The Project Gutenberg EBook of The Elements of Botany, by Asa Gray
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
Title: The Elements of Botany
For Beginners and For Schools
Author: Asa Gray
Release Date: September 18, 2010 [EBook #33757]
Language: English
Character set encoding: ISO-8859-1
*** START OF THIS PROJECT GUTENBERG EBOOK THE ELEMENTS OF BOTANY ***
Produced by Curtis Weyant, Stephen H. Sentoff and the
Online Distributed Proofreading Team at https://www.pgdp.net
GRAY'S LESSONS IN BOTANY
REVISED EDITION
THE
ELEMENTS OF BOTANY
FOR BEGINNERS AND FOR SCHOOLS
By ASA GRAY
IVISON, BLAKEMAN, AND COMPANY
NEW YORK AND CHICAGO
Copyright,
By Asa Gray.
1887.
[Pg iii]
PREFACE.
This volume takes the place of the author's Lessons in Botany and
Vegetable Physiology, published over a quarter of a century ago. It is
constructed on the same lines, and is a kind of new and much revised
edition of that successful work. While in some respects more extended,
it is also more concise and terse than its predecessor. This should the
better fit it for its purpose now that competent teachers are common.
They may in many cases develop paragraphs into lectures, and fully
illustrate points which are barely, but it is hoped clearly, stated.
Indeed, even for those without a teacher, it may be that a condensed is
better than a diffuse exposition.
The book is adapted to the higher schools, "How Plants Grow and Behave"
being the "Botany for Young People and Common Schools." It is intended
to ground beginners in Structural Botany and the principles of vegetable
life, mainly as concerns Flowering or Phanerogamous plants, with which
botanical instruction should always begin; also to be a companion and
interpreter to the Manuals and Floras by which the student threads his
flowery way to a clear knowledge of the surrounding vegetable creation.
Such a book, like a grammar, must needs abound in technical words, which
thus arrayed may seem formidable; nevertheless, if rightly apprehended,
this treatise should teach that the study of botany is not the learning
of names and terms, but the acquisition of knowledge and ideas. No
effort should be made to commit technical terms to memory. Any term used
in describing a plant or explaining its structure can be looked up when
it is wanted, and that should suffice. On the other hand, plans of
[Pg iv]structure, types, adaptations, and modifications, once understood, are
not readily forgotten; and they give meaning and interest to the
technical terms used in explaining them.
In these "Elements" naturally no mention has been made of certain terms
and names which recent cryptogamically-minded botanists, with lack of
proportion and just perspective, are endeavoring to introduce into
phanerogamous botany, and which are not needed nor appropriate, even in
more advanced works, for the adequate recognition of the ascertained
analogies and homologies.
As this volume will be the grammar and dictionary to more than one or
two Manuals, Floras, etc., the particular directions for procedure which
were given in the "First Lessons" are now relegated to those works
themselves, which in their new editions will provide the requisite
explanations. On the other hand, in view of such extended use, the
Glossary at the end of this book has been considerably enlarged. It will
be found to include not merely the common terms of botanical description
but also many which are unusual or obsolete; yet any of them may now and
then be encountered. Moreover, no small number of the Latin and Greek
words which form the whole or part of the commoner specific names are
added to this Glossary, some in an Anglicized, others in their Latin
form. This may be helpful to students with small Latin and less Greek,
in catching the meaning of a botanical name or term.
The illustrations in this volume are largely increased in number. They
are mostly from the hand of Isaac Sprague.
It happens that the title chosen for this book is that of the author's
earliest publication, in the year 1836, of which copies are rarely seen;
so that no inconvenience is likely to arise from the present use of the
name.
ASA GRAY.
Cambridge, Massachusetts,
March, 1887.
[Pg v]
CONTENTS.
[Pg 9]
Section I. INTRODUCTORY.
1. Botany is the name of the science of the vegetable kingdom in
general; that is, of plants.
2. Plants may be studied as to their kinds and relationships. This study
is Systematic Botany. An enumeration of the kinds of vegetables, as far
as known, classified according to their various degrees of resemblance
or difference, constitutes a general System of plants. A similar
account of the vegetables of any particular country or district is
called a Flora.
3. Plants may be studied as to their structure and parts. This is
Structural Botany, or Organography. The study of the organs or parts of
plants in regard to the different forms and different uses which the
same kind of organ may assume,—the comparison, for instance, of a
flower-leaf or a bud-scale with a common leaf,—is Vegetable Morphology,
or Morphological Botany. The study of the minute structure of the parts,
to learn by the microscope what they themselves are formed of, is
Vegetable Anatomy, or Histology; in other words, it is Microscopical
Structural Botany. The study of the actions of plants or of their parts,
of the ways in which a plant lives, grows, and acts, is the province of
Physiological Botany, or Vegetable Physiology.
4. This book is to teach the outlines of Structural Botany and of the
simpler parts of the physiology of plants, that it may be known how
plants are constructed and adapted to their surroundings, and how they
live, move, propagate, and have their being in an existence no less
real, although more simple, than that of the animal creation which they
support. Particularly, this book is to teach the principles of the
structure and relationships of plants, the nature and names of their
parts and their modifications, and so to prepare for the study of
Systematic Botany; in which the learner may ascertain the name and the
place in the system of any or all of the ordinary plants within reach,
whether wild or cultivated. And in ascertaining the name of any plant,
the student, if rightly taught, will come to know all about its general
or particular structure, rank, and relationship to other plants.
[Pg 10]
5. The vegetable kingdom is so vast and various, and the difference is
so wide between ordinary trees, shrubs, and herbs on the one hand, and
mosses, moulds, and such like on the other, that it is hardly possible
to frame an intelligible account of plants as a whole without
contradictions or misstatements, or endless and troublesome
qualifications. If we say that plants come from seeds, bear flowers, and
have roots, stems, and leaves, this is not true of the lower orders. It
is best for the beginner, therefore, to treat of the higher orders of
plants by themselves, without particular reference to the lower.
6. Let it be understood, accordingly, that there is a higher and a lower
series of plants; namely:—
Phanerogamous Plants, which come from seed and bear flowers,
essentially stamens and pistils, through the co-operation of which seed
is produced. For shortness, these are commonly called Phanerogams, or
Phænogams, or by the equivalent English name of Flowering Plants.[1]
Cryptogamous Plants, or Cryptogams, come from minute bodies, which
answer to seeds, but are of much simpler structure, and such plants have
not stamens and pistils. Therefore they are called in English Flowerless
Plants. Such are Ferns, Mosses, Algæ or Seaweeds, Fungi, etc. These
sorts have each to be studied separately, for each class or order has a
plan of its own.
7. But Phanerogamous, or Flowering, Plants are all constructed on one
plan, or type. That is, taking almost any ordinary herb, shrub, or
tree for a pattern, it will exemplify the whole series: the parts of one
plant answer to the parts of any other, with only certain differences in
particulars. And the occupation and the delight of the scientific
botanist is in tracing out this common plan, in detecting the likenesses
under all the diversities, and in noting the meaning of these manifold
diversities. So the attentive study of any one plant, from its growth
out of the seed to the flowering and fruiting state and the production
of seed like to that from which the plant grew, would not only give a
correct general idea of the structure, growth, and characteristics of
Flowering Plants in general, but also serve as a pattern or standard of
comparison. Some plants will serve this purpose of a pattern much better
than others. A proper pattern will be one that is perfect in the sense
of having all the principal parts of a phanerogamous plant, and simple
and regular in having these parts free from complications or disguises.
The common Flax-plant may very well serve this purpose. Being an annual,
it has the advantage of being easily raised and carried in a short time
through its circle of existence, from seedling to fruit and seed.
[Pg 11]
Section II. FLAX AS A PATTERN PLANT.
8. Growth from the Seed. Phanerogamous plants grow from seed, and their
flowers are destined to the production of seeds. A seed has a
rudimentary plant ready formed in it,—sometimes with the two most
essential parts, i. e. stem and leaf, plainly discernible; sometimes
with no obvious distinction of organs until germination begins. This
incipient plant is called an Embryo.
9. In this section the Flax-plant is taken as a specimen, or type, and
the development and history of common plants in general is illustrated
by it. In flax-seed the embryo nearly fills the coats, but not quite.
There is a small deposit of nourishment between the seed-coat and the
embryo: this may for the present be left out of the account. This embryo
consists of a pair of leaves, pressed together face to face, and
attached to an extremely short stem. (Fig. 2-4.) In this rudimentary
condition the real nature of the parts is not at once apparent; but when
the seed grows they promptly reveal their character,—as the
accompanying figures (Fig. 5-7) show.
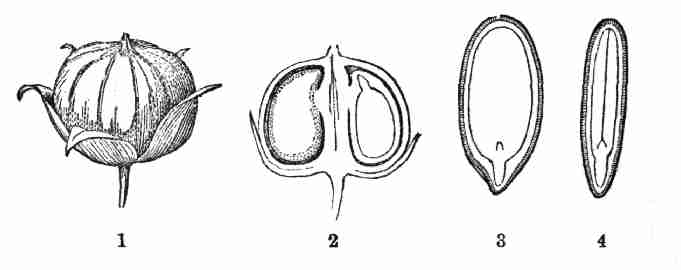
Fig. 1. Pod of Flax. 2. Section lengthwise, showing two
of the seeds; one whole, the other cut half away, bringing contained
embryo into view. 3. Similar section of a flax-seed more magnified and
divided flatwise; turned round, so that the stem-end (caulicle) of the
embryo is below: the whole broad upper part is the inner face of one of
the cotyledons; the minute nick at its base is the plumule. 4. Similar
section through a seed turned edgewise, showing the thickness of the
cotyledons, and the minute plumule between them, i. e. the minute bud on
the upper end of the caulicle.
10. Before the nature of these parts in the seed was altogether
understood, technical names were given to them, which are still in use.
These initial leaves were named Cotyledons. The initial stem on which
they stand was called the Radicle. That was because it gives rise to the
first root; but, as it is really the beginning of the stem, and because
it is the stem that produces the root and not the root that produces the
stem, it is better to name it the Caulicle. Recently it has been named
Hypocotyle; which signifies something below the cotyledons, without
pronouncing what its nature is.
[Pg 12]
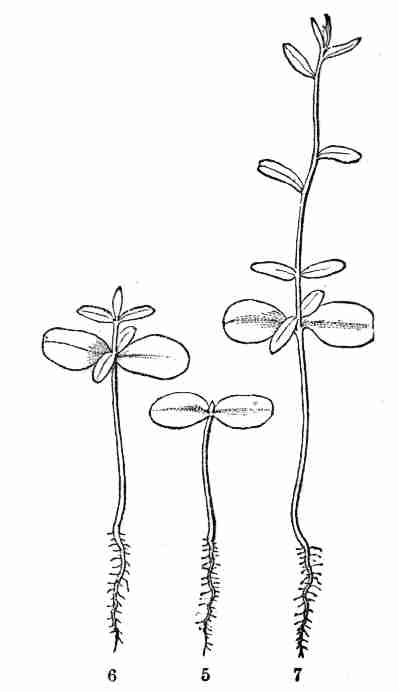
Fig. 5. Early Flax seedling; stem (caulicle), root at
lower end, expanded seed-leaves (cotyledons) at the other: minute bud
(plumule) between these. 6. Same later; the bud developed into second
pair of leaves, with hardly any stem-part below them; then into a third
pair of leaves, raised on a short joint of stem; and a fifth leaf also
showing. 7. Same still older, with more leaves developed, but these
singly (one after another), and with joints of stem between them.
11. On committing these seeds to moist and warm soil they soon sprout,
i. e. germinate. The very short stem-part of the embryo is the first
to grow. It lengthens, protrudes its root-end; this turns downward, if
not already pointing in that direction, and while it is lengthening a
root forms at its point and grows downward into the ground. This root
continues to grow on from its lower end, and thus insinuates itself and
penetrates into the soil. The stem meanwhile is adding to its length
throughout; it erects itself, and, seeking the light, brings the seed up
out of the ground. The materials for this growth have been supplied by
the cotyledons or seed-leaves, still in the seed: it was the store of
nourishing material they held which gave them their thickish shape, so
unlike that of ordinary leaves. Now, relieved of a part of this store of
food, which has formed the growth by which they have been raised into
the air and light, they appropriate the remainder to their own growth.
In enlarging they open and throw off the seed-husk; they expand, diverge
into a horizontal position, turn green, and thus become a pair of
evident leaves, the first foliage of a tiny plant. This seedling,
although diminutive and most simple, possesses and puts into use, all
the Organs of Vegetation, namely, root, stem, and leaves, each in its
proper element,—the root in the soil, the stem rising out of it, the
leaves in the light and open air. It now draws in moisture and some
food-materials from the soil by its root, conveys this through the stem
into the leaves, where these materials, along with other crude food
which these imbibe from the air, are assimilated into vegetable matter,
i. e. into the material for further growth.
12. Further Growth soon proceeds to the formation of new
parts,—downward in the production of more root, or of branches of the
main root, upward in the development of more stem and leaves. That from
which a stem with its leaves is continued, or a new stem (i. e. branch)
originated, is a Bud. The most conspicuous and familiar buds are those
of most shrubs and trees, bearing buds formed in summer or autumn, to
grow the following [Pg 13]spring. But every such point for new growth may
equally bear the name. When there is such a bud between the cotyledons
in the seed or seedling it is called the Plumule. This is conspicuous
enough in a bean (Fig. 29.), where the young leaf of the new growth
looks like a little plume, whence the name, plumule. In flax-seed this
is very minute indeed, but is discernible with a magnifier, and in the
seedling it shows itself distinctly (Fig. 5, 6, 7).
13. As it grows it shapes itself into a second pair of leaves, which of
course rests on a second joint of stem, although in this instance that
remains too short to be well seen. Upon its summit appears the third
pair of leaves, soon to be raised upon its proper joint of stem; the
next leaf is single, and is carried up still further upon its supporting
joint of stem; and so on. The root, meanwhile, continues to grow
underground, not joint after joint, but continuously, from its lower
end; and commonly it before long multiplies itself by branches, which
lengthen by the same continuous growth. But stems are built up by a
succession of leaf-bearing growths, such as are strongly marked in a
reed or corn-stalk, and less so in such an herb as Flax. The word
"joint" is ambiguous: it may mean either the portion between successive
leaves, or their junction, where the leaves are attached. For precision,
therefore, the place where the leaf or leaves are borne is called a
Node, and the naked interval between two nodes, an Internode.
 Fig.
Fig. 8. Upper part of Flax-plant in blossom.
14. In this way a simple stem with its garniture of leaves is developed
from the seed. But besides this direct continuation, buds may form and
develop into lateral stems, that is, into branches, from any node. The
proper origin of branches is from the Axil of a leaf, i. e. the angle
between leaf and stem on the upper side; and branches may again branch,
so building up the herb, shrub, or tree. But sooner or later, and
without long delay in an annual like Flax, instead of this continuance
of mere vegetation, reproduction is prepared for by
[Pg 14]
15. Blossoming. In Flax the flowers make their appearance at the end of
the stem and branches. The growth, which otherwise might continue them
farther or indefinitely, now takes the form of blossom, and is
subservient to the production of seed.
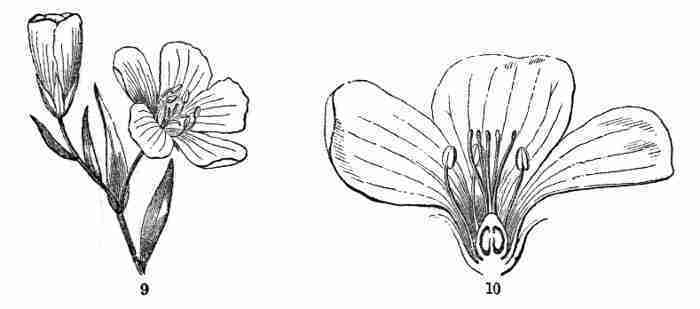
Fig. 9. Flax-flowers about natural size. 10. Section of a
flower moderately enlarged, showing a part of the petals and stamens,
all five styles, and a section of ovary with two ovules or rudimentary
seeds.
16. The Flower of Flax consists, first, of five small green leaves,
crowded into a circle: this is the Calyx, or flower-cup. When its
separate leaves are referred to they are called Sepals, a name which
distinguishes them from foliage-leaves on the one hand, and from petals
on the other. Then come five delicate and colored leaves (in the Flax,
blue), which form the Corolla, and its leaves are Petals; then a circle
of organs, in which all likeness to leaves is lost, consisting of
slender stalks with a knob at summit, the Stamens; and lastly, in the
centre, the rounded body, which becomes a pod, surmounted by five
slender or stalk-like bodies. This, all together, is the Pistil. The
lower part of it, which is to contain the seeds, is the Ovary; the
slender organs surmounting this are Styles; the knob borne on the apex
of each style is a Stigma. Going back to the stamens, these are of two
parts, viz. the stalk, called Filament, and the body it bears, the
Anther. Anthers are filled with Pollen, a powdery substance made up of
minute grains.
17. The pollen shed from the anthers when they open falls upon or is
conveyed to the stigmas; then the pollen-grains set up a kind of growth
(to be discerned only by aid of a good microscope), which penetrates the
style: this growth takes the form of a thread more delicate than the
finest spider's web, and reaches the bodies which are to become seeds
(Ovules they are called until this change occurs); these, touched by
this influence, are incited to a new growth within, which becomes an
embryo. So, as the ovary ripens into the seed-pod or capsule (Fig. 1,
etc.) containing seeds, each seed enclosing a rudimentary new plantlet,
the round of this vegetable existence is completed.
[Pg 15]
Section III. MORPHOLOGY OF SEEDLINGS.
18. Having obtained a general idea of the growth and parts of a
phanerogamous plant from the common Flax of the field, the seeds and
seedlings of other familiar plants may be taken up, and their variations
from the assumed pattern examined.
19. Germinating Maples are excellent to begin with, the parts being so
much larger than in Flax that a common magnifying glass, although
convenient, is hardly necessary. The only disadvantage is that fresh
seeds are not readily to be had at all seasons.
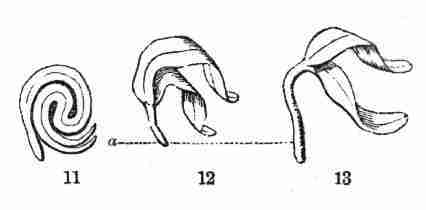
Fig. 11. Embryo of Sugar Maple, cut through lengthwise
and taken out of the seed. 12, 13. Whole embryo of same just beginning
to grow; a, the stemlet or caulicle, which in 13 has considerably
lengthened.
20. The seeds of Sugar Maple ripen at the end of summer, and germinate
in early spring. The embryo fills the whole seed, in which it is nicely
packed; and the nature of the parts is obvious even before growth
begins. There is a stemlet (caulicle) and a pair of long and narrow
seed-leaves (cotyledons), doubled up and coiled, green even in the seed,
and in germination at once unfolding into the first pair of
foliage-leaves, though of shape quite unlike those that follow.
21. Red Maple seeds are ripe and ready to germinate at the beginning of
summer, and are therefore more convenient for study. The cotyledons are
crumpled in the seed, and not easy to straighten out until they unfold
themselves in germination. The story of their development into the
seedling is told by the accompanying Fig. 14-20; and that of Sugar Maple
is closely similar. No plumule or bud appears in the embryo of these two
Maples until the seed-leaves have nearly attained their full growth and
are acting as foliage-leaves, and until a root is formed below. There is
no great store of nourishment in these thin cotyledons; so further
growth has to wait until the root and seed-leaves have collected and
elaborated sufficient material for the formation of the second internode
and its pair of leaves, which lending their help the third pair is more
promptly produced, and so on.
22. Some change in the plan comes with the Silver or Soft White Maple.
(Fig. 21-25). This blossoms in earliest spring, and it drops its large
and ripened keys only a few weeks later. Its cotyledons have not at all
the appearance of leaves; they are short and broad, and (as there is no
room to be saved by folding) they are straight, except a small fold at
the top,—a vestige of the habit of Maples in general. Their unusual
thickness is due [Pg 16]to the large store of nutritive matter they contain,
and this prevents their developing into actual leaves. Correspondingly,
their caulicle does not lengthen to elevate them above the surface of
the soil; the growth below the cotyledons is nearly all of root. It is
the little plumule or bud between them which makes the upward growth,
and which, being well fed by the cotyledons, rapidly develops the next
pair of leaves and raises them upon a long internode, and so on. The
cotyledons all the while remain below, in the husk of the fruit and
seed, and perish when they have yielded up the store of food which they
contained.
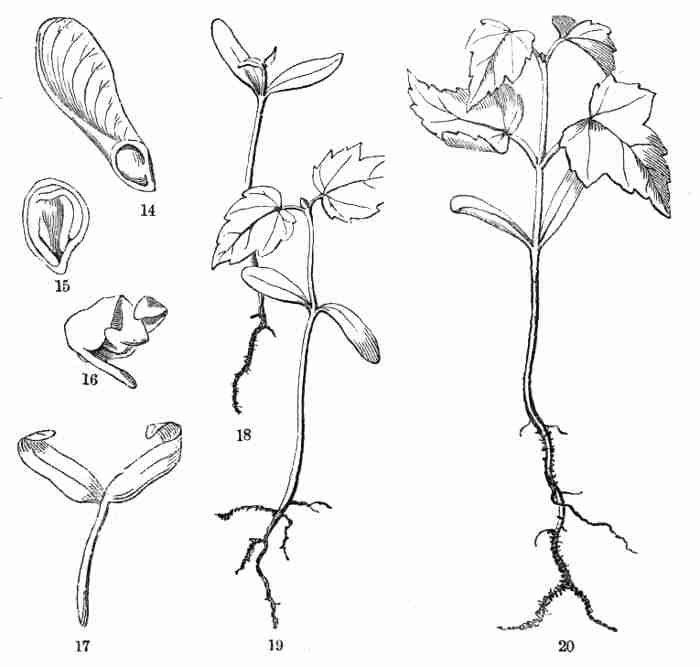
Fig. 14. One of the pair of keys or winged fruits of Red
Maple; the seed-bearing portion cut open to show the seed. 15. Seed
enlarged, and divided to show the crumpled embryo which fills it. 16.
Embryo taken out and partly opened. 17. Embryo which has unfolded in
early stage of germination and begun to grow. 18. Seedling with next
joint of stem and leaves apparent; and 19 with these parts full-grown,
and bud at apex for further growth. 20. Seedling with another joint of
stem and pair of leaves.
23. So, even in plants so much alike as Maples, there is considerable
difference in the amount of food stored up in the cotyledons by which
the growth is to be made; and there are corresponding differences in the
germination. [Pg 17]The larger the supply to draw upon, the stronger the
growth, and the quicker the formation of root below and of stem and
leaves above. This deposit of food thickens the cotyledons, and renders
them less and less leaf-like in proportion to its amount.
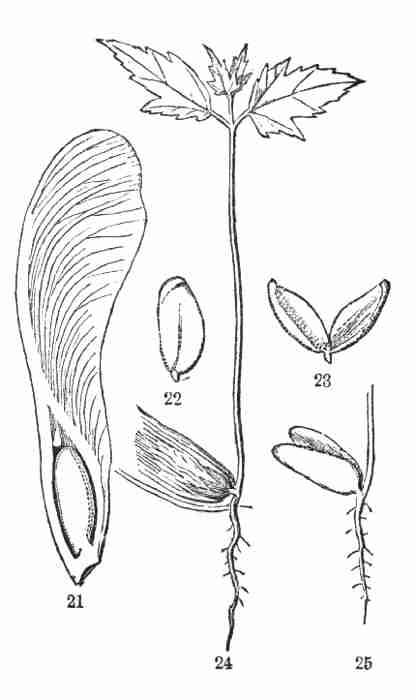
Fig. 21. Fruit (one key) of Silver Maple, Acer
dasycarpum, of natural size, the seed-bearing portion divided to show
the seed. 22. Embryo of the seed taken out. 23. Same opened out, to show
the thick cotyledons and the little plumule or bud between them. 24.
Germination of Silver Maple, natural size; merely the base of the fruit,
containing the seed, is shown. 25. Embryo of same, taken out of the
husk; upper part of growing stem cut off, for want of room.
24. Examples of Embryos with thickened Cotyledons. In the Pumpkin and
Squash (Fig. 26, 27), the cotyledons are well supplied with nourishing
matter, as their sweet taste demonstrates. Still, they are flat and not
very thick. In germination this store is promptly utilized in the
development of the caulicle to twenty or thirty times its length in the
seed, and to corresponding thickness, in the formation of a cluster of
roots at its lower end, and the early production of the incipient
plumule; also in their own growth into efficient green leaves. The case
of our common Bean (Phaseolus vulgaris, Fig. 28-30) is nearly the same,
except that the cotyledons are much more gorged; so that, although
carried up into the air and light upon the lengthening caulicle, and
there acquiring a green color, they never expand into useful leaves.
Instead of this, they nourish into rapid growth the plumule, which is
plainly visible in the seed, as a pair of incipient leaves; and these
form the first actual foliage.
25. Very similar is the germination of the Beech (Fig. 31-33), except
that the caulicle lengthens less, hardly raising the cotyledons out of
the ground. Nothing would be gained by elevating them, as they never
grow out into efficient leaves; but the joint of stem belonging to the
plumule lengthens well, carrying up its pair of real foliage-leaves.
26. It is nearly the same in the Bean of the Old World (Vicia Faba, here
called Horse Bean and Windsor Bean): the caulicle lengthens very little,
does not undertake to elevate the heavy seed, which is left below or
[Pg 18]upon the surface of the soil, the flat but thick cotyledons remaining
in it, and supplying food for the growth of the root below and the
plumule above. In its near relative, the Pea (Fig. 34, 35), this use of
cotyledons for storage only is most completely carried out. For they are
thickened to the utmost, even into hemispheres; the caulicle does not
lengthen at all; merely sends out roots from the lower end, and develops
its strong plumule from the upper, the seed remaining unmoved
underground. That is, in technical language, the germination is
hypogæous.
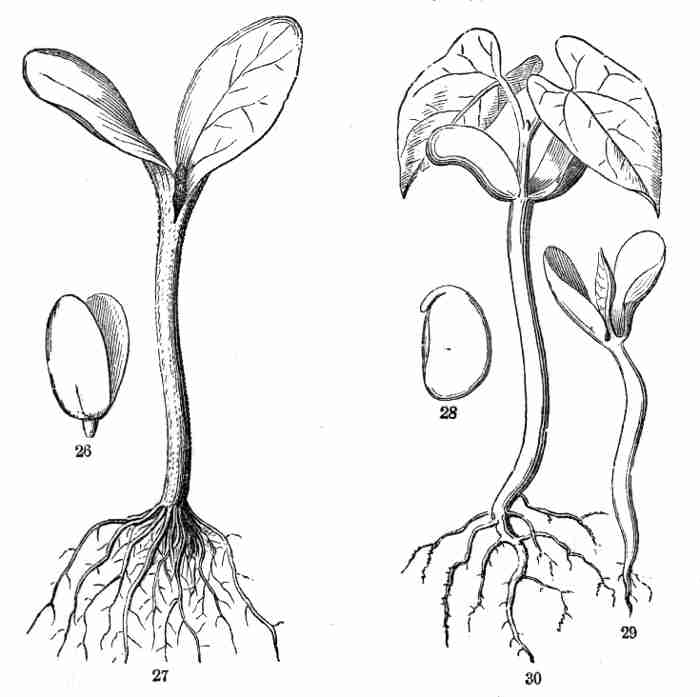
Fig. 26. Embryo of Pumpkin-seed, partly opened. 27. Young
seedling of same.
Fig. 28. Embryo of Common Bean (Phaseolus vulgaris):
caulicle bent down over edge of cotyledons. 29. Same germinating:
caulicle well lengthened and root beginning; thick cotyledons partly
spreading; and plumule (pair of leaves) growing between them. 30. Same,
older, with plumule developed into internode and pair of leaves.
27. There is sufficient nourishment in the cotyledons of a pea to make a
very considerable growth before any actual foliage is required. So it is
the stem-portion of the plumule which is at first conspicuous and
strong-growing. Here, as seen in Fig. 35, its lower nodes bear each a
useless leaf-scale instead of an efficient leaf, and only the later ones
bear leaves fitted for foliage.
[Pg 19]
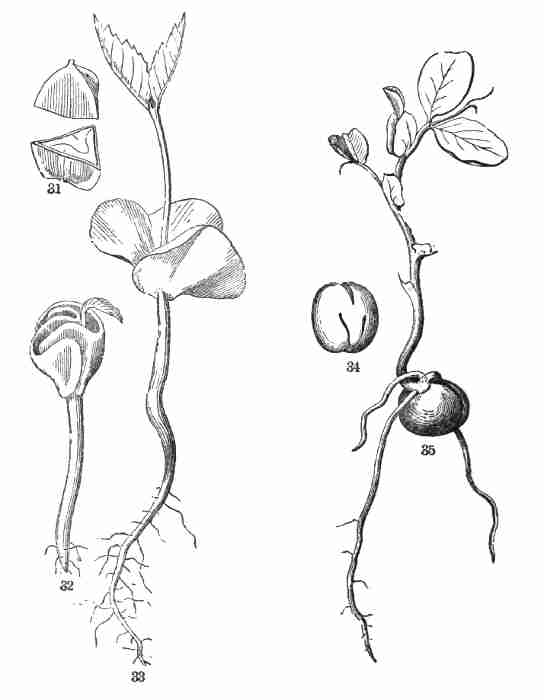
Fig. 31. A Beech-nut, cut across. 32. Beginning
germination of the Beech, showing the plumule growing before the
cotyledons have opened or the root has scarcely formed. 33. The same, a
little later, with the plumule-leaves developing, and elevated on a long
internode.
Fig. 34. Embryo of Pea, i. e. a pea with the coats
removed; the short and thick caulicle presented to view. 35. Same in
advanced germination: the plumule has developed four or five internodes,
bearing single leaves; but the first and second leaves are mere scales,
the third begins to serve as foliage; the next more so.
28. This hypogæous germination is exemplified on a larger scale by the
Oak (Fig. 36, 37) and Horse-chestnut (Fig. 38, 39); but in these the
downward growth is wholly a stout tap-root. It is not the caulicle; for
this lengthens hardly any. Indeed, the earliest growth which carries the
very short caulicle out of the shell comes from the formation of
foot-stalks to the cotyledons; above these develops the strong plumule,
below grows the stout root. The growth is at first entirely, for a long
time [Pg 20]mainly, at the expense of the great store of food in the
cotyledons. These, after serving their purpose, decay and fall away.
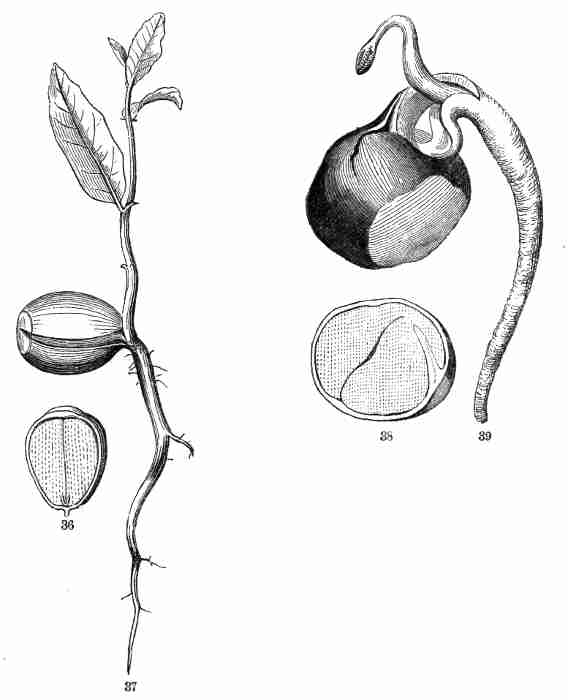
Fig. 36. Half of an acorn, cut lengthwise, filled by the
very thick cotyledons, the base of which encloses the minute caulicle.
37. Oak-seedling.
Fig. 38. Half of a horse-chestnut, similarly cut; the
caulicle is curved down on the side of one of the thick cotyledons. 39.
Horse-chestnut in germination; foot-stalks are formed to the cotyledons,
pushing out in their lengthening the growing parts.
29. Such thick cotyledons never separate; indeed, they sometimes grow
together by some part of their contiguous faces; so that the germination
seems to proceed from a solid bulb-like mass. This is the case in a
horse-chestnut.
30. Germinating Embryo supplied by its own Store of Nourishment, i. e.
the store in the cotyledons. This is so in all the illustrations thus
far, essentially so even in the Flax. This nourishment was supplied by
the mother plant to the ovule and seed, and thence taken into the embryo
during its growth. Such embryos, filling the whole seed, are
comparatively large and strong, and vigorous in germination in
proportion to the amount of their growth while connected with the parent
plant.
31. Germinating Embryo supplied from a Deposit outside of Itself. This
is as common as the other mode; and it occurs in all degrees. [Pg 21]Some
seeds have very little of this deposit, but a comparatively large
embryo, with its parts more or less developed and recognizable. In
others this deposit forms the main bulk of the seed, and the embryo is
small or minute, and comparatively rudimentary. The following
illustrations exemplify these various grades. When an embryo in a seed
is thus surrounded by a white substance, it was natural to liken the
latter to the white of an egg, and the embryo or germ to the yolk. So
the matter around or by the side of the embryo was called the Albumen,
i. e. the white of the seed. The analogy is not very good; and to avoid
ambiguity some botanists call it the Endosperm. As that means in English
merely the inwards of a seed, the new name is little better than the old
one; and, since we do not change names in botany except when it cannot
be avoided, this name of albumen is generally kept up. A seed with
such a deposit is albuminous, one with none is exalbuminous.
32. The Albumen forms the main bulk of the seed in wheat, maize, rice,
buckwheat, and the like. It is the floury part of the seed. Also of the
cocoa-nut, of coffee (where it is dense and hard), etc.; while in peas,
beans, almonds, and in most edible nuts, the store of food, although
essentially the same in nature and in use, is in the embryo itself, and
therefore is not counted as anything to be separately named. In both
forms this concentrated food for the germinating plant is food also for
man and for animals.
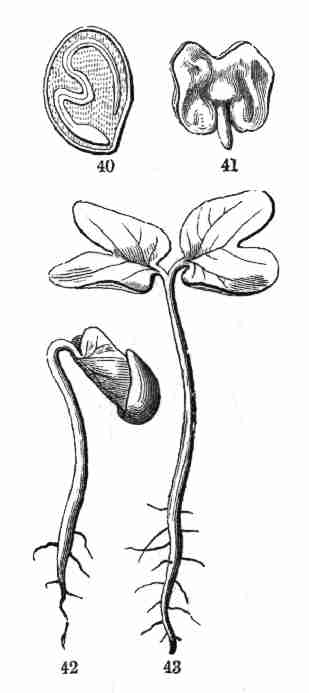
Fig. 40. Seed of Morning Glory divided, moderately
magnified; shows a longitudinal section through the centre of the embryo
as it lies crumpled in the albumen. 41. Embryo taken out whole and
unfolded; the broad and very thin cotyledons notched at summit; the
caulicle below. 42. Early state of germination. 43. Same, more advanced;
caulicle or primary stem, cotyledons or seed-leaves, and below, the
root, well developed.
33. For an albuminous seed with a well-developed embryo, the common
Morning Glory (Ipomœa purpurea, Fig. 40-43) is a convenient example,
being easy and prompt to grow, and having all the parts well apparent.
The seeds (duly soaked for examination) and the germination should be
compared with those of Sugar and Red Maple (19-21). The only essential
difference is that here the embryo is surrounded by and crumpled up in
the albumen. This substance, which is pulpy or mucilaginous in fresh and
young seeds, hardens as the seed ripens, but becomes again pulpy in
germination; and, as it liquefies, the thin cotyledons absorb it by
their [Pg 22]whole surface. It supplements the nutritive matter contained in
the embryo. Both together form no large store, but sufficient for
establishing the seedling, with tiny root, stem, and pair of leaves for
initiating its independent growth; which in due time proceeds as in Fig.
44, 45.
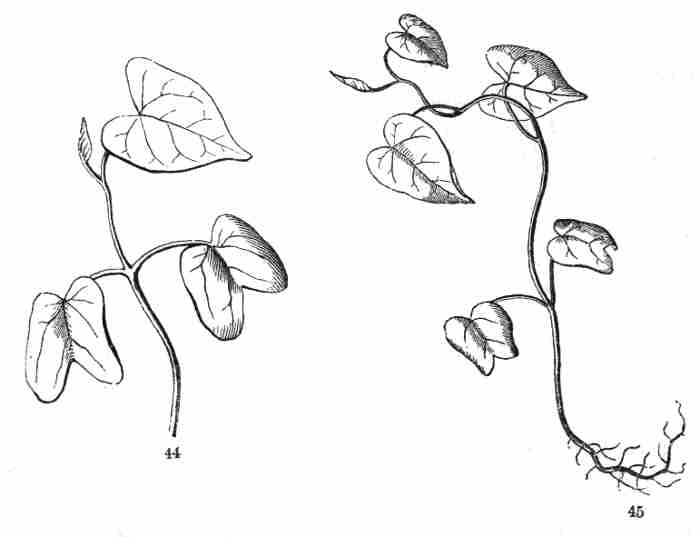
Fig. 44. Seedling of Morning Glory more advanced (root
cut away); cotyledons well developed into foliage-leaves: succeeding
internode and leaf well developed, and the next forming. 45. Seedling
more advanced; reduced to much below natural size.
34. Smaller embryos, less developed in the seed, are more dependent upon
the extraneous supply of food. The figures 46-53 illustrate four grades
in this respect. The smallest, that of the Peony, is still large enough
to be seen with a hand magnifying glass, and even its cotyledons may be
discerned by the aid of a simple stage microscope.
35. The broad cotyledons of Mirabilis, or Four-o'clock (Fig. 52, 53),
with the slender caulicle almost encircle and enclose the floury
albumen, instead of being enclosed in it, as in the other illustrations.
Evidently here the germinating embryo is principally fed by one of the
leaf-like cotyledons, the other being out of contact with the supply. In
the embryo of Abronia (Fig. 54, 55), a near relative of Mirabilis, there
is a singular modification; one cotyledon is almost wanting, being
reduced to a rudiment, leaving it for the other to do the work. This
leads to the question of the
36. Number of Cotyledons. In all the preceding illustrations, the
embryo, however different in shape and degree of development, is
evidently [Pg 23]constructed upon one and the same plan, namely, that of two
leaves on a caulicle or initial stem,—a plan which is obvious even when
one cotyledon becomes very much smaller than the other, as in the rare
instance of Abronia (Fig. 54, 55). In other words, the embryos so far
examined are all
37. Dicotyledonous, that is, two-cotyledoned. Plants which are thus
similar in the plan of the embryo agree likewise in the general
structure of their stems, leaves, and blossoms; and thus form a class,
named from their embryo Dicotyledones, or in English, Dicotyledonous
Plants. So long a name being inconvenient, it may be shortened into
Dicotyls.
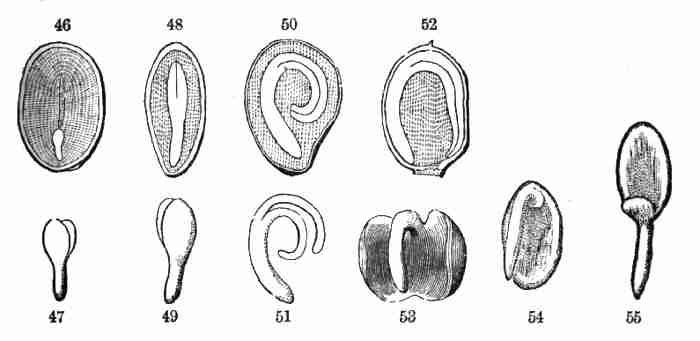
Fig. 46. Section of a seed of a Peony, showing a very
small embryo in the albumen, near one end. 47. This embryo detached, and
more magnified.
Fig. 48. Section of a seed of Barberry, showing the
straight embryo in the middle of the albumen. 49. Its embryo detached.
Fig. 50. Section of a Potato seed, showing the embryo
coiled in the albumen. 51. Its embryo detached.
Fig. 52. Section of the seed of Mirabilis or Four-o'clock,
showing the embryo coiled round the outside of the albumen. 53.
Embryo detached; showing the very broad and leaf-like cotyledons,
applied face to face, and the pair incurved.
Fig. 54. Embryo of Abronia umbellata; one of the
cotyledons very small. 55. Same straightened out.
38. Polycotyledonous is a name employed for the less usual case in which
there are more than two cotyledons. The Pine is the most familiar case.
This occurs in all Pines, the number of cotyledons varying from three to
twelve; in Fig. 56, 57 they are six. Note that they are all on the same
level, that is, belong to the same node, so as to form a circle or
whorl at the summit of the caulicle. When there are only three
cotyledons, they divide the space equally, are one third of the circle
apart. When only two they are 180° apart, that is, are opposite.
39. The case of three or more cotyledons, which is constant in Pines and
in some of their relatives (but not in all of them), is occasional among
Dicotyls. And the polycotyledonous is only a variation of the
dicotyledonous type,—a difference in the number of leaves in the whorl;
for a pair is a whorl reduced to two members. Some suppose that there
are really only [Pg 24]two cotyledons even in a Pine embryo, but these divided
or split up congenitally so as to imitate a greater number. But as
leaves are often in whorls on ordinary stems, they may be so at the very
beginning.
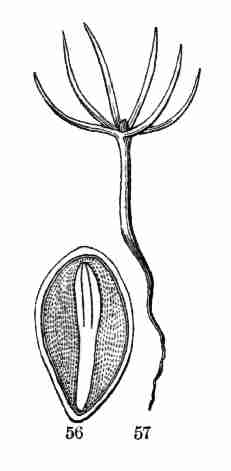
Fig. 56. Section of a Pine-seed, showing its
polycotyledonous embryo in the centre of the albumen, moderately
magnified. 57. Seedling of same, showing the freshly expanded six
cotyledons in a whorl, and the plumule just appearing.
40. Monocotyledonous (meaning with single cotyledon) is the name of the
one-cotyledoned sort of embryo. This goes along with peculiarities in
stem, leaves, and flowers, which all together associate such plants into
a great class, called Monocotyledonous Plants, or, for shortness,
Monocotyls. It means merely that the leaves are alternate from the very
first.
41. In Iris (Fig. 58, 59) the embryo in the seed is a small cylinder at
one end of the mass of the albumen, with no apparent distinction of
parts. The end which almost touches the seed coat is caulicle, the other
end belongs to the solitary cotyledon. In germination the whole
lengthens (but mainly the cotyledon) only enough to push the proximate
end fairly out of the seed; from this end the root is formed, and from a
little higher the plumule later emerges. It would appear therefore that
the cotyledon answers to a minute leaf rolled up, and that a chink
through which the plumule grows out is a part of the inrolled edges. The
embryo of Indian Corn shows these parts on a larger scale and in a more
open state (Fig. 66-68). There, in the seed, the cotyledon remains,
imbibing nourishment from the softened albumen, and transmitting it to
the growing root below and new-forming leaves above.
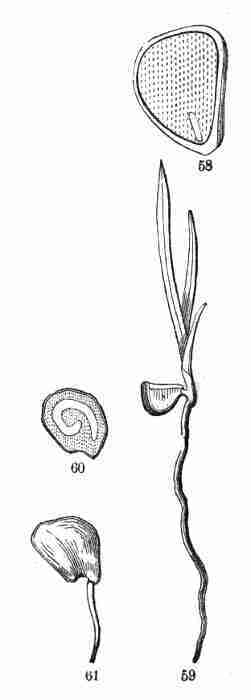
Fig. 58. Section of a seed of the Iris, or
Flower-de-Luce, enlarged, showing its small embryo in the albumen, near
the bottom. 59. A germinating seedling of the same, its plumule
developed into the first four leaves (alternate), the first one
rudimentary, the cotyledon remains in the seed.
Fig. 60. Section of an Onion seed showing the slender and
coiled embryo in the albumen, moderately magnified. 61. Seed of same in
early germination.
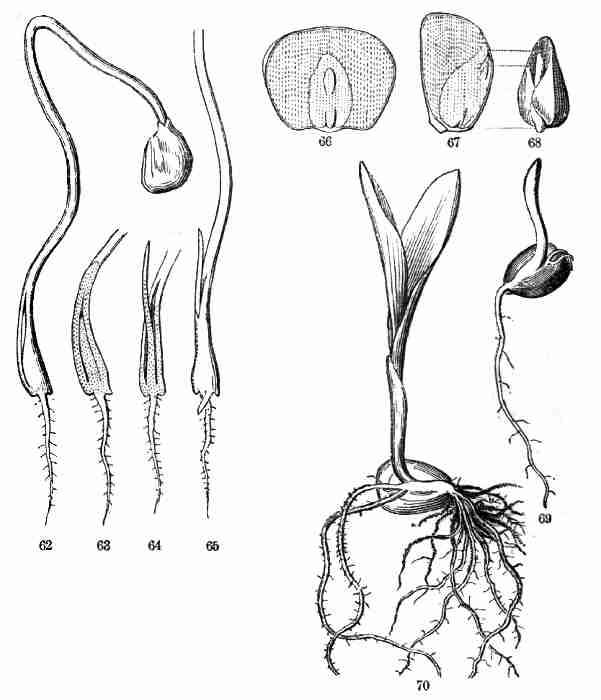
Fig. 62. Germinating Onion, more advanced, the chink at
base of cotyledon opening for the protrusion of the plumule, consisting
of a thread-shaped leaf. 63. Section of base of Fig. 62, showing plumule
enclosed. 64. Section of same later, plumule emerging. 65. Later stage
of 62, upper part cut off. 66. A grain of Indian Corn, flatwise, cut
away a little, so as to show the embryo, lying on the albumen which
makes the principal bulk of the seed. 67. A grain cut through the middle
in the opposite direction, dividing the embryo through its thick
cotyledon and its plumule, the latter consisting of two leaves, one
enclosing the other. 68. The embryo taken out whole; the thick mass is
the cotyledon, the narrow body partly enclosed by it is the plumule, the
little projection at its base is the very short radicle enclosed in the
sheathing base of the first leaf of the plumule.
Fig. 69. Grain of Indian Corn in germination, the
ascending sprout is the first leaf of the plumule, enclosing the younger
leaves within, at its base the primary root has broken through. 70. The
same, advanced; the second and third leaves developing, while the
sheathing first leaf does not further develop.
42. The general plan is the same in the Onion (Fig. 60-65), but with a
striking difference. The embryo is long, and coiled in the albumen of
the seed. To ordinary examination it shows no distinction of parts. But
germination plainly shows that all except the lower end of it is
cotyledon. For after it has lengthened into a long thread, the chink
from which the [Pg 25]plumule in time emerges is seen at the base, or near it,
so the caulicle is extremely short, and does not elongate, but sends out
from its base a simple root, and afterwards others in a cluster. Not
only does the cotyledon lengthen enormously in the seedling, but (unlike
that of Iris, Indian Corn, and all [Pg 26]the cereal grains) it raises the
comparatively light seed into the air, the tip still remaining in the
seed and feeding upon the albumen. When this food is exhausted and the
seedling is well established in the soil, the upper end decays and the
emptied husk of the seed falls away.
43. In Maize or Indian Corn (Fig. 66-70), the embryo is more developed
in the seed, and its parts can be made out. It lies against the starchy
albumen, but is not enclosed therein. The larger part of it is the
cotyledon, thickish, its edges involute, and its back in contact with
the albumen; partly enclosed by it is the well-developed plumule or bud
which is to grow. For the cotyledon remains in the seed to fulfil its
office of imbibing nourishment from the softened albumen, which it
conveys to the growing sprout; the part of this sprout which is visible
is the first leaf of the plumule rolled up into a sheath and enclosing
the rudiments of the succeeding leaves, at the base enclosing even the
minute caulicle. In germination the first leaf of the plumule develops
only as a sort of sheath, protecting the tender parts within; the second
and the third form the first foliage. The caulicle never lengthens: the
first root, which is formed at its lower end, or from any part of it,
has to break through the enclosing sheath; and succeeding roots soon
spring from all or any of the nodes of the plumule.
44. Simple-stemmed Plants are thus built up, by the continuous
production of one leaf-bearing portion of stem from the summit of the
preceding one, beginning with the initial stem (or caulicle) in the
embryo. Some Dicotyls and many Monocotyls develop only in this single
line of growth (as to parts above ground) until the flowering state is
approached. For some examples, see Cycas (Fig. 71, front, at the left);
a tall Yucca or Spanish Bayonet, and two Cocoa-nut Palms behind; at the
right, a group of Sugar-canes, and a Banana behind.
 Fig.
Fig. 71. Simple-stemmed vegetation.
[Pg 27]
Section IV. GROWTH FROM BUDS: BRANCHING.
45. Most plants increase the amount of their vegetation by branching,
that is, by producing lateral shoots.
46. Roots branch from any part and usually without definite order. Stems
normally give rise to branches only at definite points, namely, at the
nodes, and there only from the axils of leaves.
47. Buds (Fig. 72, 73). Every incipient shoot is a Bud (12). A stem
continues its growth by its terminal bud; it branches by the formation
and development of lateral buds. As normal lateral buds occupy the
axils of leaves, they are called axillary buds. As leaves are
symmetrically arranged on the stem, the buds in their axils and the
branches into which axillary buds grow partake of this symmetry. The
most conspicuous buds are the scaly winter-buds of most shrubs and trees
of temperate and cold climates; but the name belongs as well to the
forming shoot or branch of any herb.
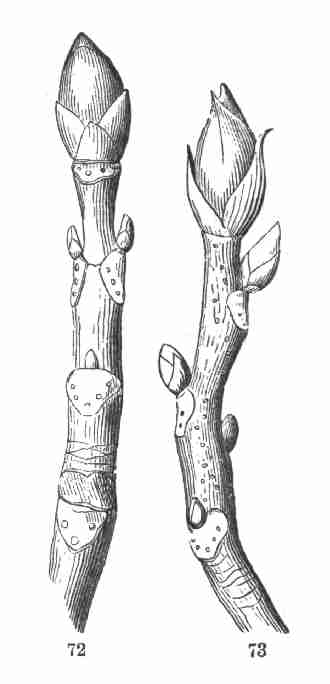
Fig. 72. Shoot of Horse-chestnut, of one year's growth,
taken in autumn after the leaves have fallen; showing the large terminal
bud and smaller axillary buds.
Fig. 73. Similar shoot of Shagbark Hickory, Carya alba.
48. The Terminal Bud, in the most general sense, may be said to exist in
the embryo,—as cotyledons, or the cotyledons and plumule,—and to crown
each successive growth of the simple stem so long as the summit is
capable of growth. The whole ascending growth of the Palm, Cycas, and
the like (such as in Fig. 71) is from a terminal bud. Branches, being
repetitions of the main stem and growing in the same way, are also
lengthened by terminal buds. Those of Horse-chestnut, Hickory, Maples,
and such trees, being the resting buds of winter, are conspicuous by
their protective covering of scales. These bud-scales, as will hereafter
be shown, are themselves a kind of leaves.
49. Axillary Buds were formed on these annual shoots early in the
summer. Occasionally they grow the same season into branches; at least,
some of them are pretty sure to do so whenever the growing terminal bud
at the end of the shoot is injured or destroyed. Otherwise they may lie
dormant until the following spring. In many trees or shrubs these
axillary buds do not show themselves until spring; but if searched for,
they may be detected, though of small size, hidden under the bark.
Sometimes, although early [Pg 28]formed, they are concealed all summer long
under the base of the leaf-stalk, which is then hollowed out into a sort
of inverted cup, like a candle-extinguisher, to cover them; as in the
Locust, the Yellow-wood, or more strikingly in the Button-wood or
Plane-tree (Fig. 74).
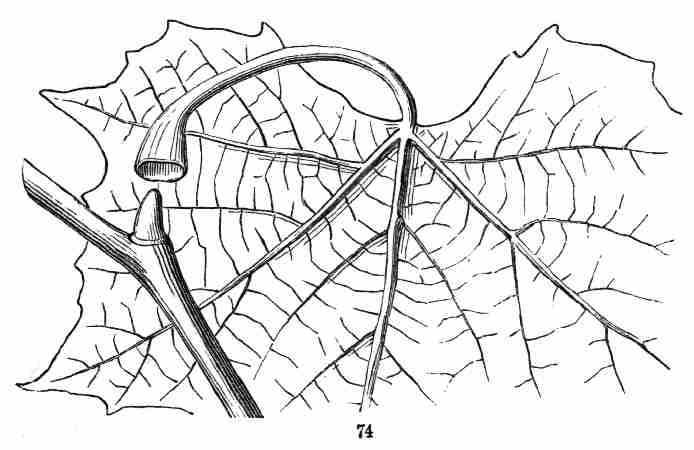
Fig. 74. An axillary bud, concealed under the hollowed
base of the leaf-stalk, in Buttonwood or Plane-tree.
50. The leaf-scars, so conspicuous in Fig. 72, 73, under each axillary
bud, mark the place where the stalk of the subtending leaf was attached
until it fell in autumn.
51. Scaly Buds, which are well represented in Fig. 72, 73, commonly
belong to trees and shrubs of countries in which growth is suspended
during winter. The scaly coverings protect the tender young parts
beneath, not so much by keeping out the cold, which of course would
penetrate the bud in time, as by shielding the interior from the effects
of sudden changes. There are all gradations between these and
52. Naked Buds, in which these scales are inconspicuous or wanting, as
in most herbs, at least above ground, and most tropical trees and
shrubs. But nearly related plants of the same climate may differ widely
in this respect. Rhododendrons have strong and scaly winter-buds; while
in Kalmia they are naked. One species of Viburnum, the Hobble-bush, has
completely naked buds, what would be a pair of scales developing into
the first leaves in spring; while another (the Snowball) has conspicuous
scaly buds.
53. Vigor of Vegetation from strong buds. Large and strong buds, like
those of the Horse-chestnut, Hickory, and the like, contain several
leaves, or pairs of leaves, ready formed, folded and packed away in
small compass, just as the seed-leaves of a strong embryo are packed
away in the seed: they may even contain all the blossoms of the ensuing
season, plainly visible as small buds. And the stems upon which these
buds rest are filled with abundant nourishment, which was deposited the
summer before in the [Pg 29]wood or in the bark. Under the surface of the
soil, or on it covered with the fallen leaves of autumn, similar strong
buds of our perennial herbs may be found; while beneath are thick roots,
rootstocks, or tubers, charged with a great store of nourishment for
their use. This explains how it is that vegetation from such buds shoots
forth so vigorously in the spring of the year, and clothes the bare and
lately frozen surface of the soil, as well as the naked boughs of trees,
very promptly with a covering of fresh green, and often with brilliant
blossoms. Everything was prepared, and even formed, beforehand: the
short joints of stem in the bud have only to lengthen, and to separate
the leaves from each other so that they may unfold and grow. Only a
small part of the vegetation of the season comes directly from the seed,
and none of the earliest vernal vegetation. This is all from buds which
have lived through the winter.
54. The Arrangement of Branches, being that of axillary buds, answers to
that of the leaves. Now leaves principally are either opposite or
alternate. Leaves are opposite when there are two from the same
joint of stem, as in Maples (Fig. 20), the two being on opposite sides
of the stem; and so the axillary buds and branches are opposite, as in
Fig. 75. Leaves are alternate when there is only one from each joint
of stem, as in the Oak, Lime-tree, Poplar, Button-wood (Fig. 74),
Morning-Glory (Fig. 45,—not counting the seed-leaves, which of course
are opposite, there being a pair of them); also in Indian Corn (Fig.
70), and Iris (Fig. 59). Consequently the axillary buds are also
alternate, as in Hickory (Fig. 73); and the branches they form
alternate,—making a different kind of spray from the other mode, one
branch shooting on one side of the stem and the next on some other. For
in the alternate arrangement no leaf is on the same side of the stem as
the one next above or next below it.
55. But the symmetry of branches (unlike that of the leaves) is rarely
complete. This is due to several causes, and most commonly to the
56. Non-development of buds. It never happens that all the buds grow. If
they did, there might be as many branches in any year as there were
leaves the year before. And of those which do begin to grow, a large
portion perish, sooner or later, for want of nourishment, or for want of
light, or because those which first begin to grow have an advantage,
which they are apt to keep, taking to themselves the nourishment of the
stem, and starving the weaker buds. In the Horse-chestnut (Fig. 72),
Hickory (Fig. 73), Magnolia, and most other trees with large scaly buds,
the terminal bud is the strongest, and has the advantage in growth; and
next in strength are the upper axillary buds: while the former continues
the shoot of the last year, some of the latter give rise to branches,
and the rest fail to grow. In the Lilac also (Fig. 75), the uppermost
axillary buds are stronger than the lower; but the terminal bud rarely
appears at all; in its place the uppermost pair of axillary buds grow,
and so each stem branches every year into two,—making a repeatedly
two-forked ramification, as in Fig. 76.
[Pg 30]
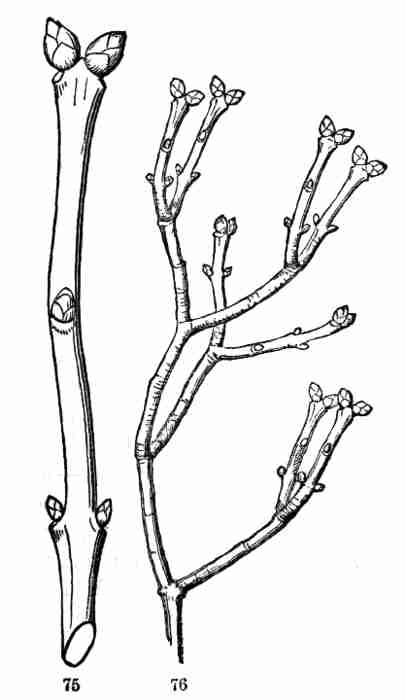
Fig. 75. Shoot of Lilac, with winter buds; the two
uppermost axillary ones strong; the terminal not developed. 76. Forking
ramification of Lilac; reduced in size.
57. Latent Buds. Axillary buds that do not grow at the proper season,
and especially those which make no appearance externally, may long
remain latent, and at length upon a favorable occasion start into
growth, so forming branches apparently out of place as they are out of
time. The new shoots seen springing directly out of large stems may
sometimes originate from such latent buds, which have preserved their
life for years. But commonly these arise from
58. Adventitious Buds. These are buds which certain shrubs and trees
produce anywhere on the surface of the wood, especially where it has
been injured. They give rise to the slender twigs which often feather
the sides of great branches of our American Elms. They sometimes form on
the root, which naturally is destitute of buds; they are even found upon
some leaves; and they are sure to appear on the trunks and roots of
Willows, Poplars, and Chestnuts, when these are wounded or mutilated.
Indeed Osier-Willows are pollarded, or cut off, from time to time, by
the cultivator, for the purpose of producing a crop of slender
adventitious twigs, suitable for basket-work. Such branches, being
altogether irregular, of course interfere with the natural symmetry of
the tree. Another cause of irregularity, in certain trees and shrubs, is
the formation of what are called
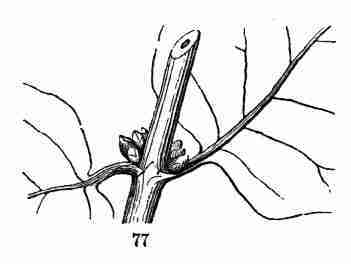 Fig.
Fig. 77. Tartarean Honeysuckle, with three accessory buds
in each axil.
59. Accessory or Supernumerary Buds. There are cases where two, three,
or more buds spring from the axil of a leaf, instead of the single one
which is ordinarily found there. Sometimes they are placed one over the
other, as in the Aristolochia or Pipe-Vine, and in the Tartarean
Honeysuckle (Fig. 77); also in the Honey-Locust, and in the Walnut and
Butternut (Fig. 78), where [Pg 31]the upper supernumerary bud is a good way
out of the axil and above the others. And this is here stronger than the
others, and grows into a branch which is considerably out of the axil,
while the lower and smaller ones commonly do not grow at all. In other
cases three buds stand side by side in the axil, as in the Hawthorn, and
the Red Maple (Fig. 79.) If these were all to grow into branches, they
would stifle each other. But some of them are commonly flower-buds: in
the Red Maple, only the middle one is a leaf-bud, and it does not grow
until after those on each side of it have expanded the blossoms they
contain.
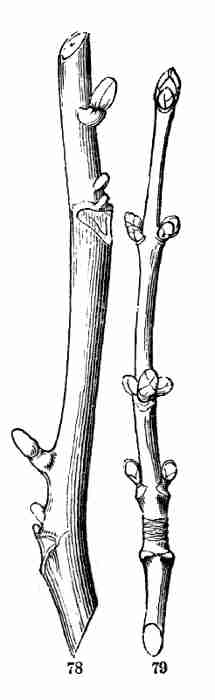
Fig. 78. Butternut branch, with accessory buds, the
uppermost above the axil.
Fig. 79. Red-Maple branch, with accessory buds placed
side by side. The annular lines toward the base in this and in Fig. 72
are scars of the bud-scales, and indicate the place of the winter-bud of
the preceding year.
60. Sorts of Buds. It may be useful to enumerate the kinds of buds which
have been described or mentioned. They are
Terminal, when they occupy the summit of (or terminate) a stem,
Lateral, when they are borne on the side of a stem; of which the
regular kind is the
Axillary, situated in the axil of a leaf. These are
Accessory or Supernumerary, when they are in addition to the normal
solitary bud; and these are Collateral, when side by side;
Superposed, when one above another;
Extra-axillary, when they appear above the axil, as some do when
superposed, and as occasionally is the case when single.
Naked buds; those which have no protecting scales.
Scaly buds; those which have protecting scales, which are altered
leaves or bases of leaves.
Leaf-buds, contain or give rise to leaves, and develop into a leafy
shoot.
Flower-buds, contain or consist of blossoms, and no leaves.
Mixed buds, contain both leaves and blossoms.
61. Definite annual Growth from winter buds is marked in most of the
shoots from strong buds, such as those of the Horse-chestnut and Hickory
(Fig. 72, 73). Such a bud generally contains, already formed in
miniature, all or a great part of the leaves and joints of stem it is to
produce, makes its whole growth in length in the course of a few weeks,
or sometimes even in a few days, and then forms and ripens its buds for
the next year's similar growth.
62. Indefinite annual Growth, on the other hand, is well marked in such
trees or shrubs as the Honey-Locust, Sumac, and in sterile shoots of
[Pg 32]the Rose, Blackberry, and Raspberry. That is, these shoots are apt to
grow all summer long, until stopped by the frosts of autumn or some
other cause. Consequently they form and ripen no terminal bud protected
by scales, and the upper axillary buds are produced so late in the
season that they have no time to mature, nor has their wood time to
solidify and ripen. Such stems therefore commonly die back from the top
in winter, or at least all their upper buds are small and feeble; so the
growth of the succeeding year takes place mainly from the lower axillary
buds, which are more mature.
63. Deliquescent and Excurrent Growth. In the former case, and wherever
axillary buds take the lead, there is, of course, no single main stem,
continued year after year in a direct line, but the trunk is soon lost
in the branches. Trees so formed commonly have rounded or spreading
tops. Of such trees with deliquescent stems,—that is, with the trunk
dissolved, as it were, into the successively divided branches,—the
common American Elm (Fig. 80) is a good illustration.

Fig. 80. An American Elm, with Spruce-trees, and on the
left Arbor Vitæ.
64. On the other hand, the main stem of Firs and Spruces, unless
destroyed by some injury, is carried on in a direct line throughout the
whole growth of the tree, by the development year after year of a
terminal bud: this forms a single, uninterrupted shaft,—an excurrent
trunk, which cannot be confounded with the branches that proceed from
it. Of such spiry or spire-shaped trees, the Firs or Spruces are
characteristic and familiar examples. There are all gradations between
the two modes.
[Pg 33]
Section V. ROOTS.
65. It is a property of stems to produce roots. Stems do not spring from
roots in ordinary cases, as is generally thought, but roots from stems.
When perennial herbs arise from the ground, as they do at spring-time,
they rise from subterranean stems.
66. The Primary Root is a downward growth from the root-end of the
caulicle, that is, of the initial stem of the embryo (Fig. 5-7, 81). If
it goes on to grow it makes a main or tap-root, as in Fig. 37, etc.
Some plants keep this main root throughout their whole life, and send
off only small side branches; as in the Carrot and Radish: and in
various trees, like the Oak, it takes the lead of the side-branches for
several years, unless accidentally injured, as a strong tap-root. But
commonly the main root divides off very soon, and is lost in the
branches. Multiple primary roots now and then occur, as in the
seedling of Pumpkin (Fig. 27), where a cluster is formed even at the
first, from the root-end of the caulicle.
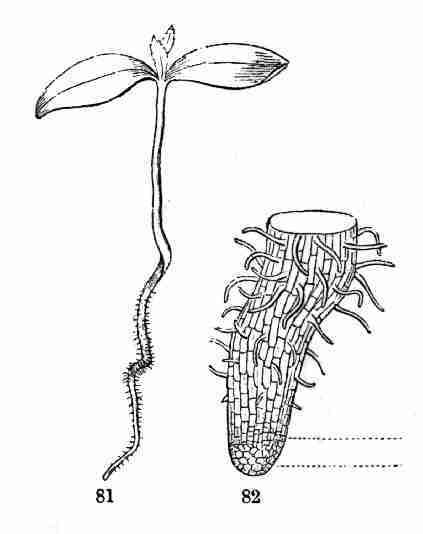
Fig. 81. Seedling Maple, of the natural size; the root
well supplied with root hairs, here large enough to be seen by the naked
eye. 82. Lower end of this root, magnified, the root seen just as
root-hairs are beginning to form a little behind the tip.
67. Secondary Roots are those which arise from other parts of the stem.
Any part of the stem may produce them, but they most readily come from
the nodes. As a general rule they naturally spring, or may be made to
spring, from almost any young stem, when placed in favorable
circumstances,—that is, when placed in the soil, or otherwise supplied
with moisture and screened from the light. For the special tendency of
the root is to avoid the light, seek moisture, and therefore to bury
itself in the soil. Propagation by division, which is so common and so
very important in cultivation, depends upon the proclivity of stems to
strike root. Stems or branches which remain under ground give out roots
as freely as roots themselves give off branches. Stems which creep on
the ground most commonly root at the joints; so will most branches when
bent to the ground, as in propagation by layering; and propagation by
cuttings equally depends upon the tendency of the cut end of a shoot
to produce roots. Thus, a piece of a plant which has stem and leaves,
either developed or in the bud, may be made to produce roots, and so
become an independent plant.
[Pg 34]
68. Contrast between Stem and Root. Stems are ascending axes; roots are
descending axes. Stems grow by the successive development of internodes
(13), one after another, each leaf-bearing at its summit (or node); so
that it is of the essential nature of a stem to bear leaves. Roots bear
no leaves, are not distinguishable into nodes and internodes, but grow
on continuously from the lower end. They commonly branch freely, but not
from any fixed points nor in definite order.
69. Although roots generally do not give rise to stems, and therefore do
not propagate the plant, exceptions are not uncommon. For as stems may
produce adventitious buds, so also may roots. The roots of the Sweet
Potato among herbs, and of the Osage Orange among trees freely produce
adventitious buds, developing into leafy shoots; and so these plants are
propagated by root-cuttings. But most growths of subterranean origin
which pass for roots are forms of stems, the common Potato for example.
70. Roots of ordinary kinds and uses may be roughly classed into
fibrous and fleshy.
71. Fibrous Roots, such as those of Indian Corn (Fig. 70), of most
annuals, and of many perennials, serve only for absorption: these are
slender or thread-like. Fine roots of this kind, and the fine branches
which most roots send out are called Rootlets.
72. The whole surface of a root absorbs moisture from the soil while
fresh and new; and the newer roots and rootlets are, the more freely do
they imbibe. Accordingly, as long as the plant grows above ground, and
expands fresh foliage, from which moisture largely escapes into the air,
so long it continues to extend and multiply its roots in the soil
beneath, renewing and increasing the fresh surface for absorbing
moisture, in proportion to the demand from above. And when growth ceases
above ground, and the leaves die and fall, or no longer act, then the
roots generally stop growing, [Pg 35]and their soft and tender tips harden.
From this period, therefore, until growth begins anew the next spring,
is the best time for transplanting; especially for trees and shrubs.
73. The absorbing surface of young roots is much increased by the
formation, near their tips, of Root-hairs (Fig. 81, 82), which are
delicate tubular outgrowths from the surface, through the delicate walls
of which moisture is promptly imbibed.
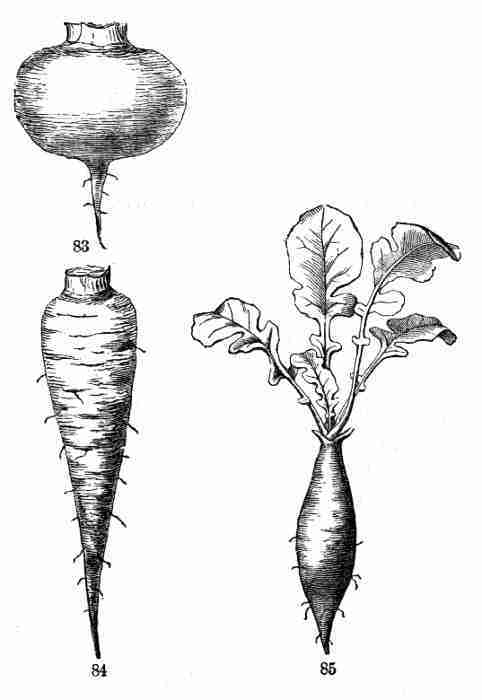 Fig.
Fig. 83-85. Forms of tap-root.
74. Fleshy Roots are those in which the root becomes a storehouse of
nourishment. Typical roots of this kind are those of such biennials as
the turnip and carrot; in which the food created in the first season's
vegetation is accumulated, to be expended the next season in a vigorous
growth and a rapid development of flowers, fruit, and seed. By the time
the seed is matured the exhausted root dies, and with it the whole
plant.
75. Fleshy roots may be single or multiple. The single root of the
commoner biennials is the primary root, or tap-root, which begins to
thicken in the seedling. Names are given to its shapes, such as
Conical, when it thickens most at the crown, or where it joins the
stem, and tapers regularly downwards to a point, as in the Parsnip and
Carrot (Fig. 84);
Turnip-shaped or napiform, when greatly thickened above, but
abruptly becoming slender below; as the Turnip (Fig. 83); and
[Pg 36]
Spindle-shaped, or Fusiform, when thickest in the middle and
tapering to both ends; as the common Radish (Fig. 85).
76. These examples are of primary roots. It will be seen that turnips,
carrots, and the like, are not pure root throughout; for the caulicle,
from the lower end of which the root grew, partakes of the thickening,
perhaps also some joints of stem above: so the bud-bearing and growing
top is stem.
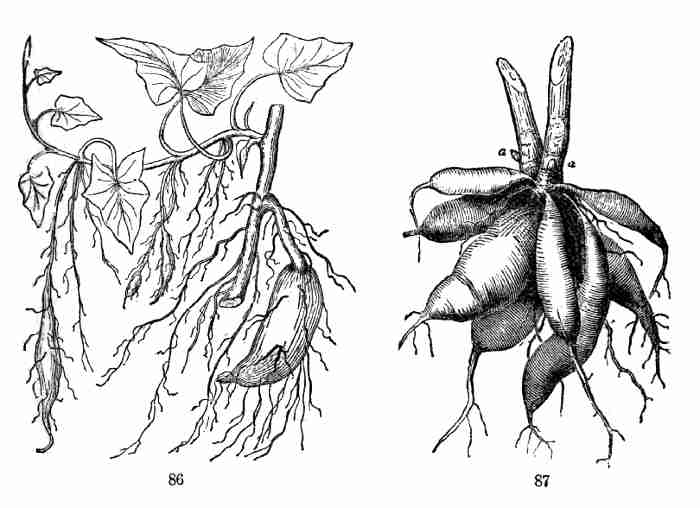
Fig. 86. Sweet-Potato plant forming thickened roots. Some
in the middle are just beginning to thicken; one at the left has grown
more; one at the right is still larger.
