical discussion can at present be gained from these topics; there either is too little actual knowledge, or there are only combinations of natural elementalities, but no elementalities of any new kind.
To sum up: we expected that a rational system might be a biological result of the future, but we could not claim at all to possess such a system. We said that transformism might be proved one day to be a true evolution, governed by one immanent principle, which then would have to be regarded as a new primary factor in nature, but we did not know the least about that principle.
Human history, on the other hand—that is, the only historical process concerned with life that is actually known to have occurred—could not teach us anything of an elemental character, since human history, at present at least, did not appear to us as a true evolution, but only as a sum of cumulations, and the singularities of this history, taken by themselves, could only be of practical or emotional interest.
Thus it is from the study of the living individual only, that we have so far gained elemental principles in biology. The analysis of individual morphogenesis and of individual inheritance has yielded us the concept of entelechy as the chief result of the first part of our lectures. We shall be able to get more proofs of the autonomy of the individual life in the beginning of the second part; indeed, the beginning of that part will bring us to a full understanding of what the living individual is, and what it is not. And then the real philosophy of life, that is, the philosophy of the individual, will occupy us for the greater half of our lectures of next summer.
Absolute, 5
Acclimatisation, 191
Acquired characters, 217, 276 f.
Adaptation (definition), 166, 171, 185
to changes from without, 172 ff.
functional, 114, 176 ff.
and Lamarckism, 272, 280
mechanical, 177 f.
morphological, 168 ff.
physiological, 184 ff.
primary and secondary, 188 f.
Adaptedness, 186 f.
Adiantum, 279
Adventitious, 55, 74, 111, 221
Albumen, 200
Allelomorphs, 231
Amphibious plants, 172 ff.
Annelids, 65, 70, 221
Answering reaction, 181
Anti-bodies, 206 f.
Antitoxins, 207 f.
A priori, 6
Aristotle, 144
Ascaris, 93
Aspergillus, 195
Assimilation, 17
Atrophy, 178
Autonomy of life, 143, 224 f., 324
Babák, 177
Baer, C. E. v., 48 f. 282
Bateson, 229 ff., 238
Bayliss, 204, 212
Begonia, 221
Bergson, 305
Berkeley, 6
Berthold, 91
Biogenetisches Grundgesetz, 248
Biology, 8 ff., 15 f.
Blaringhem, 238, 276
Blastoderm, 39
Blastomeres, 36, 59, 61 f., 79
Blastula, 37, 61, 79
Blumenbach, 26
Boirivant, 174
Bonnet, 26
Boveri, 29, 60, 95, 235 f.
Buckle, 308, 310
Bunge, v., 248
Bütschli, 91
Calcium, 97
Calkins, 33
Cambium, 120, 183, 220
Catalysis, 164, 203
Categories, 6 f.
Cause, 99 ff.
Cell, 27 f.
-division, 28 ff., 53, 94
-lineage, 58, 70
-theory, 27 f.
Chemical theory (of morphogenesis), 134 ff.
Chemistry, systematics of, 244.
Child, C. M., 180
Chromatic regulations, 197
Chromatin, 28 f.
Chromosomes, 30, 237
Chun, 66
Classification, 246 f.
Clavellina, 129, 154, 162 f.
Cleavage, 35 ff., 53, 58, 60, 63, 71, 92
Colloids, 187
Compensatory process, 112
Complex potencies, 112, 120
Conic sections, 243
Conjugation, 33
Conklin, 86
Contingency, 218, 284 ff. 304
Continuity of germ-plasm, 215, 227
Cope, 273
Correlation (of masses), 93
(of parts), 247
Correns, 228
Crampton, 70 f.
Crayfish, 105
Ctenophores, 66
Cumulation, 301 ff., 308 ff., 314, 317
Cuvier, 247
Darwin, Ch., 260 ff., 271, 283
Darwinism, 260 ff., 271, 283 ff., 293 ff., 304
Davenport, 191, 206
Delage, 32
Descent, theory of, 250 ff.
Description, 12, 50
Detto, 172
Directive stimuli, 102 ff.
Doncaster, 232
Dreyer, 92
Echinus, 27, 33 ff., 60 ff., 68, 81, 85, 87, 98, 104, 108, 111, 154, 232, 235
Ectoderm, 41, 81, 122
Egg, 31, 33 f.
Ehrlich, 207 f.
Eimer, 292
Elementary organs, 46 ff.
processes, 46 ff.
Elements of nature, 9
Embryo, 44
frog’s, 59, 65, 67
half, 59, 61, 66 ff.
whole, 61, 67 f.
Endoderm, 41, 81
Entelechy, 143 f., 224 f., 295
Entwickelungsmechanik, 57, 70, 78, 241
Enumeration, 297, 300
Enzymes, 164, 203
Epigenesis, 26, 45, 54, 72, 144, 301
Equifinality (of restitutions), 159 f.
Equipotential, 83
Eschenhagen, 195
Evolutio, 26, 45 f., 54, 59, 61, 64, 72, 144, 205, 301
Evolution, 8, 21, 46, 250, 301, 305, 311 ff., 317
Experience, 7 f., 12, 212
Experiment, 51, 56 f.
“Explaining,” 51, 309
Explicit potency, 84
Fasting, 199 f.
Ferments, 164, 203 f.
Fertilisation, 32 ff.
Fischer, 278
Foges, 107
Form, closed or open, 49
Form, organic, specific, 16 ff., 25, 92, 293 ff.
Forma accidentalis, 293
essentialis, 294 f.
Formative stimuli, 102 ff., 113, 118, 133
Francé, 158, 239
Frédéricq, 196
Frog, embryo of, 59, 65, 67
Fromm, 205
Function (mathematical), 80
Fungi, metabolism of, 201
Gaidukow, 197 f.
Galls, 101
Galton, 228, 238
Gamble and Keeble, 198
Gastrula, 41, 61, 81
Gautier, 239
Geographical distribution, 251 f.
Geometry, solid, 243
Germ-layers, 41, 44, 61
-lineage, 215
-plasm, 52, 215
Gifford, Lord, 1 ff., 322
Godlewski, 105, 155, 235
Goebel, 116
Goethe, 247
Goette, 48, 56, 214
Goltz, 181
Growth, 30, 93 f.
Gruber, 236
Haeckel, 37, 41
Half-embryo, 59, 61, 66 ff.
Haller, A. v., 26
Harmony, 107 ff., 117, 295
Hausmann, 206
Heat production, 193
Hegel, 307, 311 ff.
Herbst, 96 ff., 102, 104 ff., 172, 177, 200, 232, 236
Heredity, 21, 52
Hering, 216 f.
Hertwig, O., 60, 65
Hertwig, R., 32 f., 60, 107
His, 56, 93
History, 2, 14, 21, 250, 257, 297 ff.
of mankind, 306 ff.
Holmes, 180
Hume, 6
Hypertrophy, 112, 114
Hypertypy, 112
Idealism, 5, 7
Immunity, 204 ff.
Implicit potency, 84
Improvement (of morphogenesis), 212
Indifferent cells, 182
Inflammation, 206
Inheritance, 35, 214 ff.
of acquired characters, 217, 275 ff., 290
Irritability, 190 ff.
Jacoby, 207
Jaeger, 214
Jennings, 218
Kammerer, 176, 280
Kant, 6 f.
