
The Project Gutenberg EBook of Studies in Spermatogenesis (Part 1 of 2), by Nettie Maria Stevens This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org Title: Studies in Spermatogenesis (Part 1 of 2) Author: Nettie Maria Stevens Release Date: March 7, 2010 [EBook #31545] Language: English Character set encoding: ISO-8859-1 *** START OF THIS PROJECT GUTENBERG EBOOK STUDIES IN SPERMATOGENESIS (1 OF 2) *** Produced by Bryan Ness, Josephine Paolucci and the Online Distributed Proofreading Team at https://www.pgdp.net. (This file was produced from images generously made available by The Internet Archive/American Libraries.)
WASHINGTON, D. C.:
Published by the Carnegie Institution of Washington
September, 1905.
CARNEGIE INSTITUTION OF WASHINGTON
PUBLICATION No. 36
FROM THE PRESS OF
THE HENRY E. WILKENS PRINTING CO.
WASHINGTON, D. C.
In connection with the problem of sex determination it has seemed necessary to investigate further the so-called "accessory chromosome," which, according to McClung ('02), may be a sex determinant. This view has been supported by Sutton ('02) in his work on Brachystola magna, but rejected by Miss Wallace ('05) for the spider.
The forms selected for study have been taken from several groups of insects, and are all species whose spermatogenesis has not been previously worked out. They are (1) a California termite, Termopsis angusticollis; (2) a California sand-cricket, Stenopelmatus; (3) the croton-bug, Blattella germanica; (4) the common meal-worm, Tenebrio molitor; and (5) one of the aphids, Aphis œnotheræ.
A brief account of a chromatin element resembling the accessory chromosome in Sagitta has been added for comparison. The spermatogenesis of each form will be described in detail, and a general discussion of the results and their relation to the accessory chromosome and sex determination will follow. The spermatogenesis of the aphid has been included in another paper, but a summary of results and a few figures will be given here for reference in the general discussion.
The testes were fixed in various fluids—Flemming's strong solution, Hermann's platino-aceto-osmic, Gilson's mercuro-nitric, Lenhossek's alcoholic sublimate acetic, and corrosive acetic. Flemming's and Hermann's fluids followed by safranin gave good results in most cases. The mercuro-nitric solution and Lenhossek's fluid gave excellent fixation and were preferable to the osmic mixtures when it was desirable to stain the same material with iron-hæmatoxylin, and also with various anilin stains.
Heidenhain's iron-hæmatoxylin, either alone or with orange G or erythrosin, was used more than any other one stain. With osmic fixation safranin gave better results in some cases, because of the[Pg 4] abundance of spindle fibers and sphere substance which were stained by hæmatoxylin. The safranin-gentian combination used by Miss Wallace and others in the study of the accessory chromosome did not prove to be especially helpful with these forms. Thionin was found to be a very useful stain for distinguishing between the accessory chromosome and an ordinary nucleolus. Licht-grün was often used in combination with safranin.
In the termite it was not found to be practicable to dissect out the testes. The tip of the abdomen was therefore fixed and sectioned, young males whose wings were just apparent being used. The cells are all small, and could not be studied to advantage with less than 1500 magnification (Zeiss oil immersion 2 mm., oc. 12).
In the spermatogonium there is a very large nucleolus (plate I, fig. 1), which in the iron-hæmatoxylin preparations is very conspicuous, but does not stain like chromatin with thionin or other anilin stains, nor does it behave like an accessory chromosome during the maturation mitoses. Before each spermatogonial division it divides as in figures 2 and 3, and the same is true for each maturation mitosis. Figure 4 shows the 52 chromosomes of a spermatogonial division in metaphase. Figures 5 and 6 are young spermatocytes, showing the division of the nucleolus. Figures 8, 9, and 10 show a stage immediately following that shown in figure 6 and evidently persisting for some time. The spireme thread is very fine, stains deeply, and is wound into a dense ball, often concealing one (fig. 10) or both nucleoli (fig. 8). Figure 11 shows the next stage; the bivalent chromosomes are so disposed as to give the familiar "bouquet stage," with the loops directed away from the centrosome and sphere (c). Figures 12, 13, and 14 show the later development of the same stage, the chromatin loops becoming thicker by the concentration of the smaller granules to form the larger ones seen in figure 14. The loops now straighten out and extend in various directions across the nuclear space (figs. 15, 16, 17). In fig. 18a a longitudinal split is seen in several chromosomes. Figures 18b, 19, 20, and 21 show various stages in the contraction of these split bivalent chromosomes to form diamond-shaped tetrads, each side of which is a univalent daughter chromosome. The tetrads come into the spindle in this form (figs. 22, 23), and change to the form shown in figure 24 during the metaphase (figs. 22, 26, 28). Figures 25 and 27 show the 26 bivalent chromosomes, or tetrads, in[Pg 5] early and late metaphase, respectively, and figures 29, 30, and 31 in anaphase. This is certainly a reduction division, for the tetrads are always somewhat elongated and come into the spindle with their longer axes parallel with the axis of the spindle. The aberrant bodies in these figures are probably remains of the nucleoli; they are found only in iron-hæmatoxylin preparations. Figures 31 and 32 show exceptional cases where the cell has divided. Usually the two daughter nuclei are formed in an undivided cell. The resting-stage between the two divisions is only partial. The nucleolus appears and divides into two (figs. 33-36), and the chromosomes change into the dyad form (fig. 36), in which they come into the second maturation spindle (figs. 37, 38). The equatorial plate again shows 26 chromosomes (fig. 39). The formation of the spermatozoa is peculiar in that the original spermatocyte cell-body, as a rule, does not divide; but the four nuclei resulting from the two maturation divisions develop into sperm-heads in one cell. All have a nucleolus (fig. 41), and in a slightly later stage (fig. 42) the elongated nuclei have a distinct centrosome and sphere at the posterior end. Later stages are shown in figures 43, 44, and 45.
The points of greatest interest in the spermatogenesis of Termopsis angusticollis are, (1) the fact that no accessory chromosome is present; (2) that the method of tetrad formation and reduction are clear, despite the fact that the cells and the chromatin elements are quite small; and (3) the failure of the cell-bodies to divide and the consequent development of four spermatozoa in one cell.
The spermatogonium of Stenopelmatus contains from one to three large nucleoli, which stain much less with thionin than does the spireme (plate II, figs. 46, 47, 48). As the distinct chromosomes come into view in the prophase of mitosis, two are seen to be nearly twice as long as the others, but of equal length (figs. 48, 49, 50.) There are 46 chromosomes in the equatorial plate of a spermatogonial spindle (fig. 50). Besides the nucleolus (n), there appears in the young spermatocyte a conspicuous element (x) which stains deeply with all chromatin stains (fig. 51). It is closely applied to the nuclear membrane and is connected with an end of the spireme (figs. 51-54). At first it is quite small, and it gradually increases in size during the spireme stage. There is no "bouquet stage" in this form. Figure 55 shows the spireme segmented and split longitudinally. The segments have begun to open out at the center to give the cross which is the typical tetrad form in Stenopelmatus. Figures 56, 58, 59, and 60 show various stages in the contraction of the split segments to form crosses and[Pg 6] diamond-shaped rings. The tetrads usually remain connected by delicate linin threads, as shown in figures 57 and 60, also in figures 62 and 63, the latter taken from the metaphase of the first maturation spindle. If these linin connections persist, as they appear to do, from the segmentation of the spireme to metakinesis, the first division of the contracted tetrads must be longitudinal, corresponding to the split in the segments of figures 55, 57, 58, etc. The chromosomes in the metaphase usually appear as dumbbells (fig. 66) or elongated crosses (fig. 67), but occasionally one can be found which still shows its tetrad nature (fig. 64), so clearly indicated in the quadrivalent crosses of figure 59. In the anaphase the chromosomes are often split as in figure 68, and occasionally the two components can be seen as plainly as in figure 65. Figure 61 shows the various shapes assumed by the element x during the tetrad-stage of the chromosomes. This element x almost invariably appears in a vesicle near one pole of the spindle (figs. 67, 68); in exceptional cases it is found nearer the equatorial plate, as in figure 66, or even in the same plane with the ordinary chromosomes, but always somewhat isolated from them. In position and form this element resembles the accessory chromosomes described by Baumgartner ('04) for Gryllus domesticus; in its mode of origin it seems to differ from the other accessory chromosomes yet described.