Fig. 87. Fascicled fusiform roots of a Dahlia: a, a,
buds on base of stem.
77. A fine example of secondary roots (67), some of which remain fibrous
for absorption, while a few thicken and store up food for the next
season's growth, is furnished by the Sweet Potato (Fig. 86). As stated
above, these are used for propagation by cuttings; for any part will
produce adventitious buds and shoots. The Dahlia produces fascicled
(i. e. clustered) fusiform roots of the same kind, at the base of the
stem (Fig. 87): but these, like most roots, do not produce adventitious
buds. The buds by which Dahlias are propagated belong to the surviving
base of the stem above.
78. Anomalous Roots, as they may be called, are those which subserve
other uses than absorption, food-storing, and fixing the plant to the
soil.
Aerial Roots, i. e. those that strike from stems in the open air, are
common in moist and warm climates, as in the Mangrove which reaches the
coast of Florida, the Banyan, and, less strikingly, in some herbaceous
plants, such as Sugar Cane, and even in Indian Corn. Such roots reach
the ground at length, or tend to do so.
Aerial Rootlets are abundantly produced by many climbing plants, such
as the Ivy, Poison Ivy, Trumpet Creeper, etc., springing from the side
of stems, which they fasten to trunks of trees, walls, or other
supports. These are used by the plant for climbing.

Fig. 88. Epiphytes of Florida and Georgia, viz.,
Epidendrum conopseum, a small Orchid, and Tillandsia usneoides, the
so-called Long Moss or Black Moss, which is no moss, but a flowering
plant, also T. recurvata; on a bough of Live Oak.
79. Epiphytes, or Air-Plants (Fig. 88), are called by the former name
because commonly growing [Pg 37]upon the trunks or limbs of other plants; by
the latter because, having no connection with the soil, they must derive
their sustenance from the air only. They have aerial roots, which do not
reach the ground, but are used to fix the plant to the surface upon
which the plant grows: they also take a part in absorbing moisture from
the air.
80. Parasitic Plants, of which there are various kinds, strike their
roots, or what answer to roots, into the tissue of foster plants, or
form attachments with their surface, so as to prey upon their juices. Of
this sort is the Mistletoe, the seed of which germinates on the bough
where it falls or is left by birds; and the forming root penetrates the
bark and engrafts itself into the wood, to which it becomes united as
firmly as a natural branch to its parent stem; and indeed the parasite
lives just as if it were a branch of the tree it grows and feeds on. A
most common parasitic herb is the Dodder; which abounds in low grounds
in summer, and coils its long and slender, leafless, yellowish
stems—resembling tangled threads of yarn—round and round the stalks of
other plants; wherever they touch piercing the bark with minute and very
short rootlets in the form of suckers, which draw out the nourishing
juices of the plants laid hold of. Other parasitic plants, like the
Beech-drops and Pine-sap, fasten their roots under ground upon the roots
of neighboring plants, and rob them of their juices.
81. Some plants are partly parasitic; while most of their roots act in
the ordinary way, others make suckers at their tips which grow fast to
the roots of other plants and rob them of nourishment. Some of our
species of Gerardia do this (Fig. 89).
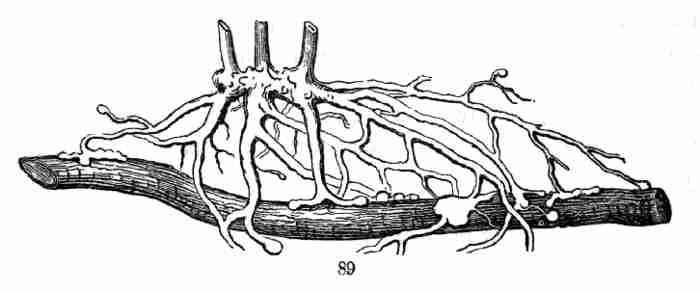
Fig. 89. Roots of Yellow Gerardia, some attached to and
feeding on the root of a Blueberry-bush.
82. There are phanerogamous plants, like Monotropa or Indian Pipe, the
roots of which feed mainly on decaying vegetable matter in the soil.
These are Saprophytes, and they imitate Mushrooms and other Fungi in
their mode of life.
83. Duration of Roots, etc. Roots are said to be either annual,
biennial, or perennial. As respects the first and second, these
terms may be applied either to the root or to the plant.
84. Annuals, as the name denotes, live for only one year, generally for
[Pg 38]only a part of the year. They are of course herbs; they spring from the
seed, blossom, mature their fruit and seed, and then die, root and all.
Annuals of our temperate climates with severe winters start from the
seed in spring, and perish at or before autumn. Where the winter is a
moist and growing season and the summer is dry, winter annuals
prevail; their seeds germinate under autumn or winter rains, grow more
or less during winter, blossom, fructify, and perish in the following
spring or summer. Annuals are fibrous-rooted.
85. Biennials, of which the Turnip, Beet, and Carrot are familiar
examples, grow the first season without blossoming, usually thicken
their roots, laying up in them a stock of nourishment, are quiescent
during the winter, but shoot vigorously, blossom, and seed the next
spring or summer, mainly at the expense of the food stored up, and then
die completely. Annuals and biennials flower only once; hence they have
been called Monocarpic (that is, once-fruiting) plants.
86. Perennials live and blossom year after year. A perennial herb, in a
temperate or cooler climate, usually dies down to the ground at the end
of the season's growth. But subterranean portions of stem, charged with
buds, survive to renew the development. Shrubs and trees are of course
perennial; even the stems and branches above ground live on and grow
year after year.
87. There are all gradations between annuals and biennials, and between
these and perennials, as also between herbs and shrubs; and the
distinction between shrubs and trees is quite arbitrary. There are
perennial herbs and even shrubs of warm climates which are annuals when
raised in a climate which has a winter,—being destroyed by frost. The
Castor-oil plant is an example. There are perennial herbs of which only
small portions survive, as off-shoots, or, in the Potato, as tubers,
etc.
Section VI. STEMS.
88. The Stem is the axis of the plant, the part which bears all the
other organs. Branches are secondary stems, that is, stems growing out
of stems. The stem at the very beginning produces roots, in most plants
a single root from the base of the embryo-stem, or caulicle. As this
root becomes a descending axis, so the stem, which grows in the
opposite direction is called the ascending axis. Rising out of the
soil, the stem bears leaves; and leaf-bearing is the particular
characteristic of the stem. But there are forms of stems that remain
underground, or make a part of their growth there. These do not bear
leaves, in the common sense; yet they bear rudiments of leaves, or what
answers to leaves, although not in the form of foliage. The so-called
stemless or acaulescent plants are those which bear no obvious stem
(caulis) above ground, but only flower-stalks, and the like.
[Pg 39]
89. Stems above ground, through differences in duration, texture, and
size, form herbs, shrubs, trees, etc., or in other terms are
Herbaceous, dying down to the ground every year, or after blossoming.
Suffrutescent, slightly woody below, there surviving from year to
year.
Suffruticose or Frutescent, when low stems are decidedly woody
below, but herbaceous above.
Fruticose or Shrubby, woody, living from year to year, and of
considerable size,—not, however, more than three or four times the
height of a man.
Arborescent, when tree-like in appearance or mode of growth, or
approaching a tree in size.
Arboreous, when forming a proper tree-trunk.
90. As to direction taken in growing, stems may, instead of growing
upright or erect, be
Diffuse, that is, loosely spreading in all directions.
Declined, when turned or bending over to one side.
Decumbent, reclining on the ground, as if too weak to stand.
Assurgent or Ascending, rising obliquely upwards.
Procumbent or Prostrate, lying flat on the ground from the first.
Creeping or Repent, prostrate on or just beneath the ground, and
striking root, as does the White Clover, the Partridge-berry, etc.
Climbing or Scandent, ascending by clinging to other objects for
support, whether by tendrils, as do the Pea, Grape-Vine, and
Passion-flower and Virginia Creeper (Fig. 92, 93); by their twisting
leaf-stalks, as the Virgin's Bower; or by rootlets, like the Ivy, Poison
Ivy, and Trumpet Creeper.
Twining or Voluble, when coiling spirally around other stems or
supports; like the Morning-Glory (Fig. 90) and the Hop.
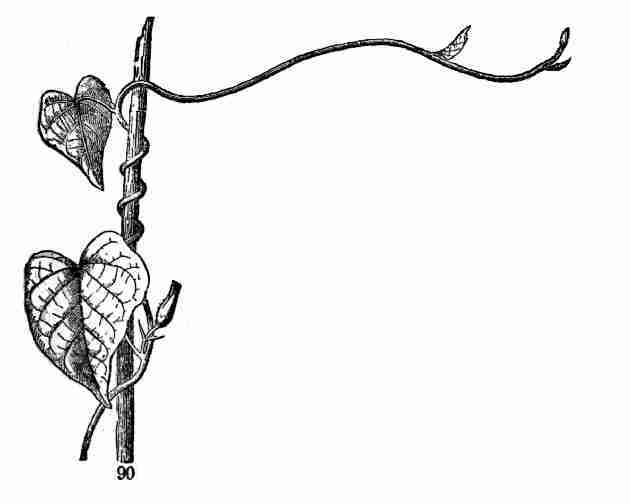 Fig.
Fig. 90. Twining or voluble stem of Morning-Glory.
91. Certain kinds of stems or branches, appropriated to special uses,
have received distinct substantive names; such as the following:
92. A Culm, or straw-stem, such as that of Grasses and Sedges.
93. A Caudex is the old name for such a peculiar trunk as a Palm-stem;
it is also used for an upright and thick rootstock.
94. A Sucker is a branch rising from stems under ground. Such are
produced abundantly by the Rose, Raspberry, and other plants said to
multiply "by the root." If we uncover them, we see at once the great
difference between these subterranean branches and real roots. They are
only creeping branches under ground. Remarking how the upright shoots
from these branches become separate [Pg 40]plants, simply by the dying off of
the connecting under-ground stems, the gardener expedites the result by
cutting them through with his spade. That is, he propagates the plant
"by division."
95. A Stolon is a branch from above ground, which reclines or becomes
prostrate and strikes root (usually from the nodes) wherever it rests on
the soil. Thence it may send up a vigorous shoot, which has roots of its
own, and becomes an independent plant when the connecting part dies, as
it does after a while. The Currant and the Gooseberry naturally multiply
in this way, as well as by suckers (which are the same thing, only the
connecting part is concealed under ground). Stolons must have suggested
the operation of layering by bending down and covering with soil
branches which do not naturally make stolons; and after they have taken
root, as they almost always will, the gardener cuts through the
connecting stem, and so converts a rooting branch into a separate plant.
96. An Offset is a short stolon, or sucker, with a crown of leaves at
the end, as in the Houseleek (Fig. 91), which propagates abundantly in
this way.
 Fig.
Fig. 91. Houseleek (Sempervivum), with offsets.
97. A Runner, of which the Strawberry presents the most familiar and
characteristic example, is a long and slender, tendril-like stolon, or
branch from next the ground, destitute of conspicuous leaves. Each
runner of the Strawberry, after having grown to its full length, strikes
root from the tip, which fixes it to the ground, then forms a bud there,
which develops into a tuft of leaves, and so gives rise to a new plant,
which sends out new runners to act in the same way. In this manner a
single Strawberry plant will spread over a large space, or produce a
great number of plants, in the course of the summer, all connected at
first by the slender runners; but these die in the following winter, if
not before, and leave the plants as so many separate individuals.
98. Tendrils are branches of a very slender sort, like runners, not
destined like them for propagation, and therefore always destitute of
buds or leaves, being intended only for climbing. Simple tendrils are
such as those of Passion-flowers (Fig. 92). Compound or branching
tendrils are borne by the Cucumber and Pumpkin, by the Grape-Vine,
Virginia Creeper, etc.
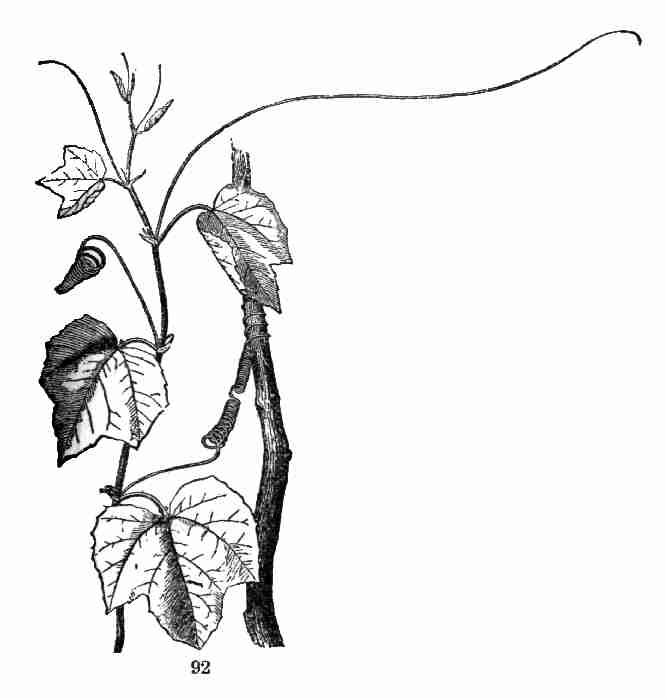
Fig. 92. A small Passion-flower (Passiflora sicyoides),
showing the tendrils.
99. A tendril commonly grows straight and outstretched until it reaches
some neighboring support, such as a stem, when its apex hooks around it
to secure a hold; then the whole tendril shortens itself by coiling up
spirally, and so draws the shoot of the growing plant nearer to the
supporting object. But the tendrils of the Virginia Creeper (Ampelopsis,
Fig. [Pg 41]93), as also the shorter ones of the Japanese species, effect the
object differently, namely, by expanding the tips of the tendrils into a
flat disk, with an adhesive face. This is applied to the supporting
object, and it adheres firmly; then a shortening of the tendril and its
branches by coiling brings up the growing shoot close to the support.
This is an adaptation for climbing mural rocks or walls, or the trunks
of trees, to which ordinary tendrils are unable to cling. The Ivy and
Poison Ivy attain the same result by means of aerial rootlets (78).
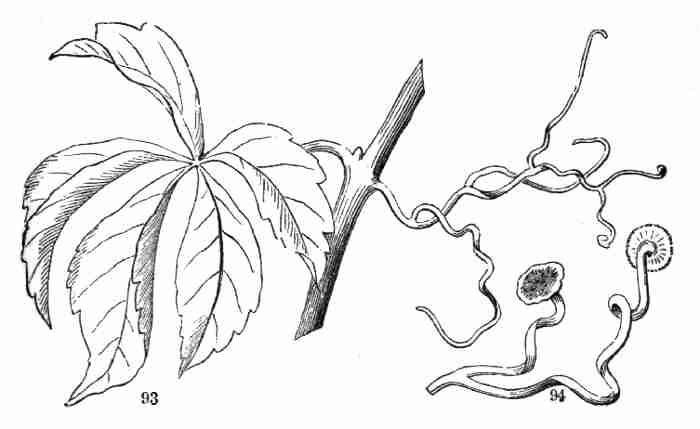
Fig. 93. Piece of the stem of Virginia Creeper, bearing a
leaf and a tendril. 94. Tips of a tendril, about the natural size,
showing the disks by which they hold fast to walls, etc.
100. Some tendrils are leaves or parts of leaves, as those of the Pea
(Fig. 35). The nature of the tendril is known by its position. A tendril
from the axil of a leaf, like that of Passion-flowers (Fig. 92) is of
course a stem, i. e. a branch. So is one which terminates a stem, as in
the Grape-Vine.
101. Spines or Thorns (Fig. 95, 96) are commonly stunted and hardened
branches or tips of stems or branches, as are those of Hawthorn,
Honey-Locust, etc. In the Pear and Sloe all gradations occur between
spines and spine-like (spinescent) branches. Spines may be reduced and
indurated leaves; as in the Barberry, where their nature is revealed by
their situation, underneath an axillary bud. But [Pg 42]prickles, such as
those of Blackberry and Roses, are only excrescences of the bark, and
not branches.
Fig. 95. A branching thorn of Honey-Locust, being an
indurated leafless branch developed from an accessory bud far above the
axil: at the cut portion below, three other buds (a) are concealed
under the petiole.
Fig. 96. Spine of Cockspur Thorn, developed from an
axillary bud, as the leaf-scar below witnesses: an accessory leaf-bud is
seen at its base.
102. Equally strange forms of stems are characteristic of the Cactus
family (Fig. 111). These may be better understood by comparison with
103. Subterranean Stems and Branches. These are very numerous and
various; but they are commonly overlooked, or else are confounded with
roots. From their situation they are out of ordinary sight; but they
will well repay examination. For the vegetation that is carried on under
ground is hardly less varied or important than that above ground. All
their forms may be referred to four principal kinds: namely, the
Rhizoma (Rhizome) or Rootstock, the Tuber, the Corm or solid
bulb, and the true Bulb.
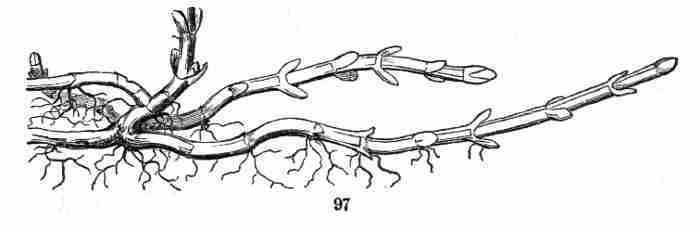
Fig. 97. Rootstocks, or creeping subterranean branches,
of the Peppermint.
104. The Rootstock, or Rhizoma, in its simplest form, is merely a
creeping stem or branch growing beneath the surface of the soil, or
partly covered by it. Of this kind are the so-called creeping,
running, or scaly roots, such as those by which the Mint (Fig. 97),
the Couch-grass, or Quick-grass, and many other plants, spread so
rapidly and widely,—"by the root," as it is said. That these are really
stems, and not roots, is evident from the way in which [Pg 43]they grow;
from their consisting of a succession of joints; and from the leaves
which they bear on each node, in the form of small scales, just like
the lowest ones on the upright stem next the ground. They also produce
buds in the axils of these scales, showing the scales to be leaves;
whereas real roots bear neither leaves nor axillary buds. Placed as they
are in the damp and dark soil, such stems naturally produce roots, just
as the creeping stem does where it lies on the surface of the ground.
105. It is easy to see why plants with these running rootstocks take
such rapid and wide possession of the soil, and why they are so hard to
get rid of. They are always perennials; the subterranean shoots live
over the first winter, if not longer, and are provided with vigorous
buds at every joint. Some of these buds grow in spring into upright
stems, bearing foliage, to elaborate nourishment, and at length produce
blossoms for reproduction by seed; while many others, fed by nourishment
supplied from above, form a new generation of subterranean shoots; and
this is repeated over and over in the course of the season or in
succeeding years. Meanwhile, as the subterranean shoots increase in
number, the older ones, connecting the successive growths, die off year
by year, liberating the already rooted side-branches as so many separate
plants; and so on indefinitely. Cutting these running rootstocks into
pieces, therefore, by the hoe or the plough, far from destroying the
plant, only accelerates the propagation; it converts one many-branched
plant into a great number of separate individuals. Cutting into pieces
only multiplies the pest; for each piece (Fig. 98) is already a
plantlet, with its roots and with a bud in the axil of its scale-like
leaf (either latent or apparent), and with prepared nourishment enough
to develop this bud into a leafy stem; and so a single plant is all the
more speedily converted into a multitude. Whereas, when the subterranean
parts are only roots, cutting away the stem completely destroys the
plant, except in the rather rare cases where the root freely produces
adventitious buds.
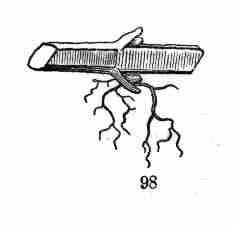
Fig. 98. A piece of the running rootstock of the
Peppermint, with its node or joint, and an axillary bud ready to grow.
106. Rootstocks are more commonly thickened by the storing up of
considerable nourishing matter in their tissue. The common species of
Iris (Fig. 164) in the gardens have stout rootstocks, which are only
partly covered by the soil, and which bear foliage-leaves instead of
mere scales, closely covering the upper part, while the lower produces
roots. As the leaves die, year by year, and decay, a scar left in the
form of a ring marks the place where each leaf was attached, that is,
marks so many nodes, separated by very short internodes.
107. Some rootstocks are marked with large round scars of a different
[Pg 44]sort, like those of the Solomon's Seal (Fig. 99), which gave this name
to the plant, from their looking somewhat like the impression of a seal
upon wax. Here the rootstock sends up every spring an herbaceous stalk
or stem, which bears the foliage and flowers, and dies in autumn. The
seal is the circular scar left by the death and separation of the base
of the stout stalk from the living rootstock. As but one of these is
formed each year, they mark the limits of a year's growth. The bud at
the end of the rootstock in the figure (which was taken in summer) will
grow the next spring into the stalk of the season, which, dying in
autumn, will leave a similar scar, while another bud will be formed
farther on, crowning the ever-advancing summit or growing end of the
stem.
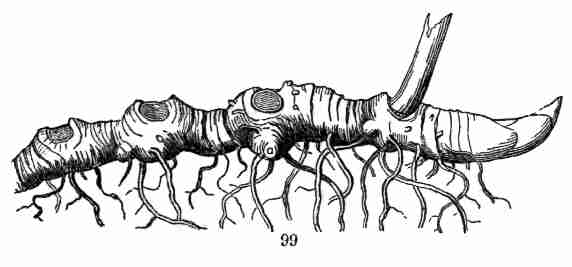
Fig. 99. Rootstock of Solomon's Seal, with the bottom of
the stalk of the season, and the bud for the next year's growth.
108. As each year's growth of stem makes its own roots, it soon becomes
independent of the older parts. And after a certain age, a portion
annually dies off behind, about as fast as it increases at the growing
end, death following life with equal and certain step, with only a
narrow interval. In vigorous plants of Solomon's Seal or Iris, the
living rootstock is several inches or a foot in length; while in the
short rootstock of Trillium or Birthroot (Fig. 100) life is reduced to a
narrower span.
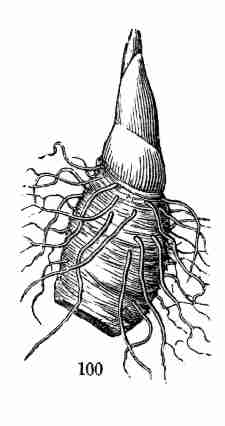
Fig. 100. The very short rootstock and strong terminal
bud of a Trillium or Birthroot.
109. An upright or short rootstock, like this of Trillium, is commonly
called a Caudex (93); or when more shortened and thickened it would
become a corm.
110. A Tuber may be understood to be a portion of a rootstock thickened,
and with buds (eyes) on the sides. Of course, there are all gradations
between a tuber and a rootstock. Helianthus tuberosus, the so-called
Jerusalem Artichoke (Fig. 101), and the common Potato, are typical and
familiar examples of the tuber. The stalks by which the tubers are
attached to the parent stem are at once seen to be different from the
roots, both in appearance and manner of growth. The scales on the tubers
are the rudiments of leaves; the eyes are the buds in their axils. The
Potato-plant [Pg 45]has three forms of branches: 1. Those that bear ordinary
leaves expanded in the air, to digest what they gather from it and what
the roots gather from the soil, and convert it into nourishment. 2.
After a while a second set of branches at the summit of the plant bear
flowers, which form fruit and seed out of a portion of the nourishment
which the leaves have prepared. 3. But a larger part of this
nourishment, while in a liquid state, is carried down the stem, into a
third sort of branches under ground, and accumulated in the form of
starch at their extremities, which become tubers, or depositories of
prepared solid food,—just as in the Turnip, Carrot, and Dahlia (Fig.
83-87), it is deposited in the root. The use of the store of food is
obvious enough. In the autumn the whole plant dies, except the seeds (if
it formed them) and the tubers; and the latter are left disconnected in
the ground. Just as that small portion of nourishing matter which is
deposited in the seed feeds the embryo when it germinates, so the much
larger portion deposited in the tuber nourishes its buds, or eyes, when
they likewise grow, the next spring, into new plants. And the great
supply enables them to shoot with a greater vigor at the beginning, and
to produce a greater amount of vegetation than the seedling plant could
do in the same space of time; which vegetation in turn may prepare and
store up, in the course of a few weeks or months, the largest quantity
of solid nourishing material, in a form most available for food. Taking
advantage of this, man has transported the Potato from the cool Andes of
Chili to other cool climates, and makes it yield him a copious supply of
food, especially important in countries where the season is too short,
or the summer's heat too little, for profitably cultivating the
principal grain-plants.
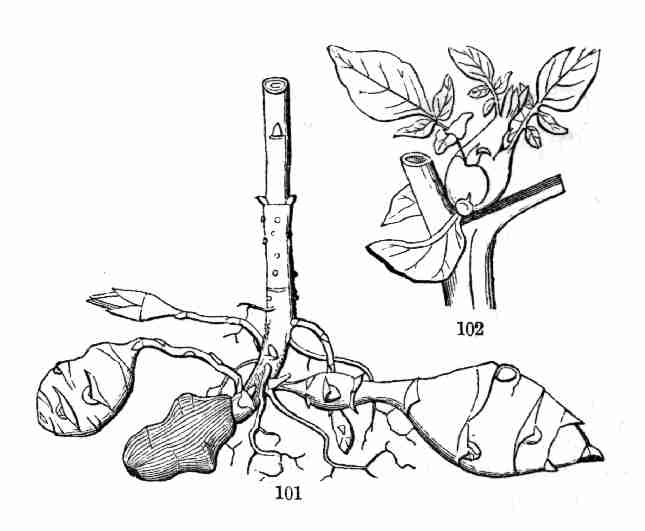
Fig. 101. Tubers of Helianthus tuberosus, called
"artichokes."
Fig. 102. Bulblet-like tubers, such as are occasionally
formed on the stem of a Potato-plant above ground.
111. The Corm or Solid Bulb, like that of Cyclamen (Fig. 103), and of
Indian Turnip (Fig. 104), is a very short and thick fleshy subterranean
stem, often broader than high. It sends off roots from its lower end, or
rather face, leaves and stalks from its upper. The corm of Cyclamen goes
on to enlarge and to produce a succession of flowers and leaves year
after year. [Pg 46]That of Indian Turnip is formed one year and is consumed
the next. Fig. 104 represents it in early summer, having below the corm
of last year, from which the roots have fallen. It is partly consumed by
the growth of the stem for the season, and the corm of the year is
forming at base of the stem above the line of roots.
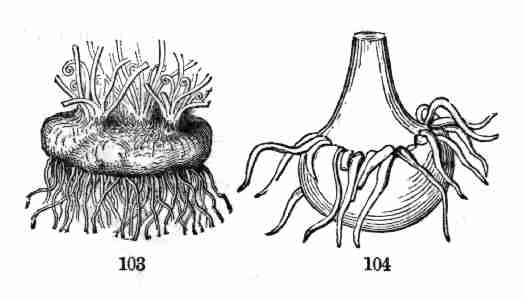
Fig. 103. Corm of Cyclamen, much reduced in size: roots
from lower face, leaf-stalks and flower-stalks from the upper.
Fig. 104. Corm of Indian Turnip (Arisæma).
112. The corm of Crocus (Fig. 105, 106), like that of its relative
Gladiolus, is also reproduced annually, the new ones forming upon the
summit and sides of the old. Such a corm is like a tuber in budding from
the sides, i. e. from the axils of leaves; but these leaves, instead of
being small scales, are the sheathing bases of foliage-leaves which
covered the surface. It resembles a true bulb in having these sheaths or
broad scales; but in the corm or solid bulb, this solid part or stem
makes up the principal bulk.
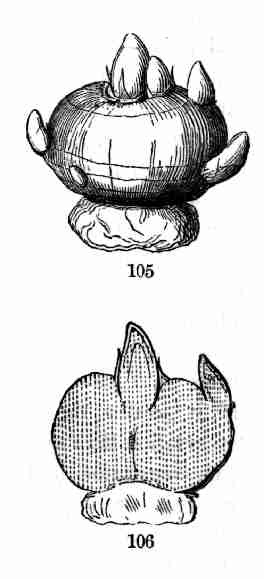
Fig. 105. Corm of a Crocus, the investing sheaths or dead
leaf-bases stripped off. The faint cross-lines represent the scars,
where the leaves were attached, i. e. the nodes: the spaces between are
the internodes. The exhausted corm of the previous year is underneath;
forming ones for next year on the summit and sides.
Fig. 106. Section of the same.
113. The Bulb, strictly so-called, is a stem like a reduced corm as to
its solid part (or plate); while the main body consists of thickened
scales, which are leaves or leaf-bases. These are like bud-scales; so
that in fact a bulb is a bud with fleshy scales on an exceedingly short
stem. Compare a White Lily bulb (Fig. 107) with the strong scaly buds of
the Hickory and Horse-chestnut (Fig. 72 and 73), and the resemblance
will appear. In corms, as in tubers and rootstocks, the store of food
for future growth is deposited in the stem; while in the bulb, the
greater part is deposited in the bases of the leaves, changing them into
thick scales, which closely overlap or enclose one another.
114. A Scaly Bulb (like that of the Lily, Fig. 107, 108) is one in which
the scales are thick but comparatively narrow.
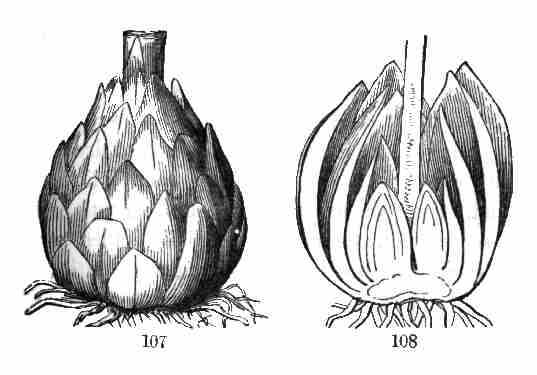
Fig. 107. Bulb of a wild Lily. 108. The same divided
lengthwise, showing two forming buds of the next generation.
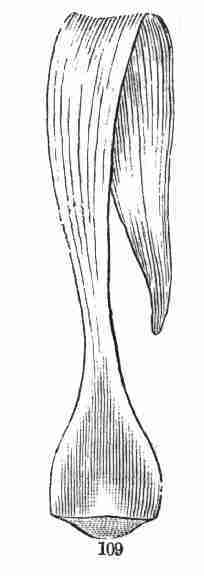
Fig. 109. A ground leaf of White Lily, its base (cut
across) thickened into a bulb-scale. This plainly shows that bulb-scales
are leaves.
115. A Tunicated or Coated Bulb is one in which the scales enwrap each
other, forming concentric coats or layers, as in Hyacinth and Onion.
[Pg 47]
116. Bulblets are very small bulbs growing out of larger ones; or small
bulbs produced above ground on some plants, as in the axils of the
leaves of the bulbiferous Lilies of the gardens (Fig. 110), and often in
the flower-clusters of the Leek and Onion. They are plainly buds with
thickened scales. They never grow into branches, but detach themselves
when full grown, fall to the ground, and take root there to form new
plants.
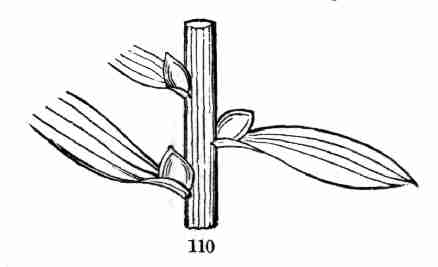
Fig. 110. Bulblets in the axils of leaves of a Tiger
Lily.
117. Consolidated Vegetation. An ordinary herb, shrub, or tree is
evidently constructed on the plan developing an extensive surface. In
fleshy rootstocks, tubers, corms, and bulbs, the more enduring portion
of the plant is concentrated, and reduced for the time of struggle (as
against drought, heat, or cold) to a small amount of exposed surface,
and this mostly sheltered in the soil. There are many similar
consolidated forms which are not subterranean. Thus plants like the
Houseleek (Fig. 91) imitate a bulb. Among Cactuses the columnar species
of Cereus (Fig. 111, b), may be likened to rootstocks. A green rind
serves the purpose of foliage; but the surface is as nothing compared
with an ordinary leafy plant of the same bulk. Compare, for instance,
the largest Cactus known, the Giant Cereus of the Gila River (Fig. 111,
in the background), which rises to the height of fifty or sixty feet,
with a common leafy tree of the same height, such as that in Fig. 89,
and estimate how vastly greater, even without the foliage, the surface
of the latter is than that of the former. Compare, in the [Pg 48]same view, an
Opuntia or Prickly-Pear Cactus, its stem and branches formed of a
succession of thick and flattened joints (Fig. 111, a), which may be
likened to tubers, or an Epiphyllum (d), having short and flat joints,
with an ordinary leafy shrub or herb of equal size. And finally, in
Melon-Cactuses, Echinocactus (c), or other globose forms (which may be
likened to permanent corms), with their globular or bulb-like shapes, we
have plants in the compactest shape; their spherical figure being such
as to expose the least possible amount of substance to the air. These
are adaptations to climates which are very dry, either throughout or for
a part of the year. Similarly, bulbous and corm-bearing plants, and the
like, are examples of a form of vegetation which in the growing season
may expand a large surface to the air and light, while during the period
of rest the living vegetable is reduced to a globe, or solid form of the
least possible surface; and this protected by its outer coats of dead
and dry scales, as well as by its situation under ground. Such are also
adapted to a season of drought. They largely belong to countries which
have a long hot season of little or no rain, when, their stalks and
foliage above and their roots beneath early perishing, the plants rest
securely in their compact bulbs, filled with nourishment and retaining
their moisture with great tenacity, until the rainy season comes round.
Then they shoot forth leaves and flowers with wonderful rapidity, and
what was perhaps a desert of arid sand becomes green with foliage and
gay with blossoms, almost in a day.

[Pg 49]
Section VII. LEAVES.
118. Stems bear leaves, at definite points (nodes, 13); and these are
produced in a great variety of forms, and subserve various uses. The
commonest kind of leaf, which therefore may be taken as the type or
pattern, is an expanded green body, by means of which the plant exposes
to the air and light the matters which it imbibes, exhales certain
portions, and assimilates the residue into vegetable matter for its
nourishment and growth.
119. But the fact is already familiar (10-30) that leaves occur under
other forms and serve for other uses,—for the storage of food already
assimilated, as in thickened seed-leaves and bulb-scales; for covering,
as in bud-scales; and still other uses are to be pointed out. Indeed,
sometimes they are of no service to the plant, being reduced to mere
scales or rudiments, such as those on the rootstocks of Peppermint (Fig.
97) or the tubers of Jerusalem Artichoke (Fig. 101). These may be said
to be of service only to the botanist, in explaining to him the plan
upon which a plant is constructed.
120. Accordingly, just as a rootstock, or a tuber, or a tendril is a
kind of stem, so a bud-scale, or a bulb-scale, or a cotyledon, or a
petal of a flower, is a kind of leaf. Even in respect to ordinary
leaves, it is natural to use the word either in a wider or in a narrower
sense; as when in one sense we say that a leaf consists of blade and
petiole or leaf-stalk, and in another sense say that a leaf is petioled,
or that the leaf of Hepatica is three-lobed. The connection should make
it plain whether by leaf we mean leaf-blade only, or the blade with any
other parts it may have. And the student will readily understand that by
leaf in its largest or morphological sense, the botanist means the
organ which occupies the place of a leaf, whatever be its form or its
function.
§ 1. LEAVES AS FOLIAGE.
121. This is tautological; for foliage is simply leaves: but it is very
convenient to speak of typical leaves, or those which serve the plant
for assimilation, as foliage-leaves, or ordinary leaves. These may first
be considered.
122. The Parts of a Leaf. The ordinary leaf, complete in its parts,
consists of blade, foot-stalk, or petiole, and a pair of
stipules.
123. First the Blade or Lamina, which is the essential part of ordinary
leaves, that is, of such as serve the purpose of foliage. In structure
it consists of a softer part, the green pulp, called parenchyma,
which is traversed and supported by a fibrous frame, the parts of which
are called ribs or veins, on account of a certain likeness in
arrangement to the veins of animals. [Pg 50]The whole surface is covered by a
transparent skin, the Epidermis, not unlike that which covers the
surface of all fresh shoots.
124. Note that the leaf-blade expands horizontally,—that is, normally
presents its faces one to the sky, the other to the ground, or when the
leaf is erect the upper face looks toward the stem that bears it, the
lower face away from it. Whenever this is not the case there is
something to be explained.
125. The framework consists of wood,—a fibrous and tough material
which runs from the stem through the leaf-stalk, when there is one, in
the form of parallel threads or bundles of fibres; and in the blade
these spread out in a horizontal direction, to form the ribs and
veins of the leaf. The stout main branches of the framework are called
the Ribs. When there is only one, as in Fig. 112, 114, or a middle one
decidedly larger than the rest, it is called the Midrib. The smaller
divisions are termed Veins; and their still smaller subdivisions,
Veinlets. The latter subdivide again and again, until they become so
fine that they are invisible to the naked eye. The fibres of which they
are composed are hollow; forming tubes by which the sap is brought into
the leaves and carried to every part.
 Fig.
Fig. 112. Leaf of the Quince:
b, blade;
p, petiole;
st, stipules.
126. Venation is the name of the mode of veining, that is, of the way in
which the veins are distributed in the blade. This is of two principal
kinds; namely, the parallel-veined, and the netted-veined.
127. In Netted-veined (also called Reticulated) leaves, the veins
branch off from the main rib or ribs, divide into finer and finer
veinlets, and the branches unite with each other to form meshes of
network. That is, they anastomose, as anatomists say of the veins and
arteries of the body. The Quince-leaf, in Fig. 112, shows this kind of
veining in a leaf with a single rib. The Maple, Basswood, Plane or
Buttonwood (Fig. 74) show it in leaves of several ribs.
128. In parallel-veined leaves, the whole framework consists of
slender ribs or veins, which run parallel with each other, or nearly so,
from the base to the point of the leaf,—not dividing and subdividing,
nor forming meshes, except by minute cross-veinlets. The leaf of any
grass, or that of the Lily of the Valley (Fig. 113) will furnish a good
illustration. Such parallel veins Linnæus called Nerves, and
parallel-veined leaves are still commonly called nerved leaves, while
those of the other kind are said to be [Pg 51]veined,—terms which it is
convenient to use, although these "nerves" and "veins" are all the same
thing, and have no likeness to the nerves and little to the veins of
animals.
129. Netted-veined leaves belong to plants which have a pair of
seed-leaves or cotyledons, such as the Maple (Fig. 20, 24), Beech (Fig.
33), and the like; while parallel-veined or nerved leaves belong to
plants with one cotyledon or true seed-leaf; such as the Iris (Fig. 59),
and Indian Corn (Fig. 70). So that a mere glance at the leaves generally
tells what the structure of the embryo is, and refers the plant to one
or the other of these two grand classes,—which is a great convenience.
For when plants differ from each other in some one important respect,
they usually differ correspondingly in other respects also.
Fig. 113. A (parallel-veined) leaf of the Lily of the
Valley. 114. One of the Calla Lily.
130. Parallel-veined leaves are of two sorts,—one kind, and the
commonest, having the ribs or nerves all running from the base to the
point of the leaf, as in the examples already given; while in another
kind they run from a midrib to the margin, as in the common
Pickerel-weed of our ponds, in the Banana, in Calla (Fig. 114), and many
similar plants of warm climates.
131. Netted-veined leaves are also of two sorts, as in the examples
already referred to. In one case the veins all rise from a single rib
(the midrib), as in Fig. 112, 116-127. Such leaves are called
Feather-veined or Penni-veined, i. e. Pinnately-veined; both terms
meaning the same thing, namely, that the veins are arranged on the sides
of the rib like the plume of a feather on each side of the shaft.
[Pg 52]
132. In the other case (as in Fig. 74, 129-132), the veins branch off
from three, five, seven, or nine ribs, which spread from the top of the
leaf-stalk, and run through the blade like the toes of a web-footed
bird. Hence these are said to be Palmately or Digitately veined, or
(since the ribs diverge like rays from a centre) Radiate-veined.
133. Since the general outline of leaves accords with the framework or
skeleton, it is plain that feather-veined (or penni-veined) leaves
will incline to elongated shapes, or at least to be longer than broad;
while in radiate-veined leaves more rounded forms are to be expected.
A glance at the following figures shows this.
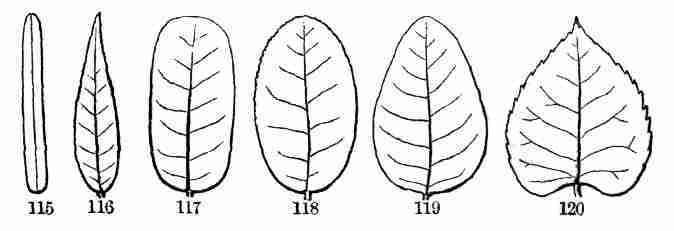
Fig. 115-120. A series of shapes of feather-veined
leaves.
134. Forms of Leaves as to General Outline. It is necessary to give
names to the principal shapes, and to define them rather precisely,
since they afford easy marks for distinguishing species. The same terms
are used for all other flattened parts as well, such as petals; so that
they make up a great part of the descriptive language of Botany. It will
be a good exercise for young students to look up leaves answering to
these names and definitions. Beginning with the narrower and proceeding
to the broadest forms, a leaf is said to be
Linear (Fig. 115), when narrow, several times longer than wide, and of
the same breadth throughout.
Lanceolate, or Lance-shaped, when conspicuously longer than wide,
and tapering upwards (Fig. 116), or both upwards and downwards.
Oblong (Fig. 117), when nearly twice or thrice as long as broad.
Elliptical (Fig. 118) is oblong with a flowing outline, the two ends
alike in width.
Oval is the same as broadly elliptical, or elliptical with the breadth
considerably more than half the length.
Ovate (Fig. 119), when the outline is like a section of a hen's egg
lengthwise, the broader end downward.
Orbicular, or Rotund (Fig. 132), circular in outline, or nearly so.
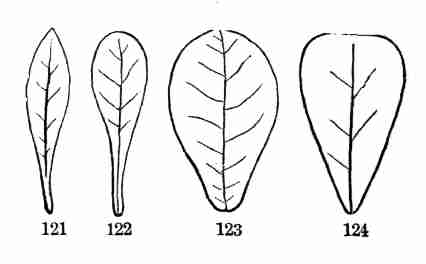
Fig. 121, oblanceolate; 122, spatulate; 123, obovate; and
124, wedge-shaped, feather-veined, leaves.
135. A leaf which tapers toward the base instead of toward the apex may
be
Oblanceolate (Fig. 121) when of the lance-shaped form, only more
tapering toward the base than in the opposite direction.
Spatulate (Fig. 122) when more rounded above, but tapering thence to a
narrow base, like an old-fashioned spatula.
[Pg 53]
Obovate (Fig. 123) or inversely ovate, that is, ovate with the
narrower end down.
Cuneate or Cuneiform, that is, Wedge-shaped (Fig. 124), broad
above and tapering by nearly straight lines to an acute angle at the
base.
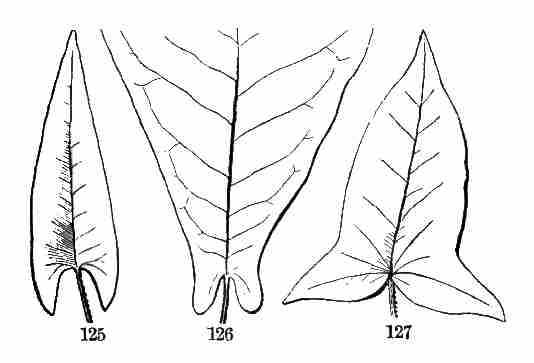
Fig. 125, sagittate; 126, auriculate; and 127,
halberd-shaped or hastate leaves.
136. As to the Base, its shape characterizes several forms, such as
Cordate or Heart-shaped (Fig. 120, 129), when a leaf of an ovate
form, or something like it, has the outline of its rounded base turned
in (forming a notch or sinus) where the stalk is attached.
Reniform, or Kidney-shaped (Fig. 131), like the last, only rounder
and broader than long.
Auriculate, or Eared, having a pair of small and blunt projections,
or ears, at the base, as in one species of Magnolia (Fig. 126).
Sagittate, or arrow-shaped, where such ears are acute and turned
downwards, while the main body of the blade tapers upwards to a point,
as in the common Sagittaria or Arrow-head, and in the Arrow-leaved
Polygonum (Fig. 125).
Hastate, or Halberd-shaped, when such lobes at the base point
outwards, giving the shape of the halberd of the olden time, as in
another Polygonum (Fig. 127).
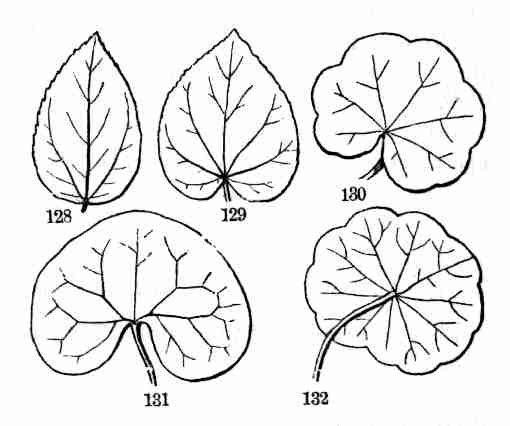 Fig.
Fig. 128-132. Various forms of radiate-veined leaves.
Peltate, or Shield-shaped (Fig. 132), is the name applied to a
curious modification of the leaf, commonly of a rounded form, where the
footstalk is attached to the lower surface, instead of the base, and
therefore is naturally [Pg 54]likened to a shield borne by the outstretched
arm. The common Watershield, the Nelumbium, and the White Water-lily,
and also the Mandrake, exhibit this sort of leaf. On comparing the
shield-shaped leaf of the common Marsh Pennywort (Fig. 132) with that of
another common species (Fig. 130), it is at once seen that a
shield-shaped leaf is like a kidney-shaped (Fig. 130, 131) or other
rounded leaf, with the margins at the base brought together and united.
137. As to the Apex, the following terms express the principal
variations:—
Acuminate, Pointed, or Taper-pointed, when the summit is more or
less prolonged into a narrowed or tapering point; as in Fig. 133.
Acute, ending in an acute angle or not prolonged point; Fig. 134.
Obtuse, with a blunt or rounded apex; as in Fig. 135, etc.
Truncate, with the end as if cut off square; as in Fig. 136.
Retuse, with rounded summit slightly indented, forming a very shallow
notch, as in Fig. 137.
Emarginate, or Notched, indented at the end more decidedly; as in
Fig. 138.
Obcordate, that is, inversely heart-shaped, where an obovate leaf is
more deeply notched at the end (Fig. 139), as in White Clover and
Wood-sorrel; so as to resemble a cordate leaf inverted.
Cuspidate, tipped with a sharp and rigid point; as in Fig. 140.
Mucronate, abruptly tipped with a small and short point, like a mere
projection of the midrib; as in Fig. 141.
Aristate, Awn-pointed, and Bristle-pointed, are terms used when
this mucronate point is extended into a longer bristle-form or slender
appendage.
The first six of these terms can be applied to the lower as well as to
the upper end of a leaf or other organ. The others belong to the apex
only.
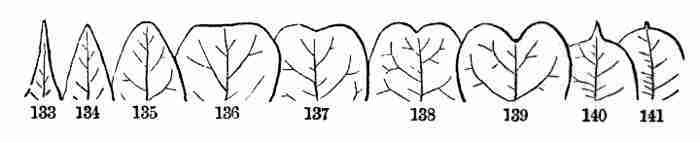 Fig.
Fig. 133-141. Forms of the apex of leaves.
138. As to degree and nature of Division, there is first of all the
difference between
Simple Leaves, those in which the blade is of one piece, however much
it may be cut up, and
Compound Leaves, those in which the blade consists of two or more
separate pieces, upon a common leaf-stalk or support. Yet between these
two kinds every intermediate gradation is to be met with.
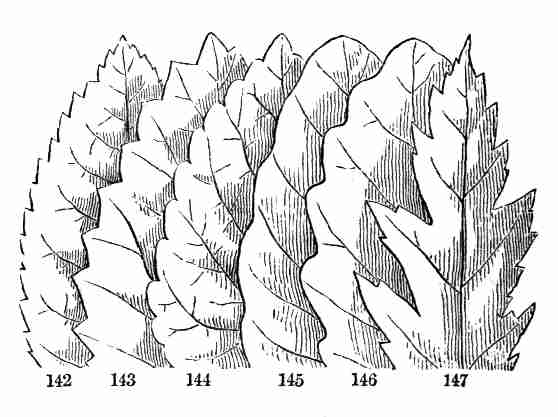 Fig.
Fig. 142-147. Kinds of margin of leaves.
139. As to Particular Outlines of Simple Leaves (and the same applies to
their separate parts), they are
[Pg 55]
Entire, when their general outline is completely filled out, so that
the margin is an even line, without teeth or notches.
Serrate, or Saw-toothed, when the margin only is cut into sharp
teeth, like those of a saw, and pointing forwards; as in Fig. 142.
Dentate, or Toothed, when such teeth point outwards, instead of
forwards; as in Fig. 143.
Crenate, or Scalloped, when the teeth are broad and rounded; as in
Fig. 144.
Repand, Undulate, or Wavy, when the margin of the leaf forms a
wavy line, bending slightly inwards and outwards in succession; as in
Fig. 145.
Sinuate, when the margin is more strongly sinuous or turned inwards
and outwards; as in Fig. 146.
Incised, Cut, or Jagged, when the margin is cut into sharp, deep,
and irregular teeth or incisions; as in Fig. 147.
Lobed, when deeply cut. Then the pieces are in a general way called
Lobes. The number of the lobes is briefly expressed by the phrase
two-lobed, three-lobed, five-lobed, many-lobed, etc., as the
case may be.
140. When the depth and character of the lobing needs to be more
particularly specified, the following terms are employed, viz.:—
Lobed, in a special sense, when the incisions do not extend deeper
than about half-way between the margin and the centre of the blade, if
so far, and are more or less rounded; as in the leaves of the Post-Oak,
Fig. 148, and the Hepatica, Fig. 152.
Cleft, when the incisions extend half way down or more, and especially
when they are sharp; as in Fig. 149, 153. And the phrases two-cleft,
or, in the Latin form, bifid, three-cleft or trifid, four-cleft
or quadrifid, five-cleft or quinquefid, etc., or many-cleft, in
the Latin form, multifid,—express the number of the Segments, or
portions.
Parted, when the incisions are still deeper, but yet do not quite
reach to the midrib or the base of the blade; as in Fig. 150, 154. And
the terms two-parted, three-parted, etc., express the number of such
divisions.
Divided, when the incisions extend quite to the midrib, as in the
lower part of Fig. 151, or to the leaf-stalk, as in Fig. 155; which
really makes the [Pg 56]leaf compound. Here, using the Latin form, the leaf is
said to be bisected, trisected (Fig. 155), etc., according to the
number of the divisions.
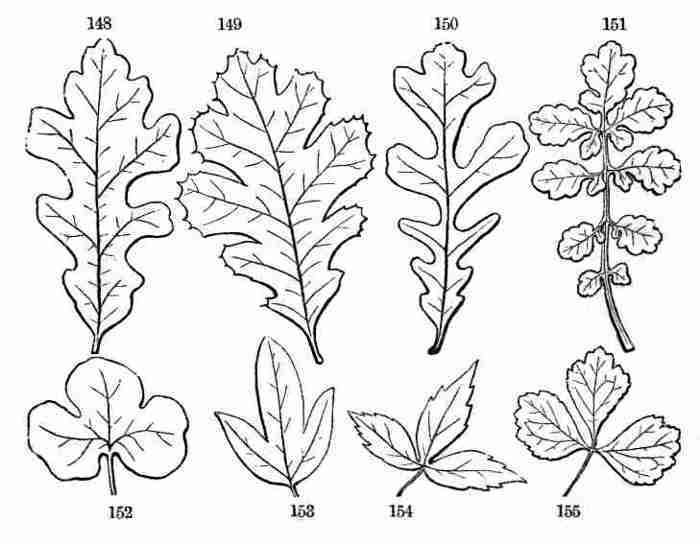
Fig. 148, pinnately lobed; 149, pinnately cleft; 150,
pinnately parted; 151, pinnately divided, leaves.
Fig. 152, palmately three-lobed; 153, palmately
three-cleft; 154, palmately three-parted; 155, palmately three-divided
or trisected, leaves.
141. The Mode of Lobing or Division corresponds to that of the veining,
whether pinnately veined or palmately veined. In the former the
notches or incisions, or sinuses, coming between the principal veins
or ribs are directed toward the midrib: in the latter they are directed
toward the apex of the petiole; as the figures show.
142. So degree and mode of division may be tersely expressed in brief
phrases. Thus, in the four upper figures of pinnately veined leaves, the
first is said to be pinnately lobed (in the special sense), the second
pinnately cleft (or pinnatifid in Latin form), the third pinnately
parted, the fourth pinnately divided, or pinnatisected.
143. Correspondingly in the lower row, of palmately veined leaves, the
first is palmately lobed, the second palmately cleft, the third
palmately parted, the fourth palmately divided. Or, in other
language of the same meaning (but now less commonly employed), they are
said to be digitately lobed, cleft, parted, or divided.
144. The number of the divisions or lobes may come into the phrase. Thus
in the four last named figures the leaves are respectively palmately
three-lobed, three-cleft (or trifid), three-parted,
three-divided, or better (in Latin form), trisected. And so for
higher numbers, as five-lobed, five-cleft, [Pg 57]etc., up to
many-lobed, many-cleft or multifid, etc. The same mode of
expression may be used for pinnately lobed leaves, as pinnately
7-lobed, -cleft, -parted, etc.
145. The divisions, lobes, etc., may themselves be entire (without
teeth or notches), or serrate, or otherwise toothed or incised; or
lobed, cleft, parted, etc.: in the latter cases making twice
pinnatifid, twice palmately or pinnately lobed, parted, or
divided leaves, etc. From these illustrations one will perceive how
the botanist, in two or three words, may describe any one of the almost
endlessly diversified shapes of leaves, so as to give a clear and
definite idea of it.
146. Compound Leaves. A compound leaf is one which has its blade in
entirely separate parts, each usually with a stalklet of its own; and
the stalklet is often jointed (or articulated) with the main
leaf-stalk, just as this is jointed with the stem. When this is the
case, there is no doubt that the leaf is compound. But when the pieces
have no stalklets, and are not jointed with the main leaf-stalk, it may
be considered either as a divided simple leaf, or a compound leaf,
according to the circumstances. This is a matter of names where all
intermediate forms may be expected.
147. While the pieces or projecting parts of a simple leaf-blade are
called Lobes, or in deeply cut leaves, etc., Segments, or
Divisions, the separate pieces or blades of a compound leaf are called
Leaflets.
148. Compound leaves are of two principal kinds, namely, the Pinnate
and the Palmate; answering to the two modes of veining in reticulated
leaves, and to the two sorts of lobed or divided leaves (141).
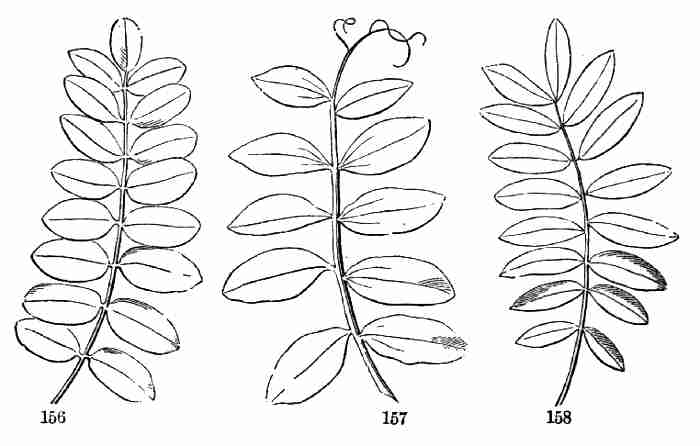
Fig. 156-158. Pinnate leaves, the first with an odd
leaflet (odd-pinnate); the second with a tendril in place of uppermost
leaflets; the third abruptly pinnate, or of even pairs.
149. Pinnate leaves are those in which the leaflets are arranged on
the sides of a main leaf-stalk; as in Fig. 156-158. They answer to the
[Pg 58]feather-veined (i. e. pinnately-veined) simple leaf; as will be
seen at once on comparing the forms. The leaflets of the former answer
to the lobes or divisions of the latter; and the continuation of the
petiole, along which the leaflets are arranged, answers to the midrib of
the simple leaf.
150. Three sorts of pinnate leaves are here given. Fig. 156 is pinnate
with an odd or end leaflet, as in the Common Locust and the Ash. Fig.
157 is pinnate with a tendril at the end, in place of the odd leaflet,
as in the Vetches and the Pea. Fig. 158 is evenly or abruptly pinnate,
as in the Honey-Locust.
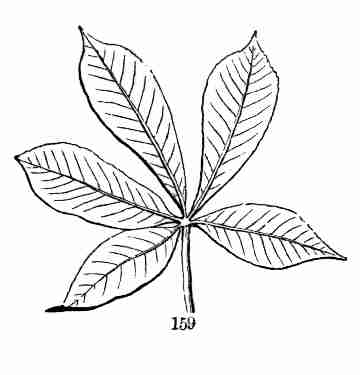
Fig. 159. Palmate (or digitate) leaf of five leaflets, of
the Sweet Buckeye.
151. Palmate (also named Digitate) leaves are those in which the
leaflets are all borne on the tip of the leaf-stalk, as in the Lupine,
the Common Clover, the Virginia Creeper (Fig. 93), and the
Horse-chestnut and Buckeye (Fig. 159). They evidently answer to the
radiate-veined or palmately-veined simple leaf. That is, the
Clover-leaf of three leaflets is the same as a palmately three-ribbed
leaf cut into three separate leaflets. And such a simple five-lobed leaf
as that of the Sugar Maple, if more cut, so as to separate the parts,
would produce a palmate leaf of five leaflets, like that of the
Horse-chestnut or Buckeye.
152. Either sort of compound leaf may have any number of leaflets; yet
palmate leaves cannot well have a great many, since they are all crowded
together on the end of the main leaf-stalk. Some Lupines have nine or
eleven; the Horse-chestnut has seven, the Sweet Buckeye more commonly
five, the Clover three. A pinnate leaf often has only seven or five
leaflets, or only three, as in Beans of the genus Phaseolus, etc.; in
some rarer cases only two; in the Orange and Lemon and also in the
common Barberry there is only one! The joint at the place where the
leaflet is united with the petiole distinguishes this last case from a
simple leaf. In other species of these genera the lateral leaflets also
are present.
153. The leaflets of a compound leaf may be either entire (as in Fig.
126-128), or serrate, or lobed, cleft, parted, etc.; in fact, may
present all the variations of simple leaves, and the same terms equally
apply to them.
154. When the division is carried so far as to separate what would be
one leaflet into two, three, or several, the leaf becomes doubly or
twice compound, either pinnately or palmately, as the case may be.
For example, while the clustered leaves of the Honey-Locust are simply
pinnate, that is, once pinnate, those on new shoots are bipinnate,
or twice pinnate, as in Fig. 160. When these leaflets are again
divided in the same way, the leaf [Pg 59]becomes thrice pinnate, or
tripinnate, as in many Acacias. The first divisions are called
Pinnæ; the others, Pinnules; and the last, or little blades
themselves, Leaflets.
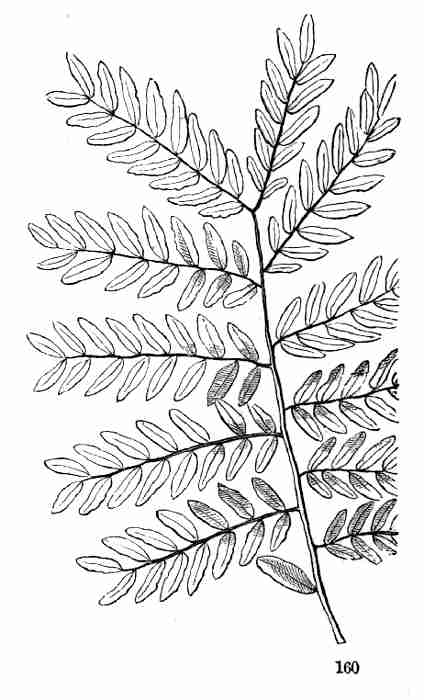 Fig.
Fig. 160. A twice-pinnate (abruptly) leaf of the
Honey-Locust.
155. So the palmate leaf, if again compounded in the same way, becomes
twice palmate, or, as we say when the divisions are in threes, twice
ternate (in Latin form biternate); if a third time compounded,
thrice ternate or triternate. But if the division goes still
further, or if the degree is variable, we simply say that the leaf is
decompound; either palmately or pinnately decompound, as the case may
be. Thus, Fig. 161 represents a four times ternately compound (in other
words a ternately decompound) leaf of a common Meadow Rue.
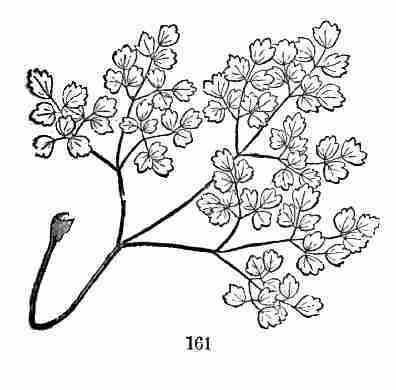 Fig.
Fig. 161. Ternately decompound leaf of Meadow Rue.
156. When the botanist, in describing leaves, wishes to express the
number of the leaflets, he may use terms like these:—
Unifoliolate, for a compound leaf of a single leaflet; from the Latin
unum, one, and foliolum, leaflet.
Bifoliolate, of two leaflets, from the Latin bis, twice, and
foliolum, leaflet.
Trifoliolate (or ternate), of three leaflets, as the Clover; and so
on.
Palmately bifoliolate, trifoliolate, quadrifoliolate,
plurifoliolate (of several leaflets), etc.: or else
Pinnately bi-, tri-, quadri-, or plurifoliolate (that is, of
two, three, four, five, or several leaflets), as the case may be: these
are terse ways of denoting in single phrases both the number of leaflets
and the kind of compounding.
157. Of foliage-leaves having certain peculiarities in structure, the
following may be noted:—
[Pg 60]
158. Perfoliate Leaves. In these the stem that bears them seems to run
through the blade of the leaf, more or less above its base. A common
Bellwort (Uvularia perfoliata, Fig. 162) is a familiar illustration. The
lower and earlier leaves show it distinctly. Later, the plant is apt to
produce some leaves merely clasping the stem by the sessile and
heart-shaped base, and the latest may be merely sessile. So the series
explains the peculiarity: in the formation of the leaf the bases,
meeting around the stem, grow together there.
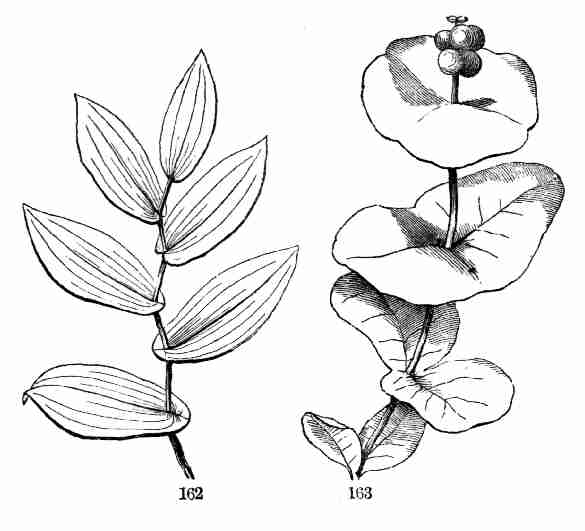
Fig. 162. A summer branch of Uvularia perfoliata; lower
leaves perfoliate, upper cordate-clasping, uppermost simply sessile.
Fig. 163. Branch of a Honeysuckle, with
connate-perfoliate leaves.
159. Connate-perfoliate. Such are the upper leaves of true Honeysuckles.
Here (Fig. 163) of the opposite and sessile leaves, some pairs,
especially the uppermost, in the course of their formation unite around
the stem, which thus seems to run through the disk formed by their
union.
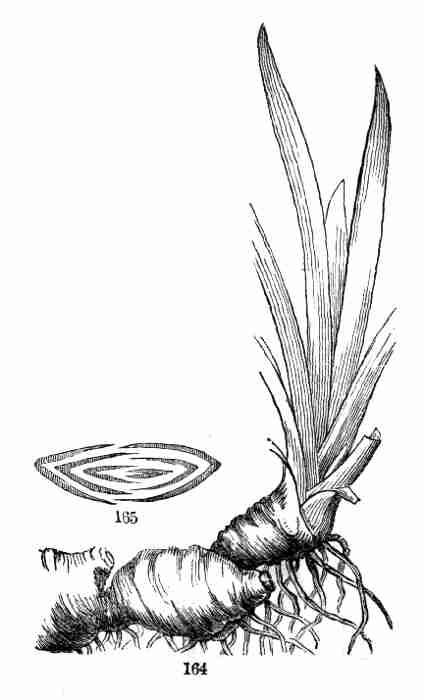
Fig. 164. Rootstock and equitant leaves of Iris. 165. A
section across the cluster of leaves at the bottom, showing the
equitation.
160. Equitant Leaves. While ordinary leaves spread horizontally, and
present one face to the sky and the other to the earth, there are some
that present their tip to the sky, and their faces right and left to the
horizon. Among these are the equitant leaves of the Iris or
Flower-de-Luce. Inspection shows that each leaf was formed as if folded
together lengthwise, [Pg 61]so that what would be the upper surface is
within, and all grown together, except next the bottom, where each leaf
covers the next younger one. It was from their straddling over each
other, like a man on horseback (as is seen in the cross-section, Fig.
165), that Linnæus, with his lively fancy, called these Equitant
leaves.
161. Leaves with no distinction of Petiole and Blade. The leaves of Iris
just mentioned show one form of this. The flat but narrow leaves of
Jonquils, Daffodils, and the cylindrical leaf of Onions are other
instances. Needle-shaped leaves, like those of the Pine, Larch, and
Spruce, and the awl-shaped as well as the scale-shaped leaves of
Junipers, Red Cedar, and Arbor-Vitæ (Fig. 166), are examples.
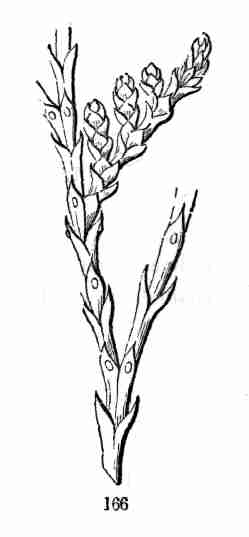
Fig. 166. Branch of Arbor-Vitæ, with awl-shaped and
scale-shaped leaves.
162. Phyllodia. Sometimes an expanded petiole takes the place of the
blade; as in numerous New Holland Acacias, some of which are now common
in greenhouses. Such counterfeit blades are called phyllodia,—meaning
leaf-like bodies. They may be known from true blades by their standing
edgewise, their margins being directed upwards and downwards; while in
true blades the faces look upwards and downwards; excepting in equitant
leaves, as already explained.
163. Falsely Vertical Leaves. These are apparent exceptions to the rule,
the blade standing edgewise instead of flatwise to the stem; but this
position comes by a twist of the stalk or the base of the blade. Such
leaves present the two faces about equally to the light. The
Compass-plant (Silphium laciniatum) is an example. So also the leaves of
Boltonia, of Wild Lettuce, and of a vast number of Australian Myrtaceous
shrubs and trees, which much resemble the phyllodia of the Acacias of
the same country. They are familiar in Callistemon, the Bottle-brush
Flower, and in Eucalyptus. But in the latter the leaves of the young
tree have the normal structure and position.
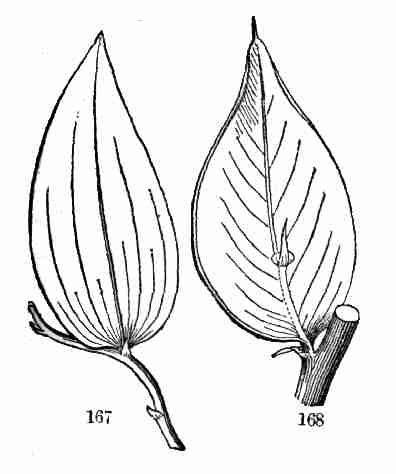
Fig. 167. The ambiguous leaf? (cladophyllum) of
Myrsiphyllum.
Fig. 168. Same of Ruscus, or Butcher's Broom.
164. Cladophylla, meaning branch-leaves. The foliage of Ruscus (the
Butcher's Broom of Europe) and of Myrsiphyllum of South Africa
(cultivated for decoration under the false [Pg 62]name of Smilax) is peculiar
and puzzling. If these blades (Fig. 167, 168) are really leaves, they
are most anomalous in occupying the axil of another leaf, reduced to a
little scale. Yet they have an upper and lower face, as leaves should,
although they soon twist, so as to stand more or less edgewise. If they
are branches which have assumed exactly the form and office of leaves,
they are equally extraordinary in not making any further development.
But in Ruscus, flowers are borne on one face, in the axil of a little
scale: and this would seem to settle that they are branches. In
Asparagus just the same things as to position are thread-shaped and
branch-like.
§ 2. LEAVES OF SPECIAL CONFORMATION AND USE.
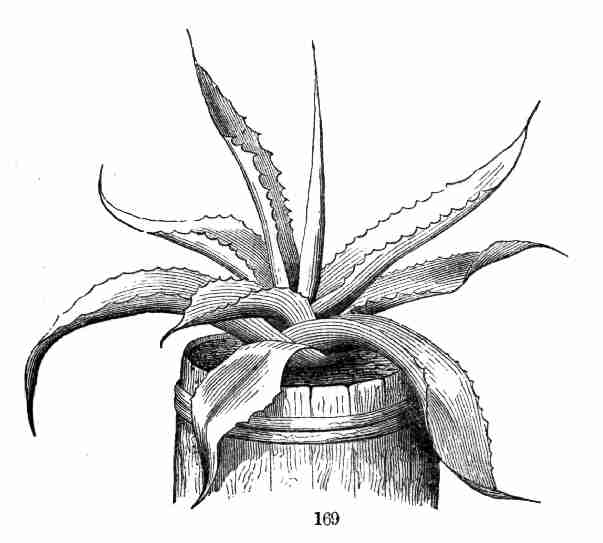
Fig. 169. A young Agave Americana, or Century-plant;
fleshy-leaved.