Kirchhoff, 50
Klebs, 96, 170, 180, 238, 276
Kölliker, 292
Korshinsky, 239
Krašan, 221, 251
Lamarck, 271 f., 291
Lamarckism, 271 f., 284 ff., 293 ff., 304
Lamprecht, 308, 310
Larva, 41 f., 44
Law of nature, 13, 16
Leibniz, 6
Lens (of eye), 105, 221
Liebmann, 256
Life, 9 f., 16, 21
Lillie, R. S., 236
Limits of regulability, 212
Lithium, 99
“Living,” 9, 16
Localisation, 101, 103, 118 ff.
Locke, 6
Loeb, J., 32, 102, 164, 179, 196, 236
Loeb, L., 208
Lyon, 87
MacDougal, 238 f., 276
Machine (definition), 139
Machine-theory of life, 138 ff., 187, 210
Maillard, 196
Manifoldness, 25 f., 30, 45
intensive, 144
Materialism, 283
Materials, transport of, 194
Matter, theory of, 8
Maturation, 31, 87
Mayenburg, v., 195
Means, of morphogenesis, 89 ff., 101, 113, 118, 228, 234
Memory, 216 f.
Mendel, 229 f.
Merrifield, 198
Mesenchyme, 39, 41, 104, 111, 151 f.
Metabolic regulations, 198 f.
Metabolism, 16, 184
of fungi, 201
Metschnikoff, 206
Micromeres, 36, 60
Miehe, 116
Mill, J. S., 57
Mimosa, 191
Minkiewicz, 198
Modification, 277
Molluscs, 70 f., 86
Morgan, T. H., 32, 66 f., 95, 107, 114 f., 162, 230
Morphaesthesia, 157
Morphogenesis, 20, 52, 76, 112, 118 f.
Morphology, 12
Movements, organic, 17
Mutations, 237 f., 276, 291
Nägeli, 266, 292
Nathansohn, 196
Natural selection, 261 f., 290
Nature, 5 ff.
Němec, 116
Newport, 57
Newt (regeneration of), 155, 221 f.
Noll, 146, 157 f.
Nomothetic, 14 f.
Normal, 78
Nuclear division, 28 f., 62, 64 f., 72, 235
Nucleus, 28, 35
rôle of nucleus in inheritance, 233 f.
Organ-forming substances, 117
Oscillariae, 197
Osmotic pressure, 93, 187, 194 f.
Overton, 196 f.
Oxidation, rôle of, 198 f.
Palaeontology, 252
Parallelism (psycho-physical), 146
Parthenogenesis, 32
Pauly, 146, 217, 273 f.
Pawlow, 204, 210, 212
Pearl, R., 212
Pfeffer, 195, 201
Phagocytosis, 206
Phenomenon, 5 f.
Philosophy, natural, 4
of nature, 4, 7, 9
of the organism, 9, 15
Phylogeny, 255, 291, 297, 304 ff.
Physiology, 12
of development (morphogenesis), 20
Planaria, 130, 155, 162 f., 200
Plants, 48 f.
Plato, 2
Pluteus, 42
Poisons, 205 ff.
Pole, 36
Polarity, 106
Potencies, complex, 112, 120
explicit, 84
implicit, 84
primary, 84, 111
prospective, 77 ff., 83, 89, 118, 125, 241
secondary, 84, 110
Poulton, 198
Precipitin, 207 f.
Pressure experiments, 63, 141
Primary potency, 84, 111
purposefulness, 146, 287
regulation, 85, 174, 188
Progress, 305
Pronuclei, 55
Prospective potency, 77 ff., 83, 89, 118, 125, 241
value, 77 f., 80, 122
Protista (Protozoa), 27, 130, 236
Protoplasm, 28, 30
morphogenetic rôle of, 67
Przibram, 112, 248
Rádl, 247
Rauber, 235
Reaction, answering, 181
Reciprocity of harmony, 156 f.
Re-differentiation, 75, 111, 163
Regeneration, 55, 74, 105, 111, 221
super-, 115 f.
Regulation, 68, 73, 85, 111, 165
defined, 166
metabolic, 198 f.
secondary, 85, 165, 188
Reinke, 146
Restitution, 21, 74, 110, 112 ff.
defined, 166
and Darwinism, 267
and Lamarckism, 286
of second order, 158
Retina, 191
Retro-differentiation, 163 f.
Rhumbler, 93
Ribbert, 114
Rickert, 315 ff.
Roux, 26, 48, 55 ff., 66 f., 76, 89, 92 f., 108, 161, 176 f., 241
Rubner, 193
Sachs, 117
Sadebeck, 279
Salamandra, 175, 281
Schneider, 146
Schultz, E., 200
Schultze, O., 67
Schwendener, 177
Science, 14, 297
natural, 1 ff.
rational, 12
Sea-urchin, see Echinus
Secondary potency, 84, 110
regulation, 85, 165, 188
Secretion, internal, 116, 200
Segmentation, 35
Selective qualities (of tissues), 186
Self-differentiation, 108
Semon, 216 f.
Sex, 107
Single, the, 315 ff.
Skeleton, 40 ff., 44, 47, 92
Spemann, 105
Spermatozoon (spermia), 32 ff.
Splitting (of hybrids), 229 f.
Stahl, 197
Standfuss, 278
Starfish, 44, 81, 122
Starling, E., 116, 204, 212
Stentor, 131
Stimuli, directive, 102 ff.
formative, 102 ff., 113, 118, 133
of restitutions, 113 f.
Structure of protoplasm, 66, 69, 72 f. 85, 88
Substance, living, 17
Sumner, 196
Super-regeneration, 115 f.
Surface-tension, 91
Sutton, 230
Symmetry, 39, 68, 70, 72, 89, 98
System, combined types of, 153 ff.
complex, 219 f.
complex-harmonious, 155
equipotential, 120
harmonious-equipotential, 121 ff., 151 f.
mixed-equipotential, 154
morphogenetic, 119 f., 163, 241
Systematics, 14 ff., 21, 243 f., 253, 264, 293, 296
Taine, 308, 310
Theology, natural, 1 ff.
Thomson, J. A., 16
Thymus, 204
Thyroid, 204
Tissue, 38
Toxins, 207 f.
Transformism, 251
Truth, 7
Tschermak, 228
Tubularia, 126 ff., 133, 158 ff.
Type, 48, 247 f., 282, 291
“Understanding” (historically), 302
Universality, postulate of, 148 f.
Universe, 5
Univocality, principle of, 161
“Values,” 317 ff.
prospective, 77 f., 80, 122
Variation, 218, 237 f., 276
Variation, fluctuating, contingent, 264 f., 273 f., 282, 290
Vernon, 232, 238
Vitalism, 143, 145 f., 210 f., 224 f., 234, 240 f., 272, 277
Vöchting, 174, 179 f., 182, 221
Volition, acts of, 274
Vries, de, 228, 238 f.
Wallace, 292
Ward, J., 8, 143
Weber, law of, 191
Weinland, 202
Weismann, 33, 52 ff., 58 f., 72, 74 f., 103, 111, 138, 214 f., 237, 277 f.
Weldon, 238
Whole, the, 28, 80, 117
-embryo, 61, 67 f.
Wigand, 255, 266, 292
Wilson, E. B., 27, 65, 70 f., 86 f., 107
Windelband, 13 f.
Winkler, 116, 221
Winterstein, 199
Wolff, C. F., 26
Wolff, G., 105, 146, 255, 266, 287 f.
Wolff, J., 177
Yung, 177
Zeleny, 112, 115, 212
Zur Strassen, 93
THE END
Printed by R. & R. Clark, Limited, Edinburgh.
BY
G. CHATTERTON HILL
Demy 8vo, Cloth, 600 pages.
