
Natural History of the Racer
Coluber constrictor
BY
HENRY S. FITCH
University of Kansas
Lawrence
1963
University of Kansas Publications
Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Theodore H. Eaton, Jr.
Published December 30, 1963
University of Kansas
Lawrence, Kansas
PRINTED BY
JEAN M. NEIBARGER, STATE PRINTER
TOPEKA, KANSAS
1963
29-7864
Natural History of the Racer
Coluber constrictor
BY
HENRY S. FITCH
| PAGE | |
| Introduction | 355 |
| Acknowledgments | 357 |
| Methods and Materials | 358 |
| Description | 362 |
| Color pattern | 362 |
| Bodily proportions | 365 |
| Lepidosis | 367 |
| Dentition | 369 |
| Hemipenis | 369 |
| Relationships | 370 |
| Range | 371 |
| Geographic Variation | 373 |
| Habitat | 375 |
| Temperature Relationships | 377 |
| Home Range and Movements | 385 |
| Food Habits | 394 |
| Methods of obtaining prey | 394 |
| Composition of food | 395 |
| Kinds of prey | 403 |
| Reproduction | 408 |
| Sexual behavior | 408 |
| Cycle of the male | 414 |
| Eggs | 416 |
| Hatching | 423 |
| Growth | 425 |
| Mortality Factors and Adaptations for Survival | 432 |
| Defense and escape | 432 |
| Natural enemies | 438 |
| Disease | 442 |
| Parasites | 442 |
| Populations | 445 |
| Composition | 445 |
| Numbers | 450 |
| Summary | 456 |
| Literature Cited | 461 |
Throughout much of the United States the racer is abundant and is one of the snakes best known to man. Its active diurnal habits and its preference for a habitat in meadows, pastures, and hayfields rather than in remote wilderness areas, result in frequent encounters with humans. The racer is a predator on many kinds of small animals, both vertebrates and invertebrates; it takes as food chiefly animals that are agricultural pests but also destroys some beneficial kinds. Yet, in general, the attitudes of rural people toward the racer are little influenced by these economic and ecologic considerations, but rather, are dominated by an unreasonable fear, despite the racer's inoffensive disposition, and inability to inflict any harm on humans.
Although an extensive literature exists regarding the racer, no thorough study of the species' natural history has been made heretofore. Obviously such study is needed. Few species of vertebrates having comparable economic bearing have been similarly neglected. In 1948, undertaking a program of ecological research on the recently created University of Kansas Natural History Reservation, I included the racer among the many common species studied to gain insight into the functioning of the local ecosystem. Live-trapping of snakes on the area was begun in 1949, and these operations were greatly intensified in the years 1957 through 1962, with efforts concentrated on the study of the racer in the 1960, 1961 and 1962 seasons. Thus my study is based upon 14 consecutive years' records on the Reservation, the northeasternmost section in Douglas County, Kansas, six and one-half miles north northeast of the University of Kansas campus at Lawrence. After the acquisition in 1956 of the 160-acre Rockefeller Tract adjacent to the Reservation on the north in Jefferson County, field work was extended to this new area, which, because it was superior habitat, in the final years of the study produced more records than the Reservation. An important but relatively minor segment of the data originated from Harvey County Park, 13 miles west of Newton, Kansas, where lines of live-traps were maintained in 1959, 1960, 1961, 1962. Smaller collections of data were obtained from the Lalouette Ranch in the Flint Hills, three miles northeast of Florence, Marion County, Kansas, and from Cedar Bluff Reservoir, 23 miles west and seven miles south of Hays in Trego County, western Kansas, where live-trapping was carried on in 1959 and 1960. Additional data were [356] obtained on numerous field trips to various collecting localities in northeastern Kansas. My first-hand knowledge of the species is also based, in part, on many years of field experience with the far western subspecies C. c. mormon in western Oregon and California, and on similar experience in 1947 and 1948 with the southern subspecies, C. c. anthicus in central Louisiana.
This varied field experience with the species at localities well scattered throughout its geographic range has added perspective to the study even though most of the records were collected within a radius of three quarters of a mile. No one locality can be regarded as entirely typical of a species' habitat over its range as a whole. According to my philosophy, the ecological niche of a species is subject to geographical variation analogous to the variation to be seen in the morphological characters of the animal itself. Different community associates, including different competitors, prey, and predators, and different physical factors enforce a somewhat different way of life on a species in geographically remote parts of its range. When analyzed these differences often turn out to have a genetic basis. Thus, limits of tolerance to heat, cold, and drought often vary geographically, and the population density, reproductive potential, seasonal cycle, and other properties of populations may be altered either by the direct effect of the environment, or through its effect on the genetic constitution, produced by natural selection.
The local population of racers studied was near the center of the species' geographic range, and is to some extent representative of the species as a whole, though differing in its ecology from other populations in proportion to their remoteness and the distinctness of their habitats. It has not been demonstrated that ecological traits of populations change in a discontinuous manner or correspond in their limits with those of named subspecies. More likely geographical variation is continuous and parallels morphological variation only in a general way. Certainly the boundaries of subspecies' ranges should not be accorded undue emphasis in an ecological study.
My investigation of the blue racer under natural conditions, combined with a compilation and analysis of published literature, has resulted in a fairly satisfactory understanding of some phases of the species' ecology and natural history, such as the food habits, the growth rate, the extent of home range and of seasonal movements. However, relatively little was learned concerning some phases of [357] the life history. Unfortunately, the traps used did not catch young of the smaller sizes. Facts concerning egg-laying, incubation, and hatching therefore are known chiefly from snakes kept in confinement. Although first-year young were captured by hand from time to time they were obtained in relatively small numbers, and little was learned regarding their population density, movements, or mortality factors. Of course, such hiatuses are to be expected; even in man such enigmas as the disparate sex ratio still challenge the investigator.
For the subspecies of racer involved in my field study the widely used vernacular "blue racer" has been adopted in this report. In general I advocate conformity with the vernacular names published by the Committee on Herpetological Common Names (1956). However, in this list, the name blue racer was assigned to Coluber constrictor foxi, an invalid subspecies of the Prairie Peninsula that has been relegated (Auffenberg, 1955:92; Smith, 1961:196) to the synonymy of C. c. flaviventris. It therefore seems appropriate that the book name "yellow-bellied racer" applied to flaviventris by the Committee should be abandoned for this subspecies, and that the name blue racer be applied officially, as it is in actual practice by both laymen and herpetologists, to all populations of this subspecies.
Financial assistance from the National Science Foundation in 1957 through 1962 is acknowledged. Although none of the three separate grants involved was made specifically for the autecological study of the racer, all three contributed to the support of the extensive program of live-trapping for snakes, which yielded most of the records upon which this report is based. Student assistants who were employed on these projects include James W. Bee, William N. Berg, Donna M. Hardy, Robert M. Hedrick, Dale Hoyt, Robert M. Packard, Robert G. Webb, and Wayne Wiens, at the Reservation; Roy Henry, Dale Horst, Dwight R. Platt, and Howard L. Schrag at Harvey County Park, and Gilbert L. Adrian at Cedar Bluff Reservoir. Dr. Edwin P. Martin, formerly of Fort Hays, Kansas State College, was helpful in planning and carrying out the field work at Cedar Bluff Reservoir. Eric Shulenberger assisted with field work and processing of data in 1962 under the National Science Foundation program for Undergraduate Research Participation. Robert Miner assisted with the examination of specimens in 1960. Mr. August Lalouette of Florence, Kansas, permitted field [358] work on his ranch and contributed information and materials to expedite this work. Mr. and Mrs. Harold Brune of Route 3, Lawrence, Kansas, kindly contributed several clutches of racer eggs found on their farm in Jefferson County, and also made available significant information accompanying them. Dr. William H. Stickel kindly made available at my request records of predation on racers from the food habits files of the U. S. Fish and Wildlife Service. Dr. William E. Duellman of the University of Kansas Museum of Natural History and Dr. Robert C. Stebbins of the University of California Museum of Vertebrate Zoology kindly permitted examination and dissection of specimens in the collections under their care. Dr. George W. Byers of the University of Kansas Department of Entomology identified numerous insects eaten by racers. My daughter, Alice V. Fitch, often assisted me with the field work and the processing of data. My wife, Virginia R. Fitch, read the manuscript critically, assisted me with the examination of museum specimens, and with typing, and helped in various other ways.
Table 1. Numbers and Distribution of Captures and Recaptures of Blue Racers on the Reservation and Rockefeller Tract
| Span of years within which each individual was captured | Number of separate years within which each individual was captured | Number of times each individual was captured | |||
| Years | Individuals | Years | Individuals | Times captured | Individuals |
| 1 | 749 | 1 | 749 | 1 | 679 |
| 2 | 137 | 2 | 197 | 2 | 181 |
| 3 | 56 | 3 | 51 | 3 | 93 |
| 4 | 32 | 4 | 15 | 4 | 31 |
| 5 | 19 | 5 | 6 | 5 | 7 |
| 6 | 13 | 6 | 1 | 6 | 14 |
| 7 | 7 | 7 | 0 | 7 | 8 |
| 8 | 2 | 8 | 1 | 8 | 4 |
| 9 | 3 | 9 | 1 | ||
| 10 | 0 | 10 | 0 | ||
| 11 | 1 | 11 | 0 | ||
| 12 | 1 | 12 | 1 | ||
| 13 | 0 | ||||
| 14 | 0 | ||||
| 15 | 0 | ||||
| 16 | 1 | ||||
This investigation was based primarily on the capture in live-traps, marking, release, and recapture of blue racers in their natural habitat. On the combined area of the Reservation and the Rockefeller [359] Experimental Tract, 1020 blue racers were recorded a total of 1688 times from August 30, 1948, to October 27, 1962. At Harvey County Park 361 blue racers were marked, and were captured a total of 467 times from May 6, 1959, to September 14, 1962, and at Cedar Bluff Reservoir 42 were captured from May 11, 1959, to June 30, 1960.
The traps used were cylinders of galvanized wire, "hardware cloth" (Fitch, 1951:77; 1960:77), having funnels opening into each end, or having a funnel at one end and a plug at the other. The traps, open at both ends, were used along hilltop rock ledges where an exposed vertical rock face provided a barrier along which a snake might travel and where it could be easily intercepted by the trap without any accessory equipment (see Pl. 21, Fig. 1). Where such natural barriers were lacking, as in level fields, barriers consisting of boards, screens or sheet metal were installed to guide the racer toward the trap and into a funnel entrance. Two such barriers at each end of a trap forming a V to guide the snake into the funnel were used in 1956 and 1957, when trapping at places away from the hibernation ledges was undertaken. Later it was found simpler and more effective to use a single barrier with a trap at each end. The barrier extended up into the funnel entrance, and usually the racer, following along the barrier on either side, would pass into the funnel and through its apex. However, it was possible for a racer to travel around the end of the barrier without entering the trap, and perhaps some did so.
The need for making the barrier and trap a tight unit impassable to the snake, was somewhat counterbalanced by the need for having the whole installation loosely constructed so that it could be easily altered, opened, adjusted, and cleaned. Since the traps were kept set in large numbers, and the task of checking them was time-consuming, speed of operation was more important than the perfect functioning of any one trap. Approximately 200 traps were kept set when operations were at their maximum. No record was kept of the number of "trap days" involved in the study, but the total was well over 100,000 for the Reservation and the Rockefeller Tract. When traps were set at both ends of a barrier, the outer end of each trap was closed with a plug. No bait was used in the traps. Occasionally small vertebrates and insects of kinds used as food by the racers may have gotten caught first and served to attract the snakes. Best catches of racers were made in the breeding season, since males were attracted by females already in the traps, and [360] several males might be captured simultaneously with one female. Occasionally as a person approached or handled a trap, a racer darted out of it, displaying a perception, acuity of vision, and skill in avoiding the inward projecting wire prongs encircling the small funnel opening, that were exceptional among the several species of snakes trapped. Doubtless many other racers that were caught in traps escaped before they were discovered. If the funnel entrance of the trap was of the same diameter as the snake itself, or only a little larger, there was little likelihood of the trapped racer escaping. However, funnel openings were usually adjusted at a diameter of approximately 11/4 inches, allowing an ample margin for even the largest racers, though inadequately small to permit ingress of a few of the largest black rat snakes, bull snakes and timber rattlers occurring locally.
Ordinarily the snakes trapped were processed in the field and released immediately. The method of marking was essentially that of Blanchard and Finster (1933:334). Two subcaudals, one on the right side and one on the left, were clipped on each snake, and when these marks healed they left permanent scars. In the racer, as in most other colubrines, the subcaudals are divided into a double series, one on the left and one on the right. Scales of the left and right sides are placed alternately. At the base of the tail one or more undersized scales usually are present on each side, and there might be some question as to precisely where the count should begin. The rule followed was to exclude from the count any small basal scales on either side that did not extend medially to contact at least one scale of the opposite side. The scale designated as "one left" (or "one right") was the first to contact one of the opposite series, regardless of whether the former was of normal size or (as was usually the case) smaller and narrower than those following it. In marking, this "number one" scale was never clipped but was left as a point of reference since a base mark was needed from which to begin the count. The marks were read from left to right, for example U 5l 2r, the "U" referring to the subcaudals or "urosteges," the "5l" indicating the fifth on the left side, and "2r" indicating the second on the right. The subcaudals clipped were the first 19 following the basal scale. When the 361 possible combinations all had been used, ending with U 20l 20r, a new series was begun duplicating the first except that on each snake the first ventral (or "gastrostege") anterior to the anal plate was clipped on the left side (G1L) to distinguish these snakes from the series [361] previously marked. Later, a third series, "G2R" was marked, and eventually a fourth series, "G3L" was started.
There were many borderline instances in which the basal scale barely contacted one of the opposite side. In such instances the formula was written U 5l ISB 2r, the subscript ISB signifying "including small basal." In other instances a basal subcaudal barely failed to contact a scale of the opposite side and this condition was indicated by the subscript NSB—"not including small basal." The condition might be so nearly equivocal that on successive occasions the same formula might be read U 5l ISB 2r and U 4l NSB 2r. Occasional misidentifications of individuals that resulted from such discrepancies were in most instances readily detected when the field records were transferred to individual file cards where the sex, size, and location of the snake at its previous captures were shown.
In some instances racers recaptured after periods of years retained conspicuous scars where scales had been clipped, but in other instances the marks had become obscure, and in fact the only trace of a mark might be a slight narrowing or notching of part of the scale originally clipped. Snakes caught and marked early in life probably retained more power of regeneration than those clipped after the attainment of maturity, but otherwise the basis for difference in extent of regeneration was not evident. In the same snake, three scales, all clipped on the same day, might show much different degrees of regeneration after the lapse of a year or more. In general, obscuring of marks by regeneration was a source of inconvenience rather than of error; only a negligible percentage of the recaptured racers had marks so obscure that their identities might have been seriously questioned, and it is doubtful that any marks were lost completely by regeneration.
Racers found in traps were removed, measured (snout-to-vent length, tail length), weighed in a cloth bag suspended from spring scales, and marked. The mouth was forced open and the snake was examined for flukes. Enamel paint of a bright color, red, green, yellow, blue or orange was smeared on the snake to gain information regarding the time of molt. The stomach was palpated for recently ingested food items, and any detected were forced up into the gullet to be identified, then were squeezed back into the stomach. The rear part of the body was palpated to detect undigested material in the gut, and if any was present, an attempt was made to squeeze out the fecal material, using only light pressure, [362] with care not to injure the racer. The inside of the trap and the ground beneath it were inspected for fecal material that might have been voided while the snake was confined. Any scatological material obtained was wrapped in a paper towel, labelled and brought back to the laboratory where it was stored. Eventually each scat was soaked for a day or more in a detergent solution, rinsed in running water in a fine gauze bag, dried, and placed with its label in a cellophane envelope for subsequent microscopical study.
Various items concerning reproductive condition were also routinely recorded. In females the ventral surface was palpated at the rear end of the body to detect the genital bursa or vagina, which in sexually mature individuals has a much thickened wall, and can be felt as a distinct lump. Males were likewise tested for sexual maturity by pipetting a small amount of fluid from the cloaca into a vial and returning it to the laboratory where it was examined microscopically for motile sperm. Also, sperm samples were often taken from males at different times throughout the season of activity, and cloacal samples from females occasionally were checked for sperm as evidence of recent copulation.
In the summer of 1962 an outdoor enclosure of 100-foot circumference was constructed, of galvanized sheet iron, with wall three feet high, set on a concrete base extending to a depth of two feet. A two-foot-deep concrete basin inside the enclosure served as a water container. The enclosure was partly shaded by a large walnut tree and the area enclosed had lush vegetation, including brome grass, various shrubs, and young trees up to 15 feet high, thus including most features of the racers' habitat, and it was situated in an area frequented by the snakes. Throughout the summer several racers were kept in the enclosure, and frequent observations on them yielded much information concerning time of activity, temperature preferences, and social and sexual behavior that could not have been obtained readily either from racers confined in small cages or from those free under natural conditions.
Hatchling racers differ much in appearance from adults; whereas the latter are of dull uniform coloration dorsally, the hatchlings have a checkered pattern of alternating blotches in several rows, including a middorsal row, with blotches much larger than those of [363] the other rows. This basic pattern is perhaps the most common one in all snakes, and is found in the young of various other genera (notably Elaphe) which lose or alter their markings during development. In these genera and in the racer, the juvenal checkered pattern may represent recapitulation of an ancestral condition. The adaptive significance of having a blotched, checkered pattern in the young, and uniform coloration in the adult is not evident. I have rarely seen the hatchlings under natural conditions except by finding them hiding beneath flat rocks. Their concealing pattern must be fully as effective as that of adults and the young themselves are more secretive than the adults.
A hatchling of 240 millimeters snout-vent length was described as follows on September 22, 1962: Ground color pale olive anteriorly, gradually darkening posteriorly, marked with chocolate blotches; middorsal blotches largest averaging about five scales long and seven or eight scales wide anteriorly; posteriorly, blotches become less distinct (tending to blend into progressively darkening ground color), and width-to-length ratio increases; gradual obscuring of blotches proceeds posteriorly, until on tail they can no longer be distinguished, and color is uniformly olive; on each side, row of lateral blotches alternates with that of middorsal blotches; lateral blotches average somewhat less than two scales long, and approximately two scales wide and are of irregular shape, sometimes subdivided; farther down on sides, lower row of lateral blotches alternates with upper lateral row; this lower lateral row, approximately same size as upper lateral row, and situated at level of first scale row, overlapping onto ventrals, where it pales to reddish brown; almost every ventral scute has one pair or more of dark reddish brown spots, tending toward semicircular shape, the arc of each projecting forward, but posteriorly on body these spots become increasingly obscure, and are indiscernible on posterior end of body and on tail; ventral surface white on chin and throat, gradually assuming suffusion of pale greenish gray posteriorly; eye dark with narrow yellowish margin around pupil; top of head grayish olive, mottled with faint and irregular dark markings; supralabials whitish, with chocolate markings, mostly wedge-shaped, in their posterior parts; rostral and internasals edged with dark posteriorly; posterior upper corner of loreal and adjacent corner of prefrontal dark; temporals having dark markings; chin almost immaculate, but with narrow black posterior edges on some of the infralabials (see Plate 19, Figs. 1 and 2, and Plate 20, Fig. 2).
The checkered pattern of the juvenile fades gradually as development proceeds. Persistence of the juvenal markings varies greatly in individuals and probably is subject to geographic variation also. By the time sexual maturity is attained the dorsal pattern often is indiscernible or represented only by faint traces. The ventral speckling is more persistent.
A female of 602 millimeters snout-vent length in mid-July 1962, at a probable age of 11 months, was described as follows: Overall dorsal color olive gray, but with remnants of juvenal pattern discernible; dark dorsal blotches have almost faded, but their edges, about one scale wide, are still distinct; low on sides, color fades to pale bluish gray, and to pale greenish blue on edges of ventrals; on anterior one-third of body midventral surface is lemon yellow; farther posteriorly it fades to ivory, almost white on tail; chin white, except for reddish brown posterior edges of last infralabials, and streak of same color on each antepenultimate infralabial; top of head olive gray with irregular scattered dark marks; preoculars pale centrally with olive brown edges; supralabials white with reddish brown triangular marks; postnasals white anteriorly, gray posteriorly; uppermost postocular brownish orange, paler centrally; two rows of semicircular spots on belly, distinct anteriorly, but fading posteriorly until indiscernible on posterior part of belly; numerous small black spots scattered irregularly over dorsal and lateral surfaces.
A male racer retained more of the juvenal pattern at an approximate age of 15 months, when he was described on November 30, 1961: Dorsal surface dark grayish brown, the large juvenal dorsal blotches (each about eight scales wide) easily discernible, but faint; top of head dark olive brown, mottled with black, paling in rostral region; supralabials white on their anterior and lower portions, marked with brown and blue-gray on their upper and posterior portions; chin white, but with rusty markings on last four infralabials; ventrals ivory-yellow with rusty spots of which the largest are approximately three-fourths of the ventrals' breadth; first row of scales bluish gray, or with greenish suffusion (in neck), most of scales having indistinct dark spots; second row of scales similarly colored but more suffused with dusky pigment, blending into the darker duller color of the dorsal surface.
A female of 720 millimeters snout-vent length, presumably about 20 months old, but not gravid, on June 13, 1962, was described as follows: Juvenal pattern no longer clearly discernible but scattered [365] traces of it remain; dorsal color predominantly grayish olive, with occasional small black spots in streaks scattered irregularly over dorsal and lateral surfaces; at anterior end of body lateral scales have bluish green edges but this shading becomes less noticeable posteriorly; head mostly olive dorsally with marking obscure; parietals have a large faint blotch; supralabials retain faint dark, brownish markings on their upper parts; dark spot on median edge of each prefrontal; supraoculars slightly darker than adjacent scutes; chin mostly white, with yellowish suffusion at edges of scales; ventral surface predominantly yellow, but fading to grayish white posteriorly; remnants of juvenal ventral spots faintly discernible as tan or whitish areas on yellow ventrals.
Munro (1950b:124) mentioned a blue racer of 749 millimeters total length (hence probably having a snout-vent length of 600 millimeters, or a little less), which retained faint juvenal markings when caught on June 23, 1948, even though it was sexually mature, since it laid eggs on the night of July 4, 1948. During several weeks of captivity this snake's markings faded perceptibly.
In fully adult blue racers, those more than three years old, the juvenal markings have become completely obliterated. In those from the area of my study, the dorsal coloration is subject to much individual variation, typically olive brown, but ranging from pale bluish gray to dark brown, dark blue, or slaty. In most, the dorsal color is uniform, but in some there are streaks and isolated scales of black. The dark dorsal color extends down the sides onto the lateral corners of the ventrals and the subcaudals. The chin is white but the remainder of the ventral surface is ivory colored.
The slender and streamlined bodily proportions of the racer are subject to change through allometric growth. The head, and especially the eyes, are relatively large in the hatchling, and become relatively smaller as growth proceeds. The relative tail length seemingly increases in the growing young and then decreases slightly in adults.
Snout-vent length and tail length were recorded in almost all the racers examined, but other measurements were recorded in relatively few. In many racers, especially the larger adults, tails had been damaged and lacked their terminal parts. Often only the tip was missing, but, of course, such individuals were not usable in the study of proportions of the tail. Table 2 summarizes information concerning relative tail length in 935 racers of both sexes and [366] various sizes, from northeastern Kansas. Nearly all measurements are from the live snakes; a few are from recently killed individuals.
In hatchlings, the proportions of the tail are not noticeably different in males and females, but data indicate that the snakes' tails are approximately seven per cent longer in males than in females; as growth proceeds, the tails become relatively longer in proportion to the body, in both sexes. The ratio reaches its maximum in young adults, having increased from approximately 28 per cent of the snout-vent length in males and 26 per cent in females, to 31 per cent in males and a little more than 28 per cent in females. In the largest racers, of both sexes, these percentages are slightly reduced. Tail-length is subject to a fairly wide range of individual variation, which tends to obscure the trends determined by sex and size.
Table 2. Relative Tail Length in Male and Female Racers of Different Size Groups
| Snout-Vent Length in Millimeters | Males | Females | ||
| Number of racers | Mean ratio of tail-length to snout-vent length | Number of racers | Mean ratio of tail-length to snout-vent length | |
| 150-200 | 7 | 27.3 ± .833 | 2 | 26.3 |
| 201-250 | 24 | 27.8 ± .490 | 23 | 25.9 ± .479 |
| 251-300 | 9 | 28.8 ± .634 | 4 | 26.6 |
| 301-350 | 20 | 29.8 ± .246 | 11 | 25.0 ± 1.060 |
| 351-400 | 8 | 28.8 ± .530 | 12 | 27.5 ± .404 |
| 401-450 | 6 | 27.8 ± .775 | 5 | 26.3 ± .357 |
| 451-500 | 12 | 29.7 ± .434 | 6 | 26.8 ± .858 |
| 501-550 | 45 | 30.5 ± .283 | 15 | 27.6 ± .284 |
| 551-600 | 76 | 31.0 ± .218 | 35 | 27.1 ± .254 |
| 601-650 | 45 | 29.6 ± .313 | 64 | 27.8 ± .163 |
| 651-700 | 50 | 30.5 ± .241 | 36 | 27.1 ± .350 |
| 701-750 | 72 | 30.5 ± .177 | 45 | 27.6 ± .373 |
| 751-800 | 45 | 30.3 ± .373 | 38 | 27.8 ± .325 |
| 801-850 | 48 | 29.7 ± .274 | 50 | 27.5 ± .205 |
| 851-900 | 18 | 29.1 ± .519 | 35 | 28.6 ± .422 |
| 901-950 | 5 | 29.8 ± .672 | 31 | 26.7 ± .252 |
| 951-1000 | 1 | 29.5 | 19 | 26.9 ± .390 |
| 1001-1050 | .... | ..... | 15 | 25.9 ± .413 |
| 1051-1100 | .... | ..... | 6 | 26.4 ± .725 |
| 1101-1150 | .... | ..... | 1 | 28.5 |
| 1151-1200 | .... | ..... | 1 | 25.0 |
In 88 racers caught in the summer of 1962 the following measurements were recorded: Head length, from tip of snout to angle of jaw; maximum head width; greatest diameter of eye; circumference of neck; circumference at mid-body; circumference at posterior end [367] of body; and circumference of tail-base. Because the measurements were small, and were made in the field on active, struggling snakes, a high degree of precision could not be attained, and the range of error was several per cent, with occasional relatively large errors. Nevertheless, ontogenetic trends are clearly indicated. Most of the racers measured were adults of small to medium size—in the range 500 to 799 millimeters, snout-vent length. Twelve females and seven males ranged from 800 to 1035 millimeters, and seven young (all females) were less than 500 millimeters. In measurements other than circumference of tail-base, significant differences could not be found between males and females of the same size group; therefore the sexes were combined to obtain larger series.
Table 3. Bodily Proportions (Expressed as Ratio of Snout-vent Length) in Racers of Different Sizes
| Size Group | Large (more than 800 millimeters) |
Medium (500 to 800 millimeters) |
Small (less than 500 millimeters) |
| Length of head | 3.61 ± .036 | 3.82 ± .025 | 5.39 |
| Width of head | 1.93 ± .049 | 2.02 ± .023 | 2.53 |
| Diameter of eye | .56 ± .008 | .63 ± .009 | 1.00 |
| Circumference of neck | 4.71 ± .082 | 5.05 ± .052 | 6.64 |
| Circumference at mid-body | 7.11 ± .238 | 7.66 ± .082 | 8.58 |
| Circumference at posterior end of body |
5.06 ± .113 | 5.03 ± .061 | 5.90 |
| Circumference of tail at base | 4.23 ± .113[1] 4.47 ± .171[2] |
4.22 ± .075[1] 4.66 ± .043[2] |
4.66[1] |
[1] Females.
[2] Males.
Table 3 shows that as compared with adults, the small young racers have stouter, stubbier bodies, relatively large heads, and, especially, large eyes. Allometric growth seems to continue throughout life and the changed proportions of the adults are accentuated in the largest and oldest individuals.
Scalation that of typical colubrid (see Pl. 19); rostral large, extending back onto dorsal surface of snout, bluntly pointed behind; paired internasals considerably wider than long, convex anteriorly, almost straight-edged posteriorly, each extends laterally to naris; [368] paired prefontals approximately twice size of internasals, and wider than long, extending laterally on each side to level of nostril; frontal convex anteriorly, concave on each side, bluntly pointed behind, nearly twice as wide anteriorly as posteriorly; parietals large; angle formed between them by frontal slightly more than 90 degrees; nostril large, situated between almost equal sized anterior nasal and posterior nasal plates; loreal slightly smaller than nasals, its anterior edge inclined forward superiorly; two rows of temporals on each side; in upper row, first one narrow and elongate, second much shortened, third intermediate in shape; in lower row all three approximately alike in size and shape; two postoculars, the lower larger; seven supralabials, first small and low, longer along upper edge than along lower, second slightly longer than high, third higher than long, contacting eye; fourth largest, contacting posterior part of eye, and lower postocular; fifth nearly as large, pointed above; sixth also large, pentagonal; seventh low and rectangular; on chin first pair of infralabials separate mental from anterior genials; second infralabial minute; third approximately twice its size; fourth much smaller, rhomboidal, fifth also large, pentagonal; sixth smaller, rhomboidal, bluntly pointed behind; seventh smaller, narrow behind; eighth small and elongate; second pair of genials longer and narrower than those of first pairs, separated from each other by smaller scales; genials in approximately five rows, but somewhat irregular in arrangement, mostly smaller and narrower than body scales; latter all smooth, arranged in 17 rows for about two-thirds of body length, then, by loss of third row on each side, reduced to 15; scales of neck region rounded and relatively small, one-third to one-fourth size of larger body scales; lowest scale row on each side largest with its scales much wider and less symmetrical than others; most of body scales of approximately hexagonal shape; on forebody they average approximately twice as long as wide, but farther posteriorly on body, width-length ratio gradually increases and some of scales, notably those of lowest row, approximately as wide as long; regularity of scale rows broken on sides just above vent by presence of many small additional scales; on tail scale rows drop out posteriorly in rapid succession, until on posterior third only four are present; ventrals strongly convex posteriorly, with free posterior edges, nearly half length of scales; anal plate divided, with diagonal suture; subcaudals in double series, those of right and left sides alternating; several minute subcaudal-like scales on each side of vent.
In the racer the maxillary, palatine, pterygoid, and dentary bones bear teeth (Fig. 1). The teeth are all much alike in size and shape, small, sharp, and recurved, typically at an angle of approximately 50 degrees. The number of teeth present is variable. Because the teeth are small and loosely attached to the jaw bones, and often are broken off in the capture and ingestion of prey, each bone usually lacks part of its complement of teeth. Even the sockets vary somewhat in number between individuals, and between the left and right sides in some snakes. Most of the skulls that I examined were not thoroughly cleaned, and the adherent dried tissues made it difficult to obtain accurate counts of the sockets. In ten skulls from Kansas and Nebraska, most frequently occurring numbers of sockets for each of the dentigerous bones were: maxillary, 15; palatine, 11; pterygoid, 18; dentary, 18.
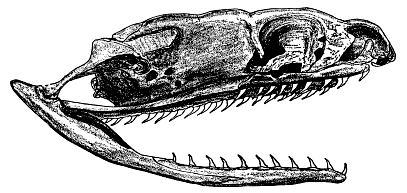
Fig. 1. Lateral view of right side of skull of adult female blue racer, × 4. University of Kansas Museum of Natural History no. 18305, from Greenwood County, Kansas.
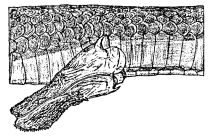
Fig. 2. Lateral view of injected and everted left hemipenis (slightly enlarged) of a blue racer from the Rockefeller Tract, Jefferson County, Kansas, showing heavy spines at base of organ, small spines of central zone and lamellae of terminal part. This hemipenis is not fully engorged.
Penial characters have proven to be useful in the classification of snakes, providing bases for separating subfamilies, genera, and species. In the racer even the subspecies have trenchant penial characters by which they may be separated in some instances. The hemipenis is roughly cylindrical, but widest at the base (Fig. 2). The sulcus spermaticus is unbranched. Approximately the basal one-third of the hemipenis has a smooth surface, broken only by the sulcus spermaticus and by three greatly enlarged spines, which form [370] hooks—one anterior, one posterior, and one dorsal. The dorsal hook is the largest of the three. Distal to the smooth part is a zone of small spines, each recurved and mounted on a fleshy tubercle. The zone of spines is poorly developed on the anterior side and is interrupted on the posterior side in the vicinity of the sulcus spermaticus but is best developed on the posterior side a short distance above and below the sulcus spermaticus. The spines are arranged in several oblique rows. Those of the proximal row are best developed, and there is rapid diminution in the size of those situated farther distally. Approximately the distal two fifths of the hemipenis forms a third zone, lacking distinct spines, but having numerous deep longitudinal grooves, alternating with lamellae which have fimbriated edges, and which fuse with each other and divide to form a reticulated pattern.