165. Leaves for Storage. A leaf may at the same time serve both ordinary
and special uses. Thus in those leaves of Lilies, such as the common
White Lily, which spring from the bulb, the upper and green part serves
for foliage and elaborates nourishment, while the thickened portion or
bud-scale beneath serves for the storage of this nourishment. The
thread-shaped leaf of the Onion fulfils the same office, and the
nourishing matter it prepares is deposited in its sheathing base,
forming one of the concentric layers of the onion. When these layers, so
thick and succulent, have given up their store to the growing parts
within, they are left as thin and dry husks. In a Houseleek, an Aloe or
an Agave, the green color of the surface of the fleshy leaf indicates
that it is doing the work of foliage; the deeper-seated white portion
within is the storehouse of the nourishment which the green surface has
elaborated. So, also, the seed-leaves or cotyledons are commonly used
for storage. Some, as in one of the Maples, the Pea, Horse-chestnut,
Oak, etc., are for nothing else. Others, as in Beech and in our common
[Pg 63]Beans, give faint indications of service as foliage also, chiefly in
vain. Still others, as in the Pumpkin and Flax, having served for
storage, develop into the first efficient foliage. Compare 11, 22-30,
and the accompanying figures.
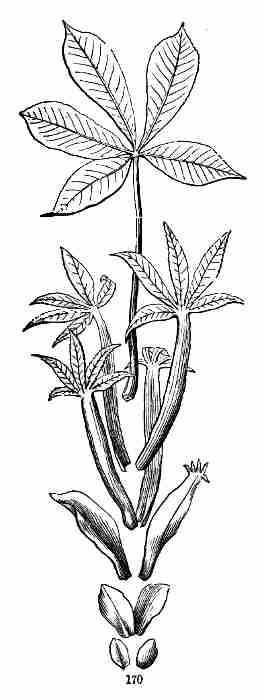
Fig. 170. Series of bud-scales and foliage-leaves from a
developing bud of the Low Sweet Buckeye (Æsculus parviflora), showing
nearly complete gradation, from a scale to a compound leaf of five
leaflets; and that the scales answer to reduced petioles.
166. Leaves as Bud-Scales serve to protect the forming parts within.
Having fulfilled this purpose they commonly fall off when the shoot
develops and foliage-leaves appear. Occasionally, as in Fig. 170, there
is a transition of bud-scales to leaves, which reveals the nature of the
former. The Lilac also shows a gradation from bud-scale to simple leaf.
In Cornus florida (the Flowering Dogwood), the four bud-scales which
through the winter protect the head of forming flowers remain until
blossoming, and then the base of each grows out into [Pg 64]a large and very
showy petal-like leaf; the original dry scale is apparent in the notch
at the apex.
Fig. 171. Shoot of common Barberry, showing transition of
foliage-leaves to spines.
167. Leaves as Spines occur in several plants. A familiar instance is
that of the common Barberry (Fig. 171). In almost any summer shoot, most
of the gradations may be seen between the ordinary leaves, with sharp
bristly teeth, and leaves which are reduced to a branching spine or
thorn. The fact that the spines of the Barberry produce a leaf-bud in
their axil also proves them to be leaves.
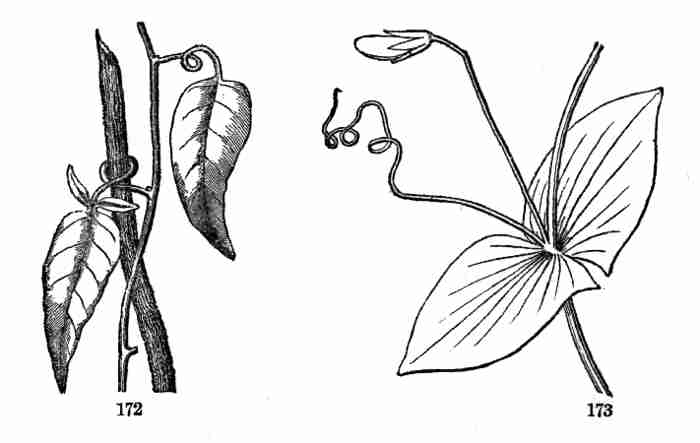
Fig. 172. Leaves of Solanum jasminoides, the petiole
adapted for climbing.
Fig. 173. Leaf of Lathyrus Aphaca, consisting of a pair
of stipules and a tendril.
168. Leaves for Climbing are various in adaptation. True foliage-leaves
serve this purpose; as in Gloriosa, where the attenuated tip of a simple
leaf (otherwise like that of a Lily) hooks around a supporting object;
or in Solanum jasminoides of the gardens (Fig. 172), and in Maurandia,
etc., where the leaf-stalk coils round and clings to a support; or in
the compound leaves of Clematis and of Adlumia, in which both the
leaflets and their stalks hook or coil around the support.
169. Or in a compound leaf, as in the Pea and most Vetches, and in
Cobæa, while the lower leaflets serve for foliage, some of the uppermost
are developed as tendrils for climbing (Fig. 167). In the common Pea
this is so with all but one or two pairs of leaflets.
170. In one European Vetch, the leaflets are wanting and the whole
petiole is a tendril, while the stipules become the only foliage (Fig.
173).
171. Leaves as Pitchers, or hollow tubes, are familiar in the common
Pitcher-plant or Side-saddle Flower (Sarracenia, Fig. 174) of our bogs.
These pitchers are generally half full of water, in which flies and
other insects are drowned, often in such numbers as to make a rich
manure for the plant. More curious are some of the southern species of
Sarracenia, which seem to be specially adapted to the capture and
destruction of flies and other insects.
Fig. 174. Leaf of Sarracenia purpurea, entire, and
another with the upper part cut off.
Fig. 175. Leaf of Nepenthes; foliage, tendril, and
pitcher combined.
[Pg 65]
172. The leaf of Nepenthes (Fig. 175) combines three structures and
uses. The expanded part below is foliage: this tapers into a tendril for
climbing; and this bears a pitcher with a lid. Insects are caught, and
perhaps digested, in the pitcher.
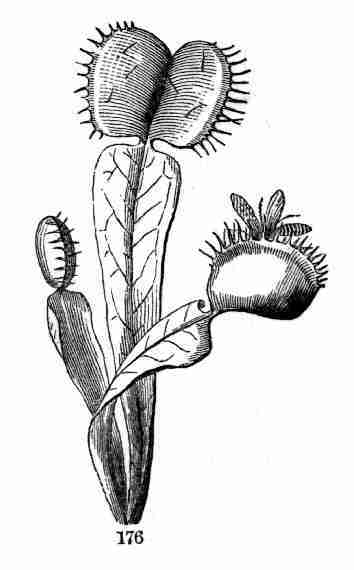
Fig. 176. Leaves of Dionæa; the trap in one of them open,
in the others closed.
173. Leaves as Fly-traps. Insects are caught in another way, and more
expertly, by the most extraordinary of all the plants of this country,
the Dionæa or Venus's Fly-trap, which grows in the sandy bogs around
Wilmington, North Carolina. Here (Fig. 176) each leaf bears at its
summit an appendage which opens and shuts, in shape something like a
steel-trap, and operating much like one. For when open, no sooner does a
fly alight on its surface, and brush against any one of the two or three
bristles that grow there, than the trap suddenly closes, capturing the
intruder. If the fly escapes, the trap soon slowly opens, and is ready
for another capture. When retained, the insect is after a time moistened
by a secretion from minute glands of the inner surface, and is digested.
In the various species of Drosera or Sundew, insects are caught [Pg 66]by
sticking fast to very viscid glands at the tip of strong bristles, aided
by adjacent gland-tipped bristles which bend slowly toward the captive.
The use of such adaptations and operations may be explained in another
place.
§ 3. STIPULES.
174. A leaf complete in its parts consists of blade, leaf-stalk or
petiole, and a pair of stipules. But most leaves have either fugacious
or minute stipules or none at all; many have no petiole (the blade being
sessile or stalkless); some have no clear distinction of blade and
petiole; and many of these, such as those of the Onion and all phyllodia
(166), consist of petiole only.
175. The base of the petiole is apt to be broadened and flattened,
sometimes into thin margins, sometimes into a sheath which embraces the
stem at the point of attachment.
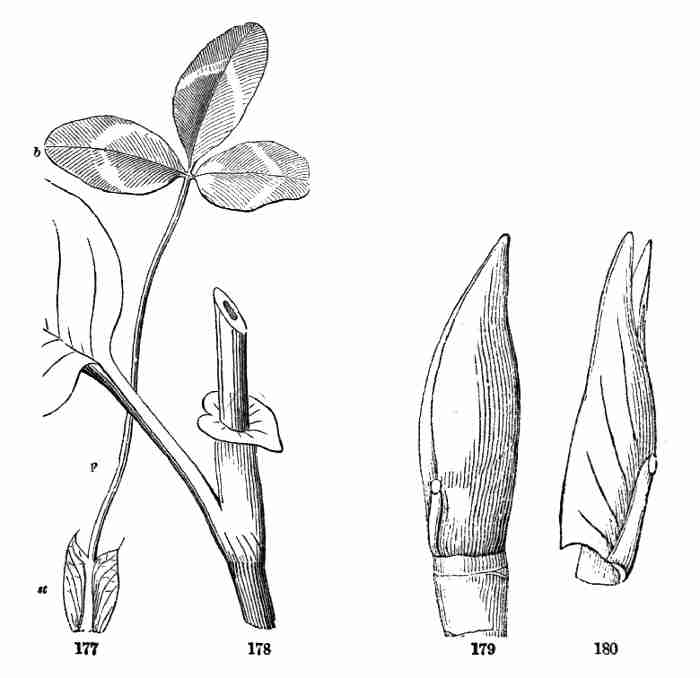
Fig. 177. Leaf of Red Clover: st, stipules, adhering to
the base of p, the petiole; b, blade of three leaflets.
Fig. 178. Part of stem and leaf of Prince's-Feather
(Polygonum orientale) with the united sheathing stipules forming a
sheath or ocrea.
Fig. 179. Terminal winter bud of Magnolia Umbrella,
natural size. 180. Outermost bud-scale (pair of stipules) detached.
176. Stipules are such appendages, either wholly or partly separated
from the petiole. When quite separate they are said to be free, as in
Fig. 112. When attached to the base of the petiole, as in the Rose and
in [Pg 67]Clover (Fig. 177), they are adnate. When the two stipules unite
and sheathe the stem above the insertion, as in Polygonum (Fig. 178),
this sheath is called an Ocrea from its likeness to a greave or
leggin.
177. In Grasses, when the sheathing base of the leaf may answer to
petiole, the summit of the sheath commonly projects as a thin and short
membrane, like an ocrea: this is called a Ligula or Ligule.
178. When stipules are green and leaf-like they act as so much foliage.
In the Pea they make up no small part of the actual foliage. In a
related plant (Lathyrus Aphaca, Fig. 173), they make the whole of it,
the remainder of the leaf being tendril.
179. In many trees the stipules are the bud-scales, as in the Beech, and
very conspicuously in the Fig-tree, Tulip-tree, and Magnolia (Fig. 179).
These fall off as the leaves unfold.
180. The stipules are spines or prickles in Locust and several other
Leguminous trees and shrubs; they are tendrils in Smilax or Greenbrier.
§ 4. THE ARRANGEMENT OF LEAVES.
181. Phyllotaxy, meaning leaf-arrangement, is the study of the position
of leaves, or parts answering to leaves, upon the stem.
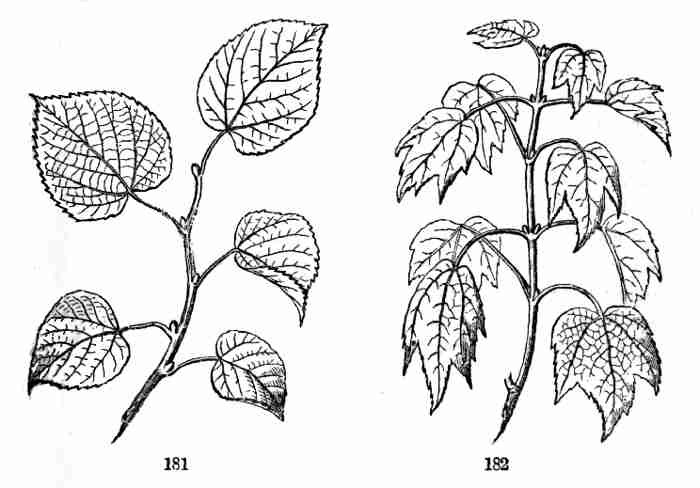
Fig. 181. Alternate leaves, in Linden, Lime-tree, or
Basswood.
Fig. 182. Opposite leaves, in Red Maple.
182. The technical name for the attachment of leaves to the stem is the
insertion. Leaves (as already noticed, 54) are inserted in three
modes. They are
Alternate (Fig. 181), that is, one after another, or in other words,
with only a single leaf to each node;
[Pg 68]
Opposite (Fig. 182), when there is a pair to each node, the two
leaves in this case being always on opposite sides of the stem;
Whorled or Verticillate (Fig. 183) when there are more than two
leaves on a node, in which case they divide the circle equally between
them, forming a Verticel or whorl. When there are three leaves in the
whorl, the leaves are one third of the circumference apart; when four,
one quarter, and so on. So the plan of opposite leaves, which is very
common, is merely that of whorled leaves, with the fewest leaves to the
whorl, namely, two.
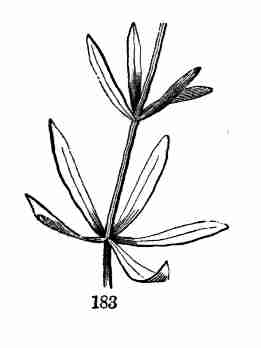 Fig.
Fig. 183. Whorled leaves of Galium.
183. In both modes and in all their modifications, the arrangement is
such as to distribute the leaves systematically and in a way to give
them a good exposure to the light.
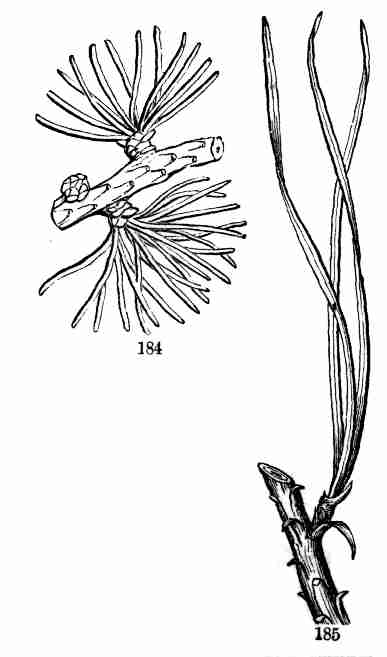
Fig. 184. A piece of stem of Larch with two clusters
(fascicles) of numerous leaves.
Fig. 185. Piece of a branch of Pitch Pine, with three
leaves in a fascicle or bundle, in the axil of a thin scale which
answers to a primary leaf. The bundle is surrounded at the base by a
short sheath, formed of the delicate scales of the axillary bud.
184. No two or more leaves ever grow from the same point. The so-called
Fascicled or Clustered leaves are the leaves of a branch the nodes
of which are very close, just as they are in the bud, so keeping the
leaves in a cluster. This is evident in the Larch (Fig. 184), in which
examination shows each cluster to be made up of numerous leaves crowded
on a spur or short axis. In spring there are only such clusters; but in
summer some of them lengthen into ordinary shoots with scattered
alternate leaves. So, likewise, each cluster of two or three
needle-shaped leaves in Pitch Pines (as in Fig. 185), or of five leaves
in White Pine, answers to a similar extremely short branch, springing
from the axil of a thin and slender scale, which represents a leaf of
the main shoot. For Pines produce two kinds of leaves,—1. primary, the
proper leaves of the shoots, not as foliage, but in the shape of
delicate scales in spring, which soon fall away; and 2. secondary, the
fascicled leaves, from buds in the axils of the former, and these form
the actual foliage.
[Pg 69]
185. Phyllotaxy of Alternate Leaves. Alternate leaves are distributed
along the stem in an order which is uniform for each species. The
arrangement in all its modifications is said to be spiral, because, if
we draw a line from the insertion (i. e. the point of attachment) of
one leaf to that of the next, and so on, this line will wind spirally
around the stem as it rises, and in the same species will always bear
the same number of leaves for each turn round the stem. That is, any two
successive leaves will always be separated from each other by an equal
portion of the circumference of the stem. The distance in height
between any two leaves may vary greatly, even on the same shoot, for
that depends upon the length of the internodes, or spaces between the
leaves; but the distance as measured around the circumference (in other
words, the Angular Divergence, or angle formed by any two successive
leaves) is uniformly the same.
186. Two-ranked. The greatest possible divergence is, of course, where
the second leaf stands on exactly the opposite side of the stem from the
first, the third on the side opposite the second, and therefore over the
first, and the fourth over the second. This brings all the leaves into
two ranks, one on one side of the stem and one on the other, and is
therefore called the Two-ranked arrangement. It occurs in all
Grasses,—in Indian Corn, for instance; also, in the Basswood (Fig.
181). This is the simplest of all arrangements, and the one which most
widely distributes successive leaves, but which therefore gives the
fewest vertical ranks. Next is the
187. Three-ranked arrangement,—that of all Sedges, and of White
Hellebore. Here the second leaf is placed one third of the way round the
stem, the third leaf two thirds of the way round, the fourth leaf
accordingly directly over the first, the fifth over the second, and so
on. That is, three leaves occur in each turn round the stem, and they
are separated from each other by one third of the circumference. (Fig.
186, 187.)
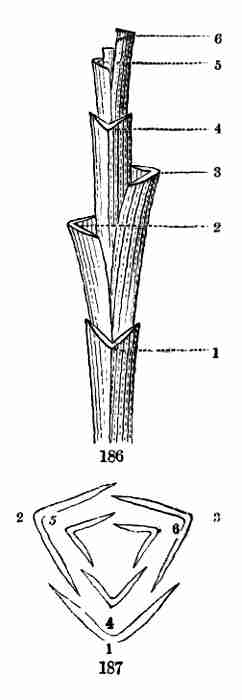
Fig. 186. Two-ranked arrangement, shown in a piece of the
stalk of a Sedge, with the leaves cut off above their bases; the leaves
are numbered in order, from 1 to 6. 187. Diagram or cross-section of the
same, in one plane; the leaves similarly numbered; showing two cycles of
three.
188. Five-ranked is the next in the series, and the most common. It is
seen in the Apple (Fig. 188), Cherry, Poplar, and the greater number of
trees and shrubs. In this case the line traced from leaf to leaf will
pass twice round the stem before it reaches a leaf [Pg 70]situated directly
over any below (Fig. 189). Here the sixth leaf is over the first; the
leaves stand in five perpendicular ranks, with equal angular distance
from each other; and this distance between any two successive leaves is
just two fifths of the circumference of the stem.

Fig. 188. Shoot with its leaves 5-ranked, the sixth leaf
over the first; as in the Apple-tree.
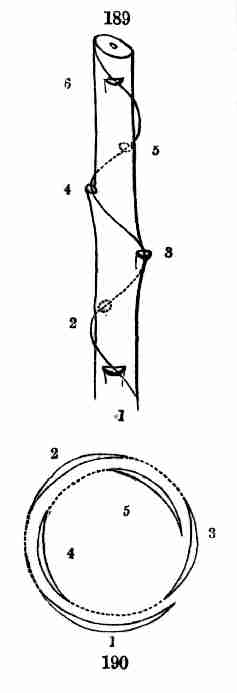
Fig. 189. Diagram of this arrangement, with a spiral line
drawn from the attachment of one leaf to the next, and so on; the parts
on the side turned from the eye are fainter.
Fig. 190. A ground-plan of the same; the section of the
leaves similarly numbered; a dotted line drawn from the edge of one leaf
to that of the next marks out the spiral.
189. The five-ranked arrangement is expressed by the fraction 2/5. This
fraction denotes the divergence of the successive leaves, i. e. the
angle they form with each other: the numerator also expresses the number
of turns made round the stem by the spiral line in completing one cycle
or set of leaves, namely, two; and the denominator gives the number of
leaves in each cycle, or the number of perpendicular ranks, namely,
five. In the same way the fraction 1/2 stands for the two-ranked mode,
and 1/3 for the three-ranked: and so these different sorts are expressed
by the series of fractions 1/2, 1/3, 2/5. Other cases follow in the same
numerical progression, the next being the
190. Eight-ranked arrangement. In this the ninth leaf stands over the
first, and three turns are made around the stem to reach it; so it is
expressed by the fraction 3/8. This is seen in the Holly, and in the
common Plantain. Then comes the
191. Thirteen-ranked arrangement, in which the fourteenth leaf is over
the first, after five turns around the stem. The common Houseleek (Fig.
191) is a good example.
192. The series so far, then, is 1/2, 1/3, 2/5, 3/8, 5/13; the numerator
and the denominator of each fraction being those of the two next
preceding ones added together. At this rate the next higher should be
8/21, then 13/34, and so on; and in fact just such cases are met with,
and (commonly) no others. These higher sorts are found in the Pine
Family, both in the leaves and the cones and in many other plants with
small and crowded leaves. But in those the number of the ranks, or of
leaves in each cycle, can only rarely [Pg 71]be made out by direct inspection.
They may be indirectly ascertained, however, by studying the secondary
spirals, as they are called, which usually become conspicuous, at least
two series of them, one turning to the right and one to the left, as
shown in Fig. 191. For an account of the way in which the character of
the phyllotaxy may be deduced from the secondary spirals, see Structural
Botany, Chapter IV.
Fig. 191. A young plant of the Houseleek, with the leaves
(not yet expanded) numbered, and exhibiting the 13-ranked arrangement;
and showing secondary spirals.
Fig. 192. Opposite leaves of Euonymus, or Spindle-tree,
showing the successive pairs crossing each other at right angles.
193. Phyllotaxy of Opposite and whorled Leaves. This is simple and
comparatively uniform. The leaves of each pair or whorl are placed over
the intervals between those of the preceding, and therefore under the
intervals of the pair or whorl next above. The whorls or pairs alternate
or cross each other, usually at right angles, that is, they decussate.
Opposite leaves, that is, whorls of two leaves only, are far commoner
than whorls of three or four or more members. This arrangement in
successive decussating pairs gives an advantageous distribution on the
stem in four vertical ranks. Whorls of three give six vertical ranks,
and so on. Note that in descriptive botany leaves in whorls of two are
simply called opposite leaves; and that the term verticillate or
whorled, is employed only for cases of more than two, unless the
latter number is specified.
194. Vernation or Præfoliation, the disposition of the leaf-blades in
the bud, comprises two things; 1st, the way in which each separate leaf
is folded, coiled, or packed up in the bud; and 2d, the arrangement of
the leaves in the bud with respect to one another. The latter of course
depends very much upon the phyllotaxy, i. e. the position and order of
the leaves upon the stem. The same terms are used for it as for the
arrangement of the leaves of the flower in the flower-bud. See,
therefore, "Æstivation, or Præfloration."
195. As to each leaf separately, it is sometimes straight and open in
vernation, but more commonly it is either bent, folded, or rolled
up. When the upper part is bent down upon the lower, as the young blade
in the Tulip-tree is bent upon the leaf-stalk, it is said to be
Inflexed or Reclined in vernation. When folded by the midrib so that
the two halves are placed face to face, it is Conduplicate (Fig. 193),
as in the Magnolia, the Cherry, and the Oak. When folded back and forth
like the plaits of a fan, it is [Pg 72]Plicate or Plaited (Fig. 194), as
in the Maple and Currant. If rolled, it may be so either from the tip
downwards, as in Ferns and the Sundew (Fig. 197), when in unrolling it
resembles the head of a crosier, and is said to be Circinate; or it
may be rolled up parallel with the axis, either from one edge into a
coil, when it is Convolute (Fig. 195), as in the Apricot and Plum; or
rolled from both edges towards the midrib,—sometimes inwards, when it
is Involute (Fig. 198), as in the Violet and Water-Lily; sometimes
outwards, when it is Revolute (Fig. 196), in the Rosemary and Azalea.
The figures are diagrams, representing sections through the leaf, in the
way they were represented by Linnæus.

Section VIII. FLOWERS.
196. Flowers are for the production of seed (16). Stems and branches,
which for a time put forth leaves for vegetation, may at length put
forth flowers for reproduction.
§ 1. POSITION AND ARRANGEMENT OF FLOWERS, OR INFLORESCENCE.
197. Flower-buds appear just where leaf-buds appear; that is, they are
either terminal or axillary (47-49). Morphologically, flowers answer
to shoots or branches, and their parts to leaves.
198. In the same species the flowers are usually from axillary buds
only, or from terminal buds only; but in some they are both axillary and
terminal.
199. Inflorescence, which is the name used by Linnæus to signify mode of
flower-arrangement, is accordingly of three classes: namely,
Indeterminate, when the flowers are in the axils of leaves, that is,
are from axillary buds; Determinate, when they are from terminal buds,
and so terminate a stem or branch; and Mixed, when these two are
combined.
200. Indeterminate Inflorescence (likewise, and for the same reason,
called indefinite inflorescence) is so named because, as the flowers
all come from axillary buds, the terminal bud may keep on growing and
prolong the stem indefinitely. This is so in Moneywort (Fig. 199).
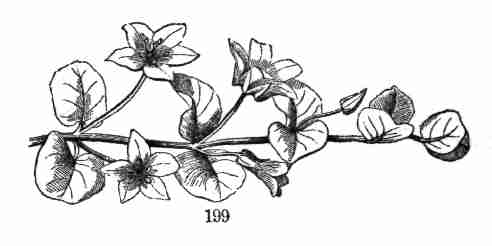
Fig. 199. Piece of a flowering-stem of Moneywort
(Lysimachia nummularia,) with single flowers successively produced in
the axils of the leaves, from below upwards, as the stem grows on.
[Pg 73]
201. When flowers thus arise singly from the axils of ordinary leaves,
they are axillary and solitary, not collected into flower-clusters.
202. But when several or many flowers are produced near each other, the
accompanying leaves are apt to be of smaller size, or of different shape
or character: then they are called Bracts, and the flowers thus brought
together form a cluster. The kinds of flower-clusters of the
indeterminate class have received distinct names, according to their
form and disposition. They are principally Raceme, Corymb, Umbel,
Spike, Head, Spadix, Catkin, and Panicle.
203. In defining these it will be necessary to use some of the following
terms of descriptive botany which relate to inflorescence. If a flower
is stalkless, i. e. sits directly in the axil or other support, it is
said to be sessile. If raised on a naked stalk of its own (as in Fig.
199) it is pedunculate, and the stalk is a Peduncle.
204. A peduncle on which a flower-cluster is raised is a Common
peduncle. That which supports each separate flower of the cluster is a
Partial peduncle, and is generally called a Pedicel. The portion of
the general stalk along which flowers are disposed is called the Axis
of inflorescence, or, when covered with sessile flowers, the Rhachis
(back-bone), and sometimes the Receptacle. The leaves of a
flower-cluster generally are termed Bracts. But when bracts of different
orders are to be distinguished, those on the common peduncle or axis,
and which have a flower in their axil, keep the name of bracts; and
those on the pedicels or partial flower-stalks, if any, that of
Bractlets or Bracteoles. The former is the preferable English name.

Fig. 200. A raceme, with a general peduncle (p),
pedicels (p'), bracts (b), and bractlets (b'). Plainly the bracts
here answer to the leaves in Fig. 199.
205. A Raceme (Fig. 200) is that form of flower-cluster in which the
flowers, each on their own foot-stalk or pedicel, are arranged along the
sides of a common stalk or axis of inflorescence; as in the Lily of the
Valley, Currant, Barberry, one section of Cherry, etc. Each flower comes
from the axil of a small leaf, or bract, which, however, is often so
small that it might escape notice, and even sometimes (as in the Mustard
Family) disappears altogether. The lowest blossoms of a [Pg 74]raceme are of
course the oldest, and therefore open first, and the order of blossoming
is ascending from the bottom to the top. The summit, never being
stopped by a terminal flower, may go on to grow, and often does so (as
in the common Shepherd's Purse), producing lateral flowers one after
another for many weeks.
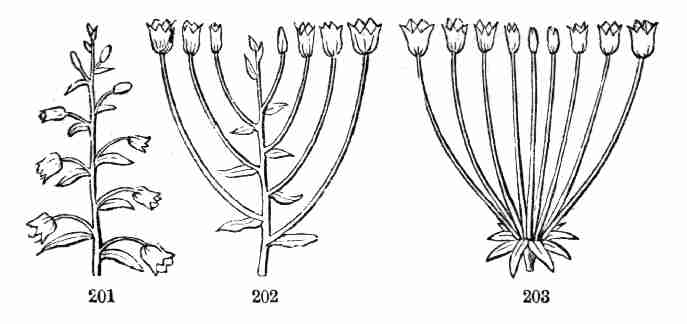 Fig.
Fig. 201. A raceme. 202. A corymb. 203. An umbel.
206. A Corymb (Fig. 202) is the same as a raceme, except that it is flat
and broad, either convex, or level-topped. That is, a raceme becomes a
corymb by lengthening the lower pedicels while the uppermost remain
shorter. The axis of a corymb is short in proportion to the lower
pedicels. By extreme shortening of the axis the corymb may be converted
into
207. An Umbel (Fig. 203) as in the Milkweed, a sort of flower-cluster
where the pedicels all spring apparently from the same point, from the
top of the peduncle, so as to resemble, when spreading, the rays of an
umbrella; whence the name. Here the pedicels are sometimes called the
Rays of the umbel. And the bracts, when brought in this way into a
cluster or circle, form what is called an Involucre.
208. The corymb and the umbel being more or less level-topped, bringing
the flowers into a horizontal plane or a convex form, the ascending
order of development appears as Centripetal. That is, the flowering
proceeds from the margin or circumference regularly towards the centre;
the lower flowers of the former answering to the outer ones of the
latter.
209. In these three kinds of flower-clusters, the flowers are raised on
conspicuous pedicels (204) or stalks of their own. The shortening of
these pedicels, so as to render the flowers sessile or nearly so,
converts a raceme into a Spike, and a corymb or an umbel into a
Head.
210. A Spike is a flower cluster with a more or less lengthened axis,
along which the flowers are sessile or nearly so; as in the Plantain
(Fig. 204).
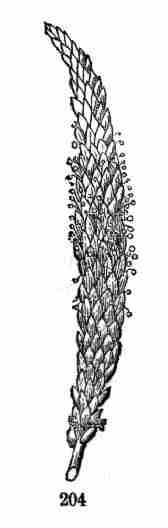 Fig.
Fig. 204. Spike of the common Plantain or Ribwort.
211. A Head (Capitulum) is a round or roundish cluster of flowers,
[Pg 75]which are sessile on a very short axis or receptacle, as in the
Button-ball, Button-bush (Fig. 205), and Red Clover. It is just what a
spike would become if its axis were shortened; or an umbel, if its
pedicels were all shortened until the flowers became sessile. The head
of the Button-bush is naked; but that of the Thistle, of the Dandelion,
and the like, is surrounded by empty bracts, which form an Involucre.
Two particular forms of the spike and the head have received particular
names, namely, the Spadix and the Catkin.
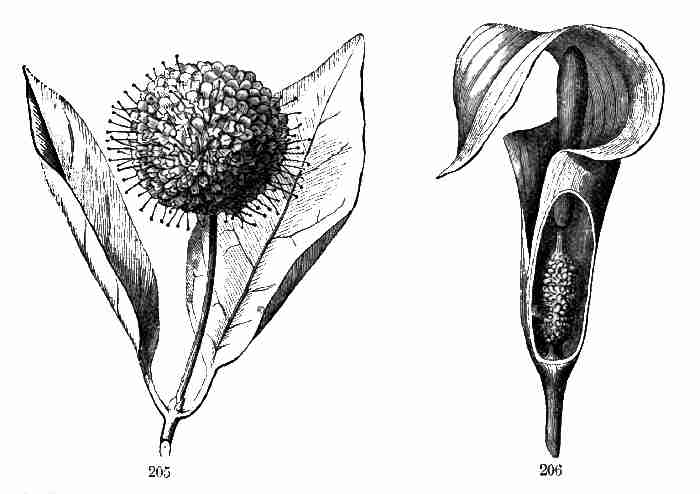
Fig. 205. Head of the Button-bush (Cephalanthus).
Fig. 206. Spadix and spathe of the Indian Turnip; the
latter cut through below.
212. A Spadix is a fleshy spike or head, with small and often imperfect
flowers, as in the Calla, Indian Turnip, (Fig. 206), Sweet Flag, etc. It
is commonly surrounded or embraced by a peculiar enveloping leaf, called
a Spathe.
213. A Catkin, or Ament, is the name given to the scaly sort of spike of
the Birch (Fig. 207) and Alder, the Willow and Poplar, and one sort of
flower-clusters of the Oak, Hickory, and the like,—the so-called
Amentaceous trees.
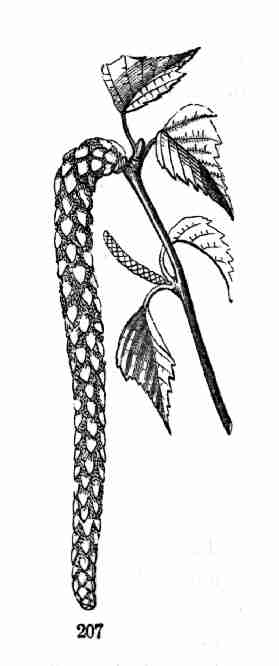 Fig.
Fig. 207. Catkin, or Ament, of Birch.
214. Compound flower-clusters of these kinds are not uncommon. When
the stalks which in the simple umbel are the pedicels of single flowers
themselves branch into an umbel, a Compound Umbel is formed. [Pg 76]This is
the inflorescence of Caraway (Fig. 208), Parsnip, and almost all of the
great family of Umbelliferous (umbel-bearing) plants.
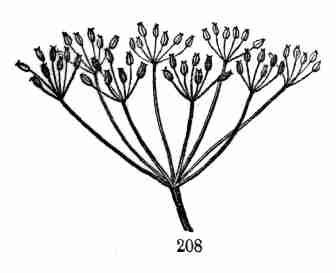 Fig.
Fig. 208. Compound Umbel of Caraway.
215. The secondary or partial umbels of a compound umbel are Umbellets.
When the umbellets are subtended by an involucre, this secondary
involucre is called an Involucel.
216. A Compound raceme is a cluster of racemes racemosely arranged, as
in Smilacina racemosa. A compound corymb is a corymb some branches of
which branch again in the same way, as in Mountain Ash. A compound
spike is a spicately disposed cluster of spikes.
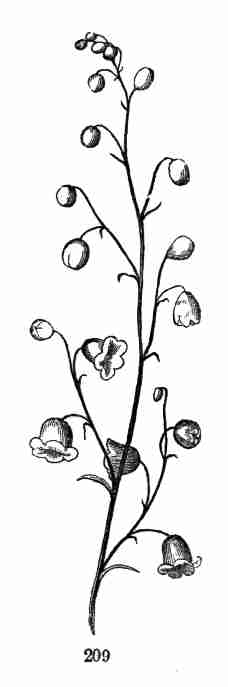 Fig.
Fig. 209. Diagram of a simple panicle.
217. A Panicle, such as that of Oats and many Grasses, is a compound
flower-cluster of a more or less open sort which branches with apparent
irregularity, neither into corymbs nor racemes. Fig. 209 represents the
simplest panicle. It is, as it were, a raceme of which some of the
pedicels have branched so as to bear a few flowers on pedicels of their
own, while others remain simple. A compound panicle is one that
branches in this way again and again.
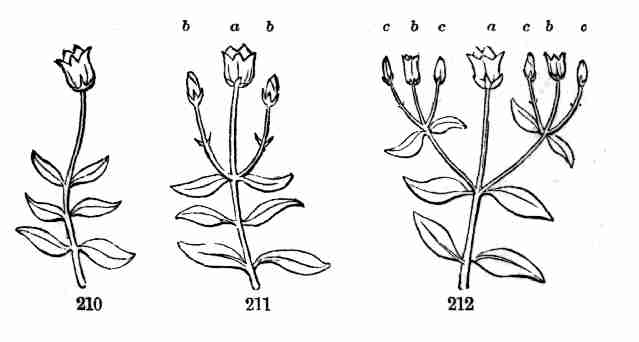
Fig. 210. Diagram of an opposite-leaved plant, with a
single terminal flower. 211. Same, with a cyme of three flowers; a,
the first flower, of the main axis; b b, those of branches. 212. Same,
with flowers also of the third order, c c.
218. Determinate Inflorescence is that in which the flowers are from
terminal buds. The simplest case is that of a solitary terminal flower,
as in Fig. 210. This stops the growth of the stem; for its terminal bud,
becoming a blossom, can no more lengthen in the manner of a leaf-bud.
Any [Pg 77]further growth must be from axillary buds developing into branches.
If such branches are leafy shoots, at length terminated by single
blossoms, the inflorescence still consists of solitary flowers at the
summit of stem and branches. But if the flowering branches bear only
bracts in place of ordinary leaves, the result is the kind of
flower-cluster called
219. A Cyme. This is commonly a flat-topped or convex flower-cluster,
like a corymb, only the blossoms are from terminal buds. Fig. 211
illustrates the simplest cyme in a plant with opposite leaves, namely,
with three flowers. The middle flower, a, terminates the stem; the two
others, b b, terminate branches, one from the axil of each of the
uppermost leaves; and being later than the middle one, the flowering
proceeds from the centre outwards, or is Centrifugal. This is the
opposite of the indeterminate mode, or that where all the flower-buds
are axillary. If flowering branches appear from the axils below, the
lower ones are the later, so that the order of blossoming continues
centrifugal or, which is the same thing, descending, as in Fig. 213,
making a sort of reversed raceme or false raceme,—a kind of cluster
which is to the true raceme just what the flat cyme is to the corymb.

Fig. 213. Diagram of a simple cyme in which the axis
lengthens, so as to take the form of a raceme.
220. Wherever there are bracts or leaves, buds may be produced from
their axils and appear as flowers. Fig. 212 represents the case where
the branches, b b, of Fig. 211, each with a pair of small leaves or
bracts about their middle, have branched again, and produced the
branchlets and flowers c c, on each side. It is the continued
repetition of this which forms the full or compound cyme, such as that
of the Laurestinus, Hobble-bush, Dogwood, and Hydrangea (Fig. 214).
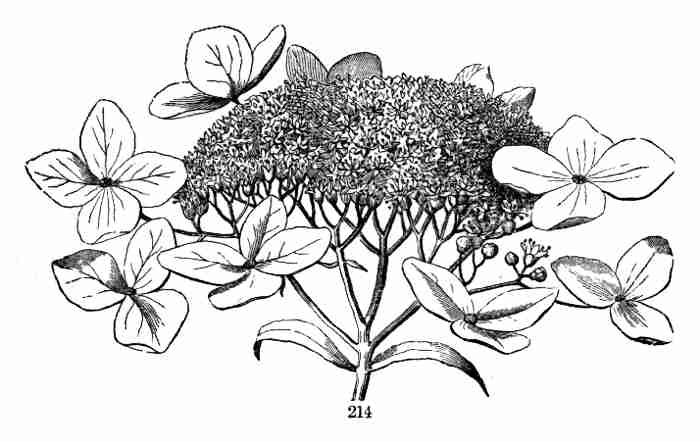
Fig. 214. Compound cyme of Hydrangea arborescens, with
neutral enlarged flowers round the circumference.
221. A Fascicle (meaning a bundle), like that of the Sweet William and
Lychnis of the gardens, is only a cyme with the flowers much crowded.
222. A Glomerule is a cyme still more compacted, so as to imitate a
head. It may be known from a true head by the flowers not expanding
centripetally, that is, not from the circumference towards the centre.
223. The illustrations of determinate or cymose inflorescence have
been taken from plants with opposite leaves, which give rise to the most
regular cymes. But the Rose, Cinquefoil, Buttercup, etc., with alternate
leaves, furnish also good examples of cymose inflorescence.
224. A Cymule (or diminutive cyme) is either a reduced small cyme of few
flowers, or a branch of a compound cyme, i. e. a partial cyme.
225. Scorpioid or Helicoid Cymes, of various sorts, are forms of
determinate inflorescence (often puzzling to the student) in which one
half of the ramification fails to appear. So that they may be called
incomplete cymes. The commoner forms may be understood by comparing a
complete [Pg 78]cyme, like that of Fig. 215 with Fig. 216, the diagram of a
cyme of an opposite-leaved plant, having a series of terminal flowers
and the axis continued by the development of a branch in the axil of
only one of the leaves at each node. The dotted lines on the left
indicate the place of the wanting branches, which if present would
convert this scorpioid cyme into the complete one of Fig. 215. Fig.
217 is a diagram of similar inflorescence with alternate leaves. Both
are kinds of false racemes (219). When the bracts are also wanting in
such cases, as in many Borragineous plants, the true nature of the
inflorescence is very much disguised.
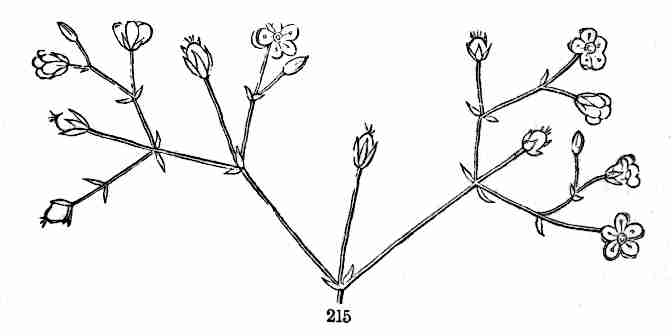
Fig. 215. A complete forking cyme of an Arenaria, or
Chickweed.
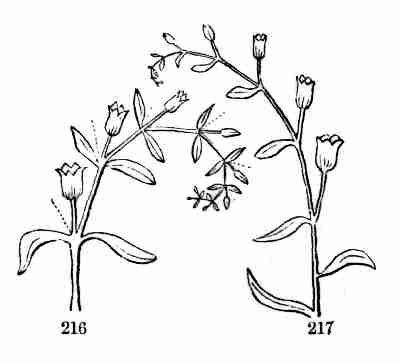
Fig. 216. Diagram of a scorpioid cyme, with opposite
leaves or bracts.
Fig. 217. Diagram of analogous scorpioid cyme, with
alternate leaves or bracts.
[Pg 79]
226. These distinctions between determinate and indeterminate
inflorescence, between corymbs and cymes, and between the true and the
false raceme and spike, were not recognized by botanists much more than
half a century ago, and even now are not always attended to in
descriptions. It is still usual and convenient to describe rounded or
flat-topped and open ramification as corymbose, even when essentially
cymose; also to call the reversed or false racemes or spikes by these
(strictly incorrect) names.
227. Mixed Inflorescence is that in which the two plans are mixed or
combined in compound clusters. A mixed panicle is one in which, while
the primary ramification is of the indeterminate order, the secondary or
ultimate is wholly or partly of the determinate order. A contracted or
elongated inflorescence of this sort is called a Thyrsus. Lilac and
Horse-chestnut afford common examples of mixed inflorescence of this
sort. When loose and open such flower-clusters are called by the general
name of Panicles. The heads of Compositæ are centripetal; but the
branches or peduncles which bear the heads are usually of centrifugal
order.
§ 2. PARTS OR ORGANS OF THE FLOWER.
228. These were simply indicated in Section II. 16. Some parts are
necessary to seed-bearing; these are Essential Organs, namely, the
Stamens and Pistils. Others serve for protection or for attraction,
often for both. Such are the leaves of the Flower, or the Floral
Envelopes.
229. The Floral Envelopes, taken together, are sometimes called the
Perianth, also Perigone, in Latin form Perigonium. In a flower which
possesses its full number of organs, the floral envelopes are of two
kinds, namely, an outer circle, the Calyx, and an inner, the Corolla.
230. The Calyx is commonly a circle of green or greenish leaves, but not
always. It may be the most brightly colored part of the blossom. Each
calyx-leaf or piece is called a Sepal.
231. The Corolla is the inner circle of floral envelopes or
flower-leaves, usually of delicate texture and colored, that is, of
some other color than green. Each corolla-leaf is called a Petal.
232. There are flowers in abundance which consist wholly of floral
envelopes. Such are the so-called full double flowers, of which the
choicer roses and camellias of the cultivator are familiar examples. In
them, under the gardener's care and selection, petals have taken the
place of both stamens and pistils. These are monstrous or unnatural
flowers, incapable of producing seed, and subservient only to human
gratification. Their common name of double flowers is not a sensible
one: except that it is fixed by custom, it were better to translate
their Latin name, flores pleni, and call them full flowers, meaning
full of leaves.
233. Moreover, certain plants regularly produce neutral flowers,
consisting of floral envelopes only. In Fig. 214, some are seen around
the margin [Pg 80]of the cyme in Hydrangea. They are likewise familiar in the
Hobble-bush and in Wild-Cranberry tree, Viburnum Oxycoccus; where they
form an attractive setting to the cluster of small and comparatively
inconspicuous perfect flowers which they adorn. In the Guelder Rose, or
Snow-ball of ornamental cultivation, all or most of the blossoms of this
same shrub are transformed into neutral flowers.

Fig. 218. A flos plenus, namely, a full double flower
of Rose.
234. The Essential Organs are likewise of two kinds, placed one above or
within the other; namely, first, the Stamens or fertilizing organs, and
second, the Pistils, which are to be fertilized and bear the seeds.
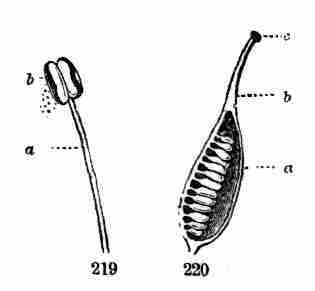
Fig. 219. A stamen: a, filament; b, anther,
discharging pollen.
Fig. 220. A pistil; with ovary, a, half cut away, to
show the contained ovules; b, style; c, stigma.
235. A Stamen consists of two parts, namely, the Filament or stalk (Fig.
219 a), and the Anther (b). The latter is the only essential part.
It is a case, commonly with two lobes or cells, each opening lengthwise
by a slit, at the proper time, and discharging a powder or dust-like
substance, usually of a yellow color. This powder is the Pollen, or
fertilizing matter, to produce which is the office of the stamen.
236. A Pistil (Fig. 220, 221) when complete, has three parts; Ovary,
Style, and Stigma. The Ovary, at base, is the hollow portion, which
contains one or more Ovules or rudimentary seeds. The Style is the
tapering [Pg 81]portion above: the Stigma is a portion of the style, usually
its tip, with moist naked surface, upon which grains of pollen may lodge
and adhere, and thence make a growth which extends down to the ovules.
When there is no style then the stigma occupies the tip of the ovary.
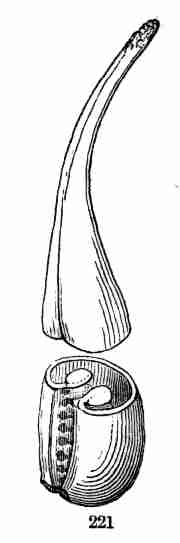
Fig. 221. Model of a simple pistil, with ovary cut across
and slightly opened ventrally, to show the ovules and their attachment.
237. The Torus or Receptacle is the end of the flower-stalk, or the
portion of axis or stem out of which the several organs of the flower
grow, upon which they are borne (Fig. 223).
Fig. 222. Flower of Sedum ternatum, a Stonecrop.
Fig. 223. Parts of same, two of each kind, separated and
displayed; the torus or receptacle in the centre; a, a sepal; b, a
petal; c, a stamen; d, a pistil.
238. The parts of the flower are thus disposed on the receptacle or axis
essentially as are leaves upon a very short stem; first the sepals, or
outer floral leaves; then the petals or inner floral leaves; then the
stamens; lastly, at summit or centre, the pistils, when there are two or
more of them, or the single pistil, when only one. Fig. 223 shows the
organs displayed, two of each kind, of such a simple and symmetrical
flower as that of a Sedum or Stonecrop, Fig. 222.
§ 3. PLAN OF FLOWER.
239. All flowers are formed upon one general plan, but with almost
infinite variations, and many disguises. This common plan is best
understood by taking for a type, or standard for comparison, some
perfect, complete, regular, and symmetrical blossom, and one as
simple as such a blossom could well be. Flowers are said to be
Perfect (hermaphrodite), when provided with both kinds of essential
organs, i. e. with both stamens and pistils.
Complete, when, besides, they have the two sets of floral envelopes,
namely, [Pg 82]calyx and corolla. Such are completely furnished with all that
belongs to a flower.
Regular, when all the parts of each set are alike in shape and size.
Symmetrical, when there is an equal number of parts in each set or
circle of organs.
240. Flax-flowers were taken for a pattern in Section II. 16. But in
them the five pistils have their ovaries as it were consolidated into
one body. Sedum, Fig. 222, has the pistils and all the other parts free
from such combination. The flower is perfect, complete, regular, and
symmetrical, but is not quite as simple as it might be; for there are
twice as many stamens as there are of the other organs. Crassula, a
relative of Sedum, cultivated in the conservatories for winter
blossoming (Fig. 224) is simpler, being isostemonous, or with just as
many stamens as petals or sepals, while Sedum is diplostemonous,
having double that number: it has, indeed, two sets of stamens.
Fig. 224. Flower of a Crassula. 225. Diagram or
ground-plan of same.
241. Numerical Plan. A certain number either runs through the flower or
is discernible in some of its parts. This number is most commonly either
five or three, not very rarely four, occasionally two. Thus the
ground-plan of the flowers thus far used for illustration is five.
That of Trillium (Fig. 226, 227) is three, as it likewise is as really,
if not as plainly, in Tulips and Lilies, Crocus, Iris, and all that
class of blossoms. In some Sedums all the flowers are in fours. In
others the first flowers are on the plan of five, the rest mostly on the
plan of four, that is, with four sepals, four petals, eight stamens (i.
e. twice four), and four pistils. Whatever the ground number may be, it
runs through the whole in symmetrical blossoms.
Fig. 226. Flower of a Trillium; its parts in threes.
Fig. 227. Diagram of flower of Trillium. In this, as in
all such diagrams of cross-section of blossoms, the parts of the outer
circle represent the calyx; the next, corolla; within, stamens (here in
two circles of three each, and the cross-section is through the
anthers); in the centre, section of three ovaries joined into a compound
one of three cells.
242. Alternation of the successive Circles. In these flowers the parts
of the successive circles alternate; and such is the rule. That is,
[Pg 83]the petals stand over the intervals between the sepals; the stamens,
when of the same number, stand over the intervals between the petals; or
when twice as many, as in the Trillium, the outer set alternates with
the petals, and the inner set, alternating with the other, of course
stands before the petals; and the pistils alternate with these. This is
just as it should be on the theory that the circles of the blossom
answer to whorls of leaves, which alternate in this way. While in such
flowers the circles are to be regarded as whorls, in others they are
rather to be regarded as condensed spirals of alternate leaves. But,
however this may be, in the mind of a morphological botanist,
243. Flowers are altered Branches, and their parts, therefore, altered
leaves. That is, certain buds, which might have grown and lengthened
into a leafy branch, do, under other circumstances and to accomplish
other purposes, develop into blossoms. In these the axis remains short,
nearly as it is in the bud; the leaves therefore remain close together
in sets or circles; the outer ones, those of the calyx, generally
partake more or less of the character of foliage; the next set are more
delicate, and form the corolla, while the rest, the stamens and pistils,
appear under forms very different from those of ordinary leaves, and are
concerned in the production of seed. This view gives to Botany an
interest which one who merely notices the shape and counts the parts of
blossoms, without understanding their plan, has no conception of.
244. That flowers answer to branches may be shown, first, from their
position. As explained in the section on Inflorescence, flowers arise
from the same places as branches, and from no other; flower-buds, like
leaf-buds, appear either on the summit of a stem, that is, as a terminal
bud, or in the axil of a leaf, as an axillary bud. And, as the plan of a
symmetrical flower shows, the arrangement of the parts on their axis or
receptacle is that of leaves upon the stem.
245. That the sepals and petals are of the nature of leaves is evident
from their appearance; they are commonly called the leaves of the
flower. The calyx is most generally green in color, and foliaceous
(leaf-like) in texture. And though the corolla is rarely green, yet
neither are proper leaves always green. In our wild Painted-cup, and in
some scarlet Sages, common in gardens, the leaves just under the flowers
are of the brightest red or scarlet, often much brighter-colored than
the corolla itself. And sometimes (as in many Cactuses, and in Carolina
Allspice) there is such a regular gradation from the last leaves of the
plant (bracts or bractlets) into the leaves of the calyx, that it is
impossible to say where the one ends and the other begins. If sepals are
leaves, so also are petals; for there is no clearly fixed limit between
them. Not only in the Carolina Allspice and Cactus (Fig. 229), but in
the Water-Lily (Fig. 228) and in a variety of flowers with more than one
row of petals, there is such a complete transition between calyx and
corolla that no one can surely tell how many of the leaves belong to the
one and how many to the other.
[Pg 84]
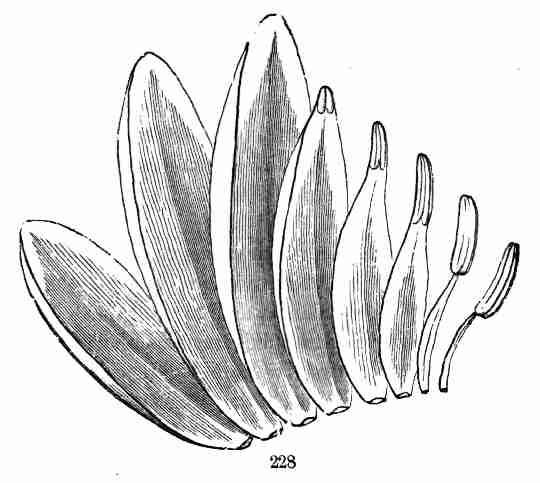
Fig. 228. Series of sepals, petals, and stamens of White
Water-Lily, showing the transitions.
 Fig.
Fig. 229. A Cactus blossom.
246. That stamens are of the same general nature as petals, and
therefore a modification of leaves, is shown by the gradual transitions
that occur between the one and the other in many blossoms; especially in
cultivated flowers, such as Roses and Camellias, when they begin to
double, that is, to change their stamens into petals. Some wild and
natural flowers show the same interesting transitions. The Carolina
Allspice and the White Water-Lily exhibit complete gradations not only
between sepals and petals, but between petals and stamens. The sepals of
our Water-Lily are green outside, but white and petal-like on the
inside; the petals, in many rows, gradually grow narrower towards the
centre of the flower; some of these are tipped with a trace of a yellow
anther, but still are petals; the next are more contracted and
stamen-like, but with a flat petal-like filament; and a further
narrowing of this completes the genuine stamen.
247. Pistils and stamens now and then change into each other in some
Willows; pistils often turn into petals in cultivated flowers; and in
the Double Cherry they are occasionally replaced by small green leaves.
Sometimes a whole blossom changes into a cluster of green leaves, as in
the "green roses" occasionally noticed in gardens, and sometimes it
degenerates into a leafy branch. So the botanist regards pistils also as
answering to leaves; that is, to single leaves when simple and separate,
to a whorl of leaves when conjoined.
[Pg 85]
§ 4. MODIFICATIONS OF THE TYPE.
248. The Deviations, as they may be called, from the assumed type or
pattern of flower are most various and extensive. The differences
between one species and another of the same genus are comparatively
insignificant; those between different genera are more striking; those
between different families and classes of plants more and more profound.
They represent different adaptations to conditions or modes of life,
some of which have obvious or probable utilities, although others are
beyond particular explanation. The principal modifications may be
conveniently classified. First those which in place of perfect
(otherwise called hermaphrodite or bisexual) flowers, give origin to
249. Unisexual, or Separated, or Diclinous Flowers, imperfect flowers,
as they have been called in contradistinction to perfect flowers; but
that term is too ambiguous. In these some flowers want the stamens,
while others want the pistils. Taking hermaphrodite flowers as the
pattern, it is natural to say that the missing organs are suppressed.
This expression is justified by the very numerous cases in which the
missing parts are abortive, that is, are represented by rudiments or
vestiges, which serve to exemplify the plan, although useless as to
office. Unisexual flowers are
Monœcious (or Monoicous, i. e. of one household), when flowers of
both sorts or sexes are produced by the same individual plant, as in the
Ricinus or Castor-oil Plant, Fig. 230.
Diœcious (or Dioicous, i. e. of separate households), when the
two kinds are borne on different plants; as in Willows, Poplars, Hemp,
and Moonseed, Fig. 231, 232.
Polygamous, when the flowers are some of them perfect, and some
staminate or pistillate only.
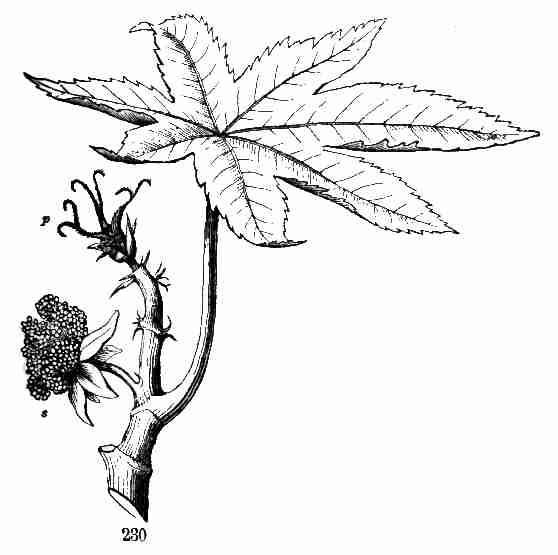
Fig. 230. Unisexual flowers of Castor-oil plant: s,
staminate flower; p, pistillate flower.
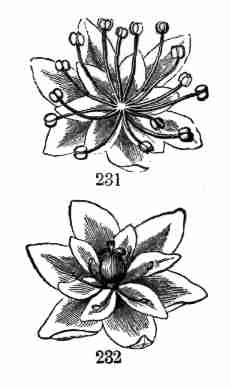
Fig. 231, staminate, and 232, pistillate flower of
Moonseed.
[Pg 86]
250. A blossom having stamens and no pistil is a Staminate or Male
flower. Sometimes it is called a Sterile flower, not appropriately,
for other flowers may equally be sterile. One having pistil but no
stamens is a Pistillate or Female flower.
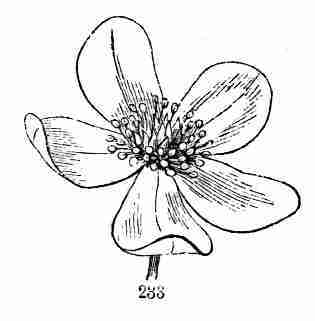
Fig. 233. Flower of Anemone Pennsylvanica; apetalous,
hermaphrodite.
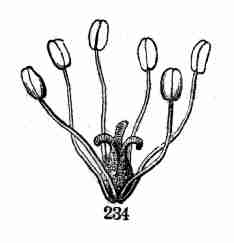
Fig. 234. Flower of Saururus or Lizard's-tail; naked, but
hermaphrodite.
251. Incomplete Flowers are so named in contradistinction to complete:
they want either one or both of the floral envelopes. Those of Fig. 230
are incomplete, having calyx but no corolla. So is the flower of Anemone
(Fig. 233), although its calyx is colored like a corolla. The flowers of
Saururus or Lizard's-tail, although perfect, have neither calyx nor
corolla (Fig. 234). Incomplete flowers, accordingly, are
Naked or Achlamydeous, destitute of both floral envelopes, as in
Fig. 234, or
Apetalous, when wanting only the corolla. The case of corolla present
and calyx wholly wanting is extremely rare, although there are seeming
instances. In fact, a single or simple perianth is taken to be a calyx,
unless the absence or abortion of a calyx can be made evident.
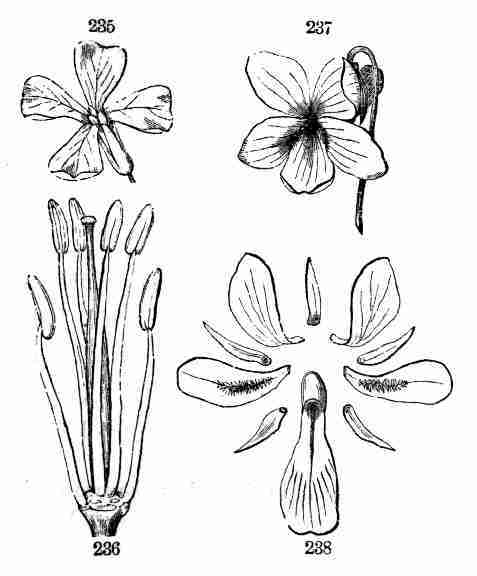
Fig. 235. Flower of Mustard. 236. Its stamens and pistil
separate and enlarged.
Fig. 237. Flower of a Violet. 238. Its calyx and corolla
displayed: the five smaller parts are the sepals; the five intervening
larger ones are the petals.
252. In contradistinction to regular and symmetrical, very many flowers
are
Irregular, that is, with the members of some or all of the floral
circles unequal or dissimilar, and
Unsymmetrical, that is, when the circles of the flower or some of them
differ in the number of their members. (Symmetrical and unsymmetrical
are used in a different sense in some recent books, but the older use
should be adhered to). Want of numerical symmetry and irregularity
commonly go together; and both are common. Indeed, few flowers are
entirely [Pg 87]symmetrical beyond calyx, corolla, and perhaps stamens; and
probably no irregular blossoms are quite symmetrical.
253. Irregular and Unsymmetrical Flowers may therefore be illustrated
together, beginning with cases which are comparatively free from other
complications. The blossom of Mustard, and of all the very natural
family which it represents (Fig. 235, 236), is regular but unsymmetrical
in the stamens. There are four equal sepals, four equal petals; but six
stamens, and only two members in the pistil, which for the present may
[Pg 88]be left out of view. The want of symmetry is in the stamens. These are
in two circles, an outer and an inner. The outer circle consists of two
stamens only; the inner has its proper number of four. The flower of
Violet, which is on the plan of five, is symmetrical in calyx, corolla,
and stamens, inasmuch as each of these circles consists of five members;
but it is conspicuously irregular in the corolla, one of the petals
being very different from the rest.
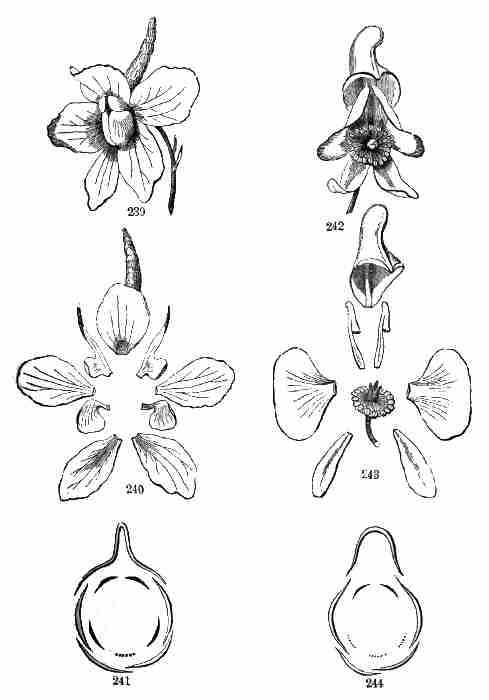
Fig. 239. Flower of a Larkspur. 240. Its calyx and
corolla displayed; the five larger parts are the sepals; the four
smaller, of two shapes, are the petals; the place of the fifth petal is
vacant. 241. Diagram of the same; the place for the missing petal marked
by a dotted line.
Fig. 242. Flower of a Monkshood. 243. Its parts
displayed; five sepals, the upper forming the hood; the two lateral
alike, broad and flat; the two lower small. The two pieces under the
hood represent the corolla, reduced to two odd-shaped petals; in centre
the numerous stamens and three pistils. 244. Diagram of the calyx and
corolla; the three dotted lines in the place of missing petals.
254. The flowers of Larkspur, and of Monkshood or Aconite, which are
nearly related, are both strikingly irregular in calyx and corolla, and
considerably unsymmetrical. In Larkspur (Fig. 239-241) the irregular
calyx consists of five sepals, one of which, larger than the rest, is
prolonged behind into a large sac or spur; but the corolla is of only
four petals (of two shapes),—the fifth, needed to complete the
symmetry, being left out. And the Monkshood (Fig. 242-244) has five very
dissimilar sepals, and a corolla of only two very small and
curiously-shaped petals,—the three needed to make up the symmetry being
left out. The stamens in both are out of symmetry with the ground-plan,
being numerous. So are the pistils, which are usually diminished to
three, sometimes to two or to one.
255. Flowers with Multiplication of Parts are very common. The stamens
are indefinitely numerous in Larkspur and in Monkshood (Fig. 242, 243),
while the pistils are fewer than the ground-plan suggests. Most
Cactus-flowers have all the organs much increased in number (Fig. 229),
and so of the Water-Lily. In Anemone (Fig. 233) the stamens and pistils
are multiplied while the petals are left out. In Buttercups or Crowfoot,
while the sepals and petals conform to the ground-plan of five, both
stamens and pistils are indefinitely multiplied (Fig. 245).
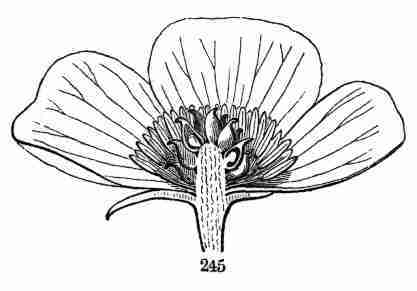
Fig. 245. Flower of Ranunculus bulbosus, or Buttercup, in
section.
256. Flowers modified by Union of Parts, so that these parts more or
less lose the appearance of separate leaves or other organs growing out
of the end of the stem or receptacle, are extremely common. There are
two kinds of such union, namely:—
Coalescence of parts of the same circle by their contiguous margins;
and
Adnation, or the union of adjacent circles or unlike parts.
257. Coalescence is not rare in leaves, as in the upper pairs of
Honeysuckles, Fig. 163. It may all the more be expected in the crowded
circles or whorls of flower-leaves. Datura or Stramonium (Fig. 246)
shows this coalescence both in calyx and corolla, the five sepals and
the five petals being thus united to near their tips, each into a tube
or long and narrow cup. These unions make needful the following terms:—
[Pg 89]
Gamopetalous, said of a corolla the petals of which are thus
coalescent into one body, whether only at base or higher. The union may
extend to the very summit, as in Morning Glory and the like (Fig. 247),
so that the number of petals in it may not be apparent. The old name for
this was Monopetalous, but that means "one-petalled;" while
gamopetalous means "petals united," and therefore is the proper term.
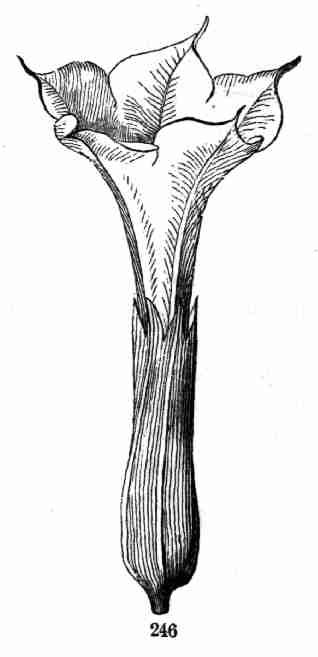
Fig. 246. Flower of Datura Stramonium; gamosepalous and
gamopetalous.
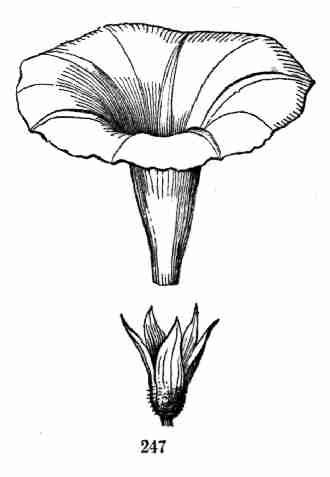
Fig. 247. Funnelform corolla of a common Morning Glory,
detached from its polysepalous calyx.
Polypetalous is the counterpart term, to denote a corolla of
distinct, that is, separate petals. As it means "many petalled," it is
not the best possible name, but it is the old one and in almost
universal use.
Gamosepalous applies to the calyx when the sepals are in this way
united.
Polysepalous, to the calyx when of separate sepals or calyx-leaves.
258. Degree of union or of separation in descriptive botany is expressed
in the same way as is the lobing of leaves (139). See Fig. 249-253, and
the explanations.
259. A corolla when gamopetalous commonly shows a distinction (well
marked in Fig. 249-251) between a contracted tubular portion below, the
Tube, and the spreading part above, the Border or Limb. The junction
between tube and limb, or a more or less enlarged upper portion of the
tube between the two, is the Throat. The same is true of the calyx.
260. Some names are given to particular forms of the gamopetalous
corolla, applicable also to a gamosepalous calyx, such as
Wheel-shaped, or Rotate; when spreading out at once, without a tube
or with a very short one, something in the shape of a wheel or of its
diverging spokes, Fig. 252, 253.
Salver-shaped, or Salver-form; when a flat-spreading border is
raised on [Pg 90]a narrow tube, from which it diverges at right angles, like
the salver represented in old pictures, with a slender handle beneath,
Fig. 249-251, 255.
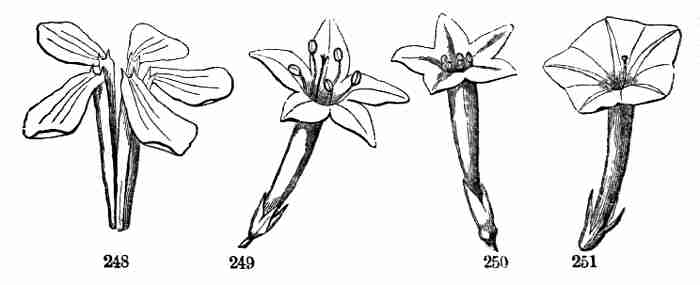
Fig. 248. Polypetalous corolla of Soapwort, of five
petals with long claws or stalk-like bases.
Fig. 249. Flower of Standing Cypress (Gilia
coronopifolia); gamopetalous: the tube answering to the long claws in
248, except that they are coalescent: the limb or border (the spreading
part above) is five-parted, that is, the petals not there united
except at very base.
Fig. 250. Flower of Cypress-vine (Ipomœa Quamoclit);
like preceding, but limb five-lobed.
Fig. 251. Flower of Ipomœa coccinea; limb almost
entire.
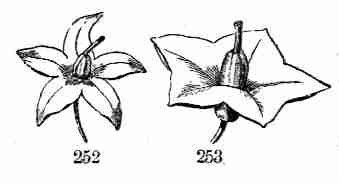
Fig. 252. Wheel-shaped or rotate and five-parted corolla
of Bittersweet, Solanum Dulcamara. 253. Wheel-shaped and five-lobed
corolla of Potato.
Bell-shaped, or Campanulate; where a short and broad tube widens
upward, in the shape of a bell, as in Fig. 254.
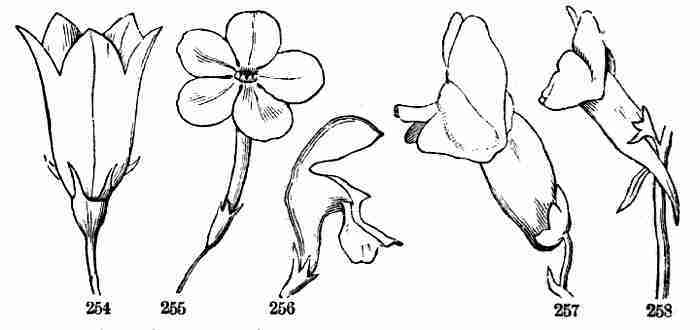
Fig. 254. Flower of a Campanula or Harebell, with a
campanulate or bell-shaped corolla; 255, of a Phlox, with salver-shaped
corolla; 256, of Dead Nettle (Lamium), with labiate ringent (or
gaping) corolla; 257, of Snapdragon, with labiate personate corolla;
258, of Toad-Flax, with a similar corolla spurred at the base.