Price 12s. 6d. net.
Post Free, Price 12s. 11d.
SOME PRESS OPINIONS
“A most praiseworthy and suggestive work—should certainly be studied by every serious thinker.”—Morning Post.
“Mr. Hill is decidedly doctrinaire, but his book is packed with scientific and sociological facts, and it gives the reader healthy intellectual exercise.”—Christian World.
“Shows wide reading, is written in a forcible and clear style, and contains much that is interesting, fresh, and acute.”—Aberdeen Free Press.
“It is a book of equal calibre with Mr. Kidd’s and goes even deeper than that remarkable production into the springs of life and conduct.”—Methodist Recorder.
“This most suggestive and valuable work, which contains abundant sociological data.”—Aberdeen Journal.
“Mr. George Chatterton Hill has written a volume of surpassing interest not alone to scientific but to theological students.”—Catholic Times.
Published by A. & C. BLACK, SOHO SQUARE, LONDON, W.
BY
W. R. BOYCE GIBSON
LECTURER IN PHILOSOPHY IN THE UNIVERSITY OF LONDON
SECOND EDITION
Crown 8vo, Cloth, with Frontispiece Portrait of Rudolf Eucken.
Price 3s. 6d. net.
Post Free, Price 3s. 10d.
SOME PRESS OPINIONS
“Mr. Gibson has given us in small compass a lucid exposition of the philosophical system of Eucken, who is Professor of Philosophy in Jena. . . . This is a most suggestive and stimulating book. In a very real sense it has brought philosophy down to earth and is deserving of serious study.”—Aberdeen Free Press.
“To it the interested reader will turn with expectation, and his expectation is likely to be more than realised. For Dr. Boyce Gibson is himself a scholar, as well as an enthusiastic lover of this great scholar.”—Expository Times.
“No reader should fail to find pleasure in a book so full of fresh and stimulating thought, expressed with great felicity of language.”—The Scottish Review.
“It is done with just the proper combination of sympathy and criticism.”—British Weekly.
“This little book on Eucken’s Philosophy is of quite exceptional interest and importance.”—The Inquirer.
“Professor Boyce Gibson . . . has performed a real service in promoting the acquaintance of English, and American students with a thinker whose distinctive views give him a special claim to their attention. . . . Professor Gibson has achieved a notable success, writing briefly, lucidly, and sympathetically.”—The New Age.
Published by A. & C. BLACK, SOHO SQUARE, LONDON, W.
1 Windelband (Geschichte und Naturwissenschaft, 3 Auflage, 1904) gives the name “nomothetic” to the whole of our “science” and calls the method of history “idiographic.” We thought it better to establish three fundamental types of all possible branches of knowledge.
2 See J. Arth. Thomson, The Science of Life, London, 1899.
3 E. B. Wilson, The Cell in Development and Inheritance, New York, Macmillan, 1896.
4 Amer. Journ. Physiol. vols. iii. and iv. 1900.
5 According to Delage (Arch. Zool. exp., 3 sér. 10, 1902), it is indifferent for the realisation of artificial parthenogenesis, whether but one, or both, or neither of the “polar bodies” has been formed. But the egg must be in the first stages of maturation to the extent that the “nuclear membrane” must be already dissolved.
6 The older theories, attributing to fertilisation (or to “conjugation,” i.e. its equivalent in Protozoa), some sort of “renovation” or “rejuvenescence” of the race, have been almost completely given up. (See Calkins, Arch. für Entwickelungsmechanik, xv. 1902). R. Hertwig recently has advocated the view, that abnormal relations between the amounts of nuclear and of protoplasmatic material are rectified in some way by those processes. Teleologically, sexual reproduction has been considered as a means of variability (Weismann), but also as a means of preserving the type!
7 The phrase “ceteris paribus” has to be added of course, as the duration of each single elementary morphogenetic process is liable to vary with the temperature and many other conditions of the medium.
8 We shall not avoid in these lectures the word “explain”—so much out of fashion nowadays. To “explain” means to subsume under known concepts, or rules, or laws, or principles, whether the laws or concepts themselves be “explained” or not. Explaining, therefore, is always relative: what is elemental, of course, is only to be described, or rather to be stated.
9 Das Keimplasma, Jena, 1892.
10 Die Bedeutung der Kernteilungsfiguren, Leipzig, 1883.
11 Unsere Körperform, Leipzig, 1875.
12 Die Entwickelungsgeschichte der Unke, Leipzig, 1875.
13 Gesammelte Abhandlungen, Leipzig, 1895. Most important theoretical papers:—Zeitschr. Biolog. 21, 1885; Die Entwickelungsmechanik der Organismen, Wien, 1890; Vorträge und Aufsätze über Entwickelungsmechanik, Heft i., Leipzig, 1905.
14 Virchow’s Archiv. 114, 1888.
15 Zeitschr. wiss. Zool. 53, 1891.
16 Zeitschr. wiss. Zool. 55, 1892.
17 In the pressure experiments I had altered the relative position of the nuclei in origine. In later years I succeeded in disturbing the arrangement of the fully formed cells of the eight-cell stage, and in getting normal larvæ in spite of that in many cases. But as this series of experiments is not free from certain complications—which in part will be understood later on (see page 73)—it must suffice here to have mentioned them. (For further information see my paper in Archiv. f. Entwickelungsmechanik, xiv., 1902, page 500.)
18 Mitteil. Neapel. 11, 1893.
19 But the elementary magnets would have to be bilateral!
20 Arch. Entw. Mech. 2, 1895.
21 Anat. Anz. 10, 1895.
22 Arch. Entw. Mech. 3, 1896.
23 It deserves notice in this connection, that in some cases the protoplasm of parts of a germ has been found to be more regulable in the earliest stages, when it is very fluid, than later, when it is more stiff.
24 Compare my Analytische Theorie der organischen Entwickelung, Leipzig, 1894, and my reviews in Ergebnisse der Anatomie und Entwickelungsgeschichte, vols. viii. xi. xiv., 1899–1905. A shorter review is given in Ergebnisse der Physiologie, vol. v., 1906. The full literature will be found in these reviews.
25 If the plane of section passes near the equator of the germ, two whole larvae may be formed also, but in the majority of cases the “animal” half does not go beyond the blastula. The specific features of the organisation of the protoplasm come into account here. See also page 65, note 17.
26 A change of the position of the cell is of course effected by each variation of the direction of the cut, which is purely a matter of chance.
27 The reader will remember (see page 65, note 17), that even the germ of Echinus is not quite equipotential along its main axis, but it is equipotential in the strictest sense around this axis. The germs of certain medusae seem to be equipotential in every respect, even in their cleavage stages.
28 Journ. Exp. Zool. 1, 1904.
29 Great caution must be taken in attributing any specific morphogenetic part to differently coloured or constructed materials, which may be observed in the egg-protoplasm in certain cases. They may play such a part, but in other cases they certainly do not (see Lyon, Arch. Entw. Mech. 23, 1907). The final decision always depends on experiment.
30 It seems that these physical conditions also—besides the real specifications in the organisation of the egg—may be different before and after maturation or (in other cases) fertilisation. (See Driesch, Archiv f. Entwickelungsmechanik, 7, p. 98; and Brachet, ibid. 22, p. 325.)