Figures 69 and 70 show the 23 bivalent chromosomes in metaphase; in figure 69 the element x is shown partly behind the large chromosome and at a different level. In figures 66 and 67 the one exceptionally large chromosome doubtless represents the two larger ones of the spermatogonia. In the anaphase the element x is sometimes as conspicuous as in figure 71; in other cases it is concealed either behind or within the polar mass of chromatin. In this form there is a distinct resting stage between the two maturation mitoses (figs. 72-75). The element x is conspicuous in one-half of the cells (figs. 72, 73); it may be included in the nucleus as in figure 72, or it may be partly or wholly outside, as in figures 74, 75, and 76. In the latter case, but not in the former, it is surrounded by its own membrane. As the chromatin begins to condense for the second mitosis, disintegration of the element x becomes apparent. This is most easily made out in cases where the element is isolated, as in figures 75 and 76; but there seems to be little doubt that it disappears before the metaphase of the second maturation mitosis. It is not possible to count the chromosomes in this stage, they are so crowded together, but it is not probable that such a conspicuous chromatin element as that seen in the first division could escape detection, even if it were in the equatorial plate among the chromosomes. No aberrant element is ever seen in these spindles;[Pg 7] and, moreover, all of the spindles and all of the spermatids appear to be exactly alike at the same stage. The chromosomes are double in the prophase (fig. 77) and always appear double in the equatorial plate (fig. 78), the paired elements corresponding to those of figure 65.
In figure 80, plate III, a pair of spermatids is shown with nuclear membrane formed and the spindle fibers twisted in a characteristic manner. Figure 81 is a slightly later stage with the spindle-remains massed against the nuclear membrane. Curiously enough there appears in the nucleus of every spermatid a body similar to the element x of the spermatocytes of the first order (figs. 82-86). This body is often applied to the nuclear membrane and connected with the spireme (figs. 84-86). It decreases in size and finally disappears (figs. 88-91). The spindle-remains divides (fig. 83), and a small part of it (a) goes to form the acrosome at the apex of the head (figs. 85-92). The larger part is probably utilized in the formation of the tail, for it gradually disappears as the tail develops.
The centrosome which, although small, is conspicuous in each mitosis, is seen in figure 83 between the two parts of the spindle-remains, applied to the outside of the nuclear membrane. In figures 85, 86, and 87 the relation of the tail (or its axial fiber) to the centrosome is shown. In figures 87 and 88, instead of the small spherical centrosome of figures 83 to 86, we have a much elongated body, at first (fig. 87) applied for its whole length to the nuclear membrane, but later lying along one side of a middle piece (m), as shown in figure 89, and in a later stage in figures 90 to 92. The mature spermatozoön with its forked anterior end appears in figure 93.
The points of especial interest in the spermatogenesis of Stenopelmatus are the development of the aberrant chromatin element x during the growth stage of the spermatocyte of the first order, its distribution to one-half of the spermatocytes of the first order, its disappearance during the rest stage between the two maturation divisions, and the development of a similar, though smaller, element in all of the spermatocytes.
Unlike the spermatogonia of Stenopelmatus, those of Blattella have both a faintly-staining nucleolus and a deeply-staining chromatin element (x), and moreover the two are always closely associated (figs. 95, 96). The number of chromatin elements in the equatorial plate of late spermatogonial mitoses is 23 (fig. 97). Later events indicate that one of the 23 is the element x, but it is impossible to distinguish it here. Figure 98 is a very early stage of the spermatocyte of the first order, showing the element x as a U-shaped body. The centrosome[Pg 8] was also conspicuous in all of the cells of this group. The spireme here, as also in figure 99, is fine and closely interwound. In figure 99 and again in figure 100 the element x is joined to the spireme as it is throughout the spireme stage. In the "bouquet" or "polarized" stage the combined nucleolus and element x are always at one side of the group of loops and down very close to the base of the figure (figs. 101, 103). In figure 102 most of the loops are cut across. The stage shown in figures 104 and 105 is a later one than that just described. Here we have again a continuous spireme connected with the element x, making it seem improbable that the bivalent chromosomes are really separated in the bouquet stage. Figure 106 gives some of the variations in form of the combined nucleolus and element x. The last of the five figures was taken from a giant cell containing at least twice the usual amount of chromatin. In one giant cell four unusually large combinations of this kind were found, and a corresponding amount of chromatin in the spireme. In figure 107 one sees the spireme divided into segments still joined by linin bridges. In figure 108 similar segments may be seen, one of them showing a longitudinal split. The element x is present, but the nucleolus has disappeared. In many cases the split, if it appears at all, closes quickly and the chromosome bends in U-shape, as in figure 109, plate IV. This figure also shows two centrosomes (c). In other cases the split persists as in figure 110 and leads to the formation of crosses of a tetrad character (figs. 111, 112, 113), as in Stenopelmatus and many other insects. Figures 114 to 117 show later stages of the U-shaped chromosomes. Perfect rings are rare. All sorts of variations are seen, broad and narrow U-shapes, rings split at one point or the opposite points, a U split at the bottom (fig. 114), pairs of parallel rods (fig. 115), and occasionally rods constricted in the middle and showing a longitudinal split in each half, as in figure 116. Figure 117 shows different views of the split rings. Apparently all of these forms straighten out so that the two components of the bivalent chromosome stand end to end as dumbbells or compressed crosses in the metaphase of the first maturation spindle (figs. 123-125). The element x remains concentrated and more or less spherical in form. Figures 118-122 are equatorial plates, with x absent in figure 120, in the same plane as the 11 other chromosomes in figure 119, far to one side in figure 118, and near one pole of the forming spindle in figure 122. It is also shown in various positions with regard to the spindle in figures 123 to 126 and 128 to 132. In figure 125 it is apparently double, and again in figure 129. In figure 130 one lagging chromosome shows the dyad nature of the products[Pg 9] of the division of the tetrad. In this form there can be no doubt that reduction occurs in the first spermatocyte division. The element x is very often concealed by the polar aggregation of chromatin, but it is sometimes as conspicuous as in figures 131 and 132. The spermatocytes of the second order go into a complete resting stage before they are completely separated, and one of a pair shows the element x, while it is lacking in the other (fig. 133). At the close of the resting stage the chromosomes appear as 11 pairs of rods of considerable length, which gradually shorten and thicken and usually bend at the center, forming U's or V's (figs. 134-138). In one stage these double U's look much like tetrads (fig. 138). The rods straighten again as they shorten still more (fig. 139), become more closely approximated, and finally form dumbbells, as in figure 141.
The element x is, of course, present in only one-half of these nuclei. In the equatorial plate, figure 142, it is absent; in figure 143 it is present, but can not be distinguished from the other chromosomes, while in figure 144 it is rendered conspicuous by its spherical form and isolated position. In only a few cases has it been possible to distinguish x in the spindle. Figures 146 and 147 show two of these cases where this element is clearly double and of different form from the other chromosomes. It is probable that it divides and so goes into one-half of all of the spermatids, as in McClung's typical cases of the accessory chromosome. Figure 145 shows the usual appearance of the other chromosomes in metaphase. The two spermatids of a pair are always alike so far as any evidence of the presence of the element x is concerned (fig. 148). Figure 149 is an exceptional case, where one chromatin element (possibly x) has evidently divided late and been left out in the cytoplasm; a smaller chromatin granule is also present in the cytoplasm of each spermatid. All of the spermatids, as in Stenopelmatus, develop a deeply-staining body, which, however, in this case is usually centrally located and often appears double (figs. 150-152).