Funnel-shaped, or Funnelform; gradually spreading at the summit of a
tube which is narrow below, in the shape of a funnel or tunnel, as in
the corolla of the common Morning Glory (Fig. 247) and of the Stramonium
(Fig. 246).
[Pg 91]
Tubular; when prolonged into a tube, with little or no spreading at
the border, as in the corolla of the Trumpet Honeysuckle, the calyx of
Stramonium (Fig. 246), etc.
261. Although sepals and petals are usually all blade or lamina (123),
like a sessile leaf, yet they may have a contracted and stalk-like base,
answering to petiole. This is called its Claw, in Latin Unguis.
Unguiculate petals are universal and strongly marked in the Pink
tribe, as in Soapwort (Fig. 248).
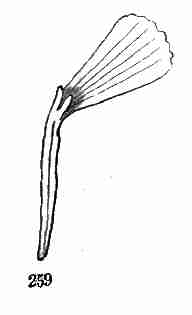
Fig. 259. Unguiculate (clawed) petal of a Silene; with a
two-parted crown.
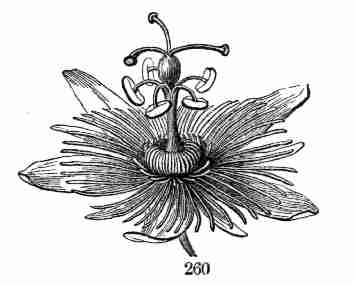
Fig. 260. A small Passion-flower, with crown of slender
threads.
262. Such petals, and various others, may have an outgrowth of the inner
face into an appendage or fringe, as in Soapwort, and in Silene (Fig.
259), where it is at the junction of claw and blade. This is called a
Crown, or Corona. In Passion-flowers (Fig. 260) the crown consists of
numerous threads on the base of each petal.
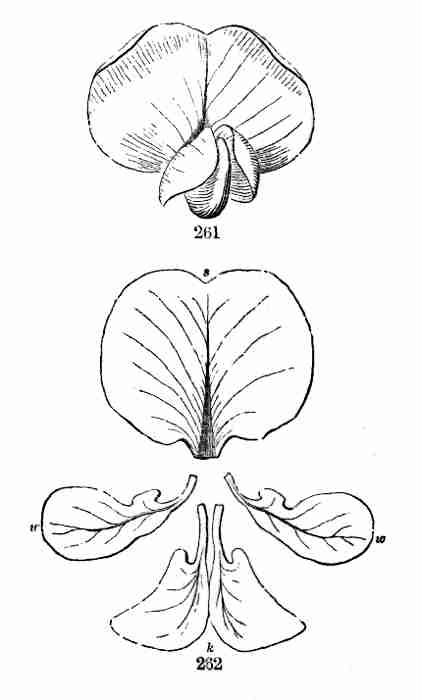
Fig. 261. Front view of a papilionaceous corolla. 262.
The parts of the same, displayed: s, Standard, or Vexillum; w,
Wings, or Alæ; k, Keel, or Carina.
263. Irregular Flowers may be polypetalous, or nearly so, as in the
papilionaceous corolla; but most of them are irregular through
coalescence, which often much disguises the numerical symmetry also. As
affecting the corolla the following forms have received particular
names:
264. Papilionaceous Corolla, Fig. 261, 262. This is polypetalous, except
that two of the petals cohere, usually but slightly. It belongs only to
the Leguminous or Pulse family. The name means butterfly-like; but the
likeness is hardly obvious. The names of the five petals of the
papilionaceous corolla are curiously incongruous. They are,
[Pg 92]
The Standard or Banner (Vexillum), the large upper petal which is
external in the bud and wrapped around the others.
The Wings (Alæ), the pair of side petals, of quite different shape
from the standard.
The Keel (Carina), the two lower and usually smallest petals; these
are lightly coalescent into a body which bears some likeness, not to the
keel, but to the prow of a boat; and this encloses the stamens and
pistil. A Pea-blossom is a typical example; the present illustration is
from a species of Locust, Robinia hispida.
265. Labiate Corolla (Fig. 256-258), which would more properly have been
called Bilabiate, that is, two-lipped. This is a common form of
gamopetalous corolla; and the calyx is often bilabiate also. These
flowers are all on the plan of five; and the irregularity in the corolla
is owing to unequal union of the petals as well as to diversity of form.
The two petals of the upper or posterior side of the flower unite with
each other higher up than with the lateral petals (in Fig. 256, quite to
the top), forming the Upper lip: the lateral and the lower similarly
unite to form the Lower lip. The single notch which is generally found
at the summit of the upper lip, and the two notches of the lower lip, or
in other words the two lobes of the upper and the three of the lower
lip, reveal the real composition. So also does the alternation of these
five parts with those of the calyx outside. When the calyx is also
bilabiate, as in the Sage, this alternation gives three lobes or sepals
to the upper and two to the lower lip. Two forms of the labiate corolla
have been designated, viz.:—
Ringent or Gaping, when the orifice is wide open, as in Fig. 256.
Personate or Masked, when a protuberance or intrusion of the base of
the lower lip (called a Palate) projects over or closes the orifice,
as in Snapdragon and Toad-Flax, Fig. 257, 258.
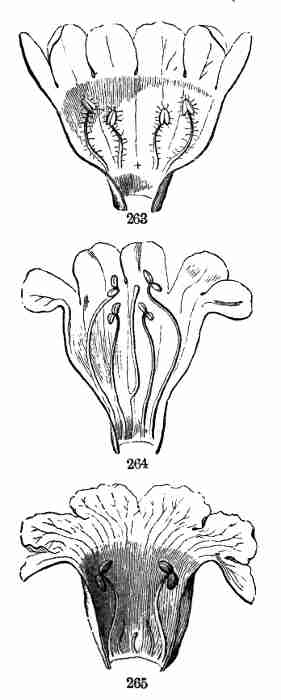
Fig. 263. Corolla of a purple Gerardia laid open, showing
the four stamens; the cross shows where the fifth stamen would be, if
present.
Fig. 264. Corolla, laid open, and stamens of Pentstemon
grandiflorus, with a sterile filament in the place of the fifth stamen,
and representing it.
Fig. 265. Corolla of Catalpa laid open, displaying two
good stamens and three abortive ones or vestiges.
[Pg 93]
266. There are all gradations between labiate and regular corollas. In
those of Gerardia, of some species of Pentstemon, and of Catalpa (Fig.
263-265), the labiate character is slight, but is manifest on close
inspection. In almost all such flowers the plan of five, which is
obvious or ascertainable in the calyx and corolla, is obscured in the
stamens by the abortion or suppression of one or three of their number.
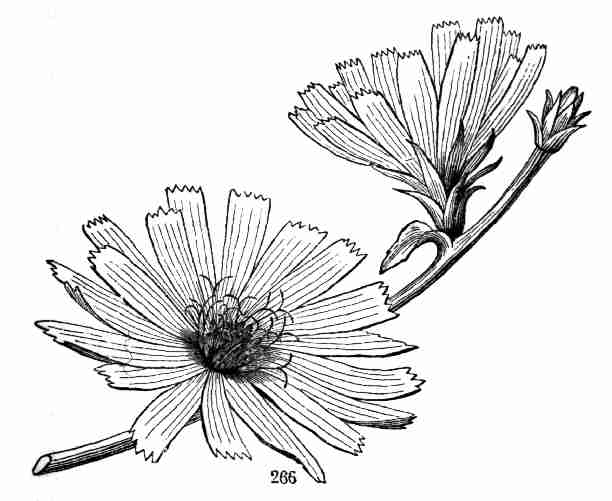 Fig.
Fig. 266. Two flower-heads of Chiccory.
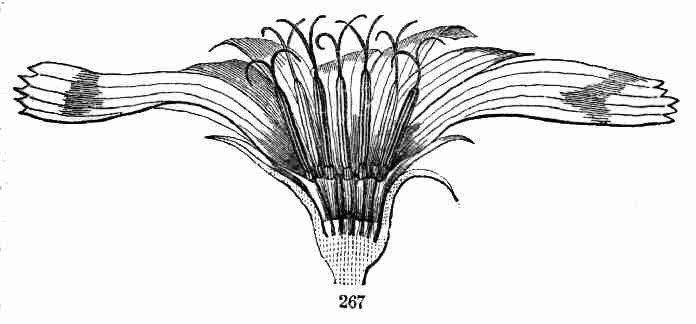
Fig. 267. One of them half cut away, better showing some
of the flowers.
267. Ligulate Corolla. The ligulate or Strap-shaped corolla mainly
belongs to the family of Compositæ, in which numerous small flowers are
gathered into a head, within an involucre that imitates a calyx. It is
best exemplified in the Dandelion and in Chiccory (Fig. 266). Each one
of these straps or Ligules, looking like so many petals, is the
corolla of a distinct flower: the base is a short tube, which opens out
into the ligule: the five minute teeth at the end indicate the number of
constituent petals. So this is a kind of gamopetalous corolla, which is
open along one side nearly [Pg 94]to the base, and outspread. The nature of
such a corolla (and of the stamens also, to be explained in the next
section) is illustrated by the flower of a Lobelia, Fig. 285.
 Fig.
Fig. 268. Head of flowers of a Coreopsis, divided
lengthwise.
268. In Asters, Daisies, Sunflower, Coreopsis (Fig. 268), and the like,
only the marginal (or Ray) corollas are ligulate; the rest (those of
the Disk) are regularly gamopetalous, tubular, and five-lobed at
summit; but they are small and individually inconspicuous, only the
ray-flowers making a show. In fact, those of Coreopsis and of
Sunflower are simply for show, these ray-flowers being not only sterile,
but neutral, that is, having neither stamens nor pistil. But in
Asters, Daisies, Golden-rods, and the like, these ray-flowers are
pistillate and fertile, serving therefore for seed-bearing as well as
for show. Let it not be supposed that the show is useless. See Section
XIII.
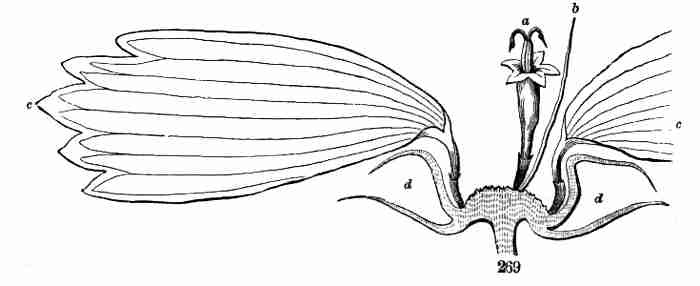
Fig. 269. A slice of the preceding more enlarged, with
one tubular perfect flower (a) left standing on the receptacle, with
its bractlet or chaff (b), one ligulate and neutral ray-flower (cc)
and part of another; dd, section of bracts or leaves of the
involucre.
269. Adnation, or Consolidation, is the union of the members of parts
belonging to different circles of the flower (256). It is of course
understood that in this (as likewise in coalescence) the parts are not
formed and then conjoined, but are produced in union. They are born
united, as the term adnate implies. To illustrate this kind of union,
take the accompanying series of flowers (Fig. 270-274), shown in
vertical section. In the first, Fig. 270, Flax-flower, there is no
adnation; sepals, petals, and stamens, are free as well as distinct,
being separately borne on the receptacle, one circle within or above the
next; only the five pistils have their ovaries coalescent. In Fig. 271,
a Cherry-flower, the petals and stamens are borne on the throat of the
calyx-tube; that is, the sepals are coalescent into a cup, and the
petals and stamens are adnate to the inner face of this; in other
[Pg 95]words, the sepals, petals, and stamens are all consolidated up to a
certain height. In Fig. 272, a Purslane-flower, the same parts are
adnate to or consolidated with the ovary up to its middle. In Fig. 273,
a Hawthorn-flower, the consolidation has extended over the whole ovary;
and petals and stamens are adnate to the calyx still further. In Fig.
274, a Cranberry-blossom, it is the same except that all the parts are
free at the same height; all seem to arise from the top of the ovary.
270. In botanical description, to express tersely such differences in
the relation of these organs to the pistil, they are said to be
Hypogynous (i. e. under the pistil) when they are all free, that is,
not adnate to pistil nor connate with each other, as in Fig. 270.
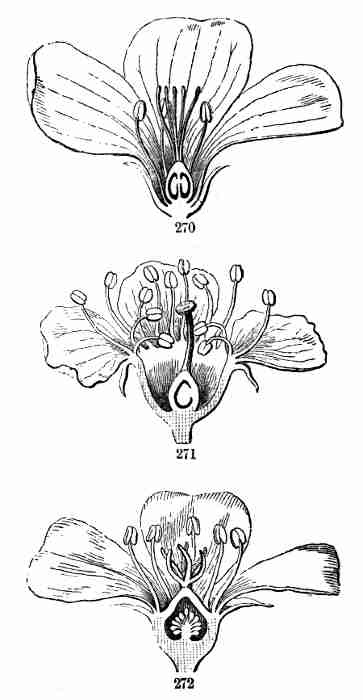
Fig. 270. Flax-flower in section; the parts all
free,—hypogynous.
Fig. 271. Cherry-flower in section; petals and stamens
adnate to tube of calyx,—perigynous.
Fig. 272. Purslane-flower in section; calyx, petals,
stamens, all adnate to lower half of ovary,—perigynous.
Perigynous (around the pistil) when connate with each other, that is,
when petals and stamens are inserted or borne on the calyx, whether as
in Cherry-flowers (Fig. 271) they are free from the pistil, or as in
Purslane and Hawthorn (Fig. 272, 273) they are also adnate below to the
ovary.
Epigynous (on the ovary) when so adnate that all these parts appear to
arise from the very summit of the ovary, as in Fig. 274. The last two
terms are not very definitely distinguished.
271. Another and a simpler form of expression is to describe parts of
the flower as being
Free, when not united with or inserted upon other parts.
Distinct, when parts of the same kind are not united. This term is the
counterpart of coalescent, as free is the counterpart of adnate. Many
writers use the term "free" indiscriminately for both; but it is better
to distinguish them.
[Pg 96]
Connate is a term common for either not free or not distinct, that
is, for parts united congenitally, whether of same or of different
kinds.
Adnate, as properly used, relates to the union of dissimilar parts.
272. In still another form of expression, the terms superior and
inferior have been much used in the sense of above and below.
Superior is said of the ovary of Flax-flower, Cherry, etc., because
above the other parts; it is equivalent to "ovary free." Or it is said
of the calyx, etc., when above the ovary, as in Fig. 273-275.
Inferior, when applied to the ovary, means the same as "calyx adnate;"
when applied to the floral envelopes, it means that they are free.
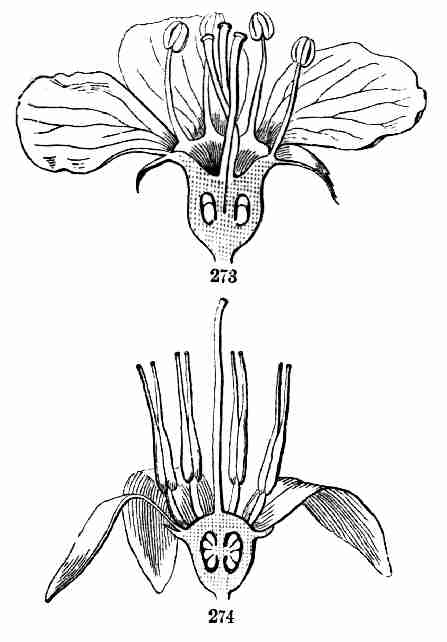
Fig. 273. Hawthorn-blossom in section; parts adnate to
whole face of ovary, and with each other beyond; another grade of
perigynous.
Fig. 274. Cranberry-blossom in section; parts epigynous.
273. Position of Flower or of its Parts. The terms superior and
inferior, or upper and lower, are also used to indicate the relative
position of the parts of a flower in reference to the axis of
inflorescence. An axillary flower stands between the bract or leaf which
subtends it and the axis or stem which bears this bract or leaf. This is
represented in sectional diagrams (as in Fig. 275, 276) by a transverse
line for the bract, and a small circle for the axis of inflorescence.
Now the side of the blossom which faces the bract is the
Anterior, or Inferior, or Lower side; while the side next the axis
is the
Posterior, or Superior, or Upper side of the flower.
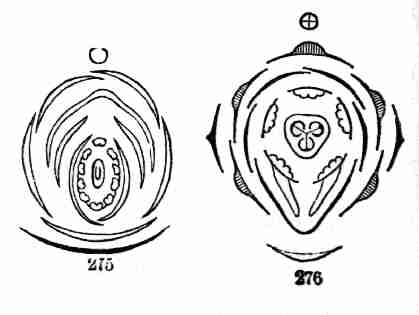
Fig. 275. Diagram of papilionaceous flower (Robinia, Fig.
261), with bract below; axis of inflorescence above.
Fig. 276. Diagram of Violet-flower; showing the relation
of parts to bract and axis.
274. So, in the labiate corolla (Fig. 256-258), the lip which is
composed of three of the five petals is the anterior, or inferior,
or lower lip; the other is the posterior, or superior, or upper
lip.
[Pg 97]
275. In Violets (Fig. 238, 276), the odd sepal is posterior (next the
axis); the odd petal is therefore anterior, or next the subtending leaf.
In the papilionaceous flower (Fig. 261, and diagram, Fig. 275), the odd
sepal is anterior, and so two sepals are posterior; consequently, by the
alternation, the odd petal (the standard) is posterior or upper, and the
two petals forming the keel are anterior or lower.
§ 5. ARRANGEMENT OF PARTS IN THE BUD.
276. Æstivation was the fanciful name given by Linnæus to denote the
disposition of the parts, especially the leaves of the flower, before
Anthesis, i. e. before the blossom opens. Præfloration, a better
term, is sometimes used. This is of importance in distinguishing
different families or genera of plants, being generally uniform in each.
The æstivation is best seen by making a slice across the flower-bud; and
it may be expressed in diagrams, as in the accompanying figures.
277. The pieces of the calyx or the corolla either overlap each other in
the bud, or they do not. When they do not overlap, the æstivation is
Valvate, when the pieces meet each other by their abrupt edges,
without any infolding or overlapping; as the calyx of the Linden or
Basswood (Fig. 277).
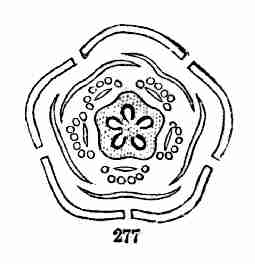
Fig. 277. Diagram of a flower of Linden, showing the
calyx valvate and corolla imbricate in the bud, etc.
Induplicate, which is valvate with the margins of each piece
projecting inwards, as in the calyx of a common Virgin's-bower, Fig.
278, or
Fig. 278. Valvate-induplicate æstivation of calyx of
common Virgin's-bower.
Fig. 279. Valvate-involute æstivation of same in
Vine-bower, Clematis Vitialla.
Involute, which is the same but the margins rolled inward, as in most
of the large-flowered species of Clematis, Fig. 279.
Reduplicate, a rarer modification of valvate, is similar but with
margins projecting outward.
Open, the parts not touching in the bud, as the calyx of Mignonette.
278. When the pieces overlap in the bud, it is in one of two ways;
either every piece has one edge in and one edge out, or some pieces are
wholly outside and others wholly inside. In the first case the
æstivation is
Convolute, also named Contorted or Twisted, as in Fig. 280, a
cross-section of a corolla very strongly thus convolute or rolled up
together, and in the corolla of a Flax-flower (Fig. 281), where the
petals only moderately overlap in this way. Here one edge of every petal
covers the next before [Pg 98]it, while its other edge is covered by the next
behind it. The other mode is the

Fig. 280. Convolute æstivation, as in the corolla-lobes
of Oleander.
Fig. 281. Diagram of a Flax-flower; calyx imbricated and
corolla convolute in the bud.
Imbricate or Imbricated, in which the outer parts cover or overlap
the inner so as to "break joints," like tiles or shingles on a roof;
whence the name. When the parts are three, the first or outermost is
wholly external, the third wholly internal, the second has one margin
covered by the first while the other overlaps the third or innermost
piece: this is the arrangement of alternate three-ranked leaves (187).
When there are five pieces, as in the corolla of Fig. 225, and calyx of
Fig. 281, as also of Fig. 241, 276, two are external, two are internal,
and one (the third in the spiral) has one edge covered by the outermost,
while its other edge covers the innermost; which is just the five-ranked
arrangement of alternate leaves (188). When the pieces are four, two are
outer and two are inner; which answers to the arrangement of opposite
leaves.
279. The imbricate and the convolute modes sometimes vary one into the
other, especially in the corolla.
280. In a gamopetalous corolla or gamosepalous calyx, the shape of the
tube in the bud may sometimes be noticeable. It may be
Plicate or Plaited, that is, folded lengthwise; and the plaits may
either be turned outwards, forming projecting ridges, as in the corolla
of Campanula; or turned inwards, as in that of Gentian Belladonna; or
Supervolute, when the plaits are convolutely wrapped round each other,
as in the corolla of Morning Glory and of Stramonium, Fig. 282.
Fig. 282. Upper part of corolla of Datura Stramonium in
the bud; and below a section showing the convolution of the plaits.
Section IX. STAMENS IN PARTICULAR.
281. Andrœcium is a technical name for the staminate system of a
flower (that is, for the stamens taken together), which it is sometimes
convenient to use. The preceding section has dealt with modifications of
the flower pertaining mainly to calyx and corolla. Those relating to the
stamens are now to be indicated. First as to
[Pg 99]
282. Insertion, or place of attachment. The stamens usually go with the
petals. Not rarely they are at base
Epipetalous, that is, inserted on (or adnate to) the corolla, as in
Fig. 283. When free from the corolla, they may be
Fig. 283. Corolla of Morning Glory laid open, to show the
five stamens inserted on it, near the base.
Hypogynous, inserted on the receptacle under the pistil or
gynœcium.
Perigynous, inserted on the calyx, that is, with the lower part of
filament adnate to the calyx-tube.
Epigynous, borne apparently on the top of the ovary; all which is
explained in Fig. 270-274.
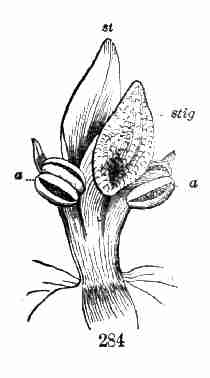
Fig. 284. Style of a Lady's Slipper (Cypripedium), and
stamens united with it; a, a, the anthers of the two good stamens;
st, an abortive stamen, what should be its anther changed into a
petal-like body; stig, the stigma.
Gynandrous is another term relating to insertion of rarer occurrence,
that is, where the stamens are inserted on (in other words, adnate to)
the style, as in Lady's Slipper (Fig. 284), and in the Orchis family
generally.
283. In Relation to each Other, stamens are more commonly
Distinct, that is, without any union with each other. But when united,
the following technical terms of long use indicate their modes of mutual
connection:—
Monadelphous (from two Greek words, meaning "in one brotherhood"),
when united by their filaments into one set, usually into a ring or cup
below, or into a tube, as in the Mallow Family (Fig. 286), the
Passion-flower (Fig. 260), the Lupine (Fig. 287), and in Lobelia (Fig.
285).
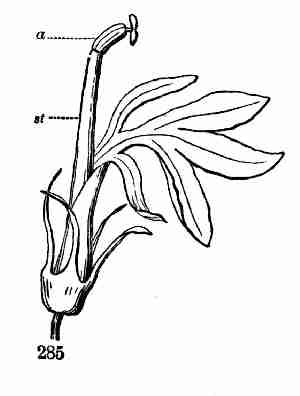
Fig. 285. Flower of Lobelia cardinalis, Cardinal flower;
corolla making approach to the ligulate form; filaments (st)
monadelphous, and anthers (a) syngenesious.
Fig. 286. Flower of a Mallow, with calyx and corolla cut
away; showing monadelphous stamens.
Fig. 287. Monadelphous stamens of Lupine. 288.
Diadelphous stamens (9 and 1) of a Pea-blossom.
Diadelphous (meaning in two brotherhoods), when united by the
filaments into two sets, as in the Pea and most of its near relatives
(Fig. 288), usually nine in one set, and one in the other.
Triadelphous (three brotherhoods), when the filaments are united in
three sets or clusters, as in most species of Hypericum.
[Pg 100]
Pentadelphous (five brotherhoods), when in five sets, as in some
species of Hypericum and in American Linden (Fig. 277, 289).
Polyadelphous (many or several brotherhoods) is the term generally
employed when these sets are several, or even more than two, and the
particular number is left unspecified. These terms all relate to the
filaments.
Syngenesious is the term to denote that stamens have their anthers
united, coalescent into a ring or tube; as in Lobelia (Fig. 285), in
Violets, and in all of the great family of Compositæ.
284. Their Number in a flower is commonly expressed directly, but
sometimes adjectively, by a series of terms which were the name of
classes in the Linnæan artificial system, of which the following names,
as also the preceding, are a survival:—
Monandrous, i. e. solitary-stamened, when the flower has only one
stamen,
Diandrous, when it has two stamens only,
Triandrous, when it has three stamens,
Tetrandrous, when it has four stamens,
Pentandrous, when it has five stamens,
Hexandrous, when with six stamens, and so on to
Polyandrous, when it has many stamens, or more than a dozen.
Fig. 289. One of the five stamen-clusters of the flower
of American Linden, with accompanying scale. The five clusters are shown
in section in the diagram of this flower, Fig. 277.
Fig. 290. Five syngenesious stamens of a Coreopsis. 291.
Same, with tube laid open and displayed.
285. For which terms, see the Glossary. They are all Greek numerals
prefixed to -andria (from the Greek), which Linnæus used for
andrœcium, and are made into an English adjective, -androus. Two
other terms, of same origin, designate particular cases of number (four
or six) in connection with unequal length. Namely, the stamens are
Didynamous, when, being only four, they form two pairs, one pair
longer than the other, as in the Trumpet Creeper, in Gerardia (Fig.
263), etc.
[Pg 101]
Tetradynamous, when, being only six, four of them surpass the other
two, as in the Mustard-flower and all the Cruciferous family, Fig. 235.
286. The Filament is a kind of stalk to the anther, commonly slender or
thread-like: it is to the anther nearly what the petiole is to the blade
of a leaf. Therefore it is not an essential part. As a leaf may be
without a stalk, so the anther may be Sessile, or without a filament.
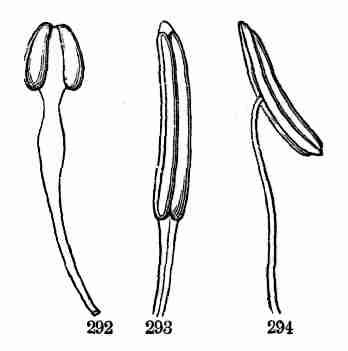
Fig. 292. Stamen of Isopyrum, with innate anther. 293. Of
Tulip-tree, with adnate (and extrorse) anther. 294. Of Evening Primrose,
with versatile anther.
287. The Anther is the essential part of the stamen. It is a sort of
case, filled with a fine powder, the Pollen, which serves to fertilize
the pistil, so that it may perfect seeds. The anther is said to be
Innate (as in Fig. 292), when it is attached by its base to the very
apex of the filament, turning neither inward nor outward;
Adnate (as in Fig. 293), when attached as it were by one face, usually
for its whole length, to the side of a continuation of the filament; and
Versatile (as in Fig. 294), when fixed by or near its middle only to
the very point of the filament, so as to swing loosely, as in the Lily,
in Grasses, etc. Versatile or adnate anthers are
Introrse, or Incumbent, when facing inward, that is, toward the
centre of the flower, as in Magnolia, Water-Lily, etc.
Extrorse, when facing outwardly, as in the Tulip-tree.
288. Rarely does a stamen bear any resemblance to a leaf, or even to a
petal or flower-leaf. Nevertheless, the botanist's idea of a stamen is
that it answers to a leaf developed in a peculiar form and for a special
purpose. In the filament he sees the stalk of the leaf; in the anther,
the blade. The blade of a leaf consists of two similar sides; so the
anther consists of two Lobes or Cells, one answering to the left, the
other to the right, side of the blade. The two lobes are often connected
by a prolongation of the filament, which answers to the midrib of a
leaf; this is called the Connective. This is conspicuous in Fig. 292,
where the connective is so broad that it separates the two cells of the
anther to some distance.
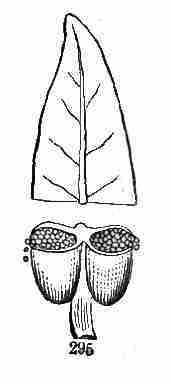
Fig. 295. Diagram of the lower part of an anther, cut
across above, and the upper part of a leaf, to show how the one answers
to the other; the filament to petiole, the connective to midrib; the two
cells to the right and left halves of the blade.
289. A simple conception of the morphological relation of an anther to a
leaf is given in Fig. 295, an ideal figure, the lower part representing
a stamen with the top of its anther cut away; the upper, the
corresponding upper part of a leaf.
[Pg 102]
290. So anthers are generally two-celled. But as the pollen begins to
form in two parts of each cell (the anterior and the posterior),
sometimes these two strata are not confluent, and the anther even at
maturity may be four-celled, as in Moonseed (Fig. 296); or rather, in
that case (the word cell being used for each lateral half of the
organ), it is two-celled, but the cells bilocellate.
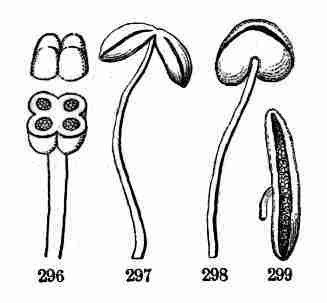
Fig. 296. Stamen of Moonseed, with anther cut across;
this 4-celled, or rather 4-locellate.
Fig. 297. Stamen of Pentstemon pubescens; the two
anther-cells diverging, and almost confluent.
Fig. 298. Stamen of Mallow; the anther supposed to answer
to that of Fig. 297, but the cells completely confluent into one.
Fig. 299. Stamen of Globe Amaranth; very short filament
bearing a single anther-cell; it is open from top to bottom, showing the
pollen within.
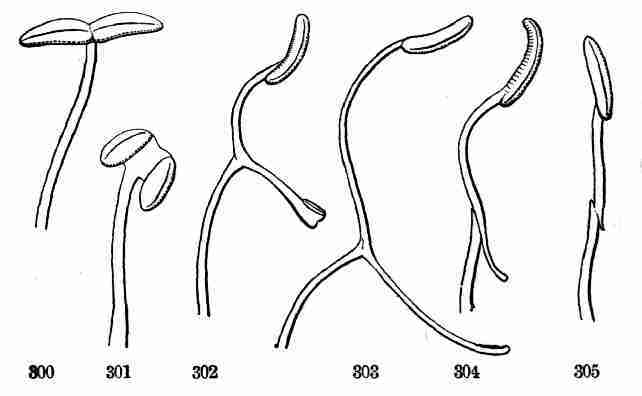
Fig. 300-305. Stamens of several plants of the Labiate or
Mint Family. Fig. 300. Of a Monarda: the two anther-cells with bases
divergent so that they are transverse to the filament, and their
contiguous tips confluent, so as to form one cell opening by a
continuous line. Fig. 301. Of a Calamintha: the broad connective
separating the two cells. Fig. 302. Of a Sage (Salvia Texana); with long
and slender connective resembling forks of the filament, one bearing a
good anther-cell; the other an abortive or poor one. Fig. 303. Another
Sage (S. coccinea), with connective longer and more thread-shaped, the
lower fork having its anther-cell wholly wanting. Fig. 304. Of a White
Sage, Audibertia grandiflora; the lower fork of connective a mere
vestige. Fig. 305. Of another White Sage (A. stachyoides), the lower
fork of connective suppressed.
291. But anthers may become one-celled, and that either by confluence
or by suppression.
292. By confluence, when the two cells run together into one, as they
nearly do in most species of Pentstemon (Fig. 297), more so in Monarda
(Fig. 300), and completely in the Mallow (Fig. 298) and all the Mallow
family.
[Pg 103]
293. By suppression in certain cases the anther may be reduced to one
cell or halved. In Globe Amaranth (Fig. 299) there is a single cell
without vestige of any other. Different species of Sage and of the White
Sages of California show various grades of abortion of one of the
anther-cells, along with a singular lengthening of the connective (Fig.
302-305).
294. The splitting open of an anther for the discharge of its pollen is
termed its Dehiscence.
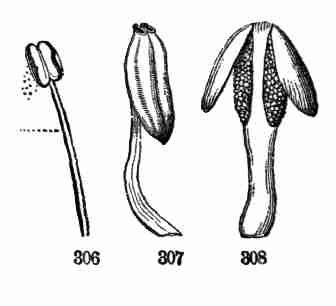
Fig. 306. Stamen with the usual dehiscence of anther down
the side of each cell.
Fig. 307. Stamen of Pyrola; cells opening by a terminal
hole.
Fig. 308. Stamen of Barberry; cells of anther each
opening by an uplifted valve.
295. As the figures show, this is commonly by a line along the whole
length of each cell, either lateral or, when the anthers are extrorse,
often along the outer face, and when introrse, along the inner face of
each cell. Sometimes the opening is only by a chink, hole, or pore at
the top, as in the Azalea, Pyrola (Fig. 307), etc.; sometimes a part of
the face separates as a sort of trap-door (or valve), hinged at the top,
and opening to allow the escape of the pollen, as in the Sassafras,
Spice-bush, and Barberry (Fig. 308).
296. Pollen. This is the powdery matter, commonly of a yellow color,
which fills the cells of the anther, and is discharged during
blossoming, after which the stamens generally fall or wither away. Under
the microscope it is found to consist of grains, usually round or oval,
and all alike in the same species, but very different in different
plants. So that the plant may sometimes be recognized from the pollen
alone. Several forms are shown in the accompanying figures.

Fig. 309. Magnified pollen of a Lily, smooth and oval;
310, of Echinocystis, grooved lengthwise; 311, of Sicyos, with bristly
points and smooth bands; 312, of Musk Plant (Mimulus), with spiral
grooves; 313, of Succory, twelve-sided and dotted.
[Pg 104]
297. An ordinary pollen-grain has two coats; the outer coat thickish,
but weak, and frequently adorned with lines or bands, or studded with
points; the inner coat is extremely thin and delicate, but extensible,
and its cavity when fresh contains a thickish protoplasmic fluid, often
rendered turbid by an immense number of minute particles that float in
it. As the pollen matures this fluid usually dries up, but the
protoplasm does not lose its vitality. When the grain is wetted it
absorbs water, swells up, and is apt to burst, discharging the contents.
But when weak syrup is used it absorbs this slowly, and the tough inner
coat will sometimes break through the outer and begin a kind of growth,
like that which takes place when the pollen is placed upon the stigma.
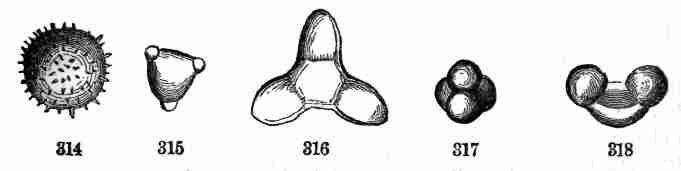
Fig. 314. Magnified pollen of Hibiscus and other
Mallow-plants, beset with prickly projections; 315, of Circæa, with
angles bearing little lobes; 316, of Evening Primrose, the three lobes
as large as the central body; 317, of Kalmia, four grains united, as in
most of the Heath family; 318, of Pine, as it were of three grains or
cells united; the lateral empty and light.
298. Some pollen-grains are, as it were, lobed (as in Fig. 315, 316), or
formed of four grains united (as in the Heath family, Fig. 317): that of
Pine (Fig. 318) has a large rounded and empty bladder-like expansion
upon each side. This renders such pollen very buoyant, and capable of
being transported to a great distance by the wind.
299. In species of Acacia simple grains lightly cohere into globular
pellets. In Milkweeds and in most Orchids all the pollen of an
anther-cell is compacted or coherent into one mass, called a
Pollen-mass, or Pollinium, plural Pollinia. (Fig. 319-322.)
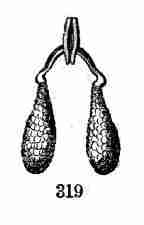
Fig. 319. Pollen, a pair of pollinia of a Milkweed,
Asclepias, attached by stalks to a gland; moderately magnified.
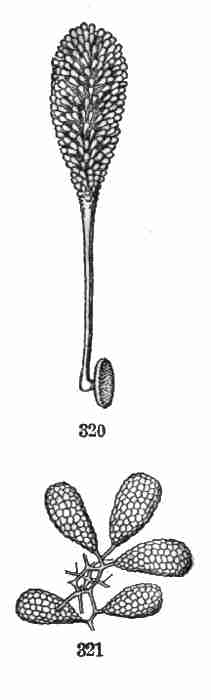
Fig. 320. Pollinium of an Orchis (Habenaria), with its
stalk attached to a sticky gland; magnified. 321. Some of the packets or
partial pollinia, of which Fig. 320 is made up, more magnified.
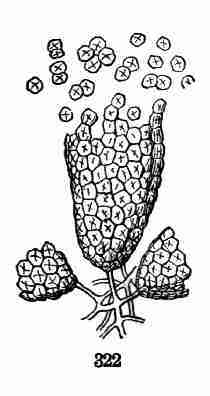
Fig. 322. One of the partial pollinia, torn up at top to
show the grains (which are each composed of four), and highly
magnified.
[Pg 105]
Section X. PISTILS IN PARTICULAR.
§ 1. ANGIOSPERMOUS OR ORDINARY GYNŒCIUM.
300. Gynœcium is the technical name for the pistil or pistils of a
flower taken collectively, or for whatever stands in place of these. The
various modifications of the gynœcium and the terms which relate to
them require particular attention.
301. The Pistil, when only one, occupies the centre of the flower; when
there are two pistils, they stand facing each other in the centre of the
flower; when several, they commonly form a ring or circle; and when very
numerous, they are generally crowded in rows or spirals on the surface
of a more or less enlarged or elongated receptacle. Their number gives
rise to certain terms, the counterpart of those used for stamens (284),
which are survivals of the names of orders in the Linnæan artificial
system. The names were coined by prefixing Greek numerals to -gynia
used for gynœcium, and changed into adjectives in the form of
-gynous. That is, a flower is
Monogynous, when it has a single pistil, whether that be simple or
compound;
Digynous, when it has only two pistils; Trigynous, when with three;
Tetragynous, with four; Pentagynous, with five; Hexagynous, with
six; and so on to Polygynous, with many pistils.
302. The Parts of a Complete Pistil, as already twice explained (16,
236), are the Ovary, the Style, and the Stigma. The ovary is one
essential part: it contains the rudiments of seeds, called Ovules. The
stigma at the summit is also essential: it receives the pollen, which
fertilizes the ovules in order that they may become seeds. But the
style, commonly a tapering or slender column borne on the summit of the
ovary, and bearing the stigma on its apex or its side, is no more
necessary to a pistil than the filament is to the stamen. Accordingly,
there is no style in many pistils: in these the stigma is sessile,
that is, rests directly on the ovary (as in Fig. 326). The stigma is
very various in shape and appearance, being sometimes a little knob (as
in the Cherry, Fig. 271), sometimes a point or small surface of bare
tissue (as in Fig. 327-330), and sometimes a longitudinal crest or line
(as in Fig. 324, 341-343), or it may occupy the whole length of the
style, as in Fig. 331.
303. The word Pistil (Latin, Pistillum) means a pestle. It came into
use in the first place for such flowers as those of Crown Imperial, or
Lily, in which the pistil in the centre was likened to the pestle, and
the perianth around it to the mortar, of the apothecary.
304. A pistil is either simple or compound. It is simple when it
answers to a single flower-leaf, compound when it answers to two or
three, or a fuller circle of such leaves conjoined.
[Pg 106]
305. Carpels. It is convenient to have a name for each flower-leaf of
the gynœcium; so it is called a Carpel, in Latin Carpellum or
Carpidium. A simple pistil is a carpel. Each component flower-leaf of
a compound pistil is likewise a carpel. When a flower has two or more
pistils, these of course are simple pistils, that is, separate carpels
or pistil-leaves. There may be only a single simple pistil to the
flower, as in a Pea or Cherry blossom (Fig. 271); there may be two such,
as in many Saxifrages; or many, as in the Strawberry. More commonly the
single pistil in the centre of a blossom is a compound one. Then there
is seldom much difficulty in ascertaining the number of carpels or
pistil-leaves that compose it.
306. The Simple Pistil, viewed morphologically, answers to a leaf-blade
with margins incurved and united where they meet, so forming a closed
case or pod (the ovary), and bearing ovules at the suture or junction of
these margins: a tapering upper portion with margins similarly inrolled,
is supposed to form the style; and these same margins, exposed at the
tip or for a portion of the length, become the stigma. Compare, under
this view, the three accompanying figures.
Fig. 323. An inrolled small leaf, such as in
double-flowered Cherry blossoms is often seen to occupy the place of a
pistil.
Fig. 324. A simple pistil (of Isopyrum), with ovary cut
across; the inner (ventral) face turned toward the eye: the ovules seem
to be borne on the ventral suture, answering to leaf-margins: the stigma
above seen also to answer to leaf-margins.
Fig. 325. Pod or simple pistil of Caltha or
Marsh-Marigold, which has opened, and shed its seeds.
307. So a simple pistil should have a one-celled ovary, only one line of
attachment for the ovules, a single style, and a single stigma. Certain
variations from this normal condition which sometimes occur do not
invalidate this morphological conception. For instance, the stigma may
become two-lobed or two-ridged, because it consists of two leaf-margins,
as Fig. 324 shows; it may become 2-locellate by the turning or growing
inward of one of the sutures, so as to divide the cavity.
308. There are two or three terms which primarily relate to the parts of
a simple pistil or carpel, and are thence carried on to the compound
pistil, viz.:—
Ventral Suture, the line which answers to the united margins of the
carpel-leaf, therefore naturally called a suture or seam, and the
ventral or inner one, because in the circle of carpel-leaves it looks
inward or to the centre of the flower.
Dorsal Suture is the line down the back of the carpel, answering to [Pg 107]the
midrib of the leaf,—not a seam therefore; but at maturity many fruits,
such as pea-pods, open by this dorsal as well as by the ventral line.
Placenta, a name given to the surface, whatever it be, which bears the
ovules and seeds. The name may be needless when the ovules grow directly
on the ventral suture, or from its top or bottom; but when there are
many ovules there is usually some expansion of an ovule-bearing or
seed-bearing surface; as is seen in our Mandrake or Podophyllum, Fig.
326.
Fig. 326. Simple pistil of Podophyllum, cut across,
showing ovules borne on placenta.
Fig. 327. Pistil of a Saxifrage, of two simple carpels or
pistil-leaves, united at the base only, cut across both above and
below.
Fig. 328. Compound 3-carpellary pistil of common St.
John's-wort, cut across: the three styles separate.
Fig. 329. The same of shrubby St. John's-wort; the three
styles as well as ovaries here united into one.
Fig. 330. Compound 3-carpellary pistil of Tradescantia or
Spiderwort; the three stigmas as well as styles and ovary completely
coalescent into one.
309. A Compound Pistil is a combination of two, three, or a greater
number of pistil-leaves or carpels in a circle, united into one body, at
least by their ovaries. The annexed figures should make it clear. A
series of Saxifrages might be selected the gynœcium of which would
show every gradation between two simple pistils, or separate carpels,
and their complete coalescence into one compound and two-celled ovary.
Even when the constituent styles and stigmas are completely coalescent
into one, the nature of the combination is usually revealed by some
external lines or grooves, or (as in Fig. 328-330) by the internal
partitions, or the number of the placentæ. The simplest case of compound
pistil is that
310. With two or more Cells and Axile Placentæ, namely, with as many
cells as there are carpels, that have united to compose the organ. [Pg 108]Such
a pistil is just what would be formed if the simple pistils (two, three,
or five in a circle, as the case may be), like those of a Pæony or
Stonecrop (Fig. 224, 225), pressed together in the centre of the flower,
were to cohere by their contiguous parts. In such a case the placentæ
are naturally axile, or all brought together in the axis or centre;
and the ovary has as many Dissepiments, or internal Partitions, as
there are carpels in its composition. For these are the contiguous and
coalescent walls or sides of the component carpels. When such pistils
ripen into pods, they often separate along these lines into their
elementary carpels.
Fig. 331, 332. Pistil of a Sandwort, with vertical and
transverse section of the ovary: free central placenta.
311. One-celled, with free Central Placenta. The commoner case is that
of Purslane (Fig. 272) and of the Pink and Chickweed families (Fig. 331,
332). This is explained by supposing that the partitions (such as those
of Fig. 329) have early vanished or have been suppressed. Indeed, traces
of them may often be detected in Pinks. On the other hand, it is equally
supposable that in the Primula family the free central is derived from
parietal placentation by the carpels bearing ovules only at base, and
forming a consolidated common placenta in the axis. Mitella and Dionæa
help out this conception.
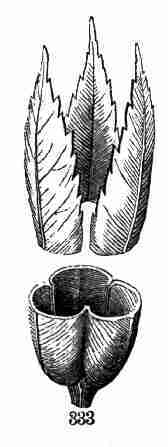
Fig. 333. Plan of a one-celled ovary of three
carpel-leaves, with parietal placentæ, cut across below, where it is
complete; the upper part showing the top of the three leaves it is
composed of, approaching, but not united.
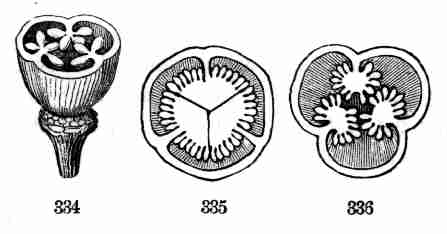
Fig. 334. Cross section of the ovary of Frost weed
(Helianthemum), with three parietal placentæ, bearing ovules.
Fig. 335. Cross section of an ovary of Hypericum
graveolens, the three large placentæ meeting in the centre, so as to
form a three-celled ovary. 336. Same in fruit, the placentæ now separate
and rounded.
312. One-celled, with Parietal Placentæ. In this not uncommon case it is
conceived that the two or three or more carpel-leaves of such a compound
pistil coalesce by their adjacent edges, just as sepal-leaves do to form
a gamosepalous calyx, or petals to form a gamopetalous corolla, and as
is shown in the diagram, Fig. 333, and in an actual cross-section, Fig.
334. Here each carpel is an open leaf, or with some introflexion,
bearing ovules along its margins; and each placenta consists of the
contiguous [Pg 109]margins of two pistil-leaves grown together. There is every
gradation between this and the three-celled ovary with the placentæ in
the axis, even in the same genus, sometimes even in different stages in
the same pistil (Fig. 335, 336).
§ 2. GYMNOSPERMOUS GYNŒCIUM.
313. The ordinary pistil has a closed ovary, and accordingly the pollen
can act upon the contained ovules only indirectly, through the stigma.
This is expressed in a term of Greek derivation, viz.:—
Angiospermous, meaning that the seeds are borne in a sac or closed
vessel. The counterpart term is
Gymnospermous, meaning naked-seeded. This kind of pistil, or
gynœcium, the simplest of all, yet the most peculiar, characterizes
the Pine family and its relatives.

Fig. 337. A pistil, that is, a scale of the cone, of a
Larch, at the time of flowering; inside view, showing its pair of naked
ovules.
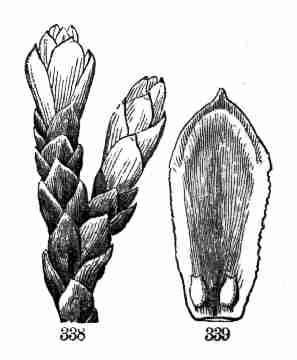
Fig. 338. Branchlet of the American Arbor-Vitæ,
considerably larger than in nature, terminated by its pistillate
flowers, each consisting of a single scale (an open pistil), together
forming a small cone.
Fig. 339. One of the scales or carpels of the last,
removed and more enlarged, the inside exposed to view, showing a pair of
ovules on its base.
314. While the ordinary simple pistil is conceived by the botanist to be
a leaf rolled together into a closed pod (306), those of the Pine, Larch
(Fig. 337), Cedar, and Arbor-Vitæ (Fig. 338, 339) are open leaves, in
the form of scales, each bearing two or more ovules on the inner face,
next the base. At the time of blossoming, these pistil-leaves of the
young cone diverge, and the pollen, so abundantly shed from the
staminate blossoms, falls directly upon the exposed ovules. Afterward
the scales close over each other until the seeds are ripe. Then they
separate that the seeds may be shed. As the pollen acts directly on the
ovules, such pistil (or organ acting as pistil) has no stigma.
315. In the Yew, and in Torreya and Gingko, the gynœcium is reduced
to extremest simplicity, that is, to a naked ovule, without any visible
carpel.
316. In Cycas the large naked ovules are borne on the margins or lobes
of an obvious open leaf. All Gymnospermous plants have other
peculiarities, also distinguishing them, as a class, from Angiospermous
plants.
[Pg 110]
Section XI. OVULES.
317. Ovule (from the Latin, meaning a little egg) is the technical name
of that which in the flower answers to and becomes the seed.
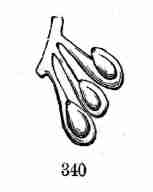
Fig. 340. A cluster of ovules, pendulous on their
funicles.
318. Ovules are naked in gymnospermous plants (as just described), in
all others they are enclosed in the ovary. They may be produced along
the whole length of the cell or cells of the ovary, and then they are
apt to be numerous, or only from some part of it, generally the top or
the bottom. In this case they are usually few or single (solitary, as
in Fig. 341-343). They may be sessile, i. e. without stalk, or they
may be attached by a distinct stalk, the Funicle or Funiculus (Fig.
340).
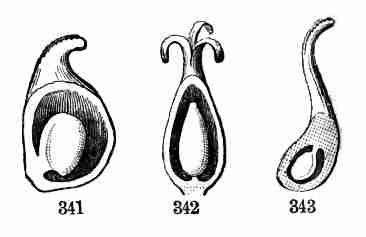
Fig. 341. Section of the ovary of a Buttercup,
lengthwise, showing its ascending ovule.
Fig. 342. Section of the ovary of Buckwheat, showing the
erect ovule.
Fig. 343. Section of the ovary of Anemone, showing its
suspended ovule.
319. Considered as to then position and direction in the ovary, they are
Horizontal, when they are neither turned upward nor downward, as in
Podophyllum (Fig. 326),
Ascending, when rising obliquely upwards, usually from the side of the
cell, not from its very base, as in the Buttercup (Fig. 341), and the
Purslane (Fig. 272),
Erect, when rising upright from the very base of the cell, as in the
Buckwheat (Fig. 342),
Pendulous, when hanging from the side or from near the top, as in the
Flax (Fig. 270), and
Suspended, when hanging perpendicularly from the very summit of the
cell, as in the Anemone (Fig. 343). All these terms equally apply to
seeds.
320. In structure an ovule is a pulpy mass of tissue, usually with one
or two coats or coverings. The following parts are to be noted, viz.—
Kernel or Nucleus, the body of the ovule. In the Mistletoe and some
related plants, there is only this nucleus, the coats being wanting.
Teguments, or coats, sometimes only one, more commonly two. When two,
one has been called Primine, the other Secundine. It will serve all
purposes to call them simply outer and inner ovule coats.
Orifice, or Foramen, an opening through the coats at the organic apex of
the ovule. In the seed it is Micropyle.
Chalaza, the place where the coats and the kernel of the ovule blend.
Hilum, the place of junction of the funiculus with the body of the
ovule.
[Pg 111]
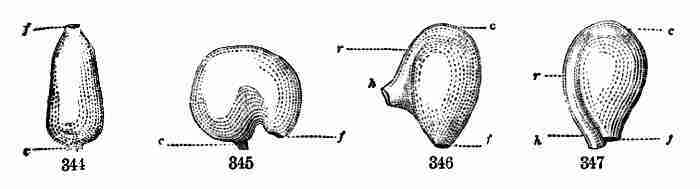
Fig. 344. Orthotropous ovule of Buckwheat: c, hilum
and chalaza; f, orifice.
Fig. 345. Campylotropous ovule of a Chickweed: c, hilum
and chalaza; f, orifice.
Fig. 346. Amphitropous ovule of Mallow: f, orifice;
h, hilum; r, rhaphe; c, chalaza.
Fig. 347. Anatropous ovule of a Violet, the parts
lettered as in the last.
321. The Kinds of Ovules. The ovules in their growth develop in three or
four different ways and thereby are distinguished into
Orthotropous or Straight, those which develop without curving or
turning, as in Fig. 344. The chalaza is at the insertion or base, the
foramen or orifice is at the apex. This is the simplest, but the least
common kind of ovule.
Campylotropous or Incurved, in which, by the greater growth of one
side, the ovule curves into a kidney-shaped outline, so bringing the
orifice down close to the base or chalaza; as in Fig. 345.
Amphitropous or Half Inverted, Fig. 346. Here the forming ovule,
instead of curving perceptibly, keeps its axis nearly straight, and, as
it grows, turns round upon its base so far as to become transverse to
its funiculus, and adnate to its upper part for some distance. Therefore
in this case the attachment of the funiculus or stalk is about the
middle, the chalaza is at one end, the orifice at the other.
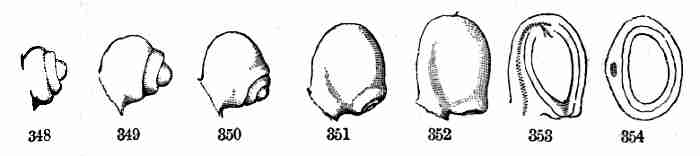
Fig. 348-350. Three early stages in the growth of ovule
of a Magnolia, showing the forming outer and inner coats which even in
the later figure have not yet completely enclosed the nucleus; 351,
further advanced, and 352, completely anatropous ovule.
Fig. 353. Longitudinal section, and 354, transverse
section of 352.
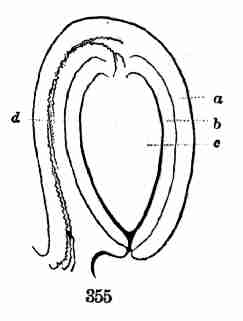
Fig. 355. Same as 353, enlarged showing the parts in
section: a, outer coat; b, inner coat; c, nucleus; d, rhaphe.
Anatropous or Inverted, as in Fig. 347, the commonest kind, so
called because in its growth it has as it were turned over upon its
stalk, to which it has continued adnate. The organic base, or chalaza,
thus becomes the apparent summit, and the [Pg 112]orifice is at the base, by
the side of the hilum or place of attachment. The adnate portion of the
funiculus, which appears as a ridge or cord extending from the hilum to
the chalaza, and which distinguishes this kind of ovule, is called the
Rhaphe. The amphitropous ovule (Fig. 346) has a short or incomplete
rhaphe.
322. Fig. 348-352 show the stages through which an ovule becomes
anatropous in the course of its growth. The annexed two figures are
sections of such an ovule at maturity; and Fig. 355 is Fig. 353
enlarged, with the parts lettered.
Section XII. MODIFICATIONS OF THE RECEPTACLE.
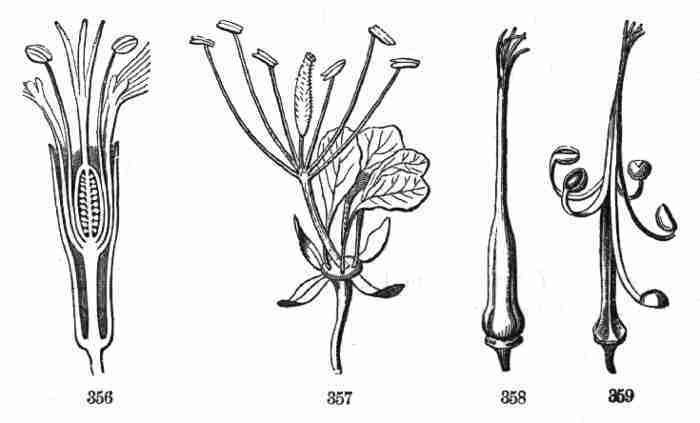
Fig. 356. Longitudinal section of flower of Silene
Pennsylvanica, showing stipe between calyx and corolla.
Fig. 357. Flower of a Cleome of the section Gynandropsis,
showing broadened receptacle to bear petals, lengthened stipe below the
stamens, and another between these and pistil.
Fig. 358. Pistil of Geranium or Cranesbill.
Fig. 359. The same, ripe, with the five carpels splitting
away from the long beak (carpophore), and hanging from its top by their
recurving styles.
323. The Torus or Receptacle of the flower (237, Fig. 223) is the
portion which belongs to the stem or axis. In all preceding
illustrations it is small and short. But it sometimes lengthens,
sometimes thickens or variously enlarges, and takes on various forms.
Some of these have received special names, very few of which are in
common use. A lengthened portion of the receptacle is called
A Stipe. This name, which means simply a trunk or stalk, is used in
botany for various stalks, even for the leaf-stalk in Ferns. It is also
applied to the stalk or petiole of a carpel, in the rare cases when
there is any, as in [Pg 113]Goldthread. Then it is technically distinguished as
a Thecaphore. When there is a stalk, or lengthened internode of
receptacle, directly under a compound pistil, as in Stanleya and some
other Cruciferæ, it is called a Gynophore. When the stalk is developed
below the stamens, as in most species of Silene (Fig. 356), it has been
called an Anthophore or Gonophore. In Fig. 357 the torus is dilated
above the calyx where it bears the petals, then there is a long
internode (gonophore) between it and the stamens; then a shorter one
(gynophore) between these and the pistil.
324. A Carpophore is a prolongation of receptacle or axis between the
carpels and bearing them. Umbelliferous plants and Geranium (Fig. 358,
359) afford characteristic examples.
Fig. 360. Longitudinal section of a young strawberry,
enlarged.
Fig. 361. Similar section of a young Rose-hip.
Fig. 362. Enlarged and top-shaped receptacle of
Nelumbium, at maturity.
325. Flowers with very numerous simple pistils generally have the
receptacle enlarged so as to give them room; sometimes becoming broad
and flat, as in the Flowering Raspberry, sometimes elongated, as in the
Blackberry, the Magnolia, etc. It is the receptacle in the Strawberry
(Fig. 360), much enlarged and pulpy when ripe, which forms the eatable
part of the fruit, and bears the small seed-like pistils on its surface.
In the Rose (Fig. 361), instead of being convex or conical, the
receptacle is deeply concave, or urn-shaped. Indeed, a Rose-hip may be
likened to a strawberry turned inside out, like the finger of a glove
reversed, and the whole covered by the adherent tube of the calyx. The
calyx remains beneath in the strawberry.
326. In Nelumbium, of the Water-Lily family, the singular and greatly
enlarged receptacle is shaped like a top, and bears the small pistils
immersed in separate cavities of its flat upper surface (Fig. 362).
 Fig.
Fig. 363. Hypogynous disk in Orange.
327. A Disk is an enlarged low receptacle or an outgrowth from it,
hypogynous when underneath the pistil, as in Rue and the Orange (Fig.
363), and perigynous when adnate to calyx-tube (as in Buckthorn, Fig.
364, 365), and Cherry (Fig. 271), or [Pg 114]to both calyx-tube and ovary, as
in Hawthorn (Fig. 273). A flattened hypogynous disk, underlying the
ovary or ovaries, and from which they fall away at maturity, is
sometimes called a Gynobase, as in the Rue family. In some Borragineous
flowers, such as Houndstongue, the gynobase runs up in the centre
between the carpels into a carpophore. The so-called epigynous disk
(or Stylopodium) crowning the summit of the ovary in flowers of
Umbelliferæ, etc., cannot be said to belong to the receptacle.
Fig. 364. Flower of a Buckthorn showing a conspicuous
perigynous disk.
Fig. 365. Vertical section of same flower.
Section XIII. FERTILIZATION.
328. The end of the flower is attained when the ovules become seeds. A
flower remains for a certain time (longer or shorter according to the
species) in anthesis, that is, in the proper state for the fulfilment
of this end. During anthesis, the ovules have to be fertilized by the
pollen; or at least some pollen has to reach the stigma, or in
gymnospermy the ovule itself, and to set up the peculiar growth upon its
moist and permeable tissue, which has for result the production of an
embryo in the ovules. By this the ovules are said to be fertilized.
The first step is pollination, or, so to say, the sowing of the proper
pollen upon the stigma, where it is to germinate.
§ 1. ADAPTATIONS FOR POLLINATION OF THE STIGMA.
329. These various and ever-interesting adaptations and processes are
illustrated in the "Botanical Text Book, Structural Botany," chap. VI.
sect. iv., also in a brief and simple way in "Botany for Young People,
How Plants Behave." So mere outlines only are given here.
330. Sometimes the application of pollen to the stigma is left to
chance, as in diœcious wind-fertilized flowers; sometimes it is
rendered very sure, as in flowers that are fertilized in the bud;
sometimes the pollen is prevented from reaching the stigma of the same
flower, although placed very near to it, but then there are always
arrangements for its transference to the stigma of some other blossom of
the kind. It is among these last that the most exquisite adaptations are
met with.
331. Accordingly, some flowers are particularly adapted to close or
self-fertilization; others to cross fertilization; some for either,
according to circumstances.
[Pg 115]
Close Fertilization occurs when the pollen reaches and acts upon a
stigma of the very same flower (this is also called self-fertilization),
or, less closely, upon other blossoms of the same cluster or the same
individual plant.
Cross Fertilization occurs when ovules are fertilized by pollen of
other individuals of the same species.
Hybridization occurs when ovules are fertilized by pollen of some
other (necessarily some nearly related) species.
332. Close Fertilization would seem to be the natural result in ordinary
hermaphrodite flowers; but it is by no means so in all of them. More
commonly the arrangements are such that it takes place only after some
opportunity for cross fertilization has been afforded. But close
fertilization is inevitable in what are called
Cleistogamous Flowers, that is, in those which are fertilized in the
flower-bud, while still unopened. Most flowers of this kind, indeed,
never open at all; but the closed floral coverings are forced off by the
growth of the precociously fertilized pistil. Common examples of this
are found in the earlier blossoms of Specularia perfoliata, in the later
ones of most Violets, especially the stemless species, in our wild Jewel
weeds or Impatiens, in the subterranean shoots of Amphicarpæa. Every
plant which produces these cleistogamous or bud-fertilized flowers bears
also more conspicuous and open flowers, usually of bright colors. The
latter very commonly fail to set seed, but the former are prolific.
333. Cross Fertilization is naturally provided for in diœcious plants
(249), is much favored in monœcious plants (249), and hardly less so
in dichogamous and in heterogonous flowers (338). Cross fertilization
depends upon the transportation of pollen; and the two principal agents
of conveyance are winds and insects. Most flowers are in their whole
structure adapted either to the one or to the other.
334. Wind-fertilizable or Anemophilous flowers are more commonly
diœcious or monœcious, as in Pines and all coniferous trees, Oaks,
and Birches, and Sedges; yet sometimes hermaphrodite, as in Plantains
and most Grasses; they produce a superabundance of very light pollen,
adapted to be wind-borne; and they offer neither nectar to feed winged
insects, nor fragrance nor bright colors to attract them.
335. Insect-fertilizable or Entomophilous flowers are those which are
sought by insects, for pollen or for nectar, or for both. Through their
visits pollen is conveyed from one flower and from one plant to another.
Insects are attracted to such blossoms by their bright colors, or their
fragrance, or by the nectar (the material of honey) there provided for
them. While supplying their own needs, they carry pollen from anthers to
stigmas and from plant to plant, thus bringing about a certain amount of
cross fertilization. Willows and some other diœcious flowers are so
fertilized, chiefly by bees. But most insect-visited flowers have the
stamens and pistils associated either in the same or in contiguous
blossoms. Even when in the same blossom, anthers and stigmas are very
commonly so situated [Pg 116]that under insect-visitation, some pollen is more
likely to be deposited upon other than upon own stigmas, so giving a
chance for cross as well as for close fertilization. On the other hand,
numerous flowers, of very various kinds, have their parts so arranged
that they must almost necessarily be cross-fertilized or be barren, and
are therefore dependent upon the aid of insects. This aid is secured by
different exquisite adaptations and contrivances, which would need a
volume for full illustration. Indeed, there is a good number of volumes
devoted to this subject.[1]
336. Some of the adaptations which favor or ensure cross fertilization
are peculiar to the particular kind of blossom. Orchids, Milkweeds,
Kalmia, Iris, and papilionaceous flowers each have their own special
contrivances, quite different for each.
337. Irregular flowers (253) and especially irregular corollas are
usually adaptations to insect-visitation. So are all Nectaries,
whether hollow spurs, sacs, or other concavities in which nectar is
secreted, and all nectariferous glands.
338. Moreover, there are two arrangements for cross fertilization common
to hermaphrodite flowers in various different families of plants, which
have received special names, Dichogamy and Heterogony.
339. Dichogamy is the commoner case. Flowers are dichogamous when the
anthers discharge their pollen either before or after the stigmas of
that flower are in a condition to receive it. Such flowers are
Proterandrous, when the anthers are earlier than the stigmas, as in
Gentians, Campanula, Epilobium, etc.
Proterogynous, when the stigmas are mature and moistened for the
reception of pollen, before the anthers of that blossom are ready to
supply it, and are withered before that pollen can be supplied.
Plantains or Ribworts (mostly wind-fertilized) are strikingly
proterogynous: so is Amorpha, our Papaws, Scrophularia, and in a less
degree the blossom of Pears, Hawthorns, and Horse-chestnut.
340. In Sabbatia, the large-flowered species of Epilobium, and
strikingly in Clerodendron, the dichogamy is supplemented and perfected
by movements of the stamens and style, one or both, adjusted to make
sure of cross fertilization.
341. Heterogony. This is the case in which hermaphrodite and fertile
flowers of two sorts are produced on different individuals of the same
species; one sort having higher anthers and lower stigmas, the other
having higher stigmas and lower anthers. Thus reciprocally disposed, a
visiting insect carries pollen from the high anthers of the one to the
high stigma of the other, and from the low anthers of the one to the low
stigma of the other. These plants are practically as if diœcious,
with the advantage that [Pg 117]both kinds are fruitful. Houstonia and
Mitchella, or Partridge-berry, are excellent and familiar examples.
These are cases of
Heterogone Dimorphism, the relative lengths being only short and long
reciprocally.
Heterogone Trimorphism, in which there is a mid-length as well as a
long and a short set of stamens and style; occurs in Lythrum Salicaria
and some species of Oxalis.
342. There must be some essential advantage in cross fertilization or
cross breeding. Otherwise all these various, elaborate, and exquisitely
adjusted adaptations would be aimless. Doubtless the advantage is the
same as that which is realized in all the higher animals by the
distinction of sexes.
§ 2. ACTION OF POLLEN, AND FORMATION OF THE EMBRYO.
343. Pollen-growth. A grain of pollen may be justly likened to one of
the simple bodies (spores) which answer for seeds in Cryptogamous
plants. Like one of these, it is capable of germination. When deposited
upon the moist surface of the stigma (or in some cases even when at a
certain distance) it grows from some point, its living inner coat
breaking through the inert outer coat, and protruding in the form of a
delicate tube. This as it lengthens penetrates the loose tissue of the
stigma and of a loose conducting tissue in the style, feeds upon the
nourishing liquid matter there provided, reaches the cavity of the
ovary, enters the orifice of an ovule, and attaches its extremity to a
sac, or the lining of a definite cavity, in the ovule, called the
Embryo-Sac.
344. Origination of the Embryo. A globule of living matter in the
embryo-sac is formed, and is in some way placed in close proximity to
the apex of the pollen tube; it probably absorbs the contents of the
latter; it then sets up a special growth, and the Embryo (8-10) or
rudimentary plantlet in the seed is the result.
Section XIV. THE FRUIT.
345. Its Nature. The ovary matures into the Fruit. In the strictest
sense the fruit is the seed-vessel, technically named the Pericarp. But
practically it may include other parts organically connected with the
pericarp. Especially the calyx, or a part of it, is often incorporated
with the ovary, so as to be undistinguishably a portion of the pericarp,
and it even forms along with the receptacle the whole bulk of such
edible fruits as apples and pears. The receptacle is an obvious part in
blackberries, and is the whole edible portion in the strawberry.
346. Also a cluster of distinct carpels may, in ripening, be
consolidated or compacted, so as practically to be taken for one fruit.
Such are raspberries, [Pg 118]blackberries, the Magnolia fruit, etc. Moreover,
the ripened product of many flowers may be compacted or grown together
so as to form a single compound fruit.
347. Its kinds have therefore to be distinguished. Also various names of
common use in descriptive botany have to be mentioned and defined.
348. In respect to composition, accordingly, fruits may be classified
into
Simple, those which result from the ripening of a single pistil, and
consist only of the matured ovary, either by itself, as in a cherry, or
with calyx-tube completely incorporated with it, as in a gooseberry or
cranberry.
Aggregate, when a cluster of carpels of the same flower are crowded
into a mass; as in raspberries and blackberries.
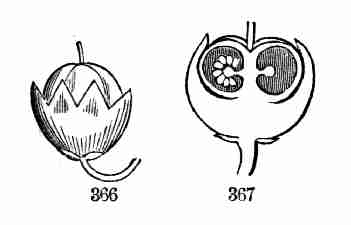
Fig. 366. Forming fruit (capsule) of Gaultheria, with
calyx thickening around its base. 367. Section of same mature, the
berry-like calyx nearly enclosing the capsule.
Fig. 368. Section of a part of a strawberry. Compare with
Fig. 360.
Fig. 369. Similar section of part of a blackberry. 370.
One of its component simple fruits (drupe) in section, showing the pulp,
stone, and contained seed; more enlarged. Compare with Fig. 375.
Accessory or Anthocarpous, when the surroundings or supports of the
pistil make up a part of the mass; as does the loose calyx changed into
a fleshy and berry-like envelope of our Wintergreen (Gaultheria, Fig.
366, 367) and Buffalo-berry, which are otherwise simple fruits. In an
aggregate fruit such as the strawberry the great mass is receptacle
(Fig. 360, 368); and in the blackberry (Fig. 369) the juicy receptacle
forms the central part of the savory mass.
Multiple or Collective, when formed from several flowers
consolidated into one mass, of which the common receptacle or axis of
inflorescence, the floral envelopes, and even the bracts, etc., make a
part. A mulberry (Fig. 408, which superficially much resembles a
blackberry) is of this multiple sort. A pine-apple is another example.
349. In respect to texture or consistence, fruits may be distinguished
into three kinds, viz.—
Fleshy Fruits, those which are more or less soft and juicy throughout;
[Pg 119]
Stone Fruits, or Drupaceous, the outer part fleshy like a berry,
the inner hard or stony, like a nut; and
Dry Fruits, those which have no flesh or pulp.
350. In reference to the way of disseminating the contained seed, fruits
are said to be
Indehiscent when they do not open at maturity. Fleshy fruits and stone
fruits are of course indehiscent. The seed becomes free only through
decay or by being fed upon by animals. Those which escape digestion are
thus disseminated by the latter. Of dry fruits many are indehiscent; and
these are variously arranged to be transported by animals. Some burst
irregularly; many are
Dehiscent, that is, they split open regularly along certain lines, and
discharge the seeds. A dehiscent fruit almost always contains many or
several seeds, or at least more than one seed.