The large genus Coluber is much in need of revision. Its many species, perhaps more than a score in all, occur in North America from southern Canada south to Guatemala, in eastern and southwestern Asia, in southern Europe, and in North Africa. All are active, slender-bodied snakes having smooth scales in few rows, and having large eyes with well developed vision. The North American species fall into two natural groups, the typical racers, and the whip snakes, often assigned to a separate genus, Masticophis (Ortenburger, 1928). The whip snakes are more specialized than the typical racers in having the eyes more enlarged, and the body form more slender and attenuate, with number of scale rows more reduced. The racers of the Old World are more diverse. Inger and Clark (1943) suggested a partitioning of the genus Coluber on the basis of the pattern by which scale rows are reduced, from the maximum number on the forebody to the minimum number at the posterior end of the body, supplemented by certain characters of the hemipenis and of the cephalic scutellation. Besides Coluber[371] and Masticophis these authors recognized within the group the genus Platyceps with several species in southern Europe and southwestern Asia; Zamenis with several species in the same region and in North Africa, and Haemorrhois, a monotypic genus of Spain, North Africa and several Mediterranean islands. Although apparently valid in principle, this arrangement has not been generally followed.
Exclusive of those species groups whose assignment to the genus Coluber are somewhat questionable, the remaining species in the genus are: C. constrictor occurring throughout most of the United States and south along a narrow Atlantic coastal strip of Mexico to Guatemala; C. oaxacae of southern Mexico; and C. spinalis of northern China. C. oaxacae is poorly known as only a few specimens have been collected, but seemingly it is a near relative and derivative of C. constrictor, especially of that species' southernmost population. C. spinalis is much more distinct, as might be expected from its geographical remoteness. It is a slender, active snake, of olive coloration dorsally with 17 scale rows and a bright yellow, black-edged dorsal stripe and yellow ventral surface. It is relatively small (up to 755 millimeters snout-vent length) and is partial to riparian habitats but is also found in forests and in dry and barren regions (Pope, 1935:224-226). It is known to feed upon lizards.
The common racer has been recorded in each of the 48 states of the mainland of the United States, also in New Brunswick, Nova Scotia, southern British Columbia, and southward through Mexico where it is limited to a narrow strip of east coast lowlands but extends as far as Guatemala. C. c. constrictor occupies the northeastern states and extends south into the Appalachian and Piedmont. C. c. priapus with its associated races paludicola, helvigularis, and anthicus has an Austroriparian distribution, occupying the Atlantic Coastal plain and the Gulf Region, and extending north in the Mississippi Valley to southern Illinois and Indiana. C. c. paludicola is localized with two disjunct populations—in the Everglades and on Cape Canaveral, Florida. C. c. helvigularis is even more restricted in range and is known only from the Appalachicola region of the Florida Panhandle and the adjacent corners of Alabama and Georgia. C. c. anthicus occupies much of central and western Louisiana and adjacent Texas. C. c. flaviventris occurs throughout the Great Plains, east in the "Prairie Peninsula" through Michigan and northern Ohio and west to the Rocky Mountains. C. c. [372] stejnegerianus occurs from Matagorda Bay in Texas southward through eastern Mexico, with a seemingly isolated population in the Sierra del Carmen region of northern Coahulia. C. c. mormon occurs in the Pacific Coast states and Great Basin.
Actually, the range limits and the continuity of distribution within the area outlined are still poorly known. The species has not been recorded from the northern parts of Maine, Vermont, New Hampshire, Michigan, Wisconsin, or Minnesota, nor from northeastern New York. It is generally absent from southwestern desert areas. Records are particularly scarce and scattered in the Rocky Mountain states, suggesting that the distribution in this area may be discontinuous. In a large area comprising all of New Mexico and Arizona, the western half of Colorado, and the southern halves of Utah and Nevada, records are so scarce as to indicate that the species is there represented by only a few well isolated relict colonies. The type locality of mormon is "Valley of the Great Salt Lake," and there are numerous records from the northern part of Utah east of Great Salt Lake (Woodbury, 1931:75), but a record from Moab is the only one known to me from the southern half of the state. The only records from western Colorado are from three miles east of Fruita and two miles west of Grand Junction, Mesa County (Maslin, 1959:56). Apparently the only valid record from Arizona is that of Shannon (1950:59) from Eagar, Apache County, in the east-central part. Shannon also recorded the racer from Boulder Dam in extreme southern Nevada. Brattstrom (1955:152) has recorded the species from the lower Pleistocene of southeastern Arizona (Curtis Ranch), bearing out the idea that the racer has partly withdrawn from a range formerly occupied in the Southwest at a time when cooler and moister climate prevailed. Other fossil occurrences are of late Pleistocene age—Vero Beach and Seminole, Florida (Brattstrom, 1953a:245) and, doubtfully, Rancho LaBrea, California (Brattstrom, 1953b:376). The range of mormon has been mapped (Wright and Wright, 1949:134) as extending east to south-central Montana on the basis of one specimen allocated on the basis of two characters. Otherwise the range of mormon seems to be entirely west of the Continental Divide, well separated from that of flaviventris by desert and mountain barriers. The conspecificity of mormon with the other subspecies needs to be more thoroughly investigated, and geographic variation within mormon also merits study.
The common racer and the several species of whip snakes (Masticophis) were revised by Ortenburger (1928). More recently with much larger series of specimens, Auffenberg (1955) again revised the classification of C. constrictor, but his study was concentrated in Florida and neighboring southeastern states with relatively little attention devoted to populations of the western and central United States. As the species occurs throughout most of the United States and south through the coastal lowlands of eastern Mexico to Guatemala, it is found over a wide range of environmental conditions. Various characters are subject to geographic variation, and some of them follow clines that are maintained over extensive areas. Such characters as the number of hemipenial spines, and the enlargement of one or more basal spines into hooks, the shape of the premaxillary bone, the number of maxillary teeth, the numbers of ventrals and caudals, color of eye, number of dorsal saddle-marks and of ventral spots in juveniles, and ratios of body proportions including tail length to total length have been used to characterize the subspecies.
Also important is the shade of coloration of adults. The subspecies constrictor, priapus and helvigularis that are characteristic of forested habitats in the eastern United States are black dorsally and have their ventral surfaces suffused to a large extent with dark or dusky coloring. Farther westward the ground color becomes progressively paler, greenish, grayish or light brown, and the ventral surface is yellow (white on the throat and neck). The same tendency appears in C. c. paludicola of the Everglades. The populations of arid climates in southern Texas and in the far western states are relatively pale colored. The species thus conforms to Gloger's Rule in changing from a pallid coloration in arid climates to a dark pattern with eumelanins predominating in a humid climate.
Perhaps the most important character that is subject to geographic variation in the racer, and certainly one of the most neglected, is body size. With information now available it is not possible to compare the sizes of different populations except in a general way. The best sources of information concerning size in several subspecies other than flaviventris, are the publications of Auffenberg (1949 and 1955). Comparison of these data with my own is not entirely satisfactory because Auffenberg did not indicate clearly differences in size between the sexes, nor indicate the [374] boundary line between young and adults. Also, his measurements are of overall length. For the purpose of comparison I have subtracted 22.5 per cent, an approximation of ratio of average tail length, from Auffenberg's figures. He stated (1955:98) that the series of specimens on which measurements were based were those "with a uniform coloration," that is to say they had lost the juvenal pattern and were probably sexually mature. Whether the same statement applied to the large series of stejnegerianus in the same author's earlier paper (1949:55) is doubtful.
C. c. constrictor: 34 New York specimens averaged 806 millimeters (Auffenberg, 1955:96).
C. c. flaviventris: 100 Kansas specimens averaged 791 millimeters (males, 746; females, 836).
C. c. priapus: 171 Florida specimens averaged 713 millimeters (Auffenberg, 1955:96).
C. c. stejnegerianus: 291 Texas specimens averaged 664 millimeters (Auffenberg, 1949:54).
C. c. mormon: 94 West Coast specimens (Museum of Vertebrate Zoology) averaged 563 millimeters (males 515, females 600).
C. c. anthicus: 35 northern Louisiana specimens estimated to average 582 millimeters (Clark, 1949:249—the author did not present individual measurements, but indicated the numbers in several size groups in his sample).
The 100 flaviventris in the above list were recorded in June, July and August, a season when the young of the preceding year are still small, and these young were not included. In a fall sample 63 males averaged 729 millimeters and 65 females averaged 886 millimeters, but with the nearly grown young (44 males and 40 females) included, the averages were changed to 651 and 768 respectively. Maximum length perhaps express differences between the subspecies as well as, or even better than, the averages listed above. The following figures indicate some of the maximum overall length measurements in inches that have been published by various authors. These measurements pertain to females unless otherwise indicated.
C. c. constrictor: 743/4, 741/4, 73, 651/2 (Auffenberg, 1955:96).
C. c. flaviventris: 71 (Pope, 1944:172), 72 (Conant, 1958:147).
C. c. priapus: 521/2 (♂) (Auffenberg, 1955:98).
C. c. stejnegerianus: 37 (Auffenberg, 1949:54), 40 (Conant, 1958:148).
C. c. mormon: 511/2 (Museum of Vertebrate Zoology).
C. c. anthicus: 70 (Conant, 1958:149).
In my own study, the largest racers recorded from the Reservation and Rockefeller Tract had the following overall lengths: 59, 571/4, 551/2, 58 ♂ (projected from stub tail).
These sets of figures show that compared with the blue racer in Kansas, with which my own study was concerned, the black racer of the northeastern states reaches a larger size, while the racers of the Southeast and especially those of the far West and of southern Texas, are dwarfed. These size differences are almost certainly correlated with behavioral and ecological differences among the snakes involved. Adaptations to exploit certain types of prey and to utilize most efficiently certain habitats and types of cover, have led to divergent trends in different parts of the range.
The racer dwells primarily in open situations, but as might be expected from its extensive geographic range, bringing it under the influence of diverse climates and habitats, its populations have diverged somewhat in adaptation to different environmental conditions. The eastern blacksnakes (subspecies constrictor and priapus) seem to prefer woodland and forest edge. In central Louisiana, anthicus occurs chiefly in an open type of woodland. The subspecies stejnegerianus is found chiefly in brushland and thorn forest. The western mormon is found in varied habitats, including moist streamside meadows, and chaparral. Published statements of herpetologists, based upon studies in limited areas, are briefly quoted below to show the trend of geographic change.
C. c. constrictor: This snake "occurs chiefly in fields" (Atkinson, 1901:148; Pennsylvania); "in more or less wooded regions and along hillsides and among bushes" (Surface, 1906:167, in Pennsylvania); "abundant, especially in wooded regions" (Hibbard, 1936:28, in Kentucky); "dry and more or less open places" (Conant, 1938:52, in Ohio); "old fields and areas about buildings" (King, 1939:572, in Tennessee).
C. c. priapus: In "drier parts of the [Okefinokee] swamp ... seems to prefer blueberries and saw palmettos" (Wright and Bishop, 1915:159 in Georgia); "common in grassy fields and in upland woods" (Allen, 1932:13, in Mississippi); "abundant along fence rows ... in dry pine-oak forest and in bottomland forest" (Trowbridge, 1937:296, in Oklahoma); "probably most abundant in open upland hammock or in old fields; limestone flatwoods" (Carr, 1950:80, in Florida); Oak and oak-hickory forest and small hill prairies in southern Illinois (Rossman, 1960:219).
C. c. paludicola: In "all parts of the freshwater Everglades, in cabbage palm hammocks, in salt marshes, and in mangrove swamps. On Key Largo ... in mesophytic hammock" (Duellman and Schwartz, 1958:296, in southern Florida).
C. c. anthicus: In "wooded areas in the vicinity of briar patches or other brushy undergrowth" (Clark, 1949:249, in northern Louisiana); "especially grassy uplands" (Fitch, 1949:88, in central Louisiana).
C. c. stejnegerianus: Of 291, 94 were in scattered brush, 92 in sparse brush, 41 in lightly wooded areas, 26 in grassy areas, 24 in heavy brush, eight in semi-arid places and six in heavily wooded situations (Auffenberg, 1949:55, in southern Texas).
C. c. mormon: In "thin brush skirting open prairie land" (Lord, 1866:304, British Columbia); cottonwood-willow and water margin habitats in prairie (Dice, 1916:310-312, in eastern Washington); "grass; amid water cress; bank of small ditch near meadow; barley field; sandy ground covered with rocks and driftwood; among sedges; in sagebrush; swimming in irrigation ditch" (Grinnell, Dixon and Linsdale, 1930:149, in northeastern California); low foothills, around the fields, and in the timber and brush along the canyon bottoms (Woodbury, 1931:75, in Utah); "open woods of Garry oak and poison oak, on grassy slopes, in chaparral, and in grain or hay fields" (Fitch, 1936:644, in southwestern Oregon); "low hot canyons where it was found to occupy areas having rather dry, rocky hills" (Ferguson, 1952:68, in northeastern Oregon).
C. c. flaviventris: Pastures, meadows, and fields (Hurter, 1911:170, in Missouri); "usually frequents dry open fields, although it is often found in bushes or cut-over land which has grown up into thickets" (Ortenburger, 1928:181); "pasture lands and on hill sides" (Peters, 1942:183, in Illinois); "along the levees in the salt marshes" (Liner, 1954:82, in southern Louisiana); "common in both prairie and woodland habitat" (Smith, 1947:34, in east-central Illinois); Flood plain, sand around sage-sumac brush, rocky slopes (Fouquette and Lindsay, 1955:411, in northwestern Texas).
Several observers have described the habitat in Kansas as follows: "grassy valleys and thinly wooded hillsides" (Burt, 1927:5); "moist and dry habitats, in wooded areas, and in prairies" (Smith, 1956:237); Oak-walnut hillside forest, cultivated field, buckbrush-sumac, prairie (Clarke, 1958:22).
Every part of the 590-acre Reservation is used to some extent by blue racers living on this area. Home ranges of most individuals are so large as to include a variety of habitats, both woodland and grassland. The habitat preferences vary according to season. In autumn some racers migrating to rock ledges to hibernate are found in mesic forest, but at that time of year leaves have begun to fall and the forest floor is less dark and humid than in summer. In spring also racers not yet back on their summer ranges are often seen either along the hilltop ledges, or moving downhill through woods toward bottomland meadows. However, in summer, the finding of a racer in mesic woodland is a rare event. Occasionally the snakes make trips across such blocks of woodland hundreds of feet wide, but they do not linger in the woodland. In decreasing order of preference the habitats used by racers on my study area may be classified as follows:
1. Tall-grass prairie, (Plate 22) either original or regenerated, dominated by native perennial grasses in stands three feet to seven feet high, including big blue-stem (Andropogon gerardi), little blue-stem (A. scoparius) Indian grass (Sorghastrum nutans), and switchgrass (Panicum virgatum).
2. Pastureland, with introduced brome grass (Bromus inermis) and associated weedy vegetation.
3. Brush, in ravines, at woodland edge, and in riparian thickets.
4. Weedy fields, dominated by such pioneer plants as ragweeds, sunflowers, horseweed, milkweed, wild lettuce, aster and goldenrod.
5. Open type of woodland dominated by such trees as honey locust and osage orange.
These habitat types are interspersed on the study area, and each racer has a wide choice of habitats without travelling beyond the limits of its own chosen area.
Grassland that has been closely grazed, mowed or burned does not provide entirely adequate food or shelter, and under such conditions clumps of brush or other dense vegetation may be of critical importance. Throughout the racer's extensive range, fields of grain and hay at times provide suitable habitat, and may support large populations, but in spring, before the young cultivated plants have made much growth, or later in the season, after they have been cut, the racer may need to depend on adjacent areas of pasture, thicket, or woodland edge and the availability of such refugia to a large extent determines the numbers of racers on cultivated areas.
In the locality of my study racers spend approximately half the year in winter dormancy. Earliest spring records and latest fall records for 13 years are shown in Table 4. The spring records in nearly all instances pertain to snakes found in the open or beneath flat rocks warmed by sunshine, usually at or near the rock ledges where hibernation occurs. Juveniles are especially well represented in these earliest spring records, and it seems that they tend to emerge a little earlier, on the average, than the adults, either because they have hibernated in more superficial and less well insulated situations or because their lesser body mass permits more rapid warming to activity than can occur in the adults. The latest fall records all pertain to racers trapped along the rock outcrops, and none was a young of the year.
Table 4. Earliest and Latest Recorded Annual Dates
When Blue Racers Were Active on the Reservation or Rockefeller Tract
| Year | Earliest spring record |
Latest fall record |
Year | Earliest spring record |
Latest fall record |
| 1950 | April 16 | November 4 | 1957 | April 30 | November 2 |
| 1951 | April 19 | November 13 | 1958 | April 25 | November 20 |
| 1952 | April 23 | November 12 | 1959 | April 6 | November 10 |
| 1953 | April 8 | November 12 | 1960 | April 22 | November 6 |
| 1954 | April 20 | November 12 | 1961 | April 19 | November 1 |
| 1955 | April 15 | November 2 | 1962 | April 23 | October 27 |
| 1956 | May 11 | November 14 |
Most of the population undoubtedly emerged somewhat later than the average date of April 16 indicated by the records in Table 4, and retired somewhat earlier than the average date of November 8. However, a small percentage of the population probably emerged even earlier each year than my records indicate, and retired into hibernation later than my records indicate. In a typical year, temperatures in April and early May are only occasionally above the level at which racers are able to become active, but are below this threshold most of the time. The same statement applies to an autumn period of late October and November. Most racers are dormant in their hibernacula during these transitional periods of spring and autumn, but some—those that have emerged early in spring, or those that have not yet retired (in fall)—retreat to temporary shelters and revert to a semi-torpid state when temperatures fall below the critical level.
Fig. 3 shows the relative extent of activity along the hilltop outcrops, as reflected by numbers of racers caught at different times during the autumn. Data from 14 years are combined, and the large composite sample indicates that in an average year there is relatively little activity along the hilltop outcrops in early September, but that activity rapidly increases to a peak in mid-October and then tapers off rapidly, usually ending in mid-November, but occasionally ending as early as late October or as late as late November.
The racers recorded in traps had, in many instances, been confined in them for from one to three days before they were found. For any one year records are not sufficiently numerous to show the trend as well as Fig. 3, but Fig. 4 shows year-to-year differences; 1958 was a fairly typical year, and also was the year in which the largest sample was obtained; in 1949 the largest catches were made earlier than usual, and the racers retired early into hibernation; in 1954 warm weather persisted until unusually late in autumn, and racers remained active beyond the time when they ordinarily would have been hibernating; in 1955 and 1961 the most concentrated activity along the outcrops, as reflected by day to day catches, came later than usual, but unseasonably cold weather ended all activity abruptly, earlier than usual.
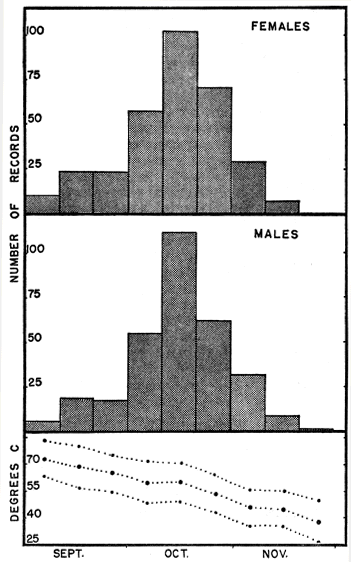
Fig. 3. Records of blue racers trapped along hilltop limestone outcrops in autumn, a composite sample of 14 years (1949 through 1962) from the Reservation and Rockefeller Tract, showing the catch grouped in ten-day intervals, beginning with September 1 to 10 and ending with November 20 to 29. Averages of the maxima, means, and minima of daily temperatures for each period are shown.
The racer's annual cycle of activity is, of course, controlled primarily by the weather, and is much delayed or accelerated in some years. But certain stabilizing factors cause the racer's annual cycle to be less variable than that of the weather. For example, in spring when persistently cool weather delays emergence from hibernation beyond the normal time, the increasing azimuth of the sun, and more intense sunshine cause the soil to warm, despite low air temperature, until emergence is finally triggered. Having once emerged, the racer is able to control its bodily temperature to a large extent by basking in sunshine to gain warmth, or by seeking shade or underground shelter to escape overheating. By such behavioral thermoregulation extremes of weather are neutralized, or at least buffered to some degree.
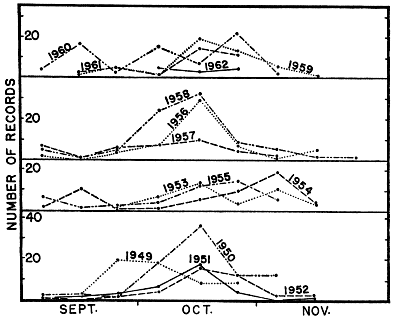
Fig. 4. Yearly variation in catch of blue racers along hilltop outcrop in autumn on the Reservation and Rockefeller Tract, grouped in same ten-day intervals indicated for Fig. 3.
In the course of my study no racers were found in their actual hibernacula. Insofar as known, these were always in deep crevices in strata of limestone near hilltops, and were inaccessible except by removal of the solid rock. The crevices where racers hibernate are known to be several feet deep in some instances, extending well below the frost line. Periodic temperature readings in two such crevices at depths of 12 inches and 30 inches, taken in the winter of 1954 (Fitch, 1956:471) showed that temperatures during dormancy are usually well within the range 0°C to 10°. Whether the [381] racers congregate in hibernating masses in regular "dens" on the Reservation has not been definitely determined, but if so, most of the hibernating groups must be small, because those trapped along the ledges are well scattered, and, in fact, give the impression of being rather uniformly distributed along them. However, ledges of northward exposure are not used as hibernation sites, so far as known, and those of full southward exposure are perhaps preferred, especially where the hilltop has a southward projecting spur, and the exposed rock face is extensive, with many cracks and fissures. I have been unable to detect differences in types of hibernation sites preferred between the racer and the copperhead, which is somewhat more numerous on the same area.
Several authors have contributed to knowledge of hibernation in the racer. Boyer and Heinze in Missouri (1934:195) noted that blue racers often were associated with copperheads in the vicinity of places chosen for hibernation. Burt (1935:329) in Kansas found blue racers emerging from dens among rocks on a prairie hillside, associating with other snakes, Diadophis punctatus, Elaphe guttata, and Pituophis melanoleucus. In the Chicago region, Pope (1944:173) reported scores of blue racers aggregating in October on and around a sand dune with oak woodland. In Ohio, Conant (1938:55) found three blue racers hibernating together about three feet underground in a small hole. One found at another locality had apparently hibernated in company with a massasauga (Sistrurus catenatus). In Maryland Cohen (1939:137) saw racers (C. c. constrictor) in the act of emerging from an old vole burrow that was a communal hibernaculum, on April 6, 8, and 10. Air temperatures at the time of emergence were 12.5° and 18.5° Centigrade. In Illinois, Schroder (1950:1-2) found seven blue racers hibernating in masses, intertwined with each other and with bull snakes at depths of 36 inches and 42 inches in an abandoned mammal burrow in a sand dune area in early February.
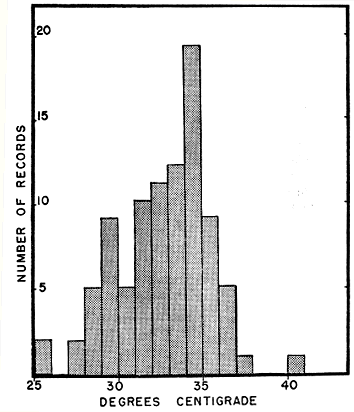
Fig. 5. Bodily temperatures of blue racers kept in a large outdoor enclosure and checked from time to time when they were active and the sun was shining, in June and July, 1962.
In the course of routine field work I often carried a Schultheis quick-reading thermometer, and from time to time I had opportunities to take the body temperatures of blue racers newly captured by hand. The trend of these records indicated the temperature range within which the snakes normally limit their activity, and the preferred temperature. In an earlier publication (Fitch, 1956:459-460) based on a few more than half the number of records of temperature now available, I discussed responses of the blue racer to temperature. The newer data bear out the trends previously revealed; of 60 records, 39 are within the six-degree range from 29° to 35°, and records are most concentrated in the one-degree range, 34° to 35°. Racers were found active at air temperatures between 15.5° and 32.4°, with the greatest concentration between 26° and 27°. Compared with most other kinds of North American snakes, the racer is remarkably tolerant of heat, and often is several degrees warmer than the level that those of other genera will normally tolerate. Racers have been seen crawling in the sunshine, or basking on days that were uncomfortably hot for humans. For example, on August 28, a large female racer released from a trap was followed and her behavior observed; after crawling some 50 feet through the grass she climbed from a ditch bank onto sunflower stalks and elm saplings, and came to rest among the stalks, in a spot strategically situated for catching grasshoppers. More than half her body was exposed to sunshine and air temperature was slightly above 34° Centigrade, yet the snake showed no sign of discomfort, and for the several minutes that she was kept under observation, did not attempt to withdraw into the shade.
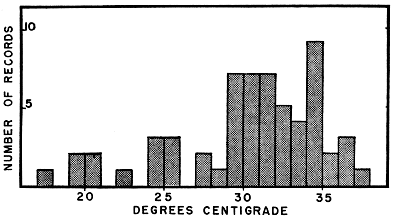
Fig. 6. Bodily temperatures of blue racers captured by hand in their natural surroundings. The preferred level is approximately the same as indicated by Fig. 5 (between 29° and 36° Centigrade), but some of the racers caught were not fully active and had lower temperatures. Some bias results from the fact that those having the lowest temperatures were the least active and hence were most easily caught.
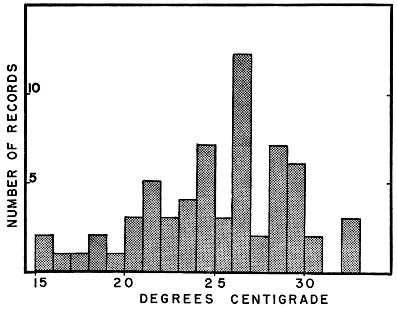
Fig. 7. Air temperatures recorded at captures of the racers whose records were used in Fig. 6. An active racer typically maintains, by basking, a bodily temperature several degrees warmer than the air.
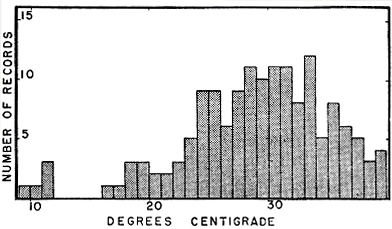
Fig. 8. Bodily temperatures of blue racers found in live-traps at Harvey County Park. Opportunity to regulate temperature by behavior was limited in these snakes in traps, which tended to match ambient temperatures.
At the Harvey County study area, bodily temperatures were recorded in many of the racers that were caught in traps. These records are much less significant than the records obtained from racers caught by hand and promptly checked for bodily temperatures. The temperatures of the trapped snakes may, to a large extent, reflect the temperatures of air and soil at the time. However, despite their confinement, the trapped racers probably were able to exercise some control over their temperatures by shifting from shade to sunshine, or from the top of the trap to its bottom, where they would be in contact with the substrate. In most of the racers removed from traps, as in those caught by hand, bodily temperatures were somewhat above air temperatures, but the difference was less in the former group of snakes. Figure 8 shows the bodily temperatures of these snakes removed from traps. Occasionally racers died in the traps from overheating. On July 2, 1960, each of two racers in traps had temperatures of 39.4°. One of these was especially vicious and frantic in its attempts to escape, but otherwise seemed unharmed. The second racer was dead, seemingly having succumbed just before it was found. Probably [385] prolonged exposure to temperature in excess of 39° would always result in death of the snake. Racers and other snakes that had become overheated in the traps and were nearing exhaustion had a characteristic limp feel when they were handled. In June, 1960, heat tolerance of a halfgrown racer was compared with that of several other snakes including a copperhead, garter snakes, and ringneck snakes. Each snake in turn was enclosed in a plastic tube plugged with cotton at one end, the snake having a quick-reading thermometer taped in place for a rectal reading. The tube was then placed in sunshine. Over periods of minutes the enclosed snake passed through a characteristic cycle. Soon it would begin to register discomfort as its temperature rose rapidly. Its struggles would become increasingly violent, then would cease abruptly. The snake would suddenly collapse, its body mostly limp, but knotted in slow contortions, its mouth gaping widely. Within a few seconds all movements would cease, but in each instance the seemingly dead snake was soon revived by holding it in cold running water. The copperhead, garter snakes, and ring-necked snakes all collapsed at temperatures near 41°. At this same temperature the racer showed signs of acute discomfort, but did not collapse even after many minutes of exposure. Probably more protracted exposure at this level would have been fatal to the racer as well as to the other kinds tested.
Blue racers that were recorded on more than one occasion were rarely caught again at the original location. For different individuals, distances between capture points ranged from zero up to a little more than three-fourths of a mile. The area of concentrated study was a mile and a half in greatest diameter; there was scant opportunity for capturing racers that moved greater distances. Even those that moved as far as a mile would have passed beyond the boundaries of the study area in most instances. Many of the marked racers that disappeared from my records probably moved beyond the limits of the study area. Nevertheless, in the great majority of instances, the distances between successive capture points for the same individual were relatively short, indicating that each racer tends to remain permanently in a restricted area.
Most captures were made in the type of grassland or brush that provides favorable habitat for the racer during the season of activity, but many other captures were made in woodland along the rock [386] ledges where the snakes come to hibernate. Four different types of movement may be recognized: 1) those in the rock ledge area where hibernation occurs; 2) those between the area where the summer is spent and the hibernation ledge—an actual small scale seasonal migration which takes place in spring and autumn—3) those within a home range, which are part of the day-to-day activities of the racer, and, 4) wandering movements by which the racer shifts its activities, perhaps permanently, from one area to another. In the records of any one snake these different types of movements cannot always be sorted with certainty. Each type will be discussed separately.
Relatively few movements along the ledges were recorded. It seems that having migrated to a ledge, the racer promptly finds its hibernaculum and retires for the winter. In spring there is equally prompt scattering of the emerging racers, which no longer find the ledge attractive. Most recorded movements along the ledges were short. Of 76 movements, nine exceeded 1000 feet, and only four others exceeded 500 feet. Most of the shorter movements were recorded within an autumn season, but several were recorded after the lapse of one or more seasons of activity. The longer movements were as follows: 1250 feet after 8 seasons (male); 1300 feet after three seasons (female); 1600 feet after one season (female); 2000 feet after one season (female); 2280 feet after seven seasons (male); 2200 feet in same season (female); 2410, 2600, and 3200 feet, each after one season (all males). The trend of these records suggests that the tendency to return year after year to the same hibernaculum is not strong; after using one for a period of years, the racer may abandon the stretch of ledge and, starting out in the opposite direction from its summer range, find a new hibernaculum as much as half a mile from the old one. Records of distances between capture points on the ledges for individual racers are shown in Fig. 12.
A total of 124 movements between summer ranges and ledges were recorded, and the distances averaged 1309 feet—approximately a quarter mile. Some racers living in hilltop fields may have had home ranges that included rock ledges, or at least were adjacent to them. In such instances no seasonal migrations would have been necessary to reach hibernacula in the autumn and summer ranges in the spring. Several short movements—100 feet, 150 feet, and 200 feet—can be explained on the basis that home ranges and hibernation ledges overlapped or were near at hand, but most [387] of the movements were longer. The longest movement was 4020 feet, after a lapse of four seasons. Twenty-four movements exceeding 2000 feet were recorded. For these the intervals between captures averaged more than double the time for the remaining movements, indicating that the longtime permanent shifts were involved in many instances.
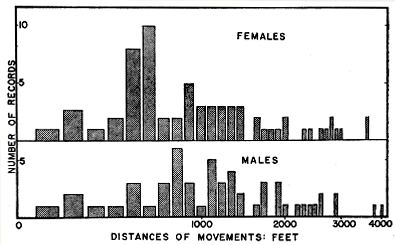
Fig. 9. Histogram of movements of blue racers between hilltop rock outcrops used for hibernation, and summer habitat on the Reservation and Rockefeller Tract. Movements of females tend to be somewhat shorter than those of males.
For all the racers living in bottomland, ranges were separated from ledges by areas of wooded hillsides averaging approximately 700 feet across. These relatively unfavorable areas had to be traversed in the course of the semi-annual migrations. Even some of the racers that lived in hilltop fields apparently crossed wooded slopes in order to reach distant hibernation ledges, or else each reached the ledge by a roundabout route although it could have found a ledge much nearer its summer range. For the 124 ledge-to-field and field-to-ledge movements, the median distance was 1030 feet. The sexes were almost equally represented in this sample but the average distance for the 55 males—1425 feet—notably exceeded that for the 69 females—1220 feet. These movements are shown in Fig. 9.