The spindle-remains (Spindelreste) forms a very conspicuous body at one side of the nucleus in the spermatids, and occasionally a mass of chromatin, probably due to imperfect mitosis, is found near the spindle-substance (fig. 150). The mass of spindle-substance at first appears structureless, but soon assumes the condition shown in figures 150 to 152. In one individual many of the spermatids had two balls of spindle-material (fig. 152), and the resulting later stages were double-tailed (fig. 153). Figure 156 shows how the spindle-substance goes into the tail and gradually disappears as the tail lengthens.[Pg 10]
The centrosome is evidently applied to the nuclear membrane, as in Stenopelmatus, and the middle-piece is developed in connection with it, as in figures 156-157, 154-155, 158-160. The element x of the spermatids gradually disappears (figs. 150, 159). An acrosome develops at the anterior end, the head condenses and lengthens, and we have the ripe spermatozoön (fig. 161). The tail is very long and is shown only in part.
Of the forms studied, Blattella alone has many degenerate spermatozoa. Some follicles have none, others a number varying perhaps from one-fourth to three-fourths of the whole number. No evidence of degeneracy was detected among the young spermatids up to the stage shown in figures 154-155, where a few like figure 162 were found. Most of the degenerate forms occur among the nearly ripe spermatozoa or in the sperm-ducts. Such are shown in figures 163 to 168. The chromatin is strangely broken up into irregular clumps, and probably no two of these degenerate sperm-heads can be found which are alike. The tails are always imperfect. The distribution and varying numbers of these degenerate spermatozoa make it impossible to interpret their condition as due to the absence of the accessory chromosome, as Miss Wallace does in the spider. The only probable explanation, it seems to me, is imperfect mitosis. Cases where more or less chromatin is left behind in the cytoplasm, especially in the first spermatocyte mitosis, are very common, and such cases as those shown in figures 149 and 150 are not rare. The giant cells, so far as I have been able to trace them, do not develop into spermatozoa.
The most important points are:
(1) The presence of the element x in the spermatogonia, closely associated with the nucleolus.
(2) The uneven number of chromatin elements in the metaphase of spermatogonial divisions.
(3) The connection of the element x with the spireme up to the stage where the spireme segments to form the bivalent chromosomes.
(4) The varied character of the tetrads, showing the first spermatocyte division to be a reducing division in the sense that it separates whole chromosomes.
(5) The fact that the element x fails to divide in the first maturation division, does divide in the second, but can not be traced beyond the equatorial plate of the latter mitosis.
(6) The similarity of all the normal spermatids, though one-half of them must contain the element x, the other half not.
(7) The varying and often large number of degenerate spermatozoa.[Pg 11]
An attempt was made to determine the somatic number of chromosomes. The dividing cells of the follicles of young eggs seemed to afford the most favorable material, but even here there was so much overlapping of the ends of the chromosomes that it was impossible to be absolutely certain of the number. In the two most favorable cases 23 were counted (fig. 94). This differs from McClung's count for similar cases among the Orthoptera, and Sutton's for Brachystola magna. The eggs have so far resisted all efforts to learn what part the odd chromosome may play in fertilization.
In the metaphase of all spermatogonial mitoses where it was possible to count accurately, 20 chromosomes were found, 19 large ones of approximately equal size, and 1 small spherical one (figs. 169, 170). There is nothing in the resting nucleus of the spermatogonia which would suggest either a nucleolus or an accessory chromosome. The chromatin stains well during the whole growth period of the spermatocytes, but it is impossible to separate the period into so definite stages as in most other forms.
In the youngest spermatocytes one finds occasionally a cyst containing cells with nuclei like those of figures 171 and 172, indicating that a brief "synapsis" or condensation stage occurs at the close of the last spermatogonial mitosis. During the greater part of the period the chromatin forms a heavy, irregular, and often segmented spireme (figs. 173, 174). Shortly before the first maturation division, such split segments as appear in figure 175 are sometimes found; some of these simulate tetrads with slender connecting bands between the paired elements. Again, one finds a few cases like figure 176, where the spireme is segmented into bivalent chromosomes, each component showing a longitudinal split. This figure also shows the small chromosome. Usually, however, the irregular and much tangled spireme (figs. 173, 174) condenses into a heavy segmented band variously disposed in the nucleus (fig. 177). This band soon separates into the bivalent chromosomes shown in figures 178 and 179, giving 9 symmetrical pairs and 1 unsymmetrical one (fig. 179 s) composed of the small chromosome and a much larger mate. In the prophase of the spindle, in rare cases, some of the chromosomes are longitudinally split and transversely constricted, forming tetrads (fig. 180), but more often they appear as in figure 181. The unequal pair appears in each figure at s. In the metaphase (fig. 182) it is the last to come into the equatorial plate, possibly because of its lack of symmetry. The smaller component of this pair is always directed toward the equator of the[Pg 12] spindle. Figure 183 shows a small tangential section of a spindle in metaphase, containing the unequal pair and one equal pair. In figure 184 a polar view of a metaphase is shown, the unequal pair, which was somewhat below the others, being indicated by stippling. Figures 184 a and 185 show that the unequal components of the unsymmetrical pair, as well as the equal components of the symmetrical pairs, are separated in metakinesis, making this clearly a reduction division. Two polar plates are shown in figures 186 and 187, one containing 10 equal elements, the other 9 equal ones and 1 small one. The telophase is shown in figure 188. There is no resting stage, but the new spindle is formed from the remains of the old one, and the spindle-shaped mass of chromatin seen in figure 188 either passes into the center of the new spindle or becomes enveloped by it. The double chromosomes separate as in figures 189 and 190. Figure 190 shows the small dyad, and figure 189 an aberrant one which may be its mate. The spindle in both divisions is peculiar in having outside of the spindle proper a dense mass of fibers which, in osmic material, stain deeply with iron hæmatoxylin. These fibers are shown in all the figures from 174 to 196. Figures 191 and 192 are equatorial plates of the two kinds of spermatocytes of the second order, figure 191 showing the small chromosome. An early anaphase appears in figures 193 and 194, which show both the small and larger chromosomes in metakinesis. Figure 195 is a later anaphase containing the divided small chromosome. In figure 196 are shown the two polar plates of a spindle corresponding to that of figure 195, and in figure 197 the polar plates of a spindle in which 10 equal chromosomes have been divided. In Tenebrio molitor the spermatids are therefore certainly of two distinct kinds, so far as the chromatin content is concerned.
In most of the young spermatids, after the nuclear membrane has formed, there appears an isolated chromatin element, which corresponds fairly well to the large or to the small component of the unsymmetrical pair, separated in the first mitosis and divided in the second. The clear portion of the nucleus containing this isolated element is at first turned toward the spindle-remains (fig. 198), but before the tail appears either the whole nucleus or its contents have rotated 180° (fig. 199). Various stages in the development of the spermatid are seen in figures 200 to 203. The clear region and the isolated element finally disappear (fig. 202 b), and the chromatin breaks up into coarser and then into finer granules within the sperm-head. In the later stages the centrosome is clearly seen at the base of the head (fig. 203).
