The Project Gutenberg EBook of Seaside Studies in Natural History, by
Elizabeth Cabot Cary Agassiz and Alexander Agassiz
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
Title: Seaside Studies in Natural History
Marine Animals of Massachusetts Bay. Radiates.
Author: Elizabeth Cabot Cary Agassiz
Alexander Agassiz
Release Date: March 5, 2011 [EBook #35490]
Language: English
Character set encoding: ISO-8859-1
*** START OF THIS PROJECT GUTENBERG EBOOK SEASIDE STUDIES IN NATURAL HISTORY ***
Produced by Bryan Ness, Tom Cosmas and the Online
Distributed Proofreading Team at https://www.pgdp.net (This
book was produced from scanned images of public domain
material from the Google Print project.)
[Pg i]
SEASIDE STUDIES
IN
NATURAL HISTORY.
BY
ELIZABETH C. AGASSIZ
AND
ALEXANDER AGASSIZ.
MARINE ANIMALS OF MASSACHUSETTS BAY.
RADIATES.
BOSTON:
JAMES R. OSGOOD AND COMPANY,
Late Ticknor & Fields, and Fields, Osgood, & Co.
1871.
[Pg ii]
Entered according to Act of Congress, in the year 1865, by
A L E X A N D E R A G A S S I Z,
in the Clerk's Office of the District Court
for the District of Massachusetts.
University Press:
Welch, Bigelow, and Company,
Cambridge.
[Pg iii]
THIS LITTLE BOOK
IS AFFECTIONATELY DEDICATED BY THE AUTHORS TO
PROFESSOR L. AGASSIZ,
WHOSE PRINCIPLES OF CLASSIFICATION HAVE BEEN THE MAIN
GUIDE IN ITS PREPARATION.
[Pg iv]
[Pg v]
PREFACE.
This volume is published with the hope of supplying a want often expressed
for some seaside book of a popular character, describing the marine animals
common to our shores. There are many English books of this kind; but they
relate chiefly to the animals of Great Britain, and can only have a general
bearing on those of our own coast, which are for the most part specifically
different from their European relatives. While keeping this object in view,
an attempt has also been made to present the facts in such a connection,
with reference to principles of science and to classification, as will give
it in some sort the character of a manual of Natural History, in the hope
of making it useful not only to the general reader, but also to teachers
and to persons desirous of obtaining a more intimate knowledge of the
subjects discussed in it. With this purpose, although nearly all the
illustrations are taken from among the most common inhabitants of our bay,
a few have been added from other localities in order to fill out this
little sketch of Radiates, and render it, as far as is possible within such
limits, a complete picture of the type.
[Pg vi]
A few words of explanation are necessary with reference to the joint
authorship of the book. The drawings and the investigations, where they are
not referred to other observers, have been made by Mr. A. Agassiz, the
illustrations having been taken, with very few exceptions, from nature, in
order to represent the animals, as far as possible, in their natural
attitudes; and the text has been written by Mrs. L. Agassiz, with the
assistance of Mr. A. Agassiz's notes and explanations.
Cambridge, May, 1865.
NOTE.
This second edition is a mere reprint of the first. A few mistakes
accidentally overlooked have been corrected; an explanation of the
abbreviations of the names of writers used after the scientific names has
been added, as well as a list of the wood-cuts. The changes which have
taken place in the opinions of scientific men with regard to the
distribution of animal life in the ocean have been duly noticed in their
appropriate place, but no attempt has been made to incorporate more
important additions which the progress of our knowledge of Radiates may
require hereafter.
Cambridge, January, 1871.
[Pg vii]
CONTENTS.
[Pg viii]
LIST OF THE WOOD-CUTS.
Unless otherwise specified, the illustrations are drawn from nature by
Alex. Agassiz.
| Fig. | | Page |
| 1. | Transverse section of an Actinia (Agassiz) | 5 |
| 2, 3, 4. | Actinia in different degrees of expansion (Agassiz) | 8 |
| 5. | Metridium marginatum fully expanded | 8 |
| 6. | Vertical section of an Actinia | 10 |
| 7. | View from above of an expanded Actinia | 11 |
| 8, 9. | Young Actiniæ | 11 |
| 10. | Rhodactinia Davisii | 13 |
| 11. | Arachnactis brachiolata | 14 |
| 12. | Young Arachnactis | 14 |
| 13. | Young Arachnactis showing the mouth | 14 |
| 14. | Bicidium parasiticum | 15 |
| 15. | Halcampa albida | 16 |
| 16. | Colony of Astrangia Danæ | 17 |
| 17. | Magnified individuals of Astrangia | 17 |
| 18. | Single individual of Astrangia | 18 |
| 19. | Lasso-cell of Astrangia | 18 |
| 20. | Limestone pit of Astrangia | 19 |
| 21. | Single individual of Halcyonium carneum | 19 |
| 22. | Halcyonium community | 20 |
| 23. | Expanded individual of Halcyonium | 20 |
| 24. | Branch of Millepora alcicornis (Agassiz) | 22 |
| 25. | Expanded animals of Millepora (Agassiz) | 22 |
| 26. | Transverse section of branch of Millepora (Agassiz) | 23 |
| 27. | Pleurobrachia rhododactyla (Agassiz) | 27 |
| 28. | The same as Fig. 27 seen in plane of tentacles (Agassiz) | 28 |
| 29. | Pleurobrachia in motion | 29 |
| 30. | Pleurobrachia seen from the extremity opposite the mouth | 30 |
| 31. | Bolina alata seen from the broad side (Agassiz) | 31 |
| 32. | Bolina seen from the narrow side (Agassiz) | 31 |
| 33. | Idyia roseola seen from the broad side (Agassiz) | 32 |
| 34. | Young Pleurobrachia still in the egg | 35 |
| 35. | Young Pleurobrachia swimming in the egg | 35 |
| 36. | Young Pleurobrachia resembling already adult | 35 |
| 37. | Young Idyia | 35 |
| 38. | Young Idyia seen from the anal pole | 36 |
| 39. | Idyia somewhat older than Fig. 37 | 36[Pg ix] |
| 40. | Idyia still older | 36 |
| 41. | Young Bolina in stage resembling Pleurobrachia | 37 |
| 42. | Young Bolina seen from the broad side | 37 |
| 43. | Young Bolina seen from the narrow side | 37 |
| 44. | Cyanea Arctica | 40 |
| 45. | Scyphistoma of Aurelia (Agassiz) | 41 |
| 46. | Scyphistoma older than Fig. 45 (Agassiz) | 41 |
| 47. | Strobila of Aurelia (Agassiz) | 41 |
| 48. | Ephyra of Aurelia (Agassiz) | 42 |
| 49. | Aurelia flavidula seen in profile (Agassiz) | 42 |
| 50. | Aurelia seen from above (Agassiz) | 43 |
| 51. | Campanella pachyderma | 44 |
| 52. | The same from below | 44 |
| 53. | Trachynema digitale | 45 |
| 54. | Haliclystus auricula | 46 |
| 55. | Lucernaria seen from the mouth side | 47 |
| 56. | Young Lucernaria | 48 |
| 57. | Hydrarium of Eucope diaphana | 50 |
| 58. | Magnified portion of Fig. 57 | 50 |
| 59. | Part of marginal tentacles of Eucope | 51 |
| 60. | Young Eucope | 51 |
| 61. | Adult Eucope, profile | 51 |
| 62. | Quarter-disk of Fig. 60 | 51 |
| 63. | Quarter-disk of Eucope older than Fig. 62 | 52 |
| 64. | Quarter-disk of adult Eucope | 52 |
| 65. | Oceania languida just escaped from the reproductive calycle | 53 |
| 66. | Same as Fig. 65 from below | 53 |
| 67. | Young Oceania older than Fig. 65 | 54 |
| | Diagram of succession of tentacles | 55 |
| 68. | Adult Oceania | 55 |
| 69. | Attitude assumed by Oceania | 56 |
| 70. | Clytia bicophora escaped from reproductive calycle | 57 |
| 71. | Somewhat older than Fig. 70 | 57 |
| 72. | Magnified portion of Hydrarium of Clytia | 57 |
| 73. | Adult Clytia | 57 |
| 74. | Zygodactyla groenlandica | 58 |
| 75. | The same seen in profile | 59 |
| 76. | Tima formosa | 61 |
| 77. | One of the lips of the mouth | 61 |
| 78. | Head of Hydrarium of Tima | 62 |
| 79. | Melicertum campanula from above (Agassiz) | 63 |
| 80. | The same seen in profile | 64 |
| 81. | Planula of Melicertum | 65 |
| 82. | Cluster of planulæ | 65 |
| 83. | Young Hydrarium | 65[Pg x] |
| 84. | Dynamena pumila | 66 |
| 85. | Magnified portion of Fig. 84 | 66 |
| 86. | Dyphasia rosacea | 67 |
| 87. | Medusa of Lafoea | 67 |
| 88. | Colony of Coryne mirabilis (Agassiz) | 68 |
| 89. | Magnified head of Fig. 88 (Agassiz) | 68 |
| 90. | Free Medusa of Coryne (Agassiz) | 68 |
| 91. | Turris vesicaria | 69 |
| 92. | Bougainvillia superciliaris | 70 |
| 93. | Hydrarium of Bougainvillia | 70 |
| 94, 95, 96. | Medusæ buds of Fig. 93 | 71 |
| 97. | Young Medusa just freed from the Hydroid | 71 |
| 98. | Tubularia couthouyi (Agassiz) | 72 |
| 99. | Cluster of Medusæ of Fig. 98 (Agassiz) | 72 |
| 100. | Female colony of Hydractinia polyclina (Agassiz) | 73 |
| 101. | Male colony of the same (Agassiz) | 73 |
| 102. | Unsymmetrical Medusa of Hybocodon prolifer (Agassiz) | 74 |
| 103. | Medusa bud of Hybocodon (Agassiz) | 74 |
| 104. | Hybocodon Hydrarium (Agassiz) | 74 |
| 105. | Dysmorphosa fulgurans | 75 |
| 106. | Proboscis of Fig. 105 with young Medusæ | 75 |
| 107. | Young Nanomia cara | 76 |
| 108. | Nanomia with rudimentary Medusæ | 76 |
| 109. | Nanomia somewhat older than Fig. 108 | 77 |
| 110. | Heart-shaped swimming bell of Nanomia | 77 |
| 111. | Cluster of Medusæ with tentacles having pendent knobs | 78 |
| 112. | Magnified pendent knob | 79 |
| 113. | Medusa with corkscrew-shaped tentacles | 79 |
| 114. | Medusa with simple tentacle | 80 |
| 115. | Adult Nanomia | 81 |
| 116. | Oil float of Nanomia | 82 |
| 117. | Physalia arethusa (Agassiz) | 83 |
| 118. | Bunch of Hydræ (Agassiz) | 84 |
| 119. | Cluster of Medusæ (Agassiz) | 84 |
| 120. | Velella mutica (Agassiz) | 84 |
| 121. | Free Medusa of Velella (Agassiz) | 84 |
| 122. | Ptychogena lactea | 86 |
| 123. | Ovary of Ptychogena | 87 |
| 124. | Synapta tenuis | 95 |
| 125. | Anchor of Synapta | 96 |
| 126. | Caudina arenata | 97 |
| 127. | Cuvieria squamata | 98 |
| 128. | Young Cuvieria | 99 |
| 129. | Cuvieria somewhat older than Fig. 128 | 99 |
| 130. | Pentacta frondosa | 100 |
| 131. | Toxopneustes drobachiensis | 102[Pg xi] |
| 132. | Portion of shell of Fig. 131 without spines (Agassiz) | 103 |
| 133. | Sea-urchin shell without spines (Agassiz) | 103 |
| 134. | Sea-urchin from the mouth side (Agassiz) | 104 |
| 135. | Magnified spine | 104 |
| 136. | Transverse section of spine | 105 |
| 137. | Pedicellaria of Sea-urchin | 105 |
| 138. | Teeth of Sea-urchin | 106 |
| 139. | Echinarachnius parma | 107 |
| 140. | Transverse section of Echinarachnius (Agassiz) | 108 |
| 141. | Ray of Star-fish, seen from mouth side (Agassiz) | 109 |
| 142. | Astracanthion berylinus | 110 |
| 143. | Single spine of Star-fish | 111 |
| 144. | Limestone network of back of Star-fish | 111 |
| 145. | Madreporic body of Star-fish | 111 |
| 146. | Cribrella oculata | 112 |
| 147. | Ctenodiscus crispatus | 114 |
| 148. | Ophiopholis bellis | 115 |
| 149. | Arm of Fig. 148, from the mouth side (Agassiz) | 116 |
| 150. | Tentacle of Ophiopholis | 116 |
| 151. | Astrophyton agassizii | 118 |
| 152. | Pentacrinus | 121 |
| 153. | Alecto meridionalis | 122 |
| 154. | Young Comatulæ | 122 |
| Figs 155, 156, 157. | Egg of Star-fish in different stages of development | 124 |
| 158. | Larva just hatched from egg | 125 |
| 159-164. | Successive stages of development of Larva | 125 |
| 165. | Larva in which arms are developing | 126 |
| 166. | Adult Star-fish Larva (Brachiolaria) | 127 |
| 167. | Fig. 166 seen in profile | 128 |
| 168-170. | Young Star-fish (Astracanthion) in different stages of development | 129 |
| 171. | Lower side of ray of young Star-fish | 130 |
| 172. | Very young Star-fish seen in profile | 130, 130 |
| 173-175. | Larvæ of Sea-urchin (Toxopneustes) in different stages of development | 130, 131 |
| 176. | Adult Larva of Sea-urchin | 132 |
| 177. | Fig. 176 seen endways | 133 |
| 178. | Sea-urchin resorbing the arms of the larva | 133 |
| 179-181. | Successive stages of young Sea-urchin | 133, 134 |
| 182. | Ophiuran which has nearly resorbed the larva | 135 |
| 183. | Larva of Ophiuran (Pluteus) | 136 |
| 184. | Young Ophiuran | 137 |
| 185. | Cluster of eggs of Star-fishes over mouth of parent | 137 |
| | Diagram of a rocky beach | 149 |
[Pg xii]
ABBREVIATIONS OF THE NAMES OF AUTHORS.
| Ag. | L. Agassiz. | | Jaeg. | Jaeger. |
| A. Ag. | A. Agassiz. | | Lam. | Lamarck. |
| Ayres | W. O. Ayres. | | Lamx. | Lamouroux. |
| Blainv. | Blainville. | | Lin. | Linnæus. |
| Bosc | Bosc. | | Lym. | Lyman. |
| Br. | Brandt. | | M. & T. | Müller and Troschel. |
| Clark | H. J. Clark. | | Mill. | Miller. |
| Cuv. | Cuvier. | | Pér. et Les. | Péron and Lesueur. |
| D. & K. | Düben and Koren. | | Sars | M. Sars. |
| Edw. | Milne-Edwards. | | Stimp. | Stimpson. |
| Forbes | Edw. Forbes. | | Til. | Tilesius. |
| Gray | J. E. Gray. | | | |
[Pg 1]
MARINE ANIMALS OF MASSACHUSETTS BAY.
ON RADIATES IN GENERAL.
It is perhaps not strange that the Radiates, a type of animals whose home
is in the sea, many of whom are so diminutive in size, and so light and
evanescent in substance, that they are hardly to be distinguished from the
element in which they live, should have been among the last to attract the
attention of naturalists. Neither is it surprising to those who know
something of the history of these animals, that when the investigation of
their structure was once begun, when some insight was gained into their
complex life, their association in fixed or floating communities, their
wonderful processes of development uniting the most dissimilar individuals
in one and the same cycle of growth, their study should have become one of
the most fascinating pursuits of modern science, and have engaged the
attention of some of the most original investigators during the last half
century. It is true that from the earliest days of Natural History, the
more conspicuous and easily accessible of these animals attracted notice
and found their way into the scientific works of the time. Even Aristotle
describes some of them under the names of Acalephæ and Knidæ, and later
observers have added something, here and there, to our knowledge on the
subject; but it is only within the last fifty years that their complicated
history has been unravelled, and the facts concerning them presented in
their true connection.
Among the earlier writers on this subject we are most indebted to Rondelet,
in the sixteenth century, who includes some account of the Radiates, in his
work on the marine animals of the Mediterranean. His position as Professor
in the University at Montpelier
[Pg 2]
gave him an admirable opportunity, of which he availed himself to the utmost,
for carrying out his investigations in this direction. Seba and Klein, two
naturalists in the North of Europe, also published at about this time
numerous illustrations of marine animals, including Radiates. But in all
these works we find only drawings and descriptions of the animals, without
any attempt to classify them according to common structural features. In 1776,
O. F. Müller, in a work on the marine and terrestrial faunæ of
Denmark, gave some admirable figures of Radiates, several of which are
identical with those found on our own coast. Cavolini also in his
investigations on the lower marine animals of the Mediterranean, and Ellis
in his work upon those of the British coast, did much during the latter
half of the past century to enlarge our knowledge of them.
It was Cuvier, however, who first gave coherence and precision to all
previous investigations upon this subject, by showing that these animals
are united on a common plan of structure expressively designated by him
under the name Radiata. Although, from a mistaken appreciation of their
affinities, he associated some animals with them which do not belong to the
type, and have since, upon a more intimate knowledge of their structure,
been removed to their true positions; yet the principle introduced by him
into their classification, as well as into that of the other types of the
animal kingdom, has been all important to science.
It was in the early part of this century that the French began to associate
scientific objects with their government expeditions. Scarcely any
important voyage was undertaken to foreign countries by the French navy
which did not include its corps of naturalists, under the patronage of
government. Among the most beautiful figures we have of Radiates, are those
made by Savigny, one of the French naturalists who accompanied Napoleon to
Egypt; and from this time the lower marine animals began to be extensively
collected and studied in their living condition. Henceforth the number of
investigators in the field became more numerous, and it may not be amiss to
give here a slight account of the more prominent among them.
Darwin's fascinating book, published after his voyage to the
[Pg 3]
Pacific, and giving an account of the Coral islands, the many memoirs of
Milne Edwards and Haime, and the great works of Quoy and Gaimard, and of
Dana, are the chief authorities upon Polyps. In the study of the European
Acalephs we have a long list of names high in the annals of science.
Eschscholtz, Péron and Lesueur, Quoy and Gaimard, Lesson, Mertens,
and Huxley, have all added largely to our information respecting these
animals, their various voyages having enabled them to extend their
investigations over a wide field. No less valuable have been the memoirs
of Kölliker, Leuckart, Gegenbaur, Vogt, and Haeckel, who in their
frequent excursions to the coasts of Italy and France have made a special
study of the Acalephs, and whose descriptions have all the vividness and
freshness which nothing but familiarity with the living specimens can give.
Besides these, we have the admirable works of Von Siebold, of Ehrenberg,
the great interpreter of the microscopic world, of Steenstrup, Dujardin,
Dalyell, Forbes, Allman, and Sars. Of these, the four latter were fortunate
in having their home on the sea-shore within reach of the objects of their
study, so that they could watch them in their living condition, and follow
all their changes. The charming books of Forbes, who knew so well how to
popularize his instructions, and present scientific results under the most
attractive form, are well known to English readers. But a word on the
investigations of Sars may not be superfluous.
Born near the coast of Norway, and in early life associated with the
Church, his passion for Natural History led him to employ all his spare
time in the study of the marine animals immediately about him, and his
first papers on this subject attracted so much attention, that he was
offered the place of Professor at Christiania, and henceforth devoted
himself exclusively to scientific pursuits, and especially to the
investigation of the Acalephs. He gave us the key to the almost fabulous
transformations of these animals, and opened a new path in science by
showing the singular phenomenon of the so-called "alternate generations,"
in which the different phases of the same life may be so distinct and
seemingly so disconnected that, until we find the relation between them, we
seem to have several animals where we have but one.
[Pg 4]
To the works above mentioned, we may add the third and fourth volumes of
Professor Agassiz's Contributions to the Natural History of the United
States, which are entirely devoted to the American Acalephs.
The most important works and memoirs concerning the Echinoderms are those
by Klein, Link, Johannes Müller, Jäger, Desmoulins, Troschel, Sars,
Savigny, Forbes, Agassiz, and Lütken, but excepting those of Forbes and
Sars, few of these observations are made upon the living specimens. It may
be well to mention here, for the benefit of those who care to know
something more of the literature of this subject in our own country, a
number of memoirs on the Radiates of our coasts, published by the various
scientific societies of the United States, and to be found in their annals.
Such are the papers of Gould, Agassiz, Leidy, Stimpson, Ayres, McCrady,
Clark, A. Agassiz, and Verrill.
One additional word as to the manner in which the subjects included in the
following descriptions are arranged. We have seen that Cuvier recognized
the unity of plan in the structure of the whole type of Radiates. All these
animals have their parts disposed around a common central axis, and
diverging from it toward the periphery. The idea of bilateral symmetry, or
the arrangement of parts on either side of a longitudinal axis, on which
all the higher animals are built, does not enter into their structure,
except in a very subordinate manner, hardly to be perceived by any but the
professional naturalist. This radiate structure being then common to the
whole type, the animals composing it appear under three distinct structural
expressions of the general plan, and according to these differences are
divided into three classes,—Polyps, Acalephs, and Echinoderms. With these
few preliminary remarks we may now take up in turn these different groups,
beginning with the lowest, or the Polyps.[1]
[Pg 5]
GENERAL SKETCH OF THE POLYPS.
[Fig. 1]
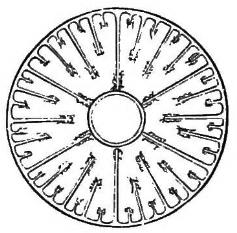 Fig. 1. Transverse section of an Actinia. (Agassiz.)
Fig. 1. Transverse section of an Actinia. (Agassiz.)
Before describing the different kinds of Polyps living on our immediate
coast, we will say a few words of Polyps in general and of the mode in
which the structural plan common to all Radiates is adapted to this
particular class. In all Polyps the body consists of a sac divided by
vertical partitions (Fig. 1.) into distinct cavities or chambers. These
partitions are not, however, all formed at once, but are usually limited to
six at first, multiplying indefinitely with the growth of the animal in
some kinds, while in others they never increase beyond a certain definite
number. In the axis of the sac, thus divided, hangs a smaller one, forming
the digestive cavity, and supported for its whole length by the six primary
partitions. The other partitions, though they extend more or less inward in
proportion to their age, do not unite with the digestive sac, but leave a
free space in the centre between their inner edge and the outer wall of the
digestive sac. The genital organs are placed on the inner edges of the
partitions, thus hanging as it were at the door of the chambers, so that
when hatched, the eggs naturally drop into the main cavity of the body,
whence they pass into the second smaller sac through an opening in its
bottom or digestive cavity, and thence out through the mouth into the
water. In the lower Polyps, as in our common Actinia for instance, these
organs occur on all the radiating partitions, while among the higher ones,
the Halcyonoids for example, they are found only on a limited number. This
limitation in the repetition of identical parts is always found to be
connected with structural superiority.
The upper margin of the body is fringed by hollow tentacles, each of which
opens into one of the chambers. All parts of the animal thus communicate
with each other, whatever is introduced at the mouth circulating through
the whole structure,
[Pg 6]
passing first into the digestive cavity, thence
through the opening in the bottom into the main chambered cavity, where it
enters freely into all the chambers, and from the chambers into the
tentacles. The rejected portions of the food, after the process of
digestion is completed, return by the same road and are thrown out at the
mouth.
These general features exist in all Polyps, and whether they lead an
independent life as the Actinia, or are combined in communities, like most
of the corals and the Halcyonoids; whether the tentacles are many or few;
whether the partitions extend to a greater or less height in the body;
whether they contain limestone deposit, as in the corals, or remain soft
throughout life as the sea-anemone,—the above description applies to them
all, while the minor differences, either in the tentacles or in the form,
size, color, and texture of the body, are simply modifications of this
structure, introducing an infinite variety into the class, and breaking it
up into the lesser groups designated as orders, families, genera, and
species. Let us now look at some of the divisions thus established.
The class of Polyps is divided into three orders,—the Halcyonoids, the
Madreporians, and the Actinoids. Of the lowest among these orders, the
Actinoid Polyps, our Actinia or sea-anemone is a good example. They remain
soft through life, having a great number of partitions and consequently a
great number of tentacles, since there is a tentacle corresponding to every
chamber. Indeed, in this order the multiplication of tentacles and
partitions is indefinite, increasing during the whole life of the animal
with its growth; while we shall see that in some of the higher orders the
constancy and limitation in the number of these parts is an indication of
superiority, being accompanied by a more marked individualization of the
different functions.
Next come the Madreporians, of which our Astrangia, to be described
hereafter, may be cited as an example. In this group, although the number
of tentacles still continues to be large, they are nevertheless more
limited than in the Actinoids; but their characteristic feature is the
deposition of limestone walls in the centre of the chambers formed by the
soft partitions, so that all the soft partitions alternate with hard ones.
The tentacles, always
[Pg7]
corresponding to the cavity of the chambers, may be
therefore said to ride this second set of partitions arising just in the
centre of the chambers.
The third and highest order of Polyps is that of the Halcyonoids. Here the
partitions are reduced to eight; the tentacles, according to the invariable
rule, agree in number with the chambers, but have a far more highly
complicated structure than in the lower Polyps. Some of these Halcyonoids
deposit limestone particles in their frame. But the tendency to solidify is
not limited to definite points, as in the Madreporians. It may take place
anywhere, the rigidity of the whole structure increasing of course in
proportion to the accumulation of limestone. There are many kinds, in which
the axis always remains soft or cartilaginous, while others, as the
so-called sea-fans for instance, well known among the corals for their
beauty of form and color, are stiff and hard throughout. Whatever their
character in this respect, however, they are always compound, living in
communities, and never found as separate individuals after their early
stages of growth. Some of those with soft axis lead a wandering life,
enjoying as much freedom of movement as if they had an individual
existence, shooting through the water like the Pennatulæ, well known on the
California coast, or working their way through the sand like the Renilla,
common on the sandy shores of our Southern States.
ACTINOIDS.
Actinia, or Sea-Anemone. (Metridium marginatum Edw.)
Nothing can be more unprepossessing than a sea-anemone when contracted. A
mere lump of brown or whitish jelly, it lies like a lifeless thing on the
rock to which it clings, and it is difficult to believe that it has an
elaborate and exceedingly delicate internal organization, or will ever
expand into such grace and beauty as really to deserve the name of the
flower after which it has been called. Figs. 2, 3, 4, and 5, show this animal in its various stages
[Pg 8]
of expansion and contraction. Fig. 2
represents it with all its external appendages folded in, and the whole
body flattened; in Fig. 3, the tentacles begin to steal out,
and the body rises slightly; in Fig. 4, the body has nearly
gained its full height, and the tentacles, though by no means fully spread, yet form
a delicate wreath around the mouth; while in Fig. 5, drawn in
life size, the
[Pg 9]
whole summit of the body seems crowned with soft, plumy fringes. We would say for the
benefit of collectors that these animals are by no means difficult to find,
and thrive well in confinement, though it will not do to keep them in a
small aquarium with other specimens, because they soon render the water
foul and unfit for their companions. They should therefore be kept in a
separate glass jar or bowl, and under such circumstances will live for a
long time with comparatively little care.
[Fig_2]
[Fig_3]
[Fig_4]
") Figs. 2,3,4. Actinia to different degrees of expansion (Agassiz)
Figs. 2,3,4. Actinia to different degrees of expansion (Agassiz)
[Fig. 5]
 fully expanded; natural size.") Fig. 5. The same Actinia (Metridium marginatum) fully expanded; natural size.
Fig. 5. The same Actinia (Metridium marginatum) fully expanded; natural size.
They may be found in any small pools about the rocks which are flooded by
the tide at high water. Their favorite haunts, however, where they occur in
greatest quantity are more difficult to reach; but the curious in such
matters will be well rewarded, even at the risk of wet feet and a slippery
scramble over rocks covered with damp sea-weed, by a glimpse into their
more crowded abodes. Such a grotto is to be found on the rocks of East
Point at Nahant. It can only be reached at low tide, and then one is
obliged to creep on hands and knees to its entrance, in order to see
through its entire length; but its whole interior is studded with these
animals, and as they are of various hues, pink, brown, orange, purple, or
pure white, the effect is like that of brightly colored mosaics set in the
roof and walls. When the sun strikes through from the opposite extremity of
this grotto, which is open at both ends, lighting up its living mosaic
work, and showing the play of the soft fringes wherever the animals are
open, it would be difficult to find any artificial grotto to compare with
it in beauty. There is another of the same kind on Saunders's Ledge, formed
by a large boulder resting on two rocky ledges, leaving a little cave
beneath, lined in the same way with variously colored sea-anemones, so
closely studded over its walls that the surface of the rock is completely
hidden. They are, however, to be found in larger or smaller clusters, or
scattered singly in any rocky fissures, overhung by sea-weed, and
accessible to the tide at high water.
[Fig. 6]
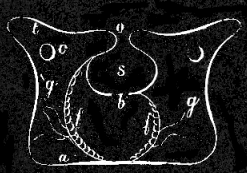 Fig. 6. Vertical section of an Actinia, showing a primary(g) and a secondary partition of g'; o mouth, t tentacles, s stomach, f f reproductive organs, b main cavity, c openings in partitions, a lower floor, or foot.
Fig. 6. Vertical section of an Actinia, showing a primary(g) and a secondary partition of g'; o mouth, t tentacles, s stomach, f f reproductive organs, b main cavity, c openings in partitions, a lower floor, or foot.
The description of Polyp structure given above includes all the general
features of the sea-anemone; but for the better explanation of the figures,
it may not be amiss to recapitulate them here in their special application.
The body of the sea-anemone may be described as a circular, gelatinous bag,
the bottom of which is flat
[Pg 10]
and slightly spreading around the margin. (Fig. 2.)
The upper edge of this bag turns in so as to form a sac within a sac.
(Fig. 6.) This inner sac, s, is the stomach or
digestive cavity, forming a simple open space in the centre of the body,
with an aperture in the bottom, b, through which the food passes
into the larger sac, in which it is enclosed. But this outer and larger
sac or main cavity of the body is not, like the inner one, a simple open
space. It is, on the contrary, divided by vertical partitions into a number
of distinct chambers, converging from the periphery to the centre. These
partitions do not all advance so far as actually to join the wall of the
digestive cavity hanging in the centre of the body, but most of them stop
a little short of it, leaving thus a small, open space between the chambers
and the inner sac. (Fig. 1.) The eggs hang on the inner
edge of the partitions; when mature they drop into the main cavity, enter
the inner digestive cavity through its lower opening, and are passed out
through the mouth.
The embryo bears no resemblance to the mature animal. It is a little
planula, semi-transparent, oblong, entirely covered with vibratile cilia,
by means of which it swims freely about in the water till it establishes
itself on some rocky surface, the end by which it becomes attached
spreading slightly and fitting itself to the inequalities of the rock so as
to form a secure basis. The upper end then becomes depressed toward the
centre, that depression deepening more and more till it forms the inner
sac, or in other words the digestive cavity described above. The open mouth
of this inner sac, which may, however, be closed at will, since the whole
substance of the body is exceedingly contractile, is the oral opening or
so-called mouth of the animal. We have seen how the main cavity becomes
divided by radiating partitions into numerous chambers; but while these
internal changes are going on, corresponding external appendages are
forming in the shape of the tentacles, which add so much to the beauty of
the animal, and play so important a part in its history. The tentacles,
[Pg 11]
at first only few in number, are in fact so many extensions of the inner
chambers, gradually narrowing upward till they form these delicate hollow
feelers which make a soft downy fringe all around the mouth. (Fig. 7.) They
do not start abruptly from the summit, but the upper margin of the body
itself thins out to form more or less extensive lobes, through which the
partitions and chambers continue their course, and along the edge of which
the tentacles arise.
[Fig. 7]
 Fig. 7. View from above of an Actinia with all its tentacles expanded; o mouth, b crescent-shaped folds at extremity of mouth, a a folds round mouth, t t t tentacles.
Fig. 7. View from above of an Actinia with all its tentacles expanded; o mouth, b crescent-shaped folds at extremity of mouth, a a folds round mouth, t t t tentacles.
The eggs are not always laid in the condition of the simple planula
described above. They may, on the contrary, be dropped from the parent in
different stages of development, sometimes even after the tentacles have
begun to form, as in Figs. 8, 9. Neither is it by means of eggs alone that
these animals reproduce themselves; they may also multiply by a process of
self-division. The disk of an Actinia may contract along its centre till
the circular outline is changed to that of a figure 8, this constriction
deepening gradually till the two halves of the 8 separate, and we have an
Actinia with two mouths, each surrounded by an independent set of
tentacles. Presently this separation descends vertically till the body is
finally divided from
[Pg 12]
summit to base, and we have two Actiniæ where there
was originally but one. Another and a far more common mode of reproduction
among these animals is that of budding like corals. A slight swelling
arises on the side of the body or at its base; it enlarges gradually, a
digestive cavity is formed within it, tentacles arise around its summit,