McCauley (1945:76) in Maryland described what seemed to be incipient territoriality in a large male racer that remained several hours in a small area, crawling about conspicuously with head [388] raised, seemingly on patrol. When an even larger male racer intruded, the first one aggressively drove him away, but neither paid any attention to a king snake that was also on the area. Other authors have noted the attachment of a racer to a small familiar area. Conant (1938:53) wrote that many of the racers he saw sought shelter in definite retreats. One of these racers was seen resting on top of a brush pile four times in a single afternoon, and each time it followed the same route to the same inaccessible spot beneath the brush.
My own observations do not bear out the idea that racers maintain regular territories, since several males may be present within a small area, even in the breeding season. Hostile behavior between males has not been observed by me under natural conditions, and in confinement has been seen only in instances of self defense. Like the racer Conant observed on a brush pile, individuals may linger in the vicinity of a favored shelter or foraging area for periods of hours, but such associations are ephemeral, and soon the snake moves on. In a uniformly favorable habitat a racer may cruise about freely in tall grass or brush. Individuals that I have attempted to follow, after flushing them or releasing them from traps, often covered distances of 100 to 300 feet within periods of a few minutes before I lost them. In such instances I maintained sufficient distance between myself and the snake so that the latter was not actively escaping. Probably the snake was not aware of pursuit in most instances, although I was able to glimpse it through the stems of grass, weeds, or shrubs, or was informed of its course by the swaying tops of grass and other vegetation.
For many of the racers captured over periods of years it was possible to plot "minimum home ranges" in the areas that they occupied. One caught 12 times in five consecutive years will serve as a typical example. There were seven locations involved; three captures were made at one point and two captures at each of two others; the other five locations were each represented by a single capture. One of the seven locations was for a capture made at a rock ledge in October, and hence can be eliminated from considerations of home range. The other six locations are based upon captures made from late May to early August, and they form a rhomboid pattern, with three locations in alignment on one side and two others inside the quadrangular figure formed by the five outlying points. Obviously such a group of records gives some idea of the location and extent of the snake's activities but the information [389] is far from complete. As shown by Odum and Kuenzler (1955), a much larger series of records, usually several dozen, with eight or more marginal locations, is necessary to illustrate even an approximation of the actual home range. Under the conditions of my study such a series of records was unattainable. Few if any of the racers recaptured had more complete series of records than the one mentioned above.
For 20 racers the records were sufficiently numerous and well distributed to permit plotting of minimum home ranges. One of these ranges was hexagonal, nine were pentagonal, eight were rhomboidal and three were triangles. In four instances the area encompassed was broken by woodland, indicating that the home range comprised two or three disjunct segments. In all instances the smaller segments were triangular. The 20 minimum home ranges averaged 6.6 acres (3.2 to 12.8). The 15 ranges of males averaged 7.3 acres, whereas the five ranges of females averaged only 4.5 acres, but the sample is too small to be relied upon for differences in the sexes.
Fig. 10. Movements of blue racers within or between areas of summer habitat on the Reservation and Rockefeller Tract. The trends are much alike for males and females.
In an earlier publication (Fitch, 1958:73) I discussed an alternative method for determining size of home range in animals that [390] move about freely within a chosen area, not having their movements restricted by attachment to a specific home base. Ordinarily any two records of the animal within its home range will be separated on the average by a distance equal to half the diameter of the area. Assuming that home ranges in general tend to have a circular shape, except as restricted by limiting environmental factors, the area can be easily computed from the average recorded movement—the home range radius. It is necessary, of course, to have a sufficiently large number of records of movements to obtain an average that is statistically reliable.
A major problem is that of recognizing movements that involve an extension of the original range or a shift away from it to a new area. A few exceptionally long movements were recorded. If these are included in the computations of home range, they greatly increase the average distance, probably introducing error. Also, the number of exceptionally short movements was greater than might have been expected if all locations of capture are at random to each other. In some instances a racer newly released may have blundered into the same trap again, or into the trap at the opposite end of its drift fence. In other instances traps may have been so strategically situated with respect to preferred travel routes that they caught the same snakes repeatedly. In still other instances, the range of an individual might have been mostly outside the study area, with only one end or corner overlapping the trap sites.
A total of 471 records for consecutive captures in field areas is available, 305 for males and 166 for females. In 20 instances successive sites of capture were the same and movement was recorded as zero. Of the 471 records, 207 involved a relatively long time span, including at least one hibernation period; the remaining 264 were based upon successive records within the same season of activity. The trends were much the same in the records involving a longer time span (up to four years) as in those records involving captures made in a single season, but for the longer periods there were some exceptionally long movements, and relatively few short movements of less than 100 feet.
Records of male racers and those of females were used for separate computations. For each series, the ten per cent of movements that were longest and the ten per cent that were shortest were eliminated from consideration in calculation of the average distance between points of capture. For the remaining 244 records of males an average movement of 595 feet was calculated, and for [391] 132 records of females an average movement of 574 feet. These distances, if accepted as typical home range radii, would represent home ranges of 26.3 acres for males and 23.8 for females. In an earlier discussion of spatial relationships in the racer (Fitch, 1958:119), based upon relatively scanty data, I estimated the home range to be approximately 23 acres in males. But with only nine records for female racers I calculated the home range to be 9.7 acres.
The disparate figures obtained from plotting minimum home range and from calculating average home range radius are not irreconcilable, since a minimum home range based on only four or five points would ordinarily include only a fraction of the actual range. Distances up to 1500 feet are included in the calculation of home range. It seems that home ranges often have a diameter of this magnitude or a little larger, although the estimated average diameter is 1140 feet. Home ranges probably most often deviate from circular shape to form an ellipse, with one diameter markedly exceeding the other. Woodland, water, roads, buildings, or cultivated fields, or other areas that are unfavorable or uninhabitable often form the boundary of a home range and influence its shape.
Many of the longer movements constituted clear-cut shifts in range. In one exceptional instance a large adult female captured in the northeastern part of the Reservation on June 22, 1950, was released 21 days later at a point 3900 feet southwest of the place of capture. On May 27, 1960 she was caught within 600 feet of the original location, seemingly having made a homing movement. Among the nine racers recorded to have made longest movements (exclusive of those movements made to or from hibernacula) four were recorded also to have made later long movements in the reverse direction, probably returning, each to its original home range, although in every instance the return movement was somewhat less than the original. A female of two-year-old size when first captured on September 2, 1957, was recaptured 3100 feet southeast on May 10, 1958. On August 7, 1959, she was recaptured again 2400 feet from the second location in the direction of the original capture. Similarly, in a three-year-old female a shift of 2730 feet was recorded at the second capture after 21 months, and at the third capture 14 months after the second, a return trip of 2360 feet had been made. A second-year female made a trip of 2640 feet between May 17 and October 1, 1960; by May 1961 she had returned 2000 feet to the vicinity of her original capture. From one year to the next an adult male shifted 2450 feet; after another year he had [392] moved back 1550 feet. Most of the longer movements recorded were those between home ranges in fields and hibernacula along ledges, but in this class of movements, distance was somewhat proportional to elapsed time. For 59 such movements exceeding 2000 feet the average was 3.1 years, whereas for 114 field-to-ledge movements of less than 2000 feet, average elapsed time was 1.6 years. This trend suggests that over periods of years a racer is likely to shift its range or its hibernaculum or both.
Fig. 11. Map showing home ranges of five blue racers, as indicated by numerous captures in successive summers, in small valley where Reservation headquarters are located, and spatial relations of their hibernacula, as represented by points of capture along hilltop limestone outcrops. In spring and autumn, traveling to and from hibernacula, the snakes migrate across wooded slopes. Each "minimum home range" is enclosed in a dotted line, and a distinctive symbol is used to show successive points of capture for each snake.
Fig. 12. Map of 600-acre area of Reservation and Rockefeller Tract, including parts where field study was most concentrated, showing movements of the 20 blue racers recorded to have shifted over the longest distances. The figure following the sex sign of each individual indicates number of months elapsed between the captures at the localities represented by the dots.
Average elapsed time between captures was 7.7 months. In the 471 field-to-field movements recorded, 53—slightly more than eleven per cent—exceeded 1500 feet and can reasonably be considered shifts of home range. The average elapsed time between captures for this group of snakes was 9.5 months. The evidence suggests [394] that, even in an area of favorable habitat, somewhat more than ten per cent of the racers in a population annually shift their home ranges somewhat, but that many stay in the same home range for periods of years or perhaps throughout life.
Shifts in range were especially noticeable where availability of suitable habitat underwent seasonal change. Along the north edge of the Reservation, prairie adjoined cultivated fields where grain or hay was grown. Until late May, the cultivated crops made little growth and the fields were almost bare. They provided insufficient shelter for the racers, which tended to keep to the prairie, where old grass of the previous year's growth furnished them with ample cover. Later in the season, crops, of oats, wheat, and alfalfa constituted suitable cover for the racers, and many of them shifted their ranges to the cultivated fields, but corn and milo crops were much less adequate for their needs. After harvesting of crops, cover in the fields was again inadequate for the racers' needs, and they tended to retreat to edge situations, or to adjacent prairie.
The racer hunts by stealth, but actively, obtaining its prey by keen eyesight and swift movements. Wright and Bishop (1915:160) wrote that because of its great speed it can catch anything that moves on the ground. As a racer moves stealthily through dense vegetation, its dull, uniform dorsal color blends well with the surface litter of dead plant material. In prowling, the snake glides along rapidly and alertly, in a jerky fashion, with frequent momentary pauses and changes of direction. Because of its inconspicuousness, it is not likely to be detected by the prey until it is close at hand. The snake is ready to dash in pursuit of any small animal that flies, jumps or runs to escape.
On August 27, 1955, my daughter observed a large racer hunting among tall weeds at the edge of the pond on the Reservation. Several times in the course of its movements, it flushed small frogs (Rana pipiens) and each time the snake darted in unsuccessful pursuit of the rapidly hopping frog. On several occasions I have been led to a blue racer by the distressed croaking of a frog that the snake had captured. In each instance, despite my cautious approach, the racer saw me before I detected it, and then darted away, abandoning its prey. On one occasion, while I was still a few yards from the racer, and before the latter had detected me, the [395] frog broke free and hopped away rapidly through tall grass and weeds, and after several leaps, hid, concealed by dense screening vegetation. The racer darted in pursuit but could not find the frog. For several minutes the snake persisted in an active search; with forebody elevated and head held high, it would turn first in one direction and then in another, with nervous, jerky movements, obviously keyed up to a high pitch of excitement. Then it became aware of my presence, lowered its head, and glided away rapidly, abandoning the search.
Although the racer depends to a large extent on sight to find its prey, scent may play some part also, as indicated by the presence in the food of young mammals taken from nests, some probably found underground. Near Garnett, Kansas, on May 4, 1952, Richard B. Loomis found a racer attacking a collared lizard (Crotaphytus collaris) beneath a large flat rock. The lizard was retaliating by biting the snake's neck. The posterior part of the snake protruded into the open, and its thrashing had directed the attention of the observer to it. Whether the racer first found the lizard under the rock, or followed it there after flushing it in the open is unknown.
An encounter between a large blue racer and an adult Great Plains skink (Eumeces obsoletus) on August 30, 1948, was described as follows: "The skink, grasped by one flank, had twisted back and seized the skin of the snake's neck in a bulldog grip, and they lay interlocked, motionless except for their rapid panting, and occasional straining of the skink to bite harder or of the snake to shift its grip and work its jaws toward the skink's head. The racer broke the skink's grip, and began to swallow it head first. When only the hind legs and tail of the skink still protruded from the racer's mouth, I lunged forward in an attempt to catch both reptiles. With a sudden movement the snake disgorged the skink, which darted away into the grass and escaped" (Fitch, 1955:78).
Many authors have contributed to knowledge of the racer's food habits. In most instances the records have been few or casual, but several intensive studies have been made, notably by Surface (1906) in Pennsylvania, Ortenburger (1928) for the species as a whole, Uhler, Cottam and Clark (1939) in Virginia, Clark (1949) in Louisiana, Auffenberg (1949) in southern Texas, Hamilton and Pollack (1956) in Georgia, and Klimstra (1959) in southern [396] Illinois. However, the findings of different authors are not strictly comparable; some have made general statements concerning the food habits but have mentioned specific items only when these were considered unusual. Certain authors have listed individual prey animals eaten; others have indicated the percentages (in bulk or in frequency) that the different kinds of prey comprised. Some writers have identified food animals only in broad categories such as "insect," "beetle" or "snake" while others have undertaken specific determinations for all the prey or for certain taxonomic groupings that were subjects of special interest.
For the eastern black racer (C. c. constrictor) the following food items have been recorded: 1 robin (Turdus migratorius, Storer, 1839:226); 1 copperhead (Agkistrodon contortrix, Verrill, 1869:158); 1 weasel (Mustela sp.—presumably the diminutive M. rixosa—Atkinson, 1901:148); 3 undetermined mammals, 1 rabbit, 1 undetermined mouse, 7 voles (2 Microtus sp., 4 M. pennsylvanicus, 1 Clethrionomys gapperi), 1 undetermined bird, 2 robin eggs, 2 garter snakes (Thamnophis sirtalis), 1 water snake (Natrix sipedon), 1 grass snake (Opheodrys vernalis), 1 green frog (Rana clamitans), 1 wood frog (R. sylvatica), 1 grasshopper (Melanoplus femur-rubrum) 2 camel crickets (Ceuthophilus sp.), 5 moths (cecropia, regal, imperial), 4 beetles, 1 currant worm, 1 ichneumonid wasp (Nematus ribesii), 1 currant worm (Surface, 1906:170); 1 ribbon snake (Thamnophis sauritus, Ditmars, 1907:282); 3 snakes (1 Liopeltis vernalis, 1 Storeria occipitomaculata, 1 undetermined), 6 white-footed mice (1 Peromyscus leucopus, 5 P. nuttalli), 1 vole (Microtus pennsylvanicus), 16 crickets (9 Gryllus pennsylvanicus, 4 G. assimilis, 2 Miogryllus verticalis, 1 Nemobius fasciatus), 2 grasshoppers (Dissosteira sp.), 1 lepidopteran, 3 elaterid beetles (Ortenburger, 1928:200). Richmond and Goin (1938:310) recorded finding the stomach of a black racer crammed with June beetles (Phyllophaga). Conant (1938:53) recorded a black racer from Ohio that had a smaller individual of its own species in its stomach. The smaller snake contained a caterpillar. Uhler, Cottam and Clark (1939:34) found food in 16 of 34 black racers from Virginia. Mammals, including a shrew (Blarina brevicauda), a mole, a flying squirrel (Glaucomys volans), a microtine, and a mouse (Peromyscus sp.) made up 26 per cent, 2 worm snakes (Carphophis amoenus), 2 ring-necked snakes (Diadophis punctatus), and 1 water snake (Natrix sipedon) made up 25.6 per cent, 5 birds including a warbler and a sparrow, made up 17.75 per cent; 2 frogs (Rana sp.) made up 9.38 per cent, 1 fence lizard (Sceloporus undulatus) made up 6.25 per cent, and insects, including cicadas (Tibicen sp.) and larval lepidopterans, made up 15.09 per cent. In Indiana, Minton (1944:457) examined 11 food-containing stomachs; there were rodents in six, snakes in five, a tree frog in one, and insects (cicadas, large grasshoppers) in four, and another black racer was found swallowing a small box turtle (Terrapene carolina). In Maryland, McCauley (1945:75) examined eight digestive tracts and recorded a shrew (Blarina brevicauda) in one, an unidentified mammal in one, 2 small cicadas in one, 2 small chickens in one, a fence lizard (Sceloporus undulatus) in one, and frogs and toads (including Hyla crucifer) in one; a ninth snake had eaten a half grown rat. In Connecticut, Finneran (1948:124) observed a large black racer eating a 21-inch garter snake (Thamnophis sirtalis). Duellman (1951:338) recorded a black racer in Greene County, Ohio, swallowing a large garter snake (Thamnophis sirtalis). In Kentucky, Barbour (1950:104) recorded remains of an unidentified snake in one stomach.
Many authors likewise have recorded food of the southern black racer (C. c. priapus). In Georgia, Wright and Bishop (1915:160) recorded finding 2 racerunners (Cnemidophorus sexlineatus), a skink (Lygosoma laterale), 4 green tree frogs (Hyla cinerea) and 1 pine woods tree frog (H. femoralis) in [397] stomachs. They also stated that the toad (Bufo lentiginosus [= terrestris]) was the most important article of food. Burt and Hoyle (1934:205) wrote that a racer from Rogers County, Oklahoma, had eaten an adult male collared lizard (Crotaphytus collaris). In Florida, Carr (1950:80) found one of these black racers eating a leopard frog (Rana pipiens). Hamilton and Pollack (1956:523) examined digestive tracts of 62 and found food in 57, comprising the following percentages by volume: Lygosoma laterale, 34.2; Eumeces fasciatus and E. egregius, 11.3; Cnemidophorus sexlineatus, 8.8; Sceloporus undulatus, 3.5; undetermined lizard, 3.5; Opheodrys aestivus, 6.6; Diadophis punctatus, 3.1; Storeria dekayi, 1.6; Coluber constrictor, 1.8; Heterodon platyrhinos, 1.8; Masticophis flagellum, 1.8; Rana sp., 5.3; Hyla cinerea, 1.8; Hyla versicolor, 1.8; Peromyscus, 1.8; undetermined rodent, 1.8; lepidopterous larva, 1.7.
In southern Illinois in an intergrading population of racers intermediate between C. c. priapus and C. c. flaviventris, Cagle (1942:188) examined several stomachs and found 1 chipmunk (Tamias striatus), 2 voles (Microtus sp.), 2 mice (Peromyscus sp.), 2 green snakes (Opheodrys sp.), 1 water snake (Natrix sipedon) and grasshoppers. From this same population Klimstra (1959:212) examined 137 digestive tracts of which 115 contained food as follows: 194 locustids, 118 gryllids, 17 undetermined beetles, 13 carabids, 6 scarabaeids, 10 lepidopterans, 9 hemipterans, 1 hymenopteran, 2 homopterans, 1 dipteran, 17 undetermined insects, 73 Peromyscus sp., 19 Microtus ochrogaster, 9 M. pinetorum, 12 Sylvilagus floridanus, 3 Scalopus aquaticus, 3 Rattus norvegicus, 4 Mus musculus, 2 Tamias striatus, 2 Synaptomys cooperi, 16 Rana pipiens, 8 Acris crepitans, 2 Rana clamitans, 2 R. palustris, 1 R. catesbeiana, 4 Hyla crucifer, 3 Pseudacris nigrita, 4 Lampropeltis calligaster, 4 Sceloporus undulatus, 4 Chrysemys picta, 1 Heterodon platyrhinos, 1 unidentified reptile, 4 Sturnella magna, 1 Otocoris alpestris, 4 unidentified birds. Percentages by volume of the various categories in this sample were: insects, 39.1; mammals, 32.9; amphibians, 10.8; reptiles, 8.3; birds, 6.3; miscellaneous, 2.6.
Food of the "buttermilk snake" (C. c. anthicus) is known only through Clark's study (1949:249). In an unstated number of examinations he found "mice" in 25, "rats" in five, lizards (Sceloporus undulatus and perhaps others) in eight, frogs (Rana pipiens) in seven, and birds in three.
The food of C. c. stejnegerianus is known only from the work of Auffenberg (1949) but his sample was based on 206 racers that had food, among the total of 291 recorded. Unfortunately, he did not present actual numbers of the various prey animals, but divided the food into seven categories and listed these as percentages. He did not indicate whether the percentages represented volumes or numbers of individual occurrences, and evidently there was some error in computation since his combined percentages totalled 111. The categories and their percentages were as follows: grasshoppers, 42.5; crickets, 13.5; miscellaneous insects, .6; earless lizards (Holbrookia sp.), 40.1; scaly lizards (Sceloporus sp.) 2.1; frogs (Rana sp.) 10.0; rodents, 2.2. Auffenberg divided his sample of racers into five size classes, and showed that the smaller snakes fed chiefly on insects whereas vertebrates were increasingly prominent in the food of the larger snakes.
The food of C. c. mormon is known chiefly through the work of Ortenburger (1928:228) who cited instances of a skink (Eumeces skiltonianus) and a young garter snake (Thamnophis sirtalis) being eaten, and listed the following items from 24 stomachs that he examined: 7 decticids, 8 acridids, 5 oedipines, 1 tryxaline, 6 Melanoplus sp., 3 M. mexicanus, 2 M. devastator, 1 M. bivittatus, 2 Dissosteira carolina, 1 Chortophaga viridis, 3 Neduba carinata, 3 Trimerotropus sp., 7 Hippiscus sp., 2 Steiroxys sp., 3 Canoula pellucida, 2 Stenopelmatus fuscus, 2 S. pictus, 4 Gryllus assimilis, 4 Ceuthophilus sp., 1 Pristoceuthophilus pacificus, 6 Gammarotettix bilobatus and 2 cicada nymphs. Grinnell, Dixon and Linsdale (1930:149) found that one of these racers had eaten a cricket. Fitch (1936:644) found another in the act of swallowing an adult vole (Microtus californicus), and recorded (1935:18) that two alligator lizards (Gerrhonotus multicarinatus) were found in the stomach of still another. [398] Woodbury (1931:75) recorded that a racer from Utah had a sagebrush scaly lizard (Sceloporus graciosus) in its stomach. Of the specimens examined in the University of California Museum of Vertebrate Zoology, no. 17256 from the Mad River, Trinity County, California, had eaten an alligator lizard (Gerrhonotus coeruleus), and no. 10120 from Yolla Bolly Mountain in the same county had eaten a bird (unidentified) and a Jerusalem cricket (Stenopelmatus sp.).
Several authors have published specific information regarding the food of C. c. flaviventris. Hurter (1911:171) caught a blue racer in the act of swallowing a copperhead (Agkistrodon contortrix). Taylor (1892:331) recorded finding garter snakes in several large racers. Pope and Dickinson (1928:53) recorded instances of blue racers feeding on racerunners (Cnemidophorus sexlineatus). Ortenburger (1928:181) examined 22 stomachs and recorded: 1 large garter snake (Thamnophis sirtalis), 1 vole (Microtus pennsylvanicus), 1 frog (Rana sp.), 31 crickets (Gryllus assimilis), 4 decticines, 2 acridids, grasshoppers (1 Hippiscus, 2 Melanoplus sp., 1 M. confusus, 1 M. differentialis, 1 Dissosteira carolina, 1 Sphargemon collare, 1 Trimerotropus sp., 1 Orphulella sp., 1 Chloealtis conspersa, 1 Chortophaga viridifasciata, 1 Omaseus sp., 1 Pedocetes sp.), and 2 caterpillars (1 noctuid, 1 sphingid). Gloyd (1928:123) recorded a hatchling glass lizard (Ophisaurus attenuatus) in the stomach of a juvenal racer. Force (1930:31) found a racer eating eggs from the nest of a cardinal (Richmondena cardinalis) and another racer eating eggs of a red-wing (Agelaius phoeniceus). Gloyd (1932:403) recorded an observation of a racer overpowering and swallowing a copperhead. Anderson (1942:210) recorded remains of crickets and grasshoppers in feces. Hudson (1942:55) recorded a racerunner (Cnemidophorus sexlineatus) in the stomach of a juvenile and recorded an earless lizard (Holbrookia maculata) 3 lizard eggs, and 14 grasshoppers (Melanoplus differentialis and others) in the stomach of another. Marr (1944:484) found a harvest mouse (Reithrodontomys montanus) in one. Breckenridge (1944:118) recorded stomach contents including a garter snake (Thamnophis sirtalis), a frog (Rana pipiens), 3 crickets and 2 moths. Mossimann and Rabb (1952:27) recorded that a racer disgorged several grasshoppers. Fouquette and Lindsay (1955:411) recorded that a blue racer had eaten a harvest mouse (Reithrodontomys sp.). Carpenter (1958:114) recorded that one blue racer had eaten a green snake (Opheodrys aestivus) and another had eaten a grasshopper and a camel cricket.
Even though the sets of data cited above are not entirely comparable, certain trends are evident. The black racers of the eastern states (especially C. c. constrictor of more northern regions) take a high proportion of vertebrates in their prey. Among these vertebrates snakes especially are well represented and the black snake would seem to be of some importance as an ophiphagous predator. The birds and mammals taken include some that are bulky (robin, cottontail, and even a weasel—the most formidable prey eaten). Presumably the rabbits that were eaten were young. In samples from the eastern United States insects made up small to insignificant parts of the food; they were lacking entirely or at least were not mentioned in the samples examined by McCauley and Wright and Bishop. In the blue racer of the central states, insects (mostly grasshoppers and crickets) are much more prominent in the food and vertebrates correspondingly less prominent. The vertebrates eaten are largely lizards, small snakes and mice. C. c. stejnegerianus is much like flaviventris in the trend of its feeding. C. c. mormon[399] is less known than these subspecies in its feeding, but indications are that it takes a higher proportion of orthopteran insects and smaller proportions of mammals and snakes than do any of the other subspecies.
In my own field study a total of 1357 food records were accumulated, one of the largest samples known for any kind of snake. Most of these records were from the small area where my population study was carried on, and studies of other kinds of animals, including those that were the racer's prey, were simultaneously in progress. Because large collections of reference materials were available, it was possible to identify to species many of the prey items found, even though they were incomplete and highly fragmented because most of them were recovered from fecal material.
The prey is, of course, swallowed entire, and the recently swallowed items squeezed from the stomachs provide the best material for the study of food habits. However, relatively few racers had detectable food items in their stomachs; digestion is rapid and often the snake was in a trap for a day or more before it was found. Therefore the greater number of records were obtained from scats. The residue in scats consisted entirely of hard and indigestible parts such as the chitin of insects' exoskeletons and the hair, feathers, scales, teeth and occasional bone fragments of the vertebrate prey. The insects eaten could usually be counted individually by sorting parts, such as heads or hind legs. With mammals, birds and reptiles the hair, feathers, or scales did not permit counting of individuals—each occurrence was assumed to represent one individual but in some instances two or more may have been present. Amphibians, lacking indigestible dermal structures were in most instances not represented at all in the scats, since their tissues were more or less completely dissolved by the digestion of the snakes. Soft-bodied larvae of insects and other invertebrates conceivably could be likewise completely digested, but such occurrences must be rare, as most of the invertebrates known to be eaten have the mouth parts, at least, heavily chitinized.
Admittedly the factors discussed above would cause some bias in the percentage composition of the food determined from scats, but I believe that the amount of error introduced was slight, because, judging from the records of items from stomachs, amphibians are not eaten frequently, and even mammals are not eaten frequently enough so that there is much chance of a snake taking two or more individuals at the same meal, unless it is robbing a nest containing a litter of young.
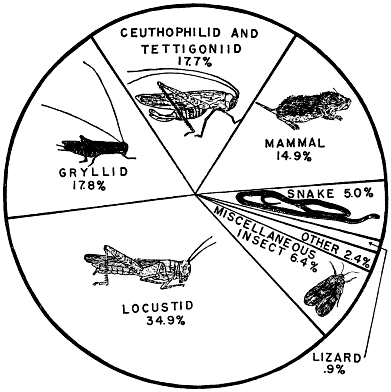
Fig. 13. Diagram showing percentage frequency of occurrence of various categories of prey in a sample of 1008 food items identified from scats and stomachs of blue racers from the Reservation and Rockefeller Tract. Insects, especially, orthopterans, made up the great majority of prey items taken.
The largest sample, based on 1008 food items, was obtained from 479 scats collected from the Reservation and Rockefeller Tract over the period 1949 through 1961. Items recorded were: 183 gryllid crickets (144 Gryllus assimilis, 36 Gryllus sp., 3 unspecified); 353 locustid grasshoppers (41 unspecified, 73 Arphia simplex, 67 Melanoplus femur-rubrum, 66 M. bivittatus, 39 M. differentialis, 17 Melanoplus sp., 15 Dissosteira carolina, 8 Chortophaga viridifasciata, 6 Syrbula admirabilis, 6 Sphargemon equale, 2 Melanoplus scudderi, 2 Schistocerca obscura, 1 S. americana); 94 camel crickets (Ceuthophilus sp.), 93 katydids (36 Neoconocephalus robustus, 15 Orchelimum vulgare, 15 O. nigripes, 6 Conocephalus sp., 4 Orchelimum sp., 2 Amblycorypha inasteca, 1 Neoconocephalus sp., 1 Daihinia brevipes); 7 cicadas (5 Tibicen sp., 1 T. pruinosa, 1 T. [401] lyrica); 45 unidentified insects; 17 beetles (including 1 Phyllophaga, 1 Calosoma scrutator, and 2 other carabids); 2 noctuid moths (Mocis latipes) and 1 caterpillar; 2 homopterans, 1 bee, 1 ant, 1 spider; 69 voles (59 Microtus ochrogaster, 9 Microtus sp., 1 M. pinetorum); 31 white-footed mice (15 Peromyscus leucopus, 14 Peromyscus sp., 1 P. maniculatus); 36 miscellaneous small mammals (6 Cryptotis parva, 4 Sigmodon hispidus, 4 Reithrodontomys megalotis, 3 Blarina brevicauda, 2 each of Scalopus aquaticus, Sylvilagus floridanus, and 1 unspecified shrew); 50 snakes (16 Coluber constrictor, 15 Diadophis punctatus, 14 Thamnophis sirtalis, 4 Elaphe obsoleta, 1 Natrix sipedon); 7 lizards (5 Eumeces fasciatus, 1 E. obsoletus, 1 Cnemidophorus sexlineatus); 3 unspecified "reptiles"; 5 birds (none identified to genus); 3 bird eggs, 1 narrow-mouthed toad (Gastrophryne olivacea).
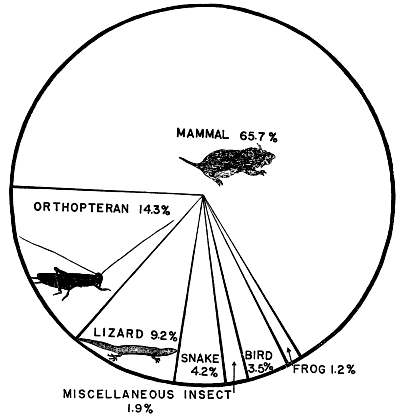
Fig. 14. Diagram showing estimated percentage by weight of various categories of prey in a sample of 1351 items, including all those represented in Fig. 13 and others from various parts of Kansas. Since the vertebrate items are on the average much bulkier than the insects eaten, vertebrates comprise most of the food, even though insects are eaten in much larger numbers.
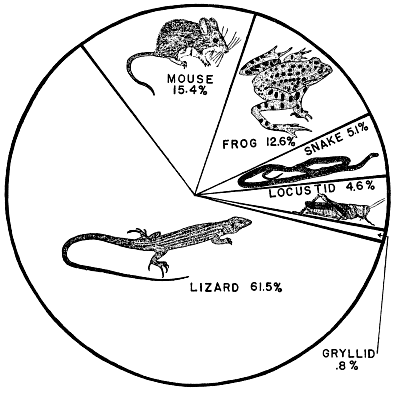
Fig. 15. Diagram showing estimated percentages by weight of various categories of prey in a sample of 69 food items squeezed out of stomachs of the blue racers captured at Harvey County Park. Most of the items were vertebrates, and lizards (Cnemidophorus) were especially prominent in the food at this locality. Samples of prey from scats (included in Figs. 13 and 14) and from stomachs show somewhat different trends, and neither is entirely representative of the actual feeding. Also, local differences in food sources are important.
Over the same period that the sample of scats was collected, a much smaller food sample of 73 prey items was collected by squeezing recently eaten food from the racers' stomachs, or by finding the snakes actually swallowing their prey. These items from stomachs are listed separately because they include relatively more vertebrates than do the items from scats. A grasshopper or cricket eaten by a large racer might have passed undetected, while a relatively [403] large item such as a vole or lizard would have produced a conspicuous bulge in the snake that ate it, and would have excited the curiosity of the investigator. A second difference is that the items from stomachs included several frogs, whereas amphibians were absent from the much larger sample from scats. A third difference is that the many insects found in stomachs were all orthopterans with the exceptions of three noctuid moths and the larva of a moth. Miscellaneous insects, such as beetles, bees and ants recorded from scats were not found in stomachs. Amphibians eaten are digested so completely that no recognizable parts of them are to be found in scats, but remains of the insects previously eaten by amphibians are to be seen in racers' scats. If not recognized as secondary items, such remains might lead to erroneous conclusions regarding the racer's food.
The items from stomachs were as follows: 21 grasshoppers (5 oedipines, 4 tryxalines, 5 Melanoplus bivittatus, 3 M. differentialis, 1 M. femur-rubrum and one each of Chortophaga viridifasciata, Dissosteira carolina, and Sphargemon equale); 8 crickets (Gryllus sp.), 3 katydids, 3 camel crickets (Ceuthophilus sp.), 3 noctuid moths, 1 larva of a moth; 10 voles (Microtus ochrogaster), 6 white-footed mice (5 Peromyscus leucopus and 1 P. maniculatus), 4 harvest mice (Reithrodontomys megalotis); 1 shrew (Cryptotis parva); 4 snakes (3 Thamnophis sirtalis, 1 Storeria dekayi); 4 lizards (2 Eumeces obsoletus, 1 Cnemidophorus sexlineatus, 1 Ophisaurus attenuatus); 4 frogs (Rana pipiens), 1 tree-frog (Hyla versicolor).