In order to determine, if possible, the value of the unsymmetrical pair of chromatin elements, very young ovaries and ovaries with egg-tubes[Pg 13] were sectioned and the chromosomes counted in the dividing cells of the egg-follicle (♀ somatic cells), and in dividing oögonia. In both cases 20 large chromosomes were found. Figure 207 is the equatorial plate from a female somatic cell of a young egg-follicle. Figure 208 a and b shows two sections of an oögonium in the prophase of mitosis. In order to determine the number and character of the chromosomes in the male somatic cells, several male pupæ were sectioned. As in the spermatogonia, 19 large chromosomes and 1 small one were found. Figure 204 shows the equatorial plate of a dividing male somatic cell, and figures 205 to 206 are daughter plates from a similar cell. (Three large chromosomes of the plate shown in figure 206 are in another section.)
From these facts it appears that the egg-pronucleus must in all cases contain 10 large chromosomes, while the spermatozoön in fertilization brings into the egg either 10 large ones or 9 large ones and 1 small one. Since the somatic cells of the female contain 20 large chromosomes, while those of the male contain 19 large ones and 1 small one, this seems to be a clear case of sex-determination, not by an accessory chromosome, but by a definite difference in the character of the elements of one pair of chromosomes of the spermatocytes of the first order, the spermatozoa which contain the small chromosome determining the male sex, while those that contain 10 chromosomes of equal size determine the female sex. This result suggests that there may be in many cases some intrinsic difference affecting sex, in the character of the chromatin of one-half of the spermatozoa, though it may not usually be indicated by such an external difference in form or size of the chromosomes as in Tenebrio. It is important that related forms should be studied in order to ascertain whether the same chromatic conditions prevail in other species of this genus or possibly in the Coleoptera in general.[A]
[A] Prof. E. B. Wilson has recently found a similar dimorphism in the spermatozoa of Lygæus and other of the Hemiptera heteroptera.
The spermatogenesis of Aphis has been fully described in another paper and will merely be briefly summarized here for the purpose of comparison with other forms.
The spermatogonia contain a large nucleolus, which gradually disappears in the prophases of mitosis (plate VII, figs. 209-211). The youngest spermatocytes closely resemble the spermatogonia (fig. 212). There is no bouquet stage and no such marked spireme stage as in[Pg 14] many other insects. The true synapsis occurs, as shown in figure 213, by pairing of like chromosomes side by side. This conjugation of like chromosomes is followed by a stage in which they are massed together at one side of the nucleus (fig. 214). In these latter stages the nucleolus has entirely faded out and nothing suggesting an accessory chromosome is present. Figures 215 and 216 are equatorial plates of the first spermatocyte mitosis. There are 5 chromosomes of different sizes and shapes, and figure 216 shows each one double. The first division of the chromosomes, though apparently longitudinal, is evidently a separation of the elements paired in a preceding stage, and is therefore a reducing division.
The anaphase of the same mitosis is shown in figures 217 and 218; it is peculiar in that one chromosome always divides more slowly than the others, the two elements hanging together at one end. In figure 219 are sister spermatocytes of the second order, the "lagging" chromosomes still connected. The second maturation division is seen in metaphase in figure 220 and in anaphase in figure 221. Figure 222 shows a young spermatid, the five chromosomes still preserving their characteristic form. Figure 223 is the equatorial plate of the first maturation division of the winter egg, showing the same form and size relations of the chromosomes as in the spermatocyte divisions. Figures 224 and 225 are equatorial plates of a polar spindle (fig. 224) and of a segmentation spindle (fig. 225) of the parthenogenetic egg, where 10 chromosomes are present, 2 of each of the sizes found in the sexual germ cells.
So far as an accessory chromosome or any other visible evidence of a sex determinant are concerned, the results are entirely negative. The conditions shown do, however, support Mendel's conception of the "purity of the germ-cells," and also afford evidence in favor of Boveri's theory of the individuality of the chromosomes.
In connection with these insect forms it is of interest to find in the spermatogenesis of Sagitta a body which stains like chromatin and behaves somewhat like the accessory chromosome. It is found in all resting stages of the spermatogonia, closely applied to the nuclear membrane (fig. 226). It divides before each spermatogonial mitosis (fig. 227) and, though not often discernible in the spindle, appears in the next generation. Figure 228 is the last spermatogonial mitosis, and figure 229 shows the element x, and the chromosomes paired at one pole of the spindle. During the various phases of the growth stage (figs. 230-232) the element x is again applied to the nuclear membrane.[Pg 15]
In the prophase of the first maturation division this element divides (figs. 233-234), and in metakinesis the two elements are found in various positions with regard to the spindle (figs. 235-237), often as conspicuous as in these figures, but sometimes concealed among the chromosomes. Before the spindle for the second division forms, this element divides again and one of the products goes into each spermatid (figs. 238-241).
As Sagitta is hermaphrodite, there would appear to be no question of sex determination by any special chromatic element. The size of the element x, its evident chromatic nature, its division before each mitosis, and its presence in mitosis and in the spermatids, with the same staining qualities as in the previous rest stages, certainly indicate some important function, either in the whole process of spermatogenesis or in the formation of the sperm-head, of which it finally becomes a part. In Sagitta this element certainly can not be regarded as a specialized spermatogonial chromosome, or as chromatin rejected from the spireme. No such element is present in the ovogenesis of Sagitta (Stevens, '03), nor has any been detected in connection with fertilization. It is certain that none is present in the first segmentation spindle of the egg.
The literature bearing on the "accessory chromosome" of McClung, the "small chromosomes" of Paulmier, and the "chromatin nucleoli" of Montgomery has been fully discussed by McClung in the paper entitled, "The accessory chromosome—sex determinant?" ('02), and will therefore be considered here only in its relation to the several forms studied. The present status of the question has been well summarized more recently by Montgomery under the heading "Heterochromosomes" in the paper, "Some observations and considerations upon the maturation phenomena of the germ cells."
Three theories as to the function of the "heterochromosomes" have been advanced: (1) That of McClung that they are sex-determinants, since in the forms which he has examined these chromatin bodies occur in only one-half of the spermatozoa, and the sex-character is the only character which divides the individuals of a species into two approximately equal groups. (2) That of Paulmier and Montgomery that they are degenerating chromatin. Montgomery regards them as "chromosomes that are in the process of disappearance[Pg 16] in the evolution of a higher to a lower chromosome number." (3) That of Miss Wallace, who suggests that in the spider only the one out of each four spermatids which contains the accessory chromosome is capable of developing into a functional spermatozoön, while the other three degenerate, as do the polar bodies given off by the egg. McClung is inclined to believe that the accessory chromosome is an element common to all of the male reproductive cells of Arthropods, and probably to vertebrate spermatocytes as well ('02).
Of the insects considered in this paper Aphis and Termopsis have no "accessory chromosome" or "heterochromosome" of any kind. The fact that no males develop from the fertilized eggs of Aphis might be offered as a reason for its absence, but such an argument would not apply to Termopsis. The sex-character may indeed be represented in the chromatin of some one of the pairs of paternal and maternal chromosomes of the spermatocytes, but there is no evident peculiarity by which one-half of the spermatozoa can be said to be different from the other half. As to McClung's statement ('02) "that if there is a cross-division of the chromosomes in the maturation mitosis, there must be two kinds of spermatozoa, regardless of the presence of the accessory chromosome," it appears to me that in a case like the aphid, where the paired elements of the five bivalent chromosomes are separated in the first maturation mitosis, there may be as many as seventeen kinds of spermatozoa instead of two. If, however, we suppose that the sex characters are segregated in the first maturation mitosis, there would, of course, be two equal classes of spermatozoa with reference to that character.