and it finally drops off from the parent and leads an independent
existence. As a number of these buds are frequently formed at once, such an
Actinia, surrounded by its little family, still attached to the parent, may
appear for a time like a compound stock, though their normal mode of
existence is individual and distinct.
[Fig. 8]
[Fig. 9]
 |
 |
| Figs. 8, 9. Young Actiniæ in different stages of growth. |
The Actinia is exceedingly sensitive, contracting the body and drawing in
the tentacles almost instantaneously at the slightest touch. These sudden
movements are produced by two powerful sets of muscles, running at right
angles with each other through the thickness of the body wall; the one
straight and vertical, extending from the base of the wall to its summit;
the other circular and horizontal, stretching concentrically around it. By
the contraction of the former, the body is of course shortened; by the
contraction of the latter, the body is, on the contrary, lengthened in
proportion to the compression of its circumference. Both sets can easily be
traced by the vertical and horizontal lines crossing each other on the
external wall of the body, as in Fig. 5. Each tentacle is in like manner
furnished with a double set of muscles, having an action similar to that
described above. In consequence of these violent muscular contractions, the
water imbibed by the animal, and by which all its parts are distended to
the utmost, is forced, not only out of the mouth, but also through small
openings in the body wall scarcely perceptible under ordinary
circumstances, but at such times emitting little fountains in every
direction.
Notwithstanding its extraordinary sensitiveness, the organs of the senses
in the Actinia are very inferior, consisting only of a few pigment cells
accumulated at the base of the tentacles. The two sets of muscles meet at
the base of the body, forming a disk, or kind of foot, by which the animal
can fix itself so firmly to the ground, that it is very difficult to remove
it without injury. It is nevertheless capable of a very limited degree of
[Pg 13]
motion, by means of the expansion and contraction of this foot-like disk.
The Actiniæ are extremely voracious; they feed on mussels and cockles,
sucking the animals out of their shells. When in confinement they may be
fed on raw meat, and seem to relish it; but if compelled to do so, they
will live on more meagre fare, and will even thrive for a long time on such
food as they may pick up in the water where they are kept.
Rhodactinia (Rhodactinia Davisii Ag.)
[Fig. 10]
 Fig. 10. Rhodactinia Davisii Ag.; natural size.
Fig. 10. Rhodactinia Davisii Ag.; natural size.
Very different from this is the bright red Rhodactinia (Fig. 10), quite
common in the deeper waters of our bay, while farther north, in Maine, it
occurs at low-water mark. Occasionally it may be found thrown up on our
sandy beaches after a storm, and then, if it has not been too long out of
its native element, or too severely buffeted by the waves, it will revive
on being thrown into a bucket of fresh sea-water, expand to its full size,
and show all the beauty of its natural coloring. It is crowned with a
wreath of thick, short tentacles (Fig. 10), and though so vivid and bright
in color, it is not so pretty as the more common Actinia marginata, with
its soft waving wreath of plume-like feelers, in comparison to which the
tentacles of the Rhodactinia are clumsy and slow in their movements.
All Actiniæ are not attached to the soil like those described above, nor do
they all terminate in a muscular foot, some being pointed or rounded at
their extremity. Many are nomadic, wandering about at will during their
whole lifetime, others live buried in the sand or mud, only extending their
tentacles beyond the limits of the hole where they make their home; while
others again lead a parasitic life, fastening themselves upon our larger
[Pg 14]
jelly-fish, the Cyaneæ, though one is at a loss to imagine what sustenance
they can derive from animals having so little solidity, and consisting so
largely of water.
Arachnactis. (Arachnactis brachiolata A. Ag.)
[Fig. 11]
 Fig. 11. Arachnactis brachiolata A. Ag., greatly magnified.
Fig. 11. Arachnactis brachiolata A. Ag., greatly magnified.
[Fig. 12]
[Fig. 13]
 |
| Fig. 12. Young Arachnactis. |
 |
| Fig. 13. Young Arachnactis seen so as to show the mouth. |
Among the nomadic Polyps is a small floating Actinia, called Arachnactis,
(Fig. 11,) from its resemblance to a spider. They are found in great plenty
floating about during the night, feeling their way in every direction by
means of their tentacles, which are large in proportion to the size of the
animal, few in number, and turned downward when in their natural attitude.
The partitions and the digestive cavity enclosed between them are short, as
will be seen in Fig. 11, when compared to the general cavity of the body
floating balloon-like above them. Around the mouth is a second row of
shorter tentacles, better seen in a younger specimen (Fig. 12). This
Actinia differs from those described above, in having two of the sides
flattened, instead of being perfectly circular. Looked at from above (as in
Fig. 13) this difference in the diameters is very perceptible; there is an
evident tendency towards establishing a longitudinal axis. In the
sea-anemone, this disposition is only hinted at in the slightly pointed
folds or projections on opposite sides of the circle formed by the mouth,
which in the Arachnactis are so elongated as to produce a somewhat narrow
slit (see
[Pg 15]
Fig. 13), instead of a circular opening. The mouth is also a
little out of centre, rather nearer one end of the disk than the other.
These facts are interesting, as showing that the tendency towards
establishing a balance of parts, as between an anterior and posterior
extremity, a right and left side, is not forgotten in these lower animals,
though their organization as a whole is based upon an equality of parts,
admitting neither of posterior and anterior extremities, nor of right and
left, nor of above and below, in a structural sense. This animal also
presents a seeming anomaly in the mode of formation of the young tentacles,
which always make their appearance at the posterior extremity of the
longitudinal axis, the new ones being placed behind the older ones, instead
of alternating with them as in other Actiniæ.
Bicidium. (Bicidium parasiticum Ag.)
[Fig. 14]
 Fig. 14. Bicidium parasiticum; natural size.
Fig. 14. Bicidium parasiticum; natural size.
The Bicidium (Fig. 14), our parasitic Actinia, is to be sought for in the
mouth-folds of the Cyanea, our common large red Jelly-fish. In any
moderate-sized specimen of the latter from twelve to eighteen inches in
diameter, we shall be sure to find one or more of these parasites, hidden
away among the numerous folds of the mouth. The body is long and tapering,
having an aperture in the extremity, the whole animal being like an
elongated cone, strongly ribbed from apex to base. At the base, viz. at the
month end, are a few short, stout tentacles. This Actinia is covered with
innumerable little transverse wrinkles (see Fig. 14), by means of which it
fastens itself securely among the fluted membranes around the mouth of the
Jelly-fish. It will live a considerable time in confinement, attaching
itself, for its whole length, to the vessel in which it is kept, and
clinging quite firmly if any attempt is made to remove it. The general
color of the body is violet or a brownish red, though the wrinkles give it
a somewhat mottled appearance.
[Pg 16]
Halcampa. (Halcampa albida Ag.)
[Fig. 15]
 Fig. 15. Halcampa albida; natural size.
Fig. 15. Halcampa albida; natural size.
Strange to say, the Actiniæ, which live in the mud, are among the most
beautifully colored of these animals. They frequently prepare their home
with some care, lining their hole by means of the same secretions which
give their slimy surface to our common Actiniæ, and thus forming a sort of
tube, into which they retire when alarmed. But if undisturbed, they may be
seen at the open door of their house with their many colored disk and
mottled tentacles extending beyond the aperture, and their mouth wide open,
waiting for what the tide may bring them. By the play of their tentacles,
they can always produce a current of water about the mouth, by means of
which food passes into the stomach. We have said, that these animals are
very brightly colored, but the little Halcampa (Fig. 15), belonging to our
coast, is not one of the brilliant ones. It is, on the contrary, a small,
insignificant Actinia, resembling a worm, as it burrows its way through the
sand. It is of a pale yellowish color, with whitish warts on the surface.
MADREPORIANS.
Astrangia. (Astrangia Danæ Ag.)
In Figure 16, we have the only species of coral
growing so far north as our latitude. Indeed, it hardly belongs in this
volume, since we have limited ourselves to the Radiates of Massachusetts
Bay,—its northernmost boundary being somewhat to the south of
Massachusetts Bay, about the shores of Long Island, and on the islands of
Martha's Vineyard Sound. But we introduce it here, though it is not included
under our
[Pg 17]
title, because any account of the Radiates, from which so important a group
as that of the corals was excluded, would be very incomplete.
[Fig. 16]
 Fig. 16. Astrangia colony; natural size.
Fig. 16. Astrangia colony; natural size.
This pretty coral of our Northern waters is no reef-builder, and does not
extend farther south than the shores of North Carolina. It usually
establishes itself upon broken angular bits of rock, lying in sheltered
creeks and inlets, where the violent action of the open sea is not felt.
The presence of one of these little communities on a rock may first be
detected by what seems like a delicate white film over the surface. This
film is, however, broken up by a number of hard calcareous deposits in very
regular form (Fig. 20), circular in outline, but divided by numerous
partitions running from the outer wall to the centre of every such circle,
where they unite at a little white spot formed by the mouth or oral
opening. These circles represent, and indeed are themselves the distinct
individuals (Fig. 17) composing the community, and they look not unlike the
star-shaped pits on a coral head, formed by Astræans. Unlike the massive
compact kinds of coral, however, the individuals multiply by budding from
the base chiefly, never rising one above the other, but spreading over the
surface on which they have established themselves, a few additional
individuals arising between the older ones. In consequence of this mode of
growth, such a community,
[Pg 18]
when it has attained any size, forms a little
white mound on the rock, higher in the centre, where the older members have
attained their whole height and solidity, and thinning out toward the
margin, where the younger ones may be just beginning life, and hardly rise
above the surface of the rock. These communities rarely grow to be more
than two or three inches in diameter, and about quarter of an inch in
height at the centre where the individuals have reached their maximum size.
When the animals are fully expanded (Fig. 18), with all their tentacles
spread, the surface of every such mound becomes covered with downy white
fringes, and what seemed before a hard, calcareous mass upon the rock,
changes to a soft fleecy tuft, waving gently to and fro in the water. The
tentacles are thickly covered with small wart-like appendages, which, on
examination, prove to be clusters of lasso-cells, the terminal cluster of
the tentacle being quite prominent. These lasso-cells are very formidable
weapons, judging both from their appearance when magnified (Fig. 19), and
from the terrible effect of their bristling lash upon any small crustacean,
or worm, that may be so unfortunate as to come within its reach.
[fig. 17]
[fig. 18]
[fig. 19]
[fig. 20]
 | |
 |
| Fig. 17. Magnified individuals of an Astrangia community in different stages of expansion. |
|
Fig. 19. Magnified lasso-cell of Astrangia. |
|
| |
 | |
 |
| Fig. 18. Single individual of Astrangia, fully expanded. |
|
Fig. 20. Limestone parts of an individual of Astrangia; magnified. |
|
The description of the internal arrangement of parts in the Actinia applies
in every particular to these corals, with the exception of the hard deposit
in the lower part of the body. As in all the Polyps, radiating partitions
divide the main cavity of the body into distinct separate chambers, and the
tentacles increasing by multiples of six, numbering six in the first set,
six in the second, and twelve in the third, are hollow, and open into the
chambers. But the feature which distinguishes them from the soft Actiniæ,
and unites them with the corals, requires a somewhat more accurate
description. In each individual, a hard deposit is formed (Fig. 20),
beginning at the base of every chamber, and rising from its floor to about
[Pg 19]
one fifth the height of the animal at its greatest extension. This lime
deposit does not, however, fill the chamber for its whole width, but rises
as a thin wall in its centre. (See Figs. 13, 17.) Thus between all the soft
partitions, in the middle of the chambers which separate them, low
limestone walls are gradually built up, uniting in a solid column in the
centre. These walls run parallel with the soft partitions, although they do
not rise to the same height, and they form the radiating lines like stiff
lamellæ, so conspicuous when all the soft parts of the body are drawn in.
The mouth of the Astrangia is oval, and the partitions spread in a
fan-shaped way, being somewhat shorter at one side of the animal than on
the other. The partitions extend beyond the solid wall which unites them at
the periphery, in consequence of which, this wall is marked by faint
vertical ribs.
HALCYONOIDS.
Halcyonium. (Halcyonium carneum Ag.)
[Fig. 21]
 Fig. 21. Single individual of Halcyonium seen from above; magnified.
Fig. 21. Single individual of Halcyonium seen from above; magnified.
We come now to the Halcyonoids, represented in our waters by the Halcyonium
(Fig. 22). In the Halcyonoids, the highest group of Polyps, the tentacles
reach their greatest limitation, which, as above mentioned, is found to be
a mark of superiority, and, connected with other structural features,
places them at the head of their class. The number of tentacles throughout
this group is always eight. They are very complicated (Fig. 21), in
comparison with the tentacles of the lower orders, being deeply lobed,
[Pg 20]
and fringed around the margin. Our Halcyonium communities (Fig. 22) usually
live in deep water, attached to dead shells, though they may occasionally
be found growing at low-water mark, but this is very rare. They have
received a rather lugubrious name from the fishermen, who call them
"dead-men's fingers," and indeed, when the animals are contracted, such a
community, with its short branches attached to the main stock, looks not
unlike the stump of a hand, with short, fat fingers. In such a condition
they are very ugly, the whole mass being somewhat gelatinous in texture,
and a dull, yellowish pink in color. But when the animals, which are
capable of great extension, are fully spread, as in Fig. 22, such a
polyp-stock has a mossy, tufted look, and is by no means an unsightly
object. When the individuals are entirely expanded, as in Fig. 23, they
become quite transparent, and their internal structure can readily be seen
through the walls of the body; we can then easily distinguish the digestive
cavity, supported for its whole length by the eight radiating partitions,
as well as the great size of the main digestive cavity surrounding it.
Notwithstanding the remarkable power of contraction and dilatation in the
animals themselves, the tentacles are but slightly contractile. This kind
of community increases altogether by budding, the individual polyps
remaining more or less united, the tissues of the individuals becoming
thicker by the deposition of lime nodules, and thus forming a massive
semi-cartilaginous pulp, uniting the whole community. In the neighborhood
of Provincetown they are very plentiful, and are found all along the shores
of our Bay in deep water.
[Fig. 22]
[Fig. 23]
 |
|
 |
| Fig. 22. Halcyonium community; natural size. |
|
Fig. 23. Individual of Halcyonium fully expanded; magnified. |
[Pg 21]
GENERAL SKETCH OF ACALEPHS.
In the whole history of metamorphosis, that wonderful chapter in the life
of animals, there is nothing more strange or more interesting than the
transformations of the Acalephs. First, as little floating planulæ or
transparent spheres, covered with fine vibratile cilia, by means of which
they move with great rapidity, then as communities fixed to the ground and
increasing by budding like the corals, or multiplying by self-division, and
later as free-swimming Jelly-fishes, many of them pass through phases which
have long baffled the investigations of naturalists, and have only recently
been understood in their true connection. Great progress has, however, been
made during this century in our knowledge of this class. Thanks to the
investigations of Sars, Dujardin, Steenstrup, Van Beneden, and many others,
we now have the key to their true relations, and transient phases of
growth, long believed to be the adult condition of distinct animals, are
recognized as parts in a cycle of development belonging to one and the same
life. As the class now stands, it includes three orders, highest among
which are the Ctenophoræ, so-called on account of their locomotive organs,
consisting of minute flappers arranged in vertical comb-like rows; next to
these are the Discophoræ, with their large gelatinous umbrella-like disks,
commonly called Jelly-fishes, Sun-fishes, or Sea-blubbers, and below these
come the Hydroids, embracing the most minute and most diversified of all
these animals.
These orders are distinguished not only by their striking external
differences, but by their mode of development also. The Ctenophoræ grow
from eggs by a direct continuous process of development, without undergoing
any striking metamorphosis; the Discophoræ, with some few exceptions, in
which they develop like the Ctenophoræ from eggs, begin life as a
Hydra-like animal, the subsequent self-division of which gives rise, by a
singular process, presently to be described, to a number of distinct
Jelly-fishes; the Hydroids include all those Acalephs which either pass the
earlier stages of their existence as little shrub-like communities,
[Pg 22]
or remain in that condition through life. These Hydroid stocks, as they are
sometimes called, give rise to buds; these buds are transformed into
Jelly-fishes, which in some instances break off when mature and swim away
as free animals, while in others they remain permanent members of the
Hydroid stock, never assuming a free mode of life. All these buds when
mature, whether free or fixed, lay eggs in their turn, from which a fresh
stock arises to renew this singular cycle of growth, known among
naturalists as "alternate generations."
The Hydroids are not all attached to the ground,—some like the Physalia
(Portuguese man-of-war), or the Nanomia, that pretty floating Hydroid of
our own waters, move about with as much freedom as if they enjoyed an
individual independent existence. As all these orders have their
representatives on our coast, to be described hereafter in detail, we need
only allude here to their characteristic features. But we must not leave
unnoticed one very remarkable Hydroid Acaleph (Fig. 24), not found in our
waters, and resembling the Polyps so much, that it has long been associated
with them. The Millepore is a coral, and was therefore the more easily
confounded with the Polyps, so large a proportion of which build coral
stocks; but a more minute investigation of its structure (Figs. 25, 26) has
recently shown that it belongs with the Acalephs.[2] This discovery is the
more important, not only as explaining the true position of this animal in
the Animal Kingdom, but as proving also the presence of Acalephs in the
earliest periods of creation, since it refers a large number of fossil
corals, whose affinities with the millepores are well understood, to that
class, instead of to the class of Polyps with which they had hitherto been
associated. But for this we should have no positive evidence of the
existence of [Pg 23]Acalephs in early geological periods, the gelatinous texture
of the ordinary Jelly-fishes making their preservation almost impossible.
It is not strange that the true nature of this animal should have remained
so long unexplained; for it is only by the soft parts of the body, not of
course preserved in the fossil condition, that their relations to the
Acalephs may be detected; and they are so shy of approach, drawing their
tentacles and the upper part of the body into their limestone frame if
disturbed, that it is not easy to examine the living animal.
[Fig. 24]
[Fig. 25]
[Fig. 26]
") |
|
") |
|
") |
| Fig. 24. Branch of Millepora alcicornis; natural size. (Agassiz.) |
|
Fig. 25. Animals of M. alcicornis expanded; magnified. a a small
Hydroid, larger Hydroid, t tentacles, m mouth. (Agassiz.) |
|
Fig. 26. Transverse section of a branch, showing pits, a a a a, of the
large Hydroids with the horizontal floors. (Agassiz.) |
The Millepore is very abundant on the Florida reefs. From the solid base of
the coral stock arise broad ridges, branching more or less along the edges,
the whole surface being covered by innumerable pores, from which the
diminutive animals project when expanded. (Fig. 25.) The whole mass of the
coral is porous, and the cavities occupied by the Hydræ are sunk
perpendicularly to the surface within the stock. Seen in a transverse cut
these tubular cavities are divided at intervals by horizontal partitions
(Fig. 26), extending straight across the cavity from wall to wall, and
closing it up entirely, the animal occupying only the outer-most open
space, and building a new partition behind it as it rises in the process of
growth. This structure is totally different from that of the Madrepores,
Astræans, Porites, and indeed, from all the polyp corals which, like all
Polyps, have the vertical partitions running through the whole length of
the body, and more or less open from top to bottom.
The life of the Jelly-fishes, with the exception of the Millepores and the
like, is short in comparison to that of other Radiates. While Polyps live
for many years, and Star-fishes and Sea-urchins require ten or fifteen
years to attain their full size, the short existence of the Acaleph, with
all its changes, is accomplished in one year. The breeding season being in
the autumn, the egg grows into a Hydroid during the winter; in the spring
the Jelly-fish is freed from the Hydroid stock, or developed upon it as the
case may be; it attains its full size in the fall, lays its eggs[Pg 24] and dies,
and the cycle is complete. The autumn storms make fearful havoc among them,
swarms of them being killed by the fall rains, after which they may be
found thrown up on the beaches in great numbers. When we consider the size
of these Jelly-fishes, their rapidity of growth seems very remarkable. Our
common Aurelia measures some twelve to eighteen inches in diameter when
full grown, and yet in the winter it is a Hydra so small as almost to
escape notice. Still more striking is the rapid increase of our Cyanea,
that giant among Jelly-fishes, which, were it not for the soft, gelatinous
consistency of its body, would be one of the most formidable among our
marine animals.
Before entering upon the descriptions of the special kinds of Jelly-fishes,
we would remind our readers that the radiate plan of structure is
reproduced in this class of animals as distinctly as in the Polyps, though
under a different aspect. Here also we find that there is a central
digestive cavity from which all the radiating cavities, whether simple or
ramified, diverge toward the periphery. It is true that the open chambers
of the Polyps are here transformed into narrow tubes, by the thickening of
the dividing partitions; or in other words, the open spaces of the Polyps
correspond to tubes in the Acalephs, while the partitions in the Polyps
correspond to the thick masses of the body dividing the tubes in the
Acalephs. But the principle of radiation on which the whole branch of
Radiates is constructed controls the organization of Acalephs no less than
that of the other classes, so that a transverse section across any Polyp
(Fig. 1), or across any Acaleph (Fig. 50), or across any Echinoderm (Fig.
140), shows their internal structure to be based upon a radiation of all
parts from the centre to the periphery.
That there may be no vagueness as to the terms used hereafter, we would
add one word respecting the nomenclature of this class, whose aliases might
baffle the sagacity of a police detective. The names Acalephs, Medusæ, or
the more common appellation of Jelly-fishes, cover the same ground, and are
applied indiscriminately to the animals they represent. The name Jelly-fish
is an inappropriate one, though the gelatinous consistency of these animals
is accurately enough expressed by it; but they have no more structural
relation to a fish than to a bird or an insect.
[Pg 25]
They have, however, received this name before the structure of animals was
understood, when all animals inhabiting the waters were indiscriminately
called fishes, and it is now in such general use that it would be difficult
to change it. The name Medusa is derived from their long tentacular appendages,
sometimes wound up in a close coil, sometimes thrown out to a great distance,
sometimes but half unfolded, and aptly enough compared to the snaky locks
of Medusa. Their third and oldest appellation, that of Acalephs,—alluding
to their stinging or nettling property, and given to them and like animals
by Aristotle, in the first instance, but afterwards applied by Cuvier in a
more limited sense to Jelly-fishes,—is the most generally accepted, and
perhaps the most appropriate of all.
The subject of nomenclature is not altogether so dry and arid as it seems
to many who do not fully understand the significance of scientific names.
Not only do they often express with terse precision the character of the
animal or plant they signify, but there is also no little sentiment
concealed under these jaw-breaking appellations. As seafaring men call
their vessels after friends or sweethearts, or commemorate in this way some
impressive event, or some object of their reverence, so have naturalists,
under their fabrication of appropriate names, veiled many a graceful
allusion, either to the great leaders of our science, or to some more
intimate personal affection. The Linnæa borealis was well named after his
famous master, by a disciple of the great Norwegian naturalist; Goethea
semperflorens, the ever-blooming, is another tribute of the same kind,
while the pretty, graceful little Lizzia, named by Forbes, is one instance
among many of a more affectionate reference to nearer friends. The
allusions of this kind are not always of so amiable a character,
however,—witness the "Buffonia," a low, noxious weed, growing in marshy
places, and named by Linnæus after Buffon, whom he bitterly hated. Indeed,
there is a world of meaning hidden under our zoölogical and botanical
nomenclature, known only to those who are intimately acquainted with the
annals of scientific life in its social as well as its professional
aspect.
[Pg 26]
CTENOPHORÆ.
The Ctenophoræ differ from other Jelly-fishes in their mode of locomotion.
All the Discophorous Medusæ, as well as Hydroids, move by a rhythmical rise
and fall of the disk, contracting and expanding with alternations so
regular, that it reminds one of the action of the lungs, and seems at first
sight to be a kind of respiration in which water takes the place of air.
The Greeks recognized this peculiar character in their name, for they
called them Sea-lungs. Indeed, locomotion, respiration, and circulation are
so intimately connected in all these lower animals, that whatever promotes
one of these functions affects the other also, and though the immediate
result of the contraction and expansion of the disk seems to be to impel
them through the water, yet it is also connected with the introduction of
water into the body, which there becomes assimilated with the food in the
process of digestion, and is circulated throughout all its parts by means
of ramifying tubes. In the Ctenophoræ there is no such regular expansion
and contraction of the disk; they are at once distinguished from the
Discophoræ by the presence of external locomotive appendages of a very
peculiar character. They move by the rapid flapping of countless little
oars or paddles, arranged in vertical rows along the surface of the disk,
acting independently of each other; one row, or even one paddle, moving
singly, or all of them together, at the will of the animal; thus enabling
it to accelerate or slacken its movements, to dart through the water
rapidly, or to diminish its speed by partly furling its little sails, or,
spreading them slightly, to poise itself with a faint, quivering movement
that reminds one of the pause of the humming-bird in the air,—something
that is neither positive motion, nor actual rest.[3]
These locomotive appendages are intimately connected with the circulating
tubes, as we shall see when we examine the structural[Pg 27] details of these
animals, so that in them also breathing and moving are in direct relation
to each other. To those unaccustomed to the comparison of functions in
animals, the use of the word breathing, as applied to the introduction of
water into the body, may seem inappropriate, but it is by the absorption of
aerated water that these lower animals receive that amount of oxygen into
the system, as necessary to the maintenance of life in them, as a greater
supply is to the higher animals. The name of Ctenophoræ or comb-bearers, is
derived from these rows of tiny paddles which have been called combs by
some naturalists, because they are set upon horizontal bands of muscles,
see Fig. 29, reminding one of the base of a comb, while the fringes are
compared to its teeth. These flappers add greatly to the beauty of these
animals, for a variety of brilliant hues is produced along each row by the
decomposition of the rays of light upon them when in motion. They give off
all the prismatic colors, and as the combs are exceedingly small, so that
at first sight one hardly distinguishes them from the disk itself, the
exquisite play of color, rippling in regular lines over the surface of the
animal, seems at first to have no external cause.
Pleurobrachia. (Pleurobrachia rhododactyla Ag.)
Among the most graceful and attractive of these animals are the
Pleurobrachia (Fig. 29), and, though not first in order, we will give it
the precedence in our description, because it will serve to illustrate some
features of the other two groups. The body of the Pleurobrachia consists of
a transparent sphere, varying, however, from the perfect sphere in being
somewhat oblong, and also by a slight compression on two opposite sides
(Figs. 27 and 28), so as to render its horizontal diameter longer in one
direction than in the other (Fig. 30). This divergence from the globular
form, so slight in Pleurobrachia as to be hardly perceptible to the casual
observer, establishing two diameters of different lengths at right angles
with each other, is equally true of the other genera. It is interesting and
important, as showing the tendency in
[Pg 28]
this highest group of Acalephs to assume a bilateral character. This
bilaterality becomes still more marked in the highest class of Radiates,
the Echinoderms. Such structural tendencies in the lower animals, hinting
at laws to be more fully developed in the higher forms, are always
significant, as showing the intimate relation between all parts of the
plan of creation. This inequality of the diameters is connected with the
disposition of parts in the whole structure, the locomotive fringes and
the vertical tubes connected with them being arranged in sets of four on
either side of a plane passing through the longer diameter, showing thus
a tendency toward the establishment of a right and left side of the body,
instead of the perfectly equal disposition of parts around a common centre,
as in the lower Radiates.
[Fig. 27]
[Fig. 28]
") |
|
") |
| Fig. 27. Pleurobrachia seen at right angles to the plane in which the tentacles are placed. (Agassiz.) |
|
Fig. 28. Pleurobrachia seen in plane of tentacles. (Agassiz.) |
The Pleurobrachia are so transparent, that, with some preparatory
explanation of their structure, the most unscientific observer may trace
the relation of parts in them. At one end of the sphere is the transverse
split (Fig. 27), that serves them as a mouth; at the opposite pole is a
small circumscribed area, in the centre of which is a dark eye-speck. The
eight rows of locomotive fringes run from pole to pole, dividing the whole
surface of the body like the ribs on a melon. (Figs. 27, 28.) Hanging from
either side of the body, a little above the area in which the eye-speck is
placed, are two most extraordinary appendages in the shape of long
tentacles, possessing such wonderful power of extension and contraction
that, while at one moment they may be knotted into a little compact mass no
bigger than a pin's head, drawn up close against the side of the body, or
hidden within it, the next instant they may be floating behind it in
various positions to a distance of half a yard and more, putting out at the
same time soft plumy fringes (Fig. 29) along one side, like the beard of a
feather. One who has never seen these animals may well be pardoned for
doubting even the most literal and matter-of-fact account of these singular
tentacles. There is no variety of curve or spiral that does not seem to be
represented in their evolutions. Sometimes they unfold gradually, creeping
out softly
[Pg 29]
and slowly from a state of contraction, or again the little
ball, hardly perceptible against the side of the body, drops suddenly to
the bottom of the tank in which the animal floating, and one thinks for a
moment, so slight is the thread-like attachment, that it has actually
fallen from the body; but watch a little longer, and all the filaments
spread out along the side of the thread, it expands to its full length and
breadth, and resumes all its graceful evolutions.
[Fig. 29]
[Fig. 30]