31 Studien über Protoplasmamechanik, Leipzig, 1886.
32 Unters. üb. mikroskopische Schäume und das Protoplasma, Leipzig, 1892.
33 Jena. Zeitschr. 26, 1892.
34 According to Zur Strassen’s results the early embryology of Ascaris proceeds almost exclusively by cellular surface-changes: the most typical morphogenetic processes are carried out by the aid of this “means.” As a whole, the embryology of Ascaris stands quite apart and presents a great number of unsolved problems; unfortunately, the germ of this form has not been accessible to experiment hitherto.
35 Rhumbler has recently published a general survey of all attempts to “explain” life, and morphogenesis in particular, in a physico-chemical way (“Aus dem Lückengebiet zwischen organismischer und anorganismischer Natur,” Ergeb. Anat. u. Entw.-gesch. 15, 1906). This very pessimistic survey is the more valuable as it is written by a convinced “mechanist.”
36 Compare the analytical discussions of Klebs, to whom we owe a great series of important discoveries in the field of morphogenetic “means” in botany. (Willkürliche Entwickelungsänderungen bei Pflanzen, Jena, 1903; see also Biol. Centralblatt, vol. xxiv., 1904, and my reply to Klebs, ibid. 23, 1903.)
37 Arch. Entw. Mech. 17, 1904.
38 Zeitschr. wiss. Zool. 55, 1902; and Mitt. Neapel. 11, 1903.
39 In certain cases part of the specific feature of the process in question may also depend on the “cause” which is localising it, e.g. in the galls of plants.
40 Herbst, “Ueber die Bedeutung die Reizphysiologie für die kausale Auffassung von Vorgängen in der tierischen Ontogenese” (Biol. Centralblatt, vols. xiv., 1894, and xv., 1895); Formative Reize in der tierischen Ontogenese, Leipzig, 1901. These important papers must be studied by every one who wishes to become familiar with the subject. The present state of science is reviewed in my articles in the Ergebnisse der Anatomie und Entwickelungsgeschichte, vols. xi. and xiv., 1902 and 1905.
41 Compare the important papers by J. Loeb, Untersuchungen zur physiologischen Morphologie der Tiere, Würzburg, 1891–2.
42 I use the word “primordia” for the German “Anlage”; it is better than the word “rudiment,” as the latter may also serve to signify the very last stage of a certain formation that is disappearing (phylogenetically).
43 A full analysis of the subject would not only have to deal with formative stimuli as inaugurating morphogenetic processes, but also with those stimuli which terminate or stop the single acts of morphogenesis. But little is actually known about this topic, and therefore the reader must refer to my other publications. I will only say here, that the end of each single morphogenetic act may either be determined at the very beginning or occur as an actual stopping of a process which otherwise would go on for ever and ever; in the first case some terminating factors are included in the very nature of the morphogenetic act itself.
44 A full account of the present state of the subject will be found in Morgan’s Experimental Zoology, New York, 1907.
45 But there certainly exist many formative relations between the real sexual organs and the so-called secondary sexual characters. Herbst has given a full analytical discussion of all that is known on this subject; but the facts are much more complicated than is generally supposed, and do not lend themselves therefore to short description. See also Foges, Pflüger’s Arch. 93, 1902.
46 It seems that in some cases (Dinophilus, certain Arthropods) the sexual products are invariably determined as “arrenogennetic” or as “thelygennetic” (Wilson, Journ. Exp. Zool. ii. and iii. 1905–6), whilst in others (Amphibia) the state of maturation or “super”-maturation determines the sex of the future organism (R. Hertwig, Verh. D. Zool. Ges. 1905–7).
47 Driesch, Die organischen Regulationen, Leipzig, 1901; Morgan, Regeneration, New York, 1901.
48 But real compensatory differentiation occurs in the cases of so-called “hypertypy” as first discovered by Przibram and afterwards studied by Zeleny: here the two organs of a pair show a different degree of differentiation. Whenever the more specialised organ is removed the less developed one assumes its form. Similar cases, which might simply be called “compensatory heterotypy,” are known in plants, though only relating to the actual fate of undifferentiated “Anlagen” in these organisms. A leaf may be formed out of the Anlage of a scale, if all the leaves are cut off, and so on.
49 For a fuller analysis compare my opening address delivered before the section of “Experimental Zoology” at the Seventh Zoological Congress, Boston, 1907: “The Stimuli of Restitutions” (see Proceedings of that Congress).
50 The problem of the stimulus of a secondary restitution as a whole must not be confused with the very different question, what the single “formative stimuli” concerned in the performance of a certain restitutive act may be. With regard to restitution as a whole these single “formative stimuli” might properly be said to belong to its “internal means”—in the widest sense of the word.
51 T. H. Morgan is very right in stating that, in regeneration, the “obstacle” itself is newly formed by the mere process of healing, previous to all restitution, and that true restitution happens all the same.
52 I merely mention here the still “simpler” one—applicable of course to regeneration proper exclusively—that for the simple reason of being “wounded,” i.e. being a surface open to the medium, the “wound” brings forth all that is necessary to complete the organism.
53 That compensatory hypertrophy cannot be due to “functional adaptation”—to be analysed later on—was proved by an experiment of Ribbert’s. Compensation may occur before the function has made its appearance, as was shown to be the case in the testicles and mammae of rabbits. (Arch. Entw. Mech. 1, 1894, p. 69.)
54 At any given time only the absolute size of the regenerated part is greater in animals which are well fed; the degree of differentiation is the same in all. Zeleny has found that, if all five arms of a starfish are removed, each one of them will regenerate more material in a given time than it would have done if it alone had been removed. But these differences also only relate to absolute size and not to the degree of differentiation. They possibly may be due in fact to conditions of nourishment, but even here other explanations seems possible (Zeleny, Journ. exp. Zool. 2, 1905).
55 For a good discussion of “super-regeneration” in the roots of plants see Němec, Studien über die Regeneration, Berlin, 1905. Goebel and Winkler have succeeded in provoking the “restitution” of parts which were not removed at all by simply stopping their functions (leaves of certain plants were covered with plaster, etc.). (Biol. Centralbl. 22, 1902, p. 385; Ber. Bot. Ges. 20, 1902, p. 81.) A fine experiment is due to Miehe. The alga Cladophora was subjected to “plasmolysis,” each cell then formed a new membrane of its own around the smaller volume of its protoplasm; after that the plants were brought back to a medium of normal osmotic pressure, and then each single cell grew up into a little plant (all of them being of the same polarity!). Two questions seem to be answered by this fact: loss of communication is of fundamental importance to restitution, and the removal of mechanical obstacles plays no part in it, for the mechanical resistances were the same at the end of the experiment as they had been at the beginning. (Ber. Bot. Ges. 23, 1905, p. 257.) For fuller analysis of all the problems of this chapter see my Organische Regulationen, my reviews in the Ergebnisse der Anatomie und Entwickelungsgeschichte, vols. viii. xi. xiv., and my Boston address mentioned above. Compare also Fitting, Ergebn. d. Physiol. vols. iv. and v.
56 The so-called “inner secretion” in physiology proper would offer a certain analogy to the facts assumed by such an hypothesis. Compare the excellent summary given by E. Starling at the seventy-eighth meeting of the German “Naturforscherversammlung,” Stuttgart, 1906.
57 The name of singular-equipotential systems might also be applied to elementary organs, the single potencies of which are awaked to organogenesis by specific formative stimuli from without; but that is not the case in the systems studied in this chapter.
58 The distance of the other boundary line from a or b would be given by the value of s.
59 A far more thorough analysis of this differentiation has been attempted in my paper, “Die Localisation morphogenetischer Vorgänge. Ein Beweis vitalistischen Geschehens,” Leipzig, 1899.