In Stenopelmatus the element x in certain stages closely resembles the "accessory chromosome" of McClung, and especially that described by Baumgartner for Gryllus domesticus, but its origin and fate are different. It first appears attached to an end of the spireme in the growth stage of the young spermatocytes, where it is much smaller than in later growth stages. It gradually increases in size, is a conspicuous element in the first maturation spindle, goes into one of each pair of spermatocytes of the second order, and there degenerates during the rest stage between the two maturation mitoses. The whole history of this element suggests that it may be rejected chromatin analogous to that observed in the ovogenesis of many forms. In Sagitta, for example, a considerable quantity of chromatin granules is given off by the chromosomes and cast out into the cytoplasm near the close of ovogenesis (Stevens, '03). Rückert ('92) has described a similar casting out of chromatin material by the chromosomes of the oöcytes of Pristiurus.[Pg 17]
The spermatogenesis of Stenopelmatus, therefore, differs from that of the other Orthoptera which have been described in having (1) a larger number of chromosomes (46), (2) an even number in the spermatogonia, (3) an accessory chromatin structure in the spermatocytes of the first order, which disappears before the second maturation division.
In Blattella we have a typical "accessory chromosome," according to McClung. It appears (1) in all resting spermatogonia closely associated with a nucleolus, (2) in the spermatogonial mitoses as an odd chromatin element, making 23 in all, (3) in the growth stage of the spermatocytes connected with an end of the spireme and also with the nucleolus. It becomes separated from the other chromatin in the tetrad-stage, remains nucleolus-like in form, and later appears in the first maturation division either among the chromosomes or in a more or less aberrant position. It passes into one of each pair of spermatocytes of the second order, persists during the rest stage, appears in the second mitosis as a dyad and then divides, going into one-half of the spermatids. The spermatids, however, as in Stenopelmatus, all have the same appearance: each has in the center—not against the nuclear membrane—a small element that stains like chromatin. Occasionally a mass of chromatin is found outside the nucleus, but this is not constant enough to support the contention of Moore and Robinson ('05) that the "nucleolus" of the related form, Periplaneta americana, is fragmented and cast out into the cytoplasm. The spermatids all appear to develop equally well for some time, but as they approach maturity a varying proportion of them become degenerate. This can not, however, be due to absence of the accessory chromosome, as Miss Wallace supposes, in the spider; for in some follicles no degenerate spermatozoa are found, and in others more than half may be degenerate. All attempts to study fertilization stages of the egg have so far failed, and the chromosomes in the female somatic cells have not proved favorable for counting. Twenty-three have been counted in several cases, but there was always some chance of error. If 23 is the somatic number in both sexes, it must be maintained by union of sex-cells containing 11 and 12 chromosomes, respectively, the same unequal number occurring in the maturated eggs as in the sperm. Under such conditions it is difficult to see how the odd chromatin element of the spermatozoa can determine sex.
The brief description of the chromatin element x in Sagitta, introduced here because it behaves like the accessory chromosome in many particulars, serves as an example of the occurrence of such an element in the spermatogenesis of a hermaphrodite form, where it can[Pg 18] not possibly be conceived of as a sex determinant. In Sagitta it is known to be confined to the male germ-cells. No such element occurs in the ovogenesis, in the sperm nucleus in the egg, or in the first segmentation spindle. Its function must, therefore, be confined to the process of spermatogenesis.
From the standpoint of sex determination, we have in Tenebrio molitor the most interesting of the forms considered in this paper. In both somatic and germ cells of the two sexes there is a difference not in the number of chromatin elements, but in the size of one, which is very small in the male and of the same size as the other 19 in the female. The egg nuclei of the female must be alike so far as number and size of chromosomes are concerned, while it is absolutely certain that the spermatids are of two equal classes as to chromatin content of the nucleus—one-half of them have 9 large chromosomes and 1 small one, while the other half have 10 large ones. Since the male somatic cells have 19 large and 1 small chromosome, while the female somatic cells have 20 large ones, it seems certain that an egg fertilized by a spermatozoön which contains the small chromosome must produce a male, while one fertilized by a spermatozoön containing 10 chromosomes of equal size must produce a female. The small chromosome itself may not be a sex determinant, but the conditions in Tenebrio indicate that sex may in some cases be determined by a difference in the amount or quality of the chromatin in different spermatozoa. This is much the most suggestive part of the work, and it will be followed up by the study of related forms.
There appears to be so little uniformity as to the presence of the heterochromosomes, even in insects, and in their behavior when present, that further discussion of their probable function must be deferred until the spermatogenesis of many more forms has been carefully worked out.
Bryn Mawr College, May 15, 1905.
Baumgartner, W. J.
'04. Some new evidences for the individuality of the chromosomes. Biol. Bull., vol. 8, no. 1.
McClung, C. E.
'99. A peculiar nuclear element in the male reproductive cells of insects. Zool. Bull., vol. 2.
'00. The spermatocyte divisions of the Acrididæ. Kans. Univ. Quart., vol. 9, no. 1.
'01. Notes on the accessory chromosomes. Anat. Anz., bd. 20, nos. 8 and 9.
'02. The accessory chromosome—Sex determinant? Biol. Bull., vol. 3, nos. 1 and 2.
'02a. The spermatocyte divisions of the Locustidæ. Kans. Univ. Quart., vol. 1, no. 8.
Montgomery, Thos. H., Jr.
'01. A study of the chromosomes of the germ-cells of Metazoa. Trans. Amer. Phil. Soc., vol. 20.
'01a. Further studies on the chromosomes of the Hemiptera heteroptera. Proc. Acad. Nat. Sci. Phila. 1901.
'04. Some observations and considerations upon the maturation phenomena of the germ-cells. Biol. Bull., vol. 6, no. 3.
Moore, J. E. S., and Robinson, L. E.
'05. On the behavior of the nucleolus in the spermatogenesis of Periplaneta americana. Quart. Jour. of Mikr. Sci., n. s., no. 192 (vol. 48, part 4).
Paulmier, F. C.
'93. Chromatin reduction in the Hemiptera. Anat. Anz., vol. 14.
'99. The spermatogenesis of Anasa tristis. Journ. of Morph., vol. 15.
Rückert, J.
'92. Zur Entwickelungsgeschichte des Ovarialeies bei Selachiern. Anat. Anz., vol. 7, no. 4 and 5.
de Sinéty, R.
'01. Recherches sur la biologie et l'anatomie des phasms. La Cellule, vol. 19.
Stevens, N. M.
'03. On the ovogenesis and spermatogenesis of Sagitta bipunctata. Zool. Jahrb., vol. 18.
Sutton, W. S.
'02. On the morphology of the chromosome group in Brachystola magna. Biol. Bull., vol. 4, no. 1.
'03. The Chromosomes in heredity. Biol. Bull., vol. 4, no. 5.
Wallace, L. B.
'00. The accessory chromosome in the spider. Anat. Anz., vol. 18.
'05. The spermatogenesis of the spider. Biol. Bull., vol. 8, no. 3.
Wilcox, E. V.
'95. Spermatogenesis of Caloptenus femur-rubrum and Cicada tibicen. Bull. Mus. Comp. Zool. Harvard Univ., vol. 27.
'96. Further studies on the spermatogenesis of Caloptenus femur-rubrum. Bull. Mus. Comp. Zool. Harvard Univ., vol. 29.
'97. Chromatic tetrads. Anat. Anz., vol. 14.
'01. Longitudinal and transverse division of chromosomes. Anat. Anz., vol. 19, no. 13.
[The figures of plates I-VI were all drawn with Zeiss oil-immersion 2 mm., oc. 12, and have been reduced one-third; those of plate VII with oc. 8, not reduced.]