Records from the Harvey County, Kansas study area include a series of 69 food items from 55 stomachs (of living snakes) and 210 food items from 113 scats. There is a relatively high proportion of vertebrates, including some frogs, in the stomachs, and with no frogs but more miscellaneous insect material in the scats. But, for the sake of brevity, the two categories of items are combined in the following list: 55 grasshoppers (12 unspecified, 1 "locustid," 31 "oedipines," 7 "tryxalines," 5 "locustines," 2 Melanoplus bivittatus and one each of M. femur-rubrum, M. scudderi, M. differentialis, and Arphia simplex); 48 crickets (31 Gryllus assimilis, 17 unspecified); 14 katydids (11 Daihinia brevipes, one each of rhadiphorine, conocephaline and Neoconocephalus sp.); 9 noctuid moths and 1 moth larva; 26 miscellaneous insects (including 13 "beetles," 1 elaterid, 1 curculionid, 1 lygaeid bug, 1 ant, 1 wasp); 1 spider, 7 mice (5 Peromyscus maniculatus, 2 unspecified), 4 unidentified mammals, 1 vole (Microtus ochrogaster), 1 shrew (Cryptotis parva), 84 lizards (77 Cnemidophorus sexlineatus, 6 Sceloporus undulatus, 1 unspecified), 6 snakes (4 "natricines," 1 Thamnophis sp., 1 Pituophis melanoleucus), 1 "reptile," 1 "bird," 9 frogs (4 unspecified, 1 Rana catesbeiana, 4 Rana pipiens, 1 Rana sp., 1 Pseudacris triseriata).
Throughout the range of the racer small mammals make up an important portion of the food, and the bulk of those eaten are voles (Microtus sp.) and white-footed mice (Peromyscus sp.). The voles being diurnal, and having habitat preferences similar to those of the racer, are especially subject to attack, but only large adult racers are capable of swallowing a full grown vole. Probably most of the voles eaten are immature. Of the white-footed mice, P. [404] maniculatus especially prefers a grassland habitat, and is usually found in situations frequented by the racer. Being mainly nocturnal and crepuscular, it is usually in hiding at times when the racer is prowling, but may be flushed from its nest in a shallow burrow or beneath a sheltering object, and overtaken by the snake. Other mammals that are important in the food are harvest mice and other mice, shrews, and young cottontails. The latter are small enough to be eaten by racers only in the early stages of their life in the nest before weaning. Rats (Rattus, Sigmodon), moles, sciurids, and weasels are less frequent prey, ordinarily too large to be eaten by racers and taken chiefly as defenseless juveniles.
Predation on birds is relatively uncommon, and in most instances it involves the eggs or nestlings, or fledglings still slow and clumsy and incapable of sustained flight, or, occasionally, injured adults. Nests that are vulnerable are chiefly those of ground nesting species, or of kinds that nest near the ground in grass or thickets. Many of the birds recorded have not been identified to species, but those identified have included a variety of small passerines and also domestic chicks.
Lizards figure prominently in most of the food samples, but only a few species, those that live on or near the ground in grassy places, have been recorded. Most of the records pertain to scaly lizards (Sceloporus undulatus and S. graciosus), earless lizards (mainly or entirely Holbrookia maculata), racerunners (Cnemidophorus sexlineatus) and skinks (Lygosoma laterale and Eumeces sp.).
Snakes are important in the racer's food in most parts of the range, but the large racers of the Northeast are those most inclined to ophiphagous habits. The common garter snake (Thamnophis sirtalis) is the species most commonly eaten. Probably this is a matter of availability rather than preference, since the garter snake is one of the commonest and most widely distributed of North American snakes, occurring throughout most of the racer's range. The green snakes (Opheodrys) also are represented frequently. The other snakes eaten are mostly medium-sized to small colubrids, of a variety of kinds. However, there are three records (from Connecticut, Missouri and Kansas) of the racer preying on the venomous copperhead. There are many records of the racer preying on smaller individuals of its own species. In my own records racer remains appeared 18 times, equalling in frequency those of the common garter snake and exceeding all other kinds. In four of these instances the scale remains were relatively few and the [405] scales were relatively large, suggesting as an alternative to actual predation that a racer may have eaten part of its own sloughed skin, or that patches of shed skin may have adhered to the scat after its deposition in the trap. However, in the remaining 14 instances the remains of racer found in scats clearly indicated cannibalism, since the scales found were small and numerous and often were associated with bone. Cannibalism seems to occur frequently enough to be a significant factor in the reduction of the first year young. Liner (1949:230) described two instances of cannibalism in a litter of blue racers hatched in captivity. In one instance two young had seized the same lizard, and one having swallowed the lizard, continued to engulf the other snake, although it was of a size approximately equal to that of the first snake. Nevertheless, swallowing was completed, with the snake eaten pressed in a series of curves. A second instance of cannibalism occurred when one young racer attempting to catch a lizard struck another racer by mistake, then retained its hold and commenced swallowing. A similar instance was observed in a brood that I kept in 1962 after hatching had occurred in the laboratory.
Hatchling turtles of two kinds (Chrysemys picta, Terrapene carolina) have been reported in the racer's food. Probably other kinds are eaten also. However, the awkward shape and almost inflexible shell of the prey on the one hand, and the slender form of the racer, with limited distensibility of the gullet on the other, would limit this type of predation to occasional instances involving an unusually large racer and a small turtle.
There seem to be no records of the racer preying on salamanders. Many kinds of frogs are eaten, chiefly ranids and hylids, and the leopard frog (Rana pipiens) is the most frequent victim. Wright and Bishop (1915:160) stated that the toad (Bufo terrestris) occupied first place in the racer's food in the region of Okefinokee Swamp, Georgia, but they mentioned no specific instances of this species being eaten. Klimstra (loc. cit.) found only four toads in his large sample of digestive tracts from Illinois. Because of their virulent dermal secretions, bufonid toads are avoided by many kinds of snakes and predation on them by the racer probably is unusual.
Most authors who have written concerning the food habits of the racer have mentioned insects as part of the diet. Statements in the literature have often seemed to imply that the racer feeds on insects in general, according to their availability. However, the large number of records now available demonstrate that the racer [406] is highly selective in choice of its insect food, that soft-bodied orthopterans, chiefly crickets, grasshoppers and katydids, are the usual insect prey, with occasional predation on moths and their larvae. Eating of other insects such as cicadas and June beetles, is a rarity, but on occasion a racer may be tempted to sample such prey when it finds the newly emerged imago before its exoskeleton has hardened. I am convinced that such rarely occurring items as carabid beetles, hemipterans, homopterans, diplopods and spiders are secondary prey items, eaten by frogs that later were eaten by the snakes, in most instances if not in all. It is noteworthy that several of the same genera of grasshoppers and crickets are prominent in the food samples collected in widely separated parts of the racer's range.
As might have been anticipated, different species of prey were not utilized by the racers to the same extent throughout the snakes' season of activity. Grasshoppers, for instance, fluctuated from a low of 25.3 per cent (frequency) in the May sample to a high of 41.4 per cent in the September sample. Availability of prey, rather than any change of preference on the part of the racer, explains this trend. Thus, the locust, Arphia simplex, which, unlike most local grasshoppers, overwinters in the adult stage, is most prominent in the food in May, represented by 15.7 per cent, but it decreases progressively to a low of 1.8 per cent in September. The common grasshoppers of the genus Melanoplus show just the opposite trend, increasing during the summer, from a low of 2.62 per cent in May (when all are nymphs and most are too small to constitute a meal worthy of a racer's attention) to a high of 31.5 per cent in September. Mammals are best represented in the food in May, when they collectively comprise nearly 30 per cent of the items taken, and they are progressively less well represented as the summer advances. Both Microtus and Peromyscus conform to this trend, but the relative numbers of Peromyscus rise again abruptly in October. The general trend may be explained by the fact that in May most small mammal populations have a high proportion of young of the year, and these young are especially vulnerable to predation by the snakes. Also, insects in general are less available in spring, and this may force the racers to utilize vertebrates to a greater extent than at other seasons. Actually, the seasonal changes in food sources are not especially striking, and it seems that each important prey species is utilized more or less throughout the season of the racer's activity.
Table 5. Distribution by Months of Various Categories of Prey Items
Recorded From Blue Racers From Kansas,
Chiefly From the Reservation
and Rockefeller Tract
| May | June | July | Aug. | Sept. | Oct. | |
| Cricket (233) | .17 | .37 | .28 | .01 | .05 | .12 |
| Ceuthophilus (94) | .01 | .19 | .30 | .14 | .13 | .23 |
| Katydid (96) | .03 | .06 | .08 | .25 | .15 | .43 |
| Melanoplus (188) | .03 | .12 | .27 | .15 | .19 | .24 |
| All grasshopper (388) | .15 | .17 | .21 | .16 | .12 | .19 |
| Microtus (79) | .45 | .32 | .13 | .02 | .07 | .01 |
| Peromyscus (39) | .41 | .18 | .08 | .08 | .05 | .20 |
| All mammal (162) | .41 | .25 | .14 | .03 | .05 | .12 |
| Lizard (70) | .07 | .46 | .29 | .15 | .01 | .02 |
| Snake (61) | .37 | .18 | .08 | .13 | .09 | .15 |
| Arphia (72) | .50 | .25 | .12 | .04 | .03 | .06 |
Table 6. Distribution of Various Common Prey Animals
in a Sample of 625 Among Racers of Different Size Groups
| Kinds of prey and their percentage frequencies in in samples |
Snout-Vent Length (mm.) of Racers in Sample | ||||||||
| 200 299 |
300 399 |
400 499 |
500 599 |
600 to 699 |
700 to 799 |
800 to 899 |
900 to 999 |
1000 to 1099 |
|
| gryllid cricket | .40 | .39 | .11 | .25 | .23 | .15 | .15 | .12 | .08 |
| Melanoplus | .09 | .14 | .14 | .14 | .15 | .20 | .27 | .25 | |
| Ceuthophilus | .20 | .11 | .13 | .06 | .07 | .12 | .09 | .04 | |
| Orchelimum | .04 | .02 | .01 | .01 | .01 | ||||
| tettigoniid | .03 | .02 | .06 | .03 | .02 | .05 | |||
| "other grasshopper" and miscellaneous orthopteran |
.30 | .25 | .10 | .09 | .03 | .12 | .07 | .09 | |
| Arphia simplex | .03 | .02 | .05 | .08 | .08 | .04 | .09 | ||
| miscellaneous insect | .04 | .04 | .09 | .07 | .04 | .08 | .05 | .04 | |
| lizard | .20 | .09 | .18 | .09 | .07 | .11 | .01 | .01 | |
| snake | .20 | .07 | .06 | .05 | .04 | .05 | .08 | .12 | |
| Microtus | .04 | .02 | .08 | .13 | .09 | .12 | .25 | ||
| Peromyscus | .09 | .02 | .05 | .06 | .03 | .05 | .04 | ||
| "other mammal" | .01 | .04 | .10 | .03 | .03 | ||||
| bird | .01 | .01 | |||||||
| Total prey items for size group |
5 | 23 | 28 | 101 | 120 | 121 | 127 | 76 | 24 |
The wide disparity in size between young and adult racers also results in utilization of different food sources to some extent. In some kinds of snakes adults and young draw their food from entirely different sources, but in the racer there is broad overlap, as shown in Table 6. The samples from the largest and smallest size groups of racers are relatively small. Two important kinds of prey—voles and grasshoppers of the genus Melanoplus—were not found at all in the smallest size groups of snakes and comprised increasing percentages in the food of the larger size groups. A large adult vole is too large to be swallowed except by an unusually large racer, and a young vole old enough to leave its nest is far too large for a hatchling racer. Grasshoppers of the genus Melanoplus are relatively large and heavily armored, and so are relatively immune to attacks from the smaller snakes. Small soft-bodied orthopterans including Gryllus, Ceuthophilus and Orchelimum, and also lizards and snakes, are best represented in the food of the smaller racers. Other types of prey showed no definite correlation with size of the racer taking them.
Many observers have published accounts of the courtship and/or mating of the racer, but all of these are, to some degree, incomplete. Because of the widely different circumstances, and the different viewpoints of the observers involved, the several accounts give much different impressions of sexual behavior in this species. Either singly or combined, the published accounts do not provide an adequate description of the process.
My own observations, made both under natural conditions and in large outdoor enclosures, are likewise somewhat incomplete, but indicate that the whole sequence of courtship and mating is divisible into the following well-defined stages: 1) the finding of a receptive female by the male; 2) the persistent following of the female by the male, who courts her by lying extended along her body and performing writhing movements, with periodic interruptions during which he momentarily leaves the female and courses rapidly through the grass around her; 3) the acceptance of the male by the female, signalled by the raising of her tail and the almost instantaneous intromission; 4) the dragging of the passive male by the female while he is firmly attached to her during the period of coitus; 5) separation of the pair and involution of the male's hemipenis.
Even in the breeding season, racers that were confined in enclosures usually were either indifferent to each other or responded with reactions of fear or hostility. In moving they tended to follow the edges, and often two moving in opposite directions would approach each other; when this occurred, one snake might strike at the other with a short jab that seemed to be mostly bluff, and then would dart away. The males, being smaller, were usually the more wary.
Sexual behavior was noticed on only a few occasions. Several large adult males were less wary than others and usually manifested curiosity or interest toward other racers. My most complete observations of sexual behavior were made on May 18, 1962, when a newly caught adult male was added to an enclosure of 100-foot circumference already containing several racers, two of which were large adult females. Within half an hour the male was found courting one of the females. She was lying in a loose coil, with the male extended along her. At my approach the female darted away in alarm for approximately three feet, and the male moved with her, so swiftly and adroitly that he maintained contact and was in approximately his original position with respect to the female when she stopped.
Spasmodic rippling movements passed down the body of the male as he lay in contact with the female. These movements lasted several seconds, increasing in intensity, alternating with longer periods of little or no movement. As each period of vigorous writhing reached its climax, the male's head jerked forward and backward several times in seeming excitement. The female's behavior was mostly passive. She seemed to be receptive, but from time to time, without any noticeable warning, she darted away for several feet as she had when the pair was first discovered. Each time the male darted forward with her, maintaining contact while she moved. These swift movements of the female seemed to be spontaneous, at least in most instances there was no evident cause for alarm. The female's movements seemed to stimulate the male's interest rather than to discourage him. In most instances the female moved only four to five feet, then stopped abruptly or turned back. She would stop in a loose resting coil, in thick grass, with the male lying over her. Often she coiled in such a way that the posterior end of her body was beneath her forebody, but this did not seem to deter the male from moving the posterior end of his body into position beside hers. After a sudden change in the [410] female's position, the rear of the male's body would perform groping movements along that of the female until his cloacal region was approximately opposite hers. The male sometimes had his chin pressed against the female's back, especially when he was moving forward along her, but more often his head was raised, and frequently was as much as 18 inches from the female's head.
At intervals averaging approximately ten minutes, during a little more than an hour of observation, the male would suddenly dart away from the female, and with unusually rapid and animated movements, he would move around her in an irregular and devious course, sometimes as far as five feet away, but usually within 18 inches. Usually on each such expedition several or many circuits were made; then the male would return to the female and would glide rapidly along her until he attained the mating position. A period of especially vigorous courting movements would follow.
At 12:55 p. m. it was necessary for me to discontinue observations, and I left the female confined in a cloth bag. Returning at 1:20 p. m. I found that the male was not displaying interest in the female confined in the bag, nor in the other female loose in the enclosure. The first female was released from the bag, and was out of sight for approximately four minutes. When relocated she was again attended by the male, who was carrying on courtship even more vigorously than he had before. At 1:35 p. m. the male achieved intromission. Although the pair was under observation at the time intromission occurred, the actual eversion of the hemipenis was not seen because the snakes were partly concealed by dense vegetation. There was a sudden flurry of movement, the male's head waving and his body thrashing. In an instant these violent movements subsided, and after a few seconds the female began to crawl forward slowly. The male had relaxed, and relinquished his contact with the female anteriorly. As she moved away he was dragged after her tail-first. He made slight backward wriggling movements that perhaps aided in maintaining sexual contact. The female's restlessness increased, and in eight minutes she dragged the male in a circuitous course a distance estimated to be between 20 and 30 feet. At 1:40 p. m. the pair was ten feet from the point where copulation had begun. The female showed increasing inclination to climb, raising her head and forebody against the trunks of saplings, and finally reaching up one to a branch 20 inches above the ground, and climbing first along the branch and then farther up the main trunk. As she progressed the [411] male was lifted from the ground, dangling limply suspended by his hemipenis and its base had become exposed. At 1:43 p. m. separation occurred and the male dropped into the grass. Semen dripped from the cloacae of both snakes. That from the female was tinged with blood. The individuals involved in this observation were kept in the enclosure subsequently but no further sexual behavior was noted.
Contrary to the popular belief that these racers have permanent mates, all available evidence indicates that they are promiscuous, and two or more males may simultaneously court the same female in the brief spring breeding season. On May 24, 1960, while I was walking in a hilltop field of brome grass, a sudden movement attracted my attention to three racers lying alongside each other. Only the posterior parts of their bodies and their tails were visible. Two were males and were performing the characteristic slow writhing movements against the body of the female from either side. Although the heads were not in view, the snakes may have been able to see me through the screening vegetation; after I had watched for approximately 20 seconds, all three suddenly took alarm, for no apparent cause, and scattered.
Further evidence of promiscuity is provided by the account of Ellicott (1880:207) who wrote regarding the eastern subspecies: "I noticed a ball of black snakes (Bascanion constrictor L) rolling slowly down a steep and stony hillside ... about two miles above Union Factory, Baltimore County, Md. ... kept together by procreative impulses." It was stated that this observation was made in early spring. "Snake balls" have often been observed, and described in the literature; usually the snakes involved were garter snakes (Thamnophis) or water snakes (Natrix). Seemingly, typical aggregations consist of a single adult female and several or many males attempting to mate with her. There is a distinct possibility that the snakes involved in Ellicott's observations were misidentified.
Sexual behavior of the racer is in most respects remarkably similar to that of the common garter snake, Thamnophis sirtalis, well known through the work of Blanchard and Blanchard (1942). In studying sexual behavior of racers, several observers have failed to differentiate between the different stages of the mating process, and have assumed that copulation was occurring when actually only the precopulatory behavior was observed. In an early description of courtship in this racer in Kansas, Brons (1882:365) stated [412] that the female "at times, seems to toy with the male, indisposed to yield to his importunities, though pressed with ardor. To avoid his suit, at times, she will dart through grass, among stones, or enter a crevice. Should he be able to reach his mate while within a hole, he is not slow in bringing her to the surface, again to be repulsed. Upon an unbroken ground the sexual union is less prolonged. Here she is unable to free herself from his quick and effectively directed moves. In case she attempts to quit him, a coil is thrown about her body, and his head laid flat upon her neck, and replaced as promptly as dislodged, evidently in the endeavor to propitiate her."
Another account probably based on courtship rather than copulation is that of Wright and Wright (1957:135), who described the behavior of a pair of C. c. priapus on Billy Island, Okefinokee Swamp in southern Georgia, on May 8, 1921, as follows: "They were stretched out, more or less coiled ... the rear parts of the bodies from the vent were entwined. The female, or smaller one seemed to have its tail around that of the male. There were contortions or quiverings from time to time.... May 8, 1921: Jackson Lee saw black snakes entwined, the male seizing the female by the top of the neck."
Blanchard and Blanchard (op. cit.) have described the dragging of the male by the female during coitus in the garter snake, and the temporarily inseparable bond formed between members of a pair by the recurved spines of the engorged hemipenis, but it has not been generally recognized that the process is much the same in other colubrines. Cottam (1937:229) described and photographed mating in a pair of C. c. mormon in Utah. The copulating racers were shown in a loose coil lying alongside each other with tails intertwined. However, when disturbed by the observers, these racers made frantic efforts to escape, crawling in a spiral course, while remaining attached and intertwined, "with no evident attempt to separate" during approximately a quarter hour of observation.
The racer is notorious for its aggressive behavior and occasional alleged attacks on humans in the breeding season. The tendency has doubtless been much exaggerated, especially in the verbal second- or third-hand accounts based on the alleged observations of eye-witnesses. Nevertheless, the supposition that large adults will sometimes pursue or attack humans when disturbed is well substantiated. In most of the instances known to me, it is the large eastern subspecies, C. c. constrictor, involved in these incidents, and [413] seemingly the smaller racers of the Middle West, far West and South are less inclined to behave aggressively. In May 1958 two pairs of large racers were confined in a semicircular wire enclosure thirty feet across and open on top, and with natural vegetation, at the Reservation headquarters. Often in approaching the cage I saw two or more racers in close association, but because of sheltering vegetation, and the snakes' timidity observation was difficult. On May 19 a pair were lying partly extended in loose coils, but immediately the female took alarm and darted away, breaking loose from the male; his hemipenis was exposed, and underwent involution and retraction in approximately 30 seconds. Unlike the female, the male on this occasion did not attempt to escape, but turned to face me with a show of aggressiveness. Probably copulation was in its final stages when the disturbance occurred.
Circling of the female racer by the male from time to time in the course of courtship has not been recognized by previous observers as a part of the mating pattern, but Pope (1944:171) described somewhat analogous behavior, probably modified by unnatural conditions of captivity and the crowding of many racers in one cage. Pope, citing earlier observations by Noble, wrote: "When sexually excited, the male blacksnakes dash wildly about before paying court to individual females. In captivity these dashes excite all specimens confined together. A male, after picking out a mate, moves his chin lightly along her back, while undulations run forward along his sides and he extends his tongue now and then. Later he throws the part of his body near his vent over the corresponding part of the female, the two tails sometimes becoming loosely intertwined."
Recorded dates of mating for the species are all in spring, but indicate a span of many weeks for the breeding season, and this spread results in part from geographical differences. Published records are as follows:
Subspecies constrictor
May 12, 1930, in Ohio (Conant, 1938:55)
Subspecies priapus
May 8, 1921, in Georgia (Wright and Wright, 1957:135)
May 9, 1921, in Georgia (Wright and Wright, 1957:135)
Subspecies flaviventris
May 3, 1931 (two pairs) in Missouri (Boyer and Heinze, 1934:195)
April 18, 1936, in Missouri (Anderson, 1942:210)
May 12, 1928, in Kansas (Gloyd, 1928:123)
Subspecies mormon
June 10, 1927, in Utah (Cottam, 1937:229)
July 7, 1938, in California (Cunningham, 1959:17)
In the course of my live-trapping, I occasionally found more than one racer in a trap. As might be expected from the low yield per trap, such double or multiple captures were relatively rare. Chance, and unusually strategic placement of certain traps were doubtless contributing factors. May and October, being the most productive months for trapping, yielded a high proportion of these combined captures. Some involved an adult and an immature snake, or two adults of the same sex. Eliminating all these, there remain 44 heterosexual captures of adults. These latter captures are significantly concentrated in their seasonal distribution and indicate a spring breeding season; 34 were in May, six were in June and four were in October. Eight of the May records and one June record each involved a trio of snakes—two males and a female in every instance. Distribution of the spring records, grouped in five-day intervals, was as follows:
| May 11 to May 15: | 14 |
| May 16 to May 20: | 5 |
| May 21 to May 25: | 8 |
| May 26 to May 30: | 13 |
| May 31 to June 4: | 1 |
| June 5 to June 9: | 0 |
| June 10 to June 14: | 2 |
| June 15 to June 19: | 2 |
| June 20 to June 24: | 0 |
| June 25 to June 30: | 1 |
Approximately 87 per cent of the records fell in the twenty-day interval, May 11 to 30, which is regarded as the main breeding season. Presumably males continue to be at the peak of breeding condition and continue to search for females after the latter have become unreceptive, partly explaining the scattering of records through most of June.
Several of the females found in traps with males in May had abundant active sperm in their cloacae and oviducts and probably had been inseminated within a few hours of the time they were checked. Others lacked sperm, but the cramped quarters inside the traps may have effectively prevented the consummation of courtship, especially when two males were confined with the same female. None of the females trapped with males in October was found to be inseminated, and it seems doubtful whether copulation ever occurs at that time of year, although males have motile sperm and seem to be in breeding condition then.
Cloacal smears indicate that males mature sexually and first produce sperm in August and September when they are a little more than a year old. Insofar as could be determined, there was no sexual activity at this time of year, and actual breeding of the [415] adolescent racers was postponed until the following May. By this time at an average age of 20 months, the snakes had made further growth.
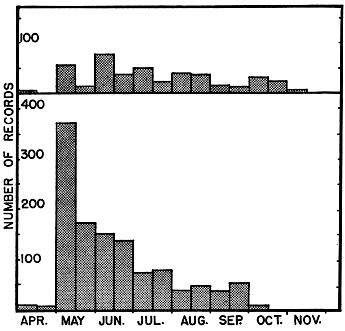
Fig. 16. Catches, in semi-monthly periods, of racers in their summer habitats, at Harvey County Park (upper) and at Reservation and Rockefeller Tract (lower). Intensive activity in spring (the breeding season), tapering off rapidly as the season advances, is well shown by the larger sample, but in the years of trapping at Harvey County Park operations usually were not fully underway until the latter part of May.
Mr. Dwight R. Platt studied the changes in the male reproductive organs during the annual cycle at Harvey County. In racers recently emerged from hibernation he found the seminiferous tubules filled with Sertoli syncytium, but containing few germ cells. Spermatogonia proliferate in May and June. During the first half of July primary spermatocytes are the dominant cells in the seminiferous tubules. By early August spermatids are dominant and the first free spermatozoa are present. In late October spermiogenesis is essentially complete and the tubules are relatively empty before the snakes hibernate. During the season of activity the seminiferous tubules increase to approximately double their minimum diameter, reaching the maximum in August. Cyclic changes [416] in size and secretory activity of the ductus deferens, ductus epididymis, and sexual segment of the renal tubules occur, with maximum size and secretory activity coinciding with the time of movement and storage of the spermatozoa. The latter are stored in both ductus deferens and ductus epididymis. Despite the short breeding season, a male racer has active sperm at all seasons.
In accounts of the racer in the humid southeastern United States, Brimley (1903:261), Wright and Bishop (1915:160) and Tinkle (1959:195) mentioned the ease with which the eggs might be found and the superficial situations in which they were sometimes deposited. Both Wright and Bishop, and Tinkle made field studies in swamps, where presumably the subsoil was saturated with moisture and too wet for the eggs. Tinkle mentioned finding one clutch beneath a discarded newspaper and another beneath a small, thin board. Surface (1906:167) stated that in Pennsylvania the eggs were to be found in loose soil, in sawdust piles, or in decaying wood of hollow logs or trees. Clark (1949:249) stated that in northern Louisiana the eggs are laid in soft, moist soil such as may be found beside decaying logs. Minton (1944:457) found two clutches under flat stones on hillsides in Indiana. In the more arid climates of the far western states the species' habits are much different in this regard. Through many years of familiarity with C. c. mormon, I have never seen its eggs. Presumably nests in this part of the range are deep underground, most often in old burrows of the pocket gopher (Thomomys), which are so abundant that in many areas the soil is riddled with them. Burrows of the ground squirrels (Spermophilus sp.) and other small digging mammals also provide potential insulated nest sites with the favorably moderate temperatures and high humidities that the eggs of snakes require.
On the morning of July 10, 1962, I was directed to the sites of two clutches recently plowed up, 11/2 miles north of the Reservation (Pl. 21, Fig. 2). The eggs were in a fallow field having a stand of sunflowers three to five feet high. The plow blades turned the soil at a depth of approximately seven inches. In each instance only a few eggs were visible. They were well scattered in the loose soil turned up by the plow; 21 were found in one clutch and 10 in the other. All the eggs were intact except two that had minute punctures from which liquid oozed. Seemingly the eggs in situ had been well above the level of the blade—at depths of four to five [417] inches. No nest cavities were discernible where the eggs were found, but elsewhere in the field tunnels of moles (Scalopus aquaticus) and prairie voles (Microtus ochrogaster) were exposed by the plow. Presumably the eggs had been in such tunnels, which had disappeared as the loose soil crumbled. Another clutch was discovered in an adjoining field on July 16. The nine eggs were at depths ranging from 61/2 to nine inches, and only the two topmost eggs had been turned up by the plow. All three clutches were within a few feet of the edges of the fields.
On the Reservation and nearby areas I have seen remains of an estimated 20 clutches that have been destroyed by predators. The remains in every instance consisted of an excavation, and the strewn torn and empty eggshells. Nests were at depths of four to eight inches in old tunnels, which most often seemed to be those of moles but also included some of the prairie vole, and perhaps some of the pine vole (Microtus pinetorum). All these nests were in open sunny places in prairie or pasture habitat.
Table 7. Numbers and Sizes of Eggs in Clutches
of the Blue Racer From Eastern Kansas
| Number of Eggs |
Lengths (mm.) |
Widths (mm.) |
Weights (grams) |
Snout-vent length of female (mm.) |
| 17 | 26.5(29-24) | 17.0(19.0-16.5) | 5.5(6.0-4.4) | 892 |
| 8 | 33.3(39-31) | 16.3(17.5-14.0) | 6.0(6.7-5.6) | 899 |
| 12 | 29.1(32-25.5) | 17.1(18.5-16.0) | 4.9(5.2-4.4) | 773 |
| 14 | 26.9(30-24) | 19.2(20-18) | 5.2(6.2-4.4) | 772 |
| 10 | 31.7(33-29) | 16.5(18-15) | 6.0(6.5-5.6) | 807 |
| 11 | 29.7(33-27) | 16.7(18-15) | 5.4(5.8-5.0) | 858 |
| 21 | 28.9(32-27) | 18.4(19.5-18) | 5.9(6.3-5.6) | 1038 |
| 13 | 30.7(34-28) | 19.3(20-18) | 6.8(7.5-6.1) | 907 |
| 18 | 29.0(30-26) | 17.9(19-16) | 4.9(5.5-4.3) | 911 |
| 12 | 30.3(34-28) | 17.8(19-15) | 5.2(6.1-3.8) | 843 |
| 14 | 30.9(36-29) | 19.4(21-18) | 6.8(7.6-6.2) | 846 |
Many observers have described the eggs of the racer, which are white, elliptical, somewhat elongate, with tough, leathery, somewhat flexible, shells, and a granular surface. Like other snake eggs, those of the racer gradually absorb moisture during incubation. They become more turgid and increase in weight and dimensions, especially in breadth, and by the time of hatching are nearly twice their size at laying. Between different clutches and even within the same clutch there is notable variation in the size of the newly laid eggs. Munro (1948:199) noted that in a small adult racer [418] kept by him, the eggs laid were larger but less numerous than those produced by a large adult. Munro noted also that shape of eggs in the two clutches differed; the smaller snake produced more elongate eggs of smaller diameter. The idea that eggs laid by the smaller females are more slender and elongate is not supported by my own data. For 11 clutches of eggs examined soon after laying, dimensions, weights, and the lengths of the females are shown in Table 7.
In a clutch of eggs beginning to hatch on September 3, 1958, dimensions and weights were as follows: length 31.8 (36-30), diameter 22.0 (24-21), weight 9.7 (10.3-9.3).
Gravid females that were kept in captivity in anticipation of their laying usually produced their clutches within a few days. The laying dates of such individuals are shown in the following list. Those with asterisks were from the Harvey County study area, others were from the Reservation and Rockefeller Tract.
June 19, 1961
June 21, 1959
June 23, 1961*
June 26, 1959
June 29-30, 1962*
July 1, 1961
July 4-5, 1962*
July 6, 1955
July 7, 1959*
July 7, 1959*
July 12, 1961
July 15, 1961*
July 18, 1961*
July 20, 1961
August 8, 1960*
A further indication of the period when laying occurs was provided by the appearance of females gravid and progressively more swollen with eggs, then their abrupt disappearance and replacement by thin and wrinkled individuals that obviously were recently parturient. The following records show the course of these events on the Reservation in the years when summer trapping was done with sufficient consistency. These dates provide a rough approximation of the time when laying occurs locally. They indicate a laying season concentrated in a period of approximately three weeks in this locality. Records from published literature also indicate that laying occurs in late June and early July at the latitude of Kansas, but somewhat earlier in the southern United States.