Fig. 371. Leafy shoot and berry (cut across) of the
larger Cranberry, Vaccinium macrocarpon.
Fig. 372, 373. Pepo of Gourd, in section. 373. One carpel
of same in diagram.
Fig. 374. Longitudinal and transverse sections of a pear
(pome).
351. The principal kinds of fruit which have received substantive names
and are of common use in descriptive botany are the following. Of fleshy
fruits the leading kind is
352. The Berry, such as the gooseberry and currant, the blueberry and
cranberry (Fig. 371), the tomato, and the grape. Here the whole flesh is
soft throughout. The orange is a berry with a leathery rind.
353. The Pepo, or Gourd-fruit, is a hard-rinded berry, belonging to
the Gourd family, such as the pumpkin, squash, cucumber, and melon, Fig.
372, 373.
354. The Pome is a name applied to the apple, pear (Fig. 374), and
quince; fleshy fruits, like a berry, but the principal thickness is
calyx, only [Pg 120]the papery pods arranged like a star in the core really
belonging to the carpels. The fruit of the Hawthorn is a drupaceous
pome, something between pome and drupe.
355. Of fruits which are externally fleshy and internally hard the
leading kind is
356. The Drupe, or Stone-fruit; of which the cherry, plum, and peach
(Fig. 375) are familiar examples. In this the outer part of the
thickness of the pericarp becomes fleshy, or softens like a berry, while
the inner hardens, like a nut. From the way in which the pistil is
constructed, it is evident that the fleshy part here answers to the
lower, and the stone to the upper face of the component leaf. The layers
or concentric portions of a drupe, or of any pericarp which is thus
separable, are named, when thus distinguishable into three portions,—
Epicarp, the external layer, often the mere skin of the fruit,
Mesocarp, the middle layer, which is commonly the fleshy part, and
Endocarp, the innermost layer, the stone. But more commonly only two
portions of a drupe are distinguished, and are named, the outer one
Sarcocarp or Exocarp, for the flesh, the first name referring to the
fleshy character, the second to its being an external layer; and
Putamen or Endocarp, the Stone, within.
Fig. 375. Longitudinal section of a peach, showing flesh,
stone, and seed.
357. The typical or true drupe is of a single carpel. But, not to
multiply technical names, this name is extended to all such fruits when
fleshy without and stony within, although of compound pistil,—even to
those having several or separable stones, such as the fruit of Holly.
These stones in such drupes, or drupaceous fruits, are called Pyrenæ,
or Nucules, or simply Nutlets of the drupe.
358. Of Dry fruits, there is a greater diversity of kinds having
distinct names. The indehiscent sorts are commonly one-seeded.
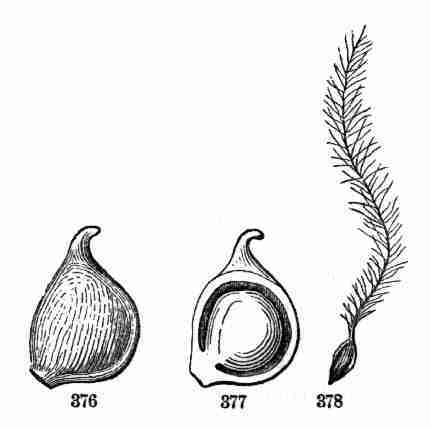
Fig. 376. Akene of a Buttercup. 377. The same, divided
lengthwise, to show the contained seed.
Fig. 378. Akene of Virgin's-bower, retaining the
feathered style, which aids in dissemination.
359. The Akene or Achenium is a small, dry and indehiscent one-seeded
fruit, often so seed-like in appearance that it is popularly taken for a
naked seed. The fruit of the Buttercup or Crowfoot is a good example,
Fig. 376, 377. Its nature, as a ripened pistil (in this [Pg 121]case a simple
carpel), is apparent by its bearing the remains of a style or stigma, or
a scar from which this has fallen. It may retain the style and use it in
various ways for dissemination (Fig. 378).
360. The fruit of Compositæ (though not of a single carpel) is also an
akene. In this case the pericarp is invested by an adherent calyx-tube;
the limb of which, when it has any, is called the Pappus. This name was
first given to the down like that of the Thistle, but is applied to all
forms under which the limb of the calyx of the "compound flower"
appears. In Lettuce, Dandelion (Fig. 384), and the like, the achenium as
it matures tapers upwards into a slender beak, like a stalk to the
pappus.
Fig. 379. Akene of Mayweed (no pappus). 380. That of
Succory (its pappus a shallow cup). 381. Of Sunflower (pappus of two
deciduous scales). 382. Of Sneezeweed (Helenium), with its pappus of
five scales. 383. Of Sow-Thistle, with its pappus of delicate downy
hairs. 384. Of the Dandelion, its pappus raised on a long beak.
361. A Cremocarp (Fig. 385), a name given to the fruit of Umbelliferæ,
consists as it were of a pair of akenes united completely in the
blossom, but splitting apart when ripe into the two closed carpels. Each
of these is a Mericarp or Hemicarp, names seldom used.
362. A Utricle is the same as an akene, but with a thin and bladdery
loose pericarp; like that of the Goosefoot or Pigweed (Fig. 386). When
ripe it may burst open irregularly to discharge the seed; or it may open
by a circular line all round, the upper part falling off like a lid; as
in the Amaranth (Fig. 387).
Fig. 385. Fruit (cremocarp) of Osmorrhiza; the two
akene-like ripe carpels separating at maturity from a slender axis or
carpophore.
Fig. 386. Utricle of the common Pigweed (Chenopodium
album).
Fig. 387. Utricle (pyxis) of Amaranth, opening all round
(circumscissile).
363. A Caryopsis, or Grain, is like an akene with the seed adhering to
the thin pericarp throughout, so that fruit and seed are incorporated
into one body; as in wheat, Indian corn, and other kinds of grain.
364. A Nut is a dry and indehiscent fruit, commonly one-celled and
one-seeded, [Pg 122]with a hard, crustaceous, or bony wall, such as the
cocoa-nut, hazelnut, chestnut, and the acorn (Fig. 37, 388.) Here the
involucre, in the form of a cup at the base, is called the Cupule. In
the Chestnut the cupule forms the bur; in the Hazel, a leafy husk.
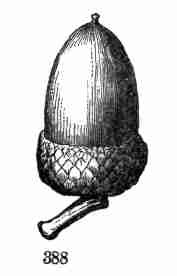
Fig. 388. Nut (acorn) of the Oak, with its cup or
cupule.
365. A Samara, or Key-fruit, is either a nut or an akene, or any other
indehiscent fruit, furnished with a wing, like that of Ash (Fig. 389),
and Elm (Fig. 390). The Maple-fruit is a pair of keys (Fig. 391).
366. Dehiscent Fruits, or Pods, are of two classes, viz., those of a
simple pistil or carpel, and those of a compound pistil. Two common
sorts of the first are named as follows:—
367. The Follicle is a fruit of a simple carpel, which dehisces down one
side only, i. e. by the inner or ventral suture. The fruits of Marsh
Marigold (Fig. 392), Pæony, Larkspur, and Milkweed are of this kind.
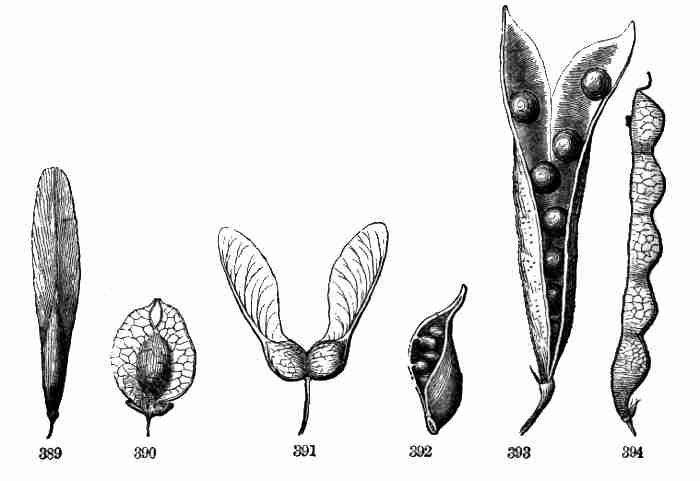
Fig. 389. Samara or key of the White Ash, winged at end.
390. Samara of the American Elm, winged all round.
Fig. 391. Pair of samaras of Sugar Maple.
Fig. 392. Follicle of Marsh Marigold (Caltha palustris).
Fig. 393. Legume of a Sweet Pea, opened.
Fig. 394. Loment or jointed legume of a Tick-Trefoil
(Desmodium).
368. The Legume or true Pod, such as the peapod (Fig. 393), and the
fruit of the Leguminous or Pulse family generally, is one which opens
along the dorsal as well as the ventral suture. The two pieces into
which it splits are called Valves. A Loment is a legume which is
constricted between the seeds, and at length breaks up crosswise into
distinct joints, as in Fig. 394.
369. The pods or dehiscent fruits belonging to a compound ovary have
several technical names: but they all may be regarded as kinds of
370. The Capsule, the dry and dehiscent fruit of any compound pistil.
The capsule may discharge its seeds through chinks or pores, as in the
[Pg 123]Poppy, or burst irregularly in some part, as in Lobelia and the
Snapdragon; but commonly it splits open (or is dehiscent) lengthwise
into regular pieces, called Valves.
Fig. 395. Capsule of Iris, with loculicidal dehiscence;
below, cut across.
Fig. 396. Pod of a Marsh St. John's-wort, with septicidal
dehiscence.
371. Regular Dehiscence in a capsule takes place in two ways, which
are best illustrated in pods of two or three cells. It is either
Loculicidal, or, splitting directly into the loculi or cells, that
is, down the back (or the dorsal suture) of each cell or carpel, as in
Iris (Fig. 395); or
Septicidal, that is, splitting through the partitions or septa, as
in St. John's-wort (Fig. 396), Rhododendron, etc. This divides the
capsule into its component carpels, which then open by their ventral
suture.
Fig. 397, 398. Diagrams of the two modes.
Fig. 399. Diagram of septifragal dehiscence of the
loculicidal type. 400. Same of the septicidal or marginicidal type.
372. In loculicidal dehiscence the valves naturally bear the partitions
on their middle; in the septicidal, half the thickness of a partition is
borne on the margin of each valve. See the annexed diagrams. A variation
of either mode occurs when the valves break away from the partitions,
these remaining attached in the axis of the fruit. This is called
Septifragal dehiscence. One form is seen in the Morning-Glory (Fig.
400).
373. The capsules of Rue, Spurge, and some others, are both loculicidal
and septicidal, and so split into half-carpellary valves or pieces.
374. The Silique (Fig. 401) is the technical name of the peculiar pod of
the Mustard family; which is two-celled by a false partition stretched
across between two parietal placentæ. It generally opens by two valves
from below upward, and the placentæ with the partition are left behind
when the valves fall off.
375. A Silicle or Pouch is only a short and broad silique, like that of
the Shepherd's Purse, Fig. 402, 403.
[Pg 124]
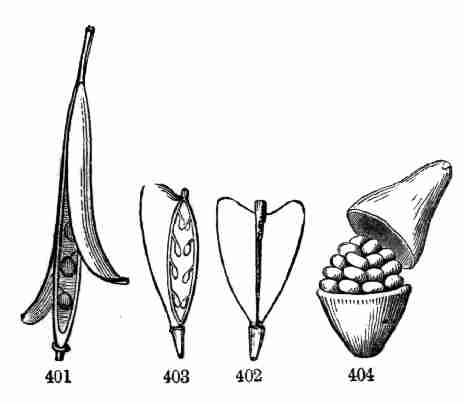
Fig. 401. Silique of a Cadamine or Spring Cress.
Fig. 402. Silicle of Shepherd's Purse.
Fig. 403. Same, with one valve removed.
Fig. 404. Pyxis of Purslane, the lid detaching.
376. The Pyxis is a pod which opens by a circular horizontal line, the
upper part forming a lid, as in Purslane (Fig. 404), the Plantain,
Henbane, etc. In these the dehiscence extends all round, or is
circumscissile. So it does in Amaranth (Fig. 387), forming a
one-seeded utricular pyxis. In Jeffersonia, the line does not separate
quite round, but leaves a portion for a hinge to the lid.
377. Of Multiple or Collective Fruits, which are properly masses of
fruits aggregated into one body (as is seen in the Mulberry (Fig. 408),
Pine-apple, etc.), there are two kinds with special names and of
peculiar structure.
Fig. 405. A fig-fruit when young. 406. Same in section.
407. Magnified portion, a slice, showing some of the flowers.
Fig. 408. A mulberry. 409. One of the grains younger,
enlarged; seen to be a pistillate flower with calyx becoming fleshy.
410. Same, with fleshy calyx cut across.
378. The Syconium or Fig-fruit (Fig. 405, 406) is a fleshy axis or
summit of stem, hollowed out, and lined within by a multitude of minute
flowers, the whole becoming pulpy, and in the common fig, luscious.
379. The Strobile or Cone (Fig. 411), is the peculiar multiple fruit of
Pines, Cypresses, and the like; hence named Coniferæ, viz.
cone-bearing [Pg 125]plants. As already shown (313), these cones are open
pistils, mostly in the form of flat scales, regularly overlying each
other, and pressed together in a spike or head. Each scale bears one or
two naked seeds on its inner face. When ripe and dry, the scales turn
back or diverge, and in the Pine the seed peels off and falls, generally
carrying with it a wing, a part of the lining of the scale, which
facilitates the dispersion of the seeds by the wind (Fig. 412, 413). In
Arbor-Vitæ, the scales of the small cone are few, and not very unlike
the leaves. In Cypress they are very thick at the top and narrow at the
base, so as to make a peculiar sort of closed cone. In Juniper and Red
Cedar, the few scales of the very small cone become fleshy, and ripen
into a fruit which closely resembles a berry.
Fig. 411. Cone of a common Pitch Pine. 412. Inside view
of a separated scale or open carpel; one seed in place: 413, the other
seed.
Section XV. THE SEED.
380. Seeds are the final product of the flower, to which all its parts
and offices are subservient. Like the ovule from which it originates, a
seed consists of coats and kernel.
Fig. 414. Seed of a Linden or Basswood cut through
lengthwise, and magnified, the parts lettered: a, the hilum or scar;
b, the outer coat; c, the inner; d, the albumen; e, the embryo.
381. The Seed-coats are commonly two (320), the outer and the inner.
Fig. 414 shows the two, in a seed cut through lengthwise. The outer coat
is often hard or crustaceous, whence it is called the Testa, or shell
of the seed; the inner is almost always thin and delicate.
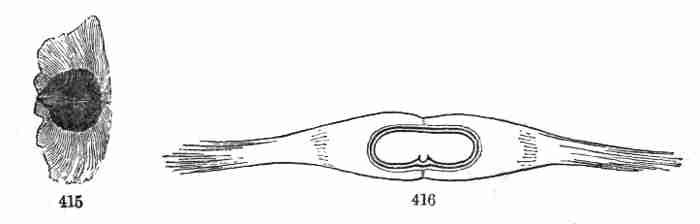
Fig. 415. A winged seed of the Trumpet-Creeper.
Fig. 416. One of Catalpa, the kernel cut to show the
embryo.
Fig. 417. Seed of Milkweed, with a Coma or tuft of long
silky hairs at one end.
382. The shape and the markings, so various in different seeds, depend
mostly on the outer coat. Sometimes this fits the kernel closely;
sometimes it is expanded into a wing, as in the Trumpet-Creeper (Fig.
415), and occasionally this wing is cut up into shreds or tufts, as in
the Catalpa (Fig. 416); or instead of a wing it may bear a Coma, or
tuft of long and soft hairs, as in the Milkweed or Silkweed (Fig. 417).
The use of wings, or downy tufts is to render the seeds buoyant [Pg 126]for
dispersion by the winds. This is clear, not only from their evident
adaptation to this purpose, but also from the fact that winged and
tufted seeds are found only in fruits that split open at maturity, never
in those that remain closed. The coat of some seeds is beset with long
hairs or wool. Cotton, one of the most important vegetable products,
since it forms the principal clothing of the larger part of the human
race, consists of the long and woolly hairs which thickly cover the
whole surface of the seed. There are also crests or other appendages of
various sorts on certain seeds. A few seeds have an additional, but more
or less incomplete covering, outside of the real seed-coats called an
383. Aril, or Arillus. The loose and transparent bag which encloses the
seed of the White Water-Lily (Fig. 418) is of this kind. So is the
mace of the nutmeg; and also the scarlet pulp around the seeds of the
Waxwork (Celastrus) and Strawberry-bush (Euonymus). The aril is a growth
from the extremity of the seed-stalk, or from the placenta when there is
no seed-stalk.
Fig. 418. Seed of White Water Lily, enclosed in its
aril.
384. A short and thickish appendage at or close to the hilum in certain
seeds is called a Caruncle or Strophiole (Fig. 419).

Fig. 419. Seed of Ricinus or Castor oil plant, with
caruncle.
385. The various terms which define the position or direction of the
ovule (erect, ascending, etc.) apply equally to the seed: so also the
terms anatropous, orthotropous, campylotropous, etc., as already defined
(320, 321), and such terms as
Hilum, or Scar left where the seed-stalk or funiculus falls away, or
where the seed was attached directly to the placenta when there is no
seed-stalk.
Rhaphe, the line or ridge which runs from the hilum to the chalaza in
anatropous and amphitropous seeds.
Chalaza, the place where the seed-coats and the kernel or nucleus are
organically connected,—at the hilum in orthotropous and campylotropous
seeds, at the extremity of the rhaphe or tip of the seed in other kinds.
Micropyle, answering to the Foramen or orifice of the ovule. Compare
the accompanying figures and those of the ovules, Fig. 341-355.
[Pg 127]
Fig. 420. Seed of a Violet (anatropous): a, hilum;
b, rhaphe; c, chalaza.
Fig. 421. Seed of a Larkspur (also anatropous); the parts
lettered as in the last.
Fig. 422. The same, cut through lengthwise: a, the
hilum; c, chalaza; d, outer seed coat; e, inner seed-coat; f,
the albumen; g, the minute embryo.
Fig. 423. Seed of a St. John's-wort, divided lengthwise;
here the whole kernel is embryo.
386. The Kernel, or Nucleus, is the whole body of the seed within the
coats. In many seeds the kernel is all Embryo; in others a large part
of it is the Albumen. For example, in Fig. 423, it is wholly embryo;
in Fig. 422, all but the small speck (g) is albumen.
387. The Albumen or Endosperm of the seed is sufficiently characterized
and its office explained in Sect. III., 31-35.
388. The Embryo or Germ, which is the rudimentary plantlet and the
final result of blossoming, and its development in germination have been
extensively illustrated in Sections II. and III. Its essential parts are
the Radicle and the Cotyledons.
389. Its Radicle or Caulicle (the former is the term long and generally
used in botanical descriptions, but the latter is the more correct one,
for it is the initial stem, which merely gives origin to the root), as
to its position in the seed, always points to and lies near the
micropyle. In relation to the pericarp it is
Superior, when it points to the apex of the fruit or cell, and
Inferior, when it points to its base, or downward.
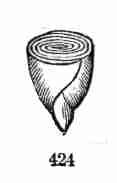
Fig. 424. Embryo of Calycanthus; upper part cut away, to
show the convolute cotyledons.
390. The Cotyledons have already been illustrated as respects their
number,—giving the important distinction of Dicotyledonous,
Polycotyledonous and Monocotyledonous embryos (36-43),—also as
regards their thickness, whether foliaceous or fleshy; and some of
the very various shapes and adaptations to the seed have been figured.
They may be straight, or folded, or rolled up. In the latter case the
cotyledons may be rolled up as it were from one margin, as in
Calycanthus (Fig. 424), or from apex to base in a flat spiral, or they
may be both folded (plicate) and rolled up (convolute), as in Sugar
Maple (Fig. 11.) In one very natural family, the Cruciferæ, two
different modes prevail in the way the two cotyledons are brought round
against the radicle. In one series they are
[Pg 128]
Accumbent, that is, the edges of the flat cotyledons lie against the
radicle, as in Fig. 425, 426. In another they are
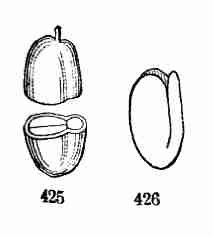
Fig. 425. Seed of Bitter Cress, Barbarea, cut across to
show the accumbent cotyledons. 426. Embryo of same, whole.
Incumbent, or with the plane of the cotyledons brought up in the
opposite direction, so that the back of one of them lies against the
radicle, as shown in Fig. 427, 428.
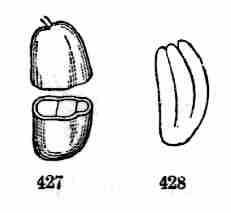
Fig. 427. Seed of a Sisymbrium, cut across to show the
incumbent cotyledons. 428. Embryo of the same, detached whole.
391. As to the situation of the embryo with respect to the albumen of
the seed, when this is present in any quantity, the embryo may be
Axile, that is occupying the axis or centre, either for most of its
length, as in Violet (Fig. 429), Barberry (Fig. 48), and Pine (Fig. 56);
and in these it is straight. But it may be variously curved or coiled in
the albumen, as in Helianthemum (Fig. 430), in a Potato-seed (Fig. 50),
or Onion-seed (Fig. 60), and Linden (Fig. 414); or it may be coiled
around the outside of the albumen, partly or into a circle, as in
Chickweed (Fig. 431, 432) and in Mirabilis (Fig. 52). The latter mode
prevails in Campylotropous seeds. In the cereal grains, such as Indian
Corn (Fig. 67) and Rice (Fig. 430a), and in all other Grasses, the
embryo is straight and applied to the outside of the abundant albumen.
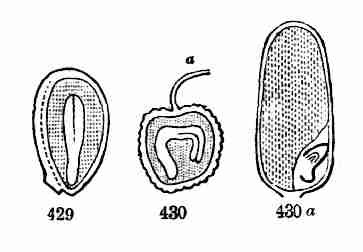
Fig. 429. Section of seed of Violet; anatropous with
straight axile embryo in the albumen. 430. Section of seed of Rock Rose,
Helianthemum Canadense; orthotropous, with curved embryo in the albumen.
430a. Section of a grain of Rice, lengthwise, showing the embryo
outside the albumen, which forms the principal bulk.
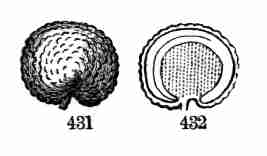
Fig. 431. Seed of a Chickweed, campylotropous. 432.
Section of same, showing slender embryo coiled around the outside of the
albumen of the kernel.
392. The matured seed, with embryo ready to germinate and reproduce the
kind, completes the cycle of the vegetable life in a phanerogamous
plant, the account of which began with the seed and seedling.
Section XVI. VEGETABLE LIFE AND WORK.
393. The following simple outlines of the anatomy and physiology of
plants (3) are added to the preceding structural part for the better
preparation of students in descriptive and systematic botany; also to
give to all learners some general idea of the life, growth, intimate
structure, and action of the beings which compose so large a part of
organic nature. Those who would extend and verify the facts and
principles here outlined will use the Physiological Botany of the
"Botanical Text Book," by Professor Goodale, or some similar book.
[Pg 129]
§ 1. ANATOMICAL STRUCTURE AND GROWTH.
394. Growth is the increase of a living thing in size and substance.
It appears so natural that plants and animals should grow, that one
rarely thinks of it as requiring explanation. It seems enough to say
that a thing is so because it grew so. Growth from the seed, the
germination and development of an embryo into a plantlet, and at length
into a mature plant (as illustrated in Sections II. and III.), can be
followed by ordinary observation. But the embryo is already a miniature
plantlet, sometimes with hardly any visible distinction of parts, but
often one which has already made very considerable growth in the seed.
To investigate the formation and growth of the embryo itself requires
well-trained eyes and hands, and the expert use of a good compound
microscope. So this is beyond the reach of a beginner.
395. Moreover, although observation may show that a seedling, weighing
only two or three grains, may double its bulk and weight every week of
its early growth, and may in time produce a huge amount of vegetable
matter, it is still to be asked what this vegetable matter is, where it
came from, and by what means plants are able to increase and accumulate
it, and build it up into the fabric of herbs and shrubs and lofty trees.
396. Protoplasm. All this fabric was built up under life, but only a
small portion of it is at any one time alive. As growth proceeds, life
is passed on from the old to the new parts, much as it has passed on
from parent to offspring, from generation to generation in unbroken
continuity. Protoplasm is the common name of that plant-stuff in which
life essentially resides. All growth depends upon it; for it has the
peculiar power of growing and multiplying and building up a living
structure,—the animal no less than the vegetable structure, for it is
essentially the same in both. Indeed, all the animal protoplasm comes
primarily from the vegetable, which has the prerogative of producing it;
and the protoplasm of plants furnishes all that portion of the food of
animals which forms their flesh and living fabric.
397. The very simplest plants (if such may specifically be called plants
rather than animals, or one may say, the simplest living things) are
mere particles, or pellets, or threads, or even indefinite masses of
protoplasm of vague form, which possess powers of motion or of changing
their shape, of imbibing water, air, and even other matters, and of
assimilating these into plant-stuff for their own growth and
multiplication. Their growth is increase in substance by incorporation
of that which they take in and assimilate. Their multiplication is by
spontaneous division of their substance or body into two or more, each
capable of continuing the process.
398. The embryo of a phanerogamous plant at its beginning (344) is
essentially such a globule of protoplasm, which soon constricts itself
into two and more such globules, which hold together inseparably in a
row; then the last of the row divides without separation in the two
other planes, to [Pg 130]form a compound mass, each grain or globule of which
goes on to double itself as it grows; and the definite shaping of this
still increasing mass builds up the embryo into its form.
Fig. 433-436. Figures to illustrate the earlier stages in
the formation of an embryo; a single mass of protoplasm (Fig. 433)
dividing into two, three, and then into more incipient cells, which by
continued multiplication build up an embryo.
399. Cell-walls. While this growth was going on, each grain of the
forming structure formed and clothed itself with a coat, thin and
transparent, of something different from protoplasm,—something which
hardly and only transiently, if at all, partakes of the life and action.
The protoplasm forms the living organism; the coat is a kind of
protective covering or shell. The protoplasm, like the flesh of animals
which it gives rise to, is composed of four chemical elements: Carbon,
Hydrogen, Oxygen, and Nitrogen. The coating is of the nature of wood
(is, indeed, that which makes wood), and has only the three elements,
Carbon, Hydrogen, and Oxygen, in its composition.

Fig. 437. Magnified view of some of a simple fresh water
Alga, the Tetraspora lubrica, each sphere of which may answer to an
individual plant.
400. Although the forming structure of an embryo in the fertilized ovule
is very minute and difficult to see, there are many simple plants of
lowest grade, abounding in pools of water, which more readily show the
earlier stages or simplest states of plant-growth. One of these, which
is common in early spring, requires only moderate magnifying power to
bring to view what is shown in Fig. 437. In a slimy mass which holds all
loosely together, little spheres of green vegetable matter are seen,
assembled in fours, and these fours themselves in clusters of fours. A
transient inspection shows, what prolonged watching would confirm, that
each sphere divides first in one plane, then in the other, to make four,
soon acquiring the size of the original, and so on, producing successive
groups of fours. These pellets each form on their surface a transparent
wall, like that just described. The delicate wall is for some time
capable of expansive growth, but is from the first much firmer than the
protoplasm within; through it the latter imbibes surrounding moisture,
which becomes a watery sap, occupying vacuities in the protoplasmic mass
which enlarge or run together as the periphery increases and distends.
When full grown the protoplasm may become a mere lining to the wall, or
some of it central, as a nucleus, this usually connected with the
wall-lining by delicate threads of the same substance. So, when full
grown, the wall with its lining—a vesicle, containing liquid or some
[Pg 131]solid matters and in age mostly air—naturally came to be named a Cell.
But the name was suggested by, and first used only for, cells in
combination or built up into a fabric, much as a wall is built of
bricks, that is, into a
401. Cellular Structure or Tissue. Suppose numerous cells like those of
Fig. 437 to be heaped up like a pile of cannon-balls, and as they grew,
to be compacted together while soft and yielding; they would flatten
where they touched, and each sphere, being touched by twelve surrounding
ones would become twelve-sided. Fig. 438 would represent one of them.
Suppose the contiguous faces to be united into one wall or partition
between adjacent cavities, and a cellular structure would be formed,
like that shown in Fig. 439. Roots, stems, leaves, and the whole of
phanerogamous plants are a fabric of countless numbers of such cells. No
such exact regularity in size and shape is ever actually found; but a
nearly truthful magnified view of a small portion of a slice of the
flower-stalk of a Calla Lily (Fig. 440) shows a fairly corresponding
structure; except that, owing to the great air-spaces of the interior,
the fabric may be likened rather to a stack of chimneys than to a solid
fabric. In young and partly transparent parts one may discern the
cellular structure by looking down directly on the surface, as of a
forming root. (Fig. 82, 441, 442).
Fig. 438. Diagram of a vegetable cell, such as it would
be if when spherical it were equally pressed by similar surrounding
cells in a heap.
Fig. 439. Ideal construction of cellular tissue so
formed, in section.
Fig. 440. Magnified view of a portion of a transverse
slice of stem of Calla Lily. The great spaces are tubular air-channels
built up by the cells.
402. The substance of which cell-walls are mainly composed is called
Cellulose. It is essentially the same in the stem of a delicate leaf or
petal and in the wood of an Oak, except that in the latter the walls are
[Pg 132]much thickened and the calibre small. The protoplasm of each living
cell appears to be completely shut up and isolated in its shell of
cellulose; but microscopic investigation has brought to view, in many
cases, minute threads of protoplasm which here and there traverse the
cell-wall through minute pores, thus connecting the living portion of
one cell with that of adjacent cells. (See Fig. 447, &c.)
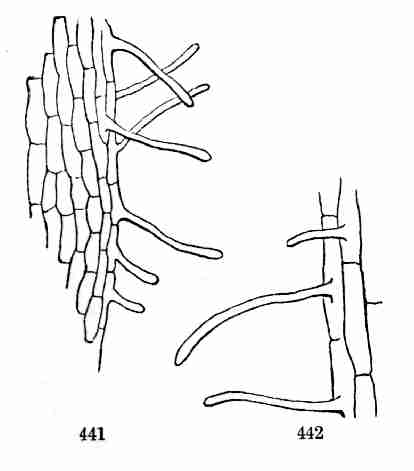
Fig. 441. Much magnified small portion of young root of a
seedling Maple (such as of Fig. 82); and 442, a few cells of same more
magnified. The prolongations from the back of some of the cells are root
hairs.
403. The hairs of plants are cells formed on the surface; either
elongated single cells (like the root-hairs of Fig. 441, 442), or a row
of shorter cells. Cotton fibres are long and simple cells growing from
the surface of the seed.
404. The size of the cells of which common plants are made up varies
from about the thirtieth to the thousandth of an inch in diameter. An
ordinary size of short or roundish cells is from 1/300 to 1/500 of an
inch; so that there may generally be from 27 to 125 millions of cells in
the compass of a cubic inch!
405. Some parts are built up as a compact structure; in others cells are
arranged so as to build up regular air-channels, as in the stems of
aquatic and other water-loving plants (Fig. 440), or to leave irregular
spaces, as in the lower part of most leaves, where the cells only here
and there come into close contact (Fig. 443).
Fig. 443. Magnified section through the thickness of a
leaf of Florida Star-Anise.
406. All such soft cellular tissue, like this of leaves, that of pith,
and of the green bark, is called Parenchyma, while fibrous and woody
parts are composed of Prosenchyma, that is, of peculiarly transformed
407. Strengthening Cells. Common cellular tissue, which makes up the
whole structure of all very young plants, and the whole of Mosses and
other vegetables of the lowest grade, even when full grown, is too
tender or too brittle to give needful strength and toughness for plants
which are to rise to any considerable height and support themselves. In
these needful strength is imparted, and the conveyance of sap through
the plant is facilitated, by the change, as they are formed, of some
cells into thicker-walled and tougher tubes, and by the running together
of some of [Pg 133]these, or the prolongation of others, into hollow fibres or
tubes of various size. Two sorts of such transformed cells go together,
and essentially form the
408. Wood. This is found in all common herbs, as well as in shrubs and
trees, but the former have much less of it in proportion to the softer
cellular tissue. It is formed very early in the growth of the root,
stem, and leaves,—traces of it appearing in large embryos even while
yet in the seed. Those cells that lengthen, and at the same time thicken
their walls form the proper Woody Fibre or Wood-cells; those of larger
size and thinner walls, which are thickened only in certain parts so as
to have peculiar markings, and which often are seen to be made up of a
row of cylindrical cells, with the partitions between absorbed or broken
away, are called Ducts, or sometimes Vessels. There are all gradations
between wood-cells and ducts, and between both these and common cells.
But in most plants the three kinds are fairly distinct.
Fig. 444. Magnified wood-cells of the bark (bast-cells)
of Basswood, one and part of another. 445. Some wood cells from the wood
(and below part of a duct); and 446, a detached wood-cell of the same;
equally magnified.
Fig. 447. Some wood cells from Buttonwood, Platanus,
highly magnified, a whole cell and lower end of another on the left; a
cell cut half away lengthwise, and half of another on the right; some
pores or pits (a) seen on the left; while b b mark sections through
these on the cut surface. When living and young the protoplasm extends
into these and by minuter perforations connects across them. In age the
pits become open passages, facilitating the passage of sap and air.
409. The proper cellular tissue, or parenchyma, is the ground-work of
root, stem, and leaves; this is traversed, chiefly lengthwise, by the
strengthening and conducting tissue, wood-cells and duct-cells, in the
form of bundles or threads, which, in the stems and stalks of herbs are
fewer and comparatively scattered, but in shrubs and trees so numerous
and crowded that in the stems and all permanent parts they make a solid
mass of wood. They extend into and ramify in the leaves, spreading out
in a horizontal plane, as the framework of ribs and veins, which
supports the softer cellular portion or parenchyma.
410. Wood-Cells, or Woody Fibres, consist of tubes, commonly between one
and two thousandths, but in Pine-wood sometimes two or three hundredths,
of an inch in diameter. Those from the tough bark of the Basswood,
[Pg 134]shown in Fig. 444, are only the fifteen-hundredth of an inch wide.
Those of Buttonwood (Fig. 447) are larger, and are here highly magnified
besides. The figures show the way wood-cells are commonly put together,
namely, with their tapering ends overlapping each other,—spliced
together, as it were,—thus giving more strength and toughness. In hard
woods, such as Hickory and Oak, the walls of these tubes are very thick,
as well as dense; while in soft woods, such as White Pine and Basswood,
they are thinner.
411. Wood-cells in the bark are generally longer, finer, and tougher
than those of the proper wood, and appear more like fibres. For example,
Fig. 446 represents a cell of the wood of Basswood of average length,
and Fig. 444 one (and part of another) of the fibrous bark, both drawn
to the same scale. As these long cells form the principal part of
fibrous bark, or bast, they are named Bast-cells or Bast-fibres.
These give the great toughness and flexibility to the inner bark of
Basswood (i. e. Bast-wood) and of Leatherwood; and they furnish the
invaluable fibres of flax and hemp; the proper wood of their stems being
tender, brittle, and destroyed by the processes which separate for use
the tough and slender bast-cells. In Leatherwood (Dirca) the bast-cells
are remarkably slender. A view of one, if magnified on the scale of Fig.
444, would be a foot and a half long.
Fig. 448. Magnified bit of a pine shaving, taken parallel
with the silver grain. 449. Separate whole wood-cell, more magnified.
450. Same, still more magnified; both sections represented: a, disks
in section, b, in face.
412. The wood-cells of Pines, and more or less of all other Coniferous
trees, have on two of their sides very peculiar disk-shaped markings
(Fig. 448-450) by which that kind of wood is recognizable.
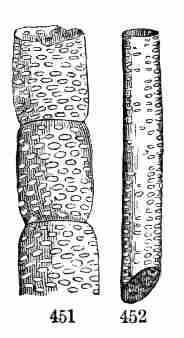
Fig. 451, 452. A large and a smaller dotted duct from
Grape-Vine.
413. Ducts, also called Vessels, are mostly larger than wood-cells:
indeed, some of them, as in Red Oak, have calibre large enough to be
discerned on a cross section by the naked eye. They make the visible
porosity of such kinds of wood. This is particularly the case with
Dotted ducts (Fig. 451, 452), the surface of which appears as if
riddled with round or oval pores. Such ducts are commonly made up of a
row of large cells more or less confluent into a tube.
Scalariform ducts (Fig. 458, 459), common in Ferns, and generally
angled by mutual pressure in the bundles, [Pg 135]have transversely elongated
thin places, parallel with each other, giving a ladder-like appearance,
whence the name.
Annular ducts (Fig. 457) are marked with cross lines or rings, which
are thickened portions of the cell-wall.
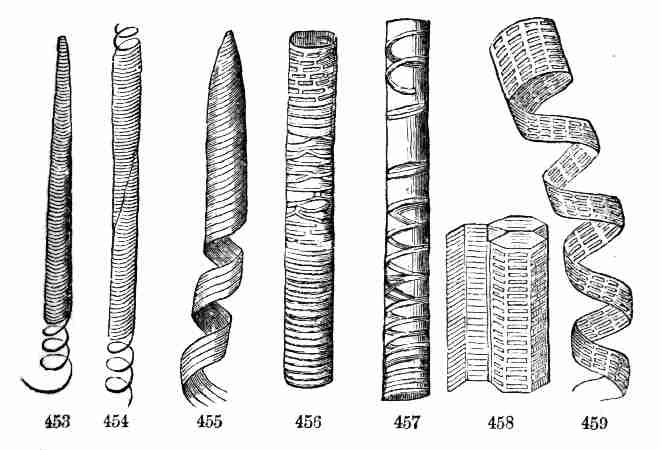
Fig. 453, 454. Spiral ducts which uncoil into a single
thread. 455. Spiral duct which tears up as a band. 456. An annular duct,
with variations above. 457. Loose spiral duct passing into annular. 458.
Scalariform ducts of a Fern; part of a bundle, prismatic by pressure.
459. One torn into a band.
Spiral ducts or vessels (Fig. 453-455) have thin walls, strengthened
by a spiral fibre adherent within. This is as delicate and as strong as
spider-web: when uncoiled by pulling apart, it tears up and annihilates
the cell-wall. The uncoiled threads are seen by gently pulling apart
many leaves, such as those of Amaryllis, or the stalk of a Strawberry
leaflet.
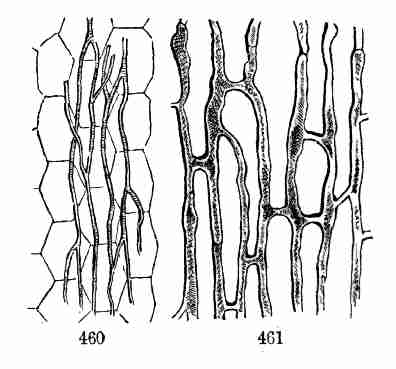
Fig. 460. Milk Vessels of Dandelion, with cells of the
common cellular tissue. 461. Others from the same older and gorged with
milky juice. All highly magnified.
Laticiferous ducts, Vessels of the Latex, or Milk-vessels are
peculiar branching tubes which hold latex or milky juice in certain
plants. It is very difficult to see them, and more so to make out their
nature. They are peculiar in branching and inosculating, so as to make a
net-work of tubes, running in among the cellular tissue; and they are
very small, except when gorged and old (Fig. 460, 461).
[Pg 136]
§ 2. CELL-CONTENTS.
414. The living contents of young and active cells are mainly protoplasm
with water or watery sap which this has imbibed. Old and effete cells
are often empty of solid matter, containing only water with whatever may
be dissolved in it, or air, according to the time and circumstances. All
the various products which plants in general elaborate, or which
particular plants specially elaborate, out of the common food which they
derive from the soil and the air, are contained in the cells, and in the
cells they are produced.
415. Sap is a general name for the principal liquid contents,—Crude
sap, for that which the plant takes in, Elaborated sap for what it
has digested or assimilated. They must be undistinguishably mixed in the
cells.
416. Among the solid matters into which cells convert some of their
elaborated sap two are general and most important. These are
Chlorophyll and Starch.
417. Chlorophyll (meaning leaf-green) is what gives the green color to
herbage. It consists of soft grains of rather complex nature, partly
wax-like, partly protoplasmic. These abound in the cells of all common
leaves and the green rind of plants, wherever exposed to the light. The
green color is seen through the transparent skin of the leaf and the
walls of the containing cells. Chlorophyll is essential to ordinary
assimilation in plants: by its means, under the influence of sunlight,
the plant converts crude sap into vegetable matter.
418. Far the largest part of all vegetable matter produced is that which
goes to build up the plant's fabric or cellular structure, either
directly or indirectly. There is no one good name for this most
important product of vegetation. In its final state of cell-walls, the
permanent fabric of herb and shrub and tree, it is called Cellulose
(408): in its most soluble form it is Sugar of one or another kind; in
a less soluble form it is Dextrine, a kind of liquefied starch: in the
form of solid grains stored up in the cells it is Starch. By a series
of slight chemical changes (mainly a variation in the water entering
into the composition), one of these forms is converted into another.
419. Starch (Farina or Fecula) is the form in which this common
plant material is, as it were, laid by for future use. It consists of
solid grains, somewhat different in form in different plants, in size
varying from 1/300 to 1/4000 of an inch, partly translucent when wet,
and of a pearly lustre. From the concentric lines, which commonly appear
under the microscope, the grains seem to be made up of layer over layer.
When loose they are commonly oval, as in potato-starch (Fig. 462): when
much compacted the grains may become angular (Fig. 463).
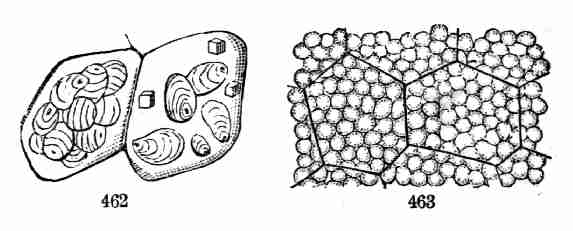
Fig. 462. Some magnified starch-grains, in two cells of a
potato. 463. Some cells of the albumen or floury part of Indian Corn,
filled with starch-grains.
420. The starch in a potato was produced in the foliage. In the soluble
form of dextrine, or that of sugar, it was conveyed through the cells of
the herbage and stalks to a subterranean shoot, and there stored up in
the [Pg 137]tuber. When the potato sprouts, the starch in the vicinity of
developing buds or eyes is changed back again, first into mucilaginous
dextrine, then into sugar, dissolved in the sap, and in this form it is
made to flow to the growing parts, where it is laid down into cellulose
or cell-wall.
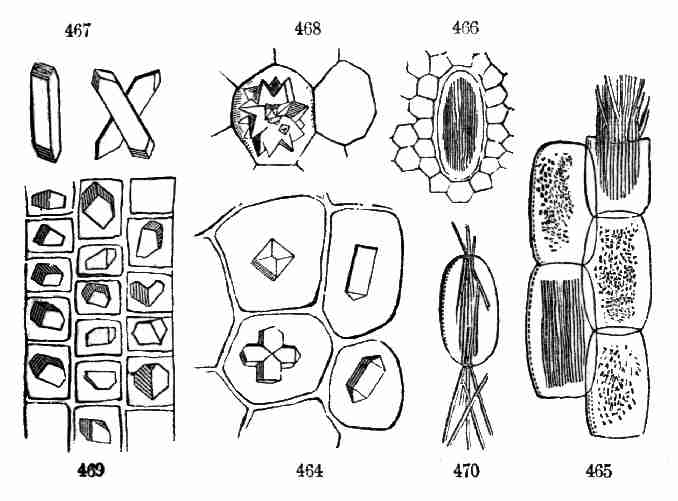
Fig. 464. Four cells from dried Onion-peel, each holding
a crystal of different shape, one of them twinned. 465. Some cells from
stalk of Rhubarb-plant, three containing chlorophyll; two (one torn
across) with rhaphides. 466. Rhaphides in a cell, from Arisæma, with
small cells surrounding. 467. Prismatic crystals from the bark of
Hickory. 468. Glomerate crystal in a cell, from Beet-root. 469. A few
cells of Locust-bark, a crystal in each. 470. A detached cell, with
rhaphides being forced out, as happens when put in water.
421. Besides these cell-contents which are in obvious and essential
relation to nutrition, there are others the use of which is
problematical. Of such the commonest are
422. Crystals. These when slender or needle-shaped are called Rhaphides.
They are of inorganic matter, usually of oxalate or phosphate or
sulphate of lime. Some, at least of the latter, may be direct
crystallizations of what is taken in dissolved in the water absorbed,
but others must be the result of some elaboration in the plant. Some
plants have hardly any; others abound in them, especially in the foliage
and bark. In Locust-bark almost every cell holds a crystal; so that in a
square inch not thicker than writing-paper there may be over a million
and a half of them. When [Pg 138]needle-shaped (rhaphides), as in stalks of
Calla-Lily, Rhubarb, or Four-o'clock, they are usually packed in
sheaf-like bundles. (Fig. 465, 466.)
§ 3. ANATOMY OF ROOTS AND STEMS.
423. This is so nearly the same that an account of the internal
structure of stems may serve for the root also.
424. At the beginning, either in the embryo or in an incipient shoot
from a bud, the whole stem is of tender cellular tissue or parenchyma.
But wood (consisting of wood-cells and ducts or vessels) begins to be
formed in the earliest growth; and is from the first arranged in two
ways, making two general kinds of wood. The difference is obvious even
in herbs, but is more conspicuous in the enduring stems of shrubs and
trees.
425. On one or the other of these two types the stems of all
phanerogamous plants are constructed. In one, the wood is made up of
separate threads, scattered here and there throughout the whole diameter
of the stem. In the other, the wood is all collected to form a layer (in
a slice across the stem appearing as a ring) between a central cellular
part which has none in it, the Pith, and an outer cellular part, the
Bark.
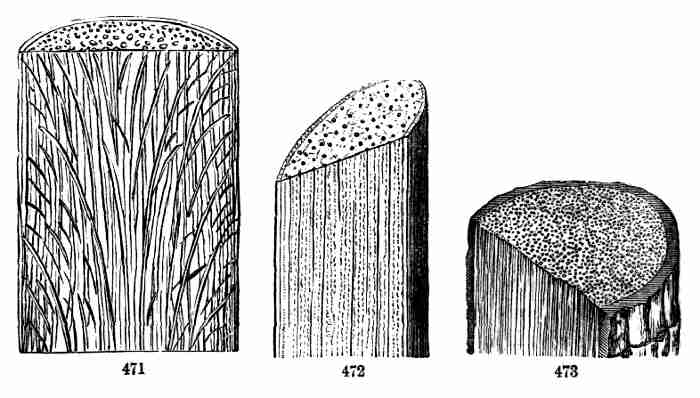
Fig. 471. Diagram of structure of Palm or Yucca. 472.
Structure of a Corn stalk, in transverse and longitudinal section. 473.
Same of a small Palm stem. The dots on the cross sections represent cut
ends of the woody bundles or threads.
426. An Asparagus-shoot and a Corn-stalk for herbs, and a rattan for a
woody kind, represent the first kind. To it belong all plants with
monocotyledonous embryo (40). A Bean-stalk and the stem of any common
shrub or tree represent the second; and to it belong all plants with
dicotyledonous or polycotyledonous embryo. The first has been called,
not very properly, Endogenous, which means inside-growing; the second,
properly enough, Exogenous, or outside-growing.
427. Endogenous Stems, those of Monocotyls (40), attain their greatest
size and most characteristic development in Palms and Dragon-trees,
therefore chiefly in warm climates, although the Palmetto and some
[Pg 139]Yuccas become trees along the southern borders of the United States. In
such stems the woody bundles are more numerous and crowded toward the
circumference, and so the harder wood is outside; while in an exogenous
stem the oldest and hardest wood is toward the centre. An endogenous
stem has no clear distinction of pith, bark, and wood, concentrically
arranged, no silver grain, no annual layers, no bark that peels off
clean from the wood. Yet old stems of Yuccas and the like, that continue
to increase in diameter, do form a sort of layers and a kind of scaly
bark when old. Yuccas show well the curving of the woody bundles (Fig.
471) which below taper out and are lost at the rind.
Fig. 474. Short piece of stem of Flax, magnified, showing
the bark, wood, and pith in a cross section.
428. Exogenous Stems, those of Dicotyls (37), or of plants coming from
dicotyledonous and also polycotyledonous embryos, have a structure which
is familiar in the wood of our ordinary trees and shrubs. It is the same
in an herbaceous shoot (such as a Flax-stem, Fig. 474) as in a
Maple-stem of the first year's growth, except that the woody layer is
commonly thinner or perhaps reduced to a circle of bundles. It was so in
the tree-stem at the beginning. The wood all forms in a cylinder,—in
cross section a ring—around a central cellular part, dividing the
cellular core within, the pith, from a cellular bark without. As the
wood-bundles increase in number and in size, they press upon each other
and become wedge-shaped in the cross section; and they continue to grow
from the outside, next the bark, so that they become very thin wedges or
plates. Between the plates or wedges are very thin plates (in cross
section lines) of much compressed cellular tissue, which connect the
pith with the bark. The plan of a one-year-old woody stem of this kind
is exhibited in the figures, which are essentially diagrams.
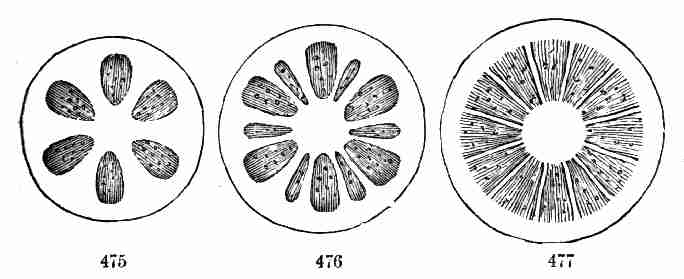
Fig. 475. Diagram of a cross section of a very young
exogenous stem, showing six woody bundles or wedges. 476. Same later,
with wedges increased to twelve. 477. Still later, the wedges filling
the space, separated only by the thin lines, or medullary rays, running
from pith to bark.
429. When such a stem grows on from year to year, it adds annually a
[Pg 140]layer of wood outside the preceding one, between that and the bark.
This is exogenous growth, or outside-growing, as the name denotes.
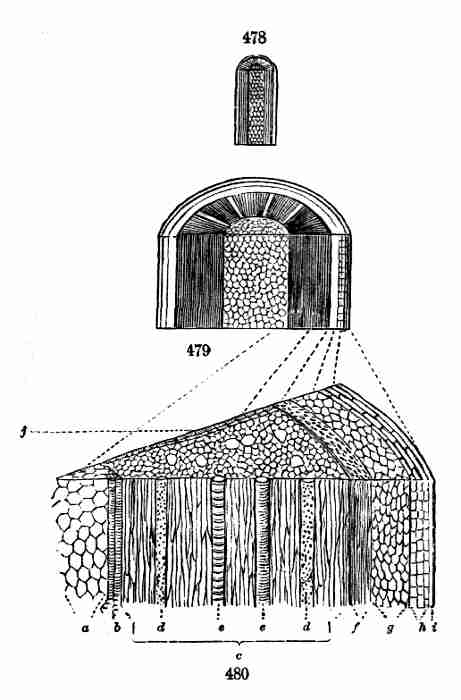
Fig. 478. Piece of a stem of Soft Maple, of a year old,
cut crosswise and lengthwise.
Fig. 479. A portion of the same, magnified.
Fig. 480. A small piece of the same, taken from one side,
reaching from the bark to the pith, and highly magnified: a, a small
bit of the pith; b, spiral ducts of what is called the medullary
sheath; c, the wood; d, d, dotted ducts in the wood; e, e,
annular ducts; f, the liber or inner bark; g, the green bark; h,
the corky layer; i, the skin, or epidermis; j, one of the medullary
rays, or plates of silver grain, seen on the cross-section.
430. Some new bark is formed every year, as well as new wood, the former
inside, as the latter is outside of that of the year preceding. The ring
or zone of tender forming tissue between the bark and the wood has been
called the Cambium Layer. Cambium is an old name of the
physiologists for nutritive juice. And this thin layer is so gorged with
rich nutritive sap when spring growth is renewed, that the bark then
seems to be loose from the wood and a layer of viscid sap (or cambium)
to be poured out between the two. But there is all the while a
connection of the bark and the wood by delicate cells, rapidly
multiplying and growing.
431. The Bark of a year-old stem consists of three parts, more or less
distinct, namely,—beginning next the wood,—
1. The Liber or Fibrous Bark, the Inner Bark. This contains some
wood-cells, or their equivalent, commonly in the form of bast or
bast-cells (411, Fig. 444), such as those of Basswood or Linden, and
among herbs those of flax and hemp, which are spun and woven or made
into cordage. It also contains cells which are named sieve-cells, on
account of numerous slits and pores in their walls, by which the
protoplasm of contiguous cells communicates. In woody stems, whenever a
new layer of wood is formed, some new liber or inner bark is also formed
outside of it.
[Pg 141]
2. The Green Bark or Middle Bark. This consists of cellular tissue
only, and contains the same green matter (chlorophyll, 417) as the
leaves. In woody stems, before the season's growth is completed, it
becomes covered by
3. The Corky Layer or Outer Bark, the cells of which contain no
chlorophyll, and are of the nature of cork. Common cork is the thick
corky layer of the bark of the Cork-Oak of Spain. It is this which gives
to the stems or twigs of shrubs and trees the aspect and the color
peculiar to each,—light gray in the Ash, purple in the Red Maple, red
in several Dogwoods, etc.
4. The Epidermis, or skin of the plant, consisting of a layer of
thick-sided empty cells, which may be considered to be the outermost
layer, or in most herbaceous stems the only layer, of cork-cells.
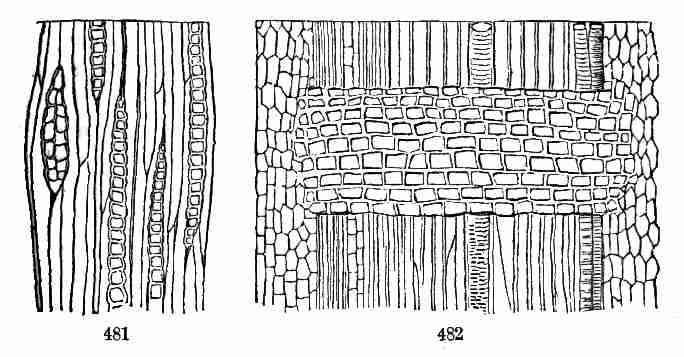
Fig. 481. Magnified view of surface of a bit of young
Maple wood from which the bark has been torn away, showing the
wood-cells and the bark-ends of medullary rays.
Fig. 482. Section in the opposite direction, from bark
(on the left) to beginning of pith (on the right), and a medullary ray
extending from one to the other.
432. The green layer of bark seldom grows much after the first season.
Sometimes the corky layer grows and forms new layers, inside of the old,
for years, as in the Cork-Oak, the Sweet Gum-tree, and the White and the
Paper Birch. But it all dies after a while; and the continual
enlargement of the wood within finally stretches it more than it can
bear, and sooner or later cracks and rends it, while the weather acts
powerfully upon its surface; so the older bark perishes and falls away
piecemeal year by year.
433. So on old trunks only the inner bark remains. This is renewed every
year from within and so kept alive, while the older and outer layers
die, are fissured and rent by the distending trunk, weathered and worn,
and thrown off in fragments,—in some trees slowly, so that the bark of
old trunks may acquire great thickness; in others, more rapidly. In
Honeysuckles and Grape-Vines, the layers of liber loosen and die when
only a year or two old. The annual layers of liber are sometimes as
distinct as those of the wood, but often not so.
[Pg 142]
434. The Wood of an exogenous trunk, having the old growths covered by
the new, remains nearly unchanged in age, except from decay. Wherever
there is an annual suspension and renewal of growth, as in temperate
climates, the annual growths are more or less distinctly marked, in the
form of concentric rings on the cross section, so that the age of the
tree may be known by counting them. Over twelve hundred layers have been
counted on the stumps of Sequoias in California, and it is probable that
some trees now living antedate the Christian era.
435. The reason why the annual growths are distinguishable is, that the
wood formed at the beginning of the season is more or less different in
the size or character of the cells from that of the close. In Oak,
Chestnut, etc., the first wood of the season abounds in dotted ducts,
the calibre of which is many times greater than that of the proper
wood-cells.
436. Sap-wood, or Alburnum. This is the newer wood, living or recently
alive, and taking part in the conveyance of sap. Sooner or later, each
layer, as it becomes more and more deeply covered by the newer ones and
farther from the region of growth, is converted into
437. Heart-wood, or Duramen. This is drier, harder, more solid, and much
more durable as timber, than sap-wood. It is generally of a different
color, and it exhibits in different species the hue peculiar to each,
such as reddish in Red-Cedar, brown in Black-Walnut, black in Ebony,
etc. The change of sap-wood into heart-wood results from the thickening
of the walls of the wood-cells by the deposition of hard matter, lining
the tubes and diminishing their calibre; and by the deposition of a
vegetable coloring-matter peculiar to each species. The heart-wood,
being no longer a living part, may decay, and often does so, without the
least injury to the tree, except by diminishing the strength of the
trunk, and so rendering it more liable to be overthrown.
438. The Living Parts of a Tree, of the exogenous kind, are only these:
first, the rootlets at one extremity; second, the buds and leaves of the
season at the other; and third, a zone consisting of the newest wood and
the newest bark, connecting the rootlets with the buds or leaves,
however widely separated these may be,—in the tallest trees from two to
four hundred feet apart. And these parts of the tree are all renewed
every year. No wonder, therefore, that trees may live so long, since
they annually reproduce everything that is essential to their life and
growth, and since only a very small part of their bulk is alive at once.
The tree survives, but nothing now living has been so long. In it, as
elsewhere, life is a transitory thing, ever abandoning the old, and
renewed in the young.
§ 4. ANATOMY OF LEAVES.
439. The wood in leaves is the framework of ribs, veins, and veinlets
(125), serving not only to strengthen them, but also to bring in the
sap, and to distribute it throughout every part. The cellular portion is
the [Pg 143]green pulp, and is nearly the same as the green layer of the bark.
So that the leaf may properly enough be regarded as a sort of expansion
of the fibrous and green layers of the bark. It has no proper corky
layer; but the whole is covered by a transparent skin or epidermis,
resembling that of the stem.
440. The cells of the leaf are of various forms, rarely so compact as to
form a close cellular tissue, usually loosely arranged, at least in the
lower part, so as to give copious intervening spaces or air passages,
communicating throughout the whole interior (Fig. 443, 483). The green
color is given by the chlorophyll (417), seen through the very
transparent walls of the cells and through the translucent epidermis of
the leaf.
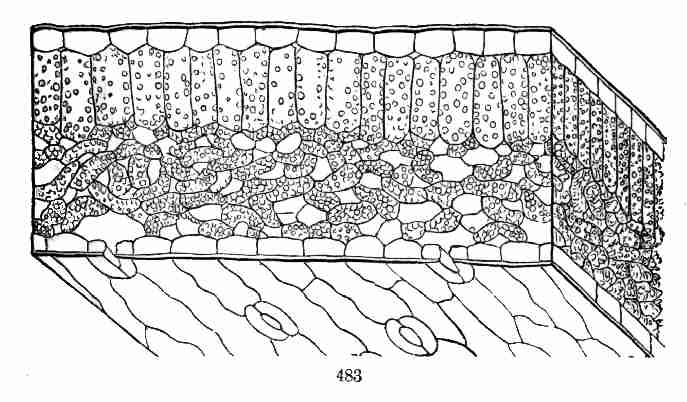
Fig. 483. Magnified section of a leaf of White Lily, to
exhibit the cellular structure, both of upper and lower stratum, the
air-passages of the lower, and the epidermis or skin, in section, also a
little of that of the lower face, with some of its stomates.
441. In ordinary leaves, having an upper and under surface, the green
cells form two distinct strata, of different arrangement. Those of the
upper stratum are oblong or cylindrical, and stand endwise to the
surface of the leaf, usually close together, leaving hardly any vacant
spaces; those of the lower are commonly irregular in shape, most of them
with their longer diameter parallel to the face of the leaf, and are
very loosely arranged, leaving many and wide air-chambers. The green
color of the lower is therefore diluted, and paler than that of the
upper face of the leaf. The upper part of the leaf is so constructed as
to bear the direct action of the sunshine; the lower so as to afford
freer circulation of air, and to facilitate transpiration. It
communicates more directly than the upper with the external air by means
of Stomates.
442. The Epidermis or skin of leaves and all young shoots is best seen
in the foliage. It may readily be stripped off from the surface of a
Lily-leaf, and still more so from more fleshy and soft leaves, such as
those [Pg 144]of Houseleek. The epidermis is usually composed of a single
layer, occasionally of two or three layers, of empty cells, mostly of
irregular outline. The sinuous lines which traverse it, and may be
discerned under low powers of the microscope (Fig. 487), are the
boundaries of the epidermal cells.
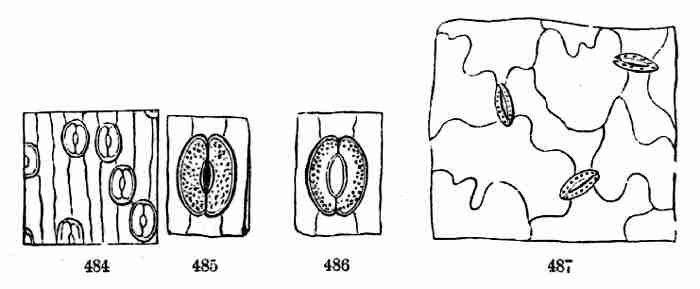
Fig. 484. Small portion of epidermis of the lower face of
a White-Lily leaf, with stomata.
Fig. 485. One of these, more magnified, in the closed
state. 486. Another stoma, open.
Fig. 487. Small portion of epidermis of the Garden
Balsam, highly magnified, showing very sinuous-walled cells, and three
stomata.
443. Breathing-pores, or Stomates, Stomata (singular, a
Stoma,—literally, a mouth) are openings through the epidermis into
the air-chambers or intercellular passages, always between and guarded
by a pair of thin-walled guardian cells. Although most abundant in
leaves, especially on their lower face (that which is screened from
direct sunlight), they are found on most other green parts. They
establish a direct communication between the external air and that in
the loose interior of the leaf. Their guardian cells or lips, which are
soft and delicate, like those of the green pulp within, by their greater
or less turgidity open or close the orifice as the moisture or dryness
varies.
444. In the White Lily the stomata are so remarkably large that they may
be seen by a simple microscope of moderate power, and may be discerned
even by a good hand lens. There are about 60,000 of them to the square
inch of the epidermis of the lower face of this Lily-leaf, and only
about 3000 to the same space on the upper face. It is computed that an
average leaf of an Apple-tree has on its lower face about 100,000 of
these mouths.
§ 5. PLANT FOOD AND ASSIMILATION.
445. Only plants are capable of originating organizable matter, or the
materials which compose the structure of vegetables and animals. The
essential and peculiar work of plants is to take up portions of earth
and air (water belonging to both) upon which animals cannot live at all,
and to convert them into something organizable; that is, into something
that, under life, may be built up into vegetable and animal structures.
All the food of animals is produced by plants. Animals live upon
vegetables, [Pg 145]directly or at second hand, the carnivorous upon the
herbivorous; and vegetables live upon earth and air, immediately or at
second hand.
446. The Food of plants, then, primarily, is earth and air. This is
evident enough from the way in which they live. Many plants will
flourish in pure sand or powdered chalk, or on the bare face of a rock
or wall, watered merely with rain. And almost any plant may be made to
grow from the seed in moist sand, and increase its weight many times,
even if it will not come to perfection. Many naturally live suspended
from the branches of trees high in the air, and nourished by it alone,
never having any connection with the soil; and some which naturally grow
on the ground, like the Live-forever of the gardens, when pulled up by
the roots and hung in the air will often flourish the whole summer long.
447. It is true that fast-growing plants, or those which produce much
vegetable matter in one season (especially in such concentrated form as
to be useful as food for man or the higher animals) will come to
maturity only in an enriched soil. But what is a rich soil? One which
contains decomposing vegetable matter, or some decomposing animal
matter; that is, in either case, some decomposing organic matter
formerly produced by plants. Aided by this, grain-bearing and other
important vegetables will grow more rapidly and vigorously, and make a
greater amount of nourishing matter, than they could if left to do the
whole work at once from the beginning. So that in these cases also all
the organic or organizable matter was made by plants, and made out of
earth and air. Far the larger and most essential part was air and water.
448. Two kinds of material are taken in and used by plants; of which the
first, although more or less essential to perfect plant-growth, are in a
certain sense subsidiary, if not accidental, viz.:—
Earthy constituents, those which are left in the form of ashes when a
leaf or a stick of wood is burned in the open air. These consist of some
potash (or soda in a marine plant), some silex (the same as
flint), and a little lime, alumine, or magnesia, iron or
manganese, sulphur, phosphorus, etc.,—some or all of these in
variable and usually minute proportions. They are such materials as
happen to be dissolved, in small quantity, in the water taken up by the
roots; and when that is consumed by the plant, or flies off pure (as it
largely does) by exhalation, the earthy matter is left behind in the
cells,—just as it is left incrusting the sides of a teakettle in which
much hard water has been boiled. Naturally, therefore, there is more
earthy matter (i. e. more ashes) in the leaves than in any other part
(sometimes as much as seven per cent, when the wood contains only two
per cent); because it is through the leaves that most of the water
escapes from the plant. Some of this earthy matter incrusts the
cell-walls, some goes to form crystals or rhaphides, which abound in
many plants (422), some enters into certain special vegetable products,
and some appears to be necessary to the well-being of the higher orders
of plants, although forming no necessary part of the proper vegetable
structure.
[Pg 146]
The essential constituents of the organic fabric are those which are
dissipated into air and vapor in complete burning. They make up from 88
to 99 per cent of the leaf or stem, and essentially the whole both of
the cellulose of the walls and the protoplasm of the contents. Burning
gives these materials of the plant's structure back to the air, mainly
in the same condition in which the plant took them, the same condition
which is reached more slowly in natural decay. The chemical elements of
the cell-walls (or cellulose, 402), as also of starch, sugar, and all
that class of organizable cell-material, are carbon, hydrogen, and
oxygen (399). The same, with nitrogen, are the constituents of
protoplasm, or the truly vital part of vegetation.
449. These chemical elements out of which organic matters are composed
are supplied to the plant by water, carbonic acid, and some combinations
of nitrogen.
Water, far more largely than anything else, is imbibed by the roots;
also more or less by the foliage in the form of vapor. Water consists of
oxygen and hydrogen; and cellulose or plant-wall, starch, sugar, etc.,
however different in their qualities, agree in containing these two
elements in the same relative proportions as in water.
Carbonic acid gas (Carbon dioxide) is one of the components of the
atmosphere,—a small one, ordinarily only about 1/2500 of its
bulk,—sufficient for the supply of vegetation, but not enough to be
injurious to animals, as it would be if accumulated. Every current or
breeze of air brings to the leaves expanded in it a succession of fresh
atoms of carbonic acid, which it absorbs through its multitudinous
breathing-pores. This gas is also taken up by water. So it is brought to
the ground by rain, and is absorbed by the roots of plants, either as
dissolved in the water they imbibe, or in the form of gas in the
interstices of the soil. Manured ground, that is, soil containing
decomposing vegetable or animal matters, is constantly giving out this
gas into the interstices of the soil, whence the roots of the growing
crop absorb it. Carbonic acid thus supplied, primarily from the air, is
the source of the carbon which forms much the largest part of the
substance of every plant. The proportion of carbon may be roughly
estimated by charring some wood or foliage; that is, by heating it out
of contact with the air, so as to decompose and drive off all the other
constituents of the fabric, leaving the large bulk of charcoal or carbon
behind.
Nitrogen, the remaining plant-element, is a gas which makes up more
than two thirds of the atmosphere, is brought into the foliage and also
to the roots (being moderately soluble in water) in the same ways as is
carbonic acid. The nitrogen which, mixed with oxygen, a little carbonic
acid, and vapor of water, constitutes the air we breathe, is the source
of this fourth plant-element. But it is very doubtful if ordinary plants
can use any nitrogen gas directly as food; that is, if they can directly
cause it to combine with the other elements so as to form protoplasm.
But when combined with hydrogen (forming ammonia), or when combined with
oxygen [Pg 147](nitric acid and nitrates) plants appropriate it with avidity.
And several natural processes are going on in which nitrogen of the air
is so combined and supplied to the soil in forms directly available to
the plant. The most efficient is nitrification, the formation of nitre
(nitrate of potash) in the soil, especially in all fertile soils,
through the action of a bacterial ferment.
450. Assimilation in plants is the conversion of these inorganic
substances—essentially, water, carbonic acid, and some form of combined
or combinable nitrogen—into vegetable matter. This most dilute food the
living plant concentrates and assimilates to itself. Only plants are
capable of converting these mineral into organizable matters; and this
all-important work is done by them (so far as all ordinary vegetation is
concerned) only
451. Under the light of the sun, acting upon green parts or foliage,
that is, upon the chlorophyll, or upon what answers to chlorophyll,
which these parts contain. The sun in some way supplies a power which
enables the living plant to originate these peculiar chemical
combinations,—to organize matter into forms which are alone capable of
being endowed with life. The proof of this proposition is simple; and it
shows at the same time, in the simplest way, what a plant does with the
water and carbonic acid it consumes. Namely, 1st, it is only in sunshine
or bright daylight that the green parts of plants give out oxygen
gas,—then they regularly do so; and 2d, the giving out of this oxygen
gas is required to render the chemical composition of water and carbonic
acid the same as that of cellulose, that is, of the plant's permanent
fabric. This shows why plants spread out so large a surface of foliage.
Leaves are so many workshops, full of machinery worked by sun-power. The
emission of oxygen gas from any sun-lit foliage is seen by placing some
of this under water, or by using an aquatic plant, by collecting the air
bubbles which rise, and by noting that a taper burns brighter in this
air. Or a leafy plant in a glass globe may be supplied with a certain
small percentage of carbonic acid gas, and after proper exposure to
sunshine, the air on being tested will be found to contain less carbonic
acid and just so much the more oxygen gas.
452. Now if the plant is making cellulose or any equivalent
substance,—that is, is making the very materials of its fabric and
growth, as must generally be the case,—all this oxygen gas given off by
the leaves comes from the decomposition of carbonic acid taken in by the
plant. For cellulose, and also starch, dextrine, sugar, and the like are
composed of carbon along with oxygen and hydrogen in just the
proportions to form water. And the carbonic acid and water taken in,
less the oxygen which the carbon brought with it as carbonic acid, and
which is given off from the foliage in sunshine, just represents the
manufactured article, cellulose.
453. It comes to the same if the first product of assimilation is sugar,
or dextrine which is a sort of soluble starch, or starch itself. And in
the plant all these forms are readily changed into one another. In the
tiny seedling, as fast as this assimilated matter is formed it is used
in growth, that is, in the formation of cell-walls. After a time some or
much of [Pg 148]the product may be accumulated in store for future growth, as
in the root of the turnip, or the tuber of the potato, or the seed of
corn or pulse. This store is mainly in the form of starch. When growth
begins anew, this starch is turned into dextrine or into sugar, in
liquid form, and used to nourish and build up the germinating embryo or
the new shoot, where it is at length converted into cellulose and used
to build up plant-structure.