60 This statement is not strictly correct for Tubularia. I found (Archiv f. Entwickelungsmechanik, ix. 1899), that a reduction of the length of the stem is always followed by a reduction of the size of the hydranth-primordium, but there is no real proportionality between them. It is only for theoretical simplification that a strict proportionality is assumed here, both in the text and the diagram. But there is an almost strict proportionality in all cases of “closed forms.”
61 One might object here that in a piece of a Tubularia stem, for instance, the tissues are in direct contact with the sea-water at the two points of the wounds only, and that at these very points a stimulus might be set up—say by a process of diffusion—which gradually decreases in intensity on its way inward. And a similar argument might apply to the small but whole blastula of Echinus, and to all other cases. But, in the first place, stimuli which only differ in intensity could hardly call forth the typical and typically localised single features realised in differentiation. On the other hand—and this will overthrow such an hypothesis completely—the dependence of the single localised effects in every case on the absolute size of the fragment or piece chosen for restoration renders quite impossible the assumption that all the singularities in the differentiation of the harmonious systems might be called forth by single stimuli originating in two fixed places in an independent way. These would never result in any “harmonious,” any proportionate structure, but a structure of the “normal” proportionality and size at its two ends and non-existent in the middle!
62 See my article in Biolog. Centralblatt, 27, 1907, p. 69. The question is rendered still more complicated by the fact that in the case of the regeneration, say, of a leg it is not the original “morphogenetic compound” which is again required for disintegration, after it has become disintegrated once already, but only a specific part of it: just that part of it which is necessary for producing the leg! On the other hand, it would be impossible to understand, on the basis of physical chemistry, how the isolated branchial apparatus of Clavellina could be transformed, by chemical processes exclusively, into a system of which only a certain part consists of that substance of which the starting-point had been composed in its completeness.
63 Besides the specified poles determined by the polar-bilateral structure of the protoplasm.
64 The pressure experiments and the dislocation experiments come into account here; for the sake of simplicity they have not been alluded to in the main line of our argument.
65 My “first proof of vitalism” was first developed in the paper, “Die Localisation morphogenetischer Vorgänge,” Leipzig, 1899. (See additional remarks in Organische Regulationem, Leipzig, 1901, and in Archiv für Entwickelungsmechanik, 14, 1902.) I cannot admit that any really serious objection has been brought forward against it. (See my articles in Biologisches Centralblatt, 22, 23, 27, and in Ergebnisse d. Anat. u. Entwickelungsgesch. 11, 14.) An historical sketch of vitalism will be found in my book, Der Vitalismus als Geschichte und als Lehre, Leipzig, 1905.
66 We are dealing here with morphogenesis and so-called vegetative physiology only; to certain psychologists, who have refuted the theory of psycho-physical parallelism, I must grant that they also have proved vitalism. (See Volume II.)
67 The eight larvae would be incomplete in some respect, but not with regard to symmetry. They would be “whole” ones, only showing certain defects in their organisation. See page 65 note 17, and page 73.
68 Reciprocal harmony may be reduced in some cases to the given proportions of one original harmonious system, from which the single constituents of the complicated system, showing reciprocal harmony, are derived. Then we have only an instance of “harmony of constellation” (see p. 109). But reciprocal harmony seems to become a problem itself, if it occurs in restitutions starting from quite a typical point, selected by the experimenter. It will be a problem of future research to give an exact formula of what happens here. Reciprocal harmony also occurs in regeneration proper. It is known that the formation of the regenerative bud and the differentiation of this bud follow each other. As the bud is composed of different elementary systems, it follows that these different systems, of which every single one is harmonious, also have to work in reciprocity to each other, in order that one whole proportionate formation may result.
69 Biol. Centralblatt. 23, 1903.
70 Certain phenomena of the physiology of growth of Geranium Robertianum, recently discussed by Francé from a vitalistic point of view (Zeitschr. Entw. lehre. 1, 1907, Heft iv.), might also belong here. I cannot see an independent proof of vitalism in these facts if taken by themselves; a pre-existing “machine” cannot be absolutely excluded here.
71 Driesch, Arch. Entw. Mech. 5, 1897.
72 Driesch, Arch. Entw. Mech. 14, 1902.
73 The root may be restored by regeneration proper, or by the production of adventitious roots, or by one of the side-roots changing its geotropism from horizontal to positive, according to the smaller or greater distance of the wound from the tip.
74 “Retro”-differentiation, of course, is not “Re”-differentiation (“Umdifferenzierung,” see p. 111), though it may help it to occur.
75 Of course such a real decay of parts may happen in other cases.
76 Certain cases of retro-differentiation occurring under conditions of strict fasting will be described in a later chapter.
77 Klebs has suppressed the reproductive phase of organisation altogether, in fungi as well as in flowering plants, or has made it occur abnormally early, merely by changing the “external conditions” and by altering the “internal” ones correspondingly. There is hardly anything like an adaptation in these cases, which, by the way, offer certain difficulties to analysis, as the boundaries between “cause” and “means” are not very sharp here.
78 Compare Herbst, Biol. Centralbl. 15, 1895; and Detto, Die Theorie der direkten Anpassung, Jena, 1904. A full account of the literature will be found in these papers.
79 Vöchting (Jahrb. wiss. Bot. 34, 1899) forced the bulbs of plants to become parts of the stem, and parts of the stem to form bulbs; in both cases the most characteristic changes in histology could be observed, being in part adaptations, but in part restitutions of the proper type. (See also my Organische Regulationen, 1901, p. 84.) A true and simple instance of a “secondary adaptation” seems to be furnished in a case described by Boirivant. In Robinia all the leaflets of a leaf-stalk were cut off: the leaf-stalk itself then changed its structure in order to assist assimilation, and also formed real stomata.
80 Arch. Entw. Mech. 17, 1904.
81 Roux, Gesammelte Abhandlungen, vol. i. 1895; in particular, Der Kampf der Teile im Organismus, Leipzig, 1881.
82 Arch. Entw. Mech. 21, 1906. By a very detailed comparative study Babák was able to prove that it is the plant proteids to which the effect of vegetable food is chiefly due; thus we have an adaptation to digestibility. Mechanical circumstances are only of secondary importance. (See also Yung.)
83 Atrophy of muscles by inactivity is not to be confused with atrophy by cutting the motor nerve; the latter is very much more complete.
84 Loeb has advocated the view that the “adaptive” growth of working muscles is simply due to the presence of a greater number of molecules in their protoplasm, muscular activity being generated by a process of chemical decomposition.
85 What has been really proved to exist by the very careful studies carried out by Child, is only certain cases of functional adaptation to mechanical conditions of the strictest kind, and relating to the general mobility only, but nothing more; such adaptations can be said to accompany restitution. See, for instance, Journ. exp. Zool. 3, 1906, where Child has given a summary of his theory.
86 Even in Vöchting’s experiments (see page 174, note 79), in which adaptations are mixed with true restitutions in the closest possible manner, a few phenomena of the latter type could most clearly be separated. The stimulus which called them forth must have been one of the hypothetic sort alluded to in a former chapter (see page 113). The best instances of true restitutions were offered in those cases, where, after the removal of all the bulbs, typical starch-storing cells were formed without the presence of any starch.
87 Beiträge zur Lehre von den Functionen der Nervencentren des Frosches, Berlin, 1869.
88 The “secondary adaptations” observed by Vöchting are too complicated and too much mingled with restitutions to allow any definite analysis of the fact of the “secondary adaptation” as such.