Fig. 29. Natural attitude of Pleurobrachia when in motion. |
| |

Fig. 30. Pleurobrachia seen from the extremity opposite the mouth. |
One word of the internal structure of these animals, to explain its
relation to the external appendages. The mouth opens into a wide digestive
cavity (Figs. 27, 28), enclosed
between two vertical tubes. Toward the
opposite end of the body these tubes terminate or unite in a single
funnel-like canal, which is a reservoir as it were for the circulating
fluid poured into it through an opening in the bottom of the digestive
cavity. The food in the digestive cavity becomes liquefied by mingling with
the water entering with it at the mouth, and, thus prepared, it passes into
this canal, from which, as we shall presently see, all the circulating
tubes ramifying throughout the body are fed. Two of these circulating
tubes, or, as they are called from the nature of the liquid they contain,
chymiferous tubes, are very large, starting horizontally and at right
angles with the digestive cavity from the point of junction between the
vertical tubes (Fig. 30) and the canal. Presently they give off two
branches, those again ramifying in two directions as they approach the
periphery, so that each one of the first main tubes has multiplied to four,
[Pg 30]
before its ramifications reach the surface, thus making in all eight
radiating tubes. So far, these eight tubes are horizontal, all diverging on
the same level; but as they reach the periphery each one gives rise to a
vertical tube, running along the surface of the body from pole to pole,
just within the rows of locomotive fringes on the outer surface, and
immediately connected with them . As in all the Ctenophoræ,
these fringes keep up a constant play of color by their rapid vibrations.
In Pleurobrachia the prevailing tint is a yellowish pink, though it varies
to green, red, and purple, with the changing motions of the animal. We have
seen that the vertical tubes between which the digestive cavity is
enclosed, start like the cavity itself from that pole of the body where the
mouth is placed, and that, as they approach the opposite pole, at a
distance from the mouth of about two thirds the whole length of the body,
they unite in the canal, which then extends to the other pole where the
eye-speck is placed. As it is just at this point of juncture between the
tubes and the canal that the two main horizontal tubes arise from which all
the others branch on the same plane (Figs. 27, 28),
it follows that they reach the periphery, not on a level with the pole
opposite the mouth, but removed from it by about one third the height of
the body. In consequence of this the eight vertical tubes arising from
the horizontal ones, in order to run the entire length of the body from
pole to pole, extend in opposite directions, sending a branch to each
pole, though the branch running toward the mouth is of course the longer
of the two. The tentacles have their roots in two sacs within the body,
placed at right angles with the split of the mouth. (Figs. 27, 30.) They
open at the surface on the opposite side from the mouth, though not
immediately within the area at which the eye-speck is placed, but
somewhat above it, and at a little distance on either side of it. The
tentacles may be drawn completely within these sacs, or be extended
outside, as we have seen, to a greater or less degree, and in every
variety of curve or spiral.
Bolina. (Bolina alata Ag.)
[Pg 31]
[Fig 31]
[Fig 32]
") |
|
") |
| Fig. 31. Bolina seen from the broad side; o eye-speck, m mouth, r
auricles, v digestive cavity, g h short rows of flappers, a f
long rows of flappers, n x t z tubes winding in the larger lobes;
about half natural size. (Agassiz.) |
|
Fig. 32. Bolina seen from the narrow side; c h short
rows of flappers, a b long rows of flappers; other letters as in Fig. 31. (Agassiz.) |
The Bolina (Fig 32), like the Pleurobrachia, is slightly oval in form, with
a longitudinal split at one end of the body, forming a mouth which opens
into a capacious sac or digestive cavity. But it differs from the
Pleurobrachia in having the oral end of the body split into two larger
lobes (Fig. 31), hanging down from the mouth. These lobes may gape widely,
or they may close completely over the mouth so as to hide it from view, and
their different aspects under various degrees of expansion or contraction
account for the discrepancies in the description of these animals. We have
seen that the Pleurobrachia moves with the mouth upward; but the Bolina, on
the contrary, usually carries the mouth downward, though it occasionally
reverses its position, and in this attitude, with the lobes spread open, it
is exceedingly graceful in form, and looks like a white flower with the
crown fully expanded. These broad lobes are balanced on the other sides of
the body by four smaller appendages, divided in pairs, two on each side
(Fig. 32), called auricles. These so-called auricles are in fact organs of
the same kind as the larger lobes, though less developed. The rows of
locomotive flappers on the Bolina differ in length from each other (Fig.
31), instead of being equal, as in the Pleurobrachia. The four longest ones
are opposite each other on those sides of the body where the larger lobes
are developed, the four short ones being in pairs on the sides where the
auricles are placed. At first sight they all seem to terminate at the
margin of the body, but a closer
[Pg 32]
examination shows that the circulating
tubes connected with the longer row extend into the lobes, where they wind
about in a variety of complicated involutions. (Fig. 32.) The movements of
the Bolina are more sluggish than those of the Pleurobrachia, and the long
tentacles, so graceful an ornament to the latter, are wanting in the
former. With these exceptions the description given above of the
Pleurobrachia will serve equally well for the Bolina. The structure is the
same in all essential points, though it differs in the size and proportion
of certain external features, and its play of color is less brilliant than
that of the Pleurobrachia. The Bolina, with its slow, undulating motion,
its broad lobes sometimes spreading widely, at other times folded over the
mouth, its delicacy of tint and texture, and its rows of vibrating fringes
along the surface, is nevertheless a very beautiful object, and well
rewards the extreme care without which it dies at once in confinement.
Idyia. (Idyia roseola Ag.)
[Fig. 33]
") Fig. 33. Idyia roseola seen from the broad side, half natural size; a anal opening, b lateral tube, c circular tube, d e f g h rows of locomotive flappers. (Agassiz.)
Fig. 33. Idyia roseola seen from the broad side, half natural size; a anal opening, b lateral tube, c circular tube, d e f g h rows of locomotive flappers. (Agassiz.)
The lowest genus of Ctenophoræ found on our coast, the Idyia (Fig. 33), has
neither the tentacles of the Pleurobrachia, nor the lobes of the Bolina. It
is a simple ovate sphere, the interior of which is almost entirely occupied
by an immense digestive cavity. It would seem that the reception and
digestion of food is intended to be the almost exclusive function of this
animal, for it has a mouth whose ample dimensions correspond with its
capacious stomach. Instead of the longitudinal split serving as a mouth, in
the Bolina and Pleurobrachia, one end of the body in the Idyia is
completely open (Fig. 33), so that occasionally some unsuspicious victim of
smaller diameter than itself may be seen to swim into this wide portal,
when suddenly the door closes behind him with a quick contraction, and he
finds himself a prisoner. The Idyia does not always obtain its food after
this indolent fashion
[Pg 33]
however, for it often attacks a Bolina or Pleurobrachia as large or even
larger than itself, when it extends its mouth to the utmost, slowly
overlapping the prey it is trying to swallow by frequent and repeated
contractions, and even cutting off by the same process such portions as
cannot be forced into the digestive cavity.
The general internal structure of the Idyia corresponds with that of the
Bolina and Pleurobrachia; it has the same tubes branching horizontally from
the main cavity, then ramifying as they approach the periphery till they
are multiplied to eight in all, each of which gives off one of the vertical
tubes connected with the eight rows of locomotive flappers. Opposite the
mouth is the eye-speck, placed as in the two other genera, at the centre of
a small circumscribed area, which in the Idyia is surrounded by delicate
fringes, forming a rosette at this end of the body. These animals are
exceedingly brilliant in color; bright pink is their prevailing hue, though
pink, red, yellow, orange, green, and purple, sometimes chase each other in
quick succession along their locomotive fringes. At certain seasons, when
most numerous, they even give a rosy tinge to patches on the surface of the
sea. Their color is brightest and deepest before the spawning season, but
as this advances, and the ovaries and spermaries are emptied, they grow
paler, retaining at last only a faint pink tint. They appear early in July,
rapidly attain their maximum size, and are most numerous during the first
half of August. Toward the end of August they spawn, and the adults are
usually destroyed by the early September storms, the young disappearing at
the same time, not to be seen again till the next summer. It is an
interesting question, not yet solved, to know what becomes of the summer's
brood in the following winter. They probably sink into deep waters during
this intervening period. The Idyia, like the Pleurobrachia, moves with the
mouth upward, but inclined slightly forward also, so as to give an oblique
direction to the axis of the body.[4]
[Pg 34]
EMBRYOLOGY OF CTENOPHORÆ.
All the Ctenophoræ are reproduced from eggs, these eggs being so
transparent that one may follow with comparative ease the changes undergone
by the young while still within the egg envelope. Unfortunately, however,
they are so delicate that it is impossible to keep them alive for any
length of time, even by supplying them constantly with fresh sea-water, and
keeping them continually in motion, both of which are essential conditions
to their existence. It is therefore only from eggs accidentally fished up
at different stages of growth that we may hope to ascertain any facts
respecting the sequence of their development. When hatched, the little
Ctenophore is already quite advanced. It is small when compared with the
size of the egg envelope, and long before it is set free, it swims about
with great velocity within the walls of its diminutive prison (Fig. 35).
The importance of studying the young stages of animals can hardly find a
better illustration than among the Ctenophoræ. Before their extraordinary
embryonic changes were understood, many of the younger forms had found
their way into our scientific annals as distinct animals, and our
nomenclature thus became burdened with long lists of names which will
disappear as our knowledge advances.
The great size of their locomotive flappers in proportion to the rest of
the body, is characteristic of the young Ctenophoræ. They seem like large
paddles on the sides of these tiny transparent spheres, and, owing to their
great power as compared with those of the adult, the young move with
extraordinary rapidity. The Pleurobrachia alone retains its quickness of
motion in after life, and although its long graceful streamers appear only
as short stumpy tentacles in the young (Fig. 34), yet
its active little body would be more easily recognized in the earlier stages
of growth
[Pg 35]
than that of the other Ctenophoræ. Figs. 34, 35, and 36 show the Pleurobrachia
at various stages of growth; Fig. 34, with its thick stunted tentacles and
short rows of flappers, is the youngest; the flappers themselves are rather
long at this age, looking more like stiff hairs than like the minute
fringes of the adult. In Fig. 35 the tentacles are already considerably
longer and more delicate; in Fig. 36 the vertical tubes are already
completed, while Figs. 27-29 present it in its adult condition.
[fig 34]
[fig 35]
[fig 36]
[fig 37]
 |
|
 |
| Fig. 34. Young Pleurobrachia still in the egg; t tentacle, e
eye-speck, c c rows of locomotive flappers, d digestive cavity;
greatly magnified. |
|
Fig. 35. Young Pleurobrachia
swimming about in the egg just before hatching; magnified. |
| | |
 |
|

|
| Fig. 36. Young Pleurobrachia resembling somewhat the adult; f funnel leading to anal opening, l lateral tubes, o o o' o' rows of locomotive flappers; magnified. |
|
Fig. 37. Young Idyia, greatly magnified; lettering as in Fig. 36; d digestive cavity. |
[fig 38]
 Fig. 38. Young Idyia seen from the anal extremity, magnified; a anal opening, other letters as in Fig. 36.
Fig. 38. Young Idyia seen from the anal extremity, magnified; a anal opening, other letters as in Fig. 36.
The Idyia differs greatly in appearance at different periods of its
development, and, indeed, no one would suspect, without some previous
knowledge of its transformations, that the young Idyia,
[Pg 36]
with its rapid gyrations, its short ambulacral tubes, like immense pouches
(Fig. 37), its large pigment spots scattered over the
surface (Fig. 38), was an earlier
stage of the rosy-hued Idyia, which glides through the water with a
scarcely perceptible motion. Figs. 37-40
represent the various stages of its growth. It will be seen how very short
are the locomotive fringes (Fig. 39) in comparison with those of the
full-grown ones (Fig. 33). It is only in the adult
Idyia that these rows attain their full height, and the tubes, ramifying
throughout the body (Fig. 40), are completed.
[fig 39]
[fig 40]
 | |
 |
| Fig. 39. Idyia somewhat older than Fig. 37, lettering as before; magnified. | | Fig. 40. Young Idyia in which the ambulacral tubes begin to ramify; magnified, letters as before. |
The Bolina, in its early condition, recalls the young Pleurobrachia. At
this period it has the same rapid motion, and when somewhat more advanced,
long tentacles, resembling those of the Pleurobrachia, make their
appearance (Fig. 41); it is only at a later period that the tentacles
become contracted, while the large lobes (Fig. 42), so characteristic of
Bolina, are formed by the elongation of the oral end of the body, the
auricles becoming more conspicuous at the same[Pg 37] time (Fig. 43). A little
later the lobes enlarge, the movements become more lazy; it assumes both in
form and habits the character of the adult Bolina.
[fig 41]
[fig 42]
[fig 43]
 |
|
 |
|
 |
| Fig. 41. Young Bolina in stage resembling Pleurobrachia; greatly magnified. |
|
Fig. 42. Young Bolina seen from the broad side, with rudimentary auricles and lobes; magnified.
|
|
Fig. 43. The same as Fig. 42,
seen from the narrow side. |
The series of changes through which the Ctenophoræ pass are as remarkable
as any we shall have occasion to describe, though not accompanied with such
absolutely different conditions of existence. The comparison of the earlier
stages of life in these animals with their adult condition is important,
not only with reference to their mode of development, but also because it
gives us some insight into the relative standing of the different groups,
since it shows us that certain features, permanent in the lower groups, are
transient in the higher ones. A striking instance of this occurs in the
fact mentioned above, that though the long tentacles so characteristic of
the adult Pleurobrachia exist in the young Bolina, they yield in importance
at a later period to the lobes which eventually become the predominant and
characteristic feature of the latter.
DISCOPHORÆ.
The disk of the Discophoræ is by no means so delicate as that of the other
Jelly-fishes. It seems indeed quite solid, and somewhat like cartilage to
the touch, and yet so large a part of its bulk consists of water, that a
Cyanea, weighing when alive about thirty-four pounds, being left to dry in
the sun for some days, was
[Pg 38]
found to have lost about 99/100 of its original weight,—only the merest
film remaining on the paper upon which it had been laid. The prominence
of the disk in this group of Jelly-fishes is well characterized by their
German name, "Scheiben quallen," viz. disk-medusæ. We shall see
hereafter that the disk, so large and seemingly solid in the
Discophoræ, thins out in many of the other Jelly-fishes, and becomes
exceedingly concave. This is especially the case in many of the Hydroid
Medusæ, where it assumes a bell-shaped form, and is constantly spoken of as
the bell. It should be remembered, however, in reading descriptions of
these animals, that the so-called bell is only a modified disk, and
perfectly homologous with that organ in the Discophoræ.
The Discophorous Medusæ are distinguished from all others by the peculiar
nature of the reproductive organs. They are contained in pouches (Fig. 50,
o, o, o, o), the contents of which are first discharged into the
main cavity, and then pass out through the mouth. Pillars support the four
angles of the digestive cavity, thus separating the lower from the upper
floor of the disk, while the chymiferous tubes (Fig. 50) branch and run
into each other near the periphery, forming a more or less complicated
anastomosing network, instead of a simple circular tube, as is the case
with the Hydroid Medusæ. (Fig. 74.)
Cyanea. (Cyanea arctica Pér. et Les.)
In our descriptions of the Discophoræ, we may give the precedence to the
Cyanea on account of its size. This giant among Jelly-fishes is represented
in Fig. 44. It is much to be regretted that these animals,
when they are not so small as to escape attention altogether, are usually
seen out of their native element, thrown dead or dying on the shore, a mass
of decaying gelatinous matter. All persons who have lived near the sea are
familiar with the so-called Sea-blubbers, sometimes strewing the sandy
beaches after the autumn storms in such numbers that it is difficult to
avoid them in walking or driving. In such a condition the Cyanea is far
from being an attractive object; to form an idea of his true appearance,
one must meet him as he swims along at midday, rather lazily withal,
his huge semi-transparent
[Pg 39]
disk, with its flexible lobed margin, glittering in the
sun, and his tentacles floating to a distance of many yards behind him.
Encountering one of those huge Jelly-fishes, when out in a row-boat one
day, we attempted to make a rough measurement of his dimensions upon the
spot. He was lying quietly near the surface, and did not seem in the least
disturbed by the proceeding, but allowed the oar, eight feet in length, to
be laid across the disk, which proved to be about seven feet in diameter.
Backing the boat slowly along the line of the tentacles, which were
floating at their utmost extension behind him, we then measured these in
the same manner, and found them to be rather more than fourteen times the
length of the oar, thus covering a space of some hundred and twelve feet.
This sounds so marvellous that it may be taken as an exaggeration; but
though such an estimate could not of course be absolutely accurate, yet the
facts are rather understated than overstated in the dimensions here given.
And, indeed, the observation was more careful and precise than the
circumstances would lead one to suppose, for the creature lay as quietly,
while his measure was taken, as if he had intended to give every facility
for the operation. This specimen was, however, of unusual size; they more
commonly measure from three to five feet across the disk, while the
tentacles may be thirty or forty feet long. The tentacles are exceedingly
numerous (see Fig. 44), arising in eight distinct bunches,
from the margin of the disk, and hanging down in a complete labyrinth.
These animals are not so harmless as it would seem, from their soft,
gelatinous consistency; it is no pleasant thing when swimming or bathing to
become entangled in this forest of fine feelers, for they have a stinging
property like nettles, and may render a person almost insensible, partly
from pain, and partly from a numbness produced by their contact, before he
is able to free himself from the network in which he is caught. The weapons
by which they produce these results seem so insignificant, that one cannot
but wonder at their power. The tentacles are covered by minute cells,
lasso-cells as they are called, (similar to those of Astrangia, Fig. 19,)
each one of which contains a whip finer than the finest thread, coiled in a
spiral within it.
[Pg 40]
[fig 44]
 Fig. 44. Cyanea arctica; greatly reduced in size.
Fig. 44. Cyanea arctica; greatly reduced in size.
Click on image to view larger sized.
[Pg 41]
These myriad whips can be thrown out at the will of the animal, and really
form an efficient galvanic battery. Behind the veil of tentacles, and
partly hidden by it, four curtains, with lobed or ruffled margins, dimly
seen in Fig. 44, hang down from the under surface of the disk. The ovaries
are formed by four pendent pouches, placed near the sides of the mouth, and
attached to four cavities within the disk. Around the circumference of the
disk are eight eye-specks, each formed by a small tube protected under a
little lappet or hood rising from the upper surface of the disk. The
prevailing color of this huge Jelly-fish is a dark brownish-red, with a
light, milk-white margin, tinged with blue, the tentacles and other pendent
appendages having a somewhat different hue from the disk. The ovaries are
flesh-colored, the curtain formed by the expansion of the lobes of the
mouth is dark brown, while the tentacles are of different colors, some
being yellow, others purple, and others reddish brown or pink.
[fig 45]
[fig 46]
[fig 47]
 |
Fig. 45. Scyphistoma of a Discophore; Aurelia flavidula. (Agassiz.)
|
|
Fig. 47. Strobila of a Discophore; Aurelia flavidula. (Agassiz.) |
|
Fig. 46. Scyphistoma, older than Fig. 45. (Agassiz.)
|
[fig 48]
") Fig. 48. Ephyra of a Discophore; Aurelia flavidula. (Agassiz.)
Fig. 48. Ephyra of a Discophore; Aurelia flavidula. (Agassiz.)
Strange to say, this gigantic Discophore is produced by a Hydroid measuring
not more than half an inch in height when full grown; could we follow the
history of any egg laid by one of these Discophoræ in the autumn, and this
has indeed been partially done, we should see that, like any other planula,
the young hatched from the egg is at first spherical, but presently becomes
pear-shaped, and attaches itself to the ground. From the upper
[Pg 42]
end tentacles project (see Fig. 45), growing more numerous, as in Fig. 46,
though they never exceed sixteen in number. As it increases in height
constrictions take place at different distances along its length, every
such constriction being lobed around its margin, till at last it looks like
a pile of scalloped saucers or disks strung together (see Fig. 47). The
topmost of these disks falls off and dies; but all the others separate by
the deepening of the constrictions, and swim off as little free disks (Fig. 48),
which eventually grow into the enormous Jelly-fish described above.
These three phases of growth, before the relation between them was
understood, have been mistaken for distinct animals, and described as such
under the names of Scyphistoma, Strobila, and Ephyra.
Aurelia. (Aurelia flavidula Pér. et Les.)
[fig 49]
") Fig. 49. Aurelia seen in profile, reduced. (Agassiz.)
Fig. 49. Aurelia seen in profile, reduced. (Agassiz.)
Another large Discophore, though by no means to be compared to the Cyanea
in size, is our common Aurelia (Figs. 49, 50) Its bluish-white disk
measures from twelve to fifteen inches in diameter, but its dimensions are
not increased by the tentacles, which have no great power of contraction
and expansion, and form a short fringe around its margin, widening and
narrowing slightly as the tentacles are stretched or drawn in. It is quite
transparent, as may be seen in Fig 49, where all the fine ramifications of
the chymiferous tubes as well as the ovaries, are seen through the vault of
the disk. Fig. 50 represents the upper surface, with
the ovaries around the mouth, occupying the same position as those of the
Cyanea, though they differ from the latter in their greater rigidity, and
do not hang down in the form of
[Pg 43]
pouches. The males and females in this kind of Jelly-fish may
be distinguished by the difference of color in the reproductive organs,
which are rose-colored in the males, and of a dull yellow in the females.
The process of development is exactly the same in the Aurelia as in the
Cyanea, though there is a very slight difference in their respective
Hydroids. They are, however, so much alike, that one is here made to serve
for both, the above figures being taken from the Hydroid of the Aurelia. It
is curious, that while, as in the case of the Aurelia and Cyanea, very
dissimilar Jelly-fishes may arise from almost identical Hydroids, we have
the reverse of the proposition, in the fact that Hydroids of an entirely
distinct character may produce similar Jelly-fishes.
[fig 50]
") Fig. 50. Aurelia flavidula, seen from above; o mouth, e e e e eyes, m m m m lobes of the mouth, o o o o ovaries, t t t t tentacles, w w ramified tubes.(Agassiz.)
Fig. 50. Aurelia flavidula, seen from above; o mouth, e e e e eyes, m m m m lobes of the mouth, o o o o ovaries, t t t t tentacles, w w ramified tubes.(Agassiz.)
The embryos or little planulæ, hatched from the Cyanea and Aurelia in the
fall, seem to be gregarious in their mode of life, swimming about together
in great numbers till they find a suitable point of attachment, and assume
their fixed Hydroid existence. The Cyaneæ, however, when adult, are usually
found singly, while the Aureliæ, on the contrary, seek each other, and
commonly herd together.
[Pg 44]
The Campanella. (Campanella pachyderma A. Ag.)
[fig 51]
 Fig. 51. Campanella seen in profile; greatly magnified.
Fig. 51. Campanella seen in profile; greatly magnified.
[fig 52]
 Fig. 52. Same, seen from below.
Fig. 52. Same, seen from below.
The Campanella (Fig. 51) is a pretty little Jelly-fish, not larger than a
pin's head, reproduced directly from eggs, without passing through the
Hydroid stage. During its early stages of growth it probably remains
attached to floating animals, thus leading a kind of parasitic existence;
but as its habits are not accurately known, this cannot be asserted as a
constant fact respecting them. The veil in this Jelly-fish is very large,
forming pendent pouches hanging from the circular canal (see Fig. 51), and
leaving but just room enough for the passage of the proboscis between the
folds. It may not be amiss to introduce here a general account of this
organ, which occurs in many of the Medusæ, though it has very different
proportions in the various kinds. It is a delicate membrane, hanging from
the circular tube, so as partially to close the mouth of the bell, leaving
a larger or smaller opening for the passage of the water, which is taken in
and forced out again by the alternate expansions and contractions of the
bell.
[Pg 45]
There are but four chymiferous tubes in the Campanella, and four stiff
tentacles, which in consequence of the peculiar character of the veil
appear, when the animal is seen in profile, to start from the middle of the
disk. The ovaries consist of eight pouches, placed near the point of
junction of the four chymiferous tubes. (Fig. 52.) This little Medusa is of
a dark yellowish color with brownish ocellated spots, scattered profusely
over the upper part of the disk.
Circe. (Trachynema digitale A. Ag.)
[fig 53]
 Fig. 53. Trachynema digitale; about twice the natural size.
Fig. 53. Trachynema digitale; about twice the natural size.
Among the Jelly-fishes, the position of which is somewhat doubtful, is the
Circe (Fig. 53), differing greatly in outline from the ordinary
Jelly-fishes. As may be seen in Figure 53, the bell forms but a small
portion of the animal; it rises in a sharp cone on the summit, thinning out
at the lower edge, to form the large cavity in which hangs the long
proboscis and the eight ovaries, four of which may be seen in Fig. 53
crowded with eggs. The Circe differs in consistency, as well as in form,
from other Jelly-fishes. It is hard and horny to the touch, and the veil,
usually so light and filmy, is here a thick folded membrane, which at every
stroke of the animal forces the water in and out of the cavity. It is very
active, moving by powerful jerks, each one of which throws it far on its
way. It advances usually in straight lines; or, if it wishes to change its
direction, it drives the water out of the veil suddenly
[Pg 46]
on one side or the other, so as to shoot off, sometimes at right angles with
its former path. Four large pedunculated eyes, hidden in the figure by the
tentacles, stand out prominently from the circular tube. When the animal is
in motion, the tentacles are carried closely curled around the edge of the
disk, as in Fig. 53, where the Circe is represented under
a magnifying power of two and a half diameters. This Jelly-fish is of a delicate
rose color, the tentacles assuming, however, a dark-purple tint at their
extremities when contracted.
Lucernaria. (Haliclystus auricula Clark.)
[fig 54]
[fig 55]