454. But that which builds plant-fabric is not the cellular structure
itself; the work is done by the living protoplasm which dwells within
the walls. This also has to take and to assimilate its proper food, for
its own maintenance and growth. Protoplasm assimilates, along with the
other three elements, the nitrogen of the plant's food. This comes
primarily from the vast stock in the atmosphere, but mainly through the
earth, where it is accumulated through various processes in a fertile
soil,—mainly, so far as concerns crops, from the decomposition of
former vegetables and animals. This protoplasm, which is formed at the
same time as the simpler cellulose, is essentially the same as the flesh
of animals, and the source of it. It is the common basis of vegetable
and of animal life.
455. So plant-assimilation produces all the food and fabric of
animals. Starch, sugar, the oils (which are, as it were, these
farinaceous matters more deoxidated), chlorophyll, and the like, and
even cellulose itself, form the food of herbivorous animals and much of
the food of man. When digested they enter into the blood, undergo
various transformations, and are at length decomposed into carbonic acid
and water, and exhaled from the lungs in respiration,—in other words,
are given back to the air by the animal as the very same materials which
the plant took from the air as its food,—are given back to the air in
the same form that they would have taken if the vegetable matter had
been left to decay where it grew, or if it had been set on fire and
burned; and with the same result, too, as to the heat,—the heat in this
case producing and maintaining the proper temperature of the animal.
456. The protoplasm and other products containing nitrogen (gluten,
legumine, etc.), and which are most accumulated in grains and seeds (for
the nourishment of their embryos when they germinate), compose the most
nutritious vegetable food consumed by animals; they form their proper
flesh and sinews, while the earthy constituents of the plant form the
earthy matter of the bones, etc. At length decomposed, in the secretions
and excretions, these nitrogenous constituents are through successive
changes finally resolved into mineral matter, into carbonic acid, water,
and ammonia or some nitrates,—into exactly or essentially the same
materials which the plants took up and assimilated. Animals depend upon
vegetables absolutely and directly for their subsistence; also
indirectly, because
457. Plants purify the air for animals. In the very process by which
they create food they take from the air carbonic acid gas, injurious to
animal respiration, which is continually poured into it by the breathing
of all animals, by all decay, by the burning of fuel and all other
ordinary combustion; and [Pg 149]they restore an equal bulk of life-sustaining
oxygen needful for the respiration of animals,—needful, also, in a
certain measure, for plants in any work they do. For in plants, as well
as in animals, work is done at a certain cost.
§ 6. PLANT WORK AND MOVEMENT.
458. As the organic basis and truly living material of plants is
identical with that of animals, so is the life at bottom essentially the
same; but in animals something is added at every rise from the lowest to
highest organisms. Action and work in living beings require movement.
459. Living things move; those not living are only moved. Plants move as
truly as do animals. The latter, nourished as they are upon organized
food, which has been prepared for them by plants, and is found only here
and there, must needs have the power of going after it, of collecting
it, or at least of taking it in; which requires them to make spontaneous
movements. But ordinary plants, with their wide-spread surface, always
in contact with the earth and air on which they feed,—the latter
everywhere the same, and the former very much so,—might be thought to
have no need of movement. Ordinary plants, indeed, have no locomotion;
some float, but most are rooted to the spot where they grew. Yet
probably all of them execute various movements which must be as truly
self-caused as are those of the lower grades of animals,—movements
which are overlooked only because too slow to be directly observed.
Nevertheless, the motion of the hour-hand and of the minute-hand of a
watch is not less real than that of the second-hand.
Fig. 488. Two individuals of an Oscillaria, magnified.
460. Locomotion. Moreover, many microscopic plants living in water are
seen to move freely, if not briskly, under the microscope; and so
likewise do more conspicuous aquatic plants in their embryo-like or
seedling state. Even at maturity, species of Oscillaria (such as in Fig.
488, minute worm-shaped plants of fresh waters, taking this name from
their oscillating motions) freely execute three different kinds of
movement, the very delicate investing coat of cellulose not impeding the
action of the living protoplasm within. Even when this coat is firmer
and hardened with a siliceous deposit, such crescent-shaped or
boat-shaped one-celled plants as Closterium or Naricula are able in
some way to move along from place to place in the water.
Fig. 489. A few cells of a leaf of Naias flexilis, highly
magnified: the arrows indicate the courses of the circulating currents.
461. Movements in Cells, or Cell-circulation, sometimes called
Cyclosis, has been detected in so many plants, especially in
comparatively [Pg 150]transparent aquatic plants and in hairs on the surface of
land plants (where it is easiest to observe), that it may be inferred to
take place in all cells during the most active part of their life. This
motion is commonly a streaming movement of threads of protoplasm,
carrying along solid granules by which the action may be observed and
the rate measured, or in some cases it is a rotation of the whole
protoplasmic contents of the cell. A comparatively low magnifying power
will show it in the cells of Nitella and Chara (which are cryptogamous
plants); and under a moderate power it is well seen in the Tape Grass of
fresh water, Vallisneria, and in Naias flexilis (Fig. 489). Minute
particles and larger greenish globules are seen to be carried along, as
if in a current, around the cell, passing up one side, across the end,
down the other and across the bottom, completing the circuit sometimes
within a minute or less when well warmed. To see it well in the cell,
which like a string of beads form the hairs on the stamens of
Spiderwort, a high magnifying power is needed.
462. Transference of Liquid from Cell to Cell, and so from place to
place in the plant, the absorption of water by the rootlets, and the
exhalation of the greater part of it from the foliage,—these and
similar operations are governed by the physical laws which regulate the
diffusion of fluids, but are controlled by the action of living
protoplasm. Equally under vital control are the various chemical
transformations which attend assimilation and growth, and which involve
not only molecular movements but conveyance. Growth itself, which is the
formation and shaping of new parts, implies the direction of internal
activities to definite ends.
463. Movements of Organs. The living protoplasm, in all but the lowest
grade of plants, is enclosed and to common appearance isolated in
separate cells, the walls of which can only in their earliest state be
said to be alive. Still plants are able to cause the protoplasm of
adjacent cells to act in concert, and by their combined action to effect
movements in roots, stems, or leaves, some of them very slow and
gradual, some manifest and striking. Such movements are brought about
through individually minute changes in the form or tension in the
protoplasm of the innumerable cells which make up the structure of the
organ. Some of the slower movements are effected during growth, and may
be explained by inequality of growth on the two sides of the bending
organ. But the more rapid changes of position, and some of the slow
ones, cannot be so explained.
[Pg 151]
464. Root-movements. In its growth a root turns or bends away from the
light and toward the centre of the earth, so that in lengthening it
buries itself in the soil where it is to live and act. Every one must
have observed this in the germination of seeds. Careful observations
have shown that the tip of a growing root also makes little sweeps or
short movements from side to side. By this means it more readily
insinuates itself into yielding portions of the soil. The root-tips will
also turn toward moisture, and so secure the most favorable positions in
the soil.
465. Stem-movements. The root end of the caulicle or first joint of stem
(that below the cotyledons) acts like the root, in turning downward in
germination (making a complete bend to do so if it happens to point
upward as the seed lies in the ground), while the other end turns or
points skyward. These opposite positions are taken in complete darkness
as readily as in the light, in dryness as much as in moisture:
therefore, so far as these movements are physical, the two portions of
the same internode appear to be oppositely affected by gravitation or
other influences.
466. Rising into the air, the stem and green shoots generally, while
young and pliable, bend or direct themselves toward the light, or toward
the stronger light when unequally illuminated; while roots turn toward
the darkness.
467. Many growing stems have also a movement of Nutation, that is, of
nodding successively in different directions. This is brought about by a
temporary increase of turgidity of the cells along one side, thus bowing
the stem over to the opposite side; and this line of turgescence travels
round the shoot continually, from right to left or from left to right
according to the species: thus the shoot bends to all points of the
compass in succession. Commonly this nutation is slight or hardly
observable. It is most marked in
468. Twining Stems (Fig. 90). The growing upper end of such stems, as is
familiar in the Hop, Pole Beans, and Morning-Glory, turns over in an
inclined or horizontal direction, thus stretching out to reach a
neighboring support, and by the continual change in the direction of the
nodding, sweeps the whole circle, the sweeps being the longer as the
stem lengthens. When it strikes against a support, such as a stem or
branch of a neighboring plant, the motion is arrested at the contact,
but continues at the growing apex beyond, and this apex is thus made to
wind spirally around the supporting body.
469. Leaf-movements are all but universal. The presentation by most
leaves of their upper surface to the light, from whatever direction that
may come, is an instance; for when turned upside down they twist or bend
round on the stalk to recover this normal position. Leaves, and the
leaflets of compound leaves, change this position at nightfall, or when
the light is withdrawn; they then take what is called their sleeping
posture, resuming the diurnal position when daylight returns. This is
very striking [Pg 152]in Locust-trees, in the Sensitive Plant (Fig. 490), and
in Woodsorrel. Young seedlings droop or close their leaves at night in
plants which are not thus affected in the adult foliage. All this is
thought to be a protection against the cold by nocturnal radiation.
470. Various plants climb by a coiling movement of their leaves or their
leaf-stalks. Familiar examples are seen in Clematis, Maurandia,
Tropæolum, and in a Solanum which is much cultivated in greenhouses
(Fig. 172). In the latter, and in other woody plants which climb in this
way, the petioles thicken and harden after they have grasped their
support, thus securing a very firm hold.
471. Tendril movements. Tendrils are either leaves or stems (98, 168),
specially developed for climbing purposes. Cobæa is a good example of
partial transformation; some of the leaflets are normal, some of the
same leaf are little tendrils, and some intermediate in character. The
Passion-flowers give good examples of simple stem-tendrils (Fig. 92);
Grape-Vines, of branched ones. Most tendrils make revolving sweeps, like
those of twining stems. Those of some Passion-flowers, in sultry
weather, are apt to move fast enough for the movement actually to be
seen for a part of the circuit, as plainly as that of the second-hand of
a watch. Two herbaceous species, Passiflora gracilis and P. sicyoides
(the first an annual, the second a strong-rooted perennial of the
easiest cultivation), are admirable for illustration both of revolving
movements and of sensitive coiling.
Fig. 490. Piece of stem of Sensitive Plant (Mimosa
pudica), with two leaves, the lower open, the upper in the closed
state.
472. Movements under Irritation. The most familiar case is that of the
Sensitive Plant (Fig. 490). The leaves suddenly take their nocturnal
position when roughly touched or when shocked by a jar. The leaflets
close in pairs, the four outspread partial petioles come closer
together, and the common petiole is depressed. The seat of the movements
is at the base of the leaf-stalk and stalklets. Schrankia, a near
relative of the Sensitive Plant, acts in the same way, but is slower.
These are not anomalous actions, but only extreme manifestations of a
faculty more or less common in foliage. In Locust and Honey-Locusts for
example, repeated jars will slowly produce similar effects.
[Pg 153]
473. Leaf-stalks and tendrils are adapted to their uses in climbing by
a similar sensitiveness. The coiling of the leaf-stalk is in response to
a kind of irritation produced by contact with the supporting body. This
may be shown by gentle rubbing or prolonged pressure upon the upper face
of the leaf-stalk, which is soon followed by a curvature. Tendrils are
still more sensitive to contact or light friction. This causes the free
end of the tendril to coil round the support, and the sensitiveness,
propagated downward along the tendril, causes that side of it to become
less turgescent or the opposite side more so, thus throwing the tendril
into coils. This shortening draws the plant up to the support. Tendrils
which have not laid hold will at length commonly coil spontaneously, in
a simple coil, from the free apex downward. In Sicyos, Echinocystis, and
the above mentioned Passion-flowers (471), the tendril is so sensitive,
under a high summer temperature, that it will curve and coil promptly
after one or two light strokes by the hand.
Fig. 491. Portion of stem and leaves of Telegraph-plant
(Desmodium gyrans), almost of natural size.
474. Among spontaneous movements the most singular are those of
Desmodium gyrans of India, sometimes called Telegraph-plant, which is
cultivated on account of this action. Of its three leaflets, the larger
(terminal) one moves only by drooping at nightfall and rising with the
dawn. But its two small lateral leaflets, when in a congenial high
temperature, by day and by night move upward and downward in a
succession of jerks, stopping occasionally, as if to recover from
exhaustion. In most plant-movements some obviously useful purpose is
subserved: this of Desmodium gyrans is a riddle.
475. Movements in Flowers are very various. The most remarkable are in
some way connected with fertilization (Sect. XIII.). Some occur under
irritation: the stamens of Barberry start forward when touched at the
base inside: those of many polyandrous flowers (of Sparmannia very
strikingly) spread outwardly when lightly brushed: the two lips or lobes
[Pg 154]of the stigma in Mimulus close after a touch. Some are automatic and
are connected with dichogamy (339): the style of Sabbatia and of
large-flowered species of Epilobium bends over strongly to one side or
turns downward when the blossom opens, but slowly erects itself a day or
two later.
476. Extraordinary Movements connected with Capture of Insects. The most
striking cases are those of Drosera and Dionæa; for an account of which
see "How Plants Behave," and Goodale's "Physiological Botany."
477. The upper face of the leaves of the common species of Drosera, or
Sundew, is beset with stout bristles, having a glandular tip. This tip
secretes a drop of a clear but very viscid liquid, which glistens like a
dew-drop in the sun; whence the popular name. When a fly or other small
insect, attracted by the liquid, alights upon the leaf, the viscid drops
are so tenacious that they hold it fast. In struggling it only becomes
more completely entangled. Now the neighboring bristles, which have not
been touched, slowly bend inward from all sides toward the captured
insect, and bring their sticky apex against its body, thus increasing
the number of bonds. Moreover, the blade of the leaf commonly aids in
the capture by becoming concave, its sides or edges turning inward,
which brings still more of the gland-tipped bristles into contact with
the captive's body. The insect perishes; the clear liquid disappears,
apparently by absorption into the tissue of the leaf. It is thought that
the absorbed secretion takes with it some of the juices of the insect or
the products of its decomposition.

Fig. 492. Plant of Dionæa muscipula, or Venus's Fly-trap,
reduced in size.
478. Dionæa muscipula, the most remarkable vegetable fly-trap (Fig. 176,
492), is related to the Sundews, and has a more special and active
apparatus for fly-catching, formed of the summit of the leaf. The two
halves of this rounded body move as if they were hinged upon the midrib;
their edges are fringed with spiny but not glandular bristles, which
interlock when the organ closes. Upon the face are two or three short
and delicate bristles, which are sensitive. They do not themselves move
when touched, but they propagate the sensitiveness to the organ itself,
causing it to close with a quick movement. In a fresh [Pg 155]and vigorous
leaf, under a high summer temperature, and when the trap lies widely
open, a touch of any one of the minute bristles on the face, by the
finger or any extraneous body, springs the trap (so to say), and it
closes suddenly; but after an hour or so it opens again. When a fly or
other small insect alights on the trap, it closes in the same manner,
and so quickly that the intercrossing marginal bristles obstruct the
egress of the insect, unless it be a small one and not worth taking.
Afterwards and more slowly it completely closes, and presses down upon
the prey; then some hidden glands pour out a glairy liquid, which
dissolves out the juices of the insect's body; next all is re-absorbed
into the plant, and the trap opens to repeat the operation. But the same
leaf perhaps never captures more than two or three insects. It ages
instead, becomes more rigid and motionless, or decays away.
479. That some few plants should thus take animal food will appear less
surprising when it is considered that hosts of plants of the lower
grade, known as Fungi, moulds, rusts, ferments, Bacteria, etc., live
upon animal or other organized matter, either decaying or living. That
plants should execute movements in order to accomplish the ends of their
existence is less surprising now when it is known that the living
substance of plants and animals is essentially the same; that the beings
of both kingdoms partake of a common life, to which, as they rise in the
scale, other and higher endowments are successively superadded.
480. Work uses up material and energy in plants as well as in animals.
The latter live and work by the consumption and decomposition of that
which plants have assimilated into organizable matter through an energy
derived from the sun, and which is, so to say, stored up in the
assimilated products. In every internal action, as well as in every
movement and exertion, some portion of this assimilated matter is
transformed and of its stored energy expended. The steam-engine is an
organism for converting the sun's radiant energy, stored up by plants in
the fuel, into mechanical work. An animal is an engine fed by vegetable
fuel in the same or other forms, from the same source, by the
decomposition of which it also does mechanical work. The plant is the
producer of food and accumulator of solar energy or force. But the
plant, like the animal, is a consumer whenever and by so much as it does
any work except its great work of assimilation. Every internal change
and movement, every transformation, such as that of starch into sugar
and of sugar into cell-walls, as well as every movement of parts which
becomes externally visible, is done at the expense of a certain amount
of its assimilated matter and of its stored energy; that is, by the
decomposition or combustion of sugar or some such product into carbonic
acid and water, which is given back to the air, just as in the animal it
is given back to the air in respiration. So the respiration of plants is
as real and as essential as that of animals. But what plants consume or
decompose in their life and action is of insignificant amount in
comparison with what they compose.
[Pg 156]
Section XVII. CRYPTOGAMOUS OR FLOWERLESS PLANTS.
481. Even the beginner in botany should have some general idea of what
cryptogamous plants are, and what are the obvious distinctions of the
principal families. Although the lower grades are difficult, and need
special books and good microscopes for their study, the higher orders,
such as Ferns, may be determined almost as readily as phanerogamous
plants.
482. Linnæus gave to this lower grade of plants the name of
Cryptogamia, thereby indicating that their organs answering to stamens
and pistils, if they had any, were recondite and unknown. There is no
valid reason why this long-familiar name should not be kept up, along
with the counterpart one of Phanerogamia (6), although organs
analogous to stamens and pistil, or rather to pollen and ovule, have
been discovered in all the higher and most of the lower grades of this
series of plants. So also the English synonymous name of Flowerless
Plants is both good and convenient: for they have not flowers in the
proper sense. The essentials of flowers are stamens and pistils, giving
rise to seeds, and the essential of a seed is an embryo (8).
Cryptogamous or Flowerless plants are propagated by Spores; and a spore
is not an embryo-plantlet, but mostly a single plant-cell (399).
483. Vascular Cryptogams, which compose the higher orders of this series
of plants, have stems and (usually) leaves, constructed upon the general
plan of ordinary plants; that is, they have wood (wood-cells and
vessels, 408) in the stem and leaves, in the latter as a frame work of
veins. But the lower grades, having only the more elementary cellular
structure, are called Cellular Cryptogams. Far the larger number of
the former are Ferns: wherefore that class has been called
484. Pteridophyta, Pteridophytes in English form, meaning
Fern-plants,—that is, Ferns and their relatives. They are mainly
Horsetails, Ferns, Club-Mosses, and various aquatics which have been
called Hydropterides, i. e. Water-Ferns.
485. Horsetails, Equisetaceæ, is the name of a family which consists
only (among now-living plants) of Equisetum, the botanical name of
Horsetail and Scouring Rush. They have hollow stems, with partitions at
the nodes; the leaves consist only of a whorl of scales at each node,
these coalescent into a sheath: from the axils of these leaf-scales, in
many species, branches grow out, which are similar to the stem but on a
much smaller scale, close-jointed, and with the tips of the leaves more
apparent. At the apex of the stem appears the fructification, as it is
called for lack of a better term, in the form of a short spike or head.
This consists of a good number of stalked shields, bearing on their
inner or under face several wedge-shaped spore-cases. The spore-cases
when they ripen open down the inner [Pg 157]side and discharge a great number
of green spores of a size large enough to be well seen by a hand-glass.
The spores are aided in their discharge and dissemination by four
club-shaped threads attached to one part of them. These are hygrometric:
when moist they are rolled up over the spore; when dry they straighten,
and exhibit lively movements, closing over the spore when breathed upon,
and unrolling promptly a moment after as they dry. (See Fig. 493-498.)
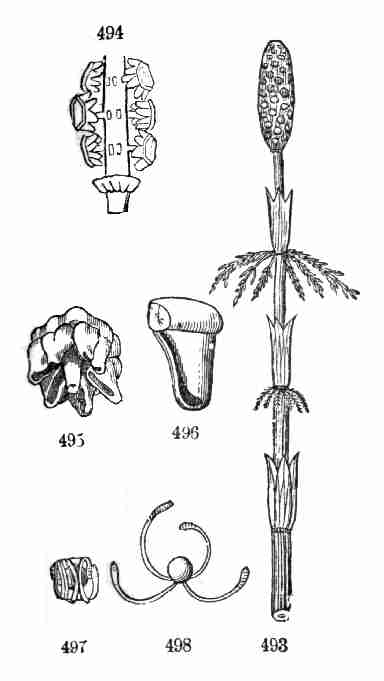
Fig. 493. Upper part of a stem of a Horsetail, Equisetum
sylvaticum. 494. Part of the head or spike of spore-cases, with some of
the latter taken off. 495. View (more enlarged) of under side of the
shield-shaped body, bearing a circle of spore-cases. 496. One of the
latter detached and more magnified. 497. A spore with the attached arms
moistened. 498. Same when dry, the arms extended.
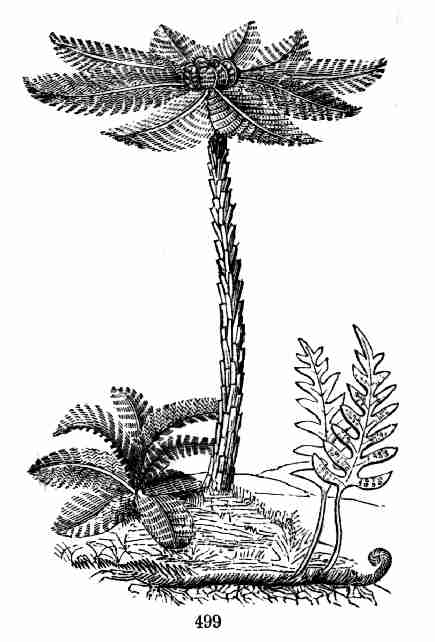
Fig. 499. A Tree-Fern, Dicksonia arborescens, with a
young one near its base. In front a common herbaceous Fern (Polypodium
vulgare) with its creeping stem or rootstock.
 Fig.
Fig. 500. A section of the trunk of a Tree-Fern.
486. Ferns, or Filices, a most attractive family of plants, are very
numerous and varied. In warm and equable climates some rise into
forest-trees, with habit of Palms; but most of them are perennial herbs.
The wood of a Fern-trunk is very different, however, from that of a
palm, or of any exogenous stem either. A section is represented in Fig.
500. The curved plates of wood each terminate [Pg 158]upward in a leaf-stalk.
The subterranean trunk or stem of any strong-growing herbaceous Fern
shows a similar structure. Most Ferns are circinate in the bud; that is,
are rolled up in the manner shown in Fig. 197. Uncoiling as they grow,
they have some likeness to a crosier.
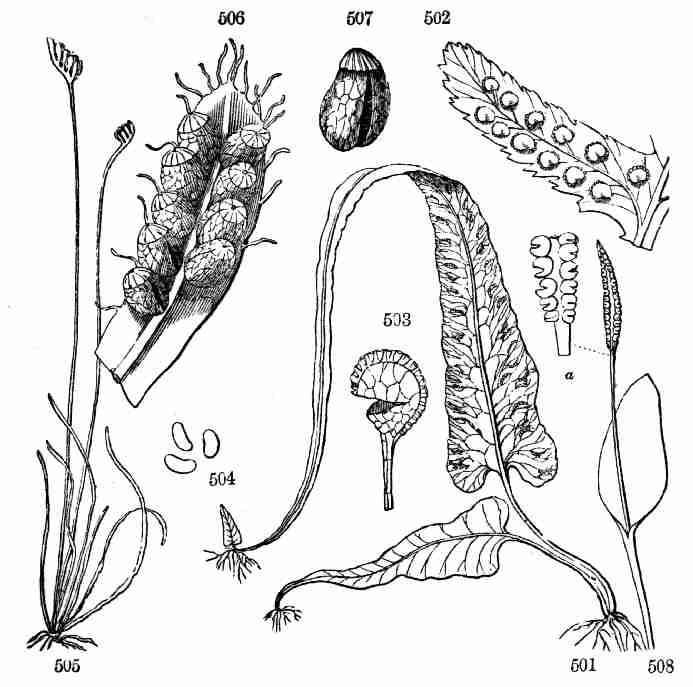
Fig. 501. The Walking-Fern, Camptosorus, reduced in size,
showing its fruit-dots on the veins approximated in pairs. 502. A small
piece (pinnule) of a Shield-Fern: a row of fruit-dots on each side of
the midrib, each covered by its kidney-shaped indusium. 503. A
spore-case from the latter, just bursting by the partial straightening
of the incomplete ring; well magnified. 504. Three of the spores of 509,
more magnified. 505. Schizæa pusilla, a very small and simple-leaved
Fern, drawn nearly of natural size. 506. One of the lobes of its
fruit-bearing portion, magnified, bearing two rows of spore-cases. 507.
Spore-case of the latter, detached, opening lengthwise. 508.
Adder-tongue, Ophioglossum; spore-cases in a kind of spike: a, a
portion of the fruiting part, about natural size; showing two rows of
the firm spore-cases, which open transversely into two valves.
487. The fructification of Ferns is borne on the back or under side of
the leaves. The early botanists thought this such a peculiarity that
they always called a Fern-leaf a Frond, and its petiole a Stipe. Usage
continues these terms, although they are superfluous. The fruit of Ferns
consists of Spore-cases, technically Sporangia, which grow out of the
veins of the leaf. Sometimes these are distributed over the whole lower
[Pg 159]surface of the leaf or frond, or over the whole surface when there are
no proper leaf-blades to the frond, but all is reduced to stalks.
Commonly the spore-cases occupy only detached spots or lines, each of
which is called a Sorus, or in English merely a Fruit-dot. In many Ferns
these fruit-dots are naked; in others they are produced under a
scale-like bit of membrane, called an Indusium. In Maidenhair-Ferns a
little lobe of the leaf is folded back over each fruit-dot, to serve as
its shield or indusium. In the true Brake or Bracken (Pteris) the whole
edge of the fruit-bearing part of the leaf is folded back over it like a
hem.
488. The form and structure of the spore-cases can be made out with a
common hand magnifying glass. The commonest kind (shown in Fig. 503) has
a stalk formed of a row of jointed cells, and is itself composed of a
layer of thin-walled cells, but is incompletely surrounded by a border
of thicker-walled cells, forming the Ring. This extends from the stalk
up one side of the spore-case, round its summit, descends on the other
side, but there gradually vanishes. In ripening and drying the shrinking
of the cells of the ring on the outer side causes it to straighten; in
doing so it tears the spore-case open on the weaker side and discharges
the minute spores that fill it, commonly with a jerk which scatters them
to the wind. Another kind of spore-case (Fig. 507) is stalkless, and has
its ring-cells forming a kind of cap at the top: at maturity it splits
from top to bottom by a regular dehiscence. A third kind is of firm
texture and opens across into two valves, like a clam-shell (Fig.
508a): this kind makes an approach to the next family.
Fig. 509. A young prothallus of a Maiden-hair, moderately
enlarged, and an older one with the first fern-leaf developed from near
the notch. 510. Middle portion of the young one, much magnified, showing
below, partly among the rootlets, the antheridia or fertilizing
organs, and above, near the notch, three pistillidia to be
fertilized.
489. The spores germinate on moistened ground. In a conservatory they
may be found germinating on a damp wall or on the edges of a
well-watered flower-pot. Instead of directly forming a fern-plantlet,
the spore grows first into a body which [Pg 160]closely resembles a small
Liverwort. This is named a Prothallus (Fig. 509): from some point of
this a bud appears to originate, which produces the first fern-leaf,
soon followed by a second and third, and so the stem and leaves of the
plant are set up.
Fig. 511. Lycopodium Carolinianum, of nearly natural
size. 512. Inside view of one of the bracts and spore-case, magnified.
Fig. 513. Open 4-valved spore-case of a Selaginella, and
its four large spores (macrospores), magnified. 514. Macrospores of
another Selaginella. 515. Same separated.
Fig. 516. Plant of Isoetes. 517. Base of a leaf and
contained sporocarp filled with microspores cut across, magnified. 518.
Same divided lengthwise, equally magnified; some microspores seen at the
left. 519. Section of a spore-case containing macrospores, equally
magnified; at the right three macrospores more magnified.
490. Investigation of this prothallus under the microscope resulted in
the discovery of a wholly unsuspected kind of fertilization, taking
place at this germinating stage of the plant. On the under side of the
prothallus two kinds of organs appear (Fig. 510). One may be likened to
an open and depressed ovule, with a single cell at bottom answering to
nucleus; the other, to an anther; but instead of pollen, it discharges
corkscrew-shaped microscopic filaments, which bear some cilia of extreme
tenuity, by the rapid vibration of which the filaments move freely over
a wet surface. These filaments travel over the surface of the
prothallus, and even to other prothalli (for there are natural hybrid
Ferns), reach and enter the ovule-like [Pg 161]cavities, and fertilize the
cell. This thereupon sets up a growth, forms a vegetable bud, and so
develops the new plant.
491. An essentially similar process of fertilization has been discovered
in the preceding and the following families of Pteridophytes; but it is
mostly subterranean and very difficult to observe.
492. Club-Mosses or Lycopodiums. Some of the common kinds, called Ground
Pine, are familiar, being largely used for Christmas wreaths and other
decoration. They are low evergreens, some creeping, all with
considerable wood in their stems: this thickly beset with small leaves.
In the axils of some of these leaves, or more commonly, in the axils of
peculiar leaves changed into bracts (as in Fig. 511, 512) spore-cases
appear, as roundish or kidney-shaped bodies, of firm texture, opening
round the top into two valves, and discharging a great quantity of a
very fine yellow powder, the spores.
493. The Selaginellas have been separated from Lycopodium, which they
much resemble, because they produce two kinds of spores, in separate
spore-cases. One kind (Microspores) is just that of Lycopodium; the
other consists of only four large spores (Macrospores), in a spore-case
which usually breaks in pieces at maturity (Fig. 513-515).
494. The Quillworts, Isoetes (Fig. 516-519), are very unlike Club Mosses
in aspect, but have been associated with them. They look more like
Rushes, and live in water, or partly out of it. A very short stem, like
a corm, bears a cluster of roots underneath; above it is covered by the
broad bases of a cluster of awl-shaped or thread-shaped leaves. The
spore-cases are immersed in the bases of the leaves. The outer
leaf-bases contain numerous macrospores; the inner are filled with
innumerable microspores.
Fig. 520. Plant of Marsilia quadrifoliata, reduced in
size; at the right a pair of sporocarps of about natural size.
495. The Pillworts (Marsilia and Pilularia) are low aquatics, which
[Pg 162]bear globular or pill-shaped fruit (Sporocarps) on the lower part of
their leaf-stalks or on their slender creeping stems. The leaves of the
commoner species of Marsilia might be taken for four-leaved Clover. (See
Fig. 520.) The sporocarps are usually raised on a short stalk. Within
they are divided lengthwise by a partition, and then crosswise by
several partitions. These partitions bear numerous delicate sacs or
spore-cases of two kinds, intermixed. The larger ones contain each a
large spore, or macrospore; the smaller contain numerous microspores,
immersed in mucilage. At maturity the fruit bursts or splits open at
top, and the two kinds of spores are discharged. The large ones in
germination produce a small prothallus; upon which the contents of the
microspores act in the same way as in Ferns, and with a similar result.
496. Azolla is a little floating plant, looking like a small Liverwort
or Moss. Its branches are covered with minute and scale-shaped leaves.
On the under side of the branches are found egg-shaped thin-walled
sporocarps of two kinds. The small ones open across and discharge
microspores; the larger burst irregularly, and bring to view globose
spore-cases, attached to the bottom of the sporocarp by a slender stalk.
These delicate spore-cases burst and set free about four macrospores,
which are fertilized at germination, in the manner of the Pillworts and
Quillworts. (See Fig. 521-526.)
Fig. 521. Small plant of Azolla Caroliniana. 522. Portion
magnified, showing the two kinds of sporocarp; the small ones contain
microspores. 523 represents one more magnified. 524. The larger
sporocarp more magnified. 525. Same more magnified and burst open,
showing stalked spore-cases. 526. Two of the latter highly magnified;
one of them bursting shows four contained macrospores; between the two,
three of these spores highly magnified.
497. Cellular Cryptogams (483) are so called because composed, even in
their higher forms, of cellular tissue only, without proper wood-cells
or vessels. Many of the lower kinds are mere plates, or ribbons, or
simple rows of cells, or even single cells. But their highest orders
follow the plan of Ferns and phanerogamous plants in having stem and
leaves for their upward growth, and commonly roots, or at least
rootlets, [Pg 163]to attach them to the soil, or to trunks, or to other bodies
on which they grow. Plants of this grade are chiefly Mosses. So as a
whole they take the name of
498. Bryophyta, Bryophytes in English form, Bryum being the Greek name
of a Moss. These plants are of two principal kinds: true Mosses
(Musci, which is their Latin name in the plural); and Hepatic Mosses,
or Liverworts (Hepaticæ).
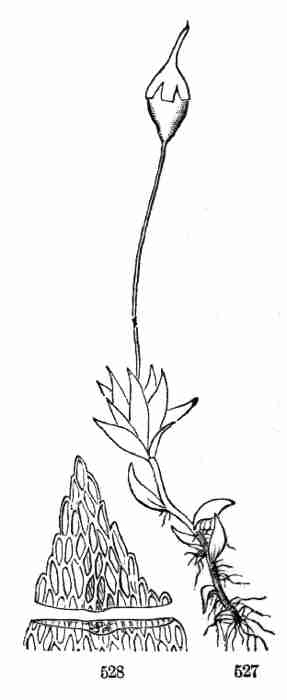
Fig. 527. Single plant of Physcomitrium pyriforme,
magnified. 528. Top of a leaf, cut across; it consists of a single layer
of cells.
499. Mosses or Musci. The pale Peat-mosses (species of Sphagnum, the
principal component of sphagnous bogs) and the strong-growing Hair-cap
Moss (Polytrichum) are among the larger and commoner representatives of
this numerous family; while Fountain Moss (Fontinalis) in running water
sometimes attains the length of a yard or more. On the other hand, some
are barely individually distinguishable to the naked eye. Fig. 527
represents a common little Moss, enlarged to about twelve times its
natural size; and by its side is part of a leaf, much magnified, showing
that it is composed of cellular tissue (parenchyma-cells) only. The
leaves of Mosses are always simple, distinct, and sessile on the stem.
The fructification is an urn-shaped spore-case, in this as in most cases
raised on a slender stalk. The spore-case loosely bears on its summit a
thin and pointed cap, like a candle-extinguisher, called a Calyptra.
Detaching this, it is found that the spore-case is like a pyxis (376),
that is, the top at maturity comes off as a lid (Operculum); and that
the interior is filled with a green powder, the spores, which are
discharged through the open mouth. In most Mosses there is a fringe of
one or two rows of teeth or membrane around this mouth or orifice, the
Peristome. When moist the peristome closes hygrometrically over the
orifice more or less; when drier the teeth or processes commonly bend
outward or recurve; and then the spores more readily escape. In Hair-cap
Moss a membrane is stretched quite across the mouth, like a drum-head,
retaining the spores until this wears away. See Figures 527-541 for
details.
500. Fertilization in Mosses is by the analogues of stamens and pistils,
which are hidden in the axils of leaves, or in the cluster of leaves at the
[Pg 164]end of the stem. The analogue of the anther (Antheridium) is a
cellular sac, which in bursting discharges innumerable delicate cells
floating in a mucilaginous liquid; each of these bursts and sets free a
vibratile self-moving thread. These threads, one or more, reach the
orifice of the pistil-shaped body, the Pistillidium, and act upon a
particular cell at its base within. This cell in its growth develops
into the spore-case and its stalk (when there is any), carrying on its
summit the wall of the pistillidium, which becomes the calyptra.
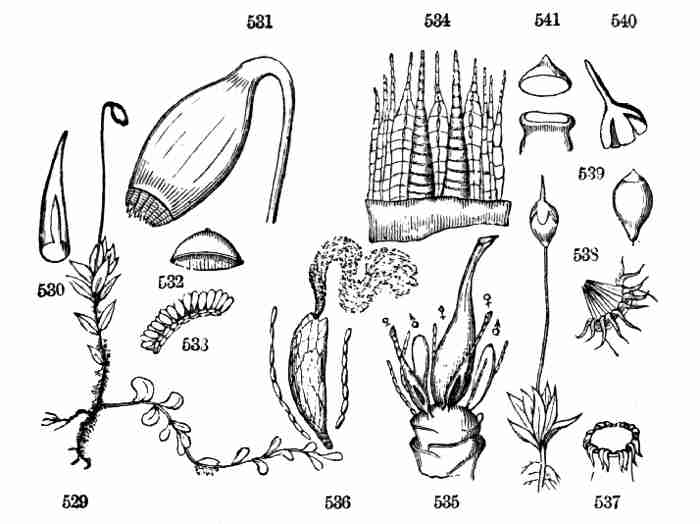
Fig. 529. Mnium cuspidatum, smaller than nature. 530. Its
calyptra, detached, enlarged. 531. Its spore-case, with top of stalk,
magnified, the lid (532) being detached, the outer peristome appears.
533. Part of a cellular ring (annulus) which was under the lid,
outside of the peristome, more magnified. 534. Some of the outer and of
the inner peristome (consisting of jointed teeth) much magnified. 535.
Antheridia and a pistillidium (the so-called flower) at end of a stem of
same plant, the leaves torn away (♂, antheridia,
♀, pistillidium), magnified. 536. A bursting antheridium, and
some of the accompanying jointed threads, highly magnified. 537. Summit
of an open spore-case of a Moss, which has a peristome of 16 pairs of
teeth. 538. The double peristome of a Hypnum. 539-541. Spore-case,
detached calyptra, and top of more enlarged spore-case and detached lid,
of Physcomitrium pyriforme (Fig. 527): orifice shows that there is no
peristome.
501. Liverworts or Hepatic Mosses (Hepaticæ) in some kinds resemble
true Mosses, having distinct stem and leaves, although their leaves
occasionally run together; while in others there is no distinction of
stem and leaf, but the whole plant is a leaf-like body, which produces
rootlets on the lower face and its fructification on the upper. Those of
the moss-like kind (sometimes called Scale-Mosses) have their tender
spore-cases splitting into four valves; and with their spores are
intermixed some slender spiral [Pg 165]and very hygrometric threads (called
Elaters) which are thought to aid in the dispersion of the spores.
(Fig. 542-544.)
Fig. 542. Fructification of a Jungermannia, magnified;
its cellular spore-stalk, surrounded at base by some of the leaves, at
summit the 4-valved spore-case opening, discharging spores and elaters.
543. Two elaters and some spores from the same, highly magnified.
Fig. 544. One of the frondose Liverworts, Steetzia,
otherwise like a Jungermannia; the spore-case not yet protruded from its
sheath.
502. Marchantia, the commonest and largest of the true Liverworts, forms
large green plates or fronds on damp and shady ground, and sends up from
some part of the upper face a stout stalk, ending in a several-lobed
umbrella-shaped body, under the lobes of which hang several thin-walled
spore-cases, which burst open and discharge spores and elaters. Riccia
natans (Fig. 545) consists of wedge-shaped or heart-shaped fronds, which
float free in pools of still water. The under face bears copious
rootlets; in the substance of the upper face are the spore-cases, their
pointed tips merely projecting: there they burst open, and discharge
their spores. These are comparatively few and large, and are in fours;
so they are very like the macrospores of Pillworts or Quillworts.
503. Thallophyta, or Thallophytes in English form. This is the name for
the lower class of Cellular Cryptogams,—plants in which there is no
marked distinction into root, stem, and leaves. Roots in any proper
sense they never have, as organs for absorbing, although some of the
larger Seaweeds (such as the Sea Colander, Fig. 553) have them as
holdfasts. Instead of axis and foliage, there is a stratum of frond, in
such plants commonly called a Thallus (by a strained use of a Greek and
Latin word which means a green shoot or bough), which may have any kind
of form, leaf-like, stem-like, branchy, extended to a flat plate, or
gathered into a sphere, or drawn out into threads, or reduced to a
single row of cells, or even reduced to single cells. Indeed,
Thallophytes are so multifarious, so numerous in kinds, so protean in
their stages and transformations, so recondite in their fructification,
and many so microscopic in size, either of [Pg 166]the plant itself or its
essential organs, that they have to be elaborately described in separate
books and made subjects of special study.
Fig. 545, 546. Two plants of Riccia natans, about natural
size. 547. Magnified section of a part of the frond, showing two
immersed spore-cases, and one emptied space. 548. Magnified section of a
spore-case with some spores. 549. Magnified spore-case torn out, and
spores; one figure of the spores united; the other of the four
separated.
504. Nevertheless, it may be well to try to give some general idea of
what Algæ and Lichens and Fungi are. Linnæus had them all under the
orders of Algæ and Fungi. Afterwards the Lichens were separated; but of
late it has been made most probable that a Lichen consists of an Alga
and a Fungus conjoined. At least it must be so in some of the ambiguous
forms. Botanists are in the way of bringing out new classifications of
the Thallophytes, as they come to understand their structure and
relations better. Here, it need only be said that
505. Lichens live in the air, that is, on the ground, or on rocks,
trunks, walls, and the like, and grow when moistened by rains. They
assimilate air, water, and some earthy matter, just as do ordinary
plants. Algæ, or Seaweeds, live in water, and live the same kind of life
as do ordinary plants. Fungi, whatever medium they inhabit, live as
animals do, upon organic matter,—upon what other plants have
assimilated, or upon the products of [Pg 167]their decay. True as these general
distinctions are, it is no less true that these orders run together in
their lowest forms; and that Algæ and Fungi may be traced down into
forms so low and simple that no clear line can be drawn between them;
and even into forms of which it is uncertain whether they should be
called plants or animals. It is as well to say that they are not high
enough in rank to be distinctively either the one or the other. On the
other hand there is a peculiar group of plants, which in simplicity of
composition resemble the simpler Algæ, while in fructification and in
the arrangements of their simple cells into stem and branches they seem
to be of a higher order, viz.:—
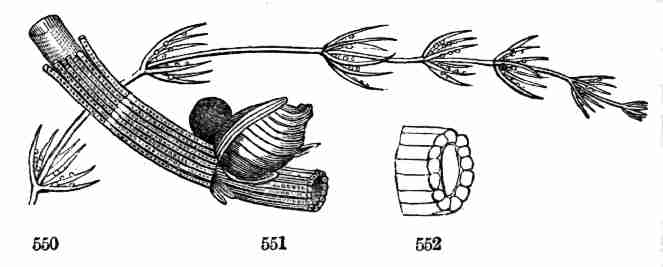
Fig. 550. Branch of a Chara, about natural size. 551. A
fruiting portion, magnified, showing the structure; a sporocarp, and an
antheridium. 552. Outlines of a portion of the stem in section, showing
the central cell and the outer or cortical cells.
506. Characeæ. These are aquatic herbs, of considerable size, abounding
in ponds. The simpler kinds (Nitella) have the stem formed of a single
row of tubular cells, and at the nodes, or junction of the cells, a
whorl of similar branches. Chara (Fig. 550-552) is the same, except that
the cells which make up the stem and the principal branches are
strengthened by a coating of many smaller tubular cells, applied to the
surface of the main or central cell. The fructification consists of a
globular sporocarp of considerable size, which is spirally enwrapped by
tubular cells twisted around it: by the side of this is a smaller and
globular antheridium. The latter breaks up into eight shield-shaped
[Pg 168]pieces, with an internal stalk, and bearing long and ribbon shaped
filaments, which consist of a row of delicate cells, each of which
discharges a free-moving microscopic thread (the analogue of the pollen
or pollen-tube), nearly in the manner of Ferns and Mosses. One of these
threads reaches and fertilizes a cell at the apex of the nucleus or
solid body of the sporocarp. This subsequently germinates and forms a
new individual.
507. Algæ or Seaweeds. The proper Seaweeds may be studied by the aid of
Professor Farlow's "Marine Algæ of New England;" the fresh-water
species, by Prof. H. C. Woods's "Fresh-water Algæ of North America," a
larger and less accessible volume. A few common forms are here very
briefly mentioned and illustrated, to give an idea of the family. But
they are of almost endless diversity.
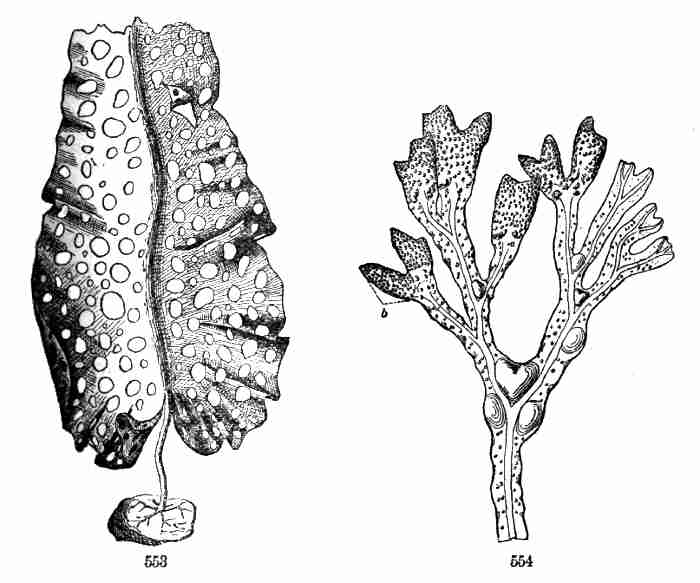
Fig. 553. Agarum Turneri, Sea Colander (so called from
the perforations with which the frond, as it grows, becomes riddled);
very much reduced in size.
Fig. 554. Upper end of a Rockweed, Fucus vesiculosus,
reduced half or more, b, the fructification.
508. The common Rockweed (Fucus vesiculosus, Fig. 554, abounding between
high and low water mark on the coast), the rarer Sea Colander (Agarum
Turneri, Fig. 553), and Laminaria, of which the larger forms are called
Devil's Aprons, are good representatives of the olive green or brownish
Seaweeds. They are attached either by a disk-like base or by root-like
holdfasts to the rocks or stones on which they grow.
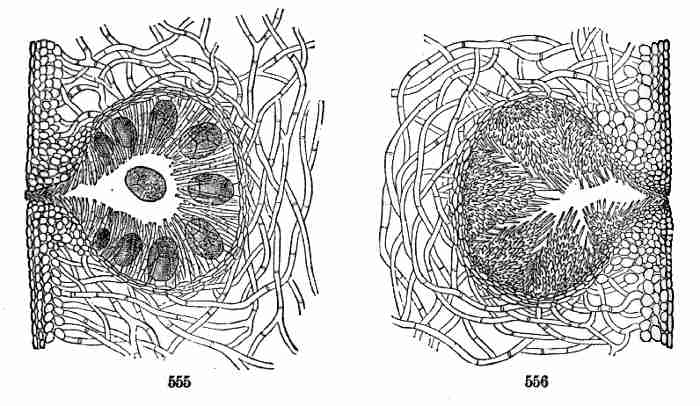
Fig. 555. Magnified section through a fertile conceptacle
of Rockweed, showing the large spores in the midst of threads of cells.
556. Similar section of a sterile conceptacle, containing slender
antheridia. From Farlow's "Marine Algæ of New England."
509. The hollow and inflated places in the Fucus vesiculosus or Rockweed
(Fig. 554) are air-bladders for buoyancy. The fructification forms in
the substance of the tips of the frond: the rough dots mark the places
where the conceptacles open. The spores and the fertilizing cells are in
different plants. Sections of the two kinds of conceptacles are given in
Fig. 555 and 556. The contents of the conceptacles are discharged
through [Pg 169]a small orifice which in each figure is at the margin of the
page. The large spores are formed eight together in a mother-cell. The
minute motile filaments of the antheridia fertilize the large spores
after injection into the water: and then the latter promptly acquire a
cell-wall and germinate.
510. The Florideæ or Rose-red series of marine Algæ (which, however, are
sometimes green or brownish) are the most attractive to amateurs. The
delicate Porphyra or Laver is in some countries eaten as a delicacy, and
the cartilaginous Chondrus crispus has been largely used for jelly.
Besides their conceptacles, which contain true spores (Fig. 560), they
mostly have a fructification in Tetraspores, that is, of spores
originating in fours (Fig. 559).
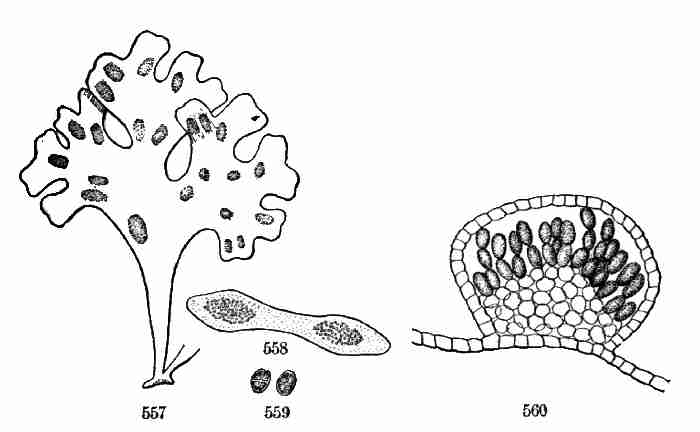
Fig. 557. Small plant of Chondrus crispus, or Carrageen
Moss, reduced in size, in fruit; the spots represent the fructification,
consisting of numerous tetraspores in bunches in the substance of the
plant. 558. Section through the thickness of one of the lobes,
magnified, passing through two of the imbedded fruit-clusters. 559. Two
of its tetraspores (spores in fours), highly magnified.
Fig. 560. Section through a conceptacle of Delesseria
Leprieurei, much magnified, showing the spores, which are single
specialized cells, two or three in a row.
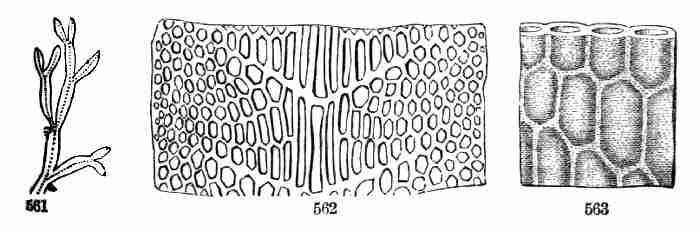
Fig. 561. A piece of the rose-red Delesseria Leprieurei,
double natural size. 562. A piece cut out and much magnified, showing
that it is composed of a layer of cells. 563. A few of the cells more
highly magnified: the cells are gelatinous and thick-walled.
511. The Grass-green Algæ sometimes form broad membranous fronds, such
as those of the common Ulva of the sea-shore, but most of them form mere
threads, either simple or branched. To this division belong almost [Pg 170]all
the Fresh-water Algæ, such as those which constitute the silky threads
or green slime of running streams or standing pools, and which were all
called Confervas before their immense diversity was known. Some are
formed of a single row of cells, developed each from the end of another.
Others branch, the top of one cell producing more than one new one (Fig.
564). Others, of a kind which is very common in fresh water, simple
threads made of a line of cells, have the chlorophyll and protoplasm of
each cell arranged in spiral lines or bands. They form spores in a
peculiar way, which gives to this family the designation of conjugating
Algæ.
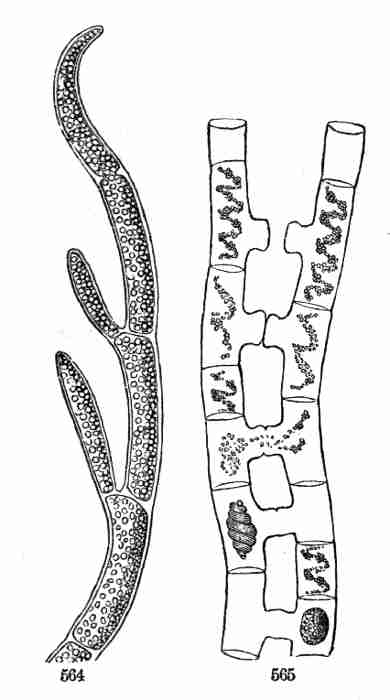
Fig. 564. The growing end of a branching Conferva
(Cladophora glomerata), much magnified; showing how, by a kind of
budding growth, a new cell is formed by a cross partition separating the
newer tip from the older part below; also, how the branches arise.
Fig. 565. Two magnified individuals of a Spirogyra,
forming spores by conjugation; a completed spore at base: above,
successive stages of the conjugation are represented.
512. At a certain time two parallel threads approach each other more
closely; contiguous parts of a cell of each thread bulge or grow out,
and unite when they meet; the cell-wall partitions between them are
absorbed so as to open a free communication; the spiral band of green
matter in both cells breaks up; the whole of that of one cell passes
over into the other; and of the united contents a large green spore is
formed. Soon the old cells decay, and the spore [Pg 171]set free is ready to
germinate. Fig. 565 represents several stages of the conjugating
process, which, however, would never be found all together like this in
one pair of threads.
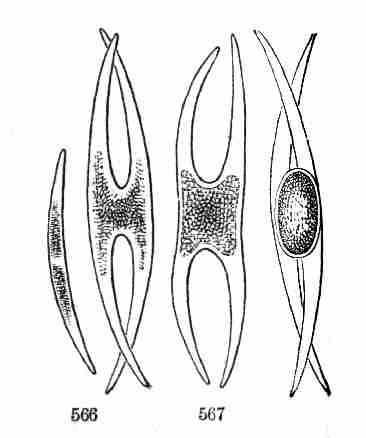
Fig. 566. Closterium acutum, a common Desmid, moderately
magnified. It is a single firm-walled cell, filled with green
protoplasmic matter.
Fig. 567. More magnified view of three stages of the
conjugation of a pair of the same.
513. Desmids and Diatomes, which are microscopic one-celled plants of
the same class, conjugate in the same way, as is shown in a Closterium
by Fig. 566, 567. Here the whole living contents of two individuals are
incorporated into one spore, for a fresh start. A reproduction which
costs the life of two individuals to make a single new one would be
fatal to the species if there were not a provision for multiplication by
the prompt division of the new-formed individual into two, and these
again into two, and so on in geometrical ratio. And the costly process
would be meaningless if there were not some real advantage in such a
fresh start, that is, in sexes.
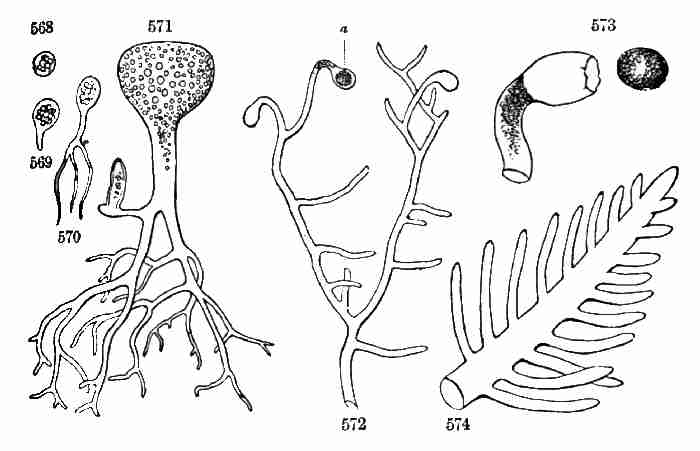
Fig. 568. Early stage of a species of Botrydium, a
globose cell. 569, 570. Stages of growth. 571. Full-grown plant,
extended and ramified below in a root-like way. 572. A Vaucheria; single
cell grown on into a much-branched thread; the end of some branches
enlarging, and the green contents in one (a) there condensed into a
spore. 573. More magnified view of a, and the mature spore escaping.
574. Bryopsis plumosa; apex of a stem with its branchlets; all the
extension of one cell. Variously magnified.
514. There are other Algæ of the grass-green series which consist of
single cells, but which by continued growth form plants of considerable
size. Three kinds of these are represented in Fig. 568-574.
515. Lichens, Latin Lichenes, are to be studied in the works of the
late Professor Tuckerman, but a popular exposition is greatly needed.
The subjoined illustrations (Fig. 575-580) may simply indicate what some
of the commoner forms are like. The cup, or shield-shaped spot, or knob,
which bears the fructification is named the Apothecium. This is mainly
[Pg 172]composed of slender sacs (Asci), having thread-shaped cells
intermixed; and each ascus contains few or several spores, which are
commonly double or treble. Most Lichens are flat expansions of grayish
hue; some of them foliaceous in texture, but never of bright green
color; more are crustaceous; some are wholly pulverulent and nearly
formless. But in several the vegetation lengthens into an axis (as in
Fig. 580), or imitates stem and branches or threads, as in the
Reindeer-Moss on the ground in our northern woods, and the Usnea hanging
from the boughs of old trees overhead.
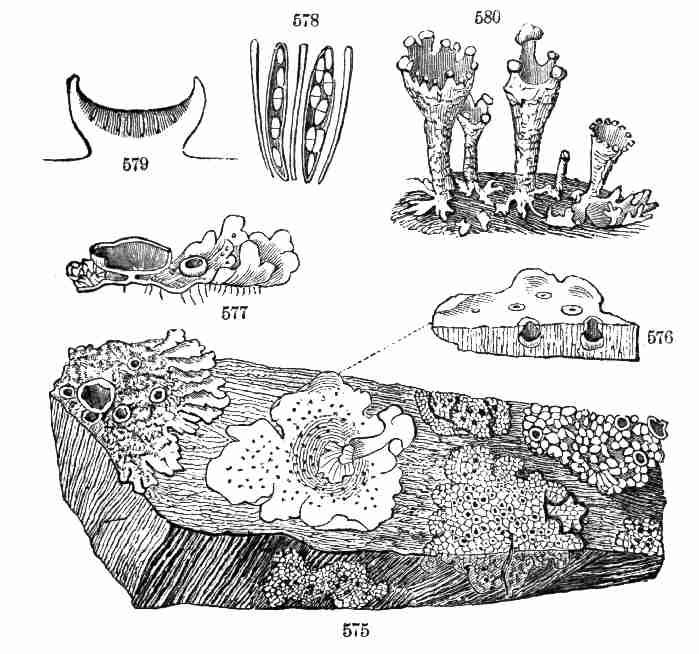
Fig. 575. A stone on which various Lichens are growing,
such as (passing from left to right) a Parmelia, a Sticta, and on the
right, Lecidia geographica, so called from its patches resembling the
outline of islands or continents as depicted upon maps. 576. Piece of
thallus of Parmelia conspersa, with section through an apothecium. 577.
Section of a smaller apothecium, enlarged. 578. Two asci of same, and
contained spores, and accompanying filaments; more magnified. 579. Piece
of thallus of a Sticta, with section, showing the immersed apothecia;
the small openings of these dot the surface. 580. Cladonia coccinea; the
fructification is in the scarlet knobs, which surround the cups.
516. Fungi. For this immense and greatly diversified class, it must here
suffice to indicate the parts of a Mushroom, a Sphæria, and of one or
two common Moulds. The true vegetation of common Fungi consists of
slender cells which form what is called a Mycelium. These filamentous
[Pg 173]cells lengthen and branch, growing by the absorption through their
whole surface of the decaying, or organizable, or living matter which
they feed upon. In a Mushroom (Agaricus), a knobby mass is at length
formed, which develops into a stout stalk (Stipe), bearing the cap
(Pileus): the under side of the cap is covered by the Hymenium, in
this genus consisting of radiating plates, the gills or Lamellæ; and
these bear the powdery spores in immense numbers. Under the microscope,
the gills are found to be studded with projecting cells, each of which,
at the top, produces four stalked spores. These form the powder which
collects on a sheet of paper upon which a mature Mushroom is allowed to
rest for a day or two. (Fig. 581-586.)
517. The esculent Morel, also Sphæria (Fig. 585, 586), and many other
Fungi bear their spores in sacs (asci) exactly in the manner of Lichens
(515).
Fig. 581. Agaricus campestris, the common edible
Mushroom. 582. Section of cap and stalk. 583. Minute portion of a
section of a gill, showing some spore-bearing cells, much magnified.
584. One of these, with its four spores, more magnified.
Fig. 585. Sphæria rosella. 586. Two of the asci and
contained double spores, quite like those of a Lichen; much magnified.
518. Of the Moulds, one of the commoner is the Bread-Mould (Fig. 587).
In fruiting it sends up a slender stalk, which bears a globular sac;
[Pg 174]this bursts at maturity and discharges innumerable spores. The blue
Cheese-Mould (Fig. 588) bears a cluster of branches at top, each of
which is a row of naked spores, like a string of beads, all breaking
apart at maturity. Botrytis (Fig. 589), the fruiting stalk of which
branches, and each branch is tipped with a spore, is one of the many
moulds which live and feed upon the juices of other plants or of
animals, and are often very destructive. The extremely numerous kinds of
smut, rust, mildew, the ferments, bacteria, and the like, many of them
very destructive to other vegetable and to animal life, are also low
forms of the class of Fungi.[1]
Fig. 587. Ascophora, the Bread-Mould. 588. Aspergillus
glaucus, the mould of cheese, but common on mouldy vegetables. 589. A
species of Botrytis. All magnified.
[Pg 175]
SECTION XVIII. CLASSIFICATION AND NOMENCLATURE.
519. Classification, in botany, is the consideration of plants in
respect to their kinds and relationships. Some system of Nomenclature,
or naming, is necessary for fixing and expressing botanical knowledge so
as to make it available. The vast multiplicity of plants and the various
degrees of their relationship imperatively require order and system, not
only as to names for designating the kinds of plants, but also as to
terms for defining their differences. Nomenclature is concerned with
the names of plants. Terminology supplies names of organs or parts, and
terms to designate their differences.
§ 1. KINDS AND RELATIONSHIP.
520. Plants and animals have two great peculiarities: 1st, they form
themselves; and 2d, they multiply themselves. They reproduce their kind
in a continued succession of
521. Individuals. Mineral things occur as masses, which are divisible
into smaller and still smaller ones without alteration of properties.
But organic things (vegetables and animals) exist as individual
beings. Each owes its existence to a parent, and produces similar
individuals in its turn. So each individual is a link of a chain; and to
this chain the natural-historian applies the name of
522. Species. All the descendants from the same stock therefore compose
one species. And it was from our observing that the several sorts of
plants or animals steadily reproduce themselves, or, in other words,
keep up a succession of similar individuals, that the idea of species
originated. There are few species, however, in which man has actually
observed the succession for many generations. It could seldom be proved
that all the White Pine trees or White Oaks of any forest came from the
same stock. But observation having familiarized us with the general fact
that individuals proceeding from the same stock are essentially alike,
we infer from their close resemblance that these similar individuals
belong to the same species. That is, we infer it when the individuals
are as much like each other as those are which we know, or confidently
suppose, to have sprung from the same stock.
523. Identity in species is inferred from close similarity in all
essential respects, or whenever the differences, however considerable,
are not known or reasonably supposed to have been originated in the
course of time under changed conditions. No two individuals are exactly
alike; a tendency to variation pervades all living things. In
cultivation, where variations are looked after and cared for, very
striking differences come to light; and if in wild nature they are less
common or less conspicuous, it is partly because they are uncared for.
When such variant forms are pretty well marked they are called
[Pg 176]
524. Varieties. The White Oak, for example, presents two or three
varieties in the shape of the leaves, although they may be all alike
upon each particular tree. The question often arises, and it is often
hard to answer, whether the difference in a particular case is that of a
variety, or is specific. If the former, it may commonly be proved by
finding such intermediate degrees of difference in various individuals
as to show that no clear distinction can be drawn between them; or else
by observing the variety to vary back again in some of its offspring.
The sorts of Apples, Pears, Potatoes, and the like, show that
differences which are permanent in the individual, and continue
unchanged through a long series of generations when propagated by
division (as by offsets, cuttings, grafts, bulbs, tubers, etc.), are not
likely to be reproduced by seed. Still they sometimes are so, and
perhaps always tend in that direction. For the fundamental law in
organic nature is that offspring shall be like parent.
Races are such strongly marked varieties, capable of coming true to
seed. The different sorts of Wheat, Maize, Peas, Radishes, etc., are
familiar examples. By selecting those individuals of a species which
have developed or inherited any desirable peculiarity, keeping them from
mingling with their less promising brethren, and selecting again the
most promising plants raised from their seeds, the cultivator may in a
few generations render almost any variety transmissible by seed, so long
as it is cared for and kept apart. In fact, this is the way the
cultivated domesticated races, so useful to man, have been fixed and
preserved. Races, in fact, can hardly, if at all, be said to exist
independently of man. But man does not really produce them. Such
peculiarities—often surprising enough—now and then originate, we know
not how (the plant sports, as the gardeners say); they are only
preserved, propagated, and generally further developed, by the
cultivator's skilful care. If left alone, they are likely to dwindle and
perish, or else revert to the original form of the species. Vegetable
races are commonly annuals, which can be kept up only by seed, or herbs
of which a succession of generations can be had every year or two, and
so the education by selection be completed without great lapse of time.
But all fruit-trees could probably be fixed into races in an equal
number of generations.
Bud-varieties are those which spring from buds instead of seed. They are
uncommon to any marked extent. They are sometimes called Sports, but
this name is equally applied to variations among seedlings.
Cross-breeds, strictly so-called, are the variations which come from
cross-fertilizing one variety of a species with another.
Hybrids are the varieties, if they may be so called,—which come from
the crossing of species (331). Only nearly related species can be
hybridized; and the resulting progeny is usually self-sterile, but not
always. Hybrid plants, however, may often be fertilized and made
prolific by the pollen of one or the other parent. This produces another
kind of cross-breeds.
525. Species are the units in classification. Varieties, although of
[Pg 177]utmost importance in cultivation and of considerable consequence in the
flora of any country, are of less botanical significance. For they are
apt to be indefinite and to shade off one form into another. But
species, the botanist expects to be distinct. Indeed, the practical
difference to the botanist between species and varieties is the definite
limitation of the one and the indefiniteness of the other. The
botanist's determination is partly a matter of observation, partly of
judgment.
526. In an enlarged view, varieties may be incipient species; and nearly
related species probably came from a common stock in earlier times. For
there is every reason to believe that existing vegetation came from the
more or less changed vegetation of a preceding geological era. However
that may be, species are regarded as permanent and essentially unchanged
in their succession of individuals through the actual ages.
527. There are, at nearly the lowest computation, as many as one hundred
thousand species of phanerogamous plants, and the cryptogamous species
are thought to be still more numerous. They are all connected by
resemblances or relationships, near and remote, which show that they are
all parts of one system, realizations in nature, as we may affirm, of
the conception of One Mind. As we survey them, they do not form a single
and connected chain, stretching from the lowest to the highest organized
species, although there obviously are lower and higher grades. But the
species throughout group themselves, as it were, into clusters or
constellations, and these into still more comprehensive clusters, and so
on, with gaps between. It is this clustering which is the ground of the
recognition of kinds of species, that is, of groups of species of
successive grades or degree of generality; such as that of similar
species into Genera, of genera into Families or Orders, of orders
into Classes. In classification the sequence, proceeding from higher
or more general to lower or special, is always Class, Order, Genus,
Species, Variety (if need be).
528. Genera (in the singular, Genus) are assemblages of closely
related species, in which the essential parts are all constructed on the
same particular type or plan. White Oak, Red Oak, Scarlet Oak, Live Oak,
etc., are so many species of the Oak genus (Latin, Quercus). The
Chestnuts compose another genus; the Beeches another. The Apple, Pear,
and Crab are species of one genus, the Quince represents another, the
various species of Hawthorn a third. In the animal kingdom the common
cat, the wild-cat, the panther, the tiger, the leopard, and the lion are
species of the cat kind or genus; while the dog, the jackal, the
different species of wolf, and the foxes, compose another genus. Some
genera are represented by a vast number of species, others by few, very
many by only one known species. For the genus may be as perfectly
represented in one species as in several, although, if this were the
case throughout, genera and species would of course be identical. The
Beech genus and the Chestnut genus would be just as distinct from the
Oak genus even if but one Beech and Chestnut were known; as indeed was
once the case.
[Pg 178]
529. Orders are groups of genera that resemble each other; that is,
they are to genera what genera are to species. As familiar
illustrations, the Oak, Chestnut, and Beech genera, along with the Hazel
genus and the Hornbeams, all belong to one order. The Birches and the
Alders make another; the Poplars and Willows, another; the Walnuts (with
the Butternut) and the Hickories, still another. The Apple genus, the
Quince and the Hawthorns, along with the Plums and Cherries and the
Peach, the Raspberry with the Blackberry, the Strawberry, the Rose,
belong to a large order, which takes its name from the Rose. Most
botanists use the names "Order" and "Family" synonymously; the latter
more popularly, as "the Rose Family," the former more technically, as
"Order Rosaceæ."
530. But when the two are distinguished, as is common in zoölogy, Family
is of lower grade than Order.
531. Classes are still more comprehensive assemblages, or great groups.
Thus, in modern botany, the Dicotyledonous plants compose one class, the
Monocotyledonous plants another (36-40).
532. These four grades, Class, Order, Genus, Species, are of universal
use. Variety comes in upon occasion. For, although a species may have no
recognized varieties, a genus implies at least one species belonging to
it; every genus is of some order, and every order of some class.
533. But these grades by no means exhaust the resources of
classification, nor suffice for the elucidation of all the distinctions
which botanists recognize. In the first place, a higher grade than that
of class is needful for the most comprehensive of divisions, that of all
plants into the two Series of Phanerogamous and Cryptogamous (6); and
in natural history there are the two Kingdoms or Realms, the
Vegetable and the Animal.
534. Moreover, the stages of the scaffolding have been variously
extended, as required, by the recognition of assemblages lower than
class but higher than order, viz. Subclass and Cohort; or lower than
order, a Suborder; or between this and genus, a Tribe; or between
this and tribe, a Subtribe; or between genus and species, a
Subgenus; and by some a species has been divided into Subspecies,
and a variety into Subvarieties. Last of all are Individuals.
Suffice it to remember that the following are the principal grades in
classification, with the proper sequence; also that only those here
printed in small capitals are fundamental and universal in botany:—
-
Series,
-
Class, Subclass, Cohort,
-
Order, or Family, Suborder, Tribe, Subtribe,
-
Genus, Subgenus or Section,
[Pg 179]
§ 2. NAMES, TERMS, AND CHARACTERS.
535. The name of a plant is the name of its genus followed by that of
the species. The name of the genus answers to the surname (or family
name); that of the species to the baptismal name of a person. Thus
Quercus is the name of the Oak genus; Quercus alba, that of the
White Oak, Q. rubra, that of Red Oak, Q. nigra, that of the
Black-Jack, etc. Botanical names being Latin or Latinized, the adjective
name of the species comes after that of the genus.
536. Names of Genera are of one word, a substantive. The older ones are
mostly classical Latin, or Greek adopted into Latin; such as Quercus
for the Oak genus, Fagus for the Beech, Corylus, the Hazel, and the
like. But as more genera became known, botanists had new names to make
or borrow. Many are named from some appearance or property of the
flowers, leaves, or other parts of the plant. To take a few examples
from the early pages of the "Manual of the Botany of the Northern United
States,"—the genus Hepatica comes from the shape of the leaf,
resembling that of the liver. Myosurus means mouse-tail. Delphinium
is from delphin, a dolphin, and alludes to the shape of the flower,
which was thought to resemble the classical figures of the dolphin.