89 General literature: Fröhlich, Das natürliche Zweckmüssigkeitsprincip in seiner Bedeutung für Krankheit und Heilung, 1894. Driesch, Die organischen Regulationen, 1901. A. Tschermak, “Das Anpassungsproblem in der Physiologie der Gegenwart,” in a collection of papers in honour of J. P. Pawlow, St. Petersburg, 1904. Bieganski, “Ueber die Zweckmässigkeit in den pathologischen Erscheinungen,” Annal. d. Naturphil. 5, 1906. Among the general text-books of physiology those by Pfeffer (Pflanzenphysiologie, 1897–1904) and von Bunge (Lehrbuch d. Phys. d. Menschen, 1901) are the fullest on the subject of “regulations.” See also different papers on general pathology by Ribbert.
90 According to investigations of the last two years, the physics of colloids seems to play as important a part in physiology as osmosis does; we here meet “means” of functioning just as we have already had “means” of organogenesis.
91 I only mention here that certain modern psychologists have assigned the true law of Weber to the sphere of judgment and not of sensation. If applied to objective reactions only, in their dependence on objective stimuli, it, of course, becomes less ambiguous, and may, in a certain sense, be said to measure “acclimatisation” with regard to the stimulus in question. The mathematical analogy of the law of Weber to the most fundamental law of chemical dynamics seems very important.
As to “acclimatisation” in the more usual meaning of the word, with regard to a change of the general faculty of resisting certain agents of the medium, “immunity” proper is to form a special paragraph of what follows, and to “acclimatisation” towards different degrees of salinity (in algae or fishes) some special remarks will also be devoted on a proper occasion. There remains only “acclimatisation” to different temperatures; but on this topic not much more than the fact is known (see Davenport, Arch. f. Entw. Mech. 2, p. 227). “Acclimatisation” does not allow of a sharp general definition; it may be the result of very different kinds of adaptations in our sense of the word.
92 I should think that the problem of the re-establishment of irritability, in principle at least, arises even when there is not a trace of so-called “fatigue” or of a “refractory period.” The process of restoring may be so rapid as not to be noticeable, nevertheless some sort of restoring is to be postulated. We may say the “irritability” of an elastic ball is re-established by its elasticity. A certain analogy to this case may perhaps be found in the muscle. But the irritability of nerves with respect to nervous conduction, and of glands with respect to secretion, or of the articulations of Mimosa may be well understood, hypothetically at least, if we assume that the ordinary course of metabolic events is apt in itself to lead to a certain state or condition of the organs in question upon which their irritability is based. Certain general conditions of functioning, as for instance the presence of oxygen for the contraction of the muscle, would better be looked upon as necessary “means” of functioning than as being part of irritability as such. “Fatigue,” of course, may also be due to the absence of such “means” or to abnormal conditions originated by functioning itself.
93 Rubner, Die Gesetze des Energieverbrauches bei der Ernährung, Leipzig u. Wein, 1902.
94 The phenomenon of fever we leave out of account here; it is regarded by some as regulation, by others as a disturbance of heat regulation. Of course, if the first view should ever prove to be the right one, fever might be classified among the real regulations of the secondary type.
95 Jahrb. wiss. Bot. 36, 1901.
96 Carbohydrates cannot be ionised, and therefore there is no doubt that in von Mayenburg’s experiments the organism itself is actively at work. As to compounds liable to ionisation, it has been noticed by Maillard that a certain regulatory character is contained simply in the physical fact that the degree of ionisation changes with concentration: decrease of concentration for instance would be followed by an increase of ionisation, and so the osmotic pressure may be preserved (C. rend. Soc. Biol. 53, 1901, p. 880).
97 In the different experiments of Nathansohn (Jahrb. wiss. Bot. 38, 1902, and 39, 1903) the salinity of the medium was changed in such a way that there was in each case either an abnormal increase or an abnormal decrease in the concentration of one single ion necessary for metabolism. The cell was found to stand these abnormal changes in such a way that in the case of the increase of the concentration of the medium it did not allow more than a certain amount of the ion in question to come in, and that in the case of the decrease it did not allow more than a certain quantity of the ion to go out. It thus seems as if the permeability of the surface were adjusted to a certain minimum and to a certain maximum of every single ion or salt, the permeability being stopped from within to without, whenever the minimum, and from without to within, whenever the maximum is reached in the cell sap; both irrespective of proper physical osmotic equilibrium (“Physiologisches Gleichgewicht”). Thus, in fact, there only would be a case of primary regulation, nothing more. It would all appear rather similar to what occurs in the kidney. Of course we do not assert that our explanation is right, but it is possible and is at the same time the most simple, and it is our general practice always to prefer the most simple hypotheses.
98 Many fishes are able to withstand great changes in the osmotic pressure of sea-water; the osmotic pressure of their body fluids, though never in a real physical equilibrium with the pressure of the medium, nevertheless may vary whenever the abnormal conditions of the latter exceed certain limits.
99 See Stahl, Naturw. Wochenschrift, N. F. 5, 1906, No. 19.
100 Arch. Anat. Phys., Phys. Abt. Suppl., 1902.
101 The adaptive phenomena discovered by Gaidukow depend upon a real alteration in the formation of pigments. In the (primary) chromatic adaptation of pupae of Lepidoptera with respect to the colour of the ground they live upon, we only have the variable effects of pre-established chromatophores (Poulton, Phil. Trans. London, 178 B, 1888; Merrifield, Trans. Ent. Soc. London, 1898). The same holds for chromatic adaptations in crabs (Gamble and Keeble, Quart. Journ. Micr. Sci. 43, 1900; Minkiewicz, Arch. Zool. exp. et gén. sér. 4, 7, notes, 1907).
102 The theory of oxidation we have shortly sketched here was developed in chapter B. 5, of my Organische Regulationen. Recent discoveries of Winterstein’s (Zeitschr. allg. Physiol. 6, 1907) have given the strongest support to my hypothetic statements, and, in fact, can be said to have brought the doctrine of organic oxidation to a critical point. There can be no doubt that oxygen not only plays the “antipoisonous” rôle I had assigned to it, but that it is not even of such great importance for the supply of functional energy as former times had assumed. No doubt it serves to drive the functional machine, but decomposition of certain chemical constituents of the organism serves this purpose even more. The latter does so in the most fundamental and original manner, so to speak, whilst oxidation only burns up its products. Almost all elemental functions, in nerve-tissue at least, go on very well in the absence of oxygen, provided that certain “poisonous” substances, resulting from this anaërobic metabolism, are constantly removed. In normal conditions that is done by oxygen, and in doing so oxygen certainly assists the supply of energy, but it does not furnish the whole of it. The difference between so-called “aërobic” and “anaërobic” life almost completely disappears under such a view, and many so-called “regulations,” of course, disappear at the same time; there is no more “intramolecular respiration.”
103 But nevertheless albumen is not to be replaced altogether in vertebrates by fat or carbohydrate; it probably serves some special function besides combustion, even in the adult.
104 Arch. Entw. Mech. 18, 1904.
105 To a physiological friend of mine I owe the suggestion that it is the permanently functioning tissues which stand hunger better than the others, at least if the sexual cells might be regarded as capable of a sécrétion interne in all cases. Then the adaptations in the state of hunger might be said to be reduced in some degree to “functional adaptation.” But it must remain an open question, it seems to me, whether such a view may indeed hold in the face of the facts observed in Planaria and infusorians.
106 In all cases where fungi of the same species are able to live on different hosts, that is, to penetrate membranes of a different chemical character, a similar objection as to the “secondary” type of such a regulation may be made.