Table 8. Dates When Parturient and Gravid Racers were Captured on
Reservation and Rockefeller Tract in Several Years, Indicating Time
of Oviposition
| Year | 1958 | 1959 | 1960 | 1961 | 1962 |
| First parturient female | June 18 | June 17 | June 22 | June 20 | June 13 |
| Last gravid female | July 8 | July 11 | July 5 | July 12 | July 16 |
Many authors have made statements regarding the size of the clutch in the racer, on the basis of those found in the field, those laid after capture, or those dissected from gravid females. Some of the statements were based upon small but unspecified samples, and are far from the mark. From records accumulated in the course of my own field work, and a summarization of those in published accounts a substantial sample is available showing the usual size of clutch in the area of my study, and the trends of geographic variation in some parts of the range.
| C. constrictor (all combined) | 151 clutches averaged 10.61 (2 to 31) eggs. |
| C. c. constrictor | 14 clutches averaged 16.80 (7 to 31) eggs. |
| C. c. priapus | 11 clutches averaged 12.60 (7 to 21) eggs. |
| C. c. mormon | 43 clutches averaged 5.79 (2 to 13) eggs. |
| C. c. flaviventris | 82 clutches averaged 11.78 (5 to 26) eggs. |
| C. c. stejnegerianus, | one clutch contained 10 eggs. |
In the foregoing list the sample of C. c. flaviventris may be divided as follows:
Reservation and vicinity: 36 clutches averaged 11.65 (6 to 21) eggs.
Harvey County study area: 21 clutches averaged 12.0 (5 to 18) eggs.
Museum specimens from Kansas: five clutches averaged 9.2 (6 to 14) eggs.
Published records (Kansas, Indiana, Iowa, Louisiana, Missouri, Oklahoma, Texas): 20 clutches averaged 12.5 (5 to 22) eggs.
Table 9. Published Records Indicating Dates of Laying
in Different Populations of Coluber constrictor
| Date of Laying |
Subspecies | Area | Authority |
| June 9, 1952 | flaviventris | SE Texas | Guidry (1953:50) |
| July 1-2, 1920 | flaviventris | Texas | Ortenburger (1928:183) |
| June 1, 1920 | flaviventris | Texas | Ortenburger (1928:183) |
| June 9, 1926 | flaviventris | Oklahoma | Force (1930:31) |
| June 24, 1926 | flaviventris | Oklahoma | Force (1930:31) |
| July 19, 1928 | flaviventris | Oklahoma | Force (1930:31) |
| July 16 | flaviventris | Oklahoma | Carpenter (1958:114) |
| July 4-5, 1948 | flaviventris | Kansas | Munro (1948:199) |
| July 4, 1948 | flaviventris | Kansas | Munro (1948:199) |
| June 26, 1930 | flaviventris | Ohio | Conant (1938:55) |
| July 3, 1961 | mormon | NW Oregon | Van de Velde, Martan and Risley (1962:212) |
| June 7, 1957 | priapus | S Illinois | Rossman (1960:219) |
| June 19, 1912 | priapus | S Georgia | Wright and Bishop (1915:160) |
| July 10, 1940 | priapus | S Illinois | Cagle (1942:187) |
| July 6 | constrictor | New York | Ditmars (1907:284) |
| June 27-July 5, 1951 | constrictor | Virginia | Werler and McCallion (1951:251) |
| June 5, 1947 | stejnegerianus | S Texas | Auffenberg (1949:54) |
Published records of clutches laid by racers, from which figures used in the foregoing account were obtained, include the following:
C. c. constrictor: 22 (Barbour, 1950:104); 21, 13 (Brimley, 1903:261); 25 (Conant, 1938:55); 8 (Ditmars, 1907:284); 7, 12, 14, 16, 19, 20, 31 (McCauley, 1945:76); 14 (Wright and Wright, 1957:136).
C. c. flaviventris: 7 (Anderson, 1942:210); 22 (Brumwell, 1951:205); 11 (Carpenter, 1958:114); 8, 9, 9 (Force, 1930:31); 10 (Guidry, 1953:50); 18 (Liner, 1949:230); 5, 8, 17 (Marr, 1944:484); 5, 14 (Munro, 1948:199); 13, 19 (Ortenburger, 1928:183); 6, 15 (Tinkle, 1959:195); 15, 19 (Wright and Wright, 1957:141).
C. c. priapus: 20, 21 (Cagle, 1942:187); 7 (Conant, 1938:55); 19 (Rossman, 1960:219); 5, 9, 11, 14 (Wright and Bishop, 1915:160); 16 (Wright and Wright, 1957:147).
C. c. mormon: 13, 9, 8, 5, 5, 5, 4, 4 (Cunningham, 1959:17); 5, 6 (Stebbins, 1954:374); 6 (Van de Velde, Martan and Risley, 1962:212); 3, 6 (Wright and Wright, 1957:144):
C. c. stejnegerianus: 10 (Auffenberg, 1949:54).
In general, the number of eggs in the clutch is proportional to the size of the female producing them. The larger and bulkier females produce more eggs. Geographic trends in number of eggs produced are perhaps controlled by differences in size between different populations; thus, the large eastern constrictor produces nearly three times as many eggs per clutch as does the small western mormon, whereas the centrally located flaviventris is somewhat intermediate in size and in numbers of eggs produced.
In most reptiles growth in length and bulk continues after attainment of sexual maturity. For many kinds including Eumeces fasciatus, Crotaphytus collaris, Cnemidophorus sexlineatus, Agkistrodon contortrix (Fitch, 1954:60; 1956:236; 1958:36; 1960:174), and Sceloporus olivaceus (Blair, 1960:94), it has been shown that the larger and older females in a population produce more offspring than do the smaller and younger individuals. This situation applies in the racer, as shown by the clutches of 52 females correlated with their sizes and presumed ages (Fig. 17, Table 10). The two-year-olds contribute a relatively small quota to the annual brood, partly because their clutches are small, but more especially because many of them fail to attain sexual maturity in time to breed. Many of the female racers that are more than two years old also fail to produce an annual clutch of eggs. The 24-day period May 28 to June 20 inclusive is judged to comprise the period when eggs have generally enlarged sufficiently to be detected in gravid females, but still have not been laid in most instances. In this period, in 1960, 1961 and 1962, ratios of gravid females to those [421] not detectably gravid in several supposed age groups arbitrarily established on the basis of size, were as follows:
| Two-year-olds | 2 gravid, 13 apparently not gravid |
| Three-year-olds | 5 gravid, 4 apparently not gravid |
| Four-year-olds | 3 gravid, 2 apparently not gravid |
| Five-year-olds | 4 gravid, 3 apparently not gravid |
| Six-year-olds (or older) | 8 gravid, 2 apparently not gravid |
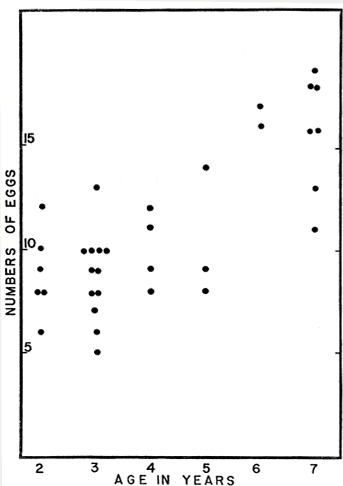
Fig. 17. Graph showing number of eggs per clutch, and correlation with supposed age (as deduced from length of body) in female blue racers from the Reservation, Rockefeller Tract, and Harvey County Park.
From the appearance of these snakes it is reasonably certain that none had already laid eggs when it was recorded, but there is some [422] possibility that a few individuals not noticeably gravid at the times they were examined, produced eggs subsequently. However, these meager data do seem to indicate that most of the two-year-old females and a minority of older individuals fail to produce clutches in the annual breeding season.
Table 10. Fecundity of Female Racers in Various Age-size Classes,
All From the Reservation and Rockefeller Tract
| Most Probable Age in Years of Females (as indicated by size) |
Number of females in sample |
Snout-vent length of females; average and extremes |
Number of eggs in clutch |
| 2 | 10 | 688 (589-748) | 9.2 (6-12) |
| 3 | 19 | 789 (756-840) | 9.9 (5-14) |
| 4 | 7 | 856 (850-861) | 10.8 (8-12) |
| 5 | 6 | 907 (892-933) | 13.0 (8-17) |
| 6 or more | 10 | 1005 (955-1088) | 15.7 (11-19) |
Under unfavorable conditions eggs can be resorbed, but probably this can occur only if initiated before ovulation. A racer in which six small eggs were palped on June 28, 1960, was kept until July 23 but did not oviposit. It no longer appeared gravid and the ova could not be detected by palpation. Another female had 13 eggs on June 21, 1960, but by July 23 when the snake was released the eggs had not been laid and no longer could be detected. Both snakes refused to feed throughout their confinement.
Like other reptilian eggs, those of the racer are dependent upon the warmth of their surroundings for incubation. They are tolerant of a wide range of environmental temperatures, but the higher the temperature the more rapidly incubation proceeds. Under natural conditions there may be much difference in hatching time in two clutches laid at the same time and in the same locality. Site of the nest—deep and well insulated, or shallow; in a well shaded situation or one exposed to maximum amounts of sunshine—would largely control rates of development. Clark (1949:249) writing of the subspecies anthicus in north-central Louisiana, stated: "eggs are laid about the first of June.... young begin to make their appearance at about ... July 1." Even for the southern states these dates of laying and hatching seem somewhat too early to reconcile with the records published by other observers, and are in need of verification, especially since they seem to be based upon vaguely remembered observations rather than upon written records. At the other extreme Surface (1906:167) wrote of constrictor in [423] Pennsylvania that hatching may occur as late as October, and that there is evidence some young may even remain in the egg over winter before hatching occurs. Several incubation periods are on record for clutches laid and hatched in captivity, as follows:
C. c. mormon, Oregon, 47 and 51 days (laid July 3, 1961, hatched August 19 and 23; Van de Velde, Martan and Risley, 1962:212).
C. c. stejnegerianus, Texas, 73 days (laid June 5, 1947, hatched August 17, Auffenberg, 1949:54).
C. c. priapus, S. Illinois, 58 and 59 days (laid July 10, 1940, hatched September 6 and 7, Cagle, 1942:187).
C. c. flaviventris, Kansas, 50 days (laid July 4 and 5, hatched August 23 and 24; Munro, 1950:124).
C. c. flaviventris, Texas, 43 days (laid June 9, 1952, hatched July 22; Guidry, 1953:50).
No incubation periods for eggs in natural nests have been recorded. In the course of my study, eggs obtained from 12 captive females were hatched in confinement, with an average incubation period of 51 days (43 to 63) as follows:
Laid July 6, 1955, hatched August 20.
Laid July 3, 1958, hatched September 3 and 4.
Laid June 21, 1959, hatched August 17.
Laid June 26, 1959, hatched August 17.
Laid July 7, 1959, hatched August 23 to 25.*
Laid June 23, 1960, hatched August 20.*
Laid June 30, 1961, hatched August 30.
Laid July 15, 1961, hatched September 1 and 2.*
Laid July 18, 1961, hatched September 2 and 3.*
Laid July 4 and 5, 1962, hatched August 15 and 16.*
Laid June 29 and 30, 1962, hatched August 14 and 15.*
Laid July 6, 1962, hatched August 16 to 20.*
In the foregoing list those entries marked with asterisks were obtained from the Harvey County study area; all others were from the Reservation and Rockefeller Tract.
Detailed observations on hatching were made on a clutch of eggs laid on June 29 and 30, 1962, by a female caught in Harvey County. The first egg in the clutch had already been laid in the trap when the female was found at 11:30 a. m., June 29. Two of the eggs were abnormal, with thin transparent shells, and were found to lack embryos when they were opened on July 7. Later, two other eggs were attacked by mold and the embryos died early in development. The clutch was kept in a can of slightly damp soil. At 2:30 p. m. on August 13, when the clutch was examined, egg no. 6 was found to have hatched. The young snake had made a 21-millimeter slit in the shell. At 12:50 a. m. on August 14, it was discovered that eggs 1, 4, 5 and 7 each had been slit. No. 4 had two parallel slits separated by a two-millimeter strip of shell, and the young racer [424] could be seen inside. At 1:05 a. m. this young snake had changed position and was lying upside down in the egg, his snout protruding slightly through one of the slits. At 1:45 a. m. he was again right side up, still in the shell. At this time each of the slit eggs showed the protruding snout of a young snake. Occasionally the viscous liquid egg white would be blown into a large bubble on the surface of the shell as the young snake exhaled. A third slit, parallel to the others, had appeared in egg no. 4. A hatchling emerged from egg no. 1 between 2:20 and 3:20 a. m., and another from no. 7 between 7:00 and 9:00 a. m. The hatchling struck vigorously many times, and vibrated his tail when he was disturbed. Egg no. 3 was first slit between 7:00 and 9:00 a. m., and three more slits appeared in it between 9:15 and 10:15 a. m. At 12:45 p. m. a hatchling was found in the act of emerging from egg no. 4, and approximately the anterior one-fourth of its body protruded. Disturbed by the movements of the observer, the little snake drew back into its shell. This hatchling began to emerge again at 12:50 and his hatching was completed at 1:00 p. m. Between 5:40 and 6:20 p. m. a hatchling emerged from egg no. 3 (for several hours this hatchling had been lying on its back inside the egg, with only its snout protruding); two slits appeared in egg no. 2 and three slits appeared in egg no. 8. At 6:50 p. m. the hatchling in the latter thrust his snout through the slit in this eggshell. This hatchling was lying on its back at first but by 10:50 it had shifted to a normal position. It emerged from the shell between 2:35 and 2:50 a. m. Egg no. 2 was the last to hatch. At 7:05 p. m. the hatchling inside made two additional small slits in the shell, and at 7:30 p. m. thrust its snout through one of them, while lying on its back. At 1:45 a. m. it was right side up, but at 3:00 a. m. had reverted to its previous position. At 4:40 a. m. it was again right side up, and it emerged from the shell at 5:55 p. m.
On August 17, at 11:00 a. m., hatchlings no. 5 and no. 6 had lost their egg teeth. All others still had their egg teeth then, but by 10:00 p. m. that of no. 8 was missing, and that of no. 4 was loose and dropped out while the snake was being handled. On August 20 at 9:00 a. m., hatchling no. 7 had lost its egg tooth; nos. 2 and 3 retained theirs only in part, and no. 1 had its egg tooth intact. By noon on August 22 no trace of an egg tooth remained on any of the hatchlings.
In the same group of hatchlings sign of impending molt was first noticed on the morning of August 17, when no. 6 was noted to have [425] its eyes clouded and milky in appearance. By evening no. 1 had attained the same stage and no. 7 was beginning to show it. On the morning of August 20, shedding had begun in no. 6, while no. 2 and no. 8 had milky eyes. The eyes had cleared in no. 1 and no. 7, and were still clear in the remaining hatchlings. On August 22 shedding had been completed by no. 1 and no. 8, and all others were in the process of shedding.
Another clutch of 14 eggs from a recently captured female was found freshly laid in a cage on July 6, 1962. Hatching of 13 occurred August 16 to 20, as shown in Table 11.
Table 11. Times of Hatching in a Clutch of Racer Eggs From Harvey County Park
| Number of Egg |
Time at which shell was slit by hatchling |
Time that hatchling emerged |
||
| Day | Hour | Day | Hour | |
| 1 | Aug. 17 | 10:50 a. m. | Aug. 18 | 3:00 to 6:00 p. m. |
| 2 | Aug. 17 | 12:15 to 12:50 p. m. | Aug. 18 | 3:25 to 4:25 p. m. |
| 3 | Aug. 17 | 1:40 p. m. | Aug. 17 | 10:45 p. m. |
| 4 | Aug. 18 | 3:00 to 6:00 p. m. | Aug. 19 | before 7:00 a. m. |
| 5 | Aug. 16 | 10:45 a. m. | Aug. 17 | 6:30 to 7:55 a. m. |
| 6 | Aug. 17 | 9:00 a. m. | Aug. 18 | 6:30 to 7:30 a. m. |
| 7 | Aug. 17 | 4:15 p. m. | Aug. 20 | 1:00 to 7:00 a. m. |
| 8 | Aug. 17 | 3:40 p. m. | Aug. 18 | 5:00 to 6:00 p. m. |
| 9 | Aug. 17 | 7:30 p. m. | Aug. 18 | 6:30 to 7:30 a. m. |
| 10 | Aug. 16 | 10:48 a. m. | Aug. 17 | 11:05 a. m. |
| 12 | ? | no record | Aug. 18 | 6:00 to 6:30 a. m. |
| 13 | Aug. 17 | 8:00 to 8:20 a. m. | Aug. 17 | 5:30 to 5:50 p. m. |
Hatching usually occurs in late August or early September, and the disparity in size between hatchlings and adults is greater than in some other kinds of snakes. In 76 young from ten clutches of eggs incubated in the laboratory, averages and extremes for measurements and weights were as follows: snout-vent length, 214.5 (186 to 244) millimeters; tail, 59.3 (44 to 73) millimeters; weight, 4.16 (2.4 to 5.8) grams. In each brood the size tended to be fairly uniform, except that there were usually one or more stunted individuals markedly smaller than the others. However, there were striking differences in size between the young of different broods. None of the young captured was as small as the average hatchling from the clutches incubated in captivity, but in the 14 years of my study only four young were captured in August. The hatchlings [426] are relatively secretive and elusive, and the lush vegetation of late summer provides them with abundant hiding places. Nevertheless it is remarkable that the hatchlings are so seldom seem, when their probable abundance is taken into account.
Probably all of those captured had already made some growth after hatching. By early November or the last week of October, racers have almost or quite completed their season of activity, and are at the hilltop ledges, preparing to hibernate, if they have not already retired into dormancy. For 25 young of the year captured in this period at the end of the growing season, measurements and weights were as follows: snout-to-vent, 327 (273 to 418) millimeters; tail 93 (72 to 114) millimeters; weight, 12.3 (7 to 19) grams. In the ten-week period between hatching and hibernation these young had already passed through their period of most rapid growth, having added, on the average, more than 50 per cent to their original lengths, and almost tripled in weight. In these young about to enter their first hibernation, variation in size and weight is much greater than in the hatchlings; some have fared much better than others, and there are significant age differences. Within any one year the time of hatching is spread over several weeks because of differences in the time of laying, and differences in nest sites, with variation in heat received, which promotes or delays the rate of incubation. Year to year differences in the trend of weather increase the dispersion as the incubation time is shortened in hot, dry summers and lengthened in those that are relatively cool and moist.
Table 12. Growth of First-year Racers
| Sex | Original record | Record(s) of recapture | ||||
| Date | Snout-vent length, millimeters |
Weight, grams |
Date | Snout-vent length, millimeters |
Weight, grams |
|
| ♂ | Aug. 17, 1959 | 223 | 5.3 | Sept. 12, 1960 | 503 | 42 |
| ♀ | Oct. 26, 1959 | 278 | 10 | Oct. 26, 1959 | 566 | 55 |
| ♀ | Nov. 2, 1959 | 340 | 18 | May 11, 1961 | 582 | 65 |
| ♀ | Nov. 10, 1959 | 348 | 13.7 | June 20, 1961 | 733 | 127 |
| ♀ | Oct. 19, 1952 | 361 | 15.3 | Oct. 9, 1953 | 620 | 68 |
| ♂ | May 9, 1953 | 368 | 17 | May 13, 1954 | 609 | 84 |
| ♀ | April 16, 1950 | 328 | 142 | Oct. 10, 1950 | 603 | 78.2 |
| ♀ | May 17, 1956 | 358 | 15.2 | Oct. 6, 1956 | 575 | 52 |
| ♂ | Nov. 1, 1953 | 330 | 11 | May 21, 1955 | 674 | 92 |
Though covering a wide size-range, the young of the year entering hibernation are still a distinct size group, not yet overlapping that of the next older group of young. Growth during the first year of life is best shown by the individuals in Table 12, all of which were marked either before their first hibernation or soon after emerging from it, and were recaptured either the following autumn, or in spring soon after emerging from a second hibernation.
These records indicate that the young racers at the time of their second hibernation have grown to a snout-vent length of well over 500 millimeters, but less than 700 millimeters, and a weight of more than 40 grams but less than 100 grams. Other racers marked in the first few weeks of life were recaptured after two or more seasons of growth, and indicate the sizes that may be expected in young adults from two to five years old, as shown in Table 13.
Table 13. Growth of Racers Marked at an Age of Less Than One Year
and Recaptured in Their Second, Third or Fourth Years
| Sex | Original record | Record(s) of recapture | ||||||
| Date | Snout-vent length in mm. |
Weight in grams |
Probable age in months |
Date | Snout-vent length mm. |
Weight in grams | Probable age in months |
|
| ♂ | Nov. 12, 1952 | 342 | .... | 21/2 | May 20, 1955 | 702 | 110 | 33 |
| ♂ | Nov. 1, 1953 | 330 | 11 | 2 | Aug. 16, 1956 | 780 | 162 | 351/2 |
| ♂ | Nov. 12, 1953 | 293 | .... | 21/2 | May 14, 1956 | 634 | 96 | 321/2 |
| Oct. 13, 1956 | 689 | 110 | 371/2 | |||||
| ♂ | April 15, 1955 | 320 | 13 | 91/2 | June 8, 1958 | 740 | 118 | 45 |
| ♂ | Oct. 2, 1955 | 348 | 18 | 1 | May 22, 1958 | 728 | 120 | 33 |
| May 21, 1960 | 795 | 130 | 57 | |||||
| ♂ | June 25, 1959 | 378 | 10 | 10 | June 30, 1960 | 590 | 60 | 22 |
| June 7, 1961 | 705 | 123 | 33 | |||||
| ♂ | Nov. 1, 1953 | 330 | 11 | 2 | Aug. 25, 1957 | 805 | 183 | 48 |
Unlike young of the year, racers in their second autumn were trapped in large numbers. By this time all were large enough to be caught in the traps of quarter-inch wire mesh, and they were the most abundant size group. Many that were marked at this stage were recaptured after intervals of months or years, showing the trend of growth. Some of these snakes in their second autumn already had overtaken the more retarded third-year individuals. [428] The two age classes cannot be separated with certainty. Selected records of individuals that were almost certainly second-year young at the time they were marked are presented in Table 14.
Table 14. Growth of Young Racers That Were Marked Near the Time of Their Second Hibernation
| Sex | Original record | Record(s) of recapture | ||||||
| Date | Snout-vent length in mm. |
Weight in grams |
Probable age in months |
Date | Snout-vent length in mm. |
Weight in grams |
Probable age in months |
|
| ♂ | Oct. 6, 1960 | 595 | 80 | 13 | July 23, 1961 | 650 | 70 | 23 |
| ♂ | Oct. 13, 1950 | 525 | 52.6 | 131/2 | Oct. 2, 1951 | 675 | 105 | 25 |
| ♂ | Nov. 2, 1950 | 545 | 55.6 | 14 | Oct. 24, 1951 | 670 | 100 | 26 |
| ♂ | Sept. 2, 1957 | 522 | 57 | 12 | Oct. 14, 1958 | 690 | 103 | 251/2 |
| ♂ | Oct. 17, 1953 | 558 | 47 | 131/2 | May 10, 1955 | 718 | .... | 32 |
| ♂ | Nov. 14, 1956 | 587 | 57 | 141/2 | May 20, 1958 | 728 | 100 | 521/2 |
| June 17, 1959 | 880 | 219 | 641/2 | |||||
| ♂ | Sept. 7, 1959 | 533 | 45 | 12 | June 6, 1961 | 740 | 112 | 33 |
| ♂ | Oct. 17, 1959 | 558 | 56 | 131/2 | July 21, 1960 | 625 | 63 | 221/2 |
| June 7, 1961 | 670 | 95 | 33 | |||||
| ♂ | Oct. 16, 1952 | 577 | 62.0 | 131/2 | May 31, 1955 | 809 | 160 | 45 |
| May 11, 1956 | 855 | 144.3 | 561/2 | |||||
| ♂ | Oct. 11, 1950 | 570 | 65.9 | 131/2 | July 11, 1956 | 820 | 193.6 | 821/2 |
| ♂ | Oct. 14, 1953 | 560 | 52 | 131/2 | June 29, 1958 | 794 | 256 | 70 |
| ♂ | Oct. 6, 1950 | 563 | 55.8 | 13 | Sept. 28, 1958 | 907 | 243 | 109 |
| ♂ | Oct. 14, 1953 | 523 | 44 | 131/2 | May 19, 1959 | 818 | 190 | 801/2 |
| May 17, 1960 | 850 | 211 | 921/2 | |||||
| ♂ | Oct. 13, 1953 | 521 | 50.1 | 131/2 | May 13, 1958 | 814 | 166.4 | 441/2 |
| June 3, 1959 | 826 | 165 | 47 | |||||
| ♂ | Nov. 5, 1953 | 512 | 34 | 14 | May 22, 1958 | 847 | 135 | 69 |
| ♀ | Aug. 11, 1953 | 534 | 39.2 | 111/2 | Sept. 18, 1954 | 670 | 143 | 241/2 |
| ♀ | Oct. 14, 1949 | 588 | 55.2 | 131/2 | Oct. 11, 1950 | 713 | 114.0 | 251/2 |
| ♀ | Oct. 6, 1950 | 570 | 60.4 | 13 | Oct. 11, 1951 | 783 | 174 | 251/2 |
| ♀ | Oct. 31, 1953 | 582 | 58 | 14 | Oct. 5, 1954 | 860 | 195 | 25 |
| ♀ | Oct. 21, 1959 | 588 | 63 | 131/2 | May 7, 1961 | 730 | 120 | 32 |
| ♀ | May 14, 1960 | 506 | 34 | 201/2 | Oct. 26, 1960 | 690 | 90 | 26 |
| ♂ | Oct. 22, 1960 | 527 | 45 | 131/2 | Oct. 7, 1961 | 620 | 74 | 25 |
| ♂ | May 3, 1960 | 530 | 48 | 20 | May 16, 1961 | 700 | 110 | 321/2 |
| ♂ | May 27, 1961 | 535 | 40 | 21 | Oct. 25, 1961 | 672 | 98 | 26 |
From the records in Table 14 and many more like them, average and extreme sizes for progressively older age groups were estimated. Even racers that were already of adult size when they were marked were tentatively identified with one or another age group, and their records of subsequent growth were used. Most of the [429] records show that the females grow more rapidly than the males, and are, on the average, larger at any given age.
Relatively few individual racers were recaptured after periods of several years. Each of the eight listed in Table 15 is among those that were captured in four or more different years, and their records are significant in revealing the trend of growth after sexual maturity has been attained. These snakes, one to three years old at the time they were marked, show well the persistent but decreasing growth, and the fluctuating weight that is characteristic of this and other species.
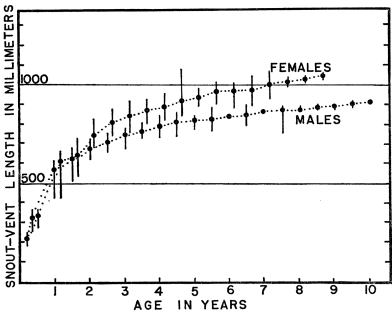
Fig. 18. Graph from records of blue racers marked early in life and recaptured, showing average and extreme snout-vent lengths for males and females of various ages. Growth is especially rapid in the first year, but continues, with gradual slowing, throughout life. In hatchlings, the sexes are of approximately equal size, but females grow to be markedly larger than males.
Some of the largest racers recorded had already reached unusually large size when they were first captured, so there was no opportunity to determine their ages. Several others, originally captured as small- or medium-sized adults, subsequently grew to a size approaching the maximum, and thus provided a basis for estimating the ages of large individuals. The records of nine such racers are listed in Table 16.
Table 15. Changes in Lengths and Weights
in Eight Racers Recaptured Repeatedly Over Periods of Years
| Date of Capture | Snout-vent length in millimeters |
Tail length length in millimeters |
Weight in grams |
Probable age in months |
| No. 1, male | ||||
| October 21, 1953 | 676 | 216 | 85 | 251/2 |
| October 12, 1954 | 733 | 238 | 128 | 371/2 |
| July 31, 1955 | 755 | 243 | 143 | 46 |
| May 20, 1956 | 785 | 250 | 153 | 551/2 |
| July 15, 1957 | 788 | 252 | 162 | 691/2 |
| June 30, 1958 | 820 | 264 | 200 | 81 |
| May 15, 1959 | 828 | 263 | 195 | 911/2 |
| May 14, 1960 | 840 | 265 | 195 | 1031/2 |
| No. 2, male | ||||
| May 9, 1955 | 678 | 212 | 105 | 32 |
| May 26, 1958 | 827 | 248 | 195 | 69 |
| May 3, 1959 | 865 | 262 | 218 | 80 |
| May 14, 1960 | 877 | 264 | 218 | 921/2 |
| July 16, 1960 | 890 | 257 | 190 | 941/2 |
| No. 3, male | ||||
| May 26, 1955 | 565 | 171 | 53 | 21 |
| August 28, 1957 | 798 | 245 | 155 | 48 |
| May 13, 1958 | 814 | 249 | 137 | 561/2 |
| May 4, 1959 | 848 | 252 | 180 | 68 |
| June 12, 1961 | 896 | 262 | 198 | 931/2 |
| No. 4, male | ||||
| May 19, 1956 | 632 | 208 | 78 | 201/2 |
| August 2, 1957 | 740 | 238 | 123 | 35 |
| May 20, 1959 | 796 | 246 | 132 | 561/2 |
| May 23, 1960 | 813 | 280 | 140 | 681/2 |
| No. 5, male | ||||
| June 30, 1956 | 594 | 188 | 73 | 22 |
| October 12, 1956 | 658 | 200 | 80 | 271/2 |
| July 14, 1957 | 690 | 220 | 102 | 341/2 |
| June 23, 1958 | 749 | 226 | 136 | 46 |
| July 24, 1959 | 751 | 239 | 133 | 59 |
| May 27, 1960 | 765 | 235 | 130 | 69 |
| No. 6, male | ||||
| October 19, 1956 | 518 | 167 | 43 | 131/2 |
| July 19, 1957 | 673 | 218 | 96 | 221/2 |
| May 5, 1960 | 780 | 250 | 175 | 56 |
| May 9, 1961 | 840 | 272 | 272 | 68 |
| No. 7, female | ||||
| May 12, 1957 | 790 | 238 | 175 | 321/2 |
| September 3, 1957 | 810 | 243 | 208 | 36 |
| October 1, 1958 | 883 | 259 | 206 | 49 |
| July 11, 1959 | 915 | 247[A] | 190 | 581/2 |
| September 12, 1960 | 930 | 249[A] | 238 | 721/2 |
| September 14, 1961 | 1012 | 268[A] | 332 | 841/2 |
| No. 8, female | ||||
| June 23, 1955 | 601 | 175 | 67 | 22 |
| August 3, 1958 | 920 | 255 | 300[B] | 60 |
| July 23, 1959 | 955 | 281 | 211 | 71 |
| May 18, 1960 | 962 | 262 | 260 | 81 |
[A] Incomplete.
[B] With food.
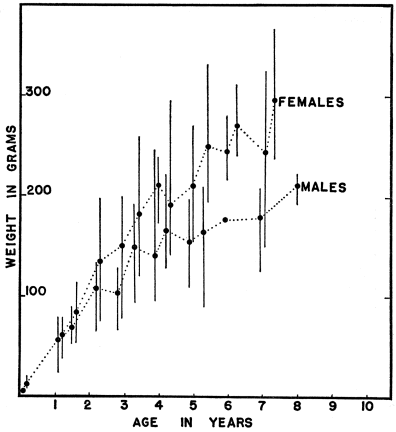
Fig. 19. Graph from records of blue racers marked early in life and recaptured, showing average and extreme weights in samples of different ages. Early in life the females outstrip males in growth and the size differential increases throughout life.
A few adult racers recaptured, including different individuals of small, medium, and large size, failed to make any measurable gain in length over periods of months, or even over several years. Most often the large individuals were those that failed to grow or made only slight gains. Some of the snakes that failed to increase in length suffered dramatic weight losses, perhaps as a result of injury, disease, or parasitism. However, other individuals that failed to gain appreciably in length did gain in weight. Doubtless both [432] genetic and environmental factors were involved. A few racers gave the impression of being stunted by adversity. All records obtained of growth (or failure to grow), throughout the period of my study were used in compiling Table 17.