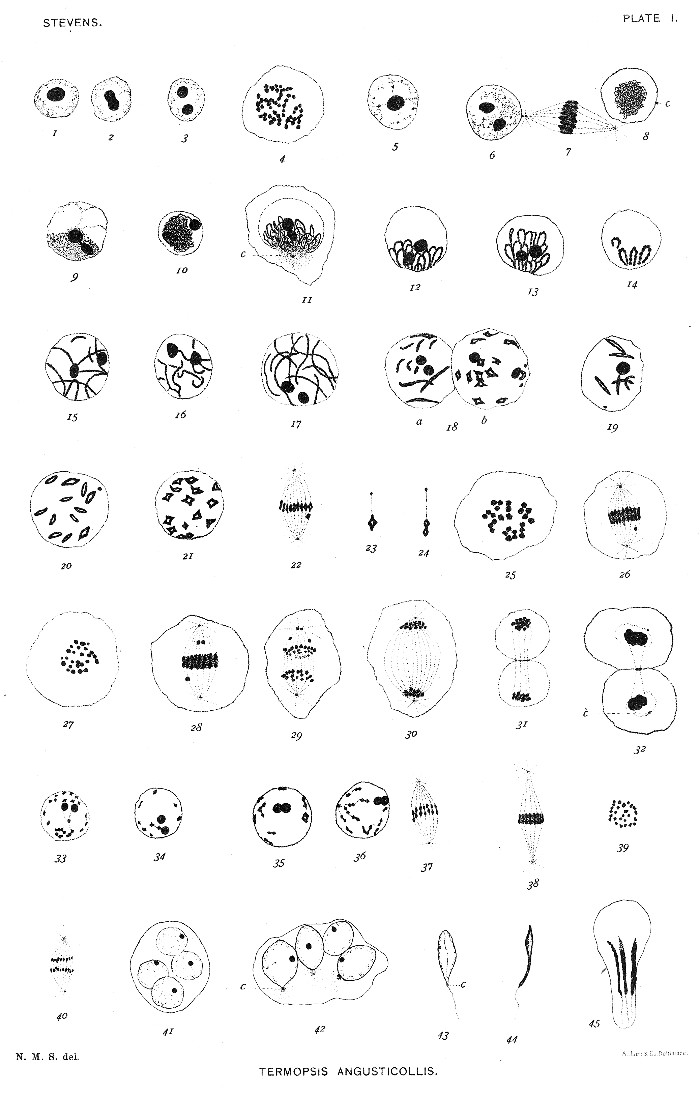
Figs. 1-3. Resting nuclei of spermatogonia, showing division of nucleolus.
4. Equatorial plate of spermatogonial mitosis, 52 chromosomes.
5-6. Young spermatocytes, showing division of nucleolus.
7. First maturation spindle, and two nuclei (6 and 8) in same cyst.
8-10. Skein-stage—so-called synapsis-stage.
11-14. Bouquet-stage, showing two nucleoli, centrosome (c) in fig. 11, and loops made up of fine, then coarser granules.
15-17. Stage following preceding; loops straightened out and extending in various directions through nucleus.
18. a, Chromosomes much shortened and longitudinally split; b, chromosomes contracted to form diamond-shaped figures.
19. Stage between 18a and 18b.
20. Stage between 19 and 18b.
21. Stage similar to 18a, one chromosome in double diamond form.
22. First maturation spindle in metaphase, chromosomes in single and double diamond shapes.
23. Chromosome in single diamond or tetrad form, as they usually come into the spindle.
24. Double diamond-form assumed before metakinesis.
25. The 26 chromosomes of an early metaphase.
26. First maturation spindle in metakinesis.
27. Equatorial plate of first maturation spindle in metakinesis.
28. Another spindle, showing three granules which are probably remains of nucleoli.
29. Anaphase of first maturation mitosis, one centrosome divided.
30. Late anaphase.
31-32. Telophase, exceptional cases of division of the cell.
33-36. Partial rest stage between first and second maturation divisions, two nucleoli present. Chromosomes in fig. 36 in form of double diamonds ready for metakinesis.
37-38. Second maturation spindle in metaphase.
39. Equatorial plate of second maturation spindle, 26 chromosomes.
40. Same in anaphase.
41. Four spermatid nuclei in one cell, each nucleus containing one nucleolus.
42. A later stage, showing elongation of nuclei, centrosome and sphere at posterior end.
43-45. Later stages in the development of the spermatozoa, nucleolus grows gradually smaller.[Pg 21]
Figs. 46-47. Nuclei of spermatogonia, showing 2 and 3 nucleoli (n).
48-49. Prophase of spermatogonial mitosis, showing two exceptionally large chromosomes of equal length.
50. Equatorial plate of spermatogonial mitosis, 46 chromosomes.
51-54. Spermatocytes in spireme stage, nucleus containing a nucleolus (n), and a chromatin element (x), which is attached to one end of spireme and gradually increases in size during growth stage of spermatocytes.
55. Spireme longitudinally split and showing the beginning of cross formation.
56. Spireme segmented, tetrads forming.
57. One split segment and a part of another connected by bands of linin.
58. More open cross and diamond forms; element x conspicuous.
59-60. More contracted cross and diamond-shaped tetrads; linin bands shown in 60, where element x is also present.
61. Different forms assumed by element x during tetrad stage (figs. 56-60).
62-63. Diamond-shaped and contracted cross-shaped tetrads from metaphase of first maturation mitosis, showing linin connections.
64. Diamond-shaped tetrad with spindle-fibers attached; a-a, probably halves of one univalent chromosome; b-b, halves of the other.
65. Dyad from anaphase of first maturation mitosis.
66-67. Metaphase of first maturation spindle, showing element x in different positions.
68. Late anaphase of same.
69-70. Equatorial plate of first maturation spindle, 23 chromosomes and element x below, in fig. 69.
71. Chromatin massed at poles of spindle; element x isolated at one pole.
72-73. Two resting spermatocytes of the second order, one containing element x, the other not.
74-76. Successive stages of breaking down of element x.
77. Prophase of second division; dyads evident, but no sign of x in this or following stages.
78. Second spermatocyte division—metakinesis.
79. Same; late anaphase.[Pg 23]

Fig. 80. Telophase of second maturation mitosis.
81. Young spermatid, showing spindle-remains at s.
82. Spermatid showing a conspicuous chromatin element in nucleus, and spindle-remains (s) elongated.
83. Spermatid, showing centrosome (c) and divided spindle-remains (s and a).
84. Older spermatid, showing centrosome (c), axial fiber of tail, and spindle-remains (s).
85. Spermatid, showing acrosome material (a) migrating to side of nucleus opposite centrosome.
86. Slightly older spermatid.
87. Later stage of spermatid, showing condensed chromatin, elongated centrosome (c), acrosome material (a), and spindle-remains (s).
88-89. Older spermatids, showing formation of acrosome (a) and middle piece (m).
90-92. More advanced stages.
93. Mature spermatozoön.
Fig. 94. Somatic cell from egg follicle, 23 chromosomes.
95. Spermatogonium, showing chromatin element (x) associated with a nucleolus (n).
96. Same, prophase of mitosis.
97. Equatorial plate of spermatogonial mitosis, 23 chromatin elements.
98. Young spermatocyte, showing centrosome (c) and U-shaped element (x).
99. Young spermatocyte, element x attached to one end of a long, fine spireme.
100. Coarser spireme stage.
101-103. Bouquet stage.
104-105. Later spireme stage.