107 The discovery of Weinland that adult dogs are able to produce “lactase” in their pancreas, whenever they are fed, quite abnormally, with milk-sugar, has recently been said to be vitiated by an analytical mistake.
108 Compare the excellent review of the subject by Bayliss and Starling in the Ergebnisse der Physiologie, 5, 1906, p. 664. The reader who misses here an analysis of the brilliant discoveries of Pawlow and his followers, relating to so-called “psychical and associative secretion,” will find these facts dealt with in another section of the book. These facts, indeed, would prove vitalism, it seems to me.
109 It would be a true secondary metabolic regulation, if after the extirpation of one gland another different one were to assume its function. Nothing is known in this respect except a few rather doubtful observations about the interchange of functions between thymus and thyroid, except also the fact that the so-called lymph-glands increase in size after the extirpation of the spleen. Even here, of course, a sort of “restitution” would be included in adaptation proper.
110 A good review is given by E. Fromm, Die chemischen Schutzmittel des Tierkörpers bei Vergiftungen, Strassburg, 1903.
111 Davenport, Arch. Entw. Mech. 2, 1895–1896, and Hausmann, Pflüger’s Arch. 113, 1906.
112 Leçons sur la pathologie comparée de l’inflammation, Paris, 1902.
113 The other steps or phases in the process of inflammation have also been regarded as adaptive: the increased quantity of body fluid for instance is said to serve to dilute poisonous substances.
114 See Jacoby, Immunität und Disposition, Wiesbaden, 1906.
115 Collected Studies on Immunity by Ehrlich and his Collaborators, translated by Ch. Bolduan, New York and London, 1906.
116 So-called genuine or innate immunity, in contrast to the immunity which is acquired, is of course a case of adaptedness only and not of adaptation. There also exists a high degree of specific adaptedness in some animals with regard to their faculty of coagulating blood. (See Leo Loeb, Biol. Bull. 9, 1905.)
117 We cannot do more than barely mention here the problem of the localisation of anti-body production. In general it seems to be true that anti-bodies are produced by those cells which require to be protected against toxins; that would agree with the general rule, that all compensation of the change of any functional state proceeds from the part changed in its function.
118 Here again I should like to except from this statement the discoveries of Pawlow. See page 204, note 108.
119 The few cases of an “improvement” of morphogenetic acts in hydroids described by myself are too isolated at present to be more than mere problems (Arch. Entw. Mech. 5, 1897). The same is true, it seems to me, with regard to certain recent discoveries made by R. Pearl on Ceratophyllum (Carnegie Inst. Wash. Publ. No. 58, 1907); and by Zeleny on a medusa (Journ. exp. Zool. 5, 1907). Pawlow’s discovery, that the enzymotic composition of the pancreatic fluid in dogs becomes more and more adapted to a specific composition of the food (either meat or bread and milk) the longer such a specific composition is offered to the individual animal, may probably be understood as a case of mere functional adaptation of the cells of the digestive glands, if it stands criticism at all (see Bayliss and Starling, Ergeb. Physiol. 5, 1906, p. 682).
120 Experiments carried out in the “Biologische Versuchsanstalt” at Vienna indeed have shown that many animal types are capable of at least a certain degree of restitution, although they had previously been denied this faculty by zoologists.
121 Ueber das Gedächtnis als eine allgemeine Function der organischen Materie, Wien, 1870. New edition in Klassiker d. exakt. Wiss., Leipzig, Engelmann.
122 Die Mneme, Leipzig, 1904.
123 Driesch, Organ. Regul. 1901.
124 The “ideal whole” is also proved to exist, if any given “Anlage,” say of a branch, is forced to give origin to a root, as has really been observed in certain plants. This case, like many other less extreme cases of what might be called “compensatory heterotypy,” are best to be understood by the aid of the concept of “prospective potency.” It is very misleading to speak of a metamorphosis here. I fully agree with Krašan about this question. See also page 112, note 48, and my Organ. Regul. pp. 77, 78.
125 Winkler has discovered the important fact, that the adventitious buds formed upon leaves may originate either from one single cell of the epidermis or from several cells together; a result that is very important with respect to the problem of the distribution of “potencies.”
126 The “regeneration” of the brain of annelids for instance is far better regarded as an adventitious formation than as regeneration proper: nothing indeed goes on here at the locality of the wound; a new brain is formed out of the ectoderm at a certain distance from it.
127 A full “analytical theory of regeneration” has been developed elsewhere (Organ. Regul. p. 44, etc.). I can only mention here that many different problems have to be studied by such a theory. The formation of the “Anlage” out of the body and the differentiation of it into the completely formed results of regeneration are two of them. The former embraces the question about the potencies not only of the regenerating body but of the elements of the Anlage also; the latter has to deal with the specific order of the single acts of regenerative processes.
128 And, of course, at the root of every new starting of certain parts of morphogenesis also, as in regeneration and in adventitious budding; these processes, as we know, being also founded upon “complex-equipotential systems,” which have had their “genesis.”
129 New edition in the “Klassiker d. exakt. Wiss.” Leipzig, Engelmann; see also Bateson, Mendel’s Principles of Heredity, Cambridge, 1902.
130 For the sake of simplicity I shall not deal here with those cases of hybridisation in which one quality is “recessive,” the other “dominant,” but only allude to the cases, less numerous though they be, where a real mixture of maternal and paternal qualities occurs.
131 This hypothesis was first suggested by Sutton and is at present held by orthodox Mendelians; but probably things are a little more complicated in reality, as seems to be shown by some facts in the behaviour of so-called “extracted recessives.” In Morgan’s Experimental Zoology, New York, 1907, a full account of the whole matter is given.
132 Arch. Entw. Mech. 21, 22, and 24, 1906–7; see also Doncaster, Phil. Trans. Royal Soc. London, B. 196, 1903. The influence of different temperature upon the organisation of the hybrids is not always quite pure, inasmuch as the paternal and the maternal forms may themselves be changed by this agent. In spite of that there exists an influence of the temperature upon the hybrid as such, at least with regard to certain features of its organisation.
133 Only the nucleus of the egg had entered its first stages of activity.
134 The first proof of vitalism, indeed, rests upon the analysis of the differentiation of an harmonious-equipotential system as a whole: this whole cannot be a machine that would relate to differentiation as a whole; the question whether there might be any machines distributed in the whole, in the form of the nuclei is of no importance at all in this argument. Moreover the pressure experiments (see page 63) prove the unimportance of such “machines” for the specificity of differentiation, and the second proof of vitalism shows that the nuclei cannot be regarded as machines accounting for differentiation in any way.
135 Boveri tried to fertilise enucleated fragments of the egg of Sphaerechinus with the sperm of Echinus. He failed to get any results in isolated experiments, but found a few small larvae of the pure Echinus type in large cultures consisting of shaken eggs. But later experiments on hybridisation in sea-urchins have shown that a full hybrid of Echinus and Sphaerechinus may be purely paternal also.
136 Surely the new results of Herbst, mentioned above, are another indication of the importance of something in the nucleus. The first stage in parthenogenesis, which he used in his experiments, is a nuclear phenomenon.
137 Boveri (Ergebn. üb. d. Konstitution etc. des Zellkerns, Jena, 1904; and “Zellen-Studien VI.” Jen. Zeitschr. 43, 1907) has made it highly probable by experiments that the different chromosomes of the nucleus of the sexual products play a different part in morphogenesis, though not in the sense of different single representatives of different single organs. This doctrine, of course, would not alter the whole problem very much: the chromosomes would only be means of morphogenesis and nothing else, no matter whether they were of equal or of different formative value. It only is with regard to the problem of the determination of sex (see page 107, note 46), that the morphogenetic singularity of one certain specific chromosome can be said to be proved.