Fig. 54. Group of Lucernaria attached to eel-grass; natural size. |
|

Fig. 55. Lucernaria seen from the mouth side. |
One of the prettiest and most graceful, as well as one of the most common
of our Jelly-fishes, is the Lucernaria (Fig. 54). It has such an
extraordinary contractility of all its parts, that it is not easy to
describe it under any definite form or position, since both are constantly
changing; but perhaps of all its various attitudes and outlines none are
more normal to it than those given in Fig. 54. It frequently raises itself
in the upright position represented here by the individual highest on the
stem, spreading itself in the form of a perfectly symmetrical cup or vase,
the margin of which is indented by a succession of inverted scallops, the
point of junction between every two scallops being crowned by a tuft of
tentacles. But watch it for a while, and the sides of this vase turn
backward, spreading completely open, till they present the whole inner
surface, with the edges even curved a little downward, drooping slightly,
and the proboscis rising in the centre. In such an attitude one may trace
with ease the shape of the mouth, the lobes surrounding it, as well as the
tubes and cavities radiating from it toward the margin. A
[Pg 47]
touch is, however, sufficient to make it close upon itself, shrinking together
in the attitude of the third individual in Fig. 54, or
even drawing its tentacles completely in, and contracting all its parts till
it looks like a little ball hanging on the stem. These are but a few of its
manifold changes, for it may be seen in every phase of expansion and contraction.
Let us now look for a moment at the details of its structure. The resemblance
to a cup or vase, as in the upper figure of the wood-cut (Fig.
54), is deceptive; for a vase is hollow, whereas the Lucernaria, though so
delicate and transparent that its upper surface, when thus stretched, seems like
a mere film, is nevertheless a solid gelatinous mass, traversed by certain channels,
cavities, and partitions, but otherwise continuous throughout. The peduncle
by which it is attached is but an extension of the floor of a gelatinous
disk, corresponding to that of any Jelly-fish. Four tubes pass through the
whole length of this peduncle, and open into four chambers, dividing the
digestive cavity above into as many equal spaces. (Fig. 55.) These spaces
are produced by folds in the upper floor of the disk, uniting it to the
lower floor at given distances, and forming so many partition-walls,
dividing the digestive sac into four distinct cavities. These lines of
juncture between the two floors, where the partitions occur, produce the
four radiating lines, running from the proboscis to the margin of the disk,
on the upper surface. (Fig. 55.) The triangular figures, running from the
mouth to each cluster of tentacles, are produced by the ovaries, which
consist of distinct pouches or bags attached to the upper surface of the
disk, and hanging down into the cavities below; every little dot within
these triangular spaces represents such a bag. Each bag is crowded with
eggs, which drop into the digestive cavity at the spawning season, and are
[Pg 48]
passed out at the mouth. The tentacles always grow in clusters, but are
nevertheless arranged according to a regular order. They are club-shaped at
their extremities, but are hollow throughout, opening into the chambers of
the digestive cavity, two of the clusters thus being connected with each
chamber. Their chief office seems to be to catch the food and convey it to
the mouth, though they may also be used as a kind of suckers, and the
animal not unfrequently attaches itself by means of these appendages.
Between every two clusters of tentacles will be observed a short, single
appendage, of an entirely different appearance. These are the so-called
auricles, and though so unlike tentacles in the adult animal, when in their
earlier stages (Fig. 56) they resemble each other closely. But as their
development goes on, the tentacles stretch out into longer, more delicate
flexible organs, while the auricles remain short and compact throughout
life. They contain a slight pigment spot representing an eye, though how
far it serves the purpose of vision remains doubtful. They are chiefly used
by the animal as a means of adhering to any surface upon which it may wish
to fasten itself; for the Lucernaria, though usually found attached to
eel-grass in shoal water, has the power of independent motion, and
frequently separates from its resting-place, floating about freely in the
water for a while, or attaching itself anew by means of the auricles and
tentacles upon some other object. The color of this pretty Acaleph varies
from a greenish hue to green, with a faint tinge of red, or to a reddish
brown. One of its commonest and most exquisite tints is that of a pale
aqua-marine. It may be found along our shores wherever the eel-grass grows,
and as far out as this plant extends. It thrives admirably in confinement,
and for this reason is especially adapted to the aquarium.
[fig 56]
 Fig. 56. Young Lucernaria; magnified.
Fig. 56. Young Lucernaria; magnified.
[Pg 49]
HYDROIDS.
Under this order, the general character of which has already been explained
in the introductory chapter on Acalephs, are included a number of groups
which, whether as Hydroid communities in their earlier phases of existence,
or as free swimming Medusæ in their farther development, challenge our
admiration, both for their beauty of form and color, and their grace of
motion. Some of them are so minute that they escape the observation of all
but those who are laboriously seeking for the hidden treasures of the
microscopic world, but the greater number are large enough to be readily
found by the most inexperienced collector, when his attention is once drawn
to them; and he may easily stock his aquarium with these pretty little
communities, and even trace the development of the Jelly-fishes upon them.
To the Hydroids belong the Campanularians, the Sertularians, and the
Tubularians. Some examples of each, as represented on our shores, will be
found under their different heads, accompanied with full descriptions.
There is another group usually considered as distinct from Hydroids, and
known as a separate order among Acalephs, under the name of Siphonophoræ,
but included with them here in accordance with the views of Vogt, Agassiz,
and others, in whose opinion they differ from the ordinary Hydroid
communities only in being free and floating, instead of fixed to the
ground. Some new facts, published here for the first time, tend to sustain
the accuracy of this classification.[5] With these few preliminary remarks
to show the connection of the order, let us now look at some of the animals
belonging to it more in detail.
Campanularians.
All the Campanularians, of which Oceania (Fig. 68),
Clytia (Fig. 73), and Eucope (Fig. 61) form a part,
belong among those little shrub-like communities of animals called Hydroids,
[Pg 50]
from which most of our Jelly-fishes are developed. They differ in one
essential feature from the Tubularians. (Fig. 93.) The whole stem, from
summit to base, is enveloped in a horny sheath, extending around both
the fertile and sterile individuals of the community, and forming a
network at the base of the stem, which serves as a kind of foundation
for the whole stock. To the naked eye such a community looks like a tiny
shrub (see Fig. 57), with the
branches growing in regular alternation on either side of the stems. The
reproductive calycles, i.e. the protecting envelopes covering the young
Medusæ, usually arise in the angles of the branches formed by a
prolongation of the sheath. These calycles or bells, as they are called,
assume a great variety of shapes,—elliptical, round, pear-shaped, or
ringed like the Clytia. (Fig. 72.) In one such bell
there may be no less than twenty or thirty Medusæ developed one below
the other; when ready to hatch, the calycle bursts and allows them to escape.
Eucope. (Eucope diaphana Ag.)
In Figs. 60 and 61 we have a representation of our little Eucope, one of
the prettiest of the Jelly-fishes belonging to this group; Fig. 57
represents the Hydroid from which it arises; a single branch with the
reproductive bell being magnified in Fig.58.
[Pg 51]
In Fig. 59 is seen a portion
of the Jelly-fish disk, with the fringe of tentacles highly magnified. The
disk of the Eucope (Fig. 60) looks like a shallow bell, of which the
proboscis often seems to form the handle; for the disk has such an
extraordinary thinness that it turns inside out with the greatest ease, so
that the inner surface may become at any moment the outer one, with the
proboscis projecting from it, as in Fig. 60, while the next movement of the
animal may reverse its whole position, and the proboscis then hangs down
from the inside, as in other Jelly-fishes. (See Fig. 61.)
[fig 57]
[fig 58]
[fig 58]
[fig 60]
[fig 61]
[fig 62]

Fig. 57. Hydrarium of Eucope; natural size.
|
|

Fig. 58. Portion of Fig. 57; magnified. |
| |
|

Fig. 59. Part of marginal tube and tentacles of Eucope, greatly magnified; e eye-speck, b base of tentacle., r reentering base of tentacle. |
|

Fig. 60. Young Eucope; magnified. |
| |
|

Fig. 61. Adult Eucope seen in profile; magnified. |
|

Fig. 62. Quarter disk of Fig. 60, seen from below; e e tentacles bearing eye-speck. |
The tentacles are solid and stiff like little hairs, and two of them, in
each quarter-segment of the disk, have small concretions at the base, which
are no doubt eye-specks. (See Fig. 62.) Along the
chymiferous tubes little swellings are developed, which increase gradually,
and become either ovaries or spermaries, according to the sex of the animal.
(Fig. 63.) In the adult the genital organs hang down,
like elongated bags, from the chymiferous
[Pg 52]
tubes. (Fig. 64.) The tentacles are numerous, multiplying
to about a hundred and ninety-two in the adult, and increasing according to
the numerical law to be explained in the description of the Oceania.
[fig 63]
[fig 64]
 between the first set (1).")
Fig. 63. Quarter-disk of young Eucope, older than Fig. 62, with a second set of tentacles (2) between the first set (1). |
|

Fig. 64. Magnified quarter-disk of adult Eucope. |
This little Jelly-fish is one of the most common in our Bay. There is not a
night or day when they cannot be taken in large numbers, from the early
spring till late in the autumn; and as the breeding season lasts during the
whole of that period, they are found in all possible stages of growth. In
consequence of this, the course of their development, and the relation
between the different phases of their existence as Hydroids, and afterwards
as Acalephs, are well known, though the successive steps of their growth
have not been traced connectedly, as in some of the other Jelly-fishes, the
Tima or Melicertum, for instance. The process is, however, so similar
throughout the class of Hydroids, that, having followed it from beginning
to end in some of the groups, we have the key to the history of others,
whose development has not been so fully traced. The eggs laid by the Eucope
in the autumn develop into planulæ, which acquire their full size as
Hydroid communities toward the close of the winter, and the development of
the young Medusæ upon them, as described above, begins with the opening
spring.
Oceania. (Oceania languida A. Ag.)[Pg 53]
The Oceania (Fig. 68) is so delicate and unsubstantial, that with the naked
eye one perceives it only by the more prominent outlines of its structure.
We may see the outline of the disk, but not the disk itself; we may trace
the four faint thread-like lines produced by the radiating tubes traversing
the disk from the summit to the margin; and we may perceive, with far more
distinctness, the four ovaries attached to these tubes near their base; we
may see also the circular tube uniting the radiating tubes, and the
tentacles hanging from it, and we can detect the edge of the filmy veil
that fringes the margin of the disk. But the substance connecting all these
organs is not to be distinguished from the element in which it floats, and
the whole structure looks like a slight web of threads in the water,
without our being able to discern by what means they are held together.
Under the microscope, however, the invisible presently becomes visible, and
we find that this Jelly-fish, like all others, has a solid gelatinous disk.
[fig 65]
[fig 66]
[fig 67]
[fig 68]

Fig. 65. Young Oceania just escaped from its reproductive calycle; magnified. |
|

Fig. 66. The same as Fig. 65, from below, still more magnified; t long tentacles., t' rudimentary tentacle, e eye-speck on each side of base of tentacles. |
| |

Fig. 67. Young Oceania, older than Fig. 65; magnified. |
|

Fig. 68. Adult Oceania; natural size. |
Let us begin with its earlier condition. When it first escapes from the
parent Hydroid stock, the Oceania is almost spherical in form. (See Fig.
65.) The disk is divided by four chymiferous tubes, running from the summit
to the margin, where they meet the circular tube in which they all unite.
At this time, it has but two well-developed tentacles, opposite each other
on the margin of the disk, just at the base of two of the chymiferous tubes
(Fig. 66), while two others are just discernible in a rudimentary
[Pg 54]
state, forming slight projections at the base of the two other tubes.
Fig. 66 gives a view of the animal from below, at this
stage of its growth, while Fig. 65 shows it in profile.
It will be seen by the latter how very spherical is the outline of the disk
at this period, while the proboscis, in which are placed the mouth and digestive
cavity, is quite long, and hangs down considerably below the lower surface of the
disk. As the animal advances in age the disk loses its spherical outline, and
becomes much flattened, as may be seen in Fig. 67. It may
be well to introduce here some explanation of the law according to which the
different sets of tentacles follow each other in successive cycles of growth,
since it is a law of almost universal application in Jelly-fishes and Polyps;
and, owing to the smaller number and simpler arrangement of the tentacles in
Oceania, it may be more easily analyzed in them than in many others, where
the number and complication of the different sets of tentacles make it very
difficult to trace their relation to each other during their successive growth.
We have seen that the Oceania begins life with only two tentacles. These form the
first set, and are marked with the number 1 in the subjoined diagram, which
gives the plan of all the different sets in their regular order. The second
set, marked 2, consists also of two, which are developed at equal distances
between the first two, i.e. at right angles with them. The third set,
however, marked 3,
[Pg 55]
consists of four, as do all the succeeding sets, and they are developed between
the first and second. The fourth set comes in between the first and third; the
fifth between the third and second; the sixth between the first and fourth; the
seventh between the fifth and second; the eighth between the third and fourth;
the ninth between the fifth and third. The ultimate number of tentacles in the
Oceania is thirty-two, or sometimes thirty-six, and the cycles always in twos
or multiples of two. But whatever be the number included in the successive
sets of tentacles, and the unit for the first set ranges from two to
forty-eight, the law in different kinds of Jelly-fishes is always the same,
the youngest set always forming between the oldest preceding set. Thus the
fourth set comes in between the first and third, and the fifth between the
second and third, the intervals occupied now by the fourth set, being
limited by the first set of tentacles on one side, and by the third set on
the other side, while the intervals occupied by the fifth set are bounded
by the second and third sets.
[fig 67b]
 Diagram of succession of tentacles.
Diagram of succession of tentacles.
The little spheres represented between the tentacles on the margin of the
disk, in Figs. 65-67, are eye-specks, and these continue to increase in
number with age; in this the Oceania differs from the Eucope, in which it
will be remembered there were but two eye-specks in each quarter-segment of
the disk throughout life. Fig. 68 represents the adult Oceania in full
size, when it averages from an inch and a half to two inches in diameter.
It is slow and languid in its movements, coming to the surface only in the
hottest hours of the summer days; at such times it basks in the sun,
turning lazily about, and dragging its tentacles after it with seeming
effort. Sometimes it remains for hours suspended in the water, not moving
even its tentacles, and offering a striking contrast to its former great
activity when young, and to the lively little
[Pg 56]
Eucope, which darts through the water at full speed, hardly stopping to rest for
a moment. If the Oceania be disturbed it flattens its disk, and folds itself up
somewhat in the shape of a bale (see Fig. 69), remaining
perfectly still, with the tentacles stretching in every direction. When the
cause of alarm is removed, it gently expands again, resuming its natural outline
and indolent attitudes. The number of these animals is amazing. At certain seasons,
when the weather is favorable, the surface of the sea may be covered with them,
for several miles, so thickly that their disks touch each other. Thus they
remain packed together in a dense mass, allowing themselves to be gently
drifted along by the tide till the sun loses its intensity, when they
retire to deeper waters. Some points, not yet observed, are still wanting
to complete the history of this Jelly-fish. By comparing such facts,
however, as are already collected respecting it, with our fuller knowledge
of the same process of growth in the Eucope, Tima, and Melicertum, we may
form a tolerably correct idea of its development. It is hatched from a
Campanularia.
[fig 69]
 Fig. 69. Attitude assumed by Oceania when disturbed.
Fig. 69. Attitude assumed by Oceania when disturbed.
Clytia. (Clytia bicophora Ag.)
In Figs. 70-73 we have the Acalephian and Hydroid stages of the Clytia
(Fig. 73), another very pretty little Jelly-fish, closely allied to the
Oceania. When first hatched, like the Oceania, it is very convex, almost
thimble-shaped (see Fig. 70), but a little later the disk flattens and
becomes more open, as in Fig. 71. In Fig. 72, we have a branch of the
Hydroid, a Campanularia, greatly magnified, with the annulated reproductive
calycle attached to it, and crowded with Jelly-fishes ready to make their
escape as soon as the calycle bursts. The adult Clytia (Fig. 73) is
somewhat smaller and more active than the Oceania, and
[Pg 57]
is easily recognized by the black base of its tentacles, at their point of
juncture with the margin of the disk. It is more commonly found at night,
than in the day-time, being nocturnal in its habits.
[fig 70]
[fig 71]
[fig 72]
[fig 73]

Fig. 70. Young Clytia just escaped from the reproductive calycle. |
|

Fig. 71. Clytia somewhat older than Fig. 70. |
| |

Fig. 72. Magnified portion of Hydrarium of Clytia. |
|

Fig. 73. Adult Clytia; twice natural size. |
Zygodactyla. (Zygodactyla groenlandica Ag.)
Little has been known, and still less published, of this remarkable genus
of Jelly-fish (Figs. 74, 75) up to the present time. The name Zygodactyla,
or Twinfinger, was given to it by Brandt, from drawings made by Mertens,
who had some opportunity of studying it in his journey around the world.
These drawings
[Pg 58]
were published in the Transactions of the St. Petersburg
Academy. In the year 1848 Professor Agassiz read a paper upon one of the
species of this genus belonging to our coast, before the American Academy,
in which he called it Rhacostoma, not being aware that it had already
received a name, and gave some account of its extraordinary phosphorescent
properties. The name Rhacostoma must of course yield to that of
Zygodactyla, which has a prior claim.
[fig 74]
 Fig. 74. Zygodactyla seen from above.
Fig. 74. Zygodactyla seen from above.
The average size of this Jelly-fish when full grown is from seven to eight
inches in diameter; sometimes it may measure even ten or eleven, but this
is rather rare. The light-violet colored disk is exceedingly delicate and
transparent, its edge being fringed with long fibrous tentacles, tinged
with darker violet at their point of juncture with the disk, and hanging
down a yard and more when fully extended, though they vary in length
according to the size of the specimen, and, in consequence of their
contractile power, may seem much shorter at some moments than at others.
The radiating tubes in this Jelly-fish are exceedingly
[Pg 59]
numerous, the whole inner surface of the disk being ribbed with them. (See
Figs. 74 and 75.) The ovaries follow the length of the tubes, though they
do not extend quite to their extremity, where they join the circular tube
around the margin of the disk; nor do they start exactly at the point where
the tubes diverge from the central cavity, but a little below it. (Fig. 74.)
Each ovary consists of a long, brownish, flat bag, split along the middle,
so closely folded together that it seems like a flat blade attached along the
length of the tube. Perhaps a better comparison would be to a pea-pod greatly
elongated, with the edges split along their line of juncture, and attached
to a tube of the same length. The ovaries are not perfectly straight, but
slightly waving, as may be seen in Fig. 74, and these undulations are
stronger when the ovaries are crowded with eggs, as is the case at the time
of spawning.
[fig 75]
 Fig. 75. Zygodactyla seen in profile.
Fig. 75. Zygodactyla seen in profile.
The large digestive cavity hangs from the centre of the under side of the
disk (Fig. 75), terminating in the proboscis, which, in this kind of
Jelly-fish, is short in proportion to the diameter of the disk, while the
opening of the mouth is very large. (Fig. 74.) It is unfortunate that a
variety of inappropriate names, likely to mislead rather than aid the
unscientific observer, have been applied to different parts of the
Jelly-fish. What we call here digestive cavity, proboscis, and mouth, are,
in fact, parts of one organ. An exceedingly delicate, transparent, filmy
membrane hangs from the under side of the disk; that membrane forms the
outer wall of the digestive cavity, which it encloses; it narrows
[Pg 60]
toward its lower margin, leaving open the circular aperture called the
mouth; this narrowing of the membrane is produced by a number of folds
in its lower part, while at its margin these folds spread out to form
ruffles around the edge of the mouth, and these ruffles again extend
into the long scalloped fringes hanging down below.
The motion of these Jelly-fishes is very slow and sluggish. Like all their
kind, they move by the alternate dilatation and contraction of the disk,
but in the Zygodactyla these undulations have a certain graceful indolence,
very unlike the more rapid movements of many of the Medusæ. It often
remains quite motionless for a long time, and then, if you try to excite it
by disturbing the water in the tank, or by touching it, it heaves a slow,
lazy sigh, with the whole body rising slightly as it does so, and then
relapses into its former inactivity. Indeed, one cannot help being
reminded, when watching the variety in the motions of the different kinds
of Jelly-fishes, of the difference of temperament in human beings. There
are the alert and active ones, ever on the watch, ready to seize the
opportunity as it comes, but missing it sometimes from too great
impatience; and the slow, steady people, with very regular movements, not
so quick perhaps, but as successful in the long run; and the dreamy,
indolent characters, of which the Zygodactyla is one, always floating
languidly about, and rarely surprised into any sudden or abrupt expression.
One would say, too, that they have their aristocratic circles; for there is
a delicate, high-bred grace about some of them quite wanting in the coarser
kinds. The lithe, flexible form of the greyhound is not in stronger
contrast to the heavy, square build of the bull dog, than are some of the
lighter, more frail species of Jelly-fish to the more solid and clumsy
ones. Among these finer kinds we would place the Tima. (Fig. 76.)
Tima. (Tima formosa Ag.)
One's vocabulary is soon exhausted in describing the different degrees of
consistency in the substance of Jelly-fishes. Delicate and transparent as
is the Tima, it has yet a certain robustness and solidity beside the
Oceania, described above. In
[Pg 61]
fact, all are gelatinous, all are more or
less transparent, and it is not easy to describe the various shades of
solidity in jelly. Perhaps they may be more accurately represented by the
impression made upon the touch than upon the sight. If, for instance, you
place your hand upon a Zygodactyla, you feel that you have come in contact
with a substance that has a positive consistency; but if you dip your
finger into a bowl where a Tima is swimming, and touch its disk, you will
feel no difference between it and the water in which it floats, and will
not be aware that you have reached it till the animal shrinks away from the
contact.
[fig 76]
[fig 77]

Fig. 76. Tima; half natural size. |
|

Fig. 77. One of the lips of the mouth at the extremity of the long proboscis; m mouth, d digestive cavity, c chymiferous tube. |
The adult Tima, represented in Fig. 76, is not more than an inch and a half
or two inches in diameter. Instead of countless tubes diverging from the
digestive cavity to the margin of the disk, as in the Zygodactyla, there
are but four. The digestive cavity in the Tima is much smaller than in the
Zygodactyla, and is placed at the end of the proboscis, which is long, and
hangs down far below the disk. This removal of the digestive cavity to the
extremity of the proboscis gives to the tubes
[Pg 62]
arising from it a very different and much sharper curve than they have
in the Zygodactyla. In the Tima they start from the end of the proboscis,
as may be seen in the wood-cut (Fig. 76), and then turn abruptly off, when
they arrive at the under surface of the disk, to reach its margin. The disk
has, as usual, its veil and its fringe of tentacles; the tentacles in the
full-grown Tima are few,—seven in all the four intermediate spaces
between the tubes, with one at the base of each tube, making thirty-two in
all. The ovaries, which are milk-white, follow the line of the tubes, as in
the Zygodactyla, and have very undulating folds when full of eggs. The tubes
meet in the digestive cavity, the margin of which spreads out to form four
ruffled edges that hang down from it. One of these ruffles, considerably
magnified, is represented in Fig. 77. In Fig. 78 we have a portion of the
Hydroid stock from which this Jelly-fish arises, also greatly magnified.
The Tima is very active, yet not abrupt in its motions; but when in good
condition it is constantly moving about, rising to the surface by the
regular pulsations of the disk, or swimming from side to side, or poising
itself quietly in the water, giving now and then a gentle undulation to keep
itself in position.
[fig 78]
 Fig. 78. Magnified head of Hydrarium of Tima.
Fig. 78. Magnified head of Hydrarium of Tima.
Though not a very frequent visitor of our shores, the appearance of the
Tima is not limited by the seasons, since they are found at all times of
the year. It is a fact, unexplained as yet, that the Tima and many other
Jelly-fishes are never seen except when full grown. What may be the haunts
and habits of these animals from the time of their hatching till they make their
[Pg 63]
appearance again in the adult condition, is not known, though it is
probable that they remain at the bottom during this period, and only come
to the surface to spawn. This impression is confirmed by the observations
made upon a very young Cyanea which was kept for a long time in
confinement; but a question of this kind cannot of course be settled by a
single experiment.[6]
[fig 79]
") Fig. 79. Melicertum campanula seen
Fig. 79. Melicertum campanula seen
from above; m mouth, o o ovaries,
t t tentacles. (Agassiz.)
Melicertum. (Melicertum campanula Pér. et Les.)
A pretty Medusa, smaller and far more readily obtained than the Tima, is
the Melicertum. (Fig. 80.) Its disk has a yellowish hue, and from its
margin hangs a heavy row of yellow tentacles, while the eight ovaries (Fig.
79) are of a darker shade of the same color. This little golden-tinted
Jelly-fish, moving through the water with short, quick throbs, produced by
the rapid rise and fall of the disk, is a very graceful object. Its bright
color, made particularly prominent by the darker undulating lines of the
ovaries, which become very marked near the spawning season, renders it more
conspicuous in the water than one would suppose from its size; for it does
not measure more than an inch in height when full grown. (See Fig. 80.)
[Pg 64]
Development of Melicertum and Tima.
[fig 80]
 Fig. 80. Melicertum seen in profile; natural size.
Fig. 80. Melicertum seen in profile; natural size.
In the Melicertum and Tima we have had the good fortune to trace the
process by which the eggs are changed into Hydroid communities. If any one
has a curiosity to follow for themselves this singular history of alternate
generations, the Melicertum is a good subject for the experiment, as it
thrives well in confinement. After keeping a number of them in a large
glass jar for a couple of days at the time of spawning, it will be found
that the ovaries, which were at first quite full of eggs, are emptied, and
that a number of planulæ; are swimming about near the bottom of the vessel.
After a day or two the outline of these planulæ, spherical at first,
becomes pear-shaped (see Fig. 81), and presently they attach themselves by
the blunt end to the bottom of the jar. (Fig. 82.) Thus their Hydroid life
begins; they elongate gradually, the horny sheath is formed around them,
tentacles arise on the upper
[Pg 65]
end, short and stunted at first, but tapering rapidly out into fine flexible
feelers, the stem branches, and we have a little Hydroid community (Fig. 83),
upon which, in the course of the following spring, the reproductive calycles
containing the Medusæ buds will be developed, as in the case of the
Eucope and Clytia. The Tima passes through exactly the same process, though
the shape of the planulæ and the appearance of the young differ from that
of the Melicertum, as may be seen in Fig. 78, where a single head of the Tima
Hydroid, greatly magnified, is represented. By combining the above observations
upon the development of the Hydroids of the Melicertum and Tima with those
previously mentioned upon the young Medusa arising from reproductive calycles in
the Eucope and Clytia, we get a complete picture of all the changes through
which any one of these Hydroid Medusæ passes, from its Hydroid condition
to the moment when it enters upon an independent existence as a free Jelly-fish.
[fig 81]
[fig 82]
[fig 83]

Fig. 81. Planula of Melicertum; magnified. |
|

Fig. 83. Young Hydrarium developed from planulæ; magnified. |

Fig. 82. Cluster of planulæ just attached to the ground. |
(Laomedea amphora Ag.)
The Medusæ of the Campanularians are not all free. On the contrary,
in many of the species they always remain attached to the Hydroid, never
attaining so high a development as the free Medusæ, and withering on
the stem after having laid their eggs.
[Pg 66]
Such is the Laomedea amphora, quite common on
all the bridges connecting Boston with the country, where, on account of
the large amount of food brought down from the sewers by the river, they
thrive wonderfully, growing to a great size, sometimes measuring from a
foot to eighteen inches in height.
[fig 84]
[fig 85]

Fig. 84. Colony of Dynamena pumila; natural size. |
|

Fig. 85. Magnified portion of Fig. 84. |
Sertularians.
The Sertularians form another group of Hydroids closely allied to the
Campanularians, though differing from them in the arrangement of the
sterile Hydræ upon the stem. Among these one of the most numerous is the
Dynamena (Dynamena pumila Lamx., Fig. 84), which hangs its yellowish
fringes from almost every sea-weed above low-water-mark. It is especially
thick and luxuriant on the fronds of our common Fucus vesiculosus. The
color is usually of a pale yellow, though sometimes it is nearly white, and
when first taken from the water it has a glittering look, such as a white
frost leaves on a spray of grass. Fig. 84 represents such a cluster in
natural size, while Fig. 85 shows a piece of the stem highly magnified,
with a reproductive calycle attached to the side of a sterile Hydra stem.
Many of these Sertularian Hydroids assume the most graceful forms, hanging
like long pendent streamers from the Laminaria, or in other instances
resembling miniature trees. One of these tree-like
[Pg 67]
Sertularians (Dyphasia rosacea Ag.), abundant on all rocks in
sheltered places immediately below low-water-mark, is represented in Fig. 86.
In both these Sertularians the Medusæ wither on the stock, never becoming
free. The free Medusæ of the Sertularians are only known in their adult
condition in a single genus, which is closely allied to Melicertum, and which
is produced from a Hydroid genus called Lafoea. Fig. 87 represents one of these
young Sertularian Medusæ (Lafoea cornuta Lamx.).
[fig 86]
[fig 87]

Fig. 86. Dyphasia rosacea, natural size. |
|

Fig. 87. Medusa of Lafoea. |
Tubularians.
In the Sertularian and Campanularian Hydroids we have found that the
communities consist generally of a large number of small individuals, so
small, indeed, that it is hardly possible at first glance to distinguish
the separate members of these miniature societies. Among the Tubularians,
on the contrary, the communities are usually composed of a small number of
comparatively large individuals; and indeed these Hydroids may even grow
singly, as in the case of the Hybocodon (Fig. 104), which attains several
inches in height. There is also another general feature in which the
Tubularians differ from both the other groups of Hydroids. In the latter,
the horny sheath which encloses the stem extends to form a protecting
calycle around the Hydra
[Pg 68]
heads. This protecting calycle is wanting round the heads of the Tubularians,
though their stems are surrounded by a sheath.
[fig 88]
[fig 89]
[fig 90]
Sarsia. (Coryne mirabilis Ag.)
Among the most common of our Tubularians is a small, mossy Hydroid (Fig.
88), covering the rocks between tides, in patches of several feet in
diameter. Fig. 89 represents a single head from this little mossy tuft
greatly magnified, in which is seen the medusa bud arising from the stem by
the process already described in the other Hydroids. In Fig. 90 we have the
little Jelly-fish in its adult condition, about the size of a small walnut,
with a wide circular opening, through which passes the long proboscis,[Pg 69]
hanging from the under surface of the disk to a considerable distance below
its margin. The four tentacles are of an immense length when compared to
the size of the animal. As a general thing, the tentacles are less numerous
in the Tubularian Medusæ than in those arising from other Hydroids; they
want also the singular limestone concretions found at the base of the
tentacles in the Campanularian Medusæ. In Fig. 91 we have one of the
Tubularian Medusæ (Turris vesicaria A. Ag.) which lifts a rather larger
number of tentacles than is usual among these Jelly-fishes. We never find
the tentacles multiplying almost indefinitely in them, as in Zygodactyla
and Eucope. The little Jelly-fish described above is known as Sarsia, while
its Hydroid is called Coryne. These names having been given to the separate
phases of its existence before their connection was understood, and when
they were supposed to represent two distinct animals. They are especially
interesting with reference to the history of Hydroids in general, because
they were among the first of these animals in whom the true relation
between the different phases of their existence was discovered. Lesson
named the Sarsia after the great Norwegian naturalist, Sars, to whom we owe
so large a part of what is at present known respecting this curious subject
of alternate generations.
[fig 91]
 Fig. 91. Turris vesicaria; natural size.
Fig. 91. Turris vesicaria; natural size.
Bougainvillia. (Bougainvillia superciliaris Ag.)
The Bougainvillia (Fig. 92), is one of our most common Jelly-fishes,
frequenting our wharves as well as our sea-shore during the spring. The
tentacles are arranged in four bunches[Pg 70] or clusters at the junction of the
radiating tubes with the circular tube, from which they may be seen
extending in every direction whenever these animals remain quietly
suspended in the water,—a favorite attitude with them, and one which they
retain sometimes for days, seeming to make no effort beyond that of gently
playing their tentacles to and fro (Fig. 92). These tentacles are capable
of immense extension, sometimes to ten or fifteen times the diameter of the
bell. The proboscis is not simple as in the Sarsia, but looks like a yellow
urn suspended at its four corners from the chymiferous tubes. The oral
opening is entirely concealed by clusters of shorter tentacles surrounding
the mouth in a close wreath, on which the eggs are supported. A highly
magnified branch of the Hydroid stock from[Pg 71] which this Medusa arises is
represented in Fig. 93. There we see the little Jelly-fishes in different
degrees of development on the stem, while in Figs. 94-97 they are given
separately and still more enlarged. In Fig. 94 the outline of the
Jelly-fish is still oval, the proboscis is but just formed, and the
tentacles appear only as round swellings or knobs. In Fig. 95 a depression
has taken place at the upper end, presently to be an opening, the proboscis
is enlarged, and the tentacles lengthened, but still turned inward. In Fig.
96 the appendages of the proboscis are quite conspicuous, the tentacles are
turned outward, and the
[Pg 72]
Jelly-fish is almost ready to break from its
attachment, having assumed its ultimate outline. Fig. 97 represents it just
after it has separated from the stem, when it has only two tentacles at
each cluster and simple knobs around the mouth, instead of the complicated
branching tentacles of the adult.
[fig 92]
[fig 93]

Fig. 92. Bougainvillia; magnified. |

Fig. 93. Hydrarium of Bougainvillia; magnified. |
[fig 94]
[fig 95]
[fig 96]
[fig 97]