Table 16. Growth in Several Racers That Were Already Adult
When Originally Captured and That Subsequently Attained Unusually Large Size
| Sex | Original record | Record(s) of recapture | ||||||
| Date | Snout-vent length in mm. |
Weight in grams |
Probable age in months |
Date | Snout-vent length in mm. |
Weight in grams |
Probable age in months |
|
| ♂ | May 14, 1956 | 815 | 191 | 561/2 | May 15, 1959 | 905 | 243 | 921/2 |
| May 16, 1961 | 945 | 287 | 1041/2 | |||||
| ♂ | May 22, 1958 | 825 | 251 | 681/2 | June 25, 1959 | 840 | 205 | 82 |
| May 27, 1960 | 880 | 210 | 93 | |||||
| Oct. 14, 1961 | 900 | 223 | 1091/2 | |||||
| ♂ | May 19, 1959 | 705 | 105 | 321/2 | May 28, 1960 | 828 | ... | 45 |
| July 3, 1961 | 900 | 265 | 581/2 | |||||
| ♀ | Nov. 6, 1954 | 823 | 190 | 38 | June 30, 1958 | 1030 | 350 | 82 |
| ♀ | June 6, 1955 | 810 | 190 | 33 | May 21, 1959 | 1087 | 345 | 801/2 |
| ♀ | Aug. 29, 1957 | 885 | 265 | 48 | Aug. 6, 1960 | 1041 | 235 | 83 |
| ♀ | May 1, 1958 | 633 | 84 | 20 | May 30, 1961 | 1088 | 320 | 57 |
| ♀ | July 20, 1958 | 862 | 203 | 461/2 | June 30, 1960 | 1020 | 238 | 70 |
| ♀ | Oct. 18, 1956 | 845 | 185 | 371/2 | May 20, 1958 | 905 | 240 | 561/2 |
| Oct. 22, 1959 | 1085 | 375 | 74 | |||||
Mortality Factors and Adaptations for Survival
The behavior patterns that are associated with defense and escape in snakes are widespread. The common racer is typical of the more generalized snakes. Almost all elements of the racer's behavior are found in snakes of other genera and perhaps, of other families. The racer differs from these only in minor details of its behavior, in the circumstances under which various behavioral traits are elicited, and their relative importance. The common racer is one of the swiftest of North American snakes, and it usually depends upon speed to avoid enemies. Racers crossing roads may detect approaching automobiles at a distance, and thereby may be stimulated to accelerate their movements and so escape onto the [433] shoulders rather than becoming traffic casualties as many individuals of most other kinds of snakes do under the same circumstances. The chances of such escape are of course much better on dirt roads that are relatively narrow and have a rough surface permitting efficient traction, than on wide smooth, paved highways. The snakes are somewhat reluctant to venture onto open expanses of pavement.
Table 17. Sizes of Racers of Different Age Groups in May and October
| Age in Years |
Males | |||||
| Snout-vent length in mm. | Weight in grams | |||||
| Average | Typical range |
Observed range |
Average | Observed range |
||
| 1+ | 539 | 432-609 | ||||
| 2- | 615 | 500-676 | 560-674 | 68.2 | 51-92 | |
| 2+ | 668 | 610-700 | 620-710 | 107.4 | 63-134 | |
| 3- | 706 | 677-730 | 648-755 | 102.1 | 65-129 | |
| 3+ | 740 | 701-754 | 667-780 | 147.0 | 93-216 | |
| 4- | 757 | 731-772 | 725-809 | 139 | 95-251 | |
| 4+ | 785 | 755-793 | 720-850 | 167.4 | 128-225 | |
| 5- | 805.5 | 773-810 | 743-855 | 152.4 | 110-198 | |
| 5+ | 810 | 801-831 | 773-858 | 163.9 | 89-211 | |
| 6- | 827 | 811-831 | 765-883 | 175.9 | 130-230 | |
| 7- | 845 | 832-850 | 788-900 | 181.2 | 125-210 | |
| 8- | 868 | 851-873 | 740-890 | 217.5 | 194-225 | |
| 8+ | 870 | 861-884 | ||||
| Females | ||||||
| 1+ | 581 | 415-658 | ||||
| 2- | 644 | 520-739 | 580-738 | 83.5 | 52-127 | |
| 2+ | 743 | 685-804 | 670-826 | 135.2 | 73-200 | |
| 3- | 810 | 740-829 | 730-880 | 149.4 | 98-219 | |
| 3+ | 836 | 804-869 | 736-915 | 181.2 | 120-268 | |
| 4- | 866 | 830-880 | 791-920 | 212.3 | 175-243 | |
| 4+ | 883 | 870-914 | 810-952 | 191.2 | 143-300 | |
| 5- | 914 | 880-929 | 833-1088 | 209.6 | 136-275 | |
| 5+ | 932 | 915-954 | 883-990 | 250.4 | 195-336 | |
| 6- | 965 | 930-961 | 892-1020 | 245.9 | 218-283 | |
| 6+ | 970 | 955-980 | 885-1003 | 271 | 243-336 | |
| 7- | 974 | 962-990 | 919-1050 | 251.3 | 150-330 | |
| 7+ | 1000 | 980-1015 | 930-1085 | 295.6 | 235-375 | |
A racer suddenly startled by close approach of a human usually flees at high speed. The first part of its course, up to 20 feet or more, is marked by a violent lateral thrashing of the body and tail, which may help the snake to gain speed, but which seem chiefly [434] designed to draw attention of a potential attacker to the spot that the snake is leaving. In any event, the mode of progression abruptly changes to a swifter and much smoother travel, and the snake may seem to vanish completely or as it glides rapidly away, its course may be marked by the slowly waving tops of tall grass. The trend of travel away from the site of disturbance is in an irregularly zig-zag course. The method of crawling, with lateral undulations of the body, supported against scattered pivot-points along the way, tends to prevent the snake from moving more than a short distance in a straight line.
In checking traps along hilltop rock ledges in October, I have often flushed racers that had come to the ledges searching for hibernation dens. In such an instance the startled racer would usually dart away down the steep slope at unusually high speed, and within a few seconds might have covered 100 feet or more, progressing with a minimum of lateral undulations, and seeming to flow or coast over the ground surface with scarcely any effort. Although coursing downhill in this manner provided an effective method of escape, racers startled at the ledges or on other sloping ground sometimes followed a course parallel to the ledge, or even turned uphill, with disastrous slowing. Often I have accomplished capture by hand, by the strategy of running downhill to get below the escaping snake, causing it to turn back up the slope, there to be slowed to the extent that it could be overtaken.
A racer that has been flushed, and has disappeared after creating a commotion by its violent thrashing, may, within a period of minutes, return surreptitiously to the vicinity, gliding back slowly and silently from the direction opposite that in which it departed. It may climb into a tree or bush where, lying stretched on outer twigs and concealed in dense foliage, it will sometimes permit close approach, rather than reveal its presence by moving. If too much disturbed however, such a snake will move away, higher, or to the opposite side of the clump, with a slow and stealthy gliding motion which is likely to escape detection. The sudden stops and abrupt changes in direction make it difficult to follow the course of a climbing racer, even though it is moving slowly. The tendency to climb to escape danger seems to be especially strong in the western subspecies, C. c. mormon. In my years of experience with the racers in the chaparral belt of western Oregon and California, I found that one of the most characteristic responses to danger was to climb into a bush or low tree and seek concealment in the thick foliage. [435] In most instances the snake was from one to eight feet above the ground; usually it did not attempt to climb high in a tree even when there was opportunity to do so. The racer's prowess as a climber is limited. In thick brush where its weight is supported by numerous stiff twigs simultaneously, it is at home and can travel rapidly, but it cannot cling effectively to rough bark of a vertical tree trunk as can the rat snakes (Elaphe sp.) and others that are more specialized in their arboreal adaptations. Chaparral-type shrubs such as Ceanothus cuneatus and Arctostaphylos viscida were those in which western yellow-bellied racers were seen climbing most frequently. Crab-apple (Pyrus ioensis) and haw (Crataegus sp.) were the shrubs most frequently utilized by blue racers on the Reservation. Carr (1950:80) noted that Florida racers were much less given to tree-climbing than were those of more northern regions. C. c. stejnegerianus of southern Texas is reputed to have arboreal tendencies highly developed (Mulaik and Mulaik, 1942:14).
The racers kept in an outdoor enclosure in the summer of 1962 spent much of their time above the ground, climbing in several walnut saplings. Usually in the warmer part of the day three or four of the five racers in the pen were climbing. In the trees a racer usually lay extended or in a loose coil among foliage in the outer twigs, at a height of five to 12 feet. Such a snake might rest for hours in almost the same position. Racers do not ordinarily enter the water voluntarily, but they are swift and efficient swimmers when the need arises. On one occasion a large female, seen near the edge of the pond on the Reservation, and approached from the landward side, took to the water without hesitation and swam strongly to the opposite shore more than 100 feet away. Other observers have recorded similar incidents.
A racer that is suddenly startled at close range may make no attempt to escape, but instead may coil and perform slow writhing movements, with the head pressed to the ground and concealed beneath part of the body. Musk, exuding from the anus is spread over the body surface as the movements continue. Only a small percentage of the racers found free in their natural habitats reacted in the manner described. In each such instance, the circumstances were such that the racer was prevented from making its usual rapid getaway, either because temperature was unusually low for activity, or because the snake was away from suitable cover. After removal from a live-trap, with handling, and especially clipping of [436] scales, racers were much more likely to behave in this passively defensive manner, which seemingly constitutes a second line of defense in snakes which have been prevented from escaping—either cornered, captured, or injured by a predator. The musk is creamlike in color and consistency, and is secreted from sausage-shaped glands in the base of the tail. Its odor is rather disagreeable, but less so than that of musks of various other genera. Also, it is less penetrating and lasting than many other musks. In handling the racers removed from traps I attempted to avoid being smeared with the musk by grasping the snake by the tail and neck and keeping its body stretched out. While the snake was being examined, and measured a droplet of musk would form at the anal orifice and would begin to flow down the snake's body. To avoid contact with the musk I would wipe away the droplet with a paper towel or leaf, but the musk droplet might be replaced several times within the few minutes that the snake was restrained.
A racer that is confronted with an object arousing its suspicion or indicating possible danger, may move away slowly with a characteristic "threat display." The forebody and neck are held rigid, well off the ground, and slightly arched, with the neck flattened in a vertical plane—causing this part of the snake to appear from lateral view larger than it actually is. The tongue is protruded frequently and waved slowly. A racer that has been disturbed and is trying to gain shelter in a direction that brings it closer to the danger usually will adopt this tense attitude, but a single threatening movement will cause it to abandon its circumspect pose, and panic in an attempt to reach the nearest shelter in the shortest possible time.
A common response to an alarm is to vibrate the tail. The tail being long and slender is vibrated much less rapidly than the short, muscular organ of a rattlesnake, or even the medium-short tail of a bull snake. The rapid twitching produces a characteristic sound in dry vegetation. A racer that vibrates its tail is fully active and aroused; the sound is heard as the snake pauses before it makes a dash for shelter. At times the sound probably functions as a decoy to distract the attention of potential predators, affording the racer an opportunity to escape.
A racer that is cornered or captured usually puts up a spirited struggle, striking vigorously and repeatedly at its tormentor. If only cornered, it will make slashing strokes at the enemy, jerking back to a coil from each stroke in a manner that causes the teeth [437] to lacerate the enemy's skin in long scratches if the stroke finds its target. The rapid recoil often causes teeth to be jerked from their sockets and left embedded in the wound inflicted by the bite. On many occasions in sustaining a bite from a racer I have received such teeth which have remained undetected for a day or more until soreness and festering led to their discovery and removal. A racer that is grasped may deliver several bites within a few seconds, chewing vigorously to imbed its teeth to the maximum at each bite.
If grasped by the tail and held clear of the ground, the racer would swing its body with a rapid whirling and twisting motion, which in a few seconds would twist off the end of the tail, unless countermeasures were promptly taken. Many racers, especially the larger and older ones, have parts of the tail missing, as a result of such escapes, and perhaps also from freezing in hibernation or from certain injuries and infections. If grasped by the body, the racer struggles with a violent lateral thrashing to break the grip of its captor, at the same time striking to bite and discharging musk, urinary wastes and feces.
Many of the racers examined bore scars from wounds that were probably inflicted by predators. A few had survived severe skeletal injuries, involving deformation of the spinal column or extensive tearing of muscle and connective tissue layers of the body wall, altering the normal body shape. A more frequent type of injury involved chiefly the integument, which had been ripped open by the teeth, claws, or bills of adversaries, despite the fact that the skin is remarkably tough and leathery. Patches of scar tissue with scales in irregular sizes, shapes and patterns characterized such injuries. The most frequently observed type of injury involved loss of part of the tail. Usually only a small terminal part was missing, but occasional stub-tailed individuals had lost as much as three-fourths of the tail.
No consistent trend of difference between the sexes in incidence of injuries to the tail was noticeable, but there was definite correlation with age. In the entire sample from the Reservation and Rockefeller Tract the percentages in each supposed age group (actually size group) lacking part of the tail were as follows: hatchlings, 2.9 per cent; one-year-olds, 9.5 per cent; two-year-olds, 15.8 per cent; three-year-olds, 14.3 per cent; four-year-olds, 15.7 per cent; five-year-olds, 23.2 per cent; six-year-olds, 30.9 per cent; seven-year-olds, 28.9 per cent; those eight years old or older, 21.1 per cent. The seeming reversal of trend in the older racers is difficult [438] to explain, but probably results from inadequate numbers in this part of the sample.
Although the racer's most characteristic response to any disturbance is to flee at high speed, certain individuals are inclined to behave aggressively under exceptional circumstances. Records of such aggressive behavior nearly all pertain to large individuals of the northeastern black racer (C. c. constrictor) in the breeding season. Cope (1900:794) wrote: "it is courageous and will sometimes attack, moving forward with the head raised from 1 to 2 feet above the ground." Ditmars (1944:13) wrote: "Occasional specimens in the breeding season ... will actually attack ... glide toward an intruder ... striking madly at one's feet or legs." Woods (1944:257) quoted a 13-year-old amateur herpetologist, Leon Gonthier, regarding the latter's encounter with an aggressive black racer on May 7, 1944, as follows: "When I ran toward the snake about 20 feet away, it turned and came for me. As I bent over to catch it, the snake grabbed me by the shirt and hung on ... held it off with a stick. It jumped twice more at me and came clear off the ground." Finneran (1948:124) describing the species' habits in Connecticut, stated: "A farmer ... constantly warned me away from Coon Ledge during the spring, saying the snakes were breeding and would 'chase you.' This very thing happened. In 1943 a blacksnake followed me for approximately ten feet, and, in 1946, a male aggressively attacked me for a period of three minutes. There was ample opportunity for escape."
Little has been published concerning the natural enemies of the common racer. The king snake (Lampropeltis getulus) is notorious for ophiphagy, and doubtless preys upon the racer at times. Wright and Bishop (1915:169) wrote of the king snake in Okefinokee Swamp, in Georgia, that "all the smaller snakes suffer, and of the larger species, the blacksnake [racer] and spreading adder are the commonest prey." However, Clark (1949:252) examined 301 stomachs of king snakes of this species in Louisiana and found no racers, although many other kinds of snakes were represented, and, collectively, made up the greater part of the food.
A rat snake (Elaphe obsoleta) found in Leon County, Florida, on June 10, 1924, had a racer in its stomach, according to the Patuxent food habits file of the U. S. Fish and Wildlife Service. A California garter snake (Thamnophis elegans terrestris) was found [439] to have a juvenal racer in its stomach (Fitch, 1940:96). A large alligator lizard (Gerrhonotus multicarinatus) was found swallowing a small racer that had been confined with it in a bag (Fitch, 1935:12).
A few records of predation on blue racers by other reptiles on the Reservation and Rockefeller Tract were obtained. In 25 scats of the prairie king snake (Lampropeltis calligaster), there were 29 vertebrate prey items of which one was a blue racer. On September 15, 1962, an adult male prairie king snake 491/2 inches in length was found to have a 27-inch yearling racer in its stomach. In 254 scats of the slender glass lizard (Ophisaurus attenuatus) vertebrate remains were rare, but there were scales and bones of one hatchling blue racer. Among 21 vertebrate prey items in 14 scats of the timber rattlesnake (Crotalus horridus) there were remains of one blue racer. Among 589 prey items of copperheads there were two juvenal blue racers (Fitch, 1960:200). In contrast to these scarce records of racers in the prey of other snakes, there were more frequent records of the blue racer preying on its own young. The 1008 food items from 479 racer scats contained remains of 16 racers, small young in most instances.
Raptorial birds are known to be important predators on snakes. Breckenridge (1944:118) reported finding remains of a blue racer in the pellet of a marsh hawk (Circus cyaneus). Many pellets of the marsh hawk collected on the Reservation were all found during the colder half of the year, and they contained no remains of reptiles. The same seasonal restrictions applied to the many pellets of four species of owls that were collected, and these also lacked remains of reptiles. One of the predators whose food habits have been most thoroughly investigated on the area is the red-tailed hawk (Buteo jamaicensis). Over the period 1955 through 1962, 1131 pellets of these hawks were collected, many from the Reservation, but more from localities scattered throughout the eastern one-fourth of Kansas. The pellets were those of nestlings and fledglings, nearly all collected from beneath the nests, in late May, June, or early July. Some 49 different nests were represented, and remains of 43 blue racers were found. In one nest which yielded a total of 191 food items, racer remains occurred 13 times, but no other nest yielded records of more than three racers, and some other species of reptiles, notably the black rat snake (Elaphe obsoleta) comprised much more important components of the food. Since only one meal in 26 contained remains of racers, it seems that a red-tailed [440] hawk would destroy only a few racers in the course of a year on its territory of perhaps half a square mile, and that its predation would not be a major factor in the racer's ecology.
That the red-tailed hawk is a natural enemy to be reckoned with throughout the racer's extensive range was demonstrated by the analysis of contents of 116 stomachs from localities well scattered over the United States and Canada. Among the 152 vertebrate prey items represented there were three racers: a juvenile C. c. constrictor from Stag Lake, New Jersey, October 16, 1927; a C. c. mormon from Weiser, Idaho, April 11, 1930, and a C. c. stejnegerianus from San Roman, Cameron County, Texas, in the spring of 1938. Because of the widely scattered geographical origin of these stomachs, a remarkably large number of species of prey were represented, and no one species predominated. Small mammals of the genera Sylvilagus, Spermophilus, and Thomomys comprised the most important component of the food; the only other reptile so well represented as the racer was the common garter snake (Thamnophis sirtalis) which likewise had three occurrences.
In June and July, 1954, a nest of broad-winged hawks (Buteo platypterus) on the Reservation yielded 71 prey items of which seven were blue racers. At least 19 species of prey were represented, although specific determinations were not possible in some instances. The prairie vole with eight records was the most frequent prey, and the racer and cardinal (nestlings) each had seven, while all other species were represented by fewer occurrences. Six of the racers were first-year young but the remaining one was thought to be an undersized second-year individual. Although an adult racer, especially a large one, would probably be an adversary too powerful to be killed and eaten by a broad-winged hawk, this hawk is perhaps one of the more important natural enemies of the first-year young in the eastern United States.
Further records of predation on racers by raptors were obtained from the U. S. Fish and Wildlife Service files through the kindness of Dr. Wm. H. Stickel. There were three records of such predation by red-shouldered hawks (Buteo lineatus) from Lunenberg, Massachusetts, May 1, 1896; Portland, Maine, March 16, 1906; and 65 miles northeast of Sarasota, Florida, June 10, 1918. There were two records of predation by broad-winged hawks from Catlettsburg, Kentucky, July 26, 1910, and Portland, Connecticut, May 18, 1912. There were four records of predation by marsh hawks on racers—Peck's Island, Maine, September 18, 1903; [441] Edgartown, Massachusetts, April 12, 1912; West [Tisbury?], Massachusetts, July 31, 1912; and Okanagan Landing, British Columbia, June 5, 1918. A sparrow hawk (Falco sparverius) from De Ranch, Wyoming, May 27, 1910, had eaten a racer, as had a barn owl (Tyto alba), from Franklin County, Kansas, November 16, 1922, and a crow (Corvus brachyrhynchos, nestling) from Onaga, Kansas, May 16, 1914.
Perhaps certain mammalian predators are even more important natural enemies than are raptorial birds, but records of predation on racers by mammals are few. Substantial samples of scats of opossums and coyotes from the Reservation have contained no remains of racers. The Fish and Wildlife Service files include a record of an opossum from Adrian, Michigan, on April 26, 1934, that had racer remains in its stomach. No racers were specifically recorded from a collection of 820 scats and 22 stomachs of raccoons (Procyon lotor) from Douglas County, Kansas, reported upon by Stains (1956:43), but occurrences of unidentified snake may have pertained in part to the racer. Skunks are probably more important natural enemies, but food habits data from the Reservation are lacking for the two kinds of skunks occurring there. Crabb (1941:356) in a study of the food habits of the spotted skunk (Spilogale putorius) in southeastern Iowa, did not report any reptiles among the many kinds of prey found in scats. Several times in many years of residence near Medford, Jackson County, Oregon, I saw remains of racers which appeared to be victims of striped skunks (Mephitis mephitis). Tracks and other sign of the skunks were often noticed along a little-used road on a hillside, passing between a pasture and a hay field, with an oak grove, high weeds, and brush, bordering the road. The racers sometimes found as victims along this road were small- or medium-sized individuals. In every instance the predator had begun eating on the tail end of the snake, and later had abandoned the remains leaving the head and part of the forebody still intact. Predation had occurred at night. Skunks foraging mostly in twilight or darkness, probably find racers inactive beneath flat rocks or in shallow burrows. The skunks would be too slow and clumsy to catch the snakes in the open when they were fully active.
The funnel traps used for catching the racers also caught many other kinds of animals. Often a racer and another kind of snake were caught together, but, ordinarily, in these instances no damage to either resulted, although racers, copperheads, king snakes, and garter snakes are all known to eat each other's young. At times, [442] large predatory ground beetles (Calosoma scrutator and Pasimachus sp.) were so abundant that a dozen or more were caught together in a trap, and several times such groups attacked and killed and partly ate young racers caught in the same traps. White-footed mice (Peromyscus leucopus), deer mice (P. maniculatus), harvest mice (Reithrodontomys megalotis), short-tailed shrews (Blarina brevicauda), and least shrews (Cryptotis parva) all were caught rather frequently in the traps, and each, on one or more occasions, gnawed and killed or severely injured a racer trapped with it. The attacks probably were motivated by hunger in all instances, the snake in some instances failing to defend itself or escape because of low temperature. Occasionally such encounters might occur even under natural conditions, the mouse or shrew finding and attacking the snake while the latter was torpid and helpless in its hibernaculum or in a more superficial temporary shelter.
Relatively few of the racers examined showed signs of disease. The most common type of affliction was an infection of the skin causing cankerlike sores, chiefly on the ventral plates, sometimes on the head or tail. These were seen most often in snakes recently emerged from hibernation, but persisted later in the season in years of exceptionally wet weather. Some of the racers most severely afflicted appeared to be in debilitated condition. The causal organism was not determined, nor was it definitely determined whether this type of disease causes mortality.
The ectoparasites of the racer are chiefly chiggers, the parasitic larvae of mites. Loomis (1956) in his study of the chigger mites of Kansas examined many of the racers captured in the early years of my field work. He checked a total of 130 racers, mostly from the Reservation, and found four different species of chiggers, all of the genus Trombicula: T. alfreddugèsi, T. lipovskyana, T. kansensis and T. sylvilagi. The common pest chigger, T. alfreddugèsi, parasitizes most of the species of reptiles, birds, and mammals occurring in Kansas, and it was by far the most common kind on the racers. The numbers per racer in different months were as follows: June, 81; July, 285; August, 432; September, 123; October, 15. Many of the racers were collected in the relatively cool and moist summers of 1950 and 1951. In a year of typical weather, heaviest infestations [443] occur in early summer, June or the first half of July, and the numbers taper off rapidly in the hot but often dry weather of late July, August, September, and October. In hot, humid weather of early summer a racer may have several hundred chiggers attached to it, filling most of the areas of exposed skin between the scales. The chiggers are conspicuous because of their bright orange color. Like T. alfreddugèsi, T. lipovskyana was also found in large numbers on racers and is found on many kinds of hosts. Loomis (op. cit.:1281) recorded it from one kind of frog, one kind of toad, one kind of turtle, two of lizards, six of snakes, 19 of birds, and nine of mammals from eastern Kansas. Five larvae of T. sylvilagi were recorded from a racer captured in October. That kind of chigger is primarily a parasite of small mammals, and perhaps cannot develop successfully when it attaches to a snake. Unlike most other kinds of chiggers, this species is most in evidence in autumn and winter. A single larva of T. kansensis was found on a racer in October. This relatively rare kind of chigger has been found on several kinds of snakes and small mammals (including pocket gophers) and is known from hot and dry rocky places. Even the racers that were heavily loaded with chiggers showed no obvious ill effects, but the chiggers are potentially vectors of various diseases.
Of the many endoparasites found in racers, the lung fluke, Neorenifer lateriporus was the only one identified and frequently observed in my study (Stewart, 1959). This is a digenetic trematode of the subfamily Reniferinae. The racer is its specific host. The life history is still unknown, but in other members of the subfamily, all of which parasitize snakes, an aquatic snail and a frog are required as hosts at different stages of the life cycle. Presumably N. lateriporus has similar requirements. The two common local water snails, Heliosoma trivolvis and Physa anatina, are both potential hosts. By far the most probable frog host is the leopard frog. In wet weather of July the recently metamorphosed leopard frogs leave the water and disperse to all habitats, probably carrying with them the parasites acquired in the tadpole stage. The racers in turn probably acquire their flukes by eating the young frogs in summer. In any case, the adult racers are nearly all parasitized, but the flukes have not been found in those racers that were less than one year old. During their first few months, the racers are too small to swallow leopard frogs, even the young. The flukes have been seen in the live racers mostly in May, when most adults are infested with the flukes. Seemingly at this season the [444] flukes migrate forward into the mouth of the host. Probably this is the time when the flukes breed and lay eggs; if so, the eggs would pass through the digestive tract of the snake and escape with its feces. The latter are usually left in terrestrial situations unfavorable for the development of an aquatic stage, but perhaps some of the eggs are washed into ponds by heavy summer rains. In late summer and fall the flukes are not to be found in the mouths of the live snakes.
Most complete records of the flukes present in racers were kept in 1959. The following table shows the numbers of racers examined and the percentage having flukes in that year.
Ortenburger (1928:182) recorded lung flukes (Renifer ellipticus) from blue racers. In Maryland, McCauley (1945:76) also recorded numerous lung flukes (Pseudorenifer sp.) in an immature racer 490 millimeters in total length. Parker (1941:34) recorded Neorenifer septicus from racers collected at Reelfoot Lake, Tennessee, Greensboro, Georgia, and Kissimmee, Florida; also he recorded N. georgianum from racers collected at Reelfoot Lake. N. septicus was recorded by the same author from the water moccasin (Agkistrodon piscivorus) and N. georgianum was also recorded from the king snake (Lampropeltis getulus).
Cloacal smears from the racers examined usually showed an abundance of ciliate protozoans, either parasites or commensals, and occasionally nematode worms. Harwood (1933:66) examined two racers from the vicinity of Houston, Texas, and found four kinds of helminths: Kalicephalus agkistrodontis, K. rectiphilus, Ophidascaris sp., and Polydelphis sp. Each parasite was found in only one of the two snakes. McCauley (loc. cit.) recorded nematodes (Physaloptera obtusissima) from black racers in Maryland, and Ortenburger (loc. cit.) recorded Physaloptera sp. from the blue racer.
Table 18. Seasonal Incidence of the Fluke Neorenifer lateriporus,
in Mouths of Blue Racers on the Reservation and Rockefeller Tract
| Apr. 15-30 |
May 1-15 |
May 15-31 |
June 1-15 |
June 15-30 |
July | Aug. | Sept. | Oct. | |
| Number of racers checked |
1 | 28 | 25 | 24 | 17 | 30 | 5 | 9 | 2 |
| Percentage of sample having flukes |
100 | 79 | 72 | 50 | 18 | 0 | 0 | 0 | 0 |
Since there is a brief annual breeding season, any local population of racers consists of a series of discrete annual age groups. The population reaches its annual maximum in early September, after undergoing sudden increase by the addition of the annual crop of hatchlings. Throughout the remainder of the year numbers of racers undergo gradual reduction as a result of the many combined mortality factors that affect them. This mortality is distributed among all the age classes, but the heaviest losses, both percentage-wise and in actual numbers, are sustained by the first-year young. Being by far the most numerous group, these young suffer more mortality than all the other age classes combined. Presumably much of this mortality is concentrated in the early weeks of life, while the young are still near their minimum size; the rate of loss is gradually reduced as larger size is attained and some of the early hazards are outgrown. In the adult age classes also, the larger and older snakes live in greater security, and the rate of mortality is higher in the smaller and younger snakes. Even before hatching, the eggs are subject to heavy losses from predators, and probably from drying, flooding, and other unfavorable climatic factors. Unfortunately it was not possible to obtain definite figures on any of these losses since the eggs were never found under natural conditions and the hatchlings were seen only in relatively small numbers.
The records obtained from trapping racers in late spring and summer in fields provided a somewhat different picture of the population from the sample obtained along the ledges in autumn. In the former sample there were 400 males to 257 females, but in the latter sample there were 355 males to 379 females. I regard the summer sex ratio as a distorted one, brought about by the greater activity of the males in the breeding season. Racers are caught most easily in May, and the fact that two or more males often were trapped with the same female, while the reciprocal combinations did not occur, demonstrates the increased activity of the males in their search for mates at this season. In autumn there is no sexual activity; both sexes probably are equally active in seeking places to hibernate when they are trapped along the hilltop outcrops. The ratio of 51.6 per cent females in my sample of 734 may indicate that in the males greater activity at other seasons results in a somewhat higher mortality. This idea is borne out by the fact that for the supposed two-, three- and four-year-olds combined, females comprise 51.2 per cent, but they comprise 55.6 per cent of those more than four years old and 61.3 per cent of those more than five years old.
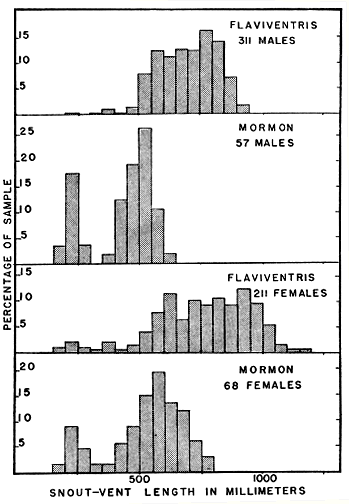
Fig. 20. Histograms comparing snout-vent length in random samples of Coluber c. flaviventris (those captured on the Reservation and Rockefeller Tract in 1960 and 1961) and C. c. mormon (specimens in the Museum of Vertebrate Zoology, University of California). In the museum sample, collected by conventional methods, the first-year young constitute a prominent and fairly distinct size group, whereas in the sample of racers from the Reservation and Rockefeller Tract, mostly caught in wire traps of quarter-inch mesh, relatively few young were secured. It is demonstrated that hatchlings are approximately the same length in both populations, but flaviventris grows much longer, and that the differences in length between the sexes is approximately the same in each population.
By assigning to each racer caught an arbitrary age, on the basis of size according to Table 17, I calculated the population (exclusive of those snakes in their first year of life) to have the composition shown in Table 19.
Table 19. Percentages of Adult Population of Blue Racers
Comprised by Each Annual Age Group
| Years of Age | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 or more |
| Percentage of population | 41.5 | 17.8 | 12.6 | 9.5 | 6.1 | 4.3 | 2.7 | 2.4 | 1.2 | 1.9 |
Figures are completely lacking to show the relative numbers of juveniles, until, already approaching adult size, the young are about to enter their second hibernation. My combined fall samples include 303 of these latter young, as against 142 racers about to enter their third hibernation. Thus, after having nearly attained adult size, these adolescent snakes sustained a loss of 53 per cent in a year. Losses must occur at an even more rapid rate in the younger and smaller snakes. It may be speculated that of the approximately 300 eggs produced by a population of 100 adult racers, 150 are lost before or during the period of incubation which lasts nearly two months. Of the 150 hatchlings emerging in early September, at least one-third probably are eliminated by the following breeding season in late May, leaving 100. The 100 survivors at this stage are still small juveniles, but by autumn they have attained adolescent size. By this time, if they had undergone a further reduction by 53 per cent, only 47 would remain—approximately the number to be expected if the population were stable from year to year.
A notable difference between the fall sample and the spring sample that I obtained was the higher proportion of large and old racers in the former sample. This difference can be attributed to the year-to-year changes in the population during the 14 consecutive years spanned by my field work. The fall sample of 734 racers represented the combined catch of the years 1949 through 1962, rather evenly distributed, but the spring sample included few snakes from the years 1949 through 1957; most were from the years 1958 [448] through 1961. In 1949 when the study was begun, the Reservation was being protected for the first time, and formerly overgrazed pastures or cultivated fields were acquiring a rank growth of grass and weedy vegetation, and thus becoming favorable habitat. The abundant new habitat promoted rapid increase in the population of racers until the newly available areas were filled to their "carrying capacity." Table 21 shows the changing trends of the different age groups. Although the separate annual samples are perhaps too small to show the composition of the population accurately, it is significant that in the fall of 1949 an unusually high proportion of the racers caught were one-year-olds, hatched in September, 1948.
Table 20 shows that in a typical group of 100 subadult and adult racers (second year and older) only a little more than one-fourth are productive females. The largest females, six years old and older, making up less than ten per cent of the adult population, contribute nearly half the total complement of eggs.