106. Various forms assumed by the combined nucleolus and element x; last figure from a giant cell.
107. Segmenting spireme.
108. Similar stage to fig. 107, one chromosome longitudinally split; element x present.[Pg 25]
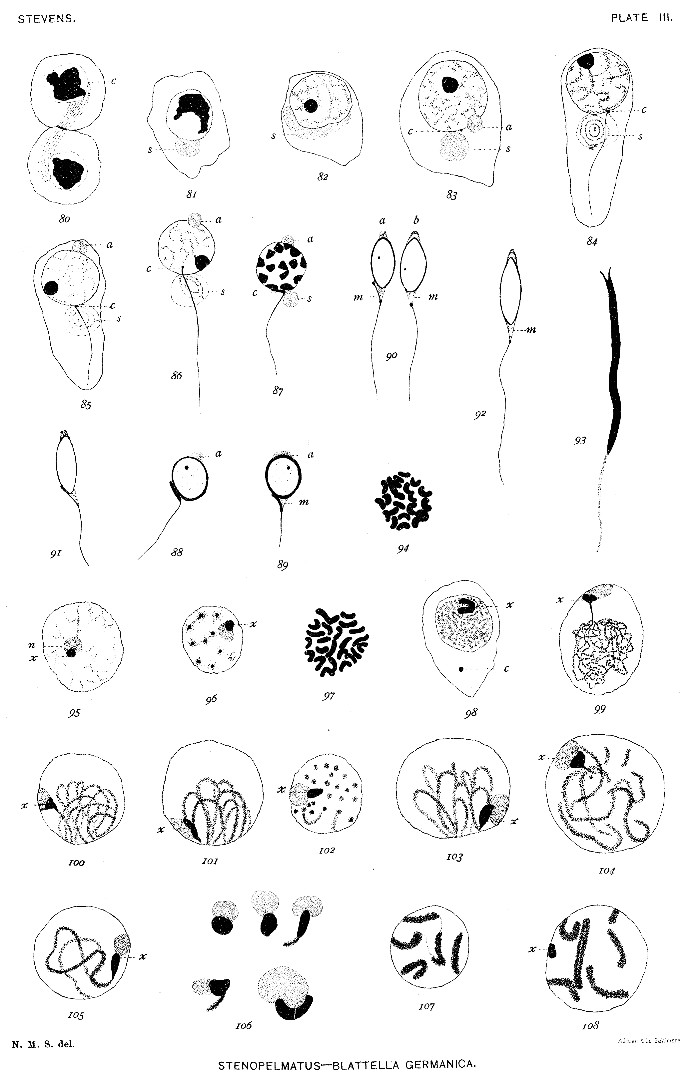
Fig. 109. Similar stage to figs. 107 and 108; chromosomes U-shaped and not longitudinally split; two centrosomes present (c).
110. Longitudinally split chromosomes.
111-113. Various stages in formation of cross-shaped tetrads.
114-117. Bent rods, U-shapes, split rings, pairs of rods, and rod-shaped tetrads (116), which are equivalent to the crosses of figs. 112-113.
118-122. Metaphase of first maturation division, showing the element x in various positions.
123-127. First maturation spindle in metaphase.
128. Same in anaphase.
129-132. Late anaphase, showing element x double in 129, and a lagging tetrad in 130.
133. Telophase, with the element x in one daughter cell.
134-136. Prophase of second maturation mitosis, showing dyads and element x.[Pg 27]
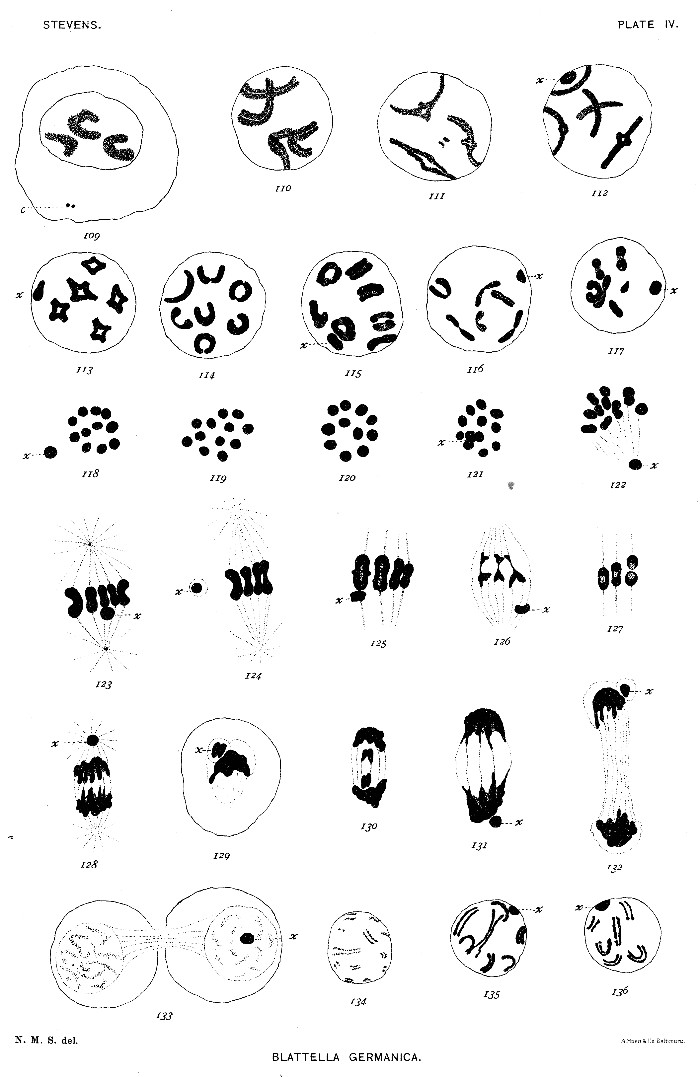
Figs. 137-141. Dyads contracting for second maturation mitosis.
142. Equatorial plate of second maturation spindle, containing 11 chromosomes.
143-144. Same, with 11 chromosomes and the element x.
145-147. Sections of second maturation spindles; element x dividing in 146 and 147.
148. Telophase of second mitosis.
149. Telophase of second mitosis, showing masses of chromatin left behind in cytoplasm.
150. Spermatid with extranuclear chromatin (a).
151. Similar stage; different view of spindle-remains (s) and of chromatin element (x2).
152-153. Spermatid with divided spindle-substance and the corresponding double-tailed form.
154-155. Stages between 156 and 158.
156-157. Older spermatids than 151, showing spindle-remains (s) and centrosome (c).
158-160. Later stages in development of sperm-head.
161. Ripe spermatozoön.
162-168. Degenerate spermatids and spermatozoa.[Pg 29]
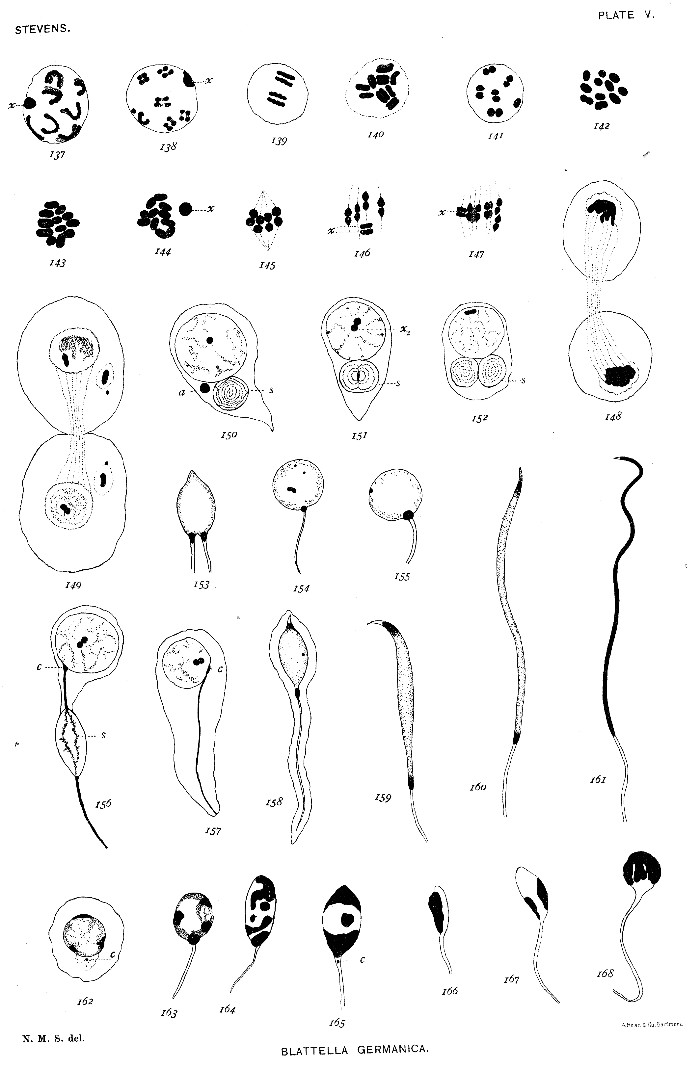
Figs. 169-170. Equatorial plates of spermatogonial mitosis, showing 19 large and 1 small chromosome.