Fig. 94 |

Fig. 95 |

Fig. 96 |

Fig. 97 |
Figs. 94, 95, 96. Medusæ buds of Fig. 93, in different
degrees of development.
Fig. 97. Young Medusa just freed from the Hydroid; magnified. |
Tubularia. (Tubularia Couthouyi Ag.)
There are several other Tubularians common in our waters which should not
be passed over without mention, although as this little book is by no means
intended as a complete text-book, but rather as a volume of hints for
amateur collectors, we would avoid as much as possible encumbering it with
many names, or with descriptions already given in more comprehensive works.
This Tubularia is interesting, however, from the fact that the Medusæ buds
are never freed from the stem, and do not develop into full-grown
Jelly-fishes, but always remain abortive. Fig. 98 represents one head of
such a Hydroid with the Medusæ buds pendent from it in a thick cluster,
while in Fig. 99 we have a few of them sufficiently magnified to show that,
though presenting the four chymiferous tubes, they are otherwise
exceedingly simple in structure, as compared with the free Jelly-fishes.
[fig 98]
[fig 99]
") Fig. 98. Tubularia; magnified. (Agassiz.) Fig. 98. Tubularia; magnified. (Agassiz.) |
|
") Fig. 99. Part of cluster of Medusæ of Fig. 98; magnified. (Agassiz.) Fig. 99. Part of cluster of Medusæ of Fig. 98; magnified. (Agassiz.) |
[Pg 73]
Hydractinia. (Hydractinia polyclina Ag.)
This is another Tubularian, covering the surface of rocks in tide-pools, or
attaching itself upon shells inhabited by hermit crabs. Indeed it was upon
these shells that the Hydractinia was first noticed, and it was long
supposed that the wanderings to which the little colony was thus subjected
were necessary for its healthy development. But subsequent observations
have shown that it attaches itself quite as frequently to the solid rock as
to these nomadic shells. It has a rosy color, and, being very small, it
looks, until one examines it closely, more like a thick red carpet of soft
moss, than like a colony of animals. These communities are distinct in sex,
the fertile individuals in each being either all male or all female. In
Fig. 100 we have a portion of a female colony, representing one fertile
head, in which the buds are crowded with Medusæ; one sterile head,
surrounded by its wreath of tentacles; and still another member of the
society whose office is not fully understood, unless it be that of a kind
of purveyor, catching food for the rest. Fig. 101 represents the
corresponding individuals taken from a male colony. The sex makes little
difference in the appearance of the reproductive heads. All the individuals
of a Hydractinia colony are connected at the base by a horny network,
rising occasionally
[Pg 74]
into points of a conical or cylindrical shape. This polymorphism among the
Tubularians is another evidence of the relation between the
Siphonophoræ, or floating Hydroids, and the fixed Hydroids.
[fig 100]
[fig 101]
 |
|
 |
Fig. 100. Female colony of Hydractinia;
a sterile individual, b fertile individual
producing female Medusæ, c fertile
individual with globular tentacles without
Medusæ, d e f g h i Medusæ in different
stages of growth, o mouth tentacles. (Agassiz.) |
|
Fig. 101. Male colony; a a sterile individuals,
b fertile individuals producing male Medusæ,
d; o globular tentacles, t slender tentacles
of sterile individual. (Agassiz.) |
Hybocodon. (Hybocodon prolifer Ag.)
Among our Medusæ derived from a Tubularian stock is the Hybocodon, viz. the
hunchbacked Medusa (Fig. 102), a singular little Jelly-fish, odd and
unsymmetrical in shape, as its name indicates, and interesting from its
relations to one of our floating communities, the Nanomia, presently to be
described. Instead of the evenly proportioned bell of the ordinary Medusæ,
the Hybocodon has a one-sided outline (Fig. 102), one large tentacle only
being fully developed, while the others remain always abortive, so that the
whole weight of the structure is thrown on one half of the bell. Upon this
large tentacle small Jelly-fishes, similar to the original, are produced by
budding, this process going on till ten or twelve such Jelly-fishes (Fig.
103) may be seen suspended from the tentacle. Up to this time it has remained
[Pg 75]
connected with the Hydroid from which it arises, a rather large
Tubularian, usually growing singly (Fig. 104), and of a deep orange-red in
color. But at this stage of its existence it frees itself, and leads an
independent life hereafter, swimming about with a quick, darting motion. In
the account of the Nanomia, the homology between its scale, or abortive
Medusa, and the Hybocodon, is traced in detail, and I need only allude to
it here. Though this Medusa is so peculiar in appearance, the Tubularian
from which it is derived is very like the Tubularia Couthouyi, already
described. This is one of the instances before alluded to, in which closely
allied forms give rise to very dissimilar ones, or, as in many cases, the
very reverse of this takes place, and closely allied forms arise from very
dissimilar ones.
[fig 102]
[fig 103]
[fig 104]
 |
Fig. 102. Unsymmetrical free Medusa of Hybocodon; r o
chymiferous tubes, v proboscis,
s circular tube, m young Medusæ at
base of long tentacle t. (Agassiz.) |
Fig. 103. Medusa bud of Hybocodon; a base of attachment,
o proboscis, c circular tube,
d young Medusæ at base of long tentacle
t. (Agassiz.) |
Fig. 104. Single head of Hybocodon Hydroid; a stem, d
Medusæ buds,
o tentacles round mouth. (Agassiz.) |
Dysmorphosa. (Dysmorphosa fulgurans A. Ag.)
Besides the budding at the base of the tentacle, as in Hybocodon, we find
another mode of development among Hydroid Medusæ, viz. that of budding from
the proboscis. One of our most common little Jelly-fishes, the Dysmorphosa
(Fig. 105), to which we owe the occasional blue phosphorescence of the sea,
so brilliant at times, buds in this manner. Fig. 105 represents an adult
Dysmorphosa, on the proboscis of which may be seen three small buds in
different stages of development. In Fig. 106 the proboscis is more
enlarged, showing one of the little Jelly-fishes similar to the parent,
just ready to drop off. We need not wonder at the
[Pg 76]
immense number of these animals, with which the sea actually swarms at times,
when we know that as fast as they are dropped, and it takes but a few days
to complete their development, they each begin the same process; so that
in the course of a week or ten days one such Medusa, supposing it to have
produced six buds only, will have given rise to forty-two Jelly-fishes,
thirty-six of which may be equally prolific in the same short period.
These Medusæ budding thus, and swimming about, carrying their
young with them, bear such a close resemblance to the floating communities
of Hydroids formerly known as Siphonophoræ, that did we not know
that some of them arise from Tubularians, it would be natural to associate
them with the Siphonophoræ.
[fig 105]
[fig 106]
 |
|
 |
| Fig. 105. Dysmorphosa seen in profile; magnified. |
|
Fig. 106. Magnified proboscis of Dysmorphosa
with young Medusæ budding from it. |
Nanomia. (Nanomia cara A. Ag.)
The Nanomia (Fig. 115), our free floating Hydroid, consists, when first
formed, of a single Hydra containing an oblong oil bubble (Fig. 107). The
whole organisation of such a Hydra is limited to a simple digestive cavity;
it has, in fact, but one organ, and one function, and consists of an
alimentary sac resembling the proboscis of a Medusa (Fig. 107); the oil
bubble is separated from it by a transverse partition, and has no
connection with the cavity. Presently, between the oil bubble and the
cavity arise a number of buds of various character (Fig. 108), which we
will describe one by one, beginning with those nearest the oil bubble,
since these upper members of the little swimming community bear a very
important part in its history. The infant community (Fig. 108) passes
rapidly into the stage represented in Fig. 109, and then through all the
stages intermediate between this and the adult, shown in its natural size
in Fig. 115. The upper buds enlarge
[Pg 77]
gradually, and soon take upon themselves a perfect Medusa structure
(Fig. 110), with the exception of the proboscis, the absence of which
is easily understood, when we find that these Medusæ, serve the
purpose of locomotion only, having no share in the function of feeding
the community, so that a digestive apparatus would be quite superfluous
for them. In every other respect they are perfect Medusæ,
attached to the Hydra as the Medusa buds always are when first formed,
having the (four) chymiferous tubes, characteristic of all Hydroid Medusæ,
radiating from the centre to the periphery; two of these tubes are very
winding, as may be seen in Fig. 110, while the other pair are straight. The
Medusæ themselves are heart-shaped in form, depressed at the centre of the
upper surface, and bulging on either side into wing-like expansions, where
they join the stem. These expansions interlock with one another, crossing
nearly at right angles. The Medusæ-like buds are the swimming bells; by
their contractions, alternately taking in and throwing out the water, they
impel the whole community forward, so that it seems rather to move like one
animal, than like a combination of individuals.
[fig 107]
[fig 108]
[fig 109]
[fig 110]

Fig. 107. Young Nanomia; magnified. |

Fig. 108. Young Nanomia with rudimentary Medusæ. |

Fig. 109. Young Nanomia, older than Fig. 108. |

Fig. 110. Heart-shaped swimming bell of Nanomia; magnified. |
[Pg 78]
[fig 111]
 Fig. 111. Cluster of Medusæ with tentacles having pendent knobs.
Fig. 111. Cluster of Medusæ with tentacles having pendent knobs.
Besides these locomotive members, the community contains three kinds of
Hydræ arising as buds from the primitive Hydra below the swimming bells,
the latter remaining always nearest the oil bubble at the top, while the
first Hydra, the founder of the community, in proportion as the new
individuals are added, is gradually pushed downward, and remains always at
the end of the string, the stem of which is formed by the elongated neck of
the primitive Hydra. All the three sets of Hydræ have certain features in
common, while they have other distinguishing characteristics marking them
as distinct individuals. They are all accompanied by triangular shields
(Fig. 111), arising with them at the same point on the parent stem, and all
are furnished with tentacles hanging down from the summit of the Hydra at
the side opposite the shield. These facts are important to remember, since
we shall presently perceive, upon analyzing their parts, that these Hydræ
have a close homology to the Hybocodon. The tentacles differ in structure
as well as in number for each kind of Hydra. Having shown in what
characters they agree, let us now take each set individually, and see what
differences they present.
In the first set which we will examine the Hydra is open-mouthed. Like the
original Hydra, it is only a digestive tube, similar in all respects to the
proboscis of a Medusa-disk. Its only function is that of feeding, and it
shows a laudable fidelity to its calling, being very constantly and
earnestly engaged in the work. Let us add, however, that in this instance
the occupation is not a wholly selfish one, since the cavity of every Hydra
communicates with that of the stem, and the food taken in at these
over-gaping mouths, is at once circulated through all parts of the
community, with the exception of the oil bubble, from which it is excluded
by the transverse partition dividing it from
[Pg 79]
all the lower members of the stock. The shields share in this general
nourishment of the compound body by means of chymiferous tubes extending
toward the outer surface, and opening into the cavity of the stem. The
mouth of this Hydra is very flexible (Fig. 111), expanding and contracting
at the will of the animal, and sometimes acting as a sucker, fastening
itself, leech-like, on the object from which it seeks to draw its
sustenance. (See Fig. 111.) The tentacles attached to this set of
Hydræ are exceedingly long and delicate. They arise in a cluster
at the upper and inner edge of the Hydra, just at its point of juncture
with the stem, and being extremely flexible and contractile, their long
tendril-like sprays are thrown out in an endless variety of attitudes.
(See Fig. 115.) Along the whole length of this kind of tentacle are
attached little pendent knobs at even distances; Fig. 112
represents such a knob greatly magnified, and absolutely paved with
lasso-cells, the inner and smaller ones being surrounded by a row of larger
ones.
[fig 112]
[fig 113]
 |
|
 |
| Fig. 112. Magnified pendent knob. |
|
Fig. 113. Medusa with corkscrew shaped tentacles. |
The second set of Hydræ (Fig. 113), are also open-mouthed, corresponding
with those described above, in everything except the tentacles, which are
both shorter and thicker, and are coiled in a corkscrew-like spiral. These
are thickly studded for their whole length with lasso-cells. (See Fig.
113.)
In the third and last set of Hydræ (Fig. 114), the mouth
[Pg 80]
is closed; they have, therefore, no share in feeding the community, but
receive their nourishment from the cavity of the stem into which they open.
They differ also from the others in having a single tentacle instead of a
cluster, and on this tentacle the lasso-cells are scattered at uneven
distances (Fig. 114). The special function of these closed Hydræ is
yet to be explained; they have oil bubbles at their upper end (see Fig. 111,
the top Hydra), and though we have never seen them drop off, it seems natural
to suppose that they do separate from the parent stock, and found new
communities similar to those from which they arise.
[fig 114]
[fig 115]

Fig. 114. Medusa with a simple thread-like tentacle. |
|

Fig. 115. Adult Nanomia, natural size, at rest. |
The intricate story of this singular compound existence does not end here.
There is still another set of individuals whose share in maintaining the
life of the community is by no means the least important. Little bunches of
buds, of a different character from any described above, may be seen at
certain distances along the lower part of the stem. These are the
reproductive individuals. They are clusters of imperfect sexual Medusæ,
resembling the rudimentary Medusæ of Tubularia (Fig. 99), which are never
freed from the parent stem, but discharge their contents at the breeding
season. Like many other compound Hydroids, the sexes are never combined, in
one of these communities; they are always either male or female, and as
those with female buds have not yet been observed, we can only judge by
inference of their probable character. Front what is already known,
however, of Hydroid communities of a like description, we suppose that the
process of reproduction must be the same in these, and that the female
stocks of Nanomia give birth to small Jelly-fishes, the eggs of which
become oil bubbles, similar to that with which our little community began.
(Fig. 108.)
[fig 116]
 Fig. 116. Oil float of Nanomia; greatly magnified.
Fig. 116. Oil float of Nanomia; greatly magnified.
By the time all these individuals have been added along the length of the
stem, the stem itself has grown to be about three inches long (Fig. 115),
though the tentacles hanging from the various members of the community give
to the whole an appearance of much greater length. The motion of this
little string of
[Pg 81]
living beings is most graceful. The oil bubble (Fig. 116)
at the upper end is their float; the swimming bells immediately below it
(Fig. 110), by the convulsive contractions of which they move along, are
their oars. The water is not taken in and expelled again by all the bells
at once, but first from all the bells on one side, beginning at the lower
one, and then from all those on the opposite side, beginning also at the
lower one; this alternate
[Pg 82]
action gives to their movements a swinging,
swaying character, expressive of the utmost freedom and grace. Whether such
a little community darts with a lightning-like speed through the water, or
floats quietly up and down, for its movements are both rapid and gentle, it
always sways in this way from side to side. Its beauty is increased by the
spots of bright red scattered along the length of the stock at the base and
tips of the Hydræ, as well as upon the tentacles. The movements and
attitudes of the tentacles are most various. Sometimes they shoot them out
in straight lines on either side, and then the aspect of the whole thing
reminds one of a tiny chandelier in which the coral drops make the
pendants, or they may be caught up in a succession of loops or floating in
long streamers; indeed, there is no end to the fantastic forms they assume,
ever astonishing you by some new combination of curves. The prevailing hue
of the whole community is rosy, with the exception of the oil bubble or
float, which looks a bright garnet color when seen in certain lights.
Let us now compare one of the Hydræ hanging from the stem (Fig. 113) with
the Hybocodon (Fig. 102). The reader will remember the unsymmetrical bell
of this singular Medusa, one half of its disk more largely developed than
the other, with the proboscis hanging from the centre, and the cluster of
tentacles from one side. Let us now split the bell so as to divide it in
two halves with the proboscis hanging between them; next enlarge the side
where there are no tentacles, and give it a triangular outline; then
contract the opposite side so as to draw up the cluster of tentacles to
meet the base of the proboscis, and what have we? The proboscis now
corresponds to the Hydra of our Nanomia, with the cluster of tentacles
attached to its upper edge (Fig. 113), while the enlarged half of the bell
represents the shield. If this homology be correct it shows that the
Nanomia is not, as some naturalists have supposed all the Siphonophores to
be, a single animal, its different parts being a mere collection of organs
endowed with special functions, as feeding, locomotion, reproduction, &c.,
[Pg 83]
but that it is indeed a community of distinct individuals corresponding
exactly to the polymorphous Hydroids, whose stocks are attached, such as
Hydractinia, and differing from them only in being free and floating.
[fig 117]
 Fig. 117. Physalia; a b air sac with crest c, m
bunches of individuals, n central tentacles, t t expanded tentacles.
(Agassiz.)
Fig. 117. Physalia; a b air sac with crest c, m
bunches of individuals, n central tentacles, t t expanded tentacles.
(Agassiz.)
The homologies of the Siphonophoræ or floating Hydroids, with many of the
fixed Hydroids, is perhaps more striking when we compare the earlier stages
of their growth. Suppose, for instance, that the planula of our Melicertum
(See Fig. 81) should undergo its development without becoming attached to
the ground,—what should we then have? A floating community (Fig. 83),
including on the same stock like the Nanomia, both sterile and fertile
Hydræ, from the latter of which Medusæ bells are developed. The little
Hydractinia community (Fig. 100), in which we have no less than four
distinct kinds of individuals, each performing a definite distinct
function, affords a still better comparison.
Physalia. (Physalia Arethusa Til.)
Among the most beautiful of the Siphonophores, is the well-known Physalia
or Portuguese man-of-war, represented in Fig. 117. The float above is a
sort of crested sac or bladder, while the long streamers below consist of a
number of individuals corresponding in their nature and functions to those
composing a Hydroid community. Among them are the fertile and sterile Hydræ
(Fig. 118), the feeders and Medusæ bells (Fig. 119). The Physalia properly
belongs to tropical waters, but sometimes floats northward, in the warm
current of the Gulf Stream, and is stranded on Cape Cod. When found so far
from their home, however, they have usually lost much of their vividness of color;
[Pg 84]
to judge of their beauty one should see them in the Gulf of Mexico,
sailing along with their brilliant float fully expanded, their crest
raised, and their long tentacles trailing after them.
[fig 118]
[fig 119]
")
Fig. 118. Bunch of Hydræ; a base of attachment, b b b single Hydræ, c c tentacles. (Agassiz.) |
|
")
Fig. 119. Bunch of Hydræ; cluster of Medusæ; b b Hydræ with tentacles, c d bunches of Medusæ (Agassiz.) |
Velella. (Velella mutica Bosc.)
Another very beautiful floating Hydroid, occasionally caught in our waters,
though its home is also far to the south, is the Velella (Fig. 120). It is
bright blue in color, and in form not unlike a little flat boat with an
upright sail. Its Medusa (Fig. 121) resembles so much that of some of our
Tubularians, that it has actually been removed on this account from the old
group of Siphonophoræ, and placed next the Tubularians; another evidence of
the close affinity between the former and the Hydroids.
[fig 120]
[fig 121]
") |
|
") |
| Fig. 120. Velella; m so-called mouth, a tentacles. (Agassiz.) |
|
Fig. 121. Free Medusa of Velella; a proboscis, b chymiferous tube, c circular tube. (Agassiz.) |
[Pg 85]
MODE OF CATCHING JELLY-FISHES.
[fig 122]
 Fig. 122. Ptychogena, natural size.
Fig. 122. Ptychogena, natural size.
Not the least attractive feature in the study of these animals, is the mode
of catching them. We will suppose it to be a warm, still morning at Nahant,
in the last week of August, with a breath of autumn in the haze that
softens the outlines of the opposite shore, and makes the horizon line a
little dim. It is about eleven o'clock, for few of the Jelly-fishes are
early risers; they like the warm sun, and at an earlier hour they are not
to be found very near the surface. The sea is white and glassy, with a
slight swell but no ripple, and seems almost motionless as we put off in a
dory from the beach near Saunders's Ledge. We are provided with two
buckets, one for the larger Jelly-fishes, the Zygodactyla, Aurelia, &c.,
the other for the smaller fry, such as the various kinds of Ctenophoræ, the
Tima, Melicertum, &c. Beside these, we have two nets and glass bowls, in
which to take up the more fragile creatures that cannot bear rough
handling. A bump or two on the stones before we are fairly launched, a
shove of the oar to keep the boat well out from the rocks along which we
skirt for a moment, and now we are off. We pull around the point to our
left and turn toward the Ledge, filling our buckets as we go. Now we are
crossing the shallows that make the channel between the inner and outer
rocks of Saunders's Ledge. Look down,—how clear the water is and how
lovely the sea-weeds, above which we are floating, dark brown and purple
fronds of the Ulvæ, and the long blades of the Laminaria with mossy green
tufts between. As we issue from this narrow passage we must be on the
watch, for the tide is rising, and may come laden with treasures, as it
sweeps through it. A sudden cry from the oarsman at the bow, not of rocks
or breakers ahead, but of "A new Jelly-fish astern!" The quick eye of the
naturalist of the party pronounces it unknown to zoölogists, un-described
by any scientific pen. Now what excitement! "Out with the net!—we have
passed him! he has gone down! no, there he is again! back us a bit." Here
he is floating close by us; now he is within the circle of the net, but he
is too delicate
[Pg 86]
to be caught safely in that way, so, while one of us moves
the net gently about, to keep him within the space enclosed by it, another
slips the glass bowl under him, lifts it quickly, and there is a general
exclamation of triumph and delight,—we have him. And now we look more
closely; yes, decidedly he is a novelty as well as a beauty. (See Fig. 122,
Ptychogena lactea A. Ag.) Those white mossy tufts for ovaries are unlike
anything we have found before (Fig. 123), and not represented in any
published figures of Jelly-fishes. We float about here for a while, hoping
to find more of the same kind, but no others make their appearance, and we
keep on our way to East Point, where there is a capital fishing ground for
Medusæ of all sorts. Here two currents meet, and the Jelly-fishes are
stranded as it were along the line of juncture, able to move neither one
way nor the other. At this spot the sea actually swarms with life; one
cannot dip the net into the water without bringing up Pleurobrachia,
Bolina, Idyia, Melicertum, &c., while the larger Zygodactyla and Aurelia
float about the boat in numbers. These large Jelly-fishes produce a
singular effect as one sees them at some depth beneath the water;
[Pg 87]
the Aureliæ, especially, with their large white disks, look like pale
phantoms wandering about far below the surface; but they constantly float upward,
and if not too far out of reach, one may bring them up by stirring the
water under them with the end of the oar.
[fig 123]
 Fig. 123. Ovary of Ptychogena; magnified.
Fig. 123. Ovary of Ptychogena; magnified.
When we have passed an hour or so floating about just beyond East Point,
and have nearly filled our buckets with Jelly-fishes of all sizes and
descriptions, we turn and row homeward. The buckets look very pretty as
they stand in the bottom of the boat with the sunshine lighting up their
living contents. The Idyia glitters and sparkles with ever-changing hues,
the Pleurobrachiæ dart about, trailing their long graceful tentacles after
them, the golden Melicerta are kept in constant motion by their quick,
sudden contractions, and the delicate transparent Tima floats among them
all, not the less beautiful because so colorless. There is an unfortunate
Idyia, who, by some mistake, has got into the wrong bucket with the larger
Jelly-fish, where a Zygodactyla has entangled it among his tentacles and is
quietly breakfasting upon it.
During our row the tide has been rising, and as we near the channel of
Saunders's Ledge, it is running through more strongly than before, and at
the entrance of the shallows a pleasant surprise is prepared us; no less
than half a dozen of our new friends (the Ptychogena as he has been
baptized), come to look for their lost companion perhaps, await us there,
and are presently added to our spoils. We reach the shore heavily laden
with the fruits of our morning's excursion.
The most interesting part of the work for the naturalist is
[Pg 88]
still to come. On our return to the Laboratory, the contents of the buckets
are poured into separate glass bowls and jars; holding them up against the
light, we can see which are our best and rarest specimens; these we dip out
in glass cups and place by themselves. If any small specimens are swimming
about at the bottom of the jar, and refuse to come within our reach, there
is a very simple mode of catching them. Dip a glass tube into the water,
keeping the upper end closed with your finger, and sink it till the lower
end is just above the animal you want to entrap; then lift your finger, and
as the air rushes out the water rushes in, bringing with it the little creature
you are trying to catch. When the specimens are well assorted, the microscope
is taken out, and the rest of the day is spent in studying the new
Jelly-fishes, recording the results, making notes, drawings, &c.
Still more attractive than the rows by day are the night expeditions in
search of Jelly-fishes. For this object we must choose a quiet night, for
they will not come to the surface if the water is troubled. Nature has her
culminating hours, and she brings us now and then a day or night on which
she seems to have lavished all her treasures. It was on such a rare
evening, at the close of the summer of 1862, that we rowed over the same
course by Saunders's Ledge and East Point described above. The August moon
was at her full, the sky was without a cloud, and we floated on a silver
sea; pale streamers of the aurora quivered in the north, and
notwithstanding the brilliancy of the moon, they too cast their faint
reflection in the ocean. We rowed quietly along past the Ledge, past Castle
Rock, the still surface of the water unbroken, except by the dip of the
oars and the ripple of the boat, till we reached the line off East Point,
where the Jelly-fishes are always most abundant, if they are to be found at
all. Now dip the net into the water. What genie under the sea has wrought
this wonderful change? Our dirty, torn old net is suddenly turned to a web
of gold, and as we lift it from the water heavy rills of molten metal seem
to flow down its sides and collect in a glowing mass at the bottom. The
truth is, the Jelly-fishes, so sparkling and brilliant in the sunshine,
have a still lovelier light of their own at night; they give out a greenish
golden light as brilliant as that of the brightest glow-worm, and on a
[Pg 89]
calm summer night, at the spawning season, when they come to the surface in
swarms, if you do but dip your hand into the water it breaks into sparkling
drops beneath your touch. There are no more beautiful phosphorescent
animals in the sea than the Medusæ; it would seem that the expression,
"rills of molten metal" could hardly apply to anything so impalpable as a
Jelly-fish, but, although so delicate in structure, their gelatinous disks
give them a weight and substance; and at night, when their transparency is
not perceived, and their whole mass is aglow with phosphorescent light,
they truly have an appearance of solidity which is most striking, when they
are lifted out of the water and flow down the sides of the net.
The various kinds present very different aspects; wherever the larger
Aureliæ and Zygodactylæ float to the surface, they bring with them a dim
spreading halo of light, the smaller Ctenophoræ become little shining
spheres, while a thousand lesser creatures add their tiny lamps to the
illumination of the ocean; for this so-called phosphorescence of the sea is
by no means due to the Jelly-fishes alone, but is also produced by many
other animals, differing in the color as well as the intensity of their
light, and it is a curious fact that they seem to take possession of the
field by turns. You may row over the same course, which a few nights since
glowed with a greenish golden light wherever the surface of the water was
disturbed, and though equally brilliant, the phosphorescence has now a pure
white light. On such an evening, be quite sure that when you empty your
buckets on your return and examine their contents you will find that the
larger part of your treasures are small crustacea (little shrimps). Of
course there will be other phosphorescent creatures, Jelly-fishes, &c.,
among them, but the predominant color is given by these little crustacea.
On another evening the light will have a bluish tint, and then the
phosphorescence is principally due to the Dysmorphosa (Fig. 105).
Notwithstanding the beauty of a moonlight row, if you would see the
phosphorescence to greatest advantage you must choose a dark night, when
the motion of your boat sets the sea on fire around you, and a long
undulating wave of light rolls off from your oar as you lift it from the
water. On a brilliant evening
[Pg 90]
this effect is lost in a great degree, and it is not until you dip your net
fairly under the moonlit surface of the sea, that you are aware how full of
life it is. Occasionally one is tempted out by the brilliancy of the
phosphorescence, when the clouds are so thick that water, sky, and land
become one indiscriminate mass of black, and the line of rocks can be
discerned only by the vivid flash of greenish golden light, when the breakers
dash against them. At such times there is something wild and weird in the
whole scene, which at once fascinates and appalls the imagination; one seems
to be rocking above a volcano, for the surface around is intensely black,
except where fitful flashes or broad waves of light break from the water under
the motion of the boat or the stroke of the oars. It was on a night like this,
when the phosphorescence was unusually brilliant, and the sea as black as ink,
the surf breaking heavily and girdling the rocky shore with a wall of fire, that
our collector was so fortunate as to find in the rich harvest he brought home
the entirely new and exceedingly pretty little floating Hydroid, described
under the name of Nanomia (Fig. 115). It was in its very infancy (Fig.
108), a mere bubble, not yet possessed of the various appendages which
eventually make up its complex structure; but it was nevertheless very
important to have seen it in this early stage of its existence, since, when
a few full-grown specimens were found in the autumn, which lived for some
days in confinement and quietly allowed their portraits to be taken (see
Fig. 115), it was easy to connect the adult animal with the younger phase
of its own life and thus make a complete history.
Marine phosphorescence is no new topic, and we have dwelt too long,
perhaps, upon a phenomenon that every voyager has seen, and many have
described. Its effect is very different, when seen from the deck of a
vessel, from its appearance as one floats through its midst, distinguishing
the very creatures that produce it, and any account of the Medusæ which did
not include this most characteristic feature would be incomplete.
[Pg 91]
ECHINODERMS.
Our illustrations and descriptions of Echinoderms are scanty in comparison
to those of the preceding class; for while, in consequence, perhaps, of the
combined influence of the Gulf Stream and the cold arctic current on the
New England shore, Acalephs are largely represented in our waters, our
marine fauna is meagre in Echinoderms. But although we have few varieties,
those which do establish themselves on a coast seemingly so ungenial for
others of their kind, such as the Echinus, and our common Star-fish, for
instance, thrive well and are very abundant. The class of Echinoderms
includes five orders, viz. Crinoids, Ophiurans, Star-fishes, Sea-urchins,
and Holothurians. The animals composing these orders differ so widely in
appearance that it was very long before their true relations were detected,
and it was seen that all their external differences were united under a
common plan. Let us compare, for instance, the worm-like Holothurians
(Figs. 124, 126, 127) with all the host of Star-fishes (Figs. 142, 146,
147) and Sea-urchins (Figs. 131, 139), or compare the radiating form of the
Star-fish, its arms spreading in every direction, with the close spherical
outline of the Sea-urchin, or the Crinoid floating at the end of a stem
(Fig. 152) with either of these, and we shall cease to wonder that
naturalists failed to find at once a unity of idea under all these
varieties of execution. And yet the fundamental structure of the class of
Echinoderms is represented as distinctly by any one of its five orders as
by any other, and is absolutely identical in all. They differ only by
trifling modifications of development.
In Echinoderms as a class, the body presents three regions differing in
structure, and on the greater or less development of these regions or
systems, as we may call them, their chief differences are based. Take, for
instance, the dorsal system, the nature of which is explained by the name,
indicating of course the back of the animal, though it does not necessarily
imply the upper side of the body, since some of the Echinoderms, as the
stemmed Crinoids, for example, carry the dorsal side downward, while
[Pg 92]
the Star-fishes and Sea-urchins carry it upward, and the Holothurians, moving
with the mouth forward, have the dorsal system at the opposite end of the
body. Whatever the natural attitude of the animal, however, and the
consequent position of the dorsal region, it exists alike in all the five
orders, though it has not the same extent and importance in each. But in
all it is made up of similar parts, bears the same relation to the rest of
the body, has the same share in the general economy of the animal. And
though when we compare the spreading back of a Star-fish with the small
area on the top of a Sea-urchin, where all the zones unite, we may not at
once see the correspondence between them, yet a careful comparison of all
their structural details shows that they are both built with the same
elements and represent the same region, though it is stretched to the
utmost in the one case, and greatly contracted in the other.
This being true of the dorsal system, let us look at another equally
important structural feature in this class. All Echinoderms have locomotive
organs peculiar to themselves, a kind of suckers which may be more or less
numerous, larger or smaller, in different species, but are always
appendages of the same character. These are variously distributed over the
body, but always with a certain regularity occupying definite spaces, shown
by investigation to be homologous in all. For instance, the rays of the
Star-fish correspond in every detail on their under side, along which the
locomotive suckers run, with the zones on the Sea-urchin, from end to end
of which the suckers are arranged; and the same is equally true of the
distribution of the suckers on the Holothurians, Ophiurans, and Crinoids,
though, as most persons are less familiar with these orders than with the
other two, it might not be so easy to point out the coincidence to our
readers. These suckers are called the ambulacra, the lines along which they
run are called the ambulacral rows or zones, while the system of locomotion
as a whole is known as the ambulacral system. Since these organs are thus
regularly distributed over the body in distinct zones or rows, it follows
that the latter must be divided by intervening spaces. These intervals are
called the interambulacral spaces; but while in some orders they are
occupied by larger plates and prominent
[Pg 93]
spines, as in the Sea-urchin and Star-fish, in others they are either
comparatively insignificant or completely suppressed, as in the Crinoids
and Ophiurans. Such are the three regions or systems which by their greater
or less development introduce an almost infinite variety of combinations into
this highest class of Radiates. It may not be amiss before proceeding further
to compare the five orders with reference to this point, and see which of these
three systems has the preponderance in each one.
Taking the orders in their rank and beginning with the lowest, we find in
the Crinoids that the dorsal system preponderates, being composed of highly
complicated plates, and developed to such a degree as to form in many
instances a stem by which the animal is attached to the ground, while the
ambulacral system is limited to a comparatively small area, and the
interambulacral system is wanting. The order of Crinoids has diminished so
much in modern geological times that we must consult its fossil forms in
order to understand fully the peculiar adaptation of the Echinoderm plan in
this group.
In the Ophiurans, the dorsal system is still large, and though it no longer
stretches out to form a stem, it folds over on the under side of the animal
so as to enclose entirely the ambulacral system, forming a kind of shield
for the arms. Here also the interambulacral system is wanting.
In the Star-fishes the dorsal system encroaches less upon the structure of
the animal. The back and oral side here correspond exactly in size, and
though the flat leathery upper surface of the animal, covered with spines,
serves as a protection to the delicate ambulacral suckers which find their
way between the rows of small plates along the under side of the arms, yet
it does not enfold them as in the Ophiurans. On the contrary, in the
Star-fishes the ambulacral rows are protected on either side by a row of
the so-called interambulacral plates, through which no suckers pass.
In the Sea-urchin, the dorsal system is contracted to a minimum, forming a
small area on the top of the animal, the rows of interambulacral plates
which are separated and lie on either side of the ambulacra in the
Star-fish being united in the Sea-urchin, and both the ambulacral and the
interambulacral systems bent
[Pg 94]
upward, meeting in the small dorsal area above, so as to form a spherical
outline. Here the ambulacral and interambulacral systems have taken a great
preponderance over the dorsal system, and the same is the case with the
Holothurians, in which the same structure is greatly elongated, the dorsal
system being thus pushed out as it were to the end of a cylinder, while the
ambulacral and interambulacral systems run along its whole length. All
Echinoderms without exception have ambulacral tubes, even though in some
there are no external ambulacral suckers connected with them.
There is one organ peculiar to the class of Echinoderms, the general
structure of which may be described here, since it is common to them all,
with the exception of the Crinoids, the anatomy of which is, however, so
imperfectly understood, that we are hardly justified in assuming that it
does not exist even in that order. This organ is known as the madreporic
body; it is a small sieve or limestone filter opening into a tube or canal;
by means of this tube, which connects with the ambulacral system, the water
from without, first filtered through the madreporic body and thus freed
from any impurities, is conveyed to the ambulacra. In the more detailed
account of the different orders we shall see what is the position of this
singular organ in each group, and how it is adapted in them all to their
special structure. The development of Echinoderms forms one of the most
wonderful chapters in the annals of Natural History. Marvellous as is the
embryonic history of the Acalephs, including all the different aspects they
assume in the cycle of their growth, it is thrown into the shade by the
transformations which Echinoderms undergo before assuming their adult
condition. This singular mode of development, although it has features
recalling the development of Jelly-fishes from Hydroids, is nevertheless
entirely distinct from it, and is known only in the class of Echinoderms.
As the whole story is given at length in the chapter on the embryology of
the Echinoderms, we need only allude to it here in general terms. We owe
the discovery of this remarkable process to Johannes Müller, one of the
greatest anatomists of this century.
[Pg 95]
HOLOTHURIANS.
[fig 124]
 Fig. 124. Synapta, natural size.
Fig. 124. Synapta, natural size.
Synapta. (Synapta tenuis Ayres.)
This is one of the most curious of the Holothurians, and easily observed on
account of its transparency, which allows us to see its internal structure.
It has a long cylindrical body (Fig. 124) along the length of which run the
five rows of ambulacra, which are in this instance closed tubes without any
projecting suckers or locomotive organs of any kind attached to them, so
that the name is retained only on account of their correspondence in
position, and not from any similarity of function to the ambulacra in
Star-fishes and Sea-urchins. But though the ambulacra in Synapta are in
fact mere water-tubes like the vertical tubes in the Ctenophoræ, by means
of which the water, first filtered through the madreporic body, circulates
along the skin, they are as organs perfectly homologous with the ambulacra
in all other Echinoderms. The mouth has a circular tube around the
aperture, and a wreath of branching tentacles encircling it. The habits of
these animals are singular. They live in very coarse mud, but they surround
themselves with a thin envelope of finer sand, which they form by selecting the
[Pg 96]
small particles with their tentacles, and making a ring around their
anterior extremity. This ring they then push down along the length of the
body, and continue this process, adding ring after ring, till they have
entirely encircled themselves with a sand tube. They move the rings down
partly by means of contractions of the body, but also by the aid of
innumerable appendages over the whole surface. To the naked eye these
appendages appear like little specks on the skin; but under the microscope
they are seen to be warts projecting from the surface, each one containing
a little anchor with the arms turned upward (Fig. 125). Around the mouth
[fig 125]