Xanthorrhiza is from two Greek words meaning yellow-root, the common
name of the plant. Cimicifuga is formed of two Latin words meaning to
drive away bugs, i. e. Bugbane, the Siberian species being used to keep
away such vermin. Sanguinaria, the Bloodroot, is named from the
blood-like color of its juice. Other genera are dedicated to
distinguished botanists or promoters of science, and bear their names:
such are Magnolia, which commemorates the early French botanist,
Magnol; and Jeffersonia, named after President Jefferson, who sent the
first exploring expedition over the Rocky Mountains. Others bear the
name of the discoverer of the plant; as, Sarracenia, dedicated to Dr.
Sarrazin, of Quebec, who was one of the first to send the common
Pitcher-plant to the botanists of Europe; and Claytonia, first made
known by the early Virginian botanist Clayton.
537. Names of Species. The name of a species is also a single word,
appended to that of the genus. It is commonly an adjective, and
therefore agrees with the generic name in case, gender, etc. Sometimes
it relates to the country the species inhabits; as, Claytonia
Virginica, first made known from Virginia; Sanguinaria Canadensis,
from Canada, etc. More commonly it denotes some obvious or
characteristic trait of the species; as, for example, in Sarracenia, our
northern species is named purpurea, from the purple blossoms, while a
more southern one is named flava, because its petals are yellow; the
species of Jeffersonia is called diphylla, meaning two-leaved, because
its leaf is divided into two leaflets. Some species are named after the
discoverer, or in compliment to a botanist who has made them known; as,
Magnolia Fraseri, named after the botanist Fraser, one [Pg 180]of the first
to find this species; and Sarracenia Drummondii, for a Pitcher-plant
found by Mr. Drummond in Florida. Such personal specific names are of
course written with a capital initial letter. Occasionally some old
substantive name is used for the species; as Magnolia Umbrella, the
Umbrella tree, and Ranunculus Flammula. These are also written with a
capital initial, and need not accord with the generic name in gender.
Geographical specific names, such as Canadensis, Caroliniana,
Americana, in the later usage are by some written without a capital
initial, but the older usage is better, or at least more accordant with
English orthography.
538. Varietal Names, when any are required, are made on the plan of
specific names, and follow these, with the prefix var. Ranunculus
Flammula, var. reptans, the creeping variety: R. abortivus, var.
micranthus, the small-flowered variety of the species.
539. In recording the name of a plant it is usual to append the name, or
an abbreviation of the name, of the botanist who first published it; and
in a flora or other systematic work, this reference to the source of the
name is completed by a further citation of the name of the book, the
volume and page where it was first published. So "Ranunculus acris,
L.," means that this Buttercup was first so named and described by
Linnæus; "R. multifidus, Pursh," that this species was so named and
published by Pursh. The suffix is no part of the name, but is an
abbreviated reference, to be added or omitted as convenience or
definiteness may require. The authority for a generic name is similarly
recorded. Thus, "Ranunculus, L.," means that the genus was so named by
Linnæus; "Myosurus, Dill.," that the Mouse-tail was established as a
genus under this name by Dillenius; Caulophyllum, Michx., that the
Blue Cohosh was published under this name by Michaux. The full reference
in the last-named instance would be, "in Flora Boreali-Americana, first
volume, 205th page,"—in the customary abbreviation, "Michx. Fl. i.
205."
540. Names of Orders are given in the plural number, and are commonly
formed by prolonging the name of a genus of the group taken as a
representative of it. For example, the order of which the Buttercup or
Crowfoot genus, Ranunculus, is the representative, takes from it the
name of Ranunculaceæ; meaning Plantæ Ranunculaceæ when written out
in full, that is, Ranunculaceous Plants. Some old descriptive names of
orders are kept up, such as Cruciferæ for the order to which Cress and
Mustard belong, from the cruciform appearance of their expanded corolla,
and Umbelliferæ, from the flowers being in umbels.
541. Names of Tribes, also of suborders, subtribes, and the like, are
plurals of the name of the typical genus, less prolonged, usually in
eæ, ineæ, ideæ, etc. Thus the proper Buttercup tribe is
Ranunculeæ, of the Clematis tribe, Clematideæ. While the Rose family
is Rosaceæ, the special Rose tribe is Roseæ.
542. Names of Classes, etc. For these see the following synopsis of the
actual classification adopted, p. 183.
[Pg 181]
543. So a plant is named in two words, the generic and the specific
names, to which may be added a third, that of the variety, upon
occasion. The generic name is peculiar: obviously it must not be used
twice over in botany. The specific name must not be used twice over in
the same genus, but is free for any other genus. A Quercus alba, or
White Oak, is no hindrance to Betula alba, or White Birch; and so of
other names.
544. Characters and Descriptions. Plants are characterized by a terse
statement, in botanical terms, of their peculiarities or distinguishing
marks. The character of the order should include nothing which is common
to the whole class it belongs to; that of the genus, nothing which is
common to the order; that of the species nothing which is shared with
all other species of the genus; and so of other divisions.
Descriptions may enter into complete details of the whole structure.
545. Terminology, also called Glossology, is nomenclature applied to
organs or parts, and their forms or modifications. Each organ or special
part has a substantive name of its own: shapes and other modifications
of an organ or part are designated by adjective terms, or, when the
forms are peculiar, substantive names are given to them. By the correct
use of such botanical terms, and by proper subordination of the
characters under the order, genus, species, etc., plants may be
described and determined with much precision. The classical language of
botany is Latin. While modern languages have their own names and terms,
these usually lack the precision of the Latin or Latinized botanical
terminology. Fortunately, this Latinized terminology has been largely
adopted and incorporated into the English technical language of botany,
thus securing precision. And these terms are largely the basis of
specific names of plants.
546. A glossary or vocabulary of the principal botanical terms used in
phanerogamous and vascular cryptogamous botany is appended to this
volume, to which the student may refer, as occasion arises.
547. Two systems of classification used to be recognized in botany,—the
artificial and the natural; but only the latter is now thought to
deserve the name of a system.
548. Artificial classifications have for object merely the ascertaining
of the name and place of a plant. They do not attempt to express
relationships, but serve as a kind of dictionary. They distribute the
genera and species according to some one peculiarity or set of
peculiarities (just as a dictionary distributes words according to their
first letters), disregarding all other considerations. At present an
artificial classification in botany is needed only as a key to the
natural orders,—as an aid in referring an unknown plant to its proper
family; and such keys are still very needful, at least for the beginner.
Formerly, when the orders themselves were not clearly made out, an
artificial classification was required to lead the [Pg 182]student down to the
genus. Two such classifications were long in vogue: First, that of
Tournefort, founded mainly on the leaves of the flower, the calyx and
corolla: this was the prevalent system throughout the first half of the
eighteenth century; but it has long since gone by. It was succeeded by
the well-known
549. Artificial System of Linnæus, which was founded on the stamens and
pistils. It consists of twenty-four classes, and of a variable number of
orders; the classes founded mainly on the number and disposition of the
stamens; the orders partly upon the number of styles or stigmas, partly
upon other considerations. Useful and popular as this system was down to
a time within the memory of still surviving botanists, it is now
completely obsolete. But the tradition of it survives in the names of
its classes, Monandria, Diandria, Triandria, etc., which are familiar in
terminology in the adjective terms monandrous, diandrous, triandrous,
etc. (284); also of the orders, Monogynia, Digynia, Trigynia, etc.,
preserved in the form of monogynous, digynous, trigynous, etc. (301);
and in the name Cryptogamia, that of the 24th class, which is continued
for the lower series in the natural classification.
550. Natural System. A genuine system of botany consists of the orders
or families, duly arranged under their classes, and having the tribes,
the genera, and the species arranged in them according to their
relationships. This, when properly carried out, is the Natural System;
because it is intended to express, as well as possible, the various
degrees of relationship among plants, as presented in nature; that is,
to rank those species and those genera, etc., next to each other in the
classification which are really most alike in all respects, or, in other
words, which are constructed most nearly on the same particular plan.
551. There can be only one natural system of botany, if by this term
is meant the plan according to which the vegetable creation was called
into being, with all its grades and diversities among the species, as
well of past as of the present time. But there may be many natural
systems, if we mean the attempts of men to interpret and express that
plan,—systems which will vary with advancing knowledge, and with the
judgment and skill of different botanists. These must all be very
imperfect, bear the impress of individual minds, and be shaped by the
current philosophy of the age. But the endeavor always is to make the
classification answer to Nature, as far as any system can which has to
be expressed in a definite and serial arrangement.
552. So, although the classes, orders, genera, etc., are natural, or as
natural as the systematist can make them, their grouping or order of
arrangement in a book, must necessarily be in great measure artificial.
Indeed, it is quite impossible to arrange the orders, or even the few
classes, in a single series, and yet have each group stand next to its
nearest relatives on both sides.
553. Especially it should be understood that, although phanerogamous
[Pg 183]plants are of higher grade than cryptogamous, and angiospermous or
ordinary phanerogamous higher than the gymnospermous, yet there is no
culmination in the vegetable kingdom, nor any highest or lowest order of
phanerogamous plants.
554. The particular system most largely used at present in the
classification of the orders is essentially the following:—
- Series I. PHANEROGAMIA: Phanerogamous or Flowering Plants.
- Class I. DICOTYLEDONES ANGIOSPERMEÆ, called for shortness in
English, Dicotyledons or Dicotyls. Ovules in a closed ovary.
Embryo dicotyledonous. Stem with exogenous plan of growth. Leaves
reticulate-veined,
- Artificial Division I. Polypetalæ, with petals mostly present
and distinct. Orders about 80 in number, Ranunculaceæ to
Cornaceæ.
- Artificial Division II. Gamopetalæ, with gamopetalous corolla.
Orders about 45, Caprifoliaceæ to Plantaginaceæ.
- Artificial Division III. Apetalæ or Incompletæ, with perianth,
when present, of calyx only. Orders about 35 in number, from
Nyctaginaceæ to Salicaceæ.
- Class II. DICOTYLEDONES GYMNOSPERMEÆ, in English Gymnosperms. No
ovary or pericarp, but ovules and seeds naked, and no proper calyx
nor corolla. Embryo dicotyledonous or polycotyledonous. Stem with
exogenous plan of growth. Leaves mostly parallel-veined. Consists
of order Gnetaceæ, which strictly connects with Angiospermous
Dicotyls, of Coniferæ, and of Cycadaceæ.
- Class III. MONOCOTYLEDONES, in English Monocotyledons or
Monocotyls. Angiospermous. Embryo monocotyledonous. Stem with
endogenous plan of growth. Leaves mostly parallel-veined.
- Division I. Petaloideæ. Perianth complete, having the
equivalent of both calyx and corolla, and all the inner series
corolline. About 18 orders.
- Division II. Calycinæ. Perianth complete (in two series) but
not corolline, mostly thickish or glumaceous. Chiefly two
orders, Juncaceæ, the true Rushes, and Palmæ, Palms.
- Division III. Spadicifloræ or Nudifloræ. Perianth none, or
rudimentary and incomplete: inflorescence spadiceous. Of five
orders, Typhaceæ and Aroideæ the principal.
- Division IV. Glumaceæ. Perianth none, or very rudimentary:
glumaceous bracts to the flowers. Orders mainly Cyperaceæ and
Gramineæ.
- Series II. CRYPTOGAMIA: Cryptogamous or Flowerless Plants.
- CLASS I. PTERIDOPHYTA, Pteridophytes (484).
- CLASS II. BRYOPHYTA, Bryophytes (498).
- CLASS III. THALLOPHYTA, Thallophytes (503).
[Pg 184]
SECTION XIX. BOTANICAL WORK.
555. Some hints and brief instructions for the collection, examination,
and preservation of specimens are added. They are especially intended
for the assistance of those who have not the advantage of a teacher.
They apply to phanerogamous plants and Ferns only, and to systematic
botany.[1]
§ 1. COLLECTION, OR HERBORIZATION.
556. As much as possible, plants should be examined in the living state,
or when freshly gathered. But dried specimens should be prepared for
more leisurely examination and for comparison. To the working botanist
good dried specimens are indispensable.
557. Botanical Specimens, to be complete, should have root or rootstock,
stem, leaves, flowers, both open and in bud, and fruit. Sometimes these
may all be obtained at one gathering; more commonly two or three
gatherings at different times are requisite, especially for trees and
shrubs.
558. In Herborizing, a good knife and a narrow and strong trowel are
needed; but a very strong knife will serve instead of a trowel or small
pick for digging out bulbs, tubers, and the like. To carry the
specimens, either the tin box (vasculum) or a portfolio, or both are
required. The tin box is best for the collection of specimens to be used
fresh, as in the class-room; also for very thick or fleshy plants. The
portfolio is indispensable for long expeditions, and is best for
specimens which are to be preserved in the herbarium.
559. The Vasculum, or Botanical Collecting-box, is made of tin, in
shape like a candle-box, only flatter, or the smaller sizes like an
English sandwich-case; the lid opening for nearly the whole length of
one side of the box. Any portable tin box of convenient size, and
capable of holding specimens a foot or fifteen inches long, will answer
the purpose. The box should shut close, so that the specimens may not
wilt: then it will keep leafy branches and most flowers perfectly fresh
for a day or two, especially if slightly moistened. They should not be
wet.
560. The Portfolio is best made of two pieces of solid binder's-board,
covered with enamel cloth, which also forms the back, and fastened by
straps and buckles. It may be from a foot to twenty inches long, from
nine to eleven or twelve inches wide. It should contain a needful
quantity of smooth but strong and pliable paper (thin so-called Manilla
paper is best), either fastened at the back as in a book, or loose in
folded sheets when not very many specimens are required. As soon as
gathered, the specimens should be separately laid between the leaves or
in the folded sheets, and kept under moderate pressure in the closed
portfolio.
[Pg 185]
561. Of small herbs, especially annuals, the whole plant, root and all,
should be taken for a specimen. Of larger ones branches will suffice,
with some leaves from near the root. Enough of the root or subterranean
part of the plant should be collected to show whether it is an annual, a
biennial, or a perennial. Thick roots, bulbs, tubers, or branches of
specimens intended to be pressed should be thinned with a knife, or cut
into slices. Keep the specimens within the length of fifteen or sixteen
inches, by folding, or when that cannot be done, by cutting into
lengths.
562. For Drying Specimens a good supply of soft and unsized paper is
wanted; and some convenient means of applying considerable pressure. To
make good dried botanical specimens, dry them as rapidly as possible
between many thicknesses of sun-dried paper to absorb their moisture,
under as much pressure as can be given without crushing the more
delicate parts. This pressure may be had by a botanical press, of which
various forms have been contrived; or by weights placed upon a
board,—from forty to eighty or a hundred pounds, according to the
quantity of specimens drying at the time. For use while travelling, a
good portable press may be made of thick binders' boards for the sides,
and the pressure may be applied by strong straps with buckles. Still
better, on some accounts, are portable presses made of wire network,
which allow the dampness to escape by evaporation between the meshes.
For herborization in a small way, a light wire-press may be taken into
the field and made to serve also as a portfolio.
563. It is well to have two kinds of paper, namely, driers of bibulous
paper, stitched into pads (or the pads may be of thick carpet-paper, cut
to size) and thin smooth paper, folded once; the specimens to be laid
into the fold, either when gathered or on returning from the excursion.
These sheets are to hold the specimens until they are quite dry. Every
day, or at first even twice a day, the specimens, left undisturbed in
their sheets, are to be shifted into fire-dried or sun-dried fresh
driers, and the pressure renewed, while the moist sheets are spread out
to dry, so as to take their turn again at the next shifting. This course
must be continued until the specimens are no longer moist to the touch.
Good and comely specimens are either made or spoiled within the first
twenty-four or thirty-six hours. After that, when plenty of driers are
used, it may not be necessary to change them so frequently.
564. Succulent plants, which long refuse to part with life and moisture,
and Spruces and some other evergreens which are apt to cast off their
leaves, may be plunged for a moment into boiling water, all but the
flowers. Delicate flowers may be encased in thin tissue paper when put
into the press. Thick parts, like the heads of Sunflowers and Thistles,
may be cut in two or into slices.
565. Dried specimens may be packed in bundles, either in folded paper or
upon single half-sheets. It is better that such paper should not be
bibulous. The packages should be well wrapped or kept in close cases.
[Pg 186]
566. Poisoning is necessary if specimens are to be permanently
preserved from the depredation of insects. The usual application is an
almost saturated solution of corrosive sublimate in 95 per cent alcohol,
freely applied with a large and soft brush, or the specimens dipped into
some of the solution poured into a large and flat dish; the wetted
specimens to be transferred for a short time to driers.
§ 2. HERBARIUM.
567. The botanist's collection of dried specimens, ticketed with their
names, place, and time of collection, and systematically arranged under
their genera, orders, etc., forms a Hortus Siccus or Herbarium. It
comprises not only the specimens which the proprietor has himself
collected, but those which he acquires through friendly exchanges, or in
other ways. The specimens of an herbarium may be kept in folded sheets
of paper; or they may be fastened on half-sheets of thick and white
paper, either by gummed slips, or by glue applied to the specimens
themselves. The former is best for private and small herbaria; the
latter for large ones which are much turned over. Each sheet should be
appropriated to one species; two or more different plants should never
be attached to the same sheet. The generic and specific name of the
plant should be added to the lower right-hand corner, either written on
the sheet, or on a ticket pasted down; and the time of collection, the
locality, the color of the flowers, and any other information which the
specimens themselves do not afford, should be duly recorded upon the
sheet or the ticket. The sheets of the herbarium should all be of
exactly the same dimensions. The herbarium of Linnæus is on paper of the
common foolscap size, about eleven inches long and seven wide. This is
too small. Sixteen and three eighths inches by eleven and a half inches
is an approved size.
568. The sheets containing the species of each genus are to be placed in
genus-covers, made of a full sheet of thick paper (such as the
strongest Manilla-hemp paper), to be when folded of the same dimensions
as the species-sheet but slightly wider: the name of the genus is to be
written on one of the lower corners. These are to be arranged under the
orders to which they belong, and the whole kept in closed cases or
cabinets, either laid flat in compartments, like "pigeon-holes," or else
placed in thick portfolios, arranged like folio volumes. All should be
kept, as much as practicable, in dust-proof and insect-proof cases or
boxes.
569. Fruits, tubers, and other hard parts, too thick for the herbarium,
may be kept in pasteboard or light wooden boxes, in a collection apart.
Small loose fruits, seeds, detached flowers, and the like may be
conveniently preserved in paper capsules or envelopes, attached to the
herbarium-sheets.
[Pg 187]
§ 3. INVESTIGATION AND DETERMINATION OF PLANTS.
570. The Implements required are a hand magnifying glass, a pocket lens
of an inch or two focus, or a glass of two lenses, one of the lower and
the other of the higher power; and a sharp penknife for dissection. With
these and reasonable perseverance the structure of the flowers and
fructification of most phanerogamous plants and Ferns can be made out.
But for ease and comfort, as well as for certainty and right training,
the student should have some kind of simple stage microscope, and under
this make all dissections of small parts. Without it the student will be
apt to fall into the bad habit of guessing where he ought to ascertain.
571. The simple microscope may be reduced to a good lens or doublet, of
an inch focus, mounted over a glass stage, so that it can be moved up
and down and also sidewise, and with (or without) a little mirror
underneath. A better one would have one or two additional lenses (say of
half and of a quarter inch focus), a pretty large stage, on the glass of
which several small objects can be placed and conveniently brought under
the lens; and its height or that of the lens should be adjustable by a
rack-work; also a swivel-mounted little mirror beneath, which is needed
for minute objects to be viewed by transmitted light.
572. For dissecting and displaying small parts on the stage of the
microscope, besides a thin-bladed knife, the only tools needed are a
good stock of common needles of various sizes, mounted in handles, and
one or more saddler's-needles, which, being triangular, may be ground to
sharp edges convenient for dissection. Also a pair of delicate-pointed
forceps; those with curved points used by the dentist are most
convenient. A cup of clean water is indispensable, with which to moisten
or wet, or in which occasionally to float delicate parts. Small flowers,
buds, fruits, and seeds of dried specimens can be dissected quite as
well as fresh ones. They have only to be soaked in warm or boiling
water.
573. The compound microscope is rarely necessary except in cryptogamic
botany and vegetable anatomy; but it is very useful and convenient,
especially for the examination of pollen. To the advanced botanist it is
a necessity, to all students of botany an aid and delight.
574. Analysis. A few directions and hints may be given. The most
important is this: In studying an unknown plant, make a complete
examination of all its parts, and form a clear idea of its floral
structure and that of its fruit, from pericarp down to the embryo, or as
far as the materials in hand allow, before taking a step toward finding
out its name and relationship by means of the keys or other helps which
the Manuals and Floras provide. If it is the name merely that is wanted,
the shorter way is to ask some one who already knows it. To verify the
points of structure one by one as they happen to occur in an artificial
key, without any preparatory investigation, is a usual but is not the
best nor the surest [Pg 188]way. It is well to make drawings or outline
sketches of the smaller parts, and especially diagrams of the plan of
the flower, such as those of Fig. 225, 227, 241, 244, 275-277. For
these, cross sections of the flower-bud or flower are to be made: and
longitudinal sections, such as Fig. 270-274, are equally important. The
dissection even of small seeds is not difficult after some practice.
Commonly they need to be soaked or boiled.
575. The right appreciation of characters and terms used in description
needs practice and calls for judgment. Plants do not grow exactly by
rule and plummet, and measurements must be taken loosely. Difference of
soil and situation are responded to by considerable variations, and
other divergences occur which cannot be accounted for by the
surroundings, nor be anticipated in general descriptions. Annuals may be
very depauperate in dry soils or seasons, or very large when
particularly well nourished. Warm and arid situations promote, and wet
ones are apt to diminish pubescence. Salt water causes increased
succulence. The color of flowers is apt to be lighter in shade, and
brighter in open and elevated situations. A color or hue not normal to
the species now and then occurs, which nothing in the conditions will
account for. A white-flowered variation of any other colored blossom
may always be expected; this, though it may be notable, no more
indicates a distinct variety of the species than an albino would a
variety of the human species. The numerical plan is subject to variation
in some flowers; those on the plan of five may now and then vary to four
or to six. Variations of the outline or lobing of leaves are so familiar
that they do not much mislead. Only wider and longer observation
suffices to prevent or correct mistakes in botanical study. But the
weighing of evidence and the balancing of probabilities, no less than
the use of the well-ordered and logical system of classification, give
as excellent training to the judgment as the search for the facts
themselves does to the observing powers.
§ 4. SIGNS AND ABBREVIATIONS.
576. For a full account of these, whether of former or actual use, see
"Structural Botany" of the "Botanical Text Book," pp. 367, 392, as also
for the principles which govern the accentuation of names. It is needful
here to explain only those used in the Manuals and Floras of this
country, for which the present volume is an introduction and companion.
They are not numerous.
577. In arranging the species, at least those of a large genus, the
divisions are denoted and graduated as follows: The sign § is prefixed
to sections of the highest rank: these sections when they have names
affixed to them (as Prunus § Cerasus) may be called subgenera. When the
divisions of a genus are not of such importance, or when divisions are
made under the subgenus itself, the most comprehensive ones are marked
by asterisks, * for the first, * * for the second, and so on.
Subdivisions are [Pg 189]marked with a prefixed +; those under this head with
++; and those under this with =, if there be so many grades. A similar
notation is followed in the synopsis of the genera of an order.
578. The interrogation point is used in botany to indicate doubt. Thus
Clematis crispa, L.? expresses a doubt whether the plant in question
is really the Clematis crispa of Linnæus. Clematis? polypetala
expresses a doubt whether the plant so named is really a Clematis. On
the other hand the exclamation point (!) is used to denote certainty
whenever there is special need to affirm this.
579. For size or height, the common signs of degrees, minutes, and
seconds, have been used, thus, 1°, 2', 3", stand respectively for a
foot, two inches, and three lines or twelfths of an inch. A better way,
when such brevity is needed, is to write 1ft. 2in. 3l.
580. Signs for duration used by Linnæus were ☉ for an annual,
♂ for a biennial, ♃ for a perennial herb,
[Symbol like numeral 5 without top bar] for a shrub or tree. DeCandolle brought in ☉ for a
plant that died after once flowering, ① if annual, ② if biennial.
581. To indicate sexes, ♂ means staminate or male plant or
blossom; ♀, pistillate or female; [Symbol like ☿ with two inverted breves], perfect or
hermaphrodite.
582. To save room it is not uncommon to use ∞ in place of
"many;" thus, "Stamens ∞," for stamens indefinitely numerous:
"∞ flora" for pluriflora or many-flowered. Still more common
is the form "Stamens 5-20," or "Calyx 4-5-parted," for stamens from five
to twenty, calyx four-parted or five-parted, and the like. Such
abbreviations hardly need explanation.
583. The same may be said of such abbreviations as Cal. for calyx,
Cor. for corolla, Pet. for petals, St. for stamens, Pist. for
pistil, Hab. for habitat, meaning place of growth, Herb. for
herbarium, Hort. for garden. Also l. c., loco citato, which avoids
repetition of volume and page.
584. "Structural Botany" has six pages of abbreviations of the names of
botanists, mostly of botanical authors. As they are not of much
consequence to the beginner, while the more advanced botanist will know
the names in full, or know where to find them, only a selection is here
appended.
[Pg 190]
ABBREVIATIONS OF THE NAMES OF BOTANISTS.
| Adans. | = | Adanson. |
| Ait. | | Aiton. |
| All. | | Allioni. |
| Andr. | | Andrews. |
| Arn. | | Arnott. |
| Aub. | | Aublet. |
| Bartr. | | Bartram. |
| Beauv. | | Palisot de Beauvois. |
| Benth. | | Bentham. |
| Bernh. | | Bernhardi. |
| Bigel. | | Jacob Bigelow. |
| Bong. | | Bongard. |
| Bonpl. | | Bonpland. |
| Br. or R. Br. | | Robert Brown. |
| Cass. | | Cassini. |
| Cav. | | Cavanilles. |
| Cham. | | Chamisso. |
| Chapm. | | Chapman. |
| Chois. | | Choisy. |
| Clayt. | | Clayton. |
| Curt. | | Curtis. |
| Curt. (M. A.) | | M. A. Curtis. |
| Darl. | | Darlington. |
| DC. | } | DeCandolle. |
| DeCand. |
| A. DC. | | Alphonse DeCandolle. |
| Desc. | | Descourtilz. |
| Desf. | | Desfontaines. |
| Desv. | | Desvaux. |
| Dill. | | Dillenius. |
| Dougl. | | Douglas. |
| Duham. | | Duhamel. |
| Dun. | | Dunal. |
| Eat. | | Eaton (Amos) or D. C. |
| Ehrh. | | Ehrhart. |
| Ell. | | Elliott. |
| Endl. | | Endlicher. |
| Engelm. | | Engelmann. |
| Engl. | | Engler. |
| Fisch. | | Fischer. |
| Frœl. | | Frœlich. |
| Gærtn. | | Gærtner. |
| Gaud. | | Gaudin. |
| Gaudich. | | Gaudichaud. |
| Ging. | | Gingins. |
| Gmel. | | Gmelin. |
| Good. | | Goodenough. |
| Grev. | | Greville. |
| Griseb. | | Grisebach. |
| Gron. | } | Gronovius. |
| Gronov. |
| Hall. | | Haller. |
| Hartm. | | Hartmann. |
| Hartw. | | Hartweg. |
| Harv. | | Harvey. |
| Haw. | | Haworth. |
| Hegelm. | | Hegelmaier. |
| Hemsl. | | Hemsley. |
| Herb. | | Herbert. |
| Hoffm. | | Hoffmann. |
| Hoffmans. | | Hoffmansegg. |
| Hook. | | Hooker. |
| Hook. f. | | J. D. Hooker. |
| Hornem. | | Hornemann. |
| Huds. | | Hudson. |
| Humb. | | Humboldt. |
| HBK. | | Humboldt, Bonpland, and Kunth. |
| Jacq. | | Jacquin. |
| Jacq. f. | | J. F. Jacquin. |
| Juss. | | Jussieu. |
| A. Juss. | | Adrien de Jussieu. |
| Kit. | | Kitaibel. |
| L. or Linn. | | Linnæus. |
| Labill. | | Labillardiere. |
| Lag. | | Lagasca. |
| Lam. | | Lamarck. |
| Ledeb. | | Ledebour. |
| Lehm. | | Lehmann. |
| Lesq. | | Lesquereux. |
| Less. | | Lessing. |
| Lestib. | | Lestibudois. |
| L'Her. | | L'Heritier. |
| Lindb. | | Lindberg. |
| Lindh. | | Lindheimer. |
| Lindl. | | Lindley. |
| Lodd. | | Loddiges. |
| Loud. | | Loudon. |
| M. Bieb. | | Marschall von Bieberstein. |
| Marsh. | | Marshall (Humphrey). |
| Mart. | | Martius. |
| [Pg 191]Mast. | = | Masters. |
| Maxim. | | Maximowicz. |
| Meisn. | } | Meisner or |
| Meissn. | Meissner. |
| Michx. or Mx. | | Michaux. |
| Michx. f. | | F. A. Michaux. |
| Mill. | | Miller. |
| Miq. | | Miquel. |
| Mitch. | | Mitchell. |
| Moç. | | Moçino. |
| Moq. | | Moquin-Tandon. |
| Moric. | | Moricand. |
| Moris. | | Morison. |
| Muell. Arg. | | J. Mueller. |
| Muell. (F.) | | Ferdinand Mueller. |
| Muhl. | | Muhlenberg. |
| Murr. | | Murray. |
| Naud. | | Naudin. |
| Neck. | | Necker. |
| Nees | } | Nees von Esenbeck. |
| N. ab E. |
| Nutt. | | Nuttall. |
| Œd. | | Œder. |
| Ort. | | Ortega. |
| P. de Beauv. | | Palisot de Beauvois. |
| Pall. | | Pallas. |
| Parl. | | Parlatore. |
| Pav. | | Pavon. |
| Pers. | | Persoon. |
| Planch. | | Planchon. |
| Pluk. | | Plukenet. |
| Plum. | | Plumier. |
| Poir. | | Poiret. |
| Radlk. | | Radlkofer. |
| Raf. | | Rafinesque. |
| Red. | | Redouté. |
| Reichenb. | | Reichenbach. |
| Rich. | | L. C. Richard. |
| Rich. f. or A. | | Achille Richard. |
| Richards. | | Richardson. |
| Ridd. | | Riddell. |
| Rœm. & Schult. | | Rœmer & Schultes. |
| Rottb. | | Rottbœll. |
| Rupr. | | Ruprecht. |
| St. Hil. | | Saint-Hilaire. |
| Salisb. | | Salisbury. |
| Schk. | | Schkuhr. |
| Schlecht. | | Schlechtendal. |
| Schrad. | | Schrader. |
| Schreb. | | Schreber. |
| Schwein. | | Schweinitz. |
| Scop. | | Scopoli. |
| Spreng. | | Sprengel. |
| Sternb. | | Sternberg. |
| Steud. | | Steudel. |
| Sull. | | Sullivant. |
| Thunb. | | Thunberg. |
| Torr. | | Torrey. |
| Tourn. | | Tournefort. |
| Trautv. | | Trautvetter. |
| Trin. | | Trinius. |
| Tuck. | | Tuckerman. |
| Vaill. | | Vaillant. |
| Vent. | | Ventenat. |
| Vill. | | Villars. |
| Wahl. | | Wahlenberg. |
| Walds. | | Waldstein. |
| Wall. | | Wallich. |
| Wallr. | | Wallroth. |
| Walp. | | Walpers. |
| Walt. | | Walter. |
| Wang. | | Wangenheim. |
| Wats. | | Sereno Watson, unless other initials are given. |
| Wedd. | | Weddell. |
| Wendl. | | Wendland. |
| Wiks. | | Wikstrom. |
| Willd. | | Willdenow. |
| Wulf. | | Wulfen. |
| Zucc. | | Zuccarini. |
| Zuccag. | | Zuccagini. |
[Pg 193]
GLOSSARY AND INDEX,
OR
DICTIONARY OF THE PRINCIPAL TERMS IN DESCRIPTIVE BOTANY, COMBINED WITH
AN INDEX.
For the convenience of unclassical students, the commoner Latin
and Greek words (or their equivalents in English form) which
enter into the composition of botanical names, as well as of
technical terms, are added to this Glossary. The numbers refer
to pages.
A, at the beginning of words of Greek derivation, commonly signifies a
negative, or the absence of something; as apetalous, without petals;
aphyllous, leafless, &c. In words beginning with a vowel, the prefix
is an; as anantherous, destitute of anther.
Abnormal, contrary to the usual or the natural structure.
Aboriginal, original in the strictest sense; same as indigenous.
Abortive, imperfectly formed, or rudimentary.
Abortion, the imperfect formation or the non-formation of some part.
Abrupt, suddenly terminating; as, for instance,
Abruptly pinnate, pinnate without an odd leaflet at the end, 58.
Acantho-, spiny.
Acaulescent (acaulis), apparently stemless; the proper stem, bearing
the leaves and flowers, being very short or subterranean.
Accessory, something additional; as Accessory buds, 30, 31;
Accessory fruits, 118.
Accrescent, growing larger after flowering.
Accrete, grown to.
Accumbent, lying against a thing. The cotyledons are accumbent when
they lie with their edges against the radicle, 128.
Acephalous, headless.
Acerose, needle-shaped, as the leaves of Pines.
Acetabuliform, saucer-shaped.
Achænium, or Achenium (plural achenia), a one-seeded, seed-like
fruit, 120.
Achlamydeous (flower), without floral envelopes, 86.
Acicular, needle-shaped; more slender than acerose.
Acinaciform, scimitar-shaped, like some bean-pods.
Acines, the separate grains of a fruit, such as the raspberry.
Acorn, the nut of the Oak, 122.
Acotyledonous, destitute of cotyledons or seed-leaves.
Acrogenous, growing from the apex, as the stems of Ferns and Mosses.
Acrogens, or Acrogenous Plants, a name for the vascular cryptogamous
plants, 156.
Aculeate, armed with prickles, i. e. aculei; as the Rose and Brier.
Aculeolate, armed with small prickles, or slightly prickly.
Acuminate, taper-pointed, 54.
Acute, merely sharp-pointed, or ending in a point less than a right
angle, 54.
[Pg 194]Adelphous (stamens), joined in a fraternity (adelphia); see
monadelphous, &c.
Aden, Greek for gland. So Adenophorous, gland-bearing.
Adherent, sticking to, or more commonly, growing fast to another body.
Adnate, literally, growing fast to, born adherent, 95. The anther is
adnate when fixed by its whole length to the filament or its
prolongation, 101.
Adnation, the state of being adnate, 94.
Adpressed or appressed, brought into contact with, but not united.
Adscendent, ascendent, or ascending, rising gradually upwards, 39.
Adsurgent, or assurgent, same as ascending, 39.
Adventitious, out of the proper or usual place; e. g. Adventitious
buds, 30.
Adventive, applied to foreign plants accidentally or sparingly
introduced into a country, but hardly to be called naturalized.
Æquilateral, equal-sided; opposed to oblique.
Aerial roots, &c., 36.
Æruginous, verdigris-colored.
Aestival, produced in summer.
Aestivation, the arrangement of parts in a flower-bud, 97.
Agamous, sexless.
Aggregate fruits, 118.
Agrestis, growing in fields.
Air-cells or Air-passages, spaces in the tissue of leaves and some
stems, 131.
Air-Plants, 36.
Akene or Akenium, 120.
Ala (plural, alæ), a wing; the side-petals of a papilionaceous
corolla, 92.
Alabastrum, a flower-bud.
Alar, situated in the forks of a stem.
Alate, winged.
Albescent, whitish, or turning white.
Albus, Latin for white.
Albumen of the seed, nourishing matter stored up with the embryo, 21,
127.
Albumen, a vegetable product, of four elements.
Albuminous (seeds), furnished with albumen, 21.
Alburnum, young wood, sap-wood, 142.
Alliaceous, with odor of garlic.
Allogamous, close fertilization.
Alpestrine, subalpine.
Alpine, belonging to high mountains above the limit of forests.
Alternate (leaves), one after another, 29, 67. Petals are alternate
with the sepals, or stamens with the petals, when they stand over the
intervals between them, 82.
Alveolate, honeycomb-like.
Ament, the scaly spike of trees like the Birch and Willow, 75.
Amentaceous, catkin-like, or catkin-bearing.
Amorphous, shapeless, without any definite form.
Amphicarpous, producing two kinds of fruit.
Amphigastrium (plural, amphigastria), a peculiar stipule-like leaf
of Liverworts.
Amphitropous ovules or seeds, 111.
Amphora, a pitcher-shaped organ.
Amplectant, embracing. Amplexicaul (leaves), clasping the stem by
the base.
Ampullaceous, swelling out like a bottle or bladder (ampulla).
Amylaceous, Amyloid, composed of starch (amylum), or starch-like.
Anandrous, without stamens.
Anantherous, without anthers.
Ananthous, destitute of flowers; flowerless.
Anastomosing, forming a net-work (anastomosis), as the veins of
leaves, 50.
Anatropous ovules or seeds, 111.
Ancipital (anceps), two-edged.
Andrœcium, a name for the stamens taken together, 98.
[Pg 195]Andro-diœcious, flowers staminate on one plant, perfect on
another.
Androgynous, having both staminate and pistillate flowers in the same
cluster.
Androphore, a column of united stamens, as in a Mallow.
Androus, or Ander, andra, andrum, Greek in compounds for male,
or stamens.
Anemophilous, wind-loving, said of wind-fertilizable flowers, 113.
Anfractuose, bent hither and thither as the anthers of the Squash, &c.
Angiospermæ, Angiospermous, with seeds formed in an ovary or
pericarp, 109.
Angular divergence of leaves, 69.
Anisos, unequal. Anisomerous, parts unequal in number.
Anisopetalous, with unequal petals. Anisophyllous, the leaves
unequal in the pairs.
Annual (plant), flowering and fruiting the year it is raised from the
seed, and then dying, 37.
Annular, in the form of a ring, or forming a circle.
Annulate, marked by rings; or furnished with an
Annulus, or ring, like that of the spore-case of most Ferns. In Mosses
it is a ring of cells placed between the mouth of the spore-case and the
lid in many species.
Annotinous, yearly, or in yearly growths.
Anterior, in the blossom, is the part next the bract, i. e. external;
while the posterior side is that next the axis of inflorescence. Thus,
in the Pea, &c., the keel is anterior, and the standard posterior,
96.
Anthela, an open paniculate cyme.
Anther, the essential part of the stamen, which contains the pollen,
14, 80, 101.
Antheridium (plural antheridia), the organ in Cryptogams which
answers to the anther of Flowering Plants, 150.
Antheriferous, anther-bearing.
Anthesis, the period or the act of the expansion of a flower.
Anthocarpus (fruits), 118.
Anthophore, a stipe between calyx and corolla, 113.
Anthos, Greek for flower; in composition, Monanthous, one-flowered,
&c.
Anticous, same as anterior.
Antrorse, directed upwards or forwards.
Apetalous, destitute of petals, 86.
Aphyllous, leafless.
Apical, belonging to the apex or point.
Apiculate, pointleted; tipped with a small point.
Apocarpous (pistils), when the several pistils of the same flower are
separate.
Apophysis, any irregular swelling; the enlargement at the base of the
spore-case of the Umbrella-Moss.
Apothecium, the fructification of Lichens, 171.
Appendage, any superadded part. Appendiculate, provided with
appendages.
Appressed, close pressed to the stem, &c.
Apricus, growing in dry and sunny places.
Apterous, wingless.
Aquatic (Aquatilis), living or growing in water; applied to plants
whether growing under water, or with all but the base raised out of it.
Arachnoid, Araneose, cobwebby; clothed with, or consisting of, soft
downy fibres.
Arboreous, Arborescent, tree-like, in size or form, 39.
Arboretum, a collection of trees.
Archegonium (plural archegonia), the organ in Mosses, &c., which is
analogous to the pistil of Flowering Plants.
Arcuate, bent or curved like a bow.
Arenose (Arenarius), growing in sand.
Areolate, marked out into little spaces or areolæ.
Argenteous, or Argentate, silvery-like.
Argillose, growing in clay.
Argos, Greek for pure white; Argophyllous or Argyrophyllous,
white-leaved, &c.
Argutus, acutely dentate.
[Pg 196]Arillate (seeds) furnished with an aril.
Arilliform, aril-like.
Arillus, or Aril, a fleshy growth from base of a seed, 126.
Aristate, awned, i. e. furnished with an arista, like the beard of
Barley, &c., 54.
Aristulate, diminutive of the last; short-awned.
Arrect, brought into upright position.
Arrow-shaped or Arrow-headed, same as sagittate, 53.
Articulated, jointed; furnished with joints or articulations, where
it separates or inclines to do so. Articulated leaves, 57.
Artificial Classification, 181.
Ascending (stems, &c.), 39; (seeds or ovules), 110.
Ascidium, a pitcher-shaped body, like leaves of Sarracenia.
Ascus (asci), a sac, the spore-case of Lichens and some Fungi.
Aspergilliform, shaped like the brush used to sprinkle holy water; as
the stigmas of many Grasses.
Asperous, rough to touch.
Assimilation, 144, 147.
Assurgent, same as ascending, 39.
Atropous or Atropal (ovules), same as orthotropous.
Aurantiacous, orange-colored.
Aureous, golden.
Auriculate, furnished with auricles or ear-like appendages, 53.
Autogamy, self-fertilization, 115.
Awl-shaped, sharp-pointed from a broader base, 61.
Awn, the bristle or beard of Barley, Oats, &c.; or any similar
appendage.
Awned or Awn-pointed, furnished with an awn or long bristle-shaped
tip, 54.
Axil, the angle on the upper side between a leaf and the stem, 13.
Axile, belonging to the axis, or occupying the axis.
Axillary (buds, &c.), occurring in an axil, 27.
Axis, the central line of any body; the organ round which others are
attached; the root and stem. Ascending and Descending Axis, 38.
Baccate, berried, berry-like, of a pulpy nature like a berry
(bacca).
Badius, chestnut-colored.
Banner, see Standard, 92.
Barbate, bearded; bearing tufts, spots, or lines of hairs.
Barbed, furnished with a barb or double hook; as the apex of the
bristle on the fruit of Echinospermum (Stickseed), &c.
Barbellate, said of the bristles of the pappus of some Compositæ when
beset with short, stiff hairs, longer than when denticulate, but shorter
than when plumose.
Barbellulate, diminutive of barbellate.
Bark, the covering of a stem outside of the wood, 138, 140.
Basal, belonging or attached to the
Base, that extremity of any organ by which it is attached to its
support.
Basifixed, attached by its base.
Bast, Bast-fibres, 134.
Beaked, ending in a prolonged narrow tip.
Bearded, see barbate. Beard is sometimes used for awn, more
commonly for long or stiff hairs of any sort.
Bell-shaped, of the shape of a bell, as the corolla of Harebell, 90.
Berry, a fruit pulpy or juicy throughout, as a grape, 119.
Bi- (or Bis), in compound words, twice; as
Biarticulate, twice-jointed, or two-jointed; separating into two
pieces.
Biauriculate, having two ears, as the leaf in fig. 126.
Bicallose, having two callosities or harder spots.
Bicarinate, two-keeled.
Bicipital (Biceps), two-headed; dividing into two parts.
[Pg 197]Biconjugate, twice paired, as when a petiole forks twice.
Bidentate, having two teeth (not twice or doubly dentate).
Biennial, of two years' continuance; springing from the seed one
season, flowering and dying the next, 38.
Bifarious, two-ranked; arranged in two rows.
Bifid, two-cleft to about the middle.
Bifoliolate, a compound leaf of two leaflets, 59.
Bifurcate, twice forked; or more commonly, forked into two branches.
Bijugate, bearing two pairs (of leaflets, &c.).
Bilabiate, two-lipped, as the corolla of Labiatæ.
Bilamellate, of two plates (lamellæ), as the stigma of Mimulus.
Bilobed, the same as two-lobed.
Bilocellate, when a cell is divided into two locelli.
Bilocular, two-celled; as most anthers, the pod of Foxglove, &c.
Binary, in twos.
Binate, in couples, two together. Bipartite, the Latin form of
two-parted.
Binodal, of two nodes.
Binomial, of two words, as the name of genus and species taken
together, 180.
Bipalmate, twice palmately divided.
Biparous, bearing two.
Bipinnate (leaf), twice pinnate, 58. Bipinnatifid, twice pinnatifid,
57.
Bipinnatisect, twice pinnately divided.
Biplicate, twice folded together.
Biserial, or Biseriate, occupying two rows, one within the other.
Biserrate, doubly serrate, as when the teeth of a leaf are themselves
serrate.
Bisexual, having both stamens and pistil.
Biternate, twice ternate; i. e. principal divisions three, each
bearing three leaflets, 59.
Bladdery, thin and inflated.
Blade of a leaf, its expanded portion, 49.
Bloom, the whitish powder on some fruits, leaves, &c.
Boat-shaped, concave within and keeled without, in shape like a small
boat.
Border of corolla, &c., 89.
Brachiate, with opposite branches at right angles to each other.
Brachy-, short, as Brachycarpous, short-fruited, &c.
Bract (Bractea), the leaf of an inflorescence. Specially, the bract
is the small leaf or scale from the axil of which a flower or its
pedicel proceeds, 73.
Bracteate, furnished with bracts.
Bracteolate, furnished with bractlets.
Bracteose, with numerous or conspicuous bracts.
Bractlet (Bracteola), or Bracteole, is a bract seated on the
pedicel or flower-stalk, 73.
Branch, Branching, 27.
Breathing-pores, 144.
Bristles, stiff, sharp hairs, or any very slender bodies of similar
appearance.
Bristly, beset with bristles. Bristle-pointed, 54.
Brunneous, brown.
Brush-shaped, see aspergilliform.
Bryology, that part of botany which relates to Mosses.
Bryophyta, Bryophytes, 163.
Bud, a branch in its earliest or undeveloped state, 27. Bud-scales,
63.
Bulb, a leaf-bud with fleshy scales, usually subterranean, 46.
Bulbils, diminutive bulbs.
Bulbiferous, bearing or producing bulbs. Bulbose or bulbous,
bulb-like in shape, &c.
Bulblets, small bulbs, borne above ground, 46.
Bulb-scales, 46.
Bullate, appearing as if blistered or bladdery (from bulla, a
bubble).
Byssaceous, composed of fine flax-like threads.
[Pg 198]Caducous, dropping off very early, compared with other parts; as the
calyx in the Poppy, falling when the flower opens.
Cæruleous, blue. Cærulescent, becoming bluish.
Cæspitose, or Cespitose, growing in turf-like patches or tufts.
Calathiform, cup-shaped.
Calcarate, furnished with a spur (calcar), 86, 87.
Calceolate or Calceiform, slipper-shaped, like one petal of the
Lady's Slipper.
Callose, hardened; or furnished with callosities or thickened spots.
Calvous, bald or naked of hairs.
Calyciflorus, when petals and stamens are adnate to calyx.
Calycine, belonging to the calyx.
Calyculate, furnished with an outer accessory calyx (calyculus) or
set of bracts looking like a calyx, as in true Pinks.
Calyptra, the hood or veil of the capsule of a Moss, 163.
Calyptrate, having a calyptra.
Calyptriform, shaped like a calyptra or candle-extinguisher.
Calyx, the outer set of the floral envelopes or leaves of the flower,
14, 79.
Cambium, Cambium-layer, 140.
Campanulate, bell-shaped, 90.
Campylotropous, or Campylotropal, curved ovules and seeds, 111.
Campylospermous, applied to fruits of Umbelliferæ when the seed is
curved in at the edges, forming a groove down the inner face; as in
Sweet Cicely.
Canaliculate, channelled, or with a deep longitudinal groove.
Cancellate, latticed, resembling lattice-work.
Candidus, Latin for pure white.
Canescent, grayish-white; hoary, usually because the surface is
covered with fine white hairs. Incanous is whiter still.
Canous, whitened with pubescence; see incanous.
Capillaceous, Capillary, hair-like in shape; as fine as hair or
slender bristles.
Capitate, having a globular apex, like the head on a pin.
Capitellate, diminutive of capitate.
Capitulum, a close rounded dense cluster or head of sessile flowers,
74.
Capreolate, bearing tendrils (from capreolus, a tendril).
Capsule, a dry dehiscent seed-vessel of a compound pistil, 122.
Capsular, relating to, or like a capsule.
Capture of insects, 154.
Carina, a keel; the two anterior petals of a papilionaceous flower,
92.
Carinate, keeled, furnished with a sharp ridge or projection on the
lower side.
Cariopsis, or Caryopsis, the one-seeded fruit or grain of Grasses,
121.
Carneous, flesh-colored; pale red. Carnose, fleshy in texture.
Carpel, or Carpidium, a simple pistil or a pistil-leaf, 106.
Carpellary, pertaining to a carpel.
Carpology, that department of botany which relates to fruits.
Carpophore, the stalk or support of a pistil extending between its
carpels, 113.
Carpos, Greek for fruit.
Cartilaginous, or Cartilagineous, firm and tough in texture, like
cartilage.
Caruncle, an excrescence at the scar of some seeds, 126.
Carunculate, furnished with a caruncle.
Caryophyllaceous, pink-like: applied to a corolla of 5 long-clawed
petals.
Cassideous, helmet-shaped.
Cassus, empty and sterile.
Catenate, or Catenulate, end to end as in a chain.
Catkin, see Ament, 75.
Caudate, tailed, or tail-pointed.
Caudex, a sort of trunk, such as that of Palms; an upright rootstock,
39, 44.
Caudicle, the stalk of a pollen-mass, &c.
Caulescent, having an obvious stem, 36.
[Pg 199]Caulicle, a little stem, or rudimentary stem (of a seedling), 11,
127.
Cauline, of or belonging to a stem, 36. Caulis, Latin name of stem.
Caulocarpic, equivalent to perennial.
Caulome, the cauline parts of a plant.
Cell (diminutive, Cellule), the cavity of an anther, ovary, &c.; one
of the anatomical elements, 131.
Cellular Cryptogams, 162.
Cellular tissue, 131.
Cellulose, 131.
Cell-walls, 130.
Centrifugal (inflorescence), produced or expanding in succession from
the centre outwards, 77.
Centripetal, the opposite of centrifugal, 74.
Cephala, Greek for head. In compounds, Monocephalous, with one head,
Microcephalous, small-headed, &c.
Cereal, belonging to corn, or corn-plants.
Cernuous, nodding; the summit more or less inclining.
Chæta, Greek for bristle.
Chaff, small membranous scales or bracts on the receptacle of
Compositæ; the glumes, &c., of grasses.
Chaffy, furnished with chaff, or of the texture of chaff.
Chalaza, that part of the ovule where all the parts grow together,
110, 126.
Channelled, hollowed out like a gutter; same as canaliculate.
Character, a phrase expressing the essential marks of a species,
genus, &c., 181.
Chartaceous, of the texture of paper or parchment.
Chloros, Greek for green, whence Chloranthous, green-flowered;
Chlorocarpous, green-fruited, &c.
Chlorophyll, leaf green, 136.
Chlorosis, a condition in which naturally colored parts turn green.
Choripetalous, same as polypetalous.
Chorisis, separation of the normally united parts, or where two or
more parts take the place of one.
Chromule, coloring matter in plants, especially when not green, or
when liquid.
Chrysos, Greek for golden yellow, whence Chrysanthous,
yellow-flowered, &c.
Cicatrix, the scar left by the fall of a leaf or other organ.
Ciliate, beset on the margin with a fringe of cilia, i. e. of hairs
or bristles, like the eyelashes fringing the eyelids, whence the name.
Cinereous, or Cineraceous, ash-grayish; of the color of ashes.
Circinate, rolled inwards from the top, 72.
Circumscissile, or Circumcissile, divided by a circular line round
the sides, as the pods of Purslane, Plantain, &c., 124.
Circumscription, general outline.
Cirrhiferous, or Cirrhose, furnished with a tendril (Latin,
Cirrhus); as the Grape-vine. Cirrhose also means resembling or
coiling like tendrils, as the leaf-stalks of Virgin's-bower. More
properly Cirrus and Cirrose.
Citreous, lemon-yellow.
Clados, Greek for branch. Cladophylla, 64.
Class, 178, 183.
Classification, 175, 183.
Clathrate, latticed; same as cancellate.
Clavate, club-shaped; slender below and thickened upwards.
Clavellate, diminutive of clavate.
Claviculate, having Claviculæ, or little tendrils or hooks.
Claw, the narrow or stalk-like base of some petals, as of Pinks, 91.
Cleistogamous (Cleistogamy), fertilized in closed bud, 115.
Cleft, cut into lobes, 55.
Close fertilization, 115.
Climbing, rising by clinging to other objects, 39, 151.
Club shaped, see clavate.
Clustered, leaves, flowers, &c., aggregated or collected into a bunch.
[Pg 200]Clypeate, buckler-shaped.
Coadunate, same as connate, i. e. united.
Coalescent, growing together. Coalescence, 88.
Coarctate, contracted or brought close together.
Coated, having an integument, or covered in layers. Coated bulb, 46.
Cobwebby, same as arachnoid; bearing hairs like cobwebs or gossamer.
Coccineous, scarlet-red.
Coccus (plural cocci), anciently a berry; now mostly used to denote
the separable carpels or nutlets of a dry fruit.
Cochleariform, spoon-shaped.
Cochleate, coiled or shaped like a snail-shell.
Cœlospermous, applied to those fruits of Umbelliferæ which have the
seed hollowed on the inner face, by incurving of top and bottom; as in
Coriander.
Coherent, usually the same as connate.
Cohort, name sometimes used for groups between order and class, 178.
Coleorhiza, a root-sheath.
Collateral, side by side.
Collective fruits, 118.
Collum or Collar, the neck or junction of stem and root.
Colored, parts of a plant which are other-colored than green.
Columella, the axis to which the carpels of a compound pistil are
often attached, as in Geranium (112), or which is left when a pod opens,
as in Azalea.
Column, the united stamens, as in Mallow, or the stamens and pistils
united into one body, as in the Orchis family.
Columnar, shaped like a column or pillar.
Coma, a tuft of any sort (literally, a head of hair), 125.
Comose, tufted; bearing a tuft of hairs, as the seeds of Milkweed,
126.
Commissure, the line of junction of two carpels, as in the fruit of
Umbelliferæ.
Complanate, flattened.
Compound leaf, 54, 57. Compound pistil, 107. Compound umbel, 75,
&c.
Complete (flower), 81.
Complicate, folded upon itself.
Compressed, flattened on opposite sides.
Conceptacle, 168.
Concinnous, neat.
Concolor, all of one color.
Conchiform, shell- or half-shell-shaped.
Conduplicate, folded upon itself lengthwise, 71.
Cone, the fruit of the Pine family, 124. Coniferous, cone-bearing.
Confertus, much crowded.
Conferruminate, stuck together, as the cotyledons in a horse-chestnut.
Confluent, blended together; or the same as coherent.
Conformed, similar to another thing it is associated with or compared
to; or closely fitted to it, as the skin to the kernel of a seed.
Congested, Conglomerate, crowded together.
Conglomerate, crowded into a glomerule.
Conjugate, coupled; in single pairs. Conjugation, 170.
Connate, united or grown together from the first formation, 96.
Connate-perfoliate, when a pair of leaves are connate round a stem,
60.
Connective, Connectivum, the part of the anther connecting its two
cells, 101.
Connivent, converging, or brought close together.
Consolidation (floral), 94.
Consolidated forms of vegetation, 47.
Contents of cells, 136.
Continuous, the reverse of interrupted or articulated.
Contorted, twisted together. Contorted æstivation, same as
convolute, 97.
Contortuplicate, twisted back upon itself.
Contracted, either narrowed or shortened.
[Pg 201]Contrary, turned in opposite direction to the ordinary.
Convolute, rolled up lengthwise, as the leaves of the Plum in
vernation, 72. In æstivation, same as contorted, 97.
Cordate, heart-shaped, 53.
Coriaceous, resembling leather in texture.
Corky, of the texture of cork. Corky layer of bark, 141.
Corm, a solid bulb, like that of Crocus, 45.
Corneous, of the consistence or appearance of horn.
Corniculate, furnished with a small horn or spur.
Cornute, horned; bearing a horn-like projection or appendage.
Corolla, the leaves of the flower within the calyx, 14, 79.
Corollaceous, Corolline, like or belonging to a corolla.
Corona, a coronet or crown; an appendage at the top of the claw of
some petals, 91.
Coronate, crowned; furnished with a crown.
Cortex, bark. Cortical, belonging to the bark (cortex).
Corticate, coated with bark or bark-like covering.
Corymb, a flat or convex indeterminate flower-cluster, 74.
Corymbiferous, bearing corymbs.
Corymbose, in corymbs, approaching the form of a corymb, or branched
in that way.
Costa, a rib; the midrib of a leaf, &c. Costate, ribbed.
Cotyledons, the proper leaves of the embryo, 11, 127.
Crateriform, goblet-shaped or deep saucer-shaped.
Creeping (stems), growing flat on or beneath the ground and rooting,
39.
Cremocarp, a half-fruit, or one of the two carpels of Umbelliferæ,
121.
Crenate, or Crenelled, the edge scalloped into rounded teeth, 55.
Crenulate, minutely or slightly crenate.
Crested, or Cristate, bearing any elevated appendage like a crest.
Cretaceous, chalky or chalk-like.
Cribrose, or cribriform, pierced like a sieve with small apertures.
Crinite, bearing long hairs.
Crispate, curled or crispy.
Croceous, saffron-color, deep reddish-yellow.
Cross-breeds, the progeny of interbred varieties, 176.
Cross fertilization, 115.
Crown, see corona. Crowned, see coronate.
Cruciate, or Cruciform, cross-shaped. Cruciform Corolla, 86.
Crustaceous, hard and brittle in texture; crust-like.
Cryptogamous Plants, Cryptogams, 10, 156.
Cryptos, concealed, as Cryptopetalous, with concealed petals, &c.
Crystals in plants, 137.
Cucullate, hooded, or hood-shaped, rolled up like a cornet of paper,
or a hood (cucullus), as the spathe of Indian Turnip, 75.
Culm, a straw; the stem of Grasses and Sedges, 39.
Cultrate, shaped like a trowel or broad knife.
Cuneate, Cuneiform, wedge-shaped, 53.
Cup-shaped, same as cyathiform or near it.
Cupule, a little cup; the cup to the acorn of the Oak, 122.
Cupular, or Cupulate, provided with a cupule.
Cupuliferous, cupule-bearing.
Curviveined, with curved ribs or veins.
Curviserial, in oblique or spiral ranks.
Cushion, the enlargement at the insertion or base of a petiole.
Cuspidate, tipped with a sharp and stiff point or cusp, 54.
Cut, same as incised, or applied generally to any sharp and deep
division, 55.
Cuticle, the skin of plants, or more strictly its external pellicle.
Cyaneous, bright blue.
Cyathiform, in the shape of a cup, or particularly of a wine-glass.
Cycle, one complete turn of a spire, or a circle, 70.
[Pg 202]Cyclical, rolled up circularly, or coiled into a complete circle.
Cyclosis, circulation in closed cells, 149.
Cylindraceous, approaching to the Cylindrical form, terete and not
tapering.
Cymbæform, or Cymbiform, same as boat-shaped.
Cyme, a cluster of centrifugal inflorescence, 77.
Cymose, furnished with cymes, or like a cyme.
Cymule, a partial or diminutive cyme, 77.
Deca- (in words of Greek derivation), ten; as
Decagynous, with 10 pistils or styles, Decamerous, of 10 parts,
Decandrous, with 10 stamens, &c.
Deciduous, falling off, or subject to fall; said of leaves which fall
in autumn, and of a calyx and corolla which fall before the fruit forms.
Declinate, declined, turned to one side, or downwards.
Decompound, several times compounded or divided, 59.
Decumbent, reclined on the ground, the summit tending to rise, 39.
Decurrent (leaves), prolonged on the stem beneath the insertion, as in
Thistles.
Decussate, arranged in pairs which successively cross each other, 71.
Deduplication, same as chorisis.
Definite, when of a uniform number, and not above twelve or so.
Definite Inflorescence, 72.
Deflexed, bent downwards.
Deflorate, past the flowering state, as an anther after it has
discharged its pollen.
Dehiscence, the regular splitting open of capsule or anther, 103, 119.
Dehiscent, opening by regular dehiscence, 119, 123.
Deliquescent, branching off so that the stem is lost in the branches,
32.
Deltoid, of a triangular shape, like the Greek capital Δ.
Demersed, growing below the surface of water.
Dendroid, Dendritic, tree-like in form or appearance.
Dendron, Greek for tree.
Deni, ten together.
Dens, Latin for tooth.
Dentate, toothed, 55. Denticulate, furnished with denticulations, or
little teeth.
Depauperate, impoverished or starved, and so below the natural size.
Depressed, flattened or as if pressed down from above.
Derma, Greek for skin.
Descending, tending gradually downwards. Descending axis, the root.
Desmos, Greek for things connected or bound together.
Determinate Inflorescence, 72.
Dextrorse, turned to the right hand.
Di- Dis (in Greek compounds) two, as
Diadelphous (stamens), united by their filaments in two sets, 99.
Diagnosis, a short distinguishing character or descriptive phrase.
Dialypetalous, same as polypetalous.
Diandrous, having two stamens, &c.
Diaphanous, transparent or translucent.
Dicarpellary, of two carpels.
Dichlamydeous (flower), having both calyx and corolla.
Dichogamous, Dichogamy, 116.
Dichotomous, two-forked.
Diclinous, having the stamens in one flower, the pistils in another,
85.
Dicoccous (fruit), splitting into two cocci or closed carpels.
Dicotyls, 23.
Dicotyledonous (embryo), having a pair of cotyledons, 23.
Dicotyledonous Plants, 23, 182.
Didymous, twin.
Didynamous (stamens), having four stamens in two pairs, 100.
Diffuse, spreading widely and irregularly.
[Pg 203]Digitate (fingered), where the leaflets of a compound leaf are all
borne on the apex of the petiole, 58.
Digynous (flower), having two pistils or styles, 105.
Dimerous, made up of two parts, or its organs in twos.
Dimidiate, halved; as where a leaf or leaflet has only one side
developed.
Dimorphism, 117. Dimorphous, Dimorphic, of two forms, 117.
Diœcious, or Dioicous, with stamens and pistils on different
plants, 85.
Dipetalous, of two petals.
Diphyllous, two-leaved.
Dipterous, two-winged.
Diplo-, Greek for double, as Diplostemonous, with two sets of
stamens.
Disciform or Disk-shaped, flat and circular, like a disk or quoit.
Discoidal, or Discoid, belonging to or like a disk.
Discolor, of two different colors or hues.
Discrete, separate, opposite of concrete.
Disepalous, of two sepals.
Disk, the face of any flat body; the central part of a head of
flowers, like the Sunflower, or Coreopsis, as opposed to the ray or
margin; a fleshy expansion of the receptacle of a flower, 113.
Disk-flowers, those of the disk in Compositæ.
Dissected, cut deeply into many lobes or divisions.
Dissepiments, the partitions of a compound ovary or a fruit, 108.
Dissilient, bursting in pieces.
Distichous, two-ranked.
Distinct, uncombined with each other, 95.
Dithecous, of two thecæ or anther-cells.
Divaricate, straddling; very widely divergent.
Divided (leaves, &c.), cut into divisions down to the base or midrib,
55.
Dodeca, Greek for twelve; as Dodecagynous, with twelve pistils or
styles, Dodecandrous, with twelve stamens.
Dodrans, span-long.
Dolabriform, axe-shaped.
Dorsal, pertaining to the back (dorsum) of an organ. Dorsal
Suture, 106.
Dotted Ducts, 148.
Double Flowers, where the petals are multiplied unduly, 79.
Downy, clothed with a coat of soft and short hairs.
Drupaceous, like or pertaining to a drupe.
Drupe, a stone-fruit, 120. Drupelet or Drupel, a little drupe.
Ducts, the so-called vessels of plants, 134.
Dumose, bushy, or relating to bushes.
Duramen, the heart-wood, 142.
Dwarf, remarkably low in stature.
E-, as a prefix of Latin compound words, means destitute of; as
ecostate, without a rib or midrib; exalbuminous, without albumen,
&c.
Eared, see auriculate, 53.
Ebracteate, destitute of bracts. Ebracteolate, destitute of
bractlets.
Eburneous, ivory-white.
Echinate, armed with prickles (like a hedgehog). Echinulate, a
diminutive of it.
Edentate, toothless.
Effete, past bearing, &c.; said of anthers which have discharged their
pollen.
Effuse, very loosely branched and spreading.
Eglandulose, destitute of glands.
Elaters, threads mixed with the spores of Liverworts, 165.
Ellipsoidal, approaching an elliptical figure.
Elliptical, oval or oblong, with the ends regularly rounded, 52.
Emarginate, notched at the summit, 54.
Embryo, the rudimentary plantlet in a seed, 11, 127.
Embryonal, belonging or relating to the embryo.
Embryo-sac, 117.
[Pg 204]Emersed, raised out of water.
Endecagynous, with eleven pistils or styles.
Endecandrous, with eleven stamens.
Endemic, peculiar to the country geographically.
Endocarp, the inner layer of a pericarp or fruit, 120.
Endochrome, the coloring matter of Algæ and the like.
Endogenous Stems, 138. Endogenous plants, an old name for
monocotyledons.
Endopleura, inner seed-coat.
Endorhizal, radicle or root sheathed in germination.
Endosperm, the albumen of a seed, 21.
Endostome, the orifice in the inner coat of an ovule.
Ennea-, nine. Enneagynous, with nine petals or styles.
Enneandrous, nine-stamened.
Ensate, Ensiform, sword-shaped.
Entire, the margins not at all toothed, notched, or divided, but even,
55.
Entomophilous, said of flowers frequented and fertilized by insects,
113.
Ephemeral, lasting for a day or less, as the corolla of Purslane, &c.
Epi-, Greek for upon.
Epicalyx, such an involucel as that of Malvaceæ.
Epicarp, the outermost layer of a fruit, 120.
Epidermal, relating to the Epidermis, or skin of a plant, 50, 141,
143.
Epigæous, growing on the earth, or close to the ground.
Epigynous, upon the ovary, 95, 99.
Epipetalous, borne on the petals or the corolla, 99.
Epiphyllous, borne on a leaf.
Epiphyte, a plant growing on another plant, but not nourished by it,
36.
Epiphytic or Epiphytal, relating to Epiphytes.
Epipterous, winged at top.
Episperm, the skin or coat of a seed, especially the outer coat.
Equal, alike in number or length.
Equally pinnate, same as abruptly pinnate, 57.
Equitant (riding straddle), 60.
Erion, Greek for wool. Erianthous, woolly-flowered. Eriophorous,
wool-bearing, &c.
Erose, eroded, as if gnawed.
Erostrate, not beaked.
Erythros, Greek for red. Erythrocarpous, red-fruited, &c.
Essential Organs of the flower, 80.
Estivation, see æstivation.
Etiolated, blanched by excluding the light, as the stalks of Celery.
Eu, Greek prefix, meaning very, or much.
Evergreen, holding the leaves over winter and until new ones appear,
or longer.
Ex, Latin prefix; privative in place of "e" when next letter is a
vowel. So Exalate, wingless; Exalbuminous (seed), without albumen,
21.
Excurrent, running out, as when a midrib projects beyond the apex of a
leaf, or a trunk is continued to the very top of a tree, 32.
Exiguous, puny.
Exilis, lank or meagre.
Eximius, distinguished for size or beauty.
Exo-, in Greek compounds, outward, as in
Exocarp, outer layer of a pericarp, 120.
Exogenous, outward growing. Exogenous stems, 139.
Exorhizal, radicle in germination not sheathed.
Exostome, the orifice in the outer coat of the ovule.
Explanate, spread or flattened out.
Exserted, protruding out of, as the stamens out of the corolla.
Exstipulate, destitute of stipules.
Extine, outer coat of a pollen-grain.
Extra-axillary, said of a branch or bud somewhat out of the axil, 31.
Extrorse, turned outwards; the anther is extrorse when fastened to the
filament on the side next the pistil, and opening on the outer side,
101.
[Pg 205]Falcate, scythe-shaped; a flat body curved, its edges parallel.
False Racemes, 78.
Family, in botany same as Order, 177.
Farina, meal or starchy matter, 136.
Farinaceous, mealy in texture. Farinose, covered with a mealy
powder.
Fasciate, banded; also applied to monstrous stems which grow flat.
Fascicle, a close cluster, 77.
Fascicled, Fasciculated, growing in a bundle or tuft, as the leaves
of Larch, 68, and roots of Peony, 35.
Fastigiate, close, parallel, and upright, as the branches of Lombardy
Poplar.
Faux (plural, fauces), the throat of a calyx, corolla, &c., 89.
Faveolate, Favose, honeycombed; same as alveolate.
Feather-veined, with veins of a leaf all springing from the sides of a
midrib, 51.
Fecula or Fæcula, starch, 136.
Female flower or plant, one bearing pistils only.
Fenestrate, pierced with one or more large holes, like windows.
Ferrugineous, or Ferruginous, resembling iron-rust; red-grayish.
Fertile, fruit-bearing, or capable of it; also said of anthers
producing good pollen.
Fertilization, the process by which pollen causes the embryo to be
formed, 114.
Fibre (woody), 133. Fibrous, containing much fibre, or composed of
fibres.
Fibrillose, formed of small fibres, or Fibrillæ.
Fibro-vascular bundle or tissue, formed of fibres and vessels.
Fiddle-shaped, obovate with a deep recess on each side.
Fidus, Latin suffix for cleft, as Bifid, two-cleft.
Filament, the stalk of a stamen, 14, 80, 101; also any slender
thread-shaped body.
Filamentose, or Filamentous, bearing or formed of slender threads.
Filiform, thread-shaped; long, slender, and cylindrical.
Fimbriate, fringed; furnished with fringes (fimbriæ).
Fimbrillate, Fimbrilliferous, bearing small fimbriæ, i. e.
fimbrillæ.
Fissiparous, multiplying by division of one body into two.
Fissus, Latin for split or divided.
Fistular, or Fistulose, hollow and cylindrical, as the leaves of the
Onion.
Flabelliform, or Flabellate, fan-shaped.
Flagellate, or Flagelliform, long, narrow, and flexible, like the
thong of a whip; or like the runners (flagellæ) of the Strawberry.
Flavescent, yellowish, or turning yellow.
Flavus, Latin for yellow.
Fleshy, composed of firm pulp or flesh.
Flexuose, or Flexuous, bending in opposite directions, in a zigzag
way.
Floating, swimming on the surface of water.
Floccose, composed of or bearing tufts of woolly or long and soft
hairs.
Flora (the goddess of flowers), the plants of a country or district,
taken together, or a work systematically describing them, 9.
Floral Envelopes, or Flower-leaves, 79.
Floret, a diminutive flower, one of a mass or cluster.
Floribund, abundantly floriferous.
Florula, the flora of a small district.
Flos, floris, Latin for flower.
Flosculus, diminutive, same as floret.
Flower, the whole organs of reproduction of Phænogamous plants, 14,
72.
Flower-bud, an unopened flower.
Flowering Plants, 10, 156. Flowerless Plants, 10, 156.
Fly-trap leaves, 65.
Fluitans, Latin for floating. Fluviatile, belonging to a river or
stream.
Foliaceous, belonging to, or of the texture or nature of, a leaf
(folium).
Foliate, provided with leaves. Latin prefixes denote the number of
leaves, as bifoliate, trifoliate, &c. Foliose, leafy; abounding in
leaves.
Foliolate, relating to or bearing leaflets (foliola);
trifoliolate, with three leaflets, &c.
[Pg 206]Folium (plural, folia), Latin for leaf.
Follicle, a simple pod, opening down the inner suture, 122.