171-175. Condensation stage, bouquet stage, spireme stage, and rather rare tetrad stage of young spermatocyte.
176. Bivalent chromosomes, with longitudinal split; small chromosome shown at s.
177. Bivalent chromosomes condensed into a close spireme.
178-179. Bivalent chromosomes separating for mitosis. The unsymmetrical pair shown in fig. 179.
180. Prophase of first maturation mitosis, showing the unsymmetrical pair and the tetrad nature of the symmetrical pairs.
181. Prophase of same mitosis, showing symmetrical and unsymmetrical pairs, as in figs. 178 and 179.
182. Metaphase, unsymmetrical pair out of the equatorial plane.
183. Tangential section of a spindle in metaphase, showing the unsymmetrical pair and one symmetrical pair.
184. Equatorial plate of same mitosis, 10 chromosomes.
184a. Early anaphase, showing separation of the elements of the unsymmetrical pair.
185. Later anaphase.
186. Polar plate, showing 9 large and 1 small chromosome.
187. Polar plate, showing 10 large chromosomes.
188. Condensation stage between the two maturation divisions.
189-190. Prophase of second maturation division, fig. 189 showing 10 equal dyads, and fig. 190, showing 9 equal and 1 small dyad.
191. Equatorial plate, showing 1 small chromosome and 9 large ones.
192. Equatorial plate, showing 10 large chromosomes.
193-194. Tangential sections of spindle in metakinesis.
195. Anaphase of same mitosis.
196. Polar plates of a spindle, showing in each 1 small chromosome and 9 large ones.
197. Polar plates of another spindle, 10 large chromosomes in each.
198. Young spermatid, showing isolated small chromosome.
199. Young spermatid, showing isolated large chromosome and rotation of nuclear contents.
200-202a, b. Older spermatids.
203. Sperm-heads, showing centrosome and granular chromatin.
204. Equatorial plate from dividing somatic cell of male pupa, showing 19 large and 1 small chromosome.
205-206. Daughter plates of a similar spindle, showing small chromosome in each; three of the large chromosomes missing in 206.
207. Equatorial plate of a dividing cell of follicle of a young egg, showing 20 large chromosomes.
208. Prophase of mitosis in a young oögonium, showing 20 large chromosomes in two sections, a and b.[Pg 31]
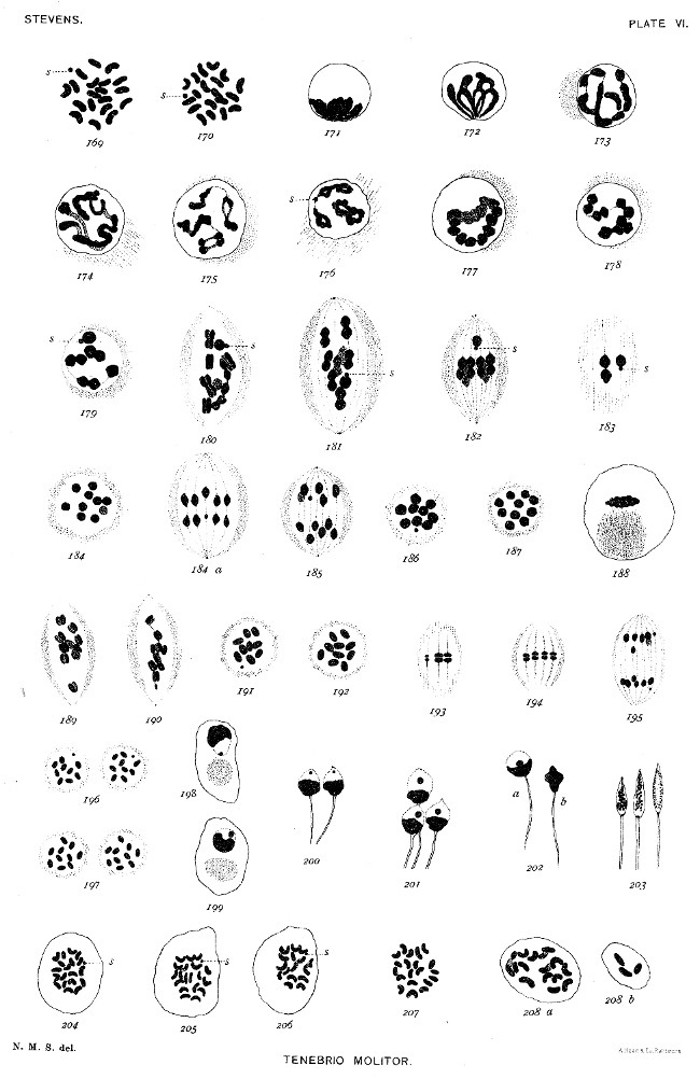
Fig. 209. Spermatogonium.
210-211. Spermatogonia in prophase of mitosis.
212. Young spermatocyte of first order.
213. Spermatocytes of first order; conjugation of the chromosomes.
214. Condensation of chromatin—spermatocytes of first order immediately before mitosis.
215. Equatorial plate of first maturation division.
216. Same, side view, showing chromosomes double.
217-218. Anaphase of same mitosis.
219. Daughter spermatocytes of second order.
220. Equatorial plate of second maturation mitosis.
221. Anaphase of same.
222. Young spermatid.
223. Equatorial plate of first polar spindle of winter egg.
224. Equatorial plate of polar spindle of parthenogenetic egg.
225. Equatorial plate of segmentation spindle of parthenogenetic egg.
Fig. 226. Resting spermatogonia.
227. Prophase of spermatogonial mitosis.
228. Last spermatogonial mitosis, metakinesis.
229. Anaphase of same, showing synapsis of chromosomes at pole of spindle, and element x.
230. Resting spermatocyte of first order.
231. Bouquet stage.
232. Later growth stage.
233. Prophase of first maturation mitosis, some of the chromosomes split longitudinally.
234. Later stage, chromosomes condensing and element x dividing.
235-237. First maturation mitosis.
238. Division of element x between the two maturation divisions.
239. Second maturation mitosis.
240. Anaphase of same, showing the element x more deeply stained than the chromosomes.
241. Young spermatids; element x still conspicuous.[Pg 33]

End of the Project Gutenberg EBook of Studies in Spermatogenesis (Part 1 of
2), by Nettie Maria Stevens
*** END OF THIS PROJECT GUTENBERG EBOOK STUDIES IN SPERMATOGENESIS (1 OF 2) ***
***** This file should be named 31545-h.htm or 31545-h.zip *****
This and all associated files of various formats will be found in:
https://www.gutenberg.org/3/1/5/4/31545/
Produced by Bryan Ness, Josephine Paolucci and the Online
Distributed Proofreading Team at https://www.pgdp.net. (This
file was produced from images generously made available
by The Internet Archive/American Libraries.)
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
https://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH F3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at https://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
https://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at https://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit https://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including including checks, online payments and credit card
donations. To donate, please visit: https://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
https://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.