Fig. 125. Anchor of Synapta; a anchor,
w plate upon which anchor is
attached; greatly magnified.
these warts are larger, but do not contain any anchors. It will be seen
hereafter that these appendages are homologous with certain organs in
other Holothurians, the warts with the anchors corresponding to the
limestone pavement covering or partially covering the surface of the
Cuvieria, for instance, while those without anchors correspond to the
so-called false ambulacra in Pentacta. By means of these appendages, though
aided also by the contractions of the body, the Synaptæ move through the
mud and collect around themselves the sand tube in which they are encased.
Their food is very coarse for animals so delicate in structure. When
completely empty of food they are white, perfectly transparent, and the
spiral tube forming the digestive cavity may be seen wound up and hanging
loosely in the centre for the whole length of the body. In such a condition
it is of a pale yellow color. But look at one that is gorged with food. The
whole length of the alimentary canal is then crowded with sand, pebbles,
and shells, distinctly seen through the transparent skin, and giving a dark
gray color to the whole body. They swallow the sand for the sake of the
nutritious substance it contains, and having assimilated and digested this,
they then eject the harder materials. The motion of the body in consequence
of its contractions
[Pg 97]
is much like that of leeches, and on this account these Synaptæ were
long supposed to be a transition type between the Radiates and worms. The
body grows to a great length, often half a yard and more, but constantly
drops large portions from its posterior part, by means of its own contractions,
or breaks itself up by the expulsion of the intestines, which are very readily
cast out. The tentacles are hollow, consisting of a central rib with branches
from either side. In the Synaptæ, as in all the Holothurians, the
madreporic body is placed near the mouth, between two of the ambulacra, and
opposite the fifth or odd one. The tube, connecting with the central tube around
the mouth, by means of which it communicates with the ambulacral tubes, is very short.
Caudina. (Caudina arenata Stimps.)
[fig 126]
 Fig. 126. Caudina arenata; natural size.
Fig. 126. Caudina arenata; natural size.
Several other Holothurians are frequently met with on our shores. Among
them is the Caudina arenata (Fig. 126), a small Holothurian, yellowish in
color, and thick in texture, by no means so pretty as the white transparent
Synapta; the tentacles are short, resembling a crown of cloves around the
mouth. It lives in the sand, and may be found in great numbers on the sandy
beaches after a storm.[Pg 98]
Cuvieria. (Cuvieria squamata D. & K.)
[fig 127]
 Fig. 127. Cuvieria; natural size.
Fig. 127. Cuvieria; natural size.
The Holothurian of our coast, excelling all the rest in beauty, is the
Cuvieria. (Fig. 127.) As it lies on the sand, a solid red lump, with
neither grace of form nor beauty of color, even the vividness of its tint
growing dull and dead when it is removed from its native element, certainly
no one could suspect that it possessed any hidden charm; but place it in a
glass bowl with fresh sea-water; the dull red changes to deep vivid
crimson, the tentacles creep out (Fig. 127) softly, and slowly, till the
mouth is surrounded by a spreading wreath, comparable for richness of tint,
and for delicate tracery, to the most beautiful sea-weeds. These tentacles,
when fully expanded, are as long as the body itself. A limestone pavement
composed of numerous pieces covers almost the whole surface of the animal;
this apparatus corresponds, as we have already mentioned, to the warts
containing anchors in the Synapta; but in the latter, the limestone
particles are smaller, whereas in the Cuvieria they are developed to a
remarkable extent. This animal is very sluggish, the ambulacral suckers,
found only on three of the tubes, being arranged in such a way as to form a
sort of sole on which they creep;
[Pg 99]
the sole is tough and leathery in texture, but free from the limestone
pavement described above. The young (Figs. 128, 129) are very common,
swimming freely about, and more readily found than the adult; they are of a
bright vermilion color, but the tentacles hardly branch at that age, nor is
the limestone pavement formed, which gives such a peculiar aspect to the
full-grown animal. The young Cuvieria, somewhat older than that represented
in Fig. 129, are found in plenty under stones at low-water mark, just after
they have given up their nomadic habits, and when the limestone pavement
begins to be developed.
[fig 128]
[fig 129]

Fig. 128. Young Cuvieria, much enlarged;
l body, g tentacles. |
|

Fig. 129. Somewhat older Cuvieria;
l body, g tentacle round mouth, g' testaete
of sole, b madreporic tentacle. |
Pentacta. (Pentacta frondosa Jäg.)
The highest of our Holothurians in structure, is the Pentacta. (Fig. 130)
It is very rare on our beaches, though occasionally found under stones at
low-water mark; farther north, in Maine, and at Grand Manan, it is very
common, covering all the rocks near low-water mark. It is a chocolate brown
in color, and
[Pg 100]
measures, when fully expanded, some fifteen to eighteen
inches in length. Unlike the Cuvieria, the ambulacral suckers are evenly
distributed and almost equally developed on all the tubes; between the five
rows of ambulacral suckers are scattered irregularly certain appendages
resembling suckers, but found on examination not to be true locomotive
suckers, and called on that account false ambulacra. These are the organs
corresponding to the warts around the mouth of the Synapta. Although the
ambulacral suckers are, as we have said, equally developed on all the
tubes, yet the Pentacta does not use them indiscriminately as locomotive
organs. In Pentacta, as well as in all Holothurians, whether provided with
ambulacral suckers, or, like the Synapta and Caudina, deprived of them, the
odd ambulacrum, viz. the one placed opposite the madreporic body, is always
used to creep upon, and forms the under surface of the animal.
[fig 130]
 Fig. 130. Pentacta frondosa; expanded about one third the natural size.
Fig. 130. Pentacta frondosa; expanded about one third the natural size.
The correspondence between the different phases of growth in the young
Pentacta, and the adult forms of the orders described above, the Synapta,
Caudina, Cuvieria, and Pentacta itself, is a striking instance of the way
in which embryonic forms illustrate the relative standing of adult animals.
In the earlier stages of its development, the ambulacral tubes alone are
developed in the Pentacta; in this condition it recalls the lower orders of
Holothurians, as the Synapta and Caudina; then a sole is formed by the
greater development of three of the ambulacra, and in this state it reminds
us of the next in order, the Cuvieria, while it is
[Pg 101]
only in assuming its adult form that the Pentacta develops its other ambulacra,
with their many suckers.
The Pentacta resembles the Trepang, so highly valued by the Chinese as an
article of food, and forms a not unsavory dish, having somewhat the flavor
of lobster.
ECHINOIDS.
Sea-urchin. (Toxopneustes drobachiensis Ag.)
[fig 131]
 Fig. 131. Toxopneustes from above, with all the appendages expanded; natural size.
Fig. 131. Toxopneustes from above, with all the appendages expanded; natural size.
Sea-urchins (Fig. 131) are found in rocky pools, hidden away usually in
cracks and holes. They like to shelter themselves in secluded nooks, and,
not satisfied even with the privacy of such a retreat, they cover
themselves with sea-weed, drawing it down with their tentacles, and packing
it snugly above them, as if to avoid observation. This habit makes them
difficult to find, and it is only by parting the sea-weed, and prying into
the most retired corners in such a pool, that one detects them. Their
motions are slow, and they are less active than either the Star-fish or the
Ophiuran, to both of which they are so closely allied.
Let us look at one first, as seen from above, with all its various organs
fully extended. (Fig. 131.) The surface of the animal is divided by ten
zones, like ribs on a melon, only that these zones differ in size, five
broad zones alternating with five narrower ones. The broad zones,
representing the interambulacral system, are composed of large plates,
supporting a number of hard projecting spines, while the narrow zones,
forming the ambulacral system, are pierced with small holes, arranged in
regular rows, (Fig. 132,) through which extend the tentacles terminating
with little cups or suckers. These zones converge towards the summit of the
animal, meeting in the small area which here represents the dorsal system;
this area is filled by ten plates, five larger ones at the extremity of the
interambulacral zones, and five smaller ones at the extremity of the
ambulacral zones. (Fig. 132.) In the five larger plates are the ovarian
openings, so-called because each
[Pg 102]
one is pierced by a small hole through which the eggs are passed out, while
in the five smaller plates are the eye-specks. The ovaries themselves consist
of long pouches or sacs, carried along the inner side of each ambulacrum; one
of these ovarian plates is larger than the others, and forms the madreporic
body, being pierced with many minute holes; here, as in the Star-fish, it is
placed between two of the ambulacral rows, and opposite the fifth or odd one.
Looked at from the under or the oral side, as seen in Fig. 134, the animal
presents the mouth, a circular aperture furnished with five teeth in its centre;
these five teeth opening
[Pg 103]
into a complicated intestine to be presently described. From the mouth, the ten
zones diverge, curving upward to meet in the dorsal area on the summit of the body.
(Fig. 133.)
[fig 132]
[fig 133]
") |
| Fig. 132. Portion of shell of Fig. 131, with spines rubbed off. (Agassiz.) |
| Fig. 133. Sea-urchin shell with all the spines removed. (Agassiz.) |
Let us now examine the appearance and functions of the various appendages
on the surface. The tentacles have a variety of functions to perform; they
are the locomotive appendages, and for this reason, as we have seen, the
zones along which they are placed are called the ambulacra, while the
intervening spaces, or the broad zones, are called the interambulacra. It
should not be supposed, however, that the locomotive appendages are the
only ones to be found on the ambulacra, for spines occur on the narrow as
well as on the broad zones, though the larger and more prominent ones are
always placed on the latter. The tentacles are also subservient to
circulation, for the water which is taken in at the madreporic body passes
into all the tentacles, sometimes called on that account water-tubes.
Beside these offices the tentacles are constantly busy catching any small
prey, and conveying it to the mouth, or securing the bits of sea-weed with
which, as has been said, these animals conceal themselves from observation.
It is curious to see their fine transparent feelers, fastening themselves
by means of the terminal suckers on such a floating piece of sea-weed,
drawing it gently down and packing it delicately over the surface of the
body. As locomotive appendages, the tentacles are chiefly serviceable on
the lower or oral side of the animal, which always moves with the mouth
downward. About this portion of the body the tentacles are numerous (Fig.
134) and large, and when the animal advances it stretches them in a given
[Pg 104]
direction, fastens them by means of the suckers on some surface, be it of
rock, or shell, or the side of the glass jar in which they are kept, and
being thus anchored it drags itself forward. The tentacles are of a violet
hue, though when stretched to their greatest length they lose their color,
and become almost white and transparent; but in their ordinary condition
the color is quite decided, and the rows along which they occur make as
many violet lines upon the surface of the body.
[fig 134]
[fig 135]
")
Fig. 134. Sea-urchin seen from the mouth side. (Agassiz.) |
|

Fig. 135. Magnified spine. |
Almost the sole function of the spines seems to be that of protecting the
animal, and enabling it to resist the attacks of its enemies, the force of
the waves, or any sudden violent contact with the rocks. The spines, when
magnified, are seen to be finely ribbed for nearly the whole length (Fig.
135), the bare basal knob serving as the point of attachment for the
powerful muscles, which move these spines on a regular ball-and-socket
joint, the ball surmounting the tubercles (seen in Fig. 132), which fit
exactly in a socket at the base of the spine. In a transverse section of a
spine (Fig. 136), we see that the ribs visible on the outside are delicate[Pg 105]
columns placed closely side by side, and connected by transverse rods
forming an exceedingly delicate pattern. Beside the tentacles and the
spines, they have other external appendages, of which the function long
remained a mystery, and is yet but partially explained; these are the
so-called pedicellariæ; they consist of a stem (s, Fig. 137), which
becomes swollen (p, Fig. 137) into a thimble-shaped knob at the end (t,
Fig. 137); this knob may seem solid and compact at first sight, but it is
split into three wedges, which can be opened and shut at will. When open,
these pedicellariæ may best be compared to a three-pronged fork, except
that the prongs are arranged concentrically instead of on one plane, and,
when closed, they fit into one another as neatly as the pieces of a puzzle.
[fig 136]
[fig 137]

Fig. 136. Transverse section of spine; magnified. |
|
Fig. 137. Pedicellariæ of Sea-urchin;
s stem, p base of fork, t fork. |
If we watch the Sea-urchin after he has been feeding, we shall learn, at
least, one of the offices which this singular organ performs in the general
economy of the animal. That part of his food which he ejects passes out at
an opening on the summit of the body, in the small area where all the zones
converge. The rejected particle is received on one of these little forks,
which closes upon it like a forceps, and it is passed on from one to the other,
down the side of the body, till it is dropped off into the water. Nothing is
more curious and entertaining than to watch the neatness and accuracy with
which this process is performed. One may see the rejected bits of food passing
rapidly along the lines upon which these pedicellariæ occur in greatest
number, as if they were so many little roads for the conveying
[Pg 106]
away of the refuse matters; nor do the forks cease from their
labor till the surface of the animal is completely clean, and free from any
foreign substance. Were it not for this apparatus the food thus rejected
would be entangled among the tentacles and spines, and be stranded there
till the motion of the water washed it away. These curious little organs
may have some other office than this very laudable and useful one of
scavenger, and this seems the more probable because they occur over the
whole surface of the body, while they seem to pass the excrements only
along certain given lines. They are especially numerous about the mouth,
where they certainly cannot have this function; we shall see also that they
bear an important part in the structure of the Star-fish, where there are
no such avenues on the upper surface, for the passage of the refuse food,
as occur on the Sea-urchin.
[fig 138]
 Fig. 138. Teeth of Sea-urchin, so-called Lantern of Aristotle.
Fig. 138. Teeth of Sea-urchin, so-called Lantern of Aristotle.
On opening a Sea-urchin, we find that the teeth (Fig. 138), which seem at
first sight only like five little conical wedges around the mouth (Fig.
134), are connected with a complicated intestine, which extends spirally
from the lower to the upper floor of the body, festooning itself from one
ambulacral zone to the next, till it reaches the summit, where it opens.
This intestine leads into the centre of the teeth, the jaws themselves,
which sustain the teeth, being made up of a number of pieces, and moved by
a complicated system of muscular bands. When the intestine is distended
with food, it fills the greater part of the inner cavity; the remaining
space is occupied in the breeding season by the genital organs. In a
section of the Sea-urchin, one may also trace the tube by which the supply
of water, first filtered through the madreporic body, is conveyed to the
ambulacra; it extends from the summit of the body to the circular tube
surrounding the mouth.
Echinarachnius. (Echinarachnius parma Gray.)
[fig 139]

Fig. 139. Echinarachnius, seen from above, with the spines on part of the shell;
a ambulacral zone, i interambulacral zone. |
Beside the Toxopneustes (Fig. 131) described above, we have another
Sea-urchin very common along our shores. Among
[Pg 107]
children who live near sandy beaches, they are well known as "sand-cakes"
(Fig. 139), and indeed they are so flat and round, that, when dried and
deprived of their bristles, they look not unlike a cake with a star-shaped
figure on its surface. (Fig. 139.) When first taken from the water they are
of a dark reddish brown color, and covered with small silky bristles. The disk
is so flat, being but very slightly convex on the upper side, that one would
certainly not associate it at first sight with the common spherical
Sea-urchin or Sea-egg, as the Toxopneustes is sometimes called. But upon
closer examination the delicate ambulacral tubes or suckers may be seen
projecting from along the line of the ambulacra, as in the spherical
Sea-urchin; and though these ambulacra become expanded near the summit into
gill-like appendages, forming a sort of rosette in the centre of the disk,
they are, nevertheless, the same organs, only somewhat more complicated.
When such a disk is dried in the sun, and the
[Pg 108]
bristles entirely removed, the lines of suture of the plates composing it, and
corresponding exactly to those of the spherical Sea-urchin, may very readily be
seen. (a and i, Fig. 139.)
[fig 140]
") Fig. 140. Transverse section of Echinarachnius; o mouth, e e ambulacra, c m ambulacral ramifications, w w interambulacra. (Agassiz.)
Fig. 140. Transverse section of Echinarachnius; o mouth, e e ambulacra, c m ambulacral ramifications, w w interambulacra. (Agassiz.)
This flat Sea-urchin or Echinarachnius, as it is called, belongs to a group
of Sea-urchins known as Clypeastroids (shield-like Sea-urchins). In a
section (Fig. 140) exposing the internal structure, one cannot but be
reminded by its general aspect of an Aurelia. Could one solidify an Aurelia
it would present much the same appearance; another evidence that all the
Radiates are built on one plan, their differences being only so many modes
of expressing the same structural idea. The teeth or jaws in this flat
Sea-urchin are not so complicated as in the Toxopneustes, being simply flat
pieces, arranged around the mouth (o, Fig. 140), without the apparatus of
muscular bands by means of which the teeth are moved in the other genus. It
is a curious fact, considered in relation to the general radiate structure
of these animals, that the teeth, instead of moving up and down like the
jaws in Vertebrates, or from right to left like those of Articulates, move
concentrically, all converging towards the centre.
STAR-FISHES.
Star-fish. (Astracanthion berylinus Ag.)
[fig 141]
") Fig. 141. Star-fish ray, seen from mouth side. (Agassiz.)
Fig. 141. Star-fish ray, seen from mouth side. (Agassiz.)
Although there is the closest homology of parts between the Star-fish and
the Sea-urchin, the arrangement of these parts, and the external appearance
of the animals, as a whole, are entirely different. The Star-fish has zones
corresponding exactly to those
[Pg 109]
of the Sea-urchin, but instead of being drawn together, and united at the summit
of the animal, so as to form a spherical outline, they are spread out on one
level in the shape of a star. This change in the general arrangement brings
the eye-specks to the extremities of the arms, and places the ovarian openings
in the angles between the arms. The madreporic body is situated on the upper
surface of the disk (Fig. 142), at the angle between two of the arms, and consequently
between two of the ambulacra, and opposite the odd one. The tube into which
it opens, runs vertically from the upper floor of the disk to the lower,
where it connects with the circular tube around the mouth, and thus
communicates with all the ambulacral rows. The ambulacral zones which, in
the Star-fish, have the shape of a furrow, run along the lower side of each
ray (Fig. 141); the interambulacral zones are divided, their plates being
arranged in rows along either side of the ambulacral furrows. The
ambulacral furrow, like the ambulacral zone in the Sea-urchin, is pierced
with numerous holes, alternating with each other in a kind of zigzag
arrangement, one hole a little in advance, the next a little farther back,
and so on, and through these holes pass the tentacles, terminating in
suckers, as in the Sea-urchins, and serving as in them for locomotive
organs. The most prominent and strongest spines are arranged upon the large
interambulacral plates on both sides of the ambulacral furrows; but the
upper surface of the animal is also completely studded with smaller spines,
scattered at various distances, apparently without any regular arrangement.
(Fig. 142.)
[fig 142]

Fig. 142. Star-fish; natural size, seen from above.
The position of the pedicellariæ is quite different from that which they
occupy in the Sea-urchin, where they are scattered singly between the
spines and tentacles, though more regularly and closely grouped along the
lines upon which the refuse food is moved off. In the Star-fish, on the
contrary, these singular organs seem to be grouped for some special purpose
around the spines, on the upper surface of the body. Every such spine
swells near its point of attachment, thus forming a spreading base
[Pg 110]
(Fig. 143), around which the pedicellariæ are arranged in a close wreath,
in the centre of which the summit of the spine projects; they differ also from
those of the Sea-urchin in having two prongs instead of three. Other
pedicellariæ are scattered independently over the surface of the animal,
but they are smaller than those forming the clusters and connected with the
spines. The function of these organs in the Star-fish remains unexplained;
the opening on the upper surface, through which the refuse food is thrown
out, is in such a position that they evidently do not serve here the same
purpose which renders them so useful to the Sea-urchin. Occasionally they
may be seen to catch small prey
[Pg 111]
with these forks, little Crustacea, for instance; but this is probably not
their only office. The Star-fish has a fourth set of external appendages in
the shape of little water-tubes. (Seen in Fig. 143.) The upper surface of the
back consists of a strong limestone network (Fig. 144), and certain openings
in this network are covered with a thin membrane through which these water-tubes
project. It is supposed that water may be introduced into the body through
these tubes; but while there can be no doubt that they are constantly filled
with water, and are therefore directly connected with the circulation through
the madreporic body (Fig. 145), no external opening has as yet been detected
in them. The fact, however, that when these animals are taken out of their native
element, the water pours out of them all over the surface of the back, so
that they at once collapse and lose entirely their fulness of outline,
seems to show that water does issue from those tubes. The ends of the arms
are always slightly turned up, and at the summit of each is a red
eye-speck. The tentacles about the eye become very delicate and are
destitute of suckers.
[fig 143]
[fig 144]
[fig 145]