The calculated number of eggs pertains to a stage before oviposition, and subsequent losses through resorption under unfavorable conditions, through inviability of embryos and through deaths of some of the gravid females, are to be expected. In the weeks of incubation further losses are sustained. Although these losses cannot be measured, they must be severe as on numerous occasions scattered and torn eggshells representing entire clutches dug out and destroyed by predators, have been found. Probably other clutches are destroyed underground by such predators as moles and egg-eating snakes, and still others by insects. Tinkle (1959:195) wrote that in a clutch of 15 eggs found under a board, four were parasitized and had small perforations. Molds destroy a high percentage of all reptilian eggs that are incubated artificially and doubtless destroy many under natural conditions also. Excessive heat or moisture, or desiccation, resulting either from climatic extremes or from poor choice of a nest site by the female, would cause further loss. In four different years, Blair (1960:108) found that losses of eggs between laying and hatching in the Texas spiny lizard (Sceloporus olivaceus) ranged from 69 per cent to 86 per cent; no other comparable study of the extent of egg losses in a species of reptile is known to me. The racer is somewhat less prolific than the spiny lizard, and potentially longer lived; the racer's eggs are larger and thicker-shelled, and they are deposited in deeper burrows. It might be expected that losses during incubation would be somewhat less in the racer than in the spiny lizard.
Table 20. Calculated Productivity in a Hypothetical Group of 100 Subadult and Adult Blue Racers
| Age-Group (years) |
Percentage of population in age-group |
Percentage of fecund females in age-group |
Percentage productive females in age-group |
Eggs per productive female |
Number of eggs produced |
| 2 | 41.5 | 51.2 | 13 | 9.2 | 26 |
| 3 | 17.8 | 51.2 | 56 | 9.9 | 51 |
| 4 | 12.6 | 51.2 | 60 | 10.8 | 42 |
| 5 | 9.5 | 55.0 | 57 | 13.0 | 39 |
| 6 and over | 18.6 | 61.3 | 80 | 15.7 | 143 |
Table 21. Percentages of Racers in Each Annual Age Group (Exclusive of Hatchlings)
in Autumnal Samples at Different Stages of the Field Study,
Showing Shift Toward Older Age-Groups in the Later Years
| Years of Age | Year or combination of years represented by each sample | ||||
| 1949 | 1950 1951 1952 |
1953 1954 1955 |
1956 1957 1958 |
1959 1960 1961 1962 |
|
| 1 | 54 | 44 | 39 | 39 | 41 |
| 2 | 14 | 24 | 21 | 19 | 19 |
| 3 | 18 | 11 | 23 | 16 | 13 |
| 4 | 4 | 10 | 10 | 11 | 9 |
| 5 | 6 | 6 | 3 | 8 | 8 |
| 6 | 2 | 1 | 3 | 1 | 3 |
| 7 | 2 | 4 | 4 | 2 | 3 |
| 8 | ...... | ...... | ...... | .5 | 2 |
| 9 | ...... | 1 | .4 | .5 | 2 |
| 10 | ...... | ...... | .4 | 2 | 1 |
| Older than 10 | ...... | ...... | ...... | .5 | ..... |
| Number in sample |
49 | 126 | 117 | 194 | 242 |
Although figures for the youngest age groups—one-year-olds and hatchlings—are missing, approximations of them may be furnished by extrapolation, from the information available regarding the productivity of the population. Some factors involved in productivity are that the sex ratio deviates from parity, slightly in favor of the females in the adolescents but more markedly in favor of the females among the older age groups; that some adult females apparently fail to produce eggs in the breeding season, but the percentage decreases in the older snakes; and that number of eggs [450] per clutch increases in proportion to the size and age of the female producing them. Too few figures are available concerning most of these factors to indicate more than the trends; nevertheless the available figures have been used in Table 20 in an attempt to estimate the productivity of a hypothetical population.
Conant (1938:178, Pl. 7) published a photograph of 106 blue racers killed in February, 1932, by farmers near Bellville, Ohio, and Pope (1944:173) mentioned that scores of blue racers aggregated in October around an oak-covered dune near Chicago. In both these instances large hibernating aggregations were involved, and the areas represented by them are unknown; nothing has been recorded regarding population densities.
The records obtained through my fall trapping, along hilltop rock outcrops, yielded no information concerning population densities, but those obtained in fields in summer did provide significant information in this regard. Even after years of trapping on the same area, the catch still consisted largely of new individuals; the method was not sufficiently effective to catch all racers present at any one time, and the total catch for a season therefore provided only a crude index of the minimum number present.
The summer trapping was carried on in three separate areas. One of these was the area of bottomland pastures and formerly cultivated fields where the Reservation headquarters are located, a block of 39 acres bounded on three sides by woodland, and on the fourth by cultivated farm land. Effective trapping in this area was carried on through the years 1955 to 1961 inclusive. A second area, of 48 acres, was one of upland fields, mostly covered with re-established prairie grasses, in the northeastern part of the Reservation. A third area, of 137 acres, also upland, was that of the Rockefeller Tract, cultivated through 1956 and sown to prairie grasses the following year, and the adjacent northwestern hilltop portion of the Reservation. Effective trapping on these two latter areas was carried on in 1958 through 1962.
For the seven years of trapping in the House Field area, the catch was as follows: 30, 33, 38, 38, 34, 24, and 20. In four years of trapping, the northeast field area yielded 42, 28, 37, 59, and 19 blue racers, and the Rockefeller Tract yielded 52, 67, 67, 126, and 106. The actual catch was hence less than one per acre in nearly all instances, but the year-to-year differences in catch are believed [451] to be caused chiefly by differences in numbers of traps used and in trapping effort, rather than by changes in the numbers of racers present.
Best index to the number of racers actually present is provided by the number of recaptures, and their ratio to first captures. The population of course, undergoes alteration from year to year, with many racers eliminated and replaced by others.
In 1955, 26 racers were caught in the May-June-July period, in the headquarters field area. In the August-September-October period of the same year five racers were caught of which only one was a member of the original 26. The five-to-one ratio indicates that the original 26 may have represented an actual population of 130, but of course the single recapture is much too small a sample to provide a reliable ratio. Some of the racers caught in May were recaptured in June, others in July, and still others not until late summer or early autumn. Somewhat different estimates can be obtained for the population depending on how the season's records are divided. For instance, in the headquarters field area in May 1955, eleven racers were caught; in the remainder of the season 20 were caught, of which two were members of the original group of eleven. The 20 to 2 ratio indicates that the 11 caught in May represented an actual population of 110. In the period May-June, 18 racers were caught, and in July-August-September-October, 15 were caught, including four of the original 18, hence indicating a population of 67. Although obviously the population underwent some change during the course of the season, the three sets of census figures apply essentially to the same population, and the divergence in them illustrates the wide range of error arising from insufficiently small samples.
Common sources of error in the censusing of natural populations of animals by the capture-recapture method ("Lincoln Index" or "Petersen Index") arise from the fact that the composition of a local population often changes between two sampling periods, or even within them. Some of the animals marked may move elsewhere, to be replaced by unmarked immigrants, or they may die and be replaced by unmarked maturing young. First-year racers that could pass through the quarter-inch mesh of the traps in spring and early summer became too large to escape in this way in the latter half of the summer, but these young were excluded from the census computations. There was doubtless some shifting of marked individuals away from the study areas and shifting of new individuals onto [452] these areas in the periods of weeks between successive samplings. Jackson (1939) has explained a method of correcting census computations based on capture-recapture ratios when there is a consistent trend of diminishing recaptures with increase in elapsed time. However, in my records no such trend is discernible; furthermore it has been demonstrated that individual racers tend to stay within the same home range area throughout most of their season of activity. Therefore, I conclude that shifts of individuals away from the study areas or into them, in the intervals between samplings constitute only a minor source of error.
A combination of the figures from the three samples listed above provides an intermediate "smoothed" figure that can be accepted with somewhat more confidence than any one of the separate censuses because it is based on more records. The combined ratios indicate a total of 105 racers in the headquarters field area. The figures obtained in the different sampling periods, and the census figures derived from their ratios are shown in tables 22 and 23. Differences from year-to-year in the census figures for any one area show no consistent trends and their variation is similar to that shown in different samples for the same season. Probably populations were fairly stable throughout the periods involved. If such stability is assumed, the samples from different years may be combined, and the composite figures derived from them may be accepted with more confidence. For the headquarters field area, for instance, 419 records of racers were gathered in all the preliminary sampling periods of the seven seasons involved; the records totalled 263 for all the secondary sampling periods, with 70 recaptures in secondary periods of the snakes recorded in the corresponding preliminary periods. A population of 75 racers is indicated—1.9 per acre. Corresponding figures for the northeast field area are: preliminary samples 453, secondary samples 163, recaptures 39, calculated number 135 (2.82 per acre). For the Rockefeller Tract the figures are as follows: Preliminary samples 807, secondary samples 476, recaptures 126, the ratio indicating a population of 153, or 1.11 per acre.
These figures represent the number of adults present in early summer when the population is near its annual low point. The first-year young, excluded from this census because they cannot be caught in representative numbers, perhaps approximate the number of adults, in May, so the figures obtained would need to be approximately doubled to be representative of the entire population. [453] By late summer the adults, and especially the yearlings, have undergone substantial reduction in numbers, but in late August and early September the hatching of a new crop of young increases the population to its annual maximum. The maximum numbers probably are about three times those obtained by censusing adults in early summer. The peak population of late summer or early autumn is estimated to consist of hatchlings, comprising somewhere near 50 per cent; adults, comprising a little more than 25 per cent; and yearlings comprising a little less than 25 per cent.
Table 22. Captures Recorded and Population Calculated From Them
on Hilltop Grassland Areas of Rockefeller Experimental Tract
and Adjacent Reservation in Four Different Years
| 1959 | 1960 | 1961 | 1962 | Four-year average | |
| First Census: | |||||
| Early May | 0 | 8 | 21 | 32 | 61 |
| Late May | 3 | 13 | 33 | 35 | 84 |
| Recaptures | 0 | 1 | 2 | 7 | 10 |
| Estimated population |
104 | 346 | 160 | 128 | |
| Second Census: | |||||
| May | 3 | 25 | 54 | 59 | 141 |
| June | 9 | 6 | 25 | 40 | 80 |
| Recaptures | 0 | 3 | 6 | 24 | 33 |
| Estimated population | 50 | 165 | 99 | 85 | |
| Third Census: | |||||
| May-June-July | 25 | 34 | 94 | 100 | 253 |
| Sept.-Oct | 8 | 13 | 20 | 13 | 54 |
| Recaptures | 1 | 3 | 5 | 4 | 13 |
| Estimated population | 200 | 182 | 376 | 324 | 263 |
| Fourth Census: | |||||
| May-June | 31 | 30 | 73 | 76 | 210 |
| July-Aug.-Sept.-Oct | 24 | 15 | 34 | 35 | 108 |
| Recaptures | 3 | 2 | 14 | 12 | 31 |
| Estimated population | 104 | 225 | 177 | 222 | 183 |
| Fifth Census: | |||||
| May | 3 | 25 | 54 | 60 | 142 |
| June to October | 23 | 20 | 67 | 40 | 150 |
| Recaptures | 0 | 5 | 14 | 20 | 39 |
| Estimated population | 100 | 258 | 120 | 162 | |
| Five Sets Combined: | |||||
| Combined first samples | 62 | 122 | 296 | 327 | 807 |
| Combined second samples | 67 | 67 | 179 | 163 | 476 |
| Recaptures | 4 | 14 | 41 | 67 | 126 |
| Estimated population | 208 | 117 | 258 | 159 | 153 |
Densities in early summer of one to three adult blue racers per [454] acre probably are typical of the better types of habitat in the region of my study. The upland field area estimated to have 2.82 racers per acre was better habitat than the other two study areas. Prior to 1948 it had been cultivated and severely eroded. In 1949 most of it was sown to seeds of prairie grasses, and by 1958 different parts of it were dominated by different species of native perennial tall grasses interspersed with areas that supported a weedy type of vegetation, and other areas that supported dense thickets of sumac, dogwood, elm saplings, or other woody plants. The abundance and diversity of dense cover and of small animals made this area especially favorable habitat for the racer.
Table 23. Captures Recorded and Populations Estimated From Them
in Headquarters Field Area of Reservation
| 1955 | 1956 | 1957 | 1958 | 1959 | 1960 | 1961 | Seven-year average |
|
| First Census: | ||||||||
| May | 11 | 21 | 7 | 24 | 17 | 13 | 6 | 99 |
| June-July-Aug. | 20 | 11 | 25 | 20 | 22 | 12 | 14 | 124 |
| Recaptures | 2 | 1 | 4 | 5 | 5 | 2 | 0 | 19 |
| Estimated population | 110 | 231 | 44 | 96 | 75 | 78 | ... | 92 |
| Second Census: | ||||||||
| May-June | 18 | 25 | 17 | 28 | 27 | 15 | 13 | 143 |
| July-Aug.-Sept.-Oct | 15 | 8 | 17 | 15 | 15 | 11 | 8 | 89 |
| Recaptures | 4 | 1 | 7 | 5 | 9 | 3 | 1 | 30 |
| Estimated population | 68 | 200 | 41 | 84 | 45 | 55 | 104 | 61 |
| Third Census: | ||||||||
| May-June-July | 26 | 28 | 22 | 31 | 31 | 20 | 19 | 177 |
| Aug.-Sept.-Oct | 5 | 7 | 13 | 12 | 6 | 5 | 2 | 50 |
| Recaptures | 1 | 2 | 7 | 5 | 3 | 1 | 1 | 20 |
| Estimated population | 130 | 128 | 47 | 87 | 63 | 75 | 148 | 63 |
| Three Sets Combined: | ||||||||
| Combined first samples | 55 | 74 | 46 | 83 | 75 | 48 | 38 | 419 |
| Combined second samples | 40 | 26 | 55 | 47 | 43 | 28 | 24 | 263 |
| Combined recaptures | 7 | 4 | 18 | 15 | 17 | 6 | 3 | 70 |
| Estimated population | 105 | 160 | 47 | 87 | 63 | 75 | 101 | 75 |
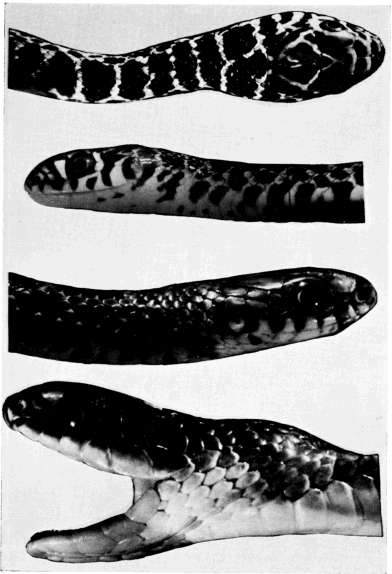
Fig. 1. Head of hatchling blue racer, dorsal view, September 1962, approximately × 3.
Fig. 2. Head of hatchling blue racer, lateral view, September 1962, approximately × 3.
Fig. 3. Head of yearling male blue racer, lateral view, August 1, 1961, a little less than twice natural size.
Fig. 4. Head of adult male blue racer, lateral view, July 16, 1961, approximately × 2. All three snakes from Rockefeller Experimental Tract, Jefferson County, Kansas.

Fig. 1. Abandoned limestone quarry on a hilltop of southward exposure on The University of Kansas Natural History Reservation, in late autumn of 1951. The crevices along the base of the ledge provided favorite hibernating sites for blue racers.

Fig. 2. Hatchling blue racer and eggshell from which it had recently emerged, in early September, 1962; × approximately 12/3.

Fig. 1. Wire funnel trap set at base of hilltop limestone outcrop in a spot strategically located for interception of blue racers searching for deep crevices in which to hibernate, October 15, 1949.

Fig. 2. Large clutch of 21 blue racer eggs, recently plowed out, at Harold Brune farm, Jefferson County, Kansas, July 10, 1962.

Fig. 1. Habitat of blue racer, blue-stem prairie on Botany Bluff at northwest corner of the University of Kansas Natural History Reservation, looking south. Trees and brush in background are along limestone outcrop at top of slope. Mowed area in foreground is southwest corner of Rockefeller Experimental Tract, a privately owned farm at the time this photograph was taken in the summer of 1951.

Fig. 2. Habitat of blue racer, blue-stem prairie on south slope of Botany Bluff, looking north along west edge of the Reservation, summer of 1951. By 1962, with exclusion of fire, and protection from mowing, prairie vegetation had largely disappeared from this slope, and had been replaced by trees and brush. As a result of these successional changes racers no longer found this slope a suitable habitat in summer, but they continued to resort to the hilltop rock outcrop to hibernate in autumn.
Some local areas probably support higher populations of racers than do areas where censuses were made, but under modern conditions, situations that offer near optimum habitat are not likely to be extensive or to persist long. On land that is capable of producing a good crop of vegetation, the crop is usually harvested either by grazing of livestock or by using the land for cultivation, with the result that the racers are, at least in some seasons, forced into marginal situations. More than 50 years ago in Missouri, Hurter (1911:170) wrote that the racer "was quite common 20 years ago in pastures, meadows and fields but as cultivation has advanced it is becoming quite rare." In 1962 the widespread and adaptable blue racer is still common in many parts of its range, including Missouri, but in most places its population densities probably are lower than formerly.
Reduction since 1911 has probably been far more drastic than the reduction that had occurred up to that time. Schmidt and Necker (1935:69), writing of the racer in the Chicago region, noted "the snakes which raise their heads and face mowing machinery tend to be exterminated in agricultural areas." They stated that in the Chicago region the racer had been exterminated by the advance of agriculture except in two extensive sand dune areas. In July 1962, Mr. V. B. Howell, a progressive farmer of the Great Bend area in central Kansas, told me that the kinds of snakes inhabiting cultivated land—blue racer, bull snakes, prairie king snakes, hog-nosed snakes, and others—had undergone great reduction in numbers during the period of his farming. He estimated that in a forty-year period the numbers had declined to perhaps five per cent of their level in the area most familiar to him, centering at his farm 11 miles northwest of Great Bend, Barton County. In accounting for this change in population density Howell pointed out the relative destructiveness to small animals of modern farm machinery as contrasted with horse-drawn equipment or that used with tractors of earlier models. Modern tractors move forward so rapidly that there is little opportunity for snakes or other small animals to avoid them, and the plows and disks cut wide swaths penetrating more deeply into the soil than did older types. On July 10, 1962, in searching the furrows of a freshly plowed small field on the Harold Brune farm, for turned-up nests of the snakes, I found two adult blue racers that had been struck and killed by the plow, possibly while they were underground. In fields that are plowed or cultivated between the times of egg-laying and hatching, the eggs are destroyed. Because of its rapid movements and alertness, the racer [456] is more likely to escape farm machines than are most other kinds of snakes. Nevertheless, it is vulnerable and survives in cultivated areas only when they are interspersed with pastures, woodlots, or streamside thickets where at least part of the population may find refuge.
Field study of the blue racer was carried on in several localities in Kansas, but chiefly at the University of Kansas Natural History Reservation (the northeasternmost section of land in Douglas County), and the adjacent 160-acre Rockefeller Experimental Tract in Jefferson County. By October 26, 1962, after 14 years of field work, a total of 1423 racers had been captured some 2197 times.
The locale of the present study was near the geographic center of the blue racer's range. The range, chiefly in the Mississippi Valley and Great Plains region, is centrally situated with respect to the other seven subspecies. An extensive but scattered literature concerning the ecology of the species as a whole, and its several geographic races, has been reviewed and utilized for comparison with my own field data.
Blue racers were caught in wire funnel traps set in prairie and pastureland habitat in summer, and along hilltop limestone outcrops in woodland in autumn. The autumn trapping along rock outcrops was carried on each year from 1949 to 1962, but effective summer trapping was carried on only in the last six years of the study. Each racer caught was individually and permanently marked by scale clipping. More than half were caught only once, but many were recorded repeatedly, with a maximum of 16 captures.
The racer occurs throughout most of the United States, and its populations are subject to much geographic variation. The snakes are largest in the northeastern part of the range, with clines of decreasing size toward the southeastern, southern and western parts of the United States. There are somewhat parallel trends in coloration; the black racer of the northeastern states grades into paler, gray or light brown subspecies in southern Florida, Texas, and the far western states. Accompanying these changes in color and size are minor morphological changes and major ecological changes. The black racers of the eastern states often inhabit forest or forest-edge habitats while the paler and smaller snakes of more southern or western areas typically inhabit scrub, chaparral, or prairie. The large, dark-colored racers of the eastern and northeastern states are especially inclined to attack larger prey including small vertebrates, [457] even weasels, rabbits, and chipmunks, whereas the smaller and paler racers of more southern and more western areas take a higher proportion of insects and rarely attack vertebrates other than small reptiles.
On the area where field work was carried on in northeastern Kansas, tall-grass prairie habitat is preferred, but fields of grain or alfalfa, grazed pasture, brush, woodland edge, groves or open woodland, and weedy fields are all utilized to some extent. The racer is strictly diurnal and largely terrestrial but it may climb through bushes or small trees in foraging or escaping.
The blue racer is a typical colubrine snake of slender build, with large eyes, and vision plays an important role in finding prey and detecting enemies. In the adult blue racer the dorsal color is variable, pale brown or gray, bluish, greenish or slaty. In the hatchling, however, there is a distinct pattern of a type widespread among colubrines and also among snakes of other groups—a series of middorsal blotches on an olive ground color, with alternating rows of smaller spots on each side. The ventral surface is pale, with dark speckling. The pattern is sharply defined on the anterior part of the body, but markings become progressively more obscure posteriorly and are scarcely discernible on the tail.
The juvenal pattern fades gradually as growth proceeds, and there is much individual variation in the rate of its loss. Some racers still retain the juvenal pattern faintly discernible after attainment of sexual maturity. There are also striking ontogenetic changes in the proportions of the head, body and tail. The diameter of the eye is approximately one per cent of the snout-vent length in hatchlings, but is only a little more than half that relative size in the largest adults. In the course of allometric growth other parts of the head also enlarge less rapidly than the body, but more rapidly than the eye. In hatchlings there is a slight average difference between the sexes in relative tail length, with males' tails the longer. Relative tail length increases slightly in both sexes up to the time of sexual maturity, and then decreases slightly with advancing age.
Racers in northeastern Kansas spend nearly half the year in hibernation, with average recorded emergence date April 16, and average date of retirement into hibernation November 8. Hibernacula are usually in crevices in hilltop limestone outcrops with south exposures. Winter temperatures within the hibernacula are usually well within the range 0 degrees to 10 degrees Centigrade. Spring [458] emergence has been recorded at an air temperature of only 12.5 degrees Centigrade. Racers bask in sunshine frequently even in warm weather, and the temperature preferendum is several degrees higher than in most other kinds of snakes. Bodily temperatures obtained from blue racers that were fully active, either under natural conditions or in a large outdoor enclosure, were concentrated in the neighborhood of 34 and 35 degrees Centigrade. For short periods racers can survive temperatures up to 45 degrees without damage, but more prolonged exposure to temperatures of slightly less than 40 degrees can be fatal. In hibernation, racers can withstand temperatures slightly below freezing, but they cannot survive being frozen solid.
Blue racers tend to limit their activities to familiar areas or home ranges; some individuals may live out their entire lives within the same home range, but others shift from time to time. Average home ranges of approximately 26 acres for males and 24 acres for females were calculated. The racers' preference for hibernacula in a habitat different from that to which summer activities are confined necessitates spring and fall migrations between the limestone outcrops where hibernation occurs and the grasslands where the snakes stay in summer. The average spring or fall migration is approximately a quarter of a mile, but an individual racer does not consistently return to the same hibernaculum. Many racers were recorded to have made movements of 2000 to nearly 4000 feet, involving shifts in home range, but some later shifted back to their original areas. Some may have made even longer shifts but their movements would not have been recorded since they would have gotten beyond the limits of the study area.
Blue racers hunt by various methods, often by coursing through dense vegetation in active search in which vision is of primary importance in locating the prey. Almost any small animal that moves nearby may be overtaken and caught with a sudden dash. From analysis of scats and prey from stomachs, a total of 1357 food items of more than 50 species was compiled. Favorite prey species were the cricket (Gryllus assimilis), grasshoppers (Arphia simplex, Melanoplus femur-rubrum, M. bivittatus, M. differentialis), camel crickets (Ceuthophilus sp.), katydid (Neoconocephalus robustus), vole (Microtus ochrogaster), white-footed mouse (Peromyscus sp.), racerunner, (Cnemidophorus sexlineatus), and leopard frog (Rana pipiens). The insects taken greatly outnumbered the vertebrates, but the vertebrates made up most of the actual bulk of prey eaten. [459] Crickets, grasshoppers, and katydids comprised most of the insect prey. Beetles, moths and cicadas were rarely taken. Vertebrate prey included miscellaneous small snakes (some of them juvenal racers), mammals, and birds. Seasonal change in the composition of the food is slight, but vertebrates figure more prominently in early summer, and insects comprise increasing percentages later in the season. Composition of the food differs according to size of the snake; gryllid and ceuthophilid crickets are best represented in the food of juveniles whereas small mammals, and grasshoppers of the genus Melanoplus are best represented in the food of large adults.
The breeding season is mainly in May. Mating is promiscuous and two or more males may court the same female simultaneously. A courting male lies on or alongside a receptive female, with spasmodic rippling abdominal movements, and with his vent adpressed to hers. At intervals in the courtship period the female moves swiftly for a few feet or a few yards shifting to a new spot, and during her activity the male strives to maintain contact with her. From time to time the male leaves the female briefly and courses rapidly around her in a devious route. Courtship is consummated when the female raises her tail in acceptance of the male and intromission is effected. During coitus, which lasts for periods of minutes, the female moves forward slowly, dragging the passive male, tail-first behind her.
Ovulation normally is in late May. Eggs from 29 to 39 millimeters in length and 14 to 21 millimeters in breadth are laid, from mid-June to early August, usually in tunnels of fossorial mammals such as voles or moles, at depths of five to 12 inches. Clutches of the blue racer average 11.8 eggs but the number is correlated with age and size of females; two-year-olds average only 9.2 eggs, whereas those females that are six years old or more average 15.7 eggs. Also, there is geographic variation in size of clutch, from only 5.8 eggs in C. c. mormon of the West Coast to 16.8 eggs in C. c. constrictor of the northeastern states. In each breeding season some females of adult size do not produce clutches. Only about 13 per cent of the two-year-olds in a small sample were fecund, but the ratio increased to 80 per cent in old adults. Incubation averages 51 (43 to 63) days.
Hatchlings usually make several longitudinal slits in the eggshell with the egg tooth before emerging, and often require a day or more to emerge after the first slit is made. Hatchlings average [460] 103/4 inches and 4.16 grams. By late October when these young are ready to enter hibernation, they have grown to 163/4 inches and 12.3 grams. Typical October lengths (overall) in inches for males and females, respectively, after successive seasons of growth, are as follows: yearlings, 273/4 and 29; two-year-olds, 341/4 and 371/4; three-year-olds, 373/4 and 413/4; four-year-olds, 40 and 443/4; five-year-olds, 411/2 and 461/4; six-year-olds, 421/4 and 481/4; seven-year-olds, 433/4 and 50; eight-year-olds, 44 and 511/4.
Judging from trends in the small samples available, sex ratio in hatchlings is approximately 1:1. In the summer trapping of blue racers, males are caught in larger numbers than females, but seemingly this is because of their greater activity. In fall along the limestone outcrops where the racers hibernate females were caught in slightly greater numbers than males among the young adults, and made up a little more than 60 per cent of the old adults. Presumably the males are eliminated more rapidly, because of their greater activity, their smaller size, or a combination of both factors. The composition by age groups of the adult racers captured was as follows: two-year-olds, 41.5 per cent; three-year-olds, 17.8 per cent; four-year-olds, 12.6 per cent; five-year-olds, 9.5 per cent; six-year-olds, 6.1 per cent; seven-year-olds, 4.3 per cent; eight-year-olds, 2.7 per cent; nine-year-olds, 2.4 per cent; ten-year-olds, 1.2 per cent; more than ten years old, 1.9 per cent.
Attempts to census blue racers on the study plots, from the capture-recapture ratios, yielded highly variable figures for different areas and even for the same area at different times. Variability is believed to result mostly from the small sizes of the samples, none of which is large enough to yield a census figure that is statistically reliable. Combined samples yielded figures indicating populations of one to three adults per acre in early summer in areas of favorable habitat. To represent the entire population in late summer—the time of its annual maximum—these figures would need to be approximately tripled.
The blue racer depends mostly on speed to escape its enemies. In escaping it often takes advantage of a downhill slope to increase its speed. A racer startled at close range often sets out with a violent thrashing that attracts attention to a given spot, then glides away so silently and rapidly that its course is not detected. The snake may circle back and approach the scene of disturbance slowly and stealthily, from the direction opposite to that taken in departing. Often a racer seeks concealment by climbing into a bush or [461] tree, and occasionally escape is effected by swimming. A racer that is caught or cornered usually makes a spirited defense by striking and biting. An alternative reaction, seen most typically when the snake is handicapped by injury or by low temperature, is to coil with the head concealed, and, with writhing movements, to smear the surface of the body with musk discharged from glands in the base of the tail.
Many natural enemies prey upon the racer, but in the present study the majority of records pertained to the red-tailed hawk. The broad-winged hawk, marsh hawk, red-shouldered hawk, sparrow hawk, and barn owl also are among the raptors that feed on this kind of snake. Among mammals the striped skunk is the only species definitely recorded to prey on the racer, feeding on the eggs as well as upon the young and adults. The common king snake, prairie king snake, timber rattlesnake, copperhead, and even the slender glass lizard have been recorded to prey on the blue racer, but probably all are of minor importance as natural enemies. Shrews (Blarina, Cryptotis) and mice (Peromyscus, Reithrodontomys) have sometimes killed and eaten racers confined in traps, and under natural conditions they possibly prey upon snakes that are immobilized when torpid from cold, in their hibernacula and temporary shelters. Chiggers are the most common ectoparasites of the racers. Four species of the chigger genus Trombicula have been recorded on racers from the Reservation. The racer is an important host of the common pest chigger, Trombicula alfreddugèsi, which often attacks humans. Most adult racers on the Reservation and nearby areas carry the fluke, Neorenifer lateriporus. In spring these flukes are conspicuous in the mouths of the racers, but in late summer they are not in evidence.
Allen, M. J.
1932. A survey of the amphibians and reptiles of Harrison County, Mississippi. Amer. Mus. Novit., 542:11-12.
Anderson, P.
1942. Amphibians and reptiles of Jackson County, Missouri. Bull. Chicago Acad. Sci., 6:203-220.
Atkinson, D. A.
1951. The reptiles of Allegheny County, Pa. Ann. Carnegie Mus., 1:145-157.
Auffenberg, W.
1949. The racer, Coluber constrictor stejnegerianus in Texas. Herpetologica, 5:53-58.
1955. A reconsideration of the racer, Coluber constrictor, in eastern United States. Tulane Studies in Zool., 2(6):89-155.
Barbour, R. W.
1950. The reptiles of Big Black Mountain, Harlan County, Kentucky. Copeia, 1950:100-107.
Baxter, G. T.
1947. The amphibians and reptiles of Wyoming. Wyoming Wild Life, August and October 1947:1-8.
Blair, W. F.
1960. The rusty lizard. Univ. Texas Press, Austin, xvi + 185 pp.
Blanchard, F. N. and Blanchard, F. C.
1942. Mating of the garter snake, Thamnophis sirtalis sirtalis (Linnaeus). Papers Michigan Acad. Sci. Arts and Letters, 27:215-234.
Blanchard, F. N. and Finster, E. B.
1933. A method of marking living snakes for future recognition with a discussion of some problems and results. Ecology, 14:334-349.
Bogert, C. M. and Cowles, R. B.
1947. Moisture loss in relation to habitat selection in some Floridian reptiles. Results of the Archbold Expeditions, No. 58; Amer. Mus. Novit., 1358:1-34.
Bonn, E. W. and McCarley, W. H.
1953. The amphibians and reptiles of the Lake Texoma area. Texas Jour. Sci., 1953(4):465-471.
Boyer, D. A. and Heinze, A. A.
1934. An annotated list of the amphibians and reptiles of Jefferson County, Missouri. Trans. Acad. Sci. St. Louis, 28(4):183-200.
Branson, E. B.
1904. Snakes of Kansas. Univ. Kansas Sci. Bull., 12(13):353-430.
Brattstrom, B. H.
1953a. Records of Pleistocene reptiles and amphibians from Florida. Quart. Jour. Florida Acad. Sci., 16(4):243-248.
1953b. The amphibians and reptiles from Rancho La Brea. Trans. San Diego Soc. Nat. Hist., 11(14):365-392.
1955. Pliocene and Pleistocene amphibians and reptiles from southeastern Arizona. Jour. Paleontology, 29(1):150-154.
Breckenridge, W. J.
1944. Reptiles and amphibians of Minnesota. Univ. Minnesota Press, 202 pp.
Brimley, C. M.
1903. Notes on the reproduction of certain reptiles. Amer. Nat., 37:261-266.
Brons, H. A.
1882. Notes on the habits of some western snakes. Amer. Nat., 16:564-567.
Brumwell, M. J.
1951. An ecological survey of the Fort Leavenworth Military Reservation. Amer. Midl. Nat., 45(1):187-231.
Burt, C. E.
1927. An annotated list of the amphibians and reptiles of Riley County, Kansas. Occas. Papers Mus. Zool. Univ. Michigan, 189:1-9.