138 H. M. Vernon, Variations in Animals and Plants, London, 1903.
139 De Vries, Die Mutationstheorie, i., 1901; and Klebs, Jahrb. wiss. Bot. 42, 1905.
140 They would not be “real exceptions” if Klebs (Arch. Entw. Mech. 24, 1907) were right in saying that both variations and mutations owe their existence to external agents. What is really proved by Klebs is the possibility of changing the type of a curve of variation and of provoking certain discontinuous varieties by external means. See also Blaringhem (Comptes rend. 1905–6, and Soc. de Biol. 59, 1905), and MacDougal (Rep. Depart. Bot. Res., 5th Year-book Carnegie Inst., Washington, 129).
141 H. de Vries, Species and Varieties: their Origin by Mutation, London, 1905. A short review of the “mutation-theory” is given by Francé in Zeitschrift f. d. Ausbau d. Entwickelungslehre, i. 1907. It is well known that Gautier, and, in the first place, Korshinsky, advocated a similar view previous to the authors named in the text.
142 Recent years have created the beginnings of a systematics based on chemical differences of metabolism and its products: such differences in fact have been found to go hand in hand with diversities of the type in some cases (v. Bunge, Przibram, etc.).
143 We prefer this unpretending definition of the theory of descent to every other. As soon as one introduces into the definition the concept of the “transmutability of species,” the term “species” would require a special definition, and that would lead to difficulties which it is unnecessary to deal with for our main purposes. It has been remarked by Krašan, (Ausichten und Gespräche über die individuelle und specifische Gestaltung in der Natur) and by several other writers, that the problem of mutability or immutability of course relates to the individuals in the first place. I should like to add to this remark that the possibility must be admitted of the individuals being transmutable, whilst the “species” are not transmutable at the same time, the line of the “species” being a fixed order, through which the “individuals” have to pass in the course of their generations. What is meant here will become clearer, when we study the different possible aspects of “phylogeny.”
144 It seems to me that my argument gives a broader logical basis to the theory of descent than does that of G. Wolff (Die Begründung der Abstammungslehre, München, 1907). Wolff starts from the concept of organic teleology, and thus finds the only reason for accepting the theory of transformism in the existence of so-called “rudimentary organs”; these organs would form an obstacle to teleology if they could not be regarded as inherited.
145 See Wigand, Der Darwinismus und die Naturforschung Newton’s und Cuvier’s, Braunschweig, 1874–7; Nägeli, Mechanisch-physiologische Theorie der Abstammungslehre, München, 1884; G. Wolff, Beiträge zur Kritik der Darwin’schen Lehre, 2nd ed. Leipzig, 1898; etc.
146 Darwinismus und Lamarckismus, München, 1905.
147 This would not be true, if the varieties of plants produced by Blaringhem, Klebs, and MacDougal by means of external agents were really “mutations” (comp. page 238, note 3).
148 Of course, the inheritance of mutations would imply a certain sort of “inheritance of acquired characters,” on the condition stated in the preceding note. But, probably, the germs of the next generation might be regarded here as being directly affected by the external agent, in a manner that will briefly be mentioned later on in the text.
149 Comp. page 238, note 2.
150 Certain English authors have applied the term “modification” to all kinds of organic properties acquired from without, whether they are adapted or not.
151 Of course the inheritance of specific values from the results of fluctuating variations, leading to new averages of variability (see p. 265), may also be understood in this manner, the conditions of nourishment acting upon the adult and upon its germs equally well.
152 Berichte üb. d. Sitzung. d. Ges. f. Bot., Hamburg, 1887, 3 Heft.
153 Quite recently Kammerer (Arch. Entw. Mech. 25, 1907, p. 7) has published very important experiments on the inheritance of “acquired” modifications with regard to the peculiarities of reproduction in Salamandra atra and S. maculosa. It seems rather improbable—though not absolutely impossible—that the germ cells were directly affected by the external modifying agent in this case.
154 We have not spoken about the hypothetic inheritance of pure physiological adaptations, for it is clear without further discussion that innate specific immunity, for instance, being a specific “adaptedness” (see p. 186) might be due to the inheritance of the results of active immunity as an adaptation, just as adaptive congenital structures might be due to such an inheritance.
155 C. E. v. Baer clearly discriminated between the type, the degree of organisation, and the histological structure. All these three topics indeed have to be taken into account separately; the third alone is of the adaptive type. All of them may be independent of each other: the Amoeba may be as adapted histologically as is a high vertebrate, but it is of much lower type; and in its own type it is of a lower degree of organisation than Radiolaria are.
156 I repeat once more that we are dealing here with dogmatic “Neo-”Lamarckism exclusively. This theory indeed claims to explain all features and properties of organic bodies on the basis of the feeling of needs and storing of contingent fulfilments and on this basis alone, just as dogmatic “Neo”-Darwinism claims to account for all those phenomena on the ground of contingent variations and natural selection. Darwin himself, as we have seen, intentionally left unexplained certain primary features of life and therefore cannot be blamed for having failed to explain them, though even then his theory remains wrong. Lamarck personally considered a real primary organisatory law of phylogeny as being of fundamental importance, and therefore he is not in the least responsible if “Neo-Lamarckism” fails as a universal theory.
157 Compare also the excellent criticism of Lamarckism lately given by G. Wolff, Die Begründung der Abstammungslehre, München, 1907.
158 It has also very often been said by Darwinians that Lamarckism is only able to explain those cases of adaptedness which relate to active functioning but not mere passive adapted characters, like “mimicry” for example. But this argument taken by itself, it seems to me, would not be fatal to Neo-Lamarckism in the special form August Pauly gave to this doctrine.
159 But nothing more. All “mutations” hitherto observed in nature or (comp. page 238, note 3) experimentally produced relate only to “varieties” and not to “species.” One could hardly say that the recent investigations about the production of mutations by external means have strengthened their importance for the general theory of transformism.
160 The word “possible” relating to originating, of course, not to surviving. It is here that natural selection may acquire its logical importance alluded to above (see page 264).
161 The discussions in the second volume of this book will show the possible significance of such an analysis. We at present are dealing with entelechy in a quasi-popular manner.
163 An immanent vitalistic phylogeny without a pre-established end has recently been advocated by H. Bergson (L’évolution créatrice, Paris, 1907).
164 In this connection the problem may be raised, whether there can be such a thing as unchangeable “species” in spite of the mutability of the individuals. Compare page 251, note 1.
165 On account of the limited size of the earth a certain final stage of human civilisation might be expected in a future time; but it would be the size of the earth which determined this end, and not the process of civilisation itself.
166 Die Grenzen der naturwissenschaftlichen Begriffsbildung, Tübingen and Leipzig, 1902.
167 The word “universality” to be understood here in quite an unpretentious quasi-popular meaning, not strictly epistemologically.
168 To avoid mistakes I wish to say here most emphatically that, according to Rickert, the method of history is regarded as completely free from subjectivity as soon as its “values” are once established. But this cannot avail to save the theory.
169 This is a rather optimistic conception of “history.” Personally, I must confess that even its emotional and practical importance seems to me to be at least diminished by the retarding effects which all sorts of “historical” considerations—in science as well as in arts and in public life—carry with them. All real progress is non-historical—and its champions almost always have become martyrs: this fact seems not to recommend history as a means of education, except for persons of a very strong character.