Fig. 143. Single spine of Star-fish, with surrounding appendages; magnified. |
|

Fig. 144. Limestone network of back of Star-fish. |
| |

Fig. 145. Madreporic body of Star-fish; magnified. |
These animals have singular mode of eating; they place themselves over
whatever they mean to feed upon, as a cockle-shell for instance, the back
gradually rising as they arch themselves above it; they then turn the
digestive sac or stomach inside out, so as to enclose their prey
[Pg 112]
completely, and proceed leisurely to suck out the animal from its shell.
Cutting open any one of the arms we may see the yellow folds of the stomach
pouches which extend into each ray; within the arms, extending along either
side of the upper surface, are also seen the ovaries, like clusters of
small yellow berries. Immediately below these, along the centre of the
lower floor of each ray, runs the ridge formed by the ambulacral furrow,
and upon either side of this ridge are placed the vesicles, by means of
which the tentacles may be filled and emptied at the will of the animal;
the rest of the cavity of the ray is filled by the liver. The mouth, which
is surrounded by a circular tube, is not furnished with teeth, as in the
Sea-urchin; but the end of each ambulacral ridge is hard, thus serving the
purpose of teeth.
Cribrella. (Cribrella oculata Forbes.)
Our coast, as we have said, is not rich in the variety of Star-fishes. We
have two large species, one of a dark-brown color (Fig. 132), the
Astracanthion berylinus, and the other, the A. pallidus, of a pinkish
tint; then there is the small Cribrella, inferior in structural rank to the
two above mentioned. (Fig. 146.) This pretty little Star-fish presents the
greatest variety of colors; some are dyed in Tyrian purple, others have a
paler shade of the same hue, some are vermilion, others a bright orange or
yellow. A glass dish filled with Cribrellæ might vie with a tulip-bed in
gayety and vividness of tints.
[fig 146]
 Fig. 146. Cribrella from above; natural size.
Fig. 146. Cribrella from above; natural size.
The disk of the Cribrella is smooth, instead of being covered, like the
larger Star-fishes, with a variety of prominent appendages. The spines are
exceedingly short, crowded like little warts over the surface. It is an
interesting fact, illustrating
[Pg 113]
again the correspondence between the adult forms of the lower orders and the
phases of growth in the higher ones, that these spines have an embryonic
character. One would naturally expect to find that these small spines of
the adult Cribrella would differ from those of the other full-grown Star-fishes
chiefly in size, that they would be a somewhat modified pattern of the same
thing on a smaller scale; but when examined under the microscope, they resemble
the spines of the higher orders in their embryonic condition; it is not, in fact,
a difference in size merely, but a difference in degree of development. The
Cribrella moves usually with two of the arms turned backward, and the three
others advanced together, the two posterior ones being sometimes brought so
close to each other as to touch for their whole length.
Hippasteria. (Hippasteria phrygiana Ag.)
Beside these Star-fishes we have the pentagonal Hippasteria (Hippasteria
phrygiana Ag.), like a red star with rounded points, found chiefly in deep
water, though it is occasionally thrown up on the beaches. It has but two
rows of large tentacles, terminating in a powerful sucking disk. The
pedicellariæ on this Star-fish resemble large two-pronged clasps, arranged
principally along the lower side. The pentagonal Star-fishes of our coast
are in striking contrast to the long-armed species we have just described;
they are edged with rows of large smooth plates, and do not possess the
many prominent spines so characteristic of the ordinary Star-fishes.
Ctenodiscus. (Ctenodiscus crispatus D. & K.)
The Ctenodiscus (Ctenodiscus crispatus D. & K., Fig. 147), an inhabitant
of more northern waters, but seeming also to be at home here occasionally,
is another pentagonal Star-fish. It lives in deep water, and frequents
muddy bottoms. The peculiar structure of their ambulacra has probably some
reference to this mode of living, for they are entirely wanting in the
sucking disks so characteristic of the other members of this class, and their
[Pg 114]
tentacles are pointed, as if to enable them to work their way
through the mud in which they make their home. The pointed tentacles of
this genus are characteristic of a large group of Star-fishes, and it is an
important fact, as showing their lower standing, that this feature, as well
as the pentagonal outline, obtains in the earlier stages of growth of our
more common Star-fishes, while in their adult condition they assume the
deeply indented star-shaped outline, and have suckers at the extremities of
the tentacles.
[fig 147]
 Fig. 147. Ctenodiscus, seen from above; natural size.
Fig. 147. Ctenodiscus, seen from above; natural size.
Solaster. (Solaster endeca Forbes.)
We find also among Star-fishes the same tendency to multiplication of parts
so common among the Polyps and Acalephs. Our Solaster (Solaster endeca/
Forbes), for instance, has no less than twelve arms; it inhabits more
northern latitudes, though sometimes found in our Bay; on the coast of
Maine it is quite common, and occurs in company with another many-rayed
species, the Crossaster papposa M. & T. The color of both of these
Star-fishes is exceedingly varied; we find in the Solaster as many
different hues as in the Cribrella, which it resembles in the structure of
its spines, while in the Crossaster bands of different tints of red and
purple are arranged concentrically, and the whole surface of the back is
spotted with brilliantly-tinged tiny wreaths of water-tubes, crowded round
the base of the different spines, which are somewhat similar to those of
the Astracanthion.
[Pg 115]
OPHIURANS.
Ophiopholis. (Ophiopholis bellis Lym.)
[fig 148]
 Fig. 148. Ophiopholis, from above; natural size.
Fig. 148. Ophiopholis, from above; natural size.
There are but two species of the ordinary forms of Ophiurans in
Massachusetts Bay; the white Amphiura (Amphiura squamata Sars), with long
slender arms, and the spotted Ophiopholis (Fig. 148), with shorter and
stouter arms, and in which the disk is less compact than in the Amphiura,
and not so perfectly circular. All Ophiurans are difficult to find, from
their exceeding shyness; they hide themselves in the darkest crevices, and
though no eye-specks have yet been detected in them, they must have some
quick perception of coming danger, for at the gentlest approach they
instantly draw away and shelter themselves in their snug retreats.
[Pg 1216
They differ from the Star-fishes in having the disk entirely distinct from
the arms; that is, the arms, instead of merging gradually into the disk,
start at once from its margin. They have no interambulacral spaces or
plates; but the whole upper surface is formed of large hard plates, which
extend from the back over the sides of the arms to their lower surface,
where they form a straight ridge along the centre. (Fig. 149.) The sides of
these plates are pierced with holes, through which the tentacles pass;
these have not, like those of the Star-fishes and Sea-urchins, a sucker at
the extremity, but are covered with little warts or tubercles (Fig. 150);
they are their locomotive appendages, and their way of moving is curious;
they first extend one of the arms in the direction in which they mean to
move, then bring forward two others to meet them, three arms being thus
usually in advance, and then they drag the rest of the body on. They move
with much more rapidity, and seem more active, than the Star-fishes;
probably owing to the greater independence of the arms from the disk. The
spines project along the margin of the arms, and not over the whole
surface, the back of the arms being perfectly free from any appendages, and
presenting only the surface of the plates. The madreporic body is formed by
a plate on the lower side of the disk, in a position corresponding
[Pg 117]
to that which it occupies in the young Star-fish; this plate is one of the large
circular shields occupying the interambulacral spaces around the mouth.
(Fig. 149.) On each side of the arms, where they join the disk, are slits
opening into the ovarian pouches. They have no teeth; but the hard ridge at
the oral end of the ambulacra, extending toward the mouth in Star-fish, is
still more distinct and sharper in the Ophiurans, approaching more nearly
the character of teeth.
[fig 149]
[fig 150]
 |
|
 |
| Fig. 149. One arm of Fig. 148; from the mouth side. |
|
Fig. 150. Ambulacral tentacle of Ophiopholis; magnified. |
Astrophyton. (Astrophyton Agassizii Stimp.)
[fig 151]
 Fig. 151. Astrophyton, Basket-fish; in a natural attitude.Click on image to view larger size.
Fig. 151. Astrophyton, Basket-fish; in a natural attitude.Click on image to view larger size.
A singular species of Ophiuran, known among fishermen as the "Basket-fish,"
(Fig. 151,) is to be found in Massachusetts Bay. Its arms are very long in
comparison to the size of the disk, and divide into a vast number of
branches. In moving, the animal lifts itself on the extreme end of these
branches, standing as it were on tiptoe (Fig. 151), so that the
ramifications of the arms form a kind of trellis-work all around it,
reaching to the ground, while the disk forms a roof. In this living house
with latticed walls small fishes and other animals are occasionally seen to
take shelter; but woe to the little shrimp or fish who seeks a refuge
there, if he be of such a size as to offer his host a tempting mouthful; he
will fare as did the fly who accepted the invitation of the spider. These
animals are exceedingly voracious, and sometimes, in their greediness for
food, entangle themselves in fishing lines or nets. When disturbed, they
coil their arms closely around the mouth, assuming at such times a kind of
basket-shape, from which they derive their name.
[Pg 118]
This Basket-fish is honorably connected with our early colonial history,
being thought worthy, by no less a personage than John Winthrop, Governor
of Connecticut, who, as he says, "had never seen the like," to be sent with
"other natural curiosities of these parts" to the Royal Society of London,
in 1670. He accompanies the specimen with a minute description, omitting
"other particulars, that we may reflect a little upon this elaborate piece
of nature." His account is as graphic as it is accurate, and we can hardly
give a better idea of the animal than by extracting some portions of it.
"This Fish," he says, "spreads itself from
[Pg 119]
a Pentagonal Root, which incompasseth the Mouth (being in the middle), into
5 main Limbs or branches, each of which, just at issuing out from the Body,
subdivides itself into two, and each of these 10 branches do again divide into two
parts, making 20 lesser branches; each of which again divide into two
smaller branches, making in all 40. These again into 80, and these into
160; and these into 320; these into 640; into 1280; into 2560; into 5120;
into 10,240; into 20,480; into 40,960; into 81,920; beyond which the
further expanding of the Fish could not be certainly trac'd";—a statement
which we readily believe, wondering only at the patience which followed
this labyrinth so far.
In a later letter, after having had an interview with the fisherman who
caught the specimen, and, as he says, "asked all the questions I could
think needful concerning it," the Governor proceeds to tell us that it was
caught "not far from the Shoals of Nantucket (which is an Island upon the
Coast of New England)," and that when "first pull'd out of the water it was
like a basket, and had gathered itself round like a Wicker-basket, having
taken fast hold upon that bait on the hook which he" (the fisherman) "had
sunk down to the bottom to catch other Fish, and having held that within
the surrounding brachia would not let it go, though drawn up into the
Vessel; until, by lying a while on the Deck, it felt the want of its
natural Element; and then voluntarily it extended itself into the flat
round form, in which it appear'd when present'd to your view." The Governor
goes on to reflect in a philosophical vein upon the purpose involved in all
this complicated machinery. "The only use," he says, "that could be
discerned of all that curious composure wherewith nature had adorned it
seems to be to make it as a purse-net to catch some other fish, or any
other thing fit for its food, and as a basket of store to keep some of it
for future supply, or as a receptacle to preserve and defend the young ones
of the same kind from fish of prey; if not to feed on them also (which
appears probable the one or the other), for that sometimes there were found
pieces of Mackerel within that concave. And he, the Fisherman, told me that
once he caught one, which had within the hollow of its embracements a very
small fish of the same kind, together with some piece or pieces of another
fish, which was judged to be of a
[Pg 120]
Mackerel. And that small one ('tis like) was kept either for its preservation
or for food to the greater; but, being alive, it seems most likely it was there
lodged for safety, except it were accidentally drawn within the net, together
with that piece of fish upon which it might be then feeding." The account
concludes by saying, "This Fisherman could not tell me of any name it hath, and
'tis in all likelihood yet nameless, being not commonly known as other Fish are.
But until a fitter English/ name be found for it, why may it not be called
(in regard of what hath been before mentioned of it) a Basket-Fish, or a
Net-Fish, or a Purs-net-Fish?" And so it remains to this day as the
Governor of Connecticut first christened it, the Basket-fish.
CRINOIDS.
The Crinoids are very scantily represented in the present creation. They
had their day in the earlier geological epochs, when for some time they
remained the sole representatives of their class, and were then so numerous
that the class of Echinoderms, with only one order, seemed as full and
various as it now does with five. The different forms they assumed in the
successive geological periods are particularly instructive; these older
Crinoids combined characters which foreshadowed the advent of the
Ophiurans, the true Star-fishes, and the Sea-urchins; and so prominently
were their prophetic characters developed, that many of them are readily
mistaken for Star-fishes or Sea-urchins.
[fig 152]
 Fig. 152. Fossil Pentacrinus.
Fig. 152. Fossil Pentacrinus.
In later times the group of Crinoids has been gradually dwindling in number
and variety. Its present representatives are the Pentacrini of Porto Rico
and the coast of Portugal, the lovely little Rhizocrinus of the Atlantic,
dredged first by the younger Sars on the coast of Norway, attached
throughout life to a stem, and the Comatula, which has a stem only in the
early stages of its growth, but is free when adult. The Pentacrinus bears
the closer relation to the more ancient Crinoids (Fig. 152), which were
always supported on a stem, while it is only in more
[Pg 121]
recent periods that we find the free Crinoids, corresponding to the Comatula.
Comatula. (Alecto meridionalis Ag.)
One large species of Comatula (Alecto Eschrichtii M. & T.) is known on
our coast, off the shores of Greenland, where it has been dredged at a
depth of about one hundred and fifty fathoms, and young specimens of the
same species have been found as far south as Eastport, Maine. The species
selected for representation here, however, (Fig. 153,) is one quite
abundant along the shores of South Carolina. It is introduced instead of
the northern one, because the latter is so rare that it is not likely to
fall into the hands of our readers. The annexed drawing (Fig. 154,
magnified from Fig. 153) represents a group of the young of the Charleston
Comatula, still attached to the parent body by their stems, and in various
stages of
[Pg 122]
development. At first sight, the Comatula, or, as it is
sometimes called, the feather-star, resembles an Ophiuran; but on a closer
examination we find that the arms are made up of short joints; and along
the sides of the arms, attached to each joint, are appendages resembling
somewhat the beards of a feather, and giving to each ray the appearance of
a plume; hence the name of feather-star. On one side the arms are covered
with a tough skin, through which project the ambulacræ, and on the same
side of the disk are situated the mouth and the anus; the latter projects
in a trumpet-shaped proboscis. On the opposite side of the disk the
Comatula is covered with plates, arranged regularly around a central plate,
which is itself covered with long cirri.
[fig 153]
[fig 154]
 seen from the back; a group of young Comatulæ attached to parent.") |
|
 |
| Fig. 153. Comatula (Living Crinoid) seen from the back; a group of young Comatulæ attached to parent. |
|
Fig. 154. Magnified view of the group of young Comatulæ of Fig. 153. |
[Pg 123]
We are indebted to Thompson for the explanation of the true relations of
the young Comatula to the present Pentacrinus and the fossil Crinoids.
Supposing these young to be full-grown animals, he at first described them
as living representatives of the genus Pentacrinus; it was only after he
had watched their development, and ascertained by actual observation that
they dropped from their stem, to lead an independent life as free Comatulæ,
that he fully understood their true connection with the past history of
their kind, as well as with their contemporaries. In Fig. 153, a faint
star-like dot (y) may be seen attached to the side of the disk by a
slight line. In Fig. 154, we have that minute dot as it appears under the
microscope, magnified many diameters; when it is seen to be a cirrus of a
Comatula, with three small Pentacrinus-like animals growing upon it, in
different stages of development. In the upper one, the branching arms and
the disk, with its many plates, are already formed; and though in the
figure the rays are folded together, they are free, and can be opened at
will. In the larger of the two lower buds, the plates of the disk are less
perfect, and the arms are straight and simple, without any ramifications,
though they are free and movable, whereas, in the smaller one, they are
folded within the closed bud.
EMBRYOLOGY OF ECHINODERMS.
All Radiates have a special mode of development, as distinct for each class
as is their adult condition, and in none are the stages of growth more
characteristic than in the Echinoderms. In the Polyps, the division of the
body into chambers, so marked a feature of their ultimate structure, takes
place early; in the Acalephs, the tubes which traverse the body are
hollowed out of its mass in the first stages of the embryonic growth, and
we shall see that in the Echinoderms also, the distinctive feature of their
structure, viz. the enclosing of the organs by separate walls, early
manifests itself. This peculiarity gives to the internal
[Pg 124]
structure of these animals so individual a character, that some naturalists,
overlooking the law of radiation, as prevalent in them as in any members of this
division, have been inclined to separate them, as a primary division of the
animal kingdom, from the Polyps and Acalephs, in both of which the
body-wall furnishes the walls of the different internal cavities, either by
folding inwardly in such a manner as to enclose them, as in the Polyps, or
by the cavities themselves being hollowed out of the general mass, as in
the Acalephs.
Star-fish. (Astracanthion.)
[fig 155]
[fig 156]
[fig 157]
 |
|
 |
|
 |
| Fig. 155. Egg of Star-fish. |
|
Fig. 156. Egg of Star-fish in which the yolk has been divided into two segments. |
|
Fig. 157. Egg in which there are eight segments of the yolk. |
The egg of the Star-fish, when first formed, is a transparent, spherical
body, enclosing the germinative vesicle and dot. (See Fig. 155.) As soon as
these disappear, the segmentation of the yolk begins; it divides first into
two portions (see Fig. 156), then into four, then into eight, and so on;
but when there are no more than eight bodies of segmentation (see Fig.
157), they already show a disposition to arrange themselves in a hollow
sphere, enclosing a space within, and by the time the segmentation is
completed, they form a continuous spherical shell. At this time the egg,
or, as we will henceforth call it, the embryo, escapes and swims freely
about. (See Fig. 158.) The wall next begins to thin out on one side, while
on the opposite side, which by comparison becomes somewhat bulging, a
depression is formed (m a, Fig. 159), gradually elongating into a loop
hanging down within the little animal, and forming a digestive cavity.
(d, Fig. 160.) At this stage it much resembles a young Actinia. The loop
spreads somewhat at its upper extremity, and at its lower end is
[Pg 125]
an opening, which at this period of the animal's life serves a double purpose,
that of mouth and anus also, for at this opening it both takes in and
rejects its food. We shall see that before long a true mouth is formed,
after which this first aperture takes its place opposite the mouth,
retaining only the function of the anus. Presently from the upper bulging
extremity of the digestive cavity, two lappets, or little pouches, project
(w/ w' Fig. 161); they shortly become completely separated from it, and
form two distinct hollow cavities (w w', Fig. 162). Here begins the
true history of the young Star-fish, for these two cavities will develop
into two water-tubes, on one of which the back of the Star-fish, that is,
its upper surface, covered with spines, will be developed, while on the
other, the lower surface, with the suckers and tentacles, will arise. At a
very early stage one of these water-tubes (w', Fig. 163) connects with a
smaller tube opening outwards, which is hereafter to be the madreporic body
(b, Fig. 163). Almost until the end of its growth, these two surfaces, as
we[Pg 126] shall see, remain separate, and form an open angle with one another; it
is only toward the end of the development that they unite, enclosing
between them the internal organs, which have been built up in the mean
while.
[fig 158]
[fig 159]
[fig 160]
[fig 161]
 |
|
 |
| Fig. 158. Larva just hatched from egg; a thickened pole. |
|
Fig. 159. Larva somewhat older than Fig. 158; m a depression at thickened pole. |

Fig. 160. Larva where the depression has become a digestive cavity d, opening at a. |
|

Fig. 161. Earlets, w w' (water-tubes), developed at the extremity of the digestive cavity d; m mouth. |
[fig 162]
[fig 163]
[fig 164]

Fig. 162. More advanced larva; a d c digestive system, v vibratile chord, m mouth. | |

Fig. 163. Profile view of larva; b madreporic opening, w' earlet, a d digestive system, m mouth, v v' vibratile chord. | |

Fig. 164. Larva showing mode of formation of mouth m, by bending of digestive cavity o |
At about the same time with the development of these two pouches, so
important in the animal's future history, the digestive cavity becomes
slightly curved, bending its upper end sideways till it meets the outer
wall, and forms a junction with it (m, Fig. 164). At this point, when the
juncture takes place, an aperture is presently formed, which is the true
mouth. The digestive sac, which has thus far served as the only internal
cavity, now contracts at certain distances, and forms three distinct,
though connected cavities, as in Fig. 163; viz. the oesophagus leading
directly from the mouth (m) to the second cavity or stomach (d), which
opens in its turn into the third cavity, the alimentary canal. Meanwhile
the water-tubes have been elongating till they now surround the digestive
cavity, extending on the other side of it beyond the mouth, where they
unite, thus forming a Y-shaped tube, narrowing at one extremity, and
dividing into two branches toward the other end. (Fig. 165.)
[fig 165]
 |
| Fig. 165. Larva in which arms are developing, lettering as before; e' e'' e''' e4 e5 e6 arms, o oesophagus. |
On the surface where the mouth is formed, and very near it on either side,
two small arcs arise, as v in Fig. 162; these are cords consisting
entirely of vibratile cilia. They are the locomotive organs of the young
embryo, and they gradually extend until they respectively enclose nearly
the whole of the upper and lower half of the body, forming two large
shields or plastrons. (Figs. 165, 166.) The corners of these shields
project, slightly at first (Fig. 165), but elongating more and more until a
number of arms are formed, stretching in various directions (Figs. 166,
167), and, by their constant upward and downward play, moving the embryo
about in the water.
[Pg 127]
[fig 166]
[fig 167]

Fig. 166. Adult larva, so-called Brachiolaria,
lettering as before; r back of young Star-fish,
t tentacles of young Star-fish, f f' brachiolar appendages.
Click on image to view larger size.

Fig. 167. Fig. 166 seen in profile, lettering as before.
[fig 168]
[fig 169]
[fig 170]

Fig. 168. Star-fish which has just resorbed the larva, seen from the back; b madreporic opening. | | 
Fig. 170. Young Star-fish which has become symmetrical, seen from the back; t' odd tentacle. |
| |

Fig. 169. Fig. 168, seen from the mouth side; m mouth, t tentacles. |
At this stage of the growth of the embryo, we have what seems quite a
complicated structure, and might be taken for a complete animal; this is
after all but the prelude to its true Star-fish existence. While these
various appendages of the embryo have been forming, changes of another kind
have taken place; on one of the two water-tubes above mentioned (w'), at
the end nearest the digestive cavity, a number of lobes are formed (t,
Fig. 166); this is the first appearance of the tentacles. In the same
region of the opposite water-tube (w) a number of little limestone rods
arise, which eventually unite to form a continuous network; this is the
[Pg 128]
beginning of the back of the Star-fish (r, Fig. 166), from which the
spines will presently project. When this process is complete, the whole
embryo, with the exception of the part where the young Star-fish is placed,
grows opaque; it fades, as it were, begins to shrink and contract, and
presently drops to the bottom, where it attaches itself by means of short
arms (f f', Fig. 166), covered with warts, which act as suckers, and
are placed just above the mouth. As soon as the Star-fish has thus secured
itself, it begins to resorb the whole external structure described above;
the water-tubes, the plastrons, and the complicated system of arms
connected with them, disappear within the little Star-fish; it swallows up,
so to speak, the first stage of its own existence; it devours its own
larva, which now becomes part and parcel of the new animal. Next the two
surfaces, the back and lower surface, on which the arms are now marked out,
while the tentacles, suckers, and spines have already assumed a certain
prominence, approach each other. At this time, however, the arms are not in
one plane; both the back and the lower surface are curved in a kind of
spiral; they begin to flatten; the arms spread out on one level,—and now
the two surfaces draw together, meeting at the circumference, and enclosing
between them the internal organs, which, as we have seen, are already
formed and surrounded by walls of their own, before the two walls of the body,
[Pg 129]
close thus over them. Fig. 168 represents the upper surface of the
Star-fish just before this junction takes place. The complicated structure
of the Brachiolaria, as the larva of the Star-fish has been called,
hitherto so essential to the life of the animal, by which it has been
supported, moved about in the water, and provided with food during its
immature condition, has made a final contribution to its further
development by the process of resorption described above, and has wholly
disappeared within the Star-fish. At this stage the rays are only just
marked out, as five lobes around the margin; Fig. 169 represents the lower
surface at the same moment, with the open mouth (m), around which the
tentacles (t) are just beginning to appear; while Fig. 170 shows us the
animal at a more advanced stage, after the two surfaces have united. It has
now somewhat the outline of a Maltese cross, the five arms being more
distinctly marked out, while the tentacles have already attained a
considerable length (Fig. 171), and the dorsal plates have become quite
distinct. Fig. 172 represents the same animal, at the same age, in profile.
This period, in which we have compared the form of the Star-fish to that of
a Maltese cross, is one of long duration; two or three years must elapse
before the arms will elongate sufficiently to give it a star-shaped form,
and before the pedicellariæ make their appearance,
[Pg 130]
and it is only then that it can be at once recognized as the young of our
common Star-fish. Even then, after it has assumed its ultimate outline,
it lacks some features of the adult, having only two rows of tentacles,
whereas the full-grown Star-fish has four.
[fig 171]
[fig 172]

Fig. 171. Lower side of ray of young Star-fish;
m mouth, b madreporic body, e eye-speck.
|

Fig. 172. Young Star-fish seen in profile;
t' odd tentacle at extremity of arm. |
[fig 173]
[fig 174]
[fig 175]

Fig. 173 |
|

Fig. 174 |

Fig. 175 |
| Fig. 173, 174, 175. Young larvæ of Toxopneustes in different stages of development; e'-eiv arms, v-v'' vibratile chord, w w' earlets (water-tubes), a o d c digestive system, r'-r''' solid rods of arms, m mouth, b madreporic opening. |
Sea-urchins.
This extraordinary process of development which we have analyzed thus at
length in the history of the Star-fish, but which is equally true of all
Echinoderms, has been hitherto described (so far as it was known) under the
name of the plutean stages of
[Pg 131]
growth. In these early stages the young, or the so-called larvæ of
Echinoderms, have received the name of Pluteus on account of their ever-changing
forms. Let us look for a moment at the plutean stages of the Sea-urchin,
as they differ in some points from those of the Star-fish. In the Pluteus
of our common Sea-urchins (see Fig. 176), the arms are supported by a
framework of solid limestone rods, which do not exist in that of the
Star-fish, and which give to the larva of the Sea-urchin a remarkable
rigidity. They are formed very early, as may be seen in Fig. 173, representing
the little Sea-urchin before any arms are discernible, though the limestone
rods are quite distinct. Figs. 173, 174, 175, may be compared with Figs. 160,
162, 165, of the young Star-fish, where it will be seen that the general
outline is very similar, though, on account of the limestone rods, the Pluteus
of the Sea-urchin seems somewhat more complicated. In
[Pg 132]
Fig. 176 the young Sea-urchin has so far encroached upon the Pluteus
that it forms the essential part of the body, the arms and rods
appearing as mere appendages. Fig. 177 shows the same animal when we
looked down upon it in its natural attitude; the Sea-urchin is carried
downward, and the arms stretch in every direction around it. In Fig. 178
the Plutens is already in process of absorption; in Fig. 179 it has wholly
disappeared; in
[Pg 133]
Figs. 180 and 181 we have different stages of the little Sea-urchin, with its spines
and suckers of a large size and in full activity. The appearance of the
Sea-urchin, as soon as this larva or Pluteus is completely absorbed, is
much more like that of the adult than is the Star-fish at the same stages,
in which, as we have seen, there is a transition period of considerable
duration.
[fig 176]

Fig. 176. Adult larva of Toxopneustes, f brachiolar appendages.
Click on image to view larger size. |
| |
|
[fig 177]

Fig. 177. Fig. 176 seen endways. |
| |
|
[fig 178]

Fig. 178. The Sea-urchin resorbing the arms of the larva. |
| |
|
[fig 179]

Fig. 179. Half a young Sea-urchin immediately after resorption of the larva;
s'' s'' spines, t' t' ambulacral tentacles. |
[Pg 134]
|
[fig 180]

Fig. 180. Young Sea-urchin older than Fig. 179; t t' tentacles, s'' s''' spines. |
| |
|
[fig 181]

Fig. 181. Still older Sea-urchin; t t tentacles, a anus, p pedicellariæ;
shell one sixteenth of an inch in diameter. |
[Pg 135]
Ophiurans.
Fig. 183 represents an Ophiuran undergoing the same process of growth, at a
period when the larva is most fully developed, and before it begins to
fail. By the limestone rods which support the arms, the Pluteus of the
Ophiuran, here represented, resembles that of the Sea-urchin more than that
of the Star-Fish, while by the character of the water-tubes and by its
internal organization it is more closely allied to the latter. It differs
from both, however, in the immense length of two of the arms; these arms
being the last signs of its plutean condition to disappear; when the young
Ophiuran has absorbed almost the whole Pluteus, it still goes wandering
about with these two immense appendages, which finally share the fate of
all the rest. Fig. 182 represents an Ophiuran at the moment when the
process of resorption is nearly completed, though the arms of the Pluteus,
greatly diminished, are still to be seen protruding from the surface of the
animal.
[Pg 136]
[fig 182]
[fig 183]
[fig 184]
[fig 185]

Fig. 182. Ophiuran which has resorbed the whole larva except the two long arms,
y y' limestone rods of young Ophiuran, r middle of back, lettering as in Fig. 183. |
| |

Fig. 183. Larva of Ophiuran; e'-eiv arms, r' riv solid rods, v v' vibratile chord, w w' water system, b madreporic body, a d digestive system. |
|

Fig. 184 Young Ophiuran which has resorbed the whole larva; r middle plate of back. |