1933. Some distributional and ecological records of Kansas reptiles. Trans. Kansas Acad. Sci., 36:186-208.
1935. Further records of the ecology and distribution of amphibians and reptiles in the Middle West. Amer. Midl. Nat., 16(3):311-336.
Burt, C. E. and Hoyle, W. L.
1934. Additional records of the reptiles of the central prairie region of the United States. Trans. Kansas Acad. Sci., 37:193-216.
Cagle, F. R.
1942. Herpetological fauna of Jackson and Union counties, Illinois. Amer. Midl. Nat., 28(1):164-200.
Carpenter, C. C.
1958. Reproduction, young, eggs and food of Oklahoma snakes. Herpetologica, 14:113-115.
Carr, A. F. Jr.
1940. A contribution to the herpetology of Florida. Univ. Florida Publ. Biol. series, 3:iv + 118 pp.
Clark, R. F.
1949. Snakes of the hill parishes of Louisiana. Jour. Tennessee Acad. Sci., 24(4):244-261.
Clarke, R. F.
1958. An ecological study of reptiles and amphibians of Osage County, Kansas. Emporia State Research Studies, 7(1):1-52.
Cohen, E.
1948. Emergence of Coluber c. constrictor from hibernation. Copeia, 1948:137-138.
Committee on Herpetological Common Names
1956. Common names for North American amphibians and reptiles. Copeia, 1956(3):172-185.
Conant, R.
1938. The reptiles of Ohio. Amer. Midl. Nat., 20(1):1-200.
1958. A field guide to reptiles and amphibians of the United States and Canada east of the 100th Meridian. Houghton-Mifflin Co., Boston xviii + 366 pp., 40 pls., 62 figs., 248 maps.
Cope, E. D.
1900. The crocodilians, lizards and snakes of North America. Rept. U. S. Nat. Mus. for 1898, pp. 153-1270.
Crabb, W. D.
1941. Food habits of the prairie spotted skunk in southeastern Iowa. Jour. Mamm., 22(4):349-364.
Crow, H. E.
1913. Some trematodes of Kansas snakes. Univ. Kansas Sci. Bull., 7:123-134.
Cunningham, J. D.
1959. Reproduction and food of some California snakes. Herpetologica, 15(1):17-19.
Curtis, L.
1949. The snakes of Dallas County, Texas. Field & Laboratory, 17:5-13.
Dice, L. R.
1916. Distribution of the land vertebrates of southeastern Washington. Univ. California Publ. Zool., 16(17):293-348.
Ditmars, R. L.
1896. The snakes found within fifty miles of New York City. Abs. Proc. Linn. Soc. New York, 8:9-24.
1907. The reptile book. Doubleday, Page and Co., New York, pp. xx + 472.
1944. Serpents of the northeastern states. New York Zool. Soc., 34 pp., 21 pls.
Duellman, W. E.
1951. Notes on the amphibians and reptiles of Greene County, Ohio. Ohio Jour. Sci., 51(6):335-341.
Duellman, W. E. and Schwartz, A.
1958. Amphibians and reptiles of southern Florida. Bull. Florida State Mus. Biol. Sci., 3(5):181-324.
Ellicott, E. L.
1880. Bundles of snakes. Am. Nat., 14:206-207.
Etheridge, R. E.
1952. The southern range of the racer Coluber constrictor stejnegerianus (Cope), with remarks on the Guatemalan species Coluber ortenburgeri Stuart. Copeia, 1952:189-191.
Ferguson, D. E.
1952. The distribution of amphibians and reptiles of Wallowa County, Oregon. Herpetologica, 8:66-68.
Finneran, L. C.
1948. Reptiles at Branford, Connecticut. Herpetologica, 4:123-126.
Fitch, H. S.
1935. Natural history of the alligator lizards. Trans. Acad. Sci. St. Louis, 29(1):1-38.
1936. Amphibians and reptiles of the Rogue River Basin, Oregon. Amer. Midl. Nat., 17(3:634-652).
1940. A biogeographical study of the ordinoides artenkreis of garter snakes (genus Thamnophis). Univ. California Publ. Zool., 44(1):1-150.
1949. Road counts of snakes in western Louisiana. Herpetologica, 5:87-90.
1951. A simplified type of funnel trap for reptiles. Herpetologica, 7:77-80.
1954. Life history and ecology of the five-lined skink, Eumeces fasciatus. Univ. Kansas Publ. Mus. Nat. Hist., 8(1):1-156.
1955. Habits and adaptations of the Great Plains skink. Ecol. Monogr., 25(3):59-83.
1956. Temperature responses in free-living amphibians and reptiles of northeastern Kansas. Univ. Kansas Publ. Mus. Nat. Hist., 8(7):417-476.
1958. Home ranges, territories and seasonal movements in vertebrates of the Natural History Reservation. Univ. Kansas Publ. Mus. Nat. Hist. 11(3):63-326.
1960. Autecology of the copperhead. Univ. Kansas Publ. Mus. Nat. Hist., 13(4):85-288.
Fitch, H. S. and Packard, R. L.
1955. The coyote on a natural area in northeastern Kansas. Trans. Kansas Acad. Sci., 58:211-221.
Fitch, H. S. and Sandidge, L. L.
1953. Ecology of the opossum on a natural area in northeastern Kansas. Univ. Kansas Publ. Mus. Nat. Hist., 7(2):305-338.
Force, E. R.
1930. The amphibians and reptiles of Tulsa County, Oklahoma, and vicinity. Copeia, 1930(2):25-39.
Fouquette, M. J. and Lindsay, H. L., Jr.
1955. An ecological survey of reptiles in parts of northwestern Texas. The Texas Jour. Sci., 7(4):402-421.
Gloyd, H. K.
1928. The amphibians and reptiles of Franklin County, Kansas. Trans. Kansas Acad. Sci., 31:115-141.
1932. The herpetological fauna of the Pigeon Lake region, Miami County, Kansas. Papers Michigan Acad. Sci. Arts and Letters, 15:389-409.
1945. The problem of too many snakes. Chicago Nat., 7(4):87-97.
Grinnell, J., Dixon, J., and Linsdale, J. M.
1930. Vertebrate natural history of a section of northern California through the Lassen Peak region. Univ. Calif. Publ. Zool., 35: v + 594 pp., 181 figs.
Guidry, E. V.
1953. Herpetological notes from southeastern Texas. Herpetologica, 9:49-56.
Hamilton, W. J., Jr., and Pollack, J. A.
1956. The food of some colubrid snakes from Fort Benning, Georgia. Ecology, 37:519-526.
Harwood, P. D.
1933. The helminths parasitic in the Amphibia and Reptilia of Houston, Texas and vicinity. Proc. U. S. Nat. Mus., 81, 17:1-71.
Hay, O. P.
1893. On the breeding habits, eggs, and young of certain snakes. Proc. U. S. Nat. Mus., 15:385-397.
Hibbard, C. W.
1936. The amphibians and reptiles of Mammoth Cave National Park Proposed. Trans. Kansas Acad. Sci., 39:277-281.
Hudson, G. E.
1942. The amphibians and reptiles of Nebraska. Nebraska Conservation Bull. no. 24, 146 pp.
Hudson, R. G.
1949. A record length milk snake. Herpetologica, 5:47.
Hurter, J. H.
1911. Herpetology of Missouri. Trans. Acad. Sci. St. Louis, 20:59-274.
Inger, F. and Clark, P. J.
1943. Partition of the genus Coluber. Copeia, 1943(3):141-145.
Jackson, C. H. N.
1939. The analysis of an animal population. Jour. An. Ecology, 8:238-246.
King, W.
1939. A survey of the herpetology of Great Smoky Mountain National Park. Amer. Midl. Nat., 21(3):531-582.
Klimstra, W. D.
1959. Food of the racer, Coluber constrictor, in southern Illinois. Copeia, 1959(3):210-214.
Langbartel, D. A.
1947. Snakes collected at Camp Eastman, Hancock County, Illinois. Herpetologica, 4:27-28.
Liner, E. A.
1949. Notes on the young of the blue racer, Coluber constrictor flaviventris. Copeia, 1949(3):230.
1954. The herpetofauna of Lafayette, Terrebonne, and Vermilion parishes, Louisiana. Proc. Louisiana Acad. Sci., 17:65-85.
1955. A herpetological consideration of the Bayou Tortue region of Lafayette Parish, Louisiana. Proc. Louisiana Acad. Sci. 18:39-42.
Linsdale, J. M.
1927. Amphibians and reptiles of Doniphan County, Kansas. Copeia, no. 164:75-81.
Loewen, S. I.
1940. On some reptilian cestodes of the genus Oochoristica (Anoplocephalidae). Trans. Amer. Microscop. Soc., 59(4):511-518.
Logier, E. B. S.
1932. Some account of the amphibians and reptiles of British Columbia. Contr. Royal Ontario Mus., Trans. Royal Canadian Inst., 18(2):311-336.
Loomis, R. B.
1956. The chigger mites of Kansas (Acarina, Trombiculidae). Univ. Kansas Sci. Bull., 37, pt. 2(19):1195-1443.
Lord, J. K.
1866. The naturalist in Vancouver Island and British Columbia. Richard Bentley, London, 375 pp.
Marr, J. C.
1944. Notes on amphibians and reptiles from the central United States. Amer. Midl. Nat., 32:478-490.
Martin, E. P.
1956. A population study of the prairie vole (Microtus ochrogaster) in northeastern Kansas. Univ. Kansas Publ. Mus. Nat. Hist., 8(6):361-416.
Maslin, T. P.
1959. An annotated check list of the amphibians and reptiles of Colorado. Univ. Colorado Studies, Biol. Series, No. 6, 98 pp.
McCauley, R. H.
1945. The reptiles of Maryland and the District of Columbia. Published by the author, Hagerstown, Maryland, 194 pp.
Minton, S., Jr.
1944. Introduction to the study of reptiles in Indiana. Amer. Midl. Nat., 32:438-477.
1949. An ectopic egg in Coluber. Herpetologica, 5:96.
Mossiman, J. E. and Rabb, G. B.
1952. The herpetology of the Tiber River area, Montana. Copeia, 1952(1):23-27.
Mulaik, S. and Mulaik, D.
1942. A neglected species of Coluber. Copeia, 1942:13-15.
Munro, D. F.
1948. Comparative sizes of two female blue racers and their eggs. Herpetologica, 4:199-200.
1949. Eating habits of young Coluber constrictor flaviventris. Herpetologica, 5:72-73.
1950a. Incubation and hatching of eggs of Coluber c. flaviventris. Herpetologica, 6:122-124.
1950b. Juvenile pattern in a mature Coluber c. flaviventris. Herpetologica, 6:124.
Neill, W. T.
1948. Hibernation of amphibians and reptiles in Richmond County, Georgia. Herpetologica, 4:107-114.
Odum, E. P. and Kuenzler, E. J.
1955. Measurement of territory and home range size in birds. Auk, 72:128-137.
Ortenburger, A. I.
1928. The whip snakes and racers, genera Masticophis and Coluber. Mem. Univ. Michigan Mus., vol. 1, pp. xviii + 247.
Owens, V.
1949. An overwintering colony of Coluber c. constrictor (Say) and Elaphe o. obsoleta (Say). Herpetologica, 5:90.
Parker, M. V.
1941. The trematode parasites from a collection of amphibians and reptiles. Jour. Tennessee Acad. Sci., 16(1):27-45.
Peters, J. A.
1942. Reptiles and amphibians of Cumberland County, Illinois. Copeia, 1942(3):182-183.
Pope, C. H.
1935. The reptiles of China. Amer. Mus. Nat. Hist., Nat. Hist. Cent. Asia, v. 10, lii + 604 pp.
1944. Amphibians and reptiles of the Chicago area. Chicago Nat. Hist. Mus. Press, 275 pp., 12 pls.
Pope, T. E. B. and Dickinson, W. E.
1928. The amphibians and reptiles of Wisconsin. Bull. Pub. Mus. City of Milwaukee, 8(1):1-138.
Richmond, N. D. and Goin, C. J.
1938. Notes on a collection of amphibians and reptiles from New Kent County, Virginia. Ann. Carnegie Mus., 27:301-310.
Rossman, D. A.
1960. Herpetological survey of the pine hills area of southern Illinois. Quart. Jour. Florida Acad. Sci. (1959), 22(4):207-225.
Schmidt, K. P. and Necker, W. L.
1935. Amphibians and reptiles of the Chicago region. Bull. Chicago Acad. Sci., 5(4):57-77.
Schroder, R. C.
1950. Hibernation of blue racers and bull snakes in western Illinois. Nat. Hist. Misc., 75:1-2.
Shannon, F. A.
1950. Coluber constrictor mormon—a state record for Arizona. Herpetologica, 6:59.
Smith, H. M.
1956. Handbook of amphibians and reptiles of Kansas. Univ. Kansas Mus. Nat. Hist. Misc. Publ., 9:357 pp.
Smith, P. W.
1947. The reptiles and amphibians of east central Illinois. Bull. Chicago Acad. Sci., 8(2):21-40.
1961. The amphibians and reptiles of Illinois. Bull. Illinois Nat. Hist. Surv., 28(1):1-298.
Stains, H. J.
1956. The raccoon in Kansas. Univ. Kansas Mus. Nat. Hist. Misc. Publ., No. 10, pp. 1-76, pls. 1-4.
Stewart, P. L.
1959. Lung flukes of Thamnophis and Coluber in Kansas. Unpublished thesis University of Kansas Library, pp. 1-62.
Stickel, W. H. and Cope, J. B.
1947. The home ranges and wandering of snakes. Copeia, 1947:127-136.
Storer, D. H.
1839. Reports on the ichthyology and herpetology of Massachusetts. Comm. Zool. and Bot. Surv. Mass., Boston, 201-258.
Surface, H. A.
1906. The serpents of Pennsylvania. Pennsylvania State Dept. Agr. Monthly Bull., Div. Zool., 4(4 and 5):113-208.
Tinkle, D. W.
1959. Observations of reptiles and amphibians in a Louisiana swamp. Amer. Midl. Nat, 62 (11):189-205.
Trowbridge, A. H.
1937. Ecological observations on amphibians and reptiles collected in southeastern Oklahoma during the summer of 1934. Amer. Midl. Nat. 18(2):285-303.
Uhler, F. M., Cottam, C. and Clarke, T. E.
1939. Food of snakes of the George Washington National Forest, Virginia. Trans. 4th Amer. Wildlife Conf., Amer. Wildlife Inst., Washington, D. C., 1939:605-632.
Van Denburgh, J.
1923. The reptiles of western North America. Occas. Papers California Acad. Sci., 10:1-1028.
Van de Velde, R. L., Martan, J. and Risley, P. L.
1962. Eggs and hatchlings of the snake Coluber constrictor mormon from Oregon. Copeia (1):212-213.
Verrill, A. E.
1869. The biter bitten. Amer. Nat., 3:158-159.
Welter, W. A. and Carr, K.
1939. Amphibians and reptiles of northeastern Kentucky. Copeia (3):128-130.
Werler, J. E. and McCallion, J.
1951. Notes on a collection of reptiles and amphibians from Princess Anne County, Virginia. Amer. Midl. Nat., 45(1):245-252.
Woodbury, A. M.
1931. A descriptive catalog of the reptiles of Utah. Bull. Univ. Utah, 21 (no. 5):x + 129 pp.
Woods, G. T.
1944. Black snake attacks a boy. Copeia, 1944(4):257.
Wright, A. H. and Bishop, S. C.
1915. A biological reconnaissance of the Okefinokee Swamp in Georgia. II. Snakes. Proc. Acad. Nat. Sci. Philadelphia, 67:139-192.
Wright, A. H. and Wright, A. A.
1957. Handbook of snakes of the United States and Canada. Comstock Publ. Associates, Cornell Univ. Press, vol. I, xviii + 564 pp.
Transmitted June 3, 1963.
29-7864
Institutional libraries interested in publications exchange may obtain this series by addressing the Exchange Librarian, University of Kansas Library, Lawrence, Kansas. Copies for individuals, persons working in a particular field of study, may be obtained by addressing instead the Museum of Natural History, University of Kansas, Lawrence, Kansas. There is no provision for sale of this series by the University Library, which meets institutional requests, or by the Museum of Natural History, which meets the requests of individuals. Nevertheless, when individuals request copies from the Museum, 25 cents should be included, for each separate number that is 100 pages or more in length, for the purpose of defraying the costs of wrapping and mailing.
* An asterisk designates those numbers of which the Museum's supply (not the Library's supply) is exhausted. Numbers published to date, in this series, are as follows:
| Vol. 1. | Nos. 1-26 and index. Pp. 1-638, 1946-1950. | ||
| *Vol. 2. | (Complete) Mammals of Washington. By Walter W. Dalquest. Pp. 1-444, 140 figures in text. April 9, 1948. | ||
| Vol. 3. | *1. | The avifauna of Micronesia, its origin, evolution, and distribution. By Rollin H. Baker. Pp. 1-359, 16 figures in text. June 12, 1951. | |
| *2. | A quantitative study of the nocturnal migration of birds. By George H. Lowery, Jr. Pp. 361-472, 47 figures in text. June 29, 1951. | ||
| 3. | Phylogeny of the waxwings and allied birds. By M. Dale Arvey. Pp. 473-530, 49 figures in text, 13 tables. October 10, 1951. | ||
| *4. | Birds from the state of Veracruz, Mexico. By George H. Lowery, Jr., and Walter W. Dalquest. Pp. 531-649, 7 figures in text, 2 tables. October 10, 1951. | ||
| Index. Pp. 651-681. | |||
| *Vol. 4. | (Complete) American weasels. By E. Raymond Hall. Pp. 1-466, 41 plates, 31 figures in text. December 27, 1951. | ||
| Vol. 5. | Nos. 1-37 and index. Pp. 1-676, 1951-1953. | ||
| *Vol. 6. | (Complete) Mammals of Utah, taxonomy and distribution. By Stephen D. Durrant. Pp. 1-549, 91 figures in text, 30 tables. August 10, 1952. | ||
| Vol. 7. | Nos. 1-15 and index. Pp. 1-651, 1952-1955. | ||
| Vol. 8. | Nos. 1-10 and index. Pp. 1-675, 1954-1956. | ||
| Vol. 9. | *1. | Speciation of the wandering shrew. By James S. Findley. Pp. 1-68, 18 figures in text. December 10, 1955. | |
| 2. | Additional records and extension of ranges of mammals from Utah. By Stephen D. Durrant, M. Raymond Lee, and Richard M. Hansen. Pp. 69-80. December 10, 1955. | ||
| 3. | A new long-eared myotis (Myotis evotis) from northeastern Mexico. By Rollin H. Baker and Howard J. Stains. Pp. 81-84. December 10, 1955. | ||
| 4. | Subspeciation in the meadow mouse, Microtus pennsylvanicus, in Wyoming. By Sydney Anderson. Pp. 85-104, 2 figures in text. May 10, 1956. | ||
| 5. | The condylarth genus Ellipsodon. By Robert W. Wilson. Pp. 105-116, 6 figures in text. May 19, 1956. | ||
| 6. | Additional remains of the multituberculate genus Eucosmodon. By Robert W. Wilson. Pp. 117-123, 10 figures in text. May 19, 1956. | ||
| 7. | Mammals of Coahulia, Mexico. By Rollin H. Baker. Pp. 125-335, 75 figures in text. June 15, 1956. | ||
| 8. | Comments on the taxonomic status of Apodemus peninsulae, with description of a new subspecies from North China. By J. Knox Jones, Jr. Pp. 337-346, 1 figure in text, 1 table. August 15, 1956. | ||
| 9. | Extensions of known ranges of Mexican bats. By Sydney Anderson. Pp. 347-351. August 15, 1956. | ||
| 10. | A new bat (Genus Leptonycteris) from Coahulia. By Howard J. Stains. Pp. 353-356. January 21, 1957. | ||
| 11. | A new species of pocket gopher (Genus Pappogeomys) from Jalisco, Mexico. By Robert J. Russell. Pp. 357-361. January 21, 1957. | ||
| 12. | Geographic variation in the pocket gopher, Thomomys bottae, in Colorado. By Phillip M. Youngman. Pp. 363-387, 7 figures in text. February 21, 1958. | ||
| 13. | New bog lemming (genus Synaptomys) from Nebraska. By J. Knox Jones, Jr. Pp. 385-388. May 12, 1958. | ||
| 14. | Pleistocene bats from San Josecito Cave, Nuevo León, México. By J. Knox Jones, Jr. Pp. 389-396. December 19, 1958. | ||
| 15. | New subspecies of the rodent Baiomys from Central America. By Robert L. Packard. Pp. 397-404. December 19, 1958. | ||
| 16. | Mammals of the Grand Mesa, Colorado. By Sydney Anderson. Pp. 405-414, 1 figure in text, May 20, 1959. | ||
| 17. | Distribution, variation, and relationships of the montane vole, Microtus montanus. By Sydney Anderson. Pp. 415-511, 12 figures in text, 2 tables. August 1, 1959. | ||
| [Pub_2] | 18. | Conspecificity of two pocket mice, Perognathus goldmani and P. artus. By E. Raymond Hall and Marilyn Bailey Ogilvie. Pp. 513-518, 1 map. January 14 1960. | |
| 19. | Records of harvest mice, Reithrodontomys, from Central America, with description of a new subspecies from Nicaragua. By Sydney Anderson and J. Knox Jones, Jr. Pp. 519-529. January 14, 1960. | ||
| 20. | Small carnivores from San Josecito Cave (Pleistocene), Nuevo León, México. By E. Raymond Hall. Pp. 531-538, 1 figure in text. January 14, 1960. | ||
| 21. | Pleistocene pocket gophers from San Josecito Cave, Nuevo León, México. By Robert J. Russell. Pp. 539-548, 1 figure in text. January 14, 1960. | ||
| 22. | Review of the insectivores of Korea. By J. Knox Jones, Jr., and David H. Johnson. Pp. 549-578. February 23, 1960. | ||
| 23. | Speciation and evolution of the pygmy mice, genus Baimoys. By Robert L. Packard. Pp. 579-670, 4 plates, 12 figures in text. June 16, 1960. | ||
| Index. Pp. 671-690 | |||
| Vol. 10. | 1. | Studies of birds killed in nocturnal migration. By Harrison B. Tordoff and Robert M. Mengel. Pp. 1-44, 6 figures in text, 2 tables. September 12, 1956. | |
| 2. | Comparative breeding behavior of Ammospiza caudacuta and A. maritima. By Glen E. Woolfenden. Pp. 45-75, 6 plates, 1 figure. December 20, 1956. | ||
| 3. | The forest habitat of the University of Kansas Natural History Reservation. By Henry S. Fitch and Ronald R. McGregor. Pp. 77-127, 2 plates, 7 figures in text, 4 tables. December 31, 1956. | ||
| 4. | Aspects of reproduction and development in the prairie vole (Microtus ochrogaster). By Henry S. Fitch. Pp. 129-161, 8 figures in text, 4 tables. December 19, 1957. | ||
| 5. | Birds found on the Arctic slope of northern Alaska. By James W. Bee. Pp. 163-211, plates 9-10, 1 figure in text. March 12, 1958. | ||
| *6. | The wood rats of Colorado: distribution and ecology. By Robert B. Finley, Jr. Pp. 213-552, 34 plates, 8 figures in text, 35 tables. November 7, 1958. | ||
| 7. | Home ranges and movements of the eastern cottontail in Kansas. By Donald W. Janes. Pp. 553-572, 4 plates, 3 figures in text. May 4, 1959. | ||
| 8. | Natural history of the salamander, Aneides hardyi. By Richard F. Johnston and Gerhard A. Schad. Pp. 573-585. October 8, 1959. | ||
| 9. | A new subspecies of lizard, Cnemidophorus sacki, from Michoacán, México. By William E. Duellman. Pp. 587-598, 2 figures in text. May 2, 1960. | ||
| 10. | A taxonomic study of the middle American snake, Pituophis deppei. By William E. Duellman. Pp. 599-610, 1 plate, 1 figure in text. May 2, 1960. | ||
| Index. Pp. 611-626. | |||
| Vol. 11. | Nos. 1-10 and index. Pp. 1-703, 1958-1960. | ||
| Vol. 12. | 1. | Functional morphology of three bats: Sumops, Myotis, Macrotus. By Terry A. Vaughan. Pp. 1-153, 4 plates, 24 figures in text. July 8, 1959. | |
| *2. | The ancestry of modern Amphibia: a review of the evidence. By Theodore H. Eaton, Jr. Pp. 155-180, 10 figures in text. July 10, 1959. | ||
| 3. | The baculum in microtine rodents. By Sydney Anderson. Pp. 181-216, 49 figures in text. February 19, 1960. | ||
| *4. | A new order of fishlike Amphibia from the Pennsylvanian of Kansas. By Theodore H. Eaton, Jr., and Peggy Lou Stewart. Pp. 217-240, 12 figures in text. May 2, 1960. | ||
| 5. | Natural history of the bell vireo. By Jon C. Barlow. Pp. 241-296, 6 figures in text. March 7, 1962. | ||
| 6. | Two new pelycosaurs from the lower Permian of Oklahoma. By Richard C. Fox. Pp. 297-307, 6 figures in text. May 21, 1962. | ||
| 7. | Vertebrates from the barrier island of Tamaulipas, México. By Robert K. Selander, Richard F. Johnston, B. J. Wilks, and Gerald G. Raun. Pp. 309-345, pls. 5-8. June 18, 1962. | ||
| 8. | Teeth of Edestid sharks. By Theodore H. Eaton, Jr. Pp. 347-362, 10 figures in text. October 1, 1962. | ||
| 9. | Variation in the muscles and nerves of the leg in two genera of grouse (Tympanuchus and Pedioecetes). By E. Bruce Holmes. Pp. 363-474, 20 figs. October 25, 1963. | ||
| 10. | A new genus of Pennsylvanian Fish (Crossopterygii, Coelacanthiformes) from Kansas. By Joan Echols. Pp. 475-501, 7 figures. October 25, 1963. | ||
| 11. | Observations on the Mississippi Kite in southwestern Kansas. By Henry S. Fitch. Pp. 503-519. October 25, 1963. | ||
| 12. | Jaw musculature of the Mourning and White-winged doves. By Robert L. Merz. Pp. 521-551, 22 figures. October 25, 1963. | ||
| More numbers will appear in volume 12. | |||
| Vol. 13. | 1. | Five natural hybrid combinations in minnows (Cyprinidae). By Frank B. Cross and W. L. Minckley. Pp. 1-18. June 1, 1960. | |
| 2. | A distributional study of the amphibians of the Isthmus of Tehuantepec, México. By William E. Duellman. Pp. 19-72, pls. 1-8, 3 figures in text. August 16, 1960. | ||
| 3. | A new subspecies of the slider tutdre (Pseudemys scripta) from Coahulia, México. By John M. Legler. Pp. 73-84, pls. 9-12, 3 figures in text. August 16, 1960. | ||
| 4. | Autecology of the copperhead. By Henry S. Fitch. Pp. 85-288, pls. 13-20, 26 figures in text. November 30, 1960. | ||
| 5. | Occurrence of the garter snake, Thamnophis sirtalis, in the Great Plains and Rocky Mountains. By Henry S. Fitch and T. Paul Maslin. Pp. 289-308, 4 figures in text. February 10, 1961. | ||
| [Pub_3] | 6. | Fishes of the Wakarusa river in Kansas. By James E. Deacon and Artie L. Metcalf. Pp. 309-322, 1 figure in text. February 10, 1961. | |
| 7. | Geographic variation in the North American cyprinid fish, Hybopsis gracilis. By Leonard J. Olund and Frank B. Cross. Pp. 323-348, pls. 21-24, 2 figures in text. February 10, 1961. | ||
| 8. | Descriptions of two species of frogs, genus Ptychohyla; studies of American hylid frogs, V. By William E. Duellman. Pp. 349-357, pl. 25, 2 figures in text. April 27, 1961. | ||
| 9. | Fish populations, following a drought, in the Neosho and Marais des Cygnes rivers of Kansas. By James Everett Deacon. Pp. 359-427, pls. 26-30, 3 figs. August 11, 1961. | ||
| 10. | Recent soft-shelled tutdres of North America (family Trionychidae). By Robert G. Webb. Pp. 429-611, pls. 31-54, 24 figures in text, February 16, 1962. | ||
| Index. Pp. 613-624. | |||
| Vol. 14. | 1. | Neotropical bats from western México. By Sydney Anderson. Pp. 1-8. October 24, 1960. | |
| 2. | Geographic variation in the harvest mouse. Reithrodontomys megalotis, on the central Great Plains and in adjacent regions. By J. Knox Jones, Jr., and B. Mursaloglu. Pp. 9-27, 1 figure in text. July 24, 1961. | ||
| 3. | Mammals of Mesa Verde National Park, Colorado. By Sydney Anderson. Pp. 29-67, pls. 1 and 2, 3 figures in text. July 24, 1961. | ||
| 4. | A new subspecies of the black myotis (bat) from eastern Mexico. By E. Raymond Hall and Ticul Alvarez. Pp. 69-72, 1 figure in text. December 29, 1961. | ||
| 5. | North American yellow bats, "Dasypterus," and a list of the named kinds of the genus Lasiurus Gray. By E. Raymond Hall and J. Knox Jones, Jr. Pp. 73-98, 4 figures in text. December 29, 1961. | ||
| 6. | Natural history of the brush mouse (Peromyscus boylii) in Kansas with description of a new subspecies. By Charles A. Long. Pp. 99-111, 1 figure in text. December 29, 1961. | ||
| 7. | Taxonomic status of some mice of the Peromyscus boylii group in eastern Mexico, with description of a new subspecies. By Ticul Alvarez. Pp. 113-120, 1 figure in text. December 29, 1961. | ||
| 8. | A new subspecies of ground squirrel (Spermophilus spilosoma) from Tamaulipas, Mexico. By Ticul Alvarez. Pp. 121-124. March 7, 1962. | ||
| 9. | Taxonomic status of the free-tailed bat, Tadarida yucatanica Miller. By J. Knox Jones, Jr., and Ticul Alvarez. Pp. 125-133, 1 figure in text. March 7, 1962. | ||
| 10. | A new doglike carnivore, genus Cynaretus, from the Clarendonian Pliocene, of Texas. By E. Raymond Hall and Walter W. Dalquest. Pp. 135-138, 2 figures in text. April 30, 1962. | ||
| 11. | A new subspecies of wood rat (Neotoma) from northeastern Mexico. By Ticul Alvarez. Pp. 139-143. April 30, 1962. | ||
| 12. | Noteworthy mammals from Sinaloa, Mexico. By J. Knox Jones, Jr., Ticul Alvarez, and M. Raymond Lee. Pp. 145-159, 1 figure in text. May 18, 1962. | ||
| 13. | A new bat (Myotis) from Mexico. By E. Raymond Hall. Pp. 161-164, 1 figure in text. May 21, 1962. | ||
| 14. | The mammals of Veracruz. By E. Raymond Hall and Walter W. Dalquest. Pp. 165-362, 2 figures. May 20, 1963. | ||
| 15. | The recent mammals of Tamaulipas, México. By Ticul Alvarez. Pp. 363-473, 5 figures in text. May 20, 1963. | ||
| More numbers will appear in volume 14. | |||
| Vol. 15. | 1. | The amphibians and reptiles of Michoacán, México. By William E. Duellman. Pp. 1-148, pls. 1-6, 11 figures in text. December 20, 1961. | |
| 2. | Some reptiles and amphibians from Korea. By Robert G. Webb, J. Knox Jones, Jr., and George W. Byers. Pp. 149-173. January 31, 1962. | ||
| 3. | A new species of frog (Genus Tomodactylus) from western México. By Robert G. Webb. Pp. 175-181, 1 figure in text. March 7, 1962. | ||
| 4. | Type specimens of amphibians and reptiles in the Museum of Natural History,
the University of Kansas. By William E. Duellman and Barbara Berg. Pp. 183-204. October 26, 1962. | ||
| 5. | Amphibians and Reptiles of the Rainforests of Southern El Petén, Guatemala. By William E. Duellman. Pp. 205-249, pls. 7-10, 6 figures in text. October 4, 1963. | ||
| 6. | A revision of snakes of the genus Conophis (Family Colubridae, from Middle America). By John Wellman. Pp. 251-295, 9 figures in text. October 4, 1963. | ||
| 7. | A review of the Middle American tree frogs of the genus Ptychohyla. By William E. Duellman. Pp. 297-349, pls. 11-18, 7 figures in text. October 18, 1963. | ||
| 8. | Natural history of the racer, Coluber constrictor. By Henry S. Fitch. Pp. 351-468, pls. 19-22, 20 figures. December 30, 1963. | ||
| More numbers will appear in volume 15. | |||
Transcriber's Note
Where tables or images split them, the text was moved to rejoin paragraphs. The List of Publications before the article was moved down to rejoin the portion after the end of the article.
Table 6 re-arranged due to use of vertically printed text.
Page 451 may have a typo "...wide range of error arising from insufficiently small samples..." should probably have been "...large samples..." (emphasis added here). This was left as is.