Follicular, resembling or belonging to a follicle.
Food of Plants, 144.
Foot-stalk, either petiole or peduncle, 49.
Foramen, a hole or orifice, as that of the ovule, 110.
Foraminose, Foraminulose, pierced with holes.
Forked, branched in two or three or more.
Fornicate, bearing fornices.
Fornix, little arched scales in the throat of some corollas, as of
Comfrey.
Foveate, deeply pitted. Foveolate, diminutive of foveate.
Free, not united with any other parts of a different sort, 95.
Fringed, the margin beset with slender appendages, bristles, &c.
Frond, what answers to leaves in Ferns, &c., 157; or to the stem and
leaves fused into one, as in Liverwort.
Frondescence, the bursting into leaf.
Frondose, frond-bearing; like a frond, or sometimes used for leafy.
Fructification, the state or result of fruiting.
Fructus, Latin for fruit.
Fruit, the matured ovary and all it contains or is connected with,
117.
Fruit-dots in Ferns; see Sorus.
Frustulose, consisting of a chain of similar pieces, or Frustules.
Frutescent, somewhat shrubby; becoming a shrub (Frutex), 39.
Fruticulose, like a small shrub, or Fruticulus. Fruticose,
shrubby, 39.
Fugacious, soon falling off or perishing.
Fulcrate, having accessory organs or fulcra, i. e. props.
Fulvous, tawny; dull yellow with gray.
Fungus, Fungi, 172.
Funicle, Funiculus, the stalk of a seed or ovule, 110.
Funnelform, or funnel-shaped, expanding gradually upwards into an
open mouth, like a funnel or tunnel, 90.
Furcate, forked.
Furfuraceous, covered with bran-like fine scurf.
Furrowed, marked by longitudinal channels or grooves.
Fuscous, deep gray-brown.
Fusiform, spindle-shaped, 36.
Galbalus, the fleshy or at length woody cone of Juniper and Cypress.
Galea, a helmet-shaped body, as the upper sepal of the Monkshood, 87.
Galeate, shaped like a helmet.
Gamopetalous, of united petals, 89.
Gamophyllous, formed of united leaves.
Gamosepalous, formed of united sepals, 89.
Geminate, twin; in pairs.
Gemma, Latin for a bud.
Gemmation, the state of budding; budding growth.
Gemmule, a small bud; the plumule, 13.
Genera, plural of genus.
Geniculate, bent abruptly, like a knee (genu), as many stems.
Generic Names, 179.
Genus, a kind of a rank above species, 177.
Germ, a growing point; a young bud; sometimes the same as embryo, 127.
Germen, the old name for ovary.
Germination, the development of a plantlet from the seed, 12.
Gerontogæous, inhabiting the Old World.
Gibbous, more tumid at one place or on one side than the other.
Gilvous, dirty reddish-yellow.
Glabrate, becoming glabrous with age, or almost glabrous.
Glabrous, smooth, in the sense of having no hairs, bristles, or other
pubescence.
[Pg 207]Gladiate, sword-shaped, as the leaves of Iris.
Glands, small cellular organs which secrete oily or aromatic or other
products; they are sometimes sunk in the leaves or rind, as in the
Orange, Prickly Ash, &c.; sometimes on the surface as small projections;
sometimes raised on hairs or bristles (glandular hairs, &c.), as in
the Sweetbrier and Sundew. The name is also given to any small
swellings, &c., whether they secrete anything or not; so that the word
is loosely used.
Glandular, Glandulose, furnished with glands, or gland-like.
Glans (Gland), the acorn or mast of Oak and similar fruits.
Glareose, growing in gravel.
Glaucescent, slightly glaucous, or bluish-gray.
Glaucous, covered with a bloom, viz. with a fine white powder of wax
that rubs off, like that on a fresh plum, or a cabbage-leaf.
Globose, spherical in form, or nearly so. Globular, nearly globose.
Glochidiate, or Glochideous, (bristles) barbed; tipped with barbs,
or with a double hooked point.
Glomerate, closely aggregated into a dense cluster.
Glomerule, a dense head-like cluster, 77.
Glossology, the department of botany in which technical terms are
explained.
Glumaceous, glume-like, or glume-bearing.
Glume; Glumes are the husks or floral coverings of Grasses, or,
particularly, the outer husks or bracts of each spikelet.
Glumelles, the inner husks of Grasses.
Gonophore, a stipe below stamens, 113.
Gossypine, cottony, flocculent.
Gracilis, Latin for slender.
Grain, see Caryopsis, 121.
Gramineous, grass-like.
Granular, composed of grains. Granule, a small grain.
Graveolent, heavy-scented.
Griseous, gray or bluish-gray.
Growth, 129.
Grumous, or Grumose, formed of coarse clustered grains.
Guttate, spotted, as if by drops of something colored.
Gymnos, Greek for naked, as
Gymnocarpous, naked-fruited.
Gymnospermous, naked-seeded, 109.
Gymnospermous gynœcium, 109.
Gymnospermæ, or Gymnospermous Plants, 183.
Gynandrous, with stamens borne on, i. e. united with, the pistil, 99.
Gynœcium, a name for the pistils of a flower taken altogether, 105.
Gynobase, a depressed receptacle or support of the pistil or carpels,
114.
Gynophore, a stalk raising a pistil above the stamens, 113.
Gynostegium, a sheath around pistils, of whatever nature.
Gynostemium, name of the column in Orchids, &c., consisting of style
and stigma with stamens combined.
Gyrate, coiled or moving circularly.
Gyrose, strongly bent to and fro.
Habit, the general aspect of a plant, or its mode of growth.
Habitat, the situation or country in which a plant grows in a wild
state.
Hairs, hair-like growths on the surface of plants.
Hairy, beset with hairs, especially longish ones.
Halberd-shaped, see hastate, 53.
Halved, when appearing as if one half of the body were cut away.
Hamate, or Hamose, hooked; the end of a slender body bent round.
Hamulose, bearing a small hook; a diminutive of the last.
Haplo-, in Greek compounds, single; as Haplostemonous, having only
one series of stamens.
[Pg 208]Hastate, or Hastile, shaped like a halberd; furnished with a
spreading lobe on each side at the base, 53.
Head, capitulum, a form of inflorescence, 74.
Heart-shaped, of the shape of a heart as painted on cards, 53.
Heart-wood, the older or matured wood of exogenous trees, 142.
Helicoid, coiled like a helix or snail-shell, 77.
Helmet, the upper sepal of Monkshood is so called.
Helvolous, grayish-yellow.
Hemi- in compounds from the Greek, half; e. g. Hemispherical, &c.
Hemicarp, half-fruit, one carpel of an Umbelliferous plant, 121.
Hemitropous (ovule or seed), nearly same as amphitropous, 123.
Hepta- (in words of Greek origin), seven; as Heptagynous, with seven
pistils or styles. Heptamerous, its parts in sevens. Heptandrous,
having seven stamens.
Herb, plant not woody, at least above ground.
Herbaceous, of the texture of an herb; not woody, 39.
Herbarium, the botanist's arranged collection of dried plants, 186.
Herborization, 184.
Hermaphrodite (flower), having stamens and pistils in the same
blossom, 81.
Hesperidium, orange-fruit, a hard-rinded berry.
Hetero-, in Greek compounds, means of two or more sorts, as
Heterocarpous, bearing fruit of two kinds or shapes.
Heterogamous, bearing two or more sorts of flowers in one cluster.
Heterogony, Heterogone, or Heterogonous, with stamens and pistil
reciprocally of two sorts, 116. Heterostyled is same.
Heteromorphous, of two or more shapes.
Heterophyllous, with two sorts of leaves.
Heterotropous (ovule), the same as amphitropous, 123.
Hexa- (in Greek compounds), six; as Hexagonal, six-angled.
Hexagynous, with six pistils or styles. Hexamerous, its parts in
sixes. Hexandrous, with six stamens. Hexapterous, six-winged.
Hibernaculum, a winter bud.
Hiemal, relating to winter.
Hilar, belonging to the hilum.
Hilum, the scar of the seed; its place of attachment, 110, 126.
Hippocrepiform, horseshoe-shaped.
Hirsute, clothed with stiffish or beard-like hairs.
Hirtellous, minutely hirsute.
Hispid, bristly, beset with stiff hairs. Hispidulous, diminutive of
hispid.
Histology, 9.
Hoary, grayish-white; see canescent, &c.
Holosericeous, all over sericeous or silky.
Homo-, in Greek compounds, all alike or of one sort.
Homodromous, running in one direction.
Homogamous, a head or cluster with flowers all of one kind.
Homogeneous, uniform in nature; all of one kind.
Homogone, or Homogonous, counterpart of Heterogone or
Homostyled.
Homologous, of same type; thus petals and sepals are the homologues of
leaves.
Homomallous (leaves, &c.), originating all round an axis, but all bent
or curved to one side.
Homorphous, all of one shape.
Homotropous (embryo), curved with the seed; curved only one way.
Hood, same as helmet or galea. Hooded, hood-shaped; see
cucullate.
Hooked, same as hamate.
Horn, a spur or some similar appendage. Horny, of the texture of
horn.
Hortensis, pertaining to the garden.
Hortus Siccus, an herbarium, or collection of dried plants, 186.
Humifuse, Humistrate, spread over the surface of the ground.
Humilis, low in stature.
[Pg 209]Hyaline, transparent, or partly so.
Hybrid, a cross-breed between two allied species, 176.
Hydrophytes, water-plants.
Hyemal, see hiemal.
Hymenium of a Mushroom, 172.
Hypanthium, a hollow flower-receptacle, such as that of Rose.
Hypo-, Greek prefix for under, or underneath.
Hypocotyle, or Hypocotyl, part of stem below the cotyledons, 11.
Hypocrateriform, properly Hypocraterimorphous, salver-shaped.
Hypogæan, or Hypogæous, produced under ground, 19.
Hypogynous, inserted under the pistil, 95, 99.
Hysteranthous, with the blossoms developed earlier than the leaves.
Icosandrous, having 20 (or 12 or more) stamens inserted on the calyx.
Imberbis, Latin for beardless.
Imbricate, Imbricated, Imbricative, overlapping one another, like
tiles or shingles on a roof, as the bud-scales of Horse-chestnut and
Hickory, 27. In æstivation, where some leaves of the calyx or corolla
are overlapped on both sides by others, 98.
Immarginate, destitute of a rim or border.
Immersed, growing wholly under water.
Impari-pinnate, pinnate with a single leaflet at the apex, 57.
Imperfect flowers, wanting either stamens or pistils, 85.
Inæquilateral, unequal-sided, as the leaf of a Begonia.
Inane, empty, said of an anther which produces no pollen, &c.
Inappendiculate, not appendaged.
Incanous, Incanescent, hoary with soft white pubescence.
Incarnate, flesh-colored.
Incised, cut rather deeply and irregularly, 58.
Included, enclosed; when the part in question does not project beyond
another.
Incomplete Flower, wanting calyx or corolla, 86.
Incrassated, thickened.
Incubous, with tip of one leaf lying flat over the base of the next
above.
Incumbent, leaning or resting upon; the cotyledons are incumbent when
the back of one of them lies against the radicle, 128; the anthers are
incumbent when turned or looking inwards.
Incurved, gradually curving inwards.
Indefinite, not uniform in number, or too numerous to mention (over
12).
Indefinite or Indeterminate Inflorescence, 72.
Indehiscent, not splitting open; i. e. not dehiscent, 119.
Indigenous, native to the country.
Individuals, 175.
Indumentum, any hairy coating or pubescence.
Induplicate, with the edges turned inwards, 97.
Induviate, clothed with old and withered parts or induviæ.
Indusium, the shield or covering of a fruit-dot of a Fern, 159.
Inermis, Latin for unarmed, not prickly.
Inferior, growing below some other organ, 96.
Infertile, not producing seed, or pollen, as the case may be.
Inflated, turgid and bladdery.
Inflexed, bent inwards.
Inflorescence, the arrangement of flowers on the stem, 72.
Infra-axillary, situated beneath the axil.
Infundibuliform or Infundibular, funnel-shaped, 90.
Innate (anther), attached by its base to the very apex of the
filament, 101.
Innovation, a young shoot, or new growth.
Insertion, the place or the mode of attachment of an organ to its
support, 95, 99.
Integer, entire, not lobed. Integerrimus, quite entire, not serrate.
[Pg 210]Intercellular Passages or Spaces, 131, 143.
Interfoliaceous, between the leaves of a pair or whorl.
Internode, the part of a stem between two nodes, 13.
Interpetiolar, between petioles.
Interruptedly pinnate, pinnate with small leaflets intermixed with
larger.
Intine, inner coat of a pollen grain.
Intrafoliaceous (stipules, &c.), placed between the leaf or petiole
and the stem.
Introrse, turned or facing inwards; i. e. towards the axis of the
flower, 101.
Intruse, as it were pushed inwards.
Inversed or Inverted, where the apex is in the direction opposite to
that of the organ it is compared with.
Involucel, a partial or small involucre, 76.
Involucellate, furnished with an involucel. Involucrate, furnished
with an involucre.
Involucre, a whorl or set of bracts around a flower, umbel, or head,
&c., 74, 75.
Involute, in vernation, 72; rolled inwards from the edges, 97.
Irregular Flowers, 86, 91.
Isos, Greek for equal in number. Isomerous, the same number in the
successive circles or sets. Isostemonous, the stamens equal in number
to the sepals or petals.
Jointed, separate or separable at one or more places into pieces, 64,
&c.
Jugum (plural Juga), Latin for a pair, as of leaflets,—thus
Unijugate, of a single pair; Bijugate, of two pairs, &c.
Julaceus, like a catkin or Julus.
Keel, a projecting ridge on a surface, like the keel of a boat; the
two anterior petals of a papilionaceous corolla, 92.
Keeled, furnished with a keel or sharp longitudinal ridge.
Kermesine, Carmine-red.
Kernel of the ovule and seed, 110.
Key, or Key-fruit, a Samara, 122.
Kidney-shaped, resembling the outline of a kidney, 53.
Labellum, the odd petal in the Orchis Family.
Labiate, same as bilabiate or two-lipped, 92.
Labiatiflorous, having flowers with bilabiate corolla.
Labium (plural, Labia), Latin for lip.
Lacerate, with margin appearing as if torn.
Laciniate, slashed; cut into deep narrow lobes or Laciniæ.
Lactescent, producing milky juice, as does the Milkweed, &c.
Lacteus, Latin for milk-white.
Lacunose, full of holes or gaps.
Lacustrine, belonging to lakes.
Lævigate, smooth as if polished. Latin, Lævis, smooth, as opposed to
rough.
Lageniform, gourd-shaped.
Lagopous, Latin, hare-footed; densely clothed with long soft hairs.
Lamellar or Lamellate, consisting of flat plates, Lamellæ.
Lamina, a plate or blade, the blade of a leaf, &c., 49.
Lanate, Lanose, woolly; clothed with long and soft entangled hairs.
Lanceolate, lance-shaped, 52.
Lanuginous, cottony or woolly.
Latent buds, concealed or undeveloped buds, 30.
Lateral, belonging to the side.
Latex, the milky juice, &c., of plants, 135.
Lax (Laxus), loose in texture, or sparse; the opposite of crowded.
Leaf, 49. Leaf-buds, 31.
Leaflet, one of the divisions or blades of a compound leaf, 57.
Leaf-like, same as foliaceous.
Leathery, of about the consistence of leather; coriaceous.
[Pg 211]Legume, a simple pod which dehisces in two pieces, like that of the
Pea, 122.
Leguminous, belonging to legumes, or to the Leguminous Family.
Lenticular, lens-shaped; i. e. flattish and convex on both sides.
Lappaceous, bur-like.
Lasio, Greek for woolly or hairy, as Lasianthus, woolly-flowered.
Lateritious, brick-colored.
Laticiferous, containing latex, 135.
Latus, Latin for broad, as Latifolius, broad-leaved.
Leaf-scar, Leaf-stalk, petiole.
Lenticels, lenticular dots on young bark.
Lentiginose, as if freckled.
Lepal, a made-up word for a staminode.
Lepis, Greek for a scale, whence Lepidote, leprous; covered with
scurfy scales.
Leptos, Greek for slender; so Leptophyllous, slender-leaved.
Leukos, Greek for white; whence Leucanthous, white-flowered, &c.
Liber, the inner bark of Exogenous stems, 140.
Lid, see operculum.
Ligneous, or Lignose, woody in texture.
Ligulate, furnished with a ligule, 93.
Ligule, Ligula, the strap-shaped corolla in many Compositæ, 93; the
membranous appendage at the summit of the leaf-sheaths of most Grasses,
67.
Limb, the border of a corolla, &c., 89.
Limbate, bordered (Latin, Limbus, a border).
Line, the twelfth of an inch; or French lines, the tenth.
Linear, narrow and flat, the margins parallel, 52.
Lineate, marked with parallel lines. Lineolate, marked with minute
lines.
Lingulate, Linguiform, tongue-shaped.
Lip, the principal lobes of a bilabiate corolla or calyx, 92.
Litoral or Littoral, belonging to the shore.
Livid, pale lead-colored.
Lobe, any projection or division (especially a rounded one) of a leaf,
&c.
Lobed or Lobate, cut into lobes, 55, 56; Lobulate, into small
lobes.
Locellate, having Locelli, i. e. compartments in a cell: thus an
anther-cell is often bilocellate.
Loculament, same as loculus.
Locular, relating to the cell or compartment (Loculus) of an ovary,
&c.
Loculicidal (dehiscence), splitting down through the back of each
cell, 123.
Locusta, a name for the spikelet of Grasses.
Lodicule, one of the scales answering to perianth-leaves in
Grass-flowers.
Loment, a pod which separates transversely into joints, 122.
Lomentaceous, pertaining to or resembling a loment.
Lorate, thong-shaped.
Lunate, crescent-shaped. Lunulate, diminutive of lunate.
Lupuline, like hops.
Lusus, Latin for a sport or abnormal variation.
Luteolus, yellowish; diminutive of
Luteus, Latin for yellow. Lutescent, verging to yellow.
Lyrate, lyre-shaped; a pinnatifid leaf of an obovate or spatulate
outline, the end-lobe large and roundish, and the lower lobes small, as
in fig. 149.
Macros, Greek for long, sometimes also used for large: thus
Macrophyllous, long or large-leaved, &c.
Macrospore, the large kind of spore, when there are two kinds, 160,
161.
Maculate, spotted or blotched.
Male (flowers or plants), having stamens but no pistil.
Mammose, breast-shaped.
Marcescent, withering without falling off.
Marginal, belonging to margin.
[Pg 212]Marginate, margined with an edge different from the rest.
Marginicidal dehiscence, 123.
Maritime, belonging to sea-coasts.
Marmorate, marbled.
Mas., Masc., Masculine, male.
Masked, see personate.
Mealy, see farinaceous.
Median, Medial, belonging to the middle.
Medifixed, attached by the middle.
Medullary, belonging to, or of the nature of, pith (Medulla); pithy.
Medullary Rays, the silver-grain of wood, 140, 141.
Medullary Sheath, a set of ducts just around the pith, 140.
Meiostemonous, having fewer stamens than petals.
Membranaceous or Membranous, of the texture of membrane; thin and
soft.
Meniscoid, crescent-shaped.
Mericarp, one carpel of the fruit of an Umbelliferous plant, 121.
Merismatic, separating into parts by the formation of partitions
across.
Merous, from the Greek for part; used with numeral prefix to denote
the number of pieces in a set or circle: as Monomerous, of only one,
Dimerous, with two, Trimerous, with three parts (sepals, petals,
stamens, &c.) in each circle.
Mesocarp, the middle part of a pericarp, when that is distinguishable
into three layers, 120.
Mesophlœum, the middle or green bark.
Micropyle, the closed orifice of the seed, 110, 126.
Microspore, the smaller kind of spore when there are two kinds, 161.
Midrib, the middle or main rib of a leaf, 50.
Milk-vessels, 135.
Miniate, vermilion-colored.
Mitriform, mitre-shaped: in the form of a peaked cap, or one cleft at
the top.
Moniliform, necklace-shaped; a cylindrical body contracted at
intervals.
Monocarpic (duration), flowering and seeding but once, 38.
Monochlamydeous, having only one floral envelope.
Monocotyledonous (embryo), with only one cotyledon, 24.
Monocotyledonous Plants, 24. Monocotyls, 24.
Monœcious, or Monoicous (flower), having stamens or pistils only,
85.
Monogynous (flower), having only one pistil, or one style, 105.
Monopetalous (flower), with the corolla of one piece, 89.
Monophyllous, one-leaved, or of one piece.
Monos, Greek for solitary or only one; thus Monadelphous, stamens
united by their filaments into one set, 99; Monandrous (flower),
having only one stamen, 100.
Monosepalous, a calyx of one piece; i. e. with the sepals united into
one body.
Monospermous, one-seeded.
Monstrosity, an unnatural deviation from the usual structure or form.
Morphology, Morphological Botany, 9; the department of botany which
treats of the forms which an organ may assume.
Moschate, Musk-like in odor.
Movements, 149.
Mucronate, tipped with an abrupt short point (Mucro), 54.
Mucronulate, tipped with a minute abrupt point; a diminutive of the
last.
Multi-, in composition, many; as Multangular, many-angled;
Multicipital, many-headed, &c.; Multifarious, in many rows or ranks;
Multifid, many-cleft; Multilocular, many-celled; Multiserial, in
many rows.
Multiple Fruits, 118, 124.
Muricate, beset with short and hard or prickly points.
Muriform, wall-like; resembling courses of bricks in a wall.
Muticous, pointless, blunt, unarmed.
Mycelium, the spawn of Fungi; i. e. the filaments from which
Mushrooms, &c., originate, 172.
[Pg 213]Naked, wanting some usual covering, as achlamydeous flowers, 86;
gymnospermous seeds, 109, 125, &c.
Names in botany, 179.
Nanus, Latin for dwarf.
Napiform, turnip-shaped, 35.
Natural System, 182.
Naturalized, introduced from a foreign country, and flourishing wild.
Navicular, boat-shaped, like the glumes of most Grasses.
Necklace-shaped, looking like a string of beads; see moniliform.
Nectar, the sweet secretion in flowers from which bees make honey, &c.
Nectariferous, honey-bearing; or having a nectary.
Nectary, the old name for petals and other parts of the flower when of
unusual shape, especially when honey-bearing. So the hollow spur-shaped
petals of Columbine were called nectaries; also the curious long-clawed
petals of Monkshood, 87, &c.
Needle-shaped, long, slender, and rigid, like the leaves of Pines.
Nemorose or Nemoral, inhabiting groves.
Nerve, a name for the ribs or veins of leaves when simple and
parallel, 50.
Nerved, furnished with nerves, or simple and parallel ribs or veins,
50.
Nervose, conspicuously nerved. Nervulose, minutely nervose.
Netted-veined, furnished with branching veins forming network, 50, 51.
Neuter, Neutral, sexless. Neutral flower, 79.
Niger, Latin for black. Nigricans, Latin for verging to black.
Nitid, shining.
Nival, living in or near snow. Niveus, snow-white.
Nodding, bending so that the summit hangs downward.
Node, a knot; the "joints" of a stem, or the part whence a leaf or a
pair of leaves springs, 13.
Nodose, knotty or knobby. Nodulose, furnished with little knobs or
knots.
Nomenclature, 175, 179.
Normal, according to rule, natural.
Notate, marked with spots or lines of a different color.
Nucamentaceous, relating to or resembling a small nut.
Nuciform, nut-shaped or nut-like.
Nucleus, the kernel of an ovule (110) or seed (127) of a cell.
Nucule, same as nutlet.
Nude, (Latin, Nudus), naked. So Nudicaulis, naked-stemmed, &c.
Nut, Latin Nux, a hard, mostly one-seeded indehiscent fruit; as a
chestnut, butternut, acorn, 121.
Nutant, nodding.
Nutlet, a little nut; or the stone of a drupe.
Ob- (meaning over against), when prefixed to words signifies
inversion; as, Obcompressed, flattened the opposite of the usual way;
Obcordate, heart-shaped, with the broad and notched end at the apex
instead of the base, 54; Oblanceolate, lance-shaped with the tapering
point downwards, 52.
Oblique, applied to leaves, &c., means unequal-sided.
Oblong, from two to four times as long as broad, 52.
Obovate, inversely ovate, the broad end upward, 53. Obovoid, solid
obovate.
Obtuse, blunt or round at the end, 54.
Obverse, same as inverse.
Obvolute (in the bud), when the margins of one piece or leaf
alternately overlap those of the opposite one.
Ocellate, with a circular colored patch, like an eye.
Ochroleucous, yellowish-white; dull cream-color.
Ocreate, furnished with Ocreæ (boots), or stipules in the form of
sheaths, 67.
Octo-, Latin for eight, enters into the composition of Octagynous,
with eight pistils or styles; Octamerous, its parts in eights;
Octandrous, with eight stamens, &c.
[Pg 214]Oculate, with eye-shaped marking.
Officinal, used in medicine, therefore kept in the shops.
Offset, short branches next the ground which take root, 40.
Oides, termination, from the Greek, to denote likeness; so
Dianthoides, Pink-like.
Oleraceous, esculent, as a pot-herb.
Oligos, Greek for few; thus Oliganthous, few-flowered, &c.
Olivaceous, olive-green.
Oophoridium, a name for spore-case containing macrospores.
Opaque, applied to a surface, means dull, not shining.
Operculate, furnished with a lid (Operculum), as the spore-case of
Mosses, 163.
Opposite, said of leaves and branches when on opposite sides of the
stem from each other (i. e. in pairs), 29, 68. Stamens are opposite the
petals, &c., when they stand before them.
Oppositifolius, situated opposite a leaf.
Orbicular, Orbiculate, circular in outline, or nearly so, 52.
Order, group below class, 178. Ordinal names, 180.
Organ, any member of the plant, as a leaf, a stamen, &c.
Organography, study of organs, 9. Organogenesis, that of the
development of organs.
Orgyalis, of the height of a man.
Orthos, Greek for straight; thus, Orthocarpous, with straight fruit;
Orthostichous, straight-ranked.
Orthotropous (ovule or seed), 111.
Osseous, of a bony texture.
Outgrowths, growths from the surface of a leaf, petal, &c.
Oval, broadly elliptical, 52.
Ovary, that part of the pistil containing the ovules or future seeds,
14, 80, 105.
Ovate, shaped like an egg, with the broader end downwards; or, in
plain surfaces, such as leaves, like the section of an egg lengthwise,
52.
Ovoid, ovate or oval in a solid form.
Ovule, the body which is destined to become a seed, 14, 80, 105, 110.
Ovuliferous, ovule-bearing.
Palate, a projection of the lower lip of a labiate corolla into the
throat, as in Snapdragon, &c.
Palea (plural paleæ), chaff; the inner husks of Grasses; the chaff
or bracts on the receptacle of many Compositæ, as Coreopsis, and
Sunflower.
Paleaceous, furnished with chaff, or chaffy in texture.
Paleolate, having Paleolæ or paleæ of a second order, or narrow
paleæ.
Palet, English term for palea.
Palmate, when leaflets or the divisions of a leaf all spread from the
apex of the petiole, like the hand with the outspread fingers, 57, 58.
Palmately (veined, lobed, &c.), in a palmate manner, 51, 56.
Palmatifid, -lobed, -sect, palmately cleft, or lobed, or divided.
Paludose, inhabiting marshes. Palustrine, same.
Panduriform, or Pandurate, fiddle-shaped (which see).
Panicle, an open and branched cluster, 81.
Panicled, Paniculate, arranged in panicles, or like a panicle.
Pannose, covered with a felt of woolly hairs.
Papery, of about the consistence of letter-paper.
Papilionaceous, butterfly-shaped; applied to such a corolla as that of
the Pea, 91.
Papilla (plural papillæ), little nipple-shaped protuberances.
Papillate, Papillose, covered with papillæ.
Pappus, thistle-down. The down crowning the achenium of the Thistle,
Groundsel, &c., and whatever in Compositæ answers to calyx, whether
hairs, teeth, or scales, 121.
Papyraceous, like parchment in texture.
Parallel-veined or nerved (leaves), 50.
[Pg 215]Paraphyses, jointed filaments mixed with the antheridia of Mosses.
Parasitic, living as a parasite, i. e. on another plant or animal, 37.
Parenchemytous, composed of parenchyma.
Parenchyma, soft cellular tissue of plants, like the green pulp of
leaves, 132.
Parietal (placentæ, &c.), attached to the walls (parietes) of the
ovary.
Paripinnate, pinnate with an even number of leaflets.
Parted, separated or cleft into parts almost to the base, 55.
Parthenogenesis, producing seed without fertilization.
Partial involucre, same as an involucel; partial petiole, a
division of a main leaf-stalk or the stalk of a leaflet; partial
peduncle, a branch of a peduncle; partial umbel, an umbellet, 76.
Partition, a segment of a parted leaf; or an internal wall in an
ovary, anther, &c.
Patelliform, disk-shaped, like the patella or kneepan.
Patent, spreading, open. Patulous, moderately spreading.
Pauci-, in composition, few; as pauciflorous, few-flowered, &c.
Pear-shaped, solid obovate, the shape of a pear.
Pectinate, pinnatifid or pinnately divided into narrow and close
divisions, like the teeth of a comb.
Pedate, like a bird's foot; palmate or palmately cleft, with the side
divisions again cleft, as in Viola pedata, &c.
Pedicel, the stalk of each particular flower of a cluster, 73.
Pedicellate, Pedicelled, borne on a pedicel.
Pedalis, Latin for a foot high or long.
Peduncle, a flower-stalk, whether of a single flower or of a
flower-cluster, 73.
Peduncled, Pedunculate, furnished with a peduncle.
Peloria, an abnormal return to regularity and symmetry in an irregular
flower; commonest in Snapdragon.
Peltate, shield-shaped; said of a leaf, whatever its shape, when the
petiole is attached to the lower side, somewhere within the margin, 53.
Pelviform, basin-shaped.
Pendent, hanging. Pendulous, somewhat hanging or drooping.
Penicillate, Penicilliform, tipped with a tuft of fine hairs, like a
painter's pencil; as the stigmas of some Grasses.
Pennate, same as pinnate. Penninerved and Penniveined, pinnately
veined, 51.
Penta- (in words of Greek composition), five; as Pentadelphous, 99;
Pentagynous, with five pistils or styles; Pentamerous, with its
parts in fives, or on the plan of five; Pentandrous, having five
stamens, 112; Pentastichous, in five ranks, &c.
Pepo, a fruit like the Melon and Cucumber, 119.
Perennial, lasting from year to year, 38.
Perfect (flower), having both stamens and pistils, 81.
Perfoliate, passing through the leaf, in appearance, 60.
Perforate, pierced with holes, or with transparent dots resembling
holes, as an Orange-leaf.
Peri-, Greek for around; from which are such terms as
Perianth, the leaves of the flower collectively, 79.
Pericarp, the ripened ovary; the walls of the fruit, 117.
Pericarpic, belonging to the pericarp.
Perigonium, Perigone, same as perianth.
Perigynium, bodies around the pistil; applied to the closed cup or
bottle-shaped body (of bracts) which encloses the ovary of Sedges, and
to the bristles, little scales, &c., of the flowers of some other
Cyperaceæ.
Perigynous, the petals and stamens borne on the calyx, 95, 99.
Peripheric, around the outside, or periphery, of any organ.
Perisperm, a name for the albumen of a seed.
Peristome, the fringe of teeth to the spore-case of Mosses, 163.
Persistent, remaining beyond the period when such parts commonly fall,
as the leaves of evergreens, and the calyx of such flowers as persist
during the growth of the fruit.
[Pg 216]Personate, masked; a bilabiate corolla with a palate in the throat,
92.
Pertuse, perforated with a hole or slit.
Perulate, having scales (Perulæ), such as bud-scales.
Pes, pedis, Latin for the foot or support, whence Longipes,
long-stalked, &c.
Petal, a leaf of the corolla, 14, 79.
Petalody, metamorphosis of stamens, &c., into petals.
Petaloid, Petaline, petal-like; resembling or colored like petals.
Petiole, a footstalk of a leaf; a leaf-stalk, 49.
Petioled, Petiolate, furnished with a petiole.
Petiolulate, said of a leaflet when raised on its own partial
leaf-stalk.
Petræus, Latin for growing on rocks.
Phalanx, phalanges, bundles of stamens.
Phænogamous, or Phanerogamous, plants bearing flowers and producing
seeds; same as Flowering Plants. Phænogams, Phanerogams, 10.
Phlœum, Greek name for bark, whence Endophlœum, inner bark,
&c.
Phœniceous, deep red verging to scarlet.
Phycology, the botany of Algæ.
Phyllocladia, branches assuming the form and function of leaves.
Phyllodium (plural, phyllodia), a leaf where the seeming blade is a
dilated petiole, as in New Holland Acacias, 61.
Phyllome, foliar parts, those answering to leaves in their nature.
Phyllon (plural, phylla), Greek for leaf and leaves; used in many
compound terms and names.
Phyllotaxis, or Phyllotaxy, the arrangement of leaves on the stem,
67.
Physiological Botany, 9.
Phytography, relates to characterizing and describing plants.
Phyton, or Phytomer, a name used to designate the pieces which by
their repetition make up a plant, theoretically, viz. a joint of stem
with its leaf or pair of leaves.
Pileus of a mushroom, 172.
Piliferous, bearing a slender bristle or hair (pilum), or beset with
hairs.
Pilose, hairy; clothed with soft slender hairs.
Pinna, a primary division with its leaflets of a bipinnate or
tripinnate leaf.
Pinnule, a secondary division of a bipinnate or tripinnate leaf, 66.
Pinnate (leaf), when leaflets are arranged along the sides of a common
petiole, 57.
Pinnately lobed, cleft, parted, divided, veined, 56.
Pinnatifid, Pinnatisect, same as pinnately cleft and pinnately
parted, 56.
Pisiform, pea-shaped.
Pistil, the seed-bearing organ of the flower, 14, 80, 105.
Pistillate, having a pistil, 85.
Pistillidium, the body which in Mosses answers to the pistil, 159,
164.
Pitchers, 64.
Pith, the cellular centre of an exogenous stem, 138.
Placenta, the surface or part of the ovary to which the ovules are
attached, 107.
Placentiform, nearly same as quoit-shaped.
Plaited (in the bud), or Plicate, folded, 72, 98.
Platy-, Greek for broad, in compounds, such as Platyphyllous,
broad-leaved, &c.
Pleio-, Greek for full or abounding, used in compounds, such as
Pleiopetalous, of many petals, &c.
Plumbeus, lead-colored.
Plumose, feathery; when any slender body (such as a bristle of a
pappus or a style) is beset with hairs along its sides, like the plume
of a feather.
Plumule, the bud or first shoot of a germinating plantlet above the
cotyledons, 13.
Pluri-, in composition, many or several; as Plurifoliolate, with
several leaflets.
Pod, specially a legume, 122; also may be applied to any sort of
capsule.
Podium, a footstalk or stipe, used only in Greek compounds, as
(suffixed) Leptopodus, slender-stalked, or (prefixed) Podocephalus,
with a stalked head, and in Podosperm, a seed stalk or funiculus.
Pogon, Greek for beard, comes into various compounds.
[Pg 217]Pointless, destitute of any pointed tip, such as a mucro, awn,
acumination, &c.
Pollen, the fertilizing powder contained in the anther, 14, 80, 103.
Pollen-growth, 117. Polleniferous, pollen-bearing.
Pollen-mass, Pollinium, the united mass of pollen, 104, as in
Milkweed and Orchis.
Pollicaris, Latin for an inch long.
Pollination, the application of pollen to the stigma, 114.
Poly-, in compound words of Greek origin, same as multi- in those of
Latin origin viz. many, as
Polyadelphous, stamens united by their filaments into several bundles,
100.
Polyandrous, with numerous stamens (inserted on the receptacle), 100.
Polycarpic, term used by DeCandolle in the sense of perennial.
Polycotyledonous, having many (more than two) cotyledons, as Pines,
23.
Polygamous, having some perfect and some unisexual flowers, 85.
Polygonal, many-angled.
Polygynous, with many pistils or styles, 105.
Polymerous, formed of many parts of each set.
Polymorphous, of several or varying forms.
Polypetalous, when the petals are distinct or separate (whether few or
many), 89.
Polyphyllous, many-leaved; formed of several distinct pieces.
Polysepalous, same as the last when applied to the calyx, 89.
Polyspermous, many-seeded.
Pome, the apple, pear, and similar fleshy fruits, 119.
Pomiferous, pome-bearing.
Porrect, outstretched.
Posterior side or portion of a flower (when axillary) is that toward
the axis, 96.
Pouch, the silicle or short pod, as of Shepherd's Purse, 123.
Præcocious (Latin, præcox), unusually early in development.
Præfloration, same as æstivation, 97.
Præfoliation, same as vernation, 71.
Præmorse, ending abruptly, as if bitten off.
Pratensis, Latin for growing in meadows.
Prickles, sharp elevations of the bark, coming off with it, as of the
Rose.
Prickly, bearing prickles, or sharp projections like them.
Primine, the outer coat of the covering of the ovule, 110.
Primordial, earliest formed; primordial leaves are the first after the
cotyledons.
Prismatic, prism-shaped; having three or more angles bounding flat
sides.
Procerous, tall, or tall and slim.
Process, any projection from the surface or edge of a body.
Procumbent, trailing on the ground, 39.
Procurrent, running through but not projecting.
Produced, extended or projecting; the upper sepal of a Larkspur is
produced above into a spur, 87.
Proliferous (literally, bearing offspring), where a new branch rises
from an older one, or one head or cluster of flowers out of another.
Propaculum or Propagulum, a shoot for propagation.
Prosenchyma, a tissue of wood-cells.
Prostrate, lying flat on the ground, 39.
Protandrous or Proterandrous, the anthers first maturing, 116.
Proteranthous, flowering before leafing.
Proterogynous or Protogynous, the stigmas first to mature, 116.
Prothallium or Prothallus, 160.
Protoplasm, the soft nitrogenous lining or contents, or living part,
of cells, 129.
Protos, Greek for first; in various compounds.
Pruinose, Pruinate, frosted; covered with a powder like hoar-frost.
Pseudo-, Greek for false. Pseudo-bulb, the aerial corms of epiphytic
Orchids, &c.
Psilos, Greek for bare or naked, used in many compounds.
Pteridophyta, Pteridophytes, 156.
Pteris, Greek for wing, and general name for Fern, enters into many
compounds.
[Pg 218]Puberulent, covered with fine and short or almost imperceptible down.
Pubescent, hairy or downy, especially with fine and soft hairs or
pubescence.
Pulverulent or Pulveraceous, as if dusted with fine powder.
Pulvinate, cushioned, or shaped like a cushion.
Pumilus, low or little.
Punctate, dotted, either with minute holes or what look as such.
Puncticulate, minutely punctate.
Pungent, prickly-tipped.
Puniceous, carmine-red.
Purpureus, originally red or crimson, more used for duller or
bluish-red.
Pusillus, weak and small, tiny.
Putamen, the stone of a drupe, or the shell of a nut, 120.
Pygmæus, Latin for dwarf.
Pyramidal, shaped like a pyramid.
Pyrene, Pyrena, a seed-like nutlet or stone of a small drupe.
Pyriform, pear-shaped.
Pyxidate, furnished with a lid.
Pyxis, Pyxidium, a pod opening round horizontally by a lid, 124.
Quadri-, in words of Latin origin, four; as Quadrangular,
four-angled; Quadrifoliate, four-leaved; Quadrifid, four-cleft.
Quaternate in fours.
Quinate, in fives. Quinque, five.
Quincuncial, in a quincunx; when the parts in æstivation are five, two
of them outside, two inside, and one half out and half in.
Quintuple, five-fold.
Race, a marked variety which may be perpetuated from seed, 176.
Raceme, a flower-cluster, with one-flowered pedicels arranged along
the sides of a general peduncle, 73.
Racemose, bearing racemes, or raceme-like.
Rachis, see rhachis.
Radial, belonging to the ray.
Radiate, or Radiant, furnished with ray-flowers, 94.
Radiate-veined, 52.
Radical, belonging to the root, or apparently coming from the root.
Radicant, rooting, taking root on or above the ground.
Radicels, little roots or rootlets.
Radicle, the stem part of the embryo, the lower end of which forms the
root, 11, 127.
Rameal, belonging to a branch. Ramose, full of branches (rami).
Ramentaceous, beset with thin chaffy scales (Ramenta), as the stalks
of many Ferns.
Ramification, branching, 27.
Ramulose, full of branchlets (ramuli).
Raphe, see rhaphe.
Ray, parts diverging from a centre, the marginal flowers of a head (as
of Coreopsis, 94), or cluster, as of Hydrangea (78), when different from
the rest, especially when ligulate and diverging (like rays or
sunbeams); also the branches of an umbel, 74.
Ray-flowers, 94.
Receptacle, the axis or support of a flower, 81, 112; also the common
axis or support of a head of flowers, 73.
Reclined, turned or curved downwards; nearly recumbent.
Rectinerved, with straight nerves or veins.
Recurved, curved outwards or backwards.
Reduplicate (in æstivation), valvate with the margins turned outwards,
97.
Reflexed, bent outwards or backwards.
Refracted, bent suddenly, so as to appear broken at the bend.
Regular, all the parts similar in shape, 82.
Reniform, kidney-shaped, 53.
[Pg 219]Repand, wavy-margined, 55.
Repent, creeping, i. e. prostrate and rooting underneath.
Replum, the frame of some pods (as of Prickly Poppy and Cress),
persistent after the valves fall away.
Reptant, same as repent.
Resupinate, inverted, or appearing as if upside down, or reversed.
Reticulated, the veins forming network, 50. Retiform, in network.
Retinerved, reticulate-veined.
Retroflexed, bent backwards; same as reflexed.
Retuse, blunted; the apex not only obtuse but somewhat indented, 54.
Revolute, rolled backwards, as the margins of many leaves, 72.
Rhachis (the backbone), the axis of a spike or other body, 73.
Rhaphe, the continuation of the seed-stalk along the side of an
anatropous ovule or seed, 112, 126.
Rhaphides, crystals, especially needle-shaped ones, in the tissues of
plants, 137.
Rhizanthous, flowering from the root.
Rhizoma, Rhizome, a rootstock, 42-44.
Rhombic, in the shape of a rhomb. Rhomboidal, approaching that
shape.
Rib, the principal piece, or one of the principal pieces of the
framework of a leaf, or any similar elevated line along a body, 49, 50.
Rimose, having chinks or cracks.
Ring, an elastic band on the spore-cases of Ferns, 159.
Ringent, grinning; gaping open, 92.
Riparious, on river-banks.
Rivalis, Latin for growing along brooks; or Rivularis, in rivulets.
Root, 33.
Root-hairs, 35.
Rootlets, small roots, or root-branches, 33.
Rootstock, root-like trunks or portions of stems on or under ground,
42.
Roridus, dewy.
Rosaceous, arranged like the petals of a rose.
Rostellate, bearing a small beak (Rostellum).
Rostrate, bearing a beak (Rostrum) or a prolonged appendage.
Rosulate, in a rosette or cluster of spreading leaves.
Rotate, wheel-shaped, 89.
Rotund, rounded or roundish in outline.
Ruber, Latin for red in general. Rubescent, Rubicund, reddish or
blushing.
Rudimentary, imperfectly developed, or in an early state of
development.
Rufous, Rufescent, brownish-red or reddish-brown.
Rugose, wrinkled; roughened with wrinkles.
Ruminated (albumen), penetrated with irregular channels or portions,
as a nutmeg, looking as if chewed.
Runcinate, coarsely saw-toothed or cut, the pointed teeth turned
towards the base of the leaf, as the leaf of a Dandelion.
Runner, a slender and prostrate branch, rooting at the end, or at the
joints, 40.
Sabulose, growing in sand.
Sac, any closed membrane, or a deep purse-shaped cavity.
Saccate, sac-shaped.
Sagittate, arrowhead-shaped, 53.
Salsuginous, growing in brackish soil.
Salver-shaped, or Salver-form, with a border spreading at right
angles to a slender tube, 89.
Samara, a wing-fruit, or key, 122.
Samaroid, like a samara or key-fruit.
Sap, the juices of plants generally, 136. Sapwood, 142.
Saprophytes, 37.
Sarcocarp, the fleshy part of a stone-fruit, 120.
[Pg 220]Sarmentaceous, Sarmentose, bearing long and flexible twigs
(Sarments), either spreading or procumbent.
Saw-toothed, see serrate, 55.
Scabrous, rough or harsh to the touch.
Scalariform, with cross-bands, resembling the steps of a ladder, 134.
Scales, of buds, 28; of bulbs, &c., 46.
Scalloped, same as crenate, 55.
Scaly, furnished with scales, or scale-like in texture.
Scandent, climbing, 39.
Scape, a peduncle rising from the ground or near it, as in many
Violets.
Scapiform, scape-like.
Scapigerous, scape-bearing.
Scar of the seed, 126. Leaf-scars, 27, 28.
Scarious or Scariose, thin, dry, and membranous.
Scion, a shoot or slip used for grafting.
Scleros, Greek for hard, hence Sclerocarpous, hard-fruited.
Scobiform, resembling sawdust.
Scorpioid or Scorpioidal, curved or circinate at the end, 77.
Scrobiculate, pitted; excavated into shallow pits.
Scurf, Scurfiness, minute scales on the surface of many leaves, as
of Goosefoot.
Scutate, Scutiform, buckler-shaped.
Scutellate, or Scutelliform, saucer-shaped or platter-shaped.
Secund, one-sided; i. e. where flowers, leaves, &c., are all turned to
one side.
Secundine, the inner coat of the ovule, 110.
Seed, 125. Seed-leaves, see cotyledons. Seed-vessel, 127.
Segment, a subdivision or lobe of any cleft body.
Segregate, separated from each other.
Semi-, in compound words of Latin origin, half; as
Semi-adherent, as the calyx or ovary of Purslane; Semicordate,
half-heart-shaped; Semilunar, like a half-moon; Semiovate,
half-ovate, &c.
Seminal, relating to the seed (Semen). Seminiferous, seed-bearing.
Sempervirent, evergreen.
Sensitiveness in plants, 149, 152.
Senary, in sixes.
Sepal, a leaf or division of the calyx, 14, 79.
Sepaloid, sepal-like. Sepaline, relating to the sepals.
Separated Flowers, those having stamens or pistils only, 85.
Septate, divided by partitions.
Septenate, with parts in sevens.
Septicidal, where dehiscence is through the partitions, 123.
Septiferous, bearing the partition.
Septifragal, where the valves in dehiscence break away from the
partitions, 123.
Septum (plural septa), a partition or dissepiment.
Serial, or Seriate, in rows; as biserial, in two rows, &c.
Sericeous, silky; clothed with satiny pubescence.
Serotinous, late in the season.
Serrate, the margin cut into teeth (Serratures) pointing forwards,
55.
Serrulate, same as the last, but with fine teeth.
Sessile, sitting; without any stalk.
Sesqui-, Latin for one and a half; so Sesquipedalis, a foot and a
half long.
Seta, a bristle, or a slender body or appendage resembling a bristle.
Setaceous, bristle-like. Setiform, bristle-shaped.
Setigerous, bearing bristles. Setose, beset with bristles or bristly
hairs.
Setula, a diminutive bristle. Setulose, provided with such.
Sex, six. Sexangular, six-angled. Sexfarious, six-faced.
Sheath, the base of such leaves as those of Grasses, which are
Sheathing, wrapped round the stem.
Shield-shaped, same as scutate, or as peltate, 53.
[Pg 221]Shrub, Shrubby, 39.
Sieve-cells, 140.
Sigmoid, curved in two directions, like the letter S, or the Greek
sigma.
Silicle, a pouch, or short pod of the Cress Family, 123.
Siliculose, bearing a silicle, or a fruit resembling it.
Silique, capsule of the Cress Family, 123.
Siliquose, bearing siliques or pods which resemble siliques.
Silky, glossy with a coat of fine and soft, close-pressed, straight
hairs.
Silver-grain, the medullary rays of wood, 139.
Silvery, shining white or bluish-gray, usually from a silky
pubescence.
Simple, of one piece; opposed to compound.
Sinistrorse, turned to the left.
Sinuate, with margin alternately bowed inwards and outwards, 55.
Sinus, a recess or bay; the re-entering angle between two lobes or
projections.
Sleep of Plants (so called), 151.
Smooth, properly speaking not rough, but often used for glabrous, i.
e. not pubescent.
Soboliferous, bearing shoots (Soboles) from near the ground.
Solitary, single; not associated with others.
Sordid, dull or dirty in hue.
Sorediate, bearing patches on the surface.
Sorosis, name of a multiple fruit, like a pine-apple.
Sorus, a fruit-dot of Ferns, 159.
Spadiceous, chestnut-colored. Also spadix-bearing.
Spadix, a fleshy spike of flowers, 75.
Span, the distance between the tip of the thumb and of little finger
outstretched, six or seven inches.
Spathaceous, resembling or furnished with a
Spathe, a bract which inwraps an inflorescence, 75.
Spatulate, or Spathulate, shaped like a spatula, 52.
Species, 175.
Specific Names, 179.
Specimens, 184.
Spermaphore, or Spermophore, one of the names of the placenta.
Spermum, Latin form of Greek word for seed; much used in composition.
Spica, Latin for spike; hence Spicate, in a spike, Spiciform, in
shape resembling a spike.
Spike, an inflorescence like a raceme, only the flowers are sessile,
74.
Spikelet, a small or a secondary spike; the inflorescence of Grasses.
Spine, 41, 64.
Spindle-shaped, tapering to each end, like a radish, 36.
Spinescent, tipped by or degenerating into a thorn.
Spinose, or Spiniferous, thorny.
Spiral Vessels or ducts, 135.
Spithameous, span-high.
Spora, Greek name for seed, used in compound words.
Sporadic, widely dispersed.
Sporangium, a spore-case in Ferns, &c., 158.
Spore, a body resulting from the fructification of Cryptogamous
plants, in them the analogue of a seed.
Spore-case (Sporangium), 158.
Sporocarp, 162.
Sport, a newly appeared variation, 176.
Sporule, same as a spore, or a small spore.
Spumescent, appearing like froth.
Spur, any projecting appendage of the flower, looking like a spur but
hollow, as that of Larkspur, fig. 239.
Squamate, Squamose, or Squamaceous, furnished with scales
(squamæ).
[Pg 222]Squamellate, or Squamulose, furnished with little scales
(Squamellæ, or Squamulæ).
Squamiform, shaped like a scale.
Squarrose, where scales, leaves, or any appendages spread widely from
the axis on which they are thickly set.
Squarrulose, diminutive of squarrose; slightly squarrose.
Stachys, Greek for spike.
Stalk, the stem, petiole, peduncle, &c., as the case may be.
Stamen, 14, 80, 98.
Staminate, furnished with stamens, 86. Stamineal, relating to the
stamens.
Staminodium, an abortive stamen, or other body in place of a stamem.
Standard, the upper petal of a papilionaceous corolla, 92.
Starch, 136, 163.
Station, the particular kind of situation in which a plant naturally
occurs.
Stellate, Stellular, starry or star-like; where several similar
parts spread out from a common centre, like a star.
Stem, 39. Stemlet, diminutive stem.
Stemless, destitute or apparently destitute of stem.
Stenos, Greek for narrow; hence Stenophyllous, narrow-leaved, &c.
Sterile, barren or imperfect.
Stigma, the part of the pistil which receives the pollen, 14, 80, 105.
Stigmatic, or Stigmatose, belonging to the stigma.
Stipe (Latin Stipes), the stalk of a pistil, &c., when it has any,
112; also of a Fern, 158, and of a Mushroom, 172.
Stipel, a stipule of a leaflet, as of the Bean, &c.
Stipellate, furnished with stipels, as in the Bean tribe.
Stipitate, furnished with a stipe.
Stipulaceous, belonging to stipules. Stipulate, furnished with
stipules.
Stipules, the appendages on each side of the base of certain leaves,
66.
Stirps (plural, stirpes), Latin for race.
Stock, used for race or source. Also for any root-like base from which
the herb grows up.
Stole, or Stolon, a trailing or reclined and rooting shoot, 40.
Stoloniferous, producing stolons.
Stomate (Latin Stoma, plural Stomata), the breathing-pores of
leaves, 144.
Stone-fruit, 119.
Storage-leaves, 62.
Stramineous, straw-like, or straw-colored.
Strap-shaped, long, flat, and narrow.
Striate, or Striated, marked with slender longitudinal grooves or
stripes.
Strict, close and narrow; straight and narrow.
Strigillose, Strigose, beset with stout and appressed, stiff or
rigid bristles.
Strobilaceous, relating to or resembling a
Strobile, a multiple fruit in the form of a cone or head, 124.
Strombuliform, twisted, like a spiral shell.
Strophiole, same as caruncle, 126. Strophiolate, furnished with a
strophiole.
Struma, a wen; a swelling or protuberance of any organ.
Strumose, bearing a struma.
Stupose, like tow.
Style, a stalk between ovary and stigma, 14, 80, 105.
Styliferous, Stylose, bearing styles or conspicuous ones.
Stylopodium, an epigynous disk, or an enlargement at the base of the
style.
Sub-, as a prefix, about, nearly, somewhat; as Subcordate, slightly
cordate; Subserrate, slightly serrate; Subaxillary, just beneath the
axil, &c.
Subclass, Suborder, Subtribe, 178.
Suberose, corky or cork-like in texture.
Subulate, awl-shaped; tapering from a broadish or thickish base to a
sharp point.
Succise, as if cut off at lower end.
Succubous, when crowded leaves are each covered by base of next above.
[Pg 223]Suckers, shoots from subterranean branches, 39.
Suffrutescent, slightly shrubby or woody at the base only, 39.
Suffruticose, rather more than suffrutescent, 37, 39.
Sulcate, grooved longitudinally with deep furrows.
Superior, above, 96; sometimes equivalent to posterior, 96.
Supernumerary Buds, 30, 31.
Supervolute, plaited and convolute in bud, 97.
Supine, lying flat, with face upward.
Supra-axillary, borne above the axil, as some buds, 31.
Supra-decompound, many times compounded or divided.
Surculose, producing suckers (Surculi) or shoots resembling them.
Suspended, hanging down. Suspended ovules or seeds hang from the very
summit of the cell which contains them.
Sutural, belonging or relating to a suture.
Suture, the line of junction of contiguous parts grown together, 106.
Sword-shaped, applied to narrow leaves, with acute parallel edges,
tapering above.
Syconium, the fig-fruit, 124.
Sylvestrine, growing in woods.
Symmetrical Flower, similar in the number of parts of each set, 82.
Sympetalous, same as gamopetalous.
Sympode, Sympodium, a stem composed of a series of superposed
branches in such a way as to imitate a simple axis, as in Grape-vine.
Synantherous or Syngenesious, where stamens are united by their
anthers, 100.
Syncarpous (fruit or pistil), composed of several carpels consolidated
into one.
Synonym, an equivalent superseded name.
Synsepalous, same as gamosepalous.
System (artificial and natural), 182, 183.
Systematic Botany, the study of plants after their kinds, 9.
Tabescent, wasting or shrivelling.
Tail, any long and slender prolongation of an organ.
Taper-pointed, same as acuminate, 54.
Tap-root, a root with a stout tapering body, 32-35.
Tawny, dull yellowish, with a tinge of brown.
Taxonomy, the part of botany which treats of classification.
Tegmen, a name for the inner seed-coat.
Tendril, a thread-shaped organ used for climbing, 40.
Terete, long and round; same as cylindrical, only it may taper.
Terminal, borne at, or belonging to, the extremity or summit.
Terminology treats of technical terms; same as Glossology, 181.
Ternate, Ternately, in threes.
Tessellate, in checker-work.
Testa, the outer (and usually the harder) coat or shell of the seed,
125.
Testaceous, the color of unglazed pottery.
Tetra- (in words of Greek composition), four; as, Tetracoccous, of
four cocci.
Tetradynamous, where a flower has six stamens, two shorter than the
four, 101.
Tetragonal, four-angled. Tetragynous, with four pistils or styles.
Tetramerous, with its parts or sets in fours. Tetrandrous, with four
stamens, 100.
Tetraspore, a quadruple spore, 169.
Thalamaflorous, with petals and stamens inserted on the torus or
Thalamus.
Thallophyta, Thallophytes, 165.
Thallus, a stratum, in place of stem and leaves, 165.
Theca, a case; the cells or lobes of the anther.
Thecaphore, the stipe of a carpel, 113.
Thorn, an indurated pointed branch, 41, 42.
Thread-shaped, slender and round or roundish, like a thread.
Throat, the opening or gorge of a monopetalous corolla, &c., where the
border and the tube join, and a little below, 89.
[Pg 224]Thyrse or Thyrsus, a compact and pyramidal panicle of cymes or
cymules, 79.
Tomentose, clothed with matted woolly hairs (tomentum).
Tongue-shaped, long and flat, but thickish and blunt.
Toothed, furnished with teeth or short projections of any sort on the
margin; used especially when these are sharp, like saw-teeth, and do not
point forwards, 55.
Top-shaped, shaped like a top, or a cone with apex downwards.
Torose, Torulose, knobby; where a cylindrical body is swollen at
intervals.
Torus, the receptacle of the flower, 81, 112.
Trachea, a spiral duct.
Trachys, Greek for rough; used in compounds, as, Trachyspermous,
rough-seeded.
Transverse, across, standing right and left instead of fore and aft.
Tri- (in composition), three; as,
Triadelphous, stamens united by their filaments into three bundles,
99.
Triandrous, where the flower has three stamens, 112.
Tribe, 178.
Trichome, of the nature of hair or pubescence.
Trichotomous, three-forked.
Tricoccous, of three cocci or roundish carpels.
Tricolor, having three colors.
Tricostate, having three ribs.
Tricuspidate, three-pointed.
Tridentate, three-toothed.
Triennial, lasting for three years.
Trifarious, in three vertical rows; looking three ways.
Trifid, three-cleft, 56.
Trifoliate, three-leaved. Trifoliolate, of three leaflets.
Trifurcate, three-forked.
Trigonous, three-angled, or triangular.
Trigynous, with three pistils or styles, 116.
Trijugate, in three pairs (jugi).
Trilobed or Trilobate, three-lobed, 55.
Trilocular, three-celled, as the pistils or pods in fig. 328-330.
Trimerous, with its parts in threes.
Trimorphism, 117. Trimorphic or Trimorphous, in three forms.
Trinervate, three-nerved, or with three slender ribs.
Triœcious, where there are three sorts of flowers on the same or
different individuals, as in Red Maple. A form of Polygamous.
Tripartible, separable into three pieces. Tripartite, three-parted,
55.
Tripetalous, having three petals.
Triphyllous, three-leaved; composed of three pieces.
Tripinnate, thrice pinnate, 59. Tripinnatifid, thrice pinnately
cleft, 57.
Triple-ribbed, Triple-nerved, &c., where a midrib branches into
three, near the base of the leaf.
Triquetrous, sharply three-angled; and especially with the sides
concave, like a bayonet.
Triserial, or Triseriate, in three rows, under each other.
Tristichous, in three longitudinal or perpendicular ranks.
Tristigmatic, or Tristigmatose, having three stigmas.
Trisulcate, three-grooved.
Triternate, three times ternate, 59.
Trivial Name, the specific name.
Trochlear, pulley-shaped.
Trumpet-shaped, tubular; enlarged at or towards the summit.
Truncate, as if cut off at the top.
Trunk, the main stem or general body of a stem or tree.
Tube (of corolla, &c.), 89.
Tuber, a thickened portion of a subterranean stem or branch, provided
with eyes (buds) on the sides, 44.
Tubercle, a small excrescence.
Tubercled, or Tuberculate, bearing excrescences or pimples.
Tubæform, trumpet-shaped.
Tuberous, resembling a tuber. Tuberiferous, bearing tubers.
Tubular, hollow and of an elongated form; hollowed like a pipe, 91.
[Pg 225]Tubuliflorous, bearing only tubular flowers.
Tunicate, coated; invested with layers, as an onion, 46.
Turbinate, top-shaped.
Turio (plural turiones), strong young shoots or suckers springing
out of the ground; as Asparagus-shoots.
Turnip-shaped, broader than high, abruptly narrowed below, 35.
Twining, ascending by coiling round a support, 39.
Type, the ideal pattern, 10.
Typical, well exemplifying the characteristics of a species, genus,
&c.
Uliginose, growing in swamps.
Umbel, the umbrella-like form of inflorescence, 74.
Umbellate, in umbels. Umbelliferous, bearing umbels.
Umbellet (umbellula), a secondary or partial umbel, 76.
Umbilicate, depressed in the centre, like the ends of an apple; with a
navel.
Umbonate, bossed; furnished with a low, rounded projection like a boss
(umbo).
Umbraculiform, umbrella-shaped.
Unarmed, destitute of spines, prickles, and the like.
Uncial, an inch (uncia) in length.
Uncinate, or Uncate, hook-shaped; hooked over at the end.
Under-shrub, partially shrubby, or a very low shrub.
Undulate or Undate, wavy, or wavy-margined, 55.
Unequally pinnate, pinnate with an odd number of leaflets, 65.
Unguiculate, furnished with a claw (unguis), 91.
Uni-, in compound words, one; as Unicellular, one-celled.
Uniflorous, one-flowered.
Unifoliate, one-leaved. Unifoliolate, of one leaflet, 59.
Unijugate, of one pair.
Unilabiate, one-lipped.
Unilateral, one-sided.
Unilocular, one-celled.
Uniovulate, having only one ovule.
Uniserial, in one horizontal row.
Unisexual, having stamens or pistils only, 85.
Univalved, a pod of only one piece after dehiscence.
Unsymmetrical Flowers, 86.
Urceolate, urn-shaped.
Utricle, a small thin-walled, one-seeded fruit, as of Goosefoot, 121.
Utricular, like a small bladder.
Vaginate, sheathed, surrounded by a sheath (vagina).
Valve, one of the pieces (or doors) into which a dehiscent pod, or any
similar body, splits, 122, 123.
Valvate, Valvular, opening by valves. Valvate, in æstivation, 97.
Variety, 176.
Vascular, containing vessels, or consisting of vessels or ducts, 134.
Vascular Cryptogams, 156.
Vaulted, arched; same as fornicate.
Vegetable Life, &c., 128. Vegetable anatomy, 129.
Veins, the small ribs or branches of the framework of leaves, &c., 49,
50.
Veined, Veiny, furnished with evident veins. Veinless, destitute
of veins.
Veinlets, the smaller ramifications of veins, 50.
Velate, furnished with a veil.
Velutinous, velvety to the touch.
Venation, the veining of leaves, &c., 50.
Venenate, poisonous.
Venose, veiny; furnished with conspicuous veins.
Ventral, belonging to that side of a simple pistil, or other organ,
which looks towards the axis or centre of the flower; the opposite of
dorsal; as the
Ventral Suture, 106.
Ventricose, inflated or swelled out on one side.
[Pg 226]Venulose, furnished with veinlets.
Vermicular, worm-like, shaped like worms.
Vernal, belonging to spring.
Vernation, the arrangement of the leaves in the bud, 71.
Vernicose, the surface appearing as if varnished.
Verrucose, warty; beset with little projections like warts.
Versatile, attached by one point, so that it may swing to and fro,
101.
Vertex, same as apex.
Vertical, upright, perpendicular to the horizon, lengthwise.
Verticil, a whorl, 68. Verticillate, whorled, 68.
Verticillaster, a false whorl, formed of a pair of opposite cymes.
Vesicular, bladdery.
Vespertine, appearing or expanding at evening.
Vessels, ducts, &c., 134.
Vexillary, Vexillar, relating to the
Vexillum, the standard of a papilionaceous flower, 92.
Villose, shaggy with long and soft hairs (Villosity).
Vimineous, producing slender twigs, such as those used for
wicker-work.
Vine, in the American use, any trailing or climbing stem; as a
Grape-vine.
Virescent, Viridescent, greenish; turning green.
Virgate, wand-shape; as a long, straight, and slender twig.
Viscous, Viscid, having a glutinous surface.
Vitta (plural vittæ), the oil-tubes of the fruit of Umbelliferæ.
Vitelline, yellow, of the hue of yolk of egg.
Viviparous, sprouting or germinating while attached to the parent
plant.
Voluble, twining; as the stem of Hops and Beans, 39.
Volute, rolled up in any way.
Wavy, the surface or margin alternately convex and concave, 55.
Waxy, resembling beeswax in texture or appearance.
Wedge-shaped, broad above, tapering by straight lines to a narrow
base, 53.
Wheel-shaped, 89.
Whorl, an arrangement of leaves, &c., in circles around the stem.
Whorled, arranged in whorls, 68.
Wing, any membranous expansion. Wings of papilionaceous flowers, 92.
Winged, furnished with a wing; as the fruit of Ash and Elm, fig. 300,
301.
Wood, 133, 142. Woody, of the texture or consisting of wood.
Woody Fibre, or Wood-Cells, 134.
Woolly, clothed with long and entangled soft hairs.
Work in plants, 149, 155.
Xanthos, Greek for yellow, used in compounds; as Xanthocarpus,
yellow-fruited.
Zygomorphous, said of a flower which can be bisected only in one plane
into similar halves.
Transcriber's Notes.
Spelling variants where it wasn't possible to determine the author's
intent were left as is. These include: "backbone" and "back-bone;"
"Buttonwood" and "Button-wood;" "cross section" and "cross-section;"
"footstalk" and "foot-stalk;" "network" and "net-work;" "Pæony" and
"Peony;" "peapod" and "pea-pod," plus plurals; "penniveined" and
"penni-veined," plus capitalized versions; "Sapwood" and "Sap-wood;"
"Snowball" and "Snow-ball;" "Verticil" and "Verticel;" "Woodsorrel" and
"Wood-sorrel."
Changed "Venation" to "Vernation" on page vi: "Vernation or
Præfoliation."
Changed "Isoëtes" to "Isoetes" on page viii: "Quillworts (Isoetes)."
Changed "liquifies" to "liquefies" on page 21: "as it liquefies."
Changed "frame-work" to "framework" on page 52: "the framework or
skeleton."
Changed "leafstalk" to "leaf-stalk" in three places: the caption to
figure 74; on page 71: "bent upon the leaf-stalk;" in the index entry
for "Petiolulate."
Changed "Honey Locust" to "Honey-Locust" in the caption to figure 95:
"branching thorn of Honey-Locust."
Page 47 has a reference to Fig. 89, which does not seem to follow
from the text. "Fig. 80" would make more sense, but the original was not
changed.
Changed "Linnaeus" to "Linnæus" on page 50: "Linnæus called Nerves."
Removed extra comma before closing parenthesis on page 51:
"the Maple (Fig. 20, 24)."
Changed "Sugar-Maple" to "Sugar Maple" on page 58: "that of the Sugar
Maple."
Changed "quadrifoliate" to "quadrifoliolate" on page 59: "trifoliolate,
quadrifoliolate, plurifoliolate."
The reference to Fig. 167 on page 64 is probably intended to refer to
Fig. 157, but was not changed.
The caption to figure 186 says that this is a two-ranked arrangement.
Although it's clear from the text and figure that this is actually a
three-ranked arrangement, the original text was preserved.
Changed colon to semi-colon in the caption to figure 219: "a, filament;
b, anther."
Changed comma to period after "Fig. 254" in its caption.
Changed "Funnel-form" to "Funnelform" on page 90: "Funnel-shaped, or
Funnelform."
Inserted closing parenthesis in caption to figure 304: "Of a Sage
(Salvia Texana)."
Changed "Butter cup" to "Buttercup" on page 120: "fruit of the
Buttercup."
Changed "carpophorse" to "carpophore" on page 121: "a slender axis or
carpophore."
Changed "cocoanut" to "cocoa-nut" on page 122: "such as the cocoa-nut."
Changed "Sepifragal" to "Septifragal" on page 123: "Septifragal
dehiscence."
Inserted missing text "(Fig." on page 128: "Rice (Fig. 430a)."
Perhaps the reference to unit 408 on page 136 should be to unit 402;
but it was not changed.
Changed "White-Pine" to "White Pine" on page 148: "White Pine and
Basswood."
Changed "discovary" to "discovery" on page 160: "discovery of a wholly
unsuspected kind."
Changed "sporo-carps" to "sporocarps" in the caption to figure 520.
Changed semi-colon to period after "microspores" in the caption to
figure 522.
Changed "Lepreiurei" to "Leprieurei" in the caption to figure 561.
Changed "Sun-flowers" to "Sunflowers" on page 187: "Sunflowers and
Thistles."
Changed "♁" to "♂" on page 189: "To indicate sexes, ♂
means staminate."
Preserved the order of the index entries, even though they aren't
strictly alphabetical.
Removed extra comma after "Amphitropous" in its index entry.
The index references for "Anemophilous" and "Entomophilous" should
probably be 115, rather than 113, but were not changed.
The index reference for "Antheridium" should probably be 159, rather
than 150, but was not changed.
Changed "short-fluited" to "short-fruited" in the index entry for
"Brachy-."
The index references for "Caulescent" and "Cauline" should probably be
38, rather than 36, but were not changed.
The index reference for "Cladophylla" should probably be 61, rather
than 64, but was not changed.
The index reference for "Dotted Ducts" should probably be 134, rather
than 148, but was not changed.
Changed "trifoliate" to "trifoliolate" in the index entry for
"Foliolate."
Changed "6" to "13" in the index entry for "Gemmule."
The index references for "Hemitropous" and "Heterotropous" should
probably be 111, rather than 123, but were not changed.
The index reference for "Hortus Siccus" was changed from 201 to 186.
The index reference for "Incised" should probably be 55, rather
than 58, but was not changed.
The index reference for "Jointed" is 64, and was not changed even though
it does not seem to make sense. Better candidates include 57 and 122.
Changed the last index reference for "Ligule" from 57 to 67;
also the reference for "Ocreate."
The index references for "Laticiferous" and "Milk-vessels" were changed
from 138 to 135.
Changed comma to semi-colon after "86" in the index entry for "Naked."
The index reference for "Panicle" should probably be 76, rather
than 81, but was not changed.
The index references for "Pentandrous" and "Triandrous" should probably
be 100, rather than 112, but were not changed.
Changed "leaf stalk" to "leaf-stalk" in its index entry, as well as the
index entry for "Petiole."
The index reference for "Pinnule" should probably be 59, rather
than 66, but was not changed.
The index reference for "Seed-vessel" should probably be 117, rather
than 127, but was not changed.
The second index reference for "Starch" is 163, and was not changed even
though it does not seem to make sense.
Changed "one" to "on" in the index entry for "Stipules": "appendages on
each side."
The index reference to page 37 for "Suffruticose" doesn't seem to make
sense, but was left as is.
The index reference for "Trigynous" should probably be 105, rather
than 116, but was not changed.
The index reference for "Unequally Pinnate" should probably be 58,
rather than 65, but was not changed.
End of the Project Gutenberg EBook of The Elements of Botany, by Asa Gray
*** END OF THIS PROJECT GUTENBERG EBOOK THE ELEMENTS OF BOTANY ***
***** This file should be named 33757-h.htm or 33757-h.zip *****
This and all associated files of various formats will be found in:
https://www.gutenberg.org/3/3/7/5/33757/
Produced by Curtis Weyant, Stephen H. Sentoff and the
Online Distributed Proofreading Team at https://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
https://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at https://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
https://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at https://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit https://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including including checks, online payments and credit card
donations. To donate, please visit: https://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
https://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.
![[Symbol like numeral 5 without top bar]](images/figsymbols.jpg){kind=link}