Fig. 185 Cluster of eggs of Star-fishes placed over the mouth of the parent. |
[Pg 137]
This mode of development, though common to all Echinoderms, appears under
very different conditions in some of them. There are certain Star-fishes,
Ophiurans, and Holothurians, passing through their development under what
is known as the sedentary process. The eggs are not laid, as in the cases
described above, but are carried in a sort of pouch over the mouth of the
parent animal, where they remain till they attain a stage corresponding to
that of Fig. 168 of the Star-fish, and having much the same cross-shaped
outline, when they escape from the pouch (as the young Ophiopholis, Fig.
184), and swim about for the first time as free animals. Fig. 185
represents a cluster of young Star-fishes of the sedentary kind at about
this period. But while this mode of growth seems at first sight so
different, we shall find, if we look a little closer, that it is
essentially the same, and that, though the circumstances under which the
development takes place are changed, the process does not differ. The
little Star-fish or Ophiuran, in the pouch, becomes surrounded by the same
plutean structure as those which are laid in the egg; it is only more
contracted to suit the narrower space in which they have to move; and the
water-tubes on which the upper and lower surfaces of the body arise, the
shields, spreading out into arms at the corners, exist, fully developed or
rudimentary, in the one as much as in the other, and when no longer
necessary to its external existence they are resorbed in the same way in
both cases. This singular process of development has no parallel in the animal
[Pg 138]
kingdom, although the growth of the young Echinoderm on the
Brachiolaria may at first sight remind us of the budding of the little
Medusa on the Hydroid stock, or even of the passage of the insect larva
into the chrysalis. But in both these instances, the different phases of
the development are entirely distinct; the Hydroid stock is permanent,
continuing to live and grow and perform its share in the cycle of existence
to which it belongs, after the Medusa has parted from it to lead a separate
life, or if the latter remains attached to the parent stock, after it has
entered upon its own proper functions. The life of the caterpillar,
chrysalis and butterfly, is also distinct and definitely marked; the moment
when the animal passes from one into the other cannot be mistaken, although
the different phases are carried on successively and not simultaneously, as
in the case of the Acalephs. But in the Echinoderms, on the contrary,
though the aspect of the Brachiolaria, or plutean stage, is so different
from that of the adult form, that no one would suppose them to belong to
the same animal, yet these two stages of growth pass so gradually into one
another, that one cannot say when the life of the larva ceases, and that of
the Echinoderm begins.
The bearing of embryology upon classification is becoming every day more
important, rendering the processes of development among animals one of the
most interesting and instructive studies to which the naturalist can devote
himself, in the present state of his science. The accuracy of this test,
not only as explaining the relations between animals now living, but as
giving the clew to their connection with those of past times, cannot but
astonish any one who makes it the basis of his investigations. The
comparison of embryo forms with fossil types is of course difficult, and
must in many instances be incomplete, for while, in the one case, death and
decay have often half destroyed the specimen, in the other, life has
scarcely stamped itself in legible characters on the new being. Yet,
whenever such comparisons have been successfully carried out, the result is
always the same; the present representatives of the fossil types recall in
their embryonic condition the ancient forms, and often explain their true
position in the animal kingdom. One of the most remarkable examples of this
in the type we are now considering,
[Pg 139]
is that of the Comatula already mentioned. Its condition in the earlier
stages of growth, when it is provided with a stem, at once shows its relation
to the old stemmed Crinoids, the earliest representatives of the class of Echinoderms.
These coincidences are still more striking among living animals, where they
can be more readily and fully traced, and often give us a key to their
relative standing, which our knowledge of their anatomical structure fails
to furnish. This is perhaps nowhere more distinctly seen than in the type
of Radiates, where the Acalephs in their first stages of growth, that is,
in their Hydroid condition, remind us of the adult forms among Polyps,
showing the structural rank of the Acalephs to be the highest, since they
pass beyond a stage which is permanent with the Polyps; while the adult
forms of the Acalephs have in their turn a certain resemblance to the
embryonic phases of the class next above them, the Echinoderms. Within the
limits of the classes, the same correspondence exists as between the
different orders; the embryonic forms of the higher Polyps recall the adult
forms of the lower ones, and the same is true of the Acalephs as far as
these phenomena have been followed and compared among them. In the class of
Echinoderms the comparison has been carried out to a considerable extent,
their classification has hitherto been based chiefly upon the ambulacral
system, so characteristic of the class, but so unequally developed in the
different orders. This places the Holothurians, in which the ambulacral
system has its greatest development, at the head of the class; next to them
come the Sea-urchins or Echinoids; then the Star-fishes; then the Ophiurans
and Crinoids, in which the ambulacral system is reduced to a minimum.
Another basis for classification in this type, which gives the same result,
is the indication of a bilateral symmetry in some of the orders. In the
Holothurians, for instance, there is a decided tendency toward the
establishment of a posterior and anterior extremity, of a right and left,
an upper and lower side of the body. In the Sea-urchins, in many of which
the mouth is out of centre, placed nearer one side than the other, this
tendency is still apparent, while in the three lower groups, the
Star-fishes, Ophiurans, and Crinoids, it is almost entirely lost, in the equal
[Pg 140]
division of identical parts radiating from a common centre. A
comparison of the embryonic and adult forms in these orders, confirms
entirely this classification based upon structural features. The
Star-fishes, in their earlier stages, resemble the mature Ophiurans, while
the Crinoids, the lowest group of all, retain throughout their whole
existence many features characteristic of the embryonic conditions of the
higher Echinoderms. In this principle of classification, already so fertile
in results, we may hope to find, in some instances, the solution of many
perplexing points respecting the structural rank of animals, the
confirmation of classifications already established; in others, an insight
into the true relations of groups which have hitherto been divided upon
purely arbitrary grounds.
[Pg 141]
DISTRIBUTION OF LIFE IN THE OCEAN.
We have seen that while our bay is rich in certain species, it is wholly
deficient or but scantily supplied with others, and that the character of
the animals inhabiting its waters is more or less directly connected with
general physical conditions. Such an area, limited though it be, gives us
some insight into the laws which, in their wider application, control the
distribution of marine life along the shores of the most extensive
continents. The coast of Massachusetts, taken as a whole, is like that of
New England generally, a rocky coast; yet it has its sandy and muddy
beaches, and though it lies for a great part open to the sea, it has
nevertheless its sheltered harbors, its quiet bays and snug recesses.
A comparison of these limited localities with far more extensive reaches of
shore, where similar physical conditions prevail, shows that they
reproduce, in fainter and less various characters of course, in proportion
to their narrower boundaries, but still with a certain fidelity, the same
combinations of animal and vegetable life. In other words, a sandy beach,
however small, gives us some idea of the nature of the animals we may look
for on any sandy coast, as, for instance, clams of various kinds,
razor-shells, quahogs, snails, &c., creatures who can penetrate the sand,
drag themselves through it or over it, leaving their winding trails as they
go, and to whom the conditions prevailing in such spots are genial. So the
narrowest mud flat on the sea-shore or muddy beach will give us the same
dead and inanimate aspect which characterizes a more extensive coast of
like character, where the gases always generated in mud are deadly to many
kinds of animals. The beings who find a home in such localities are of
closely allied species, chiefly a variety of worms, who burrow their way
into the mud, and seem to court the miasma so
[Pg 142]
fatal to other creatures. The same is true of any stony beach or rocky
shore not more than a quarter of a mile in length; it gives us an idea
of the animal population on any similar coast of greater extent.
These correspondences are of course modified by differences in climatic
conditions. The animals on a sandy beach or a rocky shore, on the coast of
Great Britain, for instance, are not absolutely identical with those of a
sandy beach or a rocky shore on the coast of New England, but they are more
or less nearly related to them. Naturalists refer to this reiteration, all
the world over, of like organic combinations under similar circumstances,
when they speak of "representative species." The aggregate result is the
same, though the individual forms are slightly modified. And here lies one
secret of the infinite variety in nature, by which the old seems ever new,
and the same thought has an eternal freshness and originality, endlessly
repeated, yet never hackneyed.
In this sense our bay presents, on a miniature scale, a variety of physical
and organic combinations, which may be compared to those more extensive
divisions in the geographical distribution of animals and plants, called by
naturalists zoölogical or botanical provinces or districts, the animal and
vegetable populations of which are technically designated as their faunæ
and floræ. Such organic realms, as we may call them, have long been
recognized on land, and the most extensive among them are easily
distinguished. No one will fail to recognize the tropical zone, with its
royal dynasty of palms and all the accompanying glories of a tropical
vegetation, its birds of brilliant plumage, its large Mammalia, lions,
tigers, panthers, elephants, and its great rivers haunted by gigantic
reptiles. Nor is the representation of vegetable and animal life less
characteristic in the temperate zone, where the oak is monarch of the
woods, with all his attendant court of elms, walnuts, beeches, birches,
maples, and the like, where birds of more sober hues, but sweeter voices,
take the place of the brilliant parrots and many-tinted humming-birds of
the tropical forest; while buffaloes, bears, wolves, foxes, and deer
represent the larger Mammalia. In the arctic zone, though marked by
peculiar and distinctive features, vegetation has dwindled to a
[Pg 143]
minimum; the birds are chiefly gulls and ducks, which go there for
the breeding season in the summer, and the reindeer and polar bears
are almost sole possessors of the snow and ice-fields; but this
meagreness in the representation of the larger land Mammalia is amply
compensated in the numbers of heavy aquatic Mammalia, the whales,
walruses, seals, and porpoises of the Arctic seas.
During the last half-century, since the geographical distribution of
animals and plants has become a subject of more careful investigation among
naturalists, these broad zones of the earth's surface, with their
characteristic populations and vegetation, have been subdivided, according
to more limited and special combinations of organic forms, into narrower
zoölogical and botanical areas. The application of these results to marine
life is however of much more recent date, and indeed it would seem at first
sight, as if the water, from its own nature, could hardly impose a barrier
so impassable as the land. The localization of the marine faunæ and floræ
is nevertheless as distinct as that of terrestrial animals and plants, and
late investigations have done much to explain the connection of this
distribution with physical conditions.
A glance at the coast of our own continent, starting from the high north
and making the circuit of its shores, from Baffin's Bay to Behring's
Straits, will show us to what a variety of physical influences the animals
who live along its shores are subjected. On the shores of Baffin's Bay,
especially on the inner coast of Greenland, where the glaciers push their
way down to the very brink of the water, and annually launch their
southward-bound icebergs, we shall hardly expect to find a very abundant
littoral fauna. On its western shore, where the ice does not advance so
far, and a greater surface of rock is exposed, the circumstances are more
favorable to the development of animal life. Here abound the winged
Mollusks (Pteropods), often swept down to the coast of Nova Scotia by the
cold current from Baffin's Bay; the "whale feed," as the fishermen call
them, because the whales devour them voraciously. Here occur also many
compound Mollusks, especially a variety of Ascidians, and the highly
colored stocks of Bryozoa. With them is found the Comatula of the
[Pg 144]
northern waters, one of the few modern Crinoids, and beside these a number of
Star-fishes, Sea-urchins, and Holothurians, not differing so essentially
from those already described as to require special mention.
Along the shore of Labrador and Newfoundland, the coast is wholly rocky,
and especially about Newfoundland it is deeply indented with bays. Here
there is ample opportunity for the growth of certain kinds of animals in
sheltered nooks. The number of species is, however, much greater along the
shores of Maine, Nova Scotia, and New Brunswick than in Labrador, owing no
doubt to the milder climate. The beautiful shore of Maine, with its
countless islands, and broken, picturesque outline, is very rich in
species. Parts of this coast are remarkable for a variety of naked
Mollusks, as well as for the great numbers of bright-colored Actiniæ, and
also for the more brilliant kinds of Holothurians, the Cuvieria, and the
like. The latter are especially abundant in the Bay of Fundy, and here also
occurs the only Northern representative on our coast of the Sea-fans or
Gorgoniæ, so common on the shores of Florida.
Farther south, from Cape Cod to Cape Hatteras, the character of the coast
changes; it becomes more sandy, and though here and there the aspect is
varied by a rocky promontory or a stony beach, yet the general character is
flat and sandy. With this new character of the shore, the fauna is also
greatly modified, and it is worthy of remark, that while thus far the
representative species have reflected the character of animals to the north
of them, they now begin to represent rather those of the Carolina shores.
South of Cape Cod come in a kind of Scallop and Periwinkle, very different
from the larger Scallops found on the coast of Maine and the British
Provinces; our Sea-urchin is replaced by the Echinocidaris, with its few
long spines, and an entirely new set of Crustacea and Worms make their
appearance on this more sandy bottom. And here we must not forget that not
only is the aspect of the animal life changed, as we pass from a rock-bound
to a sandy coast, but that of the vegetation also. The various many-tinted
sea-weeds of the rocky shore disappear almost entirely, and their place is
but poorly supplied by the long eel-grass, which is almost the only marine
plant to be found in such
[Pg 145]
a locality. Beside its more sandy character, the coast from Cape Cod to
Cape Hatteras is affected by the large amount of fresh water poured into
the sea along its whole line, greatly modifying the character of the shore
animals. The Hudson, the Delaware, the Susquehanna, the Potomac, the James,
the Roanoke, and the large estuaries connected with some of these rivers,
give a very peculiar character to the shore, and bring down, not only a vast
supply of fresh water, but also a large quantity of detritus of all sorts from
the land. Under these circumstances life would be impossible for many of the
animals which live farther north. The only locality on the North Atlantic shore,
where the conditions are somewhat similar, is at the mouth of the St. Lawrence,
that great drainage-bed through which the Canadian lakes empty their superfluous
waters into the Gulf of St. Lawrence.
The whole coast of the Carolinas, from Cape Hatteras to Florida, is a sandy
beach; but though in this respect it resembles that immediately to the
north of it, it differs greatly in other features. Comparatively little
fresh water is poured into the ocean along this shore, and its more
southerly range, instead of being protected by sand-spits like Pamlico and
Albemarle Sounds, or broken by estuaries and inlets like the coast of
Virginia, lies broadly open to the sea. On its extensive beaches we have
the large Pholas, burrowing deep below the surface, and the Cerianthus,
those long, cylindrical Actiniæ, enclosed in sheaths, with their bright
crowns of gayly-colored tentacles; the free colonies of Halcyonoids abound
also on this coast, and a new set of Sea-urchins (Spatangoids and
Clypeastroids) make their appearance.
Farther south, along the Florida coast, a new element comes in, that of the
coral reefs, enclosing shallow channels near the shore, and thus providing
sheltered harbors on their leeward side, while on their seaward side they
slope steeply to the ocean. Beside this, the reef itself affords a home for
a great variety of creatures, who bore their way into it and live in its
recesses, as some insects live in the bark of trees. Perhaps a more
favorable combination of circumstances for the development of marine life
does not exist anywhere than about the coral reefs of Florida, and
certainly nowhere is there a more rich and varied
[Pg 146]
littoral fauna, especially on their western shore within the Gulf of Mexico.
Here swims the Portuguese Man-of-War, borne gayly along on the surface of
the water by its brilliant float, here the blue Velella sets its oblique
sail to the wind, and hosts of the lighter and more brightly tinted corals
fringe the shore with a many-colored shrubbery. In these waters are also
found the blue and yellow Angel-fish, the Parrot-fish (Scarus), and the
strange Porcupine-fish (Diodon). Vegetable life is comparatively scanty
in these tropical waters, where there are scarcely any sea-weeds, except the
corallines or limestone Algæ of the reefs. The shore of the Gulf of
Mexico, as a whole, has much the same character as that of the Carolinas,
until we reach the point where the mountains and plateau of Mexico come
down to the coast. From this point to the Isthmus of Panama the coast is
again rocky.
Crossing the Isthmus and following the Pacific shore of the continent
northward, we find a sandy open shore alternating with rocky beaches as far
north as Acapulco. Along this coast there is to be found a great variety of
corals, especially Sea-fans, growing on the rocks, but no reef. The
Pocillopora, an Acalephian coral, the Pacific representative of the
Millepore of Florida, is especially abundant. On the peninsula of Lower
California we come again upon a rocky coast, with steep bluffs, extending
into the sea. Within the Gulf of California are found, on its sandy coast,
peculiar kinds of Sea-urchins, Spatangoids, and Clypeastroids, which occur
nowhere else on this coast. From Cape St. Lucas up to the Straits of Fuca,
with the exception of the large estuary forming the Bay of San Francisco,
there are scarcely a couple of harbors of any consequence. The whole shore
is most inhospitable, and the violent northwest winds in summer, and the
southeast winds in winter, render it still more bleak and difficult of
approach. In consequence of these conditions, the fauna is scanty along a
great part of the shore; the best spots for collecting are the beaches,
near the head of the peninsula, opposite the islands of Santa Barbara and
San Diego, and that within the harbor of San Francisco. On the former,
large Craw-fishes abound (Palinurus), akin to those of Florida, though
specifically different from them. In the latter, the great amount of fresh
water prevents the fauna from being exclusively marine; this harbor is,
[Pg 147]
nevertheless, the great centre of the viviparous fishes, and contains also
a large variety of peculiarly shaped Sculpins.
Farther north, between the Straits of Fuca and the island of Sitka, the
shore resembles that of Maine, with its many islands, bays, and inlets; a
succession of long, narrow islands forms a barrier along the coast,
enclosing the shore waters, so as almost to make them into an inland sea.
But little fresh water empties upon this part of the coast, and here, where
the salt water is little modified by any deposit from the land, but where
the violence of the ocean is broken by this barrier of islands, there is a
full development of marine life. The shores of the Gulf of Georgia, and
those of Vancouver's Island, seem to be especially the home of the
Star-fishes. The fauna of this locality has been but little investigated,
and yet the number of species of Star-fishes known from there is greater
than from any other region; many of them are of colossal size, measuring
some four feet in diameter. This coast seems also very favorable for the
development of Hydroids, in consequence of which its waters swarm with a
variety of Jelly-fishes. The Pennatula, that pretty compound Halcyonoid,
with its feather-like sprays, is another characteristic type of this fauna.
Beyond this, from Sitka to Behring's Straits, the same rocky coast prevails
as in Labrador and Greenland. In Behring's Straits we return again to the
forests of beautiful compound Mollusks, or rather to a variety of
"representative species," resembling the Bryozoa and Ascidians so abundant
in Baffin's Bay. The depth of the water, however, is much less here than on
the corresponding Atlantic coast, where, south of Greenland, along the
shore of Labrador, the water is very deep, while in Behring's Straits the
depth is not greater than from one hundred to one hundred and twenty
fathoms. The respective faunæ of these two shores are also affected by the
difference of temperature, the cold current from Baffin's Bay sweeping down
upon the coast of Labrador, while, through Behring's Straits, the warm
current from the Pacific pours into the Arctic Ocean.
Thus the whole coast of our continent is peopled more or less thickly with
animals. But now arises a new set of inquiries; how far into the sea do
these animals extend? how wide is their domain? Do they wander at will in
the ocean, or are they
[Pg 148]
bound by any law to keep within a certain distance
of the shore? These questions would seem to be easily answered, for
wherever we go on the surface of the sea, and as far as the eye can
penetrate into its depths, we find it full of life; and yet a closer
examination shows that all these beings have their appointed boundaries.
Along the shores, animal and vegetable life seems to be distributed in
certain definite combinations. Those who are familiar with rocky beaches
readily recognize the different bands of color produced by the various
kinds of sea-weed growing at given distances between high and
low-water-mark. First comes the olive green rockweed (the Fucus), and with
it are found barnacles and small Crustacea, myriads of which are to be seen
hopping about in this rockweed when the tide is out. Below these are the
brown crispy Rhodersperms and Melanosperms, and associated with them are
Star-fishes, Crabs, and Cockles. Next in order is the Laminarian zone. Here
we have the broad fronds of the Laminaria, the "devil's aprons," as the
fishermen call them; in this zone is the home of the Sea-urchin, and here
will be found also a few small fishes. Lastly we have the Coralline zone,
so-called on account of the lime deposit in the sea-weeds, giving them the
rigidity of corals; among these the Lobsters make their appearance, and
here are to be found also numerous clusters of Hydroids, the nurses of the
Jelly-fishes.
This distribution is not casual; these belts of animal and vegetable life
are sharply defined and so constantly associated, that they must be
controlled by the same physical laws. The first important investigations on
this subject were made by Örsted, the distinguished Danish naturalist. He
undertook a complete topographical survey of the coast near which he lived,
carrying his soundings to a depth of some twelve fathoms, and found that
both the fauna and flora of the shore were divided, according to the depth
of the water, into bands of vegetable and animal life, corresponding very
nearly with those given above. His observations were, however, limited, not
extending beyond the neighborhood of his home. It is to Edward Forbes, the
great English naturalist, whose short life was so rich in results for
science, that we owe a more complete and extensive investigation of the
whole subject.
[Pg 149]
 Diagram of a rocky beach.
Click on image to view larger size.
Diagram of a rocky beach.
Click on image to view larger size.
[Pg 150]
Aided by a friend, Captain McAndrew, who placed his yacht at his disposal,
he made a series of observations on the British, Scandinavian, and Danish
coasts, and explored also with the same object the shores of the
Mediterranean. Not content with sounding the present ocean, he sunk his
daring plummet in the seas of past geological ages, and by comparing the
nature and position of their fossil remains with those of living marine
faunæ, he measured the depths of the water along their shores. He collected
a vast amount of material, and the results of his labors have formed the
basis of all subsequent generalizations upon this subject. Nevertheless he
arrived at some erroneous conclusions, which, had he lived, he would no
doubt have been the first to correct. Dredging from low-water-mark outward,
he found that, from the Laminarian and Coralline zone, the animals began
gradually to decrease in number, and that, at a depth of two or three
hundred fathoms, the dredge always came up nearly empty. He inferred that
at a certain depth the weight of water became too great to be endured by
animals, and that the ocean beyond this line, like the land beyond the line
of perpetual snow, was barren of life. This result seemed the more probable
on account of the immense pressure to which animals are subjected, even at
a comparatively moderate depth. A column of water thirty-two feet high is
equal to one atmosphere in weight; this pressure being increased to the
same amount for every thirty-two feet of depth, it follows that a fish one
hundred and twenty-eight feet, or some twenty fathoms below the surface, is
under the pressure of almost four atmospheres plus that of the air outside.
Wherever tides run high, as in the Bay of Fundy, for instance, where an
animal is under the pressure of one atmosphere at low tide, and of three
atmospheres at high tide, we see that marine animals are uninjured by great
changes of pressure. Yet it seems natural to suppose that there is a limit
to this power of resistance; and that there must exist barren areas at the
bottom of the ocean, as destitute of life as the regions on the earth which
are above the line of perpetual snow. No doubt pressure does influence the
distribution of life in the ocean; but it would seem, from subsequent
observations, that the boundaries assigned by Forbes were far too narrow,
and that the structure of many marine animals
[Pg 151]
enables them to live under a weight, the one hundredth part of which
would be fatal to any terrestrial animal.
For some years Forbes's theory was very generally accepted, and the results
of Darwin's and Dana's investigations, showing that corals could not live
beyond a depth of fifteen fathoms, seemed to confirm it. But, quite
recently, facts derived from new and unlooked-for sources of information
have given a check to this theory. Commerce has come to the aid of science
(rewarding her for the gift first received at her hands), and the telegraph
cables, alive with the secrets of sea and land, have brought us tidings
from the deep. Dr. Wallich, the naturalist who in 1860 accompanied the
expedition to explore the bed of the Atlantic, previous to laying the
telegraphic cable, first called attention again to this subject. He brought
up various animals, highly organized, from a depth of about nineteen
hundred fathoms.
Yet, in spite of this positive evidence added to the former observations of
Ehrenberg, and to those of Sir James Ross, who, in the Antarctic Sea,
brought up an Euryale on a sounding-line from a depth of eight hundred to a
thousand fathoms, naturalists were slow to believe that the distribution of
animal life in the ocean was not limited to the shallow depths assigned by
Edward Forbes. In the Mediterranean and in the Red Sea, from depths of
eighteen hundred to two thousand fathoms, living animals have been brought
up on the telegraph wires, not of doubtful infusorial character, hovering
on the border-land between animal and vegetable life, but of considerable
size, as, for instance, one or two kinds of Crustacea, Cockles, stocks of
Bryozoa and tubes of Annelids. When the cable between France and Algiers
was taken up from a depth of eighteen hundred fathoms, there came with it
an Oyster, Cockle-shells, Annelid tubes, Bryozoa and Sea-fans. As these
animals were growing upon it, there could be no doubt that they had their
normal life and development at this depth, and since they are carnivorous,
they tell also of the existence of other animals with them on which they
feed.
The dredge, which thus far has played an important part in zoölogical
researches, is destined to revolutionize many of our
[Pg 152]
accepted theories, if we can judge of its future by the brilliant results of
the last few years.
From 1861 to the present time the Swedish government has sent several
expeditions to Spitzbergen and Greenland. They carried on dredging
operations most successfully to a depth of twenty-six hundred fathoms. For
some years past Lovén, Koren, and Danielssen, the elder and younger Sars,
and other Scandinavian naturalists, have made systematic dredgings along
the coast of Norway, which, though not extending below four hundred fathoms
or thereabouts, have yet furnished most astounding results.
The United States Coast Survey has, in connection with an exploration of
the Gulf Stream, been the first to establish a systematic series of
dredgings at great depths, continued during several years. The results have
proved conclusively that there exists everywhere, in the deep sea,
modified, of course, according to the nature of the bottom and the
temperature, a most varied fauna, totally distinct from that characteristic
of the shores and of shallower waters. Since 1867 Count Pourtales has had
charge of these investigations, first established many years ago by
Professor Bache and continued by his successor Professor Peirce. He has
dredged across the Gulf Stream between Florida and Cuba to a depth of about
seven hundred fathoms, collecting an immense number of marine animals
entirely unknown before, and characteristic of the different belts of
depth, having a most extraordinary geographical distribution, many of the
species being found in Florida, the Azores, the Faroe Islands, and the west
coast of Norway.
The English Admiralty has for two summers detailed a vessel admirably
fitted for such purposes, intrusting the scientific direction of the
expedition to Dr. Carpenter, Professor Thomson, and Mr. Jeffreys. Their
dredgings, carried on to the enormous depth of two thousand four hundred
and thirty-five fathoms, have in every respect corroborated the conclusions
drawn from the collections made by Count Pourtales and the Scandinavian
naturalists, who, not content with so thoroughly exploring their own coast,
have even sent a ship of war to dredge across the whole Atlantic.
[Pg 153]
These discoveries only show how much yet remains to be done before we shall
fully understand the laws of marine life. But we already have ample
evidence that the same beneficent order controls the distribution of
animals in the ocean as on land, appointing to all its inhabitants their
fitting home in the dim waste of waters.
[Pg 154]
SYSTEMATIC TABLE
OF THE ANIMALS DESCRIBED IN THIS VOLUME
RADIATA Cuv.
CLASS I.—POLYPI Lam.
Order I.—ACTINARIA Edw.
Metridium marginatum Edw.
Rhodactinia Davisii Ag.
Bicidiumparasiticum Ag.
Arachnactis brachiolata A. Ag.
Halcampa albida Ag.
|
|
Order II.—MADREPORIA Ag.
Astrangia Danæ Ag.
Order III.—
HALCYONARIA Edw.
Halcyonium carneum Ag.
|
CLASS II.—ACALEPHÆ Cuv.
Order I.—HYDROIDEA Johnst.
Velella mutica Bosc.
Physalia Arethusa Til.
Nanomia cara A. Ag.
Millepora alcicornis Lin.
Hydractinia polyclina Ag.
Tubularia Couthouyi Ag.
Hybocodon prolifer Ag.
Coryne mirabilis Ag.
Turris vesicaria A. Ag.
Bougainvillia superciliaris Ag.
Dysmorphosa fulguransA. Ag.
Dynamena pumila Lamx.
Dyphasia rosacea Ag.
Lafoea cornutaLamx.
Melicertum campanula Pér. et Les.
Ptychogena lactea A. Ag.
Laomedea amphora Ag.
|
|
Zygodactyla groenlandica Ag.
Tima formosa Ag.
Eucope diaphana Ag.
Clytia bicophora Ag.
Oceania languida A. Ag.
Order II.—DISCOPHORÆ Esch.
Haliclystus auricula Clark.
Trachynema digitale A. Ag.
Campanella pachyderma A. Ag.
Cyanea arctica Pér. et Les.
Aurelia flavidula Pér. et Les.
Order III.—CTENOPHORÆ Esch.
Idyia roseola/ Ag.
Pleurobrachia rhododactyla Ag.
Bolina alata Ag.
|
CLASS III.—ECHINODERMATA Klein.
Order I.—CRINOIDEA Mill.
Pentacrinus,
Alecto Eschrichtii M. & T.
Alecto meridionalis Ag.
Order II.—OPHIURIDEA Forbes.
Amphiura squamata Sars.
Ophiopholis bellis Lym.
Astrophyton AgassiziiStimp.
Order III.—ASTERIDEA Forbes.
Ctenodiscus crispatus D. & K.
Hippasteria phrygiana Ag.
Cribrellaoculata Forbes.
|
|
Solaster endeca Forbes.
Crossaster papposa M. & T.
Astracanthion pallidus Ag.
Astracanthion berylinus Ag.
Order IV.—ECHINIDEA Lam.
Toxopneustes drobachiensis Ag.
Echinarachnius parma Gray.
Order V.—HOLOTHURIDEA Brandt.
Caudina arenata Stimp.
Synapta tenuis Ayres.
Cuvieria squamata D. & K.
Pentacta frondosa Jäg/.
|
[Pg 156]
INDEX.
Abbreviations of authors' names, xii
Acalephs, 21
Actinia, 7
Actinoids, 7
Alecto Eschrichtii, 121
Alecto meridionalis, 121
Amphiura squamata, 115
Arachnactis brachiolata, 14
Astracanthion berylinus, 108
Astracanthion pallidus, 112
Astrangia Danæ, 16
Astrophyton Agassizii, 117
Aurelia flavidula, 42
Bicidium parasiticum, 15
Bolina alata, 31
Bougainvillia superciliaris, 69
Campanella pachyderma, 44
Campanularians, 49
Caudina arenata, 97
Circe, 45
Clytia bicophora, 56
Comatula, 121
Coryne mirabilis, 68
Cribrella oculata, 112
Crinoids, 120
Crossaster papposa, 114
Ctenodiscus crispatus, 113
Ctenophoræ, 26
Cuvieria squamata, 98
Cyanea aretica, 38
Development of Melicertum, 64
" " Tima, 64
Discophoræ, 37
Distribution of Life in the Ocean,
Dynamena pumila,
Dyphasia rosacea, 67
Dysmorphosa fulgurans, 75
Echinarachnius parma, 106
Echinoderms, 91
Echinoids, 101
Embryology of Astracanthion, 124
" " Ctenophoræ, 34
" " Echinoderms, 123
" " Ophiurans, 135
" " Sea-urchins, 130
" " Star-fishes, 124
Eucope diaphana, 50
Halcampa albida, 16
Halcyonium carneum, 19
Halcyonoids, 19
Haliclystus auricula, 46
Hippasteria phrygiana, 113
Holothurians, 95
Hybocodon prolifer, 74
Hydractinia polyclina, 73
Hydroids, 49
Idyia roseola, 32
Lafoea cornuta, 67
Laomedea amphora, 65
Lucernaria, 46
Madreporians, 16
Melicertum campanula, 63
Metridium marginatum, 7
Millepora alcicornis, 22
Mode of catching Jelly-fishes, 85
Nanomia cara, 76
Oceania languida, 53
Ophiurans, 115
Ophiopholis bellis, 115
Pentacrinus, 121
Pentacta frondosa, 99
[Pg 157]
Pleurobrachia rhododactyla, 27
Polyps, 52
Ptychogena lactea, 86
Physalia Arethusa, 83
Radiates, 1
Rhodactinia Davisii, 13
Sarsia, 68
Sea-urchin, 101
Sertularians, 66
Solaster endeca, 114
Star-fishes, 108
Synapta tenuis, 95
Systematic Table, 152
Tima formosa, 60
Toxopneustes drobachiensis, 101
Trachynema digitale, 45
Tubularia Couthouyi, 72
Tubularians, 67
Turris vesicaria, 69
Velella mutica, 84
Zygodactyla groenlandica, 57
THE END.
Cambridge: Stereotyped and Printed by Welch, Bigelow, & Co.
Transcriber's Note
The original spellings and gramatical constructs have been preserved
as far as possible. With the exception of the typographical corrections
detailed below, the text is that published in the original document. The
image captions were moved directly below its image from the bottom of each
page.
Images were placed in approximately the same location as in the original text. Figure 177 was rotated 90° as it fit better in this version and the lettering suggests that it may have originally been intended to be presented in this orientation. The image sizes vary from that of the original printed volume and so any notation retained from the original text regarding the same may not be accurate. Variations in the display properties of various monitors and user preferences would tend to also result in images which are not to scale with the original printed version.
Typographical Corrections
| Page xi: | Ophiopolis | => Ophiopholis |
| Page 4: | diferent | => different |
Page 21, 101,
127, 131, 148: | so called | => so-called |
End of the Project Gutenberg EBook of Seaside Studies in Natural History, by
Elizabeth Cabot Cary Agassiz and Alexander Agassiz
*** END OF THIS PROJECT GUTENBERG EBOOK SEASIDE STUDIES IN NATURAL HISTORY ***
***** This file should be named 35490-h.htm or 35490-h.zip *****
This and all associated files of various formats will be found in:
https://www.gutenberg.org/3/5/4/9/35490/
Produced by Bryan Ness, Tom Cosmas and the Online
Distributed Proofreading Team at https://www.pgdp.net (This
book was produced from scanned images of public domain
material from the Google Print project.)
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
https://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at https://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
https://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at https://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit https://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including including checks, online payments and credit card
donations. To donate, please visit: https://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
https://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.








